
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA ...Dr. Elton Lucio de Araujo Universidade...
157
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA CARACTERIZAÇÃO DE OVOS E LARVAS DE ESPÉCIES DE ANASTREPHA (DIPTERA: TEPHRITIDAE) UTILIZANDO ANÁLISES MORFOLÓGICAS E MOLECULARES VIVIAN DE SIQUEIRA DUTRA Manaus - Amazonas Janeiro - 2012
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA ...Dr. Elton Lucio de Araujo Universidade...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA
CARACTERIZAÇÃO DE OVOS E LARVAS DE ESPÉCIES DE
ANASTREPHA (DIPTERA: TEPHRITIDAE) UTILIZANDO ANÁLISES
MORFOLÓGICAS E MOLECULARES
VIVIAN DE SIQUEIRA DUTRA
Manaus - Amazonas
Janeiro - 2012

ii
VIVIAN DE SIQUEIRA DUTRA
CARACTERIZAÇÃO DE OVOS E LARVAS DE ESPÉCIES DE
ANASTREPHA (DIPTERA: TEPHRITIDAE) UTILIZANDO ANÁLISES
MORFOLÓGICAS E MOLECULARES
ORIENTADORA: DRA. BEATRIZ RONCHI TELES
CO-ORIENTADORA: DRA. JANISETE GOMES DA SILVA MILLER
Tese apresentada ao Programa de Pós
Graduação em Entomologia do Instituto
Nacional de Pesquisas da Amazônia como
parte dos requisitos para obtenção do título
de Doutor em Ciências Biológicas
(Entomologia).
Manaus - Amazonas
Janeiro - 2012

iii
BANCA EXAMINADORA
Dr. Aldo Malavasi
Biofábrica Moscamed Brasil, Juazeiro, Bahia
Dr. Elton Lucio de Araujo
Universidade Federal Rural do Semi-Árido, Mossoró, Rio Grande do Norte
Dra. Iara Sordi Joachim Bravo
Universidade Federal da Bahia, Salvador, Bahia
Dra. Keiko Uramoto
Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, São Paulo
Dr. Marco Antonio Costa
Universidade Estadual de Santa Cruz, Ilhéus, Bahia
Dr. Miguel Francisco de Souza Filho
Instituto Biológico, Campinas, São Paulo
Dr. Roberto Antonio Zucchi
Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, São Paulo

iv
Sinopse:
Este estudo descreve ovos e larvas de espécies de Anastrepha (Diptera: Tephritidae),
ampliando o conhecimento sobre esses estágios de vida. Também utilizou a análise
molecular para auxiliar na correta identificação de larvas associando-as com adultos
de sua respectiva espécie. Como informações adicionais foram registradas novas
interações entre espécies de Anastrepha, hospedeiros e parasitoides.
Palavras-chave: Entomologia Agrícola, moscas-das-frutas, imaturos, PCR-RFLP
D978 Dutra, Vivian de Siqueira
Caracterização de ovos e larvas de espécies de Anastrepha (Diptera:
Tephritidae) utilizando análises morfológicas e moleculares / Vivian de Siqueira
Dutra. --- Manaus : [s.n.], 2011.
xvi, 157 f. : il.
Tese (doutorado) --- INPA, Manaus, 2012
Orientador : Beatriz Rochi Teles
Co-orientador : Janisete Gomes da Silva
Área de concentração : Entomologia
1. Moscas-das-frutas. 2. PCR-RFLP (marcador molecular). 3. Ovos.
4. Larvas. 5. Morfologia. I. Título.
CDD 19. ed. 595.770415

v
“Faça do Senhor a sua grande alegria e Ele
dará a você os desejos do seu coração. Deixe
nas mãos do Senhor tudo o que você for fazer.
Confie Nele de todo o coração e Ele fará o que
for necessário.”
Salmos 37: 4-5

vi
AGRADECIMENTOS
A Deus, Senhor de minha vida e meu melhor amigo por tantas bênçãos e por mais uma
oportunidade de crescimento.
Ao INPA pela infraestrutura. Aos doutores do Programa de Pós Graduação em
Entomologia pelo aprendizado.
Ao CNPq pela concessão da bolsa de doutorado e financiamento do projeto.
As minhas orientadoras Dra. Beatriz Ronchi Teles e Dra. Janisete Gomes da Silva
Miller pelo incentivo, confiança e amizade.
Ao Dr. Elton Araujo e Dra. Keiko Uramoto pelos exemplares doados.
À Dra. Neusa Hamada pela disponibilidade do Laboratório de Citotaxonomia e Insetos
Aquáticos - INPA.
Aos membros da banca examinadora Dr. Aldo Malavasi, Dr. Elton Lucio de Araujo,
Dra. Iara Sordi Joachim Bravo, Dra. Keiko Uramoto, Dr. Marco Antonio Costa, Dr. Miguel
Francisco de Souza Filho e Dr. Roberto Antonio Zucchi pelas sugestões e comentários para o
enriquecimento do trabalho.
Ao meu marido Weber Novaes, companheiro e amigo em todos os momentos.
Aos meus pais Ismael e Daize Dutra, minha irmã Vanessa Dutra, meu cunhado Marco
Aurélio e minha sobrinha Ana Júlia pelo apoio e torcida à distância. Também a toda minha
família, tios, tias, primos, sogros e cunhado pelo apoio.
Aos meus amigos Danielle e Rafael Tonon, Taciane Almeida, Walter Santos, Lisiane
Dilli, Claudemir Campos, Ana Pes, Vanderly Souza, Jamile Moraes, Thiago Onari, Renata e
Felipe Gomes pelas conversas, sugestões e risadas quando eu mais precisava.
Aos amigos que ajudaram nas coletas de material Mírian Santos, Marcos Vinicuis
Garcia, Tatiana Senra, José Nilton Costa, Claudemir Campos, Ulisses Neiss, Ana Claúdia
Araújo e Felipe Gomes.
As amigas de laboratório Aline Mesquita, Lucivânia Amaral, Renata Bastos, Taciane
Almeida e Vanderly Souza. Aos amigos dos Laboratórios de Citotaxonomia e Insetos
Aquáticos (INPA) e de Genética e Biologia Molecular da Universidade Estadual de Santa
Cruz (UESC), em especial Kátia Lima. A Lenir Mota pelo apoio e amizade construída. Por
fim, a todos que torceram por mim.

vii
RESUMO
A identificação de espécies de Anastrepha Schiner (Diptera: Tephritidae) é baseada
principalmente nas características do acúleo das fêmeas. No entanto, estágios imaturos são
comumente interceptados em frutos nas barreiras quarentenárias. Assim, a identificação desse
gênero através das larvas é altamente desejável. Este estudo teve por objetivo ampliar os
conhecimentos sobre os estágios imaturos de espécies de Anastrepha por meio da
caracterização morfológica e molecular. A morfologia dos ovos de 11 espécies e das larvas de
oito espécies de Anastrepha foi descrita detalhadamente pela primeira vez. Caracteres
diagnósticos para diferenciar os ovos das onze espécies incluem ornamentação do córion;
presença ou ausência das aerópilas; posição da micrópila; formato da borda ao redor da
micrópila e presença ou ausência de apêndice respiratório. Os ovos de Anastrepha montei
Lima e Anastrepha curitis Stone possuem uma camada de superfície porosa no pólo anterior
provavelmente utilizada para respiração. Tal caractere não havia sido relatado anteriormente
para ovos de tefritídeos. Caracteres diagnósticos para diferenciar as larvas das oito espécies
estudadas incluem presença ou ausência de espinhos dorsais nos segmentos torácicos e
abdominais, número de espinhos ventrais nos segmentos torácicos e abdominais, formato do
lóbulo anal, número de cerdas no espiráculo posterior e algumas características do esqueleto
céfalo-faríngeo como comprimento total, tamanho e formato da mandíbula. Para a
caracterização molecular foi utilizada a técnica da Reação em Cadeia da Polimerase (PCR)
associada ao Polimorfismo do Comprimento dos Fragmentos de Restrição (RFLP). Foram
analisadas duas regiões do DNA mitocondrial (COI e 16S) e uma do DNA nuclear (ITS1)
para 13 espécies de Anastrepha com seis enzimas de restrição. Os melhores resultados para
diferenciação das espécies foram obtidos com os fragmentos 16S e ITS1 para Anastrepha
atrigona Hendel, A. curitis, Anastrepha manihoti Lima, Anastrepha pulchra Stone,
Anastrepha striata Schiner, Anastrepha sororcula Zucchi e Anastrepha zenildae Zucchi.
Novas associações de moscas-das-frutas com hospedeiros e parasitoides foram registrados.
Palavras-chave: moscas-das-frutas, imaturos, ornamentação do córion, espiráculo posterior,
PCR-RFLP.

viii
ABSTRACT
The identification of species of Anastrepha Schiner (Diptera: Tephritidae) is based on
the shape of the female´s aculeus. However, it is the immature stages that are commonly
intercepted at ports of entry. Thus, the correct identification of immature stages is highly
desirable. This study aimed at contributing to the identification of the immature stages of
Anastrepha species using a morphological and a molecular approach. The morphology of
eggs of 11 species and larvae of eight species of Anastrepha was described in details for the
first time. Diagnostic characters that can distinguish among the eggs of the 11 species include
chorion ornamentation; location, presence or absence of aeropyles; location of the micropyle;
pronounced rim of the chorion surrounding the micropyle and presence or absence of a
respiratory appendage. The eggs of Anastrepha montei Lima e Anastrepha curitis Stone have
a narrow patch of seemingly porous surface of the anterior pole. This character is reported for
tephritid for the first time. Diagnostic characters that can distinguish among the larvae of the
eight species studied include presence or absence of dorsal spinules on segments thoracic and
abdominal, number of ventral spinules on the segments thoracic and abdominal, shape of the
anal lobe, number of the posterior spiracular hairs and some characteristics of the
cephalopharyngeal skeleton such as total length, size and shape of the mandible. PCR-RFLP
(polymerase chain reaction associated to restriction fragment length polymorphism analysis)
was used for the molecular characterization. Two regions of the mitocondrial DNA (COI and
16S) and one region of the nuclear DNA (ITS1) were screened using six restriction enzymes
to screen 13 Anastrepha species. Interesting results to distinguish among species were
obtained for the fragments 16S and ITS1 for Anastrepha atrigona Hendel, A. curitis,
Anastrepha manihoti Lima, Anastrepha pulchra Stone, Anastrepha striata Schiner,
Anastrepha sororcula Zucchi, and Anastrepha zenildae Zucchi. New associations of fruit flies
with their hosts and parasitoids were reported for the first time.
Key words: fruit flies, immature stage, chorion ornamentation, posterior spiracular, PCR-
RFLP.

ix
SUMÁRIO
Lista de Tabelas ...................................................................................................................... xxii
Lista de Figuras ...................................................................................................................... xivi
Lista de Anexos ....................................................................................................................... xvi
Introdução Geral ....................................................................................................................... 17
Objetivo Geral .......................................................................................................................... 20
Objetivos Específicos ............................................................................................................... 20
Capítulo 1
Associação de espécies de Anastrepha (Diptera: Tephritidae) e seus parasitoides obtidos de
hospedeiros nativos e introduzidos coletados em alguns municípios brasileiros
Resumo ..................................................................................................................................... 22
Introdução ................................................................................................................................. 22
Material e Métodos ................................................................................................................... 23
Coleta de adultos ................................................................................................................... 23
Identificação dos adultos ...................................................................................................... 23
Resultados ................................................................................................................................. 24
Discussão .................................................................................................................................. 25
Conclusões ................................................................................................................................ 26
Agradecimentos ........................................................................................................................ 26
Natural Host Plants and Native Parasitoids Associated with Anastrepha pulchra and Other
Anastrepha Species (Diptera: Tephritidae) in Central Amazon, Brazil (nota científica) ......... 30
Resumo ..................................................................................................................................... 32
Host Plant of Anastrepha pulchra (Diptera: Tephritidae) in Central Amazon, Brazil -
Mistaken Identity Resolved (nota científica)............................................................................ 35
Resumo ..................................................................................................................................... 37
Capítulo 2
Egg Morphology of Anastrepha spp. (Diptera: Tephritidae) in the fraterculus Group Using
Scanning Electron Microscopy
Resumo ..................................................................................................................................... 39

x
Introdução ................................................................................................................................. 39
Material e Métodos ................................................................................................................... 39
Resultados ................................................................................................................................. 41
Discussão .................................................................................................................................. 45
Agradecimentos ........................................................................................................................ 47
Description of Eggs of Anastrepha spp. (Diptera: Tephritidae) in the spatulata Group Using
Scanning Electron Microscopy
Resumo ..................................................................................................................................... 55
Introdução ................................................................................................................................. 55
Material e Métodos ................................................................................................................... 56
Descrições ................................................................................................................................. 57
Discussão .................................................................................................................................. 59
Agradecimentos ........................................................................................................................ 62
Description of Eggs of Anastrepha curitis and Anastrepha leptozona (Diptera: Tephritidae)
Using SEM
Resumo ..................................................................................................................................... 68
Introdução ................................................................................................................................. 68
Material e Métodos ................................................................................................................... 69
Resultados ................................................................................................................................. 69
Discussão .................................................................................................................................. 71
Agradecimentos ........................................................................................................................ 73
Capítulo 3
Description of Larvae of Anastrepha spp. (Diptera: Tephritidae) in the fraterculus Group
Resumo ..................................................................................................................................... 78
Introdução ................................................................................................................................. 78
Material e Métodos ................................................................................................................... 79
Resultados ................................................................................................................................. 79
Discussão .................................................................................................................................. 93
Agradecimentos ........................................................................................................................ 95
Description of Third Instar Larvae of Five Anastrepha Species (Diptera: Tephritidae)

xi
Resumo ................................................................................................................................... 100
Introdução ................................................................................................................................. 99
Material e Métodos ................................................................................................................. 101
Resultados ............................................................................................................................... 102
Discussão ................................................................................................................................ 117
Agradecimentos ...................................................................................................................... 119
Capítulo 4
Associação de adultos e larvas de espécies de Anastrepha (Diptera: Tephritidae) utilizando a
técnica de PCR-RFLP
Resumo ................................................................................................................................... 127
Introdução ............................................................................................................................... 127
Material e Métodos ................................................................................................................. 129
Análises morfológicas ......................................................................................................... 129
Coleta de adultos ............................................................................................................. 129
Identificação dos adultos ................................................................................................. 129
Análises moleculares .......................................................................................................... 130
Extração de DNA ............................................................................................................ 130
Reação em cadeia da polimerase (PCR) ......................................................................... 130
Reação de Purificação dos fragmentos amplificados ...................................................... 131
Sequenciamento (COI) .................................................................................................... 132
PCR-RFLP ...................................................................................................................... 132
Mapas de restrição ........................................................................................................... 133
Análise dos resultados ..................................................................................................... 133
Resultados ............................................................................................................................... 133
Discussão ................................................................................................................................ 136
Conclusões .............................................................................................................................. 138
Agradecimentos ...................................................................................................................... 138
Síntese ..................................................................................................................................... 139
Referências Bibliográficas ...................................................................................................... 140

xii
LISTA DE TABELAS
Capítulo 1
Associação de espécies de Anastrepha (Diptera: Tephritidae) e seus parasitóides obtidos de
hospedeiros nativos e introduzidos coletados em alguns municípios brasileiros
Tabela 1. Famílias e espécies botânicas, peso, número de frutos, número de pupas, espécies
de Anastrepha e Braconidae.....................................................................................................26
Natural Host Plants and Native Parasitoids Associated with Anastrepha pulchra and other
Anastrepha Species (Diptera: Tephritidae) in Central Amazon, Brazil (nota científica)
Tabela 1. Espécies de Anastrepha e parasitoides coletados na Amazônia Central, Brasil......32
Capítulo 2
Egg morphology of Anastrepha spp. (Diptera: Tephritidae) in the fraterculus group using
SEM
Tabela 1. Diagnose dos ovos de seis espécies do grupo fraterculus analisadas neste
estudo........................................................................................................................................49
Description of eggs of Anastrepha spp. (Diptera: Tephritidae) in the spatulata group using
Scanning Electron Microscopy
Tabela 1. Diagnose dos ovos de três espécies de Anastrepha do grupo spatulata analisadas
neste estudo...............................................................................................................................63
Description of eggs of Anastrepha curitis and Anastrepha leptozona (Diptera: Tephritidae)
using SEM
Tabela 1. Diagnose dos ovos de Anastrepha curitis e Anastrepha leptozona analisadas neste
estudo........................................................................................................................................74
Capítulo 3
Description of Third Instar Larvae of Five Anastrepha Species (Diptera: Tephritidae)
Tabela 1. Diagnose de cinco espécies de larvas de Anastrepha analisadas neste
estudo......................................................................................................................................120
Capítulo 4

xiii
Associação de adultos e larvas de espécies de Anastrepha (Diptera: Tephritidae) utilizando a
técnica de PCR-RFLP
Tabela 1. Oligonucleotídeos iniciadores (primers) utilizados na amplificação via PCR.......129
Tabela 2. Programas específicos para amplificação de cada par de oligonucleotídeos
iniciadores (primers) para espécies de Anastrepha.................................................................130

xiv
LISTA DE FIGURAS
Capítulo 2
Egg morphology of Anastrepha spp. (Diptera: Tephritidae) in the fraterculus group using
SEM
Figura 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. antunesi. (B) A.
bahiensis. (C) A. coronilli. (D) A. distincta. (E) A. turpiniae. (F) A.
zenildae.....................................................................................................................................47
Figura 2. Vista com Microscopia Eletrônica de Varredura do ovo de A. antunesi..................48
Figura 3. Vista com Microscopia Eletrônica de Varredura dos ovos de A. bahiensis.............50
Figura 4. Vista com Microscopia Eletrônica de Varredura do ovo de A. coronilli.................50
Figura 5. Vista com Microscopia Eletrônica de Varredura do ovo de A. distincta.................51
Figura 6. Vista com Microscopia Eletrônica de Varredura dos ovos de A. turpiniae.............52
Figura 7. Vista com Microscopia Eletrônica de Varredura dos ovos de A. zenildae...............52
Description of eggs of Anastrepha spp. (Diptera: Tephritidae) in the spatulata group using
Scanning Electron Microscopy
Figura 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. manihoti. (B) A.
pickeli. (C) A. montei....................................................................................………………....62
Figura 2. Vista com MEV do ovo de A. manihoti...................................................................62
Figura 3. Vista com MEV do ovo de A. montei.......................................................................64
Figura 4. Vista com MEV do ovo de A. pickeli.......................................................................64
Figura 5. Vista com MEV da micrópila dos ovos de Anastrepha (pólo anterior à
esquerda)...................................................................................................................................65
Description of eggs of Anastrepha curitis and Anastrepha leptozona (Diptera: Tephritidae)
using SEM
Figura 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. curitis. (B) A.
leptozona...................................................................................................................................73
Figura 2. Vista com MEV do ovo de A. curitis.......................................................................73
Figura. 3. Vista com MEV do ovo de A. leptozona.................................................................75
Capítulo 3
Description of Larvae of Anastrepha spp. (Diptera: Tephritidae) in the fraterculus Group

xv
Figura 1. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. bahiensis................................95
Figura 2. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. coronilli.................................96
Figura 3. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. turpiniae................................97
Description of Third Instar Larvae of Five Anastrepha Species (Diptera: Tephritidae)
Figura 1. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. curitis...................................119
Figura 2. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para
larvas de terceiro instar (D à G) de A. pickeli.........................................................................121
Figura 3. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para
larvas de terceiro instar (D à G) de A. pulchra.......................................................................122
Figura 4. Vista com microscopia ótica para larvas de terceiro instar de A. sororcula..........123
Figura 5. Vista com Microscopia Eletrônica de Varredura (B e C) e microscopia ótica para
larvas de terceiro instar (A, D à G) de A. zenildae..................................................................124
Capítulo 4
Associação de adultos e larvas de espécies de Anastrepha (Diptera: Tephritidae) utilizando a
técnica de PCR-RFLP
Figura 1. Gel de agarose 2% com produtos de PCR do fragmento COI digerido pela enzima
VspI.........................................................................................................................................133
Figura 2. Gel de agarose 2% com produtos de PCR do fragmento 16S digerido pelas enzimas
SspI, DraI e VspI.....................................................................................................................134
Figura 3. Gel de agarose 2% com produtos de PCR do fragmento ITS1 digerido pela enzima
VspI.........................................................................................................................................135

xvi
LISTA DE ANEXOS
Anexo 1. Comprovação da publicação da nota científica intitulada “Natural Host Plants and
Native Parasitoids Associated with Anastrepha pulchra and other Anastrepha Species
(Diptera: Tephritidae) in Central Amazon, Brazil”.................................................................150
Anexo 2. Comprovação da publicação da nota científica intitulada “Host Plant of Anastrepha
pulchra (Diptera: Tephritidae) in Central Amazon, Brazil - Mistaken Identity
Resolved”................................................................................................................................152
Anexo 3. Comprovação da publicação do manuscrito intitulado “Egg morphology of
Anastrepha spp. (Diptera: Tephritidae) in the fraterculus group using SEM”.......................154
Anexo 4. Comprovação da publicação do manuscrito intitulado “Description of eggs of
Anastrepha spp. (Diptera: Tephritidae) in the spatulata group using Scanning Electron
Microscopy”............................................................................................................................155
Anexo 5. Comprovação de aceito do manuscrito intitulado “Description of Larvae of
Anastrepha spp. (Diptera: Tephritidae) in the fraterculus Group”.........................................156

17
INTRODUÇÃO GERAL
A família Tephritidae pertence à ordem Diptera, subordem Brachycera, infraordem
Muscomorpha (= Cyclorrhapha), série Schizophora, seção Acalyptratae e superfamíllia
Tephritoidea (Norrbom 2004; Zucchi 2000; Hernández-Ortiz et al. 2010). A superfamília
Tephritoidea reune as famílias Lonchaeidae, Ulidiidae (= Otitidae), Pallopteridae, Piophilidae,
Platystomatidae, Pyrgotidae, Richardiidae e Tephritidae (Norrbom 2004; Hernández-Ortiz et
al. 2010). A família Tephritidae abrange 4.448 espécies, distribuídas em 484 gêneros, sendo
considerada uma das maiores famílias de Diptera (White e Elson-Harris 1992; Norrbom
2004), e sua maior diversidade de espécies é registrada nas regiões tropical e subtropical
(Norrbom 2004).
Dentre os diversos gêneros de tefritídeos, seis apresentam espécies que podem
acarretar prejuízo econômico à fruticultura. Esses gêneros pertencem a duas subfamílias,
Dacinae e Trypetinae. Na subfamília Dacinae, Tribo Ceratitidini está o gênero Ceratitis
MacLeay, e na Tribo Dacini, os gêneros Bactrocera Macquart e Dacus Drew. Na subfamília
Trypetinae, Tribo Carpomyini está o gênero Rhagoletis Loew, e na Tribo Toxotrypanini estão
inseridos os gêneros Toxotrypana Gerstaecker e Anastrepha Schiner (Zucchi 2000; Norrbom
2000).
O gênero Anastrepha possui mais de 250 espécies descritas (Norrbom e Korytkowski
2009, 2011; Norrbom e Uchôa 2011), apresentando ampla distribuição geográfica desde a
América do Norte (Flórida, Texas e México), América Central (inclusive Ilhas do Caribe) e
América do Sul (exceto Chile e sul da Argentina) (Malavasi et al. 2000). Moscas-das-frutas
do gênero Anastrepha são encontradas em todas as regiões brasileiras e até o momento tem-se
o registro de distribuição de 112 espécies para o Brasil (Zucchi 2007, 2008). Na região Norte,
a maior diversidade de espécies é encontrada atualmente no Amazonas, com o registro de 32
espécies (Zucchi 2007, 2008; Zucchi et al. 2011b).
Dentre as espécies de Anastrepha que ocorrem no Brasil, sete causam prejuízos
econômicos à fruticultura, Anastrepha fraterculus (Wiedemann), Anastrepha grandis
(Macquart), Anastrepha obliqua (Macquart), Anastrepha pseudoparallela (Loew),
Anastrepha sororcula Zucchi, Anastrepha striata Schiner e Anastrepha zenildae Zucchi
(Zucchi 2000; Malavasi et al. 2000).
A identificação das espécies de Anastrepha é baseada em características morfológicas
das fêmeas, como padrão alar, coloração do corpo, formato e comprimento do acúleo. Dentre

18
estas características, a principal e determinante para a correta identificação é o acúleo, no
ápice do qual são observados caracteres como comprimento, formato, presença ou ausência de
constrição antes da serra, número e formato dos dentes (Zucchi 2000; Araujo e Zucchi 2006).
Há descrições de ovos para 39 espécies de Anastrepha (Emmart 1933; Lawrence 1979;
Steck e Malavasi 1988; Steck e Wharton 1988; Carroll e Wharton 1989; Murillo e Jirón 1994;
Norrbom et al. 1999; Selivon e Perondini 1999; Norrbom e Korytkowski 2009; Dutra et al.
2011a, 2011b; Figueiredo et al. 2011). Quanto às larvas, estudos sobre descrição morfológica
de espécies de Anastrepha também são escassos. Descrições de larvas de primeiro e/ou
segundo instares foram realizadas apenas para Anastrepha bistrigata Bezzi, A. grandis,
Anastrepha ludens Loew e Anastrepha suspensa (Loew) (Lawrence 1979; Steck e Malavasi
1988; Steck e Wharton 1988; Carroll e Wharton 1989). As larvas de terceiro instar, foram
descritas apenas para 13 espécies (Steck e Wharton 1988; Steck et al. 1990; Norrbom et al.
1999).
Para auxiliar na correta identificação das larvas, a técnica molecular de Reação em
Cadeia da Polimerase (PCR) associada ao Polimorfismo do Comprimento dos Fragmentos de
Restrição (RFLP) tem sido utilizada em estudos de genética de populações de Ceratitis
capitata (Wiedemann) (Gasparich et al. 1995,1997; Silva et al. 2003) e também para a
identificação de 29 espécies da tribo Ceratitidini na África (Barr et al. 2006). Baseado na
técnica PCR–RFLP, foi desenvolvido um método para identificar quatro espécies de
Rhagoletis no Chile (Salazar et al. 2002) e oito espécies de Bactrocera (Muraji e Nakahara
2002). Apesar de o gênero Anastrepha ter espécies de importância quarentenária, até o
momento apenas cinco espécies foram analisadas no estudo realizado por Armstrong et al.
(1997), A. grandis, A. ludens, A. obliqua, Anastrepha serpentina (Wiedemann) e A. striata.
Nesse estudo, os autores desenvolveram um protocolo utilizando PCR–RFLP para diferenciar
estágios imaturos de espécies de Anastrepha, Bactrocera e Ceratitis de importância
quarentenária para a Nova Zelândia. A metodologia de PCR-RFLP tem o objetivo de detectar
polimorfismos de fragmentos de DNA com rapidez e baixo custo.
O Brasil é o terceiro maior produtor mundial de frutas e em 2008 produziu mais de 43
milhões de toneladas de frutas, representando 5% da produção mundial. Apesar de todo o
sucesso neste setor, o mercado exportador brasileiro enfrenta algumas dificuldades para a
expansão competitiva, como baixo consumo de frutas comercializadas no Brasil, pouca
capacitação dos pequenos produtores e restrições quarentenárias impostas pelos países
importadores, que têm por objetivo evitar a introdução de novas pragas em seu território
(Abanorte 2009).

19
A principal forma de dispersão de espécies de Anastrepha tem sido o transporte de
frutos infestados de uma localidade para outra. Com o intuito de evitar a proliferação de novas
pragas, vários países vêm adotando medidas de proteção que exigem que os frutos importados
passem por um rigoroso tratamento quarentenário, para eliminação de diferentes estágios de
vida de possíveis novas pragas (Duarte e Malavasi 2000). Assim, a correta identificação das
larvas de Anastrepha é importante para que as estratégias de controle sejam mais específicas e
mais eficazes, pois estágios imaturos são comumente interceptados em frutos nas barreiras
quarentenárias ou pontos de entrada e é necessário aguardar a emergência de fêmeas para a
identificação da espécie. Baseado nessas informações este trabalho teve por objetivo ampliar
os conhecimentos sobre ovos e larvas de algumas espécies de Anastrepha por meio da
caracterização morfológica e molecular, visando a sua descrição e subsídios para o
desenvolvimento de um protocolo para rápida identificação e que possa ser utilizado em
programas de controle e quarentena. Vale ressaltar contudo que esse estudo não trabalhou
com as mais importantes espécies quarentenárias para os países importadores. Porém fornece
uma consistente perspectiva nesse contexto.

20
OBJETIVO GERAL
Ampliar os conhecimentos sobre ovos e larvas de espécies de Anastrepha por meio da
caracterização morfológica e molecular com a sua descrição detalhada e o fornecimento de
subsídios para o desenvolvimento de um protocolo que possa ser utilizado em programas de
defesa vegetal.
OBJETIVOS ESPECÍFICOS
1. Descrever os ovos das espécies Anastrepha antunesi Lima, Anastrepha bahiensis Lima,
Anastrepha coronilli Carrejo & González, Anastrepha curitis Stone, Anastrepha distincta
Greene, Anastrepha leptozona Hendel, Anastrepha manihoti Lima, Anastrepha montei Lima,
Anastrepha pickeli Lima, Anastrepha turpiniae Stone e A. zenildae.
2. Descrever as larvas de segundo e terceiro instares de A. bahiensis, A. coronilli, A. curitis e
A. turpiniae. Descrever larvas de terceiro instar de A. pickeli, Anastrepha pulchra Stone, A.
sororcula e A. zenildae.
3. Obter o código de barras de DNA (“DNA barcodes”) de Anastrepha antunesi, A. atrigona
Hendel, A. bahiensis, A. coronilli, A. curitis, A. distincta, A. manihoti, A. pickeli, A. pulchra,
A. striata, A. sororcula, A. turpiniae e A. zenildae por meio do sequenciamento do gene
mitocondrial COI para inclusão no banco de dados do Tephritid Barcode Initiative.
4. Obter o perfil de haplótipos de adultos e larvas das 13 espécies de Anastrepha citadas no
objetivo 3 por meio do uso de PCR-RFLP para dois genes do DNA mitocondrial (COI e 16S)
e uma região do DNA nuclear (ITS1).

21
Capítulo 1
Dutra, V.S; Ronchi-Teles, B.; Silva, J.G. Associação de
espécies de Anastrepha (Diptera: Tephritidae) e seus
parasitóides obtidos de hospedeiros nativos e introduzidos
coletados em alguns municípios brasileiros. Manuscrito
formatado para Acta Amazonica.

22
Associação de espécies de Anastrepha (Diptera: Tephritidae) e seus
parasitoides obtidos de hospedeiros nativos e introduzidos coletados em
alguns municípios brasileiros
Vivian S. DUTRA1, Beatriz RONCHI-TELES
2, Janisete G. SILVA
3
RESUMO
A dificuldade encontrada para determinar os hospedeiros para espécies de Anastrepha
ocorre devido a relatos equivocados ou incompletos, pois geralmente coletas de adultos são
realizadas com armadilhas o que impossibilita a associação com frutos hospedeiros. O
objetivo deste trabalho foi conhecer a diversidade de espécies de Anastrepha e sua relação
com hospedeiros e parasitoides obtidos de frutos nativos e introduzidos coletados em alguns
municípios brasileiros. Dos frutos coletados, foram obtidos exemplares de 19 espécies de
Anastrepha e seis espécies de parasitoides braconídeos. Dentre as espécies de moscas-das-
frutas obtidas, Anastrepha obliqua (Macquart) foi a mais abundante (17,7%) e Doryctobracon
areolatus (Szépligeti) o parasitoide mais abundante (86,0%). Também são relatados alguns
primeiros registros de associação de moscas-das-frutas com hospedeiros e parasitoides.
PALAVRAS CHAVE: moscas-das-frutas, hospedeiros, braconídeos.
INTRODUÇÃO
Os insetos da família Tephritidae, também conhecidos como moscas-das-frutas, são
consideradas pragas para a fruticultura pelo fato das fêmeas depositarem seus ovos na
superfície de frutos verdes ou em amadurecimento (Aluja 1994; Salles 2000). Dos ovos
eclodem larvas que se alimentam da polpa dos frutos, favorecendo o aparecimento de fungos
e bactérias que aceleram o apodrecimento e a queda prematura dos frutos, causando danos aos
mesmos e consequentemente prejudicando a comercialização (Aluja 1994; Duarte e Malavasi
2000; Nascimento e Carvalho 2000; Salles 2000).
A família Tephritidae compreende aproximadamente 4.500 espécies (White e Elson-
Harris 1992; Norrbom 2005). O gênero Anastrepha Schiner possui cerca de 240 espécies
descritas e algumas dessas espécies causam prejuízo à fruticultura (Zucchi 2008; Norrbom e
Korytkowski 2009). No Brasil, há o registro de ocorrência de 112 espécies de Anastrepha
1Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970 Manaus, AM, Brasil. 2Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970
Manaus, AM, Brasil. 3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil.

23
(Norrbom et al. 1999; Jesus et al. 2008; Uramoto et al. 2008; Zucchi 2007, 2008; Uramoto e
Zucchi 2010).
A dificuldade encontrada na determinação de algumas plantas hospedeiras para as
espécies de Anastrepha ocorre devido a relatos equivocados ou incompletos, pois em geral
são utilizadas apenas armadilhas para coleta de adultos, impossibilitando a associação correta
e direta com os frutos hospedeiros. Também são relativamente escassos os estudos em relação
aos hospedeiros nativos. A correta associação entre os tefritídeos e seus hospedeiros, seria
melhor registrada com a coleta de frutos de plantas introduzidas e nativas de diversas regiões
(Norrbom e Kim 1988; Hernández-Ortiz e Pérez-Alonso 1993; Aluja 1994; Hernández-Ortiz e
Aluja 1994; Zucchi 2000). Este trabalho teve como objetivo conhecer a diversidade de
espécies de Anastrepha e a associação com hospedeiros e parasitoides de frutos nativos e
introduzidos coletados em alguns municípios brasileiros.
MATERIAL E MÉTODOS
Coleta de adultos
Foram coletados frutos de hospedeiros nativos e introduzidos de espécies de
Anastrepha nos municípios de Manaus (03º06’07’’S; 60º01’30’’W), Maués (03º23’01’’S;
57º43’07’’W) e Presidente Figueiredo (02°02’04’’S; 60°01’30’’W) no estado do Amazonas;
de Porto Velho (08º45’43’’S; 63º54’14’’W) no estado de Rondônia, de Boa Vista
(02º49’11’’S; 60º40’24’’W) no estado de Roraima e de Camamu (13º56’41’’S; 39º06’14’’),
Itabuna (14º47’08’’S; 39º16’49’’W), Maracás (13º26’28’’S; 40º25’51’’W), Una
(15º17’36’’S; 39º04’31’’W) e Valença (13º22’13’’S; 39º04’23’’W) no estado da Bahia;.
As amostras de frutos foram pesadas e armazenadas individualmente em frascos
plásticos de 500 mL contendo vermiculita e cobertos por organza. Após aproximadamente dez
dias, a vermiculita foi peneirada e as pupas obtidas foram contadas e transferidas para frascos
plásticos de 30 mL contendo vermiculita úmida e cobertos com organza até a emergência dos
adultos. Os adultos foram mortos em álcool absoluto e posteriormente foram contados,
sexados e identificados. As amostras botânicas foram identificadas por Mário Terra e José
Lima.
Identificação dos adultos
A identificação das espécies de Anastrepha foi realizada através das fêmeas. O acúleo
de cada fêmea foi extrovertido e colocado entre lâmina e lamínula para análise das
características ao microscópio óptico. A identificação em nível de espécie foi baseada em

24
Steyskal (1977), Silva e Ronchi-Teles (2000), Zucchi (2000) e Zucchi et al. (2011a). Os
espécimes testemunho foram depositados na Coleção de Invertebrados do Instituto Nacional
de Pesquisas da Amazônia (INPA), Manaus, Amazonas, Brasil.
RESULTADOS
Foram coletados 74,53 kg de frutos hospedeiros de 28 espécies botânicas pertencentes
a 17 famílias (Tabela 1). No período de 2008 a 2009, as coletas foram realizadas ao longo dos
meses. Em 2010 e 2011, as coletas foram realizadas somente nos meses de janeiro a junho.
Dentre as espécies botânicas coletadas, 23 apresentaram frutos infestados. Foram coletados
3.665 frutos e obtidas 8.009 pupas, destas emergiram 3.782 adultos (1.640 machos e 2.142
fêmeas). Foram obtidos exemplares de 19 espécies de Anastrepha: Anastrepha antunesi Lima,
Anastrepha atrigona Hendel, Anastrepha bahiensis Lima, Anastrepha coronilli Carrejo &
González, Anastrepha curitis Stone, Anastrepha distincta Greene, Anastrepha fractura Stone,
Anastrepha fraterculus (Weidemann), Anastrepha leptozona Hendel, Anastrepha manihoti
Lima, Anastrepha obliqua (Macquart), Anastrepha pickeli Lima, Anastrepha pulchra Stone,
Anastrepha serpentina (Wiedemann), Anastrepha sororcula Zucchi, Anastrepha striata
Schiner, Anastrepha turpiniae Stone, Anastrepha zenildae Zucchi e uma espécie de
Anastrepha ainda não identificada (possivelmente espécie nova). Também foram obtidos 537
parasitoides da família Braconidae (Hymenoptera) das espécies Asobara anastrephae
(Muesebeck), Doryctobracon areolatus (Szépligeti), Doryctobracon brasiliensis (Szépligeti),
Opius bellus Gahan, Opius sp. e Utetes anastrephae (Viereck) (Tabela 1).
Dentre as espécies de moscas-das-frutas obtidas, A. obliqua, A. fraterculus, A. fractura
e A. bahiensis foram as mais abundantes com 17,7%, 12,9%, 11,5% e 9,5%, respectivamente.
Exemplares de A. obliqua foram obtidos em nove hospedeiros diferentes, sendo quatro
espécies botânicas da família Anacardiaceae, três de Myrtaceae, uma de Malpighiaceae e uma
de Oxalidaceae (Tabela 1). Desses nove hospedeiros, cinco são de espécies de plantas
introduzidas e quatro de plantas nativas.
Na Tabela 1, está a associação de A. striata com três espécies nativas da família
Myrtaceae; A. bahiensis e A. fraterculus ambas com duas espécies nativas de Moraceae e
Myrtaceae, respectivamente; A. manihoti e A. pickeli com uma espécie nativa de
Euphorbiaceae; A. sororcula e A. zenildae ambas com uma espécie nativa de Myrtaceae; A.
leptozona e A. serpentina ambas com uma espécie nativa de Sapotaceae; A. antunesi, A.
atrigona, A. bahiensis, A. coronilli, A. curitis, A. distincta, A. fractura e A. pulchra com uma
espécie nativa das famílias Anarcardiaceae, Loganiaceae, Moraceae, Melastomataceae,

25
Passifloraceae, Fabaceae (Mimosoideae), Celastraceae e Sapotaceae, respectivamente.
Anastrepha turpiniae foi obtida em frutos de um hospedeiro introduzido da família
Combretaceae.
Com relação aos parasitoides, a espécie mais abundante foi D. areolatus (86,0%).
NaTabela 1, estão as associações de D. areolatus parasitando as espécies A. bahiensis, A.
coronilli, A. fractura, A. fraterculus, A. obliqua, A. pickeli, A. pulchra, A. striata e A.
turpiniae; Opius sp. parasitando A. bahiensis, A. obliqua e A. fraterculus; O. bellus
parasitando A. fractura e A. manihoti; U. anastrephae parasitando A. manihoti, A. coronilli e
A. obliqua e D. brasilensis parasitando A. fractura.
DISCUSSÃO
Neste estudo, foram observadas várias associações entre moscas-das-frutas,
hospedeiros e parasitoides e algumas delas são novos registros dessas relações como A.
fractura e o hospedeiro Salacia sp. da família Celastraceae. Até então havia o registro de
apenas um hospedeiro Maquira sclerophylla (Ducke) C.C. Berg da família Moraceae para A.
fractura (Costa et al. 2009).
Também foram observadas novas associações entre moscas-das-frutas e parasitoide
para A. coronilli e U. anastrephae, A. manihoti e O. bellus, A. turpiniae e D. areolatus, A.
fractura e A. anastrephae, D. brasilensis e O. bellus. Até o momento, havia sido relatado A.
coronilli sendo parasitada por D. areolatus; A. fractura por D. areolatus e os parasitoides
figitídeos Aganaspis nordlanderi Wharton e Aganaspis pelleranoi (Brèthes); A. manihoti por
U. anastrephae e para A. turpiniae não havia sido registrado nenhum parasitoide (Zucchi
2008; Costa et al. 2009).
Foram listadas na Tabela 1, outras associações observadas entre espécies de
Anastrepha, hospedeiros e parasitoides que já haviam sido relatadas previamente (Ovruski et
al. 2000; Zucchi 2008; Costa et al. 2009; Marinho et al. 2009; Silva et al. 2010; Ronchi-Teles
et al. 2011; Zucchi et al. 2011b).
No presente estudo, foram realizados novos registros de associações entre moscas-das-
frutas, hospedeiros e parasitoides coletados em alguns municípios brasileiros, bem como
confirmadas outras associações. Estudos sobre esse tema são fundamentais para o acréscimo
de novas informações sobre as interações entre esses organismos. Estas informações tornam-
se referência para outros trabalhos a serem desenvolvidos com moscas-das-frutas e também
podendo ser úteis, por exemplo, para a elaboração de medidas para controle biológico dessas
espécies.

26
CONCLUSÕES
Neste estudo, foi registrada uma nova associação entre A. fractura e Salacia sp. da
família Celastraceae. Novas associações entre moscas-das-frutas e parasitoides braconídeos
foram registradas para A. turpiniae e D. areolatus; A. manihoti e O. bellus; A. coronilli e U.
anastrephae; A. fractura e A. anastrephae, D. brasilensis e O. bellus.
AGRADECIMENTOS
Os autores agradecem a Ana Claúdia Araújo, Claudemir Campos, Felipe Gomes, José
Nilton Medeiros da Costa, Marcos Vinicius Garcia, Mírian Santos, Neusa Hamada, Tatiana
Senra, Weber Novaes e Ulisses Neiss pelo auxílio durante as coletas dos exemplares. Esse
estudo foi financiado pelo CNPq (Conselho Nacional de Desenvolvimento Científico e
Tecnológico - projeto n°575664/2008-8), pela Coordenação de Aperfeiçoamento de Pessoal
de Nível Superior (CAPES – projeto n° 23038.037185/2008-41) e pela Rede Amazônica de
Pesquisas sobre Moscas-das-Frutas (Chamada 05/2006 Agrofuturo). Os autores também
agradecem ao CNPq pela bolsa de doutorado concedida a VSD.

27
Tabela 1. Hospedeiros e parasitoides de espécies de Anastrepha.
Famílias Espécies Hospedeiros Peso
(kg)
Frutos
(n)
Pupas
(n)
Espécie de
Anastrepha e o n. de
exemplares
Espécie de
Braconidae e o n.
de exemplares
Anacardiaceae Anacardium occidentale L. Nativo 0,9 23 15 4 A. obliqua 2 D. areolatus
Mangifera indica L. Introduzido 2,5 6 109 27 A. obliqua 5 D. areolatus
Spondias cytherea Sonn. Introduzido 0,09 8 0 0 0
Spondias mombin L. Nativo 8,2 1.055 1.481 149 A. obliqua
4 A. antunesi
87 D. areolatus
43 Opius sp.
3 U. anastrephae
Spondias purpurea L. Introduzido 1,2 213 43 7 A. obliqua 0
Celastraceae Salacia sp.♦ Nativo 0,6 14 731 181 A. fractura 60 D. areolatus
3 A. anastrephae ◘
1 D. brasiliensis ◘
1 O. bellus ◘
Combretaceae Terminalia catappa L. Introduzido 4,9 64 300 77 A. turpiniae 41 D. areolatus ◘
Duckeodendraceae Duckeoendendron cestroides Kulm. Nativo 0,04 14 0 0 0
Euphorbiaceae Manihot esculenta Crantz Nativo 5,1 88 429 114 A. pickeli
3 A. manihoti
128 D. areolatus
4 O. bellus ◘
2 U. anastrephae
Fabaceae
(Mimosoideae)
Inga edulis Mart. Nativo 2,9 17 323 111 A. distincta 0
Loganiaceae Strychnos jobertiana Baill. Nativo 0,3 12 17 8 A. atrigona 0
Malpighiaceae Malpighia glabra L. Introduzido 1,3 93 79 17 A. obliqua 21 D. areolatus
12 Opius sp.
Melastomataceae Bellucia grossularioides (L.) Nativo 3,7 476 264 50 A. coronilli 14 D. areolatus

28
Triana 1 U. anastrephae ◘
Moraceae Helicostylis scabra (Macbr.) C.C.
Berg.
Nativo 0,9 92 290 115 A. bahiensis 48 D. areolatus
Pouroma cecropiaefolia Mart. Nativo 4,6 14* 113 35 A. bahiensis 9 D. areolatus
1 Opius sp.
Myrtaceae Eugenia stipitata Mc Vaugh Nativo 3,3 37 296 56 A. obliqua 0
Psidium acutangulum D. C. Nativo 1,2 7 82 33 A. striata 0
Psidium guajava L. Nativo 10,3 343 1.140 183 A. fraterculus
61 A. striata
3 A. obliqua
2 A. sororcula
1 A. zenildae
13 D. areolatus
1 Opius sp.
Psidium guineense Swart. Nativo 2,5 207 164 21 A. fraterculus
21 A. striata
11 D. areolatus
Syzygium jambolanum (Lam.) DC. Introduzido 2,1 201 2 0 2 D. areolatus
Syzygium malaccense L. Introduzido 4,8 143 86 13 A. obliqua 1 U. anastrephae
Oxalidaceae Averrhoa carambola L. Introduzido 0,7 24 35 3 A. obliqua 3 D. areolatus
Passifloraceae Passiflora nitida Kunth. Nativo 3,4 33 205 42 A. curitis 0
Peridiscaceae Peridiscus lucidus Benth. Nativo 0,5 23 0 0 0
Rhamnaceae Ziziphus mauritiana Lam Introduzido 1,3 194 1 0 0
Sapotaceae Chrysophyllum prieurii A. DC. Nativo 0,4 55 14 6 Anastrepha sp. 0
Pouteria caimito (Ruiz & Pav.)
Radlk.
Nativo 5,2 161 1.490 105 A. serpentina
61 A. leptozona
0
Pouteria oblanceolata Pires Nativo 1,6 62 300 119 A. pulchra 20 D. areolatus
* Esse valor refere-se ao número de cachos coletados.
♦ Novo registro de hospedeiro
◘ Novo registro de parasitoide

29
Ronchi-Teles, B.; Dutra, V.S.; Tregue-Costa, A.P.;
Aguiar-Menezes, E.L.; Mesquita, A.C.A.; Silva, J.G.
2011. Natural Host Plants and Native Parasitoids
Associated with Anastrepha pulchra and other Anastrepha
Species (Diptera: Tephritidae) in Central Amazon, Brazil.
Florida Entomologist, 94(2): 347-349 (Anexo 1).

30
NATURAL HOST PLANTS AND NATIVE PARASITOIDS ASSOCIATED WITH
ANASTREPHA PULCHRA AND OTHER ANASTREPHA SPECIES (DIPTERA:
TEPHRITIDAE) IN CENTRAL AMAZON, BRAZIL
BEATRIZ RONCHI-TELES1, VIVIAN SIQUEIRA DUTRA
1, ALEXANDRA
PRISCILLA TREGUE COSTA1, ELEN DE LIMA AGUIAR-MENEZES
2, ALINE
CRISTINA ARAUJO MESQUITA1 E JANISETE GOMES SILVA
3
1Coordenação de Pesquisas em Entomologia, Instituto Nacional de Pesquisas da
Amazônia, Caixa Postal 478, 69011-970 - Manaus, Amazonas, Brasil
2Departamento de Entomologia e Fitopatologia, Universidade Federal Rural do Rio de
Janeiro, Caixa Postal 74538, 23890-000 - Seropedica, Rio de Janeiro, Brasil
3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia
Ilhéus-Itabuna km 16, 45650-000 - Ilhéus, Bahia, Brasil
A Amazônia brasileira abriga uma alta diversidade de moscas-das-frutas do
gênero Anastrepha Schiner (Diptera: Tephritidae) tendo sido registradas 53 espécies de
um total de 103 que ocorrem no Brasil. Destas, 12 espécies de Anastrepha ocorrem
exclusivamente na Amazônia brasileira (Zucchi et al. 1996; Silva & Ronchi-Teles 2000;
Zucchi 2008).
Na Amazônia Central, a maioria dos estudos sobre espécies de moscas-das-frutas
foram realizados em áreas de sistemas agroflorestais (Silva et al. 1996; Zucchi et al.
1996), utilizando armadilhas para a coleta dos exemplares (Ronchi-Teles & Silva 2005),
e apenas um estudo foi realizado em áreas de florestas não perturbadas, tendo como foco
os parasitoides (Costa et al. 2009). Assim, informações sobre a associação entre
hospedeiro / moscas-das-frutas / parasitoides ainda é limitada.
Neste estudo, foram registradas associações entre moscas-das-frutas e hospedeiros
para espécies de Anastrepha na Amazônia Central e no Brasil, e também foram
identificados parasitoides braconídeos associados com espécies de Anastrepha.

31
As coletas foram realizadas em uma área de 30 km2 da Reserva Florestal Adolpho
Ducke (RFAD) do Instituto Nacional de Pesquisas da Amazônia (INPA), localizado a
nordeste de Manaus (02º53’S, 59º59’W) no estado do Amazonas, Brasil. Essa área possui
cerca de 100 km2 de floresta primária. A média anual de temperatura é de 26,5ºC, com a
máxima mensal de 38,6 ºC (Dezembro) e mínima de 18,2 ºC (Julho) e média anual de
umidade relativa é de 82% (Araújo 1970). Foram coletados frutos maduros ou em
amadurecimento aleatoriamente no chão e na copa das árvores a cada duas semanas, no
período de outubro/2002 a junho/2003, de março a maio/2009, e de março a maio/2010.
Os frutos foram coletados em uma área de floresta de cerca de 30 km2, onde todas as
espécies de árvores já haviam sido identificadas por botânicos do Instituto Nacional de
Pesquisas da Amazônia (INPA). Os frutos coletados foram contados, pesados e
armazenados em frascos plásticos de 500 mL contendo uma camada de vermiculita e
cobertos com organza até a obtenção das pupas. As pupas obtidas foram contadas e
colocadas em frascos plásticos de 30 mL contendo uma camada de vermiculita e cobertos
com organza até a emergência dos adultos. Espécimes testemunho foram depositados na
Coleção de Invertebrados do INPA.
Foi coletado um total de 63,7 kg de frutos de 50 espécies de plantas que fazem
parte de 18 famílias botânicas. Um total de 1.398 frutos pesando 19,7 kg de 13 espécies
de plantas representado sete famílias botânicas foram infestados por moscas-das-frutas e
obtidas 880 pupas (Tabela 1). Foram registradas as associações entre Anastrepha pulchra
Stone e Mouriri collocarpa Ducke (Melastomataceae), espécie de árvore nativa e o
parasitoide Doryctobracon areolatus (Szépligeti) (Hymenoptera: Braconidae) para o
Brasil, pela primeira vez. Anastrepha pulchra foi registrada no Panamá, Venezuela e
Brasil (Amazônia) (Norrbom 2002).
Também foram registrados dois novos hospedeiros para Anastrepha atrigona
Hendel: Strychnos jobertiana Baillon (Loganiaceae) e Pouteria durlandii (Standley)
Baehni (Sapotaceae). Três espécies de parasitoides himenópteros Opius bellus Gahan,
Opius sp. (Braconidae), e Aganaspis pelleranoi (Brèthes) (Figitidae) foram observados
parasitando A. atrigona, pela primeira vez. Até o momento, A. atrigona foi registrada
apenas na Venezuela, Guiana, Suriname e Brasil (estado do Amazonas) (Norrbom et al.
1999; Zucchi 2008).

32
Foi registrado um novo hospedeiro para Anastrepha bahiensis Lima, Helicostylis
scabra (Macbride) Cornelis Christiaan Berg (Moraceae). Anastrepha bahiensis ocorre no
México e no Brasil (em vários estados) (Norrbom et al. 1999; Zucchi 2008). Neste
estudo, também foram encontradas Anastrepha bondari Lima, Anastrepha coronilli
Carrejo & González, Anastrepha obliqua Macquart e Anastrepha striata Schiner. Seus
hospedeiros listados na Tabela 1 já foram registrados anteriormente, bem como vários
outros (Norrbom 2002; Zucchi 2007, 2008). Foram observadas quatro novas espécies de
Anastrepha, ainda não descritas. Esses exemplares foram obtidos em uma espécie de
Annonaceae, uma de Bignoniaceae e duas em espécies de Sapotaceae, respectivamente
(Tabela 1).
Três espécies de braconídeos (D. areolatus, Opius sp. e O. bellus) e duas espécies
de figitídeos (Aganaspis nordlanderi Wharton e A. pelleranoi) foram associadas com
espécies de Anastrepha. Também foi realizado o primeiro registro de A. nordlanderi
parasitando A. coronilli. Neste estudo, os braconídeos e figitídeos registrados foram
encontrados previamente associados com outras espécies de Anastrepha (Canal & Zucchi
2000; Guimarães et al. 2000; Ovruski et al. 2000).
Os autores agradecem a Claudemir M. Campos e Ulisses G. Neiss pelo auxílio
durante as coletas, José Lima pela identificação das plantas, Carter R. Miller, Gary J.
Steck e dois revisores anônimos pelos comentários feitos em uma versão anterior desse
manuscrito. Esse estudo foi financiado pelo CNPq (Conselho Nacional de
Desenvolvimento Científico e Tecnológico - projeto n°575664/2008-8).
RESUMO
Um hospedeiro natural (Mouriri collocarpa) e um parasitoide (Doryctobracon
areolatus) foram registrados pela primeira vez para Anastrepha pulchra no Brasil. Foram
registrados novos hospedeiros para Anastrepha atrigona e Anastrepha bahiensis na
Amazônia brasileira. Também foram registrados parasitoides atacando A. atrigona,
Anastrepha coronilli e A. pulchra.

33
TABELA 1. ESPÉCIES DE ANASTREPHA E PARASITOIDES COLETADOS NA AMAZÔNIA CENTRAL, BRASIL.
Família Botânica Espécies Botânicas Hospedeiros N. de
frutos
Peso
(kg)
N. de
pupas
Espécies de Anastrepha
(n)
Espécies de parasitoides
(n)
Annonaceae Anonaceae (unidentified) Nativo 3 0.317 10 8 Anastrepha sp.1 0
Bignoniaceae Clytostoma sp. Nativo 2 0.096 2 1 Anastrepha sp.3
1 Anastrepha sp.4
0
Loganiaceae Strychnos jobertiana Baill. ▲ Nativo 56 1.970 17 12 A. atrigona 0
Melastomataceae Bellucia grossularioides (L.) Triana Nativo 758 5.029 68 59 A. coronilli 7 D. areolatus
1 A. nordlanderi◘
Melastomataceae Mouriri collocarpa Ducke▲ Nativo 13 0.153 108 59 A. pulchra 18 D. areolatus ◘
Moraceae Helicostylis scabra (Macbr.)
C.C.Berg. ♦
Nativo 92 0.978 290 218 A. bahiensis 48 D. areolatus
Moraceae Helicostylis tomentosa (Planch. &
Endl.) Rusby
Nativo 239 2.794 275 162 A. bahiensis 24 D. areolatus
Moraceae Naucleopsis sp. Nativo 21 0.803 9 8 A. bondari 0
Myrtaceae Eugenia patrisii Vahl. Nativo 15 0.083 16 14 A. obliqua 1 A. pelleranoi
Myrtaceae Psidium guajava L. Nativo 12 0.486 5 3 A. striata 0
Sapotaceae Chrysophyllum prieurii A.DC. Nativo 55 2.970 12 6 Anastrepha sp.4 0
Sapotaceae Pouteria durlandii (Standl.) Baehni
▲
Nativo 19 0.567 66 29 A. atrigona 10 Opius bellus ◘
2 Opius sp. ◘
2 A. pelleranoi ◘
Sapotaceae Pouteria williamii (Aubrév. &
Pellegrin) T.D. Penn.
Nativo 8 0.358 2 1 Anastrepha sp.2 0
▲ Primeiro registro de hospedeiro
♦ Novo registro de hospedeiro
◘ Novo registro de parasitoide

34
Ronchi-Teles, B.; Dutra, V.S.; Silva, J.G. 2011. Host Plant
of Anastrepha pulchra (Diptera: Tephritidae) in Central
Amazon, Brazil - Mistaken Identity Resolved. Florida
Entomologist, 94(3): 719-720 (Anexo 2).

35
HOST PLANT OF ANASTREPHA PULCHRA (DIPTERA: TEPHRITIDAE) IN
CENTRAL AMAZON, BRAZIL - MISTAKEN IDENTITY RESOLVED
BEATRIZ RONCHI-TELES1, VIVIAN SIQUEIRA DUTRA
2 E JANISETE GOMES
SILVA3
1Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal
478, 69011-970 - Manaus, Amazonas, Brasil
2Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia,
Caixa Postal 478, 69011-970 - Manaus, Amazonas, Brasil
3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus-
Itabuna km 16, 45650-000 - Ilhéus, Bahia, Brazil
Anastrepha pulchra Stone é uma das 11 espécies de Anastrepha Schiner que
pertencem ao grupo serpentina e não é considerada praga de importância econômica
(Norrbom et al. 1999; Norrbom 2002). Registros de ocorrência de A. pulchra estão restritos ao
tipo-localidade no Panamá (Stone 1942) e na Venezuela em Aragua e Bolivar (Caraballo
1981; Norrbom 2002). No Brasil, A. pulchra foi coletada em armadilhas e registrada em
Iranduba estado do Amazonas, pela primeira vez (Ronchi-Teles 2009). O conhecimento sobre
a biologia de A. pulchra é muito escasso. Caraballo (1981) fez o primeiro registro de
associação para A. pulchra com uma planta hospedeira da família Sapotaceae na Venezuela,
contudo nenhuma informação sobre essa espécie de hospedeiro foi disponibilizada. Norrbom
(2002) realizou uma revisão taxonômica do grupo serpentina e afirmou que o “único relato de
planta hospedeira” para A. pulchra “é uma espécie indeterminada de Sapotaceae” citando
Caraballo (1981) como fonte da informação. No Brasil, Ronchi-Teles (2009) registrou A.
pulchra infestando frutos da família Sapotaceae coletados na Reserva Florestal Adolpho
Ducke no estado do Amazonas, infelizmente esses frutos que não foram identificados e
nenhum gênero ou espécie do suposto hospedeiro foi conhecido.
Em recente tentativa para investigar potenciais hospedeiros e associação com
parasitoides para A. pulchra, foram coletados frutos na Reserva Ducke (RD) do Instituto
Nacional de Pesquisas da Amazônia (INPA), localizado a nordeste de Manaus (02°53’S,
59°59’W) no estado do Amazonas, Brasil. A reserva compreende uma área de 100 km2 de

36
floresta primária, e os frutos foram coletados na floresta no período de Março a Maio de 2010.
Foram obtidas 108 pupas das quais emergiram 59 adultos de A. pulchra e 18 espécimes de
Doryctobracon areolatus (Szépligeti) (Hymenoptera: Braconidae). Esses resultados sobre
associação de plantas hospedeiras e parasitoides, foram publicados juntos com informações
sobre outras espécies de Anastrepha da Amazônia Central (Ronchi-Teles et al. 2011). No
estudo de Ronchi-Teles et al. (2011), o hospedeiro natural de A. pulchra foi identificado
erroneamente como Mouriri collocarpa Ducke (localmente conhecido como “miraúba”),
espécie de árvore nativa da família Melastomataceae. Na época, a planta hospedeira M.
collocarpa foi identificada erradamente, devido a identificação ter sido baseada apenas em
frutos, pois nesse momento não havia sido possível coletar outras partes da planta. Como já
havia registro anterior para outras árvores do gênero Mouriri como hospedeiros de
Anastrepha (Zucchi 2008), foi aceito o fato de que essa espécie também poderia ser uma
planta hospedeira.
Entre abril e maio de 2011, foram coletadas mais amostras do mesmo fruto na Reserva
Biológica do Cueiras, localizada no Km 14 da ZF2, ramal que pode ser acessado pela BR 174,
em Manaus no estado do Amazonas (2°36'S, 60°07'W). Essa reserva está localizada numa
área de floresta primária contínua. Foram coletados frutos no chão da floresta e em ramos que
haviam caído da árvore. Os frutos coletados foram contados, pesados e colocados em frascos
plásticos de 500 mL contendo uma camada de vermiculita e cobertos com organza até
obtenção das pupas. As pupas foram contadas e colocadas em frascos plásticos de 30 mL
contendo uma camada de vermiculita e cobertos com organza até a emergência dos adultos.
Espécimes testemunho foram depositados na Coleção de Invertebrados do INPA.
Foram coletados 49 frutos pesando 1,41 kg e obtidos 192 pupários. Destes emergiram
um total de 113 adultos de A. pulchra (60 fêmeas e 53 machos) e dois exemplares de D.
areolatus. Neste momento, a planta hospedeira foi identificada pelos frutos, folhas e galhos
como Pouteria oblanceolata Pires (Sapotaceae) uma árvore nativa localmente conhecida
como “tuturubá” ou “abiurana-preta”. Os frutos de P. oblanceolata são amarelos, carnosos,
fibrosos e com pouco látex, possuem cerca de 4 - 4,5 cm de diâmetro e pericarpo amarelo e
liso.
Ambos os frutos de P. oblanceolata e M. collocarpa são muito similares na aparência
(formato, tamanho, cor e textura). Além disso, ambas as espécies quando plantas adultas
fazem parte do dossel da floresta Amazônica (possuindo cerca de 40 m de altura),
apresentando o tronco liso e reto. Devido a copa alta de ambas as árvores, a coleta de folhas e

37
flores torna-se muito difícil, ressaltando que essas estruturas são fundamentais para uma
correta identificação botânica.
Com o presente estudo, foi concluído com segurança que P. oblanceolata é a planta
hospedeira de A. pulchra e não M. collocarpa como relatado primeiramente em Ronchi-Teles
et al. (2011). Esses resultados corroboram com a informação prévia sobre a preferência das
espécies do grupo serpentina por plantas hospedeiras de Apocynaceae, Sapotaceae e
Myrtaceae (Norrbom 2002).
Os autores agradecem a Weber Novaes e Felipe Gomes pelo auxílio na coleta de frutos e
Mário Terra (INPA) pela identificação da planta. Os autores também agradecem a Carter R.
Miller por seus comentários sobre manuscrito. Esse estudo foi financiado pelo CNPq
(Conselho Nacional de Desenvolvimento Científico e Tecnológico - projeto n°575664/2008-
8).
RESUMO
Pouteria oblanceolata, o verdadeiro hospedeiro natural de Anastrepha pulchra, foi
registrada no estado do Amazonas, Brasil. Também foi registrado Doryctobracon areolatus
parasitando A. pulchra. Foram incluídas informações sobre o equívoco na identificação do
hospedeiro de A. pulchra.

38
Capítulo 2
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G. 2011.
Egg morphology of Anastrepha spp. (Diptera:
Tephritidae) in the fraterculus group using Scanning
Electron Microscopy. Annals of the Entomological Society
of America, 104(1): 16-24 (Anexo 3).

39
Egg Morphology of Anastrepha spp. (Diptera: Tephritidae) in the
fraterculus Group Using Scanning Electron Microscopy
VIVIAN S. DUTRA1,2BEATRIZ RONCHI-TELES
1, GARY J. STECK
2, E JANISETE G.
SILVA3
Ann. Entomol. Soc. Am. 104(1): 16-24 (2011); DOI: 10.1603/AN10105
RESUMO Com o auxílio da microscopia eletrônica de varredura (MEV), a morfologia dos
ovos de seis espécies de Anastrepha do grupo fraterculus: Anastrepha antunesi Lima,
Anastrepha bahiensis Lima, Anastrepha coronilli Carrejo & González, Anastrepha distincta
Greene, Anastrepha turpiniae Stone e Anastrepha zenildae Zucchi foi descrita
detalhadamente pela primeira vez. Os ovos foram dissecados de fêmeas capturadas em
armadilhas McPhail em Cariri no estado de Tocantins; Itacoatiara, Manaus e Presidente
Figueiredo no estado do Amazonas e Porto Velho no estado de Rondônia, na Amazônia
brasileira. Os ovos foram examinados usando-se microscópio eletrônico e óptico. Pelo menos
dez ovos de cada espécie foram examinados usando MEV. Com microscópio óptico foram
examinados entre 20 e 35 ovos por fêmea de pelo menos três fêmeas de cada espécie. Os ovos
das seis espécies são similares em seu aspecto geral. Eles são afilados em ambas as
extremidades, possuem papila no pólo anterior e a micrópila está localizada no lado dorsal do
pólo anterior. Caracteres diagnósticos para diferenciar as seis espécies incluem ornamentação
do córion, localização das aerópilas e uma pronunciada borda do córion com aparência lanosa
ao redor da micrópila. Nenhum dos ovos das espécies estudadas apresentou apêndice
respiratório.
PALAVRAS-CHAVE moscas-das-frutas, ornamentação do córion, micrópila, aerópila
Anastrepha Schiner (Diptera: Tephritidae) é um dos gêneros mais diversificados de
Tephritidae endêmicos da Região Neotropical e atualmente possui cerca de 240 espécies
descritas (Norrbom et al. 1999, Norrbom e Korytkoswski 2009). Pelo menos 50% dessas
espécies são registradas no Brasil, onde atualmente há o registro de 103 espécies de
Anastrepha descritas e várias espécies recentemente registradas mas ainda não descritas
(Zucchi 2008).
Um estudo recente baseado em caracteres morfológicos e no uso de plantas
hospedeiras propôs uma classificação filogenética com 17 grupos de espécies de Anastrepha
1Coordenação de Pesquisas em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970, Manaus, AM, Brasil. 2Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL
32614-7100. 3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil (e-mail: [email protected]).

40
(Norrbom et al. 1999). Desses grupos, o segundo maior é o grupo fraterculus com 29 espécies
descritas e amplamente distribuídas, das quais, 17 espécies ocorrem no Brasil (Norrbom et al.
1999; Zucchi 2000, 2007). O grupo fraterculus inclui algumas espécies polífagas de grande
importância econômica (Aluja 1994, Norrbom et al. 1999, Zucchi 2007).
Há dificuldades na identificação de espécies relacionadas do grupo fraterculus devido
à sobreposição de caracteres e variação no acúleo, sendo este o mais importante caracter
diagnóstico, o qual é influenciado por fatores genéticos e ambientais (Aluja 1994, Smith-
Caldas et al. 2001, Araujo e Zucchi 2006). Faz-se necessário explorar caracteres morfológicos
adicionais, tais como aqueles presentes em ovos e larvas, além de caracteres moleculares para
melhor diferenciar as espécies de Anastrepha e aumentar o entendimento sobre as relações
filogenéticas neste gênero (Norrbom et al. 1999, Smith-Caldas et al. 2001). Até o momento,
há descrição da morfologia dos ovos para apenas 25 espécies de Anastrepha e muitos desses
estudos apresentaram apenas aspectos simples da morfologia dos ovos, como comprimento e
largura visualizados em microscópio ótico (Emmart 1933, Seín 1933, Lawrence 1979, Steck e
Malavasi 1988, Steck e Wharton 1988, Carroll e Wharton 1989, Norrbom et al. 1999).
Existem estudos sobre ovos para cinco espécies do grupo fraterculus, contudo estudos
utilizando imagens em microscopia eletrônica de varredura (MEV) foram realizados apenas
para o complexo Anastrepha fraterculus (Wiedemann) (Murillo e Jirón 1994; Selivon e
Perondini 1998, 2000; Perondini e Selivon 1999; Selivon et al. 2003), Anastrepha ludens
(Loew) (Carroll e Wharton 1989, Norrbom et al. 1999), Anastrepha obliqua (Macquart)
(Murillo e Jirón 1994), Anastrepha serpentina (Wiedemann) e Anastrepha sororcula Zucchi
(Selivon e Perondini 1999).
O objetivo desse estudo foi contribuir para uma melhor identificação dos ovos de
espécies de Anastrepha e para a taxonomia do gênero através da descrição de caracteres
adicionais em MEV para algumas espécies do grupo fraterculus: Anastrepha antunesi Lima,
Anastrepha bahiensis Lima, Anastrepha coronilli Carrejo & González, Anastrepha distincta
Greene, Anastrepha turpiniae Stone e Anastrepha zenildae Zucchi.
Material e Métodos
Os ovos foram dissecados de fêmeas capturadas em armadilhas McPhail em Cariri
(11°53’27’’S; 49°09’40’’W) no estado de Tocantins; Itacoatiara (03°08’35’’S; 58°26’39’’W),
Manaus (03°06’07’’S; 60°01’30’’W) e Presidente Figueiredo (02°02’04’’S; 60°01’30’’W) no
estado do Amazonas e Porto Velho (08°45’43’’S; 63°54’14’’W) no estado de Rondônia, na

41
Amazônia brasileira. Os ovos foram preservados em microtubos plásticos de 1,5 ml contendo
etanol 80%.
As medidas de comprimento e largura dos ovos foram realizadas com micrométrica
ocular em um estereomicroscópio Wild M3C no Laboratório de Entomologia Agrícola do
Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Brasil.
Na preparação para MEV, os ovos foram transferidos para cápsulas de silicone,
desidratados em série alcoólica e no secador de ponto crítico de CO2 por três horas. Logo após
a desidratação, as amostras foram metalizadas com uma camada de ouro. Os ovos foram
examinados no microscópio eletrônico de varredura LEO 1450VP no Laboratório
Institucional de Microscopia Eletrônica de Varredura do Museu Paraense Emílio Goeldi,
Belém, Pará, Brasil e no estereomicroscópio óptico (Leica M165C) do Laboratório de
Prospecção de Bioativos de Insetos (INPA), Manaus, Amazonas, Brasil. Entre vinte e trinta
ovos por fêmea de pelo menos três fêmeas de cada espécie estudada foram examinados com
microscopia óptica. Para a visualização com MEV foram utilizados pelo menos dez ovos de
cada espécie. O pólo anterior foi definido como a extremidade do ovo que possui um pedicelo
ou uma suave projeção com micrópila e aerópilas. O pólo posterior foi definido como a
extremidade oposta ao pedicelo o qual geralmente é liso e arredondado, não possuindo
aberturas externas ou estruturas. A cabeça do embrião desenvolve-se na direção do pólo
anterior do ovo (Headrick e Goeden 1998). De acordo com a posição do embrião no interior
do ovo, o lado convexo foi considerado como o lado ventral e o lado côncavo como o lado
dorsal.
Espécimes testemunho das fêmeas e ovos foram depositados na Coleção de
Invertebrados, INPA.
Resultados
Os ovos de A. antunesi, A. bahiensis, A. coronilli, A. distincta, A. turpiniae e A.
zenildae são similares em seu aspecto geral. São alongados, afilados em ambas as
extremidades, possuem papila e micrópila localizada no pólo anterior. Caracteres diagnósticos
para diferenciar as seis espécies incluem a ornamentação do córion, localização das aerópilas
e ao redor da micrópila uma pronunciada borda no córion com a aparência lanosa. Nenhum
dos ovos das espécies estudadas apresentou apêndice respiratório.

42
Anastrepha antunesi (Figs. 1A e 2)
Material Examinado. Foram examinados 25 ovos de quatro fêmeas. As fêmeas de A.
antunesi foram coletadas em Manaus no estado do Amazonas.
Características Gerais. Os ovos apresentaram comprimento variando de 1,16-1,57
mm e largura variando de 0,18-0,22 mm. Os ovos são branco amarelados, levemente
curvados, alongados, com maior diâmetro na metade anterior do ovo e afilando-se
gradualmente em direção a ambas as extremidades (Fig. 1A).
Escultura do córion. O pólo anterior apresenta uma papila e moderada ornamentação
do córion com reticulação em arranjo poligonal. A reticulação poligonal é claramente definida
somente no lado ventral e dentro dos polígonos a superfície apresenta aparência áspera que foi
formada pela reticulação do córion. Aproximadamente 15 aerópilas estão localizadas nos
vértices dos polígonos somente no lado ventral do ovo (Fig. 2, seta b). As aerópilas
apresentam aberturas de diversos diâmetros. No lado dorsal do ovo, há uma sutil
ornamentação poligonal do córion a qual desaparece em direção ao pólo posterior, onde o
córion torna-se liso e sem reticulações. A micrópila (Fig. 2, seta a) está localizada no lado
dorsal do ovo na extremidade do pólo anterior e rodeada por uma pronunciada borda do
córion. Esta borda tem o formato de anel com um aspecto lanoso (Fig. 2). As principais
características dos ovos para diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha bahiensis (Figs. 1B e 3)
Material Examinado. Foram examinados 20 ovos de três fêmeas. Uma fêmea de A.
bahiensis foi coletada em Manaus no estado do Amazonas e as outras duas fêmeas foram
coletadas em Porto Velho no estado de Rondônia.
Características Gerais. Os ovos apresentaram comprimento variando de 1,27-1,53
mm e largura variando de 0,18-0,26 mm. Os ovos são branco amarelados, fortemente
curvados, alongados, com maior diâmetro no terço anterior do ovo e afilando-se abruptamente
na direção do pólo posterior (Fig. 1B).
Escultura do córion. O pólo anterior apresenta uma papila, uma limitada
ornamentação do córion com reticulação em arranjo poligonal (Fig. 3A) e dentro dos
polígonos a superfície possui aparência áspera (Fig. 3B). Aproximadamente 20 aerópilas estão
localizadas nos vértices dos polígonos somente no lado ventral no pólo anterior do ovo. As
aerópilas apresentam aberturas de diversos diâmetros. A reticulação é mais desenvolvida no
pólo anterior tornando-se pouco distinta em direção a porção posterior do ovo até tornar-se
completamente lisa no final do pólo posterior. A micrópila está localizada no ápice do pólo

43
anterior no lado dorsal e rodeada por uma pronunciada borda do córion. Esta borda tem o
formato de anel com um aspecto macio (Fig. 3A). As principais características dos ovos para
diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha coronilli (Figs. 1C e 4)
Material Examinado. Foram examinados 31 ovos de três fêmeas. As fêmeas de A.
coronilli foram coletadas em Manaus no estado do Amazonas.
Características Gerais. Os ovos apresentaram comprimento variando de 1,16-1,80
mm e largura variando de 0,22-0,33 mm. São creme amarelados, levemente curvados,
alongados, com maior diâmetro no terço anterior do ovo e apresentando o pólo posterior
levemente mais afilado que o pólo anterior (Fig. 1C).
Escultura do córion. O pólo anterior apresenta uma papila e cerca de 30 aerópilas
localizadas apenas no lado ventral (Fig. 4, seta b). As aerópilas apresentam aberturas de
diversos diâmetros. Toda a superfície do ovo é lisa sem nenhuma ornamentação do córion. A
micrópila está localizada no ápice do pólo anterior no lado dorsal e rodeada por uma
pronunciada borda do córion. Esta borda tem o formato de anel com uma aparência lanosa
(Fig. 4, seta a). As principais características dos ovos para diagnóstico desta espécie estão
apresentadas na Tabela 1.
Anastrepha distincta (Figs. 1D e 5)
Material Examinado. Foram examinados 37 ovos de quatro fêmeas. As fêmeas de A.
distincta foram coletadas em Itacoatiara e Presidente Figueiredo no estado do Amazonas.
Características Gerais. Os ovos apresentaram comprimento variando de 1,42-1,65
mm e largura variando de 0,22-0,26 mm. São creme amarelados, alongados, fortemente
curvados, com maior diâmetro no terço anterior do ovo e com o pólo posterior afilando-se
abruptamente (Fig. 1D).
Escultura do córion. O pólo anterior apresenta uma papila e cerca de 45 aerópilas em
toda sua extensão (Fig. 5, seta c). As aerópilas com maiores diâmetros estão concentradas no
lado ventral e aquelas com menores diâmetros estão concentradas no lado dorsal. Os ovos de
A. distincta apresentam reticulação poligonal e protuberâncias levemente desenvolvidas (Fig.
5, seta b). As reticulações possuem o formato de quadriláteros e pentágonos irregulares
próximos a micrópila, tornando-se pouco distintas e alongadas no pólo posterior do ovo. As
protuberâncias são facilmente visualizadas à medida que as reticulações tornam-se mais
alongadas. A micrópila está localizada no ápice do pólo anterior no lado dorsal e rodeada por

44
uma pronunciada borda do córion (Fig. 5, seta a). Esta borda tem o formato de anel com uma
aparência lisa e mais estreita que o anel observado nos ovos de A. bahiensis (Fig. 3A). As
principais características dos ovos para diagnóstico desta espécie estão apresentadas na Tabela
1.
Anastrepha turpiniae (Figs. 1E e 6)
Material Examinado. Foram examinados 20 ovos de duas fêmeas. Essas fêmeas
foram coletadas em Manaus no estado do Amazonas e em Cariri no estado de Tocantins.
Características Gerais. Os ovos apresentaram comprimento variando de 1,12-1,31
mm e largura variando de 0,22-0,26 mm. São creme amarelados, levemente curvados,
alongados, com maior diâmetro no terço anterior do ovo e afilando-se progressivamente em
direção a ambas as extremidades. Os ovos de A. turpiniae apresentam medidas menores de
comprimento e largura, assim parecem mais robustos que os ovos das outras espécies
descritas aqui (Fig. 1E).
Escultura do córion. O pólo anterior apresenta uma papila e cerca de 35 aerópilas
com aberturas de diâmetros diferentes. As aerópilas estão localizadas nos vértices dos
polígonos em toda extensão do pólo anterior (Fig. 6A, seta a). A ornamentação do córion é
bem desenvolvida no pólo anterior com reticulações conspícuas e distintas protuberâncias
dentro dos polígonos do córion (Fig. 6B). As reticulações consistem de quadriláteros
irregulares fortemente desenvolvidos próximos a micrópila, os quais tornam-se pouco
distintos e mais alongados em direção ao pólo posterior do ovo. A ornamentação do córion
desaparece em direção ao pólo posterior do ovo, tornando-se liso. A micrópila está localizada
no lado dorsal do ovo levemente deslocada do ápice do pólo anterior e rodeada por uma
pronunciada borda do córion (Fig. 6, seta b). Esta borda tem o formato de anel com uma
aparência lanosa similar a visualizada em A. antunesi (Fig. 2, seta a) e A. coronilli (Fig. 4, seta
a). As principais características dos ovos para diagnóstico desta espécie estão apresentadas na
Tabela 1.
Anastrepha zenildae (Figs. 1F e 7)
Material Examinado. Foram examinados 37 ovos de três fêmeas. As fêmeas de A.
zenildae foram coletadas em Cariri no estado de Tocantins.
Características Gerais. Os ovos apresentaram comprimento variando de 1,23-1,42
mm e largura variando de 0,22-0,26 mm. São creme amarelados, levemente curvados,
alongados, com maior diâmetro no terço anterior do ovo e afilando-se progressivamente em

45
direção a ambas as extremidades (Fig. 1F). Os ovos de A. zenildae são robustos como os de A.
turpiniae.
Escultura do córion. A papila está localizada no pólo anterior do ovo. A reticulação
do córion é bem desenvolvida, apresentando um número variável de protuberâncias limitadas
pelos vértices dos polígonos (Fig. 7B). Aproximadamente 40 aerópilas estão localizadas nos
vértices dos polígonos cobrindo toda extensão do pólo anterior (Fig. 7A, seta a). As aerópilas
apresentam aberturas de diversos diâmetros. As reticulações consistem em quadriláteros
irregulares fortemente desenvolvidos próximos a micrópila, as quais se tornam pouco distintas
e mais alongadas em direção ao pólo posterior do ovo. A micrópila (Fig. 7, seta b) está
localizada no lado dorsal do ovo no ápice do pólo anterior e rodeada por uma pronunciada
borda do córion com o formato de anel, apresentando uma aparência lanosa semelhante à
observada em A. antunesi (Fig. 2, seta a), A. coronilli (Fig. 4, seta a) e A. turpiniae (Fig. 6A,
seta b). As principais características dos ovos para diagnóstico desta espécie estão
apresentadas na Tabela 1.
Discussão
Neste estudo, a membrana do ovo das seis espécies de Anastrepha examinadas
correspondeu ao aspecto geral da morfologia dos ovos de Tephritidae como cor, formato e
presença ou ausência de ornamentação do córion (White e Elson-Harris 1992). Embora tendo
semelhança com outras espécies do grupo fraterculus a ornamentação do córion apresenta
características específicas para as seis espécies examinadas aqui, como descrito previamente
por Emmart (1933) e posteriormente usando MEV por Carroll e Wharton (1989), Murillo e
Jirón (1994) e Selivon e Perondini (1998).
A ornamentação do córion das seis espécies pode ser agrupada em duas amplas
categorias de ovos de Diptera Cyclorrhapha como indicado por Ferrar (1987). Anastrepha
coronilli e A. distincta pertencem à categoria de córion liso ou com pouca reticulação - a
reticulação é ausente em A. coronilli e pouco distinta em A. distincta. A ausência de
ornamentação no córion foi observada nos ovos de Anastrepha grandis (Macquart) e A.
serpentina (Emmart 1933, Steck e Wharton 1988, Selivon e Perondini 1999), e reticulação
pouco distinta observada nos ovos de A. ludens (Emmart 1933, Carroll e Wharton 1989).
Anastrepha turpiniae e A. zenildae pertencem à categoria que possuem ornamentação no
córion e ambas as espécies apresentam pronunciada reticulação. Reticulação bem
desenvolvida no córion foi observada em outras espécies de Anastrepha, Anastrepha sp. 1 aff.
fraterculus, Anastrepha sp. 2 aff. fraterculus (Selivon e Perondini 1998), A. obliqua (Murillo

46
e Jirón 1994) e A. sororcula (Selivon e Perondini 1999). A ornamentação do córion de A.
antunesi e A. bahiensis é bem específica nessas duas espécies, porque a reticulação é mais
desenvolvida no lado dorsal do pólo anterior do ovo. Assim, os resultados obtidos indicam
que essas duas espécies podem representar uma terceira categoria “reticular”, mostrando o
córion liso no lado ventral e ornamentado no lado dorsal. De acordo com Emmart (1933), a
escultura na extremidade da micrópila pode ser devido a uma deposição extra de quitina, pois
a membrana é mais espessa nessa região que no restante do ovo. Contudo, estudos recentes
indicam que a ornamentação do córion pode estar relacionada com atividade diferenciada do
epitélio folicular para adaptação no habitat (Cônsoli et al. 1999). Acredita-se que a reticulação
junto com as aerópilas desempenhem papel importante na respiração do embrião (Selivon e
Perondini 1998) e também forneçam proteção contra a dessecação (Cônsoli et al. 1999).
O pólo posterior apresenta semelhanças no padrão e estrutura para as seis espécies
estudadas e algumas diferenças específicas quanto a posição, número e estrutura das
aerópilas. As aerópilas são encontradas somente no lado ventral em A. antunesi, A. bahiensis
e A. coronilli, enquanto que em A. turpiniae, A. distincta e A. zenildae são observadas em
ambos os lados e em grande número. A variação do diâmetro das aerópilas foi verificada nos
ovos de A. bahiensis, A. coronilli e A. turpiniae. Vale ressaltar que em A. distincta as
aerópilas com diâmetros maiores estão localizadas no lado ventral, enquanto que com
diâmetros menores estão localizadas no lado dorsal. Além disso, a ausência de apêndice
respiratório foi observada nas seis espécies estudadas e também na maioria das espécies já
descritas. De acordo com a literatura, o apêndice respiratório foi descrito somente para
Anastrepha barbiellinii Lima, Anastrepha manihoti Lima, A. obliqua, Anastrepha nigrifascia
Stone e Anastrepha pittieri Caraballo (Murillo e Jirón 1994, Norrbom et al. 1999, Norrbom e
Korytkowski 2009). Acredita-se que a estrutura do córion e o número de aerópilas nos ovos
dos insetos são adaptações relacionadas ao ambiente onde eles são ovipositados. O número de
aerópilas também está relacionado com a taxa metabólica das espécies, a necessidade da troca
de gases e o controle da perda de água (Cônsoli et al. 1999).
Os ovos das seis espécies analisadas possuem uma distinta papila no pólo anterior,
contudo em A. coronilli, A. distincta, A. turpiniae e A. zenildae ela apresenta-se mais
pronunciada, semelhante ao registrado para A. grandis (Steck e Wharton 1988), A. ludens
(Carroll e Wharton 1989), Anastrepha sp. 1 aff. fraterculus (Selivon e Perondini 1998, 2000)
e A. sororcula (Selivon e Perondini 1999). De acordo com Selivon e Perondini (1998), os
ovos de Anastrepha que possuem pequena papila têm uma “morfologia intermediária” entre
os ovos com apêndices respiratórios e os ovos que não apresentam qualquer tipo de apêndice.

47
A micrópila de cinco espécies das seis estudadas está localizada no ápice do pólo
anterior do ovo como foi descrito para Anastrepha sp. 2 aff. fraterculus (Selivon e Perondini
1998, 2000; Selivon et al. 2003), A. serpentina e A. sororcula (Selivon e Perondini 1999).
Porém, para A. turpiniae (Fig. 5B) a micrópila está levemente deslocada do ápice como
descrito para Anastrepha sp. 1 aff. fraterculus (Murillo e Jirón 1994, Selivon e Perondini
1998, 2000). Em A. obliqua e A. pittieri, a micrópila está deslocada devido à presença do
apêndice respiratório (Murillo e Jirón 1994, Norrbom e Korytkowski 2009).
Os ovos analisados neste trabalho foram obtidos de fêmeas preservadas em etanol 80%
e suas características morfológicas permaneceram inalteradas. Os resultados obtidos
corroboram com os registrados por Selivon et al. (2003). Esses autores verificaram que os
ovos dissecados de fêmeas preservadas em etanol mantiveram as mesmas características da
ornamentação do córion e das estruturas internas que ovos recém ovipositados.
Em resumo, esse estudo com MEV para seis espécies de Anastrepha revelou
semelhanças com outras espécies do grupo fraterculus, cujos ovos foram descritos
previamente, bem como algumas diferenças não relatadas anteriormente. A combinação de
características exibidas por esses ovos, como a ornamentação do córion, localização das
aerópilas e da micrópila são úteis como caracteres taxonômicos. Os resultados apresentados
aqui ampliam o conhecimento sobre a morfologia da membrana do ovo para espécies de
Anastrepha do grupo fraterculus. Trabalhos semelhantes com outras espécies de Anastrepha
auxiliarão o entendimento sobre as relações filogenéticas dentro do grupo fraterculus e do
gênero como um todo.
Agradecimentos
Os autores agradecem a Hilton Tulio Costi e Rolf Junior Ferreira Silva (Museu
Paraense Emílio Goeldi) pelo auxílio na preparação do material para MEV, Carter R. Miller e
dois anônimos revisores por suas sugestões numa versão anterior desse manuscrito. Esse
projeto foi financiado pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq - projeto n° 575664/2008-8) e pela Coordenação de Aperfeiçoamento de Pessoal de
Nível Superior (CAPES - projeto n° 23038.037185/2008-41) Os autores também agradecem
ao CNPq pela bolsa de doutorado concedida a VSD.

48
Fig. 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. antunesi. (B) A.
bahiensis. (C) A. coronilli. (D) A. distincta. (E) A. turpiniae. (F) A. zenildae.

49
Fig. 2. Vista com Microscopia Eletrônica de Varredura do ovo de A. antunesi. Pólo anterior
mostrando a micrópila (seta a), ornamentação do córion e aerópila (seta b).
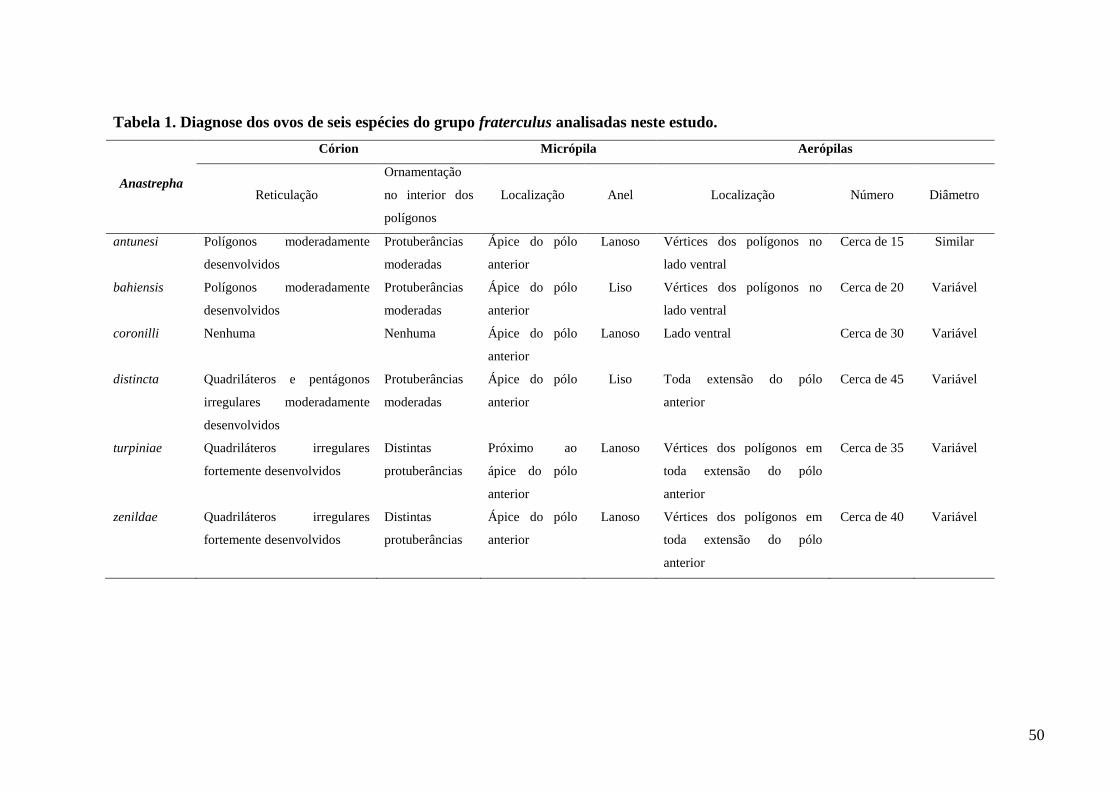
50
Tabela 1. Diagnose dos ovos de seis espécies do grupo fraterculus analisadas neste estudo.
Anastrepha
Córion Micrópila Aerópilas
Reticulação
Ornamentação
no interior dos
polígonos
Localização Anel Localização Número Diâmetro
antunesi Polígonos moderadamente
desenvolvidos
Protuberâncias
moderadas
Ápice do pólo
anterior
Lanoso Vértices dos polígonos no
lado ventral
Cerca de 15 Similar
bahiensis Polígonos moderadamente
desenvolvidos
Protuberâncias
moderadas
Ápice do pólo
anterior
Liso Vértices dos polígonos no
lado ventral
Cerca de 20 Variável
coronilli Nenhuma Nenhuma Ápice do pólo
anterior
Lanoso Lado ventral Cerca de 30 Variável
distincta Quadriláteros e pentágonos
irregulares moderadamente
desenvolvidos
Protuberâncias
moderadas
Ápice do pólo
anterior
Liso Toda extensão do pólo
anterior
Cerca de 45 Variável
turpiniae Quadriláteros irregulares
fortemente desenvolvidos
Distintas
protuberâncias
Próximo ao
ápice do pólo
anterior
Lanoso Vértices dos polígonos em
toda extensão do pólo
anterior
Cerca de 35 Variável
zenildae Quadriláteros irregulares
fortemente desenvolvidos
Distintas
protuberâncias
Ápice do pólo
anterior
Lanoso Vértices dos polígonos em
toda extensão do pólo
anterior
Cerca de 40 Variável

51
Fig. 3. Vista com Microscopia Eletrônica de Varredura dos ovos de A. bahiensis. (A) Pólo
anterior mostrando a micrópila (seta) e reticulação do córion. (B) Ampliação da área do pólo
anterior mostrando polígonos e protuberâncias do córion (seta).
Fig. 4. Vista com Microscopia Eletrônica de Varredura do ovo de A. coronilli. Pólo anterior
mostrando a micrópila (seta a) e aerópilas (seta b).

52
Fig. 5. Vista com Microscopia Eletrônica de Varredura do ovo de A. distincta. Pólo anterior
mostrando a micrópila (seta a), protuberâncias do córion (seta b) e aerópilas (seta c).

53
Fig. 6. Vista com Microscopia Eletrônica de Varredura dos ovos de A. turpiniae. (A) Pólo
anterior mostrando aerópilas (seta a) e micrópila (seta b). (B) Ampliação da área do pólo anterior
mostrando polígonos e protuberâncias do córion (seta).
Fig. 7. Vista com Microscopia Eletrônica de Varredura dos ovos de A. zenildae. (A) Pólo anterior
mostrando aerópilas (seta a) e micrópila (seta b). (B) Ampliação da área do pólo anterior
mostrando polígonos e protuberâncias do córion (seta).

54
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G. 2011.
Description of eggs of Anastrepha spp. (Diptera:
Tephritidae) in the spatulata group using Scanning
Electron Microscopy. Annals of the Entomological Society
of America, 104(5): 857-862 (Anexo 4).

55
Description of Eggs of Anastrepha spp. (Diptera: Tephritidae) in the spatulata
Group Using Scanning Electron Microscopy
VIVIAN SIQUEIRA DUTRA1, BEATRIZ RONCHI-TELES
1, GARY J. STECK
2 E JANISETE
GOMES SILVA3
Ann. Entomol. Soc. Am. 104(5): 857-862 (2011); DOI: 10.1603/AN11059
RESUMO Pela primeira vez, com microscopia eletrônica de varredura foi descrita
detalhadamente a morfologia dos ovos de três espécies de Anastrepha do grupo spatulata:
Anastrepha manihoti Lima, Anastrepha montei Lima e Anastrepha pickeli Lima. Os ovos foram
dissecados de fêmeas capturadas em armadilhas McPhail e examinados usando microscópio
eletrônico e óptico. Os ovos são afilados em ambas as extremidades, possuem uma papila no pólo
anterior e a micrópila está localizada no lado dorsal do pólo anterior. Caracteres diagnósticos para
diferenciar as três espécies incluem localização das aerópilas, presença de apêndice respiratório
em A. manihoti e A. pickeli, e uma pronunciada borda do córion com aparência lisa ao redor da
micrópila. Os ovos de A. montei possuem uma estreita camada de superfície porosa no lado
ventral do pólo anterior. Tal caractere não havia sido relatado anteriormente para ovos de
Tephritidae.
PALAVRAS-CHAVE moscas-das-frutas, micrópila, aerópila, apêndice respiratório
Anastrepha Schiner (Diptera: Tephiritidae) é um dos maiores e economicamente importantes
gêneros de moscas-das-frutas Neotropicais. Ele é endêmico das Américas, possuindo cerca de
240 espécies descritas, amplamente distribuídas pelas Américas do Sul e Central, Antilhas e sul
dos Estados Unidos (White e Elson-Harris 1992; Norrbom et al. 1999; Norrbom e Korytkowski
2009, 2011). No Brasil, tem-se o registro de 109 espécies de Anastrepha distribuídas em 12
grupos infragenéricos (Zucchi 2007, 2008; Uramoto e Zucchi 2010).
O grupo spatulata está distribuído desde o Texas até a Argentina, compreendendo dez
espécies. A maioria das espécies deste grupo infesta hospedeiros da família Euphorbiaceae ou
Olacaceae. No Brasil, tem-se o registro de ocorrência de nove espécies desse grupo (Zucchi
2007). Anastrepha manihoti Lima, Anastrepha montei Lima e Anastrepha pickeli Lima são
1Coordenação de Pesquisas em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970, Manaus, AM, Brasil. 2Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL 32614-
7100. 3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil (e-mail: [email protected]).

56
consideradas pragas de mandioca (Norrbom 2004). Anastrepha pickeli e A. montei infestam
frutos e principalmente sementes de mandioca, enquanto que A. manihoti infesta gemas terminais
e ramos jovens (Jirón e Hedström 1988, Bellotti 2008). A infestação de frutos de mandioca não
causa perda na produção, mas representa um sério problema para os produtores (Bellotti 2008).
A morfologia dos ovos do gênero Anastrepha é relativamente pouco conhecida e foram
descritas características dos ovos para apenas 34 espécies do referido gênero, até o momento. A
maioria dos estudos sobre a morfologia dos ovos utilizou a microscopia óptica para descrever
aspectos gerais como comprimento e largura (Emmart 1933, Seín 1933, Lawrence 1979, Steck e
Malavasi 1988, Steck e Wharton 1988, Norrbom et al. 1999, Norrbom e Korytkowski 2009).
Poucos estudos usaram a microscopia eletrônica de varredura (MEV) para auxiliar na descrição
dos ovos de Anastrepha (Carroll e Wharton 1989; Murillo e Jirón 1994; Selivon e Perondini
1998, 1999; Norrbom e Korytkowski 2009; Dutra et al. 2011a). A limitada literatura disponível
sugere que caracteres morfológicos, tais como os presentes em ovos e larvas, podem auxiliar no
entendimento sobre as relações filogenéticas entre as espécies de Anastrepha (Norrbom et al.
1999).
No grupo spatulata, somente descrições mínimas (dimensões e aspectos gerais) estão
disponíveis dos ovos de Anastrepha alveatoides Blanchard, Anastrepha haywardi Blanchard, A.
manihoti e A. pickeli (Norrbom et al. 1999).
Neste estudo, informações detalhadas utilizando MEV foram descritas para três espécies
do grupo spatulata: A. manihoti, A. montei e A. pickeli. Os resultados obtidos podem ser úteis na
identificação de ovos de espécies de Anastrepha e para melhor entender a taxonomia do gênero.
Material e Métodos
Os ovos examinados foram dissecados de fêmeas capturadas em armadilhas McPhail em
Manaus (03°06’07’’S, 60°01’30’’W) no estado do Amazonas; em Una (15o17’36’’S,
39o04’31’’W) no estado da Bahia e em Linhares (19°23’28’’S, 40°04’20’’W) no estado do
Espírito Santo. Os ovos foram preservados em microtubos plásticos de 1,5 ml contendo etanol
80%.
As medidas de comprimento e largura dos ovos foram realizadas com micrométrica ocular
em um estereomicroscópio Wild M3C no Laboratório de Entomologia Agrícola do Instituto
Nacional de Pesquisas da Amazônia (INPA), Manaus, Brasil.

57
Para análise com MEV, os ovos foram transferidos para cápsulas de silicone e
desidratados em série alcoólica. Em seguida, os ovos foram colocados no secador de ponto crítico
de CO2 por três horas, logo após as amostras foram metalizadas com uma camada de ouro. Os
ovos foram examinados no microscópio eletrônico de varredura LEO 1450VP no Laboratório
Institucional de Microscopia Eletrônica de Varredura do Museu Paraense Emílio Goeldi, Belém,
Pará, Brasil e no estereomicroscópio óptico (Leica M165C) do Laboratório de Prospecção de
Bioativos de Insetos do Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus,
Amazonas, Brasil. Entre dez e trinta e seis ovos por fêmea de cada espécie foram examinados
com microscopia óptica. Com MEV foram utilizados pelo menos dez ovos para visualização do
córion. O pólo anterior foi definido como a extremidade do ovo que possui um pedicelo ou uma
suave projeção com micrópila e aerópilas. O pólo posterior foi definido como a extremidade
oposta ao pedicelo o qual geralmente é liso e arredondado, não possuindo aberturas externas ou
estruturas. A cabeça do embrião desenvolve-se na direção do pólo anterior do ovo (Headrick e
Goeden 1998). Baseado na posição do embrião no interior do ovo, o lado convexo foi
considerado como o lado ventral e o lado côncavo como o lado dorsal. O termo de apêndice
respiratório foi utilizado para a estrutura responsável pela respiração dos ovos como relatado por
Murillo e Jirón (1994).
Espécimes testemunho das fêmeas e ovos foram depositados na Coleção de Invertebrados,
INPA.
Descrições
Anastrepha manihoti Lima (Figs. 1A, 2 e 5A)
Material Examinado. Foram examinados 10 ovos de uma fêmea. Essa fêmea de A.
manihoti foi coletada em Linhares no estado do Espírito Santo.
Características Gerais. Os ovos apresentaram comprimento variando de 1,76-2,02 mm,
largura variando de 0,26-0,30 mm e comprimento do apêndice respiratório variando de 0,56-0,75
mm. Os ovos são creme amarelados, levemente côncavos dorsalmente, fortemente convexos
ventralmente, alongados, com maior diâmetro na metade anterior do ovo e afilando-se
gradualmente em direção ao pólo posterior (Fig. 1A).
Escultura do córion. O pólo anterior apresenta uma pronunciada papila onde se localiza
a micrópila. Toda a superfície do ovo é lisa com reticulação hexagonal extremamente fraca.

58
Aproximadamente dez aerópilas circundam o ápice do apêndice respiratório (Fig. 2B). Foram
observadas aerópilas com vários diâmetros, variando entre 1,2 e 2,4 µm. A micrópila (Fig. 2A)
está localizada na base do apêndice respiratório no lado dorsal do pólo anterior do ovo e rodeada
por uma pronunciada borda do córion. Esta borda tem o formato de anel com um aspecto liso e
margens irregulares (Fig. 5A). As principais características dos ovos para diagnóstico desta
espécie estão apresentadas na Tabela 1.
Anastrepha montei Lima (Figs. 1C, 3 e 5C)
Material Examinado. Foram examinados 36 ovos de quatro fêmeas. As fêmeas de A.
montei foram coletadas em Una no estado da Bahia.
Características Gerais. Os ovos apresentaram comprimento variando de 1,61-2,73 mm e
largura variando de 0,11-0,15 mm. Os ovos são creme amarelados, fortemente curvados,
alongados, delgados, com maior diâmetro na metade anterior do ovo e afilando-se gradualmente
em direção ao pólo posterior (Fig. 1C).
Escultura do córion. O pólo anterior apresenta uma pronunciada papila onde se localiza
a micrópila. Toda a superfície do ovo é lisa sem nenhuma ornamentação, reticulação ou
protuberâncias. Para essa espécie não foi observado apêndice respiratório e nenhuma aerópila.
Porém, foi observada uma estreita camada de superfície porosa no lado ventral do pólo anterior
próximo a micrópila (Fig. 3, seta a). A micrópila (Fig. 3, seta b) está localizada no ápice do pólo
anterior no lado dorsal e rodeada por uma pronunciada borda do córion. Esta borda tem o formato
de anel contínuo com um aspecto liso (Fig. 5C). As principais características dos ovos para
diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha pickeli Lima (Figs. 1B, 4 e 5B)
Material Examinado. Foram examinados 30 ovos de quatro fêmeas. Duas fêmeas de A.
pickeli foram coletadas em Manaus no estado do Amazonas e duas fêmeas coletadas em Linhares
no estado do Espírito Santo.
Características Gerais. Os ovos apresentaram comprimento variando de 1,72-2,17 mm,
largura variando de 0,22-0,30 mm e comprimento do apêndice respiratório variando de 0,67-1,05
mm. Os ovos são creme amarelados, levemente côncavos dorsalmente, fortemente convexos

59
ventralmente, alongados, com maior diâmetro na metade anterior do ovo e afilando-se
gradualmente em direção ao pólo posterior (Fig. 1B).
Escultura do córion. O pólo anterior apresenta uma pronunciada papila onde se localiza
a micrópila. Toda a superfície do ovo é lisa sem nenhuma ornamentação, reticulação ou
protuberâncias. Aproximadamente 40 aerópilas estão localizadas aleatoriamente no terço apical
do apêndice respiratório (Fig. 4B). Foram observadas aerópilas com vários diâmetros, variando
entre 1,9 e 6,0 µm. A micrópila (Fig. 4A) está localizada na base do apêndice respiratório no lado
dorsal do pólo anterior do ovo e rodeada por uma pronunciada borda do córion. Esta borda tem o
formato de anel contínuo com um aspecto liso (Fig. 5A). As principais características dos ovos
para diagnóstico desta espécie estão apresentadas na Tabela 1.
Discussão
Os resultados registrados aqui estão de acordo com observações prévias sobre o padrão da
membrana de ovos de Tephritidae como a cor, formato e presença ou ausência de ornamentação
do córion (White e Elson-Harris 1992). Descrições realizadas anteriormente sobre ovos de outros
membros do grupo spatulata são breves e apenas descritivos (sem nenhuma ilustração),
descrevem o formato geral (“robusto, com longo apêndice” para A. manihoti e A. pickeli;
“robusto” para A. alveatoides; “afilado” para A. haywardi) e dimensões do ovo (comprimento,
largura e comprimento do apêndice respiratório) para A. manihoti (1,54-1,71; 0,25-0,29; e 0,62-
0,69 mm, respectivamente) e (comprimento e largura) para A. alveatoides e A. haywardi (1,35-
1,41 e 0,22 mm; 2,20-2,41 e 0,27-0,33 mm, respectivamente) (Norrbom et al. 1999). Os
resultados obtidos no presente estudo são consistentes com os apresentados acima.
Nesse estudo, utilizando MEV foram observadas diferenças qualitativas e quantitativas
que podem auxiliar a identificação das três espécies, baseada nas características do ovo.
Caracteres de diagnóstico incluem presença ou ausência, localização e número de aerópilas,
presença ou ausência e comprimento do apêndice respiratório.
As características mais notáveis dos ovos dessas três espécies de Anastrepha são presença
ou ausência de apêndice respiratório e presença, ausência ou variação no número de aerópilas. Os
ovos de A. manihoti e A. pickeli são mais largos e possuem apêndice respiratório longo e estreito
com aerópilas, enquanto que ovos de A. montei são delgados, afilados e não possuem apêndice
respiratório e aerópilas.

60
Baseado em estudos anteriores realizados com outras espécies de Anastrepha, foi sugerido
que o apêndice respiratório é utilizado para a troca de gases durante o desenvolvimento
embrionário (Murillo e Jirón 1994). De acordo com esses autores, o apêndice respiratório de
Anastrepha obliqua (Macquart) permanece na superfície dos frutos com as aerópilas para fora
(sua fig. 7). Murillo e Jirón (1994) sugerem que isso permita a não obstrução das trocas de ar
entre o ovo e a atmosfera antes da eclosão da larva. Entretanto, para A. manihoti e A. pickeli, a
localização do apêndice respiratório depois da oviposição ainda não foi registrada.
Entre as espécies de Anastrepha cuja morfologia dos ovos já foi descrita, o apêndice
respiratório foi registrado em vários grupos de espécies. No grupo fraterculus, o apêndice
respiratório está presente em Anastrepha barbiellinii Lima e A. obliqua, mas ausente em
Anastrepha amita Zucchi, Anastrepha antunesi Lima, Anastrepha bahiensis Lima, Anastrepha
coronilli Carrejo & González, Anastrepha distincta Greene, Anastrepha fraterculus
(Wiedemann), Anastrepha ludens (Loew), Anastrepha sororcula Zucchi, Anastrepha suspensa
(Loew), Anastrepha turpiniae Stone e Anastrepha zenildae Zucchi (Carroll e Wharton 1989;
Murillo e Jirón 1994, Norrbom et al. 1999, Selivon e Perondini 1999, Dutra et al. 2011a); no
grupo robusta, o apêndice respiratório está presente em Anastrepha nigrifascia Stone e
Anastrepha pittieri Caraballo (Murillo e Jirón 1994, Norrbom et al. 1999, Norrbom e
Korytkowski 2009), mas ausente em Anastrepha bella Norrbom & Korytkowski, Anastrepha
speciosa Stone (Norrbom e Korytkowski 2009) e Anastrepha cordata Aldrich (Norrbom et al.
1999, Norrbom e Korytkowski 2009); e no grupo serpentina, o apêndice respiratório está ausente
em Anastrepha serpentina (Wiedemann) (Selivon e Perondini 1999). No presente estudo, é
registrado um padrão similar para o grupo spatulata, com A. pickeli e A. manihoti possuindo
apêndice respiratório, enquanto que em A. montei é ausente. O apêndice respiratório tem sido
observado em diferentes grupos de espécies, tanto em um grande clado com muitos grupos de
espécies basais, como em clados bem maiores que incluem mais grupos de espécies derivadas,
como no cladograma apresentado por Norrbom et al. (1999). Seria interessante examinar ovos de
mais espécies basais e derivadas para mapear esses caracteres na filogenia de Anastrepha e
elucidar a evolução dessa estrutura dentro do gênero. É provável que o apêndice respiratório
tenha surgido de forma independente há muito tempo entre os vários clados de Anastrepha e de
outros Tephritidae, podendo ser uma resposta adaptativa para condições particulares do tecido
vegetal do hospedeiro onde os ovos são ovipositados.

61
A localização, número e diâmetro das aerópilas são distintos entre as três espécies.
Aerópilas foram encontradas somente no ápice do apêndice respiratório em A. manihoti e A.
pickeli, enquanto que ausente em A. montei. As aerópilas são mais largas e em maior número em
A. pickeli quanto comparada com A. manihoti. Um aspecto intrigante dos resultados obtidos foi à
ausência de apêndice respiratório em A. montei. Porém, ovos de A. montei possuem uma estreita
camada com superfície porosa no lado ventral do pólo anterior. Tal caractere não havia sido
registrado anteriormente para ovos de Tephritidae. Acredita-se que o número de aerópilas nos
ovos dos insetos seja uma adaptação relacionado com o ambiente onde eles os depositam, como
também relacionado com a taxa metabólica, necessidade de troca de gases e controle da perda de
água (Cônsoli et al. 1999), então é possível que essa aparência da superfície porosa nos ovos de
A. montei exerça a função de respiração para o ovo.
A micrópila das três espécies estudadas está localizada no ápice do pólo anterior do ovo
como previamente descrito para A. antunesi, Anastrepha bahiensis Lima, A. coronilli, A.
distincta, A. zenildae (Dutra et al. 2011a), Anastrepha sp. 2 aff. fraterculus (Selivon e Perondini
1998, 2000; Selivon et al. 2003), A. serpentina e A. sororcula (Selivon e Perondini 1999). A
micrópila está rodeada por uma pronunciada borda do córion com o formato de anel e com
aparência lisa para as três espécies. Uma importante diferença apresentada nos ovos de A.
manihoti é a aparência descontínua do anel ao redor da micrópila, a qual é útil para distinguir
essa espécie das outras estudadas aqui.
Vale salientar que as três espécies estudadas possuem córion liso sem qualquer
ornamentação distinta, reticulações ou protuberâncias. Isso poderia representar uma sinapomorfia
do grupo spatulata, podendo ser testada examinando ovos de outras espécies deste grupo. A
maioria das espécies de Anastrepha possui um padrão distinto de reticulação no córion, mas
deve-se notar que córion completamente liso também foi observado em ovos de espécies de
outros grupos de Anastrepha tais como A. coronilli [grupo fraterculus], Anastrepha grandis
(Macquart) [grupo grandis] e A. serpentina [grupo serpentina] (Steck e Wharton 1988, Selivon e
Perondini 1999, Dutra et al. 2011a).
Este estudo com MEV sobre três espécies de Anastrepha do grupo spatulata apresenta
algumas implicações importantes para questões mais amplas, como a taxonomia e filogenia
dentro do gênero. As características mostradas por esses ovos, como a presença ou ausência e
variação de apêndice respiratório e aerópilas, além da reticulação do córion são úteis como

62
caracteres taxonômicos. Além disso, esse estudo associado com o de Norrbom et al. (1999) e
Norrbom e Korytkowski (2009) mostram que informações adicionais sobre outras espécies de
Anastrepha, tanto espécies de grupos basais como derivados, auxiliarão no entendimento de
importantes aspectos das estruturas dos ovos em termos funcionais e filogenéticos.
Agradecimentos
Os autores agradecem a Keiko Uramoto, Marcos Vinicius Garcia e Tatiana Senra pelo
auxílio durante as coletas dos exemplares. Os autores também agradecem a Hilton Tulio Costi e
Rolf Junior Ferreira Silva (Museu Paraense Emílio Goeldi) pelo inestimável auxílio na
preparação do material para MEV e dois anônimos revisores por seus comentários neste
manuscrito. Esse estudo foi financiado pelo Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq), projeto n° 575664/2008-8 e pela Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES), projeto n° 23038.037185/2008-41. Os autores também
agradecem ao CNPq pela bolsa de doutorado concedida a VSD.
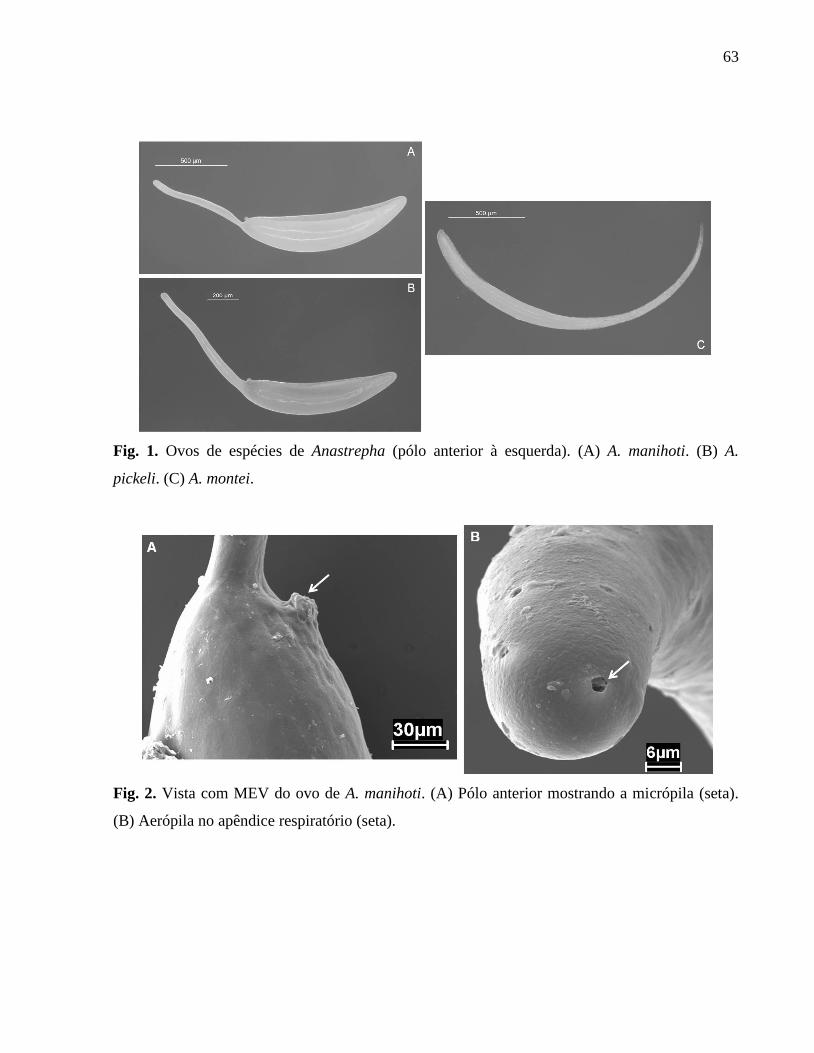
63
Fig. 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. manihoti. (B) A.
pickeli. (C) A. montei.
Fig. 2. Vista com MEV do ovo de A. manihoti. (A) Pólo anterior mostrando a micrópila (seta).
(B) Aerópila no apêndice respiratório (seta).

64
Tabela 1. Diagnose dos ovos de três espécies de Anastrepha do grupo spatulata analisadas neste estudo.
Anastrepha
Apêndice
respiratório,
comprimento
(mm)
Ornamentação
do córion
Micrópila Aerópila
Localização Anel Localização Número Diâmetro
manihoti Presente, 0,56-
0,75
Fraca reticulação Próximo a base do
apêndice respiratório
Liso com margens
irregulars
Apenas no ápice do
apêndice respiratório
Cerca de 10 Variável
montei Ausente Nenhuma Ápice do pólo anterior Liso e continuo Estreita camada de
superfície porosa
Nenhum Nenhum
pickeli Presente, 0,67-
1,05
Nenhuma Próximo a base do
apêndice respiratório
Liso e contínuo No terço apical do
apêndice respiratório
Cerca de 40 Variável

65
Fig. 3. Vista com MEV do ovo de A. montei. Pólo anterior mostrando a superfície porosa (seta a)
e a micrópila (seta b).
Fig. 4. Vista com MEV do ovo de A. pickeli. (A) Pólo anterior mostrando a micrópila (seta). (B)
Aerópilas no apêndice respiratório (seta).

66
Fig. 5. Vista com MEV da micrópila dos ovos de Anastrepha (pólo anterior à esquerda). (A) A.
manihoti. (B) A. pickeli. (C) A. montei.

67
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G.
Description of eggs of Anastrepha curitis and Anastrepha
leptozona (Diptera: Tephritidae) using SEM. Manuscrito
formatado para Annals of the Entomological Society of
America.

68
Description of Eggs of Anastrepha curitis and Anastrepha leptozona (Diptera:
Tephritidae) Using SEM
VIVIAN SIQUEIRA DUTRA1, BEATRIZ RONCHI-TELES
2, GARY J. STECK
3 E JANISETE
GOMES SILVA4
RESUMO Com o auxílio da microscopia eletrônica de varredura (MEV), neste trabalho foram
descritas as características dos ovos de Anastrepha curitis Stone e a morfologia dos ovos de
Anastrepha leptozona Hendel foi redescrita com mais detalhes. Os ovos foram dissecados de
fêmeas capturadas em armadilhas McPhail coletadas em Manaus e Presidente Figueiredo no
estado do Amazonas. Os ovos de A. curitis são muito alongados e não apresentam aerópilas, mas
uma camada de superfície porosa no lado ventral, também evidenciada em Anastrepha montei
Lima. Os ovos de A. leptozona são robustos e apresentam aerópilas apenas no lado ventral do
pólo anterior do ovo. Os ovos de ambas as espécies não possuem qualquer ornamentação no
córion, reticulação ou protuberância. Para ambas as espécies, a micrópila está localizada no ápice
do pólo anterior do ovo. Os ovos de A. curitis e A. leptozona não possuem apêndice respiratório.
As características da morfologia dos ovos são úteis para auxiliar um melhor entendimento sobre a
filogenia do gênero Anastrepha.
PALAVRAS-CHAVE moscas-das-frutas, ornamentação do córion, micrópila, aerópila
Anastrepha Schiner (Diptera: Tephiritidae) é um gênero endêmico da região Neotropical
apresentando mais de 240 espécies descritas. As espécies do referido gênero são amplamente
distribuídas pelas Américas do Sul e Central, Antilhas e sul dos Estados Unidos (White e Elson-
Harris 1992; Norrbom et al. 1999; Norrbom e Korytkowski 2009). No Brasil, tem-se o registro de
112 espécies distribuídas em 12 grupos infragenéricos de acordo com características
morfológicas e uso de hospedeiros (Norrbom et al. 1999, Zucchi 2008, Uramoto e Zucchi 2010).
O grupo leptozona compreende cinco espécies distribuídas do México ao Brasil. Para este
grupo, duas espécies têm seu hospedeiro conhecido da família Sapotaceae. Além dessa família,
Anastrepha leptozona Hendel também infesta outras cinco famílias de plantas. No Brasil, A.
1Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970, Manaus, AM, Brasil. 2Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970
Manaus, AM, Brasil. 3Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL 32614-
7100. 4Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil.

69
leptozona é amplamente distribuída e sua ocorrência já foi registrada em 15 estados (Norrbom et
al. 1999, Zucchi 2008, Frías et al. 2009).
O grupo pseudoparallela possui 20 espécies com ampla distribuição desde o Texas até a
Argentina, contudo somente duas espécies ocorrem no norte do Panamá e uma nas Antilhas. A
maioria das espécies com hospedeiro conhecido infesta frutos da família Passifloraceae (Norrbom
et al. 1999). Dez espécies deste grupo foram registradas no Brasil, entre elas Anastrepha curitis
Stone que está restrita aos estados do Amazonas e Pará na região amazônica (Zucchi 2007, 2008).
As informações sobre a morfologia dos ovos de espécies de Anastrepha ainda são
escassas, pois até o momento apenas 39 espécies foram descritas (Emmart 1933; Seín 1933;
Lawrence 1979; Steck e Malavasi 1988; Steck e Wharton 1988; Carroll e Wharton 1989;
Norrbom et al. 1999; Norrbom e Korytkowski 2009; Dutra et al. 2011a, 2011b; Figueiredo et al.
2011). Contudo, as informações registradas evidenciam que caracteres morfológicos presentes
nos estágios imaturos podem ser úteis para compreender as relações evolutivas entre as espécies
de Anastrepha (Norrbom et al. 1999).
No grupo pseudoparallela, somente ovos das espécies Anastrepha pseudoparallela
(Loew) e Anastrepha consobrina (Loew) foram medidos, suas características gerais foram
descritas e foram observados com mais detalhes em MEV (Norrbom et al. 1999, Figueiredo et al.
2011). No grupo leptozona, apenas a espécie A. leptozona teve suas características gerais
descritas.
Neste estudo, uma descrição detalhada da morfologia dos ovos foi realizada utilizando-se
MEV para A. curitis e A. leptozona. Os resultados obtidos podem ser úteis na identificação de
ovos de espécies de Anastrepha e no entendimento sobre a taxonomia desse gênero.
Material e Métodos
Ovos de A. curitis e A. leptozona foram dissecados de fêmeas capturadas em armadilhas
McPhail em Manaus (03°06’07’’S, 60°01’30’’W) e em Presidente Figueiredo (02°02’04’’S,
60°01’30’’W) no estado do Amazonas. Os ovos foram preservados em microtubos plásticos de
1,5 ml contendo etanol 80%.
As medidas de comprimento e largura dos ovos foram realizadas com micrométrica ocular
em um estereomicroscópio Wild M3C no Laboratório de Entomologia Agrícola, do Instituto
Nacional de Pesquisas da Amazônia (INPA) em Manaus, Amazonas, Brasil. Os ovos foram

70
fotografados no estereomicroscópio óptico (Leica M165C) do Laboratório de Prospecção de
Bioativos de Insetos do Instituto Nacional de Pesquisas da Amazônia (INPA), Amazonas, Brasil.
Entre dez e vinte ovos por fêmea de um total de quatro fêmeas de cada espécie foram examinados
com microscopia óptica. O pólo anterior foi definido como a extremidade do ovo que possui um
pedicelo ou uma suave projeção com micrópila e aerópilas. O pólo posterior foi definido como a
extremidade oposta ao pedicelo o qual geralmente é liso e arredondado, não possuindo aberturas
externas ou estruturas. A cabeça do embrião desenvolve-se na direção do pólo anterior do ovo
(Headrick e Goeden 1998). De acordo com a posição do embrião no interior do ovo, o lado
convexo foi considerado como o lado ventral e o lado côncavo como o lado dorsal.
Para análise com MEV, os ovos foram montados em porta amostras de alumínio e
examinados no modo ambiental no microscópio eletrônico de varredura Quanta 250 (Fei
Company) no Laboratório de Técnicas Mineralógicas da Universidade Federal do Amazonas
(UFAM) em Manaus, Amazonas. Para a MEV foram utilizados pelo menos dez ovos de cada
espécie.
Espécimes testemunho das fêmeas e ovos foram depositados na Coleção de Invertebrados,
INPA.
Resultados
Os ovos de A. curitis e A. leptozona são semelhantes em seus aspectos gerais. Eles são
alongados, afilam gradualmente em direção ao pólo posterior, possuem papila e micrópila no
pólo anterior. Pode-se diferenciar os ovos dessas duas espécies observando-se algumas
características, tais como comprimento do ovo e presença ou ausência de aerópilas no pólo
anterior. Nenhum dos ovos das espécies estudadas apresentou apêndice respiratório.
Anastrepha curitis Stone (Figs. 1A e 2)
Material Examinado. Um total de 52 ovos de quatro fêmeas foi examinado. Três fêmeas
de A. curitis foram coletadas em Manaus e uma fêmea em Presidente Figueiredo no estado do
Amazonas.
Características Gerais. Comprimento 2,13-2,58 mm e largura 0,18-0,26 mm. Os ovos
são creme amarelados, levemente curvados, muito alongados, com maior diâmetro no meio do
ovo em direção ao pólo anterior e afilando-se gradualmente em ambas as extremidades (Fig. 1A).

71
Escultura do córion. O pólo anterior apresenta uma papila na qual está localizada a
micrópila. Toda a superfície do ovo é lisa sem qualquer ornamentação do córion, reticulação ou
protuberância. Também não foi observado nenhum apêndice respiratório. Foi observada uma
camada de superfície porosa em todo pólo anterior do ovo (Fig. 2). A micrópila (Fig. 2) está
localizada no ápice do pólo anterior no lado dorsal e rodeada por uma borda do córion. Esta
borda tem o formato de anel contínuo com um aspecto liso (Fig. 2). As principais características
dos ovos para diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha leptozona Hendel (Figs. 1B e 3)
Material Examinado. Um total de 51 ovos de quatro fêmeas foi examinado. Essas
fêmeas de A. leptozona foram coletadas em Manaus no estado do Amazonas.
Características Gerais. Comprimento 1,37-1,87 mm e largura 0,25-0,31 mm. Os ovos
são creme amarelados, levemente curvados, alongados, robustos, com maior diâmetro no meio do
ovo em direção ao pólo anterior e afilando-se gradualmente em ambas as extremidades (Fig. 1B).
Escultura do córion. O pólo anterior apresenta uma papila onde se localiza a micrópila.
Toda a superfície do ovo é lisa sem qualquer ornamentação do córion, reticulação ou
protuberância (Fig. 3A). Também não foi observado nenhum apêndice respiratório. Mais de 50
aerópilas foram observadas no lado ventral do ovo próximo a micrópila (Fig. 3B). As aerópilas
apresentam vários diâmetros, variando entre 1,1 e 2,1 µm. A micrópila está localizada no ápice
do pólo anterior no lado dorsal e rodeada por uma borda do córion (Fig. 3A). Esta borda tem o
formato de anel contínuo com um aspecto liso (Fig. 3A). As principais características dos ovos
para diagnóstico desta espécie estão apresentadas na Tabela 1.
Discussão
As características gerais dos ovos de ambas as espécies estudadas aqui são semelhantes
aquelas de outras espécies de Anastrepha já descritas (White e Elson-Harris 1992, Norrbom et al.
1999). Um único estudo publicado anteriormente sobre a morfologia dos ovos de A. leptozona
descreveu as dimensões (comprimento 1,30-1,41 mm; largura 0,22-0,27 mm), formato geral do
ovo (“robusto”) e ornamentação do córion (“ausente”) (Norrbom et al. 1999). No presente estudo,
os resultados obtidos corroboram os apresentados acima.

72
Os ovos de A. curitis são delgados e muito alongados, evidenciando que essa espécie de
Anastrepha apresenta um dos ovos mais longos neste gênero. Outras três espécies também
apresentam ovos bastante longos, Anastrepha grandis (Macquart), A. montei e Anastrepha
nigrifascia Stone (Steck e Wharton 1988, Norrbom e Korytkowski 2009, Dutra et al. 2011b,
Figueiredo et al. 2011).
Ambas as espécies estudadas apresentam o córion liso sem nenhuma ornamentação,
reticulação ou protuberância. Embora a maioria das espécies de Anastrepha apresente córion com
ornamentação, a ausência de ornamentação já foi observada em outras espécies como Anastrepha
consobrina (Loew), Anastrepha coronilli Carrejo & González, A. grandis, Anastrepha manihoti
Lima, A. montei, Anastrepha pickeli Lima, Anastrepha pseudoparallela (Loew) e Anastrepha
serpentina (Wiedemann) (Emmart 1933; Steck e Wharton 1988; Selivon e Perondini 1999; Dutra
et al. 2011a, 2011b; Figueiredo et al. 2011).
Nos ovos de A. curitis, foi observada uma camada de superfície porosa semelhante à
encontrada nos ovos de A. montei (Dutra et al. 2011b), que provavelmente tem a função de
respiração para o ovo. Até o momento, apenas essas duas espécies apresentaram essa
característica. Por outro lado, nos ovos de A. leptozona foram observadas aerópilas, com diversos
diâmetros, localizadas apenas no lado ventral no pólo anterior do ovo. Esse aspecto também foi
observado em Anastrepha antunesi Lima, Anastrepha bahiensis Lima, A. coronilli e A.
serpentina (Selivon e Perondini 1999, Dutra et al. 2011a). Segundo a literatura, provavelmente o
número e o diâmetro variável das aerópilas esteja relacionado com o ambiente onde os ovos são
ovipositados, bem como a necessidade das trocas gasosas e o controle da perda de água para
evitar a desidratação (Cônsoli et al. 1999). Os ovos de ambas as espécies descritos aqui não
apresentam apêndice respiratório, como observado em algumas espécies de Anastrepha descritas
previamente tais como Anastrepha barbiellinii Lima, A. manihoti, A. nigrifascia, Anastrepha
obliqua (Macquart), A. pickeli e Anastrepha pittieri Caraballo (Murillo e Jirón 1994, Norrbom et
al. 1999, Norrbom e Korytkowski 2009, Selivon e Perondini 1999, Dutra et al. 2011b).
A micrópila das duas espécies aqui estudadas está localizada no ápice do pólo anterior do
ovo, como registrado para a maioria das espécies já descritas (Carroll e Wharton 1989; Selivon e
Perondini 1998, 1999, 2000; Norrbom e Korytkowski 2009; Dutra et al. 2011a, 2011b;
Figueiredo et al. 2011), com exceção de Anastrepha sp. 1 aff. fraterculus (Wiedemann) e
Anastrepha turpiniae (Stone) que apresentam a micrópila levemente deslocada do ápice (Murillo

73
e Jirón 1994, Selivon e Perondini 1998, 2000; Dutra et al. 2011a) e de A. barbiellinii, A.
manihoti, A. nigrifascia, A. obliqua, A. pickeli e A. pittieri que possuem apêndice respiratório no
ápice do pólo anterior (Murillo e Jirón 1994, Norrbom et al. 1999, Norrbom e Korytkowski 2009,
Selivon e Perondini 1999, Dutra et al. 2011b).
A observação detalhada da morfologia dos ovos com MEV para A. curitis e A. leptozona
mostrou diferenças que permitem a identificação dessas espécies, assim como semelhanças com
ovos de outras espécies descritas anteriormente. Características como comprimento do ovo,
presença ou ausência de aerópilas e ausência de ornamentação no córion são caracteres
taxonômicos que auxiliam o esclarecimento sobre as relações filogenéticas dentro do gênero
Anastrepha.
Agradecimentos
Os autores agradecem a Marcos Vinicius Garcia e Tatiana Senra (Embrapa Amazônia
Ocidental) pelo auxílio durante as coletas. Os autores também agradecem a Carolina Almeida e
Adriana Horbe (Universidade Federal do Amazonas) pelo auxílio nas fotografias com MEV. Esse
estudo foi financiado pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq - projeto n° 575664/2008-8), pela Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES - projeto n° 23038.037185/2008-41), pela Fundação de Amparo a Pesquisa do
Estado do Amazonas (FAPEAM - projeto nº014-017/2004) e pela Rede Amazônica de Pesquisas
sobre Moscas-das-Frutas (Chamada 05/2006 Agrofuturo). Os autores também agradecem ao
CNPq pela bolsa de doutorado concedida a VSD.

74
Fig. 1. Ovos de espécies de Anastrepha (pólo anterior à esquerda). (A) A. curitis. (B) A.
leptozona.
Fig. 2. Vista com MEV do ovo de A. curitis. Pólo anterior mostrando a micrópila (seta).

75
Tabela 1. Diagnose dos ovos de Anastrepha curitis e Anastrepha leptozona analisadas neste estudo.
Anastrepha Ornamentação do
córion
Micrópila Aerópila
Localização Anel Localização Número Diâmetro
curitis Nenhuma Ápice do pólo anterior Liso Estreita camada de
superfície porosa
Nenhum Nenhum
leptozona Nenhuma Ápice do pólo anterior Liso Lado ventral do pólo
anterior
Mais de 50 Variável
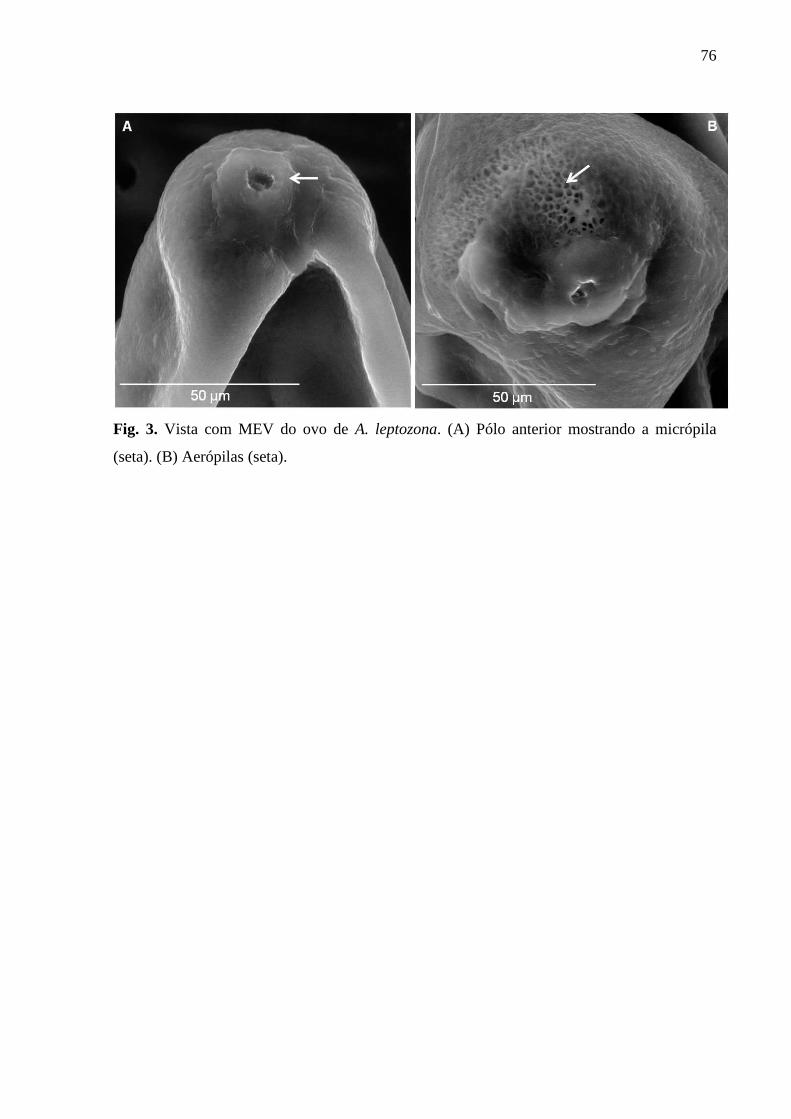
76
Fig. 3. Vista com MEV do ovo de A. leptozona. (A) Pólo anterior mostrando a micrópila
(seta). (B) Aerópilas (seta).

77
Capítulo 3
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G.
Description of Larvae of Anastrepha spp. (Diptera:
Tephritidae) in the fraterculus Group. Aceito pela Annals
of the Entomological Society of America (Anexo 5).

78
Description of Larvae of Anastrepha spp. (Diptera: Tephritidae) in the
fraterculus Group
VIVIAN S. DUTRA1,5BEATRIZ RONCHI-TELES
2, GARY J. STECK
3, E JANISETE G.
SILVA4
RESUMO A morfologia de larvas de segundo e terceiro instar de três espécies de Anastrepha
do grupo fraterculus, Anastrepha bahiensis Lima, Anastrepha coronilli Carrejo & González e
Anastrepha turpiniae Stone foram descritas e documentadas pela primeira vez. Também são
discutidos caracteres diagnósticos que podem ser utilizados para diferenciar as três espécies,
bem como a sobreposição dessas espécies com outras espécies do grupo fraterculus.
PALAVRAS-CHAVE moscas-das-frutas, morfologia, esqueleto céfalo-faríngeo
Anastrepha Schiner (Diptera: Tephiritidae) é o gênero de tefritídeos de maior importância
econômica da região Neotropical com cerca de 240 espécies descritas até o momento
(Norrbom et al. 1999, Zucchi 2008, Norrbom e Korytkowski 2009, 2011). Este gênero é
amplamente distribuído desde o sul dos Estados Unidos até o norte da Argentina e restrito aos
ambientes tropicais e subtropicais (Aluja 1994).
O Brasil apresenta um elevado número de espécies de Anastrepha e sua ocorrência já
foi relatada nos 26 estados brasileiros. Atualmente 112 espécies são registradas para o país e
deste total 33 ocorrem exclusivamente aqui (Norrbom et al. 1999; Jesus et al. 2008; Uramoto
et al. 2008; Zucchi 2007, 2008).
O grupo fraterculus compreende 29 espécies descritas e amplamente distribuídas, das
quais 17 espécies ocorrem no Brasil (Norrbom et al. 1999; Zucchi 2000, 2007). Algumas
espécies desse grupo são polífagas de grande importância econômica (Aluja 1994, Norrbom et
al. 1999, Zucchi 2007).
As larvas de Anastrepha alimentam- se do tecido vivo das plantas causando danos ao
fruto (White e Elson-Harris 1992). Larvas de moscas-das-frutas são geralmente encontradas
em frutos interceptados nos portos e aeroportos. Estudos sobre descrição morfológica de
1Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal
478, 69011-970, Manaus, AM, Brasil. 2Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970
Manaus, AM, Brasil. 3Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL
32614-7100. 4Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil.

79
larvas de terceiro instar foram realizados apenas para 13 espécies de Anastrepha e como
algumas dessas descrições são incompletas a identificação das larvas interceptadas torna-se
difícil (Steck e Wharton 1988, Carroll e Wharton 1989, Steck et al. 1990, Norrbom et al.
1999).
Neste estudo, foram descritas larvas de segundo e terceiro instar de três espécies do
grupo fraterculus, Anastrepha bahiensis Lima, Anastrepha coronilli Carrejo & González e
Anastrepha turpiniae Stone. Anastrepha bahiensis tem sido registrada do México ao Brasil
(Norrbom et al. 1999). No Brasil, essa espécie foi relatada nos estados do Amapá, Amazonas,
Bahia, Goiás, Espírito Santo, Minas Gerais, São Paulo e Santa Catarina (Zucchi 2008).
Anastrepha coronilli tem sido registrada no México, Venezuela e Brasil. No Brasil, A.
coronilli foi relatada apenas para a Amazônia brasileira (Hernandez-Ortiz 2007, Morales e
González 2007, Zucchi 2007). Anastrepha turpiniae tem sido registrada no Panamá e Brasil,
onde é encontrada em todas as regiões, com exceção da região sul (Norrbom 2004, Zucchi
2007).
Material e Métodos
Larvas de A. bahiensis, A. coronilli e A. turpiniae foram dissecadas de frutos coletados
em Manaus (03°06’07’’S; 60°01’30’’W) e Presidente Figueiredo (02°02’04’’S;
60°01’30’’W) no estado do Amazonas. Para confirmação da espécie, cerca de três larvas
foram dissecadas por fruto e as demais larvas foram deixadas nos frutos até a emergência dos
adultos. As larvas foram mortas e armazenadas em etanol 100%. De cada amostra de fruto
coletado, emergiu apenas uma espécie de Anastrepha.
Características externas como comprimento, largura, coloração e número de
segmentos foram medidas com micrométrica ocular em um estereomicroscópio Wild M3C no
Laboratório de Entomologia Agrícola do Instituto Nacional de Pesquisas da Amazônia
(INPA), Manaus, Amazonas, Brasil. As larvas foram clarificadas com hidróxido de potássio
10% em temperatura ambiente e posteriormente colocadas em lâminas escavadas em um meio
contendo álcool gel e glicerina (1:1) para visualização das características. As fileiras de
espinhos dorsais e ventrais foram contadas de acordo com as linhas médias dorsal e ventral,
respectivamente. O esqueleto céfalo-faríngeo (ECF) foi removido, colocado em glicerina e
ampliado 80X para as medições das larvas de terceiro instar e ampliado 100X para
visualização das larvas de segundo instar. Para ambos os instares, as medidas dos espiráculos
anteriores e posteriores foram feitas com objetivas de ampliação 200X e 400X. Essas
estruturas foram fotografadas usando câmera digital (Olympus DP72) acoplada ao

80
microscópio ótico (Olympus BX51) e as ilustrações do segmento caudal foram desenhadas
com câmara clara (Leica DM750) no Laboratório de Citotaxonomia e Insetos Aquáticos
(INPA). O ECF das larvas de terceiro instar foram colocados em placas de Petri com etanol
80%, fotografados, medidos e observadas às características utilizando a câmera digital (Leica
DFC420) acoplada ao estereomicroscópio (Leica M165C) do Laboratório de Prospecção de
Bioativos de Insetos (INPA).
A terminologia utilizada foi baseada em Teskey (1981), Steck e Malavasi (1988),
Steck e Wharton (1988) e Frías et al. (2009). Foram utilizados os mesmos pontos de
medições, as mesmas abreviações para os segmentos torácicos, segmentos abdominais,
espiráculo posterior e segmento caudal que os utilizados por Steck e Malavasi (1988) e Steck
e Wharton (1988). O comprimento total do esqueleto céfalo-faríngeo foi medido desde o
ápice da mandíbula até a extremidade final do corno ventral. O comprimento do corno ventral
foi medido desde a barra faringeal até extremidade final das estrias.
Na preparação para microscopia eletrônica de varredura (MEV), as larvas foram
transferidas para cápsulas de silicone, desidratadas em série alcoólica e no secador de ponto
crítico de CO2 por três horas e metalizadas com uma camada de ouro. As larvas foram
examinadas ao microscópio eletrônico de varredura LEO 1450VP no Laboratório
Institucional de Microscopia Eletrônica de Varredura do Museu Paraense Emílio Goeldi,
Belém, Pará, Brasil. Para a visualização com MEV foram utilizadas pelo menos dez larvas de
cada espécie.
Espécimes testemunho das larvas associadas com as fêmeas foram depositadas na
Coleção de Invertebrados do Instituto Nacional de Pesquisas da Amazônia (INPA).
Resultados
As larvas de A. bahiensis, A. coronilli e A. turpiniae são similares em seu aspecto
geral. Caracteres diagnósticos para diferenciar essas três espécies incluem presença ou
ausência de espinhos dorsais no terceiro segmento torácico (T3), formato do lóbulo anal,
número de troncos/extremidades das cerdas no espiráculo posterior e algumas características
do esqueleto céfalo-faríngeo.
Anastrepha bahiensis Lima (Fig. 1)
Material Examinado. Foram examinadas 32 larvas (22 larvas de terceiro instar e sete
de segundo instar) dissecadas de frutos de Helicostylis scabra (Macbr.) C.C. Berg, Moraceae
(localmente conhecida como “inharé”) e de frutos de Pouroma cecropiaefolia Moraceae

81
(localmente conhecida como “mapati”), ambas as árvores nativas da Floresta Amazônica de
terra firme.
Características das larvas de terceiro instar. As larvas possuem comprimento
variando de 5,99-10,65 mm e largura de 1,03-1,86 mm. As larvas são alongadas, cilíndricas,
afiladas nos segmentos torácicos, com segmento caudal truncado e possuem coloração
variando entre creme esbranquiçado e creme amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 2,35-3,98 µm.
O padrão de distribuição dos espinhos dorsais é T1 com 2-5 fileiras; T2 com 3-5 fileiras; T3,
A1 até A8 não apresentam fileiras de espinhos. O padrão de distribuição dos espinhos ventrais
é T1 com 4-7 fileiras; T2 com 5-8 fileiras; T3 com 3-4 fileiras; A1 com 5-7 fileiras; A2 com
10-12 fileiras, A3 com 10-15 fileiras; A4 e A5 com 12-15 fileiras; A6 e A7 com 11-14 fileiras
e A8 com 10-14 fileiras. Todos os segmentos abdominais (A1 ao A8) apresentam
protuberâncias ventrais para locomoção. Ao redor do lóbulo anal, foram visualizadas fileiras
irregulares de espinhos ventral e dorsalmente, variando entre 3-4 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 11,47-19,01 µm, botão apical com diâmetro variando de
9,23-10,54 µm e altura de 5,63-8,72 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 13,02-18,34 µm e altura de 6,71-7,33 µm. O órgão sensorial
bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 4,15-7,05 µm. As
saliências orais variam de 6-8 (Fig. 1A).
A Figura 1D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 0,84-1,28 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,15-0,24 mm, o comprimento b varia de
0,14-0,17 mm e a altura c varia de 0,12-0,18 mm. A razão entre as medidas a / c varia de 1,2-
1,3. Os dentes são curtos e pontiagudos, mas alguns deles podem ser arredondados. O
comprimento do esclerito hipofaringeal varia de 0,17-0,20 mm e a largura de 0,16-0,20 mm.
Em vista dorsal foi visualizado que o esclerito epifaringeal apresenta um lóbulo medial
direcionado anteriormente. Também em vista dorsal foi visualizado o esclerito labial em
formato de ferradura, curto, robusto e bastante esclerotinizado. A barra parastomal (= esclerito
hipostomal) é levemente arqueada e estende-se quase completamente sobre o esclerito
hipofaringeal. O esclerito ventral localiza-se abaixo do esclerito faringeal. O corno dorsal é
formado por uma área bem esclerotinizada e uma hialina, o comprimento de ambas as áreas

82
variam de 0,32-0,48 mm e 0,51-0,72 mm, respectivamente. A ponte dorsal (“dorsal bridge”)
apresenta uma projeção anterior do corno dorsal levemente esclerotinizada. O arco dorsal
possui altura variando de 0,31-0,38 mm. O esclerito anterior é esclerotinizado e possui
formato irregular. A razão entre o entalhe do corno (“cornu notch”) e corno dorsal varia de
0,6-0,7. O corno ventral é fracamente esclerotinizado. O filtro faringeal (“pharyngeal filter”) é
fracamente esclerotinizado e possui anteriormente uma barra faringeal com 8-9 cristas que
formam estrias estendendo-se por todo o comprimento do corno ventral. O corno ventral
possui comprimento variando de 0,59-0,81 mm. O corno ventral é 1,5 vezes o tamanho da
área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 11 e 14 túbulos (eventualmente 15) (Figs. 1B e E). O túbulo apresenta
comprimento entre 21,06-27,55 µm, largura distal entre 16,45-19,87 µm e largura basal entre
11,66-16,93 µm. O espiráculo anterior apresenta na linha média altura variando de 108,20-
154,82 µm; largura distal de 200,63-225,86 µm e largura basal próximo a abertura da traquéia
variando de 90,49-129,58 µm.
A Figura 1F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1
é distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente
desenvolvidos e associados com uma sensila fracamente desenvolvida. I2 é disposto
lateralmente e algumas vezes levemente ventral em relação a I1. L1 e V são pouco
desenvolvidos. D1 e I1 são associados com sensila fracamente desenvolvida. Alguns
exemplares podem ter três tubérculos ventrais (V1, V2 e V3). Os espiráculos posteriores estão
localizados acima da linha média horizontal. O lóbulo anal é bífido e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Figs. 1C e G), possuindo comprimento variando de 72,24-
98,63 µm e largura de 16,08-19,74 µm. A razão entre o comprimento e a largura das fendas
varia de 4,5-5,0. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 138,41-
168,09 µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 14-20
troncos e 20-40 extremidades; a razão entre essas extremidades / troncos varia de 1,4-2,0; esse
grupo de cerdas apresenta largura basal variando de 20,06-35,66 µm; a razão da largura basal
/ comprimento dessa fenda espiracular é de 0,2. O grupo SP-II possui 4-9 troncos e 11-22
extremidades. O grupo SP-III possui 5-12 troncos e 14-27 extremidades. O grupo SP-IV
possui 12-21 troncos e 23-41 extremidades; a razão entre essas extremidades / troncos é de

83
1,9; esse grupo de cerdas apresenta largura basal variando de 19,56-38,19 µm; a razão da
largura basal / comprimento dessa fenda espiracular é de 0,2. A média do comprimento das
cerdas é 0,4 vezes o comprimento das fendas espiraculares.
Características das larvas de segundo instar. As larvas possuem comprimento
variando de 2,13-3,78 mm e largura de 0,37-0,52 mm. As larvas de segundo instar apresentam
o mesmo formato e coloração que as larvas de terceiro instar.
Os espinhos e as fileiras de espinhos apresentam as mesmas características que as
observadas nas larvas de terceiro instar. A largura basal dos espinhos varia de 2,16-3,11 µm.
O padrão de distribuição dos espinhos dorsais é T1 e T2 com 3-4 fileiras, essas fileiras
apresentam pequenos hiatos na linha média dorsal; T3, A1 até A8 não apresentam fileiras de
espinhos. O padrão de distribuição dos espinhos ventrais é T1 com 3-5 fileiras; T2 com 4
fileiras (eventualmente 2 ou 3); T3 com 2-3 fileiras; A1 com 5 fileiras; A2 com 8-9 fileiras,
A3 com 10 fileiras; A4 e A5 com 8-9 fileiras; A6 com 8-9 fileiras (eventualmente 11); A7
com 9-11 fileiras e A8 com 8-9 fileiras (eventualmente 10). Todos os segmentos abdominais
(A1 ao A8) apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo anal
foram visualizadas três fileiras (eventualmente 4) irregulares de espinhos ventral e
dorsalmente.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 6,89-12,05 µm, botão apical com diâmetro variando de
7,20-7,62 µm e altura de 4,97-5,57 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 11,26-17,31 µm e altura de 5,30-5,47 µm. O órgão sensorial
bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 2,82-3,89 µm. As
saliências orais variam de 6-8.
O esqueleto céfalo-faríngeo possui comprimento total variando de 524,22-616,91 µm
(Fig. 1H). A mandíbula apresenta três medidas: o comprimento a varia de 102,17-122,38 µm,
o comprimento b varia de 67,44-78,43 µm e a altura c varia de 68,05-88,59 µm. A razão entre
as medidas a / c varia de 1,4-1,5. A parte apical da mandíbula é bem esclerotinizada e a parte
basal é fortemente esclerotinizada. Alguns exemplares dessa espécie possuem dois orifícios
na base da mandíbula. Os dentes são pontiagudos e possuem um dente secundário bem
desenvolvido. O comprimento do esclerito hipofaringeal varia de 90,93-114,61 µm e a largura
de 48,07-77,14 µm, apresentando a parte apical fortemente esclerotinizada e a parte basal bem
esclerotinizada. O esclerito epifaringeal, esclerito labial e a barra parastomal (= esclerito
hipostomal) apresentam as mesmas características que as larvas de terceiro instar. O corno
dorsal é formado por uma área bem esclerotinizada e uma hialina, o comprimento de ambas as

84
áreas variam de 114,58-202,43 µm e 221,54-340,55 µm, respectivamente. O arco dorsal
possui altura variando de 146,83-174,70 µm. O esclerito anterior é ausente. A razão entre o
entalhe do corno (“cornu notch”) e corno dorsal é de 0,5. O corno ventral é fracamente
esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente esclerotinizado e possui
anteriormente uma barra faringeal com 6-7 cristas que formam estrias estendendo-se por todo
o comprimento do corno ventral. O corno ventral possui comprimento variando de 270,85-
367,44 µm. O corno ventral é 1,2 vezes o tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 11 e 14 túbulos (eventualmente 9-10) (Fig. 1I). O túbulo apresenta
comprimento entre 8,85-13,19 µm, largura distal entre 9,12-11,22 µm e largura basal entre
5.88-8.97 µm. O espiráculo anterior apresenta na linha média altura variando de 73,74-96,77
µm; largura distal de 82,13-94,87 µm e largura basal próximo a abertura da traquéia variando
de 24,35-44,28 µm.
O segmento caudal com tubérculos e sensilas apresentam o mesmo padrão que as
larvas de terceiro instar. O espiráculo posterior e o lóbulo anal estão localizados da mesma
forma que nas larvas de terceiro instar. O lóbulo anal é bífido.
O espiráculo posterior apresenta três fendas cada qual com membrana estreita e poucas
trabéculas transversais, seguindo o mesmo padrão que as larvas de terceiro instar (Fig. 1J). As
fendas possuem comprimento variando de 22,38-29,91 µm e largura de 11,27-15,12 µm. A
razão entre o comprimento e a largura das fendas é de 1,9. O espiráculo posterior possui
cicatriz da ecdise aparente. A câmara onde localizam-se as fendas é arredondada e apresenta
diâmetro próximo a traquéia variando de 56,27-63,54 µm. O processo espiracular apresenta
quatro grupos de cerdas, onde SP-I possui 7-12 troncos e 14-18 extremidades; a razão entre
essas extremidades / troncos varia de 1,5-2,0; esse grupo de cerdas apresenta largura basal
variando de 6,93-15,46 µm; a razão da largura basal / comprimento dessa fenda espiracular
varia de 0,4-0,5. O grupo SP-II possui 4-6 troncos e 6-8 extremidades. O grupo SP-III possui
7-9 troncos e 10-13 extremidades. O grupo SP-IV possui 7-11 troncos e 14-16 extremidades;
a razão entre essas extremidades / troncos varia de 1,4-2,0; esse grupo de cerdas apresenta
largura basal variando de 7,75-12,33 µm; a razão da largura basal / comprimento dessa fenda
espiracular é de 0,4. A média do comprimento das cerdas é 0,5 vezes o comprimento das
fendas espiraculares.

85
Anastrepha coronilli Carrejo & González (Fig. 2)
Material Examinado. Foram examinadas 36 larvas (25 larvas de terceiro instar e 11
de segundo instar) dissecadas de frutos de Bellucia grossularioides L. Melastomataceae
(localmente conhecida como “goiaba-de-anta”), árvore nativa da Floresta Amazônica de terra
firme.
Características das larvas de terceiro instar. As larvas possuem comprimento
variando de 9,51-10,96 mm e largura de 1,55-2,37 mm. As larvas são alongadas, cilíndricas,
afiladas nos segmentos torácicos, com segmento caudal truncado e possuem coloração creme
amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 2,19-5,29 µm.
O padrão de distribuição dos espinhos dorsais é T1 com 2-3 fileiras (eventualmente 4); T2
com 3-4 fileiras (eventualmente 1-2); T3, A1 ao A8 não apresentam fileiras de espinhos. O
padrão de distribuição dos espinhos ventrais é T1 com 3-5 fileiras; T2 com 3-4 fileiras; T3
com 1-2 fileiras; A1 com 3-7 fileiras; A2 com 9-11 fileiras; A3 com 10-14 fileiras; A4 e A5
com 13-15 fileiras; A6 com 10-14 fileiras; A7 com 12-14 fileiras; A8 com 10-14 fileiras.
Todos os segmentos abdominais (A1 ao A8) apresentam protuberâncias ventrais para
locomoção. Ao redor do lóbulo anal, foram visualizadas quatro fileiras irregulares de espinhos
ventral e dorsalmente.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 17,48-25,06 µm, botão apical com diâmetro variando de
10,79-13,09 µm e altura de 8,04-13,18 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 20,18-43,47 µm e altura de 10,23-14,60 µm. O órgão
sensorial bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 4,90-8,67 µm.
As saliências orais variam de 9-10 (eventualmente 7-8) (Fig. 1A).
A Figura 1D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 1,15-1,47 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,25-0,33 mm, o comprimento b varia de
0,19-0,24 mm e a altura c varia de 0,19-0,23 mm. A razão entre as medidas a / c varia de 1,3-
1,4. Os dentes são longos e pontiagudos. O comprimento do esclerito hipofaringeal varia de
0,17-0,21 mm e a largura de 0,16-0,19 mm. Em vista dorsal foi visualizado que o esclerito
epifaringeal apresenta um lóbulo medial direcionado anteriormente. Também em vista dorsal
foi visualizado o esclerito labial em formato de ferradura, curto, robusto e bastante

86
esclerotinizado. A barra parastomal (= esclerito hipostomal) é arqueada e estende-se quase
completamente sobre o esclerito hipofaringeal. O esclerito ventral localiza-se abaixo do
esclerito faringeal. O corno dorsal é formado por uma área bem esclerotinizada e uma hialina,
o comprimento de ambas as áreas variam de 0,42-0,52 mm e 0,57-0,85 mm, respectivamente.
A ponte dorsal (“dorsal bridge”) apresenta uma projeção anterior do corno dorsal levemente
esclerotinizada. O arco dorsal possui altura variando de 0,35-0,46 mm. O esclerito anterior é
esclerotinizado e possui formato irregular. A razão entre o entalhe do corno (“cornu notch”) e
corno dorsal é de 0,6. O corno ventral é fracamente esclerotinizado. O filtro faringeal
(“pharyngeal filter”) é fracamente esclerotinizado e possui anteriormente uma barra faringeal
com 8-9 cristas que formam estrias estendendo-se por todo o comprimento do corno ventral.
O corno ventral possui comprimento variando de 0,64-0,90 mm. O corno ventral é 1,2 vezes o
tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 11-15 túbulos (eventualmente 10) (Figs. 1B e E). O túbulo apresenta
comprimento entre 17,77-37,33 µm, largura distal entre 15,87-26,76 µm e largura basal entre
12,96-21,39 µm. O espiráculo anterior apresenta na linha média altura variando de 120,81-
178,11 µm; largura distal de 193,26-271,82 µm e largura basal próximo a abertura da traquéia
variando de 85,78-133,42 µm.
A Figura 1F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1
é distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente
desenvolvidos e associados com uma sensila fracamente desenvolvida. I2 é disposto
lateralmente e algumas vezes levemente ventral em relação a I1. L1 e V são pouco
desenvolvidos. D1 e I1 são associados com sensila fracamente desenvolvida. Alguns
exemplares podem ter três tubérculos ventrais (V1, V2 e V3). Os espiráculos posteriores estão
localizados acima da linha média horizontal. O lóbulo anal é inteiro e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Figs. 1C e G), possuindo comprimento variando de 81,38-
113,57 µm e largura de 15,87-25,85 µm. A razão entre o comprimento e a largura das fendas
varia de 4,3-5,1. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 138,26-
223,37 µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 16-25
troncos (eventualmente 12 ou 30) e 45-76 extremidades (eventualmente 40); a razão entre
essas extremidades / troncos varia de 2,8-3,0; esse grupo de cerdas apresenta largura basal

87
variando de 31,60-64,03 µm; a razão da largura basal / comprimento dessa fenda espiracular
varia de 0,3-0,5. O grupo SP-II possui 5-12 troncos e 18-45 extremidades (eventualmente 16).
O grupo SP-III possui10-15 troncos (eventualmente 5-8) e 23-48 extremidades
(eventualmente 16). O grupo SP-IV possui 17-25 troncos (eventualmente 13-14) e 40-63
extremidades (eventualmente 32); a razão entre essas extremidades / troncos varia de 2,3-2,5;
esse grupo de cerdas apresenta largura basal variando de 32,44-64,90 µm; a razão da largura
basal / comprimento dessa fenda espiracular varia de 0,3-0,5. A média do comprimento das
cerdas é 0,4 vezes o comprimento das fendas espiraculares.
Características das larvas de segundo instar. As larvas possuem comprimento
variando de 4,18-5,37 mm e largura de 0,62-0,93 mm. As larvas de segundo instar apresentam
o mesmo formato e coloração que as larvas de terceiro instar.
Os espinhos e as fileiras de espinhos apresentam as mesmas características que as
observadas nas larvas de terceiro instar. A largura basal dos espinhos varia de 3,05-3,84 µm.
O padrão de distribuição dos espinhos dorsais é T1 com 3-4 fileiras; T2 com 2-3 fileiras, essas
fileiras apresentam pequenos hiatos na linha média dorsal; T3, A1 até A8 não apresentam
fileiras de espinhos. O padrão de distribuição dos espinhos ventrais é T1 com 10-11 fileiras
(eventualmente 6-7); T2 com 4-7 fileiras (eventualmente 10); T3 com 3-7 fileiras; A1 com 6-
11 fileiras (eventualmente 4-5); A2 com 10-14 fileiras; A3 com 11-14 fileiras (eventualmente
8); A4 com 10-13 fileiras; A5 com 11-14 fileiras; A6 com 12-13 fileiras (eventualmente 8);
A7 com 11-13 fileiras; A8 com 11-14 fileiras. Todos os segmentos abdominais (A1 ao A8)
apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo anal foram
visualizadas fileiras irregulares de espinhos ventral e dorsalmente, variando entre 4-5 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 12,99-18,22 µm, botão apical com diâmetro variando de
9,94-11,43 µm e altura de 7,40-9,97 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 36,73-43,39 µm e altura de 11,33-14,85 µm. O órgão
sensorial bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 5,77-8,01 µm.
As saliências orais variam de 6-7.
O esqueleto céfalo-faríngeo possui comprimento total variando de 754,71-834,01 µm
(Fig. 1H). A mandíbula apresenta três medidas: o comprimento a varia de 178,97-206,24 µm,
o comprimento b varia de 111,99-125,21 µm e a altura c varia de 132,64-163,45 µm. A razão
entre as medidas a / c varia de 1,2-1,3. A parte apical da mandíbula é bem esclerotinizada e a
parte basal é fortemente esclerotinizada. Alguns exemplares dessa espécie possuem dois
orifícios na base da mandíbula. Os dentes são pontiagudos e possuem um dente secundário

88
bem desenvolvido. O comprimento do esclerito hipofaringeal varia de 112,16-154,92 µm e a
largura de 72,21-89,78 µm, apresentando a parte apical fortemente esclerotinizada e a parte
basal bem esclerotinizada. O esclerito epifaringeal, esclerito labial e a barra parastomal (=
esclerito hipostomal) apresentam as mesmas características que as larvas de terceiro instar. O
corno dorsal é formado por uma área bem esclerotinizada e uma hialina, o comprimento de
ambas as áreas variam de 179,62-264,21 µm e 343,54-492,15 µm, respectivamente. O arco
dorsal possui altura variando de 197,97-280,86 µm. O esclerito anterior é ausente. A razão
entre o entalhe do corno (“cornu notch”) e corno dorsal é de 0,5. O corno ventral é fracamente
esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente esclerotinizado e possui
anteriormente uma barra faringeal com 7-8 cristas que formam estrias estendendo-se por todo
o comprimento do corno ventral. O corno ventral possui comprimento variando de 400,20-
488,33 µm. O corno ventral é 1,8-2,2 vezes o tamanho da área esclerotinizada do corno
dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 11-15 túbulos (Fig. 1I). O túbulo apresenta comprimento entre 10,60-17,08
µm, largura distal entre 7,44-14,03 µm e largura basal entre 6,21-11,47 µm. O espiráculo
anterior apresenta na linha média altura variando de 64,01-114,71 µm; largura distal de
105,33-128,00 µm e largura basal próximo a abertura da traquéia variando de 21,48-46,85
µm.
O segmento caudal com tubérculos e sensilas apresentam o mesmo padrão que as
larvas de terceiro instar. O espiráculo posterior e o lóbulo anal estão localizados da mesma
forma que nas larvas de terceiro instar. O lóbulo anal é inteiro.
O espiráculo posterior apresenta três fendas cada qual com membrana estreita e poucas
trabéculas transversais, seguindo o mesmo padrão que as larvas de terceiro instar (Fig. 1J). As
fendas possuem comprimento variando de 33,32-48,71 µm e largura de 11,45-18,01 µm. A
razão entre o comprimento e a largura das fendas varia de 2,7-2,9. O espiráculo posterior
possui cicatriz da ecdise aparente. A câmara onde localizam-se as fendas é oval e apresenta
diâmetro próximo a traquéia variando de 64,87-80,19 µm. O processo espiracular apresenta
quatro grupos de cerdas, onde SP-I possui 12-17 troncos e 31-40 extremidades
(eventualmente 23-26); a razão entre essas extremidades / troncos varia de 2,3-3,5; esse grupo
de cerdas apresenta largura basal variando de 15,93-30,27 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,4-0,6. O grupo SP-II possui 5-9 troncos e 12-
23 extremidades. O grupo SP-III possui 7-11 troncos (eventualmente 5) e 15-20 extremidades
(eventualmente 24). O grupo SP-IV possui 12-15 troncos (eventualmente 9-10) e 23-34

89
extremidades; a razão entre essas extremidades / troncos varia de 1,9-2,2; esse grupo de
cerdas apresenta largura basal variando de 14,08-25,62 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,4-0,5. A média do comprimento das cerdas é
0,7 vezes o comprimento das fendas espiraculares.
Anastrepha turpiniae Stone (Fig. 3)
Material Examinado. Foram examinadas 34 larvas (22 larvas de terceiro instar e 12
de segundo instar) dissecadas de frutos de Terminalia cattapa L. Combretaceae (localmente
conhecida como “castanhola”), árvore exótica introduzida no Brasil.
Características das larvas de terceiro instar. As larvas possuem comprimento
variando de 5,00-11,58 mm e largura de 1,00-2,06 mm. As larvas são alongadas, cilíndricas,
afiladas nos segmentos torácicos, com segmento caudal truncado e possuem coloração creme
amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 1,94-3,09 µm.
O padrão de distribuição dos espinhos dorsais é T1 com 4-5 fileiras (eventualmente 2-3); T2
com 4-5 fileiras (eventualmente 3); T3 com 0-2 fileiras; A1 ao A8 não apresentam fileiras de
espinhos. O padrão de distribuição dos espinhos ventrais é T1 com 4-6 fileiras (eventualmente
3 ou 8); T2 com 4-6 fileiras (eventualmente 3); T3 com 2-4 fileiras (eventualmente 5); A1
com 6-8 fileiras; A2 com 10-13 fileiras; A3 com 12-14 fileiras; A4 com 11-14 fileiras; A5
com 12-14 fileiras (eventualmente 10); A6 e A7 com 11-14 fileiras; A8 com 10-13 fileiras.
Todos os segmentos abdominais (A1 ao A8) apresentam protuberâncias ventrais para
locomoção. Ao redor do lóbulo anal, foram visualizadas fileiras irregulares de espinhos
ventral e dorsalmente variando entre 3-4 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 12,18-19,49 µm, botão apical com diâmetro variando de
9,49-13,34 µm e altura de 6,62-14,13 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 12,70-19,71 µm e altura de 6,82-12,31 µm. O órgão
sensorial bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 4,29-9,22 µm.
As saliências orais variam de 7-8 (Fig. 1A).
A Figura 1D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 0,95-1,44 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,23-0,28 mm, o comprimento b varia de

90
0,14-0,19 mm e a altura c varia de 0,15-0,20 mm. A razão entre as medidas a / c varia de 1,4-
1,5. Os dentes são longos e pontiagudos. O comprimento do esclerito hipofaringeal varia de
0,17-0,21 mm e a largura de 0,12-0,16 mm. Em vista dorsal foi visualizado que o esclerito
epifaringeal apresenta um lóbulo medial direcionado anteriormente. Também em vista dorsal
foi visualizado o esclerito labial em formato de ferradura, curto, robusto e esclerotinizado. A
barra parastomal (= esclerito hipostomal) é arqueada e estende-se quase completamente sobre
o esclerito hipofaringeal. O esclerito ventral localiza-se abaixo do esclerito faringeal. O corno
dorsal é formado por uma área bem esclerotinizada e uma hialina, o comprimento de ambas as
áreas variam de 0,29-0,48 mm e 0,40-0,74 mm, respectivamente. A ponte dorsal (“dorsal
bridge”) apresenta uma projeção anterior do corno dorsal levemente esclerotinizada. O arco
dorsal possui altura variando de 0,28-0,36 mm. O esclerito anterior é esclerotinizado e possui
formato irregular. A razão entre o entalhe do corno (“cornu notch”) e corno dorsal varia de
0,6-0,7. O corno ventral é fracamente esclerotinizado. O filtro faringeal (“pharyngeal filter”) é
fracamente esclerotinizado e possui anteriormente uma barra faringeal com 7-9 cristas que
formam estrias estendendo-se por todo o comprimento do corno ventral. O corno ventral
possui comprimento variando de 0,58-0,99 mm. O corno ventral é 2,0 vezes o tamanho da
área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 11-16 túbulos (Figs. 1B e E). O túbulo apresenta comprimento entre 19,25-
33,09 µm, largura distal entre 12,75-25,79 µm e largura basal entre 10,36-22,31µm. O
espiráculo anterior apresenta na linha média altura variando de 112,67-160,60 µm; largura
distal de 196,02-262,80 µm e largura basal próximo a abertura da traquéia variando de 97,25-
139,78 µm.
A Figura 1F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1
é distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente
desenvolvidos e associados com uma sensila fracamente desenvolvida. I2 é disposto
lateralmente e algumas vezes levemente ventral em relação a I1. L1 e V são pouco
desenvolvidos. D1 e I1 são associados com sensila fracamente desenvolvida. Alguns
exemplares podem ter três tubérculos ventrais (V1, V2 e V3). Os espiráculos posteriores estão
localizados acima da linha média horizontal. O lóbulo anal é bífido e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Figs. 1C e G), possuindo comprimento variando de 71,86-
98,86 µm e largura de 15,06-23,85 µm. A razão entre o comprimento e a largura das fendas

91
varia de 4,1-4,7. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 126,55-
174,03 µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 10-32
troncos e 13-42 extremidades; a razão entre essas extremidades / troncos é de 1,3; esse grupo
de cerdas apresenta largura basal variando de 20,76-39,86 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,2-0,4. O grupo SP-II possui 3-12 troncos e
12-24 extremidades. O grupo SP-III possui 4-10 troncos e 9-28 extremidades. O grupo SP-IV
possui 10-21 troncos (eventualmente 7-8) e 18-46 extremidades; a razão entre essas
extremidades / troncos varia de 1,8-2,1; esse grupo de cerdas apresenta largura basal variando
de 20,56-37,30 µm; a razão da largura basal / comprimento dessa fenda espiracular varia de
0,2-0,3. A média do comprimento das cerdas é 0,4 vezes o comprimento das fendas
espiraculares.
Características das larvas de segundo instar. As larvas possuem comprimento
variando de 3,50-4,50 mm e largura de 0,50-0,87 mm. As larvas de segundo instar apresentam
o mesmo formato que as larvas de terceiro instar. As larvas possuem coloração variando de
creme esbranquiçado a creme amarelado.
Os espinhos e as fileiras de espinhos apresentam as mesmas características que as
observadas nas larvas de terceiro instar. A largura basal dos espinhos varia de 1,84-3,03 µm.
O padrão de distribuição dos espinhos dorsais é T1 com 4-5 fileiras; T2 com 3-5 fileiras; T3
com 2-3 fileiras, A1 com 0-3 fileiras; A2 com 2-5 fileiras (eventualmente 0); A3 com 4-6
fileiras; A4 com 4-5 fileiras; A5 com 5 fileiras (eventualmente 0); A6 com 0-3 fileiras; A7 e
A8 não apresentam fileiras de espinhos. O padrão de distribuição dos espinhos ventrais é T1 e
T2 com 4-5 fileiras; T3 com 1-3 fileiras (eventualmente 0 ou 5); A1 com 5-10 fileiras; A2
com 10-11 fileiras (eventualmente 12); A3 com 10-12 fileiras; A4 com 11-13 fileiras; A5 com
10-12 fileiras (eventualmente 9); A6 com 11-12 fileiras (eventualmente 10 ou 13); A7 com
11-12 fileiras (eventualmente 13); A8 com 9-11 fileiras (eventualmente 12). Todos os
segmentos abdominais (A1 ao A8) apresentam protuberâncias ventrais para locomoção. Ao
redor do lóbulo anal foram visualizadas três fileiras irregulares de espinhos ventral e
dorsalmente.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um
anel basal com diâmetro variando de 8,44-11,88 µm, botão apical com diâmetro variando de
6,59-8,48 µm e altura de 4,43-5,94 µm. O órgão sensorial maxilar é cilíndrico, levemente
afilado, possui diâmetro variando 10,21-18,26 µm e altura de 6,53-7,10 µm. O órgão sensorial

92
bucal (“stomal sensory organ”) é cilíndrico e com altura variando de 3,76-5,29 µm. As
saliências orais variam de 6-8.
O esqueleto céfalo-faríngeo possui comprimento total variando de 622,04-784,97 µm
(Fig. 1H). A mandíbula apresenta três medidas: o comprimento a varia de 123,32-160,60 µm,
o comprimento b varia de 77,05-94,07 µm e a altura c varia de 97,19-122,70 µm. A razão
entre as medidas a / c varia de 1,2-1,3. A parte apical da mandíbula é bem esclerotinizada e a
parte basal é fortemente esclerotinizada. Alguns exemplares dessa espécie possuem dois
orifícios na base da mandíbula. Os dentes são pontiagudos e possuem um dente secundário
bem desenvolvido. O comprimento do esclerito hipofaringeal varia de 105,57-163,11 µm e a
largura de 72,10-83,86 µm, apresentando a parte apical fortemente esclerotinizada e a parte
basal bem esclerotinizada. O esclerito epifaringeal, esclerito labial e a barra parastomal (=
esclerito hipostomal) apresentam as mesmas características que as larvas de terceiro instar. O
corno dorsal é formado por uma área bem esclerotinizada e uma hialina, o comprimento de
ambas as áreas variam de 162,22-236,21 µm e 224,10-396,84 µm, respectivamente. O arco
dorsal possui altura variando de 129,70-199,34 µm. O esclerito anterior é ausente. A razão
entre o entalhe do corno (“cornu notch”) e corno dorsal é de 0,5. O corno ventral é fracamente
esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente esclerotinizado e possui
anteriormente uma barra faringeal com sete cristas que formam estrias estendendo-se por todo
o comprimento do corno ventral. O corno ventral possui comprimento variando de 328,55-
497,61 µm. O corno ventral é 2,0-2,1 vezes o tamanho da área esclerotinizada do corno
dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos,
variando entre 13-15 túbulos (Fig. 1I). O túbulo apresenta comprimento entre 11,50-16,52
µm, largura distal entre 8,01-11,46 µm e largura basal entre 5,00-7,87 µm. O espiráculo
anterior apresenta na linha média altura variando de 76,72-91,68 µm; largura distal de 98,49-
127,65 µm e largura basal próximo a abertura da traquéia variando de 33,27-91,68 µm.
O segmento caudal com tubérculos e sensilas apresentam o mesmo padrão que as
larvas de terceiro instar. O espiráculo posterior e o lóbulo anal estão localizados da mesma
forma que nas larvas de terceiro instar. O lóbulo anal é bífido.
O espiráculo posterior apresenta três fendas cada qual com membrana estreita e poucas
trabéculas transversais, seguindo o mesmo padrão que as larvas de terceiro instar (Fig. 1J). As
fendas possuem comprimento variando de 27,00-34,17 µm e largura de 11,58-18,34 µm. A
razão entre o comprimento e a largura das fendas varia de 1,8-2,3. O espiráculo posterior
possui cicatriz da ecdise aparente. A câmara onde localizam-se as fendas é arredondada e

93
apresenta diâmetro próximo a traquéia variando de 60,19-77,83 µm. O processo espiracular
apresenta quatro grupos de cerdas, onde SP-I possui 11-13 troncos (eventualmente 10) e 22-
29 extremidades (eventualmente 19-20); a razão entre essas extremidades / troncos varia de
2,0-2,2; esse grupo de cerdas apresenta largura basal variando de 11,42-15,49 µm; a razão da
largura basal / comprimento dessa fenda espiracular é de 0,4. O grupo SP-II possui 4-6
troncos e 8-13 extremidades (eventualmente 15). O grupo SP-III possui 6-9 troncos e 14-18
extremidades. O grupo SP-IV possui 10-12 troncos (eventualmente 8) e 19-27 extremidades
(eventualmente 37); a razão entre essas extremidades / troncos varia de 1,9-2,2; esse grupo de
cerdas apresenta largura basal variando de 10,25-14,33 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,3-0,4. A média do comprimento das cerdas é
0,5-0,7 vezes o comprimento das fendas espiraculares.
Discussão
Para as três espécies estudadas, foi observada diferença considerável no tamanho das
larvas de segundo e terceiro instares. Para todas as estruturas analisadas, as larvas de terceiro
instar possuem cerca de duas vezes o tamanho das larvas de segundo instar. Em todas as
larvas de segundo instar, foi observada a presença de um dente secundário na mandíbula, mas
essa característica não foi observada nas larvas de terceiro instar. Contudo, foi observado que
algumas estruturas como o número de saliências orais e o formato do lóbulo anal mantiveram
o mesmo padrão para ambos os instares.
Diferenças entre os instares larvais incluem o número de estrias no filtro faringeal
(“pharyngeal filter”) em A. bahiensis; para A. coronilli, maior número de fileiras de espinhos
no segmento T1 nas larvas de segundo instar, duas vezes o número de cerdas no espiráculo
posterior nas larvas de terceiro instar em relação às larvas de segundo instar. Em A. turpiniae,
larvas de segundo instar possuem fileiras de espinhos dorsais nos segmentos A1 ao A6, sendo
ausente nas larvas de terceiro instar e o número de túbulos no espiráculo anterior também
difere entre as larvas de segundo e terceiro instar para esta espécie.
As larvas de terceiro instar de A. bahiensis podem ser diferenciadas das outras duas
espécies estudadas com base no menor número de saliências orais, menor tamanho da
mandíbula e menor comprimento do ECF. A diferença entre larvas de terceiro instar de A.
coronilli e larvas das demais espécies inclui uma mandíbula maior, ECF maior, maior número
de saliências orais, maior número de cerdas no espiráculo posterior e lóbulo anal inteiro,
enquanto que nas outras duas espécies o lóbulo anal é bífido. As características que
distinguem A. turpiniae das outras espécies neste estudo são presença de espinhos dorsais no

94
segmento T3, número semelhante de fileiras de espinhos nos segmentos ventrais A4, A6 e A7,
menor razão entre troncos/extremidades no grupo de cerdas SP-I no espiráculo posterior e
menor número de estrias no filtro faringeal (“pharyngeal filter”).
Em larvas de terceiro instar de Anastrepha, a presença do esclerito ventral foi
registrado para Anastrepha leptozona Hendel por Frías et al. (2009) pela primeira vez. Estes
autores sugerem que essa estrutura “poderia ser usada para o reconhecimento dessa espécie”.
Contudo, no presente estudo foi observado esclerito ventral para algumas larvas de terceiro
instar das três espécies analisadas e de outras cinco espécies de Anastrepha (Dutra et al. dados
não publicados). Assim, essa estrutura não deve ser usada como caractere diagnóstico. Em
larvas de terceiro instar de algumas espécies já descritas, o esclerito ventral ainda não havia
sido registrado provavelmente devido à dificuldade na visualização, pois essa estrutura é
translucida e frágil.
Para as três espécies analisadas, a maioria dos caracteres descritos apresentou
sobreposição com outras cinco espécies já descritas do grupo fraterculus: Anastrepha
distincta Greene, Anastrepha fraterculus (Wiedemann), Anastrepha ludens (Loew),
Anastrepha obliqua (Macquart) e Anastrepha suspensa (Loew) (Setck et al. 1990, White e
Elson-Harris 1992, Norrbom et al. 1999). Por exemplo, para larvas de terceiro instar das três
espécies descritas, o número de saliências orais varia de 6 a 10, enquanto que em A. distincta,
A. fraterculus, A. obliqua e A. suspensa varia de 7 a 12 (Steck et al. 1990, White e Elson-
Harris 1992) e apenas A. ludens possui mais de 16 saliências orais (Carroll e Wharton 1989).
Um caractere útil para distinguir algumas espécies do grupo fraterculus seria o número de
extremidades das cerdas no grupo SP-I no espiráculo posterior, pois A. coronilli possui entre
45-76 extremidades, sendo este o maior número de extremidades descritas até o momento,
enquanto que A. ludens possui entre 17-28 extremidades e outras espécies possuem entre 10-
17 extremidades.
Várias espécies de Anastrepha do grupo fraterculus infestam Psidium (Myrtaceae)
(Norrbom et al. 1999, Araujo e Zucchi 2003). Atualmente, contudo existem informações
taxonômicas apenas para cinco espécies além das três analisadas neste estudo. Devido à
considerável sobreposição de caracteres das espécies desse grupo, faz-se necessário o
desenvolvimento de outros métodos que efetivamente permitam a distinção dessas espécies,
como análise morfométrica e/ou molecular.
É necessária a análise de um número maior de espécies do grupo fraterculus para que
suas larvas sejam descritas e assim ajudem a entender o nível de variação entre as espécies
que pertencem a este grupo. Isso provavelmente ajudaria a interpretar o nível de variação ou a

95
ausência dela quando as informações sobre as larvas fossem comparadas com outras
previamente relatadas para espécies do complexo fraterculus.
Agradecimentos
Os autores agradecem a Hilton Tulio Costi e Rolf Junior Ferreira Silva (Museu
Paraense Emílio Goeldi) pelo auxílio na preparação do material para MEV. Esse projeto foi
financiado pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq -
projeto n° 575664/2008-8), pela Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES - projeto n° 23038.037185/2008-41) e pela Fundação de Amparo a
Pesquisa do Estado Amazonas (FAPEAM - projeto n 014-017/2004). Os autores também
agradecem ao CNPq pela bolsa de doutorado concedida a VSD.

96
Fig. 1. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. bahiensis. Larvas de terceiro
instar (A) Saliências Orais. (B e E) Espiráculo Anterior. (C e G) Espiráculo Posterior. (D)
Esqueleto Céfalo-faríngeo. (F) Ilustração do Segmento Caudal. Larvas de segundo instar (H)
Esqueleto Céfalo-faríngeo. (I) Espiráculo Anterior. (J) Espiráculo Posterior. Abreviações: Ma:
comprimento da mandíbula a; Mb: comprimento da mandíbula b; Mc: altura da mandíbula c;
ASc: esclerito anterior; ES: esclerito epifaringeal; LS: esclerito labial.
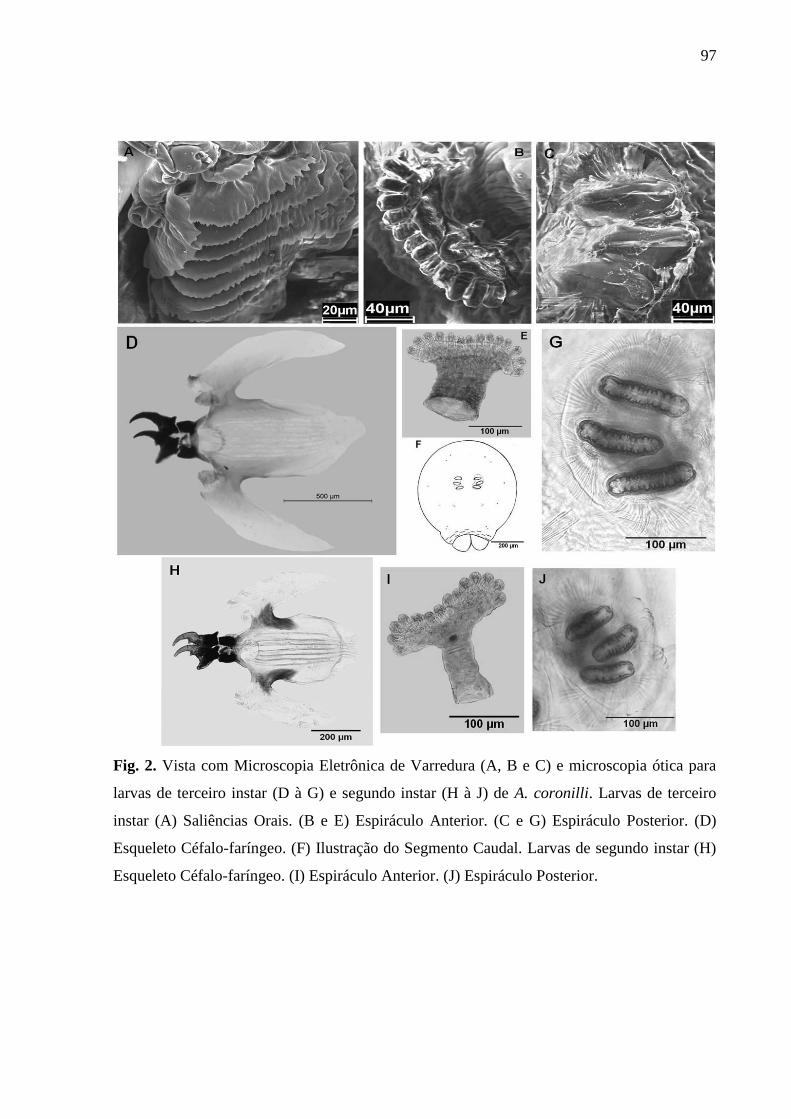
97
Fig. 2. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para
larvas de terceiro instar (D à G) e segundo instar (H à J) de A. coronilli. Larvas de terceiro
instar (A) Saliências Orais. (B e E) Espiráculo Anterior. (C e G) Espiráculo Posterior. (D)
Esqueleto Céfalo-faríngeo. (F) Ilustração do Segmento Caudal. Larvas de segundo instar (H)
Esqueleto Céfalo-faríngeo. (I) Espiráculo Anterior. (J) Espiráculo Posterior.

97
Fig. 3. Vista com Microscopia Eletrônica de Varredura (A, B e C) e microscopia ótica para larvas
de terceiro instar (D à G) e segundo instar (H à J) de A. turpiniae. Larvas de terceiro instar (A)
Saliências Orais. (B e E) Espiráculo Anterior. (C e G) Espiráculo Posterior. (D) Esqueleto
Céfalo-faríngeo. (F) Ilustração do Segmento Caudal. Larvas de segundo instar (H) Esqueleto
Céfalo-faríngeo. (I) Espiráculo Anterior. (J) Espiráculo Posterior.

99
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G.
Description of Third Instar Larvae of Five Anastrepha
Species (Diptera: Tephritidae). Manuscrito em preparação
para Annals of the Entomological Society of America.

100
Description of Third Instar Larvae of Five Anastrepha Species (Diptera:
Tephritidae)
VIVIAN SIQUEIRA DUTRA1, BEATRIZ RONCHI-TELES
2, GARY J. STECK
3 E JANISETE
GOMES SILVA4
RESUMO A morfologia de larvas de terceiro instar de cinco espécies de Anastrepha,
Anastrepha pickeli Lima, Anastrepha pulchra Stone, Anastrepha sororcula Zucchi e Anastrepha
zenildae Zucchi e a morfologia de larvas de segundo e terceiro instar de Anastrepha curitis Stone
foram descritas e documentadas pela primeira vez. Também são discutidos caracteres que podem
ser utilizados para diferenciar as cinco espécies.
PALAVRAS-CHAVE moscas-das-frutas, larva, morfologia, espiráculo posterior
As espécies de Anastrepha Schiner (Diptera: Tephiritidae) são encontradas nas regiões tropicais e
subtropicais e estão amplamente distribuídas desde o sul dos Estados Unidos até o norte da
Argentina (Aluja 1994). Este é o gênero de maior importância econômica de moscas-das-frutas
da região Neotropical com cerca de 240 espécies descritas (Norrbom et al. 1999, Zucchi 2008,
Norrbom e Korytkowski 2009). Até o momento, no Brasil foram registradas 112 espécies
(Norrbom et al. 1999; Zucchi 2007, 2008; Uramoto e Zucchi 2010).
A identificação de larvas de Anastrepha é importante para estudos de biologia básica e
evolução deste grupo e também porque a larva é o estágio geralmente encontrados em frutos
interceptados nos portos e aeroportos. Porém, estudos sobre a descrição da morfologia de larvas
de terceiro instar atualmente estão disponíveis apenas para 13 espécies de Anastrepha e algumas
dessas descrições são incompletas (Steck e Wharton 1988, Steck et al. 1990, Norrbom et al.
1999).
1Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970, Manaus, AM, Brasil. 2Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970
Manaus, AM, Brasil. 3Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL 32614-
7100. 4Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil.

101
Para complementar o conhecimento sobre os estágios imaturos de Anastrepha, o presente
estudo descreve de maneira detalhada a morfologia das larvas de terceiro instar de cinco espécies,
Anastrepha curitis Stone, Anastrepha pickeli Lima, Anastrepha pulchra Stone, Anastrepha
sororcula Zucchi e Anastrepha zenildae Zucchi e de larvas de segundo instar de A. curitis. No
Brasil, A. curitis foi registrada somente nos estados do Amazonas e do Pará, enquanto A. pickeli é
amplamente distribuída em 16 estados brasileiros, A. pulchra é restrita ao estado do Amazonas,
A. sororcula foi registrada em 20 estados das diversas regiões brasileiras e A. zenildae tem sua
ocorrência relatada em 18 estados de todas as regiões brasileiras, com exceção da região sul
(Zucchi 2007, 2008).
Material e Métodos
Larvas de A. curitis, A. pickeli, A. pulchra, A. sororcula e A. zenildae foram dissecadas de
frutos coletados em Manaus (03°06’07’’S; 60°01’30’’W) no estado do Amazonas, em Porto
Velho (08°45’43’’S; 63°54’14’’W) no estado de Rondônia, em Maracás (13°26’28’’S;
40°25’51’’W) no estado da Bahia, em Limoeiro do Norte (05°08’44’’S; 38°05’53’’W) no estado
do Ceará e em Mossoró (05°11’15’’S; 37°20’39’’W) no estado do Rio Grande do Norte. Para
confirmação da espécie, cerca de três larvas foram dissecadas dos frutos e outras foram deixadas
nos frutos até a emergência dos adultos. As larvas foram mortas e armazenadas em etanol 100%.
Características externas como comprimento, largura, coloração e número de segmentos
foram medidas com micrométrica ocular em um estereomicroscópio Wild M3C no Laboratório
de Entomologia Agrícola do Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus,
Amazonas, Brasil. As larvas foram clarificadas com hidróxido de potássio 10% em temperatura
ambiente, posteriormente as larvas foram colocadas em lâminas escavadas em um meio contendo
álcool gel e glicerina (1:1) para visualização das características. As fileiras de espinhos dorsais e
ventrais foram contadas de acordo com as linhas médias dorsal e ventral, respectivamente. O
esqueleto céfalo-faríngeo (ECF) foi removido, colocado em glicerina e ampliado 80X para as
medições das larvas de terceiro instar e ampliado 100X para visualização das larvas de segundo
instar. Para ambos os instares, as medidas dos espiráculos anteriores e posteriores foram
realizadas com objetivas de ampliação 200X e 400X. Essas estruturas foram fotografadas usando-
se câmera digital (Olympus DP72) acoplada ao microscópio ótico (Olympus BX51) e as
ilustrações do segmento caudal foram desenhadas com câmara clara (Leica DM750) no

102
Laboratório de Citotaxonomia e Insetos Aquáticos (INPA). O ECF das larvas de terceiro instar
foram colocados entre lâmina e lamínula com etanol 80% e glicerina, fotografados, medidos e
observadas as características utilizando a câmera digital (Leica DFC420) acoplada ao
estereomicroscópio (Leica M165C) do Laboratório de Prospecção de Bioativos de Insetos
(INPA).
A terminologia utilizada foi baseada em Teskey (1981), Steck e Malavasi (1988), Steck e
Wharton (1988) e Frías et al. (2009). Foram utilizados os mesmos pontos de medições, as
mesmas abreviações para os segmentos torácicos, segmentos abdominais, espiráculo posterior e
segmento caudal que aqueles utilizados por Steck e Malavasi (1988) e Steck e Wharton (1988). O
comprimento total do esqueleto céfalo-faríngeo foi medido desde o ápice da mandíbula até a
extremidade final do corno ventral. O comprimento do corno ventral foi medido desde a barra
faringeal até extremidade final das estrias.
A preparação para microscopia eletrônica de varredura (MEV) foi realizada de duas
formas, na primeira as larvas foram transferidas para cápsulas de silicone, desidratadas em série
alcoólica e no secador de ponto crítico de CO2 por três horas, sendo metalizadas com uma
camada de ouro e examinadas no microscópio eletrônico de varredura LEO 1450VP no
Laboratório Institucional de Microscopia Eletrônica de Varredura do Museu Paraense Emílio
Goeldi, Belém, Pará, Brasil.
Na segunda, as larvas foram montadas em porta amostras de alumínio e examinadas no
modo ambiental no microscópio eletrônico de varredura Quanta 250 (Fei Company) no
Laboratório de Técnicas Mineralógicas da Universidade Federal do Amazonas (UFAM) em
Manaus, Amazonas, Brasil. Para a MEV foram utilizadas pelo menos dez larvas de cada espécie.
Espécimes testemunho das larvas associadas com as fêmeas foram depositados na
Coleção de Invertebrados do Instituto Nacional de Pesquisas da Amazônia (INPA).
Resultados
As larvas de A. curitis, A. pickeli, A. pulchra, A. sororcula e A. zenildae são similares em
seu aspecto geral. Caracteres diagnósticos para diferenciar essas cinco espécies incluem ausência
de espinhos dorsais em apenas um segmento, número de espinhos ventrais nos segmentos,
formato do lóbulo anal, número de troncos/extremidades das cerdas no espiráculo posterior e

103
algumas características do esqueleto céfalo-faríngeo como comprimento total desta estrutura,
tamanho e formato da mandíbula.
As larvas de segundo e terceiro instar de A. curitis podem ser diferenciadas com base no
número de fileiras de espinhos ventrais nos segmentos (exceto nos segmentos A3 e A8), número
de saliências orais, número de cerdas no espiráculo posterior e número de cristas no filtro
faringeal (“pharyngeal filter”).
Anastrepha curitis Stone (Fig. 1)
Material Examinado. Foram examinadas 35 larvas (26 larvas de terceiro instar e nove de
segundo instar) coletadas em Manaus no estado do Amazonas. As larvas foram dissecadas de
frutos de Passiflora nitida Kunth, Passifloraceae (localmente conhecida como “maracujá-do-
mato”), uma liana típica da região Amazônica.
Características das larvas de terceiro instar. As larvas possuem comprimento variando
de 7,54-11,27 mm e largura de 1,24-1,96 mm. As larvas são alongadas, cilíndricas, afiladas nos
segmentos torácicos, com segmento caudal truncado e possuem coloração creme amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 3,83-7,24 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 8-10 fileiras (eventualmente 7); T2 com 7-8
fileiras (eventualmente 9); T3 com 6-8 fileiras; A1 com 4-6 fileiras; A2 com 4-5 fileiras
(eventualmente 3 ou 6); A3 com 2-4 fileiras; A4 e A5 com 3-4 fileiras; A6 com 1-3 fileiras; A7
com 0-1 fileiras; A8 não apresentam fileiras de espinhos. O padrão de distribuição dos espinhos
ventrais é T1 com 9-15 fileiras; T2 com 7-9 fileiras; T3 com 6-8 fileiras (eventualmente 9-10);
A1 com 7-10 fileiras (eventualmente 12); A2 com 12-16 fileiras; A3 com 13-16 fileiras; A4 com
14-18 fileiras; A5 com 14-17 fileiras (eventualmente 13); A6 com 13-17 fileiras; A7 com 14-16
fileiras (eventualmente 12 ou 18); A8 com 12-15 fileiras. O segmento torácico T3 e todos os
abdominais (A1 ao A8) apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo
anal foram visualizadas fileiras irregulares de espinhos ventral e dorsalmente, variando entre 4-5
fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 19,88-26,07 µm, botão apical com diâmetro variando de 12,37-

104
16,54 µm e altura de 9,98-14,80 µm. O órgão sensorial maxilar é cilíndrico, levemente afilado,
possui diâmetro variando 19,57-31,95 µm e altura de 9,14-13,49 µm. O órgão sensorial bucal
(“stomal sensory organ”) é cilíndrico e com altura variando de 7,40-9,67 µm. As saliências orais
variam de 8-11 (Fig. 1A).
A Figura 1D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 1,36-1,65 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,33-0,36 mm, o comprimento b varia de 0,22-
0,26 mm e a altura c varia de 0,21-0,25 mm. A razão entre as medidas a / c varia de 1,4-1,6. Os
dentes são curtos, pontiagudos e formam curvatura levemente retangular. O comprimento do
esclerito hipofaringeal varia de 0,21-0,26 mm e a largura de 0,22-0,28 mm. Em vista dorsal foi
visualizado o esclerito epifaringeal que apresenta um lóbulo medial direcionado anteriormente.
Também em vista dorsal foi visualizado o esclerito labial em formato de ferradura, curto, robusto
e bastante esclerotinizado. A barra parastomal (= esclerito hipostomal) estende-se quase
completamente sobre o esclerito hipofaringeal. O esclerito ventral localiza-se abaixo do esclerito
faringeal. O corno dorsal é formado por uma área bem esclerotinizada e uma hialina, o
comprimento de ambas as áreas variam de 0,49-0,68 mm e 0,68-0,91 mm, respectivamente. A
ponte dorsal (“dorsal bridge”) apresenta uma projeção anterior do corno dorsal levemente
esclerotinizada. O arco dorsal possui altura variando de 0,40-0,48 mm. O esclerito anterior é
esclerotinizado e possui formato irregular. A razão entre o entalhe do corno (“cornu notch”) e
corno dorsal varia de 0,6-0,7. O corno ventral é fracamente esclerotinizado. O filtro faringeal
(“pharyngeal filter”) é fracamente esclerotinizado e possui anteriormente uma barra faringeal
com 8-9 cristas (eventualmente 7) que formam estrias estendendo-se por todo o comprimento do
corno ventral. O corno ventral possui comprimento variando de 0,80-1,01 mm. O corno ventral
varia de 1,4-1,6 vezes o tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 19-25 túbulos (Figs. 1B e E). O túbulo apresenta comprimento entre 21,79-33,87 µm,
largura distal entre 15,47-34,81 µm e largura basal entre 12,20-22,11 µm. O espiráculo anterior
apresenta na linha média altura variando de 136,26-227,43 µm; largura distal de 331,69-416,63
µm e largura basal próximo a abertura da traquéia variando de 161,66-209,16 µm.
A Figura 1F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1 é

105
distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente desenvolvidos e
associados com uma sensila fracamente desenvolvida. I2 é disposto lateralmente e algumas vezes
levemente ventral em relação a I1. L1 e V são pouco desenvolvidos. D1 e I1 são associados com
sensila fracamente desenvolvida. Todos os espécimes apresentaram dois tubérculos ventrais (V1,
V2 e V3). Os espiráculos posteriores estão localizados acima da linha média horizontal. O lóbulo
anal é bífido e podendo ser protraído ou retraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Figs. 1C e G), possuindo comprimento variando de 106,21-
147,91 µm e largura de 16,91-31,49 µm. A razão entre o comprimento e a largura das fendas
varia de 4,7-6,3. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 103,31-223,44
µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 10-18 troncos
(eventualmente 20) e 25-39 extremidades (eventualmente 42); a razão entre essas extremidades /
troncos varia de 2,1-2,5; esse grupo de cerdas apresenta largura basal variando de 29,24-57,94
µm; a razão da largura basal / comprimento dessa fenda espiracular varia de 0,2-0,4. O grupo SP-
II possui 6-11 troncos e 13-26 extremidades (eventualmente 29). O grupo SP-III possui 6-11
troncos (eventualmente 13) e 15-25 extremidades (eventualmente 27). O grupo SP-IV possui 9-
15 troncos (eventualmente 7) e 23-32 extremidades (eventualmente 18); a razão entre essas
extremidades / troncos varia de 2,1-2,6; esse grupo de cerdas apresenta largura basal variando de
22,97-53,23 µm; a razão da largura basal / comprimento dessa fenda espiracular varia de 0,2-0,4.
A média do comprimento das cerdas é 0,4 vezes o comprimento das fendas espiraculares. As
principais características das larvas para diagnóstico desta espécie estão apresentadas na Tabela
1.
Características das larvas de segundo instar. As larvas possuem comprimento variando
de 6,06-6,87 mm e largura de 1,25-1,37 mm. As larvas de segundo instar apresentam o mesmo
formato e coloração que as larvas de terceiro instar.
Os espinhos e as fileiras de espinhos apresentam as mesmas características que as
observadas nas larvas de terceiro instar. A largura basal dos espinhos varia de 2,66-3,93 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 7-8 fileiras (eventualmente 6); T2 com 6-7
fileiras (eventualmente 8); T3 com 5-6 fileiras; A1 com 4-5 fileiras; A2 com 5-7 fileiras
(eventualmente 8); A3 com 6-8 fileiras (eventualmente 5); A4 com 6 ou 8 fileiras; A5 com 5-8

106
fileiras; A6 com 3-5 fileiras (eventualmente 2); A7 com 3-4 fileiras (eventualmente 0-1); A8 com
0-2 fileiras. O padrão de distribuição dos espinhos ventrais é T1 com 8-12 fileiras (eventualmente
14); T2 com 5-9 fileiras; T3 com 5-8 fileiras; A1 com 8-12 fileiras; A2 com 13-16 fileiras
(eventualmente 11); A3 com 13-16 fileiras (eventualmente 12); A4 com 13-18 fileiras; A5 com
15-17 fileiras; A6 com 14-16 fileiras (eventualmente 12); A7 com 13-16 fileiras; A8 com 12-15
fileiras. Todos os abdominais (A1 ao A8) apresentam protuberâncias ventrais para locomoção Ao
redor do lóbulo anal foram visualizadas fileiras irregulares de espinhos ventral e dorsalmente,
variando entre 4-5 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 13,28-19,57 µm, botão apical com diâmetro variando de 7,96-
10,00 µm e altura de 5,60-6,92 µm. O órgão sensorial maxilar é cilíndrico, fracamente
esclerotinizado e possui diâmetro variando 12,53-18,24 µm. O órgão sensorial bucal (“stomal
sensory organ”) é cilíndrico e com altura variando de 4,74-6,03 µm. As saliências orais variam de
6-7.
O esqueleto céfalo-faríngeo possui comprimento total variando de 627,72-930,80 µm
(Fig. 1H). A mandíbula apresenta três medidas: o comprimento a varia de 158,32-185,48 µm, o
comprimento b varia de 85,81-112,55 µm e a altura c varia de 124,19-154,13 µm. A razão entre
as medidas a / c varia de 1,2-1,3. A parte apical da mandíbula é bem esclerotinizada e a parte
basal é fortemente esclerotinizada. Alguns exemplares dessa espécie possuem dois orifícios na
base da mandíbula. Os dentes são pontiagudos e possuem um dente secundário bem
desenvolvido. O comprimento do esclerito hipofaringeal varia de 123,14-148,77 µm e a largura
de 82,36-130,60 µm, apresentando a parte apical fortemente esclerotinizada e a parte basal bem
esclerotinizada. O esclerito epifaringeal, esclerito labial e a barra parastomal (= esclerito
hipostomal) apresentam as mesmas características que as larvas de terceiro instar. O corno dorsal
é formado por uma área bem esclerotinizada e uma hialina, o comprimento de ambas as áreas
variam de 143,09-245,32 µm e 505,79-621,84 µm, respectivamente. O arco dorsal possui altura
variando de 169,06-246,78 µm. O esclerito anterior é aparentemente ausente. A razão entre o
entalhe do corno (“cornu notch”) e corno dorsal varia de 0,5-0,6. O corno ventral é fracamente
esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente esclerotinizado e possui
anteriormente uma barra faringeal com 6-7 cristas que formam estrias estendendo-se por todo o

107
comprimento do corno ventral. O corno ventral possui comprimento variando de 312,41-574,66
µm. O corno ventral varia de 2,2-2,3 vezes o tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 20-25 túbulos (Fig. 1I). O túbulo apresenta comprimento entre 13,79-19,01 µm, largura
distal entre 9,70-13,19 µm e largura basal entre 6,80-9,62 µm. O espiráculo anterior apresenta na
linha média altura variando de 93,16-111,24 µm; largura distal de 155,18-192,17 µm e largura
basal próximo a abertura da traquéia variando de 57,88-67,38 µm.
O segmento caudal com tubérculos e sensilas apresentam o mesmo padrão que as larvas
de terceiro instar. O espiráculo posterior e o lóbulo anal estão localizados da mesma forma que
nas larvas de terceiro instar. O lóbulo anal é bífido.
O espiráculo posterior apresenta três fendas cada qual com membrana estreita e poucas
trabéculas transversais, seguindo o mesmo padrão que as larvas de terceiro instar (Fig. 1J). As
fendas possuem comprimento variando de 38,79-50,78 µm e largura de 10,09-20,08 µm. A razão
entre o comprimento e a largura das fendas varia de 2,5-3,9. O espiráculo posterior possui cicatriz
da ecdise aparente. A câmara onde localizam-se as fendas é arredondada e apresenta diâmetro
próximo a traquéia variando de 86,82-97,45 µm. O processo espiracular apresenta quatro grupos
de cerdas, onde SP-I possui 9-11 troncos (eventualmente 13) e 19-24 extremidades
(eventualmente 36); a razão entre essas extremidades / troncos varia de 2,1-2,2; esse grupo de
cerdas apresenta largura basal variando de 13,77-22,14 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,4-0,5. O grupo SP-II possui 6-9 troncos e 11-16
extremidades (eventualmente 27). O grupo SP-III possui 7-9 troncos (eventualmente 11) e 11-20
extremidades. O grupo SP-IV possui 10 ou 12 troncos (eventualmente 6 ou 16) e 17-23
extremidades; a razão entre essas extremidades / troncos varia de 1,7-2,0; esse grupo de cerdas
apresenta largura basal variando de 13,05-21,48 µm; a razão da largura basal / comprimento
dessa fenda espiracular varia de 0,3-0,5. A média do comprimento das cerdas é 0,5 vezes o
comprimento das fendas espiraculares.
Anastrepha pickeli Lima (Fig. 2)
Material Examinado. Foram examinadas 33 larvas coletadas em Maracás no estado da
Bahia, em Manaus no estado do Amazonas e em Porto Velho no estado de Rondônia. As larvas
foram dissecadas de frutos de Manihot esculenta Crantz, Euphorbiaceae, um arbusto perene

108
amplamente distribuído pelo Brasil e localmente conhecido como “mandioca”, “aipim” e
“macaxeira”.
Características das larvas de terceiro instar. As larvas possuem comprimento variando
de 6,72-10,96 mm e largura de 1,34-2,17 mm. As larvas são alongadas, cilíndricas, afiladas nos
segmentos torácicos, com segmento caudal truncado e possuem coloração creme amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 3,08-5,79 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 6-8 fileiras; T2 com 6-8 fileiras
(eventualmente 5); T3 com 5-7 fileiras (eventualmente 4); A1 com 4-5 fileiras (eventualmente 3);
A2 com 2-4 fileiras; A3 com 0-3 fileiras; A4 com 0-2 fileiras; A5 com 0-1 fileiras; A6 ao A8 não
apresentam fileiras de espinhos. O padrão de distribuição dos espinhos ventrais é T1 com 8-10
fileiras (eventualmente 7); T2 com 7-9 fileiras; T3 com 6-8 fileiras; A1 com 7-10 fileiras; A2
com 13-17 fileiras (eventualmente 11); A3 com 15-17 fileiras (eventualmente 14); A4 e A5 com
15-18 fileiras (eventualmente 13-14); A6 com 15-17 fileiras (eventualmente 14); A7 com 15-16
fileiras (eventualmente 13-14); A8 com 12-14 fileiras (eventualmente 11). Todos os segmentos
abdominais (A1 ao A8) apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo
anal foram visualizadas fileiras irregulares de espinhos ventral e dorsalmente, variando entre 4-5
fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 17,75-25,08 µm, botão apical com diâmetro variando de 10,32-
13,28 µm e altura de 9,59-10,06 µm. O órgão sensorial maxilar é cilíndrico, levemente afilado,
possui diâmetro variando 18,81-24,88 µm e altura de 7,80-9,59 µm. O órgão sensorial bucal
(“stomal sensory organ”) é cilíndrico e com altura variando de 5,70-8,90 µm. As saliências orais
variam de 10-13 (Fig. 2A).
A Figura 2D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 0,97-1,31 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,19-0,30 mm, o comprimento b varia de 0,15-
0,20 mm e a altura c varia de 0,18-0,24 mm. A razão entre as medidas a / c varia de 1,2-1,4.
Alguns exemplares dessa espécie possuem dois orifícios na base da mandíbula. Os dentes são
curtos e pontiagudos, mas alguns deles podem ser arredondados. O comprimento do esclerito

109
hipofaringeal varia de 0,17-0,22 mm e a largura de 0,16-0,25 mm. Em vista dorsal foi visualizado
o esclerito epifaringeal que apresenta um lóbulo medial direcionado anteriormente. Também em
vista dorsal foi visualizado o esclerito labial em formato de ferradura, curto, robusto e bastante
esclerotinizado. A barra parastomal (= esclerito hipostomal) é levemente arqueada e estende-se
quase completamente sobre o esclerito hipofaringeal. O esclerito ventral localiza-se abaixo do
esclerito faringeal. O corno dorsal é formado por uma área bem esclerotinizada e uma hialina, o
comprimento de ambas as áreas variam de 0,41-0,54 mm e 0,56-0,96 mm, respectivamente. A
ponte dorsal (“dorsal bridge”) é levemente esclerotinizada e apresenta uma projeção anterior do
corno dorsal. O arco dorsal possui altura variando de 0,27-0,37 mm. O esclerito anterior é
esclerotinizado e possui formato irregular. A razão entre o entalhe do corno (“cornu notch”) e
corno dorsal é de 0,7. O corno ventral é fracamente esclerotinizado. O filtro faringeal
(“pharyngeal filter”) é fracamente esclerotinizado e possui anteriormente uma barra faringeal
com 8-9 cristas (eventualmente 6-7) que formam estrias estendendo-se por todo o comprimento
do corno ventral. O corno ventral possui comprimento variando de 0,55-0,81 mm. O corno
ventral varia de 1,4-1,5 vezes o tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 16-23 túbulos (Figs. 2B e E). O túbulo apresenta comprimento entre 20,88-30,83 µm,
largura distal entre 12,14-22,49 µm e largura basal entre 10,71-17,62 µm. O espiráculo anterior
apresenta na linha média altura variando de 122,98-153,01 µm; largura distal de 261,59-352,61
µm e largura basal próximo a abertura da traquéia variando de 128,87-159,67 µm.
A Figura 2F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1 é
distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente desenvolvidos e
associados com uma sensila fracamente desenvolvida. I2 é disposto lateralmente e algumas vezes
levemente ventral em relação a I1. L1 e V são pouco desenvolvidos. D1 e I1 são associados com
sensila fracamente desenvolvida. Alguns espécimes podem apresentar apenas um tubérculo
ventral (V1). Os espiráculos posteriores estão localizados acima da linha média horizontal. O
lóbulo anal é inteiro e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Fig. 2C e G), possuindo comprimento variando de 75,07-
107,11 µm e largura de 11,34-18,75 µm. A razão entre o comprimento e a largura das fendas

110
varia de 5,7-6,6. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 139,26-179,63
µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 1-7 troncos
(eventualmente 8) e 6-12 extremidades; a razão entre essas extremidades / troncos varia de 2,0-
6,0; esse grupo de cerdas apresenta largura basal variando de 4,35-9,70 µm; a razão da largura
basal / comprimento dessa fenda espiracular varia de 0,05-0,09. O grupo SP-II possui 1-7 troncos
e 3-9 extremidades. O grupo SP-III possui 1-6 troncos (eventualmente 7) e 3-9 extremidades. O
grupo SP-IV possui 1-7 troncos e 5-11 extremidades; a razão entre essas extremidades / troncos
varia de 1,5-5,0; esse grupo de cerdas apresenta largura basal variando de 3,08-9,23 µm; a razão
da largura basal / comprimento dessa fenda espiracular varia de 0,03-0,09. A média do
comprimento das cerdas é 0,4 vezes o comprimento das fendas espiraculares. As principais
características das larvas para diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha pulchra Stone (Fig. 3)
Material Examinado. Foram examinadas 24 larvas coletadas em Manaus no estado do
Amazonas. As larvas foram dissecadas de frutos de Pouteria oblanceolata Pires, Sapotaceae,
localmente conhecida como “tuturubá” ou “abiurana-preta”, árvore nativa da Floresta Amazônica
de terra firme.
Características das larvas de terceiro instar. As larvas possuem comprimento variando
de 4,43-11,68 mm e largura de 0,93-1,75 mm. As larvas são alongadas, cilíndricas, afiladas nos
segmentos torácicos, com segmento caudal truncado e possuem coloração creme amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 2,04-5,54 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 0-4 fileiras; T2 com 3-5 fileiras
(eventualmente 2); T3 nenhuma fileira (eventualmente 1); A1 ao A8 não apresentam fileiras de
espinhos. O padrão de distribuição dos espinhos ventrais é T1 com 4-5 fileiras; T2 com 3-4
fileiras (eventualmente 2); T3 com 3-4 fileiras; A1 com 5-6 fileiras; A2 e A3 com 10-13 fileiras;
A4 com 12-14 fileiras (eventualmente 8 ou 10); A5 com 10-13 fileiras (eventualmente 14); A6
com 11-14 fileiras (eventualmente 8); A7 com 8-14 fileiras; A8 com 10-13 fileiras
(eventualmente 7). O segmento torácico T3 e todos os abdominais (A1 ao A8) apresentam

111
protuberâncias ventrais para locomoção. Ao redor do lóbulo anal foram visualizadas fileiras
irregulares de espinhos ventral e dorsalmente, variando entre 3-4 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 17,97-24,87 µm, botão apical com diâmetro variando de 8,93-
13,04 µm e altura de 8,72-14,03 µm. O órgão sensorial maxilar é cilíndrico, levemente afilado,
possui diâmetro variando 13,75-29,82 µm e altura de 8,04-12,93 µm. O órgão sensorial bucal
(“stomal sensory organ”) é cilíndrico e com altura variando de 4,33-13,11 µm. As saliências orais
variam de 9-11 (Fig. 3A).
A Figura 3D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 1,08-1,40 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,24-0,30 mm, o comprimento b varia de 0,17-
0,21 mm e a altura c varia de 0,18-0,22 mm. A razão entre as medidas a / c varia é de 1,3. Os
dentes são compridos, pontiagudos e alguns exemplares dessa espécie apresentam um dente
secundário. O comprimento do esclerito hipofaringeal varia de 0,18-0,24 mm e a largura de 0,22-
0,27 mm. Em vista dorsal foi visualizado o esclerito epifaringeal que apresenta um lóbulo medial
direcionado anteriormente. Também em vista dorsal foi visualizado o esclerito labial em formato
de ferradura, curto, robusto e bastante esclerotinizado. A barra parastomal (= esclerito
hipostomal) estende-se quase completamente sobre o esclerito hipofaringeal é levemente
arqueada, mas em alguns exemplares ela pode ser reta. O esclerito ventral localiza-se abaixo do
esclerito faringeal. O corno dorsal é formado por uma área bem esclerotinizada e uma hialina, o
comprimento de ambas as áreas variam de 0,38-0,50 mm e 0,50-0,83 mm, respectivamente. A
ponte dorsal (“dorsal bridge”) é levemente esclerotinizada e apresenta uma projeção anterior do
corno dorsal. O arco dorsal possui altura variando de 0,36-0,43 mm. O esclerito anterior é
esclerotinizado e possui formato irregular. A razão entre o entalhe do corno (“cornu notch”) e
corno dorsal varia de 0,5-0,6. O corno ventral é fracamente esclerotinizado. O filtro faringeal
(“pharyngeal filter”) é fracamente esclerotinizado e possui anteriormente uma barra faringeal
com 8-9 cristas que formam estrias estendendo-se por todo o comprimento do corno ventral. O
corno ventral possui comprimento variando de 0,60-0,87 mm. O corno ventral varia de 1,6-1,7
vezes o tamanho da área esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 18-23 túbulos (Figs. 3B e E). O túbulo apresenta comprimento entre 20,79-29,31 µm,

112
largura distal entre 12,88-22,66 µm e largura basal entre 9,65-17,09 µm. O espiráculo anterior
apresenta na linha média altura variando de 122,02-177,01 µm; largura distal de 279,48-370,47
µm e largura basal próximo a abertura da traquéia variando de 141,30-194,13 µm.
A Figura 1F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1 é
distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente desenvolvidos e
associados com uma sensila fracamente desenvolvida. I2 é disposto lateralmente e algumas vezes
levemente ventral em relação a I1. L1 e V são pouco desenvolvidos. D1 e I1 são associados com
sensila fracamente desenvolvida. Alguns espécimes podem apresentar três tubérculos ventrais
(V1, V2 e V3). Os espiráculos posteriores estão localizados acima da linha média horizontal. O
lóbulo anal é bífido e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Fig. 3C e G), possuindo comprimento variando de 68,44-
101,97 µm e largura de 17,37-26,97 µm. A razão entre o comprimento e a largura das fendas
varia de 3,8-3,9. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 139,37-177,05
µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 4-5 troncos e 7-
10 extremidades (eventualmente 6); a razão entre essas extremidades / troncos varia de 1,3-1,8;
esse grupo de cerdas apresenta largura basal variando de 6,18-9,29 µm; a razão da largura basal /
comprimento dessa fenda espiracular varia de 0,08-0,09. O grupo SP-II possui 2-3 troncos
(eventualmente 1, 4 ou 5) e 3-6 extremidades (eventualmente 2). O grupo SP-III possui 4-5
troncos (eventualmente 2 ou 3) e 5-9 extremidades (eventualmente 2 ou 4). O grupo SP-IV possui
4-5 troncos (eventualmente 3 ou 7) e 5-9 extremidades (eventualmente 10); a razão entre essas
extremidades / troncos varia de 1,3-1,8; esse grupo de cerdas apresenta largura basal variando de
5,78-6,99 µm; a razão da largura basal / comprimento dessa fenda espiracular varia de 0,07-0,08.
A média do comprimento das cerdas é 0,2 vezes o comprimento das fendas espiraculares. As
principais características das larvas para diagnóstico desta espécie estão apresentadas na Tabela
1.

113
Anastrepha sororcula Zucchi (Fig. 4)
Material Examinado. Foram examinadas oito larvas coletadas em Mossoró no estado do
Rio Grande do Norte. As larvas foram dissecadas de frutos de Psidium guajava L., Myrtaceae,
localmente conhecida como “goiaba”, árvore nativa da floresta tropical.
Características das larvas de terceiro instar. As larvas possuem comprimento variando
de 8,68-10,02 mm e largura de 1,13-1,55 mm. As larvas são alongadas, cilíndricas, afiladas nos
segmentos torácicos, com segmento caudal truncado e possuem coloração creme esbranquiçada.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 2,06-3,31 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 4-5 fileiras; T2 com 4-5 fileiras
(eventualmente 3); T3 com 0-2 fileiras; A1 ao A8 não apresentam fileiras de espinhos. O padrão
de distribuição dos espinhos ventrais é T1 com 8-9 fileiras; T2 com 5-7 fileiras (eventualmente
4); T3 com 4-5 fileiras; A1 com 5-8 fileiras; A2 com 10-11 fileiras; A3 com 11-12 fileiras; A4
com 11-13 fileiras; A5, A6 e A7 com 10-12 fileiras; A8 com 10-11 fileiras. Todos os segmentos
abdominais (A1 ao A8) apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo
anal foram visualizadas fileiras irregulares de espinhos ventral e dorsalmente, variando entre 3-4
fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 17,34-20,63 µm, botão apical com diâmetro variando de 10,34-
14,61 µm e altura de 7,46-7,64 µm. O órgão sensorial maxilar é cilíndrico, levemente afilado,
possui diâmetro variando 30,39-31,61 µm e altura de 10,74-11,95 µm. O órgão sensorial bucal
(“stomal sensory organ”) é cilíndrico e com altura variando de 6,24-6,92 µm. As saliências orais
variam de 6-7 (eventualmente 8 ou 9) (Fig. 4A).
A Figura 4D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 0,95-1,23 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,21-0,27 mm, o comprimento b varia de 0,16-
0,19 mm e a altura c varia de 0,14-0,18 mm. A razão entre as medidas a / c é de 1,5. Os dentes
são compridos, pontiagudos e alguns exemplares dessa espécie apresentam um dente secundário.
O comprimento do esclerito hipofaringeal varia de 0,17-0,18 mm e a largura de 0,18-0,23 mm.
Em vista dorsal foi visualizado o esclerito epifaringeal que apresenta um lóbulo medial

114
direcionado anteriormente. Também em vista dorsal foi visualizado o esclerito labial em formato
de ferradura, curto, robusto e bastante esclerotinizado. A barra parastomal (= esclerito
hipostomal) é levemente arqueada e estende-se quase completamente sobre o esclerito
hipofaringeal. O esclerito ventral localiza-se abaixo do esclerito faringeal. O corno dorsal é
formado por uma área bem esclerotinizada e uma hialina, o comprimento de ambas as áreas
variam de 0,35-0,43 mm e 0,50-0,75 mm, respectivamente. A ponte dorsal (“dorsal bridge”) é
levemente esclerotinizada e apresenta uma projeção anterior do corno dorsal. O arco dorsal
possui altura variando de 0,34-0,43 mm. O esclerito anterior é esclerotinizado e possui formato
irregular. A razão entre o entalhe do corno (“cornu notch”) e corno dorsal é de 0,6. O corno
ventral é fracamente esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente
esclerotinizado e possui anteriormente uma barra faringeal com 8-9 cristas que formam estrias
estendendo-se por todo o comprimento do corno ventral. O corno ventral possui comprimento
variando de 0,55-0,75 mm. O corno ventral varia de 1,6-1,7 vezes o tamanho da área
esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 9-12 túbulos (Fig. 4B). O túbulo apresenta comprimento entre 19,46-27,85 µm, largura
distal entre 14,16-17,95 µm e largura basal entre 9,07-14,61 µm. O espiráculo anterior apresenta
na linha média altura variando de 119,13-160,64 µm; largura distal de 155,58-207,94 µm e
largura basal próximo a abertura da traquéia variando de 73,13-113,96 µm.
A Figura 4E ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1 é
distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente desenvolvidos e
associados com uma sensila fracamente desenvolvida. I2 é disposto lateralmente e algumas vezes
levemente ventral em relação a I1. L1 e V são pouco desenvolvidos. D1 e I1 são associados com
sensila fracamente desenvolvida. Os espiráculos posteriores estão localizados acima da linha
média horizontal. O lóbulo anal é inteiro e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Fig. 4C), possuindo comprimento variando de 71,02-92,47
µm e largura de 16,17-21,20 µm. A razão entre o comprimento e a largura das fendas é de 4,4. O
espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde localizam-se as fendas é
oval e apresenta diâmetro próximo a traquéia variando de 109,37-139,91 µm. O processo

115
espiracular apresenta quatro grupos de cerdas, onde SP-I possui 13-16 troncos e 36-48
extremidades; a razão entre essas extremidades / troncos varia de 2,7-3,0; esse grupo de cerdas
apresenta largura basal variando de 22,29-37,90 µm; a razão da largura basal / comprimento
dessa fenda espiracular varia de 0,3-0,4. O grupo SP-II possui 6-9 troncos e 13-18 extremidades
(eventualmente 21). O grupo SP-III possui 9-12 troncos e 19-30 extremidades. O grupo SP-IV
possui 12-15 troncos e 32-40 extremidades (eventualmente 47); a razão entre essas extremidades
/ troncos é de 2,6; esse grupo de cerdas apresenta largura basal variando de 21,33-32,87 µm; a
razão da largura basal / comprimento dessa fenda espiracular varia de 0,3-0,4. A média do
comprimento das cerdas é 0,5 vezes o comprimento das fendas espiraculares. As principais
características das larvas para diagnóstico desta espécie estão apresentadas na Tabela 1.
Anastrepha zenildae Zucchi (Fig. 5)
Material Examinado. Foram examinadas 19 larvas coletadas em Limoeiro do Norte no
estado do Ceará e em Mossoró no estado do Rio Grande do Norte. As larvas foram dissecadas de
frutos de Ziziphus joazeiro Mart., Rhamnaceae, conhecido localmente como “juá”, árvore do
bioma Caatinga encontrada no semi-árido nordestino brasileiro e de frutos de Psidium guajava
L., Myrtaceae, localmente conhecida como “goiaba”, árvore nativa da floresta tropical.
Características das larvas de terceiro instar. As larvas possuem comprimento variando
de 8,78-10,44 mm e largura de 1,24-1,65 mm. As larvas são alongadas, cilíndricas, afiladas nos
segmentos torácicos, com segmento caudal truncado e possuem coloração variando de creme
esbranquiçado a creme amarelado.
Todos os segmentos apresentam fileiras irregulares de espinhos distribuídas dorsal e
ventralmente. Essas fileiras de espinhos possuem comprimento variável. Os espinhos são
cônicos, simétricos, levemente curvados e possuem largura basal variando de 2,56-3,42 µm. O
padrão de distribuição dos espinhos dorsais é T1 com 5 fileiras (eventualmente 6); T2 com 4-5
fileiras; T3 com 0 ou 2 fileiras; A1 ao A8 não apresentam fileiras de espinhos. O padrão de
distribuição dos espinhos ventrais é T1 com 9 fileiras; T2 com 5-7 fileiras; T3 com 4-5 fileiras;
A1 com 6-7 fileiras (eventualmente 8); A2 e A3 com 11-12 fileiras; A4 com 12 fileiras
(eventualmente 14); A5 com 12 ou 13 fileiras; A6 com 12-13 fileiras (eventualmente 11 ou 14);
A7 com 11-13 fileiras; A8 com 11 ou 13 fileiras. Todos os segmentos abdominais (A1 ao A8)

116
apresentam protuberâncias ventrais para locomoção. Ao redor do lóbulo anal foram visualizadas
fileiras irregulares de espinhos ventral e dorsalmente, variando entre 3-4 fileiras.
O órgão sensorial da antena é cilíndrico e levemente esclerotinizado, apresenta um anel
basal com diâmetro variando de 17, 07-25,20 µm, botão apical com diâmetro variando de 8,15-
13,52 µm e altura de 6,54-7,59 µm. O órgão sensorial maxilar é cilíndrico, levemente afilado,
possui diâmetro variando 22,17-25,28 µm e altura de 8,30-15,64 µm. O órgão sensorial bucal
(“stomal sensory organ”) é cilíndrico e com altura variando de 7,44-8,37 µm. As saliências orais
variam de 9 ou 10 (Fig. 5A).
A Figura 5D apresenta o formato e o padrão de esclerotinização do esqueleto céfalo-
faríngeo (ECF). O ECF possui comprimento total variando de 1,09-1,38 mm. A mandíbula
apresenta três medidas: o comprimento a varia de 0,16-0,28 mm, o comprimento b varia de 0,17-
0,21 mm e a altura c varia de 0,17-0,21 mm. A razão entre as medidas a / c varia de 0,9-1,3. Os
dentes são fortemente esclerotinizado, longos e pontiagudos, mas em alguns exemplares podem
ser curtos e arredondados e. O comprimento do esclerito hipofaringeal varia de 0,18-0,20 mm e a
largura de 0,19-0,23 mm. Em vista dorsal foi visualizado o esclerito epifaringeal que apresenta
um lóbulo medial direcionado anteriormente. Também em vista dorsal foi visualizado o esclerito
labial em formato de ferradura, curto, robusto e bastante esclerotinizado. A barra parastomal (=
esclerito hipostomal) é reta e estende-se quase completamente sobre o esclerito hipofaringeal. O
esclerito ventral localiza-se abaixo do esclerito faringeal. O corno dorsal é formado por uma área
bem esclerotinizada e uma hialina, o comprimento de ambas as áreas variam de 0,36-0,53 mm e
0,60-0,81 mm, respectivamente. A ponte dorsal (“dorsal bridge”) é levemente esclerotinizada e
apresenta uma projeção anterior do corno dorsal. O arco dorsal possui altura variando de 0,36-
0,41 mm. O esclerito anterior é esclerotinizado e possui formato irregular. A razão entre o entalhe
do corno (“cornu notch”) e corno dorsal varia de 0,7-0,8. O corno ventral é fracamente
esclerotinizado. O filtro faringeal (“pharyngeal filter”) é fracamente esclerotinizado e possui
anteriormente uma barra faringeal com nove cristas (eventualmente 10) que formam estrias
estendendo-se por todo o comprimento do corno ventral. O corno ventral possui comprimento
variando de 0,66-0,91 mm. O corno ventral varia de 1,7-1,8 vezes o tamanho da área
esclerotinizada do corno dorsal.
O espiráculo anterior é bilobado, simétrico e possui uma única fileira de túbulos, variando
entre 11-16 túbulos (eventualmente 11) (Figs. 5B e E). O túbulo apresenta comprimento entre

117
25,12-31,38 µm, largura distal entre 11,28-18,14 µm e largura basal entre 10,80-17,49 µm. O
espiráculo anterior apresenta na linha média altura variando de 126,24-179,73 µm; largura distal
de 233,99-261,10 µm e largura basal próximo a abertura da traquéia variando de 106,17-137,07
µm.
A Figura 5F ilustra o segmento caudal com tubérculos e sensilas dorsais (D1 e D2),
intermediários (I1 e I2), lateral (L1) e ventrais (V1 e V2) bem desenvolvidas. O tubérculo D1 é
distintamente anterior ao D2. Tubérculos intermediários I1 e I2 são fortemente desenvolvidos e
associados com uma sensila fracamente desenvolvida. I2 é disposto lateralmente e algumas vezes
levemente ventral em relação a I1. L1 e V são pouco desenvolvidos. D1 e I1 são associados com
sensila fracamente desenvolvida. Os espiráculos posteriores estão localizados acima da linha
média horizontal. O lóbulo anal é inteiro e protraído.
O espiráculo posterior apresenta três fendas cada qual com membrana espessa e
numerosas trabéculas transversais (Fig. 5C e G), possuindo comprimento variando de 85,25-
108,37 µm e largura de 17,37-24,05 µm. A razão entre o comprimento e a largura das fendas
varia de 4,5-4,9. O espiráculo posterior possui cicatriz da ecdise aparente. A câmara onde
localizam-se as fendas é oval e apresenta diâmetro próximo a traquéia variando de 130,87-148,24
µm. O processo espiracular apresenta quatro grupos de cerdas, onde SP-I possui 12-16 troncos e
30-43 extremidades; a razão entre essas extremidades / troncos varia de 2,5-2,6; esse grupo de
cerdas apresenta largura basal variando de 28,53-34,40 µm; a razão da largura basal /
comprimento dessa fenda espiracular é de 0,3. O grupo SP-II possui 5-9 troncos e 11-17
extremidades. O grupo SP-III possui 5-11 troncos e 17-24 extremidades (eventualmente 29). O
grupo SP-IV possui 9-14 troncos e 26-35 extremidades; a razão entre essas extremidades /
troncos varia de 2,5-2,8; esse grupo de cerdas apresenta largura basal variando de 24,82-33,86
µm; a razão da largura basal / comprimento dessa fenda espiracular é de 0,3. A média do
comprimento das cerdas é 0,4 vezes o comprimento das fendas espiraculares. As principais
características das larvas para diagnóstico desta espécie estão apresentadas na Tabela 1.
Discussão
Larvas de terceiro instar de Anastrepha apresentam aspectos gerais semelhantes como o
formato vermiforme, a coloração e o comprimento. Contudo, foi possível distinguir entre as cinco
espécies de larvas estudadas aqui com base em alguns caracteres diagnósticos.

118
Larvas de terceiro instar de A. curitis diferem das outras quatro espécies analisadas devido
à ausência de espinhos dorsais apenas no segmento A8, esqueleto céfalo-faríngeo maior, formato
(curvatura levemente retangular) e mandíbula maior. Diferenças entre as larvas de segundo e
terceiro instar de A. curitis incluem o número de saliências orais, número de espinhos dorsais nos
segmentos A4 e A5, número de extremidades nas cerdas do grupo SP-IV e número de cristas no
filtro faringeal (“pharyngeal filter”).
As larvas de A. pickeli, A. pulchra, A. sororcula e A. zenildae podem ser diferenciadas
com base em algumas características. As larvas de A. pickeli podem ser diferenciadas das outras
espécies analisadas através da ausência de espinhos dorsais nos segmentos A6 ao A8, número
semelhante de espinhos ventrais nos segmentos A4-A5, número semelhante de troncos das cerdas
para SP-I, SP-II e SP-IV e número semelhante de extremidades das cerdas para SP-II e SP-III. As
larvas de A. pulchra podem ser diferenciadas das outras espécies estudadas devido ao mesmo
número de espinhos ventrais nos segmentos T2-T3, A2, A3, A5 e A8 e número de troncos das
cerdas para SP-III e SP-IV. As larvas de A. sororcula podem ser diferenciadas devido ao menor
número de saliências orais e mesmo número de espinhos ventrais nos segmentos A5-A7, A2 e
A8. Larvas de A. zenildae diferem das larvas das demais espécies analisadas apenas pela
semelhança no número de espinhos ventrais nos segmentos A2-A3 e A5-A6.
As larvas de A. sororcula podem ser diferenciadas de outras espécies do grupo fraterculus
descritas anteriormente com base na ausência de espinhos dorsais nos segmentos abdominais,
menor número de túbulos no espiráculo anterior e formato do lóbulo anal. Já larvas de A. zenildae
podem ser diferenciadas de outras espécies do grupo fraterculus devido à ausência de espinhos
dorsais nos segmentos abdominais e formato do lóbulo anal. Vale ressaltar que as descrições de
larvas de Anastrepha distincta Greene, Anastrepha fraterculus (Wiedemann), Anastrepha obliqua
(Macquart) e Anastrepha suspensa (Loew), espécies pertencentes ao grupo fraterculus, não
apresentaram informações sobre o esqueleto céfalo-faríngeo, fato que dificulta a adequada
diferenciação entre as diversas espécies do referido grupo.
Para as cinco espécies estudadas, as características descritas e as medições apresentaram
sobreposição com outras espécies descritas anteriormente, independentemente do grupo de
espécie no qual estão classificadas. Por exemplo, para o grupo pseudoparallela, Anastrepha
limae Stone [descrita previamente por Steck e Wharton (1988)] e A. curitis [descrita no presente
estudo] apresentam sobreposição no número de saliências orais (9-12 e 8-11, respectivamente),

119
ausência de espinhos dorsais (segmentos A3 ao A8 e A1 ao A8, respectivamente), número de
túbulos no espiráculo anterior (18-21 e 19-25, respectivamente), lóbulo anal bífido e número de
troncos das cerdas do espiráculo posterior (4-13 e 6-18, respectivamente), entretanto, estas duas
espécies apresentam diferença no comprimento do esqueleto céfalo-faríngeo, número de cristas
do filtro faringeal (“pharyngeal filter”) e no número de extremidades das cerdas do espiráculo
posterior, caracteres que podem ser úteis para a identificação das mesmas.
Neste estudo, a descrição da morfologia das larvas para cinco espécies de Anastrepha
mostrou diferenças que permitem a identificação destas, apesar da grande similaridade e
sobreposição entre elas e com espécies descritas previamente. Características como número de
fileiras de espinhos dorsais e ventrais nos segmentos, número de saliências orais, número de
cerdas no espiráculo posterior, comprimento do esqueleto céfalo-faríngeo e formato da
mandíbula são caracteres taxonômicos que podem auxiliar na identificação de espécies de
importância quarentenária e no esclarecimento sobre as relações filogenéticas dentro dos diversos
grupos de espécies e também do gênero Anastrepha.
Agradecimentos
Os autores agradecem a Marcos Vinicius Garcia e Tatiana Senra (Embrapa Amazônia
Ocidental) pelo auxílio durante as coletas; a Hilton Tulio Costi e Rolf Junior Ferreira Silva
(Museu Paraense Emílio Goeldi), Carolina Almeida e Adriana Horbe (Universidade Federal do
Amazonas) pelo auxílio nas fotografias com MEV. Esse estudo foi financiado pelo Conselho
Nacional de Desenvolvimento Científico e Tecnológico (CNPq - projeto n° 575664/2008-8), pela
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES - projeto n°
23038.037185/2008-41), pela Fundação de Amparo a Pesquisa do Estado do Amazonas
(FAPEAM - projeto nº014-017/2004) e pela Rede Amazônica de Pesquisas sobre Moscas-das-
Frutas (Chamada 05/2006 Agrofuturo). Os autores também agradecem ao CNPq pela bolsa de
doutorado concedida a VSD.

120
Fig. 1. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para larvas de
terceiro instar (D à G) e segundo instar (H à J) de A. curitis. Larvas de terceiro instar (A)
Saliências Orais. (B e E) Espiráculo Anterior. (C e G) Espiráculo Posterior. (D) Esqueleto
Céfalo-faríngeo. (F) Ilustração do Segmento Caudal. Larvas de segundo instar (H) Esqueleto
Céfalo-faríngeo. (I) Espiráculo Anterior. (J) Espiráculo Posterior. Abreviações: Ma: comprimento
da mandíbula a; Mb: comprimento da mandíbula b; Mc: altura da mandíbula c; LS: esclerito
labial.

121
Tabela 1. Diagnose de cinco espécies de larvas de Anastrepha analisadas neste estudo.
Anastrepha
Fileiras de espinhos nos
segmentos Saliências
orais
ECF
mandíbula
(mm)*
ECF
comprimento
(mm)*
N. de
túbulos
no EA**
Lóbulo
anal
Espiráculo posterior
Dorsais Ventrais Troncos Extremidades
curitis Ausente em
A8
Número diferente
em todos
8-11 0,33-0,36 1,40-1,65 19-25 Bífido Mesmo número
em SP-II e SP-III
Número
diferente para
todos
pickeli Ausente de
A6 ao A8
Mesmo número
em A4-A5
10-13 0,20-0,30 1,02-1,47 17-23 Inteiro Mesmo número
em SP-I, SP-II e
SP-IV
Mesmo
número em
SP-II e SP-III
pulchra Ausente de
A1 ao A8
Mesmo número
em T2-T3; A2,
A3, A5 e A8
9-11 0,24-0,30 1,09-1,40 17-24 Bífido Mesmo número
em SP-III e SP-
IV
Número
diferente para
todos
sororcula Ausente de
A1 ao A8
Mesmo número
em A2 e A8; A5-
A7
6-7 0,21-0,27 0,95-1,23 9-12 Inteiro Número diferente
para todos
Número
diferente para
todos
zenildae Ausente de
A1 ao A8
Mesmo número
em A2-A3 e A5-
A6.
9-10 0,16-0,28 1,09-1,38 11-16 Inteiro Número diferente
para todos
Número
diferente para
todos
*ECF = esqueleto céfalo-faríngeo
**EA = espiráculo anterior

122
Fig. 2. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para larvas de
terceiro instar (D à G) de A. pickeli. (A) Saliências Orais. (B e E) Espiráculo Anterior. (C e G)
Espiráculo Posterior. (D) Esqueleto Céfalo-faríngeo. (F) Ilustração do Segmento Caudal.

123
Fig. 3. Vista com Microscopia Eletrônica de Varredura (A à C) e microscopia ótica para larvas de
terceiro instar (D à G) de A. pulchra. (A) Saliências Orais. (B e E) Espiráculo Anterior. (C e G)
Espiráculo Posterior. (D) Esqueleto Céfalo-faríngeo. (F) Ilustração do Segmento Caudal.

124
Fig. 4. Vista com microscopia ótica para larvas de terceiro instar de A. sororcula. (A) Saliências
Orais. (B) Espiráculo Anterior. (C) Espiráculo Posterior. (D) Esqueleto Céfalo-faríngeo. (F)
Ilustração do Segmento Caudal.

125
Fig. 5. Vista com Microscopia Eletrônica de Varredura (B e C) e microscopia ótica para larvas de
terceiro instar (A, D à G) de A. zenildae. (A) Saliências Orais. (B e E) Espiráculo Anterior. (C e
G) Espiráculo Posterior. (D) Esqueleto Céfalo-faríngeo. (F) Ilustração do Segmento Caudal.

126
Capítulo 4
Dutra, V.S; Ronchi-Teles, B.; Andrade-Souza, V.; Lima,
K.M.; Steck, G.J.; Silva, J.G. Associação de adultos e
larvas de espécies de Anastrepha (Diptera: Tephritidae)
utilizando a técnica de PCR-RFLP. Manuscrito formatado
para Acta Amazonica.

127
Associação de adultos e larvas de espécies de Anastrepha (Diptera:
Tephritidae) utilizando a técnica de PCR-RFLP
Vivian S. DUTRA1,7Beatriz RONCHI-TELES
2, Vanderly ANDRADE-SOUZA
2, Kátia M.
LIMA3, Gary J. STECK
4 E Janisete G. SILVA
3
RESUMO Até o momento, a identificação de espécies de Anastrepha é baseada principalmente
nas características do acúleo das fêmeas. A identificação desse gênero através das larvas torna-se
difícil devido à grande similaridade morfológica entre as diversas espécies, sendo necessária a
observação cuidadosa para diferenciá-las. A técnica da Reação em Cadeia da Polimerase (PCR)
associada ao Polimorfismo do Comprimento dos Fragmentos de Restrição (RFLP) é útil para
auxiliar na identificação de imaturos de tefritídeos, sendo esses os estágios nos quais são
encontrados os insetos em frutas frescas exportadas ou importadas. Neste estudo, foram
analisadas duas regiões do DNA mitocondrial (COI e 16S) e uma do DNA nuclear (ITS1) para 13
espécies de Anastrepha com cinco enzimas de restrição. Os melhores resultados para
diferenciação das espécies foram obtidos com os fragmentos 16S e ITS1 para Anastrepha
atrigona Hendel, Anastrepha curitis Stone, Anastrepha manihoti Lima, Anastrepha pulchra
Stone, Anastrepha striata Schiner, Anastrepha sororcula Zucchi e Anastrepha zenildae Zucchi.
PALAVRAS-CHAVE moscas-das-frutas, imaturos, enzimas de restrição
INTRODUÇÃO
A família Tephritidae reune aproximadamente 4.500 espécies e algumas delas causam
prejuízo econômico a agricultura e fruticultura mundial, pois as larvas dos tefritídeos podem
infestar tecidos vegetais vivos como raízes, galhos jovens, folhas, flores, frutos e sementes
(Prokopy e Roitberg 1984; White e Elson-Harris 1992; Norrbom 2005). Até o momento, estão
incluídas no gênero Anastrepha Schiner cerca de 240 espécies (Zucchi 2008; Norrbom e
Korytkowski 2009) e destas, 112 são registradas no Brasil (Jesus et al. 2008; Uramoto et al.
2008; Zucchi 2007, 2008; Uramoto e Zucchi 2010).
1Programa de Pós-Graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478,
69011-970, Manaus, AM, Brasil. 2Coordenação de Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970
Manaus, AM, Brasil. 3Departamento de Ciências Biológicas, Universidade Estadual de Santa Cruz, Rodovia Ilhéus/Itabuna km 16,
45650-000, Ilhéus, BA, Brasil. 4Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL 32614-
7100.

128
A identificação das espécies de Anastrepha é baseada nas características dos adultos e a
principal estrutura analisada é o acúleo das fêmeas (Zucchi 2000; Araujo e Zucchi 2006). Quanto
às larvas, ainda não é possível identificar as mesmas com segurança, devido à grande
similaridade morfológica entre as diversas espécies de Anastrepha, sendo necessária a
observação cuidadosa para diferenciá-las. Assim, faz-se necessário a utilização de ferramentas
que tornem a identificação rápida e precisa, permitindo a utilização de quantidades mínimas de
material de qualquer estágio do inseto a ser analisado (Armstrong et al. 1997; Barr et al. 2006).
Alguns trabalhos com espécies dos gêneros Anastrepha, Ceratitis MacLeay, Bactrocera
Macquart e Rhagoletis Loew foram desenvolvidos com a técnica molecular de Reação em Cadeia
da Polimerase (PCR) associada ao Polimorfismo do Comprimento dos Fragmentos de Restrição
(RFLP) e nesses trabalhos, foi demonstrado que essa técnica é uma boa solução para auxiliar na
identificação de imaturos, pois o custo é mais baixo que as outras técnicas moleculares, o
resultado gerado também é rápido, além de ser fácil a interpretação e com menor risco de erro
(Gasparich et al. 1995,1997; Armstrong et al. 1997; Silva et al. 2003; Muraji e Nakahara 2002;
Salazar et al. 2002; Barr et al. 2006).
Este trabalho teve como objetivo obter o perfil de haplótipos de adultos e larvas de
algumas espécies de Anastrepha por meio do uso de PCR-RFLP em fragmentos de dois genes do
DNA mitocondrial (COI e 16S) e uma região do DNA nuclear (ITS1), bem como correlacionar
os perfis de haplótipos encontrados para adultos e larvas, visando estabelecer a correta
identificação das larvas com base nos perfis dos adultos. As espécies analisadas foram
Anastrepha antunesi Lima, Anastrepha atrigona Hendel, Anastrepha bahiensis Lima, Anastrepha
coronilli Carrejo & González, Anastrepha curitis Stone, Anastrepha distincta Greene,
Anastrepha manihoti Lima, Anastrepha pickeli Lima, Anastrepha pulchra Stone, Anastrepha
striata Schiner, Anastrepha sororcula Zucchi, Anastrepha turpiniae Stone e Anastrepha zenildae
Zucchi.

129
MATERIAL E MÉTODOS
Análises morfológicas
Coleta de adultos
Foram coletados frutos de hospedeiros de A. antunesi, A. atrigona, A. bahiensis, A.
coronilli, A. curitis, A. distincta, A. manihoti, A. pickeli, A. pulchra, A. striata, A. sororcula, A.
turpiniae e A. zenildae. Para A. manihoti e A. pickeli, além de frutos também foram coletados
ramos jovens de seu hospedeiro preferencial (Manihot esculenta Crantz). As coletas foram
realizadas nos municípios de Manaus, Maués e Presidente Figueiredo no estado do Amazonas, de
Camamu, Maracás e Una no estado da Bahia, de Porto Velho no estado de Rondônia e de Boa
Vista no estado de Roraima.
As amostras de frutos e ramos jovens foram pesadas e armazenadas individualmente em
frascos plásticos de 500 mL contendo vermiculita e cobertos por organza. Após
aproximadamente dez dias, a vermiculita foi peneirada e as pupas obtidas foram contadas e
transferidas para frascos plásticos de 30 mL contendo vermiculita úmida e cobertos com organza
até a emergência dos adultos. Os adultos foram mortos em etanol 100%, armazenados em freezer
-20ºC e posteriormente foram contados, sexados e identificados.
Após a pesagem dos frutos, foram retiradas algumas larvas (de uma a cinco larvas por
fruto) e outras larvas (de uma a cinco larvas por fruto) foram mantidas nos frutos até a
emergência dos adultos. As larvas foram mortas e armazenadas como descrito acima para os
adultos. Apenas em uma amostra de larvas houve a emergência de duas espécies de Anastrepha
que foram identificadas através da técnica PCR-RFLP testada neste estudo.
Identificação dos adultos
A identificação das espécies de Anastrepha foi realizada através das fêmeas. O acúleo de
cada fêmea foi extrovertido e colocado entre lâmina e lamínula para análise das características ao
microscópio óptico. A identificação ao nível de espécie foi baseada em Steyskal (1977), Silva e
Ronchi-Teles (2000), Zucchi (2000) e Zucchi et al. (2011a).
Os espécimes testemunho foram depositados na Coleção de Invertebrados do Instituto
Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazonas, e na coleção do Departamento

130
de Ciências Biológicas do Laboratório de Entomologia da Universidade Estadual de Santa Cruz
(UESC), Ilhéus, Bahia.
Análises moleculares
Extração de DNA
Foi extraído o DNA de cerca de cinco adultos de A. antunesi, A. atrigona e A. manihoti, e
cerca de dez adultos e dez larvas de A. bahiensis, A. coronilli, A. curitis, A. distincta, A. pickeli,
A. pulchra, A. striata, A. sororcula, A. turpiniae e A. zenildae, totalizando 94 adultos e 100
larvas. A extração de DNA foi realizada utilizando-se o kit de extração de tecidos animais
DNeasy Blood & Tissue (Qiagen) e o protocolo descrito em Han e McPheron (1997) que utiliza
fenol e clorofórmio, com uma etapa adicional usando proteinase K (20 mg/mL) e incubação a
55°C por 30 min.
Reação em cadeia da polimerase (PCR)
Para a reação de PCR foram utilizados primers para amplificar fragmentos dos genes
mitocondriais COI (citocromo oxidase I) e 16S, e o espaçador ITS1 (espaçador interno transcrito
1) (Tabela 1) de todas as espécies listadas acima. As reações de PCR foram realizadas nas
condições descritas em McPheron et al. (1994). Três µL de DNA extraído foram usados nessa
reação, tendo volume final de 25 µL.
Tabela 1 - Oligonucleotídeos iniciadores (primers) utilizados na amplificação via PCR.
Primer Sequência Referência
COI LCO1490: 5' GGTCAACAAATCATAAAGATATTGG 3'
HCO2198: 5' TAAACTTCAGGGTGACCAAAAAATCA 3’
Folmer et al. 1994
16S LR-J-12883: 5′ CTCCGGTTTGAACTCAGATC 3′
LR-N-13398: 5′ CGCCTGTTTATCAAAAACAT 3′
McPheron et al. 1999
ITS1 18SF: 5’ TAACTCGCATTGATTAAGTCCC 3’
5.8SR: 5’ GATATGCGTTCAAATGTCGATG 3’
Prezotto 2008
A amplificação foi realizada em termocicladores [Swift Maxi Thermal Cycler (ESCO) e
TC-512 (Techine)] com programas específicos para cada par de primers (Tabela 2).

131
Tabela 2 - Programas específicos para amplificação de cada par de oligonucleotídeos iniciadores
(primers) para espécies de Anastrepha.
Etapas COI 16S ITS1
Desnaturação inicial 94º (3 min) 95º (12 min) 94º (2 min)
Desnaturação 94º (1 min) 94º (30 seg) 94º (30 seg)
Anelamento 50º (1 min) 54º (30 seg) 54º (30 seg)
Extensão 72º (1 min) 72º (1 min) 68º (2 min)
Extensão final 72º (10 min) 72º (5 min) 72º (5 min)
Ciclos 39 40 31
Para cada reação de amplificação, foram corridos géis de agarose a 1% corados com
GelRed (10.000X) com tampão TBE 0,5X preparado de acordo com Sambrook e Russel (2001).
As amostras foram preparadas utilizando-se 2 µL de produto de PCR, 2 µL de uma solução de
azul de bromofenol 0,25%, xileno cianol 0,25% e sacarose 40% e 4µL de água milli-Q estéril.
Como marcador de peso molecular foi utilizado φX174 DNA/BsuRI digerido com Hae III. As
bandas de DNA foram visualizadas e fotografadas em fotodocumentador de luz ultravioleta
(UV).
Reação de Purificação dos fragmentos amplificados
Após a obtenção dos produtos de PCR dos exemplares de Anastrepha, as amostras foram
purificadas para a eliminação de resíduos de oligonucleotídeos iniciadores, de RNA e de di-
nucleotídeos tri-fostato que podem interferir no sequenciamento dos fragmentos de DNA que
foram amplificados. Para a purificação foram utilizadas as enzimas exonuclease I de Escherichia
coli (EXOI) e fosfatase alcalina de camarão (SAP) adicionadas ao produto de PCR.
As reações de purificação foram realizadas em termociclador (Termociclador
Mastercycler Gradiente, Eppendorf) onde foram incubados a 37°C por 30 minutos e em seguida a
80°C por 15 minutos. Para cada reação de purificação, foram corridos géis de agarose a 1,7%,
corados com brometo de etídeo (5g/mL), utilizando o tampão TBE 0,5X preparado de acordo
com Sambrook et al. (1989). Essa etapa foi realizada no laboratório de Genética e Biologia
Molecular da UESC.
As amostras foram preparadas utilizando-se 12 μL de produto de PCR na concentração de
20 ng / μL, 0,66 μL de EXOI, 0,66 μL de SAP e 0,68 μL de água Mili-Q, totalizando 20 μL de

132
solução. Os géis foram fotografados utilizando-se o sistema Image Quant 350 (GE Healthcare).
As reações que se apresentaram totalmente purificadas foram sequenciadas.
Sequenciamento (COI)
Foram obtidas três sequências de cerca de 700 pb que compreendem o tRNA cisteína, o
tRNA tirosina e um fragmento da subunidade I do gene da Citocromo Oxidase de indivíduos das
espécies A. antunesi, A. atrigona, A. bahiensis, A. coronilli, A. curitis, A. distincta, A. manihoti,
A. pickeli, A. pulchra, A. striata, A. sororcula, A. turpiniae e A. zenildae.
Os produtos de PCR foram sequenciados com os mesmos oligonucleotídeos iniciadores
utilizados na reação de amplificação. O sequenciamento dos fragmentos de DNA amplificados
foi realizado em um sequenciador automático (MegaBACETM
DNA Analysis System 1000 da GE
Healthcare) no Centro de Estudos do Genoma Humano da Universidade de São Paulo.
As sequências geradas para o gene COI foram comparadas com os genes correspondentes
de Anastrepha spp. ou de Drosophila yakuba Burla disponíveis no GenBank. O alinhamento das
sequências foi conduzido utilizando-se o programa Clustal W. As sequências serão depositadas
no GenBank. As sequências obtidas foram utilizadas para gerar mapas de restrição utilizando-se
o programa Webcutter 2.0 (Heiman 1997) que auxiliaram na escolha das enzimas testadas.
PCR-RFLP
O DNA amplificado via PCR foi digerido utilizando-se as enzimas de restrição DraI,
BsuRI, GsuI, RsaI, SspI e VspI. Pelo menos cinco adultos de cada espécie foram testados e
comparados entre si para verificar a existência de variação intrapopulacional. Logo após a
obtenção desses resultados, cinco exemplares de larvas de cada espécie também foram testados
para verificar se o padrão de bandas observadas nos adultos era o mesmo observado para as
larvas. Nos casos em que houve variação intrapopulacional, um total de 10 indivíduos de cada
estágio foram analisados. As reações de digestão foram preparadas utilizando-se 2 µL 1X
SEBuffer, 0,2 µL de enzima, 12 µL água milli-Q e 1µL do produto de PCR, correspondente a um
volume final de 15 µL. As amostras foram incubadas por períodos de 18 a 20 horas a 37ºC, com
exceção da enzima BsuRI que foi incubada a 30ºC. Após esse período foram adicionados 4 µL de
tampão de carregamento (solução de azul de bromofenol 0,25%, xileno cianol 0,25% e sacarose
40%) para a eletroforese em gel de agarose. Os produtos de digestão foram separados e

133
visualizados em gel de agarose a 2%, corados com GelRed (10.000X), com tampão TBE 0,5X
preparado de acordo com Sambrook et al. (1989), sendo visualizados e fotografados em
fotodocumentador de luz ultravioleta (UV).
Mapas de restrição
Os mapas de restrição foram gerados a partir das sequências de genes mitocondriais (COI
e 16S) e nuclear (ITS1), através do programa Webcutter 2.0 a fim de avaliar a presença de sítios
de restrição para diferentes enzimas nas três regiões que foram sequenciadas. Com base nos
mapas de restrição obtidos foram escolhidas as enzimas utilizadas para as digestões.
Análise dos resultados
Os diferentes padrões de restrição determinados pelas análises de PCR-RFLP obtidos dos
géis foram comparados entre si.
RESULTADOS
Foram analisadas duas diferentes regiões do DNA mitocondrial (COI e 16S) e uma do
DNA nuclear (ITS1) para 13 espécies de Anastrepha. Estes três fragmentos apresentaram padrões
de bandas únicos para cinco das espécies estudadas aqui, entretanto, para a maioria delas o
padrão das bandas foi similar. As espécies A. manihoti e A. pickeli, apresentaram um padrão de
bandas semelhante entre si, porém único para o grupo spatulata. Deve ser ressaltado que apenas
duas das dez espécies deste grupo foram analisadas neste estudo.
Para o fragmento COI, foram testadas as enzimas DraI, BsuRI, GsuI, SspI e VspI. O
padrão obtido com a enzima VspI diferenciou A. striata e também A. manihoti e A. pickeli das
demais espécies (Figura 1). Para a digestão com a enzima BsuRI, foram encontrados padrões de
bandas exclusivos para A. pulchra e A. striata em relação às outras espécies estudadas aqui.
Entretanto, nos testes realizados para o fragmento COI com as enzimas DraI, GsuI e SspI foi
obtido o mesmo padrão de bandas para todas as outras espécies inseridas neste trabalho.

134
Figura 1 - Gel de agarose 2% com produtos de PCR do fragmento COI digerido pela
enzima VspI. Os números acima das colunas representam as espécies: 1. A. antunesi. 2. A.
atrigona. 3. A. bahiensis. 4. A. coronilli. 5. A. curitis. 6. A. distincta. 7. A. manihoti. 8. A. pickeli.
9. A. pulchra. 10. A. sororcula. 11. A. turpiniae. 12. A. zenildae. 13. A. striata. Nas linhas das
extremidades está o marcador φX174.
Já para o fragmento 16S, foram testadas as enzimas DraI, BsuRI, SspI e VspI. Foram
obtidos resultados significativos com a enzima SspI, sendo observado um único padrão de bandas
para A. striata e outro para A. manihoti e A. pickeli (Figura 2). Foram observados padrões de
bandas semelhantes para os pares A. atrigona e A. pulchra, A. coronilli e A. zenildae, e o trio A.
bahiensis, A. sororcula e A. turpiniae. Foi possível também a diferenciação de A. sororcula de A.
zenildae (Figura 2) que são consideradas pragas que causam prejuízos econômicos a fruticultura
brasileira (Zucchi 2000; Malavasi et al. 2000). Também foi observado que A. curitis não possui
sítio de restrição para esta enzima. A digestão do fragmento 16S com DraI apresentou padrão de
bandas diferentes para A. atrigona e A. curitis (Figura 2), enquanto que as demais espécies
estudadas aqui apresentaram o mesmo padrão de bandas. A digestão do fragmento 16S com a
enzima VspI diferenciou A. manihoti das demais espécies (Figura 2). Já a enzima BsuRI não foi
eficiente para diferenciar as espécies estudadas aqui.

135
Figura 2 - Gel de agarose 2% com produtos de PCR do fragmento 16S digerido pela
enzima SspI: 1. A. bahiensis (adulto). 2. A. bahiensis (larva). 3. A. manihoti. 4. A. pickeli
(adulto). 5. A. pickeli (larva). 6. A. striata. 7. A. sororcula. 8. A. zenildae (adulto). 9. A. zenildae
(larva) Fragmento 16S digerido por DraI: 1. A. bahiensis. 2. A. atrigona. 3. A. curitis (adultos). 4
e 5. A. curitis (larva). 6. A. coronilli. Fragmento 16S digerido por VspI: 1. A. manihoti. 2. A.
pickeli. 3. A. striata. Nas linhas das extremidades está o marcador φX174.
Para o fragmento ITS1, foram testadas as enzimas DraI, RsaI e VspI. Os melhores
resultados foram obtidos com a enzima RsaI que apresentou padrões de bandas únicos para A.
atrigona e A. curitis e padrão semelhante para as demais espécies analisadas, mas essa enzima
não foi eficiente com exemplares de A. manihoti, pois em alguns não houve reconhecimento do
sítio do restrição. A digestão deste fragmento com a enzima VspI apresentou um padrão exclusivo
para A. pulchra em relação as outras espécies (Figura 3), por outro lado A. atrigona e A. curitis
não apresentaram sítio de restrição para esta enzima. As demais espécies apresentaram apenas um
sítio, sendo obtido o mesmo padrão de bandas para as mesmas. A digestão do fragmento ITS1
pela enzima DraI, não diferenciou as espécies, pois todas apresentaram o mesmo padrão de
bandas.

136
Figura 3 - Gel de agarose 2% com produtos de PCR do fragmento ITS1 digerido pela
enzima VspI: 1. A. bahiensis. 2. A. coronilli. 3. A. pulchra. 4. A. turpiniae. 5. A. zenildae. Nas
colunas das extremidades está o marcador φX174.
Foram testados produtos de PCR dos três fragmentos com as enzimas de restrição para as
larvas e elas apresentaram o mesmo padrão de bandas que os adultos de sua respectiva espécie.
As exceções foram a digestão do fragmento 16S com a enzima SspI para larvas de A. pickeli, pois
seis das doze larvas testadas não apresentaram sítio de restrição. Para A. sororcula e A. zenildae,
foram testadas 20 larvas de uma amostra onde havia a mistura de larvas de ambas as espécies,
quatro das 20 larvas testadas não apresentaram sítio de restrição, oito larvas foram identificadas
como A. sororcula e outras oito como A. zenildae.
DISCUSSÃO
No presente trabalho, foram observados padrões de bandas exclusivos para algumas
espécies diferenciando-as das demais estudadas aqui. Resultados significativos das digestões para
diferenciação das espécies foram obtidos para sete espécies de Anastrepha, A. atrigona e A.
curitis para o fragmento 16S com a enzima DraI, para A. manihoti com 16S e VspI; para A.
pulchra com ITS1 e VspI; para A. striata com COI e VspI e 16S e SspI; para A. sororcula e A.
zenildae com 16S e SspI. Entretanto, para as outras espécies estudadas o padrão de bandas foi
similar, provavelmente isso tenha ocorrido devido à proximidade filogenética destas espécies
(McPheron et al. 1999). Vale ressaltar que para A. manihoti, o fragmento ITS1 com a enzima
RsaI não reconheceu sítio de restrição em alguns exemplares testados, provavelmente devido ao
fragmento ITS1 ser um espaçador intergênico e não se manter muito conservado como, por
exemplo, o fragmento COI (Prezotto 2008).
Em um estudo realizado com PCR-RFLP para distinguir 23 espécies de tefritídeos dos
gêneros Bactrocera (16 espécies), Anastrepha (cinco espécies), Ceratitis (uma espécie) e Dirioxa

137
Walker (uma espécie), Armstrong et al. (1997) analisaram os padrões de digestão dos fragmentos
18S e 18S/ITS utilizando 23 enzimas de restrição. As espécies de Anastrepha estudadas foram
Anastrepha grandis (Macquart), Anastrepha ludens (Loew), Anastrepha obliqua (Macquart),
Anastrepha serpentina (Weidemann) e A. striata. Eles observaram que o fragmento 18S/ITS
digerido com as enzimas a RsaI e Sau3a apresentou padrões de bandas exclusivos para A.
grandis, A. obliqua, A. ludens e A. striata sendo possível a diferenciação dessas quatro espécies
entre si. Entretanto, nenhum resultado foi relatado para A. serpentina. Vale salientar que até o
momento, esse foi o único estudo realizado com PCR-RFLP para espécies de Anastrepha. No
presente estudo, as digestões realizadas com o fragmento ITS1 para A. striata não a diferenciou
das demais espécies estudadas. A diferenciação entre A. striata e as demais espécies estudadas
aqui foi obtida com os fragmentos COI e 16S como relatado acima no item sobre os resultados.
Em outro estudo utilizando a mesma técnica de PCR-RFLP, Barr et al. (2006) analisaram
30 espécies de tefritídeos dos gêneros Ceratitis (26 espécies), Trirhithrum Bezzi (uma espécie),
Capparimyia Martelli (uma espécie), Carpophthoromyia Austen (uma espécie) e Notomma Loew
(uma espécie) com três fragmentos de DNA mitocondrial (12S, 16S e ND6) digeridos com 12
enzimas de restrição (Barr et al. 2006). Os autores mostraram que a melhor enzima utilizada para
o fragmento 12S foi a SspI, para 16S foi DraI e para ND6 foi HinfI. A combinação entre esses
fragmentos e as enzimas mostrou padrões de bandas diferentes para 25 espécies, entretanto,
também mostrou alguns padrões semelhantes para quatro espécies. No presente estudo, para
espécies de Anastrepha os melhores resultados foram obtidos com o fragmento 16S combinado
com as enzimas VspI e SspI, também foram realizados testes com a enzima DraI e não foram
obtidos resultados satisfatórios para diferenciação das espécies estudadas aqui. A escolha das
enzimas DraI, VspI e SspI foi baseada nos mapas de restrição gerados anteriormente e nas
informações obtidas em estudos realizados previamente que forneceram resultados satisfatórios
com essas enzimas.
No presente estudo, foi possível diferenciar sete espécies de Anastrepha: atrigona, curitis,
manihoti, pulchra, sororcula, striata e zenildae. Essas informações são úteis na identificação de
imaturos (larvas) de espécies deste gênero que podem ser encontradas em frutas frescas
exportadas e importadas, evitando assim a dispersão de novas pragas no país.

138
CONCLUSÕES
Com a digestão do fragmento 16S com a enzima de restrição SspI foi possível diferenciar
A. striata, A. sororcula e A. zenildae. A digestão deste mesmo fragmento com a enzima DraI foi
possível diferenciar A. atrigona e A. curitis e com a enzima VspI diferenciar A. manihoti das
demais espécies estudadas aqui.
A digestão do fragmento da região ITS1 com a enzima VspI diferenciou A. pulchra das
demais espécies estudadas.
AGRADECIMENTOS
Os autores agradecem a Ana Claúdia Araújo, Claudemir Campos, Felipe Gomes, José
Nilton Medeiros da Costa, Marcos Vinicius Garcia, Mírian Santos, Tatiana Senra, Weber Novaes
e Ulisses Neiss pelo auxílio durante as coletas. Esse estudo foi financiado pelo Conselho
Nacional de Desenvolvimento Científico e Tecnológico (CNPq - projeto n° 575664/2008-8), pela
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES - projeto n°
23038.037185/2008-41), pela Fundação de Amparo a Pesquisa do Estado do Amazonas
(FAPEAM - projeto nº014-017/2004) e pela Rede Amazônica de Pesquisas sobre Moscas-das-
Frutas (Chamada 05/2006 Agrofuturo). Os autores também ao CNPq pela bolsa de doutorado
concedida a VSD.

139
SÍNTESE
No presente estudo, foram relatadas novas interações entre espécies de Anastrepha,
espécies botânicas e de parasitoides braconídeos coletados em alguns municípios brasileiros, bem
como confirmadas outras associações.
Este estudo também mostrou que a microscopia eletrônica de varredura (MEV) é uma
importante ferramenta para a descrição morfológica de ovos de 11 espécies de Anastrepha.
Revelou que existem semelhanças com outras espécies descritas anteriormente e que a
combinação das características exibidas por esses ovos, como a ornamentação do córion,
localização das aerópilas e da micrópila, presença ou ausência e variação de apêndice respiratório
são úteis como caracteres taxonômicos.
Com base na morfologia, foi possícel identificar as larvas de oito espécies de Anastrepha,
apesar da grande similaridade e sobreposição entre elas e outras espécies, cujas larvas foram
descritas previamente. Características como número de fileiras de espinhos dorsais e ventrais nos
segmentos, número de saliências orais, número de cerdas no espiráculo posterior, comprimento
do esqueleto céfalo-faríngeo e formato da mandíbula são caracteres taxonômicos que podem
auxiliar na identificação de espécies de importância quarentenária.
Com a técnica de PCR-RFLP, foi possível diferenciar sete espécies de Anastrepha
utilizando o fragmento 16S com as enzimas de restrição SspI, DraI e VspI. Todas as informações
morfológicas e moleculares sobre ovos, larvas e adultos obtidas neste estudo são úteis na
identificação de imaturos (ovos e larvas) de espécies de Anastrepha, para a elaboração de
estratégias de controle, política de defesa vegetal, além do esclarecimento sobre as relações
filogenéticas dentro dos diversos grupos de espécies de Anastrepha.

140
REFERÊNCIAS BIBLIOGRÁFICAS
Associação Central dos Fruticultores do Norte de Minas, ABANORTE, 2009. Brasil é o terceiro
maior produtor de frutas do mundo (http://www.abanorte.com.br/noticias/noticias-
principal/brasil-e-o-terceiro-maior-produtor-de-frutas-do-mundo/). Acesso em 25/10/2011.
Aluja, M. 1994. Bionomics and management of Anastrepha. Annual Review Entomology, 39:
155-178.
Araujo, E.L.; Zucchi, R.A. 2003. Moscas-das-frutas (Diptera: Tephritidae) em goiaba (Psidium
guajava L.), em Mossoró, RN. Arquivos do Instituto Biológico, 70: 73-77.
Araujo, E.L.; Zucchi, R.A. 2006. Medidas do acúleo na caracterização de cinco espécies de
Anastrepha do grupo fraterculus (Diptera: Tephritidae). Neotropical Entomology, 35: 329-
337.
Araújo, V.C. 1970. Fenologia de essências florestais amazônicas I. Manaus: INPA. Boletim do
INPA. Pesquisas Florestais.
Armstrong, K.F.; Cameron, C.M.; Frampton, E.R. 1997. Fruit fly (Diptera: Tephritidae) species
identification: a rapid molecular diagnostic technique for quarantine application. Bulletin of
Entomological Research, 87: 111–118.
Barr, N.B.; Copeland, R.S.; De Meyer, M.; Masiga, D.; Kibogo, H.G.; Billah, M.K.; Osir, E.,
Wharton, R.A.; McPheron, B.A. 2006. Molecular diagnostics of economically important
Ceratitis fruit fly species (Diptera: Tephritidae) in Africa using PCR and RFLP analyses.
Bulletin of Entomological Research, 96: 505-521.
Bellotti, A.C. 2008. Cassava pests and their management, p. 764-794. In: Capinera, J. (Ed.),
Encyclopedia of Entomology, 2nd edition, Springer Reference, The Netherlands.

141
Canal, N.A.; Zucchi, R.A. 2000. Parasitóides - Braconidae, pp. 119-126. In: Malavasi, A.;
Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no Brasil. Conhecimento
Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Caraballo, C.J. 1981. Las moscas de las frutas del género Anastrepha Schiner 1968 (Diptera:
Tephritidae) de Venezuela. Tese de Doutorado, Universidad Central de Venezuela, Maracay,
Venezuela, 210 pp.
Carroll, L.E.; Wharton, R.A. 1989. Morphology of the immature stages of Anastrepha ludens
(Diptera: Tephritidae). Annals of the Entomological Society of America, 82: 201-214.
Cônsoli, F.L.; Kitajima, E.W.; Parra, J.R.P. 1999. Ultrastructure of the natural and factitious host
eggs of Trichogramma galloi Zucchi and Trichogramma pretiosum Riley (Hymenoptera:
Trichogrammatidae). International Journal of Insect Morphology and Embryology, 28: 211-
229.
Costa, S.G.M; Querino, R.B.; Ronchi-Teles, B.; Penteado-Dias, A.M.M.; Zucchi, R.A. 2009.
Parasitoid diversity (Hymenoptera: Braconidae and Figitidae) on frugivorous larvae (Diptera:
Tephritidae and Lonchaeidae) at Adolpho Ducke Forest Reserve, Central Amazon Region,
Manaus, Brazil. Brazilian Journal of Biology, 69(2): 363-370.
Duarte, A.L.; Malavasi, A. 2000. Tratamentos Quarentenários, p. 187-192. In: Malavasi, A.;
Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no Brasil. Conhecimento
Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G. 2011a. Egg morphology of Anastrepha
spp. (Diptera: Tephritidae) in the fraterculus group using SEM. Annals of the Entomological
Society of America, 104(1): 16-24.

142
Dutra, V.S.; Ronchi-Teles, B.; Steck, G.J.; Silva, J.G. 2011b. Description of eggs of Anastrepha
spp. (Diptera: Tephritidae) in the spatulata group using Scanning Electron Microscopy.
Annals of the Entomological Society of America, 104(5): 857-862.
Emmart, E.W. 1933. The eggs of four species of fruit flies of the genus Anastrepha. Proceedings
of the Entomological Society of Washington, 35: 184-191.
Ferrar, P. 1987. A guide to the breeding habits and immature stages of Diptera Cyclorrhapha.
Entomonograph, 8: 1-907.
Figueiredo, J.V.A.; Perondini, A.L.P.; Ruggiro, E.M.; Prezzoto, L.F.; Selivon, D. 2011. External
eggshell morphology of Anastrepha fruit flies (Diptera: Tephritidae). Acta Zoologica
(Stockholm), xx: 1–9.
Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. 1994. DNA primers for amplification
of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates.
Molecular Marine Biology and Biotechnology, 3: 294-297.
Frías, D.L.; Hernández-Ortiz, V.; Munõz, L.L. 2009. Description of the Third-Instar of
Anastrepha leptozona Hendel (Diptera: Tephritidae). Neotropical Entomology, 38(4):491-496.
Gasparich, G.E.; Sheppard, W.S.; Han, H.-Y.; McPheron, B.A.; Steck, G.J. 1995. Analysis of
mitochondrial DNA and development of PCR-based diagnostic markers for Mediterranean
fruit fly (Ceratitis capitata) populations. Insect Molecular Biology, 4: 61–67.
Gasparich, G.E.; Silva, J.G.; Han, H.-Y.; McPheron, B.A.; Steck, G.J.; Sheppard, W.S. 1997.
Population genetic structure of the Mediterranean fruit fly (Diptera: Tephritidae) and
implications for worldwide colonization patterns. Annals of the Entomological Society of
America, 90: 790–797.

143
Guimarães, J.A.; Diaz, N.B.; Zucchi, R.A. 2000. Parasitóides - Figitidae (Eucoilinae), p. 127-134.
In: Malavasi, A.; Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no
Brasil. Conhecimento Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Han, H.-Y.; McPheronN, B.A. 1997. Molecular phylogenetic study of Tephritidae (Insecta:
Diptera) using partial sequences of the mitochondrial 16S ribosomal DNA. Molecular
Phylogenetics and Evolution, 7: 17–32.
Headrick, D.H.; Goeden, R.D. 1998. The biology of nonfrugivorous tephritid fruit flies. Annual
Review Entomology, 43: 217-241.
Heiman, M. 1997. Webcutter 2.0 (http://rna.lundberg.gu.se/cutter2/). Acesso em 15/12/2011.
Hernandez-Ortiz, V. 2007. Diversidad y biogeografía del género Anastrepha em Mexico, p. 53-
76. In: Hernandez-Ortiz, V. (Ed.). Moscas de la fruta en Latinoamérica (Diptera:
Tephritidae): diversidad, biologia y manejo. S y G editores, Mexico.
Hernández-Ortiz, V.; Pérez-Alonso, R. 1993. The natural host plants of Anastrepha (Diptera:
Tephritidae) in a tropical rain forest of México. Florida Entomologist, 76 (3): 447-460.
Hernández-Ortiz, V.; Aluja, M. 1994. Listado de especie del genero Neotropical Anastrepha
(Diptera: Tephritidae) com notas sobre su distribucion y plantas hospederas. Folia
Entomológica Mexicana, 88: 89-105.
Hernández-Ortiz, V.; Guillén-Aguilar, J.; López, L. 2010. Taxonomía e Identificación de Moscas
de La Fruta de Importancia Económica em América, p.49-80. In: Montoya, P.; Toledo, J.;
Hernández, E. (Eds.). Moscas de La Fruta: Fundamentos y Procedimientos para su Manejo,
S y G Editores, Distrito Federal, México.

144
Jesus, C.R.; Oliveira, M.N.; Souza Filho, M.; Silva, R.A.; Zucchi, R.A. 2008. First record of
Anastrepha parishi Stone (Diptera, Tephritidae) and its host in Brazil. Revista Brasileira de
Entomologia, 52: 135-136.
Jirón, L.F.; Hedström, I. 1988. Occurrence of fruit flies of the genera Anastrepha and Ceratitis
(Diptera: Tephritidae) and their host plant availability in Costa Rica. Florida Entomologist,
71: 62-73.
Lawrence, P.O. 1979. Immature stages of the Caribbean fruit fly, Anastrepha suspensa. Florida
Entomologist, 62: 214-219.
Malavasi, A.; Zucchi, R.A.; Sugayama, R.L. 2000. Biogeografia, p. 93-98. In: Malavasi, A.;
Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no Brasil. Conhecimento
Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Marinho, C.F.; Souza-Filho, M.F.; Raga, A.; Zucchi, R.A. 2009. Parasitóides (Hymenoptera:
Braconidae) de Moscas-das-Frutas (Diptera: Tephritidae) no Estado de São Paulo: Plantas
Associadas e Parasitismo. Neotropical Entomology, 38(3): 321-326.
McPheron, B.; Han, H.-Y.; Silva, J.G.; Norrbom, A.L. 1999. Phylogeny of Anastrepha and
Toxotrypana based upon 16S rRNA Mitochondrial DNA Sequences, p. 343-361. In: Aluja,
M.; A.L. Norrbom (Eds.). Fruit flies (Tephritidae): Phylogeny and evolution of behavior.
CRC Press, New York, New York.
McPheron, B.A.; Gasparich, G.E.; Han, H-Y.; Steck, G.J.; Sheppard, W.S. 1994. Mitochondrial
DNA restriction map for the Mediterranean fruit fly, Ceratitis capitata. Biochemical
Genetics, 32: 25–33.
Morales, P.; González, E. 2007. El género Anastrepha Schiner y su importancia económica en
frutales de Venezuela, p. 27-52. In: Hernandez-Ortiz, V. (Ed.). Moscas de la fruta en
Latinoamérica (Diptera: Tephritidae): diversidad, biologia y manejo. S y G editores, Mexico.

145
Muraji, M.; Nakahara, S. 2002. Discrimination among pest species of Bactrocera (Diptera:
Tephritidae) based on PCR– RFLP of the mitochondrial DNA. Applied Entomology and
Zoology, 37: 437–446.
Murillo, T.; Jirón, L.F. 1994. Egg morphology of Anastrepha obliqua and some comparative
aspects with eggs of Anastrepha fraterculus (Diptera: Tephritidae). Florida Entomologist, 77:
342-348.
Nascimento, A.S.; Carvalho, R.S. 2000. Manejo integrado de moscas-das-frutas, p. 169-173. In:
Malavasi, A.; Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no Brasil.
Conhecimento Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Norrbom, A.L. 2000. Fruit Fly (Diptera: Tephritidae) Classification Table
(http://www.sel.barc.usda.gov/Diptera/tephriti/Clastabl.htm). Acesso em 27/10/2011.
Norrbom, A.L. 2002. A revision of the Anastrepha serpentina species group (Diptera:
Tephritidae). Proceedings Entomological Society of Washington, 104:390-436.
Norrbom, A. 2004. Updates to Biosystematic Database of World Diptera for Tephritidae through
1999. Diptera Data Dissemination Disk 2.
Norrbom, A. 2005. The Diptera Site: nomenclator Query
(http://www.sel.barc.usda.gov:591/Diptera/names/searchNo.htm). Acesso em 14/12/2011.
Norrbom, A.L.; Kim, K.C. 1988. A list of the reported host plants of the species of Anastrepha
(Diptera: Tephritidae). U.S. Dept. Agric., Animal and Plant Health Inspection Service, Plant
Protection and Quarentine, Aphis, USDA, 114 pp.
Norrbom, A.L.; Korytkowski, C.A. 2009. A revision of the Anastrepha robusta species group
(Diptera: Tephritidae). Zootaxa, 2182: 1–91.

146
Norrbom, A.L.; Korytkowski, C.A. 2011. New species of and taxonomic notes on Anastrepha
(Diptera: Tephritidae). Zootaxa, 2740: 1–23.
Norrbom, A.L.; Uchôa, M.A. 2011. New species and records of Anastrepha (Diptera:
Tephritidae) from Brazil. Zootaxa, 2835: 61–67.
Norrbom, A.L.; Zucchi, R.A.; Henández-Ortiz, V. 1999. Phylogeny of the genera Anastrepha and
Toxotrypana (Trypetinae: Toxotrypanini) based on morphology, p. 299-342. In: M. Aluja; A.
L. Norrbom (Eds.). Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior. CRC
Press, New York, New York.
Ovruski, S.; Aluja, M.; Sivinski, J.; Wharton, R. 2000. Hymenopteran parasitoids on fruit-
infesting Tephritidae (Diptera) in Latin America and the southern United States: Diversity,
distribution, taxonomic status and their use in fruit fly biological control. Integrated Pest
Management Reviews 5: 81–107.
Perondini, A.L.P.; Selivon, D. 1999. The structure of the chorion of the Anastrepha fraterculus
(Wiedemann, 1830) eggs (Diptera, Tephritidae). Revista Brasileira de Entomologia, 43: 243-
248.
Prezotto, L.F. 2008. Análise do ITS1 do DNA ribossômico em espécies do complexo Anastrepha
fraterculus (Diptera: Tephritidae). Dissertação de Mestrado, Instituto de Biociências da
Universidade de São Paulo, São Paulo, São Paulo. 74pp.
Prokopy, R.J.; Roitberg, B.D. 1984. Foraging behavior of true fruit flies. American Scientist, 72:
41-49.
Ronchi-Teles, B. 2009. Tephritidae. p. 183-191. In: Fonseca, C.R.V.; Magalhães, C.; Rafael,
J.A.; Franklin, E. A fauna de artrópodes da Reserva Florestal Ducke: Estado atual do
conhecimento taxonômico e biológico. Editora do INPA, Manaus, Amazonas.

147
Ronchi-Teles, B.; Silva, N.M. 2005. Flutuação populacional de espécies de Anastrepha Schiner
(Diptera: Tephritidae) na região de Manaus, AM. Neotropical Entomology, 34:733-741.
Ronchi-Teles, B.; Dutra, V.S.; Tregue-Costa, A.P.; Aguiar-Menezes, E.L.; Mesquita, A.C.A.;
Silva, J.G. 2011. Natural host plants and native parasitoids associated with Anastrepha
pulchra and other Anastrepha species (Diptera: Tephritidae) in Central Amazon, Brazil.
Florida Entomologist, 94: 347-349.
Salazar, M.; Theoduloz, C.; Vega, A.; Poblete, F.; Gonzalez, E.; Badilla, R.; Meza-Basso, L.
2002. PCR–RFLP identification of endemic Chilean species of Rhagoletis (Diptera:
Tephritidae) attacking Solanaceae. Bulletin of Entomological Research, 92: 337–341.
Salles, L.A. 2000. Biologia e ciclo de vida de Anastrepha fraterculus (Wied.), p. 81-86. In:
Malavasi, A.; Zucchi, R.A. (Eds.). Moscas-das-frutas de Importância Econômica no Brasil.
Conhecimento Básico e Aplicado. Holos Editora, Ribeirão Preto, São Paulo.
Sambrook, J.; Fritsch, E.F.; Maniatis, T. 1989. Molecular Cloning: A Laboratory Manual. Cold
Spring Harbor Laboratory, Cold Spring Harbor, New York, NY. 253 p.
Sambrook, J.; Russel, D.W. 2001. Molecular Cloning: a Laboratory Manual. Cold Spring Harbor
Laboratory, Cold Spring Harbor, New York, NY. 2344 p.
Seín, F. 1933. Anastrepha (Trypetydae, Diptera) fruit flies in Puerto Rico. The Journal of the
Department of Agriculture of Puerto Rico, XVII: 183-196.
Selivon, D.; Perondini, A.L.P. 1998. Eggshell morphology in two cryptic species of the
Anastrepha fraterculus complex (Diptera: Tephritidae). Annals of the Entomological Society
of America, 91: 473-478.
Selivon, D.; Perondini, A.L.P. 1999. Description of Anastrepha sororcula and A. serpentina
(Diptera: Tephritidae) eggs. Florida Entomologist, 82: 347-353.

148
Selivon, D.; Perondini, A.L.P. 2000. Morfologia dos ovos de moscas-das-frutas do gênero
Anastrepha, p. 49-54. In: Malavasi, A.; Zucchi, R.A. (Eds.). Moscas-das-frutas de
Importância Econômica no Brasil. Conhecimento Básico e Aplicado. Holos Editora, Ribeirão
Preto, São Paulo.
Selivon, D.; Vretos, C.; Perondini, A.L.P. 2003. Evaluation of egg morphology from ethanol
preserved females of Anastrepha sp.2 aff. fraterculus (Diptera: Tephritidae). Neotropical
Entomology, 32: 527-529.
Silva, N.M.; Ronchi-Teles, B. 2000. Amapá, Amazonas, Pará, Rondônia e Roraima, p. 203-209.
In: Malavasi, A.; Zucchi, R.A. (Eds.), Moscas-das-frutas de importância Econômica no
Brasil. Conhecimento básico e aplicado. Holos-Editora, Ribeirão Preto, São Paulo.
Silva, N.M.; Silveira Neto, S.; Zucchi, R.A. 1996. The natural host plants of Anastrepha in the
State of Amazonas, Brazil, p. 353-357.In: McPheron, B.A.; Steck, G.J. (Eds.). Fruit fly pests:
a world assessment of their biology and management. St. Lucie Press, Delray Beach, Florida.
Silva, J.G.; Meixner, M.D.; McPheron, B.A.; Steck, G.J.; Sheppard, W.S. 2003. Recent
Mediterranean fruit fly (Diptera: Tephritidae) infestations in Florida - a genetic perspective.
Journal of Economic Entomology, 96: 1711–1718.
Silva, J.G.; Dutra, V.S.; Santos, M.S.; Silva, N.M.O.; Vidal, D.B.; Nink, R.A.; Guimarães, J.A.;
Araujo, E.L. 2010. Diversity of Anastrepha spp. (Diptera: Tephritidae) and associated
braconid parasitoids from native and exotic hosts in Southeastern Bahia, Brazil.
Environmental Entomology, 39(5): 1457-1465.
Smith-Caldas, M.R.B.; McPheron, B.A.; Silva, J.G.; Zucchi, R.A. 2001. Phylogenetic
relationships among species of the fraterculus group (Anastrepha: Diptera: Tephritidae)
inferred from DNA sequences of mitochondrial cytochrome oxidase I. Neotropical
Entomology, 30: 565-573.

149
Steck, G.J.; Malavasi, A. 1988. Description of immatures stages at Anastrepha bistrigata
(Diptera: Tephritidae). Annals of the Entomological Society of America, 81: 1004-1009.
Steck, J.; Wharton, R.A. 1988. Description of immature stages of Anastrepha interrupta, A.
limae, and A. grandis (Diptera: Tephritidae). Annals of the Entomological Society of America,
81: 994-1003.
Steck, G.J.; Carroll, L.E.; Celedonio-Hurtado, H.; Guillen-Aguilar. J. 1990. Methods for
identification of Anastrepha larvae (Diptera: Tephritidae), and key to 13 species. Proceedings
Entomological Society of Washington, 92: 333-346.
Steyskal, G.C. 1977. Pictorial key to species of the genus Anastrepha (Diptera: Tephritidae).
Washington, Entomological Society of Washington, 35p.
Stone, A. 1942. The fruit flies of the genus Anastrepha. Department of Agriculture Miscellaneous
Publication, United States,USA, n. 439, 112p.
Teskey, H. J. 1981. Morphology and terminology - larvae, p. 65-88. In: McAlpine, J.F.;
Vockeroth, J.R.; Wood, D.M. (Eds.). Manual of Nearctic Diptera, vol. 1 Research Branch,
Agriculture Canada. Monograph 27.
Uramoto, K.; Martins, D.S.; Lima, R.C.A.; Zucchi, R.A. 2008. Host plant record for Anastrepha
fumipennis and A. nascimentoi (Diptera, Tephritidae). Journal of Insect Science (Online), 8:
1-4.
Uramoto, K.; Zucchi, R.A. 2010. New species of Anastrepha Schiner (Diptera, Tephritidae) from
remnant area of the Atlantic Rain Forest and surroundings in the state of Espírito Santo,
Brazil. Zootaxa, 2535: 49–60.
White, I.M.; Elson-Harris, M.M. 1992. Fruit flies of economic significance: their identification
and bionomics. CAB International, Wallingford, UK, 601pp.

150
Zucchi, R.A. 2000. Taxonomia, p. 13-24. In: Malavasi, A.; Zucchi, R.A. (Eds.). Moscas-das-
frutas de Importância Econômica no Brasil. Conhecimento Básico e Aplicado. Holos Editora,
Ribeirão Preto, São Paulo.
Zucchi, R.A. 2007. Diversidad, distribución y hospederos del género Anastrepha en Brasil, p. 77-
100. In: Hernández-Ortiz, V. (Ed.), Moscas de la Fruta en Latinoamérica (Diptera:
Tephritidae): Diversidad, biologia y manejo. S y G editores, Distrito Federal, México.
Zucchi, R.A. 2008. Fruit flies in Brazil - Anastrepha species their host plants and parasitoids
(www.lea.esalq.usp.br/anastrepha/, updated on September, 2011). Acesso em 05/01/2012.
Zucchi, R.A.; Silva, N.M.; Silveira Neto, S. 1996. Anastrepha species from the Brazilian
Amazon: distribution, hosts, and lectotype designations, p. 259-264. In: McPheron, B.A.;
Steck, G.J. (Eds.). Fruit fly pests: a world assessment of their biology and management. St.
Lucie Press, Delray Beach, Florida.
Zucchi, R.A.; Silva, R.A.; Deus, E.G. 2011b. Espécies de Anastrepha e seus hospedeiros na
Amazônia brasileira, p. 53-70. In: Silva, R.A.; Lemos W.P.; Zucchi, R.A. (Eds.). Moscas-das-
frutas na Amazônia Brasileira. Diversidade, Hospedeiros e Inimigos Naturais. Embrapá
Amapá, Macapá, Amapá.
Zucchi, R.A.; Uramoto, K.; Souza-Filho, M.F. 2011a. Chave ilustrada para as espécies de
Anastrepha da região Amazônica, p. 73-90. In: Silva, R.A.; Lemos W.P.; Zucchi, R.A. (Eds.).
Moscas-das-frutas na Amazônia Brasileira. Diversidade, Hospedeiros e Inimigos Naturais.
Embrapá Amapá, Macapá, Amapá.

151
ANEXOS
(Comprovação das publicações e aceite)
Anexo 1. Comprovação da publicação da nota científica intitulada “Natural Host Plants and
Native Parasitoids Associated with Anastrepha pulchra and other Anastrepha Species (Diptera:
Tephritidae) in Central Amazon, Brazil”.

152

153
Anexo 2. Comprovação da publicação da nota científica intitulada “Host Plant of Anastrepha
pulchra (Diptera: Tephritidae) in Central Amazon, Brazil - Mistaken Identity Resolved”.

154

155
Anexo 3. Comprovação da publicação do manuscrito intitulado “Egg morphology of Anastrepha
spp. (Diptera: Tephritidae) in the fraterculus group using SEM”.

156
Anexo 4. Comprovação da publicação do manuscrito intitulado “Description of eggs of
Anastrepha spp. (Diptera: Tephritidae) in the spatulata group using Scanning Electron
Microscopy”.
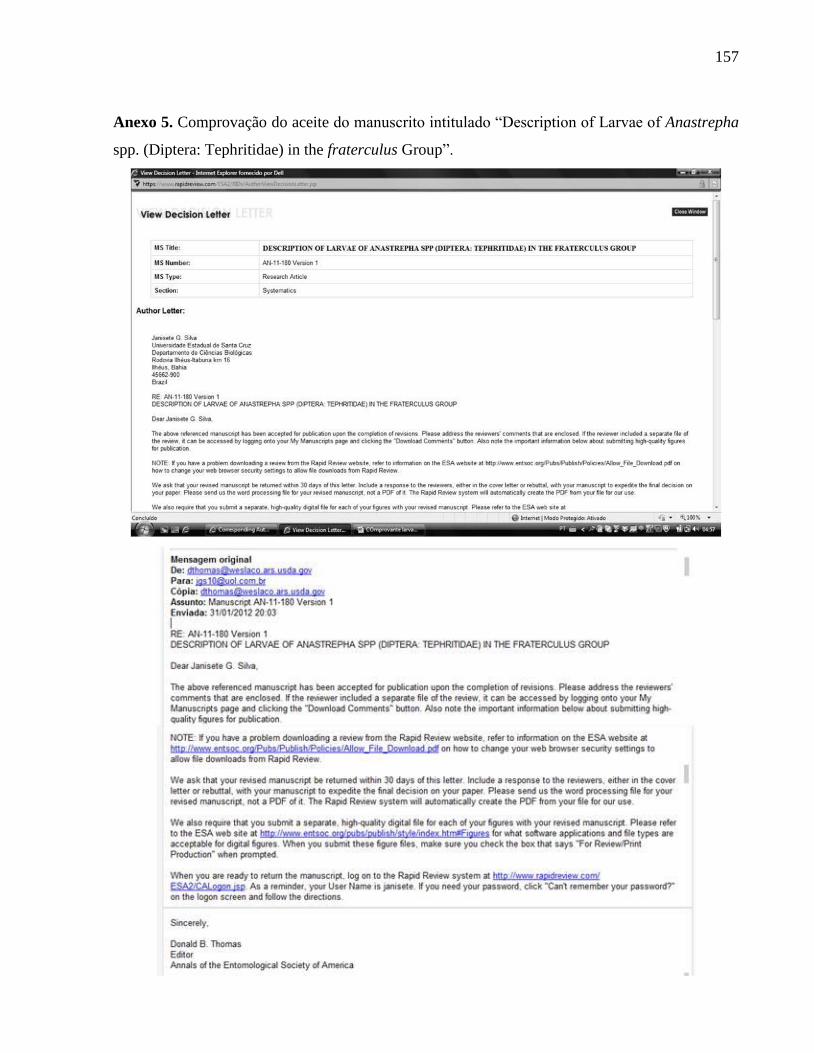
157
Anexo 5. Comprovação do aceite do manuscrito intitulado “Description of Larvae of Anastrepha
spp. (Diptera: Tephritidae) in the fraterculus Group”.








![[LIVRO] ZendFramework 2 - Elton Luís Minetto](https://static.fdocumentos.tips/doc/165x107/55cf947e550346f57ba26739/livro-zendframework-2-elton-luis-minetto.jpg)









