
MODULAÇÃO DA DEGRADAÇÃO ENZIMÁTICA DE ......2 UNIVERSIDADE DE SÃO PAULO Thalita Beatriz...
211
UNIVERSIDADE DE SÃO PAULO Thalita Beatriz Carrara da Encarnação MODULAÇÃO DA DEGRADAÇÃO ENZIMÁTICA DE GALACTOMANANO POR SUA PRÓPRIA ESTRUTURA FINA São Paulo 2012
Transcript of MODULAÇÃO DA DEGRADAÇÃO ENZIMÁTICA DE ......2 UNIVERSIDADE DE SÃO PAULO Thalita Beatriz...

UNIVERSIDADE DE SÃO PAULO
Thalita Beatriz Carrara da Encarnação
MODULAÇÃO DA DEGRADAÇÃO ENZIMÁTICA DE
GALACTOMANANO POR SUA PRÓPRIA
ESTRUTURA FINA
São Paulo
2012

2
UNIVERSIDADE DE SÃO PAULO
Thalita Beatriz Carrara da Encarnação
MODULAÇÃO DA DEGRADAÇÃO ENZIMÁTICA DE
GALACTOMANANO POR SUA PRÓPRIA ESTRUTURA FINA
MODULATION OF ENZYMATIC DEGRADATION OF
GALACTOMANNAN BY ITS FINE STRUCTURE
Orientador: Prof. Dr. Marcos Silveira Buckeridge
São Paulo
2012
Dissertação apresentada ao Instituto de Biociências da Universidade de São Paulo, para a obtenção de Título de Mestre em
Ciências Biológicas, na área de Botânica.

3
FICHA CATALOGRÁFICA
Autorizo a reprodução e divulgação total ou parcial deste trabalho, por qualquer meio
convencional ou eletrônico, para fins de estudo e pesquisa, desde que citada a fonte.
Encarnação, Thalita Beatriz Carrara
Modulação da degradação enzimática de galactomanano por sua própria
estrutura.
Orientador: Marcos Silveira Buckeridge
Dissertação (Mestrado) – Instituto de Biociências da Universidade de São
Paulo. Departamento de Botânica. 211p.
1. Galactomanano 2. α-galactosidase 3. Estrutura fina de polissacarídeos
4. Mobilização de reserva 5. Parede celular 6. Purificação enzimática
7. Sesbania virgata (CAV.) Pers. Universidade de São Paulo. Instituto de
Biociências. Departamento de Botânica.

4
Comissão Julgadora
_______________________ ______________________
Prof(a). Dr(a) Prof(a). Dr(a)
_____________________________
Prof. Dr. Marcos Silveira Buckeridge
Orientador

5
Aos meus pais, Milton e Sandra, por todo amor, carinho, dedicação e muito
apoio em todos os momentos da minha vida;
Ao meu anjo Augusto por existir em minha vida.
Dedico

6
" Voici mon secret. Il est très simple: on ne voit bien qu'avec le coeur.
L'essentiel est invisible pour les yeux."
Le Petit Prince, Antoine de Saint-Exupéry (1946).

7
Agradecimentos
Ao Instituto de Biociências da USP, principalmente ao Departamento de
Botânica;
À CAPES pela bolsa de estudo concedida;
À todo o corpo docente e aos funcionários do Departamento de Botânica por
tornarem o ambiente de trabalho muito favorável ao desenvolvimento da pós-
graduação;
Ao Professor Dr. Marcos Silveira Buckeridge pelo acolhimento em seu grupo de
pesquisa, sua orientação durante todo o desenvolvimento do Mestrado e sua
amizade;
À Dra. Sonia Dietrich agradeço pelo tempo que estive ao seu lado, aprendendo
com o seu conhecimento, me inspirando para desenvolver este trabalho, que
faz parte do seu legado, e principalmente me inspirando a ser Pesquisadora,
como ela simplesmente foi e sempre será (In memoriam);
À Professora Dra. Magdalena Rossi pela ajuda e paciência nas análises
moleculares presentes no trabalho;
Ao Professor Dr. Luis Netto e a toda equipe pela abertura do laboratório à
realização da purificação da α-galactosidase, em especial, à técnica Simone
Vidigal pela paciência, ajuda no uso do FPLC e respostas rápidas aos e-mails
de reserva do equipamento sempre feito de última hora;
À Pesquisadora Adriana Franco Paes Leme e as técnicas Romênia e Bianca
pela abertura do Laboratório de Espectrometria de Massas para a realização
do sequenciamento interno da α-galactosidase de Sesbania virgata e a grande
ajuda nesta empreitada!

8
Ao Clóvis Oliveira da Silva ou C1 ou Clovitos, como eu o chamava todo dia de
manhã, por ter aparecido do céu e me ensinado e me acompanhado durante
toda a parte experimental do meu projeto. Saiba Clovitos que sem você talvez
não tivesse chego até a metade, meu super obrigado!
À todas as meninas do Laboratório: Amanda, Dri Grandis, Bru, Dé, Egleezita,
Vivi, Gi, Cris, Eveline, por toda a ajuda em todos os quesitos, por estarem
comigo e confiarem em mim mesmo quando nem eu acreditava mais em mim!
Obrigada por absolutamente tudo!!! Aos meninos do lab também, não que eles
sejam muitos, Ivan, Maraba, Luis, Vinícius pelas ótimas risadas que demos
juntos, pelos desabafos, pelo dia-a-dia;
Ao Dr. Eduardo Aratangy, à minha terapeuta Vanessa Batista e à minha
nutricionista Fernanda Psicolaro, anjos que após muita luta apareceram em
minha vida e me ajudaram a sair da anorexia. A vocês nada do que disser será
suficiente, então só direi o meu muito obrigada por TUDO!
A todos da Academia Arena, principalmente ao Prof. José Ricardo, que
cuidaram de mim desde o primeiro dia, pegavam no meu pé e me ajudaram a
ser a Thalita novamente!
À você, Augusto, amor da minha vida, por estar ao meu lado em absolutamente
todos os momentos, por me fazer feliz, sonhar, por me fazer uma pessoa
melhor a cada dia, por me amar incondicionalmente! Obrigada pela ajuda direta
e indireta neste trabalho, obrigada por todos dias nestes últimos 7 anos e
alguns meses!
Aos meus pais, Sandra e Milton, por todo investimento em mim, por
acreditarem e me apoiarem em todas as decisões, por me amarem e estarem
absolutamente sempre ao meu lado! Saibam que este Mestrado é muito mais
de vocês do que meu, sem cada ensinamento, apoio, carinho, recebido de
vocês nada disso teria sido possível. Muito Obrigada por tudo. Amo vocês!

9
Sumário
ÍNDICE DE FIGURAS .............................................................................................................................. 12
ÍNDICE DE TABELAS .............................................................................................................................. 22
RESUMO .............................................................................................................................................. 23
ABSTRACT ............................................................................................................................................ 26
INTRODUÇÃO....................................................................................................................................... 28
1.1. SEMENTES ........................................................................................................................ 28 1.2. GERMINAÇÃO E PÓS-GERMINAÇÃO........................................................................................ 29 1.3. RESERVAS DE SEMENTES ...................................................................................................... 30 1.4. PAREDE CELULAR DE RESERVA DE SEMENTES ............................................................................. 31 1.5. POLISSACARÍDEOS DE RESERVA DE PAREDE CELULAR ................................................................... 32 1.5.1. (ARABINO)GALACTANOS ..................................................................................................... 33 1.5.2. XILOGLUCANOS ................................................................................................................. 35 1.5.3. MANANOS ....................................................................................................................... 38 1.6. GALACTOMANANOS ........................................................................................................... 39 1.6.1. ESTRUTURA QUÍMICA DOS GALACTOMANANOS ......................................................................... 40 1.6.2. PROPRIEDADES DOS GALACTOMANANOS ................................................................................. 41 1.6.3. MOBILIZAÇÃO DOS GALACTOMANANOS .................................................................................. 42 1.6.4. REGULAÇÃO DA MOBILIZAÇÃO DE RESERVA DE GALACTOMANANO ............................................... 46 1.7. CARACTERIZAÇÃO DE Α-GALACTOSIDASE .................................................................................. 47 1.8. CARACTERIZAÇÃO DA ENDO-Β-MANANASE ............................................................................... 49 1.9. ESPÉCIE SESBANIA VIRGATA .................................................................................................. 51 1.9.1. JUSTIFICATIVA PARA O USO DA ESPÉCIE.................................................................................... 51
OBJETIVOS ........................................................................................................................................... 53
CAPÍTULO 1 .......................................................................................................................................... 55
SEMIPURIFICAÇÃO E CARACTERIZAÇÃO ENZIMÁTICA DA ENZIMA Α-GALACTOSIDASE PRESENTE EM SEMENTES DE SESBANIA VIRGATA (CAV.) PERS. .................................................................................. 55
I. RESUMO ..................................................................................................................................... 56
II. INTRODUÇÃO ............................................................................................................................. 58
III. MATERIAL E MÉTODOS ............................................................................................................... 61
3.1. COLETA DO MATERIAL VEGETAL ............................................................................................ 61 3.2. DETERMINAÇÃO DA PORCENTAGEM DE GERMINAÇÃO E PICO DE ATIVIDADE ENZIMÁTICA ..................... 61 3.2.1. GERMINAÇÃO DAS SEMENTES QUIESCENTES ............................................................................. 62 3.2.2. DETERMINAÇÃO DO PESO FRESCO DO ENDOSPERMA E TEGUMENTO ............................................... 62 3.2.3. EXTRAÇÃO PROTÉICA, DETERMINAÇÃO DE PROTEÍNAS TOTAIS, DETERMINAÇÃO DA ATIVIDADE ENZIMÁTICA
DA Α-GALACTOSIDASE E ELETROFORESE EM GEL DE POLIACRILAMIDA (SDS-PAGE) .............................................. 63 3.3. PURIFICAÇÃO DA Α-GALACTOSIDASE DE SEMENTES DE SESBANIA VIRGATA (CAV.) PERS. ..................... 65 3.3.1. INCUBAÇÃO DAS SEMENTES DE SESBANIA VIRGATA PARA PURIFICAÇÃO DA Α-GALACTOSIDASE .............. 65 3.3.2. CARACTERIZAÇÃO DA TEMPERATURA ÓTIMA E PH ÓTIMO DA Α-GALACTOSIDASE EM EXTRATO BRUTO
ENZIMÁTICO 66 3.3.2.1. TEMPERATURA ÓTIMA ........................................................................................................ 66 3.3.2.2. PH ÓTIMO ....................................................................................................................... 66 3.3.3. PRIMEIRA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: PRECIPITAÇÃO COM SULFATO DE AMÔNIO ... 66 3.3.4. SEGUNDA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: CROMATOGRAFIA DE TROCA-IÔNICA COM
COLUNA ANIÔNICA (MONO-Q) .............................................................................................................. 68 3.3.5. TERCEIRA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: CROMATOGRAFIA DE GEL FILTRAÇÃO
(SUPEROSE) 69 3.3.6. TABELA DE PURIFICAÇÃO ..................................................................................................... 70

10
3.3.7. CARACTERIZAÇÃO DA Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS E DA Α-GALACTOSIDASE DE CYAMOPSIS TETRAGONOLOBA (GUAR) (MEGAZYME
®) QUANTO A TEMPERATURA ÓTIMA, PH ÓTIMO
E ASPECTOS CINÉTICOS ........................................................................................................................... 71 3.3.7.1. TEMPERATURA ÓTIMA ........................................................................................................ 71 3.3.7.2. PH ÓTIMO ........................................................................................................................ 71 3.3.7.3. ASPECTOS CINÉTICOS – KM E VMÁX .......................................................................................... 72 3.3.8. SEQUENCIAMENTO INTERNO DA Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS
72 3.4. ANÁLISES DAS SEQUÊNCIAS DE Α-GALACTOSIDASE PREVIAMENTE IDENTIFICADAS EM PLANTAS .............. 74
IV. RESULTADOS............................................................................................................................... 75
4.1. DETERMINAÇÃO DA PORCENTAGEM DE GERMINAÇÃO E PICO DE ATIVIDADE ENZIMÁTICA ..................... 75 4.2. PURIFICAÇÃO DA Α-GALACTOSIDASE DE SEMENTES DE SESBANIA VIRGATA (CAV.) PERS. ..................... 77 4.2.1. CARACTERIZAÇÃO DA TEMPERATURA ÓTIMA E PH ÓTIMO DA Α-GALACTOSIDASE EM EXTRATO BRUTO
ENZIMÁTICO 77 4.2.1.1. TEMPERATURA ÓTIMA ........................................................................................................ 77 4.2.1.2. PH ÓTIMO ........................................................................................................................ 78 4.2.2. PRIMEIRA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: PRECIPITAÇÃO COM SULFATO DE AMÔNIO .... 79 4.2.3. SEGUNDA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: CROMATOGRAFIA DE TROCA-IÔNICA COM
COLUNA ANIÔNICA (MONO-Q) ............................................................................................................... 82 4.2.4. TERCEIRA ETAPA DE PURIFICAÇÃO DA Α-GALACTOSIDASE: CROMATOGRAFIA DE GEL FILTRAÇÃO
(SUPEROSE) 85 4.2.5. TABELA DE PURIFICAÇÃO ...................................................................................................... 89 4.2.6. CARACTERIZAÇÃO DA Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS. E DA Α-GALACTOSIDASE DE CYAMOPSIS TETRAGONOLOBA (GUAR) (MEGAZYME
®) QUANTO A TEMPERATURA ÓTIMA, PH ÓTIMO
E ASPECTOS CINÉTICOS ........................................................................................................................... 90 4.2.6.1. TEMPERATURA ÓTIMA ........................................................................................................ 90 4.2.6.2. PH ÓTIMO ........................................................................................................................ 92 4.2.6.3. ASPECTOS CINÉTICOS – KM E VMÁX .......................................................................................... 93 4.2.6.4. ANÁLISE DO PERFIL PROTEICO EM GEL DE POLIACRILAMIDA SOB CONDIÇÕES DESNATURANTES DE TODAS AS
ETAPAS DE PURIFICAÇÃO ........................................................................................................................ 95 4.2.7. SEQUENCIAMENTO INTERNO DA Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS. 96 4.3. ANÁLISES DAS SEQUÊNCIAS DE Α-GALACTOSIDASES PREVIAMENTE IDENTIFICADAS EM PLANTAS ............. 98
V. DISCUSSÃO ............................................................................................................................... 100
5.1. DETERMINAÇÃO DA PORCENTAGEM DE GERMINAÇÃO E PICO DE ATIVIDADE ENZIMÁTICA .................. 100 5.2. PURIFICAÇÃO DA Α-GALACTOSIDASE DE SEMENTES DE SESBANIA VIRGATA (CAV.) PERS. ................... 101 5.3. ANÁLISES DAS SEQUÊNCIAS DE Α-GALACTOSIDASE PREVIAMENTE IDENTIFICADAS EM PLANTAS ............ 110
VI. CONCLUSÃO ............................................................................................................................. 113
VII. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................. 115
CAPÍTULO 2 ........................................................................................................................................ 120
ESTRUTURA FINA DO GALACTOMANANO E A MODULAÇÃO DA MOBILIZAÇÃO DE RESERVA EM SEMENTES DE SESBANIA VIRGATA (CAV.) PERS. ................................................................................ 120
I. RESUMO ................................................................................................................................... 121
II. INTRODUÇÃO ........................................................................................................................... 122
III. MATERIAL E MÉTODOS ............................................................................................................. 126
3.1. MATERIAL VEGETAL ......................................................................................................... 126 3.2. EXTRAÇÃO DO MANANO E GALACTOMANANO ......................................................................... 126 3.2.1. EXTRAÇÃO DO MANANO DE RESERVA DAS SEMENTES DE COFFEA ARABICA E EUTERPE EDULIS ............. 127 3.2.2. EXTRAÇÃO DO GALACTOMANANO DE RESERVA DAS SEMENTES DE SESBANIA VIRGATA, SENNA ALATA, DIMORPHANDRA MOLLIS, CYAMOPSIS TETRAGONOLOBA ............................................................................. 127 3.3. ANÁLISE DE MONOSSACARÍDEOS .......................................................................................... 128

11
3.4. HIDRÓLISE ENZIMÁTICA COM ENDO-Β-MANANASE E Α-GALACTOSIDASES...................................... 128 3.5. ANÁLISE DOS OLIGOSSACARÍDEOS DE (GALACTO)MANANO ........................................................ 130 3.5.1. ANÁLISE DOS OLIGOSSACARÍDEOS DE (GALACTO)MANANO POR HPAEC-PAD (HIGH PERFORMANCE
ANION EXCHANGE CHROMATOGRAPHY WITH PULSED AMPEROMETRIC DETECTION) .......................................... 130 3.5.2. ANÁLISE DOS OLIGOSSACARÍDEOS DE (GALACTO)MANANO POR TLC (THIN LAYER CROMATOGRAPHY) 131 3.6. ANÁLISE DA ESTRUTURA FINA DO GALACTOMANANO DE SESBANIA VIRGATA (CAV.) PERS. ATRAVÉS DE
HIDRÓLISES ENZIMÁTICAS COM ENDO-Β-MANASE DE ASPERGILUS NÍGER (MEGAZYME), Α-GALACTOSIDASE DE
CYAMOPSIS TETRAGONOLOBA (MEGAZYME) E Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS. 131 3.6.1. EXPERIMENTO I ............................................................................................................... 132 3.6.2. EXPERIMENTO II .............................................................................................................. 133 3.6.3. EXPERIMENTO III ............................................................................................................. 134 3.6.4. EXPERIMENTO IV ............................................................................................................. 135 3.6.5. EXPERIMENTO V .............................................................................................................. 136 3.7. ANÁLISE DA ESTRUTURA FINA DO (GALACTO)MANANO DE 6 ESPÉCIES COM DIFERENTES PROPORÇÕES DE
RAMIFICAÇÕES COM GALACTOSE ............................................................................................................ 137
IV. RESULTADOS............................................................................................................................. 138
4.1. ANÁLISE DA ESTRUTURA FINA DO GALACTOMANANO DE SESBANIA VIRGATA (CAV.) PERS. ATRAVÉS DE
HIDRÓLISES ENZIMÁTICAS COM ENDO-Β-MANASE DE ASPERGILUS NIGER (MEGAZYME), Α-GALACTOSIDASE DE
CYAMOPSIS TETRAGONOLOBA (MEGAZYME) E Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS. 138 4.1.1. EXPERIMENTO I ............................................................................................................... 138 4.1.2. EXPERIMENTO II .............................................................................................................. 145 4.1.3. EXPERIMENTO III ............................................................................................................. 151 4.1.4. EXPERIMENTO IV ............................................................................................................. 154 4.1.5. EXPERIMENTO V .............................................................................................................. 156 4.2. ANÁLISE DA ESTRUTURA FINA DO (GALACTO)MANANO DE 6 ESPÉCIES COM DIFERENTES PROPORÇÕES DE
RAMIFICAÇÕES COM GALACTOSE ............................................................................................................ 159
V. DISCUSSÃO ............................................................................................................................... 165
5.1. ANÁLISE DA ESTRUTURA FINA DO GALACTOMANANO DE SESBANIA VIRGATA (CAV.) PERS. ATRAVÉS DE
HIDRÓLISES ENZIMÁTICAS COM ENDO-Β-MANASE DE ASPERGILLUS NIGER (MEGAZYME®), Α-GALACTOSIDASE DE
CYAMOPSIS TETRAGONOLOBA (MEGAZYME®) E Α-GALACTOSIDASE SEMIPURIFICADA DE SESBANIA VIRGATA (CAV.) PERS.
165 5.2. ANÁLISE DA ESTRUTURA FINA DO (GALACTO)MANANO DE 6 ESPÉCIES COM DIFERENTES PROPORÇÕES DE
RAMIFICAÇÕES COM GALACTOSE ............................................................................................................ 179
VI. CONCLUSÃO ............................................................................................................................. 181
VII. BIBLIOGRAFIA ...................................................................................................................... 183
DISCUSSÃO GERAL ............................................................................................................................. 185
REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................................................... 190
ANEXO ............................................................................................................................................... 198

12
Índice de Figuras Figura 1: Estrutura e mobilização do galactano. (A) Estrutura do galactano mais
frequente composto por ligações β-(1,4) com ramificações de L-arabinofuranose a cada
16-21 resíduos da cadeia principal (não mostrada). Ainda existe um segundo tipo de
galactano formado por ligações β-(1,3)(1,6). (B) Reações envolvendo a mobilização do
polissacarídeo de reseva em Lupinus sp. β-(1,4)-D-galactano é o polímero dominante,
sendo descrito como uma ramificação do rhamnogalacturanano, por vezes ramificado
por ligações α-(1,5)-arabinano. Uma molécula de α-(1,3), (1,6)-D-galactano também
está representado embora ocorre em quantidades menores. Neste sistema de degradação,
apenas exoenzimas foram detectadas, uma das quais (exo-α-(1,4)-galactanase a partir de
Lupinus angustifolius) é capaz de libertar a maior parte (cerca 60%) das paredes
celulares intactadas da mesma espécie. In vivo, outras galactosidases e arabinosidases
parecem agir no ramos deixando apenas ramnogalacturonano. O destino dos produtos é
meramente hipotético neste esquema. Figura retirada e adaptada de Buckeridge et al.
(2000b) e Buckeridge et al. (2000c). ........................................................................... 34
Figura 2: Estrutura e mobilização de xiloglucano de reserva de sementes. (A) Estrutura
molecular do xiloglucano de reserva contendo cadeia principal de glucose unida por
ligações β-(1,4), onde ocorrem ramificações regulares com xilose ligada α-(1,6).
Algumas destas xiloses ainda podem apresentar ramificações por galactose ligadas α-
(1,2). (B) Representação esquemática da degradação de reserva de xiloglucano em
sementes. Uma sequência de XLXGXLLGXXXGXXLG é usada como um exemplo de
um segmento de um polímero de xiloglucano com a extremidade não-redutora voltada
para a esquerda. O primeiro ataque é feita por xiloglucano-endo-β-transglicosilase
(XET) e β-galactosidase (β-gal proveniente de Nasturtium apenas). Os oligossacáridos
produzidos pela ação endo da XET são atacados por β-galactosidases (Nβ-
gal=Nasturtium β-galactosidase; Cβ-gal= Copaifera β-galactosidase, o último sendo
capaz de atacar oligossacarídeos apenas em certas posições (galactoses colocadas na
ponta não-redutora). A α- xilosidase e a β-glucosidase agem em hidrólises alternadas do
final não-redutor. (*) Em leguminosas (Copaifera langsdorffii e Hymenaea courbaril),
após uma rodada de ação destas duas enzimas, a galactose estará novamente próximo do
terminal não-redutor. Desata forma, a β-galactosidase pode agora agir sobre o
oligossacarídeo XLG, libertando assim a galactose e permitindo a continuidade da ação
das outras duas exo-glicosidases. Nestas duas espécies, a galactose retirada dos
oligossacarídeos é possivelmente limitadora da degradação, pois a enzima possui um
pH ótimo de 3,2, enquanto as demais enzimas possuem um pH ótimo de 4,5. Figura
retirada e adaptada de Buckeridge et al. (2000c) e Buckeridge et al. (2000b). ............. 37
Figura 3: Estrutura molecular do polissacarídeo galactomanano e explicação para a
leitura das “abreviações” dos oligossacarídeos GalXManY. (A) Cadeia principal linear de
resíduos de D-manose unidas por ligações glicosídicas β-(1,4), na qual resíduos de D-
galactose estão unidos por ligações α-(1,6), formando ramificações simples. Figura
retirada e adaptada de Buckeridge et al. (2000a). (B) A cadeia principal de manose é
numerada a partir da extremidade redutora (à direita). O número subscrito ligado à
manose indica a quantidade de resíduos de manose que a cadeia principal do
oligossacarídeo possui, enquanto o número sobrescrito ligado a galactose indica a
posição do resíduo de galactose relativo ao resíduo de manose a partir da extremidade
redutora (McCleary et al., 1983). ................................................................................. 40

13
Figura 4: Rotas bioquímicas envolvidas no catabolismo do galactomanano somado ao
metabolismo pelo embrião dos monossacarídeos liberados na degradação da reserva.
Quando a camada de aleurona não está presente, aparentemente as hidrolases são
produzidas dentro das células endospermáticas e secretadas para fora da parede celular
onde as mesmas vias metabólicas ocorrem. Man = manose; Gal = galactose; SPS =
sacarose fostato sintase; Man-6-P = manose-6-fosfato; Fru-6-P = frutose-6-fosfato;
UDP-gal = uridina difosfatada de galactose; Suc-6-P = sacarose-6-fosfato. Figura
retirada e adaptada de Buckeridge et al. (2000b). ........................................................ 44
Figura 5: Representação esquemática dos subsítios de reconhecimento (α – ε) e ligação
da endo-β-mananase ao oligossacarídeo hipotético com 5 resíduos de manose (A-E)
ligados β-(1,4). Figura retirada de McCleary & Matheson, 1983. ................................ 50
Figura 6: Porcentagem de germinação das sementes de Sesbania virgata (cav.) Pers. ao
longo de 7 dias (168 horas) após a embebição. ............................................................ 76
Figura 7: Atividade enzimática específica da enzima α-galactosidase presente em
sementes de Sesbania virgata (Cav.) Pers. ao longo de 7 dias (168 horas) após a
embebição. .................................................................................................................. 77
Figura 8: Determinação da temperatura ótima de atividade enzimática da α-
galactosidase em extrato bruto enzimático de sementes de Sesbania virgata (Cav.) Pers.
após 4 dias de incubação em BOD. ............................................................................. 78
Figura 9: Determinação do pH ótimo de atividade enzimática da α-galactosidase em
extrato bruto enzimático de sementes de Sesbania virgata (Cav.) Pers. após 4 dias de
incubação em BOD. .................................................................................................... 79
Figura 10: Padronização da etapa de purificação da Precipitação com Sulfato de
Amônio. Determinação da saturação de sulfato de amônio responsável pela precipitação
da enzima α-galactosidase através quantificação da atividade enzimática específica da
α-galactosidase em extrato bruto e em cada precipitado ressuspendido referente à
utilização de uma porcentagem de saturação de sulfato de amônio. ............................. 80
Figura 11: Eletroforese em gel de poliacrilamida 12% do extrato bruto enzimático;
precipitados ressuspendidos de 20%, 40%, 60% e 80% de saturação de sulfato de
amônio e o descarte resultante da precipitação com sulfato de amônio, com cerca de
50µg de proteína nas amostras aplicadas. .................................................................... 81
Figura 12: Comparação da atividade enzimática da enzima α-galactosidase no: extrato
bruto (Ext.Bruto); ressuspendido do precipitado com 40% de saturação de Sulfato de
Amônio (40%); ressuspendido do precipitado com 80% de saturação de Sulfato de
Amônio (80%); sobrenadante resultante da precipitação com 80% de saturação de
Sulfato de Amônio (Descarte), após Primeira etapa de purificação: Precipitação com
Sulfato de Amônio em 40% e 80% de saturação. ......................................................... 82
Figura 13: Perfil cromatográfico da purificação em FPLC Äkta do extrato bruto
enzimático submetido à coluna aniônica Mono-Q HR 10/10 (Pharmacia Biotech).
Identificação com letras (A-C) das frações coletadas que apresentaram atividade
enzimática de α-galactosidase, onde: (A) Frações 47 a 51; (B) Frações 63 a 65; (C)

14
Frações 66 a 69. (―) Perfil proteico quantificado por UV; (―) Gradiente de NaCl
variando de 0-0.5M em 12 volumes de coluna (‖) Frações coletadas com seus
respectivos números. ................................................................................................... 83
Figura 14: Atividade enzimática da α-galactosidase expressa em absorbância a 405nm
das frações coletadas durante a purificação em FPLC Äkta do extrato bruto enzimático
submetido à coluna aniônica Mono-Q HR 10/10 (Pharmacia Biotech). (a) Frações 47 a
51; (b) Frações 63 a 65; (c) Frações 66 a 69. ............................................................... 84
Figura 15: Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue
R250 das frações reunidas 47 a 51 (pico A) após purificação em FPLC Äkta utilizando
coluna aniônica Mono-Q HR 10/10 (Pharmacia Biotech). (→) Banda proteica
concentrada de tamanho esperado para α-galactosidase. .............................................. 85
Figura 16: Perfil cromatográfico da purificação em FPLC Äkta utilizando coluna de gel
filtração Superose 6 (GE Healthcare) das frações previamente semipurificadas (frações
47 a 51) em coluna aniônica Mono-Q HR 10/10 (Pharmacia Biotech). Identificação com
letra (D) das frações coletadas que apresentaram atividade enzimática de α-
galactosidase, onde: (D) Frações 30-33. (―) Perfil proteico por quantificação por UV;
(―) Gradiente de NaCl variando de 0-0.5M em 12 volumes de coluna; (‖) Frações
coletadas com seus respectivos números. .................................................................... 86
Figura 17: Atividade enzimática da α-galactosidase expressa em absorbância a 405nm
das frações coletadas durante a purificação em FPLC Äkta, utilizando coluna de gel
filtração Superose 6 (GE Healthcare), da amostra semipurificada em coluna aniônica
Mono-Q HR 10/10 (Pharmacia Biotech). (D) Frações 30 a 33. ................................... 87
Figura 18: Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue
R250 das frações reunidas 30 a 33 (pico D) após purificação em FPLC Äkta utilizando
coluna de gel filtração Superose 6 (Pharmacia Biotech). (→) Banda proteica
concentrada de tamanho esperado para α-galactosidase. .............................................. 88
Figura 19: Determinação da temperatura ótima da α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. ...................................................................................... 91
Figura 20: Determinação da temperatura ótima da α-galactosidase de Cyamopsis
tetragonoloba (Megazyme®). ...................................................................................... 92
Figura 21: Determinação do pH ótimo da α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers. ..................................................................................................... 93
Figura 22: Determinação do pH ótimo da α-galactosidase de Cyamopsis tetragonoloba
(Megazyme). ............................................................................................................... 93
Figura 23: Curva de saturação do substrato 4-Nitrofenil-D-Galactopiranosídeo na
presença da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers. Os ensaios
enzimáticos foram conduzidos com o PNP-α-Gal nas seguintes concentrações (mM):
0.1; 0.2; 0.3; 0.5; 0.8; 1.0; 2.0; 3.0; 5.0; 8.0; 10; 13; 15; 17; 20; 25; 30. A partir dos
dados coletados foi calculado o Km e o Vmáx da α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. ...................................................................................... 94

15
Figura 24: Curva de saturação do substrato 4-Nitrofenil-D-Galactopiranosídeo na
presença da α-galactosidase de Cyamopsis tetragonoloba (Megazyme®). Os ensaios
enzimáticos foram conduzidos com o PNP-α-Gal nas seguintes concentrações (mM):
0.1; 0.2; 0.3; 0.5; 0.8; 1.0; 2.0; 3.0; 5.0; 8.0; 10; 13; 15; 17; 20; 25. A partir dos dados
coletados foi calculado o Km e o Vmáx da α-galactosidase de Cyamopsis tetragonoloba
(Megazyme®
). ............................................................................................................. 95
Figura 25: Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue
R250 de todas as etapas de purificação. (A) Extrato bruto enzimático; (B) Precipitado de
80% de Sulfato de Amônio (ressuspendido em tampão Tris-HCl 20mM ph 7,8); (C)
Alíquota das frações reunidas após purificação em FPLC Äkta utilizando coluna de
troca-iônica aniônica (Mono-Q); (D) Alíquota das frações reunidas após purificação em
FPLC Äkta utilizando coluna de gel filtração Superose 6 (GE Healthcare). ................. 96
Figura 26: Identificação com números, de 1 a 8, das bandas recortadas do gel de
poliacrilamida 12% submetidas ao sequenciamento interno para identificação das
proteínas, principalmente da α-galactosidase, juntamente com a determinação da
sequência interna da α-galactosidase semipurificada de sementes de Sesbania virgata
(Cav.) Pers. ................................................................................................................. 97
Figura 27: Análise das sequências de aminoácidos 35 de α-galactosidase de plantas
conjuntamente com a sequência dos fragmentos de α-galactosidase de Sesbania virgata
(Cav.) Pers. semipurificada determinados por Espectrometria de Massas (MALDI-
TOF). Grupos identificados por letras estão de acordo com a filogenia: (A – vermelho)
Fabaceae; (B - rosa) Fabaceae; (C – azul escuro) Fabaceae; (D - verde) Rubiaceae; (E –
roxo) Poaceae; (F – azul claro) Pinaceae. A coloração dos ramos é a mesma da árvore
de relação das 35 sequências de aminoácidos somente localizada na figura 46 do anexo,
no entanto, a identificação pelas letras é diferente, ou seja, os ramos coloridos por cada
cor são os mesmos em ambas as árvores, mas a letra que o identifica não é a mesma. (•)
Bootstrap ˃ 95, ou seja, em 95% ou mais das 1000 árvores de relação geradas os ramos
marcados com (•) ficaram agrupados como mostrado nesta árvore de relação. ............. 99
Figura 28: Análise em HPAEC-PAD (Dionex) com coluna PA-100 dos
oligossacarídeos formados após hidrólise enzimática exaustiva com Endo-β-mannanase
do galactomanano de Sesbania virgata (Cav.) Pers., visando verificar perfil dos
oligossacarídeos produto da hidrólise enzimática do galactomanano com endo-β-
mananase. (F1) dissacarídeos a decassacarídeos galactosilados ou não base do
polissacarídeo; (F2) combinação aos pares dos oligossacarídeos de F1; (F3) combinação
aos trios dos oligossacarídeos de F1. ......................................................................... 139
Figura 29: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do galactomanano de
Sesbania virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e
1U. (I) Controle (galactomanano sem endo-B-mananase); (II) perfil de oligossacarídeos
formados após hidrólise com endo-β-mananase na concentração de 0,005U; (III) perfil
de oligossacarídeos formados após hidrólise com endo-β-mananase na concentração de
0,05U; (IV) perfil de oligossacarídeos formados após hidrólise com endo-β-mananase
na concentração de 1U; (V) comparação dos perfis de oligossacarídeos formados após
hidrólise com endo-β-mananase nas concentrações de 0,005U e 0,05U; (VI)
comparação dos perfis de oligossacarídeos formados após hidrólise com endo-β-

16
mananase nas concentrações de 0,005U e 1U; (VII) comparação dos perfis de
oligossacarídeos formados após hidrólise com endo-β-mananase nas concentrações de
0,05U e 1U. (A) manobiose (M2); (B) manotriose (M3) e/ou trissacarídeo (G1M2); (C)
tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D) pentassacarídeos (manopentose –
M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4 e/ou
G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I)
oligossacarídeos >8 resíduos (G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou
G4,5
M7). Os oligossacarídeos identificados A-I são denominados oligossacarídeos F1; os
oligossacarídeos restantes são identificados como F2, quanto combinação aos pares, e
F3, quando combinação aos trios. .............................................................................. 141
Figura 30: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do manano comercial
(Megazyme®
) com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e 1U. (I) Controle
(manano sem endo-B-mananase); (II) perfil de oligossacarídeos formados após hidrólise
com endo-β-mananase na concentração de 0,005U; (III) perfil de oligossacarídeos
formados após hidrólise com endo-β-mananase na concentração de 0,05U; (IV) perfil de
oligossacarídeos formados após hidrólise com endo-β-mananase na concentração de 1U;
(V) comparação dos perfis de oligossacarídeos formados após hidrólise com endo-β-
mananase nas concentrações de 0,005U e 0,05U; (VI) comparação dos perfis de
oligossacarídeos formados após hidrólise com endo-β-mananase nas concentrações de
0,005U e 1U; (VII) comparação dos perfis de oligossacarídeos formados após hidrólise
com endo-β-mananase nas concentrações de 0,05U e 1U. (A) manobiose (M2); (B)
manotriose (M3); (D) pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F)
heptassacarídeos (manoheptose – M7 e/ou G3,4
M5). Os oligossacarídeos identificados A-
F são denominados oligossacarídeos F1. ................................................................... 144
Figura 31: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da
análise dos oligossacarídeos, produto da hidrólise enzimática com α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. dos oligossacarídeos produto da
hidrólise do galactomanano a 1% de Sesbania virgata (Cav.) Pers com endo-β-
mananase (Megazyme®
) a 0,005U, 0,05U e 1U (Experimento I). (I) Controle
(galactomanano a 1% sem enzima); (II) Controle (galactomanano a 1% com α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.; (III) Controle
(galactomanano a 1% com celulase de Trichoderma longibrachiatum (Megazyme®
));
(IV) perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia
hidrólise com endo-β-mananase a 0,005U; (V) perfil de oligossacarídeos formados após
ação da α-galactosidase sobre prévia hidrólise com endo-β-mananase a 0,05U; (VI)
perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia hidrólise
com endo-β-mananase a 1U; (VII) comparação dos perfis de oligossacarídeos formados
no Experimento I e Experimento II na concentração de 0,005U de endo-β-mannanase;
(VIII) comparação dos perfis de oligossacarídeos formados no Experimento I e
Experimento II na concentração de 0,05U de endo-β-mannanase; (IX) comparação dos
perfis de oligossacarídeos formados no Experimento I e Experimento II na concentração
de 1U de endo-β-mannanase. (A) manobiose (M2); (B) manotriose (M3); (C)
tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D) pentassacarídeos (manopentose –
M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4 e/ou
G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I)
oligossacarídeos >8 resíduos (possíveis oligossacarídeos presentes: G1,3,4
M5; G4,5
M6,

17
G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7); (K-L) octassacarídeos e
nonassacarídeos sem ramificação; (M) oligossacarídeos >10 resíduos de manose
(possivelmente resultado da desramificação do pico I). Os oligossacarídeos
identificados A-M são denominados oligossacarídeos F1; os oligossacarídeos restantes
são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos
trios........................................................................................................................... 149
Figura 32: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do galactomanano de
Sesbania virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 1U com acréscimo
ou não da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers. (I) perfil de
oligossacarídeos formados na hidrólise do galactomanano a 1% somente com endo-β-
mananase a 1U; (II) perfil de oligossacarídeos formados na hidrólise do galactomanano
a 1% com endo-β-mananase a 1U/mL e α-galactosidase de semipurificada Sesbania
virgata (Cav.) Pers. a 0,5U/mL; (III) perfil de oligossacarídeos formados na hidrólise do
galactomanano a 1% somente com α-galactosidase semipurificada Sesbania virgata
(Cav.) Pers. a 0,5U/mL; (IV) Controle (galactomanano sem enzima). (Gal) Galactose;
(A) manobiose (M2); (B) manotriose (M3); (C) tetrassacarídeos (manotetraose – M4 e/ou
G1M3); (D) pentassacarídeos (manopentose – M5 e/ou G
1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F)
heptassacarídeos (manoheptose – M7 e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos
(G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos
identificados A-I são denominados oligossacarídeos F1; os oligossacarídeos restantes
são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos
trios........................................................................................................................... 154
Figura 33: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do galactomanano de
Sesbania virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 0,005U e 1U e α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers. a 0,5U/mL. (I)
Galactomanano a 1%, 0,005U de endo-β-mananase e α-galactosidase semipurificada a
0,5U/mL; (II) Galactomanano a 0,1%, 0,005U de endo-β-mananase e α-galactosidase
semipurificada a 0,5U/mL; (III) Galactomanano a 1%, 1U de endo-β-mananase e α-
galactosidase semipurificada a 0,5U/mL; (IV Galactomanano a 0,1%, 1U de endo-β-
mananase e α-galactosidase semipurificada a 0,5U/mL. (A) manobiose (M2); (B)
manotriose(M3); (C) tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos
(manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos
(manoheptose – M7 e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos (G1,3,4
M5; G4,5
M6,
G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos identificados A-I
são denominados oligossacarídeos F1; os oligossacarídeos restantes são identificados
como F2, quanto combinação aos pares, e F3, quando combinação aos trios. ............ 155
Figura 34: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do manano comercial
(Megazyme®
) com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e 1U e α-
galactosidase de Cyamopsis tetragonoloba comercial (Megazyme®) a 0,5U/mL. (I)
perfil de oligossacarídeos formados na concentração de 0,005U de endo-β-mananase;
(II) perfil de oligossacarídeos formados na concentração de 0,05U de endo-β-mananase;
(III) perfil de oligossacarídeos formados na concentração de 1U de endo-β-mananase;

18
(IV) injeção com padrão de manose; (V) injeção com padrão de galactose. (A)
manobiose (M2); (B) manotriose (M3); (C) tetrassacarídeos (manotetraose – M4 e/ou
G1M3); (D) pentassacarídeos (manopentose – M5 e/ou G
1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F)
heptassacarídeos (manoheptose – M7 e/ou G3,4
M5). Os oligossacarídeos identificados A-
F são denominados oligossacarídeos F1. ................................................................... 158
Figura 35: Análise por Thin Layer Chromatography (TLC) dos oligossacarídeos
obtidos após hidrólise enzimática exaustiva com endo-β-mannanase (Megazyme®) das 6
espécies estudadas. (A) Euterpe edulis (150:1); (B) Coffea arabica (40:1); (C)
Dimorphandra mollis (2:1); (D) Senna alata (2:1); (E) Sesbania virgata (Cav.) Pers.
(1:1); (F) Cyamopsis tetragonoloba (1:1). (→) Oligossacarídeos exclusivo de 1 ou mais
espécies: (→1, 2) exclusivo de Cyamopsis tetragonoloba; (→3) exclusivo de Sesbania
virgata e Cyamopsis tetragonoloba; (→) Comparação dos oligossacarídeos formados
entre as espécies (A e B) com reserva de manano: (→1, 2, 3) exclusivo de Euterpe
edulis; (→4) exclusivo de Coffea arabica. (*→) Oligossacarídeos exclusivos das
espécies com reserva de manano. (Chave vermelha) Oligossacarídeos e suas
combinações em pares (F2) e em trios (F3) exclusivos de espécies com reserva de
galactomanano. ......................................................................................................... 162
Figura 36: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da
análise dos oligossacarídeos, produto da hidrólise enzimática exaustiva com endo-β-
mannanase (Megazyme®
), das 6 espécies estudadas. (A) Euterpe edulis (150:1); (B)
Coffea arabica (40:1); (C) Dimorphandra mollis (2:1); (D) Senna alata (2:1); (E)
Sesbania virgata (1:1); (F) Cyamopsis tetragonoloba (1:1). (a) manose; (b) galactose;
(c) manobiose; (d) manotriose; (f-h, j-m) oligossacarídeos ainda não-identificados; (i)
combinações aos pares e aos trios dos oligossacarídeos identificados (c-h, j-m). Os
oligossacarídeos identificados a-h e j-m são denominados oligossacarídeos F1; os
oligossacarídeos identificados por i são denominados F2, quanto combinação aos pares,
e F3, quando combinação aos trios. ........................................................................... 164
Figura 37: Explicação para a leitura das “abreviações” dos oligossacarídeos GalXMany.
A cadeia principal de manose é numerada a partir da extremidade redutora (à direita). O
número subscrito ligado à manose indica a quantidade de resíduos de manose que a
cadeia principal do oligossacarídeo possui, enquanto o número sobrescrito ligado a
galactose indica a posição do resíduo de galactose relativo ao resíduo de manose a partir
da extremidade redutora (McCleary et al., 1983). ...................................................... 177
Figura 38: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 1 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. As proteínas
realssadas pelo retângulo preto fazem referência a α-galactosidases. ......................... 198
Figura 39: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 2 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade

19
dos fragmentos gerados serem pertencentes àquela proteína ou similares. As proteínas
realssadas pelo retângulo preto fazem referência a α-galactosidases. ......................... 199
Figura 40: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 3 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 200
Figura 41: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 4 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 201
Figura 42: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 5 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 202
Figura 43: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 6 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 203
Figura 44: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 7 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 204
Figura 45: Comparação dos fragmentos gerados por espectrometria de massas
(MALDI-TOF) da BANDA 8 (gel de poliacrilamida 12% após purificação em coluna de
Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas
estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a probabilidade
dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína
realssada pelo retângulo preto faz referência a α-galactosidase. ................................. 205
Figura 46: Análise de 35 sequências de aminoácidos de α-galactosidase de plantas.
Grupos identificados por letras estão de acordo com a filogenia: (A – azul escuro)
Fabaceae; (B - verde) Rubiceae; (C - amarelo) Solanaceae; (D - vermelho) Fabaceae; (E
– azul claro) Pinaceae; (F – roxo) Poaceae. (•) Bootstrap ˃ 95, ou seja, em 95% ou mais

20
das 1000 árvores de relação geradas os ramos marcados com (•) ficaram agrupados
como mostrado nesta árvore de relação. .................................................................... 206
Figura 47: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da
análise dos oligossacarídeos, produto da hidrólise enzimática com α-galactosidase
comercial de Cyamopsis tetragonoloba (Megazyme®) dos oligossacarídeos produto da
hidrólise do galactomanano a 1% de Sesbania virgata (Cav.) Pers com endo-β-
mananase (Megazyme®
) a 0,005U, 0,05U e 1U (Experimento I). (I) Controle
(galactomanano a 1% sem enzima); (II) Controle (galactomanano a 1% com α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®); (III) Controle
(galactomanano a 1% com celulase de Trichoderma longibrachiatum (Megazyme®
));
(IV) perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia
hidrólise com endo-β-mananase a 0,005U; (V) perfil de oligossacarídeos formados após
ação da α-galactosidase sobre prévia hidrólise com endo-β-mananase a 0,05U; (VI)
perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia hidrólise
com endo-β-mananase a 1U; (VII) comparação dos perfis de oligossacarídeos formados
no Experimento I e Experimento II na concentração de 0,005U de endo-β-mananase;
(VIII) comparação dos perfis de oligossacarídeos formados no Experimento I e
Experimento II na concentração de 0,05U de endo-β-mananase; (IX) comparação dos
perfis de oligossacarídeos formados no Experimento I e Experimento II na concentração
de 1U de endo-β-mananase. (A) Manobiose (M2); (B) Manotriose (M3) e/ou
trissacarídeo G1M2; (C) tetrassacarídeos (manotetraose – M4 e/ou G
1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos
(manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos
(manoheptose – M7 e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos (possíveis
oloigossacarídeos presentes: G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou
G4,5
M7); (K-L) octassacarídeos e nonassacarídeos sem ramificação; (M)
oligossacarídeos >10 resíduos de manose (possivelmente resultado da desramificação
do pico I). Os oligossacarídeos identificados A-M são denominados oligossacarídeos
F1; os oligossacarídeos restantes são identificados como F2, quanto combinação aos
pares, e F3, quando combinação aos trios. ................................................................. 208
Figura 48: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do galactomanano de
Sesbania virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 1U com acréscimo
ou não da α-galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®). (I)
perfil de oligossacarídeos formados na hidrólise do galactomanano a 1% somente com
endo-β-mananase a 1U; (II) perfil de oligossacarídeos formados na hidrólise do
galactomanano a 1% com endo-β-mananase a 1U/mL e α-galactosidase comercial de
Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (III) perfil de oligossacarídeos
formados na hidrólise do galactomanano a 1% somente com α-galactosidase comercial
de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (IV) Controle (galactomanano
sem enzima). (Gal) Galactose; (A) manobiose (M2); (B) manotriose (M3) e/ou G1M2;
(C) tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D) pentassacarídeos
(manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6
e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou
G3,4
M5); (G-I) oligossacarídeos >8 resíduos (G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6;
G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos identificados A-I são denominados
oligossacarídeos F1; os oligossacarídeos restantes são identificados como F2, quanto
combinação aos pares, e F3, quando combinação aos trios ........................................ 209

21
Figura 49: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da
análise dos oligossacarídeos, produto da hidrólise enzimática do galactomanano de
Sesbania virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 0,005U e 1U e α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL. (I)
Galactomanano a 1%, 0,005U de endo-β-mananase e α-galactosidase comercial de
Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (II) Galactomanano a 0,1%,
0,005U de endo-β-mananase e α-galactosidase comercial de Cyamopsis tetragonoloba
(Megazyme®
) a 0,5U/mL; (III) Galactomanano a 1%, 1U de endo-β-mananase e α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (IV
Galactomanano a 0,1%, 1U de endo-β-mananase e α-galactosidase comercial de
Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL. (A) manobiose (M2); (B)
manotriose (M3) e/ou trissacarídeo G1M2; (C) tetrassacarídeos (manotetraose – M4 e/ou
G1M3); (D) pentassacarídeos (manopentose – M5 e/ou G
1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F)
heptassacarídeos (manoheptose – M7 e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos
(G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos
identificados A-I são denominados oligossacarídeos F1; os oligossacarídeos restantes
são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos
trios........................................................................................................................... 210

22
Índice de Tabelas
Tabela 1: Tabela de purificação da α-galactosidase de Sesbania virgata (Cav.) Pers.
desenvolvida após a realização das três etapas de purificação: precipitação com Sulfato
de Amônio 80%, Cromatografia de troca-iônica aniônica (Mono-Q) e Cromatografia de
gel filtração (Superose). .............................................................................................. 90
Tabela 2: Comparação da razão F1/F2 do Experimento I (hidrólise do galactomanano a
1% somente com endo-β-mananase) com a razão do Experimento II (hidrólise dos
oligossacarídeos do Experimento I somente com α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers.). .................................................................................. 147
Tabela 3: Comparação das razões Manobiose (M2)/Trissacarídeo (M3 e/ou G1M2)
(M2/M3) e Trissacarídeo (M3 e/ou G1M2)/Tetrassacarídeo (M4 e/ou G
1M3) (M3/C) dos
Experimentos I e II. ................................................................................................... 150
Tabela 4: Razão manose:galactose das 6 espécies estudadas determinada pela análise
de monossacarídeos em HPAEC-PAD (Dionex). Apresentado por ordem crescente na
quantidade de substituições de galactose: Manano (Euterpe edulis; Coffea arábica);
Galactomanano de proporção 2:1, ou seja, a cada 2 manose existe a ramificação por 1
galactose (Dimorphandra mollis; Senna alata); Galactomanano de proporção 1:1, ou
seja, teoricamente todo o polissacarídeo é substituído por galactose (Cyamopsis
tetragonoloba; Sesbania virgata) . ............................................................................ 160
Tabela 5: Oligossacarídeos produzidos na hidrólise do galactomanano de goma caroba,
razão manose:galactose 4:1 pelas endo-β-mananase de Aspergillus niger e de sementes
de Medicago sativa (alfafa). O final redutor da molécula está sempre a esquerda,
manose denominada de 1. (#) O subscrito indica a quantidade de resíduos de manose o
oligossacarídeo possui em sua cadeia principal, enquanto o sobrescrito indica a posição
da substituição da galactose na cadeia relativa ao final redutor da molécula; (*) As
estruturas dos oligossacarídeos >7 resíduos produzidos pela hidrólise da endo-β-
mananase de M.sativa não foram caracterizadas. Estes oligossacarídeos representam
26,9% do total de oligossacarídeos produzidos. (-) Oligossacarídeos não produzidos
pela hidrólise da endo-β-mananase de Aspergillus niger por esta ser mais inespecífica
em sua ação, conseguindo “triblar” a presença de ramificação com galactose diminuindo
o tamanho do oligossacarídeo. Tabela retirada e adapatada de McCleary et al. (1983)
................................................................................................................................. 168
Tabela 6: Identificação das possíveis estruturas dos oligossacarídeos de Sesbania
virgata (Cav.) Pers. componentes dos picos A-F de acordo com a comparação do tempo
de retenção real e o tempo de retenção teórico e os oligossacarídeos caracterizados e
identificados por McCleary et al. (1983) (Tabela 5). ................................................. 176

23
RESUMO
Sementes de Sesbania virgata (Cav.) Pers. acumulam suas reservas de
carbono no endosperma na forma de um polissacarídeo de parede celular, o
galactomanano. Os galactomananos são polissacarídeos constituídos de uma
cadeia principal de resíduos de D-manose ligadas β-1,4, ramificada por
resíduos de D-galactose α-1,6 ligados. A mobilização deste ocorre após a
germinação e envolve três enzimas hidrolíticas (α-galactosidase, endo-β-
mananase e exo-β-manosidase). A α-galactosidase é a primeira enzima atuar
sobre o galactomanano hidrolisando as ligações α-1,6 das galactoses
ramificadas a cadeia principal de manano (ligados β-1,4), permitindo a ação da
endo-β-mananase, que hidrolisará o polissacarídeo a oligossacarídeos, onde a
β-manosidase atuará (ligações β-1,4), transformando oligossacarídeos a
monossacarídeos a serem utilizados no desenvolvimento do embrião.
Buscando a compreensão das características da α-galactosidase e modo de
ação sobre o galactomanano, procedeu-se com a purificação, em três etapas,e
caracterização bioquímica (pH ótimo, temperatura ótima e aspectos cinéticos)
da α-galactosidase de sementes de Sesbania virgata (Cav.) Pers. Além disso,
visando evidenciar a modulação da enzima endo-β-mananase pela distribuição
de ramificações de galactose no galactomanano (estrutura fina do
galactomanano), procedeu-se com hidrólises enzimáticas do galactomanano de
Sesbania virgata (Cav.) Pers. utilizando a enzima endo-β-mananase de
Aspergillus niger (Megazyme®) somente ou em conjunto com a α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. (Capítulo 1) ou com a α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®), seguido
de análise dos oligossacarídeos por HPAEC-PAD (High Performance Anion
Exchange Chromatography with Pulsed Amperometric Detection). Também
procedeu-se com hidrólises enzimáticas de galactomananos de 6 espécies com
razão manose:galactose variando de 1:1 a 150:1 com endo-β-mananase de
Aspergillus niger (Megazyme®) e análise dos oligossacarídeos produzidos por
HPAEC-PAD. A α-galactosidase semipurificada possui, aproximadamente, 42
kDa de peso molecular em condições desnaturantes e, aproximadamente 72
kDa de peso molecular na forma nativa, sugerindo que a enzima assuma

24
estrutura quartenária. A temperatura ótima apresentada se encontra na faixa de
50°C a 55°C, pH ótimo na faixa de 4,4 a 5,4, Km= 1,8276 mM e a velocidade
máxima de 0,5024 µmolGal.min-1.mgprot-1. A espectrometria de massas gerou
os fragmentos: ALADYV-HSK-RMPGSLGHEE-QDAK-TT-GDIEDNWNSM-
TSIADS NDKW-ASYAGPGGWN-DPDMLEVGNG-GMTTEEYR-AP-LLVGCDIR-
VAVIL-WNR, estando a proteína referente a esta sequência relacionada à
mobilização de reserva. Durante a purificação e sequenciamento interno da α-
galactosidase e demais proteínas foram detectadas isoformas da α-
galactosidase de pesos moleculares variados (42 kDa a 20 kDa). Sugere-se
que estas isoformas encontradas inicialmente na purificação estejam
relacionadas com outras funções da α-galactosidase, enquanto as isoformas
encontradas após todas as etapas de purificação e identificação por
espectrometria de massas estejam relacionadas com ativação e adaptação da
α-galactosidase durante todo o processo de mobilização de reservas. Os dados
gerados das comparações dos oligossacarídeos produzidos em cada hidrólise
sugerem que as ramificações do galactomanano podem modular o
reconhecimento de sítios de clivagem pela endo-β-mananase: (1) existe a
produção de oligossacarídeos limites de digestão F1, F2 e F3 após hidrólise do
galactomanano com endo-β-mananase, como demonstrado para xiloglucanos;
(2) os oligossacarídeos F1 possuem proporções distintas quando da hidrólise
do galactomanano com endo-β-mananase em diferentes concentrações (ExP I
e EXP IV), evidenciando preferência por sítios com menor grau de
galactosilação; (3) a presença da α-galactosidase diminui a produção dos
oligossacarídeos F2 e F3, mostrando que estes não possuem resistência
intrínseca a hidrólise e que a reação atinge o equilíbrio mesmo quando ainda
existem sítios de clivagem ainda disponíveis (EXP III); (4) polissacarídeos com
estruturas diferentes, razão manose:galactose variando entre 150:1 a 1:1, são
digeridos em diferentes taxas de hidrólise pela mesma enzima, evidenciando
que a ramificação com galactose dificulta a ação da endo-β-mananase. Dessa
forma, sugere-se que a estrutura do polissacarídeo galactomanano também
contenha, pelo menos, parte da informação requerida para seu próprio
metabolismo, código para a sua degradação, estando esta informação contida
na distribuição das ramificações com resíduos de D-galactose. Sendo assim,
sugere-se que as diferentes isoformas da α-galactosidase relacionadas à

25
degradação da reserva de galactomanano de sementes de Sesbania virgata
(Cav.) Pers. seriam produto da ação proteolítica da própria enzima a fim de
melhorar a afinidade da α-galactosidase ao substrato durante o processo de
mobilização de reserva. O aumento da afinidade da α-galactosidase ao
substrato durante todo o processo de mobilização garantiria a liberação das
ramificações com galactose de forma contínua, permitindo e aumentando a
eficiência da ação da enzima endo-β-mananase aos sítios de clivagem,
garantindo a degradação do polissacarídeo a oligossacarídeos de forma
regulada, passível de bloqueio, pelo acúmulo de oligossacarídeos e galactose
livre que inibem a ação das enzimas endo-β-mananase e α-galactosidase,
respectivamente, e dificultando a ação de microorganismos, propiciando ao
embrião a maior quantidade de açúcares para o seu desenvolvimento,
aumentando as chances de sucesso no estabelecimento da plântula.

26
ABSTRACT
The seeds of Sesbania virgata (Cav.) Pers. have an endosperm which
accumulates galactomannan as a storage polysaccharide in the cell walls.
Galactomannans are composed of a linear backbone of β-(1,4)-linked D-
mannose residues with D-galactose α-(1,6)-linkages substitutions. The
galactomannans are hydrolysed after protrusion of the radicle. This process is
perfomed by three enzymes (α-galactosidase, endo-β-mannanase and exo-β-
manosidase). The α-galactosidase is the first enzyme to cleave the
polysaccharides, removing the D-galactose residues, allowing the performance
of the endo-β-mannanase, which hydrolyses the mannan backbone to mannan
oligosaccharides. The last part of the process includes exo-β-manoside, that
cleaves the mannan oligosaccharides to mannose residues, which could be
used by the embryo during growth. Aiming at understanding the function of α-
galactosidase in the process of galatomanannan degradation, we studied its
mode of action on mannans and galactomannans. The α-galactosidase of
Sesbania virgata (Cav.) Pers. was purified and characterized (pH and
temperature optimum and the enzyme kinetics). We found that the semipurified
α-galactosidase molecular weight was 42kDa at denaturating conditions, but in
native conditions was 72kDa, suggesting that the enzyme has a quaternary
structure. The enzyme optimum pH was between 4,4-5,4, optimum temperature
between 50°C-55°C, Km= 1,8276 mM and Vmáx= 0,5024 µmolGal.min-
1.mgprot-1. Mass spectrometry measures resulted the following fragments:
ALADYV-HSK-RMPGSLGHEE-QDAK-TT-GDIEDNWNSMTSIADS-NDKW-
ASYAGPGGWN-DPDMLEVGNG-GMTTEEYR-AP-LLVGCDIR-VAVIL-WNR,
being the protein from this sequence related with storage mobilization. Possible
α-galactosidase isoforms were detected during the purification, suggesting other
functions for the enzyme. The α-galactosidase isoforms detected after all
purification steps and with measured mass spectrometry (from 42kDa to 20kDa)
should be related to the storage mobilization. We suggest that the α-
galactosidase isoforms in Sesbania virgata (Cav.) Pers. seeds represents
products of the enzyme self-digestion, this process being correlated with the
enzyme/polysaccharide affinity and at last, correlated to the galactomannan
mobilization. An extract semipurified from Sesbania virgata (Cav.) Pers. and

27
enriched with α-galactosidase activity, was used along with endo-β-mannanase
from Aspergillus niger (Megazyme®) or both endo-β-mannanase and α-
galactosidase (semipurified from Sesbania virgata seeds – Chapter 1- or
commercial enzyme from Cyamopsis tetragonoloba – Megazyme®) were used
to study the fine structure of galactomannans. Hydrolysis of galactomannans
from six species with different mannose:galactose (1:1 to 150:1) ratio were
performed with endo-β-mananase from Aspergillus niger. The oligosaccharides
from all hydrolysis were analyzed by HPAEC-PAD (High Performance Anion
Exchange Chromatography with Pulsed Amperometric Detection). The
hydrolysis fragments data (HPAEC-PAD) suggest that the side-chains of the
polysaccharides can modulate the hydrolytic sites recognition on the
galactomannan by the endo-β-mannanase. This conclusion is supported by: (1)
the presence of limited digest oligosaccharides F1 and dimmers (F2) and
trimers (F3) of the F1 oligosaccharides; (2) the presence of different F1
oligosaccharides proportions after hydrolysis with endo-β-mannanase at
different concentrations, showing preference on less-branched hydrolytic sites;
(3) the α-galactosidase digestion avoided the accumulation of oligosaccharides
F2 and F3, showing that these oligosaccharides do not present intrinsic
resistance to hydrolysis and that the reaction reaches an equilibrium even when
sites of hydrolysis are still available; (4) polymers with different fine structure
(ratio mannose:galactose 1:1 to 150:1) were hydrolysed at different rates by the
endo-β-mannanase, showing that galactose branching interferes on the enzyme
action. Considering that, the branching pattern of the polysaccharide seems to
have direct influence on the interaction of the enzyme with substrate; we
suggest that the structure of the galactomannan holds part of information
required for its own degradation. The higher enzyme x substrate affinity, ensure
the galactose branches digestion, improving the endo-β-mannanase action,
ensuring the degradation of the polysaccharides to oligosaccharides. This
highly regulated degradation process prevents microorganisms predation and
increases the plantlet establishement.

28
INTRODUÇÃO
Uma das mais espetaculares inovações que surgiram durante a
evolução das plantas vasculares (Angiospermas e “Gimnospermas”) foi a
semente. Estima-se que o surgimento tenha ocorrido no Devoniano Superior há
pelo menos 365 milhões de anos. A semente é um dos principais fatores
responsáveis pela dominância das plantas na flora atual, e essa dominância
cresceu ao longo de um período de milhões de anos (Raven, 2001). Este
sucesso deve-se ao fato das sementes possuírem uma grande capacidade de
sobrevivência quando comparada com esporos livres, devido à proteção que
esta confere ao embrião, além dos recursos nela armazenados para os
estágios de germinação e estabelecimento (Raven, 2001).
1.1. Sementes
Nas Angiospermas, o surgimento da semente ocorre com a dupla
fecundação, onde uma das duas células espermáticas fecunda o óvulo
(oosfera), dando origem ao zigoto diploide e, consequentemente, o embrião;
enquanto a outra célula espermática se funde com dois núcleos polares do
saco embrionário, originando o endosperma triplóide (Raven, 2001; Beltrati &
Paoli, 2003).
A semente durante o seu desenvolvimento contém: embrião (resultado
da fusão da oosfera com uma célula espermática), endosperma (provém da
fusão de dois núcleos polares e uma célula espermática), perisperma
(resultado do desenvolvimento do nucelo) e tegumento (formado por um ou
ambos integumentos ao redor do óvulo) (Bewley & Black, 1994). Enquanto
todas as sementes maduras contêm o embrião e muitas continuam envolvidas
pelo tegumento, a persistência do endosperma ou perisperma varia de espécie
para espécie. Portanto, a persistência das partes contidas durante o
desenvolvimento da semente não é certa na semente madura (Bewley & Black,
1994; Beltrati & Paoli, 2003).

29
1.2. Germinação e Pós-Germinação
A germinação das sementes se inicia com a entrada de água na
semente, processo denominado embebição, e culmina com o início do
alongamento do eixo embrionário, normalmente a emissão da radícula (Bewley
& Black, 1994).
Durante o período compreendido entre a embebição e a emissão da
radícula, numerosos eventos ocorrem dentro da semente, como a hidratação
de proteínas, mudanças na estrutura subcelular, produção de macromoléculas
entre outros, que conjuntamente promovem a germinação (Bewley & Black,
1994).
Dessa forma, a germinação pode ser dividida em três fases: embebição,
aumento do metabolismo e iniciação do alongamento do eixo embrionário
(Bewley, 1997).
Entretanto, para que a germinação ocorra se faz necessária não só a
presença de água, mas também de outras condições ambientais favoráveis
como: temperatura adequada, presença de oxigênio, ausência de substâncias
inibidoras e algumas vezes até da luminosidade (Bewley & Black, 1994; Raven,
2001, Taiz & Zeiger, 2004).
Embora sejam necessárias condições ambientais favoráveis,
propriedades intrínsecas da semente podem impedir a sua germinação, por
não permitirem a entrada de água na semente. Nesses casos é necessário o
rompimento do tegumento da semente, pela escarificação, por exemplo, para
permitir a entrada de água e, portanto, a germinação (Labouriau, 1983).
Dessa forma, os eventos ocorridos durante esse período de germinação
são responsáveis pela reativação do metabolismo, outrora reduzido,
culminando no crescimento do embrião (Bewley & Black, 1994).
A reativação do metabolismo é suprida de energia, normalmente, por
açúcares solúveis de reserva, como a sacarose ou oligossacarídeos da série
rafinósica, pois esses podem ser rapidamente metabolizados provendo energia
para divisões e alongamentos celulares necessários para a protrusão da
radícula. No entanto, no período pós-germinativo, compreendido pelo
desenvolvimento inicial da plântula até o seu estabelecimento como organismo
autotrófico (estabelecimento da fotossíntese), a energia, bem como as

30
necessidades de carbono e nitrogênio, são supridas pela reserva de
polissacarídeos. Como as necessidades nesta fase são muito grandes a
reserva de polissacarídeos geralmente aparece em maior proporção
(Buckeridge et al., 2000b).
Assim, as sementes possuem dois tipos de reserva: açúcares solúveis e
polissacarídeos, sendo esse último melhor discutido a seguir, pois é parte do
tema principal deste trabalho.
1.3. Reservas de sementes
As Angiospermas apresentam diferentes estratégias de adaptação aos
seus respectivos ambientes, entre as quais se encontram o acúmulo de certos
compostos em suas sementes (Buckeridge et al., 2000c). As reservas das
sementes possuem basicamente duas funções que se relacionam com a
manutenção e o desenvolvimento do embrião até a formação de uma plântula
autotrófica: fonte de energia (manter processos metabólicos em
funcionamento) e fonte de matéria (carbono e nitrogênio para a construção de
tecidos vegetais que irão formar a plântula) (Buckeridge et al., 2004a;
Buckeridge et al., 2000c).
Muitos dos tecidos da semente podem funcionar como armazenadores
de reservas, por exemplo: perisperma e eixo embrionário. Entretanto, os
cotilédones e o endosperma são as principais estruturas de armazenamento
em sementes de Angiospermas (Bewley & Black, 1994). Embora alguns
endospermas contenham, principalmente, reserva de lipídeos e proteínas, a
maioria parece armazenar carboidratos, sendo eles o amido, normalmente
presente em cereais, e polissacarídeos de parede celular (por exemplo,
galactomananos) em Leguminosae (Bewley & Black, 1994).
Por outro lado, os cotilédones são relativamente mais generalistas,
podendo armazenar: proteínas, carboidratos e lipídeos (Bewley & Black, 1994).
Os produtos de degradação de lipídeos e carboidratos são utilizados na
geração de energia e produção de matéria-prima para a construção de novos
tecidos e células. Buckeridge (1993) e Buckeridge et al. (2004a) discutem que
a maioria das sementes que armazena carboidratos em grandes quantidades,

31
não armazena lipídeos, de tal forma que essas substâncias são mutuamente
exclusivas.
Neste contexto, as proteínas possuem como função o armazenamento
de nitrogênio e enxofre, elementos essenciais para a síntese de proteínas,
ácidos nucleicos e compostos secundários da plântula em crescimento
(Buckeridge et al., 2004a).
1.4. Parede celular de reserva de sementes
Os modelos mais recentes de parede celular propõem que esta seja
uma matriz complexa e espessa com diferentes papéis fisiológicos, formada,
normalmente, por três domínios independentes, mas que interagem entre si
(McCann & Roberts, 1991; Carpita & Gilbeaut, 1993; Carpita & McCann, 2000).
O domínio fundamental da parede celular é formado por microfibrilas de
celulose, envolvidas por hemiceluloses, essas últimas podendo estar
fortemente ou fracamente ligadas às fibras de celulose por pontes de
hidrogênio (Buckeridge et al., 2000c).
A interação celulose-hemicelulose está imersa no segundo domínio da
parede celular, a matriz péctica, composta de polissacarídeos ricos em ácidos
galacturônicos. Esse domínio consiste na região mais hidratada da parede
celular se comparada com o domínio celulose-hemicelulose (Buckeridge et al.,
2000c; Carpita & McCann, 2000).
O último domínio é denominado domínio protéico, este sendo composto
entre outras proteínas, por proteínas estruturais e enzimas, que operam sobre
os polímeros estruturais (Buckeridge et al., 2008). Entre as ações conhecidas,
o domínio protéico age na mobilização dos polissacarídeos de reserva da
parede celular (Carpita & McCann, 2000).
A rede formada por celulose e hemicelulose fornece força tensora,
enquanto que a rede péctica relaciona-se com a resistência a compressão
(Jarvis & McCann, 2000).
O modelo descrito acima, também pode ser aplicado à parede celular de
reserva. Entretanto, o modelo sofre uma variação, onde um domínio, ou
mesmo um de seus polissacarídeos, tenha sido depositado em maior
quantidade em relação aos demais. Aparentemente, a parede celular de

32
reserva constitui uma adaptação evolutiva da parede primária, que
desenvolveu a capacidade de armazenar quantidades apreciáveis de
carboidratos (Buckeridge et al., 2000c).
Os polímeros de reserva armazenados na parede celular podem ser de
origem péctica ou hemicelulósica. Dentre as hemicelulósicas, reconhecem-se:
mananos (mananos puros, galactomananos e glucomananos) e xiloglucanos,
enquanto os de origem péctica os arabinogalactanos (Buckeridge et al., 2000c).
A alta deposição de uma dessas substâncias nas paredes celulares de
reserva é acompanhada de pouca ou nenhuma quantidade de celulose
juntamente com os demais polissacarídeos. Essa baixa proporção de celulose
facilita o processo de mobilização das reservas, o qual seria consideravelmente
mais complexo se proporções normais de celulose estivessem presentes
(Buckeridge et al., 2000c; Buckeridge et al., 2004b).
1.5. Polissacarídeos de reserva de parede celular
Os polissacarídeos de reserva presentes na parede celular das
sementes são denominados PRPC (Buckeridge & Reid, 1996; Buckeridge et
al., 2000b). Esses polissacarídeos são mobilizados após a germinação e os
produtos de sua degradação garantem o desenvolvimento do embrião à
plântula, suprindo-a com energia e matéria-prima para a construção de órgãos
e tecidos até que a plântula se torne autotrófica (Reid, 1985a; Mayer &
Poljakoff-Mayber, 1975; Buckeridge et al., 2004b).
Os PRPC possuem características que os fazem mais convenientes
para o metabolismo celular: serem relativamente inertes ao que concerne a sua
reatividade e possuírem diferentes graus de solubilidade em água. Essas
características conferem alta compactação e baixa reatividade do polímero,
tornando possível a existência de um “compartimento celular”, a parede celular,
que permite um fluxo de água com um grau de liberdade considerável
(Buckeridge et al., 2000c).
Porém, o custo de produção desses polímeros é alto, pois, tais
compostos necessitam de um complexo sistema de biossíntese, secreção e
montagem no meio extracelular (Buckeridge et al., 2000c).

33
Em 1996, Buckeridge e Reid propuseram que os PRPC poderiam ter
passado por etapas de transferência de funções ao longo da evolução, os
quais seriam parte das transformações que levaram à formação de uma parede
de reserva a partir da parede primária. Muitas das funções primordiais da
parede primária ainda se mantêm: dureza (mananos em endospermas de
sementes de palmeiras, tomate e alface), relações hídricas (xiloglucanos em
cotilédones e galactomananos em endospermas de sementes de leguminosas)
e no controle da expansão celular (galactanos nos cotilédones de lupino e, em
menor proporção, em sementes de soja e feijão) (Buckeridge et al., 2000c).
Os polissacarídeos de parede são classificados em três grupos:
mananos, xiloglucanos e os (arabino)galactanos. O grupo dos mananos
engloba os mananos puros, glucomananos e galactomananos (Buckeridge et
al., 2000c), sendo esse último melhor discutido a seguir.
1.5.1. (Arabino)Galactanos
A primeira identificação do galactano como polissacarídeo de reserva de
semente foi realizado por Hirst et al. (1947) em sementes de Lupinus albus
(lupino), onde com base na determinação estrutural, conclui-se que o
polissacarídeo possuía cadeia principal de galactose unidas por ligações β-
(1,4).
Em um estudo mais recente realizado por Al-Kaisey & Wilkie (1992) com
o galactano de quatro espécies de lupino determinou-se a existência de dois de
galactanos: formado por ligações β-(1,3),(1,6) com algumas ligações β-(1,4), e
outro, mais frequente, composto por ligações β-(1,4) com ramificações de L-
arabinofuranose a cada 16 a 21 resíduos da cadeia principal (Figura 1).
A mobilização dos galactanos de reserva de Lupinus angustifolius foi
descrita por Crawshaw & Reid (1984) e as alterações estruturais do galactano
que acompanham o processo foram descritas por Parker (1984), onde após a
mobilização a maior parte das ramificações com galatose e arabinose é
removida da parede celular, permanecendo um material residual enriquecido
em ramnose, ácido urônico e glucose (Figura 1).
Matheson & Saini (1977) reportaram a presença de duas α-
arabinosidases e três β-galactosidases em cotilédones de Lupinus luteus, cujas

34
atividades de ambas as enzimas se mostraram relacionadas à expansão
cotiledonar.
Figura 1: Estrutura e mobilização do galactano. (A) Estrutura do galactano mais frequente
composto por ligações β-(1,4) com ramificações de L-arabinofuranose a cada 16-21 resíduos da

35
cadeia principal (não mostrada). Ainda existe um segundo tipo de galactano formado por ligações
β-(1,3)(1,6). (B) Reações envolvendo a mobilização do polissacarídeo de reseva em Lupinus sp. β-
(1,4)-D-galactano é o polímero dominante, sendo descrito como uma ramificação do
rhamnogalacturanano, por vezes ramificado por ligações α-(1,5)-arabinano. Uma molécula de α-
(1,3), (1,6)-D-galactano também está representado embora ocorre em quantidades menores. Neste
sistema de degradação, apenas exoenzimas foram detectadas, uma das quais (exo-α-(1,4)-
galactanase a partir de Lupinus angustifolius) é capaz de libertar a maior parte (cerca 60%) das
paredes celulares intactadas da mesma espécie. In vivo, outras galactosidases e arabinosidases
parecem agir no ramos deixando apenas ramnogalacturonano. O destino dos produtos é
meramente hipotético neste esquema. Figura retirada e adaptada de Buckeridge et al. (2000b) e
Buckeridge et al. (2000c).
Buckeridge & Reid (1994) isolaram uma exo-β-(1,4)-galactanase dos
cotilédones de Lupinus angustifolius e, em 1996, propuseram um modelo
hipotético de degradação dos galactanos.
Neste modelo de degradação (Buckeridge et al., 2000b), a exo-β-(1,4)-
galactanase é apontada com a enzima chave da mobilização do galactano,
sendo capaz de hidrolisar 82% das unidades de galactose da cadeia principal
do polímero, bem como cerca de 63% das galactoses presentes no polímero
em paredes celulares intactas. Os produtos da degradação, arabinose e
galactose livres podem ser utilizados na produção de sacarose para utilização
pelo embrião em desenvolvimento (Buckeridge et al., 2000b).
O galactano é uma molécula de múltiplas funções, exercendo tanto a
função de reserva de parede celular, como também, sendo um importante
elemento na modulação da expansão cotiledonar durante o desenvolvimento
da plântula.
1.5.2. Xiloglucanos
O xiloglucano é uma das principais hemiceluloses presentes em paredes
celulares de dicotiledôneas, também podendo ser acumulado como reserva de
parede celular de sementes de algumas Leguminosae-Caesalpinoidae.
Estruturalmente os xiloglucanos são compostos de uma cadeia principal
de glucose unida por ligações β-(1,4), onde ocorrem ramificações regulares
com xilose ligada α-(1,6). Algumas destas xiloses ainda podem apresentar
ramificações por galactose ligadas α-(1,2). Em parede celular primária, mas

36
não em parede celular de reserva, as ramificações por galactose ainda podem
ser ramificadas por fucose ligada α-(1,2) (Figura 2A).
O sistema de degradação do xiloglucano de reserva mais estudado foi o
componente da semente de Tropaeolum majus. Edwards et al. (1985)
acompanharam a mobilização do xiloglucano nesta espécie e notaram um
aumento da atividade enzimática da β-glucosidase, β-galactosidase,
α-xilosidase e endo-β-(1,4)-glucosidase durante a mobilização do xiloglucano.
Apesar da mobilização da reserva de xiloglucano presente em sementes
de Hymenea courbaril (jatobá) ter sido sugerida por Kooiman (1960), o
acompanhamento detalhado só foi realizado por Tiné et al. (2000) (Figura 2B).
O mecanismo de degradação proposto por Tiné & colaboradores (2000)
é dividido em três partes: transglicosilação, onde o polissacarídeo seria
parcialmente hidrolisado pela XTH (xiloglucano transglicosidase hidrolase);
desgalactosilação, etapa onde certas unidades de galactose dos produtos da
ação anterior da XTH seriam hidrolisados pela β-galactosidase; e
desmontagem, onde os oligossacarídeos remanescentes da ação da β-
galactosidase entrariam no ciclo final de degradação, onde a glucose e a xilose
seriam atacadas pela β-glucosidase e α-xilosidase, produzindo
monossacarídeos livres para a utilização do embrião no desenvolvimento
(Figura 2B).

37
Figura 2: Estrutura e mobilização de xiloglucano de reserva de sementes. (A) Estrutura molecular
do xiloglucano de reserva contendo cadeia principal de glucose unida por ligações β-(1,4), onde
ocorrem ramificações regulares com xilose ligada α-(1,6). Algumas destas xiloses ainda podem
apresentar ramificações por galactose ligadas α-(1,2). (B) Representação esquemática da

38
degradação de reserva de xiloglucano em sementes. Uma sequência de XLXGXLLGXXXGXXLG é
usada como um exemplo de um segmento de um polímero de xiloglucano com a extremidade não-
redutora voltada para a esquerda. O primeiro ataque é feita por xiloglucano-endo-β-
transglicosilase (XET) e β-galactosidase (β-gal proveniente de Nasturtium apenas). Os
oligossacáridos produzidos pela ação endo da XET são atacados por β-galactosidases (Nβ-
gal=Nasturtium β-galactosidase; Cβ-gal= Copaifera β-galactosidase, o último sendo capaz de atacar
oligossacarídeos apenas em certas posições (galactoses colocadas na ponta não-redutora). A α-
xilosidase e a β-glucosidase agem em hidrólises alternadas do final não-redutor. (*) Em leguminosas
(Copaifera langsdorffii e Hymenaea courbaril), após uma rodada de ação destas duas enzimas, a
galactose estará novamente próximo do terminal não-redutor. Desata forma, a β-galactosidase pode
agora agir sobre o oligossacarídeo XLG, libertando assim a galactose e permitindo a continuidade
da ação das outras duas exo-glicosidases. Nestas duas espécies, a galactose retirada dos
oligossacarídeos é possivelmente limitadora da degradação, pois a enzima possui um pH ótimo de
3,2, enquanto as demais enzimas possuem um pH ótimo de 4,5. Figura retirada e adaptada de
Buckeridge et al. (2000c) e Buckeridge et al. (2000b).
1.5.3. Mananos
Baseado em sua estrutura e composição, o grupo dos mananos pode
ser subdividido em três grupos: mananos puros, glucomananos e
galactmananos (Buckeridge et al., 2000a)
Glucomananos
Dentre os três subgrupos de mananos, os glucomananos são os menos
conhecidos.
A estrutura determinada por metilação apresenta uma cadeia principal
formada por um número proporcionalmente equivalente de resíduos de β-
glucopiranosídeo e β-manopiranosídeo unidos por ligações β-(1,4). Com certa
frequência pode-se detectar a presença de unidades de galactopiranosídeo
unidos α-(1,6) a cadeia principal (Buckeridge et al., 2000b).

39
Mananos Puros
Os mananos puros são definidos pelo seu conteúdo maior de 90% de
resíduos de β-manopiranosideo unidos por ligações β-(1,4) formando uma
cadeia linear sem ramificações, podendo ou não o restante da cadeia estar
ramificada com α-galactopiranosídeo (Buckeridge et al., 2000b).
Dea et al. (1986) demonstraram que mananos com taxa de ramificação
com galactose abaixo de 10% eram insolúveis e precipitavam rapidamente em
solução aquosa. Assim, os mananos são estruturalmente relacionados aos
galactomananos, apenas apresentando um grau menor de ramificações por
galactose, aumentando a interação entre as moléculas de manano, formando
cristais na parede celular, o que confere dureza à parede celular (Buckeridge et
al., 2000b).
Sachs (1862) e Keusch (1968) estudaram a germinação de sementes de
Phoenix dactylifera com reserva de manano. Determinaram a presença das
enzimas endo-β-mananase e β-manosidase responsáveis pela degradação do
manano contido nestas sementes.
Além da função de reserva de parede celular, os mananos parecem
estar relacionados à dureza da semente. Groot & Karssen (1987) e Groot et al.
(1988) evidenciaram que o manano presente da região endospérmica próxima
à extremidade da raiz de tomate, exerce papel crucial na protrusão da radícula,
pois o amolecimento do endosperma, induzido pela ação da giberelina, facilitou
a germinação.
Assim, o manano na forma de um polissacarídeo de reserva de parede
celular exerceria as funções de constritor e protetor mecânico do embrião,
como também, polissacarídeo de reserva.
1.6. Galactomananos
O primeiro estudo sobre a estrutura dos galactomananos provavelmente
foi feito por Naldeman, em 1890. Esse autor reportou a presença de galactose
e manose em várias espécies de leguminosas e também verificou que os
galactomananos se comportam como substâncias viscosas (Nadelman, 1890;
Buckeridge et al., 2000a).

40
Os galactomananos ocorrem tipicamente em endospermas de sementes
de Leguminosas, mas eles também estão presentes em sementes de espécies
de outras famílias como Compositae, Convolvulaceae, Annonaceae,
Malvaceae, Palmae, Rubiaceae e Umbelliferae (Tookey et al., 1962; Dea &
Morrison, 1975; Guzmán & Hernandez, 1982; Buckeridge et al., 2000b).
1.6.1. Estrutura química dos galactomananos
Os galactomananos são polissacarídeos compostos de uma cadeia
linear de resíduos de D-manose unidas por ligações glicosídicas β-(1,4), à qual
resíduos de D-galactose estão unidos por ligações α-(1→6), formando
ramificações simples (Buckeridge et al., 2000a) (Figura 3A).
Figura 3: Estrutura molecular do polissacarídeo galactomanano e explicação para a leitura das
“abreviações” dos oligossacarídeos GalXManY. (A) Cadeia principal linear de resíduos de D-manose
unidas por ligações glicosídicas β-(1,4), na qual resíduos de D-galactose estão unidos por ligações α-
(1,6), formando ramificações simples. Figura retirada e adaptada de Buckeridge et al. (2000a). (B)
A cadeia principal de manose é numerada a partir da extremidade redutora (à direita). O número
subscrito ligado à manose indica a quantidade de resíduos de manose que a cadeia principal do
oligossacarídeo possui, enquanto o número sobrescrito ligado a galactose indica a posição do
resíduo de galactose relativo ao resíduo de manose a partir da extremidade redutora (McCleary et
al., 1983).

41
A razão manose:galactose (M/G) e a distribuição dos resíduos de
galactose ao longo da cadeia variam de espécie para espécie, com a razão
ocorrendo entre 1:1 e 4:1. Esta propriedade é importante para estudos
quimiotaxonômicos e evolutivos (Buckeridge & Diertrich, 1990; Buckeridge et
al., 2000a).
Como dito anteriormente, os galactomananos são abundantes em
endospermas de sementes de Leguminosae (ou Fabaceae), a família com o
terceiro maior número de espécies do Reino Vegetal (Buckeridge et al., 2000a;
Buckeridge et al., 2000c). Essa família é subdividida em três subfamílias
(Caesalpinoideae, Mimosoideae e Faboideae) sendo possível classificá-las de
acordo com suas razões (M/G). A subfamília Caesalpinoideae possui a maior
razão M/G, ou seja, menor o grau de ramificação com galactose, seguida pela
Mimosoideae e finalmente Faboideae (Buckeridge et al., 2000c).
Além desse parâmetro taxonômico, visto acima, a razão M/G também
dita a solubilidade do polissacarídeo em água, e consequentemente, sua
interação com outros polissacarídeos, incluindo a celulose (Buckeridge et al.,
2000a).
1.6.2. Propriedades dos galactomananos
Embora os galactomananos sejam considerados polissacarídeos de
reserva (Buckeridge et al., 2000a), tem sido demonstrado que tal polímero
pode exercer outras funções além de reserva nas sementes.
Entre as funções, os galactomananos exercem o controle da embebição
(Reid & Bewley, 1979; Buckeridge et al., 2000c), dada sua alta solubilidade nos
primeiros estágios de germinação, esse polímero absorve grandes quantidades
de água, distribuindo-a ao redor do embrião (Buckeridge et al., 2000c).
O endosperma embebido protege o embrião contra perda d’água através
de um efeito conhecido como “tampão de água” durante os períodos de seca
após o início da germinação, protegendo-o contra estresse hídrico (Reid &
Bewley, 1979; Halmer & Bewley, 1979). Um comportamento similar também
pode ser visto em leguminosas tropicais, como em Sesbania virgata (Cav.)

42
Pers. (anteriormente publicado com o sinônimo Sesbania marginata por
Buckeridge et al., 1995a; Buckeridge & Dietrich, 1996).
Ligado à embebição, Groot & Karssen (1987) demonstraram que
distensão mecânica do endosperma, pela entrada de água, facilita a protrusão
da radícula. Sendo assim, facilitação da protrusão da radícula nas primeiras
fases da germinação também é uma função dos galactomananos.
Aparentemente as funções de reserva e embebição estão limitadas às
Leguminosas, ao passo que nas não leguminosas é aparente a existência da
função de dureza dos galactomananos. Embora, a função de dureza e proteção
parece também ser mantida em Leguminosas, como demonstrado por
Buckeridge et al. (2000c) e Lisboa et al. (2006), nas sementes de Sesbania
virgata (Cav.) Pers.
Dessa forma, fica evidente que diferenças na estrutura do
galactomanano propiciam a este polímero atuação em diversas funções
biológicas. Essas diferenças encontram-se no grau de ramificação: quanto
mais ramificado for o galactomanano, maior a possibilidade de sua função
biológica estar ligada com as relações hídricas. Em contrapartida, quanto
menos ramificado for o galactomanano, maior a sua chance de atuação com
dureza e proteção (Buckeridge et al., 2000c).
Desconhece-se qual das funções é primária entre as de reserva, de
embebição ou de dureza. Entretanto, é evidente que os galactomananos são
moléculas multifuncionais, desempenhando suas funções durante fases
distintas do crescimento e desenvolvimento das plantas (Buckeridge et al.,
2000c).
Por suas propriedades físico-químicas, os galactomananos apresentam
grande importância comercial, principalmente no que se refere a solubilidade e
a sua capacidade de formar soluções viscosas, sendo utilizados em diversos
setores industriais (Buckeridge et al., 2000a).
1.6.3. Mobilização dos galactomananos
De acordo com McCleary (1983), Buckeridge & Dietrich (1996) e
Buckeridge et al. (2000a) a mobilização do galactomanano presente no
endosperma de sementes se inicia somente após a emissão da radícula.

43
Os galactomananos são hidrolisados por três enzimas: α-galactosidase
(EC 3.2.1.22), endo-β-mananase ou β-1,4-manano-endo-hidrolase (EC
3.2.1.78) e exo-β-manosidase ou β-manosidase (EC 3.2.1.25) produzindo
galactose e manose livres ao mesmo tempo em que a glucose é produzida no
endosperma. Aparentemente essa última é disponibilizada ao embrião e servirá
como fonte de carbono e energia para o estabelecimento das plântulas
(Buckeridge et al., 2000a; Lisboa et al., 2006) (Figura 4).
A α-galactosidase é a enzima responsável pela quebra das ligações α-
(1,6) existentes entre as unidades de galactose e manose, enquanto que a
endo-β-mananase atua na cadeia principal de manano, rompendo as ligações
do tipo β-(1,4) existentes entre as unidades de manose, reduzindo o manano a
oligossacarídeos. A β-manosidase atua nas ligações β-(1,4) entre as unidades
de manose dos oligossacarídeos provenientes da ação da endo-β-mananase,
liberando manoses livres (Reid & Meier, 1972; McCleary, 1983; Buckeridge et
al., 1995a; Buckeridge & Reid, 1996; Buckeridge et al., 2000a).
De acordo com Reid & Edwards (1995) e Lisboa et al. (2006), a
degradação do galactomanano pelas enzimas requer uma ordem de atuação
das mesmas. Em sementes de leguminosas a ação da α-galactosidase é uma
condição necessária para permitir acesso à enzima endo-β-mananase a cadeia
principal do galactomanano. Apesar disso, em sementes de Lactuca sativa
(alface) a α-galactosidase só pode retirar galactose do produto de ação da
endo-β-mananase (Leung & Bewley, 1983).

44
Figura 4: Rotas bioquímicas envolvidas no catabolismo do galactomanano somado ao metabolismo
pelo embrião dos monossacarídeos liberados na degradação da reserva. Quando a camada de
aleurona não está presente, aparentemente as hidrolases são produzidas dentro das células
endospermáticas e secretadas para fora da parede celular onde as mesmas vias metabólicas
ocorrem. Man = manose; Gal = galactose; SPS = sacarose fostato sintase; Man-6-P = manose-6-
fosfato; Fru-6-P = frutose-6-fosfato; UDP-gal = uridina difosfatada de galactose; Suc-6-P =
sacarose-6-fosfato. Figura retirada e adaptada de Buckeridge et al. (2000b).
Por outro lado, a ação da β-manosidase só é possível sobre o produto
da degradação da α-galactosidase e endo-β-mananase. McCleary (1983)
demonstrou que a β-manosidase não apresenta atividade sobre o polímero
inteiro, só apresentando ação sobre oligossacarídeos de manano
desgalactosilados.

45
Apesar do conhecimento das enzimas envolvidas no processo de
mobilização do galactomanano, pouco se sabe sobre as características
bioquímicas intrínsecas das mesmas e, consequentemente, no modo de ação
dessas sobre o polissacarídeo.
Lisboa et al. (2006) purificaram e caracterizaram a enzima endo-β-
mananase de sementes de Sesbania virgata (Cav.) Pers. Neste trabalho
estimou-se seu peso molecular em 30kDa, temperatura ótima em 45°C, bem
como o pH ótimo em um intervalo entre 3,5-4,5. O ponto isoelétrico da enzima
encontra-se em pH 4,5.
Além disso, os autores demonstram que a endo-β-mananase apresenta
dois picos de atividade: um em 24horas após a embebição, e outro em 120
horas após a embebição. O primeiro pico, aparentemente, está relacionado
com a protrusão da radícula, enquanto que o segundo com a degradação do
galactomanano de reserva da semente (Lisboa et al., 2006). Esta observação
sugere que a função de amolecimento do endosperma para permitir a
protrusão da radícula esteja presente também em Leguminosae.
Em relação à mobilização do galactomanano, esses autores observaram
que a enzima endo-β-mananase, presente em sementes de Sesbania virgata
(Cav.) Pers., só possui atividade caso o galactomanano já tenha sofrido a ação
da α-galactosidase previamente (Lisboa et al., 2006).
As demais enzimas, α-galactosidase e β-manosidase, ainda não foram
caracterizadas para Sesbania virgata (Cav.) Pers., porém, os conhecimentos
das características de ambas são de suma importância para o aprofundamento
do conhecimento da mobilização de galactomananos para essa espécie, bem
como para as demais espécies com sementes que armazenam galactomanano
como reserva.
Quanto à produção dessas três hidrolases, nas sementes de Trigonella
foenum-graecum e Cyamopsis tetragonoloba as células endospérmicas não
são vivas devido à alta deposição de galactomananos nas paredes celulares
durante a maturação, assim, essas sementes possuem uma camada de
aleurona, a qual se acredita ser responsável pela produção das enzimas
hidrolíticas de mobilização (Reid, 1985a).
Por outro lado, as sementes de Ceratonia siliqua possuem as células do
endosperma vivas. Neste caso, não existe distinção clara entre o endosperma

46
e a camada de aleurona. Dessa forma, provavelmente a produção e a liberação
das enzimas hidrolíticas são feitas pelas próprias células endospérmicas
(Buckeridge et al., 2000a).
Diferentemente da última espécie, em sementes de Sesbania virgata
(Cav.) Pers. a camada de aleurona é claramente distinguida, estando presente
entre o tegumento da semente e o endosperma, ao mesmo tempo em que este
último possui corpos protéicos (Buckeridge et al., 2000a). Baseado nessas
observações, Buckerigde & Dietrich (1996) propuseram que as enzimas de
hidrólise do galactomanano poderiam ser produzidas em ambos os locais,
camada de aleurona e células endospérmicas. Tonini et al. (2007) sugerem,
ainda, que a produção dessa enzimas possa ser feita não só no endosperma,
como no endotegumento. Porém, Tonini (2008) sugere que a produção dessas
enzimas hidrolíticas só ocorra durante a maturação da semente, não existindo
produção de novo das enzimas quando a semente já se encontra madura.
1.6.4. Regulação da Mobilização de Reserva de Galactomanano
Groot & Karsen (1987) demonstraram que a giberelina regula a
germinação da semente através da indução do amolecimento do endosperma
em sementes de tomate e Spyropoulos & Reid (1985) sugeriram que o
endosperma de Leguminosae deve estar sob controle de auxina e giberelina
para a degradação das reservas contidas no endosperma.
Tonini et al. (2006) demonstraram que a inibição da mobilização dos
galactomananos em sementes de Sesbania virgata (Cav.) Pers. provavelmente
é controlada pelo tegumento da semente, pois essa contém ABA que age
negativamente na produção da enzima α-galactosidase. No entanto, Tonini et
al. (2010) sugerem que, antagonicamente ao ABA, o etileno induza a
degradação das reservas.
Além, disso o ABA tem sido descrito com um potente inibidor da
degradação do galactomanano em sementes de outras espécies: feno-grego
(Reid & Meier, 1973); caroba (Seiler, 1977); alface (Halmer & Bewley, 1979);
tomate (Nomaguchi et al., 1995) e Sesbania virgata (Cav.) Pers. (Potomati &
Buckeridge, 2002), pela interferência na atividade das enzimas de hidrólise do
galactomanano.

47
Além da regulação esperada da mobilização de reserva por
fitohormônios, foi evidenciado uma modulação do processo de degradação de
xiloglucano de reserva de sementes realizada pela estrutura do próprio
polímero de xiloglucano (Tiné et al., 2003).
Tiné et al. (2003), analisando os oligossacarídeos produzidos pela
hidrólise com celulase do xiloglucano de reserva de diferentes espécies
(Tamarindus indica, Copaifera langsdorffii e Hymenea courbaril), constituídos
de diferentes taxas de substituição por galactose, demonstraram que
xiloglucanos de estrutura diferente são hidrolisados diferentemente; que os
oligossacarídeos menos ramificados são atacados preferencialmente pela
enzima e que a presença da β-galactosidase juntamente com a celulase,
impede a formação de oligossacarídeos combinados F2 e F3, sugerindo que o
padrão das substituições por D-galactose possuem influência direta na ação
enzimática.
Dessa forma, eles concluem que a estrutura do polissacarídeo, através
de suas ramificações pela galactose, codificaria, pelo menos, uma parte da
informação para a sua própria degradação, o que eles denominaram código do
xiloglucano.
Sendo o galactomanano um polissacarídeo de reserva de parede
celular, como o xiloglucano, espera-se que a estrutura fina do polissacarídeo
também apresente, pela distribuição das ramificações de galactose, uma parte
da informação para a sua própria degradação, como o demonstrado para o
xiloglucano.
1.7. Caracterização de α-galactosidase
Alfa-galactosidases (E.C. 3.2.1.22) catalisam a hidrólise de ligações α-
(1,6) de galacto-oligossacarídeos e de polímeros de galacto-(gluco)-mananos.
Essas enzimas estão amplamente distribuídas em animais, plantas e
microorganismos (Fujimoto et al., 2003).
Em humanos, a α-galactosidase é uma exoglicosilase lisossomal que
quebra resíduos de α-D-galactose de glicolipídeos e glicoproteínas. Mutações
no gene da α-galactosidase causam a incompleta degradação de carboidratos
da globotriaosilceramida ou GL-3, um glicolipídeo, que acaba por se acumular

48
nas células e consequentemente nos tecidos e órgãos (principalmente parede
de vasos sanguíneos, coração, cérebro e rim), resultando na doença de Fabry
(Fujimoto et al., 2003).
Em plantas, o galactomanano é um dos compostos mais encontrados
como reserva de parede celular de sementes e a α-galactosidase é uma
enzima chave para a mobilização deste composto para o crescimento do
embrião (Reid, 1985). Apesar de apresentar a função de mobilização de
reserva, algumas α-galactosidases podem estar associadas a outras funções
(McCleary & Matheson, 1974).
McCleary & Matheson (1974) detectaram múltiplas formas de α-
galactosidase em sementes de Cyamopsis tetragonoloba, embora somente
uma delas estivesse associada com a degradação do galactomanano presente
no endosperma. As sementes de Lycopersicon esculentum apresentam três
isoformas da enzima α-galactosidase e, através da obtenção e sequenciamento
do cDNA da enzima, demonstraram alta similaridade desta α-galactosidase
com outras galactosidases, especialmente a de Coffea arabica (Fuertado et al.,
2001).
Atualmente, já se deduziu, a partir de genes ou cDNA, estruturas
primárias de mais de cinquenta α-galactosidases. Análises da estrutura
primária e domínios hidrofóbicos demonstraram que as α-galactosidases
podem ser classificadas em três famílias de hidrolases glicosídicas: 4, 27 e 36
(Fujimoto et al., 2003). Em geral, as α-galactosidases de eucariotos possuem
alta similaridade na sequência de aminoácidos, inclusive quanto ao seu resíduo
catalítico (aspartato), e são classificadas na família 27, enquanto α-
galactosidases de bactérias são agrupadas nas famílias 4 e 36 (Fujimoto et al.,
2003).
De acordo com o banco de dados CAZy (Carbohydrate-Active EnZymes
database) (Cantarel et al., 2008) as α-galactosidases possuem um domínio
conservado com a sequência G - [LIVMFY] - x - [LIVMFY] - x - [LIVM] - D - [DF]
- x - W - x - [RV] - [DNSF].

49
1.8. Caracterização da endo-β-mananase
As endo-enzimas como as endo-β-mananases (E.C. 3.2.1.78) catalisam
a hidrólise de ligações β-(1,4) de homoglicanos não ramificados compostos de
um único tipo de resíduo em sua cadeia principal, neste caso, manose, a
oligossacarídeos contendo no mínimo 2 a 3 resíduos (disssacarídeos e
trissacarídeos) (McCleary & Matheson, 1983).
Essas endo-enzimas que hidrolisam mananos puros vêm sendo
apontadas como importantes em eventos germinativos, antes da protrusão da
radícula, em sementes de Lycopersicon esculentum e em sementes de Lactuca
sativa, bem como, em eventos pós-germinativos de mobilização de reserva em
paredes celulares do endosperma de Lactuca sativa e Trigonella foenum-
graecum (Dirk et al., 1995).
Estudos conduzidos por McCleary & Matheson (1983) com endo-β-
mananases provenientes de fontes diversas, bactérias (Bacillus subtilis),
fungos (Aspergillus niger), animais (Helix pomatia) e plantas (Cyamopsis
tetragonoloba e Medicago sativa) evidenciaram que as endo-β-mananases
variam significativamente na habilidade de hidrolisar polissacarídeos de
galactomanano.
As endo-β-mananases de fungos prontamente hidrolisam polímeros com
alto grau de substituição por galactose e a presença de trissacarídeos 61-α-D-
galactopiranosídeo-(1,4)-β-D-manobiose no hidrolisado indica que estas
enzimas são capazes de hidrolisar alguns sítios de clivagem, onde um único
resíduo de manose não está substituído (McCleary & Matheson, 1983).
Em contraste, as endo-β-mananases de bactérias, animais e plantas
(proveniente de sementes germinadas de leguminosas) possuem uma ação
mais limitada em galactomananos com muitas substituições, onde somente
uma quantidade traço de trissacarídeo é liberada, mostrando que a ramificação
com galactose bloqueia alguns sítios disponíveis de clivagem pela endo-β-
mananase (McCleary & Matheson, 1983).
Quando estes mesmos autores compararam a diversidade e quantidade
de oligossacarídeos produzidos pela ação das endo-β-mananases de fontes
variadas sobre galactomanano de goma caroba, notaram que a enzima de
Aspergillus niger formou seis oligossacarídeos simples (F1), enquanto a de

50
Cyamopsis tetragonologa (guar) formou dez oligossacarídeos simples (F1),
evidenciando que a endo-β-mananase de plantas possui uma ação mais
limitada na clivagem do polissacarídeos altamente ramificado.
Apesar da diferença na diversidade dos oligossacarídeos, os
oligossacarídeos comuns às enzimas possuíam a mesma estrutura, indicando
que a enzima endo-β-mananase cliva os mesmos tipos de ligações, mas possui
uma especificidade diferenciada dependo da fonte (McCleary & Matheson,
1983).
As endo-β-mananases são capazes de hidrolisar oligossacarídeos de
tamanho mínimo de três resíduos de manose, no entanto, a taxa de hidrólise
aumenta consistentemente com oligossacarídeos com 5 a 6 resíduos,
indicando o número de monômeros envolvidos na afinidade máxima da enzima
pelo substrato, ou seja, a endo-β-mananase possui 5 subsítios de
reconhecimento da cadeia de manose (McCleary & Matheson, 1983).
A figura 5 representa um oligossacarídeo hipotético (cada resíduo de
manose identificado de A-E) e os 5 subsítios de reconhecimento da enzima (α-
ε), estando a extremidade redutora do oligossacarídeo a direita. De acordo,
com os dados apresentados por McCleary & Matheson (1983), a quebra do
oligossacarídeo com 5 manoses (hipotético) ocorreria sempre entre as
manoses C e D.
Figura 5: Representação esquemática dos subsítios de reconhecimento (α – ε) e ligação da endo-β-
mananase ao oligossacarídeo hipotético com 5 resíduos de manose (A-E) ligados β-(1,4). Figura
retirada de McCleary & Matheson, 1983.
A endo-β-mananase de Aspergillus niger é capaz de clivar entre as
manoses C e D mesmo havendo ramificações por galactose nas manoses A, C
ou E, porém a endo-β-mananase de Cyamopsis tetragonoloba fica bloqueada
caso exista uma substituição por galactose na manose C, novamente

51
mostrando que as enzimas de plantas possuem mais requisitos para a
clivagem e, portanto, apresentam uma maior diversidade de oligossacarídeos
(McCleary & Matheson, 1983). Além disso, evidencia a importância da co-
atuação da α-galactosidase e da endo-β-mananase para a completa
degradação do galactomanano de reserva, indicando que possivelmente as
ramificações com galactose possuam uma ação modulatória na endo-β-
mananase.
1.9. Espécie Sesbania virgata
Sesbania virgata (Cav.) Pers. está entre as espécies que acumulam
galactomanano em suas sementes. Essa espécie é pertencente à família
Leguminosae (= Fabaceae), subfamília Faboideae (= Papilionoideae), tribo
Robinieae (Kissman & Groth, 1999).
A planta é nativa do sul da América do Sul, abrangendo os seguintes
países: Paraguai, Uruguai, Argentina e Brasil. Neste último país, Sesbania
virgata (Cav.) Pers. ocorre nas regiões Sul, Sudeste e Centro-Oeste (Kissman
& Groth, 1999), normalmente associada a matas de galeria (Lisboa et al.,
2006). Esta espécie é infestante, perene, reproduzida por semente, de hábito
arbustivo, atingindo de 2 a 4 metros de altura, 25 cm de diâmetro a altura do
peito e 5 metros de copa. Possui flores pequenas e amarelas. Seus frutos
consistem em legumes indeiscentes de cor marrom opaco quando maduros,
corticosos, subarticulados, com pseudosseptos, com brusco afinamento tanto
da base, como da extremidade, sendo esta pontiaguda. Os frutos possuem de
4-8 sementes reniformes, que são lateralmente um pouco compridas, com
tegumento relativamente espesso e embrião axial curvo (Kissman & Groth,
1999; Araujo et al., 2004).
1.9.1. Justificativa para o uso da espécie
As sementes de Sesbania virgata (Cav.) Pers. são um modelo
conveniente para o estudo da mobilização de reserva de galactomananos
devido: ao grande número de sementes produzidas por indivíduo, propiciando
uma grande quantidade de material para execução de experimentos; à alta

52
viabilidade das sementes, mesmo após armazenamento por anos, podendo ser
embebidas a qualquer momento para a realização de experimentos; ao
pequeno período de tempo necessário entre a embebição ao estabelecimento
da plântula, cerca de 7 dias, permitindo a execução de muitos experimentos em
um curto período de tempo; à fácil manipulação das sementes, pois são
pequenas para a embebição e germinação em placas de Petri, ao mesmo
tempo que são grandes o suficiente para manipulação individual com mãos
nuas; à grande regularidade de comportamento observada em diversas teses e
experimentos já realizados pelo grupo; e, principalmente, à bagagem de
conhecimento desenvolvida pelo grupo na maioria dos aspectos que
concernem à mobilização de reservas: desde a localização in situ da produção
das enzimas, à purificação da endo-β-mananase e regulação do processo, até
as características do galactomanano de reserva.

53
OBJETIVOS
O presente trabalho teve como objetivo a semipurificação e a
caracterização bioquímica da enzima α-galactosidase presente em sementes
de Sesbania virgata (Cav.) Pers., associado ao estudo da estrutura fina do
polissacarídeo de reserva (galactomanano) de sementes de Sesbania virgata
(Cav.) Pers., visando determinar a interação enzima-substrato. Para tanto
buscou-se:
Avaliar a taxa de germinação das sementes, bem como, a máxima
atividade enzimática da α-galactosidase de Sesbania virgata (Cav.)
Pers. durante a germinação e estabelecimento da plântula, através de
incubação das sementes por 7 dias consecutivos em câmaras de
germinação com acompanhamento da protrusão da radícula e atividade
enzimática a cada 24 horas;
Purificar a α-galactosidase presente em extrato bruto enzimático de
sementes de Sesbania virgata (Cav.) Pers., através de precipitação com
sulfato de amônio, seguido de Cromatografia de Troca-Iônica em coluna
Aniônica (Mono-Q – Pharmacia Biotech.) em sistema FPLC e
Cromatografia de Gel Filtração (Superose 6 – GE Healthcare®) em
sistema FPLC;
Caracterizar bioquimicamente a α-galactosidase de Sesbania virgata
(Cav.) Pers., em extrato bruto e após purificação, quanto a pH e
temperatura ótimos de atividade enzimática e aspectos cinéticos,
através de ensaios enzimáticos variando o pH, temperatura e
concentração do substrato colorimétrico 4-Nitrophenil-α-D-
Galactopiranosídeo (Sigma®);
Analisar sequências gênicas de α-galactosidase de plantas depositadas
em bancos de dados, através de comparação e análise filogenética
(construção de árvore de relação);
Avaliar a estrutura fina do polissacarídeo de reserva (galactomanano),
através da análise do perfil, em HPAEC-PAD, de oligossacarídeos
produzidos após hidrólises enzimáticas do galactomanano de Sesbania

54
virgata (Cav.) Pers. com endo-β-mananase de Aspergillus niger
(Megazyme®) somente e associada à ação da α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. ou de Cyamopsis
tetragonoloba (Megazyme®).
Avaliar a estrutura fina do polissacarídeo de reserva (galactomanano),
através da comparação do perfil, em HPAEC-PAD, dos oligossacarídeos
produzidos após hidrólises enzimáticas de (galacto)mananos de reserva
de sementes de 6 espécies (proporção manose:galactose variando de
1:1 a 150:1) com endo-β-mananase de Aspergillus niger (Megazyme®).

55
CAPÍTULO 1
SEMIPURIFICAÇÃO E CARACTERIZAÇÃO ENZIMÁTICA DA ENZIMA α-GALACTOSIDASE PRESENTE EM SEMENTES DE
Sesbania virgata (CAV.) PERS.

56
I. Resumo
Sementes de Sesbania virgata (Cav.) Pers. acumulam suas reservas de
carbono no endosperma na forma de um polissacarídeo de parede celular,
o galactomanano. A mobilização deste ocorre após a germinação e envolve
três enzimas hidrolíticas (α-galactosidase, endo-β-mananase e exo-β-
manosidase). A α-galactosidase é a primeira enzima a atuar sobre o
galactomanano, hidrolisando as ligações α-(1,6) das galactoses ramificadas
à cadeia principal de manano (ligados β-1,4), permitindo a ação da endo-β-
mananase, que hidrolisará o polissacarídeo a oligossacarídeos, onde a β-
manosidase atuará (ligações β-1,4), transformando oligossacarídeos em
monossacarídeos a serem utilizados no desenvolvimento do embrião. A α-
galactosidase de Sesbania virgata (Cav.) Pers., uma leguminosa nativa,
presente no endosperma e tegumento da semente, foi semipurificada,
caracterizada quanto à temperatura ótima, pH ótimo, aspectos cinéticos (Km
e Vmáx), sequência interna, e comparada a Cyamopsis tetragonoloba (guar),
quanto as suas características bioquímicas e a 35 sequências de
aminoácidos de α-galactosidases de plantas já descritas na literatura,
quanto à similaridade na sequência de aminoácidos. Uma curva de tempo
de germinação e desenvolvimento da plântula mostrou germinação em 24
horas após embebição com atividade enzimática máxima da α-
galactosidase em 96 horas após a embebição (4° dia). A purificação da
enzima foi conduzida por 3 etapas: precipitação com Sulfato de Amônio
40% e 80%, Cromatografia de Troca-Iônica Aniônica em FPLC utilizando
coluna Mono-Q e Cromatografia de Gel Filtração em FPLC utilizando coluna
Superose 6. A α-galactosidase semipurificada possui, aproximadamente, 42
kDa de peso molecular em condições desnaturantes e, aproximadamente
72 kDa de peso molecular na forma nativa, sugerindo que a enzima assuma
estrutura quartenária. A temperatura ótima apresentada se encontra na
faixa de 50°C a 55°C, pH ótimo na faixa de 4,4 a 5,4, Km= 1,8276 mM e a
velocidade máxima de 0,5024 µmolGal.min-1.mgprot-1. Essas características
diferem daquelas encontradas para a α-galactosidase de Cyamopsis
tetragonoloba. A espectrometria de massas gerou os fragmentos:

57
ALADYV-HSK-RMPGSLGHEE-QDAK-TT-GDIEDNWNSM-TSIADSNDK
W-ASYAGPGGWN-DPDMLEVGNG-GMTTEEYR-AP-LLVGCDIR-VAVIL-
WNR, que fazem referência a 22% da proteína e possuem alta similaridade
com a sequência depositada de Cyamopsis tetragonoloba, estando
relacionada à mobilização de reserva. Durante a purificação e
sequenciamento interno da α-galactosidase foram detectadas isoformas da
α-galactosidase de pesos moleculares variados (42 kDa a 20 kDa). Sugere-
se que estas isoformas encontradas após purificação em Cromatografia de
Troca-Iônica Aniônica em FPLC utilizando coluna Mono-Q estejam
relacionadas com outras funções da α-galactosidase, enquanto as
isoformas encontradas após Cromatografia de Gel Filtração em FPLC
utilizando coluna Superose 6 e identificação por espectrometria de massas
estejam relacionadas com ativação e adaptação da α-galactosidase durante
todo o processo de mobilização de reservas. A presença de isoformas e a
possível estrutura quartenária da enzima sugerem que a α-galactosidase de
Sesbania virgata (Cav.) Pers. forme complexos enzimáticos para ativação
de α-galactosidases inativas, através de proteólise, no início da mobilização
de reserva e também forme complexos enzimáticos para a adaptação da α-
galactosidase no processo de mobilização de reserva, também por
proteólise (diminuição do peso molecular), onde as ramificações com a
galactose se tornariam raras, o que explicaria a gama de pesos moleculares
apresentados pelas diferentes isoformas (sequenciamento interno). Essas
evidências sugerem que no início da mobilização de reserva proteases
sejam transcritas e atuem sobre α-galactosidases inativas, ativando-as;
essas α-galactosidases ativas seriam as responsáveis pela ativação das α-
galactosidases inativas restantes e adaptação das α-galactosidases ativas
durante o processo de mobilização, também por proteólise, permitindo que
a enzima apresente características melhoradas para hidrolisar
eficientemente as ramificações por galactose cada vez mais raras no
decorrer do processo de mobilização. A α-galactosidase semipurificada foi
submetida, juntamente com a α-galactosidase de Cyamopsis tetragonoloba
(Megazyme®), a ensaios enzimáticos com o substrato galactomanano,
visando determinar o modo de ação da enzima e a estrutura fina do
polissacarídeo (dados apresentados no Capítulo 2).

58
II. Introdução
Uma das mais espetaculares inovações que surgiram durante a
evolução das plantas vasculares (Angiospermas e “Gimnospermas”) foi a
semente. Este sucesso deve-se ao fato das sementes possuírem uma grande
capacidade de sobrevivência quando comparada com esporos livres, devido à
proteção que esta confere ao embrião, além dos recursos nela armazenados
para os estágios de germinação e estabelecimento (Raven, 2001).
Sementes de diversas espécies de leguminosas acumulam
galactomananos na parede celular dos endospermas, sendo denominados
polissacarídeos de reserva de parede celular (PRPC) (Reid, 1985; Buckeridge
& Reid, 1996; Buckeridge et al. 2000b; Buckeridge et al., 2004a).
Os polissacarídeos de reserva, além da função de armazenar e prover
energia para o embrião, também desempenham um papel importante no
controle da embebição de água no início da germinação, absorvendo grande
quantidade de água e a distribuindo ao redor do embrião, protegendo-o contra
dessecação (Reid & Bewley, 1979; Buckeridge et al., 2000c).
Outras funções já foram descritas para os polissacarídeos de reserva de
parede celular: dureza e resistência física ao endosperma no período de
germinação, sendo importante na regulação da constrição mecânica à
protrusão da radícula (Buckeridge et al., 2000a; Lisboa et al., 2006).
Os polissacarídeos de parede são classificados em três grupos:
mananos, xiloglucanos e os (arabino)galactanos. O grupo dos mananos
engloba os mananos puros, glucomananos e galactomananos (Buckeridge et
al., 2000c).
Os galactomananos são polissacarídeos hemicelulósicos constituídos de
uma cadeia principal formada por unidades de D-manose unidas entre si por
ligações glicosídicas β-(1,4), na qual unidades de D-galactose formam
ramificações simples ligadas α-(1,6) (Moe et al., 1947; Unrau, 1961; Somme,
1968; Manzi & Cerezo, 1984).
A mobilização do galactomanano de reserva de sementes de
leguminosas já descritas se inicia após a germinação (Buckeridge et al., 2000c;
Tonini et al., 2006), sendo realizada pela ação de três enzimas hidrolíticas: α-

59
galactosidase (EC 3.2.1.22), endo-β-mananase (EC 3.2.1.78) e exo-β-
manosidase (EC 3.2.1.25), necessariamente nesta ordem (Reid & Meier, 1972;
McCleary & Matheson, 1976; Buckeridge & Dietrich, 1996), pois a presença das
ramificações de D-galactose na cadeia principal interferem na ação da endo-β-
mananase sobre a cadeia de manano (Reid & Edwards, 1995; Buckeridge et
al., 2000a; Lisboa et al, 2006).
As alfa-galactosidases (E.C. 3.2.1.22) catalisam a hidrólise de ligações
α-(1,6) de galacto-oligossacarídeos e de polímeros de galacto-(gluco)-
mananos. Essas enzimas estão amplamente distribuídas em animais, plantas e
microorganismos (Fujimoto et al., 2003).
A presença de isoformas de α-galactosidase foi primeiramente descrita
por Petek & Dong (1961) em sementes de Coffea sp e Plantago ovata. Tonini
et al. (2006) sugerem que a presença de isoformas de endo-β-mananase em
endosperma de Sesbania virgata (Cav.) Pers. seja para garantir a hidrólise do
galactomanano, mesmo com as modificações estruturais que este sofre
durante a mobilização.
Feurtado et al. (2001) sugeriram pelo menos três isoformas da α-
galactosidase durante e após a germinação de sementes de tomate e, através
da obtenção e sequenciamento do cDNA da enzima, demonstraram alta
homologia da forma ativa da α-galactosidase do tomate (40 kDa) com outras
galactosidases, especialmente com as de sementes de Coffea arabica (café)
(Zhu & Goldstein, 1994). Como apenas um gene para a α-galactosidase foi
identificado em sementes de tomate, as isoformas aparentemente se formam a
partir de modificações pós-traducionais (Feurtado et al., 2001).
McCleary & Matheson (1974) também detectaram múltiplas isoformas da
α-galactosidase, nas sementes de Cyamopsis tetragonoloba, embora apenas
uma delas (40 kDa) esteja associada com o endosperma (Overbeeke et al.
1989). DeMason & colaboradores (1992) detectaram quatro isoformas da
enzima durante o desenvolvimento do endosperma de sementes de palmeira.
Em sementes de alfafa, Cyamopsis tetragonoloba, Ceratonia siliqua, soja,
palmeira, café, feijão, Lupinus angustifolius (lupino) e Trifolium repens pelo
menos duas isoformas da α-galactosidase estão presentes, com massas
moleculares variando entre 17-57 kDa (Courtois & Petek 1966, McCleary &

60
Mathenson 1974, Williams et al. 1978, Itoh et al. 1979, Plant & Moore 1982,
Chandra Sekhar & DeMason 1990, Dhar et al. 1994).
As α-galactosidases também foram descritas relacionadas ao
amadurecimento de frutos. Soh et al. (2006) detectaram três isoformas da α-
galactosidase no fruto de Carica papaya (mamão papaia), com pesos
moleculares de 170, 57 e 29 kDa. Durante o desenvolvimento de frutos de
Cucumis melo (melão), três isoformas da α-galactosidase também foram
detectadas, uma delas com peso molecular em torno de 27 kDa (Gao &
Schaffer, 1999).
Assim, as diferentes isoformas de α-galactosidase podem estar
relacionadas tanto a diferenças estruturais do galactomanano durante a
mobilização, como por exemplo, à diminuição de substituições por galactose o
que demandaria uma α-galactosidase com maior afinidade ao substrato; como
também a diferentes funções além da degradação do galactomanano, por
exemplo, a degradação de açúcares da série rafinósica (Bewley,1997) e até o
amadurecimento de frutos (remodelamento da parede celular) (Gao & Schaffer,
1999; Soh et al., 2006).
A ação das enzimas relacionadas com a mobilização de reserva de
sementes é regulada por fito-hormônios. O ácido abscísio (ABA) é um potente
inibidor da degradação da reserva de galactomanano e está envolvido no
mecanismo de regulação dessa mobilização em sementes de Ceratonia siliqua
(Seller, 1977), Lactuca sativa (Halmer & Bewley, 1979), Lycopersicon
sculentum (Groot & Karssen, 1992), Trigonella foenum-graecum (Reid & Meier,
1973; Malek & Bewley, 1991; Kontos & Spyropoulos, 1995) e Sesbania virgata
(Cav.) Pers. (Potomati & Buckeridge, 2002; Tonini et al., 2006).
Tonini (2009) demonstrou que a mobilização de reserva de
galactomanano de sementes de Sesbania virgata (Cav.) Pers. é modulada
negativamente pela presença do ABA, enquanto o etileno é promotor da
mobilização.
Além da regulação direta realizada pelos fito-hormônios, a degradação
da reserva pode ser regulada pela ação de proteases, que normalmente são
moduladas pelos fito-hormônios (Tonini, 2009). Sabe-se que proteases são
responsáveis por modificações pós-traducionais de proteínas, com ação
proteolítica limitada a regiões específicas, necessárias para o processamento e

61
maturação de enzimas pré-formadas e controle da atividade destas (Schaller,
2004). Em sementes de Cyamopsis tetragonoloba (guar) e cevada, a enzima
α-galactosidase e β-amilase, respectivamente, podem ser ativadas através de
digestão por enzimas proteolíticas (Overbeeke et al. 1989, Guerin et al. 1992).
Tendo em vista a importância da α-galactosidase na mobilização de
reserva de galactomanano e visando a compreensão das suas características e
modo de ação sobre o galactomanano, procedeu-se com a purificação e
caracterização bioquímica da α-galactosidase de sementes de Sesbania virgata
(Cav.) Pers.
III. Material e Métodos
3.1. Coleta do Material Vegetal
As sementes de Sesbania virgata (Cav.) Pers. utilizadas na execução
dos experimentos de purificação e caracterização foram coletadas de diversos
indivíduos localizados nas imediações da Universidade de São Paulo, São
Paulo/SP, evitando, assim, vício amostral. As sementes foram separadas dos
frutos maduros e mantidas em frascos herméticos a temperatura ambiente.
As sementes coletadas foram utilizadas na preparação do extrato bruto
enzimático visando à semipurificação da enzima α-galactosidase presente nas
sementes de Sesbania virgata (Cav.) Pers.
3.2. Determinação da porcentagem de germinação e pico de
atividade enzimática
A fim de determinar a viabilidade das sementes coletadas e o momento
(em dias) após a embebição, onde se encontra a maior atividade enzimática da
α-galactosidase, procedeu-se com a embebição das sementes e
acompanhamento da germinação e desenvolvimento da plântula por 7 dias sob
luz e temperatura constantes, seguido de preparação de extrato enzimático e
mensuração da atividade enzimática.

62
3.2.1. Germinação das sementes quiescentes
Após o beneficiamento dos frutos, as sementes quiescentes foram
escarificadas mecanicamente com lixa na região abaixo do hilo e,
posteriormente, esterilizadas com uma solução de hipoclorito de sódio a 0,75%
por 15 minutos, seguido por enxágue com água destilada.
Em seguida, as sementes quiescentes foram incubadas em placas de
Petri (10cm de diâmetro) sobre papel de filtro, 10 sementes por placa e 0,5mL
de água destilada/semente (5mL), sendo mantidas sob condições de luz
constante e temperatura de 28°C, durante 7 dias (período que abrange as
fases de germinação e degradação total da reserva - galactomanano).
A cada 24 horas de incubação a partir da embebição (168 horas - 7 dias)
3 placas de Petri, contendo 10 sementes por placa, foram retiradas da
incubação e processadas individualmente.
Essas sementes coletadas, cada grupo de 10 sementes, foram
dissecadas, separando o tegumento e o endosperma do embrião (descartado)
e os tecidos submetidos à mensuração do peso fresco, extração de proteínas
mensuração da atividade enzimática, descritos a seguir.
3.2.2. Determinação do peso fresco do endosperma e tegumento
Cada grupo de 10 sementes foi pesado (peso fresco total), em seguida,
determinado a taxa de germinação através da observação da emissão da
radícula (total de sementes com emissão de radícula/total de sementes por
placa), as sementes foram dissecadas para separação do tegumento e
endosperma, os tegumentos e endospermas foram novamente pesados (peso
fresco sem embrião) e finalmente foram submetidos à extração de proteínas,
visando à determinação da atividade enzimática.

63
3.2.3. Extração protéica, determinação de proteínas totais, determinação
da atividade enzimática da α-galactosidase e eletroforese em gel de
poliacrilamida (SDS-PAGE)
Os tecidos isolados, endosperma e tegumento, das sementes coletadas
(10 sementes a cada dia, com 3 repetições) foram macerados usando
nitrogênio líquido, homogeneizados em tampão Tris-HCl 20mM pH 7.8 e
2g/100mL de Polivinilpirrolidona (PVPP) e deixados em repouso por 1 hora a
5°C. Após repouso, o extrato foi submetido à centrifugação a 10.000g, a 5°C
por 10 minutos e o sobrenadante (extrato bruto enzimático) foi submetido à
dosagem de proteínas e da atividade enzimática da α-galactosidase.
O conteúdo de proteínas totais em cada amostra foi determinado pelo
método de Bradford de quantificação de proteínas (Bradford,1976), utilizando
como padrão Albumina de Soro Bovino (BSA). Trinta microlitros de amostra
(podendo estar diluída 1:10) foram homogeneizados a 170µL de reagente de
Bradford diluído 4:1 (4 partes de reagente de Bradford para 1 parte de água
Milli-Q®). As leituras de absorbâncias foram feitas em leitor de placa Elisa no
comprimento de onda (λ) de 595nm.
O pico de atividade enzimática, dia após embebição onde se encontra a
maior atividade enzimática, foi determinado através de ensaio enzimático
utilizando as réplicas de cada coleta e o substrato enzimático 4-Nitrophenil-D-
galactopiranosídeo (Sigma Chem. Co. St. Louis, USA) (PNP-α-Gal), seguido de
leitura em espectrofotômetro a 405nm, de acordo com Alcântara et al.(1999).
As três réplicas de cada horário (24h a 168h) foram ensaiadas
separadamente, seguindo o seguinte protocolo: 10µL de tampão McIlvaine pH
5.0, 10µL de PNP-α-Gal a 50mM e 10µL de extrato bruto (proveniente de cada
réplica de cada horário). Após homogeneização, os tubos foram incubados a
45°C por 15 minutos e, em seguida, a reação foi interrompida com 2mL de
carbonato de sódio a 0,1N e a absorbância determinada a 405nm.
A atividade enzimática específica foi determinada através da seguinte
equação:

64
Onde, Constante é o coeficiente de extinção molar do PNP-α-
galactopiranosídeo; VTE é o volume total de ensaio (2,03mL); VTEB é o
volume total de extrato bruto em mL; VEE é o volume de enzima utilizada no
ensaio (0,01mL); t é o tempo do ensaio enzimático (15 minutos); TPEB é
proteína total (mg) do extrato bruto.
A análise qualitativa das proteínas foi feita através de eletroforese em
gel de poliacrilamida sob condições desnaturantes, em presença de dodecil
sulfato de sódio (SDS-PAGE), empregando-se um sistema descontínuo,
baseado no método descrito por Laemmli (1970).
As eletroforeses foram feitas em mini-géis de 0,75 mm de espessura (9
X 6cm), contendo 12% de acrilamida, montados em cuba vertical de
eletroforese modelo mini-protean (Bio-Rad). Durante a corrida foi mantida
corrente constante de 100 mA por cerca de 70 minutos, tempo necessário para
o marcador alcançar a extremidade inferior do gel.
O marcador de peso molecular utilizado foi Kaleidoscope Prestained
Standard (Bio-Rad) que contém: miosina (195 kDa), β-galactosidase (111,6
kDa), albumina de soro bovino (59,4 kDa), anidrase carbônica (30 kDa), inibidor
de tripsina de soja (24,7 kDa), lisozima (12,5 kDa) e aprotinina (6,4 kDa).
Para revelação das proteínas, após a corrida, o gel foi imerso em
solução de Coomassie Brilliant Blue R250 0,1% dissolvido em ácido
acético:metanol:água (1:4:5 - v/v/v), durante 1 hora a temperatura ambiente e
sob agitação contínua. Em seguida, o gel foi mantido em solução descorante,
ácido acético:metanol:água (1:4:5 - v/v/v), sob agitação contínua e trocas
frequentes da solução (Hames, 1990).

65
3.3. Purificação da α-galactosidase de sementes de Sesbania
virgata (Cav.) Pers.
3.3.1. Incubação das sementes de Sesbania virgata para purificação da α-
galactosidase
Com a finalidade de obter extrato bruto enzimático visando à
caracterização em extrato bruto e purificação da α-galactosidase presente em
sementes de Sesbania virgata (Cav.) Pers., aproximadamente 1000 sementes
de Sesbania virgata (Cav.) Pers. foram escarificadas mecanicamente e
esterilizadas com hipoclorito de sódio 0,75%, como descrito no item 3.2.1.
Germinação das sementes quiescentes.
As 1000 sementes foram incubadas em 4 bandejas sobre papel de filtro
por 96 horas após embebição (4 dias após embebição) sob luz constante e
temperatura de 28°C.
Ao final das 96 horas, as sementes foram pesadas (Peso Fresco Total),
dissecadas para separação do tegumento e endosperma, e estes foram
pesados para determinação do Peso fresco sem embrião.
O extrato bruto foi realizado a partir dos tegumentos e endospermas
dissecados, seguindo o mesmo protocolo anteriormente descrito no item 3.2.1.
Germinação das sementes quiescentes, resultando em um volume final de
156mL de extrato bruto disponível para purificação da α-galactosidase e
caracterização do extrato bruto.
O extrato bruto foi submetido à determinação das proteínas totais, pelo
método de Bradford (Bradford, 1976), e determinação da atividade enzimática
da α-galactosidase, ensaio enzimático com PNP-α-Gal, seguindo o mesmo
protocolo anteriormente descrito no item 3.2.1. Germinação das sementes
quiescentes.

66
3.3.2. Caracterização da temperatura ótima e pH ótimo da α-
galactosidase em extrato bruto enzimático
A fim de padronizar o ensaio enzimático da α-galactosidase para as
etapas de purificação e caracterizar esta enzima quando no extrato bruto,
procedeu-se com a determinação da temperatura ótima e pH ótimo de
atividade enzimática da α-galactosidase.
3.3.2.1. Temperatura Ótima
Para determinar a temperatura ótima da α-galactosidase em extrato
bruto, amostras de 10µL de extrato bruto foram incubadas juntamente com
10µL de PNP-α-galactopiranosídeo e 10µL de tampão McIlvaine pH 5.0 em
temperaturas crescentes: 20, 25, 30, 35, 40, 45, 50, 55 e 60°C, por 15 minutos,
seguido de acréscimo de 2mL de Na2CO3 a 0,1N e leitura a 405nm, como
descrito no item 3.2.1. Germinação das sementes quiescentes.
3.3.2.2. pH Ótimo
Para determinar o pH ótimo da α-galactosidase em extrato bruto,
amostras de 10µL de extrato bruto foram incubadas juntamente com 10µL de
PNP-α-Gal e 10µL de tampão McIlvaine variando de pH 2 a 7 (2.6; 3.0; 3.4; 4.0;
4.4; 5.0; 5.4; 6.0; 6.4 e 7.0) (McIlvaine, 1921) ou 10µL de tampão Fosfato para
pH 7.5 e 8.0, por 15 minutos, seguido de acréscimo de 2mL de Na2CO3 a 0,1N
e leitura a 405nm, de acordo com a metodologia descrita no item 3.2.1.
Germinação das sementes quiescentes.
3.3.3. Primeira Etapa de Purificação da α-galactosidase: Precipitação com
Sulfato de Amônio
Primeiramente, foi realizada uma precipitação com sulfato de amônio
com saturações de 20%, 40%, 60% e 80% visando padronizar o protocolo e
determinar a saturação, na qual a enzima α-galactosidase, é precipitada.

67
Partiu-se de 144mL de extrato bruto de tegumento e endosperma de
Sesbania virgata (Cav.) Pers., realizado como descrito no item 3.2.1.
Germinação das sementes quiescentes, acrescentou-se a massa de sulfato de
amônio referente a saturação de 20%, após homogeneização, a mistura foi
deixada a 5°C por 40 minutos, seguido de centrifugação a 13000g a 5°C por 15
minutos.
O precipitado, proteínas precipitadas com 20% de Sulfato de Amônio, foi
ressuspendido com tampão Tris-HCl 20mM pH7.8 e armazenado a -20°C até
ensaio enzimático, quantificação de proteínas pelo método de Bradford
(Bradford, 1976) utilizando BSA como padrão e eletroforese em gel de
poliacrilamida 12% (Laemmli, 1970) corado com Coomassie R250 a 0,1%
(dissolvido em ácido acético:metanol:água (1:4:5 - v/v/v)), utilizando como
padrão Kaleidoscope Prestained Standard (Bio-Rad).
Ao sobrenadante, após verificação de volume, foi acrescido massa de
sulfato de amônio referente à saturação de 40% e a mesma metodologia foi
aplicada até o acréscimo da massa referente a saturação de 80%, onde o
sobrenadante após centrifugação, denominado de descarte, foi armazenado a
-20°C até ensaio enzimático, quantificação de proteínas e eletroforese em gel
de poliacrilamida, e o precipitado de 80% também foi ressuspendido com
tampão Tris-HCl 20mM pH7.8 e armazenado a -20°C até ensaio enzimático,
quantificação de proteínas e eletroforese em gel de poliacrilamida.
Uma alíquota do extrato bruto, os precipitados de 20%, 40%, 60% e 80%
e o descarte foram submetidos a ensaio enzimático utilizando o substrato
sintético PNP-α-Gal, tampão McIlvaine pH4,4 a 45°C, como descrito no item
3.2.1. Germinação das sementes quiescentes, para determinação do
precipitado com maior atividade de α-galactosidase, e foram submetidos à
eletroforese em gel de poliacrilamida 12% (Laemmli, 1970) corado com
Coomassie R250 a 0,1% (dissolvido em ácido acético:metanol:água (1:4:5 -
v/v/v)), utilizando como padrão Kaleidoscope Prestained Standard (Bio-Rad).
Após este experimento inicial de precipitação com sulfato de amônio
seguido de ensaio enzimático optou-se por utilizar somente as saturações de
40% e 80% seguindo o protocolo descrito.
Dessa forma, os 156mL de extrato bruto, destinados a purificação, foram
submetidos a saturação de 40% de sulfato de amônio, resultando em 16,5mL

68
de precipitado ressuspendido em tampão Tris-HCl 20mM pH 7.8, que foram
congelados a -20°C até ensaio enzimático. O sobrenadante foi submetido a
saturação de 80% de sulfato de amônio resultando em 13.5mL de precipitado
ressuspendido em tampão Tris-HCl 20mM pH 7.8 e 320mL de descarte, ambos
congelados a -20°C até ensaio enzimático, quantificação de proteínas e a
próxima etapa de purificação.
Alíquotas dos precipitados de 40% e 80% ressuspendidos em tampão
Tris-HCl 20mM pH 7.8 e do descarte foram submetidas a ensaios enzimáticos
e quantificação de proteínas como descrito no item 3.2.1. Germinação das
sementes quiescentes, visando confirmar a atividade enzimática da α-
galactosidase e determinar qualquer perda nesta atividade enzimática pelo
protocolo de precipitação com sulfato de amônio, quando comparado com a
atividade enzimática determinada no extrato bruto.
Todo o volume do precipitado de 80% ressuspendido em tampão Tris-
HCl 20mM pH7.8 foi destinado a segunda etapa de purificação.
3.3.4. Segunda Etapa de Purificação da α-galactosidase: Cromatografia de
Troca-Iônica com Coluna Aniônica (Mono-Q)
Inicialmente 5 mL do precipitado de 80% ressuspendido em Tris-HCl
20mM pH7.8 foi dialisado por 24 horas, com trocas a cada 2 horas, contra o
mesmo tampão (tampão Tris-HCl 20mM pH7.8) visando retirar todo o sulfato de
amônio diluído na amostra.
Em seguida, a amostra dialisada foi concentrada em concentrador de
amostras Amicon® Ultra-4 Centrifugal Units (Millipore) e, posteriormente,
submetida à coluna de dessalinização P-10 Desalting Colunm (GE Healthcare)
para garantir completa retirada de sulfato de amônio.
Finalmente, a amostra foi submetida à Cromatografia de Troca-Iônica
Aniônica utilizando FPLC ÄKTA (GE Healthcare), com coluna Aniônica, Mono-
Q HR 10/10 (Pharmacia Biotech), tendo como eluentes: tampão Tris-HCl 20mM
pH7.8 e tampão Tris-HCl 20mM pH7.8 com 0.5M de NaCl.
A purificação da α-galactosidase ocorreu através da seguinte
metodologia em FPLC ÄKTA: equilíbrio da coluna com 5 volumes de coluna
(40mL) com tampão Tris-HCl 20mM pH7.8, seguido de injeção da amostra na

69
coluna, lavagem da coluna com 2 volumes de coluna (8mL) com tampão Tris-
HCl 20mM pH7.8 para retirar o pool de proteínas que não interagiram com a
coluna, em seguida, foi iniciado o gradiente de NaCl variando de 0M a 0.5M em
12 volumes de coluna (96mL) para liberação das proteínas gradualmente
dependendo de sua interação com a coluna, finalmente a concentração de
0.5M de NaCl foi mantida por mais 5 volumes de coluna (40mL) para limpeza
da coluna.
A atividade enzimática da α-galactosidase, nas frações de 1mL
coletadas durante o processo de purificação, foi monitorada com PNP-α-D-
galactopiranosídeo, tampão McIlvaine pH4,4 a 45°C, conforme descrito no item
3.2.1. Germinação das sementes quiescentes.
As frações que apresentaram atividade enzimática foram reunidas, uma
alíquota (50µL) sempre armazenada a -20°C e o restante submetido à
eletroforese em gel de poliacrilamida com dodecil sulfato de sódio (SDS-
PAGE), visando o acompanhamento da purificação da α-galactosidase, ou
submetido a experimentos para desenvolvimento da tabela de purificação, ou
submetido à terceira etapa de purificação.
3.3.5. Terceira Etapa de Purificação da α-galactosidase: Cromatografia de
Gel Filtração (Superose)
Primeiramente, a amostra proveniente da cromatografia aniônica foi
concentrada em concentrador de amostras Amicon® Ultra-4 Centrifugal Units
(Millipore) e, posteriormente, submetida à coluna de dessalinização P-10
Desalting Colunm (GE Healthcare) para troca do tampão Tris-HCl 20mM pH 7.8
para tampão acetato de amônio 50mM pH5.0.
A amostra foi submetida à Cromatografia de Gel filtração utilizando
FPLC ÄKTA (GE Healthcare), com coluna Superose 6 (GE Healthcare), tendo
como eluente: tampão acetato de amônio 50mM pH5.0.
A purificação da α-galactosidase ocorreu através da seguinte
metodologia em FPLC ÄKTA: equilíbrio da coluna com 4 volumes de coluna
(100mL) de água deionizada Milli-Q, seguido de equilíbrio com 2 volumes de
coluna (50mL) de tampão acetato de amônio 50mM pH5.0, em seguida, a

70
amostra foi injetada na coluna, sendo mantido um fluxo de acetato de amônio
50mM pH5.0 de 0.3mL/minuto até ausência de detecção de proteína por UV.
A atividade enzimática da α-galactosidase, nas frações de 0,5mL
coletadas durante o processo de purificação, foi monitora com PNP-α-D-Gal,
tampão McIlvaine pH 4,4 a 45°C, conforme descrito no item 3.2.1. Germinação
das sementes quiescentes.
As frações que apresentaram atividade enzimática foram reunidas e uma
alíquota (20µL) sempre foi armazenada a -20°C, sendo o restante dos 2 mL
submetidos à eletroforese em gel de poliacrilamida com dodecil sulfato de
sódio (SDS-PAGE) ou submetidos a experimentos de caracterização
enzimática e desenvolvimento da tabela de purificação ou submetidos ao
sequenciamento interno da proteína ou submetidos aos experimentos com
polissacarídeo – galactomanano (Capítulo 2).
Salienta-se que para a realização dos diversos experimentos, a
purificação foi realizada mais de uma vez, sempre partindo do mesmo volume
inicial do precipitado de 80% ressuspendido e seguindo a mesma metodologia
descrita.
3.3.6. Tabela de Purificação
Para o desenvolvimento da tabela de purificação todo o processo de
purificação, do extrato bruto à purificação em coluna Superose 6 em FPLC, foi
realizado com o armazenamento de alíquotas em cada etapa de purificação
(extrato bruto, precipitação com sulfato de amônio 80%, Mono-Q e Superose).
Cada alíquota de cada etapa foi submetida a ensaio enzimático com
PNP-α-Gal e quantificação de proteínas totais pelo método de Bradford
(Bradford, 1976), como descrito no item 3.2.1. Germinação das sementes
quiescentes.
Após coleta dos dados foram realizados cálculos para a determinação
do fator de purificação e rendimento da purificação.

71
3.3.7. Caracterização da α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers e da α-galactosidase de Cyamopsis
tetragonoloba (guar) (Megazyme®) quanto a temperatura ótima,
pH ótimo e aspectos cinéticos
A enzima α-galactosidase semipurificada de Sesbania virgata (Cav.)
Pers. proveniente de uma das purificações (seguindo os três passos de
purificação) e a α-galactosidase de guar (Megazyme®) foram caracterizadas
quanto aos aspectos de temperatura ótima, pH ótimo e aspectos cinéticos, Km
e Vmáx.
3.3.7.1. Temperatura ótima
Para determinar a temperatura ótima da α-galactosidase semipurificada
e da α-galactosidase de guar (Megazyme®), alíquotas de 5µL de amostra foram
incubadas juntamente com 5µL de PNP-α-Gal e 5µL de tampão McIlvaine pH
4,4 em temperaturas crescentes: 20, 25, 30, 35, 40, 45, 50, 55 e 60°C, por 15
minutos, seguido de acréscimo de 1mL de Na2CO3 a 0.1N e leitura a 405nm,
como descrito no item 3.2.1. Germinação das sementes quiescentes. Para
cada temperatura o ensaio enzimático foi realizado em triplicata.
3.3.7.2. pH ótimo
Para determinar o pH ótimo da α-galactosidase semipurificada e da α-
galactosidase de guar (Megazyme®), alíquotas de 5µL de amostra foram
incubadas juntamente com 5µL de PNP-α-Gal e 5µL de tampão McIlvaine
variando de pH 2 a 7 (2.6; 3.0; 3.4; 4.0; 4.4; 5.0; 5.4; 6.0; 6.4 e 7.0) (McIlvaine,
1921) por 15 minutos a 45°C, seguido de acréscimo de 1mL de Na2CO3 a 0.1N
e leitura a 405nm, de acordo com a metodologia descrita no item 3.2.1.
Germinação das sementes quiescentes. Para cada pH o ensaio enzimático foi
realizado em triplicata.

72
3.3.7.3. Aspectos Cinéticos – Km e Vmáx
Para determinar os aspectos cinéticos da α-galactosidase semipurificada
e da α-galactosidase de guar (Megazyme®), alíquotas de 5µL de amostra foram
incubadas juntamente com 5µL de tampão McIlvaine pH 4.4 e 5µL de PNP-α-
Gal variando de 0,1mM a 30mM (0.1; 0.2; 0.3; 0.5; 0.8; 1.0; 2.0; 3.0; 5.0; 8.0;
10; 13; 15; 17; 20; 25; 30mM) por 15 minutos a 45°C, seguido de acréscimo de
1mL de Na2CO3 a 0.1N e leitura a 405nm, de acordo com a metodologia
descrita no item 3.2.1. Germinação das sementes quiescentes. Para cada
concentração de substrato o ensaio enzimático foi realizado em triplicata.
3.3.8. Sequenciamento interno da α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers
O sequenciamento interno da α-galactosidase semipurificada foi
realizado no Laboratório de Espectrometria de Massas do Laboratório Nacional
de Biociências, CNPEM-ABTLuS.
Para a determinação de fragmentos da sequência interna da α-
galactosidase semipurificada, todo o processo de purificação, do extrato bruto à
purificação em coluna Superose 6 em FPLC, foi realizado com o
armazenamento de alíquotas em cada etapa de purificação (extrato bruto,
precipitação com sulfato de amônio 80%, Mono-Q e Superose).
Todo o material proveniente da purificação em Superose 6, bem como
as alíquotas de cada etapa, foram destinados à eletroforese em gel de
poliacrilamida 12% com dodecil sulfato de sódio (SDS-PAGE).
Oito bandas (proteínas) da amostra da purificação em Superose 6, entre
os pesos moleculares de 59,4 kDa e 12,5 kDa, foram recortadas do gel em
segmentos de 1mm com auxílio de um bisturi, armazenados em microtubos de
1,5 mL e submetidas a preparação para o sequenciamento interno.
Cada uma das oito bandas foi submetida a: descoloração, desidratação,
redução, alquilação, reidratação, digestão com tripsina e extração,
anteriormente à aplicação ao espectrômetro de massas. A metodologia de
cada uma das etapas de preparo está descrita a seguir.
Descoloração: os segmentos de gel contendo as proteínas foram
acrescidos de 0,5 mL da solução Destain (metanol 50%, ácido acético 2,5% em

73
água purificada) e incubados por 2 horas a temperatura ambiente. Após
descarte da solução, repetiu-se a etapa incubando somente por 1 hora.
Desidratação: a solução de Destain foi descartada e acrescentou-se aos
segmentos de gel 200 µL de acetonitrila 100%, incubou-se por 5 minutos e
descartou-se a solução. Esta etapa foi repetida e ao final o restante da
acetonitrila foi evaporado em evaporador por 2 a 3 minutos.
Redução: aos fragmentos de gel foi acrescido 30 µL de DTT 10 mM
(Ditiotreitol), incubado por 30 minutos a temperatura ambiente e descartada a
solução após centrifugação rápida (spin).
Alquilação: aos fragmentos de gel foi acrescido 30 µL de IAA 50mM
(Iodoacetamida), incubado por 30 minutos a temperatura ambiente e
descartada a solução após centrifugação rápida (spin). Os fragmentos de gel
foram lavados com 100µL de bicarbonato de amônio 100mM por 10 minutos e
descartada a solução. Os fragmentos de gel foram novamente desidratados
com 200µL de acetonitrila 100% por 5 minutos a temperatura ambiente.
Reidratação: os fragmentos de gel foram reidratados com 200µL de
bicarbonato de amônio 100mM por 10 minutos, descartada a solução após
centrifugação rápida (spin), novamente desidratados com 200µL de acetonitrila
100% por 5 minutos a temperatura ambiente. Esta última etapa foi repetida e
ao final o restante da acetonitrila foi evaporada em evaporador por 2 a 3
minutos.
Digestão das proteínas: aos fragmentos de gel for adicionados 30µL da
solução da tripsina 20ng/µL (20µg de tripsina em 1000µL de bicarbonato de
amônio 50mM gelado) e incubou-se por 30 minutos em gelo para a reidratação.
Centrifugou-se rapidamente (spin), retirou-se o excesso de tripsina, adicionou-
se 20µL de bicarbonato de amônio 50mM para cobrir o gel e incubou-se a 30°C
por 15 horas (overnight).
Extração: sem retirar a solução de bicarbonato de amônio 50mM
adicionou-se 10 µL de ácido fórmico 5% em água purificada, incubou-se por 10
minutos a temperatura ambiente e após centrifugação rápida (spin) armazenou-
se o sobrenadante em outro microtubo de 1,5 mL. Ao gel foi adicionado 12µL
ácido fórmico 5% em acetonitrila 50%, incubado por 10 minutos a temperatura
ambiente e após centrifugação rápida (spin) armazenou-se o sobrenadante no

74
tubo que fora previamente separado. Este último passo é repetido e ao final
todo o sobrenadante armazenado é evaporado até restar cerca de 5µL, o qual
é submetido ao espectrômetro de massas.
Os peptídeos de cada amostra, resultantes da digestão com tripsina,
foram analisados por LC-MSMS. Os peptídeos foram separados por gradiente
de hidrofobicidade em coluna C18 (100 µm x 100 mm) (Waters) em nano
Acquity Ultra Performance LC (Waters) acoplado a uma interface de ESI
nanospray em um espectrômetro de massa do tipo Q-Tof Ultima
(MicroMass/Waters). Volume de 4,5µL dos peptídeos foram injetados
automaticamente e os peptídeos foram eluídos da coluna com gradiente de 0-
90% de acetonitrila contendo ácido fórmico a 0,1% durante 40 min sob fluxo de
0,6 µL/min.
Os arquivos de dados gerados pelo Q-Tof foram processados em
programa Mascot Distiller v.2.3.2.0, 2009 (Matrix Science Ldt.). Para a
identificação das proteínas foi utilizado o programa Mascot Server v.2.3.01.0
(Matrix Science Ltda.), tendo como parâmetros uma clivagem perdida pela
tripsina, modificação fixa de carbamidometilação, modificação variável de
oxidação da metionina e 0,1 Da de tolerância de massas para MS e 0,1 Da de
tolerância de massas para MSMS. As buscas foram realizadas utilizando o
banco de dados Human International Protein Database (IPI) v. 3.72 (86392
sequências; 35093930 resíduos).
3.4. Análises das sequências de α-galactosidase previamente
identificadas em plantas
Visando determinar similaridades entre sequências gênicas codificantes
e de aminoácidos para α-galactosidase de plantas já depositadas em bancos
de dados públicos e a sequência interna da a-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers., foi realizada a análise das sequências de α-
galactosidase, através de alinhamento e construção de árvores de relação.
O levantamento das sequências das α-galactosidases de plantas foi
realizado por comparação através de BLAST de sequências gênicas
codificantes de Cyamopsis tetragonolobus (guar), Glycine Max (soja) e

75
Phaseolus vulgares (feijão), por serem da mesma subfamília Fabaceae que
Sesbania virgata (Cav.) Pers., contra sequências depositadas em banco de
dados públicos: GenBank (http:://www.ncbi.nlm.nih.gov) e CAZy (Carbohydrate-
Active EnZymes database) (Cantarel et al., 2008). As sequências foram
escolhidas através do valor do e-value superior a -70.
A análise das sequências por alinhamento foi feita através do programa
Mega versão 4.0 (Tamura et al., 2007), bem como a construção da árvore de
relação com bootstrap de 1000 repetições.
IV. Resultados
4.1. Determinação da porcentagem de germinação e pico de
atividade enzimática
A fim de determinar a viabilidade das sementes de Sesbania virgata
(Cav.) Pers., bem como, determinar o pico de atividade enzimática da enzima
α-galactosidase durante o desenvolvimento da plântula, foi avaliada a taxa de
germinação, através da protrusão da radícula, e a atividade enzimática da α-
galactosidase, através de ensaio enzimático com substrato enzimático 4-
Nitrophenyl-D-galactopyranoside (Sigma Chem. Co. St. Louis, USA) (PNP-α-
galactopiranosídeo), durante e após germinação, em um período de 7 dias (168
horas).
Analisando a porcentagem de germinação ao longo do período,
observou-se que a germinação completa das sementes ocorreu somente no
segundo dia após a embebição com viabilidade das sementes de 100% (Figura
6).

76
Figura 6. Porcentagem de germinação das sementes de Sesbania virgata (cav.) Pers. ao longo de 7
dias (168 horas) após a embebição.
Quanto à determinação do pico de atividade enzimática da α-
galactosidase, responsável pela retirada das substituições por galactose do
galactomanano durante a mobilização de reserva, permitindo a ação das
demais enzimas do processo de degradação (endo-β-mananase e β-
manosidase), observou-se um aumento progressivo na atividade enzimática
específica até 96 horas (4° dia após a embebição), quanto atinge seu valor
máximo, diminuindo sua atividade enzimática até 168 horas (7° dia após a
embebição), não atingindo o valor basal (Figura 7).
Sob o ponto de vista do desenvolvimento do protocolo de purificação da
enzima α-galactosidase, as sementes utilizadas no processo de purificação
foram mantidas em BOD até o 4° dia após embebição (96 horas), garantindo
máxima concentração de enzima a ser purificada.
0
20
40
60
80
100
120
0 24 48 72 96 120 144 168
% d
e s
em
en
tes
germ
inad
as
Tempo em horas após a embebição

77
Figura 7. Atividade enzimática específica da enzima α-galactosidase presente em sementes de
Sesbania virgata (Cav.) Pers. ao longo de 7 dias (168 horas) após a embebição.
4.2. Purificação da α-galactosidase de sementes de Sesbania
virgata (Cav.) Pers.
4.2.1. Caracterização da temperatura ótima e pH ótimo da α-
galactosidase em extrato bruto enzimático
Com a finalidade de padronizar o ensaio enzimático da α-galactosidase
para as etapas de purificação e caracterizar esta enzima quando em extrato
bruto, permitindo futuras comparações em condições de enzima purificada,
procedeu-se com a determinação da temperatura ótima e pH ótimo de
atividade enzimática da α-galactosidase.
4.2.1.1. Temperatura ótima
Avaliando o perfil de atividade enzimática da α-galactosidase sob
diferentes temperaturas (25° a 60°C, variando a cada 5°C), observou-se um
acréscimo progressivo na atividade enzimática de 25°C a 50°C, sendo entre
45°C e 50°C a faixa de temperatura com maior atividade enzimática, seguido
de uma queda abrupta na atividade enzimática a 60°C, valor este menor do
que a atividade enzimática a 30°C (Figura 8).
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
0 24 48 72 96 120 144 168
μm
olG
al.m
in-1
.mgp
rot-1
Tempo em horas

78
Figura 8. Determinação da temperatura ótima de atividade enzimática da α-galactosidase em
extrato bruto enzimático de sementes de Sesbania virgata (Cav.) Pers. após 4 dias de incubação em
BOD.
4.2.1.2. pH ótimo
A caracterização da atividade enzimática da α-galactosidase sob
condições variadas de pH (2,3 a 8) evidencia um aumento na atividade
enzimática entre o pH 2,3 a pH 4,4, sendo entre pH 4,4 e pH 5,4 o intervalo
com maior atividade enzimática, seguido de decréscimo mais acentuado desta
atividade enzimática entre pH 5,4 e pH 8,0, onde a atividade enzimática é
inexistente (Figura 9).
A determinação dos parâmetros de temperatura e pH ótimos (Figuras 8
e 9) em extrato bruto, nos quais a atividade enzimática é máxima, foi
considerada no desenvolvimento do protocolo do processo de purificação;
sendo assim, os ensaios enzimáticos performados, durante acompanhamento
da purificação da α-galactosidase, utilizaram PNP-αGalactopiranosídeo,
tampão McIlvaine pH 4,4 a 45°C, por 15 minutos.
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
0,16
0 10 20 30 40 50 60 70
μm
olG
al.m
in-1
.mgp
rot.
-1
Temperatura (°C)
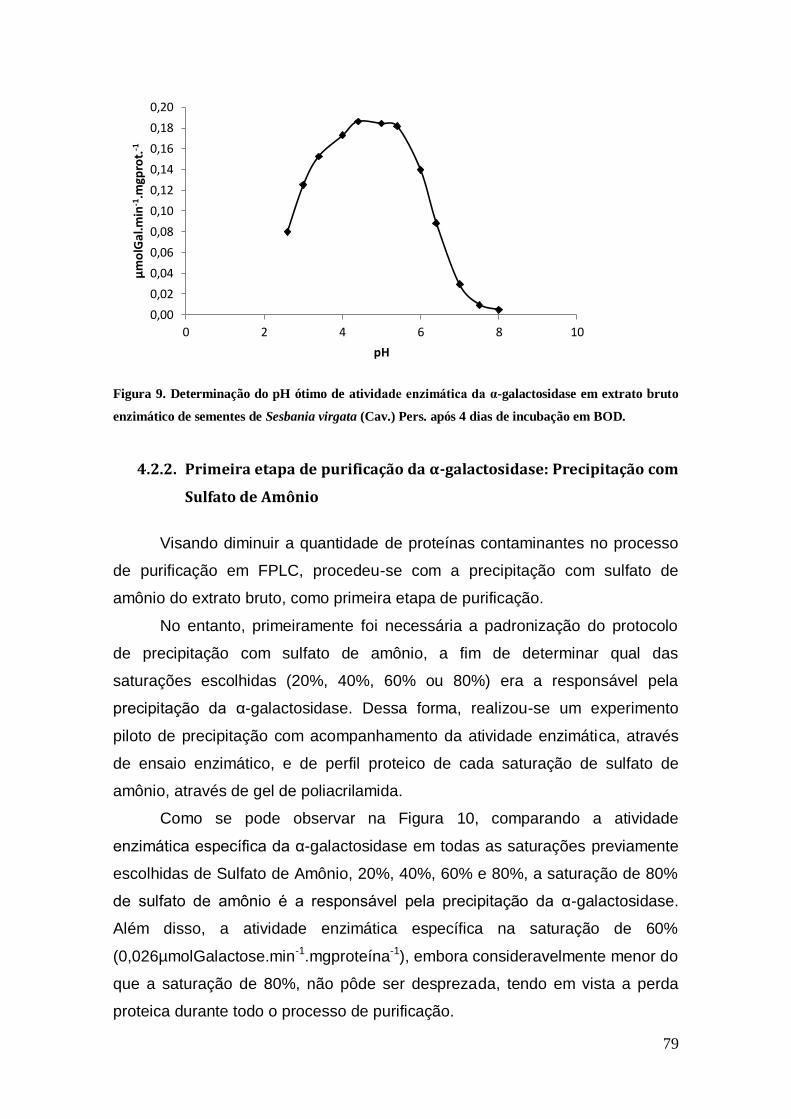
79
Figura 9. Determinação do pH ótimo de atividade enzimática da α-galactosidase em extrato bruto
enzimático de sementes de Sesbania virgata (Cav.) Pers. após 4 dias de incubação em BOD.
4.2.2. Primeira etapa de purificação da α-galactosidase: Precipitação com
Sulfato de Amônio
Visando diminuir a quantidade de proteínas contaminantes no processo
de purificação em FPLC, procedeu-se com a precipitação com sulfato de
amônio do extrato bruto, como primeira etapa de purificação.
No entanto, primeiramente foi necessária a padronização do protocolo
de precipitação com sulfato de amônio, a fim de determinar qual das
saturações escolhidas (20%, 40%, 60% ou 80%) era a responsável pela
precipitação da α-galactosidase. Dessa forma, realizou-se um experimento
piloto de precipitação com acompanhamento da atividade enzimática, através
de ensaio enzimático, e de perfil proteico de cada saturação de sulfato de
amônio, através de gel de poliacrilamida.
Como se pode observar na Figura 10, comparando a atividade
enzimática específica da α-galactosidase em todas as saturações previamente
escolhidas de Sulfato de Amônio, 20%, 40%, 60% e 80%, a saturação de 80%
de sulfato de amônio é a responsável pela precipitação da α-galactosidase.
Além disso, a atividade enzimática específica na saturação de 60%
(0,026µmolGalactose.min-1.mgproteína-1), embora consideravelmente menor do
que a saturação de 80%, não pôde ser desprezada, tendo em vista a perda
proteica durante todo o processo de purificação.
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
0 2 4 6 8 10
μm
olG
al.m
in-1
.mgp
rot.
-1
pH

80
Embora a atividade enzimática específica do descarte seja alta
(0,059µmolGalactose.min-1.mgproteína-1) comparativamente a saturação de
60%, devido ao cálculo levar em consideração o volume total de amostra, o
descarte não foi considerado por conter proteínas contaminantes (Figura 10).
Figura 10. Padronização da etapa de purificação da Precipitação com Sulfato de Amônio.
Determinação da saturação de sulfato de amônio responsável pela precipitação da enzima α-
galactosidase através quantificação da atividade enzimática específica da α-galactosidase em
extrato bruto e em cada precipitado ressuspendido referente à utilização de uma porcentagem de
saturação de sulfato de amônio.
Na Figura 11 observa-se o perfil proteico do extrato bruto, das 4
saturações de sulfato de amônio e do descarte. No perfil proteico do extrato
bruto nota-se uma ausência em proteínas de peso molecular acima de
111,6kDa, porém com grande diversidade de proteínas entre 111,6kDa e
6,4kDa.
Comparando todas as amostras, percebe-se uma diminuição gradual
das proteínas de peso molecular abaixo de 30KDa e acima de 59,4kDa a
medida que a saturação de sulfato de amônio aumenta, ao ponto de observar-
se no descarte somente 6 bandas de proteínas abaixo de 30KDa e ausência de
proteínas acima de 59,4kDa, em comparação ao extrato bruto com 11 bandas
e 2 bandas, respectivamente. Além disso, nas saturações de 60% e 80% de
sulfato de amônio existe uma concentração das proteínas de peso molecular
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Ext. Bruto 20% 40% 60% 80% Descarte
μm
olG
al.m
in-1
.mgp
rot-1

81
intermediário, entre 59,4kDa e 30kDa, intervalo de peso onde se encaixa a α-
galactosidase.
Figura 11. Eletroforese em gel de poliacrilamida 12% do extrato bruto enzimático; precipitados
ressuspendidos de 20%, 40%, 60% e 80% de saturação de sulfato de amônio e o descarte resultante
da precipitação com sulfato de amônio, com cerca de 50µg de proteína nas amostras aplicadas.
Tendo em vista os resultados do experimento piloto de precipitação com
sulfato de amônio decidiu-se por utilizar somente as concentrações de 40% e
80% de sulfato de amônio para a primeira etapa de purificação da α-
galactosidase, garantindo diminuição de proteínas contaminantes (saturação
de 40%) e máxima concentração de α-galactosidase (soma das saturações de
60% e 80%).
Na Figura 12 observa-se o perfil de atividade enzimática específica da
primeira etapa de purificação da α-galactosidase com precipitação com sulfato
de amônio nas saturações de 40% e 80% da amostra submetida à purificação,
onde se vê, novamente, a saturação de 80% como a responsável pela
precipitação da α-galactosidase, pois é a maior atividade enzimática específica
encontrada.
Padrão
195 kDa
111,6 kDa
59,4 kDa
30 kDa
24,7 kDa
12,6 kDa
Ext.Bruto 20% 40% 60% 80% Descarte

82
A fração referente a saturação de 80% foi, portanto, destinada a
segunda etapa de purificação em FPLC Äkta (GE Healthcare) utilizando coluna
de troca iônica aniônica Mono-Q (Pharmacia Biotech).
Figura 12. Comparação da atividade enzimática da enzima α-galactosidase no: extrato bruto
(Ext.Bruto); ressuspendido do precipitado com 40% de saturação de Sulfato de Amônio (40%);
ressuspendido do precipitado com 80% de saturação de Sulfato de Amônio (80%); sobrenadante
resultante da precipitação com 80% de saturação de Sulfato de Amônio (Descarte), após Primeira
etapa de purificação: Precipitação com Sulfato de Amônio em 40% e 80% de saturação.
4.2.3. Segunda etapa de purificação da α-galactosidase: Cromatografia de
troca-iônica com coluna aniônica (Mono-Q)
Com a finalidade de purificar a α-galactosidase, a amostra proveniente
da precipitação com 80% de sulfato de amônio foi submetida à cromatografia
de troca-iônica em coluna aniônica Mono-Q (Pharmacia Biotech) em sistema
de purificação FPLC Äkta (GE Healthcare) utilizando tampão Tris-HCl 20mM
pH 7.8 e gradiente salino 0 a 0.5M de NaCl. Esta purificação foi acompanhada
com ensaio enzimático de cada fração coletada durante a purificação, além da
visualização do perfil proteico por eletroforese em gel de poliacrilamida.
O cromatograma, resultado da purificação em Mono-Q, está
apresentado na Figura 13, no qual é possível verificar, anteriormente ao início
da aplicação do gradiente salino, um pico de proteínas que não interagiram
0
1
2
3
4
5
6
7
Ext.Bruto 40% 80% Descarte
µm
olG
alac
tose
.min
-1.m
gpro
t-1

83
com a coluna e saíram na lavagem da coluna com tampão Tris-HCl 20mM pH
7.8 (wash).
Além disso, pode-se verificar que os picos A, B e C, picos que
apresentaram atividade enzimática de α-galactosidase (Figura 13), deixaram de
interagir com a coluna em concentrações baixas a medianas de NaCl, entre
100mM a 250mM.
Figura 13. Perfil cromatográfico da purificação em FPLC Äkta do extrato bruto enzimático
submetido à coluna aniônica Mono-Q HR 10/10 (Pharmacia Biotech). Identificação com letras (A-
C) das frações coletadas que apresentaram atividade enzimática de α-galactosidase, onde: (A)
Frações 47 a 51; (B) Frações 63 a 65; (C) Frações 66 a 69. (―) Perfil proteico quantificado por UV;
(―) Gradiente de NaCl variando de 0-0.5M em 12 volumes de coluna (‖) Frações coletadas com seus
respectivos números.
O acompanhamento da purificação foi realizado através de ensaio
enzimático de todas as frações coletadas durante a purificação em FPLC
(Figura 14), utilizando como substrato PNP-α-Galactopiranosídeo, e
eletroforese em gel de poliacrilamida das frações reunidas que continham
atividade enzimática (Figura 15), visando determinar quais frações continham a
enzima α-galactosidase e a qualidade da purificação, respectivamente.

84
A atividade enzimática da α-galactosidase, expressa em absorbância do
PNP-α-Galactopiranosídeo a 405nm, de todas as frações coletadas na
purificação em coluna Mono-Q se encontra na Figura 14. Observam-se três
picos de atividade enzimática: frações 47 a 51 (pico A); frações 63 a 65 (pico B)
e frações 66 a 69 (pico C), evidenciando mais de uma isoforma (pelo menos
três) da enzima α-galactosidase, sendo uma delas, pico A, predominante.
Figura 14. Atividade enzimática da α-galactosidase expressa em absorbância a 405nm das frações
coletadas durante a purificação em FPLC Äkta do extrato bruto enzimático submetido à coluna
aniônica Mono-Q HR 10/10 (Pharmacia Biotech). (a) Frações 47 a 51; (b) Frações 63 a 65; (c)
Frações 66 a 69.
As frações 47 a 51 (pico A) foram reunidas e uma alíquota foi destinada
ao perfil proteico em eletroforese em gel de poliacrilamida (Figura 15), a fim de
visualizar a qualidade da purificação, através da quantidade de bandas
presentes.
O gel de poliacrilamida (Figura 15) após purificação em Mono-Q ainda
apresentou muitas proteínas contaminantes, no entanto, observou-se uma
banda (indicada pela seta) entre 59,4kDa e 30kDa. Esta banda se apresenta
mais concentrada, do que as demais, evidenciando que este passo de
purificação, apesar de não eliminar grande parte das proteínas contaminantes,
concentrou a enzima de interesse.
A
B C

85
Figura 15. Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue R250 das
frações reunidas 47 a 51 (pico A) após purificação em FPLC Äkta utilizando coluna aniônica
Mono-Q HR 10/10 (Pharmacia Biotech). (→) Banda proteica concentrada de tamanho esperado
para α-galactosidase.
4.2.4. Terceira etapa de purificação da α-galactosidase: Cromatografia de
Gel Filtração (Superose)
Com a finalidade melhorar a purificação da α-galactosidase, as frações
47 a 51 reunidas na etapa de purificação em coluna aniônica Mono-Q
(Pharmacia Biotech) foram submetidas à Cromatografia de Gel Filtração em
coluna Superose 6 (GE Healthcare) em sistema de purificação FPLC Äkta (GE
Healthcare) utilizando tampão Acetato de Amônio 50mM pH 5.0. Esta
purificação foi acompanhada com ensaio enzimático de cada fração coletada
durante a purificação somada à visualização do perfil proteico por eletroforese
em gel de poliacrilamida.
O cromatograma, resultado da purificação da α-galactosidase em coluna
Superose 6, está apresentado na Figura 16, no qual se observa a presença de
um único pico predominante. O pico identificado por D foi o único a apresentar
atividade enzimática de α-galactosidase (Figura 17).
Padrão
195 kDa
111,6 kDa
59,4 kDa
30 kDa
24,7 kDa
12,6 kDa
6,4 kDa
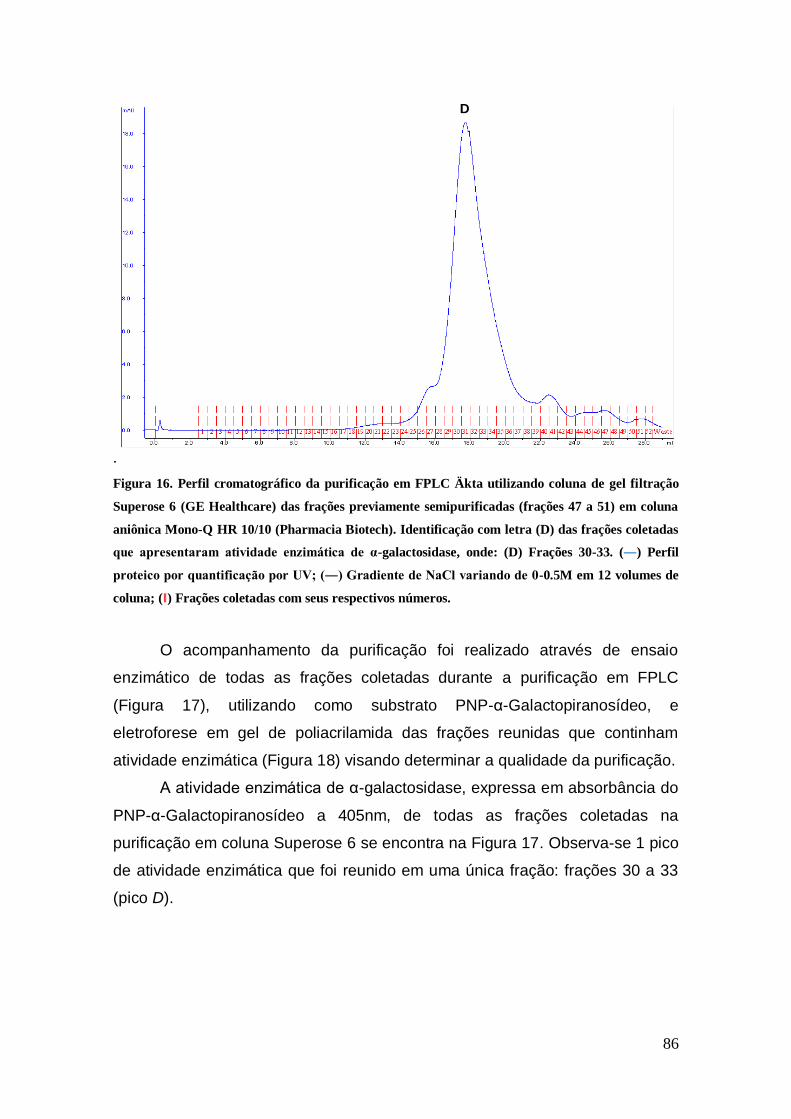
86
.
Figura 16. Perfil cromatográfico da purificação em FPLC Äkta utilizando coluna de gel filtração
Superose 6 (GE Healthcare) das frações previamente semipurificadas (frações 47 a 51) em coluna
aniônica Mono-Q HR 10/10 (Pharmacia Biotech). Identificação com letra (D) das frações coletadas
que apresentaram atividade enzimática de α-galactosidase, onde: (D) Frações 30-33. (―) Perfil
proteico por quantificação por UV; (―) Gradiente de NaCl variando de 0-0.5M em 12 volumes de
coluna; (‖) Frações coletadas com seus respectivos números.
O acompanhamento da purificação foi realizado através de ensaio
enzimático de todas as frações coletadas durante a purificação em FPLC
(Figura 17), utilizando como substrato PNP-α-Galactopiranosídeo, e
eletroforese em gel de poliacrilamida das frações reunidas que continham
atividade enzimática (Figura 18) visando determinar a qualidade da purificação.
A atividade enzimática de α-galactosidase, expressa em absorbância do
PNP-α-Galactopiranosídeo a 405nm, de todas as frações coletadas na
purificação em coluna Superose 6 se encontra na Figura 17. Observa-se 1 pico
de atividade enzimática que foi reunido em uma única fração: frações 30 a 33
(pico D).
D

87
Figura 17. Atividade enzimática da α-galactosidase expressa em absorbância a 405nm das
frações coletadas durante a purificação em FPLC Äkta, utilizando coluna de gel filtração
Superose 6 (GE Healthcare), da amostra semipurificada em coluna aniônica Mono-Q HR
10/10 (Pharmacia Biotech). (D) Frações 30 a 33.
As frações 30 a 33 (pico D) foram reunidas e uma alíquota foi destinada
ao perfil proteico em eletroforese em gel de poliacrilamida (Figura 18), a fim de
visualizar a qualidade da purificação, através da quantidade restante de
proteínas contaminantes (bandas presentes).
O perfil proteico em gel de poliacrilamida (Figura 18) após purificação
em Superose 6 ainda apresenta muitas proteínas contaminantes, no entanto,
observa-se que a banda correspondente a α-galactosidase (ver item 4.2.6.4),
banda indicada pela seta, apresenta-se mais concentrada quando comparado a
mesma banda nas etapas anteriores de purificação (figura 25).
D

88
Figura 18. Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue R250 das
frações reunidas 30 a 33 (pico D) após purificação em FPLC Äkta utilizando coluna de gel filtração
Superose 6 (Pharmacia Biotech). (→) Banda proteica concentrada de tamanho esperado para α-
galactosidase.
Como visualizado no gel de poliacrilamida (Figura 25), a purificação
proposta não foi eficiente em efetivamente purificar a α-galactosidase, no
entanto a concentrou (ver tabela de purificação) e a semipurificou.
Tendo em vista a utilização da alíquota semipurificada em experimentos
com o polissacarídeo galactomanano, procedeu-se com o sequenciamento
interno de 8 bandas (entre os pesos moleculares de 59,4 kDa e 12,6kDa) para
identificação da banda referente a α-galactosidase e determinação da
sequência interna da enzima, bem como determinar a presença ou ausência
das demais enzimas de mobilização de reserva do galactomanano (endo-β-
mananase e β-manosidase) na alíquota semipurificada.
A determinação do peso molecular da α-galactosidase foi realizado por
duas técnicas: através do gel de poliacrilamida e através da coluna Superose
(purifica por peso molecular). A diferença entre as duas técnicas permite
visualizar se a proteína assumiu estrutura quartenária com consequente
mudança no peso molecular, pois o gel de poliacrilamida desnatura as
proteínas, enquanto através da gel filtração a proteína está na forma nativa,
podendo interagir com outras.

89
O peso molecular da α-galactosidase semipurificada determinado
através do gel de poliacrilamida pelo cálculo do Rf da proteína de interesse e
sua localização na curva de calibração traçada com os valores de Rf das
proteínas marcadoras versus o log de seus pesos moleculares, conforme
Weber & Osborn (1969), foi de, aproximadamente, 42 kDa (Figura 18). Os
marcadores de peso molecular utilizados foram a miosina (195 kDa), -
galactosidase (111,6 kDa), albumina de soro bovino (59,4 kDa), anidrase
carbônica (30 kDa), inibidor de tripsina de soja (24,7 kDa), lisozima (12,5 kDa)
e aprotinina (6,4 kDa), adquiridos da Bio-Rad.
Enquanto o peso molecular da α-galactosidase determinado, através da
Gel Filtração utilizando a calibração do fabricante (Tiroglobulina 669kDa,
Ferritina 440 kDa, BSA 67 kDa e Ribonuclease 13,7 kDa), foi de
aproximadamente 72 kDa.
4.2.5. Tabela de purificação
Visando determinar o fator de purificação e o rendimento da purificação,
procedeu-se com a mensuração de proteínas totais e atividade enzimática da
α-galactosidase com PNP-αGal de alíquotas de cada etapa de purificação. Na
Tabela 1 encontram-se os valores de atividade específica, fator de purificação
e rendimento para cada etapa de purificação.
Analisando os valores da Tabela 1, nota-se um aumento de 2600% na
atividade enzimática específica da α-galactosidase após purificação em
Superose, quando comparado com a atividade enzimática específica do extrato
bruto. Além disso, o fator de purificação encontrado após as 3 etapas de
purificação é de 26,2 vezes e o rendimento da purificação de 1,28 %.

90
Tabela 1. Tabela de purificação da α-galactosidase de Sesbania virgata (Cav.) Pers. desenvolvida
após a realização das três etapas de purificação: precipitação com Sulfato de Amônio 80%,
Cromatografia de troca-iônica aniônica (Mono-Q) e Cromatografia de gel filtração (Superose).
Etapa de
Purificação
Atividade Enzimática
Específica
(μmolGal.min-1
.mgprot.-1
)
Fator de
purificação
Rendimento da
Purificação (%)
Extrato Bruto 0,041 - 100
Precipitação com
Sulfato de Amônio
80%
0,078 1,886 22,764
Mono-Q (aniônica) 0,742 17,962 3,219
Superose 6 (Gel
Filtração) 1,083 26,196 1,275
4.2.6. Caracterização da α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers. e da α-galactosidase de Cyamopsis
tetragonoloba (guar) (Megazyme®) quanto a temperatura ótima,
pH ótimo e aspectos cinéticos
A α-galactosidase semipurificada foi caracterizada bioquimicamente
quanto temperatura ótima, pH ótimo e aspectos cinéticos, Km e Vmáx.
Paralelamente a caracterização da α-galactosidase de Sesbania virgata (Cav.)
Pers. foi realizada a caracterização da α-galactosidase de Cyamopsis
tetragonoloba (guar) (Megazyme®) quanto os mesmos aspectos.
4.2.6.1. Temperatura ótima
Avaliando o perfil de atividade enzimática da α-galactosidase de
Sesbania virgata (Cav.) Pers. sob diferentes temperaturas (25° a 60°C,
variando a cada 5°C), observou-se um acréscimo progressivo na atividade
enzimática de 25°C a 50°C, sendo entre 50°C e 55°C a faixa de temperatura
com maior atividade enzimática, seguido de uma queda abrupta na atividade
enzimática a 60°C (Figura 19).

91
Figura 19. Determinação da temperatura ótima da α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers.
Avaliando o perfil de atividade enzimática da α-galactosidase
(Megazyme®) de Cyamopsis tetragonoloba sob diferentes temperaturas (25° a
60°C, variando a cada 5°C), observou-se um acréscimo progressivo na
atividade enzimática de 25°C a 40°C, sendo 40°C a temperatura com maior
atividade enzimática. Observa-se também, uma queda da atividade enzimática
a 45°C, um leve aumento na atividade enzimática a 55°C e, novamente, uma
queda na atividade enzimática a 60°C (Figura 20).
0,0000
0,2000
0,4000
0,6000
0,8000
1,0000
1,2000
0 5 10 15 20 25 30 35 40 45 50 55 60 65
Temperatura (°C)
Ati
vid
ade
Esp
ecí
fica
(µm
olG
alac
tose
.min
-1.m
gpro
t-1)

92
Figura 20. Determinação da temperatura ótima da α-galactosidase de Cyamopsis tetragonoloba
(Megazyme®).
4.2.6.2. pH ótimo
A caracterização da atividade enzimática da α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. sob condições variadas de pH
(2,3 a 7) evidenciou um aumento na atividade enzimática entre o pH 2,3 a pH
5,0, sendo entre pH 4,4 e pH 5,4 o intervalo com maior atividade enzimática,
seguido de decréscimo mais acentuado desta atividade enzimática entre pH
5,4 e pH 7,0, onde a atividade enzimática é muito baixa (Figura 21).
Enquanto a caracterização da atividade enzimática da α-galactosidase
de Cyamopsis tetragonoloba (Megazyme®) sob condições variadas de pH (2,3
a 7,0) evidenciou um aumento na atividade enzimática entre o pH 2,3 a pH 6,0,
sendo entre pH 5,4 e pH 6,4 o intervalo com maior atividade enzimática,
seguido de decréscimo acentuado desta atividade enzimática entre pH 6,4 e
pH 7,0 (Figura 22).
0,000
0,010
0,020
0,030
0,040
0,050
0,060
0,070
0 10 20 30 40 50 60 70
Temperatura (°C)
Ati
vid
ade
En
zim
átic
a (µ
mo
lGal
.min
-1)
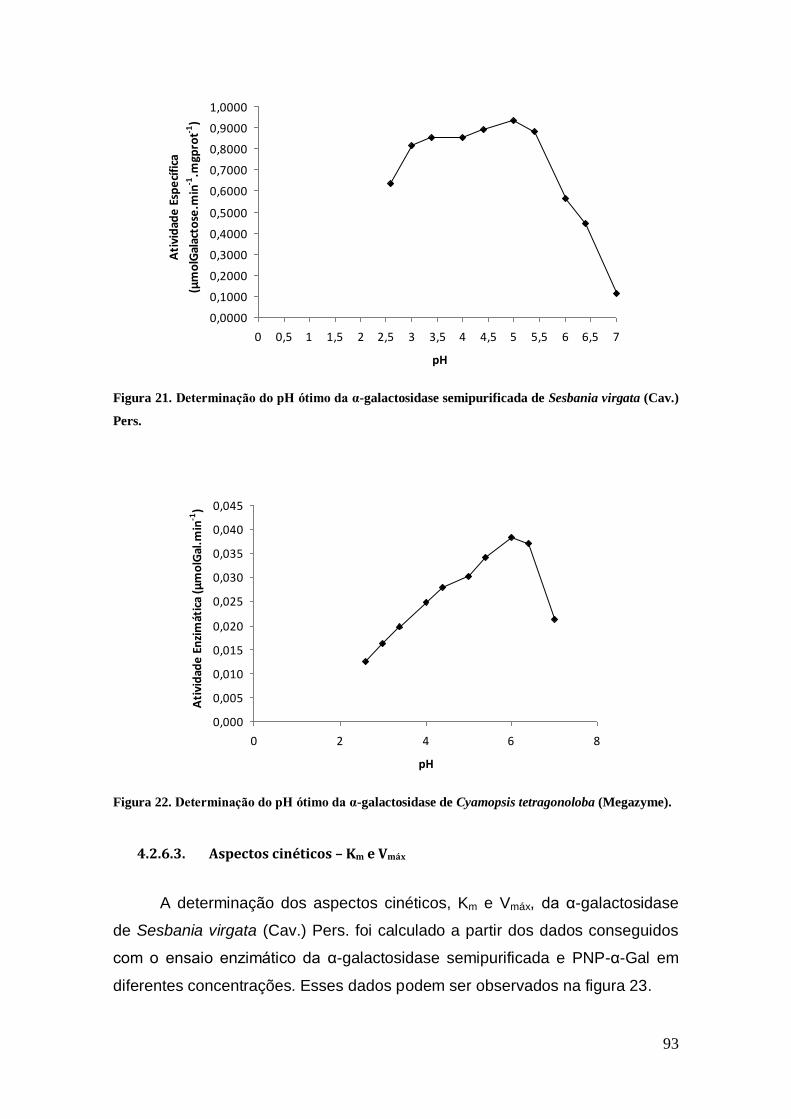
93
Figura 21. Determinação do pH ótimo da α-galactosidase semipurificada de Sesbania virgata (Cav.)
Pers.
Figura 22. Determinação do pH ótimo da α-galactosidase de Cyamopsis tetragonoloba (Megazyme).
4.2.6.3. Aspectos cinéticos – Km e Vmáx
A determinação dos aspectos cinéticos, Km e Vmáx, da α-galactosidase
de Sesbania virgata (Cav.) Pers. foi calculado a partir dos dados conseguidos
com o ensaio enzimático da α-galactosidase semipurificada e PNP-α-Gal em
diferentes concentrações. Esses dados podem ser observados na figura 23.
0,0000
0,1000
0,2000
0,3000
0,4000
0,5000
0,6000
0,7000
0,8000
0,9000
1,0000
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5 5,5 6 6,5 7
pH
Ati
vid
ade
Esp
ecí
fica
(µm
olG
alac
tose
.min
-1.m
gpro
t-1)
0,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
0,040
0,045
0 2 4 6 8
pH
Ati
vid
ade
En
zim
átic
a (µ
mo
lGal
.min
-1)

94
O Km da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
é de 1,8276mM, e o velocidade máxima é de 0,5024 µmolGal.min-1.mgproteína-
1.
Figura 23. Curva de saturação do substrato 4-Nitrofenil-D-Galactopiranosídeo na presença da
α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers. Os ensaios enzimáticos foram
conduzidos com o PNP-α-Gal nas seguintes concentrações (mM): 0.1; 0.2; 0.3; 0.5; 0.8; 1.0; 2.0; 3.0;
5.0; 8.0; 10; 13; 15; 17; 20; 25; 30. A partir dos dados coletados foi calculado o Km e o Vmáx da α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
Os aspectos cinéticos, Km e Vmáx, da α-galactosidase de Cyamopsis
tetragonoloba (Megazyme®) foram calculados a partir dos dados conseguidos
com o ensaio enzimático enzima comercial (0,250U/mL) e PNP-α-Gal em
diferentes concentrações. Esses dados podem ser observados na figura 24.
O Km da α-galactosidase de Cyamopsis tetragonoloba (Megazyme®) é de
2,398 mM e a velocidade máxima é de 0,0248 µmolGal.min-1.mgproteína-1.
0
0,1
0,2
0,3
0,4
0,5
0,6
0 5 10 15 20 25 30
Ati
vid
ad
e E
nzi
mát
ica
(µ
mo
lGal
.min
-1.m
gpro
teín
a-1)
Concentração de 4-Nitrofenil-D-Galactopiranosídeo (mM)

95
Figura 24. Curva de saturação do substrato 4-Nitrofenil-D-Galactopiranosídeo na presença da
α-galactosidase de Cyamopsis tetragonoloba (Megazyme®). Os ensaios enzimáticos foram
conduzidos com o PNP-α-Gal nas seguintes concentrações (mM): 0.1; 0.2; 0.3; 0.5; 0.8; 1.0; 2.0; 3.0;
5.0; 8.0; 10; 13; 15; 17; 20; 25. A partir dos dados coletados foi calculado o Km e o Vmáx da α-
galactosidase de Cyamopsis tetragonoloba (Megazyme®).
4.2.6.4. Análise do perfil proteico em gel de poliacrilamida sob condições
desnaturantes de todas as etapas de purificação
Para comparar qualitativamente o perfil proteico de cada etapa da
purificação: extrato bruto, precipitação com Sulfato de Amônio 80%,
cromatografias de troca-iônica (Mono-Q) e gel filtração (Superose), foi realizada
a separação das proteínas de cada etapa de purificação por eletroforese em
gel de poliacrilamida 12% (figura 25).
Analisando e comparando o perfil de proteínas do extrato bruto (figura
25 A) com os três passos de purificação (Figura 25 B, C, D), percebe-se uma
evidente diminuição na quantidade de proteínas contaminantes após
cromatografias de troca-iônica e gel filtração, no entanto, a precipitação com
sulfato de amônio 80% parece não alterar o perfil de proteínas quando
comparado com o extrato bruto.
Ao comparar-se o perfil de proteínas após as purificações em
cromatografia de troca-iônica e cromatografia de gel filtração nota-se o
desaparecimento de algumas bandas, evidenciando certa purificação, porém o
0,000
0,005
0,010
0,015
0,020
0,025
0,030
0 5 10 15 20 25
Concentração do 4-Nitrofenil-D-Galactopiranosídeo (mM)
Ati
vid
ade
Esp
ecí
fica
(µm
olG
alac
tose
.min
-1.m
gpro
t-1)

96
perfil proteico é muito parecido. Além disso, a banda de α-galactosidase
correspondente a 42KDa (entre 59,4kDa e 30kDa), evidenciada pela seta
(figura 25 D), fica mais concentrada quando compara-se o perfil proteico após
cromatografia de troca-iônica (figura 25 C) e cromatografia de gel filtração
(figura 25 D).
Dessa forma, com praticamente a manutenção do perfil proteico entre as
purificações em FPLC e concentração da α-galactosidase, considera-se que a
α-galactosidase de Sesbania virgata (Cav.) Pers. foi semipurificada.
Figura 25. Eletroforese em gel de poliacrilamida 12% corado com Coomassie Blue R250 de
todas as etapas de purificação. (A) Extrato bruto enzimático; (B) Precipitado de 80% de
Sulfato de Amônio (ressuspendido em tampão Tris-HCl 20mM ph 7,8); (C) Alíquota das
frações reunidas após purificação em FPLC Äkta utilizando coluna de troca-iônica
aniônica (Mono-Q); (D) Alíquota das frações reunidas após purificação em FPLC Äkta
utilizando coluna de gel filtração Superose 6 (GE Healthcare).
4.2.7. Sequenciamento interno da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
Inicialmente a proposta do sequenciamento interno da α-galactosidase
foi idealizada para obter-se a sequência interna da enzima purificada e
compará-la com as α-galactosidases já descritas e depositadas em bancos de
dados.

97
No entanto, após análise dos resultados da purificação, onde houve uma
semipurificação ainda com uma quantidade razoável de proteínas
contaminantes, optou-se por realizar o sequenciamento interno de oito bandas
do gel de poliacrilamida entre 59,4 kDa e 12,6 kDa (Figura 26), visando
confirmar a banda referente da α-galactosidase de Sesbania virgata (Cav.)
Pers. e, consequentemente, a sequência interna da proteína, e identificar as
demais bandas, para excluir a presença das demais enzimas da mobilização
de reservas, endo-β-mananase e β-manosidase.
Figura 26. Identificação com números, de 1 a 8, das bandas recortadas do gel de poliacrilamida
12% submetidas ao sequenciamento interno para identificação das proteínas, principalmente da α-
galactosidase, juntamente com a determinação da sequência interna da α-galactosidase
semipurificada de sementes de Sesbania virgata (Cav.) Pers.
Como o esperado, devido ao peso molecular conhecido de α-
galactosidases de plantas, a banda 1 foi identificada como uma α-
galactosidase, tendo como primeiro Hit no banco de dados a α-galactosidase
de sementes de Cyamopsis tetragonoloba (guar).
Os fragmentos identificados pela espectrometria compõem 22% da
sequência de aminoácidos da α-galactosidase, sendo a sequência de
aminoácidos apresentada abaixo:
Padrão
195 kDa
111,6 kDa
59,4 kDa
30 kDa
24,7 kDa
12,6 kDa
6,4 kDa
1 2
3 4
5 6
7
8

98
ALADYV-HSK-RMPGSLGHEE-QDAK-TT-GDIEDNWNSM-TSIADSNDK
W-ASYAGPGGWN-DPDMLEVGNG-GMTTEEYR-AP-LLVGCDIR-VAVIL-
WNR
As bandas de 2 a 8 foram sequenciadas e na Figura 39 a 45 no ANEXO
se encontram a lista de possíveis proteínas para cada uma delas, sendo que as
bandas 2, 3, 4, 5 e 8 apresentaram α-galactosidase como possível proteína
componente dessas bandas, mas não se apresentando como primeiro hit.
Ressalta-se que nenhuma dessas proteínas foram identificadas como: endo-β-
mananase ou β-manosidase, resultado importante para embasar a utilização da
alíquota semipurificada nos experimentos conduzidos com o polissacarídeo
galactomanano descrito no Capítulo 2 desta dissertação.
4.3. Análises das sequências de α-galactosidases previamente identificadas em plantas
Visando determinar similaridades entre a sequência de aminoácidos
para α-galactosidases de plantas já depositadas em bancos de dados públicos
e a sequência interna da α-galactosidase semipurificada de Sesbania virgata
(Cav.) Pers., foi realizada a análise de 35 sequências de aminoácidos de α-
galactosidases de plantas já depositadas em bancos de dados e a sequência
interna da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers.,
através de alinhamento e construção de árvores de relação.
Na Figura 27 encontra-se o resultado da construção da árvore de
relação com bootstrap de mil repetições das 35 sequências de plantas e a
sequência interna da α-galactosidase de Sesbania virgata (Cav.) Pers. Nota-se
a presença de três grandes ramos, aparentemente sem qualquer relação com a
filogenia.
Embora a relação das α-galactosidases em 3 três grandes ramos não
esteja relacionada à filogenia, alguns dos ramos mais internos, ramos
identificados de a-f estão de acordo com a filogenia: (a) Fabaceae; (b)
Fabaceae; (c) Fabaceae, (d) Rubiaceae; (e) Poaceae; (f) Pinaceae.

99
Os ramos marcados com • são ramos consistentes, visto que em 95% ou
mais das árvores geradas para a construção da árvore de consenso estes
ramos ficaram agrupados desta maneira, entre eles estão alguns dos ramos
relacionados com a filogenia: ramo b (99%), ramo d (99%) e ramo e (98%).
Os demais ramos não identificados reúnem espécies de diversas
famílias não apresentando relação com a filogenia, provavelmente apresenta
relação com a função da enzima.
Figura 27: Análise das sequências de aminoácidos 35 de α-galactosidase de plantas conjuntamente
com a sequência dos fragmentos de α-galactosidase de Sesbania virgata (Cav.) Pers. semipurificada
determinados por Espectrometria de Massas (MALDI-TOF). Grupos identificados por letras estão
de acordo com a filogenia: (A – vermelho) Fabaceae; (B - rosa) Fabaceae; (C – azul escuro)
Fabaceae; (D - verde) Rubiaceae; (E – roxo) Poaceae; (F – azul claro) Pinaceae. A coloração dos
ramos é a mesma da árvore de relação das 35 sequências de aminoácidos somente localizada na

100
figura 46 do anexo, no entanto, a identificação pelas letras é diferente, ou seja, os ramos coloridos
por cada cor são os mesmos em ambas as árvores, mas a letra que o identifica não é a mesma. (•)
Bootstrap ˃ 95, ou seja, em 95% ou mais das 1000 árvores de relação geradas os ramos marcados
com (•) ficaram agrupados como mostrado nesta árvore de relação.
V. Discussão
5.1. Determinação da porcentagem de germinação e pico de
atividade enzimática
A germinação de sementes, fase compreendida entre a embebição e
protrusão da radícula, é responsável por um conjunto de eventos fisiológicos e
bioquímicos de reativação do metabolismo embrionário, culminando com o
crescimento embrionário (Bewley & Black, 1994).
De acordo com Buckeridge & Dietrich (1996), Potomati & Buckeridge
(2002), Tonini et al. (2007) o processo germinativo em sementes de Sesbania
virgata (Cav.) Pers. ocorre entre o segundo e terceiro dias após embebição,
dado em concordância com 100% de sementes germinadas em 48 horas (2
dias) após embebição (Figura 6).
Após a protrusão da radícula, a mobilização da reserva de parede
celular de galactomanano das sementes de Sesbania virgata (Cav.) Pers. se
inicia, atingindo seu auge entre o quarto e quinto dias após a embebição
(Buckeridge & Dietrich, 1996; Tonini et al., 2006), devido ao aumento da
atividade das hidrolases, α-galactosidase, endo-β-mannanase e β-manosidase,
no endosperma (Buckeridge & Dietrich, 1996).
A máxima atividade de α-galactosidase encontrada no experimento
proposto, durante o período pós-germinativo, ocorreu, consistentemente, no
quarto dia após embebição (Figura 7), um dia atrasado ao que foi descrito por
Potomati & Buckeridge (2002), Tonini et al. (2007). No entanto, possivelmente
este atraso está relacionado às diferentes condições de embebição e
desenvolvimento da plântula (fotoperíodo e temperatura) entre ambos os
trabalhos (fotoperíodo de 12 horas e 27°C) e o experimento conduzido no
presente trabalho (claro constante e 28°C).

101
Dessa forma, as sementes Sesbania virgata (Cav.) Pers. destinadas à
purificação da α-galactosidase foram retiradas das câmaras de germinação
(BOD) no quarto dia após embebição, garantindo máxima concentração de
enzima a ser purificada.
5.2. Purificação da α-galactosidase de sementes de Sesbania
virgata (Cav.) Pers.
As α-galactosidases são enzimas que catalisam a hidrólise de resíduos
de galactose α-1,6-ligados de galacto-oligossacarídeos e polímeros galacto-
(gluco)-mananos (Fujimoto et al., 2003).
Durante a mobilização de reserva de parede celular de galactomanano
foi detectada a presença de três enzimas hidrolíticas: α-galactosidase, endo-β-
mananase e β-manosidase (Reid & Meier, 1972; McCleary & Matheson, 1976;
Buckeridge et al, 1995a; Buckeridge & Dietrich, 1996), que atuam sobre o
galactomanano, liberando manose e galactose para o metabolismo,
promovendo o crescimento da plântula (Buckeridge et al., 2000a; Lisboa et al.,
2006).
McCleary & Matheson (1975), Reid & Edwards (1995) e Lisboa et al.
(2006) evidenciaram que a degradação do galactomanano pelas três enzimas
requer uma ordem de atuação das mesmas. Em sementes de leguminosas a
ação da α-galactosidase é uma condição necessária para permitir acesso da
enzima endo-β-mananase à cadeia principal do galactomanano, uma vez que,
McCleary & Matheson (1976) demonstraram que a β-mannanase de Gleditsia
triacanthos só hidrolisa as ligações β-(1,4) entre duas manoses contínuas sem
ramificação.
Desta forma, a α-galactosidase é a primeira a atuar sobre o
polissacarídeo, hidrolisando as ligações α-(1,6) entre as unidades de manano e
galactose, em seguida, a endo-β-mananase atua nas ligações β-(1,4) da
cadeia principal de manose, hidrolisando-a a oligossacarídeos, que, por sua
vez, serão levados a monossacarídeos pela ação, nas ligações β-(1,4), da β-
manosidase nas extremidades não-redutoras dos oligossacarídeos (Reid &

102
Meier, 1972; McCleary, 1983; Buckeridge et al., 1995a; Buckeridge & Reid,
1996; Buckeridge et al., 2000a).
Neste contexto fica evidente a importância da enzima α-galactosidase no
processo de mobilização de reserva de parede celular de galactomanano e
torna-se claro que a α-galactosidase ocorre em associação com outras
hidrolases que podem dificultar o processo de purificação (Dey & Pridham,
1972).
Comumente, as técnicas mais utilizadas no processo de isolamento
desta enzima são a precipitação com sulfato de amônio, fracionamento com
solventes orgânicos, tratamento térmico, acidificação, troca-iônica, gel filtração
e focalização isoelétrica (Dey & Pridham, 1972).
O procedimento adotado para a purificação da α-galactosidase de
sementes de Sesbania virgata (Cav.) Pers. foi adaptado do protocolo
apresentado por Silva, 2001.
No presente caso, o processo de purificação não foi precedido por uma
etapa de digestão do galactomanano endógeno visando evitar a perda de
atividade enzimática durante a incubação a 30°C. Dessa forma o processo de
purificação se inicia com a precipitação em sulfato de amônio, seguido de
Cromatografia de Troca Aniônica (coluna Mono-Q – Pharmacia Biotech) em
sistema FPLC (GE Healthcare) e Cromatografia de Gel Filtração (Superose 6)
(GE Healthcare) também em sistema FPLC (GE Healthcare).
A primeira etapa de purificação: precipitação com sulfato de amônio,
inicialmente foi padronizada para se determinar a saturação necessária para a
precipitação da α-galactosidase. Conforme apresentado na Figura 10, a
saturação de 80% foi a responsável pela precipitação da enzima de interesse,
uma vez que, foi o precipitado a apresentar maior atividade enzimática.
A saturação de 60%, embora não apresentando valores de atividade
enzimática comparáveis a de 80%, foi considerada possuidora de uma fração
rica em α-galactosidase e, portanto, foi inclusa na padronização.
Diferentemente da saturação de 60%, o descarte não foi levado em
consideração, pois mesmo apresentando atividade enzimática, possuía muitas
proteínas contaminantes.
Conjuntamente aos resultados de atividade enzimática, a comparação
do perfil proteico do extrato bruto, das saturações de 20%, 40%, 60% e 80% de

103
sulfato de amônio e do descarte (Figura 11), demonstraram que a precipitação
com sulfato de amônio foi eficaz no seu objetivo, devido à gradual eliminação
de proteínas contaminantes, visualizado pelo desaparecimento de bandas.
Além disso, a presença marcada de proteínas de peso molecular
intermediário, entre 59,4kDa e 30kDa (Figura 11) nas saturações de 60% e
80%, intervalo de peso esperado para a α-galactosidase, corrobora a atividade
enzimática encontrada nestas saturações. Feurtado et al. (2001) e Lisboa et al.
(2003) demonstraram que a α-galactosidase de sementes de Lycopersicum
esculentum (tomate) e de Sesbania virgata (Cav.) Pers. possuem peso
molecular de 39.8kDa e 45kDa, respectivamente.
Assim, a padronização da precipitação com Sulfato de Amônio resultou
em um protocolo onde as saturações a serem utilizadas são de 40%, visando
retirar proteínas contaminantes e pigmentos, e de 80%, visando reunir a fração
de enzima da saturação de 60% com a da saturação de 80%, aumentando a
quantidade de enzima a ser purificada, uma vez que, o rendimento de
purificação é baixo devido a perdas a cada etapa.
A aplicação do protocolo de precipitação com sulfato de amônio na
amostra a ser submetida a todos os passos de purificação se fez necessária
para diminuir a contaminação com pigmentos e proteínas, bem como,
concentrar a α-galactosidase visando maiores rendimentos de purificação.
Na comparação dos perfis proteicos após precipitação com Sulfato de
Amônio variando de 20% a 80% (padronização) e somente nas concentrações
de 40% e 80% (etapa de purificação), nota-se que o protocolo de
padronização, as 4 concentrações, é mais eficiente na retirada de proteínas
contaminantes (figura 11 e figura 25) as custas de grande perda da enzima α-
galactosidase.
Entretanto, a precipitação com Sulfato de Amônio como etapa de
purificação (40% e 80%) apresenta praticamente o mesmo perfil proteico, mas
com um aumento na concentração da α-galactosidase, visualizado por
intensidade da banda referente à enzima (figura 25). Muito provavelmente a
saturação de 60% seria a responsável pela retirada de grande parte das
proteínas contaminantes, porém a perda enzimática é muito grande, sendo
assim, optou-se pelas saturações de 40% e 80%, incorporando algumas
proteínas contaminantes no processo de purificação.

104
A precipitação com Sulfato de Amônio é um protocolo reprodutível, pois,
como se observa nas Figuras 10 e 12, o perfil de atividade enzimática se
repete, sendo a saturação de 80% a responsável pela precipitação da α-
galactosidase.
A segunda etapa de purificação: Cromatografia de Troca-Iônica com
coluna aniônica (Mono-Q) em sistema FPLC revelou a presença de três
isoformas de α-galactosidase (Figura 13 e 14 A, B, C), sendo a isoforma A
identificada como a encontrada predominantemente às demais, devido a maior
atividade enzimática encontrada (Figura 14).
A presença de isoformas de α-galactosidase foi primeiramente descrita
por Petek & Dong (1961) em sementes de Coffea sp e Plantago ovata. Tonini
et al. (2006) sugerem que a presença de isoformas de endo-β-mananase em
endosperma de Sesbania virgata (Cav.) Pers. seja para garantir a hidrólise do
galactomanano, mesmo com as modificações estruturais que este sofre
durante a mobilização.
Por outro lado, McCleary & Matheson (1974) detectaram múltiplas
formas de α-galactosidase em sementes de Cyamopsis tetragonolobus (guar),
no entanto, somente uma delas estava relacionada ao endosperma.
Assim, as diferentes isoformas de α-galactosidase podem estar
relacionadas tanto a diferenças estruturais do galactomanano durante a
mobilização, como por exemplo, a diminuição de substituições por galactose
demandaria uma α-galactosidase com maior afinidade ao substrato; como
também a diferentes funções além da degradação do galactomanano, por
exemplo, a degradação de açúcares da série rafinósica (Bewley,1997).
Somado à demonstração da presença de isoformas, a cromatografia de
troca aniônica, concentrou a α-galactosidase (Figura 15 e Tabela 1) e, embora
o extrato semipurificado ainda apresente considerável quantidade de proteínas
contaminantes (Figura 15), a cromatografia aniônica foi eficaz na remoção de
parte desta contaminação, visto pelo pico de proteína anterior a aplicação do
gradiente salino, denominado wash, que não interagiu com a coluna (Figura 13)
e aumento de aproximadamente 18 vezes (Tabela 1) na atividade enzimática
quando comparado com o extrato bruto.
A escolha da fração 47 a 51, pico A, para dar continuação a purificação
em coluna de gel filtração foi realizada por dois motivos: a alta atividade

105
enzimática encontrada, o que possivelmente indica maior concentração de
enzima e, portanto, maior rendimento ao final da purificação; e por entender
que a presença predominante desta isoforma somado ao momento do
desenvolvimento que estas sementes foram retiradas da incubação, pico da
mobilização de reserva, indique alta probabilidade desta enzima estar
relacionada a degradação do galactomanano.
A terceira etapa de purificação: Cromatografia de Gel Filtração com
Coluna Superose 6 não foi efetiva na purificação da α-galactosidase de
Sesbania virgata (Cav.) Pers., pois, como observado na figura 18, o perfil
proteico após purificação em Superose apresentou proteínas contaminantes,
no entanto, houve uma considerável concentração da enzima, visualizado pela
intensidade da banda identificada pela seta (figura 18) e aumento de 26 vezes
na atividade enzimática da α-galactosidase quando comparado com o extrato
bruto (Tabela 1).
Sendo a Superose 6 uma coluna de gel filtração, ou seja, purifica por
peso molecular, não era esperado que um pool de proteínas de diferentes
pesos moleculares fosse agrupado em um mesmo pico. Infere-se que a α-
galactosidase possa assumir estrutura quartenária com isoformas de peso
molecular diferente, pois possui peso molecular diferente sob condições
desnaturantes, 42 kDa, e na forma nativa, 72 kDa, como também a coluna
Superose 6 apresentar problemas na separação, como por exemplo ruptura de
gel.
Embora a purificação não tenha sido efetiva na eliminação completa de
proteínas contaminantes, ela foi efetiva na concentração da α-galactosidase
(Figura 18 seta; Tabela 1), o que permitiu o sequenciamento interno da enzima
e consequente identificação como α-galactosidase.
A tabela de purificação (Tabela 1) corrobora a semipurificação da α-
galactosidase de Sesbania virgata (Cav.) Pers., evidenciado pelo aumento de
26 vezes na atividade enzimática específica, ou seja, pela atividade enzimática
específica levar em consideração a quantidade total de proteínas (em mg) na
amostra, o aumento da atividade enzimática observada faz referência a
diminuição da quantidade total de proteínas e manutenção da atividade da
enzima.

106
Além disso, a tabela de purificação (Tabela 1) mostra um rendimento de
purificação menor que 1,5%, como visto no trabalho de (Lisboa et al., 2003),
rendimento baixo para a condução dos experimentos do Capítulo 2, dado que
nos fez caracterizar também a α-galactosidase comercial de Cyamopsis
tetragonoloba (Megazyme®). Dessa forma, se ambas as enzimas possuírem as
mesmas características, os experimentos do Capítulo 2 seriam realizados
somente com a enzima comercial.
Conjuntamente aos dados da tabela de purificação (Tabela 1), os dados
da eletroforese em gel de poliacrilamida sob condições desnaturantes (SDS-
PAGE) também mostram que as etapas de purificação da α-galactosidase não
foram eficientes na efetiva purificação, no entanto havendo uma concentração
da enzima e, consequentemente, uma semipurificação (Figura 25).
Ao compararmos, na figura 25, a raia identificada como extrato bruto (A)
e a raia após precipitação em Sulfato de Amônio 80% (B) nota-se ausência de
grandes mudanças no perfil proteico, pois, como dito anteriormente, a
saturação de 60% de Sulfato de Amônio deve ser a responsável pela retirada
de grande parte das proteínas contaminantes, porém com um custo alto na
perda de enzima disponível para os próximos passos de purificação.
Por outro lado, a comparação do perfil proteico do extrato bruto (Figura
25 A) com os perfis proteicos das cromatografias em FPLC (Figura 25 C e D),
nota-se a semipurificação, pela diminuição das bandas apresentadas e
concentração da banda identificada como α-galactosidase (seta).
O mesmo pode ser dito da comparação entre as cromatografias em
FPLC, Troca-iônica aniônica Mono-Q (Figura 25 C) e Gel Filtração Superose 6
(Figura 25 D), onde o perfil proteico apresenta pequenas diferenças, com
desaparecimento e diminuição de intensidade de bandas, no entanto, os perfis
se mostram muito semelhantes, evidenciando novamente a semipurificação.
A padronização da purificação da α-galactosidase, inicialmente, levava
em consideração um quarto passo de purificação a Cromatografia de Troca-
Iônica Catiônica em FPLC utilizando a coluna CM (carboximetil). A inclusão
desta etapa anteriormente a etapa de gel filtração se mostrou efetiva na
purificação da α-galactosidase, no entanto, a coluna CM utilizada já era muito
antiga e, além de reduzir muito a atividade enzimática da α-galactosidase,
apresentou sinais evidentes de mau funcionamento, como vazamentos e

107
despressurização (bolhas de ar), o que tornou esta etapa de purificação
inviável.
Assim, as etapas de purificação propostas, Precipitação em Sulfato de
Amônio, Cromatografia de Troca-Iônica Aniônica e cromatografia de Gel
Filtração, só foram eficazes na semipurificação e concentração da enzima
(Figura 25; Tabela 1).
Embora a α-galactosidase de Sesbania virgata (Cav.) Pers. só tenha
sido semipurificada, o sequenciamento interno foi possível e eficaz no
sequenciamento de 22% da enzima.
Observando a Figura 27, nota-se que a sequência da α-galactosidase de
Sesbania virgata (Cav.) Pers. possui alta homologia com a sequência já
descrita para Cyamopsis tetragonoloba, uma vez que em 99% das árvores de
relação realizadas ambas as enzimas formaram um ramo.
O sequenciamento interno das demais bandas permitiu excluir a
presença das demais enzimas da mobilização de reserva (Endo-β-mananase e
β-manosidase), possibilitando a utilização da alíquota semipurificada nos
experimentos apresentados no Capítulo 2, pois a ausência das demais
enzimas permite a análise do modo de ação da α-galactosidase
individualmente.
Além disso, a espectrometria de massas das demais bandas corroborou
a hipótese da α-galactosidase, possivelmente, formar estrutura quartenária,
pois as bandas 2, 3, 4, 5 e 8, de peso molecular menor do que a banda 1
(identificada como α-galactosidase) podem também ser identificadas como α-
galactosidase, apesar desta possibilidade não ser o primeiro Hit, o que poderia
evidenciar que as bandas apresentadas no gel de poliacrilamida não
representam uma única proteína, sendo assim, essas bandas seriam uma
mistura de isoformas de α-galactosidase com outras proteínas (Figuras 39 - 45,
Anexo).
De forma a corroborar a hipótese da estrutura quartenária, Tonini et al.
(2010) demonstraram por Western Blot utilizando anticorpo anti α-galactosidase
e extrato bruto enzimático de Sesbania virgata (Cav.) Pers. a presença de 8
bandas variando de 78 kDa a 12 kDa reativas ao anticorpo, evidenciando a
presença de isoformas da α-galactosidase com diferentes tamanhos, que
poderiam interagir entre si formando a estrutura quartenária.

108
A interação das isoformas de α-galactosidase formando a estrutura
quartenária pode significar que a enzima necessite da formação do complexo
de uma ou mais α-galactosidase para promover a mobilização de reserva, pela
retirada das ramificações de galactose do galactomanano, de forma mais
eficiente.
No entanto, Overbeeke & colaboradores (1989), Guerin & colaboradores
(1992), demonstraram que a α-galactosidase de Cyamopsis tetragonoloba
(guar) e a β-amilase de cevada podem ser ativadas através de digestão por
enzimas proteolíticas.
Tonini & colaboradores (2010) sugeriram que haja a ação de proteases
na ativação da α-galactosidase pré-formadas (peso molecular alto) e
armazenadas na semente de Sesbania virgata (Cav.) Pers. Dessa forma, pode-
se inferir que as proteases na realidade são α-galactosidases já ativas que
reconhecem uma α-galactosidase inativa e a cliva promovendo a ativação.
Além disso, durante a mobilização de reserva o polissacarídeo
apresentaria uma mudança no padrão de ramificação com galactose, o que
demandaria uma adaptação da α-galactosidase, esta adaptação poderia ser
novamente uma proteólise por outras α-galactosidases ativas diminuindo a
ainda mais o tamanho da enzima o que explicaria a variação de pesos
moleculares encontrada.
Dessa forma, tanto a α-galactosidase poderia formar uma estrutura
quartenária para agir sobre o galactomanano, na retirada das ramificações de
galactose, como também, e mais provável, a interação entre as α-
galactosidases seria para processamento da enzima, diminuindo o seu
tamanho, ativando-a e/ou adequando-a as mudanças ao polissacarídeo.
A enzima α-galactosidase já foi descrita ocorrendo em micro-
organismos, plantas e animais (Dey & Pridham, 1972) com o pH ótimo variando
entre 3.8 a 6.7 e temperatura ótima também bastante variada (Dey & Pridham,
1972; Ademark, P. et al., 2001).
Guimarães et al. (2001), Viana et al. (2002b) demonstraram
temperaturas ótimas de α-galactosidase de sementes de Glycine max (soja)
variedade Doko e CAC-1 nas faixas de 45°C-50°C e 40°C-55°C,
respectivamente. Lima et al. (2004) apresentam a caracterização de
temperatura ótima de α-galactosidase de cotilédones de Platymiscium

109
pubensces (Tamboril-da-mata) entre 50°C-55°C, enquanto Silva (2007)
apresentou a temperatura ótima da α-galactosidase presente em cotilédones
de Schizolobium parahyba (Guapuruvu) entre 40°C-45°C. Em todos os
trabalhos, os autores relatam queda acentuada da atividade enzimática em
60°C.
Da mesma maneira, estudos de caracterização de pH ótimo de α-
galactosidase de fontes diversas demonstram ampla variação nos valores.
Guimarães et al. (2001), Viana et al. (2002b) demonstraram pH ótimo de α-
galactosidase de sementes de Glycine max (soja) variedade Doko e CAC-1 nas
faixas de pH 5.0 a 5.5 e pH 5.0 a 6.0, respectivamente. Lima et al. (2004)
apresentaram a caracterização de pH ótimo de α-galactosidase de cotilédones
de Platymiscium pubensces (Tamboril-da-mata) entre pH 4.5 a 6.0, enquanto
Silva (2007) apresentou o pH ótimo entre 5.0 e 6.0 da α-galactosidase presente
em cotilédones de Schizolobium parahyba (Guapuruvu).
Dessa forma, os resultados apresentados de temperatura ótima,
variando entre 45°C a 50°C, e pH ótimo, variando na faixa pH 4.4 e pH 5.4, da
enzima α-galactosidase de sementes de Sesbania virgata (Cav.) Pers. em
Extrato Bruto corroboram com a bibliografia (Figura 8 e 9).
O pH ótimo da enzima α-galactosidase de Sesbania virgata (Cav.) Pers.
em extrato bruto (Figura 9) e semipurificada (Figura 21) é o mesmo, estando
em ambos os casos na faixa de 4,4 a 5,4. A temperatura ótima apresenta uma
ligeira diferença (em extrato bruto: 45°C a 50°C; semipurificada 50°C a 55°C),
mas pode-se considerar praticamente a mesma faixa de temperatura ótima
(Figuras 8 e19).
No entanto, a comparação da α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. e da α-galactosidase comercial (Megazyme®)
evidencia diferenças consideráveis em todos os quesitos analisados (Figuras
19 a 24), inclusive nos aspectos cinéticos, porém os valores de temperatura
ótima, pH ótimo da α-galactosidase comercial (Megazyme®) estão dentro dos
valores descritos na bibliografia (Dey & Pridham, 1972; Viana et al., 2002b;
Lima et al. (2004).
Tendo em vista as diferenças apresentadas entre as duas enzimas,
semipurificada e a comercial, os experimentos conduzidos no Capítulo 2 foram

110
realizados com ambas as enzimas, até para determinar se essas
características influenciavam no modo de ação da enzima em seu substrato.
Como apresentado anteriormente, as características de pH, temperatura
e aspectos cinéticos são bem distintos entre as α-galactosidases já descritas
de plantas (Dey & Pridham, 1972; Ademark, P. et al., 2001), assim, as
diferenças apresentadas pelas α-galactosidases estão de acordo com o que o
já foi descrito para α-galactosidases na literatura.
5.3. Análises das sequências de α-galactosidase previamente
identificadas em plantas
Feurtado et al. (2001) analisaram a sequência genômica da α-
galactosidase de Lycopersicum esculentum (tomate) determinando o gene em
5,7Kb, com a região de codificante da enzima composta por 14 éxons
interrompidos por 14 íntrons. Além disso, resultados da análise de hibridização
do Southern com a sonda de cDNA de α-galactosidase de tomate (LeaGal)
evidenciaram 1 única cópia do gene.
O número de genes em outras espécies não foi determinado, porém em
alfafa, guar, Ceratonia siliqua, soja, Phoenix dactylifera, café, feijão, Lupinus
angustifolius e Trifolium repens, que a α-galactosidase foi purificada, foram
determinadas, pelo menos, 2 isoformas da enzima (Courtois & Petek, 1966;
McCleary & Matheson, 1974; Williams et al., 1978; Itoh et al., 1979; Plant &
Moore, 1982; Chandra Sekhar & DeMason, 1990; Dhar et al., 1994).
A α-galactosidase de tomate não foi purificada, porém, Feurtado et al.
(2001) demonstrou a presença de 3 isoformas, através da focalização
isoelétrica, e concluiu que, pela presença de apenas 1 cópia do gene de α-
galactosidase, as diferenças apresentadas pelas isoformas são provenientes
de modificações pós-transcricionais.
A análise de cDNA da α-galactosidase de tomate, performada pelo
mesmo autor, determina que a enzima madura possui 364 aminoácidos de
comprimento, 39,8kDa, pI de 4,91 e é precedida por um peptídeo sinal. A
presença de peptídeos sinais já foi reportada em café, guar, soja, feijão
(Overbeeke et al., 1989; Zhu & Goldstein, 1994; Davis et al., 1996, 1997).

111
A presença de peptídeo sinal está relacionada ao envio das proteínas a
mais de um local dentro da célula, como a parede celular, aos corpos protéicos
ou embrião, evidenciando, possivelmente diferentes funções exercidas pelas α-
galactosidases, dependendo do local para onde são enviadas (Feurtado et al.,
2001).
Feurtado et al. (2001) demonstraram, através do alinhamento da
sequência de aminoácidos de α-galactosidase de tomate com sequências de
aminoácidos de α-galactosidases de outras eudicotiledôneas, que a proteína α-
galactosidase é conservada entre as espécies, apresentando cerca de 70% de
identidade.
Além disso, a análise dessas sequências somadas as sequências de
aminoácidos de fungos e metazoários na forma de árvore de relação produziu
3 clados distintos acompanhando a taxonomia (Feurtado et al., 2001) e
totalizou 40 resíduos conservados entre as α-galactosidases de eucariotos,
entre eles a assinatura característica de α-galactosidase: G - [LIVMFY] - x -
[LIVMFY] - x - [LIVM] - D - [DF] - x - W - x - [RV] - [DNSF] (Hofman et al.,
1999).
A análise das 35 sequências de aminoácidos de α-galactosidases de
plantas, através da construção da árvore de relação (Figura 46, Anexo),
realizada no presente trabalho se mostrou bastante similar a árvore
apresentada por Feurtado et al. (2001), com alguns ramos acompanhando a
filogenia, porém a visualização não é tão clara devido a maior quantidade de
sequências analisadas. Diferentemente a Feurtado & colaboradores (2001),
neste trabalho preferiu-se a análise somente de sequências de plantas, visando
entender as relações neste clado.
Por sua vez, a análise das 35 sequências de aminoácidos de α-
galactosidases já descritas em plantas conjuntamente com a análise da
sequencia de aminoácidos da α-galactosidase de Sesbania virgata (Cav.)
Pers., através da construção da árvore de relação, quando comparada com a
árvore de relação somente das 35 sequências de aminoácidos apresenta uma
mudança na formação dos ramos e suas relações, o que aparentemente
melhorou a explicação sobre as possíveis funções das α-galactosidases
analisadas.

112
Da mesma forma que a árvore de relação das 35 sequências de
aminoácidos, análise conjunta com a α-galactosidase sequenciada também
agrupou algumas das sequências da enzima segundo a filogenia (Figura 27, A-
F), entretanto, a maioria dos ramos não acompanhou a filogenia, o que pode
ser devido a um agrupamento da enzima pela função exercida: degradação da
série rafinósica, mobilização de reserva, entre outros.
Observa-se na Figura 27, cujos ramos identificados como A, B e C são
ramos que acompanham a filogenia, família Fabaceae, que as α-
galactosidases de leguminosas não se agruparam em um único ramo, mas em
três com uma relação muito distante um do outro, corroborando a hipótese que
as α-galactosidases estão sendo agrupadas de acordo com a função,
independentemente, se acompanham a filogenia ou não.
O ramo b, ramo que agrupa as sequências de α-galactosidases de
Sesbania virgata (Cav.) Pers. e Cyamopsis tetragonolaba, muito provavelmente
agrupa as α-galactosidases responsáveis pela retirada das galactoses do
galactomanano durante a mobilização de reservas da semente, pois
sabidamente essas espécies possuem como reserva de semente o
galactomanano, reserva de parede celular (McCleary & Matheson, 1974;
Buckeridge & Dietrich, 1990).
No entanto o ramo c, que agrupa Glycine max e Phaseolus vulgaris
espécies que acumulam reserva proteica e oleogenosa, e amilácea,
respectivamente, a α-galactosidase presente seria, provavelmente, responsável
pela degradação de açúcares da série rafinósica, nos primeiros estágios da
germinação.
O ramo a, por sua vez, por estar agrupado juntamente a espécies como
Carica papaya e Cucumis sativus, espécies produtoras de frutos com
amadurecimento de fruto e, consequentemente, degradação da parede celular,
deve conter α-galactosidases responsáveis pelo remodelamento do
polissacarídeo durante a desmontagem da parede celular no amadurecimento
do fruto (Gao & Schaffer, 1999; Soh et al., 2006), e, no caso das espécies
agrupadas (Glycine Max e Pisum sativum) no processo da mínima deposição
de galactomanano existente nestas sementes.
Sugere-se que as sequências de aminoácidos de α-galactosidase que
não se agruparam de acordo com a filogenia, sejam α-galactosidases com

113
funções similares, não necessariamente relacionadas à degradação do
galactomanano, mas à degradação de oligossacarídeos da série rafinósica ou
de substratos galactosilados estocados no citoplasma, como demonstrado por
McCleary & Matheson (1974) que das múltiplas isoformas apresentadas pelo
guar, apenas uma estava relacionada à degradação de galactomanano.
A α-galactosidase sequenciada quando comparada as demais 35
sequências possui alta similaridade com a α-galactosidase de Cyamopsis
tetragonoloba (ramo consistente: 99%) e possivelmente possui a mesma
função, de mobilização de reserva, sendo também utilizada nos experimentos
conduzidos no Capítulo 2, como comparação no modo de ação da enzima.
VI. Conclusão
Baseado em todos os resultados apresentados, nota-se que as etapas
de purificação escolhidas para a purificação da α-galactosidase de Sesbania
virgata (Cav.) Pers. (precipitação com Sulfato de Amônio 80%, Cromatografia
de Troca-Iônica aniônica – Mono-Q, Cromatografia de Gel Filtração – Superose
6), não foi efetiva na purificação da α-galactosidase, mas sim em uma
semipurificação (Figura 25).
A semipurificação da α-galactosidase de Sesbania virgata (Cav.) Pers.
se mostrou livre das demais enzimas de mobilização de reserva (endo-β-
mananase e β-manosidase) e a análise da sequência da α-galactosidase
semipurificada evidenciou que esta realmente está envolvida na mobilização de
reserva.
Embora só tenha ocorrido a semipurificação da α-galactosidase de
Sesbania virgata (Cav.) Pers. foi possível a detecção de isoformas da α-
galactosidase, possivelmente estando estas isoformas relacionadas a
diferentes funções (figuras 13 e 14, B e C) e adaptações da enzima a
mudanças do polissacarídeo durante a mobilização de reservas (Figuras 26 e
27).
As duas isoformas encontradas após purificação em Mono-Q (Figuras 13
e 14, B e C) estariam relacionadas a outras funções, enquanto, as isoformas de

114
diferentes pesos moleculares encontradas após purificação em Superose
(Figuras 25 D, 38 a 42 e 45, Anexo) estariam relacionadas com a mobilização
de reserva (42kDa) e adaptações da enzima a mudanças do galactomanano
(pesos moleculares menores).
Foi possível observar uma mudança do peso molecular da enzima α-
galactosidase em condições desnaturantes (42kDa) e nativas (72kDa) (Figuras
16 e 18) possibilitando a inferência na formação de complexos enzimáticos
(estrutura quartenária) na forma nativa, sugerindo que a própria α-
galactosidase catalisaria a sua proteólise ativando α-galactosidases inativas
(manteria o peso molecular) e modificando a enzima para se adaptar as
mudanças no padrão de ramificação (diminuiria o peso molecular da enzima).
Dessa forma, é possível especular sobre a necessidade de proteases
somente no início do processo de mobilização de reserva, para a ativação das
primeiras α-galactosidases, que seriam modificadas proteoliticamente durante o
restante do processo, como proposto por Tonini et al. (2010).
A α-galactosidase de Sesbania virgata (Cav.) Pers. semipurificada e
caracterizada foi submetida a ensaios com o polissacarídeo nativo,
galactomanano, da mesma espécie, visando a compreensão do modo de ação
da enzima e, principalmente, da estrutura fina do galactomanano (Capítulo 2).

115
VII. Referências Bibliográficas
ADEMARK, P.; LARSSON, M.; TJERNELD, K.; STALBRAND, H. 2001. Multiple α-
galactosidases from Aspergillus niger: purification, characterization and substrate
specificities. Enzyme and Microbial Technology 29:441–448.
ALCÂNTARA, P.H.N., DIETRICH, S.M.C. & BUCKERIDGE, M.S. 1999. Xyloglucan mobilisation
and purification of a (XLLG/XLXG) specific β-galactosidase from cotyledons of Copaifera
langsdorffii. Plant Physiol. Biochem., 37 (9), 653-663.
BEWLEY, J. D. & BLACK, M. 1994. Seeds: Physiology of development and germination. 2ºed.
Plenum Press, New York. 445p.
BRADFORD, M.M. 1976. A rapid and sensitive method for quantification of microgan quantities
of proteins utilizing the principle of rotein-dye binding. Analytical Biochemistry 72:248-254.
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1990. Galactomannan from Brazilian seeds. Revista
Brasileira de Botânica 13:109-112.
BUCKERIDGE, M.S.; PANEGASSI, V.R. & DIETRICH, S.M.C. 1995a. Storage carbohydrate
mobilisation in seeds of Dimorphandra mollis Benth. (Leguminosae) following germination.
Revista Brasileira de Botânica 18:171-175.
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1996. Mobilisation of the raffinose family
oligosaccharides and galactomannan in germinating seeds of Sesbania marginata Benth.
(Leguminosae-Faboideae). Plant Science 117:33-43.
BUCKERIDGE, M.S. & REID, J.S.G. 1996. Major cell wall polysaccharides in legume seeds:
Structure, catabolism and biological functions. Ciência e Cultura 48:153-162.
BUCKERIDGE, M.S.; DIETRICH, S.M.C. & LIMA, D.U. 2000a. Galactomannans as the reserve
carbohydrate in legume seeds. In: GUPTA, A.K. & KAUR, N. Carbohydrate Reserves in
Plants - Synthesis and Regulation. Elsevier Science, Amsterdam. p. 283-317.
BUCKERIDGE, M.S.; SANTOS, H.P. & TINÉ, M.A.S. 2000b. Mobilisation of storage cell wall
polysaccharides in seeds. Plant Physiology and Biochemistry 38:141-156.
BUCKERIDGE, M.S.; TINÉ, M.A.S.; SANTOS, H.P. & LIMA, D.U. 2000c. Polissacarídeos de
reserva de parede celular em sementes. Estrutura, metabolismo, funções e aspectos
ecológicos. Revista Brasileira de Fisiologia Vegetal, Edição Especial, p.137-162.
BUCKERIDGE, M.S.; AIDAR, M.P.M.; SANTOS, H.P. & TINÉ, M.A.S. 2004a. Acúmulo de
reservas. In: FERREIRA, A.G. & BORGHETTI, F. Germinação: do básico ao aplicado.
Artmed, Porto Alegre. p. 31-50.
CANTAREL, B.L.; COUTINHO, P.M.; RANCUREL, C.; BERNARD, T.; LOMBARD, V. &
HENRISSAT, B. 2008. The Carbohydrate-Active EnZymes database (CAZy): an expert
resource for Glycogenomics. Nucleic Acids Research: 1–6.
CHANDRA, S.K.N. & DEMASON, D.A. 1990. Identification and immunocytochemical
localization of α-galactosidase in resting and germinating date palm (Phoenix dactylifera L.)
seeds. Planta 181:53-61.

116
COURTOIS, J.E. & PETEK, F. 1966. α-galactosidase from coffee beans. Methods in
Enzymology 8:565-670.
DAVIS, M.O.; HATA, D.J.; JOHNSON, S.A.; WALKER, J.C. & SMITH D.S. 1996. Cloning,
expression and characterization of a blood group B active recombinant α-D-galactosidase
from soybean (Glycine max). Biochemistry and Molecular Biology international 39: 471-485.
DAVIS, M.O.; HATA, D.J.; JOHNSON, S.A.; JONES, D.E.; HARMATA, M.A.; EVANS,
M.L.;WALKER, J.C. & SMITH D.S. 1997. Cloning, expression and characterization of a
blood group B active recombinant α-D-galactosidase from pinto bean (Phaseolus vulgaris).
Biochemistry and Molecular Biology international 42: 453-467.
DEY, P.M. & PRIDHAM, J.B. 1972. Biochemistry of α-galactosidases. Advan. Enzymol. 36:
91:130.
DHAR, M.; MITRA, M.; HATA, J.; BUTNARIU, O. & SMITH, D. 1994. Purification and
characterization of Phaseolus vulgaris α-D-galactosidase isozymes. Biochemistry and
Molecular Biology International 34:1055-1062.
DEMASON, D.A.; MADORE, M.A.; SEKHAR, K.N.C. & HARRIS, M.J. 1992. Role of α-
galactosidase in cell wall metabolism of date palm (Phoenix dactylifera) endosperm.
Protoplasma 166:177-186.
FEURTADO, J.A.; BANIK, M. & BEWLEY, J.D. 2001. The cloning and characterization of -
galactosidase present during and following germination of tomato (Lycopersicon esculentum
Mill.) seed. Journal of Experimental Botany 52:1239-1249.
FUJIMOTO, Z.; KANEKO, S.; MOMMA, M.; KOBAYASHI, H. & MIZUNO, H. 2003. Crystal
structure of rice -galactosidase complexed with D-galactose. J. Biol. Chem. 278 (22):
20313-20318.
GAO, Z. & SCHAFFER, A.A. 1999. A novel alkaline α-galactosidase from melon fruit with a
substrate preference for raffinose. Plant Physiology 119:979-987.
GROOT, S.P.C. & KARSSEN, C.M. 1992. Dormancy and germination of abscisic acid-deficient
tomato seeds - studies with the sitiens mutant. Plant Physiology 99:952-958.
GUERIN JR, LANCE RCM, WALLACE W. 1992 Release and activation of barley b-amylase by
malt endopeptidases. J Cereal Sci 15:5–14.
GUIMARÃES, V.M; REZENDE, S.T.; MOREIRA, M.A.; BARROS, E.G. & FELIX, C.R. 2001.
Characterization of α-galactosidase from germinating soybean seed and their use for
hydrolisis of oligossacharides. Phytochemistry 58:67-73.
HALMER, P. & BEWLEY, J.D. 1979. Mannanase production by the lettuce endosperm: control
by the embryo. Planta 144:333-340.
HAMES B. D. 1990. One-dimensional polyacrylamide gel electrophoresis. In: Hames BD,
Rickwood D (eds) Gel electrophoresis of proteins: a practical approach, 2nd edn. Oxford
University Press, New York.
HOFFMAN, K.; BUCHER, P.; FLAQUET, L.; BAIROCH, A. 1999. The PROSITE database, its
status in 1999. Nucleic Acids Research 27:215-219.

117
ITOH, T.; SHIMURA, S.; ADACHI, S. 1979. Α-galactosidase from alfafa seeds (Medicago sativa
L.). Agicultural and Biological Chemistry 43:1499-1504.
JARVIS, M.C. & MCCANN, M.C. 2000. Macromolecular biophysics of the plant cell wall:
concepts
LAEMMLI U.K. 1970. Cleavage of structural protein during the assembly of the head of
bacteriophage T4. Nature, 223, 680-685.
LIMA, E.E.; DE REZENDE, S.T.; BORGES, R.C.G. & PEREZ, S.C.J.A. 2004. Caracterização
de α´galactosidase em embrião e cotilédones de sementes de Platymiscium pubescens
Micheli, Var. Pubecens (Tamboril da mata). Revista Brasileira de Sementes, Vol. 26, nº 2,
p.82-90.
LISBOA, C.G.S. 2003. Endo-β-mananase de endosperma de Sesbania virgata (Cav.) Pers.:
purificação, caracterização e importância na germinação e desenvolvimento da plântula.
Dissertação de Mestrado. Universidade Estadual de Campinas, SP.
LISBOA, C.G.S; TONINI, P.P.; TINÉ, M.A.S. & BUCKERIDGE, M.S. 2006. Endo--mananase
from the endosperm of seeds of Sesbania virgata (Cav.) Pers. (Leguminosae): purification,
characterisation and its dual role in germination and early seedling growth. Braz. J. Plant
Physiol., 18(2):269-280.
MANZI, A.E. & CEREZO, A.S. 1984. The galactomannan-like oligosaccharides from the
endosperm of seed of Gleditsia triacanthos. Carbohydrate Research 134:115-131.
McCLEARY, B.V. & MATHESON, N.K. 1974. -D-galactosidase activity and galactomannan
and galactosylsucrose depletion in germinating legume seeds. Phytochemistry 13:1747-
1757.
MCCLEARY, B.V. & MATHESON, N.K. 1975. Galactomannan structure and β-mannanase and
β-mannosidase activity in germinating legume seeds. Phytochemistry 14:1187–1194.
McCLEARY, B.V.; MATHESON, N.K & SMALL, D.M. 1976. Galactomannans and a
galactoglucomannan in Legume seed endosperms: structural requirements for β-
mannanase hydrolysis. Phytochemistry 15:1111-1117.
McCLEARY, B.V. 1983. Enzymic interaction in the hydrolysis of galactomannan in germinating
guar: the role of exo--mannanase. Phytochemistry 22:649-658.
McIlvaine, T. C. 1921. A buffer solution for colorimetric comparison. Journal of Biological
Chemistry 49:183-189.
MOE, O.A.; MILLER, S.E. & IWEN, M.H. 1947. Investigation of the reserve carbohydrate of
leguminous seeds. I. Periodate oxidation. Journal of the American Chemical Society
69:2621-2825.
OVERBEEKE, N.; FELLINGER, A.J.; TOONEN, M.Y.; VAN WASSENAAR, D. & VERRIPS, C.T.
1989. Cloning and nucleotide sequence of the α-galactosidase cDNA from Cyamopsis
tetragonoloba (guar). Plant Molecular Biology 13:541-550.
PETEK, F. & TO DONG. 1961. Enzymologia, 23, 133.
PLANT, A.R. & MOORE, K.G. 1982. Α-D-Mannosidase and α-D-galactosidase from proteins
bodies of Lupinusangustifolius cotyledons. Phytochemistry 21:985-989.

118
POTOMATI, A. & BUCKERIDGE, M.S. 2002. Effect of abscisic acid on the mobilisation of
galactomannan and embryo development of Sesbania virgata (Cav.) Pers. (Leguminosae-
Faboideae). Revista Brasileira de Botânica 25:303-310.
RAVEN, P.H.; EVERT, R.F. & EICHHORN, S.E. 2001. Biologia Vegetal. 6ºed. Editora
Guanabara Koogan, Rio de Janeiro. 906p.
REID, J.S.G. 1985a. Cell wall storage carbohydrates in seeds. Biochemistry of the seeds gums
and hemicelluloses. Advances in Botanical Research 11:125-155.
REID, J.S.G. & BEWLEY, J.D. 1979. A dual role for the endosperm and its galactomannan
reserve in the germinative physiology of fenugreek (Trigonella foenum-graecum L.) an
endospermic leguminous seed. Planta 147:145-150.
REID, J.S.G. & EDWARDS, M. 1995. Galactomannans and other cell wall storage
polysaccharides in seeds. In: STEPHEN, A.M. Food polysaccharides and their applications.
Marcel Dekker, New York. p. 155-186.
REID, J.S.G. & MEIER, H. 1972. The function of the aleurone layer during galactomannan
mobilization in germination seeds of fenugreek (Trigonella foenum-graecum L.), crimson
clover (Trifolium incarnatum L.): a correlative biochemical and ultrastructural study. Planta
106:44-60.
SCHALLER A. 2004. A cut above the rest: the regulatory function of plant proteases. Planta
220:183–197.
SEILER, A. 1977. Galaktomannanabbau in keimenden Johanisbrotsamen (Ceratonia siliqua L.).
Planta 134:209-221.
SILVA, C.O. 2001. Papel da β-galactosidase no mecanismo de degradação do xiloglucano de
parede celular durante a mobilização de reserva em cotilédones de Copaifera lansgsdorffii.
Dissertação de Mestrado apresentada a Universidade Estadual de Campinas.
SILVA, G.A. 2007. Purificação parcial e caracterização de uma α-galactosidase de sementes
de Guapuruvu (Schizolobium parahyba). Dissertação de Mestrado pela Universidade de
Brasília.
SOH, C-P; ALI, Z.M. & LAZAN, H. 2006. Characterisation of an α-galactosidase with potential
relevance to ripening related texture changes. Phytochemistry 67:242-254.
SOMME, R. 1968. The structure of a galactomannan from Medicago Lupulina L. Acta Chemica
Scandinavica 22:537-545.
TAMURA K, DUDLEY J, NEI M & KUMAR S. 2007. MEGA4: Molecular Evolutionary Genetics
Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24:1596-1599.
TONINI, P.P.; LISBOA, C.G.S.; FRESCHI, L.; MERCIER, H.; MAZZONI-VIVEIROS, S.C. &
BUCKERIDGE, M.S. 2006. Effect of abscisic acid on galactomannan degradation and endo-
β-mannanase activity in seeds of Sesbania virgata (Cav.) Pers. (Leguminosae). Trees:
structure and function 20:669-678.
TONINI, P.P.; LISBOA, C.G.S.; SILVAI, C.O.; MAZZONI-VIVEIROS, S.C. & BUCKERIDGE,
M.S. 2007. Testa is involved in the control of storage mobilisation in seeds of Sesbania

119
virgata (Cav.) Pers., a tropical legume tree from of the Atlantic Forest. Trees: structure and
function 21:13-21.
TONINI, P. P. 2009. Conexão entre os processos de degradação das reservas de proteínas e
carboidratos e o efeito dos hormônios e açúcares em sementes Sesbania virgata de (Cav.)
Pers. Tese de Doutorado. Universidade Estadual de Campinas, SP.
TONINI, P. P.; CARRARA, T.B.; BUCKERIDGE, M. S. 2010. Storage proteins and cell wall
mobilisation in seeds of Sesbania virgata (Cav.) Pers. (Leguminosae). Trees: Structure and
Function v.24 (4): 675-684.
UNRAU, A.M. 1961. The constitution of a galactomannan from seeds of Leucaena
leucochephala. Journal of Organic Chemistry 26:3097-3101.
VIANA, S.F.; GUIMARÃES, V.M.; MOREIRA, M.A.; OLIVEIRA, M.G.A.; COSTA, N.M.B.; de
REZENDE, S.T. 2002b. Reduction of raffinose oligosaccharides in soymilk by alpha-
galactosidase from germinating soymilk seeds. In: REUNIÃO ANUAL DA SOCIEDADE
BRASILEIRA BIOQUÍMICA E BIOLOGIA MOLECULAR, 31., 2002, Caxambu. Anais
Caxambu: (SBBQ), p.95.
WEBER, K. & OSBORN, M. 1969. The Reliability of Molecular Weight Determinations by
Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis. J. Biol. Chem. 244: 4406-4412.
WILLIAMS, J.; VILLARROYA, H. & PETEK, F. 1978. Α-galactosidase II, III and IV from seeds of
Trifolium repens: Purification, physiochemical properties and mode of galactomannan
hydrolysis in vitro. Biochemical Journal 175:1069-1077.
ZHU, A. & GOLDSTEIN, J. 1994. Cloning and functional expression of DNA encoding coffe
bean α-galactosidase. Gene 140:227-231.

120
CAPÍTULO 2
ESTRUTURA FINA DO GALACTOMANANO E A MODULAÇÃO DA MOBILIZAÇÃO DE RESERVA EM SEMENTES DE Sesbania
virgata (CAV.) PERS.

121
I. Resumo
Todas as células vegetais possuem parede celular. Além das funções
típicas da parede celular primária, a parede celular ainda pode apresentar
especializações, como o acúmulo de alguns polissacarídeos específicos, em
sementes, que constituem uma reserva de carbono para o crescimento da
plântula após a germinação. O galactomanano é um dos polissacarídeos
hemicelulósicos que podem exercer este papel de reserva de parede celular.
Os galactomananos são polissacarídeos constituídos de uma cadeia principal
de resíduos de D-manose ligadas β-(1,4), ramificada por resíduos de D-
galactose α-(1,6) ligados. A mobilização deste ocorre após a germinação e
envolve três enzimas hidrolíticas: α-galactosidase, endo-β-mananase e exo-β-
manosidase. A compreensão da distribuição dos grupos de D-galactose ao
longo da cadeia principal de D-manano é determinar a estrutura fina do
polissacarídeo. Embora haja diferenças na estrutura fina de galactomananos
de diferentes espécies, demonstrou-se a correlação direta entre o conteúdo de
galactose e a viscosidade do polissacarídeo, bem como sugeriram a existência
de um requerimento mínimo de substituições por galactose para manter o
polissacarídeo solúvel. A quantidade e distribuição das substituições com D-
galactose ao longo da cadeia principal (1,4)-β-D-manano estão intima e
diretamente relacionadas à taxa de hidrólise do polissacarídeo pela
endohidrolase endo-β-mananase. Além da regulação da mobilização pelos já
esperados fitohormônios, existe a evidência em xiloglucanos de reserva que a
estrutura do polissacarídeo, através de suas ramificações pela galactose,
codificaria, pelo menos, uma parte da informação para a sua própria
degradação, o que se denominou código do xiloglucano. Visando evidenciar a
presença de um código de degradação presente na estrutura fina do
galactomanano, procedeu-se com hidrólises enzimáticas do galactomanano de
Sesbania virgata (Cav.) Pers. utilizando a enzima endo-β-mananase de
Aspergillus niger (Megazyme®) somente ou em conjunto com a α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. (Capítulo 1) ou com a α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®), seguido

122
de análise dos oligossacarídeos por HPAEC-PAD. Além disso, também
procedeu-se com hidrólises enzimáticas de galactomananos de 6 espécies com
razão manose:galactose variando de 1:1 a 150:1 com endo-β-mananase de
Aspergillus niger (Megazyme®) e análise dos oligossacarídeos produzidos por
HPAEC-PAD. Os dados obtidos das comparações dos oligossacarídeos
gerados em cada hidrólise sugerem que as ramificações do galactomanano
podem modular o reconhecimento de sítios de clivagem pela endo-β-
mananase: (1) existe a produção de oligossacarídeos limites de digestão F1,
F2 e F3 (Figura 28) após hidrólise do galactomanano com endo-β-mananase,
como demonstrado para xiloglucanos; (2) os oligossacarídeos F1 possuem
proporções distintas quando da hidrólise do galactomanano com endo-β-
mananase em diferentes concentrações (Experimento I e Experimento IV),
evidenciando preferência por sítios com menor grau de galactosilação; (3) a
presença da α-galactosidase diminui a produção dos oligossacarídeos F2 e
F3, mostrando que a estes não possuem resistência intrínseca à hidrólise e
que a reação atinge o equilíbrio mesmo quando ainda existem sítios de
clivagem ainda disponíveis (Experimento III); (4) e que polissacarídeos com
estruturas diferentes, razão manose:galactose variando entre 150:1 a 1:1, são
digeridos em diferentes taxas de hidrólise pela mesma enzima, evidenciando
que a ramificação com galactose dificulta a ação da endo-β-mananase. Através
dos resultados obtidos neste capítulo, sugere-se que a estrutura do
polissacarídeo galactomanano também contenha, pelo menos, parte da
informação requerida para seu próprio metabolismo, código para a sua
degradação, estando esta informação contida na distribuição das ramificações
com resíduos de D-galactose.
II. Introdução
Todas as células vegetais possuem parede celular. Esta estrutura é
responsável por diversas funções, entre elas: propriedades mecânicas da
parede celular, determinação da forma celular, manutenção do ambiente iônico
celular, defesa contra invasão de patógenos, sinalização e controle do

123
potencial hídrico celular (Street & Öpik, 1984; Waldron & Brett, 1990; Carpita &
Gibeaut, 1993).
A diversidade de funções exercidas pela parede celular reflete a
diversidade da composição da parede celular. Segundo Carpita & Gibeaut
(1993), a parede celular primária é composta por três domínios independentes
com funções e composição distintas: domínio celulósico-hemicelulósico,
péctico e protéico.
Além das funções típicas da parede celular primária, citadas acima, a
parede celular ainda pode apresentar especializações, entre elas: o acúmulo
de alguns polissacarídeos específicos, em sementes, que constituem uma
reserva de carbono para o crescimento da plântula após a germinação (Bewley
& Black, 1978; Buckeridge & Reid, 1996).
Entre os polissacarídeos que podem exercer este papel de reserva
estão: o xiloglucano, o galactano e o (galacto)manano. Exceto pelo galactano,
que constitui um acúmulo do domínio péctico, os demais polissacarídeos são
do domínio hemicelulósico (celulose-hemicelulose) (Buckeridge et al., 2000c).
Os galactomananos são polissacarídeos constituídos de uma cadeia
principal de resíduos de D-manose ligadas β-(1,4), ramificada por resíduos de
D-galactose α-(1,6) ligados (Buckeridge et al., 2000a). As substituições por
galactose variam de espécie para espécie, se apresentando em razões
Manose:Galactose (Man:Gal) de 1:1 a 4:1, ou seja, na proporção máxima (1:1)
teoricamente todos os resíduos de manose estão substituídos por galactose,
enquanto que na razão 4:1 a cada quatro resíduos de manose, apenas 1 é
ramificado com galactose (Buckeridge & Dietrich, 1990; Buckeridge et al.,
2000a).
A razão manose:galactose, e por consequência, a distribuição dos
resíduos de galactose, de acordo com Buckeridge & Dietrich (1990) e
Buckeridge et al. (2000a, c), são aspectos importantes do polissacarídeo,
estando relacionados à solubilidade da molécula e, consequente interação
entre os polissacarídeos.
A compreensão da distribuição dos grupos de D-galactose ao longo da
cadeia principal de D-manano é determinar a estrutura fina do polissacarídeo
(McCleary et al., 1985).

124
Diversas técnicas já foram empregadas em estudos de distribuição de
resíduos de D-galactose ao longo da cadeia principal de D-manano de
galactomananos de reserva de sementes de leguminosas (McCleary et al.,
1985).
Palmer & Ballantyne (1950) determinaram a estrutura do galactomanano
de Cyamopsis tetragonolobus (guar), como arranjo de resíduos de manose
substituídos e não-substituídos por galactose, através de raio-X. Em 1978,
Hoffman & Svensson, através de procedimento químico, determinaram que o
arranjo das substituições de D-galactose na cadeia principal de D-manano, no
galactomanano presente em guar, era aos pares e aos trios, contradizendo o
estudo de Palmer & Ballantine (1950), onde as substituições por D-galactose
foram determinadas como randômicas.
Estudos mais recentes, conduzidos por McCleary et al. (1985), utilizando
uma estratégia alternativa para análise da estrutura fina do galactomanano,
empregando enzimas particulares de degradação do galactomanano (endo-β-
mananase, principalmente) e análise da composição dos oligossacarídeos
produzidos, determinaram que a estrutura fina do galactomanano de Ceratonia
siliqua possui substituições por D-galactose não-regulares, típico de
substituições randômicas, enquanto o galactomanano de Cyamopsis
tetragonolobus (guar) possui as ramificações agrupadas aos duplos e aos trios,
não se apresentando randômica.
Embora haja diferenças na estrutura fina de galactomananos de
diferentes espécies, McCleary & colaboradores (1976) demonstraram a
correlação direta entre o conteúdo de galactose e a viscosidade do
polissacarídeo, bem como sugeriram a existência de um requerimento mínimo
de substituições por galactose para manter o polissacarídeo solúvel.
No mesmo contexto, McCleary & Matheson (1983) concluiram que a
quantidade e distribuição das substituições com D-galactose ao longo da
cadeia principal (1,4)-β-D-Manano estão intima e diretamente relacionadas à
taxa de hidrólise do polissacarídeo pela endohidrolase endo-β-mananase.
Neste mesmo trabalho, endo-β-mananases provenientes de plantas
foram ensaiadas com galactomananos altamente ramificados e o perfil de
oligossacarídeos analisado por cromatografia de gel filtração. A análise dos
perfis evidenciou, através da baixa quantidade de trissacarídeos (manotriose –

125
Man3 - e manobiose com uma galactose – Gal1Man2), que há limitação na ação
da enzima em polissacarídeos altamente ramificados.
Utilizando endo-β-mananase de fungos a taxa de hidrólise reportada ao
guar (38% de substituição com galactose) foi entre 2%-12% (Reese & Shibata,
1965; Courtois & Le Dizet, 1968; Tsujisaka et al., 1972; Emi et al., 1972). No
entanto, taxas de hidrólise para galactomananos contendo menores taxas de
substituição foram maiores: para Ceratonia siliqua (23% de substituição com
galactose) entre 15%-39% (Reese & Shibata, 1965; Courtois & Le Dizet, 1968);
para café (2% de substituição com galactose) entre 36%-58% (Tsujisaka et al.,
1972; Emi et al., 1972).
Tiné et al. (2003), analisando os oligossacarídeos produzidos pela
hidrólise com celulase de xiloglucano de reserva de diferentes espécies
(Tamarindus indica, Copaifera langsdorffii e Hymenea courbaril), constituídos
de diferentes taxas de substituição por galactose, demonstraram que
xiloglucanos de estrutura diferente são hidrolisados diferentemente; que os
oligossacarídeos menos ramificados são atacados preferencialmente pela
enzima e que a presença da β-galactosidase juntamente com a celulase,
impede a formação de oligossacarídeos complexos F2 e F3 (combinações de
oligossacarídeos menores), sugerindo que o padrão das substituições por D-
galactose possuem influencia direta na ação enzimática.
Dessa forma, eles concluem que a estrutura do polissacarídeo, através
de suas ramificações pela galactose, codificaria, pelo menos, uma parte da
informação para a sua própria degradação (Tiné et al., 2003), o que Buckeridge
(2010) denominou código do xiloglucano.
O trabalho de Tiné e colaboradores (2003) foi possível pela identificação
prévia da composição da estrutura dos oligossacarídeos produzidos após
hidrólise com celulase realizada por Buckeridge et al. (1992), pois permitiu
identificar sítios preferenciais da enzima pela presença ou ausência de
ramificação (estrutura do polissacarídeo).
Dessa forma, no presente trabalho objetivou-se compreender a
importância das ramificações de galactose presentes nos galactomananos na
atuação da endoenzima endo-β-mananase, visando determinar se a estrutura
fina do galactomanano também possui, pelo menos, parte da informação para

126
a sua própria degradação, ou seja, determinar a existência de um código em
galactomananos.
III. Material e Métodos
3.1. Material Vegetal
As sementes de Sesbania virgata (Cav.) Pers. utilizadas na execução
dos experimentos de determinação da estrutura fina foram coletadas de
diversos indivíduos localizados nas imediações da Universidade de São Paulo,
São Paulo/SP, evitando, assim, vício amostral. As sementes foram separadas
dos frutos maduros e mantidas em frascos herméticos a temperatura ambiente.
As sementes de Senna alata, Dimorphandra mollis, Cyamopsis
tetragonoloba, Coffea arabica e Euterpe edulis utilizadas na execução do
experimento de determinação e comparação das estruturas finas dos
galactomananos de diferentes proporções de galactose, foram adquiridas do
banco de sementes presente no Laboratório de Fisiologia Ecológica de Plantas
(Lafieco).
3.2. Extração do manano e galactomanano
Os polissacarídeos de reserva (manano ou galactomanano) das 6
espécies (Sesbania virgata, Senna alata, Dimorphandra mollis, Cyamopsis
tetragonoloba, Coffea arabica e Euterpe edulis) foram extraídos de suas
sementes a fim de possibilitar os experimentos de hidrólise enzimática visando
o entendimento da estrutura fina do polissacarídeo (manano ou
galactomanano) em cada espécie e a importância das ramificações de
galactose na modulação do processo de mobilização de reserva.

127
3.2.1. Extração do manano de reserva das sementes de Coffea arabica e
Euterpe edulis
A extração do manano presente nas sementes de Coffea arabica e de
Euterpe edulis foi feita, através do fracionamento da parede celular a partir do
pulverizado das sementes, método outro do utilizado para a extração do
galactomanano presente nas demais sementes, visto as diferenças de
solubilidade encontrada entre os dois polissacarídeos.
O fracionamento de parede celular inicia-se com a extração de açúcares
solúveis do pulverizado de sementes através de 4 lavagens com etanol 80% a
80°C por 20 minutos sob agitação, seguida de centrifugação a 1000g por 10
minutos e descarte do sobrenadante. Ao final das 4 etapas de extração de
açúcar solúvel o precipitado foi seco em estufa a 60°C por 24 horas.
Em seguida, realizou-se a extração de pectinas através de 3 lavagens
com oxalato de amônio 0,5% pH 7, seguido de centrifugação a 2700g por 15
minutos com descarte do sobrenadante e secagem do precipitado em estufa a
60°C por 24 horas.
Finalmente, realizou-se a extração do manano através de 3 lavagens
com hidróxido de sódio 8M, seguido de centrifugação a 3000g por 15 minutos
com descarte do precipitado, neutralização com ácido acético glacial e diálise
contra água do sobrenadante. O dialisado foi congelado a -20°C e seco em
liofilizador.
3.2.2. Extração do galactomanano de reserva das sementes de Sesbania
virgata, Senna alata, Dimorphandra mollis, Cyamopsis
tetragonoloba
A extração do galactomanano contido no endosperma das sementes de
Sesbania virgata, Senna alata, Dimorphandra mollis e Cyamopsis
tetragonoloba foi realizada segundo Buckerigde & Dietrich (1996). Partindo de
endosperma e tegumento de sementes após 12 horas de embebição, isolados,
secos e pulverizados em moinho de bolas foi extraído o galactomanano com
água quente (80°C) por 6 horas, seguido de filtração através de dupla camada
de tecido (cheesecloth). O extrato filtrado foi centrifugado a 10.000g por 30 min

128
a 5°C, seguido de precipitação do sobrenadante com 3 volumes de etanol a
5°C por 12 horas (overnight). O precipitado gerado após centrifugação a 3000g
por 5 minutos a 5°C foi coletado, seco e pesado.
3.3. Análise de monossacarídeos
A análise de monossacarídeos dos polissacarídeos extraídos das seis
espécies (Sesbania virgata, Senna alata, Dimorphandra mollis, Cyamopsis
tetragonoloba, Coffea arabica e Euterpe edulis) foi realizada através da
hidrólise ácida com H2SO4, a fim de determinar a razão manose:galactose de
cada (galacto)manano, ou seja, determinar a proporção de ramificação com
galactose.
Cinco miligramas de cada polissacarídeo foi hidrolisado a seus
monossacarídeos constituintes utilizando 72% H2SO4 – 4% H2SO4 (Saeman,
Buhl & Harris, 1945). A solução de monossacarídeos foi neutralizada com
NaOH 8M até pH entre 6 e 7, submetidas às resinas de troca iônica Dowex®
para retirada do sal e analisadas em HPAEC-PAD em coluna Carbo Pac PA-1
(DIONEX, USA) em sistema Dionex DX-500, tendo como eluentes com fluxo 1
mL/min: NaOH 20mM, água e pós-coluna contendo NaOH 500mM.
3.4. Hidrólise enzimática com Endo-β-Mananase e α-Galactosidases
Todo o estudo de determinação da estrutura fina do polissacarídeo
galactomanano, da compreensão da função das ramificações por galactose na
modulação da mobilização de reserva e determinação no modo de ação da
enzima α-galactosidase, foram realizados através de ensaios enzimáticos
utilizando galactomanano ou manano comercial (Megazyme®) como substrato,
a enzima endo-β-mananase de Aspergilus niger (Megazyme®) individualmente
ou em conjunto com as α-galactosidases (de Cyamopsis tetragonoloba –
Megazyme® – e a semipurificada de Sesbania virgata (Cav.) Pers. (Capítulo
1)).
O (galacto)manano utilizado como substrato nas hidrólises enzimáticas
foi proveniente de sementes de Sesbania virgata (Cav.) Pers. individualmente,

129
ou em conjunto com o (galacto)manano de sementes de 5 espécies (Senna
alata, Dimorphandra mollis, Cyamopsis tetragonoloba, Coffea arabica e
Euterpe edulis), enquanto o manano comercial (Megazyme®) é proveniente do
galactomanano de goma caroba submetido a tratamento enzimático e químico.
A concentração do substrato foi diferente dependo do experimento, estando na
concentração de 1% ou 0,1%.
Como o substrato, as enzimas utilizadas também variaram a
concentração dependendo do experimento. A endo-β-mananase foi utilizada
em 4 concentrações 0,005U/mL, 0,05U/mL, 0,5U/mL e 1U/mL, enquanto a α-
galactosidase comercial (Megazyme®) foi inicialmente diluída para 0,5U/mL e,
como a α-galactosidase semipurificada, foi diluída para 4 concentrações
diferentes: 1:5, 1:10, 1:100 e 1:1000.
A digestão enzimática, em si, foi preparada a partir de uma solução de
galactomanano à 1% ou 0,1% (peso/volume) em tampão acetato de amônio
50mM pH 5.0, na qual foi acrescido a endo-β-mannanase e/ou α-galactosidase
na concentração desejada.
A incubação da mistura foi realizada a 30°C por 15 horas (overnight) ou
24 horas, com posterior fervura por 5 minutos (adaptado de Cavalari, 2009).
Após a fervura, os tubos foram centrifugados à 2700g, por 5 minutos a
temperatura ambiente e o sobrenadante coletado. Ao sobrenadante foi
acrescido 3 volumes de etanol absoluto, para a precipitação do polissacarídeos
não digeridos, centrifugado à 4000g por 3 minutos, o sobrenadante coletado,
seco em concentrador de amostras (sem aquecimento) e então ressuspendido
em 1 mL de água deionizada para análise dos oligossacarídeos formados pela
digestão enzimática, por HPAEC-PAD e/ou TLC.
Os experimentos conduzidos para determinação da estrutura fina,
importância da ramificação de galactose na regulação da mobilização e o modo
de ação da α-galactosidase podem ser divididos e dois grupos. No primeiro
grupo estão cinco experimentos (Experimentos I a V) de hidrólise enzimática do
galactomanano de Sesbania virgata (Cav.) Pers. (a 1% ou 0,1%) ou manano
comercial (Megazyme) com as enzimas endo-β-mananase (0,005U/mL,
0,05U/mL e 1U/mL) e α-galactosidases (0,5U/mL, 1:5, 1:10, 1:100 e 1:1000) em
diferentes concentrações.

130
Enquanto o segundo grupo de experimentos utilizou galactomanano de
seis espécies (Sesbania virgata, Senna alata, Dimorphandra mollis, Cyamopsis
tetragonoloba, Coffea arabica e Euterpe edulis) a 1% e somente a enzima
endo-β-mananase a 0,5U/mL.
A descrição de cada experimento se encontra nos itens: 3.6. Análise da
estrutura fina do galactomanano de Sesbania virgata (Cav.) Pers. através de
hidrólises enzimáticas com Endo-β-manase de Aspergilus Níger (Megazyme),
α-Galactosidase de Cyamopsis tetragonoloba (Megazyme) e α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. e 3.7. Análise da estrutura fina
do (Galacto)manano de 6 espécies com diferentes proporções de ramificações
com galactose, respectivamente.
3.5. Análise dos oligossacarídeos de (Galacto)manano
Os oligossacarídeos produzidos pelas digestões enzimáticas com endo-
β-mananase sozinha ou em conjunto com a α-galactosidase foram analisados
por HPAEC-PAD (High Performance Anion Exchange Chromatography with
Pulsed Amperometric Detection) e/ou TLC (Thin Layer Chromatography).
3.5.1. Análise dos oligossacarídeos de (Galacto)manano por HPAEC-PAD
(High Performance Anion Exchange Chromatography with Pulsed
Amperometric Detection)
A análise por HPAEC-PAD de oligossacarídeos produto das hidrólises
enzimáticas de (galacto)mananos foi desenvolvido por Crivellari, A.C.(2009 –
dados não publicados) e performado no modelo ICS-3000 (Dionex®), utilizando
coluna CarboPac PA-100. O eluente de base utilizado foi 88mM de NaOH e o
eluente de arraste acetato de sódio 200mM em NaOH 88mM. Um gradiente da
solução de acetato de sódio 200mM em NaOH 88mM foi aplicado à amostra
visando separar os oligossacarídeos antes de sua detecção pelo detector de
pulso amperométrico.

131
3.5.2. Análise dos oligossacarídeos de (Galacto)manano por TLC (Thin
Layer Cromatography)
A análise por cromatografia em camada delgada de oligossacarídeos
produto das hidrólises enzimáticas de (galacto)mananos foi realizada como
uma segunda maneira de analisar a diferença entre os oligossacarídeos da
digestão das 6 espécies analisadas (Sesbania virgata, Senna alata,
Dimorphandra mollis, Cyamopsis tetragonoloba, Coffea arabica e Euterpe
edulis).
O material hidrolisado ressuspendido em água deionizada foi aplicado a
1 cm da base da placa de sílica (Merck®) de 20x20cm e submetido à 3 corridas
utilizando como eluente propanol:água:nitrometano na proporção 5:3:2.
A revelação da placa foi feita com a aspersão de solução etanólica
contendo 5% de ácido sulfúrico, seguido de incubação em estufa a 100°C e
fotografia do perfil de oligossacarídeos apresentados após revelação.
3.6. Análise da estrutura fina do galactomanano de Sesbania
virgata (Cav.) Pers. através de hidrólises enzimáticas com
Endo-β-manase de Aspergilus Níger (Megazyme), α-
Galactosidase de Cyamopsis tetragonoloba (Megazyme) e α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
As hidrólises enzimáticas conduzidas nos cinco experimentos descritos
abaixo visavam elucidar: a estrutura fina do galactomanano de Sesbania
virgata (Cav.) Pers., através da identificação de cada oligossacarídeo; a
importância das substituições de galactose na atuação da endoenzima endo-β-
mananase, observada através da comparação dos oligossacarídeos formados
na presença da α-galactosidase e na ausência de mesma; e a compreensão do
modo de ação da enzima α-galactosidase, através da determinação de sítios
preferenciais de ataque, visualizado pelos oligossacarídeos formados em
diferentes concentrações da endo-β-mananase.

132
3.6.1. Experimento I
As hidrólises enzimáticas conduzidas no Experimento I visaram produzir
o substrato para o Experimento II (oligossacarídeos provenientes da hidrólise
do galactomanano de Sesbania virgata (Cav.) Pers. com endo-β-mananase) e
analisá-lo quanto à proporção dos oligossacarídeos produzidos nas diferentes
concentrações da enzima. Além disso, o Experimento I visou produzir
oligossacarídeos de manano puro (sem ramificações com galactose) variando
de monossacarídeo (manose) a heptassacarídeos (oligossacarídeo de 7
manoses) de manose para identificar nas análises de HPAEC-PAD os
oligossacarídeos ramificados e não ramificados, possibilitando a determinação
da composição de cada oligossacarídeo (hidrólise do manano comercial).
Dessa forma, as digestões enzimáticas foram preparadas a partir de
uma solução de (galacto)manano de Sesbania virgata (Cav.) Pers. à 1% e
manano comercial à 1% (peso/volume) em tampão acetato de amônio 50mM
pH 5.0, na qual foi acrescido o volume equivalente a concentração final de
0,005U/mL, 0,05U/mL e 1U/mL de endo-β-mannanase de Aspergilus niger
(Megazyme®), obtendo-se um volume final de ensaio de 1 mL para cada
concentração de enzima.
As hidrólises foram mantidas a 30°C por 15 horas (overnight) e após 5
minutos de fervura foram submetidas ao protocolo de precipitação com 3
volumes de etanol, já descritas no item 3.4. hidrólise enzimática com endo-β-
mananase e α-galactosidases, para posterior análise dos oligossacarídeos
produzidos e submissão dos oligossacarídeos ao Experimento II.
A metodologia de análise dos oligossacarídeos, produtos desta hidrólise
por HPAEC-PAD, é a mesma descrita no item: 3.5. Análise dos
oligossacarídeos de (Galacto)manano.
Para cada substrato (galactomanano ou manano) em cada concentração
de endo-β-mananase (0,005U/mL, 0,05U/mL e 1U/mL) a hidrólise enzimática
foi realizada em duplicata. Além disso, dois controles foram feitos:
galactomanano de Sesbania virgata (Cav.) Pers. e manano comercial
(Megazyme®) em tampão acetato de amônio 50mM pH 5.0 sem a adição de
enzima.

133
3.6.2. Experimento II
As hidrólises enzimáticas conduzidas no Experimento II visaram produzir
a desramificação da galactose dos oligossacarídeos provenientes da digestão
do galactomanano de Sesbania virgata (Cav.) Pers. produzidos no Experimento
I e, consequente, identificação da composição desses oligossacarídeos quanto
a quantidade de manose e galactose presente, através da comparação dos
cromatogramas da análise em HPAEC-PAD do Experimento I com do
Experimento II.
Dessa forma, as digestões enzimáticas foram preparadas a partir de
200µL dos oligossacarídeos do Experimento I (provenientes da hidrólise do
galactomanano e manano) ressuspendidos em tampão acetato de amônio
50mM pH 5.0, no qual foi acrescido 10 µL de α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. ou 10 µL de α-galactosidase de Cyamopsis
tetragonoloba (Megazyme®) a 0,5U/mL.
As hidrólises foram mantidas a 30°C por 15 horas (overnight) e após 5
minutos de fervura foram submetidas ao protocolo de precipitação com 3
volumes de etanol, já descritas no item 3.4. hidrólise enzimática com endo-β-
mananase e α-galactosidases, para posterior análise dos oligossacarídeos
produzidos por HPAEC-PAD.
A metodologia de análise dos oligossacarídeos, produtos desta hidrólise
por HPAEC-PAD, é a mesma descrita no item: 3.5. Análise dos
oligossacarídeos de (Galacto)manano.
Para combinação de cada substrato (oligossacarídeos de
galactomanano ou de manano) com cada α-galactosidase (semipurificada ou
comercial) a hidrólise enzimática foi realizada em duplicata. Além disso, três
controles foram feitos: galactomanano de Sesbania virgata (Cav.) Pers. a 1% e
α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers. (duplicata),
galactomanano de Sesbania virgata (Cav.) Pers. a 1% e α-galactosidase
Comercial (Megazyme®) (duplicata) e galactomanano de Sesbania virgata
(Cav.) Pers. a 1% e celulase de Trichoderma longibachiatum (Megazyme®).

134
3.6.3. Experimento III
As hidrólises enzimáticas conduzidas no Experimento III visaram
compreender a importância das ramificações de galactose no processo de
degradação do galactomanano, através da comparação das hidrólises do
polissacarídeo galactomanano a 1% com endo-β-mananase (Megazyme®) a 1U
somente (Experimento I) e do polissacarídeo galactomanano a 1% com endo-
β-mananase (Megazyme®) a 1U conjuntamente a α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. ou a α-galactosidase de
Cyamopsis tetragonoloba (Megazyme®).
Dessa forma, a comparação da diversidade e da concentração dos
oligossacarídeos formados nas hidrólises contendo ou não a α-galactosidase
pode sugerir que a ramificação com galactose de alguma forma modula a
degradação do galactomanano.
Assim, as digestões enzimáticas foram preparadas a partir de uma
solução de galactomanano de Sesbania virgata (Cav.) Pers. à 1% em tampão
acetato de amônio 50mM pH 5.0, na qual foi acrescido 10 µL de endo-β-
mannanase de Aspergilus niger (Megazyme®) a 1U/mL somente ou
conjuntamente com 10 µL de α-galactosidase a 0,5U/mL (semipurificada de
Sesbania virgata (Cav.) Pers. ou de Cyamopsis tetragonoloba (Megazyme®)).
A concentração de 0,5U/mL só é válida para a α-galactosidase
comercial, a α-galactosidase semipurificada está na concentração após
purificação, ou seja, sem qualquer diluição.
As hidrólises foram mantidas a 30°C por 15 horas (overnight) e após 5
minutos de fervura foram submetidas ao protocolo de precipitação com 3
volumes de etanol, já descritas no item 3.4. hidrólise enzimática com endo-β-
mananase e α-galactosidases, para posterior análise dos oligossacarídeos
produzidos por HPAEC-PAD.
A metodologia de análise dos oligossacarídeos, produtos desta hidrólise
por HPAEC-PAD, é a mesma descrita no item: 3.5. Análise dos
oligossacarídeos de (Galacto)manano.
Todas as hidrólises enzimáticas foram realizadas em duplicata.

135
3.6.4. Experimento IV
As hidrólises enzimáticas conduzidas no Experimento IV visaram
compreender o modo de ação da enzima endo-β-mananase (Megazyme) em
diferentes concentrações (0,005U/mL, 0,05U/mL e 1U/mL) quando na presença
da α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers. e a α-
galactosidase comercial (Megazyme®), ambas na máxima concentração de
0,5U/mL e galactomanano nas concentrações de 1% e 0,1%.
Dessa forma, a comparação dos oligossacarídeos formados nas
diferentes concentrações da endo-β-mananase na mesma concentração da α-
galactosidase (0,5U/mL) e em concentrações distintas do galactomanano (1%
ou 0,1%) pode evidenciar preferência em alguns sítios de clivagem na cadeia
de principal de manano.
Assim, as digestões enzimáticas foram preparadas a partir de uma
solução de galactomanano de Sesbania virgata (Cav.) Pers. à 1% ou 0,1% em
tampão acetato de amônio 50mM pH 5.0, na qual foi acrescido 10 µL de endo-
β-mannanase de Aspergilus niger (Megazyme®) (0,005U/mL, 0,05U/mL e
1U/mL) e 10 µL de α-galactosidase semipurificada de Sesbania virgata (Cav.)
Pers. (0,5U/mL – concentração pós-purificação) ou 10 µL de α-galactosidase
de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL.
A concentração de 0,5U/mL só é válida para a α-galactosidase
comercial, a α-galactosidase semipurificada está na concentração após
purificação, ou seja, sem qualquer diluição.
As hidrólises foram mantidas a 30°C por 15 horas (overnight) e após 5
minutos de fervura foram submetidas ao protocolo de precipitação com 3
volumes de etanol, já descritas no item 3.4. hidrólise enzimática com endo-β-
mananase e α-galactosidases, para posterior análise dos oligossacarídeos
produzidos por HPAEC-PAD.
A metodologia de análise dos oligossacarídeos, produtos desta hidrólise
por HPAEC-PAD, é a mesma descrita no item: 3.5. Análise dos
oligossacarídeos de (Galacto)manano.
Para combinação de cada concentração da endo-β-mananase
(0,005U/mL, 0,05U/mL e 1U/mL) com cada concentração (1% ou 0,1%) do

136
substrato galactomanano e cada α-galactosidase (semipurificada ou comercial)
a 0,5U/mL, a hidrólise enzimática foi realizada em duplicata.
3.6.5. Experimento V
As hidrólises enzimáticas conduzidas no Experimento V visaram produzir
oligossacarídeos de manano puro (sem ramificações com galactose) variando
de manose a heptassacarídeos de manose para identificar nas análises de
HPAEC-PAD os oligossacarídeos ramificados e não ramificados, possibilitando
a determinação da composição de cada oligossacarídeo (hidrólise do manano
comercial).
Dessa forma, as digestões enzimáticas foram realizadas em 2 ciclos de
hidrólises, no primeiro ciclo a α-galactosidase comercial a 0,5U/mL acrescida
de uma solução de manano a 1% (Megazyme®) em tampão acetato de amônio
50mM pH 5.0, e no segundo ciclo de digestão o manano foi submetido à
hidrólise com a enzima endo-β-mananase (Megazyme®) em diferentes
concentrações (0,005U/mL, 0,05U/mL e 1U/mL).
As hidrólises foram mantidas a 30°C por 15 horas (overnight) e após 5
minutos de fervura foram submetidas a centrifugação para descarte do
sobrenadante e, em seguida, o precipitado (manano desgalactosilado) foi
submetido a novo ciclo de digestão, agora com endo-β-mananase.
Novamente as hidrólises foram mantidas a 30°C por 15 horas (overnight)
e após 5 minutos de fervura foram submetidas ao protocolo de precipitação
com 3 volumes de etanol, já descritas no item 3.4. hidrólise enzimática com
endo-β-mananase e α-galactosidases, para posterior análise dos
oligossacarídeos produzidos por HPAEC-PAD. A metodologia de análise dos
oligossacarídeos, produtos desta hidrólise por HPAEC-PAD, é a mesma
descrita no item: 3.5. Análise dos oligossacarídeos de (Galacto)manano.
Para combinação de cada concentração da endo-β-mananase
(0,005U/mL, 0,05U/mL e 1U/mL) com a α-galactosidase comercial a 0,5U/mL e
substrato galactomanano a 1%, a hidrólise enzimática foi realizada em
duplicata.

137
3.7. Análise da estrutura fina do (Galacto)manano de 6 espécies
com diferentes proporções de ramificações com galactose
Visando determinar as diferenças na estrutura fina do polissacarídeo
(manano ou galactomanano) de cada espécie (Sesbania virgata, Senna alata,
Dimorphandra mollis, Cyamopsis tetragonoloba, Coffea arabica e Euterpe
edulis) e a importância das substituições de galactose na atuação da
endoenzima endo-β-mananase, impedindo-a de hidrolisar o polissacarídeo a
oligossacarídeos, realizou-se o experimento que comparou os oligossacarídeos
(produtos da hidrólise com endo-β-mananase), das 6 espécies vegetais, que
sabidamente possuem proporções de substituição por D-galactose diferentes,
ou seja, razão manose:galactose variando de 1:1 a 500:1, através de hidrólises
enzimáticas e posterior análise por HPAEC-PAD e TLC dos oligossacarídeos
formados.
As digestões enzimáticas foram preparadas a partir de uma solução de
(galacto)manano (de cada espécie) à 1% (peso/volume) em tampão acetato de
amônio 50mM pH 5.0, na qual foi acrescido o volume equivalente a
concentração final de 0,5U de endo-β-mannanase, obtendo-se um volume final
de ensaio de 1 mL. As hidrólises foram mantidas a 30°C por 24 horas e após 5
minutos de fervura foram submetidas ao protocolo de precipitação com 3
volumes de etanol, já descritas no item 3.4. hidrólise enzimática com endo-β-
mananase e α-galactosidases, para posterior análise dos oligossacarídeos.
A metodologia de análise dos oligossacarídeos, produtos desta
hidrólise, por HPAEC-PAD e TLC, são as mesmas descritas no item: 3.5.
Análise dos oligossacarídeos de (Galacto)manano.

138
IV. Resultados
4.1. Análise da estrutura fina do galactomanano de Sesbania
virgata (Cav.) Pers. através de hidrólises enzimáticas com
Endo-β-manase de Aspergilus niger (Megazyme), α-
Galactosidase de Cyamopsis tetragonoloba (Megazyme) e α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
As hidrólises enzimáticas conduzidas nos cinco experimentos descritos
visavam elucidar: a estrutura fina do galactomanano de Sesbania virgata (Cav.)
Pers., através da identificação de cada oligossacarídeo; a importância das
substituições de galactose na atuação da endoenzima endo-β-mananase,
observada através da comparação dos oligossacarídeos formados na presença
da α-galactosidase e na ausência de mesma; e a compreensão do modo de
ação da enzima α-galactosidase, através da determinação de sítios
preferenciais de ataque desta enzima, visualizado pelos oligossacarídeos
formados após hidrólise da α-galactosidase e hidrólise da endo-β-mananase
em diferentes concentrações.
4.1.1. Experimento I
As hidrólises enzimáticas conduzidas no Experimento I visaram produzir
o substrato para o Experimento II (oligossacarídeos provenientes da hidrólise
do galactomanano de Sesbania virgata (Cav.) Pers. com endo-β-mananase) e
analisá-lo quanto à proporção dos oligossacarídeos produzidos nas diferentes
concentrações da enzima. Além disso, o Experimento I visou produzir
oligossacarídeos de manano puro (sem ramificações com galactose) variando
de monossacarídeo (manose) a decassacarídeos (oligossacarídeo de 10
manoses) de manose para identificar, nas análises de HPAEC-PAD, os
oligossacarídeos ramificados e não ramificados, possibilitando a determinação
da composição de cada oligossacarídeo (hidrólise do manano comercial).

139
A hidrólise enzimática do galactomanano de Sesbania virgata (Cav.)
Pers. resultou no perfil de oligossacarídeos apresentados na Figura 28. Os
oligossacarídeos são identificados como: oligossacarídeos F1, os quais variam
de dissacarídeos a decassacarídeos galactosilados ou não, enquanto os
oligossacarídeos identificados como F2 e F3 são oligossacarídeos F1
combinados aos pares e trios, respectivamente (Figura 28).
É possível observar nos primeiros 19 minutos de corrida um grupo de 6
picos identificados como oligossacarídeos F1, ou seja, os oligossacarídeos
base da estrutura do galactomanano.
Entre os 20 e 22 minutos de corrida, encontra-se um segundo grupo de
picos que são identificados como oligossacarídeos F2, ou seja, a combinação
aos pares dos oligossacarídeos F1. Já entre os minutos 24 e 27 da corrida,
encontra-se um terceiro grupo de picos identificados como oligossacarídeos
F3, a combinação aos trios dos oligossacarídeos F1 (Figura 28).
Ao visualizar e comparar o perfil cromatográfico apresentado na figura
28 com os demais experimentos (Experimento I a V) nota-se a junção dos
picos G e H e a separação dos oligossacarídeos F2 e F3, diferentemente do
encontrado nos Experimentos I a V, devido ao ajuste no programa do HPAEC-
PAD para justamente separar os oligossacarídeos F2 e F3.
Figura 28: Análise em HPAEC-PAD (Dionex) com coluna PA-100 dos oligossacarídeos formados
após hidrólise enzimática exaustiva com Endo-β-mannanase do galactomanano de Sesbania virgata
(Cav.) Pers., visando verificar perfil dos oligossacarídeos produto da hidrólise enzimática do
galactomanano com endo-β-mananase. (F1) dissacarídeos a decassacarídeos galactosilados ou não

140
base do polissacarídeo; (F2) combinação aos pares dos oligossacarídeos de F1; (F3) combinação aos
trios dos oligossacarídeos de F1.
Os produtos das hidrólises enzimáticas do galactomanano de Sesbania
virgata (Cav.) Pers. a 1% com endo-β-mananase de Aspergilus niger
(Megazyme) (0,005U, 0,05U e 1U) e a comparação aos pares (0,005Ux0,05U;
0,005Ux1U; 0,05Ux1U) dos oligossacarídeos formados se encontram na figura
29.
Nas três concentrações de endo-β-mananase utilizadas é possível
identificar os oligossacarídeos identificados de A-I e os oligossacarídeos
agrupados F2 e F3. Apesar da presença de todos os oligossacarídeos nas três
concentrações, a proporção entre os oligossacarídeos formados e, portanto, a
preferência na produção de cada oligossacarídeo difere em cada concentração
(Figura 29 II, III e IV).
O perfil cromatográfico comparativo da hidrólise enzimática do
galactomanano com endo-β-mananase a 0,005U e 0,05U (Figura 29 V)
evidencia a preferência da formação do pico C e G na concentração de 0,005U
de enzima, enquanto o pico A (manobiose) é preferencialmente formado em
0,05U.
O perfil cromatográfico comparativo da hidrólise enzimática do
galactomanano com endo-β-mananase a 0,005U e 1U (Figura 29 VI) apresenta
como oligossacarídeos preferencialmente produzidos na concentração de
0,005U os representados por B (manotriose), enquanto os picos A, C, E e H
são preferencialmente produzidos em 1U.
O perfil cromatográfico comparativo da hidrólise enzimática do
galactomanano com endo-β-mananase a 0,05U e 1U (Figura 29 VII) não difere
muito do comparativo a 0,005U e 1U (Figura 29 VI), onde o oligossacarídeo
preferencialmente produzido a 0,05U é a manotriose (pico B), enquanto os
oligossacarídeos formados preferencialmente por 1U são os presentes nos
picos A, C, E, G e H.

141
Figura 29: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do galactomanano de Sesbania virgata (Cav.) Pers
com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e 1U. (I) Controle (galactomanano sem endo-
B-mananase); (II) perfil de oligossacarídeos formados após hidrólise com endo-β-mananase na
concentração de 0,005U; (III) perfil de oligossacarídeos formados após hidrólise com endo-β-
mananase na concentração de 0,05U; (IV) perfil de oligossacarídeos formados após hidrólise com

142
endo-β-mananase na concentração de 1U; (V) comparação dos perfis de oligossacarídeos formados
após hidrólise com endo-β-mananase nas concentrações de 0,005U e 0,05U; (VI) comparação dos
perfis de oligossacarídeos formados após hidrólise com endo-β-mananase nas concentrações de
0,005U e 1U; (VII) comparação dos perfis de oligossacarídeos formados após hidrólise com endo-β-
mananase nas concentrações de 0,05U e 1U. (A) manobiose (M2); (B) manotriose (M3) e/ou
trissacarídeo (G1M2); (C) tetrassacarídeos (manotetraose – M4 e/ou G
1M3); (D) pentassacarídeos
(manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4 e/ou
G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I) oligossacarídeos >8
resíduos (G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos
identificados A-I são denominados oligossacarídeos F1; os oligossacarídeos restantes são
identificados como F2, quanto combinação aos pares, e F3, quando combinação aos trios.
A razão entre oligossacarídeos F1 e os oligossacarídeos F2+F3 (razão
F1/F2) representa o quanto a enzima endo-β-mananase foi capaz de reduzir o
polissacarídeo a oligossacarídeos F1 e/ou F2 e F3 e é realizada pela divisão da
somatória de todas as áreas dos picos denominados F1, pela somatória de
todas as áreas dos picos denominados F2+F3.
A razão F1/F2 encontrada para as concentrações de 0,005U, 0,05U e
1U foram 4,11; 2,85 e 5,26, respectivamente.
Os produtos das hidrólises enzimáticas do manano comercial
(Megazyme®) a 1% com endo-β-mananase de Aspergilus niger (Megazyme®)
(0,005U, 0,05U e 1U) e a comparação aos pares (0,005Ux0,05U; 0,005Ux1U;
0,05Ux1U) dos oligossacarídeos formados se encontram na figura 30.
Ao analisar os oligossacarídeos formados nas três concentrações de
enzima (0,005U, 0,05U e 1U) nota-se uma tendência na diminuição da
diversidade de oligossacarídeos formados. Nas concentrações de 0,005U e
0,05U de endo-β-mananase utilizadas é possível identificar os oligossacarídeos
identificados de A, B, D, E, H e F, picos F1, sem a detecção de qualquer pico
F2 e/ou F3. Na concentração de 1U de endo-β-mananase somente os picos A,
B e E são detectados, sem qualquer detecção de oligossacarídeos F2 e/ou F3
(Figura 30 II, III e IV).
Os oligossacarídeos comuns às três concentrações de endo-β-
mananase, A, B e E, possuem proporções distintas entre os oligossacarídeos
formados evidenciando, novamente, que a preferência na produção de cada
oligossacarídeo difere em cada concentração (Figura 30 II, III e IV).

143
O perfil cromatográfico comparativo da hidrólise enzimática do manano
com endo-β-mananase a 0,005U e 0,05U (Figura 30 V) evidencia a preferência
da formação do pico B (manotriose) na concentração de 0,005U de endo-β-
manananse, enquanto o pico A (manobiose) e pico E são preferencialmente
formados em 0,05U. Além disso, com exceção do pico D, os demais picos
possuem área total maior, indicando maior quantidade de oligossacarídeos, em
0,05U do que em 0,005U.
O perfil cromatográfico comparativo da hidrólise enzimática do manano
com endo-β-mananase a 0,005U e 1U (Figura 30 VI) apresenta como
oligossacarídeos preferencialmente produzidos na concentração de 0,005U os
representados por B (manotriose), D e F enquanto os picos A e E são
preferencialmente produzidos em 1U.
O perfil cromatográfico comparativo da hidrólise enzimática do manano
com endo-β-mananase a 0,05U e 1U (Figura 30 VII) não difere muito do
comparativo a 0,005U e 1U (Figura 30 VI), onde os oligossacarídeos
preferencialmente produzidos a 0,05U são a manotriose (pico B), pico D e pico
F enquanto o oligossacarídeo formado preferencialmente por 1U é o presente
no pico A (manobiose).
A presença de oligossacarídeos diferentes de manobiose (A) e
manotriose (B) nas hidrólises de manano com endo-β-mananase sugere a
presença remanescente de ramificações com galactose (Figura 30 V, VI, VII).
Dessa forma, realizou-se o Experimento V visando a retirada das galactoses
remanescentes, consequentemente, a hidrólise completa do manano a
manobiose e manotriose na maior concentração de endo-β-mananase (1U) e a
formação de oligossacarídeos de tamanho variado (monossacarídeo a
nonassacarídeo) sem ramificações de galactose na menor concentração
(0,005U), principalmente.
A comparação da área total dos oligossacarídeos formados nas três
concentrações de enzima permite a inferência na taxa de hidrólise do
polissacarídeo a oligossacarídeo F1 e é realizada pela soma da área de todos
os picos denominados F1.
A área total dos oligossacarídeos F1 encontrada para as concentrações
de 0,005U, 0,05U e 1U foi 308,747; 434,737 e 425,389, respectivamente.

144
Figura 30: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do manano comercial (Megazyme®) com endo-β-
mananase (Megazyme®) a 0,005U, 0,05U e 1U. (I) Controle (manano sem endo-B-mananase); (II)
perfil de oligossacarídeos formados após hidrólise com endo-β-mananase na concentração de
0,005U; (III) perfil de oligossacarídeos formados após hidrólise com endo-β-mananase na

145
concentração de 0,05U; (IV) perfil de oligossacarídeos formados após hidrólise com endo-β-
mananase na concentração de 1U; (V) comparação dos perfis de oligossacarídeos formados após
hidrólise com endo-β-mananase nas concentrações de 0,005U e 0,05U; (VI) comparação dos perfis
de oligossacarídeos formados após hidrólise com endo-β-mananase nas concentrações de 0,005U e
1U; (VII) comparação dos perfis de oligossacarídeos formados após hidrólise com endo-β-mananase
nas concentrações de 0,05U e 1U. (A) manobiose (M2); (B) manotriose (M3); (D) pentassacarídeos
(manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4 e/ou
G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5). Os oligossacarídeos
identificados A-F são denominados oligossacarídeos F1.
4.1.2. Experimento II
As hidrólises enzimáticas conduzidas no Experimento II visaram produzir
a desramificação da galactose dos oligossacarídeos provenientes da digestão
do galactomanano de Sesbania virgata (Cav.) Pers. produzidos no Experimento
I e, consequente, identificação da composição desses oligossacarídeos quanto
a quantidade de manose e galactose presentes, através da comparação dos
cromatogramas da análise em HPAEC-PAD do Experimento I com do
Experimento II.
Os produtos das hidrólises enzimáticas dos oligossacarídeos do
Experimento I (galactomanano de Sesbania virgata (Cav.) Pers. a 1% com
endo-β-mananase de Aspergilus niger (Megazyme®) a 0,005U, 0,05U e 1U)
com α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers., a
comparação do perfil de oligossacarídeos produto do Experimento I e do
Experimento II em cada concentração de endo-β-mananase e os controles da
hidrólise se encontram na figura 31.
O controle do polissacarídeo a 1% na ausência de qualquer enzima
(figura 31 I) mostra que o polissacarídeo apresenta em quantidade apreciável
os oligossacarídeos representados pelos picos C e G. Um segundo controle
com polissacarídeo a 1% e α-galactosidase semipurificada de Sesbania virgata
(Cav.) Pers. (Figura 31 II) mostra atividade esperada da α-galactosidase, pela
presença do pico de galactose, e os mesmos picos C e G apresentados pelo
controle do polissacarídeo a 1% sem qualquer enzima, evidenciando que não

146
há contaminação da enzima semipurificada por Endo-β-mananase ou β-
manosidase.
O terceiro controle com polissacarídeo a 1% e celulase de Trichoderma
longibachiatum (Megazyme®) (Figura 31 III) apresentou os mesmos picos C e
G apresentados pelo controle do polissacarídeo a 1% sem qualquer enzima,
evidenciando que o galactomanano não é digerido a oligossacarídeos pela
ação da celulase, bem como, demonstrando que o substrato de galactomanano
possui um grau de pureza considerável, sendo livre de xiloglucano, β-glucano e
celulose.
A ação da α-galactosidase semipurificada foi muito semelhante nos
oligossacarídeos produtos da hidrólise prévia com a endo-β-mananase nas três
concentrações, visto pela mesma diversidade de picos apresentados nos perfis
cromatográficos, picos estes identificados de A-M (F1) e dos oligossacarídeos
F2 e F3. (Figura 31 IV, V, VI).
Apesar da presença de todos os oligossacarídeos nas três
concentrações, existem diferenças na proporção entre os oligossacarídeos em
cada concentração de endo-β-mananase após ação da α-galactosidase
semipurificada (Figura 31 IV, V, VI).
A determinação da razão entre oligossacarídeos F1 e os
oligossacarídeos F2+F3 (razão F1/F2) após a ação da α-galactosidase
semipurificada sugere que ocorra a realocação dos oligossacarídeos F2 e F3
em picos de oligossacarídeos F1, visto que a perda das ramificações com
galactose diminuem os oligossacarídeos tornando-os F1, bem como pode
sugerir a realocação dos oligossacarídeos F1 ainda ramificados a
oligossacarídeos F1 menores. Assim, era esperado que após hidrólise com a α-
galactosidase a razão F1/F2 fosse maior da encontrada no Experimento I
(Tabela 2).
A razão F1/F2 encontrada após hidrólise com a α-galactosidase
semipurificada dos oligossacarídeos produtos da hidrólise do galactomanano
com endo-β-mananase nas concentrações de 0,005U, 0,05U e 1U foram: 4,92;
14,26 e 7,98, respectivamente, valores estes maiores do que os encontrados
após a hidrólise com endo-β-mananase no Experimento I (Tabela 2).

147
Tabela 2: Comparação da razão F1/F2 do Experimento I (hidrólise do galactomanano a 1%
somente com endo-β-mananase) com a razão do Experimento II (hidrólise dos oligossacarídeos do
Experimento I somente com α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers.).
Endo-β-Mananase
(U/mL)
Razão F1/F2
Experimento I
Razão F1/F2
Experimento II
0,005 U 4,11 4,92
0,05U 2,85 14,26
1U 5,26 7,98
O perfil cromatográfico comparativo da hidrólise enzimática do
galactomanano com endo-β-mananase a 0,005U (Experimento I) e da hidrólise
com α-galactosidase semipurificada dos oligossacarídeos produto da digestão
com endo-β-mananase na mesma concentração (Experimento II) (Figura 31
VII) evidencia o surgimento de três novos oligossacarídeos, picos K, L e M, e
desaparecimento do oligossacarídeo identificado pelo pico I, após ação da α-
galactosidase.
Além disso, após a hidrólise com α-galactosidase semipurificada dos
oligossacarídeos produtos da digestão com endo-β-mananase a 0,005U notou-
se a mudança na proporção dos picos de manobiose (A), manotriose (B),
tetrassacarídeos (C), hexassacarídeos (E) e octa e nonassacarídeos (G e H)
(Figura 31 VII). Esta mudança na proporção está explorada na Tabela 3.
Os perfis cromatográficos comparativos das hidrólises enzimáticas do
galactomanano com endo-β-mananase a 0,05U e 1U (Experimento I) e da
hidrólise com α-galactosidase semipurificada dos oligossacarídeos produtos
das digestões com endo-β-mananase nas mesmas concentrações
(Experimento II) (Figura 31 VIII e IX) apresentaram os mesmos resultados
descritos para a concentração de 0,005U: o surgimento de três novos
oligossacarídeos, picos K, L e M, desaparecimento do oligossacarídeo
identificado pelo pico I e mudança nas proporções dos picos A, B, C, E, G e H,
após ação da α-galactosidase semipurificada (Figura 31 VIII e IX).
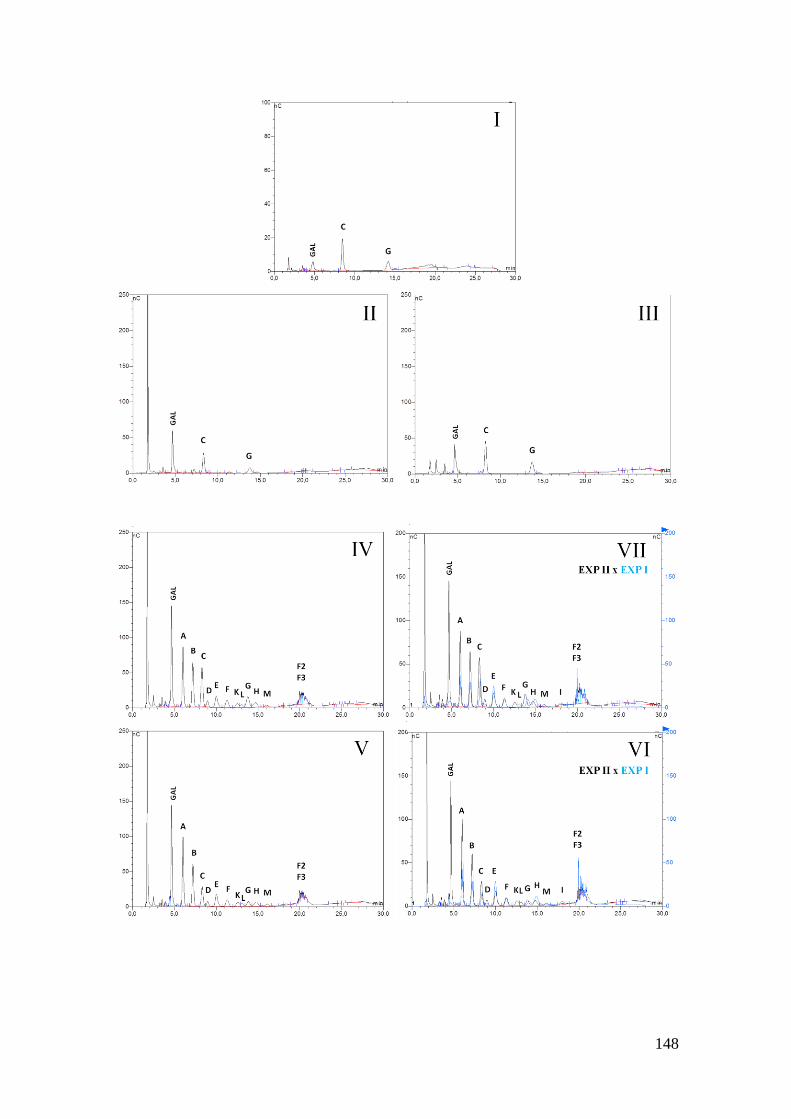
148

149
Figura 31: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da análise dos
oligossacarídeos, produto da hidrólise enzimática com α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers. dos oligossacarídeos produto da hidrólise do galactomanano a 1% de Sesbania
virgata (Cav.) Pers com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e 1U (Experimento I). (I)
Controle (galactomanano a 1% sem enzima); (II) Controle (galactomanano a 1% com α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.; (III) Controle (galactomanano a 1%
com celulase de Trichoderma longibrachiatum (Megazyme®)); (IV) perfil de oligossacarídeos
formados após ação da α-galactosidase sobre prévia hidrólise com endo-β-mananase a 0,005U; (V)
perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia hidrólise com endo-
β-mananase a 0,05U; (VI) perfil de oligossacarídeos formados após ação da α-galactosidase sobre
prévia hidrólise com endo-β-mananase a 1U; (VII) comparação dos perfis de oligossacarídeos
formados no Experimento I e Experimento II na concentração de 0,005U de endo-β-mannanase;
(VIII) comparação dos perfis de oligossacarídeos formados no Experimento I e Experimento II na
concentração de 0,05U de endo-β-mannanase; (IX) comparação dos perfis de oligossacarídeos
formados no Experimento I e Experimento II na concentração de 1U de endo-β-mannanase. (A)
manobiose (M2); (B) manotriose (M3); (C) tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6
e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I)
oligossacarídeos >8 resíduos (possíveis oligossacarídeos presentes: G1,3,4
M5; G4,5
M6, G3,4
M6;
G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7); (K-L) octassacarídeos e nonassacarídeos sem ramificação;
(M) oligossacarídeos >10 resíduos de manose (possivelmente resultado da desramificação do pico I).
Os oligossacarídeos identificados A-M são denominados oligossacarídeos F1; os oligossacarídeos
restantes são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos
trios.
McCleary et al. (1976) descreveu que a hidrólise completa de manano
com endo-β-mananase resultaria em manobiose e manotriose somente, sendo
a produção de manobiose maior do que a de manotriose, portanto, a
determinação da razão manobiose/manotriose (M2/M3) e a sua comparação
sob diferentes condições de hidrólise evidencia a taxa de conversão do

150
polissacarídeo e de oligossacarídeos grandes (F1, F2 e F3) aos produtos
máximos manobiose e manotriose.
Já a determinação e comparação da razão: manotriose/tetrassacarídeos
(pico C) (M3/C), sob diferentes condições de hidrólise permite a identificação
da mudança da proporção entre os picos, possibilitando a inferência na
estrutura do oligossacarídeo original (antes da ação da α-galactosidase) e após
a ação da α-galactosidase, bem como a inferência na realocação nos picos de
oligossacarídeos F1.
A tabela 3 reúne as razões M2/M3 e M3/C dos perfis cromatográficos
das hidrólises do galactomanano a 1% com endo-β-mananase a 0,005U, 0,05U
e 1U (Experimento I) e dos perfis cromatográficos das hidrólises com α-
galactosidase semipurificada dos oligossacarídeos produto das hidrólises do
galactomanano a 1% com endo-β-mananase a 0,005U, 0,05U e 1U
(Experimento II).
Tabela 3: Comparação das razões Manobiose (M2)/Trissacarídeo (M3 e/ou G1M2) (M2/M3) e
Trissacarídeo (M3 e/ou G1M2)/Tetrassacarídeo (M4 e/ou G
1M3) (M3/C) dos Experimentos I e II.
Razão Experimento I Experimento II
0,005U 0,05U 1U 0,005U 0,05U 1U
M2/M3 1,05 1,22 3,35 1,11 1,34 3,36
M3/C 0,74 1,53 0,39 1,01 1,91 0,48
De forma geral, todas as razões analisadas sofreram um aumento
quando houve a adição da α-galactosidase semipurificada, permitindo a
sugestão de realocação dos picos F1 B e C nos picos A e B.
A razão M2/M3 após ação da α-galactosidase aumenta
independentemente da concentração de endo-β-mananase utilizada
previamente, evidenciando que a retirada das galactoses proporciona um
aumento na digestibilidade do polissacarídeo, pelo aumento na taxa de
conversão de oligossacarídeos F1, F2 e F3 e polissacarídeo aos produtos
máximos de hidrólise: manobiose e manotriose.
Pela análise dos resultados da hidrólise dos oligossacarídeos do
Experimento I com α-galactosidase de Cyamopsis tetragonoloba (Megazyme®)

151
o modo de ação das α-galactosidases não difere, sendo o resultado muito
semelhante ao descrito para α-galactosidase semipurificada, portanto, a figura
47 expõe os resultados das digestões com α-galactosidase comercial se
encontra no Anexo.
4.1.3. Experimento III
As hidrólises enzimáticas conduzidas no Experimento III visaram
compreender a importância das ramificações de galactose no processo de
degradação do galactomanano, através da comparação das hidrólises do
polissacarídeo galactomanano a 1% com endo-β-mananase (Megazyme®) a 1U
somente (Experimento I) e do polissacarídeo galactomanano a 1% com endo-
β-mananase (Megazyme®) a 1U conjuntamente a α-galactosidase
semipurificada de Sesbania virgata (Cav.) Pers. ou a α-galactosidase de
Cyamopsis tetragonoloba (Megazyme®).
Dessa forma, a comparação da diversidade e da concentração dos
oligossacarídeos formados nas hidrólises contendo ou não a α-galactosidase
pode sugerir que a ramificação com galactose de alguma forma modula a
degradação do galactomanano.
A comparação das hidrólises do galactomanano a 1% com endo-β-
mananase a 1U na presença ou ausência da α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. está apresentada na Figura 32. Quando o
polissacarídeo galactomanano nativo foi tratado somente com α-galactosidase
semipurificada, uma pequena quantidade de galactose foi retirada e
praticamente não houve detecção de oligossacarídeo (Figura 32 III), os
oligossacarídeos C e G são os mesmos que aparecem no controle sem
qualquer enzima (Figura 32 IV).
Quando o polissacarídeo de galactomanano de Sesbania virgata (Cav.)
Pers. a 1% foi hidrolisado somente pela endo-β-mananase de Aspergillus niger
(Megazyme®) houve a produção de oligossacarídeos F1, F2 e F3, sendo a
razão F1/F2=5,26 (Figura 32 I). Além disso, a proporção de oligossacarídeos
F1 formados é praticamente a mesma de oligossacarídeos F2 e F3, mostrando
que a enzima não está sendo capaz de hidrolisar completamente o
polissacarídeo a F1, ainda mantendo intermediários F2 e F3.

152
Entretanto, a ação prolongada da combinação da endo-β-mananase de
Aspergillus niger (Megazyme®) com a α-galactosidase semipurificada de
Sesbania virgata (Cav.) Pers. sobre o galactomanano a 1% também gerou
oligossacarídeos F1, F2 e F3, porém, a proporção de oligossacarídeos F1 é
muito maior do que os oligossacarídeos F2 e F3, razão F1/F2= 11,81 (Figura
32 III), evidenciando que a presença da α-galactosidase efetivamente melhorou
a hidrólise do polissacarídeo e oligossacarídeos F2 e F3 a oligossacarídeos F1.
O Experimento III conduzido seguindo a mesma metodologia, porém
com a α-galactosidase comercial (Megazyme®) substituindo a α-galactosidase
semipurificada, apresentou os mesmos resultados, estando os cromatogramas
apresentados na figura 48 no Anexo.

153

154
Figura 32: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do galactomanano de Sesbania virgata (Cav.) Pers
com endo-β-mananase (Megazyme®) a 1U com acréscimo ou não da α-galactosidase semipurificada
de Sesbania virgata (Cav.) Pers. (I) perfil de oligossacarídeos formados na hidrólise do
galactomanano a 1% somente com endo-β-mananase a 1U; (II) perfil de oligossacarídeos formados
na hidrólise do galactomanano a 1% com endo-β-mananase a 1U/mL e α-galactosidase de
semipurificada Sesbania virgata (Cav.) Pers. a 0,5U/mL; (III) perfil de oligossacarídeos formados
na hidrólise do galactomanano a 1% somente com α-galactosidase semipurificada Sesbania virgata
(Cav.) Pers. a 0,5U/mL; (IV) Controle (galactomanano sem enzima). (Gal) Galactose; (A)
manobiose (M2); (B) manotriose (M3); (C) tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6
e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I)
oligossacarídeos >8 resíduos (G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os
oligossacarídeos identificados A-I são denominados oligossacarídeos F1; os oligossacarídeos
restantes são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos
trios.
4.1.4. Experimento IV
As hidrólises enzimáticas conduzidas no Experimento IV visaram
compreender o modo de ação da enzima endo-β-mananase (Megazyme®) em
diferentes concentrações (0,005U/mL e 1U/mL) quando na presença da α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers. e a α-
galactosidase comercial (Megazyme®), ambas na máxima concentração de
0,5U/mL e galactomanano nas concentrações de 1% e 0,1%.
Dessa forma, a comparação dos oligossacarídeos formados nas
diferentes concentrações da endo-β-mananase, na mesma concentração da α-
galactosidase (0,5U/mL) e em concentrações distintas do galactomanano (1%
ou 0,1%) pode evidenciar preferência em alguns sítios de clivagem na cadeia
de principal de manano.
Os perfis cromatográficos dos oligossacarídeos produtos das hidrólises
do galactomanano de Sesbania virgata (Cav.) Pers. a 0,1% e 1%, com endo-β-
mananase a 0,005U e 1U e α-galactosidase semipurificada a 0,5U/mL se
encontram na figura 33.

155
Independente da concentração do substrato e da enzima endo-β-
mananase a diversidade de oligossacarídeos produzida é mesma visto pela
presença dos picos A a I em todos os cromatogramas (Figura 33 I a IV).
No entanto, o aumento da concentração da enzima endo-β-mananase,
de 0,005U para 1U, ou a diminuição na concentração do substrato, de 1% para
0,1%, melhorou a eficiência da digestão, sendo o aumento da concentração da
enzima mais eficiente, devido a maior formação de oligossacarídeos F1, F2 e
F3 (Figura 33 III e IV).
A proporção dos oligossacarídeos F1 formados após hidrólise com endo-
β-mananase na menor concentração (0,005U) não reflete a proporção de
oligossacarídeos no polímero, devido a poucos fragmentos galactosilados (C-I)
aparecerem, sugerindo que os oligossacarídeos galactosilados são mais
dificilmente formados (Figura 33 I e II).
O mesmo foi visto para as hidrólises com α-galactosidase comercial de
Cyamopsis tetragonoloba (Megazyme®), dados apresentados na figura 49
presente no Anexo deste documento.
Figura 33: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do galactomanano de Sesbania virgata (Cav.) Pers

156
com endo-β-mananase (Megazyme®) a 0,005U e 1U e α-galactosidase semipurificada de Sesbania
virgata (Cav.) Pers. a 0,5U/mL. (I) Galactomanano a 1%, 0,005U de endo-β-mananase e α-
galactosidase semipurificada a 0,5U/mL; (II) Galactomanano a 0,1%, 0,005U de endo-β-mananase e
α-galactosidase semipurificada a 0,5U/mL; (III) Galactomanano a 1%, 1U de endo-β-mananase e α-
galactosidase semipurificada a 0,5U/mL; (IV Galactomanano a 0,1%, 1U de endo-β-mananase e α-
galactosidase semipurificada a 0,5U/mL. (A) manobiose (M2); (B) manotriose(M3); (C)
tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D) pentassacarídeos (manopentose – M5 e/ou
G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4 e/ou G
4M5 e/ou G
3M5); (F)
heptassacarídeos (manoheptose – M7 e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos (G1,3,4
M5;
G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos identificados A-I são
denominados oligossacarídeos F1; os oligossacarídeos restantes são identificados como F2, quanto
combinação aos pares, e F3, quando combinação aos trios.
4.1.5. Experimento V
As hidrólises enzimáticas conduzidas no Experimento V visaram produzir
oligossacarídeos de manano puro (sem ramificações com galactose) variando
de monossacarídeo (manose) a decassacarídeos (oligossacarídeo de 10
manoses) de manose para identificar, nas análises de HPAEC-PAD, os
oligossacarídeos ramificados e não ramificados, possibilitando a determinação
da composição de cada oligossacarídeo.
Os produtos das hidrólises enzimáticas do manano comercial
(Megazyme®) a 1% com endo-β-mananase de Aspergillus niger (Megazyme®)
(0,005U, 0,05U e 1U) após ação da α-galactosidase de Cyamopsis
tetragonoloba (Megazyme®) e a co-injeção de manose e galactose se
encontram na figura 34.
Ao analisar os oligossacarídeos formados nas três concentrações de
enzima (0,005U, 0,05U e 1U) nota-se uma tendência na diminuição da
diversidade de oligossacarídeos formados, mesmo após a hidrólise com α-
galactosidase em um primeiro ciclo de digestão. Nas concentrações de 0,005U
e 0,05U de endo-β-mananase utilizadas é possível identificar os
oligossacarídeos identificados de A, B, C, D, E e F, picos F1, sem a detecção
de qualquer pico F2 e/ou F3. Na concentração de 1U de endo-β-mananase
somente os picos A e B são detectados, sem qualquer detecção de
oligossacarídeos F2 e/ou F3 (Figura 34 I, II e III).

157
Os oligossacarídeos comuns às três concentrações de endo-β-
mananase, A e B, possuem proporções distintas entre os oligossacarídeos
formados podendo ser devido a diferença na concentração da enzima e não a
preferência na produção de cada oligossacarídeo em cada concentração
(Figura 34 I, II e III), visto que a concentração de 1U gerou,
preponderantemente, manobiose (A) e manotriose (B).
Nas menores concentrações de endo-β-mananase (0,005U e 0,05U) já
existe a preferência pela formação da manobiose (pico A) e, principalmente na
concentração de 0,05U, a proporção dos picos B, C, D e E, em relação ao pico
A, é maior do que a encontrada no perfil cromatográfico da concentração de 1U
de endo-β-mananase (Figura 34 I, II e III).
O perfil cromatográfico da hidrólise enzimática do manano com endo-β-
mananase a 1U (Figura 34 III) mostra a clivagem completa do manano a
manobiose e manotriose (picos A e B) com pouquíssima quantidade dos
demais oligossacarídeos (D, E e F) evidenciando ação da α-galactosidase na
retirada das galactoses restantes.
Na comparação dos perfis cromatográficos dos oligossacarídeos
formados após hidrólise com 0,05U e 1U de endo-β-mananase nota-se que os
oligossacarídeos identificados como D, E e F estão em concentração razoável
em 0,05U (em relação aos demais picos), mas a 1U eles praticamente
desaparecem, mostrando que são oligossacarídeos de manose tamanhos
diversos não galactosilados.
Os perfis cromatográficos das co-injeções com os padrões de manose e
galactose (figura 34 IV, V) mostra o tempo de retenção de 4,450 para a
manose e 4,675 para a galactose.

158
Figura 34: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do manano comercial (Megazyme®) com endo-β-
mananase (Megazyme®) a 0,005U, 0,05U e 1U e α-galactosidase de Cyamopsis tetragonoloba
comercial (Megazyme®) a 0,5U/mL. (I) perfil de oligossacarídeos formados na concentração de
0,005U de endo-β-mananase; (II) perfil de oligossacarídeos formados na concentração de 0,05U de
endo-β-mananase; (III) perfil de oligossacarídeos formados na concentração de 1U de endo-β-
mananase; (IV) injeção com padrão de manose; (V) injeção com padrão de galactose. (A)
manobiose (M2); (B) manotriose (M3); (C) tetrassacarídeos (manotetraose – M4 e/ou G1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6
e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5). Os
oligossacarídeos identificados A-F são denominados oligossacarídeos F1.

159
4.2. Análise da estrutura fina do (Galacto)manano de 6 espécies
com diferentes proporções de ramificações com galactose
Visando determinar se as substituições de galactose interferem na
atuação da endoenzima endo-β-mannanase, dificultando a ação de hidrolisar o
polissacarídeo a oligossacarídeo, realizou-se o experimento que comparou os
oligossacarídeos, produto da hidrólise com endo-β-mannanase, de
(galacto)mananos de 6 espécies vegetais (Sesbania virgata, Senna alata,
Dimorphandra mollis, Cyamopsis tetragonoloba, Coffea arabica e Euterpe
edulis, que sabidamente possuem proporções de substituição por galactose
diferentes, ou seja, razão manose:galactose variando de 1:1 a 150:1). O
método de análise dos oligossacarídeos formados foi realizado através de
HPAEC-PAD e TLC.
A razão manose:galactose das 6 espécies estudadas, apresentada na
Tabela 4, foi determinada pela análise em HPAEC-PAD dos monossacarídeos
provenientes da hidrólise ácida com ácido sulfúrico do polissacarídeo,
(galacto)manano, de cada espécie.
Pela Tabela 4 fica evidente a diferença na proporção manose:galactose
das espécies estudadas, enquanto Euterpe edulis e Coffea arabica possuem
razão manose:galactose 150:1 e 40:1, respectivamente, sendo denominados
mananos; Dimorphandra mollis e Senna Alata possuem razão 2:1 e Cyamopsis
tetragonoloba e Sesbania virgata possuem razão 1:1, e são denominados de
galactomananos.
Após identificação da razão manose:galactose dos (galacto)mananos
das 6 espécies estudadas, realizou-se a hidrólise enzimática dos mesmos
polissacarídeos com endo-β-mannanase de Aspergillus niger (Megazyme®) e o
perfil de oligossacarídeos produzidos foram analisados por Cromatografia em
Camada Delgada (TLC) (Figura 35) e por HPAEC-PAD utilizando coluna PA-
100 (Figura 36).

160
Tabela 4. Razão manose:galactose das 6 espécies estudadas determinada pela análise de
monossacarídeos em HPAEC-PAD (Dionex). Apresentado por ordem crescente na quantidade de
substituições de galactose: Manano (Euterpe edulis; Coffea arábica); Galactomanano de proporção
2:1, ou seja, a cada 2 manose existe a ramificação por 1 galactose (Dimorphandra mollis; Senna
alata); Galactomanano de proporção 1:1, ou seja, teoricamente todo o polissacarídeo é substituído
por galactose (Cyamopsis tetragonoloba; Sesbania virgata) .
Espécies Razão Manose:Galactose
Euterpe edulis 150,51 (150:1)
Coffea arabica 40,2 (40:1)
Dimorphandra mollis 1,82 (2:1)
Senna alata 1,77 (2:1)
Sesbania virgata 1,53 (1:1)
Cyamopsis tetragonoloba 1,17 (1:1)
O perfil dos oligossacarídeos revelados em TLC encontra-se na Figura
35, onde o conjunto de oligossacarídeos foi aplicado de acordo com a razão
manose:galactose da espécie. Dessa forma, em A estão os oligossacarídeos
de Euterpe edulis (150:1); em B os de Coffea arabica (40:1); em C os de
Dimorphandra mollis (2:1); em D os de Senna alata (2:1); em E os de Sesbania
virgata (1:1); e em F os de Cyamopsis tetragonoloba (1:1).
Em uma comparação inicial dos oligossacarídeos das 6 espécies nota-
se que (galacto)mananos com diferentes proporções exibem perfis de
oligossacarídeos diversos.
Quando se compara o perfil de oligossacarídeos dos galactomananos
(Figura 35 C-F) observa-se de forma imediata diferenças nas intensidades das
bandas, enquanto em C e D as bandas são fortes em sua totalidade, em E e F
somente as bandas superiores (oligossacarídeos F1) apresentam intensidade,
ainda muito mais fraca do que as das primeiras, e as bandas inferiores
(oligossacarídeos F2 e F3) estão presentes mas em baixíssima concentração.
Além disso, nota-se que Cyamopsis tetragonoloba (F) possui dois
oligossacarídeos F1 exclusivos (Figura 35 F, →1, 2) e conjuntamente com
Sesbania virgata (Cav.) Pers. possuem um oligossacarídeo F2/F3 exclusivo de
ambas as espécies (Figura 34, →3).

161
Quando se comparam as espécies com reserva de manano (Figura 35
A e B), observam-se três oligossacarídeos exclusivos da espécie A (Figura 35
A, →1, 2, 3), razão manose:galactose 150:1, e um oligossacarídeo exclusivo da
espécie B (Figura 35 B, →4), razão 40:1.
Na comparação entre as espécies com reserva de manano e as com
reserva de galactomanano, observa-se três oligossacarídeos exclusivos das
espécies com reserva de manano, oligossacarídeos identificados por *→, no
entanto, nota-se uma ausência dos oligossacarídeos de maior peso (F2/F3)
nas espécies com reserva de manano (Figura 35, chave vermelha) e presentes
em todas as espécies com reserva de galactomanano, apesar das diferenças
nas concentrações.

162
Figura 35. Análise por Thin Layer Chromatography (TLC) dos oligossacarídeos obtidos após
hidrólise enzimática exaustiva com endo-β-mannanase (Megazyme®) das 6 espécies estudadas. (A)
Euterpe edulis (150:1); (B) Coffea arabica (40:1); (C) Dimorphandra mollis (2:1); (D) Senna alata
(2:1); (E) Sesbania virgata (Cav.) Pers. (1:1); (F) Cyamopsis tetragonoloba (1:1). (→)
Oligossacarídeos exclusivo de 1 ou mais espécies: (→1, 2) exclusivo de Cyamopsis tetragonoloba;
(→3) exclusivo de Sesbania virgata e Cyamopsis tetragonoloba; (→) Comparação dos
oligossacarídeos formados entre as espécies (A e B) com reserva de manano: (→1, 2, 3) exclusivo de
Euterpe edulis; (→4) exclusivo de Coffea arabica. (*→) Oligossacarídeos exclusivos das espécies com
reserva de manano. (Chave vermelha) Oligossacarídeos e suas combinações em pares (F2) e em
trios (F3) exclusivos de espécies com reserva de galactomanano.
*
*
1
2
3
A B C D F E A A
1
3 2
4
*

163
Os perfis cromatográficos da análise por HPAEC-PAD dos
oligossacarídeos provenientes da hidrólise com endo-β-mannanase dos
polissacarídeos das 6 espécies encontram-se agrupados na Figura 36.
Da mesma maneira como no TLC (Figura 35), os perfis cromatográficos
estão identificados de forma crescente quanto à substituição por galactose,
assim: A perfil dos oligossacarídeos de Euterpe edulis (150:1); em B perfil dos
oligossacarídeos de Coffea arabica (40:1); em C perfil dos oligossacarídeos de
Dimorphandra mollis (2:1); em D perfil dos oligossacarídeos de Senna alata
(2:1); em E perfil dos oligossacarídeos de Sesbania virgata (Cav.) Pers. (1:1); e
em F perfil dos oligossacarídeos de Cyamopsis tetragonoloba (1:1).
As letras de a-m identificam cada oligossacarídeo, onde: (a) manose; (b)
galactose; (c) manobiose; (d) manotriose; (f-h, j-m) oligossacarídeos ainda não-
identificados; (i) combinações aos pares e aos trios dos oligossacarídeos
identificados c-h, j-m. Os oligossacarídeos identificados como a-h e j-m são
denominados oligossacarídeos F1; os oligossacarídeos identificados por i são
denominados F2 quando combinação aos pares e F3 quando combinação aos
trios de F1 (Figura 36).
Os picos identificados pela mesma letra (a-m) correspondem ao mesmo
oligossacarídeo, por apresentarem o mesmo tempo de retenção na coluna.
Ao analisar os perfis cromatográficos de forma crescente, quanto às
substituições por galactose, nota-se gradualmente o aparecimento dos
oligossacarídeos F2 e F3, enquanto A (150:1) não apresenta qualquer pico
denominado h e/ou i, a partir de B (40:1) estes picos estão presentes. Ainda
quanto aos picos h e i, observa-se que os perfis C e D apresentam a maior
concentração destes.
Além disso, observa-se que os perfis cromatográficos E e F possuem,
comparativamente aos demais, baixa detecção (concentração) de todos os
oligossacarídeos.
Os oligossacarídeos identificados por: a, c, d, f, g, h, i, são comuns a
todos os perfis cromatográficos, com exceção do perfil A que não possui o
oligossacarídeo identificado por g, no entanto apresenta um oligossacarídeo
exclusivo a ele, identificado por l, e um segundo oligossacarídeo, identificado
por k, presente nos perfis A, B, D e E.

164
O oligossacarídeo b parece ser exclusivo nos perfis cromatográficos B e
F, o oligossacarídeo j, apesar de baixa detecção, é exclusivo do perfil
cromatográfico de C e, por fim, o oligossacarídeo e, se apresenta em uma
quantidade razoável no perfil F, sendo exclusivo desta espécie.
Figura 36. Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da análise dos
oligossacarídeos, produto da hidrólise enzimática exaustiva com endo-β-mannanase (Megazyme®),
das 6 espécies estudadas. (A) Euterpe edulis (150:1); (B) Coffea arabica (40:1); (C) Dimorphandra
mollis (2:1); (D) Senna alata (2:1); (E) Sesbania virgata (1:1); (F) Cyamopsis tetragonoloba (1:1). (a)
manose; (b) galactose; (c) manobiose; (d) manotriose; (f-h, j-m) oligossacarídeos ainda não-
identificados; (i) combinações aos pares e aos trios dos oligossacarídeos identificados (c-h, j-m). Os
oligossacarídeos identificados a-h e j-m são denominados oligossacarídeos F1; os oligossacarídeos
identificados por i são denominados F2, quanto combinação aos pares, e F3, quando combinação
aos trios.

165
Os oligossacarídeos formados não diferem dos oligossacarídeos
identificados nos Experimentos I a V, porém a análise conduzida em HPAEC-
PAD, embora tenha seguido a mesma metodologia, este experimento foi o
primeiro a ser conduzido e na época a coluna PA-100 utilizada já estava muito
antiga, prejudicando a separação dos oligossacarídeos. Dessa forma,
consegue-se identificar a manobiose e a manotriose, mas os demais picos
possuem muitos oligossacarídeos co-eluídos e não seguem a mesma
identificação dos Experimentos I a V.
V. Discussão
5.1. Análise da estrutura fina do galactomanano de Sesbania
virgata (Cav.) Pers. através de hidrólises enzimáticas com
Endo-β-manase de Aspergillus niger (Megazyme®), α-
Galactosidase de Cyamopsis tetragonoloba (Megazyme®) e α-
galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
Diversas técnicas já foram empregadas em estudos de distribuição de
resíduos de D-galactose ao longo da cadeia principal de D-manano de
galactomananos de reserva de sementes de leguminosas (McCleary et al.,
1985).
Os estudos sobre a estrutura fina de galactomananos e glucomananos
conduzidos por McCleary foram realizados utilizando como técnicas: a hidrólise
do polissacarídeo com endo-β-mananase de Aspergillus niger, já bem
caracterizada no seu modo de ação (McCleary & Matheson, 1983), isolamento
e identificação da composição de cada oligossacarídeo por Ressonância
Magnética Nuclear (RMN) e/ou Raio-X (McCleary et al., 1976; McCleary et
al.,1983; McCleary & Matheson, 1983).
A compreensão no modo de ação da endo-β-mananase de Aspergilus
niger foi realizada pela identificação dos oligossacarídeos formados após a sua
ação sobre galactomanano de goma caroba razão manose:galactose 4:1, em
comparação com os oligossacarídeos formados após ação de endo-β-

166
mananases de bactéria (Bacillus subtilis), fungo (Irpex lacteus), animais (Helix
pomatia) e plantas (Cyamopsis tetragonoloba e Medicago sativa) sobre o
mesmo substrato (McCleary & Matheson, 1983).
De forma geral, McCleary & Matheson (1983), determinaram que endo-
β-mananases de fontes variadas possuem capacidades hidrolíticas diferentes,
requerendo um maior número de resíduos de manose sem ramificação para
conseguirem hidrolisar o polissacarídeo. A endo-β-mananase de Aspergillus
niger é a enzima menos específica no seu modo de ação, necessitando de no
mínimo de um oligossacarídeo de 3 resíduos de manose galactosilados ou não
para ainda conseguir identificar um sítio de clivagem, e com a maior
capacidade de clivar o polissacarídeo altamente galactosilado, pois possui
maior tolerância às ramificações com galactose.
Dessa forma, endo-β-mananases de fontes variadas diferem entre si na
capacidade de hidrolisar um mesmo polissacarídeo, no entanto, os
oligossacarídeos formados após a ação enzimática são os mesmos e estão
descritos na Tabela 5.
Analisando esta tabela, percebe-se que alguns oligossacarídeos são
inexistentes quando da ação da endo-β-mananase de Aspergillus niger, pois
esta enzima consegue clivar estes oligossacarídeos a fragmentos ainda
menores, exatamente pela sua maior tolerância às ramificações com galactose.
Independentemente da endo-β-mananase utilizada, McCleary &
Matheson (1983), demonstraram que quanto menor for a razão
manose:galactose de um polissacarídeo, ou seja, quanto mais ramificado for
um polissacarídeo, menor será a porcentagem de hidrólise deste dado
polissacarídeo pela endo-β-mananase.
Tiné et al. (2003) sugeriram, também através de hidrólises enzimáticas
do polissacarídeo xiloglucano e identificação dos oligossacarídeos formados,
que a estrutura do xiloglucano de reserva de sementes conteria, pelo menos,
parte da informação requerida para o seu próprio metabolismo, o que
posteriormente chamou-se de código do xiloglucano (Buckeridge, 2010).
A sugestão da presença de um código presente no polissacarídeo
contendo informações que afeta a sua hidrólise foi realizada através da
visualização de diminuição dos oligossacarídeos F2 e F3 formados na
presença da β-galactosidase, que os oligossacarídeos menos ramificados eram

167
formados preferencialmente pela enzima e que polímeros de estrutura fina
diferente possuíam porcentagem de hidrólise diferente na presença da mesma
enzima.
Assim, sugere-se, através dos resultados obtidos nesta dissertação, que
a estrutura do polissacarídeo galactomanano também contenha, pelo menos,
parte da informação requerida para seu próprio metabolismo, código para a sua
degradação, estando esta informação contida na distribuição das ramificações
com resíduos de D-galactose.
Da mesma forma que o determinado por Tiné et al. (2003), nota-se que
a hidrólise do galactomanano pela endo-β-mananase de Aspergillus niger
(Figura 28) forma oligossacarídeos F1 (base do galactomanano), bem como,
oligossacarídeos F2 e F3, evidenciando equilíbrio de reação e impedimento da
hidrólise total do polissacarídeo.
Apesar de não ter sido analisada a composição dos oligossacarídeos F2
e F3, os resultados apresentados no Experimento III (Figura 32), permitem a
sugestão que estes grupos de oligossacarídeos são mais galactosilados, pois
na presença da α-galactosidase conjuntamente com a endo-β-mananase existe
a diminuição na formação de F2 e F3 e aumento na formação de F1, sugerindo
que a ramificação com galactose parece ser a responsável pelo impedimento
da hidrólise.
A análise dos resultados do Experimento I (Figura 29) evidenciou a
preferência na formação de oligossacarídeos dependente da concentração
utilizada de endo-β-mananase. Em concentrações baixas de enzima (0,005U e
0,05U) a manotriose (M3) e/ou o trissacarídeo (G1M2) (pico B) são
preferencialmente produzidos, enquanto na concentração de 1U os
oligossacarídeos pares manobiose (M2) (pico A) e o tetrassacarídeo (G1M3) são
preferencialmente produzidos, sugerindo que a enzima, quando em baixa
concentração, prefere locais com menor quantidade de ramificações,
evidenciando que a digestão do polissacarídeo não é ao acaso.
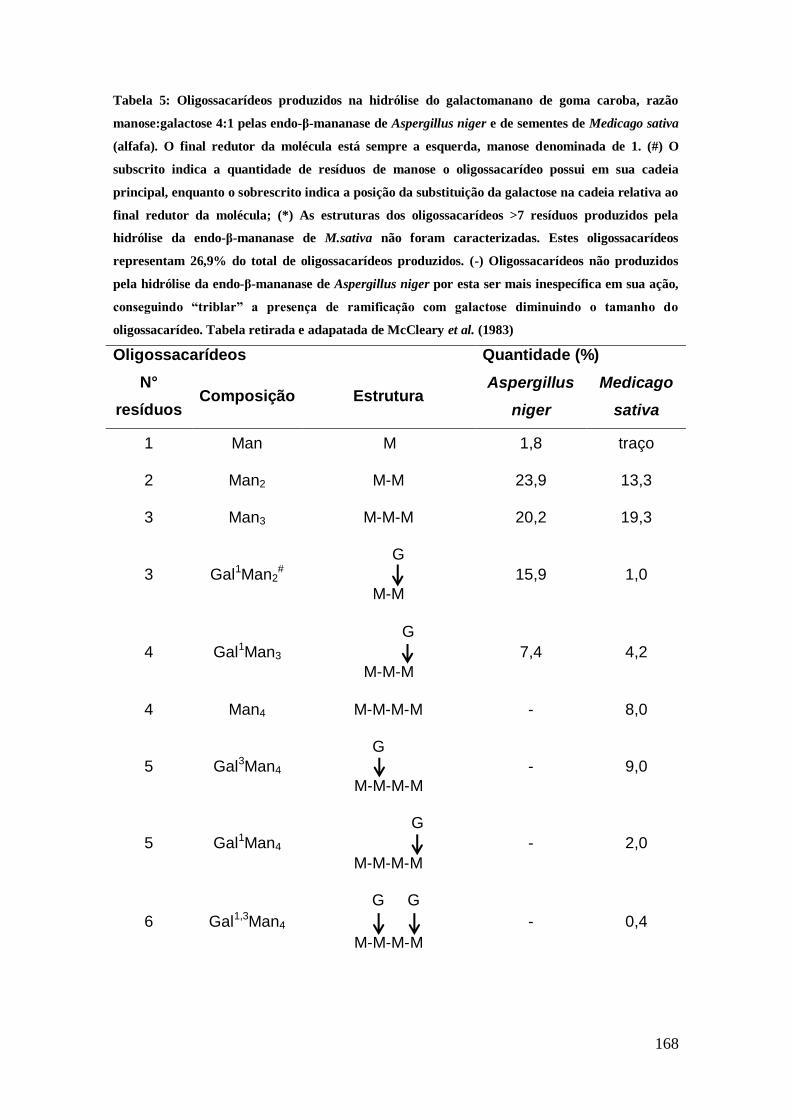
168
Tabela 5: Oligossacarídeos produzidos na hidrólise do galactomanano de goma caroba, razão
manose:galactose 4:1 pelas endo-β-mananase de Aspergillus niger e de sementes de Medicago sativa
(alfafa). O final redutor da molécula está sempre a esquerda, manose denominada de 1. (#) O
subscrito indica a quantidade de resíduos de manose o oligossacarídeo possui em sua cadeia
principal, enquanto o sobrescrito indica a posição da substituição da galactose na cadeia relativa ao
final redutor da molécula; (*) As estruturas dos oligossacarídeos >7 resíduos produzidos pela
hidrólise da endo-β-mananase de M.sativa não foram caracterizadas. Estes oligossacarídeos
representam 26,9% do total de oligossacarídeos produzidos. (-) Oligossacarídeos não produzidos
pela hidrólise da endo-β-mananase de Aspergillus niger por esta ser mais inespecífica em sua ação,
conseguindo “triblar” a presença de ramificação com galactose diminuindo o tamanho do
oligossacarídeo. Tabela retirada e adapatada de McCleary et al. (1983)
Oligossacarídeos Quantidade (%)
N°
resíduos Composição Estrutura
Aspergillus
niger
Medicago
sativa
1 Man
1,8 traço
2 Man2 23,9 13,3
3 Man3 20,2 19,3
3 Gal1Man2#
15,9 1,0
4 Gal1Man3
7,4 4,2
4 Man4 - 8,0
5 Gal3Man4
- 9,0
5 Gal1Man4
- 2,0
6 Gal1,3Man4
- 0,4
M-M-M-M
G G
M-M-M-M
G
M-M-M-M
G
M-M-M-M
M-M-M
G
M-M
G
M-M-M
M-M
M

169
6 Gal4Man5
- 3,3
6 Gal3Man5
- 3,3
7 Gal3,4Man5
11,1 9,2
8 Gal1,3,4Man5
0,6 26,9*
8 Gal4,5Man6
1,2 26,9*
8 Gal3,4Man6
4,2 26,9*
9 Gal3,4,5Man6
2,5 26,9*
9 Gal1,4,5Man6
2,3 26,9* M-M-M-M-M-M
G
G G
M-M-M-M-M-M
G
G G
M-M-M-M-M-M
G
G
M-M-M-M-M-M
G
G
M-M-M-M-M
G
G G
M-M-M-M-M
G
G
M-M-M-M-M
G
M-M-M-M-M
G

170
9 Gal1,3,4Man6
0,5 26,9*
9 Gal4,5Man7
0,7 26,9*
>9 - 7,7 26,9*
A razão F1/F2 do Experimento I (Tabela 2) corrobora a hipótese de que
existe uma preferência por sítios menos galactosilados, pois na concentração
de enzima a 1U, percebe-se uma diminuição na preferência dos sítios de
clivagem, reduzindo o polissacarídeo a oligossacarídeos F2 e F3, mas
principalmente F1, visto que a razão F1/F2(1U) é a maior das 3 concentrações.
Na menor concentração de enzima, 0,005U, aparentemente há muita
dificuldade no processamento do polissacarídeo, sendo assim, a enzima ataca
sítios de clivagem de preferência, produzindo preferencialmente F1, uma vez
que a produção de oligossacarídeos F2 e F3 requer que as enzimas lidem com
as ramificações por galactose.
Contudo, na concentração de 0,05U de enzima, parece pela menor
razão F1/F2 apresentada, que a enzima prefere hidrolisar o polissacarídeo a F2
e F3, do que reprocessar F2 e F3 a F1 pela presença das ramificações com
galactose presentes nestes oligossacarídeos, permitindo a sugestão que as
ramificações parecem dificultar a digestão pela endo-β-mananase, que não age
ao acaso, preferindo sítios menos galactosilados. Estes resultados permitem
concluir que as ramificações com galactose modulam a ação da endo-β-
mananase, possivelmente em moldes parecidos com os observados por Tiné et
al. (2003) para xiloglucanos.
Na comparação do Experimento I com o Experimento II era esperado
encontrar no perfil cromatográfico do Experimento II uma mudança no padrão
de oligossacarídeos presentes no Experimento I, pois inicialmente entendia-se
que os oligossacarídeos ramificados não eram co-eluídos com os
M-M-M-M-M-M-M
G
G
M-M-M-M-M-M
G
G G

171
oligossacarídeos não-ramificados (somente a cadeia principal de manano),
sendo assim, após o uso da α-galactosidase, os oligossacarídeos ramificados
seriam desramificados e realocados no pico referente ao oligossacarídeo de
manose, com o desaparecimento dos picos galactosilados.
Assim, o objetivo do Experimento II era observar os picos que sumiriam
para identificar os oligossacarídeos ramificados e com o aumento dos demais
picos (representantes dos oligossacarídeos de manose) inferir sobre a
composição.
Entretanto, a análise comparativa do Experimento I e do Experimento II
não evidenciou o desaparecimento de picos, com exceção do pico I, mas a
mudança na proporção dos picos (Figura 31 VII,VIII,IX) sugerindo que haja co-
eluição de oligossacarídeos com o mesmo número de resíduos, ou seja,
oligossacarídeos contendo somente manose, por exemplo manotetraose (M4)
são co-eluídos com oligossacarídeos ramificados com galactose, seguindo o
exemplo anterior, tetrassacarídeo (G1M3).
Assim, a co-eluição impossibilitou a identificação dos picos referentes a
oligossacarídeos ramificados, dificultando a identificação da composição de
cada oligossacarídeo formado após ação da endo-β-mananase de Aspergillus
niger. Sendo assim, foi necessária a utilização da descrição feita por McCleary
et al. (1983) (Tabela 5) dos oligossacarídeos formados após ação da endo-β-
mananase de Aspergillus niger juntamente com os resultados dos Experimento
I e Experimento V (Figuras 29 e 34) para inferir sobre a estrutura dos
oligossacarídeos, explicação abrangida posteriormente nesta discussão.
Embora a co-eluição tenha dificultado a identificação dos
oligossacarídeos, é inegável que a presença da α-galactosidase mudou a
proporção entre os oligossacarídeos. A razão F1/F2 de todas as concentrações
de endo-β-mananase (hidrólise anterior a ação da α-galactosidase) do
Experimento II é maior do que as razões encontradas no Experimento I,
evidenciando que a retirada da galactose propicia uma maior redução do
polissacarídeo a oligossacarídeos F1 (Tabela 2).
Aliado a isso, o aumento da razão F1/F2 encontrado no Experimento II
foi devido ao aumento na quantidade de oligossacarídeos F1 (comparação da
somatória das áreas dos picos F1 e F2-F3 de ambos os experimentos, dados
não mostrados), portanto, permite a sugestão que entre os oligossacarídeos F2

172
e F3 existem oligossacarídeos de cadeia de manose com tamanho de
oligossacarídeo F1 altamente ramificados, novamente sugerindo que a ação da
endo-β-mananase não é ao acaso, preferindo sítios de clivagem com baixa
porcentagem de ramificação.
A comparação das razões (M2/M3 e M3/C) entre os Experimentos I e II
(Tabela 3) juntamente com os oligossacarídeos identificados no trabalho de
McCleary et al. (1983) (Tabela 5) possibilitaram a inferência na composição
original dos oligossacarídeos e após a ação da α-galactosidase, pois como a
ação da endo-β-mananase foi anterior a da α-galactosidase e não
concomitante, a mudança de proporção entre os oligossacarídeos só é devido
a realocação dos oligossacarídeos desramificados.
Para a explicação da mudança das proporções dos picos utilizou-se a
descrição dos oligossacarídeos formados após ação da endo-β-mananase de
Aspergillus niger feita por McCleary et al. (1983) (Tabela 5).
Para a razão M2/M3 ter sido alterada (aumentou a quantidade de M2) só
pode ter havido a desramificação do trissacarídeo G1M2, portanto, no pico B
existe a manotriose e o oligossacarídeo ramificado G1M2.
Para a razão M3/C ter sido alterada (aumentou a quantidade de M3) só
pode ter havido a desramificação do tetrassacarídeo (G1M3), portanto no pico C
também há co-eluição de oligossacarídeos: manotetraose (M4) e
terassacarídeo G1M3.
A determinação da composição dos demais picos seguindo as
diferenças nas proporções, como realizado para os picos A, B e C, se torna
muito difícil, tendo em vista, que a desramificação de oligossacarídeos F1
maiores e dos oligossacarídeos F2 e F3 interferem na proporção dos
oligossacarídeos F1 menores impossibilitando a inferência na presença do
oligossacarídeo, às vezes ocultando ou diminuindo uma diferença na proporção
pela presença da co-eluição. Visando elucidação, toma-se como exemplo o
pico F, um heptassacarídeo, que não apresentou diferença na proporção entre
o Experimento I e Experimento II (Figura 31 VII, VIII, IX).
Sendo um heptassacarídeo (ver explicação mais abaixo), pode ser tanto
manoheptose (M7) proveniente da desramificação do nonassacarídeo (G4,5M7)
quanto um heptassacarídeo ramificado (G3,4M5). Apesar de haver uma pequena
diferença no pico D, pentassacarídeo, o pico F, heptassacarídeo, não

173
apresentou mudança na proporção, pois possivelmente recebeu por realocação
manoheptose, portanto, não é possível estabelecer quem foi o pico do
Experimento I que foi realocado para outro pico no Experimento II.
De forma geral, o Experimento II colaborou na identificação de alguns
picos e, principalmente, demonstrou que a enzima endo-β-mananase não atua
ao acaso preferindo sítios menos galactosilados, sugerindo novamente que as
ramificações com galactose modulam a sua ação.
Esta modulação pelos resíduos de galactose é também corroborada pelo
fato de que a hidrólise do manano (Megazyme®) independentemente da
concentração de enzima endo-β-mananase utilizada (0,005U, 0,05U e 1U),
Experimento I, não apresentou a geração de oligossacarídeos F2 e F3, sendo
completamente digerido a F1 (Figura 30).
O manano comercial (Megazyme®) é proveniente de tratamento
enzimático com α-galactosidase e químico com hidróxido de sódio do
galactomanano de Ceratonia siliqua (goma caroba) com razão
manose:galactose 4:1, possuindo após o tratamento 3% de galactose.
Aliado a isso, McCleary et al. (1976) afirmam que a digestão de um
manano não ramificado produz exclusivamente manobiose e manotriose, o que
foi corroborado neste trabalho.
Sendo assim, por haver mudança na proporção dos oligossacarídeos F1
formados dependente da concentração de endo-β-mananase e a presença de
oligossacarídeos diferentes de manobiose e manotriose (picos A e B) na
concentração de 1U (Figura 30), sugeriu-se que ainda houvesse ramificações
com galactose restantes no manano comercial (Megazyme®).
Dessa forma, justificou-se a execução do Experimento V, com a hidrólise
com a α-galactosidase, para garantir completa retirada das galactoses,
anteriormente à hidrólise do polissacarídeo com a endo-β-mananase nas três
concentrações (0,005U, 0,05U e 1U) (Figura 34).
Embora o objetivo do Experimento I não tenha sido atingido quanto à
produção de oligossacarídeos de manano de monossacarídeos a
heptassacarídeos, determinou-se que a baixa concentração de galactose (3%)
não é eficiente no impedimento da hidrólise (formação de F2 e F3), sugerindo
que deva existir uma quantidade de galactosilação mínima para haver

174
regulação no processo, possibilitando a explicação do porquê dos mananos
sempre possuírem um mínimo de ramificações com galactose.
As hidrólises conduzidas no Experimento V foram eficientes na retirada
das galactoses e, portanto, cumpriram com o objetivo da produção de
oligossacarídeos de manano de monossacarídeos a heptassarídeos. Isto
ocorreu porque, na concentração de 1U de endo-β-mananase houve,
preponderantemente, a produção dos oligossacarídeos manobiose (pico A) e
manotriose (pico B) e nas concentrações de 0,005U e 0,05U os
oligossacarídeos F1 diferentes de manobiose e manotriose são,
provavelmente, oligossacarídeos de manose de tamanhos diversos não
galactosilados, tendo em vista que estes picos desaparecem na concentração
de 1U, pois se fossem galactosilados, continuariam a ser produzidos (Figura
34).
Dessa forma, a partir dos dados do Experimento I X Experimento II
(Figura 31 VII, VIII, IX), dos dados de tempo de retenção dos oligossacarídeos
desgalactosilados do Experimento V (Figura 34) e da identificação dos
oligossacarídeos produtos da hidrólise de galactomanano com endo-β-
mananase de Aspergillus niger descritos por McCleary et al. (1983) (Tabela 5),
determinou-se as possíveis estruturas dos oligossacarídeos F1 das digestões
do galactomanano nos Experimentos I a V.
A partir da análise dos oligossacarídeos formados no Experimento V
com endo-β-mananase a 1U (Figura 34) determinou-se os tempos de retenção
da manobiose e manotriose, 6,084 minutos e 7,300 minutos, respectivamente.
A metodologia de análise dos oligossacarídeos por HPAEC-PAD é
isocrática somente de 0 a 11 minutos, ou seja, a concentração dos eluentes é
constante durante todo este período: 98% de NaOH 88mM e 2% de Acetato de
Sódio 200mM em NaOH 88mM. A partir dos 12 minutos até os 20 minutos a
concentração varia de 98% para 70% de NaOH 88mM e de 2% para 30% de
Acetato de Sódio 200mM em NaOH 88mM. Dos 20 minutos a 25 minutos varia-
se de 70% a 0% de NaOH 88mM e de 30% a 100% de Acetato de Sódio
200mM em NaOH 88mM. Dos 25 minutos aos 30 minutos o gradiente inicial é
reestabelecido 98% e 2% e se mantém constante até o final da corrida para o
reequilíbrio da coluna.

175
Dessa forma, criou-se a hipótese que até os 12 minutos de corrida a
cada minuto sairia um oligossacarídeo com um resíduo de manose ou
galactose a mais, assim, teoricamente aos 6 minutos um dissacarídeo
(manobiose) seria detectado. Da mesma forma, aos 7 minutos um trissacarídeo
(manotriose), aos 8 minutos um tetrassacarídeo, aos 9 minutos um
pentassacarídeo, aos 10 minutos um hexassacarídeo, aos 11 minutos um
heptassacarídeo. A partir dos 12 minutos a separação deixa de ser uniforme,
pelo aumento na concentração de acetato de sódio, e os oligossacarídeos
deixam de sair no tempo teórico e foram identificados pelas possibilidades
descritas por McCleary et al (1983) (Tabela 5).
Utilizando os tempos de retenção da digestão do galactomanano de
Sesbania virgata (Cav.) Pers. com endo-β-mananase a 1U (Experimento I –
Figura 29) para identificar os oligossacarídeos de acordo com a hipótese
formada acima, nota-se que os tempos de retenção apresentados pelos
oligossacarídeos A – F (picos) são muito próximos do tempo teórico (Tabela 6),
portanto, aceitou-se a hipótese.
Tendo em mãos o tamanho de cada oligossacarídeo em cada tempo de
retenção, utilizou-se da identificação dos oligossacarídeos formados após
hidrólise com endo-β-mananase de Aspergillus niger realizada por McCleary et
al. (1983) (Tabela 5) para identificar as possíveis estruturas dos
oligossacarídeos de Sesbania virgata (Cav.) Pers. para cada tempo de
retenção (Tabela 6).
McCleary et al. (1983) descreveram 14 oligossacarídeos formados pela
ação da endo-β-mananase de Aspergillus niger, porém no perfis
cromatográficos só observa-se 9 picos de oligossacarídeos F1, portanto,
novamente uma evidencia de co-eluição de oligossacarídeos com o mesmo
número de resíduos manose e/ou galactose, assim, cada pico identificado pode
representar mais de um oligossacarídeo.

176
Tabela 6: Identificação das possíveis estruturas dos oligossacarídeos de Sesbania virgata (Cav.)
Pers. componentes dos picos A-F de acordo com a comparação do tempo de retenção real e o tempo
de retenção teórico e os oligossacarídeos caracterizados e identificados por McCleary et al. (1983)
(Tabela 5).
Oligossacarídeo
(Picos)
Tempo de
retenção
(Real)
(minutos)
Tempo de
retenção
(Teórico)
(minutos)
Estruturas
A 5,975 6,000
B 7,159 7,000
C 8,359 8,000
D 8,909 9,000
E 9,942 10,000
F 11,200 11,000
M-M-M-M-M-M-M
M-M-M-M-M
G
G
M-M-M-M-M-M
M-M-M-M-M
G
M-M-M-M-M
G
M-M-M-M
G G
M-M-M-M-M
M-M-M-M
G
M-M-M-M
G
M-M-M
G
M-M
G
M-M
G
M-M-M
M-M

177
Vale lembrar que a manobiose e a manotriose foram identificadas
através do Experimento V e a partir delas foi idealizada a hipótese sobre os
tempos de retenção. Além disso, nas digestões do galactomanano,
diferentemente do manano tratado, houve oligossacarídeos galactosilados,
inclusive co-eluídos com a manotriose (ver Experimento II), ou seja, o pico B de
todas as hidrólises de galactomanano pode ser uma mistura de manotriose
com o oligossacarídeo de manobiose com ramificação de galactose na manose
redutora (Gal1Man2).
A leitura das “abreviações” dos oligossacarídeos GalxMany deve ser feita
da seguinte maneira: o y apresenta a quantidade de resíduos de manose na
cadeia principal do oligossacarídeo, enquanto o x apresenta em qual manose,
a partir da extremidade redutora (sempre a direita), a ramificação de galactose
está ligada. Dessa forma, o valor de y sempre será único, enquanto os valores
de x podem ser mais do que um, indicando uma ou múltiplas ramificações
(Figura 37).
Figura 37: Explicação para a leitura das “abreviações” dos oligossacarídeos GalXMany. A cadeia
principal de manose é numerada a partir da extremidade redutora (à direita). O número subscrito
ligado à manose indica a quantidade de resíduos de manose que a cadeia principal do
oligossacarídeo possui, enquanto o número sobrescrito ligado a galactose indica a posição do
resíduo de galactose relativo ao resíduo de manose a partir da extremidade redutora (McCleary et
al., 1983).
A análise comparativa da hidrólise do galactomanano exclusivamente
com endo-β-mananase (Experimento I) e da hidrólise do galactomanano com
endo-β-mananase e α-galactosidase semipurificada concomitantemente
(Experimento III) evidencia que os oligossacarídeos F2 e F3 são susceptíveis
M-M-M-M-M-M
G
G G
1 2 3 4 5 6
Gal1,3,4
Man6
Extremidade redutora

178
ao ataque da endo-β-mananase (Figura 32), pois a presença da α-
galactosidase no ensaio altera o equilíbrio da reação, produzindo mais
oligossacarídeos F1 do que F2 e F3 (ver razões F1/F2 de ambas as digestões).
Dessa forma, evidencia-se que a menor taxa de hidrólise dos
oligossacarídeos F2 e F3 nos ensaios somente com endo-β-mananase não
advém de uma possível resistência intrínseca destes fragmentos, mas
possivelmente das ramificações com galactose.
Portanto, nota-se uma importância das ramificações de galactose na
modulação do ataque da endo-β-mananase ao substrato.
Lisboa et al. (2006) demonstraram que α-galactosidases são inibidas
quando ocorre acúmulo de galactose livre. Aparentemente, o acúmulo de
oligossacarídeos galactosilados também impede a hidrólise completa da endo-
β-mananase (comparar hidrólise do manano (Experimento V) figura 34 e
hidrólise do Experimento III Figura 32). Estes fatos realçam a importância na
interação das exo-hidrolases (α-galactosidase e β-manosidase) com as endo-
hidrolases (endo-β-mananase) que ocorrem nos processos biológicos como a
degradação da reserva de sementes ou degradação da parede celular por
fungos.
Alguns autores propõem que durante a degradação do galactomanano
existe uma ordem na atuação das enzimas (Reid & Edwards, 1995; Lisboa et
al., 2006). Por outro lado, compreende-se que ambas as enzimas estejam
presentes concomitantemente durante a maior parte do processo de
degradação. Sendo assim, a α-galactosidase é capaz de alterar a afinidade da
endo-β-mananase ao substrato (galactomanano) ao retirar algumas
ramificações de galactose, dessa forma, modulando o processo de
degradação. Deste modo, a ordem de ação das enzimas se torna mais
importante quanto maior for o grau de ramificação do galactomanano.
A proporção dos oligossacarídeos F1 obtidos na digestão do
galactomanano em baixa concentração de endo-β-mananase (0,005U) é muito
diferente da proporção destes mesmos oligossacarídeos no polímero
(comparar picos A, C e E na figura 33 I e III) (Experimento IV).
A mistura dos oligossacarídeos F1 na digestão com a menor
concentração da enzima (0,005U) é rica em tetrassacarídeos (pico C) (Figura
33 I), muito embora não sendo os tetrassacarídeos os oligossacarídeos

179
presentes em maior quantidade na composição do polissacarídeo, pois na
maior concentração de enzima os oligossacarídeos preponderantemente
produzidos são a manobiose (A) e hexassacarídeos (E) (Figura 33 III) (ver
tabela 6 para visualizar as estruturas de oligossacarídeos para cada pico).
Essa diferença na proporção sugere, novamente, que o ataque da
enzima endo-β-mananase de Aspergillus niger ao polissacarídeo não é ao
acaso e as regiões menos ramificadas são preferencialmente digeridas.
McCleary et al. (1985) afirmam que as α-galactosidases não apresentam
diferenças no seu modo de ação dependente da estrutura do galactomanano,
ou seja, a quantidade de ramificação com galactose não interfere na sua ação,
sendo o seu modo de ação não randômico, apresentando uma tendência em
remover as ramificações com galactose sequencialmente de um dos lados da
cadeia de manano.
As α-galactosidases utilizadas nos experimentos, semipurificada e
comercial, muito embora apresentem características (pH ótimo, temperatura
ótima e aspectos cinéticos) diferentes, atuaram sobre o polissacarídeo ou
oligossacarídeos da mesma forma, apresentando, portanto, o mesmo modo de
ação, visto pela similaridade dos perfis de oligossacarídeos gerados
independente do experimento proposto (Experimento II, III e IV).
Sugere-se que esta similaridade esteja relacionada à importância da
mobilização de reservas no sucesso da plântula, portanto, todo o processo
seria muito conservado.
Feurtado et al. (2001) demonstraram que a sequência de aminoácidos
das α-galactosidases de plantas são bastante conservadas. Sendo assim, é
também provável que todo o processo fisiológico da mobilização de reservas
seja conservado, inclusive com uma modulação da degradação realizada pela
estrutura do polissacarídeo (distribuição das ramificações de galactoses).
5.2. Análise da estrutura fina do (Galacto)manano de 6 espécies
com diferentes proporções de ramificações com galactose
Os estudos de determinação da estrutura fina, normalmente, são
realizados comparativamente, ou seja, identificando o padrão de ação da

180
enzima sobre polissacarídeos de razão manose:galactose diversa (Emi et al.,
1972; McCleary et al., 1976; McCleary, 1983; McCleary et al., 1985).
Sendo assim, adotou-se a mesma estratégia para determinação da
estrutura fina do galactomanano de Sesbania virgata (Cav.) Pers., a fim de
compreender a ação regulatória das substituições por galactose na degradação
do polissacarídeo durante a mobilização de reserva (Tiné et al., 2003).
A análise dos oligossacarídeos provenientes da hidrólise dos
(galacto)mananos com razão manose:galactose variadas (de 150:1 a 1:1 –
Tabela 4) demonstra, como em McCleary (1983), que a quantidade e a
distribuição das ramificações com D-galactose influenciam na hidrólise do
polissacarídeo com endo-β-mananase, pois tanto a revelação do perfil dos
oligossacarídeos por TLC quanto por HPAEC-PAD, revelam a presença de
bandas únicas a determinadas razões de manose:galactose, ausência de
grupos de oligossacarídeos (F2 e F3), quando a substituição por galactose é
baixa; e proporção diversa (altura dos picos) de oligossacarídeos iguais
(identificados pela mesma letra) (Figura 35 e 36).
A ausência de oligossacarídeos F2 e F3 (Figura 35 e 36) após hidrólise
com endo-β-mananase do manano puro (150:1) de Euterpe edulis evidencia
que a baixíssima substituição por galactose não permite o equilíbrio da reação,
havendo, portanto, somente a produção de oligossacarídeos F1.
O aumento gradual da presença destes oligossacarídeos, F2 e F3, com
o aumento das substituições por D-galactose, mostra que o pequeno aumento
das ramificações por D-galactose é suficiente para levar a reação a atingir o
equilíbrio, impedindo a hidrólise mesmo de sítios disponíveis para a ação de
endo-β-mananase, assim, produzindo F1, F2 e F3.
A regulação da hidrólise, com o impedimento da hidrólise completa do
galactomanano durante a degradação de reserva (in vivo), é de extrema
importância para evitar que toda a energia contida na reserva e utilizada no
crescimento e estabelecimento da plântula seja rapidamente liberada com a
produção de monossacarídeos, os quais ficariam disponíveis para
fitopatógenos, como os fungos (Tiné et al., 2003), os quais competiriam com a
própria planta para o uso da energia armazenada no polissacarídeo de reserva.
Portanto, ambos os resultados apresentados corroboram a hipótese
formulada acima, que a presença dos oligossacarídeos F2 e F3 nas hidrólises

181
de galactomanano de Sesbania virgata (Cav.) Pers. também esteja relacionada
ao equilíbrio de reação da hidrólise, causada pelas substituições por galactose,
como demonstrado por Tiné et al. (2003).
É possível visualizar, principalmente no perfil de oligossacarídeos
revelados por HPAEC-PAD, que os galactomananos de maior substituição por
D-galactose (Figura 36 E e F) apresentam proporções de F1 (picos de a-h e j-
m) e de F2 e F3 (pico i), quando comparados com os galactomananos de razão
2:1 (Figura 36 C e D), relativamente menores, demonstrando que a enzima,
pela alta quantidade e distribuição das ramificações, não tem acesso à cadeia
principal, resultando na baixa taxa de hidrólise do polissacarídeo, como
demonstrado por Reese & Shibata (1965); Courtois & Le Dizet (1968);
Tsujisaka et al. (1972); Emi et al. (1972).
A presença de galactose, levando ao equilíbrio de reação,
aparentemente, esconde os sítios disponíveis para a ação da enzima endo-β-
mananase. Sendo assim, evidencia a importância da interação da α-
galactosidase (exo-hidrolase) com a endo-β-mananase no processo de
degradação da mobilização de reserva. Embora a exo-hidrolase não participe
diretamente na atuação da endo-β-mananase, parece haver, por parte da α-
galactosidase, uma modulação na taxa de reação, alterando a afinidade da
endo-β-mananase pelo galactomanano.
VI. Conclusão
Cpm base no conjunto de resultados apresentados, sugere-se que a
estrutura fina do galactomanano, principalmente as ramificações com
galactose, modula a ação da endo-β-mananase (de fungos ou de plantas), não
permitindo a hidrólise completa do galactomanano na ausência da α-
galactosidase.
Embora este trabalho tenha sido realizado com galactomanano, Fanuti,
Gidley & Reid (1991) e Tiné et al. (2003) encontraram efeitos similares da
ramificação de galactose no polímero de xiloglucano sobre a ação da celulase
e a xiloglucano transglicosilase hidrolase.

182
Os resultados apresentados possuem implicações para as funções
biológicas do galactomanano. Sabidamente os galactomananos exercem
funções variadas na semente, além da função de reserva de sementes
(Buckeridge et al., 2000a). Reid & Bewley, 1979; Buckeridge et al., 2000c
demonstraram que o galactomanano pela sua alta solubilidade, dada pelo alto
conteúdo de galactoses, exerce o controle da embebição, absorvendo grandes
quantidades de água, distribuindo-a ao redor do embrião e protegendo este
contra estresse hídrico (Reid & Bewley, 1979; Halmer & Bewley, 1979).
Dessa forma, o polissacarídeo altamente ramificado resulta em grande
solubilidade da molécula (Reid & Bewley, 1979; Buckeridge et al., 2000c),
assim, o polímero com apreciáveis quantidades de ramificações será
prontamente solúvel, mas de acordo com os resultados apresentados também
será hidrolisado mais lentamente.
Além da característica de tampão de água, os galactomananos, mas
principalmente os mananos, possuem a função de proteção e dureza das
sementes, protegendo-as contra patógenos e digestão dos animais
(Buckeridge et al. 2000c).
Assim, a outra implicação leva em consideração o ataque de fungos ao
galactomanano presente em sementes. A existência de ramificações com
galactose que modulam a degradação do polímero pela endo-β-mananase,
como a abrangida nesta Dissertação, pode ter sido selecionada como uma
forma de controle da degradação pela própria plântula, mas também uma
forma de impedir ou dificultar a degradação por microorganismos, denotando
um importante aspecto de coevolução das leguminosas, que possuem reserva
de galactomanano, e os fungos capazes de degradá-lo.

183
VII. Bibliografia
BEWLEY, J. D. & BLACK, M. 1978. Physiology and biochemistry of seeds in relation to
germination. Vol.1. Springer-Verlag, Berlin.
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1990. Galactomannan from Brazilian seeds. Revista
Brasileira de Botânica 13:109-112.
BUCKERIDGE, M. S.; ROCHA, D. C.; REID, J. S. G. & DIETRICH, S. M. C. 1992. Xyloglucan
structure and post-germinative metabolism in seeds ofCopaifera langsdorfii from savanna
and forest populations. Physiologia Plantarum 84:145–151.
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1996. Mobilisation of the raffinose family
oligosaccharides and galactomannan in germinating seeds of Sesbania marginata Benth.
(Leguminosae-Faboideae). Plant Science 117:33-43.
BUCKERIDGE, M.S. & REID, J.S.G. 1996. Major cell wall polysaccharides in legume seeds:
Structure, catabolism and biological functions. Ciência e Cultura 48:153-162.
BUCKERIDGE, M.S.; DIETRICH, S.M.C. & LIMA, D.U. 2000a. Galactomannans as the reserve
carbohydrate in legume seeds. In: GUPTA, A.K. & KAUR, N. Carbohydrate Reserves in
Plants - Synthesis and Regulation. Elsevier Science, Amsterdam. p. 283-317.
BUCKERIDGE, M.S.; TINÉ, M.A.S.; SANTOS, H.P. & LIMA, D.U. 2000c. Polissacarídeos de
reserva de parede celular em sementes. Estrutura, metabolismo, funções e aspectos
ecológicos. Revista Brasileira de Fisiologia Vegetal, Edição Especial, p.137-162.
CARPITA, N.C. & GIBEAUT, D.M. 1993. Structural models of primary cell walls in flowering
plants: consistency of molecular structure with the physical properties of the cell wall during
growth. The Plant Journal 3:1-30.
CAVALARI, A. A. 2009. Modificações da parede celular de frutos do mamoeiro (Carica
Papaya). Tese (Doutorado em Biologia Vegetal) - Universidade Estadual de Campinas.
COURTOIS, J.E. & LE DIZET. 1968. Bult. Soc. Chim. Biol. vol.50.
EMI, S.; FUKUMOTO, J. & YAMAMOTO, T. 1972. J. Agr. Biol. Chem. Japan 36:991.
FANUTI, C.; GIDLEY, M.J. & REID, J.S.G. 1993. Action of a pure xyloglucan endo-
transglycosylase (formly called xyloglucan-specific endo-(1,4)-β-D-glucanase) from
cotyledons of germinated nasturtium seeds. Plant Journal v.3: 691-700.
-
galactosidase present during and following germination of tomato (Lycopersicon esculentum
Mill.) seed. Journal of Experimental Botany 52:1239-1249.
HALMER, P. & BEWLEY, J.D. 1979. Mannanase production by the lettuce endosperm: control
by the embryo. Planta 144:333-340.
HOFFMAN, J. & SVENSSON, S. 1978. Carbohydrates Research 65:65-71.
LISBOA, C.G.S; TONINI, P.P.; TINÉ, M.A.S. & BUCKERIDGE, M.S. 2006. Endo- -mananase
from the endosperm of seeds of Sesbania virgata (Cav.) Pers. (Leguminosae): purification,
characterisation and its dual role in germination and early seedling growth. Braz. J. Plant
Physiol., 18(2):269-280.

184
McCLEARY, B.V.; MATHESON, N.K & SMALL, D.M. 1976. Galactomannans and a
galactoglucomannan in Legume seed endosperms: structural requirements for β-
mannanase hydrolysis. Phytochemistry 15:1111-1117.
McCLEARY, B.V. 1983. Enzymic interaction in the hydrolysis of galactomannan in germinating
guar: the role of exo-β-mannanase. Phytochemistry 22:649-658.
McCLEARY, B.V. & MATHESON, N.K. 1983. Action patterns and substrate-binding
requirements of β-D-mannanase with mannosaccharides and mannan-type
polysaccharides. Carbohydrate Research 119: 191-219.
McCLEARY, B.V.; NURTHEN, E.; TARAVEL, F.R. & JOSELEAU, J.P. 1983. Characterisation of
the oligosaccharides produced on hydrolysis of galactomannan with β-D-mannanase.
Carbohydrate Research 118: 91-109.
McCLEARY, B.V.; CLARK, A.H.; DEA, I.C.M. & REES, D.A. 1985. The fine structure of carob
and guar galactomannans. Carbohydrate Research 139:237-260.
PALMER, K.L. & BALLANTYNE, M. J. 1950. Am. Chem. Soc. 72:736-741.
REESE, E.T. & SHIBATA, Y. 1965. Can. J. Microbiol. 11:167.
REID, J.S.G. & BEWLEY, J.D. 1979. A dual role for the endosperm and its galactomannan
reserve in the germinative physiology of fenugreek (Trigonella foenum-graecum L.) an
endospermic leguminous seed. Planta 147:145-150.
REID, J.S.G. & EDWARDS, M. 1995. Galactomannans and other cell wall storage
polysaccharides in seeds. In: STEPHEN, A.M. Food polysaccharides and their applications.
Marcel Dekker, New York. p. 155-186.
SAEMAN, J.F.; BUHL, J.L. & HARRIS, E.E. 1945. Quantitative saccharification of wood and
cellulose. Ind. Eng. Chem. Anal. Ed 17: 35-37.
STREET, H.E. & ÖPIK, H. 1984. The physiology of flowering plants. Edward Arnold, Great
Britain, 3 ed. p.279.
TINÉ, M.A.S.; LIMA, D.U. & BUCKERIDGE, M.S. 2003. Galactose branching modulates the
action of cellulose on seed storage xyloglucans. Carbohydrate Polymers 52:135-141.
TSUJISAKA, Y. HIYAMA, K.; TAKENISHI, S. & FUKUMOTO, J. 1972. J. Agr. Chem. Soc.
Japan 46:155.
WALDRON, K. & BRETT, C. 1990. Physiology and biochemistry of the plant cell walls. Unwin
Hyman, London, p.193.

185
DISCUSSÃO GERAL
Os galactomananos são polissacarídeos compostos de uma cadeia
linear de resíduos de D-manose unidas por ligações glicosídicas β-(1,4), a qual
resíduos de D-galactose estão unidos por ligações α-(1,6), formando
ramificações simples (Buckeridge et al., 2000a).
O polissacarídeo galactomanano conjuntamente aos polissacarídeos
xiloglucano e (arabino)galactanos são os polímeros encontrados como
componentes de reserva de parede celular em sementes (Buckeridge et al.,
2000c).
Os galactomananos são hidrolisados por três enzimas: α-galactosidase
(EC 3.2.1.22), endo-β-mananase ou β-1,4-manano-endo-hidrolase (EC
3.2.1.78) e exo-β-manosidase ou β-manosidase (EC 3.2.1.25) produzindo
galactose e manose livres ao mesmo tempo em que a glucose é produzida no
endosperma. Aparentemente essa última é disponibilizada ao embrião e servirá
como fonte de carbono e energia para o estabelecimento das plântulas
(Buckeridge et al., 2000a; Lisboa et al., 2006).
As alfa-galactosidases (E.C. 3.2.1.22) catalisam a hidrólise de ligações
α-(1,6) de galacto-oligossacarídeos e de polímeros de galacto-(gluco)-
mananos. Essas enzimas estão amplamente distribuídas em animais, plantas e
microorganismos (Fujimoto et al., 2003).
A presença de isoformas de α-galactosidase foi primeiramente descrita
por Petek & Dong (1961) em sementes de Coffea sp. e Plantago ovata. Tonini
et al. (2006) sugerem que a presença de isoformas de endo-β-mananase em
endosperma de Sesbania virgata (Cav.) Pers. seja para garantir a hidrólise do
galactomanano, mesmo com as modificações estruturais que este sofre
durante a mobilização.
Feurtado et al. (2001) sugeriram pelo menos três isoformas da α-
galactosidase durante e após a germinação de sementes de tomate e, através
da obtenção e sequenciamento do cDNA da enzima, demonstraram alta
homologia da forma ativa da α-galactosidase do tomate (40 kDa) com outras
galactosidases, especialmente com as de sementes de Coffea arabica (café)
(Zhu & Goldstein, 1994). Como apenas um gene para a α-galactosidase foi

186
identificado em sementes de tomate, as isoformas aparentemente se formam a
partir de modificações pós-traducionais (Feurtado et al., 2001).
McCleary & Matheson (1974) também detectaram múltiplas isoformas da
α-galactosidase, nas sementes de Cyamopsis tetragonoloba, embora apenas
uma delas (40 kDa) esteja associada com o endosperma (Overbeeke et al.
1989).
Assim, as diferentes isoformas de α-galactosidase podem estar
relacionadas tanto a diferenças estruturais do galactomanano durante a
mobilização, como por exemplo, a diminuição de substituições por galactose
demandaria uma α-galactosidase com maior afinidade ao substrato; como
também a diferentes funções além da degradação do galactomanano, por
exemplo, a degradação de açúcares da série rafinósica (Bewley,1997) e até o
amadurecimento de frutos (remodelamento da parede celular) (Gao & Schaffer,
1999; Soh et al., 2006).
Tonini et al. (2010) demonstraram por Western Blot utilizando anticorpo
anti α-galactosidase e extrato bruto enzimático de Sesbania virgata (Cav.) Pers.
a presença de 8 bandas variando de 78 kDa a 12 kDa reativas ao anticorpo,
evidenciando a presença de isoformas da α-galactosidase com diferentes
tamanhos, que poderiam interagir entre si formando a estrutura quartenária.
Os autores sugerem que a interação entre as α-galactosidases seria
visando uma ação proteolítica da própria α-galactosidase para ativá-la, pois
argumentam que esta não seria transcrita de novo após a maturação da
semente, ou seja, toda a enzima necessária à degradação já estaria pronta,
mas não ativa; e para adaptar a enzima as mudanças no substrato durante o
processo de degradação da reserva.
A α-galactosidase semipurificada de Sesbania virgata (Cav.) Pers.
apresentou 6 isoformas (Capítulo I – Figura 26 e figuras 38 – 42, 45, Anexo),
sendo uma delas preponderante. No entanto, durante o início do processo de
purificação ainda separou-se mais duas isoformas (Figuras 13 e 14). Sugere-se
que as duas isoformas retiradas no início da purificação estejam relacionadas a
degradação de outros substratos, como demonstrado por Bewley (1997); Gao
& Schaffer (1999) Soh et al. (2006), na degradação de oligossacarídeos da
série rafinósica e remodelamento da parede celular, respectivamente.

187
Já as demais isoformas encontradas, sugere-se que estejam
relacionadas a ação proteolítica da própria α-galactosidase em sua ativação e
remodelamento visando o aumento da afinidade ao substrato, como
demonstrado por Tonini, et al. (2010).
A regulação da degradação de reserva de sementes já foi demonstrada
pela ação de fitohormônios. O ácido abscísio (ABA) é um potente inibidor da
degradação da reserva de galactomanano e está envolvido no mecanismo de
regulação dessa mobilização em sementes de Ceratonia siliqua (Seller, 1977),
Lactuca sativa (Halmer & Bewley, 1979), Lycopersicon sculentum (Groot &
Karssen, 1992), Trigonella foenum-graecum (Reid & Meier, 1973; Malek &
Bewley, 1991; Kontos & Spyropoulos, 1995) e Sesbania virgata (Cav.) Pers.
(Potomati & Buckeridge, 2002; Tonini et al., 2006).
Tonini (2008) demonstrou que a mobilização de reserva de
galactomanano de sementes de Sesbania virgata (Cav.) Pers. é modulada
negativamente pela presença do ABA, enquanto o etileno é promotor da
mobilização.
Além da regulação esperada por fitohormônios, Tiné et al. (2003)
sugeriram que a estrutura do xiloglucano de reserva de sementes conteria, pelo
menos, parte da informação requerida para o seu próprio metabolismo, ou seja,
o próprio polissacarídeo, através da distribuição de suas ramificações com
galactose, modularia a ação da endohidrolase, neste caso a endo-β-mananase.
Estes fatos realçam a importância na interação das exo-hidrolases (α-
galactosidase e β-manosidase) com as endo-hidrolases (endo-β-mananase)
que ocorrem nos processos biológicos como a degradação da reserva de
sementes ou degradação da parede celular por fungos (Tiné et al., 2003).
Durante a degradação do galactomanano existe uma ordem na atuação
das enzimas (Reid & Edwards, 1995 e Lisboa et al., 2006), mas compreende-
se que elas estejam presentes concomitantemente em grande parte do
processo de degradação, sendo assim, a α-galactosidase é capaz de alterar a
afinidade da endo-β-mananase ao substrato (galactomanano) ao retirar
algumas ramificações de galactose, dessa forma, modulando o processo de
degradação.
McCleary & Matheson (1983), demonstraram que quanto menor for a
razão manose:galactose de um polissacarídeo, ou seja, quanto mais ramificado

188
for um polissacarídeo, menor será a porcentagem de hidrólise de dado
polissacarídeo pela endo-β-mananase.
A partir da análise dos resultados sugere-se que as ramificações por
galactose no polissacarídeo galactomanano pode modular o reconhecimento
dos sítios de clivagem pela enzima endo-β-mananase.
Sugere-se esta modulação uma vez que: existe a produção de
oligossacarídeos limites de digestão F1, F2 e F3 (Figura 28) após hidrólise do
galactomanano com endo-β-mananase, como demonstrado para xiloglucanos
(Tiné et al., 2003); os oligossacarídeos F1 possuem proporções distintas
quando da hidrólise do galactomanano com endo-β-mananase em diferentes
concentrações (Figuras 29 e 33), evidenciando preferência por sítios com
menor grau de galactosilação; a presença da α-galactosidase diminui a
produção dos oligossacarídeos limites de digestão F2 e F3, mostrando que a
estes não possuem resistência intrínseca a hidrólise e que a reação atinge o
equilíbrio mesmo quando ainda existem sítios de clivagem ainda disponíveis
(Figura 32); e que polissacarídeos com estruturas diferentes, razão
manose:galactose variando entre 150:1 a 1:1, são digeridos em diferentes
taxas de hidrólise pela mesma enzima (Figuras 35 e 36), evidenciando que a
ramificação com galactose dificulta a ação da endo-β-mananase.
Através dos resultados obtidos nesta dissertação, sugere-se que a
estrutura do polissacarídeo galactomanano também contenha, pelo menos,
parte da informação requerida para seu próprio metabolismo, código para a sua
degradação, estando esta informação contida na distribuição das ramificações
com resíduos de D-galactose (Capítulo 2).
A modulação da degradação feita pelo próprio polissacarídeo não só
permite a regulação do processo de liberação dos açúcares, fonte de energia
para o embrião em desenvolvimento, garantindo máximo aproveitamento pelo
embrião, mas, principalmente, dificulta a ação dos microorganismos pelo
aumento da complexidade, evidenciando um aspecto importante da co-
evolução das leguminosas e fungos capazes de atacar os polissacarídeos de
reserva de sementes.
Sugere-se que as diferentes isoformas da α-galactosidase relacionadas
à degradação da reserva de galactomanano de sementes de Sesbania virgata
(Cav.) Pers. seriam produto da ação proteolítica da própria enzima a fim de

189
melhorar a afinidade da α-galactosidase ao substrato durante o processo de
mobilização de reserva. Para a ação proteolítica da α-galactosidase
possivelmente haveria a interação de duas enzimas, formando uma estrutura
quartenária, onde uma ou ambas as enzimas teriam uma parte da cadeia de
aminoácidos eliminada e, portanto, apresentariam uma diminuição do tamanho.
O aumento da afinidade da α-galactosidase ao substrato durante todo o
processo de mobilização garante a liberação das ramificações com galactose
de forma contínua, permitindo e aumentando a eficiência da ação da enzima
endo-β-mananase aos sítios de clivagem, garantindo a degradação do
polissacarídeo a oligossacarídeos de forma regulada, passível de bloqueio,
pelo acúmulo de oligossacarídeos e galactose livre que inibem a ação das
enzimas endo-β-mananase e α-galactosidase, respectivamente (Tiné et al.,
2003; Lisboa et al., 2006), e dificultando a ação de microorganismos,
propiciando ao embrião a maior quantidade de açúcares para o seu
desenvolvimento, aumentando as chances de sucesso no estabelecimento da
plântula.

190
REFERÊNCIAS BIBLIOGRÁFICAS
ADEMARK, P.; LARSSON, M.; TJERNELD, K.; STALBRAND, H. 2001. Multiple α-
galactosidases from Aspergillus niger: purification, characterization and substrate
specificities. Enzyme and Microbial Technology 29:441–448.
ALCÂNTARA, P.H.N., DIETRICH, S.M.C. & BUCKERIDGE, M.S. 1999. Xyloglucan mobilisation
and purification of a (XLLG/XLXG) specific β-galactosidase from cotyledons of Copaifera
langsdorffii. Plant Physiol. Biochem., 37 (9), 653-663.
AL-KAISEY M.T., WILKIE C.B. 1992. The polysaccharides of agricultural lupin seeds,
Carbohydr. Res. v.227: 147–161.
ARAUJO, E.C.;. MENDONÇA, A.V.R; BARROSO, D.G.; LAMÔNICA, K.R & SILVA, R.F. 2004.
Caracterização morfológica de frutos, sementes e plântulas de Sesbania virgata (Cav.)
Pers. Revista Brasileira de Sementes, vol. 26, nº1, p.105-110.
BELTRATI, C.M. & PAOLI, A.A.S. 2003. Semente. In: APEZZATO-DA-GLÓRIA, B. &
CARMELLO-GUERREIRO, S.M. Anatomia vegetal. Editora Universidade Federal de
Viçosa, Viçosa. p. 399-424.
BEWLEY, J. D. & BLACK, M. 1978. Physiology and biochemistry of seeds in relation to
germination. Vol.1. Springer-Verlag, Berlin.
BEWLEY, J. D. & BLACK, M. 1994. Seeds: Physiology of development and germination. 2ºed.
Plenum Press, New York. 445p.
BEWLEY J.D. 1997. Seed germination and dormancy. Plant Cell 9:1055-1066.
BRADFORD, M.M. 1976. A rapid and sensitive method for quantification of microgan quantities
of proteins utilizing the principle of rotein-dye binding. Analytical Biochemistry 72:248-254.
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1990. Galactomannan from Brazilian seeds. Revista
Brasileira de Botânica 13:109-112.
BUCKERIDGE, M. S.; ROCHA, D. C.; REID, J. S. G. & DIETRICH, S. M. C. 1992. Xyloglucan
structure and post-germinative metabolism in seeds ofCopaifera langsdorfii from savanna
and forest populations. Physiologia Plantarum 84:145–151.
BUCKERIGDE, M.S. 1993. A novel β-galactosidase or exo-(1→4)-β-galactanase from
cotyledons of germinated Lupinus angustifolius L. seeds, and its role in post-germinative cell
wall metabolism. Tese de Doutorado. University of Stirling.
BUCKERIDGE M.S., REID J.S.G.1994. Purification and properties of a novel â-galactosidase or
exo-(1,4)-β-D-galactanase from the cotyledons of germinated Lupinus angustifolius L.
seeds, Planta v.192: 502–511.
BUCKERIDGE, M.S.; PANEGASSI, V.R. & DIETRICH, S.M.C. 1995a. Storage carbohydrate
mobilisation in seeds of Dimorphandra mollis Benth. (Leguminosae) following germination.
Revista Brasileira de Botânica 18:171-175.

191
BUCKERIDGE, M.S. & DIETRICH, S.M.C. 1996. Mobilisation of the raffinose family
oligosaccharides and galactomannan in germinating seeds of Sesbania marginata Benth.
(Leguminosae-Faboideae). Plant Science 117:33-43.
BUCKERIDGE, M.S. & REID, J.S.G. 1996. Major cell wall polysaccharides in legume seeds:
Structure, catabolism and biological functions. Ciência e Cultura 48:153-162.
BUCKERIDGE, M.S.; DIETRICH, S.M.C. & LIMA, D.U. 2000a. Galactomannans as the reserve
carbohydrate in legume seeds. In: GUPTA, A.K. & KAUR, N. Carbohydrate Reserves in
Plants - Synthesis and Regulation. Elsevier Science, Amsterdam. p. 283-317.
BUCKERIDGE, M.S.; SANTOS, H.P. & TINÉ, M.A.S. 2000b. Mobilisation of storage cell wall
polysaccharides in seeds. Plant Physiology and Biochemistry 38:141-156.
BUCKERIDGE, M.S.; TINÉ, M.A.S.; SANTOS, H.P. & LIMA, D.U. 2000c. Polissacarídeos de
reserva de parede celular em sementes. Estrutura, metabolismo, funções e aspectos
ecológicos. Revista Brasileira de Fisiologia Vegetal, Edição Especial, p.137-162.
BUCKERIDGE, M.S.; AIDAR, M.P.M.; SANTOS, H.P. & TINÉ, M.A.S. 2004a. Acúmulo de
reservas. In: FERREIRA, A.G. & BORGHETTI, F. Germinação: do básico ao aplicado.
Artmed, Porto Alegre. p. 31-50.
BUCKERIDGE, M.S., SANTOS, H.P., TINÉ, M.A. & AIDAR, M.P.M. (2004b) Mobilização de
Reservas. In: Germinação. Do básico ao aplicado. Alfredo Gui Ferreira & Fabian Borgheti
(Eds.). Artmed, Porto Alegre pp.163-185.
BUCKERIDGE, M.S. CAVALARI, A.A. & SILVA, G.B. 2008. Parede Celular. In: Fisiologia
Vegetal, Gilberto B. Kerbauy (2 ªEdição). Guanabara Koogan, Rio de Janeiro Cap. 9: 165-
181.
BUCKERIDGE, M.S. 2010. Seed Cell Wall Storage Polysaccharides: models to understand cell
wall biosynthesis and degradation. Plant Physiology 154: 1017-1023
CANTAREL, B.L.; COUTINHO, P.M.; RANCUREL, C.; BERNARD, T.; LOMBARD, V. &
HENRISSAT, B. 2008. The Carbohydrate-Active EnZymes database (CAZy): an expert
resource for Glycogenomics. Nucleic Acids Research: 1–6.
CARPITA, N.C. & GIBEAUT, D.M. 1993. Structural models of primary cell walls in flowering
plants: consistency of molecular structure with the physical properties of the cell wall during
growth. The Plant Journal 3:1-30.
CARPITA, N.C. & McCANN, M.C. 2000. The cell wall. In: BUCHANAN, B.B.; GRUISSEM, W. &
RUSSEL, R.J. Biochemistry and molecular biology of plants. 3ºed. American Society of
Plant Physiologists, Rockville. p. 52-108.
CAVALARI, A. A. 2009. Modificações da parede celular de frutos do mamoeiro (Carica
Papaya). Tese (Doutorado em Biologia Vegetal) - Universidade Estadual de Campinas.
CHANDRA, S.K.N. & DEMASON, D.A. 1990. Identification and immunocytochemical
localization of α-galactosidase in resting and germinating date palm (Phoenix dactylifera L.)
seeds. Planta 181:53-61.
COURTOIS, J.E. & PETEK, F. 1966. α-galactosidase from coffee beans. Methods in
Enzymology 8:565-670.

192
COURTOIS, J.E. & LE DIZET. 1968. Bult. Soc. Chim. Biol. vol.50.
CRAWSHAW, L.A. & REID, J.S.G. 1984. Changes in cell-wall polysaccharides in relation to
seedling development and the mobilisation of reserves in the cotyledons of Lupinus
angustifolius Cv. Unicrop. Planta v.160:449-454.
DAVIS, M.O.; HATA, D.J.; JOHNSON, S.A.; WALKER, J.C. & SMITH D.S. 1996. Cloning,
expression and characterization of a blood group B active recombinant α-D-galactosidase
from soybean (Glycine max). Biochemistry and Molecular Biology international 39: 471-485.
DAVIS, M.O.; HATA, D.J.; JOHNSON, S.A.; JONES, D.E.; HARMATA, M.A.; EVANS,
M.L.;WALKER, J.C. & SMITH D.S. 1997. Cloning, expression and characterization of a
blood group B active recombinant α-D-galactosidase from pinto bean (Phaseolus vulgaris).
Biochemistry and Molecular Biology international 42: 453-467.
DEA, I.C.M. & MORRISON, A. 1975. Chemistry and Interactions of seed galactomannans.
Advances in Carbohydrate Chemistry and Biochemistry 31:241-312.
DEA I.C.M., CLARK A.H., MCCLEARY B.V. 1986. Effect of galactose substitution-pattern on the
interaction properties of galactomannans. Carbohydr. Res. v.147: 275–294.
DEY, P.M. & PRIDHAM, J.B. 1972. Biochemistry of α-galactosidases. Advan. Enzymol. 36:
91:130.
DHAR, M.; MITRA, M.; HATA, J.; BUTNARIU, O. & SMITH, D. 1994. Purification and
characterization of Phaseolus vulgaris α-D-galactosidase isozymes. Biochemistry and
Molecular Biology International 34:1055-1062.
DEMASON, D.A.; MADORE, M.A.; SEKHAR, K.N.C. & HARRIS, M.J. 1992. Role of α-
galactosidase in cell wall metabolism of date palm (Phoenix dactylifera) endosperm.
Protoplasma 166:177-186.
DIRK, L.M.A.; GRIFFEN, A.M.; DOWNIE, B. & BEWLEY, J.D. 1995. Multiple isozymes of endo-
β-mannanase in dry and imbibed seeds. Phytochemistry v.40: 1045-1056.
EDWARDS M., DEA I.C.M., BULPIN P.V., REID J.S.G. 1985. Xyloglucan (amyloid) mobilisation
in the cotyledons of Tropaeolum majus L. seeds following germination, Planta v.163 :133–
140.
EMI, S.; FUKUMOTO, J. & YAMAMOTO, T. 1972. J. Agr. Biol. Chem. Japan 36:991.
FANUTI, C.; GIDLEY, M.J. & REID, J.S.G. 1993. Action of a pure xyloglucan endo-
transglycosylase (formly called xyloglucan-specific endo-(1,4)-β-D-glucanase) from
cotyledons of germinated nasturtium seeds. Plant Journal v.3: 691-700.
-
galactosidase present during and following germination of tomato (Lycopersicon esculentum
Mill.) seed. Journal of Experimental Botany 52:1239-1249.
FUJIMOTO, Z.; KANEKO, S.; MOMMA, M.; KOBAYASHI, H. & MIZUNO, H. 2003. Crystal
-galactosidase complexed with D-galactose. J. Biol. Chem. 278 (22):
20313-20318.
GAO, Z. & SCHAFFER, A.A. 1999. A novel alkaline α-galactosidase from melon fruit with a
substrate preference for raffinose. Plant Physiology 119:979-987.

193
GROOT, S.P.C. & KARSSEN, C.M. 1987. Gibberellins regulate seed-germination in tomato by
endosperm weakening - a study with gibberellin-deficient mutants. Planta 171:525-531.
GROOT, S.P.C. & KARSSEN, C.M. 1992. Dormancy and germination of abscisic acid-deficient
tomato seeds - studies with the sitiens mutant. Plant Physiology 99:952-958.
GROOT, S.P.C.; KIELISZEWSKA-ROKICKIA, B.; VERMEER, E. & KARSSEN, C.M. 1988.
Giberellins-induced hydrolysis of endosperm cell walls in giberellindeficient tomato seeds
prior to radicle emergence. Planta v.174: 500-504.
GUERIN JR, LANCE RCM, WALLACE W. 1992 Release and activation of barley b-amylase by
malt endopeptidases. J Cereal Sci 15:5–14.
GUIMARÃES, V.M; REZENDE, S.T.; MOREIRA, M.A.; BARROS, E.G. & FELIX, C.R. 2001.
Characterization of α-galactosidase from germinating soybean seed and their use for
hydrolisis of oligossacharides. Phytochemistry 58:67-73.
GUZMÁN, J.M. & HERNANDEZ, G.L. 1982. Anatomía de la semilla y germinación de Turbina
corumbosa (L.) Raf., Convovulaceae. Phyton 42:1-8.
HALMER, P. & BEWLEY, J.D. 1979. Mannanase production by the lettuce endosperm: control
by the embryo. Planta 144:333-340.
HAMES B. D. 1990. One-dimensional polyacrylamide gel electrophoresis. In: Hames BD,
Rickwood D (eds) Gel electrophoresis of proteins: a practical approach, 2nd edn. Oxford
University Press, New York.
HIRST, E.L.; JONES, J.K.N.; WALDER, W.O. 1947. Pectic substances. Part 7: the constitution
of the galactan from Lupinus albus. J. Chem. Soc. 1225-1229.
HOFFMAN, J. & SVENSSON, S. 1978. Carbohydrates Research 65:65-71.
HOFFMAN, K.; BUCHER, P.; FLAQUET, L.; BAIROCH, A. 1999. The PROSITE database, its
status in 1999. Nucleic Acids Research 27:215-219.
ITOH, T.; SHIMURA, S.; ADACHI, S. 1979. Α-galactosidase from alfafa seeds (Medicago sativa
L.). Agicultural and Biological Chemistry 43:1499-1504.
JARVIS, M.C. & MCCANN, M.C. 2000. Macromolecular biophysics of the plant cell wall:
concepts
KEUSCH L. 1968. Die mobilisierung des reservemannans im keimenden Dattelsamen, Planta
v.78: 321–350.
KISSMAN, K.G. & GROTH, D. 1999. Plantas infestantes e nocivas. Tomo II. BASF, São Paulo.
P.947-950.
KOOIMAN, P.1960. On the occurence of amyloids in plant seeds. Acta Bot. Neerl. v.9: 208–
219.
KONTOS, F. & SPYROPOULOS, C.G. 1995. Production and secretion of α-galactosidase and
endo-β-mannanase activity by carob (Ceratonia siliqua L.) in the endosperm protoplast.
Journal of Experimental Botany v.46: 577-583.
LABOURIAU, L.G. 1983. A germinação das sementes. Secretaria-geral da Organização dos
Estados Americanos. Programa Regional de Desenvolvimento Científico e Tecnológico,
Washington. 173p.

194
LAEMMLI U.K. 1970. Cleavage of structural protein during the assembly of the head of
bacteriophage T4. Nature, 223, 680-685.
-galactosidase in the degradation of the
endosperm cell walls of lettuce seeds, cv. Grand Rapids. Planta 157:274-277.
LIMA, E.E.; DE REZENDE, S.T.; BORGES, R.C.G. & PEREZ, S.C.J.A. 2004. Caracterização
de α´galactosidase em embrião e cotilédones de sementes de Platymiscium pubescens
Micheli, Var. Pubecens (Tamboril da mata). Revista Brasileira de Sementes, Vol. 26, nº 2,
p.82-90.
LISBOA, C.G.S. 2003. Endo-β-mananase de endosperma de Sesbania virgata (Cav.) Pers.:
purificação, caracterização e importância na germinação e desenvolvimento da plântula.
Dissertação de Mestrado. Universidade Estadual de Campinas, SP.
LISBOA, C.G.S; TONINI, P.P.; TINÉ, M.A.S. & BUCKERIDGE, M.S. 2006. Endo-β-mananase
from the endosperm of seeds of Sesbania virgata (Cav.) Pers. (Leguminosae): purification,
characterisation and its dual role in germination and early seedling growth. Braz. J. Plant
Physiol., 18(2):269-280.
MALEK, A.E. & BEWLWY, J.D. 1991. Endo-β-mannanase activity and reserve mobilization in
excised endosperms of fenugreek is affected by volume of incubation and abscisic acid.
Seed Science Research v.1: 45-49.
MANZI, A.E. & CEREZO, A.S. 1984. The galactomannan-like oligosaccharides from the
endosperm of seed of Gleditsia triacanthos. Carbohydrate Research 134:115-131.
MATHESON N.K., SAINI H.S. 1977. α-L-arabinofuranosidases and α-D-galactosidases in
germinating-lupin cotyledons. Carbohydr. Res. v.57: 103–116.
MAYER, A.M. & POLJAKOFF-MAYBER, A. 1975. The germination of seeds. 2ºed. Pergamon
Press, Oxford. 192p.
McCANN, M.C. & ROBERTS, K. 1991. Architecture of the primary wall. In: LLOYD, C.W. The
cytoskeletal basis of plant growth and form. Academic Press, Londres. p. 109-129.
McCLEARY, B.V. & MATHESON, N.K. 1974. α-D-galactosidase activity and galactomannan
and galactosylsucrose depletion in germinating legume seeds. Phytochemistry 13:1747-
1757.
MCCLEARY, B.V. & MATHESON, N.K. 1975. Galactomannan structure and β-mannanase and
β-mannosidase activity in germinating legume seeds. Phytochemistry 14:1187–1194.
McCLEARY, B.V.; MATHESON, N.K & SMALL, D.M. 1976. Galactomannans and a
galactoglucomannan in Legume seed endosperms: structural requirements for β-
mannanase hydrolysis. Phytochemistry 15:1111-1117.
McCLEARY, B.V. 1983. Enzymic interaction in the hydrolysis of galactomannan in germinating
guar: the role of exo-β-mannanase. Phytochemistry 22:649-658.
McCLEARY, B.V. & MATHESON, N.K. 1983. Action patterns and substrate-binding
requirements of β-D-mannanase with mannosaccharides and mannan-type
polysaccharides. Carbohydrate Research 119: 191-219.

195
McCLEARY, B.V.; NURTHEN, E.; TARAVEL, F.R. & JOSELEAU, J.P. 1983. Characterisation of
the oligosaccharides produced on hydrolysis of galactomannan with β-D-mannanase.
Carbohydrate Research 118: 91-109.
McCLEARY, B.V.; CLARK, A.H.; DEA, I.C.M. & REES, D.A. 1985. The fine structure of carob
and guar galactomannans. Carbohydrate Research 139:237-260.
McIlvaine, T. C. 1921. A buffer solution for colorimetric comparison. Journal of Biological
Chemistry 49:183-189.
MOE, O.A.; MILLER, S.E. & IWEN, M.H. 1947. Investigation of the reserve carbohydrate of
leguminous seeds. I. Periodate oxidation. Journal of the American Chemical Society
69:2621-2825.
NADELMANN, H. 1890. Uber die Schleimendosperme der Leguminosen. Pringsheims Jahrb
Wissenschaft Botany 21:1-83.
NOMAGUCHI, M.; NONOGAKI, H. & YUKIO, M. 1995. Development of galactomannan-
hydrolyzing in the micropylar endosperm tip of tomato seed prior germination. Physiologia
Plantarum 94:105-109.
OVERBEEKE, N.; FELLINGER, A.J.; TOONEN, M.Y.; VAN WASSENAAR, D. & VERRIPS, C.T.
1989. Cloning and nucleotide sequence of the α-galactosidase cDNA from Cyamopsis
tetragonoloba (guar). Plant Molecular Biology 13:541-550.
PALMER, K.L. & BALLANTYNE, M. J. 1950. Am. Chem. Soc. 72:736-741.
Parker M.L. 1984. Cell wall storage polysaccharide in cotyledons of Lupinus angustifolius L. II
mobilization during germination and seedling development, Protoplasma v.120: 233–241.
PETEK, F. & TO DONG. 1961. Enzymologia, 23, 133.
PLANT, A.R. & MOORE, K.G. 1982. Α-D-Mannosidase and α-D-galactosidase from proteins
bodies of Lupinusangustifolius cotyledons. Phytochemistry 21:985-989.
POTOMATI, A. & BUCKERIDGE, M.S. 2002. Effect of abscisic acid on the mobilisation of
galactomannan and embryo development of Sesbania virgata (Cav.) Pers. (Leguminosae-
Faboideae). Revista Brasileira de Botânica 25:303-310.
RAVEN, P.H.; EVERT, R.F. & EICHHORN, S.E. 2001. Biologia Vegetal. 6ºed. Editora
Guanabara Koogan, Rio de Janeiro. 906p.
REESE, E.T. & SHIBATA, Y. 1965. Can. J. Microbiol. 11:167.
REID, J.S.G. 1985a. Cell wall storage carbohydrates in seeds. Biochemistry of the seeds gums
and hemicelluloses. Advances in Botanical Research 11:125-155.
REID, J.S.G. & BEWLEY, J.D. 1979. A dual role for the endosperm and its galactomannan
reserve in the germinative physiology of fenugreek (Trigonella foenum-graecum L.) an
endospermic leguminous seed. Planta 147:145-150.
REID, J.S.G. & EDWARDS, M. 1995. Galactomannans and other cell wall storage
polysaccharides in seeds. In: STEPHEN, A.M. Food polysaccharides and their applications.
Marcel Dekker, New York. p. 155-186.
REID, J.S.G. & MEIER, H. 1972. The function of the aleurone layer during galactomannan
mobilization in germination seeds of fenugreek (Trigonella foenum-graecum L.), crimson

196
clover (Trifolium incarnatum L.): a correlative biochemical and ultrastructural study. Planta
106:44-60.
REID, J.S.G. & MEIER, H. 1973. Enzymic activities and galactomannan mobilisation in
germinating seeds of fenugreek (Trigonella foenum-graecum L. Leguminosae). Secretion of
α-galactosidases and β-mannosidase by the aleurone layer. Planta 112:301-308.
SACHS, J. 1862. Zur Keimungsgeschichte der Dattel. Bot. Zeitung v.20: 241-246; 249-252.
SAEMAN, J.F.; BUHL, J.L. & HARRIS, E.E. 1945. Quantitative saccharification of wood and
cellulose. Ind. Eng. Chem. Anal. Ed 17: 35-37.
SCHALLER A. 2004. A cut above the rest: the regulatory function of plant proteases. Planta
220:183–197.
SEILER, A. 1977. Galaktomannanabbau in keimenden Johanisbrotsamen (Ceratonia siliqua L.).
Planta 134:209-221.
SILVA, C.O. 2001. Papel da β-galactosidase no mecanismo de degradação do xiloglucano de
parede celular durante a mobilização de reserva em cotilédones de Copaifera lansgsdorffii.
Dissertação de Mestrado apresentada a Universidade Estadual de Campinas.
SILVA, G.A. 2007. Purificação parcial e caracterização de uma α-galactosidase de sementes
de Guapuruvu (Schizolobium parahyba). Dissertação de Mestrado pela Universidade de
Brasília.
SOH, C-P; ALI, Z.M. & LAZAN, H. 2006. Characterisation of an α-galactosidase with potential
relevance to ripening related texture changes. Phytochemistry 67:242-254.
SOMME, R. 1968. The structure of a galactomannan from Medicago Lupulina L. Acta Chemica
Scandinavica 22:537-545.
-galactosidase activity and the
hydrolases of galactomannan in the endosperm of the fenugreek (Trigonella foenum-
graecum) seed. Planta 166:271-275.
STREET, H.E. & ÖPIK, H. 1984. The physiology of flowering plants. Edward Arnold, Great
Britain, 3 ed. p.279.
TAIZ, L. & ZEIGER, E. 2004. Fisiologia Vegetal. 3ºed. Artmed, Porto Alegre. 719p.
TAMURA K, DUDLEY J, NEI M & KUMAR S. 2007. MEGA4: Molecular Evolutionary Genetics
Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24:1596-1599.
TINÉ, M.A.; CORTELAZZO, A.L. & BUCKERIDGE, M.S. 2000. Xyloglucan mobilization in
cotyledons of developing plantlets of Hymenea courbaril L. (Leguminosae –
Caesalpinoideae). Plant Sci. v.154: 117-126.
TINÉ, M.A.S.; LIMA, D.U. & BUCKERIDGE, M.S. 2003. Galactose branching modulates the
action of cellulose on seed storage xyloglucans. Carbohydrate Polymers 52:135-141.
TOOKEY, H.L.; LOHMAR, R.L. & WOLFF, I.A. 1962. New sources of seed mucilages. Journal
of Agricultural and Food Chemistry v.10:131-133.
TONINI, P.P.; LISBOA, C.G.S.; FRESCHI, L.; MERCIER, H.; MAZZONI-VIVEIROS, S.C. &
BUCKERIDGE, M.S. 2006. Effect of abscisic acid on galactomannan degradation and endo-

197
β-mannanase activity in seeds of Sesbania virgata (Cav.) Pers. (Leguminosae). Trees:
structure and function 20:669-678.
TONINI, P.P.; LISBOA, C.G.S.; SILVAI, C.O.; MAZZONI-VIVEIROS, S.C. & BUCKERIDGE,
M.S. 2007. Testa is involved in the control of storage mobilisation in seeds of Sesbania
virgata (Cav.) Pers., a tropical legume tree from of the Atlantic Forest. Trees: structure and
function 21:13-21.
TONINI, P. P. 2008. Conexão entre os processos de degradação das reservas de proteínas e
carboidratos e o efeito dos hormônios e açúcares em sementes Sesbania virgata de (Cav.)
Pers. Tese de Doutorado. Universidade Estadual de Campinas, SP.
TONINI, P. P.; CARRARA, T.B.; BUCKERIDGE, M. S. 2010. Storage proteins and cell wall
mobilisation in seeds of Sesbania virgata (Cav.) Pers. (Leguminosae). Trees: Structure and
Function v.24 (4): 675-684.
TSUJISAKA, Y. HIYAMA, K.; TAKENISHI, S. & FUKUMOTO, J. 1972. J. Agr. Chem. Soc.
Japan 46:155.
UNRAU, A.M. 1961. The constitution of a galactomannan from seeds of Leucaena
leucochephala. Journal of Organic Chemistry 26:3097-3101.
VIANA, S.F.; GUIMARÃES, V.M.; MOREIRA, M.A.; OLIVEIRA, M.G.A.; COSTA, N.M.B.; de
REZENDE, S.T. 2002b. Reduction of raffinose oligosaccharides in soymilk by alpha-
galactosidase from germinating soymilk seeds. In: REUNIÃO ANUAL DA SOCIEDADE
BRASILEIRA BIOQUÍMICA E BIOLOGIA MOLECULAR, 31., 2002, Caxambu. Anais
Caxambu: (SBBQ), p.95.
WALDRON, K. & BRETT, C. 1990. Physiology and biochemistry of the plant cell walls. Unwin
Hyman, London, p.193.
WEBER, K. & OSBORN, M. 1969. The Reliability of Molecular Weight Determinations by
Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis. J. Biol. Chem. 244: 4406-4412.
WILLIAMS, J.; VILLARROYA, H. & PETEK, F. 1978. Α-galactosidase II, III and IV from seeds of
Trifolium repens: Purification, physiochemical properties and mode of galactomannan
hydrolysis in vitro. Biochemical Journal 175:1069-1077.
ZHU, A. & GOLDSTEIN, J. 1994. Cloning and functional expression of DNA encoding coffe
bean α-galactosidase. Gene 140:227-231.

ANEXO
Figura 38: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 1 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. As proteínas realssadas pelo retângulo preto fazem referência a α-
galactosidases.

199
Figura 39: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 2 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. As proteínas realssadas pelo retângulo preto fazem referência a α-
galactosidases.

200
Figura 40: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 3 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.

201
Figura 41: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 4 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.

202
Figura 42: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 5 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.

203
Figura 43: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 6 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.

204
Figura 44: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 7 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.
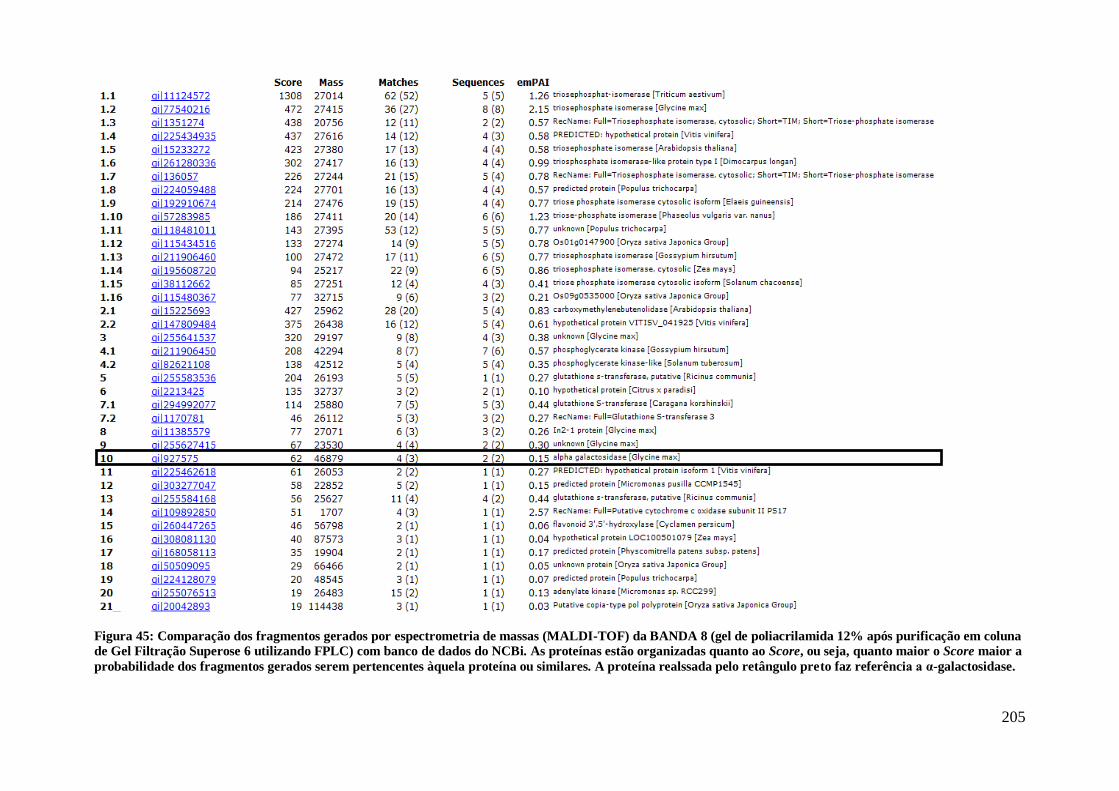
205
Figura 45: Comparação dos fragmentos gerados por espectrometria de massas (MALDI-TOF) da BANDA 8 (gel de poliacrilamida 12% após purificação em coluna
de Gel Filtração Superose 6 utilizando FPLC) com banco de dados do NCBi. As proteínas estão organizadas quanto ao Score, ou seja, quanto maior o Score maior a
probabilidade dos fragmentos gerados serem pertencentes àquela proteína ou similares. A proteína realssada pelo retângulo preto faz referência a α-galactosidase.

Figura 46: Análise de 35 sequências de aminoácidos de α-galactosidase de plantas. Grupos identificados por
letras estão de acordo com a filogenia: (A – azul escuro) Fabaceae; (B - verde) Rubiceae; (C - amarelo)
Solanaceae; (D - vermelho) Fabaceae; (E – azul claro) Pinaceae; (F – roxo) Poaceae. (•) Bootstrap ˃ 95, ou seja,
em 95% ou mais das 1000 árvores de relação geradas os ramos marcados com (•) ficaram agrupados como
mostrado nesta árvore de relação.

207

208
Figura 47: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da análise dos
oligossacarídeos, produto da hidrólise enzimática com α-galactosidase comercial de Cyamopsis tetragonoloba
(Megazyme®) dos oligossacarídeos produto da hidrólise do galactomanano a 1% de Sesbania virgata (Cav.) Pers
com endo-β-mananase (Megazyme®) a 0,005U, 0,05U e 1U (Experimento I). (I) Controle (galactomanano a 1%
sem enzima); (II) Controle (galactomanano a 1% com α-galactosidase comercial de Cyamopsis tetragonoloba
(Megazyme®); (III) Controle (galactomanano a 1% com celulase de Trichoderma longibrachiatum (Megazyme
®));
(IV) perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia hidrólise com endo-β-
mananase a 0,005U; (V) perfil de oligossacarídeos formados após ação da α-galactosidase sobre prévia hidrólise
com endo-β-mananase a 0,05U; (VI) perfil de oligossacarídeos formados após ação da α-galactosidase sobre
prévia hidrólise com endo-β-mananase a 1U; (VII) comparação dos perfis de oligossacarídeos formados no
Experimento I e Experimento II na concentração de 0,005U de endo-β-mananase; (VIII) comparação dos perfis
de oligossacarídeos formados no Experimento I e Experimento II na concentração de 0,05U de endo-β-mananase;
(IX) comparação dos perfis de oligossacarídeos formados no Experimento I e Experimento II na concentração de
1U de endo-β-mananase. (A) Manobiose (M2); (B) Manotriose (M3) e/ou trissacarídeo G1M2; (C) tetrassacarídeos
(manotetraose – M4 e/ou G1M3); (D) pentassacarídeos (manopentose – M5 e/ou G
1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7
e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos (possíveis oloigossacarídeos presentes: G1,3,4
M5; G4,5
M6, G3,4
M6;
G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7); (K-L) octassacarídeos e nonassacarídeos sem ramificação; (M)
oligossacarídeos >10 resíduos de manose (possivelmente resultado da desramificação do pico I). Os
oligossacarídeos identificados A-M são denominados oligossacarídeos F1; os oligossacarídeos restantes são
identificados como F2, quanto combinação aos pares, e F3, quando combinação aos trios.

209
Figura 48: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA- 100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do galactomanano de Sesbania virgata (Cav.) Pers com endo-β-

210
mananase (Megazyme®) a 1U com acréscimo ou não da α-galactosidase comercial de Cyamopsis tetragonoloba
(Megazyme®). (I) perfil de oligossacarídeos formados na hidrólise do galactomanano a 1% somente com endo-β-
mananase a 1U; (II) perfil de oligossacarídeos formados na hidrólise do galactomanano a 1% com endo-β-
mananase a 1U/mL e α-galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (III) perfil
de oligossacarídeos formados na hidrólise do galactomanano a 1% somente com α-galactosidase comercial de
Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (IV) Controle (galactomanano sem enzima). (Gal) Galactose;
(A) manobiose (M2); (B) manotriose (M3) e/ou G1M2; (C) tetrassacarídeos (manotetraose – M4 e/ou G
1M3); (D)
pentassacarídeos (manopentose – M5 e/ou G1M3 e/ou G
3M4); (E) hexassacarídeos (manohexose – M6 e/ou G
1,3M4
e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7 e/ou G
3,4M5); (G-I) oligossacarídeos >8 resíduos
(G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou G4,5
M7). Os oligossacarídeos identificados A-I são
denominados oligossacarídeos F1; os oligossacarídeos restantes são identificados como F2, quanto combinação
aos pares, e F3, quando combinação aos trios
Figura 49: Cromatogramas gerados por HPAEC-PAD, utilizando coluna PA-100, da análise dos
oligossacarídeos, produto da hidrólise enzimática do galactomanano de Sesbania virgata (Cav.) Pers com endo-β-
mananase (Megazyme®) a 0,005U e 1U e α-galactosidase comercial de Cyamopsis tetragonoloba (Megazyme
®) a
0,5U/mL. (I) Galactomanano a 1%, 0,005U de endo-β-mananase e α-galactosidase comercial de Cyamopsis
tetragonoloba (Megazyme®) a 0,5U/mL; (II) Galactomanano a 0,1%, 0,005U de endo-β-mananase e α-
galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (III) Galactomanano a 1%, 1U de

211
endo-β-mananase e α-galactosidase comercial de Cyamopsis tetragonoloba (Megazyme®) a 0,5U/mL; (IV
Galactomanano a 0,1%, 1U de endo-β-mananase e α-galactosidase comercial de Cyamopsis tetragonoloba
(Megazyme®) a 0,5U/mL. (A) manobiose (M2); (B) manotriose (M3) e/ou trissacarídeo G
1M2; (C) tetrassacarídeos
(manotetraose – M4 e/ou G1M3); (D) pentassacarídeos (manopentose – M5 e/ou G
1M3 e/ou G
3M4); (E)
hexassacarídeos (manohexose – M6 e/ou G1,3
M4 e/ou G4M5 e/ou G
3M5); (F) heptassacarídeos (manoheptose – M7
e/ou G3,4
M5); (G-I) oligossacarídeos >8 resíduos (G1,3,4
M5; G4,5
M6, G3,4
M6; G3,4,5
M6; G1,4,5
M6; G1,3,4
M6 e/ou
G4,5
M7). Os oligossacarídeos identificados A-I são denominados oligossacarídeos F1; os oligossacarídeos restantes
são identificados como F2, quanto combinação aos pares, e F3, quando combinação aos trios.








![Catálise Enzimática [Bioquímica I]._](https://static.fdocumentos.tips/doc/165x107/553cecca4a7959fe7f8b4a0c/catalise-enzimatica-bioquimica-i.jpg)








