
CONSEQUÊNCIAS DA ACÇÃO ANTROPOGÉNICA PARA A …
61
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA ANIMAL CONSEQUÊNCIAS DA ACÇÃO ANTROPOGÉNICA PARA A REABILITAÇÃO ECOLÓGICA DE UMA LAGOA EUTRÓFICA Tiago Manuel Barbosa da Silva MESTRADO em ECOLOGIA E GESTÃO AMBIENTAL 2008
Transcript of CONSEQUÊNCIAS DA ACÇÃO ANTROPOGÉNICA PARA A …

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
CONSEQUÊNCIAS DA ACÇÃO ANTROPOGÉNICA
PARA A REABILITAÇÃO ECOLÓGICA DE UMA
LAGOA EUTRÓFICA
Tiago Manuel Barbosa da Silva
MESTRADO em
ECOLOGIA E GESTÃO AMBIENTAL
2008

1
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
CONSEQUÊNCIAS DA ACÇÃO ANTROPOGÉNICA
PARA A REABILITAÇÃO ECOLÓGICA DE UMA
LAGOA EUTRÓFICA
Tiago Manuel Barbosa da Silva
MESTRADO em
ECOLOGIA E GESTÃO AMBIENTAL
Orientado pela Professora Doutora
Maria José L. Boavida
2008

2
Índice Pág.
Agradecimentos …………………………………………………………………………………………………… 3
Resumo …...…………………………………………………………………………………………………..…… 4
Abstract ………………………………………………………………………………………………………....… 5
Introdução ………..………………………………………………………………………………………………... 6
Materiais e Métodos …...………………………………………………………………………………………….. 9
Descrição da Lagoa de Pataias …………………………………………………………………….……. 9
Amostragem e análises …………………………………………………………………………….…... 10
Medições in-situ ………………………………………………………………………………….…….. 10
Determinações laboratoriais …………………………………………………………………….……... 10
Produtividade …………………………………………………………………………….……………. 11
Limitação pelo fósforo ………………………………………………………………………………… 12
Análise Estatística ……………………………………………………………………………………… 12
Resultados ………………………………………………………………………………………….……………. 12
Primeira parte …………………………………………………………………………….…………….. 13
Segunda parte …………………………………………………………………………….…………….. 17
Excepções da relação CHL-a – pH ………………………………………………….….…… 21
Primeira excepção ……………………………………………………….……...… 21
Segunda excepção ……………………………………………………….………... 23
Produtividade …………………………………………………………………….………..… 27
Limitação pelo fósforo ……………………………………………………………………… 28
Estatística ………………………………………………………………………….………… 28
Terceira parte ………………………………………………………………………………………….. 29
Índice do estado Trófico (TSI) ……………………………………………………………… 30
Discussão ………………………………………………………………………………………………………… 33
Primeira e Segunda partes ……………………………………………………………………………... 33
Excepções da relação CHL-a – pH ……………………………………………………….…. 35
Primeira excepção ………………………………………………………………… 35
Segunda excepção …………………………………………...………………….… 36
Relativamente ao regime de vento ……………………………………..…………………… 37
Comportamento da concentração da CHL-a ao longo da segunda parte do estudo ………… 38
Relativamente ao valor mais elevado da concentração da CHl-a a 05/09/2007…………...… 40
Produtividade primária ………………………………………………………………….…… 44
Limitação pelo fósforo ………………………………………………………………….…… 44
Estatística …………………………………………………………………………….……… 45
Terceira parte ………………………………………………………………………………….……….. 47
Conclusão ……………………………………………………………………………………………….………... 50
Bibliografia …………………………………………………………………………………………….…………. 51
Anexo I ………………………………………………………………………………………………....………… 57
Anexo II ………………………………………………………………………………………………....…….….. 60

3
Agradecimentos
À minha família, pela motivação e apoio inquestionável, desde tenra idade, na
persecução dos meus objectivos.
À Professora Doutora Maria José L. Boavida e “minha” orientadora. Sempre disponível,
no gabinete, por telefone ou internet… uma incansável e inestimável ajuda! Aprendi
muito, diria, tudo graças a si. Inequivocamente superou, ultrapassou e estilhaçou tudo e
quase todos. Palavras para quê… perfeito, perfeito era um professor assim para cada
aluno! Muito obrigado.
À Doutora Maria José Caramujo, pela ajuda e integração na equipa do laboratório.
Grande conhecedora dos meandros da ciência, sempre com uma sugestão simples e
eficiente. À imagem de todos os grandes construtores de conhecimento.
Ao Professor Doutor Dinis Duarte F. Pestana… é como, para quem anseia chegar a um
destino, e ter alguém que lhe indique o caminho. Obrigado por me ouvir.
À Eloá por partilhar comigo o tempo, conhecimento e experiências em cada saída de
campo e a todas as pessoas, que abdicaram do seu tempo e espaço para os partilharem
comigo. A todos muito obrigado.
Para o Henrique Folhas, o David Avelar, e a Catarina Miranda duas beijocas e uma
palmada, respectivamente.

4
Resumo
A lagoa de Pataias, localizada no concelho de Alcobaça, é um lago pouco
profundo (profundidade máxima 3,0m) que se encontra na fase terminal da ontogenia.
Durante décadas e até recentemente a eutroficação cultural em muito dominou e acelerou
os processos naturais.
Durante o ano de 2005 a lagoa secou completamente e os sedimentos expostos
proporcionaram a oportunidade para reduzir os nutrientes autóctones. A medida de
gestão visou a redução do pool de nutrientes autóctones, removendo a macrófita
Nymphaea alba e a camada superficial dos sedimentos numa área significativa do fundo
da lagoa.
O estudo de 2007 decorreu de Janeiro a Outubro e monitorizou várias variáveis e
parâmetros limnológicos na coluna de água (CHL-a, SRP, TP, transparência, cor,
temperatura, pH, oxigénio, condutividade, fitoplâncton e zooplâncton, combinados com
determinações da produtividade e limitação pelo fósforo) para determinar o estado trófico
da lagoa. Estes dados são comparados com os determinados no estudo de 2005,
permitindo traçar a evolução do estado trófico e, principalmente, o sucesso da aplicação
da medida de gestão que se seguiu a este estudo (Setembro de 2005). Simultaneamente,
são estudadas as flutuações dos parâmetros/variáveis objecto de monitorização, para
melhor compreender os aspectos dinâmicos do funcionamento desta lagoa.
Os resultados apontam para uma recuperação geral da lagoa com melhoria da
qualidade da água, aumento da biodiversidade e regressão do estado trófico.
Palavras-Chave: eutroficação cultural, gestão, monitorização, ontogenia, recuperação

5
Abstract
Pataias is a natural freshwater shallow lake (3 m winter maximum depth), located near
the small village of Pataias, north of Alcobaça city, in the center of Portugal. Lake
Pataias is considered, for some time now, as eutrophic, not only because of the natural
process itself (late lake ontogeny) but mainly due to human activity (cultural
eutrophication). The anthropogenic input of nutrients has by far accelerated the slow
natural process of eutrophication, dramatically depleting water quality and substantially
reducing biodiversity.
During 2005, the lake dried out completely. That was one opportunity to endure
depletion (of a nutrient rich environment) through sediment and macrophite (Nymphaea
alba) removal.
This study ran throughout 2007 (January to October) in order to determine the
lake’s trophic state by monitoring several limnological variables and parameters (CHL-a,
SRP, TP, transparency, color, temperature, pH, oxygen, conductivity, phytoplankton and
zooplankton, combined with determinations of limitation by phosphorus and lake
productivity), in order to evaluate the lake’s current trophic state. This information is
compared to the findings of the 2005 study, allowing an evaluation of the trophic state
evolution and the success of the management restoration technique (September 2005).
Simultaneously, to better understand lake Pataias dynamics, the relation among
parameters and variables (object of monitoring) was also studied in this specific
ecosystem.
Results suggest a general recovery of the lake ecosystem, with trophic state
regression, better water quality and increased biodiversity.
Keywords: eutrophication, monitoring, management, ontogeny, recovery

6
Introdução
O custo da “modernização” dos estilos de vida das populações humanas tem reclamado,
por todo o Mundo, o sacrifício do ambiente (Boavida, 1999). A percentagem de água
potável acessível à superfície é de aproximadamente 0,2 % (Wetzel, 2001) e, olhando
para os padrões de utilização das mesmas, percebe-se que caminhamos a passos largos
para a total inutilização deste recurso vital.
A partilha de recursos que, noutros tempos, despoletava conflitos entre estados no âmbito
de uma soberania absoluta do território, deu lugar a uma gestão integrada e partilhada do
mesmo. Para esta inversão de posições, em muito contribuíram a convenção de Ramsar,
as doutrinas do Direito Internacional da Água, e a transição de paradigmas. Fica assim
para trás uma perspectiva antropocêntrica, abraçando-se critérios centrados na natureza,
tendendo cada vez mais para uma harmonização de valores económicos e ecológicos
(Boavida, 1999; 2002).
No final do século XIX e até meados do século XX, os lagos eram encarados como
microcosmos, isolados do resto da paisagem. Vários investigadores salientaram as
relações funcionais dos organismos dos lagos. A concepção inovadora destes estudos
conduziu à avaliação exaustiva das interacções entre organismos e das alterações que se
verificavam nas populações em resposta às modificações das propriedades físicas,
químicas e bióticas do meio (Wetzel, 2001; Boavida, 2002). O biota está ligado de forma
inseparável à dinâmica de muitos elementos químicos, em particular à distribuição dos
nutrientes e à sua regulação biogeoquímica. De facto, foi o estudo das populações
fitoplanctónicas em lagos semelhantes, sujeitos a diferentes pressões, que caracterizou o
binómio causa/efeito. Foi a caracterização de dois lagos que sustentou o aporte de
nutrientes (devido às actividades humanas) num deles, como a diferença
comprometedora, responsável pelas elevadas quantidades de fitoplâncton. Assim,
denominou-se este, eutrófico, em oposição ao lago com poucas algas planctónicas,
denominado oligotrófico (Naumann, 1919).
Efectivamente, as provas que se têm vindo a acumular apontam, sobretudo, para a
existência de uma forte relação entre a dinâmica biológica de um lago e a concentração
de nutrientes (Wetzel, 2001). A eutroficação é um processo conhecido como
comprometedor da qualidade da água, assim como é globalmente reconhecido o papel
chave do fósforo nos “episódios” de eutroficação (Vollenweider, 1968; Wetzel, 1983;
Holtan et al., 1988).

7
O interesse do fósforo em ecologia prende-se com o papel principal que desempenha no
metabolismo biológico, uma vez que, quando comparado com a abundância natural de
outros elementos estruturalmente importantes para os biota (carbono, hidrogénio, azoto,
oxigénio, enxofre), o fósforo é o menos abundante de todos na forma disponível para os
produtores (Wetzel, 2001). Assim, a disponibilidade de fósforo frequentemente limita o
desenvolvimento das populações de fitoplâncton em lagos temperados (Schindler, 1977).
Podemos encontrar fósforo essencialmente em duas formas. O fósforo inorgânico (este é,
em geral, uma fracção menor relativamente ao orgânico) subdivide-se em duas
componentes principais: O particulado, e o dissolvido. O particulado é o que mais
representa percentualmente o fósforo total, no entanto, é a componente dissolvida do
fósforo (particularmente a parte que não se encontra complexada ou associada a iões
metálicos), que é susceptível de ser assimilada pelas comunidades planctónicas (Wetzel,
2001).
A demanda de fósforo (ortofosfato - PO43- - a única forma assimilável de fósforo) é
frequentemente superior à disponibilidade no ecossistema lacustre e embora o ortofosfato
seja tido como directamente acessível ao fitoplâncton e bacterioplâncton (Fogg, 1974), é
frequente as populações de algas subsistirem períodos mais longos, quando o ortofosfato
está presente em concentrações muito baixas (abaixo do limite de determinação: 0 – 3
µg/l) e a sobrecarga alóctone é mínima (Hutchinson, 1957). De facto, as algas conseguem
armazenar fósforo nos períodos de abundância, para depois utilizarem quando há
escassez no meio (Fitzgerald, & Nelson, 1966). Na natureza, um dos processos de
transformação do fósforo complexo em ortofosfato é a hidrólise enzimática. A maioria
das fosfatases é de largo espectro e são predominantemente fosfomonoesterases, ainda
que ocasionalmente hidrolisem polifosfatos inorgânicos (Boavida & Heath, 1988).
Tradicionalmente, a actividade da fosfatase tem sido tomada como indicador da
deficiência de fósforo, tanto em organismos (Boavida & Heath, 1988; Wynne & Ben-
Dan, 1995) como em comunidades (Rose & Axler, 1998).
Em lagos eutróficos a quantidade de biomassa é enorme, e a matéria orgânica
produzida pelas macrófitas emergentes, as macrófitas submersas e as de folha flutuante
dominam em muito a matéria orgânica de origem fitoplanctónica. Toda esta biomassa em
decomposição produz e acumula muita matéria orgânica dissolvida (DOM) (Wetzel,
1991, 1993; Boavida, 2000a) conferindo à água uma cor amarelo-acastanhada. Wetzel,
em 1991 avançou a hipótese da decomposição da matéria orgânica no geral e das
macrófitas em particular, como fonte de DOM rica em ácidos húmicos, capazes de

8
formar complexos ácidos húmicos/fosfatase, inactivando as enzimas. Esta forma de
“sequestro” pode durar bastante tempo, dando origem a um “stock” de enzimas inactivas
no ecossistema (explicando as baixas taxas de actividade enzimática da fosfatase
alcalina). Quando a circulação aproxima os complexos ácidos húmicos/fosfatase da
superfície, a radiação UV liberta as fosfatases dos complexos, retomando as fosfatases a
sua função normal (Boavida & Wetzel, 1998; Boavida, 2000a; 2000b). Portanto, é
importante ter uma “medida” da cor da água, pois para além de traduzir o ritmo de
decomposição da biomassa, também nos reporta o potencial de formação do complexo
ácido húmico/fosfatase.
São conhecidos e largamente difundidos (comunicação social) os episódios de morte de
peixes, particularmente nos Verões mais secos. O que não é tão conhecido são as
consequências para o ambiente em geral, e para nós, seres humanos em particular, da má
qualidade dessa água por mais abundante que seja. À degradação da qualidade da água
segue-se a evolução da estrutura das comunidades fito e zooplanctónicas de acordo com
o estado trófico. Representativo disso é o aumento da densidade de alguns géneros de
zooplanctontes, entre os quais: Brachionus sp. Poliarthra sp. e K. quadrata, comuns em
albufeiras caracterizadas por um aumento do estado trófico (transição de oligo para
mesotrófico) (Zurek, 1985; Pejler, 1983) e de acordo com Boavida & Caramujo (2000)
Bosmina sp. tende a dominar em ambientes mais eutróficos. Um efeito semelhante
também foi reportado por Robarts et al., (1992) e por Pinel-Alloul & Méthot (1984) em
lagos próximos do fim da sua ontogenia, sugerindo que a composição, abundância e
dinâmica das populações de zooplâncton, podem ser consequência da idade da albufeira.
Relativamente aos Rotifera e à eutroficação, tendem a aumentar a sua contribuição na
reciclagem de nutrientes ao mesmo tempo que variáveis como o oxigénio, temperatura e
a disponibilidade de alimento são factores importantes para a sua ocorrência (Mikschi,
1989) e distribuição vertical (Ruttner - Kollisko, 1980; Miracle & Vicente, 1983;
Hofmann, 1987; Baião & Boavida, 2000).
Os objectivos deste estudo foram:
1. Complementar a minha formação em Limnologia, equipamentos e técnicas.
2. Adquirir uma visão holística das interacções num lago e experimentar as
alterações/evoluções descritas em bibliografia.
3. Dar continuidade e aproveitar uma oportunidade quase única (contexto pós
biomanipulação/gestão da lagoa reportável a um estudo anterior à intervenção).
4. Ajudar a compreender e a preservar melhor um património que é de todos.

9
Materiais e Métodos Descrição da Lagoa de Pataias
A lagoa de Pataias, morfometricamente, é um lago pouco profundo com 3 m de
profundidade máxima no inverno e 0,1km2 de área, que se encontra nos estadios finais da
ontogenia. É muito importante para a biodiversidade (a segunda maior zona húmida na
área de pinhal-bravo do litoral português), e localiza-se num bosque exclusivamente
composto por Pinus pinaster (39º 40’ 39”N, 9º 0’ 12”O: http//www.earth.google.com)
adjacente à vila de Pataias, no concelho de Alcobaça (Quaresma 1999; 2001). Para além
da natural e previsível eutroficação de lagos (à medida que envelhecem – ontogenia), a
lagoa de Pataias sofreu também um processo de eutroficação cultural. O processo de
eutroficação natural é um processo lento, no entanto, com a intensificação das
actividades humanas, desenvolvimento social e empresarial não sustentado, essa natural
eutroficação foi muito acelerada.
Durante a década de noventa do século passado, a actividade de uma estufa (situada a
100 m da lagoa) e as características da bacia de drenagem facilitaram a lixiviação de
adubos e insecticidas para a lagoa, e até ao ano 2000, os esgotos da Vila de Pataias (5000
habitantes) eram directamente dirigidos para a lagoa, sem tratamento. A contínua
eutroficação cultural aumentou a sobrecarga total de nutrientes, nomeadamente fósforo e
azoto, permitiu em muito aumentar a produtividade da lagoa e acelerar o processo de
eutroficação.
Em Setembro de 2005, com a lagoa vazia e o fundo exposto, procedeu-se à remoção:
(1) da camada orgânica fruto de processos de sedimentação e rica em nutrientes, (2) de
grandes quantidades da macrófita Nymphaea alba. Esta operação de
gestão/biomanipulação visou reduzir: (1) a quantidade de nutrientes autóctones,
removendo os sedimentos (minimizando a potencial lixiviação e reciclagem de
nutrientes) e (2) os processos de decomposição e mineralização da grande biomassa de
macrófitas flutuantes, aquando do enchimento da lagoa no inverno seguinte. Com a seca,
também a ictiofauna exótica (Cyprinus carpio e Lepomis gibbosus) foi removida (os
peixes morreram, Quaresma 2006), tendo-se aproveitando a oportunidade para, em
Outubro 2006, reintroduzir a espécie autóctone Chondrostoma oligolepis (Robalo et al.,
2005) com regime alimentar com menor impacto (dieta à base de pequenos invertebrados
mas também de pequenas algas) (Maitland, 1995).

10
As macrófitas presentes na lagoa, durante este estudo, eram: Nmphaea alba, Lemna sp. e
significativa biomassa de Typha sp. , Phragmites australis e Azolla filiculoides.
Amostragem e análises
O estudo estendeu-se de Fevereiro a Outubro de 2007. As amostras foram recolhidas
quinzenalmente nos meses de Fevereiro, Março e Outubro e todas as semanas de Abril a
Setembro, sempre no mesmo sítio, o local mais profundo da lagoa.
Medições in-situ
Utilizou-se o disco de Secchi (SD) com 20 cm de diâmetro e sectores brancos e pretos,
para determinar a transparência da água. Factores ambientais, tais como a condutividade,
temperatura, oxigénio dissolvido (DO) e pH foram medidos in-situ. O pH foi medido à
superfície e a um metro com recurso ao HI8424 portable pH/mV/temperature meter –
HANNA. A temperatura, condutividade e oxigénio dissolvido foram medidos à
superfície, um metro e no fundo. Adicionalmente, e consoante a época do ano (variações
na profundidade), as três variáveis foram medidas a 1,5 e/ou 2,0 metros. A condutividade
e a temperatura foram aferidas com recurso a uma sonda multiparamétrica YSI modelo
30/10FT, enquanto o oxigénio dissolvido foi medido com uma sonda do YSI model 57,
agitando suavemente à profundidade pretendida.
Em cada dia de recolha de amostras, também o plâncton foi amostrado. A amostragem do
zooplâncton consistiu em um ou dois arrastos a filtragens a 0,5 - 1,0 - 1,5 ou 2,0 metros
(ao longo da coluna de água num movimento vertical e sempre à mesma velocidade);
o diâmetro da boca da rede de amostragem (tipo Wisconsin) é 0,12 metros e o poro é 76
µm. A fixação e preservação das amostras de zooplâncton foram feitas com uma solução
aquosa de formol saturada com açúcar, a uma concentração final de 4% (v/v).
Os aparelhos não funcionaram:
• Sonda pH de 04/05/2007 a 29/06/2007
• Sonda oxigénio de 01/06/2007 a 29/06/2007
• O SRP não foi determinado de 22/01/2007 a 15/02/2007
• A cor 29/01/2007 a 08/03/2007 e de 21/03/2007 a 11/04/2007
Determinações laboratoriais
Dentro dos parâmetros tróficos, e para além da transparência medida in situ, foram
também determinadas a clorofila a (CHL-a) e o fósforo total (TP) em amostras dos

11
primeiros 0,1 - 0,2 m da coluna de água. As amostras foram transportadas para o
laboratório a 7 - 8 ºC, em garrafas fechadas e em ambiente isolado da luz (duas horas de
viagem). No laboratório a concentração de (CHL-a) foi determinada fluorimetricamente
(excitação 420,0 nm emissão 675,0 nm: FP-750 Jasco, Japan) através de filtrados (50 –
800ml) utilizando filtros de membrana Whatman (0,45 µm) com tempos de extracção
(metanol) nunca inferiores a uma hora. Para aferir a disponibilidade de fósforo
determinou-se a concentração de SRP (soluble reactive phosphorus) e do TP (total
phosphorus) espectrofotometricamente (885 nm, Jasco VR- 530, Japan). Para o SRP
(referência à concentração de ortofosfato, ou seja, o fósforo mineralizado) utilizou-se
água da lagoa filtrada através de um filtro de membrana Whatman (0,45 µm) e o método
de Murphey & Riley (1962). Recorrendo a amostras de água da lagoa não filtradas,
determinou-se o TP depois de uma hora de hidrólise ácida com persulfato de potássio e
ácido sulfúrico concentrado, a alta temperatura e pressão (APHA, 1989).
A cor da água foi calculada após medição da absorvência num espectrofotómetro a 440
nm, de acordo com Cuthbert & Del Giogio (1992), e foi tomada como indicadora da
quantidade de compostos húmicos em solução.
A identificação das comunidades fito e zooplanctónica foi feita em laboratório
com recurso a uma lupa Nikon e microscópio binocular Leica (ampliações 40 a 1000
vezes). O zooplâncton de acordo com Ward & Whipple (1959); Scourfield & Harding
(1966); Dussart (1967) e Margaritora (1983), pelo menos até ao género. Quanto ao
fitoplâncton, este foi identificado até ao género de acordo com Lund & Lund (1995).
Produtividade
Entre Abril e Julho realizaram-se três ensaios à produtividade da lagoa. Estes testes, ao
mesmo tempo que registam a evolução da lagoa, permitem saber, para cada episódio de
recolha, a “condição geral da lagoa” à medida que se aproxima e intensifica o período
mais produtivo do ano.
O método utilizado (método das garrafas claras e escuras) consistiu em submeter metade
da amostra a um determinado período (quatro horas no presente caso) sem luz, enquanto
a outra metade foi submetida a radiação PAR (30 µEinstein m-2 s-1) pelo mesmo período.
Como resultados temos o consumo e produção de oxigénio, às escuras e com fonte
luminosa, respectivamente, através de leituras do oxigénio dissolvido (medido no início e
no fim do ensaio) e calculada a variação de oxigénio dissolvido para cada réplica. Este
método permite calcular a produtividade bruta e líquida. O ensaio foi feito para um

12
período de quatro horas com recurso a quatro réplicas para cada uma das condições
(presença/ausência de luz) e, como controlo, um réplica dupla com água desionizada e
esterilizada por UV (Odum, 1971).
Limitação pelo fósforo
O ensaio da limitação pelo fósforo realizou-se em Outubro, e é uma experiência
correspondente ao teste tipo Selenastrum modificado (Miller et al., 1978). Consistiu em
quantificar a resposta do fitoplâncton (concentração de CHL-a) na água da lagoa, quando
sujeito (em laboratório) a concentrações crescentes de fósforo (ortofosfato). A água da
lagoa foi submetida a três preparos (50, 100 e 200 µg/l – de fósforo, com três réplicas
cada um) ao longo de 14 dias, quantificando a resposta do fitoplâncton ao sétimo e
décimo quarto dias. Como controlo, utilizou-se a água da lagoa sem adição de fósforo.
A experiência desenrolou-se em ambiente controlado com recurso a radiação PAR (30
µEinstein m-2 s-1) para a totalidade dos catorze dias.
Análise Estatística
Os cálculos foram feitos recorrendo ao programa STATISTICA versão 8.0. Depois da
análise exploratória dos dados foi efectuada uma análise não paramétrica entre as várias
variáveis/parâmetros recorrendo ao coeficiente (rs) da correlação de Spearman.
Paralelamente e através de regressão múltipla (R2) também foi calculada a fitness de
algumas das correlações mais significativas.
Pareceu-me que tomar a regra de decisão “rejeitar Ho quando p < 0,05” (o que em termos
de significância é considerar P [rejeitar Ho | Ho verdadeira] < 0,05 = �) é uma forma mais
razoável de trabalhar em investigação científica (em que todas as conclusões têm que ser
equacionadas como provisórias e reversíveis) do que usar valores mais estritos para �,
por exemplo � = 0,01 ou mais estrito ainda, � = 0,001.
Resultados A exposição dos resultados está dividida em três partes. A primeira desde o começo do
estudo até 29/06/2007 (registo do primeiro bloom de algas e início do período produtivo).
A segunda, do início do período produtivo até ao fim do estudo. A terceira compara o
estado da lagoa depois da aplicação da medida de gestão (remoção de sedimentos e
macrófitas emergentes e introdução do peixe) com o estado anterior a este estudo.

13
Breve nota
No decorrer do estudo (Março) a lagoa de Pataias foi submetida à edificação (intervenção
unilateral da Junta de Freguesia) de um paredão de cimento com o objectivo de reduzir a
perda de água. Esta medida transformou um ecossistema dinâmico, num mais hermético.
A construção deste pequeno mas contundente obstáculo, permitiu não só transformar um
lago numa grande poça, como aumentar a profundidade da lagoa em, sensivelmente, um
metro.
Primeira Parte
De Janeiro a Junho registou-se visualmente (acompanhando o incremento da temperatura
e do fotoperíodo) uma sucessão na comunidade fitoplanctónica.
Nos primeiros dois meses registou-se um aumento da abundância relativa do grupo
Cladophora, o que levou ao progressivo domínio da paisagem, particularmente à
superfície, dando origem a extensos “tapetes flutuantes” de algas filamentosas. A
interferência do ensombramento (Fig. 1) produzido pelo grande desenvolvimento de
algas (Fevereiro) traduziu-se na redução da quantidade de luz que chega a maiores
profundidades. A CHL-a (um indicador da biomassa de fitoplâncton) é naturalmente
dependente da luz e sem luz não há desenvolvimento de algas. Portanto, mais
ensombramento menos biomassa e prova disso foi a progressiva redução (finais de
Fevereiro) da concentração de CHL-a (Fig. 2) coincidente com o aumento do
ensombramento, condicionando o desenvolvimento de outras algas ao longo da coluna de
água.
Figura 1: Variação da transparência de 22-01-2007 Figura 2: Variação da concentração de clorofila a
a 29-06-2007. de 22-01-2007 a 29-06-2007.
Com o início de Fevereiro e ainda com temperaturas à superfície (Fig. 3) muito baixas
(pouco menos de 10ºC), iniciou-se também a subida da concentração de CHL-a. A
15/02/2007 o acréscimo na concentração de CHL-a pode ser atribuído, não só ao
aumento da temperatura (figura 3), mas também às elevadas transparências

14
(imediatamente anteriores – Fig. 1) permitindo uma boa transmissão da radiação ao
longo da coluna de água e potenciando o desenvolvimento de algas ao longo da mesma
(verificado pela igual subtracção de sais à superfície e 1 metro - figura 4).
Figuras 3 e 4: Variação da temperatura do ar, superfície, um metro, dois metros e no fundo de 22-01-2007 a 29-06-2007 (Fig. 3, esq.).
Variação da condutividade à superfície, um metro, dois metros e no fundo de 22-01-2007 a 29-06-2007 (Fig. 4, dir.).
Depois da subida seguiu-se uma descida da concentração de CHL-a, (evidência de
decomposição indicada pelo aumento da transparência (Fig. 1) – redução do número de
algas e aumento da condutividade - Fig. 4 - senescência e aumento de sais na água)
seguido de uma ligeira subida no final de Fevereiro (Fig. 2). Em Março, e ainda que as
temperaturas tenham subido relativamente a Fevereiro, assistiu-se a uma suave descida
dos valores de CHL-a ao longo de todo o mês (Fig. 2). À medida que os valores de CHL-
a desciam, assistíamos ao aumento da abundância de algas filamentosas flutuantes
(observação pessoal), ao mesmo tempo que a A. filiculoides aparecia nesta lagoa (com
abundância sucessivamente crescente). A suave descida da concentração de CHL-a
culminou a 11/4/2007 com o valor mais baixo do ano. Assim, pode-se entender a
importância do ensombramento na dinâmica do lago. Ainda assim, o efeito deve ter sido
menor do que seria esperado, porque a concentração de oxigénio dissolvido até
aumentou. Provavelmente devido a algas filamentosas à superfície e 1 metro, algas
eucariotas unicelulares, e principalmente, as grandes quantidades da macrófita submersa
Ceratophylum (Fig. 7 e 8).
Depois do valor mais baixo do ano, seguiu-se uma subida, (um mês e meio) pois só a
25/5/2007 é que se atingiram valores iguais aos de 27/2/2007, ou seja, três meses depois
(Fig. 2). A razão da “difícil” subida dos valores de CHL-a está provavelmente uma vez
mais relacionada com o ensombramento e a flutuação da temperatura (superfície e um
metro). Aos registos mais baixos de CHL-a deste período correspondem as temperaturas
mais baixas e o inverso também é verdade. Prova disso é o aumento da Tsup em 5ºC a
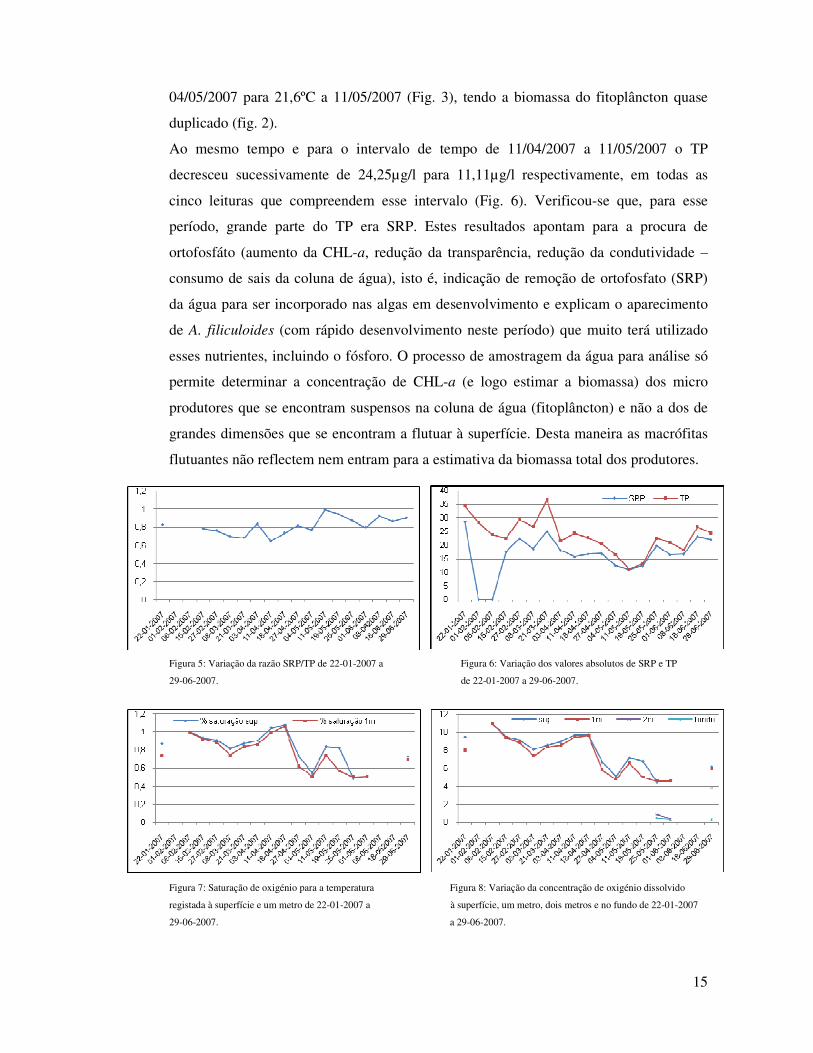
15
04/05/2007 para 21,6ºC a 11/05/2007 (Fig. 3), tendo a biomassa do fitoplâncton quase
duplicado (fig. 2).
Ao mesmo tempo e para o intervalo de tempo de 11/04/2007 a 11/05/2007 o TP
decresceu sucessivamente de 24,25µg/l para 11,11µg/l respectivamente, em todas as
cinco leituras que compreendem esse intervalo (Fig. 6). Verificou-se que, para esse
período, grande parte do TP era SRP. Estes resultados apontam para a procura de
ortofosfáto (aumento da CHL-a, redução da transparência, redução da condutividade –
consumo de sais da coluna de água), isto é, indicação de remoção de ortofosfato (SRP)
da água para ser incorporado nas algas em desenvolvimento e explicam o aparecimento
de A. filiculoides (com rápido desenvolvimento neste período) que muito terá utilizado
esses nutrientes, incluindo o fósforo. O processo de amostragem da água para análise só
permite determinar a concentração de CHL-a (e logo estimar a biomassa) dos micro
produtores que se encontram suspensos na coluna de água (fitoplâncton) e não a dos de
grandes dimensões que se encontram a flutuar à superfície. Desta maneira as macrófitas
flutuantes não reflectem nem entram para a estimativa da biomassa total dos produtores.
Figura 5: Variação da razão SRP/TP de 22-01-2007 a Figura 6: Variação dos valores absolutos de SRP e TP
29-06-2007. de 22-01-2007 a 29-06-2007.
Figura 7: Saturação de oxigénio para a temperatura Figura 8: Variação da concentração de oxigénio dissolvido
registada à superfície e um metro de 22-01-2007 a à superfície, um metro, dois metros e no fundo de 22-01-2007
29-06-2007. a 29-06-2007.

16
A génese do ortofosfato poderá estar associada a períodos de decomposição, sugeridos
pelo decréscimo da concentração de oxigénio dissolvido (18/04/2007 a 04/05/2007 e de
11/05/2007 a 01/06/2007 - Fig. 7 e 8), e também o aumento da razão SRP/TP
sensivelmente no mesmo período (Fig. 5).
O aumento da concentração de CHL-a (11/05/2007 - Fig. 2) e redução da transparência
(aumento do seston - Fig. 1) deve-se, provavelmente, à decomposição (baixas e
decrescentes concentrações de DOsup e DO1m - Fig. 7 e 8) disponibilizando não só
nutrientes (utilizados no desenvolvimento de algas, mas também e principalmente de A.
filiculoides), como também espaços livres de macrófitas à superfície (criados pela
crescente redução da abundância de algas filamentosas flutuantes) aumentando a
transmissão de radiação luminosa na coluna de água e permitindo deste modo o
desenvolvimento de algas unicelulares eucariotas (rápido desenvolvimento e tolerância a
baixas temperaturas) (Ustach & Paerl, 1982). Deste modo, a redução da transparência é,
provavelmente, o resultado da combinação do aumento da concentração de CHL-a e do
aumento de DOM e POM (decomposição).
Com cada vez melhores condições ambientais (temperatura média(Maio) perto dos 22 ºC),
à medida que os dias se tornavam mais quentes e longos, a A. filiculoides, que vinha
suplantando a população de filamentosas, também ela dominou a paisagem e a interface
ar/água. De facto, a 25/5/2007 dominou 80% da superfície da lagoa (observação pessoal).
Entretanto, a ajuda concertada entre a Câmara Municipal, a Junta de Freguesia e o
Ministério da Agricultura permitiu remover centenas de quilos de A. filiculoides.
A operação de remoção decorreu durante a última semana de Maio e inícios de Junho e
visou retirar uma grande quantidade de biomassa que, de outra maneira, após
decomposição, acabaria por devolver os nutrientes à coluna de água e/ou acumulado no
fundo, perpectuando o pool de nutrientes.
Com a remoção da A. filiculoides, reduziu-se bastante o efeito ensombramento com
implicações directas na concentração de CHL-a e biomassa (estimativa da biomassa
agora mais correcta devido à ausência de macrófitas flutuantes), tendo-se verificado
repetidas subidas depois de 25/05/2007 (Fig. 2). É de salientar a boa medida de gestão,
fundamental ao equilíbrio da lagoa.
No entanto, os dados apontam para o início da degradação da enorme biomassa de
A. filiculoides até duas semanas antes (11/05/2007) do início da remoção. Verificou-se a
alteração da cor da água (Fig. 9): ácidos húmicos dissolvidos, provenientes da
decomposição da matéria vegetal; a transparência reduziu meio metro numa semana (Fig.

17
10) decomposição: POM no seston e/ou DOM; redução da CHL-a de 11/05/2007 para
19/05/2007 (Fig. 11) aumento de SRP e TP (Fig. 12). Provavelmente, terá sido a
combinação da remoção e senescência da A. filiculoides a promover o incremento da
biomassa do fitoplâncton nos períodos subsequentes. A remoção da A. filiculoides
poderia ter sido exemplar se os cuidados a ter após remoção das plantas não tivessem
sido negligenciados, permanecendo os sacos e respectivo conteúdo disperso nas margens
da lagoa (de uma semana para a outra) repetidas vezes (observação pessoal).
Figura 9: Variação da cor (unidades Pt) ao longo da primeira Figura 10: Variação da transparência da água na
parte do estudo. A azul, a evolução das duas últimas primeira parte do estudo. A azul, a evolução das duas
amostragens antes de 25/05/2007 (indicação da degradação). últimas amostragens antes de 25/05/2007 (indicação da
degradação).
Figura 11: Variação da CHL-a na primeira parte do estudo. Figura 12: Variação da quantidade de SRP e TP na
A azul, a evolução da última amostragem até 25/05/2007 primeira parte do estudo. A azul, a evolução das duas
(indicação da degradação). últimas amostragens antes de 25/05/2007 (indicação da
degradação).
Segunda parte
A concentração de CHL-a até este período vinha apresentando valores lenta mas
progressivamente maiores. Este período destaca-se por evidenciar, para além dos
elevados valores de CHL-a (e por isso mais produtivo do que a primeira parte do estudo),
sucessivas repetições de subidas e descidas na concentração deste pigmento.

18
Figura 13: Evolução da concentração de clorofila a ao longo do estudo. Assinalada a data de separação
da primeira e segunda partes.
Depois de 29/06/2007 as consequências da sazonalidade (todas as variáveis exibiram um
padrão com maior amplitude de valores) têm efeitos transversais às variáveis e
parâmetros (concentração da CHL-a - Fig. 13; razão SRP/TP - Fig. 14; transparência -
Fig. 15; temperatura - Fig. 16; condutividade - Fig. 17; oxigénio - Fig. 18 e pH - Fig. 19),
que combinados potenciam os efeitos no ecossistema (já de si mais hermético) da lagoa.
Figura 14: Variação de SRP no TP ao longo do estudo. Figura 15: Evolução da transparência ao longo do estudo
Assinalada a data de separação da primeira e segunda Assinalada a data de separação da primeira e segunda
partes. partes.
Figura 16: Evolução da temperatura do ar, à superfície, Figura 17: Evolução da condutividade à superfície, 1 m,
1 m, 1,5 m, 2 m e fundo ao longo do estudo . Assinalada a 1,5 m, 2 m e fundo ao longo do estudo. Assinalada a
data de separação da primeira e segunda partes. data de separação da primeira e segunda partes.
29/06/2007

19
Figura 18: Concentração de oxigénio à superfície, 1 m, Figura 19: Evolução do pH à superfície e um metro
1,5 m, 2 m e fundo ao longo do estudo. Assinalada a ao longo do estudo. Assinalada a data de separação
data de separação da primeira e segunda partes. da primeira e segunda partes.
No período de 29/06/2007 a 25/07/2007 e relativamente à concentração de CHL-a, (Fig.
20) registaram-se dois “picos” e duas “depressões” sendo que as primeiras subida e
descida apresentam valores inferiores relativamente às segundas. O desenvolvimento e a
conjuntura das condições ambientais podem ser apontados como maior condicionante do
desenvolvimento do fitoplâncton.
Para se compreender a dinâmica da lagoa é importante saber a razão das constantes
subidas e descidas dos valores da clorofila a (o que se conjugou e de que maneira), para
ter havido períodos de maior desenvolvimento e outros em que esse desenvolvimento foi
menor. Nesta perspectiva é fundamental retratar a evolução de cada parâmetro ou
variável, num determinado período, e comparar com a variação da concentração de CHL-
a para o mesmo período. Este exercício ajuda, não só, a retratar correlações entre
variáveis/parâmetros, como também a interpretar a relação entre as variáveis/parâmetros,
na resposta da concentração da CHL-a.
Assim, e comparando a variação da concentração da CHL-a (Fig. 20) com a
transparência (Fig. 21), depreende-se uma relação entre as duas.
Figura 20: Evolução da CHL-a ao longo do estudo. Figura 21: Evolução da transparência ao longo do estudo.
A linha vertical assinala 29/06/2007. Setas verdes A linha vertical assinala 29/06/2007.
“picos” setas vermelhas “depressões”.
20
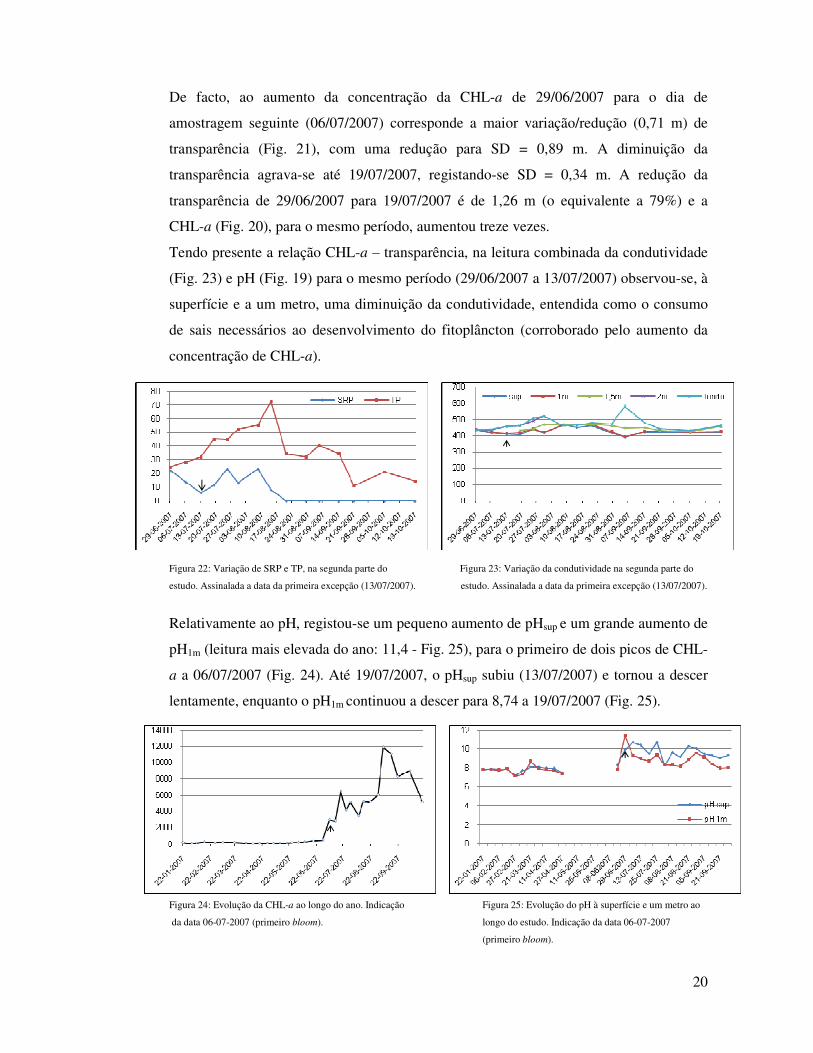
De facto, ao aumento da concentração da CHL-a de 29/06/2007 para o dia de
amostragem seguinte (06/07/2007) corresponde a maior variação/redução (0,71 m) de
transparência (Fig. 21), com uma redução para SD = 0,89 m. A diminuição da
transparência agrava-se até 19/07/2007, registando-se SD = 0,34 m. A redução da
transparência de 29/06/2007 para 19/07/2007 é de 1,26 m (o equivalente a 79%) e a
CHL-a (Fig. 20), para o mesmo período, aumentou treze vezes.
Tendo presente a relação CHL-a – transparência, na leitura combinada da condutividade
(Fig. 23) e pH (Fig. 19) para o mesmo período (29/06/2007 a 13/07/2007) observou-se, à
superfície e a um metro, uma diminuição da condutividade, entendida como o consumo
de sais necessários ao desenvolvimento do fitoplâncton (corroborado pelo aumento da
concentração de CHL-a).
Figura 22: Variação de SRP e TP, na segunda parte do Figura 23: Variação da condutividade na segunda parte do
estudo. Assinalada a data da primeira excepção (13/07/2007). estudo. Assinalada a data da primeira excepção (13/07/2007).
Relativamente ao pH, registou-se um pequeno aumento de pHsup e um grande aumento de
pH1m (leitura mais elevada do ano: 11,4 - Fig. 25), para o primeiro de dois picos de CHL-
a a 06/07/2007 (Fig. 24). Até 19/07/2007, o pHsup subiu (13/07/2007) e tornou a descer
lentamente, enquanto o pH1m continuou a descer para 8,74 a 19/07/2007 (Fig. 25).
Figura 24: Evolução da CHL-a ao longo do ano. Indicação Figura 25: Evolução do pH à superfície e um metro ao
da data 06-07-2007 (primeiro bloom). longo do estudo. Indicação da data 06-07-2007
(primeiro bloom).
21
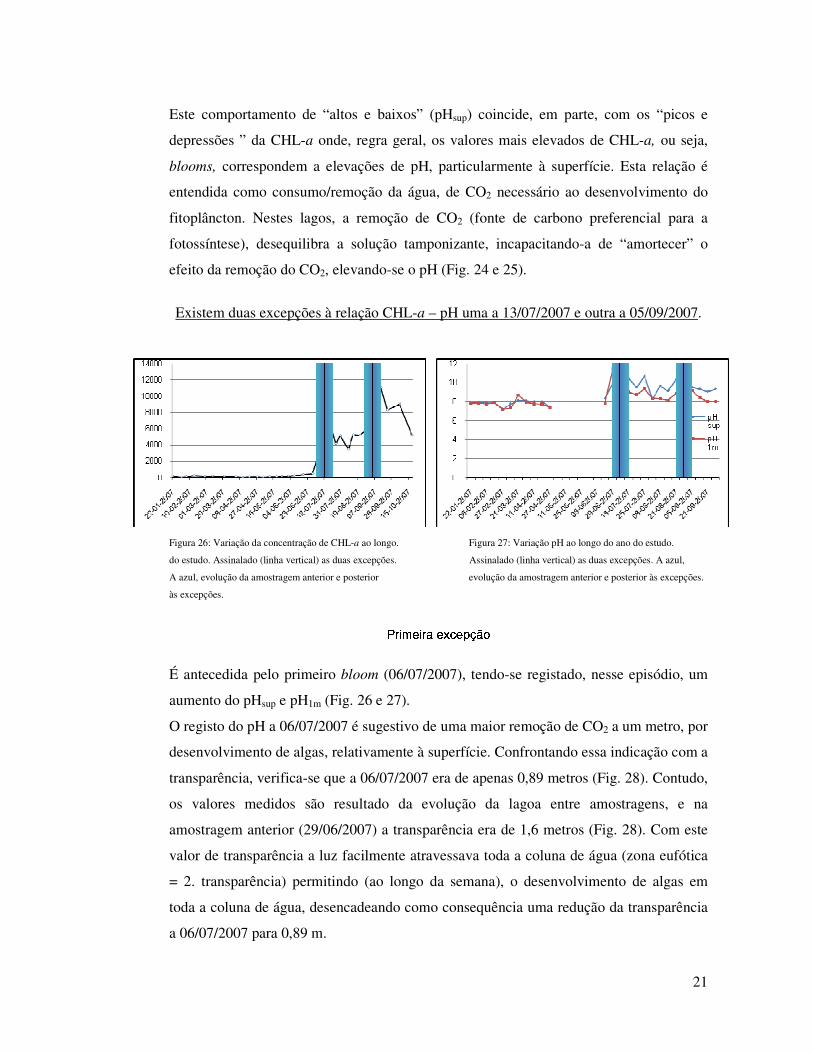
Este comportamento de “altos e baixos” (pHsup) coincide, em parte, com os “picos e
depressões ” da CHL-a onde, regra geral, os valores mais elevados de CHL-a, ou seja,
blooms, correspondem a elevações de pH, particularmente à superfície. Esta relação é
entendida como consumo/remoção da água, de CO2 necessário ao desenvolvimento do
fitoplâncton. Nestes lagos, a remoção de CO2 (fonte de carbono preferencial para a
fotossíntese), desequilibra a solução tamponizante, incapacitando-a de “amortecer” o
efeito da remoção do CO2, elevando-se o pH (Fig. 24 e 25).
Existem duas excepções à relação CHL-a – pH uma a 13/07/2007 e outra a 05/09/2007.
Figura 26: Variação da concentração de CHL-a ao longo. Figura 27: Variação pH ao longo do ano do estudo.
do estudo. Assinalado (linha vertical) as duas excepções. Assinalado (linha vertical) as duas excepções. A azul,
A azul, evolução da amostragem anterior e posterior evolução da amostragem anterior e posterior às excepções.
às excepções.
����� ����� ����� �����������
É antecedida pelo primeiro bloom (06/07/2007), tendo-se registado, nesse episódio, um
aumento do pHsup e pH1m (Fig. 26 e 27).
O registo do pH a 06/07/2007 é sugestivo de uma maior remoção de CO2 a um metro, por
desenvolvimento de algas, relativamente à superfície. Confrontando essa indicação com a
transparência, verifica-se que a 06/07/2007 era de apenas 0,89 metros (Fig. 28). Contudo,
os valores medidos são resultado da evolução da lagoa entre amostragens, e na
amostragem anterior (29/06/2007) a transparência era de 1,6 metros (Fig. 28). Com este
valor de transparência a luz facilmente atravessava toda a coluna de água (zona eufótica
= 2. transparência) permitindo (ao longo da semana), o desenvolvimento de algas em
toda a coluna de água, desencadeando como consequência uma redução da transparência
a 06/07/2007 para 0,89 m.

22
Gráfico 28: Transparência ao longo do estudo. Assinaladas as datas das duas excepções (linha vertical)
e a evolução (azul) antes e depois das datas assinaladas
Os dados da condutividade (figura 23) indicam, de 29/06/2009 a 13/07/2007, uma
sucessiva redução da condutividade da água, à superfície e a um metro, o que é sugestivo
de desenvolvimento dos produtores ao longo do primeiro metro da coluna de água.
Os dados do pH indicam a 06/07/2007 um aumento à superfície e um grande aumento a
um metro (provavelmente maior remoção de CO2 a um metro). Esta conjugação de dados
sugere que poderá ter havido um desenvolvimento “diferenciado” do fitoplâncton (maior
a um metro, do que à superfície) na coluna de água a 06/07/2007.
De 06/07/2007 para 13/07/2007:
- pHsup aumentou e o pH1m desceu (Fig. 25)
- condutividadesup desceu e a condutividade1m também desceu (Fig. 23)
- Relativamente à concentração de SRP, a procura por parte dos produtores
primários seria elevada (1º bloom a 06/07/2007) e a manutenção do bloom até à
amostragem seguinte (13/07/2007) estaria dependente da disponibilidade de
recursos. Contudo, a 13/07/2007 verificou-se uma descida na concentração de
SRP (Fig. 22) na água (relativamente a 06/07/2007) ao mesmo tempo que se
determinou uma diminuição da concentração de CHL-a (Fig. 20).
Esta conjunção de dados sugere que a água (e o seu soluto) a um metro terá chegado à
superfície. O efeito no pH, condutividade e SRP será resultado de uma mistura destas
duas massas de água (superfície e um metro), diluindo os recursos fundamentais ao
desenvolvimento de algas na zona eufótica.
Desta maneira a 13/07/2007 o desenvolvimento da comunidade de algas poderá ter sido
condicionado pela escassez de recursos.

23
� �������������������������
Neste caso e devido à evolução da transparência, não havia luz que permitisse um
desenvolvimento diferenciado para além do primeiro metro da coluna de água, ficando a
zona eufótica severamente diminuída. Com o desenvolvimento dos produtores a registar-
se na zona eufótica e a acumulação de conteúdos celulares provenientes da
decomposição, a registar-se abaixo da zona eufótica (para além de 1 metro), a superfície
ficou cada vez mais susceptível à remoção de CO2. Principalmente, nas duas amostragens
anteriores a 05/09/2007 (Fig. 26), quando se verificaram elevadas concentrações de
CHL-a e consequente remoção de sais e CO2 (como os valores de condutividadesup - Fig.
23 pHsup e pH1m - Fig. 27 sustentam), fundamentais ao desenvolvimento da comunidade
de algas.
Tendo presente a evolução da lagoa (figuras 22, 23, 27 - SRP, condutividade e pH
respectivamente), com consumo e redução de recursos nas duas amostragens anteriores,
para se registar a 05/09/2007 o valor recorde de concentração de CHL-a teria de ter
havido um “reposição” à superfície de elementos essenciais ao desenvolvimento de algas.
Observando a evolução das mesmas variáveis/parâmetros à superfície e um metro, nas
duas amostragens anteriores, observámos que:
- Em vez de o pHsup ter aumentado (em função do desenvolvimento de algas
máximo de concentração CHL-a), diminuiu ligeiramente.
- O pH1m subiu acima dos valores normais.
- E o registo da condutividade assinala uma depleção de sais à superfície, a um
metro e a um metro e meio
A figura 22 de 21/08/2007 até ao fim do estudo (19/10/2007) apontou sempre valores
abaixo do detectável (3 µg/litro). A “ausência” é interpretada como o resultado da
constante remoção de SRP por parte das algas em desenvolvimento. Deste modo, sempre
que SRP é “adicionado”, é imediatamente removido. Isto verifica-se em situações de
limitação pelo fósforo.
A 05/09/2007 para se registar o valor recorde (máximo de concentração de CHL-a), terá
de ter havido disponibilidade de recursos fundamentais ao desenvolvimento da
comunidade de algas.
Esta conjunção de dados sugere, uma vez mais, que a água (e o seu soluto) a pelo menos
um metro, terá chegado à superfície. Desta vez, a homogeneização do (pelo menos)

24
primeiro metro de coluna de água resulta no “enriquecimento” da superfície, (zona
eufótica) com substâncias fundamentais ao desenvolvimento da comunidade de algas,
reciclados na coluna de água ou fundo.
Desta maneira a 05/09/2007 o desenvolvimento da comunidade de algas poderá ter sido
potenciado pelo aumento de recursos, isto é, nutrientes. � � ����� !�"���������#���
Na zona eufótica os períodos que registaram uma diminuição da concentração de CHL-a
estão associados a processos de decomposição na coluna de água e fundo. Resultaram,
regra geral, no consumo de oxigénio, no aumento da condutividade, diminuição do pH,
aumento da transparência e aumento de SRP, isto é, o oposto do que seria de esperar para
cada uma das variáveis, quando ocorre um bloom de algas.
Quanto à concentração de oxigénio à superfície, regra geral, aumentava sempre que se
registava um bloom de algas, e reduzia entre blooms (reciclagem de nutrientes).
O processo de decomposição, como já vimos, consome oxigénio e é possível que tenha
havido períodos de anóxia junto aos sedimentos (Fig. 18):
- Exceptuando a 21/08/2007, o fundo apresentou sempre concentrações de
oxigénio dissolvido baixas ou muito baixas (média 0,4 mg O2/l)
- Particularmente entre 13/07/2007 e 25/07/2007 e a 08/08/2007, apresentou
baixas concentrações logo no primeiro metro.
- Contudo, no período de 30/08/2007 a 21/09/2007, os valores obtidos de
oxigénio dissolvido foram ligeiramente superiores (média 0,6 mg O2/l) mas
ainda assim muito baixos.
A única data com concentração de oxigénio, no fundo, acima de 0,6 mg O2/l (21/08/2007
com 3,6 mg O2/l) sugere que água rica em oxigénio terá chegado até ao fundo,
“oxigenando” toda a coluna de água no processo.
Misturar a água requer muita energia e a 21/08/2007 (duas semanas antes do máximo de
CHL-a e segunda excepção) registou-se o máximo de velocidade média do vento de todo
o estudo (25,5 km/h). Foi um episódio único com resultados singulares (concentração de
oxigénio no fundo máximo, muito acima dos valores normais) sugestivos de grande
circulação de água substanciados pelo registo das outras variáveis/parâmetros e
principalmente da velocidade do vento (Fig. 29); pH (Fig. 27); condutividade (Fig. 23);
SRP (Fig. 22); temperatura na coluna de água (Fig. 31); cor (Fig. 30).
A circulação mais intensa da água teve como consequências:

25
- descida do pHsup e do pH1m: maior taxa de hidratação do CO2 - agitação à
superfície (Fig. 27).
- homogeneização da coluna de água, tendo por isso a temperatura (Fig. 31) e a
condutividade permanecido idênticas em toda a coluna de água (Fig. 23) .
- a “remoção” de SRP: à medida que sucessivas massas de água se substituíam na
zona eufótica, os produtores primários absorviam o SRP (Fig. 22).
Os resultados sustentam a hipótese de que efectivamente terá havido circulação completa
de água a 21/08/2007 e que terá sido o vento a força motriz.
Figura 29: Evolução da velocidade do vento km/h/dia Figura 30: Evolução da cor ao longo do estudo. Indicação
na segunda parte do estudo. da data 21/08/2007.
Ambientes muito eutróficos são caracterizados pela acumulação de DOM,
frequentemente ricos em substâncias húmicas (Boavida, 2000 a).
Relativamente à cor, tal como a concentração da clorofila a, que aumentou do início ao
fim do estudo (particularmente na segunda metade – mais produtiva), a cor diminuiu
sensivelmente para metade: 120 unidades pt para 64 unidades de pt, no início e no fim do
estudo respectivamente. Uma observação criteriosa da variação da cor durante a segunda
parte do estudo evidencia uma diminuição nos períodos com elevada concentração de
oxigénio na coluna de água e fundo. O aumento da cor pode estar associado à
decomposição do fitoplâncton e a sua diminuição associada a ocorrências de blooms.
- A 21/08/2007 uma boa oxigenação da coluna de água coincidiu com o valor
mais baixo de cor (Fig. 30).

26
Figura 31: Variação da temperatura do Ar, e ao longo da coluna de água de 22/01/2007 a 19/10/2007. As setas marcam
as elevações da concentração da CHL-a (blooms).
Figura 32: Variação da concentração de CHL-a. As setas
marcam as elevações da concentração da CHL-a (blooms).
Quanto à temperatura os resultados da Fig. 31 mostraram que:
- A temperatura média: do ar; à superfície e a um metro, foram mais baixas na
primeira parte do estudo do que na segunda.
- Dias mais quentes na segunda parte do estudo resultaram numa coluna de água
mais quente do que na primeira parte do estudo.
- O aumento da temperatura do ar resultou no aquecimento da água à superfície.
- O aumento/diminuição da temperatura deu-se da superfície para o fundo.
- A temperatura pode influenciar a ocorrência de blooms. Como é exemplo,
06/07/2007 com o primeiro bloom e o registo (à data) da Tsup mais elevada
(25,3ºC).
- A ocorrência de blooms verifica-se sempre nos dias que a temperatura da água à
superfície foi mais elevada.
29/06/2007

27
O contínuo aumento da temperatura ao longo do ano levou ao gradual aquecimento da
água, da camada mais superficial para a mais profunda, e poderá ter tido um efeito
potenciador da difusão dos conteúdos das camadas de água entre si. Este diferencial de
temperatura na coluna de água poder-se-á ter traduzido (mesmo que temporariamente)
num diferencial de densidade, da água menos densa (à superfície) para a mais densa e
fria (a um metro). Com a subida da temperatura à superfície acentua-se a diferença de
densidade da água (à superfície para um metro) o que poderá ser suficiente para separar
as duas camadas, comportando-se, como duas camadas distintas. No entanto, estas
diferenças de temperatura são pequenas, assim como as variações que lhes correspondem
em densidade, não tendo, por isso, como consequência, a estratificação térmica da lagoa
durante um período significativo. Provavelmente, com o passar do tempo e trocas de
energia entre camadas subjacentes, homogeneízam-se as temperaturas, aproximando-se
as respectivas densidades, fluindo mais livremente os solutos no primeiro metro. À
medida que as trocas de energia se fazem repercutir ao longo da coluna de água, o
aquecimento que dias antes se fazia sentir à superfície, agora está mais perto do fundo.
Quanto menor for a amplitude térmica entre a superfície e o fundo, maior é a
homogeneização de solutos na coluna de água.
Produtividade
Entre Abril e Julho realizaram-se três ensaios à produtividade da lagoa (Fig. 33). Estes
testes ao mesmo tempo que registam a evolução da lagoa permitem saber (para cada
episódio de amostragem), a “condição geral da lagoa”, à medida que se aproxima e
intensifica o período mais produtivo do ano.
Figura 33: Produtividade medida em mg/O2/h em três datas distintas.

28
Nas duas primeiras amostragens não se registaram diferenças na produtividade do
fitoplâncton. A 06/07/2007 foi determinado um aumento significativo da produtividade
relativamente às duas determinações anteriores, e foi também no período entre
25/05/2007 e 06/07/2007 que se registou (a 29/06/2007) o primeiro bloom (já dentro do
período - segunda parte do estudo - mais produtivo.
Limitação pelo fósforo
Figura 34: Biomassa, medida em mg/L de CHL-a ao sétimo e décimo quarto dias.
Os resultados evidenciados pela Fig. 34 sugerem que, ao sétimo dia de experiencia, os
valores de absorvência de clorofila a são próximos e, portanto, sem grandes diferenças
entre si independentemente da quantidade de ortofosfato adicionado a cada um dos
preparos.
Quanto ao décimo quarto dia, os valores determinados revelam o incremento da
concentração de CHL-a de acordo com o aumento da concentração de ortofosfato nas
soluções. Ou seja, os resultados sugerem limitação pelo fósforo, pois quanto maior a
concentração de ortofosfato, maior é o desenvolvimento da comunidade de algas que se
obtém em resposta ao estímulo (adição de fósforo).
Estatística
Tabela 1: Coeficiente de correlação de Spearman, rs, entre variáveis/parâmetros
amostrados/determinados respectivamente com p<0,05.
Variáveis rs Temperatura ar (Nazaré) Vs. Temperatura ar (lagoa) 0,69 CHL-a Vs. SRP -0,69 CHL-a Vs SRP/TP -0,73
Box & Whisker Plot
Mean Mean±SD Mean±1,96*SD
dias 0 50 100 200 Var6 dias 0 50 100 2002
4
6
8
10
12
14
16
18
20
22
24
26
28

29
CHL-a Vs. Transparência 0,60 Temperaturaar Vs. Temperaturasup 0,68 T1m Vs. T1,5m 0,61 pHsup Vs. Condutividadesup -0,74 pH1m Vs. Condutividade1m -0,66 Condutividadesup Vs. Condutividade1m 0,97 Condutividadefundo Vs. Condutividade1,5m 0,70 Concentração Oxigéniosup Vs. pHsup 0,73 Concentração Oxigénio1m Vs. pH1m 0,53 Temperaturasup Vs. Temperatura1m 0,75 pHsup Vs. pH1m 0,79 Concentração Oxigéniosup Vs. Concentração Oxigénio1m
0,80
Concentração Oxigénio1,5m Vs. SRP/TP -0,80 Concentração CHL-a Vs. Velocidade do vento -0,76 Cor SRP 0,64
TP 0,56 SRP/TP 0,66 pHsup 0,52 pH1m 0,59 Concentração Oxigénio1,5m -0,75 Concentração Oxigéniofundo -0,81
Tabela 2: Coeficiente de correlação de Spearman, rs, entre as componentes do TSI
(PT; CHL-a; SD) com p<0,05. Correlação significativa assinalada com (*).
TSI (PT) TSI (CHL-a) TSI (SD) TSI (PT) 1,000000 0,046429 -0,021448
TSI CHL-a 0,046429 1,000000 0,604111* TSI (SD) -0,021448 0,604111* 1,000000
Com as chuvas da Primavera (2007), foi observada a movimentação de água da lagoa por
cima do recém construído paredão de cimento. Simultaneamente, verificou-se o
arrastamento da macrófita flutuante A. filiculoides que acompanhava o movimento de
saída de água via topo do paredão (observação pessoal).
Terceira parte
Breve nota
Esta parte do trabalho compreende a exposição comparativa dos resultados deste
estudo com os do estudo anterior (que decorreu de Janeiro a Agosto de 2005 – com a
lagoa completamente vazia em Agosto e antecedeu a remoção da camada mais
superficial dos sedimentos de uma área significativa do fundo da lagoa em Setembro
2005) e da macrófita N. alba. Ainda no âmbito do estudo de 2005 realizou-se uma
amostragem de monitorização (pós-reenchimento) em Abril de 2006.

30
Índice do estado Trófico (TSI) (Carlson, 1977)
Para determinar o estado trófico da lagoa foi calculado o índice do estado trófico de
Carlson (TSI) com base na concentração do TP à superfície; da clorofila a; e na
transparência de Secchi (Carlson, 1977).
Figura 35: Variação das três componentes do índice de Carlson (PT, CHL-a e SD) nos anos de 2005 e 2007. O estudo de
2005 terminou (Agosto) quando a lagoa secou (Quaresma, 2006).
Em ambos os anos, 2005 e 2007, o TSI de cada componente (TP, CHL-a e SD) foi
tendencialmente crescente ao longo do ano. À excepção dos três primeiros meses de
2005, o TSI de cada componente (TP, CHL-a e SD), foi sempre superior ao respectivo de
2007 (Fig. 35).
Dos componentes do TSI, o TP foi o que apresentou valores mais próximos (para o
conjunto dos dois anos), com indicação de melhoria e melhor qualidade de água em 2007
(Fig. 35).
Os valores (ao longo do estudo) do TSI variaram entre (PT) 37 – 66 e (SD) 47 – 80, ou
seja, entre oligotrófico e hipereutrófico.
O TSI (CHL-a) aumentou ao ponto de “sair da escala” i. e. > 100.
Figura 36: Representa os valores de TSI para cada um dos componentes
(PT, CHL-a e SD) para o mês de Abril em 2005, 2006 e 2007.
31
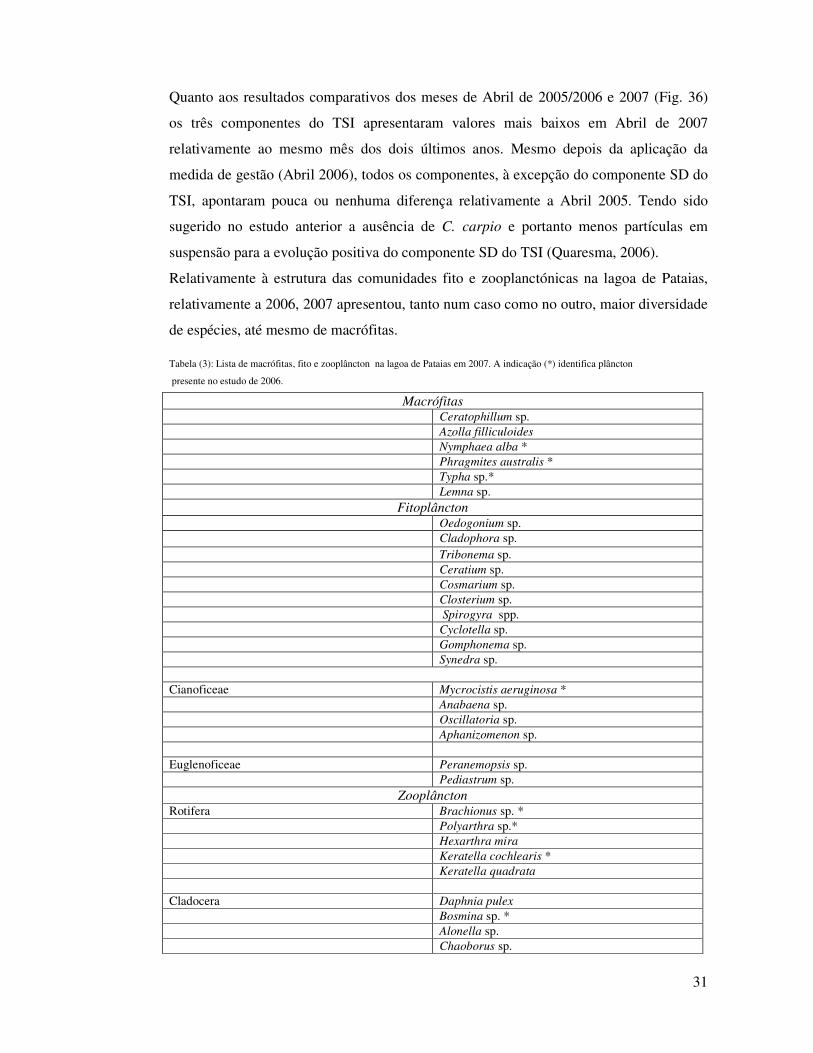
Quanto aos resultados comparativos dos meses de Abril de 2005/2006 e 2007 (Fig. 36)
os três componentes do TSI apresentaram valores mais baixos em Abril de 2007
relativamente ao mesmo mês dos dois últimos anos. Mesmo depois da aplicação da
medida de gestão (Abril 2006), todos os componentes, à excepção do componente SD do
TSI, apontaram pouca ou nenhuma diferença relativamente a Abril 2005. Tendo sido
sugerido no estudo anterior a ausência de C. carpio e portanto menos partículas em
suspensão para a evolução positiva do componente SD do TSI (Quaresma, 2006).
Relativamente à estrutura das comunidades fito e zooplanctónicas na lagoa de Pataias,
relativamente a 2006, 2007 apresentou, tanto num caso como no outro, maior diversidade
de espécies, até mesmo de macrófitas.
Tabela (3): Lista de macrófitas, fito e zooplâncton na lagoa de Pataias em 2007. A indicação (*) identifica plâncton
presente no estudo de 2006. Macrófitas
Ceratophillum sp. Azolla filliculoides Nymphaea alba * Phragmites australis * Typha sp.* Lemna sp.
Fitoplâncton Oedogonium sp. Cladophora sp. Tribonema sp. Ceratium sp. Cosmarium sp. Closterium sp. Spirogyra spp. Cyclotella sp. Gomphonema sp. Synedra sp. Cianoficeae Mycrocistis aeruginosa * Anabaena sp. Oscillatoria sp. Aphanizomenon sp. Euglenoficeae Peranemopsis sp. Pediastrum sp.
Zooplâncton Rotifera Brachionus sp. * Polyarthra sp.* Hexarthra mira Keratella cochlearis * Keratella quadrata Cladocera Daphnia pulex Bosmina sp. * Alonella sp. Chaoborus sp.
32
Copepoda Nauplii * Copidodiaptomus numidicus Acanthocyclops robustus *
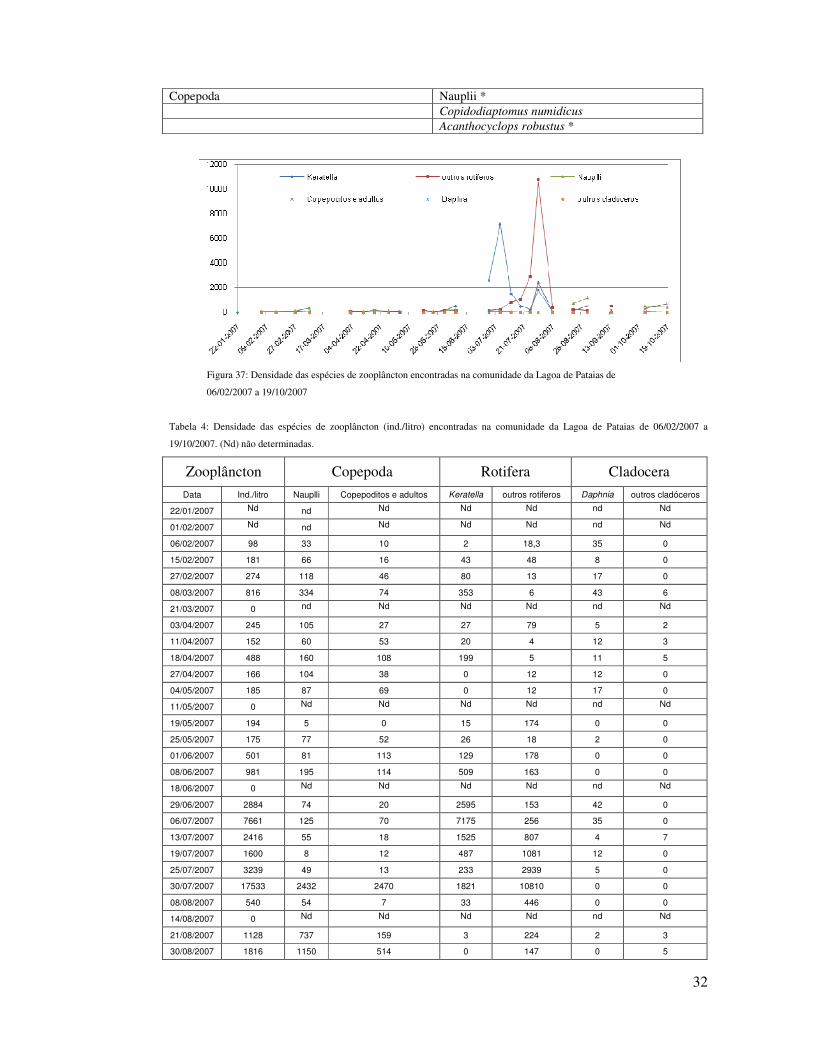
Figura 37: Densidade das espécies de zooplâncton encontradas na comunidade da Lagoa de Pataias de
06/02/2007 a 19/10/2007
Tabela 4: Densidade das espécies de zooplâncton (ind./litro) encontradas na comunidade da Lagoa de Pataias de 06/02/2007 a
19/10/2007. (Nd) não determinadas.
Zooplâncton Copepoda Rotifera Cladocera
Data Ind./litro Nauplli Copepoditos e adultos Keratella outros rotiferos Daphnia outros cladóceros
22/01/2007 Nd nd Nd Nd Nd nd Nd
01/02/2007 Nd nd Nd Nd Nd nd Nd
06/02/2007 98 33 10 2 18,3 35 0
15/02/2007 181 66 16 43 48 8 0
27/02/2007 274 118 46 80 13 17 0
08/03/2007 816 334 74 353 6 43 6
21/03/2007 0 nd Nd Nd Nd nd Nd
03/04/2007 245 105 27 27 79 5 2
11/04/2007 152 60 53 20 4 12 3
18/04/2007 488 160 108 199 5 11 5
27/04/2007 166 104 38 0 12 12 0
04/05/2007 185 87 69 0 12 17 0
11/05/2007 0 Nd Nd Nd Nd nd Nd
19/05/2007 194 5 0 15 174 0 0
25/05/2007 175 77 52 26 18 2 0
01/06/2007 501 81 113 129 178 0 0
08/06/2007 981 195 114 509 163 0 0
18/06/2007 0 Nd Nd Nd Nd nd Nd
29/06/2007 2884 74 20 2595 153 42 0
06/07/2007 7661 125 70 7175 256 35 0
13/07/2007 2416 55 18 1525 807 4 7
19/07/2007 1600 8 12 487 1081 12 0
25/07/2007 3239 49 13 233 2939 5 0
30/07/2007 17533 2432 2470 1821 10810 0 0
08/08/2007 540 54 7 33 446 0 0
14/08/2007 0 Nd Nd Nd Nd nd Nd
21/08/2007 1128 737 159 3 224 2 3
30/08/2007 1816 1150 514 0 147 0 5
33
05/09/2007 0 Nd Nd Nd Nd nd Nd
14/09/2007 856 92 188 27 542 7 0
21/09/2007 0 Nd Nd Nd Nd nd Nd
05/10/2007 840 489 282 6 57 6 0
19/10/2007 1130 417 706 0 0 7 0
Totais 15308 18192 7107 5179 282 31
33500 12286 313
Tabela 5: Densidade das espécies de zooplâncton encontradas na comunidade da Lagoa de Pataias de Janeiro a Junho de 2005.
Zooplâncton Copepoda Rotifera Cladocera
Ind./litro Total Acanthocyclops robustus Nauplii Brachionus
sp.
Polyarthra
sp.
Keratella
cochlearis
Bosmina
longirostris
Janeiro 6665 397 338 0 3090 383 2457
Fevereiro 8446 383 662 0 3002 486 3914
Abril 548 99 83 70 0 21 275
Maio 2943 309 132 0 0 316 2185
Junho 3651 171 1092 1050 0 1723 615
Totais 1359 2308 1120 6091 2929 9446
3667 10140 9446
A diferença de 2005 para 2007 quanto à biodiversidade é substancial (tabela 3).
Relativamente a 2005, em 2007 registou-se maior diversidade (tanto de fito como de
zooplâncton). No estudo de 2007 registou-se, nos períodos mais eutróficos, a ocorrência
de mais espécies (relativamente aos períodos homólogos de 2005) assim como também
se verificaram aumentos do número de espécies, nos períodos entre blooms,
relativamente aos períodos mais produtivos (2007). A abundância de rotíferos também
parece aumentar entre blooms. A comunidade zooplanctónica evidenciou alguma
variação. Regra geral, era sempre possível identificar a presença de Polyarthra sp. e de
K. quadrata. Mas não só, algumas vezes, identificava-se Brachionus sp., noutras K.
cochlearis, Polyarthra sp. e Hexarthra mira. Não se identificou a presença de todos ao
mesmo tempo, pelo menos, em quantidades significativas (observação pessoal).
As populações de cianobactérias também alternavam (observação pessoal) entre
fixadores de azoto (Anabaena sp.) e não fixadoras (M. aeruginosa).
Discussão Primeira e Segunda Partes
O comportamento cíclico da concentração da CHL-a só pode ser reflexo de combinação
ou combinações cíclicas, das condições da lagoa. Uma combinação cíclica, a influência
da CHL-a no pH, é resultado da intensa actividade fotossintética durante os blooms,
removendo grandes quantidades de CO2 da água em poucas horas (Cooke et al., 2005). A

34
remoção de grandes quantidades de CO2 (fonte preferencial de carbono inorgânico na
fotossíntese em detrimento de HCO3- e CO3
2-) por parte das espécies M. aeruginosa,
Anabaena sp., Oscillatoria sp. e Aphanizomenon sp. tem um impacto no pH,
particularmente evidente em lagos com uma fraca capacidade tamponizante (baixo
carbono inorgânico total) (Ustach & Paerl, 1982). Algumas espécies podem utilizar
bicarbonato, mas fazem-no menos eficientemente e com maior dispêndio de energia
(Cooke et al., 2005). Não é o caso destas populações de cianobactérias que apresentam
preferência por CO2 em relação a outras fontes de carbono inorgânico (Ustach & Paerl,
1982). O CO2, regra geral, é reposto na água por difusão (lenta) a partir da atmosfera,
difusão dos sedimentos ou processos de respiração e mineralização na coluna de água.
Durante períodos de elevada fotossíntese, a lenta difusão na água de CO2 da atmosfera, a
mineralização e a respiração, não conseguem eficientemente repor o CO2 (Ustach &
Paerl, 1982). Desta maneira o CO2 nos sedimentos adquire uma grande importância no
equilíbrio do carbono e consequentemente na condição ecológica da lagoa.
Portanto, na lagoa de Pataias, os períodos de grande desenvolvimento de algas mudam o
equilíbrio do carbono para os carbonatos (de fraca utilização na fotossíntese), aumentam
a concentração de oxigénio e o pH (fraca capacidade de “amortecer” o efeito da remoção
do CO2) (Cooke et al., 2005). Com a remoção do CO2, eleva-se o pH, pressionando ainda
mais a necessidade da fotossíntese por CO2, que é cada vez menos. Este comportamento
sugere o esgotamento da “capacidade de carga” da lagoa (relativamente ao pH/CO2),
quando se registam blooms.
Uma outra consequência do aumento da concentração da CHL-a é a redução da
transparência, acompanhando o desenvolvimento de algas e, a cada redução da
transparência, o aumento do ensombramento e a respectiva senescência (a ausência ou
escassez de luz dificulta o desenvolvimento de fitoplâncton e ainda desencadeia
processos de degeneração celular, libertando o conteúdo celular na água). A senescência
(seguida de decomposição), em oposição ao aumento da biomassa de fitoplâncton
(produtores), traduz-se: na redução da concentração da CHL-a, na redução da
concentração de oxigénio, no aumento de CO2, diminuição de pH, e aumento da
condutividade. Isto é, o oposto do que seria de esperar para cada uma das
variáveis/parâmetros com o aumento da concentração de CHL-a.
Somando os efeitos do pH aos do ensombramento (desencadeado pelo desenvolvimento
da comunidade de algas), temos ciclos que compreendem o desenvolvimento de algas
(aumento da concentração da CHL-a, diminuição da concentração SRP, oxigenação da

35
água, redução da transparência, aumento do pH, consumo de sais e redução da
condutividade, diminuição da cor, CO2 e nutrientes em geral), intercalados por períodos
de decomposição (diminuição da concentração de CHL-a, aumento da concentração de
SRP, redução da oxigenação, aumento da transparência, diminuição de pH, aumento da
cor, libertação de sais, libertação de nutrientes e CO2 na coluna de água), fechando o
antigo ciclo, ao mesmo tempo que gera condições para ao próximo (Fig. 38).
Figura 38: Variação de pH entre valores viáveis e menos viáveis para o
desenvolvimento da comunidade de algas.
Então, as flutuações da concentração de CHL-a observadas na segunda parte do estudo
podem ser, resultado da constante resposta a combinações de multi-estímulos. É como se
o ecossistema respondesse, de uma forma sinérgica, com um equilíbrio transitório entre a
procura e a disponibilidade de recursos. Como Antoine-Laurent Lavoisier (1789) diria…
“Nada se perde nada se ganha, tudo se transforma”.
������������ ������������
����� ����� ����� �����������
Terá sido o desenvolvimento diferenciado na coluna de água numa fase e o
desenvolvimento à superfície noutra, a determinar as excepções à relação CHL-a – pH?
Até ao primeiro bloom – 06/07/2007, as comunidades de algas que se desenvolveram em
toda a coluna de água (diluição do impacto na coluna de água), depois de 06/07/2007, só
se desenvolvem no máximo até um metro de profundidade (zona eufótica - Fig. 15).
- A 06/07/2007 registou-se um aumento do pH à superfície e um ainda maior a
um metro (figura 24 e 25).
- De 06/07/2007 a 13/07/2007 ao mesmo tempo que se verificava uma diminuição
da concentração de CHL-a (Fig. 24), também se registou uma descida da

36
concentração de SRP (Fig. 22).
- O dados da condutividade (Fig. 23) de 29/06/2009 a 13/07/2007 sugerem,
redução da condutividade da água a um metro e à superfície.
- 13/07/2007 foi a primeira amostragem com baixos valores de transparência (Fig.
28).
É possível que a 06/07/2007 tenha havido um maior desenvolvimento da comunidade de
algas a um metro do que à superfície. Ou seja, no primeiro bloom, a proximidade do
fundo e a disponibilidade de luz terão reunido melhores condições para um maior
desenvolvimento de algas (com aumento do pH, absorção de SRP e sais), na parte
inferior da coluna de água.
Então, como se interpreta a redução de SRP a 13/07/2007 (Fig. 22)?
Tratando-se de organismos fotoautotróficos, depois de 06/07/2007 a redução da zona
eufótica implica uma “concentração” numa estreita faixa da coluna de água. No entanto,
a coluna de água (de um metro de profundidade até ao fundo), encontrava-se
depauperada em nutrientes devido ao maior desenvolvimento da comunidade de algas.
A evolução do pH, condutividade e SRP de 06/07/2007 a 13/07/2007 (amostragem
seguinte ao primeiro bloom) sugere que a água (e o escasso soluto nela) abaixo de um
metro, terá chegado à superfície. Este feito terá resultado numa mistura destas duas
massas de água, traduzindo-se numa homogeneização do primeiro metro de coluna de
água, diluindo o SRP que lá se encontrava (Fig. 22).
As águas a um metro não eram pobres só em SRP, também o eram em CO2 e sais
fundamentais ao desenvolvimento das algas. Assim, as algas em desenvolvimento à
superfície (13/07/2007) terão ficado condicionadas pelos escassos recursos disponíveis,
tendo esta condição resultado no aumento do pHsup (Fig. 25); diminuição da concentração
de SRP (Fig. 22); e redução da condutividade à superfície (Fig. 23).
� �������������������������
O desenvolvimento de algas nas duas semanas anteriores (a 05/09/2007 - máximo de
clorofila a) com concentração de clorofila a na ordem das 5200 - 6000µg/l. Estas, após
senescência, ter-se-iam decomposto ainda na coluna de água (Golterman, 1975) e
acumulado os corpos celulares no fundo. Por isso as águas abaixo da zona eufótica

37
(abaixo de 0,8 m) teriam um acumular de solutos (nutrientes) e CO2 com origem na
senescência de células. Isso mesmo sugere a Fig. 18: nas duas semanas anteriores à
segunda excepção, o oxigénio que até aí havia foi consumido, possivelmente através dos
processos de decomposição.
Terá ocorrido difusão entre as camadas de água à superfície e (pelo menos) um
metro, arrastando os conteúdos na coluna de água ao misturá-la. Ter-se-á traduzido numa
diminuição do pHsup (Fig. 27) e um aumento do pH1m (Fig. 27), assim como numa
depleção de sais (Fig. 23) na coluna de água (a um metro e meio), “transportados” até à
superfície (diminuindo a condutividade a 1,5 metros) onde terão sido integrados nas
algas em desenvolvimento à superfície. Com a biomassa de algas reciclada, capacidade
tamponizante reequilibrada, mais sais e nutrientes e principalmente o CO2 reposto, as
algas à superfície desenvolveram-se bastante (Fig. 13).
Portanto, as duas excepções são resultado da mistura das camadas de água à
superfície e um metro que, num caso, o primeiro, resultou no valor de pH mais elevado, o
outro num aumento da concentração de CHL-a. Embora a génese tenha sido a mesma
para ambos os casos, diferiu no resultado final. No primeiro caso, a intrusão de água
vinda do primeiro metro da coluna de água (com poucos sais, SRP e CO2-pH elevado), na
superfície, terá comprometido a produtividade. No segundo, terá sido a intrusão de água
vinda da parte inferior da coluna de água (rica em solutos, sais, SRP e CO2-pH mais
baixo) que terá resultado na redução do pHsup, (o que terá permitido amortecer o efeito da
remoção de CO2 na água) e enriquecido a água à superfície (zona eutrófica) com
elementos essenciais ao desenvolvimento de algas, resultando nos valores máximos de
concentração de CHL-a deste estudo.
Relativamente ao regime de vento
A lagoa de Pataias é um lago pouco profundo. Portanto a circulação de toda a coluna de
água não é uma hipótese mas sim uma certeza.
Em lagos pouco profundos, o vento é um agente importante para a mistura da coluna de
água e, na lagoa de Pataias, o vento desempenhou um papel sem dúvida importante,
particularmente quando a sua energia se fez sentir a 25,5 km/h (máximo de velocidade
média). Contudo, que papel terá à velocidade média de 7,9 km/h (mínimo registado - Fig.
29)?

38
Uma vez que os dados relativos ao regime do vento são da estação meteorológica da
Nazaré, e encontrando-se a lagoa de Pataias 10 km a Este é, portanto, menos sujeita ao
efeito de costa. A topografia da bacia de drenagem e o facto de estar rodeada por um
bosque de P. pinaster, podem também contribuir para atenuar a influência do vento na
lagoa. Assim, é provável que a velocidade do vento na lagoa de Pataias seja, regra geral,
inferior à registada na Nazaré.
Poderá ter havido períodos de menor ou mesmo total ausência de mistura da coluna de
água. Nos registos da velocidade do vento, existem períodos calmos intercalados com
períodos mais agitados, e ainda, um período relativamente longo (de 30/08/2007 até ao
fim do estudo), com muito baixa velocidade do vento (a mais baixa do estudo). Estes
períodos mais calmos podem ter contribuído, por algumas horas ou poucos dias, para
uma não mistura de massas de água, nos dois metros de coluna de água. Assim, e para
estes dias, a temperatura poderá ter desempenhado um papel importante na medida em
que, directa ou indirectamente, poderá ter agido como “catalisador” fazendo a ponte entre
o CO2 na coluna de água/fundo e a superfície (substituindo o vento).
O aquecimento da água da lagoa é diferenciado, isto é, a coluna de água não aquece toda
ao mesmo tempo, primeiro aquece a camada mais superficial e esta por radiação e
condução aquece as subsequentes (Wetzel, 2001). O aumento da temperatura pode ter
desencadeado um processo dinâmico na coluna de água, uma vez que as leituras a um
metro sugerem uma fluidez entre o fundo e a superfície e vice-versa (Fig. 31).
O comportamento da concentração da CHL-a ao longo da segunda parte do estudo
Do início da segunda parte, 29/06/2007 até ao fim do estudo, os valores da concentração
da CHL-a sugerem uma crescente disponibilidade de nutrientes evidenciada pelo
aumento da biomassa fitoplanctónica, com um máximo a 05/09/2007 (Fig. 13).
O padrão crescente da concentração de CHL-a tem de ser resultado do aumento do
recurso nutrientes. A questão é saber a origem dos nutrientes, partindo do pressuposto
que existe uma quantidade finita de nutrientes na lagoa, de que parte estão nos
sedimentos (processos de sedimentação pós Setembro de 2005) e a outra parte na coluna
de água, quer na forma dissolvida, quer na particulada, ou como parte da biomassa das
comunidades do lago.
Os resultados sugerem, (1) que aos nutrientes que já faziam parte da biomassa de
fitoplâncton, ter-se-ão juntado (2) outros (crescente concentração de CHL-a - Fig. 13):

39
(1) Reciclagem de nutrientes ainda na coluna de água.
(2) Gradual mas contínua sobrecarga de nutrientes. A entrada de nutrientes
alóctones é considerada mínima (Quaresma, 2006), logo a sobrecarga de
nutrientes é autóctone e provavelmente proveniente dos sedimentos.
Considerando que:
1. Em geral, 70% ou mais do fósforo orgânico está na forma particulada (seston)
(Wetzel, 2001).
2. A hidrólise de fosfatos orgânicos dissolvidos de células senescentes é imediata
em águas quentes (Golterman, 1975).
3. O oxigénio é consumido por degradação microbiana da matéria orgânica
particulada, que sedimenta para o fundo (Wetzel, 2001).
4. Há redução da camada orgânica oxidada, nos períodos de anóxia ou muito pouco
oxigénio junto ao fundo, que serve de protecção à camada reduzida rica em
nutrientes na forma dissolvida (Mortimer, 1941).
Então, provavelmente houve:
1. Reciclagem de nutrientes (principalmente na coluna de água)
2. “Entrada” de nutrientes provenientes dos sedimentos.
2a. Há indicação de um período de dois meses e meio (25/05/2007 – 0,48 mgO2/l a
14/08/2007 – 0,5 mgO2/l) com muito pouco oxigénio no fundo e depois, de novo,
baixas concentrações de oxigénio dissolvido no fundo, na semana anterior ao
máximo de concentração de CHL-a.
Deste modo poder-se-á ter assistido à lenta mas sistemática entrada de nutrientes
(particularmente o fósforo) para a coluna de água, (com origem na camada oxidada)
contribuindo para o desenvolvimento de algas.
De acordo com Mortimer a camada oxidada pode ter 5mm a 40-50mm de espessura
(Mortimer, 1971). No entanto, para ser “reduzida” significativamente em poucos meses,
teria de ser pouco espessa. E temos razões para pensar que, de facto, era efectivamente
pouco espessa. Essas razões são:
1. Remoção de sedimentos em Setembro de 2005.
2. Os processos de hidrólise (reduzindo a acumulação no fundo), que em grande
parte decorrem na coluna de água quando quente (Golterman, 1975).
3. Nos períodos frios, a produtividade é menor

40
Logo, houve uma fraca capacidade de acumulação de matéria orgânica particulada, e
pouca acumulação desde Setembro 2005.
Sem a camada protectora (oxidada) da camada reduzida, os nutrientes são directa e
rapidamente libertados em grandes quantidades na coluna de água (Mortimer, 1971). É o
que pode ter acontecido, depois da redução diária da camada oxidada nos dois meses e
meio de baixa concentração de oxigénio dissolvido, culminando na exposição e
libertação de nutrientes reduzidos (na forma solúvel) dos sedimentos dias antes do
máximo de clorofila a (05/09/2007).
A batimetria da lagoa de Pataias também pode ter contribuído para este desfecho. Uma
vez que a distribuição da concentração de oxigénio varia com a profundidade (Fig. 18) e
o fundo da lagoa encontra-se a diferentes profundidades, a camada oxidada nos
sedimentos teve exposição diferente aos períodos de anóxia, assistindo, naturalmente, a
diferentes reduções da mesma.
Relativamente ao valor mais elevado da concentração da CHL-a a 05/09/2007
A pH 6,5 cerca de 50% do carbono inorgânico dissolvido encontra-se presente ou como
CO2 ou como H2CO3. A pH 10 o carbono inorgânico não existe nessas formas
(Hutchinson, 1957). O pH óptimo para a fotossíntese é sensivelmente 6 e a injecção de
CO2, mesmo a pH mais elevado que 10, estimula a actividade fotossintética e produção
de oxigénio (Ustach & Paerl, 1984).
A 05/09/2007 com o pHsup igual a 10,08 e o pH1m igual a 9,55 para se registar o
desenvolvimento da comunidade de algas deve ter havido disponibilidade de carbono
inorgânico na forma de CO2. Então a questão é…
De onde vem o CO2 e como chegou às algas?
O CO2 só pode ter duas origens: alóctone ou autóctone. Alóctone é extremamente difícil,
uma vez que a hidratação do CO2 na água via atmosfera é um processo lento (Stumm &
Morgan, 1970) e o regime de vento, na altura, era dos mais baixos registados ao longo do
estudo. Para além de que não existem efluentes nem afluentes na lagoa de Pataias
(Quaresma, 2006). A hipótese autóctone ganha, assim, destaque.
Se for autóctone, onde no lago teve o CO2 a sua origem?
Na coluna de água ou nos sedimentos. Provavelmente uma combinação dos dois. Algum
do CO2 com origem na coluna de água, e outro nos sedimentos através da decomposição
microbiana.
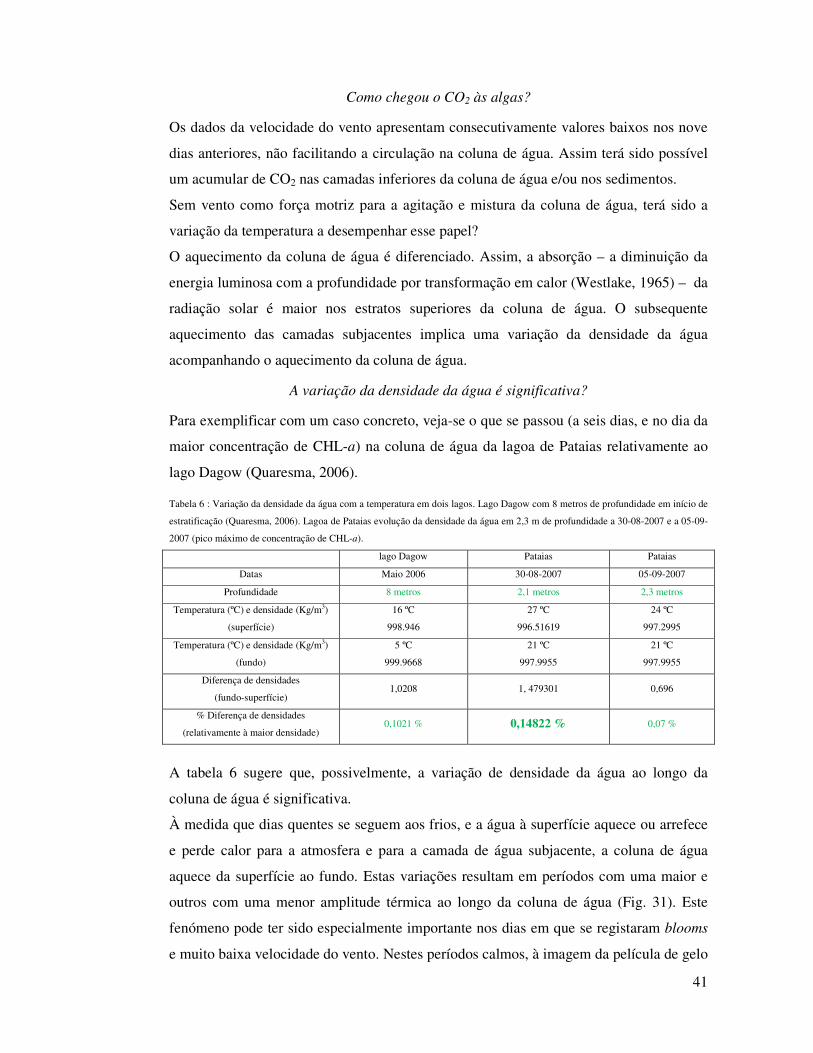
41
Como chegou o CO2 às algas?
Os dados da velocidade do vento apresentam consecutivamente valores baixos nos nove
dias anteriores, não facilitando a circulação na coluna de água. Assim terá sido possível
um acumular de CO2 nas camadas inferiores da coluna de água e/ou nos sedimentos.
Sem vento como força motriz para a agitação e mistura da coluna de água, terá sido a
variação da temperatura a desempenhar esse papel?
O aquecimento da coluna de água é diferenciado. Assim, a absorção – a diminuição da
energia luminosa com a profundidade por transformação em calor (Westlake, 1965) – da
radiação solar é maior nos estratos superiores da coluna de água. O subsequente
aquecimento das camadas subjacentes implica uma variação da densidade da água
acompanhando o aquecimento da coluna de água.
A variação da densidade da água é significativa?
Para exemplificar com um caso concreto, veja-se o que se passou (a seis dias, e no dia da
maior concentração de CHL-a) na coluna de água da lagoa de Pataias relativamente ao
lago Dagow (Quaresma, 2006).
Tabela 6 : Variação da densidade da água com a temperatura em dois lagos. Lago Dagow com 8 metros de profundidade em início de
estratificação (Quaresma, 2006). Lagoa de Pataias evolução da densidade da água em 2,3 m de profundidade a 30-08-2007 e a 05-09-
2007 (pico máximo de concentração de CHL-a).
lago Dagow Pataias Pataias
Datas Maio 2006 30-08-2007 05-09-2007
Profundidade 8 metros 2,1 metros 2,3 metros
Temperatura (ºC) e densidade (Kg/m3)
(superfície)
16 ºC
998.946
27 ºC
996.51619
24 ºC
997.2995
Temperatura (ºC) e densidade (Kg/m3)
(fundo)
5 ºC
999.9668
21 ºC
997.9955
21 ºC
997.9955
Diferença de densidades
(fundo-superfície) 1,0208 1, 479301 0,696
% Diferença de densidades
(relativamente à maior densidade) 0,1021 % 0,14822 % 0,07 %
A tabela 6 sugere que, possivelmente, a variação de densidade da água ao longo da
coluna de água é significativa.
À medida que dias quentes se seguem aos frios, e a água à superfície aquece ou arrefece
e perde calor para a atmosfera e para a camada de água subjacente, a coluna de água
aquece da superfície ao fundo. Estas variações resultam em períodos com uma maior e
outros com uma menor amplitude térmica ao longo da coluna de água (Fig. 31). Este
fenómeno pode ter sido especialmente importante nos dias em que se registaram blooms
e muito baixa velocidade do vento. Nestes períodos calmos, à imagem da película de gelo

42
em alguns lagos, o vento deixa de ser a força motriz capaz de misturar toda a coluna de
água que, nestas condições, poderá ter funcionado por camadas imiscíveis (diferentes
densidades, viscosidades, componentes solúveis - nutrientes e suspensóides) e portanto,
terem-se comportando como fluxos laminares (Wetzel, 2001).
É possível que, nos lagos pouco profundos e com muito baixas velocidades do vento, o
processo de aquecimento (que desencadeia períodos de menor ou maior “homogeneidade
térmica”), seja a força motriz capaz de misturar a coluna de água. O processo funcionaria
com base na fusão dos fluxos laminares desencadeados pela homogeneização das
respectivas densidades. Deste modo, a variação da densidade da água é o “vector” que
leva à homogeneização dos solutos na coluna de água. A aproximação de solutos ricos
em elementos fundamentais ao desenvolvimento resultaria num bloom de algas, ao passo
que a segregação de solutos na coluna de água desencadearia um esgotamento de
recursos na zona eufótica. Assim a variação da temperatura cria oportunidades para a
ocorrência de blooms.
A bibliografia (Wetzel, 2001) refere que uma velocidade do vento de apenas alguns
metros/segundo é suficiente para accionar a circulação na coluna de água. A 21/08/2007
a velocidade média do vento foi 25,5 km/h, pouco mais de 7 m/s. Terá sido suficiente
para fazer circular a coluna de água, como sustentado pelos valores da concentração do
oxigénio na coluna de água, SRP, pH, temperatura, cor e condutividade. Contudo, a
velocidade média do vento não voltou a ser tão alta nem com tanta influência directa ou
indirecta, nas outras variáveis/parâmetros. Que papel terá a velocidade média de 2,5 m/s
(mínimo registado)?
Na ausência ou muito pouco vento as próprias algas podem funcionar como
“catalisadoras” deste mecanismo?
Se parece óbvio que dias quentes implicam uma elevação da temperatura da água à
superfície, não é tão directo o aquecimento desse filme de água só em função da energia
solar. Isto é, as partículas em suspensão – seston – interferem na transmissão da radiação
solar, absorvendo parte dessa energia e aquecendo a água, ao mesmo tempo que arrefece
o fundo por ensombramento. Portanto, nos dias mais quentes aumenta a diferença de
temperatura (superfície - fundo), aumentando a diferença de densidades (separando
fluxos na coluna de água). Enquanto a radiação solar estimula o desenvolvimento, o
seston interfere na transmissão da energia solar, aumentando ainda mais a diferença de
densidades na coluna de água. O bloom leva o ecossistema ao extremo – escassez de

43
recurso(s) – ao ponto de já não conseguir manter-se, desfazendo-se. Assim, a lise celular
devolve à coluna de água os componentes utilizados no desenvolvimento das algas,
melhorando a transparência e a transmissão da radiação solar ao longo da coluna de água
enquanto os corpos celulares precipitam para os sedimentos, passando a haver uma
menor diferença de temperaturas e densidades à superfície e no fundo.
A fracção húmica da matéria orgânica dissolvida desempenha um papel importante nos
sistemas aquáticos e a cor da água é uma ferramenta simples para determinar a
concentração de substâncias húmicas (Cuthbert & Giorgio, 1992). Do início ao fim do
estudo a cor diminuiu progressivamente. A diminuição da cor nos períodos com mais
oxigénio dissolvido na coluna de água pode estar a potenciar blooms de algas; as
substâncias húmicas podem potenciar o metabolismo do plâncton fornecendo energia e
nutrientes com a mineralização (Rich & Wetzel, 1978; Benner et al., 1992).
Que papel desempenhará a oxidação dos ácidos húmicos no complexo ácido húmico/fosfatase?
Quando a circulação aproxima os complexos ácidos húmicos/fosfatase da superfície, a
radiação UV liberta as fosfatases dos complexos, retomando as fosfatases a sua função
normal (Boavida & Wetzel, 1998; Boavida, 2000a; 2000b;)
Relativamente à zona eufótica e ao ácido húmico, após a segregação do complexo ácido
húmico/fosfatase são dois os cenários possíveis:
1. Ou se forma um novo complexo (inactivando outra enzima)
2. Ou não se forma um novo complexo (oxidação/mineralização)
Num ambiente rico em oxigénio após a segregação do complexo ácido húmico/fosfatase
é possível que ocorra uma maior oxidação/mineralização do ácido húmico (não se
ligando a mais nenhuma enzima). Favorecendo a mineralização, para além da redução da
quantidade total de ácidos húmicos e da libertação da enzima, favorece principalmente
uma menor reconversão para o complexo ácido húmico/fosfatase. Isto significa, não só,
enzimas livres mas principalmente enzimas livres mais tempo. Esta condição seria
vantajosa para a comunidade de algas na lagoa, uma vez que traduzir-se-ia no aumento
do número de enzimas disponíveis, reflectindo-se num aumento da eficiência e da
eficácia da actividade enzimática, sem dispêndio de energia.
Este tipo de interacções/equilíbrio (oxidação/mineralização Vs. radiação UV e complexo
ácido húmico/fosfatase), não só parecem plausíveis como também simples, e os
processos biológicos, principalmente os mecanismos de regulação, são simples.

44
Produtividade primária
O significativo aumento da produtividade a 06/07/2007 (Fig. 33 mais de oito vezes a
média das primeiras duas determinações) também se verificou na concentração de CHL-a
pela mesma altura (Fig. 13). O aumento da concentração de clorofila a é reflexo do
aumento da biomassa de algas. À medida que a biomassa de algas aumenta, aumenta
também o número de algas que morrem e a decomposição destas células consome
oxigénio (e em alguns casos mais do que é reposto pela fotossíntese). Isso mesmo é
substanciado no cálculo da produtividade líquida ou bruta. O primeiro caso, é a diferença
entre o produzido e o consumido (garrafas claras na experiência realizada em
laboratório), o segundo, é a soma da produtividade líquida com consumo de oxigénio
(garrafas escuras), calculando-se o oxigénio total produzido.
Assim, a produtividade é reflexo da condição ecológica da lagoa.
Experiência da limitação pelo fósforo
As alterações na concentração de CHL-a verificaram-se apenas no décimo quarto dia
(Fig. 34). Para o sétimo dia e talvez devido ao período de adaptação às condições
laboratoriais, pouca ou nenhuma diferença (entre réplicas) se registou. As algas poderiam
ter reservas internas de fósforo (resultado de consumo de luxo anteriormente na lagoa) tê-
lo-ão utilizado durante os primeiros dias da experiência; a limitação pelo fósforo notou-
se assim, apenas no décimo quarto dia.
Para o décimo quarto dia os valores de absorvência sugerem um maior desenvolvimento
de acordo com as crescentes concentrações de fósforo nas soluções. Sendo assim, como o
período de duplicação da maior parte das algas é de cerca de duas semanas, os resultados
tornaram-se evidentes ao fim de catorze dias e não sete.
Pode concluir-se que, a 14/09/2007 (data da realização da experiência) a lagoa de Pataias
encontrava-se limitada pelo fósforo. Este resultado não é no entanto inesperado, uma vez
que desde 21/08/2007 as determinações de SRP resultavam em valores abaixo do
detectável (3 µg/l) e assim permaneceram até 19/10/2007 (fim do estudo) (Fig. 14 e 22).

45
Estatística
Breve nota
Em regressão linear, o coeficiente de correlação r – ou melhor ainda, o coeficiente de
determinação r2 – ajuda a precisar ideias, ainda que tenha que ser interpretado com
cuidado. É no entanto pacífico, admitindo que existe um padrão linear, considerar que
este é tanto mais forte quanto o r2 mais próximo estiver de 1. De facto, no contexto
linear, r2 x 100% é a variabilidade explicada, (1- r2) x 100% a variabilidade que o modelo
não explica. A definição de r2 pode estender-se a situações de regressão múltipla,
R2 = 1 - ,
ou melhor ainda por Ra2 (ajustado), em que o número de graus de liberdade das somas de
quadrados são usadas para moderar esta “medida de associação”. Claro que Ra2 ~ R2. De
qualquer modo, R2 (ou Ra2) ~ 1 indica que a variável resposta y é bem conhecida através
do “estímulo” x.
A proximidade da Nazaré da lagoa de Pataias é muito importante, uma vez que alguns
dados importantes são registados lá (http:// www.Windguru.com/pt). A distância não
deve ser significativa, uma vez que a temperatura do ar na Lagoa de Pataias está
significativamente correlacionada (tabela 1) com a temperatura do ar registada na
Nazaré; rs = 0,695 e a regressão linear entre as duas exibiu um R2= 0,8 ou seja, muito
semelhantes. Dito isto, considero razoável contar com os registos da Nazaré como
referência para a lagoa de Pataias (particularmente velocidade do vento).
Reflexo do aumento da temperatura ambiente é a temperatura da água à superfície; rs =
0,678 e R2= 0,644. A temperatura da água à superfície e a temperatura da água a um
metro estão correlacionadas; rs = 0,753 e o R2= 0,755 sugerem que em grande parte a
temperatura da água a um metro se deve à temperatura da água à superfície. O mesmo
acontece entre a temperatura da água a um metro e a temperatura da água a 1,5 metros,
com rs = 0,917 e R2= 0,932. Estes resultados estatísticos corroboram a interpretação feita
nos resultados relativamente às trocas de energia na coluna de água.
A correlação entre CHL-a e razão SRP/TP (quantidade de SRP no TP) também é
significativa estatisticamente; rs = -0,734 (sublinhando o inferido nos resultados), ainda
que, a razão SRP/TP não explique, inequivocamente, as flutuações da CHL-a; R2= 0,539.
A correlação; rs = 0,968 entre a condutividade à superfície e a condutividade a um metro
e R2= 0,955 mostram que o primeiro metro de água esteve, durante o tempo de estudo,

46
muito homogéneo (pelo menos relativamente à condutividade). Relativamente à
correlação, rs = 0,802 entre a concentração de oxigénio à superfície e um metro quando
comparada com a correlação rs = 0,788, entre o pH à superfície e um metro, embora
próximas, a primeira exibe um R2= 0,648 e a segunda um R2= 0,337. Isto pode significar
que o primeiro metro de coluna de água pode ter estado, pelo menos quimicamente,
estratificado. Nesse primeiro metro, também existe uma correlação entre pH a um metro
e a condutividade a um metro: rs = -0,661 e R2= 0,213. Para a mesma correlação, agora à
superfície, rs = -0,739 e R2= 0,45. As correlações corroboram a interpretação dos
resultados relativamente à remoção de sais (condutividade) aquando de blooms ao
mesmo tempo que o pH aumenta (fotossíntese e remoção de CO2). Contudo, o baixo R2 é
indicador de fraca robustez das correlações. O facto de, para além de serem interacções
indirectas das variações da CHL-a e não implicações directas da condutividade no pH,
também são interacções sinérgicas de várias variáveis/parâmetros, “diluindo a influência”
de cada um. O mesmo raciocínio aplica-se à correlação (rs = 0,732 e R2= 0,5) entre a
concentração de oxigénio à superfície e o pH à superfície e as mesmas variáveis a um
metro (rs = 0,53 e R2= 0,229).
Relativamente à velocidade do vento, esperar-se-ia que funcionasse como energia motriz,
movendo a água na lagoa, renovando a superfície com elementos essenciais ao
desenvolvimento de algas, e portanto, aumentando a concentração de CHL-a.
Incrivelmente, a correlação (rs = -0,761 e R2 = 0,487) entre a velocidade do vento e a
CHL-a é negativa. No entanto, estes dados podem ser interpretados com um
desfasamento entre a velocidade do vento e resposta do fitoplâncton, resultando numa
inferência estatística distorcida.
Quanto à cor, esta está significativamente correlacionada com SRP (rs = 0,644) e SRP/TP
(rs = 0,659). Em ambos os casos, está associada à reciclagem/decomposição da biomassa,
assim como também o está com a concentração de oxigénio a 1,5 metros e no fundo (rs =
-0,753 com R2 = 0,31 e r = -0,811 com R2 = 0,32) respectivamente, corroborando a
sugestão da associação à decomposição (junto ao fundo) e natural consumo de oxigénio
no processo.
De acordo com Carlson (1980), a correlação entre TSI (CHL-a) e TSI (SD) (tabela 2)
significa que a redução da transparência se deve às algas em suspensão.

47
Terceira parte
Os resultados da evolução anual do TSI de Carlson (Fig. 35) sugerem uma melhor
qualidade de água na lagoa no ano de 2007, à excepção dos 3 primeiros meses de 2005,
em que, de acordo com os três componentes do TSI, apresentou melhores valores de
qualidade da água.
A partir daí a água da lagoa de Pataias 2007 passou a exibir sempre melhor qualidade.
Os componentes TP e SD do TSI apresentaram valores próximos e de evolução
semelhante, sendo por isso indicadores preferenciais do estado trófico (Fig. 35). Em 2005
o componente SD do TSI sugeria uma influência das partículas em suspensão na
transparência (Quaresma, 2006). O mesmo componente (SD), em Abril de 2006 indicou
uma melhoria da qualidade da água (Fig. 36), tendo sido sugerido (ainda no estudo
anterior), que se tratou do resultado da ausência de C. carpio (Quaresma, 2006). Tal
como em 2006, em 2007 também não se registou a presença de C. carpio e,
concomitantemente, o seu impacto também não se registou. Ainda assim, e para o mesmo
componente, registou-se uma melhoria na qualidade da água em Abril de 2007
relativamente a Abril de 2006 (Fig. 36).
Quanto ao componente TP do TSI (talvez a mais correcta comparação de TSI para os
dois anos), reforçou as duas observações anteriores, dando conta que a qualidade da água
da lagoa de Pataias em 2005 foi pior que em 2007 (Fig. 35).
Relativamente à comparação do mês de Abril em 2005, 2006 e 2007: dos três, 2007 foi o
que apresentou melhores resultados, portanto melhor qualidade ecológica da água (Fig.
36). No entanto, em Abril de 2007 a lagoa apresentava pelo menos mais um metro (ou 50
%) de profundidade de água relativamente a Abril de 2006, sendo os parâmetros
considerados para o cálculo do TSI facilmente modificados por diluição. Também podem
ter sido influenciados pelo regime de chuva aquando do reenchimento da lagoa.
Ao longo de 2007, e à imagem de 2005, a evolução do TSI reflectiu a redução da
profundidade da lagoa e a influência cada vez maior do fundo na superfície, à medida
que se passava de dois metros de profundidade em Janeiro, para lagoa vazia em Agosto
(Fig. 35).

48
Maior diversidade de zooplâncton = água de melhor qualidade
Houve menos cladóceros no total (tabela 4 e 5), muito menos (o máximo de 2007 é
aproximadamente 1/5 do valor mínimo registado em 2005), e ainda que o tamanho dos
cladóceros de 2007 (até porque foram quase sempre Daphnia) seja maior do que o de
Bosmina longirostris (2005) não se sabe se os baixos valores do componente CHL-a do
TSI registados em 2007 se devem à maior capacidade filtradora dos cladóceros em 2007.
A composição, abundância e dinâmica das populações de zooplâncton são influenciadas
por várias variáveis físicas e químicas que, por sua vez, são largamente afectadas pela
amplitude das flutuações do nível da água (Wetzel, 2001). Em albufeiras, variações no
volume de água podem explicar em parte alguma abundância e a composição das
comunidades zooplanctónicas. No entanto, é plausível que outros factores como a idade
da albufeira e interacções bióticas possam também desempenhar um papel importante na
estruturação da comunidade (Pinel-Alloul & Méthot, 1984; Robarts et al., 1992). O papel
das flutuações do “volume de água prende-se com os mecanismos de disponibilização de
nutrientes, resultando, geralmente, no aumento do estado trófico. Na lagoa de Pataias,
uma vez que o volume de água não sofreu grandes variações (2007), os mecanismos que
desencadeiam um aumento da disponibilização de nutrientes serão outros. A
decomposição de algas na coluna de água e junto ao fundo serão os mecanismos de
reciclagem de nutrientes, contudo será a circulação da água o vector de transporte até à
zona eufótica. Deste modo o regime de vento e temperatura são preponderantes para a
condição ecológica da lagoa. É fácil compreender a influência do aumento da
temperatura combinada com a velocidade do vento (particularmente nos períodos mais
quentes do ano) no aumento do estado trófico em lagos pouco profundos.
Geraldes & Boavida (2004a,b) observaram que a composição do zooplâncton na
sucessão sazonal era fortemente influenciada pela temperatura. O rácio de copépodes
calanoida para ciclopoida frequentemente declina com a eutroficação (Hurlbert & Mulla,
1981; Gulati, 1984; Muck & Lampert, 1984). Herbívoros calanoida dominam em águas
oligotróficas (C. numidicus tem vantagem especialmente quando as algas se encontram
em concentrações baixas), enquanto copépodes predadores dominam em águas mais
eutróficas (o ciclopoide A. robustus apresenta taxa de desenvolvimento mais elevada em
ambientes eutróficos) (Caramujo & Boavida, 1999; Caramujo & Boavida, 2000). No
entanto, reduções na diversidade e aumento da densidade de crustáceos (cladóceros e
copépodes), também podem estar associados à idade do lago. (Popp et al., 1996).

49
Gannon & Stemberger (1978) estudaram a qualidade da água através da comunidade de
crustáceos e rotíferos e, de acordo com Baião & Boavida (2005), vários estudos foram
apontados nas últimas décadas que reportaram alterações na estrutura da comunidade ao
longo de gradientes tróficos. Nesse artigo, as autoras apontaram a substituição do
domínio dos copépodes por rotíferos, como indicação da evolução do estado trófico.
Embora usualmente considerados pouco importantes em termos de biomassa, os rotiferos
desempenham um papel importantíssimo na transferência de energia e reciclagem de
nutrientes (Park & Marshall, 2000a; Wetzel, 2001) e a sua contribuição pode aumentar
com a eutroficação (Zánkai, 1984; Park & Marshall, 2000b). E Baião & Boavida (2005)
também associaram a estados tróficos distintos alguns géneros, entre os quais:
Mesotrófico: Polyarthra sp., K. quadrata, Brachionus sp.
Eutrófico: Polyarthra sp., K. cochlearis
Parece, então, ter havido um efeito de pêndulo relativamente ao estado trófico da lagoa
de Pataias durante a segunda parte do estudo, onde se registou uma composição da
comunidade zooplanctónica (Fig. 37) de acordo com as flutuações do TSI de Carlson
(Fig. 35). Contudo, a evolução trófica da lagoa de Pataias ao longo do ano parece
favorecer a eutroficação, e à medida que vai ficando mais eutrófica, a composição da
comunidade zooplanctónica vai evoluindo também em conformidade. Assim, a lagoa de
Pataias parece padecer de envelhecimento precoce, ou então será apenas a manifestação
da flutuação do estado trófico (evolução/retrocesso) e subsequente adaptação das
populações.
Relativamente ao estudo anterior da Lagoa de Pataias (2005), a estrutura da comunidade
zooplanctónica sofreu claramente uma redução de diversidade, ao mesmo tempo que se
registou o domínio e aumento da densidade do crustáceo B. longirotris (tabela 3).
Boavida & Caramujo (2000) reportaram a presença de B. longirotris em ambientes
eutróficos. Paralelamente, os dados comparativos (figura 35) apresentaram uma melhoria
(para todos os componentes) do índice do estado trófico da lagoa relativamente a 2005.
Considero ter sido importante o escoamento de água e A. filiculoides (acompanhando o
movimento da água), via topo do paredão de cimento. Pois num ecossistema rico em
nutrientes (e à data rico em A. filiculoides) traduziu-se:
1. Na remoção de nutrientes autóctones 2. Na redução da abundância da macrófita flutuante

50
3. E na minimização da introdução (após senescência da A. filiculoides) de
compostos azotados. Considero menos positivo a não remoção (por vezes de uma semana para a outra) de
sacos cheios de A. filiculoides da margem da lagoa. Este procedimento negligente e
pouco eficiente possibilitou a lixiviação de nutrientes de volta para a lagoa, o que
segundo Wetzel (2001) acontece em poucas horas. Não terá sido preponderante para a
condição ecológica da lagoa, mas foi mais uma contribuição.
Segundo o estudo encomendado pela Comissão Temporária das Alterações Climáticas do
Parlamento Europeu, a redução da disponibilidade de água no sul da Europa será igual à
soma da quantidade de água utilizada, actualmente, na agricultura, indústria e uso de
doméstico (http://sol.sapo.pt). A confirmar-se a previsão, a maior ameaça à qualidade da
água da lagoa de Pataias é, sem dúvida, o cenário das Alterações Climáticas.
Tendo conhecimento do cenário das Alterações Climáticas e o investimento de 2005,
devem as autoridades competentes continuar a acompanhar e monitorizar (agora mais do
que nunca) a evolução da lagoa de Pataias.
Conclusão Será a água da lagoa de Pataias em 2007 melhor que a de 2005?
• Com base em todas as comparações possíveis verificou-se que, efectivamente, a
água da lagoa de Pataias em 2007 é de melhor qualidade do que a verificada, na mesma
lagoa e para os mesmos parâmetros, em 2005.
• Possivelmente já o era em Abril de 2006, no entanto, a escassez de água ou o
padrão de distribuição da precipitação (relativamente a 2007) podem ter influenciado
negativamente a avaliação da qualidade ecológica da água. Ainda assim, Abril de 2007 a
qualidade da água (relativamente a Abril 2006) foi melhor.
• Há sazonalidade dos períodos mais produtivos.
• A qualidade da água tende a agravar-se ao longo do ano.
• A qualidade da água é importante para uma boa condição ecológica da lagoa, quer
seja diluindo solutos ou reduzindo a influência do fundo na superfície. Prova disso é o
estudo de 2005, em que a lagoa em Janeiro apresentava dois metros de profundidade
(com boas indicações para a qualidade da água), degradando-se consideravelmente nos
meses seguintes, à medida que mais e mais volume de água era perdido, até se encontrar
seca oito meses depois.

51
• Na lagoa de Pataias as flutuações de densidade ao longo da coluna de água podem
separar e aproximar camadas de água adjacentes ciclicamente como resposta à variação
de temperatura. Esses períodos de aproximação e segregação, renovam, esgotam ou
impedem o desenvolvimento de fitoplâncton.
• Nos períodos classificados como eutróficos registam-se espécies características
desse estado trófico.
• A eutroficação, sazonalmente, está a acelerar a ontogenia da lagoa de Pataias.
• Num lago pouco profundo, sujeito a ventos fracos e com uma diferença
significativa da densidade da água (à superfície e no fundo), é possível que a coluna de
água funcione por fluxos laminares imiscíveis, mesmo que temporariamente.
• As Alterações Climáticas são, ao mesmo tempo, a maior ameaça/oportunidade
à/para a existência da lagoa de Pataias.
• Estes resultados sublinham, reforçam e substanciam a melhoria da qualidade
ecológica da lagoa e o sucesso da medida de gestão ambiental/biomanipulação de 2005.
Bibliografia APHA, 1989. Standart Methods for the Examination of Water and Wastewater. American Public Health Association, 1587 pp Baião, C. & Boavida, M. J., 2005. Rotifers of Portuguese reservoirs in river Tejo catchment: Relations with trophic state. Limnetica 24 (1-2): 103-114. Benner, R. & Moran, M. A. & Hodson, R. E., 1986. Biogeochemical cycling of lignocellulosic carbon in marine and freshwater ecosystems: Relative contributions of procaryotes and eukaryotes. Limnol. Oceanogr. 31: 89-100 Boavida, M. J., 1999. Wetlands: Most relevant structural and functional aspects. Limnetica, 17:57-63. Boavida, M. J., 2000a. Phosphatases in phosphorus cycling: A new direction for research on an old subject. Arch. Hidrobiol. Spec. Issues Advanc. Limnol. 55: 433-440 Boavida, M. J., 2000b. The obscure role of phosphatases in freswater lakes. Verh. Internat. Verein. Limnol. 27: 3184-3186. Boavida, M. J., 2002. Gestão e Utilização Sustentável dos Ecossistemas Internacionais de Água doce. Actaa Limnol. Bras., 14(2):51-58. Boavida, M. J. & Heath R.T. 1988. Is alkaline phosphatase always important in phosphate regeneration?. Arch. Hidrobiol. 111: 4: 507-518.

52
Boavida, M. J. & Marques R.T. 1997. Monitoring water quality in the Portuguese reservoirs of the river Tejo watershed. Verh. Internat. Verein. Limnol. 26: 740-744. Boavida, M. J. & Wetzel, R.G. 1998. Inhibition of phosphatase activity by dissolved humic substances and hydrolytic reactivation by natural UV. Freshwater Biology 40: 285-293. Caramujo, M. J. & Boavida M. J., 1999. Characteristics of the reproductive cycles and development times of Copidodiaptomus numidiscus (Copepoda: Calanoida) and Acanthocyclops robustus (Copepoda: Cyclopoida). Journal of Plancton Research. Vol.21 no. 9 pp. 1765-1778. Caramujo, M. J. & Boavida, M. J., 2000. The crustacean communities of river Tagus reservoirs. Zooplankton structure as reservoir trophic state indicator. Limnética 18:37-56. Carlson, E. R., 1977. A trophic state index for lakes. Limnol. Oceanogr. 22: 361-369 Carlson, E. R., 1980. More complications in the chlorophyll-Secchi disk relationship. Limnol. Oceanogr. 25: 379-382 Cooke, G. D. & Welch, E. B. & Peterson, S. A. & Nichols, S. A., 2005. Restoration and Management of Lakes and Reservoirs, 3rd edn. Taylor & Francis. 573pp Cuthbert, I. D. & del Giorgio, P., 1992. Towards a standard method of measuring colour in fresh water. Limnol. Oceanogr., 37: 1319-1326 Dussart, B., 1967. Les copépodes dês aux continentales D’Europe occidentale, Tome I: Calanoides et Harpacticoides. Editions N. Boubee & Cie. Fitzgerald, G. & Nelson, T., 1966. Extractive and enzymatic analysis for limiting or surplus phosphorus in algae. J. Phycologia 2: 32-37. Fogg, G. E., 1974. Algal cultures and phytoplankton ecology. Univ. Wisconsin. Gannon, J. E. & Stemberger, R. S., 1978. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. Am. Micros. Soc., 97: 16-35 Geraldes, A. M. & Boavida, M. J., 2004a. What factors affect the pelagic cladocerans of the Azibo meso-eutrophic reservoir? Ann Limnol-Int J Lim 40:101-111. Geraldes, A. M. & Boavida, M. J., 2004b. Do littoral macrophytes influence crustacean zooplankton distribution. Limnética 23:57-64. Gulati, R. D., 1984. The zooplankton and its grazing as measures of trophy in the Loosdrecht Lakes. Verh. Int. Ver. Limnol., 22: 863-867 Heath, R. T. & Cooke G.D. 1975. The significance of alkaline phosphatase in eutrophic lake. Verh. Internat. Verein. Limnol. 19: 959-965. Hutchinson, G. E. 1957. A treatise on limnology, v. 1. Wiley

53
Hofmann, W., 1987. Population Dynamics of hipolimnetic rotifers in the pluss-see (North Germany). Hidrobiology 147: 197-201. Holtan, H. L., Kamp-Nielsen, L. & Stuanes, A., 1988. Phosphorus in soil, water and sediments: na overview. Hidrobiologia 170: 19-32. http://www.windguru.com/pt http://www.earth.google.com http://sol.sapo.pt http://www.epa.gov/bioiweb1/aquatic/index.html - Aquatic Biodiversity Hurlbert, S. H. & Mulla, M. S., 1981. Impacts of mosquitofish (Gambusia affinis) predation on plankton communities. Hydrobiologia, 83: 125-151 Lavoisier, A-L., 1789. Tratado elementar da química moderna – tradução. Editora Madras. 400pp. Lund, J. W. G. & Lund, H. C., 1995. Freshwater Algae (Their microscopic world explored). Biopress Ltd. 360pp Maitland, P. S. 1995. Freshwater fish of Annexes II and IV of the EC Habitats Directive (92/43/EEC). Final report to the EC Margaritora, F., 1983. Guide per il Riconoscimento delle specile Animali delle Acque Interna Italiane. 22. Cladoceri. Stamperia Valdonega Verona 169pp. Mikschi, E., 1989. Rotifer distribution in relation to temperature and oxigem content. Hydrobiology 186/187: 209-214. Miller, W. E. & Greene, J. C. & Shiroyama, T., 1978. The Selenastrum Capricornutum Printz Algal Assay Bottle Test (Experimental Design, Application, and Data Interpretation Protocol). EPA-600/9-78-018. Miracle, M. R. & Vicente, E., 1983. Vertical distribution and rotifer concentration in the termocline of meromictic lakes. Hydrobiology 104: 259-267. Mortimer, C. H., 1971. Chemical exchanges between sediments and water in the great lakes-speculations on probable regulatory mechanisms, Limnology and Oceanography v 16(2). Muck, P. & Lampert, W., 1984. Na experimental study on the importance of food conditions for the relative abundance of calanoid copepods and cladocerans. Archiv. Für Hydrobiologie. Supplement 66: 157-179. Murphey, J.D & Riley, J.P. 1962. A modified simple solution method for the determination of phosphate in natural waters. Annal. Chem. Acta 27: 32-36

54
Naumann, E., 1919. Nagra Synpunkter angaende Limnoplanktons Oekologie; Svensk. bot. Tidskr. 13, 129–61. Odum, E. P., 1971. Fundamentals of Ecology, 3rd edition, 574 pp. Park, G. S. & Marshall, H. G., 2000a. The trophic contributions of rotifers in tidal freshwater and estuarine habitats. Estuarine, Coastal and Shelf Science 51: 729-742. Park, G. S. & Marshall, H. G., 2000b. Estuarine relationships between zooplâncton community structure and trophic gradients. J. Plankton Res. 22: 121-135. Pejler, B., 1983. Zooplanctic indicators of trophy and their food. Hydrobiologia 101 (Dev. Hidrobiol. 13): 111-114 Pinel-Alloul, B. & Méthot, G., 1984. Analyse multidimensionnelle de l’evolution du zooplâncton durant la mise en eau de trois reservoirs du Nord du Québec, Canada. Verh. Int. Verein. Limnol. 22: 1444-1455. Popp, A. & Hoagland, K. D. & Hergenrader, G. L., 1996. Zooplankton community response to reservoir aging. Hydrobiologia 339: 13-21. Quaresma, S. 1999. Contribuição para o estudo da lagoa de Pataias. Faculdade de Ciências e Tecnologia da Universidade de Coimbra, Departamento de Botânica. Quaresma, S. 2001. Plano de Gestão da Lagoa de Pataias. Faculdade de Ciências e Tecnologia da Universidade de Coimbra, Departamento de Botânica. Quaresma, S. 2006. Freshwater eutrophic ecosystems: From monitoring to management. Faculdade de Ciências da Universidade de Lisboa, Departamento de Biologia Animal. Rich, P. & Wetzel, R. G., 1978. Detritus in the lake ecosystem. Amer. Nat., 112: 57-71 Robalo J. I, Doadrio I. Almada VC & Kottelat M., 2005. Chondrostoma oligolepis, new replacement name for leuciscus macrolepidotus Steindacher, 1866 (Teleostei: Cyprinidae). Ichthyological Explorations of Freshwater. 16 (1): 47-48. Robarts, R. D. & Zahory, T. & Jarvis, A. C. & Pais-Madeira, C. M. & Sephton, L. M. & Combrink, S., 1992. Phytoplankton and zooplankton populations Dynamics and production of a recently formed african reservoir. Hydrobiologia 237: 47-60. Rose, C. & Axler, R. P., 1998. Uses of alcaline phosphatase activity in evaluating phytoplankton community phosphorus deficiency. Arch. Hydrobiol. 89: 54-87. Rutner-Kollisko, 1980. The abundace and distribuition of Filinia terminalis in various types of lakes as related to temperature, oxigem and food. Hydrobiology 73: 169-175. Schindler, D. W., 1977. Evolution of phosphorus limitation in lakes. Science 195: 260-262.

55
Scourfield, D. J. & Harding, J. P., 1966. British Freshwater cladocera. Freshwater Biological Association. Ustach, J. F. & Paerl H. W., 1982. Blue-green algal scums: Na explanation for their occurrence during freshwater blooms. Limnol. Oceanogr. 27 (2), 1982, 212-217. Vollenweider, R. A., 1969. Primary Prodution in Aquatic Environments. IBP Handbook 12, Blackwell Scientific Publishers, 213pp. Ward, H. D. & Whipple G. C., 1959. Freshwater Biology second edition. John Woley & Sons, Inc.. 1248pp Westlake, D.F., 1965. Some problems in the mesurement of radiation under water. A review. Photochem. Photobiol. 4:849-868. Wetzel, R. G., 1983. Limnology. Saunders, 767 pp. Wetzel, R. G., 1991. Extracellular enzymatic interactions: storage, redistribuition, and interspecific communications. Chróst, R. J. (eed.): Microbial Enzymes in Aquatic Environments. 6-28, Springer-Verlag. Wetzel, R. G., 1993. Humic compounds from wetlands: complexation, inactivation, and reactivation of surface bound and extracellular enzymes. Limnol. 25: 122-128. Wetzel, R.G., 2001. Limnology – Lake and River Ecosystems, 3rd edn. Academic Press. 1006 pp. Wynne, D. & Ben-Dan, T. B., 1995. The effect of light and phosphate concentrations on phosphatase activities of the photosynthetic bacterium Chlorobium spp. Can. J. Microbiology 41: 278-283. Zánkai, N. P., 1984. Predation of Cyclops vicinus (Copepoda: Cyclopoida) on small zooplankton animals in lake Balaton (Hungary). Arch. Hydrobiol. 99(3) : 360-378. Zurek, R., 1985. Species composition of zooplankton in surface waters near the Upper Silesia in the aspect of water quality. Acta Hydrobiol. 27: 339-349.

56

57
Anexo I

58
Índice de tabelas e figuras Figura 1 - pág. 13 - Variação da concentração de clorofila a de 22-01-2007 a 29-06-2007.
Figura 2 - pág. 13 - Variação da transparência de 22-01-2007 a 29-06-2007.
Figura 3 - pág. 14 - Variação da temperatura do ar, superfície, um metro, dois metros e no fundo de 22-01-2007 a
29-06-2007.
Figura 4 - pág. 14 - Variação da temperatura do ar, superfície, um metro, dois metros e no fundo de 22-01-2007 a 29-06 -
2007.
Figura 5 - pág. 15 - Variação da razão SRP/TP de 22-01-2007 a 29-06-2007.
Figura 6 - pág. 15 - Variação dos valores absolutos de SRP e TP de 22-01-2007 a 29-06-2007.
Figura 7 - pág. 15 - Saturação de oxigénio para a temperatura registada à superfície e um metro de 22-01-2007 a 29-06-2007.
Figura 8 - pág. 15 - Variação da concentração de oxigénio dissolvido à superfície, um metro, dois metros e no fundo de
22-01-2007 a 29-06-2007.
Figura 9 - pág. 17 - Variação da cor (unidades Pt) ao longo da primeira parte do estudo. A azul, a evolução das duas
últimas amostragens antes de 25/05/2007 (indicação da degradação).
Figura10 - pág. 17 - Variação da transparência da água na primeira parte do estudo. A azul, a evolução das duas
últimas amostragens antes de 25/05/2007 (indicação da degradação).
Figura 11 - pág. 17 - Variação da CHL-a na primeira parte do estudo. A azul, a evolução da última amostragem até
25/05/2007 (indicação da degradação).
Figura 12 - pág. 17 - Variação da quantidade de SRP e TP na primeira parte do estudo. A azul, a evolução das duas
últimas amostragens antes de 25/05/2007 (indicação da degradação).
Figura 13 - pág. 18 - Evolução da concentração de clorofila a ao longo do estudo. Assinalada a data de separação da
primeira e segunda partes.
Figura 14 - pág. 18 - Variação de SRP no TP ao longo do estudo. Assinalada a data de separação da primeira e segunda
partes.
Figura 15 - pág. 18 - Evolução da transparência ao longo do estudo. Assinalada a data de separação da primeira e
segunda partes.
Figura 16 - pág. 18 - Evolução da temperatura do ar, à superfície, 1 m, 1,5 m, 2 m e fundo ao longo do estudo.
Assinalada a data de separação da primeira e segunda partes.
Figura 17 - pág. 18 - Evolução da condutividade à superfície, 1 m, 1,5 m, 2 m e fundo ao longo do estudo. Assinalada a
data de separação da primeira e segunda partes.
Figura 18 - pág. 19 - Concentração de oxigénio à superfície, 1 m, 1,5 m, 2 m e fundo ao longo do estudo. Assinalada a
data de separação da primeira e segunda partes.
Figura 19 - pág. 19 - Evolução do pH à superfície e um metro ao longo do estudo. Assinalada a data de separação da
primeira e segunda partes.
Figura 20 - pág. 19 - Evolução da CHL-a ao longo do estudo. A linha vertical assinala 29/06/2007. Setas verdes “picos”
setas vermelhas “depressões”.
Figura 21 - pág. 19 - Evolução da transparência ao longo do estudo A linha vertical assinala 29/06/2007.
Figura 22 - pág. 20 - Variação de SRP e TP, na segunda parte do estudo.
Figura 23 - pág. 20 - Variação da clorofila a na segunda parte do estudo.
Figura 24 - pág. 20 - Evolução da CHL-a ao longo do ano. Indicação da data 06-07-2007 (primeiro bloom).
Figura 25 - pág. 20 - Evolução do pH à superfície e um metro ao longo do estudo. Indicação da data 06-07-2007
(primeiro bloom).
Figura 26 - pág. 21 - Variação da concentração de CHL-a ao longo do estudo. Assinalado (linha vertical) as duas
excepções. A azul, evolução da amostragem anterior e posterior às excepções.
Figura 27 - pág. 21 - Variação pH ao longo do ano do estudo. Assinalado (linha vertical) as duas excepções. A azul,
evolução da amostragem anterior e posterior às excepções.
Figura 28 - pág. 22 - Transparência ao longo do estudo. Assinaladas as datas das duas excepções (linha vertical) e a
evolução (azul) antes e depois das datas assinaladas.
Figura 29 - pág. 25 - Evolução da velocidade do vento km/h/dia na segunda parte do estudo.
Figura 30 - pág. 25 - Evolução da cor ao longo do estudo. Indicação da data 21/08/2007.

59
Figura 31 - pág. 26 - Variação da temperatura do Ar, e ao longo da coluna de água de 22/01/2007 a 19/10/2007. As
setas marcam as elevações da concentração da CHL-a (blooms).
Figura 32 - pág. 26 - Variação da concentração de CHL-a. As setas marcam as elevações da concentração da CHL-a
(blooms).
Figura 33 - pág. 27 - Produtividade medida em mg/O2/h em três datas distintas.
Figura 34 - pág. 28 - Biomassa, medida em mg/L de CHL-a ao sétimo e décimo quarto dias.
Figura 35 - pág. 30 - Variação das três componentes do índice de Carlson (PT, CHL-a e SD) nos anos de 2005 e 2007.
Figura 36 - pág. 30 - Representa os valores de TSI para cada um dos componentes (PT, CHL-a e SD) para o mês de
Abril em 2005, 2006 e 2007.
Figura 37 - pág. 32 - Densidade das espécies de zooplâncton encontradas na comunidade da Lagoa de Pataias de
06/02/2007 a 19/10/2007.
Figura 38 - pág. 35 - Variação de pH entre valores viáveis e menos viáveis para o desenvolvimento da comunidade de
algas.
Tabela 1 - pág. 28 - Coeficiente de correlação de Spearman, r, entre variáveis/parâmetros amostrados/determinados
respectivamente com p<0,05.
Tabela 2 - pág. 29 - Coeficiente de correlação de Spearman, r, entre as componentes do TSI (PT; CHL-a; SD) com
p<0,05. Correlação significativa assinalada com (*).
Tabela 3 - pág. 31 - Lista de macrófitas, fito e zooplâncton na lagoa de Pataias em 2007. A indicação (*) identifica
plâncton presente no estudo de 2006.
Tabela 4 - pág. 32 - Densidade das espécies de zooplâncton (ind./litro) encontradas na comunidade da Lagoa de
Pataias de 06/02/2007 a 19/10/2007. (Nd) não determinadas.
Tabela 5 - pág. 33 - Densidade das espécies de zooplâncton encontradas na comunidade da Lagoa de Pataias de
Janeiro a Junho de 2005.
Tabela 6 - pág. 41 - Variação da densidade da água com a temperatura em dois lagos. Lago Dagow com 8 metros de
profundidade em início de estratificação (Quaresma, 2006). Pataias evolução da densidade da
água em 2,3 m de profundidade de 30-08-2007 até 05-09-2007 (pico máximo de concentração de
CHL-a).

60
Anexo II


















