
UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE …§ão.pdf · as alterações em mecânica...
88
UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA FACULDADE DE VETERINÁRIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS JOSELITO DE OLIVEIRA NETO ALTERAÇÕES MECÂNICAS E HISTOLÓGICAS DE PULMÕES EM MODELO EXPERIMENTAL DE LESÃO AGUDA INDUZIDA PELO VENENO DA SERPENTE Crotalus durissus cascavella FORTALEZA 2015
-
Upload
truongthien -
Category
Documents
-
view
214 -
download
0
Transcript of UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE …§ão.pdf · as alterações em mecânica...

1
UNIVERSIDADE ESTADUAL DO CEARÁ
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
JOSELITO DE OLIVEIRA NETO
ALTERAÇÕES MECÂNICAS E HISTOLÓGICAS DE PULMÕES EM
MODELO EXPERIMENTAL DE LESÃO AGUDA INDUZIDA PELO
VENENO DA SERPENTE Crotalus durissus cascavella
FORTALEZA
2015

2
JOSELITO DE OLIVEIRA NETO
ALTERAÇÕES MECÂNICAS E HISTOLÓGICAS DE PULMÕES EM
MODELO EXPERIMENTAL DE LESÃO AGUDA INDUZIDA PELO
VENENO DA SERPENTE Crotalus durissus cascavella
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Veterinárias da
Faculdade de Veterinária da Universidade
Estadual do Ceará, como requisito parcial para a
obtenção do grau de Mestre em Ciências
Veterinárias.
Área de concentração: Reprodução e Sanidade
Animal.
Linha de Pesquisa: Reprodução e Sanidade de
Carnívoros, Onívoros e Aves.
Orientadora: Dra. Janaina Serra Azul Monteiro
Evangelista
FORTALEZA
2015

3

4

5
AGRADECIMENTOS
A Deus que me possibilitou a realização desse sonho tão aguardado em minha vida.
Aos meus pais José Airton de Oliveira e Maria de Fátima Bezerra Oliveira que me
apoiaram e incentivaram para a realização do curso.
À minha irmã Mayra Bezerra Oliveira, que sempre me ajudou quando precisei.
À minha esposa Raquel Horn Vasconcelos de Oliveira que esteve ao meu lado em
todos os momentos e minhas filhas Nínive e Nina que representamminha razão em conquistar
e vencer todos os desafios.
À Universidade Estadual do Ceará, que propiciou minha formação acadêmica e me
proporcionou experiências engrandecedoras e momentos memoráveis ao lado de pessoas
incríveis.
À Coordenação de Aperfeiçoamento de pessoal de Nível Superior (CAPES), pela
bolsa de mestrado concedida.
Aos professores do Programa de Pós-graduação em Ciências Veterinárias por terem
compartilhado seus conhecimentos dentro e fora das salas de aula, contribuindo para o meu
crescimento profissional.
Aos demais funcionários do PPGCV, em especial à querida Adriana, pela paciência e
orientação quanto ao funcionamento do programa.
Aos membros da banca examinadora, por aceitarem ao convite em participar e pelas
considerações feitas durante a defesa, que contribuíram para a melhora da dissertação.
À professora Dra. Janaina Serra Azul Monteiro Evangelista, por ter me acompanhado
e orientado no decorrer do curso e por ter aberto as portas de seu laboratório para que eu
pudesse crescer pessoal e profissionalmente, minha imensa gratidão.

6
Ao professor Dr.Francisco Sales Ávila Cavalcante pela parceria e utilização de
equipamentos no laboratório em que o mesmo é responsável.
Ao professor Ms. Daniel de Araujo Viana pela leitura de lâminas histológicas e pelo
laudo patológico.
A todos os alunos que fazem parte do Laboratório de Biofísica da Respiração – LBR,
da Universidade Estadual do Ceará, em especial ao aluno de doutorado Daniel Silveira Serra,
por ter me acompanhado nos experimentos de mecânica pulmonar.
A todos os alunos do Laboratório de Histologia dos efeitos causados pelos venenos de
serpentes e plantas – HISTOVESP / Universidade Estadual do Ceará (UECE).
Ao aluno de mestrado em Farmacologia da Universidade Federal do Ceará, João
Alison de Moraes Silveira, por estar sempre presente durante todas as etapas de
desenvolvimento dos experimentos.
À professora Dra. Lúcia de Fátima Lopes dos Santos da Universidade Estadual do
Ceará - (UECE), por ter participado do processo de qualificação do meu projeto.
À professora Dra. Helena Serra Azul Monteiro da Universidade Federal do Ceará –
UFC, por ter cedido seu laboratório e ter firmado sua parceria.
À professora Dra. Célia Maria de Souza Sampaio por ter-me co-orientado, estando
sempre disponível todas as vezes que a procurei e por ter participado como membro da banca
em minha defesa de mestrado.
À professora Dra. Diva Maria Borges Nojosa e à médica veterinária Roberta Rocha do
Núcleo Regional de Ofiologia (NUROF) do departamento de Biologia da Universidade
Federal do Ceará – UFC, por terem cedido gentilmente o veneno total.

7
RESUMO
Crotalus durissus cascavella (C.d.cascavella) é uma serpente encontrada na Caatinga do
Nordeste Brasileiro. Seu veneno é caracterizado em promover alterações sistêmicas e causar
óbitos em suas vítimas logo após o acidente ofídico. Esse estudo teve por objetivo investigar
as alterações em mecânica pulmonar (propriedades elásticas, resistivas e viscoelásticas) e
histopatologia em pulmões após inoculação intraperitoneal (3,0 µg/Kg) do veneno dessa
serpente (C.d.cascavella) em camundongos. Os valores médios de resistência Newtoniana
(RN), resistência tecidual(G) e elastância tecidual(H) foram significativos em todos os grupos
experimentais (1h, 3h, 6h, 12h e 24h), sendo maiores nos dois últimos grupos. A
histeresividade (η) foi significativa em 6h, 12h e 24h. Os valores médios de complacência
estática (Cst) foram significativos nos grupos de 6h, 12h e 24h sendo nesse último grupo
valores menores. A capacidade inspiratória (CI) mostrou-se significativa em 3h, 6h, 12h e
24h. Após desafio com Metacolina, RN apresentou valores médios significativos nos grupos
6h, 12h e 24h. O veneno da serpente (C.d.cascavella) induziu no tecido pulmonar, atelectasia,
enfisema, hemorragia, infiltrado inflamatório com polimorfonucleares, congestão em todos
os grupos experimentais e edema apenas no grupo de 24h. O veneno da serpente
C.d.cascavella provocou alterações mecânicas e histopatológicas no tecido pulmonar.
PALAVRAS-CHAVE: Crotalus durissus cascavella. Veneno. Mecânica Pulmonar.
Histopatologia.

8
ABSTRACT
Crotalus durissus cascavella (C. d. cascavella) is a snake found in the Brazilian Northeast
Caatinga. Its venom is featured on promoting systemic changes and cause death in his victims
after the snakebite. This study aimed to investigate the changes in lung mechanics (elastic
properties, resistive and viscoelastic) and histopathology in lungs after intraperitoneal
injection (3,0 µg/Kg) of this snake venom (C. d. cascavella) in mice. The average values of
Newtonian resistance (RN), tissue resistance (G) and tissue elastance (H) were significant in
all groups (1h, 3h, 6h, 12h and 24h) and higher in the last two groups. The histeresividade (η)
was significant in 6h, 12h and 24h. The static compliance of average values (Cst) were
significant in groups of 6h, 12h and 24h and the latter group lower values. The inspiratory
capacity (CI) was significant at 3h, 6h, 12h and 24h. After challenge with Methacholine, RN
known significant average values in groups 6h, 12h and 24h. The venom of the snake C. d.
cascavella also induced in pulmonar tissue atelectasis, emphysema, hemorrhage,
inflammatory infiltrate with polymorphonuclear cels, congestion in all experimental groups,
and edema only in the 24h group. The venom of the snake C. d. cascavellaca used mechanical
and histopathological changes in lung tissue.
Keywords: Crotalus durissus cascavella.Venom.Pulmonary Mechanics. Histopathology.

9
LISTA DE FIGURAS
Figura 1 - Famílias e gêneros de serpentes da fauna brasileira com importância médica... 16
Figura 2 - Características morfológicas de espécies de serpentes Viperidae....................... 17
Figura 3 - Fotomicrografia da parede alveolar ao final da inspiração, Aumento 400X e
coloração H.E ......................................................................................................................
28
Figura 4 - Exemplos de modelos utilizados para descrever o sistema respiratório: (A)
Modelo viscoelástico; (B) Modelo de fase constante...........................................................
39

10
LISTA DE ABREVIATURAS E SIGLAS
PH – Potencial de Hidrogênio
O2 – Gás Oxigênio
CO2 – Gás Carbônico
SINAN – Sistema de Informação de Agravos de Notificação
SINITOX/FIOCRUZ/MS - Sistema Nacional de Informações Tóxico-Farmacológicas da
Fundação Oswaldo Cruz
SIM/SUS - Sistema de Informações Hospitalares do Sistema Único de Saúde
PLA2 – Fosfolipase A2
LAO - L-amino-oxidase
FAD - Flavina adenina dinucleotídeo
MEC - Matriz Extracelular
MCh – Metacolina
Cp – complacência pulmonar
CI – Capacidade inspiratória
Cst – complacência estática
f – frequência
G – resistência tecidual
H – elastância tecidual
I – inertância
η – histeresividade

11
Patm – pressão atmosférica
Palv – pressão alveolar
Pel – pressão elástica dos pulmões
Pao – pressão na abertura das vias aéreas
PEEP – pressão positiva expiratória final
Ppl – pressão intrapleural
Raw – resistência das vias aéreas
RN – resistência Newtoniana
T – tensão superficial
V’ – fluxo
Zrs – impedância de entrada do sistema respiratório

12
SUMÁRIO
1 INTRODUÇÃO ..................................................................................................... 13
2 REVISÃO DE LITERATURA............................................................................. 15
2.1 CLASSIFICAÇÃO DAS SERPENTES DA FAUNA BRASILEIRA................. 15
2.2 ASPECTOS BIOLÓGICOS DO GÊNERO Crotalus.......................................... 18
2.2.1 Classificação e Distribuição Geográfica ........................................................... 18
2.2.2 Habitat................................................................................................................ 19
2.2.3 Hábitos Alimentares.......................................................................................... 19
2.3ASPECTOS GERAIS E EPIDEMIOLÓGICOS DO GÊNERO CROTALUS...... 19
2.4 CONSTITUINTES DO VENENO DE CROTALUS DURISSUS........................ 22
2.4.1 Crotoxina e suas subunidades............................................................................ 22
2.4.2 Crotamina.......................................................................................................... 24
2.4.3 Convulxina......................................................................................................... 24
2.4.4 Giroxina............................................................................................................. 25
2.4.5 Peptídio natriurético........................................................................................... 26
2.5 SISTEMA RESPIRATÓRIO............................................................................... 26
2.5.1 Caracterização Morfofuncional......................................................................... 26 2.5.2 Troca Gasosa..................................................................................................... 27
2.5.3 Rede de fibras do parênquima pulmonar........................................................... 29
2.5.4 Fibras de colágeno e elastina............................................................................. 30
2.5.5 Glicoproteínas.................................................................................................... 30
2.5.6 Proteoglicanos.................................................................................................... 31
2.6 MECÂNICA PULMONAR.................................................................................. 31
2.6.1 Propriedades elásticas do pulmão...................................................................... 31
2.6.2 Propriedades elásticas da parede torácica.......................................................... 32
2.6.3 Propriedades resistivas do sistema respiratório................................................. 33
2.6.4 Resistência pulmonar......................................................................................... 33
2.6.5 Resistência da parede torácica........................................................................... 34
2.6.6 Viscoelasticidade............................................................................................... 34
2.6.7 Dissipação de energia durante o ciclo respiratório............................................ 34
2.7 MODELAGEM EM MECÂNICA PULMONAR................................................ 35
2.7.1 O modelo de fase constante............................................................................... 40
2.8 PESQUISA POR HIPERRESPONSIVIDADE DE VIAS AÉREAS
MEDIANTE DESAFIO COM METACOLINA(MCH)............................................
41
3 JUSTIFICATIVA.................................................................................................. 42
4 HIPÓTESE CIENTÍFICA.................................................................................... 43
5 OBJETIVOS.......................................................................................................... 44
5.1 OBJETIVO GERAL............................................................................................. 44
5.2 OBJETIVOS ESPECÍFICOS............................................................................... 44
6 CAPÍTULO 1......................................................................................................... 45
7 CONCLUSÕES...................................................................................................... 73
8 PERSPECTIVAS................................................................................................... 74
REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 75

13
1 INTRODUÇÃO
Os venenos de serpentes são compostos de uma complexidade de substâncias
ativas, a maioria delas com estruturaproteica, que exercem um papel importante no processo
de sobrevivência desses animais. Devido às necessidades de defesa e captura de presas, as
serpentes desenvolveram evolutivamente uma grande variedade de toxinas. Essas moléculas
possuem diversos alvos biológicos, apresentando, portanto, vários efeitos importantes. A
caracterização dos efeitos específicos das frações de venenos ofídicos é de profunda
importância para o entendimento das alterações patológicas decorrentes do envenenamento
(GARCIA; LEWIS, 2003).
A composição química e as atividades biológicas dos venenos podem variar muito
entre as famílias e gêneros das serpentes (DOS-SANTOS, 2005). Segundo Barraviera e
Pereira (1999), a maior gravidade encontra-se nos acidentes causados por serpentes do gênero
Crotalus, e essas têm ganhado relevante espaço no campo científico, visto que em algumas
décadas seu veneno vem sendo alvo de pesquisadores devido a sua constituição e o efeito que
essas substâncias podem causar (PINHO; PEREIRA, 2001).
A subespécie Crotalus durissus cascavella (C. d. cascavella), encontrada na
Caatinga do Nordeste Brasileiro (BARRAVIERA,1989; PINHO; PEREIRA, 2001), produz
um veneno que conduz a alteração sistêmica, sendo responsável pela causa preliminar de
óbito após o acidente ofídico (EVANGELISTA et al., 2008).
A peçonha de Crotalus apresenta-se como um complexo tóxico-enzimático, na
qual são encontradas as enzimas fosfodiesterase, l amino oxidase, 5-nucleotidase e toxinas
como a crotoxina, convulxina, crotamina e giroxina (RANGEL-SANTOS et al., 2004).
Os animais acometidos por envenenamento crotálico manifestam as principais
complicações: insuficiência renal aguda (NISHIOKA, 2000), choque circulatório (PINHO et
al.,2000; PINHO et al., 2005), reações edematogênicas, agregação de plaquetas (MARTINS et
al., 2004) e hipotensão arterial (EVANGELISTA et al., 2008). Segundo Vital Brazil (1972)
apud Damico et al. (2005), há insuficiência respiratória aguda relacionada à paralisia
neuromuscular.
Os pulmões são os órgãos essenciais do sistema respiratório, mas não são apenas
órgãos respiratórios. Eles participam do equilíbrio térmico, pois, com o aumento da ventilação

14
pulmonar há maior perda de calor e água. Auxiliam na manutenção do pH plasmático dentro
da faixa fisiológica, regulando a eliminação de ácido carbônico (sob a forma de CO2). O
aparelho respiratório também é utilizado para outros fins como a defesa contra agentes
agressores (FRASER, 2005; ZIN; ROCCO; FAFFE, 2008). Sua função básica é de
disponibilizar oxigênio (O2) para o organismo enquanto retira gás carbônico (CO2) produzido
pelo metabolismo celular (AIRES, 1999).
Estudos realizados por Nonaka et al. (2008) e Silveira et al. (2004) analisaram,
respectivamente, a exposição ao veneno da serpente Crotalus durissus terrificus e Bothrops
jararaca ao processo de injúria pulmonar.
Nos últimos 30 (trinta) anos, houve um grande avanço na área de pesquisa com
toxinas e enzimas ofídicas, em virtude do advento das análises da estrutura bioquímica, em
especial dos venenos elapídicos, em virtude de sua maior letalidade. Contudo, novas
descobertas como das toxinas das cascavéis americanas e hipotensivas dos venenos botrópicos
geraram grande interesse para os estudos com a família Viperidae (EVANGELISTA, 2008).
O comprometimento pulmonar após envenenamento crotálico causa um quadro de
hipotensão e consequente hipoventilação, se instalando um processo de angústia respiratória,
cuja consequência será a perda do status de bem estar do animal.
Em vista disso, torna-se relevante estudar as alterações no sistema respiratório
após exposição do veneno da subespécie C. d. cascavella, buscando a um entendimento de
seus efeitos biológicos e seu comportamento agindo sob esse sistema.

15
2. REVISÃO DE LITERATURA
2.1. CLASSIFICAÇÃO DAS SERPENTES DA FAUNA BRASILEIRA
As serpentes estão classificadas no reino Animalia, filo Chordata, classe Reptilia,
subclasse Lepdosaura, ordem Squamata, subordem Serpentes. Esses animais estão presentes
praticamente em todo o mundo, constituindo o maior número de répteis viventes
(HICKMAM; ROBERTS; LARSON, 2004). Já foram descritas aproximadamente 2.300
espécies de serpentes (MELGAREJO-GIMÉNEZ, 2002). São animais vertebrados,
carnívoros, ectotérmicos e pertencem ao grupo dos répteis. (INSTITUTO BUTANTAN,
2014).
No Brasil estão presentes 75 gêneros agrupados em 10 famílias somando 371
espécies. Dessas, apenas duas famílias, Viperidae e Elapidae, são consideradas peçonhentas,
ou seja, espécies que possuem glândula produtora de veneno e aparelho inoculador, causando
intoxicações em humanos e animais (CARDOSO, 2003; BÉRNILS, 2010), constituindo um
grande risco em muitas regiões do mundo (NADUR-ANDRADE et al., 2012). O veneno
possui misturas complexas de toxinas e enzimas que mostram diferentes atividades nos
diversos sistemas biológicos, tais como citotóxica, hemorrágica, liberação debradicinina,
trombina-like, hemólise, efeitos cardiovasculares e hipotensores, necrose do tecido e efeitos
neurotóxicos (PATIÑO et al., 2013).
Nas Américas, a Família Elapidae é representada pelas serpentes corais. As 61
espécies reconhecidas estão divididas em três gêneros Micrurus com 57 espécies,
Leptomicrurus com três espécies e Micruroides com uma espécie. O gênero Micrurus ocorre
desde a Argentina até o sul dos Estados Unidos (Melgarejo, 2003). Na fauna brasileira há 22
espécies da Família Elapidae, a maioria pertencente ao gênero Micrurus (SBH, 2006).
A família Viperidae é composta pelos gêneros Bothrops, Bothropoides,
Bothriopsis, Bothrocophias, Rhinocerophis, Crotalus e Lachesis. Esses gêneros estão
distribuídos por todo o mundo e são responsáveis pelos principais acidentes ofídicos no
Brasil, tanto pela quantidade quanto pela severidade dos casos (AZEVEDO-MARQUES;
CUPO; HERING, 2003; QUEIROZ et al.,2008). O veneno contém um grande número de

16
proteínas farmacológica e bioquimicamente ativas, além de serem mais complexos que os
pertencentes à de outras famílias (DAL PAI; SANTO NETO, 1994).
O Brasil possui quatro gêneros de importância médica: Bothrops e Bothropoides,
ambos conhecidos genericamente como jararacas; Crotalus, que constituem as cascavéis; e
Lachesis, chamada popularmente de surucucus (BRASIL, 2012). A Figura 1 ilustra exemplos
de serpentes de importância clínica no Brasil.
Família Elapidae
Gênero Micrurus sp.
Família Viperidae
Gênero Bothropssp. Gênero Bothropoides sp.
Gênero Lachesis sp. Gênero Crotalus sp.
Figura 1. Famílias e gêneros de serpentes da fauna brasileira com importância médica. Fonte:
Adaptado de http://www.fiocruz.br
São utilizadas algumas características morfológicas para diferenciar serpentes
peçonhentas e não peçonhentas. O conhecimento dessas diferenças é de grande importância,

17
pois auxiliam a notificação correta dos casos, além de permitir à população identificar
possíveis riscos nas regiões onde habitam. (BRASIL, 2001).
De maneira geral, três características são analisadas para facilitar o
reconhecimento das serpentes da família Viperidae: cabeça triangular; presença de fosseta
loreal, um orifício localizado entre o olho e a narina do animal, com função termossensorial,
permitindo o movimento e a caça noturna (Figura 2a); presença de presas inoculadoras de
veneno bem desenvolvidas e móveis na região anterior do maxilar (Figura 2b); e estrutura da
cauda: Gêneros Bothrops e Bothropoides apresentam cauda lisa; Gênero Lachesis, cauda com
escamas eriçadas; e o gênero Crotalus, cauda com chocalho (Figura 2c) (BRASIL,2001).
a) b)
c)
Figura 2. Características morfológicas de reconhecimento de espécies de serpente da
Viperidae. Fonte: Adaptado de Brasil, 2001.
Legenda: a) Fosseta loreal; b) Presas inoculadoras de veneno; c) Caudas características dos gêneros Bothrops,
Bothropoides, Crotalus e Lachesis.

18
2.2. ASPECTOS BIOLÓGICOS DO GÊNERO CROTALUS
2.2.1 CLASSIFICAÇÃO E DISTRIBUIÇÃO GEOGRÁFICA
As subespécies da cascavel sul-americana, Crotalus durissus, são classificadas de
acordo com suas características morfológicas externas e distribuição geográfica, sendo
responsáveis pela alta morbidade e mortalidade representando um importante problema de
saúde pública (SANTORO et al., 1999; MARTINS et al., 2004).
No Brasil elas são representadas por apenas uma espécie, a Crotalus durissus, e
classificadas nas subespécies Crotalus durissus terrificus (encontrada nas regiões sul oriental
e meridional), Crotalus durissus collilineatus (distribuídas nas regiões secas do Centro-Oeste,
em Minas Gerais e no norte de São Paulo), Crotalus durissus cascavella (encontrada nas
áreas de caatinga da região Nordeste), Crotalus durissus ruruima (observada na região Norte),
Crotalus durissus marajoensis (observada na Ilha de Marajó). Alguns autores reconhecem
uma sexta subespécie no território brasileiro, a Crotalus durissus trigonicus, encontrada em
algumas regiões de Roraima e na savana de Rupununi na Guiana (PINHO; PEREIRA, 2001;
RATELSLANGEN, 2010; MELGAREJO, 2003; CAMILLO, 1998; AUTO, 1999).
Serpentes do gênero Crotalus são consideradas um bom modelo para estudos
relacionados à ecologia e variação geográfica, devido a sua ampla distribuição que abrange
diversos tipos de habitats, incluindo desertos, até ambientes alagados e florestais (NORMAN,
1994; BEAUPRE et al., 1998; PLACE; ABRAMSON, 2004).
O gênero é originário da América do Norte, mas se distribuindo também na
América Central e do Sul (ECHEVERRIGARAY et al, 2001). São conhecidas atualmente 30
espécies, sendo que a maior diversidade ocorre no México e nos Estados Unidos (ERNST,
1992; CAMPBELL; LAMAR, 1989).
A intensificação do processo de fragmentação de áreas florestais pode ter
facilitado àampliação na distribuição e o aumento na densidade populacional das serpentes
Crotalus durissus cascavella - cascavéis (BASTOS et al.,2005).

19
2.2.2. HABITAT
Serpentes do Gênero Crotalus são encontradas em áreas secas e pedregosas, de
baixa vegetação, sendo raramente encontradas em florestas (CAMPBELL; LAMAR, 1989;
SOERENSEN, 1990).
Apesar do número de estudos ter aumentado recentemente, ainda são escassos os
trabalhos de cunho ecológico baseados na observação de indivíduos em seu habitat natural.
Esse fato se deve em parte à dificuldade em encontrar serpentes na natureza, pois elas
apresentam densidades populacionais relativamente baixas e podem passar por longos
períodos de inatividade (SAZIMA, 1988; SECOR, 1994; BECK, 1995; OLIVEIR;
MARTINS, 2001). Apesar de estar associada preferencialmente às fitofisionomias abertas, há
registros de cascavéis em áreas com presença de mata (BASTOS et al., 2005).
2.2.3 HÁBITOS ALIMENTARES
A alimentação das serpentes Crotalus durissus compreende principalmente os
roedores, devido à grande disponibilidade ao longo do ano (SALOMÃO et al., 1995),
mamíferos não identificados, marsupiais e répteis, entretanto aves (VANZOLINI et al., 1980;
SALOMÃO et al., 1995) e lagartos teiídeos (Ameiva) podem também fazer parte da dieta
(ALMEIDA-SANTOS; GERMANO, 1996).
Aatividade de caça promovida pelas serpentes Crotalus durissus cascavella é
caracteristicamente noturna, período que coincide com o pico da atividade de suas presas,
apresentando uma estratégia de forrageamento caracterizada por permanecerem paradas
próximas a locais onde há indícios de suas presas (BURGHARDT, 1967; BROWN;
MACLEAN, 1983; REINERT et al., 1984; CHISZAR et al., 1990), aguardando por sua
passagem. O aumento da atividade de forrageio na estação seca pode estar associado a uma
maior necessidade de ingestão calórica nesse período (SALOMÃO; ALMEIDA-SANTOS,
2002).
2.3 ASPECTOS GERAIS E EPIDEMIOLÓGICOS DO GÊNERO CROTALUS
As serpentes possuem grande importância tanto na cadeia ecológica, como na área
biomédica. Pesquisas realizadas com os venenos por elas produzidos têm se intensificado,

20
uma vez que suas frações enzimático-proteicas vêm sendo usadas na fabricação de
medicamentos (SANTOS, 1995).
Destacando a anatomofisiologia da família Viperidae, estas serpentes apresentam
grande especialização no crânio, o que justifica a extrema mobilidade da maxila e outros
ossos da cabeça, conformação que favorece a efetividade nos ataques desses animais e
possibilita a inoculação do veneno durante seus botes. O veneno dessas serpentes é produzido
por duas glândulas especiais localizadas na cabeça, atrás e abaixo dos olhos, que são
glândulas salivares modificadas, onde a saliva é a própria toxina. Elas possuem reservas para
vários botes seguidos, sendo que, uma vez totalmente extraído o veneno ou secreção, as
glândulas só voltarão a estar totalmente cheias após duas semanas (SANTOS, 1995).
O veneno crotálico apresenta-se como um complexo tóxico-enzimático de várias
substâncias, dentre as enzimas podemos destacar fosfodiesterase, L-amino oxidase, 5-
nucleotidase e o fracionamento destas revela as seguintes toxinas: crotoxina, crotamina,
giroxina, convulxina e proteinases séricas, como crotalocitina, trombocitina e enzimas
trombina-like (identificadas como proteases séricas sendo capazes de degradar cadeias de
fibrinogênio, induzindo a agregação plaquetária) (RANGEL-SANTOS et al., 2004;
FONSECA, et al., 2006; SPINOSA, 2008).
Envenenamentos causados por animais peçonhentos constituem um grave
problema de saúde pública, devido a sua elevada incidência e morbimortalidade associada
(MORENO et al.,2005; PINHO;VIDAL;BURDMANN,2000; PINHO ;PEREIRA, 2001;
PINHO; OLIVEIRA; FALEIROS, 2004; PINHO;ZANETTA; BURDMANN, 2005)
principalmente na América Latina, África e Ásia (ANTUNES et al., 2010). Na Europa,
Estados Unidos e Canadá, os casos de envenenamento são raros. Na África, paralelamente, a
ocorrência desses acidentes é subnotificada, sendo desconhecida a incidência real. No
continente asiático, Paquistão, Índia e Birmânia são os países com maior frequência de casos
(CHIPPAUX,1998).
Estudos realizados sobre levantamentos epidemiológicos de acidentes ofídicos
notificados entre 1901 e 2000, observaram que o perfil de acidentes não se alterou durante
todo o século, acontecendo principalmente trabalhadores rurais do sexo masculino entre 15 e
49 anos, no início e final de cada ano (BOCHNER; STRUCHINER,2003).

21
A incidência mundial de acidentes ofídicos e sua gravidade são desconhecidas.
Estudos no ano de 2011 estimaram-se a ocorrência de 3 a 5,4 milhões de eventos por ano,
resultando em cerca de 125 a 150 mil mortes por ano (BARONE et al.,2011).
Sabe-se que os acidentes mais graves acontecem nos países pobres de regiões
tropicais e subtropicais. Entretanto, nesses países a notificação desses casos também é
subestimada, sendo por muitas vezes tratados com métodos ultrapassados e procedimentos
não efetivos (ALBUQUERQUE et al., 2004).
A sazonalidade dos acidentes ofídicos é muito variada nas diferentes regiões do
Brasil. Entretanto, há uma hipótese aceita de que o aumento dos casos esteja relacionado ao
período chuvoso e de altas temperaturas, em que as espécies residentes próximas às margens
de rios se desloquem a procura de terra firme, facilitando seu contato com o homem
(OLIVEIRA et al., 2010).
Dentre os países latinos, o Brasil ocupa a 1ª colocação em números de casos
(MINISTÉRIO DA SAÚDE, 2009), justificada pela sua grande extensão territorial. Foram
registrados 103.422 acidentes ofídicos, com 464 óbitos entre os anos de 2010 a 2013(SINAN
– Sistema de Informação de Agravos de Notificação, 2013),sendo segundo o Sinan, no ano de
2012, 19.946 casos de acidentes ofídicos em humanos no Brasil, sendo a família Viperidae
envolvida em 82,7% dos casos, e a família Elapidae, em 0,9% (BRASIL, 2012). Seguido pelo
Peru (4.500), Venezuela (2500 a 3 mil) e Colômbia (2675) (WARREL,2004).
No Brasil, os dados sobre os casos de envenenamento são coletados por meio de
alguns sistemas de notificação, alémdo Sistema de Informação de Agravos de Notificação
(SINAN/MS) há mais dois, o Sistema Nacional de Informações Tóxico-Farmacológicas da
Fundação Oswaldo Cruz (SINITOX/FIOCRUZ/MS) e o Sistema de Informações Hospitalares
do Sistema Único de Saúde (SIM/SUS). Apesar da existência desses programas, sabe-se que a
quantidade de casos é muito superior à quantidade registrada, devido, entre outros fatores, às
dificuldades de acesso aos serviços de saúde, principalmente no campo (LEMOS et al.,2009).
No Ceará, entre 2010 a 2013, foram notificados 2.328 ataques (SINAN, 2013).
Sendo em 2012, notificados 445 casos envolvendo picadas de serpentes peçonhentas, tendo
sido 68,1% atribuídos aos gêneros Bothrops sp. e Bothropoides sp., 8,3% ao gênero Crotalus
sp., 1,6% ao gênero Micrurus sp. e 0,9% ao gênero Lachesis sp. (BRASIL,2012).

22
2.4 CONSTITUINTES DO VENENO DE CROTALUS DURISSUS
2.4.1 CROTOXINA E SUAS SUBUNIDADES
A Crotoxina é o principal polipeptídeo presente no veneno crotálico e foi isolada
por Silva (1982) na subespécie Crotalus durissus terrificus, este autor destacou que este
componente correspondia a 68% do peso total do veneno. De Acordo com Hendon (1971) a
crotoxina (24 kDa) é um β-neurotoxina que tem duas subunidades, uma ácida, a crotapotina
(crotoxina A) e outra básica, a fosfolipase A2 (PLA2) (crotoxina B).
A crotapotina tem sua acidez justificada pela composição de aminoácidos: 12
resíduos de aspartato/asparagina e 14 de glutamato/glutamina versus 2 resíduos de lisina e 2
de arginina. A fosfolipase A2 crotálica contém 11 resíduos de aspartato/asparagina e 10 de
glutamato/glutamina versus 11 resíduos de lisina e 12 de arginina, caracterizando seu caráter
básico. A crotapotina e a fosfolipase A2 são ricas em cistina, contendo 7 e 8 pontes
dissulfídicas respectivamente. As duas subunidades formam a crotoxina, um complexo molar
de proporção 1:1 (BREITHAUPT et al., 1974; SPINOSA, 2008).
Variações interespecíficas e intraespecíficas foram descritas para o complexo
crotoxina. Variações intraespecíficas diferem na presença de diferentes isoformas das
subunidades A e B, as quais possuem potência e atividade biológica diferente (PEREAÑEZ et
al., 2009).
A crotapotina (crotoxina A) não possui a toxicidade e a atividade enzimática da
crotoxina, mas potencializa a sua toxicidade. Essa potencialização é explicada pelo fato de
que, quando associado ao componente B (PLA2), o componente A (crotamina) previne a
ligação da PLA2 a sítios de baixas especificidades e afinidade, direcionando a PLA2 a se ligar
a sítios de receptores de membrana (pós-sináptica) de alta especificidade e alta afinidade, o
que potencializa a toxicidade da crotoxina. Esses sítios de alta especificidade podem ser
receptores colinérgicos ou componentes da membrana fortemente associados a estes (BON et
al., 1997). Esses receptores foram denominados como tipo N (neuronal) e M (muscular). Os
receptores do tipo N foram os primeiros a serem identificados e estão presentes em grande
quantidade nas membranas de cérebro de ratos. Tais receptores ligam-se com alta afinidade às

23
PLA2s neurotóxicas. Os receptores do tipo M foram identificados em músculo esquelético de
coelhos (MATSUBARA, 2009).
A crotapotina demonstrou potencializar o edema inflamatório (induzido por
carragenina), possuir efeito antimicrobiano contra, principalmente, bactérias Gram-negativas
e aumento da pressão de perfusão renal e da resistência vascular renal em rins perfundidos
(sem, entretanto, alterar o fluxo urinário e a filtração glomerular, ao contrário da crotoxina)
(OLIVEIRA et al., 2003).
O veneno da C.d.cascavella contém pelo menos 4 isoformas de crotoxinas
formadas por diferentes combinações de crotapotinas com uma única isoforma de fosfolipase
A2 (PLA2). Ao contrário do veneno de Crotalus durissus terrificus, que contém 3 ou 4
isoformas de PLA2 (BEGHINI et al., 2000).
Os dados supracitados podem servir como indicio para sugerir as diferenças na
intensidade de atuação do veneno das diferentes subespécies.
A crotoxina é uma β-neurotoxina que possui um efeito tóxico bloqueador pós-
sináptico, diminuindo a ação da acetilcolina e impedindo a despolarização da membrana, e
pré-sináptico da liberação de acetilcolina, ou seja, ela bloqueia a transmissão neuromuscular,
impedindo a contração muscular estriada esquelética. Ela pode causar uma paralisia dos
músculos respiratórios, culminando na morte do envenenado. A crotoxina também aumenta a
permeabilidade de íons Na+ (BON et al., 1997; GOPALAKRISHNAKONE et al., 1984;
RANGEL-SANTOS et al., 2004; SPINOSA, 2008; MATSUBARA, 2009). O veneno da
Crotalus durrissus terrificus e a crotoxina B induzem a expressão protéica de cicloxigenase-2
pelas células endoteliais (MATSUBARA, 2009).
A superfamília de PLA2 consiste um amplo espectro de enzimas definidas pela
sua habilidade de catalisar a hidrólise da ligação do centro estérico (posição sn-2) de
fosfolipídios (SIX; DENNIS, 2000). Elas conferem à crotoxina uma ação miotóxica por
atuarem clivando fosfolipídios em sítios específicos de membranas de fibras musculares
estriadas esqueléticas sensíveis à PLA2, causando mionecrose, com perda de estriação das
fibras e degeneração hialina (4 a 6 horas após a injeção da crotoxina)
(GOPALAKRISHNAKONE et al., 1984).
As PLA2s induzem a um extravasamento de plasma, sendo capazes de aumentar a
permeabilidade microvascular da pele na região dorsal de ratos. Os mecanismos de formação

24
de edema induzido pelas PLA2s envolvem a ativação de fibras sensoriais C, as quais liberam
neuropeptídeos, como a substância P, que medeiam efeitos neurogênicos inflamatórios locais
(respostas vasoativas) em tecidos inervados e de mastócitos locais (pela substância P), os
quais liberam histamina e serotonina (CÂMARA et al., 2003).
Fonseca e colaboradores (2006) afirmaram então que a crotoxina apresenta duas
diferentes atividades enzimáticas (PLA2 e da protease de serina) e dois tipos diferentes de
ações biológicas (neurotóxica e coagulante).
2.4.2 CROTAMINA
A crotamina é um polipeptídeo básico, pertencente à família das miotoxinas
polipetitídicas básicas, as quais possuem a capacidade de penetração intracelular, através de
mecanismos independentes de gasto energético, por interação com proteoglicanos
(MATSUBARA, 2009).
O efeito miotóxico é causado pela crotamina, que apresenta atividade sinérgica à
da crotoxina, atua principalmente causando contrações musculares, dependentes da
despolarização da membrana das células musculares estriadas esqueléticas, através do
aumento da permeabilidade desta ao sódio. Esta ação provavelmente é exercida sobre os
canais de sódio, pela indução do influxo de cálcio (LOMONTE et al., 2003). Também foi
observado que a crotamina interage ativamente com as membranas lipídicas das células,
formando vacúolos e demonstrando uma atividade mionecrótica. Sua atividade é similar à da
alfa-toxina dos escorpiões (NICASTRO et al., 2003).
2.4.3 CONVULXINA
A convulxina é uma toxina de alto peso molecular, pertencente à família das
lectinas do tipo C, isolada de Crotalus durissus terrificus, Crotalus durissus cascavella e
Crotalus durissus collilineatus, consiste em um potente ativador plaquetário. De acordo com
Lima e colaboradores (2005) além de induzir a agregação plaquetária ao ligar-se ao receptor
GPV1 das plaquetas, ela causa convulsões, alterações respiratórias e circulatórias. É formada
por duas cadeias polipeptídicas (Cvxα e Cvxβ) covalentemente associadas em uma estrutura
trimérica (αβ)3. A indução plaquetária provocada por esta toxina consiste em uma reação
dependente de Ca+, iniciada pela ligação da convulxina à glicoproteína Ib (GPIb) (POLGAR

25
et al., 1997; JANDROT-PERRUS, 1997, KANAJI et al., 2003). Matsubara (2009) também
descreveu que esta toxina é capaz de agregar e lisar plaquetas, ligando-se com alta afinidade a
um pequeno número de sítios plaquetários, por mecanismo dependente de cálcio, fibrinogênio
e adenosina difosfato.
Leduc e Bon (1998) clonaram e sequenciaram as duas cadeias da convulxina de
Crotalus durissus terrificus. Neste estudo, observou-se uma alta similaridade entre as cadeias
α e β, com uma média de porcentagem de identidade de 74%. O sequenciamento das
subunidades resultou em cadeias compreendendo 158 e 148 resíduos de aminoácidos para
Cvxα e Cvxβ, respectivamente e um peptídeo sinal de 23 resíduos de aminoácidos idênticos
para as duas subunidades. Toyama e colaboradores (2001) isolaram e caracterizaram uma
proteína semelhante à convulxina da peçonha de Crotalus durissus collilineatus com alta
similaridade em relação à convulxina da Crotalus durissus terrificus.
2.4.4 GIROXINA
A giroxina pertence ao grupo do gênero trombina e foi isolado a partir do veneno
da serpente Crotalus durissus terrificus por Raw em 1986, por meio de três passos
cromatográficos de purificação (precipitação com sulfato de amônio, cromatografia em
Sephadex G-75 e Sepharose-1, 4-butanodiol diglycyl-p-aminobenzamidina). Após análise
eletroforética descobriu-se que a giroxina possuía um peso molecular de 34 kDa e 23 um pH
8,0, ótimo para a coagulação do sangue humano (RAW et al., 1986).
A giroxina converte o fibrinogênio em fibrina e assim aumenta o tempo de
coagulação do sangue (SANO-MARTINS et al., 2001). Além de atuar na coagulação, a
giroxina apresenta uma excelente atividade neurotóxica (BARROS et al., 2011). Fonseca e
colaboradores (2006) descreveram a giroxina apresentando duas frações principais: atividade
de trombina-like e L-amino-oxidase (LAO). A LAO é a única oxidase dependente de FAD
(Flavina adenina dinucleotídeo) encontrada em veneno de serpentes e sua toxicidade,
possivelmente, envolve a geração de peróxido de hidrogênio, produto final da oxidação de L-
aminoácidos. A LAO também induz a agregação plaquetária, envolvendo a ativação da PLA2
(TOYAMA et al., 2006).
Em estudos mais recentes verificaram que a giroxina demonstrou um elevado
nível de atividade coagulante, com uma dose mínima entre 0,015 a 0,037μg/μL (BARROS et
al.,2011).

26
Outras atividades da giroxina são a amidásica e a esterásica (MARTINS et al.,
2002). Prado-Franceschi (1990) sugere que esta toxina produz uma síndrome convulsiva
peculiar em camundongos, caracterizada por movimentos rápidos de rotação do corpo em
torno de seu eixo longitudinal.
Estudos posteriores confirmaram sua ação no sistema nervoso central, causando a
síndrome labiríntica em camundongos, corroborando os achados anteriores (MATSUBARA,
2009). O estudo desta enzima poderá servir como um modelo molecular interessante para o
desenvolvimento de novos fármacos ou agentes terapêuticos (MORAES et al., 2004;
FERRARO et al.,2005), principalmente devido a sua resistência aos inibidores de proteinases
fisiológicas (MATSUI et al., 2000;SERRANO et al., 2005).
2.4.5 PEPTÍDIO NATRIURÉTICO
Outro componente presente no veneno é o peptídio natriurético que ao ser isolado
da Crotalus durissus cascavella foi capaz de aumentar a pressão de perfusão e a resistência
vascular renal em rins perfundidos de ratos. Este componente demonstrou ser capaz de
aumentar o fluxo urinário e a filtração tubular renal e diminuir o transporte tubular de sódio e
a pressão arterial sistêmica. EVANGELISTA et al. (2008) observaram que este componente
do veneno crotálico apresentou efeitos renais e vasculares, com consequente efeito diurético e
hipotensor.
2.5 SISTEMA RESPIRATÓRIO
2.5.1 CARACTERIZAÇÃO MORFOFUNCIONAL
A traqueia divide-se em brônquios principais direito e esquerdo, os quais se
seguem dividindo em brônquios lobares e segmentares, bronquíolos terminais e respiratórios,
ductos alveolares, sacos alveolares e alvéolos. A região compreendida entre a boca ou nariz
até os bronquíolos terminais, é denominada de zona de condução, sua função é condicionar o
ar, removendo material particulado, ajustando a umidade e a temperatura, e o conduzir até os
pulmões. O epitélio de superfície dessa zona é composto por células ciliadas e secretoras, que

27
repousam sobre o tecido subepitelial, que consiste basicamente de tecido conjuntivo e
glândulas, como estas vias aéreas não contêm alvéolos e não participam das trocas gasosas,
elas constituem o espaço morto anatômico, com volume de cerca de 150 mL, em humanos. Já
a região compreendida pelos bronquíolos respiratórios, ductos alveolares e alvéolos, é
denominada de zona respiratória, por ser a região que participa das trocas gasosas (WEST,
2010; FRASER, 2005).
A traqueia, os brônquios e os bronquíolos terminais e respiratórios que não
contêm alvéolos em suas paredes, juntamente com artérias, veias, vasos linfáticos, nervos,
septo interlobular e pleura que os acompanham, formam a porção não parenquimatosa do
pulmão. Os alvéolos da zona de transição juntamente com os sacos alveolares mais distais
constituem o parênquima pulmonar propriamente dito. Há uma estimativa de que 87% do
volume pulmonar total é composto por alvéolos, o que em números equivale a cerca de 500
milhões, com área de superfície de trocas gasosas em torno de 85 m2, em humanos (WEST,
2010; FRASER, 2005).
2.5.2 TROCA GASOSA
Enquanto as células dos animais vivos liberam continuamente dióxido de carbono
e outros produtos residuais, requerem um suprimento contínuo de oxigênio e nutrientes
(BATES, 2009).
A difusão através dos tecidos é mais bem descrita pela lei de Fick, a qual afirma
que a velocidade de transferência de um gás através de uma lâmina de tecido é proporcional a
área do tecido e a diferença de pressão parcial entre os dois lados e inversamente proporcional
à espessura do tecido. A área da troca, a espessura da mesma, a solubilidade dos gases nos
tecidos, o peso molecular dos gases e a diferença de pressão entre os dois lados da membrana
de troca, são fatores que determinam a velocidade da difusão (ZIN; ROCCO, 2008).
O principal sítio de trocas gasosas a nível pulmonar é o alvéolo. Os alvéolos são
delimitados por estruturas que, vistas em corte, se assemelham a hexágonos, como observado
na Figura 3. Essa estrutura é composta por uma camada contínua de células epiteliais ao redor
de um fino interstício. Em três dimensões há alguns sólidos geométricos que se aproximam da
forma alveolar, como o dodecaedro regular.

28
Figura 3 – Fotomicrografia da parede alveolar ao final da inspiração, Aumento 400X e
coloração H.E.(Adaptado de Pavone et al., 2007).
FONTE: PAVONE, L.; ALBERT, S.; DIROCCO, J.; GATTO, L.; NIEMAN, G. Alveolar instability caused by
mechanical ventilation initially damages the nondependent normal lung. Critical Care. v. 11, p. 1-10, 2007.
A superfície alveolar é constituída principalmente por dois tipos de células:
pneumócitos tipo I e II; macrófagos alveolares também estão presentes na superfície epitelial.
O interstício contém os capilares envolvidos na troca gasosa, tecido conjuntivo e uma grande
variedade de células responsáveis por manter a forma do alvéolo e as defesas a nível alveolar
(FRASER, 2005).
O pneumócito tipo I ou célula alveolar escamosa recobre 95 % da superfície
alveolar, e apresenta pouca organela citoplasmática. As células tipo I são unidas por ligações
firmes que produzem uma barreira contra a difusão de fluidos e substâncias solúveis em água
para o interior do alvéolo (FRASER, 2005; ZIN; ROCCO; FAFFE, 2008).
O pneumócito tipo II ou célula alveolar granular tende a se localizar próximo às
junções dos alvéolos, contém muitas organelas celulares, como o retículo endoplasmático
rugoso, mitocôndrias, complexo de Golgi e numerosos grânulos eosinofílicos (corpúsculos
lamelares) que armazenam e secretam surfactante. O surfactante, que recobre a superfície
alveolar reduzindo a tensão superficial, é composto principalmente por fosfolipídios, tais
como, fosfatidilcolina saturada e insaturada, fosfatidilglicerol, fosfolípides:
fostatidiletanolamina, fosfatidilinisitol e ácido fosfatídico liso-bis, lípides neutros:
diacilglicerol e esfingomielina, colesterol. (POSSMAYER et al,1984;) e por proteínas tais
como: proteína A (SP-A), proteína B (SP-B), proteína C (SP-C) e proteína D (SP-D). Um
pequeno número de células tipo II é mitoticamente ativado e capaz de se diferenciar em
células tipo I (ZIN; ROCCO; FAFFE, 2008).

29
As membranas basais endoteliais e epiteliais são separadas por um espaço
intersticial de extensão variável que contém tecidos conectivos, como as fibras elásticas e
colágenas e uma variedade de células como os miofibroblastos e células intersticiais
contráteis. As zonas de transporte e de transição juntamente com artérias, veias, vasos
linfáticos, nervos, septo interlobular e pleura, constituem a porção não parenquimal do
pulmão. Os alvéolos das vias aéreas de transição associados com os sacos alveolares mais
distais constituem o parênquima pulmonar (FRASER, 2005).
A membrana alvéolo-capilar funciona como uma barreira entre os espaços aéreos
e o sangue. Ela é composta pelo líquido que banha os alvéolos, epitélio alveolar, membrana
basal do epitélio, estroma alveolar, membrana basal do endotélio e endotélio capilar, além
disso, o oxigênio precisa chegar à molécula de hemoglobina no interior da hemácia, logo
podemos acrescentar a barreira alvéolo capilar (o plasma, a membrana da hemácia e seu
estroma). Em alguns locais esse espaço atinge uma espessura de 0,4 μm a 0,5 μm e essa é a
principal região onde ocorrem as trocas gasosas. Nas demais regiões, a espessura varia devido
à presença de tecido conectivo (incluindo fibras de elastina e colágeno) e de uma variedade de
células, predominantemente os miofibroblastos (ZIN; ROCCO, 2008; FRASER, 2005).
2.5.3 REDE DE FIBRAS DO PARÊNQUIMA PULMONAR
Em um pulmão normal, as proteínas da matriz extracelular (MEC) são usualmente
secretadas pelo fibroblasto, e organizadas em uma rede nos espaços ao redor das células. A
MEC ocupa um volume significativo no tecido e possui informações que orientam a
migração, ligação, diferenciação e organização das células, modulando, assim, uma série de
processos (RAGHOW, 1994, JUNQUEIRA; CARNEIRO, 2008). Três grupos de
macromoléculas associam-se fisicamente para formar a matriz extracelular: as proteínas
estruturais fibrosas, como as fibras de colágeno e elastina; glicoproteínas e proteoglicanos.
Essas moléculas organizam-se em duas porções do tecido, a matriz intersticial e a membrana
basal (COTRAN et al., 1999).

30
2.5.4 FIBRAS DE COLÁGENO E ELASTINA
As fibras de colágeno são os principais constituintes da MEC. Existem vários
tipos de colágeno, sendo que os pulmões contem principalmente o colágeno tipo I e III,
envolvidos na estruturação das paredes alveolares. Sua organização se dá formando uma rede
de fibras central, que se estende desde as vias aéreas centrais aos ductos alveolares, uma
periférica partindo da pleura visceral, e um parênquima intersticial conectando as duas (SUKI
et al., 1994).
O colágeno tipo I é a principal proteína estrutural do interstício pulmonar e é
produzido principalmente durante o desenvolvimento do pulmão e nas reações fibróticas
(GOLDSTEIN, 1991), sendo adaptado para resistir às tensões e apresenta ligações covalentes
fortes entre suas fibrilas (FUNG, 1993). As fibras colágenas são mais rígidas e fortes que
qualquer outro constituinte da MEC, consequentemente, a quantidade e organização do
colágeno dentro da parede alveolar têm papel fundamental na determinação da função
pulmonar (YUAN et al, 2000).
As fibras elásticas são compostas de elastina e microfibrilas, e são sintetizadas
pelos fibroblastos e miofibroblastos e células musculares lisas do pulmão (FUNG, 1993). Elas
apresentam uma heterogeneidade estrutural e estão mecanicamente ligadas ao colágeno
através das microfibrilas e/ou proteoglicanos. O papel das microfibrilas na elasticidade do
pulmão, entretanto, ainda não foi estudado. A elastina é uma proteína hidrofóbica que se
agrega a filamentos e lâminas por ligações cruzadas, e é comumente apontada como
responsável pela elasticidade pulmonar durante a respiração em volume corrente (SUKI et al.,
1994). Entretanto, um estudo realizado comparando os efeitos da digestão de fibras de
colágeno e elastina em equações constitutivas em tiras de parênquima mostrou que o colágeno
pode ser tão importante quanto à elastina mesmo em baixos volumes (YUAN et al., 2000).
2.5.5 GLICOPROTEÍNAS
As glicoproteínas são proteínas cuja principal função é a capacidade de se ligar a
outros componentes da MEC e a proteínas específicas integrantes da membrana celular. As
principais proteínas de adesão são a fibronectina e a laminina. A fibronectina é produzida por

31
fibroblastos, monócitos, células endoteliais e outras células e sua principal função é a de
promover a ligação entre os diversos componentes da MEC, incluindo colágeno, fibrina,
proteoglicanos e às células. A laminina é a glicoproteína mais abundante nas membranas
basais e atravessa a lâmina basal, ligando-se a receptores específicos sobre a superfície das
células e componentes da MEC (SNYDER et al, 2000).
2.5.6 PROTEOGLICANOS
As moléculas de proteoglicanos formam, no tecido conjuntivo, uma substância
gelatinosa e hidratada onde se encontram as proteínas fibrosas. Os proteoglicanos fornecem
suporte mecânico ao tecido, permitindo a difusão de moléculas hidrossolúveis e migração
celular. Influenciam a formação da fibra de colágeno e interagem com várias citocinas e
fatores de crescimento (MISEROCCHI et al., 2001). Ao nível dos tecidos, encontram-se
frequentemente ligados às fibras colágenas, fibras elásticas e fibronectina, participando da
organização da MEC (ALBERTS et al., 1994). Um novo papel importante para os
proteoglicanos é o de estabilizador da rede de fibras de colágenas e elásticas, contribuindo
para a elasticidade tecidual pulmonar e estabilização em pequenos e médios volumes
(CAVALCANTE et al., 2005).
2.6 MECÂNICA PULMONAR
A difusão do ar segue a um gradiente pressórico, determinando um fluxo de ar do
local de maior pressão para o local de menor pressão. Para que isso ocorra durante a
inspiração, a pressão alveolar (Palv) tem que ser inferior à pressão atmosférica (Patm), esta
diferença de pressão é gerada pela contração dos músculos inspiratórios (ex. diafragma), os
quais aumentam a negatividade da pressão intrapleural (Ppl) distendendo as vias aéreas e os
alvéolos. Os músculos respiratórios agem de forma rítmica na parede torácica e sua ação
destina-se a vencer as impedâncias mecânicas impostas pelo sistema respiratório. Esta
impedância se refere aos componentes elásticos dos pulmões e da parede torácica, e
resistência das vias aéreas e ao atrito dos tecidos. A mecânica do sistema respiratório é
subdividida em dois sistemas elásticos: os pulmões e a parede torácica. (BATES,2009; ZIN;
ROCCO, 2009; WEST, 2010).
2.6.1 PROPRIEDADES ELÁSTICAS DO PULMÃO
A Elastância (E) é a medida do quão difícil é distender o tecido pulmonar, a fim
de se aumentar o volume que lhe rodeia, e seu valor está relacionado com a composição do

32
tecido pulmonar, a tensão superficial (T), as propriedades da parede torácica e a organização
estrutural dos constituintes do tecido. Desta maneira, podemos considerar que a pressão
associada à retração elástica (Pel, P) cresce linearmente com o volume pulmonar, de modo que
E seja a constante de proporcionalidade. (BATES, 2009; ZIN; ROCCO, 2009). É importante
ressaltar que na prática clínica é mais usual o emprego da complacência pulmonar, dada pelo
recíproco de E. Complacência pulmonar (CP) é a mudança de volume por unidade de
alteração de pressão (WEST, 2010).
Segundo a Lei de Laplace aplicada a uma superfície esférica, a pressão relaciona-
se com o raio e com a tensão superficial (T) de acordo com a equação: P = 4T/r, onde o
número quatro está associado a duas interfaces ar-líquido (interna e externa). Entretanto,
quando relacionamos esta Lei a um alvéolo esférico revestido por líquido na sua face interna,
apenas uma interface está envolvida e o numerador na Lei de Laplace tem o número 2 no
lugar de 4 (BATES, 2009;MACHADO; ZIN, 2009). De acordo com essa Lei, era de se
esperar que a tensão superficial fosse crescente à medida que os alvéolos fossem tornando-se
menores, acarretando colapsos e hiperdistensões dos espaços aéreos, entretanto, os
pneumócitos tipo II secretam o surfactante, um líquido de composição proteica e
principalmente fosfolipídios, que diminui a tensão superficial no alvéolo na medida em que
sua área diminui, evitando assim o colapso alveolar durante a expiração (ZIN; ROCCO,
2009).
2.6.2 PROPRIEDADES ELÁSTICAS DA PAREDE TORÁCICA
Da mesma forma que o pulmão possui suas propriedades elásticas intrínsecas,
assim também é a parede torácica, sendo composta pelo arcabouço torácico, o diafragma, a
parede abdominal e o mediastino. A parede torácica tende a expansão até que se consiga
atingir seu equilíbrio elástico (cerca de 75% da capacidade vital), a partir desse ponto tenderá
a retração, agindo de forma contrária a ventilação pulmonar em volumes acima desse
equilíbrio (MACHADO; ZIN, 2009).
Em indivíduos hígidos, pulmão e parede torácica movimentam-se em conjunto e
estas estruturas são separadas pelo espaço pleural. A pressão intrapleural (PPL), é a pressão no
interior do espaço pleural, e é determinada pela interação mecânica entre os pulmões e a
parede torácica. Ao final da expiração, as pressões de recolhimento elástico dos pulmões e da
parede torácica são iguais, e as forças associadas a essas pressões produzem efeitos

33
concorrentes: expansão da caixa torácica e redução do volume pulmonar, o que faz com que a
Ppl seja negativa, em torno de 3 cmH2O abaixo da pressão atmosférica. No momento da
inspiração, essa pressão torna-se mais negativa, retornando ao seu valor inicial no final da
expiração. A pressão alveolar (PAL), por sua vez, é negativa na inspiração e positiva na
expiração em relação à pressão na abertura das vias aéreas (Pao) (BERNE et al, 2004;
MACHADO; ZIN, 2009).
2.6.3 PROPRIEDADES RESISTIVAS DO SISTEMA RESPIRATÓRIO
A resistência pulmonar pode ser subdividida em dois componentes: a resistência
das vias aéreas (Raw), que influencia o fluxo de ar no interior dos pulmões, e a resistência
tecidual (Rtis), que é determinada pelas perdas energéticas geradas pela viscosidade (atrito)
pertinente à movimentação do pulmão (BATES, 2009; MACHADO; ZIN, 2009).
2.6.4 RESISTÊNCIA PULMONAR
A resistência pulmonar (RL) é obtida pela soma da resistência tecidual (Rtis) e a
resistência das vias aéreas (Raw) (WEST, 2010).
Quando o tecido pulmonar é deformado, como ocorre durante as mudanças de
volume, as fibras, células e fluidos que compõem o tecido pulmonar movem-se uns em
relação aos outros sob a ação de forças de atrito, gerando perdas energéticas, determinando a
resistência tecidual. Esse componente da resistência corresponde a cerca de 20% da
resistência pulmonar (BATES, 2009; ZIN; ROCCO; FAFFE, 2008).
Em uma abordagem simplificada e com a finalidade de descrever o escoamento de
ar ao longo das vias aéreas, estas podem ser consideradas uma série de tubos ramificados, os
quais se tornam mais estreitos, curtos e numerosos, à medida que penetram mais
profundamente nos pulmões. A resistência das vias aéreas pode ser quantificada pela
diferença de pressão entre os alvéolos e a boca, dividida pelo fluxo. Esta pode ser influenciada
pela geometria da árvore traqueobrônquica, pelo volume pulmonar, pela complacência das
vias aéreas, pela densidade e viscosidade do gás inspirado e pela musculatura lisa dos
brônquios. Baseando-se na equação de Poiseuille, seria natural pensar que a resistência das
vias aéreas tenderia a crescer dos bronquíolos (pequeno calibre) em direção aos grandes
brônquios, contudo devido ao aumento da secção transversal à medida que as vias aéreas vão

34
se tornando menores, esta resistência é maior nos brônquios de tamanho médio e muito
pequena nos bronquíolos (WEST, 2010; MACHADO; ZIN, 2009).
2.6.5 RESISTÊNCIA DA PAREDE TORÁCICA
A resistência da parede torácica é determinada pelo atrito que ocorre entre os
constituintes (caixa torácica óssea, os músculos da caixa torácica, o diafragma e o abdome) do
tecido, entre as duas pleuras e nas articulações (MACHADO; ZIN, 2009).
2.6.6 VISCOELASTICIDADE
Além dos componentes elásticos e resistivos, o sistema respiratório apresenta
propriedades viscoelásticas, que atuam no tecido pulmonar e na parede torácica (BATES et
al., 1989).
A viscoelasticidade pode ser demonstrada com materiais (ex. fios de seda) que
obedecem à lei da proporcionalidade entre a força aplicada e o alongamento resultante (Lei de
Hooke), porém, apenas por um curto período de tempo após a aplicação da força. Quando esta
força for aplicada por um período prolongado, o alongamento passa a aumentar
continuamente. Substâncias viscoelásticas quando mantidas sob deformação constante,
apresentam o chamado relaxamento de tensão (“stress relaxation”) que é uma queda de
tensão, quando o corpo é estirado. Em contrapartida, sob uma tensão constante, o corpo tende
a se deformar continuamente com o decorrer do tempo, comportamento conhecido como
“creep”. Do ponto de vista morfofuncional, a viscoelasticidade ocorre no tecido pulmonar e
na parede torácica e permite a troca de energia (pressão) entre o componente elástico e
resistivo (KOCHI, 1988).
2.6.7 DISSIPAÇÃO DE ENERGIA DURANTE O CICLO RESPIRATÓRIO
A elasticidade é uma propriedade da matéria que permite ao corporetornar à sua
forma original após sofrer deformação por uma força aplicada sobre ele. Os tecidos dos
pulmões e do tórax possuem propriedades elásticas e obedecem, dentro de certas condições, à
lei de Hooke. De acordo com essa lei, um corpo é perfeitamente elástico quando, ao sofrer
uma força distensora se deforma proporcionalmente, ou seja, a variação de comprimento (ou

35
volume) é diretamente proporcional à força (ou pressão) aplicada até que se atinja seu limite
elástico. Desta forma, quanto maior for a pressão gerada pelos músculos inspiratórios, maior
será o volume inspirado e a energia acumulada no sistema. Parte da energia gasta na
inspiração é dissipada como calor, mas parte dela é armazenada como energia potencial
elástica pelas estruturas que sofrem o estiramento, essa energia armazenada é então gasta para
realizar a expiração em volume corrente (MACHADO; ZIN, 2009; WEST, 2010; BATES,
2009).
A curva pressão-volume (PV) no sistema respiratório é obtida através de um ciclo
respiratório, sendo que o ramo da curva inspiratória não coincide com o da curva expiratória,
estes juntos formarão um loop, efeito conhecido como histerese pulmonar. Na curva PV
obtida em regime quase estático, condição em que se pode considerar desprezível as perdas
energéticas devido à viscosidade do ar, é possível quantificar a dissipação de energia sem a
contribuição da resistência das vias aéreas (APPADU; HANNING, 2003). Nessa curvaPV o
trabalho realizado durante a inspiração (trabalho positivo) é dado pela área entre o ramo
inspiratório e o eixo dos volumes, e o trabalho realizado durante a expiração é dado pela área
correspondente no ramo expiratório (trabalho negativo) e o eixo dos volumes.
Considerando o estado final do sistema, definido pelos valores de pressão e
volume ao final da expiração, como sendo igual ao estado inicial, à soma do trabalho
inspiratório e expiratório é igual à diferença entre as duas áreas acima mencionadas, ou seja, é
igual à área circunscrita pelo loop PV. Um fator importante que reduz a dissipação de energia
durante o ciclo respiratório é a camada de surfactante presente no alvéolo. Então, em
patologias onde há alteração nas propriedades ou na quantidade de surfactante, espera-se
encontrar correspondente alteração na área do loop PV (APPADU; HANNING, 2003).
2.7 MODELAGEM EM MECÂNICA PULMONAR
Esse assunto é referenciado quase que totalmente por Bates (2009).
Do ponto de vista mecânico a grande maioria dos eventos queinfluenciam a
função pulmonar ocorre no seu interior, impossibilitando a sua observação direta, deixando-
nos a intuir o que está acontecendo em seu interior a partir das relações dinâmicas observadas
entre as variáveis que podemos mensurar experimentalmente. Para isso são utilizados
modelos matemáticos. Estes nada maissão que um conjunto de equações que servem como
uma indicação precisa de nossas suposições sobre o funcionamento da mecânica pulmonar e
como um meio de se explorar as consequências dessas suposições.

36
Os testes de função pulmonar são uma ferramenta indispensável parafornecer uma
relação entre as mudanças funcionais e estruturais que ocorrem no pulmão. A resistência e a
elastância pulmonar têm sido por muito tempo as variáveis tradicionais, nos modelos animais,
para avaliar a função pulmonar, apesar de haver uma compreensão limitada da relação entre
esses parâmetros e as características estruturais do órgão.
Um exemplo de modelagem bastante simples é o chamado modelo linearde
compartimento único, constituído por um balão elástico selado ao final de um tubo, o balão
(com elastância E) representando o tecido pulmonar e o tubo (com resistência R) representa as
vias aéreas pulmonares. A equação do movimento que descreve este modelo é:
P (t) = RV’ (t) + EV (t) + P0 (1)
Onde P (t) é a pressão na entrada do modelo, V’ (t) é o fluxo de gás pelo tubo do
modelo, V (t) é o volume de gás no compartimento elástico, P0é a pressão restante aplicada e t
é o tempo.
A representação do sistema respiratório por um compartimentoúnico
uniformemente ventilado é considerada adequada desde que o fluxo fornecido ao sistema
esteja limitado a uma única frequência. Por mais que este modelo incorpore o fundamental
para o entendimento do processo da ventilação, o pulmão é muito mais complicado do que
este simples modelo nos oferece. Se o fluxo fornecido ao sistema apresentar diversas
frequências, o sistema respiratório passa a responder como sendo composto por diversos
componentes distintos, cujas contribuições relativas para o comportamento do sistema
respiratório variam com a frequência.
Com isso, podemos inferir que o ar não irá se distribuir da mesma forma, nem
aomesmo tempo para todas as unidades constituintes do pulmão. A modelagem da mecânica
do sistema respiratório que utilizaremos neste trabalho é conhecida como modelagem inversa.
Nessa abordagem considera-se que a imposição de um fluxo de ar V’ (t) na entrada das vias
aéreas corresponde à existência de uma pressão na entrada dessas vias, Pao(t). No contexto da
teoria de Sistemas Lineares, diz-se que o sistema recebeu uma entrada, dada por V’ (t), e em
resposta a ela produziu uma saída, Pao(t).
Do ponto de vista mecânico, a relação entre essa entrada e a saídadepende dos
processos físicos que ocorrem no sistema. A descrição desses processos envolve o
conhecimento das propriedades mecânicas dos constituintes internos do sistema, bem como
das interações entre esses constituintes. No sistema respiratório podem ser citados alguns

37
constituintes, como por exemplo, a rede de fibras elásticas e de colágeno, a interface ar-
líquido recobrindo alvéolos, dentre outros. As interações entre esses constituintes podem ser
exemplificadas pela transmissão das forças geradas pela tensão superficial às estruturas
sólidas do pulmão através das redes de fibras.
As propriedades mecânicas dos constituintes dificilmente são conhecidascom
precisão absoluta, e com ainda menor precisão são conhecidas às inúmeras e complexas
interações entre eles. Para contornar essas dificuldades, uma abordagem possível consiste em
identificar os constituintes com papel predominante e determinar suas propriedades e
principais interações, e buscar uma relação entre entrada (V’) e saída (Pao). Essa relação
muitas vezes é expressa por meio de equações diferenciais em que as derivadas são em
relação ao tempo. Nessas situações diz-se que essas relações matemáticas são as equações de
movimento do sistema, geralmente obtidas das leis de Newton.
Conforme foi descrito previamente, o sistema respiratório tem propriedades
elásticas e resistivas interdependentes e bastante complexas. Em mecânica Newtoniana é
muito comum abordar essa malha de interações, que são distribuídas espacialmente ao longo
de todo o sistema respiratório, por meio de descrições que agrupam as características elásticas
em um ou poucos componentes fictícios, nomeadas neste contexto como molas. O mesmo é
comumente feito para os componentes resistivos do sistema, utilizando um único componente
hipotético que tem a propriedade de dissipar energia mecânica, e cujas características
representem as propriedades combinadas dos inúmeros constituintes teciduais.
Esse constituinte dissipativo será aqui chamado de amortecedor.Para exemplificar,
a figura 4 (A) mostra um dos modelos comumente utilizados para descrever o sistema
respiratório, conhecido como modelo viscoelástico. Nessa abordagem as vias respiratórias são
representadas por um único duto cilíndrico de paredes rígidas com resistência R, que
representa o efeito combinado das resistências de todas as vias aéreas. Nessa figura o sentido
do fluxo é indicado pela seta tracejada, o que significa uma inspiração. Ao longo da via aérea
são representados os níveis de pressão alta por preenchimentos densos e baixos por pontos
rarefeitos. As regiões de trocas gasosas são representadas por uma câmara cilíndrica de
paredes rígidas e com base móvel, possibilitando variações no volume.
Note que esse modelo supõe uma distribuição perfeitamente homogênea dos gases
no pulmão. A base da câmara é conectada a uma mola, cuja ação se opõe a aumento de
volume, gerando uma pressão de recuo proporcional ao volume, medido a partir da
capacidade residual funcional. Essa constante de proporcionalidade é ET, que representa o
efeito combinado de todos os elementos elásticos do sistema respiratório. Atuando

38
mecanicamente em paralelo à mola há um amortecedor, que atuando na base do cilindro
produz uma pressão proporcional à variação do volume da câmara, supostamente igual ao
fluxo V ’. Na realidade essas quantidades (V ’ e variação do volume da câmara) são
consideradas iguais porque desprezamos a compressibilidade do ar dentro do pulmão, dentre
outros fatores. A constante de proporcionalidade entre a pressão gerada pelo amortecedor e o
fluxo é a resistência tecidual, RT, que representa o efeito combinado dos elementos resistivos
de todos os constituintes teciduais do sistema.
Uma vez estabelecido o modelo de trabalho, procede-se à estimativa dos
parâmetros do dado modelo. Com o animal conectado a um ventilador mecânico, impõe-se a
aplicação de um fluxo V’experimental ( t ) à abertura das vias aéreas e mede-se a pressão
correspondente, Pexperimental (t). Em seguida faz-se o ajuste das constantes ET e RT, de
modo que os valores de V ’(t) e PAO (t) calculados a partir das equações de movimento sejam
mais próximos possíveis doscorrespondentes valores experimentais.
Os modelos usualmente ligam as estruturas do sistema respiratório àssuas
respectivas funções. Os valores dos parâmetros do modelo devem refletir as alterações nas
estruturas do sistema associadas a patologias ou intervenções com drogas ou manobras
respiratórias. Verifica-se, por exemplo, aumento de R com o aparecimento de
broncoconstricções.
Para uma dada frequência de respiração e em pulmões sadios ou comleve grau de
injúria, esse modelo viscoelástico consegue descrever razoavelmente bem algumas
propriedades do sistema respiratório. Entretanto, há nuances presentes no sistema respiratório
que não conseguem ser descritas por esse modelo, como por exemplo, a diminuição da
resistência do sistema respiratório com o aumento da frequência respiratória.
Há muitos modelos propostos que, em melhor grau, respondem a um ou outro
aspecto da mecânica respiratória. Dentre eles, o modelo de fase constante é o mais difundido,
particularmente por descrever bem a dependência das propriedades elásticas com a frequência
e por separar, com apenas quatro parâmetros, as contribuições dos tecidos e das vias aéreas na
resistência do sistema respiratório.
Este modelo difere do modelo viscoelástico apenas pela substituição
doselementos representativos dos tecidos, ou seja, a mola e o amortecedor, conforme a Figura
4. Outra diferença é que as variáveis pressão e fluxo são tratadas no espaço das frequências.
Isso é feito a partir da transformada de Fourrier das funções V ’(t) e PAO (t), conforme as
equações abaixo:

39
No espaço das frequências, as transformadas de Fourrier da pressão e do fluxo são
relacionadas por uma simples relação de proporcionalidade, onde o fator de proporcionalidade
é uma função chamada de impedância,
Z (f): P (f) = Z (f) V’ (f) (4)
Z(f) nesse caso é chamada de impedância de entrada porque relaciona duas
quantidades medidas na entrada do pulmão.
Figura 4 - Exemplos de modelos utilizados para descrever o sistema respiratório: (A)
Modeloviscoelástico; (B) Modelo de fase constante.

40
2.7.1 O MODELO DE FASE CONSTANTE
O modelo de fase constante é um modelo avançado de mecânicapulmonar que
oferece uma distinção entre a mecânica respiratória central e periférica. Tem sido o modelo de
escolha para descrever a impedância de entrada do sistema respiratório (Zrs), que expressa
separadamente os efeitos de resistência de vias aéreas, resistência e elastância tecidual e
inertância dos gases respirados.
Para extração desses parâmetros faz-se o uso de ventilação mecânica a
frequênciaspré-definidas. O modelo de fase constante descreve a impedância utilizando a
seguinte equação:
onde RN é a resistência Newtoniana, que representa a resistência ao fluxo de ar nas vias aéreas
condutoras, e I representa a inertância do gás nas vias aéreas centrais. No modelo acima, G
reflete as propriedades dissipativas do tecido pulmonar (resistência tecidual), sendo chamado
comumente de amortecimento tecidual; H reflete o caráter elástico do tecido (elastância); f
representa a frequência, i é a raiz quadrada de -1 e α é dado por:
𝛼 =2
𝜋𝑡𝑎𝑛−1 (
𝐻
𝐺)
Outra característica importante do modelo de fase constante, além dapossibilidade
de separar as contribuições das vias aéreas e dos tecidos, diz respeito à razãoη = G/H,
conhecida como histeresividade. Duas nuances desse parâmetro merecem destaque.
A primeira é que, considerando pulmões de um mesmo grupo experimental,
verifica-se que esse parâmetro se mantém aproximadamente constante dentro do grupo. Isso
indica que os parâmetros G e H não são independentes, ou seja, um aumento da resistência
tecidual está geralmente associado a um aumento na mesma proporção do parâmetro H,
associado às características elásticas do tecido.
A segunda característica, igualmente importante, mas com maioresrepercussões
do ponto de vista fisiológico, diz respeito ao aumento de η durante broncoconstrição
(KACZKA et al., 1997). A partir de experimentos com cápsulas alveolares verificou-se que
há aumento das desigualdades regionais (heterogeneidades) nas propriedades mecânicas do
pulmão com o aumento na severidade de broncoconstricções (FREDBERG et al., 1985). Isso

41
tem levado à aceitação de que η sempre cresce à medida que o pulmão se torna
mecanicamente heterogêneo. Assim, a histeresividade pode ser usada como um indicativo
dessas heterogeneidades (BATES, 2009).
2.8PESQUISA POR HIPERRESPONSIVIDADE DE VIAS AÉREAS MEDIANTE
DESAFIO COM METACOLINA(MCH)
Estudos demonstraram que a fração do volume tecidual que éatribuído à
maquinaria contrátil é comparável nas vias aéreas, dutos alveolares e vasos sanguíneos no
parênquima pulmonar, determinando que o parênquima pulmonar, assim como as vias aéreas,
pode ser considerado um tecido (FREDBERG, 2004).
A hiperresponsividade das vias aéreas é definida como um estreitamentoexcessivo
das vias aéreas em resposta a uma variedade de estímulos químicos e físicos que têm pouco
ou nenhum efeito em indivíduos saudáveis (BRUSASCO etal., 2003). A medida da
responsividade das vias aéreas, alterada em algumas patologias, pode ser realizada através da
utilização de agonistas que provocam a broncoconstrição, com um procedimento chamado de
desafio com agonista. Este é realizado através da administração, seja por via inalatória ou
intravenosa, de um agonista da musculatura lisa das vias aéreas. Os agonistas mais
comumente utilizados são a metacolina (MCh), a acetilcolina (ACh) e o carbacol.
O termo hiperresponsividade das vias aéreas é usado para descrever afacilidade de
estreitamento das vias aéreas em resposta ao desafio com agonistas contráteis inespecíficos
(FREDBERG, 2004), é uma anormalidade funcional que pode ser desencadeada por
alérgenos, e por alguns estímulos inespecíficos, tais como poluição atmosférica, estimulantes
farmacológicos, infecções respiratórias, exercício físico e fatores emocionais (BLEASE,
2000). Alguns mecanismos quepodem estar relacionados com a hiperresponsividade
brônquica são: contratura excessiva da musculatura lisa das vias aéreas, espessura e
integridade da camada epitelial, alterações mecânicas dos brônquios, modificações no
conteúdo e no equilíbrio das proteínas da MEC e ainda regulação autonômica exacerbada.
Entretanto o percentual de participação de cada um destes fatores ainda não
étotalmente compreendido. O processo inflamatório, no qual acompanha uma vasta liberação
de mediadores inflamatórios e citocinas, tem um papel chave na hiperresponsividade
(HIZUME, 2010).

42
3 JUSTIFICATIVA
Diante da prevalência de acidentes ofídicos associados a escassos estudos com
envenenamento relacionados ao sistema respiratório, faz-se necessário um estudo mais
aprofundado e uma maior investigação dessas alterações morfofuncionais, tendo em vista o
maior esclarecimento dos mecanismos fisiopatológicos.
Essa Dissertação se enquadra na linha de pesquisa em sanidade animal visando a
um estudo morfológico com ênfase nas alterações sistêmicas que acometem a arquitetura
histológica pulmonar e vias aéreas superiores, promovidas pelo veneno desta serpente em
estudo, a fim de determinar as suas alterações histológicas e propiciar o menor sofrimento ao
animal vitimado após o acidente ofídico.
O comportamento das propriedades mecânicas do sistema respiratório, a
caracterização das estruturas pulmonares e a quantificação do processo inflamatório gerado
pela ação do veneno da serpente Crotalus durissus cascavella (C.d.cascavella)são pouco
conhecidos. Desta forma, faz-se necessário conhecer os efeitos desta possível lesão para que,
em eventual acometimento, a intervenção terapêutica seja a mais precoce e eficiente possível.

43
4 HIPÓTESE
Existe uma relação direta entre as alterações mecânicas e as alterações morfológicas
no sistema respiratório após envenenamento crotálico.

44
5 OBJETIVOS
5.1 OBJETIVO GERAL
Analisar as propriedades mecânicas e a morfologia pulmonar em modelo
experimental de lesão pulmonar aguda induzida pelo veneno da serpente Crotalus durissus
cascavella (C. d. cascavella).
5.2 OBJETIVOS ESPECÍFICOS
Estudar os efeitos biológicos do veneno bruto da serpente Crotalus durissus cascavella
(C. d. cascavella) em modelo animal.
Descrever as alterações histológicas pulmonares e das vias aéreas superiores
promovidas pela ação tóxica do veneno da serpente Crotalus durissus cascavella (C. d.
cascavella).
Observar o comportamento das propriedades mecânicas elásticas, viscosas e
viscoelásticas do sistema respiratório em resposta à lesão pulmonar aguda induzida pelo
veneno da serpente Crotalus durissus cascavella (C. d. cascavella).
Correlacionar os dados obtidos na mecânica pulmonar com a histopatologia.

45
6 CAPÍTULO 1
Mecânica Pulmonar e Histopatologia induzida pelo Veneno da
serpente Crotalus durissus cascavella
Pulmonary mechanical and Histopathology induced by Crotalus
durissus cascavella snake venom
Periódico: Acta Médica Portuguesa (Submetido em 14 de janeiro de 20015).

46
Mecânica Pulmonar e Histopatologia induzida pelo Veneno da
serpente Crotalus durissus cascavella
Pulmonary mechanical and Histopathology induced by Crotalus
durissus cascavella snake venom
Joselito de Oliveira NETO1, João Alison de Moraes SILVEIRA
2, Daniel Silveira
SERRA3, Daniel de Araújo VIANA
1, Diva Maria BORGES-NOJOSA
4, Paula Priscila
Correia COSTA2, Célia Maria de Souza SAMPAIO
5, Helena Serra Azul MONTEIRO
2,
Francisco Sales Ávila CAVALCANTE3, Janaina Serra Azul Monteiro EVANGELISTA
1
1 Laboratório deHistologia dos efeitos causados pelos venenos de serpentes e plantas –
HISTOVESP. Faculdade de Veterinária. Universidade Estadual do Ceará. Fortaleza. Brasil.
2 Faculdade de Medicina. Universidade Federal do Ceará. Fortaleza. Brasil.
3 Laboratório de Biofísica da Respiração. Centro de Ciências e Tecnologia. Universidade
Estadual do Ceará. Fortaleza. Brasil.
4 NUROF-Núcleo Regional de Ofiologia. Centro de Ciências, Departamento de Biologia
Universidade Federal do Ceará. Fortaleza. Brasil.
5 Coordenação de Ciências Biológicas. Centro de Ciências da Saúde. Universidade Estadual
do Ceará. Fortaleza. Brasil.
Correspondência: Av. Dr. Silas Munguba, 1700, Campus do Itaperi, Fortaleza-CE. CEP:
60714-903. Telefone: (85) 3101 9889. E-mail: [email protected]

47
RESUMO
Crotalus durissus cascavella (C.d.cascavella) é uma serpente encontrada na Caatinga
do Nordeste Brasileiro. Este trabalho teve por objetivo investigar as alterações em mecânica
pulmonar (propriedades elásticas, resistivas e viscoelásticas) e histopatologia em pulmões
após inoculação intraperitoneal (3,0 µg/Kg) do veneno desta serpente (C.d.cascavella) em
camundongos. Os valores médios de resistência Newtoniana (RN), resistência tecidual(G) e
elastância tecidual(H) foram significativos em todos os grupos experimentais (1h, 3h, 6h, 12h
e 24h), sendo maiores nos dois últimos grupos. A histeresividade (η) foi significativa em 6h,
12h e 24h. Os valores médios de complacência estática (Cst) foram significativos nos grupos
de 6h, 12h e 24h sendo nesse último grupo valores menores. A capacidade inspiratória (CI)
mostrou-se significativa em 3h, 6h, 12h e 24h. Após desafio com Metacolina, RN apresentou
valores médios significativos nos grupos 6h, 12h e 24h. O veneno da serpente C. d. cascavella
induziu no tecido pulmonar, atelectasia, enfisema, hemorragia, infiltrado inflamatório com
polimorfonucleares, congestão em todos os grupos experimentais e edema apenas no grupo de
24h. O veneno da C. d. cascavella provocou alterações mecânicas e histopatológicas no tecido
pulmonar, porém o mecanismo de ação continuará a ser estudado para possíveis descobertas
morfofuncionais.
PALAVRAS-CHAVE: Crotalus durissus cascavella, Mecânica Pulmonar, Histopatologia.
ABSTRACT
Crotalus durissus cascavella (C. d. cascavella) is a snake found in the Brazilian Northeast
Caatinga. This study aimed to investigate the changes in lung mechanics (elastic properties,
resistive and viscoelastic) and histopathology in lungs after intraperitoneal injection (3,0
µg/Kg) of this snake venom (C. d. cascavella) in mice. The average values of Newtonian
resistance (RN), tissue resistance (G) and tissue elastance (H) were significant in all groups

48
(1h, 3h, 6h, 12h and 24h) and higher in the last two groups. The histeresividade (η) was
significant in 6h, 12h and 24h. The static compliance of average values (Cst) were significant
in groups of 6h, 12h and 24h and the latter group lower values. The inspiratory capacity (CI)
was significant at 3h, 6h, 12h and 24h. After challenge with Methacholine, RN known
significant average values in groups 6h, 12h and 24h. The venom of the snake C. d.
cascavellaalso induced in pulmonar tissue atelectasis, emphysema, hemorrhage, inflammatory
infiltrate with polymorphonuclear cels, congestion in all experimental groups, and edema only
in the 24h group. The venom of the snake C. d. cascavellacaused mechanical and
histopathological changes in lung tissue, however the mechanism of action will continue to be
studied for possible morphological and functional findings.
Keywords: Crotalus durissus cascavella,Pulmonary Mechanics, Histopathology.
INTRODUÇÃO
A composição química e as atividades biológicas dos venenos podem variar muito
entre as famílias e gêneros das serpentes. 1A maior gravidade encontra-se nos acidentes
causados por serpentes do gênero Crotalus, e essas têm relevante espaço no campo científico,2
visto que em algumas décadas seu veneno vem sendo alvo de pesquisadores devido a sua
constituição e o efeito que essas substâncias podem causar.3
A subespécie Crotalus durissus cascavella (C.d.cascavella), encontrada na Caatinga
do Nordeste Brasileiro4,3
, produz um veneno que conduz à alteração sistêmica, sendo
responsável pela causa preliminar de óbito após o acidente ofídico.5
A peçonha de Crotalus apresenta-se como um complexo tóxico-enzimático, na qual
são encontradas as enzimas fosfodiesterase, l amino oxidase, 5-nucleotidase e toxinas como a
crotoxina, convulxina, crotamina e giroxina.6
Os animais acometidos por envenenamento crotálico manifestam como principais
complicações: insuficiência renal aguda 7, choque circulatório
8,9, reações edematogênicas,
agregação de plaquetas10
e hipotensão arterial5. Há insuficiência respiratória aguda
relacionada à paralisia neuromuscular. 11

49
O comprometimento pulmonar após envenenamento crotálico apresenta um quadro de
hipotensão e consequente hipoventilação, se instalando um processo de angústia respiratória,
cuja consequência será a perda do status de bem estar do animal acometido pelo acidente
ofídico.
Em vista disso, tornou-se relevante estudar as alterações morfofuncionais no sistema
respiratório após exposição do veneno da subespécie C. d. cascavella, buscando a um
entendimento de seus efeitos biológicos e suas ações nesse sistema.
A observação das propriedades mecânicas do sistema respiratório, a caracterização das
estruturas pulmonares e a quantificação do processo inflamatório gerado pela ação do veneno
da subespécie C. d. cascavella são pouco conhecidos. Portanto, fez-se necessário estudar os
efeitos desta possível alteração na arquitetura pulmonar para que, em eventual acometimento,
a intervenção terapêutica seja a mais precoce e eficiente possível.
Esse trabalho teve por objetivo, analisar a função pulmonar em modelo experimental
de lesão pulmonar aguda induzida pelo veneno. Para este fim, as propriedades elásticas,
resistivas e viscoelásticas foram determinadas em 1, 3, 6, 12 e 24 h após inoculação
intraperitoneal do veneno em camundongos, e os resultados foram correlacionados com dados
colhidos a partir da histopatologia pulmonar.
MATEIAL E MÉTODOS
Veneno
O veneno total foi obtido e gentilmente cedido pela professora doutora Diva Maria
Borges Nojosa e pela médica veterinária Roberta da Rocha Braga do Núcleo Regional de
Ofiologia (NUROF) do departamento de Biologia da Universidade Federal do Ceará – UFC,
Fortaleza, Brasil. O veneno foi liofilizado e diluído em solução salina (0,9%, w/v solução de
NaCl) no momento de sua utilização.
Animais
Camundongos machos Balb/C (20-30g), foram mantidos em alojamento com
temperatura controlada, umidade relativa do ar 65,3 % e 12 h claro/escuro, receberam água e
comida ad libitum. Os animais e os protocolos de pesquisa utilizados neste estudo estão de

50
acordo com as diretrizes da Comissão de Ética para uso de animais da Universidade Estadual
do Ceará – UECE, número de protocolo 12773584-4.
Protocolo Experimental
Os animais foram ventilados em padrões basais a uma frequência de 150
respirações/min, um volume corrente de 10 mL/kg, com limite pressórico de 30 cmH2O, e
uma pressão positiva expiratória final (PEEP) de 3 cmH2O.
Para a análise da função pulmonar os animais foram sedados com Diazepan
(1mg)(Novafarma Indústria) e anestesiados com Pentobarbital sódico (Hypnol 3%, Syntect)
(50 mg/mL),todos intraperitonealmente, posteriormente realizou-se procedimento cirúrgico de
traqueostomia.
Para que as variáveis da função pulmonar fossem coletadas de forma fidedigna, se faz
necessário que o animal esteja suficientemente sedado e paralisado para não haver
interferência da atividade de seus músculos respiratórios. Para tanto, após o início da
ventilação, estes foram paralisados com brometo de pancurônio (0,5 mL/kg, Pancuron®,
Cristália, Brasil), intraperitonealmente.
Antes do protocolo de análise, realizou-se duas inflações profundas (IP) com limite
pressórico de 30 cmH2O e 6s de duração, seguido por um período de 5 minutos de ventilação
nos padrões basais, para que se realizasse a padronização do histórico pulmonar. Logo após,
foi iniciado o protocolo de análise de dados, onde a impedância do sistema respiratório (Zsr)
foi medida através da técnica de oscilações forçadas de forma sequencial, em intervalos de
10s durante 2 minutos. Seguidos da realização de duas curvas pressão-volume para a coleta da
complacência estática (Cst), estimativa da capacidade inspiratória(CI) e área da curva.
A Zsr foi determinada através da medição do volume de deslocamento e pressão do
pistão no cilindro do ventilador enquanto perturbações de 3s de volume oscilatório foram
entregues para as vias aéreas. Estas perturbações eram realizadas através de 13 ondas
sinusoidais sobrepostas com variações de amplitude e frequência (1-20,5 Hz). As frequências
foram fixadas em valores mutuamente condicionadas para reduzir a distorção harmônica que
pode ocorrer em sistemas não lineares. 12
Antes do início do protocolo obtiveram-se os sinais
de calibração dinâmica necessárias para corrigir as características físicas do ventilador
mecânico em medições subsequentes de Zsr. A Zsr foi determinada através da transformada de

51
Fourier dos sinais de fluxo e a pressão do cilindro, tal como descrito anteriormente 13
. A Zsr
foi interpretada de acordo com o modelo
onde,
onde RN é a resistência Newtoniana, i é a √−1, f é a frequência (Hz), I é a inertância das vias
aéreas, e G e H caracterizam respectivamente as propriedades dissipativas e elásticas do
tecido pulmonar 12
. Outra característica importante do modelo de fase constante, diz respeito à
razão G/H, conhecida como histeresividade (η).
Hiperresponsividade do músculo liso das vias aéreas
Logo após a coleta de dados iniciais, realizaram-se novamente duas IPs seguidas por 5
minutos de ventilação com padrões basais. A hiperresponsividade do musculo liso das vias
aéreas foi avaliada através da inalação de MCh (Sigma-Aldrich®) realizada através de
aerossol produzido por um nebulizador ultrassônico (Inalasonic®, NS) acoplado na linha
inspiratória do ventilador mecânico. Para o procedimento, foram adicionados sequencialmente
4 mL de solução de MCh (0, 3, 6, 12,5, 25 e 50 mg/mL) no recipiente do nebulizador. A
nebulização foi realizada durante 30s de ventilação mecânica em padrões basais 14
e a
quantidade média de solução nebulizada foi 0,4 mL. Imediatamente após a nebulização,
realizou-se novamente a técnica de oscilações foçadas de forma sequencial em intervalos de
10s durante 2 minutos. Entre a coleta de dados das concentrações de MCh realizou-se duas
IPs seguidas por 5 minutos de ventilação com padrões basais.
Análise Morfológica
Foram obtidos fragmentos de pulmões de aproximadamente 1,5cm2 e
acondicionados em frascos contendo solução de formaldeído a 10% por um período de 48
horas. Em seguida esses fragmentos foram lavados em água corrente por 1 hora para
eliminação de resíduos de formol. Posteriormente os fragmentos foram transferidos para o
etanol a 70% permanecendo imersos nesta substância por um período de 24 horas, sendo

52
iniciado o processo de desidratação em sucessivas diluições crescentes de etanol para
posterior diafanização e em seguida cortados no micrótomo na espessura de 4 μm. Utilizou-se
Hematoxilina-Eosina (HE) para coloração e visualização em microscopia de luz (Microscópio
Nikon trinocular; ®View Software versão 7.3.1.7).
Análise Estatística
Os resultados foram apresentados como média ± desvio padrão da média, onde n
representa o número de experimentos. Foi considerada diferença estatística significativa os
resultados que apresentaram probabilidade de ocorrência da hipótese nula menor que 5% (p <
0,05). Para tanto, foi utilizado o teste t de Student, para análise de significância da diferença
entre as médias.
RESULTADOS
Efeito da exposição ao veneno C. d. cascavella na Mecânica Pulmonar
Após a inoculação do veneno, os valores médios de resistência Newtoniana (RN)
(Figura 1A), resistência tecidual(G) (Figura 1B) e elastância tecidual(H) (Figura 1C)
foramaumentados e significativos nos grupos de 1h, 3h, 6h, 12h e 24h, sendo maiores nesses
dois últimos grupos. A histeresividade (η) (Figura 1D) apresentou valores médios aumentados
e significativos nos grupos 6h,12h e 24h.
A Figura 2A mostra os valores médios de complacência estática (Cst) sendo reduzidos
e significativos nos grupos de 6h, 12h e 24h sendo nesse último grupo valores menores. A
capacidade inspiratória (CI) (Figura 2B) mostrou valores médios significativos e reduzidos
nos grupos de 3h, 6h, 12h e 24h.
Efeito da exposição ao veneno C. d. cascavella na Mecânica Pulmonar após
utilização de Metacolina
A Figura 3 apresenta a resistência Newtoniana (RN) aumentada após desafio com
Metacolina onde seus valores médios foram significativos nos grupos 6h, 12h e 24h.

53
Efeito da exposição ao veneno C. d. cascavella nos achados histopatológicos
A arquitetura histológica dos pulmões (árvore brônquica, bronquiolar e alveolar)
apresentou alteração da organização arquitetural causada por atelectasia e enfisema. A
atelectasia presente em todos os grupos apresentou intensidade moderada nos grupos 6h, 12h
e 24h (Figura 4A). O enfisema esteve presente em todos os grupos experimentais
deintensidade leve (Figura 4B). O edema foi observado apenas no grupo de 24h de
intensidade leve (Figura 4C). Em todos os grupos experimentais havia infiltração
inflamatória, embora discreta, de linfócitos e plasmócitos. Observou-se uma maior
concentração de infiltrado nos animais dos grupos de 3h e 6h (Figura 4D). Houve presença do
tecido linfoide associado ao brônquio (BALT) aumentado nos grupos 6h e 12h (Figura 4E).
A congestão foi observada em todos os grupos com intensidade leve (Figura 5).
Houve hemorragia em todos os grupos experimentais, de intensidade variável desde
leve a intensa e com maior concentração de intensidade moderada no grupo de 3h (Figura 6).
DISCUSSÃO
Os efeitos dos venenos crotálicos já são bem discutidos e elucidados na literatura, no
que se refere ao bloqueio pré-sináptico da junção neuromuscular, assim como as alterações
em rins, fígado e até mesmo sua ação local. 15,16,17,18,19
As ações deste veneno no sistema respiratório de seres humanos e animais são pouco
conhecidas, apresentadas apenas por estudos de casos e suas representações como melhores
exemplos para se estabelecer possíveis intervenções clínicas. Não foram encontradas
referências científicas a respeito da função pulmonar após inoculação desse veneno em
modelo animal.
Em estudos realizados sobre repercussões pulmonares após aplicação do veneno de
Bothrops jararaca por via intravenosa, as primeiras 48 horas seguidas do acidente são
decisivas para o estabelecimento das lesões pulmonares. 20
Em outro estudo investigando a análise das propriedades mecânicas e histopatológicas
do tecido pulmonar, encontrou-se alteração nos grupos experimentais de 3h (aumento
significativo na elastância tecidual), 6h (aumento significativo na resistência tecidual) e 12h
(aumento das propriedades viscoelásticas) após inoculação por via intramuscular do veneno
da serpente Crotalus durissus terrificus na dose de 0,6 µg/g e que após 24 horas houve

54
retorno aos valores basais. 21
Enquanto que encontramos em nosso estudo, alterações
significativas (aumento na resistência Newtoniana (RN), resistência tecidual(G) e elastância
tecidual(H)) em todos os grupos experimentais (1h, 3h, 6h, 12h e 24h), sendo maiores nos
dois últimos grupos, e nos grupos de 6h,12h e 24h, aumento na histeresividade (η), indicando
aumento na heterogeneidade do pulmão no tocante às propriedades mecânicas o que se reflete
em distribuição heterogênea das áreas ventiladas.
De modo geral, a complicação respiratória consequente ao acidente crotálico se instala
nas primeiras 24 horas, podendo surgir nos casos graves, dentro de 3 a 4 horas após a picada.
22
O infiltrado inflamatório com linfócitos e plasmócitos, presente em todos os grupos
experimentais sendo evidenciado no grupo de 6 horas é sugestivo de uma agressão tecidual
aguda do veneno C. d. cascavella. Isso provavelmente resultou em quimiotaxia, proveniente
da atividade de mediadores químicos inflamatórios liberados, como prostaglandinas,
histamina, bradicinina, ou o óxido nítrico e consequente diapedese de células inflamatórias
para o local da lesão. 24
Houve presença do tecido linfoide associado ao brônquio (BALT)
aumentado nos grupos 6h e 12h, estando relacionado com uma maior ativação do sistema
imunológico desses animais.
Estudo em que houve instilação de lipopolissacarídeo de E. coli, percebeu-se que a
lesão provocada de forma direta nos pulmões, apresentou um processo inflamatório mais
acentuado, acompanhado por aumento nos níveis de citocinas (IL-6, IL-8 e IL-10) e do
infiltrado neutrofílico.25
Em estudos com envenenamento botrópico em modelos animais, ratos e
camundongos, observou-se aparecimento de neutrófilos polimorfonucleares nas primeiras
horas. Após alcançar um pico entre 6 e 72 horas, houve substituição e aparecimento de células
mononucleares, monócitos e linfócitos.26
Outros estudos realizados com o veneno da serpente Crotalus durissus terrificus,
observou-se um aumento de células linfomononucleares no grupo de 6 horas em vias aéreas.
21 Outros estudos consideraram que em resposta à lesão aguda pulmonar ocorreu a presença
de células mononucleares. 27
A presença de hemorragia em todos os grupos experimentais, de intensidade variável
desde leve a intensa, é justificada pela existência da crotamina associada a uma alta atividade

55
de PLA2, fazendo a esse veneno ser caracterizado pela miotoxidade, podendo ser assim
responsável pelo processo hemorrágico encontrado, sendo relatado na literatura seu potencial
neurotóxico. 18,21,23
A hemorragia intensa indica extravasamento de sangue em virtude de
ruptura vascular.
A PLA2 de ação sistêmica, após a distribuição pelo corpo, se liga a receptores de alta
afinidade da membrana das células musculares em outras regiões anatômicas, induzindo a
miotoxicidade sistêmica. 28
A atividade catalítica das PLA2 irá potencializar as atividades
inflamatórias e neurotóxicas. 29
O aparecimento de edema pulmonar em um animal do grupo experimental de 24h de
exposição ao veneno retrata bem na literatura a não ocorrência frequente dessa alteração
histopatológica, podendo ser relacionado ao fator indireto da lesão gerada pelo veneno da
serpente C. d. cascavellae atribuída à ação das PLA2 sistêmico. Houve relato de uma resposta
edematogênica induzida pelo veneno da serpente Crotalus durissus terrificus onde não foi
dose-dependente e de curso rápido e transitório. 18
Há na literatura vários estudos relatando
que a PLA2 induz edema, um efeito que em alguns casos, são dependentes da habilidade das
PLA2 em se ligarem a proteínas específicas de membrana. 29
O aparecimento de edema
acentuado é encontrado em venenos de serpentes do gênero botrópico. 26
A congestão observada em todos os órgãos teve intensidade leve em todos os grupos.
Tal alteração geralmente deve estar relacionada com o processo de eutanásia dos animais. A
congestão de leitos capilares e venosos está intimamente relacionada ao desenvolvimento de
edema. A estase de sangue pouco oxigenado causa hipóxia que pode acarretar em
degeneração ou morte das células parenquimatosas, às vezes, com cicatrizes microscópicas. A
ruptura de vasos nestes locais de congestão também pode causar pequenos focos de
hemorragia. 30
A atelectasia presente em todos os grupos apresentou intensidade moderada nos
grupos 6h, 12h e 24h. Essa alteração quando ocorre em uma região segmentar do pulmão
pode distorcer o parênquima e o subsegmento adjacente e assim afetar a mecânica tecidual
local. 31
Isso é associado ao aumento da histeresividade (n).
O enfisema esteve presente em todos os grupos experimentais de intensidade leve.
Estudo em que se utilizaram animais tratados com veneno C. d. cascavella, esses
apresentaram evidente congestão vascular hepática intensa. 32
A isquemia transitória hepática

56
causa significativas alterações histológicas no fígado de rato, o que sugere o dano decorrente
da isquemia-reperfusão hepática. 33
A deficiência de alfa-1 antitripsina é um distúrbio genético
de recente descoberta e que ocorre com frequência comparável à da fibrose cística. Resulta de
diferentes mutações no gene SERPINA1 e tem diversas implicações clínicas. A alfa-1
antitripsina é produzida principalmente no fígado e atua como uma antiprotease, tendo como
principal função inativar a elastase neutrofílica, impedindo a ocorrência de dano tecidual. A
mutação mais frequentemente relacionada à doença clínica é o alelo Z, que determina
polimerização e acúmulo dentro dos hepatócitos. O acúmulo e a consequente redução dos
níveis séricos de alfa-1 antitripsina determinam, respectivamente, doença hepática e
pulmonar, sendo que esta se manifesta principalmente sob a forma de enfisema de
aparecimento precoce, habitualmente com predomínio basal. 34
Portanto, há uma hipótese de
que o veneno C. d. cascavella pode ter ocasionado lesão hepática e assim, resultar na
diminuição e/ou na deficiência da produção da alfa-1 antitripsina.
A resistência Newtoniana (RN) tem sido usada como uma boa estimativada resistência
total das vias aéreas. 35
Encontraram-se valores de significância estatística em todos os grupos
experimentais, sendo maiores nos grupos de 12h e 24h, logo estes resultados podem
representar um maior estreitamento ou um aumento na rigidez do músculo liso das vias aéreas
nestes grupos experimentais em relação ao grupo controle. A alteração no diâmetro das vias
aéreas apresenta uma dependência de alguns fatores que nãopodem ser isolados, sua
constrição depende da geração de força a partir da contração do músculo liso, 36
das
flutuações da pressão no interior das vias aéreas 37
e da natureza viscoelástica do músculo liso
e dos tecidos do parênquima ao redor. Ao reduzir o recolhimento elástico do pulmão,
rompendo fibras e destruindo anexos alveolares, ocorre uma limitação do fluxo aéreo, levando
a um aumento da resistência das vias aéreas. O aprisionamento de ar e a nova morfologia
pulmonar atuam reduzindo a superfície disponível para a troca gasosa. 38,39
A resistência de vias aéreas representa valores mais importantes quando se compara à
resistência tecidual. 40
Há correlação da variação das pressões resistivas do pulmão com a
resistência de vias aéreas. 41
A resistência tecidual (G) e elastância tecidual (H) estão relacionadas às propriedades
intrínsecas do tecido, e a análise desses parâmetros não é tão simples quanto à resistência
Newtoniana (RN). Existem várias hipóteses para explicar suas mudanças, uma delas é devido à
alteração das propriedades reológicas do tecido. 35
Outra maneira seria através da influência
que o estreitamento dasvias aéreas tem sobre estes parâmetros, estreitamento que poderia

57
resultar em uma distorção do parênquima pulmonar com fechamento de pequenas vias aéreas,
constituindo um pulmão efetivamente menor com uma elastância tecidual (H)
proporcionalmente maior.42
A resistência tecidual (G) reflete a dissipação de energia viscosa
no tecido pulmonar, parâmetro que também é alterado devido à distorção do parênquima
pulmonar que ocorre quando as vias aéreas se contraem, 42
o que pode explicar os aumentos
significantes dos valores G e H em todos os grupos experimentais, em especial nos grupos
12h e 24h, já que houve aumento no parâmetro relacionado à resistência das vias aéreas (RN).
Outra possibilidade para esse aumento é a presença de hemorragia em todos os grupos
testados, sendo de intensidade moderada no grupo de 12h e de intensidade grave no grupo de
24h, em vias aéreas de pequeno calibre, assim como a ocorrência de edema apenas no grupo
de 24h, favorecendo a oclusões dessas vias e a produção de áreas com atelectasia, presente em
todos os grupos experimentais, sendo de intensidade moderada para os grupos de 6h, 12h e
24h e no grupo controle, visto que esse processo é encontrado na prática clínica em animais
submetidos à ventilação mecânica.
Com relação ao parâmetro histeresividade (η), observou-se significância estatística nos
grupos de 6h, 12h e 24h em relação ao grupo controle. Seu valor cresce à medida que o
pulmão se torna mecanicamente heterogêneo, com isso a histeresividade pode ser usada como
um indicativo dessas heterogeneidades ventilatórias. Podemos então supor que o aumento dos
valores de η seja devido à presença de heterogeneidades ventilatórias decorrentes do aumento
da RN. A essas heterogeneidades ventilatórias podemos associar à presença de hemorragia de
intensidade moderada no grupo 12h e grave no grupo 24h, atelectasias de intensidade
moderada nos grupos 6h, 12h e 24h, enfisema de intensidade leve presente em todos os
grupos testados e edema presente no grupo 24h. A presença de secreções nas vias aéreas
periféricas pode afetar a distribuição da ventilação e assim, aumentar a inomogeneidade
mecânica. As mudanças no tônus contrátil da musculatura lisa do parênquima pulmonar não
devem ser descartadas. 31
Os parâmetros da CST e da estimativa da capacidade inspiratória (CI),obtidos através
da realização da curva PV, foram calculados a partir da equação de Salazar e Knowles, esta
equação é utilizada apenas para ajuste na metade superior do ramo expiratório da curva PV.43
A diminuição do parâmetro CSTcom significância estatística nos grupos de 6h, 12h e 24h é
reflexo do aumento da elastância tecidual (H). A diminuição da capacidade inspiratória (CI)
com significância nos grupos 3h, 6h, 12h e 24h, especialmente nesses dois últimos, estão
coerentes com o efetivo enrijecimento do tecido pulmonar indicado pelo aumento de H. Os

58
resultados desses dois parâmetros são justificados pelos mesmos achados histopatológicos
encontrados para se explicar os resultados da elastância tecidual (H).
Broncoconstrictores têm sido amplamente utilizados em pesquisas de animais com
inflamação pulmonar devido à noção de que o problema primário na asma é o encurtamento
excessivo do músculo liso das vias aéreas, quer seja por uma melhor capacidade de resposta
do músculo liso em si 44
ou por um aumento da rigidez e da espessura da parede das vias
aéreas. 45,42
Existem várias maneiras em que o pulmão pode se tornar hiperresponsivo a uma
broncoprovocação, estes incluem mecanismos pelos quais o efeito do encurtamento do
músculo liso das vias aéreas é amplificado através de efeitos geométricos 46
e os mecanismos
que envolvem o aumento do encurtamento muscular.47
Após realização de administração da Metacolina em aerossol, observou-se o aumento
com significância da resistência Newtoniana (RN) para os grupos experimentais 6h, 12h e 24h
sendo visualizado o aparecimento de tecido linfoide associado ao brônquio (BALT)
aumentado nos grupos de 6h e 12h, podendo esta hiperresponsividade estar relacionado às
condições imunológicas desses animais. A broncoconstrição causada pela Metacolina faz com
que pequenas vias aéreas se fechem por completo, 48
estas não podemmais abrir apenas com o
relaxamento da sua musculatura lisa. A capacidade residual funcional (CRF) em ratos
reportado na literatura apresentou um aumento dose dependente com a administração da
Metacolina, indicativo de aprisionamento de ar nos pulmões, sugerindo que as estruturas que
estão se fechando são vias aéreas e não alvéolos.49
Os mecanismos da Hiperresponsividade Brônquica são múltiplos, sendo associados
com inflamação aguda, crônica e com o remodelamento das vias aéreas, especialmente com o
aumento da massa de músculo liso das vias aéreas. 50,36
A maior tendência à contração pode
dever-se também a perda dos fatores que se opõem ao encurtamento do músculo liso,51
por
alterações intrínsecas ao músculo liso e por perda da retração elástica do parênquima
pulmonar.
CONCLUSÃO
O veneno de C. d. cascavella provoca alterações mecânicas e histopatológicas no
tecido pulmonar, porém este estudo não permitiu esclarecer seu mecanismo exato de ação
mecânica ehistopatológica nas vias aéreas não pôde ser esclarecido através deste estudo.
Estudos mais específicos sobre repercussões pulmonares do veneno de C. d.
cascavella se fazem necessários, a fim de se estabelecer se o processo de provocação da
injúria aguda se encontra na paralisia muscular e mionecrose sistêmica, levando a fatores

59
desencadeantes causadores de uma inflamação, ou se há uma ação direta dos fatores tóxicos
do veneno sobre o tecido pulmonar levando a alterações morfoestruturais e de propriedades
mecânicas.
REFERÊNCIAS
1. DOS-SANTOS, M. C.; ASSIS, E. B.; MOREIRA, T.D.; PINHEIRO, J.; FORTES-
DIAS, C. L., Individual venom variability in Crotalus durissus ruruima snakes, a
subspecies of Crotalus durissus from the Amazonian region. Toxicon. 2005; 46: 8:
958-61.
2. BARRAVIERA, B.; PEREIRA, P. C. M. Acidentes por serpentes do gênero Bothrops.
P.262-273. In: BARRAVIERA, B. Venenos: Aspectos clínicos e terapêuticos dos
acidentes por animais peçonhentos. Rio de Janeiro: EPUB. 1999.411p.
3. PINHO, F. M. O, PEREIRA, I. D. Ofidismo. Rev Assoc Méd Bras. 2001; 47:1: 24-9.
4. BARRAVIERA, B.Curso sobre Acidentes por animais peçonhentos: Acidentes
porSerpente do Gênero Crotalus.Arq Bras Med.1989; 64:14-20.
5. EVANGELISTA, J. S. A. M.; MARTINS, A. M. C.; NASCIMENTO, N. R. F.;
SOUSA, C.M.; ALVES, R.S.; TOYAMA, D.O.; et al. Renal and vascular effects of
the natriuretic peptide isolated from Crotalus durissus cascavellavenom. Toxicon :
official journal of the International Society on Toxinology. 2008; 52: 7:737-44.
6. RANGEL-SANTOS, A.; DOS-SANTOS, E. C.; LOPES-FERREIRA, M.; LIMA, C.;
CARDOSO, D. F.; MOTA, I. A comparative study of biological activities of crotoxina
and CB fraction of venoms from Crotalus durissus terrificus, Crotalus durissus
cascavella and Crotalus durissus collilineatus. Toxicon. 2004: 43: 7: 801-10.

60
7. NISHIOKA, S. A.; JORGE, M. T.; SILVEIRA, P. V. P.; RIBEIRO, L. A.,
SouthAmerican rattlesnake bite and soft-tissue infection: report of a case. Rev Soc
Bras de Medic Trop.2000: 3: 4:401-02.
8. PINHO, F. M.; VIDAL, E.; BURDMANN, E. A., Atualização em insuficiência
renalaguda. Insuficiência renal aguda após acidente crotálico. J Bras Nefr. 2000:
22:62-8.
9. PINHO, F. M.; ZANETTA, D. M.; BURDMANN, E. A., Acute Renal Failure after
Crotalus durissus Snakebite: A Prospective Survey on 100 Patients. Kidney Int, 2005:
67: 659-67.
10. MARTINS, A.M.C.; NOBRE, A.C.L.; ALMEIDA A.C.; BEZERRA, G.; LIMA,
A.A.M.; FONTELES, M.C, et al. Thalidomide and pentoxifylline block the renal
effects of supernatants of macrophages activated with Crotalus durissus cascavella
venom. BrazJ of Med and Bio Resear, Ribeirão Preto. 2004: 37: 10: 1525 – 30.
11. DAMICO, D. C. S.; BUENO, L. G. F.; RODRIGUES-SIMIONI, L.; MARANGONI,
S.; CRUZ-HOFLING, M. A.; NOVELLO, J. C. Neurotoxic and myotoxic actions
from Lachesis muta muta (surucucu) whole venom on the mouse and chick nerve-
muscle preparations. Toxicon. 2005: 46: 222-29.
12. HANTOS Z, DAROCZY B, SUKI B, NAGY S, FREDBERG J.J. Input impedance
and peripheral inhomogeneity of dog lungs. J of Appl Physiol. 1992:72:(1):168-78.
13. HIRAI T, MCKEOWN KA, GOMES RFM, BATES JHJ. Effects of lung volume on
lung and chest wall mechanics in rats. J Appl Physiol 1999; 86: 16-21.

61
14. XUE Z, ZHANG L, LIU Y, GUNST SJ, TEPPER R S. Chronic inflation of ferret
lungs with CPAP reduces airway smooth muscle contractility in vivo and in vitro. J Appl
Physiol, 2008: 104:610-15.
15. LUCAS, E. P. R. Estudo interlaboratorial para o estabelecimento do veneno botrópico
edo soro antibotrópico de referência nacional. [Dissertação de Mestrado].
VigilânciaSanitária. Instituto Nacional de Controle de Qualidade em Saúde, Fundação
Oswaldo Cruz - Rio de Janeiro. 104 f., 2009.
16. BARRAVIERA,B. Acidentes por animais peçonhentos. Arq BrasMed. 1990: 64: 1:
14-20.
17. BARRAVIERA, B. Estudos clínicos dos acidentes ofídicos. Revisão. J Bras Med.
1993: 65: 4:209-50.
18. SANTORO, M.C.; SOUSA-E-SILVA, M.C.; GONÇALVES, R.L.; ALMEIDA-
SANTOS, S. M.; CARDOSO, D.F.; LAPORTA-FERREIRA, I. L, et al. Comparação
das atividades biológicas em venenos de três subespécies da cascavel sul-americana
(Crotalus durissus terrificus, C. durissus cascavella e C. durissus collilineatus).
Comparative Biochemistry and Physiology. Parte C: Farm e End.1999: 122:61-73.
19. TOYAMA, M. H.; OLIVEIRA, D. G.; BERIAM, L. O. S.; NOVELLO, J. C.;
RODRIGUES-SIMIONI, L.; MARANGONI, S. Structural, enzimatic and biological
properties of new PLA2 isoform from Crotalus durissus terrificus venom.Toxicon.
2003: 41:1033-38.
20. SILVEIRA, K. S. O.; BOECHEM, N.T.; NASCIMENTO, S.M.; MURAKAMI
Y.L.B.; BARBOZA, A.P.B.; MELO, P.A, et al. Pulmonary mechanics and lung
histology in acute lung injury induced by Bothrops jararaca venom. Resp Physiol&
Neurobiol. 2004: 139:167–77.

62
21. NONAKA, P. N.; AMORIM, C. F.; PERES, A. C. P.; SILVA, C. A. M.;
ZAMUNER, S. R.; RIBEIRO, W.Pulmonary mechanic and lung histology injury
induced by Crotalus durissus terrificus snake venom. Toxicon. 2008: 51: 1158-66.
22. AMARAL, C.F.S.; MAGALHÃES, R.A.; RESENDE, N.A. Comprometimento
respiratório secundário a acidente crotálico. Rev Inst Med Trop de São Paulo. 1991:
33: 251-55.
23. BEGHINI, D. G.; TOYAMA, M H; HYSLOP, S.; SODEK, L.; NOVELLO, J. C.;
MARANGONI1, S., 5 Enzymatic characterization of a novel phospholipase A2 from
Crotalus durissus cascavella rattlesnake (Maracambóia) venom. J prot chem. 2000:
19: 8: 679-84.
24. BRAGA, M.D.M.; MARTINS, A. M. C.; AMORA, D.N.; MENEZES, D.B.;
TOYAMA, M. H.; TOYAMA, D. O, et al. Purification and biological effects of L-
amino acid oxidase isolated from Bothrops insularis venom. Toxicon, 2007: 51:199-
207.
25. MENEZES, S.L.S.; BOZZA, P. T.; NETO, H. C. C. F.; LARANJEIRA, A. P.; NEGRI, E.
M.; CAPELOZZI, V. L, et al. Pulmonary and extrapulmonary acute lung injury:
inflammatory and ultrastructural analyses. J Appl Physiol. 2005: 98: 1777-83.
26. CARDOSO, J. L. C.; JUNIOR, V. H.; FRANÇA, F. S.; MALAQUE, C. M. S.;WEN
F. H., Animais peçonhentos no Brasil: biologia, clínica e terapêutica dos acidentes. 2.
ed., São Paulo, Editora Sarvier – FAPESP, 2003, p.469.

63
27. TONG, Q.; ZHENG, L.; KANG, Q.; DODD-O, J.; LANGER, J.; LI, B, et al.
Upregulation of hipóxia-induced mitogenic factor in bacterial lipopolysaccharide-
induced acute luna injury. FEBS Letters. 2006: 580: 2207-15.
28. GUTIÉRREZ, J.M.; OWNBY,C. L. Skeletal muscle degeneration induced by venos
phospholipases A2: insigths into the mechanisms of local and systemic myotoxicity.
Toxicon. 2003: 42: 915-31.
29. IGLESIAS, C. V.; APARICIO, R.; RODRIGUES-SIMIONI, L.; CAMARGO, E. A.;
ANTUNES, E.; MARANGONI, S, et al. Effects of morin on snake venom phospholipase
A2(PLA2). Toxicon. 2005: 46: 751-58.
30. LEDUC, M. & BON, C. Cloning of subunits of convulxin, a collagen-like
plateletaggregating protein from Crotalus durissus terrificus venom. Biochem, J.
1998: 333: 389-93.
31. CORREA, F. C. F.; CIMINELLI, P.B.; FALCÃO, H.; ALCÂNTARA, B. J. C.;
CONTADOR, R. S.; MEDEIROS, A. S, et al. Respiratory mechanics and lung
histology in noemal rats anesthetized with sevoflurane. J Appl Physiol.2001: 91: 803-
10.
32. EVANGELISTA, J. S. A. M., Estudo de efeitos vasculares, renais e citotóxicos do
veneno da serpente Crotalus durissus cascavella. 2008. Tese (Doutorado) -
Universidade Federal do Ceará, Fortaleza, Ceará, 2008.
33. RHODEN, E.; TELOKEN, C.; LUCA, S.M.; RHODEN, C.; MAURI, M.; ZETTLER,
C, et al. Protective effect of allopurinol in the renal ischemia-reperfusion in
uninephrectomized rats. Gen. Pharmacol. 2000: 35: 4: 189-93.

64
34. CARMELIER, A.A.; WINTER, D.H.; JARDIM, J.R.; BARBOZA, C.E.G.; CUKIER,
A.; MIRAVITLLES, M. Alpha-1 antitrypsin deficiency: diagnosis and treatment. J
Bras Pneumo. 2008: 34(7): 514-27.
35. BATES, J.T. Lung mechanics: An inverse modeling approach. Cambridge:
Cambridge University Press, p. 150 – 187, 2009.
36. MARTIN, J.G.; DUGUET, A.; EIDELMAN, D.H. The contribution of airway
smooth muscle to airway narrowing and airway hyperresponsiveness in disease. Eur
Respir J. 2000: 16: 2: 349 – 54.
37. SALERNO D. F., WERNER R. A., ALBERS J. W., BECKER M. P.,ARMSTRONG
T.J., FRANZBLAU A. Reliability of nerve conduction studies among active workers.
Muscle Nerve. 1999: 22(10): 1372-79.
38. SAETTA, M, TURATO, G, MAESTRELLI, P, MAPP, CE, FABBRI, L. M. Cellular
and structural bases of chronic obstructive pulmonary disease. Americ J Resp
and Critic Car Med. 2001:163(6):1304-9.
39. ISHIZAWA K, KUBO H, YAMADA M, KOBAYASHI S, NUMASAKI M, UEDA
S, et al. Bone marrow-derived cells contribute to lung regeneration after elastase-
induced pulmonary emphysema. FEBS Lett. 2004:556(1-3):249-52.
40. AULER Jr., J. O.C.; W. A., CALDEIRA, M.P.; CRADOSO, W.V.; SALDIVA, P.H.
Pre- and postoperative inspiratory mechanics in ischemic and vascular heart disease.
Chest. 1987: 92: 984-90.

65
41. SALDIVA, P. H. N.; ZIN, W. A.; SANTOS, R.L.B.; EIDELMAN, D. H.; MILIC-
EMILI, J. Alveolar pressure measurement in open-chest rats. J Appl Physiol, 1992:
72(1): 302-06.
42. WAGERS, S.; LUNDBLAD, L.K.A.; EKMAN, M.; IRVIN, C.G.; BATES, J.H. The
allergic mouse model of asthma: normal smooth muscle in an abnormal lung?. JAppl
Physiol.2004: 96(6): 2019-27.
43. SALAZAR, E.; KNOWLES, J.H. An analysis of pressure-volume characteristics of
the lungs. JAppl Physiol.1964: 19(1): 97-104.
44. FREDBERG, J.J. INOUYE D.; MILLER B.; NATHAN M.; JAFARI S.; RABOUDI
S.H, et al. Airway smooth muscle, tidal stretches, and dynamically determined
contractile states. Am J Respir Crit Care Med.1997: 156(6): 1752–59.
45. MORENO, R.H.; HOGG, J.C.; PARE, P.D. Mechanics of airway narrowing. Am Ver
Respir Dis. 1986: 133(1): 1171–80.
46. BATES, J.H.; MARTIN, J.G. A theoretical study of the effect of airway smooth
muscle orientation on bronchoconstriction. J Appl Physiol. 1990: 69(3): 995–1001.
47. MACKLEM, P.T.; Bronchial hyporesponsiveness. Chest. 1987: 91(6):189–91.
48. EVANS, K.L.; BOND, R.A.; CORRY, D.B.; SHARDONOFSKY, F.R. Frequency
dependence of respiratory system mechanics during induced constriction in a murine
model of asthma. J Appl Physiol.2003: 94(1): 245-52.

66
49. LUNDBLAD, L.K.; IRVIN C.G.; ADLER A.; BATES J.H. A reevaluation of the
validity of unrestrained plethysmography in mice. J Appl Physiol. 2002: 93(4): 1198–
07.
50. BOULET, L.P, LAVIOLETTE, M, TURCOTTE, H, et al. Bronchial subepithelial
fibrosis correlates with airway responsiveness to methacholine. Chest. 1997:112:45-
51. FREDBERG, J.J. Airway obstruction in asthma: does the response to a deep
inspiration matter? Respir Res. 2001: 2:273-5.
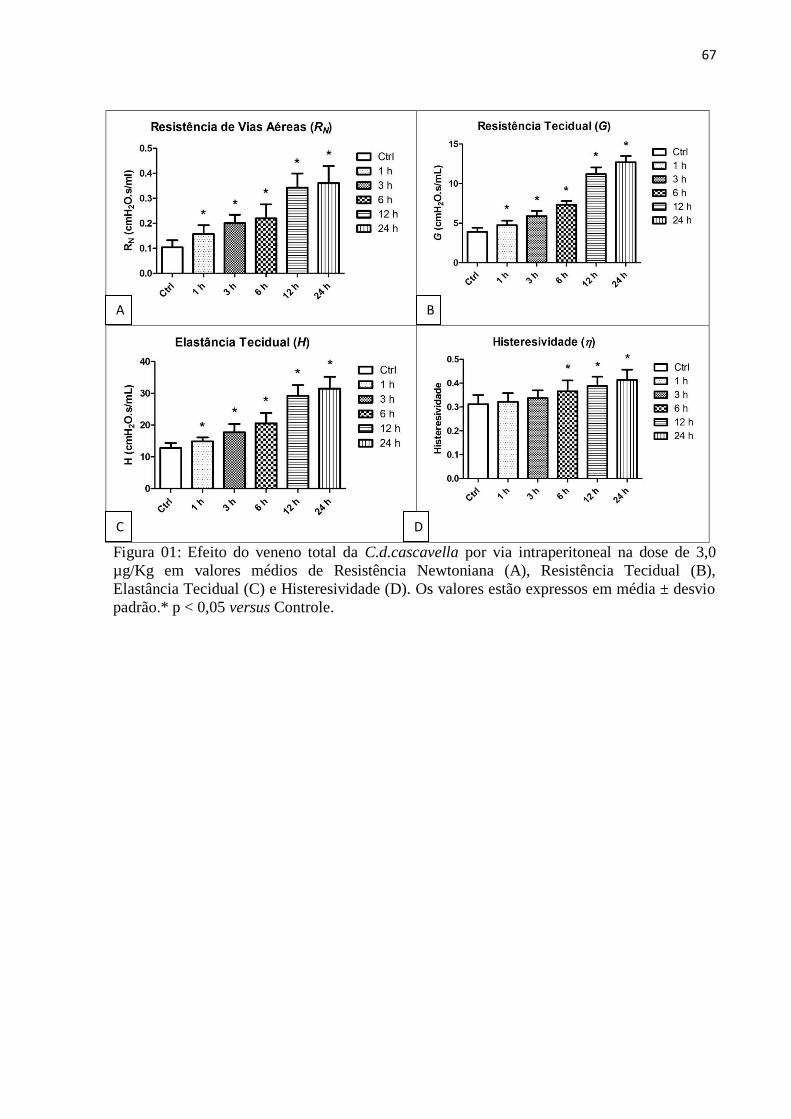
67
Figura 01: Efeito do veneno total da C.d.cascavella por via intraperitoneal na dose de 3,0
µg/Kg em valores médios de Resistência Newtoniana (A), Resistência Tecidual (B),
Elastância Tecidual (C) e Histeresividade (D). Os valores estão expressos em média ± desvio
padrão.* p < 0,05 versus Controle.
A B
C D
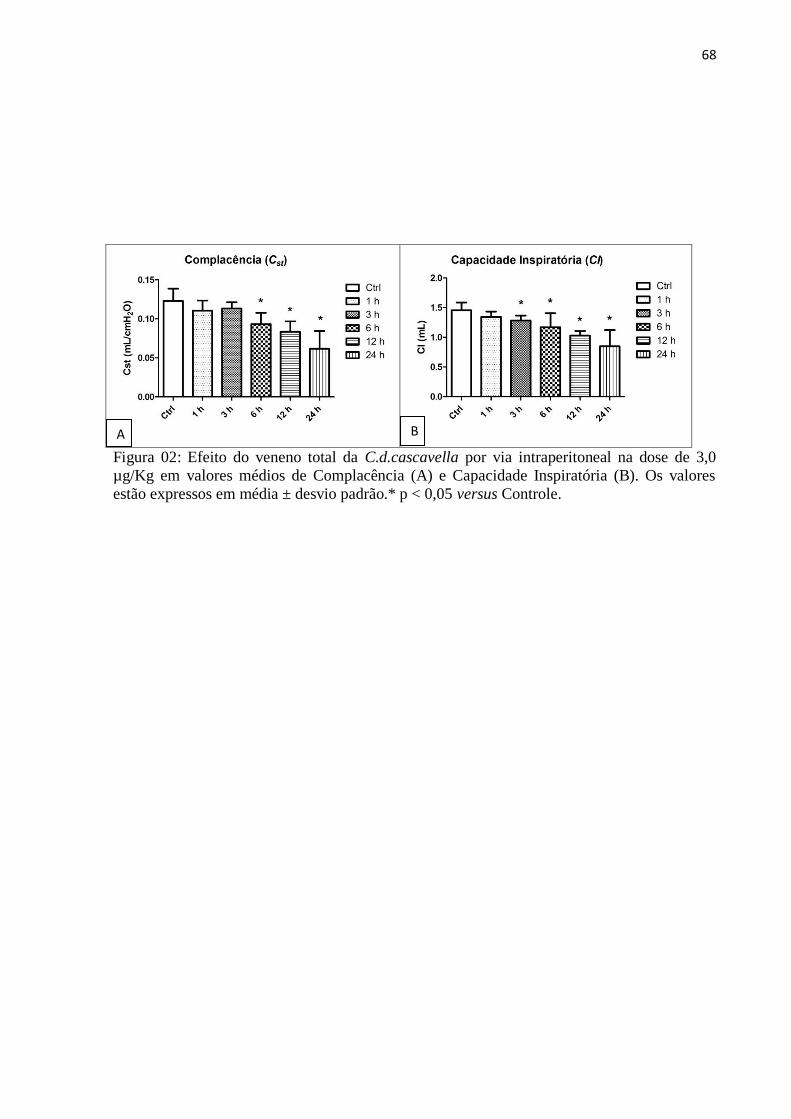
68
Figura 02: Efeito do veneno total da C.d.cascavella por via intraperitoneal na dose de 3,0
µg/Kg em valores médios de Complacência (A) e Capacidade Inspiratória (B). Os valores
estão expressos em média ± desvio padrão.* p < 0,05 versus Controle.
B A
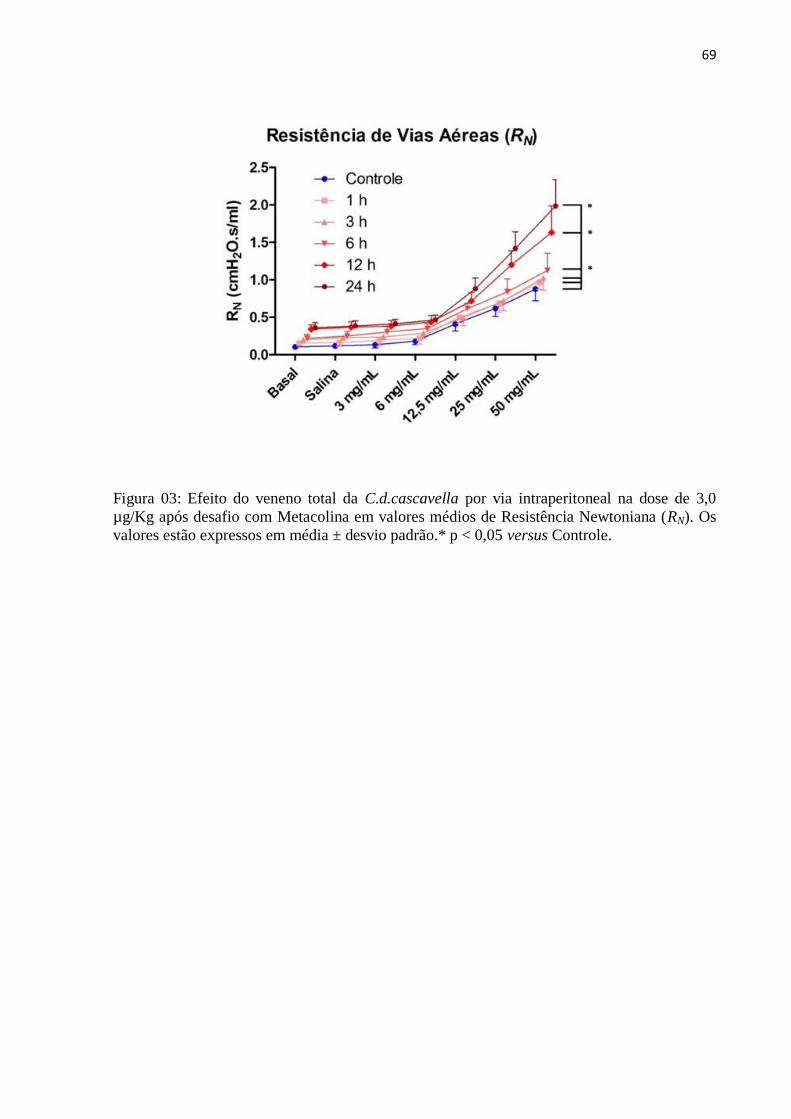
69
Figura 03: Efeito do veneno total da C.d.cascavella por via intraperitoneal na dose de 3,0
µg/Kg após desafio com Metacolina em valores médios de Resistência Newtoniana (RN). Os
valores estão expressos em média ± desvio padrão.* p < 0,05 versus Controle.

70
Figura 04. Fotomicrografias de cortes representativos de tecido pulmonar exposto ao veneno
total da C.d.cascavella por via intraperitoneal na dose de 3,0 µg/Kg (H&E, 100x). Em (A):
Atelectasia (setas) de intensidade moderada, presente nos grupos de 6h,12h e 24h; (B):
Enfisema (setas) de intensidade leve, presente nos grupos de 1h, 3h, 6h, 12h e 24h; (C):
Edema (seta) no grupo de 24h; (D): Infiltrado inflamatório (linfócitos e plasmócitos) (seta)
presente em intensidade moderada nos grupos de 3h e 6h; (E): Presença em intensidade
aumentada de Tecido Linfoide associado aos Brônquios (BALT) (seta) nos grupos de 6h e
12h. Microscópio Nikon trinocular; ®View Software – Versão 7.3.1.7.
A B
C D
E
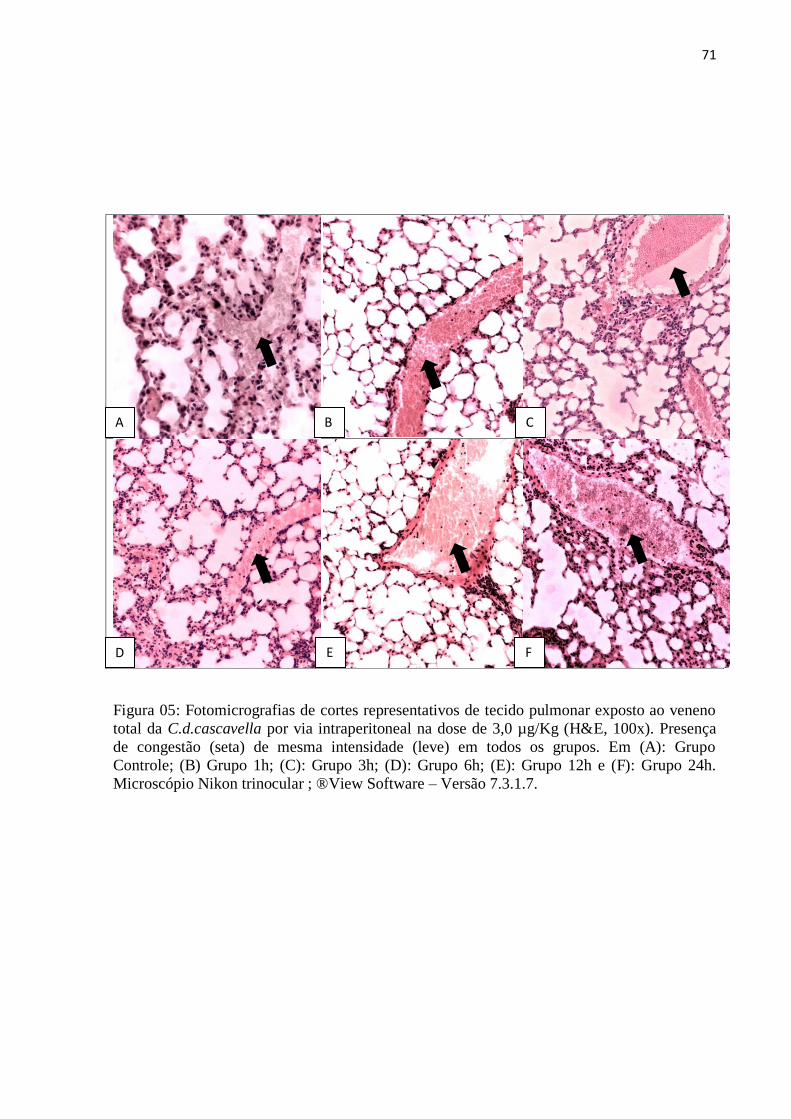
71
Figura 05: Fotomicrografias de cortes representativos de tecido pulmonar exposto ao veneno
total da C.d.cascavella por via intraperitoneal na dose de 3,0 µg/Kg (H&E, 100x). Presença
de congestão (seta) de mesma intensidade (leve) em todos os grupos. Em (A): Grupo
Controle; (B) Grupo 1h; (C): Grupo 3h; (D): Grupo 6h; (E): Grupo 12h e (F): Grupo 24h.
Microscópio Nikon trinocular ; ®View Software – Versão 7.3.1.7.
A B C
D E F
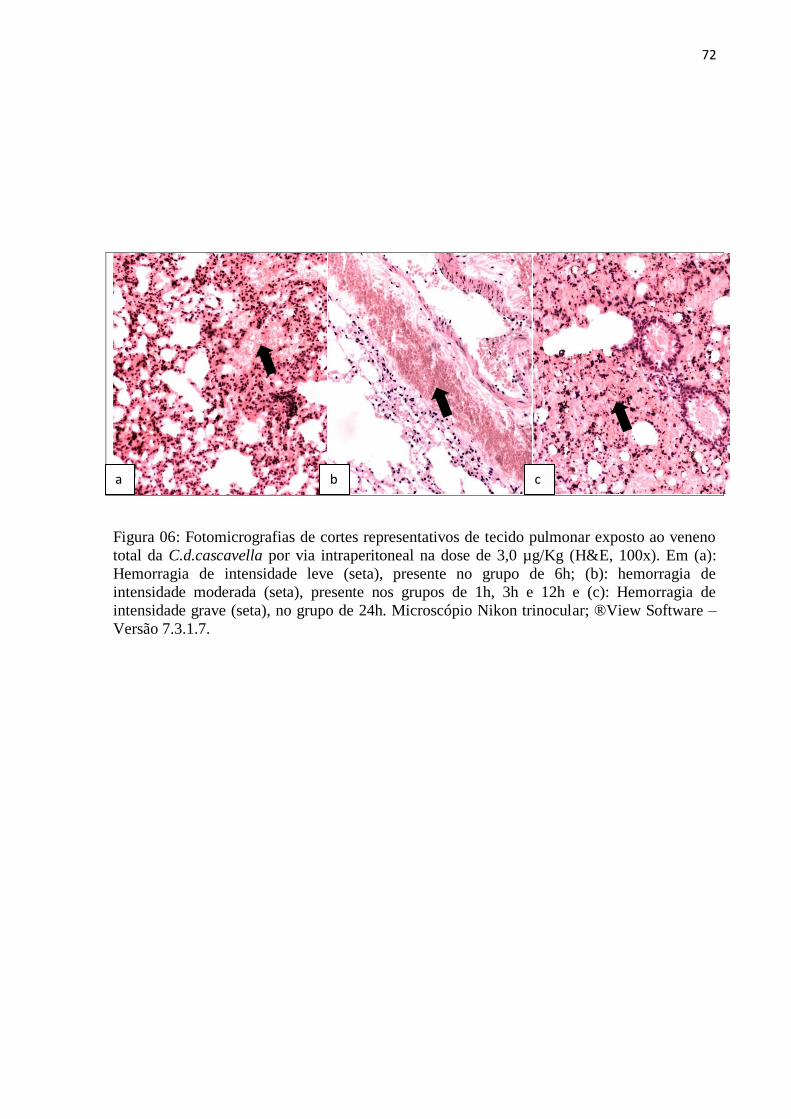
72
Figura 06: Fotomicrografias de cortes representativos de tecido pulmonar exposto ao veneno
total da C.d.cascavella por via intraperitoneal na dose de 3,0 µg/Kg (H&E, 100x). Em (a):
Hemorragia de intensidade leve (seta), presente no grupo de 6h; (b): hemorragia de
intensidade moderada (seta), presente nos grupos de 1h, 3h e 12h e (c): Hemorragia de
intensidade grave (seta), no grupo de 24h. Microscópio Nikon trinocular; ®View Software –
Versão 7.3.1.7.
a b c

73
7 CONCLUSÕES
Concluímos que o veneno de C. d. cascavella provoca alterações mecânicas e
histopatológicas no tecido pulmonar, porém seu mecanismo exato de ação mecânica
ehistopatológica nas vias aéreas não pôde ser esclarecido através deste estudo.
Estudos mais específicos sobre repercussões pulmonares do veneno de C. d.
cascavella se fazem necessários, a fim de se estabelecer se o processo de provocação da
injúria aguda se encontra na paralisia muscular e mionecrose sistêmica, levando a fatores
desencadeantes causadores de uma inflamação, ou se há uma ação direta dos fatores tóxicos
do veneno sobre o tecido pulmonar levando a alterações morfoestruturais e de propriedades
mecânicas.

74
8 PERSPECTIVAS
Utilizar os conhecimentos acerca das alterações em mecânica pulmonar, assim
como da histopatologia de tecido pulmonar promovidas pela ação farmacológica do veneno
bruto da serpente C. d. cascavella como ponto de partida para posteriores estudos com a
utilização desse veneno, assim como a utilização de substâncias isoladas contidas nele, afim
de investigações acerca de seus mecanismos fisiofarmacológicos.
Estudos posteriores serão essenciais para se determinar os fatores
desencadeadores causadores de processo inflamatório sobre o tecido pulmonar levando a
alterações morfoestruturais e de propriedades mecânicas.

75
REFERÊNCIAS BIBLIOGRÁFICAS
AIRES, M. M., Fisiologia. 2. ed., Rio de Janeiro, Guanabara Koogan, 1999.
ALBERTS, B.; BRAY, D.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WATSON, J., The cell. 3
ed. São Paulo, ed: Artes médicas sul; 8: 823-831, 1994.
ALBUQUERQUE, H. N. de.; COSTA, T. B. G. da.; CAVALCANTE, M. L. F., Estudo dos
acidentes ofídicos provocados por serpentes do gênero Bothrops no Estado da Paraíba.
Revista de Biologia e ciências da terra, v.5, n.1, 2004.
ALMEIDA-SANTOS, S. M.; GERMANO, V. J., Crotalus durissus (Neotropical
Rattlesnake). Prey.Herpetological Review, v.27, n.3, p.143, 1996.
ANTUNES, C. T.; YAMASHITA, K. M.; BARBARO, K. C.; SAIKI, M.; SANTORO. M. L.,
Compartive analysis of new Born and adult Bothrops jararaca snake venom. Toxicon. v.56,
p.1443-1458, 2010.
APPADU, B.L.; HANNING, C. D. Respiratory Physiology. In: PINNOCK, Colin; LIN,Ted;
SMITH, Tim. Fundamentals of anaesthesia. 2.ed. London: Greenwich Medical Media Ltd,
2003. p. 395-424.
AUTO, H.J.F., Animais peçonhentos. Maceió: EDUFAL, p.114, 1999.
AZEVEDO-MARQUES, M. M.; CUPO, P.; HERING, S. E. Acidentes por animais
peçonhentos: Serpentes peçonhentas. Medicina, 2003, v.36, p.480-489.
BARONE, J. M.; ALPONTI, R. F.; FREZZATTI, R.; ZAMBOTTIVILLELA, L.;
SILVEIRA, P. F., Differential efficiency of simvastatin and lipoic acid treatments on
Bothrops jararaca envenomation-induced acute kidney injury in mice. Toxicon, v.57, n.1,
p.148-156, 2011.

76
BARRAVIERA, B., Curso sobre Acidentes por animais peçonhentos: Acidentes por
Serpente do Gênero Crotalus.Arquivo Brasileiro de Medicina, v.64, p. 14-20, 1989.
BARRAVIERA, B.; PEREIRA, P. C. M. Acidentes por serpentes do gênero Bothrops. P.262-
273. In: BARRAVIERA, B. Venenos: Aspectos clínicos e terapêuticos dos acidentes por
animais peçonhentos. Rio de Janeiro: EPUB. 1999.411p.
BARROS, L C., SOARES, A.M., COSTA, F.L., RODRIGUES, V.M., FULY, A.L., GIGLIO,
J.R., GALLACCI, M., THOMAZINI-SANTOS, I.A., BARRAVIEIRA, S.R.C.S.,
FERREIRA JUNIOR, R. S., Biochemical and biological evaluation of gyroxin isolated from
Crotalus durissus terrificus venom. The Journal of Venomous Animals and Toxins including
Tropical Diseases, v.17, n.1, p.23–33, 2011.
BASTOS, E. G. M. A.; ARAÚJO, F. B.; SILVA, H. R., Records of the rattlesnakes Crotalus
durissus terrificus (Laurenti) (Serpentes, Viperidae) in the State of Rio de Janeiro,
Brazil: a possible case of invasion facilitated by deforestation. Revista Brasileira de
Zoologia, v.22, 3.ed, p.812-815, 2005.
BATES, J.H.T.; ABE, T.; ROMERO, P. V.; SATO, J. Measurement of alveolar pressure
inclosed chest dog during flow interruption. Journal of Applied Physiology, 67(1): 488-492,
1989.
BATES, J.T., Lung mechanics: An inverse modeling approach. Cambridge: Cambridge
University Press, p. 150 – 187, 2009.
BEAUPRE, S. J., DUVALL, D., O'LEILE, J., Ontogenetic variation in growth and sexual size
dimorphism in a Central Arizona population of the Western Diamond back Rattlesnake
(Crotalus atrox). Copeia, v.1, p.40-47, 1998.
BECK, D., Ecology and the energetics of tree sympatric rattlesnake species in the Sonoran
Desert. Journal of Herpetology, v.29, 2.ed, p. 211-223, 1995.
BEGHINI, D. G.; TOYAMA, M H; HYSLOP, S.; SODEK, L.; NOVELLO, J. C.;
MARANGONI1, S., 5 Enzymatic characterization of a novel phospholipase A2 from
Crotalus durissus cascavella rattlesnake (Maracambóia) venom. Journal of protein chemistry,
v. 19, n. 8, p. 679-84, 2000.

77
BERNE, R.M; LEVY, M.N; KOEPPEN, B.M; STANTON, B.A., Fisiologia. Tradução de:
Physiology, 5th ed; tradutores Nephtali Segal Grinbaum et al. Rio de janeiro: Elsevier, 2004.
BÉRNILS, R. S., Brazilian reptiles – List of species. Accessible at
http://www.sbherpetologia.org.br. Sociedade Brasileira de Herpetologia, 2010.
BOCHNER, R.; STRUCHINER, C. J., Epidemiologia dos acidentes ofídicos nos últimos 100
anos no Brasil: uma revisão. Cadernos de Saúde Pública, v.19, p.7-16, 2003.
BON, C., Snake venom and pharmacopoeia. In: Bauchot, R. (edit) Snakes a Natural
History. New York: Sterling Publishing Company, p.194-209, 1997.
BUENO, L. G. F.; RODRIGUES-SIMIONI, L.; MARANGONI, S.; CRUZ-HOFLING, M.
A.; NOVELLO, J. C. Neurotoxic and myotoxic actions from Lachesis muta muta (surucucu)
whole venom on the mouse and chick nerve-muscle preparations. Toxicon. v. 46, p.222-229,
2005.
BURGHARDT, G. M., Chemical-cue responces of inexperienced snakes: comparative
aspects. Science, v.157, p.718-721, 1967.
BLEASE, K. ; LUKACS, N.W. ; HOGABOAN, C.M. ; KUNKEL, S.L. Chemokines and
their role in airway hyper-reactivy. Respiratory Research,v. 1, n. 1, p. 54-61, 2000.
BRASIL, 2012. Epidemio-Ministério da Saúde, SVS – Sistema de Informação de Agravos de
Notificação – SINAN Net. Acesso em 15/03/2013.
BRASIL, 2001., Ministério da Saúde. Manual de diagnóstico e tratamento de acidentes por
animais peçonhentos. Brasília, 2001.
BRASIL, 2012. Epidemio-Ministério da Saúde, SVS – Sistema de Informação de Agravos de
Notificação – SINAN Net. Acesso em 15/12/2014.
BREITHAUPT, H.; RÜBSAMEN, K.; HABERMANN, E., Biochemistry and pharmacology
of the crotoxin complex. Biochemical analysis of crotapotin and the basic Crotalus
phospholipase A. European journal of biochemistry / FEBS, v. 49, n. 2, p.333-45, 1974.

78
BROWN, W. S.; MACLEAN, F. M., Conspecific scent-trailing by newborn timber
rattlesnake, Crotalus horridus. Herpetologica, v.39, p.430-436, 1983.
BRUSASCO, V.; PELLEGRINO, R. Airway Hyperresponsiveness: From Molecules to
Bedside Invited Review: Complexity of factors modulating airway narrowing in vivo:
relevance to assessment of airway hyperresponsiveness. Journal of AppliedPhysiology,v. 95,
n. 2, p. 1305–1313, 2003.
CÂMARA, P.; ESQUISATTO, L.; CAMARGO, E.; RIBELA, M.; TOYAMA, M.;
MARANGONI, S.; DE NUCCI, G.; ANTUNES, E., Inflammatory edema induced by
phospholipases A2 isolated from Crotalus durissus sp. in the rat dorsal skin: A role for mast
cells and sensory c-fibers. Toxicon, v.41, p.823-829, 2003.
CAMILLO, M.A.P., Contribuição ao estudo das giroxinas (enzimas semelhantes à trombina)
dos venenos das serpentes brasileiras Lachesis muta muta e Crotalus durissus terrificus. 1998.
75 f. Tese (Doutorado) – Instituto de Pesquisas Energéticas e Nucleares, São Paulo, São
Paulo, 1998.
CAMPBELL, J. A.; LAMAR, W. W., The venomous reptiles of Latin America. New York:
Cornell University, 1989.
CARDOSO, J. L. C.; JUNIOR, V. H.; FRANÇA, F. S.; MALAQUE, C. M. S.; WEN F.
H., Animais peçonhentos no Brasil: biologia, clínica e terapêutica dos acidentes. 2. ed., São
Paulo, Editora Sarvier – FAPESP, 2003, p.469.
CAVALCANTE, F. S.A; ITO, S.; BREWER, K.; SAKAI, H.; ALENCAR, A. M.;
ALMEIDA, M. P.; ANDRADE, J. S.; MAJUMDAR, A.; INGENITO, E. P.; SUKI, B.,
Mechanical interactions between collagen and proteoglycans: implications for the stability of
lung tissue. Journal of Applied Physiology; v.98, p. 672-679, 2005.
CHISZAR, D.; MELCER, T.; LEE, R.; RADICLIFFE, C. W.; DUVALL, D., Chemical cues
used by prairie rattlesnakes (Crotalus viridis) to follow trails of rodent prey. Journal of
Chemical Ecology, v.16,1.ed, p. 79-85, 1990.

79
CHIPPAUX, J. P., Snake-bites: apparaisal of the global situation. Bulletin World Health
Organization, v.5, n.76, p.515-524, 1998.
COTRAN, R.S.; KUMAR, V.; COLLINS, T.; ROBBINS, S.L., Pathology basis of
disease.6º ed. Philadelphia: W.B. Saunders, p. 89-112, 1999.
DAL PAI, V.; SANTO NETO, H. Ação dos venenos ofídicos sobre os tecidos animais.
In: BARRAVIERA, B. Venenos animais: uma visão integrada. Rio de Janeiro: Ed.
EPUB, 1994. p. 411.
DOS-SANTOS, M. C.; ASSIS, E. B.; MOREIRA, T.D.; PINHEIRO, J.; FORTES-DIAS, C.
L., Individual venom variability in Crotalus durissus ruruima snakes, a subspecies of
Crotalus durissus from the Amazonian region. Toxicon. v.46, n. 8, p. 958-961, 2005.
ECHEVERRIGARAY, S. ; GRAZZIOTIN, F. G. ; GRAZZIOTIN, G. G. ; AGOSTINI, G.,
Random amplified polymorphisms between two South American subspecies of rattlesnakes
(Crotalus durissus collilineatus and Crotalus durissus terrificus). Brazilian Archives of
Biology and Technology, v. 44, n.3, p. 313-317, 2001.
ERNST, C. H., Venomous Reptiles of North America. United States of America: Smithsonian
Institution Press, 1992.
EVANGELISTA, J. S. A. M.; MARTINS, A. M. C.; NASCIMENTO, N. R. F.; SOUSA,
C.M.; ALVES, R.S.; TOYAMA, D.O.; TOYAMA, M.H.; EVANGELISTA, J.J.F.;
MENEZES, D.M.; MANASSÉS, C.F.; MORAES, M.E.A.; MONTEIRO, H.S.A., Renal and
vascular effects of the natriuretic peptide isolated from Crotalus durissus cascavella venom.
Toxicon : official journal of the International Society on Toxinology, v.52, n.7, p.737-44,
2008.
EVANGELISTA, J. S. A. M., Estudo de efeitos vasculares, renais e citotóxicos do veneno da
serpente Crotalus durissus cascavella. 2008. Tese (Doutorado) - Universidade Federal do
Ceará, Fortaleza, Ceará, 2008.

80
FERRARO, G.C.; MORAES, J.R.E.; SHIMANO, A.C.; PEREIRA, G.T.; MORAES, F.R.;
BUENO DE CAMARGO, M.H., Efect of snake venom derived fbrin glue on the tendon
healing in dogs. Clinical and biomechanical study. Journal Venom Animals Toxins including
Tropical Diseases, v.11, n.3, p.261-74, 2005.
FONSECA, F. V.; ANTUNES, E.; MORGANTI, R. P.; MONTEIRO, H. S. A.; MARTINS,
A M. C.; TOYAMA, D. O.; MARANGONI, S., Characterization of a new platelet
aggregating factor from crotoxin Crotalus durissus cascavella venom. The protein journal, v.
25, n. 3, p.183–92, 2006.
FRASER, R. S., Histology and gross anatomy of the respiratory tract. In: Hamid, Q.;
Shannon, J.; Martin, J. Physiologic basis of respiratory disease. USA: PMPH, 2005, cap. 1, p.
1 – 14.
FREDBERG, J. J. Bronchospasm and its biophysical basis in airway smooth muscle.
Respiratory Research, v. 5, n. 1, p. 1 – 16, 2004.
FREDBERG, J.J.; INGRAM R.H. J.; CASTILE R.G.; GLASS G.M.; DRAZEN J.M.
Nonhomogeneity of lung response to inhaled histamine assessed with alveolar capsules.
Journal of Applied Physiology,v. 58, n. 6, p. 1914–1922, 1985.
FUNG, Y.C. Biomechanics: Mechanical Properties of Living Tissues. 2 ed: New York,
Springer-Verlag, 1993.
GARCIA, L.M.; LEWIS, R.J., Therapeutic potencial of venom peptides. Nature, v.2, p.790-
802, 2003.
GOLDSTEIN R.H., Control of type I collagen formation in the lung. The American Journal
of Physiology; v.5, p.29-40, 1991.
GOPALAKRISHNAKONE, P.; DEMPSTER, D. W.; HAWGOOD, B. J.; ELDER, H. Y.
Cellular and mitochondrial changes induced in the structure of murine skeletal muscle by
crotoxin, a neurotoxic phospholipase A2 complex. Toxicon: official jornal of the
International Society on Toxinology, v. 22, n. 1, p. 85-98, 1984.
HENDON, R.A., FRAENKEL-CONRAT, H., Biological role of the two components of
CrTX. Proceedings of the National Academy of Sciences. USA, v. 68, n. 7, 1560–1563.
1971.

81
HICKMAN, C. P. J.; ROBERTS, L. S.; LARSON, A., Princípios integrados de zoologia. 11.
ed., Rio de Janeiro, Guanabara Koogan, 2004, cap.28, p.540-541.
HIZUME, D.C. Efeitos da exposição a fumaça do cigarro sobre a inflamação, responsividade
e remodelamentos pulmonares em camundongos com inflamação pulmonar alérgica crônica.
2010. 105 f. Tese (Doutor em ciências)-Universidade de São Paulo, São Paulo, 2010.
INSTITUTO BUTANTAN. Animais Peçonhentos: Serpentes. Série Didática 5. São Paulo, SP
s/d. Disponível em: <http://www.toxnet.com.br/download/serpentes.pdf>.
Acesso em: 08 de agosto de 2014.
JANDROT-PERRUS, M.; LAGRUE, A. H.; OKUMA, M.; BON C. Adhesion and activation
of human platelets induced by convulxin involves glycoprotein VI and integrin alpha2beta1.
Journal of Biological Chemistry. V. 272, n. 43, p. 27035-27041, 1997.
JUNQUEIRA, L. C. & CARNEIRO, J. Histologia Básica. 11ª Edição. Rio de Janeiro, Editora
Guanabara Koogan. 2008.
KACZKA, D.W.; INGENITO E.P.; SUKI B.; LUTCHEN K.R. Partitioning airway and lung
tissue resistances in human: effects of bronchoconstriction. Journal of Applied Physiology, v.
82, n. 5, p. 1531–1541, 1997.
KANAJI, S.; KANAJI, T.; FURIHATA, K.; KATO, K.; WARE, J. L.; KUNICLI, T. J.
Convulxin binds to native, human glycoprotein IBα. The Journal of Biological Quemistry. v.
278, n. 41, p. 39452-39460, 2003.
KOCHI, T., OKUBO, S.; ZIN, W. A. et al. Chest wall and respiratory system mechanics in
cats: effects of flow and volume. Journal of Applied Physiology, 64: 2636-2646, 1988.
LEDUC, M. & BON, C. Cloning of subunits of convulxin, a collagen-like plateletaggregating
protein from Crotalus durissus terrificus venom. Biochemical Journal. v. 333, p.389-393,
1998.
LIMA, D.C.; ABREU, P. A.; FREITAS, C.C.; SANTOS, D.O.; BORGES, R.O.; SANTOS,
T.C.; CABRAL, L.M.; RODRIGUES, C.R.; CASTRO, H.C., Snake Venom: Any Clue for
Antibiotics and CAM? e CAM., v.2,n.1, p. 39-47, 2005.

82
LOMONTE, B.; ANGULO, Y.; CALDERÓN, L., An overview of lysine-49 phospholipase
A2 myotoxin from crotalid snake venoms and their structural determinants of myotoxic
action. Toxicon, v.42, p. 885-901, 2003.
MACHADO, M. G. R.; ZIN, W. A., Mecânica da respiração. In: ROCCO, P. R. M; ZIN, W.
A. Fisiologia respiratória aplicada. Rio de Janeiro: Guanabara Koogan, 2009, cap. 3, pág.21-
46.
MARTINS, A. M. C.; TOYAMA, M.H.; HAVT. A.; NOVELLO, J.C.; MARANGONI, S.F;
FONTELES, M.C.; MONTEIRO, H.S.A., Determination of Crotalus durissuscascavella
venom components that induce renal toxicity in isolated rat kidneys. Toxicon: official journal
of the International Society on Toxinology, v. 40, n. 8, p.1165-171, 2002.
MARTINS, A.M.C.; NOBRE, A.C.L.; ALMEIDA A.C.; BEZERRA, G.; LIMA, A.A.M.;
FONTELES, M.C.; MONTEIRO, H.S.A., Thalidomide and pentoxifylline block the renal
effects of supernatants of macrophages activated with Crotalus durissus cascavella venom.
Brazilian Journal of Medical and Biological Research, Ribeirão Preto, v. 37, n. 10,p. 1525 –
1530. 2004.
MATSUBARA, M. H. Efeitos do veneno de Crotalus durissus terrificus, da crotoxina e de
suas subunidades fosfolipase A2 e crotapotina em monocamadas de células endoteliais em
cultura. 2009. 106f. Dissertação (Mestrado em Biotecnologia) - Interunidades em
Biotecnologia, USP/Instituto Butantan/IPT, São Paulo, 2009.
MATSUI, T.; FUJIMURA, Y.; TITANI K., Snake venom proteases afecting hemostasis and
thrombosis. Biochimica et Biophysica Acta.;1477(1-2):146-56, 2000.
MELGAREJO, A. R., Serpentes Peçonhentas do Brasil. In: COSTA-CARDOSO, J.L.;
SIQUEIRA-FRANÇA, F. O.; WEN, F.H.; MÁLAQUE, C. M. S.; HADDAD, J. V.,Animais
Peçonhentos no Brasil: Biologia, Clínica e Terapêutica dos Acidentes. Sarvier-Fapesp, São
Paulo, p. 33–61, 2003.
MELGAREJO-GIMÉNEZ, A. R. Criação e Manejo de serpentes. IN: ANDRADE, A.;
PINTO, S. C.; OLICEIRA, R. S. Animais de laboratório. Criação e experimentação. Rio de
Janeiro: Editora FIOCRUZ, 2002. P. 175-198p.
MINISTÉRIO DA SAÚDE. Acidentes por animais peçonhentos: acidentes ofídicos. In: Guia
de Vigilância Epidemiológica. Secretaria de Vigilância em Saúde. Departamento de
Vigilância Epidemiológica. Brasília: Ministério da Saúde. p. 786-792, 2009.

83
MISEROCCHI G.; NEGRINI D.; PASSI A.; DE LUCA G., Development of lung edema:
interstitial fluid dynamics and molecular structure. The Journal of PhysiologicalSciences.
v.16, p. 66-71, 2001.
MORAES, J.R.E.; CORREA, P.H.A.; CAMPLESI, A.C.; MORAES, F.R., Experimental use
of fbrin glue derived from snake venom in non-pregnant canine uterus. Journal Venom
Animals Toxins including Tropical Diseases., v.10 (2):133-143, 2004.
MORENO, E.; QUEIROZ, M. A.; LIRA-DA-SILVA, R. M.; TAVARES- NETO, J., Clinical
and Epidemiological Characteristics of Snakebites in Rio Branco, Acre. Revista da Sociedade
Brasileira de Medicina Tropical, v. 38, p. 15-21, 2005.
NADUR-ANDRADE, N.; BARBOSA, A. M.; CARLOS, F. P.; LIMA, C. J.; COGO, J.
C.; ZAMUNER, S. R. Lasers In Medical Science, v. 27, p.65-70, 2012.
NICASTRO, G.; FRANZONI, L.; DE CHIARA, C.; MANCIN, A. C.; GIGLIO, J. R. &
SPISNI, A., "Solution structure of crotamine, a Na+ channel affecting toxin from Crotalus
durissus terrificus venom." European Journal of Biochemistry, v.270 (9): 1969-1979,2003.
NISHIOKA, S. A.; JORGE, M. T.; SILVEIRA, P. V. P.; RIBEIRO, L. A., South American
rattlesnake bite and soft-tissue infection: report of a case. Revista da Sociedade Brasileira de
Medicina Tropical, v.3, n.4, p.401-402, 2000.
NONAKA, P. N.; AMORIM, C. F.; PERES, A. C. P.; SILVA, C. A. M.; ZAMUNER, S. R.;
RIBEIRO, W.; COGO, J.C.; VIEIRA, R.P.; DOLHNIKOFF, M.; OLIVEIRA, L. V. F.,
Pulmonary mechanic and lung histology injury induced by Crotalus durissus terrificus snake
venom. Toxicon, v.51, p. 1158-1166, 2008.
NORMAN, D. R. Anfibios y Reptiles del Chaco Paraguayo. San José: Rare Center, v.1, 1994.
OLIVEIRA, D. G. DE; TOYAMA, MARCOS H; MARTINS, A. M.; HAVT, A.; NOBRE,
A.C.L; MARANGONI, S.; CÂMARA, P.R.; ANTUNES, E.; DE NUCCI, G.; BELIAM,
L.O.S; FONTELES, M.C.; MONTEIRO, HELENA S.A., Structural and biological
characterization of a crotapotin isoform isolated from Crotalus durissuscascavella venom.
Toxicon, v. 42, n. 1, p. 53-62, 2003.
OLIVEIRA, F. N.; BRITO, M. T.; MORAIS, I. C. O.; FOOKS, M. L.; ALBUQUERQUE, H.
N., Accidents caused by Bothrops and Bothropoides in the State of Paraiba: epidemiological
and clinical aspects. Revista da Sociedade Brasileira de Medicina Tropical, v.43, n.6, 2010.

84
OLIVEIRA, M. E.; MARTINS, M., When and where to find a pitviper: activity patterns and
habitat use of a lance head Bothrops atrox in central Amazonia, Brazil. Herpetological
Natural History, v.8, 2.ed, p. 101-110, 2001.
PAVONE, L.; ALBERT, S.; DIROCCO, J.; GATTO, L.; NIEMAN, G., Alveolar instability
caused by mechanical ventilation initially damages the nondependent normal lung. Critical
Care. v. 11, p. 1-10, 2007.
PEREAÑEZ, J.A.; NÚÑEZ, V.; HUANCAHUIRE-VEGA, S.; MARANGONI, SERGIO;
PONCE-SOTO, L. A., Biochemical and biological characterization of a PLAR (2R) from
crotoxin complex of Crotalus durissus cumanensis. Toxicon : oficial Journal of the
International Society on Toxinology, v. 53, p. 534-542, 2009.
PINHO, F. M.; OLIVEIRA, E. S.; FALEIROS, F., Snakebites in the State of Goiás, Brazil.
Revista da Associação Médica Brasileira. v.50,p.93-96,2004.
PINHO, F. M. O.; PEREIRA, I. D., Ofidismo. Revista da Associação Médica Brasileira,
v.47, n.1, p. 24-29, 2001.
PINHO, F. M.; VIDAL, E.; BURDMANN, E. A., Atualização em insuficiência renal aguda.
Insuficiência renal aguda após acidente crotálico. Jornal Brasileiro de Nefrologia, v.22, p.62-
68, 2000.
PINHO, F. M.; ZANETTA, D. M.; BURDMANN, E. A., Acute Renal Failure after Crotalus
durissus Snakebite: A Prospective Survey on 100 Patients. Kidney Int, v.67, p.659-67,2005.
POLGÁR, J.; CLEMETSON, J. M.; KEHREL, B. E.; WIEDEMANN, M.; MAGNENAT, E.
M.; WELLS, T. N.; CLEMETSON, H. J., Platelet activation and signal trans by convulxin, a
C-type lectin from Crotalus durissus terrificus (tropical rattlesnake) venom via the p62/GPVI
collagen receptor. The Journal of Biological Chemistry. v. 272, n. 21, p.13576-13583, 1997.
POSSMAYER, F.; YU, S.H.; WEBER, J.M.; HARDING, P.G., Pulmonary surfactante. Can.
The International Journal of Biochemistry & Cell Biology.v.62, p. 1121–1133, 1984.
PLACE, A. J.; ABRAMSON, C. I., A quantitative analysis of the ancestral area of
rattlesnakes. Journal of Herpetology, v.38, n.1, p.151-156. 2004.

85
PRADO-FRANCESCHI, J., On the pharmacology of convulxin and giroxin. Memórias do
Instituto Butantan, v.52, p.25-6, 1990. Suplemento.
QUEIROZ, G. P.; PESSOA, L. A.; PORTARO, F. C. V.; FURTADO, M. F. D.;
TAMBOURGI, D.V. Interspecific variation in venom composition and toxicity of brasilian
snakes from Bothrops genus. Toxicon, v. 52, p.842-851, 2008.
RANGEL-SANTOS, A.; DOS-SANTOS, E. C.; LOPES-FERREIRA, M.; LIMA, C.;
CARDOSO, D. F.; MOTA, I., A comparative study of biological activities of crotoxina and
CB fraction of venoms from Crotalus durissus terrificus, Crotalus durissus cascavella and
Crotalus durissus collilineatus. Toxicon., v.43, n.7, p. 801-810, 2004.
RAGHOW, R., The role of extracellular matrix in postinflammatory wound healing and
fibrosis. Faseb Journal, v. 8, n. 11, p. 823-831, 1994.
RATELSLANGEN. Crotalus durissus - South American rattlesnake. [2014]. Disponível em:
http://www.ratelslangen.nl/gallery/Crotalus_durissus_South_ American_rattlesnake/. Acesso
em março de 2014.
RAW I.; ROCHA M.C.; ESTEVES M.I.; KAMIGUTI A.S., Isolation and characterization of
a thrombin-like enzyme from the venom of Crotalus durissus terrifcus. Brazilian Journal of
Medical and Biological Research., v.19, n.3, p.333-338,1986.
REINERT, H. K.; CUNDALL, D.; BUSHAR, L. M., Foraging behavior of the timber
rattlesnake, Crotalus horridus. Copeia, v.4, p. 976-981, 1984.
SBH. 2014. Lista de espécies de répteis do Brasil. Sociedade Brasileira de Herpetologia
(SBH). Disponível em: http:// www.sbherpetologia.org.br/checklist/repteis.htm.
SALOMÃO, M. G.; ALMEIDA-SANTOS, S. M., The reproductive cycle in male neotropical
rattlesnake (Crotalus durissus terrificus), In J. A. CAMPBELL & E. D. BRODIE JR(eds.),
Biology of the Pit Vipers, Selva Press, Texas, p. 506-514, 2002.
SALOMÃO, M. G.; ALMEIDA-SANTOS, S. M.; PUORTO, G., Activity patterns of
Crotalus durissus (Viperidae, Crotalinae): Feeding, Reproduction and Snakebite. Studies on
Neotropical Fauna and Environment, v. 30, n. 2, p. 101-106, 1995.
SANO-MARTINS, I.S.; TOMY, S.C.; CAMPOLINA, D.; DIAS, M.B.; DE CASTRO,
S.C.B.; SOUSA-E-SILVA, M.C.C.;AMARAL, C.F.S.; REZENDE, M.A.; KAMIGUTI,

86
A.S.; WARRELL, D.A.; THEAKSTON, R.D.G. Coagulopathy following lethal and non-
lethal envenoming of humans by the South American rattlesnake in Brazil. Q J M, v.94, n.10,
p.551-559, 2001.
SANTORO, M.C.; SOUSA-E-SILVA, M.C.; GONÇALVES, R.L.; ALMEIDA-SANTOS, S.
M.; CARDOSO, D.F.; LAPORTA-FERREIRA, I. L.; SAIKI, M.; PERES, C. A.; SANO-
MARTINS, S., Comparação das atividades biológicas em venenos de três subespécies da
cascavel sul-americana (Crotalus durissus terrificus, C. durissus cascavella e C. durissus
collilineatus). Comparative Biochemistry and Physiology. Parte C: Farmacologia e
Endocrinologia, v. 122, p.61-73, 1999.
SANTOS, E., Anfíbios e répteis. 3. ed., Belo Horizonte, Itatiaia,1995, p.263.
SAZIMA, I., Um estudo de biologia comportamental da jararaca, Bothrops jararaca, com
uso de marcas naturais.Memórias do Instituto Butantan, v. 50, p. 83-99, 1988.
SECOR, S. M., Ecological significance of movements and activity range for the side
winder, Crotalus cerastes.Copeia, v.3, p.631-645, 1994.
SERRANO, S.M.; MAROUN, R.C., Snake venom serine proteinases: sequence homology vs.
substrate specifcity, a paradox to be solved. Toxicon., v.45(8), p.1115-32, 2005.
SILVA, M. G. Câncer em Fortaleza 1978-1980. Instituto e Registro de câncer do Ceará.
Fortaleza, p.135, 1982.
SILVEIRA, K. S. O.; BOECHEM, N.T.; NASCIMENTO, S.M.; MURAKAMI Y.L.B.;
BARBOZA, A.P.B.; MELO, P.A.; CASTRO, P.; MORAES V.L.G.; ROCCO, P.R.M.; ZIN,
W.A., Pulmonary mechanics and lung histology in acute lung injury induced by Bothrops
jararaca venom. Respiratory Physiology & Neurobiology. v.139, p.167–177, 2004.
SINAN, 2013. Sistema de Informação de Agravos de Notificação [documento na Internet].
Animais peçonhentos. Acesso 26 de Outubro de 2013. Disponível em:
http://dtr2004.saude.gov.br/sinanweb/index.php.
SIX, D. A.; DENNIS, E. A., The expanding superfamily of phospholipase A 2 enzymes :
classi ¢ cation and characterization. Group, v. 1488, 2000.
SOERENSEN, B., Producão de soros antipeçonhentos e características imunológicas de cada
soro. Anavenenos. On. SOERENSEN, B. Animais peçonhentos-reconhecimento, distribuição

87
geográfica, produção de soros e tratamento dos envenenamentos. Rio de Janeiro/São Paulo,
Atheneu. 1990.
SNYDER L.S.; HERTZ M.I.; HARMON K.R.; BITTERMAN P.B.; STARCHER B.C., Lung
elastin and matrix. Chest; 117:229S-34, 2000.
SPINOSA, H. S.; GÓRNIAK, S. L.; PALERMO-NETO, J., Toxicologia aplicada à
Medicina Veterinária. 1.ed. Barueri: Manole, 2008. 942p.
SUKI B.; BARABASI A-L.; LUTCHEN K. R., Lung tissue viscoelasticity: a mathematical
framework and its molecular basis. Journal of Applied Physiology, v. 76, p.2749-2759, 1994.
TOYAMA, M. H. ; CARNEIRO, Everardo M. ; MARANGONI, Sergio ; AMARAL, M. E.
C. ; VELLOSO, L. A. ; BOSCHERO, A. C. . Isolation and characterization of a Convulxin
like protein from Crotalus durisssus collilineatus venom. Journal of Protein Chemistry, NY,
USA, v. 20, n.7, p. 585-591, 2001.
TOYAMA, M.H.; TOYAMA, D.D.O.; PASSERO, L.F.D.; LAURENTI, M.D.; CORBETT,
C.E.; TOMOKANE, T.Y.; FONSECA, F.V.; ANTUNES, E.; JOAZEIRO, P.P.; BERIAM,
L.O.S.; MARTINS, M.A.C.; MONTEIRO, H.S.A; FONTELES, M.C., Isolation of a new L
amino acid oxidase from Crotalus durissus cascavella venom. Toxicon : official journal of the
International Society on Toxinology, v. 47, n. 1, p. 47- 57, 2006.
VANZOLINI, P.E.; RAMOS-COSTA, A. M. M.; VITT, L. J., Répteis das caatingas. Rio de
Janeiro, Academia Brasileira de Ciências, p.161, 1980.
VITAL BRAZIL, O. Neurotoxins from the South American rattlesnake venom. Journal of the
Formosan Medical Association. V. 71, p. 394-400, 1072. In: DAMICO, D. C. S.; BUENO, L.
G. F.; RODRIGUES-SIMIONI, L.; MARANGONI, S.; CRUZ-HOFLING, M. A.;
NOVELLO, J. C. Neurotoxic and myotoxic actions from Lachesis muta muta (surucucu)
whole venom on the mouse and chick nerve-muscle preparations. Toxicon. v. 46, p.222-229,
2005.
YUAN, H.; KONONOV, S.; CAVALCANTE, F.S.A.; LUTCHEN, K.R.; INGENITO, E.P.;
SUKI, B. Effects of collagenase and elastase on the mechanical properties of lung tissue
strips. Journal of Applied Physiology,v. 89, n. 1, p. 3-14, 2000.
ZIN, W. A.; ROCCO, P. R.M., Fisiologia Respiratória Aplicada. 1 ed. Guanabara. 2009.

88
ZIN, W. A.; ROCCO, P. R. M.; FAFFE, D. S., Fisiologia da respiração. In: Aires, M. M.
(Org.) Fisiologia. 3 ed. Rio de Janeiro: Guanabara Koogan, v. 1, p. 623-639, 2008.
WEST, J. B., Fisiologia Respiratória. 8° ed. Proto Alegre: Editora Artmed, 2010.


















