
MINISTÉRIO DA EDUCAÇÃO E DESPORTOS UNIVERSIDADE …livros01.livrosgratis.com.br/cp028280.pdf2...
49
MINISTÉRIO DA EDUCAÇÃO E DESPORTOS UNIVERSIDADE FEDERAL DE GOIÁS INSTITUTO DE PATOLOGIA TROPICAL E SAÚDE PÚBLICA Luiz Fernando Nunes Rocha Avaliação de fungicidas para isolamento in vitro de fungos patogênicos para invertebrados Orientador: Dr. W. Christian Luz Dissertação de Mestrado Goiânia-GO, 2006
Transcript of MINISTÉRIO DA EDUCAÇÃO E DESPORTOS UNIVERSIDADE …livros01.livrosgratis.com.br/cp028280.pdf2...

MINISTÉRIO DA EDUCAÇÃO E DESPORTOS
UNIVERSIDADE FEDERAL DE GOIÁS
INSTITUTO DE PATOLOGIA TROPICAL E
SAÚDE PÚBLICA
Luiz Fernando Nunes Rocha
Avaliação de fungicidas para isolamento in vitro de fungos
patogênicos para invertebrados
Orientador:
Dr. W. Christian Luz
Dissertação de Mestrado
Goiânia-GO, 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
UNIVERSIDADE FEDERAL DE GOIÁS
INSTITUTO DE PATOLOGIA TROPICAL E
SAÚDE PÚBLICA
PROGRAMA DE PÓS-GRADUAÇÃO EM
MEDICINA TROPICAL
Luiz Fernando Nunes Rocha
Avaliação de fungicidas para isolamento in vitro de fungos
patogênicos para invertebrados
Orientador:
Dr. W. Christian Luz
Dissertação submetida ao
CPGMT/IPTSP/UFG como re-
quisito parcial para obtenção de
Grau de Mestre, na área de
concentração de Parasitologia
Goiânia-GO, 2006

3
Agradecimentos
Agradeço a todos que de alguma forma colaboraram para a realização deste trabalho, em
especial:
Ao Dr. Christian Luz, Professor do Departamento de Microbiologia, Imunologia, Parasitologia e
Patologia (DMIPP), Setor de Parasitologia do Instituto de Patologia Tropical e Saúde Pública (IPTSP)
da Universidade Federal de Goiás (UFG), pela especial orientação e amizade.
Ao IPTSP e ao Programa de Pós-Graduação em Medicina Tropical da UFG, pela oportunidade.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pela bolsa
concedida.
Ao Setor de Parasitologia e ao Laboratório de Patologia de Invertebrados, em especial ao colega
Morel Cipriano de Bastos Netto, pela ajuda.
Aos meus pais e irmãos, pelo apoio, incentivo e participação.
A minha querida esposa Wanessa, pela força, compreensão e apoio.

4
Sumário
Agradecimentos............................................................................................................................. 3
Resumo ......................................................................................................................................... 5
Abstract ........................................................................................................................................ 6
Introdução ................................................................................................................................... 7
Vetores ........................................................................................................................... 7
Controle clássico de vetores .......................................................................................... 8
Controle microbiano de vetores .................................................................................... 9
Isolamento de fungos ..................................................................................................... 11
Objetivos gerais .......................................................................................................................... 12
Objetivos específicos ................................................................................................................. 12
Manuscrito 1: Utility of six fungicides for selective isolation of entomopathogenic fungi ….. 13
Manuscrito 2: In vitro susceptibility to fungicides by invertebrate-pathogenic and saprobic fungi 27
Conclusões ....................................................................................................................................... 40
Bibliografia ...................................................................................................................................... 41

5
Resumo
A alta biodiversidade de fungos patogênicos para invertebrados e seu potencial para controle de
pragas enfatiza a importância de procurar por novas espécies e linhagens eficazes. Entretanto, o
conhecimento sobre o isolamento de fungos, especialmente entomopatogênicos, com meio seletivo ou
semi-seletivo é restrito. No presente trabalho, testou-se a suscetibilidade in vitro de 41 isolados de 19
fungos patogênicos para invertebrados (6 Beauveria bassiana, B. brongniartii, Cladosporium
cladosporioides, Culicinomyces clavisporus, Duddingtonia flagrans, 6 Evlachovaea sp., Gliocladium sp.,
Hirsutella thompsonii, 8 Metarhizium anisopliae, Nomuraea rileyi, 7 Paecilomyces spp., Sporothrix
insectorum e 6 Tolypocladium cylindrosporum) e 11 contaminantes, originários de criadouros de
mosquitos no Centro-Oeste do Brasil (5 Aspergillus spp., Cunninghamella echinulata, Fusarium roseum,
Mortierella isabellina, Mucor plumbeus, Rhizopus arrhizus e Trichothecium roseum) a fungicidas
(dodine, benomyl, thiabendazole, manzate, sulfato cúprico e oxicloreto de cobre). O efeito dos fungicidas
foi avaliado na germinação de conídios e na formação de halo para os fungos B. bassiana, Evlachovaea
sp., M. anisopliae e T. cylindrosporum, e na formação de colônias e conidiogênese para todas as espécies
e isolados estudados. A germinação, formação de halo, colônias e conidiogênese em colônias de B.
bassiana, Evlachovaea sp., M. anisopliae e T. cylindrosporum variaram entre os fungicidas e suas
concentrações. Diferentes isolados destes fungos apresentaram uma distinta suscetibilidade ligada aos
fungicidas. Além disso, em concentrações críticas dos fungicidas para o desenvolvimento fúngico, uma
elevada variabilidade da suscetibilidade, entre repetições, foi observada. A maioria dos outros fungos
patogênicos e contaminantes tinham um padrão distinto de suscetibilidade a benomyl (1 mg/l), dodine (50
mg/l), manzate (100 mg/l), sulfato cúprico (200 mg/l) e thiabendazole (4 mg/l). Este último, que permitiu
desenvolvimento de um alto número de fungos patogênicos e inibiu a maioria dos contaminantes, parece
ser o fungicida mais apropriado para isolamento de fungos patogênicos para invertebrados, de substratos
com elevado teor de água e ricos em matéria orgânica. Benomyl e dodine, freqüentemente empregados
para isolamento específico de B. bassiana e M. anisopliae, foram também úteis para outras espécies.
Oxicloreto de cobre não foi apropriado para meio semi-seletivo por apresentar baixa atividade fungicida,
mesmo em concentrações elevadas de até 30 g/l. Manzate e sulfato cúprico mostraram ser menos úteis
para isolamento in vitro devido a baixa inibição dos contaminantes testados. Nenhum fungicida foi ativo,
na concentração testada, contra os fungos com crescimento rápido como C. echinulata, M. plumbeus e R.
arrhizus. Os resultados mostram a importância de se adaptar o fungicida e sua concentração para o fungo
ou os fungos alvos e possíveis contaminantes.

6
Abstract
The high biodiversity of fungi pathogenic to invertebrates and their potential for pest control
emphasize the importance to look for new effective species and strains. However, knowledge about
isolation of fungi, especially entomopathogenic, with selective or semi-selective media is restricted. In the
present study, in vitro susceptibility was tested of 41 isolates of 19 fungi pathogenic to invertebrates (6
Beauveria bassiana, B. brongniartii, Cladosporium cladosporioides, Culicinomyces clavisporus,
Duddingtonia flagrans, 6 Evlachovaea sp., Gliocladium sp., Hirsutella thompsonii, 8 Metarhizium
anisopliae, Nomuraea rileyi, 7 Paecilomyces spp., Sporothrix insectorum and 6 Tolypocladium
cylindrosporum), and 11 contaminants which originated from mosquito breeding sites in Central Brazil (5
Aspergillus spp., Cunninghamella echinulata, Fusarium roseum, Mortierella isabellina, Mucor plumbeus,
Rhizopus arrhizus and Trichothecium roseum) to fungicides (dodine, benomyl, thiabendazole, manzate,
cupric sulphate and copper oxychloride). The fungicidal effect was evaluated on conidial germination and
halo formation of B. bassiana, Evlachovaea sp., M. anisopliae and T. cylindrosporum and on colony
formation and conidiogenesis of all species and isolates studied. Germination, formation of halo, colonies
and conidiogenesis on colonies of B. bassiana, Evlachovaea sp., M. anisopliae and T. cylindrosporum
varied among fungicides and concentrations. Different isolates of these fungi showed a clear isolate-
related susceptibility to fungicides. Moreover, at critical concentrations of fungicides for fungal
development, an elevated variability of susceptibility, among replicates, was observed. Most other
pathogenic fungi and contaminants had clear patterns of susceptibility at benomyl (1 mg/l), dodine (50
mg/l), manzate (100 mg/l), cupric sulphate (200 mg/l) and thiabendazole (4 mg/l). The last fungicide,
which permitted development of a high number of pathogenic fungi and inhibited most contaminants,
seems to be the most appropriate fungicide to isolate fungi pathogenic to invertebrates from substrates
with high water contents and rich in organic material. Benomyl and dodine, often employed for specific
isolation of B. bassiana and M. anisopliae, were also useful for other pathogenic fungi. Copper
oxychloride could not be recommended for semi-selective media due to its low fungicidal activity even at
higher concentrations up to 30 g/l. Manzate and cupric sulphate showed to be less helpful for in vitro
isolation due to a low inhibition of tested contaminants. No fungicide was active, at tested concentrations,
against fast-growing C. echinulata, M. plumbeus and R. arrhizus. Results underline importance to adapt
fungicide and its concentration to target fungus or fungi and possible contaminants.

7
Introdução
Vetores
Muitos invertebrados como moluscos, ácaros e, especialmente, insetos têm grande importância na
transmissão de patógenos ou de parasitos para o homem. A classe Insecta constitui um grupo bem-
sucedido e possui o maior número de espécies em todo reino Animalia. Esta classe contém ectoparasitos e
vetores importantes, como mosquitos, outros dípteros, triatomíneos, pulgas e piolhos.
Os mosquitos (Culicidae) são insetos holometabólicos que possuem quatro estágios distintos, que
são ovo, larva, pupa e adulto. Dependendo da espécie, ocorrem em ambientes silvestres, rurais e urbanos,
sendo que somente as fêmeas são hematófagas e responsáveis pela transmissão de diversos agentes como
vírus, protozoários e helmintos. Os três gêneros de maior importância dos culicídeos são Aedes, Culex e
Anopheles.
Aedes aegypti é mundialmente conhecido como o principal vetor dos vírus da dengue e da febre
amarela urbana. No Brasil, essa espécie tem muita importância devido a sua vasta distribuição, alta
densidade e adaptação a áreas urbanas. Seus criadouros, aquáticos de pequeno porte, localizam-se em
ambientes intra e peridomiciliares. Atualmente, a dengue é uma das arboviroses mais importantes em
todo mundo por estar associada a aglomerações urbanas (Halstead 1993, WHO 1997, Gubler 2005) e
apresentar peculiaridades que dificultam a produção de vacinas e medicamentos (Yasui 1993, Khin et al.
1994). Estima-se que, a cada ano, 50 milhões de pessoas contraiam a doença em todo mundo. Dessas,
cerca de 500 mil desenvolvem a dengue hemorrágica e o número de mortos é de aproximadamente 24 mil
pessoas (WHO 2002).
Culex quinquefasciatus é encontrado em regiões tropicais e sub-tropicais. Esse mosquito é o
maior perturbador do repouso noturno humano em nosso país, além disso, transmite a Wuchereria
bancrofti, agente etiológico da filariose linfática humana, principalmente em áreas urbanas de Alagoas,
Pernambuco e Pará (Medeiros et al. 2004). A espécie é altamente sinantrópica e associada a aglomerados
urbanos e rurais, procriando, principalmente, em criadouros com água e matéria orgânica em
decomposição. As fêmeas se alimentam nas horas mais avançadas da noite, coincidindo com a presença
de microfilárias de W. bancrofti no sangue periférico. A prevalência da filariose linfática aumentou em
países de clima tropical e subtropical úmido, principalmente pela expansão não planejada da urbanização
em áreas endêmicas (Albuquerque 1993). Estima-se em cerca de 120 milhões o número de pessoas
parasitadas em todo mundo. No Brasil, esse número é de aproximadamente 49 mil pessoas e mais de 3
milhões moram em áreas de risco (Medeiros et al. 2004).
Anopheles darlingi, espécie altamente suscetível aos Plasmodium spp., é o vetor mais importante
da malária no Brasil. Reproduz, preferencialmente, em grandes coleções de águas limpas, expostas ao sol
e com muita vegetação. As fêmeas podem atacar o homem em áreas peridomiciliares, mas preferem fazê-
lo dentro das casas, principalmente ao crepúsculo vespertino e matutino. A malária é a doença parasitária
que acomete o maior número de pessoas em todo mundo, com cerca de 300-500 milhões de infectados e
1,5 a 3 milhões de mortes a cada ano (WHO 2000).

8
Os triatomíneos são insetos hemimetabólicos que exercem a hematofagia desde as ninfas até
adultos, tanto os machos como as fêmeas. Foram descritas, pelo menos 128 espécies, classificadas em 5
tribos e 17 gêneros (Schofield et al. 1999). Com exceção do gênero Linshcosteus e algumas espécies do
gênero Triatoma, todos os outros triatomíneos ocorrem exclusivamente no continente americano, desde a
Argentina até os EUA (Diotaiuti et al. 2000). Nas Américas são transmissores do Trypanosoma cruzi,
agente etiológico da doença de Chagas. Cerca de 16 a 18 milhões de pessoas estão infectadas com esse
protozoário e mais de 100 milhões vivem em áreas de risco (WHO 1999). A maioria das espécies é
silvestre, todas potencialmente transmissoras do T. cruzi, e associada a uma ampla variedade de
hospedeiros vertebrados, que servem como reservatórios do parasito. Algumas espécies adaptaram-se a
ambientes peridomiciliares ou domiciliares e têm papel importante como transmissores para o homem e
animais domésticos. No sul da América Latina, o vetor clássico intradomiciliar, T. infestans, com vasta
distribuição e densidades elevadas, é hoje, após campanhas intensas de combate, considerado erradicado
em muitas regiões do Cone Sul, inclusive no Centro-Oeste brasileiro (Dias et al. 2002). Espécies
peridomiciliares e silvestres estão, entretanto, invadindo e ocupando ambientes domiciliares e a
transmissão vetorial dessa doença, mesmo sendo baixa atualmente, não está banida. Em regiões fronteiras
entre a Argentina e Bolívia foram encontradas, recentemente, populações resistentes a piretróides,
inseticida comumente utilizado para combate desses vetores (comunicação pessoal Patrícia Juarez). Além
disso, espécies como T. sordida, T. brasiliensis, T. dimidiata, T. pseudomaculata, Panstrongylus
rufotuberculatus, Rhodnius nasutus, R. negletus, R. stali, Eratyrus mucronatus e outras espécies já foram
encontradas no interior de casas (Noireau et al. 1995; Dujardin et al. 1998, 2000; Schofield et al. 1999;
Matias et al. 2003). No estado de Goiás, T. williami e T. costalimai são espécies com alta adaptação
domiciliar comprovada (Silveira et al. 1984; Silva et al. 1992).
Controle clássico de vetores
Inseticidas sintéticos são a principal forma de combate a insetos vetores, há décadas. Os primeiros
inseticidas utilizados em campanhas para o combate de mosquitos e triatomíneos, a partir de 1945, foram
o diclorodifeniltricloretano (DDT), hexaclorocicloexano (BHC), dieldrin e outros organoclorados
(Hemingway & Ranson 2000, Aché & Matos 2001). Em 1955, a Organização Mundial de Saúde (OMS)
recomendou o uso do DDT para a erradicação global da malária através da borrifação em domicílios.
Entretanto, logo após a euforia inicial, foram registrados os primeiros casos de resistência de anofelinos
ao DDT. Mais tarde, foram encontrados outros mosquitos resistentes a inseticidas (Hemingway & Ranson
2000). Muitos organoclorados tiveram que ser retirados do mercado por afetar a saúde do homem e de
animais e por serem altamente agressivos ao meio ambiente (D’Amato et al. 2002). Novas classes de
inseticidas sintéticos, constituídas por organofosforados, carbamatos e piretróides, foram desenvolvidas
para se obterem produtos mais seguros e eficazes. A utilização indiscriminada de inseticidas, porém,
agravou o desequilíbrio ambiental e o número de casos de resistência em mosquitos, triatomíneos e outros
vetores não deixou de crescer (Zerba 1999, Vassena et al. 2000, Karunaratne & Hemingway 2001,
Alexander & Maroli 2003, Somboon et al. 2003, Audino et al. 2004).

9
A resistência de insetos a inseticidas ocorre por mecanismos comportamentais e fisiológicos
(Roberts & André 1994, Brogdon & McAllister 1998). No primeiro caso, um inseto muda de
comportamento e evita assim contato com o inseticida. A resistência fisiológica aparece com a síntese de
enzimas específicas como esterases, glutathione s- transferase ou monooxygenases, pelos insetos, que
desativam o inseticida (Hemingway et al. 2004). Essas enzimas têm sido relatadas atuando em
organoclorados, organofosforados, piretróides e carbamatos (Hemingway & Ranson 2000). A resistência
pode ocorrer também após substituição de aminoácidos chaves por outros no sítio de ligação do inseticida
o que impede a ligação específica e atuação do produto (Brengues et al. 2003).
Controle microbiano de vetores
Devido à preocupação com a resistência a inseticidas e conscientização crescente sobre o risco de
produtos químicos para o homem e o meio ambiente, microrganismos patogênicos, utilizados para
controle biológico de pragas agrícolas, estão sendo estudados em vetores e se consolidando no combate
integrado de mosquitos. Entomopatógenos são geralmente mais específicos do que inseticidas sintéticos e
apresentam baixa ou nenhuma toxidez para vertebrados, para outros insetos benéficos e para plantas
(Whiteley & Schnepf 1986). As bactérias, Bacillus thuringiensis israelensis (B.t.i.) e B. sphaericus (B.s.),
foram os primeiros microrganismos usados no controle biológico de vetores, especialmente de culicídeos.
Ambas mostraram ter ação seletiva e rápida contra larvas de culicídeos e outros dípteros de importância
em saúde pública (Federici et al. 2003, Monnerat et al. 2005). O B.t.i. produz pelo menos três diferentes
toxinas de cristal e uma toxina com atividades cito e hemolítica (Polanczyk et al. 2003). O B.s. produz
uma toxina de cristal com duas proteínas binárias e outra toxina nomeada MTX (Charles et al. 1996). A
ação conjunta e complexa destas toxinas reduz a probabilidade de induzir resistência nas larvas (Regis et
al. 2001). B.t.i. e B.s. são usados em programas de controle de mosquitos no Brasil (Regis et al. 2000,
Lima et al. 2005).
Outros microrganismos como os Baculovírus spp. estão sendo utilizados com sucesso para
combate de pragas agrícolas como Anticarsia gemmatalis que acomete plantações de soja (Moscardi &
Souza 2002). Existem poucos estudos sobre atividade de vírus em vetores (Barreau et al. 1996, Muscio et
al. 1997, 2000, Rozas-Dennis et al. 2002). Esses insetos também são acometidos por vírus, mas não se
conhecem espécies com alta virulência e potencial para combate de vetores.
O principal mecanismo de infecção de insetos por vírus e bactérias é por via oral. Larvas de
dípteros ou sifonápteros ingerem formas infectantes com o alimento. Contudo, para combate de insetos
exclusivamente hematófagos, como triatomíneos ou adultos de muitos dípteros, esses microrganismos são
inviáveis. Outros patógenos como fungos que invadem seus hospedeiros principalmente pela cutícula e
não precisam ser ingeridos parecem ter potencial para o combate integrado (Lacey & Goettel 1995).
Os fungos constituem o segundo maior grupo de organismos eucariontes do planeta, só perdendo
para os insetos (Rossman et al. 1998). Existem cerca de 1,5 milhões de espécies das quais mais de 700
são entomopatogênicas e agrupadas em 90 gêneros, causando cerca de 80% das doenças de insetos e
outros artrópodes (Glare & Milner 1991, Hawksworth 1991, Destéfano et al. 2004). A atividade de fungos

10
patogênicos para moluscos ou outros invertebrados com importância para saúde humana é pouco
conhecida. Acredita-se que menos de 5% dos fungos patogênicos para invertebrados foram descritos e
caracterizados (Hawksworth 1991). A grande biodiversidade de espécies e linhagens, como seus
metabólitos secundários tóxicos, apresentam um enorme potencial para controle de vetores e outras
pragas (Butt & Goettel 2000; Inglis et al. 2001).
A invasão do hospedeiro pelo fungo inicia-se com a adesão de formas infectantes, geralmente
conídios, à cutícula do vetor. Durante a germinação de conídios, o fungo forma um tubo germinativo e na
extremidade um apressório, que serve como apoio durante a penetração na cutícula. Quando o apressório
não é formado pode haver produção de uma massa mucilaginosa ao redor do tubo germinativo que segura
o fungo sobre a cutícula e liberando enzimas. A penetração ocorre através de processos mecânicos e
fisiológicos, como ação de enzimas e pressão mecânica das hifas. Após a penetração, o fungo forma
blastosporos que dissemina-se com a hemolinfa em todo o hospedeiro. Dependendo da virulência do
fungo e da suscetibilidade do inseto, a infecção pode levá-lo à morte. Após a morte do inseto, e em
umidade favorável, novo micélio aparece sobre o cadáver e o fungo produz conídios ou esporos que são
disseminados pelo ambiente e contaminam novos hospedeiros.
Os fungos foram utilizados entre os primeiros patógenos de insetos, mesmo sem grande sucesso,
para combater pragas agrícolas (Metchnikoff 1879). Após melhor compreensão dos mecanismos de
atuação dos fungos nos insetos o interesse para o seu uso no controle biológico cresceu bastante. No
Brasil, há mais de 60 anos, os fungos são estudados como entomopatógenos (Alves 1998). Contudo, a
grande maioria dos estudos e aplicações está relacionada ao controle de pragas agrícolas e existem poucos
trabalhos sobre fungos atuando em vetores. Os três gêneros de maior interesse para combate de estágios
aquáticos de mosquitos são, Lagenidium, Culicinomyces e Coelomomyces, porém, até hoje foi
desenvolvido um único produto à base de L. giganteum, para combate de larvas de culicídeos (Scholte et
al. 2004). Infelizmente esse produto (LaginexTM) teve registro somente por pouco tempo nos EUA e há
vários anos não está mais disponível no mercado. A principal vantagem de L. giganteum para combate é
sua alta resistência em condições de campo. Ele pode causar epizootias nos mosquitos e tem mostrado
capacidade de hibernar. Conforme Federici (1995) apenas uma única aplicação por estação é necessária
para o combate. Recentes trabalhos mostraram que outro fungo, Metarhizium anisopliae, que normalmente
não ocorre em habitats aquáticos, também tem atividade em larvas de culicídeos (Scholte et al. 2003, 2004,
Silva et al. 2004, 2005).
Os primeiros testes sobre atividade de fungos em triatomíneos foram feitos nos anos 60 (Dias &
Leão 1967). Desde então, o impacto de fatores abióticos como umidade relativa e temperatura e de fatores
bióticos como virulência ligada aos fungos e suscetibilidade ligada ao hospedeiro, foi estudado (Romaña &
Fargues 1992, Luz et al. 1994, 1998a, b, c, 1999, 2003a, b, 2004a, c, Luz & Fargues 1999, Fargues & Luz
2000, Lecuona et al. 2001). Fungos como M. anisopliae e Beauveria bassiana, foram encontrados em
habitats peridomiciliares de triatomíneos. Existem também relatos sobre ocorrência natural de B. bassiana
em triatomíneos encontrados mortos no campo na Índia e na Argentina (Parameswaran & Sankaran 1977,
Marti et al. 2005) e de outra espécie patogênica, ainda não identificada, do gênero Evlachovaea,

11
encontrada sobre uma ninfa morta de T. sordida no estado de Goiás (Luz et al. 2003b). Todos os isolados
tiveram atividade comprovada, em condições de laboratório, em triatomíneos mostrando que fungos atuam
como inimigos naturais desses insetos (Luz et al. 2004a). Em testes de campo, na proximidade de São Luís
de Montes Belos, no estado de Goiás, o número de T. sordida, em áreas peridomiciliares infestadas,
diminuiu claramente durante pelo menos 6 meses após aplicação de conídios de B. bassiana formulados
em óleo-água. Indivíduos mortos com micélio e conídios na superfície da cutícula foram detectados nesta
área (Luz et al. 2004b).
Isolamento de fungos
Até hoje, poucos trabalhos sobre a biodiversidade de fungos patogênicos e coleta de novas
espécies e isolados com potencial para controle biológico foram realizados no Brasil (Shimaxu et al.
1994, Luz et al. 2003b, 2004a). Os fungos utilizados em controle biológico são isolados diretamente de
insetos infectados vivos ou mortos, ou indiretamente de substratos contaminados utilizando insetos como
isca (Almeida & Batista-Filho 2001). Larvas de coleópteros como Tenebrio molitor, Tribolium castaneum
e Acanthocinus aedilis, de lepidópteros, Galleria mellonella e ninfas de T. infestans já foram utilizadas
como insetos-isca para isolamento de fungos de substratos (Vänninen 1995, Luz et al. 2004a). Além de
técnicas de isolamento in vivo, meios semi-seletivos ou seletivos são empregados para isolamento in vitro
e em grande escala de fungos. Fungicidas acrescidos no meio atrasam ou inibem especificamente o
crescimento de fungos não desejados e favorecem o desenvolvimento do ou dos fungos alvos. Existem
fungos contaminantes, que, na ausência de fungicida, crescem com alta velocidade e inibem o
crescimento de fungos procurados, comprometendo assim, sua detecção e isolamento. O conhecimento
sobre fungicidas e sua utilidade para isolamento de fungos patogênicos de invertebrados estão restritos a
poucos fungos e fungicidas. Dodine é um dos fungicidas mais utilizados em meio seletivo para detectar
Beauveria spp. e Metarhizium spp. (Veen & Ferron 1966; Chase et al. 1986; Sneh 1991; Liu et al.1993).
Outros produtos como sulfato cúprico, benlate e thiabendazole foram indicados para isolar Metarhizium
spp., Paecilomyces spp. e Culicinomyces clavisporus, respectivamente (Yaginuma & Takagi 1986,
Mitchell et al. 1987; Panter & Frances 2003). Existem resultados divergentes na literatura a respeito de
concentrações recomendadas dos produtos em meios seletivos para os mesmos fungos (Veen & Ferron
1966; Chase et al. 1986; Sneh 1991; Liu et al.1993). As divergências estão, provavelmente, ligadas à
qualidade dos produtos comercializados em diferentes paises e à variabilidade da suscetibilidade, entre
linhagens da mesma espécie, a um fungicida. Há poucas informações sobre o efeito específico de
fungicidas para outros fungos com patogenicidade em insetos e demais invertebrados, vetores importantes
e contaminantes, ocorrendo nos mesmos habitats. Desta forma, informações sobre a seletividade e
atividade de fungicidas em fungos patogênicos para invertebrados irão contribuir para o aprimoramento
de meios seletivos e assim permitir um aumento significativo do número de isolados e espécies com
potencial para combate de vetores e outras pragas. Existe um grande número, de fungos patogênicos para
invertebrados, desconhecidos e ainda não explorados. Além disso, bancos de germoplasma no Brasil

12
quase não dispõem de fungos específicos para vetores, como Tolypocladium cylindrosporum ou
Culicinomyces clavisporus que atacam larvas de mosquitos.
Objetivos gerais
Contribuir para o controle biológico de vetores e de outras pragas.
Contribuir para desenvolvimento de meios semi-seletivos específicos para isolamento de fungos
patogênicos para invertebrados.
Objetivos específicos
Verificar o efeito dos fungicidas benomyl, dodine, manzate, oxicloreto de cobre, sulfato cúprico e
thiabendazole sobre o desenvolvimento in vitro de B. bassiana, Evlachovaea sp., M. anisopliae e T.
cylindrosporum.
Verificar a suscetibilidade de fungos patogênicos para invertebrados como B. brongniartii,
Cladosporium cladosporioides, Culicinomyces clavisporus, Duddingtonia flagrans, Gliocladium sp.,
Hirsutella thompsonii, M. a.var. acridum, M. a. var. lepidiotum, Nomuraea rileyi, Paecilomyces spp. e
Sporothrix insectorum, aos mesmos fungicidas, exceto, oxicloreto de cobre.
Verificar o efeito do benomyl, dodine, manzate, oxicloreto de cobre, sulfato cúprico e
thiabendazole em fungos contaminantes como Aspergillus spp., Cunninghamella echinulata, Fusarium
roseum, Mortierella isabellina, Mucor plumbeus, Rhizopus arrhizus e Trichothecium roseum, coletados
em criadouros de mosquitos.

13
Utility of six fungicides for selective isolation of entomopathogenic fungi
Luiz Fernando Nunes Rocha
DMIPP, Instituto de Patologia Tropical e Saúde Pública, Universidade Federal de Goiás (UFG), CP 131,
74001-970 Goiânia, GO, Brasil
Address for correspondence: DMIPP, IPTSP, UFG, CP 131, 74001-970 Goiânia, GO, Brazil Tel: (55) 62
209 6113; Fax: (55) 62 521 1839: E-mail: [email protected]
Running title: Susceptibility of entomopathogenic fungi to fungicides
Abstract
The effect of six fungicides, dodine, benomyl, thiabendazole, manzate, cupric sulphate and copper
oxychloride, was examined in vitro upon germination and further development of Beauveria bassiana,
Evlachovaea sp, Metarhizium anisopliae and Tolypocladium cylindrosporum. Fungicidal activity depende
d on concentrations and varied among products, fungi and its strains tested. Depending on the fungicidal
concentration, germination of conidia was induced but germlings produced no or few mycelium and
conidia. M. anisopliae showed to be the most resistant species to all fungicides. Fungi were highly
resistant to copper oxychloride up to 30 g/l or susceptibility varied among replicates as shown for
Evlachovaea sp. This fungicide showed to have no use for selective medium. Considering 50% colony
forming units (CFU) grown on fungicide-added medium related to CFU grown on fungicide-free medium
a minimum for successful isolation, 1 mg/l benomyl, 200 mg/l cupric sulphate, 50 mg/l dodine, 100 mg/l
manzate and 4 mg/l thiabendazole can be recommended to isolate fungi tested in this study. There was a
good recovery of fungi from previously sterilized soils with fungicide-based medium. Thiabendazole
which is easily to obtain and can be used in low quantities showed greatest utility for selective medium of
entomopathogenic fungi.
Key words: Beauveria bassiana, Evlachovaea sp, Metarhizium anisopliae, Tolypocladium
cylindrosporum, fungicide, selective medium
Introduction
The high biodiversity of entomopathogenic fungi and their potential for insect pest control
emphasize the importance to look for new efficacious strains. If not isolated directly from infected
insects, fungi are detected in natural environments with host-baiting techniques or selective media.
Selective and semi-selective media based on antimicrobial agents contain antibiotics and other agents
with microbistatic or microbicidal activity. These agents suppress undesired microorganisms, such as
fast-growing saprobic and often antagonistic bacteria and fungi, more than the target fungus or fungi.
Many fungicides have broad spectra of activity and also affect entomopathogenic fungi. Guanidines act

14
on permeability of cellular membranes, benzimidazoles on nuclear division, while dithiocarbamates and
metallic fungicides inhibit enzymes. Selectivity of a fungicide-based medium is expected to be directly
related to the fungal susceptibility to the product and its concentration used. Knowledge about the utility
of fungicides for isolation of fungi with selective media is restricted to few entomopathogenic species.
Oxgall, cupric sulphate, dodine and benomyl were used separately or in combination to detect especially
Beauveria bassiana and Metarhizium anisopliae (Veen & Ferron 1966, Chase, Osborne & Ferguson
1986, Yaginuma & Takagi 1986, Sneh 1991, Liu et al. 1993). Benomyl was also utilized for
Paecilomyces lilacinus (Mitchell, Kannwischer-Mitchell & Dickson 1987) and thiabendazole for isolation
of Culicinomyces clavisporus (Panter & Frances 2003). However, there are considerable discrepancies
about doses recommended for the same fungicide and its use for fungal detection in literature. This is the
case of dodine and isolation of M. anisopliae from soil samples where the indicated doses of the fungicide
varied between 10 mg/l (Liu et al. 1993) and 460 mg/l (Chase, Osborne & Ferguson 1986). Another
product, benomyl was used by Mitchell, Kannwischer-Mitchell & Dickson (1987) at 50 mg/l to isolate P.
lilacinus, whereas Chase, Osborne & Ferguson (1986) recommended a much lower dose (0.38 mg/l) of
this fungicide to isolate B. bassiana and M. anispliae. Divergences about concentration-related activity,
the difficulty to obtain fungicides commonly reported in literature and the lack of information about the
effect of fungicides on other important invertebrate-pathogenic fungi underline the need to check known
data and acquire new learning about cited and other fungi and fungicides and to improve so selectivity of
media. We report on the in vitro susceptibility of B. bassiana, M. anisopliae, Tolypocladium
cylindrosporum and Evlachovaea sp, a new fungus isolated recently from Triatoma sordida (Reduviidae,
Hemiptera) with potential for biological control of triatomine vectors (Luz, Rocha & Humber 2003, Luz,
Rocha & Silva 2004), to different fungicides and a strain-related susceptibility to fungicides.
Materials and Methods
Origin of fungi
Fungi tested were obtained from different collections of entomopathogenic fungi: IPTSP, UFG,
Goiânia, GO, Brazil (IP) and USDA-ARS, Ithaca, NY, USA (ARSEF). All B. bassiana (IP 6, IP 94, IP
129, IP 132, IP 147, IP 153), Evlachovaea sp (IP 67, IP 126, IP 141, IP 142, IP 148), except IP 218,
which was found on T. sordida in Central Brazil (Luz et al. 2003), and M. anisopliae (IP 1, IP 75, IP 115,
IP 120, IP 125, IP 145), were detected in soils from Central Brazil in 2001. All ARSEF strains cited here
are T. cylindrosporum: ARSEF 705 and 1580 were isolated from Aedes sierrensis (Culicidae, Diptera),
1982 and 1984 in USA, respectively. ARSEF 1027 was found on a non-identified culicid in New Zealand
in 1982 and ARSEF 2912 on Plecia nearctica (Bibionidae, Diptera), 1973 in USA. ARSEF 962 and
ARSEF 2920 were detected in 1971 and 1970 in a soil sample in Canada and England, respectively.
Preparation of fungi
Fungi were cultured on complete medium (CM) during 15 d at 25° ± 1°C, 75 ± 5% relative
humidity and a 12 h photophase. The CM was prepared with 0.001 g FeSO4, 0.5 g KCl, 1.5 g KH2PO4,

15
0.5 g MgSO4·7 H2O, 6 g NaNO3, 0.001 g ZnSO4, 1.5 g hydrolyzed caseine, 0.5 g yeast extract, 10 g
glucose, 2 g peptone, 20 g agar and 1000 ml dist. H2O. PH was adjusted to 7. Conidia were harvested
directly from the surface of the culture by scraping. They were then suspended in 10 ml 0.1% Tween 80,
filtered, quantified based on hemacytometer counts and adjusted to final concentrations.
Fungicides
The six fungicides used in this study were one guanidine: dodine (Dodine 65W, 65% active
ingredient, Cyanamid Química do Brasil Ltda., Rio de Janeiro), two benzimidazoles: benomyl (Benlate
500, 50% active ingredient, Du Pont do Brasil, Barueri), thiabendazole (50% active ingredient, Novartis,
Rio de Janeiro), one dithiocarbamate: manzate (Manzate 800, 80% active ingredient, Du Pont do Brasil,
Barueri), and two metallic fungicides: cupric sulphate (PA, Vetec, CP Chemical Inc., Rio de Janeiro) and
copper oxychloride (Cobre Fersol, 84% active ingredient, Fersol Indústria e Comércio Ltda., São Paulo).
All fungicides were wettable powders and first dissolved in water (dodine, manzate, cupric sulphate and
copper oxychloride) or in 70% ethanol (benomyl and thiabendazole).
Evaluation of fungicidal activity
The effect of fungicides was assessed on quantitative germination and colony forming units
(CFU) with posterior production of mycelium and new conidia after inoculation of conidia on fungicide-
amended CM. Germination was tested with a single strain for each fungus: B. bassiana (IP 6),
Evlachovaea sp (IP 218), M. anisopliae (IP 120) and T. cylindrosporum (ARSEF 2920). Tests about
formation of colonies were done with the same strains except assays on strain-related susceptibility,
where 5 other strains mentioned above for each fungus were assessed. All tests were done at 25° ± 1°C
and a 12 h photophase with 4 independent replicates for each fungus, fungicide and method used.
Testing germination ten ml of fungicide-amended CM at three different concentrations of the
fungicide (Table 1) were arranged in Petri dishes (60 x 12 mm). Concentrations included an
approximately hundredfold range of values reported for entomopathogenic fungi in literature (benomyl,
cupric sulphate, dodine and thiabendazole) or the same and a tenfold inferior and superior concentration
recommended by the manufacturer for the control of phytopathogenic fungi (manzate and copper
oxychloride). Suspended conidia (500 μl) were inoculated onto the medium at a final 106 conidia/cm2.
The suspension was spread uniformly on the medium moving carefully the dishes and dried during one h
at ambient conditions. For each test conidial viability > 98% was confirmed at the same moment using
CM without fungicide. Progress of germination and development of mycelium and new conidia were
monitored daily up to 5 d post inoculation (p. i.) considering ungerminated, swollen, germination-initiated
and germinated conidia. Conidia with an elongating germ tube longer than its diameter were considered
germinated (Luz & Fargues 1997).

16
Testing formation of colonies and conidiogenesis on CFU, fungicide-amended and fungicide-free
CM arranged in Petri dishes as mentioned were inoculated with 500 μl of suspended conidia (6x102
conidia/ml) in order to obtain a final concentration of approximately 10 conidia/cm2. The suspension was
distributed equally on the surface of the medium by moving gently horizontally the dish. Growth of CFU
and conidiogenesis on the colonies were analyzed daily by microscopy (20 – 80x) up to 360 h p. i. The
total number of CFU found on fungicide-added medium was related to quantitative CFU detected at the
same moment on CM without fungicide. Initially, four concentrations of each fungicide based on
germination dynamics found in previous tests, considering also posterior formation of mycelium and new
conidia, were assayed for each fungus (Table 1). Isolation from soils was tested at the lowest
concentration that permitted growth of at least 40% of colonies of all fungi and intra-specific or generic
variability was assayed at the concentration of fungicide that permitted a emergence > 50% of colonies of
each fungus in previous tests (Table 1).
Detection of conidia in soil was carried out with red latossol. Samples were collected from an
undisturbed Cerrado region of Central Brazil. Soil showed no visible organic material and was sieved to
particles with diameters ≤ 0.6 mm. Conidia were suspended in 0.1% Tween 80 at 106 conidia/ml and then
one ml added to 10 g of previously autoclaved soil. Inoculated soil was mixed and dried for 2 h under
sterile conditions at 25°C and 75% relative humidity. One g was then suspended in 9 ml 0.1% Tween 80
in 15-ml tubes containing 2-3 dozens of glass beads (2 mm in diameter) and vortexed for three min. One
ml of the suspension was inoculated on solid fungicide amended CM in Petri dishes (100 x 20 mm) at a
final concentration of approximately 10 conidia/cm2 and growth of colonies was then evaluated as
mentioned.
Analysis of data
Arcsine-square root transformed germination and relative CFU data were examined using
analysis of variance (ANOVA), and the Student-Newman-Keuls (SNK) multiple range test of comparison
of means. Means were considered not statistically different at P > 0.05.
Results
Germination and subsequent development
Conidia of all fungi tested on fungicide-free CM started germination at 6-8 h p. i. Within 24 h
most conidia (> 98%) had germinated and first mycelium and new conidia could be detected 48 and 72 h
p. i., respectively. High germination rates ≥ 96.5%, regardless of the fungi tested, were found for benomyl
(0.5-50 mg/l), copper oxychloride (0.3-3 g/l), cupric sulphate (40-400 mg/l) and thiabendazole (0.2-20
mg/l) after 120 h incubation. At this moment only dodine, manzate and thiabendazole had a significant
effect of the concentration on the germination regardless of the fungi (F ≥ 5; df = 2, 47; P ≤ 0.01; Table
2). Germination of conidia exposed to dodine varied between 68.3% (T. cylindrosporum) and > 98% (B.
bassiana and M. anisopliae) at 50 mg/l and did not exceed 34% at 500 mg/l of the fungicide. Manzate at
2 g/l induced also a distinct variation of germination among fungi: whereas no or only few conidia of T.

17
cylindrosporum and Evlachovaea sp had swollen or germinated, an elevated germination (60.8%) was
found for M. anisopliae. For copper oxychloride and manzate at highest concentration of 30 and 20 g/l,
respectively germination could not be analyzed due to the dark coloration of the CM after adding the
fungicide. Contrarily to the other fungicides, manzate-added medium emitted a malodorous odor after
autoclaving which increased at higher concentrations and CM amended with cupric sulphate at 4 g/l did
not solidify.
Whereas mycelium and conidia of all tested fungi could be detected macroscopically on CM
added with copper oxychloride at 30 g/l, 120 h p. i., no fungal development at all was observed on cupric
sulphate or manzate-added CM at 4 g/l and 20 g/l at the same moment, respectively (Table 2). At a
tenfold lower concentration of both fungicides only M. anisopliae had produced new conidia with
manzate and using cupric sulphate and thiabendazole at highest 20 mg/l all fungi with the exception of B.
bassiana were found with new conidia. Despite a high germination rate (≥ 97.3%) of all fungi at 5 mg/l
benomyl only M. anisopliae produced few mycelium but no new conidia at this concentration. However,
distinct conidiogenesis could be observed for all fungi at a tenfold lower concentration. Few mycelium
without new conidia were found at dodine (500 mg/l) for both B. bassiana and M. anisopliae that showed
reduced germination at this concentration.
Concentration fitting of fungicides
First CFU and new conidia on colonies could be observed 48 and 72 h after inoculating conidia
on fungicide-free CM, respectively. Adding fungicide to the medium the periods until emergence of CFU
were generally retarded. The period between inoculation of conidia on the medium and appearance of
first CFU declined and the number of CFU increased with lower concentrations. At 240 h p. i. there was a
highly significant effect of the concentration on colony formation for all fungicides (F ≥ 8.4; df = 3, 63; P
< 0.001), except copper oxychloride (F = 2.3; df = 3, 63; P = 0.084). For this fungicide an elevated
number of colonies, regardless of the concentration or a high variability of CFU among replicates for
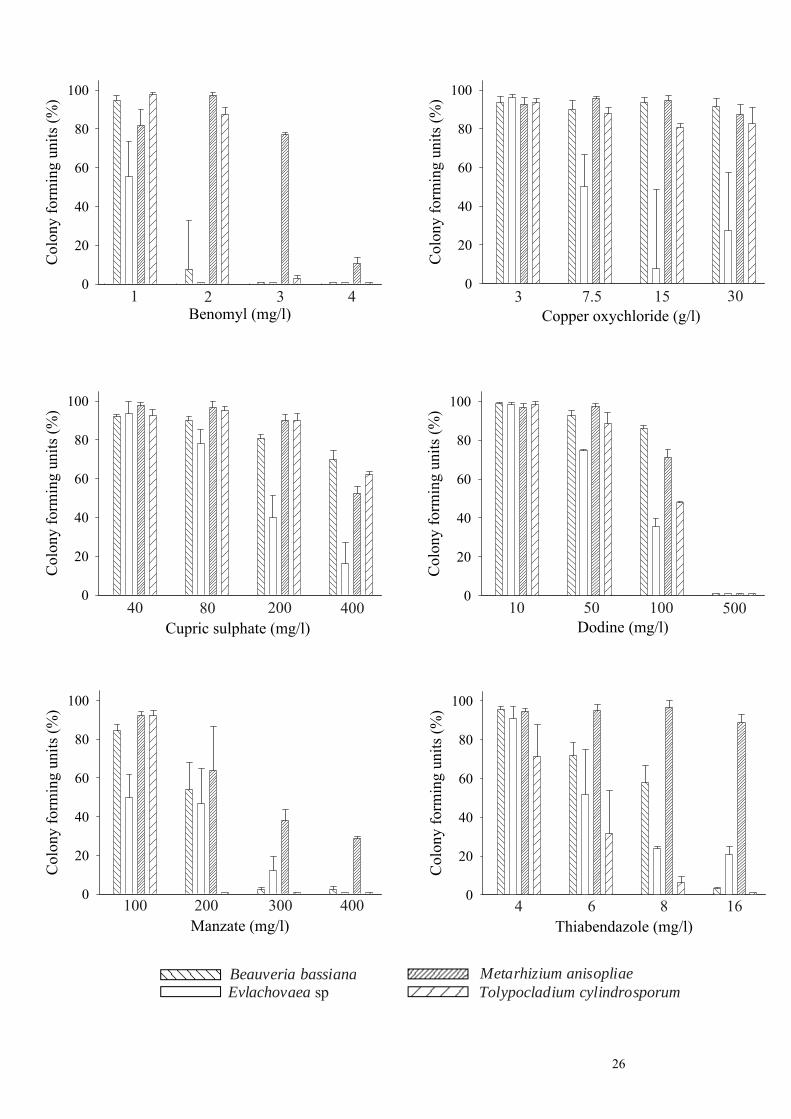
Evlachovaea sp at ≥ 7.5 g/l could be observed (Fig. 1). Cupric sulphate also permitted a distinct formation
of colonies for all fungi and concentrations although with lower numbers for Evlachovaea sp at highest
400 mg/l. No CFU at all had developed on CM amended with dodine at 500 mg/l, 240 h p. i.,
independently of the fungus tested. Similar results were found for benomyl testing B. bassiana (3 and 4
mg/l), Evlachovaea sp (2-4 mg/l) and T. cylindrosporum (4 mg/l). The last species exposed to manzate
(200-400 mg/l) and thiabendazole (16 mg/l) did not produce any colonies too and this was finally also the
case for Evlachovaea sp and manzate (400 mg/l) (Fig 1). A highly significant effect of the fungus on the
number of colonies, regardless of the concentration was found for all fungicides (F ≥ 4.1; df = 3,63; P ≤
0.01), 240 h p. i., except for dodine (F = 0.4; df = 3,63; P = 0.75). Highest values of CFU were obtained
generally for M. anisopliae, followed by B. bassiana, T. cylindrsporum and Evlachovaea sp (F = 13.8; df
3, 380; P < 0.001). First new conidia on colonies developing on fungicide-amended medium could be
observed on CFU, 72 h p. i. Advanced conidiogenesis was noticed for most fungi, 240 h p. i., even at
lower rates of CFU. However, production of new conidia was distinctly reduced when Evlachovaea sp

18
and T. cylindrosporum were exposed to cupric sulphate at 400 mg/l. Only few conidia were also found on
colonies of B. bassiana testing benomyl at 2 mg/l and thiabendazole at 16 mg/l and on colonies of T.
cylindrosporum testing the same fungicides at 3 mg/l and 6-8 mg/l, respectively.
Soil tests
All fungi produced at least 32.1% colonies and new conidia on the colonies, 360 h after
inoculating conidia mixed with soil on fungicide-amended CM, regardless of the fungicide tested (Table
3). Whereas the number of colonies did not differ among fungi tested with cupric sulphate (200 mg/l),
manzate (100 mg/l) and thiabendazole (4 mg/l) (F ≤ 0.7, df = 3, 15; P ≥ 0.58), significantly less CFU of
B. bassiana (66.2%) and Evlachovaea sp (32.1%) compared to M. anisopliae (94.7%) and T.
cylindrosporum (93.5%) were observed on benomyl (1 mg/l) added CM (F = 13.2, df = 3, 15; P < 0.001).
Testing dodine (50 mg/l) relative numbers of colonies were also distinctly reduced for Evlachovaea sp
(64.6%), compared to the other fungi where values were found at ≥ 88.8% (F = 4.5, df = 3, 15; P = 0.02).
Variability tests
For all fungal species and Evlachovaea sp a highly significant effect of the strain on quantitative
formation of colonies was noted, 240 h after exposure of conidia to fungicide-amended CM considering
all 5 fungicides tested (F ≥ 6.6; df = 4, 75; P < 0.001; Table 4). Highest variation of CFU for the same
species or genus was found at this moment for B. bassiana and Evlachovaea sp. For B. bassiana and its
strains, values of CFU varied between < 10% and > 90% when testing manzate and thiabendazole. CFU
of Evlachovaea sp found for different strains diverged between < 3% and > 67%, after inoculation of
conidia on benomyl or thiabendazole amended medium (Table 4). An elevated variation among strains
was also noted exposing M. anisopliae (20.7% up to 72%) and T. cylindrosporum (41.9% up to 98.5%) to
dodine. The same fungicide had no effect on colony formation of the B. bassiana strains. This was also
found for cupric sulphate testing Evlachovaea sp and M. anisopliae and benomyl testing T.
cylindrosporum. Highest CFU (≥ 56.6%), regardless of the fungus or strain tested, was found for cupric
sulphate.
Discussion
Results clearly showed that fungi were compatible with all fungicides tested. However, their
effect on fungal development depended on its concentration and the time of exposure. Moreover,
susceptibility to fungicides varied among fungi and tested strains. Except copper oxychloride fungicides
had different patterns of dose-related inhibition of tested fungi. Susceptibility of Evlachovaea sp to
copper oxychloride varied highly among replicates at ≥ 7.5 g/l of the product and all other fungi were
highly resistant up to 30 g/l. This fungicide showed to have no use for selective medium for the tested and
probably also for other entomopathogenic fungi. On the contrary the other products had more potential
for a specific isolation with selective media. Susceptibility of Evlachovaea sp, B. bassiana and T.
cylindrosporum to benomyl and manzate increased quickly at higher concentrations. A critical

19
concentration, where germination was almost or totally blocked up to 5 d p. i., could be observed for
Evlachovaea sp, M. anisopliae and T. cylindrosporum at 500 mg/l of dodine and for Evlachovaea sp and
T. cylindrosporum at 2 g/l manzate. M. anisopliae seemed to be more resistant to the tested fungicides
compared to the other fungi, as it was able to produce mycelium or conidia at fungicidal concentrations
that inhibited or retarded development of other fungi. This fungus was often isolated from soils using
fungicides, such as benomyl and dodine in selective media Liu et al. 1993, Keller, Kessler & Schweizer
2003, Hughes et al. 2004, Luz, Rocha & Nery 2004). We present here the first results about susceptibility
of Evlachovaea sp to fungicides. Considering 50% CFU on fungicide-added medium related to CFU
grown on fungicide-free medium as a minimum of for successful isolation, 1 mg/l benomyl, 200 mg/l
cupric sulphate, 50 mg/l dodine, 100 mg/l manzate and 4 mg/l thiabendazole can be used to isolate all
fungi tested in this study.
Tests with previously sterilized soils showed that used concentrations of fungicides permitted a
good recovery of all tested fungi, except Evlachovaea sp on benomyl-added medium. This fungus showed
an elevated variability of susceptibility among strains to this fungicide at 1 mg/l and this concentration
can hamper a successful isolation of Evlachovaea sp from soils. A lower concentration of benomyl is
probably more adequate for isolation of this genus. However, efficacy of fungicidal concentrations tested
in the present study which permitted development of at least 50% CFU on CM has to be confirmed for
isolation of fungi from unsterile soils. Under natural conditions entomopathogenic fungi occurring in the
same sample will interact between each other and fast-growing microorganisms, especially bacteria and
saprobic or phytopathogenic fungi, which may inhibit development of the target fungus or fungi. More
tests about susceptibility of potential contaminant fungi occurring in the same habitats to fungicides are
necessary. Recently Luz & Rocha (unpublished data) showed that thiabendazole (at 4 mg/l) was the most
appropriate fungicide for 12 other invertebrate-pathogenic fungi and inhibited 8 important contaminant
fungi.
Strain-related susceptibility to fungicides found in the present study and also reported by other
authors (Liu et al. 1993, Shapiro-Illan 2002) may be useful for a strain-specific isolation of fungi and
should be considered for unspecific detection of fungi of the same genus or species during prospection
activities. Results showed that an adequate concentration of a fungicide found for a specific strain can
inhibit the development of other strains of the same species or genus.
It is important to note that a successful germination is not sufficient for fungal detection and
isolation on a selective medium. Depending on the concentration of benomyl, cupric sulphate, dodine and
manzate, germination was induced but germlings produced no or few mycelium and new conidia, even
after prolonged incubation. Germlings are not visible on the medium and it is indispensable that they
develop to colonies which may be detected macroscopically and so be transferable to other media.
Formation of new conidia on colonies will be helpful for distinction between fungi. Inhibition of fungal
development after germination was especially noted for benomyl even at a lower concentration of 5 mg/l
and had also been reported for this fungicide and M. anisopliae by Milner et al. (1991).

20
In the present tests fungal susceptibility was assayed routinely with conidia. Persistence of
entomopathogenic Hyphomycetes in nature occurs mainly as conidia. It seems that entomopathogenic
fungi such as Beauveria spp develop hardly in soils and only close to mummified cadavers (Gottwald &
Tedders 1984, Studdert & Kaya 1990). If not isolated directly from those cadavers, fungi will commonly
be detected through to substrate incorporated conidia.
Fungicides cited in literature for selective media of entomopathogenic fungi such as dodine and
benomyl are not longer being manufactured or not easy to obtain. Others, manzate and thiabendazole are
restricted or banned in a few countries. Except copper oxychloride and cupric sulphate, which are
classified as slight and moderate acute toxic, respectively, other fungicides tested are toxic to humans
(Orme & Kegley 2006). Depending on the test design a suitable fungicide should be highly selective with
low risk for human beings. It should be used at a reasonable quantity, permitting a development of the
desired fungi within adequate periods of incubation on the medium. It should be autoclavable without
loosing fungicidal properties or changing consistence or color of the medium or emitting malodorous
odor, and finally be easily accessible. Thiabendazole seemed to be the fungicide that fits best to these
requisites.
Acknowledgements
The authors thank Richard A. Humber for providing T. cylindrosporum strains, identifying
Evlachovaea sp and reviewing the manuscript and the National Council of Scientific and Technological
Development (CNPq) for financial support.
References
Chase, A. R., Osborne, L. S. & Ferguson, V. M. (1986) Selective isolation of the entomopathogenic fungi
Beauveria bassiana and Metarhizium anisopliae from an artificial potting medium. Florida
Entomologist 69: 285–292.
Gottwald, T. R. & Tedders, W. L. (1984) Colonization, transmission and longevity of Beauveria bassiana
and Metarhizium ansiopliae (Deuteromycotina: Hyphomycetes) on pecan weevil larvae (Coleoptera:
Curculionidae) in the soil. Environmental Entomology 13: 557–560.
Keller, S., Kessler, P. & Schweizer, C. (2003) Distribution of insect pathogenic soil fungi in Switzerland
with special reference to Beauveria brongniartii and Metarhizium anisopliae. Biocontrol 48: 307–319.
Hughes, W. O. H., Thomsen, L., Eilenberg, J. & Boomsma, J. J. (2004) Diversity of entomopathogenic
fungi near leaf-cutting ant nests in a neotropical forest, with particular reference to Metarhizium
anisopliae var. anisopliae. Journal of Invertebrate Pathology 85: 46–53.
Liu, Z. Y., Milner, R. J., McRae, C. F. & Lutton, G. G. (1993) The use of dodine in selective media for
the isolation of Metarhizium spp. from soil. Journal of Invertebrate Pathology 62: 248–251.
Luz, C. & Fargues, J. (1997) Temperature and moisture requirements for conidial germination of an
isolate of Beauveria bassiana, pathogenic to Rhodnius prolixus. Mycopathologia 138: 117–125.

21
Luz, C., Rocha, L. F. N. & Humber, R. A. (2003) Record of Evlachovaea sp. (Hyphomycetes) on
Triatoma sordida in the State of Goiás, Brazil, and its activity against Triatoma infestans (Reduviidae,
Triatominae). Journal of Medical Entomology 40: 451–454.
Luz, C., Rocha, L. F. N., Nery, G. V. (2004) Detection of entomopathogenic fungi in peridomestic
triatomine-infested areas in Central Brazil and fungal activity against Triatoma infestans (Klug)
(Hemiptera: Reduviidae). Neotropical Entomology 33: 783–791.
Luz, C., Rocha, L. F. N. & Silva, I. G. (2004) Pathogenicity of Evlachovaea sp. (Hyphomycetes), a new
species isolated from Triatoma sordida, in Chagas disease vectors under laboratory conditions. Revista
da Sociedade Brasileira de Medicina Tropical 37: 189–191.
Milner, R. J., Huppatz, R. J. & Swaris, S. C. (1991) A new method for assessment of germination of
Metarhizium conidia. Journal of Invertebrate Pathology 57: 121–123.
Mitchell, D. J., Kannwischer-Mitchell, M. E. & Dickson, D. W. (1987) A semi-selective medium for the
isolation of Paecilomyces lilacinus from soil. Journal of Nematology 19: 255–256.
Orme, S. & Kegley, S. (2006) PAN Pesticide Database, Pesticide Action Network, North America (San
Francisco, CA), http:www.pesticideinfo.org.
Panter, C. & Frances, S. P. (2003) A more selective medium for Culicinomyces clavisporus. Journal of
Invertebrate Pathology 82: 198–200.
Shapiro-Ilan, D. I., Reilly, C. C., Hotchkiss, M. W. & Wood, B. W. (2002) The potential for enhanced
fungicide resistance in Beauveria bassiana through strain discovery and artificial selection. Journal of
Invertebrate Pathology 81: 86–93.
Sneh, B. (1991) Isolation of Metarhizium anisopliae from insects on an improved selective medium based
on wheat germ. Journal of Invertebrate Pathology 58: 269–273.
Studdert, J. P. & Kaya, H. K. (1990) Water potential, temperature, and soil type on the formation of
Beauveria bassiana soil colonies. Journal of Invertebrate Pathology 56: 380–386.
Veen, K. H. & Ferron, P. (1966) A selective medium for isolation of Beauveria tenella and Metarhizium
anisopliae. Journal of Invertebrate Pathology 8: 268–269.
Yaginuma, K. & Takagi, K. (1986) Improvement of a selective medium for isolation of Metarhizium
anisopliae (Metschnikoff) Sorokin. Japanese Journal of Applied Entomology and Zoology 30: 300–
301.
Figure 1. Relative number of colony forming units (CFU) produced by B. bassiana, Evlachovaea sp, M.
anisopliae and T. cylindrosporum on fungicide-amended medium in different concentrations and related
to the total number of CFU of fungi detected on medium without fungicide, 240 h after incubation

22
Table 1. Fungicides and concentrations used in different tests
Colony forming units
Fungicide
Unit
Germination *
Concentration fitting *
Soil tests *
Variability tests **
Benomyl
mg/l
0.5; 5; 50
1; 2; 3; 4
1
1 (E, Bb) 2 (Tc) 3 (Ma)
Copper oxychloride
g/l
0.3; 3; 30
3; 7.5; 15; 30
***
***
Cupric sulphate
g/l
0.04; 0.4; 4
0.04; 0.08; 0.2; 0.4
0.2
0.08 (E) 0.4 (Bb, Ma, Tc)
Dodine
mg/l
5; 50; 500
10; 50; 100; 500
50
50 (Tc, E) 100 (Bb, Ma)
Manzate
g/l
0.2; 2; 20
0.1; 0.2; 0.3; 0.4
0.1
0.1 (E, Tc) 0.2 (Bb, Ma)
Thiabendazole
mg/l
0.2; 2; 20
4; 6; 8; 16
4
4 (Tc), 6 (E) 8 (Bb), 16 (Ma)
* tested for all fungi ** tested for Beauveria bassiana (Bb), Evlachovaea sp (E), Metarhizium anisopliae (Ma) and Tolypocladium cylindrosporum (Tc) *** not done

23
Table 2. Cumulated germination (%) of entomopathogenic fungi and fungal development on fungicide-amended medium 120 hours after incubation * Fungicide
Concentration
Fungus tested **
F values significance ***
mg/l medium
Beauveria bassiana (IP 6)
Evlachovaea sp (IP 218)
Metarhizium anisopliae (IP 120)
Tolypocladium cylindrosporum (ARSEF 2920)
Concentration
Benomyl 50 > 98 (M-) > 98 (M-) > 98 (M-) > 98 (M-) 5 > 98 (M-) > 98 (M-) > 98 (M±) > 98 (M-) - 0.5 > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) - Copper 30000 **** (C±) **** (C+) **** (C+) **** (C+) oxychloride 3000 a > 98 (C±) > 98 (C+) > 98 (C+) > 98 (C+) - 300 a > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) - Cupric 4000 ***** (M-) ***** (M-) ***** (M-) ***** (M-) sulphate 400 a > 98 (M±) 97.3 ± 0.8 (C+) > 98 (C+) > 98 (C+) F1; 31 = 1 40 a > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) P = 0.33 Dodine 500 c 34 ± 11.5 (M±) 0 4 ± 2.5 (M±) 0 50 b > 98 (C+) 77 ± 5.1 (C+) > 98 (C+) 68.3 ± 12 (C+) F2; 47 = 145.4 5 a > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) P < 0.001 Manzate 20000 **** (M-) **** (M-) **** (M-) **** (M-) 2000 b 24.5 ± 24 (M-) 1.3 ± 1.3 (M-) 60.8 ± 13 (C±) 0 F1; 31 = 28.5 200 a 87 ± 4.8 (C+) 63.7 ± 4.7 (C+) > 98 (C+) 58.8 ± 6.9 (C+) P < 0.001 Thiabendazole 20 b 96.5 ± 0.9 (M+) 96.5 ± 0.9 (C±) > 98 (C±) > 98 (C±) 2 a > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) F2;47 = 5 0.2 a > 98 (C+) > 98 (C+) > 98 (C+) > 98 (C+) P = 0.01
* 500 μl suspended conidia were inoculated onto fungicide-amended complete medium in Petri dishes at a final 106 conidia/cm2. Dishes were then incubated at 25º ± 1ºC and 12 h photophase. Means (%) ± standard error of the mean, based on 4 replicates ** Code number, IPTSP Collection of Entomopathogenic Fungi, UFG, Brazil (IP) and the USDA-ARS Collection of Entomopathogenic Fungal Cultures, Ithaca, NY, USA (ARSEF)
*** Results were examined by analysis of variance for each fungicide and all fungi: concentrations within the same fungicide followed by different letters (a-c) showed significantly different results (P < 0.05) according to the Student-Newman-Keuls test Germination not evaluated: **** due to a dark coloration of the medium ***** as medium did not solidify at tested concentration (-) no, (±) little and (+) distinct formation of mycelium (M) and conidia (C)

24
Table 3. Percentage of colony forming units (CFU) produced by entomopathogenic fungi mixed with soil on fungicide-amended medium, 360 h after incubation *
Fungicide
Concentration
Fungi tested **
mg/l medium
Beauveria bassiana (IP 6)
Evlachovaea sp (IP 218)
Metarhizium anisopliae (IP 120)
Tolypocladium cylindrosporum (ARSEF 2920)
F values significance ***
Benomyl 1 66.2 ± 7.6 b 32.1 ± 15.7 c 94.7 ± 1.6 a 95.5 ± 1.9 a F3, 15 = 13.2
P < 0.001
Cupric sulphate 200 91.6 ± 2.1 a 84.0 ± 3.9 a 88.7 ± 4.1 a 99.3 ± 3.3 a F3, 15 = 0.2 P = 0.88
Dodine 50 93.5 ± 4.1 a 64.6 ± 8.3 b 97.5 ± 0.6 a 88.8 ± 9.2 ab F3, 15 = 4.5 P = 0.02
Manzate 100 98.6 ± 0.8 a 87.2 ± 6.4 a 94.4 ± 2.2 a 82.4 ± 6.2 a F3, 15 = 0.2 P = 0.89
Thiabendazole 4 94.5 ± 4.4 a 86.8 ± 6.2 a 89.1 ± 2.3 a 93.2 ± 6.5 a F3, 15 = 0.7 P = 0.58
* One ml of a water suspended conidia-soil mixture (5x102 conidia/ml) was inoculated onto fungicide-added and fungicide-free complete medium in Petri dishes. Dishes were then incubated at 25º ± 1ºC and 12 h photophase. CFU found on fungicide-added medium were related to the total number of CFU of each fungus detected on medium without fungicide. Means (%) ± standard error of the mean, based on 4 replicates ** Code number, IPTSP Collection of Entomopathogenic Fungi, UFG, Brazil (IP) and the USDA-ARS Collection of Entomopathogenic Fungal Cultures, Ithaca, NY, USA (ARSEF)
*** Values were examined by analysis of variance for each fungicide. Fungi followed by different letters (a-c) showed significantly different results (P < 0.05) according to the Student-Newman-Keuls

25
Table 4. Percentage of colony forming units (CFU) produced by entomopathogenic fungi on fungicide-amended medium, 240 h after incubation *
Fungicide
Concentration (mg/l)
Species or genus /Isolate
F values significance ****
Beauveria bassiana **
F4, 15
P
IP 94
IP 129
IP 132
IP 147
IP 153
Benomyl
1
97.9 ± 2.2 a
44.6 ± 2.3 c
46.9 ± 6.7 c
76.5 ± 3.4 b
86.5 ± 3.9 b
31.6
< 0.001 Cupric sulphate 400 82 ± 2.5 b 80.7 ± 3.1 b 61.8 ± 2.6 c 92.1 ± 2.7 a 85.5 ± 2.4 ab 12.2 < 0.001 Dodine 100 70.9 ± 3.4 a 80.3 ± 3.3 a 66.7 ± 3.2 a 90.4 ± 6.7 a 84.6 ± 9.2 a 2.8 0.07 Manzate 200 3.3 ± 0.7 c 91.3 ± 5.5 a 55.2 ± 1.6 b 6 ± 3.7 c 5.9 ± 2.8 c 42.1 < 0.001 Thiabendazole 8 72.8 ± 5.6 b 7.7 ± 2.1 c 8 ± 5.9 c 98.9 ± 1.1 a 85.7 ± 7.4 ab 41.7 < 0.001
Evlachovaea sp **
IP 67
IP 126
IP 141
IP 142
IP 148
Benomyl
1
2.6 ± 0.8 c
78.6 ± 3.6 a
39.6 ± 13.9 b
11.7 ± 3.3 c
20.4 ± 7.4 bc
17.2
< 0.001 Cupric sulphate 80 90.9 ± 0.8 a 86.3 ± 2.2 a 86.7 ± 2.6 a 89.6 ± 3.4 a 94.7 ± 2.4 a 2.5 0.09 Dodine 50 79.7 ± 2.8 b 95.4 ± 1 ab 94.4 ± 3.1 ab 90.5 ± 7.3 ab 99.8 ± 0.3 a 4.3 0.02 Manzate 100 72.4 ± 1.9 b 88.7 ± 2.9 ab 69 ± 7.6 b 84.8 ± 2.6 ab 94.7 ± 3.4 a 7 0.002 Thiabendazole 6 39.5 ± 4.7 b 45.8 ± 3.5 b 67.2 ± 5.8 a 41.7 ± 6.8 b 0 c 47.9 < 0.001
Metarhizium anisopliae **
IP 1
IP 75
IP 115
IP 125
IP 145
Benomyl
3
74.5 ± 4.4 b
78.3 ± 3.5 b
90.3 ± 5.2 ab
94 ± 5.5 a
80.1 ± 2.1 b
4.1
0.02 Cupric sulphate 400 72.8 ± 2.6 a 66.9 ± 3.3 a 71.3 ± 1.8 a 56.9 ± 6.3 a 64.7 ± 2.2 a 3.1 0.05 Dodine 100 20.7 ± 3.4 c 43.2 ± 5.8 b 30 ± 3.3 c 72 ± 2.2 a 64.7 ± 5.6 a 25.6 < 0.001 Manzate 200 72.5 ± 5.5 ab 79.3 ± 3.4 ab 67.1 ± 6.8 b 86 ± 5.2 a 62.8 ± 1 b 3.9 0.02 Thiabendazole 16 91.8 ± 1.3 c 99.3 ± 0.7 a 100 a 81.4 ± 2 d 96.9 ± 0.4 b 50.7 < 0.001
Tolypocladium cylindrosporum ***
ARSEF 705
ARSEF 962
ARSEF 1027
ARSEF 1580
ARSEF 2912
Benomyl
2
100 a
92.6 ± 2.5 a
96.8 ± 2.4 a
93.8 ± 2.1 a
97.2 ± 1.4 a
3
0.05 Cupric sulphate 400 56.6 ± 4.3 b 77.7 ± 1.7 b 73.5 ± 3.9 b 91.9 ± 6.1 a 73 ± 3.5 b 8.1 < 0.001 Dodine 50 41.9 ± 4.1 c 82.6 ± 1.9 b 73.9 ± 2 b 35.4 ± 7.9 c 98.5 ± 0.7 a 51.5 < 0.001 Manzate 100 38.1 ± 4.1 b 75.9 ± 8.8 a 67.7 ± 2.2 a 65.7 ± 4.7 a 77.3 ± 7.1 a 6 0.004 Thiabendazole 4 97.9 ± 0.6 ab 93.4 ± 2.1 ab 88.1 ± 3.5 b 95.5 ± 2.6 ab 99.3 ± 0.4 a 3.8 0.03
* 500 μl of 6 x 102 conidia/ml were inoculated onto fungicide-added and fungicide-free complete medium in Petri dishes. Dishes were then incubated at 25º ± 1ºC and 12 h photophase. CFU found on fungicide-added medium were related to the total number of CFU of each fungus detected on medium without fungicide. Means (%) ± standard error of the mean, based on 4 replicates ** Code number, IPTSP Collection of Entomopathogenic Fungi, UFG, Brazil *** Code number, USDA-ARS Collection of Entomopathogenic Fungal Cultures, Ithaca, NY, USA **** Values were examined by analysis of variance for each fungus and fungicide. Strains of the same species or genus followed by different letters (a - d) showed significantly different results (P < 0.05) according to the Student-Newman-Keuls test
26
Copper oxychloride (g/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
3 7.5 15 30Benomyl (mg/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
1 2 3 4
Thiabendazole (mg/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
4 6 8 16
Cupric sulphate (mg/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
40 80 200 400Dodine (mg/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
10 50 100 500
Manzate (mg/l)
Col
ony
form
ing
units
(%)
0
20
40
60
80
100
100 200 300 400
Beauveria bassianaEvlachovaea sp
Metarhizium anisopliaeTolypocladium cylindrosporum

27
In vitro susceptibility to fungicides by invertebrate-pathogenic and saprobic fungi
Luiz Fernando Nunes Rocha
Instituto de Patologia Tropical e Saúde Pública, Universidade Federal de Goiás, CP 131, 74001-970
Goiânia, GO, Brasil, [email protected]
Address for correspondene: Christian Luz, DMIPP, IPTSP, UFG, CP 131, 74001-970 Goiânia, GO,
Brasil
Tel: (55) 62 3209 6113; Fax: (55) 62 3521 1839: E-mail: [email protected]
Abstract
The effect of 5 fungicides, benomyl (1 mg/l), dodine (50 mg/l), manzate (100 mg/l), cupric sulphate (200
mg/l) and thiabendazole (4 mg/l), was tested under in vitro conditions on development of 15 isolates of
fungi pathogenic for insects and other invertebrates [Beauveria brongniartii, Culicinomyces clavisporus,
Duddingtonia flagrans, Hirsutella thompsonii, 2 Metarhizium anisopliae, Nomuraea rileyi, 7
Isaria/Paecilomyces spp, and Sporothrix insectorum], and 13 isolates of contaminant fungi [5 Aspergillus
spp, Cladosporium cladosporioides, Cunninghamella echinulata, Fusarium roseum, Gliocladium sp,
Mortierella isabellina, Mucor plumbeus, Rhizopus arrhizus, and Trichothecium roseum] originating
mostly from tree hole breeding sites of mosquitoes. Most pathogenic and contaminant fungi had clear
patterns of susceptibility or resistance to tested concentration of the fungicide. Development of both
pathogenic and contaminant fungi on fungicide-supplemented medium varied among fungi and
fungicides tested. Minimal inhibition of pathogenic fungi was found for cupric sulphate, benomyl,
dodine, thiabendazole followed by manzate. The highest inhibition of contaminants was obtained with
thiabendazole followed by benomyl and dodine > manzate and cupric sulphate. Thiabendazole was the
most appropriate fungicide to isolate fungi pathogenic to invertebrates from substrates with high water
contents and rich in organic material. The results underline the importance of adapting both a fungicide
and its concentration for a selective medium for isolating specific target fungi and while selecting against
possible contaminants.
Key words: biological control, semi-selective medium, Hyphomycetes, Zygomycetes, fungicide
Introduction
The use of fungi as biological control agents against crop pests has increased in the last few
decades. Fungi are also promising candidates for vector control, especially of mosquitoes, triatomines
and ticks [1-3]. However, of about 700 fungal species with entomopathogenic activity only a small
number are actually used for pest control. Improved fungal performance can be expected from the use of

28
new, more powerful strains and species. For this it appears to be vital to intensify the isolation and
characterization of fungi pathogenic to insects and other invertebrates. A successful isolation of these
pathogenic fungi from substrates with semi-selective media depends on antimicrobial agents. In addition
to bacteria, fast-growing fungal contaminants may inhibit their development and thus hamper the
isolation of the desired fungi. Appropriate fungicides suppress unwanted fungi and selectively favor the
development of intended fungi. Interactions between agro-chemicals (including fungicides) used have
been studied in crop protection and against the entomopathogenic fungi that either occur naturally or are
applied for use in integrated pest control [4-10], but research on semi-selective media for fungi
pathogenic to invertebrates incorporating fungicides has been restricted to relatively few fungicides and a
small number of entomopathogenic fungi. Media amended with dodine, benomyl, thiabendazole, manzate
or cupric sulphate resulted in good development and isolation of Beauveria spp, Metarhizium spp,
Paecilomyces lilacinus, Culicinomyces clavisporus, Tolypocladium cylindrosporum and Evlachovaea sp
[11-15, Rocha & Luz, personal communication]. We report on the in vitro susceptibility to five
fungicides by 15 isolates of invertebrate-pathogenic conidial fungi from different origins, and 13 isolates
of common hyphomycete or zygomycete contaminants collected in mosquito breeding sites in Central
Brazil.
Materials & Methods
Origin of fungi
Fungi tested were obtained from IPTSP (IP; Instituto de Patologia Tropical e Saúde Pública) and
CENARGEN (CG; Embrapa Recursos Genéticos e Biotecnologia, Brasília, Brazil), and from ARSEF
(USDA-ARS Plant, Soil and Nutrition Laboratory, Ithaca, NY, USA). The 15 conidial fungal pathogens
of invertebrates cited in Table 1 originated from Brazil (66.7%), Australia (13.2%), Peru (6.7%), the
Philippines (6.7%) and USA (6.7%); they were isolated from insects (60%), mites (6.6%), soils (20%),
feces (6.7%) and air (6.7%). Cladosporium cladosporioides and Gliocladium sp originated from insects
collected in Brazil and Australia, respectively (Table 1). All other fungi–5 Aspergillus spp, Fusarium
roseum, Trichothecium roseum and the zygomycetes Cunninghamella echinulata, Mortierella isabellina,
Mucor plumbeus, Rhizopus arrhizus–were isolated in 2003 from substrates collected in tree holes that are
possible breeding habitats for mosquitoes in the city of Goiânia, Brazil (Table 1). C. cladosporioides and
Gliocladium sp that are more saprobic than entomopathogenic, and the other fungi mentioned that had
not been tested for any pathogenicity before were considered as potential contaminants.
Preparation of fungi
Fungi were cultured during 15 days at 25° ± 1°C, 75 ± 5% relative humidity and a 12 h
photophase. Nomuraea rileyi was grown on potato maltose agar with yeast extract (PMYA: 170 g potato,
40 g maltose, 15 g yeast extract, 18 g agar and 1000 ml dist. H2O), C. clavisporus on Sabouraud dextrose
agar amended with yeast extract (SDAY: 10 g peptone, 40 g glucose, 2 g yeast extract, 18 g agar and
1000 ml dist. H2O), and all other fungi on complete medium (CM: 0.001 g FeSO4, 0.5 g KCl, 1.5 g

29
KH2PO4, 0.5 g MgSO4·7 H2O, 6 g NaNO3, 0.001 g ZnSO4, 1.5 g hydrolyzed casein, 0.5 g yeast extract,
10 g glucose, 2 g peptone, 20 g agar and 1000 ml dist. H2O). The pH of all media was adjusted before
autoclaving to 7. Conidia were directly harvested from the surface of cultures by scraping, suspended in
10 ml 0.1% Tween 80, and adjusted to final concentration after filtration and quantification based on
hemacytometer counts.
Fungicides
The five fungicides used in this study were a single guanidine dodine (Dodine 65W, 65% active
ingredient, Cyanamid Química do Brasil Ltda., Rio de Janeiro); two benzimidazoles benomyl (Benlate
500, 50% active ingredient, Du Pont do Brasil, Barueri) and thiabendazole (50% active ingredient,
Novartis, Rio de Janeiro); one dithiocarbamate manzate (Manzate 800, 80% active ingredient, Du Pont
do Brasil, Barueri); and one metallic fungicide cupric sulphate (PA, Vetec, CP Chemical Inc., Rio de
Janeiro). All fungicides were wettable powder formulations dissolved in water (dodine, manzate and
cupric sulphate) or in 70% ethanol (benomyl and thiabendazole). Fungicides were tested at 1 mg/l
(benomyl), 4 mg/l (thiabendazole), 50 mg/l (dodine), 100 mg/l (manzate) and 200 mg/l (cupric sulphate).
These were the maximal concentrations determined in previous studies that permitted development of six
isolates of Beauveria bassiana, Metarhizium anisopliae, T. cylindrosporum and Evlachovaea sp after
incubation up to 10 days at 25°C [Rocha & Luz, unpublished].
Evaluation of fungicide activity
The effect of fungicides was assessed on qualitative growth of mycelium after inoculation of
conidia in liquid SDY medium (Sabouraud dextrose + 1% yeast extract) for C. clavisporus and of
quantitative colony forming units (CFU) and qualitative conidiogenesis on colonies after inoculation of
conidia on fungicide-amended PMYA medium for N. rileyi or on CM for all other fungi. The
development of mycelium on solid CM was examined for fungi without distinct formation of colonies.
All fungi were also tested on fungicide-free medium. Tests were done at 25° ± 1°C and a 12 h
photophase with 4 replicates for each fungus and fungicide. Ten ml of fungicide-amended PMYA (N.
rileyi) or CM (all other fungi except C. clavisporus) were arranged in Petri dishes (60 x 12 mm), and
conidia were inoculated at a final concentration of 10 conidia/cm2. For this 0.5 ml of the suspension was
inoculated onto the medium and spread on the whole surface by shaking gently the dish. Formation of
colonies and subsequent conidiogenesis were checked daily by microscopy (20 – 80x) up to 10 days after
inoculation. The total number of CFU on fungicide-amended medium was compared to all CFU detected
on media without fungicide. Replicates with a total number of colonies found on fungicide-amended
medium exceeding the control up to 2% were considered as 100%. At higher values (> 2%) the results
were not considered and tests repeated. Qualitative development of C. clavisporus mycelium was
analyzed in liquid SDY medium. For this 150 ml medium with or without fungicide was transferred to
Erlenmeyer flasks (250 ml), inoculated with final 104 conidia/ml, and conidia then incubated at 25°C and
120 rpm for 10 days.

30
Analysis
Relative CFU data were arcsine-square root transformed and then analyzed using analysis of
variance (ANOVA) and the Student-Newman-Keuls (SNK) multiple range test of comparison of means.
Means were considered not statistically different at P > 0.05 [16].
Results
The first CFU of fungi pathogenic to invertebrates were detected between 2 and 3 days post
inoculation (p.i.) of conidia on fungicide-free media and conidiogenesis on colonies generally initiated 3
up to 5 days p.i. The total number of colonies/cm2 found on fungicide-free medium commonly did not
exceed 10 and varied between 5 and 10 CFU. Development of colonies and formation of new conidia of
these fungi on fungicide-amended media were generally somewhat retarded regardless of the product
tested. Distinct colonies and conidiogenesis could be observed for most fungi between a 3 - 6 days
exposure. The relative number of CFU at 10 days p.i. varied significantly among fungi (F12;247 = 11.7, P <
0.001) and fungicides (F4;255 = 5.1, P < 0.001), with the highest numbers of CFU for cupric sulphate,
benomyl, dodine and thiabendazole followed by manzate (Table 2). Relative numbers of CFU at this
moment on fungicide-amended media reached values between 50 and 90%, and ≥ 90% of all fungicides
and pathogenic fungi tested, except C. clavisporus and Duddingtonia flagrans for 33.9% and 53.9%,
respectively (Table 2). D. flagrans developed as diffuse mycelium without distinct CFU on fungicide-
free or fungicide-amended CM except with thiabendazole where no mycelium was detected after
incubation up to 10 days. C. clavisporus produced mycelium after exposure to SDY amended with
benomyl and cupric sulphate but no mycelium was found with dodine, manzate or thiabendazole. All
seven Isaria/Paecilomyces spp tested were found with relative CFU ≥ 65.9%, regardless of the fungicide.
High-levels of CFU (≥ 67.3%) were also observed for Sporothrix insectorum exposed to fungicides
except dodine (23.2%). No CFU of N. rileyi were detected after 10 days incubation on thiabendazole-
added medium. Manzate had the same effect on both Hirsutella thompsonii and M. anisopliae var.
acridum. Relative numbers of CFU were < 10% at the same moment for Beauveria brongniartii after
exposure to manzate, N. rileyi to dodine and H. thompsonii to thiabendazole (Table 2).
The first distinct colonies of most contaminants and conidiogenesis on colonies became visible
after 1-2 days incubation of conidia on fungicide-free or fungicide-amended medium. Relative numbers
of CFU 10 days p.i. varied significantly among fungi (F9;189 = 5.1, P < 0.001) and fungicides (F4;194 = 25,
P < 0.001) with the lowest numbers of CFU for thiabendazole > benomyl and dodine > cupric sulphate
and manzate (Table 3). No development at all up to 10 days p.i. were found for 46.2% of contaminant
fungi after exposure to one or more products. Few to no colonies were formed on benomyl or
thiabendazole by Aspergillus niger and A. terreus. Neither A. flavus and A. versicolor formed colonies on
thiabendazole-amended medium. No CFU at all could be detected after a 10 days incubation of
Cladosporium cladosporioides on benomyl, dodine or thiabendazole-added medium. M. isabellina and

31
Gliocladium sp were the only contaminants with high number (> 95.3%) of CFU after exposure to
thiabendazole. After the same period, relative number of CFU was < 1% for 80% of the species tested
with this fungicide that formed distinct colonies. Few CFU < 10% could be observed testing benomyl and
thiabendazole in T. roseum, benomyl in A. versicolor and thiabendazole in A. fumigatus. C. echinulata,
M. plumbeus and R. arrhizus developed with fast-growing diffuse mycelium and almost instantaneous
sporangiosporogenesis on fungicide-amended media regardless of fungicide tested. Figure 1 shows the
generalized effect of tested fungicides on colony formation without focusing on marked differences of
CFU for a specific fungus and a fungicide that are shown in Tables 2 and 3. Pathogenic fungi formed
distinctly more CFU than contaminants exposed to benomyl, dodine and especially thiabendazole.
Discussion
Most pathogenic and contaminant fungi proved to have a clear pattern of susceptibility and
resistance to the tested concentrations of fungicides. This is an important prior condition for specific
isolation of target fungi. Minimal CFU of 80% or higher values on media containing a fungicide, related
to numbers obtained with fungicide-free medium, which were found for 73.9% of tested pathogenic
fungi, will probably not hamper their detection and isolation. However results obtained with fungi tested
individually on fungicide-added culture medium should be interpreted carefully because of complex
interactions between target fungi and other microorganisms in the same substrate and different
susceptibility to fungicides influences isolation with semi-selective media. When a selective medium
allows only low number or total inhibition of CFU of a target fungus (is this correct English ?), lower
concentrations of the fungicide used or the adoption of another fungicide should be considered to
improve the recovery of the specific target species. In the present study thiabendazole at 4 mg/l, which in
previous studies was the maximal concentration permitting development in vitro of different strains of B.
bassiana, Evlachovaea sp, M. anisopliae and T. cylindrosporum [Rocha & Luz, unpublished] seemed to
be the most appropriate fungicide, as it favored development of a high number of fungi, pathogenic to
invertebrate but strongly inhibited most contaminants. For specific isolation of fungi inhibited by this
product, e.g., C. clavisporus, a mosquito larva-attacking fungus [19], the nematode-trapping D. flagrans
[20], H. thompsonii and N. rileyi which attacks mites [21] and noctuids [22], respectively, lower
concentrations of thiabendazole or combinations of fungicides should be tested. Little information exists
about the utility of thiabendazole for selective isolation of fungi pathogenic to invertebrates.
Thiabendazole-added to NUTNS agar medium permitted a good growth of C. clavisporus, on agar at
concentrations up to 3 mg/l whereas contaminant molds such as Cladosporium sp and Penicillium spp
were inhibited by separate or combined thiabendazole and benomyl at 1 mg/l for both fungicides [15]. In
our study C. clavisporus developed better in liquid SDY than on solid SDAY. A liquid semi-selective
medium may be preferable for isolating aquatic fungi such as C. clavisporus. A reduction of rapid growth
of a Mucor sp on the culture medium with thiabendazole (≥ 2mg/l) reported by Panter and Frances [15]
was not observed for other Zygomycetes M. plumbeus, C. echinulata and R. arrhizus at 4 mg/l of

32
thiabendazole. In addition, thiabendazole in Sabouraud medium at 10 mg/l had also a high fungicidal
activity in D. flagrans where this fungus developed well on 1 mg/l of the fungicide [20].
Benomyl and to some extent dodine, both of which are frequently used in selective media to
isolate M. anisopliae and Beauveria spp [13, 23-27], had no or little negative effect against most
pathogenic fungi. Both inhibited many contaminants at the tested concentrations and are useful for
detecting other conidial fungi pathogenic to invertebrates. Mitchell et al. [14] used a semi-selective
medium containing benomyl (50 mg/l) to quantify soil populations of P. lilacinus under field conditions.
This fungus showed to be highly resistant to benomyl. In our study, C. clavisporus, H. thompsonii, N.
rileyi and S. insectorum were inhibited by dodine. Moreover, benomyl selectively blocked the growth of
A. niger, A. terreus, A. versicolor and C. cladosporioides. This was also observed with dodine and A.
niger, A. flavus, F. roseum and C. cladosporioides.
Cupric sulphate and manzate were less appropriate for isolating of pathogenic fungi because
neither showed much effect against contaminant fungi. M. isabellina showed slight susceptibility to
manzate but this fungus was unaffected by the other fungicides tested. Moreover, manzate inhibited B.
brongniartii, C. clavisporus, H. thompsonii and M. a. var. acridum while M. a. var. lepidiotum was
highly resistant to manzate. An intra-specific variability in susceptibility to manzate, dodine and other
fungicides was also found for Metarhizium spp and B. bassiana [25, 28, Rocha & Luz, personal
communication].
C. echinulata, M. plumbeus and R. arrhizus were unaffected by any dosage levels of all
fungicides and may be problematic contaminants for isolations of pathogenic fungi. Higher doses, other
fungicides or a combined use of fungicides could be helpful but fungicidal actions against the desired
fungal target to be isolated should be tested first.
The contaminant fungi tested were all saprobes, frequently found in tree holes and in other
tropical habitats with substrates rich in organic material and with high water contents. These fungi are
also important food- and seed-spoilage molds. Aspergillus spp and Fusarium spp were also found from
mosquitoes or their breeding sites [29-32] or isolated from larval guts of Culex quinquefasciatus (Say)
[33]. Exploitation of a possible larvicidal effect of Aspergillus spp reported by different authors [34-37]
should be examined carefully because related Aspergillus spp especially A. fumigatus, but also A. flavus,
A. niger, A. versicolor and A. terreus, can be facultative pathogens of humans that cause aspergillosis or
other mycotoxin-induced pathologies [38, 39].
The results presented here showed that susceptibilities to fungicides can be both species-specific
and dependent on the product. The fungicides commonly used to isolate Beauveria spp and Metarhizium
spp are also helpful for isolating other fungi. However, dodine is no longer being manufactured, and
benomyl is not easy to acquire. Semi-selective medium based on thiabendazole will contribute best to a
successful isolation of important fungi pathogenic to invertebrates. This fungicide has been little used up
to now for isolating fungi but is registered almost worldwide and has only a low level of acute toxicity
for humans [http://www.pesticideinfo.org].

33
Acknowledgments
The authors thank Janine de Aquino Lemos, IPTSP, UFG, Goiânia, Brazil, Heloisa Frazão,
Embrapa Genetic Resources and Biotechnology, Brasília, DF, Brazil, and Richard A. Humber, USDA-
ARS, Ithaca, NY, USA, for providing fungi and the constructive review of the manuscript, and to the
National Council of Scientific and Technological Development (CNPq) for financial support.
References
1. Samish M, Rehacek J. Pathogens and predators of ticks and their potential in biological control. Ann
Rev Entomol 1999; 44: 159-182.
2. Luz C, Rocha LFN, Nery GV, Magalhães BP, Tigano MS. Activity of oil-formulated Beauveria
bassiana against Triatoma sordida in peridomestic areas in Central Brazil. Mem Inst Oswaldo Cruz
2004; 99: 211-218.
3. Scholte EJ, Knols BGJ, Samson RA, Takken W. Entomopathogenic fungi for mosquito control: a
review. J Insect Sci 2004; 4: 1-24.
4. Mietkiewski RT, Pell JK, Clark SJ. Influence of pesticide use on the natural occurrence of
entomopathogenic fungi in arable solis in the UK: field and laboratory comparisons Biocontrol Sci
Technol 1997; 7: 565-575.
5. Zermeno MAA, Barranco HN, Mier T, Toriello C. Effect of agro-chemicals on in vitro growth of the
entomopathogenic fungi Metarhizium anisoliae (Metsch) Sor and Paecilomyces fumosoroseus (Wize)
Brown & Smith. Rev Latinoam Microbiol 1999; 41: 223-229.
6. Devi PSV, Anitha C, Prasad YG. Compatibility of entomopathogenic fungus Nomuraea rileyi with
commonly used pesticides. Ind J Agricultural Sci 2002; 72: 370-372.
7. Loureiro ES, Moino A, Arnosti A, Souza GC. Efeito de produtos fitossanitários químicos utilizados
em alface e crisantemo sobre fungos entomopatogênicos. Neotrop Entomol 2002; 31: 263-269.
8. Gomez DRS, Delpin KE, Moscardi F, Nozaki MH. The impact of fungicides on Nomuraea rileyi
(Farlow) Samson epizootics and on population of Anticarsia gemmatalis Hubner (Lepidoptera:
Noctuidae), on soybean. Neotrop Entomol 2003; 32: 287-291.
9. Kouassi M, Coderre D, Todorova SI. Effects of the timing of applications on the incompatibility of
three fungicides and one isolate of the entomopathogenic fungus Beauveria bassiana (Balsamo)
Vuillemin (Deuteromycotina). J Appl Entomol 2003; 127: 421–426.
10. Er MK, Gokce A. Effects of selected pesticides used against glasshouse tomato pests on colony
growth and conidial germination of Paecilomyces fumosoroseus. Biological Control 2004; 31: 398-
404.
11. Beilhartz VC, Parberry DG, Swart HJ. Dodine: a selective agent for certain soil fungi. Trans British
Mycol Soc 1982; 79: 507-511.
12. Yaginuma K, Takagi K. Improvement of a selective medium for isolation of Metarhizium anisopliae
(Metschnikoff) Sorokin. Jap J Appl Entomol Zool 1986; 30: 300-301.

34
13. Chase AR, Osborne LS, Ferguson VM. Selective isolation of the entomopathogenic fungi Beauveria
bassiana and Metarhizium anisopliae from an artificial potting medium. Florida Entomologist 1986;
69: 285-292.
14. Mitchell DJ, Mitchell MEK, Dickson DW. A semi-selective medium for the isolation of
Paecilomyces lilacinus from soil. J Nematol 1987; 19: 255-256.
15. Panter C, Frances SP. A more selective medium for Culicinomyces clavisporus. J Invertebr Pathol
2003; 82: 198-200.
16. SAS Institute Inc. 2002. What’s New in SAS® Software for Release 8.1, Cary, NC, SAS Institute
Inc.
17. Samson RA. Paecilomyces and some allied Hyphomycetes. Stud Mycol 1974; 6: 1-119.
18. Luangsa-ard JJ, Hywel-Jones NL, Manoch L, Samson RA. On the relationships of Paecilomyces
sect. Isarioidea species. Mycol Res 2005; 109: 581-589.
19. Seif AI, Shaarawi FA. Preliminary field trials with Culicinomyces clavosporus against some
Egyptian mosquitoes in selected habitats. J Egypt Soc Parasitol 2003; 33: 291-304.
20. Paraud C, Pors I, Chartier C. Activity of Duddingtonia flagrans on Trichostrongylus colubriformis
larvae in goat feces and interaction with a benzimidazole treatment. Small Ruminant Res 2005; 55:
199-207.
21. Tanzini MR, Alves SB, Tamai MA, Moraes GJ, Ferla NJ. An epizootic of Calacarus heveae (Acari:
Eriophyidae) caused by Hirsutella thompsonii on rubber trees. Exp Appl Acarol 2000; 24: 141-144.
23. Veen KH, Ferron P. A selective medium for isolation of Beauveria tenella and Metarhizium
anisopliae. J Invertebr Pathol 1966; 8: 268-269.22. Devi KU, Murali MCH, Padmavathi J, Ramesh K.
Susceptibility to fungi of cotton boll worms before and after a natural epizootic of the
entomopathogenic fungus Nomuraea rileyi (Hyphomycetes). Biocontrol Sci Technol 2003; 13: 367-
371.
24. Sneh B. Isolation of Metarhizium anisopliae from insects on an improved selective medium based
on wheat germ. J Invertebr Pathol 1991; 58: 269-273.
25. Liu ZY, Milner RJ, McRae CF, Lutton GG. The use of dodine in selective media for the isolation of
Metarhizium spp from soil. J Invertebr Pathol 1993; 62: 248-251.
26. Keller S, Kessler P, Schweizer C. Distribution of insect pathogenic soil fungi in Switzerland with
special reference to Beauveria brongniartii and Metarhizium anisopliae. Biocontrol 2003; 48: 307-
319.
27. Hughes WOH, Thomsen L, Eilenberg J, Boomsma JJ. Diversity of entomopathogenic fungi near
leaf-cutting ant nests in a neotropical forest, with particular reference to Metarhizium anisopliae var.
anisopliae. J Invertebr Pathol 2004; 85: 46-53.
28. Ilan DIS, Reilly CC, Hotchkiss MW, Wood BW. The potential for enhanced fungicide resistance in
Beauveria bassiana through strain discovery and artificial selection. J Invertebr Pathol 2002; 86: 86-
93.

35
29. Badran RAM, Aly MZY. Studies on the mycotic inhabitants of Culex pipiens collected from fresh
water ponds in Egypt. Mycopathologia 1995; 132: 105-110.
30. Sur B, Bihri V, Sharma A, Basu SK. Survey of termite-inhabited soil and mosquito breeding sites in
Lucknow, India for potential mycopathogens of Anopheles stephensi. Mycopathologia 1999; 144: 77-
80.
31. Moraes AM, Corrado LM, Holanda VL, Costa GL, Ziccardi M, Oliveira RL, Oliveira PC.
Aspergillus from Brazilian mosquitoes - I. Genera Aedes and Culex from Rio de Janeiro State.
Mycotaxon 2001; 78: 413-422.
32. Lastra CCL, Garcia JJ, Micieli MV. Microecology of entomopathogenic fungi from aquatic
environments[ Documentos Embrapa Soja]. Londrina, Brazil: Embrapa Soja, 2002; 184: 83-85.
33. Vasanthi V, Hoti SL. Microbial flora in gut of Culex quinquefasciatus breeding in cess pits.
Southeast Asian J Trop Med Public Health 1992; 23: 312-317.
34. Vijayan V, Balarajan K. Metabolites of fungi and Actinomycetes active against mosquito larvae. Ind
J Med Res Section A, Inf Dis 1991; 93: 115-117.
35. Moraes AM, Costa GL, Barcellos MZ, Oliveira RL, Oliveira PC. The entomopathogenic potential of
Aspergillus spp in mosquitoes vectors of tropical diseases. J Basic Microbiol 2001; 41: 45-49.
36. Sundarapandian S, Sundaram MD, Tholkappian P, Balasubramanian V. Mosquitocidal properties of
indigenous fungi and Actinomycetes against Culex quinquefasciatus Say. J Biological Control 2002;
16: 89-91.
37. Govindarajan M, Jebanesan A, Reetha D. Larvicidal effect of extracellular secondary metabolites of
different fungi against the mosquito, Culex quinquefasciatus Say. Trop Biomed 2005; 22: 1-3.
38. Jayshree RS, Shafiulla M, George J, David JK, Bapsy PP, Chakrabarti A. Microscopic, cultural
and molecular evidence of disseminated invasive aspergillosis involving the lungs and the
gastrointestinal tract. J Med Microbiol 2006; 55: 961-964.
39. Segal BH, Walsh TJ. Current approaches to diagnosis and treatment of invasive aspergillosis. Am J
Respir Crit Care Medic 2006; 173: 707-717.
Figure 1. Relative median number of colony forming units (CFU) produced by 13 invertebrate-
pathogenic fungi (B. brongniartii, H. thompsonii, 2 M. anisopliae, N. rileyi, 7 Isaria/Paecilomyces spp
and S. insectorum) and 10 potentially contaminant fungi (5 Aspergillus spp, C. cladosporioides, F.
roseum, Gliocladium sp, M. isabellina, T. roseum) on fungicide-amended medium and related to the total
number of CFU of fungi detected on medium without fungicide, 10 days after incubation.

36
Table 1. Relation of tested fungi, isolated from arthropods or substrates, and informations about their origin
Species or genus
Isolate 1
Country, year of isolation
Host or substrate of origin
Aspergillus flavus 3
IP 237
Brazil, 2003
substrate in tree hole
A. fumigatus 3 IP 240 Brazil, 2003 substrate in tree hole
A. niger 3 IP 236 Brazil, 2003 substrate in tree hole
A. terreus 3 IP 242 Brazil, 2003 substrate in tree hole
A. versicolor 3 IP 241 Brazil, 2003 substrate in tree hole
Beauveria brongniartii 2 CG 619 Brazil, 1998 Thelosia camina (Lepidoptera)
Cladosporium cladosporioides 3 CG 635 Brazil, 1994 Aleurothrixus aepim (Homoptera)
Culicinomyces clavisporus 2 ARSEF 644 Australia, 1981 Anopheles amictus hilli (Diptera)
Cunninghamella echinulata 3 IP 249 Brazil, 2003 substrate in tree hole
Duddingtonia flagrans 2 CG 722 Brazil, 1997 feces
Fusarium roseum 3 IP 252 Brazil, 2003 substrate in tree hole
Gliocladium sp 3 CG 646 Australia, 1997 Inopus rubriceps (Diptera)
Hirsutella thompsonii 2 CG 524 USA, 1996 Phyllocoptruta oleivora (Acari)
Isaria amoenorosea 2,4 CG 075 Brazil, 2000 Coleoptera
I. farinosa 2,4 CG 195 Brazil, 1983 Chlosyne lacinia (Lepidoptera)
I. fumosorosea 2,4 CG 325 Philippines, 1985 Nilaparvata lugens (Homoptera)
Metarhizium anisopliae var.
acridum 2
CG 863 Peru, 2000 Schistocerca interrita (Orthoptera)
M. anisopliae var. lepidiotum 2 CG 648 Australia, 1997 Coleoptera
Mortierella isabellina 3 IP 248 Brazil, 2003 substrate in tree hole
Mucor plumbeus 3 IP 250 Brazil, 2003 substrate in tree hole
Nomuraea rileyi 2 CG 381 Brazil, 1989 Spodoptera frugiperda (Lepidoptera)
Paecilomyces carneus 2,4 CG 525 Brazil, 1995 soil
P. lilacinus 2,4 CG 362 Brazil, 1991 soil
P. marquandii 2,4 CG 190 Brazil, 1990 soil
P. variotii 2,4 CG 503 Brazil, 1995 air
Rhizopus arrhizus 3 IP 251 Brazil, 2003 substrate in tree hole
Sporothrix insectorum 2 CG 826 Brazil, 1997 Leptopharsa hevea (Hemiptera)
Trichothecium roseum 3 IP 244 Brazil, 2003 substrate in tree hole
1 Isolate number, Collection of Entomopathogenic Fungi, Embrapa Recursos Genéticos e Biotecnologia, Brasília DF, Brazil (CG), Instituto de Patologia Tropical e Saúde Pública (IP) and the USDA-ARS Collection of Entomopathogenic Fungal Cultures, Ithaca, NY, USA (ARSEF).
2 Fungal pathogens of invertebrates.
3 Potential contaminants.4 Paecilomyces sect. Isarioidea [17] is being purged for nomenclaturally and taxonomically sound reasons, and its species reassigned to other genera. Three taxa studied here are now reclassifed in Isaria while P.carneus, P. lilacinus, and P. marquandii still await reclassification, and P. variotii, the type species of Paecilomyces, remains unchanged [18].

37
Table 2. Number (%) of colony forming units (CFU) of fungi pathogenic of invertebrates grown on fungicide-amended medium and related to the
total number of CFU of each fungus detected on medium without fungicide, 10 days after incubation 1
Fungicide (mg/l) tested
F values
Species or genus
Benomyl (1)
Cupric sulphate (200)
Dodine (50)
Manzate (100)
Thiabendazole (4)
F4;15
P
Beauveria brongniartii
100 A, a
72.2 ± 8.2 B, cd
100 A, a
1.8 ± 0.8 C, e
100 A, a
138
< 0.001
Hirsutella thompsonii 71.9 ± 3.2 A, de 54.4 ± 6.2 B, d 13.9 ± 3.2 C, b 0 D, e 2.5 ± 2.5 D, c 73
< 0.001
Isaria amoenorosea 87.4 ± 2.9 A, b-e 92.6 ± 5.6 A, a-c 96.3 ± 1.4 A, a 82.7 ± 3.3 A, bc 88.9 ± 6.7 A, ab 1.4
0.28
I. farinosa 98.3 ±1.7 A, ab 85.8 ± 5.7 A, a-c 95.1 ± 4.9 A, a 93.5 ± 3.9 A, ab 87.3 ± 7.8 A, ab 1.1
0.353
I. fumosorosea 95.3 ± 1.9 A, a-c 98.6 ± 0.9 A, ab 90.5 ± 6.3 A, a 84.7 ± 3.4 A, bc 92.3 ± 4.6 A, ab 1.6
0.235
Metarhizium anisopliae var. acridum
93.5 ± 4 A, a-c 93.7 ± 3.3 A, a-c 93.5 ± 6.5 A, a 0 B, e 90.3 ± 4.9 A, a 42.1 < 0.001
M. anisopliae var. lepidiotum 88.1 ± 5.2 AB, a-d 100 A, a 95.6 ± 4.4 A, a 80.4 ± 3.6 B, c 99.3 ± 0.7 A, a 6.6
0.003
Nomuraea rileyi 93.5 ± 4.1 A, a-c 92.1 ± 3.6 A, a-c 8 ± 1.2 C, b 59.1 ± 3.2 B, d 0 D, c 88.3
< 0.001
Paecilomyces carneus 83.2 ± 5.9 A, c-e 88.6 ± 2.6 A, a-c 93.6 ± 3.7 A, a 99.4 ± 0.6 A, a 82.8 ± 13.8 A, ab 1.9
0.167
P. marquandii 99.4 ± 0.6 A, ab 95.1 ± 5 A, a-c 99.1 ± 0.9 A, a 72.5 ± 8 B, cd 91.7 ± 5.5 A, ab 5.2
0.008
P. lilacinus 94.8 ± 1.7 A, a-c 96.2 ± 3.8 A, a-c 96.2 ± 2.4 A, a 98.1 ± 2 A, a 92.4 ± 4.5 A, ab 0.6
0.705
P. varioti 76.6 ± 6.1 A, c-e 86.8 ± 6 A, a-c 92.3 ± 7.7 A, a 87.4 ± 3.8 A, bc 65.9 ± 11.7 A, b 2.2
0.116
Sporothrix insectorum 67.3 ± 5.9 B, e 79.9 ± 5.5 B, b-d 23.2 ± 5.2 C, b 96.1 ± 3.1 A, a 95.6 ± 2.5 A, ab 27.2 < 0.001
F12;39 and P values
F = 7.6 P < 0.001
F = 5 P < 0.001
F = 25.6 P < 0.001
F = 92 P < 0.001
F = 20.6 P < 0.001
1500 μl of suspended conidia were inoculated at a final concentration of 10 conidia/cm2 onto fungicide-added solid CM (complete medium) or PMYA (potato-maltose-agar) medium (N. rileyi). Dishes were incubated at 25º ± 1º C and 12 h photophase. Means (%) ± standard error of the mean, based on 4 replicates, were examined by analysis of variance (ANOVA). Mean values within the same line (effect of fungicides for one fungus) followed by capital letters (A-D) and the same column (effect of one fungicide for all fungi tested) by different small letters (a–e), respectively, are significantly different (P < 0.05) according to the Student-Newman-Keuls test.
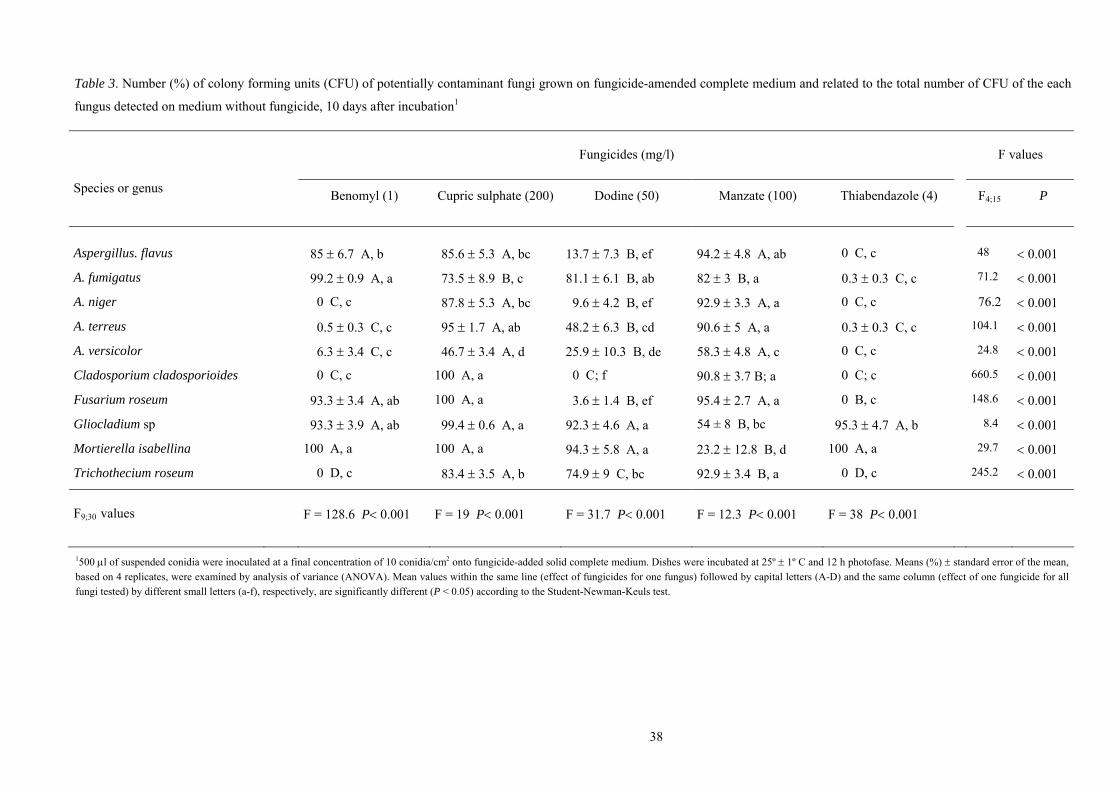
38
Table 3. Number (%) of colony forming units (CFU) of potentially contaminant fungi grown on fungicide-amended complete medium and related to the total number of CFU of the each
fungus detected on medium without fungicide, 10 days after incubation1
Fungicides (mg/l)
F values
Species or genus
Benomyl (1)
Cupric sulphate (200)
Dodine (50)
Manzate (100)
Thiabendazole (4)
F4;15
P
Aspergillus. flavus
85 ± 6.7 A, b
85.6 ± 5.3 A, bc
13.7 ± 7.3 B, ef
94.2 ± 4.8 A, ab
0 C, c
48
< 0.001
A. fumigatus 99.2 ± 0.9 A, a 73.5 ± 8.9 B, c 81.1 ± 6.1 B, ab 82 ± 3 B, a 0.3 ± 0.3 C, c 71.2 < 0.001
A. niger 0 C, c 87.8 ± 5.3 A, bc 9.6 ± 4.2 B, ef 92.9 ± 3.3 A, a 0 C, c 76.2 < 0.001
A. terreus 0.5 ± 0.3 C, c 95 ± 1.7 A, ab 48.2 ± 6.3 B, cd 90.6 ± 5 A, a 0.3 ± 0.3 C, c 104.1 < 0.001
A. versicolor 6.3 ± 3.4 C, c 46.7 ± 3.4 A, d 25.9 ± 10.3 B, de 58.3 ± 4.8 A, c 0 C, c 24.8 < 0.001
Cladosporium cladosporioides 0 C, c 100 A, a 0 C; f 90.8 ± 3.7 B; a 0 C; c 660.5 < 0.001
Fusarium roseum 93.3 ± 3.4 A, ab 100 A, a 3.6 ± 1.4 B, ef 95.4 ± 2.7 A, a 0 B, c 148.6 < 0.001
Gliocladium sp 93.3 ± 3.9 A, ab 99.4 ± 0.6 A, a 92.3 ± 4.6 A, a 54 ± 8 B, bc 95.3 ± 4.7 A, b 8.4 < 0.001
Mortierella isabellina 100 A, a 100 A, a 94.3 ± 5.8 A, a 23.2 ± 12.8 B, d 100 A, a 29.7 < 0.001
Trichothecium roseum 0 D, c 83.4 ± 3.5 A, b 74.9 ± 9 C, bc 92.9 ± 3.4 B, a 0 D, c 245.2 < 0.001
F9;30 values
F = 128.6 P< 0.001
F = 19 P< 0.001
F = 31.7 P< 0.001
F = 12.3 P< 0.001
F = 38 P< 0.001
1500 μl of suspended conidia were inoculated at a final concentration of 10 conidia/cm2 onto fungicide-added solid complete medium. Dishes were incubated at 25º ± 1º C and 12 h photofase. Means (%) ± standard error of the mean, based on 4 replicates, were examined by analysis of variance (ANOVA). Mean values within the same line (effect of fungicides for one fungus) followed by capital letters (A-D) and the same column (effect of one fungicide for all fungi tested) by different small letters (a-f), respectively, are significantly different (P < 0.05) according to the Student-Newman-Keuls test.

39
Benomyl1
Dodine50
Cupric sulphate
200
Thiabendazole4
Manzate100
Col
ony
form
ing
units
(%)
0
20
40
60
80
100 Fungi pathogenic to invertebratesContaminant fungi
Fungicides (mg/l)

40
Conclusões
A escolha adequada do fungicida e da concentração para isolamento in vitro deve considerar o/os
fungos procurados, uma variabilidade entre linhagens da mesma espécie e o tipo do substrato coletado
com possíveis contaminantes.
Thiabendazole, ainda pouco utilizado em meios seletivos, mostrou ter grande utilidade para
isolamento da maioria dos fungos patogênicos para invertebrados testados. Meios semi-seletivos com
esse fungicida têm interesse para detecção de fungos em substratos com alto teor de matéria orgânica e
água, encontrados em criadouros de mosquitos como ocos de árvores.
Benomyl e dodine, fungicidas recomendados na literatura para isolamento de B. bassiana e M.
anisopliae, tiveram também potencial para isolamento de outras espécies. Resultados contraditórios sobre
concentrações mínimas com atividade fungicida são relacionados também à variabilidade entre linhagens
da mesma espécie.
Manzate e sulfato cúprico foram menos úteis para isolamento in vitro devido à baixa inibição dos
contaminantes testados. Oxicloreto de cobre foi inapropriado para meio semi-seletivo devido à atividade
fungicida reduzida mesmo em concentrações elevadas.
Maiores concentrações ou outros produtos deveriam ser testados em contaminantes que
desenvolveram rápida e independentemente do fungicida testado, como C. echinulata, M. plumbeus e R.
arrhizus.

41
Bibliografia
Aché A, Matos AJ 2001. Interrupting Chagas disease transmission in Venezuela. Rev Inst Med Trop S Paulo
43: 37-43.
Albuquerque MFPM 1993. Urbanização, favelas e endemias: a produção da filariose no Recife, Brasil. Cad
Saúde Públ 4: 487-497.
Alexander B, Maroli M 2003: Control of phlebotomine sandflies. Med Vet Entomol 17: 1-18.
Almeida JEM, Batista-Filho A 2001. Banco de microrganismos entomopatogênicos. Rev Biotec Cienc Des
20: 77-86.
Alves SB 1998. Fungos entomopatogênicos. Controle Microbiano de Insetos. 2º ed. Piracicaba: FEALQ.
289-381.
Audino PG, Vassena C, Barrios S, Zerba E, Picollo MI 2004. Role of enhanced detoxication in a
deltamethrin-resistant population of Triatoma infestans (Hemiptera, Reduviidae) from Argentina.
Mem Inst Oswaldo Cruz 99: 335-339.
Barreau C, Jousset FX, Bergoin M 1996. Pathogenicity of the Aedes albopictus Parvovirus (AaPV), a denso-
like virus, for Aedes aegypti mosquitoes. J Invertebr Pathol 68: 299-309.
Brengues C, Hawkes NJ, Chandre F, McCarroll L, Duchon S, Guillet P, Manguin S, Morgan JC,
Hemingway J 2003. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel
mutations in the voltage-gated sodium channel gene. Med Vet Entomol 17: 87-94.
Brogdon WH, McAllister JC 1998. Insecticide resistance and vector control. Emerg Inf Dis 4: 605-613.
Butt TM, Goettel MS 2000. Bioassays of entomogenous fungi. Bioassays of entomopathogenic microbes and
nematodes. Ed. CAB International. 141-195.
Charles JF, Nielsen-Leroux C, Delécluse A 1996. Bacillus sphaericus toxins: molecular biology and mode of
action. Annu Rev Entomol 4: 451-472.
Chase AR, Osborne LS, Ferguson VM 1986. Selective isolation of the entomopathogenic fungi Beauveria
bassiana and Metarhizium anisopliae from an artificial potting medium. Florida Entomol 69: 285-292.
D`Amato C, Torres JPM, Malm O 2002. DDT (dicloro difenil tricloroetano): toxicidade e contaminação
ambiental - uma revisão. Quim Nova 25: 995-1002.
Destéfano RHR, Destéfano SAL, Messias CL 2004. Detection of Metarhizium anisopliae var. anisopliae
within infected sugarcane borer Diatraea saccharalis (Lepidoptera, Pyralidae) using specific primers.
Gen Mol Biol 27: 245-252.
Dias JCP, Leão AEA 1967. Parasitismo de fungos (Beauveria bassiana) sobre triatomíneos brasileiros
criados em laboratório. Atas SocBiol 2: 85-87.
Dias JCP, Silveira AC, Schofield CJ 2002. The impact of Chagas disease control in Latin America. A
Review. Mem Inst Oswaldo Cruz 97: 603-612.
Diotaiuti L, Pereira MH, Espínola HN 2000. Hemiptera. Parasitologia Humana. 10º ed. São Paulo: Editora
Ateneu. 292-306.

42
Dujardin JP, Schofield CJ, Panzera F 2000. Les vecteurs de la maladie de Chagas. Recherches
taxonomiques, biologiques et génétiques, Ac Roy Sci Outre-Mer, Classe Sci Nat Méd, Paris, 162 pp.
Dujardin JP, Schofield CJ, Tibayrenc M 1998. Population structure of Andean Triatoma infestans: allozyme
frequencies and their epidemiological relevance. Med Vet Entomol 12: 20-29.
Fargues J, Luz C 2000. Effect of fluctuating moisture and temperature regimes on the infection potential of
Beauveria bassiana for Rhodnius prolixus. J Invertebr Pathol 75: 202-211.
Federici BA 1995. The future of microbial insecticides as vector control agents. J Am Mosq Control Assoc
11: 260-268.
Federici BA, Park HW, Bideshi DK, Wirth MC, Johnson JJ 2003. Recombinant bacteria for mosquito
control. J Exp Biol 206: 3877-3885.
Glare TR, Milner RJ 1991. Ecology of entomopathogenic fungi. Handbook of Applied Mycology (humans,
animals e insects). Vol. 2, Ed. Arora, L. Ajello, and K. G. Mukerji. 547–612.
Gubler DJ 2005. The emergence of epidemic dengue fever and dengue hemorrhagic fever in the Americas: a
case of failed public health policy. Pan Am J Pub Health (PAHO) 17: 22-24.
Halstead SB 1993. Global epidemiology of dengue: health systems in disarray. Trop Med 35: 37-146.
Hawksworth DL 1991. The fungal dimension of biodiversity: magnitude, significance, and conservation.
Mycol Res 95: 641-655.
Hemingway J, Hawkes NJ, McCarroll L, Ranson H 2004. The molecular basis of insecticide resistance in
mosquitoes. Insect Biocherm Mol Biol 34: 653-665.
Hemingway J, Ranson H 2000. Insecticide resistance in vectors of human disease. Annu Rev Entomol 45:
371-391.
Inglis GD, Goettel MS, Butt TM, Strasser H 2001. Use of hyphomycetous fungi for managing insect pests.
Fungi as Biocontrol Agents. Ed. CAB International. 23-67.
Karunaratne SHPP, Hemingway J 2001. Malathion resistance and prevalence of the malathion
carboxylesterase mechanism in populations of mosquito vectors of disease in Sri Lanka. Bull World
Health Organization 79: 1060-1064.
Khin MN, Jirakanjanakit N, Yoksan S, Bhamarapravati N 1994. Infection, dissemination, transmission and
biological attributes of dengue-2 PDK53 candidate vaccine virus oral infection in Aedes aegypti. Am J
Trop Med Hyg 51: 864-869.
Lacey L, Goettel MS 1995. Current developments in microbial control of insects. Entomophaga 40: 202-211.
Lima JBP, Mello NV, Valle D 2005. Residual effect of two Bacillus thuringiensis var. israelensis products
assayed against Aedes aegypti (Diptera: Culicidae) in laboratory and outdoors at Rio de Janeiro,
Brazil. Rev Inst Med Trop S Paulo 47: 125-130.
Liu ZY, Milner RJ, McRae CF, Lutton GG 1993. The use of dodine in selective media for the isolation of
Metarhizium spp. from soil. J Invertebr Pathol 62: 248-251.
Lecuona RE, Edelstein MFB, Rossa FR, Arcas JA 2001. Evaluation of Beauveria bassiana (Hyphomycetes)
strains as potential agents for control of Triatoma infestans (Hemiptera: Reduviidae). J Med Entomol
38: 172-179.

43
Luz C, Fargues J 1999. Dependence of the entomopathogenic fungus Beauveria bassiana on high humidity
for infection of Rhodnius prolixus. Mycopathologia 146: 33-41.
Luz C, Fargues J, Grunewald J 1998a. The effect of fluctuating temperature and humidity on the longevity of
starved Rhodnius prolixus. J Appl Entomol 122: 219-222.
Luz C, Fargues J, Romaña CA 2003a. Influence of starvation and blood meal-induced moult on the
susceptibility of nymphs of Rhodnius prolixus Stål (Hem., Triatominae) to Beauveria bassiana (Bals.)
Vuill. infection. J Appl Entomol 127: 153-156.
Luz C, Fargues J, Romaña CA, Moreno J, Goujet R, Rougier M & Grunewald J 1994. Potential of
entomopathogenic Hyphomycetes for the control of the triatomine vectors of Chagas’ disease. Proc. 6.
Int. Coll. Invertebr. Path Microbiol Control 1: 272-276.
Luz C, Rocha LFN, Humber RA 2003b. Record of Evlachovaea sp. (Hyphomycetes) on Triatoma sordida in
the State of Goiás, Brazil, and its activity against Triatoma infestans (Reduviidae, Triatominae). J Med
Entomol 40: 451-454.
Luz C, Rocha LFN, Nery G. 2004a. Detection of entomopathogenic fungi in peridomestic triatomine-
infested areas in Central Brazil and fungal activity against Triatoma infestans (Klug) (Hemiptera:
Reduviidae). Neotrop Entomol 33: 783-791.
Luz C, Rocha LFN, Nery GV, Magalhães BP, Tigano MS 2004b. Activity of oil-formulated Beauveria
bassiana against Triatoma sordida in peridomestic areas in Central Brazil. Mem Inst Oswaldo Cruz
99: 211-218.
Luz C, Rocha LFN, Silva IG 2004c. Pathogenicity of Evlachovaea sp. (Hyphomycetes), a new species
isolated from Triatoma sordida, in Chagas disease vectors under laboratory conditions. Rev Soc Bras
Med Trop 37: 189-191.
Luz C, Silva IG, Cordeiro CMT, Tigano M 1998b. Beauveria bassiana (Hyphomycetes) as a possible agent
for biological control of Chagas disease vectors. J Med Entomol 35: 977-979.
Luz C, Silva IG, Magalhães BP, Cordeiro CMT, Tigano M 1999. Control of Triatoma infestans (Klug)
(Reduviidae: Triatominae) with Beauveria bassiana (Bals.) Vuill. preliminary assays on formulation
and application in the field. An Soc Entomol Brasil 28: 101-110.
Luz C, Tigano M, Silva IG, Cordeiro CMT, Aljanabi SM 1998c. Selection of Beauveria bassiana and
Metarhizium anisopliae isolates to control Triatoma infestans. Mem Inst Oswaldo Cruz 93: 839-846.
Matias A, Riva J, Martinez E, Torrez M, Dujardin JP 2003. Domiciliation process of Rhodnius stali
(Hemiptera: Reduviidae) in Alto Beni, La Paz, Bolivia. Trop Med Int Health 8: 264-268.
Marti GA, Scorsetti AC, Siri A, López Lastra CC 2005. Isolation of Beauveria bassiana (Bals.) Vuill.
(Deuteromycotina: Hyphomycetes) from the Chagas disease vector, Triatoma infestans (Hemiptera:
Reduviidae) in Argentina. Mycopathol 159: 389–391.
Medeiros Z 2004. A filariose bancroftiana no município de Moreno Pernambuco, Brasil. Rev Bras Epidemiol
7: 73-79.
Metchnikoff E 1879. Diseases of the larvae of the grain weevil. Insects harmful to agriculture [series]. Issue
III.

44
Mitchell DJ, Kannwischer-Mitchell ME, Dickson DW 1987. A semi-selective medium for the isolation of
Paecilomyces lilacinus. J Nematol 19: 255-256.
Monnerat RG, Dias DGS, Silva SF, Martins ES, Berry C, Falcão R, Gomes ACMM, Praça LB, Soares CMS
2005. Screening of Bacillus thuringiensis strains effective against mosquitoes. Pesq Agropec Bras 40:
103-106.
Moscardi F, Souza ML 2002. Baculovírus para o controle de pragas: panacéia ou realidade? Biotecno Ciênc
Desenvolv 24: 22-29.
Muscio OA, Bonder MA, La Torre JL, Scodeller EA 2000. Horizontal transmission of triatoma virus through
the fecal-oral route in Triatoma infestans (Hemiptera: Triatominae). J Med Entomol 37: 271-275.
Muscio OA, Torre JL, Bonder MA, Scodeller EA 1997. Triatoma vírus pathogenicity in laboratory colonies
of Triatoma infestans (Hemiptera: Reduviidae). J Med Entomol 34: 253-256.
Noireau F, Bosseno MF, Carrasco R, Telleria J, Vargas F, Camacho C, Yaksic N, Brenière F 1995. Sylvatic
triatomines (Hemiptera: Reduviidae) in Bolivia: trends toward domesticity and possible infection with
Trypanosoma cruzi (Kinetoplastida: Trypanoso-matidae). J Med Entomol 32: 594-598.
Panter C, Frances SP 2003. A more selective medium for Culicinimyces clavisporus. J Invertebr Pathol 82:
198-200.
Parameswaran G, Sankaran T 1977. Record of Beauveria bassiana (Bals.) Vuill. on Linshcosteus sp.
(Hemiptera: Reduviidae: Triatominae) in India. J Entomol Res 1: 113-114.
Polanczyk RA, Garcia MO, Alves SB 2003. Potencial de Bacillus thuringiensis israelensis Berliner no
controle de Aedes aegypti. Rev Saúde Púb 37: 813-816.
Regis L, Silva-Filha MH, Nielsen-LeRoux C, Charles JF 2001. Bacteriological larvicides of dipteran disease
vectors. Trends Parasitol 17: 377-380.
Regis L, Silva SB, Melo-Santos MA 2000. The use of bacterial larvicides in mosquito and black fly control
programmes in Brazil. Mem Inst Oswaldo Cruz 95: 207-210.
Roberts DR, Andre RG 1994. Insecticide resistance issues in vector-borne disease control. Am J Trop Med
Hyg 50: 21-34.
Romaña CA, Fargues J 1992. Relative susceptibility of different stages of Rhodnius prolixus to the
entomopathogenic hyphomycete Beauveria bassiana. Mem Inst Oswaldo Cruz 87: 363-368.
Rossman AY, Tulloss RE, Dell TEO, Thorn RG 1998. Introduction and overview. All taxa biodiversity
inventory of fungi in a Costa Rican conservation area. Ed. Parkway Publishers, Inc. 1-12.
Rozas-Dennis GS, Cazzaniga NJ, Gúerin DMA 2002. Triatoma patagonica (Hemiptera, Reduviidae), a new
host for Triatoma virus. Mem Inst Oswaldo Cruz 97: 427-429.
Schofield CJ, Diotaiuti L, Dujardin J 1999. The process of domestication in Triatominae. Mem Inst Oswaldo
Cruz 94: 375-378.
Scholte EJ, Knols BGJ, Samson RA, Takken W 2004. Entomopathogenic fungi for mosquito control: A
review. J Insect Science 4: 1-24.

45
Scholte EJ, Njiru BN , Smallegange RC, Takken W, Knols BGJ 2003. Infection of malaria (Anopheles
gambie s.s.) and filariasis (Culex quinquefasciatus) vectors with the entomopathogenic fungus
Metrhizium anisopliae. Malaria J 2: 2-8.
Shimaxu M, Teixera AR, Kishino KI 1994. Investigations on entomogenous fungi in the Cerrado region and
their utilization for microbial control of pests. Relatório técnico do Centro de Pesquisa Agropecuária
dos Cerrados, Planaltina, DF, Brasil: 202-214.
Silva IG, Silva JL, Silva HHG, Camargo M, Moura AF, Elias M, Santos AH 1992. Distribuição dos vetores
da tripossomíase americana capturados no ambiente domiciliar no estado de Goiás no período de
1984/88. An Soc Entomol 21: 139-154.
Silva RO, Silva HHG, Luz C 2004. Effect of Metarhizium anisopliae isolated from soil samples of the
central Brazilian cerrado against Aedes aegypti larvae under laboratory conditions. Rev Pat Trop 33:
207-216.
Silva RO, Silva HHG, Ulhoa CJ, Luz C 2005. Is there a relationship between N-acetyl-β-D-glucosaminidase
activity of Metarhizium anisopliae (Metschn.) Sorokin (Hyphomycetes) isolates from peridomestic
areas in Central Brazil and larvicidal effect on Aedes aegypti (L.) (Diptera, Culicidae)? J Appl Entomol
129: 158-164.
Silveira AC, Feitosa VR, Borges R 1984. Distribuição de triatomíneos capturados no ambiente domiciliar, no
período de 1975/1983, Brazil. Rev Bras Malariol Doenças Trop 36: 15-312.
Sneh B 1991. Isolation of Metarhizium anisopliae from insects on an improved selective medium based on
wheat germ. J Invertebr Pathol 58: 269-273.
Somboon P, Prapanthadara L, Suwonkerd W 2003. Insecticide susceptibility tests of Anopheles minimus s.l.,
Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus in northern Thailand. Southeast Asian J
Trop Med Pub Health 34: 87-90.
Vänninen I 1995. Distribution and occurrence of four entomopathogenic fungi in Finland: effect of
geographical location, habitat type and soil type. Mycol Res 100: 9-101.
Vassena CV, Picollo MI, Zerba EN 2000. Insecticide resistance in Brazilian Triatoma infestans and
Venezuelan Rhodnius prolixus. Med Vet Entomol 14: 51-55.
Veen KH, Ferron P 1966. A selective medium for isolation of Beauveria tenella and Metarhizium anisopliae.
J Invertebr Pathol 8: 268–26.
Whiteley HR, Schnepf HE 1986. The molecular biology of parasporal crystal body formation in Bacillus
thuringiensis. Ann Rev Microbiol 40: 549-576.
WHO 1997. Dengue haemorrhagic fever: Diagnostic, treatment, prevention and control. Genebra. 84.
WHO 1999. Chagas disease, Chile - Interruption of transmission. Weekly Epidemiological Record, 74: 9-11.
WHO 2000. WHO expert committee on malaria, WHO Technical Report Series 892. Geneva, Switzerland.
WHO 2002. Dengue and. Fact sheet Nº 117, Geneva.
Yaginuma K, Takagi K 1986. Improvement of a selective medium for isolation of Metarhizium anisopliae
(Metschnikoff) Sorokin. Japan J Appl Entomol Zool 30: 300-301.

46
Yasui K 1993. Strategies of dengue vaccine development by WHO. Using new biotechnology. Trop Med 35:
233-241.
Zerba EN 1999. Susceptibility and resistance to insecticides of Chagas disease vectors. Med Buenos Aires 59: 41-46.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















