
Alterações no comportamento de predação de Pachymerus … · 19 comportamento do predador de...
37
Alterações no comportamento de predação de Pachymerus cardo (Coleoptera: Chrysomelidae) provocado por fungos decompositores Tamires Santiago Librelon Orientador: Prof. Dr. Maurício Lopes de Faria Montes Claros Março-2015 UNIVERSIDADE ESTADUAL DE MONTES CLAROS Programa de Pós - Graduação Stricto Senso em Ciências Biológicas Área de Concentração: Biologia e Conservação DISSERTAÇÃO
Transcript of Alterações no comportamento de predação de Pachymerus … · 19 comportamento do predador de...

Alterações no comportamento de predação de Pachymerus cardo
(Coleoptera: Chrysomelidae) provocado por fungos decompositores
Tamires Santiago Librelon
Orientador: Prof. Dr. Maurício Lopes de Faria
Montes Claros
Março-2015
UNIVERSIDADE ESTADUAL DE MONTES CLAROS
Programa de Pós - Graduação Stricto Senso em Ciências Biológicas
Área de Concentração: Biologia e Conservação
DISSERTAÇÃO

TAMIRES SANTIAGO LIBRELON
Alterações no comportamento de predação de Pachymerus cardo (Coleoptera:
Chrysomelidae) provocado por fungos decompositores
Montes Claros
Março-2015
UNIVERSIDADE ESTADUAL DE MONTES CLAROS
Programa de Pós - Graduação Stricto Senso em Ciências Biológicas
Área de Concentração: Biologia e Conservação
DISSERTAÇÃO
Dissertação para defesa apresentada ao
Programa de Pós-Graduação Stricto
Sensu em Ciências Biológicas da
Universidade Estadual de Montes
Claros como requisito necessário para a
conclusão do Mestrado em Ciências
Biológicas.
Orientador: Prof. Dr. Maurício Lopes de Faria

AGRADECIMENTOS 1
2
Agradeço a Deus por ter me concedido a vida e ter me dado força, coragem e 3
sabedoria para concluir mais um ciclo da minha vida. 4
Ao meu querido orientador Maurício Lopes de Faria pela grande oportunidade, 5
pelo respeito, pelo aprendizado e paciência. 6
Ao professor Ronaldo Reis Júnior pela disponibilidade e auxilio na discussão dos 7
dados; 8
Ao professor Henrique por ceder espaço no laboratório de Microbiologia 9
Ambiental para execução de parte do meu trabalho; 10
Aos meus Pais, Paulo Roberto e Maria Marlene pela paciência e compreensão; 11
Aos minhas irmãs Roberta, Samira e Fernanda pela união, companheirismo e 12
incentivo; 13
Ao Hudson Rabelo pelo companheirismo e amor 14
Agradeço imensamente toda à equipe do LECB (Laboratório de Ecologia e 15
Controle Biológico de Inseto) pela colaboração e amizade; 16
A aqueles com as quais convivi diariamente durante o desenvolvimento deste 17
trabalho: Aline Alves, Anielle Cristina, Ane Karoline, Raquel Andrade, Gleice Mota, 18
Débora Lima; 19
Ao PROGRAMA DE FORMAÇÃO DE RECURSOS HUMANOS-PRFH225, 20
pelo provimento de minha bolsa do mestrado e ao programa de Pós-Graduação em 21
Ciência Biológicas da Universidade Estadual de Montes Claros, por todo o auxílio para 22
o desenvolvimento deste trabalho. 23
OBRIGADA!

RESUMO
RESUMO. LIBRELON, Tamires Santiago. Alterações no comportamento de predação
de Pachymerus cardo (Coleoptera: Chrysomelidae) provocado por fungos
decompositores - 2015. Dissertação Ms. Ciências Biológicas. Universidade Estadual de
Montes Claros.
1
As interações entre herbívoros e plantas hospedeiras podem ser mediadas por 2
sinais químicos emitidos por microrganismos. Os insetos, incluindo Pachymerus cardo, 3
utilizam e dependem dos compostos químicos em todo seu ciclo de vida. Tais sinais 4
químicos podem orienta-los nas respostas comportamentais. O objetivo foi analisar a 5
interferência da presença de duas espécies de fungos decompositores isolados de frutos 6
de Acrocomia aculeata no comportamento de seleção do sítio de oviposição por P. 7
cardo. Foram realizados dois bioensaios. O primeiro com fungos isolados de frutos de 8
A. aculeata. E o segundo, bioensaio foi de arena de tripla escolha, que consistiu em três 9
tratamentos (Fruto sadio, Fruto inoculado com fungo e Controle), o qual o inseto, era 10
colocado na arena central e teve igual acesso as três opções de escolha. Posteriormente 11
foi construído um etograma hierárquico para compreensão das vias comportamentais de 12
P. cardo frente aos voláteis químicos emitidos pelos frutos colonizados com fungos. Os 13
dados foram analisados através da construção de GLMs e descrições dos eventos 14
comportamentais na seleção de sítio de oviposição. O tratamento fruto sadio da arena foi 15
significativamente (P<0,05) selecionado pelos P. cardo em relação aos demais 16
tratamentos. O comportamento de Bater Asa (BA) foi associado em resposta à escolha 17
de fruto sadio, sendo P<0,05, mas as categorias comportamentais expressas por P. cardo 18
não diferiram entre os fungos testados. Dessa forma foi possível verificar que a 19
presença do fungo modifica o comportamento de seleção de sítio de oviposição de P. 20
cardo. Tais dados são relevantes para a compreensão ecológica das emissões de 21
compostos químicos por fungos como moduladores de comportamento de predadores de 22
semente. 23
24
Palavras-chaves: Interações, Microrganismo, Macaúba, Comportamento e Mediadores 25
químicos. 26
27
28

ABSTRACT 1
2
ABSTRACT: LIBRELON, Tamires Santiago. Changes in predation behavior of 3
Pachymerus cardo (Coleoptera: Chrysomelidae) caused by decomposing fungi - 2015. 4
Dissertação Ms. Ciências Biológicas. Universidade Estadual de Montes Claros. 5
6
The interactions between herbivores and host plants can be mediated by chemical 7
signals emitted by microorganisms. Insects, including Pachymerus cardo, use and 8
depend on chemical compounds throughout their lifecycle. These chemical signals can 9
guides them in behavioral responses. The aim was to evaluate the interference from the 10
presence of two species of isolated fungi decomposers in Acrocomia aculeata fruit in 11
oviposition site selection behavior by P. cardo. Two Bioassays were performed. In the 12
first, with isolates fungi of A. aculeata fruit. And the second bioassay was triple-choice 13
arena, consisting of three treatments (healthy fruit, fruit inoculated with fungi and 14
Control), which the insect was placed in the central arena and had equal access to three 15
choices. Was subsequently built a hierarchical ethogram for understanding the 16
behavioral pathways of P. cardo in front of chemical volatiles emitted by fruits 17
colonized by fungi. The data were analyzed by building GLMs and descriptions of 18
behavioral events in the selection of oviposition site. The healthy fruit treatment in 19
Arena was significantly (P <0.05) selected by P. cardo in compared to other treatments. 20
The Beat Wing behavior (BA) was associated in response to the choice of healthy fruit 21
being P <0.05, but the behavior categories expressed by P. cardo did not differ among 22
the tested fungi. Thus it was verified that the fungi presence modifies the oviposition site 23
selection behavior of P. cardo. These data are relevant to ecological understanding of 24
the emissions of chemical compounds by fungi as seed predatory behavior modulators. 25
26
Keywords: Interactions, Microorganism, Macaúba, Behavior and chemical mediators. 27
28

SUMÁRIO
1. INTRODUÇÃO ........................................................................................................... 3 1
2. MATERIAL E MÉTODOS ........................................................................................ 6 2
2.1 Sistema de estudo ................................................................................................... 6 3
2.1.1 Planta hospedeira ............................................................................................... 6 4
2.1.2 O predador da semente ...................................................................................... 7 5
2.2 Coleta de frutos de A. aculeata ............................................................................. 8 6
2.3 Obtenção dos insetos ............................................................................................. 9 7
2.4 Obtenção de fungos ............................................................................................... 9 8
2.5 Desenho experimental ......................................................................................... 10 9
2.5.1 Bioensaio com fungos ..................................................................................... 10 10
2.5.2 Bioensaio de arena ........................................................................................... 11 11
2.5.3. Elaboração do Etograma ............................................................................... 13 12
2.6 Análises de dados ................................................................................................. 14 13
3. RESULTADOS .......................................................................................................... 15 14
4. DISCUSSÃO .............................................................................................................. 21 15
5. CONCLUSÃO ........................................................................................................... 26 16
6. REFERÊNCIAS ........................................................................................................ 26 17

3
1. INTRODUÇÃO
Os sistemas ecológicos naturais são caracterizados por abranger uma vasta 1
diversidade de espécies, produzindo uma intricada rede trófica composta por indivíduos 2
que interagem entre si e com o ambiente circundante (Hastings e Powell, 1991; Irwin 3
2006; Torres-Alruiz, 2013). O consumo de plantas por insetos herbívoros representa 4
uma das interações entre espécies dominantes no ambiente, que evolutivamente 5
desempenha um papel-chave na diversificação das espécies em ambientes terrestres. 6
As interações entre herbívoros e plantas hospedeiras podem interferir no ciclo de 7
vida de alguns vegetais, principalmente quando comprometem a viabilidade das 8
sementes, interferindo diretamente no “fitness” da planta. (Panizzi e Parra 1991; 9
Almeida-Cortez, 2004). Estas interações são substancialmente mediadas por sinais 10
químicos emitidos pela planta, que permitem aos herbívoros se orientar e encontrar os 11
hospedeiros apropriados, em função da detecção de misturas características desses 12
compostos químicos (Loaiza e Céspedes 2007; Webstera et al. 2010; Martins et al. 13
2012). 14
Os compostos químicos envolvidos na comunicação dos indivíduos são 15
biossintetizados e liberados pelo emissor, produzindo uma resposta comportamental ou 16
fisiológica imediata ou de longo prazo no receptor (Paiva e Pedrosa-Marcedo 1985; 17
Nordlund e Lewis 1976). A esses sinais químicos, que são responsáveis pela 18
transmissão de informação são denominados de infoquímicos (Dicke e Sabetes 1988). 19
Os infoquímicos mediadores de interações entre dois indivíduos de espécies diferentes 20
(ação interespecífica) são designados aleloquímicos e estes compostos exercem papel 21
fundamental em todas as comunidades (Bento e Nardi, 2009; Price et al. 2011). 22

4
Os insetos herbívoros constituem o grupo de animais terrestres que mais utilizam 1
e dependem dos infoquímicos para intermediar suas interações (Vilela e Della-Lucia 2
2001). Esses compostos químicos permitem aos insetos predarem um grande número de 3
espécies florestais, sendo a predação de palmeiras tropicais muito relatadas (Scariot 4
1998; Silva et al. 2007; Fonseca 2008; Steffler et al. 2008; Donatti et al. 2009; Pereira 5
et al. 2014). 6
Em trabalho realizado em laboratório de Ecologia e controle Biológico de 7
insetos (LECB) da Universidade Estadual de Montes Claros (UNIMONTES), já foi 8
registrado maior ocorrência de predação de palmeiras da espécie Acrocomia aculeata 9
por insetos pertencentes à subfamilia Bruchinae. A interação desses herbívoros com 10
frutos geram lesões no epicarpo, tornando-os mais susceptíveis à colonização por 11
microrganismos. Essa colonização por fungos modifica o comportamento dos 12
herbívoros (Pereira et al. 2014). 13
Recentemente, a função dos microrganismos como mediadores de interações, 14
entre plantas e insetos herbívoros tem sido cada vez mais reconhecida nas pesquisas 15
ecológicas (Fonseca 2008; Jallow et al. 2008; Pereira et al. 2014). Os insetos são 16
altamente sensíveis às emissões de voláteis microbianos (Ezenwa et al. 2012), e esses 17
compostos podem mediar interações entre mais de um nível trófico, podendo atuar na 18
atratividade do fruto para os insetos (Chaudhury et al. 2010; Tomberlin et al. 2012), 19
repelência (Burkepile et al. 2006) e/ou na camuflagem de sítio de oviposição do inseto 20
(Pereira et al. 2014). 21
As substâncias químicas emitidas por microrganismos, dessa forma, produzem 22
consequências relevantes para as populações de insetos podendo afetar as respostas 23
comportamentais (Davis e Landolt 2013), bem como promover mudanças no 24

5
comportamento e história de vida das outras espécies envolvidas (Abrams 1995; Pocock 1
et al. 2012; Lewinsohn e Cagnolo 2012). 2
No presente trabalho utilizou isolados de fungos da espécie Aspergillus sp. e 3
Fusarium sp. que foram encontrados com maior frequência em frutos de A. aculeata 4
pelo grupo de pesquisadores do LECB da UNIMONTES, tais fungos foram 5
selecionados pela abundância, afim de verificar como espécies de fungos diferentes 6
interferem no comportamento de predação de bruquíneos. 7
Ambas as espécies de fungos utilizadas apresentam ampla distribuição 8
geográfica, sendo considerados cosmopolitas no ambiente. Esses fungos produzem 9
grandes quantidades de conídios, que são facilmente dispersos (Cavalcanti 2006; Fraga 10
e Pereira 2012; Oliveira et al. 2012; Prade e Matsumura 2014) e são considerados 11
fungos “oportunistas” (Cooke 1979; Pitt e Hocking 1985; Farr et al. 1989) podendo 12
atuar como saprófitos ou patógenos, dependendo da espécie (Michereff 2005; Tournas e 13
Katsoudas 2005; Gadgile e Chavan 2010). 14
Aspergillus sp. e Fusarium sp. também são importantes produtores de 15
micotoxinas que são metabólitos tóxicos naturais, que alteram o valor nutricional e 16
qualidades organolépticas dos frutos (Rodriguez-Amaya 2002; Evangelista 2005). 17
Para entendimento de como as duas espécies de fungos interferem no 18
comportamento do predador de semente foi proposta a hipótese, que espécies distintas 19
de fungos decompositores alteram o comportamento de predação de P. cardo de forma 20
diferente. 21

6
2. MATERIAL E MÉTODOS 1
2.1 Sistema de estudo
2.1.1 Planta hospedeira 2
A. aculeata (Jacq.) Lodd. ex Mart. é uma espécie pertencente à família 3
Arecaceae. É uma palmeira arbórea com porte de 10 a 15 m de altura, com estipe ereto 4
e cilíndrico. A região caulinar é revestida ou não por acúleos finos nas reminiscências 5
das bainhas (Cetec 1983; Henderson et al. 1995). As inflorescências são pequenas, 6
unissexuais, andróginas e de coloração amarela (Nucci 2007) (Fig. 1-A). 7
Fig. 1 Características morfológicas da Acrocomia aculeata. A- Visão geral e inflorescência. B- Cacho de 8
frutos maduros. C- Vista longitudinal do fruto aberto, vê-se mesocarpo em amarelo, epicarpo envolvendo 9
o mesocarpo externamente e o endocarpo constituído pela estrutura de coloração preta e a amêndoa de 10
cor branca. 11
12
Os frutos são esféricos, em forma de drupa globosa e dispostos em cachos (Fig. 13
1-B), são constituído por epicarpo cartáceo (casca); mesocarpo fino, mucilaginoso e 14
fibroso (polpa) e endocarpo duro e denso (tegumento) (Lorenzi et al. 2010). No interior 15
do endocarpo encontra-se a amêndoa, oleaginosa e comestível (Fig. 1-C). A frutificação 16
A
C
B

7
ocorre durante todo o ano e os frutos amadurecem, principalmente, entre os meses de 1
setembro a janeiro (Almeida et al. 1998; Nucci 2007; Lorenzi et al. 2010), podendo 2
permanecer sobre o solo por meses sem germinar. 3
A palmeira A. aculeata apresenta ampla distribuição geográfica, com ocorrência 4
de populações naturais em quase todo território brasileiro. Entretanto, as maiores 5
concentrações estão localizadas nos Estados de Minas Gerais, Goiás, Mato Grosso e 6
Mato Grosso do Sul (Henderson et al. 1995; Clement et al. 2005). Esta palmeira habita 7
áreas abertas do cerrado e com alta incidência solar (Lorenzi e Negrelle 2006), como 8
áreas degradadas e de pastagens (Pires et al. 2012). 9
2.1.2 O predador da semente 10
Os coleópteros pertencentes à tribo Pachymerini, família Chrysomelidae e 11
subfamília Bruchinae são predadores geralmente especialistas e utilizam quase que 12
exclusivamente palmeiras como fonte de recursos (Delobel et al. 1995; Ramirez e 13
Traveset 2010). 14
As espécies Pachymerus cardo (Fahraeus 1839) e Speciomerus revoili (Pic 15
1902) são indivíduos predadores de A. aculeata (Ramos et al. 2001; Fonseca 2008; 16
Pereira et al. 2014). Esses bruquíneos são capazes de predar até 100% das sementes 17
que permanecem sob a planta-mãe (Forget et al. 1999). 18
Os bruquíneos desenvolvem-se após a oviposição em sementes ou vagens 19
(Southgate 1979). Aproximadamente cinco dias após esse processo, as larvas eclodem e 20
penetram na semente ou vagem à procura da amêndoa (endocarpo), (Fig. 2). Geralmente 21
muitas larvas penetram no endocarpo, mas apenas um indivíduo por semente sobrevive 22
até a maturidade (Scariot 1998; Silvius e Fragoso 2002). 23
24

8
Fig. 2 Ciclo de vida do coleóptero Pachymerus cardo (Chrysomelidae, subfamília Bruchinae). A- P. 1
cardo copulando, B- Frutos de Acrocomia aculeata com ovos. C- Larva após alimentar-se de todo o 2
endosperma da semente. D- Fase pupal dos Bruchinae. F- Fruto com orifício de saída feito por P. cardo. 3
F- Individuo adulto de Bruchinae (Fonte: LECB). 4
5
2.2 Coleta de frutos de A. aculeata 6
As coletas de frutos em pré e pós-dispersão foram realizadas no município de 7
Itacambira (43° 18’ 32” W 17° 03’ 53” S) localizado no Norte de Minas Gerais e 8
levados para o laboratório de Ecologia e Controle Biológico de Insetos da Universidade 9
Estadual de Montes Claros (UNIMONTES) (Fig. 3). Os frutos coletados em pré-10
dispersão foram esterilizados por imersão durante dois minutos em solução de 11
hipoclorito de sódio a 2%, seguida de enxague com água destilada e foram colocados 12
para secar em temperatura ambiente. Posteriormente uma parte dos frutos foi utilizada 13
na inoculação de fungos e outra parte no tratamento Fruto sadio da arena, descritas no 14
tópico Bioensaio de arena. 15
16
A
E
D
C
B F

9
Fig. 3 Coletas de frutos: A - pré-dispersão e B - pós-dispersão realizadas no município de Itacambira.
2.3 Obtenção dos insetos 1
2
Para obtenção dos insetos que seriam utilizados nos bioensaios de arena, foi 3
estabelecida uma criação de bruquíneos no laboratório de Ecologia e Controle 4
Biológico de Insetos na Universidade Estadual de Montes Claros (UNIMONTES). A 5
criação foi conduzida a partir de larvas de P. cardo adquiridas pela quebra de 3.915 6
frutos de A. aculeata coletados em pós-dispersão. Cada larva foi colocada 7
separadamente em potes plásticos estéreis de 80 ml, com algodão, tampados com 8
plástico filme e mantidos em sala climatizada, sob temperatura de 25 ± 3°C, umidade 9
media relativa de 50% e fotoperíodo de 12 horas. 10
Após as larvas atingirem a idade adulta, os insetos foram transferidos para 11
aquários de vidro e alimentados com solução aquosa de 10% de mel embebido em 12
algodão. O alimento era trocado diariamente e os aquários higienizados uma vez por 13
semana. 14
2.4 Obtenção de fungos 15
Os fungos decompositores de frutos de A. aculeata utilizados nos bioensaios 16
foram obtidos de um banco de inóculo armazenado a partir do trabalho de Pereira et al. 17
(2014) que identificou 102 morfotipos de fungos, distribuídos em nove gêneros 18
A B

10
(Fusarium sp, Aspergillus sp, Acremonium, Rhizopus, Cladosporium, Memnoniella, 1
Spermospora, Penicillium, Scedosporium). As espécies Fusarium sp. e Aspergillus sp. 2
foram os mais abundantes e por isso, selecionados para os testes neste estudo (Fig. 4 e 3
5). 4
Fig. 4 Características morfológicas do gênero Fusarium sp. A. Morfologia macroscópica da colônia do 5
cultivo de Fusarium sp. B. Morfologia microscópica do microcultivo de Fusarium sp. presença de 6
maroconídios e microconídios. 7
8
9
10
11
12
13
14
15
Fig. 5 Características morfológicas do gênero Aspergillus sp. A. Morfologia macroscópica da colônia do 16
cultivo de Aspergllus sp. B. Morfologia microscópica do microcultivo de Aspergillus sp. 17
18
2.5 Desenho experimental
2.5.1 Bioensaio com fungos 19
20
Inóculos de Aspergillus sp. e Fusarium sp. foram replicados em meio de cultura 21
de BDA (Ágar Batata Dextrose) em placas de Petri de 9 mm, que foram mantidas em 22
estufa a 27 ± 3°C por um período de sete dias. Após desenvolvimento fúngico, o meio 23
A B
A B

11
foi estriado com auxílio de um bastão de vidro estéril, visando maior obtenção de 1
conídios. Esse procedimento foi realizado em câmara de fluxo laminar e as placas 2
estriadas permaneceram em temperatura ambiente por dois dias. 3
A suspensão de conídios foi obtida por meio da adição de 6 mL de água 4
destilada estéril, acrescidos com Tween 80 na proporção de 0,01 % em cada placa. Em 5
seguida, a superfície das placas de Petri com a cultura esporulada foi raspada com 6
auxílio de um bastão de vidro para a remoção dos conídios. Essas suspensões de 7
conídios foram então transferidas para um tubo Falcon, para posterior homogeneização 8
e quantificação, em Câmara de Neubauer. Após esse preparo os conídios de cada fungo 9
foram inoculados em frutos maduros, saudáveis e esterilizados (Fig. 6). 10
Fig. 6 Procedimentos do bioensaio com fungos. A Visão frontal dos materiais necessários para inoculação 11
de conídios. B Inoculação de solução de conídios em frutos maduros de A. aculeata. 12
13
Posteriormente, os frutos inoculados foram mantidos isolados em potes plásticos 14
estéreis e identificados quanto à espécie de fungo e mantidos em câmara incubadora 15
(B.O.D.), sob condições controladas, temperatura de 28 ± 2°C e umidade relativa de 16
60% durante sete dias. 17
2.5.2 Bioensaio de arena 18
Para testar o efeito direto dos compostos emitidos pelos frutos inoculados por 19
Aspergillus sp. e Fusarium sp. na escolha do sítio de oviposição das fêmeas de P. cardo, 20
A B

12
foi montada uma arena de tripla escolha semelhante à utilizada por Pereira et. al., (2014) 1
(Fig. 7 A-B). Foram utilizadas 91 fêmeas de P. cardo, das quais 41 foram testadas com 2
frutos inoculadas com Aspergillus sp. e 50 com Fusarium sp. Cada repetição foi 3
compreendida por um único e exclusivo inseto. 4
Fig. 7 Arena tripla de escolha utilizada no experimento de preferência e comportamento de oviposição. A- 5
Arena disposta dentro da Câmara de Fluxo Laminar. B- Visão de cima da arena tripla escolha. 6
7
Nesse bioensaio somente as fêmeas de P. cardo foram utilizadas. As fêmeas 8
foram selecionadas a partir observações durante o período de cópula, para aumentar a 9
probabilidade de estarem grávidas durante o bioensaio. Uma fêmea por vez foi colocada 10
na câmara central, tendo acesso equivalente aos três tratamentos: fruto de A. aculeata 11
sadio (FS), fruto inoculados com fungo (FF) e pote vazio (controle) (C). 12
Os bioensaios de arena foram conduzidos em Câmara de Fluxo Laminar entre 18 13
e 21 horas da noite, horário de maior atividade desses insetos. A fim de facilitar a 14
observação sem interferir no comportamento do inseto foi utilizada uma luz vermelha. 15
As observações ocorreram ininterruptamente por um tempo máximo de uma hora 16
para cada repetição e quando houve escolha pelas fêmeas de P. cardo por um 17
determinado tratamento foi aguardado um período de 10 minutos de permanência. 18
Durante todo tempo que o P. cardo manteve-se na arena de tripla escolha, foi registrado 19
e descrito seus repertórios comportamentais. A cada novo ensaio toda arena era trocada 20
A B

13
e ao final dos testes as arenas eram desinfetadas com álcool a 70% v/v seguida de três 1
lavagens com água para eliminar odores e possíveis esporos de fungos. 2
2.5.3. Elaboração do Etograma
Para descrição do repertório comportamental de fêmeas de P. cardo, foi elaborado um 3
Etograma, onde os comportamentos foram quantificados e qualificados em categorias, 4
levando em conta a literatura existente (Tasin et al. 2011; Lam et al. 2010; Buteler et al. 5
2009; Corrêia e Sant'Ana 2001; van Loon e Dicke 2001) e observações pessoais 6
(Tab.1). Quantificou-se a resposta pelo número de insetos testados que respondem a um 7
determinado tratamento e qualificou-se pelas atividades comportamentais relacionadas à 8
resposta motora do inseto frente à fonte de estimulo. 9
Tab. 1 Etograma comportamental de fêmeas de P. cardo submetidas ao teste de arena de tripla escolha 10
Tipo de comportamento Sigla referente ao
comportamento
Descrição do comportamento
Exploração
E
Locomoção sem direcionamento
e com antenação.
Parado Ativo Antenando
PAA
Inseto permanece parado, mas
com movimentos alternados das
antenas ou as antenas em V
Autolimpeza
AUTL
Inseto limpa partes do corpo
(patas e antenas) usando as
pernas e ou peças bucais.
Parado Inativo
PI
Inseto em repouso sem
movimento aparente.
Bater Asa
BA
Ato de estender os élitros e
desdobrar asas posteriores.
Expor Ovipositor
EO
Ato de expor o ovipositor sobre
um substrato.

14
2.6 Análises de dados
As análises foram realizadas no software R (R Core Team, 2014). A partir da 1
construção modelos lineares generalizados (GLMs), seguida de análise de resíduo para 2
verificar a adequação da distribuição de erros e o ajuste do modelo (Crawley 2002). 3
Todas essas análises foram usadas tanto para frutos inoculados com Aspergillus sp. 4
como para Fusarium sp. 5
Para analisar o efeito da presença dos fungos em relação à escolha do sítio de 6
oviposição por P. cardo, considerou-se como variável resposta a escolha de fêmeas e 7
como variável explicativa os tratamentos (FS, FF e C), esse modelo foi analisado 8
considerando a distribuição Binomial. Nessa análise foram utilizadas apenas fêmeas que 9
responderam a algum dos tratamentos. 10
Para analisar a diferença no tempo de respostas das fêmeas de P. cardo quanto à 11
escolha dos tratamentos, considerou-se como variável resposta o tempo e como variável 12
explicativa o tratamento. Esse modelo foi avaliado considerando a distribuição de 13
Poisson. 14
Para analisar a diferença no tempo de resposta em relação aos tratamentos e os 15
diferentes fungos (Aspergillus sp e Fusarium sp), assumiu-se como variável resposta o 16
tempo e como variável explicativa o tratamento e as duas espécies de fungos. O modelo 17
foi examinado considerando a distribuição de erros Quasibinomial devido à 18
sobredispersão dos dados. 19
Para verificar se algum comportamento realizado pelas fêmeas de P. cardo 20
durante o teste de arena estava associado com a escolha do tratamento fruto sadio (FS) . 21
Considerou-se como variável resposta o fruto sadio (FS) e como variável explicativa as 22

15
categorias de comportamentos E, PAA, AUTL, PI, BA e EO, esse modelo foi analisado 1
considerando a distribuição Binomial. 2
3. RESULTADOS
No experimento de preferência e comportamento de oviposição testados na arena 3
de tripla escolha, ocorreu por um período total de 2910 minutos. Do total de fêmeas 4
utilizadas, 34% não selecionaram nenhum dos tratamentos como sítio de oviposição e 5
66% testadas com Fusarium sp. e Aspergillus sp. responderam aos tratamentos. As 6
fêmeas de P. cardo que apresentaram resposta aos testes com Aspergillus sp. 7
escolheram 70% pelo tratamento FS, 20% pelo FF e 10% pelo C. Para os bioensaios 8
com Fusarium sp. as escolhas foram de 73,3% para o FS, 20% para o FF e 6,7% 9
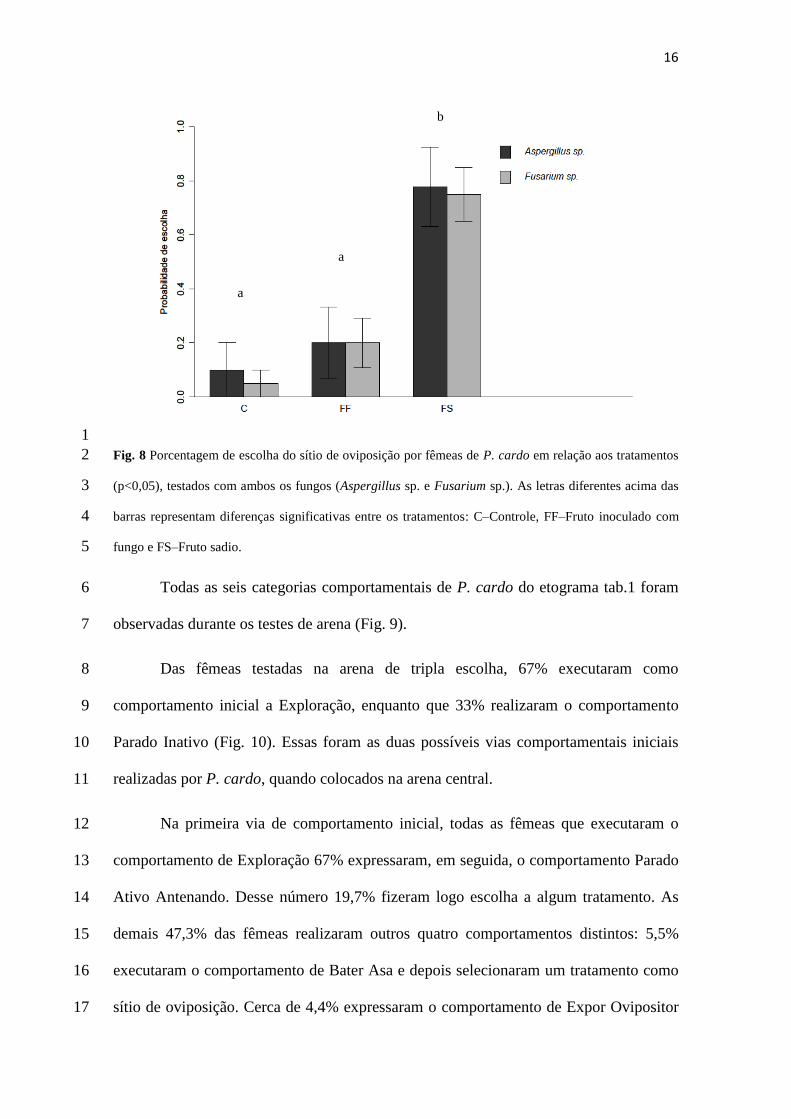
optaram pelo C (Fig.8). 10
O tempo médio de respostas das fêmeas aos tratamentos foi de 18,2 e 18,8 11
minutos para Aspergillus sp. e Fusarium sp. respectivamente. Não houve diferença 12
significativa no tempo de resposta entre os tratamentos e nem entre as espécies de 13
fungos (P>0,05). O tempo total máximo que uma fêmea levou para escolher um 14
tratamento foi de 40 minutos, enquanto o tempo mínimo foi de 1 minuto para ambas as 15
espécies de fungos. 16
Em relação à seleção de sítio de oviposição por fêmeas de P. cardo aos 17
tratamentos, a opção FS diferiu significativamente (P<0,05) dos demais tratamentos, 18
mas não houve diferença entre as espécies de fungos Aspergillus sp. e Fusarium sp. 19
testados (Fig. 8). 20
16
1
Fig. 8 Porcentagem de escolha do sítio de oviposição por fêmeas de P. cardo em relação aos tratamentos 2
(p<0,05), testados com ambos os fungos (Aspergillus sp. e Fusarium sp.). As letras diferentes acima das 3
barras representam diferenças significativas entre os tratamentos: C–Controle, FF–Fruto inoculado com 4
fungo e FS–Fruto sadio. 5
Todas as seis categorias comportamentais de P. cardo do etograma tab.1 foram 6
observadas durante os testes de arena (Fig. 9). 7
Das fêmeas testadas na arena de tripla escolha, 67% executaram como 8
comportamento inicial a Exploração, enquanto que 33% realizaram o comportamento 9
Parado Inativo (Fig. 10). Essas foram as duas possíveis vias comportamentais iniciais 10
realizadas por P. cardo, quando colocados na arena central. 11
Na primeira via de comportamento inicial, todas as fêmeas que executaram o 12
comportamento de Exploração 67% expressaram, em seguida, o comportamento Parado 13
Ativo Antenando. Desse número 19,7% fizeram logo escolha a algum tratamento. As 14
demais 47,3% das fêmeas realizaram outros quatro comportamentos distintos: 5,5% 15
executaram o comportamento de Bater Asa e depois selecionaram um tratamento como 16
sítio de oviposição. Cerca de 4,4% expressaram o comportamento de Expor Ovipositor 17
a
a
b

17
e escolheram um determinado tratamento. Um total 36,3% executaram o 1
comportamento de Autolimpeza e desses indivíduos, 19,7% fizeram escolha direta a um 2
dos tratamentos, 5,5% das que se autolimparam, realizaram o comportamento de Expor 3
Ovipositor para depois selecionar o tratamento e 11,1% após se autolimparem 4
expressaram o comportamento de Bater Asa e 1,1% expôs ovipositor para só então 5
efetuar escolha. Apenas 1,1% das fêmeas que realizou o comportamento de Parado 6
Ativo Antenando, exibiu posteriormente o comportamento Parado Inativo e não 7
respondeu aos testes. (Fig. 10). 8
Fig. 9 Representação do etograma de seleção de sítio de oviposição por P. cardo.
Na segunda via de comportamento inicial, das 33% das fêmeas de P. cardo que 9
executaram o comportamento Parado Inativo nos testes, 4,4% não escolheram um dos 10

18
tratamentos e nem expressaram outro comportamento. Em torno de 28,6%, após ficarem 1
paradas inativas realizaram o comportamento Parado Ativo Antenando e, desse valor 2
15,4% não selecionaram os tratamentos e 12,2% executaram o comportamento de 3
Autolimpeza, mas mesmo assim não houve escolha (Fig. 10). 4
O etograma hierárquico constatou que todas as vezes que o comportamento
Parado Inativo ocorreu independente da via comportamental, culminou em não escolha
dos tratamentos. E, quando os comportamentos Expor Ovipositor e Bater Asa foram
expressos sempre houve escolha de um determinado tratamento.
Com isso, distinguimos sete vias comportamentais que culminaram em escolha 5
dos tratamentos (FS, FF e C) pelas fêmeas de P. cardo (Tab. 2). Quatro dessas vias 6
(vias de escolha: 2, 3, 6 e 7), quando realizadas sempre resultaram na seleção do 7
tratamento FS, evidenciando que essas sequências comportamentais proporcionaram ao 8
inseto maior precisão e certeza de escolha do sítio de oviposição adequado. Já, quando o 9
comportamento de Autolimpeza foi executado, sem ser seguido do comportamento de 10
Bater Asa, as fêmeas se mostraram confusas quanto à escolha do sítio, visto que, 27,8% 11
e 40% delas selecionaram frutos inoculados com fungos, nas vias de escolha 4 e 5, 12
respectivamente, ao invés de frutos sadios. 13
Outra via comportamental, que elucida a confusão das fêmeas na seleção de sítio 14
de oviposição, foi quando executaram apenas dois comportamentos (via de escolha 1) e 15
logo após escolhem os tratamentos. De modo geral, os insetos selecionaram mais de 16
50% das vezes FS em todas as vias comportamentais, como preferência de sítio de 17
oviposição (Tab. 2). 18

19
Fig. 10 Etograma hierárquico de todos os comportamentos factíveis executados pelas fêmeas de P. cardo durante os
testes de arena de tripla escolha. Os dois comportamentos executados no início do teste de arena Parado inativo (PI)
ou Exploração (E). Depois de efetuar um desses comportamentos as fêmeas de P. cardo podiam efetuar outras vias
de comportamentos. Os valores em porcentagem (%) foram distribuídos dependendo da frequência dos indivíduos
que expressava o comportamento até a escolha ou a não escolha. PAA – Parado ativo antenando, AUTL –
Autolimpeza, EO – Expor ovipositor, BA – Bater asa. ESCOLHA – FF (Fruto inoculado com fungo), FS (Fruto
sadio) e C (Controle). NÃO ESCOLHA – não preferir nenhum tratamento.

20
Tab. 2 Vias comportamentais realizadas por fêmeas de P. cardo quando executaram escolha dos
tratamentos: Fruto sadio (FS), Fruto inoculado com fungo (FF) e Controle (C). As vias de escolhas são
representadas pelos comportamentos: E – Exploração, PAA – Parado Ativo Antenando, AUTL –
Autolimpeza, EO – Expor Ovipositor, BA – Bater Asa.
Via de Escolha Nº indivíduos Tratamentos (%)
FS FF C
1 E-PAA 18 55,5% 27,8% 16,7%
2 E-PAA-EO 4 100% - -
3 E-PAA-BA 5 100% - -
4 E-PAA-AUTL 18 61,1% 27,8% 11,1%
5 E-PAA-AUTL-EO 5 60% 40% -
6 E-PAA-AUTL-BA 9 100% - -
7 E-PAA-AUTL-BA-EO 1 100% - -
A sequência do comportamento de seleção do sítio de oviposição consistiu em 1
duas etapas. A primeira compõe-se da Exploração, em que os indivíduos procuraram 2
pelo sítio de oviposição e estabeleceram contato com o hospedeiro. Nessa etapa as 3
fêmeas de P. cardo desencadeava modalidades sensoriais com maior intensidade, 4
exibindo comportamentos repetitivos de contato com o hospedeiro como ficar Parado 5
Ativo Antenando, Autolimpeza e Bater Asa. E a segunda etapa consistiu em 6
reconhecimento do sítio, onde o comportamento de Parado Ativo Antenando foi 7
frequente e o de Expor Ovipositor também ocorreu. 8
Dentre todos os comportamentos expressos, o que ocorreu com maior frequência 9
foi Parado Ativo Antenando com 93% e o menos frequente foi Bater Asa com 12% para 10
os testes de arena com Aspergillus sp. Para Fusarium sp., a ocorrência do 11
comportamento Parado Ativo Antenando também foi o mais frequente com 98%, no 12
entanto, o que ocorreu menos, dentre as seis categorias de comportamentos foi Expor 13
Ovipositor com 6%. A ocorrência das categorias de comportamentos de P. cardo não 14
diferiram quanto a espécie de fungo testado (Fig. 11). 15

21
Fig. 11 Frequência de ocorrência em relação às categorias de comportamentos em testes de arena com o 1
fungos Fusarium sp. e Aspergillus sp. E – Exploração, PAA – Parado Ativo Antenando, AUTL – 2
Autolimpeza, PI – Parado Inativo, BA – Bater Asa, EO – Expor Ovipositor. 3
4
O comportamento BA realizado pelas fêmeas de P. cardo foi o único, dentre os 5
comportamentos observados, relacionado com a escolha do tratamento FS (P<0,05) 6
como preferência de sítio de oviposição. E está negativamente relacionada com a 7
escolha de FF (P <0,05). Nenhum indivíduo que BA escolheu fruto inoculado com 8
fungo. 9
4. DISCUSSÃO
A presença de Aspergillus sp. e Fusarium sp. em frutos de A. aculeata, modifica 10
o comportamento de seleção de sítio de oviposição das fêmeas de P. cardo e 11
apresentam alterações equivalentes nas categorias comportamentais e na preferência 12
pelo sítio de oviposição, o que refuta a hipótese proposta. 13
Neste mesmo sistema de estudo, Pereira et al. (2014) verificaram maior 14
preferência de seleção de sítio de oviposição de P. cardo por frutos de A. aculeata 15
íntegros, em comparação com frutos colonizados por fungos não identificados. Os 16
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
E PAA AUTL PI BA EO
Oco
rrên
cia
Comportamentos
Aspergillu sp.
Fusarium sp.

22
mesmos autores sugerem que a ação dos fungos sobre o fruto pode gerar “camuflagem” 1
química, o que dificulta a percepção pelo predador. 2
Alterações comportamentais, na seleção de sítios de oviposição na presença de 3
fungos, já foram demonstradas por outros pesquisadores, como Kruess (2001) que 4
trabalhou com Chrysomelidae da espécie Cassia rubiginosa e constatou que as fêmeas 5
preferiram, significativamente, folhas saudáveis sobre aquelas infectadas com fungos. 6
Em outro estudo, com Chrysomelidae, mas pertencente à espécie Oreina cacaliae 7
Roeder et al. (2007) verificaram que folhas sadias foram mais selecionada para 8
oviposição, do que folhas contaminadas. Rayamajhi et al. (2006), também 9
corroboraram, com o presente trabalho, visto que, a presença de fungos interferiu, 10
negativamente, no comportamento de oviposição de coleópteros Oxyops vitiosa, os 11
quais passaram a escolher, predominantemente, folhas livres de fungos, como sítio de 12
oviposição. 13
Essas alterações comportamentais são observadas para insetos pertencentes a 14
outras ordens, como, por exemplo, espécie de mariposa Lobesia botrana (Tortricidae) 15
que selecionaram, preferencialmente, sítios de oviposição com ausência de fungos, além 16
de evitar pousar em substratos contaminados por esses microrganismos (Tasin et al. 17
2011). Em experimento com moscas (Musca domestica), Lam et al. (2010) perceberam 18
que tais insetos reduziram significativamente a oviposição em fezes colonizadas por 19
Phoma e Rhizopus. Por outro lado, esses autores não verificaram interferência no 20
comportamento, quando havia a presença de Fusarium. Esses resultados diferem dos 21
observados neste estudo, uma vez que, a espécie Fusarium sp. modificou o 22
comportamento de seleção de sítio de oviposição de P. cardo. No entanto, essa 23
discordância de dados pode estar relacionada com os insetos-alvo utilizados em ambos 24
os trabalhos. 25

23
Segundo Askman e Ekbom (2006), espera-se que as consequências do 1
comportamento de seleção de sítio para oviposição sejam diferentes para insetos 2
pertencentes a grupo taxonômicos distintos, isso por haver divergência no 3
comportamento de procura, assimilação e fisiologia dos mesmos. Existem requisitos 4
específicos, na escolha de sítios de oviposição e isso pode ser determinado pelas fortes 5
associações naturais entre espécies nos diferentes habitas (Kitching 1977). 6
Em geral, a escolha dos sítios de oviposição baseia-se em vários fatores como: o 7
valor nutricional da planta (Fanara et al. 1999; Liu et al. 2008), a presença de compostos 8
químicos (Feinstein et al. 2008; Bento e Nardi 2009; Pizzamiglio-Gutierrez 2009), a 9
morfologia e idade da planta e a presença de inimigos naturais (Toscano et al. 2002; 10
Bittencourt-Rodrigues e Zucoloto 2005). 11
As fêmeas de P. cardo, possivelmente, reconhecem que os frutos colonizados 12
por fungo, não são sítios apropriados para oviposição, uma vez que poucos indivíduos 13
escolheram frutos inoculados com fungo. Os compostos químicos produzidos pelos 14
fungos podem indicar ao P. cardo, baixa qualidade do sítio ou aumento da pressão de 15
inimigos naturais, além de riscos da concorrência com competidores, assim como é para 16
outros insetos (Price et al. 2011). 17
O fato de algumas fêmeas não ter efetuado uma escolha ou ter optado pelo pote 18
vazio, pode ter ocorrido por essas fêmeas não estarem fisiologicamente prontas para 19
ovipositarem e, deste modo, não justificaria a busca por sítios. Portanto, a decisão final 20
de aceitar ou rejeitar o hospedeiro não se baseia somente em informações sensoriais, 21
mas também do estado fisiológico do inseto como: saciedade, maturidade sexual e 22
maturação dos ovos (van Loon e Dicke 2001). 23

24
Outra possível justificativa está relacionada a questões intrínsecas do indivíduo, 1
em que a capacidade de percepção e assimilação de compostos químicos sejam menos 2
aguçadas para discriminar os diferentes perfis de odor, do que em outros (Mumm e 3
Dicke 2010). Dessa forma, os compostos químicos podem estar envolvidos, no modo 4
como, os invertebrados distinguem entre frutos infectados e não infectados com fungos 5
e assim, avaliarem a adequação do sítio para seus ciclos vitais. 6
As variações das respostas dos insetos frente ao estímulo químico promovem 7
alterações nas sequências dos comportamentos de seleção do hospedeiro. Os 8
comportamentos expressos pelas fêmeas de P. cardo: Parado Ativo Antenando, 9
Autolimpeza, Bater Asa e Expor Ovipositor durante os testes de arena coincidem com 10
etapas comportamentais relatadas por van Loon e Dicke (2001), em que a seleção do 11
hospedeiro pode ser subdividida em duas fases principais e consecutivas, a primeira 12
fase de procura e a segunda de contato 13
O comportamento Parado Ativo Antenando ocorreu com maior frequência para 14
hymenópteros ao selecionarem sítios de oviposição. Esses insetos inspecionaram o 15
possível sítio, com movimentos rápidos das antenas, o que coincide com o presente 16
estudo (Buteler et al. 2009). Outro comportamento que ocorreu com frequência foi o de 17
Autolimpeza, que quando exibido nas vias comportamentais era, provavelmente, 18
indicativo que as fêmeas de P. cardo não conseguiam assimilar com precisão os sinais 19
químicos emitidos do ambiente. Isso, porque as antenas poderiam estar sujas de esporos 20
dos fungos e esse comportamento intermitente seria uma forma de limpá-las, para 21
melhorar a percepção, como já foi observado em Lepidoptera da espécie Spalgis epius, 22
que limpavam suas antenas, quando estavam revestidas de resíduos de cera (Dinesh et 23
al. 2012 e 2013). 24

25
Os comportamentos de Bater Asa e Expor Ovipositor ocorreram menos nos 1
testes de arena, entretanto, foram os que proporcionaram melhor precisão na seleção de 2
sítio de oviposição. O ato de bater asas sugere que o inseto identifica e, 3
consequentemente, reponde ao sinal químico emitido pela fonte de estímulo. Tal 4
comportamento, já foi muito verificado em espécies pertencente à ordem de 5
Hymenóptera (Askew 1968, Vinson 1972, Matthews 1975) e Lepidóptera (Buteler et 6
al., 2009, Tasin et al. 2011). 7
Os comportamentos Parado Ativo Antenando, Autolimpeza, Bater Asa e Expor 8
Ovipositor registrados para P. cardo são, provavelmente, executados em resposta a 9
mediadores químicos, encontrados, principalmente, nas antenas. (Chapman 1998, 10
Corrêia e Sant'Ana 2001, Buteler et al. 2009). As antenas dos insetos são órgãos 11
sensoriais móveis, sendo responsáveis pela percepção de odores. As moléculas de odor 12
podem ser capturadas pelas sensilas, que possuem um ou vários neurônios associados a 13
uma estrutura cuticular, essas moléculas são transportadas por proteínas solúveis 14
ligadoras de odor que converte energia química em energia elétrica, para o sistema 15
nervoso dos insetos (Nation 2002). 16
Todos as vias comportamentais e preferência de sítio de oviposição executados 17
por P. cardo, não diferiram entre as espécies de fungos Aspergillus sp. e Fusarium sp., 18
apesar de serem um grupo distinto entre si. Isso sugere que essas espécies compartilham 19
algum perfil químico, e assim o P. cardo responde ao estímulo sem mudar o padrão 20
comportamental. Dessa forma, comportamento do P. cardo pode ser desencadeado por 21
um conjunto de compostos químicos, emitidos pelo fruto ao ser infectado ou os 22
compostos do fruto deixam de ser emitidos e/ou são mascarados pela presença do 23
fungo. 24

26
5. CONCLUSÃO 1
A presença de Aspergillus sp. e Fusarium sp. em frutos de A. aculeata modifica 2
o comportamento de seleção de sítio de oviposição de P.cardo, sem alterar as categorias 3
comportamentais e a preferência de sítio de oviposição dos insetos. Dessa forma, a 4
presença de fungos intermedia as interações desses bruquíneos com a planta hospedeira. 5
Esses resultados fornecem dados relevantes sobre o papel ecológico das 6
emissões de compostos químicos, como moduladores do comportamento de seleção de 7
sítio de oviposição dos insetos. E proporcionam base para futuras investigações na 8
identificação dos perfis de voláteis dos frutos colonizados por Aspergillus sp. e 9
Fusarium sp. que promovem alterações no comportamento desses insetos. 10
6. REFERÊNCIAS
Abrams, PA (1995) Implications of dynamically variable traits for identifying, 11
classifying, and measuring direct and indirect effects in ecological communities. 12
The American Naturalist, 146: pp. 112-134. 13
Almeida, SP, Proença, CEB, Sano, SM, Ribeiro, JF (1998) Cerrado: espécies vegetais 14
úteis. Planaltina: Embrapa-CPAC: pp. 14-19. 15
Almeida-Corte, JS (2004) Dispersão e banco de sementes. In: Ferreira AG, Borghetti F 16
Germinação do básico ao aplicado. ed. Artmed: Porto Alegre: pp. 225-235. 17
Askew, RR (1968) Considerations on speciation in Chalcidoidea ( Hymenoptera ). 18
Evolution 22: pp. 642-645 19
Askmam, K e Ekbom, B (2006) Responses of ovipositing moths to host plant 20
deprivation: life history aspects and implications for intercropping. Agricultural 21
and Forest Entomology 8: pp. 213-219. 22
Bento, JMS, Nardi, C (2009) Bioecology and nutrition versus chemical ecology: The 23
multitrophic interections mediated by chemical signals. In: Panizzi, AR, Parra. 24
JRP (Org. ). Insect Bioecology and Nutrition for Integrated Pest Management. 1 25
ed. Boca Raton: CRC Press 1, pp. 163-175. 26

27
Bittencourt-Rodrigues, RS, Zucoloto, FS (2005) Effect of host age on the oviposítion 1
and performance of Ascia monuste Godart (Lepidoptera: Pieridae). Neotrop. 2
Entomol 34: pp. 169-175. 3
Burkepile, DE, Parker, JD, Woodson, CB, Mills, HJ, Kubanek, J, Sobecky, PA, Hay, 4
ME (2006) Chemically mediated competition between microbes and animals: 5
microbes as consumers in food webs. Ecology. 87: pp. 2821-2831. 6
Buteler, M, Weaver, DK, Peterson, RKD (2009) Oviposítion behavior of the wheat stem 7
sawfly when encountering plants infested with cryptic conspecifics. Environ. 8
Entomol 38: pp. 1707–15. 9
Cavalcanti, MAQ (2006). Fungos filamentosos isolados do solo em municípios na 10
região Xingó, Brasil. Acta Botanica Brasilica 20: pp. 831-837. 11
Cetec – Centro Tecnológico de Minas Gerais (1983). Produção de combustíveis líquidos 12
a partir de óleos vegetais: Estudo das oleaginosas nativas de Minas Gerais. Belo 13
Horizonte – MG. 14
Chapman, RF (1998) The Insects Structure and Function. Cambridge, Cambridge 15
University Press: pp. 770. 16
Chaudhury, M, Skoda, S, Sagel A, Welch, J (2010) Volatiles emitted from eight would-17
isolated bacteria differentially attracted gravid screw-worms 18
(Diptera:Cliporidae) to oviposit. Journal Med Entomol 47: pp. 349–354. 19
Clement, CR, Lleras, PE, Van Leeuwen, J (2005) O potencial das palmeiras tropicais no 20
Brasil: acertos e fracassos das últimas décadas. Agrociências, Montevideu 9: pp. 21
67-71. 22
Cooke, WB (1979) The ecology of fungi. pp.274. 23
Corrêia, AG, Sant'Ana, J (2001) Fundamentos da comunicação química de insetos, In: 24
Ferreira, JACP, Corrêia, AG, Vieira. Produtos naturais no controle de insetos. 25
ed. UFSCar: São Carlos: pp. 9-22. 26
Crawley, MJ (2002) Statistical computting: an introduction to data analysis using S-27
Plus. John Wiley & Sons: pp. 761. 28

28
Davis, TS, Landolt, PJ (2013) A survey of insect assemblages responding to volatiles 1
from a ubiquitous fungus in an agricultural landscape. J Chem Ecol. 39: pp. 860-2
8. 3
Delobel, A, Couturier, G, Kahn, F, Nilsson, JA (1995) Trophic relationships between 4
palms and bruchids (Coleoptera: Bruchidae: Pachymerini) in Peruvian 5
Amazonia. Amazoniana 13: pp. 209-219. 6
Dicke, M, Sabelis, MW (1988) Infochemical terminology: based on cost-benefit 7
analysis rather than origin of compounds? Functional Ecology 2: pp. 131-139. 8
Dinesh, AS, Venkatesha, MG, Ramakrishna, S (2010) Development, life history 9
characteristics and behaviour of mealybug predator, Spalgis epius (Westwood) 10
(Lepidoptera: Lycaenidae) on Planococcus citri (Risso) (Homoptera: 11
Pseudococcidae). Journal Pest Sci 83: pp. 339-345. 12
Dinesh, AZ, Venkatesha, MG (2013) A quantified ethogram for oviposition 13
behavior and oviposition preference in the hemipterophagous 14
butterfly Spalgis epius (Westwood) (Lepidoptera: Lycaenidae). Journal of 15
Ethology 31: pp. 71-77. 16
Donatti, CI, Guimarães-Jr, PR, Galetti, M (2009) Seed dispersal and predation in the 17
endemic Atlantic rainforest palm Astrocaryum aculeatissimum across a gradient 18
of seed disperser abundance. Ecology. Research 24: pp. 1187-1195. 19
Evangelista, J (2005) Contaminações em alimentos. In:Alimentos: um estudo 20
abrangente. São Paulo: Atheneu: 7 pp. 171-390. 21
Ezenwa, VO, Gerardo, NM, Inouye, DW, Medina, M, Xavier, JB (2012) Animal 22
behavior and the microbiome. Science 338: pp. 198–199. 23
Fanara, JJ, Fontdevila, A-Hasson, E (1999) Oviposiition preference and life history 24
traits in cactophilic Drosophila koepferae and D. buzzatii in association with 25
their natural host. Evolut. Ecol 13: pp. 173-190. 26
Farr, DF, Bills, GF, Chamaris, GP, Rossman, AY (1989) Fungi on plants and plant 27
products in the United States. American Phytopathology Society, (APS) Press, 28
St. Paul (Minnesota) 101: pp. 340-1990. 29

29
Feinstein, J, Purzycki, KL, Scott, M, Hequet, V, Berkov A (2008) Neotropical soldier 1
flyes (Stratiomyidae) reared from Lecythis poiteaui in French Guiana: do bat-2
pollinated flowers attract saprophiles Torrey Botc Soc 133: pp. 200-207. 3
Fonseca, FSA (2008) Padrões de ataque de insetos herbívoros em frutos de Acrocomia 4
aculeata (Jacq.) Lodd. ex. Martius (Arecaceae). Dissertação de Mestrado 5
(Ciências Biológicas) Universidade Estadual de Montes Claros, Montes Claros. 6
Forget, PM, Kitajima, K, Foster, RB (1999) Pre- and post- dispersal seed predation 7
in Tachigali versicolor (Caesalpiniaceae): effects of timing of fruiting and 8
variation among trees. Journal of Tropical Ecology 15: pp. 61- 81. 9
Fraga, ME, Pereira, MG (2012). Diversity of soil and litter isolated Trichocomaceae in 10
Atlantic Forest. Floresta Ambient 19: pp. 405-413. 11
Gadgile, DP, Chavan, AM (2010). Impact of temperature and relative humidity on 12
development of Aspergillus sp Flavus rot of mango fruit. Science Technol 3: pp. 13
48-49. 14
Hastings, A, Powell, T (1991) Chaos in a three-species food chain. Ecoloy 72: pp. 896-15
903 16
Henderson, A, Galeano, G, Bernal, R (1995) Field Guide to the Palms of the Americas. 17
New Jersey: Princepton University: pp.166-167. 18
Irwin, RE (2006) The Consequences of Direct versus Indirect Species Interactions to 19
Selection on Traits: Pollination and Nectar Robbing in Ipomopsis aggregata. The 20
American Naturalist 167: pp. 315-328. 21
Jallow, MFA, Dugassa-Gobena, D, Vidal, S (2008) Influence of an endophytic fungus 22
on host plant selection by a polyphagous moth via volatile spectrum changes. 23
Arthropod-Plant Interactions 2: pp. 53-62. 24
Kitching, RL (1977) Times resources and population dynamics in insects. Aust. Ent. 25
Mag 2: pp. 31-42 26
Kruess, A (2001) Indirect interaction between a fungal plant pathogen and a herbivorous 27
beetle of the weed Cirsium arvense. Oecologia 130: pp. 563–569. 28

30
Lam, K, Tsang, M, Labrie, A, Gries, R, Gries, G (2010) Semiochemical-Mediated 1
Oviposítion Avoidance by Female House Flies, Musca domestica, on Animal 2
Feces Colonized with Harmful Fungi. Journal Chem Ecol 36: pp. 141–147. 3
Lewinsohn, TM, Cagnoto, L (2012) keystones in a tangled bank. Science. 335: pp. 4
1449-1451. 5
Liu, Q, Tomberlin, JK, Brady, JAM, Sanford, R, YU Z (2008) Black soldier fly 6
(Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. 7
Environ. Entomol. 37: pp. 1525-1530. 8
Loaiza, JCM, Céspedes, CL (2007) Compuestos volatiles de plantas. Origen, emission 9
efectos, análisis y aplicaciones al agro. Rev. Fitotecnia Mexicana 30: pp. 327-10
351. 11
Lorenzi, GMAC, Negrelle, RRB (2006) Acrocomia aculeata (Jacq.) Lodd. ex. Mart.: 12
aspectos ecológicos, Usos e potencialidades. Visão Acadêmica, Curitiba, UFPR, 13
7(1). 14
Lorenzi, H, Kahn, F, Noblick, LR, Ferreira, E (2010) Flora Brasileira: Arecaceae 15
(Palmeiras). Instituto Plantarum. Nova Odessa, SP: pp. 382. 16
Martins, CBC, Rafael, AS, Leonardo RB, Jeffrey RA, Paulo, HGZ (2012) Volatile 17
Chemicals of Adults and Nymphs of the Eucalyptus Pest, Thaumastocoris 18
peregrinus (Heteroptera: Thaumastocoridae). Psyche: A Jounal of Entomology 19
2012: pp. 6. 20
Matthews, RW (1974) Biology of Braconidae. Annu. Rev. Entomol. 19: pp. 15-32 21
Michereff, SJ (2005) Importância dos patógenos e das doenças radiculares em solos 22
tropicais. In: MICHEREFF, S. J. et al. Ecologia e manejo de patógenos 23
radiculares em solos tropicais. Recife: UFRPE: pp. 1-18. 24
Mumm, R e Dicke, M (2010) Variation in natural plant products and the attraction of 25
bodyguards involved in indirect plant defense. Canadian Journal of Zoology 88: 26
pp.628-667. 27
Nation, JL (2002). Insect Physiology and Biochemistry. University of Florida, CRC 28
Press: pp. 485. 29

31
Nordlund, DA, Lewis, WJ (1976) Terminology of chemical-releasing stimuli in 1
intraspecific and interspecific interactions J. Chem. Ecol 2: pp. 211. 2
Nucci, SM (2007) Desenvolvimento, Caracterização e Análise da Utilidade de 3
Marcadores Microsatelites em Genética de População de Macaúba. Dissertação 4
(Mestrado em Agricultura Tropical e Subtropical) do Instituto Agronômico. 5
Campinas SP. 6
Oliveira, MV, Moura, LFWG, Ribeiro, ARC, Mota, JGSM, Freire, FCO, Magalhãe, 7
FEA (2012) Investigação da microbiota fúngica anemófila em vários setores do 8
centro vocacional tecnológico do município de Tauá-CE . Rev. de Biológia e 9
Farmácia 7: pp. 104-114. 10
Paiva, MR, Pedrosa-Macedo, JH (1985) Feromonas de Insetos. Curitiba: 11
GTZ/CONCITEC: pp. 94. 12
Panizzi, AR, Parra, JRP (1991) Ecologia nutricional de insetos e suas implicações no 13
manejo de pragas. SP Manole/CNPq: pp.359. 14
Pereira, ACF, Fonseca, FSA, Mot,a GR, Fernandes, AKC, Fagundes, M, Reis-Júnior, R, 15
Faria, ML (2014) Ecological Interactions Shape the Dynamics of Seed 16
Predation in Acrocomia aculeata (Arecaceae). Journal Plos one, 9(5): e98026. 17
Pires, TP, Souza, ES, Kuki, KN, Motoike, SY (2012) Ecophysiological traits of the 18
macaw palm: A contribution towards the domestication of a novel oil crop. 19
Industrial Crops and Products 44: pp. 200-210. 20
Pitt, JL, Hocking, AD (1997) Fungi and food spoilage. 2º ed. London: Blackie. 21
Academic and Professional. pp. 540. 22
Pizzamiglio-Gutierrez, MA (2009) Interação inseto-planta. In: Bioecologia e nutrição de 23
insetos: base para o manejo integrado de pragas. BrasíliaDF: Embrapa 24
Informação Tecnológica: pp. 211-249. 25
Pocock, MJO, Evans, DM, Memmott, J (2012) The robustness and restoralion of a 26
network of ecological networks. Science 335: pp. 973-977. 27
Prade, CA, Matsumura, ATS (2014) Comunidade de fungos do solo em uma floresta 28
ripária na região de Roca Sales, Rio Grande do Sul, Brasil. Caderno de Pesquisa, 29
série Biologia 26: pp. 36-46. 30

32
Price, PW, Denmo, RF, Eubanks, MD, Finke, DL, Kaplan, I (2011) Insect Ecology: 1
Behavior, Populations, and communities. Cambridge University Press, New 2
York: pp. 816. 3
R Core Team (2014). R: A language and environment for statistical computing. R 4
Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-5
project.org/. 6
Ramírez, N, Traveset, A (2010) Predispersal seed-predation by insects in the 7
Venezuelan Central Plain: Overall patterns and traits that influence its biology 8
and taxonomic groups. Perspectives in Plant Ecology, Evolution and Systematics 9
12: pp. 193-209. 10
Ramos, FA, Martins, I, Farias, JM, Silva, ICS, Costa, DC, Miranda AP (2001) 11
Oviposítion and Predation by Speciomerus revoili (Coleoptera, Bruchidae) on 12
seeds of Acrocomia aculeata (Arecaceae) in Brasília, DF, Brazil. Brazilian 13
Journal Biology 61: pp. 449-454. 14
Rayamajhi, MB, Van, TK, Pratt, PD, Center, TD (2006) Interactive association between 15
Puccinia psidii, and Oxyops vitiosa, two introduced natural enemies of 16
Melaleuca quinquenervia in Florida. Biol. Control 27: pp. 56–67. 17
Rodriguez-Amaya, DB, Sabino, M (2002) Pesquisa em micotoxinas no Brasil: a última 18
década em foco. Braz. J. Micróbiol., São Paulo 33: pp.1-11. 19
Roeder, G, Rahier, M, Naisbit, RE (2007) Coping with an antagonist: the impact of a 20
phytopathogenic fungus on the development and behavior of two species of 21
alpine leaf beetle. Oikos 116: pp. 1514–1523. 22
Scariot, AO (1998) Seed dispersal and predation of the palm Acrocomia aculeata. 23
Principes 42: pp. 5-8. 24
Silva, FR, Begnini, RM, Scherer, KZ, Lopes, BC, Castellani, TT (2007) Predação de 25
Sementes de Syagrus romanzoffiana (Cham.) Glassman (Arecaceae) por Insetos 26
na Ilha de Santa Catarina, SC. Rev. Brasileira de Biociências, Porto Alegre 5: 27
pp. 681-683. 28
Silvius, KM, Fragoso, JM (2002) Pulp handling by vertebrate seed dispersers increases 29
palm seed predation by bruchid beetles in the northern Amazon. Journal of 30
Ecology, 90: pp. 1024-1032. 31

33
Southgate, BJ (1979) Biology of Bruchidae. Annual Review of Entomology 24: pp. 1
449-473. 2
Steffler, CE, Donatti, CI, Galetti, M (2008) Seed Predation of Attalea dubia (Arecaceae) 3
in an Island in the Atlantic Rainforest of Brazil. Palms 52: pp. 133–140. 4
Tasin, M, Betta, E, Carlin, S, Gasperi, F, Mattivi, F, Pertot, I (2011) Volatiles that 5
encode host-plant quality in the grapevine moth. Phytochemistry 72: pp. 1999-6
2005. 7
Tomberlin, JK, Byrd, JH, Wallace, JR, Benbow, ME (2012) Assessment of 8
decomposítion studies indicates need for standardized and repeatable methods in 9
forensic entomology. Journal Forensic Research 3: pp. 147 10
Torres-Alruiz, MD, Rodríguez, DJ (2013) A topo-dynamical perspective to evaluate 11
indirect interactions in trophic webs: New indexes. Ecological Modelling 250: 12
pp. 363-369. 13
Toscano, LC, Boiça-Júnior, AL, Maruyama, W (2002) Fatores que afetam a oviposição 14
de Bemisia tabaci (Genn.) biótipo B (Hemiptera: Aleyrodidae) em tomateiro. 15
Neotrop. Entomol 31: pp. 631-634. 16
Tournas, VH, Katsoudas, E (2005). Mould and yeast flora in fresh berries, grapes and 17
citrus fruits. Int. Journal Food Microbiol 105: pp. 11-17. 18
van Loon, JJA, Dicke, M (2001) Sensory ecology of arthropods utilizing plant 19
infochemicals. In: Barth, FG, Schimidt, A (Ed.) Ecology of Sensing. Berlin: 20
Springer- Verlag: pp. 253-270. 21
Vilela, EF, Della-Lucia, TMC (2001) Introdução aos semioquímicos e Terminologia. In: 22
Vilela, EF, Della-Lucia, TMC Feromônios de insetos- Biologia, química e 23
emprego no manejo de pragas- 2º ed.. Ribeirão Preto: Ed. Holos: pp. 9-12. 24
Vinson, SB, Guillot, FS (1972) Host marking: Source of a substance that results in host 25
discrimination in insect parasitoids. Entomophaga 17: pp. 241-245 26
Webstera, B, Brucea, T, Pickett, J, Hardieb, J (2010) Volatiles functioning as host cues 27
in a blend become nonhost cues when presented alone to the black bean aphid. 28
Animal Behaviour 79: pp. 451–457. 29
30


















