







Línguas
Páginas
Legal


1
1. INTRODUÇÃO
A crescente devastação das florestas tropicais é a maior ameaça à diversidade biológica do
planeta (WILCOX & MURPHY 1985, BIERREGAARD & LOVEJOY 1989, TURNER
1996, LAURENCE & BIERREGAARD 1997, LAURENCE 1999). Com a expansão das
áreas agrícolas, urbanas e industriais, muitas matas contínuas foram reduzidas a pequenos
fragmentos florestais (HARRISON et al. 1988, WHITMORE 1997), cujos efeitos adversos
sobre as comunidades animais e vegetais têm sido demonstrados por diferentes estudos
(LAURENCE 1999, DEBINSKI & HOLT 2000, para ampla revisão). Em conjunto, esses
estudos mostraram que o processo de fragmentação do hábitat é uma das alterações
antrópicas de maior impacto sobre os ecossistemas naturais, tornando-se uma das
principais preocupações da emergente biologia da conservação (HARRIS 1984, SOULÉ
1986, MURCIA 1995, DAVIES et al. 2000).
A Teoria da Biogeografia Insular (MACARTHUR & WILSON 1967), que estabeleceu as
bases para a compreensão da composição e dinâmica das espécies de ilhas oceânicas, ao
ser aplicada a ambientes continentais, impulsionou os estudos sobre biotas de paisagens
fragmentadas terrestres (e.g. SIMBERLOFF & ABELE 1982, DIAMOND 1975, WILSON
& WILLIS 1975, TERBORGH 1976, FREEMARK & MERRIEN 1986). Dessa forma,
esperava-se que a riqueza de espécies em hábitats fragmentados estaria relacionada com o
tamanho e grau de isolamento da área, com as menores e mais isoladas apresentando
menos espécies do que as maiores e menos isoladas (DIAMOND 1975, WILSON &
WILLIS 1975, TERBORG 1976). Muitos estudos, porém, notaram tendências muito
particulares para as comunidades terrestres, concluindo que tais sistemas operam de modo
bastante diferenciado (DEBINSKI & HOLT 2000, WATSON 2002).

2
Um fragmento de hábitat pode ser entendido como uma área de vegetação delimitada no
espaço, circundada por paisagem desfavorável para populações naturais movimentarem-se
livremente para outros fragmentos de seu hábitat (WILSON & WILLIS 1975, MURCIA
1995, HANSKI & SIMBERLOFF 1997, HAILA 2002, LAURENCE 2002). Assim, entre
os fragmentos de uma paisagem existe uma área normalmente inóspita à espécie,
denominada matriz, representando uma barreira para o seu fluxo entre um e outro
fragmento (LAURENCE 1994, MALCOLM 1991, TOCHER et al. 1997, GASCON et al.
1999, DAVIES et al. 2001, LINDENMAYER & FRANKLIN 2002). Dependendo,
entretanto, da permeabilidade da matriz, algumas paisagens fragmentadas podem favorecer
o fluxo das espécies através de um sistema de metapopulações do tipo continente-ilha
(mainland-island ou source-sink), ou em mancha (patchy population), ou em desequilíbrio
(non-equilibrium metapopulation) (HARRISON 1991, HANSKI 1991, HANSKI &
SIMBERLOFF 1997).
Estudos sobre comunidades animais isoladas pela fragmentação do seu hábitat foram
conduzidos em diferentes regiões do planeta (WIEGAND et al. 2005). Entretanto, foi nas
áreas de florestas tropicais, intensamente devastadas e com alta diversidade biológica, que
tais estudos ganharam máxima expressão, com consistentes análises para os mais
diferentes grupos animais, como formigas (CARVALHO & VASCONCELOS 1999),
lepidópteros (BROWN & HUTCHINGS 1997), anfíbios (TOCHER 1998, VALLAN
2000), aves (e.g. BIERREGAARD & LOVEJOY 1989; ANJOS & BOÇON 1999,
BORGES & GUILHERME 2000), pequenos mamíferos (MALCON 1997, LYNAM &
BILLICK 1999), quirópteros (COSSON et al. 1999, BIANCONI et al. 2004) e primatas
(CHIARELLO 1999, CHIARELLO & MELO 2001, BERNARDO & GALETTI 2004).
Foi, porém, através do projeto “Dinâmica Biológica de Fragmentos Florestais” (PDBFF),
desenvolvido na Amazônia há mais de duas décadas (para uma revisão, veja LAURENCE

3
et al. 2002), que os efeitos da fragmentação florestal sobre as aves da região Neotropical
começaram a ser melhor compreendidos (e.g. LOVEJOY et al. 1983, 1984, 1986,
BIERREGAARD et al. 1992, STOUFFER & BIERREGAARD 1995, BIERREGAARD &
STOUFFFER 1997). Um dos primeiros e notáveis efeitos da fragmentação da floresta
sobre a comunidade de aves nessa região foi a constatação de um imediato aumento na
abundância de várias espécies no interior dos pequenos fragmentos recém-formados, os
quais, entretanto, rapidamente sofriam drástica perda de sua diversidade (relaxation
faunal) (DIAMOND 1975, BIERREGAARD & LOVEJOY 1989).
De modo geral, a fragmentação de hábitats dificulta o fluxo gênico entre as populações e
provoca mudanças físicas e biológicas no ambiente, como alterações microclimáticas e
aumento na taxa de predação (BIERREGAARD et al. 1992, MURCIA 1995), podendo
levar muitas espécies à extinção local (e.g. SIMBERLOFF 1994, KATTAN et al. 1994,
LAURENCE et al. 1997, RIBON et al. 2003). Porém, se os processos que causam a
extinção local também atuarem em populações de outras regiões, o resultado será a
extinção global da espécie (SIMBERLOFF 1994, TEMPLETON et al. 2001).
Os estudos sobre comunidades de aves em fragmentos florestais na região da Mata
Atlântica iniciaram-se no interior de São Paulo com WILLIS (1979), que demonstrou o
desaparecimento de muitas espécies nas matas isoladas menores, acompanhado de forte
desorganização na riqueza de espécies de algumas guildas tróficas. A partir de então,
trabalhos subseqüentes foram conduzidos em outras regiões, sobretudo nos estados de São
Paulo (e.g. ALEIXO & VIELLIARD 1995, CHIARELLO 2000, UEZU et al. 2005,
ANTUNES 2005, DEVELEY 2004), Minas Gerais (e.g. CHRISTIANSEN & PITTER
1997, D`ANGELO NETO et al. 1998, RIBON 1998, RIBON et al. 2003, RIBON 2003,
MALDONADO-COELHO & MARINI 2000), Bahia (LAPS et al. 2003) e Paraná (e.g.
ANJOS 2001, ANJOS 2004, ANJOS et al. 2004, BORNSCHEIN & REINERT 2000).

4
Embora tenham verificado resultados similares aos obtidos em São Paulo por WILLIS
(1979), tais estudos, quando analisados em conjunto, evidenciam respostas peculiares
entre as avifaunas das localidades trabalhadas, as quais podem estar relacionadas ao
histórico da fragmentação florestal a que cada uma delas foi submetida no passado.
Em seu conjunto, os estudos voltados para a Mata Atlântica representam atualmente uma
sólida compreensão sobre a resposta da avifauna à fragmentação antropogênica de seu
hábitat, equiparando-se, em linhas gerais, ao conhecimento consolidado na Amazônia
(SOUZA & BROWN 1994, MCGARIGAL & MARKS 1995, LAURENCE et al. 1998,
CARVALHO & VASCONCELOS1999, LAURENCE et al. 2000, GASCON et al. 2001).
Entre outras importantes conclusões sobre os efeitos da fragmentação florestal e da
alteração da estrutura da vegetação sobre as aves da Mata Atlântica, verificou-se, por um
lado, relativa tolerância ecológica de alguns grupos de espécies e, por outro, desolador caso
de extinções locais, o que tem estimulado constantes debates e estimativas sobre a taxa de
futuras perdas da diversidade biológica (BROOKS & BALMFORD 1996, BROOKS &
BALMFORD 1999, HARRIS & PIMM 2004).
A falta de extinções em massa na Mata Atlântica tem sido interpretada como uma pré-
adaptabilidade de seus organismos a pertubações naturais que experimentaram no passado,
em função de reduções drásticas de temperaturas durante períodos glaciais, causando
grandes mudanças na extensão e qualidade da cobertura florestal (BROWN & BROWN
1992). Contudo, outros trabalhos já demonstraram que essa idéia não é de todo verdadeira
e que a extirpação progressiva de sua fauna só ainda não ocorreu pelo fato de muitas de
suas espécies encontrarem-se no time lag da extinção, persistindo precariamente em alguns
poucos refúgios florestais, até que venham a desaparecer por completo (BROOKS &
BALMFORD 1996, BROOKS & BALMFORD 1999, ALEIXO 2001, RIBON et al. 2003,
HARRIS & PIMM 2004).

5
De fato, a identificação de várias extinções regionais (listas vermelhas estaduais) (e.g.
BERGALLO et al. 2000, STRAUBE et al. 2004) e o expressivo número de espécies
ameaçadas (CONSERVATION INTERNATIONAL DO BRASIL et al. 2000),
corroboram, por si só, a grave situação da avifauna da Mata Atlântica, ainda que somente
duas aves desse bioma tenham sido extintas na natureza (mutum-do-nordeste, Mitu mitu, e
a arara-azul-pequena, Anodorhynchus glaucus) (MMA 2003, MACHADO et al. 2005).
Ainda que os efeitos da fragmentação florestal sobre a avifauna da Mata Atlântica já sejam
relativamente bem documentados (ALEIXO 2001), a base de informações hoje disponível
parece compatível apenas com o plano genérico do conhecimento, considerando que
muitas outras de suas subregiões, distintamente peculiares em sua fisionomia vegetacional,
composição faunística e seu histórico processo de devastação, permanecem sem quaisquer
avaliações. Logo, é lícito pensar que o conhecimento até agora acumulado ainda encontra-
se em seu estágio incipiente, subjulgando a potencialidade global dos impactos provocados
sobre a biota da Mata Atlântica, ao longo de mais de 500 anos de generalizada destruição
ambiental (DEAN 1997).
Como em outras regiões do leste do Brasil, a Mata Atlântica do Estado do Espírito Santo,
que outrora recobria mais de 90% do seu território, sofreu constante e drástico processo de
devastação, restando hoje não mais do que 7% de sua cobertura vegetal original
(FUNDAÇÃO SOS MATA ATLÂNTICA 1996, IPEMA 2005b). Sem dúvida, essa
extrema modificação ambiental provocou profundas alterações na composição de
avifaunas regionais, as quais só recentemente começaram a ser notadas, seja através do
declínio de suas populações, extinções locais ou colonização de áreas por espécies
invasoras espontâneas (BAUER 1999, SIMON 2001, SIMON & CHIARELLO 2001,
WILLIS & ONIKI 2002, SIMON & LIMA 2004a, b).

6
Porém, o conhecimento sobre as comunidades de aves nas paisagens florestais
fragmentadas do Espírito Santo permanece bastante obscuro e praticamente inexplorado
quando comparado com outras regiões da Mata Atlântica. O único estudo disponível foi
realizado na região de Linhares, em matas de baixada no norte do estado, envolvendo
comparações com florestas de eucalipto, cujos resultados indicaram expressiva perda de
diversidade nos ambientes amostrados, a qual esteve relacionada com a fisionomia
simplificada da vegetação e o grau extremo de fragmentação que a região sofreu no
passado (MARSDEN et al. 2001).
A região serrana do Espírito Santo também sofreu os dramáticos impactos da ocupação
humana, logo na chegada das primeiras levas de imigrantes europeus (final do século
XIX), que promoveram a impiedosa derrubada de sua exuberante vegetação para a
exploração de terras produtivas, voltada, inicialmente, para a implantação de lavouras de
café (RUSCHI 1974, DEAN 1997). Como resultado, sua cobertura florestal original foi
extremamente reduzida, estando hoje representada apenas por porções descontínuas de
mata, concentradas, sobretudo, na porção central do estado (FUNDAÇÃO SOS MATA
ATLÂNTICA 1996, IPEMA 2005b). Apesar disso, essa região ainda é detentora de
elevada diversidade biológica (THOMAZ & MONTEIRO 1997, BAUER 1999, BAUER et
al. 2000, AZEVEDO & SANTOS 2000, BROWN & FREITAS 2000, PASSAMANI et al.
2000, SIMON 2000, VENTURINI et al. 2000, WILLIS & ONIKI 2002, RAMOS &
GASPARINI 2004), capaz, aparentemente, de persistir em um sistema fragmentado de
florestas nativas, cuja eficiência para conservação dos diferentes grupos de organismos que
ali ocorrem ainda está por ser devidamente avaliada. De fato, o único estudo que abordou o
tema de forma específica foi realizado por PASSAMANI (2003), trabalhando com
pequenos mamíferos na região de Santa Teresa, onde muitos outros grupos faunísticos
sequer foram inventariados, com exceção de lepidópteros (BROWN & FREITAS 2000),

7
aves (e.g. PARKER & GOERCK 1997, SIMON 2000, WILLIS & ONIKI 2002) e
mamíferos (PASSAMANI et al. 2000).
Embora o Município de Santa Teresa sobressaia como uma das regiões ornitologicamente
mais bem investigadas do Espírito Santo, a maioria dos estudos até hoje aí realizados
preocupou-se com a composição geral da avifauna de alguns de seus principais
remanescentes florestais, especialmente aqueles transformados em unidades de
conservação (RUSCHI 1977, FORRESTER 1993, PARKER III & GOERCK 1997,
SIMON 2000, WILLIS & ONIKI 2002). WILLIS & ONIKI (2002) foram os autores que
melhor investigaram a diversidade de aves em relação à atual situação da cobertura vegetal
da região. O estudo conduzido por eles demonstrou que a hipótese elaborada por MORRIS
(1995), segundo a qual as áreas antropicamente perturbadas contribuem para o aumento da
riqueza de espécies numa região, não é válida para ambientes ricos em espécies nativas, as
quais, quando perdidas, não são numericamente substituídas por espécies invasoras. Seus
dados mostraram que, na ausência de grandes reservas florestais, as perdas podem alcançar
até a metade da riqueza da avifauna local, sustentando a hipótese da fragilidade da biota
autóctone da Mata Atlântica (BROOKS & BALMFORD 1996, BROOKS & BALMFORD
1999, HARRIS & PIMM 2004).

8
2. OBJETIVO
O presente estudo teve como objetivo avaliar os efeitos da fragmentação florestal sobre
comunidades de aves na região serrana de Santa Teresa, Espírito Santo. Dessa forma, as
principais questões a serem respondidas sobre o problema em investigação foram:
i- qual foi o efeito do sistema de fragmentação florestal sobre a composição da avifauna
regional? Se a região sofreu perda de espécies, quantas e quais foram estas espécies?
ii- existe variação na diversidade biológica entre diferentes classes de tamanho de
fragmentos florestais? Se a variação existe, quais tipos de fragmentos detêm as maiores
riquezas de espécies, em relação aos valores de riqueza total e riqueza de espécies
dependentes do ambiente florestal?
iii- qual é a influência do estado de conservação dos fragmentos florestais sobre a riqueza
das espécies, quando analisado segundo alguns de seus descritores ambientais, tais como,
tamanho da área, grau de isolamento e permeabilidade da matriz? Alguns destes
descritores podem explicar melhor do que outros a variação da riqueza de espécies, caso
ela ocorra, entre os diferentes fragmentos florestais?
iv- as espécies que persistem na paisagem fragmentada apresentam variação de
abundância relativa entre as diferentes manchas de seu hábitat? Se sim, quais espécies
apresentam declínio ou aumento em seus índices de abundância nos fragmentos
estudados? E quais são os fragmentos que apresentam comunidades com as maiores
porcentagens de espécies com variações de abundância relativa?
v- as espécies florestais apresentam sensibilidade diferenciada ao processo de
fragmentação do seu hábitat? Se sim, seus níveis de sensibilidade são congruentes com
aqueles verificados para outras regiões da Mata Atlântica? As espécies mais afetadas

9
(maior sensibilidade) na região estudada correspondem às espécies com maior pré-
disposição à extinção na Mata Atlântica (táxons endêmicos e de alta sensibilidade
ecológica no bioma)?
vi- a composição da avifauna apresenta maior similaridade entre os fragmentos florestais
que apresentam condições semelhantes de conservação? Quais tipos de fragmentos
possuem menor e maior similaridade com a composição da avifauna original (área-
controle)? Se a composição da avifauna dos fragmentos difere da área-controle, quais são
as taxas de extinções e os táxons perdidos em cada um deles?
vii- As diferentes guildas de aves apresentam sensibilidade diferenciada ao processo de
fragmentação do seu hábitat? Se apresentam, as guildas mais sensíveis na região em
análise coincidem com aquelas de outras regiões avaliadas da Mata Atlântica? Existe
variação na composição das guildas entre os diferentes fragmentos florestais,
suficientemente capaz de apontar que os mesmos apresentam comunidades
estruturalmente modificadas em função do processo de fragmentação ocorrido na região?
Existem diferentes tipos de abordagens sobre os efeitos da fragmentação de ecossistemas
terrestes, impedindo, muitas vezes, que seus resultados favoreçam comparações ou
generalizações entre as diferentes regiões pesquisadas. Por esta razão, boa parte das
questões arroladas acima foi delineada com base nas linhas de prioridade recomendadas
para estudos desta natureza na região tropical (BIERREGAARD et al. 1997).

10
3. MATERIAIS E MÉTODOS
3.1. Área de Estudo
O Município de Santa Teresa, inserido na região centro-serrana do Estado do Espírito
Santo, ainda mantém cerca de 40% de sua cobertura florestal nativa (ca. 22.000 ha),
distribuídos sob um padrão fortemente fragmentado, resultante do desordenado processo
de desmatamento que a região sofreu no passado (Fig. 1).
Dessa sua área florestal, 22% (ca. 4.500 ha) encontram-se transformados em unidades de
conservação (Reserva Biológica Augusto Ruschi, Estação Biológica de Santa Lúcia e
Parque Natural Municipal de São Lourenço), constituídas, em sua maior parte, por
vegetação primária bem conservada. O restante (78%) corresponde a numerosos outros
remanescentes situados em propriedades privadas, cujo tamanho de sua área, grau de
isolamento, uso do solo em seu entorno, entre outras características ambientais, variam
entre eles.

11
Figura 1. Situação geográfica da área de estudo no Estado do Espírito Santo, Brasil. O mapa mais detalhado mostra a paisagem florestal fragmentada na região serrana de Santa Teresa, onde estão localizados os oito fragmentos de mata nativa estudados: FP1, FP3, FP4, PMBML; FM2, PNMSL, EBSL, RBAR (veja Tabela 1 para detalhes sobre estes fragmentos).

12
Antes de sua colonização, o Município de Santa Teresa era quase totalmentre recoberto
por matas nativas, que começaram a ser derrubadas a partir de 1875 para a implantação da
lavoura do café e culturas anuais diversas (ESTADO DO ESPÍRITO SANTO 1939,
RUSCHI 1974, MENDES & PADOVAN 2000). Entretanto, esse processo de destruição
foi atenuado algumas décadas atrás, em função da criação de unidades de conservação e
pelo seu relevo fortemente acidentado, dificultando a expansão das áreas agrícolas
(MENDES & PADOVAN 2000). Atualmente, a paisagem antrópica no entorno dos
remanescentes florestais (matriz), inclui, além das plantações de café, capoeiras, áreas de
pastagens, culturas anuais (hortaliças, principalmente) e cultura de eucalipto, mais
recentemente introduzida no município e em franca expansão em toda a região serrana do
estado (J. E. Simon, obs. pess.).
Para efeito deste estudo, delineado de acordo com a conformação atual da Mata Atlântica
de Santa Teresa, foram estabelecidas três categorias de tamanho de fragmentos florestais,
sendo elas:
a) fragmento pequeno: áreas menores que 50 ha (n = 4);
b) fragmento médio: áreas em torno de 200 ha (n = 1);
c) fragmento grande: áreas maiores que 500 ha (n = 2).
O maior número de fragmentos selecionados para a primeira categoria objetivou assegurar
a representatividade das condições locais do ambiente (córregos, taquarais, etc), para
comparação mais adequada com os fragmentos de maior tamanho. Porém, a alta
conectividade entre os remanescentes florestais da região não permitiu reconhecer nenhum
outro fragmento de médio tamanho completamente isolado dos demais pela paisagem
antrópica em seu entorno (matriz). Embora os grandes fragmentos não estivessem em
situação de perfeito isolamento, em função de algumas de suas ramificações ou franjas
florestais localizarem-se muito próximas de remanescentes vizinhos (~50 m), eles foram

13
considerados como unidades individualizadas, pelo fato de a maior parte de seu perímetro
(> 95%) estar igualmente circundado pelas paisagens antrópicas. A idade dos fragmentos
selecionados é desconhecida, mas o seu tempo de isolamento não é inferior a 30 anos,
conforme análise de fotos aéreas de 1970 disponíveis para a região (PASSAMANI 2003).
Além desses sete fragmentos, estabeleceu-se o maior fragmento ou bloco de mata nativa
da região (Reserva Biológica Augusto Ruschi) como a área-controle, perfazendo o total de
oito remanescentes florestais amostrados (Tab. 1), assim identificados [(as siglas utilizadas
para os fragmentos em propriedades privadas seguem PASSAMANI (2003)]:
- fragmento FP1 (Fig. 2a): área de 8,4 ha (19°58` S e 40°35` W), situado na localidade de
Valsugana Velha;
- fragmento FP3 (Fig. 2b): área de 2,6 ha (19°58` S e 40°35` W), situado na localidade de
Valsugana Velha;
- fragmento FP4 (não citado em PASSAMANI 2003) (Fig. 2c): 34,8 ha. (19°55` S e 40°
35`W), situado na localidade de Alto Santo Antônio;
- fragmento FM2 (Fig. 2d): 150,5 ha (19°02` S e 39°56` W), situado na localidade de
Valsugana Velha;
- fragmento PMBML (Parque do Museu de Biologia Mello Leitão) (Fig. 2e): 8,5 ha
(19°56` S e 40°35` W), situado na área urbana do município;
- fragmento PNMSL (Parque Natural Municipal de São Lourenço) (Fig. 2f): 504,8 ha (dos
quais 265,7 pertencem à área legal do parque) (19°55` S e 40° 37` W), situado na
localidade de São Lourenço/Vargem Alta;
- fragmento EBSL (Estação Biológica de Santa Lúcia) (Fig. 2g): 854,6 ha (dos quais 440
ha pertencem à área legal da estação) (19°57` S e 40°32` W) situado na localidade de
Santa Lúcia;

14
- área-controle RBAR (Reserva Biológica Augusto Ruschi): aproximadamente 4.000 ha
(dos quais 3.598 ha pertencem à área legal da reserva) (19°54` S e 40°34` W), situada na
localidade de Nova Lombardia.

15
FP1 FP3
FP4 FM2
Figura 2. Conformação espacial dos fragmentos florestais estudados (manchas escuras). No entorno de cada um deles estão representadas as matas adjacentes mais próximas (manchas claras), circunscritas num raio de 1 km (buffer representado pela linha contínua) a partir da borda do fragmento considerado (área-controle não representada).

16
PMBML
PNMSL EBSL
Figura 2 (cont.). Conformação espacial dos fragmentos florestais estudados (manchas escuras). No entorno de cada um deles estão representadas as matas adjacentes mais próximas (manchas claras), circunscritas num raio de 1 km (buffer representado pela linha contínua) a partir da borda do fragmento considerado (área-controle não representada).

17
Como o intuito principal deste estudo foi o de investigar as comunidades de aves nos
remanescentes florestais da região serrana de Santa Teresa, os mesmos foram selecionados
entre uma faixa de 600 e 900 m de altitude. A utilização dessa faixa altitudinal teve
também a preocupação de garantir a semelhança fitofisionômica entre os fragmentos,
fundamentando a comparação faunística entre eles. Foi inviável selecionar os fragmentos
aleatoriamente por diversas razões, especialmente em função da dificuldade de acesso e
conectividade da paisagem fragmentada. Com exceção do PMBML, situado na área
urbana do município, todos os outros fragmentos localizam-se na área rural, cujo tipo de
vegetação antrópica do seu entorno foi descrito anteriormente.
O clima da região, segundo a classificação de Köppen, é do tipo Cwa-CFa, marcado por
uma estação seca bem definida (abril-setembro) e uma chuvosa (outubro-março), com
precipitação média anual (sede do município) de 1.868 mm, sendo junho/julho a época
mais seca e novembro, a mais chuvosa (MENDES & PADOVAN 2000). A temperatura
média anual é de 19,9°C, com mínimas e máximas variando entre 14,3ºC e 26,2° C (dados
para a Estação Biológica de Santa Lúcia) (THOMAZ & MONTEIRO 1997).
A área do município de Santa Teresa faz parte do escudo cristalino pré-cambriano
brasileiro, mantendo, portanto, relevo representado por áreas de encostas e escarpas muito
íngremes, com típicos afloramentos rochosos e pequenas várzeas, onde o solo
predominante é do tipo latossolo vermelho-amarelo, com acidez variando de alta a muito
alta (THOMAZ & MONTEIRO 1997).
De acordo com a classificação fitogeográfica de RIZZINI (1979), a formação vegetacional
do município enquadra-se no tipo Floresta Pluvial Atlântica Montana e Sub-Montana,
distribuída entre altitudes que variam de 100 a 1.143 m (THOMAZ & MONTEIRO 1997,
MENDES & PADOVAN 2000). Contudo, as florestas das baixadas desapareceram quase
por completo, estando a maior parte da vegetação original (79%) situada acima dos 500 m,

18
coincidindo com a parte serrana do município (MENDES & PADOVAN 2000), cuja
vegetação pode ser classificada como Floresta Ombrófila Densa Montana, segundo
VELOSO & GÓES-FILHO (1982) e BRASIL (1983).
3.2. Delineamento amostral
O estudo da comunidade de aves nos fragmentos florestais selecionados foi desenvolvido
com base em amostragens padronizadas entre agosto de 2002 a fevereiro de 2004,
totalizando 18 meses de trabalhos de campo (Tab. 1). Tais amostragens foram realizadas
pelo método de transectos fixos e de distâncias limitadas (BIBBY et al. 1992), distribuídos
ao longo das trilhas pré-existentes e abertas no interior dos fragmentos. Cada transecto
correspondeu a uma seção linear de 200 m percorrida durante o tempo aproximado de uma
hora, mantendo distância mínima de 200 m entre eles e no mínimo 50 m da borda do
fragmento. Os transectos foram dispostos nas trilhas pela técnica da estratificação aleatória
(stratified random technique) (BIBBLY et al. 1998), procurando incluir porções
representativas de sua heterogeneidade ambiental. O número de transectos estabelecidos
em cada remanescente dependeu do tamanho de sua área e do tempo de amostragem do
observador, tendo variado de 1 (FP3) a 10 (PNMSL, EBSL e área-controle) (Tab. 1).
Cada amostragem foi padronizada como um esforço de campo de aproximadamente
8h/dia/mês (normalmente entre 6:00-10:00 h e 15:00-19:00 h), abrangendo todos os
transectos demarcados nos fragmentos para cada dia de trabalho. Assim, o período de cada
amostragem em áreas contendo vários transectos foi igualmente distribuído entre eles; por
exemplo, 0,8 h/transecto na área-controle. A utilização de períodos longos do dia como
unidades de tempo das amostragens tem como vantagem a possibilidade de cobrir o
período de atividade de muitas espécies da comunidade, cujos horários variam muito para a
região tropical (RALPH et al. 1997, SICK 1997, BIBBY et al. 1998).

19
O número de horas trabalhadas esteve proporcionalmente relacionado com as classes de
tamanho das áreas, do seguinte modo: fragmentos pequenos e médios - 96h (8 h/dia/mês x
12 amostragens); fragmentos grandes 144h (8 h/dia/mês x 18 amostragens); área-controle -
192h (8 h/dia/mês x 24 amostragens) (Tab. 1). Isto resultou no esforço total de campo de
aproximadamente 960 h para a caracterização da comunidade de aves na região
investigada, embora um trabalho complementar tenha sido realizado para a análise da taxa
de extinções nos fragmentos (veja item 3.8) (Tab. 1).
3.3. Parâmetros da comunidade de aves
Os efeitos da fragmentação florestal sobre a avifauna da região foram avaliados com base
em quatro parâmetros relacionados à estrutura de uma comunidade biológica, comumente
utilizados em trabalhos dessa natureza (DEBINSKY & HOLT 2000): composição da
avifauna, riqueza específica, abundância relativa das espécies e grupos ecológicos
funcionais (guildas tróficas e espaciais combinadas).
Um total de 19 grupos ecológicos funcionais (guildas) foi estabelecido para o universo das
espécies florestais (invasoras exclusas, veja item 3.6), a seguir discriminados: carnívoros
diurnos (CADI); carnívoros noturnos (CANO); grandes frugívoros de copa e/ou estrato
médio (GFCE); pequenos frugívoros de copa e/ou estrato médio (PFCE); pequenos
frugívoros do subbosque (PFSB); granívoros do solo (GRSO); granívoros de taquaral
(GRTA); insetívoros de copa e/ou estrato médio (INCE) (inclui espécies que forrageiam
no espaço aéreo próximo à copa); insetívoros do subbosque (INSB); insetívoros do solo
(INSO); insetívoros de margem de córrego (INMC); insetívoros de taquaral (INTA);
insetívoros do folhiço suspenso (aglomerado de folhas mortas acumuladas entre as lianas
ou ramagens do interior da mata) (INFS); insetívoros escaladores de tronco e\ou galho

20
(INET); insetívoros noturnos (INNO); nectarívoros (NECT); onívoros de copa e/ou estrato
médio (ONCE); onívoros do subbosque (ONSB); onívoros do solo (ONSO).
A definição desses grupos ecológicos apoiou-se na combinação da dieta preferencial,
estratos e microhábitats principais de forrageamento e período de atividade
(diurno/noturno) das espécies, a partir da consulta de ampla fonte de informações
bibliográficas (MOOJEN et al. 1941, SCHUBART et al. 1965, WILLIS 1979, MOTTA
JÚNIOR 1990, REMSEN & PARKER III 1984, PARKER et al. 1996, SICK 1997,
SOARES & ANJOS 1999, GIMENES & ANJOS 2000, LEME 2001, RIBON et al. 2003,
ANJOS et al. 2004, ANTUNES 2005, DEVELEY 2004, DURÃES & MARINI 2005).
3.4. Sensibilidade das espécies ao processo de fragmentação florestal
Várias abordagens têm sido adotadas para a determinação da sensibilidade das espécies
florestais à fragmentação do seu hábitat (RIBON 2003, ANTONGIOVANNI &
METZGER 2005, ANTUNES 2005, UEZU et al. 2005, ANJOS 2006). Uma delas baseia-
se na presença/ausência das espécies em diferentes classes de tamanho dos fragmentos
florestais, possibilitando sua classificação em três categorias de sensibilidade: baixa,
média e alta (ANJOS, 2006). Neste estudo, espécies florestais (item 3.6) presentes nos
fragmentos de maior tamanho (N = 3) (PNMSL, EBSL e área-controle), mas que
estiveram ausentes em todos os menores (N = 4) (FP1, FP3, FP4 e PMBML), foram
consideradas como de alta sensibilidade ao sistema de fragmentação regional. Todas as
demais foram consideradas como de baixa sensibilidade; porém, um único contato com a
espécie nos fragmentos pequenos durante todo o período de estudo (Índice de Abundância
Relativa = 0,08, veja item 3.5) foi desconsiderado e a espécie relacionada como ausente no
fragmento (cf. ANJOS 2006). As espécies presentes sob esta mesma condição (um único
registro) em todos os maiores fragmentos (incluindo o controle) foram desconsideradas

21
desta análise. A deficiência do número de fragmentos de tamanho médio (N=1)
inviabilizou a definição de uma categoria intermediária de sensibilidade, considerando a
importância da replicagem em experimentos amostrais (KREBS 1999, DEBINSKI &
HOLT 2000).
3.5. Cálculo do índice dos descritores ambientais dos remanescentes florestais
A maioria dos estudos com avifaunas de fragmentos de florestas tropicais analisam seus
dados com base no tamanho das áreas e o seu grau de isolamento, não considerando outros
importantes descritores do ambiente que determinam a estrutura de suas comunidades
biológicas, como a permeabilidade da matriz, conformação dos remanescentes e situação
de conservação de sua vegetação, cuja avaliação é considerada de extrema necessidade
para o estudo de biotas em paisagens fragmentadas (LAURENCE 1999, ALEIXO 2002,
METZGER 1999, RENJIFO 2001, DEVELEY 2004).
Por essa razão, este estudo estabeleceu um Índice de Conservação (IC) para a descrição da
integridade física dos fragmentos amostrados através da adaptação de alguns índices já
existentes na literatura (índices de isolamento, forma e permeabilidade), com base na
seguinte equação:
IC = (IH + IA + IS + IF + IP)/5
onde:
-IC: índice de conservação do fragmento, podendo variar de 0 (degradação extrema) a 1
(conservação plena);
-IH: índice de heterogeneidade ambiental do fragmento;
-IA: índice de área do fragmento;
-IS: índice de isolamento do fragmento;
-IF: índice de forma do fragmento;

22
-IP: índice de permeabilidade da matriz.
O índice de heterogeneidade ambiental (IH) foi calculado com base em quatro
características vegetacionais do ambiente, incluindo alguns microhábitats de
forrageamento reconhecidos para as espécies da Mata Atlântica (e.g. PARKER III et al.
1996, SICK 1997): manchas de taquara (um ou mais agrupamento de touceiras de
Chusquea, Guadua, e Merostachys e/ou espécies introduzidas), cursos d´àgua (córregos
perenes), ocorrência comum de vegetação suspensa (folhas mortas, lianas, epífitas etc) nos
estratos da floresta e predominância de vegetação primária. A presença de cada uma
dessas características correspondeu a um peso de valor igual a 0,25, fazendo IH variar de 0
(todas as características ausentes) a 1 (todas as características presentes).
O cálculo do Índice de Área (IA) apoiou-se nas três classes de tamanho dos fragmentos
(T), sendo que cada uma delas recebeu um peso de valor proporcional ao seu tamanho: T <
50 ha (peso = 0); 50 ha < T < 500 ha (peso = 0,5); T > 500 ha (peso = 1,0). As áreas
menores do que 50 ha receberam peso 0 (zero) por serem pouco efetivas na conservação
de avifaunas florestais, ao contrário das áreas maiores que 500 ha, que são capazes de
manter a quase totalidade das espécies de uma região (WILLIS 1979, D`ANGELO NETO
1998, SIMON 2000, DEVELEY 2004).
O Índice de Isolamento foi adaptado de PASSAMANI (2003), adotando-se a seguinte
equação para seu cálculo:
IS= MDM/AT
onde:
- IS: índice de isolamento;
- MDM: média das distâncias mínimas (em metros) entre o fragmento analisado e os
fragmentos vizinhos que continham pelo menos uma parte inclusa dentro de um raio de 1
km a partir do fragmento analisado;

23
- AT: área total (em ha) de todos os fragmentos vizinhos considerados [logo, o inverso do
índice de PASSAMANI (2003)], para que quanto menor o valor de IS, menor o seu grau
de isolamento e vice-versa).
O Índice de Forma foi adaptado de LAURENCE & YENSEN (1991), estabelecendo-se
para seu cálculo a seguinte equação:
IF= 200(πTA)0,5/PT
onde:
- IF: índice de forma;
- TA: área total (em hectare);
- PT: perímetro total (em metros) [logo, o inverso do índice de LAURENCE & YENSE
(1991)], para que seu valor máximo também fosse 1,0 [forma circular: melhor situação de
conservação], para mais simples aplicação no cálculo de IC.
Por fim, o Índice de Permeabilidade da matriz, desenvolvido por PASSAMANI (2003), foi
calculado com base na seguinte equação:
IP = (1xPpa + 2xPca + 3xPeu)/PT
onde:
- IP: índice de permeabilidade;
- Ppa: perímetro da pastagem;
- Pca: perímetro do café;
- Peu: perímetro do eucalipto e;
- PT: perímetro total do fragmento (perímetros em metros).
Porém, neste estudo o resultado desse índice obtido para cada fragmento foi ajustado por
uma regra-de-três, com base na variação máxima (= 2) do índice original (que,
teoricamente, pode variar de 1 a 3), para que seu novo valor máximo (= 1,0) também fosse
de mais simples aplicação no cálculo de IC.
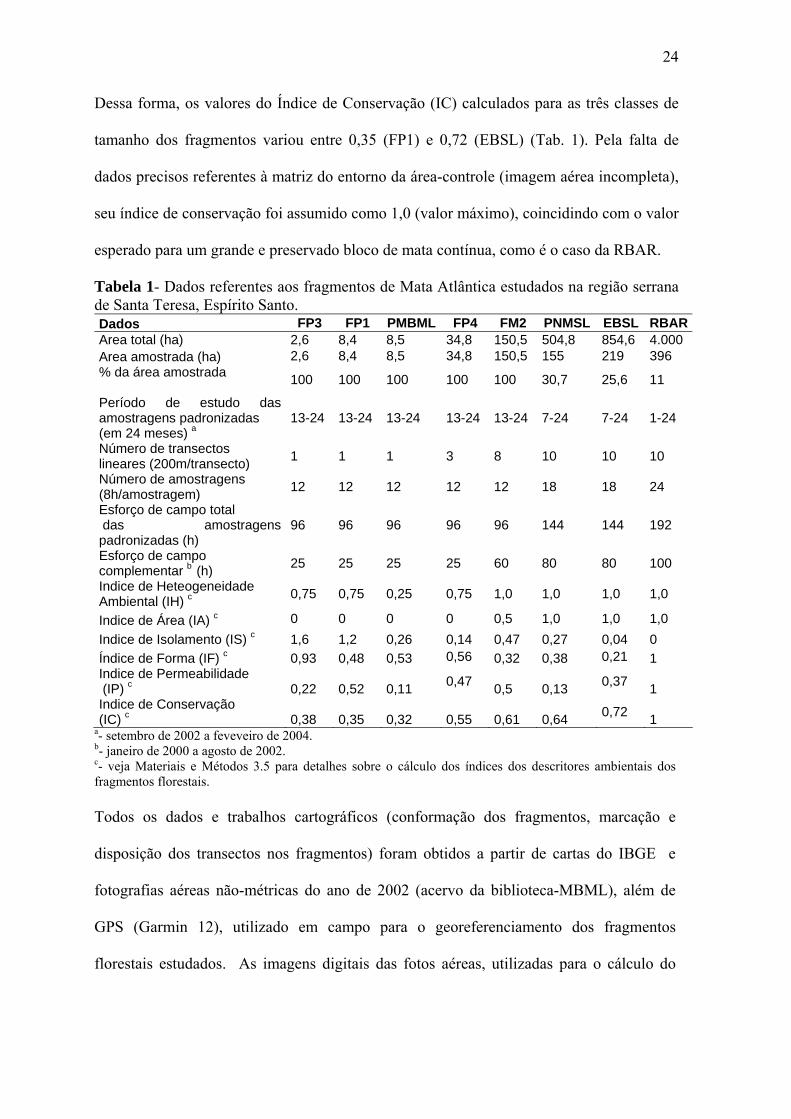
24
Dessa forma, os valores do Índice de Conservação (IC) calculados para as três classes de
tamanho dos fragmentos variou entre 0,35 (FP1) e 0,72 (EBSL) (Tab. 1). Pela falta de
dados precisos referentes à matriz do entorno da área-controle (imagem aérea incompleta),
seu índice de conservação foi assumido como 1,0 (valor máximo), coincidindo com o valor
esperado para um grande e preservado bloco de mata contínua, como é o caso da RBAR.
Tabela 1- Dados referentes aos fragmentos de Mata Atlântica estudados na região serrana de Santa Teresa, Espírito Santo. Dados FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBARArea total (ha) 2,6 8,4 8,5 34,8 150,5 504,8 854,6 4.000 Area amostrada (ha) 2,6 8,4 8,5 34,8 150,5 155 219 396 % da área amostrada 100 100 100 100 100 30,7 25,6 11
Período de estudo das amostragens padronizadas (em 24 meses) a
13-24 13-24 13-24 13-24 13-24 7-24 7-24 1-24
Número de transectos lineares (200m/transecto) 1 1 1 3 8 10 10 10
Número de amostragens (8h/amostragem) 12 12 12 12 12 18 18 24
Esforço de campo total das amostragens padronizadas (h)
96 96 96 96 96 144 144 192
Esforço de campo complementar b (h) 25 25 25 25 60 80 80 100
Indice de Heteogeneidade Ambiental (IH) c 0,75 0,75 0,25 0,75 1,0 1,0 1,0 1,0
Indice de Área (IA) c 0 0 0 0 0,5 1,0 1,0 1,0 Indice de Isolamento (IS) c 1,6 1,2 0,26 0,14 0,47 0,27 0,04 0 Índice de Forma (IF) c 0,93 0,48 0,53 0,56 0,32 0,38 0,21 1 Indice de Permeabilidade (IP) c 0,22 0,52 0,11 0,47 0,5 0,13 0,37 1 Indice de Conservação (IC) c 0,38 0,35 0,32 0,55 0,61 0,64 0,72 1
a- setembro de 2002 a feveveiro de 2004. b- janeiro de 2000 a agosto de 2002. c- veja Materiais e Métodos 3.5 para detalhes sobre o cálculo dos índices dos descritores ambientais dos fragmentos florestais. Todos os dados e trabalhos cartográficos (conformação dos fragmentos, marcação e
disposição dos transectos nos fragmentos) foram obtidos a partir de cartas do IBGE e
fotografias aéreas não-métricas do ano de 2002 (acervo da biblioteca-MBML), além de
GPS (Garmin 12), utilizado em campo para o georeferenciamento dos fragmentos
florestais estudados. As imagens digitais das fotos aéreas, utilizadas para o cálculo do

25
tamanho da área dos fragmentos, grau de isolamento, perímetro das paisagens limítrofes
dos fragmentos, etc, foram geradas e interpretadas com auxílio dos programas Idrisi 2.0,
CartaLinx 2.0 e ArcView 3.1, operados na plataforma Windows XP.
3.6. Estatística dos dados e Análises de Similaridade
A riqueza estimada de espécies nos fragmentos florestais (incluindo o controle) foi
determinada pelo método Jackknife de primeira ordem, segundo a seguinte equação
(HELTSHE & FORRESTER 1983, SMITH & BELLE 1984, KREBS 1999):
Sjack1= Sobs + Q1 (m-1/m),
onde:
- Sjack1: riqueza estimada de espécies;
- Sobs: número observado de espécies em todas as amostragens;
- Q1: número de espécies registradas em uma única amostragem;
- m: número de amostragens.
As estimativas de riqueza e as curvas de acumulação de espécies foram obtidas utilizando-
se o programa EstimateS 6.01b (COLWELL 1997), com auxílio do Excel 5.0 e Statistica
6.0, operados na plataforma Windows XP. Todos os cálculos foram realizados com base
em 50 aleatorizações (runs), considerando uma amostragem como o esforço de campo
mensal (8 h/dia) aplicado para o levantamento das espécies (12 a 24 amostragens).
O intervalo de confiança da riqueza estimada foi determinado pela seguinte equação (ZAR
1999):
IC (95%) = X ± t (α,, gl) Sx
onde:
- IC (95%): intervalo de confiança da riqueza estimada pelo método Jackknife 1;

26
- X: riqueza estimada pelo método Jackknife 1;
- t (α,, gl): valor de t da distribuição de Student (α = 5% para o teste bicaudal);
- Sx: erro padrão da riqueza estimada.
O Índice de Abundância Relativa das espécies (IAR) em cada fragmento foi determinado
pela fração entre o número de amostragens (contatos) em que a espécie foi detectada e o
número total de amostragens realizadas no fragmento em análise (n = 12 para todos os
fragmentos: março/2003 a fevereiro/2004), com valores variando de 0 (espécie não
detectada) a 1 (espécie detectada em todas as amostragens), conforme BIBBY et al.
(1998). O teste χ2(correção de Yates) foi utilizado para averiguar se o índice de abundância
relativa das espécies (testado com base no número de contatos) diferiu entre cada um dos
sete fragmentos estudados e a área-controle. O teste do χ2(correção de Yates)
também foi
utilizado para testar se as espécies mais sensíveis ao sistema de fragmentação regional
(item 3.4) estiveram numericamente mais representadas do que aquelas de baixa
sensibilidade entre o total de espécies registradas na área de estudo com maior pré-
disposição à extinção na Mata Atlântica, as quais incluem os táxons endêmicos e de alta
sensibilidade ecológica no referido bioma (item 3.7) (BROOKS et al. 1999, ALEIXO
2001, RIBON et al. 2003, HARRIS & PIMM 2004).
O tamanho da área, o grau de isolamento (IS) e a permeabilidade da matriz (IP) são
considerados como os três principais descritores ambientais (variáveis independentes)
determinantes da diversidade biológica de paisagens fragmentadas (LAURENCE 1999,
ALEIXO 2002, METZGER 1999). A análise de regressão múltipla (ZAR 1999) foi
utilizada para testar a dependência da riqueza de espécies florestais em relação à influência
mútua desses três descritores ambientais. A partir do resultado dessa análise, prosseguiu-se
com o teste de redundância das variáveis independentes, a fim de verificar qual destes

27
descritores teve maior efeito sobre a variável dependente analisada, considerando que a
mesma poderia estar distintamente relacionada com cada uma das diferentes forças do
processo de fragmentação que operam sobre a diversidade biológica.
A análise de regressão linear (ZAR 1999) foi utilizada para verificar a influência do
tamanho da área dos fragmentos (i) sobre a riqueza observada de espécies (amostragens
padronizadas), abordando distintamente a riqueza total e riqueza de espécies florestais e
(ii) sobre o conjunto das espécies que necessitam de atenção especial para sua
conservação, incluindo três grupos distintos: espécies ameaçadas de extinção no Espírito
Santo, espécies de alta sensibilidade ecológica na Mata Atlântica e endemismos deste
bioma (item 3.7). O mesmo tipo de análise foi empregado para verificar a influência do
índice de conservação dos fragmentos sobre a riqueza de espécies florestais. A regressão
linear foi ainda aplicada para avaliar alterações na riqueza de espécies das guildas em
relação ao tamanho da área dos fragmentos, a fim de verificar quais delas foram mais
afetadas com a fragmentação/redução de seu hábitat.
Quando necessário, as variáveis dependentes e independentes tiveram seus dados
ajustados por transformação logarítmica nas análises de regressão, com vistas a atender à
condição de normalidade dos dados e homoscedasticidade das variâncias, recomendada
nesse tipo de análise (ZAR 1999). Todas as análises estatísticas foram executadas no
programa Statistica 6.0 for Windows, com resultados considerados significativos quando P
< 0,05.
A variação na composição das guildas entre os fragmentos florestais foi avaliada pelo uso
da análise multivariada do tipo RA (Reciprocal Averaging) (RANDERSON 1996),
utilizando o programa PC-ORD (MCCUNE & MEFFORD 1995). Esse tipo de análise foi
aplicado para testar o grau de organização trófica-espacial da comunidade de aves entre os

28
remanescentes florestais, visando verificar o quanto os mesmos estariam segregados entre
si, em função do processo de fragmentação ocorrido na região.
O grau de similaridade biológica entre os fragmentos foi determinado com base na
composição de suas avifaunas, através da análise de agrupamento por associação de média
não-ponderada (UPGMA) (SNEATH & SOKAL 1973). Tal análise resulta em um
dendrograma de classificação hierárquica aglomerativa, cuja similaridade entre as áreas
em análise foi verificada pela distância euclidiana (MAGURRAN 1989). A elaboração do
dendrograma e o cálculo das distâncias euclidianas foram realizados com auxílio do
programa Fitopac 1.0 (SHEPHERD,1984), operado na plataforma Windows XP.
3.7. Identificação de campo, nomenclatura científica e grupos de espécies analisados
A identificação das espécies em campo apoiou-se em registros visuais e auditivos, com
auxílio de binóculo Olympus 7x35 mm e gravador Sony TCM5000-EV acoplado a
microfone Sennheiser ME-66. Quando necessária, a técnica de play-back (MARION et al.
1981) foi utilizada como ferramenta auxiliar para a identificação taxonômica das vozes
registradas, sendo essa técnica considerada de extrema importância na realização de
inventários da avifauna de florestas tropicais (JOHNSON et al. 1981, PARKER III 1991).
Vocalizações não prontamente identificadas em campo foram gravadas para posterior
confirmação em laboratório, através da comparação com arquivo de vozes das aves
disponíveis para o Brasil (VIELLIARD 1995a, b, GONZAGA & GASTIGLIONI 2001)
ou pelo envio de cópias para especialistas com ampla experiência com a avifauna da Mata
Atlântica.
O conhecimento de caracteres diagnósticos para a identificação de espécies
morfologicamente crípticas apoiou-se na bibliografia especializada (MEYER DE
SCHAUENSEE 1970, GRANTSAU 1988, RIDGELY & TURDOR 1989, 1994, SICK

29
1997, SOUZA 2002) e em consultas à coleção ornitológica do Museu de Biologia Mello
Leitão (MBML) e Museu Nacional Rio de Janeiro (MNRJ).
A seqüência sistemática e a nomenclatura científica seguem a Lista das Aves do Brasil,
organizada pelo Comitê Brasileiro de Registros Ornitológicos (2005). O estudo de
PARKER et al. (1996), combinado com SICK (1997), foi também usado para categorizar
a avifauna em elementos florestais (inclusas espécies de borda de mata) e não-florestais.
Todas as espécies registradas nos fragmentos florestais foram incluídas como membros de
suas respectivas comunidades avifaunísticas. Porém, as espécies não-florestais e as
florestais admitidas como invasoras recentes da região serrana de Santa Teresa (SIMON
2000, WILLIS & ONIKI 2002, obs. pes.) foram consideradas apenas nas análises de
riqueza total e similaridade biológica entre os fragmentos florestais.
A citação das espécies endêmicas e de alta sensibilidade ecológica da Mata Atlântica
seguiu o estudo de PARKER III et al. (1996). As espécies ameaçadas de extinção no
Estado do Espírito Santo foram citadas segundo IPEMA (2005).
3.8. Base de dados sobre a avifauna da região e taxa de extinções nos fragmentos
A região de Santa Teresa está entre as localidades mais bem inventariadas do Espírito
Santo (SIMON 2000), incluindo longo período de coleta de material ornitológico, iniciado
por volta de 1935 e prosseguindo quase ininterruptamente até os dias de hoje, sobretudo
em função dos trabalhos de Augusto Ruschi (MBML), Helmut Sick (Museu Nacional, Rio
de Janeiro - MNRJ), Olivério Pinto (Museu de Zoologia, Universidade de São Paulo -
MZUSP), Antônio Paviotti (Departamento de Zoologia, Universidade Federal de Minas
Gerais - DZUFMG) e J. E. Simon (MBML).
Além do material coletado, muitos outros estudos já foram publicados para a região
(COLLAR et al. 1992, WHITNEY & PACHECO 1995, PARKER III & GOERCK 1997,

30
SCOTT 1997, ONIKI & WILLIS 1998, WILLIS & ONIKI 1998, 2002, BAUER et al.
2000, SIMON 2000), constituindo-se em rica fonte de registros, especialmente
importantes para a compreensão dos efeitos da fragmentação florestal sobre a composição
de biotas regionais (WILLIS 1974, KATTAN et al. 1994, RENJIFO 1999, RIBON et al.
2003).
A compilação e revisão crítica das fontes de registros museológicos e bibliográficos
fizeram parte deste estudo com o intuito de subsidiar algumas conclusões (por exemplo,
extinções locais e táxons alóctones para a região) sobre a comunidade de aves na Mata
Atlântica de Santa Teresa. Com base nesta revisão, oito espécies florestais foram
admitidas como de ocorrência ou adição imprópria para a região serrana de Santa Teresa
(cf. SIMON 2000), dispensando-se, portanto, demais comentários sobre as mesmas no
escopo deste trabalho (Tab. 2).
Tabela 2. Espécies de aves consideradas como adições impróprias para a região serrana de Santa Teresa, ES. Espécies Fontes de Registro a Justificativa b Ortalis guttata 1 I-1 Pionus menstruus 2 II Crax blumenbachii 1 I-1 Aramides cajanea 2 I-2 Pyrrhura cruentata 1 I-1 Caprimulgus rufus 1 I-1 Todirostrum cinereum 3 I-2 Icterus jamacaii 1 I-1
a -1- coleção científica MBML; 2- SIMON (2000); 3- WILLIS & ONIKI (2002). b - I- espécies sem registro bibliográfico ou museológico para a região serrana do Espírto Santo, tratando-se possivelmente de espécimes provenientes de cativeiro/localidade errônea (I-1) ou equívocos de identificação (I-2); II- erro compilatório, por sua localização indevida na Tabela 1 do estudo de SIMON (2000) (Tabela 3 como sua posição correta). Todas as listagens de Augusto Ruschi (e.g. Ruschi 1969, 1977) foram excluídas dessa
compilação, em função das inúmeras inconsistências apontadas em suas publicações de
avifaunas regionais (BAUER 1999, PACHECO & BAUER 2001, SIMON 2000, WILLIS
& ONIKI 2002, SIMON 2004). O mesmo valeu para o estudo de FORRESTER (1993),
por tratar-se essencialmente de compilação de dados de terceiros (muitos observadores de

31
pássaros norte-americanos com pouca experiência com a avifauna de nosso país; cf.
WILLIS & ONIKI 2002), contendo igualmente várias adições impróprias para o
Município de Santa Teresa (SIMON 2000, WILLIS & ONIKI 2002).
A partir da revisão da base de dados secundários disponível para a região (literatura e
coleções científicas), associada aos dados de campo (veja abaixo), foi possível estabelecer
o total das espécies florestais seguramente ocorrentes na região. Com o intuito de subsidiar
algumas análises do presente estudo, tais espécies tiveram seu status de ocorrência local
(cf. SIMON 2000) avaliado, permitindo reconhecer aquelas autóctones (residentes) e
alóctones (vagante, migratória altitudinal/continental e invasora recente) para a região
serrana de Santa Teresa, a partir da base de dados acima mencionada e várias outras
bibliografias relativas ao assunto (e.g. MEYER DE SCHAUENSEE 1966, 1982, SICK
1984, 1997).
Entre janeiro de 2000 e agosto de 2002, foram realizados levantamentos assistemáticos
nos fragmentos mencionados (ca. 420 h), para aperfeiçoamento da base de dados
ornitológicos do Município de Santa Teresa e seu maior domínio das manifestações
sonoras das espécies da região (SIMON & GASPARINI 2003, SIMON 2004). Embora
não tenham sido incorporados às análises estatísticas e de similaridade acima descritas, os
registros de campo desse período são aqui fornecidos como dados complementares sobre a
composição da avifauna na área de estudo (Tab. 3). Em conjunto, os dados do período
entre janeiro de 2000 e fevereiro de 2004 (1.380 h) foram utilizados para determinar a taxa
de extinções nos fragmentos, conforme a seguinte equação:
TE= (NEP/NTR)x100
onde:
- TE: taxa de extinções;

32
- NEP: número de espécies florestais (exclusas as invasoras) perdidas no fragmento
(espécies vágeis ou inconspícuas no ambiente foram desconsideradas desta análise);
- NTR: número total de espécies florestais (exclusas as invasoras) registradas na área-
controle (veja abaixo, Tab. 3).
4. RESULTADOS
4.1. Composição da avifauna nos fragmentos florestais estudados
Um total de 261 espécies, distribuídas em 48 famílias, foram registradas nos remanescentes
de Mata Atlântica da região serrana de Santa Teresa pelas amostragens padronizadas do
presente estudo (18 meses/960h) (Tab. 3). Desse total, 187 espécies (71,6%) são florestais,
das quais 15 são consideradas como invasoras recentes na área de estudo. Do total
registrado, 37 (14,2%) e 80 (30,6%) espécies correspondem, respectivamente, a elementos
de alta sensibilidade ecológica (incluindo uma invasora, Accipiter superciliosus) e a
endemismos da Mata Atlântica (Tab. 3).
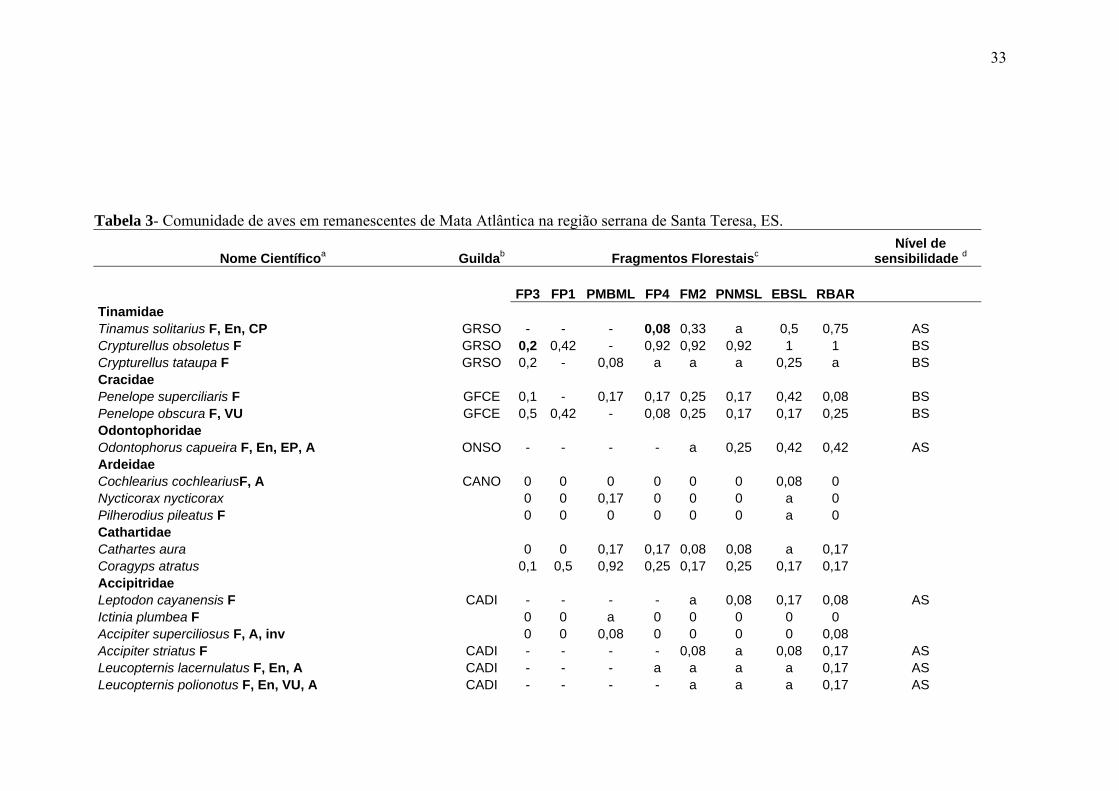
33
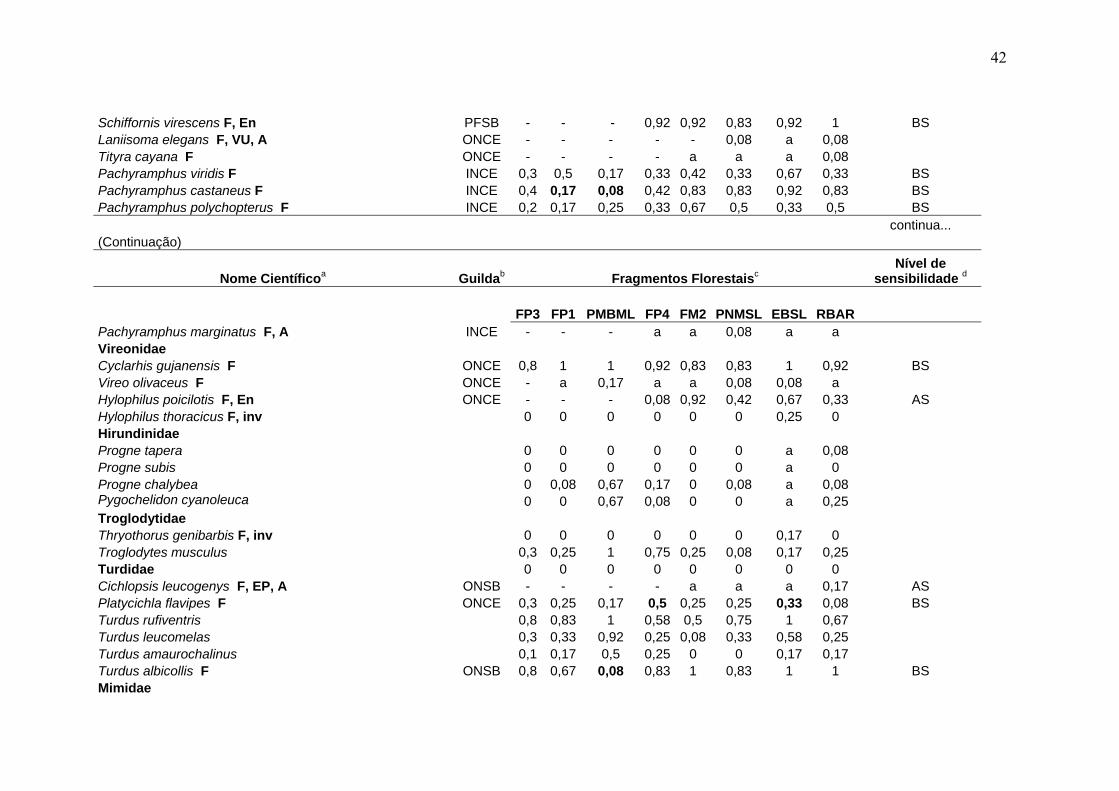
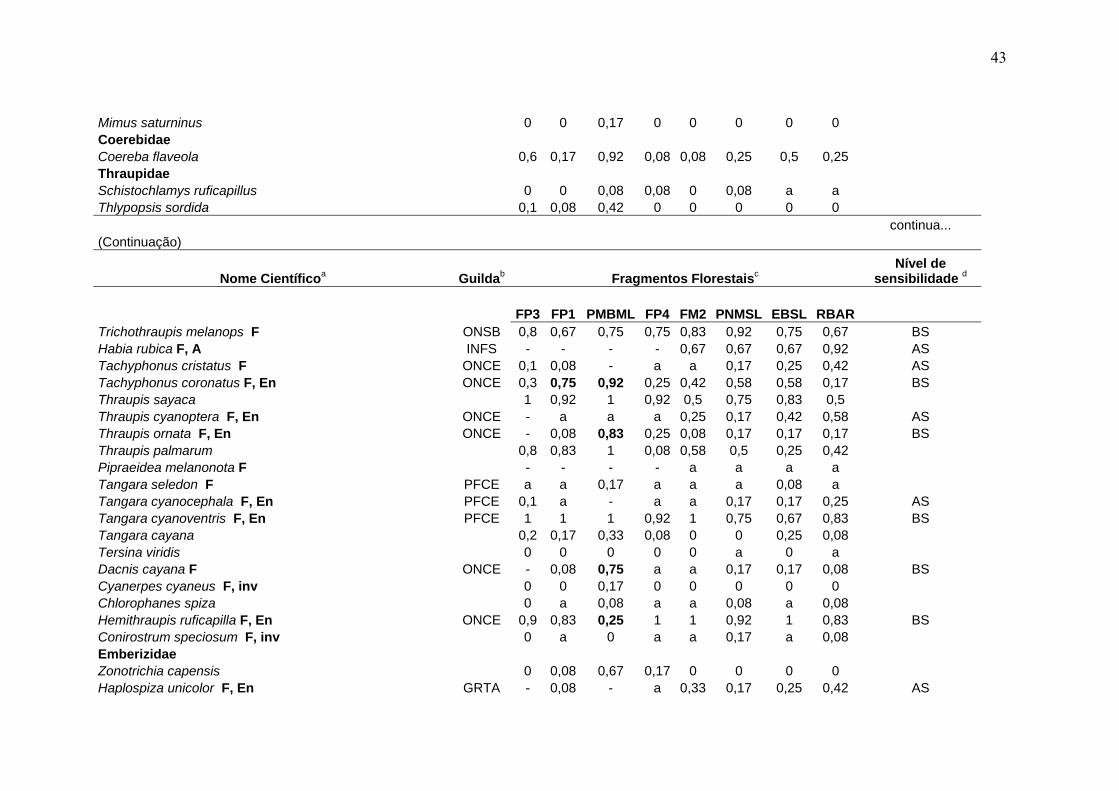
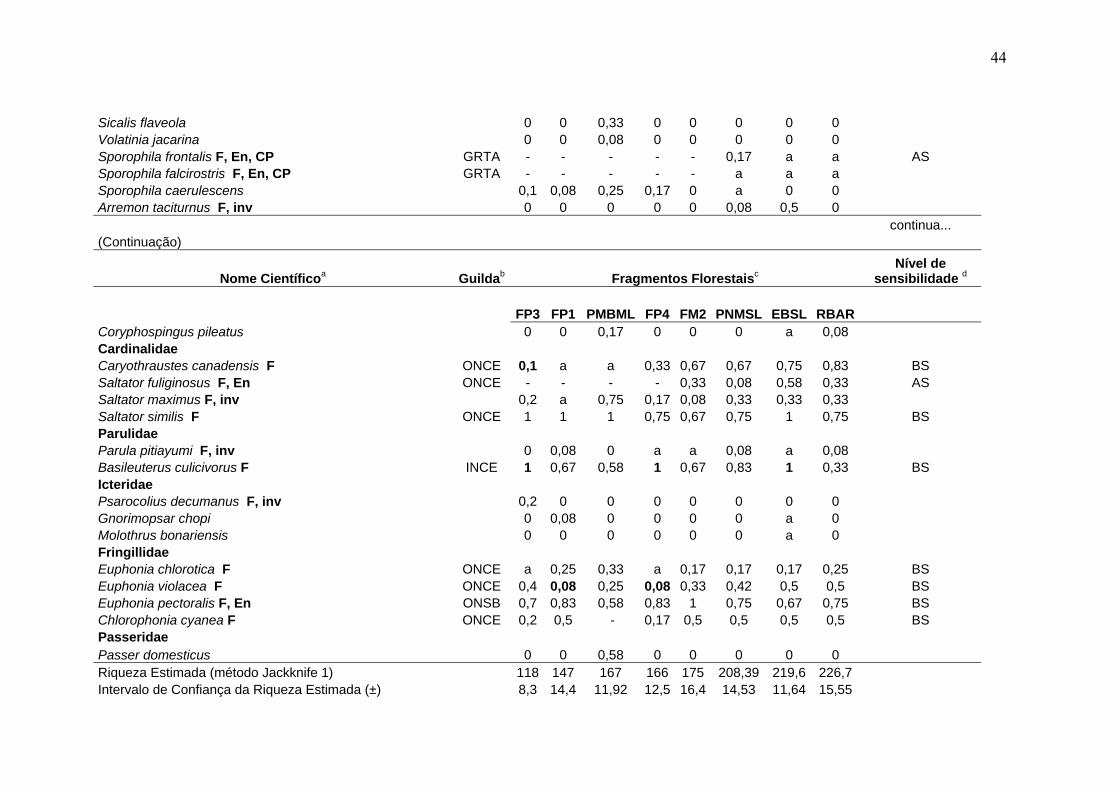
Tabela 3- Comunidade de aves em remanescentes de Mata Atlântica na região serrana de Santa Teresa, ES.
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Tinamidae Tinamus solitarius F, En, CP GRSO - - - 0,08 0,33 a 0,5 0,75 AS Crypturellus obsoletus F GRSO 0,2 0,42 - 0,92 0,92 0,92 1 1 BS Crypturellus tataupa F GRSO 0,2 - 0,08 a a a 0,25 a BS Cracidae Penelope superciliaris F GFCE 0,1 - 0,17 0,17 0,25 0,17 0,42 0,08 BS Penelope obscura F, VU GFCE 0,5 0,42 - 0,08 0,25 0,17 0,17 0,25 BS Odontophoridae Odontophorus capueira F, En, EP, A ONSO - - - - a 0,25 0,42 0,42 AS Ardeidae Cochlearius cochleariusF, A CANO 0 0 0 0 0 0 0,08 0 Nycticorax nycticorax 0 0 0,17 0 0 0 a 0 Pilherodius pileatus F 0 0 0 0 0 0 a 0 Cathartidae Cathartes aura 0 0 0,17 0,17 0,08 0,08 a 0,17 Coragyps atratus 0,1 0,5 0,92 0,25 0,17 0,25 0,17 0,17 Accipitridae Leptodon cayanensis F CADI - - - - a 0,08 0,17 0,08 AS Ictinia plumbea F 0 0 a 0 0 0 0 0 Accipiter superciliosus F, A, inv 0 0 0,08 0 0 0 0 0,08 Accipiter striatus F CADI - - - - 0,08 a 0,08 0,17 AS Leucopternis lacernulatus F, En, A CADI - - - a a a a 0,17 AS Leucopternis polionotus F, En, VU, A CADI - - - - a a a 0,17 AS

34
Rupornis magnirostris 0,7 0,67 0,75 0,92 0,83 0,33 0,67 0,42 Buteo albicaudatus 0 0,08 0 0 0,08 0 0 0 Buteo brachyurus F CADI 0 0 0,08 0 0 0 0 0 Spizaetus tyrannus F, VU CADI - - 0,08 a 0,08 0,25 0,33 0,33 AS Falconidae continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Caracara plancus 0 0 0 0 0,08 0 0 0,08 Milvago chimachima 0 0,58 a 0,42 0,17 0 0,17 0,08 Herpetotheres cachinnans F CADI - 0,08 - 0,08 0,08 a 0,17 a AS Micrastur ruficollis F CADI - - - - a 0,25 0,17 0,25 AS Falco rufigularis F, inv 0 0 0,08 0 0 0 0 0 Falco femoralis 0 0 0,08 0 0 0 0 0 Rallidae Aramides saracura F, En ONSO 0,4 0,33 0,75 0,17 0,17 0,17 0,33 0,25 BS Pardirallus nigricans 0 0 0,17 0 0 0 0 0 Charadriidae Vanellus chilensis 0 0 0,08 0 0 0 0 0 Columbidae Columbina talpacoti 0 0,17 1 0,17 0,08 0,17 a 0,25 Claravis pretiosa F GRSO - - - 0,17 a a 0,17 0,08 BS Claravis godefrida F, En, CP, A GRTA - - - - - - a 0,17 AS Patagioenas picazuro 0 0 0 0,17 0 0 0,08 0 Patagioenas cayennensis 0 0 0 0 0,08 0 0 0,08 Patagioenas plumbea F, A GFCE 0,2 - - 0,58 0,67 0,5 0,75 0,83 BS Leptotila verreauxi F GRSO 0,5 0,5 0,33 0,75 0,25 0,42 0,58 0,08 BS Leptotila rufaxilla F GRSO a 0,17 1 0,42 0,17 0,67 0,67 0,42 BS Geotrygon montana F GRSO - - - 0,17 0,25 0,25 0,42 0,33 BS Psittacidae Pyrrhura frontalis F, En GFCE 0,9 0,67 0,83 0,75 1 0,92 1 1 BS

35
Forpus xanthopterygius 0 0 0,08 0 0 0 0 0 Touit melanonotus F, En, EP, A PFCE - - - - - a a 0,08 Pionopsitta pileata F, En, VU GFCE - - - - a a 0,25 0,25 AS continua...
(Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Pionus maximiliani F GFCE 0,7 0,5 0,17 0,33 0,75 0,58 0,67 0,67 BS Amazona rhodocorytha F, En GFCE 0,5 0,08 0,08 0,33 0,33 0,17 0,08 0,17 BS Triclaria malachitacea F, En, CP GFCE - - - - a a 0,17 0,08 AS Cuculidae Piaya cayana F INCE 0,3 0,33 0,83 0,33 0,67 0,67 0,75 0,67 BS Tapera naevia 0,1 0,08 0,17 0 0,08 0 0,17 0 Tytonidae Tyto alba 0 0 a 0 0 0 0 0 Strigidae Megascops choliba 0 0 0,08 0 0 0 0,17 0 Pulsatrix koeniswaldiana F, En, A CANO - a 0,33 0,08 0,08 0,33 0,25 0,33 BS Strix virgata F CANO - - - 0,17 0,08 0,17 0,17 0,25 BS Strix huhula F, VU CANO - - - - 0,17 0,5 0,25 0,25 AS Glaucidium minutissimum F, EP CANO - - - - a 0,08 a 0,25 AS Glaucidium brasilianum F, inv 0 0 0 0 a a 0,25 a Nyctibiidae Nyctibius aethereus F, VU, A INNO - - - a 0,08 a 0,17 a AS Nyctibius griseus F, inv 0 0 0 a a a 0,25 0,08 Caprimulgidae Lurocalis semitorquatus F INNO 0 0 0 A 0,08 0,25 0,33 0,42 AS Nyctidromus albicollis F INNO 0,2 - - A a 0,17 0,33 0,17 BS

36
Nyctiphrynus ocellatus F INNO - - - - a 0,17 0,17 0,08 AS Hydropsalis torquata 0 0 0 0 0 0 0,08 0 Apodidae Streptoprocne zonaris 0 0 0,17 0 0 0 0 0 Chaetura cinereiventris F INCE - - - - 0,67 0,42 0,75 0,75 AS continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Trochilidae Ramphodon naevius F, En, EP NECT a - 0,08 0,25 0,08 0,33 0,58 0,58 BS Phaethornis squalidus F NECT 0,1 0,08 a 0,08 a 0,17 0,08 0,33 AS Phaethornis pretrei 0,2 0,42 1 0,25 0,08 0,17 0,25 0,25 Phaetornis eurynome F En NECT a 0,42 0,75 0,42 0,58 0,5 0,75 0,67 BS Eupetomena macroura 0 0 1 0 0 0,17 0,08 0,17 Aphantochroa cirrochloris En 0,1 0 1 0 0 0,08 0,25 0,08 Florisuga fusca F, En NECT a 0,42 1 0,17 0,33 0,5 0,5 0,33 BS Colibri serrirostris 0 0 1 0,08 0 0 0,17 0 Lophornis magnificus F NECT a 0,08 0,75 a a 0,17 0,17 0,17 BS Chlorostilbon aureoventris 0 0 0,08 0 0 0,08 a 0 Thalurania glaucopis F, En NECT 0,5 0,67 0,92 0,83 0,92 0,83 0,83 0,83 BS Hylocharis cyanus F NECT a - 0,17 a a a a a Leucochloris albicollis F, En NECT - 0,08 0,83 a a 0,17 0,08 0,08 BS Amazilia versicolor F NECT 0 0 0,92 0 0 0,08 0,08 0 Clytolaema rubricauda F, En NECT 0,1 0,33 0,33 0,08 0,17 0,17 0,25 0,25 BS Calliphlox amethystina 0 0 0,33 0 0 0,08 0,17 0,08 Trogonidae Trogon viridis F ONCE 0,4 0,33 a 0,25 0,58 0,75 0,5 0,75 BS Trogon surrucura F, En ONCE 0,3 0,33 0,08 0,92 0,75 0,58 0,58 0,5 BS Trogon rufus F ONSB 0,1 0,25 0,25 0,42 0,42 0,5 0,67 0,67 BS Alcedinidae

37
Ceryle torquatus 0 0 0,08 0 0 0 0 0 Chloroceryle amazona 0 0 0 0 0 0 a 0 Chloroceryle americana 0 0 0 0 0 0 0 0,08 Galbulidae Galbula ruficauda F, inv 0 0 0 0,08 a a 0,25 0 Bucconidae continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Notharchus swainsoni F, CP INCE 0 0 0 0 0 0 a 0 Malacoptila striata F, En INSB - 0,08 0,25 - 0,08 0,17 0,33 0,17 BS Chelidoptera tenebrosa F, VU, inv 0 0 0 0 a a 0,08 0,08 Ramphastidae Ramphastos vitellinus F, A GFCE 0,9 0,92 0,75 1 0,67 0,75 0,92 0,92 BS Selenidera maculirostris F GFCE 0,3 0,08 - 0,42 0,67 0,67 0,67 0,92 BS Pteroglossus bailloni F, En, A GFCE a 0,08 - a 0,33 0,17 0,42 0,17 AS Pteroglossus aracari F, inv a 0,08 0,08 0,17 0,08 0,17 0,17 a BS Picidae Picumnus cirratus 0,5 0,42 0,75 0,75 0,5 0,5 0,75 0,42 Melanerpes flavifrons F, En, VU - - - - a a a a Veniliornis maculifrons F, En INET 0,2 0,17 0,17 0,33 0,33 0,42 0,33 0,25 BS Piculus flavigula F, A INET - - - - 0,5 0,17 0,33 0,67 AS Piculus aurulentus F, En INET a 0,08 - 0,42 0,25 0,5 0,67 0,67 BS Colaptes campestris 0 0,08 0,08 0 0 0 0 0 Celeus flavescens F INET 0,1 - - 0,08 - 0,08 0,25 a AS Dryocopus lineatus F INET 0,1 0,25 a 0,17 0,17 0,25 0,17 0,33 BS Campephilus robustus F, En INET - - - - 0,25 0,17 0,17 0,17 AS Thamnophilidae Hypoedaleus guttatus F, En, A INCE - - - 0,17 0,67 0,25 0,75 0,83 BS Batara cinerea F INTA - - - - a 0,08 0,08 0,17 AS Mackenziaena severa F, En INTA 0,2 0,58 - - a 0,08 0,17 0,08 BS
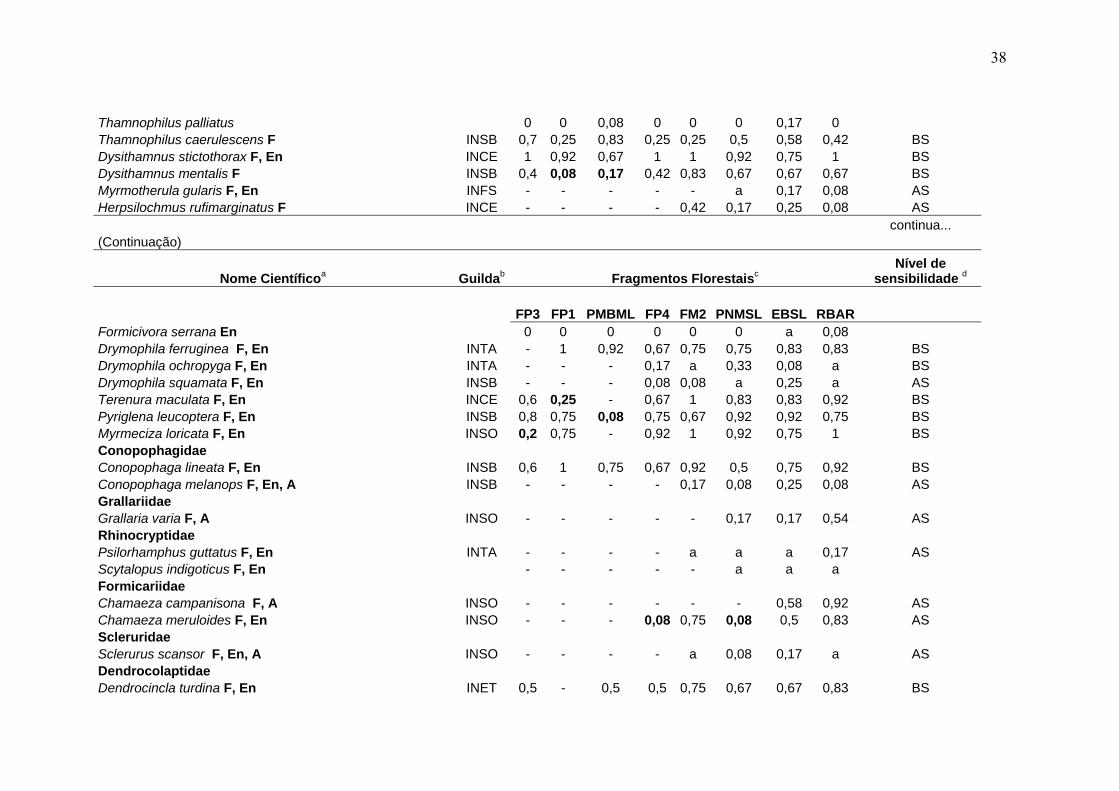
38
Thamnophilus palliatus 0 0 0,08 0 0 0 0,17 0 Thamnophilus caerulescens F INSB 0,7 0,25 0,83 0,25 0,25 0,5 0,58 0,42 BS Dysithamnus stictothorax F, En INCE 1 0,92 0,67 1 1 0,92 0,75 1 BS Dysithamnus mentalis F INSB 0,4 0,08 0,17 0,42 0,83 0,67 0,67 0,67 BS Myrmotherula gularis F, En INFS - - - - - a 0,17 0,08 AS Herpsilochmus rufimarginatus F INCE - - - - 0,42 0,17 0,25 0,08 AS continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Formicivora serrana En 0 0 0 0 0 0 a 0,08 Drymophila ferruginea F, En INTA - 1 0,92 0,67 0,75 0,75 0,83 0,83 BS Drymophila ochropyga F, En INTA - - - 0,17 a 0,33 0,08 a BS Drymophila squamata F, En INSB - - - 0,08 0,08 a 0,25 a AS Terenura maculata F, En INCE 0,6 0,25 - 0,67 1 0,83 0,83 0,92 BS Pyriglena leucoptera F, En INSB 0,8 0,75 0,08 0,75 0,67 0,92 0,92 0,75 BS Myrmeciza loricata F, En INSO 0,2 0,75 - 0,92 1 0,92 0,75 1 BS Conopophagidae Conopophaga lineata F, En INSB 0,6 1 0,75 0,67 0,92 0,5 0,75 0,92 BS Conopophaga melanops F, En, A INSB - - - - 0,17 0,08 0,25 0,08 AS Grallariidae Grallaria varia F, A INSO - - - - - 0,17 0,17 0,54 AS Rhinocryptidae Psilorhamphus guttatus F, En INTA - - - - a a a 0,17 AS Scytalopus indigoticus F, En - - - - - a a a Formicariidae Chamaeza campanisona F, A INSO - - - - - - 0,58 0,92 AS Chamaeza meruloides F, En INSO - - - 0,08 0,75 0,08 0,5 0,83 AS Scleruridae Sclerurus scansor F, En, A INSO - - - - a 0,08 0,17 a AS Dendrocolaptidae Dendrocincla turdina F, En INET 0,5 - 0,5 0,5 0,75 0,67 0,67 0,83 BS

39
Sittasomus griseicapillus F INET 0,8 0,42 0,25 0,83 0,83 0,92 0,92 1 BS Xiphocolaptes albicollis F INET 0,3 0,25 0,08 0,25 0,58 0,75 0,67 0,75 BS Dendrocolaptes platyrostris F INET - - - - 0,17 0,17 0,17 0,25 AS Xiphorhynchus fuscus F, En, A INET 1 0,92 0,33 1 1 0,92 1 1 BS Lepidocolaptes squamatus F, En, A INET 0,1 a - a 0,17 0,17 0,08 0,17 BS Campylorhamphus falcularius F, En, A INET - - - 0,17 0,08 0,58 0,67 0,67 BS continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Furnariidae Furnarius figulus 0 0 0,83 0 0 0 0 0 Furnarius rufus 0 0 a 0 0 0 0 0 Synallaxis ruficapilla F, En INSB 0,4 1 - 0,25 0,42 0,5 0,67 0,58 BS Synallaxis frontalis 0 0 0,08 0,08 0 0 0 0 Synallaxis spixi 0,1 0 a 0,17 0,08 0 0 0 Cranioleuca pallida F, En INCE 0,7 0,67 0,67 0,17 0,58 0,42 0,5 0,17 BS Anabacerthia amaurotis F, En, A - - - - a a a a Philydor lichtensteini F, En, A INFS - - - 0,08 0,67 0,5 0,42 0,67 AS Philydor atricapillus F, En, A INFS - 0,08 - 0,08 0,92 0,58 0,75 0,92 AS Philydor rufum F INCE - 0,17 0,08 0,17 0,5 0,67 0,5 0,75 BS Anabazenops fuscus F, En, A INTA 0,1 0,75 - 0,17 0,75 0,75 0,67 0,92 BS Cichlocolaptes leucophrus F, En, A INFS - - - - 0,08 0,17 0,08 0,17 AS Automolus leucophthalmus F, En INSB - 0,17 - 0,25 0,92 0,67 0,75 0,92 BS Lochmias nematura F INMC - - 0,08 - 0,67 0,75 0,5 0,58 AS Xenops minutus F INET - 0,08 - 0,08 0,08 0,5 0,33 0,17 AS Xenops rutilans F INET 0,3 0,33 0,33 0,33 0,67 0,25 0,25 0,42 BS Tyrannidae Mionectes rufiventris F, En ONSB - - - 0,08 0,33 0,42 0,33 0,17 AS Leptopogon amaurocephalus F ONCE 0,5 0,58 0,08 0,92 1 0,92 0,92 1 BS Hemitriccus diops F, En INTA 0,6 1 - 0,67 1 0,75 0,67 0,92 BS Hemitriccus nidipendulus En 0 0 0 0,08 a 0,17 a a
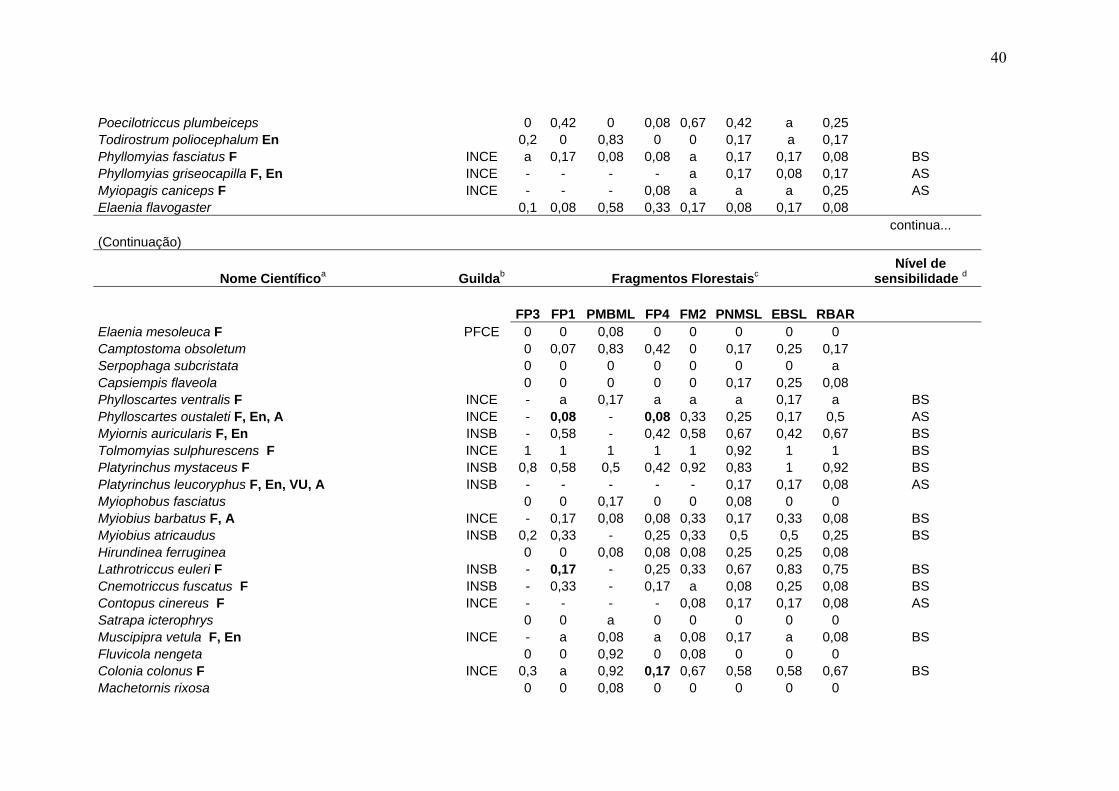
40
Poecilotriccus plumbeiceps 0 0,42 0 0,08 0,67 0,42 a 0,25 Todirostrum poliocephalum En 0,2 0 0,83 0 0 0,17 a 0,17 Phyllomyias fasciatus F INCE a 0,17 0,08 0,08 a 0,17 0,17 0,08 BS Phyllomyias griseocapilla F, En INCE - - - - a 0,17 0,08 0,17 AS Myiopagis caniceps F INCE - - - 0,08 a a a 0,25 AS Elaenia flavogaster 0,1 0,08 0,58 0,33 0,17 0,08 0,17 0,08 continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Elaenia mesoleuca F PFCE 0 0 0,08 0 0 0 0 0 Camptostoma obsoletum 0 0,07 0,83 0,42 0 0,17 0,25 0,17 Serpophaga subcristata 0 0 0 0 0 0 0 a Capsiempis flaveola 0 0 0 0 0 0,17 0,25 0,08 Phylloscartes ventralis F INCE - a 0,17 a a a 0,17 a BS Phylloscartes oustaleti F, En, A INCE - 0,08 - 0,08 0,33 0,25 0,17 0,5 AS Myiornis auricularis F, En INSB - 0,58 - 0,42 0,58 0,67 0,42 0,67 BS Tolmomyias sulphurescens F INCE 1 1 1 1 1 0,92 1 1 BS Platyrinchus mystaceus F INSB 0,8 0,58 0,5 0,42 0,92 0,83 1 0,92 BS Platyrinchus leucoryphus F, En, VU, A INSB - - - - - 0,17 0,17 0,08 AS Myiophobus fasciatus 0 0 0,17 0 0 0,08 0 0 Myiobius barbatus F, A INCE - 0,17 0,08 0,08 0,33 0,17 0,33 0,08 BS Myiobius atricaudus INSB 0,2 0,33 - 0,25 0,33 0,5 0,5 0,25 BS Hirundinea ferruginea 0 0 0,08 0,08 0,08 0,25 0,25 0,08 Lathrotriccus euleri F INSB - 0,17 - 0,25 0,33 0,67 0,83 0,75 BS Cnemotriccus fuscatus F INSB - 0,33 - 0,17 a 0,08 0,25 0,08 BS Contopus cinereus F INCE - - - - 0,08 0,17 0,17 0,08 AS Satrapa icterophrys 0 0 a 0 0 0 0 0 Muscipipra vetula F, En INCE - a 0,08 a 0,08 0,17 a 0,08 BS Fluvicola nengeta 0 0 0,92 0 0,08 0 0 0 Colonia colonus F INCE 0,3 a 0,92 0,17 0,67 0,58 0,58 0,67 BS Machetornis rixosa 0 0 0,08 0 0 0 0 0

41
Legatus leucophaius F INCE - - - - a 0,08 0,08 a Myiozetetes cayanensis 0 0 0 0,08 0 0,08 0,17 0,08 Myiozetetes similis 0,3 0,25 0,75 0,33 0,08 0,17 0,25 0,08 Pitangus sulphuratus 0,9 1 1 1 1 0,5 0,92 0,5 Myiodynastes maculatus F ONCE - a 0,25 a a 0,08 0,25 0,08 BS Megarynchus pitangua F INCE 0,8 0,58 1 0,67 0,83 0,67 1 0,67 BS continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Empidonomus varius F ONCE a a 0,25 a a a a a Tyrannus melancholicus 0,3 0,33 1 0,08 0,17 0 0,33 0,33 Rhytipterna simplex F, A INCE - 0,17 - 0,33 0,67 0,5 0,75 0,75 BS Sirystes sibilator F INCE 0,8 0,5 0,33 0,67 1 0,75 0,25 0,75 BS Myiarchus tuberculifer F INCE - - - a a 0,08 a 0,17 AS Myiarchus swainsoni F INCE 0,1 0,17 - - 0,33 a 0,08 0,17 BS Myiarchus ferox 8 0,17 0,58 0,17 0 0 0 a Attila rufus F, En ONCE 0,1 0,08 - 0,33 0,75 0,75 1 0,67 BS Oxyruncidae Oxyruncus cristatus F, A ONCE 0,3 a - 0,25 0,75 0,67 0,75 0,83 BS Cotingidae Phibalura flavirostris F, VU GFCE - - - - a a 0,08 a Carpornis cucullata F, En, A GFCE - 0,08 - - 1 0,5 0,25 0,5 AS Procnias nudicollis F, En GFCE - - a 0,17 0,33 0,17 0,5 0,58 BS Lipaugus lanioides F, En ONCE - - - - - 0,5 0,5 0,92 AS Pipridae Neopelma aurifrons F, En, VU PFSB - - - 0,08 1 0,67 0,67 1 AS Piprites chloris F, A PFCE - - - 0,08 0,08 0,25 0,17 0,33 AS Ilicura militaris F, En, A PFSB 0,4 0,42 - 1 0,75 0,58 0,67 0,75 BS Manacus manacus F PFSB - - - - a 0,42 0,58 0,17 AS Chiroxiphia caudata F, En PFSB 0,7 0,83 0,33 1 1 0,92 1 1 BS Tityridae
42
Schiffornis virescens F, En PFSB - - - 0,92 0,92 0,83 0,92 1 BS Laniisoma elegans F, VU, A ONCE - - - - - 0,08 a 0,08 Tityra cayana F ONCE - - - - a a a 0,08 Pachyramphus viridis F INCE 0,3 0,5 0,17 0,33 0,42 0,33 0,67 0,33 BS Pachyramphus castaneus F INCE 0,4 0,17 0,08 0,42 0,83 0,83 0,92 0,83 BS Pachyramphus polychopterus F INCE 0,2 0,17 0,25 0,33 0,67 0,5 0,33 0,5 BS continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Pachyramphus marginatus F, A INCE - - - a a 0,08 a a Vireonidae Cyclarhis gujanensis F ONCE 0,8 1 1 0,92 0,83 0,83 1 0,92 BS Vireo olivaceus F ONCE - a 0,17 a a 0,08 0,08 a Hylophilus poicilotis F, En ONCE - - - 0,08 0,92 0,42 0,67 0,33 AS Hylophilus thoracicus F, inv 0 0 0 0 0 0 0,25 0 Hirundinidae Progne tapera 0 0 0 0 0 0 a 0,08 Progne subis 0 0 0 0 0 0 a 0 Progne chalybea 0 0,08 0,67 0,17 0 0,08 a 0,08 Pygochelidon cyanoleuca 0 0 0,67 0,08 0 0 a 0,25 Troglodytidae Thryothorus genibarbis F, inv 0 0 0 0 0 0 0,17 0 Troglodytes musculus 0,3 0,25 1 0,75 0,25 0,08 0,17 0,25 Turdidae 0 0 0 0 0 0 0 0 Cichlopsis leucogenys F, EP, A ONSB - - - - a a a 0,17 AS Platycichla flavipes F ONCE 0,3 0,25 0,17 0,5 0,25 0,25 0,33 0,08 BS Turdus rufiventris 0,8 0,83 1 0,58 0,5 0,75 1 0,67 Turdus leucomelas 0,3 0,33 0,92 0,25 0,08 0,33 0,58 0,25 Turdus amaurochalinus 0,1 0,17 0,5 0,25 0 0 0,17 0,17 Turdus albicollis F ONSB 0,8 0,67 0,08 0,83 1 0,83 1 1 BS Mimidae
43
Mimus saturninus 0 0 0,17 0 0 0 0 0 Coerebidae Coereba flaveola 0,6 0,17 0,92 0,08 0,08 0,25 0,5 0,25 Thraupidae Schistochlamys ruficapillus 0 0 0,08 0,08 0 0,08 a a Thlypopsis sordida 0,1 0,08 0,42 0 0 0 0 0 continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Trichothraupis melanops F ONSB 0,8 0,67 0,75 0,75 0,83 0,92 0,75 0,67 BS Habia rubica F, A INFS - - - - 0,67 0,67 0,67 0,92 AS Tachyphonus cristatus F ONCE 0,1 0,08 - a a 0,17 0,25 0,42 AS Tachyphonus coronatus F, En ONCE 0,3 0,75 0,92 0,25 0,42 0,58 0,58 0,17 BS Thraupis sayaca 1 0,92 1 0,92 0,5 0,75 0,83 0,5 Thraupis cyanoptera F, En ONCE - a a a 0,25 0,17 0,42 0,58 AS Thraupis ornata F, En ONCE - 0,08 0,83 0,25 0,08 0,17 0,17 0,17 BS Thraupis palmarum 0,8 0,83 1 0,08 0,58 0,5 0,25 0,42 Pipraeidea melanonota F - - - - a a a a Tangara seledon F PFCE a a 0,17 a a a 0,08 a Tangara cyanocephala F, En PFCE 0,1 a - a a 0,17 0,17 0,25 AS Tangara cyanoventris F, En PFCE 1 1 1 0,92 1 0,75 0,67 0,83 BS Tangara cayana 0,2 0,17 0,33 0,08 0 0 0,25 0,08 Tersina viridis 0 0 0 0 0 a 0 a Dacnis cayana F ONCE - 0,08 0,75 a a 0,17 0,17 0,08 BS Cyanerpes cyaneus F, inv 0 0 0,17 0 0 0 0 0 Chlorophanes spiza 0 a 0,08 a a 0,08 a 0,08 Hemithraupis ruficapilla F, En ONCE 0,9 0,83 0,25 1 1 0,92 1 0,83 BS Conirostrum speciosum F, inv 0 a 0 a a 0,17 a 0,08 Emberizidae Zonotrichia capensis 0 0,08 0,67 0,17 0 0 0 0 Haplospiza unicolor F, En GRTA - 0,08 - a 0,33 0,17 0,25 0,42 AS
44
Sicalis flaveola 0 0 0,33 0 0 0 0 0 Volatinia jacarina 0 0 0,08 0 0 0 0 0 Sporophila frontalis F, En, CP GRTA - - - - - 0,17 a a AS Sporophila falcirostris F, En, CP GRTA - - - - - a a a Sporophila caerulescens 0,1 0,08 0,25 0,17 0 a 0 0 Arremon taciturnus F, inv 0 0 0 0 0 0,08 0,5 0 continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Coryphospingus pileatus 0 0 0,17 0 0 0 a 0,08 Cardinalidae Caryothraustes canadensis F ONCE 0,1 a a 0,33 0,67 0,67 0,75 0,83 BS Saltator fuliginosus F, En ONCE - - - - 0,33 0,08 0,58 0,33 AS Saltator maximus F, inv 0,2 a 0,75 0,17 0,08 0,33 0,33 0,33 Saltator similis F ONCE 1 1 1 0,75 0,67 0,75 1 0,75 BS Parulidae Parula pitiayumi F, inv 0 0,08 0 a a 0,08 a 0,08 Basileuterus culicivorus F INCE 1 0,67 0,58 1 0,67 0,83 1 0,33 BS Icteridae Psarocolius decumanus F, inv 0,2 0 0 0 0 0 0 0 Gnorimopsar chopi 0 0,08 0 0 0 0 a 0 Molothrus bonariensis 0 0 0 0 0 0 a 0 Fringillidae Euphonia chlorotica F ONCE a 0,25 0,33 a 0,17 0,17 0,17 0,25 BS Euphonia violacea F ONCE 0,4 0,08 0,25 0,08 0,33 0,42 0,5 0,5 BS Euphonia pectoralis F, En ONSB 0,7 0,83 0,58 0,83 1 0,75 0,67 0,75 BS Chlorophonia cyanea F ONCE 0,2 0,5 - 0,17 0,5 0,5 0,5 0,5 BS Passeridae Passer domesticus 0 0 0,58 0 0 0 0 0 Riqueza Estimada (método Jackknife 1) 118 147 167 166 175 208,39 219,6 226,7 Intervalo de Confiança da Riqueza Estimada (±) 8,3 14,4 11,92 12,5 16,4 14,53 11,64 15,55

45
Riqueza total observadae 96 117 134 136 145 180 194 196 Riqueza de espécies florestaisf 71 86 74 101 119 145 156 153 Riqueza de espécies florestais na área-controle (invasoras exclusas) (período 2000-2004) 174 Espécies florestaisf com abundância não alterada 60 69 52 84 117 142 153 continua... (Continuação)
Nome Científicoa Guildab Fragmentos Florestaisc Nível de
sensibilidade d
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Espécies florestaisf com declínio de abundância 8 14 15 14 2 3 0 Espécies florestaisf com aumento de abundância 3 3 7 3 0 0 3 Perda de espécies florestaisf 90 74 97 48 12 2 0 Taxa de extinções (%) 52 42,7 56 27,7 6,9 1,1 0
Total de espécies consideradas de alta sensibilidade ecológica na Mata Atlânticaf 7 10 5 15 23 28 29 33 Total de espécies endêmicas da Mata Atlânticaf 30 38 28 49 58 69 70 75 Total de espécies ameaçadas no Espírito Santof 1 1 2 3 6 9 12 16 Riqueza geral identificada nos fragmentos (jan/2000 - fevereiro/2004) (espécies florestais + não florestais) 108 133 156 66 196 213 236 224
a- A nomenclatura e sequência sistemática seguem a Lista das Aves do Brasil (CBRO, 2005). F- espécie florestal (do interior e borda de mata); En- endemismo da Mata Atlântica; A- espécie de alta sensibilidade ecológica no referido bioma; CP, EP, VU- status de conservação das espécies ameaçadas de extinção no Estado do Espírito Santo; inv- invasora recente na região serrana de Santa Teresa (veja Materiais e Métodos 3.7 para os detalhes de inclusão das espécies nestas categorias). b- Inclui 19 grupos ecológicos funcionais, estabelecidos a partir de guildas tróficas e espaciais combinadas (veja Materiais e métodos 3.3 para a correspondência entre a discriminação das guildas e as siglas utilizadas). c- veja Tabela 1 para tamanho da área, esforço de amostragens e outros detalhes referentes aos fragmentos estudados; os valores (0 a 1) nas colunas correspondem ao Índice de Abundância Relativa (IAR) das espécies, calculado segundo BIBBY et al. (1998): valores em negrito representam IAR com diferença significativa em relação à área-controle (χ2
(correção de Yates), p< 0,05); a letra "a" refere-se ao registro da espécie no fragmento apenas no período de jan/2000 a agosto/2002; o sinal negativo (-) refere-se à perda da espécie no fragmento. d- Nível de sensibilidade ao sistema de fragmentação regional: AS- alta sensibilidade ecológica; BS- baixa sensibilidade ecológica (veja Materiais e Métodos 3.4 para detalhes sobre o critério de inclusão das espécies nestas categorias). e- número total de espécies registradas durante as amostragens padronizadas (set/2002-fev/2004). f- número de espécies florestais (invasoras exclusas) registradas durante as amostragens padronizadas (set/2002-fev/2004).

46
Segundo a Lista Vermelha do Estado do Espírito Santo, o total mencionado ainda inclui 20
espécies (7,6%) ameaçadas de extinção, as quais encontram-se categorizadas em três status
de conservação: vulnerável (VU)- 11 espécies [Penelope obscura (Fig. 3a), Leucopternis
polionotus, Spizaetus tyrannus, Pionopsitta pileata, Strix huhula (Fig. 3b), Nyctibius
aethereus (Fig. 3c), Chelidoptera tenebrosa, Platyrinchus leucoryphus, Phibalura
flavirostris, Neopelma aurifrons e Lanisoma elegans]; em perigo (EP)- 5 espécies
[Odontophorus capueira (Fig. 3d), Touit melanonotus, Glaucidium minutissimum,
Ramphodon naevius e Cichlopsis leucogenys]; criticamente em perigo (CP)- 4 espécies
[Tinamus solitarius (Fig. 3e), Claravis godefrida, Triclaria malachitacea e Sporophila
frontalis (Fig. 3f)] (Tab. 3).
Porém, se aos dados complementares, obtidos no período anterior (jan/2000 - ago/2002),
forem somadas as amostragens padronizadas, as contagens referidas acima assumem os
seguintes valores: 277 (261 + 16) para riqueza geral registrada na área de estudo; 194 (187
+ 7) para o total de espécies florestais; 84 (80 + 4) e 38 (37 + 1), para os endemismos e
elementos de alta sensibilidade ecológica do bioma referido, respectivamente, e; 23 (20 +
3) para as ameaçadas de extinção (incluindo Notharchus swainsoni-CP; Melanerpes
flavifrons-VU e; Sporophila falcirostris-CP) (Tab. 3).

47
a b
c d
e f
Figura 3. Algumas das espécies ameaçadas de extinção no Espírito Santo registradas na área de estudo. a- Penelope obscura; b- Strix huhula; c- Nyctibius aethereus; d- Odontophorus capueira; e- Tinamus solitarius; f- Sporophila frontalis. (Fotografias: a, b, c, f - J.E.Simon; e- Lena Trindade; d- David Ribeiro).

48
4.2. Riqueza estimada de espécies nos fragmentos
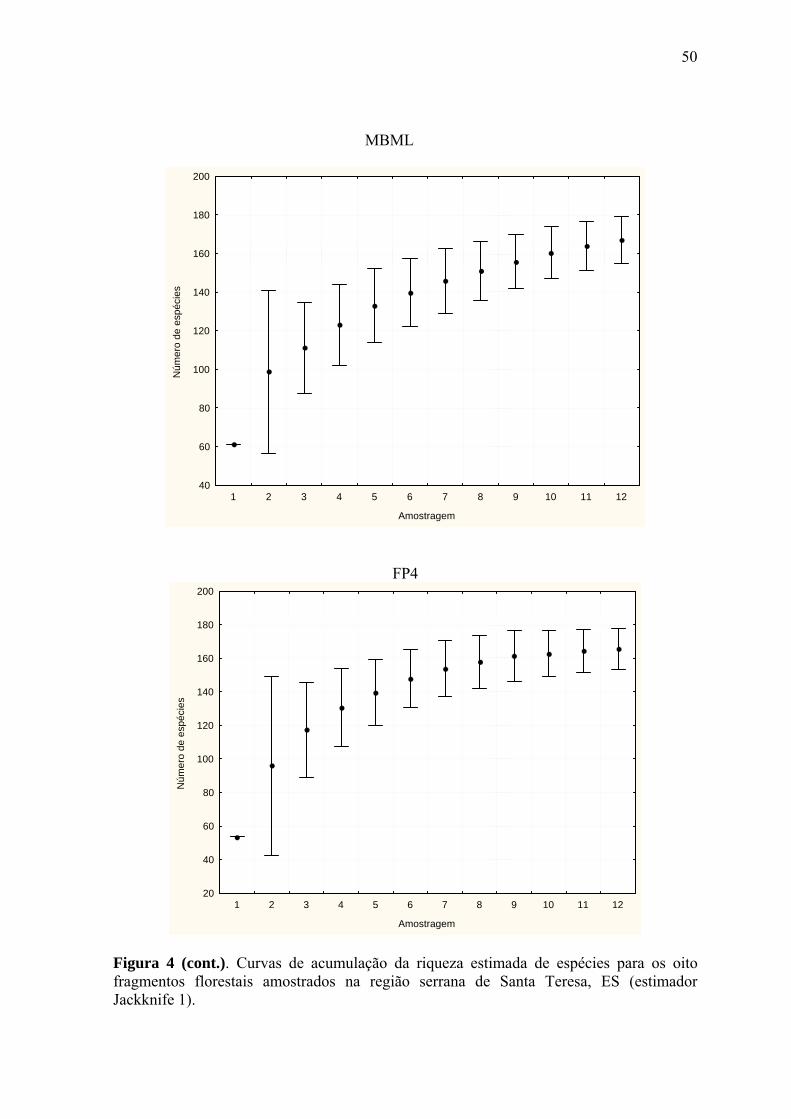
Com base no comportamento assintótico das curvas de acumulação de espécies, geradas
pelo Jackknife 1, observou-se que o método de inventário da avifauna e o esforço de
campo empregados (12 a 18 meses) foram suficientes para a determinação da riqueza de
espécies nos remanescentes selecionados (Fig. 4a-i), tornando válida a comparação entre
eles.
A riqueza estimada de espécies mostrou-se estatisticamente diferenciada entre alguns
grupos de fragmentos amostrados (Fig. 5). De acordo com o valor do seu intervalo de
confiança, os oito fragmentos puderam ser reunidos em cinco grupos distintos, a seguir
relacionados, em ordem crescente do parâmetro avaliado: FP3 (118 spp.; IC95%= ± 8,34),
FP1 (147/±14,39), PMBML/FP4/FM2 (167/±11,92; 165/±12,46; 175/±16,41,
respectivamente), PNMSL/EBSL (208/±14,53) e EBSL/RBAR (219/±11,64; 226/±15,55,
respectivamente) (Fig. 5).
Logo, todos os fragmentos estudados diferiram da área-controle, com exceção da EBSL.
Os menores fragmentos de propriedades rurais (FP1 e FP3) diferiram entre si e de todos os
demais amostrados, enquanto o PMBML, representando o único fragmento situado na área
urbana do município, não diferenciou dos fragmentos FP4 e FM2. A riqueza estimada em
PNMSL foi estatisticamente equivalente à da EBSL.

49
FP3
1 2 3 4 5 6 7 8 9 10 11 12
Amostragem
20
40
60
80
100
120
140
Núm
ero
de e
spéc
ies
FP1
1 2 3 4 5 6 7 8 9 10 11 12
Amostragem
20
40
60
80
100
120
140
160
180
Núm
ero
de e
spéc
ies
Figura 4. Curvas de acumulação da riqueza estimada de espécies para os oito fragmentos florestais amostrados na região serrana de Santa Teresa, ES (estimador Jackknife 1).
50
MBML
1 2 3 4 5 6 7 8 9 10 11 12
Amostragem
40
60
80
100
120
140
160
180
200N
úmer
o de
esp
écie
s
FP4
1 2 3 4 5 6 7 8 9 10 11 12
Amostragem
20
40
60
80
100
120
140
160
180
200
Núm
ero
de e
spéc
ies
Figura 4 (cont.). Curvas de acumulação da riqueza estimada de espécies para os oito fragmentos florestais amostrados na região serrana de Santa Teresa, ES (estimador Jackknife 1).

51
FM2
1 2 3 4 5 6 7 8 9 10 11 12
Amostragem
40
60
80
100
120
140
160
180
200
Núm
ero
de e
spéc
ies
PNMSL
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Amostragem
40
60
80
100
120
140
160
180
200
220
240
Núm
ero
de e
spéc
ies
Figura 4 (cont.). Curvas de acumulação da riqueza estimada de espécies para os oito fragmentos florestais amostrados na região serrana de Santa Teresa, ES (estimador Jackknife 1).

52
EBSL
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Amostragem
60
80
100
120
140
160
180
200
220
240
Núm
ero
de e
spéc
ies
RBAR
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Amostragem
0
20
40
60
80
100
120
140
160
180
200
220
240
260
Núm
ero
de e
spéc
ies
Figura 4 (cont.). Curvas de acumulação da riqueza estimada de espécies para os oito fragmentos florestais amostrados na região serrana de Santa Teresa, ES (estimador Jackknife 1).
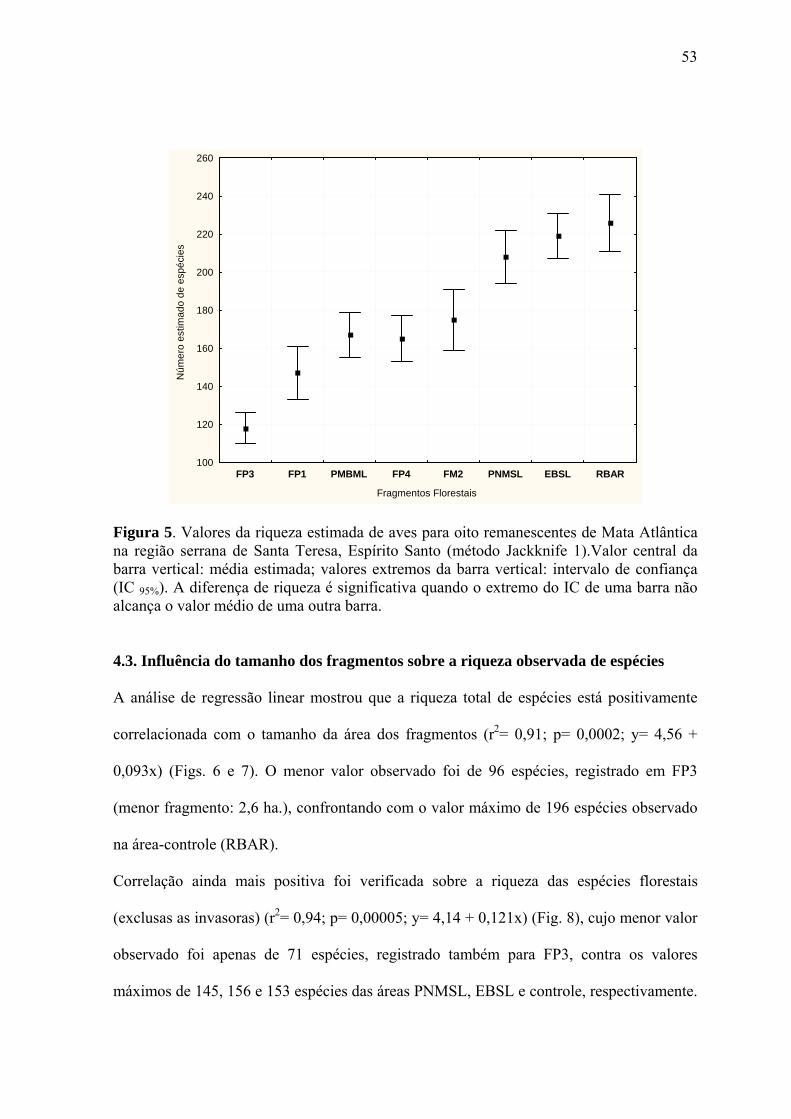
53
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Fragmentos Florestais
100
120
140
160
180
200
220
240
260
Núm
ero
estim
ado
de e
spéc
ies
Figura 5. Valores da riqueza estimada de aves para oito remanescentes de Mata Atlântica na região serrana de Santa Teresa, Espírito Santo (método Jackknife 1).Valor central da barra vertical: média estimada; valores extremos da barra vertical: intervalo de confiança (IC 95%). A diferença de riqueza é significativa quando o extremo do IC de uma barra não alcança o valor médio de uma outra barra.
4.3. Influência do tamanho dos fragmentos sobre a riqueza observada de espécies
A análise de regressão linear mostrou que a riqueza total de espécies está positivamente
correlacionada com o tamanho da área dos fragmentos (r2= 0,91; p= 0,0002; y= 4,56 +
0,093x) (Figs. 6 e 7). O menor valor observado foi de 96 espécies, registrado em FP3
(menor fragmento: 2,6 ha.), confrontando com o valor máximo de 196 espécies observado
na área-controle (RBAR).
Correlação ainda mais positiva foi verificada sobre a riqueza das espécies florestais
(exclusas as invasoras) (r2= 0,94; p= 0,00005; y= 4,14 + 0,121x) (Fig. 8), cujo menor valor
observado foi apenas de 71 espécies, registrado também para FP3, contra os valores
máximos de 145, 156 e 153 espécies das áreas PNMSL, EBSL e controle, respectivamente.
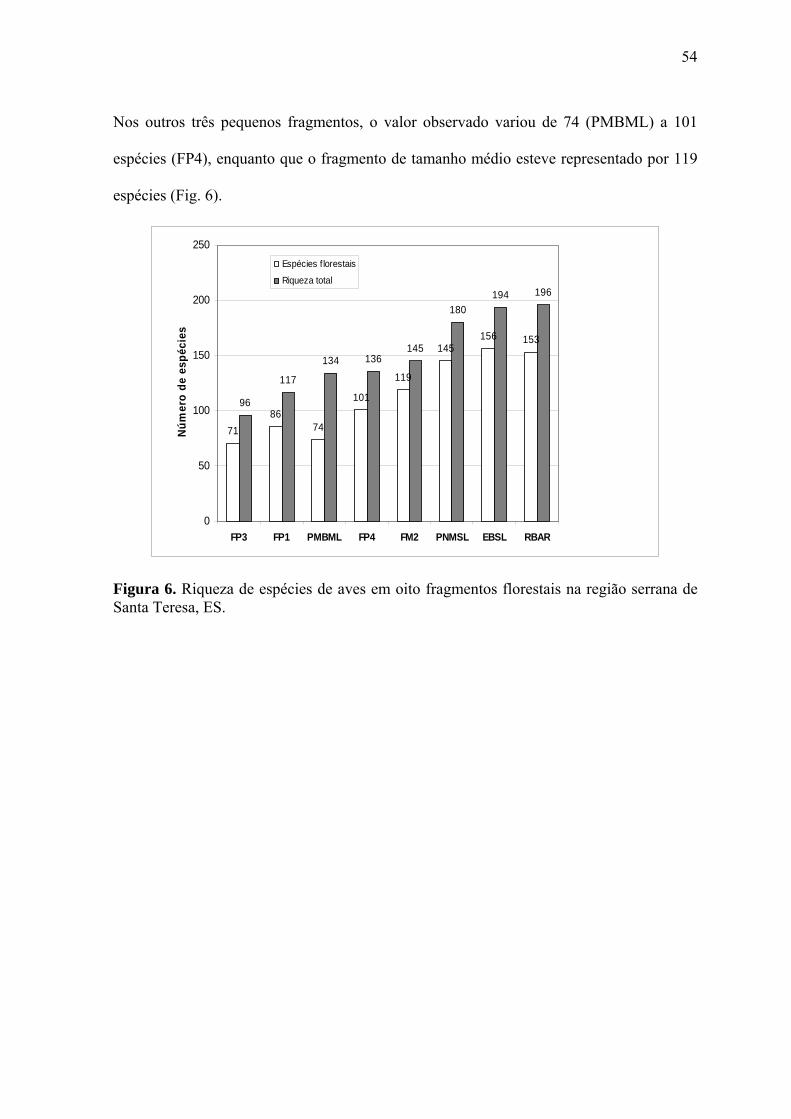
54
Nos outros três pequenos fragmentos, o valor observado variou de 74 (PMBML) a 101
espécies (FP4), enquanto que o fragmento de tamanho médio esteve representado por 119
espécies (Fig. 6).
7186
74
101
119
145156 153
96
117
134 136145
180194 196
0
50
100
150
200
250
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
Espécies f lorestais
Riqueza total
Figura 6. Riqueza de espécies de aves em oito fragmentos florestais na região serrana de Santa Teresa, ES.

55
0,95 2,12 3,54 5,01 6,22 8,18
Área (log)
4,58
4,70
4,89
4,98
5,19
5,27N
úmer
o de
esp
écie
s (lo
g)
r2 = 0,91; p = 0,0002; y = 4,58 + 0,093*x
Figura 7. Variação da riqueza total de aves em função do tamanho da área dos fragmentos florestais amostrados na região serrana de Santa Teresa, ES.
0,95 2,12 3,54 5,01 6,22 8,18
Área (log)
4,26
4,44
4,60
4,77
4,96
5,04
Núm
ero
de e
spéc
ies
(log)
r2 = 0,94; p = 0,00005; y = 4,14 + 0,12x
Figura 8. Variação da riqueza de aves florestais em função do tamanho da área dos fragmentos florestais amostrados na região serrana de Santa Teresa, ES.
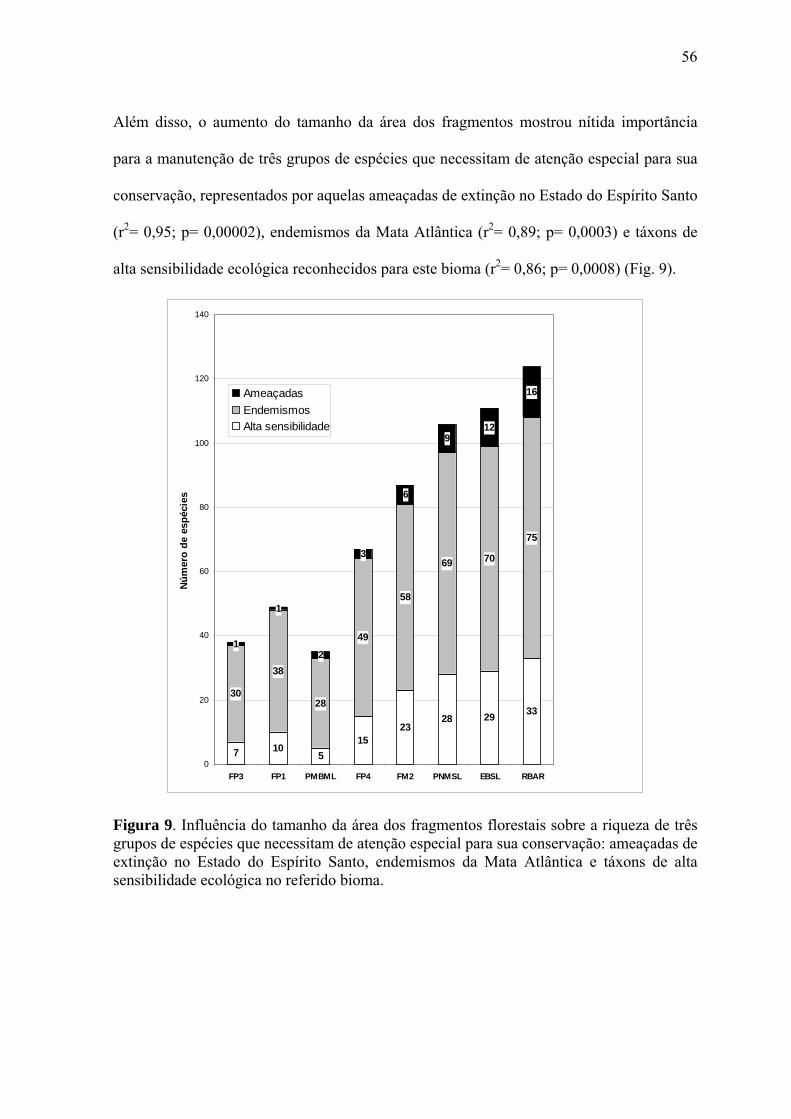
56
Além disso, o aumento do tamanho da área dos fragmentos mostrou nítida importância
para a manutenção de três grupos de espécies que necessitam de atenção especial para sua
conservação, representados por aquelas ameaçadas de extinção no Estado do Espírito Santo
(r2= 0,95; p= 0,00002), endemismos da Mata Atlântica (r2= 0,89; p= 0,0003) e táxons de
alta sensibilidade ecológica reconhecidos para este bioma (r2= 0,86; p= 0,0008) (Fig. 9).
7 105
1523
28 29 33
30
38
28
49
58
69 70
75
1
1
2
3
6
912
16
0
20
40
60
80
100
120
140
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
AmeaçadasEndemismosAlta sensibilidade
Figura 9. Influência do tamanho da área dos fragmentos florestais sobre a riqueza de três grupos de espécies que necessitam de atenção especial para sua conservação: ameaçadas de extinção no Estado do Espírito Santo, endemismos da Mata Atlântica e táxons de alta sensibilidade ecológica no referido bioma.

57
4.4. Influência dos descritores ambientais sobre a riqueza observada das espécies florestais Pela análise de regressão múltipla, verificou-se que o tamanho da área, grau de isolamento
(IS) e permeabilidade da matriz (IP), quando testados em conjunto, excerceram efeito
mútuo significativo sobre a riqueza de espécies florestais na paisagem fragmentada da
região serrana de Santa Teresa (coeficiente de determinação r2= 0,92; p= 0,0022).
Entretanto, entre esses três descritores ambientais, o tamanho da área foi o que melhor
explicou (r2= 0,70; p= 0,0023) a variação no número desse grupo de espécies nos
fragmentos estudados, sendo que os dois outros não mostraram influência individual
significativa sobre a variável dependente analisada (grau de isolamento: p= 0,20;
permeabilidade da matriz: p= 0,25).
Também verificou-se que a persistência de espécies florestais nos fragmentos foi
significativamente dependente do seu estado de conservação (r2= 0,80; p= 0,0024; y=
32,88 + 139,10x) (Fig. 10). Fragmentos com índice de conservação (IC) em torno de 0,30
(FP3, FP1 e PMBML) apresentaram riqueza média de 77 espécies, enquanto que aqueles
com índices acima de 0,60 (PNMSL e EBSL) podem sustentar uma comunidade
semelhante à de um grande bloco de mata contínua (área-controle), cuja riqueza registrada
pelas amostragens padronizadas alcançou 153 espécies (Tab. 3).
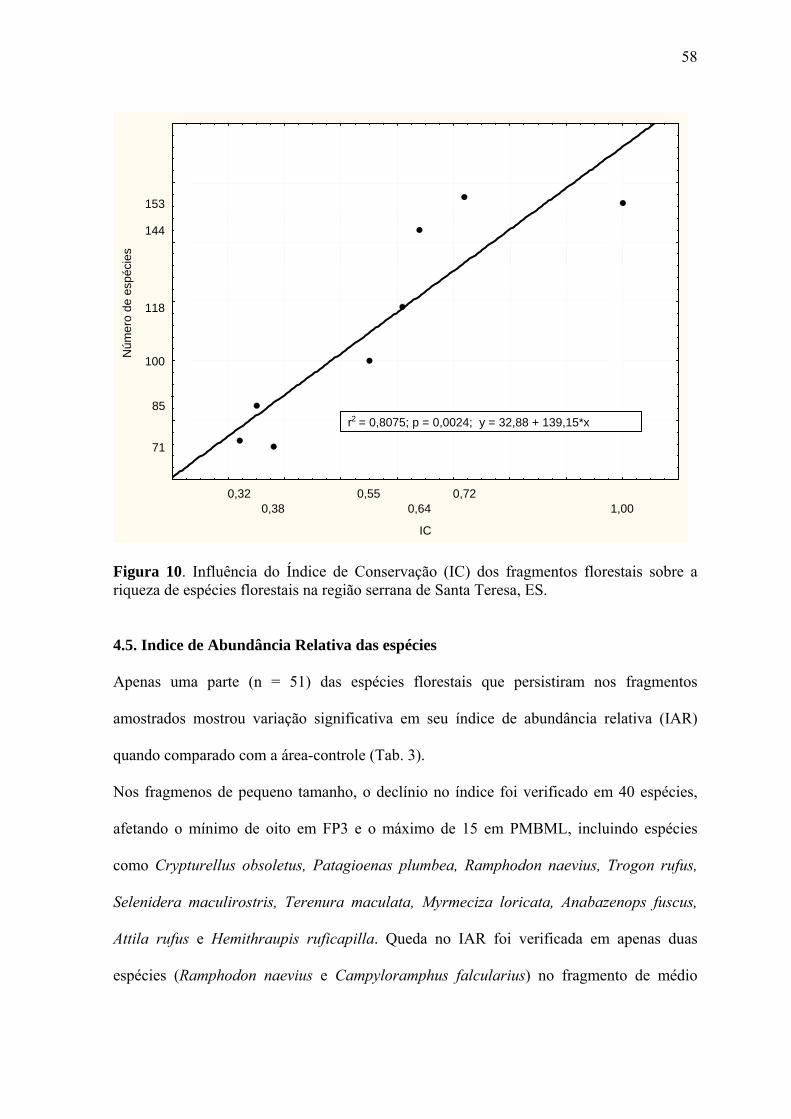
58
0,320,38
0,550,64
0,721,00
IC
71
85
100
118
144
153
Núm
ero
de e
spéc
ies
r2 = 0,8075; p = 0,0024; y = 32,88 + 139,15*x
Figura 10. Influência do Índice de Conservação (IC) dos fragmentos florestais sobre a riqueza de espécies florestais na região serrana de Santa Teresa, ES.
4.5. Indice de Abundância Relativa das espécies
Apenas uma parte (n = 51) das espécies florestais que persistiram nos fragmentos
amostrados mostrou variação significativa em seu índice de abundância relativa (IAR)
quando comparado com a área-controle (Tab. 3).
Nos fragmenos de pequeno tamanho, o declínio no índice foi verificado em 40 espécies,
afetando o mínimo de oito em FP3 e o máximo de 15 em PMBML, incluindo espécies
como Crypturellus obsoletus, Patagioenas plumbea, Ramphodon naevius, Trogon rufus,
Selenidera maculirostris, Terenura maculata, Myrmeciza loricata, Anabazenops fuscus,
Attila rufus e Hemithraupis ruficapilla. Queda no IAR foi verificada em apenas duas
espécies (Ramphodon naevius e Campyloramphus falcularius) no fragmento de médio

59
tamanho (FM2) e em três (Piculus flavigula, Hypoedaleus guttatus e Chamaeza
meruloides) para o grande fragmento PNMSL.
Por outro lado, aumento na abundância relativa foi registrado apenas em 10 espécies para o
conjunto dos fragmentos pequenos, variando entre três em FP3/FP4 e sete em PMBML,
entre elas Leptotila verreauxi, Florisuga fusca, Cranioleuca pallida, Tachyphonus
coronatus e Basileuterus culicivorus. Em relação a outras duas classes de tamanho, apenas
a ESBL (fragmento grande) apresentou espécies com IAR significativamente maior do que
a área-controle (RBAR), verificado para Leptotila verreauxi, Platycichla flavipes e
Basileuterus culicivorus (Tab. 3).
Assim sendo, as maiores porcentagens de espécies com variações em seus índices de
abundância relativa atingiram as comunidades do PMBML (30 %) e FP1 (20%),
correspondendo aos fragmentos para os quais foram atribuídos os menores valores do
índice de conservação (0,32 e 0,35, respectivamente).
4.6. Sensibilidade da avifauna florestal ao sistema de fragmentação regional
Do elenco das espécies florestais analisadas (160 espécies.) (veja Materiais e Métodos 3.4),
60 (37,5%) revelaram-se como de alta sensibilidade à fragmentação do seu hábitat, com
base em sua ausência ou inexpressiva abundância (IAR= 0,08) nos pequenos fragmentos
florestais (< 50 ha). As outras 100 (62,5%) restantes mostraram-se mais tolerantes ao
sistema de fragmentação regional, persistindo inclusive em fragmentos com áreas entre 2,6
e 34,8 ha (0,08 < IAR ≤ 1,0) (Tab. 3).
Pelo teste do χ2 verificou-se que as espécies mais sensíveis à fragmentação do seu hábitat
não estiveram numericamente mais representadas do que aquelas de baixa sensibilidade
entre o total de espécies com maior pré-disposição à extinção na Mata Atlântica, tanto em
relação aos endemismos (χ2(gl= 1, 5%)= 1,36 p= 0,19) quanto aos táxons de alta sensibilidade
ecológica (χ2(gl= 1, 5%)= 2,64 p= 0,10) do referido bioma.

60
Entre as espécies de alta sensibilidade regional, estão, por exemplo, Tinamus solitarius
(Fig. 11a), Leucopternis lacernulatus (Fig. 11b), Triclaria malachitacea (Fig. 11c),
Glaucidium minutissimum (Fig. 11d), Phaethornis squalidus (Fig. 11e), Conopophaga
melanops (Fig. 11f), Grallaria varia (Fig. 11g), Chamaeza meruloides (Fig. 11h), Philydor
atricapillus (Fig. 11i), Lochmias nematura (Fig. 11j), Platyrinchus leucoryphus (Fig. 11l),
Neopelma aurifrons (Fig. 11m), Manacus manacus (Fig. 11n) e Habia rubica (Fig. 11o)
(Tab. 3).

61
a b
c d
Figura 11. Exemplos de espécies reconhecidas como de alta sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. a- Tinamus solitarius; b-Leucopternis lacernulatus; c- Triclaria malachitacea; d- Glaucidium minutissimum (Fotografias: a, b- J. E. Simon; c, d- Pedro Lima).

62
e f
g h
Figura 11 (cont.). Exemplos de espécies reconhecidas como de alta sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. e- Phaethornis squalidus; f- Conopophaga melanops; g- Grallaria varia; h- Chamaeza meruloides. (Fotografias: J. E. Simon).

63
i j
l m
Figura 11 (cont.). Exemplos de espécies reconhecidas como de alta sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. i- Philydor atricapillus; j- Lochmias nematura; l- Xenops minutus; m- Neopelma aurifrons (Fotografias: i - l - J. E. Simon; m- Lena Trindade).

64
n o
Figura 11 (cont.). Exemplos de espécies reconhecidas como de alta sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. n- Manacus manacus; o- Habia rubica. (Fotografias: J. E. Simon).
Espécies que reveleram-se menos sensíveis à fragmentação florestal incluem, por exemplo,
Leptotila rufaxilla (Fig. 12a), Geotrygon montana (Fig. 12b), Ramphodon naevius (Fig.
12c), Thalurania glaucopis (Fig. 12d), Malacoptila striata (Fig. 12e), Dysithamnus
stictothorax (Fig. 12f), Drymophila ferruginea (Fig. 12g), Myrmeciza loricata (Fig. 12h),
Dendrocincla turdina (Fig. 12i), Anabazenops fuscus (Fig. 12j), Automolus
leucophthalmus (Fig. 12l), Platyrinchus mistaceus (Fig. 12m), Attila rufus (Fig. 12n),
Chiroxiphia caudata (Fig. 12o), Tangara cyanoventris (Fig. 12p) e Saltator similis (Fig.
12q) (Tab. 3).

65
a b
c d
Figura 12. Espécies de baixa sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. a- Leptotila rufaxilla; b- Geotrygon montana; c- Ramphodon naevius; d- Thalurania glaucopis. (Fotografias: a, b, d- J. E. Simon; c- acervo biblioteca MBML)

66
e f
g h Figura 12 (cont.). Espécies de baixa sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. e- Malacoptila striata; f- Dysithamnus stictothorax; g- Drymophila ferruginea; h- Myrmeciza loricata. (Fotografias: J. E. Simon).

67
i j l m Figura 12 (cont.). Espécies de baixa sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. i- Dendrocincla turdina; j- Anabazenops fuscus; l- Automolus leucophthalmus; m- Platyrinchus mistaceus. (Fotografias: J. E. Simon).

68
n o p q Figura 12 (cont.). Espécies de baixa sensibilidade ao sistema de fragmentação florestal na região serrana de Santa Teresa, ES. n- Attila rufus; o- Chiroxiphia caudata; p- Tangara cyanoventris; q- Saltator similis. (Fotografias: J. E Simon).

69
4.7. Análise de Similaridade Biológica entre os Fragmentos
O resultado da análise de agrupamento (UPGMA) mostrou que o PMBML foi o fragmento
mais dissimilar em sua composição faunística quando comparado com as demais áreas
amostradas (distância euclidiana: 0,66) (Fig. 13). Os fragmentos de propriedades
particulares formaram um grupo igualmente dissimilar (0,58) em relação às três reservas
florestais (EBSL, PNMSL e RBAR), com FP1/FP3 e FP4/FM2 representando dois
subgrupos com o mesmo grau de similaridade entre cada um de seus fragmentos
componentes (0,44). A maior similaridade faunística foi observada entre o fragmento
PNMSL e a área-controle (0,41), os quais, porém, tiveram forte associação com a EBSL
(0,44) (Fig. 13).
Figura 13. Dendrograma de similaridade (UPGMA) entre a composição da avifauna de oito fragmentos florestais na região serrana de Santa Teresa, ES. A escala refere-se à distância euclidiana entre os fragmentos (fragmentos florestais pequenos: FP1, F3, FP4 e MBML-urbano; fragmento médio: FM2; reservas florestais: EBSL, PNMSL e RBAR).

70
4.8. Taxa de extinções nos fragmentos
Com base no cálculo da taxa de extinções (TE), foi possível verificar que as maiores
perdas de espécies florestais ocorreram nos pequenos fragmentos, onde as taxas variaram
entre 27,7% (FP4) e 56% (PMBML) (Tab. 3).
Entre as espécies que desapareceram de um ou mais fragmentos dessa categoria, estão, por
exemplo, Tinamus solitarius (Fig. 3e, 11a), Crypturellus obsoletus, Penelope obscura
(Fig. 3a), Leptodon cayanensis (Fig. 14a), Leucopternis polionotus, Spizaetus tyrannus
(Fig. 14b), Micrastur ruficollis, Claravis pretiosa, Patagioenas plumbea, Geotrygon
montana, Pionopsitta pileata, Triclaria malachitacea (Fig. 11c), Nyctiphrynus ocellatus,
Selenidera maculirostris (Fig. 14c), Hypoedaleus guttatus, Batara cinerea, Drymophila
ochropyga, Drymophila squamata (Fig. 14d), Myrmeciza loricata (Fig. 12h),
Conopophaga melanops (Fig. 11f), Grallaria varia (Fig. 11g) Chamaeza meruloides (Fig.
11h), Sclerurus scansor, Campylorhamphus falcularius (Fig. 14e), Philydor lichtensteini,
Automolus leucophthalmus (Fig. 12l) Hemitriccus diops (Fig. 14f), Myiornis auricularis,
Carpornis cucullata, Lipaugus lanioides, Piprites chloris, Ilicura militaris, Schiffornis
virescens, Hylophilus poicilotis (Fig. 14g), Habia rubica, Haplospiza unicolor (Fig. 14h) e
Saltator fuliginosus.
A taxa de extinções no fragmento de tamanho médio alcançou 6,9%, abrangendo espécies
como Touit melanonotus, Grallaria varia (Fig.11g), Scytalopus indigoticus,
Myrmotherula gularis, Platyrinchus leucoryphus (Fig.11l), Lipaugus lanioides, Lanisoma
elegans e Sporophila falcirostris (Fig.3f). Nos maiores, foi nula (EBSL) ou alcançou o
valor de 1,1% (PNMSL), representada, neste caso, por Claravis godefrida e Chamaeza
campanisoma.

71
a b
c d
e f
g h Figura 14. Exemplo de espécies extintas em um ou mais dos pequenos fragmentos florestais estudados na região serrana de Santa Teresa, ES. a- Leptodon cayanensis; b- Spizaetus tyrannus; c- Selenidera maculirostris; d- Drymophila squamata; e- Campylorhamphus falcularius; f- Hemitriccus diops; g- Hylophilus poicilotis; h- Haplospiza unicolor (Fotografias: a, c-h: J. E Simon; b- acervo biblioteca MBML).

72
4.9. Resposta das guildas ao processo de fragmentação do seu hábitat
A aplicação da análise multivariada (RA) para o conjunto das 19 guildas analisadas
apontou forte segregação ecológica entre os fragmentos analisados, os quais ficaram
espacialmente reunidos em quatro grupos distintos (Fig. 15). O PMBML foi o mais
dissimilar entre eles, representando sozinho um dos grupos. Os demais enquadraram-se nos
três outros grupos restantes, conforme a seguinte combinação: FP1/FP3/FP4, FM2/PNMSL
e EBSL/RBAR (Fig. 15). Desta forma, os pequenos fragmentos rurais compartilharam
comunidades de grupos ecológicos funcionais estruturalmente diferenciadas daquela do
pequeno fragmento urbano (PMBML). O PNMSL assemelhou-se mais ao FM2 do que
com os outros dois grandes blocos de mata, representados pela EBSL e RBAR (área-
controle).

73
Figura 15. Dissimilaridade entre oito fragmentos florestais estudados na região serrana de Santa Teresa, ES, em função da composição de 19 grupos ecológicos funcionais avaliados (guildas tróficas espaciais combinadas) (análise de ordenação Reciprocal Averaging) (autovalor 1: 0,32; autovalor 2: 0,27). Com exceção dos nectarívoros, todas as guildas analisadas apresentaram significativa (ou
marginalmente significativa) perda de espécies com a redução do tamanho do seu hábitat
(0,47 < r2 < 0,94; 0,0002 < p < 0,057) (Tab. 4) (Fig. 16).
Quando comparadas com a área-controle, sete delas, representadas pelos carnívoros
diurnos (Fig. 16a), carnívoros noturnos (Fig. 16a), granívoros de taquaral (Fig. 16b),
insetívoros do solo (Fig. 16c), insetívoros de margem de córrego (Fig. 16c), insetívoros do
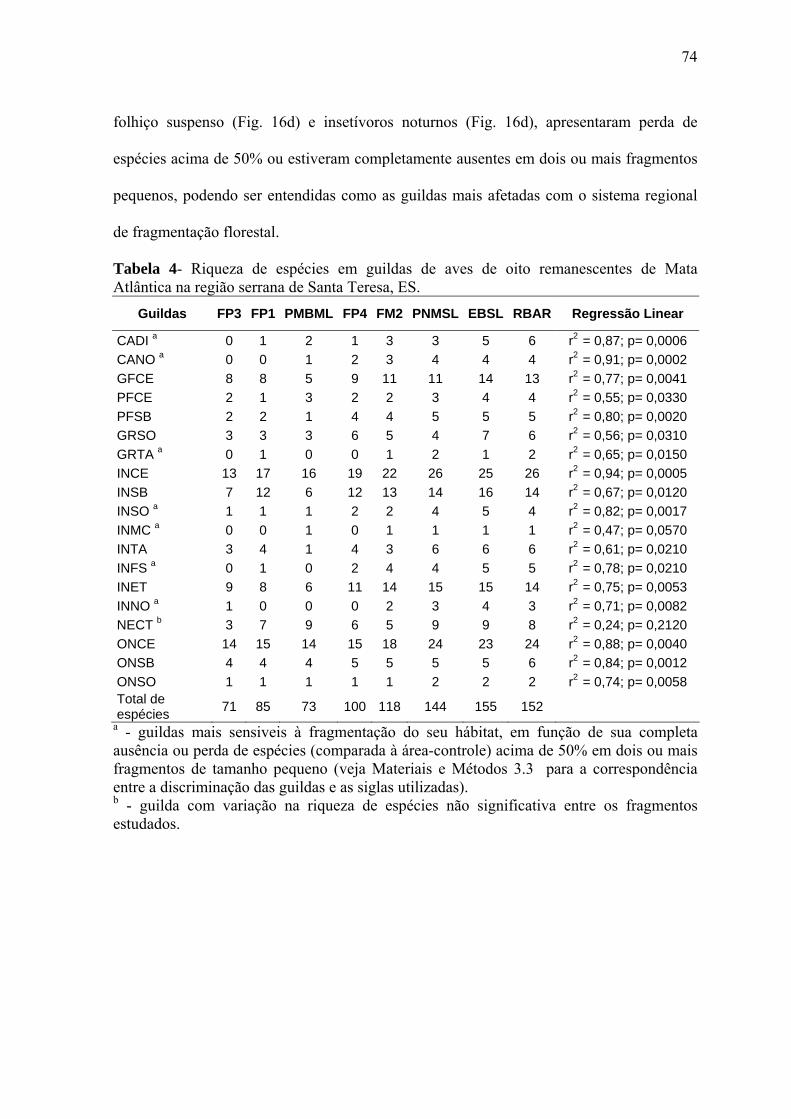
74
folhiço suspenso (Fig. 16d) e insetívoros noturnos (Fig. 16d), apresentaram perda de
espécies acima de 50% ou estiveram completamente ausentes em dois ou mais fragmentos
pequenos, podendo ser entendidas como as guildas mais afetadas com o sistema regional
de fragmentação florestal.
Tabela 4- Riqueza de espécies em guildas de aves de oito remanescentes de Mata Atlântica na região serrana de Santa Teresa, ES.
Guildas FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR Regressão Linear
CADI a 0 1 2 1 3 3 5 6 r2 = 0,87; p= 0,0006 CANO a 0 0 1 2 3 4 4 4 r2 = 0,91; p= 0,0002 GFCE 8 8 5 9 11 11 14 13 r2 = 0,77; p= 0,0041 PFCE 2 1 3 2 2 3 4 4 r2 = 0,55; p= 0,0330 PFSB 2 2 1 4 4 5 5 5 r2 = 0,80; p= 0,0020 GRSO 3 3 3 6 5 4 7 6 r2 = 0,56; p= 0,0310 GRTA a 0 1 0 0 1 2 1 2 r2 = 0,65; p= 0,0150 INCE 13 17 16 19 22 26 25 26 r2 = 0,94; p= 0,0005 INSB 7 12 6 12 13 14 16 14 r2 = 0,67; p= 0,0120 INSO a 1 1 1 2 2 4 5 4 r2 = 0,82; p= 0,0017 INMC a 0 0 1 0 1 1 1 1 r2 = 0,47; p= 0,0570 INTA 3 4 1 4 3 6 6 6 r2 = 0,61; p= 0,0210 INFS a 0 1 0 2 4 4 5 5 r2 = 0,78; p= 0,0210 INET 9 8 6 11 14 15 15 14 r2 = 0,75; p= 0,0053 INNO a 1 0 0 0 2 3 4 3 r2 = 0,71; p= 0,0082 NECT b 3 7 9 6 5 9 9 8 r2 = 0,24; p= 0,2120 ONCE 14 15 14 15 18 24 23 24 r2 = 0,88; p= 0,0040 ONSB 4 4 4 5 5 5 5 6 r2 = 0,84; p= 0,0012 ONSO 1 1 1 1 1 2 2 2 r2 = 0,74; p= 0,0058 Total de espécies 71 85 73 100 118 144 155 152
a - guildas mais sensiveis à fragmentação do seu hábitat, em função de sua completa ausência ou perda de espécies (comparada à área-controle) acima de 50% em dois ou mais fragmentos de tamanho pequeno (veja Materiais e Métodos 3.3 para a correspondência entre a discriminação das guildas e as siglas utilizadas). b - guilda com variação na riqueza de espécies não significativa entre os fragmentos estudados.
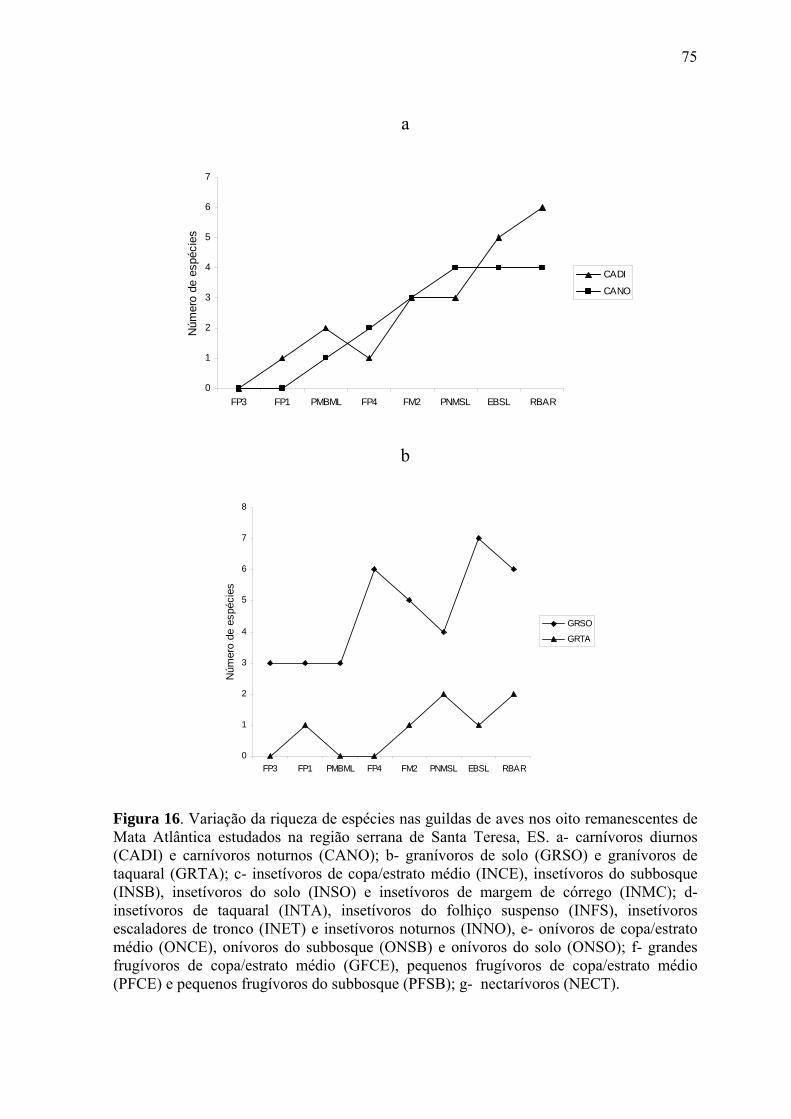
75
a
0
1
2
3
4
5
6
7
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
CADI
CANO
b
0
1
2
3
4
5
6
7
8
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
GRSO
GRTA
Figura 16. Variação da riqueza de espécies nas guildas de aves nos oito remanescentes de Mata Atlântica estudados na região serrana de Santa Teresa, ES. a- carnívoros diurnos (CADI) e carnívoros noturnos (CANO); b- granívoros de solo (GRSO) e granívoros de taquaral (GRTA); c- insetívoros de copa/estrato médio (INCE), insetívoros do subbosque (INSB), insetívoros do solo (INSO) e insetívoros de margem de córrego (INMC); d- insetívoros de taquaral (INTA), insetívoros do folhiço suspenso (INFS), insetívoros escaladores de tronco (INET) e insetívoros noturnos (INNO), e- onívoros de copa/estrato médio (ONCE), onívoros do subbosque (ONSB) e onívoros do solo (ONSO); f- grandes frugívoros de copa/estrato médio (GFCE), pequenos frugívoros de copa/estrato médio (PFCE) e pequenos frugívoros do subbosque (PFSB); g- nectarívoros (NECT).

76
c
0
5
10
15
20
25
30
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
INCE
INSB
INSO
INMC
d
0
2
4
6
8
10
12
14
16
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
INTA
INFS
INET
INNO
Figura 16 (cont.). Variação da riqueza de espécies nas guildas de aves nos oito remanescentes de Mata Atlântica estudados na região serrana de Santa Teresa, ES. a- carnívoros diurnos (CADI) e carnívoros noturnos (CANO); b- granívoros de solo (GRSO) e granívoros de taquaral (GRTA); c- insetívoros de copa/estrato médio (INCE), insetívoros do subbosque (INSB), insetívoros do solo (INSO) e insetívoros de margem de córrego (INMC); d- insetívoros de taquaral (INTA), insetívoros do folhiço suspenso (INFS), insetívoros escaladores de tronco (INET) e insetívoros noturnos (INNO), e- onívoros de copa/estrato médio (ONCE), onívoros do subbosque (ONSB) e onívoros do solo (ONSO); f- grandes frugívoros de copa/estrato médio (GFCE), pequenos frugívoros de copa/estrato médio (PFCE) e pequenos frugívoros do subbosque (PFSB); g- nectarívoros (NECT).

77
e
0
5
10
15
20
25
30
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
ONCE
ONSB
ONSO
f
0
2
4
6
8
10
12
14
16
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
GFCE
PFCE
PFSB
Figura 16 (cont.). Variação da riqueza de espécies nas guildas de aves nos oito remanescentes de Mata Atlântica estudados na região serrana de Santa Teresa, ES. a- carnívoros diurnos (CADI) e carnívoros noturnos (CANO); b- granívoros de solo (GRSO) e granívoros de taquaral (GRTA); c- insetívoros de copa/estrato médio (INCE), insetívoros do subbosque (INSB), insetívoros do solo (INSO) e insetívoros de margem de córrego (INMC); d- insetívoros de taquaral (INTA), insetívoros do folhiço suspenso (INFS), insetívoros escaladores de tronco (INET) e insetívoros noturnos (INNO), e- onívoros de copa/estrato médio (ONCE), onívoros do subbosque (ONSB) e onívoros do solo (ONSO); f- grandes frugívoros de copa/estrato médio (GFCE), pequenos frugívoros de copa/estrato médio (PFCE) e pequenos frugívoros do subbosque (PFSB); g- nectarívoros (NECT).

78
g
0
1
2
3
4
5
6
7
8
9
10
FP3 FP1 PMBML FP4 FM2 PNMSL EBSL RBAR
Núm
ero
de e
spéc
ies
NECT
Figura 16 (cont.). Variação da riqueza de espécies nas guildas de aves nos oito remanescentes de Mata Atlântica estudados na região serrana de Santa Teresa, ES. a- carnívoros diurnos (CADI) e carnívoros noturnos (CANO); b- granívoros de solo (GRSO) e granívoros de taquaral (GRTA); c- insetívoros de copa/estrato médio (INCE), insetívoros do subbosque (INSB), insetívoros do solo (INSO) e insetívoros de margem de córrego (INMC); d- insetívoros de taquaral (INTA), insetívoros do folhiço suspenso (INFS), insetívoros escaladores de tronco (INET) e insetívoros noturnos (INNO), e- onívoros de copa/estrato médio (ONCE), onívoros do subbosque (ONSB) e onívoros do solo (ONSO); f- grandes frugívoros de copa/estrato médio (GFCE), pequenos frugívoros de copa/estrato médio (PFCE) e pequenos frugívoros do subbosque (PFSB); g- nectarívoros (NECT). 4.10. Espécies florestais não detectadas em campo
Um total de 48 espécies florestais de ocorrência conhecida para a região em estudo
(literatura científica e coleções científicas, vide Materiais e Métodos 3.8) não foi registrado
pelo autor do presente estudo em seus sete anos de trabalhos de campo na região serrana de
Santa Teresa (1998-2004) (Simon 2000, Simon 2004 e dados do presente estudo/2000-
2004) (Tab. 5). Desse total, 29 espécies correspondem a elementos alóctones (vagantes,
migratórios altitudinal/continental ou invasores recentes), cujos registros para a região
mostram-se escassos nas fontes consultadas. As outras 19 espécies, tratando-se,
aparentemente, de elementos residentes, incluem 16 táxons com registros relativamente
recentes na literatura e três outras, Pipile jacutinga, Spizaetus ornatus e Orchesticus

79
abeillei, representadas apenas por registros históricos (1949-1971), a partir de material
taxidermizado do acervo ornitológico do MBML e DZUFMG. Entre estas 19 espécies,
nove estão na lista das ameaçadas de extinção no Espírito Santo (Tab. 5): Pipile jacutinga,
Harpia harpyja, Spizastur melanoleucus, Spizaetus ornatus, Touit surdus, Discosura
langsdorffi, Machaeropterus regulus, Nemosia rourei e Phaeothlypis rivularis.
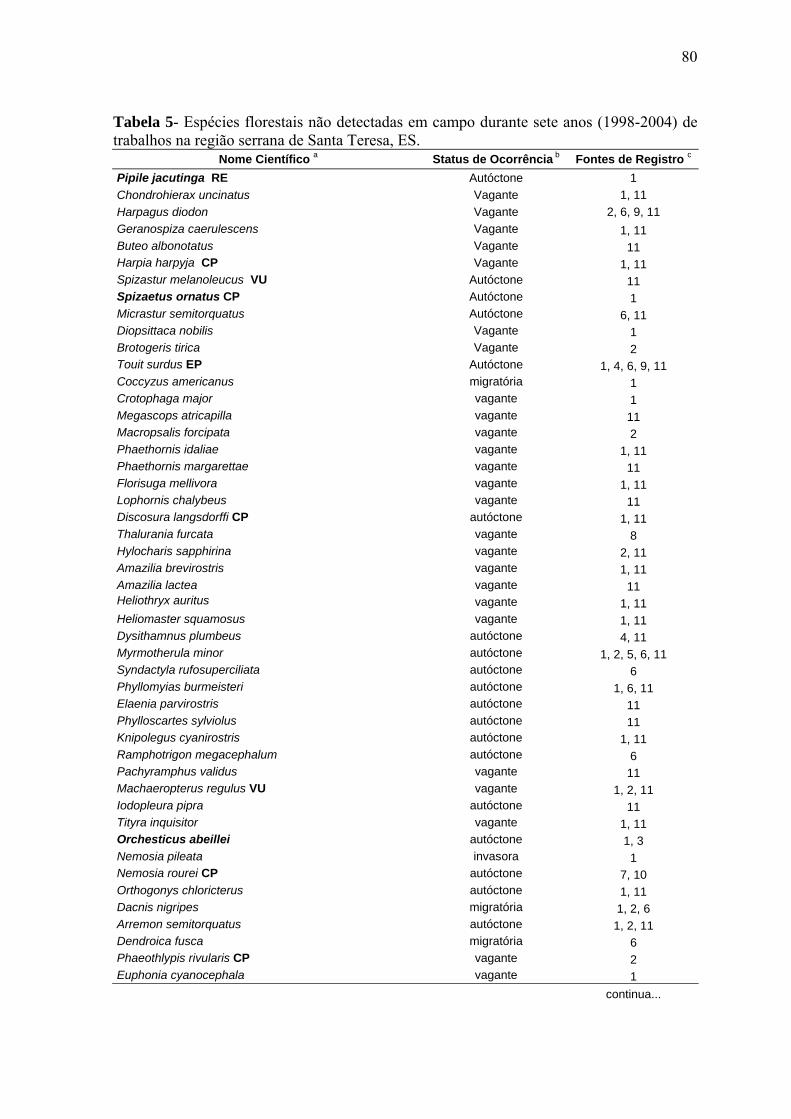
80
Tabela 5- Espécies florestais não detectadas em campo durante sete anos (1998-2004) de trabalhos na região serrana de Santa Teresa, ES.
Nome Científico a Status de Ocorrência b Fontes de Registro c Pipile jacutinga RE Autóctone 1 Chondrohierax uncinatus Vagante 1, 11 Harpagus diodon Vagante 2, 6, 9, 11 Geranospiza caerulescens Vagante 1, 11 Buteo albonotatus Vagante 11 Harpia harpyja CP Vagante 1, 11 Spizastur melanoleucus VU Autóctone 11 Spizaetus ornatus CP Autóctone 1 Micrastur semitorquatus Autóctone 6, 11 Diopsittaca nobilis Vagante 1 Brotogeris tirica Vagante 2 Touit surdus EP Autóctone 1, 4, 6, 9, 11 Coccyzus americanus migratória 1 Crotophaga major vagante 1 Megascops atricapilla vagante 11 Macropsalis forcipata vagante 2 Phaethornis idaliae vagante 1, 11 Phaethornis margarettae vagante 11 Florisuga mellivora vagante 1, 11 Lophornis chalybeus vagante 11 Discosura langsdorffi CP autóctone 1, 11 Thalurania furcata vagante 8 Hylocharis sapphirina vagante 2, 11 Amazilia brevirostris vagante 1, 11 Amazilia lactea vagante 11 Heliothryx auritus vagante 1, 11 Heliomaster squamosus vagante 1, 11 Dysithamnus plumbeus autóctone 4, 11 Myrmotherula minor autóctone 1, 2, 5, 6, 11 Syndactyla rufosuperciliata autóctone 6 Phyllomyias burmeisteri autóctone 1, 6, 11 Elaenia parvirostris autóctone 11 Phylloscartes sylviolus autóctone 11 Knipolegus cyanirostris autóctone 1, 11 Ramphotrigon megacephalum autóctone 6 Pachyramphus validus vagante 11 Machaeropterus regulus VU vagante 1, 2, 11 Iodopleura pipra autóctone 11 Tityra inquisitor vagante 1, 11 Orchesticus abeillei autóctone 1, 3 Nemosia pileata invasora 1 Nemosia rourei CP autóctone 7, 10 Orthogonys chloricterus autóctone 1, 11 Dacnis nigripes migratória 1, 2, 6 Arremon semitorquatus autóctone 1, 2, 11 Dendroica fusca migratória 6 Phaeothlypis rivularis CP vagante 2 Euphonia cyanocephala vagante 1 continua...

81
Continuação (Tab.5) a- Nome de táxons em negrito: ocorrência apenas com base em registro histórico (P. jacutinga: 1971; S. ornatus: 1949; O. abeillei: 1941, 1965). As siglas VU, EP e CP correspondem ao status de conservação da espécie no Estado do Espírito Santo (IPEMA 2005a). b- vide Materiais e Métodos (item 3.8) para a inclusão das espécies em autóctones ou alóctones (vagante, migratória ou invasora recente) para a região em estudo. c- Fontes de registro (dados secundários): 1- Coleção científica MBML; 2- Coleção científica MNRJ; 3- Coleção científica DZUFMG; 4- COLLAR et al. 1992; 5- WHITNEY & PACHECO 1995; 6- PARKER III & GOERCK 1997; 7- SCOTT 1997; 8- ONIKI & WILLIS 1998; 9- WILLIS & ONIKI 1998; 10- BAUER et al. 2000; 11- WILLIS & ONIKI 2002.

82
5. DISCUSSÃO
5.1. Composição da avifauna florestal na paisagem fragmentada de Santa Teresa
Os efeitos da fragmentação florestal sobre comunidades de aves neotropicais têm sido
intensamente explorados há pelo menos três décadas, através de estudos inicialmente
desenvolvidos na Amazônia central e na porção sudeste da Mata Atlântica, demonstrando a
resposta das espécies ao processo de redução e isolamento de seu hábitat (DEBINSKI &
HOLT 1999, LAURENCE et al. 2000, LAURENCE et al. 2002, HAGAN et al. 1996,
WATSON 2002).
Atualmente, várias regiões da Mata Atlântica já foram submetidas a esse tipo de análise
por diferentes autores, que, indistintamente, mostraram a dramática fragilidade da avifauna
à redução e alteração da qualidade do ambiente florestal, cuja reação à força do impacto,
porém, é bastante variável entre suas espécies (e.g. WILLIS 1979, ALEIXO &
VIELLIARD 1995, ALEIXO 1999, MALDONADO-COELHO & MARINI 2000, ANJOS
2001, RIBON et al. 2003, ANJOS et al. 2004, ANTUNES 2005, DEVELEY 2004).
Em Viçosa (MG), por exemplo, onde uma consistente análise foi conduzida a partir de
base histórica de registros ornitológicos e expressivo número de fragmentos florestais
amostrados por 15 anos, RIBON et al. (2003) verificaram um declínio maciço de espécies,
representando 60,7% da avifauna florestal da região estudada (28 extinções e 68 espécies
em vias de extinção). Casos semelhantes também foram verificados para o interior do
Estado de São Paulo, com o completo desaparecimento de 30 e 20 espécies para a região
de Campinas (ALEIXO & VIELLIARD 1995) e Anhembi (ANTUNES 2005),
respectivamente.
Embora vários autores tenham estudado a composição da avifauna em remanescentes
florestais do Município de Santa Teresa (PARKER III & GOERCK 1997, SIMON 2000,

83
WILLIS & ONIKI 2002, etc), os dados do presente estudo correspondem à primeira
avaliação dos efeitos da fragmentação florestal sobre as aves da região serrana do Espírito
Santo.
Com base na compilação de dados bibliográficos e museológicos disponíveis para a região
(dados secundários), foi possível verificar que 48 espécies florestais que dela constam não
foram detectadas em campo durante nossos sete anos de pesquisa na região (1998-2004)
(SIMON 2000, SIMON 2004, presente estudo e dados não publicados), a exemplo de
Pipile jacutinga, Harpagus diodon, Harpia harpyja, Dysithamnus plumbeus, Syndactyla
rufosuperciliata, Iodopleura pipra, Nemosia rourei e Arremon semitorquatus (Tabela 5).
A razão para a falta de registros dessas espécies nos levantamentos de campo do presente
estudo pode estar associada a vários fatores, como extinção local, raridade natural,
sazonalidade ou efeito de amostragem (BIBBY et al. 1998).
Muitas dessas espécies são inconspícuas em seu ambiente, podendo, de fato, não terem
sido detectadas em função do efeito de amostragem (e.g. Coccyzus americanus,
Phaethornis margarettae, Myrmotherula minor, Ramphotrigom megacephala, Orchesticus
abeillei e Phaeothlypis rivularis). Várias outras podem ser vagantes ou ocorrer em baixa
densidade, considerando alguns registros esporádicos para a região (e.g. Harpia harpyja,
Megascops atricapilla, Iodopleura pipra, Nemosia rourei e Arremon semitorquatus)
(WHITNEY & PACHECO 1995, PARKER III & GOERCK 1997, SCOTT 1997, WILLIS
& ONIKI 2002, PACHECO et al. 2003).
Das 11 espécies florestais admitidas por WILLIS & ONIKI (2002) como localmente
extintas (Pipile jacutinga, Harpia harpyja, Claravis godefrida, Melanerpes flavifrons,
Anabacerthia amaurostis, Laniisoma elegans, Pyroderus scutatus, Orthogonys
chlorocterus, Orchesticus abeillei, Cyanocompsa brissonii e Sporophila falcirostris), sete
foram por nós redescobertas durante nossos trabalhos na região (1998-2004) (C.

84
godefrida, M. flavifrons, A. amaurostis, L. elegans, P. scutatus, C. brissonii e Sporophila
falcirostris). Entre tais espécies, destacam-se dois endemismos da Mata Atlântica e
globalmente ameaçadas de extinção (PARKER III et al. 1996, MMA 2003), Claravis
godefrida e Sporophila falcirostris, e que em Santa Teresa eram conhecidos apenas por
registros históricos (C. godefrida: MBML/1943, MNRJ/1956; S. falcirostris:
MBML/1942) (SIMON 2004). Ambas foram registradas entre novembro de 2003 e março
de 2004, quando taquaras nativas frutificaram na região, chamando a atenção para a
necessidade de inventários intensivos e de longa duração para o registro de aves raras da
Mata Atlântica (SIMON 2004).
Dessa forma, talvez o único caso plausível de extinção local seja Pipile jacutinga, uma ave
de grande porte que historicamente sempre sofreu pressão de caça em toda a sua área de
distribuição geográfica, existindo relatos de dezenas de indivíduos abatidos em uma única
caçada (SICK 1997). Possivelmente, a caça pode ter sido o motivo principal de sua
extinção na região de Santa Teresa (SIMON & CAZOTTO 2002), embora a expressiva
redução do ambiente florestal do entorno do Município de Santa Teresa possa também ter
contribuído para o seu declínio populacional, por impedir que a espécie viesse a realizar
seus movimentos altitudinais, associados aos períodos de frutificação de suas plantas
preferenciais (cf. GALLETI & PIZO 1996, SICK 1997).
Se, de fato, Pipile jacutinga tratar-se da única espécie localmente extinta, o sistema de
fragmentação florestal (escala da paisagem) verificado para a região serrana de Santa
Teresa não ocasionou efeitos drásticos sobre a composição da avifauna ali ocorrente.
Dessa forma, a verificação da quase totalidade de sua avifauna original nos dias atuais
deve estar associada à condição da sua cobertura vegetal, distribuída por 40% (ca. 22.000
ha) da área do município, parte da qual representada por três unidades de conservação

85
(Estação Biológica de Santa Lúcia, Parque Natural Municipal de São Lourenço e Reserva
Biológica Augusto Ruschi) (MENDES & PADOVAN 2000, SIMON 2000).
Com base apenas nos dados de campo do presente estudo (2000-2004), os remanescentes
florestais dessa região abrangem o total de 277 espécies, das quais 194 são elementos
florestais (do interior e borda de mata), além de 84 endemismos da Mata Atlântica e 38
espécies consideradas de alta sensibilidade ecológica no referido bioma (PARKER III et
al. 1996). Nela também estão presentes 23 espécies ameaçadas no Estado do Espírito
Santo (IPEMA 2005a) (Tab. 3).
Considerando, porém, os dados secundários compilados (bibliografia e museus) (Tab. 5),
outros 16 táxons florestais residentes (excluso um caso de extinção local/Pipile jacutinga e
dois casos de desaparecimento que aguardam melhor investigação/Spizaetus
ornatus/Orchesticus abellei) ainda ocupam a região, somando 210 espécies que persistem
nas florestas serranas do Município de Santa Teresa: uma rica comunidade de aves
florestais não documentada em nenhum outro estudo realizado em paisagens fragmentadas
da Mata Atlântica (cf. WILLIS 1979, ALEIXO & VIELLIARD 1995, ANJOS et al. 1997,
CHRISTIANSEN & PITTER 1997, BORNSCHEIN & REINERT 2000, ANJOS 2001,
CHIARELLO 2000a, RIBON 1998, ANTUNES 2005, DEVELEY 2004).
Vale ressaltar, porém, que nas terras baixas do município, onde praticamente não mais
existem remanescentes representativos de mata nativa, WILLIS & ONIKI (2002)
documentaram acentuada perda de espécies de aves florestais, hoje substituídas por
avifauna típica de paisagens abertas, incluindo vários elementos invasores de zonas
desmatadas.

86
5.2. Riqueza de espécies nos fragmentos
Fundamentada na Teoria da Biogeografia de Ilhas (MACARTHUR & WILSON 1967), a
dinâmica da diversidade de espécies em remanescentes de hábitats naturais vem sendo
testada nos trabalhos sobre os efeitos da fragmentação florestal em várias regiões do
planeta. A relação da riqueza de espécies com o tamanho da área dos fragmentos tem sido
o principal foco das análises da maioria desses trabalhos, cujos resultados normalmente
evidenciam forte associação entre esse parâmetro da comunidade e a variável ambiental
considerada (BOECKLEN & GOTELLI 1984, ROSENFIELD et al. 1992, DEBINSKI &
HOLT 2000, WATSON 2002). Entretanto, várias outras características da paisagem são
dadas como determinantes da diversidade biológica em hábitats fragmentados,
especialmente a permeabilidade da matriz, efeitos de borda, grau de isolamento
(conectividade), conformação e heterogeneidade ambiental dos remanescentes florestais
(BOECKLEN 1986, LAURENCE 1999, ALEIXO 2002, METZGER 1999, RENJIFO
2001, DEVELEY 2004).
Nesse estudo, a riqueza de espécies foi analisada segundo três principais descritores
ambientais, representados pela área dos fragmentos, seu grau de isolamento e a
permeabilidade da matriz. Desses três descritores, porém, a área foi o único que explicou a
variação da riqueza observada de espécies entre os fragmentos amostrados na região de
Santa Teresa, tanto em relação à riqueza total (espécies florestais + não-florestais) quanto
em relação à riqueza das espécies dependentes do ambiente florestal (do interior e borda
de mata), para a qual a associação foi positivamente mais significativa. Embora admitido
como um importante fator para a organização das comunidades biológicas em paisagens
fragmentadas (VAN DORP & OPDAM 1987, METZGER 1999, RENJIFO 2001,
LAURENCE et al. 2001, UEZU et al. 2005), o grau de conectividade (grau de isolamento
+ permeabilidade da matriz) testado em outros estudos realizados na Mata Atlântica nem

87
sempre explicou a riqueza de aves nos fragmenos florestais (e.g. RIBON 1998, 2003,
LAPS et al. 2003, DEVELEY 2004). RIBON (1998), por exemplo, testou quatro variáveis
da paisagem, verificando que o tamanho dos fragmentos foi a que teve efeito direto sobre
a riqueza de espécies, sendo que o grau de isolamento, topografia e vegetação, ainda que
importantes, apresentaram influência menor e indireta, levando este autor a concluir que as
“ilhas de hábitat” (fragmentos florestais) funcionam como “ilhas verdadeiras” (ambientes
insulares) para a avifauna da Mata Atlântica.
A falta de relação da riqueza de espécies florestais com os dois outros descritores
ambientais testados (grau de isolamento e permeabilidade da matriz) pode ser explicada
pela disposição espacial dos fragmentos, cuja proximidade entre eles deve favorecer a
conectividade funcional para muitas espécies, em vista da porcentagem da cobertura
vegetal remanescente (ca. 40%), dando suporte à idéia de que o isolamento dos fragmentos
tem pouco efeito sobre a diversidade biológica em áreas que ainda mantêm cerca de 30%
da extensão de sua vegetação original, como verficado para a região de Viçosa, Minas
Gerais (RIBON 1998). Porém, vários estudos realizados na Mata Atlântica sugerem ou até
confirmam (ALEIXO & VIELLIARD 1995, UEZU et al. 2005, ANJOS 2006) que muitas
espécies estritamente florestais possuem pouca aptidão para atravessar os “poros da
matriz” (sensu GASCON et al. 1999), indicando que a conectividade estrutural da
paisagem nem sempre significa conectividade funcional para a avifauna do referido bioma
(UEZU et al. 2005).
Ao contrário das pesquisas na Amazônia central, onde os experimentos foram manipulados
pela criação de fragmentos com grau de isolamento, forma geométrica e escala de tamanho
planejados (1, 10, 100, 1000 ha) (LAURENCE et al. 2001), os estudos desenvolvidos na
Mata Atlântica adotaram a conformação resultante do processo histórico da fragmentação
florestal (sistema realista), muitas vezes desacompanhada de efetivo isolamento,

88
explicando porque os resultados obtidos nesta região algumas vezes não coincidem com
aqueles verificados para outros sistemas estudados. Dessa forma, a conectividade funcional
entre manchas de hábitats parece um componente bastante complexo para comparações
concludentes dos efeitos da fragmentação entre diferentes regiões, uma vez que ela
depende não somente da distribuição espacial dos fragmentos, mas também da habilidade
das espécies em deslocar-se através do ambiente do seu entorno (matriz) (SIEVING et al.
1996), sendo que as espécies aptas a passar pelos poros menores desta matriz (por
exemplo, pastagens) são as mais capacitadas a persistir em paisagens fragmentadas
(MALCOLM 1991, LAURENCE 1990, 1991, 1994).
Vários estudos realizados na região de florestas tropicais têm demonstrado uma íntima
relação entre a perda da riqueza de aves e a redução do tamanho do hábitat, incluindo os
dados da Mata Atlântica (WILLIS 1979, ANJOS & BOÇON 1999, ANJOS 2001, RIBON
1998, LAPS 2003, DEVELEY 2004). A exceção para esta região foi encontrada por
MARSDEN et al. (2001), trabalhando com fragmentos de mata de tabuleiro no norte do
Estado do Espírito Santo (Linhares/Sooretama). Entretanto, esses autores analisaram seus
dados sem distinção entre espécies florestais e não-florestais (as quais estiveram bem
representadas nos fragmentos menores), fragilizando a compreensão do efeito-área sobre a
avifauna dependente do ambiente florestal. O curto período das amostragens de campo
empregado por MARSDEN et al. (2001) (agosto-outubro/1998) pode também ter
influenciado seus resultados, uma vez que o universo das 111 espécies por eles amostradas
não incluiu muitas outras citadas para a região, para a qual são conhecidas cerca de 250
espécies (SCOTT & BROOKE 1985, STOTZ 1993, PARKER III & GOERCK 1997,
SIMÃO et al. 1999).
De fato, há informações na literatura que mostram que os resultados contrários à tendência
teórica esperada (isto é, a falta de relação entre a riqueza de espécies e o tamanho do

89
hábitat) é mais evidente entre comunidades representadas por espécies abundantes, vágeis
e generalistas de hábitat (TURNER 1996, DEBINSKI & HOLT 1999, LAURENCE 1999).
Além disso, a invasão de remanescentes de hábitats naturais por espécies oriundas das
paisagens do entorno é outra causa da equivalência de riqueza entre diferentes fragmentos
florestais, cuja composição original, porém, pode encontrar-se completamente depauperada
(ÅS 1999). Por esta razão, a riqueza de espécies, por si só, tem se mostrado pouco efetiva
para a compreensão da importância dos ambientes naturais para a conservação da sua
diversidade biológica (VIEIRA et al. 2003). De fato, o PMBML, situado na área urbana de
Santa Teresa, mostrou uma riqueza estimada de espécies semelhante a FP4 e FM2, apesar
de sua composição faunística ser extremamente diferenciada daquela verificada para a
área-controle (RBAR) (Item 5.2).
Sabe-se que grupos faunísticos respondem diferentemente aos efeitos da redução de seu
hábitat, podendo um mesmo grupo taxonômico apresentar respostas muito divergentes
entre uma região e outra, conforme já apropriadamente apontado em recente revisão feita
por DEBINSKI & HOLT (1999). Segundo estes autores, essas divergências estão
relacionadas com as diferenças existentes entre os sistemas regionais de fragmentação e os
impactos associados, como efeitos de borda, alteração nas taxas de predação no interior
dos fragmentos e conectividade da matriz, além da escala espacial analisada, que difere de
estudo para estudo.
O outro único estudo que tratou dos efeitos da fragmentação sobre a fauna de vertebrados
na região de Santa Teresa foi realizado por PASSAMANI (2003). Trabalhando com
comunidade de pequenos mamíferos (marsupiais e roedores), este autor verificou uma
riqueza de espécies semelhante (não significativa) entre as diferentes classes de tamanho
de fragmentos e as áreas-controle, diferindo, portanto, do resultado do presente estudo.
Seus resultados foram justificados com base na conectivadade funcional entre

90
remanescentes florestais, comprovada em seu estudo pela ocupação da matriz por muitas
das espécies registradas nos fragmentos florestais, que em sua maioria coincidiu com
aqueles selecionados pelo presente estudo. Logo, os resultados divergentes na riqueza de
aves e mamíferos versus o tamanho da área dos fragmentos na região de Santa Teresa
parecem relacionados com diferenças ecológicas entre estes dois grupos animais, como a
estrutura de suas comunidades e aptidão das espécies em explorar a matriz para transitar
entre os fragmentos. No caso dos pequenos mamíferos, PASSAMANI (2003) detectou o
total de 20 espécies para o conjunto dos fragmentos estudados (versus 261 espécies de
aves nas amostragens padronizadas deste estudo), muitas das quais interpretadas pelo
autor como organizadas em um sistema de metapopulações, que contribui para a
manutenção da diversidade faunística em manchas de hábitat de uma paisagem
fragmentada (HANSKI 1991, HANSKI & SIMBERLLOF 1997).
O sistema de metapopulações parece válido também para parte da comunidade de aves
florestais na região estudada, considerando que muitas de suas espécies (ca. 50%) foram
igualmente registradas em todas as classes de fragmentos, algumas das quais comumente
observadas translocando-se entre os fragmentos estudados e os seus vizinhos, por vezes
envolvendo distâncias superiores a 200 m (vide Fig. 2). Entre estas espécies, estavam, por
exemplo, Pyrrhura frontalis, Trogon viridis, Ramphastos vitellinus, Dryocopus lineatus,
Colonia colonus, Tangara cyanoventris, Caryosthraustes canadensis e Saltator similis,
que em sua maioria são típicas do estrato médio e superior da floresta e/ou com aptidão
para vôo fora do ambiente florestal.
Ao contrário, muitas outras espécies dificilmente afastam-se do interior da floresta
(fotofóbicas), mesmo em situação de proximidade com outros remanescentes florestais,
conforme demonstrado por UEZU et al. (2005) com experimentos de play-back,
reforçando o fato de que os elementos de uma comunidade biológica reagem distintamente

91
ao processo de fragmentação do seu hábitat (e.g. JOHNS 1991, SIMBERLOFF 1995,
LAURENCE et al. 1997, LAURENCE et al. 2001).
Muitas causas têm sido sugeridas para explicar a diminuição da riqueza de espécies com a
redução do hábitat. Uma das explicações mais comuns é a de que muitas espécies são
perdidas na área remanescente devido à diminuição na heterogeneidade ambiental, que
afeta a disponibilidade de recursos (fontes de alimento, microhábitats de forrageamento,
locais de nidificação, etc) ou induz o aumento na densidade de mesopredadores
(mesopredator release) e competitores potenciais (SIMBERLOFF & ABELE 1982,
SOULÉ et al. 1988, TERBORGH 1992b, BOECKLEN 1986, MALCOLM 1994,
GASCON et al. 1999, GASCON et al. 2001). Tal explicação está de acordo com os
resultados obtidos na região serrana de Santa Teresa, onde a riqueza de espécies também
esteve positivamente correlacionada com o índice de conservação dos fragmentos
amostrados, notando-se, por exemplo, que o fragmento urbano (PMBML/IC = 0,32)
apresentou menor número de espécies florestais do que um fragmento da área rural de
mesmo tamanho (FP1/IC= 0,35). Com efeito, o grau de conservação dos fragmentos
também mostrou importância sobre a riqueza de espécies na região estudada, tendo como
base alguns fatores ambientais conjuntamente analisados (por exemplo, presença de
microhábitats e forma dos fragmentos), denominados na literatura freqüentemente de
qualidade do fragmento, em oposição ao tamanho de sua área, que seria sua quantidade
(VIEIRA et al. 2003).
Entre os sete fragmentos avaliados, o mais alto valor em número absoluto de espécies
detectadas durante as amostragens padronizadas foi identificado na Estação Biológica de
Santa Lúcia (EBSL) (194 espécies), representando um bloco de aproximadamente 800 ha,
quando considerada sua área legal (440 ha) em continuidade com as florestas particulares
do seu entorno (MENDES & PADOVAN 2000, PASSAMANI 2003). Em termos de

92
riqueza total estimada pelo método Jackknife, a EBSL não diferiu da área-controle,
indicando que aquela reserva assume a mesma importância para a conservação da
diversidade regional que o maior bloco de mata contínua da região, representado pela
Reserva Biológica Augusto Ruschi (RBAR), com cerca de 4.000 ha (MENDES &
PADOVAN 2000, SIMON 2000).
Embora tenha apresentado riqueza estimada inferior à RBAR (área-controle), o Parque
Natural Municipal de São Lourenço (PNMSL, 504 ha) não diferiu da EBSL, evidenciando
que aquela unidade de conservação, ainda que de menor tamanho e associada à área urbana
do município, detém a quase totalidade da avifauna regional, incluindo 145 espécies
dependentes do ambientes florestal (versus 153 na RBAR). Logo, o fato de remanescentes
florestais com cerca de 500 ha abrigarem boa parte da diversidade regional parece explicar
porque para outras localidades serranas do Espírito Santo, também detentoras de porções
significativas de matas nativas (como Domingos Martins, Santa Maria de Jetibá e
Conceição do Castelo) (IPEMA 2005b), ainda são verificados elevados níveis de riqueza
de aves (e de outros grupos animais, veja Introdução), incluindo táxons globalmente
ameaçados (por exemplo, Cichlopsis leucogenys e Nemosia rourei) e raros endemismos da
Mata Atlântica (por exemplo, Merulaxis ater e Thripophaga macroura) (BAUER 1999,
VENTURINI et al. 2000, BAUER et al. 2001). Vale ressaltar, porém, que esta hipótese
não parece valer para outras regiões da Mata Atlântica, considerando que no sul da Bahia a
integridade da avifauna regional só é mantida em fragmentos acima de 2.000 ha (LAPS et
al. 2003).
Mesmo sob caráter de proteção integral, o Parque do Museu de Biologia Mello Leitão
(PMBML), completamente inserido na área urbana do município, apresentou riqueza de
134 espécies (amostragens padronizadas), das quais apenas 74 tratavam-se de espécies
florestais, implicando na ausência de até 51% da riqueza registrada na RBAR (153

93
espécies.). O PMBML foi também estudado por WILLIS & ONIKI (2002), que
mencionaram a mesma porcentagem de perda para este fragmento, com base em dados
secundários (coleções) e de seus levantamentos assistemáticos (1974, 1989-1997)
realizados em várias localidades de Santa Teresa. De fato, a falta de parcelas significativas
(~ 50%) da riqueza original de espécies nos pequenos remanescentes florestais (<100 ha)
tem sido frequentemente verificado nos trabalhos desenvolvidos em florestas tropicais
(TURNER 1996, TURNER & CORLETT 1996, STRATFORD & STOUFFER 1999,
PATTERSON & ATMAR 2000, LAPS et al. 2003), parecendo também ser um caso
generalizado para a avifauna da Mata Atlântica, conforme apontam vários outros estudos
desenvolvidos nesta região (WILLIS 1979, D´ANGELO NETO et al. 1998, ANJOS 2004,
RIBON 1998, RIBON 2003).
5.3. Variação no índice de abundância das espécies nos fragmentos
Prever quais espécies tendem a desaparecer mais rapidamente com a destruição de seus
hábitats é de particular interesse aos programas de conservação da biodiversidade, sendo
que flutuações no tamanho de suas populações são entendidas como um dos melhores
preditores do risco de extinção de biotas regionais (TERBORGH & WINTER 1980,
PIMM et al. 1988, DAVIES et al. 2000, ANJOS 2004).
Em seu estudo, ROBISON (2001), por exemplo, verificou a redução de até 50 % nos
valores de abundância de 37 espécies florestais 25 anos após Edwin Willis ter amostrado
as populações de aves na ilha de Barra Colorado, criada após a construção do canal do
Panamá (WILLIS 1980). Afora outras 11 espécies que se tornaram extremamente raras, 27
do total das 200 espécies residentes sofreram extinções locais, enquanto outras 15 espécies
aumentaram em 100% ou mais a sua densidade na ilha.

94
Além de ROBISON (2001), muitos outros estudos que avaliaram a abundância de aves em
fragmentos de florestas tropicais demostraram que a população de várias delas costumam
sofrer declínios abruptos provocados por causas diversas, podendo tornar-se raras ou
desaparecer por completo do local onde originalmente ocorriam (DIAMOND 1972,
SOULÉ et al. 1979¸ WILLIS 1979, LOVEJOY et al. 1984, ALEIXO & VIELLIARD
1995, LAURENCE et al. 1997, ROBISON 2001, RIBON et al. 2003, ANTUNES 2005).
Outras, por outro lado, podem se beneficiar do processo de fragmentação por um aumento
no tamanho de suas populações, modificando a estrutura da comunidade no hábitat
remanescente, um fenômeno denominado densidade compensatória (MACARTHUR et al.
1972, WRIGHT 1980).
Tais tipos de mundança na estrutura da comunidade foram também verificados para a
avifauna da região serrana de Santa Teresa, onde 51 espécies florestais registradas nos
fragmentos amostrados sofreram aumentos ou declínios em seus índices de abundância
relativa quando comparados com a área-controle. Notavelmente, os menores fragmentos
apresentaram mais espécies com alterações em seus índices do que aqueles de maior
tamanho (FM2, PNMSL, EBSL). Por exemplo, das 71 espécies florestais registradas no
menor fragmento (2,6ha), 11 (15%) sofreram variações em sua abundância relativa, das
quais 3 apresentaram índices mais elevados (Leptotila verreauxi, Cranioleuca pallida e
Basileuterus culicivorus), enquanto as demais (Crypturellus obsoletus, Patagioenas
plumbea, Trogon rufus, Myrmeciza loricata, Anabazenops fuscus, Attila rufus, Oxyruncus
cristatus e Caryosthraustes canadensis) demonstraram índíces significativamente menores
do que os da área-controle.
Mas foram os fragmentos FP1 e PMBML (para os quais foram atribuídos os menores
valores do índice de conservação) que apresentaram as comunidades com maiores
porcentagens de espécies com variações em seus índices de abundância (20% e 30%,

95
respectivamente), concordando com os resultados obtidos por ANJOS (2001) na região de
Londrina, norte do Paraná, onde o maior número de espécies com flutuações
populacionais também ocorreu em fragmentos com maior grau de comprometimento
ambiental (tamanhos pequenos e mais isolados). Em seu estudo, ANJOS (2001) verificou
alterações na abundância relativa de 63 espécies quando comparando os dados de suas
amostragens entre diferentes tamanhos de fragmentos de Mata Atlântica; desse total, 35
espécies apresentaram o efeito da densidade compensatória com o decréscimo da área dos
fragmentos, incluindo espécies como Crypturellus tataupa, Leptotila verreauxi,
Thamnophilus caerulescens, Dysithamnus mentalis, Conopophaga lineata, Lathrotriccus
euleri, Tolmomyias sulphurescens, Trichothraupis melanops e Saltator similis. Segundo
ANJOS e BOÇON (1999) e ANJOS (2004), este fenômeno é freqüente nas espécies que
persistem nos fragmentos florestais, onde podem aumentar em abundância em função do
declínio ou extinção de populações de outras espécies, em acordo com a hipótese de
apropriação de hábitat defendida por BLONDEL et al. (1988). Por outro lado,
Crypturellus obsoletus, Pyrrhura frontalis, Melanerpes flavifrons, Sittasomus
griseicapillus, Xiphocolaptes albicollis, Chiroxiphia caudata, Piprites chloris, Myiopagis
caniceps, entre outras, estiveram entre as mais afetadas com a redução do tamanho do seu
hábitat.
Comparando os dados obtidos para a Mata Atlântica por diferentes autores, incluindo o
presente estudo, é possível notar que várias espécies apresentam padrões semelhantes de
declínio ou aumento de abundância em fragmentos florestais. Embora utilizando método
diferente de amostragem, o presente estudo detectou variação no índice de abundância das
mesmas espécies que sofreram flutuações populacionais para mais (Leptotila verreauxi,
Cranioleuca pallida, Basileuterus culicivorus, Tachyphonus coronatus, etc) ou para
menos (Tinamus solitarius, Crypturellus obsoletus, Myrmeciza loricata, Automolus

96
leucophthalmus, etc) em outras regiões, demonstrando que os efeitos da fragmentação
florestal sobre avifaunas regionais são mais determinísticos do que estocásticos (CUTLER
1991).
Porém, algumas outras espécies têm mostrado respostas diferenciadas entre as regiões
analisadas, indicando que os impactos do processo de fragmentação podem operar de
maneira bastante particular sobre suas respectivas comunidades biológicas. Tomando-se
estudos que utilizaram o mesmo método de amostragem (contagens por ponto), verifica-
se, por exemplo, que Habia rubica, Dysithamnus mentalis e Chiroxiphia caudata, todas
restritas ao subbosque da mata, tiveram padrões distintos de abundância entre as regiões
de Londrina (PR) (ANJOS 2001), Viçosa (MG) (RIBON 1998) e São Paulo (SP)
(DEVELEY 2004, UEZU et al. 2005), relacionados, possivelmente, aos sistemas
particulares de fragmentação investigados. De fato, algumas das espécies evidenciaram
respostas bem particulares para as matas de Santa Teresa, em função dos valores estáveis
em seus índices entre os sete fragmentos amostrados, contrastando com os dados de outras
regiões, onde apresentaram crítico declínio em abundância mesmo em fragmentos de
maior tamanho (e.g. RIBON 1998), a exemplo de algumas espécies estritamente
dependente do ambiente florestal, como Conopophaga lineata, Thamnophilus
caerulescens e Pyriglena leucoptera (exceção para o PMBML, onde a abundância dessa
ultima espécie foi significativamente menor). Com efeito, o processo de fragmentação
ocorrido nas matas serranas de Santa Teresa não implicou em influências tão severas para
as populações de determinadas espécies que desapareceram ou tornaram-se raras em
outras paisagens fragmentadas da Mata Atlântica (Crypturellus obsoletus, Geotrygon
montana, Hypoedaleus guttatus, Grallaria varia, Philydor atricapillus, Platyrinchus
mistaceus, Lipaugus lanioides, Hemitriccus diops, etc) (ALEIXO & VIELLIARD 1995,
RIBON et al. 2003, ANTUNES 2005).

97
Além das idiossincrasias de cada espécie (raridade natural, sucesso reprodutivo,
sensibilidade a distúrbios ambientais, etc), outras causas comumente citadas para explicar
os efeitos gerados sobre as populações confinadas em fragmentos da Mata Atlântica estão
relacionadas a fatores extrínsecos (redução do hábitat e efeitos de borda) e intrínsecos
(interações ecológicas e estrutura da vegetação) dos remanescentes florestais, incluindo a
impermeabilidade da matriz, alterações microclimáticas, pressão de caça, invasão de
espécies, modificação nas taxas de competição interespecífica, apropriação de hábitat,
oferta de alimento, exclusão de territórios, perda de microhábitats de forrageamento e
nidificação (WILLIS 1979, ALEIXO & VIELLIARD 1995, CHRISTIANSEN & PITTER
1997, D`ANGELO NETO et al. 1998, RIBON 1998, ANJOS & BOÇON 1999,
MALDONADO-COELHO & MARINI 2000, ANJOS 2001, LAPS et al. 2003, RIBON
2003, RIBON et al. 2003, ANJOS 2004, ANJOS et al. 2004, DEVELEY 2004,
ANTUNES 2005, UEZU et al. 2005).
Na região de Viçosa, MG, por exemplo, a menor abundância de Campylorhamphus
falcularius e Drymophila ochropyga esteve relacionada à ausência de taquarais utilizados
no interior dos fragmentos como locais de forrageamento, assim como para Dysithamnus
mentalis e Conopophaga lineata, em relação a grotas úmidas para a construção de seus
ninhos (RIBON 2003, obs. pes.). Enquanto que o declínio de Habia rubica em fragmentos
da região de São Paulo foi relacionado à desestruturação de bandos mistos, para os quais é
considerada uma espécie nuclear (DEVELEY & PERES 2000, DEVELEY 2004).
O presente estudo não teve o objetivo de conhecer os fenômenos responsáveis pelas
variações do índice de abundância das espécies (relações causa-efeito), mas é bem
possível que as causas citadas acima possam ser aplicadas à região de Santa Teresa. Por
exemplo, o aumento na abundância de três espécies de beija-flores (Florisuga mellivora,
Lophornis magnificus e Leucochloris albicollis) no Parque do Museu de Biologia Mello

98
Leitão pode ser um resultado direto da oferta de alimento diariamente disponível no local
(bebedouros com açúcar). Da mesma maneira, a raridade de Tinamus solitarius no Parque
Natural Municipal de São Lourenço (PNMSL) deve estar intimamente associada à pressão
de caça, considerando sua diferente situação na Estação Biológica de Santa Lúcia e na
Reserva Biológica Augusto Ruschi. Além de Tinamus solitarius tratar-se de uma espécie
de grande valor cinergético no Estado do Espírto Santo (CHIARELLO 2000b, SIMON &
CHIARELLO 2001) e no próprio Município de Santa Teresa (SIMON & CAZOTTO,
2002), o parque em questão, apenas recentemente transformado em unidade de
conservação (2004), não possui até o momento qualquer sistema de fiscalização autônoma,
sendo frequente o registro de atividades de caça (poleiros, tiros, etc) neste remanescente
florestal (obs. pes.).
Outra causa igualmente importante, porém, pouco enfatizada em estudos com aves em
fragmentos da Mata Atlântica, parece ser o aumento na densidade de mesopredadores
(release predator), provacado pelo desaparecimento de consumidores de níveis mais
elevados da cadeia alimentar (por exemplo, grandes carnívoros), resultando em maiores
taxas de predação sobre outras espécies da comunidade (SOULÉ et al. 1988),
TERBORGH 1992b, MALCOLM 1994). Em recente experimento conduzido no Parque
de Biologia Mello Leitão, BARBINI & PASSAMANI (2005) verificaram queda
significativa no sucesso de sobrevivência de ninhos artificiais posicionados no solo e
estrato inferior da vegetação, cuja causa foi atribuída ao aumento da densidade do
marsupial Didelphis aurita no local estudado. Em concordância com o resultado destes
autores, J. E. SIMON (obs. pes.) observou por algumas vezes D. aurita predando ninhos
de algumas aves neste mesmo fragmento florestal (Glausis hirsuta, Leptotila rufaxilla,
Conopophaga lineata, Turdus rufiventris), incluindo o ninho de Geotrygon montana

99
(novembro de 1999), não mais ali detectada nos anos subseqüentes dos trabalhos de
campo deste estudo (2000 e 2004).
Deve-se ressaltar, entretanto, que várias das idéias mencionadas para explicar o declínio
ou aumento nas populações das espécies em fragmentos florestais da Mata Atlântica (e de
outros biomas) estão fundamentadas em observações fortuitas de autores diversos,
exigindo que as mesmas ainda sejam cientificamente testadas, como recentemente feito
com relação à dependência das espécies de Formicariidae (latu sensu) e Dendrocolaptidae
por microhábitats específicos do interior da mata (RIBON 1998, POLETTO 2004). Dessa
forma, a melhor compreensão das causas relacionadas às flutuações populacionais da
avifauna em fragmentos da Mata Atlântica ainda limita-se à extrema falta de dados sobre a
história natural das espécies, perda da diversidade genética nas populações isoladas e
intrincada rede de interações ecológicas que ocorrem entre as mesmas (ALEIXO 2001,
LAPS et al. 2003, RIBON 2003, DEVELEY 2004, ANTUNES 2005, UEZU et al. 2005).
5.4. Sensibilidade da avifauna ao processo de fragmentação florestal
O grau de sensibilidade de aves tem sido o foco de muitos estudos realizados em áreas de
florestas tropicais, em vista da crescente necessidade de se compreender a vulnerabilidade
das comunidades bióticas submetidas à intensa e contínua destruição ambiental. Em
conjunto, tais estudos demonstram que as espécies mais sensíveis são aquelas que
apresentam determinados atributos de suscetibilidade, como especificidade de
hábitat/microhábitat, baixa densidade populacional (raridade natural), distribuição restrita
(endemismo), maior tamanho corporal, territórios amplos, associação com bandos mistos,
nidificação no solo, fraca afinidade com a matriz que circunda o ambiente florestal e
exposição à pressão de caça. Em razão disso, tais espécies estão entre as primeiras a sofrer
os impactos do processo de fragmentação do seu hábitat (LECK 1979, LAURENCE 1990,

100
CAUGHLEY 1994, SISK et al. 1994, LAWTON 1995, TURNER 1996, LAURENCE et
al. 1997, SIEVING & KAR 1997, MANNE & PIMM 2001, UEZU et al. 2005).
No presente estudo, foram reconhecidas 60 espécies de alta sensibilidade para a região
serrana de Santa Teresa, todas elas caracterizadas por um ou mais atributos de
suscetibilidade mencionados na literatura, a exemplo de Tinamus solitarius (fraca
afinidade com a matriz/pressão de caça/nidificação no solo), Triclaria malachitacea
(endemismo de paisagem/baixa densidade), Grallaria varia (especificidade de
hábitat/baixa densidade), Lochmias nematura (especificidade de microhabitat) e Habia
rubica (componente de bando misto). Contudo, os mesmos atributos são encontrados em
espécies que mostraram-se mais tolerantes ao processo de fragmentação regional,
incluindo algumas registradas no fragmento menos conservado entre aqueles estudados, no
caso, o Parque do Museu de Biologia Mello Leitão (por exemplo, Pyrrhura frontalis,
Ramphodon naevius, Malacoptila striata, Dysithamnus stictothotorax, Chiroxiphia
caudata e Tangara cyanoventris).
Em seu estudo, ANJOS (2006) mostrou que alguns dos atributos de suscetibilidade a
distúrbios ambientais não estiveram claramente relacionados com os níveis de
sensibilidade em comunidade de aves na região de Londrina, norte do Paraná. Foi o caso,
por exemplo, de três espécies (Crypturellus obsoletus, Geotrygon montana e Penelope
superciliaris) com características esperadas para alta suscetibilidade (maior tamanho e/ou
hábitos terrícolas), mas que exibiram baixa sensibilidade em sua região. Enquanto que
outras três com as mesmas características foram consideradas de alta sensibilidade
(Tinamus solitarius, Crypturellus undulatus e Grallaria varia).
Comparando seus dados com aqueles disponíveis para Viçosa (RIBON et al. 2003),
ANJOS (2006) também notou que a maior parte das espécies com mesmo nível de
sensibilidade entre as duas regiões envolveu táxons tolerantes à fragmentação florestal,

101
verificando, por conseguinte, fraca congruência para espécies de alta sensibilidade. As
particularidades das metodologias aplicadas para cada região não foram consideradas por
ANJOS (2006). Entretanto, as variações nos níveis de sensibilidade das espécies entre
Londrina e Viçosa foram explicadas por este autor com base nas diferenças entre sistemas
regionais de fragmentação analisados (histórico da fragmentação, tamanho dos
remanescentes, etc), bem como pela existência de padrões diferenciados de abundância do
centro para a periferia da área de distribuição geográfica das espécies, como também
defendidas por TERBORGH & WINER (1980) e KATTAN et al. (1994).
Os critérios para a inclusão de espécies em categorias de sensibilidade para a região de
Santa Teresa foram similares aos usados em Londrina por ANJOS (2006)
(presença/ausência do táxon em diferentes classes de tamanho de fragmento). Comparando
estas duas regiões, várias congruências são observadas, quer em relação à alta (AS) ou à
baixa sensibilidade (BS), como para Tinamus solitarius (AS), Triclaria malachitacea
(AS), Geotrygon montana (BS), Grallaria varia (AS), Automolus leucophthalmus (BS) e
Pachyramphus castaneus (BS). Porém, outras espécies apresentaram diferenças em seus
níveis de sensibilidade (Herpetotheres cachinnans, Pulsatrix koeniswaldiana, Trogon
rufus, Dysithamnus sticthotorax, Campyloramphus falcularius, Habia rubica, etc),
indicando que impactos distintos operaram sobre suas respectivas comunidades
faunísticas. Logo, a tolerância diferenciada das espécies entre sistemas regionais de
fragmentação parece uma boa razão para explicar porque espécies podem estar seguras em
determinadas localidades da Mata Atlântica, mas em outras estarem em estado crítico de
conservação. Este caso pode ser ilustrado por algumas espécies de baixa sensibilidade na
paisagem de Santa Teresa (e.g. Trogon rufus, Selenidera maculirostris, Dendrocincla
turdina, Hemitriccus diops e Procnias nudicollis), porém, já extintas na região de Viçosa,
MG (RIBON et al. 2003), demonstrando que a variação geográfica nos padrões de

102
sensibilidade à fragmentação florestal assume especial relevância em planos de
conservação de comunidades bióticas regionais (ANJOS 2006).
Trabalhando com algumas espécies na região de Anhembi, São Paulo, UEZU et al. (2005)
verificaram que seus níveis de tolerância também estão intimamente associados ao grau de
conectividade da paisagem, concluindo que as respostas ao processo de fragmentação da
avifauna da Mata Atlântica são espécies-específica. Considerando, entretanto, a
importância da paisagem sobre a sensibilidade das espécies, propõe-se que a idéia sobre
suas respostas diferenciadas deveria ser ampliada para “espécies-específica-sistema”. Com
efeito, uma ave insetívora (espécie) do subbosque, com limitada capacidade de dispersão
(resposta específica) em uma matriz inóspita (sistema), parece representar bem a
abrangência deste conceito para explicar melhor a variação da tolerância das espécies em
determinada região.
O estudo de PARKER III et al. (1996) reúne um amplo banco de dados sobre as espécies
da Mata Atlântica, incluindo sua categorização em táxons sensíveis e endêmicos, os quais
normalmente são admitidos como aqueles com maior prédisposição à extinção no referido
bioma (BROOKS & BALMFORD 1996, GOERCK 1997). Em seu estudo, RIBON et al.
(2003) verificaram que as espécies pertencentes a estes dois grupos estiveram entre as
mais vulneráveis (sob algum grau de ameaça regional) na região de Viçosa. Porém,
quando testadas pelo presente estudo, tais espécies não estiveram numericamente mais
representadas entre as mais vulneráveis (alta sensibilidade) na região de Santa Teresa,
demonstrando que a mesma, além de abrigar uma elevada riqueza de espécies florestais
(item 5.1), ainda conserva um conjunto expressivo de táxons propensos à extinção na Mata
Atlântica. Contudo, tanto as espécies sensíveis como as endêmicas deste bioma
mostraram-se altamente dependentes do tamanho dos remanescentes florestais (escala do
hábitat) para sua manutenção nas matas serranas de Santa Teresa (item 5.2).

103
Como mencionado acima, vários fatores podem determinar a intolerância das espécies em
uma paisagem fragmentada, entre eles, a perda de microhábitats, baixa densidade
populacional e pressão de caça. O psitacídeo Triclaria malachitacea, por exemplo, foi
considerado sensível no interior de São Paulo, em áreas até mesmo com pequenas
alterações de seu hábitat, em função de sua raridade natural (MAGALHÃES 1999). No
presente estudo, a alta senbilidade das espécies foi determinada com base em sua ausência
nos pequenos fragmentos estudados, cujas causas podem ser resultantes de um ou mais
dos fatores conhecidos da literatura. Assim, a inexistência de córregos nos pequenos
fragmentos (exceção no MBML) explicaria as ausências pontuais do furnarídeo Lochmias
nematura e, por conseguinte, sua fraca tolerância ao processo de fragmentação regional. O
nível de alta sensibilidade foi atribuído também para os tiranídeos Mionectes rufiventris e
Platyrinchus leucoryphus, que estiveram entre algumas das espécies florestais com baixo
índice de abundância na área-controle. Da mesma maneira, Tinamus solitarius, excluso ou
raro em alguns dos fragmentos estudados (inclusive no PNMSL), destaca-se entre os casos
mais evidentes da influência da caça como fator determinante do grau de sensibilidade dos
organismos de uma região.
A seleção de hábitat é um dos principais fatores que determinam a distribuição de espécies
em fragmentos florestais, sendo a ocorrência de algumas espécies em seus microhábitats
específicos um bom indicador do seu estado de conservação ambiental (BLONDEL 1985,
STOTZ et al. 1996). A persistência de alguns insetívoros especialistas de taquarais (e.g.
Drymophila ferruginea e Hemitriccus diops) nos pequenos fragmentos florestais de Santa
Teresa, onde este ambiente esteve presente em todos eles (bambo exótico no PMBML),
sugere que a presença de microhábitats pode ser mais importante do que o tamanho de sua
área para a manutenção de parte da comunidade biótica. Esta idéia está de acordo com os

104
estudos de RIBON (1998, 2003), onde a inexistência de grotas úmidas explicou a ausência
de algumas espécies nos fragmentos da região de Viçosa, MG.
Embora aves insetívoras de taquarais sejam consideradas suscetiveis à extinção (PARKER
III et al. 1996, STOTZ et al. 1996), ANJOS (2006) verificou que a maioria destas espécies
mostrou-se tolerante (média e baixa sensibilidade) ao processo de fragmentação na região
de Londrina, em concordância com os dados de Santa Teresa e também da região de São
Paulo, onde aumentaram em abundância e riqueza nos fragmentos florestais (DEVELEY,
2004). No entanto, dependendo do grau de isolamento e da qualidade das matas
remanescentes, tais espécies podem desaparecer dos fragmentos, mesmo na presença de
taquaras no seu interior, como verificado por RIBON (1998) na região de Viçosa, MG.
Embora este autor tenha verificado algumas dessas espécies (Drymophila ferruginea e
Anabazenops fuscus) em fragmentos menores do que 3,0 ha, casos de extinção local foram
detectados nesta região, incluindo, por exemplo, Biatas nigropectus e Hemitriccus diops
(RIBON et at. 2003). Além da raridade natural e o pequeno tamanho dos fragmentos,
RIBON (1998) sugeriu a particularidade das manchas de taquaras secarem por completo
após seu período de frutificação para explicar a extinção de Biatas nigropectus em Viçosa
(o que pode ter sido válido também para Hemitriccus diops). Contudo, a persistência de
outras espécies desta guilda em pequenos fragmentos da Mata Atlântica indica que causas
mais específicas ou intrincadas devem estar envolvidas com estes tipos de extinção, sem
que nenhuma delas, entretanto, tenha até o momento sido escalerecida pela literatura. Por
isso, o estudo da história natural das espécies surge como uma das mais importantes
ferramentas para predizer com mais exatidão as respostas de comunidades bióticas ao
processo de fragmentação do seu hábitat (BIERREGAARD et al. 1992, SIMBERLOFF
1994, BIERREGAARD & STOUFFER 1997).

105
5.5. Composição da avifauna e taxa de extinções nos fragmentos florestais
Vários estudos têm demonstrado o empobrecimento faunístico em pequenos remanescentes
de vegetação natural, onde acabam por persistir apenas subconjuntos não-aleatórios da
diversidade biológica original (nested subsets) (BOLGER et al. 1991, COOK 1995,
PATTERSON & ATMAR 2000). Na região tropical, este evento tem sido explicado pela
presença de ambientes naturalmente heretogêneos, que quando deteriorados pelo processo
de fragmentação florestal, resultam em perdas não ao acaso de espécies especialistas, nem
sempre compensadas por novas taxas de colonização (ZIMMERMAN & BIERREGAARD
1986, SAUNDERS et al. 1991, TURNER 1996).
Neste trabalho, a composição da avifauna foi dissimilar entre as diferentes classes de
tamanho de fragmentos, sendo que aqueles pouco conservados (remanescentes pequenos)
foram os que mais diferiram da área-controle (Reserva Biológica Augusto Ruschi). Como
esperado, as três reservas da região (EBSL, PNMBL, RBAR) foram mais similares entre si
do que com qualquer um dos outros cinco fragmentos analisados, notando-se, porém, que o
Parque Municipal Natural de São Lourenço, apesar do seu menor tamanho, foi a reserva
faunisticamente mais relacionada com a área-controle (RBAR). Pela maior parte das matas
da EBSL situar-se em áreas de meia-encosta voltadas para a região litorânea (transectos
demarcados em altitudes de ~ 650m ), a composição de sua avifauna esteve representada
por alguns elementos considerados invasores da região serrana de Santa Teresa (e.g.
Thryothorus genibarbis, Hylophilus thoracicus e Arremon taciturnus), possivelmente
através de uma colonização recente pelo vale do rio Timbuí (SIMON 2000, WILLIS &
ONIKI 2002), justificando seu menor grau de similaridade com o PNMSL/RBAR
(transectos em ~ 800m e em posição mais interiorana).
Embora tenham apresentado as menores riquezas de espécies, os pequenos fragmentos
apresentaram comunidades faunísticas taxonomicamente semelhantes, coincidindo com o

106
modelo de distribuição de subgrupos não-aleatórios predito para hábitats de tamanho
retraído (PATTERSON 1987). As razões para a desintegração de comunidades em
fragmentos florestais, em função da perda de espécies, são amplamente citadas na
literatura, variando desde a perda da qualidade do hábitat até a desestabilização de
complexas relações ecológicas, como associações em bandos mistos e grupos mímicos em
atividades de forrageamento (BIERREGAARD et al. 1992, WIENS 1995, LAURENCE
1997, ALEIXO & VIELLIARD 1995, WILLIS 1989, DEVELEY & PERES 2000).
A estreita afinidade de espécies da região Neotropical com microhábitats específicos no
interior da mata é uma relação bem conhecida da literatura (PARKER III et al. 1996, SICK
1997, RIBON 1998, POLETTO et al. 2004). Dessa forma, é possível que a ausência de
várias espécies em alguns dos fragmentos estudados tenha sido fortemente influenciada
pela perda de sua heterogeneidade ambiental, como parece ser o caso, por exemplo, para
Neopelma aurifrons e Platyrinchus leucoryphus, ambas associadas a florestas bem
conservadas do fundo de vales úmidos (obs. pes.), conforme a exigência de alguns
Thamnophilidae estudados na região de Viçosa, MG (RIBON 1998, 2003). Da mesma
maneira, a perda de Habia rubica, considerada espécie nuclear de bandos mistos na Mata
Atlântica (DEVELEY & PERES 2000), pode explicar (por efeito cascata) a ausência de
várias outras espécies a elas associadas em alguns dos fragmentos estudados, como os
furnarídeos Philydor atricapillus e Anabazenos fuscus (DEVELEY 2004). Contudo, a
desintegração e o desaparecimento de bandos mistos não foram confirmados para alguns
fragmentos da região de Viçosa por MALDONADO-COELHO & MARINI (2000),
possivelmente em função de diferenças regionais em sua composição de espécies
(DEVELEY 2004).
Aves associadas a formigas-de-correição são muito suscetíveis à fragmentação florestal,
sobretudo aquelas denominadas “seguidoras-profissionais” destes insetos (WILLIS &

107
ONIKI 1992, BIERREGAARD & STOUFFER 1997, BORGES & GUILHERME 2000).
Embora de caráter não-obrigatório, a associação de aves da Mata Atlântica com formigas-
de-correição (Eciton burchelli e Labidus praedator) é conhecida da literatura (WILLIS
1979, WILLIS & ONIKI 1992, 1998), existindo evidências de que o seu desaparecimento
foi responsável pela perda de alguns elementos da avifauna em fragmentos do interior de
São Paulo e Minas Gerais (WILLIS 1979, CHRISTIANSEN & PITTER 1997).
No presente estudo, formigas-de-correição (Eciton sp. e Labidus sp.) foram observadas em
todos os fragmentos estudados (incluindo o fragmento urbano, PMBML), sugerindo que,
em alguns deles, a extinção de algumas aves (Dendrocincla turdina, Habia rubica, etc) que
regularmente acompanham tais insetos não esteve relacionada com este tipo de associação,
em concordância com os dados observados em Viçosa, por RIBON (1998). O registro
destas formigas nos fragmentos de Santa Teresa sugere que a matriz e a conformação
espacial das matas remanescentes da região favorecem a conectividade funcional para tais
insetos, como verificado para muitas espécies de pequenos mamíferos no estudo de
PASSAMANI (2003).
O sistema de metapopulações é muitas vezes utilizado para explicar o motivo pelo qual
várias espécies conseguem persistir em pequenos fragmentos espacialmente isolados
(HARRISON 1991, HANSKI 1991, item 5.2). Contudo, as espécies sem habilidades para
dispersão podem estar representadas em tais áreas por populações relictuais que
subsistiram à redução do seu hábitat (RIBON 1998), podendo Drymophila ferruginea
exemplificar muito bem este caso para os pequenos fragmentos (por exemplo, PMBML)
estudos na região de Santa Teresa.
A comparação da composição da avifauna antes e depois do processo de fragmentação,
muitas vezes utilizando dados históricos (coleções científicas e dados bibliográficos), é
uma maneira segura de testar a extinção local de espécies (presente estudo/item 5.1,

108
KATTAN et al. 1994, STOUFFER & BIERREGAARD 1995, RENFIJO 1999, RIBON et
al. 2003). Comparações com áreas contínuas de mata também têm fornecido importantes
conclusões, sendo que WILLIS (1979) foi o primeiro estudo a documentar extinções de
aves em fragmentos de Mata Atlântica, no interior de São Paulo, onde áreas de 21, 250 e
1.400 ha estiveram representadas por 93, 146 e 202 espécies, respectivamente.
Posteriormente, muitos outros estudos obtiveram resultados similares para outras regiões
(ALEIXO & VIELLIARD 1995, ANJOS 2004, RIBON et al. 2003), evidenciando que a
avifauna florestal deste bioma é notavelmente sensível a pertubações antrópicas, ainda que
parte de suas espécies possa persistir em hábitats de tamanho reduzido e/ou de vegetação
secundária (ALEIXO 2001, contra BROWN & BROWN 1992, PROTOMASTRO 2001).
Na região serrana de Santa Teresa, muitas espécies de aves (~50%) foram extintas nos
remanescentes menores do que 50 ha, sendo que somente aqueles com áreas em torno ou
superior a 500 ha (ESBL, PNMSL) foram capazes de manter a integridade faunística
presente na mata controle (ca. 4.000 ha). Tais resultados reforçam a opinião de
MARSDEN et al. (2001), WILLIS & ONIKI (2002) e DEVELEY (2004), em relação à
idéia de que regiões sem grandes reservas florestais mostram-se insuficientes para
conservar representativas parcelas da diversidade biológica da Mata Atlântica. Em
determinadas localidades como Santa Teresa, muitos táxons propensos à extinção
(cinergéticos, endemismos e alta sensibilidade ecológica) ainda são de comum ocorrência
(e.g. Tinamus solitarius, Chamaeza meruloides, Carpornis cucullata, Neopelma aurifrons),
sendo, por isso, muitas vezes negligenciados em análises sobre o seu status de conservação
ao longo de toda sua distribuição geográfica (SIMON et al., no prelo). De fato, algumas
espécies restritas às matas de tabuleiro do Estado do Espirito Santo (e.g. Crypturellus
noctivagus, Crax blumenbachii e Neomorphus geoffroyi) só não estão definitivamente
extintas em seu território em função de extensas áreas de florestas primárias transformadas

109
em unidades de conservação (por exemplo, Reserva Biológica de Sooretama e Reserva
Florestal da Companhia Vale do Rio Doce) (SIMON et al., no prelo).
5.6. Composição das guildas tróficas nos fragmentos florestais
A resposta das guildas de aves ao processo de fragmentação do hábitat foi analisada em
quase todos os trabalhos realizados na região da Mata Atlânica, sendo que aquelas
representadas pelos grandes frugívoros de copa, insetívoros do solo e insetívoros do
subbosque compõem o grupo mais negativamente afetado nas paisagens fragmentadas do
referido bioma (GOERCK 1997, ALEIXO 2001).
Com exceção dos nectarívoros, todas as guildas avaliadas pelo presente estudo
apresentaram perda de espécies com a redução do tamanho do seu hábitat, mostrando-se
sensíveis ao sistema de fragmentação na região de Santa Teresa. Entretanto, sete delas,
representadas pelos carnívoros diurnos, carnívoros noturnos, granívoros de taquaral,
insetívoros do solo, insetívoros de margem de córrego, insetívoros do folhiço suspenso e
insetívoros noturnos, foram as mais afetadas, chegando a desaparecer por completo de
alguns dos pequenos fragmentos florestais avaliados.
Dessa forma, a guilda dos grandes frugívoros de copa, admitida na literatura como de alta
sensibilidade na Mata Atlântica, foi pouco afetada nas matas serranas de Santa Teresa. Por
outro lado, das sete categorias mencionadas acima, quatro (carnívoros diurnos, carnívoros
noturnos, insetívoros do solo e insetívoros noturnos) mostraram resposta semelhante aos
dados obtidos no interior de São Paulo por WILLIS (1979), que usou categorias de guildas
(n= 20) semelhantes às do presente estudo (n= 19). Utilizando as mesmas categorias
adotadas por WILLIS (1979), ANJOS (2006) verificou que os insetívoros de taquarais e
grandes frugívoros de copa corresponderam aos grupos mais afetados (número elevado de

110
espécies muito sensíveis) na região de Londrina, norte do Paraná, coincidindo com o
estudo de WILLIS (1979), mas divergindo do observado para Santa Teresa.
As particularidades adotadas na categorização das guildas pelos diferentes estudos limitam
a comparação dos dados de Santa Teresa com alguns outros trabalhos realizados na Mata
Atlântica (RIBON et al. 2003, ANJOS 2004, DEVELEY 2004, ANTUNES 2005).
Contudo, tendências convergentes e divergentes na resposta de determinadas guildas
podem ser verificadas entre os dados disponíveis, evidenciando mais uma vez que cada
região é única em seu padrão de fragmentação, produzindo efeitos diferenciados em suas
respectivas comunidades biológicas. Aparentemente, a guilda dos nectarívoros (beija-
flores) é a única que mostra comportamento similar entre todas as regiões da Mata
Atlântica analisadas, não diferindo inclusive dos dados obtidos na Amazônia central
(STOUFFER & BIERREGARRD 1995). Como grupo, os nectarívoros não foram afetados
na região de Santa Teresa, mas umas poucas espécies (e.g. Ramphodon naevius e
Leucochloris albicollis) estiveram ausentes em alguns do pequenos fragmentos florestais
avaliados, em concordância com o estudo de WILLIS (1979).
Além dos esclarecimentos sobre quais são as guildas mais afetadas com a fragmentação da
Mata Atlântica, os estudos realizados nesta região têm oferecido boas explicações sobre os
motivos pelos quais declinaram em abundância ou riqueza de espécies em regiões de
florestas fragmentadas. A sensibilidade, por exemplo, dos grandes frugívoros de copa tem
sido relacionada à sua necessidade de deslocamentos sobre amplas áreas de mata contínua,
em função da variação espacial e temporal de sua fonte de alimento. Com a fragmentação
florestal, suas populações ficam sujeitas à escassez de recursos em função de seus
movimentos limitados, tornando-se susceptíveis à extinção local (WILLIS 1979,
GALLETI & PIZO 1996, GOERCK 1997). Possivelmente, em Santa Teresa, esta guilda
foi pouco afetada em função da disposição espacial e qualidade dos remanescentes

111
florestais da região, sendo que algumas espécies desta guilda (e.g. Pyrrhura frontalis,
Ramphastos vittelinus e Procnias nudicollis) observadas em vôo entre os fragmentos
florestais (incluindo o fragmento urbano, PMBML) sustentam esta hipótese.
Por sua vez, os insetívoros do solo ou do subbosque, além de possuírem fraca habilidade
de dispersão através de hábitats não florestais, normalmente estão associados a
microhábitats específicos no interior da mata (por exemplo, taquarais e grotas úmidas)
(SICK 1997, RIBON 1998). Como conseqüência, costumam apresentar distribuição em
manchas ou ocorrer em baixa densidade populacional, tornando-os igualmente vulneráveis
à fragmentação do seu hábitat (WILLIS 1979, ALEIXO & VIELLIARD 1995, GOERCK
1997, ANJOS 2001, LAPS et al. 2003, RIBON 2003, ANJOS 2006). No presente estudo, a
guilda dos insetívoros do subboque revelou-se menos vulnerável que aquela contendo
espécies do solo, evidenciando que esta última abrange espécies menos aptas a persistir em
hábitats de tamanho reduzido ou em pior estado de conservação.
Enquanto é possível que carnívoros diurnos venham a desaparecer de paisagens
fragmentadas da Mata Atlântica em função de sua baixa densidade populacional, a
extinção de insetívoros noturnos (Caprimulgidae) tem sido atribuída ao aumento nas taxas
de predação em função de seus hábitos terrícolas (WIILLIS 1979). Estas duas suposições
recebem apoio nos dados do presente estudo, considerando a raridade natural de muitas
espécies de gaviões nas matas de Santa Teresa e a presença comum de predadores
potenciais (Felis cattus, Didelphis aurita, etc) nos pequenos fragmentos amostrados (obs.
pes.). Os poucos estudos que analisaram a resposta dos carnívoros noturnos (Strigidae) em
paisagens alteradas também verificaram perdas de espécies nesta guilda (WILLIS 1979,
ANTUNES 2005, RIBON et al. 2003). Apesar de comentar sobre a necessidade de estudos
para este grupo de espécies, WILLIS (1979) sugeriu que as extinções verificadas em sua
área de estudo acompanharam o desaparecimento local de mamíferos do solo. Em seu

112
estudo, PASSAMANI (2003) não verificou mudanças na riqueza de espécies de pequenos
mamíferos entre os diferentes fragmentos analisados, significando que a sugestão de
WILLIS (1979) não parece explicar a perda dos carnívoros noturnos nos pequenos
fragmentos florestais da região de Santa Teresa. Como os carnívoros diurnos, tais aves não
são localmente abundantes, possivelmente em função da necessidade de amplos territórios
de caça, razão pela qual a guilda dos carnívoros noturnos estaria empobrecida nos
pequenos fragmentos florestais.
A definição das guildas dos granívoros de taquaral, insetívoros de margem de córrego e
insetívoros do folhiço suspenso (emaranhado de folhas mortas na ramaria do interior da
floresta) foi uma particularidade do presente estudo, dificultando sua comparação com os
demais estudos realizados na Mata Atlântica. Porém, a razão para a vulnerabilidade dessas
guildas à fragmentação na região de Santa Teresa não parece diferente das interpretações
apontadas na literatura para outras guildas de comportamento semelhante (por exemplo,
insetívoros de taquaral). A especialização destas guildas por determinados locais de
forrageio, ausentes ou reduzidos em alguns dos fragmentos estudados, pode ser a provável
causa das perdas pontuais, como exemplificada pelo desaparecimento de Lochmias
nematura nas matas destituídas de córregos em seu interior. Como este microhábitat, o
folhiço suspenso (acúmulo de folhas mortas suspensas nos estratos da floresta) também se
distribui em manchas no interior da mata, obrigando as espécies especializadas em
forragear neste tipo de substrato a realizar constantes deslocamentos em buscas de sua
fonte de alimento (REMSEN & PARKER III 1984). Dessa forma, a menor disponibilidade
desta condição nos pequenos fragmentos explicaria a exclusão pontual de outras aves
insetívoras, como Philydor spp. e Cichlocolaptes leucophrus.

113
Quase todos os estudos realizados em áreas de florestas tropicais têm demonstrado
acentuadas perdas de espécies nos hábitats remanescentes, causando drásticas mudanças na
estrutura de suas comunidades bióticas (WHITMORE 1997, ANJOS 2004).
Neste trabalho, a análise multivariada aplicada para o conjunto das 19 guildas apontou
comunidades de aves estruturalmente diferenciadas entre alguns grupos de fragmentos
estudados. As maiores dissimilaridades em relação à área-controle foram verificadas nos
fragmentos de tamanho pequeno, com destaque para o PMBML, onde muitas guildas
apresentaram significativas perdas de espécies. Porém, raras foram as guildas
completamente ausentes nesta área (granívoros de taquaral, insetívoros do folhiço suspenso
e insetívoros noturnos), sendo que algumas das espécies (e.g. Selenidera maculirostris e
Mymeciza loricata) aí extintas persistiram no menor fragmento estudado (FP3/2,6 ha).
Com efeito, o impacto negativo da fragmentação florestal sobre comunidade de aves é de
natureza complexa, podendo ser específico para cada espécie e nem sempre estar
diretamente relacionado à redução no tamanho do hábitat ou ao seu grau de isolamento na
paisagem alterada (LAURENCE et al. 1997, DEBINSKI & HOLT 2000, ANJOS 2001,
ANTUNES 2005).
A principal limitação ao se tentar fazer generalizações sobre a resposta das comunidades
de aves na região da Mata Atlântica reside no fato de que cada paisagem é única em seu
padrão e histórico de fragmentação. Porém, a falta de sistematização dos protocolos de
coleta e análise dos dados é outra séria limitação, por impedir a efetiva comparação dos
resultados obtidos entre as diferentes regiões deste bioma. Dessa forma, o grande problema
com a compreensão das abordagens sobre a fragmentação do hábitat é que o conhecimento
até agora reunido ainda se encontra, paradoxalmente, bastante fragmentado (WATSON
2002).

114
5.7. Implicações e ações para a conservação da avifauna florestal
Ainda que áreas perturbadas de matas tropicais possam suportar porções da diversidade
biológica, dramáticas perdas de espécies podem ocorrer quando o processo de
fragmentação florestal é intenso (TURNER & CORLETT 1996, WATSON 2002).
Comparada a outras paisagens fragmentadas da Mata Atlântica, a região serrana de Santa
Teresa, ainda que destituída de 60% de suas florestas nativas, encontra-se em situação
privilegiada de conservação, testemunhada por sua elevada diversidade faunística e
florística recentemente registrada na literatura (THOMAZ & MONTEIRO 1997, BROWN
& FREITAS 2000, PASSAMANI et al. 2000, SIMON 2000, WILLIS & ONIKI 2002,
PASSAMANI 2003, presente estudo). Sem dúvida, a rica comunidade de aves florestais
verificada para a região de estudo está relacionada à extensão e qualidade de sua vegetação
remanescente, em parte representada por três unidades de conservação, que totalizam cerca
de 5.000 ha de matas nativas em bom estado de conservação. Uma parcela da avifauna
registrada nestas reservas é rara ou não persiste em remanescentes de propriedades
privadas, os quais normalmente são menores do que 150 ha e circundados por ambientes
antrópicos desfavoráveis para muitas espécies. Com efeito, são individualmente poucos
efetivos para abrigar comunidades de aves mais complexas, contrastando, portanto, com os
resultados encontrados para pequenos mamíferos por PASSAMANI (2003), que verificou
diversidade semelhante entre pequenos e grandes fragmentos florestais, a qual foi, em
parte, interpretada pelo autor como uma resposta direta à permeabilidade da matriz na
paisagem estudada. Em oposição também aos dados obtidos para pequenos mamíferos na
região de Santa Teresa (PASSAMANI 2003), a perda em torno de 50% da avifauna
florestal nos pequenos fragmentos sugere fraca habilidade de muitas espécies em
atravessar a matriz de hábitats antrópicos, impedindo que as mesmas venham ali a se
restabelecer por intermédio da imigração de indivíduos de fragmentos vizinhos. Em

115
conjunto, porém, os fragmentos situados em terras particulares representam uma
expressiva área de vegetação nativa, favorável a um amplo espectro de condições
ambientais, igualmente importante para a conservação da diversidade biológica na escala
da paisagem. De fato, vários estudos já demonstraram que pequenos fragmentos (~ 100 ha)
podem funcionar como refúgios para muitas espécies animais e vegetais mesmo após
décadas de seu isolamento, fornecendo uma oportunidade para a restauração de biotas
regionais em paisagens fragmentadas (CHIARELLO 2000a, TURNER & CORLLET
1996).
As ações para a conservação das espécies de mamíferos nas matas serranas de Santa
Teresa foram muito bem discutidas por PASSAMANI et al. (2000) e PASSAMANI
(2003), podendo muito bem ser aplicadas para a avifauna da região. Como suas principais
recomendações, aqui apresentadas de forma sistematizada, tais autores mencionaram:
- necessidade de proteção voltada também para os remanescentes florestais situados em
propriedades privadas, pelo papel que desempenham na manutenção da diversidade
regional e conectividade estrutural da paisagem (stepping stones), a qual pode favorecer o
sistema de metapopulações para muitas espécies;
- aumento da permabilidade do hábitat-matriz, através do manejo adequado dos ambientes
do entorno dos remanescentes florestais, priorizando a recomposição natural da vegetação
nativa e o uso do solo baseado em culturas arbustivas/arbóreas;
- revegetação de áreas de solo degradado, inclusive com a utilização de plantios de
eucalipto, desde que adequadamente manejado (cultivo sem capina do subbosque e
colheita parcelada), visando atenuar os impactos físicos e biológicos ligados à borda dos
fragmentos;
- melhor fiscalização contra as atividades antrópicas de degradação do ambiente florestal;

116
- implementação de planos de manejo dos remanescentes florestais preocupados com o
desenvolvimento econômico e social da região;
- realização de novos estudos para obtenção de outras importantes informações para
subsidiar os trabalhos de conservação da Mata Atlântica de Santa Teresa.
Entretanto, entre estas recomendações, vale ressaltar que a cultura do café, a mais
intensamente praticada na região e desejável para a conectividade funcional da paisagem
para pequenos mamíferos (PASSAMANI 2003), tem pouco valor para as aves, uma vez
que poucas são as espécies florestais (Crypturellus tataupa, Leptotila verreauxi,
Nyctidromus albicollis, entre outras) capazes de ocupar esta cultura como extensão do seu
hábitat (obs. pes.), seja para obtenção de recursos (fontes de alimento, locais de
nidificação, etc) ou meio de transposição para outras manchas de seu hábitat.
Dessa forma, a taxação de impostos sobre terras improdutivas e lavouras de café, como
sugerida por WILLIS & ONIKI (2002) para áreas ou produções de “luxo”, poderia ser
uma prática interessante a favor de culturas mais indicadas para melhor qualidade do
hábitat-matriz. Entretanto, em Santa Teresa prevalece a agricultura dos pequenos
produtores, desacompanhada de tecnologias relacionadas ao aumento da produtividade de
suas plantações, parecendo impraticável que a conservação da diversidade biológica venha
justificar a adotação de tal medida (pelo menos a curto e médio prazo) no município.
Na região de Santa Teresa, muitas pessoas ainda praticam a exploração ilegal de recursos
florestais (caça, madeira, palmito, orquídeas, taquaras, etc), muitas vezes difícil de ser
coibida com base apenas nos trabalhos de fiscalização dos órgãos responsáveis. Assim, a
implementação de um programa de inclusão de moradores locais estimulados e aptos a
trabalhar com conservação ambiental é também recomendável para a manutenção da
integridade biológica das florestas nativas da região.

117
Por fim, vale ressaltar que os dados obtidos para aves e mamíferos na região de Santa
Teresa, aliados a outros estudos realizados na Mata Atlântica, evidenciam que o problema
da fragmentação das florestas deste bioma é muito mais complexo do que se pode prever e
que generalizações sobre suas consequências são muito difíceis de serem feitas. Cada
paisagem parece única no padrão de fragmentação, estrutura da vegetação, uso do solo,
tipo de matriz e composição das espécies. Portanto, estratégias de manejo de ambientes
fragmentados não devem limitar-se apenas em preceitos e modelos científicos pré-
definidos. Na verdade, devem estar fundamentadas na interpretação dos dados obtidos na
região em análise, a fim de que as mesmas possam ser mais efetivas na conservação da
diversidade biológica (PASSAMANI 2003).

118
6. CONCLUSÕES
Com base nos resultados discutidos na secção anterior, as seguintes conclusões podem ser
apresentadas como respostas às questões que nortearam o desenvolvimento do presente
estudo (veja Objetivos):
6.1. (questão i) Em muitas regiões do leste do Brasil, a fragmentação da Mata Atlântica
afetou drasticamente a comunidade de aves dependente do ambiente florestal, levando
várias espécies à raridade ou extinção local. Na escala da paisagem, o processo de
fragmentação na região serrana de Santa Teresa (onde ainda restam cerca de 40 % da
cobertura florestal nativa) não implicou em sérias ameaças para a diversidade regional,
tendo causado possivelmente a extirpação apenas de Pipile jacutinga, uma ave de grande
porte que sempre foi historicamente caçada em toda sua área de distribuição geográfica.
6.2. (questão ii) Pequenos fragmentos (< 100 ha) de Mata Atlântica costumam apresentar
riqueza de espécies de aves semelhante a fragmentos maiores e mais conservados, porém,
são dissimilares em sua composição faunística, o que reforça a idéia de que este parâmetro
da comunidade, por si só, é pouco útil para a tomada de decisões relativas à conservação de
biotas regionais.
Na região de Santa Teresa, os pequenos fragmentos (<50 ha), mesmo situados em uma
paisagem com boa conectividade estrutural (numerosos fragmentos e proximidade entre
eles), possuem avifaunas depauperadas, normalmente 50 % daquelas conhecidas das
reservas florestais do município. O fato de remanescentes florestais de Santa Teresa com
cerca de 500 ha abrigarem expressivas parcelas da diversidade regional parece explicar
porque algumas outras localidades serranas do Espírito Santo, com matas de tamanho
similar, ainda mantêm elevados níveis de riqueza de aves, incluindo táxons globalmente
ameaçados e raros endemismos da Mata Atlântica.

119
6.3. (questão iii) O índice de conservação dos fragmentos, aplicado pela primeira vez na
análise de paisagens fragmentadas da Mata Atlântica, com base na combinação de cinco
diferentes descritores ambientais (heterogeneidade, grau de isolamento, forma e área dos
fragmentos, além da permeabilidade da matriz), mostrou-se útil para explicar a persistência
das espécies nos remanescentes florestais da região serrana de Santa Teresa, ES.
Fragmentos para os quais foram atribuídos índices de conservação em torno de 0,30
(pequenos fragmentos) mantêm, em média, apenas 50% da comunidade de aves florestais
de um grande bloco de mata contínua (área-controle), enquanto que aqueles com índices
acima de 0,60 (PNMSL e EBSL) podem sustentar a quase (ou mesma) totalidade da
avifauna regional.
Quando testados em conjunto, o tamanho da área, grau de isolamento e permeabilidade da
matriz excerceram um efeito significativo sobre a riqueza de espécies florestais na região
serrana de Santa Teresa (r2= 0,92). Porém, entre estes três descritores ambientais, o
tamanho da área foi o que melhor explicou (r2= 0,70) a variação no número de espécies
nos fragmentos estudados, coincidindo com o resultado de outras análises conduzidas na
Mata Atlântica.
A falta de relação da riqueza de espécies florestais com os dois outros descritores
ambientais (grau de isolamento e permeabilidade da matriz) pode ser explicada pela
disposição espacial dos fragmentos, cuja proximidade entre eles deve favorecer a
conectividade funcional para muitas espécies, em vista da porcentagem da cobertura
vegetal remanescente (ca. 40%), dando suporte à idéia de que o isolamento dos fragmentos
tem pouco efeito sobre a diversidade biológica em áreas que ainda mantêm cerca de 30%
da extensão de sua vegetação original, como verificado para a região de Viçosa, Minas
Gerais. Porém, os dados reunidos para a Mata Atlântica demonstram que muitas espécies
estritamente florestais possuem pouca aptidão para atravessar os “poros da matriz”

120
(hábitats antrópicos), tornando-se altamente dependentes da área dos remanescentes
florestais para sua manutenção na paisagem fragmentada do referido bioma.
Todas as análises sobre os efeitos da fragmentação da Mata Atlântica adotaram a
conformação histórica da fragmentação florestal (sistema realista), podendo explicar
porque os resultados obtidos nesta região algumas vezes não coincidem com aqueles
verificados para outros sistemas estudados, como o da região da Amazônia central, onde
os experimentos foram manipulados pela criação de fragmentos com grau de isolamento,
forma geométrica e escala de tamanho planejados (1, 10, 100, 1000 ha).
6.4. (questão iv) Parte da avifauna (51 spp.) que persistiu nos fragmentos florestais da
região serrana de Santa Teresa apresentou variações em seus índices de abundância
relativa, quando comparados com a área-controle. Os dados da literatura, combinados
com aqueles obtidos pelo presente estudo, sugerem que várias espécies apresentam
padrões semelhantes de declínio e aumento de abundância, demonstrando que os efeitos da
fragmentação são mais determinísticos do que estocásticos para a avifauna da Mata
Atlântica. Porém, algumas espécies mantiveram índices estáveis entre diferentes classes de
tamanho de fragmento, indicando que o processo de fragmentação na região estudada não
foi tão severo quanto outras regiões do leste do Brasil, onde o declínio populacional fez
com que muitas espécies se tornassem raras ou viessem a desaparecer por completo.
As comunidades com as maiores porcentagens de espécies com variações em seus índices
de abundância foram encontradas nos fragmentos menos conservados (Parque de Biologia
Mello Leitão e FP1), concordando, por exemplo, com os resultados obtidos na região de
Londrina, norte do Paraná, onde o maior número de espécies com flutuações populacionais
também ocorreu em fragmentos com maior grau de comprometimento ambiental.
Além das idiossincrasias das espécies, muitas causas relacionadas a fatores extrínsecos e
intrínsecos dos remanescentes florestais têm sido utilizadas para explicar as variações

121
populacionais em avifaunas confinadas nas paisagens fragmentadas da Mata Atlântica (por
exemplo, alterações microclimáticas e modificações nas taxas de competição
interespecífica). Muitas delas, porém, ainda não foram cientificamente testadas e, portanto,
a melhor compreensão das causas relacionadas às flutuações populacionais da avifauna em
paisagens fragmentadas da Mata Atlântica ainda limita-se à extrema falta de dados sobre a
história natural das espécies, perda de diversidade genética nas populações isoladas e a
intrincada rede de interações ecológicas que ocorrem entre as mesmas.
6.5. (questão v) No presente estudo, 60 espécies foram reconhecidas como de alta
sensibilidade para a região serrana de Santa Teresa, enquanto outras 99 mostraram-se mais
tolerantes ao sistema de fragmentação regional, ocorrendo em remanescentes florestais
menores do que 50 ha, como o Parque do Museu de Biologia Mello Leitão (8,5 ha), situado
na área urbana do município.
Independente de sua categoria de sensibilidade, muitas destas espécies são caracterizadas
por um ou mais atributos de suscetibilidade mencionados na literatura (por exemplo,
especificidade por microhábitat, maior peso corporal e baixa densidade populacional),
apoiando a idéia de que os níveis de tolerância de comunidades biológicas estão
intimamente relacionados às particularidades da paisagem fragmentada (qualidade da
vegetação, porcentagem de matas, conformação dos fragmentos, qualidade das matas, grau
de conectividade, etc).
Os dados de Santa Teresa, quando comparados com os de outras regiões que utilizaram
metodologias semelhantes, também apontam incongruências nos níveis de sensibilidade
das espécies, indicando que impactos distintos operaram sobre suas respectivas
comunidades faunísticas. Logo, a tolerância diferenciada das espécies entre sistemas
regionais de fragmentação parece uma boa razão para explicar porque espécies podem

122
estar seguras em determinadas localidades da Mata Atlântica, mas em outras estarem em
estado crítico de conservação.
O presente estudo verificou também que espécies consideradas propensas à extinção na
Mata Atlântica (táxons endêmicos e de alta sensibilidade ecológica) não estiveram
numericamente mais representadas entre as mais vulneráveis (alta sensibilidade) na região
de Santa Teresa, demonstrando que a mesma, além de abrigar uma elevada riqueza de
espécies florestais, ainda conserva um conjunto expressivo de táxons de interesse para a
conservação da diversidade biológica do referido bioma.
6.6. (questão vi) Com base em intensivos levantamentos de campo (12-18 meses),
verificou-se que a composição da avifauna foi dissimilar entre as diferentes classes de
tamanho de fragmentos, sendo que aqueles pouco conservados (remanescentes pequenos:
< 50 ha) foram os que mais diferiram da área-controle (Reserva Biológica Augusto
Ruschi).
Como esperado, as três reservas da região (EBSL, PNMBL, RBAR) foram mais similares
entre si do que com qualquer um dos outros cinco fragmentos analisados. Apesar do seu
menor tamanho (~ 500 ha) (versus ~ 800 ha/EBSL), o Parque Municipal Natural de São
Lourenço foi a reserva faunisticamente mais relacionada com a área-controle (RBAR).
Embora tenham apresentado as menores riquezas de espécies, os pequenos fragmentos
apresentaram comunidades faunísticas taxonomicamente semelhantes, coincidindo com o
modelo de distribuição de subgrupos não-aleatórios (subset nested) predito para hábitats
de tamanho reduzido. É possível que a notável desintegração das comunidades de aves em
alguns dos fragmentos estudados tenha sido fortemente influenciada pela perda de sua
heterogeneidade ambiental. Entretanto, causas mais intrincadas relacionadas a interações
interespecíficas (por exemplo, desestruturação de bandos mistos) podem ter igualmente

123
contribuído para a exclusão de espécies no interior destes fragmentos, como já sugerido
por alguns outros estudos realizados na Mata Atlântica.
Na região estudada, os fragmentos florestais menores do que 50 ha chegaram a perder até
50% de suas espécies, sendo que somente aqueles com áreas em torno ou superior a 500 ha
(ESBL, PNMSL) foram capazes de manter a mesma (ou semelhante) integridade faunística
de uma grande área de mata contínua (~ 4.000 ha). Tais resultados reforçam a opinião de
que regiões sem grandes reservas florestais mostram-se insuficientes para conservar
representativas parcelas da diversidade biológica da Mata Atlântica, muitas vezes
negligenciada porque em determinadas regiões detentoras de expressivas porções de
florestas nativas (como Santa Teresa), muitos táxons propensos à extinção ainda são de
comum ocorrência, interferindo em análises sobre o status de conservação das espécies ao
longo de toda sua distribuição geográfica.
6.7. (questão vii) Apesar de atingirem quase todas as guildas analisadas (menos os
nectarívoros), as extinções verificadas com a redução do hábitat na região de Santa Teresa
foram mais acentuadas em sete delas, representadas pelos carnívoros diurnos, carnívoros
noturnos, granívoros de taquaral, insetívoros do solo, insetívoros de margem de córrego,
insetívoros do folhiço suspenso e insetívoros noturnos.
A guilda dos grandes frugívoros de copa, admitida como de alta sensibilidade na Mata
Atlântica, foi pouco afetada nas matas serranas de Santa Teresa. Por outro lado, das sete
guildas mencionadas acima, quatro (carnívoros diurnos, carnívoros noturnos, insetívoros
do solo e insetívoros noturnos) mostraram resposta semelhante aos dados obtidos no
interior de São Paulo, onde as categorias de guildas analisadas foram semelhantes às do
presente estudo. Tais resultados, aliados a outras comparações interregionais, reforçam a
idéia de que cada região é única em seu padrão de fragmentação, produzindo efeitos
diferenciados em suas respectivas comunidades biológicas.

124
As comunidades de aves, analisadas segundo seus grupos ecológicos funcionais (guildas
tróficas e espaciais combinadas), mostaram-se estruturalmente diferenciadas entre alguns
grupos de fragmentos estudados. As maiores dissimilaridades em relação à área-controle
foram verificadas nos fragmentos de tamanho pequeno, onde muitas guildas apresentaram
perdas significativas espécies.
Além das paisagens com padrões e históricos particulares de fragmentação, a falta de
sistematização dos protocolos de coleta e análise dos dados é outra séria limitação ao se
tentar fazer generalizações sobre a resposta das comunidades de aves na região da Mata
Atlântica. Dessa forma, o grande problema com a compreensão das abordagens sobre a
fragmentação do hábitat é que o conhecimento até agora reunido na literatura ainda se
encontra bastante fragmentado.

125
7. BIBLIOGRAFIA ALEIXO, A. Effects of selective logging on a bird community in the Brazilian Atlantic Forest. The Condor, v.101, p.537-548. 1999. ALEIXO, A. Conservação da avifauna da Floresta Atlântica: efeitos da fragmentação e a importância de florestas secundárias. In: ALBUQUERQUE, J. L. B.; CÂNDIDO JR, J.F.; STRAUBE, F. C.; ROOS, A. L. (Org.). Ornitologia e conservação: da ciência às estratégias. Tubarão: Unisul, 2001. p.199-206.
ALEIXO, A. Molecular systematics and the role of the "várzea"-"terra-firme" ecotone in the diversification of Xiphorhynchus woodcreepers (Aves: Dendrocolaptidae). The Auk, v.119, p.621-640. 2002.
ALEIXO, A.; VIELLIARD, J. M. E. Composição e dinâmica da avifauna da mata de Santa Genebra, Campinas, São Paulo, Brasil. Revista Brasileira de Zoologia, v.12, no.3, p.493-511. 1995.
ANJOS, L. Bird communities in five Atlantic forest fragments in southern Brazil. Ornitologia Neotropical, v. 12, p.11-27. 2001.
ANJOS, L. Species richness and relative abundance of birds in natural and anthropogenic fragments of Brazilian Atlantic forest. Anais da Academia Brasileira de Ciências, v.76, no.2, p.429-434. 2004.
ANJOS, L. Bird species sensitivity in a fragmented landscape of the Atlantic forest in southern Brazil. Biotropica, v.38, no.2, p.1-6. 2006.
ANJOS, L.; SCHUCHMANN, K. L.; BERNDT, R. Avifauna composition, species richness, and status in the Tibagi river basin, Paraná State, southern Brazil. Ornitologia Neotropical, v.8, p.145-173. 1997.
ANJOS, L.; BOÇON, R. Bird communities in natural forest patches in southern Brazil. Wilson Bulletin, v.111, no.3, p.397-414. 1999.
ANJOS, L.; ZANETTE, L.; LOPES, E. V. Effects of fragmentation on the bird guilds of the Atlantic forest in north Paraná, southern Brazil. Ornitologia Neotropical, v.15, p.137-144. 2004.
ANTONGIOVANNI, M.; METZGER, J. P. Influence of matrix habitats on the occurence of insectivorous bird species in Amazonian forest fragments. Biological Conservation, v.122, p. 441-451. 2005.
ANTUNES, A. Z. Alterações na composição da comunidade de aves ao longo do tempo em um fragmento florestal no sudeste do Brasil. Revista Brasileira de Ornitologia, v.13, no.1, p.47-61. 2005.
ÅS S. Invasion of matrix species in small habitat patches. Conservation Ecology [on line], v.3, no. 1, p.1-13. 1999.

126
AZEVEDO, C. O.; SANTOS, H. S. Perfil da fauna de himenópteros parasitóides (Insecta, Hymenoptera) em uma área de Mata Atlântica da Reserva Biológica de Duas Bocas, Cariacica, ES, Brasil. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.117-126. 2000.
BARBINI, I. G.; PASSAMANI, M. Pequenos mamíferos e a predação de ninhos artificiais no Museu de Biologia Prof. Mello Leitão (ES). Revista Natureza [on line], v.1, no. 2, p. 56-61. 2005.
BAUER, C. Padrões atuais de distribuição de aves florestais na região sul do Estado do Espírito Santo, Brasil.1999. Dissertação de mestrado, UFRJ, Museu Nacional. Rio de Janeiro, 1999.
BAUER, C.; PACHECO, J. F.; VENTURINI, A. C.; WHITNEY, B. M. Rediscovery of the Cherry-throated Tanager Nemosia rourei in southern Espírito Santo, Brazil. Bird Conservation International, v.10, p.93-104. 2000.
BERNARDO, C. S. S.; GALETTI, M. Densidade populacional de primatas em um fragmento florestal no sudeste do Brasil. Revista Brasileira de Zoologia, v.21, no.4, p.827-832. 2004.
BLONDEL, J. Habitat selection in istand versus mainland birds. In: CODY, M. L. (ed.). Habitat selection in birds. Academic Press. San Diego. 558p. 1985.
BLONDEL, J.; CHESSEL, D.; FROCHOT, B. Birds impoverishment, niche expansion, and density inflation in mediterranean island habitats. Ecology v.69: p. 1899-1917. 1988.
BIANCONI, G. V.; MIKICH, S. B.; PEDRO, W. A. Diversidade de morcegos (Mammalia, Chiroptera) em remanescentes florestais do município de Fênix, noroeste do Paraná, Brasil. Revista Brasileira de Zoologia, v.21, no.4, p.943-954. 2004.
BIBBY, C.; BURGUESS, N. D.; HILL, D. A. Bird census techniques. Londres: Academic Press, 1992.
BIBBY, C.; JONES, M.; MARSDEN, S. J. Expedition field techniques: bird surveys. Expedition Advisory Centre, 137p. 1998.
BIERREGAARD, R. O. J.; LOVEJOY, T. E. Birds of Amazonian forest fragments: effects of insularixation. Prec. XIX Intern. Ornithol. Cong., p. 1564-1579. 1986.
BIERREGAARD, R. O. J.; LOVEJOY, T. E. Effects of forest fragmentation on Amazonian understory bird communities. Acta Amazonica, v.19, p.215-241. 1989.
BIERREGAARD, R. O. J.; LOVEJOY, T. E.; KAPOS, V.; SANTOS, A. A.; HUTCHINGS, R. W. The biological dynamics of tropical rainforest fragments. Bioscience, v.42, no.11, p.859-866. 1992.
BIERREGAARD, R. O. J.; STOUFFER, P. C. Understory birds and dynamic habitat mosaics in Amazonian rainforests. In: LAURENCE, W.F.; BIERREGAARD, R.O.J. (Org.). Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago: University of Chicago Press, 1997. p.138-155.

127
BOECKLEN, W. J. E. GOTELLI, N. J. Island biogeographic theory and conservation practice: species area or specious-area relationships? Biological Conservation, v.29, p.63-80. 1984.
BOLGER, D.T.; ALBERTS, A.C.; SOULÉ, M.E. Occurrence patterns of bird species in habitat fragments: sampling, extinction, and nested species subsets. The American Naturalist, v.137, p.155-166. 1991
BORGES, S. H.; GUILHERME, E. Comunidade de aves em um fragmento florestal urbano em Manaus, Amazonas, Brasil. Revista Brasileira de Ornitologia, v.8, no.1, p.17-23. 2000.
BORNSCHEIN, M. R.; REINERT, B. L. Aves de três remanecentes florestais do norte do Estado do Paraná, sul do Brasil, com sugestões para a conservação e manejo. Revista Brasileira de Zoologia, v.17, no.3, p.615-636. 2000.
BRASIL. Departamento Nacional de Produção Mineral. Projeto RADAM. v.32. Folhas SF 23/24 Rio de Janeiro/Vitória. Rio de Janeiro. 780p. 1983.
BROOKS, T. M.; BALMFORD, A. Atlantic forest extinctions. Nature, v.380, p.115. 1996.
BROOKS, T. M.; TOBIAS, J.; BALMFORD, A. Deforestation and bird extinction in the Atlantic forest. Animal Conservation, v.2, p.211-222. 1999.
BROWN, K. S.; BROWN, G. G. Habitat alteration and species loss in Brazilian forest. In: WHITMORE, T.C.; SAYER, J.A. (Org.). Tropical deforestation and species extinction. Londres: Chapmam and Hall, 1992. p.119-142.
BROWN, K. S. & FREITAS, A. V. L. Diversidade de Lepidoptera em Santa Teresa, Espirito Santo. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.71-116. 2000.
BROWN, K. S.; HUTCHINGS, R. W. Disturbance, fragmentation, and the dynamics of diversity in Amazonian forest butterflyes. In: LAURENCE, W. F.; BIERREGAARD, R. O. J. (Org.). Tropical forest remnants: ecology, management, and conservation of fragmented communities. The University of Chigaco Press: 1997. p.91-110.
CARVALHO, K. S.; VASCONCELOS, H. L. Forest fragmentation in central Amazonia and effects on litter-dwelling ants. Biological Conservation, v.91, p.151-157. 1999.
CHIARELLO, A. G. Effects of fragmentation of the Atlantic forest on mammal communities in south-eastern Brazil. Biological Conservation, v.89, p.71-82. 1999.
CHIARELLO, A. G. Conservation value of a native forest fragment in a region of extensive agriculture. Revista Brasileria de Biologia, v.60, no.2, p.237-247. 2000a.
CHIARELLO, A. G. Influência da caça ilegal sobre mamíferos e aves das matas de tabuleiro do norte do estado do Espírito Santo. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.229-247. 2000b.

128
CHIARELLO, A. G. Density and populations size of mammals in remnants of Brazilian Atlantic Forest. Conservation Biology, v.14, no.6, p.1649-1657. 2000c.
CHIARELLO, A. G.; MELLO, F. R. Primate population densities and sizes in Atlantic forest remnants of northern Espirito Santo, Brasil. International Journal of Primatology, Londres, v.22, no.3, p.379-396. 2001.
CHRISTIANSEN, M. B.; PITTER, E. Species loss in a forest bird community near Lagoa Santa in Southeastern Brazil. Biological Conservation, v.80, p.23-32. 1997.
COLLAR, N. J.; GONZAGA, L. P.; KRABBE, N.; MADROÑO NIETO, A.; NARANJO, L. G.; PARKER III, T. A.; WEGE, D. C. Threatened Birds of the Americas: the ICBP/IUCN Read Data Book. Cambridge, U.K: International Council of Bird Preservation, 1150 p. 1992.
COLWELL, R. K. EstimateS: Statistical estimation of species richness and shared species from samples. 1997. Disponível: http://viceroy.eeb.uconn.edu/estimates [Acessado em 26 de agosto de 2004]. COOK, R. R. The relationship between nested subsets, habitat subdivision, and species diversity. Oecologia, v. 101,p. 204-210.1995. CONSERVATION INTERNATIONAL DO BRAZIL; FUNDAÇÃO SOS MATA ATLÂNTICA; FUNDAÇÃO BIODIVERSITAS; INSTITUTO DE PESQUISAS ECOLÓGICAS; SECRETARIA DO MEIO AMBIENTE DO ESTADO DE SÃO PAULO; INSTITUTO ESTADUAL DE FLORESTAS - MG. Avaliação e ações prioritárias para a conservação da biodiversidade da Mata Atlântica e Campos Sulinos. Brasília: Ministério do Meio Ambiente, Secretaria de Biodiversidade e Florestas, 40 p. 2000.
COSSON, J. F.; PONS, J. M.; MASSON, D. Effects of forest fragmentation on frugivorous and nectarivorous bats in French Guiana. Journal of Tropical Ecology, v.15, p.515-534. 1999.
CULLEN, L., BODMER, R. E.; PADUA, C. V. Effects of hunting in habitat fragments of the Atlantic Forest, Brazil. Biological Conservation, v.95, p.49-56. 2000.
CUTLER, A. Nested faunas and extinction in fragmented habitats. Conservation Biology, v.5, no.4, p.495-505.1991.
DAVIES, K. F.; MARGULES, C.; LAWRENCE, J. W. Which traits of species predict population decline in experimental forest fragments? Ecology, v.2, p.1450-1461. 2000.
D'ANGELO NETO, S.; VENTURIN, N.; FILHO, A. T. O.; COSTA, F. F. Avifauna de quatro fisionomias florestais de pequeno tamanho (5-8 ha) no campus da UFLA. Revista Brasileira de Biologia, v.58, no.3, p.463-472. 1998. DEBINSKI, D. M.; HOLT, R. D. A survey and overview of habitat fragmentation experiments. Conservation Biology, v.14, no.2, p.342-355. 2000.
DEAN, W. A Ferro e Fogo: a História e a Devastação da Mata Atlântica Brasileira. São Paulo: Companhia das Letras, 484p. 1997.

129
DEVELEY, P. F. Efeitos da Fragmentação e o Estado de Conservação da Floresta na Diversidade de Aves de Mata Atlântica. 2004. Tese de Doutorado, Instituto de Biociência da Universidade de São Paulo. São Paulo. 2004. DEVELEY, P. F.; PERES, C. A. Resource seasonality and the structure of mixed species bird flocks in a coastal Atlantic forest of southeastern Brazil. Journal of Tropical Ecology, v.16, no.1, p.33-53. 2000. DIAMOND, J. M. The island dilemma: lessons of modern biogeographic studies for the design of natural reserves. Biological Conservation, v.7, p.129-147. 1975.
ESTADO DO ESPÍRITO SANTO. Município de Santa Teresa. Estatística, Corografia e História. Rio de Janeiro: Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, 99p. 1939. FISZION, J. T.; MARCHIORO, N. P. X.; BRITEZ, R. M.; CABRAL, D. C.; CAMELY, N. C.; CANAVESI, V.; CASTELLA, P. R.; CASTRO, E. B. V.; JUNIOR, L. C.; CUNHA, M. B. S.; FIGUEIREDO, E. O.; FRANKE, I. L.; GOMES, H.; GOMES, L. J.; HREISEMNOU, V. H. V.; LANDAU, E. C.; LIMA, S. M. F.; LOPES, A. T. L.; NETO, E. M.; MELLO, A. L.; OLIVEIRA, L. C.; ONO, K. Y.; PEREIRA, N. W. V.; RODRIGUES, A. S.; RODRIGUES, A. A. F.; RUIZ, C. R.; SANTOS, L. F. G. L.; SMITH, W. S.; SOUZA, C. R. Causas antrópicas. In: RAMBALDI, D. M.; OLIVEIRA, D. A. S. D. (Org.). Fragmentação de ecossistemas: causas, efeitos sobre a biodiversidade e recomendações de políticas públicas. Brasília: MMA/SBF, 2003. p. 65-99.
FORMAN, R. T. T.; GALLI, A. E.; LECK, C. F. Forest size and avian diversity in New Jersey woodlots with some land use implications. Oecologia, v.26, no.9, p.1-8. 1976.
FREEMARK, K. E.; MERRIAN, H. G. Importance of area and habitat heterogeneity to bird assemblages in temperate forest fragments. Biological Conservation, v.36, p.115-141. 1986.
FUNDAÇÃO SOS MATA ATLÂNTICA; INPE. Evolução dos remanescentes florestais e ecossistemas associados do domínio da Mata Atlântica no período de 1985 – 1990. São Paulo. 1993. GALETTI, M.; PIZO, M. A. Fruit eating by birds in a forest fragment in southeastern Brazil. Revista Brasileira de Ornitologia, 4: 71-79. 1996.
GALETTI, M.; ZIPARRO, V. B.; MORELLATO, P. C. Fruiting phenology and frugivory on the Palm Euterpe edulis in a lowland Atlantic forest of Brazil. Ecotropica, v.5, p.115-122. 1999.
GASCON, C., LOVEJOY, T. E., BIERREGAARD, R. O. J., MALCOLM, J. R., STOUFFER, P. C., VASCONCELOS, H. L., LAURENCE, W. F., ZIMMERMAN, B. L., TOCHER, M. D.; BORGES, S. H. Matrix habitat and species richness in tropical forest remnants. Biological Conservation, v. 91, p.223-229. 1999.

130
GASCON, C.; LAURENCE, W. F.; LOVEJOY, T. E. Fragmentação florestal e biodiversidade na Amazônia Central. In: GARAY, I. E. G.; DIAS, B. F. S. (Org.). Conservação da biodiversidade em ecossistemas tropicais: avanços conceituais e revisão de novas metodologias de avaliação e monitoramento. Petrópolis: Editora Vozes, 2001 p. 112-127.
GIMENES, M. R.; ANJOS, L. D. Distribuição espacial de aves em um fragmento florestal do campus da Universidade Estadual de Londrina, norte do Paraná, Brasil. Revista Brasileira de Zoologia, v.17, no.1, p.263-271. 2000.
GOERCK, J. M. Patterns of rarity in the birds of Atlantic Forest of Brazil. Conservation Biology, v.11, no.1, p.112-118. 1997.
GONZAGA, L. P., CASTIGLIONI, G. D. A. Aves das montanhas do sudeste do Brasil. Sonopress Rimo da Amazônia Indústria e Comércio Ltda. 2001.
GRANTSAU, R. Os Beija-flores do Brasil: uma Chave de Identificação para todas Formas de Beija-flores do Brasil. Rio de Janeiro: Expressão e Cultura, 233 p. 1988.
GUEVARA, E.; GUEVARA, J. Dados preliminares sobre la diversidad y estructura de la comunidad de aves del bosque integral otongachi, Pichincha, Ecuador. Ornitologia Neotropical, v.16, p.129-136. 2005.
HANSKI, I. E.; GILPIN, M. E. Metapopulation dynamics: brief history, and conceptual domain. In: HANSKI, I.; GILPI, M. E. (Org.) Metapopulation dynamics: empirical and theoretical investigations. Academic Press, London, 1991. p.3-16.
HANSKI, I. E; SIMBERLOFF, D. S.C. The metapopulation approach, its history, conceptual domain, and application to conservation. In: HANSKI, I. e GILPIN, M. E. (Org.). Metapopulation biology: ecology, genetics and evolution. San Diego: Academic Press, 1997. p.5-12. HAGAN, J. M.; HAEGEN, W. M. V.; MCKINLEY, P. S. The early development of forest fragmentation effects on birds. Conservation Biology, v.10, no.1, p.188-202. 1996. HAILA, Y. A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological Applications, v.12, p.321-334. 2002.
HARRIS, L. D. The Fragmented Forest. Chicago: University of Chicago Press. 1984.
HARRIS, L. D. The Fragmented Forest: Island Biogeography Theory and Preservation of Biotic Diversity. Chicago: University of Chicago Press. 1984.
HARRIS, L. D. Edge effects and conservation of biotic diversity. Conservation Biology, v.2, p.330-332. 1988. HARRISON, S. Local extinction in a metapopulation context: an empirical avaliation. In: GILPIN, M. E.; HANSKI, L. (Org.) Metapopulation dynamics: empirical and theoretical investigations. London: Academic Press, 1991.

131
HARRIS, G. M.; PIMM, S. L. Bird species's tolerance of secondary forest habitats and its effects on extincton. Conservation Biology, v.18, no.6, p.1607-1616. 2004.
HELTSHE, J. F. E.; FORRESTER N. E. Estimating species richness using the Jackknife procedure. Biometrics, v.39, p.1-11. 1983. HUMPHEREYS, W. F.; KITCHENER, D. J. The effect of habitat utilization on species-area curves: implication for optimal reserve area. Journal of Biogeography, v.9, p.391-396. 1982. IPEMA. Lista da Fauna e Flora Ameaçadas de Extinção no Estado do Espírito Santo. Disponível: http://www.ipema-es.org.br/hp/faunameacada.htm 2005a [capiturado em16 out. 2005]. IPEMA. Conservação da Mata Atlântica no Estado do Espírito Santo: Cobertura Florestal e Unidades de Conservação. IPEMA: Vitória, ES. 142p. 2005b. JOHNSON, R. R.; BROWN, B. T.; HAIGHT, L. T.; SIMPSON, J. M. Playback recordings as a special avian censusing technique. Studies in Avian Biology, v.6, p.68-75. 1981. KARR, J. R. Surveying birds in the tropics. Studies in Avian Biology, v.6, p.533-548. 1981a.
KARR, J. R. Surveying birds with mist nets. In: RALPH, C. J.; SCOTT, J. M. (Org.) Estimating numbers of terrestrial birds, p. 62-67. 1981b.
KATTAN, G. R.; ALVAREZ - LOPEZ, H.; GERALDO, M. Forest fragmentation and bird extinctions: San Antonio eighty years later. Conservation Biology, v.8, no.1, p.138-146. 1994. KLEIN, B. C. Effects of forest fragmentation on dung and carrion beetle communities in central Amazonian. Ecology, 70, p.1715-1725. 1989. LAMBECK, R. Focal species: a multi species umbrella for nature conservation. Conservation Biology, v.11, p.849-856. 1997. LAPS, R.; CORDEIRO, P.H.C.; RIBON, R.; RODRIGUES, A.A.F.; UEJIMA, A. Aves. In: RAMBALDI, D.M.; OLIVEIRA, D.A.S.D. (Org.). Fragmentação de ecossistemas: causas, efeitos sobre a biodiversidade e recomendações de políticas públicas. Brasília: MMA/SBF, 2003. p. 153-181.
LAURENCE, W. F. Comparative responses of five arboreal marsupials to tropical forest fragmentation. Journal of Mammalogy, v.71, p.641-653. 1990. LAURENCE, W. F. E YENSEN, E. Predicting the impacts of edge effects in fragmented habitats. Conservation Biology, v.55, no.1, p.77-92. 1991.

132
LAURENCE, W. F. Edge effects in tropical forest fragments: application of a model for the design of nature reserves. Biological Conservation, v.57, p.205-219. 1991. LAURENCE, W. F. Rainforest fragmentation and the structures of small mammal communities in tropical Queensland. Biological Conservation, v.69, p.23-32. 1994. LAURENCE, W. F. Ecology and management of fragmented tropical landscapes. Biological Conservation, v.91, p.2-3. 1999a. LAURENCE, W. F. Introduction and synthesis. Biological Conservation, v.91, p.101-107. 1999b. LAURENCE, W. F.; VASCONCELOS, H. L.; LOVEJOY, T. E. Forest loss and fragmentation in the Amazon: implications for wildlife conservation. Oryx, v.34, p.39-45. 2000. LAURANCE, W. F. Hyperdynamism in fragmented habitats. Veg. Sci., v.13, p.595-602. 2002.
LAURENCE, W. F.; BIERREGAARD, R. O. J.; GASCON, C.; DIDHAM, R. K.; SMITH, A. P.; LYNAM, A. J.; VIANA, V. M.; LOVEJOY, T. E.; SIEVING, K. E.; SITES, J. W.; ANDERSEN, M.; TOCHER, M. D.; KRAMER, E. A.; RESTREBO, C.; MORITZ, C. Tropical forest fragmentation: synthesis of a diverse and dynamic discipline. In: LAURENCE, W. F. & BIERREGAARD, R. O. J. (Org.) Tropical forest remnants: ecology, management, and conservation of fragmented communities, The University of Chigaco Press. 1997. p. 502-513.
LAURENCE, W. F.; FERREIRA, L. V.; RANKIN-de-MERONA, J. M.; LAURENCE, S. G. Rain forest fragmentation and the dynamics of Amazonian tree communities. Ecology, v.79, p.2032-2040. 1998. LAURENCE, W. F.; LOVEJOY, T. E.; VASCONCELOS, H. L.; BRUNA, E. M.; DIDHAM, R. K.; STOUFFER, P. C.; GASCON, C.; BIERREGAARD, R. O. J.; LAURENCE, W. F.; SAMPAIO, E. Ecosystem decay of Amazonian forest fragments: a 22-years investigation. Conservation Biology, v.16, no.2, p.605-618. 2002. LAWTON, J. H. Population dynamic principles. In: LAWTON, J. H.; MAY, R. M. (Org.). Extinction rates. Oxford: Oxford University Press, 1995. p.145-163. LECK, C. F. Avian extinction in an insulated tropical wet-forest preserve, Equador. The Auk 96, 343-352. 1979. LEME, A. Foraging substrate selection by ochrerumped antbird Drymophila ochropyga. Revista Brasileira de Ornitologia, v.9, no.1, p.7-11. 2001. LINDENMAYER, D. B.; FRANKLIN, J. F. Conserving Forest Biodiversitu. A Comprehensive Multiscale Approach. Washington: Island Press. 2002. LOVEJOY, T. E.; BIERREGAARD, R. O. J.; RANKIN, J. M.; SCHUBART, H. O. R. Ecological dynamics of tropical forest fragments. In: SUTTON, S. L.; WHITMORE, T. C.;

133
CHADWICK, A. C. (Org) Tropical rain forest: ecology and management. Oxford: Blackwell Scientific Plubication. 1983.
LOVEJOY, T. E.; RANKIN, J. M.; BIERREGAARD Jr., R. O.; BROWN Jr., K.S.; EMMONS, M. E.; VAN DER VOORT. Ecosystem decay of Amazon forest remnants. In: NITECKI, M. H. (Org.) Extinctions. Chicago: University of Chicago Press, 1984. p.295-325.
LOVEJOY, T. E.; BIERRENGAARD Jr, R. O.; RYLANDS, A. B.; MALCOM, J. R.; QUINTELA, C. E.; HARPER, L. H.; BROWN Jr., K. S.; POWELL, G. V. N., SCon \HUBART, H. O. R.; HAYS, M. B. Edge effects and other effects of isolation on Amazon frest fragment. In: SOULÉ, M. E. (Org.). Conservation biology: the science of scarcity and diversity. Sinauer Associates, Sunderland, Massachusetts. p. 257-285.
MACARTHUR, R. H. E.; WILSON, E. O. The Theory of Island Biogeography. Princeton: Princeton University Press, 1967.
MACHADO, A. B. M.; MARTINS, C. S.; DRUMMOND, G. M. (Org.) Lista da Fauna Brasileira Ameaçada de Extinção. Incluindo as Listas das Espécies Quase Ameaçadas e Deficientes de Dados. Belo Horizonte: Fundação Biodiversitas. 2005. MAGALHÃES, F. C. R. As Aves na Fazenda Barreiro Rico. Plêiade: São Paulo. 1999. MAGURRAN, A. E. Ecological Diversity and its Measurement. Princeton: Princeton University, 1988. MALCOLM, J. R. The small mammals of Amazon forest fragments: pattern and process. PhD. Thesis. University of Florida, USA. 1991. MALCOLM, J. R. Biomass and diversity of small mammals in Amazonian forest fragments. In: LAURENCE, W. F.; BIERREGAARD, R. O. J. (Org) Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago: University of Chicago Press, 1997. p. 207-221.
MALDONADO-COELHO, M.; MARINI, M. A. Effects of forest fragment size and sucessional slage on mixed-species bird flocks in southeastern Brazil. The Condor, v.102, p.585-594. 2000. MARION, W. R.; O'MEARE, T. E.; MAEHR, D. S. Use of playback recordings in sampling elusive or secretive birds. Studies in Avian Biology,v. 6, p.81-85. 1981. MARSDEN, S. J.; WHIFFIN, M.; GALETTI, M. Bird diversity and abundance in forest fragments and Eucalypus plantations around an Atlantic forest reserve, Brazil. Biodiversity and Conservation, v.10, p.737-751. 2001. MCCUNE, B. E.; MEFFORD, M. J. PC-ORD - Multivariate Analysis of Ecological Data, version 2.0. Gleneden Beach: MjM Soft Design, 1995. MINISTÉRIO DO MEIO AMBIENTE. Instrução Normativa MMA de 27 de maio de 2003. Disponível em http://www.mma.gov.br.

134
MENDES, S. L.; PADOVAN, M. D. P. A Estação Biológica de Santa Lúcia, Santa Teresa, Espírito Santo. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.7-34. 2000. MANNE,L.; L., PIMM, S. L. Beyond eight forms of rarity: which species are threatened and which wil be next?. Animal Conservation, v.4, p.221-229. 2001. METZGER, J. P. Estrutura da paisagem e fragmentação: análise bibliográfica. Anais da Academia Brasileira de Ciências, v.71, p.445-463. 1999.
MEYER DE SCHAUENSEE, R. A Guide to the Birds of South America. Academy of Natural Scienses of Philadelphia: Ed. Livingston, 470p. 1970.
MITTERMEIER, R. A.; FONSECA, G. A. B.; RYLANDS, A. B.; MITTERMEIER, C. G. Atlantic forest. In: MITTERMEIER, R. A.; MYERS, N.; GIL, P. R.; MITTERMEIER, C. G. (Org.) Hotspots: earth's biologically richest and endangered terrestrial ecoregions. Cidade do México: Conservation International e Cemex, 1999. p. 137-147.
MITTERMEIER, R. A.; MYERS, N.; THOMSEN, J. B.; FONSECA, G. A. B.; OLIVIERI, S. Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation priorities. Conservation Biology, v.12, p.516-520. 1998. MOOJEN, J.; CARVALHO, J. C.; LOPES, H. S. Observações sobre o conteúdo gástrico das aves brasileiras. Memórias do Instituto Oswaldo Cruz, v.36, p.404-444. 1941.
MORRIS, J. The truth in the balance. Nature, v.375, p.115-116. 1995.
MOTTA-JUNIOR, J. C. Estrutura trófica e composição das avifaunas de três hábitats terrestres na região central do estado de São Paulo. Revista Brasileira de Ornitologia, v.1, p.65-71. 1990. MURCIA, C. Edge Effects in fragmented forest: implications for conservation. Tree, v.10, no.2, p.58-62. 1995. NEWMARK, W. D. Tropical forest fragmentation and the local extinction of understory birds in the Eastern Usambara mountains, Tanzania. Conservation Biology, v.5, p.67-78. 1991. ONIKI, Y.; WILLIS, E. O. Building and incubation at a nest of Frilled Coquettes, Lophornis magnifica (Trochilidae). Ornitologia Neotropical, v.9, no.1, p.77-80. 1998a. ONIKI, Y.; WILLIS, E. O. Nesting of Yellow-Fronted Woodpeckers, Melanerpes flavifrons (Picidae). Ornitologia Neotropical, v.9, no.1, p.81-85. 1998b.
PACHECO, J. F.; BAUER, C. A lista de aves do Espírito Santo de Augusto Ruschi (1953): uma análise crítica. In: ALBUQUERQUE, J. L. B.; CÂNDIDO Jr, J. F.; STRAUBE, F. C.; ROOS, A. L. (Org.) Ornitologia e conservação: da ciência às estratégias. Tubarão: Unisul, 2001a. p.261-278.

135
PACHECO, J. F.; BAUER, C. As aves do Espírito Santo do Príncipe Maximiliano de Wied. Atualidades Ornitológicas, v.99, p.6. 2001b.
PACHECO, J. F.; FONSECA, P. S. M.; PARRINI, R. Coletânia cronológica de registros recentes de Harpia harpyja (L.) para os estados do Rio de Janeiro e Espírito Santo. Atualidades Ornitológicas, v.111, no.7. 2003. PARKER III, T. A. On the use of tape recorders in avifaunal surveys. The Auk, v.108, no.2, p.443-444. 1991.
PARKER III, T. A.; GOERCK, J. M. The importance of national parks and biological reserves to birds conservation in the Atlantic forest region of Brazil. In: REMSEN, J. V. J. (Org.) Studies in neotropical ornithology honoring Ted Parker. Washington: The American Ornitologistis' Union, 1997. p. 527-541.
PARKER III, T. A.; STOTZ, D. F.; FITZPATRICK, J. W. Ecological and distributional databases. In: STOTZ, D. F.; FITZPATRICK, J. W.; PARKER III, T. A.; MOSKOVITS, D. K. (Org.) Neotropical birds: ecology and conservation. Chicago: The University of Chicago Press, 1996. p. 113-436.
PASSAMANI, M.; MENDES, S. L.; CHIARELLO, A.G. Non-volant mammlas of the Estação Biológica de Santa Lúcia and adjacents areas of Santa Teresa, Espírito Santo, Brazil. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.201-214. 2000.
PASSAMANI, M. O Efeito da Fragmentação da Mata Atlântica Serrana sobre a Comunidade de Pequenos Mamíferos de Santa Teresa, Espírito Santo. 2003. Tese de Doutorado, UFRJ. Rio de Janeiro. 2003. PATTERSON, B. D. The principle of nested subsets and its implications for biological conservation. Conservation Biology v. 1, p. 323-334. 1987. PATTERSON, B. D.; ATMAR, W. Analyzing species composition in fragments. Bonn. Zool. Monogr. v. 6, p.9-24. 2000. PRICE, O. F.; WOINARSKI, J. C. Z.; ROBINSON, D. Very large area requirements for frugivorous birds in monsoon rainforests of the Northern Territory, Australia. Biological Conservation, v.91, p.169-180. 1999. PIMM, S. L.; JONES, H. L.; DIAMOND, J. On the risk of extinction. American Naturalist. v.132, p.757-785. 1988 POLETTO, F.; ANJOS, L.; LOPES, E. V.; FAVARO, F. L.; VOLPATO, G. H.; SERAFINI, P. P. Caracterização do microhabitat e vulnerabilidade de cinco espécies de arapaçus (Aves: Dendrocolaptidae) em um fragmento florestal do norte do estado do Paraná, sul do Brasil. Revista Brasileira de Ornitologia, v.2, p.89-96. 2004. RALPH, C. J. Et al. Manual de métodos de campo en ornitologia. USDA Forest Service. 1997.

136
RAMOS, A. D.; GASPARINI, J. L. Anfíbios do Goiapaba-Açu, Fundão, Estado do Espírito Santo. Fundão: Santo Antônio, 75 p. 2004. RANDERSON, P. F. Ordination. In: FRY, J. C. (Org.) Biological data analysis: a practical approach. New York: Oxford University Press. 1996. p. 173-209. REMSEN, J. V. J.; PARKER III, T. A. Arboreal Dead-leaf-searching birds of the Neotropics. The Condor, v.86, p.36-41. 1984. RENJIFO, L. M. Composition changes in a subandean avifauna after long-term forest fragmentation. Conservation Biology, v.13, p.1124-1139. 1999.
RENJIFO, L. M. Effects of natural and anthropogenic landscape matrices on the abundance of subandean bird species. Ecological Applications, v.11, p.14-31. 2001.
RIBON, R. Fatores que Influenciam a Distribuição da Avifauna em Fragmentos de Mata Atlântica nas Montanhas de Minas Gerais. 1998. Dissertação de Mestrado, UFMG. Belo Horizonte, Minas Gerais. 1998 RIBON, R. Aves em Fragmentos de Mata Atlântica do Sudeste de Minas Gerais: Incidência, Abundância e Associação à Topografia. 2003. Tese de Doutorado, UFMG. Belo Horizonte, Minas Gerais. 2003. RIBON, R.; SIMON, J. E.; MATTOS, G. T. Bird extinctions in Atlantic forest fragments of the Viçosa region, southeastern Brazil. Conservation Biology, v.17, no.6, p.1827-1839. 2003. RIDGELY, R. S.; TUDOR, G. The Birds of South America, 1. Austin : University of Texas Press. 516p. 1989.
RIDGELY, R. S.; TUDOR, G. The Birds of South America, 2. Austin: Universidade of Texas Press. 814p. 1994.
RIZINI, C. T. Tratado de Fitogeografia do Brasil. Aspectos Sociológicos e Florísticos. São Paulo: Editora da Universidade de São Paulo. 1979.
ROSENFIELD, R. N.; MORASKY, C. M.; BIELEFELDT, J. J.; LOOPE, W. L. Forest Fragmentation and Island Biogeography: a Summary and Bibliography. Denver: United States Depart. of the Interior. Nat. Park Serv. Natur. Res. Publ. Office, 51p. 1992.
RUSCHI, A. O café e o estado do Espírito Santo. Boletim do Museu de Biologia Mello Leitão, v.42, p.1-51. 1974. RUSCHI, A. A ornitofauna da Estação Biológica do Museu Nacional. Boletim do Museu de Biologia Mello Leitão, v.88, p.1-10. 1977. RUSCHI, A. As aves do recinto da sede do Mus. Biol. M. Leitão, em S. Teresa, observadas durante os anos 1968-1969. Boletim do Museu de Biologia Mello Leitão, série Proteção à Natureza, v.31, p.1-14. 1969.

137
SAUNDERS, D. A.; HOBBS, R. J.; MARGULES, C. Biological consequences of ecosystem fragmentation: a review. Conservation Biology, v.7, p.18-32. 1991. SCHUBART, O.; AGUIRRE, A. C.; SICK, H. Contribuição para o conhecimento da alimentação das aves brasileiras. Arquivos de Zoologia, v.12, p.95-249. 1965. SCOTT, D. A. A possible re-sighting of the Cherry-throated Tanager Nemosia rourei in Espírito Santo, Brazil. Cotinga, v.7, p.61-63. 1997.
SCOTT, D. A.; BROOKE, M. L. The endangered avifauna of southeastern Brazil: a report on the BOU/WWF expeditions of 1980/81 and 1981/82. In: DIAMOND, A. W.; LOVEJOY, T. E. (Org) Conservation of Tropical Forest Birds. Cambridge: International Council for Birds Preservation, 1985. p. 115-139.
SHAFFER, M. L. Minimum population sizes for species conservation. Bioscience, v.31, no.2, p.131-134. 1981.
SHEPHERD, G. J. Fitopac 1.0: Manual do usuário. Campinas: Unicamp. 88p. 1984.
SICK, H. Migração e Aves na América do Sul Continental. Brasília: Centro de Estudos de Migração de Aves (Publicação Técnica 2). 86p. 1984.
SICK, H. Ornitologia Brasileira. Edição revista e ampliada por José Fernando Pacheco. Rio de Janeiro: Nova Fronteira. 862p. 1997.
SIEVING, K. E.; WILLSON, M. F.; de SANTO, T. L. Habitat barriers to movement of understory birds in fragmented south temperate rainforest. The Auk, v.113, no.4, p.944-949. 1996.
SIEVING, K. E.; KARR, J. R. Avian extinction and persistence mechanisms in lowland Panana. In: LAURENCE, W. F.; BIERREGAARD, R. O. J. (Org) Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago: The University of Chigaco Press. 1997. p. 156-170.
SIMÃO, I.; SANTOS, F. A. M.; PIZO, M. A. Vertical stratification and diet of psittacids in a tropical lowland forest of Brazil. Revista Brasileira de Ornitologia, v.5, no.2, p.169-174. 1997.
SIMBERLOFF, D. Habitat fragmentation and population extinction of birds. Ibis, v.137, p.105-111. 1995. SIMBERLOFF, D.; ABALE, L. G. Refuge design and island biogeographyc theory effects of fragmentation. The American Naturalist, v.20, no.1, p.41-51. 1982.
SIMON, J. E. Composição da avifauna da Estação Biológica de Santa Lúcia, Santa Teresa - ES. Boletim do Museu de Biologia Mello Leitão, v.11/12, p.149-170. 2000.
SIMON, J.E. Status de conservação de Ara chloroptera (Psittacidae) no Estado do Espírito Santo. Livro de Resumos do IX Congresso Brasileiro de Ornitologia.Curitiba: PUC, 2001. p.364-365.

138
SIMON, J. E. Registro de aves raras com a frutificação de taquaras na região de Santa Teresa, Espírito Santo, Brasil. Resumos do XII Congresso Brasileiro de Ornitologia, p.381. 2004. Universidade Regional de Blumenau. 2004. SIMON, J. E.; CHIARELLO, A. G. Status de conservação dos Tinamidae do Estado do Espírito Santo. Resumos do IX Congresso Brasileiro de Ornitologia, p.366-367. 2001. Curitiba, Paraná, PUC. Resumos do IX Congresso Brasileiro de Ornitologia. 2001.
SIMON, J. E.; CAZOTTO, R. L. Pressão de caça sobre a avifauna da região de Santa Teresa. Resumos do X Congresso Brasileiro de Ornitologia. p.107-108. 2002. Fortaleza, Ceará. 2002.
SIMON, J. E.; GASPARINI, J. L. Descrição da vocalização de Phyllodytes kautskyi Peixoto e Cruz, 1988 (Amphibia, Anura, Hylidae). Boletim do Museu de Biologia Mello Leitão, v.16/12, p.5-54. 2003. SIMON, J. E.; LIMA, S. R. Diversidade da avifauna na região da APA do morro Mestre Álvaro, município da Serra, Espírito Santo. Resumos do XII Congresso Brasileiro de Ornitologia, p.383-384. 2004. Universidade Regional de Blumenau. 2004a. SIMON, J. E.; LIMA, S. R. Inventário preliminar da avifauna da região da APA do morro Mestre Álvaro, município da Serra, Espírito Santo. Resumos do XII Congresso Brasileiro de Ornitologia , p.382. 2004. Universidade Regional de Blumenau. 2004b. SMITH, E. P.; BELLE, V. Non parametric estimation of species richness. Biometrics, v.40, p.119-129. 1984.
SNEATH, P. H. E.; SOKAL, R. R. Numerical Taxonomy. San Francisco: W.H. Freeman and Company, 573p. 1973.
SOARES, E. S.; ANJOS, L. Efeito da fragmentação florestal sobre aves escaladoras de tronco e galho na região de Londrina, norte do Estado do Paraná, Brasil. Ornitologia Neotropical, v.10, p.61-68. 1999. SOULÉ. M. E.; SIMBERLOFF, D.S. What do genetics and conservation tell us about the design of nature reserves? Biological Conservation, v.35, p.19-40. 1986.
SOUZA, D. All the Birds of Brazil: an Identification Guide. Salvador: DALL, 356p. 2002.
SOULÉ, M. E.; WILCOX, B. A.; HOLTBY, C. Bening neglect: a model of faunal collapse in the game reserves of east Africa. Biological Conservation, v.15, p.259-272. 1979.
SOLUÉ, M. R.; BOLGER, D. T.; ALBERTS, A. C.; SAUVAJOT, R. S.; WRIGHT, J.; SORICE, M.; HILL, S. Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conservation Biology, v.2, p.75-92. 1988.

139
STRATFORD, J.A.; STOUFFER, P.C. Local extinctions of terrestrial insectivorous birds in a fragmented landscape near Manaus, Brazil. Conservation Biology, v.13, no.6, p.1416-1423. 1999.
STRAUBE, C. S.; URBEN-FILHO, A.; KAJIWARA, D. Aves: In: MIKICH, S. B.; BÉRNILS, R. S. (Org.). Livro Vermelho da Fauna Ameaçada no Estado do Paraná. Curitiba: Instituto Ambiental do Paraná, 2004. p.145-496. STOTZ, D. F. Geographic variation in species composition of mixed species flocks in lowland humid forests in Brazil. Papéis Avulsos de Zoologia, v.38, no.4, p.61-75. 1993.
STOTZ, D. F.; FITZPATRICK, J. W.; PARKER III, T. A.; MOSKOVITS, D. K. Neotropical birds: Ecology and conservation. Chicago: The University of Chicago Press. 1996.
STOUFFER, P. C.; BIERREGAARD, R. O. J. Effects of forest fragmentation on understory hummingbirds in Amazonian Brazil. Conservation Biology, v.9, p.1094. 1995a. STOUFFER, P. C.; BIERREGAARD, R. O. J. Use of Amazonian forest fragments by understory insectivorous birds: effects of fragment size surrounding secondary vegetation, and time since isolation. Ecology, v.76, no.8, p.2429-2445. 1995b.
SUTHERLAND, W. J. The Conservation Handbook: Research, Management and Policy. Oxford: Blackwell Science. 2000.
TEMPLETON, A. R.; ROBERTSON, R. J.; BRISSON, J.; STRASBURG, J. Disrupting evolutionary processes: the effect of fragmentation on Collared Lizards in the Missouri Ozarks. Proceedings of the National Academy of Science, v.98, p.5426-5432. 2001.
TERBORGH, J. Island biogegraphy and conservation: strategy and limitations. Science, v.193, p.1029-1030. 1976.
TERBORGH, J.; WINTER, B. Some causes of extinction. In: SOULÉ, M. E.; WILCOX, B. A. (Org.) Conservation biology: an evolutionary-ecological perspective. Sunderland: Sinauer Associates. 1980. p.119-133.
TERBORGH, J. Maintenance of diversity in tropical forests. Biotropica, v.24, p.283-292. 1992. THOMAZ, L. D.; MONTEIRO, R. Composição florística da Mata Atlântia de encosta da Estação Biológica de Santa Lúcia, município de Santa Teresa, ES. Boletim do Museu de Biologia Mello Leitão, v.7, p.3-48. 1997.
TOCHER, M. D. Diferenças na composição de espécies entre três tipos de florestas e campo de pastagem na Amazônia Central. In: GASCON, C.; MOUTINHO, P. (Org.) Floresta Amazônica: Dinâmica, Regeneração e Manejo. Manaus: Ministério de Tecnologia e Ciência, INPA, 1998. p. 219-232.
TOCHER, M. D.; GASCON, C.; ZIMMERMAN, B. Fragmentation effects on a central Amazonian frog communiy: a ten-year study. In: LAURANCE, W. F.; BIERREGAARD,

140
R. O. (Org.) Tropical Forest Remnants: Ecology, Management and Conservation of Fragmented Communities. Chicago: University of Chicago Press, 1997. p.124-137
TURNER, I. M. Species loss in fragments of tropical rain forest: a review of the evidence. Journal of Applied Ecology, v.33, p.200-209. 1996. TURNER, I. M.; CORLETT, R. T. The conservation value of small, isolated fragments of lowland tropical rain forest. Trends in Ecology and Evolution, v.11, no.8, p.330-333. 1996. UEZU, A.; METZGER, J. P.; VIELLIARD, J. M. E. Effects on structural and functional connectivity and patch size on the abundance of seven Atlantic Forest bird species. Biological Conservation, In Press. 2005. VALLAN, D. Influence of forest fragmentation on amphibian diversity in the nature reserve of Ambohitantely, highland Madagascar. Biological Conservation, v.96, p.31-43. 2000. VAN DORP, D.; OPDAM, P. F. M. Effects of patch size, isolation and regional abundance on forest bird communities. Landscape Ecology, v.1, p.59-73. 1987. VELOSO, H. P.; GÓES FILHO, L. Fitogeofrafia brasileira: classificação fisionômica-economica da vegetação neotropical. Boletim Técnico do Projeto RADAMBRASIL, v.1, p.3-79. 1982.
VENTURINI, A. C.; REHEN, M. P.; PAZ, P. R.; CARMO, L. P. Contribuição ao conhecimento das aves da região centro-serrana do Espírito Santo: Municípios de Santa Maria de Jetibá e Itarana. Atualidades Ornitológicas, v.98, p.11-12; v.99, p.12. 2000.
VIEIRA, M. V.; FARIA, D. M.; FERNANDEZ, F. A. S.; FERRARI, S. F.; FREITAS, S. R.; GASPAR, D. A.; MOURA, R. T.; PARDINI, R.; PIRES, A. S.; RAVETTA, A.; MELLO, M. A. R.; RUIZ, C. R.; SETZ, E. Z. F. Mamíferos. In: RAMBALDI, D. M.; OLIVEIRA, D. A. S. D. (Org.). Fragmentação de ecossistemas: causas, efeitos sobre a biodiversidade e recomendações de políticas públicas. Brasília: MMA/SBF, 2003. p. 125-151. VIELLIARD, J. M. E. Guia sonoro das aves do Brasil. [CD 1]. Campinas: Universidade Estadual de Campinas. 1995a.
VIELLIARD, J.M. Canto de aves do Brasil. CD. Sociedade Brasileira de Ornitologia. 1995b.
WATSON, D. M. A conceptual framework for studying species composition in fragments, islands and other patchy ecosystems. Journal of Biogeography, v.29, p.823-834. 2002. WHITMORE, T. C. Tropical forest disturbance, disappearence, and species loss. In: LAURENCE, W.F.; BIERREGAARD, R.O.J. (Org.). Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago: Univesity of Chicago Press, p. 3-12. 1997.

141
WHITNEY, B. M.; PACHECO, J. F. Distribution and conservation status of four Myrmotherula antwrens (Formicariidae) in the Atlantic Forest of Brazil. Bird Conservation International, v.5, p.295-313. 1995. WIEGAND, S. S. K.; HENLE, K. The conservation biology of a specialist and a generalist gecko in the fragmented landscape of the western Austrilian Wheatbelt. Sarreetal, p.39-51. 2005. WIENS, J. A. Population responses to patchy enviroments. Annual Review of Ecology and Systematics, v.7, p.81-120. 1976.
WIENS, J. A. Habitat fragmentation: island vs. Landscape perspectives on bird conservation. Ibis, v. 137, no.S97, p. 104.1995
WILCOX, B. A.; MURPHY D. D. Conservation strategy: the effects of fragmentation on extinctions. American Naturalist, v.125, p.879-887. 1985. WILLIS, E. O. Populations and local extinctions of birds on Barro Colorado Island, Panama. Ecological Monographs, v.44, p.153-169. 1974. WILLIS, E. O. The composition of avian communities in remanescent woodlots in southern Brazil. Papéis Avulsos de Zoologia, v.33, no.1, p.1-25. 1979. WILLIS, E. O. Ecological roles of migratory and resident birds on Barro Colorado Island, Panama. In: KEAST, A.; MORTON, E. S. (Org.) Migrant birds in the Neotropics: ecology, behavior, distribution and conservation. Washington: Smithsonian, 1980, p.205-225 WILLIS, E. O. Mimicry in bird flocks of cloud forests in southeastern Brazil. Revista Brasileira de Biologia, v.49, p.615-619. 1989. WILLIS, E. O.; ONIKI, Y. As aves e as formigas de correição. Boletim do Museu Paranaense Emílio Goeldi, Série Zoologia, v. 8, no.1, p. 123-151.1992
WILLIS, E. O.; ONIKI, Y. Na trilha das formigas carnívoras. Ciência Hoje, v.8, no.47, p.26-32. 1998a. WILLIS, E. O.; ONIKI, Y. One-parent nesting in Cinnamon-vented Pihas (Lipaugus lanioides, Cotinginae, Tyrannidae). Ornitologia Neotropical, v.9, p.129-159. 1998b.
WILLIS, E.O. & ONIKI, Y. Birds of Santa Teresa, ES, Brazil: Do humans add or subtract species? Papéis Avulsos de Zoologia, v.42, p.193-264. 2002 WILSON, M. E.; SANTO, T. L., SABAG, C.; ARMESTO, J. J. Avian communities of fragmented south-temperate rain forests of Chile. Conservation Biology, v.8, p.508-520. 1994.

142
WILSON, E. O.; WILLIS, E. O. Applied biogeography. In: CODY, M. L.; DIAMOND, J. M. (Org.) Ecology and evolution of communities. Cambridge: Belknap Press of Havard Univ, 1975. p.523-534.
ZAR, J. H. Biostatistical Analysis. Prentice Hall, New Jersey, 663p. 1999. ZIMMERMAN, B. L.; BIERREGAARD, R. O. J. Relevance of the equilibrium theory of island biogeography and species-area relations to conservation with a case from Amazonia. Journal of Biogeography, v.13, p.133-143. 1986.