
Variabilidade do domínio IIS6 do gene do canal de sódio...
117
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMAS DE PÓS-GRADUAÇÃO DO INPA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA EVOLUTIVA Ana Paula Barbosa da Silva MANAUS – AMAZONAS Março, 2014 Variabilidade do domínio IIS6 do gene do canal de sódio, associada à resistência aos inseticidas piretróides, em populações de Anopheles darlingi e Anopheles marajoara da Amazônia brasileira
Transcript of Variabilidade do domínio IIS6 do gene do canal de sódio...

i
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMAS DE PÓS-GRADUAÇÃO DO INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA EVOLUTIVA
Ana Paula Barbosa da Silva
MANAUS – AMAZONAS
Março, 2014
Variabilidade do domínio IIS6 do gene do canal
de sódio, associada à resistência aos inseticidas
piretróides, em populações de Anopheles darlingi
e Anopheles marajoara da Amazônia brasileira

ii
Ana Paula Barbosa da Silva
ORIENTADORA: Joselita Maria Mendes dos Santos, Dra.
CO-ORIENTADOR: Ademir de Jesus Martins Júnior, Dr.
Tese apresentada ao Programa de Pós-
Graduação do INPA como parte dos
requisitos para obtenção do título de Doutor
em Genética, Conservação e Biologia
Evolutiva.
MANAUS – AMAZONAS
Março, 2014
Variabilidade do domínio IIS6 do gene do canal de
sódio, associada à resistência aos inseticidas
piretróides, em populações de Anopheles darlingi e
Anopheles marajoara da Amazônia brasileira

iii
S586 Silva, Ana Paula Barbosa da
Variabilidade do domínio IIS6 do gene do canal de sódio, associada à resistência aos inseticidas piretróides, em populações de Anopheles darlingi e Anopheles marajoara da Amazônia brasileira / Ana Paula Barbosa da Silva. --- Manaus: [s.n], 2014.
xvii, 117 f. : il. color. Tese (doutorado) --- INPA, Manaus, 2014.
Orientador : Joselita Maria Mendes dos Santos.
Co-orientador : Ademir de Jesus Martins Júnior.
Área de concentração : Genética, Conservação e Biologia Evolutiva.
1. Malária. 2. Anofelinos. 3. Resistência kdr. 4. Inseticidas. 5. Canal de sódio I. Título.
CDD 616.9362
Sinopse:
Anopheles darlingi e Anopheles marajoara são duas importantes espécies vetoras de
malária no Brasil. O domínio IIS6 do gene do canal de sódio desses anofelinos
neotropicais foi sequenciado de amostras oriundas de seis localidades da Amazônia
brasileira, para investigar a presença de mutações kdr (knockdow resistance). Para A.
darlingi foram obtidos cinco sítios polimórficos, que agruparam as amostras em cinco
distintos haplótipos. Para A. marajoara foram obtidos quatro sítios polimórficos, que
agruparam as amostras em quatro distintos haplótipos. Todas as substituições encontradas
foram sinônimas e a clássica mutação kdr não foi encontrada. Não foi possível
correlacionar a distribuição dos haplótipos encontrados com a perda de susceptibilidade à
deltametrina, evidenciada em algumas populações.
Palavras-chave: malária, haplótipos, deltametrina, anofelinos, canal de sódio.

iv
DEDICATÓRIA
Aos meus pais, Raimundo e Raimunda, pelo amor,
carinho e incentivo. Aos meus irmãos, por toda a
ajuda e companheirismo. E em especial ao meu filho
amado
CESAR TASSO

v
AGRADECIMENTOS
A Deus, por estar sempre ao meu lado em todos os momentos da minha vida, protegendo-me
e guiando-me...
Ao meu filho Cesar Tasso, por ter tornado minha vida muito mais alegre e colorida.
Aos meus pais, pelo amor, companheirismo, carinho e apoio incondicional.
Aos meus irmãos, pelo carinho, apoio e incentivo.
Ao Instituto Nacional de Pesquisas da Amazônia (INPA) - Coordenação de Sociedade,
Ambiente e Saúde (CSAS) e ao Laboratório de Vetores da Malária e Dengue pela
oportunidade de desenvolver este projeto.
À Coordenação do GCBEv, em especial à Dra. Vera Scarpassa e Dra. Eliana Feldberg, pelo
incentivo e apoio financeiro.
À CAPES, pela concessão da bolsa.
Ao CNPq pelos recursos financeiros por meio do Edital no 55/2008 – CT/AMAZÔNIA.
À Secretaria Municipal de Educação (SEMED), por meio do Projeto Qualifica, por
proporcionar minha dedicação exclusiva ao doutorado.
À Dra. Joselita Maria Mendes dos Santos, minha orientadora, pelos ensinamentos, dedicação
e confiança demonstrados durante toda a realização deste trabalho.
Ao Dr. Ademir de Jesus Martins Júnior, meu co-orientador, pelos ensinamentos, por todo o
apoio logístico, pelo carinho, dedicação e pelas brincadeiras que tornaram o desenvolvimento
desse trabalho super divertido.
Ao Dr. José Bento Pereira Lima, pela importante colaboração e incentivo.
Ao Dr. Wanderli Pedro Tadei pelo apoio logístico no desenvolvimento deste projeto.
À Dra. Denise Valle, pelo apoio logístico.

vi
Ao Mestre Juracy de Freitas Maia pela valiosa colaboração, apoio técnico e esclarecimentos
sempre que solicitado e pela amizade.
Ao colega Carlos Eduardo Freitas Lemos, pelo incentivo e ajuda no desenho dos mapas.
À minha querida amiga Mirian dos Santos, por todo carinho a mim dispensado.
Ao meu mais que amigo e compadre Rosinaldo Clemente, pelo suporte, carinho e incentivo.
Aos colegas Wallison e Noêmia, pelo apoio técnico e companheirismo, principalmente,
quando eu estava grávida.
À equipe do Laboratório de Vetores da Malária e Dengue, em especial Adelina Loureiro,
Maria do Desterro, Zilá Oliveira, Bastos, Cristóvão, Carlos Praia, pela colaboração técnica,
solidariedade, boa vontade, amizade e carinho durante o período em que estive neste
laboratório.
À equipe do Laboratório de Fisiologia e Controle de Artrópodes Vetores da FIOCRUZ de
Manguinhos, em especial Ignez, Luiz, Felipe, Oscar, Sandrine e Luciana, pelos ensinamentos
e pela convivência divertida.

vii
EPÍGRAFE
“Costumávamos pensar que nosso futuro estava nas estrelas.
Agora sabemos que ele está nos nossos genes”.
(James Watson)

viii
RESUMO
Mutações não sinônimas no domínio IIS6 do gene do canal de sódio (NaV) têm sido
frequentemente associadas à resistência aos piretróides em diversas espécies de anofelinos.
Neste trabalho, foi realizada a primeira investigação da diversidade nucleotídica dessa região
em duas importantes espécies vetoras de malária humana no Brasil, Anopheles darlingi e
Anopheles marajoara. Uma região parcial do domínio IIS6 foi sequenciada de fêmeas adultas,
capturadas dentro e fora dos domicílios, das cidades de Manaus e Coari (AM), e larvas mais e
menos susceptíveis à deltametrina, das cidades de Manaus, São Gabriel da Cachoeira e
Iranduba, no estado do Amazonas, Rio Branco, no Acre, e Macapá, no Amapá. Além disso,
um bioensaio simplificado de knockdown usando larvas foi adaptado para avaliar o status de
susceptibilidade ou tolerância dos anofelinos neotropicais. Com base nesse bioensaio foram
evidenciadas perdas de susceptibilidade nas populações do estado do Amazonas (A. darlingi:
Manaus; A. marajoara: Iranduba). A análise da diversidade nucleotídica do NaV de A.
darlingi revelou cinco sítios polimórficos, que agruparam as amostras em cinco distintos
haplótipos (A, B, C, D e I). Em A. marajoara foram evidenciados quatro sítios polimórficos,
que agruparam as amostras em quatro haplótipos (E, F, G e H). Todas essas substituições
foram sinônimas e mutações kdr (knockdown resistance) no sítio 1014 não foram detectadas,
o que sugere a ocorrência de outros mecanismos de resistência relacionados com a perda de
susceptibilidade à deltametrina, como por exemplo, resistência metabólica. Muito embora
mutações kdr não tenham sido encontradas, é importante ressaltar que sua ocorrência é
permissiva, uma vez que o códon do sítio 1014 dessas duas espécies é formado por TTA, e
uma única mutação (TTT ou TCA) pode mudar o aminoácido para fenilalanina ou serina,
conforme observado em outros insetos. A sequência de aminoácidos traduzidos da região
gênica analisada revelou total homologia com a de outros anofelinos dispostas nos bancos de

ix
dados públicos e as diferenças nucleotídicas observadas estiveram de acordo com a filogenia
das espécies. Não foi possível correlacionar a frequência de alguns dos haplótipos com a
perda de susceptibilidade ao inseticida ou com as fêmeas capturadas dentro ou fora dos
domicílios. A presente análise da região parcial do domínio IIS6 do gene NaV de A. darlingi e
A. marajoara representa uma oportunidade ímpar para a compreensão da diversificação
evolutiva de um gene diretamente relacionado com resistência aos piretróides. Além disso,
descrevemos pela primeira vez a padronização de um teste de susceptibilidade rápido, simples
e de baixo custo, em larvas de anofelinos. Todas essas informações poderão ser usadas pelos
órgãos competentes para direcionar as medidas estratégicas a serem adotadas para combater
os vetores da malária.

x
ABSTRACT
Non synonymous mutations in the IIS6 domain of the voltage-gated sodium channel gene
(NaV) have been frequently associated with resistance to pyrethroids in several species of
anophelines. In this work, the first investigation of the nucleotide diversity of this region was
carried out in Anopheles darlingi and Anopheles marajoara, two main human malaria vectors
in Brazil. A partial region of the IIS6 domain was sequenced from adults females collected
indoor and outdoor from Manaus and Coari (AM), and most and least susceptible larvae to
deltamethrin from Manaus, São Gabriel da Cachoeira, and Iranduba, in the Amazonas State,
Rio Branco, Acre, and Macapá, Amapá. In addition, a simplified knockdown bioassay using
larvae was adapted to assess the status of susceptibility or tolerance of Neotropical
anophelines. The results of the bioassays revealed low susceptibility in the populations from
Amazonas State (A. darlingi: Manaus; A. marajoara: Iranduba). The analysis of the
nucleotide diversity in the NaV of A. darlingi showed five polymorphic sites that assembled
the samples into five haplotypes (A, B, C, D and I). In A. marajoara four polymorphic sites
were detected that assembled the samples into four haplotypes (E, F, G and H). All these
substitutions were synonymous and the kdr (knockdown resistance) mutations at 1014 site
were not detected, suggesting the occurrence of other resistance mechanisms related to the
loss of susceptibility to deltamethrin, such as metabolic resistance. Although kdr mutations
have not been found, it is important that its occurrence is permissive, once the codon 1014 of
these two species is formed by TTA and a single mutation (TTT or TCA) can change the
amino acid to phenylalanine or serine as observed in other insects. The amino acid translation
of the region analyzed showed complete homology with other anophelines placed in public
databases and the nucleotide differences observed were in agreement with the phylogeny of

xi
the species. It was not possible to correlate the frequency of some haplotypes with the loss of
susceptibility to the insecticide or females captured indoor or outdoor. Our analysis of the
IIS6 domain of the A. darlingi and A. marajoara’s NaV represents a unique opportunity for
understanding the evolutionary diversification of a gene directly associated with pyrethroid
resistance. Furthermore, we describe for the first time the padronization of a simple, quick and
cost effective susceptibility test in anophelines larvae. All this information can be used by the
competent institutions to direct the strategic measures to be adopted to combat malaria
vectors.

xii
SUMÁRIO
1 INTRODUÇÃO ............................................................................................................... 18
1.1 A situação global da malária ...................................................................................... 18
1.2 Os vetores da malária ................................................................................................. 22
1.3 Considerações gerais sobre Anopheles darlingi ........................................................ 23
1.4 Considerações gerais sobre Anopheles marajoara .................................................... 25
1.5 Estratégias para o combate aos vetores da malária .................................................... 27
1.6 Modo de ação dos piretróides .................................................................................... 30
1.7 Mecanismos de resistência aos piretróides ................................................................ 31
1.8 Mutações kdr como mecanismo de resistência .......................................................... 34
1.9 Associação entre ITN e mutação kdr ......................................................................... 43
1.10 Ferramentas moleculares para o diagnóstico de mutações kdr .................................. 45
2 HIPÓTESES .................................................................................................................... 47
3 OBJETIVOS .................................................................................................................... 48
3.1 Objetivo Geral ............................................................................................................ 48
3.2 Objetivos Específicos ................................................................................................ 48
4 MATERIAL E MÉTODOS ............................................................................................ 49
4.1.1 Área de estudo .................................................................................................... 49
4.1.2 Obtenção das amostras ....................................................................................... 51
4.1.3 Bioensaio simplificado de knockdown ............................................................... 53
4.1.4 Extração de DNA das larvas ............................................................................... 56
4.1.5 Extração de DNA dos adultos ............................................................................ 57
4.1.6 Quantificação do DNA ....................................................................................... 58
4.1.7 Amplificação, clonagem e sequenciamento do domínio IIS6 do gene do canal de
sódio usando a técnica de primers degenerados ............................................................... 59
4.1.8 Amplificação, clonagem e sequenciamento do domínio IIS6 do gene do canal de
sódio usando primers específicos ...................................................................................... 64
4.1.9 Análises dos dados ............................................................................................. 66
5 RESULTADOS E DISCUSSÃO .................................................................................... 67

xiii
5.1 Bioensaio simplificado de knockdown ....................................................................... 67
5.2 Variação no domínio IIS6 do gene NaV de Anopheles darlingi e Anopheles
marajoara ............................................................................................................................. 71
6 CONCLUSÕES ............................................................................................................... 82
7 REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 84

xiv
LISTA DE FIGURAS
Figura 1. Distribuição da malária no mundo. ........................................................................... 20
Figura 2. Total de casos de malária diagnosticados no Brasil de 1959 até 2012. .................... 21
Figura 3. Mapa da distribuição global das espécies de anofelinos. .......................................... 23
Figura 4. Anopheles darlingi, principal vetor da malária na região amazônica. ...................... 24
Figura 5. Anopheles marajoara, um importante vetor no estado do Amapá............................ 26
Figura 6. Representação esquemática do canal de sódio inserido na membrana celular,
evidenciando seus quatro domínios homólogos (I-IV), cada um composto por seis segmentos
hidrofóbicos (S1-S6). ............................................................................................................... 31
Figura 7. Distribuição das mutações kdr ao redor do mundo: (a) América, (b) África, (c) Ásia
.................................................................................................................................................. 42
Figura 8. Mapa do Brasil, enfatizando as localidades onde as populações de Anopheles
darlingi e Anopheles marajoara foram coletadas. ................................................................... 50
Figura 9 - (a) Copos telados acondicionados em caixa de poliestireno usados para o transporte
das fêmeas de Anopheles darlingi e Anopheles marajoara; (b) copos plásticos com filó usados
para oviposição; (c) cubas esmaltadas onde as larvas eram criadas. ........................................ 52
Figura 10. Alimentação usada para criação de larvas de anofelinos. ....................................... 53
Figura 11 - Bioensaio simplificado de knockdown, aplicado a anofelinos neotropicais. ......... 55
Figura 12. Larvas de 4º estádio de Anopheles darlingi, usadas nos bioensaios de knockdown.
.................................................................................................................................................. 56
Figura 13. Perfil eletroforético em gel de acrilamida (10%) da amplificação da região IIS6 do
gene NaV de Anopheles darlingi, usando os primers descritos por Enayati et al. (2003). 1 a
14= Amostras de DNA da população de Manaus; M= Marcador Molecular. .......................... 61
Figura 14. Perfil eletroforético em gel de agarose da amplificação da região IIS6 do NaV de
Anopheles darlingi, usando os primers descritos por Enayati et al. (2003). M1= Marcador
Molecular Ladder 50 pb; 1= produto amplificado; 2= produto amplificado; C= Controle
negativo; M2= Marcador Molecular Gene Ruler. .................................................................... 62

xv
Figura 15. Perfil eletroforético em gel de agarose da amplificação da região IIS6 do NaV de
Anopheles darlingi, usando os primers descritos por Enayati et al. (2003), enfatizando o
recorte das bandas. M1= Marcador Molecular Ladder 50 pb; 1= produto amplificado; 2=
produto amplificado; C= Controle. ........................................................................................... 63
Figura 16. Placa de Petri evidenciando o crescimento das colônias de Escherichia coli com os
clones da região IIS6 do gene NaV de Anopheles darlingi. ..................................................... 63
Figura 17. Perfil eletroforético em gel de acrilamida (10%) da amplificação da região IIS6 do
gene NaV de Anopheles darlingi, usando primers específicos. 1 a 6= Amostras de DNA da
população de Coari; M= Marcador Molecular. ........................................................................ 65
Figura 18. Perfil de susceptibilidade à deltametrina em populações naturais de Anopheles
darlingi e Anopheles marajoara da Amazônia brasileira. Os tempos medianos de knockdown
de cada população (KT50) foram calculados para 0,1 e 0,4 ppm de inseticida. SI = índice de
susceptibilidade, SGC = São Gabriel da Cachoeira. ................................................................ 69
Figura 19. Uma árvore de Neighbor-Joining mostrando as relações filogenéticas dos
haplótipos encontrados no domínio IIS6 do gene NaV de Anopheles darlingi e Anopheles
marajoara com as sequências de outros anofelinos (detalhes na Tabela 4). Os valores entre os
ramos indicam a percentagem de vezes que um nó foi suportado no teste de bootstrap (1000
repetições) (Tamura et al., 2011). ........................................................................................... 74
Figura 20. Diversidade nucleotídica da região parcial do domínio IIS6 do gene do canal de
sódio de anofelinos. Os nucleotídeos representados por letra maiúscula e minúscula
correspondem às regiões codificantes e ao íntron, respectivamente. O códon homólogo àquele
onde ocorre a clássica mutação kdr (1014) está sublinhado. Os sítios monomórficos estão
representados por pontos (.), as substituições pelos respectivos nucleotídeos e deleções por
traços (-). AdNaV_haplo_A, AdNaV_haplo_B, AdNaV_haplo_C, AdNaV_haplo_D e
AdNaV_haplo_I são os haplótipos de Anopheles darlingi ao passo que AmNaV_haplo_E,
AmNaV_haplo_F, AmNaV_haplo_G e AmNaV_haplo_H são os de Anopheles marajoara aqui
evidenciados. AdarC2 corresponde à sequência obtida do projeto genoma de A. darlingi
(Scaffold_542). As outras sequências foram obtidas do GenBank e VectorBase (Tabela 5). . 76
Figura 21. Relações genealógicas entre os haplótipos da região parcial do domínio IIS6 do
gene do canal de sódio de Anopheles darlingi, reveladas por estatística de parsimônia
(Clement et al., 2000). AdNaV_haplo_A corresponde ao haplótipo A, AdNaV_haplo_B ao

xvi
haplótipo B, AdNaV_haplo_C ao haplótipo C, AdNaV_haplo_D ao haplótipo D e
AdNaV_haplo_I ao haplótipo I. As linhas que conectam as formas representam os passos
mutacionais entre os haplótipos. Nelas estão contidas as indicações das mutações ................ 77
Figura 22. Relações genealógicas entre os haplótipos da região parcial do domínio IIS6 do
gene do canal de sódio de Anopheles marajoara, reveladas por estatística de parsimônia
(Clement et al., 2000). AmNaV-haplo_E corresponde ao haplótipo E, AmNaV-haplo_F ao
haplótipo F, AmNaV-haplo_G ao haplótipo G e AmNaV-haplo_H ao haplótipo H. As linhas
que conectam as formas representam os passos mutacionais entre os haplótipos. Nelas estão
contidas as indicações das mutações. ....................................................................................... 81

xvii
LISTA DE TABELAS
Tabela 1. Relação das 41 espécies vetoras de malária humana dominantes na África, Ásia e
Américas. .................................................................................................................................. 22
Tabela 2. Lista das espécies de anofelinos nas quais a mutação kdr já foi detectada, bem
como, as localidades e referências bibliográficas. .................................................................... 37
Tabela 3. Métodos moleculares usados para a detecção das mutações kdr. ............................. 46
Tabela 4. Detalhes das sequências do domínio IIS6 das espécies de anofelinos usadas no
alinhamento nucleotídico. ......................................................................................................... 78
Tabela 5. Distribuição dos haplótipos do domínio IIS6 do gene do canal de sódio de
Anopheles darlingi. ................................................................................................................... 78
Tabela 6. Distribuição dos haplótipos do domínio IIS6 do gene do canal de sódio de
Anopheles marajoara. .............................................................................................................. 81

18
1 INTRODUÇÃO
1.1 A situação global da malária
A malária é uma doença parasitária causada por protozoários do gênero
Plasmodium, que podem ser transmitidos ao homem pela picada do mosquito fêmea do
gênero Anopheles. Na sua forma típica, caracteriza-se por acessos de febre com intervalos de
24, 48 ou 72 horas (Forattini, 2002).
A doença é também conhecida por febre terçã, febre quartã, febre palúdica, sezão,
impaludismo, febre palustre, maleita, paludismo, febre dos pântanos, febre do brejo e febre
romana. Vale ressaltar que o nome malária surgiu por se achar que fosse decorrente do ar
contaminado – “mal ar”, pois no início do aparecimento da doença não se sabia da existência
do agente etiológico, que foi descoberto somente em 1880 por Charles Alphonse Laveran, e
nem do papel dos anofelinos como vetores, descoberto oito anos mais tarde por Patrick
Manson e Ronald Ross (Neves, 1988; Litsios, 1996).
Plasmodium falciparum e Plasmodium vivax são as principais espécies de agentes
etiológicos da malária que afetam os seres humanos. A forma mais grave da doença é causada
por P. falciparum, que se não for tratada pode ser fatal em indivíduos não imunes. A outra, P.
vivax, não é usualmente letal, mas o perigo está na reincidência do parasita, que pode se
manifestar no mesmo indivíduo após meses ou até mesmo anos de aquisição da infecção, por
causa das formas persistentes (hipnozoítos) presentes no fígado (WHO, 2009; Lacerda et al.,
2012).
De todas as afecções consideradas endêmicas, nenhuma marcou tanto a vida
humana como a malária, fazendo vítimas ao longo dos séculos, até mais que todas as grandes

19
epidemias de peste, de cólera e de varíola reunidas. De acordo com a Organização Mundial de
Saúde (OMS), aproximadamente 207 milhões de casos foram reportados em 2012, com uma
estimativa de 627.000 mortes (WHO, 2013).
Dentre os fatores que contribuem para esse panorama podemos destacar a ausência
de uma vacina antimalárica efetiva, a distribuição de Plasmodium resistentes a drogas, o
desenvolvimento de resistência aos inseticidas nos mosquitos vetores, dentre outros (WHO,
2011).
Ela ocorre em maior incidência nos países subdesenvolvidos e em desenvolvimento
(Figura 1), principalmente, dos continentes africano (80%), asiático (15%) e americano (14%)
(WHO, 2011). Nas Américas, 21 países são afetados pela malária, dos quais, o Brasil
apresenta o maior número de casos (50,7%), com aproximadamente um milhão deles, sendo
diagnosticados na região amazônica (99% dos casos) (SIVEP-Malaria, 2013), habitada por
mais de 26 milhões de pessoas, representando cerca de 12% da população brasileira (IBGE,
2013).

20
Figura 1. Distribuição da malária no mundo. Fonte: WHO (2011).
Dados sobre os casos de malária diagnosticados no Brasil desde 1959 até 2012
evidenciaram que até meados de 1988 P. vivax e P. falciparum ocorriam com a mesma
intensidade (Figura 2). Após esse período, houve um aumento na ocorrência de malária por P.
vivax, possivelmente, resultante de adaptações naturais que possam estar aumentando à
susceptibilidade das pessoas a esse tipo de parasita, adicionado às dificuldades de se controlar
a doença.
Em toda a região amazônica, muitos fatores interferem na dinâmica da transmissão
da malária, tais como: fatores sócioeconômicos (migração, habitação, densidade populacional,
renda); ambientais (hidrologia, clima, topografia, vegetação); biológicos (ciclo de vida dos
vetores e os agentes patológicos, imunidade da população) e médicossanitários (efetividade
do sistema de saúde) (Tadei et al., 1998; FUNASA, 2002). Apesar disso, existe um consenso

21
mundial que a luta antivetorial continua a ser o meio mais eficiente para tentar interromper a
transmissão da malária em áreas endêmicas (WHO, 2009).
Figura 2. Total de casos de malária diagnosticados no Brasil de 1959 até 2012.
Fonte: SIVEP-Malária (2013).

22
1.2 Os vetores da malária
Os vetores da malária fazem parte do gênero Anopheles, que inclui, aproximadamente,
484 espécies, distribuídas em sete subgêneros (Harbach, 2004), das quais, 70 apresentam
competência vetorial para a malária humana (Service e Townson, 2002), com 41 delas sendo
consideradas espécies vetoras dominantes (Sinka et al., 2012) (Erro! Fonte de referência não
encontrada.). A distribuição geográfica das espécies desse gênero é ampla, podendo ocorrer nos mais
diversos ambientes, indo desde os desertos até as florestas tropicais (Figura 3) (Kiszewski et al., 2004).
Tabela 1. Relação das 41 espécies vetoras de malária humana dominantes na África, Ásia e Américas.
Continente
Espécies
África
Anopheles arabiensis, Anopheles funestus, Anopheles gambiae, Anopheles melas, Anopheles
merus, Anopheles moucheti e Anopheles nili
Ásia
Anopheles barbirostris, Anopheles lesteri, Anopheles sinensis, Anopheles aconitus, Anopheles
annularis, Anopheles balabacensis, Anopheles culicifacies, Anopheles dirus, Anopheles farauti,
Anopheles flavirostris, Anopheles fluviatilis, Anopheles koliensis, Anopheles leucosphyrus,
Anopheles maculatus, Anopheles minimus, Anopheles punctulatus, Anopheles stephensi,
Anopheles subpictus e Anopheles sundaicus
Américas Anopheles freeborni, Anopheles pseudopunctipennis, Anopheles quadrimaculatus, Anopheles
albimanus, Anopheles albitarsis, Anopheles aquasalis, Anopheles darlingi, Anopheles
marajoara e Anopheles nuneztovari
Compilado a partir de Sinka et al. (2010a), Sinka et al. (2010b), Sinka et al. (2011) e (Sinka et al., 2012).
No Brasil, o subgênero Nyssorhynchus é considerado um dos mais importantes, por
conter as principais espécies vetoras de malária (por exemplo, Anopheles darlingi, Anopheles
marajoara, Anopheles aquasalis, Anopheles albitarsis).

23
Figura 3. Mapa da distribuição global das espécies de anofelinos.
Fonte: Kiszewski et al. (2004)
1.3 Considerações gerais sobre Anopheles darlingi
Anopheles darlingi Root, 1926 é uma espécie de anofelino que apresenta grande
relevância epidemiológica, por ser o principal vetor da malária em quase toda a sua área de
ocorrência (Consoli e Lourenço-de-Oliveira, 1994; Olano et al., 2001; Forattini, 2002). Os fatores
que contribuem para isso são sua elevada antropofilia/endofagia e também sua susceptibilidade
frente aos agentes etiológicos P. falciparum e P. vivax (Forattini, 2002; Grieco et al., 2005).

24
Os indivíduos adultos desta espécie apresentam pequeno ou médio porte. A veia costa das
asas apresenta a mancha pré-umeral de tamanho grande, com envergadura correspondente a três ou
mais do que a da mancha umeral. Os tarsos III a V das patas posteriores são completamente brancos
(Figura 4) ou apresentam pequeno anel escuro basal nos tarsômeros (Faran, 1980).
Figura 4. Anopheles darlingi, principal vetor da malária na região amazônica. Fonte: Galardo, 2010.
Esse mosquito possui ampla distribuição geográfica, ocorrendo desde o sul do México
(Chiapas) até a Nicarágua na América Central, sendo encontrado também na Colômbia e na porção
leste da Cordilheira dos Andes (Sinka et al., 2012). No Brasil, ocorre em quase todos os estados,
com exceção de Santa Catarina e Rio Grande do Sul (Consoli e Lourenço-de-Oliveira, 1994).
A. darlingi utiliza corpos d´água onde existe pouca correnteza para o desenvolvimento de
suas larvas e pupas. Seus criadouros são de águas profundas, limpas, pouco turvas e ensolaradas ou
parcialmente sombreadas, onde suas formas imaturas habitam as margens, escondidas entre a
vegetação emergente ou flutuante e os detritos vegetais caídos na superfície líquida. Estes
criadouros são utilizados, indiscriminadamente, durante todo o ano e, por serem permanentes,

25
funcionam como focos de resistência durante a estação mais seca. Contudo, durante a estação
chuvosa, essa espécie pode empregar uma grande variedade de coleções líquidas de tamanho e
profundidade menores, tais como: valas, poças e impressões de patas de animais (Forattini, 2002).
Por causa de sua importância epidemiológica, muitos estudos envolvendo a biologia e a
estrutura genética das populações de A. darlingi têm sido realizados, a fim de conhecer melhor a
composição genética da espécie e entender os mecanismos envolvidos na dinâmica de transmissão
da malária (Freitas-Sibajev et al., 1995; Calvo et al., 2004; Conn et al., 2006; González et al.,
2007; Scarpassa e Conn, 2007; Mirabello et al., 2008; Silva et al., 2008; Silva et al., 2010).
1.4 Considerações gerais sobre Anopheles marajoara
Anopheles marajoara Galvão e Damasceno, 1942 (Figura 5) é outro importante vetor da
malária, principalmente, no estado do Amapá (Brasil), e em Orinoquía (Colômbia), onde é
incriminado como vetor primário (Conn et al., 2002; Brochero et al., 2005).
Esse mosquito é um membro do complexo Anopheles albitarsis, que inclui mais cinco
espécies: Anopheles albitarsis sensu strictu, Anopheles deaneorum, Anopheles janconnae (chamado
anteriormente de espécie “E”), Anopheles oryzalimnetes (chamada anteriormente de espécie “B”) e
espécie “F”. Essas espécies são isomórficas, mas apresentam diferenças quanto à distribuição
geográfica, ecologia, capacidade vetorial e preferências por hospedeiros (Galvão e Damasceno,
1942; Linthicum, 1988; Rosa-Freitas, 1989; Rosa-Freitas et al., 1990; Wilkerson et al., 1995a;
Wilkerson et al., 1995b; Lehr et al., 2005; Brochero et al., 2007; Motoki et al., 2009).

26
Figura 5. Anopheles marajoara, um importante vetor no estado do Amapá. Fonte: Galardo (2010)
As espécies deste complexo desenvolvem–se em vários tipos de criadouros, temporários
ou permanentes, naturais ou artificiais. Normalmente, ocorrem durante o ano todo, porém são mais
abundantes durante a estação chuvosa (Consoli e Lourenço-de-Oliveira, 1994).
A distribuição geográfica de A. marajoara vai desde a Costa Rica, na América Central,
até a Bolívia, na América do Sul (Wilkerson et al., 1995a; Wilkerson et al., 1995b; Rubio-Palis et
al., 2003). No Brasil, ela tem sido registrada na região amazônica e também na região sudeste do
país (Wilkerson et al., 1995a; Sinka et al., 2012).
Devido a sua importância epidemiológica, muitos estudos sobre a ecologia, morfologia, e
análises moleculares já foram realizados em populações de A. marajoara, bem como, nas outras
espécies que compõem o complexo A. albitarsis, principalmente, com o intuito de elucidar o status
taxonômico de cada uma delas (Galvão e Damasceno, 1942; Rubio-Palis et al., 2003; Li et al.,
2005; Posso et al., 2006; Galardo, 2010).

27
1.5 Estratégias para o combate aos vetores da malária
As ferramentas estratégicas para o combate à malária enquadram-se dentro de dois
domínios principais: prevenção, por meio do controle de mosquitos vetores, e gerenciamento de
casos, através do diagnóstico e tratamento da malária, sendo o primeiro considerado o mais efetivo
(WHO, 2009).
As formas de controle de mosquitos vetores são didaticamente classificadas como:
mecânicas (eliminação de criadouros), biológicas (uso de predadores ou parasitas) ou químicas
(aplicação de inseticidas sintéticos) (Consoli e Lourenço-de-Oliveira, 1994; Rose, 2001).
O desenvolvimento de inseticidas químicos, que permanecem ativos por longos períodos,
foi um dos mais relevantes avanços ocorrido no século XX (Mellanby, 1992) e ainda hoje tem
papel fundamental no controle de vetores de doenças e pragas na agricultura.
Existem quatro grupos principais de inseticidas, classificados de acordo a sua natureza
química e modos de ação. São eles: organoclorados, organofosforados, carbamatos e piretróides.
O primeiro inseticida empregado no controle de anofelinos foi o DDT, um organoclorado
que teve seu uso iniciado em Nápoles no ano de 1944 para combater uma epidemia de tifo
(Mellanby, 1992). Em 1955, a OMS propôs a erradicação global da malária por meio da borrifação
domiciliar desse inseticida. De baixo custo, altamente eficiente, conseguiu dizimar populações de
vetores em escala global. No entanto, o desenvolvimento de problemas ambientais e sanitários, aliado
ao surgimento de resistência, resultou na proibição do produto em muitos países (Govere et al., 2002).
Contudo, após a “Convenção de Estocolmo sobre Poluentes Orgânicos Persistentes”, realizada em 2007,
o seu uso foi restabelecido em áreas restritas com alta transmissão de malária, como por exemplo, na
África (WHO, 2007a).

28
Os organofosforados (malathion, temephos, fenitrotion, etc.), foram desenvolvidos na década
de 1940 e desde então tem sido usados como inseticidas, herbicidas e reguladores de crescimento em
plantas. Apesar de serem biodegradáveis e não cumulativos, apresentam como desvantagens
instabilidade química e elevada toxicidade em vertebrados (Morales-Rojas e Moss, 2002).
Os carbamatos, também referidos como metilcarbamatos por serem derivados do ácido
metilcarbâmico (Fukuto, 1990), são compostos usados como inseticidas, nematicidas e acaricidas.
Apresentam baixa persistência ambiental, sendo menos tóxicos para os seres vivos do que os
organoclorados. Devido ao amplo uso na agricultura têm sido incriminados como agentes contaminantes
de alimentos, água e ar, com efeitos adversos em seres humanos e animais (Gupta, 1994).
Por volta dos anos de 1970 foi lançada uma classe de inseticida considerada mais eficiente e
menos tóxica que as demais: os piretróides sintéticos. Esses inseticidas chamaram a atenção por
apresentarem maior capacidade letal contra insetos, bastando pequenas doses do produto para um efeito
satisfatório (Omoto, 2000). Por conta disso, eles praticamente substituíram ou suplementaram o uso
das outras classes em diversas áreas de controle de pragas e representam aproximadamente 23% do
mercado de inseticidas químicos, mais de um quarto do mercado mundial (Schleier III e Peterson,
2011).
Os piretróides são análogos sintéticos dos inseticidas ésteres do ácido crisântemo
(piretrinas I) e do ácido pirétrico (piretrinas II), naturalmente encontrados nas folhas de
Chrysanthemum cinerafolis. São classificados como tipo I, compostos que não apresentam o grupo
alfa-ciano, como a permetrina, e tipo II, compostos que apresentam o grupo alfa-ciano, como a
deltametrina (Davies et al., 2007b). São inseticidas biodegradáveis, não cumulativos e raramente
provocam intoxicações agudas em aves e mamíferos (Palchick, 1996).

29
Atualmente o controle da malária está basicamente dependente dessa classe de inseticida,
que tem sido amplamente empregada em campanhas de borrifação intradomiciliar (Indoor Residual
Spraying-IRS) e também para controlar pestes agrícolas no mundo todo. Além disso, é a única
classe aprovada pelo Programa de Avaliação de Pesticidas da OMS (World Health Organization
Pesticide Scheme-WHOPES) para impregnação de telas e mosquiteiros (Insecticide Treated Nets-
ITN) (Phillips-Howard et al., 2003; WHO, 2006a; WHO, 2007b; WHOPES, 2014), devido à sua
rápida atuação contra insetos e baixa toxicidade em mamíferos (Hemingway e Ranson, 2000).
IRS é um método no qual inseticidas residuais são aplicados nas superfícies das paredes e
dos tetos das residências (WHO, 2006b). A intenção dessa estratégia é de que os mosquitos, após a
ingestão de sangue, irão repousar nessas superfícies e permanecer por tempo suficiente para
absorver uma dose letal do inseticida. ITN é um mosquiteiro que repele, incapacita ou mata os
mosquitos que entram em contato com o inseticida impregnado no material da rede, sendo assim
uma barreira química e ao mesmo tempo uma barreira física contra os insetos. Existem duas
categorias de ITNs: os mosquiteiros convencionais e os de longa duração (Long Lasting Insecticide
Treated Nets-LLINs) (Hill et al., 2006; WHO, 2006b).
O sucesso de algumas dessas estratégias causou euforia no mundo todo, fazendo surgir o
otimismo de que a eliminação da malária como um problema de saúde pública seria possível, por
meio do extermínio de seus vetores. Entretanto, hoje elas estão seriamente ameaçadas devido ao
surgimento de populações de vetores resistentes a esses compostos químicos. Como novas classes
de inseticidas alternativos e igualmente interessantes ainda não estão disponíveis no mercado, a
seleção para a resistência tende a continuar aumentando, a menos que efetivas estratégias de
gerenciamento de resistência sejam implementadas (Jones et al., 2013).

30
1.6 Modo de ação dos piretróides
Os piretróides, assim como o DDT e seus análogos, pertencem ao grupo de neurotoxinas
que compartilham um modo de ação similar. Esses compostos têm como alvo o canal de sódio
regulado por voltagem, presente nas células do sistema nervoso central e periférico (neurônios,
miócitos, células endócrinas e ovários), alterando a cinética de propagação de impulsos nervosos
(Schleier III e Peterson, 2011).
Estruturalmente, o canal de sódio é uma proteína transmembranar integral, composta por
quatro domínios homólogos (I-IV), cada um deles composto por seis hélices (S1-S6) conectadas por
alças de ligação (loops) (Figura 6). Os segmentos S5, S6 e a alça de ligação S5-S6 formam um poro
aquoso central, e as hélices S1-S4 de cada domínio unem-se para formar quatro domínios
independentes sensíveis à voltagem (Catterall et al., 2003; Davies et al., 2007a).
A subunidade α do canal de sódio de A. gambiae é composta por 6.417 nucleotídeos,
apresentando uma ORF (Open Reading Frame) que codifica 2.139 aminoácidos, com massa
molecular calculada de 240 kDa. O gene NaV desse mosquito, localizado no loco para (paralysis)
do cromossomo X, tem aproximadamente 74 kpb em comprimento, composto por 35 exons,
incluindo dois exons duplicados. Além disso, apresenta 32 introns, que podem sintetizar diferentes
RNAs mensageiros (RNAm) por meio de splicing alternativo (Davies et al., 2007a).
Os efeitos dos piretróides são estereoespecíficos e dois diferentes sítios de ligação no
canal de sódio foram identificados. O primeiro foi proposto por O'reilly et al. (2006), no qual as
hélices IIS5 e IIIS6 teriam papel fundamental na interação com a molécula do inseticida e que a
ligação adicional na alça de ligação IIS4-S5 poderia explicar a maior potência dos piretróides em
comparação com o DDT. O segundo foi proposto por Du et al. (2013), no qual o sítio de ligação
seria uma espécie de “bolso” formado pela alça de ligação IS4-S5 e pelas hélices IS5 e IIS6. Para

31
ambos os modelos, o efeito seletivo dos inseticidas sobre os insetos seria explicado pela não
conservação dos resíduos de aminoácidos dessas regiões entre os artrópodes e não artrópodes.
Figura 6. Representação esquemática do canal de sódio inserido na membrana celular, evidenciando seus quatro domínios homólogos (I-IV), cada um composto por seis segmentos hidrofóbicos (S1-S6). Fonte: Martins e Valle (2012).
1.7 Mecanismos de resistência aos piretróides
A resistência pode ser definida como a habilidade de indivíduos de uma dada espécie
suportar doses de substâncias tóxicas, que seriam letais para a maioria dos indivíduos da população
normal (Subramanyam e Hagstrum, 1996). Sendo, portanto, um marco na mudança da composição
genética de uma determinada população, em resposta à pressão de seleção. Trata-se de um caso
típico de Seleção Natural, que consiste no aumento da frequência relativa de alguns indivíduos
"pré-adaptados", presentes em uma população, resultante da aplicação constante de um mesmo
produto químico (Crow, 1957).
O uso intensivo e extensivo de piretróides, assim como aconteceu para outras classes de
inseticidas vem selecionando populações resistentes a esses compostos, gerando um fator limitante
aos esforços para combater populações de vetores (Dong, 2007). O fenômeno de resistência tem

32
sido observado em mais de 500 espécies de insetos ao redor do mundo, entre as quais mais de 50
são anofelinos (Hemingway e Ranson, 2000). Dois principais mecanismos são incriminados como
responsáveis pela resistência aos piretróides: resistência metabólica e insensibilidade do sítio-alvo
(Ranson et al., 2011; Martins e Valle, 2012).
Resistência metabólica ocorre quando elevada atividade de uma ou mais enzimas resulta
em uma porção suficiente de inseticida, sendo sequestrado ou detoxificado antes de atingir o seu
alvo e de promover o efeito desejado (Ranson et al., 2011). Isso ocorre devido ao aumento no
número de moléculas disponíveis (amplificação gênica ou superativação da expressão) ou por
mutação na porção gênica codificadora da enzima, produzindo a metabolização mais eficiente do
inseticida (Hemingway, 2000; Hemingway e Ranson, 2000; Karaağaç, 2012).
Esse mecanismo de resistência é altamente complexo, muito embora recentes avanços
venham caracterizando os principais genes das enzimas responsáveis pela detoxificação, abrindo
caminho para o desenvolvimento de marcadores moleculares para a identificação e/ou
acompanhamento da resistência em determinadas populações (Hemingway et al., 2004). Três
principais superfamílias de enzimas estão envolvidas no processo de detoxificação: Esterases,
Oxidases de Função Mista (OFM, ou simplesmente P450) e Glutationa S-Transferases (GST)
(Hemingway e Ranson, 2000).
Ensaios bioquímicos colorimétricos têm sido amplamente empregados para a detecção de
alterações na atividade de enzimas detoxificantes. Nesse teste, a atividade enzimática de uma
população natural é comparada a de linhagens controle, com o emprego de substratos específicos
para cada família enzimática (Brogdon e Mcallister, 1998b).
Já em nível transcricional, mais recentemente, a técnica de microarray ganhou destaque
na investigação da resistência metabólica. Nela, os detox chips (detoxification chips) comparam a

33
expressão de virtualmente todos os genes das famílias relacionadas com o metabolismo de
inseticidas (GSTs, OFMs, Esterases, Carboxiesterases), entre mosquitos resistentes e susceptíveis.
Além destes, constam ainda outros genes, como por exemplo, os do metabolismo redox, envolvidos
na proteção contra radicais livres (David et al., 2005).
A análise da expressão gênica por meio de detox chip em A. gambiae evidenciou elevada
atividade dos genes de GST (GSTE2), P450 (CYP6Z1 e CYP325) e Peroxidases, em mosquitos
resistentes a DDT (David et al., 2005). Em um estudo realizado por Müller et al. (2008a), genes
com função antioxidante (Superóxido-dismutase, GST, Peroxidase e P450) foram diferencialmente
expressos em populações de A. arabiensis de Camarões tolerantes à deltametrina. Elevada
expressão de CYP6P3, um gene da família P450, foi observada em populações de A. gambiae
resistentes à permetrina (Müller et al., 2008b). Expressão diferencial também foi observada em A.
funestus, cujos genes de P450 (CYP6P9, CYP6M7) e COI (do sistema redox) foram mais
expressivos nos indivíduos resistentes (Christian et al., 2011).
A resistência por insensibilidade do sítio alvo ocorre quando há uma alteração das
moléculas que diretamente interagem com o inseticida, tornando-o menos tóxico ou ineficiente
(Brogdon e Mcallister, 1998b; Hemingway et al., 2004; Pittendrigh et al., 2008). Uma vez que os
alvos de inseticidas são moléculas estruturais do sistema nervoso, altamente conservadas ao longo
da evolução, poucas mudanças são permissivas sem a perda de suas funções fisiológicas (Ffrench-
Constant et al., 1998). Desta forma, é comum que a seleção de mutações que conferem resistência
ocorra em sítios homólogos entre diferentes espécies de insetos (Martinez-Torres et al., 1998;
Dong, 2007; Davies et al., 2007a).
Vale destacar que a insensibilidade do sítio alvo, por ser uma característica que atribui a
maior parte da variação genética relacionada à resistência a um único gene (Wondji et al., 2007), é

34
o mecanismo melhor compreendido. Com isto, diagnósticos moleculares para sua detecção fazem
parte integrante das estratégias de monitoramento de resistência aos inseticidas em muitos
programas de controle da malária (Corbel e N'guessan, 2013).
1.8 Mutações kdr como mecanismo de resistência
Vários estudos evidenciaram que a resistência ao efeito knockdown dos piretróides sobre
várias espécies de insetos está associada com mutações pontuais no NaV, sua molécula-alvo. Por
definição, efeito knockdown é a perda da coordenação e paralisia provocada pelo inseticida,
frequentemente, acompanhada por espasmos e tremores (Schleier III e Peterson, 2011).
Este mecanismo de resistência foi primeiramente observado na mosca Musca domestica,
no qual a substituição de um aminoácido leucina por fenilalanina no segmento hidrofóbico IIS6
(L1014F) resultou em moderado aumento da resistência ao DDT, ficando conhecida como
resistência/mutação kdr (knockdown resistance) (Williamson et al., 1996).
Em anofelinos a mutação kdr L1014F foi identificada pela primeira vez em linhagens de
A. gambiae resistentes a piretróides (Martinez-Torres et al., 1998) e desde então tem sido detectada
em uma série de outras espécies (Reimer et al., 2005; Girod et al., 2006; Awolola et al., 2007;
Dabiré et al., 2009; Balkew et al., 2010; Coetzee e Koekemoer, 2013). Ainda no sítio 1014, outra
substituição, leucina por serina (L1014S), foi identificada em A. gambiae e também associada ao
fenótipo kdr (Ranson et al., 2000).
As mutações L1014F e L1014S foram inicialmente observadas em populações de A.
gambiae da África ocidental e oriental, respectivamente. Portanto, a primeira é algumas vezes
referida como kdr-w (kdr west), e a segunda como kdr-e (kdr east) (Pinto et al., 2007). Em todo

35
caso, nota-se que a distribuição destas mutações está fortemente relacionada com as espécies
crípticas do complexo A. gambiae (Santolamazza et al., 2008).
Em populações asiáticas de A. sinensis, em adição às substituições L1014F/L1014S,
foram reportadas as mutações L1014C e L1014W, que mudam o aminoácido leucina para cisteína e
triptofano, respectivamente. Além disso, no sítio imediatamente anterior ao da clássica mutação
kdr, ocorre a substituição N1013S, que muda o aminoácido aspargina para serina (Kim et al., 2007;
Tan et al., 2012a). Em populações de A. culicifacies da Índia, também em adição às substituições
L1014F/L1014S, foi descrita uma nova mutação no sítio 1010, substituindo valina por leucina
(V1010L) (Singh et al., 2010).
Outra mutação, localizada na posição 918 da alça de ligação IIS4-S5 do gene do canal de
sódio de Musca domestica, que substitui uma metionina por treonina, sinérgica à clássica mutação
L1014F, foi associada a elevados níveis de resistência a DDT e piretróides, sendo, portanto,
chamada de mutação super kdr (Ingles et al., 1996). Situação análoga foi observada em outras
espécies de insetos, tais como, na mosca do chifre Haematobia irritans (Jamroz et al., 1998), no
pulgão do pessegueiro Myzus persicae (Anstead et al., 2007; Eleftherianos et al., 2008), na tripes
da cebola Thrips tabaci (Toda e Morishita, 2009) e na traça Tuta absoluta (Haddi et al., 2012).
Porém, ainda não há registros de substituição homóloga em anofelinos.
Com base nas técnicas moleculares existentes atualmente, foi possível identificar e
mapear a distribuição das mutações kdr entre as mais diversas espécies de anofelinos ao redor do
mundo. Desde que foi descrita pela primeira vez em 1998 (Martinez-Torres et al., 1998), as
alterações no NaV das espécies desse gênero têm sido amplamente monitoradas, de forma que
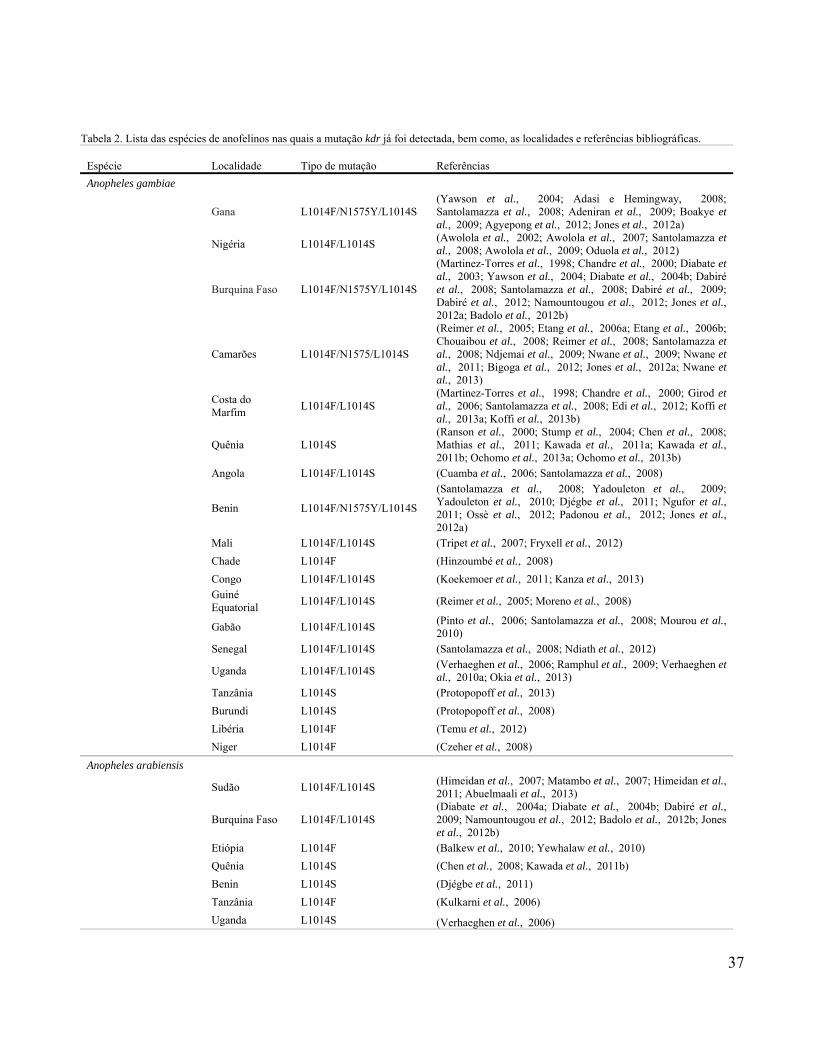
contabilizamos 98 referências publicadas até o final de 2013 (Tabela 2).

36
Até o presente momento, mutações no NaV foram descritas em pelo menos 13 diferentes
espécies de anofelinos. A. gambiae é de longe a mais estudada (62 registros), apresentando três
variantes mutacionais (L1014F, L1014S e N1575Y), detectadas em 20 dos 54 países da África
(Figura 7). Em seguida vem outro anofelino africano, A. arabiensis (17 registros), apresentando
duas variantes (L1014F e L1014S), detectadas em sete países.
37
Tabela 2. Lista das espécies de anofelinos nas quais a mutação kdr já foi detectada, bem como, as localidades e referências bibliográficas.
Espécie Localidade Tipo de mutação Referências Anopheles gambiae
Gana L1014F/N1575Y/L1014S (Yawson et al., 2004; Adasi e Hemingway, 2008; Santolamazza et al., 2008; Adeniran et al., 2009; Boakye et al., 2009; Agyepong et al., 2012; Jones et al., 2012a)
Nigéria L1014F/L1014S (Awolola et al., 2002; Awolola et al., 2007; Santolamazza et al., 2008; Awolola et al., 2009; Oduola et al., 2012)
Burquina Faso L1014F/N1575Y/L1014S
(Martinez-Torres et al., 1998; Chandre et al., 2000; Diabate et al., 2003; Yawson et al., 2004; Diabate et al., 2004b; Dabiré et al., 2008; Santolamazza et al., 2008; Dabiré et al., 2009; Dabiré et al., 2012; Namountougou et al., 2012; Jones et al., 2012a; Badolo et al., 2012b)
Camarões L1014F/N1575/L1014S
(Reimer et al., 2005; Etang et al., 2006a; Etang et al., 2006b; Chouaibou et al., 2008; Reimer et al., 2008; Santolamazza et al., 2008; Ndjemai et al., 2009; Nwane et al., 2009; Nwane et al., 2011; Bigoga et al., 2012; Jones et al., 2012a; Nwane et al., 2013)
Costa do Marfim L1014F/L1014S
(Martinez-Torres et al., 1998; Chandre et al., 2000; Girod et al., 2006; Santolamazza et al., 2008; Edi et al., 2012; Koffi et al., 2013a; Koffi et al., 2013b)
Quênia L1014S (Ranson et al., 2000; Stump et al., 2004; Chen et al., 2008; Mathias et al., 2011; Kawada et al., 2011a; Kawada et al., 2011b; Ochomo et al., 2013a; Ochomo et al., 2013b)
Angola L1014F/L1014S (Cuamba et al., 2006; Santolamazza et al., 2008)
Benin L1014F/N1575Y/L1014S
(Santolamazza et al., 2008; Yadouleton et al., 2009; Yadouleton et al., 2010; Djégbe et al., 2011; Ngufor et al., 2011; Ossè et al., 2012; Padonou et al., 2012; Jones et al., 2012a)
Mali L1014F/L1014S (Tripet et al., 2007; Fryxell et al., 2012) Chade L1014F (Hinzoumbé et al., 2008) Congo L1014F/L1014S (Koekemoer et al., 2011; Kanza et al., 2013)
Guiné Equatorial L1014F/L1014S (Reimer et al., 2005; Moreno et al., 2008)
Gabão L1014F/L1014S (Pinto et al., 2006; Santolamazza et al., 2008; Mourou et al., 2010)
Senegal L1014F/L1014S (Santolamazza et al., 2008; Ndiath et al., 2012)
Uganda L1014F/L1014S (Verhaeghen et al., 2006; Ramphul et al., 2009; Verhaeghen et al., 2010a; Okia et al., 2013)
Tanzânia L1014S (Protopopoff et al., 2013) Burundi L1014S (Protopopoff et al., 2008) Libéria L1014F (Temu et al., 2012) Niger L1014F (Czeher et al., 2008) Anopheles arabiensis
Sudão L1014F/L1014S (Himeidan et al., 2007; Matambo et al., 2007; Himeidan et al., 2011; Abuelmaali et al., 2013)
Burquina Faso L1014F/L1014S (Diabate et al., 2004a; Diabate et al., 2004b; Dabiré et al., 2009; Namountougou et al., 2012; Badolo et al., 2012b; Jones et al., 2012b)
Etiópia L1014F (Balkew et al., 2010; Yewhalaw et al., 2010) Quênia L1014S (Chen et al., 2008; Kawada et al., 2011b) Benin L1014S (Djégbe et al., 2011) Tanzânia L1014F (Kulkarni et al., 2006) Uganda L1014S (Verhaeghen et al., 2006)

38
Anopheles sinensis
China L1014F/L1014S/L1014C/L1014W/N1013S
(Tan et al., 2012a; Tan et al., 2012b; Chang et al., 2013; Zhong et al., 2013)
Coréia L1014F/L1014C (Kang et al., 2012) Laos L1014S (Verhaeghen et al., 2010b) Cambodja L1014S (Verhaeghen et al., 2010b) Vietnam L1014S (Verhaeghen et al., 2010b) Anopheles stephensi
Dubai L1014F (Enayati et al., 2003)
Índia L1014F/L1014S (Gayathri e Murthy, 2006; Singh et al., 2011)
Anopheles subpictus
Sri Lanka L1014F (Karunaratne et al., 2007)
Indonésia L1014F (Syafruddin et al., 2010)
Anopheles albimanus
México L1014F (Lol et al., 2013)
Nicarágua L1014C (Lol et al., 2013)
Costa Rica L1014C (Lol et al., 2013)
Anopheles sacharovi
Turquia L1014F/L1014S (Lüleyap et al., 2002)
Anopheles culicifacies
Índia L1014F/L1014S/V1010L (Singh et al., 2009; Singh et al., 2010)
Anopheles sundaicus
Indonésia L1014F (Syafruddin et al., 2010)
Anopheles aconitus
Indonésia L1014F (Syafruddin et al., 2010)
Anopheles vagus
Indonésia L1014F (Syafruddin et al., 2010)
Laos L1014S (Verhaeghen et al., 2010b)
Cambodja L1014S (Verhaeghen et al., 2010b)
Vietnam L1014S (Verhaeghen et al., 2010b)
Anopheles paraliae
Laos L1014S (Verhaeghen et al., 2010b)
Cambodja L1014S (Verhaeghen et al., 2010b)
Vietnam L1014S (Verhaeghen et al., 2010b)
Anopheles peditaeniatus
Laos L1014F/L1014S (Verhaeghen et al., 2010b)
Cambodja L1014F/L1014S (Verhaeghen et al., 2010b)
Vietnam L1014F/L1014S (Verhaeghen et al., 2010b)

39
A. sinensis é a terceira, com seis registros. Porém, foi a que, surpreendentemente,
apresentou o maior número de variantes kdr (L1014F, L1014S, L1014C, L1014W e N1013S),
distribuídas em cinco dos 50 países da Ásia, com a maioria delas sendo detectada na China.
Segundo Kang et al. (2012), esse fato está relacionado com o elevado tamanho populacional e
a ampla distribuição geográfica da espécie, que tendem a aumentar a variabilidade genética.
Entre as demais espécies asiáticas, A. stephensi apresentou três registros de duas
variantes (L1014F e L1014S), detectadas em Dubai e na Índia. A.subpictus (L1014F), A.
culicifacies (L1014F, L1014S e V1010L) e A. vagus (L1014F) apresentaram dois registros.
Enquanto que, A. sacharovi (L1014F/L1014S), A. sundaicus (L1014F), A. aconitus (L1014F),
A. paraliae (L1014S) e A. peditaeniatus (L1014F/L1014S) tiveram somente um registro.
A presença da mutação kdr nas Américas foi observada em apenas uma espécie de
anofelino, A. albimanus, que apresentou as variantes L1014F e L1014C em populações
oriundas do México, Nicarágua e Costa Rica (Tabela 2).
Um levantamento da distribuição geográfica da mutação kdr em populações de A.
gambiae no continente africano, realizado por Pinto et al. (2007), detectou a presença da
variante L1014F nos países do Oeste (kdr-w), desde a Nigéria até o Senegal, a presença da
L1014S (kdr-e) no leste (Quênia), e ambas as mutações ocorrendo no Centro-Oeste,
compreendendo Angola, Gabão, Guiné Equatorial, e Camarões. Esse mesmo padrão de
distribuição foi reportado um ano depois por Santolamazza et al. (2008). Atualmente, a
ocorrência de ambas as mutações, em simpatria, tem sido observada na maioria dos países
africanos, exceto em Niger, Etiópia, Chade e Libéria que reportaram somente a presença da
L1014F, e Burundi e Quênia com somente a L1014S (Figura 7).

40
É preciso destacar que o complexo A. gambiae compreende sete espécies crípticas:
A. gambiae s.s., A. arabiensis, A. melas, A. merus, Anopheles quadriannulatus espécie A, A.
quadriannulatus espécie B e Anopheles bwambae. Elas são indistinguíveis morfologicamente,
porém, são classificadas de acordo com inversões cromossômicas fixas e polimórficas
(Slotman et al., 2005). Há ainda as formas moleculares Savannah, Mopti, Bamako, Forest e
Bissau, classificadas de acordo com inversões paracêntricas do segundo cromossomo de A.
gambiae s.s. (Coluzzi et al., 1979).
A mutação L1014F foi primeiramente descrita em populações de A. gambiae da
forma Savannah, também conhecida como forma S, e até meados de 1999 essa mutação não
ocorria em simpatria com a forma Mopti (M) (Chandre et al., 1999). Entretanto, estudos
posteriores identificaram a presença dessa mutação também na forma M, possivelmente,
resultante de introgressão a partir da forma S (Weill et al., 2000; Diabate et al., 2003).
Introgressão também foi sugerida por Tripet et al. (2007) ao detectar o alelo kdr na forma
Bamako. Por outro lado, um novo evento mutacional independente (de novo phenomeno)
poderia explicar a emergência da mutação kdr em A. arabiensis (Diabate et al., 2004a).
Mesmo em uma única espécie a mesma mutação pode ocorrer várias vezes, de
forma independente. A análise da sequência parcial do domínio IIS6 do NaV de populações de
A. gambiae de várias localidades africanas, sugeriu que os alelos kdr surgiram através de no
mínimo quatro eventos mutacionais independentes, com dois deles (TTT/TCA), a partir de
um único passo mutacional sobre o ancestral comum (TTA), resultando nas duas diferentes
substituições de aminoácidos (fenilalanina/serina) previamente descritas (Pinto et al., 2007).
Apesar dos 15 anos de pesquisa, algumas dúvidas ainda surgem com relação à
mutação kdr, principalmente, quando o assunto é associação genótipo e fenótipo. Uma das

41
técnicas adotadas para testar tal associação é realização de bioensaios com inseticidas (cones
da OMS, Teste de garrafas, ITN, LLIN) seguidos da genotipagem da mutação kdr entre os
anofelinos mortos e sobreviventes ao teste. Em outras palavras, através de bioensaios,
seguidos da genotipagem dos indivíduos detectados como resistentes e susceptíveis, busca-se
testar a hipótese de que a mutação é observada em maior frequência entre os resistentes.
Dos 98 trabalhos aqui analisados, 63 (64,3%) realizaram bioensaio seguido de
genotipagem e encontraram associação entre a presença da mutação e a resistência aos
inseticidas. Destes, aproximadamente seis detectaram o envolvimento de mais de um
mecanismo de resistência (kdr e metabólica) (Awolola et al., 2009; Namountougou et al.,
2012; Chang et al., 2013; Nwane et al., 2013; Zhong et al., 2013; Ochomo et al., 2013a) e
dois apenas sugeriram a ocorrência desse fenômeno (Verhaeghen et al., 2010a; Temu et al.,
2012).

42
Figura 7. Distribuição das mutações kdr ao redor do mundo: (a) América, (b) África, (c) Ásia

43
Por outro lado, dos 98, somente em seis trabalhos (9,5%) não foi possível associar o
genótipo (kdr) com o fenótipo (resistência) (Cuamba et al., 2006; Matambo et al., 2007;
Moreno et al., 2008; Balkew et al., 2010; Verhaeghen et al., 2010b; Jones et al., 2012b).
Nestes casos, a não associação mutação versus resistência foi sugerida devido ao baixo
tamanho amostral (Cuamba et al., 2006), mutação similarmente distribuída entre
sobreviventes e mortos ao bioensaio com inseticida (Matambo et al., 2007; Balkew et al.,
2010; Verhaeghen et al., 2010b; Jones et al., 2012b), ou mutação entre os indivíduos
susceptíveis, muito embora, neste último exemplo, os bioensaios tenham sido
reconhecidamente realizados fora dos padrões recomendados pela OMS (Moreno et al.,
2008).
Por fim, em 28 trabalhos (28,6%) foi realizada somente a genotipagem das
amostras do campo, considerando a presença da mutação como indício suficiente de
resistência.
1.9 Associação entre ITN e mutação kdr
O uso de ITNs/LLINs tem sido uma importante ferramenta usada para tentar
reduzir a morbidade e a mortalidade geradas pela malária (WHO, 2007b). De acordo com um
levantamento realizado por Lengeler (2004), a implementação dessa estratégia na África Sub-
Saariana, entre 1986 e 2003, foi capaz de reduzir a taxa de incidência da morbidade em 50% e
a mortalidade infantil em 17%. Entretanto, a manutenção dessa eficiência ainda é hoje uma
questão controversa.

44
A implementação de ITNs foi capaz de prevenir a mortalidade de crianças em uma
área com elevada transmissão de malária no Quênia (Phillips-Howard et al., 2003).
Resultados similares foram obtidos na Nigéria (Adeogun et al., 2012), onde as LLINs foram
eficientes em matar ou reduzir a alimentação sanguínea de populações de A. gambiae
sabidamente portadoras da mutação kdr ou de resistência metabólica, em Mali (Fryxell et al.,
2012), em Benin (Corbel et al., 2004; Ngufor et al., 2011) e na Uganda (Okia et al., 2013).
O uso de mosquiteiros também foi eficiente contra populações de A. arabiensis oriundas do
Zâmbia, muito embora tenha sido reportado que o alelo kdr não estava presente nessa
população (Norris e Norris, 2011).
Por outro lado, redução na susceptibilidade das populações de A. gambiae quando
expostas a ITNs foi observada na Uganda (John et al., 2008). Além disso, foram
evidenciados aumentos na frequência kdr para essa mesma espécie após a distribuição de
LLINs no Quênia (Stump et al., 2004), em Niger (Czeher et al., 2008), no Senegal (Ndiath et
al., 2012) e em Benin (Aïzoun et al., 2014). Vale ressaltar que na região de Oueme (Benin),
apesar do aumento na frequência da mutação kdr após três anos de implementação do
programa de distribuição de ITNs e IRS em larga escala, esta não foi associada à pressão
seletiva dos mosquiteiros (Padonou et al., 2012).
A mais recente atualização da WHOPES continua indicando apenas piretróides
(deltametrina, alfacipermetrina, permetrina e uma combinação de deltametrina ou permetrina
e butóxido de piperonila - PBO) para LLINs (WHOPES, 2014). Contudo, frente à
possibilidade da perda de efetividade ocasionada pela resistência, o desenvolvimento de
mosquiteiros impregnados com outras classes de inseticidas é uma alternativa promissora. Em
um estudo realizado com mosquiteiros impregnados com clorpirifós-metil (organofosforado)
e lambdacialotrina (piretróide), foi evidenciado que, sozinhos ou em combinações, eles foram

45
eficientes em matar ou reduzir a alimentação sanguínea de A. gambiae da Costa do Marfim,
mesmo em áreas com alta frequência das mutações kdr e ace.IR. Esta última mutação, que
ocorre no gene da acetilcolinesterase, confere resistência a organofosforados (Asidi et al.,
2005).
1.10 Ferramentas moleculares para o diagnóstico de mutações kdr
O fenômeno de resistência pode ser estudado em vários níveis, indo desde os
ensaios biológicos e análises bioquímicas, até a caracterização molecular de genes que
conferem resistência. Atualmente, busca-se o desenvolvimento de ferramentas que possam
fazer um screening genético de populações naturais em larga escala, capazes de predizerem a
predisposição daquelas populações à resistência ao inseticida. Assim sendo, identificar e
localizar marcadores genéticos associados à resistência passou a constar entre as prioridades
do Plano Global para Gerenciamento de Resistência aos Inseticidas da OMS (GPIRM)
(WHO, 2012).
Nesse sentido, a busca por novos marcadores genéticos que identifiquem o genótipo
kdr e o associem com o fenótipo de resistência em populações naturais de insetos de maneira
simples e rápida impõe grandes desafios. Alguns métodos baseados na genotipagem de DNA
estão atualmente disponíveis para detectar a mutação kdr em populações do campo e sob
condições de laboratório. Essas técnicas têm como principais vantagens a alta sensibilidade e
a capacidade de distinguir entre indivíduos homozigotos e heterozigotos (Hemingway e
Ranson, 2000). Na Tabela 3 estão listados os principais métodos empregados na detecção das
mutações kdr, enfatizando os equipamentos requeridos para cada uma delas.

46
Tabela 3. Métodos moleculares usados para a detecção das mutações kdr.
Método
Equipamentos necessários
Mutação
Referências
Reação em cadeia da polimerase alelo especifica (AS-PCR) Termociclador, equipamentos para eletroforese e transiluminador
L1014F/S/C (Martinez-Torres et al., 1998; Ranson et al., 2000)
Ensaio de ligação de nucleotídeos (HOLA) Termociclador, leitor de placa ELISA L1014F/S (Lynd et al., 2005) Sonda de oligonucleotídeos com sequências especificas (SSOP-ELISA)
Termociclador, shaker e leitor de placa ELISA L1014F/S (Kulkarni et al., 2006)
PCR por sonda de oligonucleotídeos com sequências especificas (PCR-Dot Blot)
Termociclador, shaker e membrane de nylon L1014F/S (Kolaczinski et al., 2000)
Transferência de energia por ressonância fluorescente (FRET)/Análise de curva melt (MCA)
PCR em Tempo Real L1014F/S (Verhaeghen et al., 2006)
Alongamento de PCR com fluorescência Termociclador e equipamentos para eletroforese
L1014F/S (Tripet et al., 2006)
Melt de alta resolução (HRM) PCR em Tempo Real L1014F/S (Liew et al., 2004) Loop alelo específico mediado por amplificação isotermal (AS-LAMP)
Turbidímetro e banho maria L1014F (Badolo et al., 2012a)
PCR – Ensaio de polimorfismo usando enzimas de restrição (PCR-RFLP)
Termociclador L1014F/C (Tan et al., 2012b)
Ensaio de PCR usando primer para análise de restrição (PIRA-PCR)
Termociclador, equipamentos para eletroforese e transiluminador
L1014F/S (Janeira et al., 2008)
Ensaio de Multiplex PCR usando primer para análise de restrição (mPIRA-PCR) �
Termociclador e equipamentos para eletroforese
L1014F/S (Kazanidou et al., 2009)
Sistema de amplificação de mutação refratária (ARMS) � Termociclador, equipamentos para eletroforese e transiluminador
L1014F (Singh et al., 2009)

47
2 HIPÓTESES
H0: Não existem diferenças genéticas na região IIS6 do gene do canal de sódio entre
populações de Anopheles darlingi e Anopheles marajoara da Amazônia brasileira;
H1: Existem diferenças genéticas na região IIS6 do gene do canal de sódio entre populações
de Anopheles darlingi e Anopheles marajoara da Amazônia brasileira;
H0: Populações naturais de A. darlingi e A. marajoara não apresentam mutações não
sinônimas na região IIS6 do gene do canal de sódio;
H1: Populações naturais de A. darlingi e A. marajoara apresentam mutações não sinônimas na
região IIS6 do gene do canal de sódio;
H0: Não existe correlação entre a presença de mutações e resistência aos inseticidas
piretróides em anofelinos neotropicais;
H1: Existe correlação entre a presença de mutações e resistência aos inseticidas piretróides em
anofelinos neotropicais.

48
3 OBJETIVOS
3.1 Objetivo Geral
• Isolar, caracterizar e analisar a região parcial do domínio IIS6 do gene do canal de
sódio em populações de Anopheles darlingi e Anopheles marajoara da Amazônia
brasileira.
3.2 Objetivos Específicos
• Comparar as sequências parciais da região IIS6 do gene do canal de sódio de A.
darlingi e A. marajoara com as sequências de outros anofelinos dispostas nos bancos
de dados genômicos;
• Avaliar o grau de polimorfismo da região parcial do domínio IIS6 do gene do canal de
sódio de A. darlingi e A. marajoara;
• Investigar a presença de mutações não sinônimas na região IIS6 do gene do canal de
sódio e analisar suas frequências em populações naturais de A. darlingi e A.
marajoara;
• Correlacionar a presença de mutações não sinônimas com a resistência à deltametrina
por meio de testes de susceptibilidade.

49
4 MATERIAL E MÉTODOS
4.1.1 Área de estudo
Foram estudadas populações procedentes de seis municípios da região Norte do
Brasil: Manaus, São Gabriel da Cachoeira (SGC), Coari e Iranduba, no estado do Amazonas;
Macapá, no estado do Amapá; e Rio Branco, no estado do Acre (Erro! Fonte de referência
não encontrada.).
O estado do Amazonas possui uma área absoluta de 1.559.159,148 km2 e uma
população de 3.807.921 habitantes, sendo o segundo estado menos populoso
(aproximadamente 2,23 hab./km2) (IBGE, 2013). Apresenta clima equatorial úmido, com
temperaturas médias anuais sempre acima de 22°C e duas estações bem definidas: chuvosa
(dezembro a maio) e seca ou menos chuvosa (junho a novembro) (Bonetti, 1999). Manaus
(3°07'07,78"S; 60°01'18.07"O) e Iranduba (3°08'44,48"S; 60°15'26,63"O) estão localizadas
na mesorregião denominada Centro Amazonense e são duas das oito cidades que compõem a
Região Metropolitana de Manaus. São Gabriel da Cachoeira (SGC) (0°33'29,66"S;
68°08'35,31"O) está localizada na mesorregião Norte Amazonense, noroeste do estado,
distando cerca de 850 km da capital Manaus. Coari (4°05'06"S; 63°08'27"O) está localizada
na mesorregião Centro Amazonense, distante cerca de 363 km da capital do estado (IBGE,
2013).
O estado do Amapá possui uma área absoluta de 142.828,512 km2 e uma
população de 734.996 habitantes (4,69 hab./km2). Apresenta clima equatorial úmido, com
temperaturas médias anuais de 27°C e duas estações bem definidas: verão (julho a dezembro)
e inverno (janeiro a junho) (IBGE, 2013). Macapá (0°24'29,53"N; 50°55'44,13"O), a capital

50
do estado, está situada a sudoeste, pertencendo à mesorregião Sul do Amapá e à microrregião
Macapá (IEPA, 2014).
O estado do Acre possui uma área absoluta de 164.123,040 km2 e uma população
de 776.463 habitantes (4,47 hab./km2). Apresenta clima equatorial úmido, com temperaturas
médias anuais variando de 24 a 32ºC e duas estações bem definidas: chuvosa (novembro a
abril) e seca (maio a outubro). Rio Branco (9°58'29"S; 67°48'36"O), a capital do estado,
pertence à mesorregião Vale do Acre e à microrregião Rio Branco (IBGE, 2013).
Figura 8. Mapa do Brasil, enfatizando as localidades onde as populações de Anopheles
darlingi e Anopheles marajoara foram coletadas.

51
4.1.2 Obtenção das amostras
Para a obtenção das amostras, mosquitos adultos e larvas foram coletados no
período compreendido entre dezembro de 2009 até julho de 2013. Os espécimes adultos
foram capturados a cada 60 minutos, das 17:00 até as 24:00 horas, com auxílio de um
aspirador manual movido à pilha, empunhados por dois capturadores que retêm todos os
mosquitos que se aproximam. Essas coletas foram realizadas dentro e fora dos domicílios, e
também nas paredes dos currais, seguindo os critérios e as recomendações da Organização
Mundial de Saúde (OMS) (WHO, 1975).
Posteriormente, eles foram transferidos para copos telados acondicionados em uma
caixa térmica de poliestireno (Figura 9a) e transportados para o Laboratório de Malária e
Dengue do Instituto Nacional de Pesquisas da Amazônia (INPA). Algumas fêmeas foram
imediatamente colocadas em microtubos eppendorf (1,5 mL) devidamente etiquetados,
contendo isopropanol absoluto, e mantidas até as análises moleculares. Outras, após o repasto
sanguíneo, que foi feito usando um hamster (Mesocricetus auratus), foram isoladas em copos
telados para postura individual (Figura 9b).
A coleta das larvas foi feita nos criadouros usando o método de imersão, com
auxílio de conchas, pipetas e bacias plásticas, das 09:00 até as 12:00 horas e das 14:00 até as
17:00 horas (WHO, 1975). As larvas de 1º até 3º estádio foram conduzidas ao laboratório de
Malária e Dengue do INPA e mantidas até o 4º estádio larval.
A identificação dos adultos, dos ovos e das larvas foi feita utilizando as chaves
taxonômicas de Gorham et al. (1967), Faran e Linthicum (1981) e Consoli e Lourenço-de-
Oliveira (1994).

52
Na manutenção dos mosquitos em laboratório, foi utilizado o método descrito em
Santos et al. (1981). Cada desova foi colocada em cubas esmaltadas de 13 cm e 16 cm de
diâmetro, por 6 cm de altura (Figura 9c). Em cada cuba, foram adicionados 450 mL e 500 mL
de água destilada, respectivamente, e a troca de água era realizada a cada dois dias.
Figura 9 - (a) Copos telados acondicionados em caixa de poliestireno usados para o transporte das fêmeas de
Anopheles darlingi e Anopheles marajoara; (b) copos plásticos com filó usados para oviposição; (c) cubas
esmaltadas onde as larvas eram criadas.
A alimentação das larvas foi à base de ração para peixes moída (Tetramin e
Goldfish, na proporção 2:1) (Figura 10). Os mosquitos foram criados em um insetário com
temperatura constante de 26ºC ± 1ºC e umidade relativa entre 80% e 90%, até atingirem o 4º
estádio larval.

53
Figura 10. Alimentação usada para criação de larvas de anofelinos.
4.1.3 Bioensaio simplificado de knockdown
Para avaliar a resistência ao efeito knockdown provocado pelos piretróides, um
bioensaio simplificado de larvas foi padronizado, a partir da metodologia descrita em Kawada
et al. (2009). De acordo com esse bioensaio, larvas de 4º estádio foram individualmente
colocadas em copos plásticos contendo 20 mL da solução de inseticida, sendo acompanhado o
tempo de knockdown a cada 5 minutos, por até 30 minutos (Figura 11).
O tempo médio de knockdown (KT50), ou seja, o tempo necessário para que 50%
das larvas de uma dada população sofram o efeito do inseticida, foi contabilizado em seis
categorias: 1 – de 0 a 5 minutos; 2 – de 6 a 10 minutos; 3 – de 11 a 15 minutos; 4 – de 16 a 20
minutos; 5 – de 21 a 30 minutos; 6 - acima de 30 minutos.
Por definição, efeito knockdown é a perda da coordenação e paralisia provocada
pelo inseticida, frequentemente, acompanhada por espasmos e tremores (Schleier III e

54
Peterson, 2011). Assim sendo, as larvas que apresentavam tremores, que permaneciam no
fundo do copo ou eram incapazes de nadar e flutuar, ou até mesmo que ficavam paralisadas,
foram consideradas sob o efeito do inseticida (Figura 12).
O inseticida usado foi deltametrina grau-técnico (Sigma) diluída em acetona pura
(Pureza Analítica/P.A.), conforme recomendações do fabricante, a partir do qual foram
testadas sete distintas concentrações (0,05 ppm; 0,1 ppm; 0,2 ppm; 0, 3ppm; 0,4 ppm; 0,5
ppm e 0,6 ppm) para a determinação das doses diagnósticas desse inseticida sobre as
populações de anofelinos. Entendem-se aqui como doses diagnósticas a menor e a maior
concentração capaz de atingir o KT50 das larvas da população susceptível (referência).
Os experimentos foram realizados em triplicata, usando 20 larvas por experimento,
sendo um controle, no qual foi inserido somente o solvente (acetona P.A), para avaliar se esse
composto químico estava exercendo algum efeito danoso sobre as larvas.
Após a obtenção das doses discriminantes, foi calculado o índice de
susceptibilidade, ou seja, o produto do KT50 da menor dose pelo KT50 da maior dose. O
índice de susceptibilidade varia de um a 36, assim sendo, a população menos tolerante terá
valor próximo de um e a mais tolerante terá valor próximo de 36. Finalizado os experimentos,
as larvas foram inseridas, individualmente, em microtubos devidamente etiquetados, contendo
isopropanol absoluto (100%) e mantidas à temperatura ambiente até as análises moleculares.
Uma das dificuldades na realização de bioensaios em anofelinos amazônicos é a
falta de colonização de muitas das espécies sob condições de laboratório. Assim sendo, não
existem colônias que possam ser usadas como referência para avaliação da susceptibilidade a
inseticidas. No entanto, existe um consenso de que populações coletadas em áreas com pouca
ou nenhuma utilização de inseticidas possam ser usadas para esse fim. Dessa forma, a

55
população de São Gabriel da Cachoeira foi utilizada com esse propósito, pois as amostras
foram coletadas em um sítio na BR-307, a cerca de 9 km de distância do centro da cidade
(00o08’48.6”S; 66º57’29.5”W), que por estar longe de áreas urbanas, não recebe aplicação
direta de inseticidas.
Figura 11 - Bioensaio simplificado de knockdown, aplicado a anofelinos neotropicais.

56
Figura 12. Larvas de 4º estádio de Anopheles darlingi, usadas nos
bioensaios de knockdown.
4.1.4 Extração de DNA das larvas
Foram realizadas extrações individuais de DNA genômico, seguindo o protocolo
descrito em Wilkerson et al. (1995a). O número total de indivíduos analisados foi de 240
larvas de A. darlingi das populações de Manaus, SGC e Macapá (40 mais susceptíveis de
Manaus, 40 menos susceptíveis de Manaus, 40 mais susceptíveis de SGC, 40 menos
susceptíveis de SGC, 40 mais susceptíveis de Macapá, 40 menos susceptíveis de Macapá) e
240 larvas de A. marajoara das populações de Iranduba/AM, Macapá/AP e Rio Branco/AC
(40 mais susceptíveis de Iranduba, 40 menos susceptíveis de Iranduba, 40 mais susceptíveis
de Macapá, 40 menos susceptíveis de Macapá, 40 mais susceptíveis de Rio Branco, 40 menos
susceptíveis de Rio Branco), ambas submetidas ao bioensaio com deltametrina.
Etapa 1: Foram realizadas extrações individuais de larvas de 4º estádio, onde cada
larva foi homogeneizada em 50 μL de Tampão de Lise - TL (SDS 1%, Tris-HCl 50 mM, pH
8.0, EDTA 25 mM, pH 8.0 e NaCl 52 mM) com auxílio de um bastão de acrílico, em

57
microtubos do tipo Eppendorf de 1,5 mL. Em seguida, foram adicionados 100 μL de TL para
remover resíduos das paredes do tubo e do bastão. As amostras foram encubadas a 65ºC por
30 minutos em banho-maria. Posteriormente, foram submetidas à centrifugação a 14000 rpm
por 30 segundos e então foram adicionados 100 μL de Acetato de Potássio em cada tubo. As
amostras foram misturadas em vórtex e mantidas em gelo por 1 hora no refrigerador. Após
esse período, foram centrifugadas novamente a 14000rpm por 10 minutos e os sobrenadantes
foram transferidos para novos tubos Eppendorf de 1,5 mL. Então foram adicionados 500 μL
de Etanol (100%) gelado e as amostras foram mantidas a -20ºC por 1 hora (ou “overnight”).
Etapa 2: Posteriormente as amostras foram centrifugadas a 14000 rpm por 30
minutos e descartado o sobrenadante. Adicionou-se ao precipitado 500 μL de Etanol (70%)
gelado e centrifugou-se a 14000 rpm por 10 minutos. Foi retirado novamente o sobrenadante
e secado o precipitado em fluxo laminar. Após a evaporação do Etanol, os precipitados foram
ressuspendidos em 15 μL de água Milli-Q autoclavada e estocado em temperatura a -20ºC
para posterior quantificação.
4.1.5 Extração de DNA dos adultos
Foi extraído DNA genômico de 240 indivíduos adultos de A. darlingi, coletados
dentro e fora dos domicílios nas cidades de Manaus e Coari (Manaus: 60 intradomiciliares e
60 peridomiciliares; Coari: 60 intradomiciliares e 60 peridomiciliares), seguindo a técnica de
fenol-clorofórmio descrita em Sambrook e Russell (2001).
Etapa 1: Foram realizadas extrações individuais de mosquitos adultos, onde cada
mosquito foi homogeneizado em 50 μL de TL, com auxílio de um bastão de acrílico, em

58
microtubos do tipo Eppendorf de 1,5 mL. Em seguida, foram adicionados 100 μL de TL para
remover resíduos das paredes do tubo e do bastão e 15 μL de solução de Proteinase K. As
amostras foram encubadas a 50ºC por 1 hora em banho-maria. Posteriormente foram
adicionados 150 μL de Fenol em TE (Tris-EDTA). Os tubos foram misturados em vórtex e
centrifugados a 14000 rpm por 1 minuto. Em seguida, as fases aquosas (superiores) foram
transferidas para novos tubos e foram adicionados 300 μL de Fenol-clorofórmio-álcool-
isoamílico. Os tubos foram misturados novamente em vórtex e centrifugados a 14000 rpm por
1 minuto. Novamente, as fases aquosas foram transferidas para novos tubos, e estes
receberam 600 μL de Clorofórmio-álcool-isoamílico. Os tubos foram misturados em vórtex,
centrifugados a 14000 rpm por 1 minuto e as fases aquosas transferidas para novos tubos.
Adicionou-se 120 μL de Acetato de Sódio e 1200 μL de Etanol (100%) gelado. Em seguida,
as amostras foram mantidas a -20ºC por 1 hora (ou “overnight”).
Etapa 2: Nessa etapa foram realizados os mesmos procedimentos descritos na
etapa 2 do protocolo anterior.
4.1.6 Quantificação do DNA
Após a extração foi realizada a quantificação para verificar a concentração do
DNA das amostras, pois normalmente as bases aminadas encontradas nos ácidos nucléicos
são cromóforos muito fortes que permitem uma rápida e eficiente quantificação desses
polímeros em solução. As quatro bases encontradas no DNA absorvem entre 250 a 270 nm,
de forma que é observado um pico em torno de 260 nm em DNA de alta massa molecular
(Azevedo et al., 2003).

59
A quantificação foi realizada em espectrofotômetro (Gene Quant Pro-Pharmacia)
com os seguintes procedimentos: Adicionou-se 68 μL de água Milli-Q autoclavada e 2 μL do
DNA (fator de diluição 35) na cubeta. Em seguida foram realizadas as leituras. Para a
amplificação do DNA a concentração considerada ótima foi entre 100 a 3000 ng/μL e razão
de 1400 a 2000, conforme Caccone et al. (1996).
Após a quantificação, as amostras foram diluídas em água Milli-Q autoclavada
para a obtenção de amostras com uma concentração final de 10 ng/μL de solução.
Posteriormente, tanto as amostras puras (DNA estoque) quanto às diluídas foram mantidas em
freezer a -20ºC até o início das análises moleculares.
4.1.7 Amplificação, clonagem e sequenciamento do domínio IIS6 do gene do canal de
sódio usando a técnica de primers degenerados
Durante o desenvolvimento deste trabalho, o genoma de A. darlingi ainda não
havia sido publicado. Por isso, em uma fase inicial, para a amplificação de uma região parcial
de 200 pb foi utilizada a técnica de primers degenerados, usando os primers Gen1F (5’-GGA
GAR TGG ATY GAR TCN AT GTG GGA-3’) e Gen1R (5’-TTN GAC AAA AGC AAG
GCT AAG AA-3’), previamente desenhados para Anopheles stephensi (Enayati et al., 2003).
Nessa estratégia, os DNAs extraídos foram agrupados em 10 distintos pools
gênicos, conforme segue: para A. darligi - (1) intradomicílio de Manaus, (2) peridomicílio de
Manaus, (3) larvas mais susceptíveis de Manaus, (4) larvas menos susceptíveis de Manaus,
(5) larvas mais susceptíveis de Macapá, (6) larvas menos susceptíveis de Macapá, (7) larvas

60
mais susceptíveis de SGC, (8) larvas menos susceptíveis de SGC; para A. marajoara - (9)
larvas mais susceptíveis de Macapá, (10) larvas menos susceptíveis de Macapá.
Em seguida foram realizadas as etapas de amplificação, clonagem, purificação e
sequenciamento, conforme metodologia descrita em Martins et al. (2009). Foram utilizadas as
amostras diluídas do DNA genômico em tubo tipo Eppendorf de 0,2 mL com a seguinte
composição: 2 µL de DNA; 1,3 U de Pfu Polymerase (Biotools) (0,025 U/μL); 5,0 µL de
Buffer 10X com MgCl2 (1X); 1,0 µL de dNTP (0,2 mM); 5,0 µL Gen1F (forward) (1 µM);
5,0 µL de Gen1R (reverse) (1 µM); 30,8 µL de água Milli-Q autoclavada.
O volume total da reação foi de 50 μL. Os tubos foram levados ao termociclador
(marca Veriti 384-well, Applied Biosystems), programado para realizar 30 ciclos com os
seguintes perfis de temperatura: 3 minutos a 94ºC, para desnaturação inicial; 30 segundos a
94ºC, para desnaturação; 30 segundos a 54ºC, para anelamento dos primers; 45 segundos a
72ºC, para alongamento das fitas sintetizadas; 7 minutos a 72ºC, para o ciclo de alongamento
final.
O produto amplificado foi submetido à corrida eletroforética (170 V por 40
minutos), em gel de acrilamida (10%) (TBE 1X), para avaliar a qualidade do material obtido.
Em cada poço do gel, foram aplicados 4,0 μL do produto purificado + 1,0 μL de 6X Orange
DNA Loading Dye (Fermentas), 4,0 μL de Marcador Molecular O’Gene Ruler de 100 pb
(Fermentas) + 1 μL de 6X Orange DNA Loading Dye (Fermentas).

61
Figura 13. Perfil eletroforético em gel de acrilamida (10%) da amplificação da região IIS6 do gene NaV de Anopheles darlingi, usando os primers descritos por Enayati et al. (2003). 1 a 14= Amostras de DNA da população de Manaus; M= Marcador Molecular.
Com o sucesso da amplificação (Figura 13), o restante do produto (45 μL) foi
submetido à corrida eletroforética em gel de agarose (1,5%) (TBE 0,5X) (120 V por 60
minutos). Em cada poço foram aplicados 45 μL do produto amplificado de cada reação + 4,5
μL de Blue Green Loading Dye I (LGC Biotecnologia), 1 μL de Marcador Molecular (Ladder
50 pb) + 1 μL de Blue Green Loading Dye I (LGC Biotecnologia) + 8 μL de tampão 0,5X, e 5
μL de Gene Ruler Ultra Low Range (Fermentas) + 1 μL de Blue Green Loading Dye I (LGC
Biotecnologia) + 4 μL de tampão 0,5X.
200 pb

62
Figura 14. Perfil eletroforético em gel de agarose da amplificação da região IIS6 do NaV de Anopheles darlingi, usando os primers descritos por Enayati et al. (2003). M1= Marcador Molecular Ladder 50 pb; 1= produto amplificado; 2= produto amplificado; C= Controle negativo; M2= Marcador Molecular Gene Ruler.
As bandas foram recortadas do gel (Figura 15) e purificadas, usando o Kit Illustra
TM GFXTM PCR DNA and Gel Band Purification (GE Healthcare), com TAE e TBE,
conforme recomendações do fabricante. Como o produto purificado apresentou boa
qualidade, foi dado início a ligação e posterior transformação. A etapa de clonagem foi
realizada usando o kit CloneJet PCR Cloning kit (Fermentas). O produto ligado foi então
transformado em células competentes de E. coli DH5α, por meio de choque térmico a 42º C e
inseridas em placas de Petri contendo meio de cultura LB sólido com ampicilina (100 μL/mL)
(Figura 16). Cerca de 100 colônias de cada pool foram inoculadas em meio líquido rico em
nutrientes (Circle Growth, Gibco) em placas de 96 poços fundos, por 22 horas sob 200 rpm de
rotação a 37º C.
200 pb

63
Figura 15. Perfil eletroforético em gel de agarose da amplificação da região IIS6 do NaV de Anopheles darlingi, usando os primers descritos por Enayati et al. (2003), enfatizando o recorte das bandas. M1= Marcador Molecular Ladder 50 pb; 1= produto amplificado; 2= produto amplificado; C= Controle.
Figura 16. Placa de Petri evidenciando o crescimento das colônias de Escherichia coli com os clones da região IIS6 do gene NaV de Anopheles darlingi.
Após esse período, o DNA plasmidial foi extraído por meio da técnica de lise
alcalina, usando Acetato de Potássio 3 M, GET (Glicose, EDTA, Tris-HCl) e NaOH 0,2N +
M1 1 2 C M2
200 pb

64
SDS 1%, seguida de filtração em placas Millipore, precipitação e lavagem em isopropanol e
etanol (Sambrook e Russell, 2001). Os plasmídeos então purificados foram submetidos à
reação de sequenciamento nucleotídico, usando o kit Big Dye 3.1 em sequenciador ABI 377
(Applied Biosystems) da plataforma de sequenciamento de DNA PDTIS, da Fundação
Oswaldo Cruz.
4.1.8 Amplificação, clonagem e sequenciamento do domínio IIS6 do gene do canal de
sódio usando primers específicos
Logo após a publicação dos dados do genoma de A. darlingi (Marinotti et al.,
2013), um novo par de primers, 42F (5’-TCG TGT TTT ATG CGG AGA ATG G-3’) e 422R
(5’-CAC GGA CGC AAT TTG ACT TGT-3’), agora específico para A. darlingi, foi
desenhado, e isso permitiu a amplificação de uma região genômica maior (400 pb).
Nessa estratégia foram analisadas as populações de A. darlingi de Coari (40
intradomiciliares; 40 peridomiciliares) e A. marajoara de Rio Branco (40 mais susceptíveis e
40 menos susceptíveis).
Os DNAs foram individualmente amplificados em tubo tipo Eppendorf de 0,2 mL
com a seguinte composição: 1 µL de DNA; 0,5 U de Pfu Polymerase (Biotools) (0,02 U/μL);
2,5 µL de Buffer 10X com MgCl2 (1X); 0,5 µL de dNTP (0,2 mM); 1,5 µL de 42F (forward)
(0,6 µM); 1,5 µL de 422R (reverse) (0,6 µM); 17,5 µL de água Milli-Q autoclavada.
O volume total da reação foi de 25 μL. Os tubos foram levados ao termociclador
(marca Veriti 384-well, Applied Biosystems), programado para realizar 35 ciclos com os
seguintes perfis de temperatura: 3 minutos a 94ºC, para desnaturação inicial; 30 segundos a

65
94ºC, para desnaturação; 30 segundos a 60ºC, para anelamento dos primers; 60 segundos a
72ºC, para alongamento das fitas sintetizadas; 5 minutos a 72ºC, para o ciclo de alongamento
final.
O produto amplificado foi submetido à corrida eletroforética (170 V por 40
minutos), em gel de acrilamida (10%) (TBE 1X), para avaliar a qualidade do material obtido.
Em cada poço do gel, foram aplicados 4,0 μL do produto purificado + 1,0 μL de 6X Orange
DNA Loading Dye (Fermentas), 4,0 μL de Marcador Molecular (Ladder 100 pb) (Invitrogen)
+ 1 μL de 6X Orange DNA Loading Dye (Fermentas).
Figura 17. Perfil eletroforético em gel de acrilamida (10%) da amplificação da região IIS6 do gene NaV de Anopheles darlingi, usando primers específicos. 1 a 6= Amostras de DNA da população de Coari; M= Marcador Molecular.
Com o sucesso da amplificação (Figura 17), o restante do produto (20 μL) foi
purificado usando o kit IllustraTM GFXTM PCR and Gel Band Purification (GE Healthcare),
de acordo com as instruções do fabricante. Os produtos purificados foram diretamente
submetidos à reação de sequenciamento nucleotídico de ambas as fitas do DNA, usando o kit

66
Big Dye 3.1 em sequenciador ABI 377 (Applied Biosystems) da plataforma de
sequenciamento de DNA PDTIS, da Fundação Oswaldo Cruz.
4.1.9 Análises dos dados
As sequências obtidas foram alinhadas e corrigidas manualmente usando o
programa BioEdit 7.0.5.3 (Hall, 1999). O programa Mega 5 (Tamura et al., 2011) foi usado
para tradução das sequências de nucleotídeos, análise de sítios polimórficos e para construção
da árvore filogenética. O programa DnaSP 5.10 (Librado e Rozas, 2009) foi usado para
determinar o número de haplótipos, considerando como distintos somente aqueles
representados por, no mínimo, duas sequências encontradas. O programa TCS 1.13 (Clement
et al., 2000) foi usado para a análise genealógica entre os haplótipos. As comparações
estatísticas dos números de haplótipos em cada grupo foi conduzida no programa BioEstat 5.0
(Ayres et al., 2007).

67
5 RESULTADOS E DISCUSSÃO
5.1 Bioensaio simplificado de knockdown
Existem poucos trabalhos na literatura atual que descrevem o status de
susceptibilidade das populações de anofelinos neotropicais frente aos inseticidas piretróides.
De acordo com Santos et al. (2007), isso ocorre principalmente, devido a não colonização de
algumas das espécies em laboratório, o que dificulta a padronização dos bioensaios.
Entre os bioensaios mais empregados na detecção de resistência em anofelinos
constam: testes com papéis (WHO, 2006b) ou garrafas de vidro (Brogdon e Mcallister,
1998a), impregnados com inseticidas. Essas metodologias, embora eficientes em alguns
países, apresentam algumas dificuldades técnicas quanto a sua aplicabilidade no Brasil para
algumas espécies de vetores.
Os testes padrões da OMS com papéis impregnados, embora úteis e eficazes,
tornaram-se caros ao longo dos anos, requerem uma grande quantidade de insetos a serem
testados, não estão disponíveis para alguns inseticidas e as dosagens disponíveis não são
aplicáveis a todas as espécies de vetores (Brogdon e Mcallister, 1998a).
As provas de garrafa, muito embora sejam de baixo custo, também requerem uma
grande quantidade de insetos adultos. Além disso, o avaliador pode se contaminar ao
introduzir ou retirar os insetos de dentro das garrafas impregnadas (Brogdon e Chan, 2013).
Assim sendo, descrevemos aqui pela primeira vez a padronização de uma
metodologia simples, rápida e de baixo custo, capaz de determinar o índice de
susceptibilidade de uma dada população de anofelino ainda no estágio larval.

68
Essa metodologia foi inicialmente desenvolvida por Kawada et al. (2009) para
testar o nível de susceptibilidade das larvas de Aedes aegypti frente aos piretróides. Esses
autores concluíram que o novo método foi eficiente quando comparado aos testes tradicionais,
principalmente, quando o objetivo é mapear resistência tipo kdr, que se manifesta durante
todos os estágios do ciclo de vida do mosquito.
No entanto, para um panorama mais completo, os testes com papéis (WHO,
2006b) ou garrafas de vidro (Brogdon e Mcallister, 1998a) impregnados com inseticidas
devem ser considerados para mosquitos adultos, uma vez que os mecanismos de resistência
metabólica podem se expressar, especificamente, em determinados estágios do
desenvolvimento dos insetos (WHO, 2013).
Devido a não existência de colônias susceptíveis (ou de referência) para essas
espécies, foram comparados os índices de susceptibilidade (SI) entre populações naturais
(Figura 18). Com base na população de SGC, foram identificadas as concentrações de 0,1 e
0,4 ppm, como sendo as doses diagnósticas de deltametrina para as populações de A. darlingi
e A. marajoara.
Assim sendo, para A. darlingi, SGC foi a que exibiu menor tolerância (SI=4). Esse
resultado é esperado, uma vez que as coletas realizadas nessa localidade foram feitas em um
sítio na BR-307, que por estar longe de áreas urbanas, não recebe aplicação direta de
inseticidas.
Baixa tolerância (SI=6) também foi obtida na população de Macapá. Já a maior
tolerância foi evidenciada em Manaus (SI=15). Em comparação com Macapá, a cidade de
Manaus é maior e mais populosa, apresentando maiores índices de malária e dengue, tendo,
portanto, maior aplicação de piretróides. Assim sendo, a pressão de seleção sofrida pelos
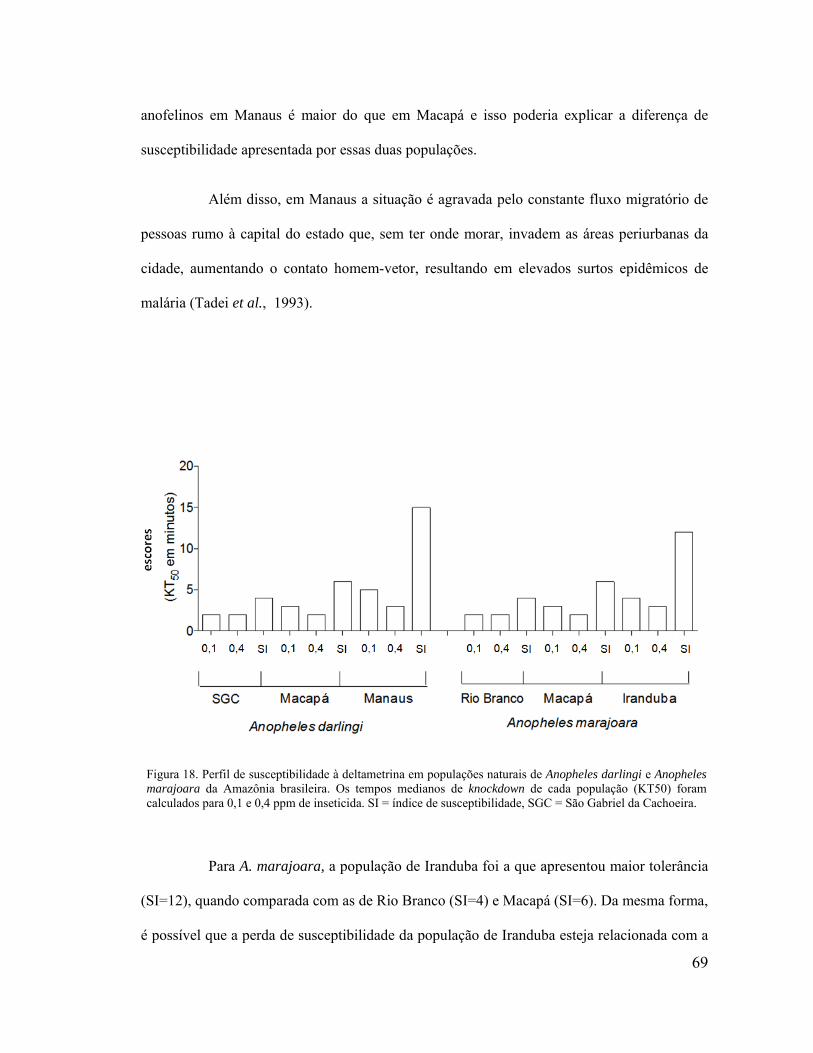
69
anofelinos em Manaus é maior do que em Macapá e isso poderia explicar a diferença de
susceptibilidade apresentada por essas duas populações.
Além disso, em Manaus a situação é agravada pelo constante fluxo migratório de
pessoas rumo à capital do estado que, sem ter onde morar, invadem as áreas periurbanas da
cidade, aumentando o contato homem-vetor, resultando em elevados surtos epidêmicos de
malária (Tadei et al., 1993).
Figura 18. Perfil de susceptibilidade à deltametrina em populações naturais de Anopheles darlingi e Anopheles marajoara da Amazônia brasileira. Os tempos medianos de knockdown de cada população (KT50) foram calculados para 0,1 e 0,4 ppm de inseticida. SI = índice de susceptibilidade, SGC = São Gabriel da Cachoeira.
Para A. marajoara, a população de Iranduba foi a que apresentou maior tolerância
(SI=12), quando comparada com as de Rio Branco (SI=4) e Macapá (SI=6). Da mesma forma,
é possível que a perda de susceptibilidade da população de Iranduba esteja relacionada com a

70
sua proximidade da capital Manaus. Somente 20 km separam esses dois municípios, que
estão, atualmente, conectados pela Ponte Rio Negro. Portanto, as medidas estratégicas de
combate aos anofelinos adotadas em Iranduba, podem ser um reflexo daquelas adotadas em
Manaus.
Muito embora a malária seja um grave problema de saúde pública no Brasil,
existem poucos estudos avaliando a resistência aos inseticidas nas populações de seus vetores.
Isso ocorre, principalmente, devido à falta de colonização desses mosquitos, que dificulta a
execução e padronização de bioensaios entre grupos, sendo, portanto, um empecilho para a
obtenção de um perfil bem definido do real status de susceptibilidade das populações de
anofelinos da Amazônia (Santos et al., 1999).
Mesmo assim, resistência fisiológica intermediária ao DDT foi detectada três
décadas atrás em populações de A. albitarsis do Mato Grosso (Brown e Pal, 1973) e em
populações de A. galvoi e A. rondoni de São Paulo (Brown, 1986). No entanto, um estudo
mais recente, conduzido pela Secretaria de Vigilância em Saúde do Ministério da Saúde, tem
evidenciado populações brasileiras de A. darlingi, A. aquasalis, A. nuneztovari e A. albitarsis
susceptíveis à cipermetrina, seguindo os padrões descritos para o gênero (Santos et al., 2007).
Em países vizinhos, o primeiro registro de populações de A. darlingi resistentes a
inseticidas foi em amostras coletadas em Quibdo, Chocó, nordeste da Colômbia (Quiñones et
al., 1987; Suarez et al., 1990). Nesse mesmo país, recentemente foi identificada resistência
cruzada entre DDT e lambdacialotrina em populações oriundas de Medio Atrato, Chocó
(Fonseca-González et al., 2009). Também na Colômbia, foram identificadas populações de
A. marajoara resistentes a DDT (Quiñones e Suarez, 1990).

71
A utilização constante de uma única classe de inseticida além de aumentar a
pressão de seleção, aumenta a frequência dos genótipos resistentes. A detoxificação
enzimática (Esterases e Oxidases de Função Mista) pode ser o principal mecanismo reduzindo
a susceptibilidade das populações brasileiras de A. darlingi e A. marajoara, conforme
evidenciado por Fonseca-González et al. (2009) na Colômbia.
De acordo com Omoto (2000), uma boa estratégia para o manejo da resistência ou
da perda de susceptibilidade dos insetos é a rotação e mistura de inseticidas. Essa estratégia
consiste na utilização rotacionada de inseticidas estruturalmente distintos e com diferentes
modos de ação, baseado no fato de que a frequência da resistência a um produto tende a
diminuir quando produtos com diferentes modos de ação e detoxificação são utilizados.
Colocamos aqui, como sugestão, que essa medida seja adotada no estado do Amazonas, com
o intuito de prevenir problemas mais graves de resistência. Outra sugestão seria a distribuição
de mosquiteiros impregnados com diferentes tipos de inseticidas.
5.2 Variação no domínio IIS6 do gene NaV de Anopheles darlingi e Anopheles
marajoara
O canal de sódio regulado por voltagem é o alvo dos inseticidas piretróides, que
são amplamente usados para controlar insetos vetores de doenças e pragas na agricultura.
Estudos prévios evidenciaram que um dos principais mecanismos de resistência, mutações
tipo kdr, ocorre no domínio IIS6 desse canal em diversos insetos, inclusive em mosquitos do
gênero Anopheles (Martinez-Torres et al., 1998; Ranson et al., 2000; Awolola et al., 2007;
Bass et al., 2007; Kim et al., 2007; Adasi e Hemingway, 2008; Singh et al., 2009; Kang et
al., 2012; Lol et al., 2013). Portanto, a avaliação da diversidade molecular desta região

72
genômica em anofelinos neotropicais tem o potencial de informar sobre a predisposição
genética para esse tipo de resistência em populações naturais.
O resultado do sequenciamento nucleotídico dos fragmentos de 200 pb e 400 pb,
tanto de A. darlingi quanto de A. marajoara, a partir de primers degenerados previamente
desenvolvidos para o vetor da malária asiático A. stephensi (Enayati et al., 2003) e
específicos para A. darlingi evidenciou conter região homóloga aos exons 20, 21, incluindo o
íntron entre eles, correspondente ao domínio IIS6 do NaV de outros anofelinos, dispostos nos
bancos de dados genômicos (GenBank e VectorBase) (Davies et al., 2007a; Tan et al.,
2012a; Lol et al., 2013).
Ao todo foram obtidas 90 sequências de boa qualidade para A. marajoara e 305
para A. darlingi. A tradução dedutível em aminoácidos do fragmento sequenciado revelou
total similaridade, com aqueles obtidos em outros anofelinos. Tal similaridade é esperada,
uma vez que, como o canal de sódio é essencial no processo de transmissão e propagação de
impulsos nervosos, mudanças de aminoácidos poderiam comprometer o papel fisiológico da
proteína (Ffrench-Constant et al., 1998).
Comparações entre as regiões codificantes das sequências dos genes AdNaV e
AmNaV com as de outros anofelinos, revelaram identidade variando de 73,6% (Anopheles
minimus) a 99,7% (Anopheles darlingi - AdarC2) (Tabela 4). Na Figura 19 são apresentadas
as prováveis relações filogenéticas entre elas. A se considerar pelo alto valor de bootstrap, as
sequências foram divididas em dois grandes grupos, sendo o primeiro formado pelos
anofelinos do Velho Mundo (África e Ásia), e o segundo pelos anofelinos do Novo Mundo
(América), seguindo um grau de divergência dentro do esperado para a filogenia destas
espécies (Harbach, 2013).

73
Sequências de aminoácidos do canal de sódio de A. gambiae comparada com as de
outras espécies do grupo Neoptera também evidenciaram grau de divergência dentro do
esperado para a filogenia das espécies, sugerindo que este canal está mais fortemente sujeito à
pressão de seleção purificadora do que outras proteínas ortólogas naqueles organismos
(Davies et al., 2007a).
Ao comparar as sequências nucleotídicas entre as espécies de anofelinos
evidenciou-se que muitas das variações encontradas ocorreram no íntron (59), mostrando
alguns pontos de mutação e eventos de inserção/deleção (indels) (Figura 20). Foram
observados 29 sítios polimórficos no exon 20 e 66 no exon 21.
Além de A. darlingi e A. marajoara, o único anofelino neotropical, cuja sequência
do NaV está disponível nos bancos de dados públicos, é A. albimanus. Alto índice de
conservação foi evidenciado entre essas espécies. No entanto, é interessante destacar que em
A. albimanus recentemente foi descrita a presença de mutações kdr em populações naturais
(L1014F no México, L1014C na Nicarágua e Costa Rica) (Lol et al., 2013).
Ainda entre essas espécies, observou-se que A. darlingi possui o íntron maior do
que A. marajoara e A. albimanus, que apresentaram uma deleção de duas bases nas mesmas
posições (153 e 154), sendo que para esta última espécie foi evidenciada uma deleção
adicional na posição 179.

74
Figura 19. Uma árvore de Neighbor-Joining mostrando as relações filogenéticas dos haplótipos encontrados no domínio IIS6 do gene NaV de Anopheles darlingi e Anopheles marajoara com as sequências de outros anofelinos (detalhes na Tabela 4). Os valores entre os ramos indicam a percentagem de vezes que um nó foi suportado no teste de bootstrap (1000 repetições) (Tamura et al., 2011).
Tem sido proposto que diferenças no tamanho do íntron possam ser,
potencialmente, usadas como identificação taxonômica das espécies de vetores da malária nos
Neotrópicos (Lol et al., 2013). Nossos resultados reforçam essa proposta.
De acordo com Davies et al. (2007a), as diferenças observadas, tanto na
composição de bases dos exons, quanto na composição e tamanho do íntron, podem ser
resultantes de pequenas adaptações ou ajustes finos de funcionalidade do canal de sódio, que
podem ser desenvolvidos entre diferentes espécies.
Nas populações de A. darlingi aqui avaliadas, foram obtidos cinco sítios
polimórficos, que resultaram em cinco haplótipos, cujas relações genealógicas estão

75
representadas na Figura 21. Apenas uma substituição ocorreu no íntron, além de outras quatro
nos exons, porém todas foram sinônimas.
Assim sendo, a mutação kdr, no sítio 1014, não foi detectada, sugerindo a
ocorrência de outros mecanismos envolvidos na perda de susceptibilidade de algumas
populações à deltametrina. No entanto, vale ser destacado que sua ocorrência é permissiva,
uma vez que o códon nesse sítio é formado por TTA e uma única mutação, por exemplo, TTT
(Martinez-Torres et al., 1998) ou TCA (Ranson et al., 2000), pode mudar o aminoácido de
leucina para fenilalanina ou serina, conforme já ocorreu em outras espécies de anofelinos,
como em Anopheles gambiae s.s.(Martinez-Torres et al., 1998; Ranson et al., 2000),
Anopheles funestus (Morgan et al., 2010), Anopheles stephensi (Enayati et al., 2003),
Anopheles culicifacies (Hoti et al., 2006; Singh et al., 2009), Anopheles sinensis (Kim et al.,
2007; Tan et al., 2012a), Anopheles sacharovi (Luleyap et al., 2002), Anopheles vagus
(Verhaeghen et al., 2010b), inclusive em A. albimanus (Lol et al., 2013).
A população de Manaus apresentou quatro haplótipos (A, B, C e D), SGC três (B,
C e D) e Macapá dois (B e D). Curiosamente, apesar da proximidade com Manaus, Coari
apresentou somente, e com exclusividade, o haplótipo I. Coincidentemente, este é similar à
sequência do Projeto Genoma de A. darlingi (AdarC2), cujas amostras também foram
procedentes daquela localidade (Marinotti et al., 2013).

76
Figura 20. Diversidade nucleotídica da região parcial do domínio IIS6 do gene do canal de sódio de anofelinos. Os nucleotídeos representados por letra maiúscula e minúscula correspondem às regiões codificantes e ao íntron, respectivamente. O códon homólogo àquele onde ocorre a clássica mutação kdr (1014) está sublinhado. Os sítios monomórficos estão representados por pontos (.), as substituições pelos respectivos nucleotídeos e deleções por traços (-). AdNaV_haplo_A, AdNaV_haplo_B, AdNaV_haplo_C, AdNaV_haplo_D e AdNaV_haplo_I são os haplótipos de Anopheles darlingi ao passo que AmNaV_haplo_E, AmNaV_haplo_F, AmNaV_haplo_G e AmNaV_haplo_H são os de Anopheles marajoara aqui evidenciados. AdarC2 corresponde à sequência obtida do projeto genoma de A. darlingi (Scaffold_542). As outras sequências foram obtidas do GenBank e VectorBase (Tabela 4).
Exon 21
Exon 20

77
Figura 21. Relações genealógicas entre os haplótipos da região parcial do domínio IIS6 do gene do canal de sódio de Anopheles darlingi, reveladas por estatística de parsimônia (Clement et al., 2000). AdNaV_haplo_A corresponde ao haplótipo A, AdNaV_haplo_B ao haplótipo B, AdNaV_haplo_C ao haplótipo C, AdNaV_haplo_D ao haplótipo D e AdNaV_haplo_I ao haplótipo I. As linhas que conectam as formas representam os passos mutacionais entre os haplótipos. Nelas estão contidas as indicações das mutações
A genealogia em rede apresentada sugere que o haplótipo B seja o ancestral
(Figura 21). De fato, este foi o mais frequente nos grupos do intra e peridomicílio e nas larvas
mais e menos susceptíveis de Manaus, SGC e Macapá (Tabela 5). Para esta última localidade,
exceção foi observada no grupo de larvas menos susceptíveis, onde o D foi o mais frequente.
O haplótipo A foi, exclusivamente, encontrado nas amostras intra e peridomiciliares de
Manaus.
A rede de haplótipos no formato estrela de A. darlingi sugere diferenciação
genética combinada com expansão geográfica recente. Resultados similares foram obtidos ao
analisar a variabilidade do íntron I do NaV de populações africanas de A. gambiae (Gentile et
al., 2004).
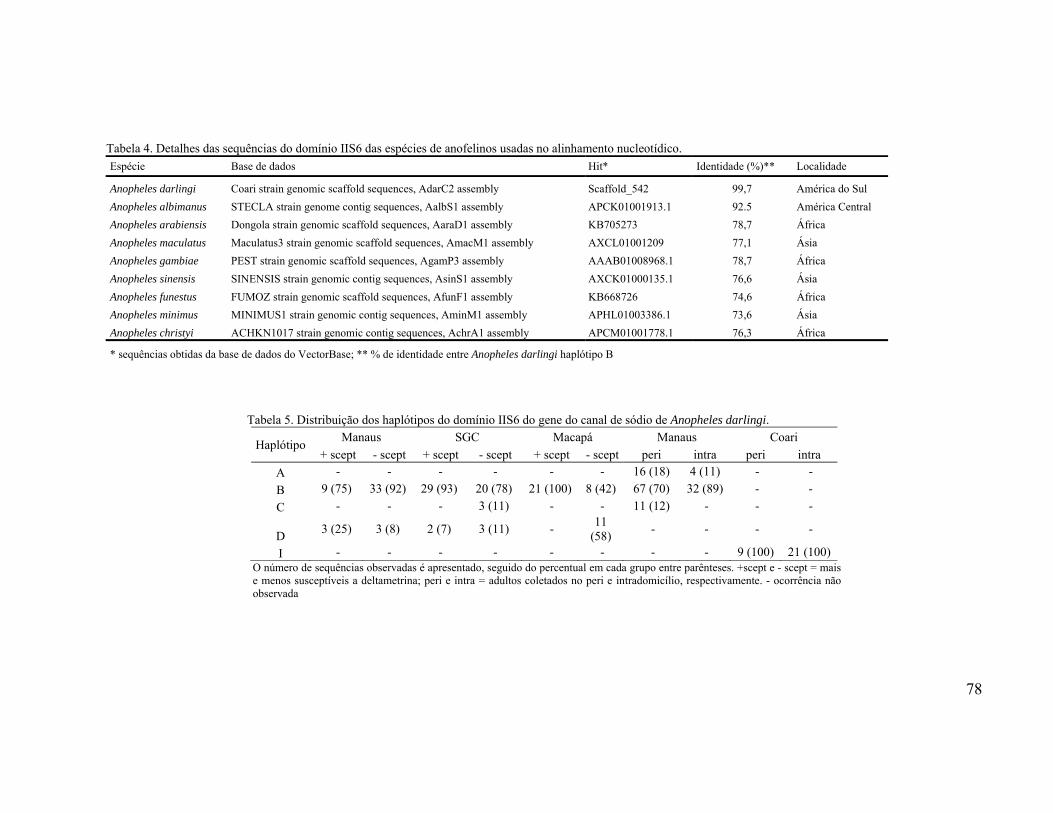
78
Tabela 4. Detalhes das sequências do domínio IIS6 das espécies de anofelinos usadas no alinhamento nucleotídico. Espécie Base de dados Hit* Identidade (%)** Localidade
Anopheles darlingi Coari strain genomic scaffold sequences, AdarC2 assembly Scaffold_542 99,7 América do Sul Anopheles albimanus STECLA strain genome contig sequences, AalbS1 assembly APCK01001913.1 92.5 América CentralAnopheles arabiensis Dongola strain genomic scaffold sequences, AaraD1 assembly KB705273 78,7 África Anopheles maculatus Maculatus3 strain genomic scaffold sequences, AmacM1 assembly AXCL01001209 77,1 Ásia Anopheles gambiae PEST strain genomic scaffold sequences, AgamP3 assembly AAAB01008968.1 78,7 África Anopheles sinensis SINENSIS strain genomic contig sequences, AsinS1 assembly AXCK01000135.1 76,6 Ásia Anopheles funestus FUMOZ strain genomic scaffold sequences, AfunF1 assembly KB668726 74,6 ÁfricaAnopheles minimus MINIMUS1 strain genomic contig sequences, AminM1 assembly APHL01003386.1 73,6 Ásia Anopheles christyi ACHKN1017 strain genomic contig sequences, AchrA1 assembly APCM01001778.1 76,3 África
* sequências obtidas da base de dados do VectorBase; ** % de identidade entre Anopheles darlingi haplótipo B
Tabela 5. Distribuição dos haplótipos do domínio IIS6 do gene do canal de sódio de Anopheles darlingi.
Haplótipo Manaus SGC Macapá Manaus Coari + scept - scept + scept - scept + scept - scept peri intra peri intra
A - - - - - - 16 (18) 4 (11) - - B 9 (75) 33 (92) 29 (93) 20 (78) 21 (100) 8 (42) 67 (70) 32 (89) - - C - - - 3 (11) - - 11 (12) - - -
D 3 (25) 3 (8) 2 (7) 3 (11) - 11 (58) - - - -
I - - - - - - - - 9 (100) 21 (100) O número de sequências observadas é apresentado, seguido do percentual em cada grupo entre parênteses. +scept e - scept = mais e menos susceptíveis a deltametrina; peri e intra = adultos coletados no peri e intradomicílio, respectivamente. - ocorrência não observada

79
A. marajoara apresentou quatro sítios polimórficos, gerando quatro haplótipos
distintos (E, F, G e H), todos com substituições sinônimas. Macapá apresentou todos estes,
tendo ainda exclusividade para o haplótipo E. O mais frequente, tanto em Macapá como
Iranduba, foi o haplótipo F, que, como sugere a rede de haplótipos (Figura 22), deve ser o
haplótipo mais ancestral entre eles. Embora nossa amostragem não tenha sido grande o
suficiente para discriminar diferenças marcantes interpopulacionais, vale notar a ocorrência de
duas linhagens de A. marajoara, recentemente descritas por Mckeon et al. (2010).
Os inseticidas borrifados dentro ou fora das residências apresentam diferenças
quanto à pressão de seleção que exercem sobre a população alvo. Quando aplicados no
intradomicílio, por exemplo, os piretróides persistem em substratos como paredes e pisos
(efeito residual), que pode perdurar por até três meses. Dessa forma, eles continuam a
exterminar os insetos que entram em contato com esses substratos mesmo após dias de
aplicação do inseticida. O mesmo não acontece no peridomicílio, onde a dispersão e a perda
da efetividade desses compostos são mais rápidas (Santos et al., 2007).
Um estudo prévio revelou maior variabilidade genética em populações de A.
darlingi coletadas no intra do que no peridomicílio, também de amostras procedentes de
Manaus e Coari (Silva et al., 2010). De certa forma, aquele resultado nos encorajou a
investigar se havia alguma relação similar, considerando o gene NaV, que estaria,
possivelmente, sob pressão de seleção. Contudo, no presente trabalho, a diferença observada
na frequência dos haplótipos das amostras de A. darlingi coletadas no intra e peridomicílio em
Manaus não foi significativa (χ2= 5,864; gl= 2; p= 0,0533).
É possível que não vejamos substituições não sinônimas em um alelo selecionado
para resistência. Isto porque a mutação de fato relacionada pode estar em uma região ainda

80
não analisada do gene, mas que esteja ligada a algum outro polimorfismo, mesmo que neutro,
no domínio aqui sequenciado.
De fato, em populações de A. funestus da República do Malawi, embora não
tenham sido detectadas mudanças na sequência de aminoácidos do canal de sódio, revelaram-
se haplótipos com substituições sinônimas em indivíduos resistentes (Wondji et al., 2007).
Portanto, mesmo não apresentando mudança de aminoácidos, vale investigar a frequência dos
haplótipos entre os mais e menos susceptíveis. Neste sentido, foram encontradas diferenças
altamente significativas para A. darlingi (χ2= 16,77; gl= 1; p< 0,0001) e A. marajoara (χ2=
11,56; gl= 3; p= 0,009), ambos de Macapá.
No caso de A. marajoara de Macapá, os haplótipos G e H apareceram somente
nas larvas menos tolerantes (mais susceptíveis), o que levou àquela diferença. No entanto,
estes haplótipos também foram observados nas larvas mais tolerantes de Iranduba (Tabela 6),
que apresentou um alto índice de susceptibilidade ou maior tolerância à deltametrina (Figura
18). Portanto, não é possível concluir sobre a ligação de algum haplótipo desta espécie com
aumento de tolerância à piretróide.
Em A. darlingi a diferença significativa nas frequências entre mais e menos
tolerantes (susceptíveis) é explicada pela presença do haplótipo D apenas entre as larvas mais
tolerantes e em alta frequência (58%). Como Macapá apresentou um índice de
susceptibilidade parecido ao de SGC (Figura 18), localidade provavelmente, livre de pressão
de seleção com inseticidas, não estamos seguros em afirmar que haplótipo D de A. darlingi
esteja ligado a algum fator relacionado à resistência. Ensaios adicionais com novas coletas
seriam importantes para elucidar melhor estas questões, além de exploração para além da
região IIS6 do NaV.

81
Figura 22. Relações genealógicas entre os haplótipos da região parcial do domínio IIS6 do gene do canal de sódio de Anopheles marajoara, reveladas por estatística de parsimônia (Clement et al., 2000). AmNaV-haplo_E corresponde ao haplótipo E, AmNaV-haplo_F ao haplótipo F, AmNaV-haplo_G ao haplótipo G e AmNaV-haplo_H ao haplótipo H. As linhas que conectam as formas representam os passos mutacionais entre os haplótipos. Nelas estão contidas as indicações das mutações.
Tabela 6. Distribuição dos haplótipos do domínio IIS6 do gene do canal de sódio de Anopheles marajoara.
Haplótipo Macapá Iranduba
+ scept - scept + scept - scept
E 1 (4) 4 (36) - -
F 16 (55) 7 (64) 14 (100) 28 (78)
G 8 (27) - - 5 (14)
H 4 (14) - - 3 (8) O número de sequências observadas é apresentado, seguido do percentual em cada grupo entre parênteses. +scept e - scept = mais e menos susceptíveis à deltametrina; peri e intra = adultos coletados no peri e intradomicílio, respectivamente. - ocorrência não observada.

82
6 CONCLUSÕES
• Houve sucesso na padronização do bioensaio simplificado de knockdown em A.
darlingi e A. marajoara. Essa metodologia, por ser simples e rápida, pode ser inclusive
usada sob condições de campo;
• Nos bioensaios com deltametrina, foram evidenciadas perdas de susceptibilidade nas
populações desses anofelinos do estado do Amazonas, possivelmente resultantes de
pressão de seleção por inseticidas;
• O sequenciamento da região parcial do domínio IIS6 do gene do canal de sódio revelou
a presença de mutações sinônimas que agruparam as amostras em cinco distintos
haplótipos (A, B, C, D e I) para A. darlingi e quatro (E, F, G e H) para A. marajoara,
respectivamente;
• Mutações do tipo kdr não foram evidenciadas, sugerindo o envolvimento de outros
mecanismos de resistência para explicar a perda de susceptibilidade dos anofelinos do
estado do Amazonas (Manaus e Iranduba).
• Muito embora mutações kdr não tenham sido encontradas, sua ocorrência é permissiva
uma vez que o códon do sítio 1014 dessas espécies é formado por TTA e a presença de
uma única mutação (TTT/TCA) possibilita a mudança de aminoácido;
• A tradução dedutível em aminoácidos da região analisada revelou total homologia com
as de outros anofelinos dispostas nos bancos de dados públicos e as diferenças
nucleotídicas observadas estiveram de acordo com a filogenia desses anofelinos;
• Não foi possível correlacionar a distribuição de alguns dos haplótipos aqui obtidos com
a perda de susceptibilidade das populações à deltametrina;

83
• A presente análise da região parcial do domínio IIS6 do NaV de A. darlingi e A.
marajoara representa uma oportunidade ímpar para a compreensão da diversificação
evolutiva de um gene diretamente relacionado com resistência aos piretróides;
• Além disso, descrevemos pela primeira vez, a padronização de um teste de
susceptibilidade rápido, simples e de baixo custo, em larvas de anofelinos;
• Todas essas informações poderão ser usadas pelos órgãos competentes para direcionar
as medidas estratégicas a serem adotadas para combater os vetores da malária na
região amazônica.

84
7 REFERÊNCIAS BIBLIOGRÁFICAS
Abuelmaali, S.A.; Elaagip, A.H.; Basheer, M.A.; Frah, E.A.; Ahmed, F.T.A.; Elhaj, H.F.A.;
Seidahmed, O.M.E.; Weetman, D.; Hamid, M.M.A. 2013. Impacts of agricultural
practices on insecticide resistance in the malaria vector Anopheles arabiensis in
Khartoum State, Sudan. PLoS ONE, 8(11): e80549.
Adasi, K.; Hemingway, J. 2008. Susceptibility to three pyrethroids and detection of
knockdown resistance mutation in Ghanaian Anopheles gambiae sensu strictu. Journal
of Vector Ecology, 33(2): 255-262.
Adeniran, T.T.; Brown, C.A.; Rogers, W.; Wilson, M.D.; Appawu, M.A.; Boakye, D.A. 2009.
Susceptibility status of Anopheles gambiae sensu stricto (Diptera: Culicidae) to
pyrethroid and carbamate insecticides in the Greater Accra region of Ghana, West
Africa. International Journal of Tropical Insect Science, 29(3): 124-129.
Adeogun, A.O.; Olojede, J.B.; Oduola, A.O.; Awolola, T.S. 2012. Village-scale evaluation of
PermaNet 3.0: an enhanced efficacy combination long-lasting insecticidal net against
resistant population of Anopheles gambiae s.s. Malaria Chemotherapy, Control &
Elimination, 1: 1-9.
Agyepong, N.; Mak-Mensah, E.E.; Brown, C.A. 2012. Prevalence of Anopheles gambiae s.s.
and their pyrethroid knock down resistance pattern in five selected communities in
kumasi metropolis using polymerase chain reaction (PCR). European Journal of
Experimental Biology, 2(2): 304-310.
Aïzoun, N.; Aikpon, R.; Akogbéto, M. 2014. Evidence of increasing L1014F kdr mutation
frequency in Anopheles gambiae s.l. pyrethroid resistant following a nationwide
distribution of LLINs by the Beninese National Malaria Control Programme. Asian
Pacific Journal of Tropical Biomedicine, 4(3): 239-243.

85
Anstead, J.A.; Mallet, J.; Denholm, I. 2007. Temporal and spatial incidence of alleles
conferring knockdown resistance to pyrethroids in the peach-potato aphid, Myzus
persicae (Hemiptera: Aphididae), and their association with other insecticide
resistance mechanisms. Bulletin of Entomological Research, 97(3): 243-252.
Asidi, A.N.; N'Guessan, R.; Koffi, A.A.; Curtis, C.F.; Hougard, J.M.; Chandre, F.; Corbel, V.;
Darriet, F.; Zaim, M.; Rowland, M.W. 2005. Experimental hut evaluation of bednets
treated with an organophosphate (chlorpyrifos-methyl) or a pyrethroid
(lambdacyhalothrin) alone and in combination against insecticide-resistant Anopheles
gambiae and Culex quinquefasciatus mosquitoes. Malaria Journal, 4: 25.
Awolola, T.S.; Brooke, B.D.; Hunt, R.H.; Coetzee, M. 2002. Resistance of the malaria vector
Anopheles gambiae s.s. to pyrethroid insecticides, in south-western Nigeria. Annals of
Tropical Medicine and Parasitology, 96(8): 849-852.
Awolola, T.S.; Oduola, A.M.J.; Strode, C.; Koekemoer, L.L.; Brooke, B.; Ranson, H. 2009.
Evidence of multiple pyrethroid resistance mechanisms in the malaria vector
Anopheles gambiae sensu stricto from Nigeria. Transactions of the Royal Society of
Tropical Medicine and Hygiene, 103(11): 1139-1145.
Awolola, T.S.; Oduola, A.O.; Oyewole, I.O.; Obansa, J.B.; Amajoh, C.N.; Koekemoer, L.L.;
Coetzee, M. 2007. Dynamics of knockdown pyrethroid insecticide resistance alleles in
a field population of Anopheles gambiae s.s. in southwestern Nigeria. Journal of
Vector Borne Diseases, 44: 181-188.
Ayres, M.; Ayres-Júnior, M.; Ayres, D.L.; Santos, A.A. 2007. BIOESTAT - Aplicações
estatísticas nas áreas das ciências bio-médicas. ONG Mamirauá: Belém.
Azevedo, M.O.; Felipe, M.S.S.; Brigido, M.M.; Maranhão, A.Q.; De-Souza, M. 2003.
Técnicas básicas em Biologia Molecular. Ed. Universidade de Brasília: Brasília. 211
pp.

86
Badolo, A.; Okado, K.; Guelbeogo, W.M.; Aonuma, H.; Bando, H.; Fukumoto, S.; Sagnon,
N.F.; Kanuka, H. 2012a. Development of an allele-specific, loop-mediated, isothermal
amplification method (AS-LAMP) to detect the L1014F kdr-w mutation in Anopheles
gambiae s.l. Malaria Journal, 11: 227.
Badolo, A.; Traore, A.; Jones, C.M.; Sanou, A.; Flood, L.; Guelbeogo, W.M.; Ranson, H.;
Sagnon, N.F. 2012b. Three years of insecticide resistance monitoring in Anopheles
gambiae in Burkina Faso: resistance on the rise? Malaria Journal, 11: 232.
Balkew, M.; Ibrahim, M.; Koekemoer, L.L.; Brooke, B.D.; Engers, J.; Aseffa, A.; Michael,
T.G.; Elhassen, I. 2010. Insecticide resistance in Anopheles arabiensis (Diptera:
Culicidae) from villages in central, northern and south west Ethiopia and detection of
kdr mutation. Parasites & Vectors, 3: 40.
Bass, C.; Nikou, D.; Donelly, M.J.; Williamson, M.S.; Ranson, H.; Ball, A.; Vontas, J.; Field,
L.A. 2007. Detection of knockdown resistance (kdr) mutations in Anopheles gambiae:
a comparison of two new high-throughput assays with existing methods. Malaria
Journal, 6: 111.
Bigoga, J.D.; Ndangoh, D.N.; Ambene, P.H.A.; Patchoké, S.; Fondjo, E.; Leke, R.G.F. 2012.
Pyrethroid resistance in Anopheles gambiae from the rubber cultivated area of Niete,
South Region of Cameroon. Acta Tropica, 124: 210-214.
Boakye, D.A.; Adasi, K.; Appawu, M.; Brown, C.A.; Wilson, M.D. 2009. Pattern of
household insecticide use and pyrethroid resistance in Anopheles gambiae sensu
stricto (Diptera: Culicidae) within the Accra metropolis of Ghana. African
Entomology, 17(2): 125-130.

87
Bonetti, J.C. 1999. Pressupostos bioclimáticos de conforto térmico para uma arquitetura dos
trópicos úmidos. Dissertação (Mestrado em Ciências Ambientais), Universidade
Federal do Amazonas, Manaus. 80 pp.
Brochero, H.H.L.; Li, C.; Wilkerson, C. 2007. A newly recognized species in the Anopheles
(Nyssorhynchus) albitarsis complex (Diptera: Culicidae) from Puerto Carreño,
Colombia. American Journal of Tropical Medicine and Hygiene, 76(6): 1113-1117.
Brochero, H.L.; Rey, G.; Buitrago, L.S.; Olano, V.A. 2005. Biting activity and breeding sites
of Anopheles species in the municipality Villavicencio, Meta, Colombia. Journal of
American Mosquito Control Association, 21: 182-186.
Brogdon, W.G.; Chan, A. Diretrizes para avaliar a resistência a inseticida em vetores usando o
bioensaio com garrafa do CDC. Disponível em: http://www.cdc.gov/malaria. Data de
acesso: 28 de Janeiro de 2013.
Brogdon, W.G.; McAllister, J.C. 1998a. Simplification of adult mosquito bioassays through
use of time-mortality determinations in glass bottles. Journal of American Mosquito
Control Association, 14(2): 159-164.
Brogdon, W.G.; McAllister, J.C. 1998b. Insecticide resistance and vector control. Emerging
infectious Diseases, 4(4): 605-613.
Brown, A.W.A. 1986. Insecticide resistance in mosquitoes: a pragmatic review. Journal of
American Mosquito Control Association, 2: 123-140.
Brown, A.W.A.; Pal, R. 1973. Resistance des arthropods aux insecticides. World Health
Organization: Geneve. 541 pp.
Caccone, A.; Garcia, B.A.; Powell, J.R. 1996. Evolution of the mitochondrial DNA control
region in the Anopheles gambiae complex. Insect Molecular Biology, 5: 51-69.

88
Calvo, E.; Andersen, J.; Francischetti, I.M.; Capurro, M.; deBianchi, A.G.; James, A.A.;
Ribeiro, J.M.C.; Marinotti, O. 2004. The transcriptome of adult female Anopheles
darlingi salivary glands. Insect Molecular Biology, 13(1): 73-88.
Catterall, W.A.; Chandy, K.G.; Clapham, D.E.; Gutman, G.A.; Hofmann, F.; Harmar, A.J.;
Abernethy, D.R.; Spedding, M. 2003. International Union of Pharmacology:
Approaches to the nomenclature of voltage-gated ion channels. Pharmacological
Reviews, 55(4): 573-574.
Chandre, F.; Brengues, C.; Dossou-Yovo, J.; Ma, G.S.; Darriet, F.; Diabate, A.; Carnevale, P.;
Guillet, P. 1999. Current distribution of a pyrethroid resistance gene (kdr) in
Anopheles gambiae complex from West Africa and further evidence for reproductive
isolation of the Mopti form. Parassitologia, 41: 319-322.
Chandre, F.; Darriet, F.; Duchon, S.; Finot, L.; Manguin, S.; Carnevale, P.; Guillet, P. 2000.
Modifications of pyrethroid effects associated with kdr mutation in Anopheles
gambiae. Medical and Veterinary Entomology, 14: 81-88.
Chang, X.L.; Xue, Y.Q.; Zhang, A.D.; Zhu, G.D.; Fang, Q. 2013. Deltamethrin resistance,
metabolic detoxification enzyme and kdr mutation in Anopheles sinensis in region
along Huaihe River in Anhui Province. Zhongguo Xue Xi Chong Bing Fang Zhi Za
Zhi, 25(3): 263-267.
Chen, H.; Githeko, A.K.; Githure, J.I.; Mutunga, J.; Zhou, G.; Yan, G. 2008. Monooxygenase
levels and knockdown resistance (kdr) allele frequencies in Anopheles gambiae and
Anopheles arabiensis in Kenya. Journal of Medical Entomology, 45(2): 242-250.
Chouaibou, M.; Etang, J.; Brévault, T.; Nwane, P.; Hinzoumbé, C.K.; Mimpfoundi, R.;
Simard, F. 2008. Dynamics of insecticide resistance in the malaria vector Anopheles
gambiae s.l. from an area of extensive cotton cultivation in Northern Cameroon.
Tropical Medicine and International Health, 13(4): 476-486.

89
Christian, R.N.; Strode, C.; Ranson, H.; Coetzer, N.; Coetzee, M.; Koekemoer, L.L. 2011.
Microarray analysis of a pyrethroid resistant African malaria vector, Anopheles
funestus, from southern Africa. Pesticide Biochemistry and Physiology, 99: 140-147.
Clement, M.; Posada, D.; Crandall, K.A. 2000. TCS: a computer program to estimate gene
genealogies. Molecular Ecology, 9: 1657-1659.
Coetzee, M.; Koekemoer, L.L. 2013. Molecular systematics and insecticide resistance in the
major African malaria vector Anopheles funestus. Annual Review of Entomology, 58:
393-412.
Coluzzi, M.; Sabatini, A.; Petrarca, V.; Di Deco, M.A. 1979. Chromosomal differentiation
and adaptation to human environments in the Anopheles gambiae complex.
Transactions of the Royal Society of Tropical Medicine and Hygiene, 73: 483-497.
Conn, J.E.; Vineis, J.H.; Bollback, J.P.; Onyabe, D.Y.; Wilkerson, R.C.; Póvoa, M.M. 2006.
Population structure of the malaria vector Anopheles darlingi in a malaria-endemic
region of Eastern Amazonian Brazil. American Journal of Tropical Medicine and
Hygiene, 74(5): 798-806.
Conn, J.E.; Wilkerson, C.; Nazaré, M.; Segura, O.; Raimundo, T.; De Souza, L. 2002.
Emergence of a new neotropical malaria vector facilitated by human migration and
changes in land use American Journal of Tropical Medicine and Hygiene, 66: 18-22.
Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. 1994. Principais mosquitos de importância
sanitária no Brasil. Fiocruz: Rio de Janeiro. 228 pp.
Corbel, V.; Chandre, F.; Brengues, C.; Akogbéto, M.; Lardeux, F.; Hougard, J.M.; Guillet, P.
2004. Dosage-dependent effects of permethrin-treated nets on the behaviour of
Anopheles gambiae and the selection of pyrethroid resistance. Malaria Journal, 3: 22.

90
Corbel, V.; N'Guessan, R. 2013. Distribution, mechanisms, impact and management of
insecticide resistance in malaria vectors: a pragmatic review. In: Manguin, S., (Ed.)
Anopheles mosquitoes - New insights into malaria vectors, pp. 579-633. InTech,
Disponível em: http://www.intechopen.com/books/anopheles-mosquitoes-new-
insights-into-malaria-vectors.
Crow, J.F. 1957. Genetics of insecticide resistance to chemicals. Annual Review of
Entomology, 2: 227-246.
Cuamba, N.; Choi, K.S.; Townson, H. 2006. Malaria vectors in Angola: distribution of
species and molecular forms of the Anopheles gambiae complex, their pyrethroid
insecticide knockdown resistance (kdr) status and Plasmodium falciparum sporozoite
rates. Malaria Journal, 5: 2.
Czeher, C.; Labbo, R.; Arzika, I.; Duchemin, J.B. 2008. Evidence of increasing Leu-Phe
knockdown resistance mutation in Anopheles gambiae from Niger following a
nationwide long-lasting insecticide-treated nets implementation. Malaria Journal, 7:
189.
Dabiré, K.R.; Diabaté, A.; Djogbenou, L.; Ouari, A.; N'Guessan, R.; Ouédraogo, J.B.;
Hougard, J.M.; Chandre, F.; Baldet, T. 2008. Dynamics of multiple insecticide
resistance in the malaria vector Anopheles gambiae in a rice growing area in South-
Western Burkina Faso. Malaria Journal, 7: 188.
Dabiré, K.R.; Diabaté, A.; Namountougou, M.; Toé, K.H.; Ouari, A.; Kengne, P.; Bass, C.;
Baldet, T. 2009. Distribution of pyrethroid and DDT resistance and the L1014F kdr
mutation in Anopheles gambiae s.l. from Burkina-Faso (West Africa). Transactions of
the Royal Society of Tropical Medicine and Hygiene, 103(11): 1113-1120.
Dabiré, R.K.; Namountougou, M.; Sawadogo, S.P.; Yaro, L.B.; Toé, H.K.; Ouari, A.;
Gouagna, L.C.; Simard, F.; Chandre, F.; Baldet, T.; Bass, C.; Diabaté, A. 2012.

91
Population dynamics of Anopheles gambiae s.l. in Bobo-Dioulasso city: bionomics,
infection rate and susceptibility to insecticides. Parasites & Vectors, 5: 127.
David, J.P.; Strode, C.; Vontas, J.; Nikou, D.; Vaughan, A.; Pignatelli, P.M.; Louis, C.;
Hemingway, J.; Ranson, H. 2005. The Anopheles gambiae detoxification chip: a
highly specific microarray to study metabolic-based insecticide resistance in malaria
vectors. PNAS, 102(11): 4080-4084.
Davies, T.G.E.; Field, L.M.; Usherwood, P.N.; Williamson, M.S. 2007a. A comparative study
of voltage-gated sodium channels in the Insecta: implications for pyrethroid resistance
in Anopheline and other Neopteran species. Insect Molecular Biology, 16(3): 361-375.
Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. 2007b. DDT, pyrethrins,
pyrethroids and insect sodium channels. Life, 59(3): 151-162.
Diabate, A.; Baldet, T.; Chandre, C.; Dabiré, K.R.; Kengne, P.; Guiguemde, T.R.; Simard, F.;
Guillet, P.; Hemingway, J.; Hougard, J.M. 2003. kdr mutation, a genetic marker to
assess events of introgression between the molecular M and S forms of Anopheles
gambiae (Diptera: Culicidae) in the Tropical Savannah Area of West Africa. Journal
of Medical Entomology, 40(2): 195-198.
Diabate, A.; Baldet, T.; Chandre, E.; Dabiré, K.R.; Simard, F.; Ouedraogo, J.B.; Guillet, P.;
Hougard, J.M. 2004a. First report of a kdr mutation in Anopheles arabiensis from
Burkina Faso, West Africa. Journal of the American Mosquito Control Association,
20: 195-196.
Diabate, A.; Brengues, C.; Baldet, T.; Dabiré, K.R.; Hougard, J.M.; Akogbéto, M.; Kengne,
P.; Simard, F.; Guillet, P.; Hemingway, J.; Chandre, F. 2004b. The spread of the Leu-
Phe kdr mutation through Anopheles gambiae complex in Burkina Faso: genetic
introgression and de novo phenomena. Tropical Medicine and International Health,
9(12): 1267-1273.

92
Djégbe, I.; Boussari, O.; Sidick, A.; Martin, T.; Ranson, H.; Chandre, F.; Akogbéto, M.;
Corbel, V. 2011. Dynamics of insecticide resistance in malaria vectors in Benin: first
evidence of the presence of L1014S kdr mutation in Anopheles gambiae from West
Africa. Malaria Journal, 10: 261.
Dong, K. 2007. Insect sodium channels and insecticide resistance. Departament of
Entomology, 7: 17-30.
Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. 2013.
Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel.
PNAS, 110(29): 11785-11790.
Edi, C.V.A.; Koudou, B.G.; Jones, C.M.; Weetman, D.; Ranson, H. 2012. Multiple-
insecticide resistance in Anopheles gambiae mosquitoes, Southern Côte d'Ivoire.
Emerging infectious Diseases, 18(9): 1508-1511.
Eleftherianos, I.; Foster, S.P.; Williamson, M.S.; Denholm, I. 2008. Characterization of the
M918T sodium channel gene mutation associated with strong resistance to pyrethroid
insecticides in the peach-potato aphid, Myzus persicae (Sulzer). Bulletin of
Entomological Research, 98: 183-191.
Enayati, A.A.; Vatandoost, H.; Ladonni, H.; Townson, H.; Hemingway, J. 2003. Molecular
evidence for a kdr-like pyrethroid resistance mechanism in the malaria vector
mosquito Anopheles stephensi. Medical and Veterinary Entomology, 17: 138-144.
Etang, J.; Fondjo, E.; Chandre, E.; Morlais, I.; Brengues, C.; Nwane, P.; Chouaibou, M.;
Ndjemai, H.; Simard, F. 2006a. First report of knockdown mutations in the malaria
vector Anopheles gambiae from Cameroon. American Journal of Tropical Medicine
and Hygiene, 74(5): 795-797.
Etang, J.; Fondjo, E.; Chandre, F.; Morlais, I.; Brengues, C.; Nwane, P.; Chouaibou, M.;
Ndjemai, H.; Simard, F. 2006b. Short report: first report of knockdown mutations in

93
the malaria vector Anopheles gambiae from Cameroon. American Journal of Tropical
Medicine and Hygiene, 74(5): 795-797.
Faran, M.E. 1980. Mosquito studies (Diptera: Culicidae). XXXIV. A revision of the
Albimanus section of the subgenus Nyssorhynchus of Anopheles. Contributions of the
American Entomological Institute (Ann Arbor), 15: 1-215.
Faran, M.E.; Linthicum, K.J. 1981. A handbook of the Amazon species of Anopheles
(Nyssorhynchus) (Diptera: Culicidae). Mosquitoes Systematic, 13: 1-81.
Ffrench-Constant, R.H.; Pittendrigh, B.; Vaughan, A.; Anthony, N. 1998. Why are there so
few resistance-associated mutations in insecticide target genes? Philosophical
Transactions of the Royal Society B: Biological Science, 353(1376): 1685-1693.
Fonseca-González, I.; Quiñones, M.L.; McAllister, J.; Brogdon, W.G. 2009. Mixed-function
oxidases and esterases associated with cross-resistance between DDT and lambda-
cyhalothrin in Anopheles darlingi Root 1926 populations from Colombia. Memórias
do Instituto Oswaldo Cruz, 104(1): 18-26.
Forattini, O.P. 2002. Culicidologia Médica. EDUSP: São Paulo. 860 pp.
Freitas-Sibajev, M.G.R.; Conn, J.E.; Mitchell, S.E.; Cockburn, A.F.; Seawright, J.A.; Momen,
H. 1995. Mitochondrial DNA and morphological analysis of Anopheles darlingi
populations from Brasil (Diptera: Culicidae). Systematic Mosquitoes, 27(2): 78-99.
Fryxell, R.T.T.; Seifert, S.N.; Lee, Y.; Sacko, A.; Lanzaro, G.; Cornel, A. 2012. The
knockdown resistance mutation and knockdown time in Anopheles gambiae collected
from Mali evaluated through a bottle bioassay and a novel insecticide-treated net
bioassay. Journal of the American Mosquito Control Association, 28(2): 119-122.
Fukuto, T.R. 1990. Mechanism of action of organophosphorus and carbamate insecticides.
Environmental Health Perspectives, 87: 245-254.

94
FUNASA. Plano de intensificação das ações de controle da malária na Amazônia Legal.
Disponível em: http://www.funasa.gov.br/epi/malaria/malaria0.htm. Data de acesso:
02 de março de 2002.
Galardo, A.K.R. 2010. A importância dos Anopheles darlingi Root, 1926 e Anopheles
marajoara Galvão e Damasceno, 1942 na transmissão da malária no município de
Macapá/AP - Brasil. Biologia de Agentes Infecciosos - UFPA, Universidade Federal
do Pará, Belém-PA. 147 pp.
Galvão, A.L.A. ; Damasceno, R.G. 1942. Sobre um novo anofelino da Ilha Marajó, Anopheles
(Nyssorhynchus) marajoara n. sp. (Diptera: Culicidae). Folia Clinical Biology, 14: 60-
66.
Gayathri, V.; Murthy, P.B. 2006. Reduced susceptibility to deltamethrin and kdr mutation in
Anopheles stephensi Liston, a malaria vector in India. Journal of American Mosquito
Control Association, 22(4): 678-688.
Gentile, G.; Santolamazza, F.; Fanello, C.; Petrarca, V.; Caccone, A.; Torre, A.D. 2004.
Variation in an intron sequence of the voltage-gated sodium channel gene correlates
with genetic differentiation between Anopheles gambiae s.s. molecular forms. Insect
Molecular Biology, 13(4): 371-377.
Girod, R.; Orlandi-Pradines, E.; Rogier, C.; Pages, F. 2006. Malaria transmission and
insecticide resistance of Anopheles gambie (Diptera: Culicidae) in the French Military
Camp of Port- Bouet, Abidjan (Côte d'Ivoire): implications for vector control Journal
of Medical Entomology, 43(5): 1082-1087.
González, R.; Wilkerson, R.; Súarez, M.F.; García, F.; Gallego, G.; Cárdenas, H.; Posso, C.E.;
Duque, M.C. 2007. A population genetics study of Anopheles darlingi (Diptera:
Culicidae) from Colombia based on Random Amplified Polymorphic DNA-

95
Polymerase Chain Reaction and Amplified Fragment Lenght Polymorphism markers.
Memórias do Instituto Oswaldo Cruz, 102(3): 255-262.
Gorham, J.R.; Stojanovich, C.J.; Scott, H.G. 1967. Clave ilustrada para los mosquitos
anofelinos de Sudamérica Oriental. Comunicable Disease Center/United States Public
Health Service, Atlanta. 64 pp.
Govere, J.M.; Durrheim, D.N.; Kunene, S. 2002. Malaria trends in South Africa and
Swaziland and the introduction of synthetic pyrethroids to replace DDT for malaria
control. South African Journal of Science, 98(1-2): 19-21.
Grieco, J.P.; Achee, N.L.; Roberts, D.R.; Andre, R.G. 2005. Comparative susceptibility of
three species of Anopheles from Belize, Central America, to Plasmodium falciparum.
Journal of American Mosquito Control Association, 21: 279-290.
Gupta, R.C. 1994. Carbofuran toxicity. Journal of Toxicology and Environmental Health,
43(4): 383-418.
Haddi, K.; Berger, M.; Bielza, P.; Cifuentes, D.; Field, L.M.; Gorman, K.; Rapisarda, C.;
Williamson, M.S.; Bass, C. 2012. Identification of mutations associated with
pyrethroid resistance in the voltage-gated sodium channel of the tomato leaf miner
(Tuta absoluta). Insect Biochemistry and Molecular Biology, 42: 506-513.
Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41: 95-98.
Harbach, R.E. 2004. The classification of genus Anopheles (Diptera: Culicidae): a working
hypothesis of phylogenetic relationships. Bulletin of Entomological Research, 95: 537-
553.
Harbach, R.E. The phylogeny and classification of Anopheles. In: Manguin, S., (Ed.)
Anopheles mosquitoes - New insights into malaria vectors, pp. 3-55. InTech,

96
Disponível em: http://www.intechopen.com/books/anopheles-mosquitoes-new-
insights-into-malaria-vectors.
Hemingway, J. 2000. The molecular basis of two contrasting metabolic mechanisms of
insecticide resistance Insect Biochemistry and Molecular Biology, 30: 1009-1015.
Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. 2004. The molecular basis of
insecticide resistance in mosquitoes. Insect Biochemistry and Molecular Biology, 34:
653-665.
Hemingway, J. ; Ranson, H. 2000. Insecticide resistance in insect vectors of human disease.
Annual Review of Entomology, 45: 371-391.
Hill, J.; Lines, J.; Rowland, M. 2006. Insecticide-treated nets. Advances in Parasitology, 61:
77-128.
Himeidan, Y.E.; Chen, H.; Chandre, F.; Donnelly, M.J.; Yan, G. 2007. Permethrin and DDT
resistance in the malaria vector Anopheles arabiensis from Eastern Sudan. American
Journal of Tropical Medicine and Hygiene, 77(6): 1066-1068.
Himeidan, Y.E.; Hamid, M.M.A.; Jones, C.M.; Ranson, H. 2011. Extensive permethrin and
DDT resistance in Anopheles arabiensis from eastern and central Sudan. Parasites &
Vectors, 4: 154.
Hinzoumbé, C.K.; Péka, M.; Nwane, P.; Gouni, I.D.; Etang, J.; Ekobo, A.S.; Simard, F. 2008.
Insecticide resistance in Anopheles gambiae from south-western Chad, Central Africa.
Malaria Journal, 7: 192.
Hoti, S.L.; Vasuki, V.; Jambulingam, P.; Sahu, S.S. 2006. Kdr allele-based PCR assay for
detection of resistance to DDT in Anopheles culicifacies sensu lato Giles population
from Malkangiri District, Orissa, India. Current Science, 91(5): 658-661.
IBGE. 2013. Censo demográfico 2013. Disponível em:
www.ibge.gov.br/estadosat/perfil.php?sigla=am. Data de acesso: 04 de abril de 2013.

97
IEPA. Instituto de Pesquisas Cientificas e Tecnológicas do estado do Amapá. Disponível em:
www.iepa.org.br. Data de Acesso: 11 de janeiro de 2014.
Ingles, P.J.; Adams, P.M.; Knipple, D.C.; Soderlund, D.M. 1996. Characterization of voltage-
sensitive sodium channel gene coding sequences from insecticide-susceptible and
knockdown-resistant house fly strains. Insect Biochemistry and Molecular Biology,
26: 319-326.
Jamroz, R.C.; Guerrero, F.D.; Kammlah, D.M.; Kunz, S.E. 1998. Role of the kdr and super-
kdr sodium channel mutations in pyrethroid resistance: correlation of allelic frequency
to resistance level in wild and laboratory populations of horn flies (Haematobia
irritans). Insect Biochemistry and Molecular Biology, 28(12): 1031-1037.
Janeira, F.; Vicente, J.L.; Kanganje, Y.; Moreno, M.; Rosário, V.E.; Cravo, P.; Pinto, J. 2008.
A Primer-Introduced Restriction Analysis-Polymerase Chain Reaction Method to
detect knockdown resistance mutations in Anopheles gambiae. Journal of Medical
Entomology, 45(2): 237-241.
John, R.; Ephraim, T.; Andrew, A. 2008. Reduced susceptibility to pyrethroid insecticide
treated nets by the malaria vector Anopheles gambiae s.l. in western Uganda. Malaria
Journal, 7: 92.
Jones, C.M.; Haji, K.A.; Khatib, B.O.; Bagi, J.; Mcha, J.; Devine, G.J.; Daley, M.; Kabula,
B.; Ali, A.S.; Majambere, S.; Ranson, H. 2013. The dynamics of pyrethroid resistance
in Anopheles arabiensis from Zanzibar and an assessment of the underlying genetic
basis. Parasites & Vectors, 6: 343.
Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donelly, M.J.;
Wilding, C.S. 2012a. Footprints of positive selection associated with a mutation
(N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. PNAS,
109(17): 6614-6619.

98
Jones, C.M.; Toé, H.K.; Sanou, A.; Namountougou, M.; Hughes, A.; Diabaté, A.; Dabiré, R.;
Simard, F.; Ranson, H. 2012b. Additional selection for insecticide resistance in urban
malaria vectors: DDT resistance in Anopheles arabiensis from Bobo-Dioulasso,
Burkina Faso. PLoS ONE, 7(9): e45995.
Kang, S.; Jung, J.; Lee, S.; Hwang, H.; Kim, W. 2012. The polymorphism and the
geographical distribution of the knockdown resistance (kdr) of Anopheles sinensis in
the Republic of Korea. Malaria Journal, 11: 151.
Kanza, J.P.B.; Fahime, E.E.; Alaoui, S.; Essassi, E.M.; Brooke, B.; Malafu, A.N.; Tezzo,
F.W. 2013. Pyrethroid, DDT and malathion resistance in the malaria vector Anopheles
gambiae from the Democratic Republic of Congo. Transactions of the Royal Society
of Tropical Medicine and Hygiene, 107: 8-14.
Karaağaç, S.U. 2012. Insecticide Resistance. In: Perveen, F., (Ed.). Insecticides - Advances in
Integrated Pest Management, pp. 469-478. InTech. Disponível em:
http://www.intechopen.com/books/insecticides-advances-in-integrated-pest-manage
ment/insecticide-resistance.
Karunaratne, S.H.; Hawkes, N., J.; Perera, M.D.; Ranson, H.; Hemingway, J. 2007. Mutated
sodium channel genes and elevated monooxygenases are found in pyrethroid resistant
populations of Sri Lankan malaria vectors. Pesticide Biochemistry and Physiology,
88(1): 108-113.
Kawada, H.; Dida, G.O.; Ohashi, K.; Komagata, O.; Kasai, S.; Tomita, T.; Sonye, G.;
Maekawa, Y.; Mwatele, C.; Njenga, S.M.; Mwandawiro, C.; Minakawa, N.; Takagi,
M. 2011a. Multimodal pyrethroid resistance in malaria vectors, Anopheles gambiae
s.s., Anopheles arabiensis, and Anopheles funestus s.s. in western Kenya. PLoS ONE,
6(8): e22574.

99
Kawada, H.; Futami, K.; Komagata, O.; Kasai, S.; Tomita, T.; Sonye, G.; Mwatele, C.;
Njenga, S.M.; Mwandawiro, C.; Minakawa, N.; Takagi, M. 2011b. Distribution of a
knockdown resistance mutation (L1014S) in Anopheles gambiae s.s. and Anopheles
arabiensis in Western and Southern Kenya. PLoS ONE, 6(9): e24323.
Kawada, H.; Higa, Y.; Nguyen, Y.T.; Tran, S.H.; Nguyen, H.T.; Takagi, M. 2009. Nationwide
investigation of the pyrethroid susceptibility of mosquito larvae collected from used
tires in Vietnam. PLOS Neglected tropical Diseases, 3(3): 1-8.
Kazanidou, A.; Nikou, D.; Grigoriou, M.; Vontas, J.; Skavdis, G. 2009. A Multiplex PCR
Assay for simultaneous genotyping of kdr and ace-1 loci in Anopheles gambiae.
American Journal of Tropical Medicine and Hygiene, 80(2): 236-238.
Kim, H.; Baek, J.H.; Lee, W.J.; Lee, S.H. 2007. Frequency detection of pyrethroid resistance
allele in Anopheles sinensis populations by real-time PCR amplification of specific
allele (rtPASA). Pesticide Biochemistry and Physiology, 87(1): 54-61.
Kiszewski, A.; Mellinger, A.; Spielman, A.; Malaney, P.; Sachs, S.E.; Sachs, J. 2004. A
global index representing the stability of malaria transmission. American Journal of
Tropical Medicine and Hygiene, 70: 486-498.
Koekemoer, L.L.; Spillings, B.L.; Christian, R.N.; Lo, T.C.M.; Kaiser, M.L.; Norton, R.A.I.;
Oliver, S.V.; Choi, K.S.; Brooke, B.D.; Hunt, R.H.; Coetzee, M. 2011. Multiple
insecticide resistance in Anopheles gambiae (Diptera: Culicidae) from Pointre Noire,
Republic of the Congo. Vector-Borne and Zoonotic Diseases, 11(8): 1193-1200.
Koffi, A.A.; Alou, L.P.A.; Adja, M.A.; Chandre, F.; Pennetier, C. 2013a. Insecticide
resistance status of Anopheles gambiae s.s. population from M'Bé: a WHOPES-
labelled experimental hut station, 10 years after the political crisis in Côte d'Ivoire.
Malaria Journal, 12: 151.

100
Koffi, A.A.; Alou, L.P.A.; Kabran, J.P.; N'Guessan, R.; Pennetier, C. 2013b. Re-visiting
insecticide resistance status in Anopheles gambiae from Côte d'Ivoire: a nation-wide
informative survey. PLoS ONE, 8(12): e82387.
Kolaczinski, J.H.; Fanello, C.; Hervé, J.P.; Conway, D.J.; Carnevale, P.; Curtis, C.F. 2000.
Experimental and molecular genetic analysis of the impact of pyrethroid and non-
pyrethroid insecticide impregnated bednets for mosquito control in an area of
pyrethroid resistance. Bulletin of Entomological Research, 90(02): 125-132.
Kulkarni, M.A.; Rowland, M.; Alifrangis, M.; Mosha, F.W.; Matowo, J.; Malima, R.; Peter,
J.; Kweka, E.; Lyimo, I.; Magesa, S.; Salanti, A.; Rau, M.E.; Drakeley, C. 2006.
Occurrence of the leucine-to-phenylalanine knockdown resistance (kdr) mutation in
Anopheles arabiensis populations in Tanzania, detected by a simplified high-
throughput SSOP-ELISA method. Malaria Journal, 5: 56.
Lacerda, M.V.G.; Mourão, M.P.G.; Alexandre, M.A.A.; Siqueira, A.M.; Magalhães, B.M.L.;
Martinez-Espinosa, F.E.; Santana Filho, F.S.; Brasil, P.; Ventura, A.M.R.S.; Tada,
M.S.; Couto, V.S.C.D.; Silva, A.R.; Silva, R.S.U.; Alecrim, M.G.C. 2012.
Understanding the clinical spectrum of complicated Plasmodium vivax malaria: a
systematic review on the contributions of the Brazilian literature. Malaria Journal, 11:
12.
Lehr, M.A.; Kilpatrick, C.W.; Wilkerson, R.C.; Conn, J.E. 2005. Cryptic species in Anopheles
(Nyssorhynchus) albitarsis (Diptera: Culicidae) complex: incongruence between
Random Amplified Polymorphic DNA-Polymerase Chain Reaction identification and
analysis of Mitochondrial DNA COI gene sequences. Annals of the Entomological
Society of America, 98(6): 908-917.
Lengeler, C. 2004. Insecticide-treated bed nets and curtains for preventing malaria. Cochrane
Database of Systematic Reviews, (2): Art. No.: CD000363.

101
Li, C.; Wilkerson, C.; Fonseca, M. 2005. Isolation of polymorphic microsatellite markers
from the malaria vector Anopheles marajoara (Diptera: Culicidae). Molecular ecology
notes, 5: 65-67.
Librado, P.; Rozas, J. 2009. DnaSP v5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics, 25: 1451-1452.
Liew, M.; Pryor, R.; Palais, R.; Meadows, C.; Erali, M.; Lyon, E.; Wittwer, C. 2004.
Genotyping of single-nucleotide polymorphisms by High-Resolution Melting of small
amplicons. Clinical Chemistry, 50: 1156-1164.
Linthicum, K.J. 1988. A revision of the Argyritarsis section of the subgenus Nyssorhynchus
of Anopheles. Mosquitoes Systematic, 20: 98-271.
Litsios, S. 1996. The tomorrow of malaria. Pacific/Ecotrends. World Health Organization:
Geneva. 181 pp.
Lol, J.C.; Castellanos, M.E.; Liebman, K.A.; Lenhart, A.; Pennington, P.M.; Padilla, N.R.
2013. Molecular evidence for historical presence of knock-down resistance in
Anopheles albimanus, a key malaria vector in Latin America. Parasites & Vectors, 6:
268.
Lüleyap, H.P.; Alptekin, D.; Kasap, H.; Kasap, M. 2002. Detection of knockdown resistance
mutations in Anopheles sacharovi (Diptera: Culicidae) and genetic distance with
Anopheles gambiae (Diptera: Culicidae) using cDNA sequencing of the voltage-gated
sodium channel gene. Journal of Medical Entomology, 39(6): 870-874.
Lynd, A.; Ranson, H.; McCall, P.J.; Randle, N.P.; Black IV, W.C.; Walker, E.D.; Donnelly,
M.J. 2005. A simplified high-throughput method for pyrethroid knock-down
resistance (kdr) detection in Anopheles gambiae. Malaria Journal, 4: 16.

102
Marinotti, O.; Cerqueira, G.C.; Almeida, L.G.P.; Ferro, M.I.T.; Loreto, E.L.S.; Zaha, A.;
Teixeira, S.M.R.; Wespiser, A.R.; Silva, A.A.; Schlindwein, A.D.; Pacheco, A.C.L.;
Silva, A.L.C.; Graveley, B.R.; Walenz, B.P.; Lima, B.A.; Ribeiro, C.A.G.; Nunes-
Silva, C.G.; Carvalho, C.R.; Soares, C.M.A.; Menezes, C.B.A.; Matiolli, C.; Caffrey,
D.; Araújo, D.A.M.; Oliveira, D.M.; Golenbock, D.; Grisard, E.C.; Fantinatti-
Garboggini, F.; Carvalho, F.M.; Barcellos, F.G.; Prodoscimi, F.; May, G.; Azevedo-
Junior, G.M.; Guimarães, G.M.; Goldman, G.H.; Padilha, I.Q.M.; Batista, J.S.; Ferro,
J.A.; Ribeiro, J.M.C.; Fietto, J.L.R.; Dabbas, K.M.; Cerdeira, L.; Agnez-Lima, L.F.;
Brocchi, M.; Carvalho, M.O.; Teixeira, M.M.; Maia, M.M.D.; Goldman, M.H.S.;
Schneider, M.P.C.; Felipe, M.S.S.; Hungria, M.; Nicolás, M.F.; Pereira, M.; Montes,
M.A.; Cantão, M.E.; Vincentz, M.; Rafael, M.S.; Silverman, N.; Stoco, P.H.; Souza,
R.C.; Vicentini, R.; Gazzinelli, R.T.; Neves, R.O.; Silva, R.; Astolfi-Filho, S.; Maciel,
T.E.F.; Ürményi, T.P.; Tadei, W.P.; Camargo, E.P.; Vasconcelos, A.T.R. 2013. The
genome of Anopheles darlingi, the main neotropical malaria vector. Nucleic Acids
Research, 17: 1-14.
Martinez-Torres, D.; Chandre, E.; Williamson, M.S.; Darriet, F.; Bergé, J.B.; Devonshire,
A.L.; Guillet, P.; Pasteur, N.; Pauron, D. 1998. Molecular characterization of
pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae
s.s. Insect Molecular Biology, 7(2): 179-184.
Martins, A.J.; Lins, R.M.M.A.; Linss, J.G.B.; Peixoto, A.A.; Valle, D. 2009. Voltage-gated
sodium channel polymorphism and metabolic resistance in pyrethroid-resistant Aedes
aegypti from Brazil. American Journal of Tropical Medicine and Hygiene, 81(1): 108-
115.
Martins, A.J.; Valle, D. The pyrethroid knockdown resistance. In: Soloneski, S. and
Larramendy, M., (Eds.). Insecticides - basic and other applications. InTech.

103
Disponível em: http://www.intechopen.com/books/insecticides-basic-and-other-
applications/the-pyrethroid-knockdown-resistance.
Matambo, T.S.; Abdalla, H.; Brooke, B.D.; Koekemoer, L.L.; Mnzava, A.; Hunt, R.H.;
Coetzee, M. 2007. Insecticide resistance in the malarial mosquito Anopheles
arabiensis and association with the kdr mutation. Medical and Veterinary
Entomology, 21: 97-102.
Mathias, D.K.; Ochomo, E.; Atieli, F.; Ombok, M.; Bayoh, M.N.; Olang, G.; Muhia, D.;
Kamau, L.; Vulule, J.M.; Hamel, M.J.; Hawley, W.A.; Walker, E.D.; Gimnig, J.E.
2011. Spatial and temporal variation in the kdr allele L1014S in Anopheles gambiae
s.s. and phenotypic variability in susceptibility to insecticides in Western Kenya.
Malaria Journal, 10: 10.
McKeon, S.N.; Lehr, M.A.; Wilkerson, R.C.; Ruiz, J.F.; Sallum, M.A.; Lima, J.B.; Povoa,
M.M.; Conn, J.E. 2010. Lineage divergence detected in the malaria vector Anopheles
marajoara (Diptera: Culicidae) in Amazonian Brazil. Malaria Journal, 9: 271.
Mellanby, K. 1992. The DDT story. British Crop Protection Council: United Kingdom. 115
pp.
Mirabello, L.; Vineis, J.H.; Yanoviak, S.P.; Scarpassa, V.M.; Póvoa, M.M.; Padilla, N.;
Achee, N.L.; Conn, J.E. 2008. Microsatellite data suggest significant population
structure and differentiation within the malaria vector Anopheles darlingi in Central
and South America. BMC Ecology, 8(3): 1-15.
Morales-Rojas, H.; Moss, R.A. 2002. Phosphorolytic reactivity of o-Iodosylcarboxylates and
related nucleophiles. Chemical Reviews, 102(7): 2497-2522.
Moreno, M.; Vicente, J.L.; Cano, J.; Bersoza, P.J.; Lucio, A.; Nzambo, S.; Bobuakasi, L.;
Buatiche, J.N.; Ondo, M.; Micha, F.; Rosario, V.E.; Pinto, J.; Benito, A. 2008.

104
Knockdown resistance mutations (kdr) and insecticide susceptibility to DDT and
pyrethroids in Anopheles gambiae from Equatorial Guinea. Tropical Medicine and
International Health, 13(3): 430-433.
Morgan, J.C.; Irving, H.; Okedi, L.M.; Steven, A.; Wondji, C.S. 2010. Pyrethroid resistance
in an Anopheles funestus population from Uganda. PLoS ONE, 5(7): e11872.
Motoki, M.T.; Wilkerson, R.C.; Sallum, M.A.M. 2009. The Anopheles albitarsis complex
with the recognition of Anopheles oryzalimnetes Wilkerson and Motoki, n.sp. and
Anopheles janconnae Wilkerson and Sallum, n.sp. (Diptera: Culicidae). Memórias do
Instituto Oswaldo Cruz, 104(6): 823-850.
Mourou, J.R.; Coffinet, T.; Jarjaval, F.; Pradines, B.; Amalvict, R.; Rogier, C.; Kombila, M.;
Pagès, F. 2010. Malaria transmission and insecticide resistance of Anopheles gambiae
in Libreville and Port-Gentil, Gabon. Malaria Journal, 9: 321.
Müller, P.; Chouaibou, M.; Pignatelli, P.; Etang, J.; Walker, E.D.; Donelly, M.J.; Simard, F.;
Ranson, H. 2008a. Pyrethroid tolerance is associated with elevated expression of
antioxidants and agricultural practice in Anopheles arabiensis from an area of cotton
fields in Northern Cameroon. Molecular Ecology, 17: 1145-1155.
Müller, P.; Warr, E.; Stevenson, B.J.; Pignatelli, P.M.; Morgan, J.C.; Steven, A.; Yawson,
A.E.; Mitchell, S.N.; Ranson, H.; Hemingway, J.; Paine, M.J.; Donnelly, M.J. 2008b.
Field-caught permethrin-resistant Anopheles gambiae overexpress CYP6P3, a P450
that metabolises pyrethroids. PLOS Genetics, 4(11): e1000286.
Namountougou, M.; Simard, F.; Baldet, T.; Diabaté, A.; Ouédraogo, B.; Martin, T.; Dabiré,
R.K. 2012. Multiple insecticide resistance in Anopheles gambiae s.l. populations from
Burkina Faso, West Africa. PLoS ONE, 7(11): e48412.
Ndiath, M.O.; Sougoufara, S.; Gaye, A.; Mazenot, C.; Konate, L.; Faye, O.; Sokhna, C.;
Trape, J.F. 2012. Resistance to DDT and pyrethroids and increased kdr mutation

105
frequency in Anopheles gambiae after the implementation of permethrin-treated nets
in Senegal. PLoS ONE, 7(2): e31943.
Ndjemai, H.N.M.; Patchoké, S.; Atanga, J.; Etang, J.; Simard, F.; Bilong, C.F.B.; Reimer, L.;
Cornel, A.; Lanzaro, G.C.; Fondjo, E. 2009. The distribution of insecticide resistance
in Anopheles gambiae s.l. populations from Cameroon: an update. Transactions of the
Royal Society of Tropical Medicine and Hygiene, 103: 1127-1138.
Neves, D.P. 1988. Parasitologia Humana. Atheneu: São Paulo. 462 pp.
Ngufor, C.; N'Guessan, R.; Boko, P.; Odjo, A.; Vigninou, E.; Asidi, A.; Akogbéto, M.;
Rowland, M. 2011. Combining indoor residual spraying with chlorfenapyr and long-
lasting insecticidal bed nets for improved control of pyrethroid-resistant Anopheles
gambiae: an experimental hut trial in Benin. Malaria Journal, 10: 343.
Norris, L.C.; Norris, D.E. 2011. Efficacy of long-lasting insecticidal nets in use in Macha,
Zambia, against the local Anopheles arabiensis population. Malaria Journal, 10: 254.
Nwane, P.; Etang, J.; Chouaibou, M.; Toto, J.C.; Hinzoumbé, C.K.; Mimpfoundi, R.;
Ambene, H.P.F.; Simard, F. 2009. Trends in DDT and pyrethroid resistance in
Anopheles gambiae s.s. populations from urban and agro-industrial settings in
southern Cameroon. BMC Infectious Diseases, 9: 163.
Nwane, P.; Etang, J.; Chouaibou, M.; Toto, J.C.; Koffi, A.; Mimpfoundi, R.; Simard, F. 2013.
Multiple insecticide resistance mechanisms in Anopheles gambiae s.l. populations
from Cameroon, Central Africa. Parasites & Vectors, 6: 41.
Nwane, P.; Etang, J.; Chouaibou, M.; Toto, J.C.; Mimpfoundi, R.; Simard, F. 2011. kdr-based
insecticide resistance in Anopheles gambiae s.s. populations in Cameroon: spread of
the L1014F and L1014S mutations. BMC Research Notes, 4: 463.

106
O'Reilly, A.O.; Khambay, B.P.S.; Williamson, M.S.; Field, L.A.; Wallace, B.A.; Davies,
T.G.E. 2006. Modelling insecticide-binding sites in the voltage-gated sodium channel.
Biochemical Journal, 396: 255-263.
Ochomo, E.; Bayoh, M.N.; Brogdon, W.G.; Gimnig, J.E.; Ouma, C.; Vulule, J.M.; Walker,
E.D. 2013a. Pyrethroid resistance in Anopheles gambiae s.s. and Anopheles arabiensis
in western Kenya: phenotypic, metabolic and target site characterizations of three
populations. Medical and Veterinary Entomology, 27: 156-164.
Ochomo, E.O.; Bayoh, N.M.; Walker, E.D.; Abongo, B.O.; Ombok, M.O.; Ouma, C.;
Githeko, A.K.; Vulule, J.; Yan, G.; Gimnig, J.E. 2013b. The efficacy of long-lasting
nets with declining physical integrity may be compromised in areas with high levels of
pyrethroid resistance. Malaria Journal, 12: 368.
Oduola, A.O.; Idowu, E.T.; Oyebola, M.K.; Adeogun, A.O.; Olojede, J.B.; Otubanjo, O.A.;
Awolola, T.S. 2012. Evidence of carbamate resistance in urban populations of
Anopheles gambiae s.s. mosquitoes resistant to DDT and deltamethrin insecticides in
Lagos, South-Western Nigeria. Parasites & Vectors, 5: 116.
Okia, M.; Ndyomugyenyi, R.; Kirunda, J.; Byaruhanga, A.; Adibaku, S.; Lwamafa, D.K.;
Kironde, F. 2013. Bioefficacy of long-lasting insecticidal nets against pyrethroid-
resistant populations of Anopheles gambiae s.s. from different malaria transmission
zones in Uganda. Parasites & Vectors, 6: 130.
Olano, V.A.; Brochero, H.L.; Saénz, R.; Quiñones, M.L.; Molina, J.A. 2001. Mapas
preliminares de la distribución de especies de Anopheles vectores de malaria en
Colombia. Biomédica, 21: 402-408.
Omoto, C. Modo de ação dos inseticidas e resistência de insetos a inseticidas. In: Guedes,
J.C., Costa, I.D. and Castiglioni, E., (Eds.). Bases e técnicas de manejo de insetos, p.
248. Universidade Federal de Santa Maria, Santa Maria.

107
Ossè, R.; Gnanguenon, V.; Sèzonlin, M.; Aïkpon, R.; Padonou, G.; Yadouléton, A.;
Akogbéto, M.C. 2012. Relationship between the presence of kdr and Ace-1 mutations
and the infection with Plasmodium falciparum in Anopheles gambiae s.s. in Benin.
Journal of Parasitology and Vector Biology, 4(3): 31-39.
Padonou, G.G.; Sezonlin, M.; Ossè, R.; Aïzoun, N.; Agbo, F.O.; Oussou, O.; Gbédjissi, G.;
Akogbéto, M. 2012. Impact of three years of large scale indoor residual spraying
(IRS) and insecticide treated nets (ITNs) interventions on insecticide resistance in
Anopheles gambiae s.l. in Benin. Parasites & Vectors, 5: 72.
Palchick, S. 1996. The biology of disease vectors. University Press of Colorado: Colorado.
220 pp.
Phillips-Howard, P.A.; Nahlen, B.L.; Kolczack, M.S.; HIghtower, A.W.; Kuile, F.O.; Alaii,
J.A.; Gimnig, J.E.; Arudo, J.; Vulule, J.M.; Odhacha, A.; Kachur, S.P.; Schoute, E.;
Rosen, D.H.; Sexton, J.D.; Oloo, A.J.; Hawley, W.A. 2003. Efficacy of permethrin-
treated bed nets in the prevention of mortality in young children in an area of high
perennial malaria transmission in western Kenya. American Journal of Tropical
Medicine and Hygiene, 68: 23-29.
Pinto, J.; Lynd, A.; Elissa, N.; Donnelly, M.J.; Costa, C.; Gentile, C.; Caccone, A.; Rosário,
V.E. 2006. Co-occurrence of East and West African kdr mutations suggest high levels
of resistance to pyrethroid insecticides in Anopheles gambiae from Libreville, Gabon.
Medical and Veterinary Entomology, 20: 27-32.
Pinto, J.; Lynd, A.; Vicente, J.L.; Santolamazza, F.; Randle, N.P.; Gentile, G.; Moreno, M.;
Simard, F.; Charlwood, J.D.; Rosário, V.E.; Caccone, A.; Torre, A.D.; Donnelly, M.J.
2007. Multiple origins of knockdown resistance mutations in the Afrotropical
mosquito vector Anopheles gambiae. PLoS ONE, 2(11): e1243.

108
Pittendrigh, B.R.; Margam, V.M.; Sun, L.; Huesing, J.E. 2008. Resistance in the Post-
Genomics age. In: Onstad, D.W., (Ed.) Insect resistance management: biology,
economics and prediction, p. 305. Elsevier Ltd., Illinois.
Posso, C.E.; González, R.; Cárdenas, H.; Tascón, R. 2006. Estructura genética de Anopheles
darlingi Root, An. nuneztovari Gabaldon y An. marajoara Galvão & Damasceno de
Colombia mediante RAPD-PCR. Revista Colombiana de Entomología, 32(1): 49-56.
Protopopoff, N.; Matowo, J.; Malima, R.; Kavishe, R.; Kaaya, R.; Wright, A.; West, P.A.;
Kleinschmidt, I.; Kisinza, W.; Mosha, F.W.; Rowland, M. 2013. High level of
resistance in the mosquito Anopheles gambiae to pyrethroid insecticides and reduced
susceptibility to bendiocarb in north-western Tanzania. Malaria Journal, 12: 149.
Protopopoff, N.; Verhaeghen, K.; Bortel, W.V.; Roelants, P.; Marcotty, T.; Baza, D.;
D'Alessandro, U.; Coosemans, M. 2008. A significant increase in kdr in Anopheles
gambiae is associated with an intensive vector control intervention in Burundi
highlands. Tropical Medicine and International Health, 13(12): 1479-1487.
Quiñones, M.L.; Suarez, A.F. 1990. Indoor resting heights of some anophelines in Colombia.
Journal of American Mosquito Control Association, 6: 602-604.
Quiñones, M.L.; Suarez, A.F.; Fleming, G.A. 1987. Estado de la susceptibilidad al DDT de
los principales vetores de malaria en Colombia y su implicación epidemiológica.
Biomédica, 7: 81-86.
Ramphul, U.; Boase, T.; Bass, C.; Okedi, L.M.; Donelly, M.J.; Müller, P. 2009. Insecticide
resistance and its association with target-site mutations in natural populations of
Anopheles gambiae from eastern Uganda. Transactions of the Royal Society of
Tropical Medicine and Hygiene, 103: 1121-1126.
Ranson, H.; Jensen, B.; Vulule, J.M.; Wang, X.; Hemingway, J.; Collins, F.H. 2000.
Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan

109
Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect
Molecular Biology, 9(5): 491-497.
Ranson, H.; N'Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. 2011. Pyrethroid
resistance in African anopheline mosquitoes: what are the implications for malaria
control? Trends in Parasitology, 27(2): 91-98.
Reimer, L.; Fondjo, E.; Patchoké, S.; Diallo, B.; Lee, Y.; Ng, A.; Ndjemai, H.M.; Atanga, J.;
S.F., T.; Lanzaro, G.; Cornel, A.J. 2008. Relationship between kdr mutation and
resistance to pyrethroid and DDT insecticides in natural populations of Anopheles
gambiae. Journal of Medical Entomology, 45(2): 260-266.
Reimer, L.J.; Tripet, F.; Slotman, M.; Spielman, A.; Fondjo, E.; Lanzaro, G.C. 2005. An
unusual distribution of the kdr gene among populations of Anopheles gambiae on the
Island of Bioko, Equatorial Guinea. Insect Molecular Biology, 14(6): 683-688.
Rosa-Freitas, M.G. 1989. Anopheles (Nyssorhynchus) deaneorum, a new species in the
albitarsis complex (Diptera: Culicidae). Memórias do Instituto Oswaldo Cruz, 84(4):
535-543.
Rosa-Freitas, M.G.; Deane, L.M.; Momen, H. 1990. A morphological, isoenzymatic and
behavioral study of ten populations of Anopheles (Nyssorhynchus) albitasis Lynch-
Arribálzaga, 1878 (Diptera: Culicidae) including from de type-locality - Baradero,
Argentina. Memórias do Instituto Oswaldo Cruz, 85(3): 275-289.
Rose, R.I. 2001. Pesticides and public health: integrated methods of mosquito management.
Emerging infectious Diseases, 7(1): 17-23.
Rubio-Palis, Y.; Wilkerson, R.C.; Guzman, H. 2003. Morphological characters of adult
Anopheles (Nyssorhynchus) marajoara in Venezuela. Journal of American Mosquito
Control Association, 19: 107-114.

110
Sambrook, J.; Russell, D.W. 2001. Molecular Cloning: a laboratory manual. Cold Spring
Harbor Laboratory Press, New York. 724 pp.
Santolamazza, F.; Calzetta, M.; Etang, J.; Barrese, E.; Dia, I.; Caccone, A.; Donnelly, M.J.;
Petrarca, V.; Simard, F.; Pinto, J.; Torre, A.D. 2008. Distribution of knock-down
resistance mutations in Anopheles gambiae molecular forms in west and west-central
Africa. Malaria Journal, 7: 74.
Santos, J.B.; Santos, F.; Macêdo, V. 1999. Variação da densidade anofélica com o uso de
mosquiteiros impregnados com deltametrina em uma área endêmica de malária na
Amazônia brasileira. Caderno de Saúde Pública do Rio de Janeiro, 15(2): 281-292.
Santos, J.M.M.; Contel, E.P.B.; Kerr, W.E. 1981. Ciclo biológico, postura e estádios larvais
de Anopheles darlingi Root 1926 (Diptera: Culicidae) da Rodovia Manaus-Boa Vista.
Acta Amazonica, 11(4): 789-797.
Santos, R.L.; Fayal, A.S.; Aguiar, A.E.F.; Vieira, D.B.R.; Póvoa, M.M. 2007. Avaliação do
efeito residual de piretróides sobre anofelinos da Amazônia Brasileira. Revista de
Saúde Pública, 41(2): 276- 283.
Scarpassa, V.M.; Conn, J.E. 2007. Population genetic structure of the major malaria vector
Anopheles darlingi (Diptera: Culicidae) from the Brazilian Amazon, using
microsatellite markers. Memórias do Instituto Oswaldo Cruz, 102(3): 319-327.
Schleier III, J.J.; Peterson, R.K.D. Pyrethrins and pyrethroid insecticides. In: López, O. and
Fernández-Bolaños, J.G., (Eds.) Green trends in insect control, pp. 94-131. Royal
Society of Chemistry.
Service, M.W.; Townson, H. The Anopheles vector. In: Gilles, H.M. and Warrell, D.A., (Eds.)
Essential Malariology, pp. 59-84. Arnold, London.

111
Silva, A.P.B.; Tadei, W.P.; Santos, J.M.M. 2008. Análisis de polimorfismo en poblaciones de
Anopheles darlingi de la Amazonía Brasileña. Revista Latinoamericana de Genética,
1: 228-234.
Silva, A.P.B.; Tadei, W.P.; Santos, J.M.M. 2010. Variabilidade genética em populações de
Anopheles darlingi (Diptera: Culicidae) e relação ao comportamento da atividade de
picar, analisada por RAPD. Acta Amazonica, 40(3): 585-590.
Singh, O.P.; Bali, P.; Hemingway, J.; Subbarao, S.K.; Dash, A.P.; Adak, T. 2009. PCR-based
methods for the detection of L1014 kdr mutation in Anopheles culicifacies sensu lato.
Malaria Journal, 8: 154.
Singh, O.P.; Bali, P.; Hemingway, J.; Subbarao, S.K.; Dash, A.P.; Adak, T. 2009. PCR-based
methods for the detection of L1014 kdr mutation in Anopheles culicifacies sensu lato.
Malaria Journal, 8: 154-161.
Singh, O.P.; Dykes, C.L.; Das, M.K.; Pradhan, S.; Bhatt, R.M.; Agrawal, O.P.; Adak, T.
2010. Presence of two alternative kdr-like mutations, L1014F and L1014S, and a
novel mutation, V1010L, in the voltage gated Na+ channel of Anopheles culicifacies
from Orissa, India. Malaria Journal, 9: 146.
Singh, O.P.; Dykes, C.L.; Lather, M.; Agrawal, O.P.; Adak, T. 2011. Knockdown resistance
(kdr)-like mutations in the voltage-gated sodium channel of a malaria vector
Anopheles stephensi and PCR assays for their detection. Malaria Journal, 10: 59.
Sinka, M.E.; Bangs, M.J.; Manguin, S.; Chareonviriyaphap, T.; Patil, A.P.; Temperley, W.H.;
Gething, P.W.; Elyazar, I.R.F.; Kabaria, C.W.; Harbach, R.E.; Hay, S.I. 2011. The
dominant Anopheles vectors of human malaria in the Asia-Pacific region: occurrence
data, distribution maps and bionomic précis. Parasites & Vectors, 4: 89.
Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil,
A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; Okara, R.M.; Boeckel, T.V.;

112
Godfray, H.C.J.; Harbach, R.E.; Hay, S.I. 2010a. The dominant Anopheles vectors of
human malaria in Africa, Europe and the Middle East: occurrence data, distribution
maps and bionomic précis. Parasites & Vectors, 3: 117.
Sinka, M.E.; Bangs, M.J.; Manguin, S.; Rubio-Palis, Y.; Chareonviryiyaphap, T.; Coetzee,
M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.;
Kabaria, C.W.; Burkot, T.R.; Harbach, R.E.; Hay, S.I. 2012. A global map of
dominant malaria vectors. Parasites & Vectors, 5: 69.
Sinka, M.E.; Rubio-Palis, Y.; Manguin, S.; Patil, A.P.; Temperley, W.H.; Gething, P.W.;
Boeckel, T.V.; Kabaria, C.W.; Harbach, R.E.; Hay, S.I. 2010b. The dominant
Anopheles vectors of human malaria in the Americas: occurrence data, distribution
maps and bionomic précis Parasites & Vectors, 3: 72.
SIVEP-Malaria. Notificações de casos de malária. Disponível em:
http://portalweb04.saude.gov.br/sivep_malaria/. Data de acesso: 24 de abril de 2013.
Slotman, M.A.; della Torre, A.; Calzetta, M.; Powell, J.R. 2005. Differential introgression of
chromosomal regions between Anopheles gambiae and An. arabiensis. American
Journal of Tropical Medicine and Hygiene, 73(2): 326-335.
Stump, A.D.; Atieli, F.K.; Vulule, J.M.; Besansky, N.J. 2004. Dynamics of the pyrethroid
knockdown resistance allele in western Kenyan populations of Anopheles gambiae in
response to insecticide treated bed net trials. American Journal of Tropical Medicine
and Hygiene, 70(6): 591-596.
Suarez, A.F.; Quiñones, M.L.; Palacios, J.D.; Carrillo, A. 1990. First record of DDT
resistance in Anopheles darlingi Journal of American Mosquito Control Association,
6: 72-74.

113
Subramanyam, B.H.; Hagstrum, D.W. Resistance measure and management. In:
Subramanyam, B.H. and Hagstrum, D.W., (Eds.) Integrated management of insects in
stored products. Marcel Dekker Inc., New York.
Syafruddin, D.; Hidayati, A.P.N.; Asih, P.B.S.; Hawley, W.A.; Sukowati, S.; Lobo, N.F.
2010. Detection of 1014F kdr mutation in four major Anopheline malaria vectors in
Indonesia. Malaria Journal, 9: 315.
Tadei, W.P.; Santos, J.M.M.; Scarpassa, V.M.; Rodrigues, I.B. 1993. Incidência, distribuição
e aspectos ecológicos de espécies de Anopheles (Diptera: Culicidae), em regiões
naturais e sob impacto ambiental da Amazônia brasileira. In: Ferreira, E.J.G., Santos,
G.M., Leão, E.L.M. and Oliveira, L.A., (Eds.) Bases científicas para estratégias de
preservação e desenvolvimento da Amazônia, vol. 2, pp. 167-196. Instituto Nacional
de Pesquisas da Amazônia, Manaus.
Tadei, W.P.; Thatcher, B.D.; Santos, J.M.M.; Scarpassa, V.M.; Rodrigues, I.B.; Rafael, M.S.
1998. Ecologic observations on anopheline vectors of malaria in the Brazilian
Amazon. American Journal of Tropical Medicine and Hygiene, 59(2): 325-335.
Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. 2011. MEGA5:
Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary
Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution, 28:
2731-2739.
Tan, W.L.; Li, C.X.; Wang, Z.M.; Liu, M.D.; Dong, Y.D.; Feng, X.Y.; Wu, Z.M.; Guo, X.X.;
Xing, D.; Zhang, Y.M.; Wang, Z.C.; Zhao, T.Y. 2012a. First detection of multiple
knockdown resistance (kdr)-like mutations in voltage-gated sodium channel using
three new genotyping methods in Anopheles sinensis from Guangxi Province, China.
Journal of Medical Entomology, 49: 1012-1020.

114
Tan, W.L.; Wang, Z.M.; Li, C.X.; Chu, H.L.; Xu, Y.; Dong, Y.D.; Wang, Z.C.; Chen, D.Y.;
Liu, H.; Liu, D.P.; Liu, N.; Sun, J.; Zhao, T. 2012b. First report on co-occurrence
knockdown resistance mutations and susceptibility to beta-cypermethrin in Anopheles
sinensis from Jiangsu Province, China. PLoS ONE, 7(1): e29242.
Temu, E.A.; Maxwell, C.; Munyekenye, G.; Howard, A.F.V.; Munga, S.; Avicor, S.W.;
Poupardin, R.; Jones, J.J.; Allan, R.; Kleinschmidt, I.; Ranson, H. 2012. Pyrethroid
resistance in Anopheles gambiae, in Bomi County, Liberia, compromises malaria
vector control. PLoS ONE, 7(9): e44986.
Toda, S.; Morishita, M. 2009. Identification of three point mutations on the sodium channel
gene in pyrethroid-resistant Thrips tabaci (Thysanoptera: Thripidae). Journal of
Economic Entomology, 102: 2296-2300.
Tripet, F.; Wright, J.; Cornel, A.; Fofana, A.; Mcabee, R.; Meneses, C.; Reimer, L.; Slotman,
M.; Thiemann, T.; Dolo, G.; Traoré, S.; Lanzaro, G. 2007. Longitudinal survey of
knockdown resistance to pyrethroid (kdr) in Mali, West Africa, and evidence of its
emergence in the Bamako form of Anopheles gambiae s.s. American Journal of
Tropical Medicine and Hygiene, 76(1): 81-87.
Tripet, F.; Wright, J.; Lanzaro, G. 2006. A new high-performance PCR diagnostic for the
detection of pyrethroid knockdown resistance kdr in Anopheles gambiae. American
Journal of Tropical Medicine and Hygiene, 74(4): 658-662.
Verhaeghen, K.; Bortel, W.V.; Roelants, P.; Backeljau, T.; Coosemans, M. 2006. Detection of
the East and West African kdr mutation in Anopheles gambiae and Anopheles
arabiensis from Uganda using a new assay based on FRET/Melt Curve analysis.
Malaria Journal, 5: 16.
Verhaeghen, K.; Bortel, W.V.; Roelants, P.; Okello, P.E.; Talisuna, A.; Coosemans, M.
2010a. Spatio-temporal patterns in kdr frequency in permethrin and DDT resistant

115
Anopheles gambiae s.s. from Uganda. American Journal of Tropical Medicine and
Hygiene, 82(4): 566-573.
Verhaeghen, K.; van Bortel, W.; Trung, H.D.; Sochantha, T.; Koeokenchanh, K.; Coosemans,
M. 2010b. Knockdown resistance in Anopheles vagus, An. sinensis, An. paraliae and
An. peditaeniatus populations of the Mekong region. Parasites & Vectors, 3: 59.
Weill, M.; Chandre, F.; Brengues, C.; Manguin, S.; Akogbéto, M.; Pasteur, N.; Guillet, P.;
Raymond, M. 2000. The kdr mutation occurs in the Mopti form of Anopheles gambiae
s.s. through introgression. Insect Molecular Biology, 9(5): 451-455.
WHO. 1975. Manual on practical entomology in malaria. Part II. Methods and Techniques.
World Health Organization (WHO Offset Publication 13): Geneva.
WHO. 2006a. The World Health Report 2006. Disponível em:
http://www.who.int/whr/2006/en/. Data de acesso: 29 de outubro de 2006.
WHO. 2006b. Guidelines for testing mosquito adulticides for indoor residual spraying and
treatment of mosquito nets. World Health Organization: Geneva. 60 pp.
WHO. 2007a. The use of DDT in malaria vector control. World Health Organization: Geneva
WHO. 2007b. Insecticide-treated mosquito nets: a WHO position statement. Worl Health
Organization: Geneva.
WHO. 2009. Global malaria control and elimination: report of a technical review. World
Health Organization: Geneva.
WHO. 2011. Malaria report. In: WLC-i-P, D., (Ed.) WHO Global Malaria Programme, p.
259. World Health Organization: Geneva.
WHO. 2012. Global plan for insecticide resistance management in malaria vectors (GPIRM).
World Health Organization: Geneva

116
WHO. 2013. Malaria. Disponível em: http://www.who.int/mediacentre/factsheets/fs094/en/,
Data de acesso: 04 de dezembro de 2013.
WHO. 2013. Test procedures for insecticide resistance monitoring in malaria vector
mosquitoes. World Health Organization: Geneva. 30 pp.
WHOPES. WHO recommended long-lasting insecticidal nets. Disponível em:
http://www.who.int/whopes/Long_lasting_insecticidal_nets_06_Feb_2014.pdf?ua=1.
Data de acesso: 06 de fevereiro de 2014.
Wilkerson, R.C.; Gaffigan, T.V.; Lima, J.B. 1995a. Identification of species related to
Anopheles (Nyssorhynchus) albitarsis by random amplified polymorphic DNA-
polymerase chain reaction (Diptera: Culicidae). Memórias do Instituto Oswaldo Cruz,
90(6): 721-732.
Wilkerson, R.C.; Parsons, T.J.; Klein, T.A.; Gaffigan, T.V.; Bergo, E.; Consolim, J. 1995b.
Diagnosis by random amplified polymorphic DNA polymerase reaction of four cryptic
species related to Anopheles (Nyssorhynchus) albitarsis (Diptera: Culicidae) from
Paraguay, Argentina and Brazil. Journal of Medicine and Entomology, 32(5): 697-
704.
Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. 1996. Identification of
mutations in the housefly para- type sodium channel gene associated with knockdown
resistance (kdr) to pyrethroids insecticides. Mol. Gen. Genet., 252: 51-60.
Wondji, C.S.; Morgan, J.; Coetzee, M.; Hunt, R.H.; Steen, K.; Black IV, W.C.; Hemingway,
J.; Ranson, H. 2007. Mapping a Quantitative Trait Locus (QTL) conferring pyrethroid
resistance in the African malaria vector Anopheles funestus. BioMed Central, 8: 34-47.
Yadouleton, A.W.; Padanou, G.; Asidi, A.; Moiroux, N.; Banganna, S.B.; Corbel, V.;
N'Guessan, R.; Gbenou, D.; Yacoubou, I.; Gazard, K.; Akogbéto, M.C. 2010.

117
Insecticide resistance status in Anopheles gambiae in southern Benin. Malaria
Journal, 9: 83.
Yadouleton, A.W.M.; Asidi, A.; Djouaka, R.F.; Braïma, J.; Agossou, C.D.; Akogbéto, M.C.
2009. Development of vegetable farming: a cause of the emergence of insecticide
resistance in populations of Anopheles gambiae in urban areas of Benin. Malaria
Journal, 8: 103.
Yawson, A.E.; McCall, P.J.; Wilson, M.D.; Donnelly, M.J. 2004. Species abundance and
insecticide resistance of Anopheles gambiae in selected areas of Ghana and Burkina
Faso. Medical and Veterinary Entomology, 18: 372-377.
Yewhalaw, D.; Bortel, W.V.; Denis, L.; Coosemans, M.; Duchateau, L.; Speybroeck, N.
2010. First evidence of high knockdown resistance frequency in Anopheles arabiensis
(Diptera: Culicidae) from Ethiopia. American Journal of Tropical Medicine and
Hygiene, 83(1): 122-125.
Zhong, D.; Chang, X.; Zhou, G.; He, Z.; Fu, F.; Yan, Z.; Zhu, G.; Xu, T.; Bonizzoni, M.;
Wang, M.H.; Cui, L.; Zheng, B.; Chen, B.; Yan, G. 2013. Relationship between
knockdown resistance, metabolic detoxification and organismal resistance to
pyrethroids in Anopheles sinensis. PLoS ONE, 8(2): e55475.


















