
UNIVERSIDADE DE SÃO PAULO ESCOLA DE ENGENHARIA DE...
49
UNIVERSIDADE DE SÃO PAULO ESCOLA DE ENGENHARIA DE LORENA DANIELLE UCHIMURA PASCOLI APLICAÇÃO DE CELULASES TERMOACIDÓFILAS NA HIDRÓLISE DA CELULOSE VISANDO À PRODUÇÃO DE ETANOL DE SEGUNDA GERAÇÃO Lorena 2016
Transcript of UNIVERSIDADE DE SÃO PAULO ESCOLA DE ENGENHARIA DE...

UNIVERSIDADE DE SÃO PAULO
ESCOLA DE ENGENHARIA DE LORENA
DANIELLE UCHIMURA PASCOLI
APLICAÇÃO DE CELULASES TERMOACIDÓFILAS NA HIDRÓLISE DA
CELULOSE VISANDO À PRODUÇÃO DE ETANOL DE SEGUNDA GERAÇÃO
Lorena
2016

DANIELLE UCHIMURA PASCOLI
APLICAÇÃO DE CELULASES TERMOACIDÓFILAS NA HIDRÓLISE DA
CELULOSE VISANDO À PRODUÇÃO DE ETANOL DE SEGUNDA GERAÇÃO
Trabalho de Conclusão de Curso
de Graduação apresentado à
Escola de Engenharia de Lorena da
Universidade de São Paulo como
requisito parcial para conclusão do
curso de Engenharia Bioquímica.
Orientador: Prof. Dr. Júlio César
dos Santos
Lorena
2016

AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHO, POR QUALQUER MEIOCONVENCIONAL OU ELETRÔNICO, PARA FINS DE ESTUDO E PESQUISA, DESDE QUE CITADA A FONTE
Ficha catalográfica elaborada pelo Sistema Automatizadoda Escola de Engenharia de Lorena,
com os dados fornecidos pelo(a) autor(a)
Pascoli, Danielle Uchimura Aplicação de celulases termoacidófilas na hidróliseda celulose visando à produção de etanol de segundageração / Danielle Uchimura Pascoli; orientador JúlioCésar dos Santos. - Lorena, 2016. 48 p.
Monografia apresentada como requisito parcialpara a conclusão de Graduação do Curso de EngenhariaBioquímica - Escola de Engenharia de Lorena daUniversidade de São Paulo. 2016Orientador: Júlio César dos Santos
1. Bioetanol. 2. Hidrólise enzimática. 3. Enzimastermoacidófilas. I. Título. II. dos Santos, JúlioCésar, orient.

Aos meus pais, por sempre estarem ao
meu lado e terem me ensinado o valor e
importância da educação.

AGRADECIMENTOS
Em primeiro lugar, à Deus, por todas as bênçãos e oportunidades que me
concedeu e por estar sempre me guiando em meus caminhos.
Aos meus pais, Pedro e Elizabeth, por sempre acreditarem em mim, me
incentivarem e me apoiarem durante toda a minha vida. Por terem me ensinado o
valor da educação, do respeito e humildade. Toda e qualquer conquista é graças
ao amor e dedicação de vocês.
À toda a minha família, que está sempre ao meu lado me apoiando em todos
os momentos.
Aos meus amigos da escola Natural Vivência, por todos os momentos de
alegria e por me mostrarem que tempo e distância não enfraquecem amizades
verdadeiras.
Às minhas amigas da faculdade, Pamela e Karina, por terem feito parte de
um momento tão importante da minha vida. Obrigada por tornarem meus anos de
faculdade tão maravilhosos e pela amizade tão sincera e especial.
À minha amiga Bruna, pelo companheirismo durante a Iniciação Científica e
pela amizade leve e divertida que foi fundamental durante meus anos de faculdade.
Ao meu namorado, Gabriel, por sempre me apoiar em minhas decisões, por
acreditar em mim e me incentivar a dar sempre o meu melhor. Obrigada por todo o
amor, companheirismo, amizade e paciência.
Ao meu professor orientador, Dr. Júlio César dos Santos, por partilhar seu
conhecimento e por toda a atenção e tempo dedicados à realização deste trabalho.
A todos os professores e funcionários da Escola de Engenharia de Lorena -
USP, por terem me proporcionado, de alguma maneira, a oportunidade de vivenciar
experiências únicas nesta escola durante os últimos anos.

RESUMO
PASCOLI, D. U. Aplicação de celulases termoacidófilas na hidrólise da
celulose visando à produção de etanol de segunda geração. 2016. 48f.
Monografia (Graduação) – Escola de Engenharia de Lorena, Universidade de São
Paulo, Lorena, 2016.
Biocombustíveis têm sido estudados como alternativas aos combustíveis fósseis,
visto que são fontes renováveis de energia e não agridem o meio ambiente. O
etanol de segunda geração, ou bioetanol, é um biocombustível que vem ganhando
bastante atenção nos últimos anos. Um procedimento comum para a produção
deste biocombustível consiste em processos de pré-tratamento ácido e hidrólise
enzimática da biomassa lignocelulósica, a fim de liberar os carboidratos presentes
em sua estrutura, seguido de fermentação por micro-organismos específicos.
Porém, este procedimento é encarecido por ajustes físico-químicos feitos no meio
obtido após o pré-tratamento para permitir a atuação de enzimas. Neste contexto,
o presente trabalho teve como objetivo avaliar o uso de celulases termoacidófilas
durante a hidrólise enzimática da celulose, visando ao favorecimento da viabilidade
econômica da produção industrial de etanol de segunda geração. Neste estudo,
enzimas termoacidófilas foram produzidas por micro-organismos geneticamente
modificados e empregadas na hidrólise da celulose presente no slurry obtido após
pré-tratamento ácido de switchgrass. Os resultados deste trabalho mostraram que
o uso destas enzimas pode ser bastante promissor, visto que ambas foram capazes
de hidrolisar o substrato e liberar açúcares redutores sob condições extremas.
Desta maneira, o emprego de enzimas termoacidófilas pode representar uma
redução de custos na produção industrial de bioetanol, visto que permite integrar o
pré-tratamento ácido da biomassa e subsequente hidrólise enzimática, sem
necessidade de resfriamento ou neutralização prévios.
Palavras-chave: Bioetanol, hidrólise enzimática, enzimas termoacidófilas.

ABSTRACT
PASCOLI, D. U. Application of thermoacidophilic cellulases in cellulose
hydrolysis aiming to second generation ethanol production. 2016. 48f.
Monografia (Graduação) – Escola de Engenharia de Lorena, Universidade de São
Paulo, Lorena, 2016.
Biofuels have been studied as an alternative to fossil fuels since they are renewable
energy sources and do not harm the environment. Second generation ethanol, or
bioethanol, is a biofuel that is gaining a lot of attention over the past years. A
common procedure for bioethanol production consists in pretreatment and
enzymatic hydrolysis of lignocellulosic biomass to release the carbohydrates
present in its structure, followed by fermentation by specific microorganisms.
However, this procedure gets more expensive due to physicochemical adjustments
necessary in the medium obtained after the pretreatment aiming to allow the action
of the enzymes. In this context, the objective of this work was to evaluate the use of
thermoacidophilic cellulases in pretreated medium, focusing on an economically
viable large-scale second-generation ethanol production. In this study,
thermoacidophilic enzymes were produced by genetically modified microorganisms
and used during enzymatic hydrolysis of the cellulose present in the slurry produced
after acid pretreatment of switchgrass. The results of this investigation
demonstrated that the use of these enzymes could be very promising since both
could hydrolysate the substrate and release reducing sugars under extreme
conditions. This way, the use of thermoacidophilic enzymes can result in cost
reduction in industrial production of bioethanol since they allow the possibility of
integrating acid biomass pretreatment with the subsequent enzymatic hydrolysis,
without the requirement of previous cooling or neutralization.
Keywords: Bioethanol, enzymatic hydrolysis, thermoacidophilic enzymes.

LISTA DE TABELAS
Tabela 1 - Variação da composição da biomassa lignocelulósica (em % base seca)
de acordo com a espécie do vegetal................................................................ 21
Tabela 2 - Distribuição das amostras para determinação de atividade de β-
glucosidase nos extratos pré-purificados obtidos a partir do cultivo de E. coli
transformada com gene de S. solfataricus ....................................................... 31
Tabela 3 - Volumes de cada componente presente nos tubos de microcentrífuga
correspondentes aos experimentos de hidrólise enzimática de celulose de
switchgrass empregando celulase comercial de T. reesei 1 μg/mL .................... 33
Tabela 4 - Volumes de cada componente presente nos tubos de microcentrífuga
correspondentes aos experimentos de hidrólise enzimática de celulose de
switchgrass empregando endoglunacase......................................................... 33
Tabela 5 - Volumes de cada componente presente nos tubos de microcentrífuga
correspondentes aos experimentos de hidrólise enzimática de celulose de
switchgrass empregando β-glucosidase .......................................................... 34
Tabela 6 - Volumes de cada componente presente nos tubos de microcentrífuga
correspondentes aos experimentos de hidrólise enzimática de celulose de
switchgrass empregando coquetel de enzimas................................................. 34
Tabela 7 - Resultados de absorbância obtidos para as amostras de β-glucosidase
dos extratos pré-purificados obtidos a partir do cultivo de E. coli transformada com
gene de S. solfataricus ................................................................................... 38

LISTA DE FIGURAS
Figura 1 - Diferentes tipos de energia utilizadas no mundo em 2014.................. 15
Figura 2 - Esquema simplificado da produção de etanol a partir de biomassa
lignocelulósica ............................................................................................... 17
Figura 3 - Quantidade estimada da redução na emissão de gases estufa ao
substituir gasolina comum por etanol derivado de switchgrass, para cada ano de
colheita de switchgrass. Mínima (cinza), média (azul) e máxima (verde) ............ 18
Figura 4 - Representação esquemática das associações entre celulose (fibrila
elementar), hemicelulose e lignina na parede celular vegetal ............................ 19
Figura 5 - Representação esquemática da estrutura da celulose ....................... 20
Figura 6 - Principais açúcares monoméricos constituintes da porção hemicelulósica
da parede celular ........................................................................................... 20
Figura 7 - Unidades de construção da lignina: álcool p-cumarílico, álcool coniferílico
e álcool sinapílico ........................................................................................... 21
Figura 8 - Representação esquemática da produção de etanol a partir de biomassa
lignocelulósica ............................................................................................... 23
Figura 9 - Esquema mostrando o efeito da etapa de pré-tratamento na estrutura da
parece celular ................................................................................................ 23
Figura 10 - Sítio de ação das diferentes enzimas do complexo celulolítico ......... 25
Figura 11 - Eletroforese em gel SDS-PAGE para o extrato enzimático de β-
glucosidase pré-purificado. Os resultados correspondem ao precipitado (1) e
sobrenadante (2) obtidos após sonicação e ao precipitado (3) e sobrenadante (4)
obtidos após aquecimento em banho maria. Na Figura, M corresponde aos padrões
de peso molecular .......................................................................................... 37
Figura 12 - Curva padrão de albumina bovina sérica (BSA) obtida para o método
BCA .............................................................................................................. 40
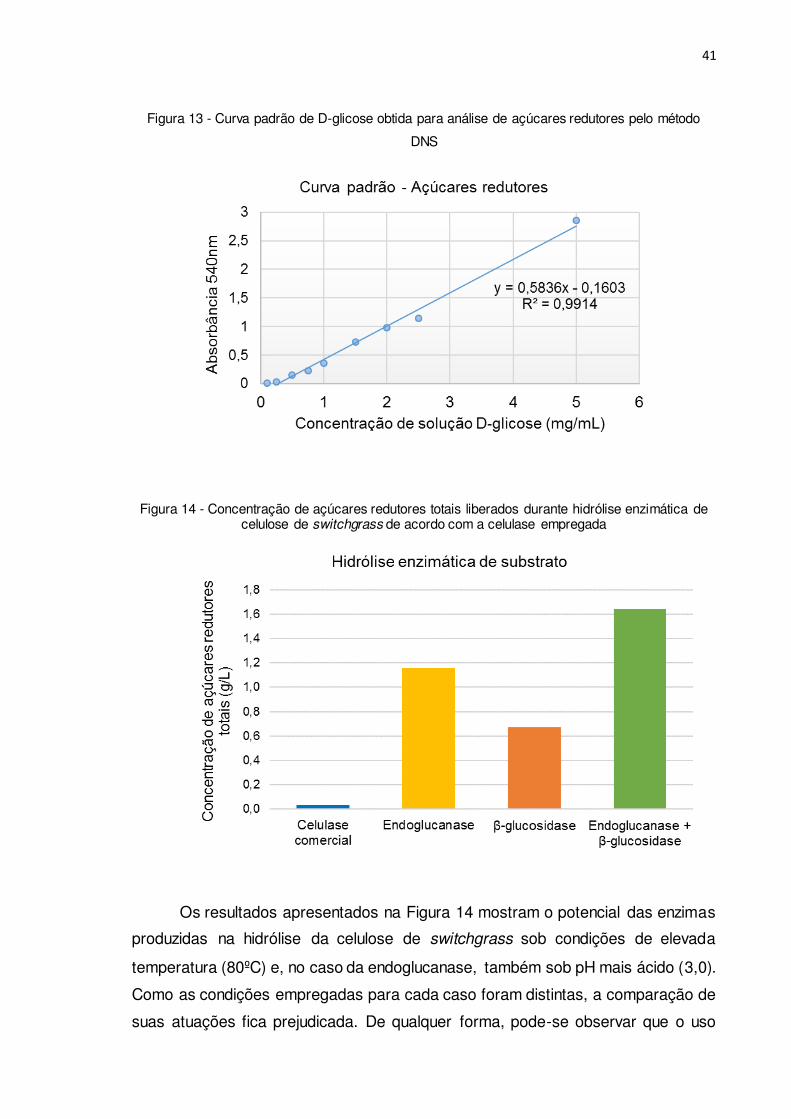
Figura 13 - Curva padrão de D-glicose obtida para análise de açúcares redutores
pelo método DNS ........................................................................................... 41

Figura 14 - Concentração de açúcares redutores totais liberados durante hidrólise
enzimática de celulose de switchgrass de acordo com a celulase empregada .... 41

LISTA DE ABREVIATURAS E SIGLAS
ART Açúcares Redutores Totais
BCA Ácido Bicinconínico
BSA Albumina Bovina Sérica
LB Luria-Bertani
PNP p-nitrofenol
SD Synthetic Dropout
SDS-PAGE Sodium Dodecyl Sulfate - PolyAcrylamide Gel
Electrophoresis
S-WR Standard Working Reagent
β -PNPG p-nitrofenil-β-D-glucopiranosídeo

SUMÁRIO
1. INTRODUÇÃO......................................................................................... 12
2. OBJETIVOS ............................................................................................ 14
2.1 Geral .................................................................................................... 14
2.2 Específicos ........................................................................................... 14
3. REVISÃO DE LITERATURA ..................................................................... 15
3.1 Biocombustíveis: etanol de segunda geração derivado da biomassa switchgrass ................................................................................................. 15
3.2 Características dos materiais lignocelulósicos ......................................... 18
3.3 Aspectos da conversão biotecnológica da biomassa lignocelulósica a etanol .................................................................................................................. 22
3.4 A hidrólise enzimática da celulose e o potencial das enzimas termoacidófilas .................................................................................................................. 25
4. METODOLOGIA ...................................................................................... 27
4.1 Matéria-prima e pré-tratamento .............................................................. 27
4.2 Manipulação genética de micro-organismos ............................................ 27
4.3 Produção dos extratos enzimáticos ......................................................... 28
4.3.1. Recuperação e pré-purificação dos extratos enzimáticos .................. 28
4.4 Caracterização do extrato enzimático de β-glucosidase ........................... 29
4.4.1 Eletroforese SDS-PAGE .................................................................. 29
4.4.2 Método β-PNPG .............................................................................. 30
4.4.3 Método BCA (ácido bicinconínico) .................................................... 31
4.5 Hidrólise enzimática ............................................................................... 32
5. RESULTADOS E DISCUSSÃO................................................................. 36
5.1 Caracterização do extrato enzimático de β-glucosidase ........................... 37
5.1.1 Eletroforese SDS-PAGE .................................................................. 37
5.1.2 Análise de atividade pelo método β-PNPG ........................................ 38
5.1.3 Determinação da quantidade de proteínas pelo método BCA ............. 39
5.2 Hidrólise enzimática e avaliação da atuação do coquetel enzimático......... 40
6. CONCLUSÕES........................................................................................ 43
7. REFERÊNCIAS ....................................................................................... 44

12
1. INTRODUÇÃO
O consumo elevado e crescente de combustíveis fósseis tornou-se
insustentável a ponto de intensificar a busca por fontes alternativas de energia em
todo o mundo. Os combustíveis fósseis não são renováveis, o que faz com que a
dependência energética a partir deles seja um problema recorrente. Além disso, a
queima destes combustíveis libera gases como o dióxido de carbono, o qual é
apontado como o principal causador do efeito estufa. Sendo assim, fontes
alternativas de energia estão sendo avaliadas, garantindo uma opção mais
sustentável e menos agressiva ao meio ambiente (HIMMEL et al., 2007). Uma fonte
alternativa muito estudada atualmente é o etanol de segunda geração, ou bioetanol,
que é proveniente de biomassa lignocelulósica.
O bioetanol obtido por via biotecnológica tem se mostrado muito promissor,
pois utiliza matérias primas disponíveis em abundância e não compete com a
cadeia de produção de alimentos (ABRIL; ABRIL, 2009). A biomassa
lignocelulósica é proveniente de subprodutos agrícolas e florestais e é composta
principalmente por celulose e hemicelulose, que correspondem a polímeros de
carboidratos, e pela lignina, uma macromolécula aromática. Estas três frações
encontram-se intimamente associadas, dando origem a uma estrutura recalcitrante
na parede celular vegetal (GÍRIO et al., 2010).
De maneira geral, o processo de obtenção de bioetanol se dá pela realização
de um pré-tratamento da biomassa, usualmente químico, que favorece a liberação
dos açúcares presentes na parede celular vegetal. A etapa subsequente é a
hidrólise enzimática, que promove a liberação dos açúcares monoméricos que
compõem os polissacarídeos presentes no material. Após este processo, os
açúcares obtidos são submetidos ao processo de fermentação, empregando-se
micro-organismos específicos, para obtenção do etanol como produto final
(DEMIRBAS, 2005).
O pré-tratamento pode ser feito de diversas formas, sendo comum uma
alternativa que emprega condições de alta temperatura e pressão, na presença de
catalisador ácido diluído (CARDONA et al., 2010). Sendo assim, ao final deste
processo, é obtido uma mistura com pH ácido e temperatura elevada. Estas

13
condições são inapropriadas para o uso de enzimas, pois estas seriam inativadas.
Neste caso, para se empregar a hidrólise enzimática após o pré-tratamento,
tornam-se necessários ajustes físico-químicos (neutralização e resfriamento) da
mistura obtida. Em escala industrial, estes ajustes acabam encarecendo o
processo, pois o volume de meio é muito grande, exigindo muitos equipamentos e
reagentes.
Neste contexto, o presente trabalho explora uma alternativa que pode reduzir
a necessidade de ajustes físico-químicos do meio obtido após o pré-tratamento.
Para isso, este trabalho avalia o potencial do emprego de celulases termoacidófilas
na hidrólise enzimática logo após a etapa de pré-tratamento, visto que estas
enzimas são termorresistentes e podem atuar sob pH ácido. Deste modo, a
aplicação de celulases termoacidófilas na produção de bioetanol pode oferecer um
elevado potencial de redução no custo de produção em larga escala.

14
2. OBJETIVOS
2.1 Geral
Avaliar o uso de celulases termoacidófilas durante a hidrólise enzimática da
celulose de Panicum virgatum (switchgrass) pré-tratada com ácido diluído,
visando ao favorecimento da viabilidade econômica da produção industrial
de etanol celulósico.
2.2 Específicos
Produção do extrato enzimático contendo endoglucanase de Sulfolobus
solfataricus expresso em Saccharomyces cerevisiae;
Produção e caracterização do extrato enzimático contendo β-glucosidase de
Sulfolobus solfataricus expresso em Escherichia coli;
Avaliação do potencial de uso em separado ou combinado das enzimas
endoglucanase e β-glucosidase produzidas na hidrólise da celulose presente
no slurry de switchgrass sob condições de elevada temperatura e/ou pH.

15
3. REVISÃO DE LITERATURA
3.1 Biocombustíveis: etanol de segunda geração derivado da biomassa
switchgrass
Desde a Revolução Industrial, os combustíveis fósseis representam a fonte
energética mais utilizada no mundo. Em 2014, 78,4% da energia total consumida
no planeta ainda era proveniente de combustíveis fósseis, tais como petróleo,
carvão e gás natural (Figura 1) (MILANO et al., 2016). O consumo acelerado destes
combustíveis acaba gerando elevadas emissões de dióxido de carbono (CO2), o
que tem provocado mudanças climáticas severas no nosso planeta, intensificando
o efeito estufa (DEMAIN, 2009). Deve-se considerar também que, por se tratarem
de fontes não renováveis, esperam-se sérias limitações no fornecimento destes
combustíveis em um futuro próximo.
Figura 1 - Diferentes tipos de energia utilizadas no mundo em 2014
Fonte: Adaptado de MILANO et al., 2016.
Deste modo, fontes alternativas de energia têm sido avaliadas a fim de
reduzir a dependência em recursos não renováveis. Em 2014, apenas 19% da
energia consumida no mundo era do tipo renovável, sendo 10% desta considerada
energia renovável moderna e 9% proveniente de biomassa tradicional. A energia

16
renovável moderna consiste em 3,8% de energia hidrelétrica, 5,4% de outros tipos
de energia (como solar, eólica, geotérmica e de biomassa) e apenas 0,8% de
biocombustíveis (Figura 1) (MILANO et al., 2016). Apesar de constituírem uma
pequena porção da energia utilizada no mundo atualmente, as energias renováveis
representam uma alternativa muito promissora aos combustíveis fósseis, pois
oferecem uma geração de energia mais limpa e sustentável, além de reduzir a
emissão de gases estufa na atmosfera e contribuir com o meio ambiente.
Entre os biocombustíveis mais produzidos no mundo está o etanol, cuja
tecnologia de primeira geração consiste na produção a partir de matérias-primas
sacaríneas ou amiláceas, como cana-de-açúcar, milho, trigo, mandioca, entre
outros (RODRIGUES, 2011). Porém, o uso de matérias-primas alimentícias para
fins energéticos pode representar um problema adicional. Assim, começou-se a
desenvolver o etanol de segunda geração, que consiste na sua produção a partir
de materiais lignocelulósicos (Figura 2), como, por exemplo, bagaço de cana-de-
açúcar, espiga de milho, gramíneas, etc. (BAI; LUO; VAN DER VOET, 2010;
MISHRA et al., 2015; RODRIGUES, 2011)
Enquanto a produção de etanol de primeira geração é largamente estudada
e possui tecnologias maduras, o aprimoramento dos processos envolvidos na
produção do etanol de segunda geração ainda se encontra em fase de pesquisa e
desenvolvimento. Estas novas tecnologias têm sido direcionadas à viabilização da
produção de etanol e outros produtos dentro do conceito de uma biorrefinaria.
Em uma biorrefinaria são empregados diversos processos químicos e
biotecnológicos a fim de transformar a biomassa lignocelulósica em diferentes
produtos. Primeiramente, ocorre a desconstrução da biomassa através de
diferentes tipos de pré-tratamento químicos, físicos e bioquímicos, resultando nas
frações de celulose, hemicelulose e lignina (Figura 2), em sua forma polimérica,
oligomérica ou monomérica. Depois, estes componentes são convertidos em uma
variedade de produtos através de processos como extração, hidrólise,
transesterificação, fermentação, pirólise, entre outros (STÖCKER, 2008; WYMAN,
1994).

17
Figura 2 - Esquema simplificado da produção de etanol a partir de biomassa lignocelulósica
Biomassalignocelulósica
Pré-tratamentoPré-hidrólise
Hidrólise ácida ou enzimática
Fermentação da glicose
Recuperação do produto
ETANOL
Fermentação da xilose
Processamento da lignina
Químicos, reforçadores ou
combustível para caldeiras
Químicos, reforçadores ou
combustível para caldeiras
Hemicelulose
Lignina
Celulose
Fonte: Adaptado de WYMAN, 1994.
Diferentes matérias-primas têm sido avaliadas para a obtenção etanol de
segunda geração e sua escolha depende da sua disponibilidade em cada região.
Uma planta que vem recebendo destaque em alguns estudos é a switchgrass, uma
gramínea nativa das Planícies Centrais na América do Norte cujo nome científico é
Panicum virgatum. Atualmente ela é estudada como uma alternativa ao milho como
matéria-prima na produção de etanol celulósico (BOUTON, 2007). De fato, cerca
de 90% do etanol produzido e utilizado nos Estados Unidos é derivado de milho
(DIAS et al., 2005). O milho representa o principal produto agrícola do país norte-
americano, com uma colheita de mais de 90 milhões de acres no ano de 2013
(NATIONAL CORN GROWERS ASSOCIATION, 2014). Apesar do custo da
produção de etanol derivado de milho ser menor em relação ao derivado de
switchgrass (PIMENTEL; PATZEK, 2005), o uso do milho para produção de etanol
compete diretamente com a indústria alimentícia, acarretando no aumento do preço
dos alimentos e reduzindo seus benefícios econômicos.
Assim, estudos vêm sendo realizados a fim de avaliar a utilização de switchgrass
como biomassa na produção de bioetanol. A switchgrass é uma das espécies
vegetais predominantes encontradas nas pradarias de grama alta, e devido à sua
característica de raiz profunda, esta gramínea apresenta bom crescimento em
diversos tipos de ambiente, inclusive em locais com solo de qualidade relativamente
baixa. Outras vantagens desta planta consistem na proteção do solo contra erosão,

18
aumento da concentração de carbono no solo por “sequestro” deste em faixas mais profundas e menor utilização de fertilizantes e pesticidas (BOUTON, 2007;
WRIGHT, 2007).
De acordo com U.S. Department of Energy (2016), 1 tonelada de switchgrass
pode produzir 366 litros de etanol, enquanto 1 tonelada de milho pode produzir
cerca 470 litros. Apesar dos números estarem a favor da utilização do milho, a
switchgrass apresenta um saldo energético positivo, pois gera muito mais energia
do que gasta em seu plantio e bioprocesso, representando assim uma ótima
relação custo-benefício (VITAL, 2007). Além disso, Schmer et al. (2008) afirmam
que o uso de etanol proveniente da gramínea resulta na liberação de cerca de 94%
menos gases estufa que a gasolina, sustentando o fato de que switchgrass é uma
alternativa promissora aos combustíveis fósseis (Figura 3).
Figura 3 - Quantidade estimada da redução na emissão de gases estufa ao substituir gasolina comum por etanol derivado de switchgrass, para cada ano de colheita de switchgrass. Mínima
(cinza), média (azul) e máxima (verde)
Fonte: Adaptado de SCHMER et al., 2008.
3.2 Características dos materiais lignocelulósicos
A biomassa lignocelulósica é composta principalmente por celulose,
hemicelulose e lignina (GÍRIO et al., 2010; MAMMAN et al., 2008; PÉREZ et al.,
2002; RODRIGUES, 2011), frações macromoleculares que estão intimamente

19
associadas na parede celular e que de modo geral correspondem a mais de 75%
da biomassa vegetal (Figura 4) (ABRIL; ABRIL, 2009). Uma série de outras
substâncias, tais como óleos vegetais, proteínas e cinzas, correspondem à fração
restante (WYMAN, 1994).
Figura 4 - Representação esquemática das associações entre celulose (fibrila elementar), hemicelulose e lignina na parede celular vegetal
Fonte: CARVALHO et al., 2009.
A celulose, considerada o principal constituinte da parede celular vegetal, é
um homopolímero não-ramificado e linear, constituído por unidades de D-glicose
ou anidroglucopiranose unidos por ligações glicosídicas β-(1→4). A estrutura da
celulose contém ligações de hidrogênio intramoleculares e intermoleculares,
contribuindo para sua estrutura cristalina e formação da fibrila elementar (Figura 5)
(DUTTA; PAL, 2014). Esta molécula corresponde a 38-50% dos tecidos vegetais,
porcentagem que pode variar de acordo com a matéria-prima utilizada (MAMMAN
et al., 2008).
A hemicelulose é um heteropolímero ramificado que, em geral, corresponde
a cerca de 23-32% da biomassa vegetal (MAMMAN et al., 2008). É constituída por
macromoléculas de pentoses (β-D-xilose e α-L-arabinose), hexoses (β-D-manose,
β-D-glicose e α-D-galactose) e/ou ácidos urônicos (ácido α-D-glucurônico, ácido α-
D-4-O-metilgalacturônico e ácido α-D-galacturônico) (GÍRIO et al., 2010; PÉREZ et
al., 2002). Sendo assim, a hemicelulose se distingue da celulose por ser um
polímero de carboidratos de composição variada de açúcares, cadeias menores,
além de apresentar ramificações (Figura 6) (FENGEL; WEGENER, 1989).

20
Figura 5 - Representação esquemática da estrutura da celulose
Fonte: STOCKER, 2008.
Figura 6 - Principais açúcares monoméricos constituintes da porção hemicelulósica da parede
celular
Fonte: Adaptado de MAMMAN et al., 2008.
A lignina é a macromolécula aromática mais abundante do planeta e
corresponde a aproximadamente 15-25% do tecido vegetal (MAMMAN et al.,
2008). Possui uma estrutura complexa, hidrofóbica e amorfa e é constituída por
unidades de hidroxifenilpropano (LEE, 1997). É sintetizada a partir de três
radicais: álcool coniferílico, álcool p-cumarílico e álcool sinapílico (Figura 7),
resultando em uma estrutura heterogênea. A lignina fornece impermeabilidade,

21
resistência e suporte às plantas contra o ataque microbiológico e ao estresse
oxidativo (MAMMAN et al., 2008; PÉREZ et al., 2002; RODRIGUES, 2011).
Figura 7 - Unidades de construção da lignina: álcool p-cumarílico, álcool coniferílico e álcool
sinapílico
Fonte: Adaptado de FENGEL; WEGENER, 1989.
A composição e porcentagem relativa destes componentes na parede
celular varia de acordo com a espécie do vegetal (PÉREZ et al., 2002; ROSA;
GARCIA, 2009), como mostrado na Tabela 1.
Tabela 1 - Variação da composição da biomassa lignocelulósica (em % base seca) de acordo com a espécie do vegetal
Eucalipto Pinheiro Switchgrass Bagaço
Celulose 49,5% 44,5% 32,0% 37,0%
Hemicelulose 13,0% 22,0% 25,0% 28,0%
Lignina 27,5% 28,0% 18,0% 21,0%
Outros 10,0% 5,5% 25,0% 14,0%
Fonte: Adaptado de ROSA; GARCIA, 2009.

22
3.3 Aspectos da conversão biotecnológica da biomassa lignocelulósica a etanol
O processo convencional de produção de etanol celulósico abrange,
basicamente, quatro etapas: o pré-tratamento, responsável pelo rompimento da
matriz lignocelulósica (Figura 8); a hidrólise enzimática, que promove a
despolimerização da celulose em glicose por meio da ação de enzimas celulolíticas;
a fermentação, que é a conversão dos açúcares a etanol geralmente realizada por
leveduras; e a destilação, retificação e desidratação, que separam e purificam o
produto final (Figura 9) (DEMİRBAŞ, 2005; MARGEOT et al., 2009; SANTOS et al.,
2012). A hemicelulose também pode ser empregada na produção de etanol e,
dependendo do tipo de pré-tratamento utilizado, esta já é hidrolisada em açúcares
monoméricos (pentoses e hexoses) nesta etapa. A fermentação das pentoses, no
entanto, depende de micro-organismos específicos ou geneticamente modificados,
tais como cepas recombinantes de Escherichia coli, Zymomonas mobilis e
Saccharomyces cerevisiae (CARDONA; QUINTERO; PAZ, 2010).
Figura 8 - Esquema mostrando o efeito da etapa de pré-tratamento na estrutura da parece celular
Fonte: Adaptado de KUMAR et al., 2009.
A etapa de pré-tratamento é fundamental devido à forte associação da
celulose e da hemicelulose com a lignina na parede celular. Esta parte do processo
tem o objetivo de aumentar a área de superfície interna do substrato, romper a
matriz lignocelulósica e aumentar a porosidade da biomassa, favorecendo a
hidrólise enzimática posterior (Figura 8) (CARDONA; QUINTERO; PAZ, 2010;

23
DEMAIN, 2009; KUMAR et al., 2009). Para isso, podem ser utilizados diversos
métodos físicos, químicos e biológicos, tais como moinho de bolas, hidrólise ácida
e alcalina, explosão a vapor, pirólise, tratamento com solventes, peróxido de
hidrogênio, entre outros (PANDEY et al., 2000; SANTOS et al., 2012).
Figura 9 - Representação esquemática da produção de etanol a partir de biomassa lignocelulósica
BIOMASSA LIGNOCELULÓSICA
PRÉ-TRATAMENTO
CELULOSE
GLICOSE
HIDRÓLISE ENZIMÁTICA
VinhoVinho
FERMENTAÇÃO
ETANOL HIDRATADO
ETANOL HIDRATADO
DESTILAÇÃO/RETIFICAÇÃO
ETANOL ANIDRO
ETANOL ANIDRO
DESIDRATAÇÃO
Fonte: Adaptado de SANTOS et al., 2012.
A hidrólise ácida diluída é um dos métodos mais estudados atualmente - ela
é conduzida em reator sob temperatura e pressão elevadas, e o tempo de reação
pode variar (CARDONA; QUINTERO; PAZ, 2010; DEMİRBAŞ, 2005). O ácido

24
sulfúrico é o catalisador mais utilizado, embora o ácido clorídrico e o ácido nítrico
também possam ser empregados. De acordo com Cardona, Quintero e Paz (2010),
condições ótimas do pré-tratamento ácido com ácido sulfúrico foram obtidas com
H2SO4 5%, a 121ºC, por 60 minutos. A mistura de sólidos e líquidos obtida após
pré-tratamento ácido é comumente chamada de “slurry”, e esta pode conter
compostos tóxicos e inibitórios que são produzidos durante o tratamento com ácido.
Estes produtos, como por exemplo derivados de furano (furfural e hidroxi-metil-
furfural), são tóxicos aos micro-organismos empregados na fermentação. Além
disso, estes compostos também podem inibir a ação de enzimas durante a hidrólise
enzimática (KIM et al., 2011).
Após o pré-tratamento, é necessária a hidrólise das cadeias de celulose para
obtenção dos açúcares fermentescíveis. Na hidrólise enzimática, são adicionadas
ao meio fermentativo enzimas capazes de quebrar a celulose em monômeros de
glicose, que serão posteriormente fermentados a etanol por leveduras. As enzimas
que hidrolisam a celulose são denominadas celulases (endoglucanase,
celobiohidrolase e β-glucosidase), que agem de forma sinérgica e possuem
diferentes sítios de ligação (CASTRO; PEREIRA JR., 2010; LEE, 1997; WYMAN,
1994). Primeiramente, a endoglucanase hidrolisa as ligações glicosídicas no
interior da cadeia, quebrando-a em posições randômicas e criando novas
terminações de cadeia. Então, a celobiohidrolase quebra as ligações glicosídicas
das extremidades da cadeia, liberando moléculas de celobiose (dímero de glicose).
Por fim, a celobiose é hidrolisada pela enzima β-glucosidase, liberando assim
monômeros de glicose individuais (Figura 10) (CARVALHO et al., 2009; CASTRO;
PEREIRA JR., 2010; HIMMEL et al., 2007; KUMAR et al., 2009; KUMAR; SINGH;
SINGH, 2008; STÖCKER, 2008). Porém, este processo requer atenção a
características específicas do complexo enzimático, pois algumas enzimas podem
sofrer inibição pelo seu próprio produto (CASTRO; PEREIRA JR., 2010).
A etapa seguinte do processo é a fermentação, na qual são empregados
micro-organismos (leveduras, bactérias ou fungos filamentosos) capazes de
transformar os açúcares fermentescíveis presentes no hidrolisado lignocelulósico
em produtos de interesse, como por exemplo xilitol, manitol e etanol (GÍRIO et al.,
2010). Para a produção de etanol, são utilizadas leveduras para fermentar a glicose
presente no meio e transformá-la em etanol e gás carbônico, sendo a levedura S.

25
cerevisiae utilizada há milhares de anos para este fim (CARDONA; QUINTERO;
PAZ, 2010; ROSA; GARCIA, 2009).
Figura 10 - Sítio de ação das diferentes enzimas do complexo celulolítico
Fonte: Adaptado de KUMAR et al., 2008.
3.4 A hidrólise enzimática da celulose e o potencial das enzimas termoacidófilas
Celulases podem ser produzidas por uma variedade de micro-organismos, e
os fungos filamentosos do gênero Trichoderma (em especial a espécie Trichoderma
reesei) têm recebido atenção comercial graças à sua capacidade de secretar
grandes quantidades da enzima (CASTRO; PEREIRA JR., 2010; WYMAN, 1994;
ZHANG; LYND, 2004). A reação de hidrólise enzimática ocorre geralmente em
condições mais brandas de pressão, temperatura e pH, pois enzimas sofrem
alterações em sua conformação estrutural e tornam-se inativas quando são
expostas a condições de meio extremas. De acordo com Castro e Pereira Jr.
(2010), as propriedades cinéticas e físico-químicas das celulases dependem do
micro-organismo que a produziu e do tipo da enzima. Considerando o micro-
organismo Trichoderma sp., o pH ótimo de atuação pode variar entre 4,8 e 5,1 e a
temperatura entre 45ºC e 60ºC. Por isso, são necessários ajustes físico-químicos
no hidrolisado obtido após o pré-tratamento ácido antes deste passar para a etapa
de hidrólise enzimática, visto que ele foi tratado com ácido forte sob altas

26
temperaturas – condições inapropriadas para o uso de enzimas. Assim, o
resfriamento e neutralização do meio são etapas necessárias após o pré-
tratamento ácido, representando custos extras na produção de etanol de segunda
geração.
A fim de oferecer uma alternativa a este cenário, foi estudado a possível
aplicação de enzimas termoacidófilas durante a etapa de hidrólise enzimática. A
Archaea da espécie Sulfolobus solfataricus é uma bactéria termoacidófila que
cresce aerobicamente sob condições extremas, com condições ótimas de
temperatura por volta de 80ºC e pH 3 (BROCK et al., 1972; MILLER; BLUM, 2010;
SHE et al., 2001). Foi então descoberto que a Archaea S. solfataricus apresenta
genes que codificam a produção de celulases, em especial uma endoglucanase
extracelular. Esta enzima apresenta as mesmas características de resistência a
altas temperaturas e pH ácido provenientes da Archaea, condições semelhantes às
utilizadas durante a etapa de pré-tratamento da biomassa lignocelulósica, podendo
então atuar sob condições que normalmente desnaturariam outras enzimas. Por
isso, esta enzima pode ser uma alternativa interessante, pois favorece a redução
da necessidade de uso de álcalis e possibilita o uso da enzima antes do
resfriamento total da mistura oriunda do pré-tratamento (LALITHAMBIKA et al.,
2012). Além disso, a Archaea S. solfataricus também possui um gene que expressa
a produção de β-glucosidase, outra celulase termoestável com atividade ótima a
85ºC (AGUILAR et al., 1997).

27
4. METODOLOGIA
Todos experimentos foram realizados no laboratório do grupo de pesquisas do
Dr. Paul Blum, localizado no departamento Nebraska Center for Energy Sciences
Research (University of Nebraska - Lincoln), no estado de Nebraska, Estados
Unidos.
4.1 Matéria-prima e pré-tratamento
A biomassa lignocelulósica utilizada foi a gramínea switchgrass disponível no
laboratório do Dr. Paul Blum na University of Nebraska. Esta foi pré-tratada com
base nas condições descritas por Zhang (2003), utilizando ácido sulfúrico 1% em
autoclave (pH 3, 121ºC por 10 minutos). Depois todo o material, incluindo sólido e
líquido (slurry), foi armazenado a 4°C para uso posterior.
4.2 Manipulação genética de micro-organismos
Os micro-organismos produtores de enzimas estão disponíveis no
laboratório do Dr. Paul Blum, no qual fez-se a manipulação genética destes,
conforme descrito no artigo de Lalithambika et al. (2012). Em especial, o gene
Sso1354 presente no genoma da Sulfolobus solfataricus apresentou grande
atividade enzimática da endoglucanase extracelular. Então, procedeu-se à
manipulação genética isolando o gene Sso1354 da S. solfataricus e inserindo-o no
genoma da levedura Saccharomyces cerevisiae. Esta levedura apresenta
condições de crescimento brandas (30ºC), ao contrário da Archaea (+80ºC). Assim,
esta manipulação genética acabou tornando a produção da endoglucanase
termoacidófila mais facilmente executável em laboratório.
Um procedimento semelhante foi realizado com o gene Sso1353 da S.
solfataricus, responsável pela codificação da produção de outra celulase, a β-
glucosidase. Este gene foi então expressado na bactéria Escherichia coli,
apresentando bons rendimentos. A enzima β-glucosidase também se mostrou

28
termorresistente, permanecendo ativa sob temperaturas altas. Porém, diferente da
endoglucanase, esta não se mostrou resistente a meios com pH ácido. Além disso,
a enzima β-glucosidase é intracelular, fazendo-se necessários procedimentos de
rompimento celular para obtenção da enzima isolada.
4.3 Produção dos extratos enzimáticos
A endoglucanase foi produzida pelo cultiva da levedura S. cerevisiae
geneticamente manipulada. Para isto, empregou-se o meio seletivo SD-leu-ura
(Synthetic Dropout), composto por acetato de potássio 1%, extrato de levedura
0,1% e dextrose 0,05%, sem leucina e uracila. Uma alçada de células foi transferida
para um frasco Erlenmeyer de 125 mL contendo 50 mL de meio previamente
esterilizado em autoclave a 121ºC por 15 minutos. As células foram cultivadas em
incubadora com movimento rotatório a 200 rpm e 30ºC até que fosse atingida a
fase estacionária.
A β-glucosidase foi produzida pelo cultivo de E. coli geneticamente
manipulada. Neste caso, empregou-se o meio LB (Luria-Bertani) contendo triptona
1%, extrato de levedura 0,5% e cloreto de sódio 1%. Uma alçada de células foi
transferida para um frasco Erlenmeyer de 2 L contendo 1 L de meio esterilizado em
autoclave a 121ºC por 15 minutos, sendo em seguida adicionados 100 μg/mL de ampicilina. As células foram cultivadas em incubadora com movimento rotatório a
200 rpm e 37ºC até atingirem fase estacionária.
Em ambos os casos, o tempo de fermentação no qual a fase estacionária foi
atingida foi verificado pela constância nos valores de densidade óptica medida a
600 nm, utilizando o espectrofotômetro da marca Biotek, modelo Synergy H1 Hybrid
Multimode Reader.
4.3.1. Recuperação e pré-purificação dos extratos enzimáticos
Após a fermentação para produção de endoglucanase, o meio contendo as
células de S. cerevisiae foi centrifugado a 3000 xg por 5 minutos. O sobrenadante

29
contendo a endoglucanase extracelular foi mantido refrigerado a 4ºC e o precipitado
foi descartado.
A recuperação e pré-purificação da β-glucosidase obtida no cultivo de E. coli foi
feita com base nas condições descritas por Haseltine et al. (1999), conforme
brevemente descrito a seguir: a cultura de 1 L contendo células de E. coli foi
centrifugada a 7000 xg por 10 minutos, e o precipitado contendo as células foi
ressuspenso em solução tampão de fosfato de sódio 80 mM. As células então foram
rompidas através de processo de sonicação em gelo a 4ºC por 20 minutos. As
células rompidas foram clarificadas por centrifugação a 13000 xg por 15 minutos a
4ºC, e alíquotas de 1 mL foram retiradas do precipitado e do sobrenadante. O
sobrenadante foi ressuspenso em solução tampão de fosfato de sódio 80 mM e
aquecido em banho-maria a 85ºC por 30 minutos. Posteriormente o mesmo foi
reclarificado por centrifugação e foram retiradas novamente alíquotas de 1 mL do
precipitado e sobrenadante. O sobrenadante final foi mantido refrigerado a 4ºC e o
precipitado foi descartado. As quatro amostras retiradas foram utilizadas na etapa
de identificação da β-glucosidase por eletroforese, detalhada na seção 4.4.1 deste
trabalho.
4.4 Caracterização do extrato enzimático de β-glucosidase
4.4.1 Eletroforese SDS-PAGE
A pureza e peso molecular da enzima β-glucosidase foram determinadas por
eletroforese SDS-PAGE seguindo a metodologia descrita por Haseltine et al.
(1999). Para determinação do peso molecular, utilizou-se como referência um
marcador padrão de peso molecular da marca Bio-Rad (Precision Plus Protein™
Dual Color Standards). Após a eletroforese, o gel foi mantido em corante Comassie
Blue R250 durante toda a noite para que as bandas proteicas se tornassem visíveis.

30
4.4.2 Método β-PNPG
A atividade enzimática da β-glucosidase foi determinada por monitoramento
da liberação de p-nitrofenol (PNP) a partir da reação de p-nitrofenil- β-D-
glucopiranosídeo (PNPG). O procedimento foi realizado com base nas condições
descritas por Patchett, Daniel e Morgan (1987). Para este ensaio, foram preparadas
três soluções: a) solução 100 mM de acetado de sódio (NaAc) pH 4,5, b) solução
contendo 10 mM de β-PNPG em 100 mM de NaAc pH 4,5, e c) solução 1 M de
bicarbonato de sódio (NaHCO3). Foram preparadas também diluições 1:10 e 1:100
do extrato enzimático de β-glucosidase utilizando 10mM Tris-Cl pH 7 como
solvente.
As amostras foram distribuídas em tubos de microcentrífuga de acordo com
a Tabela 2, sendo realizadas em duplicata. Os tubos foram incubados a 80ºC por
30 minutos e a reação foi interrompida adicionando-se 500μL da solução 1 M
NaHCO3. A absorbância de PNP liberado foi medida a 420 nm utilizando
espectrofotômetro.
Os valores de absorbância foram utilizados para o cálculo de atividade
enzimática, o qual foi realizado conforme Equação 1, sendo feita uma média dos
valores obtidos com cada diluição do extrato.
� = ×� × �×[ − �.�⁄ ]� (Eq. 1)
Z = Atividade enzimática em U/mL de extrato
x = Soma da média das absorbâncias dos controles (tubos 2 e 3) e da média
das absorbâncias dos tubos com enzimas e NaAc (tubo 4 para diluição 1:10, e
tubos 8 e 9 para diluição 1:100)
y = Média entre absorbâncias dos tubos com enzimas e substrato β-PNPG +
NaAc (tubos 6 e 7 para diluição 1:10, e tubos 10 e 11 para diluição 1:100)
t = tempo de incubação (30 minutos)
d = diluição do extrato enzimático
Ɛ = absortividade molar do PNP (18500 M-1.cm-1)

31
b = caminho óptico da cubeta (1 cm)
10 = fator para correção da diluição da amostra no tubo de incubação (100 µL
foram diluídos para 1 mL)
106 = fator para transformação de mol de PNP em µmol.
Uma unidade de β-glucosidase foi definida como a quantidade de enzima
necessária para liberar 1 μmol de PNP por minuto.
Tabela 2 - Distribuição das amostras para determinação de atividade de β-glucosidase nos extratos pré-purificados obtidos a partir do cultivo de E. coli transformada com gene de S.
solfataricus
Tubo Amostra β-PNPG + NaAc NaAc
1 (branco) - - 1mL
2 (controle) - 1mL -
3 (controle) - 1mL -
4 (1:10) 100μL extrato enzimático diluído 10 vezes - 1mL
5 (1:10) 100μL extrato enzimático diluído 10 vezes - 1mL
6 (1:10) 100μL extrato enzimático diluído 10 vezes 1mL -
7 (1:10) 100μL extrato enzimático diluído 10 vezes 1mL -
8 (1:100) 100μL extrato enzimático diluído 100 vezes - 1mL
9 (1:100) 100μL extrato enzimático diluído 100 vezes - 1mL
10 (1:100) 100μL extrato enzimático diluído 100 vezes 1mL -
11 (1:100) 100μL extrato enzimático diluído 100 vezes 1mL -
4.4.3 Método BCA (ácido bicinconínico)
O ensaio de BCA foi realizado a fim de determinar a concentração de
proteínas obtida durante o processo. O método BCA baseia-se na redução de Cu2+
para Cu+ pelas ligações peptídicas das proteínas. Os íons Cu2+ produzidos se ligam
à molécula de BCA e, ao fazê-lo, formam um complexo de coloração púrpura que
absorve luz a 562 nm. A quantidade de íons Cu2+ reduzidos é diretamente
proporcional à concentração de proteína presente, e esta pode ser estimada pela
comparação com uma proteína padrão, como a albumina bovina sérica (BSA).

32
A metodologia utilizada neste trabalho foi descrita por Smith et al. (1985). O
reagente S-WR (Standard Working Reagent) utilizado neste método consiste na
mistura de 100 mL do reagente A e 2 ml do reagente B. O reagente A é uma solução
aquosa de BCA-Na2 1%, Na2CO3•H2O 2%, tartarato de sódio 0,16%, hidróxido de
sódio 0,4% e NaHCO3 0,95%. O reagente B consiste em CuSO4•5H2O 4% em água
deionizada.
Primeiramente, foi construída uma curva padrão de BSA utilizando soluções
com as seguintes concentrações (μg/mL): 0 (branco), 100, 250, 500, 750 e 1000.
Em tubos de ensaio, foram adicionados 100 μL de cada solução padrão de BSA e
2 mL de reagente S-WR. Os tubos foram encubados a 37ºC por 30 minutos e, ao
término da reação, as absorbâncias foram lidas a 562 nm contra branco utilizando
espectrofotômetro da marca Biotek, modelo H1 Hybrid Multimode Plate Reader.
Em seguida, a mesma reação foi realizada com o extrato enzimático de β-
glucosidase. Foi feita uma diluição 1:10 do extrato enzimático utilizando água
destilada como solvente e 100 μL desta diluição foi adicionado em um tubo de
ensaio, em duplicata. Em seguida, 2 mL de reagente S-WR foram adicionados em
cada tubo, e estes foram incubados a 37ºC por 30 minutos. Ao término do período
de incubação, as absorbâncias das amostras foram lidas a 562 nm contra o branco.
4.5 Hidrólise enzimática
Os experimentos de hidrólise enzimática foram realizados em tubos de
microcentrífuga (Eppendorf) de 1 mL, em quatro abordagens diferentes: a)
utilizando celulase comercial de T. reesei, b) utilizando somente a endoglucanase,
c) utilizando somente a β-glucosidase, e d) utilizando coquetel enzimático de
endoglucanase e β-glucosidase. O preparado comercial de T. reesei foi usado
como controle para melhor avaliar dos resultados da hidrólise enzimática com as
enzimas produzidas.
As reações foram realizadas a 80ºC por 30 minutos, exceto para a
preparação de Trichoderma reesei, para a qual a temperatura empregada foi de
37ºC e o tempo de reação foi de 4 horas. Com relação ao pH, os experimentos
foram realizados na presença de solução tampão citrato-fosfato 50 mM. Para a
enzima de Trichoderma reesei, utilizou-se pH 4,5, sendo os ensaios com

33
endoglucanase e β-glucosidase realizados em pH 3 e 5, respectivamente. Para os
ensaios com o coquetel contendo as duas enzimas (endoglucanase e β-
glucosidase), a reação ocorreu a pH 5.
Para cada ensaio foram preparadas soluções branco contendo água
destilada ao invés do extrato enzimático. Após o período de incubação, a glicose
liberada no processo foi medida por método DNS. Os volumes dos componentes
empregados em cada tudo de microcentrífuga correspondentes aos ensaios de
hidrólise enzimática estão apresentados nas Tabelas 3, 4, 5 e 6. O substrato
utilizado foi o slurry de switchgrass, que corresponde ao meio contendo as frações
sólida e líquida obtidas após pré-tratamento ácido da biomassa (seção 4.1). Antes
do uso nas reações de hidrólise enzimática, o slurry foi diluído 100 vezes em
tampão citrato-fosfato 50 mM pH 4,5.
Tabela 3 - Volumes de cada componente presente nos tubos de microcentrífuga correspondentes aos experimentos de hidrólise enzimática de celulose de switchgrass empregando celulase
comercial de T. reesei 1 μg/mL
Branco pH 4,5 Celulase comercial
100 μL solução tampão
citrato-fosfato 50 mM pH 4,5
100 μL substrato*
100 μL água destilada
100 μL solução tampão
citrato-fosfato 50 mM pH 4,5
100 μL substrato*
100 μL celulase comercial
*Slurry de switchgrass diluído 100 vezes.
Tabela 4 - Volumes de cada componente presente nos tubos de microcentrífuga correspondentes aos experimentos de hidrólise enzimática de celulose de switchgrass empregando endoglunacase
Branco pH 3 Endoglucanase
100 μL solução tampão
citrato-fosfato 50 mM pH 3
100 μL substrato*
100 μL água destilada
100 μL solução tampão citrato-fosfato 50 mM
pH 3
100 μL substrato*
100 μL extrato enzimático de endoglucanase
*Slurry de switchgrass diluído 100 vezes.

34
Tabela 5 - Volumes de cada componente presente nos tubos de microcentrífuga correspondentes aos experimentos de hidrólise enzimática de celulose de switchgrass empregando β-glucosidase
Branco pH 5 β-glucosidase
100 μL solução tampão
citrato-fosfato 50 mM pH 5
100 μL substrato*
100 μL água destilada
100 μL solução tampão citrato-fosfato 50
mM pH 5
100 μL substrato*
100 μL extrato enzimático de β-glucosidase
*Slurry de switchgrass diluído 100 vezes.
Tabela 6 - Volumes de cada componente presente nos tubos de microcentrífuga correspondentes aos experimentos de hidrólise enzimática de celulose de switchgrass empregando coquetel de
enzimas
Branco pH 5 Endoglucanase + β-glucosidase
100 μL solução tampão citrato-
fosfato 50 mM pH 5
100 μL substrato*
100 μL água destilada
100 μL solução tampão citrato-fosfato
50 mM pH 5
100 μL substrato*
50 μL de cada extrato enzimático
*Slurry de switchgrass diluído 100 vezes.
A quantificação dos açúcares redutores produzidos foi feita por método DNS,
com base nas condições descritas por Bernfeld (1955). Para este ensaio foi
utilizado o reagente DNS, que é composto por 18,2 g de tartarato de sódio e
potássio (NaK), 1 g de hidróxido de sódio (NaOH), 1 g de ácido dinitrosalicílico
(DNS), 0,2 g de fenol e 0,05 g de sulfato de sódio (Na2SO4), além de água destilada
q.s.p. 100 mL. Foi preparada também uma solução 40% de tartarato de sódio e
potássio.
Primeiramente, foi construída uma curva padrão utilizando soluções de D-
glicose com as seguintes concentrações (mg/mL): 0; 0,1; 0,25; 0,5; 0,75; 1,0; 1,5;
2,0; 2,5 e 5,0. Então, foram adicionados, em tubos de microcentrífuga, 100 μL de
cada solução padrão de glicose, 200 μL de solução tampão citrato-fosfato 50mM
(pH 5) e 300 μL de reagente DNS. Os tubos foram aquecidos em banho-maria com
água fervente por 15 minutos, e, logo após retirar os tubos da água, foram

35
adicionados 100 μL de solução 40% tartarato de sódio e potássio para encerrar a
reação. Após o resfriamento dos tubos em temperatura ambiente, as absorbâncias
foram lidas a 540 nm utilizando espectrofotômetro.
Em seguida, para as amostras de hidrólise enzimática, o método DNS foi
empregado em todos os ensaios a fim de determinar a concentração de açúcares
redutores produzidos durante as reações. Para isso, foram adicionados 300 μL de
reagente DNS em todos os tubos de microcentrífuga e estes foram então colocados
em banho-maria com água fervente por 15 minutos. Ao final desta etapa, foram
adicionados 100 μL da solução 40% tartarato de sódio e potássio aos tubos ainda
quentes para encerrar a reação. Após resfriamento em temperatura ambiente, as
absorbâncias foram lidas a 540 nm utilizando espectrofotômetro.

36
5. RESULTADOS E DISCUSSÃO
Neste trabalho foram produzidas duas celulases, endoglucanase e β-
glucosidase, ambas provenientes de micro-organismos geneticamente
manipulados. A endoglucanase apresentou característica extracelular, enquanto a
β-glucosidase apresentou característica intracelular. Considerando-se a
experiência prévia do grupo de pesquisa no qual o trabalho foi desenvolvido, já
estavam disponíveis informações que indicavam a possibilidade de uso da
endoglucanase neste processo.
Por outro lado, a β-glucosidase é intracelular e haviam poucas informações
no grupo de pesquisa relacionadas à sua produção e atividade. Desta forma, foram
necessários procedimentos mais elaborados de separação para que o extrato
enzimático fosse obtido. Após sua obtenção, fez-se análise de eletroforese SDS-
PAGE para identificar a presença da enzima no extrato enzimático, procedendo-se
também a uma análise com β-PNPG para determinar a atividade da β-glucosidase,
além de análise da concentração de proteínas presentes no extrato por método
BCA.
O coquetel enzimático empregado neste trabalho contendo as duas
celulases, endoglucanase e β-glucosidase, foi baseado nas conclusões da
pesquisa de Lalithambika et al. (2012) utilizando a Archaea Sulfolobus solfataricus.
Naquele trabalho, foi comprovado que a endoglucanase da Archaea seria capaz de
hidrolisar em oligômeros a celulose presente no meio reacional, comprovando ser
esta uma enzima extracelular. Ainda de acordo com Lalithambika et al. (2012), uma
enzima transportadora possibilitou a entrada dos oligômeros no citoplasma da
Archaea, onde uma subsequente hidrólise foi realizada pela enzima β-glucosidase,
a qual é intracelular. Com base nos resultados destes autores, propôs-se que a β-
glucosidase fosse extraída das células e empregada em conjunto com a
endoglucanase no meio reacional, conforme será relatado adiante.

37
5.1 Caracterização do extrato enzimático de β-glucosidase
5.1.1 Eletroforese SDS-PAGE
A eletroforese SDS-PAGE foi realizada com intuito de observar a presença da
enzima β-glucosidase no extrato enzimático. O resultado é apresentado na Figura
11, sendo que as alíquotas retiradas do precipitado e sobrenadante após a
sonicação mostram o efeito da lise celular, enquanto as alíquotas do precipitado e
sobrenadante após o aquecimento em banho-maria mostram o efeito da
desnaturação proteica devido à exposição a alta temperatura (80ºC).
Figura 11 - Eletroforese em gel SDS-PAGE para o extrato enzimático de β-glucosidase pré-purificado. Os resultados correspondem ao precipitado (1) e sobrenadante (2) obtidos após
sonicação e ao precipitado (3) e sobrenadante (4) obtidos após aquecimento em banho maria. Na Figura, M corresponde aos padrões de peso molecular
A análise do gel indicou a presença de diversas proteínas no sobrenadante pós-
sonicação (poço 2, Figura 11), as quais foram produzidas pelo micro-organismo
hospedeiro e incluem a enzima de interesse. Esta grande quantidade de diferentes
proteínas é explicada pelo fato de que, com o rompimento celular causado pela
sonicação, proteínas intracelulares foram liberadas e passaram a fazer parte do

38
meio. Como a enzima de interesse é termorresistente (Lalithambika et al., 2012),
procedeu-se ao procedimento de purificação pela precipitação de proteínas a 80°C.
A análise do efeito do aquecimento em banho-maria a 80ºC mostrou que a
maioria das proteínas presentes sofreu desnaturação e precipitou após serem
expostas a altas temperaturas (poço 3), com exceção da β-glucosidase e outras
moléculas que permaneceram no sobrenadante (poço 4) (Figura 11). A β-
glucosidase foi identificada com base em seu peso molecular próximo de 50-60
kDa, conforme experiência prévia do grupo de pesquisa do Dr. Paul Blum.
5.1.2 Análise de atividade pelo método β-PNPG
A reação β-PNPG foi realizada e as absorbâncias obtidas para cada
amostra são mostradas na Tabela 7.
Tabela 7 - Resultados de absorbância obtidos para as amostras de β-glucosidase dos extratos
pré-purificados obtidos a partir do cultivo de E. coli transformada com gene de S. solfataricus
Tubo* Densidade óptica a 420nm
1 (branco) 0
2 (controle) 0,0095
3 (controle) 0,0185
4 (1:10) 0,0576
5 (1:10) -0,0009
6 (1:10) 2,1778
7 (1:10) 2,2353
8 (1:100) 0,0028
9 (1:100) 0,0012
10 (1:100) 0,2234
11 (1:100) 0,2502
*detalhes descritos na Tabela 2

39
Conforme pode ser observado na Tabela 7 e conforme descrição da
composição dos tubos da Tabela 2, as medições incluíram um tubo empregado
como branco (para zerar o espectrofotômetro), além de tubos controle, em
duplicata, para que fossem descontadas nos cálculos de atividade as absorbâncias
devidas aos reagentes empregados (tubos 2 e 3) e aos componentes do
extrato enzimático (tubos 4 e 5 para o extrato diluído 10 vezes, e tubos 8 e 9 para
o extrato diluído 100 vezes). Os tubos 6 e 7 correspondem a uma análise em
duplicata para o extrato diluído 10 vezes e os tubos 10 e 11 correspondem ao
extrato diluído 100 vezes. A leitura de absorbância do tubo 5 foi desconsiderada
por ser um valor discrepante.
Com base nos resultados da Tabela 7 e Equação 1, foi possível o cálculo da
atividade de β-glucosidase do extrato pré-purificado obtido. O resultado
correspondeu a 0,394 U/mL.
5.1.3 Determinação da quantidade de proteínas pelo método BCA
A curva padrão de albumina bovina sérica (BSA) foi obtida a partir das
diferentes concentrações da solução padrão e suas respectivas absorbâncias,
tendo sido linear até 1000 µg/mL, conforme ajuste mostrado na Figura 12.
As leituras obtidas para os extratos enzimáticos foram realizadas em
duplicata e resultaram em valores de absorbância iguais a 0,1957 e 0,1943.
Utilizando a equação da curva padrão (Figura 12), foi possível calcular as
correspondentes concentrações de enzima por regressão linear, obtendo-se os
valores 211,78 e 210,22 μg/mL. Foi calculada a média entre estes valores (211
μg/mL), sendo esta multiplicada por 10 (fator de diluição). Assim, o ensaio resultou
em uma concentração de proteínas de 2110 μg/mL. Este valor foi então multiplicado
por 25 mL (volume total de extrato enzimático obtido no processo), e assim foi
obtida a massa total de proteínas produzidas, de 52,75 mg.

40
Figura 12 - Curva padrão de albumina bovina sérica (BSA) obtida para o método BCA
5.2 Hidrólise enzimática e avaliação da atuação do coquetel enzimático
Para avaliação da eficiência da hidrólise enzimática da celulose pelas
enzimas produzidas, a quantidade de açúcares redutores totais (ART) liberada no
meio foi determinada pelo método de DNS. Com este fim, inicialmente, uma curva
padrão de D-glicose foi obtida a partir das diferentes concentrações da solução
padrão de glicose e suas respectivas absorbâncias, sendo o ajuste linear
apresentado na Figura 13.
Procedeu-se então às reações de hidrólise da celulose presente no slurry de
switchgrass com os extratos enzimáticos de endoglucanase e β-glucosidase em
separado ou combinados. O total de açúcares redutores obtidos é apresentado na
Figura 14, juntamente com o resultado obtido empregando preparação comercial
de celulase.
41
Figura 13 - Curva padrão de D-glicose obtida para análise de açúcares redutores pelo método
DNS
Figura 14 - Concentração de açúcares redutores totais liberados durante hidrólise enzimática de celulose de switchgrass de acordo com a celulase empregada
Os resultados apresentados na Figura 14 mostram o potencial das enzimas
produzidas na hidrólise da celulose de switchgrass sob condições de elevada
temperatura (80ºC) e, no caso da endoglucanase, também sob pH mais ácido (3,0).
Como as condições empregadas para cada caso foram distintas, a comparação de
suas atuações fica prejudicada. De qualquer forma, pode-se observar que o uso

42
dos extratos enzimáticos de endoglucanase e β-glucosidase resultou em maior
concentração de açúcares redutores em comparação ao uso da preparação
comercial. De fato, com base na experiência prévia do grupo de pesquisas no qual
o trabalho foi realizado, a preparação comercial de celulase foi encubada durante
4h e, mesmo assim, resultou em uma concentração de ART mais baixa. Este baixo
desempenho pode ser atribuído à sensibilidade da celulase comercial de T. reesei
às substâncias presentes no slurry, as quais podem atuar como inibidoras da
atividade enzimática (KIM et al., 2011). Por outro lado, pode-se observar também
que, mesmo no slurry, as enzimas endoglucanase e β-glucosidase foram capazes
de hidrolisar a celulose de switchgrass em condições extremas.
Cabe comentar ainda que a liberação de açúcares redutores na reação
catalisada pelas enzimas do extrato de β-glucosidase sugere a presença de outras
enzimas do complexo celulolítico. Seja como for, quando, nas mesmas condições,
fez-se o uso combinado dos dois extratos enzimáticos (endoglucanase+ β-
glucosidase), a liberação de ART foi cerca de 2,5 vezes superior ao do uso apenas
do extrato de β-glucosidase.

43
6. CONCLUSÕES
A metodologia empregada para produção e pré-purificação das enzimas
endoglucanase e β-glucosidase possibilitou a obtenção de extratos enzimáticos
capazes de atuar na hidrólise da celulose presente no slurry de switchgrass obtido
por pré-tratamento ácido diluído. A β-glucosidase intracelular foi extraída das
células e o extrato pré-purificado apresentou atividade de 0,394 U/mL na hidrólise
de β-PNPG a 80ºC. O uso deste mesmo extrato na hidrólise da celulose de
switchgrass presente no slurry resultou na liberação de açúcares redutores,
indicando a possibilidade de que outras enzimas do complexo celulolítico
estivessem presentes. A endoglucanase obtida foi capaz de atuar em condições
termoacidófilas (80ºC, pH 3,0), atuando também em conjunto com a β-glucosidase
sob temperatura de 80ºC a pH 5,0. O potencial das enzimas foi demonstrado,
indicando a possibilidade de uso de hidrolases do complexo celulolítico em
condições que podem ser encontradas em correntes materiais oriundas de pré-
tratamento ácido de biomassa, sem necessidade de extensivo resfriamento ou
neutralização.

44
7. REFERÊNCIAS
ABRIL, D.; ABRIL, A. Ethanol from lignocellulosic biomass. Ciencia e Investigacion Agraria, v. 36, n. 2, p. 177–190, 2009.
AGUILAR, C. F. et al. Crystal structure of the beta-glycosidase from the hyperthermophilic archeon Sulfolobus solfataricus: resilience as a key factor in thermostability. Journal of molecular biology, v. 271, n. 5, p. 789–802, 1997.
BAI, Y.; LUO, L.; VAN DER VOET, E. Life cycle assessment of switchgrass-derived ethanol as transport fuel. International Journal of Life Cycle Assessment, v. 15, n. 5, p. 468–477, 2010.
BERNFELD, P. α-and β-Amylases. Methods in Enzymology, v. 1, p. 149-158, 1955.
BOUTON, J. H. Molecular breeding of switchgrass for use as a biofuel crop. Current Opinion in Genetics & Development, v. 17, p. 553–558, 2007.
BROCK, T. D. et al. Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. Archiv für Mikrobiologie, v. 84, n. 1, p. 54–68, 1972.
CARDONA, C. A.; QUINTERO, J. A.; PAZ, I. C. Production of bioethanol from sugarcane bagasse: Status and perspectives. Bioresource Technology, v. 101, n. 13, p. 4754–4766, 2010.
CARVALHO, W. et al. Uma visão sobre a estrutura, composição e biodegradação da madeira. Quimica Nova, v. 32, n. 8, p. 2191–2195, 2009.
CASTRO, A. M. DE; PEREIRA JR., N. Produção, propriedades e aplicação de celulases na hidrólise de resíduos agroindustriais. Quimica Nova, v. 33, n. 1, p. 181–188, 2010.
DEMAIN, A. L. Biosolutions to the energy problem. Journal of Industrial Microbiology and Biotechnology, v. 36, n. 3, p. 319–332, 2009.
DEMİRBAŞ, A. Bioethanol from Cellulosic Materials: A Renewable Motor Fuel from

45
Biomass. Energy Sources, v. 27, n. 4, p. 327–337, 2005.
DIAS, M. E. et al. Ethanol as Fuel : Energy , Carbon Dioxide Balances , and Ecological Footprint. v. 55, n. 7, p. 593–602, 2005.
DUTTA, S.; PAL, S. Promises in direct conversion of cellulose and lignocellulosic biomass to chemicals and fuels: Combined solvent-nanocatalysis approach for biorefinary. Biomass and Bioenergy, v. 62, p. 182–197, 2014.
FENGEL, D.: WEGENER, G. Wood: chemistry, ultrastructure reactions. New York: W. De Gruyter, p. 613, 1989.
GÍRIO, F. M. et al. Hemicelluloses for fuel ethanol: A review. Bioresource Technology, v. 101, n. 13, p. 4775–4800, 2010.
HASELTINE, C. et al. Coordinate Transcriptional Control in the Hyperthermophilic Archaeon Sulfolobus solfataricus Coordinate Transcriptional Control in the Hyperthermophilic Archaeon Sulfolobus solfataricus. Journal of Bacteriology, v. 181, n. 13, p. 3920–3927, 1999.
HIMMEL, M. E. et al. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science (New York, N.Y.), v. 315, n. 5813, p. 804–7, 2007.
KIM, Y. et al. Soluble inhibitors/deactivators of cellulase enzymes from lignocellulosic biomass. Enzyme and Microbial Technology, v. 48, n. 4–5, p. 408–415, 2011.
KUMAR, P. et al. Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Industrial and Engineering Chemistry Research, v. 48, n. 8, p. 3713–3729, 2009.
KUMAR, R.; SINGH, S.; SINGH, O. V. Bioconversion of lignocellulosic biomass: Biochemical and molecular perspectives. Journal of Industrial Microbiology and Biotechnology, v. 35, n. 5, p. 377–391, 2008.
LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, v. 227, p. 680–685, 1970.

46
LALITHAMBIKA, S. et al. Carbohydrate hydrolysis and transport in the extreme thermoacidophile Sulfolobus solfataricus. Applied and Environmental Microbiology, v. 78, n. 22, p. 7931–7938, 2012.
LEE, J. Biological conversion of lignocellulosic biomass to ethanol. Journal of Biotechnology, v. 56, n. 1, p. 1–24, 1997.
MAMMAN, A. S. et al. Furfural: Hemicellulose/xylose- derived biochemical. Biofuels, Bioproducts and Biorefining, v. 6, n. 3, p. 246–256, 2008.
MARGEOT, A. et al. New improvements for lignocellulosic ethanol. Current Opinion in Biotechnology, v. 20, n. 3, p. 372–380, 2009.
MILANO, J. et al. Microalgae biofuels as an alternative to fossil fuel for power generation. Renewable and Sustainable Energy Reviews, v. 58, p. 180–197, 2016.
MILLER, P. S.; BLUM, P. H. Extremophile-inspired strategies for enzymatic biomass saccharification. Environmental technology, v. 31, n. 8–9, p. 1005–1015, 2010.
MISHRA, A. et al. Lignocellulosic ethanol production employing immobilized Saccharomyces cerevisiae in packed bed reactor. Renewable Energy, v. 98, p. 57–63, 2015.
NATIONAL CORN GROWERS ASSOCIATION. World of Corn. p. 12, 2014.
PANDEY, A. et al. Biotechnological potential of agro-industrial residues. I: Sugarcane bagasse. Bioresource Technology, v. 74, n. 1, p. 69–80, 2000.
PATCHETT, M. L.; DANIEL, R. M.; MORGAN, H. W. Purification and properties of a stable beta-glucosidase from an extremely thermophilic anaerobic bacterium. Biochem. J., v. 243, p. 779–787, 1987.
PÉREZ, J. et al. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: An overview. International Microbiology, v. 5, n. 2, p. 53–63, 2002.
PIMENTEL, D.; PATZEK, T. Ethanol production using corn, switchgrass, and wood; Biodiesel production using soybean and sunflower. Natural Resources Research,

47
v. 14, n. 1, p. 65–76, 2005.
RODRIGUES, J. A. R. Do engenho à biorrefinaria. A usina de açúcar como empreendimento industrial para a geração de produtos bioquímicos e biocombustíveis. Quimica Nova, v. 34, n. 7, p. 1242–1254, 2011.
ROSA, S. DA; GARCIA, J. O etanol de segunda geração: limites e oportunidades. Revista do BNDES, p. 117–155, 2009.
SANTOS, F. A et al. Potencial da palha de cana-de-açúcar para produção de etanol. Quimica nova, v. 35, n. 5, p. 1004–1010, 2012.
SCHMER, M. R. et al. Net energy of cellulosic ethanol from switchgrass. Proceedings of the National Academy of Sciences, v. 105, n. 2, p. 464–469, 2008.
SHE, Q. et al. The complete genome of the crenarchaeon Sulfolobus solfataricus P2. Proceedings of the National Academy of Sciences of the United States of America, v. 98, n. 14, p. 7835–7840, 2001.
SMITH, P. K. et al. Measurement of protein using bicinchoninic acid. Analytical Biochemistry, v. 150, n. 1, p. 76–85, 1985.
STÖCKER, M. Biofuels and biomass-to-liquid fuels in the biorefinery: Catalytic conversion of lignocellulosic biomass using porous materials. Angewandte Chemie - International Edition, v. 47, n. 48, p. 9200–9211, 2008.
U.S. DEPARTMENT OF ENERGY. Ethanol Feedstocks. Department of Energy Biomass Program, 2016. Disponível em: < http://www.afdc.energy.gov/fuels/ethanol_feedstocks.html>. Acesso em: 09 Ago 2016.
VITAL, N. A planta que pode salvar o mundo. Dinheiro Rural, ed. 37, 2007. Disponível em <http://revistadinheirorural.terra.com.br/secao/agrotecnologia/a-planta-que-pode-salvar-o-mundo> Acesso em: 09 Ago 2016.
WYMAN, C. E. Ethanol from lignocellulosic biomass: Technology, economics, and opportunities. Bioresource Technology, v. 50, n. 1, p. 3–15, 1994.

48
WRIGHT, L. Historical perspective on how and why switchgrass was selected as a "model” high-potential energy crop. Oak Ridge National Laboratory, Environmental Sciences Division, Report ORNL/TM-2007/109, 2007.
ZHANG, Y. H. P.; LYND, L. R. Cellodextrin preparation by mixed-acid hydrolysis and chromatographic separation. Analytical Biochemistry, v. 322, n. 2, p. 225–232, 2003.
ZHANG, Y. H. P.; LYND, L. R. Toward an aggregated understanding of enzymatic hydrolysis of cellulose: Noncomplexed cellulase systems. Biotechnology and Bioengineering, v. 88, n. 7, p. 797–824, 2004.


















