
UFRRJ INSTITUTO DE BIOLOGIA CURSO DE PÓS-GRADUAÇÃO ... - Verônica... · ufrrj instituto de...
83
UFRRJ INSTITUTO DE BIOLOGIA CURSO DE PÓS-GRADUAÇÃO MULTICÊNTRICO EM CIÊNCIAS FISIOLÓGICAS DISSERTAÇÃO INFLUENCIA DE DIFERENTES DOSES DE CIPIONATO DE ESTRADIOL NAS ALTERAÇÕES HIDROELETROLÍTICAS DE RATAS OVARIECTOMIZADAS VERONICA CRISTINA LOPES MENEZES Seropédica, RJ 2015
Transcript of UFRRJ INSTITUTO DE BIOLOGIA CURSO DE PÓS-GRADUAÇÃO ... - Verônica... · ufrrj instituto de...

UFRRJ
INSTITUTO DE BIOLOGIA
CURSO DE PÓS-GRADUAÇÃO MULTICÊNTRICO EM
CIÊNCIAS FISIOLÓGICAS
DISSERTAÇÃO
INFLUENCIA DE DIFERENTES DOSES DE CIPIONATO
DE ESTRADIOL NAS ALTERAÇÕES
HIDROELETROLÍTICAS DE RATAS
OVARIECTOMIZADAS
VERONICA CRISTINA LOPES MENEZES
Seropédica, RJ
2015

2
UFRRJ
INSTITUTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO MULTICÊNTRICO EM CIÊNCIAS
FISIOLÓGICAS
INFLUÊNCIA DE DIFERENTES DOSES DE CIPIONATO DE
ESTRADIOL NAS ALTERAÇÕES HIDROELETROLÍTICAS DE RATAS
OVARIECTOMIZADAS
VERONICA CRISTINA LOPES MENEZES
Sob orientação do Professor
Luis Carlos Reis
Dissertação apresentada ao Programa
Multicêntrico de Pós-graduação em
Ciências Fisiológicas da Universidade
Federal Rural do Rio de Janeiro como
requisito parcial para obtenção do título de
Mestre em Ciências Fisiológicas. Área de
concentração: Fisiologia.
Seropédica, RJ
Julho de 2015

3

4

5
Dedicatória
À minha família

6
Agradecimentos
Neste momento em que estou a caminho de concluir mais uma etapa da minha formação
aproveito para agradecer primeiramente a minha família, especialmente meus pais Angela e Jorge
por terem me apoiado em tudo desde sempre e por acreditarem junto comigo no meu sonho de
me formar e seguir na vida acadêmica. Foram diversas vezes em que minha família me trouxe até
à Rural nos fins de semana e feriados e sempre me apoiaram em todos os momentos.
Gostaria de agradecer ao meu orientador Prof. Luis Carlos Reis, por ser um orientador
sempre presente, com que podemos contar e que se preocupa muito com seus alunos. Gostaria de
agradecer também, Mestre, pelo senhor ter acreditado no meu potencial desde a Iniciação
Científica e ter passado muito de sua experiência para nós, além de se empenhar tanto para que
nós tivéssemos um bom começo na vida acadêmica e especialmente poder começar da forma
certa. Sempre vou lembrar com carinho de todos os momentos e das reuniões em sua casa,
sempre regadas com boa conversa e boa companhia e também do respeito pelos alunos.
Agradeço também ao Prof. André Mecawi pelas idéias e discussões no decorrer do
trabalho, pelas sugestões que surgiram, pelo suporte quando nos ajudou na bancada na coleta do
material, por trazer animais de Ribeirão Preto quando precisávamos, pelas importantes
considerações na discussão dos dados e pelos momentos divertidos durante os experimentos!
É muito legal olhar para os últimos cinco anos de DCF e ver o quanto meus amigos
também ajudaram na minha formação. Lembro que cheguei ao laboratório muito tímida e logo fui
bem recebida por todos os colegas e professores e hoje vejo o quanto que isso me chamou
atenção e me fez querer continuar no laboratório.
Gostaria de agradecer ao Prof. Wellington pela amizade e pela ajuda nos cálculos de
doses e também pela ajuda na padronização do fotômetro de chama. Agradeço também aos
Professores Fábio, Emerson, Frederico, Magda e Michele com quem tive aulas durante a
graduação e pude aprender muito com seus conhecimentos. Agradeço também à Profa. Alba ,
chefe do Departamento, pelo apoio no dia-a-dia e pela amizade.
Aproveito também para agradecer aos colegas de laboratório Bruno Paes Leme, Livia
Monteiro, Raoni dos Santos, Silvana do Nascimento e Evandro Valentim por toda ajuda durante
o trabalho, por serem ótimos colegas e mais ainda serem amigos em que posso confiar. Obrigada
a vocês pelos vários bons momentos divertidos no laboratório, pela companhia durante os
experimentos. Agradeço também aos colegas por me ajudarem a ver os meus defeitos com
carinho e a melhorar na minha forma de ser. Vocês sempre terão um lugar especial nas minhas
lembranças.
Gostaria de agradecer também à Fabrícia por ter me ensinado a ser mais sagaz com os
experimentos...lembro muito dela falando “Agilidade, Verônica” (risos), também pela orientação
durante a Iniciação Científica e pelo aprendizado dos protocolos de gaiola metabólica, por vários
momentos engraçados e pela deliciosa culinária mineira nas reuniões na casa do Mestre.
Gostaria de agradecer a Iracema Araújo pela amizade, ajuda em diversos momentos no
laboratório desde a Iniciação Científica, pelas discussões nos seminários e momentos divertidos.
Gostaria de agradecer ao Danilo Lustrino pela confiança, e por me apresentar no
laboratório e me passar o seu projeto de Iniciação Científica para que desse continuidade. Você
não imagina como foi legal o experimento do pCPA!
Gostaria de agradecer também a Marissa, Claudinho, Phelipe Fontanezi e gostaria de
agradecer muito ao Roberto Laureano pelo apoio desde a Iniciação Científica. Lembro que o
Roberto me apoiou e ajudou muito quando estava começando no laboratório e com quem pude
aprender os experimentos de labirinto em cruz elevado e campo aberto e que além disso, diversas

7
vezes foi me buscar no laboratório após às 23 horas quando estava fazendo o experimento do
pCPA .
Gostaria de agradecer à Nayana por ter me recebido na sua casa em Vargem Grande para
que pudesse frequentar a disciplina de Fisiopatologia da Tireóide. Foi muito bom fazer a
disciplina com você e agradeço pela ajuda na discussão do artigos.
Agradeço também a Güinever Eustaquio pela colaboração nos seminários na disciplina
de Receptores Nucleares da Profa. Carmem na UFRJ. Adorei a semana em que fiquei como
agregada no LEM e conhecendo outros colegas.
Agradeço também aos demais colegas Fernanda Kohn, Poliana, Raquel Teixeira, Carla
Franzini, Carla Myrra, Julia Vilas Boas e Bruna Sbano por todos os momentos divertidos , pelo
apoio e conversas no corredor.
Também agradeço aos técnicos Maria Rita, Vicente, Raquel Nascimento, Ipojucan e aos
secretários Dione e Franklin pelo apoio no dia-a-dia e pela amizade.
Gostaria de agradecer aos órgãos de fomento CNPq no período de 2011 a 2013 em que fui
bolsista de Iniciação Científica por dois anos e à CAPES no período de 2013 a 2015 em que fui
bolsista de Mestrado , respectivamente.

8
RESUMO
MENEZES, Veronica Cristina Lopes – Influência de diferentes doses de cipionato de estradiol
nas alterações hidroeletrolíticas de ratas ovariectomizadas. - Dissertação (Mestrado em
Ciências Fisiológicas, Fisiologia). Instituto de Biologia, Departamento de Ciências
FisiológicasUniversidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2015.
Palavras-chave: estrógeno, ovariectomia, apetite ao sódio, ingestão alimentar, sistema
serotoninérgico
A distribuição de receptores estrogênicos em estruturas centrais envolvidas na regulação da
homeostase hidroeletrolítica como o órgão vasculoso da lâmina terminal, núcleo subfornicial,
núcleo dorsal da rafe indica que o estradiol pode atuar nessas estruturas em resposta a alterações
nos fluidos corporais. Nosso objetivo foi verificar se a reposição hormonal pode influenciar de
maneira concentração-dependente o status hidroeletrolítico e neuroendócrino de ratas castradas
com reposição hormonal em diferentes doses de forma comparativa. Ratas Wistar (~230 g) foram
previamente adaptadas, por 5 dias, em gaiolas metabólicas, com acesso ad libitum aos
bebedouros volumétricos de água e salina hipertônica e ao alimento, sendo mantidas sob ciclo
claro-escuro de 12 horas em sala com temperatura controlada em 22º±2 ºC. Ao final da
adaptação, as ratas previamente anestesiadas com cetamina (75 mg/kg) e xilazina (5 mg/kg)
foram submetidas à cirurgia de ovariectomia bilateral. Os animais foram divididos em 4 grupos:
OVX, reposição com óleo de milho), repostos com óleo de milho cipionato de estradiol (E2) 2,5
µg/kg (E2 2,5), 10,0 µg/kg (E2 10,0) e 40 µg/kg (E2 40,0). O tratamento de reposição foi feito
pela via subcutânea, diariamente durante 7 dias tendo sido iniciado no dia seguinte à cirurgia.
Foram realizados três protocolos experimentais: avaliação sob condições basais, depleção de íons
sódio e reapresentação de fluidos. Neste estudo o estradiol apresentou efeito dose dependente nos
seguintes parâmetros sob condições basais: peso corporal diário, volume urinário diário, ingestão
de alimento diário. Após depleção de sódio não houve diferença em relação ao volume urinário
de 2 e de 24 horas após o experimento. No entanto após a reapresentação dos fluidos houve efeito
dose-dependente no comportamento ingestivo de água e de salina hipertônica tanto nos animais
depletados de sódio quanto nos animais controles.Os dados suportam que o estradiol modula o
comportamento ingestivo dos animais sob condições basais e após a depleção de sódio.

9
ABSTRACT
MENEZES, Veronica Cristina Lopes – Influence of different doses of estradiol cipionate on
the hidroelectrolytic challenges of female ovariectomized rats. - Dissertation (Master in
Physiological Sciences, Physiology). Institute of Biology, Department of Physiological Sciences,
Federal Rural University of Rio de Janeiro,Seropédica, RJ, 2015.
Key-words: estrogen, ovariectomy, sodium apettite , food intake, serotonergic system.
The distribution of estrogen receptors in brain structures that are envolved in the hidroelectrolyte
balance such as the lamina terminalis (LT), subfornical organ (SFO) and dorsal raphe nucleus
(DRN) demonstrated that estradiol can modulate important responses in body fluids. In the
literature experimental data support that estrogen can increase the triptofan-hydroxilase type 2 ,
the main enzyme for the serotonin synthesis. The primary question here is whether or not
differences in the baseline or stimulated intake are a function of different levels of circulating
gonadal hormones in female ovariectomized rats. Female Wistar intact rats (~230 g) were
previously aclimated in metabolic cages during 5 days and ad libitum access to water and
hypertonic saline (1.8%) bottles and food. Room temperature was maintained at 22±2 ºC with
12:12 h light-dark cycle (lights off at 19:00). Rats were anesthetized with intraperitoneal
injections of a mixture of ketamine (75 mg/kg) and xylazine (5 mg/kg) and then bilaterally
ovariohysterectomized. There were four experimental groups: OVX (replaced with corn oil), 2,5
µg/kg (E2 2,5), 10,0 µg/kg (E2 10,0) and 40,0 µg/kg (E2 40,0), daily during seven days, s.c.
After 24 h of the surgery the hormonal replacement initiated (estradiol cypionate, EC, Pfizer,
Animal Health). We did three experimental protocols: baseline evaluations, sodium depletion and
fluid replacement. The estrogen replacement exibitted a dose dependent effect in the following
parameters under basal conditions: daily body weight, daily urinary volume and daily food
intake. After sodium depletion there were no difference in the urinary volume after 2 and 24
hours of the experiment. But after fluid reposition we observed a dose dependent effect in the
ingestive behaviour of water and hypertonic saline intake in sodium depleted and control animals.
Our data support that estradiol can alter the natriorexigenic and dipsogenic responses especially
after sodium depletion depending of the estrogenic status.

10
Lista de abreviaturas
Ace- amídala central
ADH- hormônio antidiurético
AEs- amídala estendida
ANP – peptídeo natriurético atrial
ANG 1- angiotensina tipo 1
ANG 2 – angiotensina tipo 2
AP- área postrema
ADN- ácido desóxi-ribonucléico
ARNm - ácido-ribonucleico mensageiro
AVP- arginina-vasopressina
AV3V- região antero-ventral do terceiro ventrículo
BNP- peptídeo natriurético cerebral
CRH- hôrmonio liberador de corticotrofina
ENaC- canais para sódio epiteliais
ER α – receptor estrogênico tipo alfa
ER β – receptor estrogênico tipo beta
ER’s- receptores estrogênicos
E2- 17 β-estradiol, estradiol
ECA- enzima conversora de angiotensina
HVM- hipotálamo ventro-medial
LCR- líquido céfalo-raquidiano
LEC- líquido extracelular
LT- lamina terminalis
NLET- núcleo leito da estria terminal
NPOM – núcleo mediano pré-óptico
NaCl- cloreto de sódio
NPV- núcleo paraventricular
NPB- núcleo parabraquial
NOS- núcleo supra-óptico
NDR- núcleo dorsal da rafe
NPY- neuropeptídeo Y
NTS- núcleo do trato solitário
OCV’s – órgãos circunventriculares
OSF-órgão subfornicial
OT- ocitocina
OVX – rata ovariectomizada tratada com veículo
OVX-EC– rata ovariectomizada tratada com estrógeno
OVLT- órgão vasculoso da lâmina terminal
OTr- receptor para ocitocina
POMC- pró-ópiomelanocortina
SNC- sistema nervoso central
SRAA- sistema renina-angiotensina-aldosterona
5-HT 1Ar – receptor serotoninérgico tipo 1 A
TPH 2 – triptofano-hidroxilase tipo 2
T3- tri-iodo-tironina; T4- tetra-iodo-tironina

11
SUMÁRIO
1. Introdução...........................................................................................................................página 12
1.1 Revisão de Literatura.........................................................................................................página 12
1.2 Justificativa........................................................................................................................ página 27
2. Objetivos.............................................................................................................................página 29
2.1 Objetivos gerais..................................................................................................................página 29
2.2 Objetivos específicos...........................................................................................................página 29
3.0 Material e métodos..............................................................................................................página 29
3.0.1 Animais e Manutenção....................................................................................................página 29
3.0.2 Procedimentos cirúrgicos................................................................................................página 30
3.0.2.1 Ovariectomia e seu aspecto translacional...................................................................página 30
3.0.3 Protocolo de reposição hormonal...................................................................................página 31
3.0.4 Tratamentos hormonais e doses ....................................................................................página 31
3.0.5 Tratamento com cipionato de estradiol.........................................................................página 32
3.0.5.1 Preparação das alíquotas de estrógeno.......................................................................página 32
3.0.6 Avaliação das ingestões de água e salina hipertônica (NaCl 1,8%)............................página 33
3.0.7 Avaliação do volume urinário e osmolalidade urinária e plasmática.........................página 34
3.0.8 Avaliação do sódio plasmático........................................................................................página 34
3.1 Avaliação do índice uterino e hipofisário.........................................................................página 35
4.0 Protocolo experimental......................................................................................................página 35
4.1 Experimento 1- Efeito do tratamento com estradiol sobre o apetite ao sódio, sede em ratas
OVX controle e sob reposição hormonal................................................................................página 35
4.1.2 Desenho experimental.....................................................................................................página 36
5.0 Análise estatística ...............................................................................................................página 36
6.0 Resultados e discussão........................................................................................................página 37
7.0 Conclusões...........................................................................................................................página 66
7.1 Conclusão geral....................................................................................................................página 66
7.2 Conclusões específicas.........................................................................................................página 66
8.0 Considerações finais e perspectivas ................................................................................página 67
9.0 Referências bibliográficas..................................................................................................página 67
10.0 Anexo..................................................................................................................................página 83

12
1. Introdução
1.1 Revisão de literatura
1.1.1 Regulação do equilíbrio hidroeletrolítico
A ingestão de água e salina hipertônica são comportamentos motivados importantes para a
homeostase dos indivíduos. A ingestão de água ocorre em situações em que há desidratação
celular, perda de volume sanguíneo e hiperosmolalidade plasmática. Hoje sabemos por
evidências da literatura que em condições de hiperosmolalidade, não estão ativados apenas os
mecanismos de sede e de redução da excreção urinária de água mediada pela vasopressina, mas
também mecanismos que inibem o consumo adicional de soluto e que induzem a excreção de
sódio mediada pela ocitocina e pelo peptídeo natriurético atrial (ANTUNES-RODRIGUES, et al.
1997; JOHNSON & THUNHORST, 1997).
Os estudos de Verney (1947) apresentaram o conceito de osmolalidade e propuseram a
existência de um mecanismo osmorreceptor relacionado à liberação de vasopressina em resposta
ao aumento de osmolalidade. Posteriormente, Anderson & Mccann (1955), postularam a
existência de um osmorreceptor que seria um sensor para sódio localizado em regiões cerebrais
desprovidas de barreira hematoencefálica, que poderiam estar envolvidas na regulação do apetite
ao sódio e sua excreção em resposta a variações da concentração desse soluto no líquido
cefalorraquidiano (LCR).Atualmente é bem conhecido que os osmorreceptores são neurônios
especializados capazes de traduzir variações da pressão osmótica no líquido extracelular (LEC)
em sinais elétricos que ativam áreas do sistema nervoso central envolvidas no controle da
ingestão e excreção de sal e água pela liberação de acetilcolina ou angiotensina.
Essas células sensíveis às variações de osmolalidade ou concentração de sódio do LEC
estão localizadas nos órgãos circunventriculares (OCV’s) localizadas principalmente no terceiro
ventrículo. Além de sua localização no sistema nervoso central (SNC), receptores para sódio
também estão localizados perifericamente nas terminações nervosas aferentes e adjacentes aos
vasos hepáticos, renais e intestinais. Por exemplo, os receptores para sódio localizados em vasos
hepáticos são estimulados pelo aumento na concentração de sódio na veia porta e que por
aferências vagais ativam neurônios do núcleo do trato solitário (NTS) que é um importante centro

13
de integração os quais, então, estimulam estruturas neurais do bulbo, que induzem o aumento da
natriurese e inibição da absorção intestinal de sódio.
E recentemente, o estudo de Noda (2007) verificou que canais para sódio sensíveis às
concentrações de sódio são localizados em células ependimárias e que esses canais atuam como
sensores dos níveis de sódio dos fluidos corporais.
Quando estudamos as vias de entrada para o sistema nervoso central temos que considerar
que fatores humorais (osmolalidade, concentração de sódio,hormônios) e mecânicos (volemia)
agem como sinais-chave para o aparecimento da sede e do apetite ao sódio. (SCHULKIN,1991;
OLDFIELD & MCKINLEY, 1995); JONHSON & THUNHORST, 1997; MCKINLEY et al.
2001). Uma via envolve fatores humorais, que pelo plasma ou LCR, sinalizam aos OCV’s
prosencefálicos como órgão vasculoso da lâmina terminal (OVLT) e órgão subfornicial (OSF) e
romboencefálicos como a área postrema (AP). A outra via é constituída de aferências vagais e
glossofaríngeas provenientes de receptores viscerais (osmossodiorreceptores hepáticos ou renais,
barorreceptores renais ou cardiovasculares) que se dirigem para núcleos integradores do tronco
encefálico, como o núcleo do trato solitário (NTS) e núcleo parabraquial (NPB).
Na literatura há o modelo hodológico da sede extracelular que foi proposto por
JONHSON & THUNHORST,1997; MCKINLEY et al. 2001. De acordo com este modelo os
sinais que chegam ao OSF e ao OVLT são, em seguida integrados primeiramente no núcleo
mediano pré-óptico (NPOM) por meio de conexões mediadas por angiotensina II. Do NPOM
partem sinais ativando neurônios dos núcleos paraventricular e supra-óptico hipotalâmicos que
controlam a secreção de vasopressina e ocitocina, e a ele chegam projeções do tronco encefálico
(AP, NTS, NPB e núcleos bulbares noradrenérgicos) com os quais o NPOM apresenta conexões
recíprocas (OLDFIELD & MCKINLEY, 1995; JONHSON & THUNHORST, 1997;
MCKINLEY et al. 2001).
O NTS, por sua vez, não apenas se conecta ao NPB, mas também a estruturas
telencefálicas subcorticais chaves para o processamento da informação envolvida com o controle
da sede e do apetite ao sódio, tais como a amígdala estendida (AEs)-que forma um contínuo
envolvendo o núcleo leito da estria terminal (NLET). Essas áreas telencefálicas corticais, a área
insular agranular (córtex gustatório primário) e o córtex límbico pré-frontal estariam envolvidas
na integração e coordenação das informações endócrinas e autonômicas (viscerossensorial e

14
somatossensorial) com o comportamento motivado requerido para a sede e apetite ao sódio
(ALBEID et al. 1995).
O controle exercido pelo núcleo parabraquial, particularmente pela sua porção lateral
envolve serotonina e peptídeos (colecistocinina) e pode ser importante para evitar ingestão
excessiva de sódio quando o apetite ao sódio é ativado durante a desidratação. O NTS e o NPB
possuem conexões diretas com o córtex gustatório, além de projeções para o tálamo gustatório de
onde partem colaterais para amídala central (Ace) e ao BNST (NOGREN, 1995).
O controle da homeostase hidroeletrolítica conta ainda com um importante sistema de
controle homeostático do volume e da tonicidade dos fluidos corporais. Os sistemas renina
angiotensina aldosterona (SRAA), o sistema ocitocinérgico, juntamente com o peptídeo atrial
natriurético (ANP) e a inervação simpática renal representam mecanismos reguladores da
reabsorção renal de sódio que atuam de modo integrado. A renina é uma enzima secretada pelas
células justaglomerulares presentes na arteríola aferente e participa dessa reação em resposta à
hiponatremia, alteração no volume de fluidos extracelular e hipotensão. A renina converte o
angiotensinogênio em angiotensina I (ANG I) que , posteriormente, por ação da enzima
conversora de angiotensina (ECA) é a responsável pela conversão da angiotensina I em
angiotensina II (ANG II). A ANG II possui efeito vasoconstritor, promotor da reabsorção renal
de sódio e indutor do comportamento dipsogênico. A angiotensina II, por sua vez poderá atuar na
região cortical da glândula adrenal, estimulando a síntese e liberação de mineralocorticóides,
principalmente a aldosterona, um hormônio que auxilia na retenção corpórea de íons sódio.
A ingestão de água e de sódio é ativada por ação da ANG II nos núcleos da lâmina
terminal OSF,OVLT e NPOM e se reduz quando há lesão nesses núcleos (THUNHORST et al.
1990). É na lâmina terminal (LT), particularmente nas estruturas pré-óptico-periventriculares da
parte anterior e ventral do terceiro ventrículo (AV3V), que reside o sinergismo entre a
angiotensina e a aldosterona para produzir apetite ao sódio. Lesões na região do AV3V, que
inclui o OVLT e NPOM inibem a ingestão de sódio induzida pela ANG II, mas não aquela
induzida por ação mineralo-corticoide (FITTS et al. 1990; DE LUCA JR. et al. 1992). O órgão
subfornicial (OSF) por sua vez, possui conexões bidirecionais com outros dois núcleos da lâmina
terminal e envia projeções diretas ao NTS, ao complexo da amígdala estendida e ao córtex pré-
frontal, que por sua vez, estabelece fortes conexões com o córtex gustatório primário
(OLDFIELD & MCKINLEY, 1995).

15
Com relação ao controle neuroendócrino emsituações de hipernatremia há um aumento na
atividade do SRAA, aumento na secreção de ANP e na pressão arterial. Há também o estiramento
dos cardiomiócitos e aumento na secreção da ocitocina que possui efeito natriurético, e desse
modo reforça os estímulos para a excreção renal de sódio (FITZSIMONS, 1998) e parece que a
ocitocina influencia também a inibição do apetite ao sódio, mas essa questão ainda está em
estudo.A ocitocina plasmática aumenta em resposta a uma sobrecarga osmótica, e esse aumento é
revertido em resposta à diluição do líquido extracelular durante a ingestão de água que precede o
apetite ao sódio em animais hipovolêmicos (BLACKBURN et al. 1995, VERBALIS et al. 1995).
Considerando que a injeção central, e não periférica, de ocitocina inibe a ingestão de sódio, foi
sugerido que a ocitocina plasmática é um marcador da ocitocina produzida e liberada no cérebro.
A ocitocina central agiria então inibindo os circuitos cerebrais que induzem apetite ao sódio. Isso
aconteceria concomitantemente à ativação de mecanismos facilitadores do apetite ao sódio
(angiotensina II, aldosterona, desativação dos receptores de volume) nas fases iniciais de
hipovolemia, quando esse apetite está inibido. A ocitocina cerebral elevada inibiria a ingestão de
sódio enquanto a sede é produzida. A água ingerida dilui os líquidos corporais, o que inibiria a
liberação cerebral de ocitocina. A ação dos sinais facilitadores estaria então liberada para
produzir o apetite ao sódio. Assim surgiu uma explicação para o apetite ao sódio ocorrer atrasado
em relação à sede em animais hipovolêmicos, atraso que evita o acréscimo de uma desidratação
intracelular à hipovolemia já instalada: a ocitocina participaria de mecanismos inibidores da
ingestão de sódio, que seriam desativados durante a redução da osmolaridade extracelular
liberando, então, a ação dos mecanismos facilitadores (BLACKBURN et al. 1995; VERBALIS et
al. 1995).
O hormônio antidiurético (ADH) ou vasopressina (AVP) também é importante nesse
mecanismo de regulação. Em condição de hipovolemia ou desidratação e hiperosmolalidade
plasmática os sistemas de controle homeostáticos reconhecem esse erro e sinalizam para a
neurohipófise promover a liberação da vasopressina, que estimula a reabsorção renal de sódio nos
túbulos renais e reabsorção de água pela inserção de canais de aquaporina tipo 2 na membrana
basolateral dos túbulos coletores renais. A vasopressina possui ainda um efeito vasoconstritor
levando ao aumento da pressão arterial. Todos esses mecanismos podem de algum modo ser
influenciados pela reposição hormonal com estrógeno.

16
Outro mecanismo de regulação ocorre pela atividade dos canais de sódio epiteliais
(ENaC) encontrados nos túbulos renais em sua porção cortical e são também responsáveis pela
reabsorção de sódio. Nesse contexto, a serotonina poderia atuar como um inibidor endógeno dos
canais ENaC, e desse modo podemos dizer que em condições de déficit serotoninérgico de
algum modo a redução ou mesmo ausência da serotonina poderá estimular a reabsorção renal de
sódio e com isso estaria diminuindo a sua excreção. Frindt & Palmer (2012) evidenciaram que a
aldosterona é capaz de aumentar os níveis protéicos da subunidade alfa dos canais para sódio
epiteliais (ENaC α), presentes nos canais de sódio epiteliais renais, modulando positivamente
esses canais. Assim um outro possívelmecanismo seria a modulação do canal pela aldosterona,
que aumenta a permeabilidade da barreira apical dos epitélios, aumentando assim a absorção de
Na+ no intestino grosso e a reabsorção do íon no ducto coletor, o que reduz sua excreção urinária.
1.1.2 Efeitos da modulação estrogênica na homeostase hidromineral
Os primeiros estudos que demonstraram que os hormônios gonadais estão envolvidos no
controle da homeostase hidroeletrolítica observaram que fêmeas bebiam menos água que os
machos sob as mesmas condições (RICHTER & BRAILEY,1929). O estudo mais antigo que
diretamente demonstrou que a ingestão de água se alterava como consequência de flutuações no
ciclo ovulatório foi realizado na espécie Macaca nemestrina em que foi encontrado que a
ingestão diária de água diminuía durante a fase periovulatória ou durante o momento em que os
autores chamavam de “odor sexual na pele” que foi observado durante dois ou três ciclos
menstruais (KROHN & ZUCCKERMAN, 1937). Posteriormente,estudos realizados em ratas por
Antunes-Rodrigues & Covian (1963) demonstram primeiramente que há uma diminuição na
ingestão de sódio durante o estro e elevada ingestão de sódio durante o diestro quando baixos
níveis plasmáticos de estrogênio são encontrados. Danielsen & Buggy (1980) também
encontraram dados similares em ratas intactas durante a fase do estro.
Um estudo de Vijande et al. (1978) avaliou o efeito dipsogênico da angiotensina II que foi
administrada pela via subcutânea a ratos machos e fêmeas. Foi evidenciado que as fêmeas adultas
beberam mais água do que os machos e também demonstrou que as fêmeas demonstravam maior
sensibilidade a ANG II durante o proestro. Os machos e as fêmeas castradas aos nascimento e
fêmeas androgenizadas ao nascimento bebiam volumes similares de água após a injeção de ANG

17
II. O padrão de ingestão induzida por estímulo é diferente em ambos os sexos, mas parece
depender do estágio de desenvolvimento dos animais.
Estudos de reposição hormonal nas décadas seguintes demonstraram que o estradiol é
suficiente para diminuir a ingestão de fluidos em roedores ovariectomizados (STRICKER et al.
1991). Neste estudo os autores evidenciaram que machos e fêmeas gonadectomizados ingeriram
menor volume de salina assim como os machos intactos, mas o consumo era muito menor quando
as fêmeas eram tratadas com estrógeno. Esses resultados indicam que um robusto apetite por
sódio pode ser produzido em ratos por prolongada privação de sódio e que o estrógeno pode
diminuir esse comportamento. Enquanto que o tratamento com progesterona não exerce efeito na
ingestão de água sozinho ou em combinação com E2 (THRASHER & FREGLY, 1977).
Recentemente,Antunes-Rodrigues et al. (2013)em um estudo de revisão reportou que o
estradiol participa do controle do balanço hidroeletrolítico pelos seguintes mecanismos:
aumentando a responsividade de neurônios vasopressinérgicos e ocitocinérgicos em neurônios do
núcleo paraventricular (NPV) e do núcleo supra-óptico (NOS) bem como a secreção de
vasopressina (AVP) e ocitocina (OT); aumentando a liberação de fatores natriuréticos como
peptídeo natriurético atrial (ANP) e OT em resposta à estimulação osmótica e outras alterações
sistêmicas; diminuindo a responsividade à ANG II central, mediada por receptores
angiotensinérgicos tipo 1 (AT1), e aumentando a atividade serotoninérgica no núcleo dorsal da
rafe (NDR) que conseqüentemente inibe o apetite ao sódio. Santollo & Daniels (2015) também
revisitaram o tema do papel do estrógeno na regulação da homeostase hidroeletrolítica.
Por outro lado, o sistema serotoninérgico também desempenha um papel importante e
parece influenciar em muitas vias de integração cerebral (Azmitia & Segal, 1978), sendo que o
estrógeno exerce uma influência direta neste sistema. Tal suposição foi confirmada recentemente
por Susuki et al. (2013) que evidenciou que o receptor estrogênico tipo beta (ERβ) é
predominantemente expresso no NDR e sugeriram que agonistas de ERβ poderiam ser utilizados
como tratamento para a depressão ao invés da terapia com estradiol.
Foi evidenciado também que o mesmo sistema participa ativamente do controle da
homeostase hidroeletrolítica - em especial o núcleo dorsal da rafe mesencefálica, rico em
populações de neurônios serotoninérgicos e que foi descrito por Reis (2007) é uma das principais
estruturas responsáveis pelo controle do apetite ao sódio.

18
Além disso, diversos estudos apontaram que os níveis serotoninérgicos bem como a
atividade neural desse sistema podem ser alterados durante o ciclo estral em função das
flutuações hormonais (KUENG et al. 1976; BIEGON et al. 1980; DESAN et al. 1988).
Na literatura estudos suportam a idéia de que os distúrbios decorrentes do déficit da
atividade serotoninérgica em ratas ovariectomizadas (OVX) seriam resultantes de uma supra-
regulação dos autorreceptores serotoninérgicos subtipo 1 A (5-HT1A) somatodendríticos do
(NDR) da rafe mesencefálica ou aumento da expressão dos autorreceptores serotoninérgicos
subtipo 1B (5-HT1B) em sua sub-região ventromedial, ambas são revertidas pela reposição
estrogênica (LU & BETHEA, 2002). No núcleo hipotalâmico ventromedial os receptores 5-
HT1A parecem estar supra-regulados durante o estro, ou seja, a expressão desse receptor parece
estar aumentada quando os níveis estrogênicos estão elevados. Esse seria mais um dos
mecanismos que contribuem para a inibição do apetite ao sódio durante o estro. Ainda neste
contexto um estudo de Creech et al. (2012) demonstrou que o estrógeno é capaz de levar a
dessensibilização dos receptores 5-HT1A no núcleo paraventricular do hipotálamo (NPV).
Sabemos por evidências da literatura que os inibidores de recaptação de serotonina (SSRIs)
podem induzir a dessensibilização dos receptores 5-HT1A no NPV. Neste experimento o
estrógeno foi capaz de produzir uma dessensibilização mais rápida. Os autores verificaram que a
dose de 10 µg/kg de benzoato de estradiol, duas vezes ao dia durante dois dias foi o mínimo
necessário para ocorrer uma dessensibilização parcial de aproximadamente 40%.
Somponpun (2007) descreveu possíveis mecanismos pelos quais o estrógeno atua na
manutenção do equilíbrio hidroeletrolítico, com especial atenção para os hormônios AVP e OT.
ERα está expresso em áreas osmosensitivas da lâmina terminal enquanto que em neurônios
magnocelulares hipotalâmicos do núcleo supra-óptico (NOS) e paraventricular (NPV) o receptor
ERβ está predominantemente expresso (SLADEK & SOMPONPUN 2007, 2003; SHUGHRUE
& MERCHENTHALER, 2001). Cada subtipo de receptor é diferentemente regulado em resposta
a alterações na osmolalidade plasmática, sugerindo uma importância funcional para esses
receptores. Essa heterogeneidade no padrão de expressão dos receptores estrogênicos permite que
o estrógeno possa desempenhar funções específicas em diferentes tipos celulares e dessa forma
desempenhar seu papel na osmorregulação. Kisley e colaboradores (2000) realizando estudo
imunoistoquímico com ratas ovariectomizadas (OVX) após reposição com E2 evidenciaram
aumentada expressão de proteína c-Fos em neurônios magnocelulares do núcleo paraventricular.

19
A retenção de líquidos corporais é comumente observada em mulheres que possuem
níveis plasmáticos elevados de estrógeno como no momento da ovulação ou pela administração
de estrógeno exógeno. Os níveis plasmáticos de vasopressina são encontrados elevados em
mulheres jovens durante a fase folicular do ciclo menstrual em paralelo com elevadas
concentrações de E2 (SOMPONPUN, 2007). Adicionalmente, a liberação de vasopressina basal
ou por estímulos está também elevada em mulheres na pós-menopausa, que recebem a terapia
estrogênica. Num estudo realizado por MAFFEI e colaboradores (2001) mulheres na pós-
menopausa e que utilizam a reposição hormonal apresentaram aumento nos níveis plasmáticos de
ANP e peptídeo natriurético cerebral (BNP).
O estrógeno pode ainda alterar o balanço entre inputs inibitórios ou estimulatórios para os
neurônios magnocelulares. Isso pode trazer efeitos genômicos indiretos que podem interferir ao
nível do núcleo da célula, alterando a expressão gênica e efeitos não – genômicos diretos através
dos quais o estrógeno pode se ligar aos receptores alfa e beta (Hartley et al. 2004) e produzir
diversos efeitos biológicos.
Dalmasso et al. (2011) realizaram um estudo com ratas intactas no estro e no diestro e
ovariectomizadas com ou sem reposição estrogênica e submetidas à depleção de sódio. Sua
equipe demonstrou o papel modulatório do estrógeno na inibição do apetite por sódio que pode
envolver a participação de neurônios excitatórios do OVLT e neurônios serotonérgicos no NDR
inibindo a resposta natriorexigênica. A atividade de neurônios do OVLT em ratas
ovariectomizadas com reposição estrogênica é praticamente ausente após depleção de sódio,
enquanto que a atividade dos neurônios do NDR aumenta ou permanece inalterada.
Assim como na ingestão de água o estrógeno reduz a ingestão de salina hipertônica na
maioria dos experimentos. Essas observações diferem da ingestão de água porque a resposta não
parece ter dimorfismo. Ao observarem os efeitos do E2 na ingestão de água que só ocorre em
fêmeas, parece que tanto machos quanto fêmeas diminuem a sua ingestão de salina quando
tratados com E2 (STRICKER et al. 1991). Neste estudo os autores argumentaram que não há
dimorfismo sexual quanto ao efeito organizacional do E2. Por exemplo, a combinação do déficit
de sódio e do diurético furosemida estimula o apetite ao sódio em machos e fêmeas. Essa
ingestão é resultado do aumento nos níveis plasmáticos de angiotensina II (ANG II) e da perda de
sódio causada pelo diurético. Apesar do efeito ser encontrado em ambos os sexos, a ingestão de
salina é duas vezes maior em fêmeas do que machos (WOLF, 1982).Como após a depleção de

20
sódio a ingestão de salina hipertônica é muito maior em fêmeas do que em machos isso nos leva a
pensar que em fêmeas, de algum modo o estrógeno sensibiliza mais os neurônios osmossensitivos
aos efeitos dipsogênicos e da angiotensina II e natriorexigêncos da aldosterona.
1.1.3- Efeitos da ação estrogênica na secreção, na regulação osmótica e na expressão gênica
vasopressina e ocitocina
De acordo com os estudos de Verney (1947) e de Sladeck & Somponpun (2008) o
principal estímulo para a liberação de vasopressina e ocitocina é o aumento da osmolalidade
plasmática.
Sabemos por evidências da literatura que elevados níveis plasmáticos de estrógeno e
progesterona podem levar à retenção de fluidos (Davidson et al. 1984; Stachenfeld, et al. 1998),
hipertensão (Oelkers, 1996) e edema (Oian et al. 1997, Tollan et al.1993).
O aumento do risco de hipertensão no período pré-menopausa em mulheres e sua
prevenção pela terapia estrogênica envolvem mecanismos que ainda não foram completamente
elucidados e a redução abrupta dos níveis estrogênicos, como ocorre na menopausa em situações
fisiológicas ou experimentalmente, como na cirurgia de ovariectomia é capaz de alterar muitos
sistemas biológicos. Mais especificamente, o estrógeno influencia processos fisiológicos relativos
ao balanço dos fluidos corporais, como a regulação da pressão arterial, os níveis basais de
vasopressina (AVP), bem como a liberação de vasopressina estimulada pelo aumento da pressão
osmótica. Desse modo, a elevação dos níveis de AVP e o aumento da reabsorção renal de sódio
contribuem para a retenção de água encontrada em altos status estrogênicos. No entanto, o
mecanismo central pelo qual o estrógeno induz o aumento na secreção de vasopressina ainda não
está completamente elucidado(STACHENFELD, et al. 1998).
A participação do estrógeno na regulação osmótica de liberação de vasopressina pode
ocorrer provavelmente via sistema nervoso central. O estrógeno atua diretamente em neurônios
vasopressinérgicos no hipotálamo (Akaishi et al. 1990; Barron et al. 1986; Sar & Stumpf, 1980).
Adicionalmente, receptores estrogênicos são encontrados na neurohipófise e em neurônios
produtores de vasopressina no núcleo supraóptico do hipotálamo (SAR & STUMPF, 1980). Há
evidencias de que a inervação vasopressinérgica nos núcleos paraventricular e supra-óptico seja

21
influenciada pelo estrógeno (STONE et al. 1982). Além disso, o estrógeno pode afetar a liberação
de vasopressina indiretamente via catecolaminas (CROWLEY et al. 1978).
Estudos que utilizaram o modelo de infusão de salina hipertônica demonstraram que um
baixo limiar osmótico para liberação de vasopressina é encontrado quando os níveis plasmáticos
de estrógeno e progesterona estão elevados como na fase luteal do ciclo menstrual (Vokers et al.
1998) e na gravidez (DAVIDSON, et al. 1984).
Um outro hormônio gonadal que não exerce efeito sobre a regulação osmótica induzida
pela vasopressina é a progesterona (CALZONE, et al. 2001). Neste estudo mulheres receberam
uma infusão de salina 3% e dois tipos de anticoncencionais por via oral: um que apresentava
somente estrógeno e outro que apresentava estrógeno mais progesterona. O objetivo foi comparar
esses anticoncepcionais e avaliar se somente o estrógeno ou o estrógeno mais a progesterona
poderiamalterar o limiar osmótico para secreção de vasopressina. Os autores verificaram que a
progesterona não alterou a osmolalidade plasmática basal, e provavelmente o estrógeno sozinho
seria capaz de alterar esse parâmetro.
1.1.4 – Modulação estrogênica sobre o sistema renina-angiotensina-aldosterona, função
renal e cardiovascular
O risco de doença cardiovascular tende a ser elevado nas mulheres na pré-menopausa ou
nas mulheres submetidas à menopausa induzida cirurgicamente. Nesta fase da vida são freqüentes
os casos de hipertensão pós-menopausa, bem como de osteoporose em decorrência da perda de
densidade óssea, que pode ser influenciada também pelo déficit estrogênico.
Sabemos por evidências da literatura que os hormônios estrogênicos desempenham um
papel protetor no sistema cardiovascular e acredita-se que esse efeito seja mediado pelo receptor
de estrógeno tipo alfa (ERα). Esquerda et al. (2007) reportou que a ativação do ERα tem sido
considerada na mediação do relaxamento vascular e pode desempenhar um papel protetor
enquanto o ERβ é demonstrado como importante nas respostas inflamatórias à injúria vascular.
Com relação à pressão arterial um estudo de Laragh (1976) mencionou que as pílulas
anticoncepcionais que contém estrógeno podem produzir alterações no sistema renina-
angiotensina aldosterona, principalmente por aumentar a atividade de renina plasmática e
excreção de aldosterona e também retenção de sódio, o que pode predispor à hipertensão.

22
Os hormônios gonadais também parecem exercer uma influência sobre a função renal,
inclusive com dimorfismo sexual em seus efeitos. Enquanto que os androgênios aumentam a
reabsorção tubular proximal de sódio e a pressão intraglomerular, os estrogênios levam à
vasodilatação renal e diminuição da fração de filtração ( PECHERE-BERTSCHI et al. 2007).
Gracelli et al. 2013 avaliaram os efeitos dos hormônios gonadais na função renal em
modelo de desnervação renal em ratas ovariectomizadas com reposição de estrógeno ou
progesterona. Os autores verificaram se haveria algum efeito desses hormônios na reabsorção
renal de sódio e cloreto, bem como na liberação de catecolaminas pelo rim. Foi relatado que o
estrógeno aumenta a liberação de catecolaminas pelo rim, ou seja exerce uma influência positiva
sobre o sistema nervoso autônomo simpático, que é um dos fatores que contribuem para a
retenção de sódio.
Um outro mecanismo de regulação ocorre pela influência do estrógeno na expressão de
subunidades dos canais ENaC (Gambling et al. 2004), que são canais de sódio epiteliais
encontrados nos túbulos renais proximais e distais (VERLANDER et al. 1998; BRUNETTE et al.
2001).
Além disso, estrógeno e progesterona atuam em tecidos renais modulando a NA-K-
ATPase (principalmente no córtex renal) por mecanismos genômicos e esses efeitos poderiam ser
importantes para balanço de Na+, Cl
- e água (GRACELLI et al. 2012). Além disso, o estrógeno,
in vitro, aumenta a expressão do co-transportadorNa-K-ATPase nas células renais dos túbulos
distais em cultura por mecanismos não genômicos (VERLANDER, et al. 1998). Investigações
mostraram que o rim controla a expressão de receptores estrogênicos nucleares (Erα, ERβ) e
transmembranares como o GPR30 (GAVA et al. 2011; LEMALE, et al. 2008). Esse efeito nos
túbulos proximais poderiam ser mediados diretamente via seus receptores ou poderiam também
atuar indiretamente via sistema renina angiotensina-aldosterona (NASCIMENTO, et al. 2008).
1.1.5 – Influência estrogênica sobre o peso corporal e sobre o comportamento
alimentar
Sinchak & Wagner (2012) reportaram em um estudo de revisão alguns mecanimos
genômicos pelos quais o estrógeno regula o metabolismo energético.

23
É também bastante conhecido na literatura que o estrógeno diminui o apetite em roedores
e primatas (BUTERA et al. 1984, CZAJA, et al. 1974). Em ratos o agonista ER α (PPT) diminui
a ingestão de alimento e o tamanho da refeição, enquanto que o agonista ER β diarilpropionitrila
(DPN) não teve efeito (SANTOLLO et al. 2007). Em camundongos o silenciamento do ERα no
hipotálamo ventro-medial (HVM) leva a hiperfagia, obesidade e intolerância à glicose
(MUSATOV, et al. 2007). Em porquinho da guiné o estradiol também diminuiu a ingestão de
alimento e a frequência das refeições (ROEPKE, et al. 2010). Assim, as propriedades do estradiol
em reduzir o apetite podem ser mediadas através de uma sinalização rápida iniciada via mER
envolvendo ERα e mER-Gαq mas não o ERβ. Em humanos a ingestão energética de mulheres é
mais baixa durante a fase periovulatória, quando os níveis estrogênicos estão no seu pico
máximo, já a ingestão mais alta ocorre durante a fase luteal em que predominam os níveis de
progesterona (JOHNSON, et al. 1994). Essas alterações cíclicas na ingestão energética são
associadas com diminuição no consumo de carbohidratos próximo ao período da ovulação, e
aumento no consumo de gordura durante a fase luteal (JOHNSON, et al. 1994).
De acordo com Geary & Asarian (1999, 2002) a dose de 2 µg/rata foi capaz de reduzir o
peso dos animais e o tamanho da refeição em ratas ovariectomizadas, sendo que as ratas tratadas
com essa dose apresentaram maior frequência na ingestão.
Silva et al. (2010)evidenciaram que ratas ovariectomizadas e tratadas com 10 µg/kg de
cipionato de estradiol por 8 dias apresentavam hipofagia. Neste trabalho foi demonstrado que tal
efeito poderia ser mediado por neuropeptídios orexigênicos como neuropeptídeo Y (NPY),
peptídeo agouti relativo (AgRP) e orexina A, cuja expressão estava reduzida e pela expressão
aumentada de mediadores anorexigênicos como melanocortina tipo 4 (MC4), leptina e hormônio
liberador de corticotrofina (CRH).
Em muitos aspectos, os efeitos do estrógeno na homeostase energética estão relacionados
com mecanismos periféricos que se relacionam com os níveis de leptina. ERs e receptor para
leptina (OB-R) possuem quase que os mesmos mecanismos de transdução de sinal que envolve a
via STAT 3 e PI3K (GAO et al. 2007; JEFFERY et al. 2011; ROEPKE et al. 2007).
A deleção de ERα também abole a supra-regulação na expressão de pró-
ópiomelanocortina (POMC) induzida pela leptina em camundongos AKITA diabéticos
(HIROSAWA et al. 2008). Além disso, a queda de OB-R em neurônios POMC de fêmeas de
camundongos diminui a expressão de ER α no hipotálamo e camundongas ovariectomizadas com

24
déficit de POMC apresentam uma tendência em acumular mais gordura do que as fêmeas
controles (SHI et al. 2010). No entanto, o aumento assimétrico da formação de sinapses induzida
pelo estrógeno no pericário de neurônios POMC é independente da sinalização de leptina como
esse efeito foi observado em camundongos ob/ob e db/db (GAO et al. 2007). O estrogênio
também aumenta agudamente a concentração de leptina circulante em mulheres (Lin et al. 2005)
e estimula a liberação de leptina em cultura de tecido adiposo de fêmeas e não de machos
(CASABIELL et al. 1998). A expressão de ácido ribonucleico mensageiro (ARNm) de leptina
também é mais alta em mulheres na pré-menopausa do que em menopausa (FAJARDO et al.
2004). Convencionalmente, em camundongos machos e ratas intactas a anorexia causada pela
implantação crônica de estradiol é associada com níveis séricos, plasmáticos e no fluido cérebro-
espinhal de leptina (ROCHA et al. 2001,TRITOS et al. 2004). Por outro lado, agonistas seletivos
de ER α cronicamente administrados a ratas ovariectomizadas reduz os níveis circulantes de
leptina associada ao peso corporal e gordura visceral (WEIGT et al. 2012). Esse poderia
representar um efeito genômico seguido de ativação ER α ou alternativamente uma forma de
compensar as alterações em face da prolongada exposição ao hormônio.
Um estudo de Fungfuang et al. (2013) evidenciou que existe um aumento significativo na
expressão de ARNm para leptina no tecido adiposo durante o proestro comparado com o diestro.
Esses achados sugerem que o aumento dos níveis séricos de leptina pelo estrógeno durante o
proestro, pode ser um dos fatores que regulam o comportamento alimentar.
O sistema serotoninérgico também participa do controle do comportamento alimentar e
pode ser influenciado pelo estradiol. Rivera et al. (2012) demonstraram que receptores
serotoninérgicos tipo 2C (5-HT2C) pós-sinápticos desempenham importante papel em mediar os
efeitos anorexigênicos do estradiol no sistema serotoninérgico. Os autores mencionaram que o
tratamento agudo de 1 µg/0,1 ml de 17 β-estradiol pela via subcutânea durante dois dias foi capaz
de reduzir o consumo de alimento durante a fase de escuro e a associação do estrógeno com
mCPP (metacloro-fenilpiperazina), um agonista dos receptores 5-HT2C intensificou a resposta
anorexigênica e ainda evidenciou que o estrógeno aumenta o conteúdo proteico desse receptor no
tronco cerebral caudal, mas não no hipotálamo de ratas ovariectomizadas. Também é conhecido
que o estrógeno dessensibiliza o receptor 5HT1A no PVN e em parte essa resposta ocorre pela
ativação do GPR 30 ( ROSSI et al. 2010; XU et al. 2009).

25
Em outro trabalho Santollo et al. (2011) monitoraram a ingestão de alimento por 48 horas
seguidas de microinfusões agudas de veículo em doses diárias de estradiol (0,25 a 10,0 µg) em
diferentes regiões encefálicas como núcleo mediano pré-óptico e núcleo arqueado e o núcleo
dorsal da rafe em modelo de ratas ovariectomizadas. Os autores apontam que ainda não se sabe se
o estradiol exerce efeito modulatório no núcleo paraventricular no contexto da ingestão de
alimento, mas evidenciaram que o hipotálamo ventromedial não está envolvido nesta regulação.
Neste estudo as infusões no núcleo mediano pré-óptico anterior (MPOA) com a dose mais alta
(2,5 µg) de E2 foi suficiente para diminuir a ingestão de alimento. E também a administração de
E2 no núcleo dorsal da rafe resultou em diminuição da ingestão de alimento por ser esta uma
região rica em neurônios serotoninérgicos.
1.1.6 Alguns efeitos da terapia estrogênica em outros sistemas biológicos
O estrógeno também é capaz de aumentar a atividade mitótica nos lactotrofos da
adenohipófise e conseqüentemente aumentar o peso da glândula hipofisária. Um aumento dose-
dependente no índice mitótico foi observado após 7 dias do início do tratamento, representando
uma aceleração da atividade mitótica de 1,7 % por dia em ratas ovariectomizadas na ausência de
reposição estrogênica e 3,7% por dia na presença de uma dose farmacológica de 50 µg/rata/dia ou
230 µg/kg/ dia, sendo esta uma dose suprafisiológica (LONAN & LEVY, 2009).
Thomas et al. (1986) verificou que hormônios gonadais como o estrógeno e progesterona
poderiam alterar os níveis séricos de glicose, insulina, tri-iodotironina (T3) e tetra-iodotironina
(T4) em ratas ovariectomizadas. A glicemia foi reduzida em ratas ovariectomizadas e repostas
com estrógeno, mas foi aumentada pelos tratamentos de estrógeno com progesterona. A insulina
foi reduzida pela ovariectomia e normalizada pela reposição estrogênica e aumentada pelo
tratamento do estrógeno mais progesterona. A concentração plasmática de T3 foi reduzida pela
ovariectomia e foi normalizada pela reposição estrogênica e aumentada pela associação dos
tratamentos (estrógeno mais progesterona). Já a concentração plasmática de T4 só foi afetada
pela reposição com estrógeno, sendo que sua concentração estava reduzida.
O estrógeno também é capaz de alterar o metabolismo da glicose. Liu et al. (2013) em
modelo de desnervação hepática evidenciou que ratas ovariectomizadas apresentaram 17 % de
redução da glicose plasmática, que foi completamente restaurada pela reposição hormonal
sistêmica. Além disso, a administração de estrógeno por microdiálise no núcleo paraventricular

26
do hipotálamo (NPV) ou no hipotálamo ventro-medial (HVM) restaurou a glicose plasmática.
Adicionalmente, esse estudo demonstrou que a desnervação simpática no fígado bloqueou o
efeito do estrógeno no HVM e a sensibilidade hepática à insulina, ou seja, a administração intra-
hipotalâmica de estrógeno regula a sensibilidade periférica à insulina via sistema simpático .
1.1.7 Um breve comentário sobre ações genômicas e não-genômicas para estrógenos
Até o momento encontram-se descritos dois subtipos de ER: o ER α e o ER β. Além
desses dois subtipos, são conhecidas variantes de ambas as isoformas, que podem ter o ARNm
incompleto ou conter maior ou menor número de bases em seus exons.
Dentre os efeitos genômicos do estrógeno sabemos que os receptores de estrógeno
promovem transcrição gênica por um mecanismo que consiste na dimerização do receptor e na
ligação do dímero a uma sequência do ácido desóxi-ribonucléico (ADN), chamada de elemento
responsivo ao estrógeno (ERE), a qual está presente nas regiões promotoras dos genes
responsivos a esses receptores. No entanto, a transcrição gênica também pode ser ativada por
meio de outras porções da molécula de DNA, como os sítios AP1 e Sp1.
O estrógeno pode ainda alterar o balanço entre inputs inibitórios ou estimulatórios para os
neurônios magnocelulares. Isso pode trazer efeitos genômicos indiretos que podem interferir ao
nível do núcleo da célula, alterando a expressão gênica e efeitos não – genômicos diretos
através dos quais o estrógeno pode se ligar aos receptores alfa e beta (Hartley et al. 2004) e
produzir diversos efeitos biológicos.
Já nas ações não-genômicas do hormônio existe grande número de relatos de ações
rápidas dos estrógenos alterando as propriedades elétricas de neurônios em questão de minutos.
Essas ações tem sido denominadas de não-genômicas e parecem ser decorrentes da atuação direta
dos estrógenos na membrana celular, modulando a ativação de proteína G e segundos
mensageiros; ainda não se sabe ao certo quais os tipos de receptores envolvidos nesse modo de
ação; uma aventada é a existência de receptores de membrana para estrógenos. Wang et al. 2015
evidenciou a existência do receptor estrogênico alfa tipo 36 (Er α 36) que é principalmente
expresso no citoplasma e na membrana plasmática ER-α36 está envolvido com a sinalização
rápida do estrógeno, no entanto o mesmo receptor inibe a sinalização genômica do hormônio.

27
Mohamed & Rahman (2000) mencionaram que a regulação gênica do hormônio
estrogênico é tecido específica, tendo avaliado a expressão de ERs em rim, fígado, útero e outros
tecidos.
1.2 Justificativa
Na literatura verificamos muitas diferenças metodológicas em muitos estudos quanto à
duração da reposição hormonal, o período de tempo em que o tratamento é realizado, a dose
utilizada do hormônio, a sua composição química (benzoato de estradiol, 17-β estradiol,
etinilestradiol e outros), biodisponibilidade, e linhagem de animais de laboratório (Long Evans,
Wistar, Sprague Dawley). A literatura também reporta que há diferenças na expressão dos
receptores estrogênicos cerebrais nas diferentes linhagens e também é importante considerar se
houve ou não intervalo entre a cirurgia e o início da reposição hormonal, pois tudo isso influencia
na expressão dos receptores estrogênicos.
Assim, uma das intenções de nosso estudo é auxiliar na compreensão dos efeitos de
tratamentos hormonais com estrogênio em alguns parâmetros de avaliação considerando doses
infrafisiológicas, fisiológicas e suprafisiológicas do hormônio para estudar se há efeito dose-
dependente relativo aos mecanismos fisiológicos estudados e compreender como o estrógeno age
alterando a expressão comportamental dos animais.
Um outro ponto a ser considerado é que na literatura encontramos alguns trabalhos sobre
os efeitos organizacionais e ativacionais dos hormônios sexuais, considerando o dimorfismo. No
entanto, a literatura suporta muito mais informações sobre como o estrógeno regula a ingestão de
água,mas não define a discussão sobre de que maneira o estrógeno regula o apetite ao sódio. Essa
discussão da ingestão de sódio modulada por estrógeno é bastante complexa.
Adicionalmente é bastante discutida a influência do estrógeno sobre o sistema
serotoninérgico e a maioria dos autores acredita que o estrógeno exerce uma modulação sobre
este sistema pelo aumento da expressão da enzima triptofano-hidroxilase tipo 2 (TPH2), sendo
esta encontrada principalmente em tecidos cerebrais (DONNER & HANDA, 2009). Contudo, em
grande parte desses experimentos os cientistas utilizam baixas doses de estrógeno como foi
utilizado 20µg/animal no estudo de Dalmasso et al. (2011). Nosso grupo, todavia, utiliza doses
muito superiores do hormônio (40 - 80µg/Kg). Esta dose suprafisiológica mimetiza os níveis de
estrógeno circulantes em fêmeas no último terço de gestação.

28
Dados não publicados de nosso laboratório com a dose de 80µg/Kg demonstraram uma
redução da expressão da enzima TPH2 a partir do NDR extraído dos encéfalos de ratas
ovariectomizadas tratadas com estrógeno durante 14 dias sob condições basais. Ao contrário do
esperado não observamos um aumento da expressão de ARNm da TPH2 em animais tratados
com estrógeno mas sim uma diminuição significativa quando comparamos os resultados dos
animais tratados com óleo.
Ao final da gestação verificamos que os níveis de estrógeno e ocitocina aumentam e que
os níveis de progesterona diminuem. De acordo com Murata et al. (2003) o estrógeno estimula a
expressão de ARNm de receptores de ocitocina (OTr) no útero em ratas ovariectomizadas e,
aparentemente o mesmo ocorre no SNC aumentando a liberação de ocitocina.
Rossi et al. (2010) demonstraram que o uso de agonista dos receptores ERβ é capaz de
levar a dessensibilização do autoreceptor 5-HT1A aumentando desta forma a mobilização e
liberação de serotonina (5-HT). Popova et al. (2010) verificaram que a ativação a longo prazo
dos receptores 5-HT1A com agonista seletivo leva a uma considerável queda do ARNm da TPH2
no mesencéfalo de camundongos. No entanto, não existem pesquisas que mostrem quais
mecanismos ocorrem após estimulação serotonérgica a longo prazo e com altas doses de
estrógeno. Com estudos futuros pretendemos compreender se esse seria um mecanismo de
controle homeostático específico do sistema serotoninérgico como uma tentativa de diminuir a
liberação do neurotransmissor, mediante redução da expressão de ARNm para TPH2.
Há também outros estudos que reportam que o estrógeno é um fator de proteção
cardiovascular e alguns estudos que relatam casos de hipertensão pós-menopausa, já que em
condições de déficit estrogênico é comum que mulheres no climatério mudem seus hábitos
alimentares, dentre eles, a adição de mais sódio ao alimento. Nosso estudo também pretende
contribuir com experimentos que ajudem na compreensão dos mecanismos envolvidos entre
homeostase hidroeletrolítica, déficit estrogênico, terapia estrogênica e pressão arterial.
2.0 Objetivos
2.1 Objetivo geral:
a) Avaliar o efeito da administração de diferentes doses de cipionato de estradiol nas
alterações hidroeletrolíticas de ratas Wistar ovariectomizadas.

29
2.2 Objetivos específicos:
a) Avaliar o efeito do estradiol sobre o apetite ao sódio, sede, natriurese, sódio plasmático,
volume urinário, sódio urinário e índice uterino condições basais e após depleção de sódio;
b) Avaliar a ingestão alimentar diária e o peso corporal diário em condições basais e verificar se a
reposição estrogênica em diferentes doses pode alterar esses parâmetros.
c) Realizar a dosagem dos níveis de estradiol plasmáticos e relacioná-los com a dose de hormônio
administrada em ratas ovariectomizadas.
3.0 Material e métodos
3.0.1 Animais e Manutenção
Este trabalho foi conduzido conforme o “Guide for the Care and Use of Laboratory
Animals” (NIH Publicação Nº 85-23, revisado 1996) e os protocolos experimentais avaliados e
aprovados pela Comissão de Ética em Experimentação Animal da Universidade Federal Rural do
Rio de Janeiro em consonância com a Lei AROUCA 11794 de 8 de outubro de 2008. ( Processo
nº 23083.011699/2013-99).
Foram utilizadas 120 ratas da linhagem Wistar, adultas (~230 g, ± 60 dias), e
provenientes do Biotério Experimental do Departamento de Ciências Fisiológicas da
Universidade Federal Rural do Rio de Janeiro. Destes animais, 64 fêmeas foram utilizadas nos
protocolos de avaliação em gaiolas metabólicas e 56 fêmeas foram utilizadas nos protocolos de
coleta de sangue e de cérebro. Os animais foram mantidos em caixas coletivas (5 animais por
caixa), aclimatados à temperatura controlada (23 ± 2ºC), sob regime de luz com ciclo claro-
escuro de 12 horas (luz 7:00-19:00h). Os animais tiveram acesso à água e alimento ad libitum. Os
experimentos foram realizados no período da manhã, entre 09:00 – 12:00 horas.Para avaliação de
ingestão de água e de solução salina hipertônica (NaCl 1,8%), ingestão de ração moída,volume
urinário, excreção renal de sódio, os animais foram transferidos para gaiolas metabólicas, nas
quais foram aclimatados por 5 dias antes do experimento.
Todos os esforços foram feitos para minimizar o sofrimento dos animais, seguindo o
“princípio dos 3R” para reduzir o número de animais utilizados no estudo e otimizar os
protocolos experimentais de modo a obter o máximo de dados de cada animal testado.

30
3.0.2 Procedimentos cirúrgicos
Todas as cirurgias foram realizadas sob plano anestésico induzido cetamina (Vetbrands,
Brasil , 60 mg/kg, i.p.) combinada com xilazina (Bayer, Brasil, 7,5 mg/kg, i.p.) sendo seguidas de
doses profiláticas de pentabiótico veterinário (0,2 mL/rata , i.m., Fort Dodge) para evitar
possíveis infecções.
3.0.2.1 O modelo experimental de ovariectomia e seu aspecto translacional
A menopausa é definida como um período em que a síntese de hormônios gonadais está
reduzida, bem como a produção de folículos ovarianos, sendo que na mulher esse processo se dá
por volta dos 51 anos.
Em modelos experimentais não podemos dizer que as ratas possuem a fase de menopausa,
pois elas não possuem ciclo menstrual, e sim ciclo estral. Então, desse modo, só poderemos
denominar tal fase como período de déficit na síntese de hormônios gonadais ou insuficiência
ovariana primária, que ocorre nos modelos de estudo da senescência natural, que na rata ocorre
por volta de um ano e meio a dois anos de vida. O termo “ menopausa” é referido em
experimentação com o objetivo de evidenciar o aspecto translacional do modelo.
De modo artificial , reproduzimos em laboratório o modelo de insuficiência gonadal,
mediante a cirurgia de ovariectomia, em que os ovários são retirados para reduzir os níveis
plasmáticos de estrógeno endógeno e com isso poderemos avaliar os efeitos da reposição
hormonal em diversos parâmetros de interesse.
Em nosso protocolo experimental a ovariectomia bilateral (OVX) foi realizada em ratas
adultas, por um acesso para-costal e em condições assépticas. Após indução anestésica com a
perda dos reflexos medulares, os animais foram submetidos à tricotomia e, em seguida, foi
realizada uma incisão bilateral da pele entre a última costela e a coxa. Posteriormente, o tecido
subcutâneo e a musculatura foram divulsionados com auxílio de pinça e tesoura cirúrgicas, para
visualização dos ovários. Os ovários foram exteriorizados e foi realizada a ligadura das trompas

31
uterinas, artérias e veias ovarianas, seguida da remoção dos ovários. Por fim, foi realizada a
sutura da musculatura e pele.
3.0.3 Protocolo de reposição hormonal
3.0.4 Tratamentos hormonais e doses
Com o intuito de verificar se há efeito dose – dependente nos parâmetros avaliados
utilizamos neste estudo diferentes doses de cipionato de estradiol ( Pfizer, Saúde Animal) para
um estudo comparativo entre doses infra-fisiológicas (2,5 µg/kg), fisiológicas (10 µg/kg) e
supra-fisiológicas (40 µg/kg) em ratas ovariectomizadas com e sem reposição hormonal pelo
período de 7 dias de tratamento. Sierra-Ramirez, et al. (2011) avaliaram a farmacocinética e a
farmacodinâmica do cipionato de estradiol sob duas diferentes vias de administração ( subcutânea
e intramuscular). No entanto, a informação sobre a meia-vida do hormônio é citada na espécie
humana, mas não está muito clara neste artigo.
De acordo com a literatura, a dose de 2 µg/kg (Geary & Asarian, 1999) é capaz de
produzir níveis plasmáticos de estradiol similares aos níveis de pico encontrados durante o ciclo
ovariano de ratas intactas, sendo capaz de diminuir a ingestão de alimento e retomar o peso
corporal à normalidade (BUTCHER et al., 1974; CECCHINI, et al., 1983; LU et al., 1985).
Dados publicados na literatura suportam evidências que 10 μg/kg (2,5 μg/rata) de
estrógeno (Liu et al., 2013; Vilhena-Franco et al., 2011; Silva et al. 2010)é uma dose suficiente
para reposição estrogênica em condições fisiológicas similares à concentração de estradiol
encontrada em ratas durante o proestro. Esse dado foi confirmado por Smith et al. (1975) que
verificou que a dose de 2,0 μg/rata a cada 4 dias é a dose de reposição em que os níveis de
estradiol são encontrados na manhã do proestro. Mecawi et al. (2011) verificou que ratas
ovariectomizadas tratadas com estrógeno com doses de 10 µg/kg e 40 µg/kg e que foram
submetidas à hemorragia apresentaram um aumento nos níveis plasmáticos de ocitocina e
vasopressina.

32
3.0.5 Tratamento com cipionato de estradiol
No dia da cirurgia de ovariectomia os animais foram pesados previamente para
homogeneização dos grupos experimentais. O grupo tratado recebeu diferentes doses de
estrógeno que foram administradas de acordo com o peso corporal, sendo que o volume
administrado não excedeu 0,13 ml para as ratas mais pesadas, sendo que o volume mínimo
administrado era de 0,1 ml. Todas as administrações foram feitas pela via subcutânea e realizadas
diariamente, entre 9:00 e 12:00 horas, tendo início 24 horas após a ovariectomia, e prosseguindo
durante 7 dias, com o objetivo de mimetizar a terapia estrogênica da mulher em menopausa. O
tratamento com estrógeno foi realizado com cipionato de estradiol (ECP, Pfizer Saúde Animal)
diluído em óleo de milho filtrado. Previamente às injeções os animais foram pesados na balança
analítica (modelo Helmac 1000) e seus pesos foram divididos por 2, já que a concentração da
solução preparada era o dobro da dose. Após a divisão do peso o valor resultante sofria uma
aproximação matemática.
Dessa forma teremos quatro grupos experimentais: OVX-óleo, E2 2,5, E2 10, E2 40.
3.0.5.1 Preparação das alíquotas de estrógeno
Considerando que o frasco original de cipionato de estradiol ( Pfizer, Saúde Animal)
contém 2 mg de estradiol em cada ml, objetivamos preparar 3 alíquotas que correspondem às
seguintes doses administradas: 40 μg/kg (solução 1), 10 μg/kg (solução 2) e 2,5 μg/kg (solução 3)
ou administração de veículo (óleo de milho filtrado). As soluções de estradiol foram preparadas
para o dobro da concentração desejada para que o volume administrado aos animais fosse
reduzido a aproximadamente 0,1 ml por animal.
Realizamos as diluições a partir do frasco original, utilizando a seguinte fórmula, em que:
C1- concentração do frasco inicial (µg/ml)
V1- volume do frasco inicial (ml)
C2- concentração do frasco final (µg/ml)
C1.V1= C2.V2

33
V2- volume do frasco final (ml)
Primeira etapa - Preparo da Solução 1 – 40 µg/kg ( 10 µg/rata)
Colocamos 0,4 ml do estrógeno do frasco original (Pfizer , Saúde Animal) e adicionamos
9,6 ml de óleo de milho filtrado, totalizando 10 ml.
Segunda etapa – Preparo da Solução 2 – 10 µg/kg (2,5 µg/rata)
Colocamos 0,1 ml do estrógeno do frasco original ( Pfizer, Saúde Animal) e adicionamos
9,9 ml de óleo de milho filtrado, totalizando 10 ml.
Terceira etapa – Preparo da Solução 3 – 2,5 µg/kg (0,625 µg/rata)
Colocamos 0,1 ml do estrógeno do frasco original ( Pfizer, Saúde Animal) e adicionamos
39,9 ml de óleo de milho filtrado, totalizando 40 ml.
3.0.6 Avaliação das ingestões de água e solução salina hipertônica (NaCl 1,8%)
Ratas OVX normovolêmicas e normohidratadas foram previamente adaptadas às gaiolas
metabólicas por cinco dias. Os animais receberam em suas gaiolas dois bebedouros volumétricos
individuais graduados em 0,1 ml, um contendo água e o outro, NaCl 1,8%, e comedouros
contendo ração moída ad libitum ,previamente pesada,até o final do experimento. A ingestão dos
fluidos foi avaliada diariamente, por 7 dias, em ciclos de 24 horas. Posterior a esta avaliação, foi
calculado o índice de preferência por sódio: (ingestão de NaCl 1,8% / ingestão de água + ingestão
de NaCl 1,8%) x 100 (FRANKMANN et al., 1986).
Para avaliação do efeito da terapia estrogênica no apetite e na saciedade ao sódio e à água
utilizamos o protocolo da furosemida que consiste num protocolo de depleção de volume e de
depleção corpórea de íons sódio através da administração do diurético de alça , furosemida (20
mg/Kg, s.c.) mais dieta pobre em sódio. Durante as 24 horas anteriores ao experimento os
animais tiveram acesso à água destilada e dieta pobre em NaCl e condicionados em gaiolas

34
metabólicas. No dia seguinte , após 24 horas de depleção de sódio foi realizada a reapresentação
dos fluidos colocando bebedouros individuais contendo NaCl 1,8% e água filtrada que foram
oferecidos aos animais depletados de sódio. Em seguida, a ingestão dos fluidos foi avaliada
cumulativamente por 300 minutos. Nos protocolos sob condições basais, depleção de sódio e de
reapresentação de fluidos foram utilizados os mesmos animais em sequência.
3.0.7 Avaliação do volume urinário e osmolalidade urinária e plasmática
O volume urinário foi avaliado diariamente sob condições basais a partir do primeiro dia
da terapia estrogênica. Após a depleção de sódio com furosemida o volume urinário foi avaliado
2 horas após a depleção de sódio, 24 horas após a depleção e 2 horas após início da avaliação de
ingestão de fluidos.
Após as coletas das amostras de urina realizadas em gaiolas metabólicas. Também foi
coletado o plasma no 8º dia do experimento 24 horas após depleção de sódio. A osmolalidade das
amostras de sangue e urina foi determinada por meio do osmômetro. Para uniformizar os
resultados, e para comparação entre médias dos diferentes grupos, os dados obtidos foram
expressos em mOsm/kg H2O/ para osmolalidade.
3.0.8 Avaliação do sódio plasmático
O conteúdo de sódio das amostras de plasma 24 horas após depleção de sódio foi
determinado por meio de fotometria de chama (Modelo Micronal B462). Para uniformizar os
resultados, e para comparação entre médias dos diferentes grupos, os dados obtidos foram
expressos em μEq/L para excreção de sódio. Os dados de sódio urinário diário, 24 horas após a
depleção de sódio e 2 horas após reapresentação de fluidos estão em análise. A diluição das
amostras de plasma foi 1:100 e das amostras de urina foi 1:200 num volume total de 3 ml em
cada leitura.

35
3.1 Avaliação do índice hipofisário e uterino
Após a eutanásia dos animais, as hipófises e os úteros foram coletados inteiros para
obtenção do peso úmido desses tecidos e assim realizar a determinação dos seus respectivos
índices absolutos para validação das cirurgias e reposições hormonais.
4.0 Protocolos experimentais
4.1 Experimento 1 – Efeito do tratamento com estradiol sobre o apetite ao sódio, sede,
índice uterino e hipofisário em ratas OVX controle e ratas OVX tratadas com estrógeno
Ratas submetidas à cirurgia de ovariectomia foram avaliadas diariamente em gaiolas
metabólicas quanto à ingestão de água e de salina hipertônicas que foram ofertadas aos animais
em buretas volumétricas individuais. Para avaliação da influência estrogênica nos
comportamentos dispsogênicos e natriorexigênicos sob condições basais, após depleção de sódio
com furosemida e após reapresentação de água e salina hipertônica (1,8%). No último dia de
reposição hormonal os animais foram submetidos ao protocolo de depleção de íons sódio e no dia
seguinte os mesmos animais passaram pelo protocolo de reapresentação dos fluidos para
avaliação das ingestões cumulativas. Ao final das avaliações os animais foram decapitados e
coletados os úteros e a hipófise para avaliação do índice uterino e hipofisário, respectivamente.
Essa avaliação permite verificar a eficácia dos tratamentos hormonais, sendo realizada pela
divisão do peso do órgão pelo peso do animal.

36
4.1.2 Desenho experimental
Experimento 1
Ovariectomia
___________________________________________________________________________
0 4d 7d 8d
Avaliações diárias
OVX+O / OVX+EC
Coleta de sangue
e encéfalos (Experimento 2)
Depleção de sódio Reapresentação dos fluidos
Avaliações cumulativas
_________________________________________________________________________
8d 9d 9d
2h 24h 0 120 180 240 300 min
Coleta do
Volume
Urinário Coleta do
Volume urinário
5.0 Análise estatística
Os resultados estão apresentados como médias ± erro padrão da média (EPM). Os dados
foram analisados usando o programa Graphpad Prism 6.0 (San Francisco, USA) e submetidos ao
teste de Grubbs que identifica pelo menos um outlier. Posteriormente foram analisados pelo teste
de normalidade Shapiro-Wilk. Após verificar que os dados apresentavam distribuição normal foi
realizado o teste ANOVA de medidas repetidas, ANOVA two way ou ANOVA one-way, seguido
de pos-teste de Bonferroni. O nível de significância adotado foi de 5% (α = 5%). Consideraremos
o efeito dose dependência sempre que houver diferença do grupo tratado comparado ao controle.
37
6.0 Resultados e discussão
6.1 Avaliação da influência da ovariectomia e reposição hormonal sobre o peso corporal
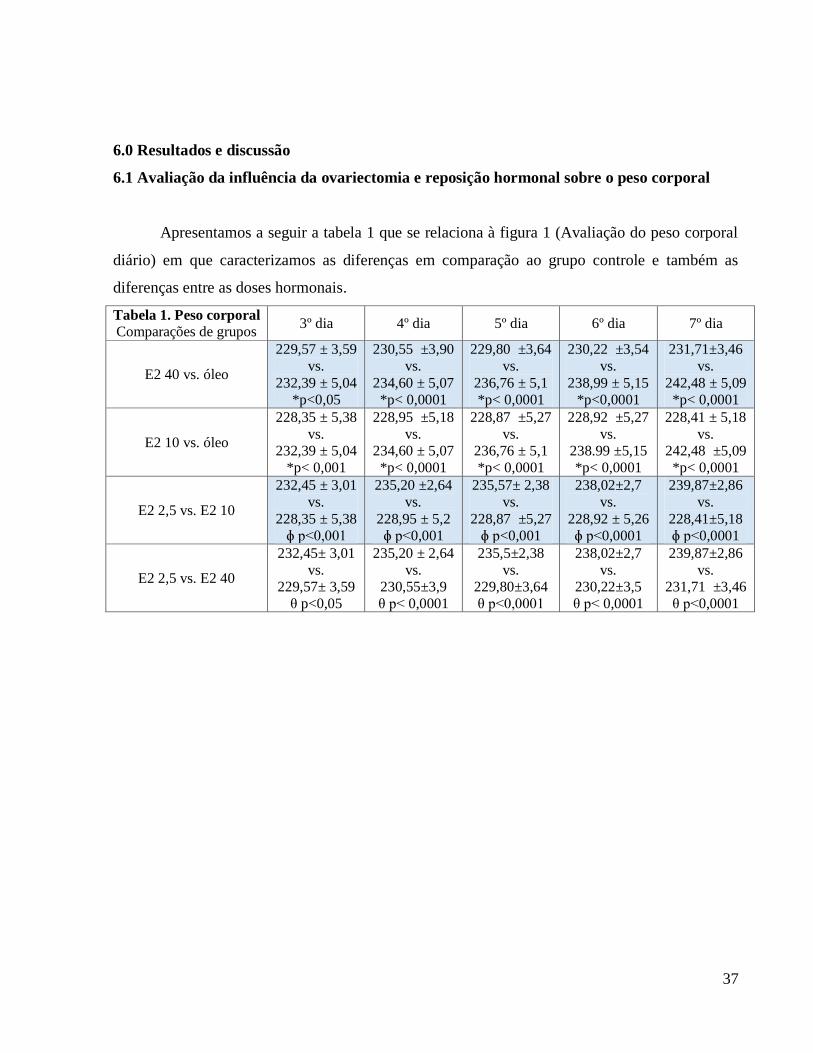
Apresentamos a seguir a tabela 1 que se relaciona à figura 1 (Avaliação do peso corporal
diário) em que caracterizamos as diferenças em comparação ao grupo controle e também as
diferenças entre as doses hormonais.
Tabela 1. Peso corporal
Comparações de grupos 3º dia 4º dia 5º dia 6º dia 7º dia
E2 40 vs. óleo
229,57 ± 3,59
vs.
232,39 ± 5,04
*p<0,05
230,55 ±3,90
vs.
234,60 ± 5,07
*p< 0,0001
229,80 ±3,64
vs.
236,76 ± 5,1
*p< 0,0001
230,22 ±3,54
vs.
238,99 ± 5,15
*p<0,0001
231,71±3,46
vs.
242,48 ± 5,09
*p< 0,0001
E2 10 vs. óleo
228,35 ± 5,38
vs.
232,39 ± 5,04
*p< 0,001
228,95 ±5,18
vs.
234,60 ± 5,07
*p< 0,0001
228,87 ±5,27
vs.
236,76 ± 5,1
*p< 0,0001
228,92 ±5,27
vs.
238.99 ±5,15
*p< 0,0001
228,41 ± 5,18
vs.
242,48 ±5,09
*p< 0,0001
E2 2,5 vs. E2 10
232,45 ± 3,01
vs.
228,35 ± 5,38
ɸ p<0,001
235,20 ±2,64
vs.
228,95 ± 5,2
ɸ p<0,001
235,57± 2,38
vs.
228,87 ±5,27
ɸ p<0,001
238,02±2,7
vs.
228,92 ± 5,26
ɸ p<0,0001
239,87±2,86
vs.
228,41±5,18
ɸ p<0,0001
E2 2,5 vs. E2 40
232,45± 3,01
vs.
229,57± 3,59
θ p<0,05
235,20 ± 2,64
vs.
230,55±3,9
θ p< 0,0001
235,5±2,38
vs.
229,80±3,64
θ p<0,0001
238,02±2,7
vs.
230,22±3,5
θ p< 0,0001
239,87±2,86
vs.
231,71 ±3,46
θ p<0,0001
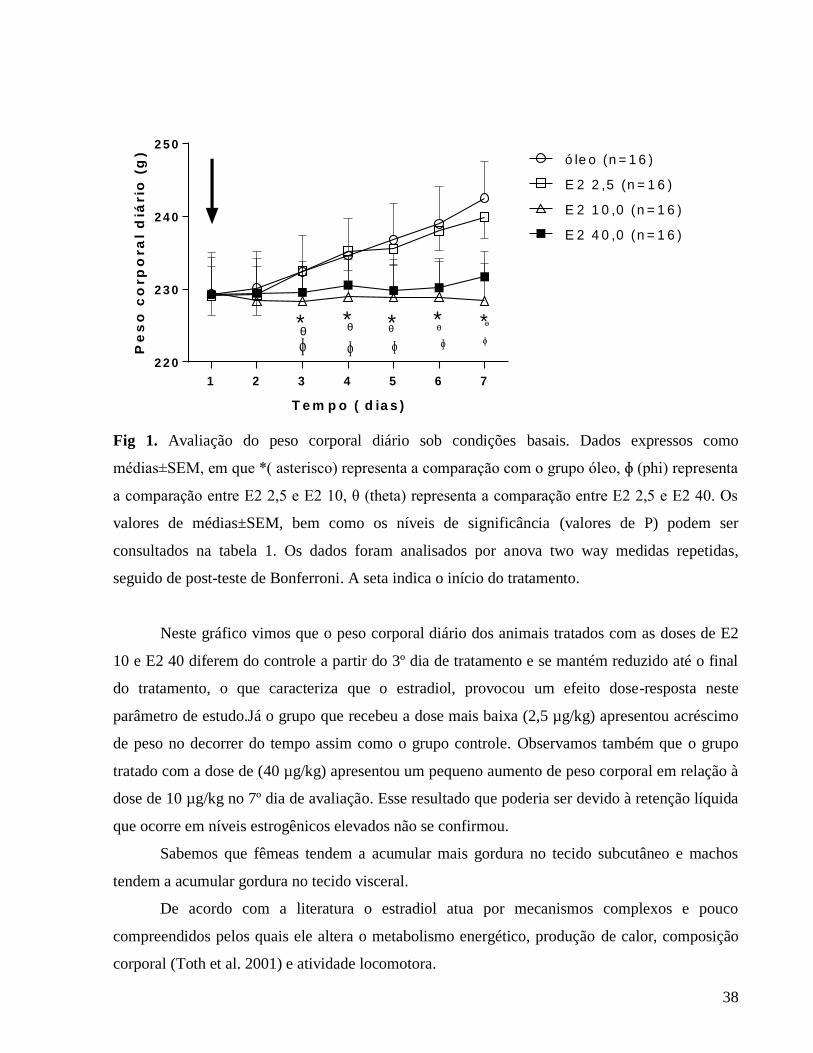
38
T e m p o ( d ia s )
Pe
so
co
rp
ora
l d
iário
(g
)
1 2 3 4 5 6 7
2 2 0
2 3 0
2 4 0
2 5 0
ó le o (n = 1 6 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 6 )
* * * * *ɸ ɸ ɸ ɸ ɸ θ θ θ θ
θ
Fig 1. Avaliação do peso corporal diário sob condições basais. Dados expressos como
médias±SEM, em que *( asterisco) representa a comparação com o grupo óleo, ɸ (phi) representa
a comparação entre E2 2,5 e E2 10, θ (theta) representa a comparação entre E2 2,5 e E2 40. Os
valores de médias±SEM, bem como os níveis de significância (valores de P) podem ser
consultados na tabela 1. Os dados foram analisados por anova two way medidas repetidas,
seguido de post-teste de Bonferroni. A seta indica o início do tratamento.
Neste gráfico vimos que o peso corporal diário dos animais tratados com as doses de E2
10 e E2 40 diferem do controle a partir do 3º dia de tratamento e se mantém reduzido até o final
do tratamento, o que caracteriza que o estradiol, provocou um efeito dose-resposta neste
parâmetro de estudo.Já o grupo que recebeu a dose mais baixa (2,5 µg/kg) apresentou acréscimo
de peso no decorrer do tempo assim como o grupo controle. Observamos também que o grupo
tratado com a dose de (40 µg/kg) apresentou um pequeno aumento de peso corporal em relação à
dose de 10 µg/kg no 7º dia de avaliação. Esse resultado que poderia ser devido à retenção líquida
que ocorre em níveis estrogênicos elevados não se confirmou.
Sabemos que fêmeas tendem a acumular mais gordura no tecido subcutâneo e machos
tendem a acumular gordura no tecido visceral.
De acordo com a literatura o estradiol atua por mecanismos complexos e pouco
compreendidos pelos quais ele altera o metabolismo energético, produção de calor, composição
corporal (Toth et al. 2001) e atividade locomotora.

39
O estrógeno influencia o gasto energético modulando a temperatura corporal e a
termogênese. Além disso pode atuar sobre a mobilização de gordura, modificando também o
perfil lipídico (níveis de colesterol e de triglicerídeos), além de aumentar o estresse oxidativo
mitocondrial.
Gao et al. (2008) mencionarm que o estrogênio e a leptina podem interagir via estatina 3
(STAT 3), regulando genes em comum em regiões hipotalâmicas relacionadas a homeostase
energética e reprodução.
Assim, a perda de peso promovida pelos hormônios gonadais é elaborada por um amplo
conjunto de fatores envolvendo queda na expressão de neuropeptídios orexigênicos e aumento
dos anorexigênicos (SILVA et al. 2011).Além disso Donner & Handa (2009) evidenciaram que
aumento da TPH2 e consequentemente da síntese de serotonina é mediada pelo receptor
estrogênico beta (Erβ). A ativação desse sistema também é um estímulo para saciedade. E
também o aumento do consumo de oxigênio (GAO et al. 2007) e do estresse oxidativo (PUERTA
et al. 1998), aumento da mobilização de gordura e um dos mecanismos seria por β-oxidação.
Todos esses fatores somados resultam em saciedade e perda de peso.
6.2 Avaliação do efeito da ovariectomia e da reposição hormonal sobre o peso uterino e o
peso hipofisário
A seguir apresentamos a tabela 2 que se relaciona com a figura 2 (avaliação do peso
uterino). Nesta tabela inserimos as diferenças em relação ao grupo controle, caracterizando assim
um fenômeno de dose-dependência.
Tabela 2: Peso uterino
E2 40 vs. óleo 0,69 ± 0,19 vs. 0,22 ± 0,04g,
*p<0,0001
E2 10 vs. óleo
0,69 ± 0,15 vs. 0,21 ± 0,04 g,
*p<0,0001
E2 2,5 vs. óleo 0,61 ± 0,1 vs. 0,21 ± 0,04 g ,
*p<0,001
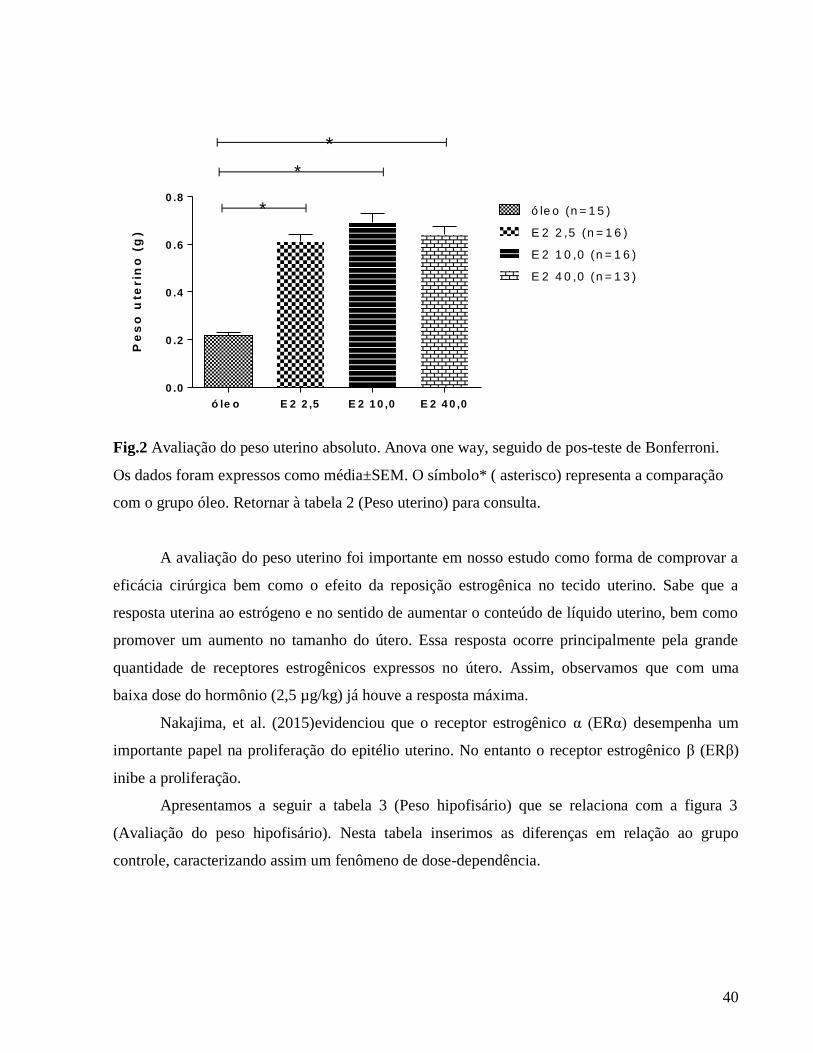
40
Pe
so
ute
rin
o (
g)
ó le o E 2 2 ,5 E 2 1 0 ,0 E 2 4 0 ,0
0 .0
0 .2
0 .4
0 .6
0 .8
ó le o (n = 1 5 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 3 )
*
*
*
Fig.2 Avaliação do peso uterino absoluto. Anova one way, seguido de pos-teste de Bonferroni.
Os dados foram expressos como média±SEM. O símbolo* ( asterisco) representa a comparação
com o grupo óleo. Retornar à tabela 2 (Peso uterino) para consulta.
A avaliação do peso uterino foi importante em nosso estudo como forma de comprovar a
eficácia cirúrgica bem como o efeito da reposição estrogênica no tecido uterino. Sabe que a
resposta uterina ao estrógeno e no sentido de aumentar o conteúdo de líquido uterino, bem como
promover um aumento no tamanho do útero. Essa resposta ocorre principalmente pela grande
quantidade de receptores estrogênicos expressos no útero. Assim, observamos que com uma
baixa dose do hormônio (2,5 µg/kg) já houve a resposta máxima.
Nakajima, et al. (2015)evidenciou que o receptor estrogênico α (ERα) desempenha um
importante papel na proliferação do epitélio uterino. No entanto o receptor estrogênico β (ERβ)
inibe a proliferação.
Apresentamos a seguir a tabela 3 (Peso hipofisário) que se relaciona com a figura 3
(Avaliação do peso hipofisário). Nesta tabela inserimos as diferenças em relação ao grupo
controle, caracterizando assim um fenômeno de dose-dependência.
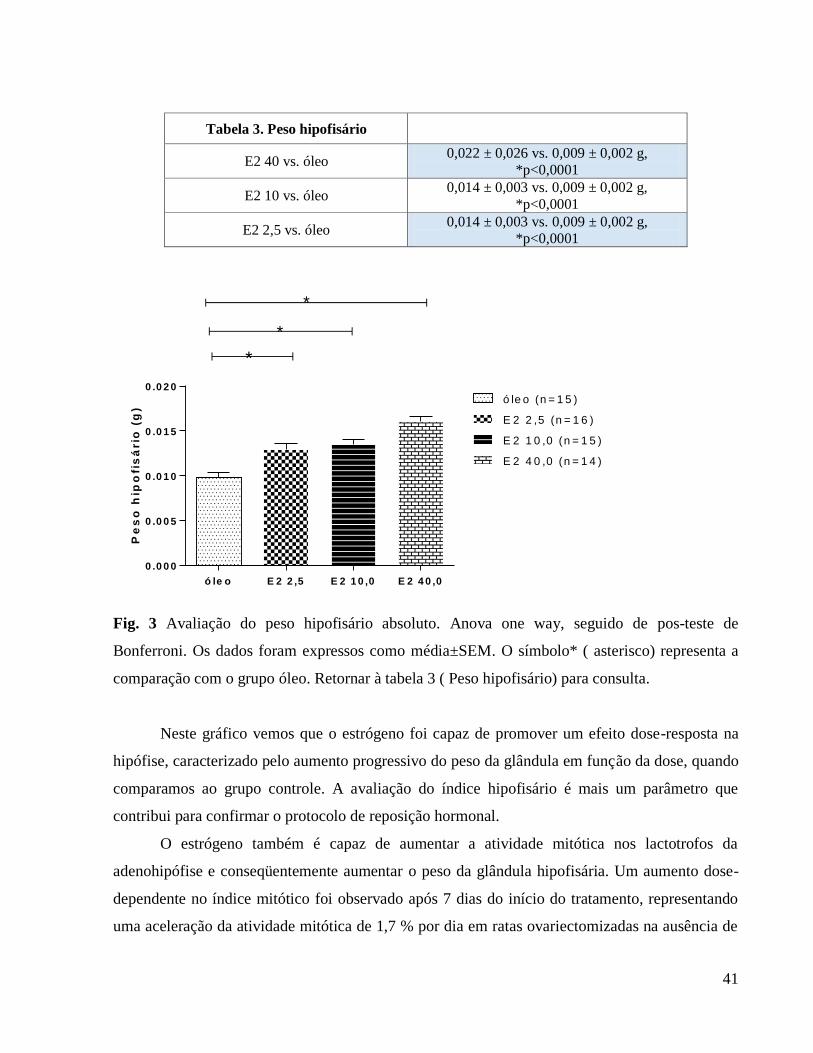
41
Tabela 3. Peso hipofisário
E2 40 vs. óleo 0,022 ± 0,026 vs. 0,009 ± 0,002 g,
*p<0,0001
E2 10 vs. óleo 0,014 ± 0,003 vs. 0,009 ± 0,002 g,
*p<0,0001
E2 2,5 vs. óleo 0,014 ± 0,003 vs. 0,009 ± 0,002 g,
*p<0,0001
Pe
so
hip
ofi
sá
rio
(g
)
ó le o E 2 2 ,5 E 2 1 0 ,0 E 2 4 0 ,0
0 .0 0 0
0 .0 0 5
0 .0 1 0
0 .0 1 5
0 .0 2 0
ó le o (n = 1 5 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 5 )
E 2 4 0 ,0 (n = 1 4 )
*
*
*
Fig. 3 Avaliação do peso hipofisário absoluto. Anova one way, seguido de pos-teste de
Bonferroni. Os dados foram expressos como média±SEM. O símbolo* ( asterisco) representa a
comparação com o grupo óleo. Retornar à tabela 3 ( Peso hipofisário) para consulta.
Neste gráfico vemos que o estrógeno foi capaz de promover um efeito dose-resposta na
hipófise, caracterizado pelo aumento progressivo do peso da glândula em função da dose, quando
comparamos ao grupo controle. A avaliação do índice hipofisário é mais um parâmetro que
contribui para confirmar o protocolo de reposição hormonal.
O estrógeno também é capaz de aumentar a atividade mitótica nos lactotrofos da
adenohipófise e conseqüentemente aumentar o peso da glândula hipofisária. Um aumento dose-
dependente no índice mitótico foi observado após 7 dias do início do tratamento, representando
uma aceleração da atividade mitótica de 1,7 % por dia em ratas ovariectomizadas na ausência de

42
reposição estrogênica e 3,7% por dia na presença de uma dose farmacológica de 50 µg/rata/dia ou
230 µg/kg/ dia, sendo esta uma dose suprafisiológica (LONAN & LEVY, 2009).
6.3 Avaliação do comportamento ingestivo de água e salina hipertônica sob condições basais
em função da dose hormonal administrada
Não foi observada dose dependência na avaliação de ingestão de água em relação ao
controle,logo não há representação de dados significativos em relação ao controle, no entanto
observamos diferença entre doses de estradiol que pode ser observada a seguir na tabela 4.
Tabela 4. Ingestão basal de água
Comparações de grupos 2º dia 3º dia
E2 10 vs. E2 40
8,27±0,59
vs.
6,52 ± 0,56
Ψ p<0,01
9,96 ±0,54
vs.
8,31 ±0,45
Ψ p< 0,01
E2 2,5 vs. E2 40
7,88± 0,52
v.s
.6,52 ±0,56 ,θ p<0,05
_______
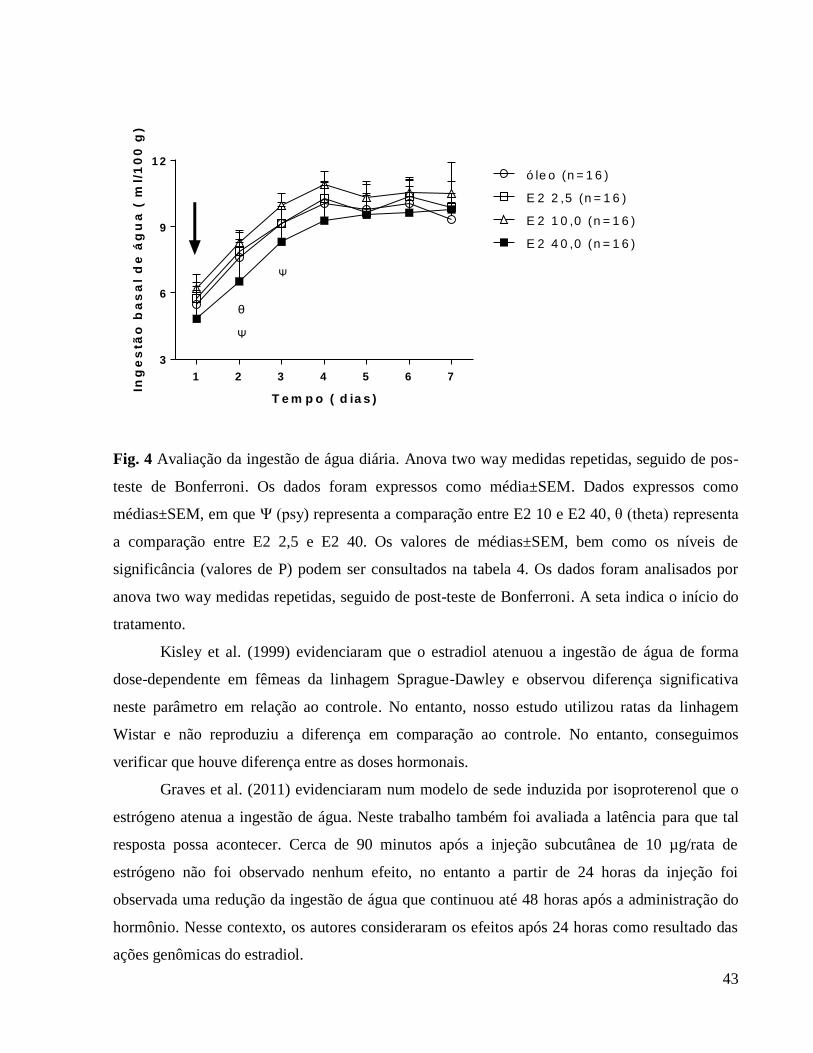
43
T e m p o ( d ia s )
Ing
es
tão
ba
sa
l d
e á
gu
a (
ml/
10
0 g
)
1 2 3 4 5 6 7
3
6
9
1 2
ó le o (n = 1 6 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 6 )
θ
Ψ
Ψ
Fig. 4 Avaliação da ingestão de água diária. Anova two way medidas repetidas, seguido de pos-
teste de Bonferroni. Os dados foram expressos como média±SEM. Dados expressos como
médias±SEM, em que Ψ (psy) representa a comparação entre E2 10 e E2 40, θ (theta) representa
a comparação entre E2 2,5 e E2 40. Os valores de médias±SEM, bem como os níveis de
significância (valores de P) podem ser consultados na tabela 4. Os dados foram analisados por
anova two way medidas repetidas, seguido de post-teste de Bonferroni. A seta indica o início do
tratamento.
Kisley et al. (1999) evidenciaram que o estradiol atenuou a ingestão de água de forma
dose-dependente em fêmeas da linhagem Sprague-Dawley e observou diferença significativa
neste parâmetro em relação ao controle. No entanto, nosso estudo utilizou ratas da linhagem
Wistar e não reproduziu a diferença em comparação ao controle. No entanto, conseguimos
verificar que houve diferença entre as doses hormonais.
Graves et al. (2011) evidenciaram num modelo de sede induzida por isoproterenol que o
estrógeno atenua a ingestão de água. Neste trabalho também foi avaliada a latência para que tal
resposta possa acontecer. Cerca de 90 minutos após a injeção subcutânea de 10 µg/rata de
estrógeno não foi observado nenhum efeito, no entanto a partir de 24 horas da injeção foi
observada uma redução da ingestão de água que continuou até 48 horas após a administração do
hormônio. Nesse contexto, os autores consideraram os efeitos após 24 horas como resultado das
ações genômicas do estradiol.

44
Findlay et al. (1979) realizaram um estudo em ratas ovariectomizadas e tratadas com 20
µg de benzoato de estradiol no modelo de sede induzida por isoprenalina e também pelo aumento
da hipertonicidade com injeção subcutânea de salina hipertônica (2%). Neste trabalho foi
demonstrado que a sede de origem extracelular ativa o sistema renina-angiotensina-aldosterona
que responde a diferentes mecanismos induzidos por desidratação. Os autores evidenciaram que
somente o comportamento ingestivo causado pela ativação de mecanismos extracelulares parece
ser mais sensível às variações do ciclo ovariano e aos hormônios ovarianos.
No entanto, em condições basais, embora sem alterações significativas, observamos que a
dose intermediária (10 µg/kg) apresentou um aumento não significativo na ingestão de água
durante todo o período de avaliação. Já o grupo que recebeu a dose alta (40 µg/kg) apresentou um
aumento não significativo. Desse modo, podemos supor que em níveis estrogênicos altos poderia
haver downregulation dos receptores AT1, o que impossibilitaria os efeitos dipsogênicos da
angiotensina II.
Em relação à ingestão basal de salina hipertônica não observamos efeito dose-resposta
neste parâmetro, mas no grupo que recebeu a dose de 10 µg/kg verificamos que houve uma
redução não significativa o consumo de salina. Esse dado já e bem conhecido na literatura.
A seguir apresentamos a tabela 5 que se refere à figura 5 (Ingestão basal de salina
hipertônica). Não observamos dose dependência em relação ao grupo controle, mas houve
diferença quanto à ingestão de salina entre as doses do hormônio.
Tabela 5. Ingestão basal de salina (1,8%)
Comparações de grupos 2º dia 4º dia 5º dia
E2 10 vs. E2 40
1,29 ± 0,16 vs.
3,01±0,42
Ψ p<0,01
1,35±0,33
vs.
3,7±0,65
Ψ p<0,0001
1,29 ±0,31
vs.
2,79 ± 0,57
Ψ p< 0,05
E2 2,5 vs. E2 10
___________
2,9 ± 0,58
v.s
1,35 ± 0,3
ɸ p<0,05
_________
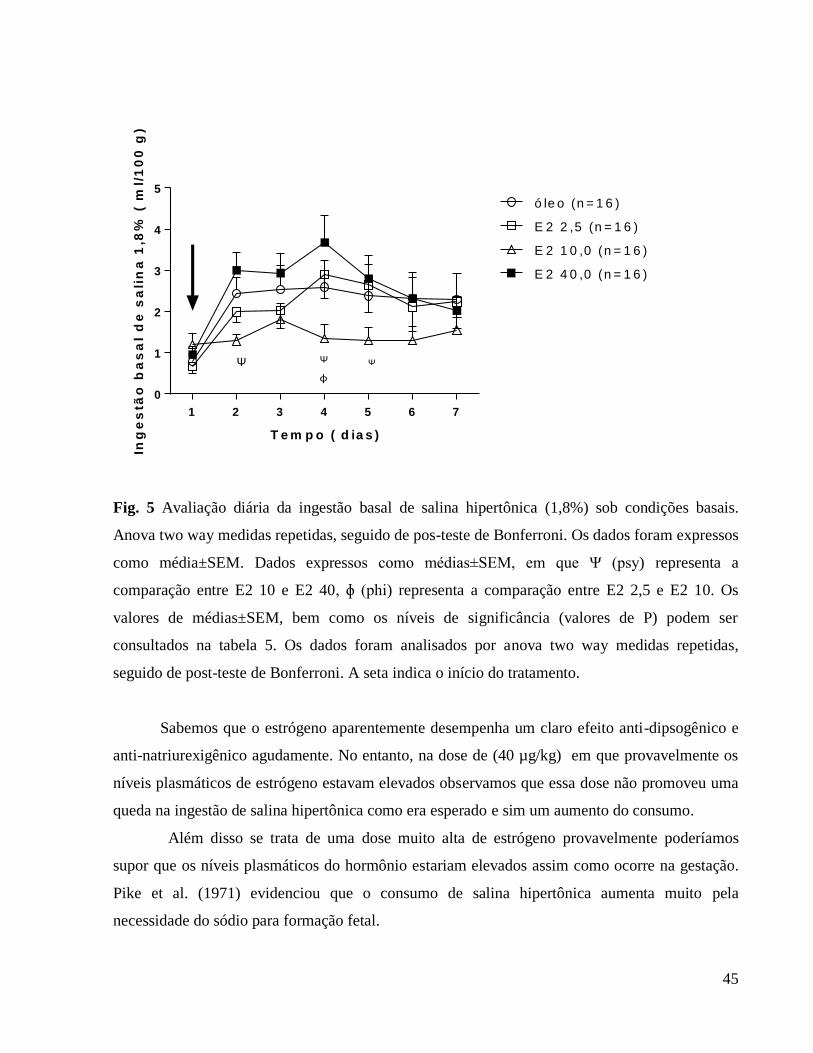
45
T e m p o ( d ia s )
Ing
es
tão
ba
sa
l d
e s
ali
na
1,8
% (
ml/
10
0 g
)
1 2 3 4 5 6 7
0
1
2
3
4
5
ó le o (n = 1 6 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 6 )
Ψ Ψ Ψ
ɸ
Fig. 5 Avaliação diária da ingestão basal de salina hipertônica (1,8%) sob condições basais.
Anova two way medidas repetidas, seguido de pos-teste de Bonferroni. Os dados foram expressos
como média±SEM. Dados expressos como médias±SEM, em que Ψ (psy) representa a
comparação entre E2 10 e E2 40, ɸ (phi) representa a comparação entre E2 2,5 e E2 10. Os
valores de médias±SEM, bem como os níveis de significância (valores de P) podem ser
consultados na tabela 5. Os dados foram analisados por anova two way medidas repetidas,
seguido de post-teste de Bonferroni. A seta indica o início do tratamento.
Sabemos que o estrógeno aparentemente desempenha um claro efeito anti-dipsogênico e
anti-natriurexigênico agudamente. No entanto, na dose de (40 µg/kg) em que provavelmente os
níveis plasmáticos de estrógeno estavam elevados observamos que essa dose não promoveu uma
queda na ingestão de salina hipertônica como era esperado e sim um aumento do consumo.
Além disso se trata de uma dose muito alta de estrógeno provavelmente poderíamos
supor que os níveis plasmáticos do hormônio estariam elevados assim como ocorre na gestação.
Pike et al. (1971) evidenciou que o consumo de salina hipertônica aumenta muito pela
necessidade do sódio para formação fetal.

46
Sabemos que o núcleo dorsal da rafe possui um importante papel no controle do apetite ao
sódio (Franchini et al. 2002; Badaue-Passos et al. 2007; Reis, 2007) e também há dados da
literatura que suportam a possibilidade de o estrogênio dessensibilizar receptores 5-HT1A no
núcleo paraventricular (Creech et al. 2012). Não sabemos se essa dessensibilização também
poderia estar ocorrendo em neurônios do NDR.
Além disso, estudos translacionais em mulheres demosntraram que a hipertensão é duas
vezes mais prevalente em mulheres na pós-menopausa e que essa prevalência é reduzida pela
terapia hormonal (SCUTERI, et al. 2001).
Em nosso estudo observamos que níveis estrogênicos altos (40 µg/kg) levam ao aumento
no consumo de salina hipertônica e não em sua redução. Com isso, podemos supor que a médio e
longo prazo o uso de terapias de reposição hormonal pode ser um fator predisponente à
hipertensão. Assim como o uso prolongado de contraceptivos poderia levar ao mesmo efeito.
O mecanismo pelo qual o estrógeno aumenta a ingestão espontânea de salina hipertônica
pode estar associado à vasodilatação promovida pelo estrógeno. O estrogênio pode influenciar a
atividade de células endoteliais via mecanismos genômicos e não genômicos. Ao se ligar ao ERα,
estrogênios aumentam a produção de agentes vasodilatadores como óxido nítrico, prostaciclinas e
prostaglandina E2 (DARBLADE et al. 2002; VEGETO et al. 2003). Desse modo, em doses altas
do hormônio haveria uma queda na pressão arterial e como mecanismo compensatório os animais
aumentariam o consumo de salina hipertônica.
A seguir apresentamos a Tabela 6 ( ingestão de alimento) que se relaciona com a figura 6
(Avaliação da ingestão alimentar).Caracterizamos a seguir as diferenças em relação ao grupo
controle, o que evidenciou um efeito de dose-dependência neste parâmetro.
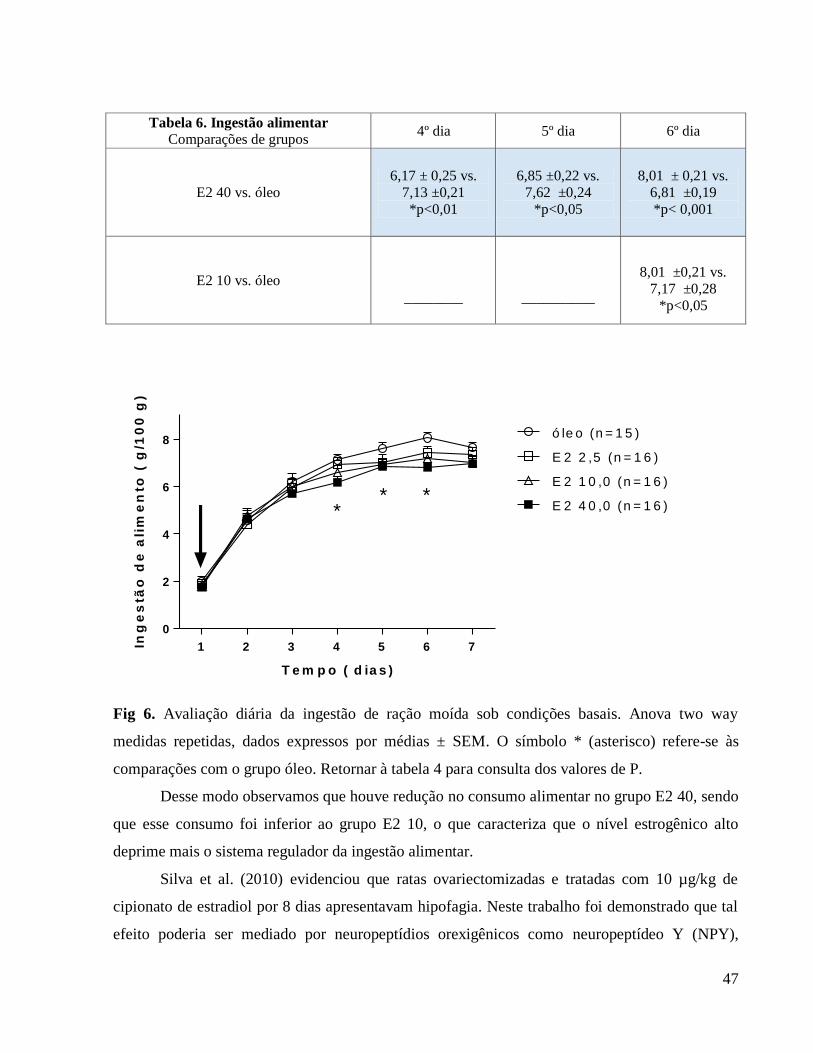
47
Tabela 6. Ingestão alimentar
Comparações de grupos 4º dia 5º dia 6º dia
E2 40 vs. óleo
6,17 ± 0,25 vs.
7,13 ±0,21
*p<0,01
6,85 ±0,22 vs.
7,62 ±0,24
*p<0,05
8,01 ± 0,21 vs.
6,81 ±0,19
*p< 0,001
E2 10 vs. óleo
________
__________
8,01 ±0,21 vs.
7,17 ±0,28
*p<0,05
T e m p o ( d ia s )
Ing
es
tão
de
ali
me
nto
( g
/10
0 g
)
1 2 3 4 5 6 7
0
2
4
6
8ó le o (n = 1 5 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 6 )**
*
Fig 6. Avaliação diária da ingestão de ração moída sob condições basais. Anova two way
medidas repetidas, dados expressos por médias ± SEM. O símbolo * (asterisco) refere-se às
comparações com o grupo óleo. Retornar à tabela 4 para consulta dos valores de P.
Desse modo observamos que houve redução no consumo alimentar no grupo E2 40, sendo
que esse consumo foi inferior ao grupo E2 10, o que caracteriza que o nível estrogênico alto
deprime mais o sistema regulador da ingestão alimentar.
Silva et al. (2010) evidenciou que ratas ovariectomizadas e tratadas com 10 µg/kg de
cipionato de estradiol por 8 dias apresentavam hipofagia. Neste trabalho foi demonstrado que tal
efeito poderia ser mediado por neuropeptídios orexigênicos como neuropeptídeo Y (NPY),

48
peptídeo agouti relativo (AgRP) e orexina A, cuja expressão estava reduzida e pela expressão
aumentada de mediadores anorexigênicos como melanocortina 4 (MC4), leptina e CRH.
De acordo com a literatura fêmeas em estro são mais sensíveis do que machos ou do que
fêmeas intactas em diestro ao efeito anorexigênico de agonistas serotoninérgicos como a
fenfluramina (RIVERA, et al. 2005). Além disso, a administração exógena de E2 aumenta o
efeito anorexigênico em ratas OVX (ECKEL, et al. 2005), sugerindo que os efeitos
anorexigênicos da fenfluramina relacionados ao estro também podem ser mediados pelo
estrógeno endógeno. Além disso, o estrógeno aumenta a expressão de Pet-1 e de transportadores
de serotonina , que poderiam estar implicados na regulação de neurônios serotoninérgicos no
DRN durante o mesmo período em que o E2 diminui a ingestão de alimento e o peso corporal em
ratas OVX (RIVERA, et al. 2005). O sistema serotoninérgico também participa do controle do
comportamento alimentar e pode ser influenciado pelo estradiol. Rivera et al. (2012)
demonstraram que receptores serotoninérgicos tipo 2C (5-HT2C) pós-sinápticos desempenham
importante papel em mediar os efeitos anorexigênicos do estradiol no sistema serotoninérgico.
Dados prévios da literatura demonstraram que E2 modula neuropeptídios relativos ao
controle da ingestão alimentar nas regiões cerebrais supra-citadas, incluindo NPY, fator de
liberação de corticotrofina (CRF), POMC e neurotransmissores como a serotonina (RIVERA, et
al. 2005; ECKEL, et al. 2005). De fato, receptores para estrógeno e leptina estão co-localizados
em regiões que controlam o metabolismo e a função gonadal como núcleo arqueado (ARC),
hipotálamo ventromedial (HVM), núcleo pré-óptico anterior (POA).
Outro estudo documentou que o estrógeno não apresenta efeito nos níveis de ghrelina
circulantes em mulheres que estejam normalmente ciclando ou em menopausa (DAFOPOULOS
et al. 2010).
6.4 Avaliação da reposição hormonal sobre o volume urinário e a osmolalidade urinária sob
condições basais
A seguir apresentamos a tabela 7 que se relaciona com a figura 7 ( Avaliação do volume
urinário). Demonstramos nesta tabela que houve dose-dependência em relação ao volume
urinário.
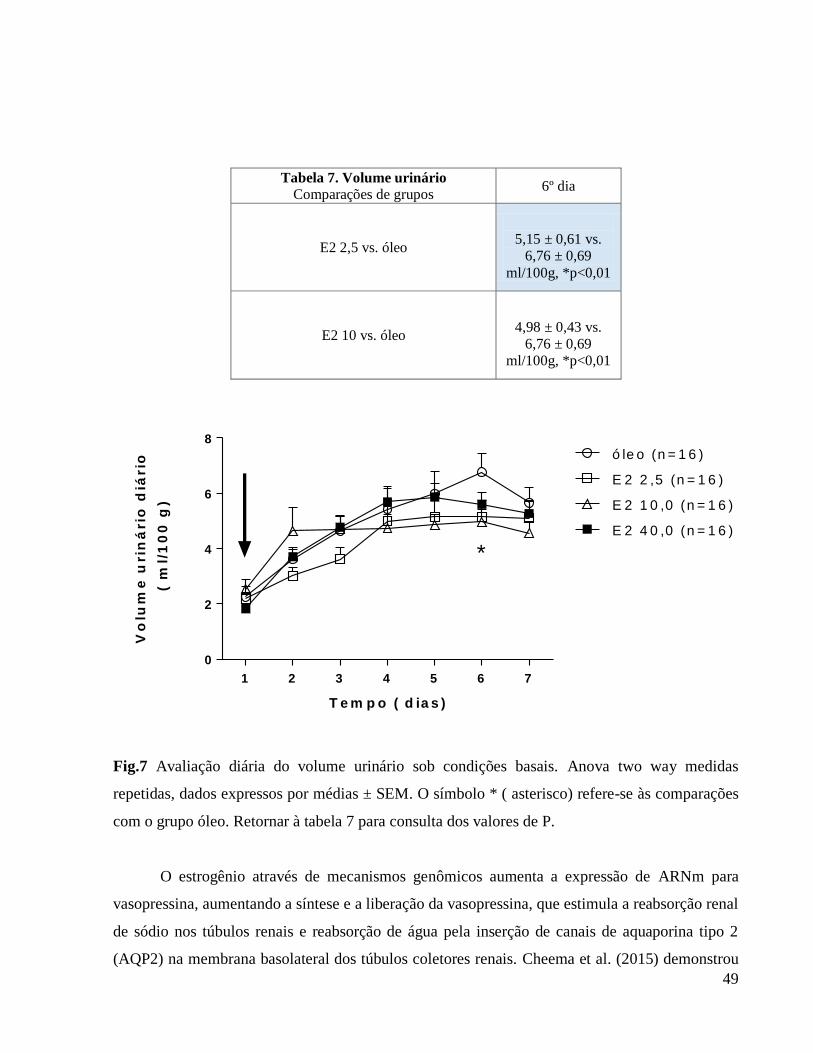
49
Tabela 7. Volume urinário
Comparações de grupos 6º dia
E2 2,5 vs. óleo
5,15 ± 0,61 vs.
6,76 ± 0,69
ml/100g, *p<0,01
E2 10 vs. óleo
4,98 ± 0,43 vs.
6,76 ± 0,69
ml/100g, *p<0,01
T e m p o ( d ia s )
Vo
lum
e u
rin
ário
diá
rio
( m
l/1
00
g)
1 2 3 4 5 6 7
0
2
4
6
8
ó le o (n = 1 6 )
E 2 2 ,5 (n = 1 6 )
E 2 1 0 ,0 (n = 1 6 )
E 2 4 0 ,0 (n = 1 6 )
*
Fig.7 Avaliação diária do volume urinário sob condições basais. Anova two way medidas
repetidas, dados expressos por médias ± SEM. O símbolo * ( asterisco) refere-se às comparações
com o grupo óleo. Retornar à tabela 7 para consulta dos valores de P.
O estrogênio através de mecanismos genômicos aumenta a expressão de ARNm para
vasopressina, aumentando a síntese e a liberação da vasopressina, que estimula a reabsorção renal
de sódio nos túbulos renais e reabsorção de água pela inserção de canais de aquaporina tipo 2
(AQP2) na membrana basolateral dos túbulos coletores renais. Cheema et al. (2015) demonstrou
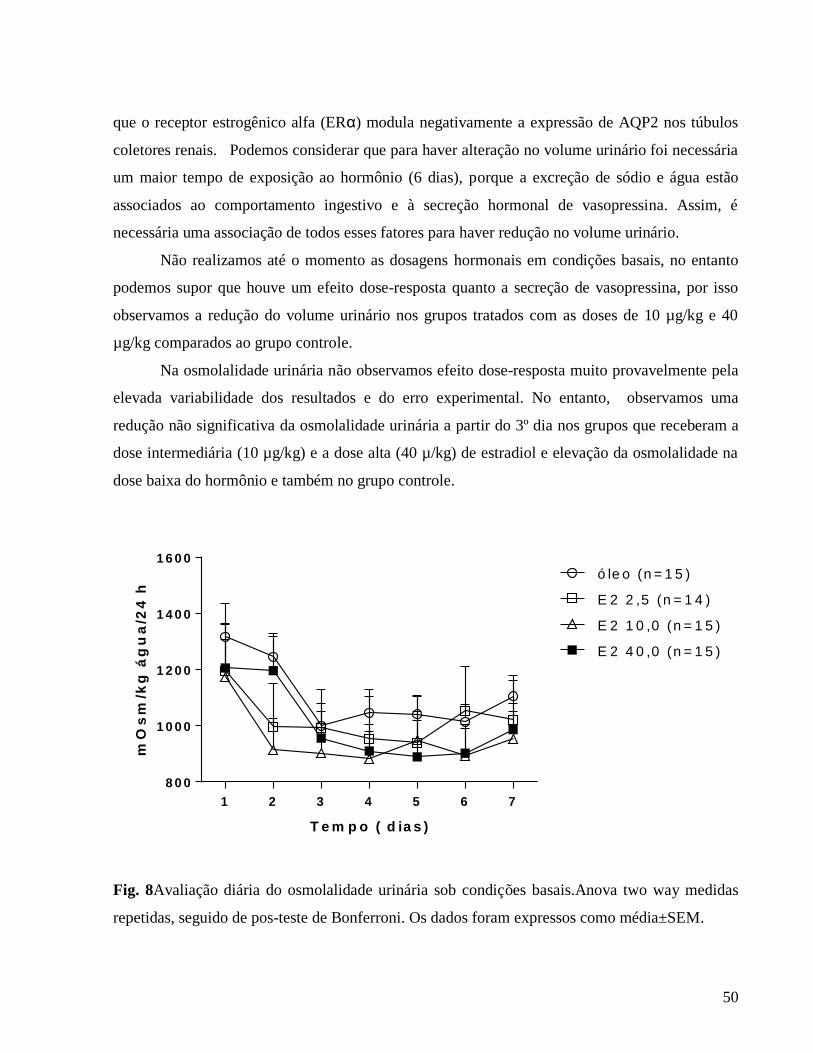
50
que o receptor estrogênico alfa (ERα) modula negativamente a expressão de AQP2 nos túbulos
coletores renais. Podemos considerar que para haver alteração no volume urinário foi necessária
um maior tempo de exposição ao hormônio (6 dias), porque a excreção de sódio e água estão
associados ao comportamento ingestivo e à secreção hormonal de vasopressina. Assim, é
necessária uma associação de todos esses fatores para haver redução no volume urinário.
Não realizamos até o momento as dosagens hormonais em condições basais, no entanto
podemos supor que houve um efeito dose-resposta quanto a secreção de vasopressina, por isso
observamos a redução do volume urinário nos grupos tratados com as doses de 10 µg/kg e 40
µg/kg comparados ao grupo controle.
Na osmolalidade urinária não observamos efeito dose-resposta muito provavelmente pela
elevada variabilidade dos resultados e do erro experimental. No entanto, observamos uma
redução não significativa da osmolalidade urinária a partir do 3º dia nos grupos que receberam a
dose intermediária (10 µg/kg) e a dose alta (40 µ/kg) de estradiol e elevação da osmolalidade na
dose baixa do hormônio e também no grupo controle.
T e m p o ( d ia s )
mO
sm
/kg
ág
ua
/24
h
1 2 3 4 5 6 7
8 0 0
1 0 0 0
1 2 0 0
1 4 0 0
1 6 0 0
ó le o (n = 1 5 )
E 2 2 ,5 (n = 1 4 )
E 2 1 0 ,0 (n = 1 5 )
E 2 4 0 ,0 (n = 1 5 )
Fig. 8Avaliação diária do osmolalidade urinária sob condições basais.Anova two way medidas
repetidas, seguido de pos-teste de Bonferroni. Os dados foram expressos como média±SEM.

51
O estradiol poderia ativar mecanismos que ajudam na retenção de sódio quando os níveis
estrogênicos estão altos e por isso observamos a redução da osmolalidade urinária.
É conhecido que o aumento de volume extracelular na mulher ocorre durante a fase pré-
ovulatória do ciclo menstrual quando os níveis estrogênicos estão aumentando (STEPHENSON
et al. 1988). Além disso, a retenção de água e sódio ocorre durante a gestação (STACHENFELD
et al. 2005) e em mulheres na pós-menopausa que recebem estrogênio (DIGNAM, et al. 1956). A
dose mais alta de estrógeno (40 µ/kg) provavelmente favoreceu ainda mais a retenção de sódio
por ser uma dose que se assemelha os níveis estrogênicos durante a fase final da gestação. Esses
achados poderiam estar associados com ações do estrógeno e da progesterona na função
renal,levando a retenção de água e sódio (STACHENFELD et al. 2005). No entanto, nosso
estudo não avaliou o efeito da reposição com progesterona em nos animais testados. O estradiol
estimula o aumento da expressão de ARNm do ENaC no rim de ratas (GAMBLING, et al. 2004),
principalmente nos túbulos proximais e distais (BRUNETTE, et al. 2001), favorecendo a retenção
de íons sódio.
Um outro possível mecanismo é a ação estrogênica sobre o sistema nervoso autônomo
simpático, já que a inervação simpática renal contribui para a retenção de sódio. O estrógeno
exerce efeitos diretos na inervação simpática renal de acordo com GRACELLI et al. 2013 ).
Neste estudo foi utilizado o modelo desnervação renal em ratas castradas com reposição de
estrogênio (2,0 mg/kg) ou progesterona. Foi observado que a reposição estrogênica normalizou
os níveis plasmáticos de catecolaminas após a desnervação renal.
6.5 Avaliação da influência da reposição hormonal sobre o volume urinário após a depleção
de sódio
Avaliamos também o volume urinário após a depleção de sódio com furosemida para
confirmação da eficácia do modelo. Assim observamos nas duas primeiras horas um aumento da
diurese, como era esperado. No entanto, não foi observado efeito em função da dose do hormônio
estrogênico mas somente em função do tempo de avaliação.
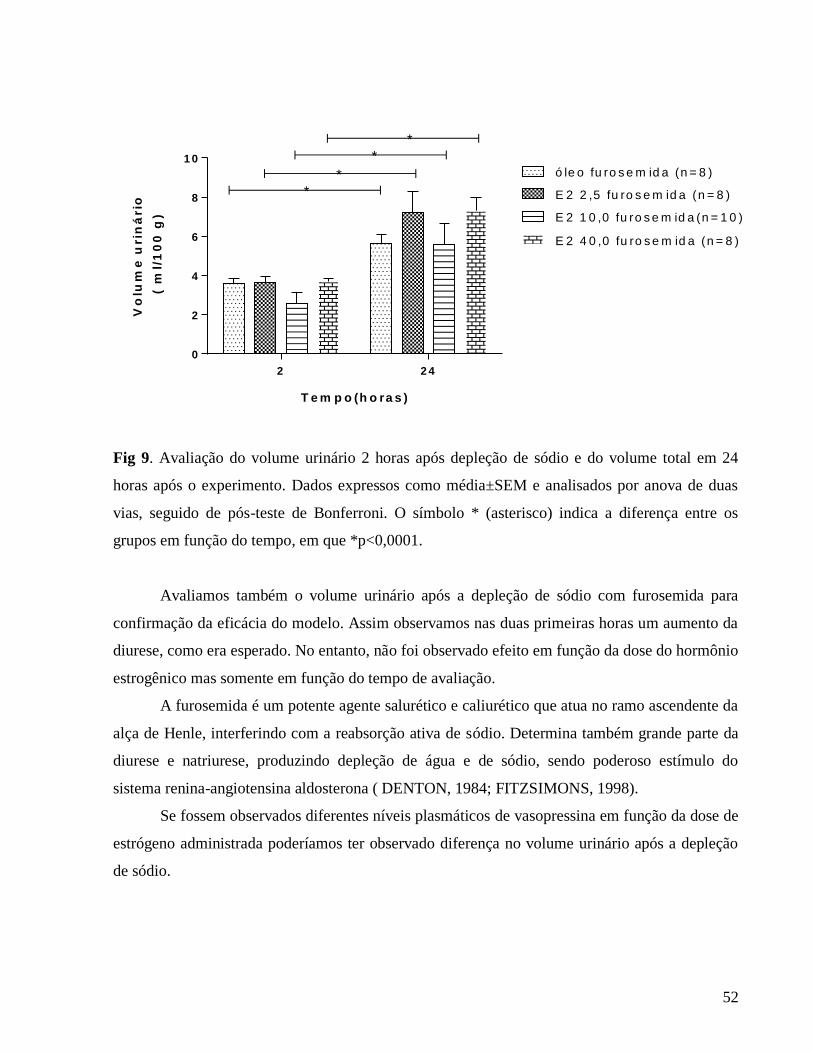
52
T e m p o (h o ra s )
Vo
lum
e u
rin
ário
( m
l/1
00
g)
2 2 4
0
2
4
6
8
1 0
ó le o fu ro s e m id a (n = 8 )
E 2 2 ,5 fu ro s e m id a (n = 8 )
E 2 1 0 ,0 fu ro s e m id a (n = 1 0 )
E 2 4 0 ,0 fu ro s e m id a (n = 8 )
**
**
Fig 9. Avaliação do volume urinário 2 horas após depleção de sódio e do volume total em 24
horas após o experimento. Dados expressos como média±SEM e analisados por anova de duas
vias, seguido de pós-teste de Bonferroni. O símbolo * (asterisco) indica a diferença entre os
grupos em função do tempo, em que *p<0,0001.
Avaliamos também o volume urinário após a depleção de sódio com furosemida para
confirmação da eficácia do modelo. Assim observamos nas duas primeiras horas um aumento da
diurese, como era esperado. No entanto, não foi observado efeito em função da dose do hormônio
estrogênico mas somente em função do tempo de avaliação.
A furosemida é um potente agente salurético e caliurético que atua no ramo ascendente da
alça de Henle, interferindo com a reabsorção ativa de sódio. Determina também grande parte da
diurese e natriurese, produzindo depleção de água e de sódio, sendo poderoso estímulo do
sistema renina-angiotensina aldosterona ( DENTON, 1984; FITZSIMONS, 1998).
Se fossem observados diferentes níveis plasmáticos de vasopressina em função da dose de
estrógeno administrada poderíamos ter observado diferença no volume urinário após a depleção
de sódio.
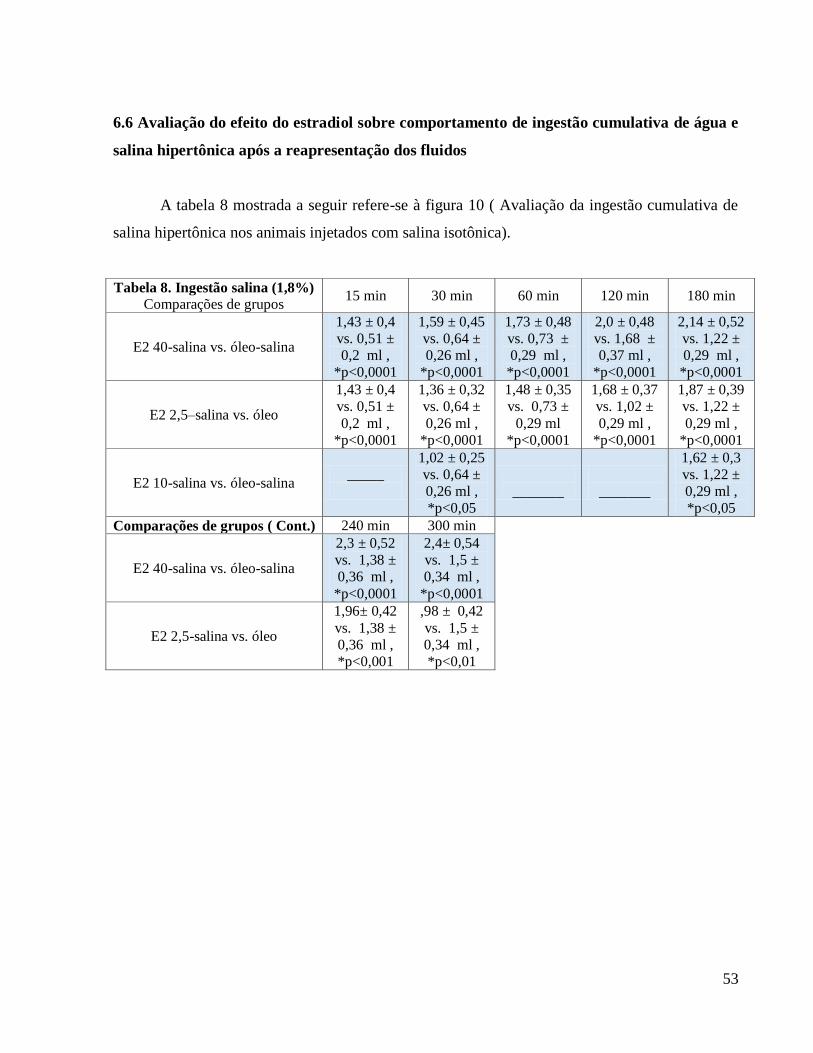
53
6.6 Avaliação do efeito do estradiol sobre comportamento de ingestão cumulativa de água e
salina hipertônica após a reapresentação dos fluidos
A tabela 8 mostrada a seguir refere-se à figura 10 ( Avaliação da ingestão cumulativa de
salina hipertônica nos animais injetados com salina isotônica).
Tabela 8. Ingestão salina (1,8%)
Comparações de grupos 15 min 30 min 60 min 120 min 180 min
E2 40-salina vs. óleo-salina
1,43 ± 0,4
vs. 0,51 ±
0,2 ml ,
*p<0,0001
1,59 ± 0,45
vs. 0,64 ±
0,26 ml ,
*p<0,0001
1,73 ± 0,48
vs. 0,73 ±
0,29 ml ,
*p<0,0001
2,0 ± 0,48
vs. 1,68 ±
0,37 ml ,
*p<0,0001
2,14 ± 0,52
vs. 1,22 ±
0,29 ml ,
*p<0,0001
E2 2,5–salina vs. óleo
1,43 ± 0,4
vs. 0,51 ±
0,2 ml ,
*p<0,0001
1,36 ± 0,32
vs. 0,64 ±
0,26 ml ,
*p<0,0001
1,48 ± 0,35
vs. 0,73 ±
0,29 ml
*p<0,0001
1,68 ± 0,37
vs. 1,02 ±
0,29 ml ,
*p<0,0001
1,87 ± 0,39
vs. 1,22 ±
0,29 ml ,
*p<0,0001
E2 10-salina vs. óleo-salina _____
1,02 ± 0,25
vs. 0,64 ±
0,26 ml ,
*p<0,05
_______
_______
1,62 ± 0,3
vs. 1,22 ±
0,29 ml ,
*p<0,05
Comparações de grupos ( Cont.) 240 min 300 min
E2 40-salina vs. óleo-salina
2,3 ± 0,52
vs. 1,38 ±
0,36 ml ,
*p<0,0001
2,4± 0,54
vs. 1,5 ±
0,34 ml ,
*p<0,0001
E2 2,5-salina vs. óleo
1,96± 0,42
vs. 1,38 ±
0,36 ml ,
*p<0,001
,98 ± 0,42
vs. 1,5 ±
0,34 ml ,
*p<0,01
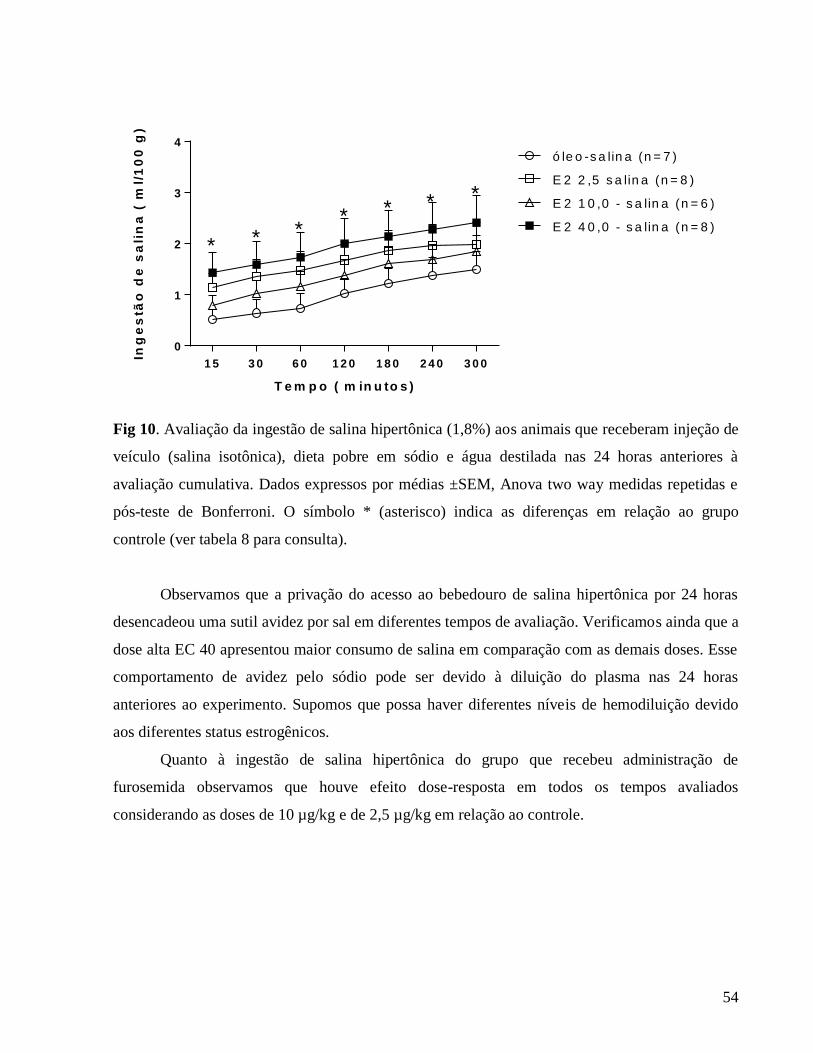
54
T e m p o ( m in u to s )
Ing
es
tão
de
sa
lin
a (
ml/
10
0 g
)
1 5 3 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0
0
1
2
3
4
ó le o -s a lin a (n = 7 )
E 2 2 ,5 s a lin a (n = 8 )
E 2 1 0 ,0 - s a lin a (n = 6 )
E 2 4 0 ,0 - s a lin a (n = 8 )* *
*
*
** *
Fig 10. Avaliação da ingestão de salina hipertônica (1,8%) aos animais que receberam injeção de
veículo (salina isotônica), dieta pobre em sódio e água destilada nas 24 horas anteriores à
avaliação cumulativa. Dados expressos por médias ±SEM, Anova two way medidas repetidas e
pós-teste de Bonferroni. O símbolo * (asterisco) indica as diferenças em relação ao grupo
controle (ver tabela 8 para consulta).
Observamos que a privação do acesso ao bebedouro de salina hipertônica por 24 horas
desencadeou uma sutil avidez por sal em diferentes tempos de avaliação. Verificamos ainda que a
dose alta EC 40 apresentou maior consumo de salina em comparação com as demais doses. Esse
comportamento de avidez pelo sódio pode ser devido à diluição do plasma nas 24 horas
anteriores ao experimento. Supomos que possa haver diferentes níveis de hemodiluição devido
aos diferentes status estrogênicos.
Quanto à ingestão de salina hipertônica do grupo que recebeu administração de
furosemida observamos que houve efeito dose-resposta em todos os tempos avaliados
considerando as doses de 10 µg/kg e de 2,5 µg/kg em relação ao controle.
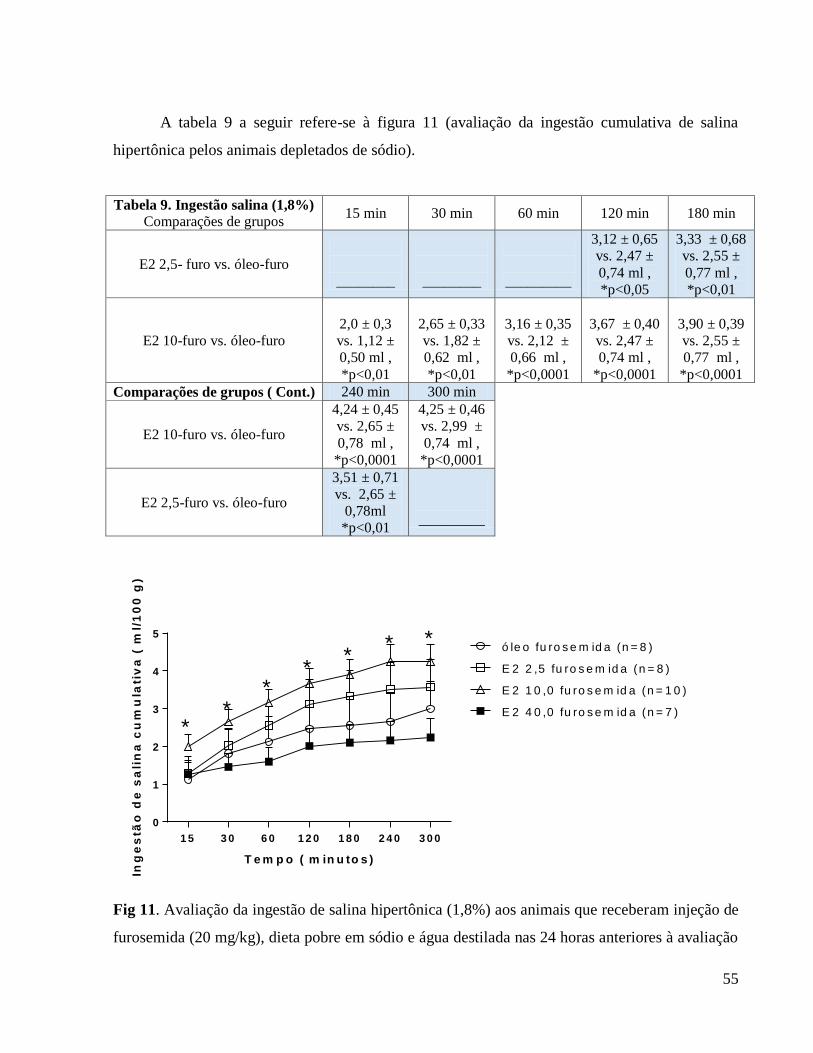
55
A tabela 9 a seguir refere-se à figura 11 (avaliação da ingestão cumulativa de salina
hipertônica pelos animais depletados de sódio).
Tabela 9. Ingestão salina (1,8%)
Comparações de grupos 15 min 30 min 60 min 120 min 180 min
E2 2,5- furo vs. óleo-furo
________
________
_________
3,12 ± 0,65
vs. 2,47 ±
0,74 ml ,
*p<0,05
3,33 ± 0,68
vs. 2,55 ±
0,77 ml ,
*p<0,01
E2 10-furo vs. óleo-furo
2,0 ± 0,3
vs. 1,12 ±
0,50 ml ,
*p<0,01
2,65 ± 0,33
vs. 1,82 ±
0,62 ml ,
*p<0,01
3,16 ± 0,35
vs. 2,12 ±
0,66 ml ,
*p<0,0001
3,67 ± 0,40
vs. 2,47 ±
0,74 ml ,
*p<0,0001
3,90 ± 0,39
vs. 2,55 ±
0,77 ml ,
*p<0,0001
Comparações de grupos ( Cont.) 240 min 300 min
E2 10-furo vs. óleo-furo
4,24 ± 0,45
vs. 2,65 ±
0,78 ml ,
*p<0,0001
4,25 ± 0,46
vs. 2,99 ±
0,74 ml ,
*p<0,0001
E2 2,5-furo vs. óleo-furo
3,51 ± 0,71
vs. 2,65 ±
0,78ml
*p<0,01
_________
T e m p o ( m in u to s )
Ing
es
tão
de
sa
lin
a c
um
ula
tiv
a (
ml/
10
0 g
)
1 5 3 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0
0
1
2
3
4
5
ó le o fu ro s e m id a (n = 8 )
E 2 2 ,5 fu ro s e m id a (n = 8 )
E 2 1 0 ,0 fu ro s e m id a (n = 1 0 )
E 2 4 0 ,0 fu ro s e m id a (n = 7 )
**
**
** *
Fig 11. Avaliação da ingestão de salina hipertônica (1,8%) aos animais que receberam injeção de
furosemida (20 mg/kg), dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação

56
cumulativa. Dados expressos por médias ±SEM, Anova two way medidas repetidas e pós-teste de
Bonferroni.O símbolo *(asterisco) representa as comparações em relação ao grupo controle.
Retornar à tabela 9 para verificar os dados estatísticos.
Após o desafio hidroeletrolítico percebemos que o grupo tratado com a dose de 40 µg/kg
apresentou menor consumo de salina hipertônica sob estímulo. Isso poderia ser explicado pela
elevada ingestão de salina já em condição basal e desse modo podemos supor que os níveis
plasmáticos de sódio já estavam elevados anteriormente, e supostamente a necessidade de busca
pelo sódio sob estímulo seria menor. Além disso, é bastante provável que nesse grupo os
mecanismos que favorecem a retenção de íons sódio e também de água estejam mais
sensibilizados para restauração de volume sanguíneo, pois a reposição com a dose alta (40 µg/kg)
provavelmente resultaria em hipernatremia. Por isso, quando os níveis estrogênicos estão altos
não há um significativo apetite pelo sódio.
Já o grupo tratado com a dose de 10 µg/kg apresentou significativo apetite ao sódio, pois
já em condições basais a ingestão espontânea de sal era baixa, logo seus níveis plasmáticos de
sódio também eram possivelmente mais baixos. Assim, após o desafio houve uma ativação
significativa do sistema-renina angiotensina aldosterona, desencadenando o apetite pelo sódio.
Na ingestão cumulativa de água do grupo que recebeu salina houve efeito dose-resposta
no tempo de 300 min entre os grupos E2 10 vs. Óleo (0,63 ± 0,13 vs. 0,38± 0,20 g, *p<0,05).
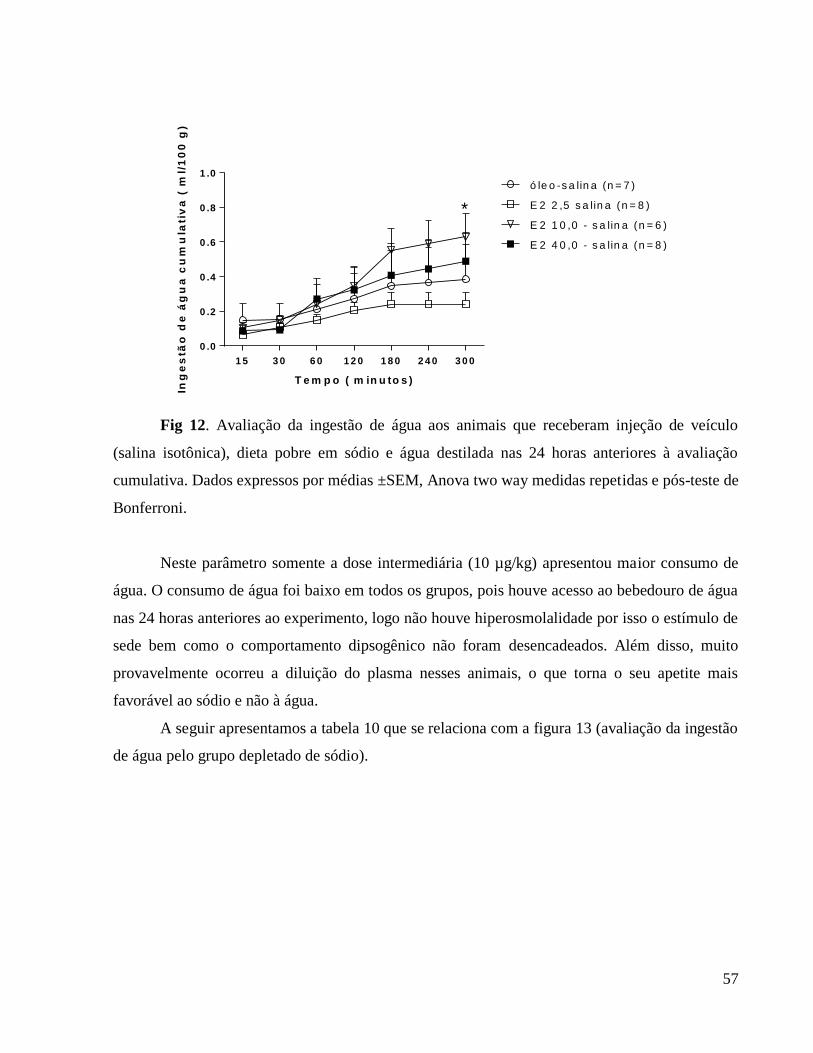
57
T e m p o ( m in u to s )
Ing
es
tão
de
ág
ua
cu
mu
lati
va
( m
l/1
00
g)
1 5 3 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
ó le o -s a lin a (n = 7 )
E 2 2 ,5 s a lin a (n = 8 )
E 2 1 0 ,0 - s a lin a (n = 6 )
E 2 4 0 ,0 - s a lin a (n = 8 )
*
Fig 12. Avaliação da ingestão de água aos animais que receberam injeção de veículo
(salina isotônica), dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação
cumulativa. Dados expressos por médias ±SEM, Anova two way medidas repetidas e pós-teste de
Bonferroni.
Neste parâmetro somente a dose intermediária (10 µg/kg) apresentou maior consumo de
água. O consumo de água foi baixo em todos os grupos, pois houve acesso ao bebedouro de água
nas 24 horas anteriores ao experimento, logo não houve hiperosmolalidade por isso o estímulo de
sede bem como o comportamento dipsogênico não foram desencadeados. Além disso, muito
provavelmente ocorreu a diluição do plasma nesses animais, o que torna o seu apetite mais
favorável ao sódio e não à água.
A seguir apresentamos a tabela 10 que se relaciona com a figura 13 (avaliação da ingestão
de água pelo grupo depletado de sódio).
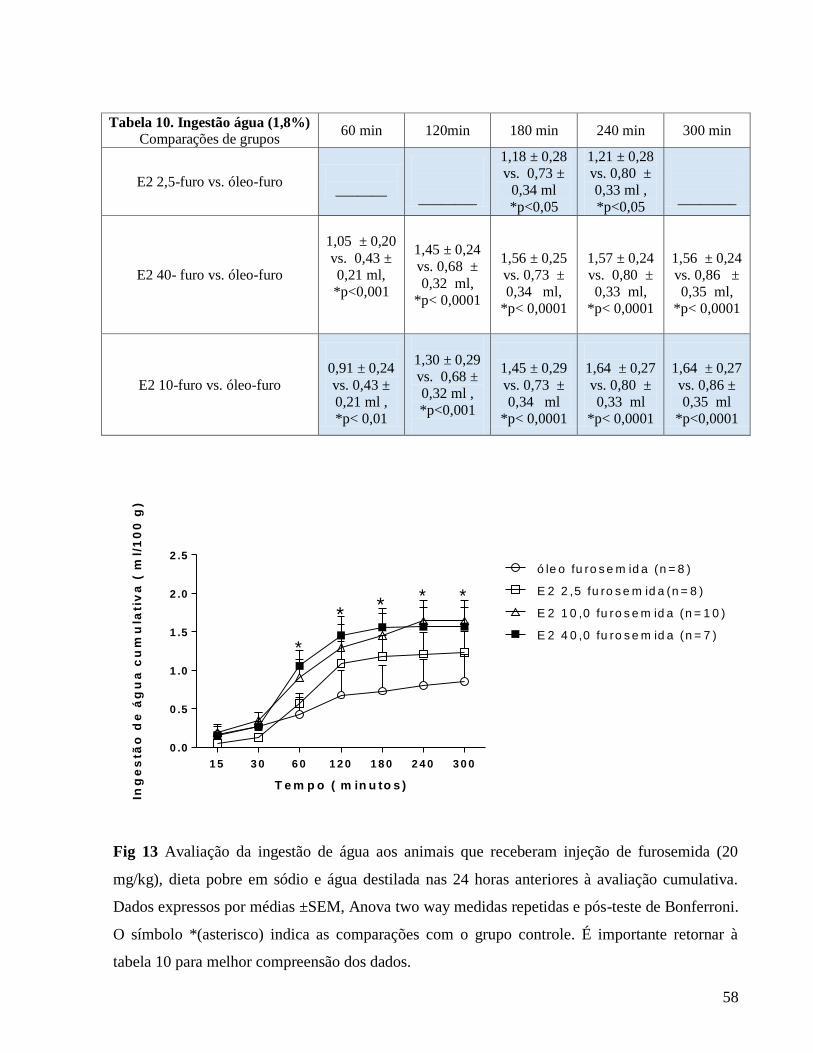
58
Tabela 10. Ingestão água (1,8%)
Comparações de grupos 60 min 120min 180 min 240 min 300 min
E2 2,5-furo vs. óleo-furo
_______
________
1,18 ± 0,28
vs. 0,73 ±
0,34 ml
*p<0,05
1,21 ± 0,28
vs. 0,80 ±
0,33 ml ,
*p<0,05
________
E2 40- furo vs. óleo-furo
1,05 ± 0,20
vs. 0,43 ±
0,21 ml,
*p<0,001
1,45 ± 0,24
vs. 0,68 ±
0,32 ml,
*p< 0,0001
1,56 ± 0,25
vs. 0,73 ±
0,34 ml,
*p< 0,0001
1,57 ± 0,24
vs. 0,80 ±
0,33 ml,
*p< 0,0001
1,56 ± 0,24
vs. 0,86 ±
0,35 ml,
*p< 0,0001
E2 10-furo vs. óleo-furo
0,91 ± 0,24
vs. 0,43 ±
0,21 ml ,
*p< 0,01
1,30 ± 0,29
vs. 0,68 ±
0,32 ml ,
*p<0,001
1,45 ± 0,29
vs. 0,73 ±
0,34 ml
*p< 0,0001
1,64 ± 0,27
vs. 0,80 ±
0,33 ml
*p< 0,0001
1,64 ± 0,27
vs. 0,86 ±
0,35 ml
*p<0,0001
T e m p o ( m in u to s )
Ing
es
tão
de
ág
ua
cu
mu
lati
va
( m
l/1
00
g)
1 5 3 0 6 0 1 2 0 1 8 0 2 4 0 3 0 0
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
ó le o fu ro s e m id a (n = 8 )
E 2 2 ,5 fu ro s e m id a (n = 8 )
E 2 1 0 ,0 fu ro s e m id a (n = 1 0 )
E 2 4 0 ,0 fu ro s e m id a (n = 7 )
*
**
* *
Fig 13 Avaliação da ingestão de água aos animais que receberam injeção de furosemida (20
mg/kg), dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação cumulativa.
Dados expressos por médias ±SEM, Anova two way medidas repetidas e pós-teste de Bonferroni.
O símbolo *(asterisco) indica as comparações com o grupo controle. É importante retornar à
tabela 10 para melhor compreensão dos dados.

59
Avaliamos neste gráfico o papel do estrógeno em modular o comportamento dispsogênico
sob estímulo de depleção de sódio. Lembramos que neste modelo há tanto perda de água quanto
perda de sódio, ocorrendo primeiramente o apetite ao sódio seguida da ingestão de água.
Observamos que o modelo ocasionou em hiperosmolalidade plasmática, sendo esse um
importante estímulo para sede e liberação de vasopressina.
Assim podemos supor que as três doses hormonais provocaram diferentes níveis de
hemodiluição e consequentemente diferentes necessidades de reposição de água ou sódio. Sabe-
se que o estrógeno diminui o limiar osmótico para liberação de vasopressina, logo é provável que
um elevado nível estrogênico seja capaz de ocasionar a liberação de AVP mais facilmente. Além
disso, os neurônios osmossensitivos podem ser mais sensibilizados aos efeitos dipsogênicos da
ANG II, por isso também o consumo de água é maior em níveis estrogênicos altos sob estímulo
osmótico.
Não encontramos diferença significativa entre os grupos com relação à preferência por
sódio, no entanto percebemos que a dose de 10 µg/kg e de 40 µg/kg apresentaram um percentual
de preferência levemente menor que a dose baixa 2 µg/kg e que o controle.
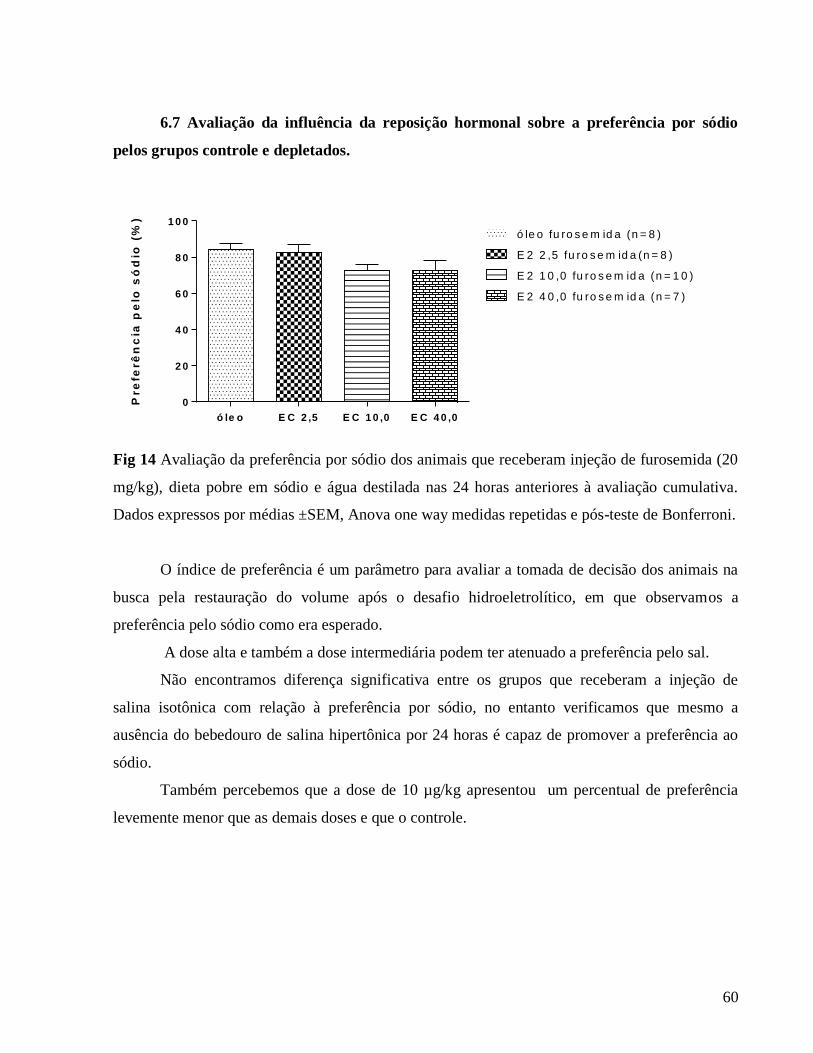
60
6.7 Avaliação da influência da reposição hormonal sobre a preferência por sódio
pelos grupos controle e depletados.
Pre
ferê
nc
ia p
elo
só
dio
(%
)
ó le o E C 2 ,5 E C 1 0 ,0 E C 4 0 ,0
0
2 0
4 0
6 0
8 0
1 0 0
ó le o fu ro s e m id a (n = 8 )
E 2 2 ,5 fu ro s e m id a (n = 8 )
E 2 1 0 ,0 fu ro s e m id a (n = 1 0 )
E 2 4 0 ,0 fu ro s e m id a (n = 7 )
Fig 14 Avaliação da preferência por sódio dos animais que receberam injeção de furosemida (20
mg/kg), dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação cumulativa.
Dados expressos por médias ±SEM, Anova one way medidas repetidas e pós-teste de Bonferroni.
O índice de preferência é um parâmetro para avaliar a tomada de decisão dos animais na
busca pela restauração do volume após o desafio hidroeletrolítico, em que observamos a
preferência pelo sódio como era esperado.
A dose alta e também a dose intermediária podem ter atenuado a preferência pelo sal.
Não encontramos diferença significativa entre os grupos que receberam a injeção de
salina isotônica com relação à preferência por sódio, no entanto verificamos que mesmo a
ausência do bebedouro de salina hipertônica por 24 horas é capaz de promover a preferência ao
sódio.
Também percebemos que a dose de 10 µg/kg apresentou um percentual de preferência
levemente menor que as demais doses e que o controle.
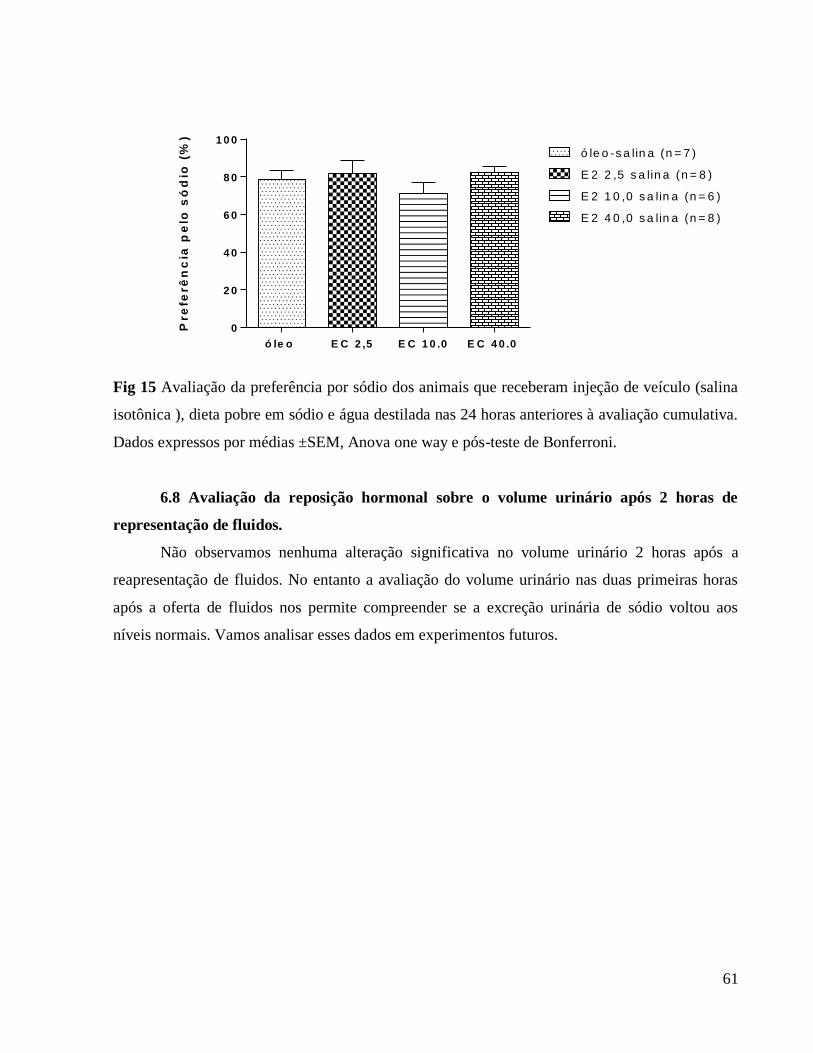
61
Pre
ferê
nc
ia p
elo
só
dio
(%
)
ó le o E C 2 ,5 E C 1 0 .0 E C 4 0 .0
0
2 0
4 0
6 0
8 0
1 0 0
ó le o -s a lin a (n = 7 )
E 2 2 ,5 s a lin a (n = 8 )
E 2 1 0 ,0 s a lin a (n = 6 )
E 2 4 0 ,0 s a lin a (n = 8 )
Fig 15 Avaliação da preferência por sódio dos animais que receberam injeção de veículo (salina
isotônica ), dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação cumulativa.
Dados expressos por médias ±SEM, Anova one way e pós-teste de Bonferroni.
6.8 Avaliação da reposição hormonal sobre o volume urinário após 2 horas de
representação de fluidos.
Não observamos nenhuma alteração significativa no volume urinário 2 horas após a
reapresentação de fluidos. No entanto a avaliação do volume urinário nas duas primeiras horas
após a oferta de fluidos nos permite compreender se a excreção urinária de sódio voltou aos
níveis normais. Vamos analisar esses dados em experimentos futuros.
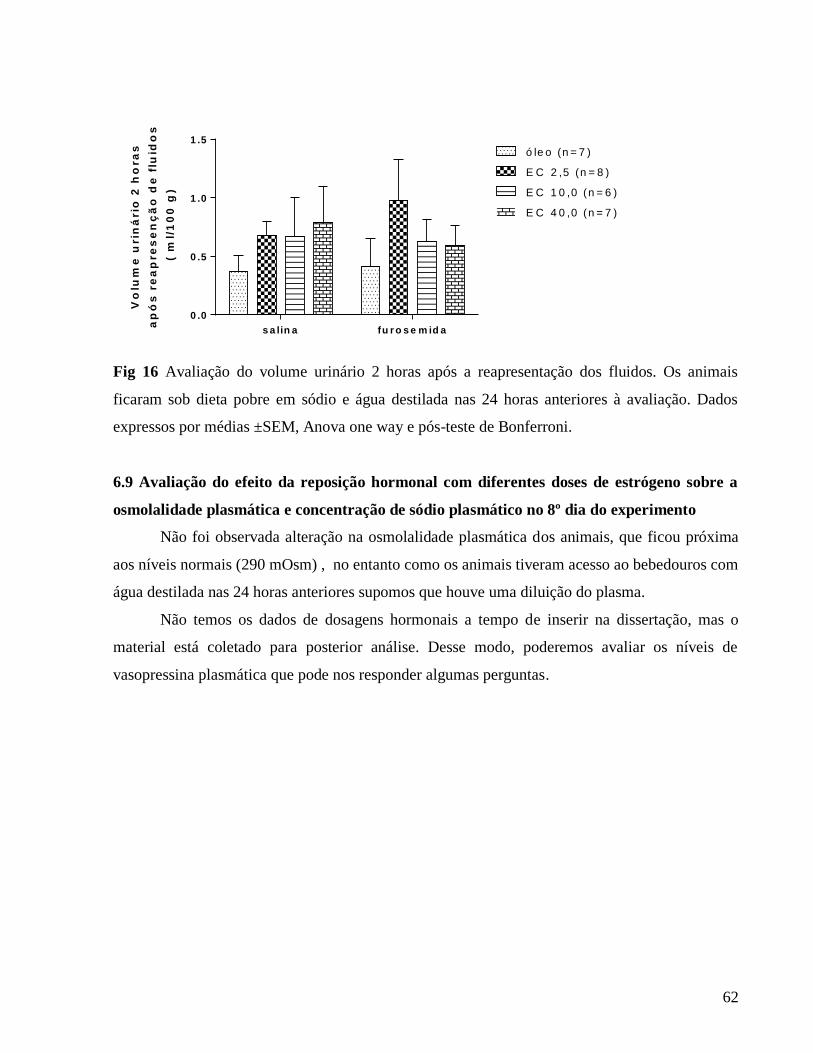
62
Vo
lum
e u
rin
ário
2 h
ora
s
ap
ós
re
ap
re
se
nç
ão
de
flu
ido
s
( m
l/1
00
g)
s a lin a fu r o s e m id a
0 .0
0 .5
1 .0
1 .5
ó le o (n = 7 )
E C 2 ,5 (n = 8 )
E C 1 0 ,0 (n = 6 )
E C 4 0 ,0 (n = 7 )
Fig 16 Avaliação do volume urinário 2 horas após a reapresentação dos fluidos. Os animais
ficaram sob dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação. Dados
expressos por médias ±SEM, Anova one way e pós-teste de Bonferroni.
6.9 Avaliação do efeito da reposição hormonal com diferentes doses de estrógeno sobre a
osmolalidade plasmática e concentração de sódio plasmático no 8º dia do experimento
Não foi observada alteração na osmolalidade plasmática dos animais, que ficou próxima
aos níveis normais (290 mOsm) , no entanto como os animais tiveram acesso ao bebedouros com
água destilada nas 24 horas anteriores supomos que houve uma diluição do plasma.
Não temos os dados de dosagens hormonais a tempo de inserir na dissertação, mas o
material está coletado para posterior análise. Desse modo, poderemos avaliar os níveis de
vasopressina plasmática que pode nos responder algumas perguntas.
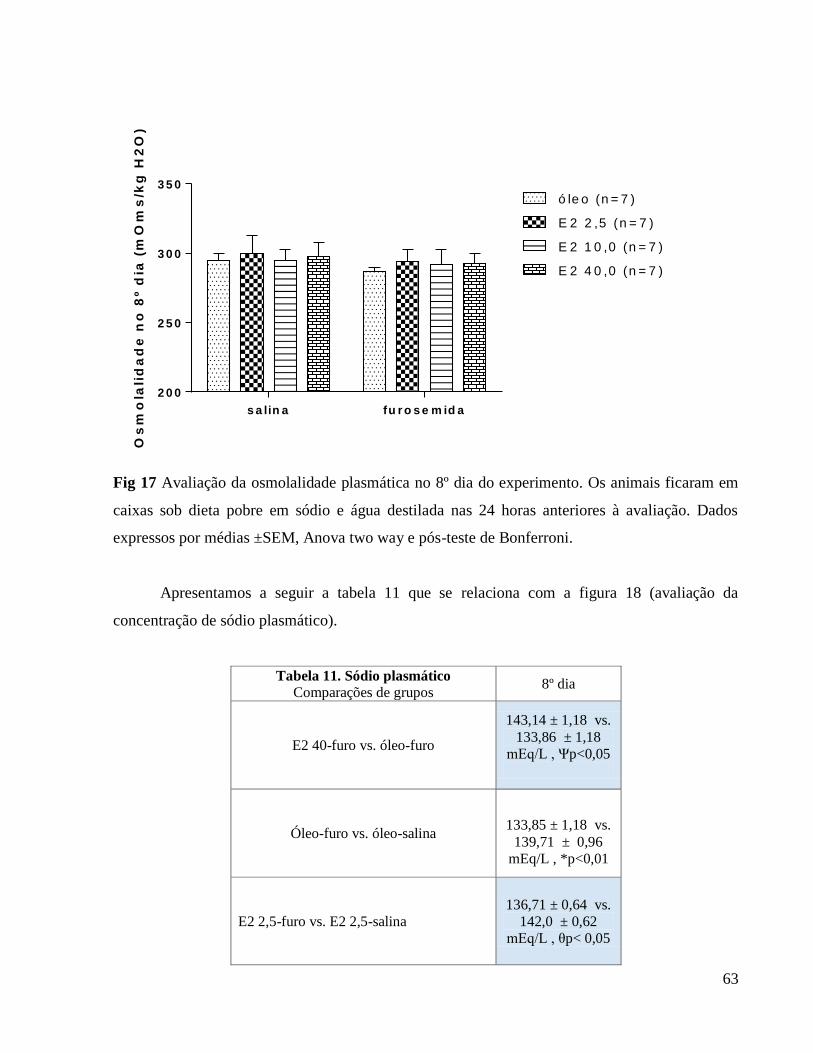
63
Os
mo
lali
da
de
no
8º d
ia (
mO
ms
/kg
H2
O)
s a lin a fu r o s e m id a
2 0 0
2 5 0
3 0 0
3 5 0
ó le o (n = 7 )
E 2 2 ,5 (n = 7 )
E 2 1 0 ,0 (n = 7 )
E 2 4 0 ,0 (n = 7 )
Fig 17 Avaliação da osmolalidade plasmática no 8º dia do experimento. Os animais ficaram em
caixas sob dieta pobre em sódio e água destilada nas 24 horas anteriores à avaliação. Dados
expressos por médias ±SEM, Anova two way e pós-teste de Bonferroni.
Apresentamos a seguir a tabela 11 que se relaciona com a figura 18 (avaliação da
concentração de sódio plasmático).
Tabela 11. Sódio plasmático
Comparações de grupos 8º dia
E2 40-furo vs. óleo-furo
143,14 ± 1,18 vs.
133,86 ± 1,18
mEq/L , Ψp<0,05
Óleo-furo vs. óleo-salina
133,85 ± 1,18 vs.
139,71 ± 0,96
mEq/L , *p<0,01
E2 2,5-furo vs. E2 2,5-salina
136,71 ± 0,64 vs.
142,0 ± 0,62
mEq/L , θp< 0,05
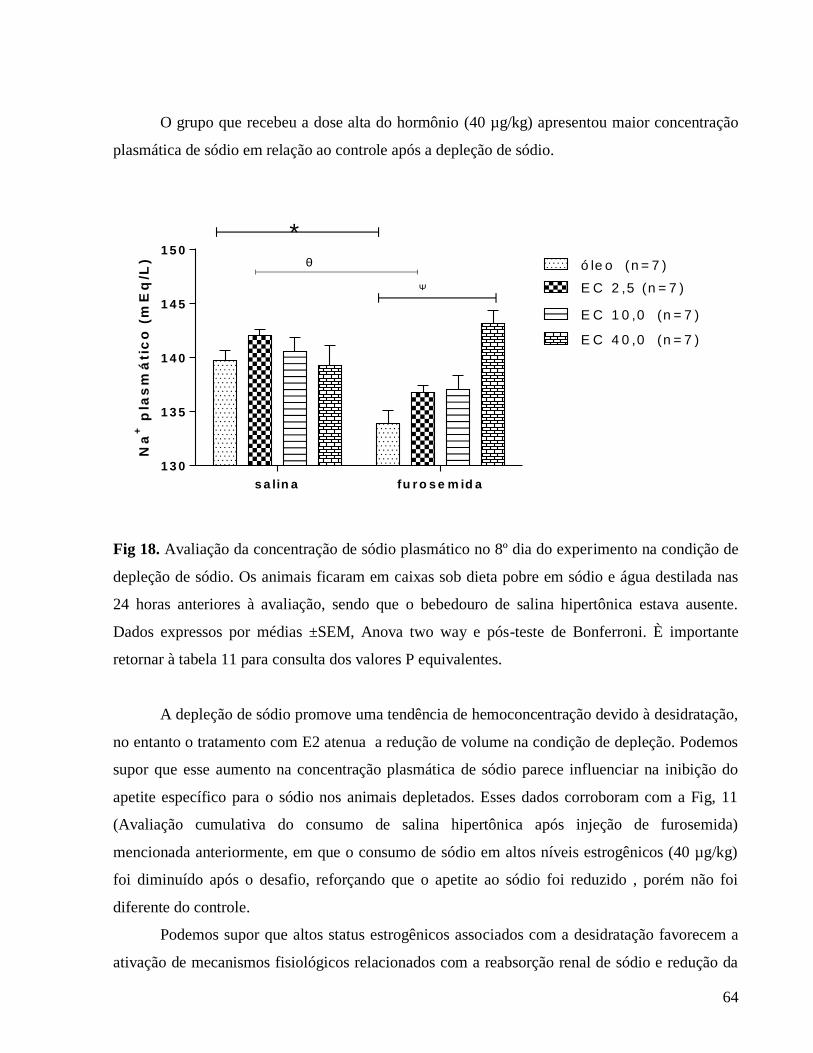
64
O grupo que recebeu a dose alta do hormônio (40 µg/kg) apresentou maior concentração
plasmática de sódio em relação ao controle após a depleção de sódio.
Na
+p
las
má
tic
o (
mE
q/L
)
s a lin a fu r o s e m id a
1 3 0
1 3 5
1 4 0
1 4 5
1 5 0
ó le o (n = 7 )
E C 2 ,5 (n = 7 )
E C 1 0 ,0 (n = 7 )
E C 4 0 ,0 (n = 7 )
*θ
Ψ
Fig 18. Avaliação da concentração de sódio plasmático no 8º dia do experimento na condição de
depleção de sódio. Os animais ficaram em caixas sob dieta pobre em sódio e água destilada nas
24 horas anteriores à avaliação, sendo que o bebedouro de salina hipertônica estava ausente.
Dados expressos por médias ±SEM, Anova two way e pós-teste de Bonferroni. È importante
retornar à tabela 11 para consulta dos valores P equivalentes.
A depleção de sódio promove uma tendência de hemoconcentração devido à desidratação,
no entanto o tratamento com E2 atenua a redução de volume na condição de depleção. Podemos
supor que esse aumento na concentração plasmática de sódio parece influenciar na inibição do
apetite específico para o sódio nos animais depletados. Esses dados corroboram com a Fig, 11
(Avaliação cumulativa do consumo de salina hipertônica após injeção de furosemida)
mencionada anteriormente, em que o consumo de sódio em altos níveis estrogênicos (40 µg/kg)
foi diminuído após o desafio, reforçando que o apetite ao sódio foi reduzido , porém não foi
diferente do controle.
Podemos supor que altos status estrogênicos associados com a desidratação favorecem a
ativação de mecanismos fisiológicos relacionados com a reabsorção renal de sódio e redução da

65
natriurese como aumento dos níveis de aldosterona, diminuição dos níveis de OT e ANP, o que
auxiliaria na retenção de sódio. Mais estudos são necessários para avaliar esse parâmetro.
Ao compararmos a concentração plasmática de sódio nos animais injetados com a salina
isotônica não observamos alteração em função da dose de estradiol. E ao observar o consumo de
salina hipertônica sob condições basais (Fig.5) verificamos que o mesmo já estava mais elevado
no grupo EC 40 porém sem alteração significativa. Esse dado sugere que a diluição do plasma
pode ter influenciado neste parâmetro, supostamente porque os animais tiveram acesso ao
bebedouro de água nas 24 horas anteriores ao experimento. No entanto não medimos o consumo
de água durante esse período.
Verificamos que a furosemida promoveu uma significativa redução nos níveis de sódio
plasmáticos nos grupos óleo-furosemida e E2 2,5 furosemida ao compararmos com os seus
respectivos controles que receberam a administração de salina. Essa resposta era esperada. Desse
modo, podemos inferir que a baixa dose do hormônio (2,5 µg/kg) não é suficiente para acionar
mecanismos que favorecem a retenção de sódio.
Além disso, um estudo recente de Hofmeister et al. (2015) demonstrou que a pele
também possui reservatórios para sódio e que este, por sua vez, poderia estar ligado aos
glicosaminoglicanos existentes no músculo esquelético e na epiderme.
A partir da hipótese anterior podemos supor que o estrógeno também poderia aumentar a
síntese dos glicosaminoglicanos em outros reservatórios de sódio e também poderia ser uma
forma de reter mais sódio.
7.0 Conclusões
7.1 Conclusão geral
Nosso estudo sugere que provavelmente o hormônio estrogênico também exerce um efeito
dose dependente em parâmetros comumente avaliados em estudos de regulação hidroeletrolítica e
exerce um efeito modulatório sobre a expressão comportamental dos animais seletivamente na
ingestão alimentar e no volume urinário sob condições basais e nos comportamentos
natriorexigênicos e dipsogênicos após depleção de sódio. Há possibilidade de o estradiol
desempenhar efeitos neuroendócrinos e também genômicos. Tais dados poderiam auxiliar a
elucidar os resultados comportamentais.

66
7.2 Conclusões específicas
1) Os dados sugerem que o estrógeno sensibiliza os neurônios osmosentitivos para
desencadear respostas dipsogênicas e natriurexigênicas da angiotensina II em regiões
cerebrais envolvidas com o controle da homeostase hidroeletrolítica. Um dos possíveis
mecanismos é a influência estrogênica sobre a expressão gênica dos receptores AT1 em
órgãos circunventriculares.
2) A resposta natriorexigênica desencadeada por diferentes níveis hormonais é intensificada
quando há um desafio hidroeletrolítico cuja origem do estímulo é extracelular como
ocorre no modelo de depleção de sódio, ou seja, depende do paradigma experimental
utilizado.
3) Possivelmente, após a depleção de sódio haveria diferentes níveis de diluição do plasma a
partir de diferentes status estrogênicos, no entanto nosso estudo não aferiu essa resposta.
4) O efeito-dose dependente reflete a intensidade com que o sistema renina-angiotensina
aldosterona é sensibilizado.
5) Nossa suposição é que poderia existir um cross-talk intracelular envolvendo diferentes
fatores de transcrição e mecanismos de super-expressão ou de silenciamento gênico entre
sistema renina- angiotensina, estrógeno e níveis hormonais de AVP, OT em neurônios
osmossensitivos para ajuste da osmolalidade - tonicidade dos fluidos corporais.
8.0 Considerações finais e perspectivas
Em experimentos futuros seria interessante investigar os mecanismos intracelulares pelos
quais o estradiol ativa os neurônios osmossensitivos e indiretamente o sistema renina
angiotensina aldosterona após estímulo osmótico pode ser uma maneira - em associação com
dados que já existem na literatura- para explicar o papel do estrógeno em inibir o apetite ao
sódio.Há muitos estudos de comportamento, imunoistoquímicos e mesmo análises hormonais,
mas falta conhecer o que o estrógeno ativa ou inibe em cascatas intracelulares importantes
para ativação dos neurônios osmossensitivos.
Uma outra possibilidade para estudos futuros seria realizar o mesmo protocolo de
reposição hormonal e alterar o paradigma experimental, acrescentando novos desafios como

67
sobrecarga salina, protocolo furocap, oferta de dieta hipossódica ou hiperssódica e avaliar se
há outras respostas possíveis neste contexto.
9.0 Referências bibliográficas
ALHEID, G. DE OLMOS, J.; BELTRAMINO, C. Amygdala and extended amygdala, In: Paxinos, G.
The rat nervous system, 2nd ed, San Diego, Academic Press,p 495-578, i 1995.
ANTUNES-RODRIGUES, J.; RUGINSK, S.G. ; MECAWI, A.S. MARGATHO, L.O.; CRUZ, J.C.;
VILHENA-FRANCO, T.; REIS, W.L.; VENTURA, R.R.; REIS, L.C.; VIVAS, L.M. ; ELIAS,
L.L.K. Mapping and signaling of neural pathways involved in the regulation of hydromineral homeostasis
Braz J Med Biol Res 46(4), 2013.
ANTUNES-RODRIGUES, J.; FAVARETTO, A.L.; GUTKOWSKA,J.; MCCANN, S.M.
The neuroendocrine control of atrial natriuretic peptide release. Mol Psychiatry, 2(5):359-67, 1997.
ANTUNES-RODRIGUES, J.; COVIAN, M.R.; Hypothalamic control of sodium chloride and water
intake. Acta Physiol Lat Am 13: 94-100, 1963.
AKAISHI,T.: SAKUMA Y. Estrogen-induced modulation of hypothalamic osmoregulation in female
rats. Am J Physiol RegulatoryIntegrative Comp Physiol258: R924–R929, 1990.
ASARIAN, L.; GEARY, N.Cyclic Estradiol Treatment Normalizes Body Weight and Restores
Physiological Patterns of Spontaneous Feeding and Sexual Receptivity in Ovariectomized RatsHormones
and Behavior42, 461–471 ,2002.
AZMITIA, E.C; SEGAL, M. An autoradiographic analysis of the differential ascending projections of
the dorsal and median raphe nuclei in the rat. J Comp Neurol, 179 (3), 641-668, 1978.
BADAUÊ-PASSOS, D.; GODINO, A.; JOHNSON, A.K.; VIVAS, L. ANTUNES-RODRIGUES, J.
Dorsal raphe nuclei integrate allostatic information evoked by sodium depletion-induced ingestion. Exp
Neurol 206:86-94, 2007.

68
BARRON,W.M.; SCHREIBER,J.; LINDHEIMER, MD. Effect of ovarian sex steroids on
osmoregulation and vasopressin secretion in the rat. Am J Physiol Endocrinol Metab250: E352–E361,
1986.
BLACKBURN, R.E., SAMSON W.K. FULTON, R.J. STRICKER, E.M. VERBALIS, J.G. Central
oxytocin and ANP receptors mediate osmotic inhibition of salt apetite in rats. Am J Physiol, 269R245-
R251,1995.
BIEGON, A.; BERCOVITZ, SAMUEL, D. Serotonin receptor concentration during the estrous cycle of
the rat. Brain Res. 187:221–225, 1980.
BOTELHO, L.M.; BLOCK, C.H.; KHOSLA, M.C.; SANTOS, R.A. Plasma angiotensin (1-7)
immunoreactivity is increased by salt load, water deprivation, and hemorrhage. Pept, 15(4):723-9, 1994.
BRUNETTE, M.G.; LECLERC, M. Effect of estrogen on calcium and sodium transport by the nephron
luminal membranes. J Endocrinol 170: 441-450, 2001.
BUTCHER, R.L.; COLLINS, W.E.; FUGO, N.W. Plasma concentration of LH, FSH, prolactin,
progesterone and estradiol-17b throughout the 4-day estrous cycle of the rat. Endocrinology 94:1704–
1708, 1974.
BUTERA, P.C.; CZAJA, J.A. Intracranial estradiol in ovariectomized guinea pigs: effects on ingestive
behaviors and body weight. Brain research. 322:41–48, 1984.
CALZONE, W.L.; SILVA, C.; KEEFE, D.L.; STACHENFELD, N.S. Progesterone does not alter
osmotic regulation of AVPAm J Physiol Regulatory Integrative Comp Physiol281: R2011–R2020, 2001.
CASABIELL, X.; PINEIRO, V.; PEINO, R.; LAGE, M.; CAMINA, J.; GALLEGO, R.; VALLEJO,
L.G.; DIEGUEZ, C.; CASANUEVA, F.F. Gender differences in both spontaneous and stimulated leptin
secretion by human omental adipose tissue in vitro: dexamethasone and estradiol stimulate leptin release
in women, but not in men. The Journal of clinical endocrinology and metabolism 83:2149–2155,
1998.

69
CECCHINI, D.J.; CHATTORAJ, S.C. ; FANOUS, A.S. ; PANDA, S.K.; BRENNAN, T.F.;
EDELIN, K.C. Radioimmunoassay of 2-hydroxyestrone in plasma during the estrous cycle of the rat:
interrelationships with estradiol, progesterone, and the gonadotropins. Endocrinology 112:1122–1126;
1983.
CHEEMA, M.U.; IRSIK, D.L.; WANG, Y.; MILLER-LITTLE, W.; HYNDMAN, K.A.; MARKS,
E.S.; FROKIAER, J.; BOESEN, E.; NORREGAARD, R. Estradiol regulates AQP2 expression in the
collectin duct – a novel inhibitory for estrogen receptor alpha Am J Renal Physiol ( junho, 2015) doi:
10.1152/ajprenal.00685.2014.
CREECH, R.D.; Q LI, G.A. CARRASCO, L.D. VAN DE KAR, N.A. Estradiol induces partial
desensitization of serotonin 1A receptor signaling in the paraventricular nucleus of the hypothalamus and
alters expression and interaction of RGSZ1 and Gaz Neuropharmacology 62: 2040-2049, 2012.
CROWLEY, R.S. ; AMICO, J.A. Gonadal steroid modulation of oxytocin and vasopressin gene
expression in the hypothalamus of the osmotically stimulated rat. Endocrinology133: 2711– 2718, 1993.
CROWLEY,W.R. ; O’DPNOHUE, T.L.; GEORGE, J.M.; JACOBOWITZ,D.M. Changes in
pituitary oxytocin and vasopressin during theestrous cycle and after ovarian hormones: evidence for
mediation by norepinephrine. Life Sci23: 2579–2586, 1978.
CURTIS, K.S. Estrogen and the central control of body fluid balance. Physiology & Behavior97 :180–
192, 2009.
CZAJA, J.A; GOLDFOOT, D.A.; KARAVOLAS, H.J. Comparative facilitation and inhibition of
lordosis in the guinea pig with progesterone, 5-alpha-pregnane-3,2-dione, or 3-alpha-hydroxy-5-
alphapregnan- 20-one. Hormones and behavior. 5:261–274, 1974.
DAFOPOULOS, K.; CHALVATZAS, N.; KOSMAS,G.; KALLITSARIS, A.; POURNARAS, S.;
MESSINIS, I.E. The effect ofestrogens on plasma ghrelin concentrations in women. Journal of
endocrinological investigation 33:109–112, 2010.
DALMASSO, C.; AMIGONE, J.L. ; VIVAS, L.Serotonergic system involvement in the inhibitory
action of estrogen on induced sodium appetite in female rats. Physiol Behav, 1;104(3):398-407, 2011.

70
DANIELSEN, J.; BUGGY, J. Depression of ad lib and angiotensin-induced sodium intake at oestrus.
Brain Res Bull. 5, 501–504, 1980.
DARBLADE, B.; PENDARIES, C.; KRUST, A. Estradiol alters nitric oxide production in the mouse
aorta through the alpha-, but not beta-, estrogen receptor. Circ Res. 90(4):413–419, 2002.
DAVISON, J.M.; GILMORE, E.A.; DURR, J.; ROBERTSON, G.L.; LINDHEIMER, M.D. Altered
osmotic thresholds for vasopressin secretion and thirst in human pregnancy. Am J Physiol Renal
FluidElectrolyte Physiol246: F105–F109, 1984.
DENTON, D.A; COGHLAN, J.P.; FEI, D.T.; MCKINLEY, M.; NELSON, J.; SCOGGINS.;
TARJAN, E.; TREGEAR, G.W.; TRESHAM, J.J.; WEISINGER, R. Stress, ACTH, salt intake and
high blood pressure. Clin Exp Hypertens A.6(1-2):403-15, 1984.
DESAN, P.H.;WOODMANSEE, W.W.; RYAN, S.M.; SMOCK, T.K.; MAIER, S.F. Monoamine
neurotransmitters and metabolites during the estrous cycle, pregnancy, and the postpartum period.
Pharmacol. Biochem.Behav. 30:563–568, 1988.
DIGNAM, W.S.; VOSKIAN, J. ; ASSALI, N.S. Effects of estrogens on renal hemodynamics and
excretion of electrolytes in human subjects. J Clin Endocrinol Metab 16: 1032-1042, 1956.
DONNER, N. & HANDA, J.R. Estrogen receptor beta regulates the expression of tryptophan-
hidroxylase 2 mRNA within serotonergic neurons of the rat dorsal raphe nuclei. Neuroscience, 163:705-
718, 2009.
ELIAS, P.C.L.; ELIAS, L.L.K,.; MOREIRA, A.C. Padronização do teste de infusão de salina
hipertônica para o diagnóstico de diabetes insípido com dosagem da vasopressina plasmática. Arq Bras
Endocrinol Metabol, 42:198-204, 1998.
ESQUERDA, M.E.D.; CRAIG, T.; HINOJOSA-LABORDE, C.Effect of Ovariectomy on Renal
Estrogen Receptor α and Estrogen Receptor βin Young Salt-Sensitiveand -Resistant Rats.
Hypertension,50:768-772,2007.

71
ECKEL, L.A.; HOUPT, T.A.; GEARY, N. Estradiol treatment increases CCK-induced c-Fos expression
inthe brains of ovariectomized rats. American journal of physiology. Regulatory, integrative and
comparative physiology. 283:R1378–1385, 2002.
FAJARDO, M.E; MALACARA, J.M.; MARTINEZ-RODRIGUEZ, H.G.; BARRERA-
SALDANHA, H.A. Hormone and metabolic factors associated with leptin mRNA expression in pre- and
postmenopausal women. Steroids. 69:425–430, 2004.
FITTS, D.A, TJEPKES, D.S.; BRIGHT R.O. Salt apetite and lesions of the ventral part of the ventral
median preoptic nucleus. Behav Nerosci, 104:818-27, 1990.
FITZSIMONS, J.T. Angiotensin,thirst and sodium apetite. Phisiol Rev, 78:583-686,1998.
FRANKMANN, S.P.; DORSA, P.M.; SAKAI, R.R.; SIMPSON, J.B. A single experience with
hyperoncotic colloid dialysis persistently alters water and sodium intake.In:De-Caro G, Epstein AN, Massi
M (Eds.), The Physiology of Thirst and Sodium Appetite, Plenum Press, New York, pp. 115-121, 1986.
GAMBLING, L.; DUNFORD, S.; WILSON, C.A.; MCARDLE, H. L.; BAINES, D.L. Estrogen and
progesterone regulate alpha, beta, and gamma ENaC subunit mRNA levels in female rat kidney Kidney
Int 65: 1774-1781, 2004.
GAO, Q.; MEZEI, G.; NIE, Y.; RAO, Y.; CHOI, C.S.; BECHMANN, I.;LERANTH, C.;
ALLERAND, D.; TORAN-PRIEST, C.A.; ROBERTS, J.L.; GAO, X.B.; MOBBS,C.; SHULMAN,
G.I.; DIANO, S.; HORVATH, T.L. Anorectic estrogen mimics leptin’s effect on the rewiring of
melanocortin cells and Stat3 signaling in obese animals Nature medicine. 13:89–94, 2007.
GAVA, A.L.; FREITAS, F.P.;MEYRELLES, S.S.; GRACELLI, J.B. Gender-dependent effects of
aging on the kidney Braz J Med Biol Res 44: 905-913, 2011.
GEARY, N.; ASARIAN, L. Cyclic Estradiol Treatment Normalizes Body Weight and Test Meal Size in
Ovariectomized Rats Physiology & Behavior, Vol. 67, No. 1, pp. 141–147, 1999

72
GRACELLI, J.B.; SOUZA-MENEZES, J.; BARBOSA, C.M.; ORNELLAS, F.S.; TAKIYA, C.M. ;
ALVES, L.M. Role of estrogen and progesterone in the modulation of CNG-A1 and Na/K+- ATPase
expression in the renal cortex. Cell Physiol Biochem 30: 160-172 ,2012.
GRACELI, J.B.; CICLINI, M.A.; BISSOLI, G.R.; MOYSES, M.R. Roles of estrogen and
progesterone in modulating renal nerve function in the rat kidney, Brazilian Journal of Medical and
Biological Research 46: 521-527 2013.
GRAVES, N.S.; HAYES, H.; FAN, L.; CURTIS, K.S. Time course of behavioral, physiological and
morphological changes after estradiol treatment of ovariectomized rats. Physiol Behav, 103(0):261-
267,2011.
GUTKOWSKA, J.; THIBAULT, G.; JANUSZEWICZ, P.; CANTIN, M.; GENEST, J. Direct
radioimmunoassay of atrial natriuretic factor. Biochem Biophys ResCommun, 122(2):593-601, 1984.
FINDLAY, A.L.; FITZSIMONS, J.T. ; KUCHARZYK, J. Dependence of spontaneous and
angiotensin-induced drinking in the rat upon the oestrous cycle and ovarian hormones. J
Endocrinol. 82(2):215-25, 1979.
FRANCHINI, L.F.; JOHNSON, A.K.; DE OLMOS, J.; VIVAS, L. Sodium apetite and Fos activation
in serotonergic neurons, Am J Physiol Regulatory Integrative Comp Physiol 282:235-243, 2002.
FRINDT, G.; PALMER, L.G. Regulation of epithelial Na+ channels by adrenal steroids:
mineralocorticoid and glucocorticoid effects. Am J Physiol Renal Physiol, 302(1): 20–26, 2012.
FITZSIMONS JT. Angiotensin, thirst, and sodium appetite. Physiol Rev 78: 583– 686, 1998.
FUNGFUANG, W.; NAKADA, T.; NAKAO, N.; TERADA, M.; YOKOSUKA, M.;GIZURARSON,
S.; HAU, S.; MOON, C.~SAITO, T.R. Serum leptin concentrations, leptin mRNA expression, and food
intake during the estrous cycle in rats Lab Anim Res 29(1):1-6, 2013.

73
HAANWINCKEL, M.A.; ELIAS, L.K.; FAVARETTO, A.L.; GUTKOWSKA, J.; MCCANN, S.M.;
ANTUNES-RODRIGUES, J. Oxytocin mediates atrial natriuretic peptide release and natriuresis after
volume expansion in the rat. Proc Natl Acad SciUSA, 92(17):7902-7906, 1995.
HARTLEY, D.E.; DICKSON, S.L.; FORSLING, M.L. Plasma vasopressin concentrations and Fos
protein expression in the supraoptic nucleus following osmotic stimulation or hypovolaemia in the
ovariectomized rat: effect of oestradiol replacement. J Neuroendocrinol16: 191–197, 2004.
HIROSAWA, M.; MINATA, M.; HARADA, K.H.; HITOMI, T.; KRUST, A.; KOIZUMI, A.
Ablation of estrogen receptor alpha (ERalpha) prevents upregulation of POMC by leptin and insulin.
Biochemical and biophysical research communications. 371:320–323, 2008.
HOFMEISTER, L.H.; PERISIC, S.; TITZE, J. Tissue sodium storage: evidence for kidney-like
extrarenal countercurrent systems? Pflugers Arc, 467(3):551-558, 2015.
JEFFERY, G.S.; PENG, K.C.; WAGNER, E.J. The role of phosphatidylinositol-3-kinase and AMP-
activated kinase in the rapid estrogenic attentuation of cannabinoid-induced changes in energy
homeostasis. Pharmaceuticals 4:630–651, 2011.
JOHNSON, A.K.; THUNHORST, R.L. The neuroendocrinology of thirst and salt appetite: visceral
sensory signals and mechanisms of central integration. Front Neuroendocrinol.18(3):292-353, 1997.
JOHNSON, W.G.; CORRIGAN, S.A.; LEMMON, C.R.; BERGERON, K.B. CRUSCO, A.H. Energy
regulation over the menstrual cycle. Physiology & Behavior. 56:523–527, 1994.
KISLEY, L. R.; SAKAI, R. R.; FLANAGAN-CATO, L. M. & FLUHARTY, S. J. Estrogen increases
angiotensin II-induced c-Fos expression in the vasopressinergic neurons of the paraventricular nucleus in
the female rat. Neuroendocrinology, 72:306-17, 2000.
KROHN, P.L. ; ZUCKERMAN, S. Water metabolismo in relation to the menstrual cycle. J. Physiol. 88,
369–387,1937.

74
KUENG, W.; WIRZ-JUSTICE,A.; MENZI, R.; CHAPPUIS-ARNDT, E. Regional brain variations
of tryptophan, monoamines, monoamine oxidase activity, plasma free and total tryptophan during the
estrous cycle of the rat. Neuroendocrinology21:289–296, 1976.
LARAGH, J.H. Oral contraceptives - induced hypertension - nine years later. Am J ObstetGynecol.
1;126(1):141-7, 1976.
LEMALE, J.; BLOCH-FAURE, M.; GRIMONT, A.; ABIDA, B. IMBERT-TEBOUL, M.
CRAMBERT, G. Membrane progestin receptors alpha and gamma in renal epithelium. Biochim Biophys
Acta 1783: 2234-2240, 2008.
LIN, K.C.; SAGAWA, N.; YURA, S.; ITOH, H.; FUJII, S. Simultaneous increases of leptin and
gonadotropinreleasing hormone following exogenous estrogen administration in women with normally
menstrual cycle. Endocrine jornal 52:449–454, 2005.
LU, J.K.; LAPOLT, P.S.; NASS, T.E.; MATT, D.W.; JUDD, H.J. Relationof circulating estradiol and
progesterone to gonadotropin secretion and estrous cyclicity in aging female rats. Endocrinology
116:1953–1959; 1985.
LU, Z. N. & BETHEA, L. C. Ovarian Steroid Regulation of 5-HT1A Receptor Binding and G protein
Activation in Female Monkeys. Neuropsychopharmacology, 27: 12-24, 2002.
MAFFEI, S.; DEL RY, S.; PRONTERA, C.; CLERICO, A. Increase in circulating levels of cardiac
natriuretic peptides after hormone replacement therapy in postmenopausal women Clinical Science, 101,
447–453, 2001.
MARSTON, O.J.; GARFIELD, A.S.; HEISLER, L.K. Role of central serotonin and melanocortin
systems in the control of energy balance. European journal of pharmacology. 660:70–79, 2011.
MCKINLEY, M.J.; ALLEN, A.M.; MAY,C.N.; MCALLEN, R.M..; OLDFIELDB.J.; SY,D.;
MEMDELSOHNN, F.A. Neural pathways from the lamina terminalis influencing cardiovascular and
body fluid homeostasis. Clin Exp Pharmacol Physiol, 28:990-2, 2001.

75
MECAWI, A.S.; VILHENA-FRANCO, T.; FONSECA, F.V.; REIS, L.C.; ELIAS, L.L.; ANTUNES-
RODRIGUES, J. The role of angiotensin II on sodium appetite after low-sodium diet.J
Neuroendocrinol, 25(3):281-91,2013 .
MECAWI, A. S.; LEPLETIER, A.; ARAUJO, I.G. FONSECA, F.V.; REIS, L.C. Oestrogenic
influence on brain AT1 receptor signalling on thirst and salt appetite in osmotic-stimulated and sodium-
depleted rats. Exp. Physiol. 93, 1002–1010. 2008.
MIYATA, S.; TSUJIOKA, H.; MASANOBU, I.; MATSUNAGA,W.; KURAMOTO,H.;
KIYOHARA,T.; Time course of Fos e Fras expression in the hypothalamic supraoptic neurons during
chronic osmotic stimulation Molecular Brain Research , 90:39-47, 2001.
MOHAMED, M.K.; RAHMAN, A.A. Effect of long-term ovariectomy and estrogen replacement on the
expression of estrogen receptor gene in female ratsEuropean Journal of Endocrinology 142 307–314,
2000.
MURATA, T.; NARITA, K.; HONDA, K.; MATSUKAWA, S.; HIGUCHI, T.Differential regulation
of estrogen receptor alpha and beta mRNAs in the rat uterus during pregnancy and labor: possible
involvement of estrogen receptors in oxytocin receptor regulation. Endocrine Journal, 50(5):579-587,
2003.
MUSATOV, S.; CHEN, W.; PTAFF, D.W.; MOBBS, C.V.; YANG, X.J.; CLEGG, D.J.; KAPLIII,
M.G.; OGAWA, S. Silencing of estrogen receptor alpha in the ventromedial nucleus of hypothalamus
leads to metabolic syndrome. Proceedings of the National Academy of Sciences of the United States of
America. 2007.
NAKAJIMA, T.; TANIMOTO, Y.; TANAKA, M.; CHAMBON, P.; WATANABE, H.; IGUSHI, T.;
SATO, T. Neonatal estrogen receptor β is important in the permanent inhibition of epithelial cell
proliferation in the mouse uterus. Endocrinology. May 28:en20151012. [Epub ahead of print]
NASCIMENTO, D.S. ; REIS, C.U.; GOLDENBERG, R.C.; ORTIGA-CARVALHO, T.M. PAZOZ-
MOURA,C.C.; GUGGINO, S.E. Estrogen modulates ClC-2 chloride channel gene expression in rat
kidney. Pflugers Arch 446: 593-599, 2003.

76
NODA, M. Hydromineral neuroendocrinology: mechanism of sensing sodium levels in the mammalian
brain Exp Physiol92.3:513–522,2007.
NOLAN, L.A. ; LEVY, A. Prolonged oestrogen treatment does not correlate with a sustained increase in
anterior pituitary mitotic index in ovariectomized Wistar rats Journal of Endocrinology 200, 301–309,
2009.
NORGREN, R. Gustatory system. In: Paxinos, G. (ed). The rat Nervous System, 2nd
. Ed. San Diego,
Academic Press, p.751-71, 1995.
OELKERS, W.K.H. Effects of estrogen and progestogens on the renin-aldosterone system and blood
pressure. Steroids61: 166– 171, 1996.
OIAN, P. TOLLAN, A.; FADNES, H.O. NODDELAND, H.; MALTAU, J.M. Transcapillary fluid
dynamics during the menstrual cycle. Am J Obstet Gynecol 156: 952–955, 1987.
OLDFIELD, B.J.; MCKINLEY, M.J. Circunventricular organs. In: Paxinos, G. (ed). The rat Nervous
System, 2 nd.ed. San Diego, Academic Press, p. 391-403, 1995.
PECHERE-BERTSCHI, A.; BURNIER, M. Gonadal steroids, salt-sensitivity and renal function Curr
Opin nephrol hypertens. 16(1):16-21, 2007.
PIKE, R.L.; YAO, C. Increased sodium chloride appetite during pregnancy in the rat. J Nutr.101(2):169-
75, 1971.
POPOVA, N.K. ;NAUMENKO, V.S.; CYBKO, A.S.; BAZOVKINA, D.V.Receptor-genes cross-talk:
effect of chronic 5-HT(1A) agonist 8-hydroxy-2-(di-n-propylamino) tetralin treatment on the expression
of key genes in brain serotonin system and on behavior. Neuroscience. 2010 Aug 11;169(1):229-35.
PUERTA, M.; ROCHA, M.; GONZALEZ-COVADELA,S.; MCBENNETT, S.M.; ANDREWS, J.F.
Changes incytochrome oxidase activity in brown adipose tissue during oestrous cycle in the rat. European
journal of endocrinology / European Federation of Endocrine Societies.139:433–437,1998.

77
REIS, L.C. Papel do sistema serotoninérgico na regulação do equilíbrio hidroeletrolítico. Tese de
Doutorado, Faculdade de Medicina da USP de Ribeirão Preto, 265p, 1993.
RICHTER, C.P.; BRAILEY, M.E. Water-intake and its relation to the surface área of the body. Proc.
Natl. Acad. Sci. USA15, 570–578, 1929.
RIVERA, H.M.; SANTOLLO, J.; NIKONOVA, L.V.; ECKEL, L.A. Estradiol increases the anorexia
associated with increased 5-HT2C receptor activation in ovariectomized ratsPhysiol Behav.18; 105(2):
188–194, 2002.
RIVERA, H.M.; ECKEL, L.A. The anorectic effect of fenfluramine is increased by estradiol treatmentin
ovariectomized rats. Physiology & Behavior. 2005; 86:331–337, 2005.
ROCHA, M.;GRUESO, E.; PUERTA, M. The anorectic effect of oestradiol does not involve changes in
plasma and cerebrospinal fluid leptin concentrations in the rat. The Journal of endocrinology. 171:349–
354, 2001.
ROEPKE, T.A.; MALYALA, A. BOSCH, M.A.; KELLY, M.J. RONNKLEIV, O.K. Estrogen
regulation of genes important for K+ channel signaling in the arcuate nucleus. Endocrinology. 148:4937–
4951, 2007.
ROEPKE, T.A.; BOSCH, M.A.; RICK, E.A.; LEE,B.; WAGNER, E.J.; SEIDLOVA-WUTTKE, D.
WUTTKE,W.; SCANLAN, T.S. Contribution of a membrane estrogen receptor to the estrogenic
regulation of body temperature and energy homeostasis. Endocrinology. 151:4926–4937, 2010.
ROSSI, D.V.; DAI, Y.; THOMAS, P. ; CARRASCO, G.A. ; DON CARLOS, L.L.; MUMA, N.A.;
LI, Q.Estradiol-induced desensitization of 5-HT1A receptor signaling in the paraventricular nucleus of the
hypothalamus is independent of estrogen receptor-beta. Psychoneuroendocrinology, 35(7):1023-33,
2010.
SANTOLLO, J.; WILEY, M.D.; ECKEL, L.A. Acute activation of ER alpha decreases food intake,
meal size, and body weight in ovariectomized rats. American journal of physiology. Regulatory,
integrative and comparative physiology. 293:R2194–2201, 2007.

78
SANTOLLO, J.; TORREGROSSA, A.M. ECKEL, L.A. Estradiol acts in the medial preoptic area,
arcuate nucleus, and dorsal raphe nucleus to reduce food intake in ovariectomized rats. Horm
Behav.60(1):86-93, 2011.
SANTOLLO, J.; DANIELS, D. Control of fluid intake by estrogens in the female rat: role of the
hypothalamus. Front Syst Neurosci, 4;9:25, 2015.
SAR, M.; STUMPF, W.E. Simultaneous localization of [3H]estradiol and neurophysin I or arginine
vasopressin in hypothalamic neurons demonstrated by a combined technique of drymount autoradiography
and immunohistochemistry. NeurosciLett17: 179–184, 1980.
SCHULKIN, J. Sodium hunger: the search for a salty taste. Cambridge, Cambridge University Press,
1991.
SCUTERI, A.; BOS, A.J.; BRANT, L.J.; TALBOT, L.; LAKATTA, E.G.; FLEG, J.L. Hormone
replacement therapy and longitudinal changes in blood pressure in postmenopausal women. Ann Intern
Med.135(4):229–238, 2001.
SHI, H.; SORRELL, J.E.; CLEGG, D.J.; WOODS, S.C.; SEELEY, R.J. The roles of leptin receptors
on POMC neurons in the regulation of sex-specific energy homeostasis. Physiology & Behavior
100:165–172, 2010.
SHUGHRUE, P.J; MERCHENTHALER, I. Distribution of estrogen receptor beta immunoreactivity in
the rat central nervous system. J Comp Neurol 436: 64–81, 2001.
SIERRA-RAMIREZ. J.F.; LARA-RICALDEA, R.; LUJANC, M.; VELÁZQUEZ-RAMÍREZA, N.;
GODÍNEZ- VICTORIA, M.; HERNÁDEZ-MUNGUÍA.; PADILLAB,A.; GARZA-FLORESA,
J.Comparative pharmacokinetics and pharmacodynamics after subcutaneousand intramuscular
administration of medroxyprogesterone acetate (25 mg)and estradiol cypionate (5 mg) Contraception 84:
565–570, 2011.
SILVA, L.E.C.M; CASTRO, M.; AMARAL, F.C.; ANTUNES-RODRIGUES, J.; ELIAS, L.L.K.
Estradiol-induced hypophagia is associated with the differential mRna expression of hypothalamic
neuropeptides Braz J Med Biol Res, 43(8) 759-766, 2010.

79
SINCHAK, K.; WAGNER, E.J. Estradiol signaling in the regulation of reproduction and
energy balance. Front Neuroendocrinol. 33(4):342-63, 2012.
SLADEK, C.D.; SOMPONPUN, S.J. Estrogen receptors: their roles in regulation of vasopressin release
for maintenance of fluid and electrolyte homeostasis. Front Neuroendocrinol, 29 (1): 114-127, 2008.
SMITH, M.S.; FREEMAN, M.E.; NEILL, J.D. The control of progesterone secretion during estrous
cycle and early pseudopregnancy in the rat: prolactin, gonadotropin and steroid levels associated with
rescue of the corpus luteum of pseudopregnancy. Endocrinology. 96(1):219-26, 1975.
SOMPOPUN, S.J.; SLADEK, C.D. Osmotic regulation of estrogen receptor-beta in rat vasopressin and
oxytocin neurons. J Neurosci 23: 4261–4269, 2003.
SOMPONPUN, J. S. Neuroendocrine regulation of fluid and electrolyte balance by ovarian steroids:
contributions from central oestrogen receptors. J Neuroendocrinol, 19(10):809-18, 2007.
STACHENFELD, N.S.; KEEFE, D.L.; TAYLOR, H.S.Responses to a saline load in gonadotropin-
releasing hormone antagonistpretreated premenopausal women receiving progesterone or estradiol-
progesterone therapy. J Clin Endocrinol Metab 90: 386-394, 2005.
STACHENFELD, N.S. DIPIETRO, L.; PALTER, S.F.; NADEL, E.R.Estrogen influences osmotic
secretion of AVPand body water balance in postmenopausal women Am J Physiol. 274(1 Pt 2):R187-95,
1998.
STEPHENSON, L.A.; KOLKA, M.A. Plasma volume during heat stress and exercise in women. Eur J
Appl Physiol Occup Physiol 57: 373-381, 1988.
STONE, J.D.; CROFTON, J.T. SHARE, L. Sex differences in central cholinergic and angiotensinergic
control of vasopressin release. Am J Physiol Regulatory Integrative Comp Physiol 263: R1030–R1034,
1992.
STRICKER, E.M.; THIELS, E.; VERBALIS, J.G. Sodium apetite in rats after prolonged dietary
sodium deprivation:a sexually dimorphic phenomenon. Am. J. Physiol. 260, R1082–R1088, 1991.

80
SUSUKI, H.; BARROS, R.P.A.; SUGIYAMA, N.;KRISHNAN, B.C.;YADEN, B.C.; KIM, H.J.;
WARNER, M.; GUSTAFSSON, J.A. Involvement of estrogen receptor b in maintenance of serotonergic
neurons of the dorsal raphe Molecular Psychiatry 18: 674 -680, 2013.
THRASHER, T.N.; FREGLY, M.J. Effect of chronic treatment with an estrogen-progestogen
combination on beta adrenergic-induced thirst. Pharmacol Biochem Behav. 8(2):177-83, 1978.
THUNHORST, R.L.; EHRLICH, K.J.; SIMPSON, J.B. Subfornical organ participates in salt apetite.
Behav Neurosci, 104:637-42, 1990.
TOLLAN, A.; OIAN, P.; FADNES, H.O.; MALTAU, J.M. Evidence for altered transcapillary fluid
balance in women with the pré-menstrual syndrome. Acta Obstet Gynecol Scand72: 238–242, 1993.
TOTH, M.J.; TCHERNOF, A.; SITES, C.K; POEHLMAN, E.T. Effect of menopausal status on body
compositionand abdominal fat distribution. Int J Obes Relat Metab Disord. 24:226–231, 2000.
TRITOS, N.A.; SEGAL-LIEBERMAN, G. VEZERIDIS, P.S.; MARATOS-FLIER, E. Estradiol-
induced anorexia is independent of leptin and melanin-concentrating hormone. Obesity research. 12:716–
724, 2004.
VECSEI, P. Glucocorticoids: cortisol, corticosterone and compounds. Methods of hormone
radioimmunoassay, Acad Press, M:767-792, 1979.
VEGETO, E.; BELCREDITO, S.; ETTERI, S. Estrogen receptor-alpha mediates the brain
antiinflammatory activity of estradiol. Proc Natl Acad Sci100(16):9614–9619, 2003.
VERBALIS, J.G. BLACKBURN, R.E. HOFFMAN, G.E.; STRICKER, E.M. Establishing behavioral
and physiological functions of central oxytocin: insights from studies of oxytocin and ingestive
behaviours. Adv Exp Med Biol, 395:209-25, 1995.

81
VERLANDER, J.; TRAN, T.M.; ZHANG, L.;KAPLAN, M.R. ; HEBERT, S.C. Estradiol enhances
thiazide-sensitive NaCl cotransporter density in the apical plasma membrane of the distal convoluted
tubule in ovariectomized rats. J Clin Invest 101: 1661-1669, 1998.
VERNEY EB. The antidiuretic hormone and the factors which determine its release. Proc R Soc Lond B
Biol Sci. 135(878):25-106,1947.
VIJANDE, M.; COSTALES, M.; SCHIAFFINI, O. MARIN, B. Angiotensin-induced drinking: sexual
differences. Pharmacol Biochem Behav. 8(6):753-5, 1978.
VILHENA-FRANCO,T.; MECAWI, A.S.; ELIAS, L.L.K; ANTUNES-RODRIGUES,J. Oestradiol
potentiates hormone secretion and neuronal activation in response to hypertonic extracellular volume
expansion in ovariectomised rats. J Neuroendocrinol, 23, 481-489,2011.
VOKES, T.J.; WEISS, N.M., SCHREIBER, J.; GASKILL, M.B.; ROBERTOSON, G.L.
Osmoregulation of thirst and vasopressin during normal menstrual cycle. Am J Physiol Regulatory
IntegrativeComp Physiol254: R641–R647, 1988.
WANG, Z.Y.; YIN, L. Estrogen receptor alpha-36 (ER-α36): A new player in human breast cancer. Mol
Cell Endocrinol. doi: 10.1016/j.mce.2015.04.017. [Epub ahead of print]
WEIGT, C.; HERTRAMPF, T.; ZOTH, N.; FRITZMEIER, K.H.; DIEL, P. Impact of estradiol. ER
subtype specific agonists and genistein on energy homeostasis in a rat model of nutrition induced obesity,
Molecular and cellular endocrinology 351:227–238, 2012.
WOLF ,G. Refined salt apetite methodology for rats demonstrated by assessing sex differences. J.
Comp.Physiol.Psychol. 96, 1016–1021, 1982.
XU, H.; QIN, S.; CARRASCO, G.A.; DAI, Y. FILARDO E.J.; PROSSNITZ, E.R.; BATTAGLIA,
G. DONCARLOS, L.L. MUMA, N.A. Extra-nuclear estrogen receptor GPR30 regulates serotonin
function in rat hypothalamus. Neuroscience. 158:1599–1607, 2009.
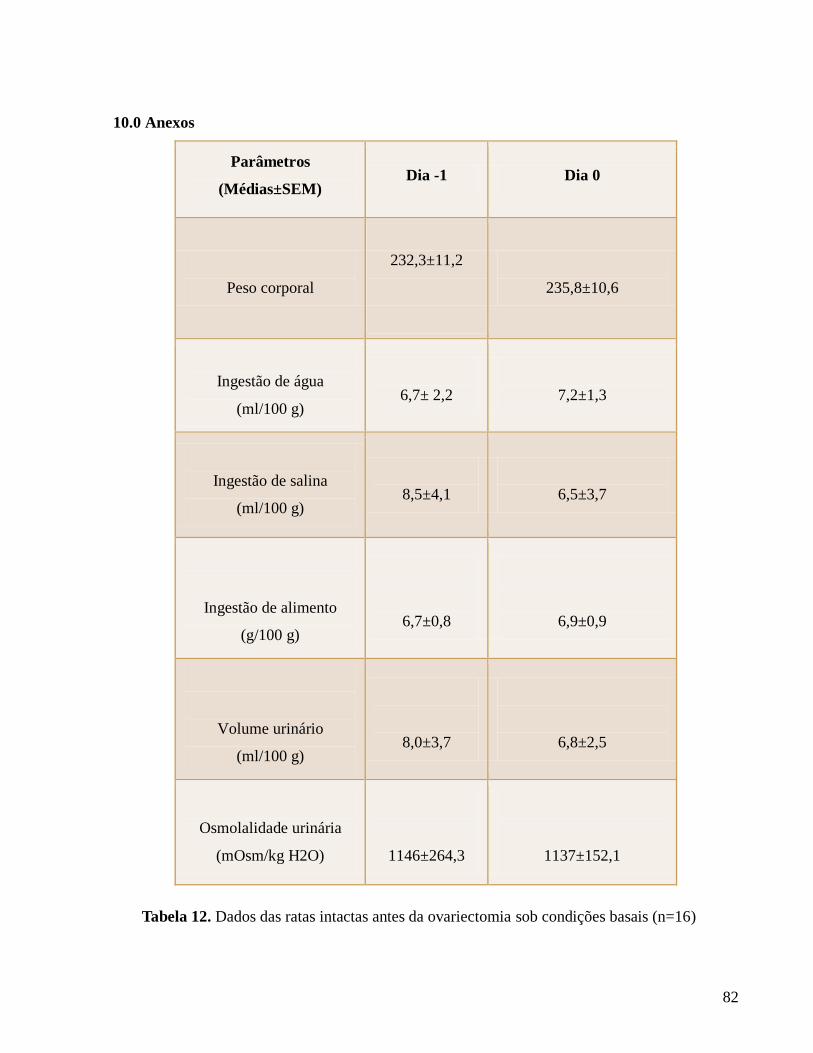
82
10.0 Anexos
Parâmetros
(Médias±SEM) Dia -1 Dia 0
Peso corporal
232,3±11,2
235,8±10,6
Ingestão de água
(ml/100 g)
6,7± 2,2
7,2±1,3
Ingestão de salina
(ml/100 g)
8,5±4,1
6,5±3,7
Ingestão de alimento
(g/100 g)
6,7±0,8
6,9±0,9
Volume urinário
(ml/100 g)
8,0±3,7
6,8±2,5
Osmolalidade urinária
(mOsm/kg H2O)
1146±264,3
1137±152,1
Tabela 12. Dados das ratas intactas antes da ovariectomia sob condições basais (n=16)

83


















