
Paulo Augusto de Lima Filho - repositorio.ufrn.br
92
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DIVERSIFICAÇÃO EVOLUTIVA DE GOBÍDEOS NO LITORAL BRASILEIRO: PADRÕES CITOGENÉTICOS E ECOMORFOLÓGICOS Paulo Augusto de Lima Filho Natal-RN 2011
Transcript of Paulo Augusto de Lima Filho - repositorio.ufrn.br

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
DIVERSIFICAÇÃO EVOLUTIVA DE GOBÍDEOS NO LITORAL BRASILEIRO:
PADRÕES CITOGENÉTICOS E ECOMORFOLÓGICOS
Paulo Augusto de Lima Filho
Natal-RN
2011

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
DIVERSIFICAÇÃO EVOLUTIVA DE GOBÍDEOS NO LITORAL BRASILEIRO:
PADRÕES CITOGENÉTICOS E ECOMORFOLÓGICOS
Paulo Augusto de Lima Filho
Dissertação apresentada ao Programa de Pós-
Graduação em Ecologia da Universidade
Federal do Rio Grande do Norte, como parte
dos requisitos para a obtenção do título de
mestre em Ecologia.
Orientador: Dr. Wagner Franco Molina
Natal-RN
2011

Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Lima Filho, Paulo Augusto de.
Diversificação evolutiva de gobídeos no litoral brasileiro: padrões citogenéticos e
ecomorfológicos / Paulo Augusto de Lima Filho. – Natal, RN, 2011.
90 f. : Il.
Orientador: Prof. Dr. Wagner Franco Molina.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências.
Programa de Pós-Graduação em Ecologia
1. Gobiidae – Dissertação 2. Evolução cariotípica – Dissertação. 3. Morfometria geométrica –
Dissertação. I. Molina, Wagner Franco. II. Universidade Federal do Rio Grande do Norte. IV. Título.
RN/UF/BSE-CB CDU 597.556.33

“Aprender é uma coisa
que nuca se cansa,
nunca se tem medo e
nunca se arrepende”
(Leonardo da Vinci)

“Dedico aos meus pais, familiares,
amigos e colegas, pelo apoio,
confiança e paciência”

AGRADECIMENTOS
Não só nesta trajetória, mas durante toda minha vida, agradeço a Deus, pois
me presenteou com a vida, a força e a coragem.
Agradeço a minha família pelo apoio, compreensão, carinho e paciência
demonstrado. Maior gratidão devo aos meus pais, Paulo Augusto de Lima, Maria da
Conceição Dutra de Andrade e minha “boa drasta” Geovana da Silva Nascimento,
pela palavra sempre oportuna.
À meus tios e tias dedico gratidão adicional por ter me apoiado neste sonho,
Dailva, Hugo, Dedé, Neide e Gracinete me acolhendo e auxiliando durante toda esta
trajetória no Rio Grande do Norte.
Agradeço também aos meus irmãos, Henrique, Ricardo, Isaura e Tatiane que
me deram forte base familiar, meus sobrinhos e primos.
A minha querida Dayse, agradeço pela ajuda neste projeto, pelo carinho,
apoio e compreensão estando sempre presente ao meu lado.
Agradeço aos meus antigos amigos e aos novos, que conquistei nesta
jornada, desde o Cursinho da Poli, CEDAP, graduação e pós-graduação na
Universidade Federal do Rio Grande do Norte, colegas residentes das republicas
que fiz parte, especialmente a Rogerson Rodrigues, grande amigo que me apoiou e
incentivou durante toda minha peregrinação acadêmica.
Agradeço a todas as pessoas que realmente me ajudaram no
desenvolvimento deste trabalho, os quais considero como amigos e colegas de
ofício. A meu ver, verdadeiramente praticaram ciência, democratizando o
conhecimento e trabalhando em equipe.
Sou grato aos meus amigos de trabalho no laboratório, pelo ambiente que
promoveram e pelos ensinamentos que melhoraram decisivamente minha pesquisa:
Clóvis, Pantera, Gideão, Amanda, Emanuel, Uedson, Calado, Pablo, Eurico,
Washington, Layse, Alyson e Guinga.
Agradeço ao meu orientador, professor Dr. Wagner Franco Molina, por
acompanhar e contribuir no meu desenvolvimento acadêmico, orientando-me com
liberdade e exigência, permitindo o meu desenvolvimento científico e a realização
deste trabalho.
Agradeço ao programa de Pós-Graduação em Ecologia, pela oportunidade de
mestrado, sua equipe de funcionários e docentes pelo apoio e conhecimentos
adquiridos durante o decorrer deste trabalho.

Agradeço a Universidade Federal do Rio Grande do Norte (UFRN), ao Centro
de Biociências (CB) e ao Departamento de Biologia Molecular e Genética (DBMG).
Agradeço a CAPES (Coordenação de Aperfeiçoamento de Pessoal de nível
Superior), pela bolsa de estudos oferecida. Ao CNPq (Conselho Nacional de
Desenvolvimento Científico e Tecnológico) pelo financiamento deste trabalho, sob
processo de número: Nº556793/2009-9 e ao IBAMA pela autorização de coleta das
espécies (Processo Nº 19135/1).

SUMÁRIO
Lista de Figuras VIII
Lista de Tabelas XI
Lista de Abreviaturas XII
Resumo XIII
Abstract XIV
1. INTRODUÇÃO 1
1.1. Principal componente vertebrado marinho 1
1.2. Aspectos citogenéticos da família Gobiidae 3
1.3. A evolução refletida na morfologia 5
2. OBJETIVOS 8
3. MATERIAL E MÉTODOS 9
3.1. Material 9
3.2. Métodos 11
3.2.1. Técnica de estimulação mitótica 11
3.2.2. Técnica de obtenção de cromossomos mitóticos 11
3.2.3. Preparação das lâminas 12
3.2.4. Análises cromossômicas 12
3.2.5. Detecção de regiões organizadoras de nucléolos (RONs) 12
3.2.6. Detecção da heterocromatina constitutiva (Banda-C) 14
3.2.7. Coloração com fluorocromos base-específico 14
3.2.8. Análises por morfometria geométrica (MG) 14
4. CAPÍTULOS 15
4.1. Caracterização da diversidade na família Gobiidae por meio da citogenética e morfometria geométrica
15
4.2. Análises citogenéticas e morfométricas interpopulacionais no peixe marinho Bathygobius soporator (Gobiidae - Perciformes)
44
5. CONCLUSÕES 61
6. REFERÊNCIAS BIBLIOGRÁFICAS 71

LISTA DE FIGURAS
Figura 1. Desenho morfológico esquemático de Gobiidae (gênero Bathygobius). Em destaque (a) nadadeiras dorsais separadas, primeira nadadeira dorsal com raios, olhos grandes, ausência de linha lateral; (b) nadadeiras pélvicas fusionadas presentes na maioria das espécies
2
Figura 2. Mapa com pontos de coleta dos exemplares da família Gobiidae. Litoral do Maranhão (MA), Rio Grande do Norte (RN), Bahia (BA) e Atol das Rocas.
9
Figura 3. Espécies analisadas no presente estudo: a. Ctenogobius smaragdus, b. Ctenogobius boleosoma, c. Bathygobius soporator, d. Bathygobius
mystacium, e. Coryphopterus glaucofraenum.
10
CAPÍTULO I
Figura 1. Imagem digitalizada de Coryphopterus glaucofraenum retratando os 11 landmarks utilizados nas análises morfológicas interespecífica em Gobiidae. Descrição esquemática dos landmarks. 1. Ponto distal do osso pré-maxilar; 2. Origem da primeira nadadeira dorsal; 3. Término da primeira nadadeira dorsal; 4. Origem da segunda nadadeira dorsal; 5. Término da segunda nadadeira dorsal; 6. Término da nadadeira anal; 7. Origem da nadadeira anal; 8. Origem da nadadeira pélvica; 9. Margem anterior ocular; 10. Margem posterior ocular; 11. Base superior da nadadeira peitoral. Barra=0,5cm.
20
Figura 2. Cariótipo de Ctenogobius smaragdus a partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
22
Figura 3. Cariótipo de Ctenogobius boleosoma a partir de coloração convencional (a). Em destaque (b) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
22
Figura 4. Metáfase de C. smaragdus (a) e C. boleosoma (b), exibindo marcações CMA3
+ na maior parte dos cromossomos em posição centromérica e terminal. Barra=5µm.
22
Figura 5. Cariótipo de Bathygobius soporator a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (4º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
23
Figura 6. Cariótipo de Bathygobius mystacium a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (5º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
23

Figura 7. Cariótipo de Coryphopterus glaucofraenum a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
24
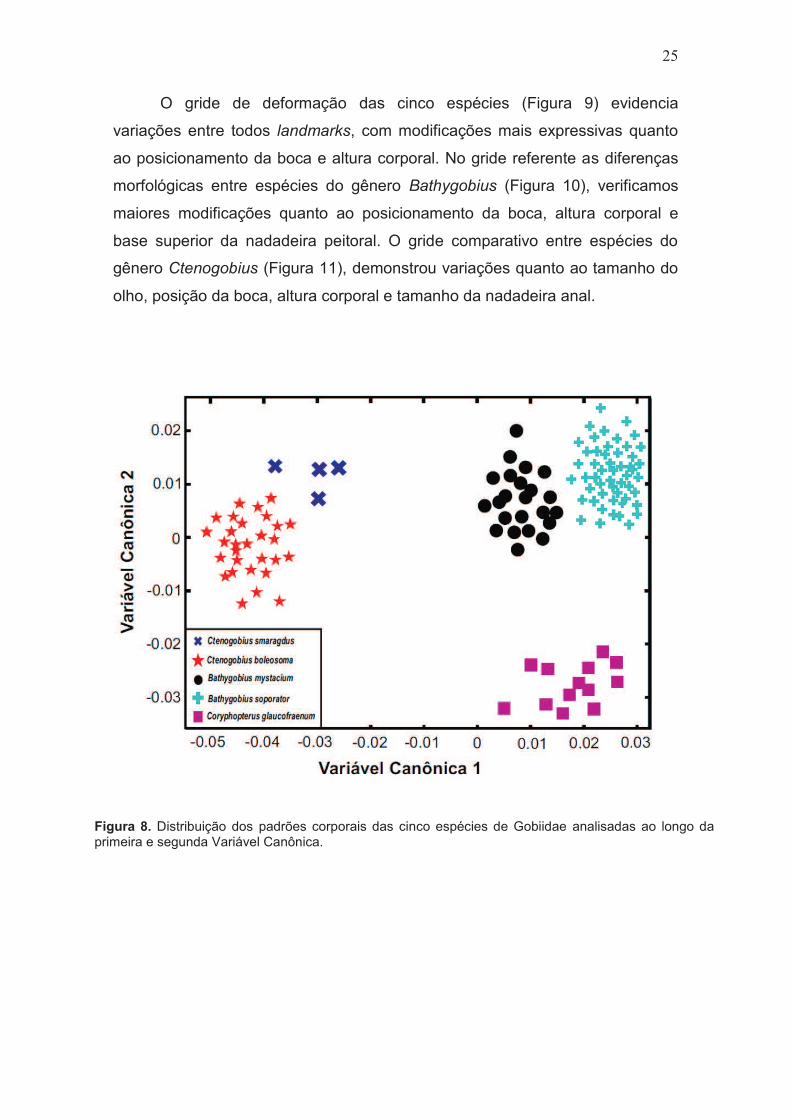
Figura 8. Distribuição dos padrões corporais das cinco espécies de Gobiidae analisadas ao longo da primeira e segunda Variável Canônica.
25
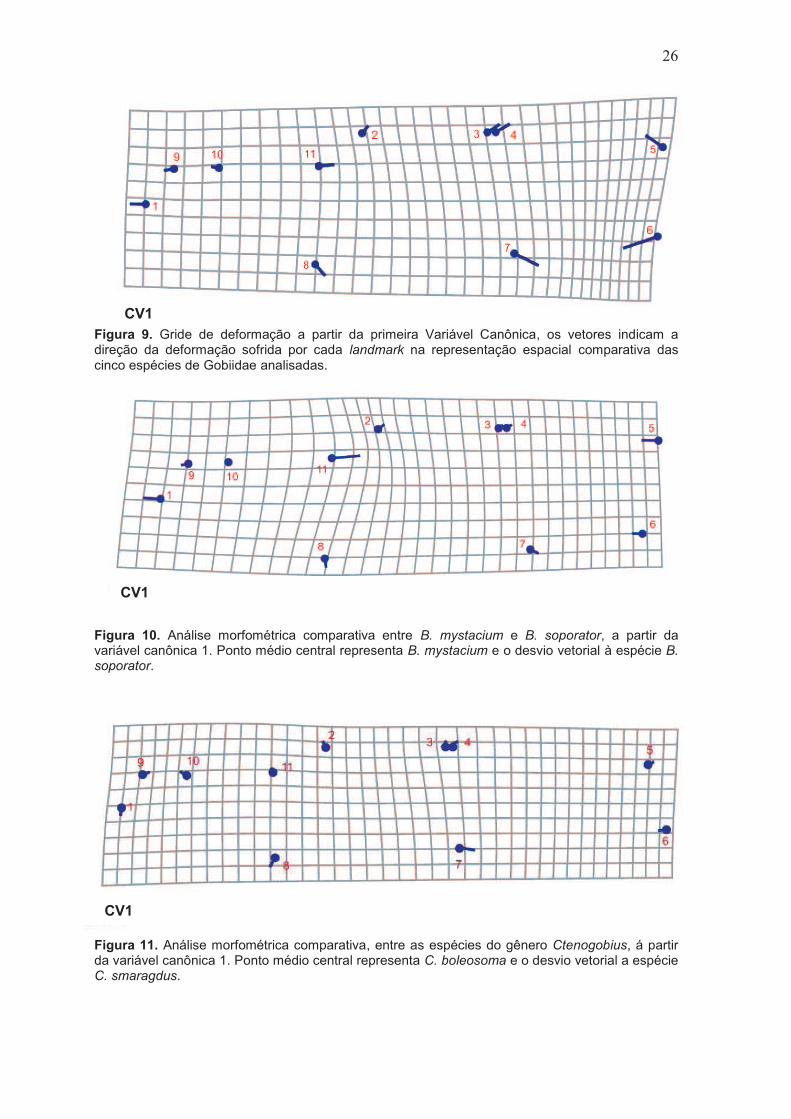
Figura 9. Gride de deformação a partir da primeira Variável Canônica, os vetores indicam a direção da deformação sofrida por cada landmark na representação espacial comparativa das cinco espécies de Gobiidae analisadas.
26
Figura 10. Análise morfométrica comparativa entre B. mystacium e B. soporator, a partir da variável canônica 1. Ponto médio central representa B.
mystacium e o desvio vetorial à espécie B. soporator.
26
Figura 11. Análise morfométrica comparativa, entre as espécies do gênero Ctenogobius, à partir da variável canônica 1. Ponto médio central representa C. boleosoma e o desvio vetorial a espécie C. smaragdus.
26
CAPÍTULO II
Figura 1. Imagem digitalizada de B. soporator retratando os 09 landmarks definidos para análises morfológicas interpopulacionais. Descrição esquemática dos landmarks. 1. Ponto distal do osso pré-maxilar; 2. Origem da primeira nadadeira dorsal; 3. Término da primeira nadadeira dorsal; 4. Origem da segunda nadadeira dorsal; 5. Término da segunda nadadeira dorsal; 6. Término da nadadeira anal; 7. Origem da nadadeira anal; 8. Origem da nadadeira pélvica; 9. Base infra-ocular medial. Barra=1,5cm.
49
Figura 2. Cariótipo de B. soporator para populações contientais (MA, RN e BA) à partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (4º) corado com impregnação argêntea, CMA3 e DAPI. Barra= 5µm.
50
Figura 3. Cariótipo de B. soporator população do Atol das Rocas, a partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra= 5µm.
50
Figura 4. Distribuição das amostras de B. soporator ao longo da primeira e segunda variável canônica (63,5% e 30,5% da variância respectivamente), a partir das variáveis morfométricas.
52
Figura 5. Distribuição das populações continentais amostradas de B. soporator ao longo da primeira e segunda variável canônica (76,2% e 23,5% da variância respectivamente). Os símbolos em destaque indicam a média morfométrica de cada população.
52

Figura 6. Gride comparativo entre população continental e insular de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população do Atol das Rocas.
53
Figura 7. Gride comparativo entre populações continentais de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população do Maranhão.
53
Figura 8. Gride comparativa entre populações continentais de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população da Bahia.
54

LISTA DE TABELAS
CAPÍTULO I
Tabela 1. Número de indivíduos utilizados para estudos morfométricos e citogenéticos.
19
Tabela 2. Dados citogenéticos das cinco espécies de Gobiidae analisadas. 21
Tabela 3. Resultados post-hoc ANOVA a partir de procrustes, classificadas com a média do centróide, efeito individual, para todos os espécimes em estudo e espécies dos gêneros Bathygobius e Ctenogobius.
24
Tabela 4. Dados citogenéticos disponíveis da família Gobiidae (Perciformes). 28
CAPÍTULO II
Tabela 1. Número de indivíduos coletados, seus respectivos pontos de coleta e sua utilização para estudos morfométricos e citogenéticos.
48
Tabela 2. Porcentagem de indivíduos alocados para cada população e suas interfaces baseados na CVA.
51
Tabela 3. Resultados post-hoc ANOVA a partir de procrustes, classificadas com a média do centróide, efeito individual, entre as populações continentais.
51

LISTA DE ABREVIATURAS
a – Acrocêntrico
AgNO3 – Nitrato de prata
Ag-RONs – Regiões organizadoreas de nucléolo evidenciadas pela impregnação com nitrato de prata
ANOVA – Análise de variância
AR – Atol das Rocas
AT – Adenina e Timina
BA – Bahia
BC – Bandamento C
BRA - Brasil
CMA3 – Cromomicina A3
CVA – Análise de variáveis canônicas
DAPI – 4`4’,6-diamidino-2-fenilindol
DNA – Ácido desoxirribonucléico
EUA – Estado Unidos da América
GC - Guanina e Citosina
LGRM – Laboratório de Genética de Recursos Marinhos
m – Metacêntrico
MA – Maranhão
MANOVA – Análise multivariada de variância
MG – Morfometria geométrica
NE – Nordeste
NF – Número de braços cromossômicos
N-S – Norte Sul
p – Probabilidade estatística do teste
RN – Rio Grande do Norte
sm - Submetacêntrico
st – Subtelocêntrico
UFRN – Universidade Federal do Rio Grande do Norte
VC – Variável canônica

RESUMO
Os peixes da ordem Perciformes caracterizam-se como um modelo importante para o entendimento da estrutura genética de populações marinhas, pois apresentam tanto exemplos de conservação cromossômica como de diversificação cariotípica para alguns grupos. Gobiidae é a família mais especiosa no ambiente marinho. Entre seus representantes, muitas espécies fazem parte de uma fauna críptica pouco percebida e estudada, apresentando ampla distribuição com características comportamentais e reprodutivas que as tornam propícias à ação de barreiras biogeográficas. Morfologicamente esta família apresenta estruturas corporais reduzidas através de simplificações e regressões. Dados citogenéticos, apesar da importância em inferências evolutivas, são incipientes frente sua diversidade de espécies, sobretudo para espécies do Atlântico Ocidental. Visando estimar a diversidade evolutiva em Gobiidae foram desenvolvidas análises citogenéticas e dos padrões corporais, através de morfometria geométrica, em cinco espécies presentes no litoral brasileiro, Coryphopterus glaucofraenum, Bathygobius mystacium, B.
soporator, Ctenogobius smaragdus e C. Boleosoma. Os dados obtidos demonstram marcante diversidade cariotípica e morfológica entre as espécies. As inversões pericêntricas e fusões desempenham papel importante na evolução cromossômica desta família, ocasionando diferenças cariotípicas estruturais e numéricas em todas as espécies. Comparações cariotípicas e morfológicas entre amostras geográficas de B. soporator provenientes do litoral do Maranhão, Rio Grande do Norte e Bahia evidenciaram padrões citogenéticos comuns, mas padrões morfológicos diferentes. Uma amostra proveniente do Atol das Rocas revelou conspícua diferenciação cariotípica e morfológica das demais populações continentais, confirmando a presença de uma nova espécie insular. As abordagens empregadas revelam diversificações compatíveis com um grupo com características de baixa vagilidade e amplamente sujeito a seleção ambiental decorrente de exigências ecológicas peculiares.
Palavras-Chave: Gobiidae, evolução cariotípica, morfometria geométrica,
Coryphopterus, Bathygobius, Ctenogobius.

ABSTRACT
The fishes of the order Perciformes are characterized as an important model for understanding the genetic structure of marine populations, because besides they present examples of conservation chromosomal, also they present the karyotype diversification for some groups. Gobiidae family is the most specious in the marine environment. Among its representatives, many species are part of a cryptic fauna little noticed and studied, a wide distribution with behavioral and reproductive characteristics, that make them conducive to the action of biogeographical barriers. Morphologically this family presents reduced body structures through simplification and regressions. Despite their importance in evolutionary inferences, cytogenetics data are incipient facing their species diversity, especially with western Atlantic species. In order to estimate the evolutionary diversity in Gobiidae, it were developed cytogenetic analysis and the standards body, through geometric morphometrics in five species on the Brazilian coast, Coryphopterus glaucofraenum, Bathygobius
mystacium, B. soporator, Ctenogobius smaragdus e C. Boleosoma. The data show significant karyotype and morphological diversity among the species. The pericentric inversions and mergers play an important role in chromosomal evolution of this family, causing karyotypic structural and numerical differences in all species. Karyotypic and morphological comparisons among geographic samples of B. soporator from the coast of Maranhão, Rio Grande do Norte and Bahia showed cytogenetics patterns commons, but different morphological patterns. A sample from the Atol das Rocas revealed conspicuous morphological and karyotypic differentiation of another continental populations, confirming the presence of a new island species. The approaches done reveal diversification consistent with characteristics of a group of low vagile and largely able to environmental selection due from peculiar ecological requirements.
Keywords: Gobiidae, karyotype evolution, geometric morphometry, Coryphopterus,
Bathygobius, Ctenogobius.

1
1. INTRODUÇÃO
1.1 Principal componente vertebrado marinho
Os peixes constituem um grupo polifilético com certa de 28.000
espécies, o que corresponde a mais da metade das formas vivas de
vertebrados. Deste total, cerca de 60% vive no ambiente marinho e a sua
maioria habita as zonas costeiras (Nelson, 2006). Representam o grupo de
vertebrados com maior êxito em termos de diversificação devido à grande
variabilidade de habitats presentes no meio marinho e dulcícola.
A ordem Perciformes com 10.033 espécies é a mais diversificada de
todas as ordens de peixes, dominantes nos oceanos e em muitos ambientes
dulcícolas tropicais e subtropicais. Dentre os Perciformes a subordem
Gobioidei possui um número estimado de aproximadamente 2.211 espécies,
distribuídas em 270 gêneros, correspondendo a aproximadamente 22% dos
Perciformes (Pezold, 1984; Nelson, 2006). Nesta subordem, a família Gobiidae
é a que apresenta maior número de espécies (Robertson, 1998; Nelson, 2006),
considerada monofilética, possui 210 gêneros e um número estimado de 1.950
espécies (Pezold, 1984; Tracker, 2003; Nelson, 2006), o que corresponde a
cerca de 88% da subordem Gobioidei (Pezold, 1993).
Gobiidae compartilha características crípticas, tanto morfológicas,
biológicas e comportamentais, que aliadas a grande quantidade de espécies e
ao número insuficiente de informações biológicas existentes, contribui para a
imprecisão em algumas caracterizações taxonômicas.
Anatomicamente apresentam suas nadadeiras pélvicas fundidas
formando um disco de sucção que auxilia o animal em seu posicionamento no
ambiente, além disso, exibem nadadeiras dorsais separadas, primeira
nadadeira dorsal com espinhos, olhos grandes e ausência de linha lateral
(Nelson, 2006) (Figura 1). A maior parte das espécies é bentônica, vivendo em
tocas ou esconderijos em ambientes marinhos e estuarinos. Contudo, algumas
formas exibem inúmeras adaptações ecológicas, como por exemplo, formas
nectônicas ou habilidade para respirar o oxigênio atmosférico (Robertson,

2
1998; Tracker, 2003).
Quanto aos aspectos biogeográficos, este grupo demonstra-se
amplamente distribuído podendo ser encontrado ao longo do Atlântico, Pacífico
e Índico em áreas tropicais, subtropicais e temperadas (Menezes & Figueiredo,
1985; Mendes, 2000; Tracker, 2003; Nelson, 2006). Apresenta representante
em habitats estuarinos e dulcícolas, freqüentemente sendo os mais abundantes
em água doce no interior de ilhas oceânicas (Nelson, 2006).
Figura 1. Desenho morfológico esquemático de Gobiidae (gênero Bathygobius). Em destaque (a) nadadeiras dorsais separadas, primeira nadadeira dorsal com raios, olhos grandes, ausência de linha lateral; (b) nadadeiras pélvicas fusionadas presentes na maioria das espécies (adaptado de Nelson, 2006).
Os Gobídeos representam elementos dominantes em habitats
bentônicos nos recifes tropicais, podendo ser morfologicamente crípticos em
relação a membros da própria família e outras como Labrisomidae, Blenniidae
(Sale, 1991; Galvão, 2006) e Eleotridae, em áreas estuarinas. A maior parte
das espécies apresenta tamanho reduzido, não ultrapassando 10 cm de
comprimento, seu maior representante Glossogobius giuris, atinge cerca de 50
cm e o menor Trimmatom nanus corresponde ao menor vertebrado existente,
uma fêmea madura atinge 8-10mm (Nelson, 2006).
Seus hábitos alimentares são diversos, sendo em sua maioria carnívoros
e onívoros, com alguns representantes herbívoros. Muitas espécies que vivem
próximas a costa, se alimentam principalmente de invertebrados bentônicos e
algas. Em contraste a esta situação, espécies em habitats recifais podem
apresentar hábitos alimentares mais especializados. Diversas espécies são
semi-pelágicas e se alimentam de copépodas planctônicos. Espécies
b a

3
dulcícolas muitas vezes se alimentam de diatomáceas e microalgas
(Emmanuel & Ajibola, 2010). É comum uma estreita associação entre
diferentes espécies de gobídeos com outros animais, como esponjas, moréias,
camarões e ouriços do mar. Espécies de pequeno porte podem estar
associadas a peixes maiores, alimentando-se de ectoparasitas e realizando a
limpeza destes (Colin, 1975).
Os representantes de Gobiidae apresentam considerável importância
ecológica e comercial, sua elevada abundância em determinadas localidades
os tornam um componente fundamental na cadeia alimentar (De Sylva, 1975).
Várias espécies são pescadas e comercializadas de forma significativa, onde
são utilizados como iscas na pesca de peixes de maior porte (Emmanuel &
Ajibola, 2010). Em diversos países Ocidentais devido suas cores exuberantes,
comportamentos peculiares e tamanhos apropriados, algumas espécies são
intensamente comercializadas no segmento da aquariofilia, compondo de 5 a 7
% das vendas no mercado mundial de peixes ornamentais marinhos (Wabnitiz
et al., 2003). No Brasil a espécie Elacatinus figaro já foi uma das mais
importantes no comércio de exportação de peixes ornamentais marinhos do
país (Gasparini et al., 2005), mas atualmente encontra-se na lista de espécies
ameaçadas de extinção, resguardada do extrativismo (IBAMA, 2004).
A diversidade de espécies, amplo padrão de distribuição, características
ecológicas e comportamentais peculiares, evidenciadas por representes da
família Gobiidae demonstram a grande capacidade adaptativa dos peixes que a
compõe, cuja carência de informações existentes para o grupo, os torna
importante modelo para análises evolutivas no ambiente marinho.
1.2 Aspectos citogenéticos da família Gobiidae
Os Perciformes apresentam tanto exemplos de conservação
cromossômica, compartilhada por muitas espécies, quanto de diversificação
cariotípica em alguns grupos (Brum & Galetti, 1997), dos quais, polimorfismos
cromossômicos numéricos e estruturais, além de vários sistemas sexuais
recorrentes (Galetti et al., 2000). Aproximadamente 8% das espécies de
Perciformes foram cariotipadas (Galetti et al., 2000; Afonso & Galetti, 2005;

4
Galetti et al., 2006; Molina & Bacurau, 2006). Apesar da maioria dos seus
representantes serem marinhos, neste ambiente apenas 2% das espécies
apresentam dados citogenéticos (Almeida-Toledo et al., 1993; Brum, 1996).
Cerca de 60% dos Perciformes estudados até então, apresentam um
cariótipo caracterizado por 48 cromossomos monobraquiais, o que faz acreditar
que este seja um caráter plesiomórfico para o grupo (Galetti et al., 2000). As
informações cariotípicas disponíveis para Perciformes marinhos têm
confirmado esta hipótese (Klinkhardt et al., 1995; Brum & Galetti, 1997), o que
demonstra para estas espécies que a especiação em alguns grupos, parece
não ter sido seguida por diferenciação cariotípica significante.
A família Gobiidae destaca-se por apresentar grande diversidade
cariotípica (Galetti et al., 2000), com cariótipos variando de 2n=30, em
Neogobius euricephalus (Ene, 2003), a 2n=52, em Gobius niger (Vitturi &
Catalano, 1989). Apesar da grande diversidade cariotípica, a família exibe com
maior freqüência espécies que apresentam valores diploides com 44, 46 e 48
cromossomos (Ene, 2003). Dados cromossômicos estão disponíveis para
aproximadamente 4,5% das espécies da família Gobiidae (Klinkhardt et al.,
1995), sendo a sua maioria restrito a identificação dos valores diplóides e da
macroestrutura cariotípica (Caputo, 1998). No litoral brasileiro são encontradas
cerca de 40 espécies de Gobiidae (Menezes et al., 2003), destas apenas duas
possuem dados citogenéticos, Bathygobius soporator, com 2n=48 e Awaous
strigatus, com 2n=46 (Brum et al.,1996; Souza et al., 1998; Cipriano et al.,
2002).
Para representantes da ordem Perciformes, um dos mecanismos que
permite explicar a existência de diversificação cromossômica estrutural, é a
ocorrência de inversões pericêntricas, que constituem os rearranjos
cromossômicos mais frequentes nessa ordem (Cano et al., 1982; Galetti et al.,
2000; Ueno & Takai, 2000; Molina & Galetti, 2004). A variabilidade numérica
derivada de processos de fissão e fusão tem sido relatada em algumas famílias
como Pomacentridae, Labridae, Apogonidae, entre outras (Molina & Galetti,
2002; Sena & Molina, 2007; Araújo et al., 2010). Em Gobiidae rearranjos
Robertsonianos estão entre os principais mecanismos de diferenciação
cromossômica (Thode et al., 1988; Amores et al., 1990; Vasil’ev & Grigoryan,
1993; Caputo et al., 1997; Galetti et al., 2000; Ene, 2003).

5
A presença de variabilidade cariotípica intra e interpopulacional foram
constatadas em algumas espécies da família como, por exemplo, Aphia minuta
que apresentou cinco citótipos distintos em uma população de onze indivíduos
presentes na costa de Ortona, Itália (Caputo et al., 1999). Espécies do gênero
Gobius, como G. niger e G. paganellus, exibem populações cariotipicamente
monomórficas (Klinkhardt, 1992; Vasil'ev & Grigoryan, 1992; Caputo et al.,
1997) e populações com polimorfismo acentuado, envolvendo vários tipos de
rearranjos cromossômicos (Vitturi & Catalano, 1989; Amores et al., 1990;
Caputo et al., 1997).
As modificações frequentes e complexas na estrutura cariotípica dos
Gobídeos parecem estar ligadas ao padrão evolutivo deste grupo (Caputo et
al., 1997). Embora os estudos cromossômicos em gobídeos estejam longe de
ser completos, sua abrangência as espécies do oeste do Atlântico contribuirá
substancialmente para o conhecimento da diversidade cariotípica e seus
mecanismos regentes nesta região, o que até então é desconhecido.
1.3 A evolução refletida na morfologia
Pesquisas relacionadas ao formato corporal dos organismos assumem
importante papel em estudos biológicos, uma vez que podem refletir
experiências biológicas e evolutivas (Bookstein, 1991; Zelditch et al., 2004),
tendo como base o conceito de que o fenótipo apresentado por um organismo
decorre da interação existente entre um dado genótipo e o meio ambiente em
que se encontra (Ricklefs & Miles, 1994) e que a evolução de um mesmo
fenótipo em populações naturais pode ser evidência de seleção natural e
adaptação (Mcguigan et al., 2005).
Em contrapartida, variações morfológicas podem afetar o
comportamento e interação ecológica dos organismos, principalmente se
levarmos em conta características físicas e químicas do ambiente (Webb,
1984). Assim, o formato corporal pode ter relevância evolutiva e ecológica para
um dado organismo (Klingenberg et al., 2003). Diante disto, análises de
estruturas morfológicas como as morfométricas podem ser de grande valia

6
(Schluter, 1993), sendo utilizada em distintas abordagens biológicas durante
anos (Monteiro et al., 2002; Rosenberg, 2002).
A morfometria é a análise da forma do corpo em relação ao tamanho
através de métodos numéricos, propiciando a interpretação e comparação dos
padrões de variação de caracteres quantitativos (Blackith & Reyment, 1971;
Cavalcanti & Lopes, 1993).
Historicamente, o estudo da variação corporal consistia em medidas
relativas de estruturas anatômicas que eram analisadas por meio de arranjos
canônicos, componentes principais e outros recursos estatísticos multivariados,
configurando a técnica denominada de morfometria tradicional (Monteiro et al.,
2002).
A morfometria geométrica (MG) é um conjunto de técnicas
computacionais e estatísticas, que no estudo da forma de estruturas biológicas
possibilita, além de analisar e quantificar, visualizar graficamente a variação de
forma da estrutura estudada, considerando simultaneamente várias
características de uma estrutura corporal complexa, em que são utilizados
marcos anatômicos ou landmarks (Rohlf, 1993; Monteiro & Reis, 1999). A
utilização de landmarks comuns às amostras analisadas permite identificar as
variações de forma entre as mesmas estruturas morfológicas nos diferentes
exemplares em estudo.
Na superposição de marcos anatômicos, a variação de forma é estudada
nas próprias coordenadas dos mesmos, após a retirada dos efeitos de
tamanho, orientação e posição. Logo, se as configurações estão centradas em
um ponto comum, proporcionalizadas de forma que tenham o mesmo tamanho
e rotacionadas, as coordenadas encontradas localizam um ponto no espaço de
forma (Shape Space), chamado de espaço da forma de Kendall (Kendall,
1984;1989; Dryden & Mardia, 1998; Rohlf, 1999).
Esta nova abordagem permite a descrição da variabilidade usando uma
análise estatística robusta bem como a utilização de Thin-plate spline (TPS)
para ilustrar e descrever os resultados em termos de deformações. Uma das
principais inovações da MG foi a reformulação das idéias de D’Arcy Thompson,
que visualizava as formas biológicas como distorções de um sistema de
coordenadas geométricas (Thompson, 1917).

7
O estudo da morfometria tem interessado diversas áreas do
conhecimento mostrando-se de grande importância na compreensão da
biologia evolutiva dos organismos (Blackith & Reyment, 1971). É considerada
uma ferramenta muito útil na sistemática, descrição de estruturas
populacionais, identificação de estoques pesqueiros, dimorfismo sexual e
fornecimento de conhecimento sobre o grau de variação intra e interespecífico
(Fonteles Filho,1989; Peres, et al., 1995; Ehliger, 1991; Vidales et al., 1997;
Fairbairn, 1997; Baras, 1999; Silva, 2000; Fulford & Rutherford, 2000; Rapp Py-
Daniel & Cox Fernandes, 2005; Aguirre & Shervette, 2005).
No caso dos peixes da família Gobiidae, que representam um
excepcional exemplo de especiação e diversificação, com alto índice de
endemismos nos recifes tropicais, o conhecimento do grau de variação
morfológico, seja ele interespecífico, intrapopulacional e interpopulacional,
associada a ferramentas citogenéticas e de genética molecular, pode nos
auxiliar na identificação dos mecanismos evolutivos atuantes na diversificação
desta família.

8
2. OBJETIVOS
2.1 Objetivo geral
Diante das informações disponíveis o presente trabalho se propôs
investigar aspectos de diversificação na família Gobiidae, analisando espécies
do Atlântico, por meio de análises citogenéticas resolutivas, aliadas a análises
dos padrões corporais existentes em níveis inter e intraespecíficos a fim de
identificar a presença de processos evolutivos neste peculiar grupo de peixes.
2.2 Objetivos específicos
· Analisar citogeneticamente as espécies Bathygobius mystacium, B.
soporator, Ctenogobius smaragdus, C. boleosoma e Coryphopterus
glaucofraenum, utilizando coloração convencional, bandamento C,
impregnação por nitrato de prata e coloração com fluorocromos base-
específicos (CMA3 e DAPI);
· Estabelecer uma comparação dos padrões morfológicos das espécies
Bathygobius mystacium, B. soporator, Ctenogobius smaragdus, C.
boleosoma e Coryphopterus glaucofraenum, por meio da técnica de
morfometria geométrica, com fins de aferição ecomorfológica,
taxonômica e sistemática;
· Analisar possíveis estruturações populacionais na espécie Bathygobius
soporator ao longo do litoral brasileiro (MA, RN e BA) e Atol das Rocas,
através da associação de características citogenéticas entre as
populações dessas localidades e os padrões corporais de cada
população.

9
3. MATERIAL E MÉTODOS
3.1 Material
Os exemplares da família Gobiidae, foram coletados ao longo do litoral
brasileiro no litoral dos estados do Rio Grande do Norte, praia de Búzios
(5°41'57.41"S, 35°11'34.71"O) e Barra do Cunhaú (6°19'15.50"S,
35°2'29.31"O), da Bahia, cidade de Salvador (13° 0'39.61"S, 38°31'27.21"O),
do Maranhão, cidade de São Luís (2°28'14.06"S, 44°15'49.15"O) e Atol das
Rocas (3º51’’S, 33º49’’W) (Figura 2), com o auxílio de rede de confecção
própria e sacos plásticos.
Figura 2. Mapa com pontos de coleta dos exemplares da família Gobiidae. Litoral do Maranhão (MA), Rio Grande do Norte (RN), Bahia (BA) e Atol das Rocas.

10
As espécies utilizadas nas análises foram Bathygobius mystacium
(Ginsburg, 1947), n=23; Bathygobius soporator (Valenciennes, 1837), n=170;
Ctenogobius smaragdus, (Valenciennes, 1837), n=04; Ctenogobius boleosoma,
(Jordan e Gilbert, 1882), n=29 e Coryphopterus glaucofraenum (Gill, 1863),
n=13 (Figura 2). As mesmas foram identificadas a partir da chave de
classificação de Bohlke & Chaplin (1993) e Hoese & Murdy (2002).
Figura 3. Espécies analisadas no presente estudo. a. Ctenogobius smaragdus, b. Ctenogobius boleosoma, c. Bathygobius soporator, d. Bathygobius mystacium, e. Coryphopterus glaucofraenum. Barra=1 cm.
Os espécimes utilizados para estudos morfológicos e citogenéticos
foram mantidos em aquários aerados até serem fotografados e submetidos ao
processo de estimulação mitótica.
Exemplares testemunho de cada espécie se encontram preservados em
formol 5% no Laboratório de Genética de Recursos Marinhos, para posterior
depósito no Museu de Ciências Morfológicas da UFRN.
a b
c
d
e

11
3.2 Métodos
3.2.1 Técnica de estimulação mitótica
Os exemplares foram estimulados mitoticamente através da técnica
preconizada por Molina (2001) e Molina et al. (2010), que faz uso de complexos
de antígenos do ar, bacterianos e fúngicos (Munolan® e Aminovac®). Este
procedimento consiste na inoculação intramuscular de solução do composto (2
comprimidos/1ml de água destilada); na proporção de 1ml/50g de peso
corporal, por um período de 24 a 48 horas. Decorrido este tempo, os
exemplares foram anestesiados com óleo extraído de cravo (Syzygium
aromaticum) (Eugenol) e posteriormente sacrificados para extração do rim
cefálico.
3.2.2 Técnica de obtenção de cromossomos mitóticos
A técnica de obtenção de cromossomos mitóticos adotada seguiu o
método preparação in vitro descrito por Gold et al. (1990). Os exemplares
tiveram o rim anterior (cefálico) e posterior removidos e utilizados para
obtenção de suspensão celular. Estes foram colocados em 9 ml de meio de
cultura RPMI 1640 e dissociados com ajuda de seringas de vidro, transferindo
o material logo em seguida para tubos falcon de 15ml, completando o volume
até 10ml do mesmo. Foi adicionada solução de colchicina 0,025% por 30
minutos, para cessar a divisão mitótica, deixando agir em temperatura
ambiente. Em seguida o material foi centrifugado por 10 minutos a 900rpm.
Após o excedente ser removido acrescentamos 9 ml de solução hipotônica de
KCl (0,075M), por 28 a 30 minutos em temperatura ambiente, ao término
novamente centrifugado por 10 minutos. Solução fixadora de metanol e ácido
acético (3:1) foi utilizada para fixação do material em três ciclos de
centrifugação por 10 minutos, sendo o material conservado à -20ºC em tubos
de 1,5ml com tampa para análises posteriores.

12
3.2.3 Preparação das lâminas
Um total de três ou quatro gotas de suspensão celular foram gotejadas
sobre uma lâmina recoberta com um filme d’água destilada aquecida à 60oC,
após secar ao ar, foi corada com solução Giemsa 5%, diluído em tampão
fosfato pH 6,8, por um período de 08 minutos, posteriormente lavada com água
destilada e seca ao ar.
3.2.4 Análises cromossômicas
Lâminas previamente preparadas com as suspensões celulares de cada
indivíduo foram analisadas através de microscópio óptico, sob o aumento de
1000 vezes. Uma média de 30 metáfases foi analisada para cada exemplar,
visando o estabelecimento do valor diploide modal para cada espécie e a
definição dos tipos cromossômicos. As melhores metáfases foram fotografadas
em fotomicroscópio de epifluorescência (OlympusTM BX50), sob aumento de
1000X, por meio de sistema digital de captura (DP70, Olympus), com o uso dos
software DPController, v. 1.2.1.108 (Olympus Optical Co. Ltd.) e utilizadas na
definição da fórmula cromossômica e construção do cariótipo. Os
cromossomos foram definidos quanto à posição dos centrômeros em
metacêntricos (m), com a razão entre o braço maior e menor (RB) variando de
1,00 a 1,70; submetacêntricos (sm), RB = 1,71 – 3,00; subtelocêntricos (st), RB
= 3,01 – 7,00; e acrocêntricos (a), RB > 7,01 (Levan et al., 1964).
3.2.5 Detecção das Regiões Organizadoras de Nucléolos (RONs)
As regiões organizadoras de nucléolo (RONs) foram evidenciadas de
acordo com a técnica descrita por Howell e Black (1980), com pequenas
modificações. Uma solução gelatinosa contendo 1g de gelatina incolor
acrescido de 50 ml de água e 0,5ml de ácido fórmico foi depositada na lâmina,
misturada a quatro gotas de nitrato de prata (Ag-NO3) 50%, cobrindo-a em
seguida com lamínula. Logo após foi incubada em estufa a 60ºC entre cinto a
oito minutos, até que uma coloração âmbar aparecesse. Ao término do

13
processo a lâmina foi lavada com água destilada, seca ao ar e analisada ao
microscópio.
3.2.6 Detecção de Heterocromatina Constitutiva (Banda-C)
A observação de regiões de heterocromatina constitutiva foi realizada de
acordo com o método desenvolvido por Summer (1972) com pequenas
alterações, visando uma melhor qualidade das preparações. Inicialmente as
lâminas foram imersas em HCl 0,2 N à temperatura ambiente, por 14 minutos,
lavando-a logo em seguida em água destilada e seca ao ar. Após isto, as
lâminas foram incubadas numa solução saturada de Ba(OH)2.8H2O, à 5%,
durante um período de 1 à 2 minutos à 42ºC. Após incubação foram expostas à
HCl 0,2N rapidamente, lavadas com água destilada e mantidas em solução de
2xSSC à 60ºC por uma hora, sendo posteriormente coradas com solução à 5%
de Giemsa diluído em tampão fosfato, pH 6,8 por 6 minutos e visualizadas ao
microscópio.
3.2.7 Coloração com Fluorocromos base-específicos
Para a dupla coloração com os fluorocromos CMA3 e DAPI, as lâminas
foram envelhecidas por três dias, coradas com 30 µL de CMA3 0,5 mg/mL por 2
horas, e em seguida coradas com 30 µL de DAPI 2 µL/mL por 30 minutos,
segundo metodologia preconizada por (Carvalho et al., 2005), com algumas
modificações. Posteriormente as lâminas foram montadas em tampão glicerol-
McIlvaine pH 7,0 (1:1), preservadas em câmara escura, por três dias e então
analisadas. As metáfases foram fotografadas em fotomicroscópio de
epifluorescência (OlympusTM BX50) com filtros apropriados em aumento de
1000X.
3.2.8 Análises por Morfometria Geométrica
Para a análise da morfometria geométrica, imagens no formato JPEG
com dimensão 3264x2448, foram obtidas da face lateral esquerda dos

14
espécimes, com escala métrica, utilizando-se uma câmera digital Sony H10
(8,1 megapixels) acoplada a um tripé VF WT3540.
O programa tpsUtil foi utilizado para ordenar as imagens dos espécimes
num único arquivo com o formato TPS, possibilitando o acesso às imagens
pelos demais programas da série. O software tpsDig2 (Rohlf, 2006) foi utilizado
para localizar os landmarks ou marcos anatômicos nas imagens, que
posteriormente foram utilizadas nas análises morfométricas. Os landmarks
foram escolhidos segundo a classificação de Bookstein (1991), sendo nove
utilizados para estudos interpopulacionais e onze para distinção de espécies.
As coordenadas referentes ao posicionamento espacial de cada landmark
foram sobrepostas utilizando-se o software CoordGen6.
Com a utilização do software CVAGen6, análises das variáveis
canônicas (VC) e MANOVA permitiram comparações morfométricas entre as
espécies, populações e testes de alocação. O programa MorphoJ 1.02b
possibilitou a quantificação do discernimento morfológico através da distância
de Mahalanobis, ANOVA e a formação dos grides de deformação à partir da
variável canônica que se mostrou mais significativa.

15
4.1 Capítulo I
Caracterização da diversidade na família Gobiidae por
meio da citogenética e morfometria geométrica
Lima Filho, P. A. & Molina, W. F.
Resumo
A família Gobiidae é a mais especiosa no ambiente marinho, apresenta características morfológicas com simplificações e perdas em estruturas corpóreas. Tamanha diversidade parece ter sido acompanhada por alterações cromossômicas significativas quando comparada a outros representantes da Ordem Perciformes com elevado conservadorismo cromossômico. Poucas informações citogenéticas estão disponíveis para espécies sul-americanas. Neste trabalho cinco espécies de gobídeos do Atlântico foram caracterizadas citogeneticamente por meio de coloração convencional com Giemsa, bandamento C, identificação de cístrons ribossomais pela técnica de Ag-RONs, coloração com os fluorocromos CMA3/DAPI e comparadas morfologicamente por meio da morfometria geométrica. Identificou-se uma marcante diversidade cariotípica entre as espécies Coryphopterus glaucofraenum (2n=40; 2sm+38a; NF=42), Ctenogobius smaragdus (2n=48; 2sm+42a; NF=50), Ctenogobius
boleosoma (2n=44; 2sm+42a; NF=46), Bathygobius mystacium (2n=48; 2m+4st+42a; NF=54) e Bathygobius soporator (2n=48; 2m+6st+40a; NF=56). Quanto aos padrões morfológicos todas as cinco espécies apresentam perfis morfológicos distintos, contudo exibem maior proximidade morfológica entre espécies cogenéricas. Os dados cromossômicos ressaltam o papel das inversões pericêntricas, fusões e fissão na evolução cariotípica do grupo. A diversidade cariotípica e morfológica presente nesta família sugerem uma dinâmica na evolução compatível com grupos dulcícolas sujeitos a intensas fragmentações populacionais, aliado a diversificações morfológicas adequadas a exploração de habitats peculiares.
Palavras-chave: Gobiidae; diversidade cariotípica; morfometria geométrica;
citogenética de peixes.

16
Characterization of the diversity of Gobiidae family by
cytogenetics and geometric morphometrics
Lima Filho, P. A. & Molina, W. F.
Abstract
Gobiidae family is the most specious in the marine environment, it shows morphologic features with simplifications and losses on body structure. Such diversity seems to have been accompanied by chromosomal changes significantly when compared to others representatives of the order Perciformes with high chromosomal conservatism. Few cytogenetic informations are available for South American species. In this study five species of Atlantic gobídeos were characterized cytogenetically by conventional Giemsa staining, C banding, identification of ribosomal cistrons by the technique of Ag-NORs, staining with the fluorochromes CMA3/DAPI, and also compared morphologically and by means of geometric morphometry. It was identified a remarkable karyotypic diversity among species Coryphopterus glaucofraenum
(2n=40; 2sm+38a; NF=42), Ctenogobius smaragdus (2n=48; 2sm+42a; NF=50), Ctenogobius boleosoma (2n=44; 2sm+42a; NF=46), Bathygobius
mystacium (2n=48; 2m+4st+42a; NF=54) e Bathygobius soporator (2n=48; 2m+6st+40a; NF=56). Against to the morphologic patterns, all five species have distinct morphological profiles, but exhibit greater proximity among morphological species co-generics. Chromosomes data underscore the role of pericentric inversions, merges and fission in karyotype evolution of the group. The karyotypic and morphological diversity present in this family suggests a dynamic evolution compatible with freshwater groups vulnerable to intense population fragmentation, morphological diversification with appropriate exploration of peculiar habitats.
Keywords: Gobiidae, karyotypic diversity, geometric morphometrics,
cytogenetics of fishes.

17
Introdução
Os peixes da família Gobiidae estão distribuídos nas regiões tropical,
subtropical e temperada em habitats costeiros e dulcícolas. Com cerca de
1.950 espécies, representam a maior família de peixes marinhos (Nelson,
2006), componente importante e predominante da fauna de peixes em
ambientes recifais. Devido à ocorrência de espécies morfologicamente crípticas
e de difícil amostragem, a biologia deste grupo permanece pouco estudada
(Thacker, 2003). Na costa brasileira existem cerca de 40 espécies (Menezes et
al., 2003), sendo a sua maioria compartilhada com a fauna caribenha.
Contrastando com outros grupos de Perciformes marinhos, a família
Gobiidae apresenta elevada diversidade cariotípica, o que a torna um modelo
particularmente importante para o entendimento da estrutura genética de
populações marinhas (Brum & Galetti, 1997). No que diz respeito aos aspectos
citogenéticos, a maioria das informacões para este grupo estão resumidas
prioritariamente a descrições numéricas e de morfologia cromossômica que
apesar da pouca ajuda no esclarecimento de relações filogenéticas (Ene, 2003;
Caputo et al., 1997), contribui substancialmente para o conhecimento da sua
diversidade para família (Vasil’ev & Grigoryan, 1993). Estudos citogenéticos
demonstraram uma tendência à ocorrência de translocações Robertsonianas
para a família, com demonstrações de variações cromossômicas intra e inter-
populacionais (Ene, 2003).
A dificuldade em classificar e identificar representates da família
Gobiidae, ocorre devido às características morfológicas escassas e marcante
variação morfológica interpopulacional (Springer, 1983; Winterbottom, 1984),
gerando para muitas espécies confusões taxonômicas e de nomenclatura
(Kottelat, 1997). No entanto, o uso de análises por morfometria geométrica já
utilizada com sucesso em diversos grupos de peixes (e. Ibañez et al., 2007;
Maderbacher et al., 2008; Chizinski et al., 2010), que permite identificar
variações geométricas da forma, com base numa configuração de pontos
anatômicos com redução dos efeitos inerentes ao tamanho, a localização e a
posição estrutural ocupada (Monteiro et al., 2002; Parsons et al., 2003), não foi
aplicada em espécies desta família com vistas a estabelecer comparações
ecomorfológicas e identificar diferenciações entre populações e espécies.

18
Em alguns grupos de peixes a estrutura cariotípica parece ter sofrido
poucas mudanças evolutivas quando comparadas às mudanças morfológicas
(Molina et al., 2002; Motta Neto & Molina, 2010). Não está claro se entre os
Gobiidae, diferentemente de inúmeras outras famílias de Perciformes (Molina,
2006), existem divergências nas taxas evolutivas entre os padrões
cromossômicos e morfológicos.
Neste sentido, a fim de se obter melhor compreensão da diversificação
cromossômica e mofológica desta família, aqui são apresentados os primeiros
registros citogenéticos de cinco espécies do Oeste do Atlântico, cujos padrões
são comparados citogeneticamente em associação com seus caracteres
ecomorfométricos.
Material e métodos
Foi coletado um total de 119 exemplares de cinco diferentes espécies de
Gobiidae (Tabela 1) provenientes do litoral do Estado do Rio Grande do Norte
(RN) (Praia de Búzios - 5º 46’ S, 35º 12’ W) e estuário de Barra do Cunhaú
(6°19'15.50"S, 35°2'29.31"O), NE do Brasil. Deste total apenas a espécie
Ctenogobius smaragdus foi coletada no estuário de Barra do Cunhaú e as
demais proveniente da praia de Búzios. Os espécimes foram capturados com
uso de rede de confecção própria, transferidos às dependências do laboratório
de Genética de Recursos Marinhos na UFRN e mantidos em aquários aerados
até serem fotografados para análises morfológicas e se procedesse à
realização de preparações cromossômicas.
Análises citogenéticas
Precedente aos métodos de obtenção de cromossomos metafásicos, os
exemplares foram submetidos à estimulação mitótica, “in vivo” com duas
aplicações intramuscular/peritoneal de Aminovac® (complexo de antígenos
bacterianos e fúngicos) na razão de um comprimido diluído em 0,5 ml de água
na proporção de 100 g de peso do animal (Molina et al., 2010).
Os cromossomos mitóticos foram obtidos a partir de células do rim
anterior, segundo Gold et al. (1990). Cerca de trinta metáfases foram

19
analisadas de cada exemplar para se estabelecer o número modal, sendo que,
as melhores metáfases foram fotografadas em um microscópio OlympusTM
BX50, (1000x), acoplado à uma câmera digital (Olympus, modelo DP70), e
utilizadas na confecção do cariótipo. A morfologia cromossômica foi
determinada de acordo com Levan et al. (1964). As regiões heterocromáticas
foram evidenciadas a partir da metodologia desenvolvida por Sumner (1972),
enquanto que a detecção das Ag-RONs foi realizada conforme a técnica
descrita por Howell & Black (1980). A coloração com os fluorocromos CMA3 e
DAPI seguiu o protocolo de Carvalho et al. (2005).
Tabela 1. Número de indivíduos utilizados para estudos morfométricos e citogenéticos.
Espécies Análises
citogenéticas
Análises
morfométricas
Coryphopterus glaucofraenum (05♀, 03♂) 13
Ctenogobius boleosoma (02♀, 03♂) 29
Ctenogobius smaragdus (02♀, 02♂) 04
Bathygobius mystacium (02♀, 02♂) 23
Bathygobius soporator (09♀, 11♂) 50
Total 41 109
Análises morfológicas
Para as análises por morfometria geométrica (MG), imagens no formato
JPEG com dimensão 3264x2448, foram obtidas da face lateral esquerda dos
espécimes, com escala de métrica, utilizando-se uma câmera digital Sony H10
(8,1 megapixels) acoplada a um tripé VF WT3540.
O programa tpsUtil foi utilizado para ordenar as imagens dos espécimes
num único arquivo com o formato TPS. O software tpsDig2 (Rohlf, 2006) foi
utilizado para localizar 11 landmarks nas imagens (Figura 1), escolhidos
segundo a classificação de Bookstein (1991). As coordenadas referentes ao
posicionamento espacial de cada landmark foram sobrepostas utilizando-se o
software CoordGen6.

20
Com a utilização do software CVAGen6, análises das variáveis
canônicas (VC) e MANOVA permitiram comparações morfométricas entre as
espécies, testes de alocação e agrupamentos. Para realizar o teste estatístico
ANOVA, D2 (distância de Mahanalobis) e gerar matrizes (Gride) de deformação
comparativa, utilizou-se o programa MorphoJ 1.02b®.
Figura 1. Imagem digitalizada de Coryphopterus glaucofraenum retratando os 11 landmarks utilizados nas análises morfológicas interespecífica em Gobiidae. Descrição esquemática dos landmarks. 1. Ponto distal do osso pré-maxilar; 2. Origem da primeira nadadeira dorsal; 3. Término da primeira nadadeira dorsal; 4. Origem da segunda nadadeira dorsal; 5. Término da segunda nadadeira dorsal; 6. Término da nadadeira anal; 7. Origem da nadadeira anal; 8. Origem da nadadeira pélvica; 9. Margem anterior ocular; 10. Margem posterior ocular; 11. Base superior da nadadeira peitoral.Barra=0,5cm.
Resultados
As análises citogenéticas realizadas nos gobídeos C. glaucofraenum, C.
boleosoma, C. smaragdus, B. mystacium e B. soporator, indicaram um número
diplóide que variou de 40 a 48 cromossomos, com diferentes números
fundamentais, e cariótipos constituídos por elementos metacêntricos,
submetacêntricos, subtelocêntricos e na sua maioria acrocêntricos (Tabela 2).
Em todas as espécies analisadas pôde-se observar Ag-RONs simples
em posição terminal no braço curto de um dos maiores pares do cariótipo. A
única exceção foi observada em B. mystacium, cujos sítios ribossomais
ocupam posição intersticial. A coloração com os fluorocromos evidenciou um
padrão CMA3+/DAPI- verificando-se a presença de regiões ricas em bases GC
nas porções correspondentes às regiões organizadoras de nucléolos para
todas as espécies. Regiões heterocromáticas ricas em GC (CMA3+/DAPI-)

21
estavam presentes na maioria dos cromossomos nas regiões centroméricas e
terminais, das duas espécies do gênero Ctenogobius (Figura 4).
O bandamento C revelou a presença de blocos heterocromáticos
evidentes nas espécies C. smaragdus, B. soporator e B. mystacium e
reduzidos em C. glaucofraenum, preferencialmente localizados na região
centromérica da maioria dos cromossomos, com a presença de alguns blocos
em regiões terminais para todas as espécies e em posição pericentroméricas
em alguns cromossomos de B. mystacium.
A espécie C. glaucoraenum apresentou seus primeiros pares
acrocêntricos com tamanhos relativamente superiores (Figura 7), que configura
a seu cariótipo uma redução desequilibrada de tamanho cromossômico,
diferente do padrão de redução proporcional apresentado pelas demais
espécies analisadas.
Tabela 2. Dados citogenéticos das cinco espécies de Gobiidae analisadas.
Espécies 2n Fórmula
Cariotípica NF Ag-RONs Banda
C CMA3
+
DAPI-
Coryphopterus glaucofraenum 40 2sm+38a 42 Simples (1º T) C e T 1º T
Ctenogobius boleosoma 44 2sm+42a 46 Simples (1º T) - Múltiplas
Ctenogobius smaragdus 48 2sm+46a 50 Simples (1º T) C e T Múltiplas
Bathygobius mystacium 48 2m+4st+42a 54 Simples (5º I) C, T e I 5º I
Bathygobius soporator 48 2m+6st+40a 56 Simples (4º T) C e T 4º T
Tipos cromossômicos, m=metacêntrico, sm=submetacêntrico, st=subtelocêntrico, a=acrocêntrico; T=terminal, C=centromérica, I=intersticial, NF= número fundamental, - técnica não disponível.

22
Figura 2. Cariótipo de Ctenogobius smaragdus a partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
Figura 3. Cariótipo de Ctenogobius boleosoma a partir de coloração convencional (a). Em destaque (b) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
Figura 4. Metáfase de C. smaragdus (a) e C. boleosoma (b), exibindo marcações CMA3+ na maioria
dos cromossomos em posição centromérica e terminal. Barra=5µm.
CMA3
1º
DAPI
a b c
a b
1º
CMA3
DAPI
b a

23
Figura 5. Cariótipo de Bathygobius soporator a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (4º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
Figura 6. Cariótipo de Bathygobius mystacium a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (5º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
a b c
DAPI
CMA3
4º
a b c
DAPI
CMA3
5º

24
Figura 7. Cariótipo de Coryphopterus glaucofraenum a partir de coloração convencional (a) e bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra=5µm.
O teste de MANOVA utilizando as variáveis canônicas demonstraram
alto poder discriminante onde as variáveis 1 e 2 explicaram a maior parte da
variação morfológica observada (53%) para as espécies. A variação
morfológica foi significante (Wilks’ Lambda=0.0002; X2=496.9974; df=72;
p<0,001; distância de Mahalanobis >6.0) entre as espécies, permitindo ainda
agrupar com total precisão todos representantes das cinco espécies (Figura 8).
Obteve-se maior proximidade morfológica entre as espécies cogenéricas
pertencentes aos gêneros Bathygobius e Ctenogobius (Mahalanobis
equivalente a 8,36 e 14,24, respectivamente), no entanto, significativamente
distintas para Bathygobius. As espécies destes dois gêneros são discriminadas
entre si ao longo da variável canônica 1. Os exemplares de C. Glaucofraenum
são perfeitamente discriminados de B. soporator e B. mystacium pela variável
canônica 2.
Tabela 3. Resultados post-hoc ANOVA a partir de procrustes, classificadas com a média do centróide, efeito individual, para todos os espécimes em estudo e espécies dos gêneros Bathygobius e Ctenogobius.
Grupos p F df
Todos os espécimes <0,0001 23,55 4
Bathygobius <0,0001 47,58 1
Ctenogobius 0,36* 0,88 1
* Valor não significativo.
a b c
CMA3
1º
DAPI
25
O gride de deformação das cinco espécies (Figura 9) evidencia
variações entre todos landmarks, com modificações mais expressivas quanto
ao posicionamento da boca e altura corporal. No gride referente as diferenças
morfológicas entre espécies do gênero Bathygobius (Figura 10), verificamos
maiores modificações quanto ao posicionamento da boca, altura corporal e
base superior da nadadeira peitoral. O gride comparativo entre espécies do
gênero Ctenogobius (Figura 11), demonstrou variações quanto ao tamanho do
olho, posição da boca, altura corporal e tamanho da nadadeira anal.
Figura 8. Distribuição dos padrões corporais das cinco espécies de Gobiidae analisadas ao longo da primeira e segunda Variável Canônica.
26
Figura 9. Gride de deformação a partir da primeira Variável Canônica, os vetores indicam a direção da deformação sofrida por cada landmark na representação espacial comparativa das cinco espécies de Gobiidae analisadas.
Figura 10. Análise morfométrica comparativa entre B. mystacium e B. soporator, a partir da variável canônica 1. Ponto médio central representa B. mystacium e o desvio vetorial à espécie B. soporator.
Figura 11. Análise morfométrica comparativa, entre as espécies do gênero Ctenogobius, á partir da variável canônica 1. Ponto médio central representa C. boleosoma e o desvio vetorial a espécie C. smaragdus.
CV1
CV1
CV1

27
Discussão
Os gobídeos analisados demonstraram marcante diversidade cariotípica,
tanto numérica exibido por C. boleosoma (2n=44) e C. glaucofraenum (2n=40),
quanto estrutural entre as espécies C. smaragdus, B. mystacium e B.
soporator, que apresentam mesmo número cromossômico (2n=48), mas
divergências quanto ao número de elementos bibraquiais (NF=50, 54 e 56).
As espécies já analisadas citogeneticamente da família Gobiidae
demonstram cariótipos altamente variáveis (Galetti et al., 2000). Através do
levantamento de dados citogenéticos disponíveis para o grupo (Tabela 4),
encontram-se descrições citogenéticas de 81 espécies, o que corresponde a
4,15%, referente ao total de espécies da família, destas os números diplóides
mais frequentes correspondem a 2n=44 e 46, ambos com frequência
aproximada de 33%, seguido por 2n=48, com cerca de 10%. Acompanhado da
diversificação numérica, identifica-se notável variação na macroestrutura
cariotípica identificada por largos valores de variação para o número de braços
cromossômicos (NF= 38 a 98), com valor modal de 24,7% para espécies com
NF=46.
Os estudos cromossômicos em gobídeos estiveram concentrados nas
décadas de 70-90, desenvolvidos basicamente no continente europeu e
asiático. Diante das metodologias vigentes na maioria dos casos se
restringiram ao estabelecimento dos valores diplóides e macroestrutura
cariotípica, oferecendo informações limitadas ao uso de abordagens
filogenéticas (Caputo, 1998). Apesar disso, os dados disponíveis sugerem
diferentes mecanismos envolvidos na diferenciação cariotípica deste peculiar
grupo de peixes. Os principais mecanismos são as inversões pericêntricas,
fissões e fusões cêntricas, comum a muitos grupos de peixes (Amores et al.,
1990; Phillips & Rab, 2001), estes últimos responsáveis pelas intensas
divergências numéricas em Gobiidae (Cataudella et al., 1973; Thode et al.,
1985; Amores et al., 1990, Klinkhardt, 1992; Caputo et al., 1996, 1997).
Entre os rearranjos cromossômicos, as fusões robertsonianas têm sido
apontadas como um dos mecanismos mais comuns envolvidos na
diferenciação cariotípica deste táxon (Thode et al.,1988; Galetti et al., 2000),
responsável por modificações no número e estrutura cromossômica, relatado

28
para várias espécies de Gobiidae presentes no Mar Negro e Mediterrâneo
(Ene, 2003).
Tabela 4. Dados citogenéticos disponíveis para a família Gobiidae (Perciformes).
Subfamília Espécie 2n Fórmula cariotípica NF Referências Amblyopinae Odontamblyopus rubicundus** (=Gobioides rubicundus)
46 2m+26sm+10st+8a 84 Manna & Prasad (1974)
O. rubicundus 46 4m+16sm+26st/a 66 Arai & Sawada (1975) Trypauchen vagina 46 12m+6sm+10st+18a 74 Khuda-Bukhsh (1978)
Oxudercinae Apocryptes bato
46
24m+10sm+12a
80
Nayak & Khuda-Bukhsh (1987)
Boleophthalmus boddarti 46 46m/sm 92 Subrahmanyan (1969) B. glaucus 46 12m+20sm+2st+12a 80 Manna & Prasad (1974) B. pectinirostris 46 46st/a 46 Arai & Sawada (1975) Periophthalmus modestus** (=Periophthalmus cantonensis)
46 18m+12sm+16st/a 76 “
Pseudapocryptes elongatus** (=Apocryptes lanceolatus)
38 14m+22sm+2st 76 Nayak & Khuda-Bukhsh (1987)
Sicydiinae Sicyopterus japonicus
44
10m+10sm+24a
64
Arai & Fujiki (1979)
Gobiinae Acentrogobius pflaumi
50
48m/sm+2st/a
98
Nogusa (1960)
Amblygobius albimaculatus 44 2m+42st/a 46 Nishikawa et al. (1974) Aphia minuta
1 44 44a 44 Caputo et al. (1999)
“ 43 42a+1st 42 “ “ 42 1m+1st+40a 44 “ “ 42 1M+1m+40a 44 “ “ 41 2M+1st+38a 44 “ Bathygobius fuscus 48 48a 48 Arai & Sawada (1975) B. mystacium 48 2m+4st+42a 54 Presente estudo B. soporator 48 2m+46a 50 Brum et al. (1996) “ “ 2m/sm+46a 50 Cipriano et al. (2002) B. soporator 48 2m+6st+40a 56 Presente estudo Coryphopterus glaucofraenum 40 2sm+38a 42 Presente estudo Elacatinus macrodon** (=Gobiosoma macrodon)
38 38a 38 Musammil (1974)
E. zebrella** (=Gobiosoma zebrella)
38 38a 38 “
Glossogobius fasciatopunctatus 44 10m+28sm+2st+4a 84 Fei & Tao (1987) G. giuris 46 46a 46 Rishi & Singh (1982) Gobiodon citrinus 44 2m+42st/a 46 Arai & Sawada (1974) “ 43 1m+42st/a 44 “ G. quinquestrigatus 44 44a 44 Arai & Fujiki (1979) G. rivulatus 44 44a 44 “ Gobius bucchichi 44 2sm+42a 46 Thode & Alvarez (1983) G. cobitis 46 46a 46 Caputo et al. (1997) G. cruentatus 46 2st+44a 48 Thode & Alvarez (1983) G. fallax
1 38 8m/sm+30a 46 Thode et al. (1988) “ 39 7m/sm+32a 46 “ “ 40 6m/sm+34a 46 “ “ 40 7m/sm+33a 47 “ “ 41 5m/sm+36a 46 “ “ 42 4m/sm+38a 46 “ “ 43 3m/sm+40a 46 “ G. niger
1 52 2m+4sm+16st+30a 74 Vitturi & Catalano (1989)
“ 51 3m+4sm+16st+28a 74 Caputo et al. (1997) “ 50 4m+4sm+16st+26a 74 “ “ 49 5m+4sm+16st+24a 74 “ G. paganellus
1 48 2sm+46a 50 “
Gobiusculus flavescens 46 6m/sm+40a 52 Klinkhardt (1992) Mesogobius batrachocephalus 30 16m+14a 46 Ivanov (1975) Neogobius melanostomus** (=N. cephalarges)
46 46a 46 Vasil’ev (1985)
N. fluviatilis 46 46a 46 “
N. gymnotrachelus1 46 46a 46
Vasil’ev & Grogoryan(1992)
N. melanostomus 46 46a 46 Vasil’ev (1985) N. rhodioni 46 46a 46 Vasil’ev & Vasilieva (1994)

29
Padogobius bonelli** (= P. martensi)
46 1m+3sm+2st+40a 52 Cataudella et al. (1973)
Pomatoschistus lozanoi 37 3m+12sm+10st+12a 62 Webb (1980) P. microps 46 4m+16sm+20st+6a 86 Klinkhardt (1989) P. minutus 46 4m+16sm+16st+10a 82 “ “ “ 18sm+18st+10a 82 “ P. norvegicus 32 10m+10sm+8st+4a 60 Webb (1980) P. pictus 46 22m/sm+12st+12a 80 Klinkhardt (1992) Ponticola constructor** (=Neogobius constructor)1 42 4m/sm+38a 46 Vasil’ev & Vasilieva (1994)
P. cyrius** (=Neogobius cyrius)1
36 -- -- “
P. eurycephalus** (=Neogobius eurycephalus)1
32 12m+2sm+18a 46 Ene (2003)
“ 31 13m+2sm+16a 46 Ene (2003) “ 30 14m+2sm+14a 46 “ P. kessleri** (=Neogobius kessleri)
46 46a 46 Vasil’ev (1985)
Proterorhinus marmoratus 46 46a 46 Rab (1985) Rhinogobius similis** (=Gobius similis)
44 -- -- Nogusa (1960)
Zosterisessor ophiocephalus (=Gobius ophiocephalus)1
46 2m/sm+44a 48 Caputo et al. (1996)
Z. ophiocephalus
46 46a 46 Vasil’ev (1985)
“ 45 1st+45a 47 “ Gobionellinae Acanthogobius lactipes** (=Aboma lactipes)
40 40a 40 Arai & Sawada (1974)
A. flavimanus 44 44st/a 44 “ “ “ 36st+8a 80 Arai & Kobayashi (1973) “ “ 10m/sm/st+34a 54 Arai & Sawada (1975)
Awaous grammepomus 46 46st/a 46 Khuda-Bukhsh & Barat (1987)
A. strigatus 46 (♀) X1X1X2X2 -- Souza et al. (1998) “ 45 (♂)X1X2Y -- “
A. tajasica 46 46a 46 Stange & Passamani (1986)
Chaenogobius annularis 44 18sm+26st/a 62 Arai & Sawada (1975) “ “ 36m/sm+8a 80 Arai et al. (1974) “ “ 44a 44 Nogusa (1960) Gymnogobius castaneus** (=Chaenogobius castaneus)
44 36m/sm/st+8a 80 Nishikawa et al. (1974)
G. isaza**(=Chaenogobius isaza)
44 12sm+32st/a 56 Arai & Sawada (1975)
G. urotaenia** (=Chaenogobius urotaenia)
44 -- -- Nogusa (1960)
“ 42 14sm+28a 56 Yamada (1967) Chaenogobius annularis** (=Chasmichthys dolichognatus)
44 44st/a 44 Arai & Sawada (1975)
Chasmichthys gulosus 44 44st/a 44 “ “ “ 16m/sm/st+28a 60 Nishikawa et al. (1974) Ctenogobius boleosoma 44 2sm+42a 46 Presente estudo C. smaragdus 48 2sm+46a 50 Presente estudo C. shufeldti** (=Gobionellus shufeldti)
48 48a (♀) 48 Pezold (1984)
“ 47 46a+1m (♂) 48 “ Mugilogobius abei** (=Gobius abei)
46 -- -- Nogusa (1960)
Gillichthys mirabilis 44 12sm+32a 56 Chen & Ebeling (1971) Luciogobius grandis 44 -- -- Arai (1981) L. guttatus 44 -- -- Arai & Kobayashi (1973) Pterogobius elapoides 44 14sm+30st 88 “ P. zonoleucus 44 14sm+30st 88 Arai & Sawada (1975) Quietula guaymasiae 42 6m+4sm+32a 52 Cook (1978) Q. y-cauda 42 42a 42 “ Rhinogobius brunneus 44 44a 44 Nishikawa et al. (1974) R. flumineus 44 44a 44 Arai & Kobayashi (1973) R. flumineus** (=Tukugobius flumineus)
44 44a 44 Nadamitsu (1974)
R. giurinus 44 44a 44 Nishikawa et al. (1974) Tridentiger obscurus 44 10m/sm+34a 54 Arai et al. (1974) T. trigonocephalus 44 28m/sm/st+16a 72 Arai et al. (1973)

30
**Nomenclatura válida para a espécie; 1Espécies que apresentam polimorfismos cromossômicos decorrentes de fusão Robertsoniana.
Os rearranjos cromossômicos aparentemente envolvidos na
diversificação de Gobiinae (e. B. soporator, B. mystacium e C. glaucofraenum)
e Gobionellinae (e. C. boleosoma e C. smaragdus) são as inversões
pericêntricas, mecanismo de diversificação mais frequente em Perciformes
(Molina & Galetti, 2004), e fusões em tandem ou robertsonianas.
Sugere-se um cariótipo basal para Gobiidae caracterizado por 2n=46
cromossomos acrocêntricos, a partir do qual o aumento do número de
cromossomos bi-braquiais representaria cariótipos mais derivados (Vasil’ev &
Grigoryan, 1993). Apesar de C. glaucofraenum apresentar 2n=40, número
inferior ao considerado basal, os primeiros pares acrocêntricos apresentam
tamanho superior de forma desproporcional em relação aos demais, indicando
a ocorrência de fusão em tandem para a formação dos mesmos, mecanismo
comum observado na diversificação cariotípica de outros gobídeos como G.
paganellus (Amores et al., 1990).
Apresentando cariótipos caracterizados por 2n=48, as espécies C.
smaragdus, B. mystacium e B. soporator, possuem padrões cariotípicos
considerados derivados para família, com possibilidade de ação de diferentes
processos à estruturação cromossômica dessas espécies. É provável que
inversões pericentroméricas seguidas de fissões cromossômicas, tenham
fornecido um passo crucial no estabelecimento do valor diplóide destas
espécies e influenciado à formação dos valores diplóides presentes nas
espécies Yongeichthys criniger (Arai & Sawada, 1974), Ctenogobius shufeldti
(Pezold, 1984), Gobius niger (Vitturi & Catalano, 1989) e G. paganellus (Caputo
et al., 1997).
As divergências na macroestrutura cariotípica entre B. mystacium
(2n=48; 2m+4st+42a) e B. soporator (2n=48; 2m+6st+40a), podem ser
explicadas pela ocorrência de uma inversão pericentromérica, envolvendo as
regiões portadoras dos sítios ribossomais. A presença de um número maior de
elementos acrocêntricos no cariótipo de B. mystacium (NF=54) poderia indicar
a ocorrência de um cariótipo basal em relação ao de B. soporator (NF=56).
“ 46 16sm+6st+24a 68 Fei & Tao (1987) Yongeichthys criniger** (=Ctenogobius criniger) 50 34m/sm+6st+10a 90 Arai & Sawada (1974)

31
Entretanto a presença de RONs intersticiais em um par cromossômico
acrocêntrico em B. mystacium (5o par) indica que inversões pericêntricas
ocorreram tanto em cromossomos acrocêntricos como, neste caso, podem ter
ocorrido em cromossomos bibraquiais.
Outras espécies de gobídeos têm demonstrado marcações
cromossômicas de Ag-RONs intercaladas a regiões heterocromáticas ricas em
bases GC (Mandrioli et al., 2001; Ene, 2003), fato observado para as espécies
C. glaucofraenum, C. boleosoma, C. smaragdus, B mystacium e B. soporator,
podendo favorecer a ocorrência de rearranjos envolvendo os pares portadores
destas regiões (Vicari et al., 2003).
É provável que o cariótipo mais basal dentre as espécies analisadas,
seja o de C. boleosoma (NF=46), o qual apresenta RONs em cromossomos bi-
braquiais, como as demais apresentam cariótipos considerados derivados para
o grupo, é possível que esta localização de RONs seja uma característica
plesiomórfica entre as espécies analisadas.
Registros de RONs disponíveis para gobídeos são escassos, entretanto
indicam diferentes padrões de disposição (Ene, 2003). Caputo (1998)
identificou quatro fenótipos de RONs para gobídeos: (1) posição terminal-
centromérica em braços curtos de um par de cromossomos
acrocêntricos/subtelocêntricos em G. fallax (Thode et al., 1983),
Potamoschistus minutus (Klinkardt, 1992; Caputo, 1998), P. pictus (Klinkardt,
1992), P. marmoratus (Caputo, 1998); (2) braços curtos de dois pares de
cromossomos submetacêntricos em Gobiusculus flavescens (Klinkardt, 1992) (3) porção terminal no braço longo em Zosterisessor ophiocephalus (Caputo,
1998) e (4) intersticial/pericêntrica em um par de cromossomos acrocêntricos
Gobius cobitis (Caputo, 1998). Ene (2003) aponta mais dois possíveis
fenótipos, que corresponderia: (5) região pericentromérica em um par
acrocêntrico Proterohinus marmoratus (Ra´b, 1985) e (6) posição
pericentromérica em cromossomo metacêntrico N. eurycephalus (Ene, 2003).
Apesar da constatação de RONs múltiplas em Gobius fallax (Thode et
al.,1983), G. paganellus (Caputo, 1998), Gobiusculus flavescens (Klinkardt,
1992) e Gobius niger (Mandrioli et al., 2001) o padrão encontrado caracterizado
por marcações do tipo simples nas espécies analisadas é considerado um
caráter comum não apenas aos teleósteos, como também para a maioria dos

32
vertebrados (Ameniya & Gold, 1986).
Mesmo com total distinção entre as espécies, a partir da macroestrutura
cariotípica, o mapeamento de regiões heterocromáticas, heterocromatinas GC-
ricas e cístrons ribossomais, fornece indícios da existência em algum grau de
homeologias cromossômicas entre as espécies. A presença destas regiões
associadas configurando marcações simples em cromossomos de tamanho e
configuração semelhante pode ser indício de uma provável manutenção de
regiões cromossômicas sintênicas ao longo da diferenciação de linhagens
filéticas para família Gobiidae.
A presença de grandes blocos heterocromáticos nas porções terminais
dos cromossomos como nos cariótipos das espécies de Gobiidae, poderia ter
um papel facilitador em eventos de fusões em tandem ou robertosonianas.
Nesta família, heterocromatinas heterogêneas tem sido identificadas em
diversas espécies, como G. paganellus, G. niger, G. cobitis e Zosterisessor
ophiocephalus (Caputo et al., 1997), o que associa a variabilidade
cromossômica encontrada no grupo às características estruturais e
composicionais das heterocromatinas centroméricas e teloméricas.
A existência de heterocromatinas GC-ricas localizadas em posição
centromérica e terminal dos cromossomos das espécies do gênero
Ctenogobius, C. boleosoma e C. smaragdus, é similar aquelas encontradas na
maioria dos cromossomos mono e bi-braquiais de Neogobius eurycephalus,
que exibe cariótipos com 2n=30, 31, 32 e mesmo número de braços
cromossômicos (NF=46), resultado de múltiplas fusões entre elementos mono-
braquiais (Ene, 2003).
Para os peixes marinhos, onde o isolamento geográfico é menos comum
que em sistemas de água doce, o surgimento de cromossomos sexuais pode
ter desempenhado um papel decisivo no processo de especiação (Galetti et al.,
2000), sendo gerados frequentemente por eventos robertsonianos (Almeida &
Toledo, 1984; Brum et al., 1992). Em outras espécies do gênero Ctenogobius,
como C. shufeldti o mecanismo de translocação robertsoniana está envolvida
na origem de um sistema sexual múltiplo X1X1X2X2\X1X2Y (Pezold, 1984). De
forma geral a presença de heterocromatinas ricas em bases GC
equilocalmente à sítios ribossomais, têm sido também apontadas como
precursoras de rearranjos em outras espécies (Vicari et al., 2003).

33
Diferentemente de outros grupos Perciformes (Molina, 2006), em
Gobiidae existe uma estreita relação entre rearranjos cromossômicos e
processo de especiação. Um fator decisivo para grande diversidade
cromossômica que a família Gobiidae apresenta, parece estar relacionado à
sua diversidade ecológica e evolutiva que propiciou ocupação de múltiplos
habitats. A maioria dos membros desta família são bentônicos, habitantes de
recifes de corais cuja alta produtividade (Fraser & Currie, 1996), elevada
complexidade espacial (Lingo & Szedlmayer, 2006) e complexidade ecológica
tem sido implicada nos altos níveis de diversidade (Bellwood & Wainwright,
2002). Estes fatores associados levam a uma alta taxa de cladogênese que
aliado a eventos vicariantes paleogeográficos favorecem a especiação dos
peixes residentes (Alfaro et al., 2007).
Adaptações ecológicas se traduzem em modificações morfológicas
substanciais. Muitas espécies de peixes apresentam grande variação
morfológica, entretanto possuem cariótipos crípticos (Molina et al., 2005).
Análises citogenéticas e de morfometria geométrica em cinco espécies da
família Haemulidae, demonstraram elevada constância citogenética, e
completa diversidade morfológica relacionada a adaptações ecomorfológicas
funcionais para cada espécie (Motta Neto & Molina, 2010). Associadas,
técnicas genéticas e de comparações morfológicas vêm sendo crescentemente
utilizadas na família Gobiidae (Taylor & Hellberg, 2003; Tornabene et al., 2010),
auxiliando na compreensão dos processos envolvidos na diversificação
filogenética de diferentes espécies.
A perfeita distinção dos padrões corporais entre as cinco espécies,
associada a cariótipos conspicuamente diversos indica que o processo de
especiação na família atrelou processos adaptativos a eventos alopátricos em
populações capazes de fixar novos rearranjos. Esta diversidade cariotípica
presente associada à morfológica, sugerem uma dinâmica na evolução
cariotípica compatível com grupos dulcícolas (Cipriano et al., 2008) sujeitos a
intensas fragmentações populacionais, aliado a diversificações morfológicas
adequadas a exploração de habitats peculiares (Pouilly et al., 2003; Freitas et
al., 2005; Ready et al., 2006; Concheiro-Pérez et al., 2007).
Morfologicamente as modificações encontradas entre as espécies estão
relacionadas à altura corporal, posicionamento da boca e posição da nadadeira

34
peitoral. Alterações no posicionamento da nadadeira peitoral e aumento da
altura, refletem adaptações relacionadas ao processo de natação, com
implicações diretas sobre o uso de habitat (Wainwright et al., 2002) e
interações bióticas (Werner, 1977). As demais adaptações, referentes ao
posicionamento ou tamanho da boca, indicam diferentes graus de
especialização alimentar para as espécies, podendo estar relacionado ao
tamanho potencial da presa (Piorski et al., 2005). Espécies do mesmo gênero
demonstraram maior proximidade morfológica como B. soporator/ B. mystacium
e C. smaragdus/ C. boleosoma, sendo as últimas não significantemente
discriminadas entre si.
Os perfis ecomorfológicos para Bathygobius e Ctenogobius, sugerem
diferenças no padrão alimentar, devido a modificações no posicionamento da
boca e características natatórias, com diferenças no posicionamento das
nadadeiras peitorais, responsáveis pela elevação da parte anterior do corpo por
estar à frente do centro de gravidade (Gosline, 1971) e altura corporal menor,
garantindo a B. mystacium e C. boleosoma maior maneabilidade, agilidade
(Breda et al., 2005) e estabilidade local (Horn & Gibson, 1988).
Os representantes da família Gobiidae estudados demonstraram
diversidade cariotípica e morfológica, evidência para o grupo de relação entre
processo especiativo e modificação cromossômica. Devido a grande
diversidade de espécies e imprecisões taxonômicas, a utilização de
ferramentas citogenéticas e de morfometria geométrica associadas, podem
auxiliar a identificação da composição real desta família e processos evolutivos
que permitiram tamanha diversificação.

35
Referências
Alfaro, M.E., Santini, F., Brck, C.D. (2007). Do reffs drive diversification in marine
Teleosts? Evidence from the pufferfishes and their allies (Order Tetraodontiformes). Evolution, 61-9: 2104–2126.
Almeida-Toledo, L. F., Foresti, F., Toledo-Filho, S. A. (1984). Complex sex
chromosome system. In: Eigenmannia sp. (Pisces, Gymnotiformes). Genetica, 64, 3, 165-169.
Amemiya, C. T. Gold, J. R. (1990). Cytogenetic studies in North American minnows
(Cyprinidae). XVII. Chromosomal NOR phenotypes of 12 species, with comments on cytosystematic relationship among 50 species. Hereditas, 112:231-247.
Amemiya, C.T., Gold, J.R. (1986). Chromomycin A3 stains nucleolus organizer regions
of fish chromosomes. Copeia, 1986:226–231. Amores, A., Giles, V., Thode, G. (1990). Adaptive charater of a Robertsonian fusion in
chromosomes of the fish Gobius paganellus (Pises, Perciformes). Heredity, 65: 151-155.
Amores, A., Giles, V., Thode, G. (1990). Adaptive charater of a Robertsonian fusion in
chromosomes of the fish Gobius paganellus (Pises, Perciformes). Heredity, 65: 151-155.
Arai, R. (1981). Fishes of the genus Luciogobius e Inu (Gobiidae) from peninsula Idzu,
Central Japanese Memoirs National Science Museum, 14: 151-166. Arai, R., Fujiki, S. (1979). Chromosomes of Japanese gobioid fishes (IV). Bulletin
National Science Museum, 5: 153-159. Arai, R., Katsuyama, I., Sawada, Y. (1974). Chromosomes of Japanese gobioid fishes
(II). Bulletin National Science Museum, 17: 269-279. Arai, R., Kobayasi, H. (1973). A chromosome study on thirteen species of Japanese
gobiid fishes. Japan Journal Ichthyology, 20: 1-6. Arai, R., Sawada, Y. (1974). Chromosomes ofJapanese gobioid fishes. Bull. Nail. Sci.
Mus. Tokyo, 17(2): 97`105. Arai, R., Sawada, Y. (1975). Chromosomes of Japanese gobioid fishes (III). Bulletin
National Science Museum, 1: 225-232. Bellwood, D.R., Wainwright, P.C. (2002). The history and biogeography of fishes on
coral reefs. Pp. 5–32 in P. F. Sale, ed. Coral reef fishes. Academic Press, San Diego, CA.

36
Bookstein, F. L. (1991). Morphometric tools for landmark data. Cambridge University Press, Cambridge. New York, 435p.
Breda, L., Oliveira, E. F., Goulart, E. (2005). Ecomorfologia de locomoção de peixes
com enfoque para espécies neotropicais. Acta Scientiarum: Biological Sciences, 27(4): 371-381.
Brum, M. J. I., Corrêa, M. M. O., Purcell, C. A.; Ribeiro, V. P., Muratori, R. S. (1996).
Análise cromossômica em Xenomelaniris brasiliensis (Atherinidae), Micropogonias furnieri (Scianidae) e Bathygobius soporator (Gobiidae) do litoral do estado do Rio de Janeiro. In: VI Simpósio de Citogenética Evolutiva e aplicada de Peixes Neotropicais, Universidade Federal de São Carlos. São Carlos, Brasil. 101p.
Brum, M.J.I., Galetti Jr., P.M. (1997). Teleostei plan ground karyotype. Journal of
Computational Biology, 2: 91-102. Brum, M.J.I., P.M. Galetti Jr., M.M.O. Corrêa, Aguilar, C.T. (1992). Multiple sex
chromosomes in South Atlantic fish Brevoortia aurea, Clupeidae. Brazilian Journal of Genetics, 15: 547.533.
Caputo, V. (1998). Nucleolar organizer (NOR) location and cytotaxonomic implications
in six species of gobiid fishes (Perciformes, Gobiidae). Italy Journal Zoology, 65: 93-99.
Caputo, V., Vitturi, R., Odierna, G., Cano, J., Olmo, E., Colomba, M.S. (1996).
Characterization of mitotic chromosomes in the Gobiid fish Zosterissor
ophiocephalus (Pallas, 1811) (Perciformes, Gobiidae). Biol. Zent. bl. 115: 328-336.
Caputo, V., Marchegiani, F., Sorice, M., Olmo, E. (1997). Heterochromatin
heterogeneity and chromossome variability in four species of gobiid fishes (Perciformes: Gobiidae). Cytogenetics and Cell Genetics, 79: 266-271.
Caputo, V., Sorice, M., Vitturi, R., Magistrelli, H., Olmo, E. (1998). Cytogenetic studies
in some species of Scorpaeniformes (Teleostei: Percomorpha). Chromosomal Research, 6: 255–262.
Caputo, V., Caniglia, M.L., Machella, N. (1999). The chromosomal complement of
Aphia minuta, a paedomorphic goby. Journal of Fish Biology, 55: 455-458. Carvalho, R., Soares Filho, W.S., Brasileiro-Vidal, A.C., Guerra, M. (2005). The
relationships among lemons, limes and citron: A chromosomal comparison. Cytogenetic Genome Research, 109:276-282.
Cataudella, S., Civitelli, M.V., Capanna, E. (1973). The chromosomes of some
mediterranean teleosts: Scorpaenidae, Serranidae, Labridae, Blenniidae, Gobiidae (Pisces: Scorpaeniformes, Perciformes). Bolletino di Zoologia, 40(3-4):

37
385-389. Chen, T.R., Ebeling, A.W. (1971). Chromosomes of the gobi fishes in the genus
Gilichtys. Copeia, 1971: 171-174. Cheverud, J.M. (1989). A comparative analysis of morphological variation patterns in
the papionins. Evolution, 43:1737–1747. Chizinski, C. J., Pope, K. L., Wilde, G. R., Strauss, R. E. (2010). Implications of
stunting on morphology of freshwater fishes. Journal of Fish Biology, 76 , 564–
579. Cipriano, R. R., Cestari, M. M., Fenocchio, A. S. (2002). Levantamento citogenético de
peixes marinhos do litoral do Paraná. In: Simpósio de citogenética e genética de peixes, IX. Universidade Estadual de Maringá. Paraná, Brasil. 111p.
Cipriano, R. R., Fenocchio, A. S., Artoni, R. F., Molina, W. F., Noleto, R. B., Kantek, D.
L. Z., Cestari, M. M. (2008). Chromosomal Studies of Five Species of the Marine Fishes From the Paranaguá Bay and the Karyotypic Diversity in the Marine Teleostei of the Brazilian Coast. Brazil archive biology technology, v.51 n.2: pp.303-314.
Concheiro-Pérez, G. A. C., Líban, O., Ortí, G., Bermingham, E., Doadrio, I., Zardoya,
R. (2007). Phylogeny and biogeography of 91 species of heroine cichlids (Teleostei: Cichlidae) based on sequences of the cytochrome b gene. Molecular Phylogenetics and Evolution, 43: 91–110.
Cook, P.C. (1978). Karyotypic analysis of the gobiid fish genus Quietula Jordan and
Evermann. Journal Fish Biology, 12: 173-179. Doebley, J., Stec, A. (1993). Inheritance of the morphological differences between
maize and teosinte: comparison of results for two F2 populations. Genetics, 134: 559-570.
Ene, A.C. (2003). Chromossomal polymorphism in the goby Neogobius eurycephalus
(Perciformes: Gobiidae). Marine Biology, 142: 583-588. Fei, Z., Tao, R. (1987). Studies on the karyotypes of four species in Gobiodae and
compared with other fishes in there. Journal Zhejiang College of Fisheries, 6: 127-131.
Fraser, R.H., Currie, D.J. (1996). The species richness-energy hypothesis in a system
where historical factors are thought to prevail: coral reefs. American Natural, 148:138–159.
Freitas, C . E . C ., Petrere, M., Barrella, W. (2005). Natural and artificially-induced
habitat complexity and freshwater fish species composition. Fisheries Management and Ecology, 12, 63–67.

38
Galetti Jr., P. M., Aguilar, C. T., Molina, W. F. (2000). An overview on marine fish
cytogenetics. Hydrobiologia, 420: 55.62. Gold, J.R., Li, C., Shipley, N.S., Powers, P.K. (1990). Improved methods for working
with fish chromosomes with a review of metaphase chromosome banding. Journal Fish Biology, 37: 563-575.
Gosline, W. A. (1971). Functional morphology and classification of teleostean fishes.
Honolulu: University of Hawaii, 208 p. Horn, M.H., Gibson, R.N. (1988). Intertidal fishes. Sei. Amer. 258 (1): 54-60. Howell, W.M., Black, D.A. (1980). Controlled silver staining of nucleolus organizer
region with protective colloidal developer: a 1-step method. Experientia, 36: 1014-1015.
Ibanez, A., Cowx, I.G., O’Higgins, P. (2007). Geometric morphometrics analysis of fish
scales for identifying genera, species and local populations within Mugilidae. Canadian Journal of Fisheries and Aquatic Sciences, 64: 1091-1100.
Ivanov, V.N. (1975). Chromosomes of the Black sea Gobiidae - Gobius melanostomus
(Pallas) and Gobius batrachocephalus (Pallas) Tsitologiya i Genetika. Cytology and Genetics, 9: 551-552.
Khuda-Bukhsh, A.R. (1978). Somatic chromosomes of an estuarine fish, Trypauchen
vagina (Fam.: Gobiidae) from Sagar Island, West Bengal, India. Current Science, 47: 137-138.
Khuda-Bukhsh, A.R., Barat, A. (1987). Chromosomes in fifteen species of Indian
teleost (Pisces). Caryologia, 40: 131-144. King, M. (1993). Species evolution. The role of chromosome change. Cambridge:
Cambridge University Press. Klinkhardt, M. (1989). Untersuchungen zur Karyologie und Fruchtbarkeit von
Pomatoschistus microps (Kroyer) und P. minutus (Pallas) (Teleostei, Gobiidae). Aus einem inneren Küstengewässer der südwestlichen Ostsee. Zoologischer Anzeiger, 222: 177-190.
Klinkhardt, M. B. (1992). Chromosome structures of four Norwegian gobies (Gobiidae,
Teleostei) and a hypothetical model of their karyo-evolution. Chromatin, 1: 169–
183. Kottelat, M. (1997). European freshwater fishes: an heuristic checklist of the freshwater
fishes of Europe (exclusive of former USSR), with an introduction for non-systematists and comments on nomenclature and conservation. Biology, (Bratislava), 52 (Supplement 5), 1-271.

39
Larson, A. (1998). The comparison of morphological and molecular data in
phylogenetic systematics. In R. DeSalle and B. Schierwater. Molecular approaches to ecology and evolution, 275–296.
Levan, A., Fredga, K., Sandeberg, A. A. (1964). Nomenclature for centromeric position
on chromosomes. Hereditas, 52: 201-220. Lingo, M.E., Szedlmayer, S.T. (2006). The influence of habitat complexity on reef fish
communities in the northeastern Gulf of Mexico. Environment Biological Fishes, 76:71–80.
Maderbacher, M., Bauer, C., Herler, J., Postl, L., Makasa, L., Sturmbauer, C. (2008)
Assessment of traditional versus geometric morphometrics for discriminating populations of the Tropheus moorii species complex (Teleostei: Cichlidae), a Lake Tanganyika model for allopatric speciation. Journal of Zoological
Systematics and Evolutionary Research, 46:153–161.
Madrioli, M., Manicardi, G.C., Machella, N., Caputo, V. (2001). Molecular and cytogenetic analysis of the goby Gobius niger (Teleostei, Gobiidae). Genetica, 110: 73-78.
Manna, G.K., Prasad, R. (1974). Chromosome analysis in three species of fishes
belonging to family Gobiidae. Cytologia, 39: 609-618. Menezes, N. A., Buckup, P. A., Figueiredo, J. L., Moura, R. L. (2003). Catálogo das
espécies de peixes marinhos do Brasil. São Paulo, Museu de Zoologia da Universidade de São Paulo. 160p.
Molina, W. F., Alves, D. E. O., Araújo, W. C., Martinez, P. A., Silva, M. F. M., Costa, G.
W. W. F. (2010). Performance of human immunostimulating agents in the improvement of fish cytogenetic preparations. Genetic Molecular Research, 9 (3): 1807-1814.
Molina, W. F., Lima, L. C. B., Vasconcelos, A. J. M., Accioly, I. (2005). Extensive
karyotypic similarity in marine fishes of the family Haemulidae, Caryologia. Molina, W.F. (2006). Chromosome changes and stasis in marine fishes groups. In: Eva
Pisano, Catharine Ozouf-Costaz, Fausto Foresti, B. G. Kapoor. Fish Cytogenetics. 1ed. Enfield: Science Publisher, v 1, p. 1-52.
Molina, W.F., Galetti, J.P.M. (2004a). Karyotypic changes associated to the dispersive
potential on Pomacentridae (Pisces, Perciformes). Journal of Experimental
Marine Biology and Ecology, 309: 109 – 119.

40
Molina, W.F., Galetti, J.P.M. (2004b). Multiple pericentric inversions and chromosome divergence in the reef fishes Stegastes (Perciformes, Pomacentridae). Genetics and Molecular Biology, 27: 543.548.
Molina. W.F., Galetti, P.M. (2002). Robertsonian rearrangements in the reef fish
Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genetic Molecular Biology, 25:373-377.
Monteiro, L. R., Diniz-Filho, A. F., Reis, S. F., Araújo, E. D. (2002). Geometric
estimates of heritability in biological shape. Evolution, 56 (3): 563-572. Motta Neto, C. C., Molina, W. F. (2010). Haemulidae, modelo cariotípico de estase
evolutiva. Dissertação de mestrado. PPG Ecologia. UFRN, Natal, RN. 114p. Musammil, A. (1974). A chromosome study of twospecies of Gobiosoma from
Venezuela (Gobiidae, Teleostei). Bulletin Institute Oceanography University Oriente, 13: 11-16.
Nadamitsu, S. (1974). Karyological studies of two types of Gobioid fish, "Yoshinobori".
Bulletin Hiroshima Women’s University, 9: 3-9. Nayak, K., Khuda-Bukhsh, A.R. (1987). Chromosomes of two species of fishes
Apocryptes (Gobiidae: Pisces). Environmental Ecology, 5: 371-373. Nelson, J.S. (2006). Fishes of the world. 4 ed. John Willey & Sons, New Jersey. Nishikawa, S., Amaoka, K., Nakanishi, K. (1974). A comparative study of
chromosomes of twelve species of gobioid fish in Japan. Japan Journal Ichthyology, 21: 61-71.
Nogusa, S. (1960). A comparative study of the chromosomes of fishes with particular
considerations on taxonomy and evolution. Memoirs Hyogo University Agricultural, 3: 1-62.
Oliveira, C., Foresti, F., Hilsdorf, A.W.S. (2009).Genetics of neotropical fish: from
chromosomes to populations. Fish Physiology and Biochemistry, 35:81–100. Parsons, K. J., Beren, W. R., Hrbek, T. (2003). Getting into shape: An empirical
comparison of traditional truss-based morphometric methods with a newer geometric method applied to New World cichlids. Environmental Biology of Fishes, 67: 417–431.
Pezold, F. (1984). Evidence for multiple sex chromosomes in the freshwater goby,
Gobionellus shufeldti (Pisces: Gobiidae). Copeia, 1984: 235-238. Phillips, R., Ráb, P. (2001). Chromosome evolution in the Salmonidae (Pisces): An
update. Biological Review, 76: 1-25.

41
Piorski, N. M., Alves, J. R., Machado, M. R. D., Correia, M. M. V. (2005). Alimentação e ecomorfologia de duas espécies de piranhas (Characiformes: Characidae) do lago de Viana, estado do Maranhão, Brasil. Acta Amazônica, 35(1): 63-70.
Pouilly, M., Lino, F., Bretenoux, J. G., Rosales, C. (2003). Dietary-morphological
relationships in a fish assemblage of the Bolivian Amazonian floodplain. Journal of Fish Biology, 62(5):1137-1158.
Rab, P. (1985). Karyotype of the Danude goby, Proterorhinus marmoratus (Pisces,
Gobiidae). Folia Zoologica, 34: 329-334. Rapp Py-Daniel, L. H., Cox Fernandes, C. (2005). Dimorfismo sexual em Siluriformes e
Gymnotiformes (Ostariophysi) da Amazônia. Acta Amazônica, 35 (1): 97-110. Ready, J. S., Ferreira, E. J. G., Kullander, S. O. (2006). Discus fishes: mitochondrial
DNA evidence for a phylogeographic barrier in the Amazonian genus Symphysodon (Teleostei: Cichlidae). Journal of Fish Biology, 69: 200–211.
Rishi, K.K., Singh, J. (1982). Karyological studies on five estuarine fishes. Nucleus, 25:
178-180. Rocha, L.A., Robertson, D.R., Roman, J., Bowen, B.W. (2005). Ecological speciation in
tropical reef fishes. Proceedings of the Royal Society of London. Series B, Biological Sciences, 272, 573–579.
Rohlf, F. J. (2006). tpsDig version 2.10. Ecology and Evolution, Suny at Stony Brook.
http://life.bio.sunysb.edu/morph/. Souza, I.L., Moreira-Filho, O. Bertollo, L.A.C. (1998). Contribuição citogenética em
Gobiidae. VII Simpósio de Citogenética Evolutiva Aplicada de Peixes Neotropicais, p. 26. Londrina, Paraná, Brazil.
Springer, V.G. (1983). Tyson belos, new genus and species of western Pacific fish
(Gobiidae, Xenistminae), with discussion of gobioid osetology and classification. Smithson. Control Zoology, 390: 1–40.
Stange, E.A.R., Passamani, M. (1986). Caracterização cromossômica de Awaous
tajasica (Lichtenstein, 1822) (Pisces, Gobiidae). In: I Simpósio de Citogenetica Evolutiva Aplicada a Peixes Neotropicais. São Carlos, São Paulo. p. 59.
Subrahmanyam, K. (1969). A karyotypic study of the estuarinefish Boleophthalmus
boddaerti (Pallas) with calcium treatment. Current Science, 38: 437-439. Sumner, A.T. (1972). A simple technique for demonstrating centromeric
heterochromatin. Experimental Cell Research, 75: 304. Taylor, M.S., Hellberg, M.E. (2003). Genetic evidence for local retention of pelagic
larvae in a Caribbean reef fish. Science, 299, 107– 109.

42
Thode, G., Alvarez, M.C. (1983). The chromosome complements of two species of Gobius (Teleostei, Perciformes). Experientia, 39: 1312-1314.
Thode, G., Giles, V., Alvarez, M. C. (1985). Multiple chromosome polymorphism in
Gobius paganellus (Teleostei, Perciformes). Heredity, 54: 3-7. Thode, G., Martinez, G., Ruiz, J. L., Lopez, J. R. (1988). A complex chromosomal
polymorphism in Gobius fallax (Gobiidae, Perciformes). Genetica, 76: 65–71. Tornabene, L., Baldwin, C., Weigt Lee, A., Pezold, F. (2010). Exploring the diversity of
western Atlantic Bathygobius (Teleostei: Gobiidae) with cytochrome c oxidase-I, with descriptions of two new species. Aqua, International Journal of Ichthyology, vol. 16 no. 4.
Tracker, C.E. (2003). Molecular phylogeny of the gobioid fishes (Teleostei:
Perciformes: Gobioidei). Molecular Phylogenetics and Evolution, 26: 354-368.update. Biological Reviews, 76: 1.26.
Vasil’ev, V.P., Grigoryan, K.A. (1993). Karyology of the Gobiidae. Journal Ichthyology,
33:1–16. Vasil'ev, V.P. (1985). The evolutionary karyology of fishes. Evoljuzionnaja kariologiya
ryb. Nauka, Moscow. p. 300. Vasil'ev, V.P., Grogoryan, K.A. (1992). Karyology of fishes from the family Gobiidae.
Voprosy Ikhtiologii, 32: 27-40. Vasil'ev, V.P., Vasil'yeva, E.D. (1994). The new systematics of freshwater Caucasian
gobies: three distinct species within former N. Platyrostris constructor (Gobiidae). In: Abstract. VIII Congress Soc. Europ. Ichthyology Oviedo, p. 66.
Vicari, M.R. Artoni, R.F., Bertollo, L.A.C. (2003). Heterochromatin polymorphism
associated with 18S rDNA: a differential pathway among Hoplias malabaricus fish populations. Cytogenetics Genome Research, 1001:24-28.
Vitturi, R., Catalano, E. (1989). Multiple chromosome polymorphism in the gobiid fish
Gobius niger jozo L. 1758 (Pisces, Gobiidae). Cytologia, 54: 231-235. Wainwright, P. C., Collar, D.C., Alfaro, M.E. (2007). Integrated diversification of
locomotion and feeding in labrid fishes. Biology Letters, 4, 84–86. Webb, C.J. (1986). Karyology of the Indo-Pacific Parioglossus raoi (Herre) (Teleostei:
Gobioidei) from Fiji. Australian Journal Marine and Freshwater Researsh, 37: 347-351.
Werner, E. E. (1977). Species packing and niche complementarity in three sunfishes.
American Naturalist, 111:553–579.

43
Winterbottom, R. (1984). A review of the gobiid fish genus Trimma from the Chagos Archipelago, central Indian Ocean, with the description of seven new species. Canadian Journal Zoology, 62: 695–715.
Yamada, J. (1967). An observation of the chromosomes in the embryonic cells of a
goby, Chaenogobius urotaenia (Hilgendorf). Bulletin of the Faculty of Fisheries Hokkaido University, 18: 183-187.

44
4.2 Capítulo II
Análises citogenéticas e morfométricas interpopulacionais no
peixe marinho Bathygobius soporator (Gobiidae - Perciformes)
Lima Filho, P. A. & Molina, W. F.
Resumo
Bathygobius soporator, também conhecido como amboré, espécie bentônica de baixa vagilidade, habitante de zonas intertidais apresenta uma extensa distribuição anfi-Atlântica. Diante de condições propícias ao fracionamento populacional e indícios de ausência de coesão genética em toda sua área de distribuição, persistem dúvidas sobre o real status desta espécie. Aqui são comparadas por meio de técnicas citogenéticas e de morfometria geométrica quatro populações geograficamente distribuídas em largo espaço oceânico, ao longo do litoral brasileiro, no Maranhão (MA), Rio Grande do Norte (RN), Bahia (BA) e Atol das Rocas (AR), único atol do Atlântico. Exceto para a amostra do AR, todas as amostras exibiram um cariótipo similar, com 2n=48 cromossomos, composto por 2m+6st+40a (NF=56). As Ag-RONs são simples localizadas em posição terminal no braço curto do 4o par (st). Blocos heterocromáticos estão distribuídos em regiões centroméricas e terminais da maioria dos pares cromossômicos. A população do AR exibe uma fórmula cariotípica diferenciada (2n=48; 28st+20a; NF=76) com ausência do par metacêntrico característico de outras populações da espécie. Os sítios ribossomais nesta amostra se apresentam sobre o braço curto do 1o par cromossômico (st). Morfologicamente as populações continentais (MA, RN e BA) apresentam médias significativamente distintas, com alguma sobreposição em parte da amostra, entretanto exibem total distinção com a população do AR. A combinação de dados citogenéticos e morfométricos sugere que as populações continentais aparentemente se encontram respondendo a regimes seletivos distintos, mas sem indicações de divergências cromossômicas dentro do nível de análise empregado, por outro lado, os padrões conspicuamente diversos na população do Atol das Rocas, parecem confirmar a existência de uma nova espécie.
Palavras-chave: Gobiidae; citogenética de peixes; Atol das Rocas;
morfometria geométrica; nova espécie.

45
Cytogenetics and morphometric analyses interpopulation in
marine fish Bathygobius soporator (Gobiidae – Perciformes)
Lima Filho, P. A. & Molina, W. F.
Abstract
Bathygobius soporator, also known as Amboró, benthic specie of low vagile, it lives in intertidal zones and it has an extensive distribution amphi-Atlantic. Given the conditions conducive to population fragmentation and lack of evidence of genetic cohesiveness throughout its distribution area, there remain doubts about the real status of this species. Here, they are compared by means of cytogenetic techniques and geometric morphometric four populations geographically distributed in wide ocean, along the Brazil's coast: Maranhao (MA), Rio Grande do Norte (RN), Bahia (BA) and Atol das Rocas (AR), the only atoll in the Atlantic. Except for the sample from AR, all samples showed a similar karyotype with 2n=48 chromosomes, consisting by 2m +6 st +40 a (FN=56). As Ag-RONs are simply located in terminal position on short arm of 4o pair (st). Heterochromatic blocks are distributed in centromeric regions and terminals of the most of the chromosomes pairs. The population from AR displays dinstinctive karyotype (2n=48; 28st+20a; NF=76) without metacentric pair, characteristic of the populations from the specie. The ribosomal sites in this sample are presented on the short arm from chromosome first pair (st). Morphologically, the continental populations (MA, RN and BA) have significantly different averages, with some overlap in the sample, however they show total distinction from AR population. The combination of cytogenetic and morphometric data suggests that continental populations are apparently responding to different selective regimes, but there are not indications of chromosomal differences within the analysis level considered, on the other hand, conspicuously different patterns in the population from Atol das Rocas , seem to confirm the new specie existence.
Keywords: Gobiidae; fishes cytogenetics; Atol das Rocas, geometric morphometrics, new specie.

46
Introdução
A família mais especiosa presente no ambiente marinho, Gobiidae é
composta por indivíduos de pequeno porte presentes em todos os mares
tropicais e subtropicais, sendo constituída por cerca de 1.950 espécies (Nelson,
2006). Dentre os gobídeos a espécie Bathygobius soporator (Valenciennes,
1837), apresenta distribuição anfi-atlântica, no oeste do Atlântico distribui-se da
Flórida (EUA) ao Rio Grande do Sul (BRA) (Cervigo´n, 1966; Robins & Ray,
1986; Miller & Smith, 1989; Burns et al., 2010), e no Atlântico leste, pode ser
encontrado na costa da África, do Senegal a Angola (Cervigo´n, 1966; Miller &
Smith, 1989; Afonso et al., 1999).
Apesar da ampla distribuição geográfica, B. soporator apresenta
deslocamento limitado, é uma espécie bentônica sedentária, euritérmica,
eurialina (Fanta, 1997). Litorâneo residente (Demartini, 1999), pode ser
encontrado em poças de maré na zona intertidal de regiões costeiras e ilhas
oceânicas. Apresenta acentuado comportamento de homing (Gibson &
Yoshiyama, 1999) e provavelmente quando em habitat adequado se limite a
uma área reduzida por toda sua vida, uma vez que os adultos exibem
fidelidade à poça ou toca (Martins et al., 2006).
A princípio, tamanha distribuição geográfica pode ser explicada pela
difícil caracterização de barreiras físicas absolutas no ambiente marinho, que
associado a fase larval livre natante e influência de correntes marinhas, altas
taxas de dispersão seriam promovidas (Imron et al., 2007), ocasionando
conectividade entre as populações marinhas e possibilitando processos
genéticos evolutivos interdependentes (Luttikhuizen, 2003). No entanto, com o
deslocamento limitado de B. soporator associado a características reprodutivas
como ovos adesivos, curto período pelágico larval (Tavolga, 1953; Peters,
1983) e características ambientais locais, sua capacidade de dispersão e
efetivação em fluxo gênico torna-se questionável e o isolamento de
populações, seguido de especiação, possível.
Diante de condições propícias ao fracionamento populacional, não pode
ser descartada a possibilidade que B. soporator constitua um complexo de
espécies, como muitas outras marinhas, que a princípio detinham de grande
distribuição geográfica, e foram consideradas sob um exame minucioso,

47
complexos morfologicamente similares, mas geneticamente distintas
(Knowlton, 2000).
Uma vez que a família Gobiidae apresenta elevada diversidade
cariotípica (Galetti et al., 2000), e que especiações são frequentemente
associadas a rearranjos cromossômicos (Mandrioli et al., 2001), visando
identificar possíveis diferenciações populacionais cariotípicas e morfológicas, o
presente trabalho compara populações de B. soporator geograficamente
distribuídas no atlântico sul, ao longo do litoral brasileiro e no Atol das Rocas,
por meio de técnicas citogenéticas e de morfometria geométrica.
Material e métodos
Foram coletados um total de 170 exemplares de Bathygobius soporator
provenientes de quatro áreas da costa nordeste brasileira, praia do Araçagi, em
São Luís (MA) (2°28'14.06"S, 44°15'49.15"O), praia do Farol da Barra, em
Salvador (BA) (13°0'39.61"S, 38°31'27.21"O), praia de Búzios, em Nísia
Floresta (RN) (5°41'57.41"S, 35°11'34.71"O) e Atol das Rocas (3º51’’S,
33º49’’W) (Tabela 1). Os espécimes foram capturados com uso de rede de
confecção própria, submetidos aos protocolos de obtenção de cromossomos
mitóticos e fotografados em campo, ou acondicionados em sacos plásticos com
água e oxigênio e transportados às dependências do Laboratório de Genética
de Recursos Marinhos na UFRN, sendo mantidos em aquários aerados até
serem fotografados para análise morfológica e se procedesse à realização de
preparações cromossômicas. Para identificação taxonômica utilizou-se a chave
de classificação de Bohlke & Chaplin (1993).
Preparações cromossômicas
Precedente aos métodos de obtenção de cromossomos metafásicos, os
exemplares foram submetidos à estimulação mitótica, “in vivo” com duas
aplicações intramuscular/peritoneal de Aminovac® (complexo de antígenos
bacterianos e fúngicos) na razão de um comprimido diluído em 0,5 ml de água
na proporção de 100g de peso do animal (Molina et al., 2010).
Os cromossomos mitóticos foram obtidos a partir de células do rim
48
anterior, segundo Gold et al. (1990). Cerca de trinta metáfases foram
analisadas para cada exemplar para se estabelecer o número modal, sendo
que, as melhores metáfases foram fotografadas em um microscópio OlympusTM
BX50 (1000x), acoplado à uma câmera digital (Olympus, modelo DP70), e
utilizadas na confecção do cariótipo. A morfologia cromossômica foi
determinada de acordo com Levan et al. (1964).
As regiões heterocromáticas foram definidas segundo Sumner (1972), a
detecção das marcações Ag-RONs foi realizada conforme a técnica descrita
por Howell e Black (1980). A coloração com os fluorocromos CMA3 e DAPI
seguiu o protocolo de Carvalho et al. (2005).
Tabela 1: Número de indivíduos coletados, seus respectivos pontos de coleta e sua utilização para estudos morfométricos e citogenéticos.
Pontos de Coleta Análises
morfométricas Análises
citogenéticas Maranhão 50 14 (09♀ / 05♂)
Rio Grande do Norte 50 20 (09♀ / 11♂)
Bahia 50 16 (10♀ / 06♂)
Atol das Rocas 20 10 (06♀ / 04♂)
Total 170 60
*Todos os espécimes coletados foram utilizados para análises morfométricas.
Análises morfológicas
Para as análises por morfometria geométrica (MG), imagens no formato
JPEG com dimensão 3264x2448, foram obtidas da face lateral esquerda dos
espécimes, com escala métrica, utilizando-se uma câmera digital Sony H10
(8,1 megapixels) acoplada a um tripé VF WT3540.
O programa tpsUtil foi utilizado para ordenar as imagens dos espécimes
num único arquivo com o formato TPS. O software tpsDig2 (Rohlf, 2006) foi
utilizado para localizar nove landmarks nas imagens (Figura 1), escolhidos
segundo a classificação de Bookstein (1991). As coordenadas referentes ao
posicionamento espacial de cada landmark foram sobrepostas utilizando-se o
software CoordGen6.
Com a utilização do software CVAGen6, análises das variáveis
canônicas (VC) e MANOVA permitiram comparações morfométricas entre as

49
populações, testes de alocação e agrupamentos. Para realizar o teste
estatístico ANOVA, D2 (distância de Mahanalobis) e gerar matrizes (Gride) de
deformação comparativa utilizou-se o programa MorphoJ 1.02b®.
Figura 1. Imagem digitalizada de B. soporator retratando os nove landmarks definidos para análises morfológicas interpopulacionais. Descrição esquemática dos landmarks: 1. Extremidade distal do osso pré-maxilar; 2. Origem da primeira nadadeira dorsal; 3. Término da primeira nadadeira dorsal; 4. Origem da segunda nadadeira dorsal; 5. Término da segunda nadadeira dorsal; 6. Término da nadadeira anal; 7. Origem da nadadeira anal; 8. Origem da nadadeira pélvica; 9. Base infra-ocular medial. Barra=1,5cm.
Resultados
As populações de B. soporator provenientes da costa brasileira (litoral do
MA, RN e BA) apresentaram características citogenéticas semelhantes, com
2n=48, e fórmula cariotípica consistindo de 2m+6st+40a (NF=56).
Sítios Ag-RONs foram localizados no braço curto do quarto par
cromossômico subtelocêntrico (Figura 2). Estas regiões se mostraram
heterocromáticas apresentando um padrão CMA3+ DAPI-. Além destas, o
bandamento C evidenciou segmentos heterocromáticos em regiões
centroméricas na maioria dos cromossomos e terminal em alguns, estendendo-
se nos braços curtos de cromossomos subtelocêntricos.
Os espécimes provenientes do Atol das Rocas demonstraram 2n=48,
estrutura cariotípica de 28st+20a (NF=76) e padrões diferenciados quando
submetidos aos diferentes bandamentos, comparados aos apresentados pelas
demais populações. A impregnação com nitrato de prata revelou sítios Ag-
RONs em posição terminal no braço curto do 1º par cromossômico (Figura 3),
intercalado a blocos heterocromáticos ricos em GC (CMA3+/DAPI-). O

50
bandamento C revelou a presença de conspícuos blocos heterocromáticos em
posição centromérica em grande parte dos cromossomos, de forma mais
evidente em braços curtos em todos os cromossomos subtelocêntricos.
Figura 2. Cariótipo de B. soporator para populações contientais (MA, RN e BA) à partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (4º) corado com impregnação argêntea, CMA3 e DAPI. Barra= 5µm.
Figura 3. Cariótipo de B. soporator população do Atol das Rocas, a partir de coloração convencional (a) bandamento C (b). Em destaque (c) o par organizador nucleolar (1º) corado com impregnação argêntea, CMA3 e DAPI. Barra= 5µm.
a b c
4º
CMA3
DAPI
a c
1º
CMA3
DAPI
b

51
As populações continentais exibiram médias morfométricas
significativamente diferentes (Figura 5), embora apresentem indivíduos com
morfologia intermediária entre os grupos (Tabela 2). A população do Atol das
Rocas por sua vez, apresentou total distinção morfológica (distância de
Mahalanobis >4,9) em relação a todas as populações continentais (Figura 4).
Tabela 2. Porcentagem de indivíduos alocados para cada população de B. soporator e suas interfaces baseados na CVA.
Origem da População Identidade comum* Intermediários** N
Pop. MA 76% 24% 50
Pop. RN 90% 10% 50
Pop. BA 82% 18% 50
Pop. Atol das rocas 100% 0% 20
*Porcentagem de indivíduos da população que compartilham um morfotipo comum; **morfótipos correspondentes a outras populações.
Através de MANOVA, as variáveis canônicas 1 e 2 utilizadas para o
estudo mostraram-se significativas para F (p<0,0001), explicando cerca de
63,5% e 30,5% respectivamente, da variação morfológica observada para as
populações. Os resultados da ANOVA a partir de análises procrustes
confirmaram a distinção morfológica das populações continentais (Tabela 3).
Tabela 3. Resultados post-hoc ANOVA a partir de procrustes, classificadas com a média do centroide, efeito individual, entre as populações continentais.
População p F df
RN x MA < 0, 0001 25,69 1
RN x BA < 0, 0001 33,15 1
MA x BA < 0, 0001 390,24 1

52
Figura 4. Distribuição das amostras de B. soporator ao longo da primeira e segunda variável canônica (63,5% e 30,5% da variância respectivamente), a partir das variáveis morfométricas.
Figura 5. Distribuição das populações continentais amostradas de B. soporator ao longo da primeira e segunda variável canônica (76,2% e 23,5% da variância respectivamente). Os símbolos em destaque indicam a média morfométrica de cada população.
Ao verificarmos os grides de deformação comparativos, a partir da
primeira variável canônica entre as populações, verificamos modificações em
todos os landmarks. O gride comparativo formado para populações do RN x

53
Atol das Rocas (Figura 6) demonstra um morfotipo para espécies do AR com
significativa redução no conjunto de nadadeiras dorsais e anal estabelecendo
modificações no seu posicionamento, aumento corporal e da região anterior.
Entre as populações do RN x MA (Figura 7) observa-se para a população do
MA redução na altura corporal e modificações no posicionamento da boca e
base infra-ocular medial. A comparação entre as populações RN x BA (Figura
8) aponta modificações expressivas na altura corporal, região anterior mais
fusiforme e alterações no posicionamento da nadadeira anal e pélvica para
população da BA.
Figura 6. Gride comparativo entre população continental e insular de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população do Atol das Rocas.
Figura 7. Gride comparativo entre populações continentais de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população do Maranhão.

54
Figura 8. Gride comparativa entre populações continentais de B. soporator, a partir da variável canônica 1. Ponto médio central representa população do RN e o desvio vetorial população da Bahia.
Discussão
Os dados citogenéticos para as populações continentais monstraram-se
coincidentes, com 2n=48 e fórmula cariotípica consistindo de 2m+6st+40a
(NF=56). No entanto a população do Atol das Rocas com 2n=48 apresentou
estrutura caríotipica 28st+20a (NF=76) e total distinção morfológica (distância
de Mahalanobis >4,9) com as populações continentais, estes novos dados
associados à distinção molecular constatada (Lima et al., 2005), deixa claro a
existência de uma nova espécie para o gênero Bathygobius presente no oeste
do Atlântico.
Teleósteos marinhos apresentam baixa variabilidade no número de
cromossomos e alta uniformidade na macro-estrutura cariotípica (Cipriano et
al., 2008), entretanto representantes da família Gobiidae demonstram
cariótipos altamente variáveis (Caputo, 1998; Ene, 2003) e uma estreita relação
entre rearranjos cromossômicos e processo de especiação (Cap. I), que auxilia
a identificação de espécies crípticas através de caracteres cromossômicos e
marcadores citotaxonômicos.
As diferenças encontradas na estrutura cariotípica das populações
continentais e AR podem ser atribuídas a diferentes mecanismos de
modificação cromossômica como eventos de inversões pericêntricas frequente

55
em Perciformes (Molina & Galetti, 2004), fissão cromossômica e fusões
Robersonianas, são importante fonte de diversificação cariotípica não apenas
para família Gobiidae (Cataudella et al., 1973; Thode et al., 1985; Vitturi &
Catalano, 1989; Amores et al., 1990; Klinkhardt, 1992; Caputo et al., 1996;
1997), como para Pomacentridae (Ojima & Kashiwagi, 1981) e Cichlidae
(Martins et al., 1995).
O gênero Bathygobius apresenta 26 espécies (Eschmeyer, 2010),
destas apenas quatro possuem descrição citogenética B. mystacium (Cap. I),
B. soporator litoral brasileiro (Brum et al., 1996; Cipriano et al., 2002), B.
soporator presente no AR e B. fuscus (Arai & Sawada, 1975) costa japonesa.
Estas espécies compartilham 2n=48 cromossomos, caracterizado como padrão
aos Perciformes (Galetti et al., 2000), indicativo de derivação cariotípica para
representantes da família Gobiidae, que considera 2n=46 cromossomos
(NF=46) cariótipo ancestral/basal (Vasil’ev & Grigoryan, 1993). Este aumento
no número cromossômico pode ser decorrente de inversões pericêntricas
seguidos de fissão cromossômica, processos aparentemente comum ao
gênero e ocasional a família, presentes em Yongeichthys criniger (Arai &
Sawada, 1974), Ctenogobius shufeldti (Pezold, 1984), C. smaragdus (Cap. I),
Gobius niger (Vitturi & Catalano, 1989), G. paganellus (Caputo et al., 1997).
A quantidade de heterocromatina, assim como seus aspectos
composicionais possuem um papel importante na diversificação cariotípica de
muitos peixes (Caputo et al., 1997; Canapa et al., 2002; Molina et al., 2002;
Molina & Galetti, 2002). A análise da heterocromatina constitutiva presente no
cariótipo da população do AR aponta blocos conspícuos, deflagrando a
ocorrência de um cariótipo mais sujeito à macro mudanças cariotípicas. É
provável que a ocorrência de inversões pericêntricas tenha configurado tais
mudanças, justificado pelo elevado número de cromossomos bi-braquiais com
braços curtos completamente heterocromáticos, que normalmente é apontado
como resultado de inversões cromossômicas (King, 1993).
A heterocromatina constituiva apresentada em populações continentais
dispõe de posição centromérica e terminal em alguns pares cromossômicos,
com blocos na sua maioria pouco heterocromáticos, distintos do padrão
apresentado pela população AR. Entretanto seu posicionamento levanta
indícios da ocorrência de fusão Robertsoniana à formação dos cromossomos

56
metacêntricos, devido sua ocupação em posição centromérica evidente e
diferencial aos demais blocos heterocromáticos. Estudos realizados nos
gobídeos G. niger e N. eurycephalus com polimorfismo cromossômico,
apontam características estruturais e de composição da heterocromatina
centromérica GC-ricas, como facilitadoras a ocorrência de fusões
Robertsonianas (Caputo et al.,1997; Ene, 2003).
O uso de Ag-RONs constitui um efetivo marcador citotaxonômico para
peixes (Affonso, 2000). Sua utilização tem ajudado a formular hipóteses
filogenéticas em alguns grupos (Cross et al., 2006) e na identificação de fauna
críptica, como aqui apresentado. A presença de único par portador de sítios
compartilhado às populações continentais e AR, localizadas em posição
terminal-centromérica em braços curtos de cromossomos
submetacêntricos/subtelocêntricos é comum a diferentes espécies de Gobiidae
como Potamoschistus minutus (Klinkardt, 1992; Caputo, 1998), P. pictus
(Klinkardt, 1992), P. marmoratus (Caputo, 1998) e idicativo de possível
característica plésiomórfica às espécies presentes no litoral brasileiro (Cap. I).
A associação entre RONs e heterocromatina CMA3+ observada nas
populações analisadas, é freqüentemente descrita em peixes (Ameniya & Gold,
1986; Almeida-Toledo et al., 1996), sendo as regiões GC-ricas propensas a
uma alta taxa de recombinação que pode favorecer processos de fusões ou
fissões cêntricas (Redi et al., 1990), e quando adjacentes, sobrepostas ou
intercaladas às RONs pode favorecer rearranjos aos pares portadores (Vicari et
al., 2003).
Apesar da proximidade do AR com a costa brasileira, sua colonização
pela espécie B. soporator é proveniente da região caribenha e parece ter
ocorrido após as espécies da costa brasileira e caribenha terem se separado
(Lima et al., 2005). As diferenças apresentadas na macroestrutura cariotípica,
posicionamento dos sítios Ag-RONs equilocais à heterocromatinas ricas em
bases GC evidenciadas pela dupla coloração apresentando um padrão
CMA3+/DAPI- e blocos heterocromáticos, entre populações continentais e AR
são significativas e podem ser utilizadas como indicativo carioevolutivo à
espécies do gênero Bathygobius, presentes na província biogeográfica
caribenha e brasileira.

57
Estudos citogenéticos realizados em populações localizadas na baía de
Paranaguá, Paraná e litoral do Rio de Janeiro apresentaram cariótipos com
2n=48 cromossomos, 2m+46a (NF=50) (Brum et al., 1996; Cipriano et al.,
2002). Apesar da diferença ao padrão encontrado no presente trabalho, a não
detecção de cromossomos submetacêntricos pode ser atribuída ao menor grau
de condensação cromossômica e não utilização de técnicas citogenéticas para
detecção das regiões organizadoras de nucléolos (Ag-RONs), heterocromatina
constitutiva (Banda C) e coloração com os fluorocromos CMA3 e DAPI.
No ambiente marinho barreiras físicas entre populações de peixes são
difíceis de caracterizar (Joyeux et al., 2001), a fase larval pelágica garante
potencial à dispersão (Palumbi, 1994; Mora & Sale, 2002) e pode promover
conectividade entre populações. Entretanto, a identificação de quebra
filogeográfica entre populações de gobídeos no mar do Caribe, com retenção
local de larvas ao redor das ilhas (Taylor & Hellberg, 2005; 2006) traz indícios
da possibilidade de especiação na presença de barreiras geográficas
transitórias ou fracas.
Análises moleculares em diferentes populações continentais de B.
soporator demonstraram alta estruturação populacional, sugerindo níveis
limitados no fluxo gênico desta espécie ao longo da costa brasileira (Lima et al.,
2005). A presença de deslocamento limitado associado a características
reprodutivas como ovos adesivos, curto período pelágico larval (Tavolga, 1953;
Peters, 1983) e eventos vicariantes históricos, podem ter contribuído com este
cenário de redução do fluxo gênico.
Espécies do gênero Bathygobius presentes na costa atlântica americana
são consideradas altamente conservadas morfologicamente, o que dificulta a
determinação do número de espécies válidas para o gênero, usando apenas
caracteres morfológicos (Tornabene et al., 2010). Caracteres merísticos
tradicionais usados em chaves taxonômicas não são capazes de distinguir as
amostras populacionais de B. soporator presente no Atol das Rocas e da costa
brasileira (Lima et al., 2005). Entretanto, a análise utilizando morfometria
geométrica evidenciou total distinção morfológica entre estas duas populações,
o que demonstra eficácia e sensibilidade à técnica para o reconhecimento de

58
fauna críptica e aplicabilidade a resolução de problemas taxonômicos presente
para o gênero Bathygobius.
A associação de técnicas genéticas e morfométricas vêm auxiliando
extensivamente no reconhecimento de espécies crípticas de gobídeos e seus
mecanismos evolutivos atuantes. Evidências de especiação simpátrica
(ecológica) foram demonstradas no gênero Gobiodon, onde a separação em
nível de espécie não seguia a geografia, mas a mudanças no coral anfitrião
(Munday et al., 2004). Taylor & Hellberg (2003) analisando o gobídeo
Elacatinus evelyna, evidenciaram uma abrupta descontinuidade genética entre
populações separadas por apenas 20 km, com diferenças nos padrões de
coloração por habitat, os quais teriam desempenhado papel importante em sua
diversificação.
Análises morfométricas apontaram distinção à população AR de B.
soporator, que demonstrou apresentar estrutura corporal mais fusiforme com
região anterior e altura corporal maiores, modificações no posicionamento da
boca e do olho e redução nas nadadeiras anal e dorsais, em relação à
população continental do RN. Estas modificações morfológicas em conjunto
podem garantir maior velocidade de locomoção (Breda et al., 2005),
estabilidade na natação (Bond, 1979) e especialização alimentar (Piorski et al.,
2005), adaptações estas que podem estar relacionadas às condições
ecológicas peculiares dessa região insular.
Morfologicamente as populações continentais de B. soporator
apresentam médias corporais significantemente distintas. A principal variação
morfológica relaciona-se a altura corporal, que demonstra elevação progressiva
conforme o aumento da latitude. Embora a variação térmica entre as
populações distribuídas não seja visivelmente drástica (média anual 57 anos:
MA 26,3ºC e BA 25,2ºC) (EMBRAPA, 2003), ela pode estar relacionada ao
aumento da altura média do corpo das populações distribuídas no sentido
Norte-sul. Esta distribuição clinal se adequa a regra de Bergman que associa o
aumento da latitude/diminuição da temperatura com um aumento do volume
corporal do organismo (Gaston et al., 2008), descrita para diversos grupos de
vertebrados (Lomolino et al., 2006) inclusive peixes (Macpherson & Duarte,
1994; Smith & Brown, 2002) na costa brasileira, como pomacentrídeo
Abudefduf saxatilis (Molina et al., 2006).

59
O gobídeo B. soporator é uma espécie encontrada principalmente em
áreas intertidais habitando poças de maré, com alta capacidade de reação a
mudanças rápidas, desenvolvendo ao longo de sua evolução estratégias
fisiológicas para sobreviver em ambientes variáveis (Fanta, 1997). Estas
características evitam que a espécie tenha que se deslocar, o que associado
ao comportamento reprodutivo e limitação no fluxo gênico, sugerem a seleção
de morfótipos melhores adaptados a cada área.
Em função de características ambientais, muitas espécies de peixes
apresentam considerável plasticidade fenotípica que as permite aumentar sua
aptidão em determinado ambiente (Mérona et al., 2009), sugere-se que a
capacidade de um padrão genotípico, produzir múltiplos fenótipos em resposta
a variações no meio ambiente (Pfennig et al., 2010), seja uma importante
estratégia de adaptação à populações que enfrentam ambientes varáveis
(Stearns, 1989; Scheiner, 1993), desempenhando um papel importante na sua
diversificação (West-Eberhard, 1989).
As variações intrapopulacionais e interpopulacionais identificadas na
espécie são comuns em peixes. A variação morfológica intrapopulacional é
normalmente relacionada ao habitat dos indivíduos (Smith & Skúlason, 1996),
onde a competição intraespecífica atua na correlação entre a morfologia e
escolha do habitat (Wilson & Turelli, 1986). Variações morfológicas do tipo
interpopulacionais normalmente estão relacionadas à disponibilidade de
recursos e competição com outras espécies (Lindsey, 1981; Lavin & McPhail,
1985, 1986; Magnan, 1988; Mittelbach et al., 1992; Walker, 1997).
Apesar das variações no posicionamento da boca nas populações
continentais de B. soporator, análises alimentares em três populações
distribuídas ao longo da costa brasileira, Pará (Norte), Bahia (Nordeste) e São
Paulo (Sudeste), evidenciaram semelhanças tróficas caracterizando um hábito
onívoro, com consumo principal de crustáceos e algas (Lopes & Oliveira-Silva,
1998; Corrêa & Uieda, 2007; Oliveira-Raiol et al., 2011), discernível ao padrão
exibido pela espécie na costa da Nigéria, Atlântico Oriental (Lawson & Thomas,
2010). Estas características ecológicas diferenciadas ao longo da extensa
distribuição geográfica de B. soporator, quando associadas ao reduzido fluxo
gênico apresentado por suas populações ao longo da costa brasileira, sugerem
uma distinção marcante de B. soporator nas porções Leste e Oeste do

60
Atlântico. Estudos genéticos e morfométricos são indicados para auxiliar esta
possível imprecisão taxonômica.
A combinação de variação genética e morfológica tem sido aplicada no
esclarecimento do padrão evolutivo de gênero Bathygobius. Análises utilizando
ambas as abordagens realizadas em representantes da costa da Flórida (EUA)
à Venezuela e áreas insulares, como Bahamas e Belize, permitiram identificar
duas novas espécies para o gênero, Bathygobius geminatus e Bathygobius
antilliensis, presentes no oeste do Atlântico (Tornabene et al., 2010).
A diversidade encontrada na família Gobiidae tem sido explicada
principalmente por processos vicariantes levando à alopatria (Huyse et al.,
2004). Os dados cromossômicos aliados as análises morfológicas parecem
corroborar um cenário evolutivo de fragmentações populacionais e adaptação a
novos habitats como promotores da extrema diversidade presente no grupo.

61
5. CONCLUSÕES
· Os gobídeos analisados demonstram marcante diversidade cariotípica e morfológica, evidenciando estreita relação entre rearranjos cromossômicos e processo de especiação para família;
· Rearranjos cromossômicos envolvidos na diversificação de B. soporator, B. mystacium, C. glaucofraenum, C. boleosoma e C. smaragdus são as inversões pericêntricas, mecanismos fissão e fusão cromossômica;
· A diversidade cariotípica e morfológica sugere uma dinâmica na evolução cariotípica, compatível com grupos dulcícolas sujeitos a intensas fragmentações populacionais;
· A utilização da citogenética e morfometria geométrica demonstra eficiência à identificação das espécies da família Gobiidae, mesmo quando crípticas como B. soporator e B. mystacium;
· Os dados citogenéticos e morfométricos para populações continentais e AR de B. soporator confirmam a existência de uma nova espécie para o gênero Bathygobius presente no oeste do Atlântico;
· As populações de B. soporator provenientes da costa brasileira (litoral do MA, RN e BA) apresentam morfotipos distintos. A principal variação morfológica relaciona-se a altura corporal e se adequa a regra de Bergman;
· Os dados cromossômicos aliados as análises morfológicas corroboram um cenário evolutivo de fragmentações populacionais e adaptação a novos habitats como promotores da extrema diversidade presente na família.

62
Referências
Affonso, P. R. A. M. (2000). Caracterização citogenética de peixes de corais da
Família Pomacanthidae (Perciformes). Masters thesis, Universidade Federal de São Carlos, São Paulo, Brasil.
Afonso, P., Porteiro, F. M., Santos, R. S., Barreiros, J. P., Worms, J., Wirtz, P. (1999).
Coastal marine fishes of São Tomé Island (Gulf of Guinea). Life Marine Science, 17, 65– 92.
Almeida Toledo, L.F. (1998). Cytogenetic markers in neotropical freshwater fishes. In:
Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M.S., Lucena, C.A.S. (eds). Phylogeny and Classification of Neotropical Fishes. Edipucrs, Porto Alegre, Brasil. p. 583-588.
Almeida-Toledo, L.F., Stocker, A.J., Foresti, F., Almeida-Toledo Filho, S. (1996).
Fluorescence in situ hybridization with rDNA probes on chromosomes of two nucleolus organizer region phenotypes of a species of Eigenmannia (Pisces, Gymnotoidei, Sternopygidae). Chromosome Research, 4: 301-305.
Amemiya, C.T., Gold, J.R. (1986). Chromomycin A3 stains nucleolus organizer regions
of fish chromosomes. Copeia, 1986:226–231. Amores, A., Giles, V., Thode, G. (1990). Adaptive charater of a Robertsonian fusion in
chromosomes of the fish Gobius paganellus (Pises, Perciformes). Heredity, 65: 151-155.
Arai, R., Katsuyama, I., Sawada, Y. (1974). Chromosomes of Japanese gobioid fishes
(II). Bulletin National Science Museum, 17: 269-279. Arai, R., Sawada, Y. (1975). Chromosomes of Japanese gobioid fishes. (III) Bull. Tókio,
Japan. National Science Museum, 1: 225-232. Bertollo, L. A. C., Oliveira, C., Molina, W. F., Margarido, V.P., Fontes, M. S., Pastori
M.C., Falcão, J. N., Fenocchio, A. S. (2004). Chromosome evolution in the erythrinid fish, Erythrinus erythrinus (Teleostei, Characiformes). Heredity, 93: 228.233.
Bohlke, J.E., Chaplin, C.C.G. (1993). Fishes of the Bahamas and adjacent tropical
waters. University of Texas Press, Texas. 857 pp. Bond, C.E. (1979). Biology of fishes. Philadelphia: Saunders College Publishing. Bookstein, F. L. (1991). Morphometric tools for landmark data. Cambridge University
Press, Cambridge, New York, 435p.

63
Breda, L., Oliveira, E. F., Goulart, E. (2005). Ecomorfologia de locomoção de peixes com enfoque para espécies neotropicais. Acta Scientiarum: Biological Sciences, 27(4): 371-381.
Brum, M. J. I. (1995). Correlações entre a Filogenia e a Citogenética de Peixes
Teleósteos. Revista Brasileira Genética, Série Monografias, 2: 5-42. Brum, M. J. I., Corrêa, M. M. O., Purcell, C. A., Ribeiro, V. P., Muratori, R. S. (1996).
Análise cromossômica em Xenomelaniris brasiliensis (Atherinidae), Micropogonias furnieri (Scianidae) e Bathygobius soporator (Gobiidae) do litoral do estado do Rio de Janeiro. In: VI Simpósio de Citogenética Evolutiva e aplicada de Peixes Neotropicais, Universidade Federal de São Carlos, São Carlos, p. 101.
Brutlag, D.L. (1980). Molecular arrangement and evolution of heterochromatic DNA.
Annual Review Genetics, 14:121-44. Burns, M. D. M., Garcia, A. M., Vieira, J. P. (2010). Pisces, Perciformes, Gobiidae,
Ctenogobius stigmaticus (Poey, 1860): new species record at Patos Lagoon estuary, state of Rio Grande do Sul, Brazil. Check List, Journal of Species List and Distribution, ISSN 1809-127X.
Canapa, A., Cerioni, P. N., Barucca, M., Olmo, E., Caputo, V. (2002). A centromeric
satellite DNA may be involved in heterochromatin compactness in gobiid fishes. Chromosome Research, 10: 297.304.
Caputo, V. (1998). Nucleolar organizer (NOR) and cytotaxonomic implications in six
species of gobiid fishes (Perciformes, Gobiidae). Italy Journal Zoology, 65:93–99. Caputo, V., Marchrgiani, F., Sorice, M., Olmo, E. (1997). Heterochromatin
heterogeneity and chromosome variability in four species of gobiid fishes (Perciformes: Gobiidae). Cytogenetics and Cell Genetic, 79: 266–271.
Caputo, V., Vitturi, R., Odierna, G., Cano, J., Olmo, E., Colomba, M.S. (1996).
Characterization of mitotic chromosomes in the Gobiid fish Zosterissor
ophiocephalus (Pallas, 1811) (Perciformes, Gobiidae). Biol. Zent. bl. 115: 328-336.
Carvalho, R., Soares Filho, W.S., Brasileiro-Vidal, A.C., Guerra, M. (2005). The
relationships among lemons, limes and citron: A chromosomal comparison. Cytogenetic Genome Research, 109:276-282.
Cataudella, S., Civitelli, M.V., Capanna, E. (1973). The chromosomes of some
mediterranean teleosts: Scorpaenidae, Serranidae, Labridae, Blenniidae, Gobiidae (Pisces: Scorpaeniformes, Perciformes). Bolletino di Zoologia, 40(3-4): 385-389.
Cervigo´n, F. M. (1966). Los peces marinos de Venezuela. Fondo de Cultura

64
Científica, Caracas. 951 pp. Cipriano, R. R., Cestari, M. M., Fenocchio, A. S. (2002). Levantamento citogenético de
peixes marinhos do litoral do Paraná. In: Simpósio de citogenética e genética de peixes, IX. Paraná. Universidade Estadual de Maringá, 2002. p.111.
Cipriano, R. R., Fenocchio, A. S., Artoni, R. F., Molina, W. F., Noleto, R. B., Kantek, D.
L. Z., Cestari, M. M. (2008). Chromosomal Studies of Five Species of the Marine Fishes From the Paranaguá Bay and the Karyotypic Diversity in the Marine Teleostei of the Brazilian Coast. Brazil archive biology technology, v.51 n.2: pp.303-314.
Corradini, N., Rossi, F., Giordano, E., Caizzi, R., Vern, F., Dimitri, P. (2007) Drosophila
melanogaster as a model for studying protein-encoding genes that are resident in constitutive heterochromatin. Heredity, 98:3-12.
Corrêa, M.O. D. A., Uieda, V. S. (2007). Diet of the ichthyofauna associated with
marginal vegetation of a mangrove forest in southeastern Brazil. Iheringia, Série Zoologia, Porto Alegre. 97(4):486-497.
Cross, I, Merlo, A., Machado, M., Infante, C., Canãvate, J.P., Rebordinos, L. (2006).
Cytogenetic characterization of the sole Solea senegalensis (Teleostei: Pleuronectiformes: Soleidae): Ag-NOR, (GATA)n, (TTAGGG)n and ribosomal genes by one-color and two-color FISH. Genetica, 128: 253– 259.
Demartini, E.E. (1999). Intertidal spawning, p. 143-164 In: Horn, M.H., Martin, K.L.M., &
Chotkowski, M.A. (Eds). Intertidal Fishes: life in two worlds. San Diego, Academic Press. 399p.
EMBRAPA. (2003). Empresa Brasileira de Pesquisa Agropecuária. Banco de dados
climáticos. Estimativa de 57 anos. Disponível: http://www.bdclima.cnpm.embrapa.br/metodo.php. Acesso em 10/02/2010.
Emmanuel, O. L., Ajibola, E. T. (2010). Food and feeding habits and reproduction in
Frillfin goby, Bathygobius soporator (Cuvier and Valenciennes, 1837) in the Badagry Creek, Lagos, Nigeria. International Journal of Biodiversity and Conservation, 2(12), 414-421.
Ene, A.C. (2003). Chromosomal polymorphism in the goby Neogobius eurycephalus
(Perciformes: Gobiidae). Marine Biology, 142: 583.588. Eschmeyer, W. N. (2010). Catalog of Fishes electronic version (6 May 2010).
http://research.calacademy.org/ichthyology/ catalog/fishcatmain.asp. Fanta, E. (1997). Behaviour and circadian rhythm of the fish Bathygobius soporator
Valenciennes (Gobiidae) under the influence of environmental salinity and
temperature. Revista brasileira de Zoologia, v. 14, n.1, p.221-224.

65
Ferreira, C.E.L., Floeter, S.R., Gasparini, J.L., Ferreira, B.P., Joyeux, J.C. (2004). Trophic structure patterns of Brazilian reef fishes: a latitudinal comparison. Journal Biogeography, 31, 1093–1106.
Floeter, S. R., Gasparini, J. L. (2000). The sothwestern Atlantic reef fish fauna:
composition and zoogeographic patterns. Journal of Fish Biology, 56, 1099 -1114.
Galetti Jr., P. M., Aguilar, C. T., Molina, W. F. (2000). An overview on marine fish
cytogenetics. Hydrobiologia, 420: 55.62. Gaston, K.J., Chown, .SL., Evans, K.L. (2008). Ecogeographical rules: elements of a
synthesis. Journal Biogeography, 35:483–500. Gibson, R.N., Yoshiyama, R.M. (1999). Intertidal fish communities, p. 264-296. In:
Horn, M.H., Martin, K.L.M., Chotkowski, M.A. (Eds). Intertidal fishes: life in two worlds. San Diego, Academic Press, 399p.
Gold, J.R., Li, C., Shipley, N.S., Powers, P.K. (1990). Improved methods for working
with fish chromosomes with a review of metaphase chromosome banding. Journal Fish Biology, 37: 563-575.
Horn, M.H., Gibson, R.N. (1988). Intertidal fishes. Sei. Amer. 258 (1): 54-60. Howell, W.M., Black, D.A. (1980). Controlled silver staining of nucleolus organizer
region with protective colloidal developer: a 1-step method. Experientia, 36: 1014-1015.
Huyse, T., Houdt, J.V., Volckaert, F.A. (2004). Paleoclimatic history and vicariant
speciation in the "sand goby" group (Gobiidae, Teleostei). Molecular Phylogenetic Evolution,32(1):324-36.
Imron, J. B., Hale, P., Degnan, B. M., Degnan, S. M. (2007). Pleistocene isolation and
recent gene flow in Haliotis asinina, an Indo-Pacific vetigastropod with limited dispersal capacity. Molecular Ecology, 16: 289-304.
Joyeux, J. C., Floeter, S. R., Ferreira, C. E. .L., Gasparini, J. L. (2001). Biogeography
of tropical reef fish: the South Atlantic puzzle. Journal of Biogeography, 28, 831–
841. King, M. (1993). Species evolution. The role of chromosome change. Cambridge.
Cambridge University Press. Klinkhardt, M. B. (1992). Chromosome structures of four Norwegian gobies (Gobiidae,
Teleostei) and a hypothetical model of their karyo-evolution. Chromatin, 1: 169–
183.

66
Knowlton, N. (2000). Molecular genetic analyses of species boundaries in the sea. Hydrobiologia, 420, 73–90.
Lavin, P.A., McPhail, J.D. (1986). Adaptive divergence of trophic phenotype among
freshwater populations of the threespine stickleback (Gasterosteus aculeatus). Canadian Journal of Fisheries and Aquatic Science, 43:2455-2463.
Lavin, P.A., McPhail, J.D. (1985). The evolution of freshwater diversity in the
threespine stickleback (Gasterosteus aculeatus): site-specific differentiation of trophic morphology. Canadian Journal of Zoology, 63:2632-2638.
Levan, A., Fredga, K., Sandeberg, A.A. (1964). Nomenclature for centromeric position
on chromosomes. Hereditas, 52: 201-220. Lima, D., Freitas, J. E. P., Solé-Cava, A. M. (2005). Genetic detection of cryptic
species in the frillfin goby Bathygobius soporator. Journal Experimental Marine Biology and Ecology, 320:211–223.
Lindsey, C.C. (1981). Stocks are chameleons: Plastisity in gill rakers of coregonid
fishes. Canadian Journal of Fisheries and Aquatic Science, 38:1497-1506. Lomolino, M. V., Riddle, B. R., Brown, J. H. (2006). Biogeography. Sinauer, 3ª ed.
Massachusetts, 845p. Lopes, P.R.D., Oliveira-Silva, J.T. (1998). Alimentação de Bathygobius soporator
(Valenciennes, 1837) (Actinopterygii: Gobiidae) na localidade de Cacha Pregos (Ilha de Itaparica), Bahia, Brasil. Biotemas, 11(1): 81-92.
Luttikhuizen, P. C. (2003). Spatially structure genetic variation in a broadcast-spawning
bivalve: quantitative versus molecular traits. Journal Evolution Biology, 16, 260–
272. Macpherson, E., Duarte, C.M. (1994). Patterns in species richness, size, and latitudinal
range of East Atlantic fishes. Ecography, 17:242–248. Madrioli, M., Manicardi, G.C., Machella, N., Caputo, V. (2001). Molecular and
cytogenetic analysis of the goby Gobius niger (Teleostei, Gobiidae). Genetica, 110: 73-78.
Magnan, P. (1988). Interactions between Brook charr, Salvelinus fontinalis, and
nonsalmonid species: ecological shift, morphological shift, and their impact on zooplankton communities. Canadian Journal of Fisheries and Aquatic Science, 45:999-1009.
Martins, I.C., Portella-Castro, L.B., Julio Jr., H.F. (1995). Chromosome analysis of
Cichlidae family (Pisces - Perciformes) from the Parana river. Cytologia, 60:223-231.

67
Martins, R. F., Nomura, M. N., Hadel, V. F. (2006). Fidelidade de Bathygobius
soporator (Valenciennes, 1837) (Pisces: Gobiidae) à poça de maré do Istmo do Baleeiro, São Sebastião (SP). In: XXVI Congresso Brasileiro de Zoologia, Resumos. Londrina.
Mérona, B., Mol, J., Vigouroux, R., Chaves, P. T. (2009). Phenotypic plasticity in fish
life-history traits in two neotropical reservoirs: Petit-Saut Reservoir in French Guiana and Brokopondo Reservoir in Suriname. Neotropical Ichthyology, 7(4):683-692.
Miller, P.J., Smith, R.M. (1989). The West African species of Bathygobius (Teleostei:
Gobiidae) and their affinities. Journal Zoology, 218, 277– 318. Mittelbach, G.G., Osenberg, C.W., Wainwright, P.C. (1992). Variation in resource
abundance affects diet and feeding morphology in the pumpkinseed sunfish (Lepomis gibbosus). Oecologia, 90:8-13.
Molina, W. F., Alves, D. E. O., Araújo, W. C., Martinez, P. A., Silva, M. F. M., Costa, G.
W. W. F. (2010). Performance of human immunostimulating agents in the improvement of fish cytogenetic preparations. Genetic Molecular Research, 9 (3): 1807-1814.
Molina, W. F., Galetti Jr., P. M. (2004), Karyotypic changes associated to the
dispersive potential on Pomacentridae (Pisces, Perciformes). Journal of Experimental Marine Biology and Ecology, 309, 109-119.
Molina, W. F., Maia-Lima, F. A., Affonso, P. R. A. M. (2002). Divergence between
karyotypical pattern and speciation events in Serranidae fish (Perciformes). Caryologia, 55: 299.305.
Molina, W. F., Shibatta, O. A., Galetti Jr., P. M. (2006). Multivariate morphological
analyses in continental and island populations of Abudefduf saxatilis (Linnaeus) (Pomacentridae, Perciformes) of Western Atlantic. Pan-American Journal of Aquatic Sciences, 1 (2): 49-56.
Molina, W.F., Galetti Jr., P.M. (2002). Robertsonian rearrangements in the reef fish
Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genetic and Molecular Biology, 25: 373.377.
Mora, C., Sale, P.F. (2002). Are populations of coral reef fish open or closed. Trends in
Ecology and Evolution, 17: 422-428. Munday, P.L., Herwerden, L.v., Dudgeon, C.L. (2004). Evidence for sympatric
speciation by host shift in the sea. Current Biology, 14:1498–1504. Nelson, J.S. (2006). Fishes of the world. 4 ed. John Willey & Sons, New Jersey.

68
Ojima,Y., Kashiwagi, E. (1981). Chromosomal evolution associated with Robertsonian fusion in the genus Drascyllus (Chrominae, Pisces). Proc. Japan Academic, 57(Ser. B.) 368’370.
Oliveira-Raiol, R. D., Soares, B. E., Montag, L. F. A. (2011). Ecologia alimentar do
peixe muré, Bathygobius soporator (valenciennes, 1837) em poças de maré na zona costeira amazônica: dados preliminares. XIX Encontro Brasileiro de Ictiologia, Manaus, Amazonas, Brasil. 41.
Palumbi, S.R. (1994). Genetic divergence, reproductive isolation, and marine
speciation. Annual Review of Ecology and Systematics, 25: 547–572. Peters, K.M., (1983). Larval and early juvenile development of the frillfin goby,
Bathygobius soporator (Perciformes: Gobiidae). Northeast Gulf Science, 6, 137– 153.
Pezold, F. (1984). Evidence for multiple sex chromosomes in the freshwater goby,
Gobionellus shufeldti (Pisces: Gobiidae). Copeia, 1984: 235-238. Pfennig, D. W., Wund, M. A., Snell-Rood, E. C., Cruickshank, T., Schlichting, C. D.,
Moczek, A. P. (2010). Phenotypic plasticity’s impacts on diversification and speciation. Trends in Ecology and Evolution, 25 459–467.
Piorski, N. M., Alves, J. R., Machado, M. R. D., Correia, M. M. V. (2005). Alimentação
e ecomorfologia de duas espécies de piranhas (Characiformes: Characidae) do lago de Viana, Maranhão, Brasil. Acta Amazônica, 35(1): 63-70.
Redi, C.A., Garagna, S., Zuccotti, M. (1990). Robertsonian chromosome formation and
fixation: the genomic scenario. Biological Journal Linnean Society, 41: 235-255. Robins, C.R., Ray, G.C. (1986). A field guide to Atlantic coast fishes of North America.
Houghton Mifflin Company, Boston, USA. 354p. Rocha, L.A. (2003). Patterns of distribution and processes of speciation in Brazilian
reef fishes. Journal Biogeography, 30, 1161– 1171. Rohlf, F. J. (2006). tpsDig version 2.10. Ecology and Evolution, Suny at Stony Brook.
http://life.bio.sunysb.edu/morph/. Rosa, R. S., Rosa, l. L., Rocha, L. A. (1997). Diversidade da ictiofauna de poças de
maré da praia do Cabo Branco, João Pessoa, Paraíba, Brasil. Revista brasileira de Zoologia, 14 (1): 201 – 212.
Scheiner, S.M. (1993). Genetics and evolution of phenotypic plasticity. Annual Review
of Ecology and Systematics, 24:35-68.

69
Smith, K.F., Brown, J.H. (2002). Patterns of diversity, depth range and body size among pelagic fishes along a gradient of depth. Global Ecology Biogeography, 11:313–322.
Smith, T.B., Skúlason, S. (1996). Evolutionary significance of resource polymorphism
in fishes, amphibians, and birds. Annual Review of Ecology and Systematics, 27:111-133.
Stearns, S.C. (1989). The evolutionary significance of phenotypic plasticity.
BioScience, 39:436-445. Sumner, A.T. (1972). A simple technique for demonstrating centromeric
heterochromatin. Experimental Cell Research, 75: 304. Tavolga, W.N. (1953). Spawning and embryonic development in the gobiid fish,
Bathygobius soporator. Anat. Rec. 117, 596–597. Taylor, M.S., Hellberg, M.E. (2003). Genetic evidence for local retention of pelagic
larvae in a Caribbean reef fish. Science, 299, 107– 109. Taylor, M.S., Hellberg, M.E. (2005). Marine radiations at small geographic scales:
speciation in Neotropical reef gobies (Elacatinus). Evolution, 59, 374–385. Taylor, M.S., Hellberg, M.E. (2006). Comparative phylogeography in a genus of coral
reef fishes: biogeographical and genetical concordance in the Caribbean. Molecular Ecology, 15: 695-707.
Thode, G., Giles, V., Alvarez, M. C. (1985). Multiple chromosome polymorphism in
Gobius paganellus (Teleostei, Perciformes). Heredity, 54: 3-7. Tornabene, L., Baldwin, C., Weigt Lee, A., Pezold, F. (2010). Exploring the diversity of
western Atlantic Bathygobius (Teleostei: Gobiidae) with cytochrome c oxidase-I, with descriptions of two new species. Aqua, International Journal of Ichthyology, vol. 16 no. 4.
Vasil’ev, V.P., Grigoryan, K.A. (1993). Karyology of the Gobiidae. Journal Ichthyology,
33:1–16. Vicari, M.R. Artoni, R.F., Bertollo, L.A.C. (2003). Heterochromatin polymorphism
associated with 18S rDNA: a differential pathway among Hoplias malabaricus fish populations. Cytogenetics Genome Research, 1001:24-28.
Vitturi, R., Catalano, E. (1989). Multiple chromosome polymorphism in the gobiid fish
Gobius niger jozo L. 1758 (Pisces, Gobiidae). Cytologia, 54: 231-235. Walker, J.A. (1997). Ecological morphology of lacustrine threespine stickleback
Gasterosteus aculeatus (Gasterosteidae) body shape. Biological Journal of the Linnean Society, 61:3-50.

70
West-Eberhard, M. J. (1989). Phenotypic plasticity and the origins of diversity. Annual
Review of Ecology and Systematics, 20:249-278. Wilson, D.S., Turelli, M. (1986). Stable underdominance and the evolutionary invasion
of empty niches. American Naturalist, 127:835-850.

71
6. REFERÊNCIAS BIBLIOGRÁFICAS
Affonso, P.R.A.M., Galetti, P.M. (2005). A diversificação cromossômica de peixes recifais do gênero Centropyge (Perciformes, Pomacanthidae). Genetica 123:227-233.
Aguirre, W.E., Shervette, V. R. (2005). Morphological diversity of the Cynoscion group (Perciformes: Sciaenidae) in the Gulf os Guayaquil, Ecuador (Perciformes:
Sciaenidae), a comparative approach. Environmental Biology of Fishes, 73: 403- 413.
Almada, F., Alamada, V.C., Guillemaud, T., Wirtz, P. (2005). Phylogenetic relationships of the north-eastern Atlantic and Mediterranean blenniids. Biological Journal of the Linnean Siciety, 86:283-295.
Almeida-Toledo, L. F., Coutinho-Barbosa, A. C., Rosenberg, C., Daniel, M. F. Z., Toledo-Filho, S. A. (1993). Hibridação in situ com sondas de rDNA em cromossomos de Piaractus mesopotamicus, Colossoma macropomum e de seus híbridos. Brazilian Journal Genetica, 16 (Suplement), 141.
Amores, A., Giles, V., Thode, G. (1990). Adaptive charater of a Robertsonian fusion in chromosomes of the fish Gobius paganellus (Pises, Perciformes). Heredity, 65: 151-155.
Amores, A., Martinez, I.A.G., Reina, J., Alvarez, M. C. (1993). Karyotype, C-banding, and Ag-NOR analysis in Diplodus bellottii (Sparidae, Perciformes). Intra-individual poly-morphism involving heterochromatic regions. Genome, 36, 672-675.
Araujo, W. C., Martinez, P. A., Molina, W.F. (2010). Mapping of Ribosomal DNA by FISH, EcoRI Digestion and Replication Bands in the Cardinalfish Apogon
americanus (Perciformes). Cytologia, 75, 1109-117.
Baras, E. (1999). Functional implications of early sexual growth dimorphism in vudu. Journal of Fish Biology, 54 (1): 119.
Bezerra, D. D. (2002). Estudos citogenéticos populacionaisem Hoplerythrinus
unitaeniatus (Pisces – Erythrinidae). Análise da biodiversidade. Dissertação (Mestrado em genética e evolução). Universidade Federal de São Carlos – São Carlos.
Blackith, R.E. Reyment, R.A. (1971). Multivariate morphometrics. London, Academic Press, 412p.
Bohlke, J.E., Chaplin, C.C.G. (1993). Fishes of the Bahamas and adjacent tropical waters. University of Texas Press, Texas. 857 pp.
Bookstein, F. L. (1991). Morphometric tools for landmark data. Cambridge University

72
Press, Cambridge, New York, 435p.
Brum, M. J. I. (1996). Cytogenetic Studies of Brazilian Marine Fish. Brazilian Journal of Genetetics, 19 (3): 421-427.
Brum, M. J. I., Corrêa, M. M. O., Purcell, C. A., Ribeiro, V. P., Muratori, R. S. (1996). Análise cromossômica em Xenomelaniris brasiliensis (Atherinidae), Micropogonias furnieri (Scianidae) e Bathygobius soporator (Gobiidae) do litoral do estado do Rio de Janeiro. In: VI Simpósio de Citogenética Evolutiva e aplicada de Peixes Neotropicais, Universidade Federal de São Carlos, São Carlos, p. 101.
Brum, M.J.I., Galetti Jr., P.M. (1997). Teleostei plan ground karyotype. Journal of Computational Biology, 2: 91-102.
Cano, J., Thode, G., Alvarez, M.C. (1982). Karyoevolutive consideration in 29 mediterranea teleost fishes. Vie Milieu, 32: 21-24.
Caputo, V. (1998). Nucleolar organizer (NOR) location and cytotaxonomic implications in six species of gobiid fishes (Perciformes, Gobiidae). Italy Journal Zoology, 65: 93-99.
Caputo, V., Caniglia, M.L., Machella, N. (1999). The chromosomal complement of Aphia minuta, a paedomorphic goby. Journal Fish Biology, 55:455–458.
Caputo, V., Marchrgiani, F., Sorice, M., Olmo, E. (1997). Heterochromatin heterogeneity and chromosome variability in four species of gobiid fishes (Perciformes: Gobiidae). Cytogenetic Cell Genetetic, 79: 266–271.
Caputo, V., Sorice, M., Vitturi, R., Magistrelli, H., Olmo, E. (1998). Cytogenetic studies in some species of Scorpaeniformes (Teleostei: Percomorpha). Chromosomal Research, 6: 255–262.
Carvalho, R., Soares Filho, W.S., Brasileiro-Vidal, A.C., Guerra, M. (2005). The relationships among lemons, limes and citron: A chromosomal comparison. Cytogenetic Genome Research, 109:276-282.
Cavalcanti, M. J., Lopes, P. R. D. (1993). Análise morfométrica multivariada de cinco espécies de Serranidae (Teleostei: Perciformes). Acta Biologica Leopoldensia, v.15, n.1, p.53-64.
Cipriano, R. R., Cestari, M. M., Fenocchio, A. S. (2002). Levantamento citogenético de peixes marinhos do litoral do Paraná. In: Simpósio de citogenética e genética de peixes, IX. Paraná. Universidade Estadual de Maringá, 2002. p.111.
Colin, P. (1975). The neon gobies: The comparative biology of the gobies of the genus Gobiosoma, subgenus Elacatinus (Pisces: Gobiidae) in the tropical western North Atlantic Ocean. Neptune City, NJ: T.F.H.

73
De Sylva, D.P. (1975). Nektonic food webs in estuaries. In: Cronin (ed). Estuarine research, New York: Academic Press, 1: 420- 447,587.
Dryden, I. L., Mardia, K. V. (1998). Statistical shape analysis. John Wiley & Sons, New York.
Ehlinger, T.J. (1991). Allometry and analysis of morphometric variation in the bluegill, Lepomis macrochirus. Copéia, 2: 347-57.
Emmanuel, O. L., Ajibola, E. T. (2010). Food and feeding habits and reproduction in Frillfin goby, Bathygobius soporator (Cuvier and Valenciennes, 1837) in the Badagry Creek, Lagos, Nigeria. International Journal of Biodiversity and Conservation, 2(12), 414-421.
Ene, A.C. (2003). Chromosomal polymorphism in the goby Neogobius eurycephalus (Perciformes: Gobiidae). Marine Biology, 142: 583.588.
Fairbairn, D.J. (1997). Allometry for sexual size dimorphism: pattern and process in the coevolution of body size in males and females. Annual Review, of Ecology, Evolution and Systematics, 28: 659-87.
Fonteles Filho, A. A. (1989). Recursos pesqueiros: biologia e dinâmica populacional. Ed. Imprensa Oficial do ceará, Fortaleza. 296 p.
Fulford, R.S. Rutherford, D.A. (2000). Discrimination of larval Morone geometric shape differences with landmark morphometrics. Copeia, 4: 965-972.
Galetti Jr., P. M., Aguilar, C. T., Molina, W. F. (2000). An overview on marine fish cytogenetics. Hydrobiologia, 420: 55.62.
Galetti Jr., P.M., Molina, W.F., Affonso, P.R.A.M. and Aguilar, C.T. (2006). Assessing genetic diversity of Brazilian reef fishes by chromosomal and DNA markers. Genetica, 126: 161-177.
Galetti Jr., P.M., Molina, W.F., Affonso, P.R.A.M., Aguilar, C.T. (2006). Assessing genetic diversity of Brazilian reef fishes by chromosomal and DNA markers. Genetica, 126: 161-177.
Galvão, T. B. (2006). Caracterização citogenética em peixes recifais: Ophioblennius
atlanticus, Scartella cristata, Labrisomus nuchipinnis e Bathygobius soporator. Dissertação. Universidade Federal do Rio Grande do Norte. Pós-graduação em genética e biologia molecular. Natal, RN.
Gasparini, J. L., Floeter, S.R., Ferreira, C. E. L. (2005). Marine Ornamental Trade in Brazil. Biodiversity Conservation, 14, 2883-2899.
Gold, J.R., Li, C., Shipley, N.S., Powers, P.K. (1990). Improved methods for working with fish chromosomes with a review of metaphase chromosome banding. Journal Fish Biology, 37: 563-575.

74
Grigoryan, K.A., Vasil’ev, V.P. (1993). Karyotypes of five species of goby (Gobiidae)
from the basins of the Black and Caspian Seas. Journal Ichthyology, 33:137–
143.
Hoese, D.F., Murdy, E.O. (2003). Perciformes: Gobioidei: Gobiidae. In: Western Central Atlantic. Carpenter, K. E. Department of Biological Sciences Old Dominion University Norfolk, Virginia, USA.v.3, p.1781.
Howell, W.M., Black, D.A. (1980). Controlled silver staining of nucleolus organizer region with protective colloidal developer: a 1-step method. Experientia, 36: 1014-1015.
IBAMA (2004). Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis. Instrução normativa n 5, de 21 de maio de 2004. 10 p.
Kendall, D. G. (1984). Shape-manifolds, procrustean metrics and complex projective spaces. Bulletin of the London Mathematical. London Mathematical Society, 16: 81–121.
Kendall, D. G. (1989). A Survey of the Statistical Theory of Shape. Statistical Science, 4(2): 87–120.
Klingenberg, C. P., Barluenga, M., Meyer, A. (2003). Body shape variation in cichlid fishes of the Amphilophus citrinellus species coplex. Biological Journal of the Linnean Society, 80: 397-408.
Klinkhardt, M. B. (1992). Chromosome structures of four Norwegian gobies (Gobiidae, Teleostei) and a hypothetical model of their karyo-evolution. Chromatin, 1: 169–
183.
Klinkhardt, M.B., Tesche, M., Greven, H. (1995). Database of fish chromosomes. Westarp Wissenschaften, Magdeburg, Germany. 273 pp.
Levan, A., Fredga, K., Sandeberg, A.A. (1964). Nomenclature for centromeric position on chromosomes. Hereditas, Lund, 52: 201-220.
McGuigan, K., Chenoweth, S. F., Blows, M. W. (2005). Phenotypic Divergence along Lines of Genetic Variance. The American Naturalist, 165 (1): 32-43.
Mendes, L. F. (2000). História natural, biologia alimentar, repartição espacial, densidades populacionais e ecomorfologia do Gobióides e Blenióides (Perciformes) do Arquipélago de Fernando de Noronha, PE. Tese de Doutorado, USP,SP, Brasil.
Menezes, N. A., Buckup, P. A., Figueiredo, J. L., Moura, R. L. (2003). Catálogo das espécies de peixes marinhos do Brasil. São Paulo, Museu de Zoologia da Universidade de São Paulo. 160p.
Menezes, N. A., Figueiredo, J. L. (1985). Manual de peixes marinhos do sudeste do Brasil. V. Teleostei(4). São Paulo, Museu de Zoologia da Universidade de São

75
Paulo. 105p.
Molina, W. F., Alves, D. E. O., Araújo, W. C., Martinez, P. A., Silva, M. F. M., Costa, G. W. W. F. (2010). Performance of human immunostimulating agents in the improvement of fish cytogenetic preparations. Genetic Molecular Research, 9 (3): 1807-1814.
Molina, W. F., Galetti Jr., P. M. (2004), Karyotypic Changes Associated To The Dispersive Potential On Pomacentridae (Pisces, Perciformes). Journal of Experimental Marine Biology and Ecology, 309, 109-119.
Molina, W.F. (2001). An alternative method for mitotic stimulation in fish cytogenetics. Chromosome Science, 5:149-152.
Molina, W.F., Bacurau, T.O. (2006) Structural and numerical chromosomal variation in marine Perciformes (Priacanthidae and Gerreidae). Cytologia, Japão, 71(3): 237-242.
Molina, W.F., Galetti Jr., P.M. (2002). Robertsonian rearrangements in the reef fish Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genetic and Molecular Biology, 25: 373.377.
Monteiro, L. R., Diniz-Filho, A. F., Reis, S. F., Araújo, E. D. (2002). Geometric estimates of heritability in biological shape. Evolution, 56 (3): 563-572.
Monteiro, L. R., Reis, S. F. (1999). Princípios de Morfometria Geométrica. Ribeirão Preto, Ed. Holos, 198 p.
Muss, A., Robertson, D.R., Stepien, C.A., Wirtz, P., Bowen, B.W. (2001). Phylogeography of Ophioblennius: The role of ocean currents and geography in reef fish evolution. Evolution, 55:561-572.
Nelson, J.S. (2006). Fishes of the world. 4 ed. John Willey & Sons, New Jersey.
Oliveira, C., Foresti, F., Hilsdorf, A.W.S. (2009).Genetics of neotropical fish: from chromosomes to populations. Fish Physiology and Biochemistry, 35:81–100.
Peres, P. R., Valentin, J. L., Fernandez, F. (1995). Tópicos em tratamentos de dados biológicos. Rio de Janeiro: Universidade Federal do Rio de Janeiro.
Pezold, F. (1984). Evidence for multiple sex chromosomes in the freshwater goby, Gobionellus shufeldti (Pisces: Gobiidae). Copeia, 1984(1):235-238.
Pezold, F. (1993). Evidence for a monophyletic Godiidae. Copeia, 1993(3):634-643.
Randall, J.E., Lobel, P.S., Kennedy, C.W. (2005). Comparative ecology of the gobies Nes longus and Ctenogobius saepepallens, both symbiotic with the snapping shrimp Alpheus floridanus. Environmental Biology of Fishes, 74, 119-127.
Rapp Py-Daniel, L. H., Cox Fernandes, C. (2005). Dimorfismo sexual em Siluriformes e

76
Gymnotiformes (Ostariophysi) da Amazônia. Acta Amazônica, 35 (1): 97-110.
Ricklefs, R. E., Miles, D. B. (1994). Ecological and evolutionary inferences from morphology: An ecological perspective. p.13-41. In: Wainwright, P.C., Reilly, S.M. Ecological morphology, University of Chicago Press, Chicago.
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs, 17:179–186.
Rohlf, F. J. (1999). Shape Statistics: Procrustes Superimpositions and Tangent Spaces. Journal of Classification, 16:197-223.
Rohlf, F. J. (2006). tpsDig version 2.10. Ecology and Evolution, SUNY at Stony Brook. http://life.bio.sunysb.edu/morph/.
Rohlf, F. J., Marcus, L. F. (1993). A revolution in morphometrics. Trends in Ecology Evolution, 8: 129-132.
Rosenberg, M.S. (2002). Fiddler crab claw shape variation: A geometric morphometric analysis across the genus Uca. Biological Journal of the Linnean Society, 75(2):147-162.
Sale, P.F. (1991). The ecology of fishes on coral reefs. Academic Press, London, UK.
Schluter, D. (1993). Adaptative radiation in sticklebacks: size, shape, and habitat use efficiency. Ecology, 74: 699-709.
Sena, D.C.S., Molina, W.F. (2006). Robertsonian rearrangements and pericentric inversions in Scaridae fish (Perciformes). Genetics and Molecular Research, 6 (3): 575-580.
Silva, E. L. (2000). Utilização de Técnicas Multivariadas no Estudo Morfométrico da albacora laje Thunnus albacares (Bonnaterre, 1788) do Atlântico. Dissertação de Mestrado. Universidade Federal Rural de Pernambuco, 64p.
Souza, I.L., Moreira-Filho, O., Bertollo, L. A. C. (1998). Contribuição citogenética em Gobiidae. VII Simpósio Citogenetica Evolutiva Aplicada Peixes Neotropicais. Resumo, p.26.
Sumner, A.T. (1972). A simple technique for demonstrating centromeric heterochromatin. Experimental Cell Research, 75: 304.
Thode, G., Martinez, G., Ruiz, J. L., Lopez, J. R. (1988). A complex chromosomal polymorphism in Gobius fallax (Gobiidae, Perciformes). Genetica, 76: 65–71.
Thompson, D. W. (1917). On growth and form. Cambridge Univ. Press, London, 753p.
Tracker, C.E. (2003). Molecular phylogeny of the gobioid fishes (Teleostei: Perciformes: Gobioidei). Molecular Phylogenetics and Evolution, 26: 354-368.update. Biological Reviews, 76: 1.26.

77
Ueno, K., Takai, A. (2000). Chromosome evolution involving Robertsonian rearrangements in Xirichthys fish (Labridae, Perciformes). Cytobios, 103: 7-15.
Vasil’ev, V.P., Grigoryan, K.A. (1993). Karyology of the Gobiidae. Journal Ichthyology,
33:1–16.
Vasil'ev, V.P., Grogoryan, K.A. (1992). Karyology of fishes from the family Gobiidae. Voprosy Ikhtiologii, 32: 27-40.
Vidales, K., Markakis, G., Tsimenides, N. (1997). Discrimination between populations of picare Spicara smaris (Linneus, 1758) in the Aegean Sea, using multivariate analysis of phenetic characters. Fisheries Research, 30: 191 - 197.
Vitturi, R., Catalano, E. (1989). Multiple chromosome polymorphism in the gobiid fish Gobius niger jozo (Linneus 1758) (Pisces, Gobiidae). Cytologia, 54: 231–235.
Wabnitiz, C., Taylo, M., Green, E., Razak, T. (2003). From ocean to aquarium. UNEP-WCMC, Cambridge, UK. 66p.
Webb, P. W. (1984). Body form, locomotion and foraging in aquatic vertebrates. American Zoologist, 24: 107-120.
Zelditch, M. L., Swiderski, D. L., Sheets, H. D., Fink, W. L. (2004). Geometric morphometrics for biologists. Part II: Analysis shape variables. Elsevier Academy Press, 153-290.


















