
O USO DO HABITAT POR MORCEGOS EM SISTEMAS … · tendências gerais em resposta ao desflorestamento...
57
Universidade de Lisboa Faculdade de Ciências Departamento de Biologia Animal O USO DO HABITAT POR MORCEGOS EM SISTEMAS NATURAIS E AGROFLORESTAIS NA SAVANA BRASILEIRA SÓNIA MARISA SOARES PINA MESTRADO EM ECOLOGIA E GESTÃO AMBIENTAL 2011
Transcript of O USO DO HABITAT POR MORCEGOS EM SISTEMAS … · tendências gerais em resposta ao desflorestamento...

Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Animal
O USO DO HABITAT POR MORCEGOS EM SISTEMAS
NATURAIS E AGROFLORESTAIS NA SAVANA
BRASILEIRA
SÓNIA MARISA SOARES PINA
MESTRADO EM ECOLOGIA E GESTÃO AMBIENTAL
2011

2
Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Animal
O USO DO HABITAT POR MORCEGOS EM SISTEMAS
NATURAIS E AGROFLORESTAIS NA SAVANA
BRASILEIRA
SÓNIA MARISA SOARES PINA
Dissertação orientada por:
Dr. Christoph Meyer (CBA/FCUL)
Dr. Marlon Zortéa (LZE/UFG)
2011

3
Resumo
O processo de conversão de sistemas naturais em áreas para pastagens,
agricultura e silvicultura no Brasil tem sido cada vez mais acelerado e são poucos
os estudos que abordam o impacto destas transformações nas comunidades de
morcegos. Visando contribuir neste aspecto, realizou-se entre Dezembro de 2010
e Abril de 2011 capturas com redes de neblina mensalmente, durante cinco
meses, em oito fragmentos de dois tipos de habitat: 4 plantações de eucalipto
(Eucalyptus) e 4 fragmentos de floresta semidecidual no Cerrado brasileiro, para
avaliar as diferenças de diversidade e abundância e testar o efeito das
características da paisagem e variáveis locais nas comunidades de morcegos.
Dos 209 indivíduos capturados 75 foram em fragmentos de eucalipto e 134 em
áreas de floresta nativa. Destes, 90% eram frugívoros e 97% pertencentes à
família Phyllostomidae. A espécie mais capturada foi Carollia perspicillata (92
indivíduos) seguida por Artibeus planirostris (39) e Platyrrhinus lineatus (21). As
áreas nativas apresentaram uma maior riqueza de espécies, menor dominância e
maior diversidade e equitabilidade comparativamente às plantações de eucalipto.
Além disso, áreas florestadas apresentam uma fauna mais completa e com
espécies mais sensíveis como alguns morcegos da subfamília Phyllostominae.
Através da ordenação não métrica multidimensional (NMDS) verificou-se uma
fraca separação entre os habitats amostrados embora uma análise de
similaridades (ANOSIM) indicou uma ausência de diferenças significativas na
composição de espécies. A proporção de cobertura florestal (Cerrado e
Eccalyptus) foi a única variável de paisagem com influência significativa na
composição de espécies. Pelo contrário, a composição de espécies não
apresenta qualquer relação com as distâncias geográficas entre os fragmentos
florestais. Aplicando o modelo linear generalizado de efeitos mistos (GLMM), para
as três espécies com maior abundância, verificou-se que o factor Habitat teve
influência apenas na abundância de Artibeus planirostris, a espécie mais
abundante em Cerrado. Os resultados demonstram o empobrecimento das
comunidades de morcegos em plantações de eucalipto e a necessidade de se
perceber a resposta de cada espécie a esta conversão do habitat nativo.
Palavras-chave: morcegos; Cerrado; eucaliptos; diversidade; habitat; Chiroptera;
Eucalyptus

4
Abstract
The conversion of natural systems into areas for cattle grazing, agriculture, and
silviculture in Brazil has been increasing rapidly, and there are few studies on the
impact of these changes on bat assemblages. Aiming to contribute towards filling
this gap, I compared bat assemblages in semideciduous forest fragments in
Brazilian Cerrado with those of eucalyptus plantations (Eucalyptus sp.).
Specifically, I assessed differences in species richness, diversity, abundance, and
species composition between habitat types and evaluated bat responses with
respect to a variety of landscape characteristics and local-scale variables. For this,
I sampled bats with mist nets in 4 replicates each of eucalyptus plantations and
fragments of native Cerrado over the course of 5 months, between December
2010 and April 2011. Of the total of 209 individuals captured, 75 were caught in
eucalyptus fragments and 134 in native forest areas. Of these, 90% were
frugivores and 97% belonged to the family Phyllostomidae. The most frequently
captured species were Carollia perspicillata (92 individuals) followed by Artibeus
planirostris (39) and Platyrrhinus lineatus (21). Native areas had greatest species
richness, lower dominance and greatest diversity and evenness compared to
eucalypt plantations. Gleaning animalivorous species of the subfamily
Phyllostominae seem to be most sensitive to plantation forests as they were not
captured in this habitat. Non-metric multidimensional scaling (NMDS) indicated no
strong separation between habitats types and there were no significant differences
in species composition based on an analysis of similarities (ANOSIM). The
proportion of forest cover (both Cerrado and Eucalyptus) in the landscape was the
only landscape-scale variable which had a significant influence on species
composition. In contrast, species composition was unrelated to geographical
distance between forest fragments. A generalized linear mixed effects model
(GLMM) for the three most abundant species only showed significant differences
in abundance for Artibeus planirostris, the species being more abundant in native
Cerrado. This results highlights the impoverishment of bat assemblages in
eucalypt plantations and how is important understand the bat sepecies-specific
responses to this native habitat conversion.
Keywords: bats; Cerrado; eucalyptus; diversity; habitat; Chiroptera; Eucalyptus

5
Índice
Resumo 3
Abstract 4
Índice 5
1. Introdução 6
• Objectivos 9
2. Materiais e Métodos 10
2.1. Área de estudo 10
2.2. Recolha de dados 12
2.3. Tratamento dos dados e análise estatística 14
2.3.1. Variáveis métricas locais e de paisagem 14
2.3.2. Riqueza, abundância, diversidade e composição de espécies 15
2.3.3. Modelação da fauna de morcegos com ambiente 17
2.3.4. Autocorrelação espacial 18
3. Resultados 18
3.1. Riqueza, abundância, diversidade e composição de espécies 18
3.2. Modelação da fauna de morcegos com o ambiente 25
3.3. Autocorrelação espacial 26
4. Discussão 26
4.1. Enquadramento da composição taxionómica obtida, na região e bioma em estudo 26
4.2. Riqueza, dominância, diversidade e composição de espécies 29
4.3. Modelação da fauna de morcegos com o ambiente 36
5. Considerações finais 39
6. Agradecimentos 42
7. Bibliografia 43
Anexo I 56
Anexo II 56
Anexo III 57

6
1. Introdução
O Cerrado e os sistemas agroflorestais
A eco-região de savanas do Brasil, também conhecida como Cerrado,
distribui-se de forma contínua em 10 estados da federação, ocupando cerca de 2
milhões de km2 das regiões Centro-Oeste, Sudeste e Nordeste do país, além de
encraves na porção norte brasileira (Eiten, 1994). É o segundo maior ecossistema
Neotropical, revestindo 20 a 25% da área total do território brasileiro (Machado et
al., 2004; Zortéa & Alho, 2008; Ferreira et al., 2009).
O Cerrado é caracterizado por uma vegetação estratificada em espécies
gramíneas, arbustivas e arbóreas, com flora e fauna bem adaptadas ao clima
sazonal, isto é, uma estação chuvosa que dura de Outubro a Março, seguida por
uma estação seca de Abril a Setembro (Klink & Machado, 2005; Klink & Machado,
2005; Ferreira et al., 2009). Este ecossistema é extremamente heterogéneo e
estudos têm indicado até 25 unidades fisiográficas distintas e mais de 70 sistemas
de terras diferentes para a região (Felfili et al., 2004). Tais características,
associadas a uma topografia suave torna este ecossistema altamente atractivo
para actividades agro-pecuárias, principalmente por monoculturas comerciais e
bovinocultura (Ferreira et al., 2009). Segundo um estudo realizado em 2002, 55%
do Cerrado já foram desmatados ou transformados pela acção humana (Machado
et al., 2004).
A perda de biodiversidade e a fragmentação da floresta são as duas
principais consequências da intensa pressão humana experiênciada nos trópicos
nas últimas décadas (Bernard & Fenton, 2007). A fragmentação e o isolamento
dos habitats naturais constituem a principal ameaça à biodiversidade a nível
regional e global (Bianconi et al., 2006; Klingbeil & Willig, 2009). O
reflorestamento, para a produção de carvão vegetal, papel e celulose, tem
incrementado a formação de grandes maciços florestais utilizando espécies do
género Eucalyptus, que têm substituído a vegetação nativa do Cerrado (Pereira &
Oliveira, 2005). Particularmente, no Estado de Goiás, localizado na região Centro-
Oeste brasileira, o plantio de eucalipto tem, recentemente, substituído algumas
áreas de pastagens e de lavouras temporárias. Em 2010 foram estimadas pela
ABRAF (2011) o plantio de quase 60 mil hectares de eucalipto no Estado.

7
Os quirópteros – diversidade e conservação
No mundo existem mais de 1150 espécies de morcegos (Schipper et al.,
2008), encontrando-se a maior diversidade de espécies na região neotropical,
com 83 géneros e cerca de 300 espécies (Simmons, 2005; Bianconi et al., 2006).
No Brasil são conhecidos 64 géneros de morcegos pertencentes a nove famílias
(Reis et al., 2007) e o Cerrado abriga mais de 100 espécies de morcegos, o que
corresponde a 60% da fauna brasileira de morcegos, e mais de 40% do total da
fauna de morcegos da América do sul (Aguiar & Zortéa, 2008). No Cerrado, tal
como em biomas como a Mata Atlântica, a Floresta Amazónica e outros, são
encontradas muitas espécies da família Phyllostomidae, as quais são importantes
dispersoras de sementes e polinizadoras de plantas (Lobova et al., 2009), como
por exemplo, as espécies do género Artibeus Leach, Sturnira Gray, Glossophaga
E. Geoffroy e Lonchophylla Thomas. Neste ecossistema, encontram-se também
espécies com outros hábitos alimentares, tais como as espécies insectívoras
como Lophostoma spp., hematófagas como Desmodus rotundus (E. Geoffroy,
1810), as carnívoras como Chrotopterus auritus (Peters, 1856) e as piscívoras
como Noctilio leporinus (Linnaeus, 1758).
Os morcegos são considerados um excelente bioindicador das alterações
humanas, provocadas na qualidade do habitat (Jones et al., 2009). Nas florestas
Neotropicais, os Chiroptera podem representar um taxon chave, devido à sua
participação na polinização e dispersão de sementes (Willig et al., 2007). Além
disso, os morcegos compõem um grupo modelo ideal para avaliar respostas à
fragmentação do habitat, uma vez que estes são altamente diversos e móveis,
com o potencial de se movimentarem ao longo de extensas áreas fragmentadas
(Meyer & Kalko, 2008).
Estudos realizados sobre o efeito da fragmentação em morcegos
neotropicais têm produzido resultados equívocos levando a que as respostas
destes à fragmentação não estejam bem compreendidas (Faria, 2006; Klingbeil &
Willig, 2009). Perceber e prever como as populações de quirópteros vão
responder à ruptura de habitat é um grande desafio para os biólogos. Foi
demonstrado por vários autores (e.g. Faria, 2006; Gorresen & Willig, 2004; Willig
et al., 2007) que uma comunidade de morcegos tende a ser menos diversa e
dominada por menos espécies em habitats fragmentados. Por outro lado, os

8
morcegos neotropicais respondem à fragmentação do habitat de um modo
específico consoante a espécie (Klingbeil & Willig, 2009) sendo evidente duas
tendências gerais em resposta ao desflorestamento e à fragmentação do habitat.
Usualmente, os filostomídeos insectívoros e carnívoros decrescem em
abundância e riqueza de espécies enquanto, em contraste, os filostomídeos
frugívoros e nectarívoros, exibem frequentemente abundâncias maiores em
florestas que foram sujeitas ao desflorestamento e à fragmentação do habitat
(Clarke et al., 2005; Castro-Arellano et al., 2007; Willig et al., 2007; Bobrowiec &
Gribel, 2010).
A fragmentação do habitat é uma característica dominante da paisagem
moderna (Ewers & Didham, 2006) e uma vez que, a resposta dos morcegos à
fragmentação do habitat nos neotrópicos é equívoca, torna-se cada vez mais
importante perceber a relação entre o grau de fragmentação e a variação em
aspectos da biodiversidade (Klingbeil & Willig, 2009). Assim é necessário a
realização de estudos que tentem perceber, para além de questões como a
qualidade dos habitats (e.g. estudos de habitats perturbados versus não
perturbados, Fenton et al., 1992), o efeito das variáveis locais (e.g. área e
tamanho dos fragmentos em estudo, distância entre os fragmentos, entre outras)
e da estrutura da paisagem (e.g. densidade de fragmentos, cobertura florestal,
comprimento de borda, entre outras) nas respostas das populações de morcegos
e também específicas de cada espécie à fragmentação (Klingbeil & Willig, 2009).
Para Gorresen & Willig (2004), as características da paisagem como
tamanho do fragmento, cobertura florestal e densidade de fragmentos, foram
atributos importantes relacionados com a resposta dos filostomídeos, em
paisagens fragmentadas, tanto a nível das espécies como da comunidade.
Existe pouca informação disponível sobre o valor da conservação de
plantações de árvores comerciais (Zurita et al., 2006) e as paisagens de produção
(“production landscapes”) são raramente consideradas como áreas prioritárias
para a conservação nos trópicos (Barlow et al., 2007).
Os estudos de biodiversidade desenvolvidos em plantações de eucalipto e
em outros tipos de plantações são muito menos extensivos e mais recente do que
a pesquisa desenvolvida em plantações de pinheiros (Danielsen & Heegaard,
1995; Bernhard-Reversat, 2001; Borsboom et al., 2002; Lindenmayer et al., 2003;
Barlow et al., 2007). Tem sido demonstrado que as comunidades de vertebrados

9
e invertebrados são menos diversas do que aquelas presentes em vegetação
nativa, onde esta redução na diversidade deve-se muito à relativa simplicidade
estrutural destas plantações (Lindenmayer et al., 2003). Barlow et al. (2007b)
concluíram, segundo um estudo realizado com borboletas em plantações de
Eucalyptus, que estas podem ajudar a conservar um número limitado de espécies
de floresta.
Muitos dos trabalhos desenvolvidos em plantações, estudaram
comunidades de aves (Marsden et al., 2001; Zurita et al., 2006; Lantschner et al.,
2008; Gabriel et al., 2009; Filloy et al., 2010; Hsu et al., 2010) e poucos são os
trabalhos que se focaram nas comunidades de morcegos (Harvey & Villalobos,
2007; Borkin & Parson, 2010).
Segundo o estudo desenvolvido por Harvey & Villalobos (2007) na
Costa Rica, as plantações de cacau e banana, mantém comunidades de
morcegos semelhantes aquelas observadas em áreas de florestas nativas. Por
outro lado, num estudo realizado no Brasil sobre morcegos, foi observado
uma menor riqueza de espécies e uma estrutura da comunidade
simplificada em plantações de Araucária comparativamente à floresta nativa
(Reis et al., 2006).
Uma vez que são poucos os estudos realizados em plantações de
eucaliptos e sobre as comunidades de morcegos, vê-se cada vez mais acrescida
a necessidade de realizar-se estudos comparativos de morcegos em
plantações de Eucalyptus.
Objectivos
Sendo assim, este trabalho teve como objectivo investigar as diferenças de
diversidade e abundância entre plantações de eucalipto (Eucalyptus spp.) e
fragmentos de floresta nativa (Cerrado), ao nível das espécies e da comunidade.
Para alcançar este objectivo pretendeu-se:
- Comparar a riqueza de espécies, diversidade, equitabilidade e
abundância entre os vários fragmentos de Cerrado e plantações de Eucalyptus,
verificando se há diferenças significativas entre os dois tipos de habitat, partindo
do pressuposto que habitats naturais apresentariam uma maior riqueza e
abundância;
10
- Testar o efeito, sobre a composição de espécies e abundância, dos
seguintes atributos locais e características da paisagem: área do fragmento e
comprimento de borda, distância entre fragmentos, tamanho médio do fragmento,
densidade de fragmentos e cobertura florestal. Previu-se encontrar diferenças na
estrutura e composição da comunidade de morcegos entre as plantações de
eucalipto e os fragmentos de floresta, apresentando esta última uma estrutura de
comunidade mais simplificada. Esperou-se ainda, que a riqueza de espécies,
diversidade e abundância fosse dependente das características da paisagem.
2. Materiais e Métodos
2.1 Área de estudo
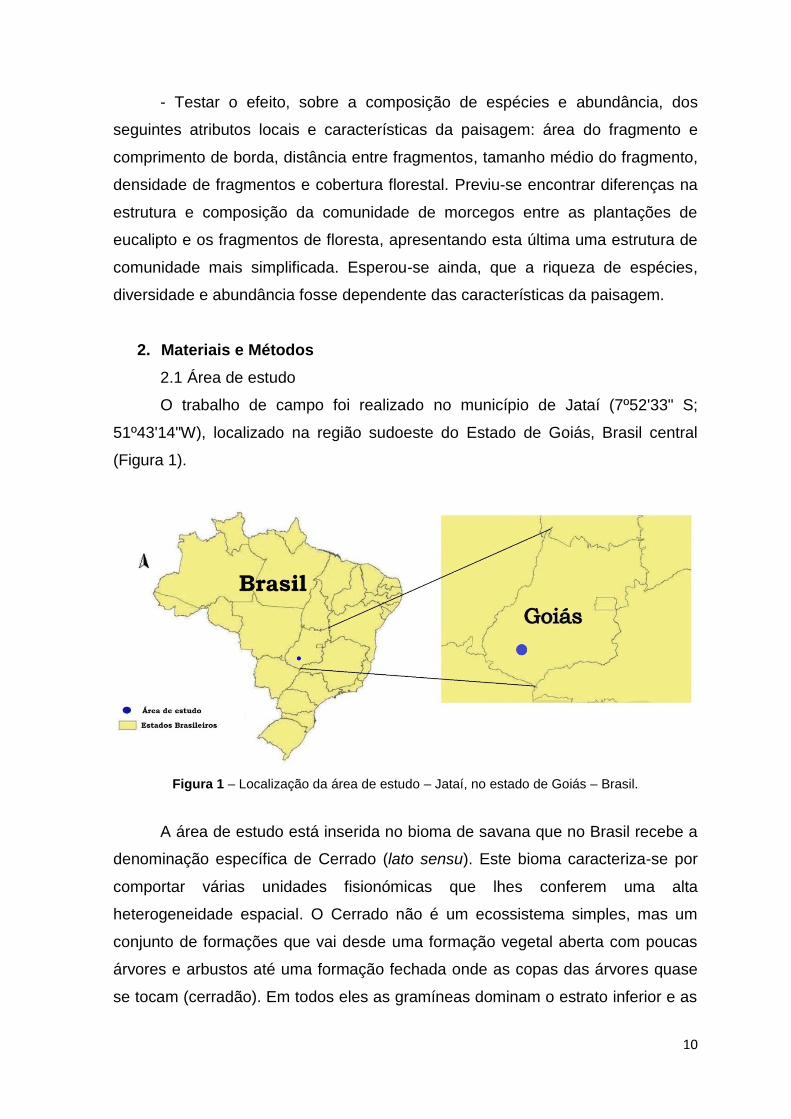
O trabalho de campo foi realizado no município de Jataí (7º52'33" S;
51º43'14"W), localizado na região sudoeste do Estado de Goiás, Brasil central
(Figura 1).
Figura 1 – Localização da área de estudo – Jataí, no estado de Goiás – Brasil.
A área de estudo está inserida no bioma de savana que no Brasil recebe a
denominação específica de Cerrado (lato sensu). Este bioma caracteriza-se por
comportar várias unidades fisionómicas que lhes conferem uma alta
heterogeneidade espacial. O Cerrado não é um ecossistema simples, mas um
conjunto de formações que vai desde uma formação vegetal aberta com poucas
árvores e arbustos até uma formação fechada onde as copas das árvores quase
se tocam (cerradão). Em todos eles as gramíneas dominam o estrato inferior e as

11
árvores são relativamente baixas, geralmente tortuosas e com folhas espessas
(Felfili et al., 2004; Salgado-Labouriau, 2005). O clima é tipicamente sazonal, com
Invernos secos e Verões chuvosos. A chuva ocorre maioritariamente entre
Outubro e Abril (Felfili et al., 2005).
O município de Jataí localiza-se na Microrregião Sudoeste de Goiás,
dominada por sedimentos antigos e faixas de derrames basálticos. Apresenta um
relevo plano e suavemente ondulado com altitudes que variam de 696 a 700
metros. As temperaturas vão de 18.2ºC (Julho) a 24.4ºC (Outubro) e possui uma
variação pluviométrica que oscila de 1000 mm a 1600 mm, distribuídos
irregularmente durante o ano (Silva et al., 2009). O clima é classificado como Aw,
Tropical Húmido de Savana, conforme a classificação de KÖppen (1948). A
vegetação dominante nesta zona é floresta semidecidual.
A região sudoeste de Goiás engloba aproximadamente 27 milhões de ha,
representando cerca de 16% do total da área do estado. A maioria desta área
está altamente fragmentada, devido à conversão do Cerrado em terras para
produção agrícola (Alho, 2005). Os sistemas agro-florestais são ainda raros na
área, no entanto, a plantação de exóticas como Eucalyptus está a aumentar,
tornando-se um importante elemento destas paisagens fragmentadas.
As áreas seleccionadas, para a realização deste trabalho foram de dois
tipos: plantações de Eucalyptus e fragmentos de floresta semidecidual (Figura 2).
Figura 2 – Fotografia aérea das áreas de estudo a) Plantação de Eucalyptus; b) Fragmento de
floresta semidecídual. (Fonte: Fabiano R. Melo).
a) b)

12
As plantações de Eucalyptus são áreas sem sub-coberto vegetal. Os
fragmentos de savana brasileira seleccionados têm como vegetação dominante a
floresta semidecidual.
Junto à maioria destas áreas existe a passagem de uma rodovia. Estas
estão também, na sua maioria, envolvidas por plantações de soja, de cana-de-
açúcar e pastagem de gado.
2.2 Recolha de dados
O trabalho de campo foi realizado entre Dezembro de 2010 e Abril de 2011.
Foram seleccionados 4 fragmentos de cada tipo de habitat em estudo: plantações
de Eucalyptus e fragmentos de floresta semidecidual (Figura 3). Estes foram
amostrados mensalmente totalizando 8 noites de captura por mês. Todas as
capturas foram realizadas na semana de lua nova, para reduzir potenciais
tendências no sucesso de captura, devido ao comportamento de fobia lunar dos
morcegos (e.g. Morrison, 1978).
A selecção dos locais de estudo foi dependente da disponibilidade de
plantações de Eucalyptus adequadas, presentes na área de estudo. Estes
fragmentos têm diferentes áreas e formas irregulares.
Figure 3 – Áreas de amostragem (Euca = Plantações de Eucalyptus; F – Fragmentos de floresta).
Fonte: Google Earth (Dezembro de 2010).
5 km

13
Os morcegos foram capturados usando 10 redes de neblina por noite, com
dimensões de 10 a 14m x 2.5 m e malha de 16 mm, dispostas ao longo de um
transecto linear em cada fragmento (Figura 4).
As redes foram montadas sempre no mesmo e único transecto, ao longo
de 5 meses, dentro de cada fragmento em estudo (Figura 4), sendo abertas ao
pôr-do-sol e verificadas de 20 a 30 minutos durante 4 a 5 horas/noite.
Os morcegos capturados foram retirados manualmente das redes e
transferidos para sacos individuais de algodão. Os animais foram manipulados no
campo onde foi realizada a biometria e identificação (Figura 5).
Figura 4 – Redes de neblina armadas em a) Plantação de Eucalyptus; b) Fragmento de Floresta
semidecidual.
Figura 5 – Análise biométrica dos morcegos capturados.
a) b)

14
Foi registado numa Ficha de Campo a espécie, o sexo, o estado
reprodutor, o peso corporal (g) e o tamanho do antebraço (mm). Utilizou-se uma
balança digital, com resolução de 0,1 g, para aferir o peso e um paquímetro com
0,01 mm, para medir o antebraço dos morcegos. Estes foram identificados
usando Vizotto & Taddei (1973) e Gardner (2007). Para alguns taxa, foi
necessário consultar literatura adicional, por exemplo: Platyrrhinus (Ferrell &
Wilson, 1991); pequenos Artibeus (Handley, 1987); Sturnira (Davis, 1980); Myotis
(Laval, 1973).
Ao fim da noite, logo após o encerramento dos trabalhos, os animais foram
libertados no mesmo local de captura. Alguns exemplares foram levados ao
laboratório para o procedimento de fixação e conservação (Licença número
35128167) para uma posterior correcta identificação. Estes foram posteriormente
guardados junto à colecção de morcegos do Laboratório de Zoologia da
Universidade Federal de Goiás.
2.3 Tratamento dos dados e análise estatística
As análises realizadas foram restritas aos morcegos capturados
pertencentes à família Phyllostomidae, uma vez que estes são os morcegos que
podem seguramente revelar uma abundância realística quando utilizadas as
redes de neblina ao nível do solo (Zortéa et al., 2010). Além disto, esta é uma
família muito rica em espécies e numericamente dominante nas comunidades de
morcegos neotropicais (e.g. Kalko, 1998).
2.3.1 Variáveis métricas locais e de paisagem
Para cada um dos oito fragmentos foram calculadas variáveis locais e
medidas ao nível da paisagem, usando o software ImageJ 1.45j e algumas
ferramentas do Google Earth.
As variáveis locais medidas foram: a área (Hc) e o comprimento da borda
(m) de cada fragmento e a distância mais próxima em linha recta entre o vértice
de cada fragmento e entre o centro de cada fragmento (m).
Foram calculadas as seguintes medidas que caracterizam a composição e
configuração da paisagem, cercando cada área por um círculo para uma escala

15
focal de 3 km, com o raio de 1,5 km centrado em cada fragmento: tamanho médio
do fragmento, número de fragmentos de eucalipto (n.frag.euca), de floresta
(n.frag.mata) e o total (n.frag.total), densidade de fragmentos de eucalipto
(dens.euca), de floresta (dens.mata) e o total (dens.total) e a proporção de
cobertura florestal de eucalipto (p.cober.euca), de floresta (p.cober.mata) e o total
(p.cober.total).
A escala focal foi escolhida tendo em conta as usadas em Klingbeil & Willig
(2009) (raio de 1 Km; 3 Km e 5 Km) e Meyer & Kalko (2008) (raio de 0,5 Km; 1
Km e 1,5 Km) e a adaptação ao trabalho realizado. Neste caso, optou-se pela
escala focal de 3 Km, para minimizar sobreposição espacial dos fragmentos
vizinhos e portanto a autocorrelação espacial entre os locais de amostragem.
2.3.2 Riqueza, abundância, diversidade e composição de espécies
Para avaliar a riqueza estimada para cada tipo de habitat usou-se a curva
de acumulação de espécies baseada em amostras ao acaso (1000
randomizações) (Gotelli & Colwell, 2001). Assim avaliou-se o número de espécies
esperadas de ocorrer em cada local usando um estimador de riqueza de espécies
não paramétrico. Escolheu-se o estimador de Jackknife de primeira ordem
(Jack1), uma vez que tem em consideração a heterogeneidade de movimentos de
animais móveis como os morcegos, baseado no trabalho desenvolvido por Brose
& Martinez (2004). Os cálculos foram realizados usando o software EstimateS
8.2.0 (Colwell, 2009).
Para comparar a riqueza de espécies rarefaccionadas, a dominância e a
equitabilidade (índice PIE de Hurlbert “Probability of interspecific encounter”) entre
os diferentes tipos de habitat, usou-se o software EcoSim, com 1000 iterações. A
rarefacção permite a comparação da riqueza de espécies ao nível da abundância
menor, através da transformação das abundâncias, obtidas para cada tipo de
habitat, para a mesma escala, que é definida pela menor abundância. Neste caso
o nível de abundância comum era de 75 indivíduos. A equitabilidade refere-se ao
quão similar as espécies estão representadas numa comunidade, podendo variar
de 0 (dominância de espécies únicas) a 1 (abundância igual para todas as
espécies) e a dominância permite aferir a fracção da população que é
representada pelas espécies mais comuns. O Hurlbert‟s PIE é um índice de

16
equitabilidade que calcula, numa comunidade, a probabilidade de dois indivíduos
amostrados ao acaso representarem duas espécies diferentes (Stirling & Wilsey,
2001; Gotelli & Entsminger, 2001; Melo, 2008). Em cada caso, a significância
estatística foi determinada segundo os intervalos de confiança de 95% simulados,
gerados pelo programa (Gotelli & Entsminger, 2001).
O esforço de captura foi calculado segundo Straube & Bianconi (2002),
como m2rede h. Este foi de 25960 h m2 e 25530 h.m2, para Eucalipto e Floresta,
respectivamente, o que é relativamente semelhante. Para cada tipo de habitat, foi
calculada a abundância relativa para cada espécie e tipo de habitat como:
Abundância relativa = Abundância Absoluta / Esforço de Captura.
Determinou-se a diversidade segundo o Índice de Shannon-Wiener (H’)
usando-se o Software PAST. Apesar do índice de Shannon-Wiener ter sido várias
vezes questionado (Lande, 1996; Magurran, 2004) foi também usado
anteriormente por muitos autores, o que torna possível comparar os resultados
obtidos neste trabalho com outros estudos publicados (Aguirre, 2002; Faria et al.,
2006; Rex et al., 2008).
Usou-se o coeficiente de Bray-Curtis para descrever a dissimilaridade em
termos de composição de espécies entre os dois tipos de habitat (Magurran,
2004), através do Software EstimateS 8.2.0 (Colwell, 2009). Aplicou-se depois
uma escala não métrica multidimensional (NMDS) para ordenar os locais e avaliar
as diferenças inter-locais na composição de espécies. O NMDS é um teste
robusto, uma técnica de ordenação não linear considerado como um método
efectivo para analisar dados ecológicos da comunidade (McCune & Grace, 2002).
As diferenças na composição de espécies entre os dois tipos de habitat
foram avaliadas através de uma análise de similaridades (ANOSIM; e.g. McArdle
& Anderson, 2001), que é um procedimento não métrico de permutação
multivariada, muito semelhante a uma típica ANOVA univariada, que testa por
diferenças entre grupos primeiramente definidos da amostra da comunidade,
baseados numa matriz de (dis)similaridade (Clarke, 1993). A medida de distância
usada foi a de Bray-Curtis. Estes cálculos foram realizados com o Software PAST.

17
2.3.3 Modelação da fauna de morcegos com ambiente
Para além das variáveis métricas locais e de paisagem calculadas foram
ainda consideradas as distâncias obtidas na matriz de NMDS de Bray-Curtis, que
permite representar as diferenças na composição de espécies entre fragmentos
(Anexo I).
Uma vez que se possuem dados de apenas quatro fragmentos para cada
tipo de habitat (floresta e eucalipto) e um elevado número de variáveis, teve-se de
realizar pares de comparações das variáveis preditivas de modo a obter as
variáveis significativamente correlacionadas. Isto resultou num total de seis
variáveis explicativas. Como variáveis a escala local foi considerado a área e o
perímetro (comprimento de borda) e em termos de paisagem a proporção de
cobertura de mata, proporção de cobertura de eucalipto, número de fragmentos
de mata e número de fragmentos de eucalipto. Estas foram standardizadas, ou
seja transformadas na mesma escala de grandeza de valores, pois existem
valores de grandeza muito diferente, como o caso das áreas e dos perímetros.
Realizou-se uma análise multivariada de permutações da variância usando
matrizes de distâncias (ADONIS) que relaciona variáveis ambientais com a
composição de espécies. Esta análise de permutações é feita dentro de cada tipo
de habitat e não inclui permutações entre tipos de habitat. A ADONIS é usada em
ecologia quando se tem um número limitado de amostras de indivíduos (Zapala &
Schork, 2006) e permitiu verificar quais das seis variáveis tem efeito significativo
na composição de espécies.
Para verificar se existem diferenças significativas na abundância de cada
espécie entre os dois tipos de habitat (eucalipto e floresta) usaram-se modelos
lineares generalizados de efeitos mistos (GLMM). Uma vez que a abundância de
espécies são contagens, usou-se a distribuição de Poisson. O factor fixo nestes
modelos foi a variável habitat e os factores aleatórios os fragmentos e as noites
de captura, ajustado pelo método de estimação dos parâmetros de máxima
verossimilhança (Zuur et al., 2007).
Como o número de capturas para a maioria das espécies era muito baixa,
apenas 3 espécies (Carollia perspicillata; Artibeus planirostris; Platyrrhinus
lineatus) foram consideradas na realização deste teste, por serem as únicas com
dados suficientes. Neste trabalho foram seleccionados quatro fragmentos para
cada um dos dois tipos de habitat (eucalipto X floresta), que foram amostrados

18
mensalmente em apenas um ponto de colecta em diferentes noites cada um,
durante cinco meses. Considerou-se assim as noites aleatórias e aninhadas aos
fragmentos. Para a realização deste procedimento recorreu-se ao pacote de R
lme4 (Bates et al., 2011). Para analisar as relações da fauna de morcegos com o
ambiente, realizou-se a selecção do modelo usando likelihood ratio test e tendo
em conta o valor criterioso de Akaike (AIC). Para cada espécie é calculado o
modelo com e sem o factor fixo (Habitat) e depois aplicado o Likelihood ratio test,
que compara os dois modelos e permite concluir sobre a influência do factor fixo
na abundância de cada espécie em análise (Zuur et al., 2009).
Todas estas análises foram realizadas usando o pacote estatístico R
(versão 2.2.0, R Development Core Team, 2006).
2.3.4 Autocorrelação espacial
Os dados de abundância estão muitas vezes autocorrelacionados
espacialmente. Para perceber esta correlação foi construída uma matriz de
distâncias ecológicas e geográficas (Anexo II). Realizou-se um Teste de Mantel
para testar se existe correlação entre as distâncias ecológicas e geográficas.
Foram realizadas 5000 permutações, aplicando-se o índice de Bray-Curtis para a
matriz de abundância das espécies (Anexo III) e as distâncias geográficas do
utilizador, que são Euclídicas, para a matriz de distâncias geográficas. Estas
análises foram realizadas usando o pacote estatístico R (versão 2.2.0, R
Development Core Team, 2006).
3. Resultados
3.1 Riqueza, abundância, diversidade e composição de espécies
Foram capturados 209 morcegos, pertencentes a duas famílias, 9 géneros
e 13 espécies (Tabela I), ao longo de 40 noites de amostragem. Os morcegos da
Família Phyllostomidae representaram a maioria das capturas (97%) e a maioria
das espécies (92%). A outra família registada foi a Vespertilionidae com sete
capturas de uma única espécie.

19
Tabela I – Relação de espécies e respectivas abundâncias registadas em áreas de floresta nativa
e de monocultura de eucalipto no sudoeste de Goiás, Brasil.
Familia Subfamilia Espécies Eucalipto Floresta Grupo
Funcional
Phyllostomidae Phyllostominae Chrotopterus auritus 0 1 Carnívoro
Phyllostomus hastatus 3 4 Omnívoro
Lophostoma brasiliense 0 3 Insectívoro
Stenodermatinae Artibeus planirostris 4 35 Frugívoro
Artibeus lituratus 5 10 Frugívoro
Artibeus cinereus 1 4 Frugívoro
Sturnira lilium 5 9 Frugívoro
Sturnira tildae 0 1 Frugívoro
Platyrrhinus lineatus 8 13 Frugívoro
Platyrrhinus helleri 0 1 Frugívoro
Desmodontinae Desmodus rotundus 1 2 Hematófago
Carolliinae Carollia perspicillata 48 44 Frugívoro
Vespertilionidae Myotinae Myotis nigricans 0 7 Insectívoro
Total 75 134
A espécie mais abundante foi Carollia perspicillata com 92 capturas, o
equivalente a 44% do total, seguida de Artibeus planirostris com 39 capturas
(19%), Platyrrhinus lineatus com 21 capturas (10%), Artibeus lituratus com 15
capturas (10%) e Sturnira lilium com 14 capturas (7%). As restantes espécies
totalizaram juntas apenas 14% das capturas.
Nas áreas de Eucalipto foram capturados 75 indivíduos. A espécie mais
abundante foi Carollia perspicillata, com 48 indivíduos, seguida de Platyrrhinus
lineatus, com oito indivíduos. Em floresta nativa capturaram-se 134 indivíduos,
sendo a espécie Carollia perspicillata (44) novamente a mais capturada, seguida
de Artibeus planirostris (35) e Platyrrhinus lineatus (13).
Nas áreas de vegetação nativa observou-se a ocorrência das 13 espécies
registadas no estudo, incluindo cinco espécies exclusivas (Chrotopterus auritus,
Lophostoma brasiliense, Sturnira tildae, Platyrrhinus helleri e Myotis nigricans), de
modo que nas áreas de eucalipto anotou-se apenas oito espécies (Tabela I,
Figura 6).
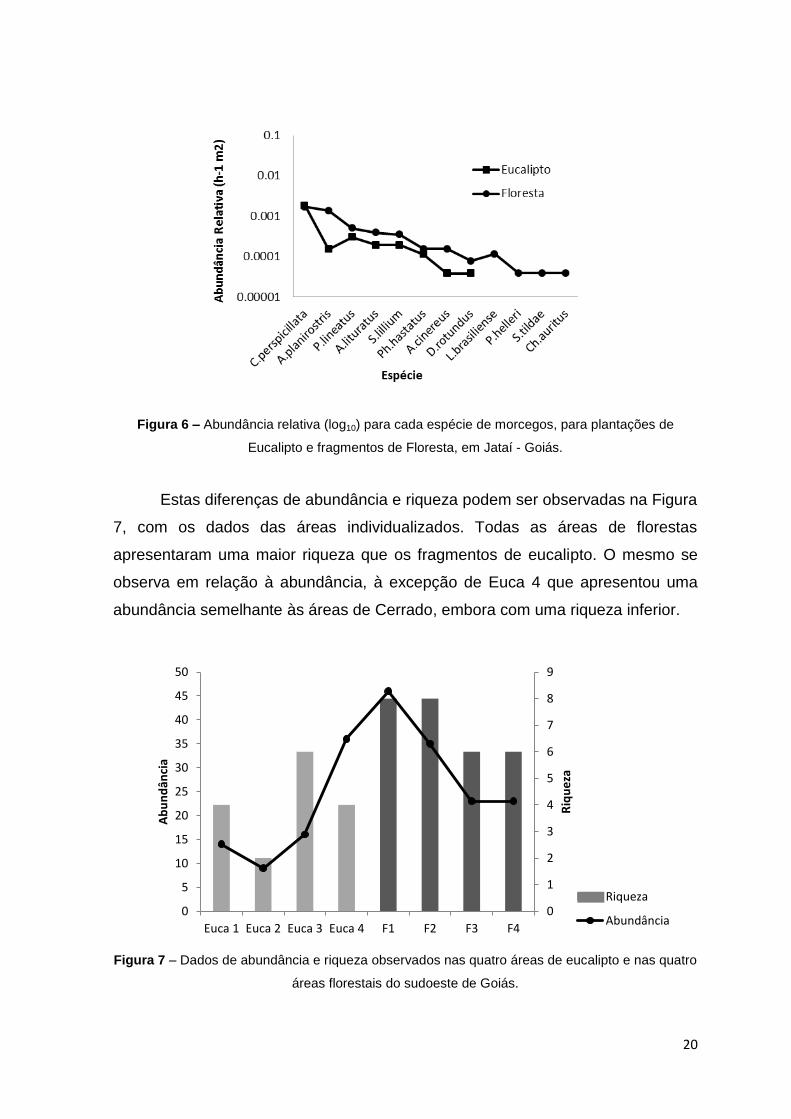
A distribuição da abundância relativa segundo as espécies apresenta
valores mais elevados para a Floresta comparativamente ao Eucalipto, à
excepção de uma espécie, Carollia perspicillata (Figura 6).
20
Figura 6 – Abundância relativa (log10) para cada espécie de morcegos, para plantações de
Eucalipto e fragmentos de Floresta, em Jataí - Goiás.
Estas diferenças de abundância e riqueza podem ser observadas na Figura
7, com os dados das áreas individualizados. Todas as áreas de florestas
apresentaram uma maior riqueza que os fragmentos de eucalipto. O mesmo se
observa em relação à abundância, à excepção de Euca 4 que apresentou uma
abundância semelhante às áreas de Cerrado, embora com uma riqueza inferior.
Figura 7 – Dados de abundância e riqueza observados nas quatro áreas de eucalipto e nas quatro
áreas florestais do sudoeste de Goiás.
0
1
2
3
4
5
6
7
8
9
0
5
10
15
20
25
30
35
40
45
50
Euca 1 Euca 2 Euca 3 Euca 4 F1 F2 F3 F4
Riq
ue
za
Ab
un
dân
cia
Riqueza
Abundância

21
As diferenças detectadas nos dois tratamentos (floresta nativa X eucalipto)
como a baixa riqueza e abundância e alta dominância observada na monocultura
de eucalipto expressou uma diversidade mais baixa neste ambiente (H’ = 1.286)
quando comparada com a área florestal nativa (H’ = 1.830).
Quanto aos grupos funcionais, 90% dos indivíduos capturados eram
frugívoros (Tabela II). Os insetívoros e carnívoros foram capturados
exclusivamente em áreas de floresta nativa.
Tabela II – Riqueza de espécies (S), número de capturas (N) e abundância relativa (%) para cada
grupo funcional de morcegos, para as áreas de Eucalipto e Floresta.
Eucalipto Floresta Total
Grupo Funcional S N % S N % S N %
Frugívoro 6 71 95 8 117 87 8 188 90.0
Insectívoro 0 0 0 2 10 7 2 10 4.8
Omnívoro 1 3 4 1 4 3 1 7 3.3
Hematófago 1 1 1 1 2 2 1 3 1.4
Carnívoro 0 0 0 1 1 1 1 1 0.5
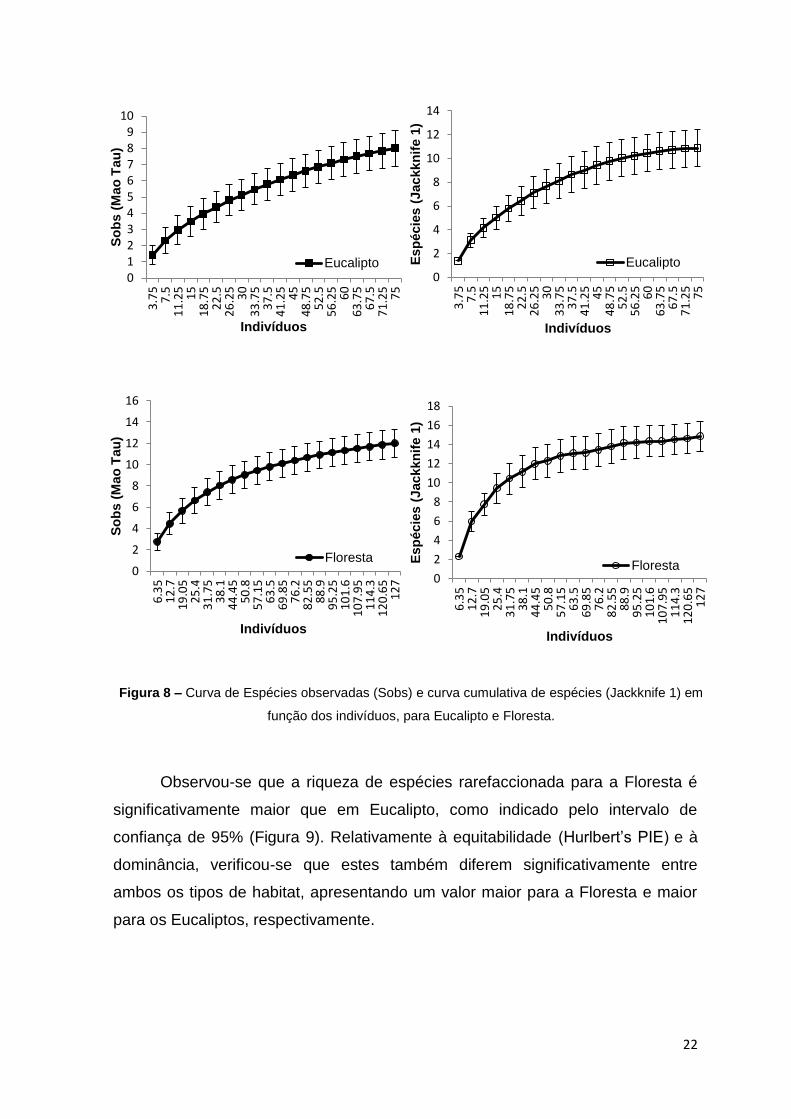
Uma vez que não se observou explicitamente uma estabilização da curva
cumulativa de espécies, principalmente no caso do Eucalipto em que não é
perceptível a aproximação a uma relação assimptótica, pode-se dizer apenas que
o número de espécies acumulado é satisfatório para o tamanho da amostra.
Conseguiu-se um nível de inventariação de espécies de 80.8% para Floresta e
73.73% para Eucalipto, relativamente ao que seria esperado segundo o estimador
usado – Jack1 (Eucalipto - 10.85 espécies; Floresta – 14.85 espécies) (Figura 8).
22
0
2
4
6
8
10
12
14
16
18
6.3
51
2.7
19
.05
25
.43
1.7
53
8.1
44
.45
50
.85
7.1
56
3.5
69
.85
76
.28
2.5
58
8.9
95
.25
10
1.6
10
7.9
51
14
.31
20
.65
12
7
Esp
écie
s (
Jackkn
ife 1
)
Indivíduos
Floresta
0123456789
10
3.7
57
.51
1.2
51
51
8.7
52
2.5
26
.25
30
33
.75
37
.54
1.2
54
54
8.7
55
2.5
56
.25
60
63
.75
67
.57
1.2
57
5
So
bs (
Mao
Tau
)
Indivíduos
Eucalipto0
2
4
6
8
10
12
14
3.7
57
.51
1.2
51
51
8.7
52
2.5
26
.25
30
33
.75
37
.54
1.2
54
54
8.7
55
2.5
56
.25
60
63
.75
67
.57
1.2
57
5
Esp
écie
s (
Jackkn
ife 1
)
Indivíduos
Eucalipto
0
2
4
6
8
10
12
14
16
6.3
51
2.7
19
.05
25
.43
1.7
53
8.1
44
.45
50
.85
7.1
56
3.5
69
.85
76
.28
2.5
58
8.9
95
.25
10
1.6
10
7.9
51
14
.31
20
.65
12
7
So
bs (
Mao
Tau
)
Indivíduos
Floresta
Figura 8 – Curva de Espécies observadas (Sobs) e curva cumulativa de espécies (Jackknife 1) em
função dos indivíduos, para Eucalipto e Floresta.
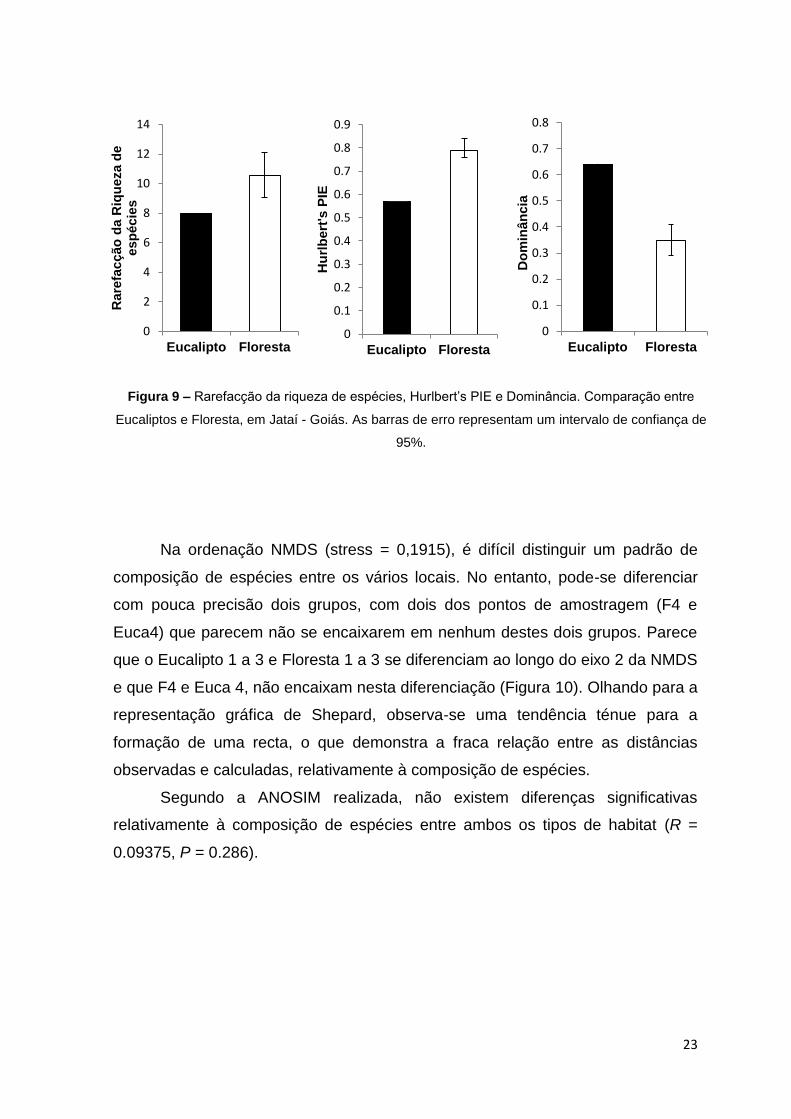
Observou-se que a riqueza de espécies rarefaccionada para a Floresta é
significativamente maior que em Eucalipto, como indicado pelo intervalo de
confiança de 95% (Figura 9). Relativamente à equitabilidade (Hurlbert‟s PIE) e à
dominância, verificou-se que estes também diferem significativamente entre
ambos os tipos de habitat, apresentando um valor maior para a Floresta e maior
para os Eucaliptos, respectivamente.
23
0
2
4
6
8
10
12
14
Eucalipto Floresta
Rare
facção
da R
iqu
eza d
e
esp
écie
s
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Eucalipto Floresta
Hu
rlb
ert
's P
IE
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Eucalipto Floresta
Do
min
ân
cia
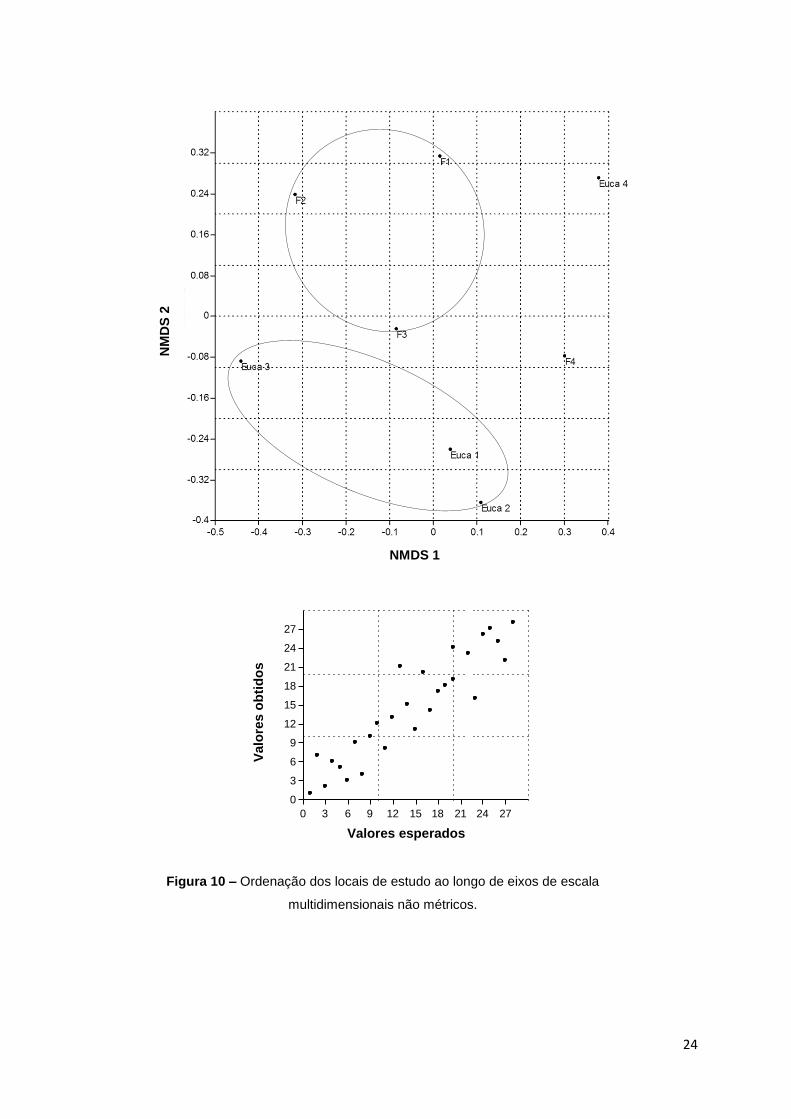
Na ordenação NMDS (stress = 0,1915), é difícil distinguir um padrão de
composição de espécies entre os vários locais. No entanto, pode-se diferenciar
com pouca precisão dois grupos, com dois dos pontos de amostragem (F4 e
Euca4) que parecem não se encaixarem em nenhum destes dois grupos. Parece
que o Eucalipto 1 a 3 e Floresta 1 a 3 se diferenciam ao longo do eixo 2 da NMDS
e que F4 e Euca 4, não encaixam nesta diferenciação (Figura 10). Olhando para a
representação gráfica de Shepard, observa-se uma tendência ténue para a
formação de uma recta, o que demonstra a fraca relação entre as distâncias
observadas e calculadas, relativamente à composição de espécies.
Segundo a ANOSIM realizada, não existem diferenças significativas
relativamente à composição de espécies entre ambos os tipos de habitat (R =
0.09375, P = 0.286).
Figura 9 – Rarefacção da riqueza de espécies, Hurlbert‟s PIE e Dominância. Comparação entre
Eucaliptos e Floresta, em Jataí - Goiás. As barras de erro representam um intervalo de confiança de
95%.
24
0 3 6 9 12 15 18 21 24 27
Target rank
0
3
6
9
12
15
18
21
24
27
Obta
ined r
ank
Figura 10 – Ordenação dos locais de estudo ao longo de eixos de escala
multidimensionais não métricos.
NM
DS
2
NMDS 1
Valores esperados
Valo
res o
bti
do
s
25
3.2 Modelação da fauna de morcegos com o ambiente
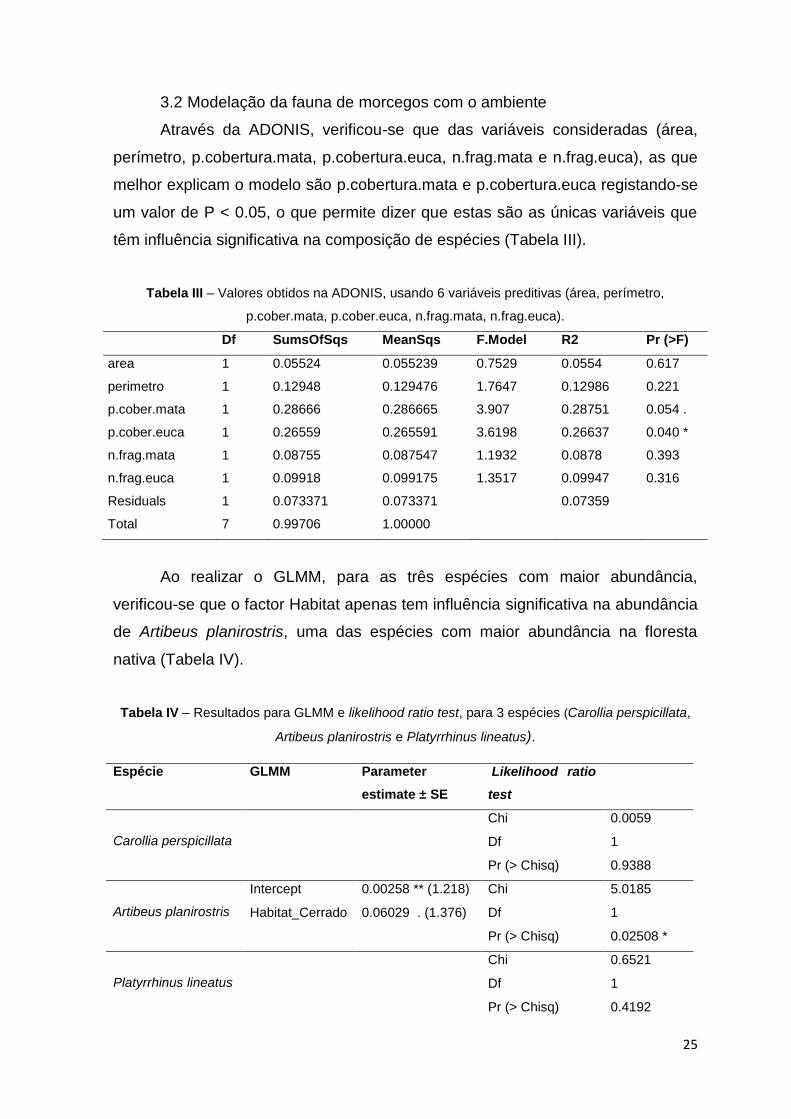
Através da ADONIS, verificou-se que das variáveis consideradas (área,
perímetro, p.cobertura.mata, p.cobertura.euca, n.frag.mata e n.frag.euca), as que
melhor explicam o modelo são p.cobertura.mata e p.cobertura.euca registando-se
um valor de P < 0.05, o que permite dizer que estas são as únicas variáveis que
têm influência significativa na composição de espécies (Tabela III).
Tabela III – Valores obtidos na ADONIS, usando 6 variáveis preditivas (área, perímetro,
p.cober.mata, p.cober.euca, n.frag.mata, n.frag.euca).
Df SumsOfSqs MeanSqs F.Model R2 Pr (>F)
area 1 0.05524 0.055239 0.7529 0.0554 0.617
perimetro 1 0.12948 0.129476 1.7647 0.12986 0.221
p.cober.mata 1 0.28666 0.286665 3.907 0.28751 0.054 .
p.cober.euca 1 0.26559 0.265591 3.6198 0.26637 0.040 *
n.frag.mata 1 0.08755 0.087547 1.1932 0.0878 0.393
n.frag.euca 1 0.09918 0.099175 1.3517 0.09947 0.316
Residuals 1 0.073371 0.073371
0.07359
Total 7 0.99706 1.00000
Ao realizar o GLMM, para as três espécies com maior abundância,
verificou-se que o factor Habitat apenas tem influência significativa na abundância
de Artibeus planirostris, uma das espécies com maior abundância na floresta
nativa (Tabela IV).
Tabela IV – Resultados para GLMM e likelihood ratio test, para 3 espécies (Carollia perspicillata,
Artibeus planirostris e Platyrrhinus lineatus).
Espécie GLMM Parameter
estimate ± SE
Likelihood ratio
test
Carollia perspicillata
Chi 0.0059
Df 1
Pr (> Chisq) 0.9388
Artibeus planirostris
Intercept 0.00258 ** (1.218) Chi 5.0185
Habitat_Cerrado 0.06029 . (1.376) Df 1
Pr (> Chisq) 0.02508 *
Platyrrhinus lineatus
Chi 0.6521
Df 1
Pr (> Chisq) 0.4192

26
3.3 Autocorrelação espacial
Verificou-se, pelo Teste de Mantel que não existe relação entre as
distâncias geográficas (distância entre centros (R = 0.0817, P = 0.6608) e
distância entre o vértice mais próximo (R = 0.0812, P = 0.3284), dos vários
fragmentos em estudo e a composição de espécies obtida.
4. Discussão
4.1 Enquadramento da composição taxionómica obtida, na região e bioma
em estudo
A composição taxionómica de morcegos obtida neste trabalho apresenta
características semelhantes ao padrão das comunidades neotropicais obtido em
outros trabalhos (e.g. Lim & Engstrom, 2000; Kalko & Handley, 2001; Estrada &
Coates-Estrada, 2002; Esbérard, 2003; Bernard & Fenton, 2007; Zortéa et al.,
2010). Este padrão pode ser explicado pela dinâmica das competições
interespecíficas, onde espécies mais adaptadas e sem fortes pressões
competitivas apresentam maiores densidades, enquanto espécies com mais
pressões competitivas exibem abundâncias menores (Stevens & Willig, 2000).
A composição taxionómica de morcegos na região Neotropical é em geral,
dominada por espécies da família Phyllostomidae. Na verdade, esta família é a
mais rica em número de espécies no Brasil, com 90 espécies (Reis et al., 2007).
Esta dominância numérica e ecológica da família Phyllostomidae, é um padrão
comum em estudos com comunidades de morcegos na região Neotropical (e.g.
Estrada & Coates-Estrada, 2002; Bernard & Fenton, 2007; Zortéa et al., 2010),
também porque estes são os morcegos que podem seguramente ser capturados
usando as redes de neblina ao nível do solo (e.g. Kalko, 1998).
Esta predominância foi verificada na área em estudo, quer em número de
espécies quer em número de indivíduos, tendo-se capturado apenas sete
indivíduos de uma espécie (Myotis nigricans), pertencente a uma outra família -
Vespertilionidae. De todos os morcegos capturados neste estudo, 97% pertencem
à família Phyllostomidae. O mesmo valor foi observado num outro trabalho
realizado numa área próxima aos sítios do presente estudo (Zortéa & Alho 2008).

27
Segundo os estudos realizados em Cerrado, a riqueza de espécies de
morcegos para uma comunidade local pode variar de 15 a 25 espécies, no caso
em que são usadas redes de neblina ou quando a inventariação é feita através de
capturas em grutas (Zortéa & Alho, 2008). Esta riqueza de espécies está também
documentada para a região de Goiás, através de vários trabalhos realizados. Por
exemplo, Zortéa et al. (2010) registaram 21 espécies na Bacia Hidrográfica do Rio
Corumbá e Rodrigues et al. (2002) obtiveram 19 espécies no Parque Nacional
das Emas.
Neste trabalho a riqueza foi baixa (N = 13) e uma das explicações
possíveis poderia ser a presença de factores que influenciam a abundância e
diversidade de morcegos, como provavelmente a escassez de abrigos, a falta de
habitat nativo e uma composição florística modificada, uma vez que esta se
encontra altamente alterada pelo Homem. A riqueza observada neste estudo
corresponde apenas a 12,6% das espécies conhecidas registadas para o
Cerrado, segundo a lista compilada por Aguiar & Zortéa (2008). Seria importante
a realização de outros métodos de amostragem, como por exemplo, capturas
diurnas em abrigos e também a amostragem acústica usando-se detectores de
ultra-sons (Kunz & Parsons, 2009) e um aumento no esforço de amostragem, de
modo a capturar-se/detectar-se mais espécies de morcegos na área,
nomeadamente de famílias como os vespertilionídeos, os molossídeos, os
emballonurídeos e os mormoopídeos.
As famílias não amostradas mas esperadas para a região são também, por
outro lado, mais difíceis de amostrar, ou por ocorrerem em baixas densidades ou
por apresentarem comportamento de forrageio a grandes alturas, o que dificulta a
interceptação destes morcegos nas redes de neblina armadas ao nível do solo,
que foi a metodologia usada.
Segundo uma visão zoogeográfica, todas as espécies observadas neste
estudo já foram registadas para o Cerrado (Marinho-Filho, 1996; Aguiar & Zortéa,
2008; Zortéa & Alho, 2008). No entanto existem duas espécies que parecem estar
menos descritas para este bioma: Lophostoma brasiliense (Zortéa & Alho, 2008) e
Sturnira tildae (Gregorin et al., 2011).
Lophostoma brasiliense é uma espécie predominantemente insectívora e
está aparentemente associada a habitats de floresta primária e secundária, a
fragmentos florestais, pastos e pomares e no cerrado a veredas e outras áreas

28
húmidas. Abriga-se em ocos de árvores e casas, tendo sido frequentemente
encontrada em cupinzeiros arbóreos (Mangolin et al., 2007; Nogueira et al., 2007).
A espécie Sturnira tildae deve ocorrer na maioria dos estados brasileiros
apesar de ter sido registada em apenas 10 deles (Zortéa, 2007). Para o cerrado
apenas Marinho-Filho (1996), faz referência na sua lista de espécies brasileiras
sem referenciar de onde o dado foi obtido. Só bem mais recentemente Gregorin et
al. (2011) citaram esta pela primeira vez, no bioma Cerrado num trabalho
realizado no Estado do Tocantins que fica imediatamente no norte de Goiás. A
espécie possui hábito frugívoro e está associada a sub-bosque de áreas
florestais, podendo ser também encontrada em habitats alterados como clareiras
(Zortéa, 2007).
As espécies dominantes variam geograficamente, no entanto espécies
como C. perspicillata e Artibeus spp. têm sido citadas como as mais comuns na
região Neotropical (Zortéa & Alho, 2008). A espécie Carollia perspicillata foi a
mais comum neste estudo com 44% das capturas, seguida por Artibeus
planirostris e Platyrrhinus lineatus. Estas três espécies são todas frugívoras e
como apontado por Aguiar & Zortéa (2008) estão entre os morcegos mais comuns
do Cerrado.
Carollia perspicillata é uma espécie muito abundante e um dos morcegos
mais comuns em levantamentos em diversas áreas da região Neotropical
como a Amazônia (Bernard et al., 2001; Bernard, 2002; Marques-Aguiar et al.,
2009), o Cerrado (Gonçalves & Gregorin, 2004) e a Mata Atlântica (Baptista &
Mello, 2001; Dias & Peracchi, 2008). É uma espécie considerada generalista
(Cloutier & Thomas, 1992; Giannini & Kalko, 2004) e alguns autores sugerem
ainda, que esta possui uma maior eficiência em adaptar-se aos processos de
fragmentação e/ou modificação do habitat (Brosset et al., 1996; Wilson et al.,
1996; Estrada & Coates-Estrada, 2002) podendo a sua flexibilidade estar
relacionada com o facto de esta alimentar-se de plantas pioneiras, como por
exemplo do género Piper, que costumam ser abundantes nas margens de
fragmentos florestais e em floresta secundária.
Artibeus planirostris e Platyrrhinus lineatus foram as duas espécies mais
capturadas depois de C. perspicillata, padrão observado também por Rodrigues et
al. (2002). Todas as demais dez espécies tiveram 15 ou menos capturas.

29
A ocorrência do morcego-vampiro Desmodus rotundus era esperada,
devido à forte conversão dos habitats marginais em pastagens que abrigam o
gado, um dos seus principais alimentos (Greenhall et al., 1983). Num município
vizinho a Jataí, D. rotundus foi a segunda espécie mais abundante dentre 25
táxons amostrados (Zortéa & Alho, 2008). É importante destacar o registo da
espécie Chrotopterus auritus, uma carnívora que embora largamente distribuida
pelo Brasil (Nogueira et al., 2007) é pouco abundante.
4.2 Riqueza, dominância, diversidade e composição de espécies
Estudos abordando a conservação da biodiversidade em plantações de
eucaliptos e outras plantações são relativamente recentes (Danielsen &
Heegaard, 1995; Bernhard-Reversat, 2001; Borsboom et al., 2002; Lindenmayer
et al., 2003; Barlow et al., 2007) e mais escassos quando comparados aos
realizados em plantações de pinheiros (Lindenmayer et al., 2003). São vários os
trabalhos existentes que escrevem sobre a conservação da biodiversidade para
diferentes plantações florestais, como de palmeiras e borracha (Danielsen &
Heegaard, 1995), cacau (Faria et al., 2006), cacau e banana (Harvey & Villalobos,
2007), Eucalyptus (Borsboom et al., 2002; Barlow et al., 2007b) e eucaliptos e
coníferas (Lindenmayer et al., 2003). A maioria dos trabalhos existentes referem-
se a comunidades de aves (Marsden et al., 2001; Zurita et al., 2006; Lantschner et
al., 2008; Gabriel et al., 2009; Filloy et al., 2010; Hsu et al., 2010). Existem alguns
trabalhos que incidem noutros grupos, como borboletas (Barlow et al., 2007b),
pequenos mamíferos (Umetsu & Pardini, 2007) e morcegos (Harvey & Villalobos,
2007; Borkin & Parson, 2010).
Marsden et al. (2001) estudaram comunidades de aves dentro da Reserva
de Sooretama/Linhares e em fragmentos de floresta e plantações de Eucalyptus
presentes num raio de 7 Km em redor da reserva, no Estado do Espírito Santo,
Brasil. Estes autores concluíram que a riqueza de espécies para plantações de
Eucalyptus > 30 anos é extremamente baixa e muito menor que a registada para
outras plantações no Brasil. Eles sugerem que um sub-bosque pobre em
vegetação e disperso é a principal razão para a ausência da maioria das
espécies. A riqueza de espécies observada nas plantações de Eucalyptus diferiu
significativamente da obtida em fragmentos de floresta e na reserva. Verificaram

30
ainda que a riqueza de espécies nos fragmentos de floresta não diminui com o
tamanho do fragmento, distância à reserva ou qualidade da floresta nativa.
Zurita et al. (2006) estudaram comunidades de aves em floresta nativa e
em plantações comerciais de árvores exóticas (Pinus spp.) e nativas (Araucaria
angustifolia), na Argentina. Aqui verificou-se que a riqueza de espécies é 50%
menor nas plantações comparativamente à floresta nativa. Observaram ainda que
as comunidades de aves nas plantações comerciais eram maioritariamente
compostas por espécies de floresta generalistas. Assim, concluíram que a maior
alteração na riqueza de espécies de aves e na sua composição está associada à
substituição da floresta nativa pelas plantações de árvores comerciais sob as
condições de gestão correntes.
Lantschner et al. 2008 analisaram o efeito das plantações de Pinus
ponderosa nas comunidades de aves, considerando diferentes práticas de gestão
(cobertura de árvores densa e esparsa) e diferentes contextos de paisagem onde
estão inseridas (Floresta de Austrocedros chilensis e estepes), na Patagónia –
Argentina. Concluíram que as plantações de pinheiro podem providenciar habitat
para um número substancial de espécies nativas de aves, que está dependente
com as práticas de gestão e do contexto da paisagem das áreas de florestamento
onde ocorrem.
Gabriel et al. (2009) tinham como um dos objectivos comparar a riqueza e
abundância de espécies de avifauna encontradas em talhões de eucalipto e em
remanescentes de vegetação nativa de cerradão numa fazenda de produção
comercial de madeira. Verificaram que a riqueza de espécies era
significativamente menor nas plantações de eucalipto relativamente ao cerradão.
A densidade do sub-bosque foi a única variável que influenciou, na riqueza e
abundância de espécies observadas nas plantações de eucalipto.
Filloy et al. (2010) examinaram a composição da comunidade de aves e o
número de espécies nativas em dois tipos de uso do solo (campos de soja e
plantações de eucalipto) em dois diferentes biomas (floresta e savana), no
Paraguai e na Argentina. Dentro de cada bioma, a composição da comunidade de
aves diferiu entre os campos de soja e as plantações de eucalipto. A composição
da comunidade de aves entre o mesmo uso do solo (soja ou eucalipto), mas em
biomas diferentes, foi similar. A composição das comunidades locais em
ecossistemas antropizados parece ser extremamente influenciado pelas

31
condições locais provenientes do tipo de uso do solo. As plantações de eucalipto
e os campos de soja são menos prejudiciais tanto para as aves de floresta como
de savana, em contextos dominados por floresta e savana, respectivamente.
Hsu et al. (2010) investigaram o valor relativo de conservação de
plantações de eucalipto através da avaliação da riqueza de avifauna, abundância
e composição de espécies, em cinco tipos de habitat (floresta, floresta ripária,
plantações, plantações ripárias, pastos ripários), na Austrália. A riqueza de
espécies e a abundância foram similares para todos os tipos de habitat à
excepção das plantações, que suportaram menos espécies e em menores
números.
Barlow et al. (2007b) examinaram os factores que influenciam o valor das
plantações de Eucalyptus em larga escala para as borboletas tropicais que se
alimentam de frutas, na Amazónia Brasileira. Focaram-se na avaliação da
importância dos factores de paisagem versus nível-local que determinam a
diversidade e composição da comunidade de borboletas. Verificaram que a
estrutura da vegetação a nível local e os factores composicionais são mais
importantes do que o contexto da paisagem, para as borboletas que se alimentam
de frutas.
Umetsu & Pardini (2007) avaliaram o uso da matriz da paisagem por
pequenos mamíferos, comparando a distribuição de espécies entre os fragmentos
de floresta e os habitats adjacentes predominantes (vegetação nativa em estádios
iniciais de regeneração, plantações de eucaliptos, áreas de agricultura e áreas
rurais com construções), no Brasil. Encontraram uma forte dissimilaridade nas
comunidades de pequenos mamíferos entre a vegetação nativa e os habitats
antrópicos.
Para avaliarem a importância relativa dos sistemas agro-florestais para a
conservação da biodiversidade, Harvey & Villalobos (2007) caracterizaram as
comunidades de aves e morcegos que ocorrem em floresta, em 2 tipos de
sistemas agro-florestais (cacau e banana) e em plantações de monoculturas, na
reserva de Talamanca, Costa Rica. Os sistemas agro-florestais mantêm a
comunidade de morcegos, que era tão rica em espécies, abundante e diversa
quanto a existente em floresta. Apresentava o mesmo padrão de espécies
dominantes, mas continha mais morcegos nectarívoros que as florestas. O
mesmo foi verificado para as aves, no entanto a composição de espécies da

32
comunidade apresentava-se mais modificada, com menos espécies dependentes
de floresta, mais espécies de áreas abertas e espécies dominantes diferentes.
Por último, Borkin & Parsons (2010) reviram o conhecimento actual sobre a
utilização das plantações florestais de exóticas, pelo morcego de cauda longa
(Chalinolobus tuberculatus) e reportaram pela primeira vez todo o tipo de
plantações de exóticas que estes morcegos usam na Nova Zelândia.
Tal como era esperado, a substituição da floresta nativa por plantações de
eucalipto influenciou a riqueza e diversidade das espécies de morcegos. No
presente trabalho, as diferenças na riqueza de espécies foram significativas, com
as áreas de floresta nativa a apresentarem uma maior riqueza de espécies que as
monoculturas de eucalipto.
A substituição da floresta nativa por plantações de monoculturas de árvores
resulta na simplificação da estrutura e composição da população vegetal, o que
pode influênciar grandemente uma comunidade (Marsden et al., 2001).
Verificou-se que os fragmentos de floresta suportam uma comunidade de
morcegos muito mais rica que as plantações de eucalipto. O mesmo parece
acontecer em diferentes florestas subtropicais e para outros grupos faunísticos,
nomeadamente as aves. Estes resultados parecem também ser independentes
das espécies de árvores plantadas e foi demonstrado com aves em plantações de
eucalipto no Brasil em Floresta Atlântica (Marsden et al., 2001) e em cerradão
(Gabriel et al., 2009), em plantações de Pinus na Floresta Atlântica na Argentina
(Zurita et al., 2006) e com morcegos em plantações de borracha e de palmeiras
na Indonésia (Danielsen & Heegaard, 1995).
Normalmente, espécies mais dependentes de floresta são afectadas pela
substituição da floresta nativa, e as comunidades em plantações comerciais são
constituídas maioritariamente por espécies de floresta e de borda generalistas
(Zurita et al., 2006).
O observado declínio das espécies de morcegos pode estar relacionado
com a dependência das espécies pelos habitats florestais nativos, onde as
espécies dependentes de floresta são as mais sensíveis á substituição das
florestas nativas. É importante assim destacar que as áreas de eucalipto não
suportaram cinco dos táxons observados e foram utilizados pelas espécies mais
comuns, enquanto as áreas florestais além de maior riqueza de espécies
apresentaram também uma comunidade mais diversa em relação aos hábitos

33
alimentares, incluindo espécies insectívoras (Lophostoma brasiliense e Myotis
nigricans) e uma carnívora (Chrotopterus auritus), que estiveram ausentes nas
plantações de eucaliptos.
No estudo realizado por Harvey & Villalobos (2007) sobre comunidades de
morcegos, em florestas e sistemas agro-florestais de banana e cacau, foi
verificado que a riqueza de espécies foi similar em ambos os habitats, no entanto
foram detectadas algumas diferenças subtis na comunidade, como um maior
número de morcegos nectarívoros nos sistemas agro-florestais comparativamente
às florestas. Foram vários os motivos apresentados, que combinados poderiam
justificar os altos níveis de biodiversidade nos sistemas agro-florestais, como as
copas estruturalmente similares às de floresta, a abundância de recursos
alimentares (frutos, néctar e insectos) e a grande proximidade entre os
fragmentos de floresta e os sistemas agroflorestais.
Também para Bernard & Fenton (2007), ao compararem habitats de
floresta e fragmentos cercados por vegetação do tipo savana na Amazónia
Central, a matriz de savanas com árvores e arbustos dispersos e a relativa curta
distância entre os fragmentos e a floresta primária, parecem facilitar a
conectividade entre estes habitats, conduzindo a comunidades estatisticamente
semelhantes mas com visíveis diferenças de estrutura e composição.
Deve-se destacar neste trabalho, que apesar de algumas das áreas de
eucalipto e de floresta estarem relativamente próximas ou até conectadas, como é
o caso da plantação de eucalipto 3 e do fragmento de floresta 4 (ver Figura 3),
obteve-se uma diferença significativa na riqueza de espécies para os dois tipos de
habitat.
Relativamente à dominância, verificou-se que esta foi significativamente
maior para as plantações de eucalipto. A menor diversidade verificada, o declínio
na riqueza de espécies e o aumento na proporção de espécies dominantes nas
plantações de eucalipto, conduziram a uma comunidade de morcegos mais pobre
em espécies e menos diversa nestas plantações. Assim, as espécies mais
sensíveis e com mais funções ecológicas, como Lophostoma brasiliense,
Platyrrhinus helleri, Sturnira tildae e Chrotopterus auritus, apenas estiveram
presentes nos fragmentos de floresta.
Danielsen & Heegaard (1995) obtiveram também uma maior dominância
para as comunidades de morcegos, num estudo de biodiversidade que realizaram

34
na Indonésia, em áreas desflorestadas e em plantações de borracha e plantações
de palmeiras. No entanto, eles salientam que resultados obtidos numa área
podem não ser necessariamente usados para prever os efeitos específicos das
espécies resultantes de perturbações de outra comunidade de morcegos.
Observou-se ainda que, espécies especialistas tendem a estar apenas
presentes nos fragmentos de floresta nativa (Cerrado) enquanto espécies
generalistas estão também presentes em plantações de eucaliptos. Assim, notou-
se a ausência de espécies florestais ecologicamente exigentes nas áreas de
plantações de eucalipto, como Lophostoma brasiliense, insectívoro que captura
as suas presas em folhas da copa ou no chão, no meio das árvores (Smith, 2008).
Barlow et al. (2007) salientam que plantações de eucaliptos falham como
promotores de conectividade para espécies especialistas de aves, quando estas
necessitam de cruzar extensas áreas. No entanto, segundo Gabriel et al. (2009) a
oferta de itens alimentares (insectos e néctar de Eucalyptus spp.), pelas
plantações aumenta a frequência de algumas espécies de aves, o que pode
aumentar a permeabilidade da matriz na paisagem fragmentada.
Existem vários factores que podem influenciar a presença das diferentes
espécies de morcegos, mamíferos, aves e insectos em plantações florestais como
o tempo da plantação, o clima e tipo de bioma onde se encontra inserida e a
presença ou ausência de sub-bosque.
Segundo Marsden et al. (2001) a riqueza de espécies de aves numa
plantação de Eucalyptus de 30 anos foi muito menor do que a encontrada para
uma plantação de 80 anos, onde a riqueza vegetal do sub-bosque existente era
maior.
Para Bernhard-Reversat (2001) o crescimento da vegetação rasteira em
plantações como as de eucalipto é favorecido pela presença de chuva elevada
em países como o Congo (precipitação média anual de 1250 mm), permitindo a
colonização por espécies que usem vegetação arbustiva. Por outro lado, Marsden
et al. (2001) afirmam que a pobreza de espécies em Eucalyptus pode ser
resultado de uma limpeza intensiva da vegetação do sub-coberto. Em plantios de
eucaliptos adjacentes à reserva de Sooretama (Espírito Santo), Marsden et al.
(2001) registaram apenas oito espécies de aves, justificando esse baixo número
pelo intenso manejo, que não permitiria o desenvolvimento de sub-bosque.
Portanto, o sub-bosque torna as plantações menos hostis, sendo atractivo para

35
diversas espécies, que podem utilizá-lo não apenas para deslocamento, mas
também como área de alimentação e abrigo.
Muitos estudos têm também ressaltado o valor de conservar fragmentos de
floresta nativa dentro das plantações. Em geral, quanto maior o fragmento de
floresta nativa presente na plantação, mais espécies de vertebrados, podem
suportar. Contudo, estes fragmentos nem sempre têm de ser grandes para serem
eficientes na conservação da biodiversidade (Lindenmayer et al., 2003). As
plantações de árvores têm o potencial de possibilitar um serviço de conservação
em muitos dos trópicos húmidos, desde que estes aumentem em tamanho
rapidamente e apresentem um contraste estrutural menor com a vegetação
nativa, que muitos outros usos do solo agrícolas mais intensivos (Barlow et al.,
2007b).
Apesar de ténue, a diferenciação na composição de espécies entre
Eucalipto e Floresta obtida era previsível, uma vez que se espera que as
comunidades de morcegos presentes em plantações de eucaliptos sejam mais
pobres em espécies e menos abundantes que as existentes em fragmentos de
floresta nativa. Era suposto que Floresta 4 e Eucalipto 4, se encaixassem nesta
diferenciação, não se tendo encontrado motivos aparentes para tal situação. No
entanto, segundo a ANOSIM realizada, não existem diferenças significativas
relativamente à composição de espécies entre ambos os tipos de habitat em
estudo. Esta ausência de diferenças significativas contradiz o esperado neste
trabalho, uma vez que se pressupõe que as comunidades de morcegos em
plantações de eucaliptos fossem mais afectadas.
Bernard & Fenton, (2007), que comparam a composição de espécies entre
os habitats de floresta e fragmentos cercados por vegetação do tipo savana na
Amazónia Central - Brasil, também não obtiveram uma separação evidente entre
os locais amostrados segundo a MDS, concluindo que as savanas parecem não
funcionar como uma barreira ecológica para os morcegos. Contudo, verificaram
que a composição de espécies difere significativamente entre a savana e as áreas
florestais, indicando que os morcegos poderam reconhecer estas como habitats
distintos de forrageio.

36
4.3 Modelação da fauna de morcegos com o ambiente
A fragmentação do habitat é uma característica dominante da paisagem
moderna (Ewers & Didham, 2006) e a resposta das espécies à fragmentação da
floresta pode variar fortemente de acordo com os seus requisitos ecológicos,
características da paisagem e variáveis locais (Bernard & Fenton, 2007).
Através da análise multivariada de permutações da variância (ADONIS)
verificou-se que das variáveis em análise (área, perímetro, proporção de
cobertura de mata, proporção de cobertura de eucalipto, número de fragmentos
de mata e número de fragmentos de eucalipto), a proporção de cobertura florestal
(eucalipto e floresta) é a única com influência na composição de espécies. Aliás,
esta era uma das variáveis que se esperava que tivesse influência na composição
de espécies, uma vez que se pressupõe que os habitats naturais comportam
comunidades de morcegos mais ricas, diversas e abundantes.
Wethered & Lawes (2005) que estudaram comunidades de aves em
florestas fragmentadas concluíram que é perigoso ignorar a influência da
interveniente matriz nos planos de conservação da biodiversidade. Para eles a
influência da matriz da paisagem, estrutura e área do habitat na riqueza de
espécies e na comunidade, é complexa e dependente de cada espécie.
Relativamente à variável área, os resultados são diferentes para vários
trabalhos e observa-se que estão muito dependentes de variáveis como o habitat
da matriz envolvente. Por exemplo, segundo o trabalho realizado por Fischer &
Lindenmeyer (2002a) na Austrália, não existe uma relação significativa entre as
espécies de aves e a área de fragmentos de plantações de eucaliptos numa
matriz de plantações de pinho (“radiata pine” Pinus radiata), mas é evidente uma
relação significativa entre as espécies e a área dos fragmentos numa matriz de
pasto. Estas diferenças podem-se dever à conectividade entre os habitats
permitida pela matriz da paisagem, como Estrada & Coates-Estrada (2002)
escrevem num trabalho realizado com morcegos no Brasil, onde verificaram que
os morcegos evitam uma matriz composta apenas por pasto.
À semelhança com outro trabalho (Faria, 2006), neste estudo também não
foi encontrado um efeito significativo do tamanho dos fragmentos na composição
de espécies. Faria (2006) escreve que muitas espécies de morcegos parecem
persistir à forte fragmentação da paisagem, devido à sua facilidade de explorar o
mosaico dos habitats modificados, na matriz e realizar movimentos de longa

37
distância. No entanto, Faria (2006) acrescenta que as espécies podem reagir de
diferentes formas às diversas componentes da matriz. Ao analizar a abundância
das espécies, Faria (2006) verificou que uma das espécies dominantes de floresta
(Artibeus obscurus) foi significativamente menos comum em pequenos
fragmentos comparativamente com os de maiores dimensões.
Apesar de neste trabalho não se ter verificado uma influência da variável
número de fragmentos na composição de espécies, para Zurita et al. (2006) a
presença de floresta nativa perto de plantações pode conduzir a um aumento da
riqueza de espécies, neste caso de aves, nas plantações arbóreas. No entanto,
olhando a paisagem da nossa área de estudo verifica-se um número de vários
pequenos e médios fragmentos que poderiam estar a contribuir para a riqueza
observada nos eucaliptais. Além disso, os morcegos que possuem boa
capacidade de locomoção e dispersão, poderam usar estas áreas de eucalipto
como rotas de vôo, já que estas são caracterizadas por um subosque limpo que
facilita estes deslocamentos.
A partir do citado anteriormente, pode-se deduzir que a ausência de
influência significativa das variáveis locais de paisagem na composição de
espécies pode dever-se à conectividade e proximidade entre os habitats
amostrados permitida pela matriz da paisagem. No entanto, deve-se realçar que
as espécies podem reagir de diferentes formas às diferentes componentes da
matriz, tendo-se verificado neste caso, uma menor diversidade e riqueza de
espécies em plantações de eucalipto.
Para Klingbeil & Willig (2009) a estrutura da paisagem pode ser
problemática, uma vez que é composta por características que escondem
respostas individuais e podem potencialmente apenas reflectir o padrão das
espécies dominantes.
Verificou-se que das três espécies analisadas, Carollia perspicillata,
Platyrrhinus lineatus e Artibeus planirostris, o factor habitat, apenas tem um efeito
significativo na última espécie mencionada. Esta diferença deve-se prender a
semelhanças vs diferenças morfológicas e padrões ecológicos das espécies em
questão, como alimentação, escolha de abrigos e tipo de habitat preferencial.
O uso do habitat por morcegos varia muito entre espécies, dependendo do
seu tamanho corporal e modo de forrageio. Geralmente espécies maiores são
esperadas de procurarem comida em habitats abertos enquanto espécies mais

38
pequenas são associadas a habitats mais densos. No entanto, o uso dos
diferentes habitats varia muito consoante a espécie (Loeb & O‟Keefe, 2006). Por
outro lado, espécies consideradas grandes como A. planirostris (Antebraço = 62-
73 mm; Peso = 55 g) provavelmente viajam distâncias maiores (>2,5 Km) durante
a noite em busca de alimento quando comparadas com espécies, por exemplo, do
género Carollia (Antebraço = 38-44 mm; Peso = 18 g), que realizam pequenos
voos em busca de alimento (Klingbeil & Willig, 2009).
A espécie Artibeus planirostris é abundante em habitats tipo cerrado, com
árvores dispersas e arbustos e uma componente de erva difundida, o cerradão,
constituído por grupos de muitas árvores, alguns arbustos, e pouca erva. A sua
presença parece associada a áreas florestadas, abrigando-se em árvores. No
cerrado, alimenta-se de frutos de Vismia, o que constituiu grande parte da sua
alimentação. Apesar de se alimentarem maioritariamente de frutos, podem ainda
consumir alguns artrópodes (Hollis, 2005; Zortéa, 2007).
A espécie Carollia perspicillata pode ser encontrada desde florestas
húmidas sempre verdes a florestas secas decíduas, sendo no entanto mais
comum em habitats de crescimento secundário (Cloutier & Thomas, 1992; Passos
et al., 2003). Trata-se de um frugívoro de sub-bosque, que se alimenta
primariamente de plantas de sucessão secundária como os representantes dos
géneros Piper, Ficus, Cecropia e Solanum. É considerada como uma espécie
generalista e no Brasil alimenta-se de pelo menos 28 espécies de frutos (Cloutier
& Thomas, 1992). Grande parte dos frutos da sua dieta são produzidos em
árvores baixas e arbustos, atribuindo-se uma classificação a este morcego de
frugívoro ao nível do solo. A sua presença parece associada a águas paradas e
áreas abertas. É dos morcegos mais comuns capturados ao nível do solo, onde
forrageia numa larga variedade de frutos, algumas flores e insectos e pode
abrigar-se em buracos de árvores (Cloutier & Thomas, 1992; Filho et al., 2007).
A espécie Platyrrhinus lineatus embora predominantemente frugívora,
alimenta-se também de insectos, néctar, pólen e folhas (Zortéa, 1993; Zortéa,
2007). Já foi observada a comer frutos dos géneros Vismia, Cecropia, Ficus e
Carica. Abriga-se em grutas e também sob a folhagem densa da floresta,
incluindo folhas de palmeira e outras plantas (Willig & Hollander, 1987; Reis et al.,
2007).

39
Por ser uma espécie de grande plasticidade ecológica e amplamente
distribuída, percebe-se que os tipos de habitat em estudo não tenham um grande
efeito na abundância de Carollia perspicillata. Pelo contrário, parece que as
plantações de eucaliptos não possuem as características necessárias para uma
maior presença de Artibeus planirostris. As plantações de eucaliptos são áreas
menos densas e com uma estrutura de vegetação mais simplificada,
comparativamente aos fragmentos de florestas, com menos recursos, de alimento
e abrigo, para esta espécie. Pensa-se que estas áreas de eucalipto sirvam, para
as espécies de morcegos capturadas e outras, de corredores ecológicos entre os
seus abrigos e as áreas de alimentação.
Segundo, Barros & Rui (2008), os dados obtidos através de detectores de
ultra-sons no sul do Brasil, indicam que a vegetação arbórea de grande porte
favorece a presença e actividade de quirópteros, mesmo quando se trata de
espécies vegetais exóticas.
O Teste de Mantel apontou que não existe relação entre as distâncias
geográficas dos vários fragmentos em estudo e a composição de espécies obtida.
No estudo de Klingbeil & Willig (2009), que comparam a abundância de espécies
entre os locais de amostra e as distâncias em linha recta entre os centros
geográficos destes locais, também não foi encontrada uma relação significativa.
Apesar destes resultados, acredita-se que plantações de eucaliptos em que
estejam presentes fragmentos de floresta nativa, aumente a riqueza de espécies,
como sugerido por Zurita et al. (2006).
5. Considerações Finais
Este estudo representa uma primeira tentativa de identificar as respostas
das comunidades de morcegos à conversão do habitat em plantações de
eucalipto e às características da paisagem daí resultantes, em cerrado no
sudoeste de Goiás.
Alguns dos resultados obtidos, através das várias análises estatísticas
efectuadas, são menos conclusivos relativamente ao que era esperado. Talvez
esta diferença se deva aos poucos indivíduos capturados no total do trabalho,
uma vez que das treze espécies amostradas apenas três apresentaram mais de

40
20 capturas. No entanto, é possível destacar que em termos de composição de
espécies verificamos importantes diferenças entre o hábitat natural (Cerrado) e as
monoculturas de eucalipto, com a floresta nativa com mais espécies e uma
comunidade mais complexa, mais diversa e menos dominante que o eucalipto.
Foi possível observar que as comunidades presentes em plantações de
eucaliptos apresentam uma estrutura/composição algo diferente daquelas
presentes em fragmentos de floresta nativa, uma vez que, como é de esperar, são
dominadas por espécies mais generalistas e de fácil adaptação a habitats
alterados. No entanto, supõe-se que estas plantações florestais poderiam ser
mais ricas se forem tidos em conta alguns detalhes, como a manutenção de um
subcoberto e a presença de fragmentos de floresta de adequado tamanho e
distância nestas plantações.
Apesar das plantações de Eucalyptus estarem longe de poderem substituir
a floresta nativa, é importante referir que as plantações de eucaliptos em estudos
não são “desertos biológicos”. De toda maneira, estudos complementares
deveriam ser conduzidos para verificar como os morcegos estão a utilizar as
áreas de eucalipto para se referir a estas monoculturas como habitats adequados
a estes.
As medidas de biodiversidade das comunidades de morcegos para os dois
tipos de habitat, não responderam por si só às características da paisagem,
tornando-se evidente a necessidade de perceber a resposta de cada espécie a
esta alteração do habitat nativo.
Os resultados apresentados aqui fornecem evidências do empobrecimento
das comunidades de morcegos em habitats antropizados relacionados com a
deflorestação e a criação de plantações arbóreas de eucalipto. Sugere-se que as
medidas de conservação deveriam ser preferencialmente voltadas para a
preservação do habitat a nível local e em segundo plano focarem-se na matriz da
paisagem.
Como trabalho para um futuro próximo, sugeria-se realizar colectas na
estação seca (Maio a Setembro) e aumentar o esforço de amostragem. Isto é
importante para avaliar se a sazonalidade tem alguma influência nos padrões
observados neste estudo realizado na estação chuvosa. Além disso, o aumento
do esforço iria gerar mais dados para examinar as respostas de diferentes
espécies, entre as diferentes estações e perceber o que acontece neste tipo de

41
habitat e consequentemente permitirem produzir linhas de gestão e acções de
conservação. Seria ainda importante perceber a resposta de cada espécie à
conversão do habitat nativo em plantações de eucalipto.

42
6. Agradecimentos
Primariamente quero agradecer aos meus pais, Rosa Muñoz e José Pina,
que como sempre me apoiaram e encorajaram na realização deste trabalho.
Quero agradecer aos meus orientadores, Christoph Meyer pelo material,
atenção e ajuda despendida no trabalho, e Marlon Zortéa por ter primariamente
aceite ser meu orientador no Brasil e por ter facilitado todas as condições para a
minha chegada, adaptação, estadia, trabalho e partida do Brasil.
Não posso deixar de agradecer também, à Universidade Federal de Goiás
e ao Professor Fabiano Melo que disponibilizou juntamente com o orientador
Marlon Zortéa os recursos para a realização do trabalho de campo.
Agradeço aos proprietários das fazendas pela permissão de utilização das
áreas de estudo.
Aos colegas e amigos Vinycio Carrijo, Diego Afonso, Daniel Santos, Paola
Da Mata, Wagner Filho, Felipe Zenha, Marluci Fachi, Karen Almeida entre muitos
outros, que me ajudaram no campo e me receberam de braços abertos em Jataí,
aqui fica um muito obrigado.
Quero ainda agradecer à D. Maria De Carvalho e seus familiares que me
acolheram em sua casa e à minha amiga Karla Gomes que tanto me incentivou
para este desafio.
Por último agradeço a uma pessoa muito especial pelo seu apoio, carinho,
atenção e mão-de-obra, Tiago Dores.

43
7. Bibliografia
Aguiar, L. M. S. & Zortéa, M. 2008. A diversidade de morcegos conhecida
para o cerrado. IX Simpósio Nacional Cerrado. II Simpósio Internacional Savanas
Tropicais. Brasília.
ABRAF. 2011. Anuário estatístico da ABRAF ano base 2010. ABRAF –
Associação Brasileira de Produtores de Florestas Plantadas. Brasília, 130 p.
Aguirre, L. F. 2002. Structure of a Neotropical savana bat community.
Journal of Mammalogy, 83: 775 - 784.
Barlow, J., Araujo, I. S., Overal, W. L., Gardner, T. A., Mendes, F. S., Lake,
I. R. & Peres, C. A. 2007b. Diversity and composition of fruit-feeding butterflies in
tropical Eucalyptus plantations. Biodiversity Conservation. Springer.
Barlow, J., Gardner, T. A., Araujo, I. S., Ávila-Pires, T. C., Bonaldo, A. B.,
Costa, J. E., Esposito, M. C., Ferreira, L. V., Hawes, J., Hernandez, M. I. M.,
Hoogmoed, M. S., Leite, R. N., Lo-Man-Hung, N. F., Malcolm, J. R., Martins, M.
B., Mestre, L. A. M., Miranda-Santos, R., Nunes-Gutjahr, A. L., Overal, W. L.,
Parry, L., Peters, S. L., Ribeiro-Junior, M. A., da Silva, M. N. F., da Silva Motta, C.
& Peres C. A. 2007. Quantifying the biodiversity value of tropical primary,
secondary, and plantation forests. PNAS, 104 (47): 18555 - 18560.
Baptista, M. & Mello, M. A. R. 2001. Preliminary inventory of the bat species
of the Poço das Antas Biological Reserve, Rio de Janeiro. Chiroptera Neotropical,
7 (1-2): 133 - 135.
Barros, M. A. S. & Rui, A. M. 2008. Monitoramento de Quirópteros, pp: 2-
31. In: Magalhães, R. G. (Coordenador Geral). Relatório Final do Monitoramento
Ambiental Pré-implantação do Parque Eólico de Palmares. Palmares do Sul (RS),
159 p.

44
Bates, D., Maechler, M. & Bolker, B. 2011. Lme4: Linear mixed-effects
models using S4 classes. R package version 0.999375-39. http://CRAN.R-
project.org/package=lme4
Bernard, E. 2002. Diet, activity and reproduction of bat species (Mammalia,
Chiroptera) in Central Amazonia, Brasil. Revista Brasileira de Zoologia, 19 (1): 173
- 188.
Bernard, E., Albernaz, A. L. K. M. & Magnusson, W. E. 2001. Bat species
composition in three localities in the Amazon Basin. Studies on Neotropical Fauna
and Environment, 36 (3): 177 -184.
Bernard, E. & Fenton, M. B. 2007. Bats in a fragmented landscape: species
composition, diversity and habitat interactions in savanas of Santarém, Central
Amazonia, Brazil. Biological Conservation, 134: 332 - 343.
Bernhard-Reversat, F. 2001. Effect of exotic tree plantations on plant
diversity and biological soil fertility in the Congo Savanna: with special reference to
Eucalypts. Center for International Forestry Research, Indonesia.
Bianconi, G. V., Mikich, S. B. & Pedro, W. A. 2006. Movements of bats
(Mammalia, Chiroptera) in Atlantic Forest remnants is southern Brazil. Revista
Brasileira de Zoologia, 23 (4): 1199 – 1206.
Bobrowiec, P. E. D. & Gribel, R. 2010. Effects of different secondary
vegetation types on bat community composition in Central Amazonia, Brazil.
Animal Conservation, 13: 204-216.
Borkin, K. M. & Parsons, S. 2010. The importance of exotic plantation forest
for the New Zealand long-tailed bat (Chalinolobus tuberculatus). New Zealand
Journal of Zoology, 37 (1): 35 – 51.
Borsboom, A. C., Lees, J. W. N., Mathieson, M. & Hogan, L. 2002.
Measurement and integration of fauna biodiversity values in Queensland

45
agroforestry systems. A report for the RIRDC/ Land and Water Australia/
FWPRDC Joint Venture Agroforestry Program Suppoted by the Natural Heritage
Trust. Rural Industries Research and Development Corporation, 02 (044).
Brose, U. & Martinez, N. D. 2004. Estimating the richness of species with
variable mobility. Oikos, 105: 292 - 300.
Brosset, A., Charles-Dominique, P., Cockle, A., Cosson, J. F. & Masson, D.
1996. Bat communities and deforestation in French Guiana. Canadian Journal of
Zoology, 74: 1974 - 1982.
Castro-Arellano, I., Presley, S. J., Saldanha, L. N., Willig, M. R. & Wunderle
Jr., J. M. 2007. Effects of reduced impact logging on bat biodiversity in terra firme
forest of lowland Amazonia. Biological Conservation, 138: 269-285.
Clarke, K. R. 1993. Non-parametric multivariate analyses of changes in
community structure. Australian Journal of Ecology, 18: 117 - 143.
Clarke, F. M., Rostant, L. V. & Racey, P. A. 2005. Life after logging: post-
logging recovery of a neotropical bat community. Journal of Applied Ecology, 42:
409 - 420.
Cloutier, D. & Thomas, D. W. 1992. Carollia perspicillata. Mammalian
Species, 417: 1 - 9.
Colwell, R. K. 2009. EstimateS: Statistical estimation of species richness
and shared species from samples. Version 8.2. User's Guide and application
published at: ttp://purl.oclc.org/estimates.
Danielsen, F. & Heegaard, M. 1995. Impact of logging and plantation
development on species diversity: a case study from Sumatra. Pg. 73-92. In:
Sandbukt (ed.) Management of tropical forests: towards an integrated perspective.
Oslo: Centre for Development and the Environment, University of Oslo.

46
Davis, W.B. 1980. New Sturnira (Chiroptera: Phyllostomidae) from Central
and South America, with key to currently recognized species. Occasional Papers,
Museum of Texas, Tech University, 70: 1 - 5.
Dias, D. & Peracchi, A. L. 2008. Quirópteros da Reserva Biológica do
Tinguá, Estado do Rio de Janeiro, sudeste do Brasil (Mammalia: Chiroptera).
Revista Brasileira de Zoologia, 25 (3): 401-407.
Eiten, G. 1994. Vegetação do Cerrado. Pp. 17-73. In: Ferreira, L. G.,
Ferreira, M. E., Rocha, G. F., Nemayer, M., Ferreira, N. C. 2009. Dinâmica
agrícola e desmatamentos em áreas de cerrado: uma análise a partir de dados
censitários e imagens de resolução moderada. Revista Brasileira de Cartografia,
61 (02): 117-127.
Esbérard, C. E. L. 2003. Diversidade de morcegos em área de Mata
Atlântica regenerada no Sudeste do Brasil. Revista Brasileira de Zoociências, 5
(2): 189-211.
Estrada, A. & Coates-Estrada, R. 2002. Bats in continuous forest, forest
fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico.
Biological Conservation, 103: 237 - 245.
Ewers, R. M. & Didham, R. K. 2006. Confounding factors in the detection of
species responses to habitat fragmentation. Biological Reviews, 81: 117 - 142.
Faria, D. 2006. Phyllostomid bats of a fragmented landscape in the north-
eastern Atlantic forest, Brazil. Journal of Tropical Ecology, 22: 531-542.
Faria, D., Laps, R. R., Baumgarten, J. & Cetra, M. 2006. Bat and bird
assemblages from forests and shade cacao plantations in two contrasting
landscapes in the Atlantic Forest of Southern Bahia, Brazil. Biodiversity and
Conservation, 15: 587 – 612.

47
Felfili, J. M., Júnior, M. C. S., Sevilha, A. C., Fagg, C. W., Walter, B. M. T.,
Nogueira, P. E. & Rezende, A. V. 2004. Diversity, floristic and structural patterns
of cerrado vegetation in central Brazil. Plant Ecology, 175: 37- 46.
Felfili, J. M., Sousa-Silva, J. C. & Scariot, A. 2005. Biodiversidade, ecologia
e conservação do Cerrado: avanços no conhecimento. In: Scariot, A., Sousa-
Silva, J. C. & Felfili, J. M. (eds). Cerrado: ecologia, biodiversidade e conservação.
Ministério do Meio Ambiente, Brasília. Pp. 25 -44.
Fenton, M. B., Acharya, L., Audet, D., Hickey, M. B. C., Merriman, C.,
Obrist, M. K. & Syme, D. M. 1992. Phyllostomid bats (Chiroptera: Phyllostomidae)
as indicators of habitat disruption in the Neotropics. Biotropica, 24: 440 – 446.
Ferreira, L. G., Ferreira, M. E., Rocha, G. F., Nemayer, M. & Ferreira, N. C.
2009. Dinâmica agrícola e desmatamentos em áreas de cerrado: uma análise a
partir de dados censitários e imagens de resolução moderada. Revista Brasileira
de Cartografia, 61 (02): 117-127.
Ferrel, C. S. & Wilson, D. E. 1991. Platyrrhinus helleri. Mammalian Species,
372: 1-5.
Filho, H. O., Lima, I. P. & Fogaça, F. N. O. 2007. Subfamília Carolliinae. In:
Reis, N. R., Peracchi, A. L., Pedro, W. A. & Lima I. P. (eds.). Morcegos do Brasil.
Londrina. Pp. 99-105.
Filloy, J., Zurita, G. A., Corbelli, J. M., Bellocq, M. I. 2010. On the similarity
among bird communities: Testing the influence of distance and land use. Acta
Oecologica, 36: 333 – 338.
Fischer, J. & Lindenmayer, D. B. 2002a. Small patches can be valuable for
biodiversity conservation: two case studies on birds in southeastern Australia.
Biological Conservation, 106: 129 – 136.

48
Gabriel, V. A., Ranieri, S. B. L. & de Abreu Bovo, A. A. 2009. Avifauna em
plantios de eucalipto e em fragmentos de cerradão no Município de Três Lagoas,
Mato Grosso do Sul, Brasil. Anuais do III Congresso Latino Americano de
Ecologia, São Lourenço – MG.
Gardner, A.L. 2007. Mammals of South America. Vol. 1- Marsupials
Xenarthrans Shrews and Bats. The University of Chicago Press, Chicago and
London, 669 p.
Giannini, N. P. & Kalko, E. K. V. 2004. Trophic structure in a large
assemblage of phyllostomid bats in Panama. Oikos, 105 (2): 209-220.
Gonçalves, E. & Gregorin, R. 2004. Quirópteros da estação ecológica da
Serra das Araras, Mato Grosso do Sul, Brasil, com o primeiro registro de Artibeus
gnomus e A. anderseni para o cerrado. International Journal of Biodiversity, 1
(5):143 - 149.
Gorresen, P. M. & Willig, M. R. 2004. Landscape responses of bats to
habitat fragmentation in atlantic forest of Paraguay. Journal of Mammalogy, 85 (4):
688 – 697.
Gotelli, N. J. & Colwell, R. K. 2001. Quantifying biodiversity: procedures and
pitfalls in the measurement and comparison of species richness. Ecology
Letters, 4: 379 - 391.
Gotelli, N. J. & Entsminger, G. L. 2001. EcoSim: Null models software for
ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. Available at:
http://homepages.together.net/ ~gentsmin/ecosim.htm.
Greenhall, A. M., Joermann, G., Schmidt, U. & Seidel, M. R. 1983.
Desmodus rotundus. Mammalian Species, 202: 1 - 6.
Gregorin, R., Gonçalves, E., Aires, C. C. & Carmignotto., A. P. 2011.
Morcegos (Mammalia: Chiroptera) da Estação Ecológica Serra Geral do

49
Tocantins: composição específica e considerações taxonômicas. Biota
Neotropica, 11 (1): 299 – 311.
Handley, C.O., Jr. 1987. New species of mammals from northern South
America: fruit-eating bats, genus Artibeus Leach. Fieldiana Zoology, Field
Museum of Natural History, 39: 163-172.
Harvey, C. A. & Villalobos, J. A. G. 2007. Agroforestry systems conserve
species-rich but modified assemblages of tropical birds and bats. Biodiversity and
Conservation, 16: 2257 - 2292.
Hollis, L. 2005. Artibeus planirostris. Mammalian Species, 775: 1 - 6.
Hsu, T., French, K. & Major, R. 2010. Avian assemblages in eucalypt
forests, plantations and pastures in northern NSW, Australia. Forest Ecology and
Management, 260: 1036 – 1046.
Jones, G., Jacobs, D. S., Kunz, T. H., Willig, M. R. & Racey, P. A. 2009.
Carpenoctem: the importance of bats as bioindicators. Endangered Species
Research, 8: 93 - 115.
Kalko, E. K. V. 1998. Organisation and diversity of tropical bat communities
through space and time. Zoology, 101: 281 - 297.
Kalko, E. K. V. & Handley, C. O. Jr. 2001. Neotropical bats in the canopy:
diversity, community structure, and implications for conservation. Plant Ecology,
153: 319 – 333.
Klingbeil, B. T. & Willig, M. R. 2009. Guild-specific responses of bats to
landscape composition and configuration in fragmented Amazonian rainforest.
Journal of Applied Ecology, 46: 203 - 213.
Klink, C. A. & Machado, R. B. 2005. A conservação do Cerrado brasileiro.
Megadiversidade, 1 (1): 147-155.

50
Klink, C. A. & Machado, R. B. 2005. Conservation of the Brazilian Cerrado.
Conservation Biology, 19 (3): 707-713.
KÖppen, W. 1948. Climatologia: um estudo dos climas da Terra. México.
Fundo de Cultura Económica. 478 p.
Kunz, T. H. & Parsons, S. 2009. Ecological and behavioral methods for the
study of bats. 2nd edn. The Jones Hopkins University Press. 533 p.
Lande, R. 1996. Statistics and partitioning of species diversity, and similarity
among multiple communities. Oikos, 76: 5 – 13.
Lantschner, M. V., Rusch, V. & Peyrou, C. 2008. Bird assemblages in pine
plantations replacing native ecosystems in NW Patagonia. Biodiversity and
Conservation, 17: 969 – 989.
Laval, R. K. 1973. A revision of the Neotropical bats of the genus Myotis.
Bulletin of Natural History Museum, Los Angeles County, 15: 1 - 54.
Lim, B. K. & Engstrom, M. D. 2000. Preliminary survey of bats from the
upper Mazaruni of Guyana. Chiroptera Neotropical, 6 (1 - 2): 119-128.
Lindenmayer, D. B., Hobbs, R. J. & Salt, D. 2003. Plantation forests and
biodiversity conservation. Australian Forestry, 66 (1): 62 - 66.
Lobova, T.A., Geiselman, C.K. & Mori, S. A. 2009. Seed dispersal by bats in
the Neotropics. New York Botanical Garden Press, New York.
Loeb, S. C. & O‟Keefe, J. M. 2006. Habitat use by forest bats in South
Carolina in relation to local, stand, and landscape characteristics. Journal of
Wildlife Management, 70 (5): 1210 – 1218.

51
Machado, R. B., Neto, M. R., Pereira, P. P., Caldas, E. F., Gonçalves, D.
A., Santos, N. S., Tabor, K. & Steininger, M. 2004. Estimativas de perda da área
do Cerrado brasileiro. Conservação Internacional. Brasília. Brasil, 23 p.
Magurran, A. E. 2004. Measuring biological diversity. Blackwell Publishing.
Oxford, UK.
Mangolin, R., Motta, A. G., Esbérard, E. L. & Bergallo, H. G. 2007. Novos
resgistros de Lophostoma brasiliensis Peters para o sudoeste do Brasil
(Mammalia, Chiroptera, Phyllostomidae). Revista Brasileira de Zoociências, 9 (1):
225 - 228.
Marinho-Filho, J. S. 1996. The Brazilian Cerrado Bat Fauna and its
conservation. Chiroptera Neotropical, 2 (1): 37 – 39.
Marques-Aguiar, S. A., Aguiar, G. F. S., Saldanha, L. N., Rocha, M. M. B.,
Fonseca, R. T. A., Reis-Filho, V. O. 2009. Diversidade de morcegos (Mammalia:
Chiroptera) no Baixo Rio Xingu, Estado do Pará. Anais do IX Congresso de
Ecologia do Brasil, São Lourenço – MG.
Marsden, S. J., Whiffin, M. & Galetti, M. 2001. Bird diversity and abundance
in forest fragments and Eucalyptus plantations around an Atlantic forest reserve,
Brazil. Biodiversity and Conservation, 10: 737 – 751.
McArdle, B.H. & M.J. Anderson. 2001. Fitting multivariate models to
community data: A comment on distance-based redundancy analysis. Ecology, 82:
290 – 297.
McCune, B. & Grace, J. B. 2002. Analysis of ecological communities. MjM
Software, Gleneden Beach, OR.
Melo, A. S. 2008. O que ganhamos „confundido‟ riqueza de espécies e
equabilidade em um índice de diversidade?. Biota Neotropica, 8 (3): 21 - 27.

52
Meyer, C. F. J. & Kalko, E. K. V. 2008. Assemblage-level responses of
phyllostomid to tropical forest fragmentation: land-bridge islands as a model
system. Journal of Biogeography, 35: 1711 - 1726.
Morrison, D. W. 1978. Lunar phobia in a neotropical bat, Artibeus
jamaicensis (Chiroptera: Phyllostomidae). Animal Behaviour, 26: 852 - 855.
Nogueira, M. R., Peracchi, A. L. & Moratelli, R. 2007. Subfamília
Phyllostominae. In: Reis, N. R., Peracchi, A. L., Pedro, W. A. & Lima I. P. (eds.).
Morcegos do Brasil. Londrina. Pp. 61-97.
Passos, F. C., Silva, W. R., Pedro, W. A. & Bonin, M. R. 2003. Frugivoria
em morcegos (Mammalia, Chiroptera) no Parque Estadual Intervales, sudeste do
Brasil. Revista Brasileira de Zoologia, 20: 511 - 517.
Pereira, J. A. A. & Oliveira, C. A. 2005. Efeitos do Eucalyptus
camaldulensis sobre a colonização mecorrízica e a nodulação em Dimorphandra
mollis e Stryphnodendron adstringens, em Brasilândia, Minas Gerais. Cerne, 11
(4): 409-415.
Reis, N. R., Peracchi, A. L., Pedro, W. A. & Lima, I. P. 2007. Morcegos do
Brasil. Londrina, Brasil. 253p.
Reis, N. R., Peracchi, A. L., Lima, I. P. & Pedro, W. A. 2006. Riqueza de
espécies de morcegos (Mammalia, Chiroptera) em dois diferentes habitats, na
região centro-sul do Paraná, sul do Brasil. Revista Brasileira de Zoologia, 23 (3):
813-816.
Rex, K., Kelm, D. H., Wiesner, K., Kunz, T. H. & Voigt, C. C. 2008. Species
richness and structure of three Neotropical bat assemblages. Biological Journal of
Linnean Society, 94: 617 – 619.
Rodrigues, F. H., Silveira, L., Jácomo, A. T., Carmignotto, A. P., Bezerra, A.
M., Coelho, A. C., Garbogini, H., Pagnozzi, J. & Hass, A. 2002. Composição e

53
caracterização da fauna de mamíferos do Parque Nacional das Emas, Goiás,
Brasil. Revista Brasileira de Zoologia, 19 (2): 589 - 600.
Salgado-Labouriau, M. L. 2005. Alguns aspectos sobre a Paleoecologia
dos Cerrados. In: Scariot, A., Sousa-Silva, J. C. & Felfili, J. M. (eds). Cerrado:
ecologia, biodiversidade e conservação. Ministério do Meio Ambiente, Brasília.
Pp. 109 - 118.
Schipper, J., Chanson, J.S., Chiozza, F., Cox, N.A. & Hoffmann, M. 2008.
The status of the World's land and marine mammals: diversity, threat, and
knowledge. Science, 322: 225 - 230.
Silva, F. B., Assis, M. L., Rocha, I. R., Lima, O. M., Carvalho, L. A. A. 2009.
Urbanização e impermeabilização do solo: etapas de um processo erosivo nas
proximidades da Mata do Açude, Jataí/GO. XI – EREGEO – Simpósio Regional
de Geografia. Jataí.
Simmons, N. B. 2005. Order Chiroptera. Mammal Species of the World: A
Taxonomic and Geographic Reference. D.E. Wilson & D.M. Reeder (eds). Johns
Hopkins University Press. Pp. 312 - 529.
Smith, P. 2008. Pygmy round-eared bat Lophostoma brasiliense. Mammals
of Paraguay. 28: 1 – 6.
Stevens, R. D. & Willig, M. R. 2000. Community structure, abundance, and
morphology. OIKOS, 88: 48 - 56.
Stirling, G. & Wilsey, B. 2001. Empirical relationships between Species
Richness, Eveness, and Proportional Diversity. The American Naturalist, 158 (3):
286 - 299.
Straube, F. C. & Bianconi, G. V. 2002. Sobre a Grandeza e a unidade
utilizada para estimar esforço de captura com utilização de redes-de-neblina.
Chiroptera Neotropical, 8 (1-2): 150 - 152.

54
Umetsu, F. & Pardini, R. 2007. Small mammals in a mosaico of forest
remnants and anthropogenic habitats – evaluating matrix quality in na Atlantic
forest landscape. Landscape Ecology, 22: 517 – 530.
Vizotto, L. D. & Taddei, V.A. 1973. Chave para determinação de
quirópteros brasileiros. Publicação da Faculdade de Filosofia, Ciências e Letras
de São José do Rio Preto, São José do Rio Preto, 72 p.
Wethered, R. & Lawes, M. J. 2005. Nestedness of bird assemblages in
fragmented Afromontane forest: the effect of plantation forestry in the matrix.
Biological Conservation, 123: 125 – 137.
Willig, M. R. & Hollander, R. R. 1987. Vampyrops lineatus. Mammalian
Species, 275: 1 - 4.
Willig, M. R., Presley, S. J., Bloch, C. P., Hice, C. L., Yanoviak, S. P., Díaz,
M. M., Chauca, L. A., Pacheco, V. & Weaver, S. C. 2007. Phyllostomid bats of
Lowland Amazonia: effects of habitat alteration on abundance. Biotropica, 39 (6):
737 - 746.
Wilson, D. E., Ascorra, C. F., Solari, S. 1996. Bats as indicators of habitat
disturbance, pp. 613 - 625. In: Wilson, D. E. & Sandoval, A. (eds). Manu: The
biodiversity of southeastern Peru. Washington, Office of biodiversity programs.
National Museum of Natural History, Smithsonian Institution, 365p.
Zapala, M. A. & Schork, N. J. 2006. Multivariate regression analysis of
distance matrices for testing associations between gene expression patterns and
related variables. Proceedings of the National Academy of Sciences of the United
States of America, 103 (51): 19430 – 19435.
Zortéa, M. 1993. Folivory in Platyrrhinus (Vampyrops) lineatus (Chiroptera:
Phyllostomidae). Bat Research News, 39 (3 - 4): 59-60.

55
Zortéa, M. 2007. Subfamília Stenodermatinae. In: Reis, N. R., Peracchi, A.
L., Pedro, W. A. & Lima, I. P. (eds.). Morcegos do Brasil. Londrina. Pp. 107-127.
Zortéa, M. & Alho, C. J. R. 2008. Bat diversity of a Cerrado habitat in central
Brazil. Biodiversity Conservation. Springer.
Zortéa, M., de Melo, F. R., Carvalho, J. C. & Rocha, Z. D. 2010. Morcegos
da Bacia do rio Corumbá, Goiás. Chiroptera Neotropical, 16 (1): 611 - 617.
Zuur, A. F., Ieno, E. N. & Smith, G. M. 2007. Analysing Ecological Data.
Springer. 672 p.
Zuur, A. F., Ieno, E. N., Walker, N. J., Saveliev, A. A. & Smith, G. M. 2009.
Mixed effects models and extensions in ecology with R. Springer, New York.
Zurita, G. A., Rey, N., Varela, D. M., Villagra, M. & Bellocq, M. I. 2006.
Conversion of the Atlantic Forest into native and exotic tree plantations: Effects on
bird communities from the local and regional perspectives. Forest Ecology and
Management, 235: 164 - 173.
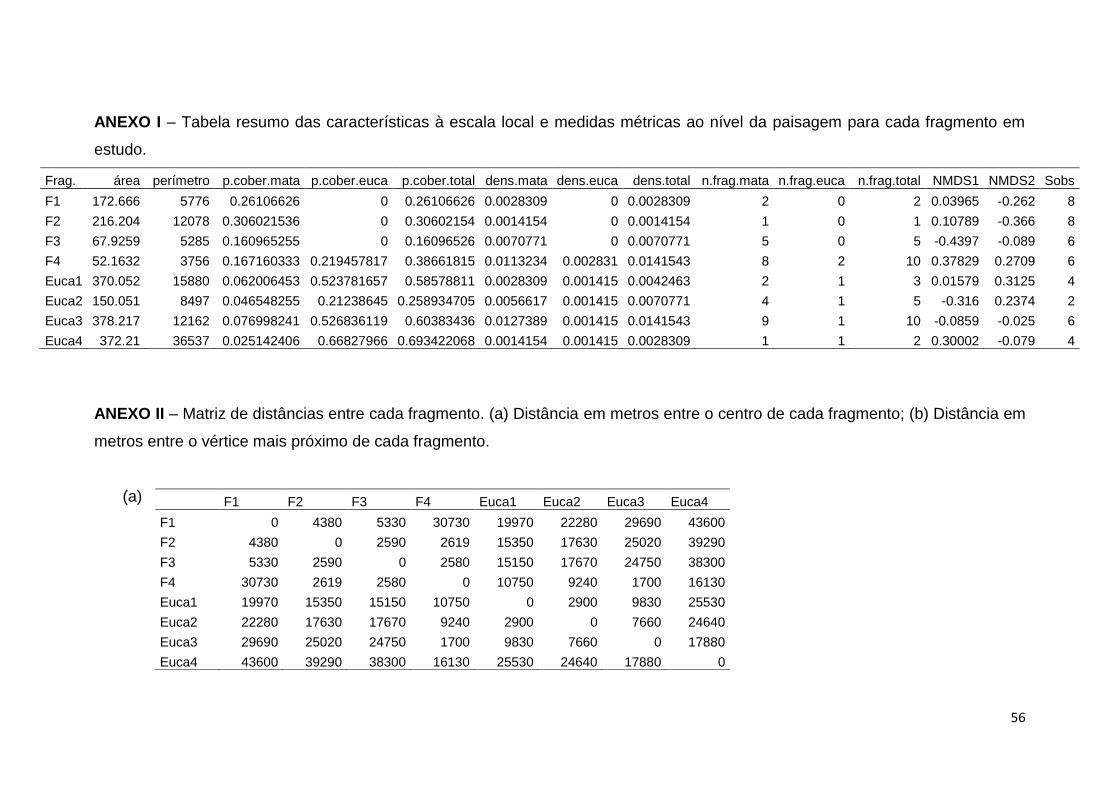
56
ANEXO I – Tabela resumo das características à escala local e medidas métricas ao nível da paisagem para cada fragmento em
estudo.
Frag. área perímetro p.cober.mata p.cober.euca p.cober.total dens.mata dens.euca dens.total n.frag.mata n.frag.euca n.frag.total NMDS1 NMDS2 Sobs
F1 172.666 5776 0.26106626 0 0.26106626 0.0028309 0 0.0028309 2 0 2 0.03965 -0.262 8
F2 216.204 12078 0.306021536 0 0.30602154 0.0014154 0 0.0014154 1 0 1 0.10789 -0.366 8
F3 67.9259 5285 0.160965255 0 0.16096526 0.0070771 0 0.0070771 5 0 5 -0.4397 -0.089 6
F4 52.1632 3756 0.167160333 0.219457817 0.38661815 0.0113234 0.002831 0.0141543 8 2 10 0.37829 0.2709 6
Euca1 370.052 15880 0.062006453 0.523781657 0.58578811 0.0028309 0.001415 0.0042463 2 1 3 0.01579 0.3125 4
Euca2 150.051 8497 0.046548255 0.21238645 0.258934705 0.0056617 0.001415 0.0070771 4 1 5 -0.316 0.2374 2
Euca3 378.217 12162 0.076998241 0.526836119 0.60383436 0.0127389 0.001415 0.0141543 9 1 10 -0.0859 -0.025 6
Euca4 372.21 36537 0.025142406 0.66827966 0.693422068 0.0014154 0.001415 0.0028309 1 1 2 0.30002 -0.079 4
ANEXO II – Matriz de distâncias entre cada fragmento. (a) Distância em metros entre o centro de cada fragmento; (b) Distância em
metros entre o vértice mais próximo de cada fragmento.
(a)
F1 F2 F3 F4 Euca1 Euca2 Euca3 Euca4
F1 0 4380 5330 30730 19970 22280 29690 43600
F2 4380 0 2590 2619 15350 17630 25020 39290
F3 5330 2590 0 2580 15150 17670 24750 38300
F4 30730 2619 2580 0 10750 9240 1700 16130
Euca1 19970 15350 15150 10750 0 2900 9830 25530
Euca2 22280 17630 17670 9240 2900 0 7660 24640
Euca3 29690 25020 24750 1700 9830 7660 0 17880
Euca4 43600 39290 38300 16130 25530 24640 17880 0
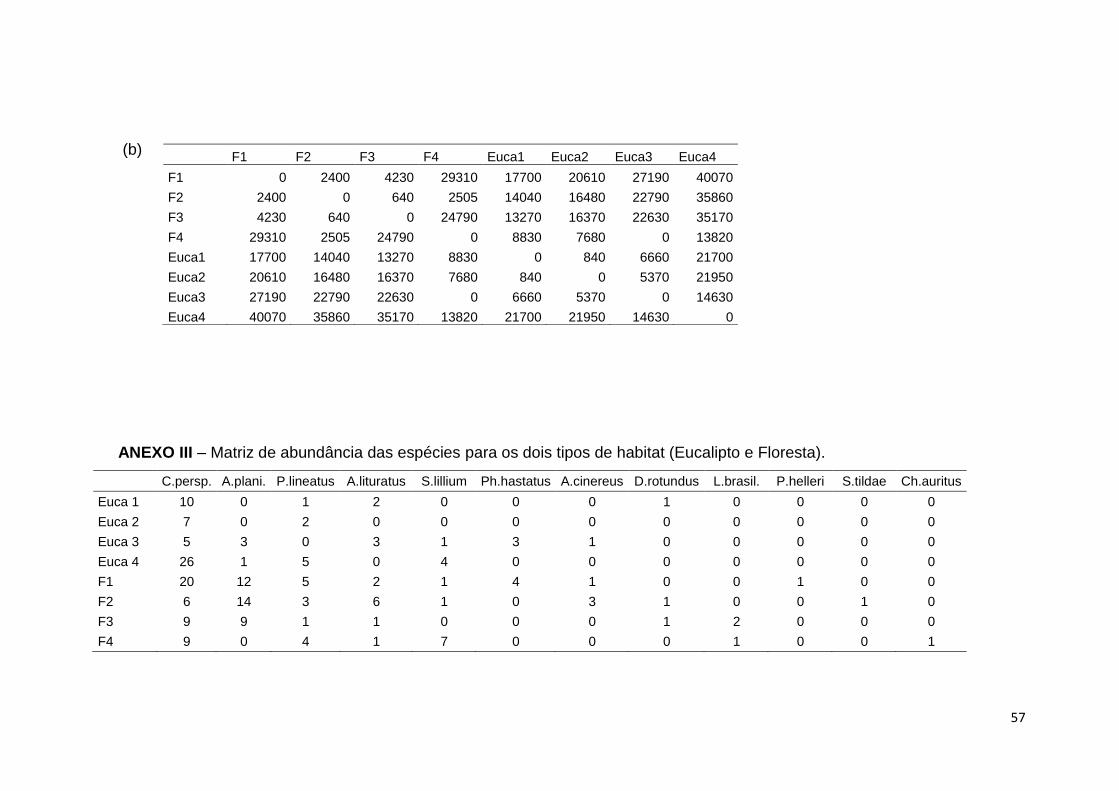
57
(b)
ANEXO III – Matriz de abundância das espécies para os dois tipos de habitat (Eucalipto e Floresta).
C.persp. A.plani. P.lineatus A.lituratus S.lillium Ph.hastatus A.cinereus D.rotundus L.brasil. P.helleri S.tildae Ch.auritus
Euca 1 10 0 1 2 0 0 0 1 0 0 0 0
Euca 2 7 0 2 0 0 0 0 0 0 0 0 0
Euca 3 5 3 0 3 1 3 1 0 0 0 0 0
Euca 4 26 1 5 0 4 0 0 0 0 0 0 0
F1 20 12 5 2 1 4 1 0 0 1 0 0
F2 6 14 3 6 1 0 3 1 0 0 1 0
F3 9 9 1 1 0 0 0 1 2 0 0 0
F4 9 0 4 1 7 0 0 0 1 0 0 1
F1 F2 F3 F4 Euca1 Euca2 Euca3 Euca4
F1 0 2400 4230 29310 17700 20610 27190 40070
F2 2400 0 640 2505 14040 16480 22790 35860
F3 4230 640 0 24790 13270 16370 22630 35170
F4 29310 2505 24790 0 8830 7680 0 13820
Euca1 17700 14040 13270 8830 0 840 6660 21700
Euca2 20610 16480 16370 7680 840 0 5370 21950
Euca3 27190 22790 22630 0 6660 5370 0 14630
Euca4 40070 35860 35170 13820 21700 21950 14630 0


















