
Jardim Botânico do Rio de Janeiro - Byrsonima sericea ......fragmentos de diferentes tamanhos na...
52
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical Biologia reprodutiva de Byrsonima sericea (Malpighiaceae), em fragmentos de diferentes tamanhos na Restinga de Massambaba, Arraial do Cabo, Rio de Janeiro Bernardo Souza Dunley 2006
Transcript of Jardim Botânico do Rio de Janeiro - Byrsonima sericea ......fragmentos de diferentes tamanhos na...

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Biologia reprodutiva de Byrsonima sericea (Malpighiaceae),
em fragmentos de diferentes tamanhos na Restinga de Massambaba, Arraial do Cabo, Rio de Janeiro
Bernardo Souza Dunley
2006

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Biologia reprodutiva de Byrsonima sericea (Malpighiaceae), em fragmentos de diferentes tamanhos na Restinga de
Massambaba, Arraial do Cabo, Rio de Janeiro
Bernardo Souza Dunley
Dissertação apresentada ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Botânica. Orientador: Dr. Leandro Freitas
Rio de Janeiro 2006

Biologia reprodutiva de Byrsonima sericea (Malpighiaceae), em fragmentos de diferentes tamanhos na Restinga de Massambaba,
Arraial do Cabo, Rio de Janeiro
Bernardo Souza Dunley
Dissertação submetida ao corpo docente da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários para a obtenção do grau de Mestre.
Aprovada por: Prof. Dr. Leandro Freitas (Orientador) ______________________ Profa. Dra. Dorothy Sue Dunn de Araujo ______________________ Prof. Dr. Marcelo Trindade Nascimento ______________________ em __/__/2006
Rio de Janeiro 2006

Dunley, Bernardo Souza.
D921 Biologia reprodutiva de Byrsonima sericea (Malpighiaceae) em fragmentos de diferentes tamanhos na Restinga de Massambaba, Arraial do Cabo, Rio de Janeiro / Bernardo Souza Dunley. – Rio de Janeiro, 2006.
xi, 41 f. : il. ; 28 cm. Dissertação (mestrado) – Instituto de Pesquisas Jardim Botânico
do Rio de Janeiro/Escola Nacional de Botânica Tropical, 2006. Orientador: Leandro Freitas. Banca examinadora: Dorothy Sue Dunn de Araujo, Marcelo
Trindade Nascimento. Bibliografia.
1. Byrsonima sericea. 2. Malpighiaceae. 3. Fragmentação de habitat. 4. Biologia reprodutiva. 5. Restinga de Massambaba. 6. Galhas. 7. Abelhas solitárias 8. Arraial do Cabo (RJ). 9. Rio de Janeiro (Estado). I. Título. II. Escola Nacional de Botânica Tropical.
CDD 583.79
iv

Para Yasmin, em sua longa estrada...
Para Carmen, meus pais e irmãos, os
“construtores” da estrada.
v

AGRADECIMENTOS
À Maria Helena Campos Baeta Neves e Flávio da Costa Rodrigues (IEAPM) pelo
estímulo e impulso inicial nesta jornada.
A Leandro Freitas (JBRJ) pela orientação científica e filosófica durante todo o
trabalho, fornecendo as bases essenciais para a concretização deste trabalho.
À Dorothy Sue Dunn de Araujo e Ricardo Ferreira Monteiro (UFRJ), Cyl Farney
Catarino de Sá (JBRJ), Maria Célia Rodrigues Correia (MN-UFRJ) e Isabel Cristina
Sobreira Machado (UFPE) por suas prestimosas contribuições bibliográficas e valorosos
comentários.
À Tania Ocimoto Oda e Luiz Antônio de Souza (IEAPM) pela contribuição dos
dados climatológicos.
À Maria Cristina Gaglianone (UENF) pela identificação das abelhas.
Em especial, ao amigo Sávio Henrique Calazans, pelo importante auxílio com a
literatura sobre fragmentação de ecossistemas.
Às colegas Jakeline Prata de Assis Pires (ENBT-JBRJ) e Denise Espellet Klein
(UENF) pelo auxílio nos experimentos de tubo polínico.
A meus pais, Edson e Marilda, e irmãos, Leonardo e Lilian, pelo indispensável
apoio e amizade, e minha esposa e filha, Carmen e Yasmin, pelo carinho e paciência, sem
reclamar das “pedras” no caminho.
A todos os amigos e colegas de classe, professores e todos os profissionais da Escola
Nacional de Botânica Tropical, pela competência e parceria, pelos auxílios e trocas de
conhecimento, fundamentais ao sucesso de todos.
Ao Instituto de Pesquisas Jardim Botânico do Rio de Janeiro e ao PROAP/CAPES
pelo apoio financeiro e estrutural ao longo do curso, e ao PMA/Petrobrás pelo auxílio
financeiro para participação no 56º Congresso Nacional de Botânica.
E aos meus amigos de longa estrada, por ainda continuarem sendo verdadeiramente
amigos mesmo em minha “ausência”.
vi

RESUMO
A fragmentação de habitats é considerada uma das mais poderosas forças de modificação
dos ecossistemas, em grande parte como resultado das atividades humanas. Dentre os
vários efeitos, a alteração dos processos associados a interações inseto-planta pode
conduzir a estados de endogamia em suas populações. A Restinga de Massambaba perdeu
uma parte significativa de sua cobertura vegetal, em processos intensificados nas últimas
seis décadas, que ainda não cessaram. Byrsonima sericea possui flores produtoras de óleo,
polinizadas principalmente por abelhas solitárias de médio porte. Com o objetivo de
avaliar os efeitos da fragmentação de habitat sobre a biologia reprodutiva de B. sericea
foram estudadas: (1) densidade e cobertura lineares, (2) fenologia reprodutiva, (3)
composição e freqüência da fauna de polinizadores, (4) produção de frutos, e (5)
ocorrência de galhas entomógenas, em seis fragmentos (três pequenos - FP - e três maiores
- FM) em formação arbustiva aberta na restinga. Foi também analisado o sistema
reprodutivo de B. sericea nas populações estudadas. Não houve diferenças entre FP e FM
para as fenofases reprodutivas na maioria dos meses e, também, para densidade e cobertura
lineares, e ocorrência de galhas caulinares. A freqüência de visita de polinizadores e a
produção de frutos foram maiores em FP. Galhas florais foram mais numerosas em FM.
Esta espécie é auto-incompatível com discreta autocompatibilidade. A maior freqüência de
visitas de abelhas em FP refletiu em aumento na produção de frutos, o que pode estar
relacionado à auto-incompatibilidade de B. sericea combinada com a permanência de um
tamanho de população ainda suficiente para as polinizações cruzadas nestas áreas. No atual
estágio, a fragmentação produz pouco efeito direto sobre a biologia reprodutiva e
distribuição populacional de B. sericea, ou mesmo é favorecida por este processo. Tais
efeitos são mais evidentes sobre a dinâmica de seus polinizadores e galhadores florais. É
possível que este desequilíbrio nestas interações possa conduzir a um futuro
empobrecimento na diversidade genética destas populações causado por taxas de
endogamia crescentes.
vii

ABSTRACT
Habitat fragmentation is considered one of the most powerful modification forces of
ecosystems, mostly as result of human activities. Among several effects, changes in
interactions between insects and plants, may increase endogamy of populations. The
Massambaba restinga lost a significant part of its original vegetation cover due to
anthropogenic impacts, which were intensified in the last six decades and still persist.
Byrsonima sericea bears oil-producing flowers, pollinated mainly by medium-sized
solitary bees. To evaluate the effects of habitat fragmentation on B. sericea populations, I
studied: (1) linear density and coverage, (2) reproductive phenology, (3) composition and
frequency of pollinators, (4) fruit production, and (5) entomogenous gall occurrence, in six
fragments (three small - FP - and three large - FM) in open restinga scrub. I also analyzed
the reproductive system of B. sericea in these populations. No differences were found
between FP and FM as regards reproductive phenology in most months, linear density and
coverage, and stem gall occurrence. Frequency of pollinator visits and fruit production
were higher in FP. Floral galls were more numerous in FM. This species is self-
incompatible with slight self-compatibility. The higher frequency of bee visits in FP leads
to an increase in fruit production, which may be related to the self-incompatibility of
B. sericea combined with a population size that permits cross-pollination in these areas.
Nowadays, fragmentation exerts a small negative effect on B. sericea reproductive biology
and population distribution, or it may even be favored by this process. Its effects are more
evident on pollinators and floral-galler dynamics. It is possible that disturbance of these
interactions may lead to a future reduction in genetic diversity of populations caused by
increased endogamy rates.
viii

SUMÁRIO
Lista de Figuras .................................................................................................................. x
Lista de Tabelas ................................................................................................................ xi
INTRODUÇÃO GERAL .................................................................................................. 1
Sucesso reprodutivo de Byrsonima sericea (Malpighiaceae) em fragmentos de restinga
de diferentes tamanhos ............................................................................................ 6
INTRODUÇÃO .............................................................................................................. 7
MATERIAIS E MÉTODOS ......................................................................................... 9
ÁREA DE ESTUDO E CARACTERIZAÇÃO DOS FRAGMENTOS ............................................ 9
ESPÉCIE ESTUDADA ..................................................................................................... 12
DENSIDADE E COBERTURA DE B. SERICEA NOS FRAGMENTOS ....................................... 14
FENOLOGIA REPRODUTIVA, AGENTES POLINIZADORES E PRODUÇÃO DE FRUTOS ......... 15
REGISTRO DE GALHAS ENTOMÓGENAS ......................................................................... 16
ANÁLISES ESTATÍSTICAS .............................................................................................. 18
RESULTADOS ............................................................................................................. 18
DISTRIBUIÇÃO DE INDIVÍDUOS DE B. SERICEA NOS FRAGMENTOS ................................ 18
FENOLOGIA REPRODUTIVA E PRODUÇÃO DE FRUTOS ................................................... 19
VISITAS DE POLINIZADORES E OCORRÊNCIA DE GALHAS ENTOMÓGENAS ..................... 20
DISCUSSÃO ................................................................................................................. 26
LITERATURA CITADA ............................................................................................ 32
CONCLUSÕES GERAIS ................................................................................................ 38
REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 39
ix

LISTA DE FIGURAS
Figura 1. Simulação dos eventos de fragmentação em parte da Restinga de
Massambaba ........................................................................................................... 4
Figura 2. Disposição dos fragmentos na Restinga de Massambaba ............................ 12
Figura 3. Morfoespécies de galhas entomógenas em Byrsonima sericea ..................... 17
Figura 4. Fenologia reprodutiva de Byrsonima sericea e médias de temperatura ..... 20
Figura 5. Entradas de ninhos de abelhas solitárias ...................................................... 22
Figura 6. Visitantes florais de Byrsonima sericea ......................................................... 23
Figura 7. Número total de visitas de polinizadores de Byrsonima sericea .................. 24
Figura 8. Composição relativa de morfoespécies de galhas por fragmento ............... 25
Figura 9. Galhas florais predadas .................................................................................. 29
Figura 10. Hipótese de forrageamento de uma abelha solitária ................................. 31
x

LISTA DE TABELAS
Tabela 1. Localização, tamanho, distância mínima para o fragmento vizinho e
distância mínima entre indivíduos dos fragmentos de restinga estudados ....... 11
Tabela 2. Caracterização dos fragmentos de restinga estudados ................................ 13
Tabela 3. Distribuição espacial de Byrsonima sericea nos fragmentos, por intercepto
de linhas .................................................................................................................. 19
Tabela 4. Espécies de abelhas visitantes das flores de B. sericea ................................. 21
Tabela 5. Quantidade de galhas entomógenas em B. sericea ....................................... 25
xi

INTRODUÇÃO GERAL
A fragmentação de habitats pode ser definida como o processo onde habitats contínuos
são divididos ao longo do tempo em áreas ou manchas menores, de diferentes tamanhos, e
mais ou menos isoladas entre si (Rambaldi & Oliveira 2003). Tal processo pode ser resultado
de fenômenos naturais (vulcanismo, terremotos, tsunamis, incêndios naturais, tempestades,
etc.), ou, mais freqüentemente, por perturbações antrópicas diversas (desmatamento para
habitações em várias escalas, exploração madeireira e pecuária, agricultura, etc.). Os efeitos
negativos sobre as populações animais e vegetais decorrentes da fragmentação de habitat
estão bem documentados por diversas pesquisas realizadas nos últimos anos (ver Rambaldi &
Oliveira 2003). Entre tais efeitos, destacam-se o aumento na endogamia e a redução da
variabilidade genética das populações, que conseqüentemente aumentam o risco de sua
extinção (Aizen et al. 2002a, b; Jacquemyn et al. 2002).
A desestabilização causada pela fragmentação de habitat e a perda da conectividade
entre os ecossistemas podem afetar as interações entre as plantas e seus polinizadores,
competidores ou inimigos naturais. O aumento dos níveis de consangüinidade das populações
afetadas pode alterar a persistência das espécies individualmente e a sua resistência, ou
tolerância, a inimigos naturais (Hoffmeister et al. 2005). Assim, a biologia reprodutiva, em
particular sob a ótica da interação inseto-planta e do sucesso reprodutivo, pode constituir um
elemento auxiliar na elucidação destes efeitos.
Mais de 80% das angiospermas viventes dependem, em diferentes graus, de animais
para sua polinização e reprodução sexual (Bawa 1990, Buchmann & Nabhan 1996). A
dependência dessas relações mutualísticas para a reprodução pode ampliar a suscetibilidade
da planta à fragmentação ou outras formas de perturbação do ambiente. A degradação do
mutualismo planta-animal pode provocar consideráveis reduções na qualidade e no número de
sementes e freqüentemente constituem o primeiro passo para o colapso demográfico de várias
populações vegetais. Existem exemplos de plantas que beiram à extinção em decorrência da
deficiência de sua reprodução sexual associada à perda de seus agentes polinizadores
(Buchmann & Nabhan 1996, Renner 1998, Cox & Elmqvist 2000, Paton 2000).
As plantas com flores produtoras de óleo representam um número considerável de
espécies de angiospermas de alguns grupos tropicais. Tais plantas ofertam o óleo como
recompensa aos polinizadores, em lugar do néctar. O óleo floral é usado pelos insetos, no
caso, grupos especializados de abelhas, para alimentar suas larvas e possivelmente para
revestimento das paredes dos ninhos (Neff & Simpson 1981, Vogel 1990). As abelhas
1

misturam o óleo coletado ao pólen de outras espécies vegetais, obtendo, assim, alimento de
alto teor energético para suas larvas (Simpson & Neff 1981). As flores produtoras de óleo
estão representadas em pelo menos oito famílias de angiospermas: Malpighiaceae,
Krameriaceae, Cucurbitaceae, Primulaceae, Solanaceae, Scrophulariaceae, Iridaceae e
Orchidaceae (Endress 1994).
O gênero Byrsonima (murici), com cerca de 150 espécies, distribui-se pelas Antilhas e
Américas do Sul e Central (Mabberley 1993). A espécie escolhida para este trabalho reflete
um exemplo de planta com considerável dependência do mutualismo planta-polinizador.
Byrsonima sericea DC. é uma espécie produtora de óleo floral, através de glândulas
especializadas, os elaióforos, nas sépalas, embora em estudos sobre sua biologia reprodutiva
em uma floresta perenifólia de Pernambuco (Silva 1990, Teixeira & Machado 2000), nem
todos os indivíduos desta espécie apresentaram os elaióforos nas suas flores. Segundo os
critérios propostos por Hoffmeister et al. (2005), B. sericea seria uma possível espécie-chave
para estudos dos efeitos de fragmentação de habitat. Dados sobre a freqüência e abundância
desta espécie em outras restingas, bem como de sua distribuição geográfica, foram incluídos
nesta análise (Silva 1990, Araujo et al. 1998, Menezes & Araujo 1999, Araujo 2000, Teixeira
& Machado 2000, Pereira et al. 2001, Sá 2002, Rodrigues 2002, Araujo et al. 2004). Entre os
trabalhos que tratam da biologia reprodutiva de B. sericea, são de especial interesse os de
Silva (1990), Teixeira & Machado (2000) e Rodrigues (2002).
Além de áreas de mata perenifólia na Mata Atlântica, B. sericea também ocorre em
formações arbustivas de restinga, onde adquire menor porte e maior ramificação de suas
partes aéreas, tornando por vezes difícil o reconhecimento do caule principal, e distribuídas
em populações de freqüência relativa baixa a moderada (Silva 1990, Araujo et al. 1998,
Menezes & Araujo 1999, Araujo 2000, Teixeira & Machado 2000, Pereira et al. 2001,
Sá 2002, Rodrigues 2002, Araujo et al. 2004). Também são encontradas outras espécies de
Malpighiaceae produtoras de óleo floral coexistindo nas mesmas formações vegetais da
restinga, destacando-se Peixotoa hispidula A. Juss. e Stigmaphyllon paralias A. Juss. (Araujo,
2000).
Uma outra forma de interação entre insetos e plantas, e que também interfere direta ou
indiretamente em seu sucesso reprodutivo, são as cecídias, mais comumente conhecidas como
galhas. Em geral, há uma interação muito íntima do inseto (larva) com a planta, levando a
uma maior especificidade se comparada a outras formas de interações inseto-planta (Redfern
& Askew 1992). Alguns trabalhos relacionam os efeitos da fragmentação de habitat com a
dinâmica de interações entre insetos fitófagos e seus parasitóides. Em geral, predadores e
2

parasitóides de herbívoros sofrem com a fragmentação (Hoffmeister et al. 2005), ou, por
outro lado, são favorecidos, ainda que indiretamente pela maior disponibilidade de presas nas
áreas perturbadas (Bolger et al. 2000).
Existem alguns dados sobre pesquisas de fragmentação na Amazônia que vêm
indicando o aumento da riqueza de alguns insetos, associado à redução do fragmento
(R. F. Monteiro, comun. pess.). Em um levantamento de vários trabalhos realizados no Parque
Nacional da Restinga de Jurubatiba (Macaé, RJ), Monteiro et al. (2004) apresentam as
seguintes espécies e grupos de insetos indutores de galhas (segundo seus tipos florais) em
Byrsonima sericea: botão floral, Bruggmanniella byrsonimae Maia & Couri (Cecidomyiidae,
Diptera) e Lepidoptera não identificada; folha circular, Oligotrophini (Cecidomyiidae,
Diptera); caulinar, Cecidomyiidae (Diptera).
A restinga de Massambaba está contida nos municípios de Arraial do Cabo, Araruama e
Saquarema, todos componentes da chamada Região dos Lagos, no estado do Rio de Janeiro,
e, juntamente com outros ecossistemas da região, está inserida no domínio da Mata Atlântica.
Considerada um dos quatorze Centros de Diversidade Vegetal (CDV) no Brasil (Araujo
1997), a Mata Atlântica é detentora de um considerável grau de endemismo e está seriamente
ameaçada, sendo, por isso, considerada um “hotspot” (Conservation International 2006)
Historicamente, a restinga de Massambaba vem sofrendo sucessivos ciclos de impactos
antrópicos que contribuem para a perda da cobertura vegetal original, iniciada pela atividade
salineira, seguida da urbanização com discreta industrialização regional, e, mais atualmente,
intensificação da especulação imobiliária, principalmente ilegal nas áreas remanescentes de
restinga, aliada ao modesto, mas crescente, incremento no turismo. Nos distritos do município
de Arraial do Cabo conhecidos como Monte Alto e Figueira vêm ocorrendo de forma
acelerada, nos últimos 20-30 anos, a fragmentação da estreita faixa de restinga entre a praia
Grande e a lagoa de Araruama, em decorrência de loteamentos (Figura 1).
Os primeiros núcleos urbanos conhecidos na região têm início com a feitoria de
Américo Vespúcio em 1503 (Praia dos Anjos, Arraial do Cabo) e, mais adiante, com a
fundação de Cabo Frio, em 1616. A atividade salineira começa em 1823, e por volta de 1930
já existiam 2.100 ha de salinas, distribuídas pelos esporões da Lagoa de Araruama e em áreas
de restinga adjacentes a esta, em Cabo Frio (Palmer, Pontas do Costa e Ambrósio), na
Massambaba (Figueira) e no nascente vilarejo de operários de salinas de Monte Alto
(Bidegain & Bizerril 2002, Prado 2002). Nesta época, a vegetação de restinga, que se estendia
ao longo da Praia Grande até o Morro de Nazaré, em Saquarema, deveria ser contínua ou com
poucas interrupções. Uma ferrovia – Estrada de Ferro Maricá – cortava o que até então
3

deveria ser o maior núcleo não impactado de restinga, delimitado pelo Canal do Itajurú e
Palmer (Cabo Frio), Lagoa de Araruama, Praia do Foguete (Cabo Frio) e a Praia Grande, no
Arraial do Cabo (Pinto 1947), terminando no porto da Praia dos Anjos, em Arraial do Cabo,
entre 1937 e 1962, quando foi desativada (Bidegain & Bizerril 2002).
FIGURA 1. Simulação dos eventos de fragmentação em parte da Restinga de Massambaba.
Os quatro diagramas correspondem à porção leste da Restinga de Massambaba
(aproximadamente os primeiros doze quilômetros). Em preto, as formações vegetais de
restinga; em cinza, praias, áreas urbanas e salinas; em branco, fábrica, oceano, lagoa e demais
corpos hídricos; área estriada, afloramentos rochosos (compilado com informações de Pinto
1947, Bidegain & Bizerril 2002, Prado 2002, Semads 2002).
Em 1940 tem início o maior ciclo de desenvolvimento na região com a inauguração da
rodovia RJ-106 (Bidegain & Bizerril 2002). Na década de 50 ocorre o primeiro desmatamento
em larga escala da restinga, com o início da construção da Cia. Nacional de Álcalis (CNA)
com suas áreas de salinas para suprir a produção de carbonato de sódio (barrilha), o parque
fabril, a vila de operários, áreas externas para deposição de efluentes e resíduos industriais,
áreas adjacentes à fábrica para plantio de casuarinas e eucaliptos, possivelmente para geração
4

de energia (S. Marchi, comun. pess.), e construção de um canal com 5,5 km de extensão, ao
longo da restinga, interligando a fábrica com a Lagoa de Araruama (Bidegain & Bizerril
2002). Com o início de suas atividades, em 1960, o fluxo de moradores e a demanda de
serviços aumentam gradativamente, e, em igual proporção, a ocupação de mais áreas de
restinga, principalmente em Monte Alto e Figueira, e na estrada para Cabo Frio (Prado 2002).
É possível que com a chegada da CNA, as salinas que já estavam na região ampliaram a
sua produção (e, assim, mais áreas sobre a restinga) diante da nova demanda oferecida pela
fábrica (o cloreto de sódio é uma de suas principais matérias-primas), pelo menos até o início
da década de 80, quando a atividade salineira começa a entrar em declínio (Bidegain &
Bizerril 2002). Com isto, um novo ciclo de impactos surge, de menor expressão territorial,
mas ininterrupto, que é a exploração de áreas de restinga e de salinas desativadas para a
especulação imobiliária, seja em grande escala (condomínios e grandes empreendimentos) ou
pequena (imóveis particulares, isolados ou agregados).
Durante algum tempo a posse de terras em nome da CNA funcionou como um “efeito-
tampão”, evitando as ocupações nas áreas remanescentes de restinga. Recentemente este
cenário tem dado sinais de redução de seu efeito, com a construção do aeroporto de Cabo Frio
sobre uma área de 300 ha de restinga (em 1998), novos condomínios previstos sobre áreas de
restinga, ou adjacentes a estas, e as ocupações ilegais isoladas não dão sinais de que vão
cessar frente à atual pressão ambiental. Em uma das áreas estudadas durante este trabalho, na
localidade de Monte Alto, cerca de 50% de sua vegetação foi suprimida em favor de novas
construções populares.
Os estudos na região vêm aumentando significativamente nos últimos anos,
especialmente nos campos da fitossociologia e conservação (Araujo, D. S. D. 2000, Sá, C. F.
C. 2002, Cordeiro, S. Z., 2005), etnobotânica (Fonseca-Kruel, V. S. & Peixoto, A. L. 2004) e
ecologia (Scarano, F. R. 2002, Rocha, C. F. D. et al. 2005), e a biologia reprodutiva pode
auxiliar na definição de padrões e compreensão dos processos que possam alterar o
funcionamento deste ecossistema. Estes esforços poderão ser importantes para quantificar e
qualificar os impactos antrópicos, delineando, assim, os limites aceitáveis das perturbações e
viabilizando o apoio às decisões de implantação de políticas públicas de manejo ambiental
sustentável (Rambaldi & Oliveira 2003).
O objetivo geral deste trabalho é investigar se a fragmentação de habitat na Restinga de
Massambaba está afetando a biologia reprodutiva de B. sericea pela redução na freqüência de
visitas de polinizadores e produção de frutos, ou alterando suas fenofases reprodutivas ou seu
sistema reprodutivo, e, ainda, interferindo na incidência de insetos galhadores nesta espécie.
5

Sucesso reprodutivo de Byrsonima sericea (Malpighiaceae) em
fragmentos de restinga de diferentes tamanhos 1
ABSTRACT
Habitat fragmentation frequently affects ecosystem functionality and may increase endogamy
of populations by changing mutualistic relationships between plants and their pollinators. The
aim of this study was evaluate the effects of habitat fragmentation on Byrsonima sericea DC.
populations in open restinga scrubs (three small patches up to 0.3 ha (FP) and three large
patches from 14.0 to 99.0 ha (FM)) were studied with emphasis on reproductive biology and
interaction with insects (pollinators and gallers). No differences were found between FP and
FM linear density and coverage, reproductive phenology in most months, and stem gall
occurrence. The frequency of pollinator visits and fruit production was higher in FP, while
floral galls were more numerous in FM. Results indicate little effect of habitat fragmentation
on B. sericea reproductive biology and spatial distribution at the present dimensional and
temporal scale. These effects increased pollinator frequency and reduced floral gall
establishment, which favored B. sericea higher fruit set. Nevertheless, higher flower offering
to bees and reduced galler attack within the small remnant areas may result in more bee visits
to a few individuals of B. sericea in the area, and consequently to increased endogamy in the
future.
RESUMO
A fragmentação de habitat freqüentemente compromete a funcionalidade de um ecossistema,
podendo elevar as taxas de endogamia de suas populações, pela alteração da relação
1 Manuscrito preparado seguindo as normas da revista Biotropica.
6

mutualística entre as plantas e seus polinizadores. O objetivo deste estudo foi avaliar os
efeitos da fragmentação de habitat sobre populações de Byrsonima sericea DC. em áreas de
restinga (três fragmentos pequenos com até 0.3 ha (FP) e três maiores com 14.0 a 99.0 ha
(FM)), com ênfase em sua biologia reprodutiva e interação com insetos (polinizadores e
galhadores). Não houve diferenças entre FP e FM para densidade e cobertura lineares,
fenofases reprodutivas na maioria dos meses e ocorrência de galhas caulinares. A freqüência
de visitas de polinizadores e a produção de frutos foram maiores em FP, enquanto que a
ocorrência de galhas florais foi maior em FM. Os resultados indicam pouco efeito da
fragmentação de habitat sobre a biologia reprodutiva e densidade populacional de B. sericea
na atual escala dimensional e temporal. Tais efeitos resultaram em aumento na freqüência de
visitas de polinizadores e redução do estabelecimento de galhas florais, o que favoreceu a
maior produção de frutos de B. sericea em um primeiro momento. Entretanto, é possível que,
no futuro, a maior oferta de flores às abelhas e o menor ataque de galhas dentro das áreas
remanescentes reduzidas resultem em mais visitas das abelhas em poucos indivíduos de
B. sericea na área e, conseqüentemente, a um aumento da endogamia.
Key words: Byrsonima sericea; habitat fragmentation; pollination; solitary bees; insect
galls; Brazil; restinga.
A FRAGMENTAÇÃO DE HABITATS FREQÜENTEMENTE INTERFERE DE FORMA NEGATIVA NO
SUCESSO REPRODUTIVO DAS ANGIOSPERMAS (Bawa 1990, Aizen & Feinsinger 1994a, b,
Aizen et al. 2002a, b, Jacquemyn et al. 2002). O aumento dos níveis de consangüinidade nas
populações remanescentes de áreas fragmentadas pode alterar a persistência destas espécies,
bem como sua resistência a inimigos naturais (Hoffmeister et al. 2005). Entretanto, não são
poucos os estudos demonstrando que a fragmentação apresenta aspectos positivos em relação
a tais sistemas (Ghazoul & Shaanker 2004), tais como maior produção de sementes e maior
7

carga de pólen nos estigmas (Bosch et al. 2002), maior produção de flores nos fragmentos
(Johnson et al. 2004, Somanathan et al. 2004), ou ainda, a ausência de efeitos aparentes em
certas plantas (Quesada et al. 2004).
A despeito da complexidade das interações e variabilidade nas respostas, é inegável
que a cada dia novos estudos têm, sob uma ótica integradora, contribuído para a elucidação
destes fenômenos (Ghazoul & Shaanker 2004) e para apontar quais são as diretrizes
necessárias para minimizar os impactos sobre os ecossistemas (Rambaldi & Oliveira 2003).
Entre as características reprodutivas que podem determinar o grau de sensibilidade das plantas
à fragmentação de habitat, estão o tipo de sistema reprodutivo, o grau de especialização do
sistema de polinização (Aizen et al. 2002a), o tamanho e comportamento do polinizador e o
tipo de dispersão (Ghazoul & Shaanker 2004).
As flores de Byrsonima sericea (Malpighiaceae) são polinizadas preferencialmente por
fêmeas de abelhas de médio porte que buscam óleo e pólen para alimentação de suas larvas e,
possivelmente, para a impermeabilização dos ninhos (Teixeira & Machado 2000,
Rodrigues 2002). As principais abelhas polinizadoras de B. sericea compreendem espécies de
médio porte da família Apidae (Centridini), de hábito solitário, com particular destaque para
os gêneros Centris e Epicharis. Estas abelhas caracteristicamente nidificam no solo escavando
buracos (Roubik 1989). Embora sejam solitárias, para algumas espécies os ninhos constituem
grandes agregações (Silveira et al. 2002).
As fêmeas destas espécies desenvolveram estruturas especializadas para coleta e
transporte de óleo e pólen, principalmente escopas bem desenvolvidas nas pernas posteriores
(Ramalho & Silva 2002, Silveira et al. 2002). A oferta de óleo e a atração de abelhas
Centridini têm sido interpretadas como um sistema de recompensa muito específico para estes
animais, representando um evento evolucionário independente (Fenster et al. 2004),
responsável pelo sucesso das Malpighiaceae nas Américas (Anderson 1979, Vogel 1990). Já o
8

sistema reprodutivo de B. sericea tem sido interpretado como auto-incompatível (Teixeira &
Machado 2000) a discretamente autocompatível (Silva 1990, Rodrigues 2002).
Outro tipo de interação planta-animal que pode interferir negativamente no sucesso
reprodutivo desta espécie, por exemplo, parasitando suas flores, diz respeito aos insetos
galhadores. São reconhecidas algumas espécies de insetos que produzem galhas (cecídias) em
flores, folhas ou caule de B. sericea em áreas de restinga (Flinte et al. 2003, Monteiro et al.
2004). A dinâmica de interações entre insetos fitófagos e seus parasitóides comumente é
alterada pelos efeitos causados pela fragmentação de habitat, tanto positiva como
negativamente (Bolger et al. 2000, Hoffmeister et al. 2005).
Com o objetivo de verificar os efeitos da fragmentação no sucesso reprodutivo de
B. sericea, os seguintes parâmetros foram comparados entre fragmentos de duas classes de
tamanho em uma área de restinga: (1) densidade e cobertura linear das populações;
(2) fenologia reprodutiva (floração e frutificação), (3) composição, freqüência e
comportamento da fauna de polinizadores, (4) produção de frutos e sementes (i.e.,
fecundidade), (5) ocorrência e tipos de galhas.
MATERIAIS E MÉTODOS
ÁREA DE ESTUDO E CARACTERIZAÇÃO DOS FRAGMENTOS.—Este estudo foi realizado em áreas
fragmentadas de formações arbustivas das restingas de Massambaba e de Cabo Frio,
respectivamente nos municípios de Arraial do Cabo e Cabo Frio (22°55’– 22°59’S, 42°02’–
42°08’W), no estado do Rio de Janeiro, sudeste do Brasil. Foram selecionados seis
fragmentos de restinga, distribuídos em duas classes de tamanho (três fragmentos pequenos
(FP), entre 0.07 e 0.28 ha; três fragmentos maiores (FM), entre 14.0 e 99.0 ha).
As áreas de restinga estudadas correspondem a remanescentes de um sistema contínuo
de formações arbustivas distintas, que vem sendo impactado há aproximadamente seis
9

décadas: um primeiro ciclo, em grande escala e pontual, de urbanização e industrialização
(salinas e indústria química) e, um segundo ciclo que teve início na década de 1980 com a
queda da atividade salineira e pressão para a especulação imobiliária, que, embora tenha se
dado em menor escala do que o primeiro, continua até os dias atuais (Pinto 1947, Bidegain &
Bizerril 2002, Prado 2002).
Os critérios para escolha destas áreas levaram em conta: (1) respeito a uma ordem de
grandeza distinta entre as duas classes de tamanho, (2) grau de isolamento satisfatório entre os
fragmentos, (3) presença de um número suficiente de indivíduos de B. sericea para os
experimentos e observações, e (4) maior correspondência possível entre as formações vegetais
e características do terreno.
Os elementos delimitadores dos fragmentos incluídos neste trabalho abrangem tanto
áreas matriciais (salinas, condomínios, núcleos urbanos, pastagens e lagoas) como linhas de
interrupção (ruas, estradas asfaltadas ou não pavimentadas e canais artificiais). Para os
fragmentos que apresentaram distâncias mínimas entre outro(s) fragmento(s) inferiores a
100 m, todas estas áreas foram somadas e interpretadas como “sistemas de fragmentos” ou
“fragmentos compostos”, e uma nova distância mínima, a partir destes sistemas, foi calculada
(Tabela 1). Os fragmentos maiores, em particular, apresentam desenhos muito variados, mais
ou menos extensos, inclusive sujeitos a diferentes regimes de exposição eólica e marinha
(Figura 2).
A formação vegetal predominante nas áreas de estudo foi do tipo arbustiva aberta
(Tabela 2). As designações para tais formações aqui apresentadas seguem as propostas por
Araujo et al. (1998), contudo tais formações para a Restinga de Massambaba são
relativamente diferentes de suas correspondentes em outras restingas do estado do Rio de
Janeiro, necessitando de uma classificação mais específica com base em novos estudos
florísticos e fitossociológicos (D. S. D. Araujo, comun. pess.).
10
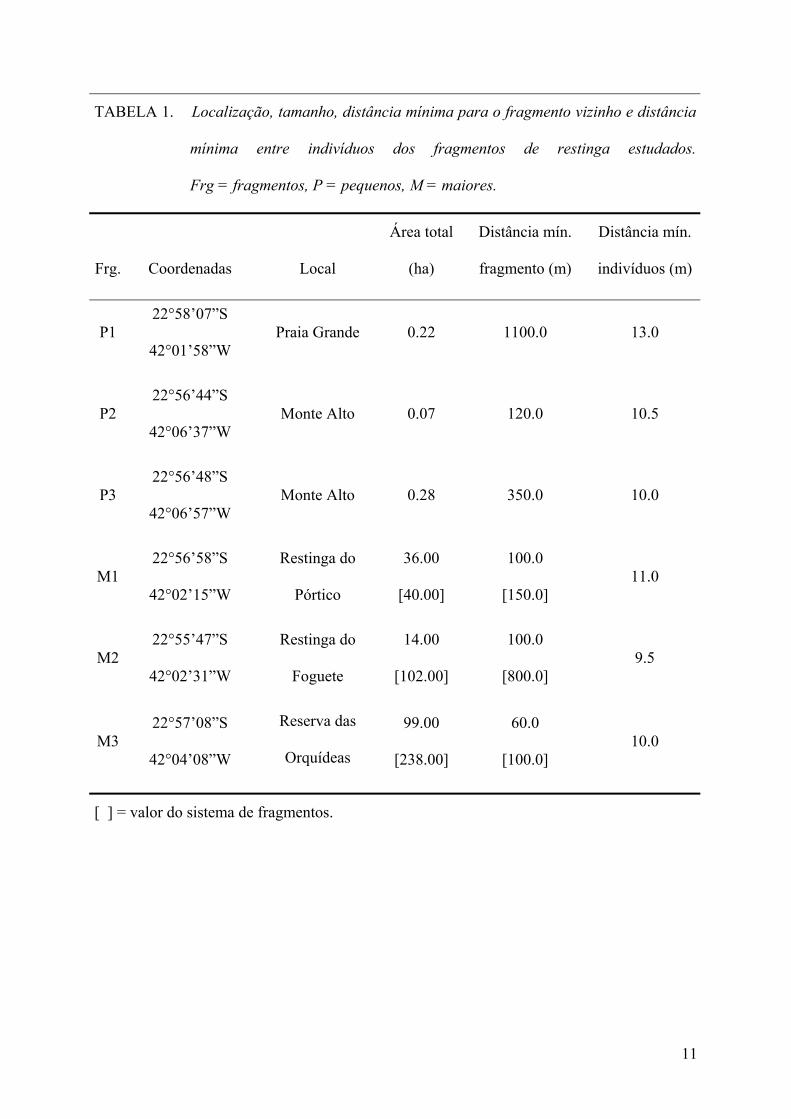
TABELA 1. Localização, tamanho, distância mínima para o fragmento vizinho e distância
mínima entre indivíduos dos fragmentos de restinga estudados.
Frg = fragmentos, P = pequenos, M = maiores.
Frg. Coordenadas Local
Área total
(ha)
Distância mín.
fragmento (m)
Distância mín.
indivíduos (m)
P1 22°58’07”S
42°01’58”W Praia Grande 0.22 1100.0 13.0
P2 22°56’44”S
42°06’37”W Monte Alto 0.07 120.0 10.5
P3 22°56’48”S
42°06’57”W Monte Alto 0.28 350.0 10.0
M1 22°56’58”S
42°02’15”W
Restinga do
Pórtico
36.00
[40.00]
100.0
[150.0] 11.0
M2 22°55’47”S
42°02’31”W
Restinga do
Foguete
14.00
[102.00]
100.0
[800.0] 9.5
M3 22°57’08”S
42°04’08”W
Reserva das
Orquídeas
99.00
[238.00]
60.0
[100.0] 10.0
[ ] = valor do sistema de fragmentos.
11

FIGURA 2. Disposição dos fragmentos na Restinga de Massambaba. As manchas escuras
compreendem formações arbustivas distintas, e os trechos de formação halófila-psamófila
foram omitidos. O centro dos círculos tracejados indica o local do fragmento (P – pequenos;
M – maiores) onde foram concentrados os experimentos, e suas coordenadas estão indicadas
na Tabela 1. A estrela indica a localização da estação meteorológica do IEAPM. Note que P2
apresenta um estreito corredor de conectividade com o sistema de fragmentos M3 (Adaptado
de Semads 2002).
ESPÉCIE ESTUDADA.—Byrsonima sericea apresenta, em áreas de restinga aberta, hábito
predominantemente arbustivo (com cerca de 0.5 a 3.0 m de altura), com o sistema aéreo muito
ramificado desde a base, sendo eventualmente difícil reconhecer o caule principal, mas
também podem ocorrer formas subarborescentes, com um tronco mais definido e alto (até
4.0 m de altura média). Esta espécie também ocorre no cerrado (região Centro-Oeste do
Brasil) e em trechos litorâneos da Mata Atlântica, nas regiões SE e NE (Silva 1990, Teixeira
12

& Machado 2000, Araujo 2000, Rodrigues 2002). As flores são arranjadas em racemos
terminais, que se abrem em padrão cornucópia (muitas flores durante várias semanas) (sensu
Gentry 1974), são zigomorfas, andróginas, pentâmeras, vivamente amarelas, apresentando dez
elaióforos (i.e., glândulas secretoras de óleo) nas sépalas, que às vezes podem estar ausentes
(Teixeira & Machado 2000).
TABELA 2. Caracterização dos fragmentos de restinga estudados. Frg = fragmentos,
P = pequenos, M = maiores.
Frg. Formação vegetal * Topografia Matriz circundante Anos
P1 arbustiva fechada de
pós-praia
dunas {7-9 m},
montículos
urbano, pasto,
morro 50-55
P2 arbustiva aberta de
Clusia dunas {6-8 m} urbano, ocupações 20-30
P3 arbustiva aberta de
Clusia plano, montículos urbano, ocupações 20-30
M1 arbustiva aberta de
Clusia plano, montículos
urbano, salinas,
rodovia 50-55
M2 arbustiva aberta de
Ericaceae plano, montículos
urbano, salinas,
rodovia 50-55
M3 arbustiva aberta de
Clusia plano, montículos
pasto, salinas,
rodovia 50-55
cf. Araújo et al., 1998, [ ] = valor do sistema de fragmentos, { } altura média das dunas,
montículos = dunas pequenas de até 2.0 m de altura.
13

Nas populações estudadas, não foi encontrado nenhum indivíduo sem elaióforos nas
sépalas, incluindo outros indivíduos não utilizados no estudo reprodutivo. O androceu possui
dez estames, contendo grãos de pólen secos. O gineceu é composto por ovário súpero,
tricarpelar e trilocular com um óvulo por lóculo, apresentando três estiletes que terminam em
uma diminuta área estigmática. Esta é revestida por uma membrana (opérculo) que deve ser
rompida para que a polinização possa ser efetuada.
Investigações sobre o sistema reprodutivo têm indicado a espécie como auto-
incompatível (Teixeira & Machado 2000) ou discretamente autocompatível (Silva 1990,
Rodrigues 2002). Foram realizados testes de polinizações manuais na restinga de
Massambaba, os quais indicaram seu sistema como preferencialmente auto-incompatível, com
pequena autocompatibilidade (sucesso para alogamia = 24,8% e autopolinização manual =
2,9%, B. S. Dunley, dados não publ.). Foram realizados testes de receptividade do estigma
após fricção do opérculo em diferentes estágios pré-antésicos e pós-antésicos, com solução
peróxido de hidrogênio (Zeisler 1938) a fim de determinar a condição mais adequada para a
condução dos experimentos. Exsicatas de B. sericea foram depositadas no Herbário do Jardim
Botânico do Rio de Janeiro (RB) (B. S. Dunley s/n, RB 413002, RB 413003, RB 413004).
DENSIDADE E COBERTURA DE B. SERICEA NOS FRAGMENTOS.—Para determinação e
quantificação da população de B. sericea foi adotado o método de intercepto de linha
(Brower & Zar 1984, Pereira et al. 2001) em transects de 400 m para cada fragmento
(20 linhas de 20 m, espaçadas a 10 m entre si), com exceção do fragmento P2, onde só foi
possível delimitar o transect com 200 m, respeitando o mesmo padrão de distribuição sobre o
terreno (“grid”) adotado nos demais fragmentos. Foram determinados e comparados os
índices de densidade linear (ID) e de cobertura linear (IC) para todos os fragmentos, segundo
as fórmulas apresentadas em Brower & Zar (1984).
14

FENOLOGIA REPRODUTIVA, AGENTES POLINIZADORES E PRODUÇÃO DE FRUTOS.—Foram
selecionados três indivíduos adultos de B. sericea, em cada fragmento, para monitoramento e
registro das fenofases reprodutivas (produção de botões, flores e frutos imaturos e maduros)
através do percentual de intensidade de Fournier, que consiste em uma escala semiquantitativa
de cinco categorias (0 a 4), a intervalos de 25% (Fournier 1974, Bencke & Morellato 2002a,
b). Os indivíduos para estudo foram escolhidos ao acaso dentre os que apresentavam
semelhança em relação à altura e cobertura (área) de copa. Os registros fenológicos foram
realizados mensalmente, entre maio de 2004 e junho de 2005, sendo que os registros foram
quinzenais durante os meses de maior intensidade de floração e frutificação (outubro de 2004
a fevereiro de 2005). Os registros de temperatura (médias mensais) foram obtidos de uma
estação meteorológica pertencente ao Instituto de Estudos do Mar Almirante Paulo Moreira –
IEAPM, situada em área próxima ao fragmento M3 (Figura 2).
As observações dos visitantes florais foram efetuadas em três indivíduos por
fragmento, em plena floração e diferentes daqueles em que foram conduzidos os experimentos
do sistema reprodutivo. Quando a floração nestes indivíduos cessava, eram escolhidos outros
indivíduos próximos, em plena floração, até encerar o período de observações, entre outubro
de 2004 e fevereiro de 2005. Assim, cerca de seis indivíduos por fragmento foram
monitorados para a determinação da freqüência de visitas e composição dos polinizadores.
Outros trabalhos indicam o período da manhã como o de maior intensidade de visitas de
polinizadores (Teixeira & Machado 2000, Rodrigues 2002) e em função destes dados, as
observações foram efetuadas entre 05:30h e 13:00h.
Foram registrados a visita, o comportamento e o horário do evento, sendo que cada
sessão de observação durou 20 minutos. No ato da captura, registro e acondicionamento do
inseto visitante, a contagem do tempo era interrompida até que se reiniciasse o ciclo de
observações. Os ciclos de observações dos agentes polinizadores foram distribuídos
15

eqüitativamente (quantidade de observações e faixa de horários) entre os seis fragmentos,
totalizando 167 minutos de observações em cada fragmento. Foram realizados registros
fotográficos de visitas às flores e de espécimes capturados e fixados.
A produção de frutos foi quantificada em flores expostas aos polinizadores em três
racemos por cada um de três indivíduos, por cada fragmento, totalizando 39 racemos e
822 flores tratadas ou marcadas. Foram contados todos os botões e flores de cada
inflorescência, as quais foram isoladas em sacos de “voile” após esta contagem, durante 7-10
dias. Os sacos foram removidos após 40-50 dias e os frutos produzidos foram contados.
Frutos muito pequenos (menos de 6 mm) e soltos dentro dos sacos não foram considerados na
contagem.
REGISTRO DE GALHAS ENTOMÓGENAS.—Foi registrada e quantificada a presença de galhas
entomógenas (cecídias) em três indivíduos por fragmento (à exceção do fragmento P2, que foi
ocupado por habitações no decorrer do trabalho), em novembro de 2005. As galhas foram
classificadas em dois tipos gerais: florais e caulinares. Apesar de ocorrer mais de um subtipo
de galha floral ou caulinar, para os propósitos deste estudo foram consideradas apenas estas
duas categorias gerais.
Foram quantificadas, por contagem direta ou estimativa para indivíduos com número
de galhas maior ou igual a cinco (média de galhas em 15 racemos, multiplicada pelo total de
racemos), todas as galhas presentes nos indivíduos amostrados, tanto as florais como as
caulinares, sendo também categorizadas segundo a morfoespécie (Figura 3). Foram
denominadas três morfoespécies para os dois tipos gerais de galhas: floral indumentosa
(botões inchados e fechados inicialmente, formando um padrão de estrela com as sépalas e
elaióforos, que ao abrir-se mostra grande desenvolvimento de indumento em seu interior),
floral esferóide lisa (receptáculo inchado como um “falso-fruto”) e floral esferóide rugosa
16

(padrão “falso-fruto”, mais intumescido e com a superfície parda e crestada); caulinar calóide
(padrão de intumescência caulinar “estourada”), caulinar tuberculóide (padrão de “tubérculo”
esferóide, freqüentemente próximo das regiões de inserções dos ramos) e caulinar apical (os
ápices dos ramos apresentam-se intumescidos, e as folhas jovens, próximas aos meristemas,
tornam-se reduzidas ou atrofiadas).
FIGURA 3. Morfoespécies de galhas entomógenas em Byrsonima sericea. Galhas florais
(a, indumentosa; b, esferóide rugosa; c, esferóide lisa) e caulinares (d, calóide;
e, tuberculóide; f, apical). (barras = 5 mm).
17

Ainda não está confirmado se a morfoespécie floral indumentosa é de fato uma galha,
mas os indícios morfológicos e a presença de larvas em seu interior, assim como a
modificação na anatomia e histologia do botão atacado, parecem confirmar esta hipótese.
Foram escolhidos indivíduos semelhantes quanto ao porte e estado fenológico.
ANÁLISES ESTATÍSTICAS.—Foram comparadas as médias e erros-padrão dos parâmetros
analisados entre as duas classes de tamanho de fragmentos, menores e maiores, através do
teste-t e do teste do qui-quadrado (para dados percentuais). Nos casos em que os dados não
apresentaram normalidade, ou nos dados de percentual de intensidade de Fournier, foi
aplicado o teste de Mann-Whitney (teste-U). Todas as análises estatísticas foram geradas pelo
programa NCSS & PASS (Hintze 2004).
RESULTADOS
DISTRIBUIÇÃO DE INDIVÍDUOS DE B. SERICEA NOS FRAGMENTOS.—O número médio de
indivíduos amostrados em FM foi aproximadamente o dobro do valor em FP (Tabela 3), mas
não foi observada diferença significativa entre FP e FM para os índices de densidade linear
(ID) e de cobertura linear (IC) ( x ± SE, ID—FP = 0.031 ± 0.004, FM = 0.059 ± 0.020,
t = –2.51, P = 0.066; IC—FP = 0.081 ± 0.01, FM = 0.11 ± 0.05, t = –0.98, P = 0.38). Durante
os trabalhos de campo a área P2 foi intensamente perturbada pela população de seu entorno
para desmatamentos e loteamentos. Ao término da coleta de dados no campo, quatro dos sete
indivíduos interceptados na amostragem em P2 foram cortados para dar lugar a construções.
18
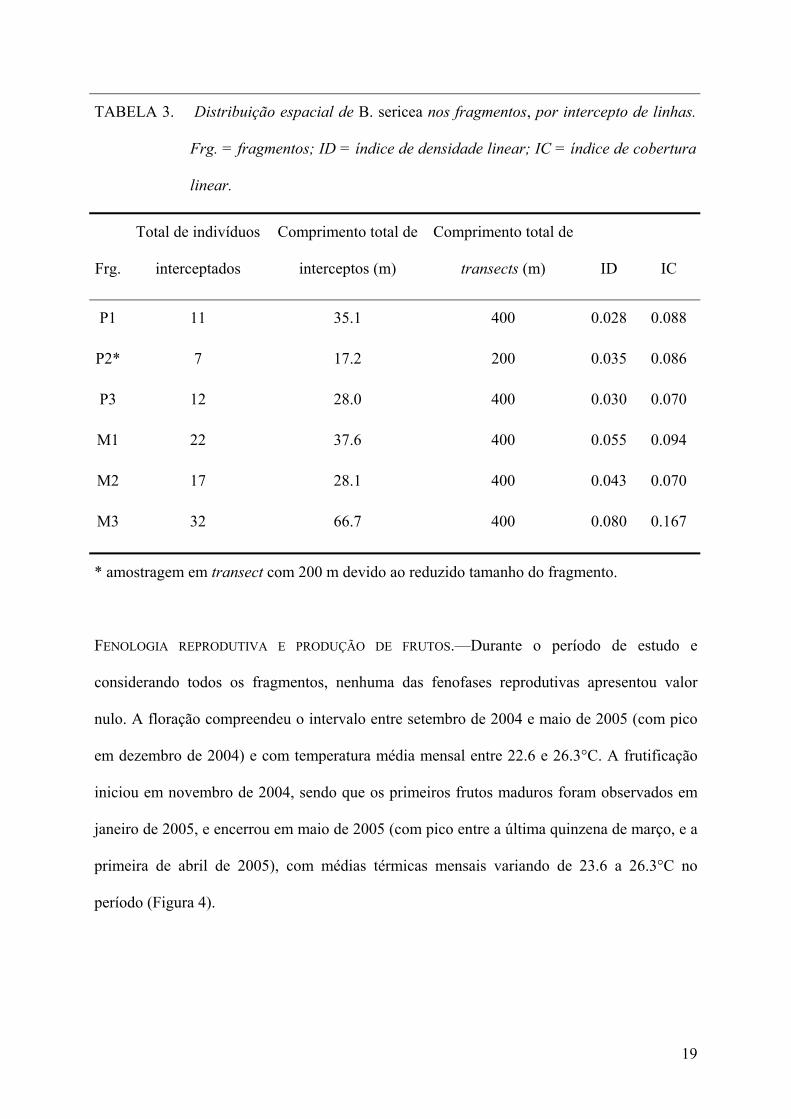
TABELA 3. Distribuição espacial de B. sericea nos fragmentos, por intercepto de linhas.
Frg. = fragmentos; ID = índice de densidade linear; IC = índice de cobertura
linear.
Frg.
Total de indivíduos
interceptados
Comprimento total de
interceptos (m)
Comprimento total de
transects (m) ID IC
P1 11 35.1 400 0.028 0.088
P2* 7 17.2 200 0.035 0.086
P3 12 28.0 400 0.030 0.070
M1 22 37.6 400 0.055 0.094
M2 17 28.1 400 0.043 0.070
M3 32 66.7 400 0.080 0.167
* amostragem em transect com 200 m devido ao reduzido tamanho do fragmento.
FENOLOGIA REPRODUTIVA E PRODUÇÃO DE FRUTOS.—Durante o período de estudo e
considerando todos os fragmentos, nenhuma das fenofases reprodutivas apresentou valor
nulo. A floração compreendeu o intervalo entre setembro de 2004 e maio de 2005 (com pico
em dezembro de 2004) e com temperatura média mensal entre 22.6 e 26.3°C. A frutificação
iniciou em novembro de 2004, sendo que os primeiros frutos maduros foram observados em
janeiro de 2005, e encerrou em maio de 2005 (com pico entre a última quinzena de março, e a
primeira de abril de 2005), com médias térmicas mensais variando de 23.6 a 26.3°C no
período (Figura 4).
19
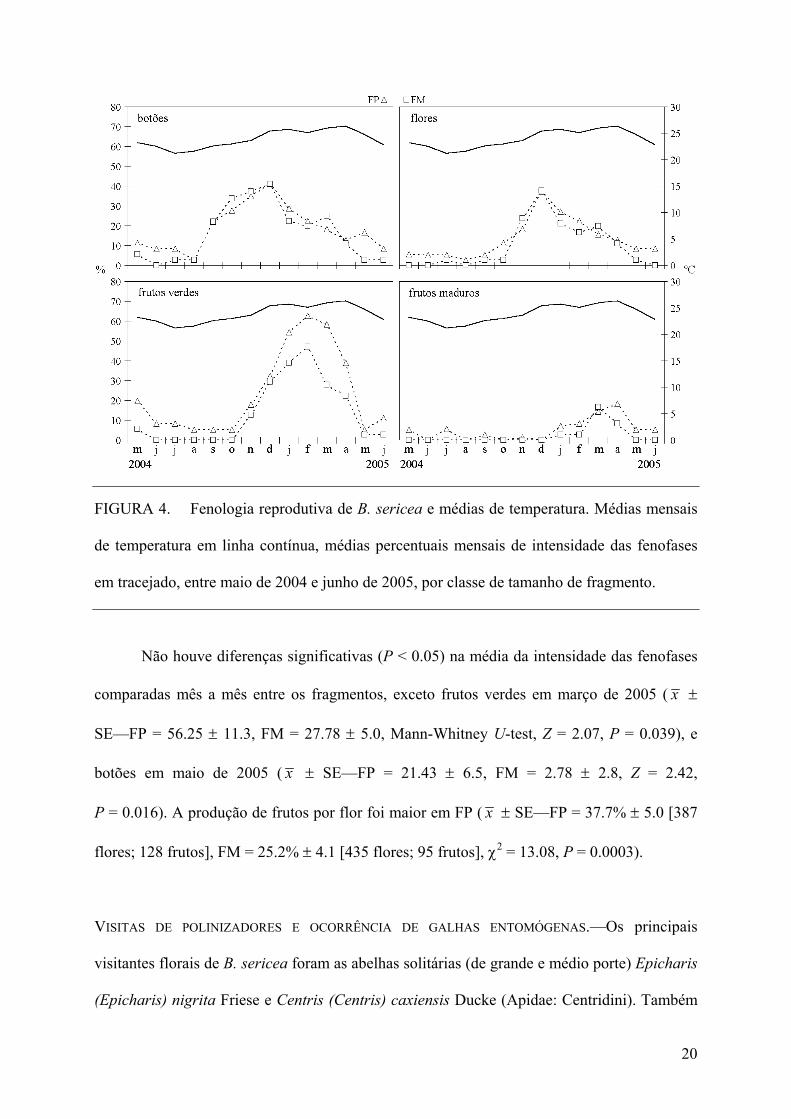
FIGURA 4. Fenologia reprodutiva de B. sericea e médias de temperatura. Médias mensais
de temperatura em linha contínua, médias percentuais mensais de intensidade das fenofases
em tracejado, entre maio de 2004 e junho de 2005, por classe de tamanho de fragmento.
Não houve diferenças significativas (P < 0.05) na média da intensidade das fenofases
comparadas mês a mês entre os fragmentos, exceto frutos verdes em março de 2005 ( x ±
SE—FP = 56.25 ± 11.3, FM = 27.78 ± 5.0, Mann-Whitney U-test, Z = 2.07, P = 0.039), e
botões em maio de 2005 ( x ± SE—FP = 21.43 ± 6.5, FM = 2.78 ± 2.8, Z = 2.42,
P = 0.016). A produção de frutos por flor foi maior em FP ( x ± SE—FP = 37.7% ± 5.0 [387
flores; 128 frutos], FM = 25.2% ± 4.1 [435 flores; 95 frutos], χ2 = 13.08, P = 0.0003).
VISITAS DE POLINIZADORES E OCORRÊNCIA DE GALHAS ENTOMÓGENAS.—Os principais
visitantes florais de B. sericea foram as abelhas solitárias (de grande e médio porte) Epicharis
(Epicharis) nigrita Friese e Centris (Centris) caxiensis Ducke (Apidae: Centridini). Também
20
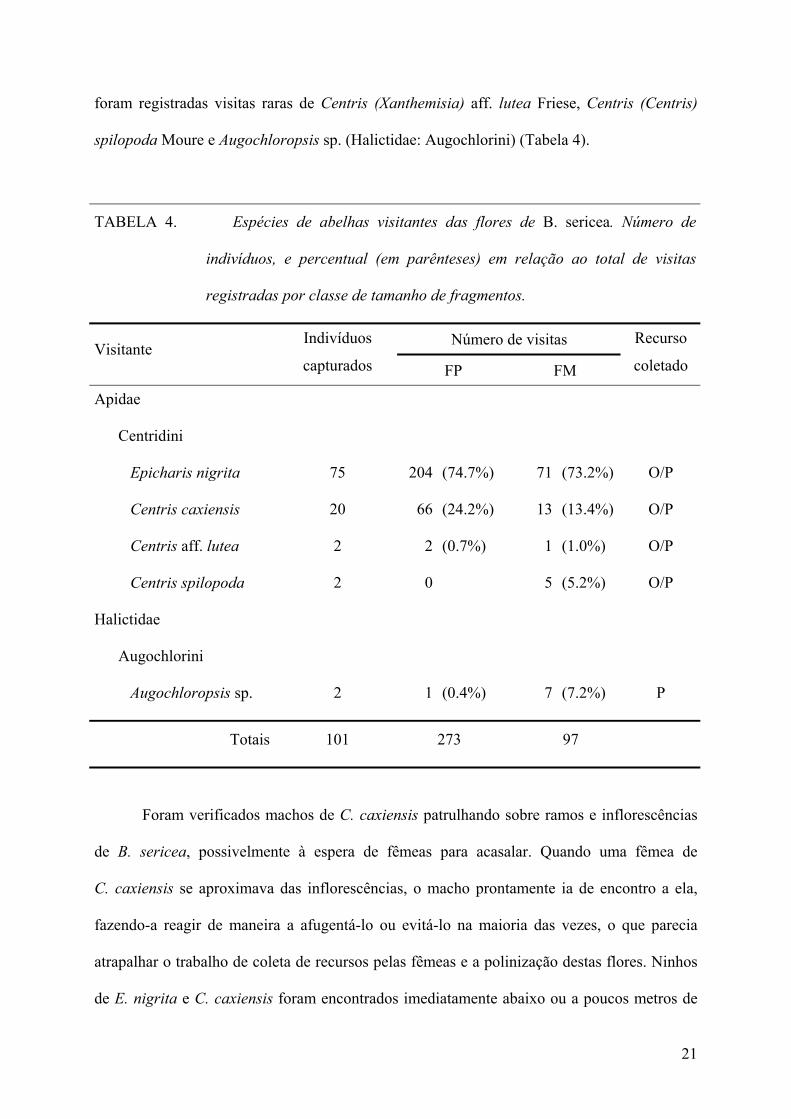
foram registradas visitas raras de Centris (Xanthemisia) aff. lutea Friese, Centris (Centris)
spilopoda Moure e Augochloropsis sp. (Halictidae: Augochlorini) (Tabela 4).
TABELA 4. Espécies de abelhas visitantes das flores de B. sericea. Número de
indivíduos, e percentual (em parênteses) em relação ao total de visitas
registradas por classe de tamanho de fragmentos.
Número de visitas Visitante Indivíduos
capturados FP FM
Recurso
coletado
Apidae
Centridini
Epicharis nigrita 75 204 (74.7%) 71 (73.2%) O/P
Centris caxiensis 20 66 (24.2%) 13 (13.4%) O/P
Centris aff. lutea 2 2 (0.7%) 1 (1.0%) O/P
Centris spilopoda 2 0 5 (5.2%) O/P
Halictidae
Augochlorini
Augochloropsis sp. 2 1 (0.4%) 7 (7.2%) P
Totais 101 273 97
Foram verificados machos de C. caxiensis patrulhando sobre ramos e inflorescências
de B. sericea, possivelmente à espera de fêmeas para acasalar. Quando uma fêmea de
C. caxiensis se aproximava das inflorescências, o macho prontamente ia de encontro a ela,
fazendo-a reagir de maneira a afugentá-lo ou evitá-lo na maioria das vezes, o que parecia
atrapalhar o trabalho de coleta de recursos pelas fêmeas e a polinização destas flores. Ninhos
de E. nigrita e C. caxiensis foram encontrados imediatamente abaixo ou a poucos metros de
21

distância de alguns indivíduos de B. sericea, tanto em FP como em FM (Figura 5).
FIGURA 5. Entradas de ninhos de abelhas solitárias. Esquerda, entrada (seta) de ninho de
Centris caxiensis, situada na lateral de uma pequena duna de areia contendo vegetação em P1;
centro, entrada (seta) de ninho de Epicharis nigrita em trecho arenoso plano (em P1),
imediatamente embaixo de um indivíduo de Byrsonima sericea; direita, entrada (cone
arenoso) de ninho de abelha solitária não identificada, em M1 embaixo de um indivíduo de
Allagoptera arenaria (Arecaceae) (barras = 1 cm).
A composição e a freqüência relativa dos polinizadores em FP e FM foi similar,
excetuando que Centris spilopoda foi observada apenas em FM (Tabela 4). Porém, a
freqüência total de visitas foi maior em FP que em FM (visitas totais/167 min. x ± SE—FP =
91.0 ± 9.1, FM = 32.3 ± 12.7, t = 3.77, P = 0.02). Epicharis nigrita e C. spilopoda realizaram
vibração dos músculos de vôo para coleta de pólen, comportamento que não foi verificado
para C. caxiensis, C. aff. lutea e Augochloropsis sp. Também foram observadas algumas
visitas de diminutas abelhas do gênero Dialictus (Halictidae: Halictini) (Figura 6) e uma
espécie de Vespidae, entretanto não foi possível determinar o tipo de forrageamento e suas
visitas parecem não ter importância para a polinização.
22

FIGURA 6. Visitantes florais de Byrsonima sericea. a. Centris aff. lutea ♀;
b. Epicharis nigrita ♀; c. Centris caxiensis ♀; d. Augochloropsis sp; e. Dialictus sp;
f. Centris spilopoda ♀ (note a escopa bem desenvolvida, em primeiro plano, na perna
posterior esquerda; g. fêmea de Epicharis nigrita coletando óleo dos elaióforos em flores
abertas e também pólen por vibração; h. fêmea de Centris caxiensis coletando óleo em botões
(barras = 5 mm).
23
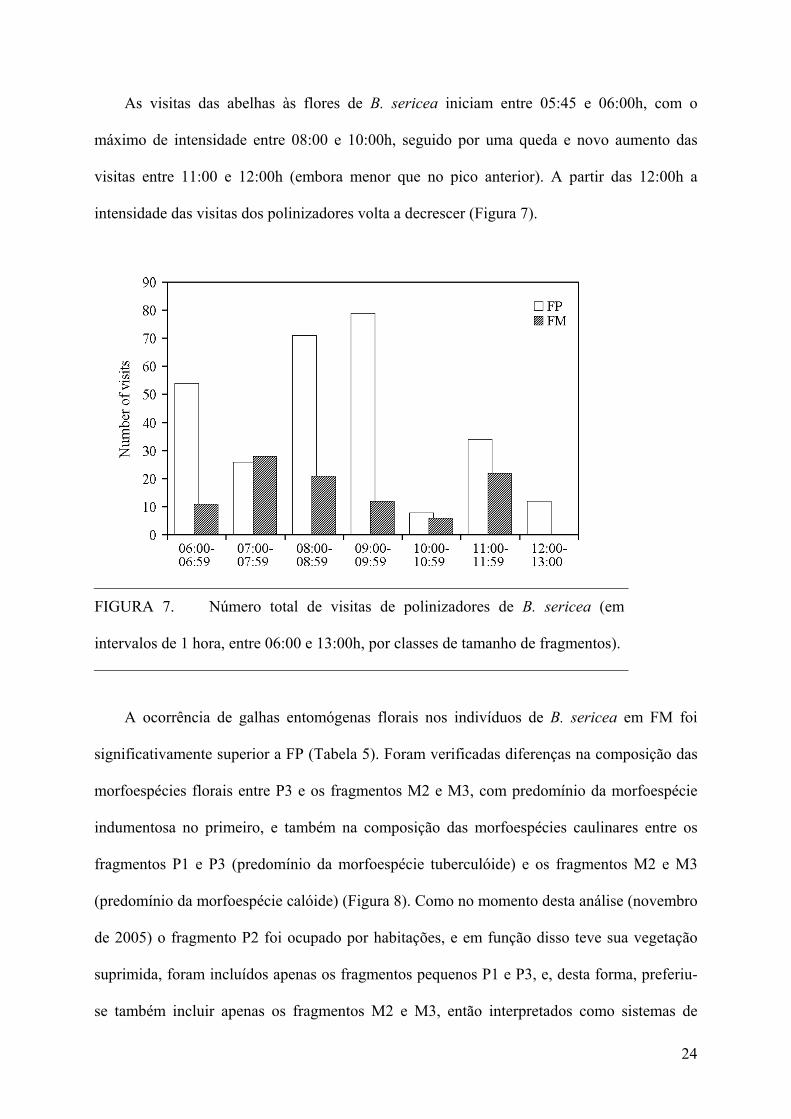
As visitas das abelhas às flores de B. sericea iniciam entre 05:45 e 06:00h, com o
máximo de intensidade entre 08:00 e 10:00h, seguido por uma queda e novo aumento das
visitas entre 11:00 e 12:00h (embora menor que no pico anterior). A partir das 12:00h a
intensidade das visitas dos polinizadores volta a decrescer (Figura 7).
FIGURA 7. Número total de visitas de polinizadores de B. sericea (em
intervalos de 1 hora, entre 06:00 e 13:00h, por classes de tamanho de fragmentos).
A ocorrência de galhas entomógenas florais nos indivíduos de B. sericea em FM foi
significativamente superior a FP (Tabela 5). Foram verificadas diferenças na composição das
morfoespécies florais entre P3 e os fragmentos M2 e M3, com predomínio da morfoespécie
indumentosa no primeiro, e também na composição das morfoespécies caulinares entre os
fragmentos P1 e P3 (predomínio da morfoespécie tuberculóide) e os fragmentos M2 e M3
(predomínio da morfoespécie calóide) (Figura 8). Como no momento desta análise (novembro
de 2005) o fragmento P2 foi ocupado por habitações, e em função disso teve sua vegetação
suprimida, foram incluídos apenas os fragmentos pequenos P1 e P3, e, desta forma, preferiu-
se também incluir apenas os fragmentos M2 e M3, então interpretados como sistemas de
24
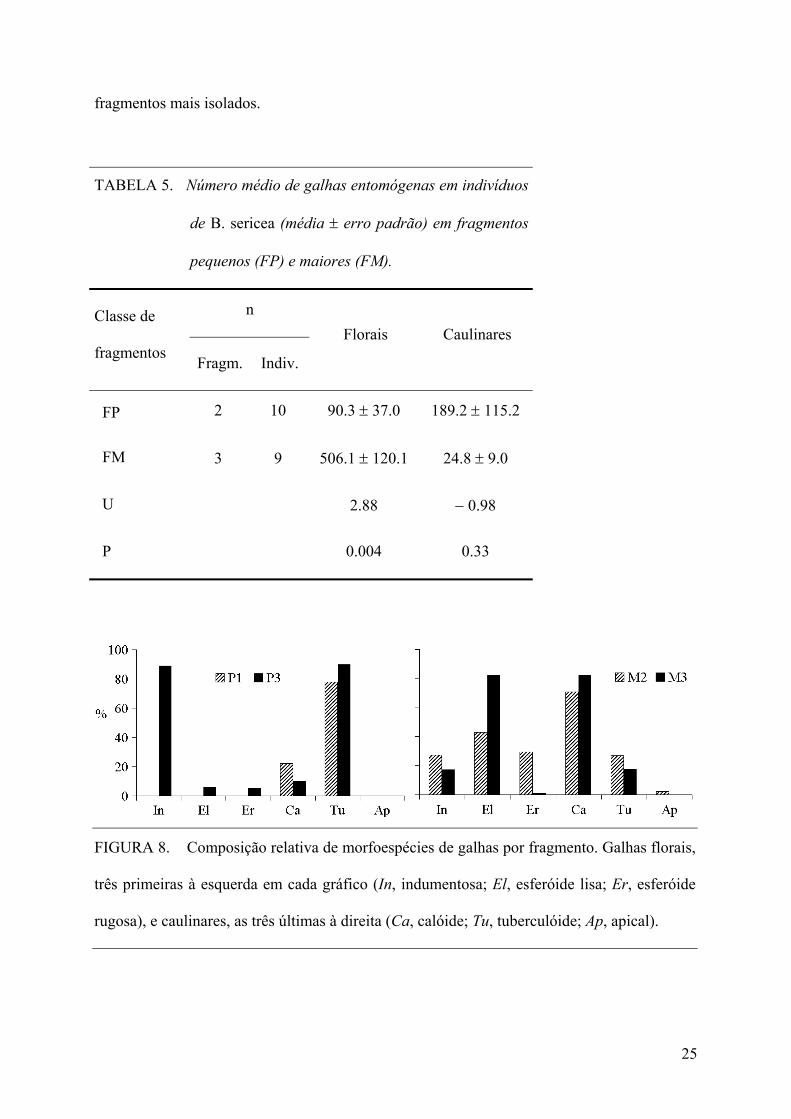
fragmentos mais isolados.
TABELA 5. Número médio de galhas entomógenas em indivíduos
de B. sericea (média ± erro padrão) em fragmentos
pequenos (FP) e maiores (FM).
n Classe de
fragmentos Fragm. Indiv.
Florais Caulinares
FP 2 10 90.3 ± 37.0 189.2 ± 115.2
FM 3 9 506.1 ± 120.1 24.8 ± 9.0
U 2.88 − 0.98
P 0.004 0.33
FIGURA 8. Composição relativa de morfoespécies de galhas por fragmento. Galhas florais,
três primeiras à esquerda em cada gráfico (In, indumentosa; El, esferóide lisa; Er, esferóide
rugosa), e caulinares, as três últimas à direita (Ca, calóide; Tu, tuberculóide; Ap, apical).
25

DISCUSSÃO
Os eventos que iniciaram o processo de isolamento das seis áreas analisadas neste estudo
podem ser considerados relativamente recentes (cinco a seis décadas para alguns, e duas a três
para outros). Estes fragmentos também apresentam diferentes características dentro de cada
uma das duas classes de tamanho, tais como área e formato, grau de isolamento, topografia,
entre outros. Os resultados dos experimentos e observações indicam pouca influência dos
efeitos da fragmentação de habitat sobre as populações de B. sericea, entretanto estes parecem
afetar parte da fauna entomológica associada a elas, tais como polinizadores e galhadores.
Alguns trabalhos mostram padrões diferentes, com aumento na densidade e
dominância de B. sericea, em áreas de restinga quando suas populações são submetidas a
impactos mais intensos e encontram-se em regeneração, tais como desbastes de vegetação (Sá
2002) ou queimadas (Araújo & Peixoto 1977), o que sugere que são favorecidas por tais
perturbações. Em situações semelhantes, porém em estágio mais climácico, sua densidade e
dominância são baixas (Assumpção & Nascimento 2000) sugerindo que esta espécie pertence
à categoria de pioneira tardia na sucessão ecológica.
Como a natureza dos impactos envolvidos no processo de fragmentação da Restinga
de Massambaba é diferente das experimentadas nestas outras áreas, não se tratando de
regeneração mas de áreas remanescentes que foram isoladas pela supressão da vegetação
contínua em seu entorno, a densidade e a cobertura linear de B. sericea nos fragmentos
menores não chegou a ser alterada. É possível que o aumento, nestas áreas, do trânsito de
pessoas e animais domesticados (pisoteio) dificulte o estabelecimento de novos indivíduos, ou
intervenham diretamente em plantas já estabelecidas, cortando-as ou queimando-as
seletivamente para diversos fins. Alguns dos experimentos conduzidos nos fragmentos
menores tiveram parte dos ramos tratados (ou mesmo o indivíduo todo) cortados ou
queimados para loteamentos ilegais, ou possivelmente como forma de hostilizar uma possível
26

intervenção em favor da desocupação das referidas áreas.
Durante as incursões de campo, foi possível perceber visualmente uma tendência no
aumento do número de flores abertas e frutos produzidos nos fragmentos menores, o que foi
detectado pelo método de Fournier apenas em duas ocasiões, cada uma em um dado mês
(março e maio de 2005 respectivamente, para frutos verdes e botões). Certas espécies vegetais
submetidas a efeitos deletérios decorrentes da redução da área natural (alteração do
microclima e dos regimes hídrico e trófico) amplificam a produção de flores por indivíduo
(Johnson et al. 2004), o que parece ser o caso de B. sericea em certas áreas de restinga (C. F.
C. Sá, comun. pess.), embora estudos sistemáticos neste sentido não tenham sido realizados.
Diferenças nas fenofases reprodutivas de B. sericea que não foram detectadas pelo percentual
de intensidade de Fournier poderiam ser evidenciadas pela contagem (ou outra estimativa)
direta de frutos produzidos em função do número de flores nos racemos de cada indivíduo
analisado, ou, de botões emitidos ou flores abertas.
A maior produção de frutos em FP está relacionada à maior freqüência de visitas de
polinizadores nestas áreas, associada ao sistema reprodutivo auto-incompatível de
B. sericea e também à presença nestas populações de uma quantidade de indivíduos ainda
satisfatória para a efetivação de polinizações cruzadas. A observação de ninhos das principais
abelhas polinizadoras em FP indica que estas abelhas não encontram maiores dificuldades em
se estabelecerem nestas áreas. Para abelhas nativas a fragmentação de habitat pode resultar
em substituição por espécies exóticas (Aizen & Feinsinger 1994b) e os efeitos negativos sobre
as primeiras estão mais ligados à qualidade dos fragmentos do que à sua área ou grau de
isolamento (Gathmann et al. 1994, Tscharntke et al. 1998). A mobilidade dos polinizadores é
menos suscetível ao arranjo dos fragmentos na paisagem do que para outros grupos de insetos
(Cane 2001), e este parece ser o caso para as populações de B. sericea e seus polinizadores
nas áreas estudadas na restinga de Massambaba.
27

A raridade e a variabilidade populacional podem estar fortemente relacionadas (Kruess
& Tscharntke 1994, Zabel & Tscharntke 1998), e esta hipótese poderia explicar em parte a
ausência de Centris spilopoda em FP, de rara ocorrência em FM. O maior número de visitas
de abelhas nos fragmentos menores foi conseqüência do menor ataque dos galhadores às
partes florais, uma vez que a quantidade de flores visitadas tende a estar relacionada
positivamente ao número de flores abertas (Bosch et al. 2002). Como a área em um fragmento
menor foi significativamente reduzida, é possível que as abelhas forrageiem mais
intensamente em busca de recursos (óleo e pólen), em um mesmo indivíduo, ou em poucos
indivíduos, o que conduziria a um aumento na freqüência de visitas, porém não
necessariamente a um aumento no fluxo de pólen.
A riqueza de abelhas polinizadoras em B. sericea encontrada neste estudo foi similar
àquela da restinga de Maricá (Rodrigues 2002). Na floresta úmida perenifólia (Mata
Atlântica) de Pernambuco (Teixeira & Machado 2000), a riqueza de abelhas foi bem superior
(19 espécies) e duas espécies foram comuns a este trabalho: E. nigrita e C. spilopoda.
Contudo, a composição das espécies de abelhas polinizadoras nas áreas estudadas foi
diferente daquela registrada na mesma época do ano (outubro a fevereiro) em Maricá e no
Parque Ecológico Chico Mendes, no Rio de Janeiro, não havendo nenhuma espécie em
comum (Rodrigues 2002). Isso mostra que menores distâncias geográficas não
necessariamente refletem maior similaridade entre duas áreas distintas, e que a perda de áreas
naturais regionalmente próximas pela ocupação humana pode extinguir importantes relações
específicas locais.
A menor ocorrência de galhadores florais nos fragmentos menores poderia estar
relacionada, por exemplo, com uma maior ocorrência de insetos parasitóides ou dificuldade
de estabelecimento dos galhadores adultos nestas áreas. Parasitóides compreendem insetos,
geralmente euritomídeos (Hymenoptera: Eurytomidae), que são inimigos naturais de
28

galhadores (Monteiro et al. 2004). Foram observados ataques de possíveis himenópteros
parasitóides em galhas florais da morfoespécie esferóide-lisa, em um dos fragmentos grandes
(Figura 9). Deve-se considerar, ainda, que a freqüência da ocorrência de galhas em um
fragmento de restinga pode estar associada ao tipo de formação vegetal, entretanto, tais
relações ecológicas ainda não estão bem esclarecidas e demandam maior número de pesquisas
neste sentido (Monteiro et al. 2004).
FIGURA 9. Galhas florais predadas. Estas galhas, da
morfoespécie esferóide lisa, foram possivelmente submetidas a
ataques de himenópteros parasitóides (barra = 5 mm).
A qualidade da matriz circundante e do próprio fragmento pode influenciar no
movimento de alguns insetos e em sua habilidade em colonizar plantas hospedeiras (Jonsen et
al. 2001). De um “ponto de vista alado” (Hunter 2002), em se tratando de insetos, a
conectividade entre ambientes mais ou menos isolados é mais um aspecto da biologia das
espécies envolvidas do que propriamente da estrutura da paisagem (Haslet 2001, Hunter
2002). Assim, de modo geral para os insetos, e mais especificamente para os fitófagos
29

(galhadores) na Restinga de Massambaba, outros efeitos, associados (i.e., alteração do
microclima e da cadeia trófica, influência da borda e da matriz, aumento do trânsito de
pessoas e animais domesticados) ou não (i.e., níveis relativos de exposição eólica e marinha,
topografia do terreno, disponibilidade relativa do lençol freático, tipo de formação vegetal),
que não apenas o isolamento relativo e o desenho dos fragmentos, mas que definam mais a
qualidade do habitat, interfiram no estabelecimento destes insetos em função de sua biologia
intrínseca e inter-relações.
A desestabilização de processos mutualísticos, antagonísticos e em níveis tróficos
causados pela fragmentação de habitat pode afetar as interações entre as plantas e seus
polinizadores, competidores e inimigos naturais (Hoffmeister et al. 2005). Os ciclos de
fragmentação na Restinga de Massambaba são relativamente recentes (cerca de cinco a seis
décadas), o que pode ainda representar um curto espaço de tempo para possíveis alterações na
diversidade genética das populações de B. sericea ou das espécies de abelhas que as
polinizam. Entretanto, a supressão de galhadores florais nos fragmentos menores e a redução
de indivíduos de B. sericea em função da própria redução destas áreas, e que produziriam
relativamente mais flores, poderiam conduzir a um aumento na taxa de endogamia para esta
espécie. Uma hipótese a ser futuramente investigada seria a possibilidade destes fatores
alterarem os padrões de campanhas de forrageio intrafragmentos das abelhas resilientes nestas
áreas reduzidas, que poderiam visitar mais flores em cada vez menos indivíduos para
satisfazer suas necessidades de recursos florais (Figura 10).
Parece razoável supor que os fragmentos maiores sejam mais semelhantes funcional e
estruturalmente a um sistema maior não perturbado (Rambaldi & Oliveira 2003), como seria a
restinga contínua, há cerca de seis décadas. Assim, o padrão de ocorrência das galhas florais
nos fragmentos maiores seria o mais próximo de um sistema contínuo e, ainda, pode significar
um atributo ecológico em que ao reduzir a quantidade de flores disponíveis por indivíduo em
30

B. sericea, obrigaria os polinizadores a forragear em mais indivíduos diferentes por unidade
de área.
FIGURA 10. Hipótese de forrageamento de uma abelha solitária – em uma mesma área,
em momentos distintos: antes (A) e após (B) sofrer os efeitos da fragmentação de habitat.
Os círculos correspondem a indivíduos de B. sericea, sendo que a produção de flores,
suprimida por galhadores em “A”, está representada por diâmetros diferentes. Em “B”,
contornos contínuos indicam a situação atual do tamanho do fragmento e dos indivíduos
existentes; em tracejado, a situação anterior. A estrela indica um ninho de uma abelha
solitária, que parte para uma campanha de forrageio, seguindo o trajeto pontilhado (com
setas). Note que em “A”, a abelha precisou forragear 12 vezes, em mais indivíduos de
B. sericea, ficando menos tempo em cada um; em “B”, a abelha forrageou mais vezes em
menos indivíduos, o que resulta em aumento na freqüência de visitas, ou, ainda, o tempo
de forrageio em um mesmo indivíduo poderia ser ampliado.
31

Ainda neste raciocínio, como se tratam de abelhas de médio porte, o que lhes
conferiria uma maior autonomia de vôo, ou seja, maior potencial para forragear em indivíduos
mais separados espacialmente (Ghazoul & Shaanker 2004) é provável que o papel de tais
abelhas na variabilidade genética populacional em B. sericea seja realmente importante,
considerando que B. sericea possui sistema reprodutivo totalmente (Teixeira & Machado
2000), ou preferencialmente (Silva 1990, Rodrigues 2002) auto-incompatível e cuja
autopolinização também é dificultada pela presença do opérculo protegendo a superfície
estigmática antes e após a antese (Rodrigues 2002).
LITERATURA CITADA
AIZEN, M.A., ASHWORTH, L. E GALETTO, L. 2002a. Reproductive success in fragmented
habitats: do compatibility systems and pollination specialization matter? Journal of
Vegetation Science 13(6): 885-892.
______, E FEINSINGER, P. 1994a. Forest fragmentation, pollination, and plant reproduction in
a Chaco dry forest, Argentina. Ecology 75(2): 330-351.
______, E ______. 1994b. Habitat fragmentation, native insect pollinators, and feral
honeybees in argentine “Chaco Serrano”. Ecological Applications 4(2): 378-392.
______, VÁZQUEZ, D.P., SMITH-RAMÍREZ, C. 2002b. Historia natural y conservación de los
mutualismos planta-animal del bosque templado de Sudamérica austral. Revista Chilena
de Historia Natural 75: 79-97.
ANDERSON, W.R. 1979. Floral conservatism in Neotropical Malpighiaceae. Biotropica 11:
219-223.
ARAUJO, D.S.D. 2000. Análise florística e fitogeográfica das restingas do Estado do Rio de
Janeiro. Tese de Doutorado, Curso de Pós-graduação em Ecologia. Instituto de Biologia,
Universidade Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, Estado do Rio de Janeiro,
32

176p.
______ & PEIXOTO, A.L. 1977. Renovação de uma comunidade vegetal de restinga após uma
queimada. Trabalhos no XXVI Congresso Nacional de Botânica. Academia Brasileira de
Ciências. P. 1-17.
______, SCARANO, F.R., SÁ, C.F.C.DE, KURTZ, B.C., ZALUAR, H.T., MONTEZUMA, R.C.M.,
OLIVEIRA, R. C. DE. 1998. Comunidades vegetais do Parque Nacional da Restinga de
Jurubatiba. In: Esteves, F. A. (ed.). Ecologia das lagoas costeiras do Parque Nacional da
Restinga de Jurubatiba e do município de Macaé (RJ). Núcleo de Pesquisas Ecológicas de
Macaé (NUPEM) – Universidade Federal do Rio de Janeiro, 464p.
ASSUMPÇÃO, J. & NASCIMENTO, M.T.N. 2000. Estrutura e composição florística de quatro
formações vegetais de restinga no complexo lagunar Grussaí/Iquipari, São João da Barra,
RJ, Brasil. Acta Bot. Bras. 14(3): 301-315.
BAWA, K.S. 1990. Plant-pollinator interactions in tropical rain forests. Annual Review of
Ecology and Systematics 21: 399-422.
BENCKE, C.S.C. & MORELLATO, L.P.C. 2002a. Comparação de dois métodos de avaliação da
fenologia de plantas, sua interpretação e representação. Revista Brasil. Bot. 25(3): 269-
275.
______. 2002b. Estudo comparativo da fenologia de nove espécies arbóreas em três tipos de
floresta atlântica no sudeste do Brasil. Revista Brasil. Bot. 25(2): 237-248.
BIDEGAIN, P. & BIZERRIL C. (orgs.) 2002. Lagoa de Araruama: perfil ambiental do maior
ecossistema lagunar hipersalino do mundo. Rio de Janeiro: Semads. 160 p. il.
BOLGER, D.T., SUAREZ, A.V., CROOKS, K.R., MORRISON, S.A., CASE, T.J. 2000. Arthropods
in urban habitat fragments in southern California: area, age, and edge effects. Ecological
Applications 10(4): 1230-1248.
33

BOSCH, M., SIMON, J., ROVIRA, A.M., MOLERO, BLANCHÉ, C. 2002. Pollination ecology of
the pre-Pyrenean endemic Petrocoptis montsicciana (Caryophyllaceae): effects of
population size. Biological Journal of the Linnean Society 76: 79-90.
BROWER, J.E. & ZAR, J.H. 1984. Field & laboratory methods for general ecology. 2 ed., Wm.
Brown Co., Dubuque. 226p.
CANE, J.H. 2001. Habitat fragmentation and native bees: a premature veredict? Conservation
Ecology 5: 149-161.
FENSTER, C.B., ARMBRUSTER, W.S., WILSON, P., DUDASH, M.R., THOMSON, J.D. 2004.
Pollination syndromes and floral specialization. Annual Review of Ecological Evolution
and Systematics 35: 375-403.
FLINTE, V., ARAUJO, C.O., MACEDO, M.V. & MONTEIRO, R.F. 2003. Comunidade de insetos
associada a Byrsonima sericea (Malpighiaceae) na Restinga de Jurubatiba, RJ:
observações preliminares. In: VI Congresso de Ecologia do Brasil, Fortaleza.
FOURNIER, L.A. 1974. Un método cuantitativo para la medición de características fenológicas
en árboles. Turrialba 24: 422-423.
GATHMANN, A., GREILER, H.J., TSCHARNTKE, T. 1994. Trap-nesting bees and wasps
colonizing set-aside fields: succession and body size, management by cutting and sowing.
Oecologia 98: 8-14.
GENTRY, A.H. 1974. Flowering phenology and diversity in tropical Bignoniaceae. Biotropica
6(1): 64-68.
GHAZOUL, J. & SHAANKER, R.U. 2004. Sex in space: a synthesis. Biotropica 36(2): 180-183.
HASLET, J. R. 2001. Biodiversity and conservation of Diptera in heterogeneous land mosaics:
a fly’s eye view. Journal of Insect Conservation 5: 71-75.
HINTZE, J. 2004. NCSS-PASS. Number Cruncher Statistical System-Power Analysis and
Sample Size. Kaysville, Utah.
34

HOFFMEISTER, T.S., VET, L.E.M., BIERE, A. HOLSINGER, K., FILSER, J. 2005. Ecological and
evolutionary consequences of biological invasion and habitat fragmentation. Ecosystems
8: 657-667.
HUNTER, M.D. 2002. Landscape structure, habitat fragmentation, and the ecology of insects.
Agricultural and Forest Entomology 4: 159-166.
JACQUEMYN, H., BRYS, R. & HERMY, M. 2002. Patch occupancy, population size and
reproductive success of a forest herb (Primula elatior) in a fragmented landscape.
Oecologia (Berl.) 130: 617-625.
JOHNSON, S.D., COLLIN, C.L., WISSMAN, H.J., HALVARSSON, E., ÅGREN, J. 2004. Factors
contributing to variation in seed production among remnant populations of the endangered
daisy Gerbera aurantiaca. Biotropica 36(2): 148-155.
JONSEN, I.D., BOURCHIER, R.S. & ROLAND, J. 2001. The influence of matrix habitat on
Aphthona flea beetle immigration to leafy spurge patches. Oecologia 127: 287-294.
KRUESS A. & TSCHARNTKE, T. 1994. Habitat fragmentation, species loss, and biological
control. Science 264: 1581-1584.
MONTEIRO, R.F., ODA, R.A.M., NARAHARA, K.L. & CONSTANTINO, P.A.L. 2004. Galhas:
diversidade, especificidade e distribuição. In: Rocha, C.F.D., Esteves, F.A. & Scarano,
F.R. (orgs.). 2004. Pesquisas de longa duração na Restinga de Jurubatiba: ecologia,
história natural e conservação. São Carlos: RiMa, 376p.
PEREIRA, M.C.A., ARAUJO, D.S.D., PEREIRA, O.J. 2001. Estrutura de uma comunidade
arbustiva da restinga de Barra de Maricá - RJ. Revista Brasil. Bot. 24(3): 273-281.
PINTO, M.S. 1947. A indústria de álcalis no Brasil. Ministério da Agricultura. DNPM,
Boletim nº 28. Rio de Janeiro. 154 p. il.
PRADO, S.M. 2002. Da anchova ao salário mínimo: uma etnografia sobre injunções de
mudança social em Arraial do Cabo. Niterói: EdUFF. 145 p. il.
35

QUESADA, M., STONER, K.E., LOBO, J.A., HERRERÍAS-DIEGO, Y., PALACIOS-GUEVARA, C.,
MUNGUÍA-ROSAS, M.A., O.-SALAZAR, K.A., ROSAS-GUERRERO, V. 2004. Effects of forest
fragmentation on pollinator activity and consequences for plant reproductive success and
mating patterns in bat-pollinated bombacaceous trees. Biotropica 36(2): 131-138.
RAMALHO, M. & SILVA, M. 2002. Flora oleífera e sua guilda de abelhas em uma comunidade
de restinga tropical. Sitientibus série Ciências Biológicas 2 (1/2): 34-43.
RAMBALDI, D.M. & OLIVEIRA, D.A.S. (orgs.). 2003. Fragmentação de ecossistemas: Causas,
efeitos sobre a biodiversidade e recomendações de políticas públicas. Brasília:
MMA/SBF, 510p.
RODRIGUES, L.F.M. 2002. Biologia floral e sistema de reprodução de Byrsonima sericea DC.
(Malpighiaceae). Dissertação de Mestrado, Curso de Ciências Biológicas (Botânica).
Museu Nacional, Universidade Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, Estado
do Rio de Janeiro, 82p.
ROUBIK, D. W. 1989. Ecology and natural history of tropical bees. Cambridge: Cambridge
University Press, 514p.
SÁ, C.F.C.DE. 2002. Regeneração de um trecho de floresta de restinga na Reserva Ecológica
Estadual de Jacarepiá, Saquarema, Estado do Rio de Janeiro: II – Estrato arbustivo.
Rodriguésia 53(82): 5-23.
SEMADS. 2002. Mapa ambiental da lagoa de Araruama. Rio de Janeiro: Projeto Planágua. 1
mapa, color., 94 cm. Escala 1: 50.000.
SILVA, S.I. 1990. Floração e frutificação de duas variedades de Byrsonima sericea DC.
Dissertação. Curso de Mestrado da Universidade Federal Rural de Pernambuco. Recife-
PE.
SILVEIRA, F.A., MELO, G.A.R., ALMEIDA, E.A.B. 2002. Abelhas brasileiras: sistemática e
identificação. 1. ed. Belo Horizonte: MMA, PROBIO-PNUN. Fundação Araucária. 253 p.
36

il.
SOMANATHAN, H., BORGES, R.M., CHAKRAVARTHY, V.S. 2004. Does neighborhood floral
display matter? Fruit set in carpenter bee-pollinated Heterophragma quadriloculare and
beetle-pollinated Lasiosiphon eriocephalus. Biotropica 36(2): 139-147.
TEIXEIRA, L.A.G. & MACHADO, I.C. 2000. Sistema de polinização e reprodução de
Byrsonima sericea DC (Malpighiaceae). Acta. Bot. Bras. 14(3): 347-357.
TSCHARNTKE, T., GATHMANN, A., STEFFAN-DEWENTER, I. 1998. Bioindication using trap-
nesting bees and wasps and their natural enemies: community structure and interactions.
Journal of Applied Ecology 35: 708-719.
VOGEL, S. 1990. History of the Malpighiaceae in the light of pollination ecology. Memories
of the New York Botanical Garden 55: 130-142.
ZABEL, J. & TSCHARNTKE, T. 1998. Does fragmentation of Urtica habitats affect
phytophagous and predatory insects differentially? Oecologia 116: 419-425.
ZEISLER, M. 1938. Über die Abgrenzung der eigentlichen Narbenflüche mit Hilfe von
Reaktionen. Beiheft Botanisches Zentralblatt 58: 308-318.
37

CONCLUSÕES GERAIS
Considerando a escala espacial do presente trabalho, determinada pelo desenho dos
fragmentos nas áreas estudadas e a escala de tempo relativamente recente dos processos
antrópicos envolvidos no isolamento destas áreas, a fragmentação de habitat surtiu pouco
efeito sobre as populações de Byrsonima sericea em formação arbustiva de restinga, e
influenciou mais diretamente na dinâmica de seus principais polinizadores e
estabelecimento de insetos galhadores de suas flores. Tais efeitos resultaram em maior
freqüência de visitas dos polinizadores às flores e redução da ocorrência de galhas florais
nas populações de B. sericea em fragmentos menores, que, combinadamente, resultaram
em maior produção de frutos nestas áreas mais fragmentadas.
Assim, a fragmentação de habitat favoreceu B. sericea no atual estado em que se
encontra. Entretanto, em estágios mais avançados da fragmentação em curso, ou
analisando outros aspectos não investigados neste trabalho (sucesso na dispersão de
sementes e estabelecimento de plântulas, dinâmica de nutrientes no sistema, etc.), é
possível que ocorram efeitos negativos sobre estas populações.
A imensa complexidade que o tema da fragmentação de habitat demanda, reflete a
complexidade das interações entre os componentes de um ecossistema (Rambaldi &
Oliveira 2003), ainda que consideremos apenas os componentes bióticos. Hoffmeister et al.
(2005) propõem que estudos sobre efeitos da fragmentação de habitat devem integrar
“elementos estáticos” (i.e., alterações na cadeia trófica e na abundância e riqueza de
espécies) com “elementos dinâmicos” (i.e., processos evolucionários sendo modificados
pela plasticidade fenotípica dos indivíduos e mudança de genótipo nas populações
envolvidas). A ampliação de abordagens de estudos para o sistema envolvendo a
fragmentação da restinga de Massambaba e Byrsonima sericea certamente fornecerá
elementos para melhor compreensão dos efeitos da fragmentação, tanto no seu componente
teórico como para subsídio para ações de conservação.
38

REFERÊNCIAS BIBLIOGRÁFICAS
Aizen, M.A., Ashworth, L. & Galetto, L. 2002a. Reproductive success in fragmented habitats: do compatibility systems and pollination specialization matter? Journal of Vegetation Science 13(6): 885-892.
______, Vázquez, D.P., Smith-Ramírez, C. 2002b. Historia natural y conservación de los
mutualismos planta-animal del bosque templado de Sudamérica austral. Revista Chilena de Historia Natural 75: 79-97.
Araujo, D.S.D. 1997. Cabo Frio Region. In: Davis, S.D., Heywood, V.H., Herrera-
MacBryde, O., Villa-Lobos, J. & Hamilton, A.C. (eds). Centers of Plant Diversity: a guide and strategy for their conservation. v.3, The Americas, WWF/IUCN, Oxford: 373-375.
______. 2000. Análise florística e fitogeográfica das restingas do Estado do Rio de Janeiro.
Tese de Doutorado, Curso de Pós-graduação em Ecologia. Instituto de Biologia, Universidade Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, Estado do Rio de Janeiro, 176p.
______, Pereira, M.C.A., Pimentel, M.C.P. 2004. Flora e estrutura de comunidades
na Restinga de Jurubatiba – síntese dos conhecimentos com enfoque especial para formação aberta de Clusia. In: Rocha, C.F.D., Esteves, F.A., Scarano, F.R. (orgs.). Pesquisas de longa duração na Restinga de Jurubatiba: ecologia, história natural e conservação. São Carlos: RiMa, 376p.
______, Scarano, F.R., Sá, C.F.C.de, Kurtz, B.C., Zaluar, H.T., Montezuma, R.C.M.,
Oliveira, R.C.de. 1998. Comunidades vegetais do Parque Nacional da Restinga de Jurubatiba. In: Esteves, F. A. (ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do município de Macaé (RJ). Núcleo de Pesquisas Ecológicas de Macaé (NUPEM) – Universidade Federal do Rio de Janeiro, 464p.
Bawa, K.S. 1990. Plant-pollinator interactions in tropical rain forests. Annual Review of
Ecology and Systematics 21: 399-422. Bidegain, P. & Bizerril C. (orgs.) 2002. Lagoa de Araruama: perfil ambiental do maior
ecossistema lagunar hipersalino do mundo. Rio de Janeiro: Semads. 160 p. il. Bolger, D.T., Suarez, A.V., Crooks, K.R., Morrison, S.A., Case, T.J. 2000. Arthropods in
urban habitat fragments in southern California: area, age, and edge effects. Ecological Applications 10(4): 1230-1248.
Buchmann, S.L. & Nabhan, G.P. 1996. The forgotten pollinators. Island Press, Covelo,
CA. Conservation International 2006. Biodiversity hotspots. Disponível em:
http://www.conservation.org/ImageCache/news/content/press_5freleases/2005/february/hotspots2_5fkit/hotspots2map_2epdf/v1/hotspots2map.pdf. Acesso em: 13 jan. 2006.
39

Cordeiro, S. Z. 2005. Composição e distribuição da vegetação herbácea em três áreas com fisionomias distintas na Praia do Peró, Cabo Frio, RJ, Brasil. Acta Botanica Brasilica 19(4): 679-693.
Cox, P.A. & Elmqvist, T. 2000. Pollinator extinction in the Pacific Islands. Conservation
Biology 14(5): 1237-1239. Endress, P.K. 1994. Diversity and evolutionary biology of tropical flowers. Cambridge:
Cambridge University Press: 162-166. Fonseca-Kruel, V. S. da & Peixoto, A. L. 2004. Etnobotânica na Reserva Extrativista
Marinha de Arraial do Cabo, RJ, Brasil. Acta Botanica Brasilica 18(1): 177-190. Hoffmeister, T.S., Vet, L.E.M., Biere, A. Holsinger, K., Filser, J. 2005. Ecological and
evolutionary consequences of biological invasión and habitat fragmentation. Ecosystems 8: 657-667.
Jacquemyn, H., Brys, R. & Hermy, M. 2002. Patch occupancy, population size and
reproductive success of a forest herb (Primula elatior) in a fragmented landscape. Oecologia (Berl.) 130: 617-625.
Mabberley, D.J. 1993. The plant-book: a portable dictionary of the higher plants.
Cambridge University Press. 4ed. New York. Menezes, L.F.T. & Araujo, D.S.D. 1999. Estrutura de duas formações vegetais do cordão
externo da Restinga de Marambaia, RJ. Acta Botanica Brasilica 13(2): 223-235. Monteiro, R.F., Oda, R.A.M., Narahara, K.L. & Constantino, P.A.L. 2004. Galhas:
diversidade, especificidade e distribuição. In: Rocha, C.F.D., Esteves, F.A. & Scarano, F.R. (orgs.). 2004. Pesquisas de longa duração na Restinga de Jurubatiba: ecologia, história natural e conservação. São Carlos: RiMa, 376p.
Neff, J.L. & Simpson, B.B. 1981. Oil-collecting structures in the Anthophoridae
(Hymenoptera): morphology, function and use in systematics. Journal of the Kansas Entomological Society 54: 95-123.
Paton, D.C. 2000. Disruption of bird-plant pollination systems in southern Australia.
Conservation Biology 14: 1232-1234. Pereira, M.C.A., Araujo, D.S.D., Pereira, O.J. 2001. Estrutura de uma comunidade
arbustiva da restinga de Barra de Maricá - RJ. Revista Brasileira de Botânica 24(3): 273-281.
Pinto, M.S. 1947. A indústria de álcalis no Brasil. Ministério da Agricultura. DNPM,
Boletim nº 28. Rio de Janeiro. 154 p. il. Prado, S.M. 2002. Da anchova ao salário mínimo: uma etnografia sobre injunções de
mudança social em Arraial do Cabo. Niterói: EdUFF. 145 p. il.
40

Rambaldi, D.M. & Oliveira, D.A.S. (orgs.). 2003. Fragmentação de ecossistemas: Causas, efeitos sobre a biodiversidade e recomendações de políticas públicas. Brasília: MMA/SBF, 510p.
Redfern, M. & Askew, R.R. 1992. Plant galls. Richmond Publishing, England. Renner, S.S. 1998. Effects of habitat fragmentation of plant pollinator interactions in the
tropics. In: Newbery, D.M., Prins, H.H.T. & Brown, N.D. (eds.) Dynamics of tropical communities. Blackwell Science, London, UK: 339-360.
Rocha, C. F. D., Van Sluys, M., Bergallo, H. G. & Alves, M. A. S. 2005. Endemic and
threatened tetrapods in the restingas of the biodiversity corridors of Serra do Mar and of the Central da Mata Atlântica in eastern Brazil. Brazilian Journal of Biology 65(1): 159-168.
Rodrigues, L.F.M. 2002. Biologia floral e sistema de reprodução de Byrsonima sericea
DC. (Malpighiaceae). Dissertação de Mestrado, Curso de Ciências Biológicas (Botânica). Museu Nacional, Universidade Federal do Rio de Janeiro (UFRJ), Rio de Janeiro, Estado do Rio de Janeiro, 82p.
Sá, C.F.C.de. 2002. Regeneração de um trecho de floresta de restinga na Reserva
Ecológica Estadual de Jacarepiá, Saquarema, Estado do Rio de Janeiro: II – Estrato arbustivo. Rodriguésia 53(82): 5-23.
Scarano, F. R. 2002. Structure, function and floristic relationships of plant communities in
stressful habitats marginal to the Brazilian Atlantic Rainforest. Annals of Botany 90(4): 517-524.
Semads. 2002. Mapa ambiental da lagoa de Araruama. Rio de Janeiro: Projeto Planágua.
1 mapa, color., 94 cm. Escala 1: 50.000. Silva, S.I. 1990. Floração e frutificação de duas variedades de Byrsonima sericea DC.
Dissertação. Curso de Mestrado da Universidade Federal Rural de Pernambuco. Recife-PE.
Simpson, B.B. & Neff, J.L. 1981. Floral rewards: alternatives to pollen and nectar. Annals
of the Missouri Botanical Garden 68: 301-322. Teixeira, L.A.G. & Machado, I.C. 2000. Sistema de polinização e reprodução de
Byrsonima sericea DC (Malpighiaceae). Acta Botanica Brasilica 14(3): 347-357. Vogel, S. 1990. History of the Malpighiaceae in the light of pollination ecology. Memoirs
of the New York Botanical Garden 55: 130-142.
41






![galeria nara roesler bruno dunley//no lugar em que já estamos...Hamish Fulton’s first solo show in South America, curated by Alexia Tala; and Cães sem plumas [prólogo] curated](https://static.fdocumentos.tips/doc/165x107/5f4c19e860bad950872bf5e3/galeria-nara-roesler-bruno-dunleyno-lugar-em-que-j-estamos-hamish-fultonas.jpg)









