
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA...
57
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA Programa de Pós-graduação em Botânica – (PPG Botânica) GERMINAÇÃO, DESENVOLVIMENTO E RESPOSTA AO ALAGAMENTO DE Macrolobium acaciifolium (Benth.) Benth. (FABACEAE) DE POPULAÇÕES DE VÁRZEA E IGAPÓ DA AMAZÔNIA CENTRAL LUCÉLIA RODRIGUES SANTOS Manaus, Amazonas Março de 2016
-
Upload
truongdiep -
Category
Documents
-
view
212 -
download
0
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
Programa de Pós-graduação em Botânica – (PPG Botânica)
GERMINAÇÃO, DESENVOLVIMENTO E RESPOSTA AO ALAGAMENTO DE
Macrolobium acaciifolium (Benth.) Benth. (FABACEAE) DE POPULAÇÕES DE VÁRZEA
E IGAPÓ DA AMAZÔNIA CENTRAL
LUCÉLIA RODRIGUES SANTOS
Manaus, Amazonas
Março de 2016

i
LUCÉLIA RODRIGUES SANTOS
GERMINAÇÃO, DESENVOLVIMENTO E RESPOSTA AO ALAGAMENTO DE
Macrolobium acaciifolium (Benth.) Benth. (FABACEAE) DE POPULAÇÕES DE VÁRZEA
E IGAPÓ DA AMAZÔNIA CENTRAL
Orientador: Dr. Florian Karl Wittmann
Dissertação apresentada ao Instituto Nacional
de Pesquisas da Amazônia, como parte dos
requisitos para a obtenção do título de Mestre
em Botânica.
Manaus, Amazonas
Março de 2016

ii
S237 Santos, Lucélia Rodrigues
Germinação, desenvolvimento e resposta ao alagamento de Macrolobium acaciifolium (Benth.)
Benth. (Fabaceae) de populações de várzea e igapó da Amazônia Central/ Lucélia Rodrigues
Santos. --- Manaus: [s.n.], 2016.
57 f. : il. color.
Dissertação (Mestrado) --- INPA, Manaus, 2016.
Orientador : Florian Karl Wittmann.
Área de concentração: Botânica.
1. Germinação. 2. Macrolobium acaciifolium. 3. Biomassa.
I. Título.
CDD 583.323
Sinopse
Foram investigados o padrão de germinação, a biomassa de plântulas e as respostas morfológicas ao alagamento
de plântulas de Macrolobium acaciifolium (Benth.) Benth.de populações de várzea e igapó, com o objetivo de
observar se essas características sofrem influência do tipo de solo da localidade onde as populações em estudo
estão localizadas. Foram analisadas a porcentagem de germinação, o índice de velocidade de germinação, a
biomassa e crescimento, o surgimento de alterações morfológicas externas decorrentes do alagamento do
substrato, a quantidade de nutrientes das folhas e do substrato. Os resultados mostraram haver correlação entre o
tipo de substrato e algumas das variáveis estuadas.
Palavras chaves: Germinação, Biomassa, Alagamento do solo, Nutrientes.

iii
À Joaquina Rodrigues
In memorian

iv
AGRADECIMENTOS
Ao Instituto Nacional de Pesquisas da Amazônia pelo apoio oferecido para a
realização do curso de Pós-graduação em Botânica.
Ao Conselho Nacional de Pesquisa e Desenvolvimento Científico e Tecnológico –
CNPq pela concessão da bolsa de estudo.
Ao Projeto MAUÁ/Max Planck pelo apoio logístico.
Ao meu orientador Dr. Florian Wittmann pela orientação e sobretudo, pela paciência
ao indicar quais os caminhos a seguir e me possibilitar conhecer o que há de fascinante nos
ecossistemas alagáveis.
À Dra. Astrid Wittmann pela colaboração com comentários e correções.
Aos professores do curso de Botânica pelo conhecimento compartilhado.
Ao professor/coordenador Mike Hopkins pela disponibilidade e gentileza.
À minha turma de 2014, Yuri Pastor e minhas “cumplices” Marcília Freitas, Raquel
Chaves e Dariene Santos, obrigado pela amizade que espero levar por toda vida.
Aos alunos da turma de 2013 pela acolhida.
Às secretárias, Léia, Helcineide, Jéssica Yara e Carminha pelo auxílio junto à
coordenação de Botânica.
Aos colegas do projeto Max Planck, Mário Picanço, Valdeney, Beth, Kelvin, Celso e
Wallace por toda a ajuda prestada.
A meus irmãos Francisca, Maria Lúcia e Osvaldo pelo apoio que me deram nesses
dois anos, cada um a seu modo.
Aos meus pais Maria e Adão e ao tio José, meu segundo pai, por tudo.
À dona Jacira e seu filho Israel pela amizade que me dedicaram durante minha
permanência em Manaus.
Ao Fábio Nogueira pela disponibilidade de me ajudar em pleno fim de semana,
obrigada pela companhia, as conversas e risadas.

v
“A sabedoria da natureza é tal que não produz nada de supérfluo ou inútil.”
Nicolau Copérnico

vi
RESUMO
As áreas alagáveis amazônicas representam 16% dos 5.000.000 km² da Amazônia Brasileira.
Os principais tipos de áreas inundáveis são as várzeas e os igapós. Ecossistemas estes com
características bem distintas quanto à físico-química da água e do solo; enquanto a várzea é
inundada por águas brancas ricas em nutrientes, o igapó sofre inundação de águas pretas com
uma menor quantidade nutrientes dissolvidos, diferença esta que se reflete no solo dessas
áreas. As espécies vegetais que habitam áreas alagáveis lançam mão mecanismos fisiológicos
específicos para tolerar o estresse da redução de oxigênio durante a inundação. Entretanto,
existem limitações impostas também pela físico-química desses dois ambientes de forma que
poucas espécies podem ser encontradas na várzea e no igapó, como é o caso do Macrolobium
acaciifolium (Benth.) Benth., espécie escolhida para investigar a influência do solo de várzea
e igapó na germinação, crescimento e desenvolvimento de plântulas em casa de vegetação.
Foram analisadas as características biométricas das sementes, a evolução da germinação em
relação ao tempo, a área foliar específica, a quantidade de nutrientes encontrados nas folhas e
a quantidade de biomassa nas plântulas e indivíduos das populações de várzea e igapó em seu
solo de origem e em vermiculita. Considerando a origem, as sementes variam apenas no peso
e na espessura, tendo as sementes de várzea apresentado maiores médias de peso e as de igapó
as maiores médias de espessura. Não houve diferença significativa na porcentagem de
germinação, as médias de comprimento e peso da parte aérea foram maiores nas plântulas de
igapó do que nas de várzea. Não houve diferença nas médias de peso da raiz, no crescimento e
na biomassa. Não houve diferença nas alterações morfológicas externas e nas taxas de
sobrevivência com relação ao alagamento. Não houve diferença significativa com relação ao
comprimento e peso da parte aérea. A menor média de diâmetro foi observada nos indivíduos
de várzea que cresceram em solo de igapó, as médias de área foliar específica não diferiram
estatisticamente entre si. A quantidade de macro e micronutrientes nas folhas de M.
acaciifolium de várzea e igapó variou nos dois tipos de solo e com relação à procedência,
sendo que os indivíduos de igapó em solo de igapó de modo geral exibiram mais nutrientes
nas folhas comparados aos indivíduos de várzea. Os resultados sugerem que os indivíduos de
M. acaciifolium de igapó apresentam uma maior plasticidade adaptativa às condições do meio
do que os indivíduos de M. acaciifolium de várzea.
Palavras chaves: Germinação, Biomassa, Nutrientes.

vii
ABSTRACT
Amazon floodplains represent 16% of all the 5.000.000 km² of the Brazilian Amazon, where
the major floodable areas are várzeas and flooded forests (igapós). These two ecosystems
feature very different characteristics in terms of physical-chemical water and soil: the várzea
is flooded by nutrient-rich white waters, while the igapó is flooded by black waters with fewer
nutrients dissolved. This difference is reflected in the soil of these areas, and in order to
tolerate the stress caused by the decrease of oxygen during flooding times, plant species in
floodplains developed specific physiological mechanisms. However, there are also limitations
imposed by the physical and chemical of these two ecosystems, so that few species can be
found in várzea and flooded forests, such as Macrolobium acaciifolium (Benth.) Benth., the
specie by which we investigated the influence of várzea soil and igapó germination, growth
and development of seedlings in greenhouse. It was analyzed the biometric characteristics of
the seeds, the evolution of germination over time, the specific leaf area, the amount of
nutrients in leaves and the amount of biomass in seedlings and individuals of várzea and
igapó populations on its soil of origin and on vermiculite. In what concerns the origin of the
seed, there was a difference only in thickness since the igapó seeds presented higher
averages, while lowland seeds presented higher averages in weight, but there was no
significant difference in the percentage of germination. The average length and weight of the
aerial part and in the length of the root were higher in seedlings in the igapó than in várzea.
No significant difference in the averages of root weight, growth and biomass was registered,
nor in external morphology and survival rate with respect to flooding. There was no
significant difference regarding the length and weight of the aerial part. The lowest average
diameter was observed in floodplain individuals developed in igapó soil, and the average of
the specific leaf area was not statistically different from each other. The amount of macro and
micro-nutrients in the leaves of M. acaciifolium in a floodplain or igapó varied in both soil
types and with respect to the origin, since the igapó individuals in an igapó soil, in general,
presented more nutrients in the leaves compared to floodplain individuals. The results
suggest that M. acaciifolium of igapó individuals are more plastic-prone to adapt to
environmental conditions than individuals of M. acaciifolium in várzea.
Key-words: Germination, Biomass, Nutrients.

viii
SUMÁRIO
FICHA CATALOGRÁFICA ...................................................................................
........
ii
SINOPSE ......................................................................................................... ii
DEDICATÓRIA ...................................................................................................... iii
AGRADECIMENTO
S
......................................................................................... iv
EPÍGRAFE ........................................................................................................ v
RESUMO ...................................................................................................... vi
ABSTRACT ........................................................................................................ vii
INTRODUÇÃO .............................................................................................
...
13
OBJETIVO GERAL...................................................................................................... 16
Objetivos específico ....................................................................................................... 16
REFERÊNCIAS.............................................................................................................
....
17
Capítulo I ............................................................................................................ 19
Germinação, Biometria de sementes e Biomassa de Plântulas de Macrolobium
acaciifolium (Benth.) Benth. provenientes de populações de áreas de várzea
igapó da Amazônia Central.....................................................................................
19
Resumo........................................................................................................................... 20
Abstratct........................................................................................................................... 20
1 Introdução............................................................................................................. 21
2 Material e Métodos.......................................................................................................... 22
3 Resultados e Discussão.................................................................................................... 27
4 Conclusão......................................................................................................................... 33
5 Referências.......................................................................................................................
.
34
Capítulo II.................................................................................................................... 37
Crescimento, desenvolvimento e alterações morfológicas de plântulas de
Macrolobium acaciifolium (Benth.) Benth. em solos alagados de várzea e igapó
da Amazônia Central............................................................................................... 37
Resumo.......................................................................................................................... 38
Abstract.......................................................................................................................... 38
1 Introdução...................................................................................................................... 40
2 Material e Métodos........................................................................................................ 41
3 Resultados e Discussão.............................................................................................
.....................................................................
44
4
.
1
Conclusão......................................................................................................................
.
49
5
.
2
Referências....................................................................................................................
.
50
SÍNTESE GERAL....................................................................................................... 52
APÊNDICES.......................................................................................................... 53

ix
LISTA DE FIGURAS
CAPÍTULO 1
Figura 1 Localização das áreas de estudo: Igarapé Tarumã-mirim e Ilha da
Marchantaria.....................................................................................
23
Figura 2 Análise da fertilidade do solo usado como substrato. Laboratório de
Análises de Solo e de Plantas – LASP.
EMBRAPA....................................................................................... 25
Figura 3 Sementes de M. acaciifolium provenientes de ambientes de várzea e
igapó da Amazônia Central................................................................
28
Figura 4 Polígonos de frequência germinativa de sementes de Macrolobium
acaciifolium de área de várzea plantado em solo de
várzea...................................................................................... 30
Figura 5 Polígonos de frequência germinativa de sementes de Macrolobium
acaciifolium de área de várzea plantado em
vermiculita.................................................................................... 30
Figura 6 Polígonos de frequência germinativa de sementes de Macrolobium
acaciifolium de área de igapó plantado em solo de
igapó............................................................................................. 30
Figura 7 Polígonos de frequência germinativa de sementes de Macrolobium
acaciifolium de área de igapó plantado em
vermiculita.................................................................................... 30

x
CAPÍTULO 2
Figura 1 Análise da fertilidade do solo usado como substrato. Laboratório de
Análises de Solo e de Plantas – LASP. EMBRAPA.............................. 42
Figura 2 Formação de raízes adventícias e lenticelas em M. acaciifolium em
condição de alagamento parcial............................................................ 44
LISTA DE TABELAS
CAPÍTULO 1
Tabela 1 Análise Granulométrica do solo usado como substrato. Laboratório
de Análises de Solo e de Plantas – LASP. EMBRAPA........................
26
Tabela 2 Resumo da análise de variância para o comprimento (mm), peso (g),
largura (mm) e espessura (mm) das sementes de Macrolobium
acaciifolium de amostras de populações de várzea e igapó da
Amazônia Central............................................................................ 27
Tabela 3 Médias de porcentagem e velocidade de germinação de sementes de
Macrolobium acaciifolium de várzea e igapó da Amazônia
Central............................................................................................... 29
Tabela 4 Médias de biomassa seca de plantas jovens de M. acaciifolium em
diferentes substratos....................................................................... 31
CAPÍTULO 2
Tabela 1 Distribuição dos tratamentos.............................................................
41
Tabela 2 Análise Granulométrica do solo usado como substrato. Laboratório
de Análises de Solo e de Plantas – LASP. EMBRAPA........................ 42

xi
Tabela 3 Peso da raiz e parte aérea de M. acaciifolium. T1 = plantas de várzea
em solo de igapó, T2 = plantas de várzea em solo de várzea, T3 =
plantas de igapó em solo de várzea, T4 = plantas de igapó em solo de
igapó................................................................................................ 45
Tabela 4 Área foliar específica. T1 = plantas de várzea em solo de igapó, T2 =
plantas de várzea em solo de várzea, T3 = plantas de igapó em solo
de várzea, T4 = plantas de igapó em solo de
igapó............................................................................................ 46
Tabela 5 Quantidade de macronutrientes em folhas de M. acaciifolium 90 dias
após alagamento em solos de várzea e igapó.
T1 = plantas de várzea em solo de igapó, T2 = plantas de várzea em
solo de várzea, T3 = plantas de igapó em solo de várzea, T4 = plantas
de igapó em solo de igapó...................................................................
47
Tabela 5 Quantidade de micronutrientes em folhas de M. acaciifolium 90 dias
após alagamento em solos de várzea e igapó.
T1 = plantas de várzea em solo de igapó, T2 = plantas de várzea em
solo de várzea, T3 = plantas de igapó em solo de várzea, T4 = plantas
de igapó em solo de igapó................................................................. 48

12
INTRODUÇÃO GERAL
A Amazônia Brasileira possui cerca de 5.000.000 km², dos quais 800.000 km² são
constituídos por áreas úmidas, entre as quais destacam-se as várzeas e os igapós (Melack e
Hess, 2010; Junk et al., 1989). As várzeas abrangem aproximadamente 500.000 km² das áreas
alagáveis amazônicas, sofrem inundação periódica do Rio Amazonas e seus afluentes de
águas barrentas com alta carga de sedimentos provenientes dos Andes e encostas pré-andinas,
possuindo pH próximo da neutralidade (Melack e Hess, 2010; Junk et al., 2010; Junk e
Piedade, 2005; Furch, 1984).
As áreas de igapó representam 300.00 km² das áreas alagáveis na Amazônia (Melack e
Hess, 2010). Ao contrário das várzeas, são inundadas por rios de águas pretas ou claras com
pH ácido. Essas águas são pobres em nutrientes por serem provenientes dos antigos escudos
das Guianas e do Brasil Central (Prance, 1979; Sioli, 1984; Lowe-Mcconnell, 1999; Junk,
2010). A diferença do teor nutricional das águas reflete-se no solo. As concentrações em
toneladas/ha dos nutrientes P, K, Ca e Mg em substrato de várzea chegam a ser de 5 a 15
vezes mais alta do que em solos de igapó (Furch, 1997).
O ecossistema de várzea é considerado o que apresenta maior produção de biomassa
devido à deposição de nutrientes, a cada ano, a camada de deposição de sedimentos pode
atingir de 0,3 a 1 metro. Essa diferença é explicada pelo fato de que as espécies de várzea
contam com uma maior disponibilidade nutricional (Piedade et al., 2005; Almeida et al.,
2004).
Por ser rico em nutrientes, o solo de várzea permite uma maior produtividade natural;
a produção anual de folhas novas e a taxa de rotatividade foliar nas florestas sobre solo de
várzea é superior à encontrada em florestas sobre solo de igapó (Ferreira, 1997; Kubitzki,
1989).
Árvores que crescem em solos de várzea possuem uma concentração média de P, K.
Ca e Mg quatro vezes maior do que as árvores de igapó e a produtividade acima do solo em
florestas de várzea equivale a quase o dobro da produtividade em florestas de igapó (Furch,
1997; Malhi et al., 2004).
O padrão sazonal de variação do nível da água, o chamado “pulso de inundação”,
permitiu que as espécies vegetais que colonizam esses ecossistemas desenvolvessem

13
diferentes tipos de características adaptativas fenológicas, fisiológicas e morfológicas que
garantissem o sucesso do estabelecimento das espécies (Parolin et al, .2004, Junk et al.,
1989).
A altura e a duração da inundação periódica induzem o surgimento de alterações no
comportamento ecofisiológico das árvores que colonizam as áreas alagáveis, tornando
possível sua adaptação às condições de escassez de oxigênio durante longos períodos
(Wittmann et al., 2006). Essas espécies arbóreas sobrevivem em um estado de dormência e
podem também apresentam um vigoroso crescimento na fase inundada (Parolin et al., 2004).
Schöngart et al., (2005) monitorando indivíduos de Macrolobium acaciifolium
(Benth.) Benth. constataram que na várzea as árvores desta espécie apresentaram um
incremento radial duas vezes superior ao daquelas árvores de igapó. Também é postulado que
a maior disponibilidade de nutrientes dos ambientes de várzea possibilita que algumas
espécies tenham mais sucesso competitivo se comparadas a outras provenientes de ambiente
de igapó onde a disponibilidade nutricional é mais baixa (Ferreira et al., 2005).
Desta forma, este estudo propõe investigar se as diferenças entre as condições
nutricionais dos solos de várzea e dos solos de igapó exercem influência significativa sobre a
germinação e crescimento inicial de plântulas da espécie Macrolobium acaciifolium (Benth.)
Benth. de ocorrência comum nos dois ambientes, assim como as suas respostas
morfoanatômicas quando submetidas à condição de alagamento em sua fase de
desenvolvimento inicial.
A espécie em estudo, M. acaciifolium, é classificada como clímax quanto ao tipo de
sucessão ecológica (Schöngart et al., 2002; Félix-da-Silva et al., 2013). Os frutos são do tipo
legume com sementes relativamente grandes contendo elevado teor de água, sendo, portanto,
recalcitrantes, a germinação é do tipo fanerocotiledonar hipógea com cotilédones
armazenadores (Maia et al., 2005). Apesar da literatura afirmar que cada fruto possui apenas
uma semente, neste trabalho durante a abertura dos frutos foram encontrados alguns contendo
duas sementes. A espécie possui folhas compostas, pinadas, paripinadas, multifolioladas com
18-20 pares, margem superior verde escura e inferior mais opaca, as flores são brancas com
estames vermelhos e exalam odor agradável. Sementes ovoides, achatadas, esverdeadas
assumindo tons amarronzados quando maduras (Souza, 2012).
O presente trabalho foi dividido em dois capítulos. No primeiro investigou-se a
influência do substrato na germinação e crescimento inicial de plantas de Macrolobium
acaciifolium de populações de várzea e igapó plantadas em seu solo de origem. No segundo

14
foi feito um experimento cruzado, onde mudas de igapó foram plantadas em solo de igapó e
em solo de várzea e mudas de várzea foram plantadas em solo de várzea e em solo de igapó
para observar as respostas de crescimento e desenvolvimento da espécie nos dois substratos
em condição de alagamento parcial.

15
OBJETIVO GERAL
O presente estudo tem como objetivo correlacionar as condições nutricionais e físicas
dos solos de várzea e igapó com a germinação, o crescimento e o desenvolvimento de plantas
jovens da espécie Macrolobium acaciifolium (Benth.) Benth., comum aos dois ambientes,
bem como observar as respostas morfológicas ao alagamento nas duas espécies.
Objetivos específicos
Realizar a análise granulométrica e química do solo de várzea e igapó dos locais de
coleta das sementes.
Analisar a interferência do tipo de substrato no comportamento germinativo e
produção de biomassa dos indivíduos das populações de M. acaciifolium provenientes de
populações de igapó e várzea.
Observar as alterações morfológicas, a sobrevivência, crescimento e a biomassa de
plântulas de Macrolobium acaciifolium de populações de várzea e igapó em diferentes
condições nutricionais nos substratos de várzea e igapó em alagamento parcial.

16
REFERÊNCIAS
Almeida, S. S. de; Amaral, D. D. do; Silva, A. S. L. da. 2004. Análise florística e estrutura de
florestas de várzea no estuário amazônico. Acta Amazonica 34(4): 513 - 524.
Félix-da-Silva, M.M.; Bastos, M.N.C.; Gurgel, E.S.C. 2013. Macrolobium Schreb.
(Leguminosae, Caesalpinioideae) na Floresta Nacional de Caxiuanã, Pará, Brasil. Boletim do
Museu Paraense Emílio Goeldi, Ciências Naturais 8: 75-93.
Ferreira, L. V.; Almeida, S. S.; Amaral, D. D.; Parolin, P. 2005. Riqueza e composição de
espécies da floresta de igapó e várzea na Estação Científica Ferreira Pena: subsídios para o
plano de manejo da Floresta Nacional de Caxiuanã. Pesquisas, Botânica, (56): 103-116.
Ferreira, L. 1997. Effects of the duration of flooding on species richness and floristic
composition in three hectares in the Jaú National Park in floodplain forests in central
Amazonia. Biodiversity and Conservation, 6(10), 1353-1363.
Furch, B. 1984. Unterschungen zur Ünberschwemmungstoleranz der Várzea unde ders Igapó:
Blattpigmente. Biogeographica (19):77- 83.
Junk, W.J. 1989. Flood tolerance and tree distribution in Central Amazonian floodplains. In:
Nielsen, L.B.; Nielsen, I.C.; Baisley, H. (Eds). Tropical Forests:
Botanical Dynamics, Speciation and Diversity Academic Press, London. p. 47-64.
Junk W., Piedade Mtf, Wittmann F, Schöngart J, Parolin P. 2010. Central Amazonian
floodplain forests: ecophysiology, biodiversity and sustainable management. Springer Verlag,
Berlin. 615 pp.
Prance, G. T. 1979. Notes on the vegetation of Amazonia. III. Terminology of Amazonian
Forest types subjected to inundation. Brittonia. 31: 26-38.
Kubitzki K. 1989. The ecogeographical differentiation of Amazonian inundation forests. Plant
Systematics and Evolution. 162 p. 285–304.
Lowe-Mcconnell, R. H. 1999. Estudos ecológicos de comunidades de peixes tropicais.
EDUSP, São Paulo. 534 pp.
Maia, L.A., Maia, S.; Parolin, P. 2005. Seedling Morphology of Non-pioneer Trees in Central
Amazonian Várzea Floodplain forests. Ecotropica, 11:1–8.
Maguire, J.D. 1962. Speed of germination: aid in selection and evaluation for seedling
emergence and vigor. Crop Science. 2(2):176-177.
Malhi, Y.; Baker, T.R.; Phillips, O.L.; Almeida, S.; Alvarez, E.; Arroyo, L.; Chave, J.;
Czimczik, C.I.; Di Fiori, A.; Higuchi, N.; Killeen, T.J.; Laurance, S.G.; Laurance, W.F.;
Lewis, S.L.; Montoya, L. M. M.; Monteagudo, A.; Neill, D.A.; Vargas, P.N.; Patiño, S.;
Pitman, N.C.A.; Quesada, C.A.; Salomão, R.; Silva, J.N.M.; Lezama, A.T.; Martínez, R.V.;
Terborgh, J.; Vinceti, B.; Lloyd, J. 2004. The above-ground coarse wood productivity of 104
Neotropical forest plots. Global Change Biology, (10): 563-591.

17
Melack, J. M.; Hess, L. L. 2010. Remote sensing of the distribution and extent of wetlands in
the Amazon basin. In: Junk, W. J., Piedade, M. T. F.; Wittmann, F. K.; Schöngart, J.. Central
Amazonian floodplain forests: ecophysiology, biodiversity and sustainable management. New
York: Springer Verlag, Dordrecht-Heidelberg-London. p. 43-59.
Parolin, P. 2004. Floristic study of an igapo floodplain forest in Central Amazonia, Brazil
(Taruma-Mirim, Rio Negro). Amazoniana, 18(2), 29-47.
Piedade, M. T. F.; Schöngart, J.; Junk, W. J. 2005. O manejo sustentável das áreas alagáveis
da Amazônia Central e as comunidades herbáceas aquáticas. Uakari, 1(1):43-55.
Schöngart, J.; Piedade, M. T. F.; Ludwigshausen, S.; Horna, V.; Worbes, M. 2002. Phenology
and stem-growth periodicity of tree species in Amazonian floodplain forests. Journal of
Tropical Ecology, 18:4, 581-597.
Schöngart, J.; Piedade, M.T.F.; Wittmann, F.; Junk, W.J.; Worbes, M. 2005. Wood growth
patterns of Macrolobium acaciifolium (Benth.) (Fabaceae) in Amazonian black-water and
white-water floodplain forests. Oecologia, (145): 454-461
Sioli, H. 1984. The Amazon: limnology and landscape ecology a mighty tropical river and its
basin. Dr. W. Junk Publisher, Dordrecht, The Netherlands. 763 pp.
Souza, L. A. G. de. 2012. Guia da biodiversidade de Fabaceae do Alto Rio Negro. FINEP,
118p.
Wittmann, F.; Schöngart; Parolin, P.; Worbes, M.; Piedade, M. T.F.; Junk, W. J. 2006. Wood
specific gravity of trees in Amazonian white-water forests in relation to flooding. IAWA
Journal, 27(3): 255-266.

18
Capítulo I
Santos, L. R.; Wittmann, F. K. & Almeida, M. C.
Germinação, Biometria de sementes e Biomassa de
Plântulas de Macrolobium acaciifolium (Benth.)
Benth. provenientes de populações de áreas de
várzea e igapó da Amazônia Central
Formatado de acordo com as normas da Revista Biota Neotropica.

20
Germinação, biometria de sementes e biomassa de plântulas de
Macrolobium acaciifolium (Benth.) Benth. provenientes de populações de
áreas de várzea e igapó da Amazônia Central
Resumo - Este estudo teve como objetivo verificar o efeito dos substratos de várzea e igapó
na formação de sementes, na germinação e na biomassa de plântulas da espécie Macrolobium
acaciifolium (Benth.) Benth., de ocorrência comum nesses dois ecossistemas. Foi realizada a
análise biométrica de sementes de indivíduos de M. acaciifolium de população de várzea e de
igapó Amazônia Central, sendo que foram medidas 100 sementes de cada ambiente. O teste
de germinação foi conduzido em casa de vegetação, os tratamentos foram divididos em:
sementes de igapó em solo de igapó, sementes de igapó em vermiculita, sementes de várzea
em solo de várzea e sementes de várzea em vermiculita. Cada tratamento contou com 100
sementes, sendo cinco repetições de 20 sementes. Na análise biométrica as sementes de
várzea apresentaram maior média de peso do que as sementes de igapó, que por sua vez
apresentaram a maior média de largura. O índice de velocidade de germinação em solo de
várzea foi inferior a observado nos outros substratos. As plântulas de sementes oriundas de
várzea e plantadas em solo de várzea apresentaram menores médias de comprimento da raiz e
de comprimento e peso da parte aérea.
Palavras chaves: Germinação, Biometria de sementes, Biomassa.
Germination, seeds biometrics and seedlings biomass of Macrolobium acaciifolium
(Benth.) Benth. from várzea and igapó populations in Central Amazon
Abstratct - The present study is aimed at verifying the effect of floodplain and igapó
substrates in seed formation, germination and biomass of seedlings of the species
Macrolobium acaciifolium (Benth.) Benth., which are common occurrences in these two
ecosystems. It was conducted the biometric analysis of seeds of M. acaciifolium from the
population of várzea and igapó in Central Amazon, where 100 seeds of each environment
were measured. The germination test was conducted in a greenhouse, the treatments were
divided into igapó seeds in igapó soil, seeds in vermiculite, várzea seeds in várzea soils, and
várzea seeds in vermiculite. Each treatment had 100 seeds, in five repetitions of 20 seeds. In
the biometric analysis, várzea seeds showed the highest weight average than igapó seeds,
which in turn had the highest width average. The germination rate in várzea soil was lower
than observed in other substrates. Seedlings from seeds from várzea and planted in várzea soil
had the lowest average of root length and of aerial part weight and length.
Keywords: Germination, Seeds biometrics, Biomass.

21
1 Introdução
A germinação das sementes é uma fase crítica no processo de estabelecimento da
planta em seu ambiente natural, portanto, informações a cerca do comportamento biológico da
semente estão entre as principais ferramentas para o manejo das populações de plantas
(Vázquez-Yanes & Orozco-Segovia, 1993). Do ponto de vista fisiológico, entende-se como
germinação a retomada do crescimento embrionário após a embebição de água pela semente
finalizando com a protusão da radícula (Carvalho & Nakagawa, 2012).
O tempo de germinação das sementes é uma das características relacionadas com as
estratégias de estabelecimento das espécies em determinado local, assim, juntamente com o
estudo de outras particularidades como a morfologia inicial das plântulas pode fornecer
importantes informações que sirvam de base para o conhecimento dos processos de
regeneração natural e conservação das espécies (Ng, 1978, Santos et al., 2005).
Os aspectos fisiológicos e morfológicos de espécies que colonizam as planícies
inundáveis podem mudar de acordo com o tipo de área onde se encontram. Parolin et al.
(2003) verificaram que espécies de igapó tendem a apresentar sementes maiores e mais
pesadas do que espécies de várzea. Os autores observaram ainda que a porcentagem de
germinação, comprimento das folhas e altura das plantas eram significativamente maiores nas
espécies de áreas de igapó comparadas com as espécies de várzea, a despeito de a várzea
apresentar solos mais ricos em nutrientes do que os solos de igapó.
A quantidade de nutrientes disponíveis nos solos alagáveis da Amazônia Central age
significativamente sobre a formação das sementes, em solos pobres observa-se que as
sementes possuem mais reservas para nutrir a plântula por mais tempo e tentar garantir o
sucesso do seu estabelecimento (Parolin, 2000). O crescimento inicial de plântulas pode sofrer
forte influência da quantidade de reservas de nutrientes das sementes, a mobilização dos
principais compostos de reserva nos tecidos de armazenamento (carboidratos, lipídios e
proteínas) ocorre após a conclusão da germinação para fornecer nutrientes à plântula até que
ela se torne autotrófica (Buckeridge, et al., 2004, Ferreira et al., 2009; Bewley et al., 2013).
Singh e Arunachalan (2002) ao avaliarem o tratamento para quebra de dormência de cinco
espécies de leguminosas arbóreas observaram uma correlação positiva entre o tamanho das
sementes e a taxa de germinação, indicando que sementes com maior quantidade de reserva
de nutrientes germinaram mais rápido.

22
É sabido ainda que a germinação de sementes também é influenciada por caraterísticas
do substrato como aeração, capacidade de retenção de água, estrutura entre outros fatores que
podem favorecer ou prejudicar a germinação (Kanashiro, 1999).
De acordo com Moreira & Moreira (1996), 59,5% das espécies que colonizam
florestas alagáveis apresentam germinação do tipo hipógea enquanto que na terra firme corre
o inverso, 54,5% das espécies possuem germinação do tipo epígea. Segundo estes autores as
sementes com germinação do tipo hipógea possuem teor de reserva mais elevado.
Considerando que a espécie M. acaciifolium colonizou áreas com particularidades tão
distintas quanto a várzea e o igapó, a proposta deste estudo foi avaliar os aspectos do processo
germinativo e características morfométricas de indivíduos de populações dos dois ambientes
sendo que as informações obtidas podem servir de base para trabalhos de manejo florestal
incluindo recuperação de matas ciliares.
Material e Métodos
Os frutos de populações várzea foram coletados na região da Ilha da Marchantaria (03º
13’ S 59º 58’ W) e proximidades em áreas que sofrem inundação periódica pelas águas do
Rio Solimões. Os frutos de populações de igapó foram coletados próximo ao igarapé Tarumã-
mirim (3º 01’ 38” S 60º 10” 45’W) que faz parte da bacia do rio Negro, situado a noroeste da
cidade de Manaus, no quilômetro 21 da BR 174 (Figura 1).
As coletas foram realizadas entre os meses de abril e maio de 2015. Foram coletados
apenas frutos maduros, que após levados para a sede do Grupo MAUÁ/Projeto Max Planck,
no Instituto Nacional de Pesquisas da Amazônia, tiveram suas sementes retiradas
manualmente. As sementes passaram foram desinfectadas por imersão em solução de
hipoclorito de sódio a 10% por dez minutos e em seguida foram lavadas em água corrente.

23
Figura 1 – Localização das áreas de estudo: Lago Tarumã-mirim e Ilha da Marchantaria.
Fonte: Google Maps, 2015.
Escala: 1:5000
Com auxílio de um paquímetro digital foram medidos comprimento, largura e
espessura de 200 sementes (divididas em dois grupos, 100 sementes de várzea e 100 sementes
de igapó). Para determinar a massa foi utilizada balança analítica com precisão de quatro
casas decimais.
As sementes foram postas para germinar em bandejas plásticas de 20 cm x 60 cm,
dispostas em bancadas na casa de vegetação do Grupo MAUÁ/Max Planck, localizada nas
dependências do Instituto Nacional de Pesquisas da Amazônia, Campus I. O experimento foi
conduzido em temperatura ambiente, que no período variou de 27 ºC a 39 ºC na cidade de
Manaus (INPE, 2015). Cada tratamento contou com cinco repetições de 20 sementes,
totalizando 100 sementes por tratamento (Oliveira et al., 2011). As regas foram feitas com
água de poço artesiano. As sementes foram plantadas em seu solo de origem de acordo com
sua procedência e em vermiculita, totalizando 2 tratamentos para cada espécie: M.
acaciifolium de igapó + solo de igapó e M. acaciifolium de igapó + vermiculita; M.
acaciifolium de várzea + solo de várzea e em vermiculita.
Cada bandeja foi considerada uma unidade amostral e cada tratamento contou com
cinco repetições de 20 sementes.
Lago Tarumã-mirim
Ilha da Marchantaria
N

24
O início do experimento se deu no dia 2 de junho de 2015 e foi encerrado dia 3 de
agosto do mesmo ano, quando a germinação estabilizou-se. Foi considerado como germinada
a plântula que apresentou emergência do caulículo.
A porcentagem de germinação, o tempo médio de germinação e a frequência
germinativa foram calculados segundo as fórmulas propostas por Labouriau & Agudo (1987).
O índice de velocidade de germinação foi calculado de acordo com Maguire (1962).
Para a medição do crescimento retirou-se vinte plantas com altura mínima de 30 cm
dentro de cada tratamento, a parte aérea foi separada e cortada da raiz com tesoura, as duas
partes foram medidas com trena. A parte aérea foi medida desde a gema apical até o colo e as
raízes do final do colo até a extremidade final da raiz primária.
A determinação da biomassa seca foi efetuada após realizar as medições as partes das
plantas, tendo estas sido acondicionadas em sacos de papel kraft e levadas para a estufa a 80º
C por 72 horas até atingirem peso constante (Lopes et al. 2008). Após a secagem o material
foi pesado em balança analítica com precisão de quatro casas decimais.
O delineamento experimental utilizado foi o inteiramente casualizado (DIC). Os dados
foram submetidos ao teste de normalidade de Kolmogorov-Smirnov e posteriormente
realizou-se a análise de variância, as médias foram comparadas pelo teste de Tukey a 5% de
probabilidade. O programa utilizado para as análises foi o Assistat 7.7 Beta.

25
Quadro 1 - Análise da fertilidade do solo usado como substrato. Laboratório de Análises de Solo e de Plantas – LASP. EMBRAPA.
Amostra pH C M.O. P K Na Ca Mg Al H+Al SB t T V m Fe Zn Mn Cu
(H2O) (g/kg) mg/dm3 cmolc/dm
3 % mg/dm
3
Solo de
Igapó 4,69 14,84 25,53 4 12 3 0,06 0,06 1,77 3,83 0,16 1,93 3,99 4,10 91,53 406 0,76 0,50 0,66
Solo de
várzea 6,16 2,95 5,08 120 58 19 4,76 1,24 0,0 0,66 6,23 6,23 6,89 90,42 0,00 464 5,71 128 3,71
pH em água relação 1:2,5
P, Na, K, Fe, Zn, Mn e Cu – Extrator Mehlich-1
Ca, Mg – Extrator KCl 1mol/L
H + Al – Extrator de acetato de cálcio 0,5 mol/L
SB – Soma de bases
CTC (T) – Capacidade de troca catiônica efetiva
CTCt (t) – Capacidade de troca catiônica a pH 7,0
V – índice de saturação por bases
m – Índice de saturação por alumínio
Matéria orgânica (M.O.) – (C orgânico) x 1,724 – Walkley-Black

26
Tabela 1 - Análise Granulométrica do solo usado como substrato. Laboratório de Análises de Solo e de Plantas – LASP. EMBRAPA.
Solo Areia Grossa Areia Fina Areia Total Silte Argila
2,00 – 0,20
(mm)
0,20 – 0,05
(mm)
2,00 - 0,05
(mm)
0,05 - 0,002
(mm)
< 0,002
(mm)
(g/kg)
Várzea 10,87 651,60 662,47 171,95 188,5
Igapó 632,59 143,97 776,55 149,03 51,50

27
Resultados e Discussão
O resultado da análise biométrica de sementes mostrou que as sementes de populações
do ambiente de várzea apresentaram maiores valores de peso enquanto que as sementes
coletadas no igapó apresentaram maior média de espessura. As médias de largura e
comprimento não apresentaram diferença estatística (Tabela 2).
Tabela 2 – Resumo da análise de variância para o comprimento (mm), peso (g) e espessura (mm) das
sementes de Macrolobium acaciifolium de amostras de populações de várzea e igapó da Amazônia Central.
Ambientes Comprimento
(mm)
Peso
(g)
Largura
(mm)
Espessura
(mm)
Várzea 38,35±4,047a 5,90±1,363a 32,17±2,870a 6,44±1,053a
Igapó 39,080±3,59a 4,72±1,315b 31,10±3,10a 7,50±1,162b
Médias seguidas por letras distintas diferem entre si ao nível de 1% de probabilidade de erro pelo Teste de
Tukey. (p < 0.05). Média ± desvio padrão, n=100.
As sementes de igapó apesar de terem maior espessura estavam mais leves do que as
sementes de várzea, isso pode ter ocorrido por estas últimas se apresentarem mais largas e
fazendo com que sua massa ficasse mais distribuída.
Feitoza et al. (2014) em estudo realizado com 50 sementes coletadas na floresta de
várzea da ilha do Combu em Belém, encontrou médias de comprimento para sementes de M.
acaiifolium variando de 23,25 mm e 32,28 mm, largura de 2,66 mm a 40,44 mm e espessura
de 4,95 mm a 7,18 mm. Castro (2012) encontrou valores similares para a biometria de 100
sementes coletadas em floresta secundária de várzea inundada pelo rio Solimões, média de
comprimento igual a 36,95 mm, peso 5,32 mm, largura 32,37 mm e espessura igual a 6,09
mm. Os valores encontrados por Castro (2012) foram aproximados dos encontrados no
presente estudo.
Parolin (2000) em um estudo estimando a massa de 31 sementes de espécies de várzea
e 27 sementes de espécies de igapó, encontrou uma média global de peso de 7,1 g para
sementes de igapó e de apenas 1,2 g para sementes de várzea. Segundo a autora espécies que
se desenvolvem em solos mais pobres em nutrientes tendem a investir mais na quantidade de
reservas de nutrientes das sementes, sendo, portanto, comum estas apresentarem maior
biomassa. No entanto, ao comparar características biométricas de sementes de Hevea
spruceana de populações de várzea e igapó, Liberato (2010) observou que as sementes

28
diferiam significativamente entre si em comprimento, largura e espessura, sendo que as
sementes de várzeas apresentaram maiores valores destas duas últimas variáveis. No caso
deste estudo, não houve diferença significativa entre comprimento e largura das sementes,
apenas as variáveis peso e espessura foram estatisticamente diferentes.
Segundo Piña-Rodrigues & Aguiar (1993) o tamanho e o peso das sementes de
determinada espécie são características de elevada plasticidade, podendo variar de ano para
ano, entre indivíduos ou mesmo dentro de um indivíduo. As sementes de M. acaciifolium
apresentaram pequenas diferenças no formato (Figura 3).
Figura 3 – Sementes de M. acaciifolium provenientes ambientes de várzea e igapó da Amazônia
Central.
A qualidade fisiológica das sementes de Macrolobium acaciifolium foi avaliada por
meio da porcentagem de germinação, índice de velocidade de germinação e desempenho de
plântulas em diferentes substratos.
A germinação foi estabilizada ao 52º dia após a semeadura, não houve diferença
significativa na taxa de germinação para os substratos testados (Tabela 3). No índice de
velocidade de germinação, no entanto, o tratamento onde sementes de várzea foram plantadas
em solo de várzea apresentou média inferior aos demais.
Sementes de igapó
Sementes de várzea

29
Tabela 3 – Médias de porcentagem e velocidade de germinação de sementes de
Macrolobium acaciifolium de várzea e igapó da Amazônia Central.
Origem das sementes Tipo de substrato Germinação
(%)
IVG
Várzea
Solo de várzea 56±13,20a 1,39±0,248b
Vermiculita 83±16,04a 1,54±0,059a
Igapó Solo de igapó 68±18,40a 1,57±0,001a
Vermiculita 85±15,81a 1,57±0,002a
ns não significativo (p >= 0.05). As médias seguidas pela mesma letra não diferem estatisticamente
entre si. Foi aplicado o Teste de Tukey. Média ± desvio padrão.
Maia et al. (2005) encontraram valores de taxa de germinação de 90% para um lote de
500 sementes de M. acaciifolium coletadas em área de várzea do Rio Solimões. As autoras
utilizaram como substrato areia lavada. No presente estudo observou-se que as sementes de
várzea apresentaram valores mais elevados de taxa de germinação e velocidade de
germinação na vermiculita, podendo-se supor que substratos mais porosos favorecem o
processo germinativo da semente desta espécie. As sementes de igapó tiveram maior taxa
germinativa no substrato de igapó, mas esse valor não diferiu estatisticamente do encontrado
na germinação em vermiculita. Esse resultado contrasta com a afirmação de Reis et al. (2002),
pois segundo esses autores em solos arenosos a tendência é que haja uma baixa taxa de
germinação devido a reduzida capacidade que esse tipo de substrato tem de manter a umidade
pois, as partículas de maior diâmetro possuem menos eficiência para reter moléculas de água
por apresentarem menos tensão superficial do que as partículas de diâmetros menores que
predominam em solos mais argilosos. A rápida drenagem do substrato prejudicaria o processo
de embebição da semente diminuindo a porcentagem de germinação (Reis, et al., 2002). No
presente estudo, o solo argiloso não implicou em uma maior taxa de germinação, em
compensação o índice de velocidade de germinação foi menor nesse substrato.
O período de germinação da espécie foi inferior a 60 dias, sendo considerado rápido
em todos os tratamentos (Vázquez-Yanes & Orozco-Segovia, 1982). O início da germinação
das sementes coletadas no igapó se deu no quinto dia, tanto em solo de igapó quanto em
vermiculita. Das sementes coletadas na várzea, o início da germinação na vermiculita foi
registrado no sexto dia em solo de várzea no nono dia.
A frequência de germinação exibiu valores polimodais tanto para os lotes de sementes
de várzea quanto os lotes de sementes de igapó (Figuras 4, 5,6 e 7).

30
Figura 4 - Polígonos da frequência germinativa de sementes de
Macrolobium acaciifolium de área de várzea plantado em solo de
várzea.
Figura 5 - Polígonos da frequência germinativa de sementes de
Macrolobium acaciifolium de área de várzea plantado em vermiculita.
Figura 6 - Polígonos da frequência germinativa de sementes de
Macrolobium acaciifolium de área de igapó plantado em solo de igapó.
Figura 7 - Polígonos da frequência germinativa de sementes de
Macrolobium acaciifolium de área de igapó plantado em solo de
vermiculita.

31
Observou-se que tanto nas sementes coletadas no igapó como nas sementes coletadas
na várzea e plantadas em vermiculita os intervalos entre o registro das germinações foi mais
curto do que nas sementes de várzea plantadas em solo de várzea. Parolin et al. (2001) em um
estudo com mudas de outras espécies constatou que na várzea a porcentagem de germinação e
a duração foram menores do que no igapó. O substrato com partículas maiores, como é o caso
do solo de igapó e da vermiculita em comparação com o solo de várzea onde predominam
partículas mais finas, pode facilitar a emergência da plântula.
Maia et al. (2005) em seu estudo registrou o início da germinação de M. acaciifolium
de várzea como tendo ocorrido ao décimo terceiro dia e foi estabilizada ao quadragésimo
nono dia, sendo este último valor bem aproximado aos encontrados no presente trabalho
(início entre o quinto e sétimo dias e término por volta do quinquagésimo).
O tempo médio de germinação das sementes de M. acaciifolium nos diferentes
substratos variou de 27,95 a 30,54 dias. Considerando que esta espécie ocorre nas menores
cotas de inundação, a germinação agrupada em um intervalo curto de tempo pode servir de
estratégia para o aproveitamento da fase terrestre, que nessas áreas fica restrito a poucos
meses do ano (Junk et al., 1989, Wittmann et al., 2010). Parolin e Junk (2002) em um estudo
com 12 espécies de áreas alagáveis constataram que a taxa de germinação foi maior em
sementes de condição de não-alagamento e em algumas espécies, mesmo tendo iniciado a
germinação, a semente apodreceu logo depois.
A análise da biomassa seca de plântulas de M. acaciifolium apontou diferenças
significativas quanto ao peso e comprimento da parte aérea e para o comprimento da raiz
(Tabela 4).
Tabela 4 – Médias de Biomassa seca de plantas jovens de Macrolobium acaciifolium em
diferentes substratos.
Compartimento e
medida analisados
Origem das sementes e substrato
Igapó Várzea
Solo de igapó Vermiculita Solo de várzea Vermiculita
Peso da raiz 0,762±0,09a 0,7063±0,07a 0,668±0,14a 0,683±0,11a
Peso da parte aérea 2,62±0,51a 2,473±0,37a 1,722±0,59b 2,234±0,40a
Comprimento da
raiz 18,82±4,44a 21,85±3,23a 12,99±2,85b 20,29±3,94a
Comprimento da
parte aérea 52,175±8,12a 55,57±6,704a 39,95±9,17b 49,475±6,81a
Médias seguidas por letras distintas na horizontal diferem entre si ao nível de 5% de probabilidade de erro pelo
Teste de Tukey. (p < 0.01), n=20. Media ± desvio padrão.
As plântulas oriundas das sementes coletadas na área de igapó apresentaram maiores
valores de peso e comprimento com exceção do peso da raiz onde não houve diferença

32
estatística significativa. Observou-se que as raízes das plantas de várzea apesar de mais curtas
formavam um emaranhado mais denso, enquanto as raízes das plantas de igapó se mostravam
mais longas e menos ramificadas.
As raízes das plantas que se desenvolveram em solo de várzea apresentaram a menor
média de comprimento, não investindo tanto no crescimento radicular provavelmente por
estarem em um meio mais rico em nutrientes (Quadro 1), não necessitando expandir tanto a
superfície de contato quanto as plantas em vermiculita e em substrato de igapó.
Neves et al. (2005) sugerem que além das características genéticas que determinam a
morfologia da raiz, atributos físico-químicos do solo agem de forma significativa em sua
formação. De acordo com Beutler & Centurion (2003) a porosidade do solo influencia no
crescimento das raízes e na absorção de nutrientes e água de acordo com a quantidade de
macroporos (poros grandes, predominam em solos mais arenosos) e microporos (poros
menores, comuns em solos mais argilosos). Solos com predominância de microporos
apresentam tendência a indução do crescimento lateral das raízes, cujo diâmetro é reduzido
sendo mais fácil penetrar em espaços menores (Beutler & Centurion, 2003), como foi
constatado neste estudo com as raízes em substrato de várzea.
As maiores médias de crescimento aéreo foram observadas nas plantas de igapó, mas
essas médias não diferiram estatisticamente da média de comprimento encontrada nas plantas
de várzea plantadas em vermiculita. O substrato de igapó é reconhecidamente mais pobre em
nutrientes, sugerindo que para a espécie em estudo, uma taxa mais elevada de nutrientes não
implicou em um maior incremento de biomassa na parte aérea nesta fase.
Esses resultados corroboram com aqueles encontrados por Parolin et al. (2003), que os
justificam pelo fato de que as espécies que ocorrem nas cotas mais altas de inundação do
igapó tendem a desenvolver-se mais rapidamente em altura do que as espécies que ocorrem na
várzea. Aparentemente, esse comportamento também ocorre entre indivíduos de M.
acaciifolium quando em ambientes diferentes.
Entretanto, foi observado que quando plantadas em vermiculita, um substrato inerte, as
plantas jovens de M. acaciifolium de várzea tiveram maior média de crescimento do que
sementes do mesmo lote que germinaram e se desenvolveram em substrato de várzea.
Restando a hipótese da influência granulométrica do substrato no desenvolvimento inicial das
mudas. Tanto a vermiculita quanto o substrato de igapó são mais porosos do que o substrato
de várzea, por este conter mais argila. Os poros maiores do substrato são preenchidos por ar e
água, e os poros menores apenas por água, fazendo com que a textura exerça considerável

33
influência na aeração (Zanetti et al. 2003) o que pode explicar os resultados encontrados neste
estudo.
As raízes respiram, consumindo oxigênio e expelindo gás carbônico. Para o adequado
funcionamento de seu metabolismo é necessário que o processo de trocas gasosas seja
eficiente e este processo está intimamente associado com a aeração do solo (Letey, 1985). O
substrato mais compactado reduz o estoque de oxigênio e pode interferir nas trocas gasosas
entre a raiz e o meio, podendo limitar a atividade metabólica radicular, um dos principais
hormônios vegetais do crescimento, a citosina, é produzido em outras partes da planta, mas é
fabricado em maior quantidade nas raízes. Limitações no metabolismo radicular podem
reduzir a produção de hormônios e refletir no crescimento total do vegetal (Mapfumo et al.,
1998, Ishaq et al., 2001).
Seriam necessários mais estudos enfocando o metabolismo de plântulas de M.
acaciifolium para definir até que ponto o substrato influencia em seu desenvolvimento.
Conclusão
A caracterização biométrica das sementes de M. acaciifolium não indicou diferenças
significativas nas variáveis, comprimento e largura em relação aos ambientes de origem. A
média do peso das sementes de várzea foi maior, mas a espessura das sementes de igapó
exibiu maior média.
Não houve diferença significativa na porcentagem de germinação para as amostras de
diferentes ecossistemas alagáveis.
O índice de velocidade de germinação para o tratamento de sementes de várzea
plantadas em solo de várzea mostrou-se inferior aos demais tratamentos.
As plântulas originadas a partir de sementes coletadas na área de igapó apresentaram
maiores médias de comprimento e peso da parte aérea e comprimento da raiz quando
comparadas com as médias das plântulas de sementes de várzea.
Não houve diferença significativa quanto às médias do peso da raiz.

34
Referências
BEUTLER A. N.; CENTURION, J. F. 2003. Efeito do conteúdo de água e da compactação do
solo na produção de soja. Pesquisa Agropecuária Brasileira 38:849-856.
BEWLEY, JD; BRADFORD KJ; HILHORST HWM; NONOGAKI H. 2013. Seeds:
Physiology of development, germination and dormancy, 3rd
ed. Springer. New York 392p.
BUCKERIDGE, M. S.; AIDAR, M. P.; SANTOS, H. P.; TINÉ, M. A. S. 2004. Acúmulo de
Reservas. In: FERREIRA, A. G. e BORGHETTI, F. (eds.) Germinação: do básico ao
aplicado. p. 31-50.
CARVALHO, N. M.; NAKAGAWA, J. 2012. Sementes: ciência, tecnologia e produção.
5.ed. FUNEP. Jaboticabal.
CASTRO, D. J. L. de. 2012. Características funcionais de espécies arbóreas de diferentes
estádios sucessionais na várzea da Amazônia Central. Dissertação de mestrado em Ecologia.
Instituto Nacional de Pesquisas da Amazônia. Manaus.
FEITOZA, G. V.; SANTOS, J. M. U. dos; GURGEL, E. S. C.; OLIVEIRA, D. M. T. 2014.
Morphology of fruits, seeds, seedlings and saplings of three species of Macrolobium Schreb.
(Leguminosae, Caesalpinioideae) in the Brazilian Amazon floodplain. Acta Botanica Brasilica
28:422-433.
FERREIRA, C. da S.; PIEDADE, M. T. F.; TINE, M. A. S.; ROSSATO, D. R.; PAROLIN, P.
BUCKERIGDE, M. S. 2009. The role of carbohydrates in seed germination and seedling
establishment of Himatanthus sucuuba, an Amazonian tree with populations adapted to
flooded and non-flooded conditions. Annals of Botany 104:1111-1119.
ISHAQ, M.; IBRAHIM, M.; HASSAN, A.; SAEED, M.; LAL, R. 2001. Subsoil compaction
effects on crops in Punjab, Pakistan: II. Root growth and nutrient uptake of wheat and
sorghum. Soil and Tillage Research 60:153-161.
JUNK, W.J. 1989. Flood tolerance and tree distribution in Central Amazonian floodplains. In:
Nielsen, L.B.; Nielsen, I.C.; Baisley, H. (Eds). Tropical Forests: Botanical Dynamics,
Speciation and Diversity Academic Press, London. p. 47-64.
KANASHIRO, S. 1999. Efeito de diferentes substratos na produção da espécie Aechemea
fasciata (Lindley) Baker em vasos. Dissertação (Mestrado)– Escola Superior de Agricultura
"Luiz de Queiroz", Piracicaba.
LABOURIAU, L.G.; AGUDO, M. 1987.On the physiology of seeds germination in Salvia
hispanica L. I. Temperature effects. Anais da Academia Brasileira de Ciências 59:37-56.
LETEY, J. 1985. Relationship between soil physical properties and crop production.
Advances in Soil Science 1:277-294.

35
LOPES, I. L. M.; JARDIM, M. A. G.; MEDEIROS, T. D. S. 2008. Germinação e
desenvolvimento morfológico de plantas oleaginosas de floresta de várzea. Botânica Aplicada
– Pesquisa, 89: 252-254.
LIBERATO, M. A. R. 2010. Metabolismo energético e emissão de compostos orgânicos por
Hevea spruceana (Benth.) Müll. Arg em diferentes ecossistemas inundáveis da Amazônia
Central. Tese de doutorado em Botânica. Instituto Nacional de Pesquisas da Amazônia.
Manaus.
MAGUIRE, J.D. 1962. Speeds of germination-aid selection and evaluation for seedling
emergence and vigor. Crop Science 2:176-177.
MAIA, L.A., MAIA, S.; PAROLIN, P. 2005. Seedling Morphology of Non-pioneer Trees in
Central Amazonian Várzea Floodplain forests. Ecotropica, 11:1–8. Maguire, J.D. 1962.
Speed of germination: aid in selection and evaluation for seedling emergence and vigor. Crop
Science 2:176-177.
MAPFUMO, E.; CHANASYK, D. S.; BARON, V. S. 1998. Forage growth and yield
components as influenced by subsurface compaction. Agronomy Journal 90:805-812.
MOREIRA, M. F.; MOREIRA, F. W. 1996. Características da germinação de sementes de 64
espécies de leguminosas florestais nativas da Amazônia em condições de viveiro. Acta
Amazonica 26: 3-16.
NEVES, O. S. C.; BENEDITO, D. da S.; MACHADO, R. V.; CARVALHO, J. de G.
Crescimento. 2005. Produção de matéria seca e acúmulo de N, P, K, Ca, Mg e S na parte
aérea de mudas de andiroba (Carapa guianensis Aubl.) cultivadas em solo de várzea, em
função de diferentes doses de fósforo. Revista Árvore 28:343-349.
NG, F. S. P. 1978. Strategies of Establishment in Malayan forest trees. In: P. B. Tomlinson
and M.H. Zimmermann (eds.) Tropical trees as Living Systems. Cambrigde University,
Cambrigde, p. 129-162.
OLIVEIRA, A. K. M. de,RIBEIRO, J. F. W., PEREIRA, K. C. L., SILVA, C. A. A.
Germinação de sementes de Aspidosperma tomentosum Mart. (Apocynaceae)em diferentes
temperaturas. Revista Brasileira de Biociências 9:392-397.
PAROLIN, P. 2000. Seed mass in Amazonian floodplain forests with contrasting nutrient
supplies. Journal of Tropical Ecology 16:417-428.
PAROLIN, P. 2001. Seed germination and early establishmentin 12 tree species from
nutrient-rich and nutrient-poor Central Amazonian floodplains. Aquatic Botany 70:89–103.
PAROLIN, P.; JUNK, W. J. 2002. The effect of submergence on seed germination in trees
from amazonian floodplains. Boletim do Museu Paraense Emílio Goeldi Série Botanica
18:321-329.

36
PAROLIN, P.; FERREIRA, L. V.; JUNK, W. J. 2003. Germination characteristics and
establishment of trees from central Amazonian flood plains. Tropical Ecology 44: 157-169.
PIÑA-RODRIGUES, F. C. M.; AGUIAR, I. B. de. Maturação e dispersão de sementes. 1993.
In: Sementes Florestais Tropicais. AGUIAR, I. B. de, PIÑA-RODRIGUES, F. C. M;
FIGLIOLIA, M. B. ABRATES, Brasília, 350pp.
REIS, E. F.; SCHAEFER, C. E. G. R.; VIEIRA, L. B.; SOUZA, C. M.; Fernandes, H. C.
2002. Avaliação do contato solo-semente em um solo argiloso sob plantio direto, em
diferentes teores de água do solo. Engenharia na Agricultura 10:31-39.
SANTOS, D. L.; SUGAHARA, V. Y.; TAKAKI, M. 2005. Efeitos da luz e da temperatura na
germinação de sementes de Tabebuia serratifolia (Vahl) Nich, Tabebuia chrysotricha (Mart.
Ex DC.) Standl e Tabebuia roseo-alba (Ridl) Sand Bignoniaceae. Ciência Florestal 15:87-92.
SINGH, N.D.; ARUNACHALAN, A. 2002. Effect of pre-sowing treatments and seed size on
germination in five leguminous tree species. Range Manegement and Agroforestry 2:139-143.
VÁZQUEZ-YANES, C.; OROZCO-SEGOVIA A. 1993. Patterns of Seed Longevity and
germination in the tropical rainforest. Annual Review of Ecology and Systematics 24: 69-87.
ZANETTI, M., FERNANDES, C.; CAZETTA, J. O.; CORÁ, J. E.; MATTOS JUNIOR, D.
2003. Caracterização física de substratos para a produção de mudas e porta-enxertos cítricos
sob telado. Revista Laranja 24:519-530.
WITTMANN, F.; SCHÖNGART, J.; JUNK, W. J. 2010. Phytogeography, species diversity,
community structure and dynamics of central Amazonian floodplain forests. In: JUNK, W. J.,
PIEDADE, M. T. F.; WITTMANN, F. K.; SCHÖNGART, J. Central Amazonian floodplain
forests: ecophysiology, biodiversity and sustainable management. Dordrecht Heidelberg
London-New York, p. 61-102

37
Capítulo II ___________________________________________________________________________
Santos, L. R. & Wittmann, F. K. Crescimento,
desenvolvimento e alterações morfológicas de
plântulas de Macrolobium acaciifolium (Benth.)
Benth. em solos alagados de várzea e igapó da
Amazônia Central
Formatado de acordo com as normas da Revista Biota Neotropica.

38
Crescimento, desenvolvimento e alterações morfológicas de plântulas de
Macrolobium acaciifolium (Benth.) Benth. em solos alagados de várzea e
igapó da Amazônia Central
Resumo – A espécie Macrolobium acaciifolium (Benth.) Benth. ocorre em áreas de
maiores cotas de inundação de várzea e igapó da Amazônia Central, tendo se adaptado às
condições edáficas distintas destes dois ambientes. No presente estudo analisaram-se as
respostas morfológicas ao alagamento parcial, o crescimento, acúmulo de biomassa e o
conteúdo de nutrientes das folhas de plântulas de M. acaciifolium em substrato de várzea e
igapó. As sementes de M. acaciifolium foram coletadas em áreas de várzea e igapó e
germinadas em vermiculita em casa de vegetação. As plântulas produzidas foram
transplantadas para vasos contendo solo de várzea e solo de igapó, totalizando quatro
tratamentos: plântulas de várzea em solo de várzea e em solo de igapó e plântulas de igapó em
solo de igapó e em solo de várzea. Cada tratamento contou com 40 plântulas. Após uma
semana as plântulas foram submetidas a alagamento parcial do solo. Aos 90 dias encerrou-se
o experimento. Em todos os tratamentos observaram-se alterações na morfologia externa dos
indivíduos e ausência de mortalidade relacionada ao alagamento. Os indivíduos de ambas as
populações apresentaram menor peso e comprimento da raiz quando plantados em solo de
várzea. Não houve diferença significativa com relação ao comprimento e peso da parte aérea.
A menor média de diâmetro foi observada nos indivíduos de várzea que cresceram em solo de
igapó. As médias de área foliar específica não diferiram estatisticamente entre si. Os
indivíduos de várzea e igapó plantados em solo de igapó apresentaram as menores médias de
diâmetro comparadas às médias dos demais tratamentos. A quantidade de macro e
micronutrientes nas folhas de M. acaciifolium de várzea e igapó variou nos dois tipos de solo
e com relação à procedência, sendo que os indivíduos de igapó em solo de igapó de modo
geral exibiram mais nutrientes nas folhas comparados aos indivíduos de várzea, sugerindo que
indivíduos das duas populações possuem estratégias diferentes quanto ao uso de nutrientes.
Palavras chaves: Biomassa, Alagamento do solo, Nutrientes.
Growth, development and morphological changes in seedlings of Macrolobium acaciifolium (Benth.) Benth. in várzea and igapó soils in Central Amazon
Abstratct -The specie Macrolobium acaciifolium (Benth.) Benth. occurs in areas of higher flood levels of floodplain and igapó in Central Amazon, since it adapted to different soil conditions of these two environments. In the present study we analyzed the morphological responses to partial flooding, growth, biomass accumulation, and nutrient content in the seedlings leaves of M. acaciifolium in várzea and igapó substrate. Seeds of M. acaciifolium were collected in várzea and igapó areas, and germinated in vermiculite in a greenhouse. Seedlings produced were transplanted into pots containing várzea and igapó soils, in a total of four treatments: várzea seedlings in várzea and igapó soils, and igapó seedlings in igapó and in várzea soils. Each treatment had 40 seedlings, and after a week the seedlings were subjected to partial flooding. After 90 days the experiment was concluded. In all treatments it was observed changes in the external morphology of individuals and no mortality related to flooding. Individuals of both populations had lower weight and length of root when planted in lowland soil. There was no significant difference regarding the length and weight of the aerial

39
part. The lowest average diameter was observed in lowland and igapó individuals grown in igapó soil. The averages of specific leaf area were not statistically different from each other. Várzea and igapó Individuals planted in igapó soil showed the lowest average diameter compared to the average of the other treatments. The amount of macro and micro-nutrients in the leaves of várzea and igapó M. acaciifolium varied in both soil types and origin, while igapó individuals in igapó soil showed, in general, more nutrients in the leaves compared to várzea and igapó individuals, suggesting that both populations have different strategies for the use of nutrients.
Keywords: Biomass, Soil flooding, Nutrients.

40
1. Introdução
Nos ecossistemas alagáveis de várzea e igapó a quantidade de minerais contida nos
sedimentos transportados pela água age significativamente sobre a qualidade do solo (Worbes
et al., 1992). Os solos de igapó são formados por sedimentos provenientes de rochas bastante
erodidas do período Pré-cambriano (3,1 milhões de anos) e que formam amplas regiões de
areia branca pobres em minerais (Furch, 1997, Junk, 1998). Por outro lado, os solos de várzea
são formados por rochas sedimentares alcalinas desde o Período Terciário até o recente, que
ainda estão sob processo erosivo produzindo, portanto, uma quantidade elevada de sedimentos
ricos em minerais como Ca, Mg, Na e K (Furch, 1997, Junk, 1993).
Os solos de várzea por serem relativamente ricos em nutrientes possibilitam uma alta
produtividade, de forma que algumas árvores de várzea perdem as folhas no período de
inundação para substituí-las por uma folhagem nova quando as águas baixam.
Árvores de igapó por sua vez, por se desenvolverem em solos pobres
nutricionalmente, investem em manter suas folhas no período de cheia para evitar o gasto
energético da reposição de folhagem. Esse mecanismo pode explicar as diferenças na
produção de serrapilheira, que é muito superior nas florestas de várzea (10,3 t/ha) comparada
com a produção em solos de igapó (6,7 t/ha) (Kubitzki, 1989, Ferreira, 1997). Segundo Furch
(1997) a diferença entre a razão solo/nutrientes entre igapó e várzea é inferior às diferenças
quanto ao conteúdo de minerais (nitrogênio, fósforo, potássio, cálcio e magnésio), o que
sugere que as plantas dos dois ambientes possuem estratégias diferentes no que diz respeito ao
uso desses nutrientes.
Ao exercerem influência sobre os parâmetros químicos, biológicos e físicos do solo, as
inundações periódicas que ocorrem na várzea e no igapó (Pezeshki, 1994 e 2001) agem,
consequentemente, sobre o comportamento da comunidade vegetal fazendo com que haja
diferenças nas formações florestais dos dois ambientes (Prance, 1979, Ferreira, 1997).
Algumas plantas sob condições de alagamento podem apresentar estratégias para criar
tolerância à anoxia, e assim garantir seu estabelecimento no ambiente (Pimenta et al., 1998).
Este é o caso, por exemplo, de algumas espécies que na época das cheias tendem a investir na
formação de raízes adventícias, hipetrofia de lenticelas, redução do número de folhas entre
outras estratégias de sobrevivência (Parolin, 2012).

41
Deste modo, o presente trabalho buscou investigar as respostas ao alagamento e o
incremento de biomassa de duas populações da espécie Macrolobium acaciifolium (Benth.)
Benth. provenientes de várzea e igapó. Considerando as condições nutricionais dos solos
desses dois ambientes.
Material e Métodos
Os frutos de M. acaciifolium de população de várzea foram coletadas nas
proximidades da ilha da Marchantaria (03º 13’ S 59º 58’ W), na bacia pertencente ao rio
Solimões. Os frutos de populações de igapó foram coletados próximo ao Lago Tarumã-mirim
(3º 01’ 38” S 60º 10” 45’W) que faz parte da bacia do rio Negro.
As coletas foram realizadas entre os meses de abril e maio de 2015. Foram coletados
apenas frutos maduros, que após levados para a sede do Grupo MAUÁ/Projeto Max Planck,
no Instituto Nacional de Pesquisas da Amazônia, tiveram suas sementes retiradas
manualmente. As sementes passaram foram desinfectadas por imersão em solução de
hipoclorito de sódio a 10% por dez minutos e em seguida foram lavadas em água corrente.
A semeadura ocorreu em junho de 2015. As sementes foram plantadas em bandejas
contendo vermiculita de textura média e mantidas em temperatura ambiente em casa de
vegetação telada. Após 30 dias decorridos da germinação, as plântulas foram transplantadas
para vasos de polietileno com capacidade para 3,5 kg contendo substrato de igapó ou várzea,
conforme os tratamentos na Tabela 1.
Tabela 1 – Distribuição dos tratamentos
Origem da semente Substrato Repetições
Várzea Várzea 4 x 10
Igapó 4 x 10
Igapó Igapó 4 x 10
Várzea 4 x 10
Após uma semana do transplante as mudas foram submetidas a alagamento parcial. Os
vasos foram colocados em tanques de pvc com capacidade para 500 litros ficando com raiz e
parte do caule submersos por uma coluna de água de aproximadamente 10 cm acima do solo.
A água utilizada foi proveniente de poço artesiano do Instituto Nacional de Pesquisas da
Amazônia; a cada 15 dias os tanques foram esvaziados para troca da água para evitar
possíveis interferências em decorrência de alterações nas suas qualidades químicas e físicas.

42
O solo utilizado como substrato foi coletado em áreas próximas ao local de coleta das
sementes, uma vez que o local ao redor das matrizes ainda se encontrava inundado. Para
realizar análise foram coletadas dez sub amostras de solo a 30 cm de profundidade em cada
área, profundidade esta recomendada por Furch (1997) para solos alagáveis.
Aos 30 e 90 dias de experimento foi realizada a contagem do número de indivíduos
vivos dentro dos quatro tratamentos e também observadas a ocorrência de alterações
morfológicas externas.
Para a análise de crescimento em altura, a cada quinze dias o comprimento de dez
plantas de cada tratamento foi aferido com auxílio de fita métrica.
Para a análise de crescimento em diâmetro, em igual intervalo de tempo, foi medida a
circunferência do caule de dez plantas por tratamento a cinco centímetros do solo utilizando
paquímetro digital.
No final do experimento, para a determinação da área foliar mediu-se a área foliar de
50 folíolos por tratamento usando o medidor de área foliar Leaf Area Meter, Delta-T Devices.
Após a medição os foliolos foram secos em estufa a 40 graus até atingirem peso constante. A
partir dos valores de área e peso foi calculada a área foliar específica pela fórmula:
AFE = AF/MS
Em que:
AFE = área foliar específica
AF= área foliar
MS = massa seca dos folíolos
Para a aferição da biomassa seca, as mesmas dez plantas selecionadas para medição da
altura e diâmetro, foram retiradas dos vasos aos 90 dias, lavadas e seccionadas com tesoura,
separando-se raiz e parte aérea. Os compartimentos foram acondicionados em sacos de papel,
levados à estufa a 80º por 72 horas, quando alcançaram peso constante, e pesados em balança
analítica com precisão de quatro casas decimais (Piedade, 1998 adaptado por Ferreira, 2006).
Selecionou-se ao acaso folhas adultas com aspecto fitossanitário saudável dentro de
todos os tratamentos para determinação do teor de nutrientes. As análises foram realizadas
pelo Laboratório de Análises de Solo e de Plantas – LASP. EMBRAPA. Foram avaliadas as
quantidades de macro e micronutrientes.
O delineamento experimental utilizado foi o inteiramente casualizado (DIC). Os dados
de peso bem como os de altura da raiz e da parte aérea foram submetidos ao teste de

43
normalidade de Kolmogorov-Smirnov e posteriormente realizou-se a análise de variância, as
médias foram comparadas pelo teste de Tukey a 5% de probabilidade.
Os dados referentes ao diâmetro e área foliar específica foram submetidos ao teste de
Kolmogorov-Smirnov e não apresentaram distribuição normal, tendo sido então aplicado o
teste de Student-Newman-Keuls (SNK) para comparação de médias.
O programa estatístico utilizado foi o Assistat 7.7 Beta.

42
Figura 1 - Análise da fertilidade do solo usado como substrato. Laboratório de Análises de Solo e de Plantas – LASP. EMBRAPA.
Amostra pH C M.O. P K Na Ca Mg Al H+Al SB t T V m Fe Zn Mn Cu
(H2O) (g/kg) mg/dm3 cmolc/dm
3 % mg/dm
3
Solo de
Igapó 4,69 14,84 25,53 4 12 3 0,06 0,06 1,77 3,83 0,16 1,93 3,99 4,10 91,53 406 0,76 0,50 0,66
Solo de
várzea 6,16 2,95 5,08 120 58 19 4,76 1,24 0,0 0,66 6,23 6,23 6,89 90,42 0,00 464 5,71 128 3,71
pH em água relação 1:2,5
P, Na, K, Fe, Zn, Mn e Cu – Extrator Mehlich-1
Ca, Mg – Extrator KCl 1mol/L
H + Al – Extrator de acetato de cálcio 0,5 mol/L
SB – Soma de bases
CTC (T) – Capacidade de troca catiônica efetiva
CTCt (t) – Capacidade de troca catiônica a pH 7,0
V – índice de saturação por bases
m – Índice de saturação por alumínio
Matéria orgânica (M.O.) – (C orgânico) x 1,724 – Walkley-Black

43
Tabela 2 - Análise Granulométrica do solo usado como substrato. Laboratório de Análises de Solo e de Plantas – LASP. EMBRAPA.
Solo Areia Grossa Areia Fina Areia Total Silte Argila
2,00 – 0,20
(mm)
0,20 – 0,05 (mm) 2,00 - 0,05
(mm)
0,05 - 0,002
(mm) < 0,002 (mm)
(g/kg)
Várzea 10,87 651,60 662,47 171,95 188,5
Igapó 632,59 143,97 776,55 149,03 51,50

44
Resultados e Discussão
Ao final do experimento com plântulas de Macrolobium acaciifolium, aos 90 dias não
foi observada a mortalidade de plantas. Em todos os tratamentos foram registradas alterações
morfológicas ligadas à submersão parcial, como raízes adventícias e formação de lenticelas no
caule que surgiram a partir do oitavo dia de alagamento (Figura 2).
Figura 2- Formação de raízes adventícias e lenticelas em Macrolobium acaciifolium em condição de
alagamento parcial em solo de igapó.
Foto: Santos, 2015.
Não foi registrado clorose ou perda de folhas. A ausência de mortalidade na fase
inicial de desenvolvimento dessa espécie sob alagamento e a formação de estruturas
adaptativas faz parte do sucesso de sua adaptação ao pulso de inundação das áreas onde
ocorre (Parolin et al., 2002), não sendo afetado pela qualidade nutricional do solo. Este
resultado diverge do encontrado por Santiago e Paoli (2003) para as espécies Adelia
membranifolia (Müll. Arg.) Pax & K. Hoffm (Euphorbiaceae) e Peltophorum dubium
(Spreng.) Taub. (Fabaceae) submetidas ao alagamento em substrato agrícola Plantamax, de
acordo com os autores as alterações morfológicas devido ao alagamento foram encontradas
apenas nos indivíduos que se desenvolveram em substrato com mais nutrientes. Os mesmos
autores registraram o mesmo comportamento para Genipa americana L., onde algumas
plantas não desenvolveram respostas morfológicas sob estresse por déficit nutricional quando
em substrato alagado (Santiago e Paoli, 2007).
Raízes adventícias
Lenticelas
hipertrofiadas
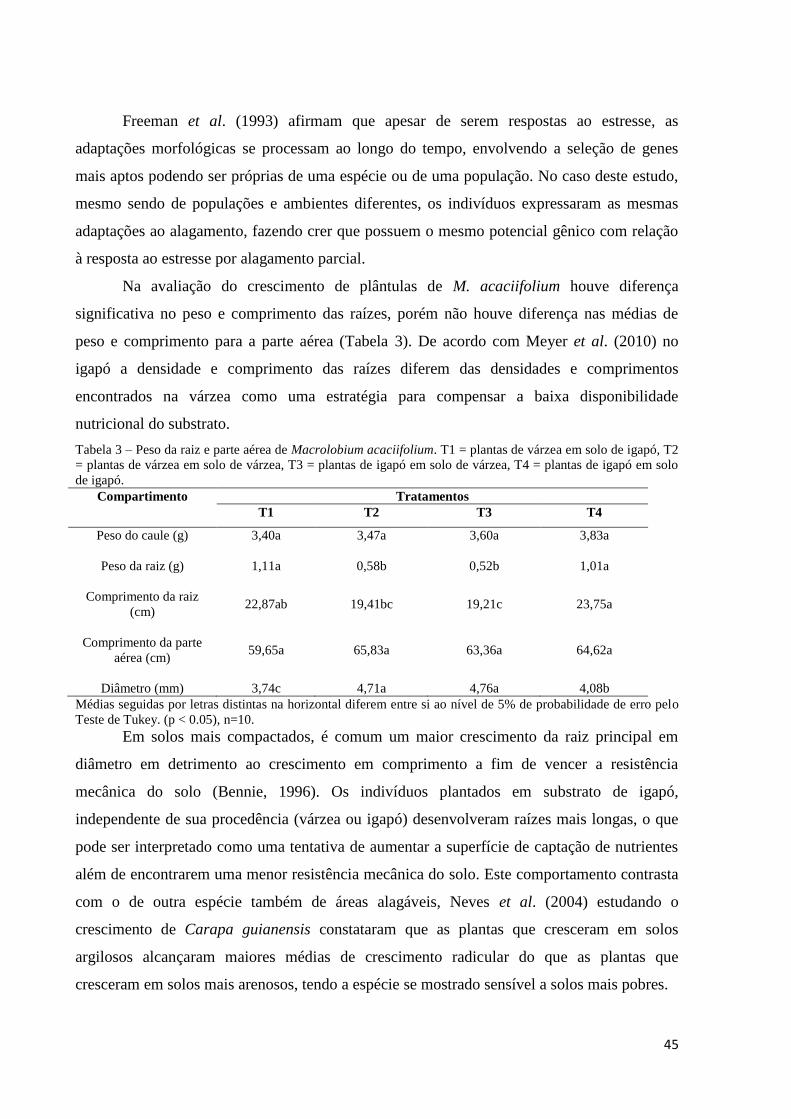
45
Freeman et al. (1993) afirmam que apesar de serem respostas ao estresse, as
adaptações morfológicas se processam ao longo do tempo, envolvendo a seleção de genes
mais aptos podendo ser próprias de uma espécie ou de uma população. No caso deste estudo,
mesmo sendo de populações e ambientes diferentes, os indivíduos expressaram as mesmas
adaptações ao alagamento, fazendo crer que possuem o mesmo potencial gênico com relação
à resposta ao estresse por alagamento parcial.
Na avaliação do crescimento de plântulas de M. acaciifolium houve diferença
significativa no peso e comprimento das raízes, porém não houve diferença nas médias de
peso e comprimento para a parte aérea (Tabela 3). De acordo com Meyer et al. (2010) no
igapó a densidade e comprimento das raízes diferem das densidades e comprimentos
encontrados na várzea como uma estratégia para compensar a baixa disponibilidade
nutricional do substrato.
Tabela 3 – Peso da raiz e parte aérea de Macrolobium acaciifolium. T1 = plantas de várzea em solo de igapó, T2
= plantas de várzea em solo de várzea, T3 = plantas de igapó em solo de várzea, T4 = plantas de igapó em solo
de igapó.
Compartimento Tratamentos
T1 T2 T3 T4
Peso do caule (g) 3,40a 3,47a 3,60a 3,83a
Peso da raiz (g) 1,11a 0,58b 0,52b 1,01a
Comprimento da raiz
(cm) 22,87ab 19,41bc 19,21c 23,75a
Comprimento da parte
aérea (cm) 59,65a 65,83a 63,36a 64,62a
Diâmetro (mm) 3,74c 4,71a 4,76a 4,08b
Médias seguidas por letras distintas na horizontal diferem entre si ao nível de 5% de probabilidade de erro pelo
Teste de Tukey. (p < 0.05), n=10.
Em solos mais compactados, é comum um maior crescimento da raiz principal em
diâmetro em detrimento ao crescimento em comprimento a fim de vencer a resistência
mecânica do solo (Bennie, 1996). Os indivíduos plantados em substrato de igapó,
independente de sua procedência (várzea ou igapó) desenvolveram raízes mais longas, o que
pode ser interpretado como uma tentativa de aumentar a superfície de captação de nutrientes
além de encontrarem uma menor resistência mecânica do solo. Este comportamento contrasta
com o de outra espécie também de áreas alagáveis, Neves et al. (2004) estudando o
crescimento de Carapa guianensis constataram que as plantas que cresceram em solos
argilosos alcançaram maiores médias de crescimento radicular do que as plantas que
cresceram em solos mais arenosos, tendo a espécie se mostrado sensível a solos mais pobres.

46
As maiores médias de diâmetro foram encontradas nos indivíduos plantados em solo
de várzea, independente da origem das sementes. Schöngart et al. (2005), em um estudo
dendrocronológico constataram que indivíduos de Macrolobium acaciifolium (Benth.) Benth.
de várzea apresentaram um incremento radial duas vezes superior ao encontrado nas árvores
de mesma espécie que estavam em área de igapó. Sugerindo que para a variável diâmetro o
solo é um fator limitante.
Os valores de área foliar específica variaram de 109,22 cm2/g
a 177,17 cm
2/g
e
diferença entre os tratamentos não foi significativa (Tabela 4).
Tabela 4 – Área foliar específica (AFE). T1 = plantas de várzea em solo de igapó, T2 = plantas de várzea em
solo de várzea, T3 = plantas de igapó em solo de várzea, T4 = plantas de igapó em solo de igapó.
Tratamento Área foliar específica (cm2/g)
T1 141,25a
T2 177,17a
T3 134,26a
T4 109,22a
Médias seguidas por mesma letra não diferem entre si ao nível de 1% de probabilidade de erro pelo Teste SNK.
(p < 0.01), n=50.
As folhas são os órgãos que mais respondem às variações de disponibilidade de
recursos, expressando isso em características estruturais (Smith et al., 1997). A área foliar
específica também é um atributo relacionado à captura de luminosidade, sendo associada à
capacidade fotossintética da planta (Evans & Seemann, 1989). Contudo, há estudos apontando
que a disponibilidade de água e minerais exerce mais influência na alocação de massa foliar
do que a luz (Kwesiga & Grace, 1986). Uma alta taxa de interceptação de luminosidade pode
não significar uma maior produtividade pois, podem ocorrer variações na quantidade de
nutrientes e carbono em nas zonas da folha responsáveis pela interceptação (Pooter et al.,
2009).
Neste estudo, as folhas do tratamento T1 e T4, que cresceram em um substrato mais
pobre em nutrientes não tiveram, estatisticamente, maiores médias de AFE comparadas com
as do tratamento T2 e T3 que se desenvolveram em substrato mais rico nutricionalmente.
Como as condições de luminosidade foram iguais em todos os tratamentos, concluiu-
se que de modo geral para a variável AFE os indivíduos de M. acaciifolium das duas
populações estudadas, mesmo em diferentes condições nutricionais, apresentaram respostas
plásticas semelhantes.
Na análise nutricional das folhas observou-se que as quantidades de macronutrientes
variaram dentro dos tratamentos (Tabela 5). A quantidade de nutrientes disponível no

47
substrato de várzea foi superior à encontrada em solo de igapó, mas, isso não implicou
necessariamente em uma maior quantidade de macronutrientes nas folhas, o que pode ter
ocorrido pela interação entre os próprios macronutrientes e o conteúdo de matéria orgânica e
pH dos substratos.
A amostra de solo de várzea apresentou menor quantidade de matéria orgânica (5,08 g
kg-1
) e maior pH (6,1) comparada à amostra de solo de igapó (matéria orgânica igual a 25,53 g
kg-1
e pH 4,69) (Figura 1). De acordo com Pavinato et al., (2008) a decomposição de material
orgânico e a liberação de compostos gerados nesse processo exerce influência sobre a
disponibilidade de nutrientes do solo. Segundo esses autores esta influência se dá pela
transformação de um íon simples em um íon complexo ou, pela adsorção de íons que
apresentam antagonismo inibindo a ação de alguns grupos funcionais.
Tabela 5 - Quantidade de macronutrientes em folhas de M. acaciifolium 90 dias após alagamento em
solos de várzea e igapó. T1 = plantas de várzea em solo de igapó, T2 = plantas de várzea em solo de
várzea, T3 = plantas de igapó em solo de várzea, T4 = plantas de igapó em solo de igapó.
Tratamentos Macronutrientes
g kg-1
N P K Ca Mg S
T1 22,57 1,12 8,82 4,33 8,46 0,85
T2 20,72 1,19 8,21 6,69 8,02 0,95
T3 20,97 1,19 7,84 8,08 8,64 1,02
T4 22,51 1,23 8,12 6,94 8,88 0,69
A distribuição mais uniforme foi a do nitrogênio, seguido do fósforo e do magnésio
em todos os tratamentos a maior quantidade de nitrogênio encontrada foi nos tratamentos T1 e
T4, cujo substrato foi solo de igapó, o nitrogênio está associado à quantidade de matéria
orgânica, sendo que quase a totalidade de nitrogênio no solo se encontra na forma orgânica
(Black, 1968). A maior absorção de nitrogênio foi nas plântulas plantadas em solo de igapó,
onde havia a maior quantidade de matéria orgânica no solo. A quantidade de fósforo
encontrado nas folhas aparentemente não apresentou nenhuma correlação com a quantidade
deste nutriente no solo. Segundo Velloso et al. (1993) após o alagamento, a solubilidade do
fósforo aumenta causando uma maior eficiência na sua utilização pela planta, a quantidade
desse nutriente foi semelhante em todos os tratamentos.
Nos Tratamentos T1 e T2, onde as plantas eram provenientes de sementes de várzea,
foi encontrado mais K nas folhas comparado aos demais tratamentos e uma menor quantidade
de magnésio apesar da sua maior disponibilidade no substrato de várzea do que no de igapó.

48
Pode ocorrer antagonismo entre elementos durante a absorção de minerais, fazendo com que a
presença de um iniba a absorção do outro, relação essa observada entre o potássio e o
magnésio e também entre o potássio e o cálcio (Malavolta,1980). Nos tratamentos T3 e T4,
onde foram registradas as maiores quantidades de cálcio e magnésio as quantidades de
potássio foram menores, podendo indicar reação de antagonismo entre esses minerais.
No caso da quantidade de enxofre, o elemento foi encontrado em maior quantidade nas
folhas de indivíduos plantados em solo de várzea, independente da origem das sementes.
Segundo Lefroy et al., (1993) quanto maior o pH maior a disponibilidade de enxofre. Neste
estudo no substrato de várzea o pH ficou em 6,16 e no substrato de igapó em 4,69 sugerindo
que o a maior absorção de S pode ter sido influenciada pelo pH mais neutro encontrado no
solo de várzea.
A análise dos micronutrientes está sumarizada na Tabela 5 a seguir.
Tabela 6 – Quantidade de micronutrientes em folhas de M. acaciifolium 90 dias após alagamento em solos de
várzea e igapó. T1 = plantas de várzea em solo de igapó, T2 = plantas de várzea em solo de várzea, T3 = plantas
de igapó em solo de várzea, T4 = plantas de igapó em solo de igapó
Tratamentos Micronutrientes
mg k-1
B Cu Fe Mn Zn
T1 23,15 4,33 440,55 24,31 14,71
T2 29,47 4,02 294,19 42,13 15,72
T3 30,70 4,16 281,06 36,76 35,80
T4 36,68 4,83 349,69 30,27 29,19
As quantidades de boro e zinco foram maiores nas folhas de plantas de área de igapó,
independente do substrato, demonstrando que os indivíduos de igapó tendem a absorver mais
desse nutriente do que os indivíduos de várzea.
O cobre foi encontrado em maior quantidade nas folhas dos tratamentos T1 e T4,
ambos com substrato de igapó. A disponibilidade do cobre diminui com o aumento do pH do
solo, o pH entre 6 e 7 reduz a solubilidade do cobre e este fica mais preso à argila e à matéria
orgânica (Potafos, 1996). Os substratos dos tratamentos T1 e T4 eram formados por solos de
igapó, que possuíam pH mais baixo, menos argila e matéria orgânica do que o solo de várzea
usado nos tratamentos T2 e T3 (Figura 1).
Foi observado que ao contrário do que aconteceu com o zinco e o boro, a maior
disponibilidade de Mn no solo implicou numa maior quantidade deste elemento nas folhas
(T2 e T3). Este elemento é associado à síntese de clorofila e a ativação de enzimas, sendo
importante para o crescimento vegetal (Larcher, 2000).

49
A absorção de ferro apresentou valores mais elevados, nos tratamentos com solo de
igapó, a despeito deste substrato possuir uma quantidade de ferro inferior a encontrada no
substrato de várzea. Em solos inundados há o aumento os níveis da forma reduzida do ferro
(Fe2+
), facilmente absorvida pelas plantas, o que também é induzido pela maior quantidade de
matéria orgânica. A quantidade de matéria orgânica no substrato de igapó foi
aproximadamente, cinco vezes superior à encontrada no substrato de várzea, o que pode ter
influenciado na absorção de ferro pelas plantas nos tratamentos T1 e T4. Observou-se ainda
que quanto maior a quantidade de ferro menor a de Mn. Malavolta (1980) associa esse efeito à
inibição competitiva que o íon Fe+2
exerce sobre o Mn+2
.
De modo geral, observou-se que o comportamento da espécie Macrolobium
acaciifolium em solos de várzea e igapó apresenta algumas diferenças quanto à utilização de
nutrientes independente da quantidade do nutriente no substrato.
Conclusão
As plantas de populações de várzea quando plantadas em solo de igapó apresentaram
menor diâmetro do que nos demais tratamentos.
Houve diferenças nas quantidades de macro e micronutrientes nas folhas de M.
acaciifolium de várzea e igapó nos dois tipos de solo, no entanto, os valores encontrados
estavam dentro dos limites necessários para o funcionamento do metabolismo vegetal, não
sendo observados sinais de deficiência nutricional.

50
Referências
BENNIE, A.T.P. 1996. Growth and mechanical impedance. In: Waisel,Y.; Eshel, A.; Kafkafi,
U. (eds.) Plants roots: the hiddenhalf. 2.ed.Marcel Dekker, New York.
FREEMAN, D.C., GRAHAM, J.H.; EMLEN, J.M. 1993. Developmental stability in plants:
Symmetries, stress and epigenesis. Genetica 9:97-119.
FURCH, K. 1997. Chemistry of várzea and igapó soils and nutrient inventory of their
floodplain forests. In: Junk, W. J. (ed.). The Central Amazon floodplains. Ecology of a
Pulsing System. Berlin. p. 47-64.
EVANS, J. R.; SEEMANN, J. R. 1989. The allocation of protein nitrogen in the
photosynthetic apparatus: costs, consequences and control. In: Briggs, W. R. (ed.)
Photosynthesis. New York. p.183-205.
JUNK, W.J. 1998. A várzea do Rio Solimões-Amazonas: Conceitos para o aproveitamento
sustentável e seus recursos. Anais do IV Simpósio de Ecossistemas Brasileiros. Águas de
Lindóia. p. 1-24.
JUNK, W.J. 1993. Wetlands of Tropical South America. In: Whigham, D.; Hejny, S.;
Dykyjova, D. (Eds). Wetlands of the world. Dr. W.J. Junk Publishers, Dordrecht, Boston. p.
679-739.
BLACK, C. A. 1968. Soil plant relationships. John Wiiley and sons (eds.). Wiley, 792 p.
LARCHER, W. 2000. Ecofisiologia Vegetal. RIMA, São Carlos.
MALAVOLTA, E. 1980. O reino da nutrição mineral de planta. In: Malavolta, E. Elementos
de nutrição mineral de plantas. Ceres, São Paulo. p. 9-15.
POTAFOS. 1996 Nutri-Fatos:Informação agronômica sobre nutrientes para as culturas.
Piracicaba, SP – Arquivo do agrônomo Nº 10.
POOTER, H.; NIINEMETS, Ü.; POOTER, L.; WRIGHT, I. J.; VILLAR, R. 2009. Causes
and consequences of variation in leaf mass per area (LMA): a meta-analysis. New
Physiologist 182:556-588.
MEYER, U.; JUNK, W. J.; LINCK, C. 2010. Fine roots and mycorrhizal associations in two
Amazonian Central inundation forests: igapó and várzea. In: Amazonian floodplains forests:
ecophysiology, biodiversity and sustainable management. Junk, W. J.; Piedade, M. T. F.;
Wittmann, F. K.; Schöngart, J. (eds.), New York: Springer Verlag, Dordrecht-Heidelberg-
London. p. 163-178.

51
PAROLIN, P.; JUNK, W. J. 2002. The effect of submergence on seed germination in trees
from amazonian floodplains. Boletim do Museu Paraense Emílio Goeldi Série Botanica
18:321-329.
LEFROY, R. D. B.; SAMOSIR, S. S. R.; BALIR, G. J. 1993. The dynamics of sulphur,
phosphorus and iron in flooded soils as affected by changes in Eh and pH. Australian Journal
of Soil Research. 31:493-508.
PAVINATO, P. S.; ROSOLEM, C. A. Disponibilidade de nutrientes do solo e decomposição
e liberação de compostos orgânicos. 2008. Revista Brasileira de Ciência do Solo 32:911-920.
SCHÖNGART, J.; PIEDADE, M.T.F.; WITTMANN, F.; JUNK, W.J.; WORBES, M. 2005.
Wood growth patterns of Macrolobium acaciifolium (Benth.) (Fabaceae) in Amazonian black-
water and white-water floodplain forests. Oecologia 145: 454-461
VELLOSO, A. C. X.; OLIVEIRA, C.; LEAL, J. R. 1993. Processos redox em glei húmico do
Estado do Rio de Janeiro: III. Variações das concentrações de Fe II. e fosfato. Revista
Brasileira de Ciência do Solo 17:27-33.
KWESIGA, F.; GRACE, J. 1986. The role of the red-fair ratio in the response of tropical tree
seedlings to shade. Annals of botany 57: 283-290.
SMITH, W. K.; VOGELMANN T. C.; DELUCIA E. H.; BELL D. T.; SHEPHERD K. A.
1997. Leaf Form and Photosynthesis: do leaf structure and orientation interact to regulate
internal light and carbon dioxide? BioScience 47:785-793.
SOUZA, L. A. G. de. 2012. Guia da biodiversidade de Fabaceae do Alto Rio Negro. Manaus.
Projeto Fronteiras. 118 p.
NEVES, O. S. C.; BENEDITO, D. da S.; MACHADO, R. V.; CARVALHO, J. de G.
Crescimento. 2005. Produção de matéria seca e acúmulo de N, P, K, Ca, Mg e S na parte
aérea de mudas de andiroba (Carapa guianensis Aubl.) cultivadas em solo de várzea, em
função de diferentes doses de fósforo. Revista Árvore 28:343-349.
SANTIAGO, E.F.; PAOLI. A. A.S. 2003. O aumento em superfície em Adelia membranifolia
(Müll. Arg.) Pax & K. Hoffm. e Peltophorum dubium (Spreng.) Taub., em resposta ao
estresse por deficiência nutricional e alagamento do substrato. Revista Brasileira de Botânica.
26:503-513.
SANTIAGO, E.F.; PAOLI. A. A.S. 2007. Respostas morfológicas em Guibourtia hymenifolia
(Moric) J. Leonard (Fabaceae) e Genipa americana L. (Rubiaceae), submetidas ao estresse
por deficiência nutricional e alagamento do substrato. Revista Brasileira de Botânica, 30: 131-
140.

52
SÍNTESE GERAL
Ausência de sinais de subdesenvolvimento de plantas mesmo quando da diminuição
de nutrientes disponíveis no substrato ou exposição ao alagamento.
Indivíduos de população de várzea quando transplantados para solo de igapó
apresentaram uma diminuição considerável do crescimento em diâmetro.
As relações de absorção de alguns nutrientes nas duas populações independem da
quantidade disponível no substrato.
Por fim, chegou-se a conclusão de que são necessários mais estudos a cerca do
comportamento da espécie com relação à absorção e estratégias de uso de nutrientes.

53
APÊNDICES

54

55


















