
Influência das Plantas de Sapal na Biogeoquímica do...
156
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e Guadiana Ricardo Manuel Pires Mendes Dissertação de Mestrado em Química Tecnológica Especialização em Química Tecnológica 2012
Transcript of Influência das Plantas de Sapal na Biogeoquímica do...

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
Influência das Plantas de Sapal na
Biogeoquímica do Mercúrio e Metilmercúrio:
Sapais do Tejo e Guadiana
Ricardo Manuel Pires Mendes
Dissertação de
Mestrado em Química Tecnológica
Especialização em Química Tecnológica
2012


UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
Influência das Plantas de Sapal na
Biogeoquímica do Mercúrio e Metilmercúrio:
Sapais do Tejo e Guadiana
Ricardo Manuel Pires Mendes
Dissertação de
Mestrado em Química Tecnológica
Especialização em Química Tecnológica
Orientadores:
Prof. Doutor Manuel Luís Matos Lopes (DQB/FCUL) e
Doutor João Alfredo Vieira Canário (Investigador no Instituto Português do Mar e da
Atmosfera)
2012


Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
i
Agradecimentos
Ao Doutor João Canário, orientador deste projeto, por todo o apoio, bom humor e
excelente pessoa que é.
Ao Professor Doutor Manuel Matos Lopes e pela possibilidade de efectuar este estágio
sob a sua orientação e pelas suas correcções e opiniões.
Ao Instituto Português do Mar e da Atmosfera pela disponibilidade de todos os meios
necessários à realização deste trabalho.
Ao LNEG, pelo apoio prestado e pela disponibilidade de equipamento na determinação de
alúminio.
À Doutora Helena Lourenço e Susana Gonçalves, pelo apoio prestado e pela
disponibilidade de equipamento na determinação de ferro e mangânes.
À Doutora Marta Nogueira, pelo apoio prestado na determinação dos potenciais redox e
pH.
Á Mestre Bárbara Anes, pelos ensinamentos e ajuda prestada no tratamento de dados.
Um muito Obrigado!
Ao Doutor Miguel Caetano, pelos primeiros ensinamentos, conselhos e saída de campo
Ao Doutor Nelson O'Driscoll pelas leituras de determinação de metilmercúrio.
Ao Mestre Pedro Brito, pelo companheirismo e ajuda durante os trabalhos de campo e à
Mestre Rute Cesário, pela simpatia e pelo ensino das primeiras técnicas
Aos restantes colegas do IPMA. Joana Raimundo, Patrícia Pereira, Rui, Maria João e
Teresa pela amizade e companheirismo durante todo o estágio.
Ao Denis Rodrigues, pelo companheirismo nestes anos de faculdade, e pela capacidade
de trabalho sempre demonstrada e com que eu me identifico. Se nestes anos, alguma vez
fui incorreto, as minhas sinceras e sentidas desculpas.
Ao André Ferreira, um amigo de vida, pelo apoio e ajuda sempre demonstrada e á
Margarida Murta, por ser a amiga e a pessoa que é.
Á Rita Santos, o meu grande amor, que sempre me ajudou, apoiou e esteve sempre ao
meu lado, mesmo nas alturas mais complicadas e dificeis do estágio.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
ii
Ao tio Vitor, tia Helena, primos Diogo, António e Pedro, perdoêm-me as ausências e os
silêncios neste ano.
E a referência mais importante, à minha Mãe, patrocinadora deste mestrado, pela
educação, amor e carinho, e por nada me ter faltado. Um exemplo de vida. Se há
“vencidos da vida” ela é uma “vencedora da vida”.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
iii
Resumo
O presente trabalho incide num estudo realizado sobre a variação sazonal
(inverno/primavera) of biogeoquímica do Hg em sedimentos, águas e plantas em
sapais com Hg de distintas origens naturais (origem antropogénica – sapal do Rosário
e origem geológica – sapal de Castro Marim)
Durante as amostragens foram recolhidas partes aereas de plantas, águas
sobrenadantes, cores de sedimentos de zonas colonizadas por plantas (colonizados
por S. fruticosa e S. maritima) e de zonas não colonizadas.
Foram determinadas a Humidade (%), LOI (%),Eh, pH, %Biomassa (só no caso dos
cores com plantas), e concentrações de SO42-, Cl-, HS-, AVS, Hg, Al, Fe, Mn, Hg e
MeHg.
Os resultados mostram que os teores de Hgsed e MeHgsed do sedimento no sapal do
Rosário são uma ordem de grandeza mais elevados do que no sapal de Castro Marim,
o que demonstra a contaminação a que o sapal do Rosário foi sujeito.
No sapal do Rosário os teores de Hg e MeHg dissolvidos, são maiores no inverno do
que na primavera, mas no sapal de Castro Marim, os teores de Hg e MeHg dissolvidos
são maiores na primavera, uma vez que podem reter o mercúrio na fração dissolvida.
Concluímos também que foram nas raízes das plantas onde houve maiores
percentagens de MeHg face ao Hg total, na maioria dos casos, durante a primavera.
De uma maneira geral, verificou-se que os teores de Hg e MeHg na biomassa são
muito superiores aos teores de mercúrio no sedimento o que é devido ao facto da
biomassa subterrânea absorver/adsorver Hg e MeHg existentes no sedimento,
acumulando nas raizes.
Os teores de Hg e MeHg nas partes aéreas são inferiores aos teores na biomassa
subterrânea, tendo um fraco transporte das raízes para as partes aéreas.
Palavras-chave:mercúrio, metilmercúrio, sapais, variação sazonal, diagénese

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
iv
Abstract
Our present work focus on a study on the seasonal variation (winter/spring) of mercury
biogeochemistry in salt marshes, with the mercury from distinct natural origins
(anthropogenic origin - Rosário’s salt marshes, and geologic origin - Castro Marim’s
marshes).
During the sampling period we also collected aerial portions of local plants, overlaying
water samples and sediment cores from colonized (colonized by S. fruticosa and S.
maritima) and non-colonized areas.
In the laboratory, we determined the Humidity (%), LOI (%), Eh, pH, % of biomass (only
in the case of the cores obtained from colonized areas), and were analysed
concentrations of Cl-, SO42, HS-, AVS, Al, Fe, Mn, Hg, MeHg.
Our results show that the concentrations of Hg and MeHg determined from the solid
fraction of the sediment obtained from the Rosário’s salt marshes are of a greater order
of magnitude then those determined from the Castro Marim’s salt marshes. These
results reveal the degree of contamination the Rosário’s salt marshes underwent.
In Rosário’s salt marshes the concentrations of dissolved Hg and MeHg are greater
during the winter than in the spring, but in Castro Marim’s salt marshes they are greater
in the spring, since they can retain Hg in the dissolved fraction.
We also concluded that a greater percentage of MeHg is found in the plants’ roots
when compared with the total Hg present, in the majority of cases during the spring.
We could generally verify that the concentrations of Hg and MeHg in the biomass are
much higher than the concentrations of Hg in the sediment which is due to the fact that
the subterranean biomass can absorb/adsorb Hg and MeHg existing in the sediment,
collecting them in the plants’ roots.
Concentrations of Hg and MeHg in the aerial portions of the plants than in the
subterranean biomass, having a weak rate of transport from the roots to the aerial
portions.
Keywords:mercury, metylmercury,salt marshe, sazonal variation, diagenesys

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
v
Índice
Agradecimentos ............................................................................................................. i
Resumo ......................................................................................................................... iii
Abstract ........................................................................................................................ iv
Índice ............................................................................................................................. v
Índice de Figuras ......................................................................................................... viii
Índice de Tabelas ........................................................................................................ xiii
Lista de Acrónimos ...................................................................................................... xiv
I. Introdução .................................................................................................................. 1
1. Sapais .................................................................................................................... 1
1.1. Caracteristicas gerais dos sapais .................................................................... 1
1.2. Flora nos sapais .............................................................................................. 2
2. O mercúrio ............................................................................................................. 3
2.1. O ciclo biogeoquímico do mercúrio ................................................................. 3
2.2. Propriedades físico-químicas do mercúrio ....................................................... 5
2.3. Toxicidade do mercúrio e metilmercúrio .......................................................... 7
2.4. Formação de metilmercúrio ............................................................................. 8
2.5. Mercúrio em águas naturais .......................................................................... 10
2.6. Mercúrio em sedimentos ............................................................................... 12
3. Reações de diagénese nos sapais ...................................................................... 15
3.1. Comportamento geral .................................................................................... 15
3.2. Mercúrio em sapais ....................................................................................... 18
4. Objetivos deste trabalho ...................................................................................... 19
II. Caracterização da área de estudo ........................................................................... 21
1. Estuário do Tejo ................................................................................................... 22
2. Estuário do Guadiana .......................................................................................... 24
III. Materiais e Métodos ............................................................................................... 26
1. Descontaminação do material .............................................................................. 26
2. Trabalho de Campo ............................................................................................. 27
3. Processamento das amostras .............................................................................. 28
3.1 Sedimentos .................................................................................................... 28
3.1.1. Águas intersticiais .................................................................................. 28
3.1.2 Fração sólida .......................................................................................... 28
3.1.3. Procedimento especial para sulfuretos dissolvidos e AVS ..................... 29
3.2. Águas sobrenadantes.................................................................................... 29

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
vi
3.3 Plantas ........................................................................................................... 30
3.3.1 Biomassa subterrânea ............................................................................ 30
3.3.2 Biomassa aérea ...................................................................................... 30
4. Análise das amostras ........................................................................................... 32
4.1. Análise das águas ......................................................................................... 32
4.1.1. Determinação de ferro e manganês totais dissolvidos ........................... 32
4.1.2. Determinação de cloretos dissolvidos .................................................... 32
4.1.3. Determinação de sulfatos ...................................................................... 33
4.1.4. Determinação de sulfuretos dissolvidos totais ....................................... 33
4.1.5. Determinação de mercúrio total dissolvido ............................................. 34
4.1.6. Determinação de metilmercúrio dissolvido ............................................. 35
4.2. Análise à fração sólida do sedimento ............................................................ 36
4.2.1. Teor de humidade .................................................................................. 36
4.2.2. Teor em matéria orgânica sedimentar ................................................... 36
4.2.3. Determinação do pH e EH ...................................................................... 37
4.2.4. Determinação da concentração de ferro, manganês e alumínio total .... 37
4.2.5. Determinação da concentração mercúrio total ....................................... 39
4.2.6. Determinação de metilmercúrio ............................................................. 39
4.2.7. Determinação de sulfuretos insolúveis ................................................... 40
4.3 Análise das amostras vegetais ....................................................................... 41
4.3.1 Determinação da percentagem de biomassa subterrânea ...................... 41
4.3.2. Determinação de mercúrio total ............................................................. 41
4.3.3. Determinação de metilmercúrio ............................................................. 42
5. Materiais Certificados de Referência ................................................................... 43
IV. Resultados e Discussão ......................................................................................... 45
1. Caracterização do sedimento .............................................................................. 45
1.1. Fração sólida dos sedimentos ....................................................................... 45
1.1.1. Teores de humidade, LOI, potencial redox e pH .................................... 45
1.1.2. Alumínio, ferro e manganês totais .......................................................... 51
1.1.3. Sulfuretos Ácidos Voláteis ..................................................................... 59
1.1.4. Biomassa subterrânea ........................................................................... 61
1.2. Águas intersticiais ......................................................................................... 63
1.2.1. Clorinidade ............................................................................................. 63
1.2.2. Sulfatos e Sulfuretos dissolvidos totais .................................................. 65
1.2.3. Ferro e manganês dissolvidos ............................................................... 73

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
vii
2. Águas sobrenadantes .......................................................................................... 78
3. Mercúrio e metilmercúrio...................................................................................... 79
3.1 Fração sólida do sedimento ........................................................................... 79
3.1.1. Mercúrio ................................................................................................. 79
3.1.2. Metilmercúrio ......................................................................................... 84
3.2 Águas intersticiais .......................................................................................... 87
3.2.1. Mercúrio ................................................................................................. 87
3.2.2. Metilmercúrio ......................................................................................... 89
3.3 Plantas ........................................................................................................... 93
3.3.1. Biomassa subterrânea ........................................................................... 93
3.3.1.1. Mercúrio ........................................................................................... 93
2.3.1.2. Metilmercúrio .................................................................................... 95
3.3.2. Biomassa aerea ..................................................................................... 99
3.3.2.1. Mercúrio ........................................................................................... 99
3.3.2.2. Metilmercúrio .................................................................................. 100
3.4. Distribuição de mercúrio e metilmercúrio entre sedimentos e biomassa
subterrânea ........................................................................................................ 103
3.5. Correlações entre mercúrio e metilmercúrio ................................................ 109
3.6. Cálculo dos stocks de Hg e MeHg ............................................................... 113
3.7 Cálculo dos fluxos difusivos ......................................................................... 115
3.8. Comparação dos valores de mercúrio e metilmercúrio ................................ 117
V. Conclusões Gerais ................................................................................................ 119
Relatório de ambiente e segurança ........................................................................... 120
Análise de custos ...................................................................................................... 122
Apêndice ................................................................................................................... 123
1.1 Técnicas de análise ...................................................................................... 123
1.2.1 espectroscopia de Absorção Atómica por Chama ................................ 123
1.2.2. espectroscopia de Absorção Atómica Eletrotérmica ............................ 124
1.2.3. espectroscopia de Fluorescência Atómica – Vapor Frio com Injeção de
Fluxo .............................................................................................................. 125
1.2.4. espectroscopia de Fluorescência Atómica – Cromatografia Gasosa ... 126
1.2.5. espectroscopia Molecular Ultravioleta - Visível .................................... 127
1.2.6. Polarografia ......................................................................................... 127
Glossário ................................................................................................................... 129
Referências ............................................................................................................... 130

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
viii
Índice de Figuras
Figura 1 – Ciclo Biogeoquímico do Mercúrio (adaptado de Monteiro (2010) e Canário (2004a)).4
Figura 2 - Variação de cloro- e hidroxi-complexos de Hg2+
em solução aquosa, consoante a concentração de Cloro e o pH. ....................................................................................... 11
Figura 3- Esquema explicativo da diagenética em sedimentos não colonizados por plantas e em sedimentos colonizados por plantas (adaptado de Otte & Jacob, 2003). ................ 16
Figura 4 – Estuário do Tejo, com destaque para a zona do sapal do Rosário, onde se realizou a amostragem. .................................................................................................................. 23
Figura 5 - Estuário do Guadiana. Destaque para a zona do sapal de Castro Marim, onde se realizou a amostragem. .................................................................................................. 24
Figura 6 – Fotografia de um core de sedimentos num corer metálico, e fotografia do local de amostragem no sapal do Rosário, onde se pode ver solos colonizados por plantas e solos não colonizados. ................................................................................................... 27
Figura 7- Imagens das plantas em sapal, juntamente com desenhos do seu perfil. Em cima temos a S. fruticosa e em baixo a S. marítima (adaptado de Duarte, 2010 e Antunes Dias & Marques, 1999). ................................................................................................. 31
Figura 8 - Esquema da montagem para a extração do AVS e SEM (Adaptado de Canário,(2000). .............................................................................................................. 40
Figura 9 - Perfis verticais dos teores de Humidade (%), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ................................................................................................................... 45
Figura 10 - Perfis verticais dos teores de Humidade (%) nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ....................................................................................................................................... 46
Figura 11 - Perfis verticais dos teores de LOI (%), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ................................................................................................................... 47
Figura 12 - Perfis verticais dos valores de LOI (%), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ....................................................................................................................................... 47
Figura 13 – Perfis verticais dos potenciais redox, nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. 48
Figura 14 – Perfis verticais dos potenciais redox, nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ....................................................................................................................................... 49
Figura 15 – Perfis verticais dos pH, nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ................... 50
Figura 16 – Perfis verticais dos pH, nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ................... 50

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
ix
Figura 17 - Perfis verticais dos teores de alumínio (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 51
Figura 18 - Perfis verticais dos teores de alumínio (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ................................................................................................................... 52
Figura 19 - Perfis verticais dos teores de ferro (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 53
Figura 20 - Razões dos teores de Fe/Al dos sedimentos recolhidos no Sapal de Castro Marim. ....................................................................................................................................... 53
Figura 21 - Perfis verticais dos teores de ferro (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ....................................................................................................................................... 54
Figura 22 - Razões dos teores de Fe/Al dos sedimentos recolhidos no sapal do Rosário. ........ 55
Figura 23 - Perfis verticais dos teores de manganês (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados. ............................................................................................................ 56
Figura 24 - Razões dos teores de (Mn/Al)×10-2
dos sedimentos recolhidos no sapal de Castro Marim ............................................................................................................................. 56
Figura 25 - Perfis verticais dos teores de manganês (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 57
Figura 26 - Razões dos teores de (Mn/Al)×10-2 dos sedimentos recolhidos no Sapal do Rosário. .......................................................................................................................... 58
Figura 27 - Perfis verticais dos teores de AVS (µmol g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 59
Figura 28 - Perfis verticais dos teores de AVS (μmol g-1
), nos sedimentos recolhidos no sapal do Rosário em sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 60
Figura 29 - Perfis verticais em profundidade da Biomassa (%) nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima. .... 61
Figura 30 - Perfis verticais da percentagem de Biomassa (%) nos sedimentos recolhidos no sapal do Rosário em sedimentos colonizados por S. fruticosa e S. marítima. ............... 62
Figura 31 - Perfis verticais dos teores de cloretos (g L-1
) nas águas intersticiais dos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. .......................................................................................... 63
Figura 32- Perfis verticais dos teores de cloretos (g L-1
) nas águas intersticiais dos sedimentos recolhidos no Sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. .......................................................................................... 64
Figura 33 - Perfis verticais dos teores de sulfatos (mM) nas águas intersticiais dos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. .......................................................................................... 65

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
x
Figura 34 - Razões dos teores de (SO42-
/Cl-) nas águas intersticiais dos sedimentos recolhidos
no sapal de Castro Marim. ............................................................................................. 66
Figura 35 - Perfis verticais dos teores de Sulfatos (mM) nas águas intersticiais dos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados............................................................................................ 66
Figura 36 - Perfis das razões dos teores de (SO42-
/Cl-) dos sedimentos recolhidos no Sapal de
Rosário. .......................................................................................................................... 67
Figura 37 - Perfis verticais dos teores de sulfuretos dissolvidos (HS-) (µM) nas águas
intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim. ........................................................ 68
Figura 38 - Perfis verticais dos teores de Sulfuretos dissolvidos (µM) dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. .......................................................................................................................... 68
Figura 39 - Perfis dos teores de SO42-
(mM) e HS- (μM) nas águas intersticiais dos sedimentos
recolhidos no sapal de Castro Marim. ............................................................................ 70
Figura 40 - Perfis dos teores de SO42-
(mM) e HS-(μM) nas águas intersticiais dos sedimentos
recolhidos no sapal do Rosário. ..................................................................................... 71
Figura 41 - Perfis verticais dos teores de ferro (mg L-1
) nas águas intersticiais dos sedimentos para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim. ............................................................................ 73
Figura 42 – Perfis verticais dos teores de ferro dissolvido (mg L-1
) nas águas intersticiais dos sedimentos para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. ................................................................ 74
Figura 43 - Perfis verticais dos teores de manganês (mg L-1
) nas águas intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim. .................................................................................................. 76
Figura 44 – Perfis verticais dos teores de manganês dissolvido (mg L-1
) nas águas intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. ..................................................................................... 77
Figura 45 - Perfis verticais dos teores de mercúrio (μg g-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim. ......... 79
Figura 46 - Perfis das razões dos teores de ([Hg]/[Al]) e ([Hg])/(%LOI) dos sedimentos recolhidos no sapal de Castro Marim. ............................................................................ 80
Figura 47 - Perfis verticais dos teores de mercúrio total (µg g-1
), em sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. ............. 81
Figura 48 - Perfis das razões dos teores de ([Hg]/[Al]) e ([Hg])/(%LOI) dos sedimentos recolhidos no sapal do Rosário. ..................................................................................... 82
Figura 49 - Perfis verticais dos teores de metilmercúrio (ng g-1
), nos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim. ....................................................................................................................................... 84

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
xi
Figura 50 - Perfis verticais dos teores de metilmercúrio total (ng g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados. ................................................................................................................... 85
Figura 51 - Perfis percentagens de MeHg em relação à proporção de mercúrio total dos sedimentos recolhidos no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.). ... 86
Figura 52 - Perfis verticais dos teores de mercúrio (ng L-1
) para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim. ......... 87
Figura 53 - Perfis verticais dos teores de mercúrio total (ng L-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. ....... 88
Figura 54 - Perfis verticais dos teores de metilmercúrio (ng L-1
) para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim. ....................................................................................................................................... 89
Figura 55 - Perfis verticais dos teores de metilmercúrio (ng L-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário. ....... 90
Figura 56 - Perfis percentagens de MeHg em relação à proporção de mercúrio dissolvido nos sedimentos recolhidos no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.). ... 92
Figura 57 - Perfis verticais do teor de mercúrio total (μg g-1
) na biomassa subterrânea nos sedimentos colonizados por S. fruticosa e S. marítima recolhidos no sapal de Castro Marim. ............................................................................................................................ 93
Figura 58 – Perfis verticais dos teores de mercúrio total (μg g-1
) na biomassa subterrânea de S. fruticosa e S. maritima, recolhida no sapal do Rosário. ................................................. 94
Figura 59 - Perfis verticais do teores de metilmercúrio total (ng g-1
) na biomassa subterrânea nos sedimentos colonizados por S. fruticosa e S. maritima, recolhidos no sapal de Castro Marim. ................................................................................................................. 95
Figura 60 - Perfis verticais dos teores de metilmercúrio total (ng g-1
) na biomassa subterrânea da S. fruticosa e S. marítima recolhida no sapal do Rosário,. ........................................ 96
Figura 61 - Perfis percentagens de MeHg em relação à proporção de mercúrio dissolvido na biomassa subterrânea recolhida no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.). ................................................................................................................................ 97
Figura 62 - Valores dos teores de mercúrio total (μg g-1
) nas partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima, no sapal de Castro Marim. ... 99
Figura 63 - Teores de mercúrio total (μg g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal do Rosário. ................................... 100
Figura 64 - Valores dos teores de metilmercúrio total (ng g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal de Castro Marim. ..................................................................................................................................... 100
Figura 65 - Teores de metilmercúrio (ng g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal do Rosário. ................................... 101
Figura 66 – Percentagem de metilmercúrio em função do teor de mercúrio total nas partes fotossintéticas e não fotossintéticas para a S. fruticosa e S. marítima, recolhidas no sapal de Castro Marim ................................................................................................. 101

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
xii
Figura 67 - Percentagem de metilmercúrio em função do teor de Mercúrio total nas partes fotossintéticas e não fotossintéticas para a S. fruticosa e S. marítima, recolhidas no sapal do Rosário. ......................................................................................................... 101
Figura 68 - Perfis dos teores de Hg na biomassa subterrânea e nos sedimentos colonizados, recolhidos no sapal de Castro Marim. .......................................................................... 103
Figura 69 - Perfis dos teores de Hg na biomassa subterrânea e nos sedimentos colonizados, recolhidos no sapal do Rosário. ................................................................................... 104
Figura 70 - Perfis dos teores de MeHg na biomassa subterrânea e nos sedimentos colonizados recolhidos no sapal de Castro Marim. .......................................................................... 105
Figura 71 - Perfis dos teores de MeHg na biomassa subterrânea e nos sedimentos colonizados recolhidos no sapal do Rosário. ................................................................................... 106
Figura 72 - Correlações entre os teores de Hg na fração sólida e na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal de Castro Marim. ................................... 109
Figura 73 - Correlações entre os teores de MeHg na fração sólida e MeHg na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal de Castro Marim. ........ 110
Figura 74 – Correlações entre os teores de MeHg e Hg na biomassa subterrânea nos sedimentos colonizados recolhidos no sapal de Castro Marim. ................................... 110
Figura 75 – Correlações entre os teores de MeHg e Hg na biomassa subterrânea nos sedimentos colonizados recolhidos no sapal do Rosário. ............................................ 111
Figura 76 - Correlações entre os teores de MeHg na fração sólida e MeHg na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal do Rosário. ................. 111
Figura 77 - Correlações entre os teores de Hg na fração sólida e na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal do Rosário. ............................................ 112
Figura 78 – Esquema de funcionamento do “Advanced Mercury Analyser 254” (adaptado de Costley et al., 2000). .................................................................................................... 124

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
xiii
Índice de Tabelas
Tabela 1 – Espécies de mercúrio mais comuns (adaptado de IUPAC (1998)). ........................... 6
Tabela 2 – Espécies principais de mercúrio presentes na fração sólida e nas águas intersticiais dos sedimentos (Canário, 2004a). ................................................................................. 14
Tabela 3 - Condições operatórias usadas para análise de Fe e Mn por EAA-Chama. .............. 32
Tabela 4 – Condições operatórias para a determinação de mercúrio total dissolvido por Fluorescência Atómica ................................................................................................... 35
Tabela 5 – Condições operatórias usadas para análise de Fe, Al, e Mn por EAA-Chama ........ 38
Tabela 6 - Concentrações e incertezas associadas dos padrões PACS-2, MESS-3, HISS-1, 1646a, BCR-60 e BCR-62 para Hg, Al, Mn, Si, e Fe ...................................................... 43
Tabela 7 – Concentrações médias, desvios padrões e erros de método associados para o mercúrio presente nas plantas e na fracção sólida do sedimento. ................................. 44
Tabela 8 – Teores de inverno e de primavera dos teores das espécies químicas analisadas para as águas sobrenadantes, recolhidas no sapal de Castro Marim. ........................... 78
Tabela 9 - Teores de inverno e de primavera dos teores das espécies químicas analisadas para as águas sobrenadantes, recolhidas no sapal de Rosário. ............................................ 78
Tabela 10 – Valores dos stocks obtidos para o sapal de Castro Marim e do Rosário.............. 113
Tabela 11 - Valores dos fluxos difusivos com a indicação dos respetivos sentidos, para o HgD nos sedimentos e HgD nas águas sobrenadantes ........................................................ 116
Tabela 12 - Teores mercúrio e metilmercúrio nacionais e internacionais no sedimento e dissolvidos.................................................................................................................... 117
Tabela 13 – Teores mercúrio e metilmercúrio nacionais em sedimentos colonizados por plantas halófitas ........................................................................................................................ 118
Tabela 14 – Lista de material mais consumido e respetivos custos ......................................... 122
Tabela 15 – Custos das análises, com nos valores apresentados em Diário da Républica, 2ª série- Nº 223 – 20 de Novembro de 2006, Despacho nº23597/2006 ........................... 122

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
xiv
Lista de Acrónimos
AVS - Sulfuretos Ácidos Voláteis ou Monosulfuretos metálicos (do inglês, Acid volatile
sulphides)
CH2O – Fórmula geral dos compostos de carbono
CVAFS - espectroscopia de Fluorescência Atómica por Vapor - Frio (do Inglês Could
Vapor – Atomic Fluorescence Spectroscopy)
DPCSV – Polarografia Diferencia de Redissolução catódica
DPP – Polarografia Diferencial por Impulsos
EH – Potencial redox em relação ao elétrodo de hidrogénio
HgD – Mercúrio na fração dissolvida do sedimento
Hgraiz – Mercúrio absorvido pela biomassa subterrânea
Hgsed – Mercúrio na fração sólida do sedimento
HMDE – Elétrodo de mercúrio de gota suspensa
L.D. – Limite de deteção
LOI - Matéria Orgânica Sedimentar (do inglês, Lost on Ingnition)
MeHg – Metilmercúrio
p – valor da probabilidade estatística
r – coeficiente de correlação
– Tortuosidade do sedimento
- Porosidade do sedimento

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 1
I. Introdução
1. Sapais
1.1. Caracteristicas gerais dos sapais
Os sapais são ambientes específicos de estuários, deltas, ou lagos, e são zonas de
transição entre ecossistemas marinhos e terrestres. São caracterizados por serem
plataformas de sedimentos finos (por deposição fluvial ou por erosão e transporte de
ondas) e de matéria orgânica. São alagados consoante as marés e colonizados por
plantas, que apresentam características que lhes permitem desenvolver-se neste tipo
de ambientes (Mitsh & Gosselink, 2000; Caçador & Vale, 2001; Davidson-Arnott, et al.,
2002).
Os sapais são ecossitemas de extrema importância para o ambiente devido à elevada
produção biológica primária, proteção à erosão costeira, fluxos hidrológicos regulares,
ciclos biogeoquímicos de nutrientes e metais, e habitats com uma grande variedade de
espécies, onde se incluem insetos, anfíbios, peixes e aves (Mitsh & Gosselink, 2000;
Caçador & Vale, 2001; Válega, et al., 2008a; Canário, et al., 2010).
Os sapais podem atuar como fontes, sumidouros ou transformadores de espécies e
elementos químicos, dependendo de vários parâmetros tais como o pH, condições
redox, oxigénio dissolvido, matéria orgânica e interações entre microrganismos (Mitsh
& Gosselink, 2000; Otte & Jacob, 2003).
Podem ainda reter nutrientes e contaminantes (Válega, et al., 2008a), sendo os
nutrientes mais comuns, em geral, compostos de azoto, fósforo e ferro (Mitsh &
Gosselink, 2000).
De uma maneira geral, estas zonas sofrem grandes perturbações de origem
antropogénica devido à proximidade a cidades e indústrias, ocupação para fins
agrícolas e construção, tendo sido por isso afetados por contaminações provenientes
dos efluentes que têm, muitas vezes, elevado o teor em metais pesados.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
2 Ricardo M.P.Mendes
1.2. Flora nos sapais
Os sapais de Portugal são colonizados por espécies halófitas, sendo as mais comuns
a Halimione portulacoides, Sarcocornia fruticosa, Spartina marítima e Juncos
marítimos (Antunes Dias & Marques, 1999; Caçador & Vale, 2001; Canário, et al.,
2007a; Caetano, et al., 2008; Duarte, et al., 2010).
Estas plantas podem imobilizar ou mobilizar metais nos sedimentos da rizosfera,
consoante a excreção de oxigénio das raízes para o sedimento, influenciando assim
os processos redox e ainda alterando o pH (Caçador & Vale, 2001; Otte & Jacob,
2003; Weis & Weis, 2004). Conseguem suportar períodos em que ficam submersas
por várias horas e suportar elevada salinidade. Permitem que se depositem mais
sedimentos, que são estabilizados pelas suas raízes e onde é produzida e acumulada
mais matéria orgânica (Mitsh & Gosselink, 2000; Davidson-Arnott, et al., 2002).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 3
2. O mercúrio
2.1. O ciclo biogeoquímico do mercúrio
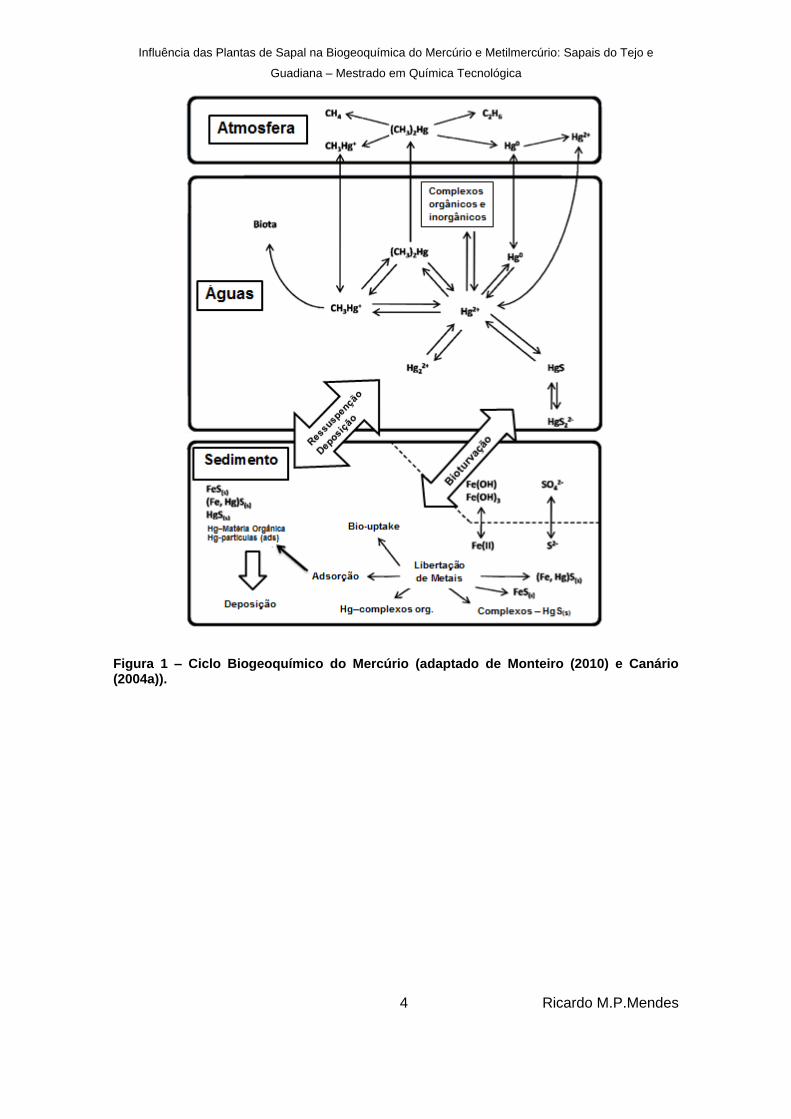
Na figura 3 encontra-se representado o ciclo biogeoquímico do mercúrio que resume a
especiação e forma química do mercúrio no meio ambiente (meio atmosférico, meio
aquático e meio sedimentar).
O ciclo, de uma forma geral, centra-se nas reações que envolvem as seguintes espécies:
Hg0 ↔ Hg2+ ↔ CH3Hg+
O mercúrio (II) encontra-se fundamentalmente em sedimentos e águas na forma de
compostos simples ou compostos mais complexos, podendo sofrer dois processos
distintos: ser reduzido a mercúrio elementar através de processos de fotoredução,
passando para a atmosfera (Canário & Vale, 2004b) ou através de redução (Mason, et al.,
1995) encontrando-se então como gás dissolvido na água ou como vapor de mercúrio na
atmosfera; ser biologicamente metilado através de micro-organismos (Baldi, 1997).
O teor destas três espécies de mercúrio referidas num determinado meio depende de
diversos fatores mas fundamentalmente dos equilíbrios entre as reações em que as
referidas espécies se encontrem envolvidas.
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
4 Ricardo M.P.Mendes
Figura 1 – Ciclo Biogeoquímico do Mercúrio (adaptado de Monteiro (2010) e Canário (2004a)).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 5
2.2. Propriedades físico-químicas do mercúrio
O desenvolvimento industrial conduziu ao aparecimento, ou ao aumento de
substâncias químicas que até então não existiam ou existiam em muito pequenas
quantidades no ambiente. A libertação destas substâncias mais ou menos reativas
afetam os sistemas em que estão inseridas. Entre os poluentes, destacam-se os
metais pesados, onde se inclui o mercúrio (Hg).
O mercúrio é um poluente global sendo que as suas formas químicas apresentam um
elevado grau de toxicidade para o biota, sendo por isso, considerado um dos metais
pesados mais tóxicos (Pacyna, et al., 2008). O mercúrio é o único metal líquido à
temperatura ambiente (Horvat, 1996), com uma densidade de 13,5 g cm-3, a sua
temperatura de fusão é de -38,9ºC (considerado o ponto de fusão mais baixo de todos
os metais), e um ponto de ebulição é a 357,25ºC (Andrew & Nriagu, 1979; O’Neil,
1985 ).
É caracterizado por uma baixa resistividade elétrica, elevada tensão superficial e
elevada condutividade térmica na gama líquida (Andrew & Nriagu, 1979). A pressão de
vapor é de 0,0703 Pa a 10ºC e 0,17 Pa a 20ºC, com a concentração de saturação de
vapor de mercúrio no ar a variar entre 5,5 mg m-3 a 10ºC para 12 mg m-3 a 20ºC
(Andrew & Nriagu, 1979).
Devido á sua volatilidade, é facilmente transportado por grandes distâncias atravéz do
transporte atmosférico. Por conseguinte, conduz a contaminação de locais que se
encontram a grandes distâncias das fontes antropogénicas (Pirrone, et al., 2008).
O mercúrio possui três os estados de oxidação, sendo eles o mercúrio elementar
(Hg0), o ião mercuroso (Hg+) e ião mercúrico (Hg2+), que podem formar espécies mais
ou menos estáveis. O mercúrio elementar (Hg0) e o ião mercúrico (Hg2+) são os
estados de oxidação mais comuns na natureza (Horvat, 1996).
Como referido anteriormente o mercúrio elementar (Hg0) é bastante volátil,
relativamente não reativo e por isso eficientemente transportado, como um vapor na
atmosfera por longas distâncias, apresentando baixa solubilidade (IUPAC, 1998;
Ullrich, et al., 2001). A sua oxidação a HgO pelo oxigénio é termodinamicamente lenta
e este composto é facilmente decomposto por ação da luz em Hg0 e O2 (NOAA, 1996;
Morel, et al., 1998; Jackson, 1998).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
6 Ricardo M.P.Mendes
A espécie Hg+ não é termodinamicamente estável, sendo encontrada na forma de
catião diatómico Hg22+. Esta espécie não forma complexos estáveis e transforma-se
rapidamente em Hg0 + Hg2+, onde o Hg2+ forma complexos estáveis na presença de
ligandos (NOAA, 1996; Morel, et al., 1998; Jackson, 1998).
A espécie inorgânica Hg2+, forma complexos estáveis com os iões cloreto, sulfato,
nitrato, hidróxido e com ligandos orgânicos, como – CH3 ou – COOH. Por ser um ácido
de Lewis fraco, tem maior afinidade com ligandos considerados bases de Lewis fracas,
como os sulfuretos, sendo por isso, a formação de HgS altamente favorável (NOAA,
1996; Morel, et al., 1998; Jackson, 1998).
Os compostos em que o mercúrio está ligado a um radical orgânico e outro grupo são
solúveis em água, dissociando-se no catião R-Hg+ e no anião X-. Os aniões mais
comuns são o Cl-, o OH-, o NO3- e o SO4
2-. Os ligandos de iões de halogéneos (Cl-, I- e
Br-) conferem à ligação um caráter covalente linear (R-Hg-X) (Jackson, 1998).
Na natureza o mercúrio encontra-se fundamentalmente na forma de sulfureto de
mercúrio (HgS), usualmente designado por cinábrio.
A Tabela 1 apresenta um resumo geral das espécies químicas em que o Hg se
encontra.
Tabela 1 – Espécies de mercúrio mais comuns (adaptado de IUPAC (1998)).
Mercúrio elementar Hg0
Espécies inorgânicas de mercúrio Ião mercúrico Hg2+
Ião mercuroso Hg+
Sulfureto de mercúrio HgS
Espécies orgânicas de mercúrio Metilmercúrico CH3Hg+
Etilmercúrio C2H5Hg+
Dimetilmercúrio (CH3)2Hg
As fontes antropogénicas de mercúrio, são bastantes diversificadas, resultando, de
maneira geral, de indústrias de extração e purificação de metais preciosos, indústrias
cloro-alcalinas, e da queima de combustíveis fosseis como carvão, turfa e madeira em
centrais termoelétricas.
O mercúrio teve aplicações em, instrumentos de medição de temperatura e pressão,
na produção de lâmpadas e baterias, no processamento de madeiras (antifúngico) e

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 7
em amálgamas para a manufatura eletrolítica de cloretos e hidróxido de sódio (NOAA,
1996; Morel, et al., 1998; IUPAC, 1998; Ullrich, et al., 2001). Atualmente a utilização de
mercúrio encontra-se proibida na EUA e Canadá.
2.3. Toxicidade do mercúrio e metilmercúrio
O Hg elementar é pouco reativo, por isso, a toxicidade deste está fundamentalmente
associada às suas formas orgânicas e inorgânicas.
O metilmercúrio é a forma de Hg mais abundante, sendo considerado cancerígeno e
teratogénico (WHO, 1990). A sua elevada toxicidade deve-se à solubilidade lipídica e
propriedades iónicas que apresenta, o que faz com que seja um composto que
atravessa facilmente as membranas celulares dos organismos vivos por transporte
passivo, inibindo a síntese proteica (Baldi, 1997).
O metilmercúrio funciona como uma potente neurotoxina, que se bioacumula nos
organismos, devido a taxas de eliminação muito inferiores ás taxas de acumulação e
por isso, a sua concentração amplifica ao longo da cadeia trófica: é absorvido por
seres unicelulares (fitoplâncton e zooplâncton) sendo esses seres ingeridos por peixes
pequenos, que por sua vez são ingeridos por peixes maiores. Consequentemente a
concentração do metal vai aumentando ao longo da hierarquia predatória. Verifica-se a
sua acumulação em peixes, invertebrados, mamíferos (especialmente organismos
marinhos) e plantas aquáticas (Kumar & Tate, 1982; WHO, 1990; NOAA, 1996; Morel,
et al., 1998; Ullrich, et al., 2001) constituindo um sério risco para a saúde pública. Os
teores em metilmercúrio, existente no peixe, podem variar entre 60 a 90% do mercúrio
total (O’Neil, 1985 ).
No organismo humano os alvos da toxicidade do metilmercúrio são o cérebro e o
sistema nervoso central, sendo afetadas as funções sensoriais, visuais e auditivas
(WHO, 1990). Quanto a mecanismos de toxicidade, o mercúrio estabelece fortes
ligações com os grupos que contêm enxofre (grupos tiol) em proteínas e enzimas,
danificando ou corrompendo os mecanismos da sua síntese e atividade (O’Neil, 1985 ;
WHO, 1990). Em termos de consequências de envenenamento o metilmercúrio pode
atravessar a placenta e afetar o feto.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
8 Ricardo M.P.Mendes
Têm existido vários acidentes devido a alimentos contaminados por metilmercúrio. O
acidente da baía de Minamata (Japão), que decorreu nos anos de 1950, foi o mais
mediático. Contudo também houve outros, como nos anos de 1971/72 no Iraque,
devido à ingestão de grão com um fungicida feito à base de metilmercúrio (O’Neil,
1985 ; Ullrich, et al., 2001).
2.4. Formação de metilmercúrio
A formação de metilmercúrio no ambiente pode ocorrer em três compartimentos
ambientais: na coluna de água, nos sedimentos e no biota, sendo a biometilação o
processo biogeoquímico mais frequente na formação de metilmercúrio (Morel, et al.,
1998; Ullrich, et al., 2001).
A biometilação depende de variáveis ambientais como a disponibilidade de iões de Hg
inorgânico e a população microbiológica metilante (destacando-se as bactérias sulfato-
redutoras) (NOAA, 1996). O balanço entre os processos de metilação e desmetilação,
influenciam a quantidade de metilmercúrio produzida (Morel, et al., 1998; Ullrich, et al.,
2001; Merrit & Amirbahman, 2009).
De uma maneira geral, as taxas de metilação são mais elevadas em condições
anóxicas. Estas estão igualmente dependentes de parâmetros como os teores de
matéria orgânica, nutrientes, concentração de sulfatos e sulfuretos, cloretos (efeito da
salinidade), temperatura, pH e potencial redox, que estimulam ou não, o crescimento
microbiano favorecendo as reações de metilação (NOAA, 1996; Ullrich, et al., 2001;
Miranda, et al., 2007).
A variação sazonal destes parâmetros, faz com que existam períodos do ano com
taxas de metilação mais elevadas (Canário, et al., 2005; Canário, et al., 2007a;
Canário, et al., 2007b). Vários estudos apontam para um aumento dos processos de
metilação durante os meses do verão, sendo por isso a temperatura um dos fatores
determinantes (Canário, et al., 2005; Canário, et al., 2007a). As condições redox do
sedimento também são outro fator que influencia a metilação do mercúrio, pois baixos
potenciais redox, associados a ambientes anóxicos favorecem as taxas de metilação
(Compeau & Bartha, 1984; Ullrich, et al., 2001). A presença de matéria orgânica nos
sedimentos também favorece a metilação, pois pressupõe a existência de mais
carbono disponível para ser utilizado pelas bactérias metiladoras (Furutani & Rudd,
1980)

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 9
A influência direta do pH nos processos de metilação não é consensual, apenas se
sabe que o pH influencia a mobilidade do mercúrio e metilmercúrio (Ullrich, et al.,
2001). Condições de elevada salinidade tornam o metilmercúrio menos estável
(Compeau & Bartha, 1984).
Aparentemente as condições para as quais se verificam taxas de metilação maiores
de mercúrio, são pH baixo, baixa salinidade e presença de matéria orgânica em
decomposição em condições redutoras (Ullrich, et al., 2001).
Os micro-organismos podem produzir metilmercúrio utilizando mercúrio armazenado
em sedimentos por contaminação de efluentes industriais e domésticos nos estuários
dos rios, podendo através de reações diagenéticas (definida na página 9), ficar
disponivel para processos de metilação (Compeau & Bartha, 1984; Gilmoure, et al.,
1992; Canário, et al., 2003a). As bactérias sulfato redutoras, são os principais agentes
metiladores de mercúrio em meios anóxicos de ambientes lacustres e estuarinos
(NOAA, 1996; Merrit & Amirbahman, 2009), sendo o processo geral o seguinte:
A disponibilidade de nutrientes, abundância de aceitadores de eletrões, atividade
bacteriana, distribuição das populações sulfaturedutoras (sendo que nem todas as
bactérias sulfato-redutoras são responsáveis pela metilação do mercúrio), são
condições que afetam os principais agentes metiladores (Nagase, et al., 1982; Choi &
Bartha, 1994; King, et al., 2000; Guimarães, et al., 2006).
O enxofre é um elemento extremamente importante no controlo da metilação. Como
anteriormente referido, as bactérias sulfato-redutoras são os principais agentes
metiladores (Miranda, et al., 2007). Em condições redutoras e elevadas concentrações
de sulfuretos, a metilação é inibida devido à formação de sulfuretos de mercúrio e a
sua consequente precipitação, que levam a uma diminuição do mercúrio disponível
para ser metilado. (Morel, et al., 1998), Ks (HgS) = 10-58
Hg2+ + S2- HgS
A metilação abiótica pode ocorrer na doação de grupos metil, provenientes de
materiais húmicos, ao Hg2+ (Nagase, et al., 1982), ou ocorrer fotoquimicamente,
quando há transferência de grupos metil na presença de radiação ultravioleta
(Siciliano, et al., 2005). A seguinte reação é um exemplo de transmutação abiótica do

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
10 Ricardo M.P.Mendes
CH3Hg(II) em (CH3)2Hg é que pode assumir um papel importante na redução de
metilmercúrio dos sedimentos (Gagnon, et al., 1997):
2CH3Hg+ + H2S (CH3Hg)2S + 2H+ (CH3)2Hg (volátil) + HgS + 2H+
2.5. Mercúrio em águas naturais
O mercúrio é introduzido nos sistemas aquáticos por processos naturais, como a
deposição atmosférica ou por fontes antropogénicas, sendo adsorvido por partículas
finas existentes em suspensão, ficando uma pequena porção na forma solubilizada
(Jackson, 1998).
Nas águas naturais, o mercúrio dissolvido varia ao longo da coluna de água, existindo
gradientes geoquímicos horizontais e verticais que fazem variar a especiação e
distribuição do mercúrio (Morel, et al., 1998). A sua distribuição entre fração dissolvida
ou particulada, depende da salinidade, concentração e natureza da matéria orgânica
(Canário, et al., 2008b).
A fração particulada é definida, operacionalmente, como sendo constituída por todas
as partículas que ficam retidas num filtro de 0,45 μm de porosidade, e incluem
partículas inorgânicas e biológicas.
A fração dissolvida, está distribuída pelas seguintes espécies químicas: Hg0 (volátil e
não reativo) com concentrações mais elevadas na interface água/atmosfera; espécies
inorgânicas de Hg2+ e mercúrio orgânico (principalmente MeHg+, Me2Hg e EtHg), com
concentrações mais elevadas na interface água-sedimento (Morel, et al., 1998).
Em águas naturais, o Hg2+ não se encontra como ião livre, mas sim complexado com
outras espécies como hidróxidos (Hg(OH)+, Hg(OH)2, Hg(OH)3-, cloretos (HgCl-, HgCl2,
HgCl3, HgCl42-) ou ambos (HgClOH), dependendo, a sua especiação, do pH e da
salinidade (Morel, et al., 1998). Na água do mar, predominam as espécies de mercúrio
complexadas com cloretos, principalmente, HgCl3- e HgCl4
2-, enquanto que em
ambientes estuarinos, predominam as formas Hg(OH)+, HgCl-, HgClOH. Nos rios,
contrariamente, predominam complexos formados com hidróxidos.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 11
O diagrama de fases (Figura 2), ilustra a
variação das espécies de Hg, em função do
pCl (-log[Cl-])e do pH (Morel, et al., 1998).
Cerca de 95% de Hg2+ nas águas encontra-se
ligado a matéria orgânica dissolvida, podendo
ser de espécies organometálicas de mercúrio
(como o MeHg), que se encontra na forma de
cloro- ou hidroxo-complexos (CH3HgCl ou
CH3HgOH) (Meili, 1997; Morel, et al., 1998). A
proporção de metilmercúrio face ao mercúrio
dissolvido é usualmente mais elevada na
coluna de água do que nos sedimentos, e é
mais alta em águas fluviais, do que em águas
estuarinas. Em águas estuarinas e marinhas o
metilmercúrio pode ser inferior a 5% do
mercúrio total, mas a proporção sobe para cerca de 30% do total em colunas de água
de rios e lagos (Ullrich, et al., 2001), podendo haver casos em que a proporção é
maior, especialmente em águas anóxicas e ricas em sulfuretos (Canário, 2004a).
O Hg apresenta elevada afinidade com sulfuretos, sendo a espécie mais importante o
cinábrio (HgS). Os sulfuretos controlam a solubilidade do Hg nas águas, devido à
baixa solubilidade dos sulfuretos de mercúrio (Morel, 1998). Os oxi-hidróxidos de ferro
e de manganês, também são importantes na mobilidade do mercúrio pelas águas,
devido à sua elevada capacidade de adsorver e coprecipitar Hg2+, e de o libertar após
a sua dissolução. A formação e dissolução dos oxi-hidróxidos de ferro e manganês
são controlados pelas propriedades redox e quantidade de oxigénio, podendo assim
influenciar a metilação (Gagnon, et al., 1997; Meili, 1997; Ullrich, et al., 2001).
Os compostos resultantes da especiação de mercúrio podem ser classificados em
águas naturais de duas categorias (Cossa & Couran, 1988; IUPAC, 1998):
Espécies reativas: (Hg2+, Hg2+ formando complexos inorgânicos, e complexos
lábeis de Hg com a matéria orgânica);
Figura 2 - Variação de cloro- e hidroxi-complexos de Hg
2+ em
solução aquosa, consoante a concentração de Cloro e o pH.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
12 Ricardo M.P.Mendes
Espécies não reativas: como compostos que mantêm ligações covalentes, tipo
C-Hg, tais como: (CH3Hg+) Hg(CN)2: espécies ligadas ao enxofre, tais como:
HgS.
2.6. Mercúrio em sedimentos
Os sedimentos são considerados como sendo constituídos pela fração sólida, águas
intersticiais e por gases gerados em reações no seu interior, interatuando com a
coluna de água e servindo de substrato à fauna e flora (Berner, 1980; Stumm &
Morgan, 1996).
Existem vários tipos de interações entre os metais, os constituintes dos sedimentos e
a coluna de água sobrenadante, que podem ser responsáveis pelas diferenças entre
as espécies químicas nos sedimentos (Schulz, 2000; Ramalhosa, 2002; Canário,
2004a), sendo estas:
Precipitação/solubilização dos carbonatos, sulfuretos e hidróxidos dos metais.
Adsorção/desadsorção física e química dos metais nos materiais argilosos, nas
substâncias húmicas e resíduos orgânicos.
Adsorção física, química e coprecipitação dos metais com oxi-hidróxidos de
ferro e manganês.
Ao conjunto de reações que envolvem todos os parâmetros e processos físico-
químicos atrás referidos, designam-se por reações diagenéticas.
Os oxi-hidróxidos de ferro e manganês são elementos presentes nos sedimentos e
permitem a adsorção de elementos vestigiários na sua estrutura, influenciando de
forma relevante a distribuição dos metais nos sistemas aquáticos (Förstner &
Wittmann, 1981). A adsorção dos iões metálicos por estes óxidos pode ser
interpretada como sendo devido à formação de complexos, ou à troca iónica em H+ ou
com outros catiões, à medida que os iões metálicos são adsorvidos nos óxidos.
Verificou-se que sedimentos em meios óxicos têm uma capacidade de adsorção de
metais, superior à de sedimentos em meios anóxicos, pois a redução do ferro e
manganês presentes nos óxidos hidratados provoca uma libertação instantânea dos
metais adsorvidos (Förstner & Wittmann, 1981). Estes factos contribuem para apoiar a

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 13
hipótese de que o ferro e o manganês podem influenciar a distribuição e transporte de
contaminantes em sistemas aquáticos (Förstner & Wittmann, 1981).
O mercúrio em sedimentos resulta da deposição da matéria particulada em suspensão
da coluna de água, sendo a sua adsorção e retenção controladas por parâmetros
físico-químicos, tais como o pH, a força iónica, o potencial redox, a área superficial dos
constituintes do sedimento, os teor em matéria orgânica (Schuster, 1991), sulfuretos,
ferro, manganês, alumínio, silício e ação de organismos (Schulz, 2000). Os
sedimentos constituem a maior reserva de Hg em ambientes aquáticos (NOAA, 1996;
Ullrich, et al., 2001) e se contaminados, podem contribuir como fonte de mercúrio para
o sistema aquático, mesmo após as descargas de efluentes terem cessado (Covelli, et
al., 1999).
Como referido anteriormente, em sedimentos óxicos e subóxicos, o Hg encontra-se
associado a oxi-hidróxidos de ferro e manganês e a matéria orgânica (ácidos húmicos
e compostos orgânicos contendo enxofre), que de uma forma geral constituem uma
percentagem elevada do sedimento (NOAA, 1996; Jackson, 1998; Canário, et al.,
2003a; Canário, et al., 2003b). Se esses sedimentos se tornarem anóxicos, ocorre a
redução do sulfato a sulfureto e o Hg coprecipita com sulfureto de ferro e/ou reage
com o ião sulfureto, formando HgS que precipita (Jackson, 1998).
São exemplos de processos físico-químicos a erosão, a dragagem, a diagénese e a
bioturbação nos sedimentos, os quais podem aumentar a presença de compostos de
mercúrio orgânico na coluna de água afetando o ambiente à escala local (Válega, et
al., 2008a). No entanto, a desadsorção do mercúrio existente no sedimento para as
águas intersticiais e consequentemente para a coluna de água, é um processo lento,
ocorrendo maioritariamente sob a forma de HgCl2 e Hg(OH)2, sendo essa desadsorção
dependente do pH, força iónica e teor em cloretos do meio (Jackson, 1998).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
14 Ricardo M.P.Mendes
A variação das espécies químicas de Hg consoante a capacidade oxidante e redutora
dos sedimentos (na fração sólida e nas águas intersticiais), está presente na Tabela 2,
Tabela 2 – Espécies principais de mercúrio presentes na fração sólida e nas águas intersticiais dos sedimentos (Canário, 2004a).
Fração Sólida Água intersticial
Condições óxicas
Hg-óxidos de ferro Hg-óxidos de manganês
Hg-matéria orgânica
Hg0, HgCl2, Hg(OH)2, HgCl4
2-, HgCl3
-
HgHS2-, HgS2
2-
Condições anóxicas
HgS ou adsorvido à superfície de sulfuretos minerais
Complexos com diversos ligandos orgânicos

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 15
3. Reações de diagénese nos sapais
3.1. Comportamento geral
A vegetação dos sapais influência a dinâmica dos ecossistemas estuarinos e retém
eficientemente metais resultantes de descargas para o sistema (Caçador & Vale,
2001). Estas retenções ou libertação de metais ocorrem através das reações
diagenéticas.
Os parâmetros como potencial redox (EH), pH e oxigénio dissolvido, são importantes
na disponibilidade de metais vestigiais (como o Hg, Pb, Cd), e na sua partição entre as
frações sólida e dissolvida. A variação das condições de Eh e pH, induz variações na
especiação e solubilidade do metal, podendo resultar na alteração da difusão das
espécies nas águas intersticiais e a sua incorporação nas plantas. Estes parâmetros
dependem do oxigénio que o sistema de raízes bombeia para a rizosfera, o que pode
indicar se a planta está ativa ou não (Otte & Jacob, 2003; Weis & Weis, 2004;
O’Driscoll, et al., 2011).
No esquema A da Figura 3, estão representadas três camadas (I, II e III) com
propriedades redox distintas. A camada I é a mais superficial e a mais oxidada, sendo
designada por óxica. A camada II é uma camada intermédia, sendo moderadamente
reduzida e designada por camada subóxica. A camada III é a camada reduzida, sendo
por isso considerada como a camada anóxica. Acima da camada I encontram-se as
águas sobrenadantes.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
16 Ricardo M.P.Mendes
Figura 3- Esquema explicativo da diagenética em sedimentos não colonizados por plantas e em sedimentos colonizados por plantas (adaptado de Otte & Jacob, 2003).
Num sedimento sem vegetação ou com outras perturbações (esquema A e B), as
camadas sedimentares estão químicamente bem definidas.
Na camada óxica (camada I) o ferro existe predominantemente na forma de ião férrico
(Fe(III)), enquanto o enxofre está presente como ião sulfato (SO42-). Na camada
anóxica (camada III), predominam as espécies mais reduzidas como o ferro na forma
de ião ferroso (Fe(II)) e sulfuretos (S2-). Na camada subóxica (camada II), as espécies
variam conforme a sua proximidade à camada óxica ou anóxica.
Nas camadas I e III, devido à remobilização dos metais, o ferro e outros metais podem
ser relativamente imobilizados. O ferro precipita como óxidos, e os metais têm
afinidade de adsorção elevada ou estão altamente imobilizados, devido à ligação com
sulfuretos. Na camada II, o ferro na forma Fe(II) e outros metais ganham relativa
mobilidade. É esperado que a concentração de metais aumente na direção da camada
I, devido ao gradiente de difusão causado pela libertação de ferro e outros metais dos
Mobilidade dos
metais ( Fe, Me)
Espécies
dominantes
Concentrações totais
de metais no sedimento
Baixo Alta

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 17
sulfuretos na camada anóxica, e subsequente coprecipitação na camada oxidada (Otte
& Jacob, 2003).
No esquema C, coloca-se a hipótese de o sedimento estar colonizado por plantas.
Ora, neste caso, as plantas halófitas de sapal libertam oxigénio na zona da rizosfera
ocorrendo que a camada óxica pode aumentar e alargar as suas propriedades com a
oxidação de metais de sulfuretos noutras camadas com diferentes estados de
oxidação na rizosfera (Caçador & Vale, 2001; Otte & Jacob, 2003). Nesta camada, o
oxigénio bombeado pelas plantas oxida os sulfuretos presentes nos sedimentos. O
ferro, o manganês e o enxofre interagem repetidamente em reações redox, alterando a
mobilidade e disponibilidade dos metais, tornando-os mais disponíveis, e permitindo a
sua captura pelas raízes das plantas, que são transportados para os tecidos
superiores destas (Marins, et al., 1997; Weis & Weis, 2004; Canário, et al., 2007a;
Válega, et al., 2008a; O’Driscoll, et al., 2011)
No esquema D, após o desenvolvimento das raízes, há um acumular de matéria
orgânica retida pela rede de raízes no seu máximo desenvolvimento. Como efeito, a
matéria orgânica vai reagir com o oxigénio, sendo oxidada, e consequentemente há
uma diminuição das espécies oxidadas diminuindo assim a camada óxica, levando a
um aumento da camada subóxica com propriedades redox intermédias (Caçador &
Vale, 2001; Otte & Jacob, 2003).
No esquema E, com a mudança sazonal de estação do ano, a atividade das raízes
declina. As raízes ao degradarem-se fazem aumentar a matéria orgânica nas camadas
sedimentares, levando a um maior consumo de oxigénio e consequentemente a uma
diminuição da camada óxica, aumentando a camada anóxica (Caçador & Vale, 2001;
Otte & Jacob, 2003).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
18 Ricardo M.P.Mendes
3.2. Mercúrio em sapais
As concentrações de mercúrio e a proporção de MeHg face à percentagem de Hg total
em sapais, pode variar com a sua localização e com a espécie ou planta que o
coloniza o tipo de planta que o coloniza (Canário, et al., 2007a). Vários estudos
demonstraram que as concentrações de Hg e MeHg são mais elevadas nos
sedimentos colonizados por plantas do que nos sedimentos não colonizados.
No caso dos sedimentos colonizados por plantas foram observados maiores
concentrações de Hg e MeHg nas camadas sedimentares da rizosfera (zona de
sedimtos sob influencia da raíz), uma fraca mobilidade do Hg das partes inferiores das
plantas para as partes superiores, baixa acumulação nas partes superiores das
plantas, e fraca incorporação de Hg nas folhas via atmosfera (Weis & Weis, 2004;
Canário, et al., 2007a; Válega, et al., 2008a; Canário, et al., 2010).
São vários os estudos que indicam a existência e influência de bactérias na rizosfera
como agentes metiladores. Em Canário, et al., (2010), foi observada uma ausência de
correlação entre a concentração de Hg total e os correspondentes níveis de MeHg e
sugerindo que a metilação depende unicamente de fatores ambientais e
microbiológicos, podendo a transformação biológica de Hg para MeHg aumentar a
toxicidade dos sedimentos colonizados por plantas.
Guimarães, et al., (2000), verificaram que nas raízes de várias macrófitas tinham uma
ordem de grandeza mais elevada de metilação nas suas raízes do que nos restantes
sedimentos não colonizados.
Sendo a metilação atribuída a micro-organismos junto às raízes das plantas e
associados a sólidos. Como explicação desta observação, foi proposto que as raízes
expelem o que é considerado uma fonte de carbono que aumenta o carbono dissolvido
e estimula a atividade microbiana. Souza, et al., (1999), observaram que aplicando
antibióticos junto de plantas de sapais, as concentrações de Hg acumuladas na
rizosfera diminuíram significativamente pois o antibiótico inibia as bactérias existentes
nessa camada.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 19
4. Objetivos deste trabalho
O principal objetivo deste trabalho foi identificar a influência das plantas de sapal e da
variação sazonal nos processos biogeoquímicos responsáveis pela partição e
transporte do mercúrio e metilmercúrio em sapais utilizando-se os sapais do Rosário e
se Castro Marim como modelos.
Os objectivos específicos são os seguintes:
1. Estudar a variação das concentrações de mercúrio e metilmercúrio em
sedimentos, águas intersticiais e plantas (partes aéreas e raízes) de duas
espécies de halófitas em dois sistemas com mercúrio de diferentes origens.
2. Identificar os principais processos biogeoquimicos responsáveis pela partição e
transporte do mercúrio e metilmercúrio nos sapais estudados
3. Estudar a variação sazonal das concentrações de mercúrio e metilmercúrio nos
diversos compartimentos ambientais analisados
4. Estimar os stocks e fluxos difusivos de mercúrio e metilmercúrio nos dois
sistemas e para cada espécie de halófita.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
20 Ricardo M.P.Mendes

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 21
II. Caracterização da área de estudo
O critério para a escolha das áreas de estudo foi a origem da fonte de mercúrio:
origem natural ou antropogénica. Com uma fonte de Hg de origem natural, foi
escolhido o estuário do Guadiana (sapal de Castro Marim), por ser pouco estudado,
pouco poluído e por atravessar a faixa pírítica ibérica (considerada fonte natural de
Hg) (Caetano, et al., 2008). O sapal do Rosário, no estuário do Tejo, foi escolhido por
estar nas proximidades de uma zona considerada fonte antropogénica de Hg, devido à
zona industrial e urbana aí existente (Caçador & Vale, 2001; Duarte, et al., 2010).
A vegetação nos sapais é constituída por espécies de plantas, sujeitas a inundações
periódicas, que se desenvolvem nas margens dos estuários em condições de baixo
hidrodinamismo (Antunes Dias & Marques, 1999).
Segundo Catarino e Caçador (1981), os sapais do Estuário do Tejo e Guadiana têm,
de modo geral, vegetação em comum, sendo os géneros Halimione, Sarcocornia e
Spartina os mais importantes na composição, estrutura e dinâmica das formações
vegetais superiores (Antunes Dias & Marques, 1999).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
22 Ricardo M.P.Mendes
1. Estuário do Tejo
O estuário do rio Tejo tem uma extensão longitudinal de 80 Km, (desde S. Julião da
Barra até à secção de Muge no Ribatejo) correspondendo esta última ao limite da
maré dinâmica. A largura do estuário é variável, 0,7 Km no extremo montante, 15 Km
na parte central e 2 Km no canal de saída.
O estuário ocupa uma área total de 320 Km2 e em condições de baixa-mar e marés
vivas 40% fica emersa, apresentando uma importante zona entre marés (Ferreira,
1988; Antunes Dias & Marques, 1999). Esta zona encontra-se periodicamente em
contacto com o ar atmosférico e sujeita por isso as transformações fisico-químicas daí
resultantes (Canário & Vale, 2004b).
No estuário, a principal fonte de água doce é o rio Tejo. O caudal médio anual do rio é
de 400 m3.s-1, mas as variações anuais podem ir desde 60 m3s-1 a 2200 m3.s-1. O
tempo de residência da água doce no estuário oscila entre 65 dias para caudais na
ordem dos 100 m3 s-1 e 6 dias para caudais de cheia da ordem dos 2211 m3.s-1
(Sirgado, 1995). Em termos de circulação da água no estuário, verifica-se que esta é
essencialmente influenciada pela ação das marés, às quais se adiciona, sobretudo nas
zonas de montante do estuário, o efeito de correntes induzidas pelos caudais fluviais
do rio Tejo.
O estuário do Tejo é um estuário poluído por descargas diretas provenientes de
indústrias e efluentes, estando a contaminação por metais distribuida ao longo de duas
zonas: a Cala do Norte e o Barreiro, consideradas como fontes antropogénicas de
metais (Antunes Dias & Marques, 1999; Caçador & Vale, 2001; Canário, 2004a;
Canário, et al., 2005).
No estuário existem sapais de proporções consideráveis na margem sul do Tejo: os
sapais de Corroios, Talaminho, Sarilhos Pequenos, Paio Pires, Rosário, Alcochete,
Barco e Pancas (Caçador & Vale, 2001). Existem ainda pequenos sapais mais a
montante do estuário (Cala do Norte, Alverca, Vila Franca de Xira), mas de dimensões
muito reduzidas. Os sapais de Alcochete e Pancas são sapais contidos na Reserva
Natural do Estuário do Tejo.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 23
Figura 4 – Estuário do Tejo, com destaque para a zona do sapal do Rosário, onde se realizou a amostragem.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
24 Ricardo M.P.Mendes
2. Estuário do Guadiana
O estuário do Guadiana é um estuário longo e estreito, do tipo de um rio em vale
encaixado. A largura média do estuário na zona da embocadura é cerca de 2 km,
diminuindo progressivamente para montante, sendo a largura média da ordem de 1 km
e a profundidade média da ordem dos 3 metros.
Possui regiões de sapal tanto na margem Portuguesa (sapal de Castro Marim, na
Figura 5) como na margem espanhola (sapal de Ayamonte). O limite de penetração da
água salgada no interior do estuário depende do caudal do rio, sendo que, no verão o
limite de propagação situa-se na região de Alcoutim (a 40 km da foz) podendo os
efeitos das marés serem sentidos em Mértola (a 65 km da foz). As estruturas
reguladoras do rio aí existentes impedem a sua propagação mais para montante.
Figura 5 - Estuário do Guadiana. Destaque para a zona do sapal de Castro Marim, onde se realizou a amostragem.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 25
O rio Guadiana possui um caudal muito variável tanto na escala sazonal como
interanual, como consequência do clima e da natureza do solo, muito impermeável e
da sua bacia hidrográfica. Em situações de cheia o caudal pode exceder os 10000 m3
s-1, mas de verão são frequentes situações de caudal praticamente nulo. O caudal
médio é da ordem dos 80 m3 s-1 (INAG, 2012).
A maioria da região do sapal de Castro Marim na costa da margem Portuguesa e o de
Ayamonte na margem Espanhola são zonas protegidas com estatutos de reservas
naturais. O sapal de Castro Marim compreende uma área formada por sapais, salinas
e esteiros, com zonas secas de xistos, grés vermelho, areias e arenitos. O sapal foi
sendo destruído para terrenos agrícolas, ou salinas. Contudo, a passagem a Reserva
Natural da maioria da área de sapal, permitiu que se tenham desenvolvido programas
de recuperação dos sapais.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
26 Ricardo M.P.Mendes
III. Materiais e Métodos
1. Descontaminação do material
O material utilizado, para recolha e pré-tratamento das amostras, foi previamente
descontaminado. O processo de descontaminação envolveu as seguintes etapas:
i) Lavagem com água destilada (Elix, Millipore)
ii) Imersão em ácido nítrico (HNO3 65%, Panreac) a 20% (v/v) durante 2
iii) Lavagens repetidas com água ultrapura (Milli-Q, Millipore)
iv) Imersão em ácido clorídrico (HCl 37%, Panreac) a 20% (v/v) durante 2 dias
v) Lavagem (3 vezes) com água ultrapura (Milli-Q, Millipore)
vi) Secagem do material numa câmara de fluxo laminar (Microflow Laminar Flow
Workstation) no interior de uma sala limpa, de modo a evitar contaminações por
deposição de poeiras (Caetano, 1998)
vii) Colocação do material descontaminado em sacos de plástico selados até
utilização.
O processo de descontaminação das bombas de teflon para digestão de sedimentos,
envolveu as seguintes etapas:
i) Lavagem com água destilada (Elix, Millipore)
ii) Imersão do material 24h em detergente Extran
iii) Lavagem (3 vezes) com água ultrapura (Milli-Q, Millipore)
iv) Imersão em ácido nítrico (HNO3 65%, Panreac) a 30% (v/v) durante dois dias.
v) Lavagem repetidas vezes com água ultrapura (Milli-Q, Millipore).
Seguidamente todo o material foi seco em câmara de fluxo laminar (Microflow Laminar
Flow Workstation) no interior de uma sala limpa.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 27
2. Trabalho de Campo
Os trabalhos de campo foram realizados no sapal de Castro Marim e no sapal do
Rosário, nos estuários do Guadiana e Tejo, respetivamente. Com o auxílio de um
corer metálico, foram recolhidos vários cores de sedimentos colonizados por
Sacocornia fruticosa e colonizados por Spartina marítima e de sedimento não
colonizado (ver figura 6). Estes sedimentos foram secionados em camadas de 2 em 2
cm até 0 aos 20 cm, e de 5 em 5 cm dos 20 aos 30 cm.
Figura 6 – Fotografia de um core de sedimentos num corer metálico, e fotografia do local de amostragem no sapal do Rosário, onde se pode ver solos colonizados por plantas e solos não colonizados.
Para a análise de sedimentos (fração sólida e águas intersticiais, as amostras
seccionadas foram colocadas em tubos de policarbonato de 250 cm3 (previamente
descontaminados), consoante a sua profundidade, sendo completamente cheios e
hermeticamente fechados com tampas de rosca de polipropileno. Para análise a
sulfuretos, as camadas seccionadas foram colocadas em tubos de polipropileno de 50
cm3 (Digitubes, SCP Science) e imediatamente congelados.
Foram recolhidas cores de sedimentos colonizados planta e de sedimentos não
colonizados, que foram seccionados para sacos de polietileno, respetivamente
identificados consoante a sua profundidade. As partes aéreas das plantas foram
recolhidas com o auxílio de uma faca, e colocadas em sacos de polietileno
devidamente identificados.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
28 Ricardo M.P.Mendes
As águas sobrenadantes (na superfície dos sedimentos) foram recolhidas por
aspiração com o auxílio de uma seringa, numa zona não colonizada por plantas e
processadas in situ. Todas as amostras foram transportadas para o laboratório em
malas térmicas refrigeradas.
3. Processamento das amostras
3.1 Sedimentos
3.1.1. Águas intersticiais
Os tubos de policarbonato (250 cm3) contendo as amostras de sedimento foram
centrifugados (centrifuga KUBOTA 6800) a 6000 rpm, durante 45 minutos, a uma
temperatura de 4ºC, de modo a separar a fração sólida dos sedimentos e as águas
intersticiais. As águas intersticiais, resultantes da centrifugação, foram filtradas com
filtros (Whatman, FP30/ 0,45CA), com uma porosidade de 0,45 μm, tendo as amostras
sido acidificadas a 0,5% com HCl 37% (livre de mercúrio, Panreac) em frascos de
vidro, para determinação de mercúrio total e metilmercúrio dissolvidos. Cerca de 10
mL da amostra de águas intersticiais foram colocados em tubos de polipropileno e
acidificadas a HNO3 65% (livre de mercúrio, Panreac) para determinação de Fe, Mn,
Cl- e SO42- dissolvidos.
As amostras para determinação de metilmercúrio foram congeladas e as amostras
para determinação de Fe, Mn, Cl- e SO42- foram armazenadas no frigorífico a uma
temperatura de 4ºC, até à análise.
3.1.2 Fração sólida
Os sedimentos com raízes das plantas foram lavados com água ultrapura (18,2 MΩ
cm, Milli-Q, Millipore), num peneiro de malha de 0,500 mm (Retch), de modo a separar
as raízes do restante sedimento. As partículas de sedimento que foram arrastadas
pelas águas de lavagens e passaram através do peneiro foram recolhidas para o
interior de caixas. As águas de lavagens são então deixadas em repouso para que as
partículas em suspensão se depositem. Após um dia, quando se verificou que grande
parte das partículas estão depositadas, separam-se as águas de lavagem. As
partículas de sedimentos depositadas no fundo da caixa foram então postas a secar

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 29
numa estufa (Heraeus) a 40ºC (temperatura máxima que se pode atingir de modo que
o mercúrio não volatilize).
Das camadas recolhidas para sacos de polietileno de sedimentos não colonizados, foi
retirado uma pequena porção a qual foi posta diretamente a secar em estufa
(Memmert) a 40ºC.
Todos os sedimentos (colonizados e não colonizados) após estarem secos, foram
moídos num moinho homogeneizador (Retch, type: vibro).
3.1.3. Procedimento especial para sulfuretos dissolvidos e
AVS
Os tubos com as amostras de sedimentos para análise de sulfuretos dissolvidos e
AVS, foram descongelados, abertos e uma pequena amostra de sedimento húmido foi
retirada para extração dos AVS. De seguida os tubos são centrifugados (Centrífuga de
Bancada Excelsa®3 Modelo 28) a 3000 rpm durante 10 minutos. As águas resultantes
da centrifugação foram filtradas, por filtros (Whatman, FP30/ 0,45CA), com uma
porosidade de 0,45 μm. Todo este processo foi realizado numa câmara de luvas em
atmosfera de azoto (é necessário atmosfera de azoto de modo a que os sulfuretos não
se oxidem).
3.2. Águas sobrenadantes
As águas sobrenadantes recolhidas in situ foram imediatamente filtradas com filtros
(Whatman, FP30/ 0,45CA) com uma porosidade de 0,45 μm, que se colocaram na
ponta da seringa. Após a filtração foram acidificadas a 0,5% com HCl 37% (livre de
mercúrio, Panreac) introduzidos em frascos de vidro e cerca de 10 mL da amostra,
colocadas em tubos de polipropileno e acidificadas a 0,1% com HNO3 65% (livre de
mercúrio, Panreac).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
30 Ricardo M.P.Mendes
3.3 Plantas
3.3.1 Biomassa subterrânea
As raízes foram obtidas a partir da lavagem dos sedimentos colonizados por plantas
com água ultrapura (18,2 MΩ cm, Milli-Q, Millipore), num peneiro de malha de 0,500
mm, tendo o cuidado de se retirar todo o sedimento. Caso existam grãos de areia
estes foram, juntamente com a biomassa existente, secos em estufa (Memmert) a
40ºC (Caetano, et al., 2008; Caetano, et al., 2009; Duarte, et al., 2010). Após uma
semana a biomassa foi retirada da estufa, e com o auxílio de uma lupa separam-se
grãos de areia existentes. A biomassa é então homogeneizada num moinho (Retch,
tipo MM200, frequência: 30).
3.3.2 Biomassa aérea
A biomassa aérea que consiste na parte superior das plantas, foi lavada com água
ultrapura (18,2 MΩ cm, Milli-Q, Millipore) de modo a remover partículas de sedimentos.
Após a lavagem separou-se o material em órgãos fotossintéticos e não fotossintéticos.
Sendo que a S. fruticosa não possui um verdadeiro sistema de folhas e caules bem
definidos as suas componentes fotossintéticas são mais “carnudas” (Antunes Dias &
Marques, 1999), e as componentes não-fotossintéticas, de aspeto seco, com pouca
água.
A S. marítima é uma planta constituída por um talo (órgão não fotossintético) que se
separa em folhas (órgãos fotossintéticos) (Duarte, et al., 2010; Caetano, et al., 2008;
Caetano, et al., 2009).
Com o auxílio de uma tesoura, separam-se as componentes das partes aéreas das
duas plantas, sendo posteriormente colocadas a secar em estufa (Memmert) a 40ºC.
Ao fim de uma semana de secagem são homogeneizadas num moinho Retch (tipo
MM200, frequência: 30). As respetivas plantas podem ser observadas na figura 7.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 31
Figura 7- Imagens das plantas em sapal, juntamente com desenhos do seu perfil. Em cima temos a S. fruticosa e em baixo a S. marítima (adaptado de Duarte, 2010 e Antunes Dias & Marques, 1999).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
32 Ricardo M.P.Mendes
4. Análise das amostras
4.1. Análise das águas
Estes procedimentos foram realizados para as águas intersticiais e águas
sobrenadantes.
4.1.1. Determinação de ferro e manganês totais dissolvidos
A determinação dos teores de ferro e manganês nas águas intersticiais foi efetuada
diretamente das amostras acidificadas, por espectroscopia de absorção atómica com
atomização à chama, num aparelho Spectra AA 55B da Varian. Quantificaram-se os
metais através do método curva de calibração, com padrões de 0; 0,5; 1,0; 2,0; 2,5; e
5,0 mg L-1, para o ferro e com padrões de 0; 0,1; 0,3; 0,5; 1,0; 1,5 mg L-1, para o
manganês.
Tabela 3 - Condições operatórias usadas para análise de Fe e Mn por EAA-Chama.
Elemento Comprimento de onda(nm) Tipo de Chama (Comb/Oxi.)
Fe 248,3 C2H2/Ar
Mn 279,5 C2H2/Ar
4.1.2. Determinação de cloretos dissolvidos
Para determinação do teor em cloreto nas amostras de água intersticial foi utilizado o
método de Mohr descrito pela APHA (1995). O método consiste na utilização do ião
cromato como indicador da titulação do cloreto com ião prata, que ao ficar em excesso
reagirá com o indicador formando espécie Ag2CrO4 de cor avermelhada. O
fundamento do método por ser descrito pelas seguintes reações:
Ag+ + Cl- → AgCl (s) Branco pKS = 9,96
2Ag+ + CrO42- → Ag2CrO4 (s) Vermelho acastanhado pKS = 12,2
De uma ampola de Titrisol (Merck) preparou-se uma solução titulante de AgNO3 0,1 M.
Após este procedimento pipetou-se 1 cm3 de água intersticial para um erlenmeyer de
titulação e adicionaram-se algumas gotas do indicador K2CrO4 0,25M. Titulou-se com
a solução de nitrato de prata até o aparecimento da cor vermelho acastanhado.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 33
4.1.3. Determinação de sulfatos
O teor em sulfato nas águas intersticiais foi determinado por turbidimetria segundo o
método descrito em APHA (1995).
O método tem por base a reação do anião sulfato (SO42-) com o ião bário (Ba2+) ,
formando o sulfato de bário (BaSO4), num meio que conduz à formação de uma
suspensão coloidal, com o glicerol.
Para isso, o ião sulfato (SO42-) é precipitado em meio ácido por um excesso de cloreto
de bário (BaCl2) formando cristais de sulfato de bário, altamente insolúvel:
Ba2+ + SO42- BaSO4
A adição de glicerol, etanol e cloreto de sódio origina a formação de uma emulsão,
facilitando a permanência do (BaSO4) em suspensão, permitindo assim a quantificação
da radiação dispersa pelas partículas deste em relação à radiação incidente
recorrendo à técnica de espectrofotometria de UV-Visível (Apêndice).
A 200 μL de amostra foram adicionados 5 cm3 de uma solução contendo 50 cm3 de
glicerol 87% (Merck), 30 cm3 HCl 37% (Panreac), 300 cm3 de água ultrapura (18,2 MΩ
cm, Milli-Q, Millipore), 100 cm3 de etanol a 90% e 75 g de NaCl (p.a., MERCK).
A mistura foi agitada durante uns segundos com um agitador magnético e adicionou-
se BaCl2 dihidratado (Merck) em excesso. Cronometrou-se imediatamente um minuto
de agitação, a velocidade constante, e mediu-se a absorvância ao comprimento de
onda de 420 nm num espectrofotómetro Jasco V-630, com células de 1 cm de
percurso ótico. A quantificação dos sulfatos foi realizado através do método da curva
de calibração com padrões de 50, 35, 25, 10 e 5 mM de Sulfato de Sódio anidro (p.a.,
Merck)
4.1.4. Determinação de sulfuretos dissolvidos totais
O teor em sulfuretos inorgânicos dissolvidos totais, [HS-]T (Σ [H2S]+[HS-]+[S2-] +[Sx2-])
foi determinado por análise direta das água intersticiais, pipetaram-se diretamente
100μL que se verteram numa célula polarográfica contendo 9,9 cm3 de uma solução
aquosa de NaCl 37% a 36 g dm-3 até pH 10-12 (aferido com NaOH 1 M), previamente
desarejada durante 5 minutos. Recorreu-se ao método de Voltametria Diferencial de
Redissolução Catódica (DPCSV) (ver Apêndice). (Luther, et al., 1985; Luther, et al.,

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
34 Ricardo M.P.Mendes
1989). A análise foi efetuada num polarógrafo Metrohm equipado com um processador
VA 693 e um posto VA 694, onde se utilizou um elétrodo de mercúrio de gota
suspensa (HMDE), um elétrodo de referência Ag/AgCl/NaCl (36‰) e um elétrodo de
platina como auxiliar. Em DPCVS fez-se a deposição do sulfureto durante 60 s a um
potencial de -400mV tendo-se feito um varrimento catódico entre -400 e -900 mV com
uma velociadade de 6,0 mV s-1. O potencial do pico de sulfureto foi cerca de -700 mV
em relação ao elétrodo de referência.
O método de quantificação foi a curva de calibração padrão, tendo-se adicionado
volumes de 100μL de padrões, uma solução resultante da diluição de uma solução
mãe de sulfureto, previamente padronizada por iodometria de 500 mg dm-3 de
Na2S.H2O anidro em água Milli-Q. A quantificação do sinal foi feita através da altura do
pico observado ao potencial referido.
4.1.5. Determinação de mercúrio total dissolvido
A metedologia para a determinação do mercúrio total dissolvido, baseou-se com o
método da EPA (2002). O método inicia-se com a oxidação prévia de todo o Hg
dissolvido presente na amostra a Hg2+ com BrCl, (EPA, 2002; Leopold, et al., 2008).
Para eliminar o excesso de BrCl existente na solução, adiciona-se NH2OH.HCl,
seguida da adição de uma solução de 2% de SnCl2 em HCl a 10% (reagentes livres de
mercúrio), para redução do Hg2+ a Hg0, sendo este passo de extrema importância, pois
garante que o Hg passe á sua forma de vapor, sendo posteriormente arrastado por
uma corrente de árgon.
A determinação do mercúrio total dissolvido é realizada a partir de amostras
acidificadas. Foi transferido uma alíquota de 30-40 mL de amostra (água intersticial e
sobrenadante) para um balão volumétrico de 50 mL, onde foram adicionado 7,5 mL de
HCl 33% (livro de Hg), e 1mL de solução KBr/KBrO3, e perfez-se o volume a 50 mL
com água ultrapura (18,2 MΩ cm, Milli-Q, Millipore). Deixou-se repousar as amostras
durante 30 minutos e como a cor amarela não persistiu durante esse tempo,
adicionou-se 1 mL de solução KBr/KBrO3 e aguardou-se mais 30 minutos. Preparou-se
um branco com os mesmos reagentes.
O Hg0 é purgado da solução através de um separador gás-líquido, sendo o árgon
utilizado como gás de arraste, e pré-concentrado numa amálgama de ouro; (Method,
2003; Leopold, et al., 2008; Leopold, et al., 2009; EPA, 2002). Após este processo, o
Hg0 é desadsorvido termicamente (450ºC) da amálgama de ouro, e transportado

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 35
diretamente para a célula de Fluorescência Atómica (CVAFS) para determinação
analítica (ver apêndice) (EPA, 2002; Leopold, et al., 2009).
O equipamento utilizado neste trabalho é constituído por um detetor de fluorescência
atómica, detetor Merlin Mercury mod 10.025S (PS Analytical Ltd), com sistema de
injeção de fluxo conduzida por uma bomba peristáltica multicanal e separador gás-
líquido (Gómes-Ariza, et al., 2004). Para a quantificação do sinal utilizou-se a medida
da altura máxima do pico de fluorescência máxima do mercúrio, depois convertido em
concentração a partir da curva de calibração constituída por todas as leituras dos
padrões.
Tabela 4 – Condições operatórias para a determinação de mercúrio total dissolvido por Fluorescência Atómica
Condições Operatórias
Comprimento de onda (nm) 253,7
Volume amostra injetada (cm3) 8
Tempo de purga (s) 20
Tempo de reação (s) 30
Tempo de leitura (s) 30
Temperatura da célula (ºC) 60
4.1.6. Determinação de metilmercúrio dissolvido
Para a determinação de metilmercúrio (MeHg) aplicou-se o método de determinação
do MeHg em águas por destilação, etilação aquosa, purga, captura e deteção por
CVAFS, com acoplamento da Cromatografia Gasosa com Pirólise à espectroscopia de
Fluorescência Atómica (Bings, et al., 2010), descrito por EPA (EPA, 1998)
O método consiste numa destilação prévia da amostra, a 125ºC sob fluxo de azoto.
Após a destilação, o pH do destilado é ajustado a 4,9 com um tampão de Acetato e a
amostra é etilada (para converter todo o mercúrio em metiletilmercúrio, mais volátil)
num vaso fechado de purga através da adição de Tetraetil Borato de Sódio (NaBEt4).
O metiletilmercúrio é separado da solução por purga com N2 para um coletor. O
metiletilmercúrio capturado é termicamente desadsorvido do coletor e conduzido por
um gás inerte de arraste, através de uma coluna cromatográfica pirolítica de

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
36 Ricardo M.P.Mendes
decomposição, aquecida a 800ºC, para converter as formas orgânicas de mercúrio em
mercúrio elementar (Hg0), sendo este conduzido para deteção numa célula de
Fluorescência Atómica (EPA, 1998; Diez, et al., 2009). O equipamento utilizado
consiste num Detetor (Merlin Mercury mod. 10.023S, PS Analytical) utilizando um
pirolisador (PS Analytical 10.565) (Evans, et al., 1999; Gómes-Ariza, et al., 2004) e um
cromatógrafo (Agillent, 6890 Series, GC System). A quantificação dos sinais foi
realizada através do método de adição padrão, sendo o mesmo feito para sedimentos
e plantas.
4.2. Análise à fração sólida do sedimento
4.2.1. Teor de humidade
Para determinar o teor de humidade do sedimento (percentagem da massa total do
sedimento (%H), colocaram-se porções das amostras de sedimentos em cadinhos de
porcelana, que foram colocados em estufa (Memmer) a 105ºC, durante 24horas.
Decorrido este tempo, os cadinhos são retirados da estufa e arrefecidos num
exsicador até à temperatura ambiente. Os cadinhos foram pesados (Mettler Tolledo
AG 245) vazios, com o sedimento fresco, e após arrefecerem. A %H de cada camada
é determinada através da seguinte expressão:
onde msed(fresco) representa a massa de sedimento antes de secar em estufa a
105ºC, e msed (seco) representa a massa de sedimento após a secagem a 105ºC.
4.2.2. Teor em matéria orgânica sedimentar
O teor de matéria orgânica sedimentar (percentagem da massa total do sedimento
(%LOI - Lost oy Ingnition), é estimada através de um método termogravimétrico. Este
método termogravimétrico é baseado na destruição de matéria orgânica por
aquecimento do sedimento previamente seco.
Os cadinhos de porcelana contendo amostras de sedimentos secos a 105ºC, são
colocados numa mufla (Nabertherm Labotherm) a 450ºC durante duas horas. Após

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 37
duas horas, os cadinhos são pesados. A expressão que determina a %LOI de cada
camada é determinada através da seguinte expressão:
onde msed(seco) representa a massa de sedimento após a secagem a 105ºC, e
msed(450ºC) representa a massa de sedimento após estar sujeita a 450ºC.
4.2.3. Determinação do pH e EH
A determinação do pH e EH, foram realizados pela introdução dos elétrodo de pH e de
EH diretamente nos sedimentos através de uma sonda WTW multiline p3 pH/LF. O
elétrodo calibrado com dois tampões (4.00 e 7.00±0,02 a 20ºC) e para o potencial
redox, foram calibrados com uma solução que estabelece na interface um potencial de
220±5 mVvs Ag/AgCl a 25ºC. Os valores medidos para o potencial redox, foram
depois convertidos para potenciais normais de hidrogénio (EH).
4.2.4. Determinação da concentração de ferro, manganês e
alumínio total
Procedeu-se à determinação dos teores de Al, Fe e Mn recorrendo-se a uma digestão
prévia do sedimento e posterior análise por espectroscopia de absorção atómica
(EAA-Chama) para quantificação (Apêndice A).
A digestão das amostras de sedimento é feita por ataque ácido de acordo com o
método descrito por Rantala e Loring (1975). Este método consiste na digestão de
uma porção de sedimento, com água régia e ácido fluorídrico, seguida da
estabilização do produto final com ácido bórico. A água régia, que é mistura de ácido
clorídrico e nítrico (destilado) na proporção 3:1, permite uma solubilização dos metais
existentes na amostra de sedimento através do forte poder oxidante do ácido nítrico e
do cloro livre (que se liberta quando da mistura dos dois ácidos), bem como da
complexação dos iões resultantes pelo ião cloreto resultante da ionização do ácido
clorídrico (Lee, 1980). Os silicatos e aluminosilicatos presentes no sedimento são

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
38 Ricardo M.P.Mendes
destruídos com a adição de ácido fluorídrico, sendo adicionado ácido bórico de modo
a evitar a precipitação de fluoretos devido à formação de ácido fluorbórico, segundo a
seguinte reação química:
H3BO3 + 4HF → HBF4 + 3H2O
Foram introduzidos 100 mg de amostra e de materiais certificados de referência de
sedimento seco e homogeneizado em autoclaves de teflon, e adicionou-se 1 cm3 de
água régia e 6 cm3 de ácido fluorídrico 40% (MERCK, p.a.).
As autoclaves foram colocadas numa estufa a 100 ºC durante uma hora e após este
período de tempo, o conteúdo foi transferido para tubos de polipropileno de 50 cm3
(Digitubes, SCP Science), os quais já continham 5,6 g de ácido bórico (p.a., MERCK).
Depois de aferir a solução com água desionizada (18,2 MΩ cm, Milli-Q, Millipore), para
o volume total de 50 cm3, os tubos são agitados, para homogeneizar as soluções.
A determinação do teor dos elementos maioritários (Al, Fe e Mn) é efetuada num
espectrómetro de absorção atómica, num aparelho Spectra AA 55B da Varian para as
determinações dos teores de Fe e Mn e num aparelho SOLLAR 969 AA Spectrometer
com atomização à chama para determinação dos teores de Al (ver Apêndice), nas
condições operatórias indicadas na tabela 5.
Tabela 5 – Condições operatórias usadas para análise de Fe, Al, e Mn por EAA-Chama
Elemento Comprimento de onda
(nm)
Tipo de Chama
(Comb/Oxi.)
Fe 248,3 C2H2/Ar
Mn 279,5 C2H2/Ar
Al 309,3 C2H2/N2O
Os teores de metais nas amostras foram determinados pelo método da curva de
calibração, com padrões de 0; 0,5; 1,0; 2,0; 2,5; e 5,0 mg/L, para o ferro, com padrões
de 0; 0,1; 0,3; 0,5; 1,0; 1,5 mg/L; para o manganês, e 0; 1; 2; 5; 10; 15; 20 mg/L.
Para verificar a exatidão das determinações, foram utilizados materiais de referência
de concentração certificada: 1646a, corresponde a sedimentos estuarinos, e PACS-2,
MESS-3, HISS-1, correspondem a sedimentos marinhos.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 39
4.2.5. Determinação da concentração mercúrio total
O teor Hg total na fração sólida dos sedimentos foi determinado recorrendo à
espectroscopia de Absorção Atómica com Decomposição Eletrotérmica, num
autoanalisador de mercúrio LECO AMA-254 (ver Apêndice).
Para esta análise as amostras de sedimento, previamente secas e moídas, de massa
conhecida foram colocadas numa barquinha que é automaticamente introduzida no
AMA-254. Para verificar a exatidão das determinações foram utilizados materiais de
referência de concentração certificada, sendo determinados os teores de amostras de
sedimento com concentrações de mercúrio de materiais de referência certificados:
PACS-2 e MESS-3, que correspondem a sedimentos marinhos.
4.2.6. Determinação de metilmercúrio
Para se determinar o MeHg presente na fração sólida dos sedimentos, procedeu-se a
uma digestão alcalina das amostras de sedimento previamente seco, com uma mistura
de KOH/MeOH, de modo a destruir a matriz orgânica e solubilizar o MeHg.
Após a digestão procedeu-se a uma extração orgânica em diclorometano (DCM), de
modo a separar os componentes orgânicos dos restantes componentes dissolvidos
pela digestão alcalina.
Seguiu-se uma pré-concentração em solução aquosa com Na2S, de modo a separar-
se o MeHg dos componentes orgânicos, graças à boa afinidade do Hg com os
sulfuretos.
Por fim ocorreu uma reextracção para DCM, para se poder se poder determinar o
MeHg por GC-AFS (O’Driscoll, et al., 2011).
A quantificação por GC-AFS, ocorreu como descrito pela EPA (1998) ( ver Apêndice
1.2.4). Foram usados os seguintes materiais de referência: IAEA-405 e CRM-580
(sedimentos estuarinos).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
40 Ricardo M.P.Mendes
4.2.7. Determinação de sulfuretos insolúveis
Os sulfuretos reativos insolúveis, denominados AVS, “Acid Volatile Sulphides” são
extraídos do sedimento através de uma solução de HCl 1 M. Juntamente com os
sulfuretos são extraídos também os metais associados denominados por SEM,
“Simultaneously Extracted Metals” (Madureira, 1997) bem como alguns óxidos e
carbonatos de ferro e manganês.
O processo consiste na passagem dos sulfuretos precipitados à forma de à
temperatura ambiente H2S, utilizando uma solução de HCl.
MSn + 2HCl H2S ↑ + MCl2
onde M representa o metal ligado ao ião sulfureto pelo que a estequiometria pode não
ser rigorosamente MS. O H2S formado é arrastado para uma solução alcalina que o
retém na forma HS-.
H2S + OH- HS- + H2O
Cerca de 50 mg de sedimento húmido foi pesado em pequenos pedaços de parafilme
que foram introduzidos rapidamente em frascos contendo 10 cm3 de HCl 1 M (livre de
Hg, Panreac). A mistura ficou a reagir durante 30 minutos em atmosfera de azoto,
sendo o H2S produzido recolhido em 20 cm3 de NaOH 1M.
Figura 8 - Esquema da montagem para a extração do AVS e SEM (Adaptado de Canário,(2000).
Solução de HCl Solução de NaOH

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 41
4.3 Análise das amostras vegetais
A análise às plantas consiste na determinação da percentagem de biomassa
encontrada nos sedimentos recolhido e na determinação da concentração de mercúrio
nas partes aéreas e nas partes subterrâneas (biomassa subterrânea).
4.3.1 Determinação da percentagem de biomassa subterrânea
A percentagem de biomassa subterrânea de cada segmento, foi determinada através
da seguinte equação:
Onde: mraiz seca, corresponde à massa da raíz seca encontrada em cada segmento e
m(raíz+sed)seco, corresponde à massa do sedimento com as raízes, seco.
Ora, como não temos possibilidade de determinar diretamente m(raíz+sed)seco, este é
calculado pela soma entre: mraiz seca (determinado através da pesagem das raízes
secas) e msed seco (massa do sedimento seco).
Como também não é possível determinar diretamente msed seco, este é calculado
através da expressão:
Onde se subtrai à m(raíz+sed) humidos, a massa de água existente em cada segmento
segmento de sedimento e mraiz seca
4.3.2. Determinação de mercúrio total
O teor de Hg total é determinado em todas as partes aéreas e subterrâneas,
recorrendo à espectroscopia de Absorção Atómica com Decomposição Eletrotérmica,
num autoanalisador de mercúrio LECO AMA-254 (ver Apêndice).
Para esta análise as amostras de partes aéreas e subterrâneas, previamente secas e
moídas, de massa conhecida, foram colocadas numa barquinha que é
automaticamente introduzida no AMA-254. Juntamente com as amostras de sedimento

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
42 Ricardo M.P.Mendes
são determinadas as concentrações de mercúrio de materiais de referência
certificados: BCR-60 e BCR-62, para material biológico de origem vegetal.
4.3.3. Determinação de metilmercúrio
Para a determinação do MeHg presente nos tecidos vegetais procedeu-se a uma
digestão ácida das amostras de previamente secas, com uma solução HBr
concentrado (Merck) saturada com CuSO4 de acordo com o métido descrito em
Canário et al (2006). O HBr concentrado possibilita adegradação da matriz de celulose
e os iões Cu2+ encontram-se descritos como eficientes na manuntenção dos iões
HgCH3+ em solução (Canário, et al., 2004c).
Após a digestão procedeu-se à extração do MeHg para a fase orgânica com
diclorometano (DCM). Seguidamente, ao extracto orgânico foi adicionada uma solução
de Na2S de modo a extrair o MeHg da fase orgânica purificando o analito devido à
afinidade entre o Hg e o enxofre. Finalmente, foi adicionado ao extracto aquoso 2 mL
de diclorometano e 1 mL de HCl concentrado (livre de Hg). O HCl possiblita que ocorra
a reacão:
S2- + 2H3O+ H2S + 2H2O
O H2S como é gasoso é retirado da solução através de um fluxo de azoto deixando o
MeHg livre para passar à fase orgânica. O MeHg é finalmente quantificado por GC-
AFS. (Canário, et al., 2006; O’Driscoll, et al., 2011). Neste procedimento foi utilizado o
seguinte material de referência: AIEA-140/TM (planta marinha).
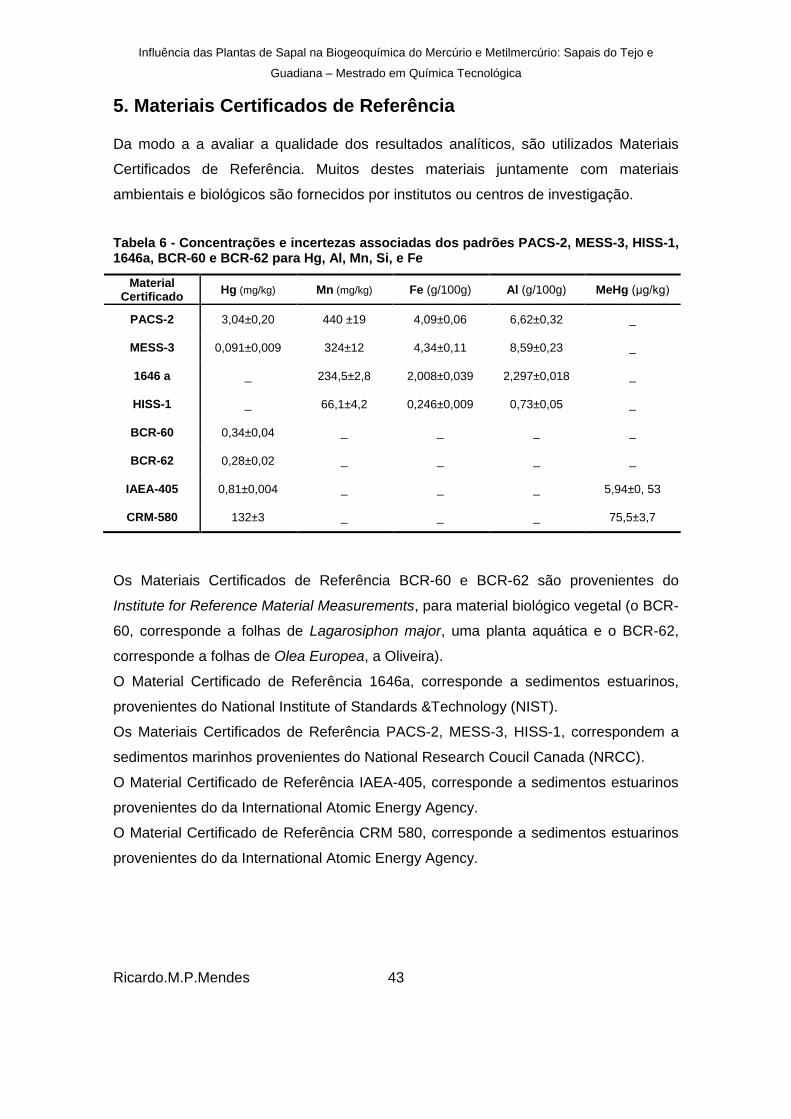
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 43
5. Materiais Certificados de Referência
Da modo a a avaliar a qualidade dos resultados analíticos, são utilizados Materiais
Certificados de Referência. Muitos destes materiais juntamente com materiais
ambientais e biológicos são fornecidos por institutos ou centros de investigação.
Tabela 6 - Concentrações e incertezas associadas dos padrões PACS-2, MESS-3, HISS-1, 1646a, BCR-60 e BCR-62 para Hg, Al, Mn, Si, e Fe
Material Certificado
Hg (mg/kg) Mn (mg/kg) Fe (g/100g) Al (g/100g) MeHg (μg/kg)
PACS-2 3,04±0,20 440 ±19 4,09±0,06 6,62±0,32 _
MESS-3 0,091±0,009 324±12 4,34±0,11 8,59±0,23 _
1646 a _ 234,5±2,8 2,008±0,039 2,297±0,018 _
HISS-1 _ 66,1±4,2 0,246±0,009 0,73±0,05 _
BCR-60 0,34±0,04 _ _ _ _
BCR-62 0,28±0,02 _ _ _ _
IAEA-405 0,81±0,004 _ _ _ 5,94±0, 53
CRM-580 132±3 _ _ _ 75,5±3,7
Os Materiais Certificados de Referência BCR-60 e BCR-62 são provenientes do
Institute for Reference Material Measurements, para material biológico vegetal (o BCR-
60, corresponde a folhas de Lagarosiphon major, uma planta aquática e o BCR-62,
corresponde a folhas de Olea Europea, a Oliveira).
O Material Certificado de Referência 1646a, corresponde a sedimentos estuarinos,
provenientes do National Institute of Standards &Technology (NIST).
Os Materiais Certificados de Referência PACS-2, MESS-3, HISS-1, correspondem a
sedimentos marinhos provenientes do National Research Coucil Canada (NRCC).
O Material Certificado de Referência IAEA-405, corresponde a sedimentos estuarinos
provenientes do da International Atomic Energy Agency.
O Material Certificado de Referência CRM 580, corresponde a sedimentos estuarinos
provenientes do da International Atomic Energy Agency.
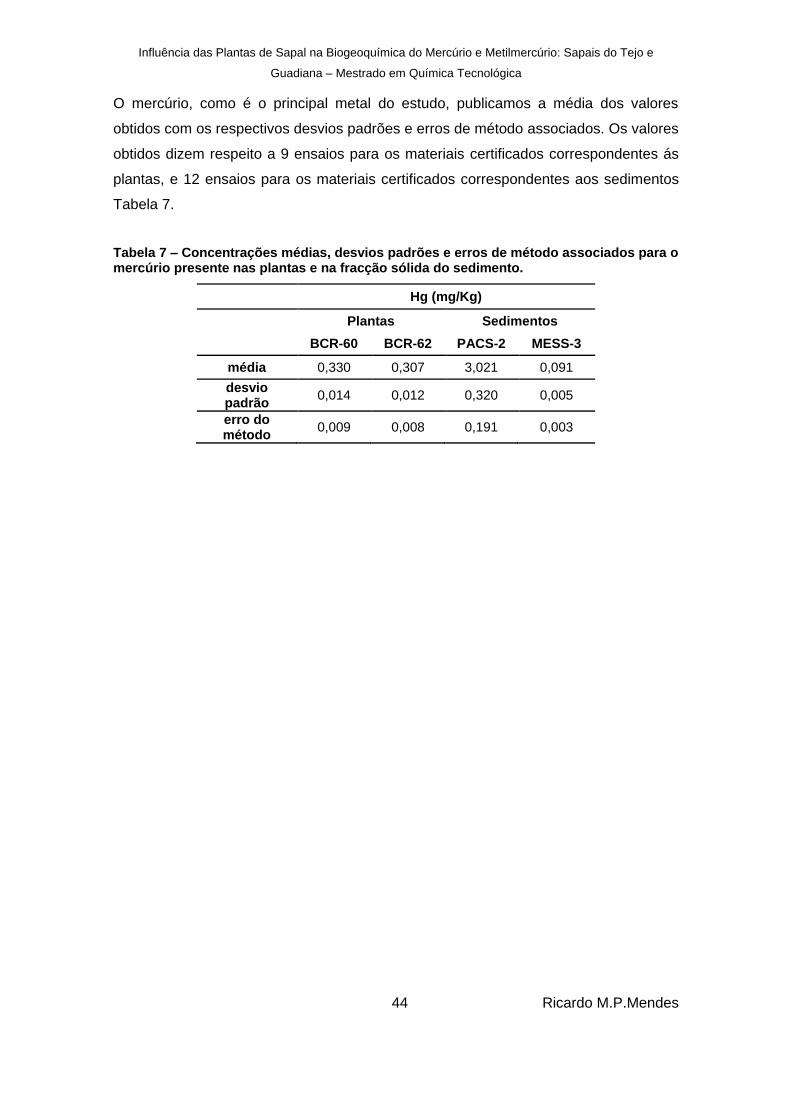
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
44 Ricardo M.P.Mendes
O mercúrio, como é o principal metal do estudo, publicamos a média dos valores
obtidos com os respectivos desvios padrões e erros de método associados. Os valores
obtidos dizem respeito a 9 ensaios para os materiais certificados correspondentes ás
plantas, e 12 ensaios para os materiais certificados correspondentes aos sedimentos
Tabela 7.
Tabela 7 – Concentrações médias, desvios padrões e erros de método associados para o mercúrio presente nas plantas e na fracção sólida do sedimento.
Hg (mg/Kg)
Plantas Sedimentos
BCR-60 BCR-62 PACS-2 MESS-3
média 0,330 0,307 3,021 0,091
desvio padrão
0,014 0,012 0,320 0,005
erro do método
0,009 0,008 0,191 0,003
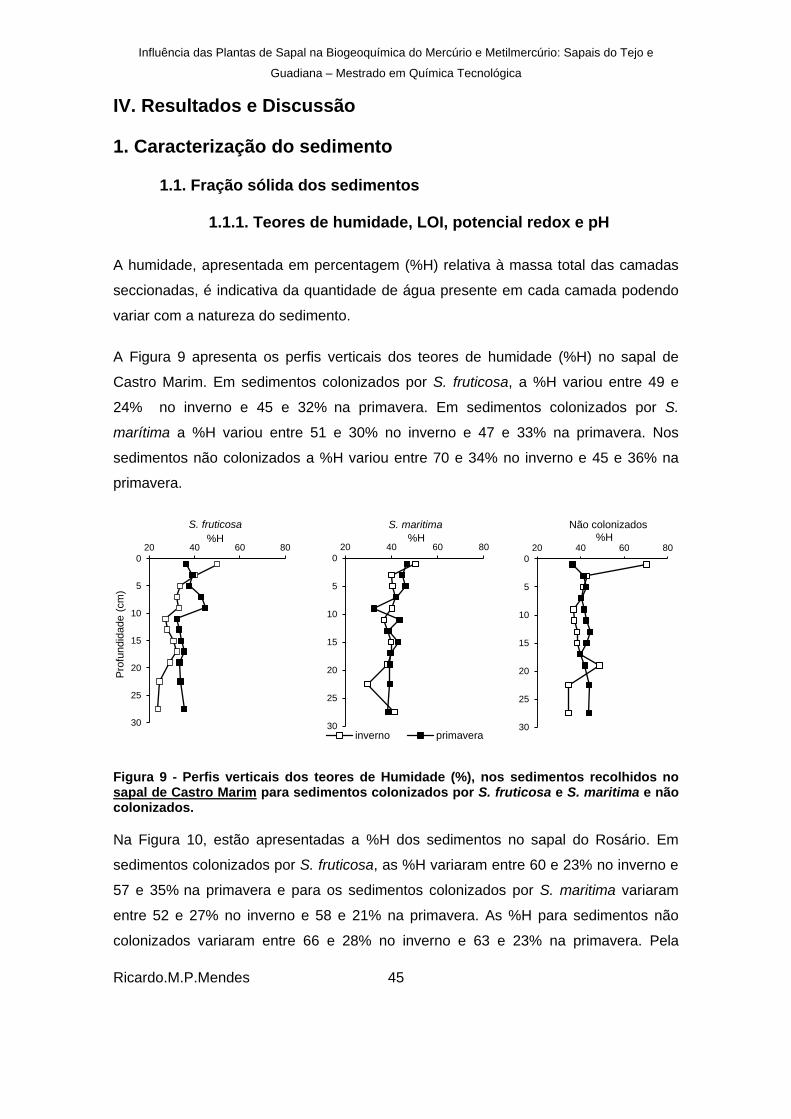
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 45
IV. Resultados e Discussão
1. Caracterização do sedimento
1.1. Fração sólida dos sedimentos
1.1.1. Teores de humidade, LOI, potencial redox e pH
A humidade, apresentada em percentagem (%H) relativa à massa total das camadas
seccionadas, é indicativa da quantidade de água presente em cada camada podendo
variar com a natureza do sedimento.
A Figura 9 apresenta os perfis verticais dos teores de humidade (%H) no sapal de
Castro Marim. Em sedimentos colonizados por S. fruticosa, a %H variou entre 49 e
24% no inverno e 45 e 32% na primavera. Em sedimentos colonizados por S.
marítima a %H variou entre 51 e 30% no inverno e 47 e 33% na primavera. Nos
sedimentos não colonizados a %H variou entre 70 e 34% no inverno e 45 e 36% na
primavera.
Figura 9 - Perfis verticais dos teores de Humidade (%), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
Na Figura 10, estão apresentadas a %H dos sedimentos no sapal do Rosário. Em
sedimentos colonizados por S. fruticosa, as %H variaram entre 60 e 23% no inverno e
57 e 35% na primavera e para os sedimentos colonizados por S. maritima variaram
entre 52 e 27% no inverno e 58 e 21% na primavera. As %H para sedimentos não
colonizados variaram entre 66 e 28% no inverno e 63 e 23% na primavera. Pela
0
5
10
15
20
25
30
20 40 60 80
Pro
fundid
ade (
cm
)
%H
S. fruticosa
0
5
10
15
20
25
30
20 40 60 80%H
S. maritima
inverno primavera
0
5
10
15
20
25
30
20 40 60 80%H
Não colonizados
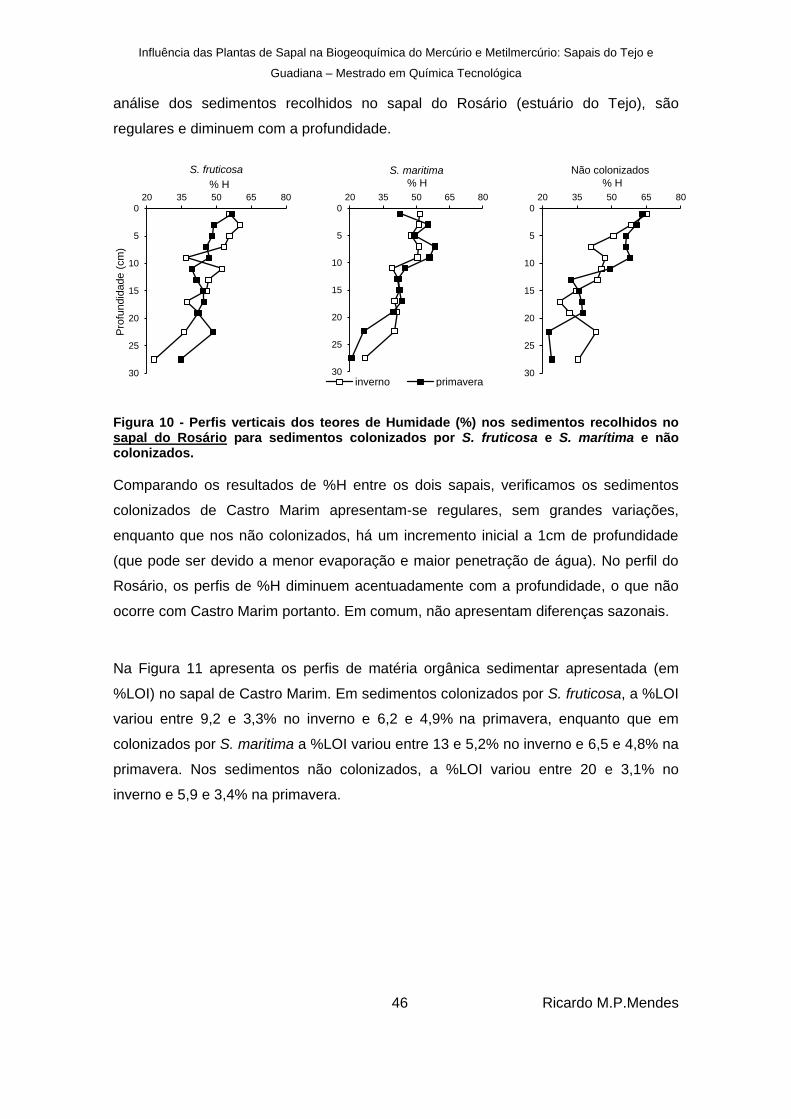
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
46 Ricardo M.P.Mendes
análise dos sedimentos recolhidos no sapal do Rosário (estuário do Tejo), são
regulares e diminuem com a profundidade.
Figura 10 - Perfis verticais dos teores de Humidade (%) nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Comparando os resultados de %H entre os dois sapais, verificamos os sedimentos
colonizados de Castro Marim apresentam-se regulares, sem grandes variações,
enquanto que nos não colonizados, há um incremento inicial a 1cm de profundidade
(que pode ser devido a menor evaporação e maior penetração de água). No perfil do
Rosário, os perfis de %H diminuem acentuadamente com a profundidade, o que não
ocorre com Castro Marim portanto. Em comum, não apresentam diferenças sazonais.
Na Figura 11 apresenta os perfis de matéria orgânica sedimentar apresentada (em
%LOI) no sapal de Castro Marim. Em sedimentos colonizados por S. fruticosa, a %LOI
variou entre 9,2 e 3,3% no inverno e 6,2 e 4,9% na primavera, enquanto que em
colonizados por S. maritima a %LOI variou entre 13 e 5,2% no inverno e 6,5 e 4,8% na
primavera. Nos sedimentos não colonizados, a %LOI variou entre 20 e 3,1% no
inverno e 5,9 e 3,4% na primavera.
0
5
10
15
20
25
30
20 35 50 65 80
Pro
fundid
ade (
cm
)
% H
S. fruticosa
0
5
10
15
20
25
30
20 35 50 65 80
% H
S. maritima
inverno primavera
0
5
10
15
20
25
30
20 35 50 65 80
% H
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 47
Figura 11 - Perfis verticais dos teores de LOI (%), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
A Figura 12 apresenta as %LOI nos sedimentos no sapal do Rosário. Nos sedimentos
colonizados por S. fruticosa, as %LOI variaram entre 9,6 e 1,8% no inverno e 8,1 e
2,8% na primavera e para os sedimentos colonizados por S. marítima estes variaram
entre 9,9 e 2,4% no inverno e 10,3 e 1,1% na primavera. Em sedimentos não
colonizados variaram entre 8,1 e 2,0% no inverno e 8,5 e 1,5% na primavera.
Figura 12 - Perfis verticais dos valores de LOI (%), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Analisando os perfis de percentagem de matéria orgânica sedimentar para o sapal de
Castro Marim, verificamos que nos perfis dos sedimentos colonizados por S. fruticosa
apresentam-se regulares. O mesmo ocorre com os outros perfis, com a excessão de
0
5
10
15
20
25
30
0 5 10 15 20
Pro
fundid
ade (
cm
) %LOI
S. fruticosa
0
5
10
15
20
25
30
0 5 10 15 20
%LOI S. maritima
inverno primavera
0
5
10
15
20
25
30
0 5 10 15 20
%LOI
Não colonizados
0
5
10
15
20
25
30
0 2 4 6 8 10 12
Pro
fundid
ade (
cm
)
% LOI
S. fruticosa
0
5
10
15
20
25
30
0 2 4 6 8 10 12
% LOI
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 2 4 6 8 10 12
% LOI
Não colonizados
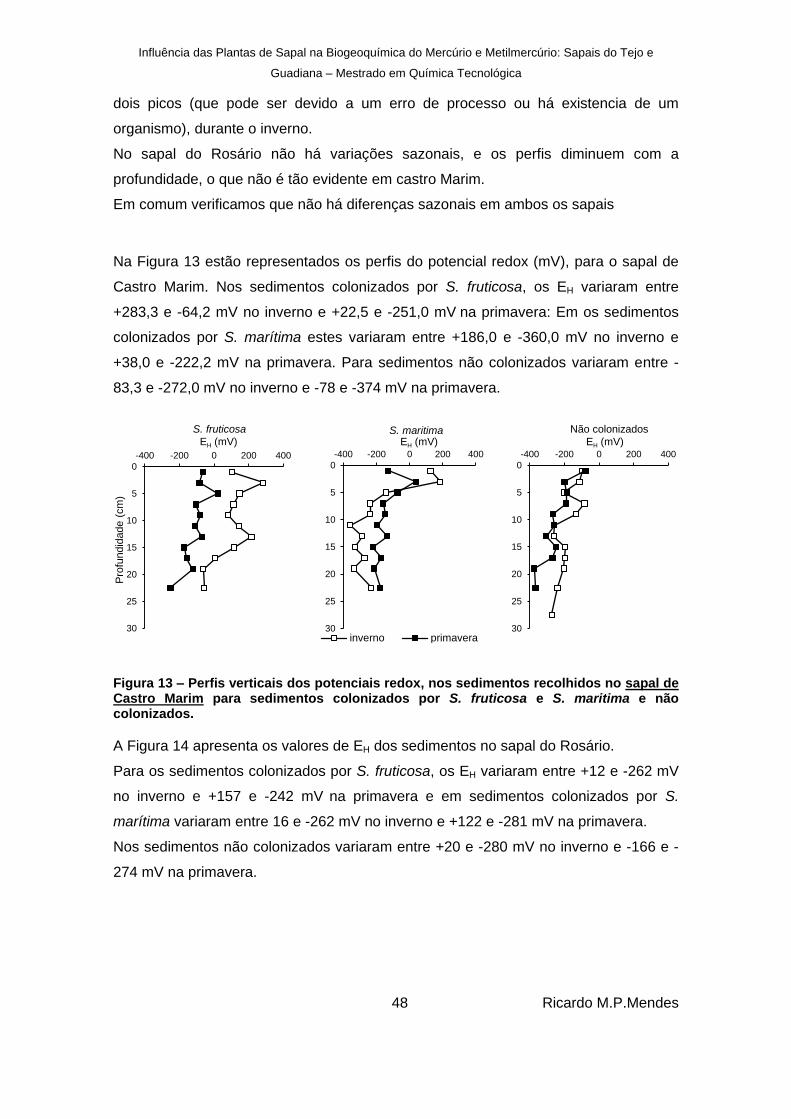
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
48 Ricardo M.P.Mendes
dois picos (que pode ser devido a um erro de processo ou há existencia de um
organismo), durante o inverno.
No sapal do Rosário não há variações sazonais, e os perfis diminuem com a
profundidade, o que não é tão evidente em castro Marim.
Em comum verificamos que não há diferenças sazonais em ambos os sapais
Na Figura 13 estão representados os perfis do potencial redox (mV), para o sapal de
Castro Marim. Nos sedimentos colonizados por S. fruticosa, os EH variaram entre
+283,3 e -64,2 mV no inverno e +22,5 e -251,0 mV na primavera: Em os sedimentos
colonizados por S. marítima estes variaram entre +186,0 e -360,0 mV no inverno e
+38,0 e -222,2 mV na primavera. Para sedimentos não colonizados variaram entre -
83,3 e -272,0 mV no inverno e -78 e -374 mV na primavera.
Figura 13 – Perfis verticais dos potenciais redox, nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
A Figura 14 apresenta os valores de EH dos sedimentos no sapal do Rosário.
Para os sedimentos colonizados por S. fruticosa, os EH variaram entre +12 e -262 mV
no inverno e +157 e -242 mV na primavera e em sedimentos colonizados por S.
marítima variaram entre 16 e -262 mV no inverno e +122 e -281 mV na primavera.
Nos sedimentos não colonizados variaram entre +20 e -280 mV no inverno e -166 e -
274 mV na primavera.
0
5
10
15
20
25
30
-400 -200 0 200 400
Pro
fundid
ade (
cm
)
EH (mV)
S. fruticosa
0
5
10
15
20
25
30
-400 -200 0 200 400
EH (mV) S. maritima
inverno primavera
0
5
10
15
20
25
30
-400 -200 0 200 400
EH (mV)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 49
Figura 14 – Perfis verticais dos potenciais redox, nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
Comparando sapais, verificamos que no sapal de Castro Marim, em sedimentos
colonizados por S. fruticosa é de notar que os valores de EH são positivos durante o
inverno (podendo sugerir que o sedimento é mais óxico), enquanto os valores na
primavera são na generalidade negativos. Em sedimentos colonizados por S.
maritima, os valores de EH entre os 0 e 5 cm apresentam um incremento positivo
nestas camadas, contudo os restantes valores são negativos. Os valores são de uma
maneira geral, regulares.
No sapal do Rosário, nos perfis de sedimentos colonizados, os valores de EH são
muito irregulares, com valores alternados positivos e negativos ao longo da
profundidade (principalmente durante a primavera) em várias camadas. Os valores no
inverno são negativos.
Em sedimentos não colonizados, no sapal de Castro Marim os valores são sempre
negativos, e sem grandes variações, enquanto que no sapal do os valores são sempre
negativos, com os valores na primavera a apresentarem-se mais irregulares que os
valores de inverno.
Sendo o intervalo entre -150 e 200 mV considerado o intervalo onde a grande maioria
dos metais tem mobilidade (Jackson, 1998), verificamos que a maior parte dos valores
em ambos os sapais, se encontra dentro deste intervalo, ou seja, os metais possuem
mobilidade.
0
5
10
15
20
25
30
-400 -200 0 200
Pro
fundid
ade (
cm
) EH (mV)
S. fruticosa
0
5
10
15
20
25
30
-400 -200 0 200
EH (mV)
S. maritima
inverno primavera
0
5
10
15
20
25
30
-400 -200 0 200
EH (mV) Não colonizados
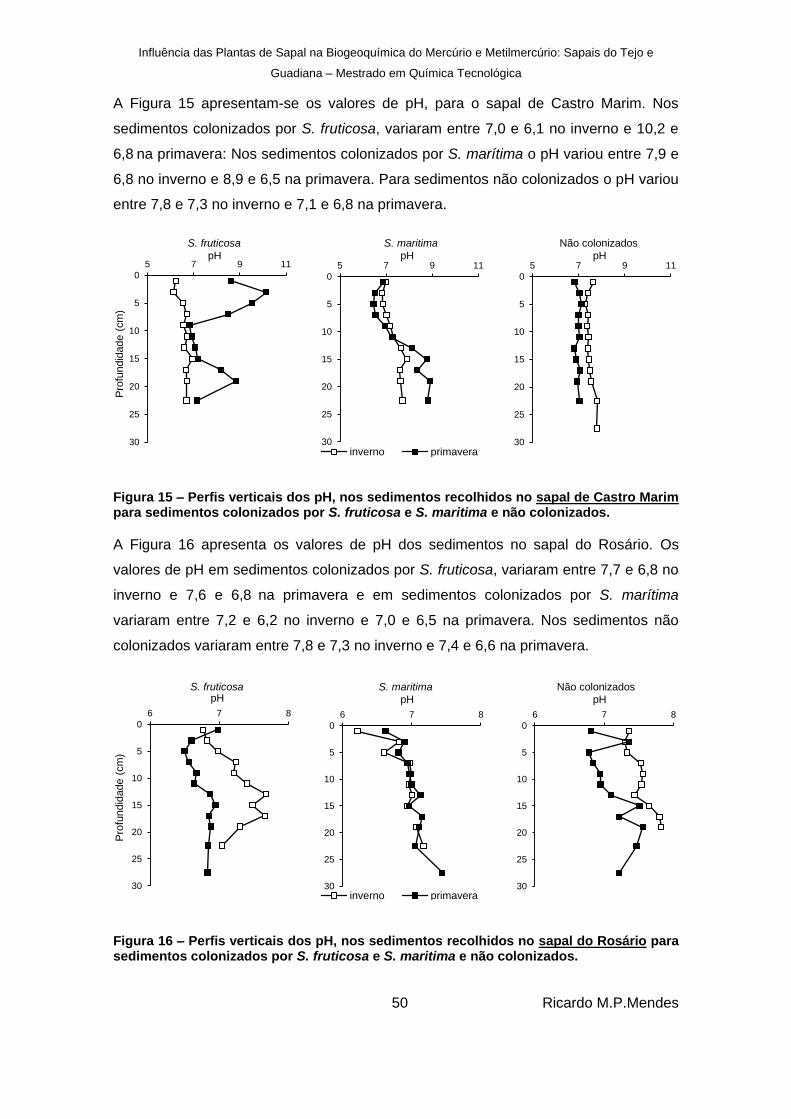
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
50 Ricardo M.P.Mendes
A Figura 15 apresentam-se os valores de pH, para o sapal de Castro Marim. Nos
sedimentos colonizados por S. fruticosa, variaram entre 7,0 e 6,1 no inverno e 10,2 e
6,8 na primavera: Nos sedimentos colonizados por S. marítima o pH variou entre 7,9 e
6,8 no inverno e 8,9 e 6,5 na primavera. Para sedimentos não colonizados o pH variou
entre 7,8 e 7,3 no inverno e 7,1 e 6,8 na primavera.
Figura 15 – Perfis verticais dos pH, nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
A Figura 16 apresenta os valores de pH dos sedimentos no sapal do Rosário. Os
valores de pH em sedimentos colonizados por S. fruticosa, variaram entre 7,7 e 6,8 no
inverno e 7,6 e 6,8 na primavera e em sedimentos colonizados por S. marítima
variaram entre 7,2 e 6,2 no inverno e 7,0 e 6,5 na primavera. Nos sedimentos não
colonizados variaram entre 7,8 e 7,3 no inverno e 7,4 e 6,6 na primavera.
Figura 16 – Perfis verticais dos pH, nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
0
5
10
15
20
25
30
5 7 9 11
Pro
fundid
ade (
cm
)
pH
S. fruticosa
0
5
10
15
20
25
30
5 7 9 11pH
S. maritima
inverno primavera
0
5
10
15
20
25
30
5 7 9 11pH
Não colonizados
0
5
10
15
20
25
30
6 7 8
Pro
fundid
ade (
cm
)
pH S. fruticosa
0
5
10
15
20
25
30
6 7 8
pH
S. maritima
inverno primavera
0
5
10
15
20
25
30
6 7 8
pH
Não colonizados
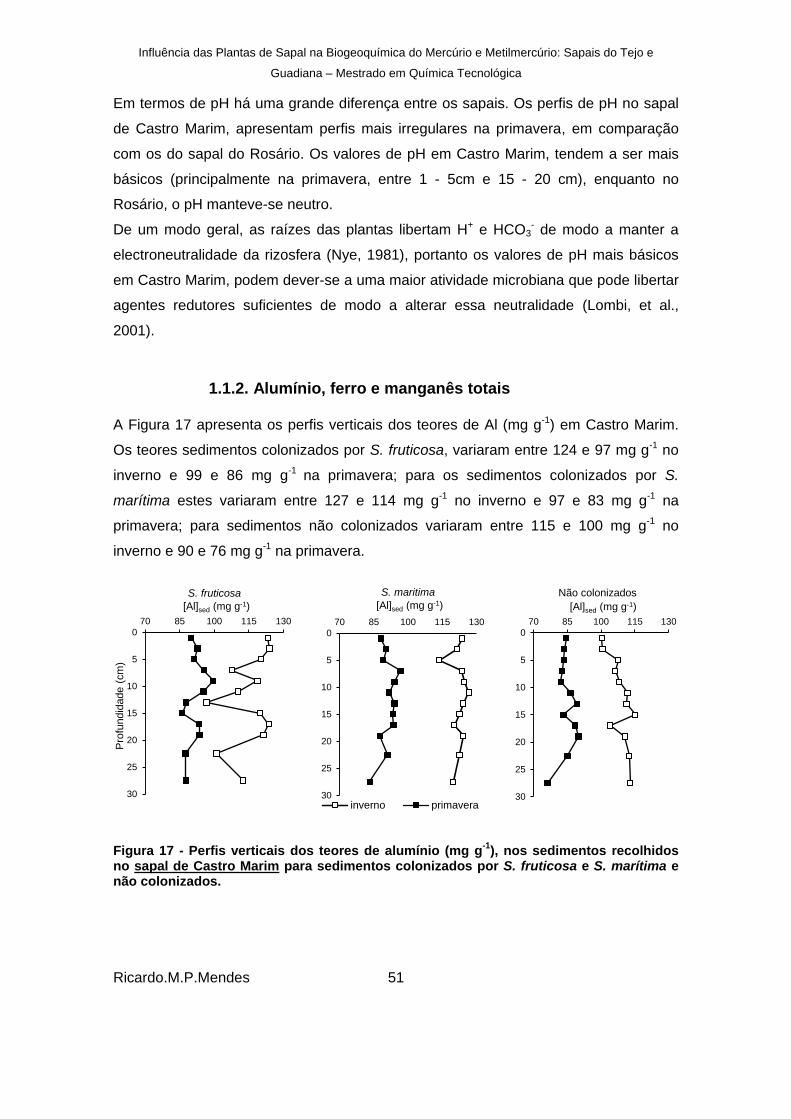
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 51
Em termos de pH há uma grande diferença entre os sapais. Os perfis de pH no sapal
de Castro Marim, apresentam perfis mais irregulares na primavera, em comparação
com os do sapal do Rosário. Os valores de pH em Castro Marim, tendem a ser mais
básicos (principalmente na primavera, entre 1 - 5cm e 15 - 20 cm), enquanto no
Rosário, o pH manteve-se neutro.
De um modo geral, as raízes das plantas libertam H+ e HCO3- de modo a manter a
electroneutralidade da rizosfera (Nye, 1981), portanto os valores de pH mais básicos
em Castro Marim, podem dever-se a uma maior atividade microbiana que pode libertar
agentes redutores suficientes de modo a alterar essa neutralidade (Lombi, et al.,
2001).
1.1.2. Alumínio, ferro e manganês totais
A Figura 17 apresenta os perfis verticais dos teores de Al (mg g-1) em Castro Marim.
Os teores sedimentos colonizados por S. fruticosa, variaram entre 124 e 97 mg g-1 no
inverno e 99 e 86 mg g-1 na primavera; para os sedimentos colonizados por S.
marítima estes variaram entre 127 e 114 mg g-1 no inverno e 97 e 83 mg g-1 na
primavera; para sedimentos não colonizados variaram entre 115 e 100 mg g-1 no
inverno e 90 e 76 mg g-1 na primavera.
Figura 17 - Perfis verticais dos teores de alumínio (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
0
5
10
15
20
25
30
70 85 100 115 130
Pro
fundid
ade (
cm
)
[Al]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
30
70 85 100 115 130
[Al]sed (mg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
70 85 100 115 130
[Al]sed (mg g-1)
Não colonizados
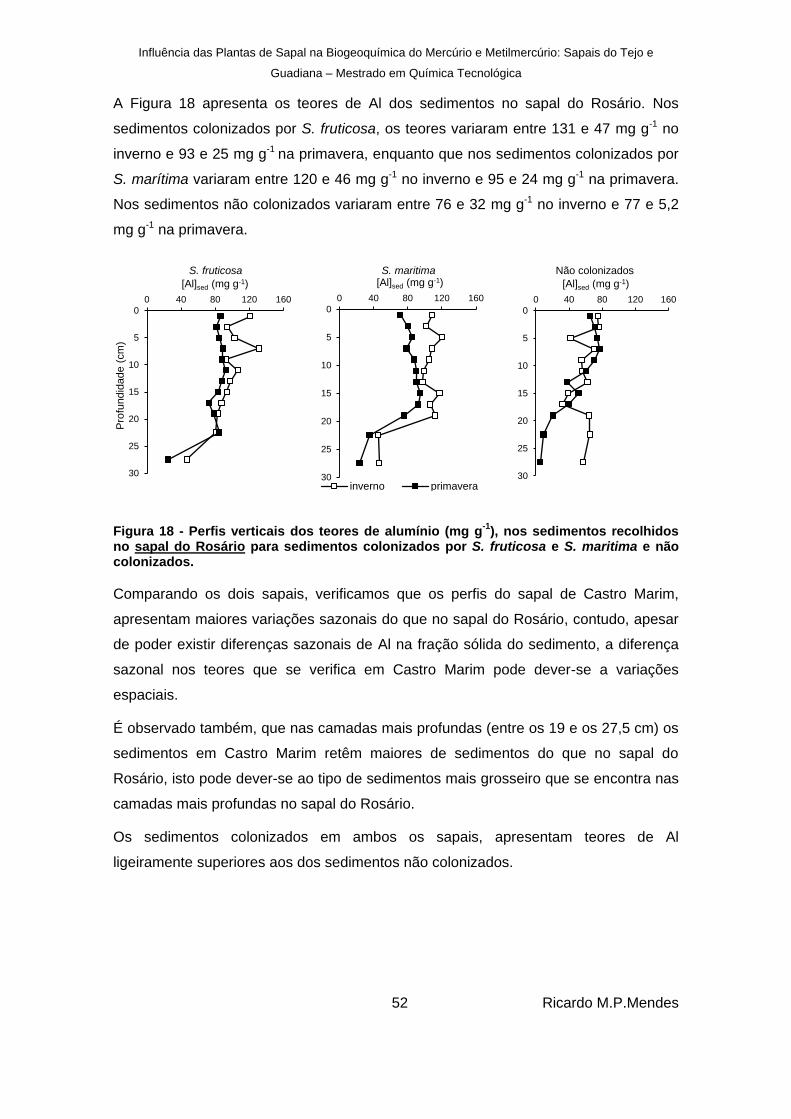
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
52 Ricardo M.P.Mendes
A Figura 18 apresenta os teores de Al dos sedimentos no sapal do Rosário. Nos
sedimentos colonizados por S. fruticosa, os teores variaram entre 131 e 47 mg g-1 no
inverno e 93 e 25 mg g-1 na primavera, enquanto que nos sedimentos colonizados por
S. marítima variaram entre 120 e 46 mg g-1 no inverno e 95 e 24 mg g-1 na primavera.
Nos sedimentos não colonizados variaram entre 76 e 32 mg g-1 no inverno e 77 e 5,2
mg g-1 na primavera.
Figura 18 - Perfis verticais dos teores de alumínio (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
Comparando os dois sapais, verificamos que os perfis do sapal de Castro Marim,
apresentam maiores variações sazonais do que no sapal do Rosário, contudo, apesar
de poder existir diferenças sazonais de Al na fração sólida do sedimento, a diferença
sazonal nos teores que se verifica em Castro Marim pode dever-se a variações
espaciais.
É observado também, que nas camadas mais profundas (entre os 19 e os 27,5 cm) os
sedimentos em Castro Marim retêm maiores de sedimentos do que no sapal do
Rosário, isto pode dever-se ao tipo de sedimentos mais grosseiro que se encontra nas
camadas mais profundas no sapal do Rosário.
Os sedimentos colonizados em ambos os sapais, apresentam teores de Al
ligeiramente superiores aos dos sedimentos não colonizados.
0
5
10
15
20
25
30
0 40 80 120 160
Pro
fundid
ade (
cm
)
[Al]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
30
0 40 80 120 160
[Al]sed (mg g-1) S. maritima
inverno primavera
0
5
10
15
20
25
30
0 40 80 120 160
[Al]sed (mg g-1)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 53
A Figura 19 apresenta os perfis verticais dos teores de Fe (mg g-1), do sapal de Castro
Marim. Os teores de sedimentos colonizados por S. fruticosa, variaram entre 68 e 41
mg g-1 no inverno e 37 e 42 mg g-1 na primavera, enquanto que em sedimentos
colonizados por S. marítima variaram entre 57 e 50 no inverno e 40 e 33 mg g-1 na
primavera. Os teores dos sedimentos não colonizados variaram entre 52 e 42 mg g-1
no inverno e 35 e 31 mg g-1na primavera.
Figura 19 - Perfis verticais dos teores de ferro (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Os perfis dos teores de Fe são normalizados com os perfis dos teores de alumínio, e
servem para indicar diferenças na fração alumino-silicatada do sedimento (Loring,
1991).
Figura 20 - Razões dos teores de Fe/Al dos sedimentos recolhidos no Sapal de Castro Marim.
0
5
10
15
20
25
30
15 30 45 60 75
Pro
fundid
ade (
cm
)
[Fe]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
30
15 30 45 60 75
[Fe]sed (mg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
15 30 45 60 75
[Fe]sed (mg g-1)
Não colonizados
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8
Pro
fundid
ade (
cm
)
([Fe]/[Al])sed
S. fruticosa
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8
([Fe]/[Al])sed
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8
([Fe]/[Al])sed
Não colonizados
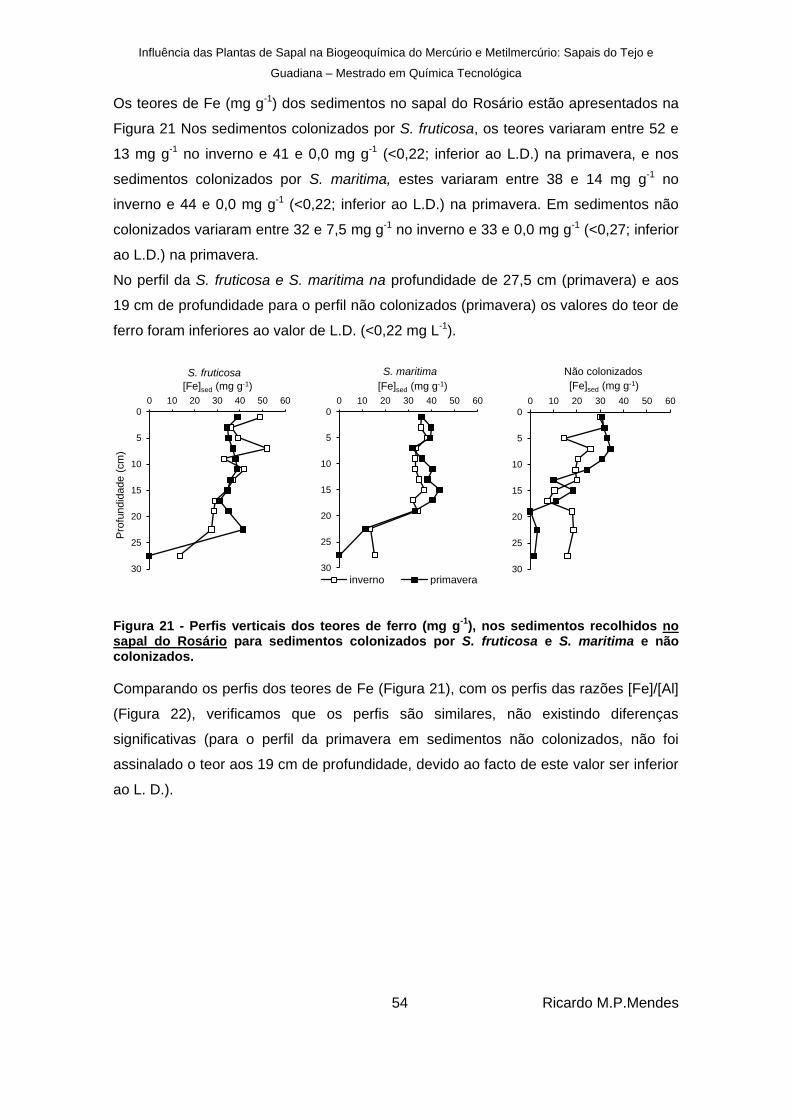
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
54 Ricardo M.P.Mendes
Os teores de Fe (mg g-1) dos sedimentos no sapal do Rosário estão apresentados na
Figura 21 Nos sedimentos colonizados por S. fruticosa, os teores variaram entre 52 e
13 mg g-1 no inverno e 41 e 0,0 mg g-1 (<0,22; inferior ao L.D.) na primavera, e nos
sedimentos colonizados por S. maritima, estes variaram entre 38 e 14 mg g-1 no
inverno e 44 e 0,0 mg g-1 (<0,22; inferior ao L.D.) na primavera. Em sedimentos não
colonizados variaram entre 32 e 7,5 mg g-1 no inverno e 33 e 0,0 mg g-1 (<0,27; inferior
ao L.D.) na primavera.
No perfil da S. fruticosa e S. maritima na profundidade de 27,5 cm (primavera) e aos
19 cm de profundidade para o perfil não colonizados (primavera) os valores do teor de
ferro foram inferiores ao valor de L.D. (<0,22 mg L-1).
Figura 21 - Perfis verticais dos teores de ferro (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
Comparando os perfis dos teores de Fe (Figura 21), com os perfis das razões [Fe]/[Al]
(Figura 22), verificamos que os perfis são similares, não existindo diferenças
significativas (para o perfil da primavera em sedimentos não colonizados, não foi
assinalado o teor aos 19 cm de profundidade, devido ao facto de este valor ser inferior
ao L. D.).
0
5
10
15
20
25
30
0 10 20 30 40 50 60
Pro
fundid
ade (
cm
)
[Fe]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40 50 60
[Fe]sed (mg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40 50 60
[Fe]sed (mg g-1)
Não colonizados
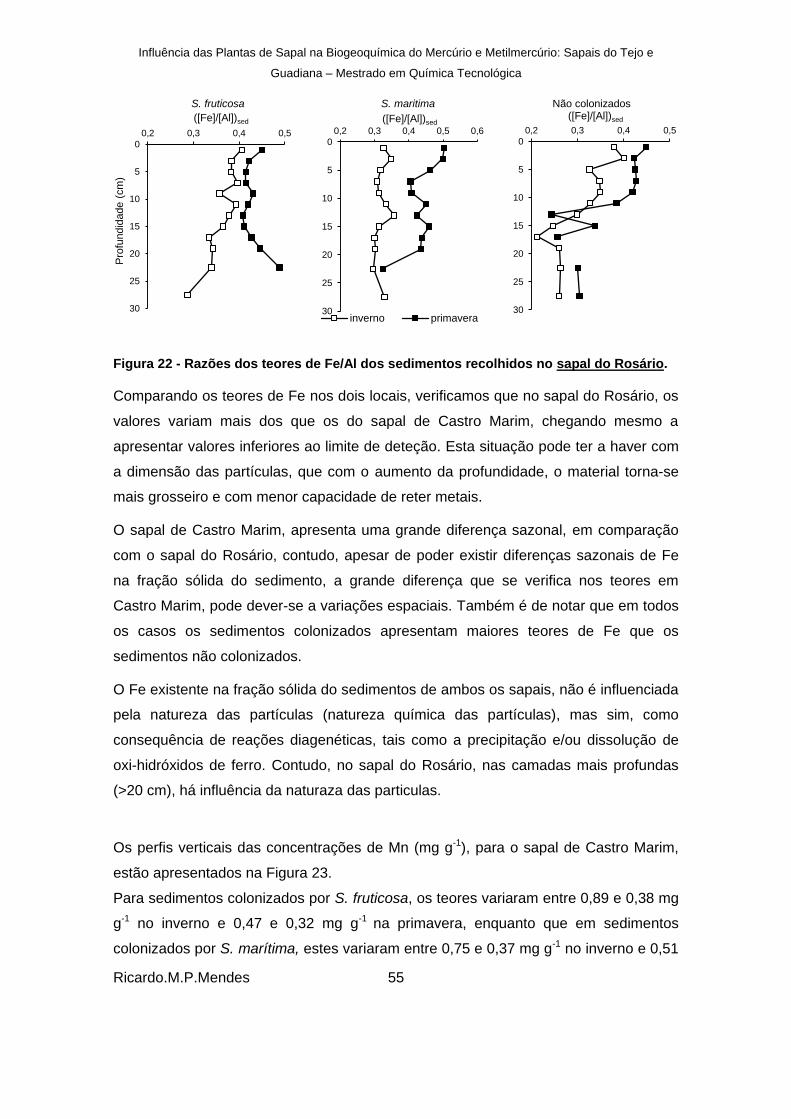
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 55
Figura 22 - Razões dos teores de Fe/Al dos sedimentos recolhidos no sapal do Rosário.
Comparando os teores de Fe nos dois locais, verificamos que no sapal do Rosário, os
valores variam mais dos que os do sapal de Castro Marim, chegando mesmo a
apresentar valores inferiores ao limite de deteção. Esta situação pode ter a haver com
a dimensão das partículas, que com o aumento da profundidade, o material torna-se
mais grosseiro e com menor capacidade de reter metais.
O sapal de Castro Marim, apresenta uma grande diferença sazonal, em comparação
com o sapal do Rosário, contudo, apesar de poder existir diferenças sazonais de Fe
na fração sólida do sedimento, a grande diferença que se verifica nos teores em
Castro Marim, pode dever-se a variações espaciais. Também é de notar que em todos
os casos os sedimentos colonizados apresentam maiores teores de Fe que os
sedimentos não colonizados.
O Fe existente na fração sólida do sedimentos de ambos os sapais, não é influenciada
pela natureza das partículas (natureza química das partículas), mas sim, como
consequência de reações diagenéticas, tais como a precipitação e/ou dissolução de
oxi-hidróxidos de ferro. Contudo, no sapal do Rosário, nas camadas mais profundas
(>20 cm), há influência da naturaza das particulas.
Os perfis verticais das concentrações de Mn (mg g-1), para o sapal de Castro Marim,
estão apresentados na Figura 23.
Para sedimentos colonizados por S. fruticosa, os teores variaram entre 0,89 e 0,38 mg
g-1 no inverno e 0,47 e 0,32 mg g-1 na primavera, enquanto que em sedimentos
colonizados por S. marítima, estes variaram entre 0,75 e 0,37 mg g-1 no inverno e 0,51
0
5
10
15
20
25
30
0,2 0,3 0,4 0,5
Pro
fundid
ade (
cm
) ([Fe]/[Al])sed
S. fruticosa
0
5
10
15
20
25
30
0,2 0,3 0,4 0,5 0,6
([Fe]/[Al])sed
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,2 0,3 0,4 0,5
([Fe]/[Al])sed Não colonizados
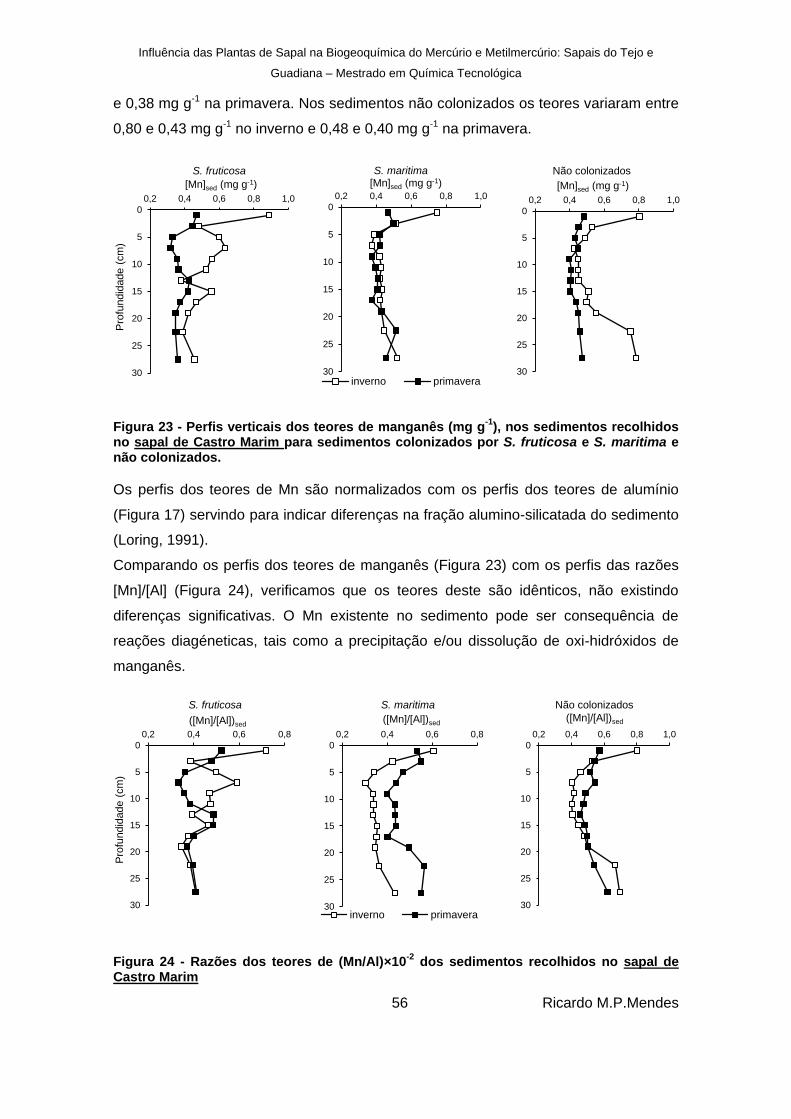
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
56 Ricardo M.P.Mendes
e 0,38 mg g-1 na primavera. Nos sedimentos não colonizados os teores variaram entre
0,80 e 0,43 mg g-1 no inverno e 0,48 e 0,40 mg g-1 na primavera.
Figura 23 - Perfis verticais dos teores de manganês (mg g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
Os perfis dos teores de Mn são normalizados com os perfis dos teores de alumínio
(Figura 17) servindo para indicar diferenças na fração alumino-silicatada do sedimento
(Loring, 1991).
Comparando os perfis dos teores de manganês (Figura 23) com os perfis das razões
[Mn]/[Al] (Figura 24), verificamos que os teores deste são idênticos, não existindo
diferenças significativas. O Mn existente no sedimento pode ser consequência de
reações diagéneticas, tais como a precipitação e/ou dissolução de oxi-hidróxidos de
manganês.
Figura 24 - Razões dos teores de (Mn/Al)×10-2
dos sedimentos recolhidos no sapal de Castro Marim
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8 1,0
Pro
fundid
ade (
cm
)
[Mn]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8 1,0
[Mn]sed (mg g-1) S. maritima
inverno primavera
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8 1,0
[Mn]sed (mg g-1)
Não colonizados
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8
Pro
fundid
ade (
cm
)
([Mn]/[Al])sed
S. fruticosa
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8
([Mn]/[Al])sed
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,2 0,4 0,6 0,8 1,0
([Mn]/[Al])sed
Não colonizados
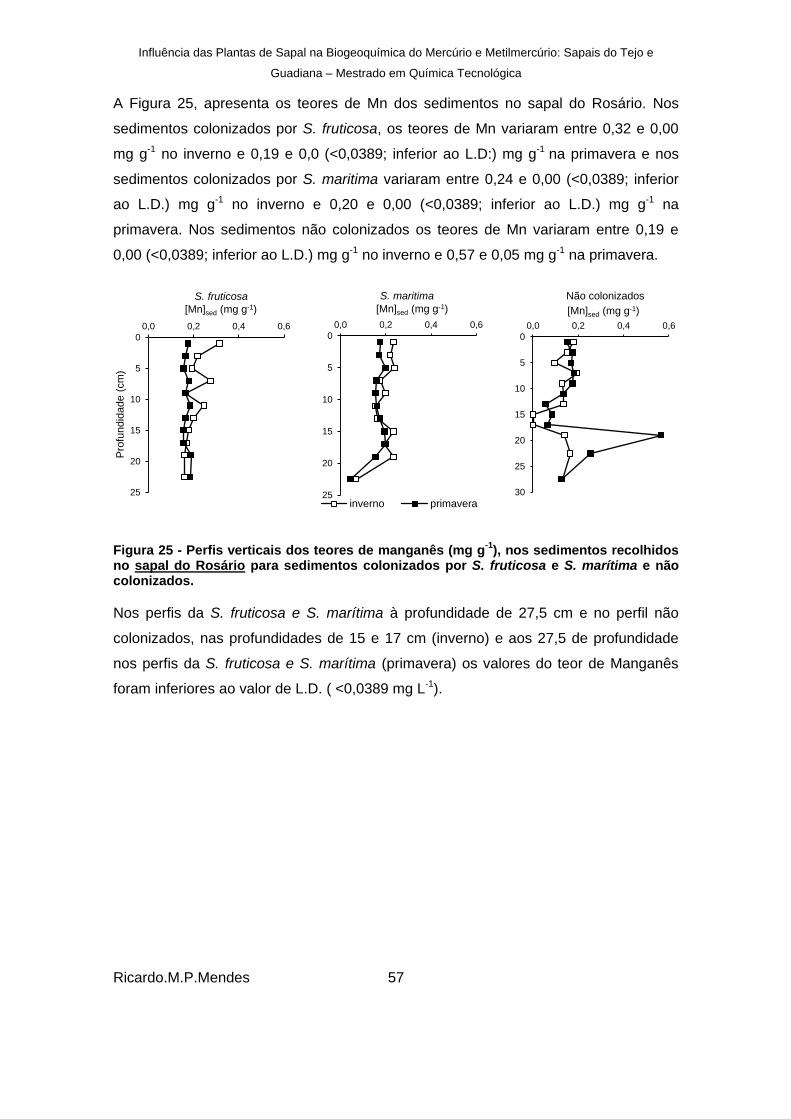
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 57
A Figura 25, apresenta os teores de Mn dos sedimentos no sapal do Rosário. Nos
sedimentos colonizados por S. fruticosa, os teores de Mn variaram entre 0,32 e 0,00
mg g-1 no inverno e 0,19 e 0,0 (<0,0389; inferior ao L.D:) mg g-1 na primavera e nos
sedimentos colonizados por S. maritima variaram entre 0,24 e 0,00 (<0,0389; inferior
ao L.D.) mg g-1 no inverno e 0,20 e 0,00 (<0,0389; inferior ao L.D.) mg g-1 na
primavera. Nos sedimentos não colonizados os teores de Mn variaram entre 0,19 e
0,00 (<0,0389; inferior ao L.D.) mg g-1 no inverno e 0,57 e 0,05 mg g-1 na primavera.
Figura 25 - Perfis verticais dos teores de manganês (mg g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Nos perfis da S. fruticosa e S. marítima à profundidade de 27,5 cm e no perfil não
colonizados, nas profundidades de 15 e 17 cm (inverno) e aos 27,5 de profundidade
nos perfis da S. fruticosa e S. marítima (primavera) os valores do teor de Manganês
foram inferiores ao valor de L.D. ( <0,0389 mg L-1).
0
5
10
15
20
25
0,0 0,2 0,4 0,6
Pro
fundid
ade (
cm
)
[Mn]sed (mg g-1)
S. fruticosa
0
5
10
15
20
25
0,0 0,2 0,4 0,6
[Mn]sed (mg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,0 0,2 0,4 0,6
[Mn]sed (mg g-1)
Não colonizados
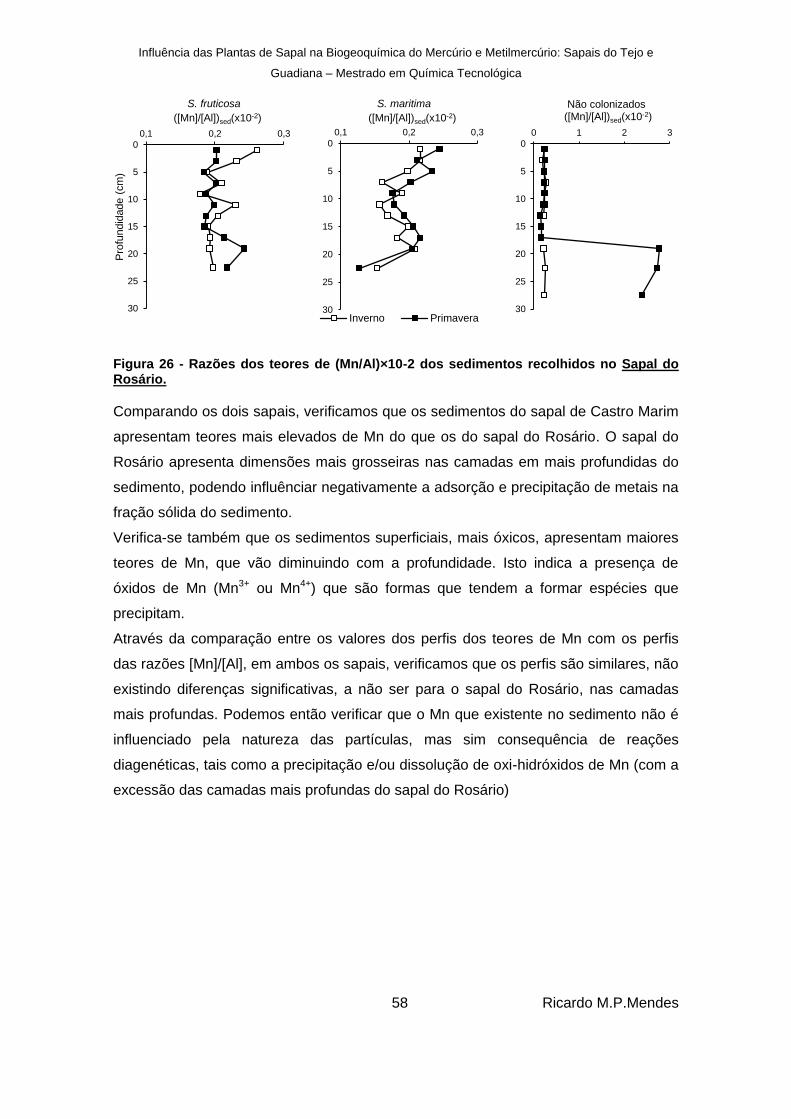
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
58 Ricardo M.P.Mendes
Figura 26 - Razões dos teores de (Mn/Al)×10-2 dos sedimentos recolhidos no Sapal do Rosário.
Comparando os dois sapais, verificamos que os sedimentos do sapal de Castro Marim
apresentam teores mais elevados de Mn do que os do sapal do Rosário. O sapal do
Rosário apresenta dimensões mais grosseiras nas camadas em mais profundidas do
sedimento, podendo influênciar negativamente a adsorção e precipitação de metais na
fração sólida do sedimento.
Verifica-se também que os sedimentos superficiais, mais óxicos, apresentam maiores
teores de Mn, que vão diminuindo com a profundidade. Isto indica a presença de
óxidos de Mn (Mn3+ ou Mn4+) que são formas que tendem a formar espécies que
precipitam.
Através da comparação entre os valores dos perfis dos teores de Mn com os perfis
das razões [Mn]/[Al], em ambos os sapais, verificamos que os perfis são similares, não
existindo diferenças significativas, a não ser para o sapal do Rosário, nas camadas
mais profundas. Podemos então verificar que o Mn que existente no sedimento não é
influenciado pela natureza das partículas, mas sim consequência de reações
diagenéticas, tais como a precipitação e/ou dissolução de oxi-hidróxidos de Mn (com a
excessão das camadas mais profundas do sapal do Rosário)
0
5
10
15
20
25
30
0,1 0,2 0,3
Pro
fundid
ade (
cm
) ([Mn]/[Al])sed(x10-2)
S. fruticosa
0
5
10
15
20
25
30
0,1 0,2 0,3
([Mn]/[Al])sed(x10-2)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 1 2 3
([Mn]/[Al])sed(x10-2) Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 59
1.1.3. Sulfuretos Ácidos Voláteis
Os sulfuretos ácidos voláteis, denominados AVS, (“Acid Volatile Sulfide”, Ex: FeS), e
correspondem a compostos de sulfuretos que se encontram precipitadas na fração
sólida do sedimento.
A Figura 27 apresenta os teores de AVS para o sapal de Castro Marim, que variaram
para os sedimentos colonizados por S. fruticosa, entre 52 e 0,9 µmol g-1 no inverno e
5,5 e 1,1 µmol g-1na primavera, enquanto que, em sedimentos colonizados por S.
marítima estes variaram entre 3,4 e 0,7 µmol g-1no inverno e 2,7 e 0,3 µmol g-1 na
primavera. Nos sedimentos não colonizados variaram entre 85 e 5,9 µmol g-1no
inverno e 53 e 1,1 µmol g-1na primavera.
Figura 27 - Perfis verticais dos teores de AVS (µmol g-1
), nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
A Figura 28 apresenta, os teores de AVS nos sedimentos no sapal do Rosário. Em
sedimentos colonizados por S. fruticosa os teores de AVS variaram entre 37 e 1,2
μmol g-1 no inverno e 3,5 e 0,5 μmol g-1 na primavera e para os sedimentos
colonizados por S. marítima, variaram entre 57 e 2,3 μmol g-1 no inverno e 5,2 e 1,4
μmol g-1 na primavera. Nos sedimentos não colonizados variaram entre 65 e 2,4 μmol
g-1 no inverno e 37 e 1,4 μmol g-1 na primavera.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90
Pro
fundid
ade (
cm
)
[AVS] (μmol g-1)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90
[AVS] (μmol g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90
[AVS] (μmol g-1)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
60 Ricardo M.P.Mendes
Figura 28 - Perfis verticais dos teores de AVS (μmol g-1
), nos sedimentos recolhidos no sapal do Rosário em sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
No sapal de Castro Marim e do Rosário, verificamos que existe uma diferença nos
teores de AVS entre sedimentos colonizados e não colonizados.. Os teores de AVS,
em Castro Marim e no Rosário, na primavera em sedimentos colonizados apresentam-
se muito baixos comparativamente aos teores na primavera em sedimentos não
colonizados.
As diferenças teores de AVS no inverno, deve-se há influencia das raizes.
Em Castro Marim, os teores de AVS da S. fruticosa, sao dimensionalmente superiores
a partir do momento em que não foram encontradas raizes, na S. maritima os teores
foram muito baixos, tanto no inverno como na primavera.
No Rosário, os teores de AVS dos sedimentos colonizados no inverno foram da
mesma dimensão que sedimentos não colonizados, o que não ocorre na primavera.
Verificamos assim que as raizes parecem funcionar de maneira diferente entre os
sapais. Enquanto em Castro Marim, os teores só atingem valores elevados quando
não há raizes, tanto no inverno como na primavera, no sapal do Rosário há a
indicação que nas raizes não funcionam no inverno.
Contudo, podemos concluir que a atividade das raízes influência os teores de AVS no
sedimento.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
Pro
fundid
ade (
cm
) [AVS] (μmol g-1) S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
[AVS] (μmol g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
[AVS] (µM) Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 61
1.1.4. Biomassa subterrânea
A Figura 29 apresenta a biomassa expressa em percentagem (%Bio) em sedimentos
colonizados, no sapal de Castro Marim. Em sedimentos colonizados por S. fruticosa,
que variaram entre 0,22 e 0,08 % no inverno e 1,1 e 0,27 % na primavera. Nos
sedimentos colonizados por S. marítima variaram entre 3,5 e 0,52 % no inverno e 1,0
e 0,10 % na primavera.
Figura 29 - Perfis verticais em profundidade da Biomassa (%) nos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima.
A Figura 30 representa os perfis de %Bio em sedimentos colonizados, no sapal de
Rosário. Nos sedimentos colonizados por S. fruticosa, as percentagens de biomassa
variaram entre 3,4 e 0,06% no inverno e 0,86 e 0,05% na primavera; para os
sedimentos colonizados por S. marítima estes variaram entre 1,71 e 0,03% no inverno
e 4,0 e 0,01% na primavera.
0
5
10
15
20
25
30
0 1 2 3 4
Pro
fundid
ade (
cm
)
% Biomassa Subterrânea
S. fruticosa
0
5
10
15
20
25
30
0 1 2 3 4
% Biomassa subterrânea S. maritima
Inverno Primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
62 Ricardo M.P.Mendes
Figura 30 - Perfis verticais da percentagem de Biomassa (%) nos sedimentos recolhidos no sapal do Rosário em sedimentos colonizados por S. fruticosa e S. marítima.
No sapal de Castro Marim, a percentagem de biomassa (%Bio), foi diferente entre as
espécies e entre as estações do ano.
Para a S. fruticosa, a %Bio durante o inverno atingiu um máximo de 2,2% a uma
profundidade média de 3 cm, mas não foram encontradas mais raízes abaixo dos 13
cm de profundidade média. Na primavera, foram encontradas raízes ao longo de todo
o core, o que indica que houve um crescimento das raízes durante esta estação,
resultante de uma maior atividade desta planta.
Para a S. maritima, foram encontradas raízes ao longo de todo o core, com maior
%Bio nas camadas superficiais (máximo de 3,5% aos 1 cm de profundidade média),
sendo a %Bio durante o inverno muito superior à da primavera, o que sugere que
houve uma degradação de raízes do inverno para a primavera.
No sapal do Rosário a percentagem de biomassa para a S. fruticosa no inverno atingiu
um máximo de 3,4% aos 2 cm de profundidade média, não ultrapassando os 11 cm de
profundidade. Na primavera os valores de %Bio não ultrapassaram os 1% e atingiu a
profundidade de 22,5 cm.
Comparando com o sapal do Rosário com o sapal de Castro Marim, verificamos que
nos resultados para a S. maritima, há maiores %Bio na primavera do que no inverno
(no sapal de Rosário ocorre o contrário). Também se observa maiores %Bio nos
primeiros 10 cm de sedimento.
0
5
10
15
20
25
30
0 1 2 3 4 5
Pro
fundid
ade (
cm
)
% Biomassa subterrânea S. fruticosa
0
5
10
15
20
25
30
0 1 2 3 4 5
%Biomassa subterrânea S. maritima
inverno primavera
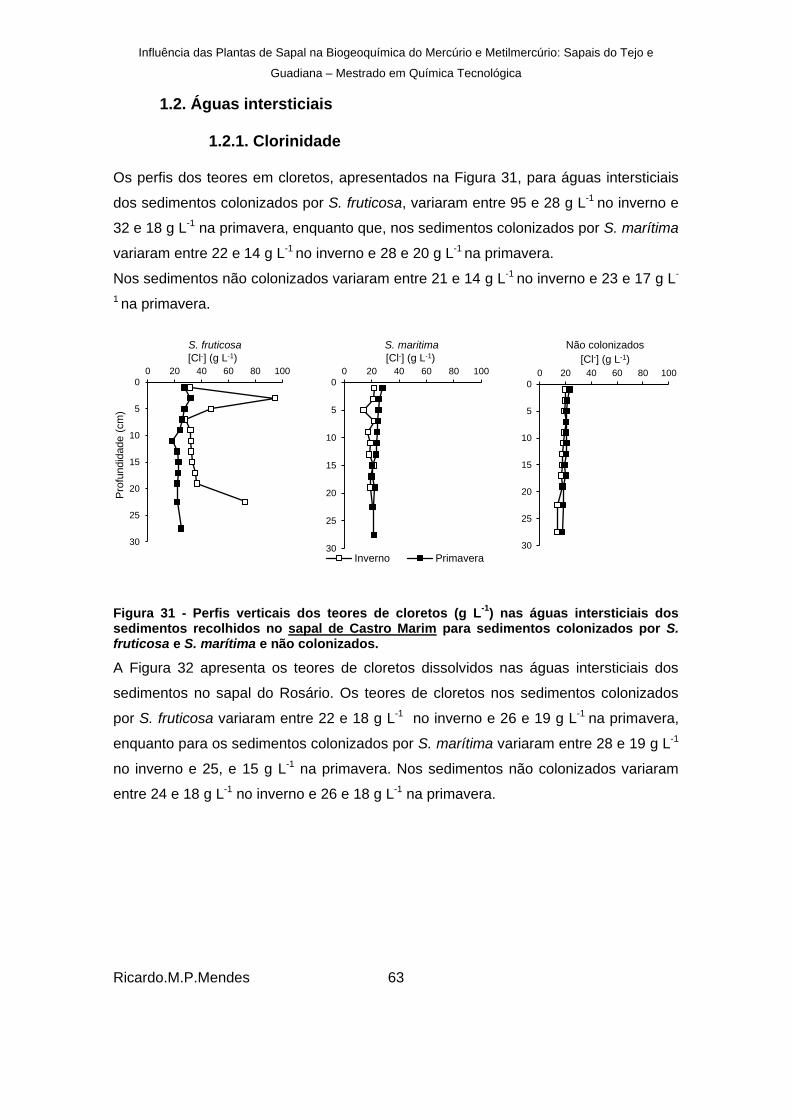
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 63
1.2. Águas intersticiais
1.2.1. Clorinidade
Os perfis dos teores em cloretos, apresentados na Figura 31, para águas intersticiais
dos sedimentos colonizados por S. fruticosa, variaram entre 95 e 28 g L-1 no inverno e
32 e 18 g L-1 na primavera, enquanto que, nos sedimentos colonizados por S. marítima
variaram entre 22 e 14 g L-1 no inverno e 28 e 20 g L-1 na primavera.
Nos sedimentos não colonizados variaram entre 21 e 14 g L-1 no inverno e 23 e 17 g L-
1 na primavera.
Figura 31 - Perfis verticais dos teores de cloretos (g L-1
) nas águas intersticiais dos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
A Figura 32 apresenta os teores de cloretos dissolvidos nas águas intersticiais dos
sedimentos no sapal do Rosário. Os teores de cloretos nos sedimentos colonizados
por S. fruticosa variaram entre 22 e 18 g L-1 no inverno e 26 e 19 g L-1 na primavera,
enquanto para os sedimentos colonizados por S. marítima variaram entre 28 e 19 g L-1
no inverno e 25, e 15 g L-1 na primavera. Nos sedimentos não colonizados variaram
entre 24 e 18 g L-1 no inverno e 26 e 18 g L-1 na primavera.
0
5
10
15
20
25
30
0 20 40 60 80 100
Pro
fundid
ade (
cm
)
[Cl-] (g L-1)
S. fruticosa
0
5
10
15
20
25
30
0 20 40 60 80 100
[Cl-] (g L-1)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 20 40 60 80 100
[Cl-] (g L-1)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
64 Ricardo M.P.Mendes
Figura 32- Perfis verticais dos teores de cloretos (g L-1
) nas águas intersticiais dos sedimentos recolhidos no Sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Pela análise ao teor de cloretos (mg L-1), no sapal de Castro Marim, verificamos que á
excessão do inverno para a S. fruticosa, os perfis são regulares ao longo
profundidade, sem variações de teores entre estações. Na situação da S. fruticosa,
verificamos que no inverno o perfil é irregular e os teores são superiores aos dos
outros perfis, apresentando um pico nos primeiros 5 cm de profundidade e outro pico
aos 22,5 cm de profundidade.
No sapal do Rosário, os teores de cloretos nos perfis dos sedimentos colonizados por
S. Fruticosa são regulares, apresentando maiores teores na primavera do que no
inverno. O mesmo se verifica para os sedimentos não colonizados, apesar de não
terem perfis tão regulares. Na situação para a S. marítima, verifica-se que não há
grandes diferenças entre inverno e a primavera. Em sedimentos não colonizados, as
concentrações de cloretos, variam em certas camadas.
Comparando os dois sapais, verificamos que os teores de cloretos não variam muito,
estando dentro da mesma gama de valores, à excessão das concentrações no sapal
de Castro Marim, onde no perfil da S. fruticosa. A variação sazonal é minima, contudo
há maiores teores de Cloretos na primavera.
0
5
10
15
20
25
30
10 15 20 25 30
Pro
fundid
ade (
cm
) [Cl-] (g L-1)
S. fruticosa
0
5
10
15
20
25
30
10 15 20 25 30
[Cl-] (g L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
10 15 20 25 30
[Cl-] (g L-1)
Não colonizados
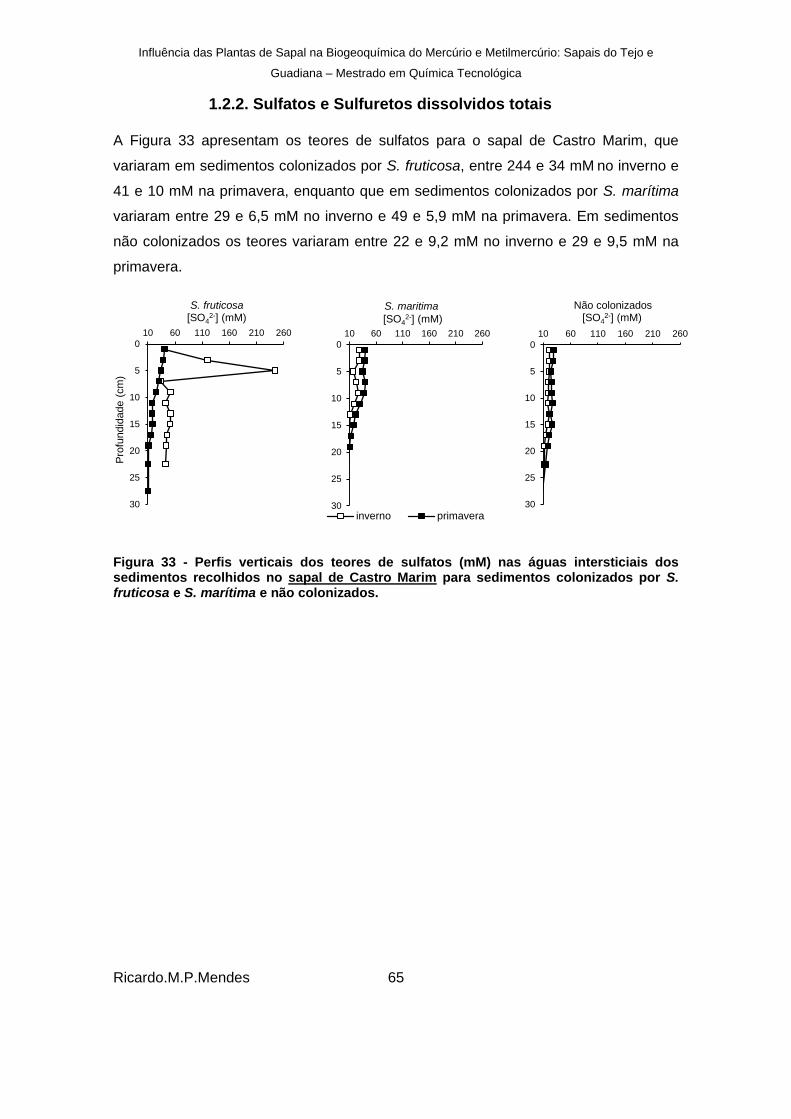
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 65
1.2.2. Sulfatos e Sulfuretos dissolvidos totais
A Figura 33 apresentam os teores de sulfatos para o sapal de Castro Marim, que
variaram em sedimentos colonizados por S. fruticosa, entre 244 e 34 mM no inverno e
41 e 10 mM na primavera, enquanto que em sedimentos colonizados por S. marítima
variaram entre 29 e 6,5 mM no inverno e 49 e 5,9 mM na primavera. Em sedimentos
não colonizados os teores variaram entre 22 e 9,2 mM no inverno e 29 e 9,5 mM na
primavera.
Figura 33 - Perfis verticais dos teores de sulfatos (mM) nas águas intersticiais dos sedimentos recolhidos no sapal de Castro Marim para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
0
5
10
15
20
25
30
10 60 110 160 210 260
Pro
fundid
ade (
cm
)
[SO42-] (mM)
S. fruticosa
0
5
10
15
20
25
30
10 60 110 160 210 260
[SO42-] (mM)
S. maritima
inverno primavera
0
5
10
15
20
25
30
10 60 110 160 210 260
[SO42-] (mM)
Não colonizados
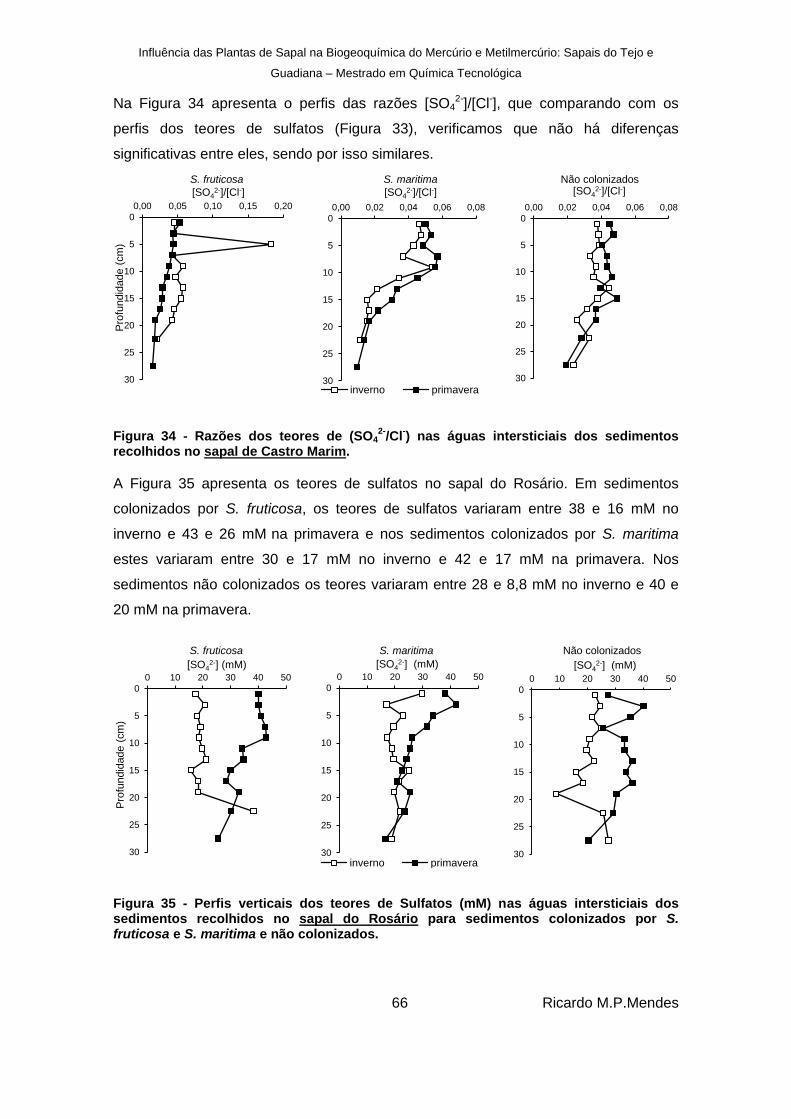
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
66 Ricardo M.P.Mendes
Na Figura 34 apresenta o perfis das razões [SO42-]/[Cl-], que comparando com os
perfis dos teores de sulfatos (Figura 33), verificamos que não há diferenças
significativas entre eles, sendo por isso similares.
Figura 34 - Razões dos teores de (SO42-
/Cl-) nas águas intersticiais dos sedimentos
recolhidos no sapal de Castro Marim.
A Figura 35 apresenta os teores de sulfatos no sapal do Rosário. Em sedimentos
colonizados por S. fruticosa, os teores de sulfatos variaram entre 38 e 16 mM no
inverno e 43 e 26 mM na primavera e nos sedimentos colonizados por S. maritima
estes variaram entre 30 e 17 mM no inverno e 42 e 17 mM na primavera. Nos
sedimentos não colonizados os teores variaram entre 28 e 8,8 mM no inverno e 40 e
20 mM na primavera.
Figura 35 - Perfis verticais dos teores de Sulfatos (mM) nas águas intersticiais dos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. maritima e não colonizados.
0
5
10
15
20
25
30
0,00 0,05 0,10 0,15 0,20
Pro
fundid
ade (
cm
)
[SO42-]/[Cl-]
S. fruticosa
0
5
10
15
20
25
30
0,00 0,02 0,04 0,06 0,08
[SO42-]/[Cl-]
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,00 0,02 0,04 0,06 0,08
[SO42-]/[Cl-]
Não colonizados
0
5
10
15
20
25
30
0 10 20 30 40 50
Pro
fundid
ade (
cm
)
[SO42-] (mM)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40 50
[SO42-] (mM)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40 50
[SO42-] (mM)
Não colonizados
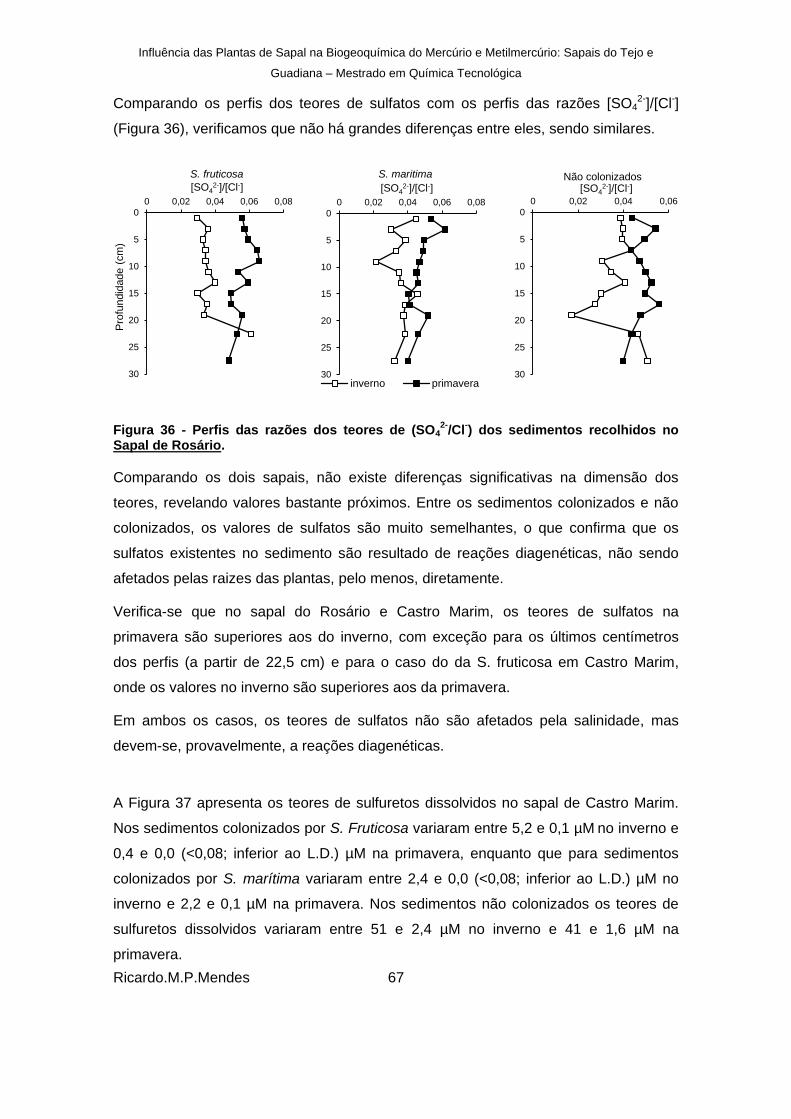
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 67
Comparando os perfis dos teores de sulfatos com os perfis das razões [SO42-]/[Cl-]
(Figura 36), verificamos que não há grandes diferenças entre eles, sendo similares.
Figura 36 - Perfis das razões dos teores de (SO42-
/Cl-) dos sedimentos recolhidos no
Sapal de Rosário.
Comparando os dois sapais, não existe diferenças significativas na dimensão dos
teores, revelando valores bastante próximos. Entre os sedimentos colonizados e não
colonizados, os valores de sulfatos são muito semelhantes, o que confirma que os
sulfatos existentes no sedimento são resultado de reações diagenéticas, não sendo
afetados pelas raizes das plantas, pelo menos, diretamente.
Verifica-se que no sapal do Rosário e Castro Marim, os teores de sulfatos na
primavera são superiores aos do inverno, com exceção para os últimos centímetros
dos perfis (a partir de 22,5 cm) e para o caso do da S. fruticosa em Castro Marim,
onde os valores no inverno são superiores aos da primavera.
Em ambos os casos, os teores de sulfatos não são afetados pela salinidade, mas
devem-se, provavelmente, a reações diagenéticas.
A Figura 37 apresenta os teores de sulfuretos dissolvidos no sapal de Castro Marim.
Nos sedimentos colonizados por S. Fruticosa variaram entre 5,2 e 0,1 µM no inverno e
0,4 e 0,0 (<0,08; inferior ao L.D.) µM na primavera, enquanto que para sedimentos
colonizados por S. marítima variaram entre 2,4 e 0,0 (<0,08; inferior ao L.D.) µM no
inverno e 2,2 e 0,1 µM na primavera. Nos sedimentos não colonizados os teores de
sulfuretos dissolvidos variaram entre 51 e 2,4 µM no inverno e 41 e 1,6 µM na
primavera.
0
5
10
15
20
25
30
0 0,02 0,04 0,06 0,08
Pro
fundid
ade (
cm
)
[SO42-]/[Cl-]
S. fruticosa
0
5
10
15
20
25
30
0 0,02 0,04 0,06 0,08
[SO42-]/[Cl-]
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 0,02 0,04 0,06
[SO42-]/[Cl-]
Não colonizados
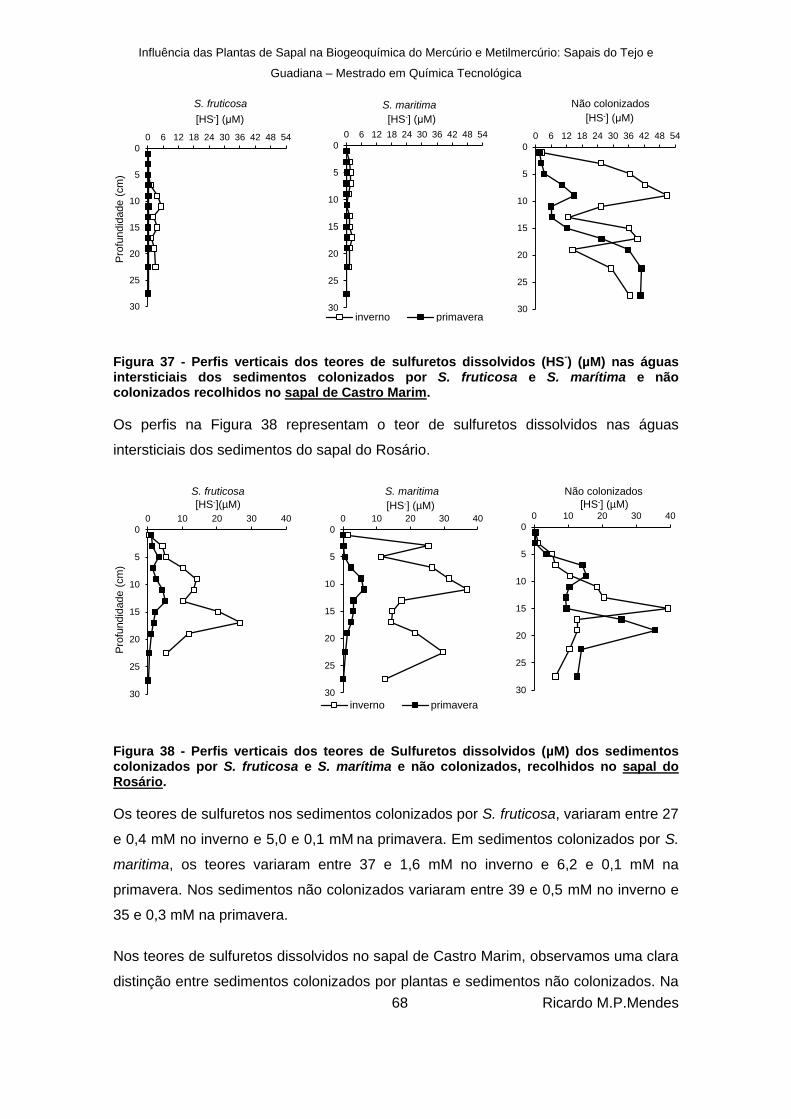
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
68 Ricardo M.P.Mendes
Figura 37 - Perfis verticais dos teores de sulfuretos dissolvidos (HS-) (µM) nas águas
intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim.
Os perfis na Figura 38 representam o teor de sulfuretos dissolvidos nas águas
intersticiais dos sedimentos do sapal do Rosário.
Figura 38 - Perfis verticais dos teores de Sulfuretos dissolvidos (µM) dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
Os teores de sulfuretos nos sedimentos colonizados por S. fruticosa, variaram entre 27
e 0,4 mM no inverno e 5,0 e 0,1 mM na primavera. Em sedimentos colonizados por S.
maritima, os teores variaram entre 37 e 1,6 mM no inverno e 6,2 e 0,1 mM na
primavera. Nos sedimentos não colonizados variaram entre 39 e 0,5 mM no inverno e
35 e 0,3 mM na primavera.
Nos teores de sulfuretos dissolvidos no sapal de Castro Marim, observamos uma clara
distinção entre sedimentos colonizados por plantas e sedimentos não colonizados. Na
0
5
10
15
20
25
30
0 6 12 18 24 30 36 42 48 54
Pro
fundid
ade (
cm
) [HS-] (μM)
S. fruticosa
0
5
10
15
20
25
30
0 6 12 18 24 30 36 42 48 54
[HS-] (μM)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 6 12 18 24 30 36 42 48 54
[HS-] (μM)
Não colonizados
0
5
10
15
20
25
30
0 10 20 30 40
Pro
fundid
ade (
cm
)
[HS-](µM)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40
[HS-] (µM)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40
[HS-] (µM)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 69
situação de sedimentos colonizados por plantas, as concentrações de sulfuretos são
muito baixas, com um ligeiro aumento no inverno (quando não há raízes).
Na Sapal do Rosário, Verificamos também que nos sedimentos colonizados, durante o
inverno há maiores teores de sulfuretos do que na primavera.
Em ambos os sapais verifica-se que nos sedimentos colonizados a influência das
raizes durante a primavera (comparando com os sedimentos não colonizados), e a
ausência de variação sazonal nos sedimentos nos não colonizados.
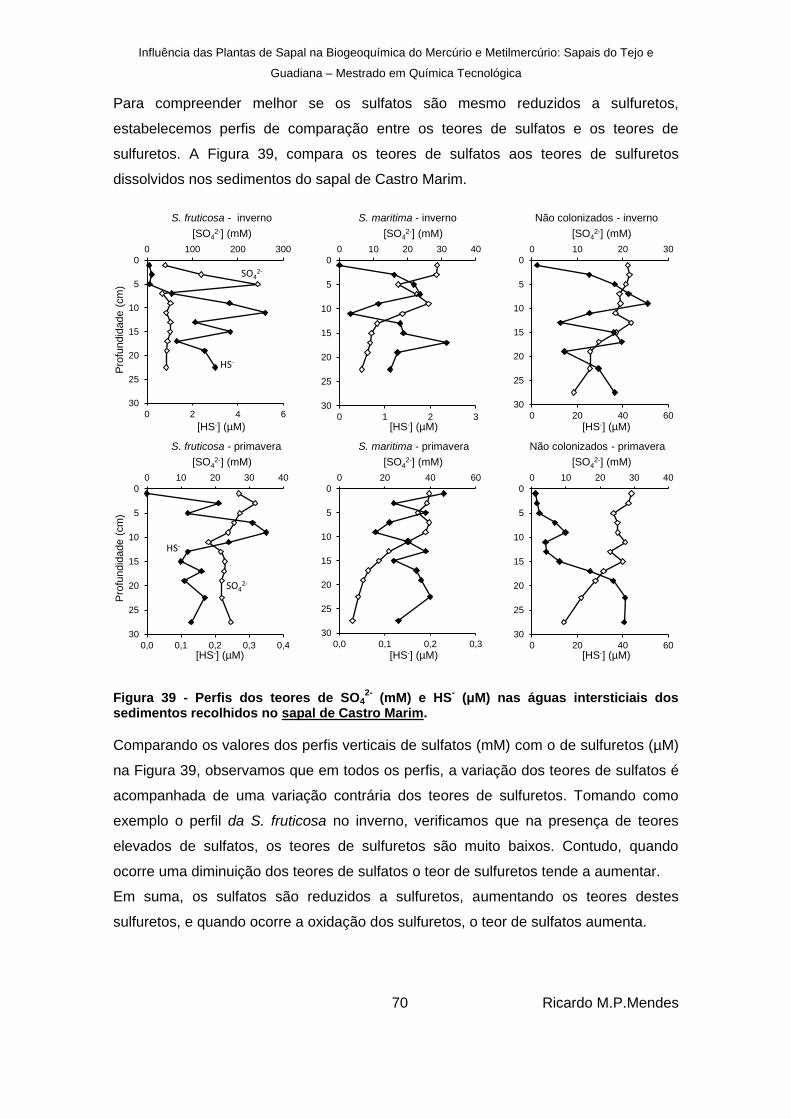
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
70 Ricardo M.P.Mendes
Para compreender melhor se os sulfatos são mesmo reduzidos a sulfuretos,
estabelecemos perfis de comparação entre os teores de sulfatos e os teores de
sulfuretos. A Figura 39, compara os teores de sulfatos aos teores de sulfuretos
dissolvidos nos sedimentos do sapal de Castro Marim.
Figura 39 - Perfis dos teores de SO42-
(mM) e HS- (μM) nas águas intersticiais dos
sedimentos recolhidos no sapal de Castro Marim.
Comparando os valores dos perfis verticais de sulfatos (mM) com o de sulfuretos (µM)
na Figura 39, observamos que em todos os perfis, a variação dos teores de sulfatos é
acompanhada de uma variação contrária dos teores de sulfuretos. Tomando como
exemplo o perfil da S. fruticosa no inverno, verificamos que na presença de teores
elevados de sulfatos, os teores de sulfuretos são muito baixos. Contudo, quando
ocorre uma diminuição dos teores de sulfatos o teor de sulfuretos tende a aumentar.
Em suma, os sulfatos são reduzidos a sulfuretos, aumentando os teores destes
sulfuretos, e quando ocorre a oxidação dos sulfuretos, o teor de sulfatos aumenta.
0 2 4 6
0
5
10
15
20
25
30
0 100 200 300
[HS-] (µM)
Pro
fundid
ade (
cm
)
[SO42-] (mM)
S. fruticosa - inverno
SO42-
HS-
0 1 2 3
0
5
10
15
20
25
30
0 10 20 30 40
[HS-] (µM)
[SO42-] (mM)
S. maritima - inverno
0 20 40 60
0
5
10
15
20
25
30
0 10 20 30
[HS-] (µM)
[SO42-] (mM)
Não colonizados - inverno
0,0 0,1 0,2 0,3 0,4
0
5
10
15
20
25
30
0 10 20 30 40
[HS-] (µM)
Pro
fundid
ade (
cm
)
[SO42-] (mM)
S. fruticosa - primavera
SO42-
HS-
0,0 0,1 0,2 0,3
0
5
10
15
20
25
30
0 20 40 60
[HS-] (µM)
[SO42-] (mM)
S. maritima - primavera
0 20 40 60
0
5
10
15
20
25
30
0 10 20 30 40
[HS-] (µM)
[SO42-] (mM)
Não colonizados - primavera
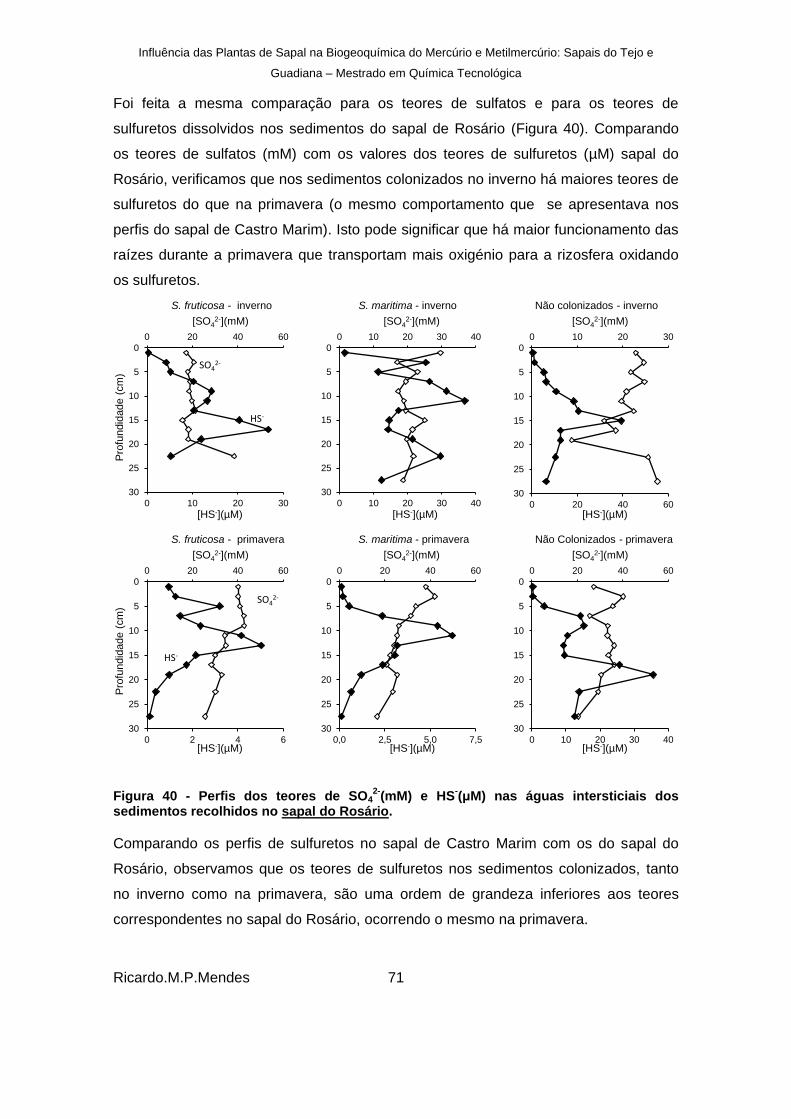
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 71
Foi feita a mesma comparação para os teores de sulfatos e para os teores de
sulfuretos dissolvidos nos sedimentos do sapal de Rosário (Figura 40). Comparando
os teores de sulfatos (mM) com os valores dos teores de sulfuretos (µM) sapal do
Rosário, verificamos que nos sedimentos colonizados no inverno há maiores teores de
sulfuretos do que na primavera (o mesmo comportamento que se apresentava nos
perfis do sapal de Castro Marim). Isto pode significar que há maior funcionamento das
raízes durante a primavera que transportam mais oxigénio para a rizosfera oxidando
os sulfuretos.
Figura 40 - Perfis dos teores de SO42-
(mM) e HS-(μM) nas águas intersticiais dos
sedimentos recolhidos no sapal do Rosário.
Comparando os perfis de sulfuretos no sapal de Castro Marim com os do sapal do
Rosário, observamos que os teores de sulfuretos nos sedimentos colonizados, tanto
no inverno como na primavera, são uma ordem de grandeza inferiores aos teores
correspondentes no sapal do Rosário, ocorrendo o mesmo na primavera.
0 10 20 30
0
5
10
15
20
25
30
0 20 40 60
[HS-](µM)
Pro
fundid
ade (
cm
)
[SO42-](mM)
S. fruticosa - inverno
SO42-
HS-
0 10 20 30 40
0
5
10
15
20
25
30
0 10 20 30 40
[HS-](µM)
[SO42-](mM)
S. maritima - inverno
0 20 40 60
0
5
10
15
20
25
30
0 10 20 30
[HS-](µM)
[SO42-](mM)
Não colonizados - inverno
0 2 4 6
0
5
10
15
20
25
30
0 20 40 60
[HS-](µM)
Pro
fundid
ade (
cm
)
[SO42-](mM)
S. fruticosa - primavera
SO42-
HS-
0,0 2,5 5,0 7,5
0
5
10
15
20
25
30
0 20 40 60
[HS-](µM)
[SO42-](mM)
S. maritima - primavera
0 10 20 30 40
0
5
10
15
20
25
30
0 20 40 60
[HS-](µM)
[SO42-](mM)
Não Colonizados - primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
72 Ricardo M.P.Mendes
Este facto sugere, que os sedimentos do sapal do Rosário são mais permeáveis, ou
então, as raízes das plantas do sapal de Castro Marim são menos ativas que as raízes
das plantas no sapal do sapal do Rosário.
Também se verifica que nos dois sapais, há maiores teores de sulfuretos no inverno
do que na primavera. Estas diferenças podem estar relacionada com uma maior
atividade das raízes durante a primavera que bombeiam mais oxigénio para a rizosfera
oxidando os sulfuretos. Este comportamento é observado em todos os perfis tanto os
do sapal de Castro Marim e do Rosário.
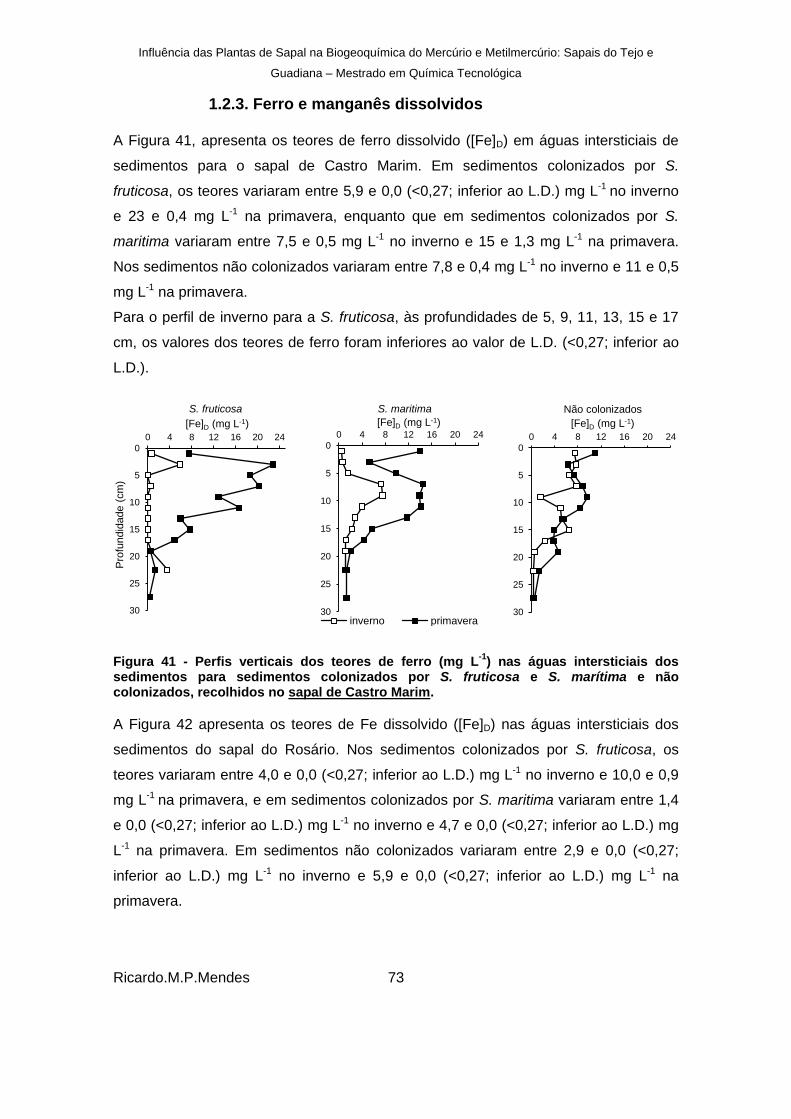
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 73
1.2.3. Ferro e manganês dissolvidos
A Figura 41, apresenta os teores de ferro dissolvido ([Fe]D) em águas intersticiais de
sedimentos para o sapal de Castro Marim. Em sedimentos colonizados por S.
fruticosa, os teores variaram entre 5,9 e 0,0 (<0,27; inferior ao L.D.) mg L-1 no inverno
e 23 e 0,4 mg L-1 na primavera, enquanto que em sedimentos colonizados por S.
maritima variaram entre 7,5 e 0,5 mg L-1 no inverno e 15 e 1,3 mg L-1 na primavera.
Nos sedimentos não colonizados variaram entre 7,8 e 0,4 mg L-1 no inverno e 11 e 0,5
mg L-1 na primavera.
Para o perfil de inverno para a S. fruticosa, às profundidades de 5, 9, 11, 13, 15 e 17
cm, os valores dos teores de ferro foram inferiores ao valor de L.D. (<0,27; inferior ao
L.D.).
Figura 41 - Perfis verticais dos teores de ferro (mg L-1
) nas águas intersticiais dos sedimentos para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim.
A Figura 42 apresenta os teores de Fe dissolvido ([Fe]D) nas águas intersticiais dos
sedimentos do sapal do Rosário. Nos sedimentos colonizados por S. fruticosa, os
teores variaram entre 4,0 e 0,0 (<0,27; inferior ao L.D.) mg L-1 no inverno e 10,0 e 0,9
mg L-1 na primavera, e em sedimentos colonizados por S. maritima variaram entre 1,4
e 0,0 (<0,27; inferior ao L.D.) mg L-1 no inverno e 4,7 e 0,0 (<0,27; inferior ao L.D.) mg
L-1 na primavera. Em sedimentos não colonizados variaram entre 2,9 e 0,0 (<0,27;
inferior ao L.D.) mg L-1 no inverno e 5,9 e 0,0 (<0,27; inferior ao L.D.) mg L-1 na
primavera.
0
5
10
15
20
25
30
0 4 8 12 16 20 24
Pro
fundid
ade (
cm
)
[Fe]D (mg L-1)
S. fruticosa
0
5
10
15
20
25
30
0 4 8 12 16 20 24
[Fe]D (mg L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 4 8 12 16 20 24
[Fe]D (mg L-1)
Não colonizados
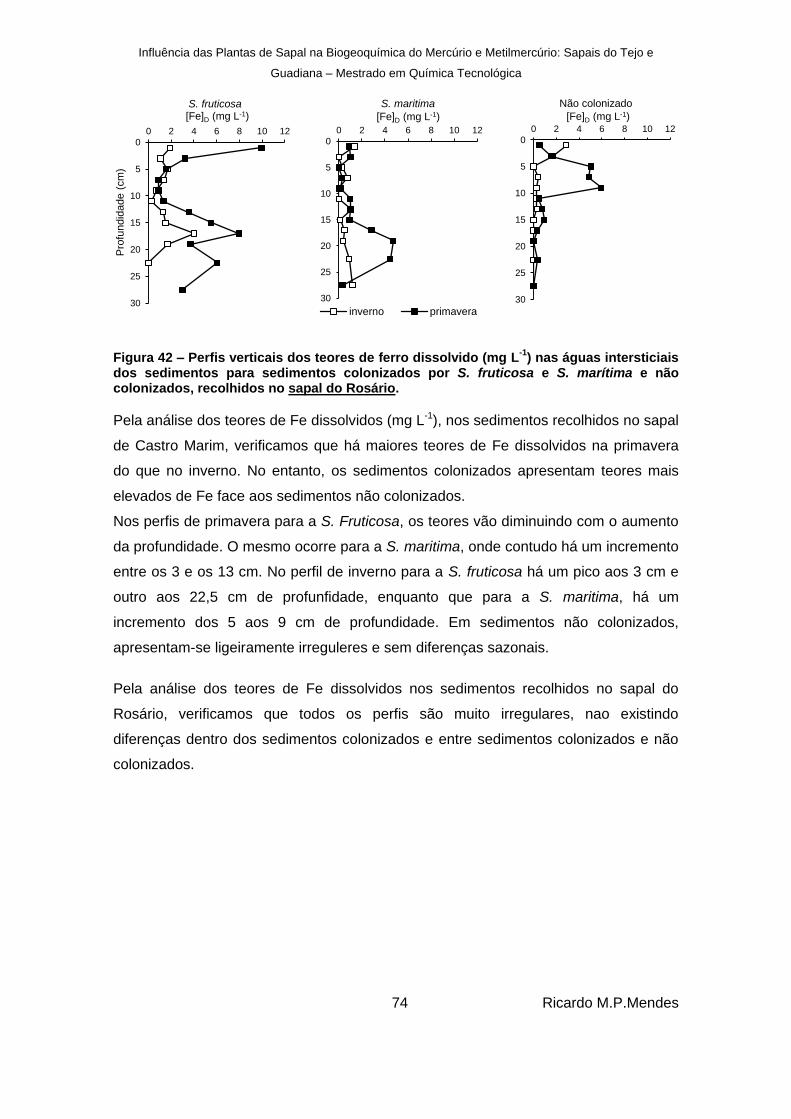
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
74 Ricardo M.P.Mendes
Figura 42 – Perfis verticais dos teores de ferro dissolvido (mg L-1
) nas águas intersticiais dos sedimentos para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
Pela análise dos teores de Fe dissolvidos (mg L-1), nos sedimentos recolhidos no sapal
de Castro Marim, verificamos que há maiores teores de Fe dissolvidos na primavera
do que no inverno. No entanto, os sedimentos colonizados apresentam teores mais
elevados de Fe face aos sedimentos não colonizados.
Nos perfis de primavera para a S. Fruticosa, os teores vão diminuindo com o aumento
da profundidade. O mesmo ocorre para a S. maritima, onde contudo há um incremento
entre os 3 e os 13 cm. No perfil de inverno para a S. fruticosa há um pico aos 3 cm e
outro aos 22,5 cm de profunfidade, enquanto que para a S. maritima, há um
incremento dos 5 aos 9 cm de profundidade. Em sedimentos não colonizados,
apresentam-se ligeiramente irreguleres e sem diferenças sazonais.
Pela análise dos teores de Fe dissolvidos nos sedimentos recolhidos no sapal do
Rosário, verificamos que todos os perfis são muito irregulares, nao existindo
diferenças dentro dos sedimentos colonizados e entre sedimentos colonizados e não
colonizados.
0
5
10
15
20
25
30
0 2 4 6 8 10 12
Pro
fundid
ade (
cm
) [Fe]D (mg L-1) S. fruticosa
0
5
10
15
20
25
30
0 2 4 6 8 10 12
[Fe]D (mg L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 2 4 6 8 10 12
[Fe]D (mg L-1)
Não colonizado

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 75
Comparando sapais, concluímos que em sedimentos colonizados, no sapal de Castro
Marim os teores de FeD são superiores aos FeD no sapal do Rosário. Esta observação
é acompanhada com uma diminuição dos teores de Fe na fração sólida do sedimento
(Figura 19, ). Ora, de maneira geral, baixos valores de pH, induzem uma maior
mobilidade dos metais (Jackson, et al., 1993), podendo resultar em reações de
adsorção/desasorção, envolvendo o Fe e o Mn (Schindler & Sposito, 1991):
Fe-OM+ + H+ -Fe-OH + M2+ , onde M é o metal em questão.
Contudo, no sapal de Castro Marim, os valores de pH em sedimentos colonizados são
tendêncialmente básicos na primavera (Figura 15).
Esta situação pode dever-se a reações de redução do ferro, mediadas por bactérias
redutoras de ferro, traduzidas nas seguintes reacções (Haese, 2000), onde são
consumidos iões H+, com o consequente aumento do pH, e de ferro na forma solúvel:
4Fe(OH)3 + CH2O + 7H+ 4Fe2+ + HCO3- + 10H2O
4FeOOH + CH2O + 7H+ 4Fe2+ + HCO3- + 6H2O
2Fe3O4 + CH2O + 11H+ 6Fe2+ + HCO3- + 6H2O
Em suma, o aumento das concentrações de FeD no sapal de Castro Marim, através da
redução de óxi-hidróxidos de Ferro, pode levar a um aumento do Hg disponivel para
metilação, que devido á sua origem geogénica não se encontra tão disponivel.
Desta maneira, e tendo em conta os teores de sulfatos e de sulfuretos, verificamos
que o sedimento em Castro Marim apresenta um caráter subóxico, com todas as
caracteristicas que lhes são inerentes (Figura 3).
No sapal do Rosário, não há grandes diferenças nos teores de Fe na fração sólida
entre inverno e primavera, contudo para a fração dissolvida isso varia com
profundidade de certas camadas dos sedimentos. Tendo também em conta os sulfatos
e os sulfuretos dissolvidos, este sedimento pode ser qualificado como subóxico.
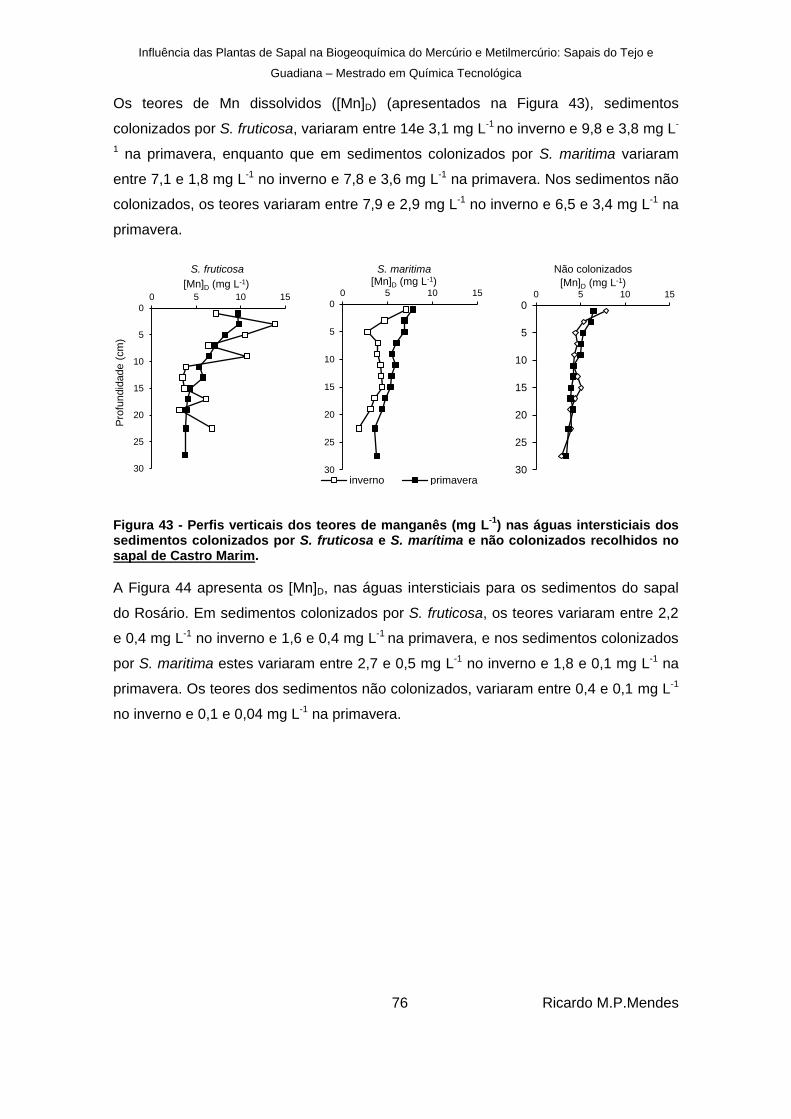
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
76 Ricardo M.P.Mendes
Os teores de Mn dissolvidos ([Mn]D) (apresentados na Figura 43), sedimentos
colonizados por S. fruticosa, variaram entre 14e 3,1 mg L-1 no inverno e 9,8 e 3,8 mg L-
1 na primavera, enquanto que em sedimentos colonizados por S. maritima variaram
entre 7,1 e 1,8 mg L-1 no inverno e 7,8 e 3,6 mg L-1 na primavera. Nos sedimentos não
colonizados, os teores variaram entre 7,9 e 2,9 mg L-1 no inverno e 6,5 e 3,4 mg L-1 na
primavera.
Figura 43 - Perfis verticais dos teores de manganês (mg L-1
) nas águas intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim.
A Figura 44 apresenta os [Mn]D, nas águas intersticiais para os sedimentos do sapal
do Rosário. Em sedimentos colonizados por S. fruticosa, os teores variaram entre 2,2
e 0,4 mg L-1 no inverno e 1,6 e 0,4 mg L-1 na primavera, e nos sedimentos colonizados
por S. maritima estes variaram entre 2,7 e 0,5 mg L-1 no inverno e 1,8 e 0,1 mg L-1 na
primavera. Os teores dos sedimentos não colonizados, variaram entre 0,4 e 0,1 mg L-1
no inverno e 0,1 e 0,04 mg L-1 na primavera.
0
5
10
15
20
25
30
0 5 10 15
Pro
fundid
ade (
cm
)
[Mn]D (mg L-1)
S. fruticosa
0
5
10
15
20
25
30
0 5 10 15
[Mn]D (mg L-1) S. maritima
inverno primavera
0
5
10
15
20
25
30
0 5 10 15
[Mn]D (mg L-1)
Não colonizados
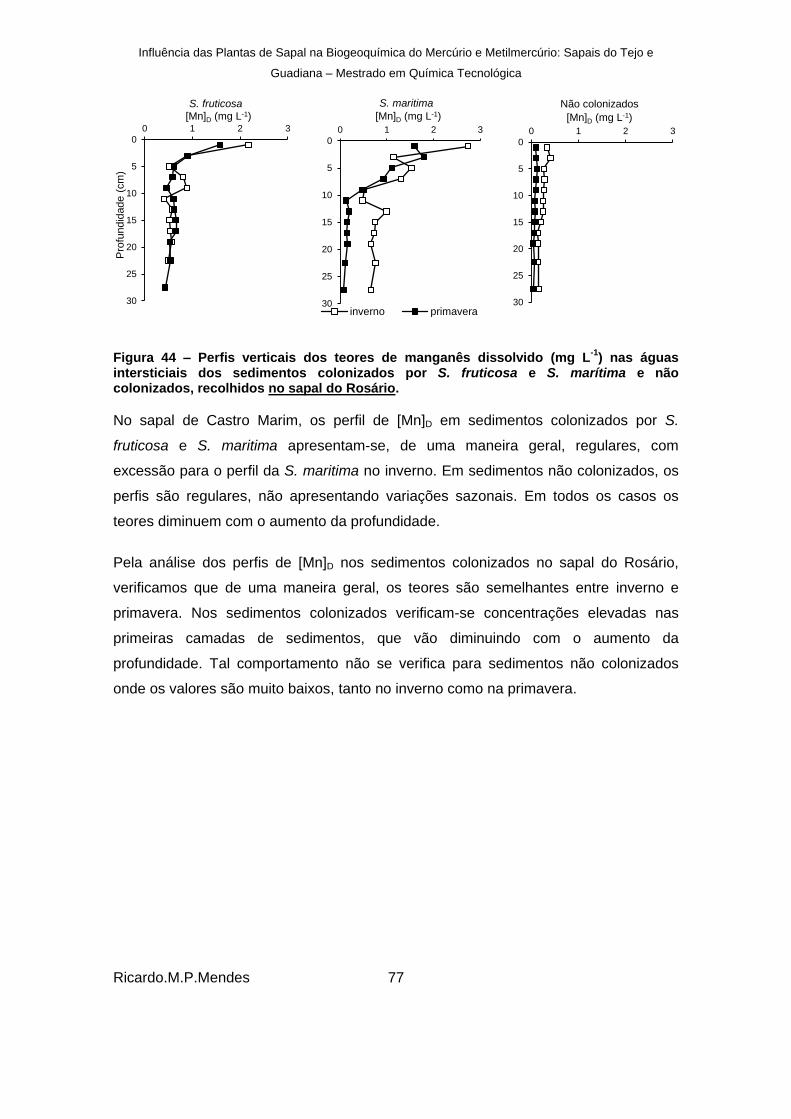
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 77
Figura 44 – Perfis verticais dos teores de manganês dissolvido (mg L-1
) nas águas intersticiais dos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
No sapal de Castro Marim, os perfil de [Mn]D em sedimentos colonizados por S.
fruticosa e S. maritima apresentam-se, de uma maneira geral, regulares, com
excessão para o perfil da S. maritima no inverno. Em sedimentos não colonizados, os
perfis são regulares, não apresentando variações sazonais. Em todos os casos os
teores diminuem com o aumento da profundidade.
Pela análise dos perfis de [Mn]D nos sedimentos colonizados no sapal do Rosário,
verificamos que de uma maneira geral, os teores são semelhantes entre inverno e
primavera. Nos sedimentos colonizados verificam-se concentrações elevadas nas
primeiras camadas de sedimentos, que vão diminuindo com o aumento da
profundidade. Tal comportamento não se verifica para sedimentos não colonizados
onde os valores são muito baixos, tanto no inverno como na primavera.
0
5
10
15
20
25
30
0 1 2 3P
rofu
ndid
ade (
cm
) [Mn]D (mg L-1)
S. fruticosa
0
5
10
15
20
25
30
0 1 2 3
[Mn]D (mg L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 1 2 3
[Mn]D (mg L-1)
Não colonizados
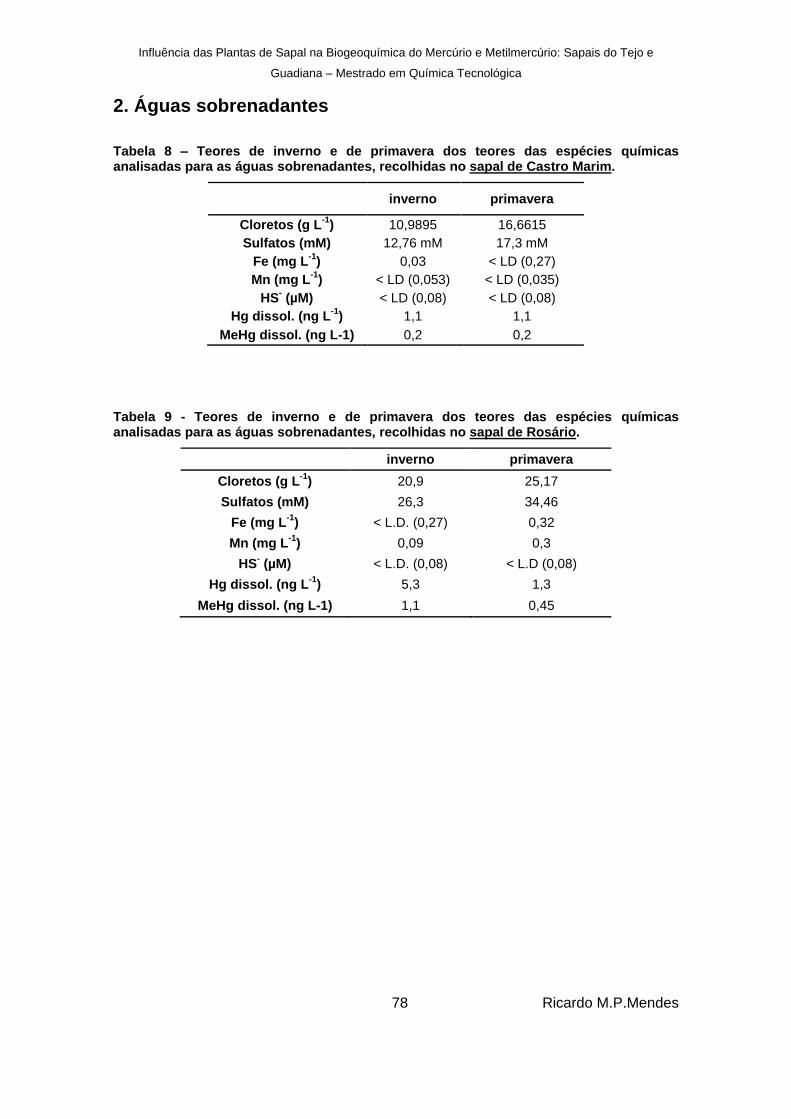
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
78 Ricardo M.P.Mendes
2. Águas sobrenadantes
Tabela 8 – Teores de inverno e de primavera dos teores das espécies químicas analisadas para as águas sobrenadantes, recolhidas no sapal de Castro Marim.
inverno primavera
Cloretos (g L-1
) 10,9895 16,6615
Sulfatos (mM) 12,76 mM 17,3 mM
Fe (mg L-1
) 0,03 < LD (0,27)
Mn (mg L-1
) < LD (0,053) < LD (0,035)
HS- (µM) < LD (0,08) < LD (0,08)
Hg dissol. (ng L-1
) 1,1 1,1
MeHg dissol. (ng L-1) 0,2 0,2
Tabela 9 - Teores de inverno e de primavera dos teores das espécies químicas analisadas para as águas sobrenadantes, recolhidas no sapal de Rosário.
inverno primavera
Cloretos (g L-1
) 20,9 25,17
Sulfatos (mM) 26,3 34,46
Fe (mg L-1
) < L.D. (0,27) 0,32
Mn (mg L-1
) 0,09 0,3
HS- (µM) < L.D. (0,08) < L.D (0,08)
Hg dissol. (ng L-1
) 5,3 1,3
MeHg dissol. (ng L-1) 1,1 0,45
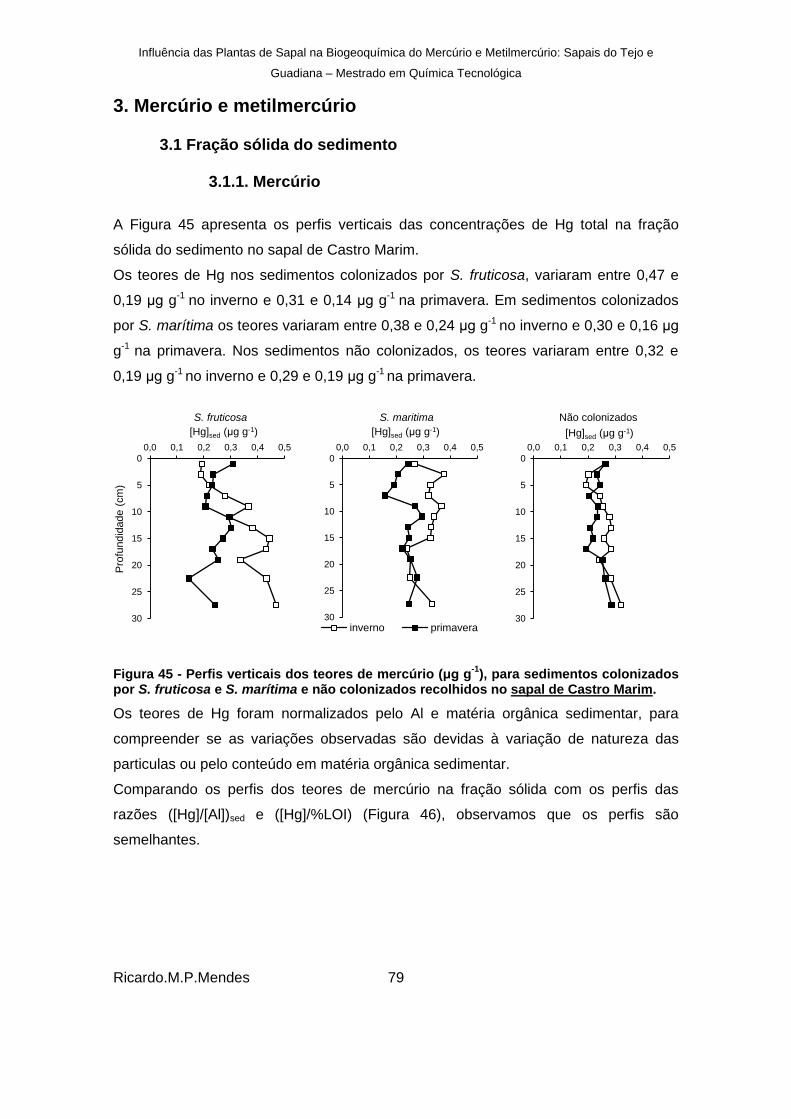
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 79
3. Mercúrio e metilmercúrio
3.1 Fração sólida do sedimento
3.1.1. Mercúrio
A Figura 45 apresenta os perfis verticais das concentrações de Hg total na fração
sólida do sedimento no sapal de Castro Marim.
Os teores de Hg nos sedimentos colonizados por S. fruticosa, variaram entre 0,47 e
0,19 μg g-1 no inverno e 0,31 e 0,14 μg g-1 na primavera. Em sedimentos colonizados
por S. marítima os teores variaram entre 0,38 e 0,24 μg g-1 no inverno e 0,30 e 0,16 μg
g-1 na primavera. Nos sedimentos não colonizados, os teores variaram entre 0,32 e
0,19 μg g-1 no inverno e 0,29 e 0,19 μg g-1 na primavera.
Figura 45 - Perfis verticais dos teores de mercúrio (μg g-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim.
Os teores de Hg foram normalizados pelo Al e matéria orgânica sedimentar, para
compreender se as variações observadas são devidas à variação de natureza das
particulas ou pelo conteúdo em matéria orgânica sedimentar.
Comparando os perfis dos teores de mercúrio na fração sólida com os perfis das
razões ([Hg]/[Al])sed e ([Hg]/%LOI) (Figura 46), observamos que os perfis são
semelhantes.
0
5
10
15
20
25
30
0,0 0,1 0,2 0,3 0,4 0,5
Pro
fundid
ade (
cm
)
[Hg]sed (μg g-1)
S. fruticosa
0
5
10
15
20
25
30
0,0 0,1 0,2 0,3 0,4 0,5
[Hg]sed (μg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,0 0,1 0,2 0,3 0,4 0,5
[Hg]sed (μg g-1)
Não colonizados
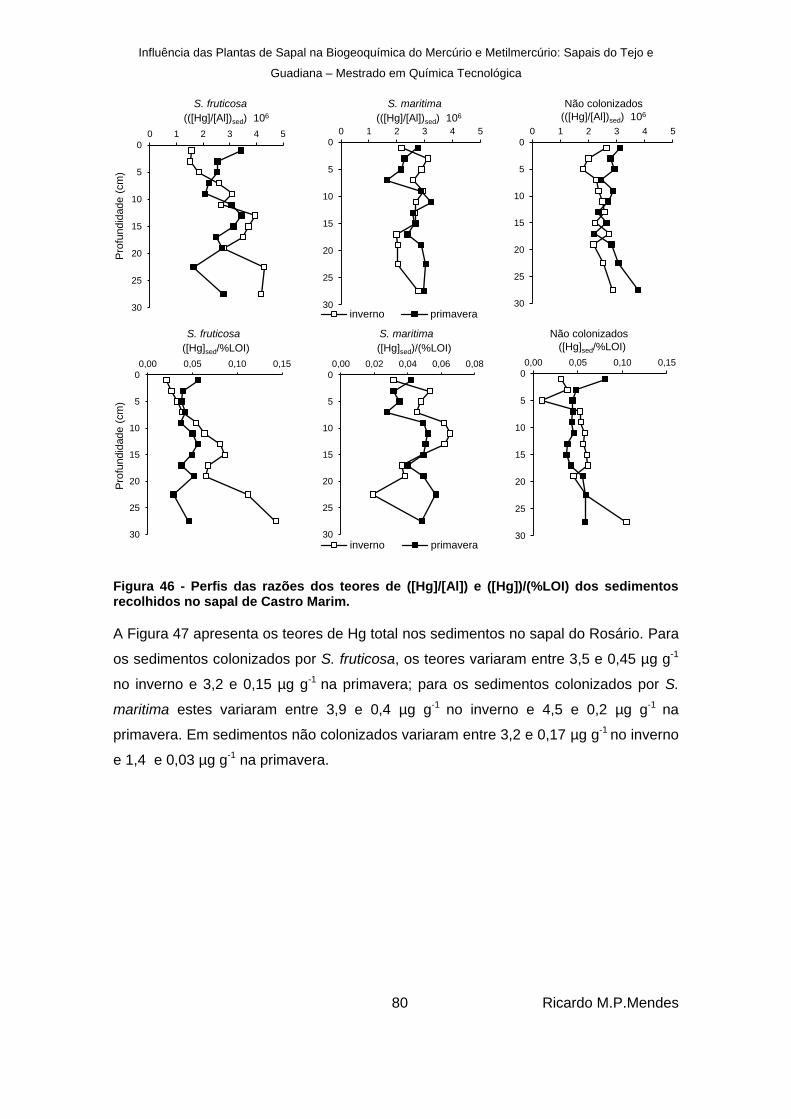
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
80 Ricardo M.P.Mendes
Figura 46 - Perfis das razões dos teores de ([Hg]/[Al]) e ([Hg])/(%LOI) dos sedimentos recolhidos no sapal de Castro Marim.
A Figura 47 apresenta os teores de Hg total nos sedimentos no sapal do Rosário. Para
os sedimentos colonizados por S. fruticosa, os teores variaram entre 3,5 e 0,45 µg g-1
no inverno e 3,2 e 0,15 µg g-1 na primavera; para os sedimentos colonizados por S.
maritima estes variaram entre 3,9 e 0,4 µg g-1 no inverno e 4,5 e 0,2 µg g-1 na
primavera. Em sedimentos não colonizados variaram entre 3,2 e 0,17 µg g-1 no inverno
e 1,4 e 0,03 µg g-1 na primavera.
0
5
10
15
20
25
30
0 1 2 3 4 5
Pro
fundid
ade (
cm
) (([Hg]/[Al])sed)
106
S. fruticosa
0
5
10
15
20
25
30
0 1 2 3 4 5
(([Hg]/[Al])sed)
106
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 1 2 3 4 5
(([Hg]/[Al])sed)
106
Não colonizados
0
5
10
15
20
25
30
0,00 0,05 0,10 0,15
Pro
fundid
ade (
cm
)
([Hg]sed/%LOI)
S. fruticosa
0
5
10
15
20
25
30
0,00 0,02 0,04 0,06 0,08
([Hg]sed)/(%LOI)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,00 0,05 0,10 0,15
([Hg]sed/%LOI)
Não colonizados
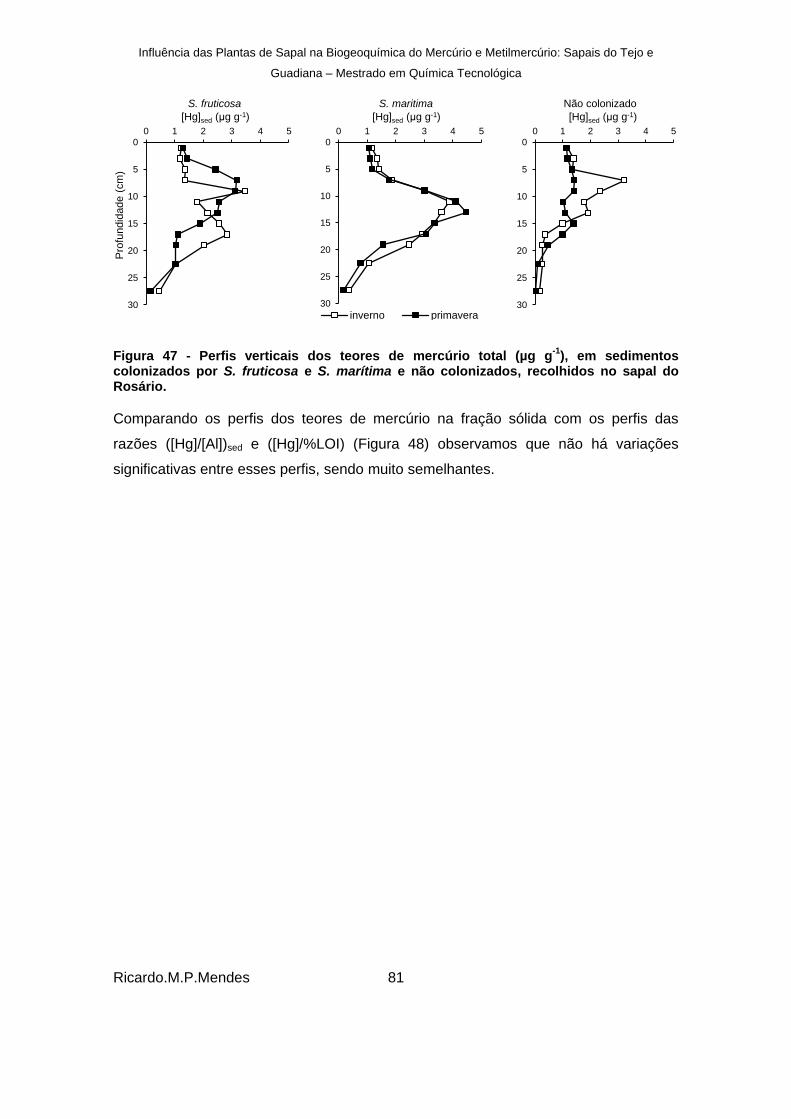
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 81
Figura 47 - Perfis verticais dos teores de mercúrio total (µg g-1
), em sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
Comparando os perfis dos teores de mercúrio na fração sólida com os perfis das
razões ([Hg]/[Al])sed e ([Hg]/%LOI) (Figura 48) observamos que não há variações
significativas entre esses perfis, sendo muito semelhantes.
0
5
10
15
20
25
30
0 1 2 3 4 5
Pro
fundid
ade (
cm
) [Hg]sed (μg g-1)
S. fruticosa
0
5
10
15
20
25
30
0 1 2 3 4 5
[Hg]sed (μg g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 1 2 3 4 5
[Hg]sed (μg g-1)
Não colonizado
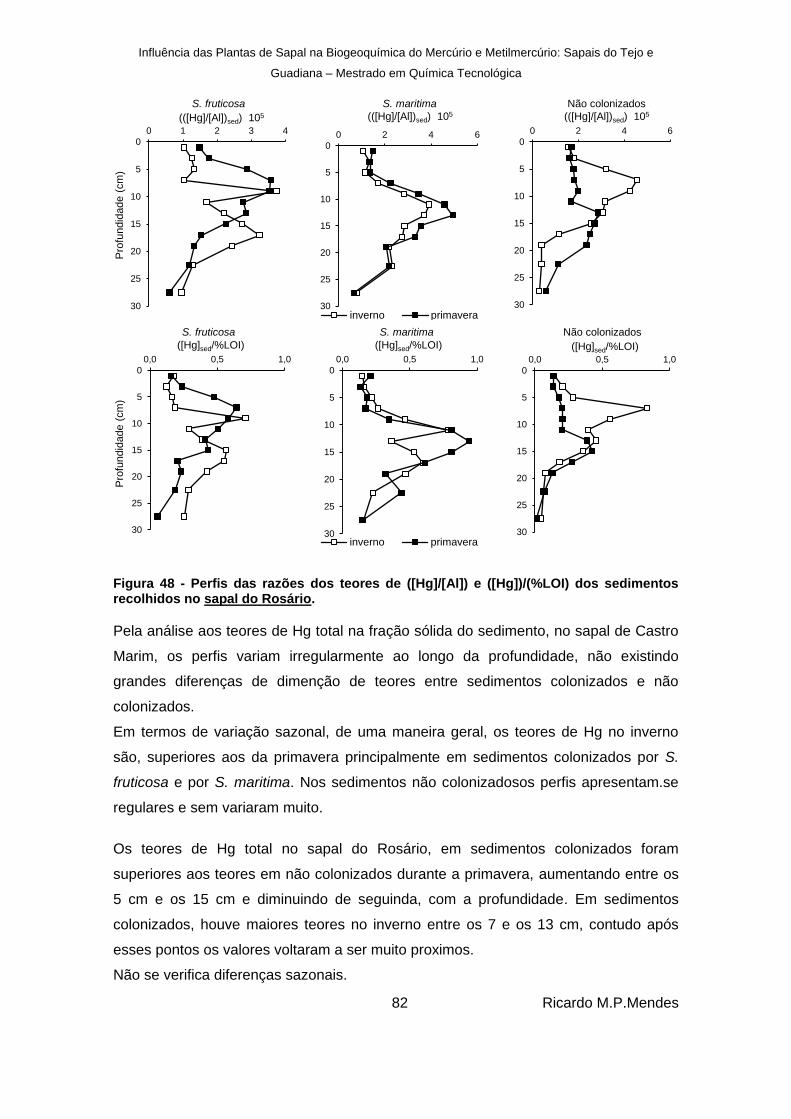
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
82 Ricardo M.P.Mendes
Figura 48 - Perfis das razões dos teores de ([Hg]/[Al]) e ([Hg])/(%LOI) dos sedimentos recolhidos no sapal do Rosário.
Pela análise aos teores de Hg total na fração sólida do sedimento, no sapal de Castro
Marim, os perfis variam irregularmente ao longo da profundidade, não existindo
grandes diferenças de dimenção de teores entre sedimentos colonizados e não
colonizados.
Em termos de variação sazonal, de uma maneira geral, os teores de Hg no inverno
são, superiores aos da primavera principalmente em sedimentos colonizados por S.
fruticosa e por S. maritima. Nos sedimentos não colonizadosos perfis apresentam.se
regulares e sem variaram muito.
Os teores de Hg total no sapal do Rosário, em sedimentos colonizados foram
superiores aos teores em não colonizados durante a primavera, aumentando entre os
5 cm e os 15 cm e diminuindo de seguinda, com a profundidade. Em sedimentos
colonizados, houve maiores teores no inverno entre os 7 e os 13 cm, contudo após
esses pontos os valores voltaram a ser muito proximos.
Não se verifica diferenças sazonais.
0
5
10
15
20
25
30
0 1 2 3 4
Pro
fundid
ade (
cm
) (([Hg]/[Al])sed)
105
S. fruticosa
0
5
10
15
20
25
30
0 2 4 6
(([Hg]/[Al])sed)
105
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 2 4 6
(([Hg]/[Al])sed)
105
Não colonizados
0
5
10
15
20
25
30
0,0 0,5 1,0
Pro
fundid
ade (
cm
)
([Hg]sed/%LOI)
S. fruticosa
0
5
10
15
20
25
30
0,0 0,5 1,0
([Hg]sed/%LOI)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,0 0,5 1,0
([Hg]sed/%LOI)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 83
Por fim, comparando os teores de Hg total entre o sapal de Castro Marim e o sapal do
Rosário, observamos que os teores de Hg no sapal do Rosário são uma ordem de
grandeza superiores aos do sapal de Castro Marim. Esta diferença é justificada com a
contaminação antropogénica existente no sapal do Rosário.
Foi observado também observado que em nenhum dos dois sapais houve diferenças
sazonais, pois os valores dos dois sapais não apresentaram grandes variações.
O facto de a Comparação entre os perfis dos teores de mercúrio na fração sólida com
os perfis das razões ([Hg]/[Al])sed e ([Hg]/%LOI), ter mostrado são semelhantes em
ambos os sapais, permite-nos concluir que a distribuição de Hg total resulta da
deposição natural, e/ou de reações diagenéticas posteriores à sua deposição

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
84 Ricardo M.P.Mendes
3.1.2. Metilmercúrio
No sapal de Castro Marim, os perfis verticais dos teores de MeHg na fração sólida
(apresentados na Figura 49) nos sedimentos colonizados por S. fruticosa, variaram
entre 7,3 e 0,0 (inferior ao L.D.) ng g-1 no inverno e 16 e 0,11 ng g-1 na primavera,
enquanto que para sedimentos colonizados por S. marítima estes variaram entre 22 e
2,1 ng g-1 no inverno e 16 e 1,2 ng g-1 na primavera. Nos sedimentos não colonizados,
os teores de MeHg variaram entre 0,87 e 0,0 (inferior ao L.D.) ng g-1 no inverno e 1,3 e
0,0 (inferior ao L.D.) ng g-1 na primavera.
Os perfis de teores de MeHg da S. fruticosa são irregulares, e dos 19 aos 27,5 cm de
profundidade os valores foram inferiores ao L.D.. Os perfis dos teores de MeHg da S.
maritima são irregulares e os sedimentos não colonizados os valores dos 11 aos 27,5
cm são inferiores ao L.D..
Figura 49 - Perfis verticais dos teores de metilmercúrio (ng g-1
), nos sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim.
A Figura 50 apresenta os teores de metilmercúrio na fração sólida dos sedimentos do
sapal do Rosário. Nos sedimentos colonizados por S. fruticosa, os teores variaram
entre 57e 0,0 (inferior ao L.D.) ng g-1 no inverno e 202 e 1,3 ng g-1 na primavera. Em
sedimentos colonizados por S. marítima estes os teores variaram entre 430 e 12 ng g-1
no inverno e 212 e 1,3 ng g-1 na primavera. Nos sedimentos não colonizados os teores
de MeHg variaram entre 26 e 0,0 (inferior L.D.) ng g-1 no inverno e 3,5 e 0,0 (inferior ao
L.D.) ng g-1 na primavera.
0
5
10
15
20
25
30
0 5 10 15 20 25
Pro
fundid
ade (
cm
)
[MeHg]sed (ng g-1)
S. fruticosa
0
5
10
15
20
25
30
0 5 10 15 20 25
[MeHg]sed (ng g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 5 10 15 20 25
[MeHg]sed (ng g-1)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 85
Figura 50 - Perfis verticais dos teores de metilmercúrio total (ng g-1
), nos sedimentos recolhidos no sapal do Rosário para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados.
Pela análise aos teores de MeHg no sapal de Castro Marim, verificamos que nos
sedimentos colonizados, os teores são mais elevados do que os teores em
sedimentos não colonizados. Em sedimentos não colonizados, para além de não
haver teores muito elevados, não foi detetado MeHg a partir dos 10 cm.
Nos sedimentos colonizados, os teores de MeHg têm um incremento até aos 11-13 cm
(depende do perfil), e diminuem gradualmente com o aumento da profundidade. Os
sedimentos colonizados por S. marítima foi onde foram detetados maiores teores de
MeHg.
Pela análise aos teores de metilmercúrio nos sedimentos do sapal do Rosário,
verificamos que os teores em sedimentos colonizados por S. fruticosa são mais
elevados na primavera do que no inverno.
Em sedimentos colonizados por S. maritima, os teores de inverno são mais elevados
do que na primavera. Há portanto uma diferença entre as espécies de plantas na
dimensão do teor.
Verifica-se também que em sedimentos não colonizados os teores de MeHg, foram
muito menores comparativamente aos teores nos sedimentos colonizados. Confirma-
se assim uma diferença na produção de metilmercúrio entre sedimentos colonizados e
sedimentos não colonizados.
Comparando os dois sapais verificou-se diferenças entre os teores dos sedimentos
colonizados com o dos não colonizados, o que demonstra o efeito das raízes no
favorecimento da produção de metilmercúrio. As duas espécies de plantas,
0
5
10
15
20
25
30
0 100 200 300 400 500
Pro
fundid
ade (
cm
) [MeHg]sed (ng g-1)
S. fruticosa
0
5
10
15
20
25
30
0 100 200 300 400 500
[MeHg]sed (ng g-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 100 200 300 400 500
[MeHg]sed (ng g-1)
Não colonizados
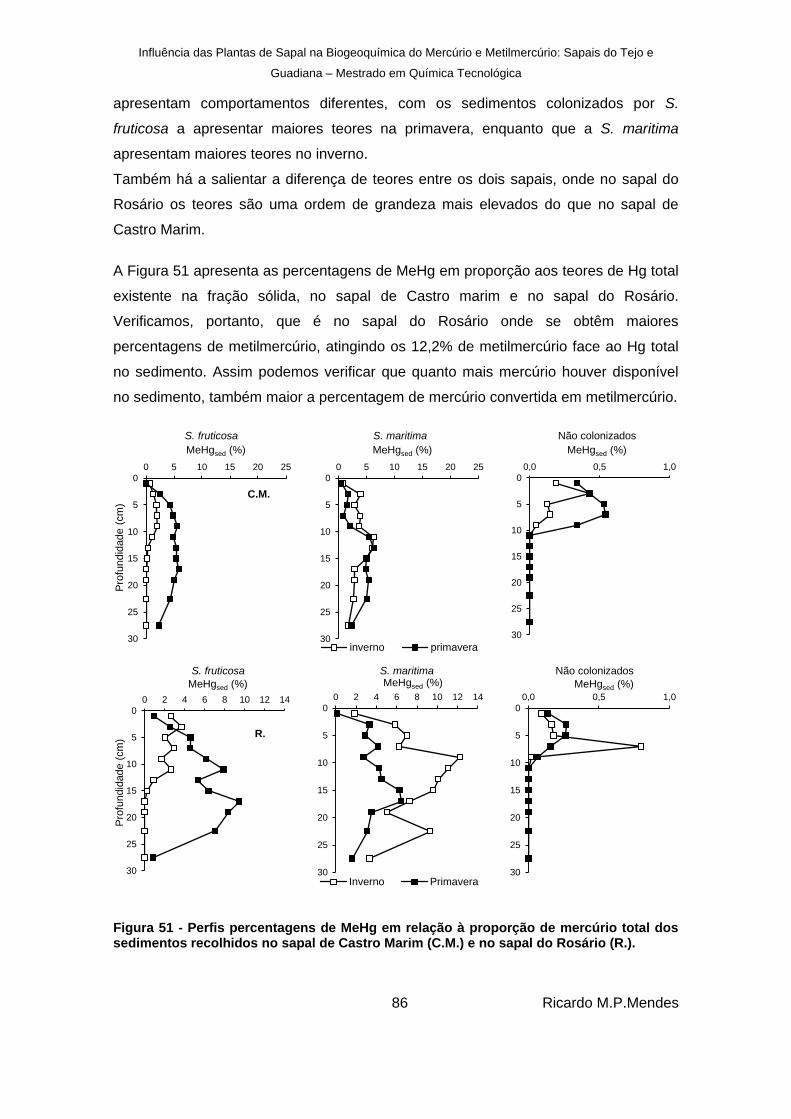
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
86 Ricardo M.P.Mendes
apresentam comportamentos diferentes, com os sedimentos colonizados por S.
fruticosa a apresentar maiores teores na primavera, enquanto que a S. maritima
apresentam maiores teores no inverno.
Também há a salientar a diferença de teores entre os dois sapais, onde no sapal do
Rosário os teores são uma ordem de grandeza mais elevados do que no sapal de
Castro Marim.
A Figura 51 apresenta as percentagens de MeHg em proporção aos teores de Hg total
existente na fração sólida, no sapal de Castro marim e no sapal do Rosário.
Verificamos, portanto, que é no sapal do Rosário onde se obtêm maiores
percentagens de metilmercúrio, atingindo os 12,2% de metilmercúrio face ao Hg total
no sedimento. Assim podemos verificar que quanto mais mercúrio houver disponível
no sedimento, também maior a percentagem de mercúrio convertida em metilmercúrio.
Figura 51 - Perfis percentagens de MeHg em relação à proporção de mercúrio total dos sedimentos recolhidos no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.).
0
5
10
15
20
25
30
0 5 10 15 20 25
Pro
fundid
ade (
cm
)
MeHgsed (%)
S. fruticosa
0
5
10
15
20
25
30
0 5 10 15 20 25
MeHgsed (%)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0,0 0,5 1,0
MeHgsed (%)
Não colonizados
0
5
10
15
20
25
30
0 2 4 6 8 10 12 14
Pro
fundid
ade (
cm
)
MeHgsed (%)
S. fruticosa
R.
0
5
10
15
20
25
30
0 2 4 6 8 10 12 14
MeHgsed (%) S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0,0 0,5 1,0
MeHgsed (%)
Não colonizados
C.M.
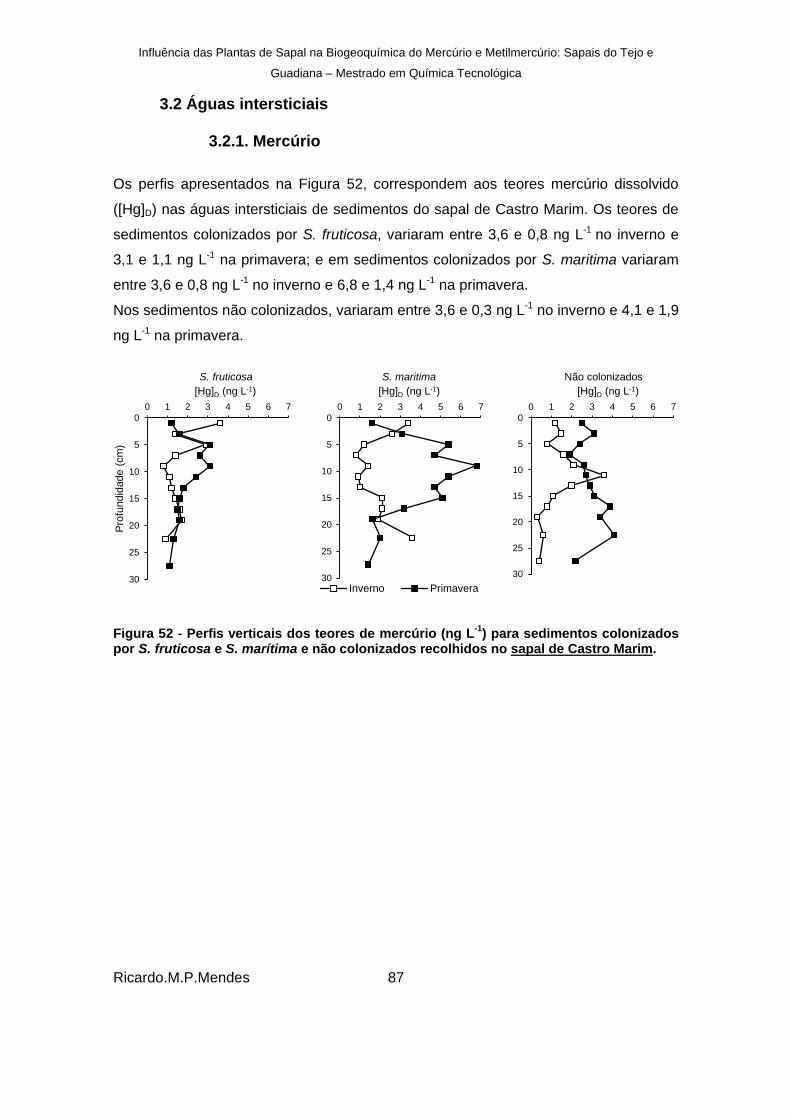
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 87
3.2 Águas intersticiais
3.2.1. Mercúrio
Os perfis apresentados na Figura 52, correspondem aos teores mercúrio dissolvido
([Hg]D) nas águas intersticiais de sedimentos do sapal de Castro Marim. Os teores de
sedimentos colonizados por S. fruticosa, variaram entre 3,6 e 0,8 ng L-1 no inverno e
3,1 e 1,1 ng L-1 na primavera; e em sedimentos colonizados por S. maritima variaram
entre 3,6 e 0,8 ng L-1 no inverno e 6,8 e 1,4 ng L-1 na primavera.
Nos sedimentos não colonizados, variaram entre 3,6 e 0,3 ng L-1 no inverno e 4,1 e 1,9
ng L-1 na primavera.
Figura 52 - Perfis verticais dos teores de mercúrio (ng L-1
) para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados recolhidos no sapal de Castro Marim.
0
5
10
15
20
25
30
0 1 2 3 4 5 6 7
Pro
fundid
ade (
cm
)
[Hg]D (ng L-1)
S. fruticosa
0
5
10
15
20
25
30
0 1 2 3 4 5 6 7
[Hg]D (ng L-1)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 1 2 3 4 5 6 7
[Hg]D (ng L-1)
Não colonizados
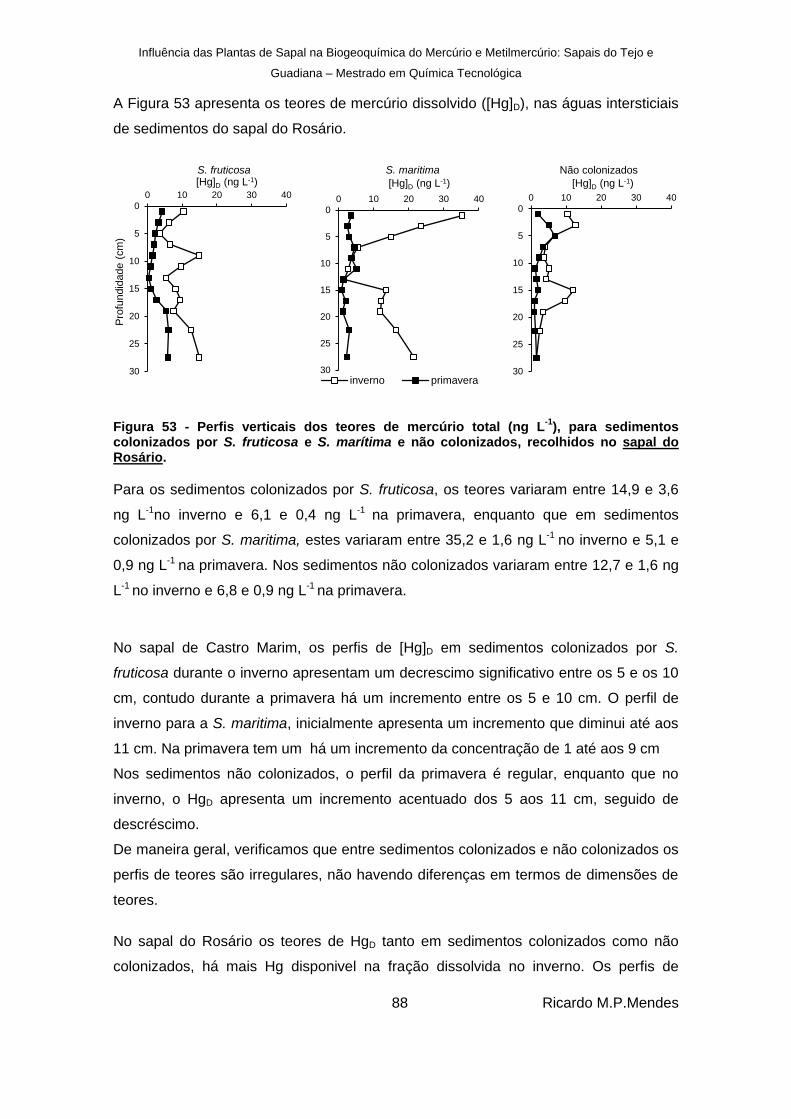
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
88 Ricardo M.P.Mendes
A Figura 53 apresenta os teores de mercúrio dissolvido ([Hg]D), nas águas intersticiais
de sedimentos do sapal do Rosário.
Figura 53 - Perfis verticais dos teores de mercúrio total (ng L-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
Para os sedimentos colonizados por S. fruticosa, os teores variaram entre 14,9 e 3,6
ng L-1no inverno e 6,1 e 0,4 ng L-1 na primavera, enquanto que em sedimentos
colonizados por S. maritima, estes variaram entre 35,2 e 1,6 ng L-1 no inverno e 5,1 e
0,9 ng L-1 na primavera. Nos sedimentos não colonizados variaram entre 12,7 e 1,6 ng
L-1 no inverno e 6,8 e 0,9 ng L-1 na primavera.
No sapal de Castro Marim, os perfis de [Hg]D em sedimentos colonizados por S.
fruticosa durante o inverno apresentam um decrescimo significativo entre os 5 e os 10
cm, contudo durante a primavera há um incremento entre os 5 e 10 cm. O perfil de
inverno para a S. maritima, inicialmente apresenta um incremento que diminui até aos
11 cm. Na primavera tem um há um incremento da concentração de 1 até aos 9 cm
Nos sedimentos não colonizados, o perfil da primavera é regular, enquanto que no
inverno, o HgD apresenta um incremento acentuado dos 5 aos 11 cm, seguido de
descréscimo.
De maneira geral, verificamos que entre sedimentos colonizados e não colonizados os
perfis de teores são irregulares, não havendo diferenças em termos de dimensões de
teores.
No sapal do Rosário os teores de HgD tanto em sedimentos colonizados como não
colonizados, há mais Hg disponivel na fração dissolvida no inverno. Os perfis de
0
5
10
15
20
25
30
0 10 20 30 40
Pro
fundid
ade (
cm
)
[Hg]D (ng L-1) S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40
[Hg]D (ng L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 10 20 30 40
[Hg]D (ng L-1)
Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 89
primavera são regulares em todas as situações, enquanto que no inverno apresentam-
se irregulares, com grandes variaçoes de teores entre camadas.
Entre sapais, verificamos que no sapal de Castro Marim há maiores concentrações de
HgD durante a primavera, e no sapal do Rosário onde há maiores concentrações de
HgD durante o inverno. Os teores HgD total no Rosário são superiores aos teores no
sapal de Castro Marim.
3.2.2. Metilmercúrio
Os seguintes perfis representam os teores de metilmercúrio ([MeHg]D) dissolvido nas
águas intersticiais.
Figura 54 - Perfis verticais dos teores de metilmercúrio (ng L-1
) para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal de Castro Marim.
Os teores de metilmercúrio dissolvido [MeHg]D apresentados na Figura 54, nas águas
intersticiais de sedimentos colonizados por S. fruticosa, variaram entre 0,9 e 0,0
(inferior ao limite de deteção) ng L-1 no inverno e 1,0 e 0,2 ng L-1 na primavera,
enquanto que para os sedimentos colonizados por S. maritima variaram entre 1,1 e 0,2
ng L-1 no inverno e na 1,7 e 0,3 ng L-1 na primavera.
Nos sedimentos não colonizados os teores variaram entre 0,4 e 0,0 (inferior L.D.) ng L-
1 no inverno e 0,9 e 0,0 (inferior ao L.D.) ng L-1 na primavera.
No perfil da S. fruticosa para o inverno, os teores dos 19 aos 22,5 cm de profundidade
foram inferiores ao limite de deteção. Para o perfil dos sedimentos não colonizados, a
0
5
10
15
20
25
30
0,0 0,5 1,0 1,5 2,0
Pro
fundid
ade (
cm
)
[MeHg]D (ng L-1)
S. fruticosa
0
5
10
15
20
25
30
0,0 0,5 1,0 1,5 2,0
[MeHg]D (ng L-1)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0,0 0,5 1,0 1,5 2,0
[MeHg]D (ng L-1)
Não colonizados
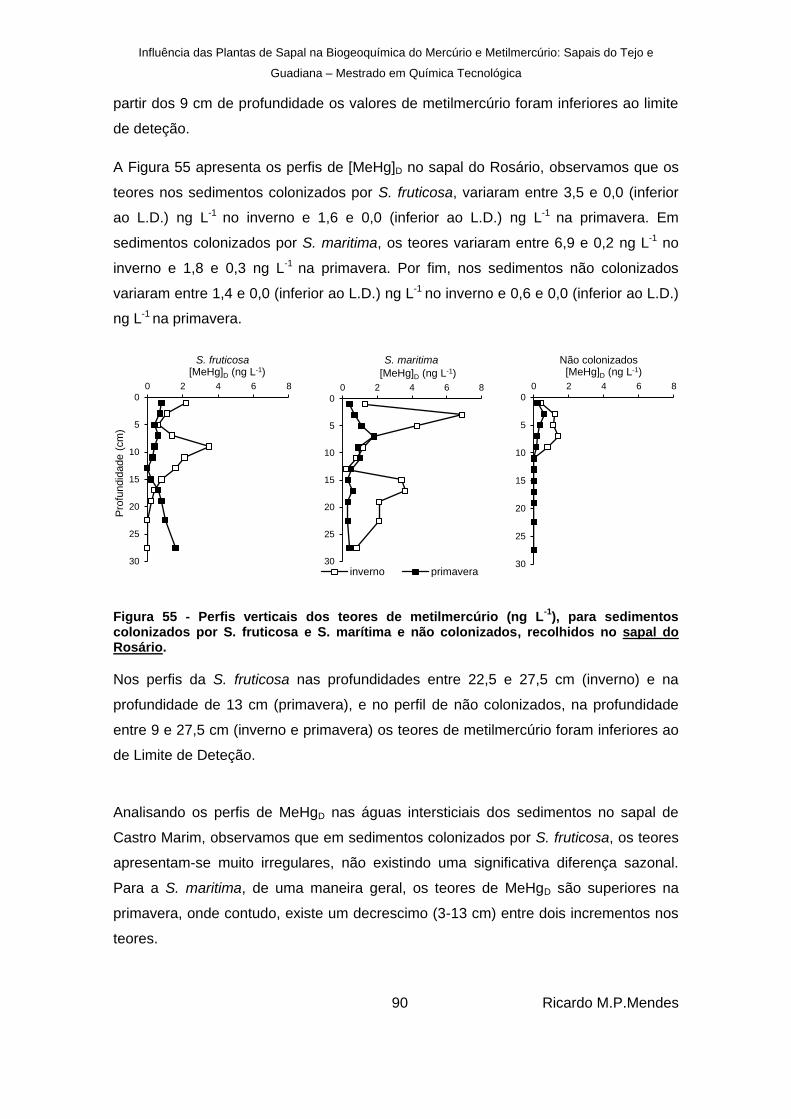
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
90 Ricardo M.P.Mendes
partir dos 9 cm de profundidade os valores de metilmercúrio foram inferiores ao limite
de deteção.
A Figura 55 apresenta os perfis de [MeHg]D no sapal do Rosário, observamos que os
teores nos sedimentos colonizados por S. fruticosa, variaram entre 3,5 e 0,0 (inferior
ao L.D.) ng L-1 no inverno e 1,6 e 0,0 (inferior ao L.D.) ng L-1 na primavera. Em
sedimentos colonizados por S. maritima, os teores variaram entre 6,9 e 0,2 ng L-1 no
inverno e 1,8 e 0,3 ng L-1 na primavera. Por fim, nos sedimentos não colonizados
variaram entre 1,4 e 0,0 (inferior ao L.D.) ng L-1 no inverno e 0,6 e 0,0 (inferior ao L.D.)
ng L-1 na primavera.
Figura 55 - Perfis verticais dos teores de metilmercúrio (ng L-1
), para sedimentos colonizados por S. fruticosa e S. marítima e não colonizados, recolhidos no sapal do Rosário.
Nos perfis da S. fruticosa nas profundidades entre 22,5 e 27,5 cm (inverno) e na
profundidade de 13 cm (primavera), e no perfil de não colonizados, na profundidade
entre 9 e 27,5 cm (inverno e primavera) os teores de metilmercúrio foram inferiores ao
de Limite de Deteção.
Analisando os perfis de MeHgD nas águas intersticiais dos sedimentos no sapal de
Castro Marim, observamos que em sedimentos colonizados por S. fruticosa, os teores
apresentam-se muito irregulares, não existindo uma significativa diferença sazonal.
Para a S. maritima, de uma maneira geral, os teores de MeHgD são superiores na
primavera, onde contudo, existe um decrescimo (3-13 cm) entre dois incrementos nos
teores.
0
5
10
15
20
25
30
0 2 4 6 8
Pro
fundid
ade (
cm
)
[MeHg]D (ng L-1) S. fruticosa
0
5
10
15
20
25
30
0 2 4 6 8
[MeHg]D (ng L-1)
S. maritima
inverno primavera
0
5
10
15
20
25
30
0 2 4 6 8
[MeHg]D (ng L-1) Não colonizados

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 91
No caso de sedimentos não colonizados, os teores de metilmercúrio são ligeiramente
superiores na primavera mas a partir dos 9 cm não foram detetados valores para o
metilmercúrio, tanto de inverno como na primavera
No sapal do Rosário, observamos que para sedimentos colonizados e não
colonizados, há maiores concentrações de MeHgD no inverno. Para os sedimentos
colonizados por S. maritima, há incremento onde se atingiu-se um valor máximo de 6,9
ng L-1 aos 3 cm, contudo segue-se um decréscimo acentuado dos 3 aos 13 cm,
seguido de um novo incremento. No caso dos sedimentos não colonizados, verifica-se
que a partir dos 9 cm não se conseguiu detetou mais metilmercúrio.
Entre sapais, observamos que no sapal de Castro Marim, há teores mais elevados de
MeHgD na primavera do que no inverno, ocorrendo o contrário no sapal do Rosário.
No sapal do Rosário há muito mais metilmercúrio dissolvido que no sapal de Castro
Marim, tanto no inverno como na primavera, por haver maior contaminação de Hg
provenientes de fontes antropogenicas.
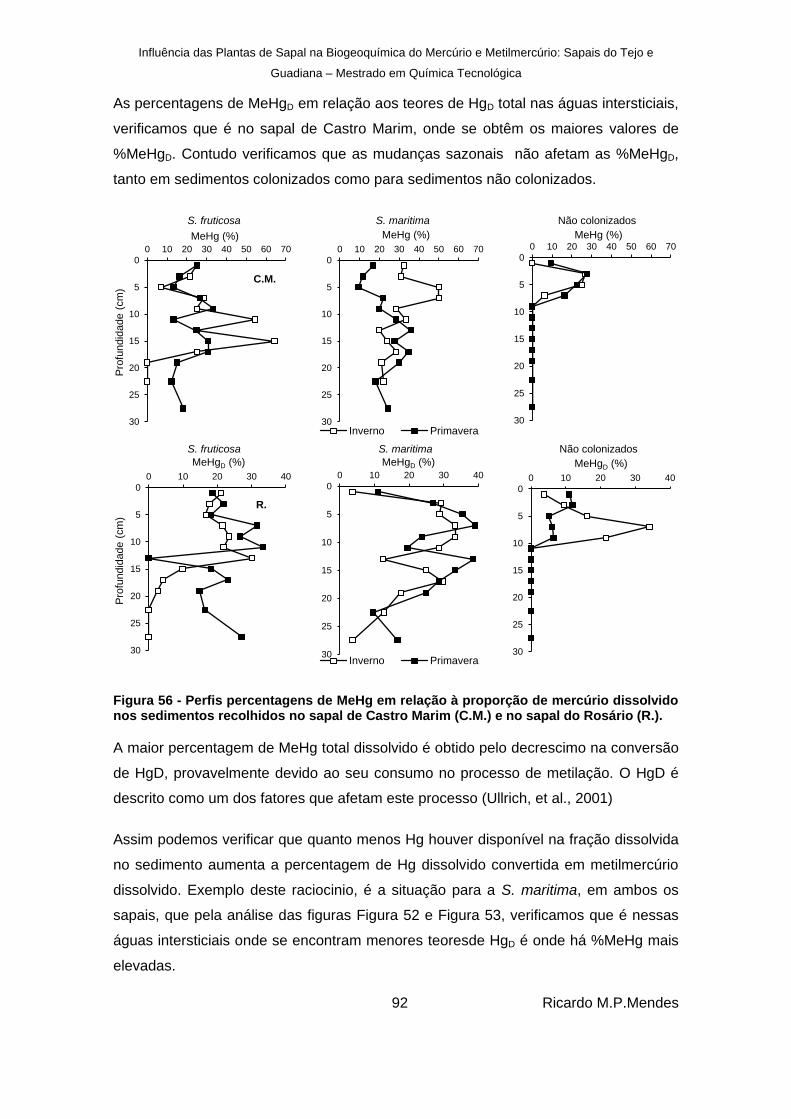
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
92 Ricardo M.P.Mendes
As percentagens de MeHgD em relação aos teores de HgD total nas águas intersticiais,
verificamos que é no sapal de Castro Marim, onde se obtêm os maiores valores de
%MeHgD. Contudo verificamos que as mudanças sazonais não afetam as %MeHgD,
tanto em sedimentos colonizados como para sedimentos não colonizados.
Figura 56 - Perfis percentagens de MeHg em relação à proporção de mercúrio dissolvido nos sedimentos recolhidos no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.).
A maior percentagem de MeHg total dissolvido é obtido pelo decrescimo na conversão
de HgD, provavelmente devido ao seu consumo no processo de metilação. O HgD é
descrito como um dos fatores que afetam este processo (Ullrich, et al., 2001)
Assim podemos verificar que quanto menos Hg houver disponível na fração dissolvida
no sedimento aumenta a percentagem de Hg dissolvido convertida em metilmercúrio
dissolvido. Exemplo deste raciocinio, é a situação para a S. maritima, em ambos os
sapais, que pela análise das figuras Figura 52 e Figura 53, verificamos que é nessas
águas intersticiais onde se encontram menores teoresde HgD é onde há %MeHg mais
elevadas.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
Pro
fundid
ade (
cm
)
MeHg (%)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
MeHg (%)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
MeHg (%)
Não colonizados
0
5
10
15
20
25
30
0 10 20 30 40
Pro
fundid
ade (
cm
)
MeHgD (%)
S. fruticosa
0
5
10
15
20
25
30
0 10 20 30 40
MeHgD (%)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 10 20 30 40
MeHgD (%)
Não colonizados
C.M.
R.
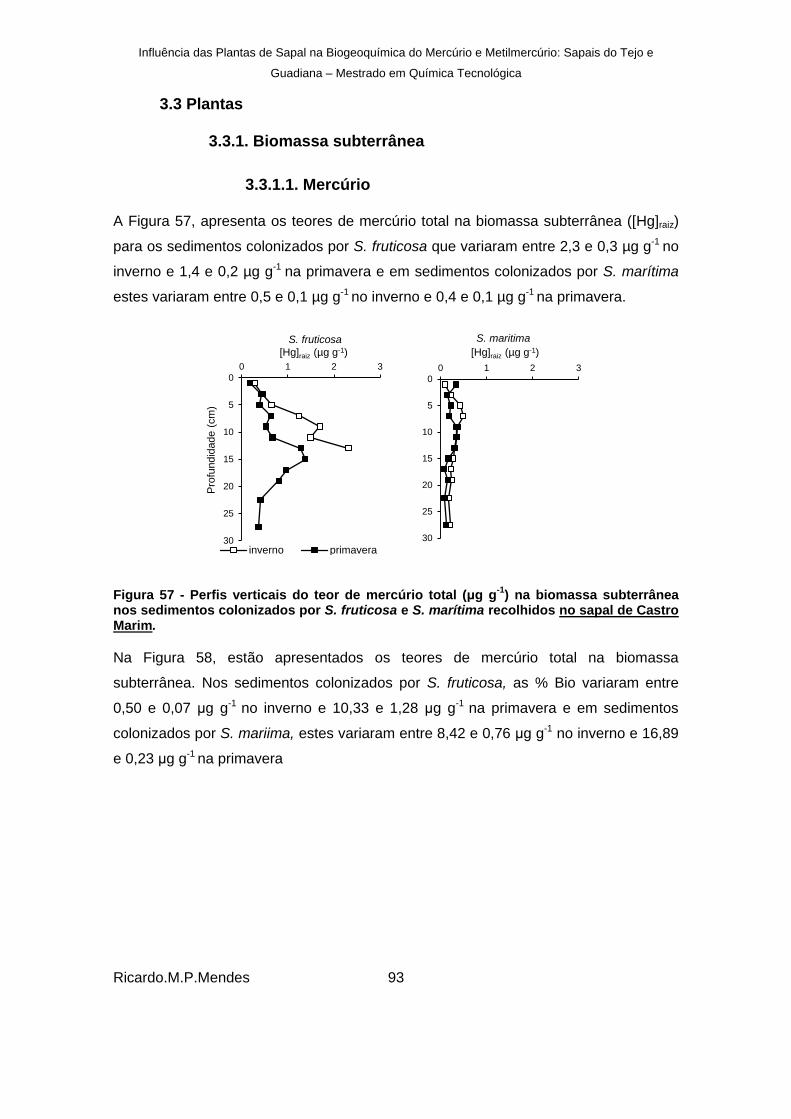
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 93
3.3 Plantas
3.3.1. Biomassa subterrânea
3.3.1.1. Mercúrio
A Figura 57, apresenta os teores de mercúrio total na biomassa subterrânea ([Hg]raiz)
para os sedimentos colonizados por S. fruticosa que variaram entre 2,3 e 0,3 µg g-1 no
inverno e 1,4 e 0,2 µg g-1 na primavera e em sedimentos colonizados por S. marítima
estes variaram entre 0,5 e 0,1 µg g-1 no inverno e 0,4 e 0,1 µg g-1 na primavera.
Figura 57 - Perfis verticais do teor de mercúrio total (μg g-1
) na biomassa subterrânea nos sedimentos colonizados por S. fruticosa e S. marítima recolhidos no sapal de Castro Marim.
Na Figura 58, estão apresentados os teores de mercúrio total na biomassa
subterrânea. Nos sedimentos colonizados por S. fruticosa, as % Bio variaram entre
0,50 e 0,07 μg g-1 no inverno e 10,33 e 1,28 μg g-1 na primavera e em sedimentos
colonizados por S. mariima, estes variaram entre 8,42 e 0,76 μg g-1 no inverno e 16,89
e 0,23 μg g-1 na primavera
0
5
10
15
20
25
30
0 1 2 3
Pro
fundid
ade (
cm
)
[Hg]raiz (µg g-1)
S. fruticosa
inverno primavera
0
5
10
15
20
25
30
0 1 2 3
[Hg]raiz (µg g-1)
S. maritima

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
94 Ricardo M.P.Mendes
Figura 58 – Perfis verticais dos teores de mercúrio total (μg g-1
) na biomassa subterrânea de S. fruticosa e S. maritima, recolhida no sapal do Rosário.
Os teores de Hg total presentes na biomassa subterrânea (Hgraiz) do sapal de Castro
Marim, para a S. fruticosa são maiores no inverno do que na da primavera, atingindo
esta planta o valor máximo encontrado. Na S. maritima os teores são muito
semelhantes entre estações, embora menores do que na S. fruticosa.
No sapal do Rosário, os teores de Hgraiz da S. fruticos são maiores na primavera face
ao inverno. Na S. maritima os teores são muito semelhantes entre estações, contudo
entre os 9 e 11 cm os teores de Hgraiz têm um incremento, onde excedem claramente
os teores no inverno.
Comparando sapais, verificamos que no sapal do Rosário os teores Hgraiz são maiores
que os encontrados no sapal de Castro Marim. Também os perfis de teores mercúrio
são muito diferentes entre as plantas.
Uma vez dentro nas raizes o Hg pode ser deslocado para as partes fotossintéticas das
plantas (Castro, et al., 2009), variando de acordo com o tipo de planta (Weis & Weis,
2004).
0
5
10
15
20
25
30
0 5 10 15 20
Pro
fundid
ade (
cm
)
[Hg]raiz (µg g-1)
S. fruticosa
0
5
10
15
20
25
30
0 5 10 15 20
[Hg]raiz (µg g-1) S. maritima
inverno primavera
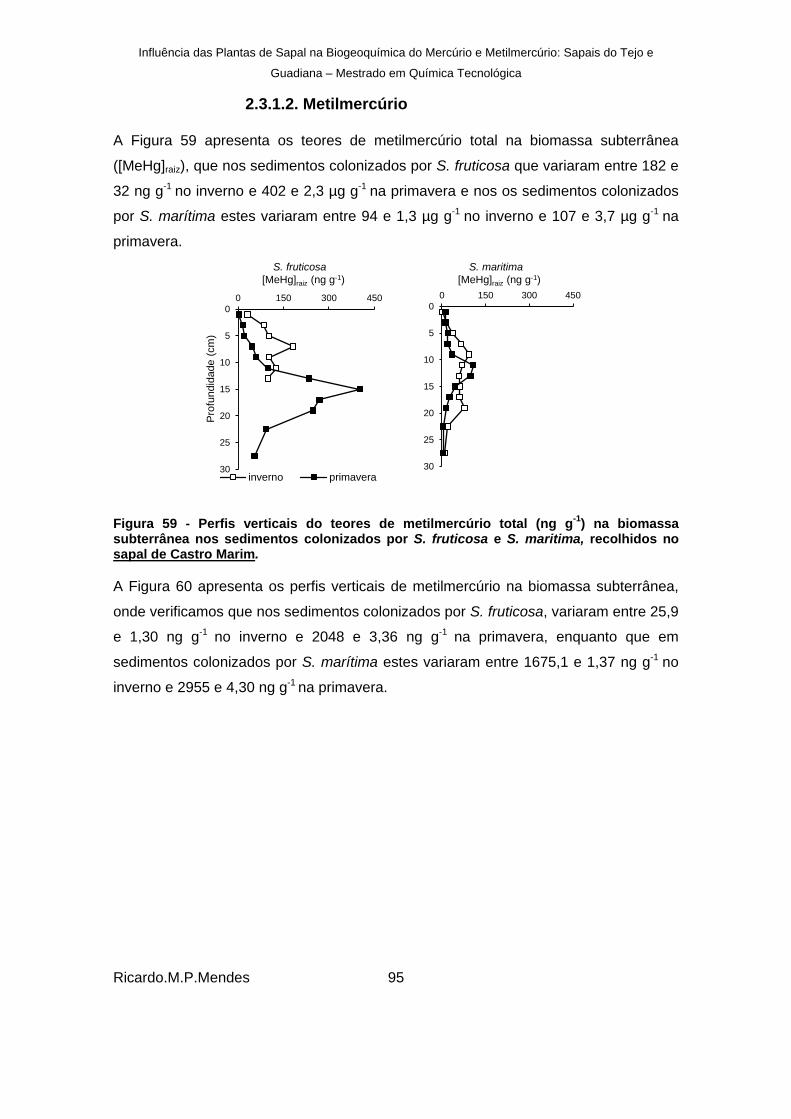
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 95
2.3.1.2. Metilmercúrio
A Figura 59 apresenta os teores de metilmercúrio total na biomassa subterrânea
([MeHg]raiz), que nos sedimentos colonizados por S. fruticosa que variaram entre 182 e
32 ng g-1 no inverno e 402 e 2,3 µg g-1 na primavera e nos os sedimentos colonizados
por S. marítima estes variaram entre 94 e 1,3 µg g-1 no inverno e 107 e 3,7 µg g-1 na
primavera.
Figura 59 - Perfis verticais do teores de metilmercúrio total (ng g-1
) na biomassa subterrânea nos sedimentos colonizados por S. fruticosa e S. maritima, recolhidos no sapal de Castro Marim.
A Figura 60 apresenta os perfis verticais de metilmercúrio na biomassa subterrânea,
onde verificamos que nos sedimentos colonizados por S. fruticosa, variaram entre 25,9
e 1,30 ng g-1 no inverno e 2048 e 3,36 ng g-1 na primavera, enquanto que em
sedimentos colonizados por S. marítima estes variaram entre 1675,1 e 1,37 ng g-1 no
inverno e 2955 e 4,30 ng g-1 na primavera.
0
5
10
15
20
25
30
0 150 300 450
Pro
fundid
ade (
cm
)
[MeHg]raiz (ng g-1)
S. fruticosa
inverno primavera
0
5
10
15
20
25
30
0 150 300 450
[MeHg]raiz (ng g-1)
S. maritima
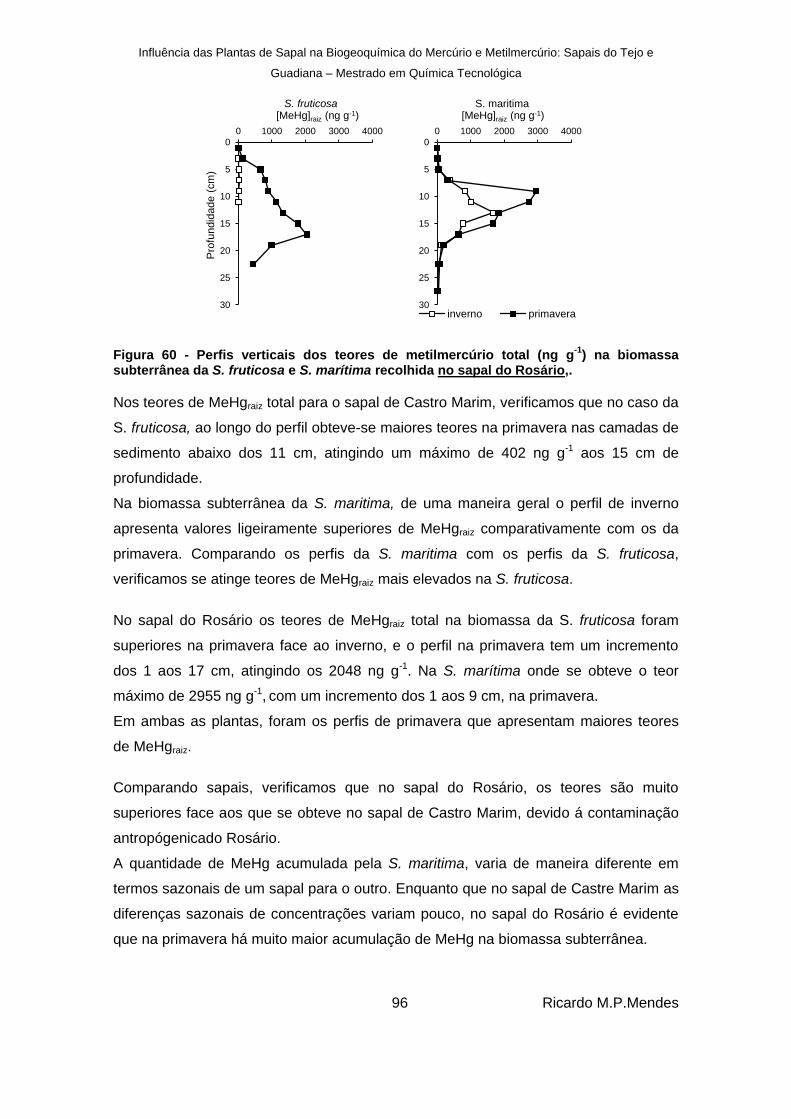
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
96 Ricardo M.P.Mendes
Figura 60 - Perfis verticais dos teores de metilmercúrio total (ng g-1
) na biomassa subterrânea da S. fruticosa e S. marítima recolhida no sapal do Rosário,.
Nos teores de MeHgraiz total para o sapal de Castro Marim, verificamos que no caso da
S. fruticosa, ao longo do perfil obteve-se maiores teores na primavera nas camadas de
sedimento abaixo dos 11 cm, atingindo um máximo de 402 ng g-1 aos 15 cm de
profundidade.
Na biomassa subterrânea da S. maritima, de uma maneira geral o perfil de inverno
apresenta valores ligeiramente superiores de MeHgraiz comparativamente com os da
primavera. Comparando os perfis da S. maritima com os perfis da S. fruticosa,
verificamos se atinge teores de MeHgraiz mais elevados na S. fruticosa.
No sapal do Rosário os teores de MeHgraiz total na biomassa da S. fruticosa foram
superiores na primavera face ao inverno, e o perfil na primavera tem um incremento
dos 1 aos 17 cm, atingindo os 2048 ng g-1. Na S. marítima onde se obteve o teor
máximo de 2955 ng g-1, com um incremento dos 1 aos 9 cm, na primavera.
Em ambas as plantas, foram os perfis de primavera que apresentam maiores teores
de MeHgraiz.
Comparando sapais, verificamos que no sapal do Rosário, os teores são muito
superiores face aos que se obteve no sapal de Castro Marim, devido á contaminação
antropógenicado Rosário.
A quantidade de MeHg acumulada pela S. maritima, varia de maneira diferente em
termos sazonais de um sapal para o outro. Enquanto que no sapal de Castre Marim as
diferenças sazonais de concentrações variam pouco, no sapal do Rosário é evidente
que na primavera há muito maior acumulação de MeHg na biomassa subterrânea.
0
5
10
15
20
25
30
0 1000 2000 3000 4000
Pro
fundid
ade (
cm
)
[MeHg]raiz (ng g-1) S. fruticosa
0
5
10
15
20
25
30
0 1000 2000 3000 4000
[MeHg]raiz (ng g-1) S. maritima
inverno primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 97
Assim verificamos, de uma maneira geral, que no inverno a biomassa subterrânea
apresenta pequenas concentações de MeHg quando comparados com a primavera,
sugerindo que a atividade das plantas é responsável pela captura e acumulação de
MeHg na biomassa subterrânea (Weis & Weis, 2004).
A Figura 61 apresenta, a percentagem de MeHgraiz em relação ao mercúrio total na
biomassa subterrânea (Hgraiz). Verificamos que no sapal de Castro Marim, obteve-se
valores de percentagem de MeHg mais elevados para a S. marítima, apesar de ter
menores teores de mercúrio total e metilmercúrio.
No sapal do Rosário, as percentagem MeHgraiz foram semelhantes aos do sapal de
Castro Marim.
Figura 61 - Perfis percentagens de MeHg em relação à proporção de mercúrio dissolvido na biomassa subterrânea recolhida no sapal de Castro Marim (C.M.) e no sapal do Rosário (R.).
0
5
10
15
20
25
30
0 10 20 30 40
Pro
fundid
ade (
cm
)
MeHgraiz (%)
S. fruticosa
C.M.
0
5
10
15
20
25
30
0 10 20 30 40
MeHgraiz (%)
S. maritima
Inverno Primavera
0
5
10
15
20
25
30
0 10 20 30
Pro
fundid
ade (
cm
)
MeHgraiz (%)
S. fruticosa
R.
0
5
10
15
20
25
30
0 20 40 60
MeHgraiz (%)
S. maritima
Inverno Primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
98 Ricardo M.P.Mendes
Comparando sapais, é de salientar que as percentagens de MeHgraiz são diferentes
entre a S. fruticosa e a S. maritima, sendo que na S. fruticosa as percentagens de
MeHgraiz , evoluem de maneira semelhante, principalmente durante a primavera. Na S.
maritima, tanto de inverno como durante a primavera, a evolução das percentagens de
MeHgraiz são semelhantes.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 99
3.3.2. Biomassa aerea
3.3.2.1. Mercúrio
A Figura 62 apresenta os teores de mercúrio nas componentes aéreas (partes
fotossintéticas e não fotossintéticas) da S. fruticosa e S. marítima.
Figura 62 - Valores dos teores de mercúrio total (μg g-1
) nas partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima, no sapal de Castro Marim.
Pela análise aos resultados dos teores de Hg total na biomassa aerea das plantas do
sapal de Castro Marim, observamos que nas partes fotossintéticas foram obtidos
maiores teores face ás partes não fotossintéticas. Também é de assinalar diferenças
sazonais como maiores teores de Hg no inverno do que na primavera.
Isto pode ser interpretado com o transporte de Hg para as partes fotossintéticas da
planta, com a sua consequente acumulação durante o inverno devido a uma menor
atividade das mesmas. Durante a primavera, há uma maior atividade da planta, por
causa do aumento da temperatura e do tempo de insulação, pode levar que estas a
libertem mais mercúrio, diminuindo assim o Hg na planta (Weis & Weis, 2004).
A Figura 63 apresenta os teores de mercúrio total (µg g-1) determinados na biomassa
aérea (partes fotossintéticas e partes não fotossintéticas) para o saal do Rosário.
Os teores de Hg total que foram determinados nas partes aéreas do sapal do Rosário,
verificamos que os teores entre as partes fotossintéticas e não fotossintéticas, variam
consoante a planta.A S. maritima apresenta maiores teores nas partes fotossintéticas
do que nas partes não fotossintéticas, sendo contrário ao comportamento da S.
fruticosa.
0,00
0,04
0,08
0,12
0,16
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticas
S. maritima
[Hg
] (µ
g g
-1)
inverno primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
100 Ricardo M.P.Mendes
Figura 63 - Teores de mercúrio total (μg g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal do Rosário.
3.3.2.2. Metilmercúrio
A Figura 64, apresenta os teores de MeHg nas partes fotossintéticas e não
fotossintéticas da S. fruticosa e S. marítima: Os valores de teores de MeHg na
primavera são inferiores ao limite de deteção (<0,10 ng g-1), em todas as partes.
Figura 64 - Valores dos teores de metilmercúrio total (ng g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal de Castro Marim.
A Figura 65 apresenta os teores de metilmercúrio (ng g-1) determinados na biomassa
aérea (partes fotossintéticas e partes não fotossintéticas), para o sapal do Rosário. Os
teores de metilmercúrio para a primavera são inferiores ao L.D. (<0,10 ng g-1), em
todas as partes.
0,00
0,01
0,02
0,03
0,04
0,05
0,06
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticaS. maritima
[Hg
] (µ
g g
-1)
inverno primavera
0
2
4
6
8
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticas
S. maritima
[MeH
g]
(ng g
-1)
inverno

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 101
Figura 65 - Teores de metilmercúrio (ng g-1
) para as partes fotossintéticas e não fotossintéticas para as plantas S.fruticosa e S. marítima no sapal do Rosário.
A Figura 66 e a Figura 67 apresentam as percentagem de metilmercúrio face ao
mercúrio total, observamos que nos dois sapais estudados. Na primavera não foi
detetado MeHg.
Figura 66 – Percentagem de metilmercúrio em função do teor de mercúrio total nas partes fotossintéticas e não fotossintéticas para a S. fruticosa e S. marítima, recolhidas no sapal de Castro Marim
Figura 67 - Percentagem de metilmercúrio em função do teor de Mercúrio total nas partes fotossintéticas e não fotossintéticas para a S. fruticosa e S. marítima, recolhidas no sapal do Rosário.
Os teores de metilmercúrio determinados permitem verificar que, tanto para o sapal do
Rosário como para o sapal de Castro Marim, não foi determinado metilmercúrio na
0
2
4
6
8
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticaS. maritima
[Me
Hg
] (n
g g
-1)
inverno
0
4
8
12
16
20
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticas
S. maritima
MeH
g (
%)
inverno
0
4
8
12
16
20
Partesfotossintéticas
S. fruticosa
Partesfotossintéticas
S. maritima
Partes nãofotossintéticas
S. fruticosa
Partes nãofotossintéticaS. maritima
MeH
g (
%)
inverno

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
102 Ricardo M.P.Mendes
primavera. Tal facto interpretado com o aumento das temperaturas e do tempo de
insolação, e uma maior atividade das plantas, leva a acelerar os mecanismos de
desmetilação do mercúrio.
De um modo geral, tal como se pode verificar neste estudo, as plantas de sapal
acumulam diferentes quantidades de metais entre as partes aereas e subterrâneas,
apresentando maior acumulação de mercúrio e metilmercúrio total na biomassa
subterrânea. Estes resultados estão de acordo com outros artigos publicados que
sujerem existir uma baixa translocação de Hg entre as raizes e as partes aereas das
plantas (Ernst, 1990; Otte, 1991; Matheus, et al., 2004; Canário, et al., 2010)
Com o intuito de sobreviveram em sedimentos contaminados com metais, as plantas
de sapal desenvolvem vários mecanismos que regulam a concentração deste, tais
como, mecanismos de restrição de absorção e restrição do metal para os orgãos
aereos, com a excessão de plantas que hiper acumulam Hg (Vesk, et al., 1999;
MacFarlane & Burchett, 2000; Porchenriedes, 2003). A quantidade de Hg que é
excretada das plantas através das folhas, varia de acordo com as espécies de plantas
e variam sazonalmente (Weis & Weis, 2004)
O processo de translocação do Hg detetado nestas plantas, do sistema radicular para
a parte aerea vegetal é efetuada por proteinas transportadoras (Marschner, 1995). Já
nas partes aereas, os metais são armazenados em compartimentos celulares, onde
tenham uma interferência reduzida com processos celulares vitais (Carvalho, 2009).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 103
3.4. Distribuição de mercúrio e metilmercúrio entre sedimentos e
biomassa subterrânea
No sapal de Castro Marim, os teores de Hg total existentes nos sedimentos e na
biomassa subterrânea, verificamos que nos perfis da S. fruticosa, tanto de inverno
como na primavera, a biomassa subterrânea contem teores de mercúrio superiores
aos encontrados nos sedimentos
Nos perfis da S. marítima, já não ocorre o mesmo que nos perfis da S. fruticosa, aqui
os perfis são muito irregulares, isto pode dever-se ao facto dos baixos teores de Hg
disponíveis no sedimento, estando a disponibilidade relacionada com a origem
geológica deste (Canário, et al., 2007a).
Figura 68 - Perfis dos teores de Hg na biomassa subterrânea e nos sedimentos colonizados, recolhidos no sapal de Castro Marim.
No sapal do Rosário, as plantas apresentam um comportamento diferente ao
apresentado no sapal de Castro Marim. A S. fruticosa apresenta muito pouco mercúrio
na biomassa subterrânea durante o inverno, sugerindo estar menos ativa, o que pode
0
5
10
15
20
25
30
0 1 2 3
Pro
fundid
ade (
cm
)
[Hg] (µg g-1)
S. fruticosa - inverno
Sedimento
Biomassa Subterrânea
0
5
10
15
20
25
30
0,0 0,2 0,4 0,6
[Hg] (µg g-1)
S. maritima - inverno
0
5
10
15
20
25
30
0,0 0,5 1,0 1,5
Pro
fundid
ade (
cm
)
[Hg] (µg g-1)
S. fruticosa - primavera
0
5
10
15
20
25
30
0,0 0,1 0,2 0,3 0,4
[Hg] (µg g-1)
S. maritima - primavera
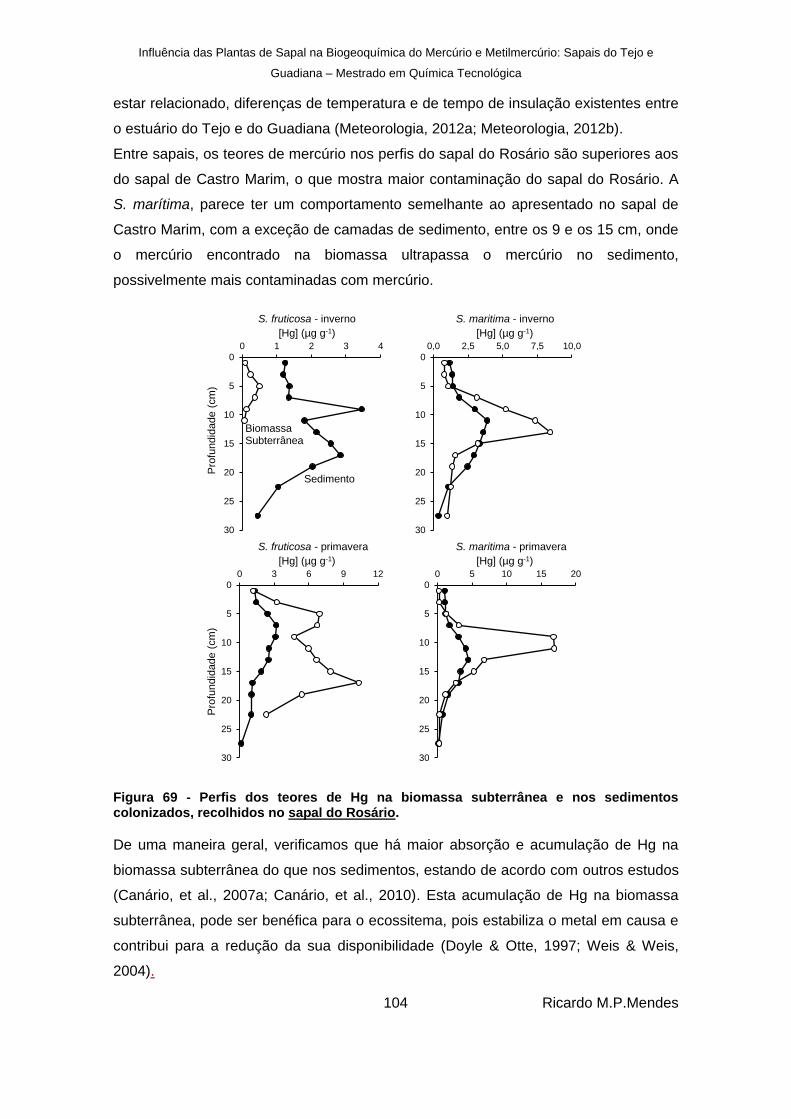
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
104 Ricardo M.P.Mendes
estar relacionado, diferenças de temperatura e de tempo de insulação existentes entre
o estuário do Tejo e do Guadiana (Meteorologia, 2012a; Meteorologia, 2012b).
Entre sapais, os teores de mercúrio nos perfis do sapal do Rosário são superiores aos
do sapal de Castro Marim, o que mostra maior contaminação do sapal do Rosário. A
S. marítima, parece ter um comportamento semelhante ao apresentado no sapal de
Castro Marim, com a exceção de camadas de sedimento, entre os 9 e os 15 cm, onde
o mercúrio encontrado na biomassa ultrapassa o mercúrio no sedimento,
possivelmente mais contaminadas com mercúrio.
Figura 69 - Perfis dos teores de Hg na biomassa subterrânea e nos sedimentos colonizados, recolhidos no sapal do Rosário.
De uma maneira geral, verificamos que há maior absorção e acumulação de Hg na
biomassa subterrânea do que nos sedimentos, estando de acordo com outros estudos
(Canário, et al., 2007a; Canário, et al., 2010). Esta acumulação de Hg na biomassa
subterrânea, pode ser benéfica para o ecossitema, pois estabiliza o metal em causa e
contribui para a redução da sua disponibilidade (Doyle & Otte, 1997; Weis & Weis,
2004).
0
5
10
15
20
25
30
0 1 2 3 4
Pro
fundid
ade (
cm
)
[Hg] (µg g-1)
S. fruticosa - inverno
Sedimento
Biomassa Subterrânea
0
5
10
15
20
25
30
0,0 2,5 5,0 7,5 10,0
[Hg] (µg g-1)
S. maritima - inverno
0
5
10
15
20
25
30
0 3 6 9 12
Pro
fundid
ade (
cm
)
[Hg] (µg g-1)
S. fruticosa - primavera
0
5
10
15
20
25
30
0 5 10 15 20
[Hg] (µg g-1)
S. maritima - primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 105
Os teores de metilmercúrio nos perfis referentes ao sapal de Castro Marim mostram a
rizosfera influenciam claramente a metilação de mercúrio, tal com noutros estudos
(Canário, et al., 2007a; Castro, et al., 2009; Canário, et al., 2010).
A dimensão da de biomassa, parece influenciar a quantidade total de metilmercúrio
produzida. Na primavera existe picos de mercúrio que excedem claramente os valores
obtidos no inverno, contudo no inverno há uma distribuição muito superior de
metilmercúrio nas raízes.
Figura 70 - Perfis dos teores de MeHg na biomassa subterrânea e nos sedimentos colonizados recolhidos no sapal de Castro Marim.
No sapal do Rosário Os teores de metilmercúrio nos perfis mostra dois
comportamentos distintos. Para a S. fruticosa, durante o inverno, os teores de MeHg
são maiores no sedimento do que na biomassa subterrânea, situação que se inverte
na primavera. Nos perfis da S. marítima, verifica-se o há elevados teores de
0
5
10
15
20
25
30
0 50 100 150 200
Pro
fundid
ade (
cm
)
[MeHg] (ng g-1)
S. fruticosa - inverno
Sedimento
Biomassa Subterrânea
0
5
10
15
20
25
30
0 25 50 75 100
[MeHg] (ng g-1)
S. maritima - inverno
0
5
10
15
20
25
30
0 200 400 600
Pro
fundid
ade (
cm
)
[MeHg] (ng g-1)
S. fruticosa - primavera
0
5
10
15
20
25
30
0 20 40 60 80 100 120
[MeHg] (ng g-1)
S. maritima - primavera
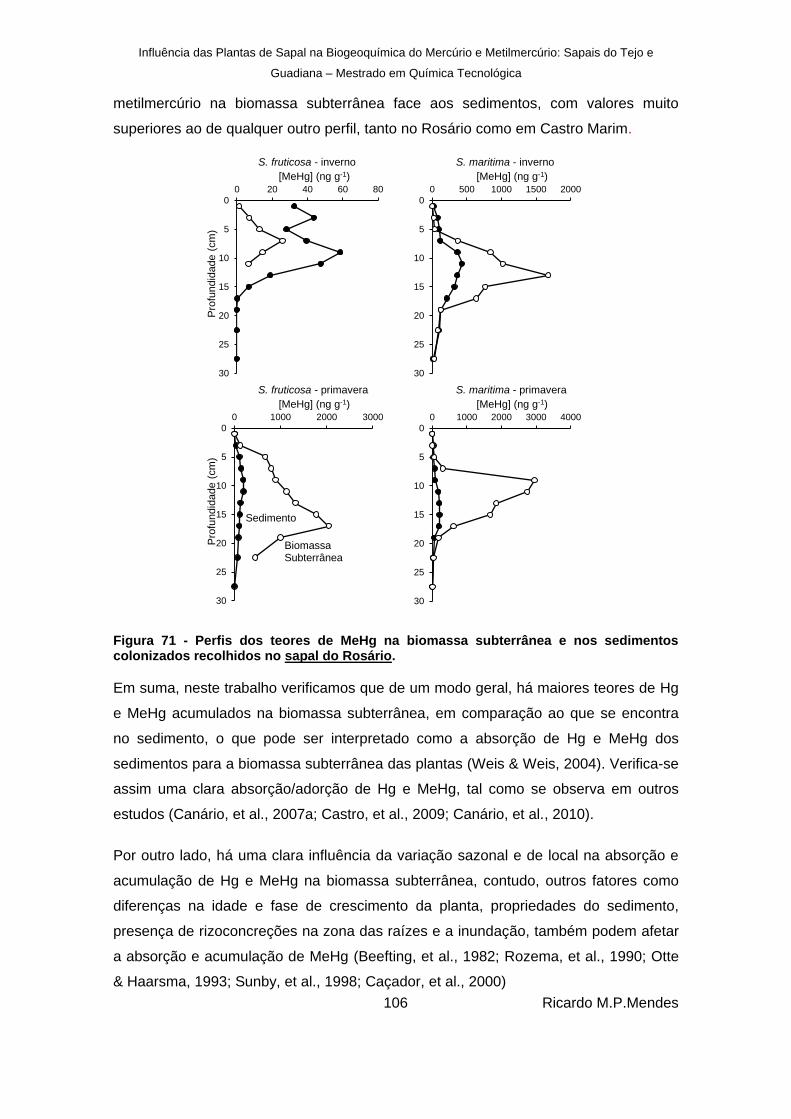
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
106 Ricardo M.P.Mendes
metilmercúrio na biomassa subterrânea face aos sedimentos, com valores muito
superiores ao de qualquer outro perfil, tanto no Rosário como em Castro Marim.
Figura 71 - Perfis dos teores de MeHg na biomassa subterrânea e nos sedimentos colonizados recolhidos no sapal do Rosário.
Em suma, neste trabalho verificamos que de um modo geral, há maiores teores de Hg
e MeHg acumulados na biomassa subterrânea, em comparação ao que se encontra
no sedimento, o que pode ser interpretado como a absorção de Hg e MeHg dos
sedimentos para a biomassa subterrânea das plantas (Weis & Weis, 2004). Verifica-se
assim uma clara absorção/adorção de Hg e MeHg, tal como se observa em outros
estudos (Canário, et al., 2007a; Castro, et al., 2009; Canário, et al., 2010).
Por outro lado, há uma clara influência da variação sazonal e de local na absorção e
acumulação de Hg e MeHg na biomassa subterrânea, contudo, outros fatores como
diferenças na idade e fase de crescimento da planta, propriedades do sedimento,
presença de rizoconcreções na zona das raízes e a inundação, também podem afetar
a absorção e acumulação de MeHg (Beefting, et al., 1982; Rozema, et al., 1990; Otte
& Haarsma, 1993; Sunby, et al., 1998; Caçador, et al., 2000)
0
5
10
15
20
25
30
0 20 40 60 80
Pro
fundid
ade (
cm
)
[MeHg] (ng g-1)
S. fruticosa - inverno
0
5
10
15
20
25
30
0 500 1000 1500 2000
[MeHg] (ng g-1)
S. maritima - inverno
0
5
10
15
20
25
30
0 1000 2000 3000
Pro
fundid
ade (
cm
)
[MeHg] (ng g-1)
S. fruticosa - primavera
Sedimento
Biomassa Subterrânea
0
5
10
15
20
25
30
0 1000 2000 3000 4000
[MeHg] (ng g-1)
S. maritima - primavera

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 107
As reações de redução de SO42- para S2- e Fe (III) para Fe(II) são mediadas por micro-
organismos designadas por as bactérias redutoras de sulfatos e redutoras de ferro,
que estão envolvidas em reações de complexação e metilação de metais,
principalmente de mercúrio (Kelley & Tuovinen, 1988; Choi & Bartha, 1994; Otte &
Jacob, 2003; Avramescu, et al., 2011).
Estas bactérias estão associadas à rizosfera, sendo influenciadas pela atividade das
raízes (Weis & Weis, 2004; Avramescu, et al., 2011). No periodo da primavera, ocorre
o aumento da temperatura e do tempo de insulação (Meteorologia, 2012a;
Meteorologia, 2012b). Como consequência as plantas oxidam os sedimentos que
proporciona a remobilização e aumento de disponibilidade de contaminantes metálicos
(o que ocorre neste trabalho, pelo aumento da disponibilidade de matais na fração
dissolvida , diminuição dos teores de sulfuretos dissolvidos e um ligeiro aumento dos
teores de sulfatos), ocorrendo também a libertação de compostos orgânicos por parte
das raizes, estimulando a atividade bacterial (Weis & Weis, 2004). Isto demonstra a
complexidade da biogeoquímica dos sapais, e a interdependência de várias variáveis
que influênciam a metilação, e contribuem para a sua variabilidade (Canário, et al.,
2010).
As bactérias na rizosfera, utilizam o Hg que se encontra na fração dissolvida para a
acumulação de Hg na biomassa subterranea das plantas através de processos
complexos que envolvem a produção de compostos lábeis nos exudados das raízes
(De Souza, et al., 1999). A acumulação de Hg e MeHg nas raízes varia com a
profundidade, em todas as plantas analisadas. Isto indica a existência de camadas
preferenciais para a retenção e com maior eficiência na remoção de Hg e MeHg dos
sedimentos para a biomassa subterrânea, tal como noutros estudos (Canário, et al.,
2007a; Canário, et al., 2010).
De acordo com vários estudos (Weis & Weis, 2004; Wang & Greger, 2004; Skinner, et
al., 2007) as raízes conseguem capturar Hg e MeHg através da ligação química Hg-S
a duas classes de peptidos ricos em cisteína: as metalotioninas e as fitoquelatinas
(Wang & Greger, 2004; Zenk, 1996). Os mecanismos envolvem a ligação que se
estabelece entre o Hg, com grupos de tiois (R-SH) na cisteina destes peptidos, que
reagem com a glutationa e são transportados para vacuolos (Brouwer, et al., 1993;
Zenk, 1996).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
108 Ricardo M.P.Mendes
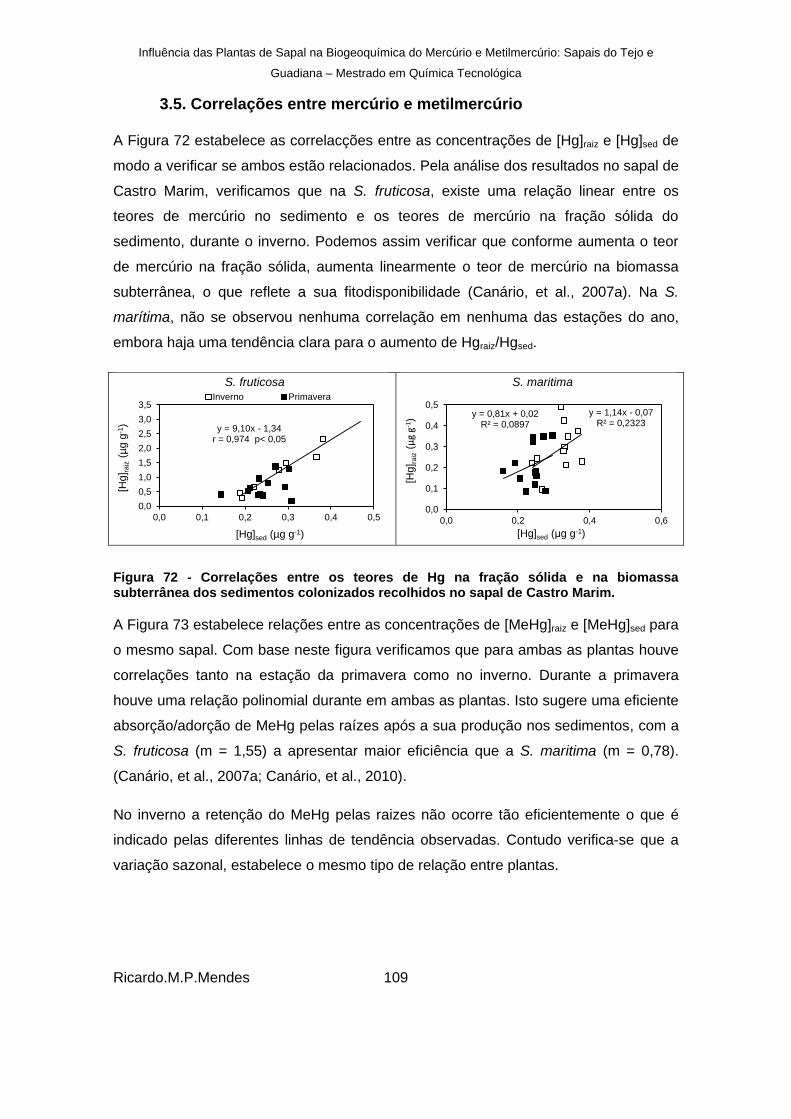
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 109
3.5. Correlações entre mercúrio e metilmercúrio
A Figura 72 estabelece as correlacções entre as concentrações de [Hg]raiz e [Hg]sed de
modo a verificar se ambos estão relacionados. Pela análise dos resultados no sapal de
Castro Marim, verificamos que na S. fruticosa, existe uma relação linear entre os
teores de mercúrio no sedimento e os teores de mercúrio na fração sólida do
sedimento, durante o inverno. Podemos assim verificar que conforme aumenta o teor
de mercúrio na fração sólida, aumenta linearmente o teor de mercúrio na biomassa
subterrânea, o que reflete a sua fitodisponibilidade (Canário, et al., 2007a). Na S.
marítima, não se observou nenhuma correlação em nenhuma das estações do ano,
embora haja uma tendência clara para o aumento de Hgraiz/Hgsed.
Figura 72 - Correlações entre os teores de Hg na fração sólida e na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal de Castro Marim.
A Figura 73 estabelece relações entre as concentrações de [MeHg]raiz e [MeHg]sed para
o mesmo sapal. Com base neste figura verificamos que para ambas as plantas houve
correlações tanto na estação da primavera como no inverno. Durante a primavera
houve uma relação polinomial durante em ambas as plantas. Isto sugere uma eficiente
absorção/adorção de MeHg pelas raízes após a sua produção nos sedimentos, com a
S. fruticosa (m = 1,55) a apresentar maior eficiência que a S. maritima (m = 0,78).
(Canário, et al., 2007a; Canário, et al., 2010).
No inverno a retenção do MeHg pelas raizes não ocorre tão eficientemente o que é
indicado pelas diferentes linhas de tendência observadas. Contudo verifica-se que a
variação sazonal, estabelece o mesmo tipo de relação entre plantas.
y = 9,10x - 1,34 r = 0,974 p< 0,05
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
0,0 0,1 0,2 0,3 0,4 0,5
[Hg] r
aiz (
µg g
-1)
[Hg]sed (µg g-1)
S. fruticosa Inverno Primavera
y = 1,14x - 0,07 R² = 0,2323
y = 0,81x + 0,02 R² = 0,0897
0,0
0,1
0,2
0,3
0,4
0,5
0,0 0,2 0,4 0,6
[Hg] r
aiz (
µg
g-1)
[Hg]sed (μg g-1)
S. maritima

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
110 Ricardo M.P.Mendes
Figura 73 - Correlações entre os teores de MeHg na fração sólida e MeHg na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal de Castro Marim.
Na Figura 74 verificamos que há uma correlação linear entre as concentrações de
MeHg e Hg na biomassa subterrânea, durante a primavera para ambas as plantas. O
declive da S. fruticosa é superior ao declive da S. maritima, o que siginifica que o
armazenamento de MeHg e Hg na raiz é proporcional, o que significa que a metilação
do mercúrio não diminui a capacidade que essa planta tem em extrair o Hg dos
sedimentos. O delive da S. fruticosa (m = 312,2) é superior ao da S. maritima (m =
209,8), o que significa que a primeira é mais eficiente na captura de Hg e MeHg por
parte da raiz.
Figura 74 – Correlações entre os teores de MeHg e Hg na biomassa subterrânea nos sedimentos colonizados recolhidos no sapal de Castro Marim.
Verifica-se que no sapal do Rosário não existe (Figura 75) relação entre [Hg]raiz e
[Hg]sed para a S. fruticosa, tanto no inverno como na primavera. No entanto, para a S.
maritima, verifica-se uma tendência exponencial durante o inverno, o que indica uma
relação entre o Hg no sedimento e o Hg capturado pela raiz.
y = 10,962x + 64,82 r = 0,5418 p< 0,05
y = 1,55x2 - 7,82x + 17,67 r = 0,753 p< 0,05
0
100
200
300
400
500
0 5 10 15 20
[MeH
g] r
aiz (
ng
g-1)
[MeHg]sed (ng g-1)
S. fruticosa Inverno Primavera
y = 26,96ln(x) - 13,67 r = 0,584 p< 0,05
y = 0,78x2 - 9,79x + 36,21 r = 0,723 p< 0,05
0
20
40
60
80
100
120
0 5 10 15 20 25
MeH
gra
iz (
ng
g-1)
[MeHg]sed (ng g-1)
S. maritima
y = -72,79x2 + 208,20x - 6,01 r = 0,831 p< 0,05
y = 312,19x - 80,39 r = 0,912 p< 0,05
0
100
200
300
400
500
0,0 0,5 1,0 1,5 2,0 2,5
[MeH
g] r
aiz (
ng g
-1)
[Hg]raiz(µg g-1)
S. fruticosa Inverno Primavera
y = 209,83x - 11,45 r = 0,626 p< 0,05
0
20
40
60
80
100
120
0,0 0,2 0,4 0,6
[MeH
g] r
aiz
(ng g
-1)
[Hg]raiz(µg g-1)
S. maritima

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 111
Figura 75 – Correlações entre os teores de MeHg e Hg na biomassa subterrânea nos sedimentos colonizados recolhidos no sapal do Rosário.
Na Figura 76 verificamos que no perfil para a S. fruticosa durante o inverno não há
qualquer tipo de correlação entre o [MeHg]raiz e o [MeHg]sed, contudo na primavera há
correlação que se pode traduzir na forma de uma função exponencial, ou seja há
estabelece-se uma ligação entre o metrilmercúrio do sedimento e a sua captura para a
raiz.
Na S. marítima não há correlação durante a primavera, contudo durante o inverno há
uma correlação linear, indicando que conforme aumenta o teor de metilmercúrio no
sedimento, aumenta linearmente o teor de metilmercúrio na raíz da planta.
Figura 76 - Correlações entre os teores de MeHg na fração sólida e MeHg na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal do Rosário.
Na figura 77, verificamos que para a S. fruticosa e para a S. maritima há uma
correlação linear na primavera, que está fortemente relacionada com o teor de
mercúrio total encontrado na raiz. O que indica que o armazenamento de MeHg e Hg
na raiz é proporcional, significando que a metilação do mercúrio não diminui a
capacidade que essa planta tem em extrair o Hg dos sedimentos.
O delive da S. fruticosa (m = 215,8) é superior ao da S. maritima (m = 177,2), o que
significa que a primeira é mais eficiente na captura de Hg e MeHg por parte da raiz.
0
2
4
6
8
10
12
0 1 2 3 4
[Hg] r
aiz (
µg
g-1)
[Hg]sed(µg g-1)
S. fruticosa Inverno Primavera
y = 0,51e0,63x r = 0,841 p< 0,05
0
4
8
12
16
20
0 1 2 3 4 5
[Hg] r
aiz
(µ
g g-1
)
[Hg]sed (μg g-1)
S. maritima
y = 0,10x1,90 r = 0,918 p< 0,05
0
500
1000
1500
2000
2500
3000
0 50 100 150 200 250
[MeH
g] r
aiz (
ng
g-1)
[MeHg]sed (ng g-1)
S. fruticosa Inverno Primavera
y = 3,2x - 136,48 r= 0,893 p< 0,05
0
500
1000
1500
2000
2500
3000
3500
0 100 200 300 400 500
[MeH
g] r
aiz (
ng
g-1)
[MeHg]sed (ng g-1)
S. maritima
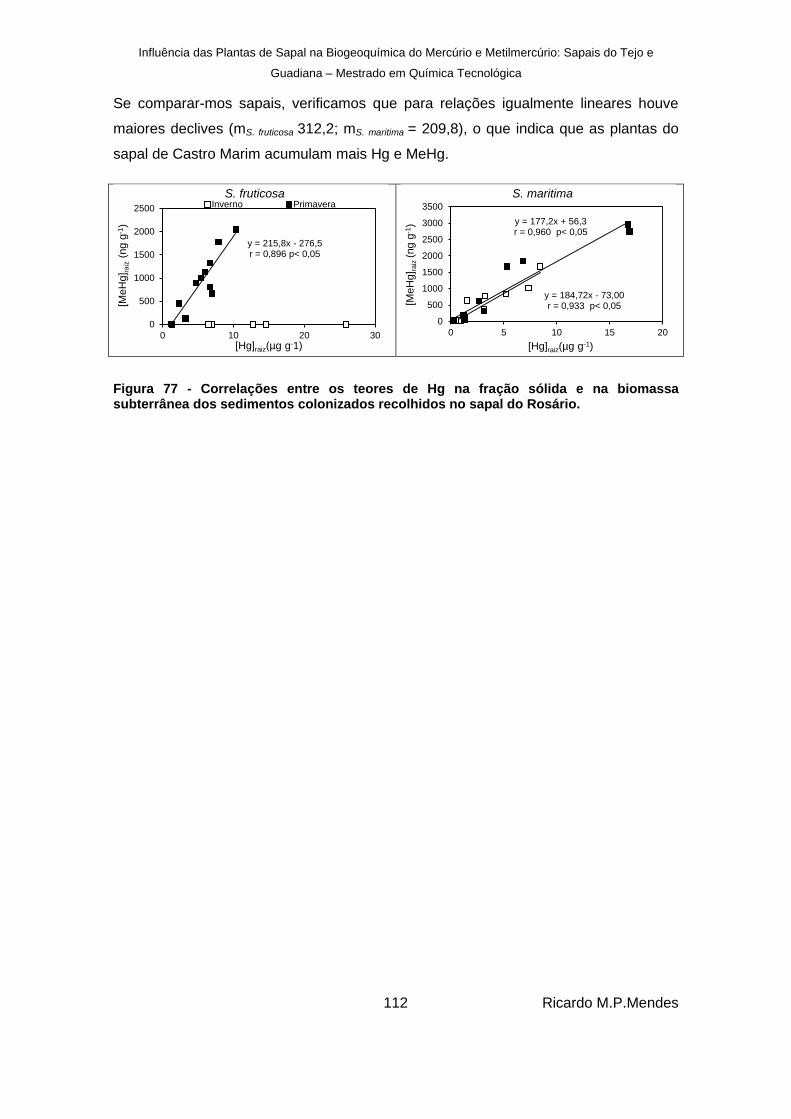
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
112 Ricardo M.P.Mendes
Se comparar-mos sapais, verificamos que para relações igualmente lineares houve
maiores declives (mS. fruticosa 312,2; mS. maritima = 209,8), o que indica que as plantas do
sapal de Castro Marim acumulam mais Hg e MeHg.
Figura 77 - Correlações entre os teores de Hg na fração sólida e na biomassa subterrânea dos sedimentos colonizados recolhidos no sapal do Rosário.
y = 215,8x - 276,5 r = 0,896 p< 0,05
0
500
1000
1500
2000
2500
0 10 20 30
[MeH
g] r
aiz (
ng g
-1)
[Hg]raiz(µg g-1)
S. fruticosa Inverno Primavera
y = 184,72x - 73,00 r = 0,933 p< 0,05
y = 177,2x + 56,3 r = 0,960 p< 0,05
0
500
1000
1500
2000
2500
3000
3500
0 5 10 15 20
[MeH
g] r
aiz
(ng g
-1)
[Hg]raiz(µg g-1)
S. maritima
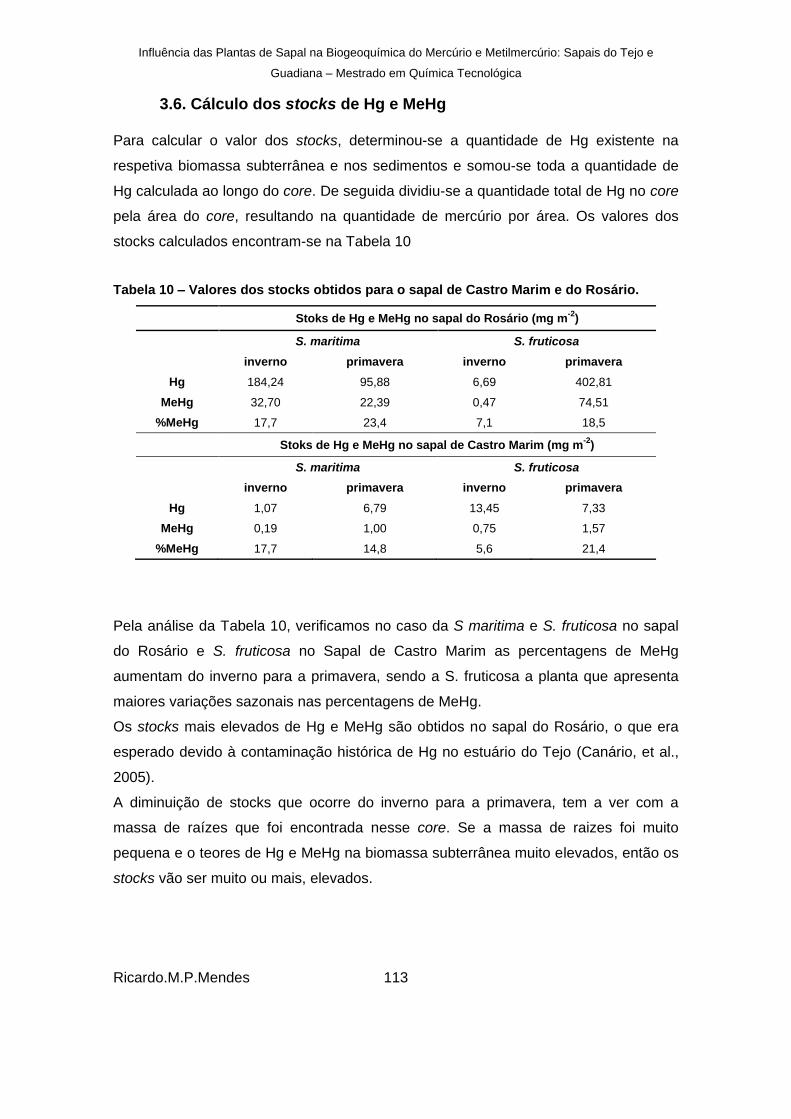
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 113
3.6. Cálculo dos stocks de Hg e MeHg
Para calcular o valor dos stocks, determinou-se a quantidade de Hg existente na
respetiva biomassa subterrânea e nos sedimentos e somou-se toda a quantidade de
Hg calculada ao longo do core. De seguida dividiu-se a quantidade total de Hg no core
pela área do core, resultando na quantidade de mercúrio por área. Os valores dos
stocks calculados encontram-se na Tabela 10
Tabela 10 – Valores dos stocks obtidos para o sapal de Castro Marim e do Rosário.
Stoks de Hg e MeHg no sapal do Rosário (mg m
-2)
S. maritima S. fruticosa
inverno primavera inverno primavera
Hg 184,24 95,88 6,69 402,81
MeHg 32,70 22,39 0,47 74,51
%MeHg 17,7 23,4 7,1 18,5
Stoks de Hg e MeHg no sapal de Castro Marim (mg m
-2)
S. maritima S. fruticosa
inverno primavera inverno primavera
Hg 1,07 6,79 13,45 7,33
MeHg 0,19 1,00 0,75 1,57
%MeHg 17,7 14,8 5,6 21,4
Pela análise da Tabela 10, verificamos no caso da S maritima e S. fruticosa no sapal
do Rosário e S. fruticosa no Sapal de Castro Marim as percentagens de MeHg
aumentam do inverno para a primavera, sendo a S. fruticosa a planta que apresenta
maiores variações sazonais nas percentagens de MeHg.
Os stocks mais elevados de Hg e MeHg são obtidos no sapal do Rosário, o que era
esperado devido à contaminação histórica de Hg no estuário do Tejo (Canário, et al.,
2005).
A diminuição de stocks que ocorre do inverno para a primavera, tem a ver com a
massa de raízes que foi encontrada nesse core. Se a massa de raizes foi muito
pequena e o teores de Hg e MeHg na biomassa subterrânea muito elevados, então os
stocks vão ser muito ou mais, elevados.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
114 Ricardo M.P.Mendes
Canário, et al, (2010), determinou os stocks de Hg e MeHg nas plantas S. fruticosa e
S. maritima, para o sapal de Alcochete na primavera. Os valores de stocks de Hg e
MeHg para a S. fruticosa foram de 930 mg m-2 e 74 mg m-2, respetivamente. Para a S.
maritima, os stocks de Hg e MeHg foram 220 mg m-2 e 28 mg m-2, respetivamente.
Comparando estes stocks com os stocks presentes neste estudo, verificamos que no o
sapal de Alcochete foram encontrados maiores quantidades de Hg por m2, contudo os
stocks de MeHg são muito próximos.
Devidos às baixas concentrações de Hg e MeHg nas partes aereas das plantas e nas
a contribuição destas é muito pequena, e por isso, não foram consideradas.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 115
3.7 Cálculo dos fluxos difusivos
Para examinar de que forma a difusão do HgD e de MeHgD existente nas àguas
intersticiais do sedimento, que contribui para a contaminação das águas
sobrenadantes, calcularam-se os fluxos difusivos do Hg e do MeHg.
Os fluxos difusivos (J), são estimados a partir da 1ª Lei de Fick (Schulz, 2000):
Onde representa a porosidade do sedimento, o quadrado a tortuosidade, Ds o
coeficiente de difusão (dm2 s-1) para o mercúrio iónico em água do mar, dC/dx a
variação da concentração de mercúrio iónico (mol dm-3) em função da coordenada
espacial x (dm).
O valor da porosidade pode ser calculado para cada camada através da expressão
(Berner, 1980; Caetano, 1998) :
Onde mA representa a massa de água intersticial (g) e mS a massa de sedimento seco
(g).
Os valores para o cálculo da porosidade, foram obtidos com base na determinação da
humidade do sedimento.
O cálculo do valor da tortuosidade, foi feito através da Lei de Boudreau (Boudreau,
1997):
Esta lei é usada para calcular o quadrado da tortuosidade para vários valores de
porosidade. Os coeficientes de difusão para o Hg e MeHg, foram obtidos por valores
tabelados (Schulz, 2000; Gill & Bruland, 1990).
As concentrações de de Hg e MeHg dissolvido nas águas sobrenadantes foram
retirados da Tabela 8 e Tabela 9.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
116 Ricardo M.P.Mendes
A Tabela 11, apresenta os valores dos fluxos difusivos com a indicação dos respetivos
sentidos entre HgD e MeHgD presente na camada de sedimentos entre 0 e 2 cm de
sedimento e o HgD e MeHgD presente nas águas sobrenadantes.
Pelos valores e sentidos dos fluxos, verificamos que quanto ao mercúrio, todos os
sentidos foram negativos, o que indica que há uma difusão de mercúrio do sedimento
para as àguas sobrenadantes, resultando assim numa contaminação do meio
aquático.
Quanto ao MeHg verificamos que apresenta valores de fluxos difusivos positivos e
valores negativos, o que significa que há diferentes sentidos de fluxo.
Isto significa que a presença de maiores teores de MeHg em águas sobrenadantes do
que nos sedimentos, faz com haja difusão de MeHg da coluna de água para os
sedimentos (valor de fluxo positivo, sentido descendente). A presença de maiores
teores de MeHg em sedimentos do que nas águas sobrenadantes, faz com haja
difusão de MeHg dos sedimentos para a coluna de água (valor de fluxo negativo,
sentido ascendente).
Tabela 11 - Valores dos fluxos difusivos com a indicação dos respetivos sentidos, para o HgD nos sedimentos e HgD nas águas sobrenadantes
Sapal Tipo de
sedimento Estação do ano
JHg(ng m-2
s-1
) Sentido do Fluxo
de Hg JMeHg(ng dm
2 s
-1)
Sentido do Fluxo
de Hg
Ro
sá
rio
S. fruticosa inverno -7,83E-05
-5,55E-06
primavera -5,46E-05 -1,81E-06
S. maritima inverno -5,57E-04 -1,22E-06
primavera -2,95E-05 1,82E-07
Não colonizados
inverno -7,12E-05 3,21E-06
primavera -1,87E-05 1,30E-06
Ca
str
o M
ari
m
S. fruticosa inverno -3,36E-05 -3,09E-06
primavera -1,05E-06 -2,99E-07
S. maritima inverno -3,13E-05 -4,03E-06
primavera -7,17E-06 -2,85E-07
Não colonizados
inverno -2,03E-06 1,33E-06
primavera -1,48E-05 -1,20E-07
Resultados identicos tem sido observados no estuário do Tejo, para o MeHg (Cesário
et. al., dados não publicados), e que podem ser explicados, com a maior ou menor
degradação do MeHg na coluna de água, devido a inumeros factores ambientais, nos
quais se destacam a concnetração de oxigénio dissolvido, o pH, e o potencial redox.
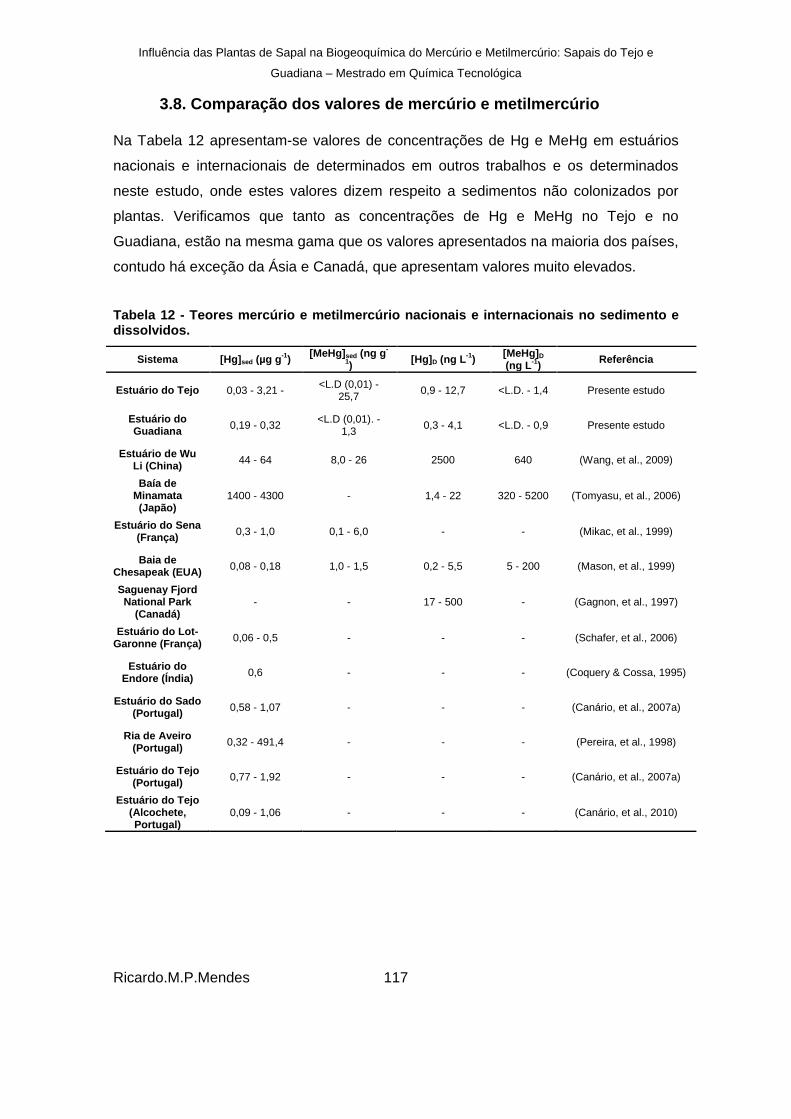
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 117
3.8. Comparação dos valores de mercúrio e metilmercúrio
Na Tabela 12 apresentam-se valores de concentrações de Hg e MeHg em estuários
nacionais e internacionais de determinados em outros trabalhos e os determinados
neste estudo, onde estes valores dizem respeito a sedimentos não colonizados por
plantas. Verificamos que tanto as concentrações de Hg e MeHg no Tejo e no
Guadiana, estão na mesma gama que os valores apresentados na maioria dos países,
contudo há exceção da Ásia e Canadá, que apresentam valores muito elevados.
Tabela 12 - Teores mercúrio e metilmercúrio nacionais e internacionais no sedimento e dissolvidos.
Sistema [Hg]sed (µg g-1)
[MeHg]sed (ng g-
1)
[Hg]D (ng L-1
) [MeHg]D (ng L
-1)
Referência
Estuário do Tejo 0,03 - 3,21 - <L.D (0,01) -
25,7 0,9 - 12,7 <L.D. - 1,4 Presente estudo
Estuário do Guadiana
0,19 - 0,32 <L.D (0,01). -
1,3 0,3 - 4,1 <L.D. - 0,9 Presente estudo
Estuário de Wu Li (China)
44 - 64 8,0 - 26 2500 640 (Wang, et al., 2009)
Baía de Minamata (Japão)
1400 - 4300 - 1,4 - 22 320 - 5200 (Tomyasu, et al., 2006)
Estuário do Sena (França)
0,3 - 1,0 0,1 - 6,0 - - (Mikac, et al., 1999)
Baia de Chesapeak (EUA)
0,08 - 0,18 1,0 - 1,5 0,2 - 5,5 5 - 200 (Mason, et al., 1999)
Saguenay Fjord National Park
(Canadá) - - 17 - 500 - (Gagnon, et al., 1997)
Estuário do Lot-Garonne (França)
0,06 - 0,5 - - - (Schafer, et al., 2006)
Estuário do Endore (Índia)
0,6 - - - (Coquery & Cossa, 1995)
Estuário do Sado (Portugal)
0,58 - 1,07 - - - (Canário, et al., 2007a)
Ria de Aveiro (Portugal)
0,32 - 491,4 - - - (Pereira, et al., 1998)
Estuário do Tejo (Portugal)
0,77 - 1,92 - - - (Canário, et al., 2007a)
Estuário do Tejo (Alcochete, Portugal)
0,09 - 1,06 - - - (Canário, et al., 2010)
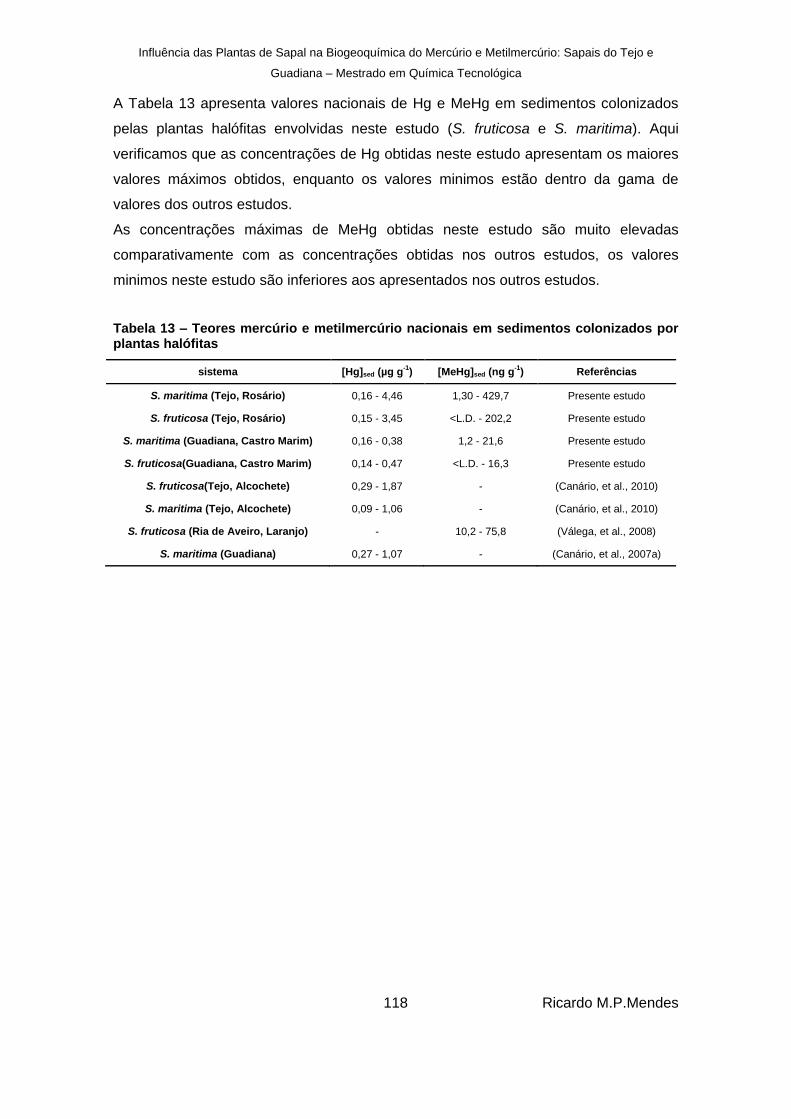
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
118 Ricardo M.P.Mendes
A Tabela 13 apresenta valores nacionais de Hg e MeHg em sedimentos colonizados
pelas plantas halófitas envolvidas neste estudo (S. fruticosa e S. maritima). Aqui
verificamos que as concentrações de Hg obtidas neste estudo apresentam os maiores
valores máximos obtidos, enquanto os valores minimos estão dentro da gama de
valores dos outros estudos.
As concentrações máximas de MeHg obtidas neste estudo são muito elevadas
comparativamente com as concentrações obtidas nos outros estudos, os valores
minimos neste estudo são inferiores aos apresentados nos outros estudos.
Tabela 13 – Teores mercúrio e metilmercúrio nacionais em sedimentos colonizados por plantas halófitas
sistema [Hg]sed (µg g-1) [MeHg]sed (ng g
-1) Referências
S. maritima (Tejo, Rosário) 0,16 - 4,46 1,30 - 429,7 Presente estudo
S. fruticosa (Tejo, Rosário) 0,15 - 3,45 <L.D. - 202,2 Presente estudo
S. maritima (Guadiana, Castro Marim) 0,16 - 0,38 1,2 - 21,6 Presente estudo
S. fruticosa(Guadiana, Castro Marim) 0,14 - 0,47 <L.D. - 16,3 Presente estudo
S. fruticosa(Tejo, Alcochete) 0,29 - 1,87 - (Canário, et al., 2010)
S. maritima (Tejo, Alcochete) 0,09 - 1,06 - (Canário, et al., 2010)
S. fruticosa (Ria de Aveiro, Laranjo) - 10,2 - 75,8 (Válega, et al., 2008)
S. maritima (Guadiana) 0,27 - 1,07 - (Canário, et al., 2007a)

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 119
V. Conclusões Gerais
Deste trabalho resultaram as seguintes observações:
O sapal de Castro Marim apresenta maiores teores e maior variação sazonal
nos teores de Fe, Mn e Al, face sapal do Rosário. Os sedimentos colonizados
retêm maiores teores destes metais comparativamente aos sedimentos não
colonizados.
Concluímos que a atividade das raízes influência significativamente os teores
de AVS e de sulfatos dissolvidos, pois comparativamente com os teores dos
sedimentos não colonizados, os teores de AVS e de sulfatos dissolvidos em
sedimentos colonizados, foram muito baixos. Durante a primavera, quando as
plantas apresentam maior atividade, os teores de AVS e de sulfatos dissolvidos
diminuiram face ao inverno.
Verificamos que os teores de Hg e MgHg na fração sólida do sedimento no
sapal do Rosário são uma ordem de grandeza mais elevados do que no sapal
de Castro Marim, o que demonstra a contaminação a que o sapal do Rosário
está sujeito.
No sapal do Rosário os teores de Hg e MeHg dissolvidos, são maiores no
inverno do que na primavera. No sapal de Castro Marim, os teores de Hg e
MeHg dissolvidos são maiores na primavera.
Os teores de Hg e MeHg nas partes aéreas, são mais baixos que os teores na
biomassa subterrânea das plantas. Deste modo, há uma fraca distribuição de
Hg e MeHg das raízes para as partes aéreas, possivelmente por bloqueio da
passagem de Hg e MeHg das raízes para as partes aéreas das plantas. Os
baixos teores também podem estar associados a mecanismo remoção de Hg e
MeHg, por redução a Hg0 e produção de dimetilmercúrio.
De uma maneira geral, verificou-se que os teores de Hg e MeHg na biomassa
subterrânea são muito superiores aos teores de Hg e MeHg no sedimento, e
que as plantas tendem a acumular a maioria do Hg e MeHg na biomassa
subterrânea

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
120 Ricardo M.P.Mendes
Relatório de ambiente e segurança
Neste relatório estão presentes as situações de maior exigência em termos de
segurança assim como as substâncias que exigiram maiores cuidados de
manuseamento.
Durante a realização do projeto houve procedimentos que exigram maiores situações
cuidados e atenções. A descontaminação do material foi realizada numa sala
apropriada para o efeito, onde como medidas de seguranças teve-se de recorrer a
mascára, devido aos vapores dos àcidos nitrícos e cloridrico.
Ácido Nítrico
símbolo de perigosidade
No. CAS : 7697-37-2 Frase(s) - R R 8 Favorece a inflamação de matérias combustíveis. R35 Provoca queimaduras graves. Frase(s) - S S23 Não respirar os gases/vapores/fumos/aerossóis. S26 Em caso de contacto com os olhos, lavar imediata e abundantemente com água e consultar um especialista. S36 Usar vestuário de protecção adequado. S45 Em caso de acidente ou de indisposição, consultar imediatamente o médico (se possível mostrar-lhe o rótulo).
Ácido clorídrico
Frase(s) - R R 34 Provoca queimaduras. R 37 Irritante para as vias respiratórias. Frase(s) - S S 26 Em caso de contacto com os olhos, lavar imediata e abundantemente com água e consultar um especialista. S 45 Em caso de acidente ou de indisposição, consultar imediatamente o médico (se possível mostrar-lhe o rótulo).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 121
No processo de digestão, houve especial cuidado com a utilização do àcidos
fluoridrico. Todo o procedimento foi realizado na hotte com várias de luvas sobre as
mãos.
Os desperdicios da resultantes da titulação para determinação de cloretos, devido á
sua perigosidade parao meio aquático, foram armazenados em frascos e guardados
para recolha de residuos. O mesmo procedimento ocorreu para os desperdicios para
as soluçoes mistura ne determinação de sulfatos.
Ácido Fluorídrico
No. CAS : 7664-39-3
Frase(s) - R R 26/27/28 Muito tóxico por inalação, em contacto com a pele e por ingestão. R 35 Provoca queimaduras graves. Frase(s) - S S 7/9 Manter o recipiente bem fechado em local bem ventilado. S 26 Em caso de contacto com os olhos, lavar imediata e abundantemente com água e consultar um especialista. S 36/37 Usar vestuário de protecção e luvas adequadas. S 45 Em caso de acidente ou de indisposição, consultar imediatamente o médico (se possível mostrar-lhe o rótulo).
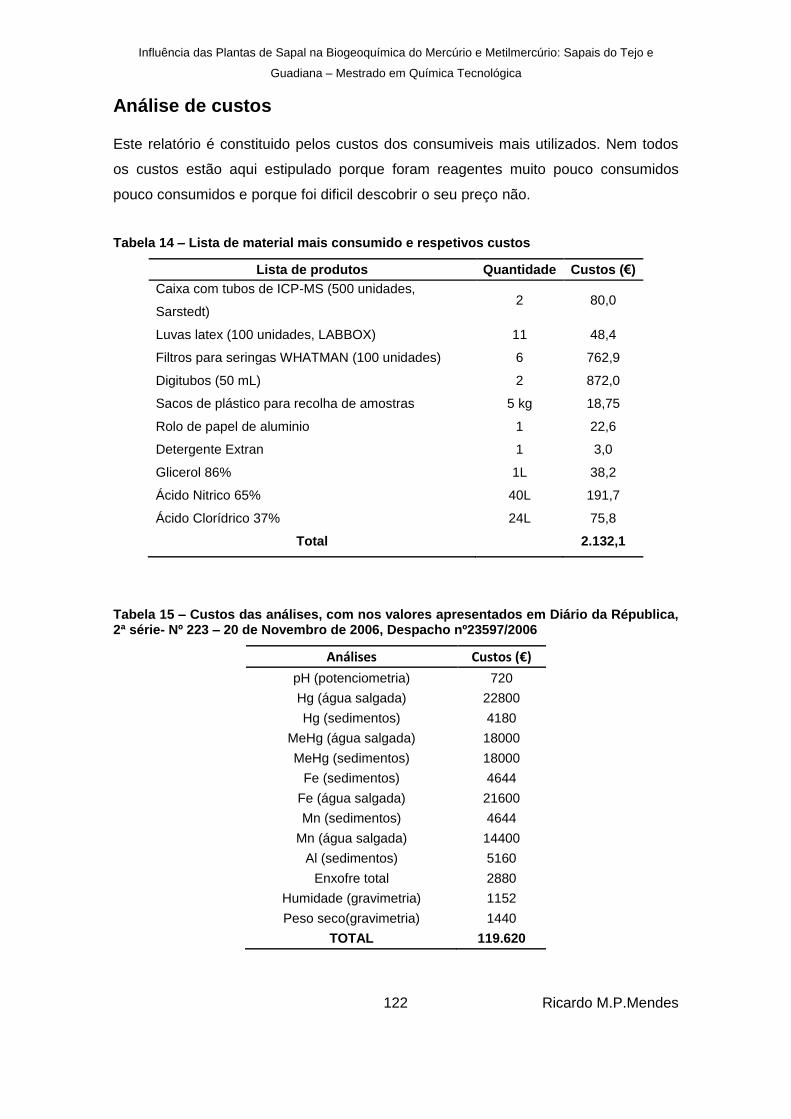
Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
122 Ricardo M.P.Mendes
Análise de custos
Este relatório é constituido pelos custos dos consumiveis mais utilizados. Nem todos
os custos estão aqui estipulado porque foram reagentes muito pouco consumidos
pouco consumidos e porque foi dificil descobrir o seu preço não.
Tabela 14 – Lista de material mais consumido e respetivos custos
Lista de produtos Quantidade Custos (€)
Caixa com tubos de ICP-MS (500 unidades,
Sarstedt) 2 80,0
Luvas latex (100 unidades, LABBOX) 11 48,4
Filtros para seringas WHATMAN (100 unidades) 6 762,9
Digitubos (50 mL) 2 872,0
Sacos de plástico para recolha de amostras 5 kg 18,75
Rolo de papel de aluminio 1 22,6
Detergente Extran 1 3,0
Glicerol 86% 1L 38,2
Ácido Nitrico 65% 40L 191,7
Ácido Clorídrico 37% 24L 75,8
Total
2.132,1
Tabela 15 – Custos das análises, com nos valores apresentados em Diário da Républica, 2ª série- Nº 223 – 20 de Novembro de 2006, Despacho nº23597/2006
Análises Custos (€)
pH (potenciometria) 720
Hg (água salgada) 22800
Hg (sedimentos) 4180
MeHg (água salgada) 18000
MeHg (sedimentos) 18000
Fe (sedimentos) 4644
Fe (água salgada) 21600
Mn (sedimentos) 4644
Mn (água salgada) 14400
Al (sedimentos) 5160
Enxofre total 2880
Humidade (gravimetria) 1152
Peso seco(gravimetria) 1440
TOTAL 119.620

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 123
Apêndice
1.1 Técnicas de análise
A elaboração do projeto de estágio a decorrer no IPMA I.P., tem como objetivo o
estudo de processos biogeoquímicos que envolvem o mercúrio (Hg) e o metilmercúrio
(MeHg) em sapais. Para o estudo desses processos é necessário caracterizar
quimicamente o sedimento, através da quantificação de elementos importantes para o
estudo da biogeoquímica do Hg como o ferro (Fe), alumínio (Al), manganês (Mn). Para
esta quantificação utiliza-se geralmente a espectroscopia de Absorção Atómica com
atomização à chama (EAA).
1.2.1 espectroscopia de Absorção Atómica por Chama
Esta técnica foi utilizada para determinar as concentrações totais de ferro, manganês,
alumínio e silício em sedimentos.
O modelo de funcionamento consiste nos seguintes procedimentos: o
espectrofotómetro aspira uma amostra líquida que é conduzida através de um capilar,
sendo nebulizada e misturada com um gás combustível, onde tem de passar por
defletores de modo a recolher pequenas gotas de líquido existente. A mistura amostra,
gás combustível e gás oxidante são conduzidos ao queimador onde são lançados
numa chama devidamente alinhada a um feixe de luz (emitido por uma lâmpada
especial) com o comprimento de onda de interesse, sendo assim possível medir a
quantidade de radiação absorvida pelos átomos. (Skoog & Leary, 1992; Gonçalves,
2001))
O tipo de chama que se utiliza depende do tipo de metal, pois a maioria dos elementos
tem uma temperatura apropriada para a sua atomização. O tipo de chama mais
utilizado é a de Ar/Acetileno, cuja gama de temperatura varia entre os 2100-2400ºC
(considerada moderadamente quente e rica em gás combustível), sendo este tipo de
chama que determinará as quantidades de Fe e Mn. Para o Al e Si, devido à facilidade
que estes elementos têm em formar óxidos muito refratários é necessário um tipo de
chama mais quente e por isso utiliza-se uma chama Ar/Acetileno e Protóxido de Azoto
(Óxido Nitroso) (Gonçalves, 2001).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
124 Ricardo M.P.Mendes
1.2.2. espectroscopia de Absorção Atómica Eletrotérmica
Este método foi utilizado para determinar a concentração de Hg total no sedimento.
O método de atomização eletrotérmica permite medir diretamente amostras sólidas,
sendo a calibração um problema especial (Bings, et al., 2010). Esta técnica permite
assim, determinar diretamente a concentração de Hg existente em amostras sólidas. O
equipamento AMA-254 (Advanced Mercury Analyser) da LECO Instruments serve para
análise direta de Hg de amostras sólidas (Costley, et al., 2000), podendo ser
analisadas massas entre os 30 e 300 mg.
O modo de funcionamento consiste numa amostra sólida que é colocada numa barca
de níquel, sendo esta barca introduzida num tubo de decomposição constituído por
quartzo, contendo um catalisador (oxalato de cobalto com uma mistura de óxidos de
manganês, cobalto e acetato de cálcio). A amostra é inicialmente seca, a 120ºC,
seguindo-se a decomposição térmica a 750ºC numa atmosfera rica em oxigénio (gás
de arraste). O mercúrio libertado na forma de Hg0 é capturado e concentrado numa
amálgama de ouro, sendo separado de outros produtos da decomposição. A
amálgama é aquecida a 900ºC para libertar o Hg que é transportado para a célula de
leitura (sistema de deteção), que se encontra a uma temperatura de 120ºC, para a
análise por EAA, a um comprimento de onda específico de 253,7 nm, por incidência de
uma fonte de luz.
Figura 78 – Esquema de funcionamento do “Advanced Mercury Analyser 254” (adaptado de Costley et al., 2000).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 125
O tempo total da análise é de 5 minutos para amostras de sedimentos e biológicas,
distribuídos pelas etapas na seguinte forma: tempo de secagem, 10 segundos; tempo
de decomposição, 150 segundos e tempo de espera 45 segundos ( (Costley, et al.,
2000; LECO, 2012). As vantagens desta técnica são significativas na medida em que
se consegue realizar uma análise ao Hg total significativamente rápida (5 min); não
necessita de preparação prévia da amostra ou de um longo processo de digestão, isto
é, de processos para extrair os metais da amostra e dissolve-los numa solução (ao
contrário do que acontece com a absorção atómica onde é necessário a digestão das
amostras) (Costley, et al., 2000).
Como desvantagens, não permite uma correção do ruído de fundo, e pode ter
problemas com as amostras (sólidas ou líquidas) que contenham significativas
concentrações de compostos orgânicos. A eficiência e seletividade da amálgama
ainda não foram totalmente investigadas, podendo a amálgama sofrer com o “efeito de
memória”. O limite de deteção foi calculado em 0,12 mg/g e os intervalos de
concentração entre 0-50 e 0-1600 ng (Costley, et al., 2000).
1.2.3. espectroscopia de Fluorescência Atómica – Vapor Frio
com Injeção de Fluxo
O método para determinar o Hg total dissolvido em águas intersticiais e sobrenadantes
por oxidação, purga, captura em através de sistema de espectroscopia de
Fluorescência Atómica por Vapor Frio com Injeção de Fluxo (em inglês FI-CV-AFS). É
um método que apresenta um limite de deteção de 0,2 ng/L, quando não há
interferências significativas presentes, para um intervalo entre os 0,3 – 100 ng/L (EPA,
2002). É utilizado um sistema automático de Injeção de Fluxo em todo o sistema
(Leopold, et al., 2008; Leopold, et al., 2009).
O método inicia-se fora do aparelho, onde todo o Hg dissolvido presente na amostra é
oxidado a Hg2+ com BrCl, (EPA, 2002; Leopold, et al., 2008). Seguidamente adiciona-
se NH2OH.HCl, para eliminar o excesso de BrCl existente na solução. Pela adição de
SnCl2, o Hg2+ é reduzido a Hg0, sendo este passo de extrema importância, pois
garante que o Hg passe à sua forma mais volátil. O Hg0 é purgado da solução através
de um separador gás-líquido, sendo o Árgon utilizado como gás de arraste, e pré-
concentrado numa amálgama de ouro; (Method, 2003; Leopold, et al., 2008; Leopold,

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
126 Ricardo M.P.Mendes
et al., 2009; EPA, 2002). Após este processo, o Hg0 é desadsorvido termicamente
(450ºC) da amálgama de ouro, e transportado diretamente para a célula de
Fluorescência Atómica para determinação analítica (EPA, 2002; Leopold, et al., 2009).
O equipamento, utilizado neste trabalho, consiste num Merlin Mercury Detetor mod
10.025S, PS Analytical Ltd, com sistema de injeção de fluxo conduzida por uma
bomba peristáltica multicanal e separador gás-liquido (Gómes-Ariza, et al., 2004).
1.2.4. espectroscopia de Fluorescência Atómica –
Cromatografia Gasosa
Para determinar os compostos orgânicos de Hg, principalmente o metilmercúrio
(MeHg) acoplou-se a Cromatografia Gasosa com Pirólise à espectroscopia de
Fluorescência Atómica (CG-Pi-EFA, em inglês representada pelas siglas CG-Py-AFS)
(Bings, et al., 2010), através do método de determinação do MeHg em águas por
Destilação, Etilação aquosa, Purga, Captura e deteção por EFA, da EPA (EPA, 1998).
Esta técnica tem a vantagem de ter um intervalo de deteção entre 0,02 – 5ng/L (Diez,
et al., 2009).
O método consiste numa destilação prévia da amostra, a 125ºC sob fluxo de Azoto.
Após a destilação, o pH do destilado é ajustado a 4,9 com um tampão de Acetato e
etilada (para converter todo o MeHg em metiletilmercúrio, mais volátil) num vaso
fechado de purga através da adição de Tetraetil Borato de Sódio (NaBEt4). O
metiletilmercúrio é separado da solução por purga com N2 para um coletor (Carbontrap
®) de carbono grafítico ( (EPA, 1998)). O metiletilmercúrio capturado é termicamente
desadsorvido do Carbontrap®, para um gás inerte que conduz o metiletilmercúrio
libertado, através de uma coluna cromatográfica pirolítica de decomposição, aquecida
a 800ºC, para converter as formas orgânicas de mercúrio em mercúrio elementar (Hg0)
e então este é conduzido para a célula de Fluorescência Atómica para deteção (EPA,
1998; Diez, et al., 2009).
O equipamento utilizado consiste num Merlin Mercury Detetor mod. 10.023S, PS
Analytical utilizando um pirolisador PS Analytical 10.565 (Evans, et al., 1999; Gómes-
Ariza, et al., 2004) e um cromatógrafo Agillent, 6890 Series, GC System.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 127
1.2.5. espectroscopia Molecular Ultravioleta - Visível
Quando um fotão de radiação incide numa partícula ocorre a absorção, caso a energia
do fotão for igual à diferença exata de energia do estado fundamental e o estado de
energia mais elevado. A energia do fotão é transferida para a partícula, passando esta
a encontrar-se num estado de energia mais elevado (estado excitado). A absorção de
radiação ultravioleta e visível por parte das moléculas ocorre em uma ou mais bandas
de absorção, em que cada uma das quais é composta por inúmeras níveis discretos
de energia bastante próximos um dos outros. Cada uma destes níveis é originado pela
transição de um eletrão do estado eletrónico fundamental para um dos muitos níveis
de energia rotacional e vibracional do estado eletrónico excitado.
No espectrofotómetro de UV-Visível a energia é originada por uma lâmpada de
tungsténio que emite radiação na gama do ultravioleta-visível. Este género de
equipamentos está apetrechado com filtros, em que cada um transmite uma diferente
porção do estado. A solução do analíto absorve a radiação da sua cor complementar,
sendo esta a cor selecionada pelo filtro. A energia com o comprimento de onda
selecionado passa pelo filtro e de seguida pela solução amostra, sendo então
registada por uma fotocélula. A radiação transmitida é então medida por comparação
com uma célula de solvente em que percentagem de transmitância é 100. (Skoog, et
al., 1996)
1.2.6. Polarografia
A polarografia é um caso particular de voltametria, cuja resposta é originada pela
combinação do transporte de massa por difusão e convecção. Na polarografia linear é
utilizado um elétrodo de mercúrio gotejante, DME (“dropping mercury electrode”) e é
efetuado um varrimento linear do potencial. Uma representação da corrente em função
da diferença de potencial demonstra as oscilações de corrente originadas pelas gotas
de mercúrio que caem do capilar. As concentrações de sulfuretos inorgânicos
dissolvidos nas amostras de água são, em geral, determinadas com um elétrodo de
mercúrio, aproveitando a facilidade de oxidação do mercúrio devido à precipitação do
sulfureto.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
128 Ricardo M.P.Mendes
Usando as técnicas de polarografia diferencial com impulsos (DPP) e de voltametria
de redissolução catódica (DPCSV), determina-se o teor em sulfuretos ficando o
precipitado adsorvido à superfície da gota de mercúrio num passo prévio, consoante a
concentração da amostra (Madureira, 1997).
Na voltametria diferencial de redissolução catódica, o analito é primeiro adsorvido no
minielétrodo durante um determinado período de tempo, formando um composto
insolúvel, a um potencial mais positivo do que o valor de equilíbrio elétrodo/composto
insolúvel/anião em solução.
A análise da solução contendo HS-, fez-se por Polarografia Diferencial por Impulsos
(DPP), onde se utilizou um elétrodo getejante de mercúrio (DME), onde ocorrem as
seguintes reações:
Os iões HS- reagem com o mercúrio do elétrodo formando sulfureto de mercúrio que é
adsorvido a este. A deposição do sulfureto é feita durante 60s a um potencial
constante de -400 mV.
HS- + Hg HgS (precipita) + 2e-
É então feito um varrimento catódico entre -400 e 900 mV com uma velocidade de 5
mV s-1. Nesta fase, o sulfureto de mercúrio é “redissolvido” passando o ião sulfureto à
solução e regenerando o mercúrio do elétrodo.
2e- + HgS (precipitado) Hg + S2-
Tal como na técnica de DPP, os iões HS- reagem com o mercúrio do elétrodo,
formando o sulfureto de mercúrio, que é adsorvido a este. Numa segunda fase o
sulfureto de mercúrio é redissolvido, regenerando-se o mercúrio do elétrodo e o ião
sulfureto passa à solução (Canário, 2000).

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 129
Glossário
Meios Anóxicos Meios com maior potencial redutor e elevado teor em espécies
reduzidas.
Meios Óxicos Meios com maior potencial oxidante e elevado teor em espécies
oxidadas.
Meios Subóxicos Meios compreendidos entre os meios óxicos e anóxicos com
propriedades e espécies intermédias dos dois meios.
Sapal De uma forma geral, é como uma pradaria de halófitas em sedimentos
aluviais na fronteira de corpos de água salgada cujo o nível varia com as marés.
Estuário Localização geral das zonas onde os rios encontram o mar, e onde a água
dos rios se mistura com a água do mar.
Rizosfera Camada sedimentar que é diretamente influenciada pelas raízes das
plantas, sendo importante na determinação de processos na mobilidade de metais e a
sua disponibilidade para reações físico-químicas.
Corer Peça metálica com uma extremidade em forma semicilíndrica utilizada para
retirar amostras de sedimentos
Core Amostras de sedimentos obtidas através do corer, em forma cilíndrica.
Tortuosidade Medida de difusão das águas intersticiais entre os poros do
sedimentos.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
130 Ricardo M.P.Mendes
Referências
Andrew, A. & Nriagu, J., 1979. The global cycle of mercury. In: J. Nriagu, ed. The biogeochemistry of mercury in the environment. s.l.:Elsevier, North Holland Biomedical Press, pp. 1 - 21.
Antunes Dias, A. & Marques, J., 1999. Estuários. Estúário do Tejo: o seu Valor e um pouco da sua História. Alcochete: Reserva Natural do Estuário do Tejo.
APHA, 1995. Standard methods for the examination of water and waste water. 19ª Ed ed. Washington DC: American Public Health Association.
Avramescu, M.-L.et al., 2011. Biogeochemical factors influencing net mercury methylation in contaminated freshwaters sedimens from the St. Lawrence River in Conrwall, Ontario, Canada. Science of the total Environment, Volume 409, pp. 968-978.
Baldi, F., 1997. Metal ions in Biological Systems. New York: Marcel Dekker, Inc.
Beefting, W., Nieuwenhuize, J., Stoppler, M. & Mohl, C., 1982. Heavy metal accumulation in salt marshes from the Western and Eastern Scheldt. Science of the Total Environment, Volume 25, pp. 199-223.
Berner, R., 1980. Early Diagenesis. A theorical Approach, USA: Princeton University Press.
Bings, N. H., Bogaerts, A. & Broekaert, J. A., 2010. Atomic Spectroscopy: A review. Analytical Chemistry, Volume 82, pp. 4653-4681.
Boudreau, B., 1997. Diagenetic models and their impletation: modelling trasport and reactions in aquatic sediments. Berlin, Heidelberg: Springer.
Brouwer, M., Hoexum-Brouwer, T. & Cashon, R., 1993. a putative gluthatione-binding site in CdZn-metallothionein identified by equilibrium binding and molecular-modellig studies. Biochem. J., Volume 1993, pp. 219-225.
Caçador, I. & Vale, C., 2001. Salt Marshes. In: M. Dekker, ed. Metals in the Environment. Basel(New York): Marcel Dekker, Inc.
Caçador, M., Vale, C. & Catarino, F., 2000. Seasonal variation of Zn, Pb, Cu, and Cd concentrations in the root-system os Spartina maritima and Hamilione portulacoides from Tagus estuary salt marshes. Mar. Environ. Res., Volume 49, pp. 279-290.
Caetano, M., 1998. Biogeoquímica do manganês, ferro, cobre e cádmio em sedimentos da Ria Formosa. Disssertação apresentada à Universidade do Algarve para a obtenção do Grau de Doutor em Ciências do Mar, s.l.: s.n.
Caetano, M., Caçador, I., Duarte, B. & Vale, C., 2009. Stock and losses of trace metals from salt marsh plants. Marine Environmental Research, Volume 67, pp. 75-82.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 131
Caetano, M., Vale, C., Cesário, R. & Fonseca, N., 2008. Evidence for preferential depths retention in roots of salt marsh plants. Science of the Total Environment, Volume 390, pp. 466-474.
Canário, J., 2000. Mercúrio em Sedimentos Contaminados e Águas Intersticiais da Cala do Norte do Estuário do Tejo, Lisboa: Faculdade de Ciências e Tecnologias.
Canário, J., 2004a. Mercúrio e Monometilmercúrio na cala do norte do estuário do Tejo – Diagénese, trocas com a coluna de água e interacção com o biota, s.l.: s.n.
Canário, J., Antunes, P., Lavrado, J. & Vale, C., 2004c. Simple method for monomethylmercury determination in estuarine sediments. Trends in Analytical Chemistry, Volume 23, pp. 798-805.
Canário, J., Branco, V. & Vale, C., 2007b. Seasonal variation of monomethylmercury concentration in surface sediments of the Tagus Estuary (Portugal);. Environmental Pollution, Volume 148, pp. 380-383.
Canário, J., Caetano, M. & Vale, C., 2006. Validation and application of an analytical method for monomethylmercury quantification in aquatic plant tissues. Analytical Chimica Acta, Volume 580, pp. 258-262.
Canário, J. & Vale, C., 2004b. Rapid release of mercury from intertidal sediments exposed to solar radiation: A field experiment. Environmental Science & Technology, Volume 38, pp. 3901-3907.
Canário, J., Vale, C. & Caetano, M., 2003a. Mercury in sediments and pore waters at contaminated site in the Tagus. Ciencias Marinas, 29(4).
Canário, J., Vale, C. & Caetano, M., 2005. Distribution of Monomethylmercury and mercury in surface sediments of the Tagus Estuary (Portugal). Marine pollution Bulletin, Volume 50, pp. 1121-1145.
Canário, J., Vale, C., Caetano, M. & Cesário, R., 2007a. Evidence for elevated production of Methylmercuryin Salt Marshes;. Environmental Science Technology, Volume 41, pp. 7376-7382.
Canário, J., Vale, C., Caetano, M. & Madureira, M. J., 2003b. Mercury in contaminated sediments and pore waters enriched in sulphate (Tagus Estuary, Potugal). Environmental Pollution, Volume 126, pp. 425-433.
Canário, J., Vale, C. & Nogueira, M., 2008b. The pathway of mercury in contaminated waters determined by association with organic carbon (Tagus Estuary, Portugal). Applied Geochemistry, Volume 148, pp. 380 - 383.
Canário, J. et al., 2010. Mercury in sediments and vegetation in a moderately contaminated salt marsh (Tagus Estuary, Portugal). Journal of Environmental Sciences, Volume 22, pp. 1151-1157.
Carvalho, A., 2009. Mecanismos de resposta a mercúrio em plantas de sapal, Aveiro: Universidade de Aveiro.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
132 Ricardo M.P.Mendes
Castro, R. et al., 2009. Accumulation, distribution and cellular partitioning of mercury in several halophytes of a contaminated salt marsh. Chemosphere, Volume 76, pp. 1348-1355.
Catarino, F. & Caçador, I., 1981. Produção de Biomassa e estratégia de Desenvolvimento em Spartina Maritima e outros Elementos da Vegetação dos Sapais do Estuátio do Tejo. Boletim da Sociedade Broteriana, 34(Série 2).
Choi, S. & Bartha, R., 1994. Cobalamin-mediated mercury methylation by Desulfovibrio desulfuricans L.S.. Apllied and Environmental Microbiology, Volume 59, pp. 290 - 295.
Compeau, G. & Bartha, R., 1985. Sulphate-reducing bacteria: principal methylators of mercury in anoxic estuarine sediment. Applied and Environmental Microbiology, Volume 50, pp. 498 - 502.
Compeau, G. C. & Bartha, R., 1984. Methylation and demethylation of mercury under controlled redox, pH and salinity conditions. Applied and Environmental Microbiology, Volume 50, pp. 1203 - 1207.
Coquery, M. & Cossa, D., 1995. Mercury speciation is surface sediments of the North Sea. Netherlands Journal of Sea Research, 34(4), pp. 245-257.
Cossa, D. G. C. & Couran, P., 1988. Dissolved mercury behavior in the saint Lawrence Estuary. Estuarine, Coastal and Shelf Science, Volume 26, pp. 227 - 230.
Costley, C. T. et al., 2000. Determination of mercury in environmental and biological samples using pyrolysis atomic absorption spectrometry with gold amalgamation;. Analytica Chimica Acta, Volume 405.
Covelli, S., Faganeli, J., Horvart, M. & Bramati, A., 1999. Porewater distribution and benthic flux measurements of mercury and methylmercury in the Gulf of Triest (Northern Adriatic Sea). Estuarine, Coastal and Shelf Science, Volume 48, pp. 415-428.
Davidson-Arnott, R. G. D., Proosdij, D., Ollerhead, J. & Schostak, L., 2002. Hydrodynamics and sedimentation in salt marshes: examples from a macrotidal marsh, Bay of Fundy. Geomorphology, Volume 48, pp. 209-231.
De Souza, M. P., Huang, C. P., Chee, N. & Terry, N., 1999. Rhizospherebactéria enhance the accumulation of selenium and mercury in wetland plants. Planta, Volume 209.
Diez, S., Carrasco, L. & Bayona, J. M., 2009. Simultaneous determination of methyl- and ethyl- mercury by solid-phase microextraction followed by gas chromatography atomic fluorescence detection. Journal of Chromatography A, Volume 1216.
Doyle, M. & Otte, M., 1997. Organism-induced accumulation of Fe, Zn and As in wetland soil. Environ Pollut, 96(1), pp. 1-11.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 133
Duarte, B. et al., 2010. Accumulation and biological cycling of heavy metal in four march species, from Tagus estuary (Portugal). Environmental Pollution, Volume 158, pp. 1661-1668.
Dyrssen, D. & Wedborg, M., 1991. The sulfur-mercury(II)system in natural waters. Water Air Soil Pollut., Volume 56, pp. 507-519.
EPA, 1998. Method 1630: Methylmercury in Water by Distilation, Aqueous Ethylation, Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry, Washington: United States Environmental Protection Agency.
EPA, 2002. Method 1631, Revision E: Mercury in Water by Oxidation, Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry, Washington: United States Environmental Protection Agency.
Ernst, W., 1990. Ecophysiology of plants in waterlogged and flooded environments.. Aqua Bot, Volume 38, pp. 73-90.
Evans, E. et al., 1999. Comparison of AFS and ICP-MS detection coupled with gaschromatograpgy for the determination of methylmercury in marine samples. Analytica Chimica Acta, Volume 390, pp. 245-253.
Ferreira, J. G., 1988. Mercúrio em Algas Macrófitas no Estuário do Tejo; Dissertação apresentada à Faculdade de Ciencias e Tecnologia da Universidade Nova de Lisboa para a obtenção do Grau de Doutor em Ciencias do Ambiente, Lisboa: s.n.
Förstner, U. & Wittmann, G., 1981. Metal Pollution in the Aquatic Environment. Berlim: Springer-Verlag.
Furutani, A. & Rudd, J., 1980. Measurement of Mercury methylation in lake water and sediment samples. Appl. Environ. Microbiol., Volume 40, pp. 770-776.
Gagnon, C., Pelletier, E. & Mussi, A., 1997. Behaviour of anthropogenic mercury in coastal marine sedimens. Marine Chemistry, Volume 59, pp. 159-176.
Gill, G. & Bruland, K., 1990. Mercury speciation in surface freshwaters systems in California and other areas. Environmental Science Technology, 24(9), pp. 1392-1400.
Gilmoure, C. C., Henry, E. A. & Mitchell, R., 1992. Sulfate stimulation of mercury methylation in freshwater sediment. Environmental Science Technology, Volume 26, pp. 2281-2287.
Gómes-Ariza, J., Lorenzo, F., Garcia-Barrera, T. & Sánchez-Rodas, D., 2004. Analytical approach for routine methylmercury determination in seafood using gas chromatography – atomic fluorescence spectrometry;. Analytica Chimica Acta, Volume 511, pp. 165-173.
Gonçalves, M. d. L. S., 2001. Métodos Instrumentais para Análise de Soluções - Analise Quantitativa. 4ª Edição ed. s.l.:s.n.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
134 Ricardo M.P.Mendes
Guimarães, J. R. et al., 2000. Mercury net methylation in five tropical flood plain regions of Brasil: high in the root zone of floating macrophyte mats but low in surface sediments and flooded soils. Science of the Total Environment, Volume 261, pp. 99-107.
Guimarães, J. et al., 2006. Hg methylation and the microbial consortium in the sediment and in periphyton of tropical macrophytes: effect of different inhibitors. Wisconsin, EUA, Proceedings 8th International Conference Mercury as a Global pollutant.
Haese, R., 2000. The Reactivity of Iron. In: H. Schulz & M. Zabel, edits. Marine Geochemistry. Heidelberg: Springer, pp. 233-261.
Horvat, M., 1996. Mercury analysis and speciation in environmental samples.. In: W. Baeyens, R. Ebinghaus & O. Vasiliev, edits. Regional and global mercury cycles: sources, fluxes and mass balances.. Dordrecht: Kluweer, pp. 1 - 31.
INAG, 2012. www.inag.pt/estuarios/Inicio/frame_page.htm. [Online].
IUPAC, 1998. The determination of Mercury species in environmental and biological Samples. Pure & Applied Chemistry, Volume 70, 8, pp. 1585 -1615.
Jackson, L., Kalff, J. & Rasmussen, J., 1993. Sediment pH and redox potencial affect the bioavailability of Al, cu, Fe, Mn and Zn to rooted aquatic macrophytes. Can. J. Fish. Aquat. Sci., Volume 50, pp. 143-148.
Jackson, T. A., 1998. Mercury in Aquatic Ecosystems . In: Metal Metabolism in Aquatic Enviroments. London: Chapman & Hall, pp. 77-739.
Kelley, B. & Tuovinen, O., 1988. Microbiological oxidations of minerals in mine tailings. In: W. Salomons & U. Forstner, edits. Chemistry and biology of Solid waste - Dredged Material and mine Tailings. New York: Springer-Verlag, pp. 33-53.
King, J., Kostka, J., Fischer, M. & Saunders, F., 2000. Sulfate-reducing bacteria methylare mercury at variables rates in pure culture and marine sediments. Applied Environmental Microbiology, Volume 66, pp. 2430-2437.
Kumar , V. & Tate, B., 1982. Mercury. Journal of Chemical Education, Volume 11, 59, pp. 971-972.
LECO, 2012. http://www.leco.com/products/organic/ama254/ama_254.html. [Online].
Lee, J., 1980. Química Inorgânica. 3ª Ed. ed. S. Paulo: s.n.
Leopold, K., Foulke, M. & Worsfol, P. J., 2009. Gold-Coated Silica as a Preconcentration Phase for the Determination of Total Dissolved Mercury in Natural Waters Using Atomic Fluorescence Spectrometry. Analytical Chemistry, Volume 81, (9), pp. 3421-3428.
Leopold, K., Harwardt, . L., Schuster, . M. & Schlemmer, G., 2008. A new fully automated on-line digestion system for ultra trace analysis of mercury in natural waters by means of FI-CV-AFS. Talanta, Volume 76, pp. 383-388.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 135
Lombi, E., Wenzel, W., gobran, G. & Adriano, D., 2001. Dependency of Phytoavailability of metals on Indigenous and Induced Rhizorphere Processes: a review. In: G. Gobran, W. Wenzel & E. Lombi, edits. Trace elements in the Rhizosphere. Boca Raton: CRC Press LLC, pp. 3-24.
Loring, D., 1991. Normalization of heavy-metal data from estuarine and coastal sediments. ICES Journal os Marine Science , Volume 48, pp. 101-115.
Luther, G., Ferdelman, T., Kostka, J. T. E. & Church, T., 1989. Concentrations and form of dissolves sulfide in the oxic water column of the ocean. Mar. Chem., Volume 27, pp. 165-177.
Luther, G., Giblin, A. E. & Varsolona, R., 1985. Polarographic analysis of sulphur species in marine porewaters. Limnol. Oceanog.l, Volume 30, pp. 727-736.
MacFarlane, G. & Burchett, M., 2000. Celullar distribution of copper, lead, and zinc in the grey mangrove Avicennia marina (Forsk.) Vierh. Aqua. Bot., Volume 68, pp. 45-59.
Madureira, M. J., 1997. Biogeoquímica do Enxofre em Sedimentos de Sapais. Efeitos na Química do Ferro e Manganês., Lisboa: Instituto Superior Técnico.
Marins, R., Lacerda, L., Gonçalves, G. & Paiva, E., 1997. Effects of root metabolism on the post-depositional mobilization of mercury in salt marsh soil. Bulletin of Environmental Contamination and Toxicology, Volume 58, pp. 733-738.
Marschner, H., 1995. Mineral Nutrition of higher Plants. Second Ed. ed. Londos: Academic Press Limited.
Mason, R., Fitzgerald, W. & Morel, F., 1995. The Role of Microoganisms in Elemental Mercury Formation in Natural Waters. Water, air and soil Pollutant, Volume 80, pp. 775-787.
Mason, R. P. et al., 1999. Mercury in the Chesapeake Bay. Marine Chemistry, Volume 65, pp. 77-96.
Matheus, D., Moran, B., McCabe, P. & Otte, M., 2004. Zinc tolerance, uptake, accumulation and distribution in plant and protoplasts of five european populations of the wetland grass Glyceria fluitans. Aqua Bot, Volume 80, pp. 39-52.
Meili, M., 1997. Mercury in lakes and rivers. In: M. Dekker, ed. Metal Ions in Biological Systems. Mercury and its effects on Environment and Biology. New York: s.n., pp. 21-51.
Merrit, K. A. & Amirbahman, A., 2009. Mercury methylation dynamics in estuarine and coastal marine environments – a critical review;. Earth-Science Reviews, Volume 96, pp. 54-66.
Meteorologia, I. d., 2012a. Boletim climatológico mensal - Março de 2012, s.l.: s.n.
Meteorologia, I. d., 2012b. Boletim climatológico mensal - Maio de 2012, s.l.: s.n.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
136 Ricardo M.P.Mendes
Method, M. M., 2003. Millenium Merlin Method for total mercury in drinking, surface, ground, industrial & domestic waste waters and saline waters, Kent: s.n.
Mikac, N., Niessen, S., Ouddane, B. & Wartel, M., 1999. Speciation os mercury in sediments of the Seine estuary (France). Applied Organometallic Chemistry, Volume 13, pp. 715-725.
Miranda, M. et al., 2007. Mercúrio em sistemas aquáticos: Factores ambientais que afectam a metilação. Oecologia Brasiliensis, 11(2), pp. 240 - 251.
Mitsh, W. & Gosselink, J., 2000. Wetlands. 3ª ed. s.l.:John Wiley & Sons Inc.
Monteiro, C. E. S. S., 2010. Mercúrio e Meteilmercúrio em cores de sedimento do Estuário do Tejo, Aveiro: Universidade de Aveiro.
Morel, F., Kraepiel, A. M. L. & Amyot, M., 1998. The chemical cycle and bioaccumulation of mercury. Annual Review of Ecology and Systematics, Volume 29, pp. 543-566.
Nagase, H., Ose, Y., Sato, T. & Isikawa, T., 1982. Methylation of Mercury by humic substances in aquatic environment. Science of the Total Environment, Volume 24, pp. 133 - 142.
NOAA, 1996. Contaminants in aquatic habitats at hazardous waste sites: Mercury;, Seattle: s.n.
Nye, P., 1981. pH changes across the rizosphere induced by roots. Plant and Soil, Volume 61, pp. 7-26.
O’Driscoll, N., Canário, J., Crowell, N. & Webster, T., 2011. Mercury Speciation and distribution on coastal wetlands and tidal mudflats. Water, Air, & Soil Pollution, Volume 220, pp. 313-326.
O’Neil, P., 1985 . Environmental Chemistry. London: George Allen & UnWin.
Otte, M., 1991. contamination of coastal wetlands with heavy metals: factors affecting uptake of heavy metals by salt marsh plants.. In: L. Rozema & J. Verkleij, edits. Ecological responses to Environmental Stress. s.l.:Kluwer academic Publisher, pp. 126-133.
Otte, M. & Haarsma, M. B. R. R. J., 1993. Relation between heavy metal concentration in salt marsh plants and soil. Environ. pollut, Volume 82, pp. 13-22.
Otte, M. L. & Jacob, D. L., 2003. Conflicting processes in the wetland plant rhizosphere: metal retention or mobilization?. Water, Air and Soil Pollution, Volume 3, pp. 91 - 104.
Pacyna, J. M., Pacyna, E. G., Styeenhuisen, F. & Wilson, S., 2008. Global anthropogenic mercury emission of mercury to the atmosphere. In: C. J. Cleveland, ed. Enciclopedia of Earth. s.l.:s.n.
Pereira, M. E. et al., 1998. Tidal export of particlate mercury from the most contaminated area of Aveiro's laggon, Portugal. Science of the Total Environment, Volume 213, pp. 157-163.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
Ricardo.M.P.Mendes 137
Pirrone, N. et al., 2008. Global Mercury Emissions to the atmosphere from Natural and Antropogenic Sources. Rome, Italy, s.n.
Porchenriedes, C. B. J., 2003. Estrés por Metales Pesados. In: M. Reigosa, N. Pedrol & A. Sanchez, edits. Ecofisiologia Vegetal. Madrid: Paraninfo.
Pyzik, A. & Sommer, S., 1981. Sedimentary iron monosulfides: kinetics ena mechanism of formation. Geochimica et Cosmochimica Acta, Volume 45, pp. 687-698.
Ramalhosa, E., 2002. Mercúrio na Ria de Aveiro: Associações, Reactividade e Especiação, s.l.: s.n.
Rantala, R. & Loring, D., 1975. Multi-element analysis of silicate rocks and marine sediments by atomic absorption spectrophotometry. Atomic Absorption Newsletter, Volume 14, pp. 117-120.
Rozema, J., Bijwaard, P., Prast, G. & Broekman, R., 1990. Ecophysiological adaptations of coastal halophytes from foredunes and salt marshes. Vegetatio, Volume 62, pp. 499-521.
Schafer, J. et al., 2006. mercury in the Lot-Garonne river system (France): sourcer, fluxes and anthropogenic componenet. Applied Geochemistry, Volume 21, pp. 515-527.
Schindler, P. & Sposito, G., 1991. Surface complexation at (hydr)oxide surfaces. In: G. Bolt, ed. Interactions at the Soil Colloid-Soil Solution Interface. Dordrecht: Kluwer Academic Publishers, pp. 115-145.
Schulz, H. D., 2000. Quantification of Early diagenesis: Dissolved constituents in Marine Pore Waters. In: H. D. Schulz & M. Zabel, edits. Marine Geochemistry. s.l.:Springer, pp. 85 - 128.
Schuster, E., 1991. The Behaviour of Inorganic Mercury in the Soil With Special Emphasis on Complexation and Adsorption Processes – An Overview. Water, Air and Soil Pollution, Volume 56, pp. 667-680.
Siciliano, S. et al., 2005. Abiotic production of methylmercury by solar radiation. Environmental Science Technology, Volume 39, pp. 1071-1077.
Sirgado, P., 1995. Contaminação de Sedimentos da Cala do Norte do Estuário do Tejo, s.l.: DCEA/FCT.
Skinner, K., Wright, N. & Porter-Goff, E., 2007. Mercury uptake and accumulation by four apecies of aquatic plants. Environ. Pollut, Volume 145, pp. 234-237.
Skoog, D. & Leary, J., 1992. Principles of Instrumental Analysis. 4ª ed. New York: Saunders College Publishing.
Skoog, D., West, D. & Holer, F., 1996. Fundamentals of Analytical Chemistry. 7ª ed. s.l.:Saunders College Publishing.

Influência das Plantas de Sapal na Biogeoquímica do Mercúrio e Metilmercúrio: Sapais do Tejo e
Guadiana – Mestrado em Química Tecnológica
138 Ricardo M.P.Mendes
Skoog, D., West, D. & Holler, F., 1992. Fundamentals of Analytical Chemistry. 6ª ed. s.l.:Saunders College Publishing.
Stumm, W. & Morgan, J., 1996. Aquatic Chemistry: Chemical Equilibria and Rates in Natural Waters. Canada: John Wiley & Sons.
Sunby, B., Vale, C. C. M. C. F., Madureira, M. & Caetano, M., 1998. Metal- rich concretions on the roots of salt marsh plants. Limn Ocean, Volume 43, pp. 245-252.
Tomyasu, T. et al., 2006. Spatial variation os mercury in sediment of Minamata Bay, japan. Science of the Total Environment, 368(1), pp. 283 - 290.
Ullrich, S. M., Tanton, T. W. & Abdrashitova, S. A., 2001. Mercury in the Aquatic Environment: A review of factors affecting methylation. Critical Reviews in Environmental Science and Technology, Volume 31, 3, pp. 241-293.
Válega, M. et al., 2008a. Mercury mobility in a salt marsh colonized by Halimione portulacoides. Chemosphere, Volume 72, p. 1607 – 1613.
Válega, M. et al., 2008b. Mercury mobility in a salt marsh colonized by Halimione portulacoides. Chemosphere, Volume 73, pp. 1224-1229.
Válega, M. et al., 2008. Assessment of methylmercury production in a temperate salt marsh (Ria de Aveiro Lagoon, Portugal). Marine Pollution Bulletin, 56(1), pp. 136-162.
Vesk, P., Nockolds, C. & Allaway, W., 1999. Mechanisms of metal tolerance in higher plants. In: A. Shaw, ed. Heavy Metal Tolerance in Plants: Evolutionary Aspects. s.l.:s.n., pp. 179-194.
Wang, S. et al., 2009. Total mercury and monomethylmercury in water, sediments, and hydrophytes from the rivers, estuary, and bay along the Bohai sea coast, northeastern china. Applied Geochemistry, 24(9), pp. 1702 - 1711.
Wang, Y. & Greger, M., 2004. Plant and environment interections. J. Environ. Qual., Volume 33, pp. 1779-1785.
Weis, J. & Weis, P., 2004. Metal uptake, transport and release by wetland plants: implication for phytoremediation. Environmental International, Volume 30, pp. 685-700.
WHO, 1990. Methylmercury. Environmental Health Criteria 101, Geneve,: s.n.
Zenk, M., 1996. Heavy metal detoxification in higher plants. Gene, Volume 179, pp. 21-30.


















