
FERNANDO ROCHA DOS SANTOS NÍVEIS SÉRICOS DE … · 2011-08-09 · positiva das medianas do NO...
87
1 FERNANDO ROCHA DOS SANTOS NÍVEIS SÉRICOS DE ÓXIDO NÍTRICO E EXPRESSÃO DA ÓXIDO NÍTRICO SINTASE INDUZÍVEL NO BAÇO DE CÃES NATURALMENTE INFECTADOS COM Leishmania (Leishmania) chagasi . Dissertação apresentada ao Programa de Pós- Graduação em Ciências Biológicas da Universidade Vale do Rio Doce, como requisito parcial para obtenção do título de Mestre em Ciências Biológicas (Área de concentração: Imunopatologia de Doenças Infecciosas e Parasitárias). Orientador: Dr. Luiz Cosme Cotta Malaquias NpIMUNO/UNIVALE Co-Orientadores: Dr. Alexandre Barbosa Reis Dra. Cláudia Carneiro Martins NUPEB/ICEB/UFOP. Governador Valadares 2008
Transcript of FERNANDO ROCHA DOS SANTOS NÍVEIS SÉRICOS DE … · 2011-08-09 · positiva das medianas do NO...

1
FERNANDO ROCHA DOS SANTOS
N ÍVE IS SÉRICOS DE ÓXIDO N ÍTRICO E EXPRESSÃO DA ÓXIDO
N ÍTRICO S INTASE INDUZÍVE L NO BAÇO DE CÃES NATURALMENTE
INFECTADOS COM Leishmania (Le ishmania ) chagas i.
D isser tação ap resen tada ao P rograma de Pós -Graduação em C iênc ias B io lóg icas da Un ive rs idade Va le do R io Doce, como requ is i to pa rc ia l pa ra ob tenção do t í t u l o de Mest re em C iênc ias B io lóg i cas (Á rea de concent ração : Imunopa to log ia de Doenças In fecc iosas e Paras i tá r ias ) .
Or ien tado r :
D r . Lu iz Cosme Cot ta Ma laqu ias
Np IMUNO/UNIVALE
Co -Or ien tado res :
D r . A lex andre Barbosa Re is
D ra . C láud ia Carne i ro Mar t ins
NUPEB/ ICEB/UFOP.
Governador Va ladares
2008

2
FERNANDO ROCHA DOS SANTOS
N ÍVE IS SÉRICOS DE ÓXIDO N ÍTRICO E EXPRESSÃO DA ÓXIDO
N ÍTRICO S INTASE INDUZÍVE L NO BAÇO DE CÃES NATURALMENTE
INFECTADOS COM Leishmania (Le ishmania ) chagas i.
D isser tação ap resen tada ao P rograma de Pós -Graduação em C iênc ias B io lóg icas da Un ive rs idade Va le do R io Doce, como requ is i to pa rc ia l pa ra ob tenção do t í t u l o de Mest re em C iênc ias B io lóg i cas (Á rea de concent ração : Imunopa to log ia de Doenças In fecc iosas e Paras i tá r ias ) .
Governador Va ladares , 08 de Ju lho de 2008 .
Banca examinadora:
P ro f . D r . Lu iz Cosme Co t ta Malaqu ias
Un ive rs idade Va le do R io Doce
P ro f . D r . Wanderson Gera ldo L ima
Un ive rs idade Federa l de Ouro P re to
P ro fa . Dra . Vanessa Peruh ype Maga lhães Pascoa l
Cent ro de Pesqu isas René Rachou

3
Ded ico es te t raba lho aos meus f i l hos Rhuan e
M yrna . Espero que se i nsp i rem no meu
es forço , dete rminação e perseverança e
avancem mais do que eu avance i no
conhec imento c ien t í f i co . Lembrando que
buscar com humi ldade o conhec imento ,
p r inc ipa lmente das ve rdades in te rnas , nos
ap rox imará do Cr iado r .

4
AGRADECIMENTOS
Aos meus o r ien tado res Dr . Lu iz Cosme Cot ta Malaqu ias , Dr . A lexandre
Barbosa Reis e Dra C láud ia Mar t i ns Carne i ro pe la pac iênc ia ao me
ens ina rem a rac ioc inar c i ênc ia . Agradeço po r i rem além da c iênc ia e
t ransmi t i rem a fe to em mu i tos momentos .
À Me Pau la Me lo Abreu V ie i ra por me aux i l i a r nos exper imentos de
imuno-h is toquím ica e na aná l i se de imagens com ded icação , pac iênc ia e
humi ldade.
À Cr is t i ane , m inha esposa , pe lo ca r i nho nos momentos impor tan tes e
dec is i vos de minha v ida , aos meus f i l hos po r compreenderem m inhas
ausênc ias , mesmo quando eu es tava f i s i camen te p resente , à Rose l y,
m inha i rmã, pe lo apo io e ao Breno, meu sob r inho, pelo supor te d ig i t a l .
À Técn i ca Mar ia Amél i a Gonça lves e aos B ioquímicos Ana E l i e te da
Conce ição Te les e Va ld iney J . O l i ve i ra po r me subs ti t u í rem no t raba lho .
A todos do NUPEB/ ICEB/UFOP que de a lguma fo rma me ajudaram,
espec ia lmente à Técn i ca Mar ia Chaves do Labora tór io de
Imunopato log ia , por seu apo io nas técn i cas de h i s tolog ia p r inc ipa lmente
na mic ro tomia dos tec idos .
À Coordenação de Aper fe i çoamen to de Pessoa l de N ível Super ior
(CAPES) e UNIVALE pe la bo lsa de es tudos . À P re fe i tura Mun ic ipa l de
I t a ipé pe lo apo io .
Ao Deus -Cr iado r po r t udo que me perm i t i u aprender nessa caminhada . . .

5
“Deus tem seu a l ta r no se io da c r i ação , e o
t em também em cada coração humano . No
p r imei ro o f i c i am as po tênc ias cósmicas ; no
segundo, a consc iênc ia i nd iv i dua l . ”
Car los Bernardo Gonzá les Peco tche
(Raumso l ) – O Espí r i t o

6
RESUMO
Este es tudo ob je t i vou ava l i a r os n íve i s sé r i cos de óx ido n í t r i co (NO) , a exp ressão esp lên i ca da enz ima Óx ido N í t r i co S in tase induz íve l ( iNOS) por método imuno-h i s toquímico ( IHQ) em cães natu ra lmen te i n fec tados (C I) com Le ishman ia (L . ) chagas i compara t i vamen te a cães não i n fec tados (CNI), po is o baço é um ó rgão l in fó ide re lac ionado com a de fesa i na ta e adap ta t i va f requentemen te acomet ido po r es ta paras i t ose . Os an imais fo ram agrupados de acordo com a fo rma c lí n i ca em cães ass in tomát i cos (CA) , o l i goss in tomát i cos (CO) e s in tomát i cos (CS) e de aco rdo com a carga paras i t á r i a em an ima is com ba ixo (BP) , méd io (MP) e a l t o pa ras i t i smo (AP) . A lém d i sso , fo ram rea l i zadas co r re lações dos resu l t ados de NO e iNOS com a exp ressão esp lên i ca das c i t oc inas IL-10 , IFN- γ e TNF-α e com imunog lobu l i nas sé r i cas IgG1 , IgG2 e IgG to ta l an t i -Leishmania . Observou -se ma io r ca rga paras i t á r i a em CS em re lação a CA (p<0,01 ) . A med iana dos n í ve is sé r i cos de NO dos C I fo i s i gn i f i ca t i vamente maio r quando comparada à med iana dos n í ve is sé r i cos de NO em CNI (p<0 ,05 ) . Quando subd iv id idos de aco rdo com a fo rma c l í n i ca , a med iana de NO em CS fo i s i gn i f i ca t i vamente maio r que a med iana de NO em CNI (p<0,01 ) . Observou -se também uma co r re lação pos i t i va das med ianas do NO sér ico com a dens idade paras i t á r i a esp lên i ca (p<0,0001 , r=0 ,6444) . A aná l i se de co r re lação most rou -se pos i t i va en t re n í ve i s sér i cos de NO e exp ressão esplên i ca de TNF-α (p=0,005, r=0 ,589 ) , com IgG2 (p=0 ,019 , r=0 ,466 ) e IgG to ta l (p=0,047, r=0 ,400 ) . Foram observadas fo r tes marcações da enz ima iNOS em C I e ausênc ia dessa marcação em CNI. Os t i pos de marcação imuno-h is toquímica fo ram categor izados como d i fusa , foca l e pon tua l . Na ava l i ação semi -quan t i t a t i va , as marcações fo ram mais f requentemente observadas em CS, segu ida po r CO e CA. Quant i ta t i vamente , os resu l t ados da IHQ demonst ram que a med iana da expressão de iNOS fo i maio r em CS (p<0,01) e CO (p<0,05) comparada à med iana em CNI. Es tes resu l t ados sugerem que o agravamento c l í n i co es tá assoc iado à maio r exp ressão de iNOS. A assoc iação en t re a exp ressão da iNOS com a ca rga paras i t á r i a esp lên i ca não apresen tou va lo r s ign i f i ca t i vo (p=0 ,055) , mas observou-se um p redomín io de maio r exp ressão de iNOS em cães com a l to pa ras i t i smo, embora i s to não tenha se to rnado uma regra . Aná l i ses de co r re lações en t re iNOS e c i t oc inas esp lên i cas TNF-α , IFN-γ e IL-10 ou com IgG1 , IgG2 e IgG to ta l não fo ram s ign i f i ca t i vas (p>0,05 ) . A t ravés des te es tudo fo i poss íve l suger ir que oco r re um aumento dos n í ve is de NO com o agravamento c l ín i co e com o aumento da dens idade paras i t á r i a em Le i shmaniose V iscera l Can ina (LVC) . Conc lu i -se haver exp ressão esp lên i ca da enz ima iNOS em cães com LVC e sugere -se que a ex p ressão es tá aumen tada com o agravamento c l in i co . A lém d isso , os resu l t ados perm i tem in fe r i r que o NO sér i co pode se r usado como fe r ramen ta prognós t i ca em LVC. Pa lavras chave: Le i shmaniose V iscera l Can ina . Óx ido n í t r i co . Óx ido n í t r i co s in tase induz íve l ( iNOS) . C i toc inas . Imunoglobu l inas.

7
ABSTRACT
This s tud y a imed to eva lua te ser i c l eve ls o f n i t r i c ox ide (NO) , sp leen ex press ion o f the enzyme induc ib le n i t r i c oxide s yn thase ( iNOS) b y immunoh is tochemis t ry i n dogs na tura l l y i n fec ted w i th Leishmania (L . ) chagas i ( ID ) compared to non - in fec ted dogs (N ID) because sp leen is an o rgan l ympho id re la ted to i nna te and adapt i ve defense a f fec ted by the paras i t e . The an imals were grouped acco rd ing to c l in i ca l fo rm in as ymptomat i c (AD) , o l i gos yn tomat i c (OD) and s ymptomat i c dogs (SD) and i n acco rdance w i th t he paras i t e l oad in an ima ls w i th l ow (LP) , med ium (MP) and h igh paras i t i sm (HP) . Add i t i ona l l y, da ta f rom NO and iNOS exp ress ion were co r re la ted w i th t he sp len ic exp ress ion o f cytok ines IL-10 , IFN-γ , TNF-α and w i th se ra immunog lobu l ins IgG1 , IgG2 and IgG to ta l an t i -Leishmania. In SD the re was h igher paras i t e l oad i n re la t ion to AD (p<0,01 ) . The med ian sera l eve ls o f NO in ID was s ign i f i can t l y h igher than the med ian found in N ID (p<0 ,05 ) . When assessed acco rd ing to c l i n i ca l fo rm, the med ian o f SD was s ign i f i can t l y h igher t han the med ian found i n N ID (p<0,01 ) . There was a lso a pos i t i ve co r re la t ion between o f med ian se r ic NO w i th sp len i c pa ras i t e dens i t y (p<0,0001 , r=0 ,6444) . The ana l ys i s o f co r re la t ion showed a pos i t i ve co r re la t ion between se r i c l eve ls o f NO and sp len i c exp ress ion o f TNF-α (p=0,005, r=0 ,589 ) and IgG2 (p=0 ,019 , r=0 ,466 ) and to ta l IgG (p=0 ,047 , r= 0 ,400) . Semiquant i t a t i ve l y ana lys i s o f the IHC showed h igher ex p ress ion o f t he enz yme iNOS in ID and l ack o f express ion i n N ID . The iNOS labe l ing were ca tego r ized as d i f fuse , foca l and punc tua l . F rom qua l i t a t i ve po in t o f v i ew, the s t ronges t and p rominen t l abe l i ng was observed in SD, fo l l owed b y OD and AD. Quan t i t a t i vel y ana l ys i s showed ex p ress ion o f iNOS was h igher in SD (p<0.01 ) and OD (p<0.05 ) compared to N ID (p=0 ,001) . These resu l ts sugges t tha t t he c l in i ca l de ter io ra t i on i s assoc ia ted w i th inc reased ex press ion o f iNOS. The assoc ia t ion between the ex press ion o f iNOS w i th the sp len i c paras i t e l oad was no t s i gn i f i can t (p=0,055) , bu t t here was a tendency o f h igher ex p ress ion o f iNOS in dogs w i th h igh paras i t i sm, a lt hough th is was no t a ru le . Ana l ys i s o f co r re la t ion between iNOS and exp ress ion sp len i c cytok ines IL-10 , TNF-α , IFN-γ o r w i th immunog lobu l i ns IgG1 , IgG2 and IgG to ta l was no t s i gn i f i can t (p>0.05) . These resu lts sugges t t ha t ser i c NO may be assoc ia ted w i th t he c l i n i ca l worsen ing and inc reased sp len i c pa ras i t e load i n Can ine V iscera l Le i shmanias is (CVL) . In conc lus ion , i t was observed the exp ress ion o f the enz yme iNOS in the sp leen i n CVL and sugges ts t h is exp ress ion inc rease w i th the c l ini ca l worsen ing o f the d isease . A lso , t he resu l t s sugges t tha t l eve ls o f se r i c NO may be used as p rognos t i c t oo l i n CVL. Key words : Can ine V iscera l Le i shmanias is . N i t r i c oxide . Induc ib le n i t r i c ox ide syn thase ( iNOS) . Cytok ines . Immunog lobu l ins .

8
L ISTA DE FIGURAS
Figu ra 1 - Dens idade paras i t á r i a esp lên i ca (LDU – “Le i shmania Donovan Un i ts ” ) po r fo rma c l ín ica de l e ishmaniose v i scera l can ina . Cães ass in tomát i cos (CA; n= 10 ) , cães o l i goss in tomát i cos (CO; n= 10 ) e cães s in tomát i cos (CS; n= 15) . D i fe rença s ign i f i ca t i va rep resentada po r p< 0 ,01 ; Tes te de Kruska l Wal l i s e pós- tes te de Dunn . .. . . . . . . . . . . . . . . . . . . . . . . . . . .37 F igu ra 2 - N íve is sér i cos de NO (NO2
- e NO3- ) em cães não in fec tados
(CNI; n=10) , comparados a cães natu ra lmen te i n fec tados com L. (L . ) chagas i (C I ; n=35) . D i fe renças s ign i f i ca t i vas representadas po r (p<0,05 ) ; Tes te de Mann Wh i tney. . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 F igu ra 3 - N íve is sé r i cos de NO (NO2
- e NO3- ) de cães não i n fec tados
(CNI; n= 10 ) , comparados a cães ass in tomát i cos (CA; n=10) , cães o l i goss in tomát i cos (CO; n=10) e cães s in tomát i cos (CS; n=15) . D i fe renças s ign i f i ca t i vas rep resentadas po r (p<0,01) ; Tes te de Kruska l Wal l i s e pós - tes te de Dunn . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .40 F igu ra 4 - N íve is sé r i cos de NO (NO2
- e NO3- ) de cães in fec tados com
Leishmania (L . ) chagas i (C I ; n=35) subd i v id idos em an imais ap resentando ba ixo paras i t i smo (BP; LDU en t re 0 e 10 ) , méd io paras i t i smo (MP; LDU en t re 11 e 250 ) e a l t o pa ras i ti smo (AP; LDU en t re 251 e 2564) . D i fe renças s ign i f i ca t i vas representadas por p<0,05 (en t re BP e MP) e p<0,001 (en t re BP e AP) . Tes te de Kruskal Wal l i s e pós -tes te de Dunn . B) Cor re lação de Spearman en t re NO sér i co e dens idade paras i t á r i a esp lên i ca de C I (p<0001 , r=0 ,6444, n=35) . . . . . . . . . . . . . . . . . . . . . . . . 41 F igu ra 5 - Aná l i ses de co r re lação dos n í ve is sér icos de NO (NO2
- e NO3- )
em cães i n fec tados com Leishmania (L . ) chagas i (C I , n=21) com ex p ressão esp lên i ca de IL-10 (A ; p=0 ,971) , IFN-γ (B ; p=0,181) e TNF-α (C ; p=0 ,005 , r=0 ,589 ) . Cor re lação de Spearman . . . . . .. . . . . . . . . . . . . . . . . . . . . . . 42 F igu ra 6 - Aná l i se Aná l i se de co r re lação dos n íve i s sé r i cos de NO (NO2
- e NO3
- ) cães natu ra lmente i n fec tados com Le ishman ia (L . ) chagas i (C I ; n=24) com n íve is sé r i cos de IgG to ta l an t i -Leishmania (A ; p=0 ,047 , r=0 ,400 ) , IgG1 (B ; p=0 ,964) e IgG2 (C; p=0 ,019 , r=0 ,466 ) . Co r re lação de Spearman. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42 F igu ra 7 - Fo tomic rogra f i as de de co r te h i s to lóg i co esp lên i co de cães na tu ra lmente i n fec tados com Le i shmania (L. ) chagas i submet ido a reação imuno-h is toquím ica an t i - iNOS. A ) Ausênc ia de marcação no grupo cont ro le (CNI) ; B ) Exp ressão d i sc re ta em cão ass in tomát i co (CA) ; C ) moderada em cão o l i goss in tomát ico (CO) ; D) acen tuada em cão s in tomát i co (CS) . 600X. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 F igu ra 8 - Ava l i ação quant i t a t i va da ex p ressão de Óx ido Ní t r i co S in tase i nduz íve l ( iNOS) no baço de cães não in fec tados (CNI) e na tu ra lmente

9
i n fec tados com Le ishmania (L . ) chagas i. Imuno-h i s toquím ica an t i -iNOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47 F igu ra 9 - Ava l i ação quant i t a t i va da ex p ressão de Óx ido Ní t r i co S in tase i nduz íve l ( iNOS) no baço de cães não i n fec tados (CNI, n=8 ) e na tu ra lmente in fec tados com Leishman ia (L . ) chagas i, c l ass i f i cados po r fo rmas c l ín i cas de LVC (CA; n=8,CO; n=8e CS ; n=13) . D i fe rença s ign i f i ca t i va en t re CNI e CO rep resentada po r p<0,05 e en t re CNI e CS rep resentada po r p<0 ,01 . K ruska l Wal l i s e pós - tes te de Dunn. Imuno-h is toquímica an t i - iNOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47 F igu ra 10- Ava l i ação quant i ta t i va da exp ressão de Óx ido N í t r i co S in tase i nduz íve l ( iNOS) no baço de cães não i n fec tados (CNI, n=8 ) e na tu ra lmente in fec tados com Leishman ia (L . ) chagas i, c l ass i f i cados po r dens idade paras i t á r i a esp lên i ca (Ba ixo paras i t i smo : LDU en t re 0 e 10 ) , méd io paras i t i smo: LDU en t re 11 e 250 e a l t o pa ras it i smo : LDU en t re 251 e 2564) . p>0 ,05 . Tes te de Kruska l Wa l l i s e pós -tes te de Dunn . . . . . . . 48 F igu ra 11- Cor re lação de Spearman da exp ressão esp lên i ca de iNOS em cães natura lmen te i n fec tados com Leishmania (L . ) chagas i (C I ; n=21) com a exp ressão esp lên i ca de IL-10 (A ; p=0,110) , IFN-γ (B; p=0 ,697 e TNF-α (C ; p=0 ,205) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . 49 F igu ra 12- Cor re lação de Spearman da exp ressão esp lên i ca de iNOS em cães natura lmen te i n fec tados com Leishmania (L . ) chagas i (C I ; n=24) com as imunoglobu l i nas IgG1 , IgG2 e IgG to ta l an t i -Leishmania (A ; p=0,076) , IgG1 (B ; p=0,999) , IgG2 (C; p=0 ,203) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50

10
L ISTA DE TABELAS
Tabe la 1 - F reqüênc ia de graus de paras i t i smo esp lêni co assoc iado a fo rmas c l í n i cas em cães natu ra lmente in fec tados com Leishmania (L . ) chagas i. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38 Tabe la 2 - Méd ia percentua l de marcação da exp ressão de iNOS por g rupo c l í n i co quanto ao t ipo e in tens idade nos f ragmentos esp lên i cos de cães in fec tados com Leishmania (L . ) chagas i. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46

11
L ISTA DE ABREVIATURAS
ABC Complexo Av id ina B io t ina
AMG Aminoguan id ina
AP A l to Paras i t i smo
BP Ba ixo Paras i t i smo
CA Cães Ass in tomát i cos
CD4+ “ Clus ter o f d i fe ren t i a t ion ” 4 pos i t i vo (Grupo de d i fe renc iação)
CD8+ “ Clus ter o f d i fe ren t i a t ion ” 8 pos i t i vo (Grupo de d i fe renc iação)
CCZ Cent ro de Cont ro le de Zoonoses
C I Cães na tu ra lmente i n fec tados
CNI Cães Não In fec tados
cNOS Óx ido Ní t r i co S in tase cons t i tu t i va
CO Cães Ol i goss in tomát i cos
CS Cães S in tomát i cos
DAB D iaminobenz id ina
eNOS Óx ido Ní t r i co S in tase endote l i a l
HE Hematox i l i na Eos ina
H IV V í rus da Imunode f ic i ênc ia Humana
IFN- γ In te r fe ron gama
Ig Imunog lobu l i na
IL -10 In te r l euc ina 10
iNOS Óx ido Ní t r i co S in tase induz íve l
LDU “Le i shman Donovan Un i ts ”
LPG Lipo fos fog l i canos
LPS L ipopo l i ssacár ides
LT Le i shman iose Tegumenta r
LT- α1β2 L in fo tox ina-α1β2
LV Le i shman iose V iscera l
LVC Le i shman iose V iscera l Can ina
RNAm Ác ido R ibonuc lé i co mensage i ro
NK Célu las “na tu ra l k i l l e r ”
nNOS Óx ido Ní t r i co S in tase neurona l
NO Óx ido Ní t r i co

12
PALM Ba inha l i n fó ide per i -a r te r i o la r
PALMS Ba inhas l in fó ides per i -a r ter io la res
PB Po lpa Branca
PBS “Phospha te Bu f fe red Sa l ine ” (So lução sa l i na tamponada com
fos fa to )
PV Po lpa Verme lha
R IF I Reação de Imuno f luo rêscenc ia Ind i re ta
RT-PCR Reação em Cadeia da Po l imerase com Transc r i ção Reversa
SRD Sem Raça De f in ida
TGF-β Fato r de C resc imento T rans fo rman te be ta
Th1 L in fóc i t os T “he lper ” t ipo 1
Th2 L in fóc i t os T “he lper ” t ipo 2
TNF-α Fa to r de Necrose Tumora l a l fa
ZM Zona marg ina l

13
SUMÁRIO
1 INTRODUÇÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 1 .1 ASPECTOS GERAIS DAS LE ISHMANIOSES. . . . . . . . . . . . .. . . . . . . . . . . . . . . . . 15 1 .2 FORMAS CLÍN ICAS DA LE ISHMANIOSE V ISCERAL CANIA. . . . . . 17 1 .3 ASPECTOS IMUNOPATOLÓGICOS DA LVC. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . 18 1 .4 BAÇO: ESTRUTURA E FUNÇÃO. . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . 21 1 .5 B IOLOGIA DO ÓXIDO N ÍTR ICO: SUA PARTIC IPAÇÃO NA RESPOSTA IMUNE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 2 OBJETIVOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 2 .1 OBJ ETIVO GERAL. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 2 .2 OBJ ETIVOS ESPECÍF ICOS. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 2 .2 .1 Dete rmina r a ca rga pa ras i tá r i a no baço de cães na tu ra lmen te in f ec tados com L . (L . ) ch ag as i po r tado res de d i f e ren tes f o rmas c l í n i cas de l e i shman iose e def in i r os g raus de paras i t i smo esp lên i co . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 2 .2 .2 De te rmina r os n í ve is do óx ido n í t r i co sé r i co de cães na tu ra lmen te in f ec tados com L . (L . ) chagas i e associa r os resu l tados com as fo rmas c l ín i cas e g raus de paras i t i smo esp lên i co . . . . . . . . . . . . . . . . . 30 2 .2 .3 Ava l i a r a exp ressão da enz ima iNOS em baço de cães na tu ra lmen te in f ec tados com L . ( L . ) c h ag as i e assoc ia r os resu l tados com as fo rmas c l ín i cas e g raus de paras i t i smo esp lên i co . . . . . . . . . . . . . . . . . 30 2 .2 .4 Assoc ia r os per f i s de exp ressão de iNOS em baço e os n íve i s de NO sér i cos com a p rodução das c i toc inas INF- γ , TNF-α e IL -10 . . . . . . . 30 2 .2 .5 Assoc ia r os per f i s de exp ressão de iNOS em tec ido esp lên ico e os n íve i s sé r i cos de NO com os n í ve is de IgG1, IgG2 e IgG to ta l an t i -L e i shm an ia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30 3 MATERIAIS E MÉTODOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . 31 3 .1 ANIMAIS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 3 .2 AVALIAÇÃO CLÍN ICA. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .32 3 .3 ANÁLISE PARASITOLÓGICA. . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . 32 3 .4 QUANTIF ICAÇÃO SÉRICA DE NO. . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . 33 3 .5 REAÇÃO IMUNO-H ISTOQUÍM ICA PARA DETECÇÃO DE INOS. . 34 3 .6 ANÁLISES ESTATÍST ICAS. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36 4 RESULTADOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 4 .1 DETERMINAÇÃO DA CARGA PARASITÁRIA . . . . . . . . . . . . .. . . . . . . . . . . . . . 37 4 .1 .1 Carga pa ras i tá r i a po r fo rma c l ín i ca . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . 37 4 .1 .2 Ca tegor i zação po r g raus de pa ras i t i smo esp lênico . . . . . . . . . . . . . . . . . . 38 4 .2 N ÍVE IS SÉRICOS DE ÓXIDO N ÍTRICO. . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . 38 4 .2 .1 Dosagem de NO sér i co em cães não in f ec tados comparados a cães i n fec tados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 4 .2 .2 De terminação dos n í ve i s sér i cos de NO comparada po r carga pa ras i tá r i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40 4 .3 ANÁLISE DE CORRELAÇÃO ENTRE OS NÍVE IS SÉRICOS DE NO COM EXPRESSÃO ESPLÊNICA DE IL-10 , IFN-γ E TNF-α E COM OS N ÍVE IS SÉRICOS DE IgG1 , IgG2 E IgG TOTAL ANTI-LEISHMANIA. . 41 4 .4 EXPRESSÃO ESPLÊNICA DE iNOS. . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . 43 4 .4 .1 Aná l i se semi -quan t i ta t i va de iNOS. . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 4 .4 .1 .1 Marcação D i fusa (MD) . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45

14
4 .4 .1 .2 Marcação Foca l (MF) . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 4 .4 .1 .3 Marcação Pontua l (MP) . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 4 .4 .2 Aná l i se quan t i ta t i va de iNOS. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 4 .4 .2 .1 Expressão esp lên i ca de iNOS comparados po r fo rma c l í n i ca . . . . . 46 4 .4 .2 .2 Expressão esp lên i ca de iNOS comparada po r ca rga paras i tá r ia . . 48 4 .4 .2 .3 Aná l i se de co r re lação en t re a exp ressão da iNOS com n íve i s esp lên i cos de IL-1 0 , IFN-γ e TNF-α e com n íve is de IgG1 , IgG2 e IgG to ta l an t i -Leishmania. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . 48 5 DISCUSSÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51 6 CONCLUSÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61 7 REFERÊNCIAS B IBLIOGRÁFICAS. . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . 62

15
1 INTRODUÇÃO
1 .1 ASPECTOS GERAIS DAS LE ISHMANIOSES
As Le i shmanioses são um con jun to de doenças em sua ma io r ia
zoonoses causadas por p ro tozoár ios da o rdem K inetop las t i da, f amí l ia
Trypanosomat idae e gênero Leishmania (LA INSON e t a l ., 1987) . O
gênero Le ishman ia f o i desc r i t o pe la p r imei ra vez po r Ross em 1903,
agrupando espéc ies de p ro tozoár ios un i ce lu la res , d igenét i cos que
hab i tam o t ra to d iges t i vo de hospede i ros inver teb rados em suas fo rmas
f l age ladas (p romas t i go ta e paramas t i go ta ) . Es tes paras i tos também
in fec tam cé lu las do s is tema fagoc í t i co mononuc lear (SFM) de
ve r teb rados em sua fo rma sem f l age los ex te r io r i zados (amas t i go ta ) . A
t ransmissão ocor re pe la p i cada da fêmea do f l ebo tomíneo Lutzomya
l ong ipa lp i s, que tem s ido reg i s t rada em ambien tes ru ra i s e u rbanos ,
p róx imos a an imais domést i cos e hab i tações humanas (MARZOCHI e t
a l . , 1985; LA INSON e t a l ., 1987; LA INSON & RANGEL, 2005; S ILVA
e t a l ., 2007) .
A fo rma in fec tan te , p romas t i go ta metac íc l i ca , p resen te no inseto
ve to r é a i nocu lada na derme jun to à sa l i va du ran te o repas to sangu íneo.
Com base no desenvo l v imento das fo rmas p romast igo tas no tubo
d iges t i vo do inse to ve to r , LA INSON & SHAW (1988) p ropuseram um
agrupamento das espéc ies do gênero Leishmania em do is subgêneros:
pa ras i t os que têm seu desenvo l v imento l im i tado ao es tômago do veto r
( sup rap i l á r i a ) per tencem ao subgênero Le ishman ia , enquan to os paras i t os
que se aderem às paredes do p i lo ro ou í l eo a t ravés do f lage lo
(Per ip i l á r i a ) per tencem ao subgênero Viann ia.
As l e i shmanioses podem ser d iv idas em do is g rupos cl í n i cos:
Le i shman iose Tegumenta r (LT ) e Le i shmaniose V isceral (LV) .
As d i fe ren tes man i fes tações c l í n i cas das l e i shmanioses es tão
envo lv idas com a espéc ie do paras i to , espéc ie do vetor e es tado imune do
hospede i ro den t re ou t ros aspectos (MARZOCHI e t a l ., 1985) .

16
A LT pode se man i fes tar em d i ve rsas fo rmas c l ín i cas ta i s como:
cu tânea, cu tânea mucosa , e cu tânea d i fusa (GONTIJO & CARVALHO,
2003) .
A LV humana, t ambém chamada de ca lazar , também man ifes ta -se
em d i fe ren tes fo rmas c l í n i cas , va r i ando desde a fo rma c l í n i ca
ass in tomát i ca ou sub -c l í n i ca , a té as fo rmas c l í n i cas (aguda e c rôn i ca )
(BADARO e t a l ., 1986) .
A LV é uma doença de cará te r c rôn i co que a t i nge órgãos i n te rnos ,
p r inc ipa lmente o f í gado, baço, medu la óssea e l i n fonodos . A in fecção
pode se r ass in tomát i ca ou pode evo lu i r ap resentado feb re reco r ren te e
mal -es ta r segu ido de p rogress i va p io ra , anemia e hepa to-esp lenomega l i a ,
l i n fadenopat i a , l eucopen ia , pe rda de apet i te , fad iga e es tado de
deb i l idade p rogress i vo . Se não t ra tada l eva à mor te dent ro de 2 a 3 anos
(MALLA & MAHAJAN, 2006) .
O agente e t i o lóg ico da LV no ve lho mundo ( Índ ia e les te da
Á f r i ca ) é a Leishmania (L . ) donovan i; na Ch ina , Ás ia Cen t ra l , sudes te da
Europa e Med i te r râneo é a Leishmania (L . ) i n fan tum e no Novo Mundo
(Amér i ca La t ina ) é a Leishmania (L . ) chagas i (LA INSON, 1970 ;
LA INSON & SHAW, 1972) .
Cor ren temente as le i shmanioses a fe tam 12 mi l hões de ind i v íduos
em todo mundo com oco r rênc ia ap rox imada de 500.000 novos casos de
LV po r ano. C inco países l i de ram a p reva lênc ia da doença: Índ ia ,
Bang ladesh, Nepa l , Sudão e Bras i l (ENGWERDA e t a l ., 2004) . Com a
ex pansão da A IDS , pac ien tes co - i n fec tados com Le ishman ia e H IV j á são
no t i f i cados , o que ocas iona aumento nos d i s tú rb ios imuno lóg i cos , uma
vez que a i n fecção pe lo H IV l eva à dep leção de cé lulas T CD4+ ,
f avo recendo o desenvo lv imento de fo rmas graves de le i shmanioses , ou
po r ou t ro lado l evam ao aumen to da rep l i cação do v írus (DESJEUX &
ALVAR, 2003; S INHA e t a l ., 2005; KUMAR e t a l ., 2007) .
Nos ú l t imos anos a LV , que e ra t ip icamen te c i r cunscr i t a à zona
ru ra l , vem u rban izando-se e a tua lmente a LV es tá p resente em 19 dos 27
es tados b ras i l e i ros . Nos anos 90 as reg iões Nor te , Sudes te e Cent ro -
Oeste passaram a cont r i bu i r e levando a i nc idênc ia da LV no Bras i l . Nos
ú l t imos c inco anos oco r re ram 3500 casos novos , a maio r i a na reg ião

17
Nordes te . Es te p rocesso de expansão e u rban ização es tá l i gado à
p rox im idade en t re as hab i tações , a l t a dens idade popu lac iona l e à
suscet ib i l i dade da popu lação (GONTIJO & MELO, 2004).
Dados da l i t e ra tu ra re la tam que o aparec imento de casos humanos
de le ishman iose é an teced ido pe lo achado de cães i nfec tados
(PALATNIK-DE-SOUZA e t a l. , 2001) . No en tan to , a inda ex is tem
cont rovérs ias sob re es ta ques tão , que j us t i f i ca a rea l i zação de novos
es tudos , po is a e l im inação dos cães so ropos i t i vos nas á reas endêmicas
não tem imped ido o aparec imento de casos humanos, embora ba ix em a
p reva lênc ia da Le i shmaniose V i scera l Can ina (LVC) (BRAGA e t a l .,
1998) . Out ros pesqu isadores demonst ra ram a poss ib i li dade de in fecção
do i nseto ve tor pe lo homem, suger indo a poss ib i l i dade des te func ionar
como reservató r io da l e ishmaniose (COSTA e t a l ., 2000) . En t re tan to ,
es te mecan ismo é pouco ace i t o e cont rad iz t odo o parad igma desc r i t o na
l i t e ra tu ra du ran te a década de 50 , quando fo ram de fi n idos os pape is do
homem, da raposa e do cão no c i c lo ep idemio lóg i co da LV no Bras i l
(DEANE & DEANE, 1954) .
1 .2 FORMAS CLÍN ICAS DA LE ISHMANIOSE V ISCERAL CANINA
A LVC assume grande impor tânc ia no es tudo das l e i shmanioses ,
po i s o cão tem s ido responsab i l i zado como o p r i nc ipa l reservató r i o
domést i co da LV , encont rando-se nos i nquér i tos ep idemio lóg icos de
reg iões endêmicas , a l t a i nc idênc ia de cães in fec tados . Es tes achados
oco r rem p rovave lmente dev ido a sua suscep t ib i l i dade à i n fecção e
p r inc ipa lmente pe lo e levado paras i t i smo cu tâneo comumente encon t rado ,
poss ib i l i t ando a manutenção do c i c lo b io lóg ico do paras i to ao i n fec ta r o
i nseto ve to r no amb ien te domés t i co (DEANE & DEANE, 1954 ; KEENAN
e t a l ., 1984a ; ABRANCHES e t a l ., 1991b; BRAGA e t a l ., 1998 ; COSTA
e t a l ., 1999; REIS e t a l ., 2006b) .
A lém d i s to , o i n tenso paras i t i smo cu tâneo mesmo em cães
ass in tomát i cos fo r ta lece sua impor tânc ia como fon te de i n fecção para

18
f l ebo tomíneos (MOLINA e t a l ., 1994) . Out ro impor tan te aspecto que se
deve ressa l ta r é o fa to de que o cão é um ex ce len te modelo exper imenta l
pa ra o en tend imento da LV humana bem como para tes tes de drogas e
vac inas (MORENO & ALVAR, 2002; ALVAR e t a l ., 2004) .
A LVC pode se r ca tego r izada em t rês d is t in tas fo rmas c l ín i cas :
Cães Ass in tomát icos (CA) – Cães que ap resentam apenas
so ro log ia pos i t i va , sem s ina i s c l ín i cos .
Cães Ol igoss in tomát i cos (CO) - Observam-se no máx imo t rês
s ina i s c l ín i cos , podendo aparecer i so ladamen te ou combinados de fo rma
b randa e i nespecí f i ca : a lopec ia loca l i zada, moderado emagrec imento ,
adenopat i a , esp lenomegal i a l eve, o lhos opacos .
Cães S in tomát i cos (CS) – Os an imais ap resen tam uma
combinação de ma is de t rês s in tomas assoc iados . Podem apresenta r
a lopec ia loca l ou genera l i zada, desp igmentação , es fo l i ação , nódu los ,
fo rmação de pús tu las , l esões cu tâneas u l ce radas , gera lmen te no foc inho,
o re lha e ex t remidades , con jun t i v i t e , i r i t e e uveí te, ce ra to -con jun t i v i t e ,
on i cogr i fose, aumen to dos l in fonodos , hepatomega l i a, esp lenomega l i a e
deb i l idade gera l . Ou t ros s ina i s também podem ocor rer como d ia r ré ia,
apa t i a ge ra l , hemorrag ia i n tes t ina l , pares ia do t rem pos ter i o r , bem como
emagrec imento progress i vo evo lu indo para caquex ia e ób i t o (POZIO e t
a l . , 1981; TRYPHONAS e t a l ., 1977; MANCIANTI e t a l ., 1988;
BARROUIN-MELO e t a l ., 2006; REIS e t a l ., 2006a) .
1 .3 ASPECTOS IMUNOPATOLÓGICOS DA LVC
A h i s tó r i a natura l da LVC é um tóp i co amplamente i nves t i gado po r
d ive rsos au to res oco r rendo uma sér i e de man i fes tações
imunopa to lóg i cas , sendo o acúmulo de cé lu las fagocít i cas mononuc leares
nos tec idos l in fó ides uma carac ter í s t i ca fundamen tal . A lém d is to ,
observa-se h ipe rp las ia ce lu la r nos órgãos a fe tados : f í gado , baço, medu la
óssea , mucosa do in tes t ino de lgado e l in fonodos (TAFURI e t a l ., 2001;
ENGWERDA e t a l ., 2004) .

19
No contex to h i s topato lóg ico observa-se no f í gado de cães
na tu ra lmente ou exper imenta lmen te i n fec tados (C I) com Leishmania (L . )
chagas i reações granu lomatosas , h iperp las ia e h ipe r t ro f ia das cé lu las de
Kup f fe r , das qua i s mu i tas se ap resentam paras i t adas por fo rmas
amas t i go tas . Re la ta -se também a p resença de i n f i l t rado
l in foh i s t i op lasmoc i tá r i o acentuado no compar t imento hepá t i co . No baço
d ive rsos t raba lhos na l i t e ra tu ra desc revem a reação in f lamató r i a c rôn i ca
e d i fusa, podendo oco r re r t ambém a presença de granu lomas c i r cundando
macró fagos rep le tos de paras i t os (OLIVE IRA e t a l ., 1993; TAFURI e t
a l . , 1996) e em modelo mur ino c i t a -se a perda da m ic roarqu i t e tu ra
esp lên i ca com o agravamento da LV (ENGWERDA e t a l ., 2004) .
Os l i n fonodos apresentam aumen to no tamanho e número dos
fo l í cu los l i n fó ides , com marcada h iperp las ia , h ipert ro f ia medu la r e
macró fagos medu la res paras i t ados , fenômeno es te i n timamen te
re lac ionado com a l i n fadenopat i a f reqüentemente observada na LVC
(L IMA e t a l ., 2004) .
Na medu la óssea observam-se fo rmas amast i go tas no in ter io r dos
macró fagos , podendo oco r re r anemia e t romboc i topen ia nos an imais
i n fec tados (TAFURI e t a l ., 2001) .
A in tens idade do paras i t i smo no baço , f í gado e medula óssea
encon t ra -se assoc iada à fo rma c l ín ica da doença e ac red i t a -se que a
d isseminação na pe le do an imal oco r ra após a v isce ra l i zação do paras i t o
p r inc ipa lmente após sua chegada à medu la óssea (G IUNCHETTI e t a l .,
2006; REIS e t a l ., 2006a; G IUNCHETTI e t a l ., 2008) .
D i ve rsos es tudos cons ide rando as d i fe ren tes fo rmas c l ín i cas da
LVC demonst raram que CA apresen tam paras i t i smo cu tâneo sendo ,
po r tan to , fon te de i n fecção aos f l ebo tomíneos , embora CO e CS
ap resentem maio r in fec t i v i dade (MOLINA e t a l ., 1994; GUARGA e t a l .,
2000; TRAVI e t a l ., 2001) .
O quadro b ioquím ico c láss ico na LVC ava l i ado pe la ele t ro fo rese
de p ro te ínas sé r i cas most ra i nversão das tax as de albumina /g lobu l ina ,
l evando a h ipe rgamaglobu l i nemia em ap rox imadamente 70% dos casos .
Es te quadro é marcante e exp l i ca a in tensa a t i v i dade po l i c l ona l de
cé lu las B , com e levada p rodução de imunog lobu l inas ( Ig ) o r i g inando os

20
quadros mais g raves com desenvo l v imento de l esões rena is com
g lomeru lone f r i t e mesang iop ro l i f e ra t i va dev ido à depos i ção de
imunocomp lexos no g loméru lo de Malp igh i (TRYPHONAS e t a l ., 1977 ;
TAFURI e t a l ., 1989 ; POLI e t a l ., 1991) .
Es ta i n tensa a t i v idade po l i c lona l na LVC leva à p rodução de
d ive rsas c lasses e subc lasses de Ig p roduz idas du ran te a evo lução da
doença . A lguns au to res admi tem que a IgG1 es ta re lac ionada com
p rogressão enquanto ou t ros most ram o cont rá r io (BORJA-CABRERA e t
a l . , 2004 ; REIS e t a l ., 2006c; RODRIGUEZ-CORTES e t a l ., 2007) .
REIS e t a l . (2006c) demonst ra ram que cães natu ra lmente
i n fec tados c lass i f i cados como o l i go e s in tomát i cos são capazes de
p roduz i r a l t os n íve i s de an t i co rpos de d i ve rsas c lasses ( IgG, IgM, IgA ,
IgE) , e most raram uma assoc iação de a l t os t í t u l os de IgG2 com a l ta
ca rga paras i t á r i a , p r inc ipa lmente na pe le e baço . Es tes au to res também
mos t ra ram que o paras i t i smo de medu la óssea e baço es tá pos i t i vamente
co r re lac ionado com a grav idade da doença .
Os mecan ismos re lac ionados com a res i s tênc ia à in fecção po r L .
chagas i a inda não es tão c la ros na i n fecção natu ra l can ina . A LVC
s in tomát i ca tem s ido assoc iada com mudanças imuno lóg i cas envo lvendo
cé lu las T (P INELL I e t a l ., 1995) , es tando re lac ionada com depressão da
imun idade ce lu la r , conseqüentemen te com perda da capac idade de de fesa
imune , com queda na produção de c i toc inas do T ipo 1, favo recendo a
d isseminação e reprodução dos paras i tos (ABRANCHES e t a l ., 1998) .
A lguns t raba lhos mos t ram assoc iação en t re a t i vação de cé lu las
p rodutoras de IFN-γ , IL-12 , IL-2 e TNF-α e a fo rma ass in tomát i ca da
LVC. Por ou t ro l ado , observa-se que paras i t i smo esplên i co es tá
assoc iado com maio res n í ve is de exp ressão de IL-4 e IL -10 (P INELL I e t
a l . , 1995; LAGE e t a l ., 2007) i nd i cando uma p rováve l re lação en t re es ta
c i t oc ina e a fo rma c l í n i ca da doença.

21
1 .4 BAÇO: ESTRUTURA E FUNÇÃO
O baço é um ó rgão l i n fó ide secundár io comp lexo e com d ive rsas e
impor tan tes funções para o o rgan ismo, a tuando a t i vamen te no
metabo l i smo do fer ro , des t ru i ção de hemácias ve lhas (e r i t ro fagoc i tose
esp lên i ca) , a lém de se r um reserva tór io de sangue em s i tuações de
es forço . Na fase embr ionár ia o baço tem a função hematopo ié t i ca , que é
parc ia lmen te perd ida após o nasc imen to . Possu i reg iões compos tas por
cé lu las de de fesa responsáve is po r fagoc i t ose de o rgan ismos es t ranhos,
ap resentação de an t ígenos , p rodução de an t i co rpos e cé lu las de memór ia .
Sua o r i gem embr io lóg i ca são as cé lu las p rogen i to ras mesodermais
(BRENDOLAN e t a l. , 2007) .
O baço é c i r cundado po r uma espessa cápsu la de tec ido
con jun t i vo , que é reves t i da po r ep i t é l i o pav imentoso s imp les . Os sep tos
de tec ido con jun t i vo pene t ram no parênqu ima des te órgão, conduz indo os
vasos sanguíneos para o in te r i o r . É cons t i tu ído pe la po lpa b ranca (PB) e
po lpa verme lha (PV) . A PB tem um ar ran jo de ba inha c i l í nd r i ca
c i r cu lando a a r té r ia cen t ra l , composta po r t rês sub-compar t imentos : A
ba inha l in fó ide per i -a r ter io la r (PALM) , a zona margina l (ZM) e os
fo l í cu los . A cápsu la compõe-se de tec ido conec t i vo , f i b roso , f ib ras
e lás t i cas e muscu la tu ra l i sa . A camada ex te rna da cápsu la esp lên i ca é
composta de cé lu las mesote l i a i s , que podem não se r ev identes no co r te
h is to lóg ico . Da par te i n te rna da cápsu la o r i g inam-se t rabécu las de
muscu la tu ra l i sa e t ec ido f ib ro -e lás t i co em d i reção ao parênqu ima
esp lên i co , con tendo também vasos sangu íneos , l i n fá ti cos e nervos , sendo
os vasos l in fá t i cos do t ipo e fe ren tes , a t ravés dos qua is l i n fóc i t os migram
para os nodos l in fá t i cos esp lên i cos (CESTA, 2006) .
O sangue chega ao baço a t ravés da a r té r i a esp lên i ca a fe ren te , que
se d iv ide em ar tér i as t rabecu la res que penet ram no parênqu ima, se
rami f i ca em pequenas a r te r ío las que en t ram na PV, onde se to rnam
ar te r ío las cent ra i s . Ou t ras a r te r ío las menores o r i ginam cap i l a res da PB.
As a r te r ío las são c i r cundadas po r tec ido l in fó ide , fo rmando as PALMS,
composta po r cé lu las T , cé lu las dendr í t i cas e cé lu las es t romais . Os

22
f o l í cu los são fo rmados po r cé lu las B fo l i cu la res , cé lu las dendr í t i cas e
cé lu las es t roma is , podendo oco r rer expansão c lona l de cé lu las B , quando
a t i vadas , l evando a h iper t ro f i a fo l i cu la r (ENGWERDA e t a l ., 2004) .
A ZM s i t ua-se en t re a PB e PV, cons is t indo num se io reves t i do
po r cé lu las endo te l ia is que permi te um f luxo ráp ido e um pon to de
en t rada de l eucóc i tos na PB. Es ta reg ião esp lên i ca é composta po r
macró fagos meta lo f ín i cos marg ina is , macró fagos da zona marg ina l ,
cé lu las B e T , sendo espec ia l i zada em fagoc i tose de par t í cu las es t ranhas
ao o rgan ismo, como p ro tozoár ios , bac té r ias e v í rus (OLIVER e t a l .,
1997; MARTIN & KEARNEY, 2002 ; P ILLA I e t a l ., 2005) .
Os fa to res que a tuam na migração de l i n fóc i tos B , T, cé lu las
dendr í t i cas no in ter i o r esp lên i co são cont ro lados po r c i toc inas , como a
l in fo tox ina -α1β2 (LT-α1β2) e fa to r de nec rose tumora l α (TNF-α ) , es tes
po r sua vez con t ro lam a exp ressão de qu imioc ina l i gan te CXC13
(CXCL13) , responsáve l pe la a t ração de l i n fóc i t os B aos fo l í cu los de
cé lu las B . A qu imioc ina l i gan te CC19 (CCL19) e qu imioc ina l i gan te CC
21 (CCL21) a tuam d i r i g i ndo cé lu las T à zona de l in fóc i tos T ,
conseqüentemen te o per fe i t o func ionamento do baço e a manutenção de
sua arqu i t e tu ra é dependente da harmon ia de d i ve rsas qu imioc inas e do
TNF-α , que so f rem a l te rações na sua produção quando há paras i tose
esp lên i ca em modelo mur ino (ENGWERDA e t a l ., 2002) .
As respos tas adap ta t i vas e i na tas podem ser e f i c ientemen te
mon tadas no baço, fazendo des te um impor tan te ó rgão na homeos tase
imune . Enquanto a PV es tá res t r i t a à respos ta adapta t i va , a ZM es tá
envo lv ida em ambos os t ipos de respos ta , a t ravés de macró fagos e
l in fóc i tos , es ta é uma região c ruc ia l onde pa tógenos da co r ren te
sanguínea são seqües t rados . Cé lu las B e macró fagos da ZM podem
fo rmar uma in teração en t re imun idade i na ta e adap tat i va (MEBIUS e t a l . ,
2004) .
A PV cons is te de s inusó ides que fo rmam lab i r i n tos at ravés de
con jun tos ce lu la res conhec idos como co rdões esp lên icos ou de B i l ro th ,
es tes , po r sua vez , são fo rmados po r f ib ras re t i cu lares com cé lu las
re t i cu la res , macró fagos assoc iados e hemác ias . São cons ideradas cé lu las
re t i cu la res os mio f i b rob las tos que tem a função de cont ração do ó rgão .

23
As f i b ras re t i cu lares são compos tas de co lágeno e fi b ras e lás t i cas ,
m io f i b r i l as , l âmina basa l de cé lu las re t i cu la res e f i b ras nervosas
ad renérg i cas não mie l in izadas (MEBIUS & KRAAL, 2005) .
A lguns t raba lhos mos t ram que no baço mur ino , cé lu las T CD4+
aumentam suas percen tagens após a in fecção com Leishmania i n fan tum
superando o número de cé lu las T CD8+ (GOMES-PEREIRA e t a l ., 2004) .
Ou t ros exper imentos reve la ram que o número de cé lu las T CD4+ e
T CD8+ aumentavam no baço após in fecção com Leishmania in fan tum,
mos t rando que es tas par t i c i pam em con jun to na respos ta imune ce lu la r
con t ra o paras i t o (TSAGOZIS e t a l ., 2003) .
1 .5 B IOLOGIA DO ÓXIDO N ÍTR ICO: SUA PARTIC IPAÇÃO NA
RESPOSTA IMUNE
A e l im inação de mic roo rgan i smos i n t rac i t op lasmát icos ocor re
a t ravés da p rodução de espéc ies reat i vas do ox igên io e do n i t rogên io ,
sendo um des tes p rodutos o óx ido n í t r i co (NO) , uma molécu la
responsáve l po r numerosos fenômenos f i s io lóg i cos (BOGDAN 1997,
2001) .
O NO é um gás inco lo r à t emperatu ra ambien te , pouco so lúve l em
água , a l t amen te reat i vo , com meia -v ida de 5 a 30 segundos . É s in te t i zado
pe la ação da enz ima Óx ido N í t r i co S in tase (NOS) sobre o subs t ra to L-
a rg in ina, conver tendo-a em L-c i t ru l ina e l ibe rando NO. Es te pode ser
ex c re tado a t ravés do a r exp i rado em sua fo rma in i c ia l ou pe lo suo r ,
sa l i va e fezes em sua fo rma ox idada n i t r i to ou n i t ra to . A ex creção
p r inc ipa l ocor re pe la u r ina (BAYLIS & VALLANCE, 1998; ELLIS e t a l . ,
1998) .
Mu i tos exper imentos têm s ido fe i t os para se ava l i a r a par t i c ipação
do NO l i gada ao cont ro le da pressão a r te r i a l , neu rot rasmissão ,
re laxamento muscu lar , per is ta l t i smo in tes t i na l , coagu lação sanguínea,
adesão e agregação p laquetá r i a (ADAMS, 1996 ; JORES e t a l ., 1992;
FAHEY et a l ., 1996 ; WALDECK et a l ., 1998; RADOMSKI e t a l ., 1990;

24
FA INT e t a l ., 1991 ; LORENTE et a l ., 1996) , bem como ver i f i ca r sua
par t i c ipação na b io log ia de tumores (AMBER et a l ., 1988; STUEHR &
NATHAN, 1989) , reações i n f l amató r ias au to - imunes (SINGH e t a l .,
2000) , no choque sépt i co (TRZECIAK e t a l . , 2008) e em doenças
i n f l amató r i as (GRABBE & SCHUWARZ, 1998) .
A l i t e ra tu ra desc reve t rês i soenz imas de Óx ido Ní t ri co S in tases
(NOS) p resen tes no c i t oso l , sendo duas cons t i t u t i vas (eNOS e nNOS) e
uma induz íve l ( iNOS) (CERQUEIRA & YOSHIDA, 2002) ,
es t ru tu ra lmen te semelhan tes , mas regu ladas de d i fe ren tes fo rmas . As
cNOS oco r rem em duas i so fo rmas , neu rona l (nNOS) e endote l i a l (eNOS),
a nNOS es tá envo lv ida com p rocessos homeos tá t i cos como
neuro t ransmissão, pe r i s ta l t i smo e a eNOS com cont role imed ia to da
p ressão a r te r i a l e vasod i l a tação (F ILHO & ZILBERSTEIN , 2000) .
As cNOS têm peso molecu lar de 133 kDa ap resentando-se nas
fo rmas monomér i cas , são regu ladas por um mecan ismo Ca2 +/ ca lmodu l ina
que a t i va e ina t i va as enz imas, es tas têm s ín tese basa l e p roduzem
concent rações p i comola res de NO. Ao con t rá r i o , Óx ido N í t r i co S in tase
i nduz íve l ( iNOS) necess i t a da Ca2 +/ ca lmodu l ina apenas para sua
a t i vação , não oco r rendo ina t i vação quando a concen tração de cá l c io ca i .
A iNOS é expressa quando há indução da s ín tese do RNAm por
endo tox inas bac te r i anas , l i popo l i sacár i des (LPS) , ou c i t oc inas
re lac ionadas com respos ta do T ipo 1 , como IFN-γ , TNF-α , IL -1 β. Esta
enz ima t em peso molecu lar de 130 kDa e ap resen ta-se sob a fo rma de
monômero e te t râmero . A at i vação da iNOS leva a concent rações
nanomola res de NO (1000 vezes maio r que as cNOS) e a ação dessa
enz ima requer n i co t i namida aden ina d inuc leo t ídeo fos fa to (NADPH) ,
t e t rah id rob iop te r ina (BH4) , f l av ina aden ina d inuc leo t ídeo (FAD) , f l av ina
mononuc leo t ídeo (FMN) , O2 e heme como co fa to res ( IGNARRO, 1996; L I
& POULOS, 2005; TE IXE IRA e t a l ., 2002) .
A loca l i zação imuno-h is toquím ica de iNOS no baço de ra tos ,
i nduz ida po r Prop ion ibacte r ium acnes e LPS fo i demonst rada em
macró fagos , a lguns l in fóc i t os , neut ró f i los e eos inóf i l os , suger indo ass im
a ex pressão da enz ima em vár ios t ipos ce lu la res (FÖRSTERMANN &
DUN, 1996) .

25
A geração de NO é ca rac te r ís t i ca de cé lu las do s is tema imune ta is
como: cé lu las dendr í t i cas , NK, monóc i tos , macró fagos , m ic róg l i a ,
cé lu las de Kupf fe r , eos inó f i l os e neu t ró f i l os . A lém d is to , ou t ros t ipos
ce lu la res como: cé lu las endo te l i a i s , ep i t e l i a is , célu las da muscu la tu ra
l i sa , f ib rob las tos , condróc i t os , hepatóc i tos , cé lu las mesang ia i s e cé lu las
de Schwann também são po tenc ia i s p roduto ras de NO (MONCADA e t a l .,
1991) .
O p roduto da ox idação do NO gerado pe las cé lu las o ri g ina n i t r i t os
e n i t ra tos (MARLETTA, 1988; MARLETTA e t a l ., 1988 ; MAYER e t a l .,
1989) e sua a t i v i dade an t im icrob iana tem s ido demonst rada po r uma sér ie
de exper imentos , sendo que há ev idênc ias demonst rando que a p rodução
de NO é par te i n tegran te da respos ta do hospede i ro à in fecção
(MACMICKING e t a l. ,1997) .
A l t a p rodução de NO por macró fagos depende de L-a rgin ina
ex t race lu la r em quan t idades adequadas (HEVEL & MARLETTA, 1992) .
Na maio r i a dos t ipos ce lu la res , o t ranspor te da a rgin ina oco r re v ia
s is tema ca t i ôn i co i ndependente do pH e Na+ , enquan to a a t i v i dade é
med iada po r uma famí l i a de p ro te ínas t ranspor tado ras de aminoác idos
ca t i ôn i cos (CAT1 , CAT2A, CAT2B) (N ICHOLSON e t a l ., 2001) . O
per fe i t o func ionamento desse s is tema é en tão necessár io pa ra que o
s is tema de de fesa in t race lu la r a tue .
A NADPH ox idase e a iNOS são enz imas envo lv idas na p rodução
de i n te rmed iá r ios do ox igên io e espéc ies rea t i vas do n i t rogên io
respec t i vamente . A fusão dessas mo lécu las a l t amente reat i vas pode gera r
pe rox in i t r i to (OONO-) , S -n i t ro t io is (RSNO), óx ido de n i t rogên io I I I
(N 2O3) , t e t róx ido de d in i t rogên io IV (N2O4) e complexos d in i t ros i l - fe r ro
(CDNF) (CERQUEIRA & YOSHIDA, 2002; PACHER e t a l ., 2007) ,
l evando à fo rmação de vá r i as mo lécu las com poder ant im ic rob iano.
O NO pode es ta r re lac ionado tan to na defesa do hospede i ro
(BRANDONIS IO e t a l. , 2001; PANARO e t a l. , 2001) e com a v i ru lênc ia
de p ro tozoár ios (HERTZ & MANSFIE LD, 1999 ; GENESTRA et a l .,
2006a) .
Pesqu isadores re la tam a p resença de mo lécu las p ro teto ras
p resentes na membrana ce lu lar des tes paras i tos , t a is como: g l i co l íp ides ,

26
g l i co inos i to l fos fo l íp ides e l ipo fos fog l i cano (LPG) , os ú l t imos com
potenc ia l capac idade in ib ido ra de s ín tese de NO, quando macró fagos
a inda não fo ram at i vados pe lo IFN-γ . Es tes es tudos re la tam que LPG
podem in ib i r a s ín tese de IL -12 e en tão b loquear i nd i re tamente a indução
da iNOS (L IEW e t a l ., 1997) .
A p rodução de NO e a t i v idade an t i -Le ishmania fo i de tec tada em
l inhagem de macró fagos can inos i n fec tados após incubação com IFN-γ ,
TNF-α e IL -2 (P INE LLI e t a l ., 2000) . Ou t ros pesqu isadores
demonst ra ram a impor tânc ia da geração de NO como m ic rob ic ida , em
LVC, usando cu l t u ra de cé lu las mononuc leares do sangue per i fé r i co de
cães exper imenta lmente i n fec tados com L. (L . ) chagas i, nas qua i s o
i n i b ido r especí f i co da iNOS, aminoguan id ina (AMG), l evou ao aumento
da ca rga paras i t á r ia e d im inu i ção dos n í ve is de NO em sobrenadan te de
cu l tu ra (RODRIGUES e t a l ., 2007) .
A impor tânc ia l e ishmanic ida do NO v ia iNOS em cu l tura de
macró fagos mur inos , t ambém usando i n ib ido res específ i cos da enz ima
mos t ra ram que a a t i v i dade da iNOS é c ruc ia l para con t ro le da
pers is tênc ia da Leishmania em hospede i ros imunocompetentes após a
reso lução da in fecção p r imár ia . D ive rsos t raba lhos demonst ram que a
fa lha em mante r iNOS at i va pode l evar a uma reat i vação de in fecção
l a ten te causada po r o rgan ismos sens íve i s ao NO, duran te a fase de
imunossupressão (L IEW e t a l ., 1990a; STENGER e t a l ., 1996) .
O NO e a sua capac idade l e i shmanic ida em macró fagos de cães
vac inados com promast i go tas de Le ishman ia (L . ) in fan tum ap resenta ram
assoc iação en t re a p rodução de NO induz ido po r IFN-γ (BACELLAR e t
a l . , 2000 ; PANARO e t a l ., 2001) . Refo rçando a impor tânc ia do NO como
m ic rob ic ida , demonst rou -se um aumen to da expressão de iNOS em
cu l tu ra de macró fagos can inos in fec tados com Leishman ia (L . ) in fan tum
es t imu lados com IFN-γ . Naque le es tudo, a maio r exp ressão de iNOS
levou ao aumento na p rodução de NO (S ISTO e t a l ., 2001) .
A iNOS tem s ido assoc iada a res is tênc ia à in fecção por
Leishmania ma jo r em camundongos C57BL/6 , nos qua i s t em s ido
demonst rada a ex pressão des ta enz ima desde o in í c io da i n fecção na

27
pe le . Cont ra r i amente , macró fagos de camundongos BALB/c , suscep t íve i s
à i n fecção , ap resen tam n íve i s menores de exp ressão de iNOS dev ido à
i n i b i ção causada por TGF-β e d im inu i ção da respos ta ao IFN-
γ (STENGER e t a l ., 1994; BOGDAN, 1997; 2001) .
GANTT e t a l ., (2003) ava l i ando os mecan ismos de de fesa das
l e i shmanias “ i n v i t ro ” mos t ra ram que es tas podem l ibera r p ro tease
ca teps ina B , capazes de a t i var TGF-β l a ten te e ass im a t i va r a a rg inase . O
TGF-β a tua r i a de fo rma a pro longar o pa ras i t i smo , i n ib indo o s i s tema de
de fesa dos macró fagos e des ta fo rma d iminu indo a p rodução de NO.
Re la ta -se que a po la r i zação da respos ta T ipo 1 (Th1) é e f i c i en te
na geração de NO e es tá assoc iada com doenças i n f l amató r i as e
e l im inação de patógenos in t race lu la res v ia iNOS. Em con t ras te , quando
p redominam no me io ex t race lu la r c i t oc inas como IL-4, IL -5 , IL-10 e
TGF-β ocor re i n i b i ção da produção de NO ( respos ta T ipo 2 ou Th2)
(NAGELKERKEN e t a l ., 1993; BACELLAR e t a l ., 2000) . Exper imentos
demonst ra ram que o p redomín io de cé lu las do T ipo 2 , com aumentada
p rodução de IL-4 , ca rac te r i zam-se po r não se rem p rote to ras con t ra as
l e i shmanioses , ocor rendo exacerbação dos s in tomas (LOHOFF e t a l .,
1997) .
Os e fe to res ce lu la res Th1 e Th2 se or i g inam a par t ir de cé lu las
p recurso ras chamadas Thp. Es tas quando são es t imu ladas pe la IL -12 dão
o r i gem a cé lu las T ipo 1 e quando es t imu ladas pe la IL -4 o r i g inam cé lu las
T ipo 2 (MOSMANN & COFFMAN, 1989) . En t re tan to , são mui tos os
fa to res que in f l uenc iam na po la r i zação para um dos t i pos de respos ta
imune f ren te a paras i tos do gênero Leishmania spp . Recentemente fo i
desc r i t o que a lém das popu lações e fe to ras der i vadas de LT aux i l i a res
desc r i t as ac ima, a ex is tênc ia de cé lu las T regu la tór i as (T CD4+CD25+ )
poder iam favo recer a i n fecção levando uma modu lação da imun idade
p ro te to ra ao paras i to (L IU e t a l ., 2003; XU e t a l ., 2003) .
Es tudos re la ta ram que cães natura lmente i n fec tados com
Leishmania (L . ) chagas i, s in tomát i cos , apresenta ram ma iores n íve is de
c i t oc inas IL-10 e TGF-β (BELKA ID e t a l ., 2001; CORREA e t a l ., 2007) ,
suger indo que a ex i s tênc ia de cé lu las regu la tó r i as poder ia i n f luenc ia r a

28
i n te ração paras i to -hospede i ro e des ta fo rma favo recer a man i fes tação de
fo rmas c l ín i cas graves na LVC.
A lém da sua par t i c i pação na imun idade a i n fecções , o NO pode
es ta r envo lv ido em p rocessos imunopato lóg i cos , po is sua a l t a reat i v idade
pode ocas ionar danos ce lu la res acar re tando um e fe i to tóx i co indese jáve l
no compar t imento ce lu la r . Há consenso de que o e fe it o c i t o tóx ico do NO
oco r re dev ido à s inerg ia com o íon superóx ido , p r i nc ipa lmente pe lo
subprodu to perox in i t r i to ( ISCHIROPOULOS, 2004; NORMAN & L IEW,
1995) . Es te p rodu to do NO é também um fo r te ox idante e pode reag i r
d i re tamen te com ampla gama de molécu las p resen tes no meio ce lu la r ,
p r inc ipa lmente os g rupos r i cos em e lé t rons , sua me ia v ida é cu r ta (~10 –
20 ms) , mas su f i c i en te para c ruzar a membrana ce lu lar , d i fund indo -se
uma a duas cé lu las de d iâmet ro (DENICOLA e t a l. , 1998) . Seus
p r inc ipa is a lvos são p ro te ínas , sendo i nc lus i ve capaz de i na t i var a iNOS
po r mod i f i cação ox idat i va de seu grupo heme (HUHMER e t a l. , 1997) ,
aminoác idos , p r inc ipa lmente ox idação das c i s te inas (SH IBATA e t a l. ,
1996) e po r mecan ismos ind i re tos de reações qu ímicas l eva a n i t ração de
t i ros inas , a fe tando a es t ru tu ra e função p ro té i ca , resu l tando na geração
de ep i t opos an t i gên i cos , mudança na a t i v idade de enz imas , a l t e rada
o rgan ização do c i to -esque le to e a l t e rações impor tantes na t ransdução de
s ina i s (SCHOPFER e t a l. , 2003) . J á se demonst rou seu po tenc ia l
ox idat i vo também sobre os aminoác idos t r i p to fano , met ion ina e
h is t id ina , a lém de reag i r com l i p íd ios e ác idos nuclé icos (PACHER et
a l . , 2007) .
Ev idênc ias es tão se acumulando no sen t ido de admi t ir que o NO
cont r i bua para a lgumas cond ições pa to lóg i cas como asma, a r t r i te
reumató ide, l esões a te rosc le ró t i cas , t ubercu lose, esc le rose múl t ip l a ,
A lzhe imei r e gas t r i te induz ida por Hel i cobacter py lo r i (DUSSE et a l. ,
2003) .
A ava l i ação da expressão da enz ima iNOS em tec ido esp lên i co de
cães natu ra lmente in fec tados Le ishmania (L . ) chagas i, bem como a
quan t i f i cação sé r ica de NO é um tóp i co pouco inves ti gado . A lém d isso , a
assoc iação en t re esses b iomarcadores , as d i fe ren tes fo rmas c l ín i cas , os
g raus de paras i t i smo esp lên i co e suas co r re lações com o per f i l de

29
c i t oc inas e subc lasses de an t i co rpos pode aux i l i a r no en tend imento da
imunopa togênese da LVC. Nes te sent ido , o p resente traba lho , buscou
fo rnecer i n fo rmações ad i c iona i s à e luc idação do complexo mecan ismo de
i n te ração paras i t o -hospede i ro na LVC cons iderando o compar t imen to
esp lên i co como um impor tan te ó rgão l i n fó ide a t ing ido pe lo paras i t o .

30
2 OBJET IVOS
2 .1 OBJ ETIVO GERAL
Cor re lac ionar n í ve is sé r i cos de NO e a exp ressão da enz ima iNOS
no baço com per f i l de c i t oc inas e subc lasses de Ig de cães na tu ra lmente
i n fec tados com Leishmania (Le i shmania ) chagas i , apresentando
d i fe ren tes fo rmas c l ín i cas e g raus de paras i t i smo esp lên i co .
2 .2 OBJ ETIVOS ESPECÍF ICOS
2 .2 .1 De te rminar a ca rga paras i t á r i a no baço de cães natu ra lmente
i n fec tados com L . (L . ) chagas i po r tado res de d i fe ren tes fo rmas c l ín icas
de l e i shmaniose e de f in i r os g raus de paras i t i smo esp lên i co
2 .2 .2 De te rminar os n í ve is do óx ido n í t r i co sé r i co de cães
na tu ra lmente i n fec tados com L. (L . ) chagas i e assoc ia r os resu l t ados com
as fo rmas c l í n i cas e g raus de paras i t i smo esp lên i co.
2 .2 .3 Ava l i a r a exp ressão da enz ima iNOS em baço de cães
na tu ra lmente i n fec tados com L. (L . ) chagas i e assoc ia r os resu l t ados com
as fo rmas c l í n i cas e g raus de paras i t i smo esp lên i co.
2 .2 .4 Assoc ia r os per f i s de exp ressão de iNOS em baço e os n íve is de
NO sér i cos com a produção das c i toc inas INF-γ , TNF-α e IL -10 .
2 .2 .5 Assoc ia r os per f i s de ex pressão de iNOS em tec ido esp lên i co e
os n íve is sé r icos de NO com os n í ve is de IgG1 , IgG2 e IgG to ta l an t i -
Leishmania.

31
3 MATERIAIS E MÉTODOS
Este es tudo é f ru to de uma co laboração en t re o Núc leo de
Pesqu isa em Imuno log ia da UNIVALE e o Labora tó r io de Imunopato log ia
do NUPEB/UFOP.
O banco de dados d i sponíve l pa ra c i t oc inas e an t i corpos des tes
an imais poss ib i l i t ou aná l i ses de co r re lações en t re os n í ve is sér i cos de
NO e ex pressão esp lên i ca de iNOS com a exp ressão esp lên i ca de IL-10 ,
IFN- γ , TNF-α e n íve is sé r i cos de Ι gG1, IgG2 e IgG to ta l an t i–
Leishmania .
3 .1 ANIMAIS
Soro e f ragmen tos de baço de 35 cães sem raça de f i nida (SRD) de
ambos os sexos , com idade es t imada en t re 2 a 6 anos, p rocedentes do
cen t ro de con t ro le de zoonoses (CCZ) de Be lo Hor izon te – MG, fo ram
se lec ionados para es te es tudo. O uso ex per imenta l des tes fo i ap rovado
pe lo comi tê de é t i ca da UFMG (CETEA) . Os an imais hav iam s ido
p rev iamente t ra tados com ant i -he lmín t i cos de l a rgo espect ro (Enda l
p lus®) e imun izados cont ra parvov i rose, l ep tosp i rose , pa ra in f luenza e
hepat i t e (P f i zer®) . Os cães inser i dos nes te es tudo ap resentavam tes tes
so ro lóg i cos pe la Reação de Imuno f l uo rêscenc ia Ind i re ta (R IF I) e ELISA
pos i t i vos (C I) .
Dez cães com t í tu los in fe r io res a 1 :40 (R IF I) , ex ames
paras i t o lóg i cos nega t ivos e c l i n i camen te sad ios fo ram cons iderados não
i n fec tados e i nc lu ídos no grupo cont ro le (CNI) .

32
3 .2 AVALIAÇÃO CLÍN ICA
Os cães so ropos i t i vos po r R IF I fo ram c l in icamente class i f i cados
de aco rdo com presença ou ausênc ia de s ina is c l ín icos em: Ass in tomát i co
(CA, n=10) , quando não hav ia s ina i s ou s in tomas da doença;
O l i goss in tomát i co (CO, n=10) , com no máx imo t rês s ina i s c l ín i cos
i nc lu indo có rnea opaca e /ou a lopec ia e /ou moderada perda de peso;
S in tomát i co (CS , n=15) , apresentando s ina i s c l ín i cos de l e i shmaniose
v isce ra l , com córneas opacas , severa perda de peso , on icogr i fose, l esões
cu tâneas , apat ia e ce ra to -con jun t i v i t e . A ava l i ação c l ín ica fo ra rea l i zada
po r um méd ico veter i ná r io t re inado em LVC.
3 .3 ANÁLISE PARASITOLÓGICA
Após nec rops ia , amos t ras do baço dos cães so ropos i ti vos fo ram
submet idas a exames paras i t o lóg icos para pesqu isa de fo rmas
amas t i go tas de Leishmania po r meio de es f regaços e c lass i f i cados de
aco rdo com a ca rga paras i t á r i a . Nes te t raba lho fo i usado o índ i ce de
ca rga paras i tá r ia ob t i do pe la aná l i se do tec ido esplên i co .
Amost ras do baço fo ram co le tadas e impressões fo ram rea l i zadas
em duas l âm inas m ic roscóp i cas que após secas ao a r fo ram f i xadas em
metano l , co radas com Giemsa e examinadas ao mic roscóp io óp t i co . Fo i
con tado o número de fo rmas amast i go tas po r 1000 cé lu las nuc leadas e os
resu l t ados ex pressos como “Le i shman Donovan Un i ts ” (LDU) de aco rdo
com STAUBER (1955) mod i f i cado po r REIS e t a l . (2006 a ,b ,c ) .
O paras i t i smo esp lên i co fo i ca tegor izado como ba ixo (BP) , méd io
(MP) e a l t o paras i t i smo (AP) con fo rme os va lo res de LDU
es ta t i s t i camente catego r izados em terc i s da segu in te fo rma: BP (0 – 10 ) ,
MP (11– 250) e AP (251 – 2564) de aco rdo com LAGE et a l . (2007) .

33
3 .4 QUANTIF ICAÇÃO SÉRICA DE NO
A ava l i ação da concen t ração de NO sér i co fo i rea l izada pe la med ida
de n i t r i t os (NO2- ) e n i t ra tos (NO3
- ) , de aco rdo com a reação de Gr iess
(GREEN e t a l ., 1982; GUTMAN & HOLLYWOOD, 1992) . Os so ros dos
cães fo ram cod i f i cados an tes da rea l i zação da dosagem e decod i f i cados
após , com pos te r i o r aná l i se dos dados ob t i dos .
Para ava l i ação dos n íve i s sér i cos de NO, a 100 µ L de so ro de cada
cão d i lu ído 1 :4 em água des t i l ada fo ram ad i c ionados 10 µL de n i t ra to
redutase (1U/mL) (S igma-A ld r i ch Co. , S t . Lou i s , MO, USA) , 10 µL de
NADPH (6mM) (S igma-A ld r i ch Co. , S t . Lou is , MO, USA) e 10 µL de
FAD (200 mM) (S igma-A ld r i ch Co . , S t . Lou is , MO, USA) . Es te
p roced imento fo i necessár io pa ra reduz i r n i t ra to (NO3- ) em n i t r i t o (NO2) .
Após um per íodo de incubação po r 12 horas a 37oC, as amost ras fo ram
submet idas ao t ra tamento com 1 /20 de su l fa to de z inco (300g/L) (S igma-
A ldr i ch Co . , S t Lou is , MO, USA) . Subseqüen temente , as amost ras fo ram
cen t r i f ugadas a 10000g po r 15 minu tos . Após cent r i fugação , 100 µL de
cada amos t ra fo ram ad i c ionadas em dup l i ca ta em p lacas de 96 poços de
fundo p lano . Pos ter i o rmente , fo ram ad i c ionados 100 µ L de reagente de
Gr iess (0 .1% naphth yle thylend iam ine d ihyd roch lo r ide, 1%
su lphanylam ide , 5% phosphor i c ac id ) (S igma-A ld r i ch Co . , S t Lou is , MO,
USA) às amost ras p rocessadas de so ro . Após i ncubação em câmara
escu ra po r 10 minu tos , as amost ras fo ram submet idas ao l e i t o r
au tomát i co de mic rop lacas e a abso rbânc ia fo i ava l iada no compr imento
de onda de 570 nm. A concent ração de n i t r i t o das amost ras fo i
de terminada pe la ex t rapo lação da cu rva -padrão de d ilu ição da so lução de
n i t r i t o de sód io (com var i ação l inear de 0 ,78 -100 µmol /L) poss ib i l i t ando
a iden t i f i cação de va lo res de cada amost ra . Os resul t ados (em µM) fo ram
ex p ressos como méd ia da concen t ração de NO sér i co .

34
3 .5 REAÇÃO IMUNO-H ISTOQUÍM ICA PARA DETECÇÃO DE INOS
Co r tes h i s to lóg i cos p rocessados e para f in izados foram submet idos à
m ic ro tomia para a ob tenção de 2 cor tes com espessu ra de 4 µm e es tes
fo ram f i xados em lâminas ge la t i n izadas . A segu i r , os co r tes fo ram
despara f in izados em x i lo l (do i s banhos de 15 minu tos cada) , h id ra tados
em concent rações dec rescentes de á l coo l (100 , 90 , 80 e 70%) em banhos
de 5 m inu tos cada . Em segu ida , os co r tes fo ram lavados em água
co r ren te po r 5 minu tos e en tão se p rocedeu ao b loque io da perox idase
endógena (180ml de á l coo l met í l i co /20mL de peróx ido de h id rogên io
30%) por 30 minu tos à tempera tu ra ambien te . Segu iu -se a l avagem em
PBS com t rês banhos de 5 minu tos cada .
Para a recuperação an t i gên i ca fo i u t i l i zado o tampão c i t ra to pH
6 .0 em fo rno de mic roondas po r 10 minu tos . Depo is de re t i radas do fo rno
de m icroondas , os co r tes fo ram de ixados à temperatura amb ien te para
oco r re r um res f r iamento p rogress i vo dos mesmos a inda imersos no
tampão c i t ra to pH 6 .0 . Só en tão fo ram lavados em PBS, em t rês banhos
de 5 minu tos cada .
Após a secagem das bo rdas dos co r tes com pape l absorvente ,
ap l i cou -se o soro no rmal de cava lo (R .T .U . Vectas tain Un iversa l E l i t e
ABC; Vecto r Labora to r ies , Inc . ) po r 30 minu tos em câmara úmida a
37°C, com o ob je t i vo de b loquear s í t i os i nespec í f i cos . Em segu ida ,
secou-se o excesso de so ro no rma l de cava lo e os cor tes fo ram incubados
po r 18 ho ras em câmara úm ida a 4°C com o an t i co rpo po l i c lona l IGG de
coe lho an t i - iNOS (Rabb i t an t i -Mouse iNOS, SC-651 , Santa C ruz
B io techno logy, INC. ) d i l u ído 1 :200 em so lução d i l uen te de an t i co rpo
com componentes redutores de “background” (DakoCytomat ion , USA) .
Pos te r i o rmen te , os co r tes fo ram lavados em PBS e i ncubados com
o an t i co rpo secundár io b io t in i l ado (R .T .U. Vectas tain Un ive rsa l E l i t e
ABC; Vecto r Labora to r ies , Inc . ) po r 30 minu tos em câmara úmida a
37°C. Segu i ram-se t rês banhos de 5 m inu tos cada, em PBS. Os co r tes
fo ram en tão incubados com o complexo ABC (R .T .U. Vec tas ta in
Un ive rsa l E l i t e ABC; Vector Labora tor i es , Inc . ) po r 30 m inu tos em

35
câmara úmida a 37°C . Segu iu -se a lavagem em PBS, t rês banhos de 5
m inu tos cada. A reve lação da reação da perox idase fo i ob t i da a t ravés da
i ncubação em so lução de DAB (50mg de d iaminobenz id ina em 250 mL de
PBS e 500 µL de Peróx ido de H id rogên io 30%) du ran te 5 minu tos . No
i n tu i t o de i n te r romper a reve lação, os co r tes fo ram lavados em água
co r ren te po r c inco m inu tos .
Para rea l i za r a con t ra -co lo ração , fo i u t i l i zado Hematox i l i na de
Har r i s por 10 segundos à temperatu ra amb ien te segu ida pe la l avagem dos
co r tes por 5 minu tos em água co r ren te . O mate r i a l fo i des id ra tado em
á l coo l 100% e seco em es tu fa 56°C. Após a secagem as l âminas fo ram
mon tadas com res ina (P ró -C i to ) e l amínu la .
Como cont ro le da reação fo ram inc lu ídos co r tes de pe le ob t idos de
cão inocu lado com ant ígeno de Leishmania (V iann ia ) b ras i l i ens is e
sapon ina, que j á hav ia most rado a l t a ex pressão de iNOS, du ran te a fase
de padron ização da reação imuno-h is toquímica . Como con t ro le nega t ivo ,
omi t iu -se o an t i corpo an t i - iNOS, que fo i subs t i t u ído po r PBS em uma
lâm ina . Os tec idos ass im obt idos fo ram u t i l i zados para ana l i sa r a
ex p ressão de iNOS no baço. Es te exper imento fo i real i zado em dup l i ca ta
em d ias d i fe ren tes e em cada ba ter i a hav ia 1 cont role pos i t i vo e 1
cont ro le nega t i vo . As l âminas fo ram submet idas à aná l i se qua l i t a t i va
usando-se mic roscóp io óp t i co e pos te r io rmente à anál i se quant i t a t i va da
ex p ressão de iNOS a t ravés de mic roscóp io Le i ca DM 5000/Le i ca CTR
5000 , p rocessadas com o aux í l i o do so f tware Le i ca Qwin P lus ,
ana l i sando-se 20 campos a leató r i os de cada l âm ina e os resu l t ados
ex p ressos em µm2 . Seqüenc ia lmen te os d i fe ren tes grupos c l í n i cos fo ram
assoc iados aos resu l tados de exp ressão de iNOS.
Fo ram u t i l i zados d i fe ren tes parâmet ros na observação da
ex p ressão da iNOS, a saber : d i fusa (quando a marcação oco r r i a em toda
ex tensão do campo mic roscóp io ana l i sado) , foca l (quando os focos de
marcação f i cavam de l im i tados em áreas bem de f in idas) e pontua l
(quando cé lu las marcadas i so ladamen te e ram observadas ) .

36
3 .6 ANÁLISES ESTATÍST ICAS
Rea l i za ram-se aná l i ses es ta t ís t i cas u t i l i zando-se tes tes espec í f i cos
para compara r os resu l t ados das dosagens de NO em so ro e exp ressão de
iNOS esp lên i ca com o apo io ins t rumenta l do so f tware PRISM 4 .0
adotando-se n í ve l de s ign i f i cânc ia a 5%. Os dados fo ram submet idos aos
tes tes não paramét r i cos . Para aná l i se da ca rga paras i t á r i a po r g rupos
c l í n i cos , dos n í ve is sé r icos de NO por g rupos c l í n icos , n íve i s sé r i cos de
NO po r carga paras i t á r i a , ava l i ação quan t i t a t i va de iNOS po r g rupos
c l í n i cos e ava l iação quan t i t a t i va de iNOS po r ca rga paras i t á r i a fo i usado
o tes te de Kruska l Wal l i s . Os d i fe ren tes graus de paras i t i smo esp lên i co
fo ram es t imados po r t e rc i s . Para aná l i se dos n í ve i s de NO ent re an imais
i n fec tados e não i n fec tados usou-se o tes te de Mann-Whi tney. Para
aná l i ses de co r re lação en t re n í ve is sér i cos de NO e per f i l sé r i co de Ig ,
co r re lação en t re n íve is sé r i cos de NO e dens idade paras i tá r ia esp lên i ca ,
co r re lação en t re n íve is sé r i cos de NO e per f i l de ci t oc inas esp lên i cas ,
co r re lação en t re exp ressão esp lên i ca de iNOS e per fi l sé r i co de Ig e
co r re lação en t re exp ressão esp lên i ca de iNOS e per fi l de c i t oc inas
esp lên i cas usou-se co r re lação de Spearman .

37
4 RESULTADOS
4 .1 DETERMINAÇÃO DA CARGA PARASITÁRIA
4 .1 .1 Carga pa ras i tá r ia po r fo rma c l í n i ca
A carga paras i t á r ia esp lên i ca dos d i fe ren tes grupos c l í n i cos de
cães catego r izados em CA, CO e CS ap resentou med iana de LDU igua i s a
2 , 20 e 298 respec t i vamente , havendo d i fe rença s igni f i ca t i va en t re os
g rupos (p<0 ,01 ) . Ver i f i cou -se aumento da ca rga paras i tá r i a assoc iada à
maio r g rav idade da LVC, como demons t rado na F igu ra 1 .
F i g u r a 1 - D e n s i d a d e p a r a s i t á r i a e s p l ê n i c a ( L D U – “ Le i s h ma n i a D o n o v a n U n i t s ” ) p o r f o r ma c l í n i c a d e l e i s h ma n i o s e vi s c e r a l c a n i n a . C ã e s a s s i n t o má t i c o s ( C A ; n = 1 0 ) , c ã e s o l i g o s s i n t o mát i c o s ( C O ; n = 1 0 ) e c ã e s s i n t o má t i c o s ( C S ; n = 1 5 ) . D i f e r e n ç a s i g n i f i ca t i v a r e p r e s e n t a d a p o r p < 0 , 0 1 ; T e s t e d e K r u s k a l W a l l i s e p ó s - t e s t e d e D u n n .

38
4 .1 .2 Ca tegor i zação por g raus de paras i t i smo esp lêni co
O paras i t i smo esp lên i co fo i ca tegor izado como ba ixo (BP) , méd io
(MP) e a l t o paras i t i smo (AP) con fo rme os va lo res de LDU
es ta t i s t i camente catego r izados em terc i s da segu in te fo rma: BP (0 – 10 ) ,
MP (11– 250) e AP (251 – 2564) . Es ta c lass i f i cação permi t iu es t imar o
percen tua l de grau paras i t i smo por g rupo c l ín ico . Fo i observado que o
g rupo CS ap resen tava 53 ,3% de AP , 33 ,3% de MP e apenas 13 ,3% de BP;
O grupo CO apresen tava 20% de AP, 50% de MP e 30% de BP; Enquanto
o g rupo CA ap resen tava 30% de MP e 70% de BP (Tabe la 1 ) .
T a b e l a 1 - F r e q ü ê n c i a d e g r a u s d e p a r a s i t i s mo e s p l ê ni c o a s s o c i a d o a f o r ma s c l í n i c a s e m c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i
G R A U D E
P A R A S I T I S M O F O R M A S C L Í N I C A S
C A % C O % C S %
B P ( n = 1 2 ) 7 0 3 0 1 3 , 3
M P ( n = 1 3 ) 3 0 5 0 3 3 , 3
A P ( n = 1 0 ) 0 2 0 5 3 , 3
T o t a l % 1 0 0 1 0 0 1 0 0
C A – C ã e s a s s i n t o má t i c o s ; C O – C ã e s o l i g o s s i n t o má t ic o s ; C S – C ã e s s i n t o má t i c o s ; B P – B a i x o p a r a s i t i s mo ; M P - M é d i o p a r a s i t i s mo ; A P – A l t o p a r a s i t i s mo . ( n = 3 5 ) .
4 .2 N ÍVE IS SÉRICOS DE ÓXIDO N ÍTRICO
A dete rm inação dos n í ve is sé r i cos de óx ido n í t r i co fo i rea l i zada
de acordo com a redução de n i t ra tos a n i t r i tos , es tes reve lados pe la
reação de Gr iess e os resu l t ados assoc iados às fo rmas c l ín i cas dos cães ,
bem como ao grau de paras i t i smo esp lên i co .
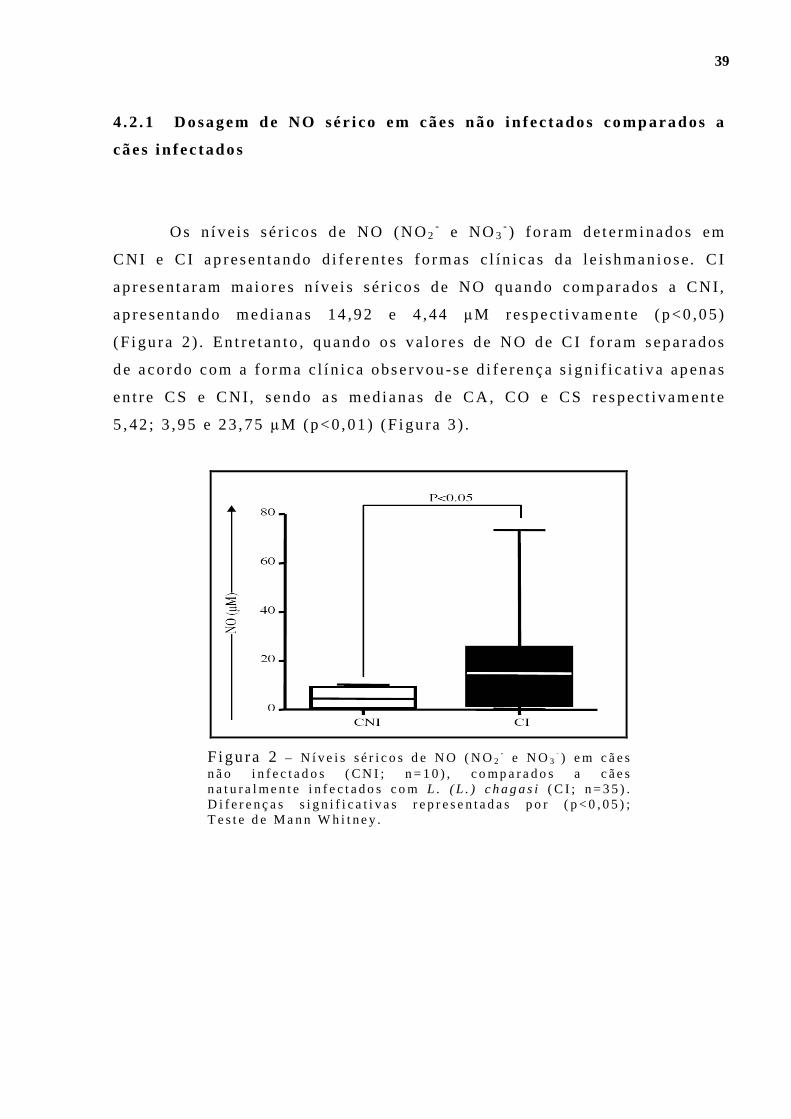
39
4 .2 .1 Dosagem de NO sér i co em cães não i n fec tados comparados a
cães i n fec tados
Os n í ve i s sé r i cos de NO (NO2- e NO3
- ) f o ram dete rminados em
CNI e C I ap resentando d i fe ren tes fo rmas c l ín icas da le i shmaniose. C I
ap resenta ram maiores n íve i s sé r i cos de NO quando comparados a CNI,
ap resentando med ianas 14 ,92 e 4 ,44 µM respect i vamente (p<0 ,05)
(F igu ra 2 ) . En t re tan to , quando os va lo res de NO de C I fo ram separados
de acordo com a fo rma c l í n i ca observou -se d i fe rença s ign i f i ca t i va apenas
en t re CS e CNI, sendo as med ianas de CA, CO e CS respec t i vamente
5 ,42 ; 3 ,95 e 23 ,75 µM (p<0,01 ) (F igu ra 3 ) .
F igu ra 2 – N í v e i s s é r i c o s d e N O ( N O2- e N O3
- ) e m c ã e s n ã o i n f e c t a d o s ( C N I ; n = 1 0 ) , c o mp a r a d o s a c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L . ( L . ) c h a g a s i ( C I ; n = 3 5 ) . D i f e r e n ç a s s i g n i f i c a t i v a s r e p r e s e n t a d a s p o r ( p < 0 , 0 5) ; T e s t e d e M a n n W h i t n e y .

40
F i g u r a 3 - N í v e i s s é r i c o s d e N O ( N O2- e N O3
- ) d e c ã e s n ã o i n f e c t a d o s ( C N I ; n = 1 0 ) , c o mp a r a d o s a c ã e s a s s i n t o má t i c o s ( C A ; n = 1 0 ) , c ã e s o l i g o s s i n t o má t i c o s ( C O ; n = 1 0 ) e c ã e s s i n t o má t i c o s (C S ; n = 1 5 ) . D i f e r e n ç a s s i g n i f i c a t i v a s r e p r e s e n t a d a s p o r ( p < 0 , 0 1) ; T e s t e d e K r u s k a l W a l l i s e p ó s - t e s t e d e D u n n . .
4 .2 .2 De te rminação dos n í ve i s sé r i cos de NO comparada por ca rga
pa ras i tá r i a
Fo ram ava l i ados também os n í ve is sé r i cos de NO de aco rdo com o
grau de paras i t i smo esp lên i co . Observa-se que a mediana de NO do grupo
com BP (1 ,49 µM) é s ign i f i ca t i vamen te menor que a med iana dos grupos
com MP (18 ,84 µM) e AP (26 ,37 µM) (p<0,05 ) (F igu ra 4A) . Es te
resu l t ado fo i re fo rçado pe la aná l i se de co r re lação en t re es tas va r i áve is
que reve la ram ex is t i r uma fo r te co r re lação en t re a ca rga paras i t á r i a e os
n íve i s sé r i cos de NO (p<0,0001 , r= 0 ,6444) (F igura 4B) .
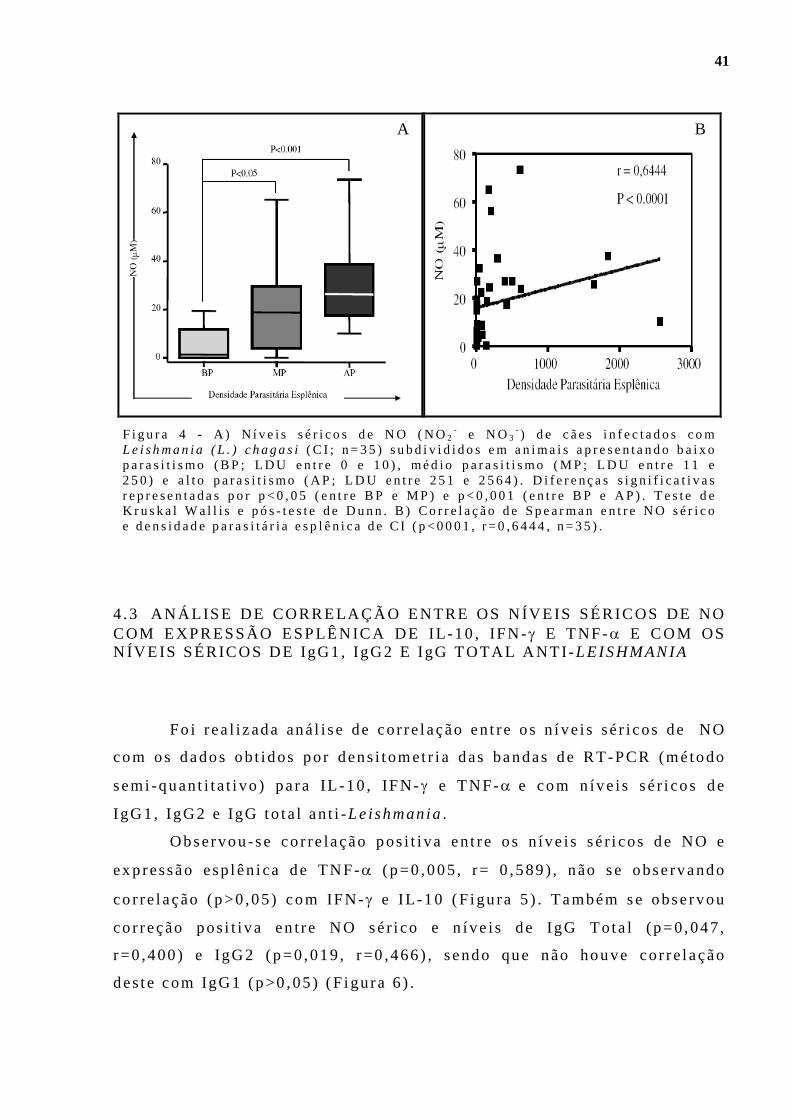
41
F i g u r a 4 - A ) N í v e i s s é r i c o s d e N O ( N O2
- e N O3- ) d e c ã e s i n f e c t a d o s c o m
L e i s h m a n i a ( L . ) c h a g a s i ( C I ; n = 3 5 ) s u b d i v i d i d o s e m a n i ma i s a p r e s e n t a n d o b ai x o p a r a s i t i s mo ( B P ; L D U e n t r e 0 e 1 0 ) , mé d i o p a r a s i t i smo ( M P ; L D U e n t r e 1 1 e 2 5 0 ) e a l t o p a r a s i t i s mo ( A P ; L D U e n t r e 2 5 1 e 2 5 6 4 ) . D i f e r e n ç a s s i g n i f i c a t i v a s r e p r e s e n t a d a s p o r p < 0 , 0 5 ( e n t r e B P e M P ) e p < 0 , 0 0 1 ( e n t r e B P e A P ) . T e s t e d e K r u s k a l W a l l i s e p ó s - t e s t e d e D u n n . B ) C o r r e l a ç ã o de S p e a r ma n e n t r e N O s é r i c o e d e n s i d a d e p a r a s i t á r i a e s p l ê n i c a d e C I ( p < 0 0 0 1 , r = 0 , 6 4 4 4 , n = 3 5 ).
4 .3 ANÁLISE DE CORRELAÇÃO ENTRE OS NÍVEIS SÉRICOS DE NO COM EXPRESSÃO ESPLÊNICA DE IL-10 , IFN-γ E TNF-α E COM OS N ÍVE IS SÉRICOS DE IgG1 , IgG2 E IgG TOTAL ANTI-LEISHMANIA
Fo i rea l i zada aná l i se de cor re lação en t re os n í ve is sé r icos de NO
com os dados ob t idos por dens i t omet r i a das bandas de RT-PCR (método
semi -quant i t a t i vo ) pa ra IL -10 , IFN-γ e TNF-α e com n íve is sé r i cos de
IgG1 , IgG2 e IgG to ta l an t i -Le ishmania.
Observou -se co r re lação pos i t i va en t re os n íve is sé ri cos de NO e
ex p ressão esp lên i ca de TNF-α (p=0 ,005, r= 0 ,589 ) , não se observando
co r re lação (p>0 ,05) com IFN-γ e IL-1 0 (F igu ra 5 ) . Também se observou
co r reção pos i t i va en t re NO sér i co e n í ve is de IgG Tota l (p=0,047,
r=0 ,400 ) e IgG2 (p=0,019, r=0 ,466 ) , sendo que não houve cor re lação
des te com IgG1 (p>0 ,05 ) (F igura 6 ) .
B A

42
F i g u r a 5 – A n á l i s e s d e c o r r e l a ç ã o d o s n í v e i s s é r i c o s d e N O ( N O2
- e N O3- ) e m
c ã e s i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i ( C I , n = 2 1 ) c o m e x p r e s s ã o e s p l ê n i c a d e I L - 1 0 ( A ; p = 0 , 9 7 1 ), I F N -γ ( B ; p = 0 , 1 8 1 ) e T N F -α ( C ; p = 0 , 0 0 5 , r = 0 , 5 8 9 ) . C o r r e l a ç ã o d e S p e a r ma n .
F i g u r a 6 - A n á l i s e d e c o r r e l a ç ã o d o s n í v e i s s é r i c o s d e N O ( N O2
- e N O3- ) d e
c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i ( C I ; n = 2 4 ) c o m n í v e i s s é r i c o s d e I g G t o t a l a n t i -L e i s h m a n i a ( A ; p = 0 , 0 4 7 , r = 0 , 4 0 0 ) , I g G 1 ( B ; p = 0 , 9 6 4 ) e I g G 2 ( C ; p = 0 , 0 1 9 , r = 0 , 4 6 6 ) . C o r r e l a ç ã o d e S p e a r ma n.
B
0 .0 0 .2 0 .4 0 .6 0 .80
2 0
4 0
6 0
8 0
IgG 2
NOµM
p = 0 ,0 19 r = 0 ,4 66
C
1 .50 .0 0 .5 1 .00
2 0
4 0
6 0
8 0
IgG 1
NO
(µM
) p = 0 ,9 64
0 .00 0 .15 0 .30 0 .45 0 .600
2 0
4 0
6 0
8 0
IgG T o ta l
NO
(µM
)
p = 0 ,0 47 r = 0 ,4 00
A
0 1 0 2 0 3 0 0
2 0
4 0
6 0
I F N - γ
NO
(µ
M)
p = 0 , 1 8 1
0 1 0 0 0 2 0 0 0 3 0 0 00
2 0
4 0
6 0
8 0
I L - 1 0
8 0N
O (µ
M) p = 0 , 9 7 1
A B
0 1 0 2 0 3 0
r = 0 , 5 8 9 p = 0 , 0 0 5
C
NO
(µ
M)
8 0
6 0
4 0
2 0
0
T N F -α

43
4.4 EXPRESSÃO ESPLÊNICA DE iNOS
4 .4 .1 Aná l i se semi -quan t i ta t i va de iNOS
Cor tes h is to lóg i cos de f ragmentos de baço dos cães natura lmente
i n fec tados com L . chagas i fo ram submet idos à reação imuno-
h is toquímica i nves t i gando-se a expressão da enz ima iNOS (F igu ra 7) .
Fo ram u t i l i zados d i fe ren tes parâmet ros na observação da exp ressão da
iNOS, a saber : d i fusa (quando a marcação oco r r i a em toda ex tensão do
campo m icroscóp io ana l i sado) , foca l (quando os focos de marcação
f i cavam de l im i tados em áreas bem def i n idas ) e pontua l (quando cé lu las
marcadas i so ladamente e ram observadas ) (Tabe la 2 ) . Observou -se
também marcação nas t rabécu las e cápsu las , que fo i i nc lu ída como
marcação d i fusa.

44
F igu ra 7 – F o t o m i c r o g r a f i a s d e c o r t e h i s t o l ó g i c o e s p l ê n i c o d e c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i s u b me t i d o a r e a ç ã o i mu n o - h i s t o q u í m i c a a n t i - i N O S . A ) A u s ê n c i a d e ma r c a ç ã o n o g r u p o c o n t r o le ( C N I ) ; B ) E x p r e s s ã o d i s c r e t a e m c ã o a s s i n t o má t i c o ( C A ) ; C ) mo d e r a d a e m c ã o o l i g o s s i n t o má t i c o ( C O ) ; D ) a c e n t u a d a e m c ã o s i n t o má t i c o ( C S ) . 6 0 0 X .
A B
C D

45
4 .4 .1 .1 Marcação D i fusa (MD)
No grupo CS , 53% dos an imais apresentaram marcação d i fusa
acentuada (MDA) , 30 ,8% marcação d i fusa moderada (MDM) e 16 ,2% não
ap resenta ram MD. 45% dos CO ap resen taram MDA, 18 ,2% MDM, 18 ,2%
marcação d i fusa l eve (MDL) e 18 ,6% não ap resenta ram MD. CA
ap resenta ram MDL em 57,1% dos an ima is e 42 ,9% de les não
ap resenta ram MD (Tabe la 2 ) .
4 .4 .1 .2 Marcação Foca l (MF)
Apresenta ram marcação foca l acentuada (MFA) 30 ,8% dos an imais
do grupo CS , 7 ,7% marcação foca l moderada (MFM), 5 ,5% marcação
foca l l eve (MFL) e 56% não ap resenta ram MF. 18 ,2% de CO
ap resenta ram MFA, 18 ,2% MFM, 9 ,1% MFL e 54 ,5% não apresentavam
MF. 14 ,2% de CA ap resen ta ram MFL e 85 ,8% não apresentavam MF
(Tabe la 2 ) .
4 .4 .1 .3 Marcação Pontua l (MP)
No grupo CS 7 ,7% dos an imais apresentaram marcação pon tual
acentuada (MPA) , 15 ,4% marcação pon tua l moderada (MPM) , 15 ,4%
marcação pon tua l leve (MPL) e 61 ,5% não ap resen ta ram MP. 9 ,1% de CO
ap resenta ram MPM, 18 ,2% MPL e 72 ,7% não ap resenta ram MP. 14 ,3% de
CA ap resen taram MPM, 14 ,3% MPL e 71 ,4% não ap resenta ram MP
(Tabe la 2 ) .

46
T a b e l a 2 – M é d i a p e r c e n t u a l d e ma r c a ç ã o d a e x p r e s s ão d e i N O S p o r g r u p o c l í n i c o q u a n t o a o t i p o e i n t e n s i d a d e n o s f r a g me n t o s e s p l ê n ic o s d e c ã e s i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i
Forma
clínica Marcação Difusa Marcação Focal Marcação Pontual
AUS. LEVE MOD. ACEN. AUS. LEVE MOD. ACEN. AUS. LEVE MOD. ACEN.
CA
(n=8) 42,9 57,1 0 0 85,8 14,2 0 0 71,4 14,3 14,3 0
CO
(n=8) 18,6 18,2 18,2 45 54,5 9,1 18,2 18,2 72,7 18,2 9,1 0
CS
(n=13) 16,2 0 30,8 53 56 5,5 7,7 30,8 61,5 15,4 15,4 7,7
C A – C ã e s a s s i n t o má t i c o s ; C O – c ã e s o l i g o s s i n t o má t ic o s ; C S – C ã e s S i n t o má t i c o s . A U S = a u s e n t e , M O D . = mo d e r a d a ; A C E N . = a c e n t u a d a .
4 .4 .2 Aná l i se quan t i ta t i va de iNOS
4 .4 .2 .1 Expressão esp lên i ca de iNOS comparados po r fo rma c l í n i ca
A aná l i se quant i t a t i va de iNOS em f ragmentos esp lênicos de 8
CNI e 29 C I fo i rea l i zada usando o ana l i sador de imagens Le i ca Qwin
P lus fo togra fando-se a lea to r i amente 20 campos mic roscóp i cos de cada
co r te h is to lóg i co ca l cu lando-se a méd ia de do is exper imentos . CNI não
ap resenta ram marcação detec táve l , po r tan to não fo i poss íve l quan t i f i car
iNOS nes te g rupo. A quan t i f i cação da exp ressão da iNOS em C I
ap resentou grande d ispersão dos dados , va r i ando de 0 a 1 .051 .769 µm2
com med iana 30 .561 µm2 (F igu ra 8 ) . As aná l i ses en t re as d i fe ren tes
fo rmas c l ín icas de C I com CNI most ra ram d i fe rença sign i f i ca t i va en t re
CS e CNI (p<0 ,01) e en t re CO e CNI (p<0,05 ) (F igu ra 9 ) .

47
F i g u r a 8 – A v a l i a ç ã o q u a n t i t a t i v a d a e x p r e s s ã o d e Óx i d o N í t r i c o S i n t a s e i n d u z í v e l ( i N O S ) n o b a ç o d e c ã e s n ã o i n f e c t a d o s ( C NI ) e n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i. I mu n o - h i s t o q u í m i c a a n t i - i N O S .
F i g u r a 9 – A v a l i a ç ã o q u a n t i t a t i v a d a e x p r e s s ã o d e Ó x i d o N í t r i co S i n t a s e i n d u z í v e l ( i N O S ) n o b a ç o d e c ã e s n ã o i n f e c t a d o s ( C NI , n = 8 ) e n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i, c l a s s i f i c a d o s p o r f o r ma s c l í n i c a s d e L V C ( C A ; n = 8 , C O ; n = 8 e C S ; n = 1 3 ) . D i f e r e n ç a s i g n i f i ca t i v a e n t r e C N I e C O r e p r e s e n t a d a p o r p < 0 , 0 5 e e n t r e C N I e C S r e p r e s e n ta d a p o r p < 0 , 0 1 . K r u s k a l W a l l i s e p ó s - t e s t e d e D u n n . I mu n o - h i s t o q u í m i c a a n t i- i N O S

48
4 .4 .2 .2 Expressão esp lên i ca de iNOS comparada po r ca rga paras i tá r ia
Não fo ram encont radas d i fe renças s ign i f i ca t i vas quando os
va lo res da exp ressão de iNOS fo ram agrupados po r carga paras i t á r ia
(p=0,055) . As med ianas de iNOS em cães com BP, MP e AP fo ram 4302,
21523, e 171524 µm2 respec t i vamente . Os resu l t ados demonst ram que
50% dos cães com BP não exp ressam n íve is detec táve is da enz ima, no
en tan to , em 100% dos cães com AP a ex p ressão oco r re (F igu ra 10) .
F i g u r a 1 0 – A v a l i a ç ã o q u a n t i t a t i v a d a e x p r e s s ã o d e Ó x i d o N í t r i co S i n t a s e i n d u z í v e l ( i N O S ) n o b a ç o d e c ã e s n ã o i n f e c ta d o s ( C N I , n = 8 ) e n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i, c l a s s i f i c a d o s p o r d e n s i d a d e p a r a s i t á r i a e s p l ê n i c a ( B a i x o p a r a s i t i s mo : L D U e n t r e 0 e 1 0 ) , mé d i o p a r a s i t i s mo : L D U e n t r e 1 1 e 2 5 0 e a l t o p a r a s it i s mo : L D U e n t r e 2 5 1 e 2 5 6 4 ) . p > 0 , 0 5 . T e s t e d e K r u s k a l W a l l i s e p ó s -t e s t e d e D u n n .
4 .4 .2 .3 Aná l i se de co r re lação en t re a exp ressão da iNOS com n íve is
esp lên i cos de IL-1 0 , IFN-γ e TNF-α e com n í ve is de IgG1 , IgG2 e IgG
to ta l an t i -Leishmania
Fo ram rea l i zadas aná l i ses de co r re lação de Spearman en t re os
dados quant i t a t i vos da ex p ressão de iNOS em C I com os n í ve is de IL -10 ,

49
IFN- γ e TNF-α, não sendo encont rados resu l t ados s ign i f i ca t i vos (p>
0 ,05 ) (Figura 11 ) . As aná l i ses de co r re lações en t re os dados
quan t i t a t i vos da exp ressão de iNOS em C I com os n í ve i s de IgG To ta l
an t i – Leishmania, IgG1 e IgG2 também não ap resentaram resu l t ados
s ign i f i ca t i vos (p>0,05) (Figu ra 12 ) .
F i g u r a 1 1 – C o r r e l a ç ã o d e S p e a r ma n d a e x p r e s s ã o e s p l ê n i c a d e i NO S e m c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i ( C I ; n = 2 1 ) c o m a e x p r e s s ã o e s p l ê n i c a d e I L - 1 0 ( A ; p = 0 , 1 1 0 ) , I F N -γ ( B ; p = 0 , 6 9 7) e T N F -α ( C ; p = 0 , 2 0 5 ) .

50
0.0 0.2 0.4 0.60
50
100
150
IgG Total
0.0 0.5 1.0 1.50
50
100
150
IgG 1
0.00 0.25 0.50 0.750
50
100
150
IgG 2
Áre
a m
arca
da p
ara
iNO
S (µm
2 ) x
104
p = 0,776 p= 0,966
p= 0,203
A
C
B
Áre
a m
arca
da p
ara
iNO
S (µm
2 ) x
104
Áre
a m
arca
da p
ara
iNO
S (µm
2 ) x
104
F i g u r a 1 2 – C o r r e l a ç ã o d e S p e a r ma n d a e x p r e s s ã o e s pl ê n i c a d e i N O S e m c ã e s n a t u r a l me n t e i n f e c t a d o s c o m L e i s h m a n i a ( L . ) c h a g a s i ( C I ; n = 2 4 ) c o m a s i mu n o g l o b u l i n a s I g G 1 , I g G 2 e I g G t o t a l a n t i -L e i s h m a n i a ( A ; p = 0 , 0 7 6 ) , I g G 1 ( B ; p = 0 , 9 9 9 ) , I g G 2 ( C ; p = 0 , 2 0 3 ) .

51
5 D ISCUSSÃO
A LV ap resenta ampla d is t r i bu i ção ep idemio lóg i ca em áreas
t rop i ca i s e subt rop i ca is do g lobo (TESH, 1995) . É uma das mais
re levantes e emergen tes doenças paras i t á r i as de todo o mundo , podendo
l evar a 98% de mor ta l idade se não t ra tada . No Bras i l , a impor tânc ia da
LV es tá no fa to da sua a l t a inc idênc ia e amp la d is tr i bu ição geográ f i ca ,
seu p rocesso de expansão , u rban ização e na poss ib i li dade de a t ing i r
fo rmas graves assoc iadas à desnut r i ção e in fecções concomi tan tes . Na
maio r i a das á reas endêmicas 80% dos casos reg i s t rados oco r rem em
c r i anças com menos de 10 anos , sendo no t i f i cados aprox imadamente
1980 casos po r ano. Des tes , 90% s i t uam-se na reg ião no rdes te (GONTIJO
& MELO, 2004; S ILVA e t a l ., 2001).
Do pon to de v is ta ep idemio lóg i co a LVC é apontada como mais
impor tan te que a doença humana dev ido a sua ma io r preva lênc ia e pe lo
fa to de CA e CS serem in fecc iosos ao inse to ve to r e p r inc ipa lmente
dev ido à i n fecção can ina an teceder o aparec imen to de casos humanos
(DEANE & DEANE, 1954; KEENAN e t a l ., 1984b ; ABRANCHES e t a l .,
1991a; MOLINA e t a l ., 1994) .
A suscep t i b i l i dade can ina à i n fecção e o s ta tus c l ín i co
desenvo lv ido es tão re lac ionados com d ive rsos mecan ismos de i n te ração
paras i t o -hospede i ro . Para cont r i bu i r com os avanços na e luc idação des tes
mecan ismos em LVC fo ram rea l i zados nes te es tudo , a ava l i ação da ca rga
paras i t á r i a esp lên ica , quant i f i cação dos n íve is séri cos de NO e a
ex p ressão da enz ima iNOS, assoc iando os resu l tados ob t i dos com n íve is
da exp ressão de IFN-γ , TNF-α , IL -10 , IgG1 , IgG2 e IgG to ta l an t i -
Leishmania .
Os resu l t ados na F igu ra 1 e Tabe la 1 most ram aumen to da ca rga
paras i t á r i a em cães s in tomát i cos , ou se ja , com maior compromet imento
c l í n i co . Es tes resu l tados se ap rox imam do expos to po r a lguns au tores
que re la ta ram haver co r re lação en t re a ca rga paras itá r i a do baço e
l in fonodos e o agravamento c l ín ico da LVC (SANCHEZ e t a l ., 2004;
G IUNCHETTI e t a l ., 2006) . Os resu l t ados es tão também de aco rdo com o

52
ex pos to po r REIS et a l. (2006a) que ap resenta ram uma co r re lação en t re o
s ta tus c l í n i co na LVC e grau de paras i t i smo most rado em esca la
dec rescente , re f le t ido pe la o rdem: medu la óssea, baço , l i n fonodos , pe le e
f í gado .
A quan t i f i cação de NO tem s ido rea l i zada po r vá r i os
pesqu isadores , dev ido ao seu po tenc ia l c i t o tóx ico re lac ionado com
mecan ismos de de fesa ina ta em vár i as doenças i n fecciosas (MACHADO
e t a l. , 2004) . Na Le i shmaniose, PANARO e t a l ., (1999) demonst ra ram o
po tenc ia l m ic rob i c ida do NO em macró fagos humanos in fec tados com L.
i n fan tum es t imu lados com IFN-γ e LPS bacte r i ano. An ter io rmente ,
VOULDOUKIS e t a l ., (1996) hav iam demons t rado a capac idade
m ic rob ic ida do NO ao re la ta r o aumento sér i co de L-c i t ru l ina , metabó l i to
da L-a rg inase, ao so f re r ação da iNOS, em cães i n fec tados com
Leishmania in fan tum submet idos a t ra tamento com ant imon ia l
pentava len te (g lucan t ime) .
En fa t i zando a impor tânc ia do NO como agen te l e ishmanic ida
pesqu isadores demonst ra ram que macró fagos can inos , an te r io rmente
vac inados com p romas t i go tas mor tas de Leishman ia in fan tum,
ap resentam ma io r capac idade de p rodução de NO associada à ma io r
capac idade de mata r o pa ras i t o (PANARO e t a l ., 2001) . P INELL I e t a l .,
(2000) ap resenta ram aumento dos n íve is de NO em sobrenadante de
cu l tu ra de macró fagos can inos in fec tados com Leishmania i n fan tum
a t i vados com IFN-γ , TNF-α e IL -2 e LPS . Out ros t raba lhos most ra ram a
impor tânc ia dessas c i toc inas na a t i vação da exp ressão de iNOS e
conseqüentemen te da p rodução de NO (L IEW e t a l . , 1990a ,b ) .
PANARO e t a l . (2008) , em exper imentos “ in v i t ro ” , suger i ram que
a p rodução de NO es tá envo l v ida na p ro teção e no p rognós t i co de cães
na tu ra lmente i n fec tados po r L . in fan tum, most rando que a adoção de
l in fóc i tos au tó logos reduz ia a p rodução de NO po r macró fagos de
an imais s in tomát icos e ass in tomát icos à med ida que a i n fecção se
to rnava mais c rôn ica .
Nes te t raba lho , os resu l tados demonst ram que C I ap resen taram a
med iana dos n íve is sé r i cos de NO s ign i f i ca t i vamente maiores que a
med iana do grupo de CNI (p<0 ,05 ) (F igu ra 2 ) . En t re os C I , os n í ve is de

53
NO em CS fo ram s ign i f i ca t i vamen te maiores quando comparados aos
n íve i s ob t idos em CNI (p<0 ,01 ) (F igu ra 3 ) . Quando ava l i ados os n íve i s
sé r i cos de NO em re lação à ca rga paras i t á r i a encon tra ram-se n í ve i s
s i gn i f i can temen te menores de NO em cães com BP em re lação a MP
(p<0,05 ) e AP (p<0 ,001) . Re fo rçando es tes resu l t ados a aná l i se de
co r re lação en t re NO sér i co e ca rga paras i t á r i a revelou uma fo r te
co r re lação pos i t i va (p=0 ,0001, r= 0 ,6444) (F igura 4A e B) .
O aumento da p rodução de NO em cães na tu ra lmente infec tados
com a l to pa ras i t i smo e s in tomát i cos poder ia se r i nduz ido po r mo lécu las
do paras i t o como o LPG (L IEW e t a l . ,1997) . Naque le t raba lho , os
au to res re la ta ram que fos fog l i canos (PG) l ibe rados pe lo paras i t o são
capazes de i n ib i r ou i nduz i r a s ín tese de NO po r macró fagos mur inos
es t imu lados com IFN-γ e LPS . No en tan to , es tes são aspectos
cont roversos , po is exper imentos rea l i zados reve la ram que molécu las
p resentes na Leishmania como LPG podem in ib i r a p rodução de IL-12 ,
que é o p r inc ipa l i ndu tor de respos ta do T ipo 1 (PROUDFOOT et a l. ,
1996; L IEW et a l . ,1997) .
Fo i encon t rada uma assoc iação en t re os n íve is sé r icos de NO e a
ex p ressão esp lên i ca da c i toc ina TNF-α (p=0 ,005) e en t re a p rodução de
IgG2 (p=0 ,019) e IgG to ta l (p=0,047) (F igu ras 5 e 6) . É sab ido que o
TNF-α é uma das c i t oc inas i nduto ras da a t i vação da exp ressão de iNOS
(L IEW e t a l. 1990b) e es ta c i toc ina pode se r p roduz ida po r macró fagos ,
cé lu las Th1 , Th2 e mastóc i tos (BEUTLER & CERAMI, 1989) .
A co r re lação pos i t i va com os i so t ipos de IgG sugere uma
po la r i zação da respos ta para o per f i l Th2. An ima is suscept íve i s
desenvo lvem uma respos ta t i p i camen te Th2 e uma respos ta humora l
ex uberan te (HONORÉ e t a l. , 1998; ROLÃO e t a l. , 2007) . Níve is maio res
de IgG2 têm s ido reg is t rados re lac ionados a mau p rognós t i co
(BOURDOISEAU e t a l ., 1997; REIS e t a l ., 2006c) bem como a bom
p rognós t i co na LV (DEPLAZES e t a l ., 1995; N IETO e t a l ., 1999) ,
havendo cont rovérs ias en t re os au tores , necess i t ando-se i nves t i gações
ad i c iona is .

54
Deve-se cons ide rar t ambém o fa to des te es tudo te r si do rea l i zado
com cães natu ra lmente i n fec tados . A p rodução de NO pode te r s ido
impor tan te na fase aguda da doença, de f i n indo qua l se r ia o p rognós t i co
dos cães . MANNA e t a l. , (2006) re la ta ram em es tudo de “coo r te ” que
cães ass in tomát icos natura lmen te i n fec tados com L . in fan tum que
ap resenta ram expressão de IFN-γ no tempo i n i c i a l do es tudo
permaneceram ass in tomát i cos ou evo lu í ram com poucos s in tomas , mas
an imais que não ap resenta ram es ta ex p ressão evo lu í ram para fo rma
c l i n i ca s in tomát i ca . A exp ressão de iNOS em modelo mur ino , na fase
aguda , t em demonst rado que o NO gerado p romove o con t ro le de
i n fecções po r L. ma jo r, sendo ex pressa na pe le e l in fonodos d renantes da
l esão em camundongos res is ten tes C57BL/6 l evando à d im inu i ção no
número de paras i t os . Já em camundongos suscept íve i s, que não se cu ram
espontaneamente (BALB/c ) , es ta expressão é s ign i f i ca t i vamente menor e
não oco r re redução da ca rga paras i t á r ia (STENGER et a l. , 1994) .
Ou t ro impor tan te aspecto que pode exp l i ca r em par te a p resença
de maio res n í ve is de NO em cães ap resentando a l to pa ras i t i smo é a
poss ib i l i dade de par te da p rodução do NO te r s ido sin te t i zada pe las
p róp r i as le ishman ias . A lguns au to res demonst ra ram a capac idade de
l e i shmanias p roduz i rem NO e re lac ionaram es te aspecto d i re tamente à
capac idade i n fecc iosa do paras i to . A p rodução de NO po r fo rmas
p romas t i go tas e amast i go tas axên i cas de L . amazonens i s fo i
demonst rada , oco r rendo d im inu i ção quando se emprega o uso de
i n ib ido res de NOS como o L-NAME (NG-n i t ro -L-a rg in ina met i l es te r ) .
Es tes au to res suger i ram a inda que o NO gerado pe lo paras i t o possa
d im inu i r a respos ta m ic rob ic ida dos macró fagos reduz indo a a t i v idade da
iNOS, poss ive lmente po r a tua r em mecan ismos pós - t ransc r i c iona i s da
enz ima, in ib indo a p rodução de NO ou re ta rdando a apoptose da cé lu la
pa ras i t ada . Também demons t ra ram que a capac idade de p roduz i r NO es tá
p resente em vár ias espéc ies como L . amazonens i s , L . b ras i l i ens is e L .
chagas i em mecan ismos dependentes do Ca2 +(GENESTRA e t a l. , 2006
a ,b ; 2003) . Os mecan ismos de regu lação da s ín tese de NO têm s ido
amplamente inves t i gados e sua p rodução pode se r in ib ida por um

55
mecan ismo de “ feedback ” nega t i vo , onde o NO regu la sua p róp r i a
p rodução (L IEW, 1994) .
Os resu l t ados sugerem que maiores n íve is de NO, na fase c rôn i ca
da LVC, es tão assoc iados a mau prognós t i co , re lac ionados com a l t o
pa ras i t i smo, podendo-se in fe r i r que o NO pode se r usado como
b iomarcador da progressão c l ín i ca da doença. No en tan to , se r i am
necessár ios t raba lhos ad i c iona is pa ra se es tabe lecer uma a f i rmação, po i s
ex is tem l im i tações metodo lóg i cas que devem ser conside radas na
dosagem sér i ca do NO pe lo método de Gr iess cu jos resu l tados podem não
re f l e t i r apenas os aspec tos re lac ionados com a doença , mas também
in te r fe rênc ia pe la d ie ta ou p rodução de n i t r i t os pela f l o ra in tes t i na l
(MARZINZIG e t a l . , 1997) . É impor tan te ressa l ta r que a va l i dade da
de terminação das concent rações de n i t r i t o e n i t ra to como verdade i ro
í nd ice da p rodução de NO endógeno poder ia ge ra r cont rovérs ias dev ido a
fa to res como: i n terações qu ím icas in v i vo (KNOWLES & MONCADA,
1994; BENJAMIN & VALLANCE, 1994) . No en tan to , o método de Gr iess
tem s ido va l idado po r d i versos au tores que most ram que as
concent rações de n i t r i to e n i t ra to a l t e ram-se como resu l tado de d i ve rsas
doenças (FARREL e t a l ., 1992; M IDDLETON e t a l . , 1993; BALDEWEG
e t a l . , 1996) .
A aná l i se qua l i t a t i va da exp ressão da iNOS é apresen tada na
F igu ra 7 e Tabe la 2 . Podem ser observados d i fe ren tes t ipos de marcação
imuno-h is toquím ica . O grupo de CNI não ap resentou marcação
de tec táve l . Quando os an imais fo ram ava l i ados de aco rdo com a fo rma
c l í n i ca ve r i f i cou -se que a marcação em CA var i ou de l eve a moderada e
não se encon t rou nes tes an ima is marcação acentuada em nenhum dos
c r i t é r i os adotados . Po r sua vez , os g rupos CO e CS apresentaram uma
maio r f reqüênc ia e i n tens idade da ex p ressão de iNOS, sendo mais
p roeminente no ú l t imo . Es ta marcação pode es ta r re lac ionada com os
eventos in f l amató r i os observados na LVC. O t i po de marcação d i fusa
pode es ta r re lac ionado com a i n tensa h ipe rp las ia de macró fagos no baço
de cães com LV (TAFURI e t a l. , 2001) . A marcação foca l pode es ta r
re lac ionada com a marcação pr inc ipa lmente nas PALMS, ZM ou
fo l í cu los , po is macró fagos e cé lu las dendr í t i cas estão p resentes nessas

56
es t ru tu ras e podem es ta r a t i vadas e p roduz i r iNOS. J á a marcação
pontua l re lac iona-se com marcação em cé lu las i so ladas . Quanto à
marcação nos vasos esp lên icos , MONCADA e t a l . (1991) c i t am vár i as
cé lu las capazes de ex p ressar iNOS, en t re e las as célu las endo te l i a i s que
são poss i ve lmente as marcadas nes te exper imento .
Uma poss íve l exp l i cação para a exp ressão da iNOS por C I
con fo rme a F igura 8 pode es ta r oco r rendo v ia recepto res do t i po To l l
(TRLs ) , uma impor tan te famí l i a de recepto res p resentes em cé lu las
imunes de mamí fe ros que reconhecem mot ivos conservados de pa tógenos
(pad rão molecu la r assoc iado à pa tógenos – PAMPs) . Os TRLs p resentes
nas membranas dos macró fagos ou l in fóc i tos de cães podem es ta r sendo
a t i vados por PAMPs p resentes nas le i shmanias (LPG) (BECKER e t a l .,
2003; L IESE e t a l ., 2007) . Es ta a t i vação l eva a p rodução de c i t oc inas
p ró - in f lamató r i as dependente da a t i vação do fa to r nuc lea r kB (NFkB) , o
qua l d i spara mecan ismos e fe to res m ic rob ios tá t i cos /mic rob i c idas por
an imais res is ten tes em in fecções po r Trypanosoma cruz i , T rypanosoma
b ruce i , Le ishmania sp., Toxop lasma gond i i e P lasmodium sp . (TAKEDA
e t a l ., 2003; GAZZINE LLI e t a l ., 2004) .
Observou -se c la ramente que hav ia uma ma io r exp ressão de iNOS
assoc iada à g rav idade da doença cons iderando-se aval i ação semi -
quan t i t a t i va (Tabe la 2 ) e es ta fo r te marcação es tava assoc iada aos cães
ap resentando a l t o pa ras i t i smo esp lên i co , embora es ta observação não
tenha se to rnado uma regra e tenham s ido encon t rados cães com médio
ou ba ixo paras i t i smo com a l t a exp ressão de iNOS, bem como CS ou com
a l t o pa ras i t i smo com ba ix a exp ressão de iNOS.
O aumen to da expressão de iNOS es ta r assoc iado com a fo rma
c l í n i ca , cons ide rando-se as aná l i ses semi -quant i ta ti va e quan t i t a t i va de
iNOS, (mais i n tenso em CS e CO) pode es ta r re lac ionado ao maio r
número de cães com AP nes tes grupos (Tabe la 2 , F igura 9 ) . Quanto a
CNI não ap resen tar marcação detec táve l , es tá de acordo com resu l t ados
de LAGE e t a l. (2007) cu jo t raba lho reve lou que es te g rupo não
ap resenta exp ressão esp lên i ca de IFN-γ , que po r sua vez tem s ido
demonst rado es ta r re lac ionado com a a t i vação da ex pressão de iNOS

57
(HEVEL & MARLETTA, 1992; HOLAN et a l ., 2006; NIEDBALA e t a l. ,
2002; P INE LLI e t a l. 2000) .
Apesar de não te r s ido observado d i fe rença s ign i f i ca t i va quan to à
ex p ressão esp lên i ca de iNOS por dens idade paras i t á ria esp lên i ca , do
ponto de v i s ta semi -quant i t a t i vo (Tabe la 1 e 2 ) os resu l t ados sugerem
que a maior ex p ressão de iNOS não es tá necessar iamente l i gada à
redução da ca rga paras i t á r i a no tec ido esp lên i co ou que es ta expressão
u l t rapassa os n íve is idea i s pa ra o c lea rence esp lêni co e to rna-se tóx i co
ao hospede i ro poss i ve lmente pe la geração de perox ini t r i t o (PACHER et
a l . , 2007) . ANDERSON et a l. , (2005) mos t ra ram que camundongos
C57BL/6 gera lmente res i s ten tes , quando exper imenta lmen te in fec tados
po r cepas v i ru len tas de L. ma jor , fa l haram em e l im inar os paras i t os,
apesar de exp ressarem a l tos n íve is de iNOS na lesão e de ap resenta rem
uma respos ta po lar i zada de Th1 .
Deve-se cons ide ra r a inda que a exp ressão de iNOS e a p rodução
de NO são apenas par te da respos ta imune na LVC. REIS et a l. , (2006b)
demonst ra ram que C I ap resentam d iminu i ção nas taxas de cé lu las T
CD4+ /CD8+ no sangue per i fé r i co comparados a CNI e que CA apresentam
s ign i f i ca t i vamente maio r quan t idade cé lu las T CD8+ e CD4+ que CS, bem
como ma io r exp ressão MHC c lasse I I po r l i n fóc i tos ci r cu lan tes . Ass im, a
queda na taxa de cé lu las T CD4+ /CD8+ , bem como a queda de l i n fóc i t os
T CD8+ e CD4+ , impor tan tes fon tes de IFN-γ poder iam levar os
macró fagos de CS e CO a es ta rem ine f i c i en tes em p romoverem a
e l im inação dos paras i tos , po r uma a t i vação i ncomp leta dev ido à
i nsu f i c i en te p rodução de IFN-γ . De fo rma semelhante CARRERA e t a l .,
(1996) sugerem que a p rodução de IL -12 é f i rmemente regu lada em
macró fagos mur inos po r p romast i go tas de l e ishmanias du ran te a
i n fecção. Ass im, o cont ro le dessa impor tan te c i t oc ina a t i vadora de
respos ta do T ipo 1 pode es ta r assoc iada, de fo rma ind i re ta , às
i nsu f i c i en tes p roduções de IFN-γ e ex p ressão de iNOS que se r i am
necessár ios para promover a mor te dos paras i tos em C I .
Há mui tas ev idênc ias c l í n i cas e ex per imenta i s i nd icando que a
reso lução da i n fecção l e i shmania l é dependente de cé lu las T e

58
macró fagos a t i vados po r c i toc inas der i vadas de cé lulas T , havendo
t raba lhos demons t rando a impor tânc ia do IFN-γ no cont ro le das
l e i shmanioses (CARVALHO e t a l ., 1985; L IEW e t a l ., 1982; SACKS e t
a l . , 1987) . Out ros es tudos re la taram cães ass in tomát icos com LVC
ap resentando aumen to na p rodução de IL -2 , TNF-α e IFN-γ (CABRAL e t
a l . , 1992; P INELL I e t a l ., 1994a,b ; P INELL I e t a l ., 1995) .
Con t ra r i amente , a IL -10 tem s ido assoc iada à sup ressão de produção de
c i t oc inas do T ipo 1 e à modu lação da a t i v i dade le i shmanic ida de
macró fagos (BOGDAN e t a l ., 1991; MOSMANN & MOORE, 1991) . A
ex p ressão de RNAm do IFN-γ es tá d im inu ída no in íc i o de in fecção
ex per imenta l de cães i n fec tados com Leishmania in fan tum. Num per íodo
p ré -pa ten te subseqüente oco r re aumento do IFN-γ e pos te r io r dec résc imo
quando aparecem os s ina i s c l ín i cos da doença, havendo também
ex p ressão de RNAm para IL-10 em cu l t u ra de cé lu las mononuc leares do
sangue per i fé r i co (SANTOS-GOMES e t a l ., 2002) . CORREA et a l. ,
(2007) demonst ra ram que cães natu ra lmente in fec tados po r Le ishman ia
(L . ) chagas i ap resen tam maio res n íve i s de IL-10 e TGF-β e es tas tem
s ido assoc iadas com fo r te in ib i ção da capac idade m ic rob ic ida de
macró fagos . GAZZINE LLI e t a l ., (1992) mos t ram que a IL -10 e TGF-
β também in ib i ram a mor te de fo rmas t r i pomast i gos tas do Trypanosoma
c ruz i po r macró fagos mur inos a t i vados po r IFN-γ em mecan ismo
dependente de NO. Por sua vez , resu l tados ob t i dos po r ANDERSON et
a l . (2005) most ra ram que em camundongos C57BL/6 gera lmente
res i s ten tes à i n fecção po r L . ma jo r quando i nocu lados com cepas do
paras i t o res i s ten te ao t ra tamento não oco r r i a a cu ra da l esão na derme
mesmo com uma po lar i zação de respos ta do T ipo 1 , havendo aumen to da
ca rga paras i t á r i a , demonst rando que a p rodução de IL-10 es tava
re lac ionada com o aumento da suscept i b i l i dade .
Apesar de não te r s i do observada co r re lação en t re a iNOS e as
c i t oc inas IL -10 , IFN-γ , e TNF-α , e os an t i co rpos IgG1 , IgG2 ou IgG
to ta l an t i–Leishman ia (p>0,05 ) (F iguras 11 e 12 ) , LAGE e t a l . (2007)
demonst ra ram uma co r re lação en t re IL -10 e o s ta tus c l ín ico severo e a l ta

59
carga paras i t á r i a ao desc rever que 100% dos C I com AP ap resentavam
ex p ressão de RNAm de IL-10 .
A suscept i b i l i dade à i n fecção pode ser dev ido ao estabe lec imento
de c i r cu i t os imunor regu la tó r i os , onde a maior i ndução de c i toc inas
regu la tó r i as como o TGF-β e IL -10 poder ia render os macró fagos
i ne f i c i en tes no c lea rence dos paras i tos (BELKA ID e t a l. , 2001; GANTT
e t a l, 2003 , ANDERSON et a l . , 2005 ; CORREA et a l. , 2007) .
Também podemos supor que em LVC possa oco r re r a pa rt i c ipação
de cé lu las T regu la tó r i as (T reg) an te r i o rmen te es tudadas no mode lo
mur ino (XU e t a l ., 2003) , como as cé lu las T CD4+CD25+ que poder iam
es ta r e levando os n íve i s esp lên i cos de IL-10 imped indo a po la r i zação
para respos ta do T ipo 1 , modu lando a p rodução de IFN- γ , iNOS, NO e
conseqüentemen te p ro tegendo o paras i to con t ra os mecan ismos
m ic rob ic idas dos macró fagos (RONCAROLO e t a l ., 2006 ; ANDERSON et
a l . , 2005) . Resu l tados opos tos fo ram ap resentados po r NYLEN e t a l .,
(2007) , que descar tam es tas cé lu las como pr inc ipa i s fon tes de IL-10 na
fase in ic i a l da l e ishmaniose humana.
A impor tânc ia da par t i c ipação da IL-10 na pers i s tênc ia do
paras i t o é re fo rçada po r a lguns au to res que demonstraram em
camundongos nocau tes para IL-10 ou uso de an t i corpos b loqueadores
an t i - IL-10 l evava a cu ra es té r i l quando in fec tados com L . ma jor. No
en tan to , oco r r i a l a tênc ia da le ishman iose quando hav ia exp ressão de IL-
10 . Es tes au to res demonst raram a inda que a IL -10 p roduz ida po r cé lu las
T reg ocor re r i a na tu ra lmente , e e ra c ruc ia l pa ra pers is tênc ia da in fecção
po r L . ma jo r em le i shmaniose cu tânea em modelo mur ino (BELKA ID e t
a l . , 2002) .
A supos ta par t i c i pação de cé lu las regu la tó r i as merece
i nves t i gação em fu tu ros t raba lhos , p r i nc ipa lmente re lac ionando sua
p resença com a fo rma c l ín i ca , po is render iam in fo rmações que poder iam
e luc ida r aspectos igno rados da imunopa to log ia em LVC.
A f a lha na e l im inação dos paras i tos mesmo quando o baço can ino
es tá exp ressando a l t os n íve i s de iNOS, pode es ta r li gado a mecan ismo de
escape dos paras i tos pe la exp ressão de molécu las por le i shmanias como
as perox idox inas LcPx n-1 e LcPx n-2 ou superóx ido d ismutase que

60
con ferem p ro teção aos paras i t os con t ra os mecan ismos m icrob i c idas
(OLIV IER e t a l ., 2005) .
A inda en t re os poss íve i s mecan ismos de escape presentes no
gênero Leishmania, re la ta -se a sua capac idade de l ibe ra r ca theps ina B ,
uma p ro tease capaz de c l i va r TGF-β l a ten te à fo rma a t i va
imunossupresso ra capaz de p ro longar a sob rev ivênc ia do paras i t o em
macró fagos (WILSON et a l. , 2005) .
A lém dos mecan ismos de escape dos p ro tozoár ios deve-se
cons ide rar o fa to de te rmos t raba lhado com cães natu ra lmente in fec tados ,
es tes poder iam es tar pa ras i t ados po r d i fe ren tes cepas de L. (L . ) chagas i e
es tas poder iam ap resenta r ma io r ou menor patogen ic idade e v i ru lênc ia
sendo mais ou menos res is ten tes ao NO gerado pe la cé lu la hospede i ra
(NATAMI e t a l ., 2000) .
A de te rminação da fo rma c l í n i ca que se es tabe lece no an imal pode
es ta r re lac ionada com d i fe renças gené t i cas (HOWARD e t a l. , 1980;
FOWELL & LOCKSLEY, 1999 ; L IPOLDOVÁ e t a l, 2000 , 2002) ,
i n fecções concomi tan tes (BARRAL-NETTO e t a l. , 1995; HASSAN et a l .,
2006; RODRÍGUEZ-SOSA e t a l. , 2006) , número de p icadas e e fe i t o da
sa l i va de f l ebó tomos i n fec tados (MBOW e t a l. , 1998; DE MOURA et a l. ,
2007; OLIVE IRA e t a l. , 2008) e fa to res nu t r i c iona is (HOLZMULLER et
a l . , 2006 ; ANSTEAD et a l . 2001; 2003) .

61
6 CONCLUSÃO
Tomados em con jun to , os resu l tados sugerem que em cães
na tu ra lmente in fec tados po r L. chagas i a ex acerbação das fo rmas c l ín i cas
es tá assoc iado ao aumento da ca rga paras i t á r i a , aumen to do n í ve is
sé r i cos de NO e da ex pressão esp lên i ca de iNOS. O es tabe lec imen to de
c i r cu i tos imunor regu la tó r i os com a p rodução de c i toc inas regu la tó r i as
poder ia t o rnar os macró fagos dos cães in fec tados i ne fe t i vos na
e l im inação dos paras i tos e a p rodução de NO poder ia es ta r assoc iada aos
mecan ismos imunopa to lóg i cos na LVC.
.

62
7 REFERÊNCIAS BIBL IOGRÁFICAS :
ABRANCHES, P . ; CAMPINO, L e SANTOS-GOMES G. M. Can ine l e i shmanias is . New concepts o f ep idemio logy and immunopa tho logy: t he i r impac t in the cont ro l o f human v isce ra l l e i shmanias is . Acta Med Por t , v .11 , n .10 , p .871-5 . Oc t , 1998. ABRANCHES, P . ; S ILVA-PEREIRA, M. C . ; CONCEICAO-S ILVA F . M . ; SANTOS-GOMES, G. M. ; JANZ, J . G . Can ine l e ishmaniasis : pa tho log i ca l and eco log i ca l fac to rs in f l uenc ing t ransmiss ion o f i n fec t ion . Jou rna l Pa ras i to logy, v .77 , n .4 , p .557-61 . Aug, 1991a. ABRANCHES, P . ; SANTOS-GOMES G. ; RACHAMIM N. ; CAMPINO L. ; SCHNUR, L. F . ; JAFFE, C . L . An exper imenta l model fo r can ine v i scera l l e i shmanias is . Paras i te immuno logy, v .13 , n .5 , p .537-50 . Sep , 1991b. ADAMS, H. R . Ph ys io log ic , pa thoph ys io log i c , and therapeut i c imp l i ca t i ons fo r endogenous n i t r i c ox ide. J Am Vet Med Assoc. V .209, n7 , p 1297-1302. 1996. ANDERSON, C . F . ; MENDEZ, S . ; SACKS, D . L . Nonhea l ing i n fec t ion desp i te Th1 po la r i za t i on p roduced by a s t ra in o f Lei shmania major in C57BL/6 mice. The Jou rna l o f Immuno logy, 174 : 2934-2941.2005 . ANSTEAD, G. M. ; CHANDRASEKAR, B . ; ZHAO, W. ; YANG J .; PEREZ L. E . ; MELBY, P . C . Ma lnu t r i t i on a l te rs the innate immune response and i ncreases ear l y v i sce ra l i za t i on fo l l owing Le i shman ia donovan i i n fec t ion . I n fec t i on and immun i t y , v .69 , n .8 , p .4709-18. Aug, 2001 . ANSTEAD G. M. ; CHANDRASEKAR B. ; ZHANG Q. ; MELBY P . C. Mu l t inu t r i en t undernut r i t i on d ys regu la tes the res iden t macrophage p ro in f l ammato ry cytok ine network , nuc lea r fac to r -kappaB act i va t ion , and n i t r i c ox ide produc t ion . J Leukoc B io l; v .74 (6 ) , p .982 -91 . Dec, 2003. APPAY, V . ; N IXON, D. F . ; DONAHOE, S . M . ; G ILLESPIE , G . M . ; DONG, T . ; K ING, A . ; OGG, G. S . ; SP IEGE L, H . M . ; CONLON, C . ; SP INA, C . A . ; HAVLIR , D . V . ; R ICHMAN, D. D. ; WATERS, A. ; EASTERBROOK, P . ; MCMICHAEL, A . J . ; ROWLAND-J ONES, S. L . . H IV-spec i f i c CD8(+) T ce l l s p roduce an t i v i ra l c ytokines bu t are impai red i n cyto l yt i c func t i on . The Jou rna l o f Exper imen ta l Med ic ine, v .192, n .1 , p .63 -75 . Ju l , 2000 .

63
ALVAR, J . ; CANAVATE, C . ; MOLINA, R . ; MORENO, J . ; N IETO, J . . Can ine l e i shmanias i s . Advances in Paras i to logy, v .57 , p .1 -88 . 2004 . AMBER, L. J . ; H IBBS, J . B . JR. ; TAINTOR, R. R . ; VAVRIN, Z . The L-a rg in ine dependen t e f fec to r mechan ism i s i nduced in mur ine adenocarc inoma ce l l s b y cu l t u re supernatan t f rom c it o tox ic ac t i va ted macrophages . J . Leukoc. B io l ., v .43 ,p .187 -192 . 1988 . BACELLAR, O. ; D 'OLIVE IRA, A . ; JERONIMO J R. S . ; CARVALHO E. M . IL-10 and IL-12 a re the ma in regu la to ry cytok ines in v i scera l l e i shmanias is . Cytok ine , v .12 , n .8 , p .1228-31. Aug, 2000 . BADARO, R. ; JONES, T . C . ; CARVALHO, E . M . ; SAMPAIO, D . ; REED, S . G. ; BARRAL, A . ; TE IXE IRA, R . ; J OHNSON, JR. W. D. New perspect i ves on a subc l in i ca l fo rm o f v isce ra l l e ishmanias is . The Jou rna l o f In fec t ious D iseases, v .154 , n .6 , p .1003-11. Dec , 1986. BALDEWEG, T . ; SOORANNA, S . ; DAS, I . ; CATALAN, J . ; GAZZARD. B . Serum n i t r i t e concen t ra t ion sugges t a ro le fo r ni t r i c ox ide i n A IDS . AIDS , v .10 , n .4 , p .451-452. 1996. BARRAL-NETTO M, DA S ILVA JS, BARRAL A , REED S. Up- regu la t ion o f T he lpe r 2 and down-regu la t ion o f T he lper 1 cytok ines du r i ng mur ine re t rov i rus - induced immunode f i c i ency s ynd rome enhances suscept ib i l i t y o f a res i s tan t mouse s t ra in to Le i shmania amazonensi s . Am J Pa tho l. 146(3 ) :635 -42 . Mar , 1995 . BARROUIN-MELO, S . M. ; LARANJEIRA, D . F . ; ANDRADE F ILHO, F . A . DE; TR IGO, J . ; J ULIAO, F . S . ; FRANKE, C . R . ; PALIS AGUIAR, P . H . ; CONRADO DOS-SANTOS, W. L. ; PONTES-DE-CARVALHO, L. Can sp leen asp i ra t ions be sa fe l y used fo r t he paras i to log ica l d iagnos is o f can ine v isce ra l l e i shmanios is? A s tudy on assymptomat i c and po l ys ymptomat i c an imals . Vete r i na ry Jou rna l , v .171, n .2 , p .331 -9 . Mar , 2006 . BAYLIS C . ; VALLANCE P. Measurement o f n i t r i t e and ni t ra te l eve l s in p lasma and ur ine - -what does th ismeasure te l l us about the ac t i v i t y o f the endogenous n i t r i c ox ide sys tem? Curr Op in Nephro l Hyper tens . V .7 (1) , p .59 -62 . Jan , 1998.

64
BECKER, I . ; SALA IZA, N . ; AGUIRRE, M. ; DELGADO, J . ; CARRILLO-CARRASCO, N. ; KOBEH, L. G . ; RUIZ, A . ; CERVANTES, R .; TORRES, A . P . ; CABRERA, N . ; GONZALEZ, A . ; MALDONADO, C . ; ISIBAS I, A . Le i shman ia l ipophosphog l ycan (LPG) ac t i va tes NK ce ll s th rough to l l -l i ke recepto r -2 . Molecu la r and B iochemica l Pa ras i to logy, v .130 , n .2 , p .65 -74 . Aug, 2003. BELKA ID, Y . ; HOFFMANN, K. F . ; MENDEZ, S . ; KAMHAWI, S . ; UDEY, M. C . ; WYNN , T . A . ; SACKS D. L. The ro le o f in ter leuk in ( IL) -10 i n the pers is tence o f Le i shmania majo r in the skin a f t e r hea l i ng and the the rapeut i c po ten t i a l o f an t i - IL-10 receptor ant i body fo r s ter i l e cu re . The Jou rna l o f Exper imen ta l Med ic ine, v .194 , n .10 , p .1497-506 . Nov, 2001 . BELKA ID, Y . ; P ICCIR ILLO, C . A . ; MENDEZ, S . ; SHEVACH, E . M . ; SACKS, D . L . CD4+CD25+ regu la to ry T ce l l s con t ro l Le i shmania majo r pe rs is tence and immun i t y. Nature , v .420, n .6915, p .502 -7 . Dec , 2002 . BENJ AMIN, N . ; VALLANCE, P . P lasma n i t r i t e as a marker o f n i t r i c ox ide p roduct i on . Lancet, v . 344 , n .8927, p .960 . 1994 . BEUTLER B, CERAMI A .The b io logy o f cachect i n /TNF- -a p r imary med ia to r o f the hos t response. Annu Rev Immuno l . ;7 :625-55 . Rev iew. 1989 BOGDAN, C . O f m ic robes , macrophages and n i t r i c ox ide . Behr ing Ins t i tu te Mi t te i l ungen , n .99 , p .58-72 . Mar , 1997. BOGDAN, C . N i t r i c ox ide and the immune response . Natu re immuno logy, v .2 , n .10 , p .907-16 . Oct , 2001 . BOGDAN, C. ; VODOVOTZ, Y . ; NATHAN, C. Macrophage deact i va t ion b y i n te r l euk in 10 . The Jou rna l o f Exper imenta l Med ic ine, v .174 , n .6 , p .1549-55. Dec 1 , 1991 . BORJA-CABRERA, G . P . ; CRUZ MENDES, A. ; PARAGUAI DE SOUZA, E . ; HASHIMOTO OKADA, L. YKAWASAKI. ; , J . K . ; DE A . TR IVE LLATO F . A . ; COSTA A. C . ; REIS , A . B . ; GENARO, O. ; BATISTA, L. M . ; PALATNIK , M . ; PALATNIK-DE-SOUSA, C. B . E f fec t i ve immuno therapy aga ins t can ine v isce ra l l e ishman ias i s w i th t he FML-vacc ine. Vacc ine, v .22 , n .17 -18 , p .2234-43. Jun 2 , 2004.

65
BOURDOISEAU, G. , C . BONNEFONT, E . HOAREAU, C . BOEHRINGER, T . STOLLE E L. CHABANNE. Spec i f i c IGG1 and IGG2 an tibod y and l ymphocyte subset l eve l s i n na tu ra l l y Le i shmania infan tum- in fec ted t reated and un t rea ted dogs . Vete r ina ry immuno logy and immunopa tho logy, v .59 , n .1 -2 , p .21 -30 . Oct 6 , 1997 . BRAGA, M. D. M . ; COELHO, I . C . B. ; POMPEU, M. M . L. ; EVANS, T . G . ; MACAULLIFE, I . T . ; TE IXE IRA M. J . ; L IMA, J . W. D . O. Cont ro le do ca lazar can ino : comparação dos resu l t ados de um programa de e l im inação ráp ida de cães so ro r reagen tes po r ensa io imuno-enz imát i co com out ro de e l im inação ta rd ia de cães so ro r reagentes po r t es te de imuno f l uo rescênc ia ind i re ta de e luato de pape l de fi l t ro . Rev is ta da Soc iedade B ras i l e i ra de Med ic ina T rop i ca l, v .31 , n .5 , p .419 -424 . 1998 . BRANDONIS IO O. ; PANARO M. A. ; S ISTO M. ; ACQUAFREDDA A . ; FUMAROLA L. ; LEOGRANDE D. ; MITOLO V . N i t r i c ox ide produc t ion b y Le i shmania - i n fec ted macrophages and modu la t ion by c ytok ines and p ros tag land ins. Parass i to log ia . v . 43 Supp l 1 , p .1 -6 . 2001 . BRANDONIS IO, O . ; SP INE LL I R . ; PEPE M. Dendr i t i c ce ll s i n Le i shman ia i n fec t ion . Microbes and i n fec t i on / Ins t i tu t Pas teu r, v .6 , n .15 , p .1402-9 . Dec , 2004. BRENDOLAN, A . ; ROSADO, M. M. ; CARSETTI, R . ; SELLERI, L . ; DEAR, T . N. Deve lopmen t and func t ion o f the mammal ian sp leen. Bioessays, v .29 , n .2 , p .166 -77. Feb , 2007. CABRAL, M . ; O 'GRADY, J . ; ALEXANDER, J . Demonst ra t i on o f Le i shman ia spec i f i c ce l l med ia ted and humora l immuni t y i n as ymptomat i c dogs . Paras i te immuno logyogy, v .14 , n .5 , p .531 -9 . Sep , 1992 . CARRERA, L. ; GAZZINE LL I , R . T . ; BADOLATO, R. ; H IENY, S . ; MULLER, W. ; KUHN, R . ; SACKS, D . L . Le i shmania p romast i go tes se lec t i ve l y i nh ib i t i n te r l euk in 12 induc t ion i n bone mar row-der i ved macrophages f rom suscept i b le and res is tan t m ice . The Jou rna l o f Exper imen ta l Med ic ine, v .183 , n .2 , p .515 -26. Feb 1 , 1996 . CARVALHO, E. M . ; BADARO, R . ; REED, S . G. ; J ONES, T. C . ; JOHNSON, JR. W. D . Absence o f gamma in te r fe ron and i n te r l euk in 2 p roduct i on du r i ng ac t i ve v i scera l l e ishmanias is . The Jou rna l o f c l in i ca l inves t i ga t i on, v .76 , n .6 , p .2066-9 . Dec , 1985.

66
CERQUEIRA, N . F . ; YOSHIDA, W. B . Óx ido n í t r i co : Revi são. Ac ta C i rú rg i ca B ras i l e i ra , v .17 , n .6 , p .417-23. 2002. CESTA, M. F . Normal s t ruc tu re , func t ion , and h is to logy o f t he sp leen. Tox ico log i ca l Pa tho logy, v .34 , n .5 , p .455-65 . 2006 . CORREA, A. P . ; DOSSI, A . C . ; OLIVE IRA VASCONCELOS, R. DE; MUNARI, D . P . ; DE L IMA, V . M. Eva luat i on o f t rans format ion growth fac to r beta1 , i n te r leuk in -10 , and in te r fe ron-gamma in male symptomat i c and asymptomat i c dogs natu ra l l y i n fec ted by Le i shmania (Le i shmania ) chagas i . Vete r ina ry Paras i to logy, v .143 , n .3 -4 , p .267 -74. Feb 28 , 2007 . COSTA, C . H. ; GOMES, R . B . ; S ILVA, M. R . ; GARCEZ, L. M . ; RAMOS, P . K . ; SANTOS, R. S . ; SHAW, J . J . ; DAVID, J . R . ; MAGUIRE, J . H . Competence o f the human hos t as a reservo i r fo r Le ishmania chagas i . The Jou rna l o f I n fec t ious D iseases, v .182 , n .3 , p .997 -1000 . Sep, 2000. COSTA, C . H . ; PEREIRA, H . F . ; PEREIRA, F . C . ; TAVARES, J . P . ; ARAUJ O M. V . ; GONCALVES, M. J . Is t he househo ld dog a r i sk fac to r fo r Amer i can v isce ra l l e i shmanias is i n Braz i l? Transac t ions o f the Roya l Soc ie ty o f T rop i ca l Med ic ine and Hyg iene, v .93 , n .5 , p .464 . Sep , 1999 . DE MOURA T . R . ; OLIVE IRA F . ; NOVAIS F . O . ; MIRANDA J . C . ; CLARÊNCIO J . ; FOLLADOR I . ; CARVALHO E . M. ; VALENZUELA J . G . ; BARRAL-NETTO M. ; BARRAL A . ; BRODSKYN C. ; DE OLIVE IRA C . I . Enhanced Le i shman ia b raz i l i ens i s In fec t i on Fol l ow ing P re -Ex posure to Sandf ly Sa l i va . PLoS Neg l T rop D is. v .28 , n .2 , p .84 . Nov , 2007 . DEANE, L. M. ; DEANE M. P . Dogs natu ra l l y i n fec ted by Le i shmania donovan i in Ceara. Hosp i ta l (R io J . ), v .45 , n .6 , p .703 -7 . Jun , 1954 . DENICOLA A . ; SOUZA J . M. ; RADI R . D i f fus ion o f pe rox yn i t r i t e ac ross
e r yth rocyte membranes . Proc Na t l Acad Sc i USA, v .95 , p .3566-3571.
1998 .
DEPLAZES, P . ; SMITH, N . C . ; ARNOLD, P . ; LUTZ, H . ; ECKERT, J . Spec i f i c IGG1 and IGG2 ant ibod y responses o f dogs to Le i shmania

67
i n fan tum and o ther pa ras i t es . Paras i te immuno logy, v .17 , n .9 , p .451 -8. Sep, 1995. DESJEUX, P . ; ALVAR, J . Le i shman ia /H IV co - in fec t ions: ep idemio logy i n Eu rope. Anna ls o f t rop i ca l med ic ine and pa ras i to logy, v .97 Supp l 1 , p .3 -15 . Oct , 2003 . DUSSE, L. M . S . A . ; V IE IRA, L. M . ; CARVALHO, M. G. ; Rev isão sob re óx ido n i t r i c . Jo rna l B ras i le i ro de Pa to log ia e Med ic ina Labora to ri a l , v .39 , n .4 , p .343 -350 . 2003. ELL IS G. ; ADATIA I . ; YAZDANPANAH M. ; MAKELA S. K . N i t r i t e and n i t ra te ana l yses : a c l i n i ca l b iochemis t r y pe rspect ive .A Cl in B iochem, v . 31 , n .4 , p .195-220. Jun ; 1998 . ENGWERDA, C . R . ; ATO, M. ; COTTERELL, S . E . ; MYNOTT, T . L . ; TSCHANNERL, A . ; GORAK-STOLINSKA, P . M. ; KAYE, P . M . A ro le fo r tumor nec ros is fac to r -a lpha i n remodel i ng the sp len i c marg ina l zone du r i ng Le i shman ia donovan i i n fec t ion . The Amer ican Journa l o f Pa tho logy, v .161, n .2 , p .429 -37. Aug, 2002. ENGWERDA, C. R . ; ATO, M. ; KAYE, P . M. Macrophages , patho logy and paras i t e pe rs is tence i n exper imenta l v isce ra l l e i shmanias is . Trends in Pa ras i to logy, v .20 , n .11 , p .524 -30. Nov , 2004. FA INT, R . W. ; MACKIE , I . J . ; MACHIN, S . J . P la te le t aggrega t i on is i nh ib i t ed by a n i t r i c ox ide - l i ke fac to r re leased f rom human neut roph i ls i n v i t ro . Br. J . Haemato l . , v .77 , p .539 -545. 1991. FARRELL, A . J . ; BLAKE, D. R . ; PALMER, R. M . J . ; MONCADA, S . Inc reased concen t ra t i on o f n i t r i t e in s ynov ia l f l u id an se rum samp les sugges t i nc reased n i t r i c ox ide s yn thes is in rheumatic d i seases . Ann . Reum. D is. , v . 51 , p .1219-1222. 1992. FAHEY, T . J . ; YOSHIOKA, T . ; SHIRES, G. T . ; FANTIN I , G . A . The ro le o f t umor nec ros i s fac to r and n i t r i c ox ide i n the acu te ca rd iovascu la r response to endotox in . Ann . Surg . V .223, n1 , p .63-69 , 1996. F ILHO, R . F . ; Z ILBERSTEIN, B . Óx ido n í t r i co : o s imples mensage i ro pe rcor rendo a comp lex idade . Metabo l i smo, s ín tese e funções . Ass Med B ras, v .46 , n .3 , p .265-71 . 2000 .

68
FOWELL DJ , LOCKSLEY RM. Le i shmania major in fec t i on o f inbred m ice : unmask ing gene t i c dete rm inants o f in fec t ious d iseases .B ioessays . 21 (6 ) :510 -8 . Jun , 1999. FÖRSTERMANN, U ; DUN, N. J . Im munoh is tochemica l l ocal i za t ion o f n i t r i c ox ide s yn thases . Methods in Enzymology, v .268 , p . 510–515 . 1996 . GANTT, K . R . ; SCHULTZ-CHERRY, S . ; RODRIGUEZ, N . ; JERONIMO, S . M. ; NASCIMENTO, E. T . ; GOLDMAN, T . L . ; RECKER, T. J . ; M ILLER, M. A. ; W ILSON, M. E . Ac t i va t i on o f TGF-be ta by Le i shmania chagas i : impor tance fo r pa ras i t e su rv iva l i n macrophages . Jou rna l o f Immuno logy , v .170 , n .5 , p .2613-20. Mar 1 , 2003 . GAZZINE LL I , R . T . ; OSWALD, I . P . ; H IENY, S . ; JAMES, S . L . ; SHER, A . The m ic rob ic ida l ac t i v i t y o f i n ter fe ron -gamma- t reated macrophages aga ins t Tr ypanosoma c ruz i i nvo l ves an L-a rg in ine -dependen t , n i t rogen ox ide-med ia ted mechan ism inh ib i tab le by i n te r l euk in-10 and t rans fo rm ing growth fac to r -be ta . European jou rna l o f immuno logy, v .22 , n .10 , p .2501-6 . Oct , 1992 . GAZZINE LL I , R . T . ; ROPERT, C. ; CAMPOS, M. A . Ro le of t he To l l / i n te r l euk in -1 recepto r s i gna l i ng pathway i n hos t res is tance and pa thogenes i s du r i ng i n fec t i on w i th p ro tozoan paras ites . Immuno log i ca l Rev iews, v .201, p .9 -25 . Oct , 2004 . GENESTRA, M. ; SOUZA, W. J . ; GUEDES-S ILVA, D . ; MACHADO, G. M . C . ; CYSNE-FINKELSTEIN, L. ; BEZERRA, R . J . S . ; MONTEIRO, F . ; LEON, L. L . N i t r i c ox ide b iosyn thes i s by Le i shman ia amazonens i s p romas t i go tes conta in ing a h igh percen tage o f metacyc l i c fo rms . Arch ives o f Mic rob io logy , v .185 , p .348–354 . 2006a. GENESTRA, M. ; GUEDES-S ILVA, D . ; SOUZA, W. J . S . ; CYSNE-F INKE LSTEIN, L. ; SOARES-BEZERRA, R . J . ; MONTEIRO, F. P . ; LEONA, L. L . N i t r i c Ox ide Syn thase (NOS) Charac te r iza t i on in Le i shman ia amazonens is Ax en ic Amast i go tes . Arch ives o f Med ica l Research , v .37 , p .328–333. 2006b . GENESTRA, M; SOUZA, W. J . S . ; CYSNE-FINKE LSTEIN, L.; LEON, L. L . Comparat i ve ana l ys i s o f the n i t r i c ox ide p roducti on b y Leishmania sp . Medica l M ic rob io logy and Immuno logy . v .192, p .217–223 .2003.

69
G IUNCHETTI, R . C . ; MAYRINK, W. ; GENARO, O. ; CARNEIRO, C . M . ; CORREA-OLIVEIRA, R . ; MARTINS-F ILHO, O. A . ; MARQUES, M. J . ; TAFURI, W. L . ; REIS , A . B . Re la t ionsh ip be tween can ine v isce ra l l e i shmanios is and the Leishmania (Le ishmania ) chagas i bu rden i n dermal i n f l ammato ry foc i. Journa l o f Compara t i ve Pa tho logy l, v .135, n .2 -3 , p .100-7 . Aug, 2006. G IUNCHETTI, R . C . ; MAYRINK, W. ; CARNEIRO, C . M . ; CORREA-OLIVE IRA, R . ; MARTINS-F ILHO, O . A . ; MARQUES, M. J .; TAFURI, W. L. ; REIS , A . B . H is topa tho log i ca l and immunoh is tochemica l i nves t i ga t i ons o f the hepa t i c compar tmen t assoc ia ted w i th pa ras i t i sm and se rum b iochemica l changes i n can ine v isce ra l l e ishman ias is . Research i n Ve te r ina r y Sc ience, v . 84 , p . 269-277. 2008. GOMES-PEREIRA, S . ; RODRIGUES, O . R . ; SANTOS-GOMES, G. M . Dynamics o f CD62L/CD45RB CD4+ and CD8+ l ymphocyte subsets i n hepat i c and sp len i c t i ssues du r i ng mur ine v isce ra l l e i shmanias is . Immuno logy Le t ters , v .95 , n .1 , p .63 -70 . Aug 15 , 2004 . GONTIJ O, B . ; DE CARVALHO M. L. Amer i can cu taneous le i shmanias is . Rev is ta da Soc iedade B ras i le i ra de Med ic ina T rop i cal , v .36 , n .1 , J an - p .71 -80 . Feb , 2003 . GONTIJ O, C . M. F . ; MELO, M. N . Le i shmaniose v isce ral no Bras i l : quadro a tua l , desa f i os e perspec t i vas . Rev is ta B ras i l e i ra de Ep idemio log ia, v .7 , n .3 , p .338 -49. 2004 . GRABBE, S . ; SCHUWARZ, T . Immunnoregu la to ry mechan isms invo lved i n e l i c i t a t ion o f a l l e rg i c con tac t hyp ersens i t i v i t y. Immuno logy Today, v .19 , p .37 -42 . 1998. GREEN, L. C . ; WAGNER, D. A . ; GLOGOWSKI, J . ; SK IPPER, P . L . ; W ISHNOK, J . S . ; TANNENBAUM, S . R . Ana l ys i s o f n i t rate , n i t r i te , and [15N]n i t ra te i n b io log ica l f lu i ds . Ana ly t i ca l B iochemis t ry , v .126 , n .1 , p .131-8 . Oc t , 1982. GUARGA, J . L . ; LUC IENTES, J . ; PERIBANEZ, M. A. ; MOLINA, R . ; GRACIA M. J . ; CASTILLO, J . A . Exper imenta l in fec t ion o f Ph lebo tomus pern i c i osus and de termina t ion o f t he natu ra l i n fec ti on ra tes o f Le i shman ia in fan tum in dogs . Acta Trop i ca , v .77 , n .2 , p .203-7 . Nov 2 , 2000 .

70
GUTMAN, S . I . ; HOLLYWOOD, C . A . S imp le , rap id method fo r de termin ing n i t ra tes and n i t r i tes i n b io log i ca l f l uids . Cl in i ca l Chemis t ry , v .38 , n .10 , p .2152. Oct , 1992. HASSAN MF, ZHANG Y, ENGWERDA CR, KAYE PM, SHARP H, B ICKLE QD. The Sch is tosoma manson i hepa t i c egg granu loma p rov ides a favo rab le mic roenv i ronment fo r sus ta ined growth of Le i shmania donovan i . Am J Patho l. v .169 , n .3 , p .943-53 . Sep, 2006. HERTZ, C . J . ; MANSFIE LD, J . M . IFN-gamma-dependent n i t r i c ox ide p roduct i on i s no t l i nked to res is tance in exper imenta l A f r i can t r ypanosomias i s . Cel lu la r immuno logy , v .192 , n .1 , p .24 -32 . Feb 25, 1999 . HEVEL, J . M. ; MARLETTA, M. A. Macrophage n i t r i c ox ide syn thase: re la t i onsh ip between enzyme-bound te t rahyd rob iop te ri n and syn thase ac t i v i t y. Biochemis t ry, v .31 , n .31 , p .7160-5 . Aug 11 , 1992 . HOLAN, V . , P INDJAKOVA, J . ; KRULOVA, M. ; NEUWIRTH, A. ; FR IC, J . ; ZAJ ICOVA, A. P roduct ion o f n i t r i c ox ide du r i ng gra f t re jec t ion i s regu la ted b y the Th1/Th2 ba lance, t he a rg inase ac t iv i t y , and L-a rg in ine metabo l i sm. Transp lan ta t i on , v .81 , n .12 , p .1708-15 . Jun 27 , 2006. HOLZMULLER, P . ; H IDE, M . ; SERENO, D. ; LEMESRE, J . L. Le i shman ia in fan tum amas t i go tes res is tan t t o n i t r i c ox ide cyto tox i c i t y: Impac t on i n v i t ro pa ras i t e deve lopmen ta l cyc le and metabo l i c enz yme ac t i v i t i es . In f ec t ion , genet i cs and evo lu t i on : j ou rna l o f mo lecu la r ep idemio logy and evo lu t i ona ry gene t i cs i n in f ec t ious d iseases, v .6 , n .3 , p .187 -97. May, 2006 . HONORÉ S , GARIN YJ , SULAHIAN A , GANGNEUX JP , DEROUIN F . In f l uence o f the hos t and paras i t e s t ra in in a mouse model o f v i sce ra l Le i shman ia i n fan tum in fec t ion . FEMS Immuno l Med Mic rob io l . 21 (3 ) :231 -9 . Ju l ; 1998.
HOWARD JG, HALE C , CHAN-LIE W WL. Immuno log i ca l regula t ion o f ex per imenta l cu taneous l e ishmanias i s . 1 . Immunogenet i c aspects o f suscept i b i l i t y t o Le i shmania t rop i ca in m ice. Parasi t e Immuno l Win te r ; 2 (4 ) :303 -14 . 1980 .

71
HUHMER A. F . ; N ISHIDA C . R . ; ORTIZ DE MONTELLANO P . R . ; SCHONEICH C . Inac t i va t ion o f t he i nduc ib le n i t r i c ox ide syn thase by perox yn i t r i t e . Chem Res Tox i co l, v .10 , p .618–626 . 1997. IGNARRO L. J. Phys io logy and pathoph ys io logy o f n i t r i c ox ide. K idney In t Supp l , v .6 , n .55 , p .2 -5 . Jun , 1996. JORES, K. ; YU, J . ; MCCORMACK, D . G. Cap i l l a r y an a rte r i o la r responses to loca l vasod i l a to rs a re impa i red i n a ra t mode l o f seps is . J . App l . Phys io l. V . 84 , n .3 , p .837 -844 .1992 . KEENAN, C. M. ; HENDRICKS, L. D . ; L IGHTNER, L. ; WEBSTER, H . K . ; JOHNSON A. J . V iscera l l e ishmanias is in the German shepherd dog. I . In fec t i on , c l i n i ca l d isease , and c l i n i ca l pa tho logy. Vete r ina ry Pa tho logy, v .21 , n .1 , p .74-9 . Jan , 1984a . KEENAN, C . M . ; HENDRICKS, L. D . ; L IGHTNER, L. ; JOHNSON, A. J . V iscera l l e i shmanias is in t he German shepherd dog. I I . Patho logy. Ve te r i na ry Pa tho logy, v .21 , n .1 , p .80 -6 . Jan , 1984b . KNOWLES, R. E . ; MONCADA, S . N i t r i c ox ide s yn thases i n mammal . Biochem. J. v . 298, p . 249 -258 . 1994 . KUMAR, P . ; SHARMA, P . K . ; JA IN , R . K . ; GAUTAM, R. K . ; BHARDWAJ, M. ; KAR, H. K . Ora l u l ce r as an unusua l fea tu re o f v isce ra l l e ishmanias is i n an A IDS pa t i en t . Ind ian Journa l o f Med ica l Sc iences, v .61 , n .2 , p .97 -101 . Feb, 2007 . LAGE, R . S . ; OLIVE IRA, G . C . ; BUSEK, S . U . ; GUERRA, L. L . ; G IUNCHETTI, R . C . ; CORREA-OLIVEIRA, R . ; REIS , A . B . Ana l ys i s o f t he cytok ine p ro f i l e in sp leen ce l l s f rom dogs natura l l y i n fec ted b y Le i shman ia chagas i . Vete r ina ry immuno logy and immunopa tho logy, v .115, n .1 -2 , p .135-45. Jan 15 , 2007 . LA INSON, R. Summary o f recen t abs t rac ts . V . Le i shmanias is . CDT D iges t, v .1 , n .2 , p .605-11 . Oct , 1970 . LA INSON, R . ; RANGEL E . F . Lu tzomyia l ong ipa lp is and the eco-ep idemio logy o f Amer ican v i scera l l e ishmanias i s , w ith pa r t i cu la r re fe rence to Braz i l : a rev iew. Memór ias do Ins t i tu to Oswa ldo Cruz, v .100, n .8 , p .811-27 . Dec , 2005 .

72
LA INSON, R. ; SHAW, J . J . Le i shman ias i s o f the New Wor ld : t axonomic p rob lems . Br i t i sh med ica l bu l l e t in , v .28 , n .1 , p .44-8 . Jan , 1972 . LA INSON, R. ; SHAW, J . J . Observat ions on the deve lopmen t o f Le i shman ia (L. ) chagas i Cunha and Chagas i n t he midgut o f t he sand f l y vec to r Lu tzomyia long ipa lp i s (Lu tz and Ne iva ) . Anna les de Paras i to log ie Huma ine e t Comparee, v .63 , n .2 , p .134-45 . 1988. LA INSON, R. , J . J . SHAW, F . T . S i l ve i ra e R . R . Braga . Amer i can v isce ra l l e i shmanias is : on the o r ig in o f Le i shman ia (Le i shmania ) chagas i . Transac t ions o f the Roya l Soc ie ty o f T rop i ca l Med icine and Hyg iene, v .81 , n .3 , p .517 . 1987 . L I H . ; POULOS T . L . S t ruc tu re- funct i on s tud ies on ni t r i c ox ide s yn thases . J Ino rg B iochem, v .99 , n .1 , p .293-305. Jan , 2005. L IESE, J . ; SCHLEICHER, U . ; BOGDAN, C . TLR9 s igna l ing i s essent ia l f o r the i nnate NK ce l l response in mur ine cu taneous l e ishmanias i s . European Jou rna l o f Immuno logy. V . 37 , Issue 12 , P . 3424 – 3434.2007 . L IEW F. Y . Regu la t i on o f n i t r i c ox ide syn thes is in i n fec t i ous and au to immune d i seases . Immuno l Le t t , v .43 (1 -2) , p .95 -98 . Dec , 1994 . L IEW, F . Y . ; HALE, C . ; HOWARD, J . G. Immuno logi c regu la t i on o f ex per imenta l cu taneous le ishman ias i s . V . Characte r iza t i on o f e f fec to r and spec i f i c suppresso r T ce l l s. Journa l o f Immuno logy, v .128, n .4, p .1917-22. Ap r , 1982 . L IEW, F . Y . ; MILLOTT, S . ; PARKINSON, C . ; PALMER, R. M. ; MONCADA, S . Macrophage k i l l i ng o f Le i shmania paras it e i n v i vo i s med ia ted by n i t r i c ox ide f rom L-a rg in ine. Jou rna l o f Immuno logy , v .144, n .12 , p .4794-7 . Jun 15 , 1990a . L IEW, F . Y . , Y . L I , S . M ILLOTT. Tumor nec ros is fac to r -a s ynerg i zes w i th IFN-g i n med ia t i ng k i l l i ng o f Leishmania ma jo r th rough the i nduct i on o f n i t r i c ox ide. J . Immuno l . 145 :4306 . 1990b.

73
LIEW, F . Y . ; WEI, X . Q . ; PROUDFOOT, L. C ytok ines and n i t r i c ox ide as e f fec to r mo lecu les aga ins t paras i t i c in fec t ions . Ph i l osoph ica l t ransac t i ons o f the Roya l Soc ie ty o f London. Ser ies B , B io log i ca l sc iences, v .352 , n .1359 , p .1311-5 . Sep 29 , 1997 . L IMA, W. G . ; M ICHALICK, M . S . ; DE MELO, M. N . ; TAFURI, W. L. ; TAFURI W. L. Can ine v isce ra l l e i shmanias is : a h is topatho log i ca l s tudy o f l ymph nodes. Ac ta T rop i ca , v .92 , n .1 , p .43 -53 . Sep , 2004. L IPOLDOVÁ M, SVOBODOVÁ M, KRULOVÁ M, HAVELKOVÁ H, BADALOVÁ J , NOHÝNKOVÁ E , HOLÁN V , HART AA, VOLF P , DEMANT P . Suscep t ib i l i t y t o Le i shmania majo r in fec ti on i n m ice: mu l t ip l e l oc i and hete rogene i t y o f immunopatho log ica l phenot ypes . Genes Immun. 1 (3 ) :200-6 . Feb , 2000 . L IPOLDOVÁ M, SVOBODOVÁ M, HAVELKOVÁ H, KRULOVÁ M, BADALOVÁ J , NOHÝNKOVÁ E, HART AA, SCHLEGEL D, VOLF P , DEMANT P. Mouse genet i c model fo r c l in i ca l and immuno log i ca l he terogene i t y o f l e i shmanias is . Immunogene t i cs. 54(3 ) :174 -83. Jun , 2002 . L IU , H . ; HU, B . ; XU, D . ; L IEW, F . Y . CD4+CD25+ regula to ry T ce l l s cu re mur ine co l i t i s : t he ro le o f IL-10 , TGF-be ta , and CTLA4 . Jou rna l o f Immuno logy , v .171 , n .10 , p .5012-7 . Nov 15 , 2003 . LOHOFF, M. ; FERRICK, D . ; M ITTRUCKER, H. W. ; DUNCAN, G. S . ; B ISCHOF, S . ; ROLL INGHOFF, M. ; MAK, T . W. In te r fe ron regu la to r y fac to r -1 i s requ i red fo r a T he lpe r 1 immune response in v i vo . Immun i t y , v .6 , n .6 , p .681-9 . Jun , 1997 . LORENTE, L. ; ALLER, M. A. ; AR IAS , J . L . ; ALONSO, M. S . ; AR IAS , J . C l in ica l b io logy o f n i t r i c ox ide . Br i t i sh Jou rna l o f Su rgery , v .83 , p .1010-1019. 1996. MACHADO, P . R . L . ; ARAÚJO, M. I . A . S . ; CARVALHO, L. ; CARVALHO, E . M. Immune response mechan isms to in fect i ons . Anais B ras i le i ros de Dermato log ia, R io de J ane i ro , v . 79 , n . 6 , 2004 MACMICKING, J . ; X IE Q.W. ; NATHAN C . N i t r i c ox ide and macrophage funct i on . Annua l Rev iew Immuno logy, v . 15 , p .323 -50. Ap r i l , 1997.

74
MALLA, N . ; MAHAJAN, R. C . Pathophys io logy o f v i sce ral l e i shmanias is - some recent concepts . The Ind ian Journa l o f Med ica l Research, v .123, n .3 , p .267-74 . Mar , 2006 . MANCIANT I, F . ; GRAMICCIA , M . ; GRADONI, L . ; P IERI, S. S tud ies on can ine l e i shmanias is con t ro l . 1 . Evo lu t i on o f i n fect i on o f d i f f e ren t c l i n i ca l fo rms o f can ine le ishman ias i s fo l lowing ant imon ia l t rea tment . Transact ions o f the Roya l Soc ie ty o f T rop i ca l Med icine and Hyg iene, v .82 , n .4 , p .566 -7 . 1988 . MANNA L. ; REALE S . ; V IOLA E . ; V ITALE F . ; MANZILLO V . F . ; M ICHELE P . L. ; CARACAPPA S. ; GRAVINO A . E . Le i shmania DNA load and cytok ine ex p ress ion l eve ls in as ymptomat i c natu ra l l y i n fec ted dogs . Vet Pa ras i to l, v .20 , n .142 (3 -4 ) , p .271 -280 . Dec, 2006. MARZINZIG M. ; NUSSLER, A . K . ; STADLER, J . ; MARZINZIG, E . ; BARTHLEN, W. ; NUSSLER, N.C . ; BEGER, H. G . ; MORRIS , S . M. ; BRUCKNER, U. B . Ana l yt i ca l Methods : Improved methods to measure en p roducts o f n i t r i c ox ide i n b io log ica l f lu ids : N i t ri t e , n i t ra te and s -n i t roso th io l s . Ni t r i c Ox ide : B io logy and chemis t ry. Vo l . 1 , n .2 , p . 177 -189. Ap r i l , 1997 . MARLETTA, M. A. Mammal ian s yn thes is o f n i t r i t e , n i tra te , n i t r i c ox ide , and N-n i t rosat ing agen ts . Chemica l Research in Tox i co logy, v .1 , n .5 , p .249-57 . Sep-Oc t , 1988. MARLETTA, M. A . ; YOON, P . S . ; IYENGAR, R . ; LEAF, C . D. ; W ISHNOK, J . S . Macrophage ox idat ion o f L-a rg in ine to n i t r i t e and n i t ra te : n i t r i c ox ide i s an i n te rmed ia te . Biochemis t ry, v .27 , n .24 , , p .8706-11. Nov 29 , 1988. MARTIN, F . ; KEARNEY, J . F . Marg ina l -zone B ce l l s . Na tu re Rev iews . Immuno logy , v .2 , n .5 , p .323 -35. May, 2002 . MARZOCHI, M . C . ; COUTINHO, S . G. ; DE SOUZA, W. J . ; DE TOLEDO, L. M . ; GRIMALDI J UNIOR, G. ; MOMEN, H . ; PACHECO RDA, S . ; SABROZA, P . C . ; DE SOUZA, M. A . ; RANGEL J UNIOR, F . B . Can ine v i sce ra l l e ishmanias i s i n R io de Jane i ro , Braz i l . C l in i ca l , pa ras i t o log i ca l , t herapeut i ca l and ep idemio log ica l f ind ings (1977-1983) . Memór ias do Ins t i tu to Oswaldo Cruz, v .80 , n .3 , p .349 -57. Ju l -Sep , 1985 .

75
MAYER, B . ; SCHMIDT, K . ; HUMBERT, P . ; BOHME, E. B iosyn thes is o f endo the l ium-der ived re lax ing fac to r : a cytoso l i c enzyme in po rc ine ao r t i c endothe l i a l ce l l s Ca2+-dependen t l y conver t s L-a rg in ine in to an ac t i va to r o f so lub le guanyl y l c yc lase. B iochemica l and B iophys i ca l Research Communica t ions, v .164 , n .2 , p .678 -85. Oc t 31 , 1989 . MBOW ML, BLEYENBERG J A, HALL LR , T ITUS RG. Ph lebotomus papatas i sand f l y sa l i va r y g land l ysa te down- regu lates a Th1 , bu t up -regu la tes a Th2, response i n mice in fec ted w i th Le ishmania majo r . J Immuno l , v .161, n .10 , p .5571-5577. 1998. MEBIUS, R . E . ; KRAAL, G . S t ruc tu re and funct ion o f the sp leen. Na tu re Rev iews. Immuno logy , v .5 , n .8 , p .606 -16 . Aug, 2005 . MEBIUS, R . E . ; NOLTE, M. A . ; KRAAL, G . Deve lopmen t and funct ion o f t he sp len i c marg ina l zone. Cr i t i ca l Rev iews in Immuno logy, v .24 , n .6 , p .449 -64. 2004. M IDDLETON, S . J . ; SHORTHOUSE, M. ; HUNTER, J . O. Increaded n i t r i c ox ide s yn thes is in u lce ra t i ve co l i t es . Lancet, v . 341, p .25 -38 . 1993 . MOLINA, R . ; AMELA, C . ; N IETO, J . ; SAN-ANDRES, M. ; GONZALEZ, F . ; CASTILLO, J . A . ; LUCIENTES, J . ; ALVAR J . In fec ti v i t y o f dogs na tu ra l l y i n fec ted w i th Le i shmania in fan tum to co lon ized Ph lebotomus pern i c i osus . Transac t i ons o f the Roya l Soc ie ty o f T rop i ca l Med icine and Hyg iene, v .88 , n .4 , p .491-3 . Ju l -Aug, 1994. MONCADA, S . , R . M . PALMER E E . A . H IGGS. N i t r i c ox ide: ph ys io logy, pathoph ys io logy, and pharmaco logy. Pharmaco l Rev, v .43 , n .2 , p .109 -42. Jun , 1991 . MORENO, J . ; ALVAR, J . Can ine l e i shmanias is : ep idemio log ica l r i sk and the exper imenta l model . Trends in Pa ras i to logy, v .18 , n .9 , p .399 -405 . Sep, 2002. MOSMANN TR; COFFMAN RL. TH1 and TH2 ce l l s : d i f fe rent pat terns o f l ymphok ine sec re t ion l ead to d i f fe ren t func t iona l proper t ies . Annu Rev Immuno l . ;7 :145 -173 .1989 .

76
MOSMANN, T . R . ; MOORE, K . W. The ro le o f IL-10 i n cross regu la t ion o f TH1 and TH2 responses . Immuno logy Today, v .12 , n .3 , p .A49-53. Mar , 1991 . NATAMI, A . ; SAHIB I , H . ; LASRI, S . ; BOUDOUMA, M. ; GUESSOUSS-IDRRISSI N . ; RHALEM, A . Sero log i ca l , c l in ica l and his topa tho log i ca l changes in na tu ra l l y i n fec ted dogs w i th Le i shmania in fan tum in t he Khemisse t p rov ince , Morocco. Vete r ina ry Research, v .31 , n .3 , p .355 -63 . May-J un , 2000 . NAGELKERKEN, L. ; GOLLOB, K . J . ; T IE LEMANS, M. ; COFFMAN, R. L . Ro le o f t rans fo rm ing growth fac to r -beta in t he pre fe ren t i a l i nduct i on o f T he lpe r ce l l s o f t ype 1 by s taphylococca l en te ro tox in B . European jou rna l o f immuno logy , v .23 , n .9 , p .2306-10 . Sep, 1993. N ICHOLSON, B. ; MANNER, C . K . ; KLEEMAN, J . ; MACLEOD, C . L . Sus ta ined n i t r i c ox ide p roduc t ion i n macrophages requ i res the arg in ine t ranspor te r CAT2 . J B io l Chem, v .276 , n .19 , p .15881-5 . May 11 , 2001. N IEDBALA, W. ; WEI, X . Q . ; CAMPBELL, C . ; THOMSON, D .; KOMAI-KOMA, M. ; L IEW, F . Y . N i t r i c ox ide p re fe ren t ia l l y induces t ype 1 T ce l l d i f f e ren t ia t i on b y se lec t i ve l y up - regu la t i ng IL -12 recepto r beta 2 ex p ress ion v ia cGMP. Proceed ings o f the Na t i ona l Academy o f Sc iences o f the Un i ted S ta tes o f Amer i ca, v .99 , n .25 , p .16186-91 . Dec 10 , 2002 . N IETO, C . G. ; GARCIA-ALONSO, M. ; REQUENA, J . M . ; M IRON, C . ; SOTO, M. ; ALONSO, C. ; NAVARRETE, I . Ana l ys i s o f the humora l immune response aga ins t t o ta l and recombinan t an t i gens o f Le i shmania i n fan tum: co r re la t ion w i th d isease progress ion in can ine exper imenta l l e i shmanias is . Vete r ina ry immuno logy and immunopa tho logy, v .67, n .2 , p .117 -30. Feb 1 , 1999. NORMAN J . E . ; L IEW F. Y . N i t r i c ox ide. Mol Med Today . v .1 , n .8 , p .358. Nov , 1995 . NYLEN, S . ; MAURYA, R. ; E IDSMO, L. ; MANANDHAR, K . D .; SUNDAR, S . ; SACKS, D. Sp len i c accumula t ion o f IL-10 RNAm in T ce l l s d i s t i nc t f rom CD4+CD25+ (Foxp3) regu la to r y T ce l l s i n human v isce ra l le ishman ias is . The Journa l o f Exper imen ta l Med ic ine, v .204, n .4 , p .805 -17. Ap r 16 , 2007 .

77
OLIVE IRA, G. G. ; SANTORO, F . ; SADIGURSKY, M. The subc l i n i ca l fo rm o f exper imen ta l v i sce ra l l e i shmanias is i n dogs. Memór ias do Ins t i tu to Oswaldo Cruz , v .88 , n .2 , p .243-8 . Ap r -Jun , 1993 . OLIVE IRA F , LAWYER PG, KAMHAWI S , VALENZUELA J G. Immuni t y t o D i s t i nc t Sand F l y Sa l i va r y P ro te ins P r imes the An t i -Le i shmania Immune. Response towards P ro tec t ion o r Exacerba t ion o f D i sease. PLoS Neg l T rop D is. ; 2 (4 ) :e226 . Ap r 16 , 2008 OLIVER, A. M. ; MARTIN, F . ; GARTLAND, G. L. ; CARTER, R . H . ; KEARNEY, J . F . Marg ina l zone B ce l l s ex h ib i t un ique ac t i va t ion , p ro l i f e ra t i ve and immunog lobu l i n sec re to r y responses . European jou rna l o f immuno logy , v .27 , n .9 , p .2366-74 . Sep, 1997 . OLIV IER, M . ; GREGORY, D . J . ; FORGET, G. Subvers ion mechan isms b y wh ich Le i shman ia paras i t es can escape the hos t immune response: a s igna l i ng po in t o f v i ew. C l in M icrob io l Rev , v .18 , n .2 , p .293-305. Ap r , 2005 . PALATNIK-DE-SOUZA, C. B . ; SANTOS, W. R . ; FRANÇA-S ILVA, J . C . ; COSTA, R. T . ; REIS , A . B . ; PALATNIK , M . ; MAYRINK, W. ; GENARO, O. Impac t o f can ine con t ro l i n t he ep idemio logy o f can ine and human v isce ra l l e ishman ias is in Braz i l . The Amer i can jou rna l o f t rop ica l med ic ine and hyg iene, v .65 , n .5 , p .510 -517.2001 . PANARO, M. A . ; ACQUAFREDDA, A . ; L IS I , S . ; LOFRUMENTO, D. D . ; M ITOLO, V . ; S ISTO, M. ; FASANELLA, A. ; TROTTA, T . ; BERTANI, F . ; CONSENTI, B . ; BRANDONIS IO, O . N i t r i c ox ide p roduc t ion by macrophages o f dogs vacc inated w i th k i l l ed Le i shmania in fan tum p romas t i go tes . Comp Immuno l Mic rob io l In f ec t D is , v .24 , n .3 , p .187 -95 . Ju l , 2001 . PANARO, M. A . ; ACQUAFREDDA, A . ; L IS I , S . ; LOFRUMENTO, D. D . ; TROTTA, T . ; SATALINO, R . ; SACCIA , M . ; M ITOLO, V . ; BRANDONIS IO, O . Induc ib le n i t r i c ox ide syn thase and n i t r i c ox ide p roduct i on i n Le i shmania in fan tum- in fec ted human macrophages s t imu la ted w i th in ter fe ron -gamma and bacter i a l l i popo l ysacchar ide . I n te rna t iona l Journa l o f C l in i ca l & Labora tory Rese arch , v .29 , n .3 , p .122-7 . 1999. PANARO M. A. ; BRANDONIS IO O. ; DE CAPRARI IS D . ; CAVALLO P . ; C IANCIULL I A . ; M ITOLO V . ; OTRANTO D. Can ine l e i shmanias is in Southern I t a l y: a ro le fo r n i t r i c ox ide re leased f rom ac t i va ted

78
macrophages i n as ymptomat i c i n fec t i on? Paras i t Vec to rs , v .9 , n .1 , p .10 . 2008 . PACHER, P . ; BECKMAN, J . S . ; L IAUDET, L. N i t r i c ox ide and perox yn i t r i t e in hea l th and d isease . Phys io l Rev, v .87 , n .1 , p .315-424. Jan , 2007. PARKER, S . J . ; ROBERTS, C. W. ; ALEXANDER, J . CD8+ T ce l l s a re the majo r l ymphocyte subpopu la t i on i nvo l ved i n t he pro tec t i ve immune response to Tox op lasma gond i i i n m ice . Cl in i ca l and Exper imen ta l Immuno logy , v .84 , n .2 , p .207-12 . May, 1991. P ILLA I , S . ; CARIAPPA, A . ; MORAN, S . T . Marg ina l zone B ce l l s . Annu Rev Immuno l , v .23 , p .161 -96. 2005. P INE LLI , E . ; BOOG, C. J . ; RUTTEN, V . P . ; VAN DIJ K, B . ; BERNADINA, W. E . ; RUITENBERG, E . J . A can ine CD8+ cyto tox i c T -ce l l l i ne spec i f i c fo r Le i shmania in fan tum- in fec ted macrophages . Tissue An t igens, v .43 , n .3 , p .189-92 . Mar , 1994a. P INE LLI , E . ; GEBHARD, D. ; MOMMAAS, A. M . ; VAN HOEIJ, M . ; LANGERMANS, J . A . ; RUITENBERG, E . J . ; RUTTEN, V . P . In fec t ion o f a can ine macrophage ce l l l i ne w i th le i shmania i n fantum: dete rm inat ion o f n i t r i c ox ide p roduc t ion and an t i - l e i shmania l ac ti v i t y. Vete r inary Paras i to logy, v .92 , n .3 , p .181-9 . Oc t 1 , 2000. P INE LLI , E . ; GONZALO, R. M. ; BOOG, C . J . ; RUTTEN, V. P . ; GEBHARD, D . ; DEL REAL G. ; RUITENBERG, E . J . Le i shmania i n fan tum-spec i f i c T ce l l l i nes der i ved f rom as ymptomat i c dogs that l yse i n fec ted macrophages in a ma jo r h is tocompat ib i l i t y comp lex - res t r i c ted manner . European jou rna l o f immuno logy, v .25 , n .6 , p .1594-600 . Jun , 1995 . P INE LLI , E . ; K ILL ICK-KENDRICK, R . ; WAGENAAR, J . ; BERNADINA, W. ; DEL REAL G. ; RUITENBERG, E. J . Ce l l u l a r and humora l immune responses in dogs ex per imenta l l y and na tu ra l l y i n fec ted w i th Le i shman ia i n fan tum. I n fec t i on and immun i t y , v .62 , n .1 , p .229-35 . Jan , 1994b . POLI , A . ; ABRAMO, F. ; MANCIANTI, F . ; N IGRO, M. ; P IERI, S . ; B IONDA, A . Rena l invo l vemen t i n can ine l e ishmanias is . A l i gh t -m ic roscop i c , immunoh is tochemica l and e lec t ron -mic roscop i c s tudy. Nephron, v .57 , n .4 , p .444-52 . 1991 .

79
POZIO, E . ; GRADONI, L . ; BETTIN I , S . ; GRAMICCIA , M . Le i shmanias is i n Tuscany ( I t a l y) : V I . Can ine l e ishmanias is in the focus o f Monte Argen tar io (Grosseto ) . Acta T rop i ca , v .38 , n .4 , p .383 -93. Dec, 1981. PROUDFOOT L, N IKOLAEV AV, FENG GJ , WEI WQ, FERGUSON MA, BRIMACOMBE JS, L IEW FY. Regu la t ion o f the exp ress ion o f n i t r i c ox ide s yn thase and l e i shmanic ida l ac t i v i t y b y g l ycocon juga tes o f Le i shman ia l ipophosphog l ycan i n mur ine macrophages . Proc Na t l Acad Sc i U S A. v .1 ;93 (20 ) :10984-9 . 1996. RADOMSKI, M . W. ; PALMER, R . M . J . ; MONCADA, S . An L-a rg in ine: n i t r i c ox ide pathway p resent in human p la te le ts regu la tes aggrega t ion , Proc . Na t l . Acad . Sc i . USA, v .87 , p .5193-5197 . 1990 . REIS , A . B . ; MARTINS-F ILHO, O. A . ; TE IXE IRA-CARVALHO, A . ; CARVALHO, M. G . ; MAYRINK, W. ; FRANCA-S ILVA, J . C . ; G IUNCHETTI, R . C . ; GENARO, O. ; CORREA-OLIVE IRA, R . Paras i t e dens i t y and impai red b iochemica l /hemato log ica l s ta tus a re assoc ia ted w i th severe c l i n i ca l aspects o f can ine v isce ra l l e ishmanias is . Research in Vete r i na ry Sc ience, v .81 , n .1 , p .68-75 . Aug, 2006a . REIS , A . B . ; TE IXE IRA-CARVALHO, A . ; G IUNCHETTI, R . C . ; GUERRA, L. L . ; CARVALHO, M. G . ; MAYRINK, W. ; GENARO, O. ; CORREA-OLIVE IR A, R . ; MARTINS-F ILHO, O. A . Phenot yp ic fea tu res o f c i rcu la t ing l eucocytes as immuno log i ca l markers fo r c l in i ca l s ta tus and bone marrow paras i t e dens i t y i n dogs na tu ra l l y i n fec ted by Le i shman ia chagas i. C l in i ca l and Exper imenta l Immuno logy, v .146 , n .2 , p .303 -11. Nov , 2006b . REIS , A . B . ; TE IXEIRA-CARVALHO, A . ; VALE, A. M . ; MARQUES, M. J . ; G IUNCHETTI, R . C . ; MAYRINK, W. ; GUERRA, L. L . ; ANDRADE, R . A . ; CORREA-OLIVE IRA R . ; MARTINS-F ILHO, O. A . Isot ype pa t te rns o f immunog lobu l i ns : ha l lmarks fo r c l in i ca l s ta tus and t i ssue paras i t e dens i t y i n Braz i l i an dogs na tu ra l l y i n fec ted b y Le i shmania (Le i shmania ) chagas i . Vete r ina ry immuno logy and immunopa tho logy, v .112, n .3 -4 , p .102-16. Aug 15 , 2006c. RODRIGUES, C . A . ; BATISTA, L. F . ; TE IXE IRA, M . C . ; PEREIRA, A . M . ; SANTOS, P . O . ; DE SA OLIVE IRA, G . G . ; DE FREITAS, L. A . ; VERAS, P . S . Per iphera l b lood mononuc lear ce l l supernatan ts f rom as ymptomat i c dogs immunized and exper imenta l l y cha ll enged w i th Le i shman ia chagas i can s t imu la te can ine macrophages to reduce

80
i n fec t ion i n v i t ro . Vete r i na ry Paras i to logy, v .143, n .3 -4 , p .197 -205. Feb 28 , 2007 . RODRÍGUEZ-SOSA M, R IVERA-MONTOYA I , ESPINOZA A , ROMERO-GRIJ ALVA M, LÓPEZ-FLORES R , GONZÁLE Z J , TERRAZAS L I. Acu te cys t i ce rcos is favou rs rap id and more severe les ions caused b y Le i shman ia ma jo r and Le i shmania mex i cana in fec t ion , a ro le fo r a l t e rnat i ve l y ac t i va ted macrophages . Cel l Immuno l . 242 (2 ) :61 -71 . Aug, 2006 . RODRIGUEZ-CORTES, A . ; FERNANDEZ-BELLON, H . ; RAMIS , A . ; FERRER, L. ; ALBEROLA, J . ; SOLANO-GALLEGO, L. Le i shmania -spec i f i c i so t ype l eve ls and the i r re la t i onsh ip w i th spec i f i c ce l l -med ia ted immuni t y pa rameters i n can ine l e ishmanias is . Vete r inary immuno logy and immunopatho logy, v .116 , n .3 -4 , p .190-8 . Ap r 15 , 2007. ROLÃO N, CORTES S , GOMES-PEREIRA S, CAMPINO L. Le i shmania i n fan tum: mixed T -he lper -1 /T -he lper -2 immune response i n ex per imenta l l y i n fec ted BALB/c mice . Exp Paras i to l. Mar ;115(3 ) :270-6 . 2007 . RONCAROLO, M. G . ; GREGORI, S . , BATTAGLIA , M . ; BACCHETTA, R . ; FLE ISCHHAUER, K . ; LEV INGS, M. K. In te r l euk in -10-sec re t i ng t ype 1 regu la to r y T ce l l s i n rodents and humans . Immuno log i ca l Rev iews, v .212, p .28-50 . Aug, 2006. SACKS, D . L . ; LAL, S . L . ; SHRIVASTAVA, S . N. ; BLACKWELL, J . ; NEVA, F . A . An ana l ys i s o f T ce l l respons iveness in Ind ian ka la-azar . Jou rna l o f Immuno logy , v .138, n .3 , p .908-13 . Feb 1 , 1987 . SANCHEZ, M. A. ; D IAZ, N . L . ; ZERPA, O. ; NEGRON, E. ; CONVIT , J . ; TAP IA , F . J . Organ-spec i f i c immun i ty i n can ine v i scera l l e ishman ias i s : ana l ys i s o f s ymptomat i c and as ymptomat i c dogs na tu ra l l y i n fec ted w i th Le i shman ia chagas i . The Amer i can jou rna l o f t rop i ca l med ic ine and hyg iene, v .70 , n .6 , p .618 -24. Jun , 2004 . SANTOS-GOMES, G. M. ; ROSA, R . ; LEANDRO, C. ; CORTES, S . ; ROMÃO, P . ; S ILVEIRA, H . C ytok ine ex p ress ion du r ing the ou tcome o f can ine exper imen ta l i n fec t ion b y Le i shman ia in fan tum. Vete r ina ry immuno logy and immunopa tho logy, v .88 , n .1 -2 , p .21 -30 . Sep 6 , 2002 .

81
SCHOPFER F . J . ; BAKER P. R . ; FREEMAN B. A. NO-dependent p ro te in n i t ra t i on : a ce l l s igna l i ng even t o r an ox ida t i ve in f l ammato ry response? Trends B iochem Sc i , v .28 , p .646–654. 2003 SHARMA, S . K . ; DUBE, A . ; NADEEM, A. ; KHAN, S . ; SALEEM, I . ; GARG, R. ; MOHAMMAD, O. Non PC l i posome ent rapped p romast i go te an t i gens e l i c i t pa ras i t e spec i f i c CD8+ and CD4+ T -ce l l immune response and p ro tec t hamsters aga ins t v isce ra l l e i shmanias is. Vacc ine, v .24 , n .11 , p .1800-10. Mar 10 , 2006. SHIBATA M. ; ARAKI N . ; HAMADA J . ; SASAKI T . ; SHIMAZU K . ; FUKUUCHI Y . Bra in n i t r i t e p roduct ion dur ing g loba l i schemia and reper fus ion : an i n v ivo m icrod ia l ys i s s tud y. Bra in Res, v .734, p .86–90 . 1996 . S ILVA, E . S . ; GONTIJ O, C . M . ; PACHECO, R. S . ; F IUZA, V . O. ; BRAZIL R . P . V iscera l l e i shmanias is in the Met ropo li t an Reg ion o f Be lo Hor izon te , S ta te o f M inas Gera is , Braz i l . Memór ias do Ins t i tu to Oswaldo Cruz, v .96 , n .3 , p .285-91 . Ap r , 2001 . S ILVA, J . G. ; WERNECK, G . L. ; CRUZ MDO, S . ; COSTA, C . H . ; DE MENDONÇA, I . L . Na tu ra l i n fec t ion o f Lu tzomyia l ongipa lp is by Le i shman ia sp . in Teres ina , P iau i S ta te , Braz i l . Cad Saúde Pub l i ca, v .23 , n .7 , p .1715-20 . Ju l , 2007. S INGH, V . K . ; MEHROTRA, S . ; NARAYAN, P . ; PANDEY, C . M . ; AGARWAL, S . S . ; Modu la t i on o f au to immune d iseases by n i t r i c ox ide . Immuno l . Res ., v .22 , n .1 , p .1 -19 . 2000 . S INHA, P . K . ; PANDEY, K. ; BHATTACHARYA, S . K . D iagnos i s & management o f l e i shmania /H IV co - in fec t ion . The Ind ian Jou rna l o f Med ica l Research, v .121 , n .4 , p .407 -14 . Ap r , 2005 . S ISTO, M. ; BRANDONIS IO, O . ; PANARO, M. A . ; ACQUAFREDDA, A. ; LEOGRANDE, D. ; FASANELLA, A . ; TROTTA, T . ; FUMAROLA, L. ; M ITOLO, V . Induc ib le n i t r i c ox ide syn thase exp ress ion i n Le i shmania -i n fec ted dog macrophages. Comp Immuno l M ic rob io l In f ec t D i s , v .24 , n .4 , p .247 -54. Oc t , 2001. STAUBER, L. A . Le i shman ias i s i n the hamste r . In : Cole , W.H. (Ed . ) , Some Ph ys io log i ca l Aspec ts and Consequence o f Parasi t i sm. Rugers Un ivers i ty Press , New Brunsw ick. p . 77-99 . 1955 .

82
STENGER, S . ; DONHAUSER, N. ; THURING, H. ; ROLLINGHOFF, M. ; BOGDAN, C. Reac t i va t i on o f l a ten t l e ishman ias i s by i nh ib i t ion o f i nduc ib le n i t r i c ox ide syn thase. The Jou rna l o f Exper imen ta l Med ic ine, v .183, n .4 , p .1501-14 . Ap r 1 , 1996 . STENGER, S . ; THURING, H . ; ROLLINGHOFF M. ; BOGDAN, C. T i ssue ex p ress ion o f induc ib le n i t r i c ox ide s yn thase i s c lose l y assoc ia ted w i th res i s tance to Le i shmania ma jo r. The Jou rna l o f Exper imen ta l Med ic ine, v .180, n .3 , p .783-93 . Sep 1 , 1994 . STUEHR, D . J . ; NATHAN, C . F . N i t r i c ox ide : a macrophage p roduc t respons ib le fo r cytos tas is an resp i ra to r y i nh ib i t ion in t umor ta rge t ce l l s . The Jou rna l o f Exper imen ta l Med ic ine, v .169, p .1543. 1989 . TAFURI, W. L. ; TAFURI, W. L. ; BARBOSA, A . J . ; M ICHALICK, M. S . ; GENARO, O. ; FRANCA-S ILVA, J . C . ; MAYRINK, W. ; NASCIMENTO, E . H is topatho logy and immunocytochemica l s tudy o f type 3 and t ype 4 complement recepto rs i n t he l i ve r and sp leen o f dogs na tura l l y and ex per imenta l l y i n fec ted w i th Le i shmania (Le i shman ia) chagas i . Rev Ins t Med T rop Sao Pau lo, v .38 , n .2 , p .81 -9 . Mar-Apr , 1996 . TAFURI, W. L. ; DE OLIVE IRA, M . R . ; MELO, M. N . ; TAFURI, W. L. Can ine v i sce ra l l e ishmanios i s : a remarkab le h is topatho log i ca l p i c tu re o f one case repo r ted f rom Braz i l . Vete r ina ry Paras i to logy, v .96 , n .3, p .203-12 . Apr 2 , 2001. TAFURI, W. L. ; MICHALICK, M . S . ; D IAS , M . ; GENARO, O. ; LE ITE, V . H. ; BARBOSA, A . J . ; BAMBIRRA, E . A . ; DA COSTA, C. A . ; MELO, M. N. ; MAYRINK, W. Op t i ca l and e lec t ron m icroscop i c s tud y o f t he k idney o f dogs natu ra l l y and exper imenta l l y i n fec ted w i th Le i shmania (Le i shmania ) chagas i . Rev Ins t Med T rop Sao Pau lo, v .31 , n .3 , p .139 -45 . May-J un , 1989 . TAKEDA, K . ; KA ISHO T. ; AK IRA, S . To l l - l i ke recepto rs . Annua l Rev iew o f Immuno logy, v .21 , p .335 -76 . 2003. TE IXE IRA, S . A . ; CASTRO, G. M . ; PAPES, F . ; MARTINS, M. L. ; ROGERIO, F . ; LANGONE, F. ; SANTOS, L. M . ; ARRUDA, P .; DE NUCCI G. ; MUSCARA, M. N. Express ion and ac t i v i t y of n i t r i c ox ide s yn thase i so fo rms i n ra t b ra in du r ing the deve lopment o f exper imental

83
a l l e rg i c encepha lom ye l i t i s . Bra in Res Mo l B ra in Res, v .99 , n .1 , p .17-25 . Feb 28 , 2002 . TESH, R . B. Cont ro l o f zoonot i c v isce ra l l e i shmanias is : i s i t t ime to change s t ra teg ies? The Amer i can jou rna l o f t rop i ca l med ic ine and hyg iene, v .52 , n .3 , p .287 -92. Mar , 1995 . TRAVI, B . L . ; TABARES, C . J . ; CADENA, H. ; FERRO, C.; OSÓRIO, Y . CANINE v iscera l l e i shmanias is in Co lomb ia : re la t ionsh ip between c l i n i ca l and paras i to log i c s ta tus and i n fec t i v i t y fo r sand f l i es . The Amer i can jou rna l o f t rop i ca l med ic ine and hyg iene, v .64 , n .3 -4 , p .119-24 . Mar -Apr , 2001. TRYPHONAS, L. ; ZAWIDZKA, Z . ; BERNARD, M. A. ; JANZEN, E . A . V iscera l l e ishmanias is in a dog: c l i n ica l , hemato log i ca l and patho log i ca l observat i ons . Can J Comp Med, v .41 , n .1 , p .1 -12 . J an , 1977. TRZECIAK S . ; C INEL I . ; PHILL IP DELLINGER R. ; SHAPIRO N. I . ; ARNOLD R . C . ; PARRILLO J . E . ; HOLLENBERG S. M. ; Resusc i ta t i ng the m ic roc i r cu la t ion i n seps i s : the cen t ra l ro le o f n i t r i c ox ide, emerg ing concepts fo r nove l t he rap ies , and cha l l enges fo r c lin i ca l t r i a ls . Acad Emerg Med. V 15 , n5 , p . 399 -413 . May, 2008. TSAGOZIS , P . ; KARAGOUNI, E . ; DOTSIKA, E . CD8(+) T ce l l s w i th pa ras i t e -spec i f i c cyto tox i c ac t i v i t y and a Tc1 p ro fi l e o f cytok ine and chemok ine sec re t ion deve lop i n exper imenta l v i sceral le ishman ias i s . Paras i te immuno logy, v .25 , n .11 -12 , p .569-79 . Nov -Dec, 2003 . VOULDOUKIS , I . ; DRAPIER, J . C . ; NUSSLER, A. K . ; TSELENT IS , Y . ; DA S ILVA, O . A . ; GENTIL IN I , M . ; MOSSALAYI, D . M . ; M ONJOUR, L. ; DUGAS, B. Can ine v isce ra l l e i shmanias is : success fu l chemotherap y i nduces macrophage an t i l e i shmania l ac t i v i t y v i a the L-a rg in ine n i t r i c ox ide pathway. Ant imic rob Agen ts Chemother, v .40 , n .1 , p .253 -6 . J an , 1996 . WALDECK,K. ; NY, L. ; PERSSON, K. E . ; Med ia to rs and mechan isms o f re la t i on i n rabb i t u re tha l smooth musc le . B r . J . Pharmaco l. V .123 , n .4 , p . 617 -624 . 1998 . W ILSON, M. E . ; JERONIMO, S . M . ; PEARSON, R . D. Immunopa thogenes i s o f i n fec t ion w i th the v i sce ra l i zi ng Le i shmania spec ies . Microb Pathog , v .38 , n .4 , Apr , p .147 -60 . 2005 .

84
XU, D. ; L IU , H . ; KOMAI-KOMA, M. ; CAMPBELL, C . ; MCSHARRY, C. ; ALEXANDER, J . ; L IEW, F . Y . CD4+CD25+ regu la to r y T ce l l s suppress d i f fe ren t i a t ion and func t ions o f Th1 and Th2 ce l l s , Le i shmania major i n fec t ion , and co l i t i s i n m ice . Journa l o f Immuno logy , v .170 , n .1 , p .394-9 . Jan 1 , 2003 .

85
UNIVERSIDADE VALE DO R IO DOCE
FACULDADE DE C IÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM C IÊNCIAS BIOLÓGICAS
Fe rnando Rocha dos San tos
NÍVEIS SÉRICOS DE ÓXIDO NÍTRICO E
EXPRESSÃO DA ÓXIDO NÍTRICO SINTASE
INDUZÍVEL NO BAÇO DE CÃES
NATURALMENTE INFECTADOS COM
Leishmania (Leishmania) chagasi
Governador Va ladares
2008

86
Santos, Fernando Rocha dos Níveis séricos de óxido nítrico e expressão da óxido nítrico Sintase induzível no baço de cães naturalmente infectados com Leishmania (Leishmania) chagasi /, Fernando Rocha dos Santos. --2008. 82f. Dissertação (mestrado) – Universidade Vale do Rio Doce, Programa de Pós-Graduação em Ciências Biológicas, Governador Valadares, MG, 2008. Orientador: Luiz Cosme Cotta Malaquias. 1. Leishmaniose visceral canina. 2. Óxido nítrico – Níveis Séricos. 3. Parasitologia. I. Malaquias, Luiz Cosme Cotta II. Universidade Vale do Rio Doce. III. Título.
CDD 636.089696

87


















