
Faculdade de Engenharia Qufmica de Lorena Departamento de...
86
. - . ·--·----··--·----------------- --- -- Faculdade de Engenharia Qufmica de Lorena Departamento de Biotecnologia Pós-Graduação em Biotecnologia Industrial Avaliação ln Vivo dos Níveis de Atividade de Invertase em Leveduras Este exemplar corresponde a versão final da dissertação de mestrado aprovada pela banca examinadora. ' / I , i Lorena - SP - Brasil 1998
Transcript of Faculdade de Engenharia Qufmica de Lorena Departamento de...

. - . ·--·----··--·----------------- --- --
Faculdade de Engenharia Qufmica de Lorena Departamento de Biotecnologia
Pós-Graduação em Biotecnologia Industrial
Avaliação ln Vivo dos Níveis de Atividade de Invertase em Leveduras
Este exemplar corresponde a versão final da dissertação de mestrado aprovada pela banca examinadora.
' / I , i
Lorena - SP - Brasil 1998

-----. -------- ··--· . ·-- -- --------· .. --· ------· ------------ -····· -----------------------------------
Ficha Catalográf"1e2 Elaborada pela Biblioteca do Departamento de
Biotecnologia da F AENQUIL
CARVALHO. Patrícia Maria Barroso de Avaliação in vivo dos níveis de anvidade de invertase em leveduras/
C:; 3 1 a Patrícia Maria Barroso de Carvalho. -Lorena. 1998. ~p '
Dissertação (Mestrado) • Faculdade de Engenharia Química de Lorena.! Depanamento de Biotecnologia
Orientadora: Medeiros. Maria Bemadete
1. Biotecnologia 2. Leveduras 3. lnvertase 4. Método GOD-PAP. l. Título ll. Medeiros. Maria Bemadete, orientadora
574.6 CDU

·--- ------ ··- -- -·-- ··---··-···--· ·- ·-·-----------··-- - -· ··-·· -~--· -·--- ···- .....
i
ÍNDICE
Páginas
LISTA DE FIGURAS iii LISTA DE TABELAS V
RESUMO vi ABSTRACT vii
- 1. INTRODUÇAO ...•............................•.•.............•..................•....•.•.•.......•••.• 1 1.1. BIOTECNOLOGIA 1 1.2. LEVEDURAS 4 1.3. INvERTASE .. 7 1.4. APLICAÇÃO INDUSTRIAL DA INVERT ASE 13 1.5. MÉTODOGOD-PAP 15
2. OWETIVOS: 16
, 3 . .MA. TERIAL E ME TODOS 17
3.1. AVALIAÇÃO DOS NÍVEIS DE PRODUÇÃO DE INVERTASE 17 3 .1.1. Leveduras 17 3 .1.2. Ativação das células 17 3. 1.3. Obtenção de massa celular 18 3. 1. 4. Cultivo para a produção de in vertase . . . . . .. . .. . . . . . .. . . . .. .. .. . . .. .. .. .. . .. . .. . 18 3.1.5. Ensaio enzimático 19 3.1.5. l. Preparo das células para a dosagem de invertase 19 3.1.5.2. Reação enzimática 19 3.1.6. Curvas de crescimento e concentração de glicose no meio 20
3.2.DOSAGEM DE GLICÍDIOS 20 3.3. DETERMINAÇÃO DOS PARÂMETROS ENVOLVIDOS NO AUMENTO DA ABSORBÂNCIA DURAi~ A DETERMINAÇÃO DA GLICOSE PELO MÉTODO GOD-PAP 22 3 .3 .1. Medida do pH das soluções de glicídios adicionadas da solução de reação do método GOD-P AP 22 3.3.2. Influência do aquecimento a temperatura de fervura da água na inversão da sacarose 22 3.3.3. Efeito da solução de reação do método GOD-PAP sobre diferentes glicídios 23 3.3.4. Efeito das enzimas do método GOD-PAP sobre a sacarose 23

--------·-··-·-- - - - - --- -- - -- - - - ii
3.3.S. Cinética da reação de hidrólise da sacarose pelo reagente GOD-PAP 23 3.3.6. Determinação da variação da absorbância para diversas concentrações de glicose na presença de sacarose 24
- 4. RESULTADOS E DISCUSSAO e ••••••••••••••••••••••• 25 4.1. IDENTIFICAÇÃO DAS VARIÁ VEIS RELACIONADAS À INADEQUAÇÃO DO MÉTODO GOD-PAP PARA DOSAGEM DA GLICOSE EM MISTURA COM SACAROSE 26 4.1.1. Influência do pH 27 4.1.2. Influência do aquecimento da solução de sacarose 28 4.1.3. Reação do método GOD-PAP com diferentes glicídios 32 4.2. ClNÉTICA DA REAÇÃO DE HIDRÓLISE DA SACAROSE PELA SOLUÇÃO DE REAÇÃO DO M:Él'ODO GOD-PAP 35 4.3. ADAPTAÇÃO DO MÉ'TOOO ·································································· 37 4.3.1. Determinação da variação da absorbância para diversas concentrações de glicose na presença de sacarose 3 7 4 .4. PRODUÇÃO DE INVERT ASE 40 4.5. CRESCIMENTO CELULAR E CONCENTRAÇÃO DE GLICOSE NO MEIO 44
5. CONCLUSOES 58
6. REFERÊNCIAS BIBLIOGRAFICAS 59
7. APÊ~DICES

.. - ·-·· ·-····--·--···---------- ili
ÍNDICE DE FIGURAS
Página
FIGURA 1: CINÉTICA DA REAÇ.ÃO DE HIDRÓLISE DA SACAROSE PELA SOLUÇÃO DE REAÇÃO DO MÉTODO GOD-PAP 35
FIGURA 2: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE DEBARYOMYCES HANSENJJ (50059) 45
FIGURA 3: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE RHOOOTHORUU RUBRA (50596) 45
FIGURA 4: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE SA.CCHAROMYCESCEREVJSUE(50613) 46
FIGURA 5: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE PICHJA BECKJ { 5053 7) 4 7
FIGURA 6: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE PICHIA Of{}...fERJ (50808) 47
F'IGUR.\ 7: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DEAUREOBASID!lJM PULUL4NS(51102) 48
FIGl"R.\ 8: Cor--.;CENTR..AÇ.i.O DE GUCOSE NO \{EIO DE CUL rrvo E CRESCIMENTO CELL1.AR DE TORCL4SPORA PRETORJE.VSJS (50811 ) 49
flGl'R.\ 9: CONCENTRAÇAO DE GLICOSE NO ~{EIO DE CUL nvo E CRESCIMF\'TO CELULAR DE WIUJOPSISCAllFORNJCA (50420) 50
FIGUR.\ 10: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE CRYPTOCOCCUS ATER (51032) 51
FIGURA 11: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE CRYPTOCOCCUS MACERANS (50361 ) 51
FIGURA 12: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE SACCHAROMYCES KLVYVERY (50629) 52

FIGURA 13: CONCENTRAÇÃO DE GLICOSE NO MEIO DE cu.nvo E CRESCIMENTO CELULAR DE ZYGOSSA.CHAROMYCESSP(50965) 53
FIGURA 14: CONCENTIUÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE DEIWUOMICF.S SP (50385) 53
FIGURA 15: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE SÃCCJUROMYCES CF.REJIISIAE (IR2) 54
FIGURA 16: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE SÃCC™ROMTCF.S CEREVJSUE (ATCC 9763) 55
FIGURA 17: CONCENTRAÇÃO DE GLICOSE NO MEIO DE CULTIVO E CRESCIMENTO CELULAR DE PJCHM CARSONJ (50809) 55

------------------------------- ----------
V
ÍNDICE DE TABELAS
Página
TABELA 1: VALORES DO PH DAS SOLUÇÕES DE GLICÍDIOS ADICIONADAS À SOLUÇÃO DE REAÇÃO 00 MÉTOOO GOD-PAP 27
TABELA 2: INFLUÊNCIA 00 1EMPO DE AQUECIMENTO A 96°C DA SOLUÇÃO DE SACAROSE 40 MM SOBRE O AUMENTO DA CONCENTRAÇÃO DE GLICOSE 29
T ADE.LA 3: INFLUÊNOA 00 TEMPO DE AQUECIMENTO A 96°C DAS SOLUÇÕES DE SACAROSE l O E 20 MM SOBRE O AUMENTO DA CONCENTRAÇÃO DE GLICOSE 29
TABELA 4: INFLUÊNCIA DO TEMPO DE AQUECIMENTO A 96°C DA SOLUÇÃO DE SACAROSE 10 MM DILUÍDA EM ÁGUA DESTILADA SOBRE O AUMENTO DA CONCENTRAÇÃO DE GLICOSE 30
TABELA S: INFLUÊNCIA DO TEMPO AQUECIMENTO A 96°C DE SOLUÇÕES DE SACAROSE l O MM SOBRE O AUMENTO DA CONCENTRAÇÃO DE GLICOSE - .. 3 l
TABELA 6: REAÇÃO DE XILOSE, FRUTOSE, TREALOSE, CELOBIOSE E RAFINOSE 40 MM COM A SOLUÇÃO DE REAÇÃO DO MÉTODO GOD-PAP 33
TABELA 7: EFEITO DA ENZIMA GLICOSE OXIDASE SOBRE A SOLUÇÃO DE
SACAROSE 10 MM ·-···-········-···········································-··············35
TABELA 8: CONCENTRAÇÃO DE AÇÚCARES REDUTORES TOTAIS DA SOLUÇÃO DE SACAROSE 10 MM - 36
TABELA 9: VARIAÇÃO EM FUNÇÃO DO TEMPO DE LEITURA DOS VALORES DE CONCENTRAÇÃO DE GLICOSE DETERMlNADOS PELO MÉTODO GOD-P AP A PARTIR DE DE SOLUÇÕES DE GLICOSE NAS CONCENTRAÇÕES DE 5 A 180 µGIML DILUÍDAS EM SACAROSE 10 MM 38
TABELA 10: NÍVEIS DE ATIVIDADE DA INVERTASE EM DIFERENTES LEVEDURAS 42

RESUMO· · --
Avaliação ln Vivo dos Níveis de Atividade de lnffrtase em Leveduras. Patricia Maria Barroso de Carvalho. Dissertação de Mestrado. Programa de Pós Graduação em Biotecnologia Industrial, Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena. Orientador: Ora. Maria Bemadete Medeiros (Departamento de Biotecnologia, FAENQUIL, CP 116, 12.000-000, Lorena I SP-Brasil). Banca examinadora: Dra. Denise Maria Mano Pessoa e Dr. Arnaldo Márcio Ramalho Prata. Setembro de 1998.
A avaliação in vivo dos nrvers de atividade de inverta.se (13-frutofuranosideo frutoidrolase E.C. 3.2.1.26) é de fundamental importância no estudo do potencial de linhagens de .le~ para aplicação biotecnológica. A maior parte dos processos industriais que fazem uso dessa enzima emprega células vivas de leveduras.
Em sua maioria, os métodos descritos para a quantificação da invertase correlacionam a atividade dessa enzima com a quantidade de glicose ou açúcares redutores totais ( AR T) liberados após a hidrólise enzimática da sacarose em condições pré determinadas. O método da Glicose Oxidase-Peroxidase (GOD-PAP - Merck 8443) é um método colorimétrico utilizado na quantificação da glicose na faixa de concentração de O a 200 µg/mL.
No presente trabalho, foi feita uma avaliação dos níveis de atividade de invertase em 16 linhagens de leveduras pertencentes a 9 diferentes gêneros. Dentre as linhagens testadas, 31,25% não apresentaram atividade enzimática nas condições experimentais enquanto 68, 7 5°/o apresentaram atividade em níveis que variaram de 0,8 a 18 U (U=µg de ART liberados por mio).
O método GOD-P AP forneceu leituras crescentes de absorbância à 51 O nm, sempre que foi utilizado para determinar a concentração de glicose liberada após o ensaio enzimático. Esse resultado foi atribuído à hidrólise subseqüente e contínua da sacarose por algum reagente( s) não enzimático(s) presente(s) na solução de reação do método. O estudo realizado sobre a cinética dessa reação demonstrou que o método GOD-P AP só pode ser utilizado para a determinação da concentração de glicose em presença de sacarose mediante adaptação: trabalhar com concentração inicial de glicose entre 40 e 100 µglmL e efetuar a leitura de absorbância com o tempo de 21 min de reação.

ABSTRACT
ln Vivo Evaluation of Invertase Activity Leveis in Y easts. Patricia Maria Barroso de Carvalho. Dissertação de Mestrado. Programa de Pós Graduação em Biotecnologia Industrial, Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena. Orientador: Ora. Maria Bernadete Medeiros (Departamento de Biotecnologia, F AENQUIL, CP 116, 12.000-000, Lorena I SP-Brasil). Banca examinadora: Ora. Denise Maria Mano Pessoa e Dr. Arnaldo Márcio Ramalho Prata. Setembro de 1998.
The in vivo evaluation of invertase @-fructofmanoside fructohydrolase E.C. 3.2.1.26) activity leveis is a subject of major concem to the study of yeasts strains potential for biotecnological application. Most of the industrial processes which makes use of this enzyme employs whole yeasts cells.
ln general, methods described for estimating invertase activity correlate this enzyme activity with either the amount of glucose or total reducing sugars (TRS) produced after sucrose enzymatic hydrolysis under standardized conditions. The Glucose Oxidase-Peroxidase Method (GOD-PAP - Merck) is a colorimetric method used to determine the amount of glucose in a concentration range from O to 200 µg/mL.
ln the present work, we performed an evaluation of the invertase activity levels in 16 strains belonging to 9 different genera. Amongst the tested strains, 31.25°/o did not show invertase activity under experimental conditions, whereas 68.75% showed activity in leveis ranging between 0.8 and 18 U (U=µg ofTRS produced per min).
At 510 nm, the GOD-PAP Method yielded increasing mesurements of absorbance whenever it was employed to determine the glucose concentration produced after the enzymatic assay. We regarded this result as being due to the subsequent and continuous hydrolysis of sucrose by some non-enzymatic reagent(s) included in the reaction solution of the method. The study performed on the kinetics of this reaction showed that the GOD-PAP Method can only be used for the assessment of glucose concentration in the presence of sucrose, providing that some assumptions be made, that is, to work with an initial glucose concentration from 40 to 100 µg/mL, and to accomplish absorbance mesurements 21 min after the beginning of the reaction.

-·· --------- ---·-. --···----------- 1
1. INTRODUÇÃO
1.1. Biotecnologia
A biotecnologia surgiu como um campo de atividades
interrelacionadas das ciências da biologia, química e engenharia. Segundo
Buli (1995) pode ser definida como a aplicação de princípios científicos e da
engenharia no processamento de materiais por agentes biológicos para a
obtenção de bens e serviços. A biotecnologia tem suas origens na
microbiologia aplicada à medicina, agricultura e indústria e, no seu início,
tinha por ferramentas linhagens naturais isoladas em meio seletivo e
purificadas. A moderna biotecnologia, no entanto, permite ainda a alteração
da estrutura genética das células com a conseqüente criação de novos
microrganismos e metabólitos.
As leveduras provavelmente representam os primeiros orgamsmos
explorados pelo homem como agentes biotecnológicos (Kirsop, 1988). A
utilização de leveduras em processos para a produção de bebidas e
alimentos remonta aos primórdios da história da humanidade. Tais processos
desenvolveram-se quase por acaso, ainda antes de que fossem conhecidas
suas bases científicas e mesmo sem a noção de que neles as leveduras
executavam funções específicas. A fermentação de sucos de frutas e de
extratos de cereais por populações mistas de leveduras provavelmente
constituiu o primeiro uso desses microrganismos pelo homem, resultando em
tipos de bebidas alcoólicas que podem ser consideradas como precursoras
dos modernos vinhos e cervejas. Como resultado da longa história das
leveduras a serviço do homem, muito se foi aprendendo sobre a
manipulação e o controle da atividade das leveduras, tendo esse
conhecimento permitido a evolução da primitiva produção doméstica para a

escala industrial.
O interesse no potencial biotecnológico das leveduras foi
impulsionado pelo reconhecimento de características tais como a facilidade
de manuseio e de obtenção de grandes quantidades de massa celular, a
habilidade de crescer em uma variedade de substratos, faixas de pH e taxas
de oxigênio. As células das leveduras crescem de modo estável tanto como
haplóides quanto como diplóides e políplóides, sendo a obtenção de
mutantes mais fácil na fase haplóide.
Além disso, as leveduras são apropriadas como objeto para estudos
experimentais como modelo de células eucariotas, pois podem ser, de
maneira semelhante às bactérias, cultivadas e manipuladas com facilidade e,
dessa forma contnbuem, também, para a elucidação de processos
bioquímicos, metabólicos e genéticos das células eucarióticas (Stewart &
Russell, 1988).
Assim como outros organismos, as leveduras utilizam os açúcares
como fontes de carbono e energia nos processos biossintéticos necessários
ao crescimento. A contribuiçàodas leveduras, notadamente da
Saccharomyces cerevisiae e das espécies intimamente relacionadas a ela,
para o progresso da humanidade, tem sido principalmente baseada em sua
capacidade de efetuar rápida e eficientemente a conversão de açúcares em
dióxido de carbono e etanol usado tanto como bebida como para propósitos
industriais e combustível.
As indústrias farmacêuticas e de alimentos têm tido uma longa e
lucrativa associação com as leveduras as quais constituem-se em ricas fontes
de enzimas de importância comercial que podem ser produzidas a partir de
açúcares simples e nitrogênio amoniacal. Como exemplo cita-se a
p-galactosidase (E.C. 3.2.1.23) produzida por Kluyveromyces fragilis e
Kluyveromyces lactis, a triacilglicerol lipase (E.C. 3.1. l.3) produzida por

Saccharomyces lipolytica e Candida lipolytica e a (Wlllllase (E.C. 3.2.1.L)
produzida por Schwanniomyces alluvius, Schwanniomyces castelli e
Saccharomyces diastaticus.
Uma grande variedade de espécies de leveduras é explorada na
biotransformação, biorremediação e na produção de flavorizantes, óleos,
gorduras e proteínas heterólogas (Roberts & Wildman, 1995). A replicação
e segregação dos cromossomos e a síntese de macromoléculas ocorrem nas
leveduras e nos eucariotos superiores de forma similar, tomando promissor
o seu uso para a clonagem e expressão de genes heterólogos para, por
exemplo, a produção de interferon, hormônio do crescimento humano e
insulina (Williamson, 1988).
Segundo Stewart & Russel ( 1988), o uso industrial das leveduras
pode ser classificado em três grupos de acordo com a relação do produto
com a bioquímica do microrganismo. No primeiro está o uso dos
constituintes celulares das leveduras para a produção de alimentos e ração
na forma de proteína unicelular, e de componentes macromoleculares como
lipídeos, proteínas, peptideos, enzimas, coenzimas, vitaminas e ácidos
nucléicos. No segundo, o uso de seus produtos de excreção na obtenção de
cerveja, vinho, cidra, etanol industrial, glicerol e dióxido de carbono para
bebidas carbonadas e na fermentação de massas de pães e bolos. E, no
terceiro, o uso em processos que exploram a interação enzima-substrato,
como por exemplo, a utilização da p-galactosidase para a hidrólise da
lactose na produção de laticínios destinados a pessoas com intolerância a
esse dissacarídeo.

1.2. Levedaras
Leveduras e orgarusmos leveduriformes são microfungos que
diferenciam-se dos fungos filamentosos por não produzirem esporos
assexuais ( conídias) em estruturas aéreas distintas ( conidióforos) e por
apresentarem-se predominantemente ou exclusivamente sob forma
unicelular. As células são geralmente esféricas, ovais, ou cilíndricas. Como
células simples crescem e reproduzem-se mais rapidamente que os fungos
filamentosos. Em contraste com os fungos filamentosos, o estágio sexual das
leveduras não ocorre em corpos de frutificação. Sua reprodução vegetativa
faz-se geralmente por gemulação, fissão ou gemulação-fissão que é
característico das leveduras com gemulação bipolar. Não possuem clorofila
e são portanto incapazes de suprir suas necessidades orgânicas via
fotossíntese ou quimiossíntese, devendo, portanto, viver como saprófitas,
simbiontes ou parasitas.
As leveduras são organismos ubíquos. Muitas são encontradas na
superficie das plantas onde são capazes de explorar os nutrientes
naturalmente exudados e exudados mais copiosos oriundos de lesões ou de
senescência de parte das plantas. São mais abundantes em folhas e frutas e
no néctar das flores. Outras são encontradas na superficie externa e no trato
digestivo de animais, especialmente de insetos. Poucas são patógenos de
plantas e de animais de sangue quente, incluindo o homem. Ocorrem
também com freqüência nos solos e nos corpos d'água (CarWe &
Watkinson, 1996).
Taxonomicamente não constituem um grupo definido de
microrganismos, sendo dificil fornecer uma lista de características
específicas que abranja todas as leveduras. Muitas têm ciclos de vida
complexos e algumas podem apresentar uma fase filamentosa. Possuem

s
parede celular rfg1da, de espessura quo varia de 1 SO a 400 um.em moção da
espécie e das condições da cultura. A parede é composta de glucana,
manana, proteínas, lipídeos e quitina que, dependendo da espécie, estão
presentes em diferentes proporções ou mesmo ausentes. O núcleo é bem
organizado com membrana nuclear, mas não possuem qualquer meio de
locomoção, o que as encaixa na divisão Eumycota.
A classificação taxonômica das leveduras é feita com base no seu
modo de reprodução o qual pode ser sexuado ou assexuado. A propriedade
de formar esporos sexuais dentro de ascos, ou produzí-los externamente em
basídios, colocam algumas leveduras na sub-divisão Ascomycotina e outras
na Basidiomycotina, respectivamente. As espécies para as quais não se
conhece um estágio sexual perfeito são consideradas como sendo fungos
imperfeitos e estão incluídas na sub-divisão Deuteromycotina.
Além das divergências apresentadas quanto aos ciclos de vida sexual
e assexual, as espécies de leveduras também não formam um grupo
homogêneo no que concerne ao metabolismo energético. Há espécies que
usam preferencialmente vias fennentativas enquanto outras preferem as vias
oxidativas (Gonzáles Siso et al., 1996). Apresentam uma variação no
número de cromossomos que vai de 3 a mais de 20. Essa diversidade indica
que levedura é uma estrutura morfológica que foi favoravelmente
selecionada e que apareceu recorrentemente no curso da evolução (Benitéz
et ai., 1996).
Nas leveduras classificadas na sub-divisão Ascomycotina, uma célula
diplóide sofre meiose e forma quatro a oito esporos ( ascósporos) encerrados
dentro do asco. Os ascósporos são células haplóides; células haplóides de
diferentes organismos se combinam para formar um novo organismo
diplóide. A essa sub-divisão pertencem numerosas espécies de leveduras
entre as quais encontram-se as principais leveduras de importância industrial
- --

6
como: Saccharomyces cernisiae e Saecharoa,yces.,~~~
utilizadas para a produção de etanol e de proteína unicelular, Hansenula
polymorpha e Hansenula anomala, utilizadas para a produção de triptofano,
L-alanina, L-lisina , L-valina e outros aminoácidos, e Saccharomyces
elipsoideus, utilizada para a produção de vinho.
A sub-divisão Basidiomycotina não tem fornecido leveduras de
importância industrial. Esses organismos reproduzem-se formando esporos
assexuais externos em uma estrutura chamada basídio ou em um promicélio
originado a partir de um teliósporo.
A sub-divisão Deuteromycotina inclui as leveduras que reproduzem-
se geralmente por gemulação; muitas entretanto produz.em esporos
assexualmente. Essa sub-divisão compreende umas poucas espécies
relevantes do ponto de vista industrial, dentre as quais destacam-se as
pertencentes ao gênero Cândida: C. unlis é usada na produção de ração
animal na forma de proteína unicelular, C. ltpolitica, na produção de ácido
cítrico e C. boidinii, C. tropicalis e C. gutlliermondii, na produção de
xilitol.
Organismos leveduriformes podem ser fungos superiores dimórficos
que crescem como as leveduras por meio de gemulação, fissão ou
conídiações sucessivas ou, podem ser organismos que possuem corpos de
frutificação simples mas que, por alguma razão, não são considerados como
leveduras. Algumas espécies leveduriformes são bastante úteis em processos
biotecnológicos, como por exemplo a produção de riboflavina por
Ashbya gossypii e de lipase por Geotrichum candidum.

7
u. lnvertase
A invertase (f3-frutofuranosideo frutoidrolase, E.C. 3.2.1.26) catalisa a
hidrólise da rafinose e da sacarose. A ação da invertase sobre a sacarose
resulta numa mistura equimolar de glicose e frutose - açúcar invertido
(Wiseman & Woodward, 1975). A invertase também realiza a transferência
de grupos frutosil produzindo fruto-oligossacarideos. A atividade de
transfrutosilação da invertase aumenta em concentrações de sacarose
superiores a 120 g/L (Vitolo & Yassuda, 1991).
A atividade da invertase em leveduras foi observada por Bertholet, em
1833, sendo a enzima isolada por ele em 1866. Essa enzima, desde então,
tem sido extensivamente estudada e, questões como sua localização na
célula da levedura e a permeabilidade da membrana à sacarose, foram temas
de prolongada discusão.
Wickerham ( 1958) forneceu evidências de que a maiona das
leveduras que fermentam a sacarose secreta quantidades variáveis de
invertase durante o seu crescimento. Esse autor observou ainda, que na
maioria das espécies por ele estudadas o nível de produção de invertase
extracelular é uma caracteristica da linhagem. Islam & Lampem ( 1962)
demonstraram, que a fermentação da sacarose por protoplastos de
Saccharomyces cerevistae consiste de uma secreção inicial de invertase no
meio, seguida da hidrólise da sacarose e da assimilação e fermentação das
hexoses acumuladas externamente.
A síntese da invertase por leveduras não necessita de um indutor
específico (Zimmerman & Schell, 1977) mas sofre repressão catabólica pela
presença de glicose no meio de cultura (Moreno et ai., 1979). Da mesma
forma, quando a levedura cresce em sacarose, a síntese é reprimida assim
que a concentração das hexoses produzidas pela hidrólise desse dissacarídeo

g
chega a uma ccmceatraçlo critica (Mouteneeourt e1 ai., 1973).
A partir da década de 60, ficou constatado que há diferentes formas
moleculares de invertase. Em S. cerevisiae, a espécie mais estudada quanto
a produção dessa enzima, há a produção de duas diferentes formas de
invertase: uma que é glicosilada e que está localizada externamente à
membrana celular, retida no espaço periplasmático e em associação com a
parede celular, e outra que não é glicosilada e que permanece no citoplasma
(Zárate & Belda, 1996). A forma extracelular é uma glicoproteína com
massa molecular de 270 kDa que contém 50% de fosfomanana e 3% de
glicosamina. A forma interna é livre de carboidratos, corresponde à metade
da enzima extracelular e tem massa molecular de 135 kDa. Como a
membrana celular é impermeável à sacarose, somente a invertase externa
está disponível para a reação esse dissacarídeo (Grossmann & Zimmermann,
1979~ Mizunaga et ai., 1981).
A invertase externa nativa é oligomérica ocorrendo principalmente
como dímero, tetrâmero e octâmero (Zech & Gõrish, 1995). Para ser
excretada, a invertase é inserida co-traducionalmente no retículo
endoplasmático, onde é glicosilada, assume a estrutura quartemária
oligomérica, sendo então transportada para o aparelho de Golgi, empacotada
em vesículas secretoras e, finalmente, liberada no espaço periplasmático
(Esmon et ai., 1987). A glicosilação estabiliza a enzima com respeito à
desnaturação térmica e é necessária para o arranjo em tetrâmeros e
octâmeros.
As quantidades relativas das duas formas da enzima ( externa e
intracelular) dependem das condições em que as células foram cultivadas
(Moreno et ai., 1975). A enzima externa é quantitativamente mais afetada
pela repressão por glicose, podendo estar presente em níveis baixos (menos
de l Oo/o do total) em células reprimidas, ou seja, quando cultivadas em meio

9
com concentração de glicose superior a O,OS%, ·,Oll podo.-, a ·fonna
predominante ( mais de 95% do total) em células desreprimidas (Gascón &
Ottolenghi, 1972). Segundo Elorza et ai. (1977), em células de
S. cerevisiae, a repressão da síntese da invertase pela glicose é exercida em
três níveis: em nível da transcrição, em nível da tradução e em nível da
estabilidade do RNA mensageiro. A presença de glicose não tem efeito
sobre os processos de glicosilação e secreção e não interfere na atividade
catalítica da enzima.
As isoenzimas não aparentam ser interconversíveis e diferem na
cinética da sua síntese durante o ciclo celular e na composição de
aminoácidos (Perlman & Halvorson, 1981). Porém, de acordo com
Ottolenghi (1972), essa diferença na composição de aminoácidos
aparentemente não tem significância para a função da enzima em nível
molecular.
Pela análise genética de S. cerevisiae foi estabelecida a existência de
uma família de seis genes estruturais, não alélicos, denominados SUC, que
codificam diferentes isoenzirnas. Qualquer um desses genes leva à produção
das duas formas de invertase e ao consumo de sacarose e rafinose
(Rodriguez et ai., 1981). Naumov et ai. (1996) analisaram 91 linhagens
diferentes de S. cerevisiae e verificaram que na maioria das linhagens um
único gene SUC2 estava presente, mas em algumas linhagens foram
encontrados dois ou três genes SUC.
O gene SUC2 de S. cerevisiae foi o mais amplamente estudado
( Carlson et ai., 1983; Perlman et al., 1984; Sarokin & Carlson, 1985;
Perlman et al.; 1986; Sarokin & Carlson, 1986). Codifica as duas formas da
enzima sintetizadas a partir de códons de iniciação diferentes dando origem
a dois RNAs mensageiros distintos. Um, de 1,9 kb, codifica um precursor da
forma externa glicosilada, cuja produção é regulada pela repressão por

10
da proteína formada) uma seqüência responsável pela produção de um
peptídio sinal. E outro, de 1,8 kb, codifica a forma não glicosilada
intracelular que não possui o peptídeo sinal e é produzida constitutivamente
em níveis baixos, sem função fisiológica aparente (Carlson et al., 1983).
Invertases de linhagens do gênero Saccharomyces produzidas pelos
genes SUCl a SUC5 apresentaram comportamento muito similar entre si.
Todas sofrem repressão pela glicose e as constantes cinéticas para a
hidrólise da sacarose por essas enzimas parcialmente purificadas são
semelhantes entre si, porém não idênticas (Rodriguez et al., 1981). Também
não se observou diferenças discerníveis quanto ao comportamento das
formas externa e interna de invertase produzidas por qualquer um desses
genes ( Ottolenghi, 1972).
Mais recentemente, além do gênero Saccharomyces, outros gêneros
de levedura têm recebido crescente atenção quanto à produção de invertase,
devido tanto à diversidade metabólica desses gêneros quanto à possibilidade
de se realizar modificações genéticas em algumas espécies. As técnicas de
biologia molecular permitiram o rápido avanço na compreensão de muitas
leveduras pertencentes a outros gêneros que não Saccharomyces.
As propriedades da invertase de Torulaspora pretoriensis são
similares às da invertase externa de S. cerevisiae. A enzima purificada por
Oda & Tonomura ( 1994) é uma glicoproteína tetramérica cujo conteúdo de
carboidrado representa 50% da sua massa molecular total. A massa
molecular estimada para os monômeros e para a proteína nativa é de 130 e
530 kDa, respectivamente. A atividade da invertase foi detectada no espaço
periplasmático e/ou na parede celular. Portanto, a hidrólise da sacarose dá-
se externamente à membrana celular. O pH ótimo para atividade encontra-se
na faixa de 4,5-5,0 e a temperatura é de 45ºC, respectivamente. Em pH 4,5,

-----··-----------------------------:--=----- 11
os valores de Km calculados para essa enmnas com sacarose e rafiRose como substrato, foram de 48,3 e 120 mM, respectivamente.
A invertase de Candida utilis purificada por Chávez et ai. (1997) é
uma glicoproteína dimérica composta de dois monômeros idênticos com
massa molecular aparente de 150 kDa. A enzima possui atividade em
sacarose e em rafinose apresenta 64% da atividade observada em sacarose.
É estável em pH na faixa de 3 - 6, sendo que o pH ótimo para a sua
atividade é 5,5. A temperatura ótima encontrada foi de 7<fC. Os valores de
Km observados para a enzima de C. utilis tendo a sacarose como substrato
foram cerca de 400/o menores que os encontrados para a de S. cerevisiae,
indicando uma maior afinidade da invertase de C. utilis pela a sacarose.
A invertase de Schizosaccaromyces pombe é também uma
glicoproteína cujo conteúdo de carboidrato é de 67o/o. Está associada à
parede celular e contém resíduos terminais de galactose e manose. A enzima
é estável em temperaturas inferiores a 60ºC e o pH ótimo para a sua
atividade é 4,0. O valor de Km para a sacarose é 22 mM ( Zárate & Belda,
1996)
Rodriguez et ai. ( 1995) purificaram e caracterizaram a invertase de
Pichia anomaia. Esses autores verificaram que a maioria das características
da enzima analisada por eles era similar às das invertases produzidas por
outras leveduras. A invertase foi encontrada quase que exclusivamente em
associação com a parede celular. A enzima purificada era uma glicoproteína
oligomérica composta de 3 ou 4 subunidades de tamanho idêntico
(86,5 kDa). A presença de carboidrato ligado à porção N-terminal da
proteína contribui para aproximadamente 30% da massa total da molécula,
porção essa que consiste de oligossacarídeo de manose. Também, da mesma
forma que para S. cerevisiae, a síntese da invertase por P. anomala é
influenciada pela concentração da fonte de carbono no meio; concentrações

12
supenores A
decréscimo na atividade foi observado para valores de pH inferiores a 4, O e
superiores a 6,5.
Em um trabalho posterior Pérez et ai. (1996) clonaram e
seqüenciaram o gene INVI que codifica a invertase em P. anomala. A
seqüência de 5 50 aminoácidos prevista a partir da seqüência de nucleotídeos
apresentou homologia significativa com as invertases das espécies
S. cerevisiae e Schizosaccharomyces occidentalts, na relação de 37,8% e
45,6%, respectivamente. A maior semelhança foi encontrada na porção N-
terminal da proteína madura a qual apresentou várias regiões altamente
conservadas, uma das quais contendo três dos dez sítios potenciais para
glicosilação identificados na seqüência da invertase de P. anomala.
A caracterização da invertase heteróloga, obtida da expressão do gene
SUC2 de S. cerevisiae em S. pombe, forneceu informações sobre a
habilidade dessa levedura no processamento glicoproteínas heterólogas,
indicando que ela pode ser capaz de galactosilar não somente as suas
próprias glicoproteínas mas também glicoproteinas heterólogas codificadas
por genes heterólogos e introduzidos nessa levedura por transformação.
Essa habilidade pode colocar S. pombe em posição de vantagem em relação
a S. cerevisiae na expressão de glicoproteínas de eucariotos superiores com
oligossacarídeos do tipo complexo ou h.tbrido, os quais além de manose
possuem galactose e ácidos siálicos (Zárate & Belda, 1996).

13
1.4. Aplicaçio illd9Strial da â1Yei tase
A frutose é o monossacarídeo mais doce que se conhece, tem duas
vezes o poder adoçante da sacarose e, portanto, os processos para a
produção de frutose são de considerável valor comercial. A descoberta por
Bruyn & Eckensteín, em 1895, da interconversão da glicose em frutose, em
condições alcalinas, forneceu bases químicas para a produção de frutose a
partir de glicose. Porém, não houve produção industrial decorrente dessa
descoberta devido à formação de sub-produtos não metabolizáveis e
indesejáveis para o consumo humano. Uma alternativa para a indústria de
alimentos é a produção de frutose a partir da hidrólise enzimática da
sacarose utilizando para isso, a invertase de leveduras. A frutose está
disponível no mercado em solução com a glicose, como açúcar invertido,
derivado da hidrólise enzimática da sacarose de cana de açúcar ou
beterraba, como também pura e cristalizada ou como xarope.
A invertase purificada pode ser utilizada, também, na produção de
marzipan e na manufatura do recheio cremoso de bombons; em produtos
para a higiene oral para prevenção e remoção de placas dentais; na hidrólise
da rafinose para a produção de melibiose; na estocagem de tâmaras para
evitar a cristalização da sacarose; e em biossensores, na forma imobilizada,
para a determinação de sacarose (Ribeiro, 1977).
As leveduras que produzem a invertase são utilizadas in vivo na
produção de massas para pães e bolos, de álcool industrial e de bebidas
alcoólicas (Kretzschmar, 196 l ). Para a produção de proteína unicelular e de
complexos vitamínicos, as leveduras são propagadas em matéria-prima
contendo como substrato a sacarose (Crueger & Crueger, 1989).
A invertase é usada comercialmente no processo de produção de
fruto-oligossacarídeos (FOS) a partir da sacarose pela transferência de

-------·---- ------ 14
grupos iiutosil (Hayasbi et ai., 1994; Cllen,; 199S; <>a,, 1996}. Os FOS
possuem a metade do poder adoçante da sacarose e não são susceptíveis à
decomposição pelas enzimas digestivas do homem e de animais e, portanto,
são açúcares não cariogênicos. Também exibem efeito estimulante sobre o
crescimento da bifidobacteria intestinal, a qual é favorável à saúde e
nutrição humanas sendo, por isso, no Japão, correntemente usados em uma
variedade de alimentos (Cheng et ai., 1996). Dentre os usos potenciais dos
FOS está incluído seu emprego como flavorizante em iorgutes (West, 1996).
Um fator critico para a utilização de enzimas em escala industrial é a
redução de custos; as enzimas, em geral são solúveis e instáveis e podem ser
usadas apenas uma vez em um determinado processo. O desenvolvimento de
técnicas de imobilização permitiu a redução da quantidade de enzima
utilizada na conversão de substratos com consequente diminuição de custos.
A imobilização consiste na ligação, por meios fisicos ou químicos, da
molécula da enzima a um suporte inerte com retenção da atividade catalítica.
A enzima pode, dessa forma, ser usada continuamente. Diferentes materiais
têm sido estudados como suportes para imobilizar a invertase produzida por
leveduras, tais como poliestireno (Gemeiner et ai., 1995), Concanavalina A
(Mutlu et ai., 1996), Dowex (Ribeiro, 1997). Em todos os casos, obtem-se
maior estabilidade aliada à possibilidade de uso continuo e repetido da
invertase, ampliando assim o interesse para a sua aplicação industrial.

JS
1.5. Método GOD-P AP '"' . ;:,,· ·:-,· :-" ~-~ "'- ..,. ·r <\ ,. .,. J, .. - •.•• :
Os métodos colorimétricos usados para quantificar a atividade da
invertase são geralmente baseados na dosagem de açúcares redutores totais
(ART) ou da glicose liberados após a hidrólise enzimática da sacarose.
Desde fins dos anos 50, o método baseado no uso da enzima glicose
oxidase tem sido modificado e usado para a determinação de glicose no
sangue (Raabo & Terkilasen, 1960). Posteriormente, o uso desse método
que combina especificidade e simplicidade foi extendido para a
quantificação da atividade de enzimas que hberam glicose a partir de seus
substratos, como a maltase, P- galactosidase, u-amilase, P-glicosidase e
invertase.
Nesse método, na presença da enzima glicose oxidase, a glicose é
transformada em glucanolactona a qual reage com água para formar ácido
glucônico e peróxido de hidrogênio. Na presença de peroxidase, elétros são
transferidos do peróxido de hidrogênio formado para um aceptor gerando
cor, com aceptores como a o-dianisidina e o 2 ,2 '-azino-di( 3-
etilbenzotiazolina-6-sulfonato (ABTS), ou fluorescência com o ácido
hidroxifenilacético. Dependendo do aceptor de elétrons diferenres faixas de
sensibilidade são obtidas (Baker, 1991 ).
O Método GOD-P AP (Merck 8443) foi desenvolvido para a dosagem
de glicose no sangue e tem sensibilidade para a faixa de concentração de O a
200 µg/mL. Nesse caso, o peróxido de hidrogênio reage na presença de
peroxidase com a 4-aminofenazona e o 2,4-diclorofenol formando a
antipirilquinooimina. A quantidade desse corante formada é proporcional à
concentração de glicose na amostra.

16
2. OBJETIVOS:
O presente trabalho teve como objetivos:
• Avaliar os níveis de atividade de invertase em leveduras que assimilam ou
fermentam a sacarose.
• Avaliar a utilização do método GOD-PAP (Merck 8443) para a
quantificação da glicose formada a partir da hidrólise enzimática da
sacarose.
• Avaliar a influência da concentração de glicose na repressão da síntese da
invertase em diferentes linhagens de levedura.

17
3. MATERIAL E MÉTODOS
3.1. Avaliação dos níveis de produção de invertase
Para avaliar os níveis de produção de invertase em leveduras, foi
desenvolvida a metodologia que se segue:
3.1.1. Leveduras
Foram utiliz.adas linhagens procedentes da coleção de cultura de
leveduras do Instituto de Microbiologia Dr. Paulo de Goes da Universidade
Federal do Rio de Janeiro e duas linhagens de Saccharomyces cerevisiae,
ATCC 9763 e IR2 procedente do Fermentation Research Institute de
T sukuba, Japão. As linhagens, previamente identificadas, foram
selecionadas segundo a capacidade de fermentar e/ou assimilar a sacarose.
As culturas estoque foram mantidas sob refrigeração em tubos com meio
G YMP (glicose 20 g/L; extrato de levedura 5 g/L; extrato de malte 1 O g/L;
fosfato de sódio dibásico 2 g/L; ágar 20 g/L - esterilizado em autoclave a
12 1 ºC por 15 min).
3.1.2. Ativação das células
Uma porção de células das culturas estoque foi tranferida para tubos
de ensaio contendo 3 mL de meio líquido (glicose 20 g/L; extrato de
levedura 5 g/L; fosfato de sódio dibásico 2 g/L; pH 4,5 - esterilizado em
autoclave a 12lºC por 15 min). Os tubos foram incubados a 30ºC por 16 h
sob agitação de 200 rpm. Após o período de incubação, as células foram
colhidas por centrifugação, lavadas duas vezes e resuspensas em 3 mL de

18
água destilada est«Jl.
3.1.3. Obtenção de massa celular
A suspensão de células do item anterior foi vertida em um erlenmeyer
de 125 mL contendo 20 mL de meio de cultura (Y east Nitrogen Base -
Difco) 6, 7 g/L, extrato de levedura 1 O g/L, glicerol 3 g/l, - esterilizado por
vapor fluente por 15 min). Os erlenmeyers foram incubados sob agitação de
200 rpm a 30ºC por 24 h. As células foram colhidas por centrifugação,
lavadas duas vezes e resuspensas em l O mL de água destilada estéril. As
células foram usadas como ínóculo.
3.1.4. Cultivo para a produção de invertase
As leveduras foram cultivadas em quatro erlenmeyers de 250 mL
contendo 50 mL de meio (YNB 6, 7 g/L, extrato de levedura l O g/L, glicose
1 g/L - esterilizado por vapor fluente por 15 min) nos quais foi inoculado um
volume determinado da suspensão celular, de forma a se obter uma
absorbância inicial a 600 nm de O, 1 no sistema inoculado. Dois erlenmeyers
foram utilizados para avaliar os níveis de atividade da invertase nas células
após 4 e 8 h de cultivo e os outros dois, para a elaboração de curva de
crescimento celular.

9
3.1.S. Euaio euimitico
3.1.5.1. Preparo das células para a dosagem de invertase
As células foram preparadas centrifugando-se e lavando duas vezes
com água destilada estéril todo o conteúdo dos erlenmeyers. A massa celular
foi resuspensa em 4 mL de água destilada estéril. Foi feita a leitura da
absorbância dessa suspensão de células a 600 nm em espectrofotómetro
Shimadzu UV-150-02 e, o volume dela ser utilizado na reação enzimática
foi calculado de modo a fornecer no volume final da reação enzimática
(3000 µL) uma absorbância de 2 e 4 para os tempos de 4 e 8 h de
incubação, respectivamente (Rodriguez et ai., 1995).
3.1.5.2. Reação enzimática
Os ensaios foram conduzidos em tubos de centrifuga contendo
1500 µL de uma solução de sacarose 1 O mM em tampão acetato de sódio
O, 1 M, pH 5,0. Após a adição da suspensão de células, preparada como
descrito acima, completou-se o volume para 3000 µL com tampão acetato
de sódio O, 1 ~ pH 5,0.
Em seguida, os tubos foram deixados em banho termostatizado a 30ºC
por 15 min. Após esse período, para paralisar a reação enzimática, os tubos
foram aquecidos à temperatura de fervura da água (96ºC) por 2 min. Para
cada ensaio foi feito wn branco interrompendo-se a reação imediatamente
após a adição da suspensão de células. Os tubos foram centrifugados e no
sobrenadante foi dosada a concentração de glicose.
A atividade da enzima foi definida em termos de µg de AR T liberados

20
por min de reação a 30°C. A atividade normalizada foi expressa como µg de ART.min-1/00 (Chávez et al., 1997).
3.1.6. Curvas de crescimento e concentração de glicose no meio
Durante o cultivo das leveduras, foram retiradas amostras de 2 mL em
intervalos de 2 h por 1 O horas. As Células foram centrifugadas, lavadas duas
vezes em água destilada estéril e resuspensas em 2 mL de água. A leitura
das absorbâncias foi feita em duplicata, em um espectrofotômetro Shimadzu
UV-150-02 a 600 nm. As curvas foram obtidas graficando-se o logarítimo
da absorbância em função do tempo de incubação (h).
A concentração de glicose no meio foi calculada mediante a seguinte
fórmula:
% glicose= [(Gi - Gt)/Gi].100,
onde, Gi é a concentração inicial de glicose (1 g/L) e Gt é a concentração de
glicose dosada em um tempo determinado. A glicose foi dosada pelo método
GOD-PAP.
3.2. Dosagem de glicídios
Foram utilizados dois métodos para dosagem de glicídios redutores:
um, GOD-PAP, específico para dosagem de glicose e outro, de Somogy,
para dosagem de açúcares redutores totais (ART) (Somogy, 1952), ambos
com sensibilidade para a faixa de concentração de O a 200 µglmL .
•

21
Método GOD-PAP (Merc:k 8"3)
Reagentes:
1. Mistura tampão-enzima:
Tampão fosfato
Tampão Tris
pH da solução tampão
0,1 mmol/L
0,05 mmol/L
8,0
Glicose oxídase (GOD)
Peroxídase (POD)
7KU/L
1,4 KU/L
2. Corantes:
4-Aminofenazona
2,4-Diclorofenol
0,25 mmol/L
0,3 mmol/L
A solução de reação foi feita dissolvendo-se a mistura tampão-enzima
e os corantes em 500 mL de água bidestilada. Essa solução foi mantida sob
refrigeração a l OºC.
Para a dosagem de glicose, adicionou-se a 100 µL de amostra 1 mL
da solução de reação. Após 15-20 min de reação a 3 7ºC, foi feita a leitura
da absorbância a 51 O nm em um espectro fotômetro Shimadzu UV-150-02.
Foi feita uma curva de calibração utilizando-se soluções de glicose em
concentrações de l O - 200 µg/mL e, para o cálculo da concentração de
glicose, utilizou-se a equação obtida dessa curva.

3.3. Determiaaçio dos parimetros envolvidas ao aumento da
absorbância durante a dosagem da glkose pelo método GOD-P AP
3.3.1. Medida do pH das soluções de glicídios adicionadas da
solução de reação do método GOD-P AP
Foram medidos o pH da mistura da solução de reação do método
GOD-P AP com as soluções de glicose 40 ug/ml, e de sacarose 1 O mM
(3 420 µg/mL) em tampão acetato de sódio 0,1 M, pH 5,0 utilizando-se
potenciômetro Micronal 83 7 4.
Um volume de 20 mL das soluções dos glicídios foi diluído em 20 mL
de tampão acetato 0,1 mM, pH 5,0 antes da medida ser efetuada, simulando-
se, dessa forma, a diluição e o pH da dosagem de glicose após o ensaio
enzimático.
3.3.2. Influência do aquecimento a temperatura de fervura da
água na inversão da sacarose
A 5 mL das soluções de sacarose 1 O, 20 e 40 mM acrescentou-se
5 mL de tampão acetato O, 1 mM, pH 5,0 ou em água destilada, quando
indicado, e, em seguida, colocou-se l ml de cada diluição em três tubos de
ensaio. O primeiro não foi aquecido, o segundo foi aquecido por 1 min à
temperatura de fervura da água (96°C), e o último foi aquecido à mesma
temperatura por 2 min, Após o aquecimento, foi feita pelo método
GOD-PAP a dosagem da glicose formada.

23
3.3.3. Efeito da sohaçle de reaçio do método GOD-P AP sobre
diferentes glicídios
Foram feitas soluções 40 µg/mL de x.ilose, frutose, trealose, rafinose e
celobiose. Foi efetuada a reação, em triplicara, destas soluções de glicídios
com o reagente GOD-PAP seguindo-se a metodologia sugerida pela Merck
para a dosagem de glicose por meio do reagente em questão.
3.3.4. Efeito das enzimas do método GOD-P AP sobre a sacarose
Foi feita a reação de l 00 µL das soluções de sacarose l O mM e de
glicose 40 µg/m.L com l mL da mistura tampão-enzima do método
GOD-PAP. Após 15 min de incubação a 37ºC, a dosagem de ART formados
foi efetuada pelo método de Somogy.
3.3.5. Cinética da reação de hidrólise da sacarose pelo reagente
GOD-PAP
Foi feita uma diluição de 1 mL da solução de sacarose 1 O mM em
1 mL de tampão acetato 0,1 M, pH 5,0. A cinética da reação dessa solução
com o reagente GOD-P AP) foi acompanhada em um espectrofotômetro
Hitachi U 200 l por 2 h.
O tempo zero da reação correspondeu a 17 min após a adição do
reagente GOD-P AP. Sendo desse tempo, 15 min correspondentes à
incubação a 37ºC, conforme recomendação da Merck, e 2 min
correspondentes ao tempo de ajustar o aparelho às condições desejadas.
Foi feito o gráfico da concentração de glicose em função do tempo de

reação.
3.3.6. Determinação da variação da absorbância para diversas
concentrações de glicose na presença de sacarose
Foram feitas diluições de l mL de soluções de glicose 10, 20, 40, 80,
120, 140, 200, 240, 280, 320, e 360 µg/ml em l mL de sacarose 10 mM. A
cinética da hidrólise de cada uma das soluções de sacarose pela solução de
reação do método GOD-PAP foi acompanhada por 30 min em um
espectrofotômetro Hitachi U-2001. Os ensaios foram feitos em triplicata.
Como no item anterior, o tempo zero das reações correspondeu a
17 min após a adição do reagente GOD-PAP.

4. RESULTADOS E DISCUSSÃO
A determinação in vivo da invertase periplasmática de leveduras é de
grande interesse, pois, o emprego dessa enzima na indústria alimentícia, de
bebidas alcoólicas e do álcool industrial dá-se pela utilização de células
vivas (Silveira et ai., I 996).
Os métodos descritos para a determinação da atividade da invertase
periplasmática são baseados na medida da glicose ou dos açúcares
redutores totais (ART) formados pela hidrólise enzimática da sacarose sob
condições pré-estabelecidas de pH, temperatura, massa celular e tempo de
reação.
A literatura cita amplamente a utilização da enzima glicose oxidase
para a dosagem da glicose, na forma de "kits" de reagentes ou da
elaboração dos reagentes em laboratório e por meio de biossensores
(Cheetham, I 995).
Não obstante, no presente trabalho, a metodologia proposta para
avaliar os níveis de produção de invertase mostrou-se inadequada na etapa
da quantificação (pelo método GOD-P AP) da glicose formada pela ação
dessa enzima numa solução contendo sacarose 1 O mM em tampão acetato
0,1 M, pH 5,0. Por essa metodologia, não foi possível determinar a
quantidade de invertase produzida pelas leveduras ou mesmo determinar se
houve ou não síntese da enzima, pois as dosagens de glicose em solução
contendo sacarose forneciam, em todas as determinações, leituras de
absorbância crescentes.
Vitolo & Borzani (1983) verificaram que a interrupção da reação
enzimática em um dado momento é um ponto crítico na determinação da
atividade da invertase em células intactas de leveduras; a parede celular
protege a enzima e impede a rápida inativação da invertase.

26
As leituras de absorbância crescentes no entanto somente eram obtidas quando a glicose formada durante a reação enzimática era dosada
pelo método GOD-PAP. Quando a determinação dos ART era feita pelo
método de Somogy, esse resultado não se repetia. Além disso, após a
reação enzimática, as células eram separadas do meio de reação por
centrifugação. Dessa forma, mesmo que a reação enzimática não tivesse
sido o interrompida pelo aquecimento, no momento da dosagem dos
açúcares não havia invertase presente no meio de reação.
Investigou-se o que poderia estar afetando a adequação do método
GOD-PAP para a dosagem de glicose nas condições experimentais
estabelecidas para o presente trabalho.
4.1. Identificação das variáveis relacionadas à inadequação do método
GOD-P AP para dosagem da glicose em mistura com sacarose
O aumento das leituras de absorbância nas dosagens da glicose
formada após os ensaios enzimáticos poderia estar relacionado à
contaminação da sacarose com glicose ou com a hidrólise da sacarose. A
hidrólise da sacarose poderia estar ocorrendo em função do pH da reação
enzimática, do aquecimento à temperatura de fervura da água (96°C) para a
interrupção dessa reação ou da combinação desses dois fatores. Outra
hipótese seria a inversão química ou enzimática da sacarose pelos
componentes da solução de reação do reagente GOD-PAP.

-····---------
4. 1.1. hdluâlda do pH
A sacarose é facilmente hidrolisada em meio ácido. O açúcar
invertido industrial é obtido por hidrólise ácida em pH 3,5, e o mel de
abelha, que é um açúcar invertido natural, tem pH 4,0 (Alais & Linden,
1991).
A solução de reação do método GOD-PAP tem pH 8,0. Porém, como
nos ensaios enzimáticos foi utilizado tampão acetato 0,1 M, pH 5,0, e como
a reação da glicose com a enzima glicose oxidase resulta na formação de
ácido glucônico, pensou-se na possibilidade de desvio do pH para a faixa
ácida, o que poderia resultar em uma hidrólise ácida da sacarose. Foi, então,
feita a determinação do pH da mistura.
Os valores determinados de pH na faixa de 6,0-7,7 não sugerem uma
hidrólise ácida da sacarose (Tabela 1). A formação de ácido glucônico não
altera significativamente o pH do sistema de reação. O desvio do pH de 8,0
para 6,0, nos ensaios com tampão acetato O, 1 M, pH 5,0 e sacarose, devem
ser atribuídos a influência do pH do tampão, uma vez que a solução de
sacarose foi feita em tampão acetato O, 1 M, pH 5,0.
Tabela 1: Valores do pH das soluções de glicídios adicionadas à solução
de reação do método GOD-PAP.
Soluções pH
Glicose+ Solução de reação
Sacarose + Solução de reação Tampão acetato + Solução de reação
7,67
6,03
6,00

--------·· ------
4.1.2. Influência do aquecimento da ~o de sacarose
Esses experimentos foram realizados com o objetivo de verificar se a
sacarose utilizada na reação enzimática, estava sendo hidrolisada com o
aquecimento por 2 min a 96ºC, utilizado para interromper a reação
enzimática.
Na Tabela 2, as leituras apresentadas para os tempos l e 2 min de
aquecimento e sem aquecimento da reação enzimática correspondem à
primeira leitura obtida após a adição do reagente GOD-P AP seguido de
incubação por 15-20 mina 37°C, conforme recomendação da Merck. Após
esse período, no entanto, esses valores aumentavam progressivamente
sendo impossível determinar qual o real valor da concentração de glicose
presente na solução.
Por esse experimento, verificou-se que o aquecimento a 96ºC por
2 min não é responsável pela hidrólise da sacarose. A obtenção de
concentrações de glicose em torno de 170 ug/ml, com e sem aquecimento, a
princípio indicava que a sacarose estivesse contaminada com glicose e que
essa reagia lentamente com a glicose oxidase fornecendo, dessa forma,
valores sempre crescentes de leitura de absorbância. Em vista disto, optou-
se pelo uso de uma solução de sacarose mais diluída que, nesse caso, daria
menor interferência nos ensaios enzimáticos. Para verificar esta hipótese, o
mesmo ensaio foi repetido com soluções de sacarose 1 O e 20 mM.

Tabela 2: JnfWncia do tempo de aqueci•nento a 96°C da solução de
sacarose 40 mM sobre o aumento da concentração de glicose.
TempG de aquecimento Leitura da absorbância Concentração de glicose
(min) (510 mn) (µg/mL)
2 0,553 170,58
l 0,548 168,97
sem aquecimento 0,549 169,29
A Tabela 3, apresenta as leituras iniciais de absorbância das soluções
10 e 20 mM com e sem aquecimento. Obteve-se resultado idêntico ao
anterior: leituras que aumentavam progressivamente sem que houvesse
influência do tempo de aquecimento das soluções a 96°C.
Tabela 3: Influência do tempo de aquecimento a 96ºC das soluções de
sacarose 1 O e 20 mM sobre o aumento da concentração de
glicose.
Concentração da Tempo de Leitura da Concentraçâo
solução de sacarose aquecimento absorbância de glicose
(mM) (min) (510 nm) (µg/mL)
2 0,108 27,03
10 1 0,102 25,10
sem aquecimento 0,106 26,39
2 0,198 56,06
20 1 0,204 58,00
sem aquecimento 0,201 57,03

30
Como a reaç.ão eomnática era realizada em tampão acdato de sódio
O, 1 M, pH 5,0, para esses ensaios as soluções de sacarose foram diluídas
nesse tampão, simulando, assim, as condições da reação enzimática. Para
eliminar a possibilidade de que a combinação desse valor de pH com a
temperatura de 96ºC promovesse a hidrólise da sacarose, esse ensaio foi
repetido com a solução de sacarose 1 O mM, substituindo-se na diluição o
tampão por água destilada (Tabela 4 ).
Tabela 4: Influência do tempo de aquecimento a 96ºC da solução de
sacarose 1 O mM diluída em água destilada sobre o aumento
da concentração de glicose.
Tempo de aquecimento Leitura da absorbância Concentração de glicose
(min) (510nm) (µglmL)
2 0,097 23,48
1 0,091 21,55
sem aquecimento 0,093 22,19
Os resultados obtidos foram semelhantes aos dos ensaios realizados
com o tampão acetato 0,1 M, pH 5,0. Permanecia, portanto, a conclusão de
que a presença de glicose em concentrações em tomo de 22 µg/mL, era
devido à contaminação da sacarose por glicose.
Na tentativa de se encontrar uma sacarose que não fosse contaminada
ou que fosse menos contaminada por glicose, foram testadas outras quatro
amostras de sacarose de fabricantes diferentes, sendo que de um fabricante
foram utilizadas amostras de 2 lotes. Foram então preparadas soluções

31
1 O mM dessas amostras de sacarose e o ensaio foi repetido fa7.endo.se as
diluições em tampão acetato O, 1 M, pH 5,0 (Tabela 5).
Tabela 5: Influência do tempo de aquecimento a 96ºC de soluções de
sacarose I O mM sobre o awnento da concentração de glicose.
Tempo de Leitura da Concentração
Amostra aquecimento absorbância de glicose
(mio) (510nm) (µg/mL)
2 0,094 22,52
I 1 0,085 19,61
sem aquecimento 0,094 22,52
2 0,131 34,45
2 1 0,120 30,90
sem aquecimento 0,103 25,42
2 O, 107 26,71
3 1 0,091 21,55
sem aquecimento O, 110 27,68
2 0,059 11,23
4 l 0,062 12, 19
sem aquecimento 0,061 11,87
Para as quatro amostras de sacarose obteve-se, quanto ao aumento
das leituras de absorbância, um resultado idêntico aos dos ensaios
anteriores.

32
4.1.3. Reaçio do método GOD-P AP cem difeftlltes gliddiGs
Para explicar os aumentos observados nas leituras de absorbância,
levantou-se a hipótese de que se a sacarose estivesse contaminada com
glicose e frutose livres, a glicose oxidase estaria reagindo inicialmente com
a glicose e em seguida com a frutose que, de alguma forma, estaria sendo
convertida em uma aldose. Por outro lado, a ligação glicosídica da molécula
de sacarose ( do tipo J3- l ,2) poderia estar sendo rompida pelas enzimas ou
reagentes cromógenos presentes na solução de reação do método
GOD-PAP, hberando desta forma os monômeros de glicose e frutose.
Se assim fosse, a especificidade do reagente seria duvidosa, podendo
haver reação com outros glicídios que não a glicose. E, em misturas de
açúcares, após reação com a solução de reação do método GOD-P AP, a
formação de antipirilquinonimina não corresponderia necessariamente à
quantidade de glicose. As informações obtidas no presente trabalho até
então, indicavam que pelo método GOD-P AP obtinha-se uma leitura de
absorbância pontual para dosagem de glicose e crescente para sacarose.
Para verificar a possibilidade da solução de reação do método
GOD-P AP reagir com outros glicídios e, no caso de estar havendo hidrólise
da sacarose, assim como para conferir se havia especificidade para um
determinado tipo de ligação, foi elaborado um ensaio utilizando-se soluções
40 mM de xilose ( ai dose), frutose ( cetose ), trealose ( dímero de glicose,
ligação n-I, 1 ), celobiose ( dímero de glicose, ligação J3- l, 4) e rafinose
(trissacarídeo composto de galactose, glicose e frutose, ligações a-1,6 e
J3- l ,2).
Nenhum dos glicídios testados reagiu com a solução de reação do
método GOD-PAP (Tabela 6). Não se obteve leitura de absorbância com
xilose, frutose e rafinose. As leituras de absorbância obtidas com a

33
celobiose e com a trealose foram coostaotes e correspondem a uma
concentração de glicose de cerca de 2 µg/mL, podendo ser atribuídas a
traços de glicose presentes na solução desses dissacarídeos ou, como esses
valores estão fora da curva de calibração, podem ser considerados sem
significado. Com esses resultados concluiu-se que os reagentes e/ou
enzimas presentes na solução de reação do método GOD-PAP não eram
capazes de reagir com outros açúcares ou de converter frutose em glicose.
Permanecia, no entanto, a dúvida quanto ao porquê da reação contínua com
a sacarose; se esse efeito era ocasionado pela hidrólise desse dissacarídeo
seguida de reação com a glicose formada ou pela reação direta da enzima
glicose oxidase com a sacarose.
Tabela 6: Reação de xilose, frutose, trealose, celobiose e rafinose
40 µg/mL com a solução de reação do método GOD-PAP. .. - -
Glicídio Leitura da absorbância Concentração de glicose
(510nm) (µg/mL)
Xilose 0,000 0,00
Glicose O, 157 40,00
Frutose 0,000 0,00
Celobiose 0,009 2,19
Trealose 0,008 2,04
Rafinose 0,000 0,00
Como a enzima glicose oxidase é específica para a D-glicose,
reagindo com o carbono 1 da molécula e convertendo-a em ácido glucônico

e na molécula de sacarose é este carbono o que está ligado à fiutose, a hipótese de reação direta com a sacarose era, portanto, menos provável.
Concluiu-se que havia de fato hidrólise da sacarose e que essa, uma
vez que não era causada pelo aquecimento a 96ºC e/ou pela alteração do pH
da reação enzimática, deveria ser promovida por alguma substância ( outra
enzima ou reagente) presente na solução de reação do método GOD-P AP.
Pela mesma razão acima exposta, se a hidrólise fosse enzimática, não
seria realizada pela glicose oxidase. A outra enzima que está presente nesse
reagente é a mutarotase bacteriana, que acelera a interconverção de
açúcares anoméricos e que também não causaria esta hidrólise. Restava a
possibilidade de que a hidrólise fosse química e ocasionada pelos outros
reagentes.
Para eliminar definitivamente a possibilidade de uma hidrólise
enzimática da sacarose, foi feito um ensaio com as enzimas do método
GOD-PAP livres dos reagentes cromógenos. A dosagem de ART formados
foi efetuada pelo método químico colorimétrico de Somogy.
Não ocorreu inversão da solução de sacarose pela enzima glicose
oxidase (Tabela 7), como evidenciado pela ausência de ART. Dessa forma,
concluiu-se que as enzimas da solução de reação do método GOD-PAP não
invertem a sacarose.
33
Tabela 7: Efeito da enzima glicose oxidase sobre a solução de sacarose lOmM.
Glicídio Concentração Leitura da absorbância
(µg/mL) (510nm)
Sacarose 3 420
40
0,0
0,105 Glicose
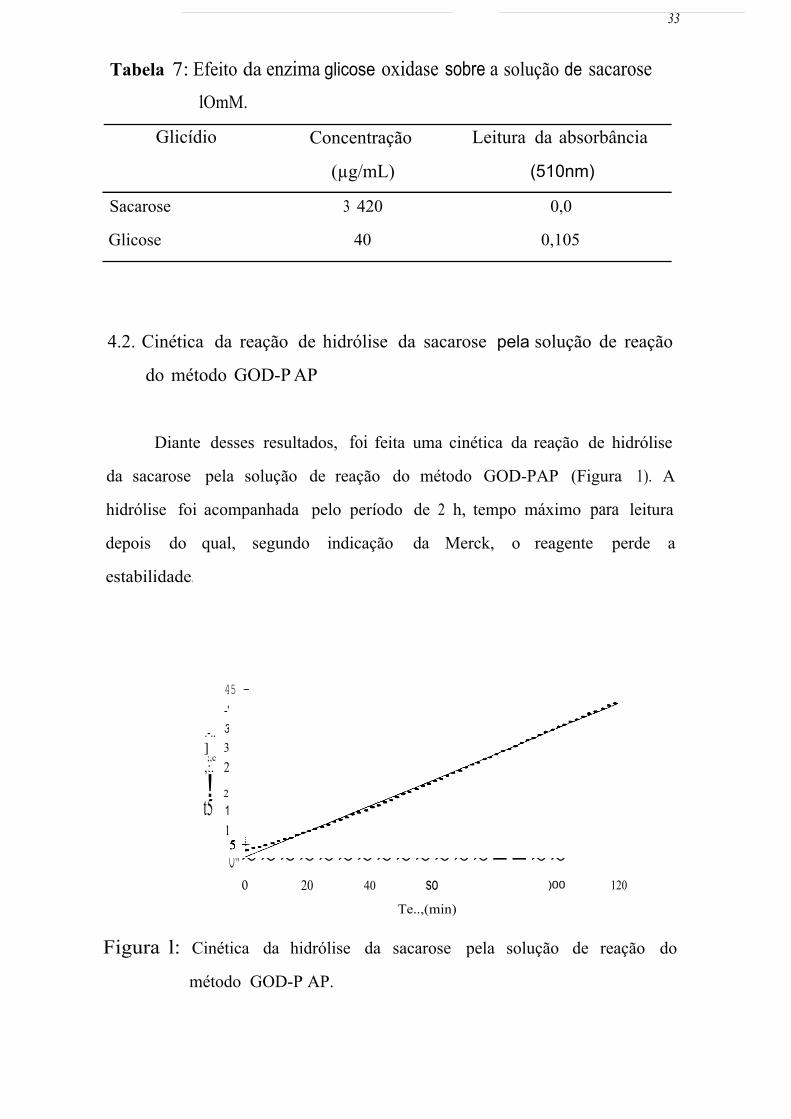
4.2. Cinética da reação de hidrólise da sacarose pela solução de reação
do método GOD-P AP
Diante desses resultados, foi feita uma cinética da reação de hidrólise
da sacarose pela solução de reação do método GOD-PAP (Figura 1). A
hidrólise foi acompanhada pelo período de 2 h, tempo máximo para leitura
depois do qual, segundo indicação da Merck, o reagente perde a
estabilidade.
45 - -'O T
35-:- .-.. . ] 30 -L- ;,c ,:. 25 -f-
! 2() t t5 15 t
10 + y =0,3272x+ l,8189 R; =0,9982
o~•~~~~~~~~~~~~~--~~ o 20 40 so
Te..,(min)
)00 120
Figura l: Cinética da hidrólise da sacarose pela solução de reação do
método GOD-P AP.

------------------------ 36
Para confirmar que a reação observada era provocada pela hidrólise
da sacarose e não pela presença de AR T contaminantes, foi determinada,
pelo método de Somogy, a concentração de ART presente na solução de
sacarose l O mM utilizada no experimento descrito acima.
Pelo resultados apresentados nas Tabelas 7 e 8, concluiu-se que a
solução de sacarose não possui glicose e frutose livres. Por isso, as leituras
obtidas pelo método GOD-PAP não podem ser atnbuídas à contaminação
da sacarose pelos monossacarídeos que a compõem mas sim, à hidrólise
química da sacarose. Portanto, em primeira análise, não é possível a
utilização do método GOD-P AP para a dosagem de glicose na presença de
sacarose. Portanto, foi feita uma proposta de adaptação do método.
Tabela 8: Concentração de açúcares redutores totais da solução de
sacarose l O mM_
Glicídio Concentração
(µg/mL)
Leitura da absorbância
(510nm)
Sacarose
Glicose
3 420
40
0,0
0,106

- ----------- 31
4.3. Adaptaçio do método
4.3.1. Determinação da variação da absorbincia para diversas
concentrações de glicose na presença de sacarose
A Tabela 9, apresenta a variação das concentrações das soluções de
glicose em presença de sacarose. Nessa tabela, o tempo zero de leitura
corresponde a 17 min após a adição da solução de reação ( 15 min de
incubação a 3 7ºC recomendados pela Merck e mais 2 min consumidos para
ajustar o espectrofotômetro ). Não foi observada influência da concentração
inicial de glicose livre na hidrólise da sacarose. Durante os 30 min de
leitura, a cinética da reação de hidrólise da sacarose pela solução de reação
do método GOD-PAP pôde ser descrita por um modelo linear em todos os
casos. Para verificar o ajuste desses modelos foram feitas tabelas de análise
de variância (ANOVA). Os gráficos com as equações das retas e as tabelas
de ANOVA estão apresentadas em apêndice.

-···· ·--· ···--····· ······ ----·
• ~ e,
1 o o o o
"O o - ~ e o ü Q. cn o
"O
~ = = ·- e o u - - ~ u
"O o ~ ta o o () ~
~ e u u
"O ~ o "O = , ... ~ = - .... tJ "O
= 1 u () = o o o Q) 00
"O cn = u V) 1- o Q) ca "O > IJl 'Jl Q)
o IO '-> "O ee = t:
1-o e ::1 Q) - ':.) 1.J e: o u e»
"O cri cu o e: Q.
~ e u o - u ·- o "5o "O o Q)
"O ~ cn o- ] ~ g- E - o Q) cn o u
K,:$ "O ()o 1- cu -~ -~ > Q. .. ~ = - ~ .e = E--
i -
s -
-lf'l-OOQ\NO'\O'l'CM'CQ\'CM'C lf'lNOOMOONl"-Olf'lQ\Ml"--lt'lQ\..-, :,: ô o· -· ..: M M .....,· M .....,· • ~ vi vi vi '° 0- ...... ------------ ....... -- ----- ........................ _.._
~~OO'Cin-•1"-0~MOMl"-01"-
8 O,-... N_OO.M_ -O.-_ .o. •.oo.N.l"-.O • • • ·gg--NNMMM .... ~ot'I - I"- 00 °' °' O O O O O O O O O °' e- 0"-0\_.. .............. _ ............................... - (ll
~ ~ e (,:!
Õb () N Q\ -o O'l'CNO'I N O\ -e ..... oi .... - ·- 1 - o "' ..... ~ "' O'\ M O\ N ,..._ º· .... 00 N °' ..... Q. - ,;ti ~ ô 0 .. _;,.... .. N ... ,....;- ..... ~ .... - ~ ~ X) N M ,r, ·e: e, ,... ,... ac:i 00 00 00 X) :ic 00 00 X) X) 00 X) ee X) - W}
o u ~ "O e 00
(,:!
N .... ..... °' 00 - 00 "' X) - X) ...., O' N ,r, ;.a s 00 ""· °' •. °' ..... °'· N ~ o ,r, Xl. N. '°· - ,r, -V
~ - - - N N ...... ,.... -i -i ..,. v-. ..,..., -6 -o E .,., -c -c -c se -e -c -c -o -o -c -c -o -..e -c -o E ~ - - ;3 o:
~ "'· or. _, ...... ...... ...,. r-- '° M ...... r::: ...... ,.:; ...... ,.... V r-- N ...... r-: ...... - or. ~ ..,. X) N - ,r, ~ ...,. 1-o
;;: .... ;;: ;;: N N ,...; r-. r-: ..,. ..,. .,.., "'· .,., ::;: a. ,..... .,,. ..,. ..,. ..,. ..,. ..,. ..,. ..,. ..,. ..,. ..,. ..,. ~ ... .E= V
.D (,:! .... .... ("') N -o - .... 00 r-- o o ,r, NN-0 - o OV'l º· ..., °' .,., °' ..... X) ...., 00 ,...., t'-N !"- N (,:!
N ,...;,...; .... ,..., ..... • ~vi ,r, -.() -6 ........ ........ ,;ti oô °' 00 N N N N N N N N N N N N N N N N rJl
V e (ll o
~-~-~~- ...... \O Q\ N °' °' '° V'> N V"l °' '° ] o ...... ,... - '° °' .... 00 ...... ,... - V") °' ~ ~...,·vi vi '6 '6 ................ ,;ti ,;ti e
N M ("">t"'l 00 u - - - - - - - - - - - - - - - - r:.n u 1- Q. (,:!
Vl
N ...... 00 NI"- N M .... 00 °' o V") 8.~ - N ~ ,..._ '°· °' V") ,... o N .,., ,... OM '° ~ - "' o ........ oô oô oô oô e,.."~ e,,." . ....., ....., o - - - ..... °' _, - ~ - - - - - - - >
(ll o º- rJl
e. = o N ..,. '° 00 N .... '° 00 o .D e·- o N .... '° 00 N o e- e - - - - - N N N N .... E-- -

39
A primeira leitura de absorbincia das soluções S, l O e 20 P!VJDl correspondem, respectivamente a concentrações de glicose 54,44, 26,60, e
10,21 % superiores à concentração esperada. Na faixa entre 40 e 100 µg/ml,
a primeira leitura de absorbância fornece valores de concentração de glicose
muito próximos aos que estavam presentes na solução. E, apesar de
prosseguir aumentando, essas leituras fornecem concentrações equivalentes
às esperadas nos 4 primeiros minutos para as concentrações de 40 e
60 µg/ml, entre 2 e 8 min para a concentração de 80 µgim] e entre 4 e
1 O min para a concentração de 100 µg/ml. Na faixa de 120 a 180 µg/ml, as
leituras iniciais fornecem concentrações de glicose de 9 a 13% menores que
as concentrações reais sendo que, a última leitura, efetuada 30 min após a
primeira, corresponde a concentrações de 3 a 9,32% menores que as
esperadas.
Para a utilização do método GOD-P AP para a dosagem de glicose em
presença de sacarose, se faz necessário observar duas condições: que a
concentração de glicose esteja na faixa entre 40 e 100 µg/ml e que o tempo
decorrido entre o momento de adição da solução de reação e a leitura da
absorbância seja rigorosamente o mesmo para todas as leituras. Conforme
os resultados apresentados, a leitura da absorbància deve ser feita
preferencialmente 21 min após a adição do reagente.
A concentração de glicose presente no sistema de reação ao final do
ensaio enzimático esteve abaixo da faixa de calibração de 40-100 µg/ml em
todos os ensaios preliminares cujas concentrações de AR T foram
determinadas pelo método de Somogy. Por isso, optou-se por dar
seguimento ao trabalho utilizando-se apenas o método de Somogy para a
dosagem de AR T.

40
4.4. Produçlo de invenue
Os meios usados para o cultivo de microrganismos devem conter
todos os elementos mnna forma apropriada para a síntese das substâncias
celulares e para a produção de produtos metabólicos.
O meio Y east Nitrogen Base (YNB) foi desenvolvido por Wickerham
( 1958). Contém vários elementos traço, 9 vitaminas, 3 aminoácidos -
histídina, metionina e triptofano, e os principais sais minerais - fosfato de
potássio, sulfato de magnésio, cloreto de sódio e cloreto de cálcio. A fonte
de nitrogênio é o sulfato de amônio, que pode ser usado pela maioria das
leveduras conhecidas. A fonte de carbono deve ser acrescentada (Phaff et
al., 1978).
Em ensaios preliminares, a levedura Saccharomyces cerevisiae
(50613) foi cultivada em YNB acrescido de 0,1% de glicose e, como
resultado, não se obteve crescimento celular. Acrescentou-se ao meio O, lo/o
de extrato de levedura, tendo sido então obtido crescimento celular.
Resultados idênticos foram obtidos para seleção de leveduras produtoras de
xilitol (Barbosa et ai., 1988). Os extratos de levedura contém 18
aminoácidos os quais podem estimular o crescimento de certas leveduras
fastidiosas.
A fonte de carbono a ser uti1izada foi escolhida mediante comparação
do cultivo dessa mesma levedura em YNB com 0,1 % de extrato de levedura
e 0,1 % de glicídio na forma de sacarose ou glicose. Como o crescimento
celular em sacarose foi escasso, concluiu-se que as linhagens que
apresentassem baixa atividade de invertase apresentariam pouco
crescimento em meio contendo sacarose.
Além disso, Martínez-Pastor & Estruch (1996) observaram que
quando as células de levedura foram inoculadas em um meio, visando a

.. ----------------··-·----··------- -------- 41
desrepressão da invertase, cuja fonte de cmbono alo era a glicose, a expressão dessa enzima foi retardada. A atividade da invertase foi detectada
nas primeiras horas de cultivo somente em meio contendo glicose como
fonte de carbono. Segundo os mesmos autores, a ausência de atividade da
invertase nessa condição não representa uma falha na transcrição dos genes
reprimíveis pela glicose, e sim uma completa inibição da tradução na
ausência de fontes de carbono rapidamente utilizáveis, como a glicose. A
capacidade de tradução só é retomada após várias horas de incubação.
Portanto, quando a indução transcricional é medida por meio da
atividade enzimática, há que se considerar as condições de cultivo
favoráveis à tradução, como a presença de glicose, uma vez que a ausência
de atividade enzimática não é sempre sinônimo de ausência de trancrição.
A principal condição para a obtenção de leveduras ricas em invertase
é o cultivo em meio contendo glicose em concentração inferior a 0,5 g/L
(Ribeiro, 1997). Essa condição foi considerada como critério para
desrepressão da síntese da invertase no presente trabalho.
Diante do exposto, foi proposto cultivar as leveduras por 1 O h em
meio contendo concentração inicial de l g/L de glicose, monitorar a
concentração de glicose no meio e fazer duas determinações da atividade da
invertase, com 4 e 8 h de cultivo.
Na Tabela l O estão apresentadas as atividades e as atividades
normaliz.adas da invertase em diferentes leveduras.

·-·---···-----·-·· --····-··--·--------- ·'2
Tabela 10: Níveis de atividade da invertase em diferentes leveduras.
Atividade da lnvertase
LEVEDURAS ug.ml." µg.mL-1/00
4h 8b 4h 8b
S. cerevisiae (IR2) 1,00 2,00 0,50 0,50
S. cerevisiae (ATCC 9763) 1,06 2,34 0,53 0,58
S. cerevisiae ( 50613) 0,00 0,00 0,00 OJ>O
S. kluyve,y(50629) 1,15 1,73 0,57 0,43
Zygossacharomyces sp (50965) 0,22 0,22 0,11 0,05
P. becki ( 50808) 0,00 0,00 0,00 0,00
P. carsoni(50809) 0,22 0,36 0,11 0,09
P. ohmeri (50537) 0,00 0,00 0,00 0,00
W califomtca (50420) 0,00 0,57 0,00 0,14
D. hanseni (50059) 0,00 0,00 0,00 0,00 ..
Debaryomyces sp (50385) 0,08 0,04 0,04 0,01 ·-
R. rubra (50596) 0,00 0,00 0,00 0,00
T pretoriensis (50811) 0,00 0,47 0,00 0,12
e. macerans ( 50361) 4,94 4,85 2,47 2,21
C. ater (51032) 1,28 l, 70 0,64 0,42
A. pululans (51102) 0,08 0,00 0,04 0,00
Considerando-se os resultados obtidos nas condições experimentais,
as leveduras estudadas podem ser agrupadas, quanto à síntese de invertase
nos tempos de 4 e 8 h de cultivo, como se segue:
• não produziram a enzima - D. hanseni (50059), R. rubra (50596),
S. cerevisiae (50613), P. becki (50808) e P. ohmeri (50537)~
• produziu somente no tempo 4 h -A. pululans (51102)

43
• produzi•am somae no tempo 8 h - W. colifomica (S0420) e T. pretoriensis ( 50811);
• produziram a enzima nos doi.s tempos - C. ater (51032),
C. macerans (50361), Zygossacharomyces sp (50965), P. carsoni
(50809), S. cerevisiae ATCC 9763, S. cerevisiae IR2 e Debaryomyces
sp (50385), S. kluyvery (50629).
As leveduras procedentes da coleção de culturas do Instituto de
Microbiologia da UFRJ, utili~das no presente trabalho, foram escolhidas
conforme o critério de assimilar ou fermentar a sacarose, como evidenciado
nos testes bioquímicos utilizados nos trabalhos de identificação dessas
leveduras. A ausência de atividade da invertase em algmnas linhagens
(50059, 50537, 50596, 50613, 50808) deve refletir mais a dificuldade de se
determinar a atividade da invertase em células vivas do que a incapacidade
dessas leveduras produzirem a enzima.
A quantificação da atividade das enzimas do espaço periplasmático
em suspensão de células de leveduras, está sujeita a erros quantitativos por
uma variedade de razões. De maior importância é a relação entre a
velocidade de formação do produto pela enzima que está sendo monitorada
e a velocidade dos processos que dissipam o produto. Se a velocidade de
assimilação, for relativamente alta a concentração do produto pode ser
subestimada ou não quantificada (Arnold, 1991).
Silveira et ai. ( 1996) não detectaram atividade da invertase em
células vivas de S. cerevisiae seguindo as metodologias descritas na
literatura para o ensaio enzimático e atribuíram o fato ao consumo de
glicose e frutose produzidas pela ação da enzima. Os níveis reais de
atividade somente foram evidenciados após inibição da enolase ( enzima da

-------- ---- --· ----- - - ---------------------- - ---- -- -- :.w---··-·-· ···-
via glicolítica que converte o 2-fosfoglicerato ·em fosfoenolpiruvato) durante
o ensaio enzimático, pela adição de fluoreto de sódio 50 mM.
4.5. Crescimento celular e concentração de glicose no meio de cultivo
A concentração inicial de glicose presente no meio de cultivo para a
síntese de invertase era de l g/L. A presença de glicose em concentração
inferior a 0,5 g/L foi considerada como condição para desrepressão da
síntese da enzima. Seguindo-se esse critério, as leveduras estariam
desreprimidas quando o consumo ultrapassasse 50% da glicose inicial.
Não foi detectada a produção de invertase pelas linhagens
D. hanseni (50059) e R. rubra (50596) (Figuras 2 e 3) por meio da
metodologia proposta. D. hanseni apresentou pouco crescimento celular e
baixo consumo de glicose. A concentração de glicose no meio de cultivo
permaneceu acima de 0,5 g/L durante o crescimento celular. R. rubra
apresentou melhor crescimento do que D. hanseni mas também não havia
consumido 50% da glicose presente no meio com 8 h de cultivo.
Em ambos os experimentos as células encontravam-se sob condição
de repressão da síntese da invertase nos tempos nos quais foi medida a
atividade enzimática. Portanto, é possível que em condições de
desrepressão, essas duas leveduras produzam a enzima em questão.

--· - -·------------·--· --· -·-----------·----------;is-----··· --·
100 90-r ~l
li i
! 70, ~ 60 + -;,
50 + -8 40 + $. 30 .i
20 j_
10 + o • o
e =
• 2
-+-cresc. celular
4 6 Temp,(11)
8 10
Figura 2: Concentração de glicose no meio de cultivo e crescimento celular
de Debaryomyces hansenii (50059).
100 ------------ 90 - ~------- ! 70 _ , ---- % de glicose
~ 60 - · --+- cresc. celular ";'.: :() - ---·--- ~ 40 - '/!. 30 -
20 - !O - O~------------------- 0.1
8 !O
- 10
e = - - 1 -:
-; ~ "" -
O 2 4 6 Temp> (b)
Figura 3: Concentração de glicose no meio de cultivo e crescimento celular
de Rhodothorula rubra (50596).

Seguindo-se a metodologia proposta também não se detectou
produção de invertase pelas leveduras S. cerevisiae (50613), P. becki
(50808) e P. ohmeri (50537) (Figuras 4 a 6). Com 4 h de cultivo essas
leveduras não haviam consumido 50~ô da glicose presente no meio de
cultura, estando, portanto, sob condição de repressão da síntese de
invertase. Com 8 h de cultivo, no entanto, quase 100°/o da glicose já havia
sido consumida, proporcionando condições para a síntese da enzima.
A não detecção de atividade de invertase para essas leveduras após
oito horas de cultivo pode significar a existência de baixos níveis de
atividade da enzima associada à rápida assimilação da glicose e frutose
formadas a partir da hidrólise da sacarose.
. -· - -........,..,
100 - !O <)'.) - ~- s ;,, 70 - -----------
~ ------ % de gh;;-Ose = - ] 60 - - - ";'.: 50 - -+-- cresc . .xkllar ,,: - Ji -e 40 - 11
~ 30 - li= • ~ 20 - !O T o o
o 2 4 6 8 10 Teqio (h)
Figura 4: Concentração de glicose no meio de cultivo e crescimento celular
de Saccharomyces cerevisiae ( 50613 ).

·-···--·-··· --·····----------------
o 2 4 6 8 10
Te-.,()a)
Figura S: Concentração de glicose no meio de cultivo e crescimento celular
de Pichia becki (50808).
- 10
~ e s =
-= =": - 1 ,.c
~ ,. "i
~ :t o .s
o o 1 4 6 8 10 ..
Te~(b)
Figura 6: Concentração de glicose no meio de cultivo e crescimento celular
de Pichia ohmeri (50537).

----·-·--··-- ----
A. pululans (51102) (Figura 7) apresentou pouco crescimento e COllSUDlO
inferior a 20% da glicose do meio de cultura com 1 O h de cultivo. Portanto,
nos tempos 4 e 8 h de cultivo encontrava-se sob condição de repressão da
síntese de invertase. Foi verificada atividade da invertase presente com 4 h
(0,08 ug.ml,") e ausente com 8 h de cultivo. Com 4 h de cultivo, a atividade
normalizada obtida foi de 0,04 ug.ml," /00.
A atividade detectada com 4 h de cultivo, quando ainda não tinha
havido consumo da glicose do meio, indica a existência de condição de
desrepressão da síntese da invertase, em presença de concentrações de
glicose superiores a 0,5 g/L. Para P. anomala a síntese de invertase só é
reprimida em concentrações de glicose superiores a 2 g/L (Rodrigues et ai.,
1995).
A não detecção da atividade com 8 h pode ser devido à assimilação
da glicose e frutose formadas, considerando-se que as células colhidas com
8 h de cultivo estariam com o metabolismo mais ativo do que aquelas
colhidas com 4 h.
100 • • • - 10 1 • • 90 -----.
~...;. '1 70.,. _.._%de glicose e ! = ~ 60 +
1-+-cresc. ceillar ~ ~ 1 \(> 50 + ~ 1 -1 -8 40 + __.------+ ~ 30 + j •
20 + L 10 .; o. • 0,1
o 2 4 6 8 10
Te~(h)
Figura 7: Concentração de glicose no meio de cultivo e crescimento celular
de Aureobasidium pululans (51102).
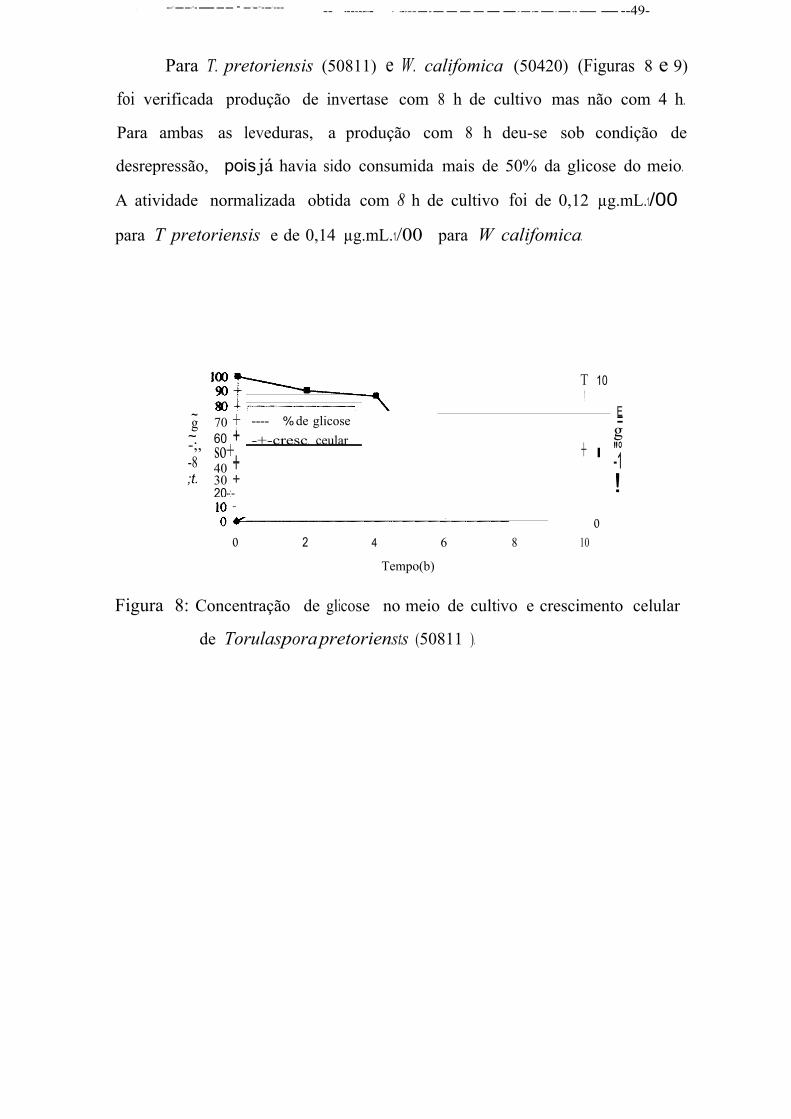
" --···---·----·-·-··--·-·-- -- --------- - ----------------------------49-
Para T. pretoriensis (50811) e W. califomica (50420) (Figuras 8 e 9)
foi verificada produção de invertase com 8 h de cultivo mas não com 4 h.
Para ambas as leveduras, a produção com 8 h deu-se sob condição de
desrepressão, pois já havia sido consumida mais de 50% da glicose do meio.
A atividade normalizada obtida com 8 h de cultivo foi de 0,12 µg.mL.1/00
para T pretoriensis e de 0,14 µg.mL.1/00 para W califomica.
T 10 i !
E ~ 70 + ---- % de glicose = g 1 g ~ 60 + -+-cresc. ceular -;, "° so+ + l -1 -8 40 + ;t. 30 + ! 20--:--
o o 2 4 6 8 10
Tempo(b)
Figura 8: Concentração de glicose no meio de cultivo e crescimento celular
de Torulaspora pretoriensts (50811 ).

····--· ··--· ··----· -·-· -···· ·-·--···-··-· . ----··-- --·------------r,r---···----
100 10 ex) - e ~ ro -
-----% de gbcose j 1
= ! 70 - g ..:! (iO - i ";'e 50 ~ -+-cresc. cehllarj t l ,,e
-8 40 + -1 ~ 30 +
1 ! 20 - 10 - o o
o 2 4 6 8 10
Teq,o(h)
Figura 9: Concentração de glicose no meio de cultivo e crescimento celular
de Williopsis ca/ifomica (50420).
Com 4 e 8 h de cultivo de C. ater (51032) (Figura 10) a concentração
de glicose no meio era superior a 50o/o e portanto as células encontravam-se
sob condição de repressão. A atividade da invertase foi de 4, 73 e de
6,30 ug.ml," com 4 e 8 h de cultivo, respectivamente. A atividade
normalizada da invertase obtida com 4 h (0,64 µg.mL.1/00) foi maior do
que aquela apresentada com 8 h (0,42 µg.mL-1/00).
Esperava-se, para todas as leveduras estudadas, que a atividade
verificada com 8 h fosse o dobro da obtida com 4 h, considerando-se que
para o ensaio enzimático realizado com 8 h, foi empregado o dobro de
células. Esse resultado corrobora a suposição de que as células
metabolicamente mais ativas estariam assimilando mais rapidamente os
produtos da hidrólise da sacarose e dessa forma os valores detectados
estariam sub-estimados.
Para C. macerans (50361) (Figura 11) as atividades normalizadas
verificadas com 4 e 8 h foram semelhantes, 2,47 e 2,21 µg.mL-1/DO,
respectivamente.

Também para essas leveduras, a repressão da síntese da invertase
deve ocorrer em concentrações de glicose superiores a 0,5 g/L.
100 T lO ~T 1
i
so + ---- % de gli;:ose e ~ :± ! r:: ~ -+- cresc, cem lar g -;, l,C
-8 :i -1 ~ 30 + .s 1
20 + 10 o
o 2 4 6 8 lO Teq,o(h)
Figura 1 O: Concentração de glicose no meio de cultivo e crescimento
celular de Cryptococcus ater (51032).
: -. - --
100 90 - ~- ·------- e " 70 - ---- °'o de gbcose e !:
] 60 - -+- cresc ce lu lar -;: 50 - ...:;
-8 40-:- ~ ~ 30' i' • ~ 20 .;
10 - o 0.1
o 2 4 6 8 10
Tempo(h)
Figura 11: Concentração de glicose no meio de cultivo e crescimento
celular de Cryptococcus macerans (50361).

·----------- -· -·-- ---- - --- --- ----------·- ----,.,51..--····--- . -·····
Para S. kluyvery (50629), Zygossacbaromyces sp (S096S) e Debaryomyces sp (50385) (Figuras 12 a 14) foi verificada produção de
invertase em duas condições distintas, sob repressão com 4 h e sob
desrepressão com 8 h. Para Zygossacharomyces sp (50965) a atividade
normalizada obtida com 4 h (0,11 µg.mL-1/00) foi o dobro da obtida com
8 h (0,05 µg.mL-1/00). Para Debaryomyces sp (50385) a atividade
normalizada verificada com 4 h (0,04 µg.mL-1/00) foi 4 vezes maior do que
a apresentada com 8 h (0,01 µg.mL-1,'DO).
Esse resultado, embora contrário ao esperado, corrobora a hipótese de
que as células metabolicamente mais ativas estariam assimilando mais
rapidamente os produtos da hidrólise da sacarose e, dessa forma, os valores
de atividade apresentados com 8 h estariam sub-estimados.
100 - 10 <X) - &) - ------ e ~ 70 - -%de glicose e
J;
J fi) - -.-..:resc. celular ~ :,e 50 - - 1 ~ 40 - ~ ~ 30 - r • ""\
20 - - 10 - o o
o 2 4 6 8 !O
Temp>(b)
Figura 12: Concentração de glicose no meio de cultivo e crescimento
celular de S.kluyvery (50629).

100 1 - lO
90 + i
V ~t ã ! ~ i =v= g ~ 'ic '° -8 40 + 1-+-cresc. ceklltr: -1 ~ 30+ .! •
ro + o . o o 2 4 6 8 10
Tempo(h)
Figura 13: Concentração de glicose no meio de cultivo e crescimento
celular de Zygossacharomyces sp (50965).
100 - 10 'X) -
~- 41 70 - ----- % de g bco se E s = w- - ~ ~ cres e ,;e ln lar :::: x ~- - l ~ ~ ~) - ~ ~ 30 - iC • "'\
20 - - IO ~ o o
o 2 4 6 8 10 Tempo(h)
Figura 14: Concentração de glicose no meio de cultivo e crescimento
celular de Debaryomyces sp ( 50385).
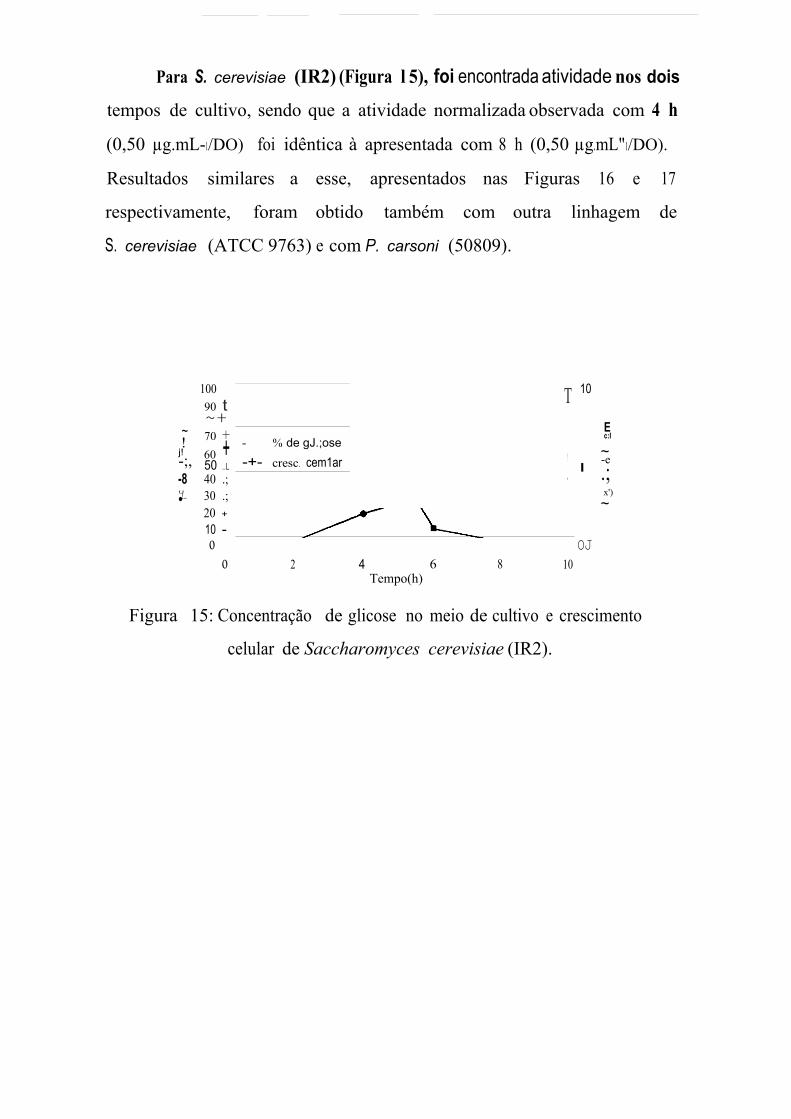
Para S. cerevisiae (IR2) (Figura l 5), foi encontrada atividade nos dois
tempos de cultivo, sendo que a atividade normalizada observada com 4 h
(0,50 µg.mL-1/DO) foi idêntica à apresentada com 8 h (0,50 µg.mL"1/DO).
Resultados similares a esse, apresentados nas Figuras 16 e 17
respectivamente, foram obtido também com outra linhagem de
S. cerevisiae (ATCC 9763) e com P. carsoni (50809).
100 T 10 90 t ~+ E ~ 70 + ! - % de gJ.;ose
c:I
j! 60 + ! ~ -;, 50 ..L -+- cresc. cem1ar l -e
-8 40 .; ' .; ':!.. 30 .; x') • ~
20 +
10 - o OJ
o 2 4 6 8 10 Tempo(h)
Figura 15: Concentração de glicose no meio de cultivo e crescimento
celular de Saccharomyces cerevisiae (IR2).
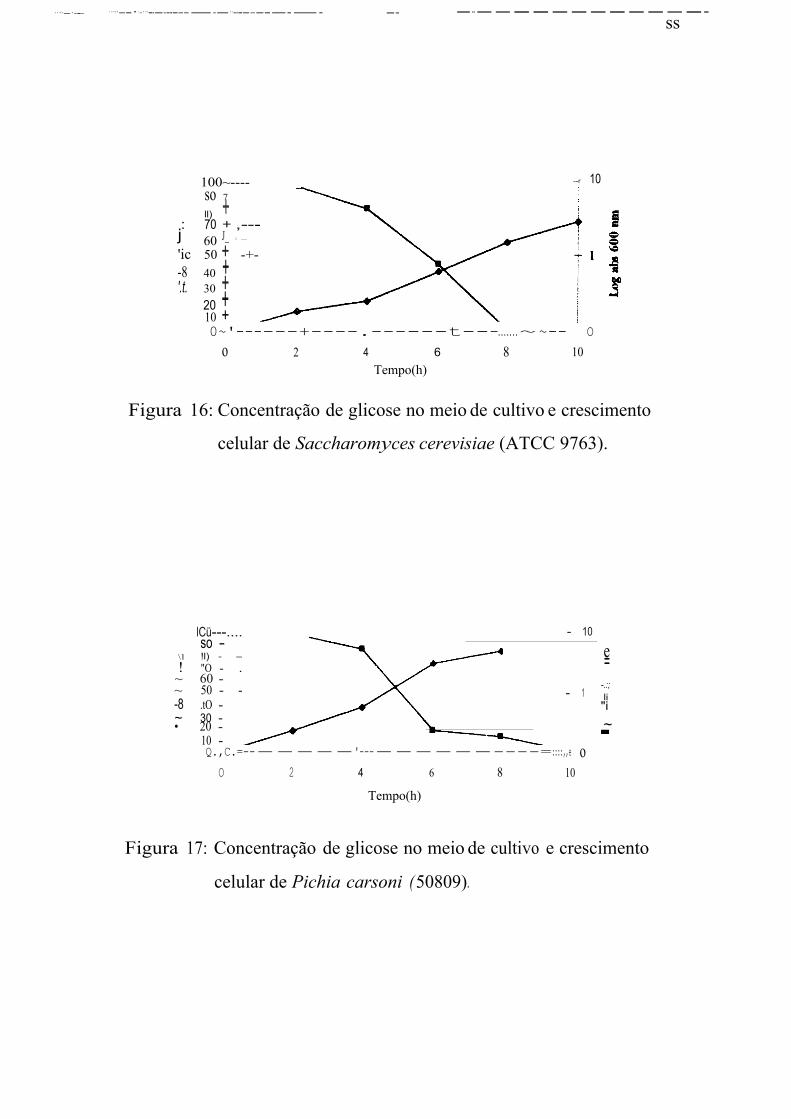
······-·--- ······--··-···------------··----------- -- . ---------------- ss
100~---- so 7
ll) + : 70 + ,---""-d-~--~ j 60 J_ 1, ---- 70 e gill,AJse 'ic 50 + -+- cresc. cekllar -8 40 + '.t. 30 +
20 + 10 + 0~'------+----.------t--- ....... ~~-- O
--r 10
o 2 4 6 Tempo(h)
8 10
Figura 16: Concentração de glicose no meio de cultivo e crescimento
celular de Saccharomyces cerevisiae (ATCC 9763).
ICü---...._ so -
\1 !l) - ------ ! "O - ......_º·ode l!OCOSe ~ 60 - - ~ 50 - -+- -:resc .::e lular -8 .tO - ~ 30 - • 20 -
10 - Q.,C.=-------'-------------=::::,,e
- 10
e = - 1
-.:; li "i ~ -
o 4 6 8 10
Tempo(h)
O 2
Figura 17: Concentração de glicose no meio de cultivo e crescimento
celular de Pichia carsoni ( 50809).

A dificuldade de se comparar as atividades enzimáticas obtidas no
presente trabalho com dados de literatura reside no fato de que os vários
autores utilizam em seus trabalhos unidades definidas de diversos modos.
Silveira et ai. ( 1996) definem l unidade como sendo 1 µmol de glicose
produzida por minuto de reação por mg de células. Cbávez et ai. (1997)
definem 1 unidade como a quantidade de enzima que hidroliz.a 1 mM de
sacarose por min de reação. Para Vitolo & Borzani (1983) a atividade da
invertase é definida como sendo a quantidade, expressa em g, de ART
liberada por min de reação por g de peso seco de células.
Além disso, as metodologias empregadas para o cultivo das células e
as condições utilizadas para os ensaios enzimáticos variam de acordo com
os autores. A isso acrescente-se que, no ensaio enzimático, alguns autores
trabalham com células vivas íntegras enquanto outros utilizam protoplastos
(Kuo & Lampem, 1971) ou frações celulares (Zárate & Belda, 1996).
No presente trabalho, os melhores resultados foram obtidos com
C macerans (50361 ). Pelo fto dessa levedura, até o presente momento, não
Ter aplicação industrial, não foi encontrado na literatura nenhum dado sobre
a atividade da invertase nesse gênero.
Com relação às demais linhagens utilizadas, por comparação com
C. macerans (50361), o gênero Saccharomyces e a espécie C. ater (51032)
apresentaram níveis médios de atividade ( de 1,00 a 2,34 ug.ml,") E, as
demais linhagens que produziram a invertase foram classificadas como
produtoras de baixos níveis de atividade ( de 0,04 a 0,57 ug.ml,").
Muitas aplicações industriais de leveduras produtoras de invertase, no
entanto, envolvem condições em que altos níveis de atividade não são
desejáveis. Há evidências de que a habilidade de S. cerevisiae fermentar
altas concentrações de sacarose, incluindo aquelas presentes na massa para

5. CONCLUSÕES
• A utilização do método GOD-PAP (Merck 8443) para a dosagem de
glicose em presença de sacarose requer cautela. Os reagentes do método
hidrolisam a sacarose durante o tempo de reação. A glicose formada é
quantificada pelo método GOD-PAP levando à obtenção de leituras de
absorbância crescentes.
• Para que o método GOD-PAP (Merck 8443) seja utilizado para dosagem
de glicose em presença de sacarose é necessário que a concentração
inicial de glicose esteja na faixa de 40 - l 00 µg/m.L e que o tempo de
leitura seja o mesmo para todos as amostras. Esse tempo deve ser
preferencialmente de 21 min após a adição do reagente.
• Do universo de linhagens de leveduras testadas, quanto a produção de
invertase, 68)5'% produziram a enzima e 3 l ,25o/o não a produziram nas
condições experimentais.
• A concentração de glicose que reprime a síntese da invertase varia de
acordo com a linhagem; 50o/o das leveduras testadas produziram invertase
em concentrações de glicose superiores a 0,5 g/L.
• A linhagem que apresentou melhor resultado quanto a produção de
invertase foi C. macerans (50361).

59
A ,
6. REFERENCIAS BIBLIOGRAFICAS
- ALAIS, C., LINDEN, G. Simple carbohydrates and derived products. In:-----
Food and Biochemistry. New York: Ellis Horwood, 1991, cap. 2,
p. 18-37.
- ARNOLD, W. N. Periplasmic space. ln: ROSE, A. H., HARRISON, J.S. eds.
Toe yeasts. London: Academic Press, 1991. v, 4, p. 279-295.
- BAKER, W. L. Glucose oxidase reaction for estimation of glucose without
horseradish peroxidase. Some microbiological and fermentation
application. J. Inst. Brew., v. 97, p.457-462, 1991.
- BARBOS~ M. F. S., MEDEIROS, M. B., MANCILM I. M.,
SCHNEIDER, H., LEE, H. Screening of yeasts for production of xylitol
from D-xylose and some factors which affect xylitol yield in Candida
gutlltermondii. J. lnd. Microbiol., v. 3, p. 241-251, 1988.
- BENÍTEZ, T., GASENT-RA~Z, J. M.; CASTREJÓN, F., CODÓN, A.
Development of new strains for the food industry. Biotechnol. Prog.,
v.12,p.149-163, 1996.
- BULL, A. T. Introduction. ln: MOO-YOUNG, M. ed. Compreensive
Biotecnology. ed. London: Pergamon Press, 1985. v. 1, p. 3 - 5.

--·-----~·--·---------·--
- CARLILE, M. J., WATKINSON, S. C. Fungai diversity. ln:- The fungi.
London: Academic Press., 1996, cap. 2, p. 9-75.
- CARLSON, M., TAUSSIG, R., KUSTU, S., BOTSTEIN. Toe secreted form
of invertase in Saccharomyces cerevisiae is sinthesized from m.RNA
encoding a signal sequence. Mol. Cell. Biol., v. 3, n. 3, p. 439-447,
1983.
- CHÁ VEZ, F. P., RODRIGUEZ, L., DÍAZ, J., DELGADO, J. M.,
CREMA TA, J. A. Purification and caracterization of an invertase from
Candida utilis: comparison with natural and recombinant yeast
invertases. J. Biotechnol., v. 53, p. 67-74, 1997.
- CHEN, W. C. í3-frutofuranosidase production by Aspergillus Japonicus in
shaking batch cultures as affected by initial sucrose concentration.
Biotechnol. Lett., v. 18, n 1, p. 69-72, 1996.
- CHEN, W. C. Production of '3-frutofuranosidase by Aspergillus japonicus in
batch and fed-batch cultures. Biotechnol. Lett., v. l 7, n. 12,
p. 1291-1924, 1995.
- CHENG, C. Y., DUAN, K. J., SHEU, D. C., LIN, C. T., LI, S. Y. Production
of frutooligosaccharides by immobilized mycelium of Aspergillus
japonicus. J. Chem. Tech. Biotechnol., v. 66, p. 135-138, 1996.

61
- CHEETHAM, P. S. J. Principies of industrilal biocatalysis and bioprocessing.
ln: WISEMAN, A, ed. Handbook of Enzyme Biotechnology. 3 ed.
London: Ellis Horwood, 1995. cap. 4 p. 83-233.
- CRUEGER, W., CRUEGER A. Proteína de origen unicelular (SCP). In:----
eds. Biotecnologia: manual de microbiologia industrial. Zaragoza:
Editorial Acnbia, 1989. cap. 16, p. 353-364.
- ELO~ M. V., LOSTAU, C. M., VILLANUEVA, J. R., SENTADREAU,
R., SÁNCHEZ, E. Invertase mesenger ribonucleic acid in
Saccharomyces cerevisiae. Biochim. Biophys. Acta., v. 475, p. 651-655,
1977.
ESMON, P. C., ESMON, B., SCHAUER, I. E., TAYLOR, A.,
SCHEKMAN, R. Structure, assembly, and secretion of octameric
invertase. J. Biol. Chem., v. 262, p. 4387-4394, 1987.
.. . -. ,.._
- GASCÓN, S., OTTOLENGHl, P. Influence of glucose concentrarion of the
medium on the invertase content of a strain of Saccharomyces bearing
the SUC2 gene. Compt. Rend. Trav. Lab. Carlsberg, v. 39, n. 2,
p. 15-24, 1972.

62
- GEMEINER, P, DOCOLOMANSKY, P., NAHÁLKA, J., STEFUC" V.,
DA.i'\ITELSSON, 8. New approaches for verification of kinetic
parameters of immobilized Concanavalin A: Invertase preparations
investigated by flow microcalorimetry. Biotechnol. Bioeng .v. 49,
p. 26-35, 1995.
- GONZÁLES SISO, M. I., RAMIL, E., CERDÁN, M. E., FREIRE-PICOS,
M. A. Respirofermentative metabolism in Kluyveromyces lactis: ethanol
production and crabtree effect. Enzyme and Microbial Tecnology. V.18,
p. 585-591,1996.
- GROSSMANN, M. K., ZIM:MERMANN, F. K. Toe structural genes of
internai invertases in Saccharomyces cerevisiae. Molec. Gen. Genet.,
V. 175, p. 223-229, 1979.
- HA Y ASHI, S., TOBOUSHI, M., T AKASAKI, Y., rMADA, K. Long-Term
continuous reaction of immobilized ~-frutofuranosidase. Biotechnol.
Lett., v. 16, p. 22 7-228, 1994.
- ISLAM, M. F., LAMPEN, J. O. Invertase secretion and sucrose fermentation
by Saccharomyces cerevisiae protoplasts. Biochim. Biophys. Acta.,
V. 58,p.294-302, 1962.
- KIRSOP, B. E. Resource Centers. ln: KIRSOP, B.E., KURTZMAN, C.P.
eds. Yeasts - Living Resources for Biotechnology. Cambridge:
Cambridge University Press, 1988. cap. 1, p. 1-35

. 63
- KRETZSCHMAR, H. Tratamiento, mejora y aplicaciones de la levadura. ln: -
- ed. Levaduras y alcoholes., 1955. cap. 14, p. 393-455.
- KUO, S. C., LAMPEN, J. O. Osmotic Regulation of invertase Formation and
secretion by protoplasts of Saccharomyces. J. Bacteriol. v. 101,
p. 183-191, 1971.
- MARTÍNEZ-PASTOR, M. T., ESTRUCR F. Sudden depletion of carbon
source blocks translation but not transcription, in the yeast
Saccharomyces cerevisiae. FEBS Letters, v. 390, p. 319-322, 1996.
- MIZUNAG1' T., TKACZ, J. S., RODRJGUEZ, L., HACKEL, R.,
LAMPEN, J. O. Temperature-sensitive forms of large and small
invertase in a mutant derived from a SUC l strain of Saccharomyces
cerevisiae . Mol. Cell. Biol., v. 1, p. 460-468, 1981.
- MONTENECOlJRT, B. S., KUO, S. ,CLAMPEN, J. O. Saccharomyces
mutants with invertase formation Resistant to repression by hexoses.
J. Bacteriol., v. 114, p. 233-238, 1973.
- MORENO, F., HERRERO, P., P~ F., GASCÓN, S. Invertase and
a-galactosidase sinthesis by yeast. Cell. Mol. Biol., v. 25, p. 1-6, 1979.

- MORENO, F., OCHO~ A. G., GASCÓN, S., Vll.LANUEV ~ J. R.
Molecular forms of yeast invertase. Eur. J. Biochem.,v. 50, p. 571-579,
1975.
- MUTLU, M.; TURHAN, N., ERCAN, M. T.; ÔWURAL, A. R., ÇELEBI,
S. S. Kinetics of invertase immobilised on poly (phe-lys) coated
polystyene beads. Biotechnology Techniques, v. 10, n 2, p. 71-76, 1996.
- MYERS, D. K., LAWLO~ D. T. M., ATTFIELD, P. V. Influence of
invertase activity and glicerol synthesis and retention on fennentation of
media with a high sugar concentration by Saccharomyces cerevisiae.
Appl. Environ. Microbiol., v. 63, n. 1, p. 145-150, 1997.
- NAUMOV, G. l., NAUMOV~ E. S., SANCHO, E. D., KORHOL~ M. P.
Polymeric SUC genes in natural population of Saccharomyces
cerevisiae. FEMS Microbiol. Lett., v. 135, p. 31-35, 1996.
- ODA, Y , TONO~ K. Purification and caracterization of invertase from
Torulaspora pretoriensis YK-1. Biosci. Biotech. Biochem., v. 58, n. 6,
p. 1155-1157, 1994.
OTTOLENGHI, P. Some properties of five non allelic
J3-D-Fructofuranosidases (invertases) of Saccharomyces. Compt. Rend.
Trav. Lab. Carlsberg, v. 38, n. 13, p. 213-221, 1972.

6.S
- PÉREZ, J. A., RODRÍGUEZ, J., RODRÍGUEZ, L., RUIZ T. Cloning and
sequence analysis of the invertase gene INV 1 from the yeast Pichia
anomala. Curr. Genet., v. 29, p. 234-240, 1996.
- PERLMAN, D., HAL VORSON, H. O. Distinct repressible mRNA for
cytoplasmic and secreted yeast invertase are encoded by a single gene.
Cell, v. 25, p. 525-536, 1981.
- PERLMAN, D., RANEY, P., HAL VORSON, H. O. Cytoplasmic and
secreted Saccharomyces cerevisiae invertase mRNAs encoded by one
gene can be differentially or coordinately regulated. Mol. Cell. Biol.,
V. 4, p. }682-1688, 1984.
- PERLMAN, D., RANEY, P., HAL VORSON, H. O. Mutations affecting the
signal sequence alter synthesis and secretion of yeast invertase. Proc.
Natl. Acad. Sei. US& v. 83, p. 5033-503 7, 1986.
- PHAFF, H. J. Micorganismos Industriales. lnvestigacion v Ciencia, v. 62,
p. 52-65, 1981.
- RAABO, E., TERKILDSEN, T. C. On the determination of blood glucose.
Scandinav. J. Clin. & Lab., v. 12, p. 402-407, 1960.
- RIBEIRO, R. R. Caracterização cinética da invertase imobilizada em Dowex-
l X8-50. São Paulo: USP/ Faculdade de Ciências Farmacéuticas ,1997.
(Tese de Mestrado).

--·----·-~-----
- ROBERTS, I. N., WILDMAN, H. G. The diverse potencial of yeasts.
Bio/technology, v. 13, p. 1246, 1995.
- RODRIGUEZ, L., LAMPEN, MACKA Y, V. L. SUCl gene of
Saccharomyces: a structural gene for the large (glycoprotein) and Small
( carbohydrate-free) form of invertase. Mol. Cell. Biol., v. l, n. 5, p. 469-
474, 1981.
- RODRÍGUEZ, J., PÉREZ, J. A., RUIZ, T., RODRÍGUEZ, L.
Characterization of the invertase from Pichia anomala. Biochem. J.,
V. 306,p.235-239, 1995.
SAROKIN, L., CARLSON, M. Upstream of the SUC2 gene confers
regulated expression to a heterologous gene in Saccharomyces
cerevisiae. Mol. Cell. Biol., v. 5, n.10, p. 2521-2526, 1985.
. -, ,..
- SAROKIN, L., CARLSON, M. Short repeated elements in the upstream
rezulatorv rezion of the SUC2 gene of Saccharomvces cerevisiae. Mol. '-" •· - ..... .. -
Cell. Biol., v.ó, n. 7, p. 2324-2333, 1986.
- SILVE~ M. C. F., CAR V AJAL, E., BON, E. P. S. Assay for in vivo
invertase using NaF. Anal. Biochem., v. 238, p. 26-28, 1996.
- SOMOGY, M. Notes on sugar determination. J. Biol. Chem., v. 195,
p. 19 _23, 1952.

68
- WICKERHAM, L. J. Evidence ofthe Production of extracellular invertase by
certain strains of yeasts. Arch. Biochem. Biophys., v. 76, p. 439-448,
1958.
- WILLIAMSON, D. H. Yeast clonning and biotechnology. ln: WALKER, J.
M., GINGOLD, E. B, eds. Molecular Biology and Biotechnology.
London:The Royal Society of Chemistry, 1988, cap. 5, p. 85-102.
- WISEMAN, A., WOODW ARD, J. Industrial yeast Invertase stabilization.
Process Biochem. v. July/August, 1975.
- ZÁRA TE, V., BELDA, F. Caracterization of the heterologous invertase
produced by Schizosaccaromyces pombe from the SUC2 gene of
Saccharomyces cerevisiae. J. Appl. Biotechnol., v. 80, p. 45-52, 1996.
- ZECH, M., GôRISH, H. Invertase from Saccharomyces cerevisiae.
reversible inactivation by components of industrial molasses media.
Ennme and Microbial Technology, v. l 7, p. 41-46, 1995.
- ZIMMERMANN, F. K., SCHEEL, I. Mutants of Saccharomyces cerevisiae
resistant to carbon catabolite repression. Molec. Gen. Genet., v. 154,
p. 75-82, 1977.
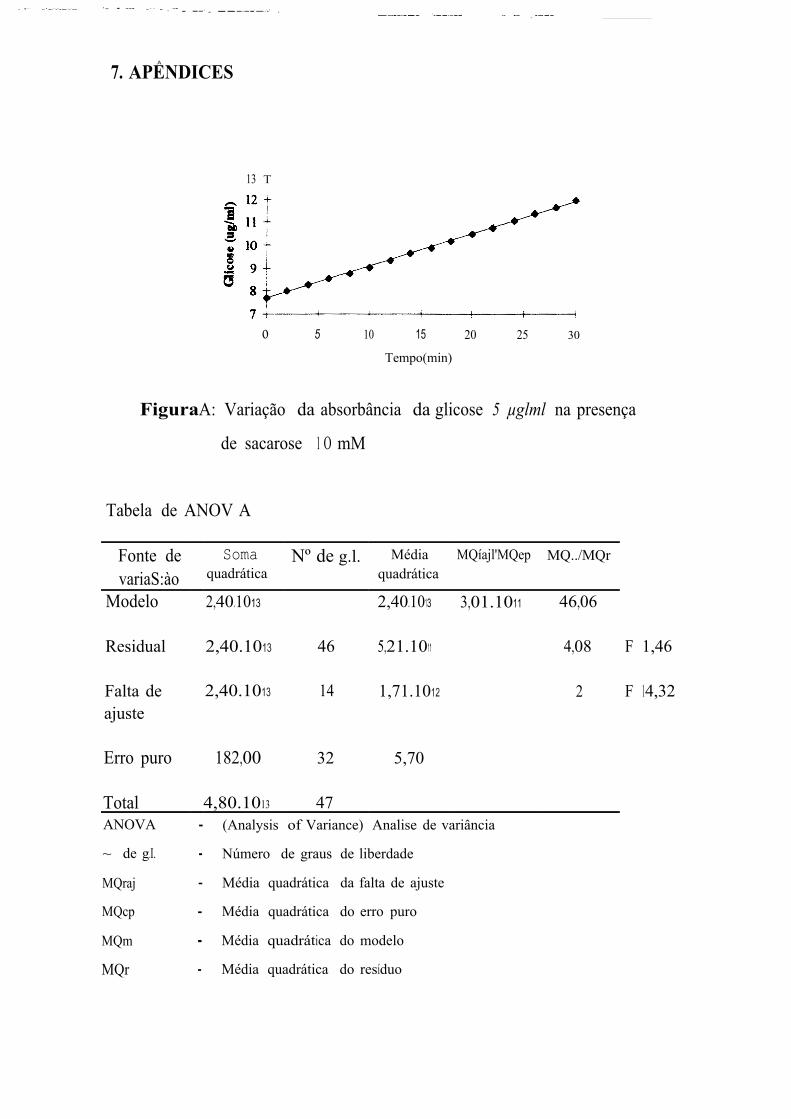
- •···· --·--···-··--··-··--- ·- - --- -·· - . --- - --· - ------·--·-· - . -----·--·--- ·------- -- -- . ----
A
7. APENDICES
13 T
y = O, 1399x + 7,6873 R2 =0,99'J3
o 5 10 15
Tempo(min)
20 25 30
Figura A: Variação da absorbância da glicose 5 µglml na presença
de sacarose 1 O mM
Tabela de ANOV A
Fonte de Soma Nº de g.l. Média MQíajl'MQep MQ../MQr
variaS:ào quadrática quadrática
Modelo 2,40.1013 2,40.1013 3,01.1011 46,06
Residual 2,40.1013 46 5,21.1011 4,08 F 1,46
Falta de 2,40.1013 14 1,71.1012 2 F 14,32 ajuste
Erro puro 182,00 32 5,70
Total 4,80.1013 47 ANOVA - (Analysis of Variance) Analise de variância
~ de g.I. - Número de graus de liberdade
MQraj - Média quadrática da falta de ajuste
MQcp - Média quadrática do erro puro
MQm - Média quadrática do modelo
MQr - Média quadrática do resíduo

20 T 19 t
::- 18 - _,ê 17 -:- ~ .:- 16 .... i. 15 .; ! 14 l a 13 !
12 r 11-:~~~~~~~~~-+-~-+-~------ o
y = Q,210lx + 12,681 R2 =0,9997
5 10 15
Tempo(min)
20 25 30
Figura B: Variação da absorbância da glicose 10 µg/ml na presença
de sacarose 1 O mM
Tabela de ANOVA
Fonte de Soma N2 de g.l. Média MQfajlMQcp MQ.JMQr varia~ào quadrática quadrática
Modelo 180, 1 O 1 180, 1 O 0,0 l 467,89
Residual 17,71 46 0,38 4,08 F 1,46
Falta de 0,06 14 0,00 2 F 14,32 3JUSte
Erro puro 17,65 32 0,55
Total 197 ,81 47

15
Tempo(m)
Figura C: Variação da absorbância da glicose 10 µg/mJ na presença
30 -e-
29-:- ,.... 28 - e 27 - ~ 26 i ::, -t-
';' 25 t ! 24 + ~ 23 I C5 : 22
21 + 20 +-'. --+---+----+---+----+-----,
o
y = 0.2369" + rum R2 = 0,9998
5 10 20 25 30
de sacarose 20 mM
Tabela de ANOVA
Fonte de Sorna Nº de g.l. Média MQraJMQ.,,, MQml'MQr variação quadrática quadrática
Modelo 228,98 228,98 0,0 l 808,69 .. - -a
Residual 11,78 46 0,26 4,08 F 1,46
Falta de 0,05 14 0,00 2 F 14,32 ajuste
Erro puro 11, 73 "'I ., 0,37 s :
Total 252,53 47
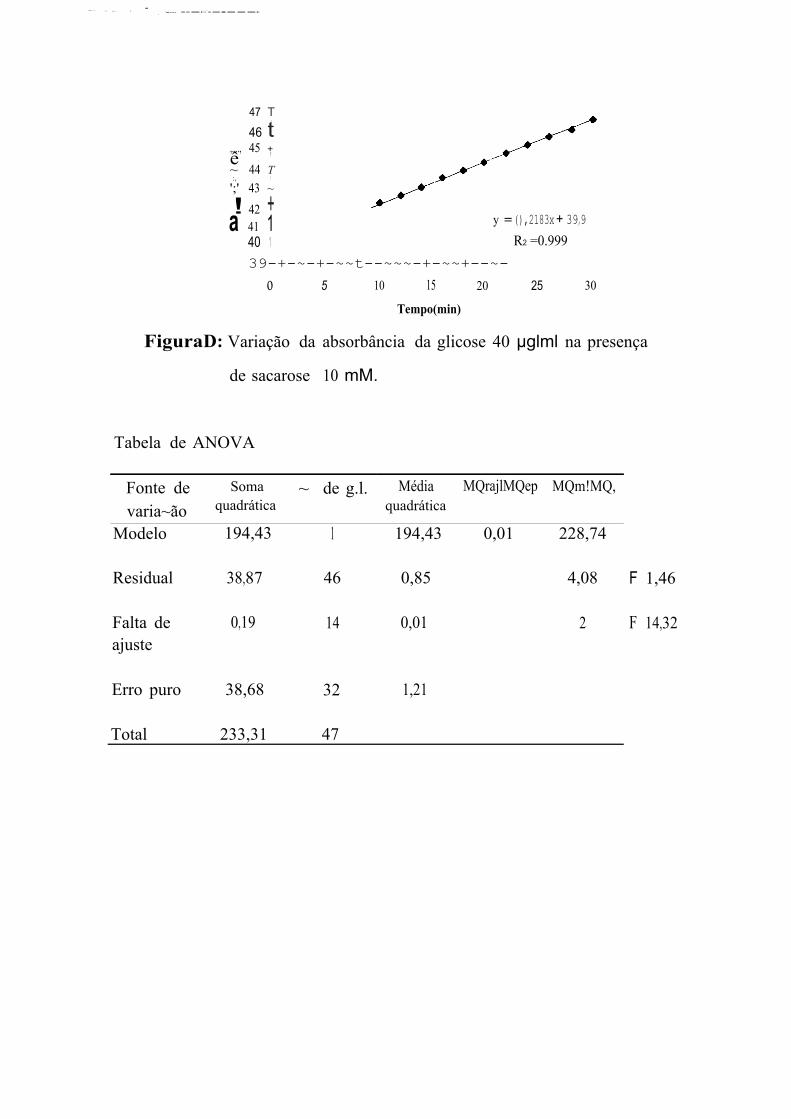
·-·-- .. ... . -- . ·---- ----·--·--------
47 T 1
46 t ,...., 45 + ê ' ~ 44 T :, 1
';' 43 ~ ! 42 + a 41 1
40 1 39-+-~-+-~~t--~~~-+-~~+--~-
y = (),2183x + 39,9 R2 =0.999
o 5 10 15 Tempo(min)
20 25 30
Figura D: Variação da absorbância da glicose 40 µglml na presença
de sacarose 10 mM.
Tabela de ANOVA
Fonte de Soma ~ de g.l. Média MQrajlMQep MQm!MQ, varia~ão quadrática quadrática
Modelo 194,43 1 194,43 0,01 228,74
Residual 38,87 46 0,85 4,08 F 1,46
Falta de 0,19 14 0,01 2 F 14,32 ajuste
Erro puro 38,68 32 1,21
Total 233,31 47
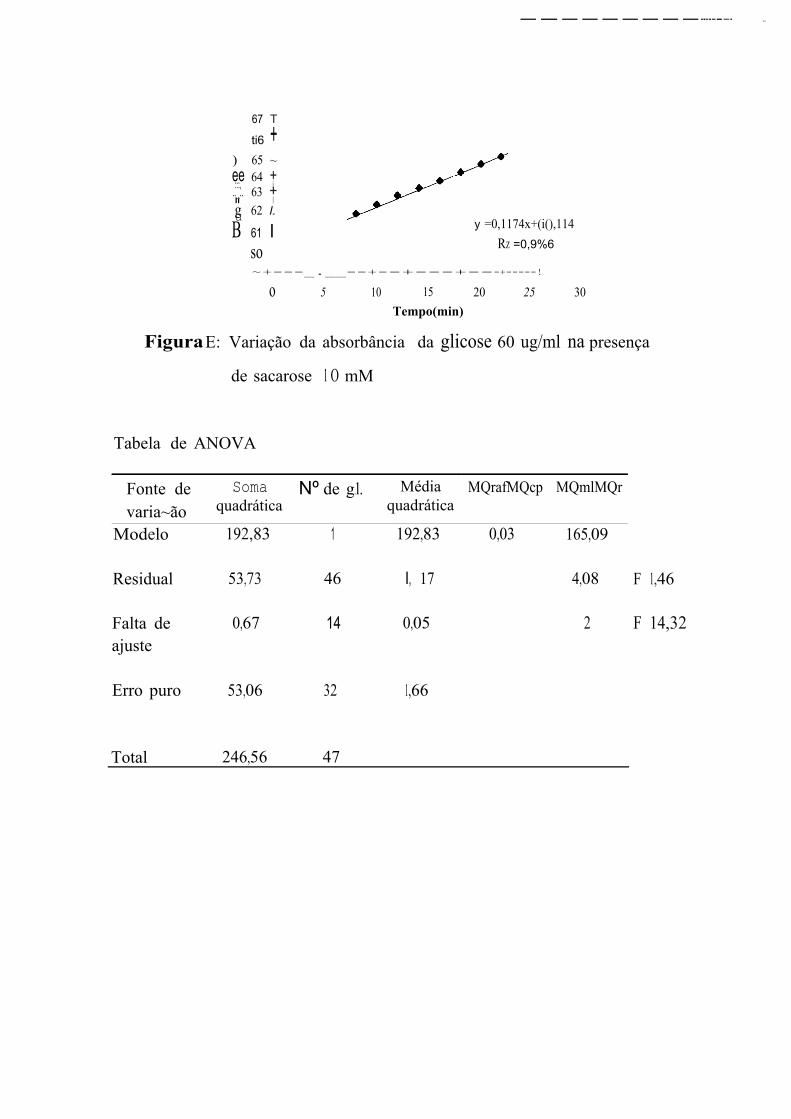
----------------- ·--
67 T
ti6 + ) 65 ~ ee 64 + :::, i .._.. 63 + li I g 62 l. u ' B 61 l
so ~+---_.__--+--+---+---+-----!
y =0,1174x+(i(),114 Rz =0,9%6
15 Tempo(min)
Figura E: Variação da absorbância da glicose 60 ug/ml na presença
o 5 10 20 25 30
de sacarose 1 O mM
Tabela de ANOVA
Fonte de Soma Nº de g.l. Média MQrafMQcp MQmlMQr varia~ão quadrática quadrática
Modelo 192,83 1 192,83 0,03 165,09
Residual 53,73 46 l, 17 4,08 F 1,46
Falta de 0,67 14 0,05 2 F 14,32 ajuste
Erro puro 53,06 32 1,66
Total 246,56 47
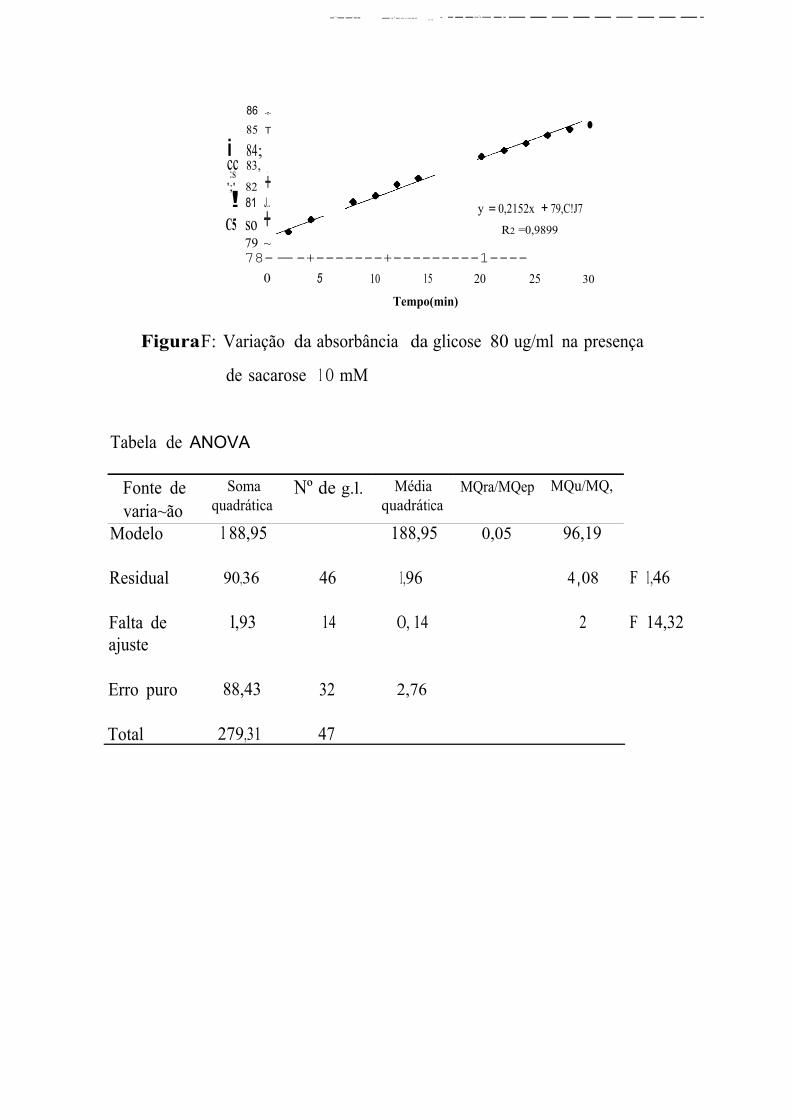
-··--·-- --·------- .. - -----·--------------
86 -e-
85 T
i 84; cc 83, :s : ';' 82 + ! 81 J..
C5 so + 79 ~ 78-·--+-------+---------1----
0
•
y = 0,2152x + 79,C!J7 R2 =0,9899
5 10 15
Tempo(min)
20 25 30
Figura F: Variação da absorbância da glicose 80 ug/ml na presença
de sacarose 1 O mM
Tabela de ANOVA
Fonte de Soma Nº de g.l. Média MQra/MQep MQu/MQ, varia~ão quadrática quadrática
Modelo l 88,95 188,95 0,05 96,19
Residual 90,36 46 1,96 4 08 F 1,46 '
Falta de l,93 14 O, 14 2 F 14,32 ajuste
Erro puro 88,43 32 2,76
Total 279,31 47

-·-···· ...• - .. --· . ·-·-- ·-------------. - ------------ -·-· -------- ------·----
1~ - 105 +
i ,.._ 104 , ] 103 -t- • i .:, 102 + ! 101 + ;; 100 f '..,; 99 +
98 ~ <n -.-!--+----.-----,....---------
º
y = 0,2386x + 9&, 164 ' R- =0,9911
5 10 15
Tempo(min)
20 25 30
Figura G: Variação da absorbância da glicose 100 µglml na presença
de sacarose 10 mM.
Tabela de ANOVA
Fonte de Soma N2 de g.l. Média MQfa/MQq, MQn/MQr vana~ão quadrática quadrática
Modelo 43430,21 1 43430,21 1757 ,84 46, 19
Residual 43256,20 46 940,35 4,08 F 1,46
Falta de 43200,03 14 3085,72 ') F 14,32 ajuste
Erro puro 56, 17 32 1,76
Total 86686,41 47

ll7 - ll6 ~
i 115 - :ili 114..,... ~ :
-; 1)3 + ! 112 + a 111 +
llO ' 109~·~~--~---~----~~--~~~---; o
y = Q,2199x+ 109,92 R2 =0,9949
5 10 15
Tempo(min)
20 25 30
Figura H: Variação da absorbância da glicose 120 µg!ml na presença
de sacarose 10 mM.
Tabela de ANOVA
Fonte de Soma ~ de g.l. Média MQrajMQcp MQu/MQr vana~ão quadrática quadrática
Modelo 197,29 197,29 0,14 501,58
Residual 18,09 46 0,39 4,08 F 1,46
Falta de l ,02 14 0,07 2 F 14,32 ajuste
Erro puro 17 ,08 32 0,53
Total 215,39 47

. . . " . . ------------------------- -
129 - 128..;...
.-. ": P7 - ~ - ~ t 126- ~___.,~
! 125 + ~ ã 124 t ~ Y =0,1932x+ 122,66 123 ~. R2 = 0,9953
1n~•~~--~-+-~---~~-----~--+-~---< o 5 10 15 20 25 30
Tempo(min)
Figura 1: Variação da absorbância da glicose 140 µg/ml na presença
de sacarose 1 O mM
Tabela de ANOVA
Fonte de Soma N2 de g.l. Média MQra_/MQ.., MQnlMQr varia~ão quadrática quadrática
Modelo 152,29 1 152,29 0,04 158,71
Residual 44, 14 46 0,96 4,08 F 1,46
Falta de 0,72 14 0,05 2 F 14,32 ajuste
Erro puro 43,42 32 1,36
Total 196,43 47

147 -
146- ~ :::, 145 - -· s -+- ? :~ = - ~_._-.-- j 142 r ~ y=~1Sx+140,t9 0 14 t - __. Ri = 0.9933
140 .----. 139+---------r----+---+-----i
O 5 10 15 20 25 30
Tempo(min)
Figura J: Variação da absorbância da glicose 160 µg/ml na presença
de sacarose 1 O mM.
Tabela de ANOVA
Fonte de Soma Nº de g.l. Média MQra/MQci, MQnlMQr varia~ào quadrática quadrática
Modelo 165,66 1 165,66 0,09 254,42
Residual 29,95 46 0,65 4,08 F 1,46
Falta de 1, 12 14 0,08 2 F 14,32 ajuste
Erro puro 28,83 32 0,90
Total 195,61 47

------ ---·------- -- ·- - - - -- ---- ------
164 - -. 163- ~ e I ez - ____..---- 1 l 6 t - ,__ _,_____..--- - ";' ieo - ~- ! t59 -. ~ ij 158 -
157 ..:: 156 t •
O 5
y=~+l57.15 R"" = 0.9&3 l
10 15 20 25 30
Tempo(min)
Figura L: Variação da absorbância da glicose 180 µg/ml na presença
de sacarose 10 mM.
Tabela de ANOV A
Fonte de Soma Nº de g.l. Média MQrajMQcp MQolMQr variaS:ãO quadrática quadrática
Modelo 176,35 1 176,35 0,03 31,65
Residual 256,33 46 5,57 4,08 F 1,46
Falta de 3,04 14 0,22 2 F 14,32 ajuste
Erro puro 253,29 32 7,92
Total 432,68 47


















