
Expressão de Transtirretina no Sistema Gastrointestinal...
55
Escola Superior de Tecnologia da Saúde do Porto Instituto Politécnico do Porto Susete Marlene Machado Costelha Expressão de Transtirretina no Sistema Gastrointestinal do Murganho Dissertação de Mestrado em Tecnologia Bioquímica em Saúde Setembro de 2011
Transcript of Expressão de Transtirretina no Sistema Gastrointestinal...

Escola Superior de Tecnologia da Sauacutede do Porto Instituto Politeacutecnico do Porto
Susete Marlene Machado Costelha
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
Dissertaccedilatildeo de Mestrado em Tecnologia Bioquiacutemica em Sauacutede
Setembro de 2011
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
ii
ESCOLA SUPERIOR DE TECNOLOGIA DA SAUacuteDE
DO PORTO
INSTITUTO POLITEacuteCNICO DO PORTO
Susete Marlene Machado Costelha
Expressatildeo de Transtirretina no Sistema
Gastrointestinal do Murganho
Dissertaccedilatildeo submetida agrave Escola Superior de Tecnologia da Sauacutede do Porto para cumprimento
dos requisitos necessaacuterios agrave obtenccedilatildeo do grau de Mestre em Tecnologia Bioquiacutemica em Sauacutede
realizada sob a orientaccedilatildeo cientiacutefica da Professora Doutora Maria Joatildeo Saraiva (Professora
Catedraacutetica Aacuterea das Neurociecircncias Instituto de Ciecircncias Biomeacutedicas Abel Salazar da
Universidade do Porto) e sob co-orientaccedilatildeo da Profordf Doutora Ceciacutelia Santos (Professora
Associada Aacuterea das Ciecircncias da Sauacutede Faculdade de Ciecircncias da Sauacutede da Universidade da
Beira Interior) e Profordm Doutor Ruacuteben Fernandes (Professor Adjunto Aacuterea de Bioquiacutemica
Unidade de Mecanismos Moleculares da Doenccedila do Centro de Investigaccedilatildeo em Sauacutede e
Ambiente - ESTSP e Centro de Farmacologia e Biopatologia Quiacutemica ndash FMUP)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iii
Agradecimentos
Gostaria de agradecer a todos os que de alguma forma contribuiacuteram para a realizaccedilatildeo deste
trabalho Em especial
Agrave Professora Maria Joatildeo Saraiva por me ter possibilitado os meios para a execuccedilatildeo desta
tese e por toda a orientaccedilatildeo e atenccedilatildeo que dedicou agrave mesma
Agrave Professora Ceciacutelia pela orientaccedilatildeo e por todo o apoio durante todo este tempo Pela boa
disposiccedilatildeo e enorme optimismo em relaccedilatildeo a tudo
Ao Professor Ruacuteben e Professora Cristina por terem organizado este mestrado e estarem
sempre disponiacuteveis para tudo o que fosse necessaacuterio
A todos os meus amigos do laboratoacuterio (vocecircs sabem quem satildeo) o meu muito obrigado
pelo apoio boa disposiccedilatildeo (que seria de mim sem os nossos almoccedilos) Por terem tornado
mais faacutecil todo este processo de aprendizagem e por serem sobretudo uns amigos muito
especiais
Um agradecimento especial agrave Anabela e ao Paul pela paciecircncia e dedicaccedilatildeo com que me
ajudaram sempre que necessaacuterio Eacute tudo mais faacutecil quando existem pessoas assim
Um obrigado especial agrave Paula Francisco Tacircnia Doutora Rosaacuterio Almeida e Doutora
Isabel Cardoso por toda a ajuda e apoio
Aos meus pais e avoacutes por serem as pessoas mais especiais deste mundo natildeo seria nada
sem vocecircs
Agrave minha irmatilde por ter dado um significado ainda mais especial agrave minha vida e por garantir
que todos os dias satildeo uma animaccedilatildeo Eacutes sem duacutevida uma estrela
Ao Filipe por todo o apoio dedicaccedilatildeo e amor por estar sempre ao meu lado por ser a
minha fonte de optimismo
A todos os meus amigos o meu obrigado por estarem comigo em todas as fases da minha
vida esta natildeo eacute excepccedilatildeo O meu agradecimento especial a todos voacutes
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iv
Resumo
A transtirretina (TTR) eacute uma proteiacutena plasmaacutetica constituiacuteda por quatro subunidades
idecircnticas de aproximadamente 14KDa e de massa molecular de 55 KDa (Blake et al
1978)
A TTR eacute responsaacutevel pelo transporte de tiroxina (T4) (Andrea et al 1980) e retinol
(vitamina A) neste uacuteltimo tipo de transporte atraveacutes da ligaccedilatildeo agrave proteina de ligaccedilatildeo ao
retinol (RBP) (Kanai et al 1968) Eacute sintetizada principalmente pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) e tambeacutem sintetizada pelas ceacutelulas epiteliais do
plexo coroacuteide e secretada para o liacutequido cefaloraquidiano (LCR) (Aleshire et al 1983)
Existem outros locais que expressam TTR mas em menor quantidade nomeadamente a
retina do olho (Martone et al 1988) o pacircncreas (Kato et al 1985) o saco vitelino
visceral (Soprano et al 1986) o intestino (Loughna et al 1995) o estocircmago coraccedilatildeo
muacutesculo e baccedilo (Soprano et al 1985) A TTR eacute uma proteiacutena do ponto de vista
filogeneacutetico extremamente conservada o que jaacute de si eacute um indicador da sua importacircncia
bioloacutegica (Richardson 2009)
O objectivo deste trabalho foi avaliar a expressatildeo de transtirretina ao longo do sistema
gastrointestinal do murganho nos seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon e
tambeacutem bexiga com cerca de 3 meses de idade O segundo objectivo foi identificar as
ceacutelulas responsaacuteveis por essa expressatildeo nos oacutergatildeos em estudo
Foi possiacutevel verificar que apenas o estocircmago apresenta valores de expressatildeo normalizada
de TTR diferente de zero expressatildeo essa muito inferior agrave do fiacutegado tal como se esperava
Por imunohistoquiacutemicaimunofluorescecircncia foi possiacutevel determinar que as ceacutelulas que
expressam TTR satildeo pouco abundantes e estatildeo presentes na regiatildeo glandular do estocircmago
do murganho e tambeacutem do humano Para aleacutem disto verificou-se que a TTR co-localiza
com somatostatina e que as ceacutelulas que sintetizam TTR correspondem agraves ceacutelulas D
responsaacuteveis pela secreccedilatildeo de somatostatina
Palavras-chave Transtirretina Polineuropatia Familiar Amiloidoacutetica Sistema
Gastrointestinal
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
v
Abstract
Transthyretin (TTR) is a plasma protein composed of four identical subunits of
approximately 14 KDa and molecular mass of 55 KDa (Blake et al 1978)
TTR is responsible for the transport of thyroxine (T4) (Andrea et al 1980) and retinol
(vitamin A) by binding to the retinol binding protein (RBP) (Kanai et al 1968) It is
synthesized primarily by the liver and secreted into the blood (Murakami et al 1987) and
also synthesized by epithelial cells of the choroid plexus and secreted into the
cerebrospinal fluid (CSF) (Aleshire et al 1983) There are other sites that express TTR
but in smaller quantities namely the retina of the eye (Martone et al 1988) pancreas
(Kato et al 1985) visceral yolk sac (Soprano et al 1986) intestine (Loughna et al
1995) stomach heart muscle and spleen (Soprano et al 1985)
TTR is a highly conserved protein which from a phylogenetic standpoint is itself an
indicator of its biological importance (Richardson 2009)
The aim of this study was to evaluate the expression of transthyretin along the
gastrointestinal tract of mice (3 months old) ndash esophagus stomach duodenum colon and
bladder as well The second aim was to identify the cells responsible for this expression in
the organs under study
It was possible to verify that only the stomach showed values of normalized expression
different from zero This expression was lower than in the liver as expected
By immunohistochemistry immunofluorescence it was determined that cells expressing
TTR are present in the glandular region of the stomach of mice and also in human
stomach It was also determined that D cells responsible for secretion of somatostatin
synthesize TTR
Key-words Transthyretin Familial Amyloid Polyneuropathy Gastrointestinal Tract
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
ii
ESCOLA SUPERIOR DE TECNOLOGIA DA SAUacuteDE
DO PORTO
INSTITUTO POLITEacuteCNICO DO PORTO
Susete Marlene Machado Costelha
Expressatildeo de Transtirretina no Sistema
Gastrointestinal do Murganho
Dissertaccedilatildeo submetida agrave Escola Superior de Tecnologia da Sauacutede do Porto para cumprimento
dos requisitos necessaacuterios agrave obtenccedilatildeo do grau de Mestre em Tecnologia Bioquiacutemica em Sauacutede
realizada sob a orientaccedilatildeo cientiacutefica da Professora Doutora Maria Joatildeo Saraiva (Professora
Catedraacutetica Aacuterea das Neurociecircncias Instituto de Ciecircncias Biomeacutedicas Abel Salazar da
Universidade do Porto) e sob co-orientaccedilatildeo da Profordf Doutora Ceciacutelia Santos (Professora
Associada Aacuterea das Ciecircncias da Sauacutede Faculdade de Ciecircncias da Sauacutede da Universidade da
Beira Interior) e Profordm Doutor Ruacuteben Fernandes (Professor Adjunto Aacuterea de Bioquiacutemica
Unidade de Mecanismos Moleculares da Doenccedila do Centro de Investigaccedilatildeo em Sauacutede e
Ambiente - ESTSP e Centro de Farmacologia e Biopatologia Quiacutemica ndash FMUP)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iii
Agradecimentos
Gostaria de agradecer a todos os que de alguma forma contribuiacuteram para a realizaccedilatildeo deste
trabalho Em especial
Agrave Professora Maria Joatildeo Saraiva por me ter possibilitado os meios para a execuccedilatildeo desta
tese e por toda a orientaccedilatildeo e atenccedilatildeo que dedicou agrave mesma
Agrave Professora Ceciacutelia pela orientaccedilatildeo e por todo o apoio durante todo este tempo Pela boa
disposiccedilatildeo e enorme optimismo em relaccedilatildeo a tudo
Ao Professor Ruacuteben e Professora Cristina por terem organizado este mestrado e estarem
sempre disponiacuteveis para tudo o que fosse necessaacuterio
A todos os meus amigos do laboratoacuterio (vocecircs sabem quem satildeo) o meu muito obrigado
pelo apoio boa disposiccedilatildeo (que seria de mim sem os nossos almoccedilos) Por terem tornado
mais faacutecil todo este processo de aprendizagem e por serem sobretudo uns amigos muito
especiais
Um agradecimento especial agrave Anabela e ao Paul pela paciecircncia e dedicaccedilatildeo com que me
ajudaram sempre que necessaacuterio Eacute tudo mais faacutecil quando existem pessoas assim
Um obrigado especial agrave Paula Francisco Tacircnia Doutora Rosaacuterio Almeida e Doutora
Isabel Cardoso por toda a ajuda e apoio
Aos meus pais e avoacutes por serem as pessoas mais especiais deste mundo natildeo seria nada
sem vocecircs
Agrave minha irmatilde por ter dado um significado ainda mais especial agrave minha vida e por garantir
que todos os dias satildeo uma animaccedilatildeo Eacutes sem duacutevida uma estrela
Ao Filipe por todo o apoio dedicaccedilatildeo e amor por estar sempre ao meu lado por ser a
minha fonte de optimismo
A todos os meus amigos o meu obrigado por estarem comigo em todas as fases da minha
vida esta natildeo eacute excepccedilatildeo O meu agradecimento especial a todos voacutes
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iv
Resumo
A transtirretina (TTR) eacute uma proteiacutena plasmaacutetica constituiacuteda por quatro subunidades
idecircnticas de aproximadamente 14KDa e de massa molecular de 55 KDa (Blake et al
1978)
A TTR eacute responsaacutevel pelo transporte de tiroxina (T4) (Andrea et al 1980) e retinol
(vitamina A) neste uacuteltimo tipo de transporte atraveacutes da ligaccedilatildeo agrave proteina de ligaccedilatildeo ao
retinol (RBP) (Kanai et al 1968) Eacute sintetizada principalmente pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) e tambeacutem sintetizada pelas ceacutelulas epiteliais do
plexo coroacuteide e secretada para o liacutequido cefaloraquidiano (LCR) (Aleshire et al 1983)
Existem outros locais que expressam TTR mas em menor quantidade nomeadamente a
retina do olho (Martone et al 1988) o pacircncreas (Kato et al 1985) o saco vitelino
visceral (Soprano et al 1986) o intestino (Loughna et al 1995) o estocircmago coraccedilatildeo
muacutesculo e baccedilo (Soprano et al 1985) A TTR eacute uma proteiacutena do ponto de vista
filogeneacutetico extremamente conservada o que jaacute de si eacute um indicador da sua importacircncia
bioloacutegica (Richardson 2009)
O objectivo deste trabalho foi avaliar a expressatildeo de transtirretina ao longo do sistema
gastrointestinal do murganho nos seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon e
tambeacutem bexiga com cerca de 3 meses de idade O segundo objectivo foi identificar as
ceacutelulas responsaacuteveis por essa expressatildeo nos oacutergatildeos em estudo
Foi possiacutevel verificar que apenas o estocircmago apresenta valores de expressatildeo normalizada
de TTR diferente de zero expressatildeo essa muito inferior agrave do fiacutegado tal como se esperava
Por imunohistoquiacutemicaimunofluorescecircncia foi possiacutevel determinar que as ceacutelulas que
expressam TTR satildeo pouco abundantes e estatildeo presentes na regiatildeo glandular do estocircmago
do murganho e tambeacutem do humano Para aleacutem disto verificou-se que a TTR co-localiza
com somatostatina e que as ceacutelulas que sintetizam TTR correspondem agraves ceacutelulas D
responsaacuteveis pela secreccedilatildeo de somatostatina
Palavras-chave Transtirretina Polineuropatia Familiar Amiloidoacutetica Sistema
Gastrointestinal
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
v
Abstract
Transthyretin (TTR) is a plasma protein composed of four identical subunits of
approximately 14 KDa and molecular mass of 55 KDa (Blake et al 1978)
TTR is responsible for the transport of thyroxine (T4) (Andrea et al 1980) and retinol
(vitamin A) by binding to the retinol binding protein (RBP) (Kanai et al 1968) It is
synthesized primarily by the liver and secreted into the blood (Murakami et al 1987) and
also synthesized by epithelial cells of the choroid plexus and secreted into the
cerebrospinal fluid (CSF) (Aleshire et al 1983) There are other sites that express TTR
but in smaller quantities namely the retina of the eye (Martone et al 1988) pancreas
(Kato et al 1985) visceral yolk sac (Soprano et al 1986) intestine (Loughna et al
1995) stomach heart muscle and spleen (Soprano et al 1985)
TTR is a highly conserved protein which from a phylogenetic standpoint is itself an
indicator of its biological importance (Richardson 2009)
The aim of this study was to evaluate the expression of transthyretin along the
gastrointestinal tract of mice (3 months old) ndash esophagus stomach duodenum colon and
bladder as well The second aim was to identify the cells responsible for this expression in
the organs under study
It was possible to verify that only the stomach showed values of normalized expression
different from zero This expression was lower than in the liver as expected
By immunohistochemistry immunofluorescence it was determined that cells expressing
TTR are present in the glandular region of the stomach of mice and also in human
stomach It was also determined that D cells responsible for secretion of somatostatin
synthesize TTR
Key-words Transthyretin Familial Amyloid Polyneuropathy Gastrointestinal Tract
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iii
Agradecimentos
Gostaria de agradecer a todos os que de alguma forma contribuiacuteram para a realizaccedilatildeo deste
trabalho Em especial
Agrave Professora Maria Joatildeo Saraiva por me ter possibilitado os meios para a execuccedilatildeo desta
tese e por toda a orientaccedilatildeo e atenccedilatildeo que dedicou agrave mesma
Agrave Professora Ceciacutelia pela orientaccedilatildeo e por todo o apoio durante todo este tempo Pela boa
disposiccedilatildeo e enorme optimismo em relaccedilatildeo a tudo
Ao Professor Ruacuteben e Professora Cristina por terem organizado este mestrado e estarem
sempre disponiacuteveis para tudo o que fosse necessaacuterio
A todos os meus amigos do laboratoacuterio (vocecircs sabem quem satildeo) o meu muito obrigado
pelo apoio boa disposiccedilatildeo (que seria de mim sem os nossos almoccedilos) Por terem tornado
mais faacutecil todo este processo de aprendizagem e por serem sobretudo uns amigos muito
especiais
Um agradecimento especial agrave Anabela e ao Paul pela paciecircncia e dedicaccedilatildeo com que me
ajudaram sempre que necessaacuterio Eacute tudo mais faacutecil quando existem pessoas assim
Um obrigado especial agrave Paula Francisco Tacircnia Doutora Rosaacuterio Almeida e Doutora
Isabel Cardoso por toda a ajuda e apoio
Aos meus pais e avoacutes por serem as pessoas mais especiais deste mundo natildeo seria nada
sem vocecircs
Agrave minha irmatilde por ter dado um significado ainda mais especial agrave minha vida e por garantir
que todos os dias satildeo uma animaccedilatildeo Eacutes sem duacutevida uma estrela
Ao Filipe por todo o apoio dedicaccedilatildeo e amor por estar sempre ao meu lado por ser a
minha fonte de optimismo
A todos os meus amigos o meu obrigado por estarem comigo em todas as fases da minha
vida esta natildeo eacute excepccedilatildeo O meu agradecimento especial a todos voacutes
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iv
Resumo
A transtirretina (TTR) eacute uma proteiacutena plasmaacutetica constituiacuteda por quatro subunidades
idecircnticas de aproximadamente 14KDa e de massa molecular de 55 KDa (Blake et al
1978)
A TTR eacute responsaacutevel pelo transporte de tiroxina (T4) (Andrea et al 1980) e retinol
(vitamina A) neste uacuteltimo tipo de transporte atraveacutes da ligaccedilatildeo agrave proteina de ligaccedilatildeo ao
retinol (RBP) (Kanai et al 1968) Eacute sintetizada principalmente pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) e tambeacutem sintetizada pelas ceacutelulas epiteliais do
plexo coroacuteide e secretada para o liacutequido cefaloraquidiano (LCR) (Aleshire et al 1983)
Existem outros locais que expressam TTR mas em menor quantidade nomeadamente a
retina do olho (Martone et al 1988) o pacircncreas (Kato et al 1985) o saco vitelino
visceral (Soprano et al 1986) o intestino (Loughna et al 1995) o estocircmago coraccedilatildeo
muacutesculo e baccedilo (Soprano et al 1985) A TTR eacute uma proteiacutena do ponto de vista
filogeneacutetico extremamente conservada o que jaacute de si eacute um indicador da sua importacircncia
bioloacutegica (Richardson 2009)
O objectivo deste trabalho foi avaliar a expressatildeo de transtirretina ao longo do sistema
gastrointestinal do murganho nos seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon e
tambeacutem bexiga com cerca de 3 meses de idade O segundo objectivo foi identificar as
ceacutelulas responsaacuteveis por essa expressatildeo nos oacutergatildeos em estudo
Foi possiacutevel verificar que apenas o estocircmago apresenta valores de expressatildeo normalizada
de TTR diferente de zero expressatildeo essa muito inferior agrave do fiacutegado tal como se esperava
Por imunohistoquiacutemicaimunofluorescecircncia foi possiacutevel determinar que as ceacutelulas que
expressam TTR satildeo pouco abundantes e estatildeo presentes na regiatildeo glandular do estocircmago
do murganho e tambeacutem do humano Para aleacutem disto verificou-se que a TTR co-localiza
com somatostatina e que as ceacutelulas que sintetizam TTR correspondem agraves ceacutelulas D
responsaacuteveis pela secreccedilatildeo de somatostatina
Palavras-chave Transtirretina Polineuropatia Familiar Amiloidoacutetica Sistema
Gastrointestinal
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
v
Abstract
Transthyretin (TTR) is a plasma protein composed of four identical subunits of
approximately 14 KDa and molecular mass of 55 KDa (Blake et al 1978)
TTR is responsible for the transport of thyroxine (T4) (Andrea et al 1980) and retinol
(vitamin A) by binding to the retinol binding protein (RBP) (Kanai et al 1968) It is
synthesized primarily by the liver and secreted into the blood (Murakami et al 1987) and
also synthesized by epithelial cells of the choroid plexus and secreted into the
cerebrospinal fluid (CSF) (Aleshire et al 1983) There are other sites that express TTR
but in smaller quantities namely the retina of the eye (Martone et al 1988) pancreas
(Kato et al 1985) visceral yolk sac (Soprano et al 1986) intestine (Loughna et al
1995) stomach heart muscle and spleen (Soprano et al 1985)
TTR is a highly conserved protein which from a phylogenetic standpoint is itself an
indicator of its biological importance (Richardson 2009)
The aim of this study was to evaluate the expression of transthyretin along the
gastrointestinal tract of mice (3 months old) ndash esophagus stomach duodenum colon and
bladder as well The second aim was to identify the cells responsible for this expression in
the organs under study
It was possible to verify that only the stomach showed values of normalized expression
different from zero This expression was lower than in the liver as expected
By immunohistochemistry immunofluorescence it was determined that cells expressing
TTR are present in the glandular region of the stomach of mice and also in human
stomach It was also determined that D cells responsible for secretion of somatostatin
synthesize TTR
Key-words Transthyretin Familial Amyloid Polyneuropathy Gastrointestinal Tract
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
iv
Resumo
A transtirretina (TTR) eacute uma proteiacutena plasmaacutetica constituiacuteda por quatro subunidades
idecircnticas de aproximadamente 14KDa e de massa molecular de 55 KDa (Blake et al
1978)
A TTR eacute responsaacutevel pelo transporte de tiroxina (T4) (Andrea et al 1980) e retinol
(vitamina A) neste uacuteltimo tipo de transporte atraveacutes da ligaccedilatildeo agrave proteina de ligaccedilatildeo ao
retinol (RBP) (Kanai et al 1968) Eacute sintetizada principalmente pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) e tambeacutem sintetizada pelas ceacutelulas epiteliais do
plexo coroacuteide e secretada para o liacutequido cefaloraquidiano (LCR) (Aleshire et al 1983)
Existem outros locais que expressam TTR mas em menor quantidade nomeadamente a
retina do olho (Martone et al 1988) o pacircncreas (Kato et al 1985) o saco vitelino
visceral (Soprano et al 1986) o intestino (Loughna et al 1995) o estocircmago coraccedilatildeo
muacutesculo e baccedilo (Soprano et al 1985) A TTR eacute uma proteiacutena do ponto de vista
filogeneacutetico extremamente conservada o que jaacute de si eacute um indicador da sua importacircncia
bioloacutegica (Richardson 2009)
O objectivo deste trabalho foi avaliar a expressatildeo de transtirretina ao longo do sistema
gastrointestinal do murganho nos seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon e
tambeacutem bexiga com cerca de 3 meses de idade O segundo objectivo foi identificar as
ceacutelulas responsaacuteveis por essa expressatildeo nos oacutergatildeos em estudo
Foi possiacutevel verificar que apenas o estocircmago apresenta valores de expressatildeo normalizada
de TTR diferente de zero expressatildeo essa muito inferior agrave do fiacutegado tal como se esperava
Por imunohistoquiacutemicaimunofluorescecircncia foi possiacutevel determinar que as ceacutelulas que
expressam TTR satildeo pouco abundantes e estatildeo presentes na regiatildeo glandular do estocircmago
do murganho e tambeacutem do humano Para aleacutem disto verificou-se que a TTR co-localiza
com somatostatina e que as ceacutelulas que sintetizam TTR correspondem agraves ceacutelulas D
responsaacuteveis pela secreccedilatildeo de somatostatina
Palavras-chave Transtirretina Polineuropatia Familiar Amiloidoacutetica Sistema
Gastrointestinal
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
v
Abstract
Transthyretin (TTR) is a plasma protein composed of four identical subunits of
approximately 14 KDa and molecular mass of 55 KDa (Blake et al 1978)
TTR is responsible for the transport of thyroxine (T4) (Andrea et al 1980) and retinol
(vitamin A) by binding to the retinol binding protein (RBP) (Kanai et al 1968) It is
synthesized primarily by the liver and secreted into the blood (Murakami et al 1987) and
also synthesized by epithelial cells of the choroid plexus and secreted into the
cerebrospinal fluid (CSF) (Aleshire et al 1983) There are other sites that express TTR
but in smaller quantities namely the retina of the eye (Martone et al 1988) pancreas
(Kato et al 1985) visceral yolk sac (Soprano et al 1986) intestine (Loughna et al
1995) stomach heart muscle and spleen (Soprano et al 1985)
TTR is a highly conserved protein which from a phylogenetic standpoint is itself an
indicator of its biological importance (Richardson 2009)
The aim of this study was to evaluate the expression of transthyretin along the
gastrointestinal tract of mice (3 months old) ndash esophagus stomach duodenum colon and
bladder as well The second aim was to identify the cells responsible for this expression in
the organs under study
It was possible to verify that only the stomach showed values of normalized expression
different from zero This expression was lower than in the liver as expected
By immunohistochemistry immunofluorescence it was determined that cells expressing
TTR are present in the glandular region of the stomach of mice and also in human
stomach It was also determined that D cells responsible for secretion of somatostatin
synthesize TTR
Key-words Transthyretin Familial Amyloid Polyneuropathy Gastrointestinal Tract
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
v
Abstract
Transthyretin (TTR) is a plasma protein composed of four identical subunits of
approximately 14 KDa and molecular mass of 55 KDa (Blake et al 1978)
TTR is responsible for the transport of thyroxine (T4) (Andrea et al 1980) and retinol
(vitamin A) by binding to the retinol binding protein (RBP) (Kanai et al 1968) It is
synthesized primarily by the liver and secreted into the blood (Murakami et al 1987) and
also synthesized by epithelial cells of the choroid plexus and secreted into the
cerebrospinal fluid (CSF) (Aleshire et al 1983) There are other sites that express TTR
but in smaller quantities namely the retina of the eye (Martone et al 1988) pancreas
(Kato et al 1985) visceral yolk sac (Soprano et al 1986) intestine (Loughna et al
1995) stomach heart muscle and spleen (Soprano et al 1985)
TTR is a highly conserved protein which from a phylogenetic standpoint is itself an
indicator of its biological importance (Richardson 2009)
The aim of this study was to evaluate the expression of transthyretin along the
gastrointestinal tract of mice (3 months old) ndash esophagus stomach duodenum colon and
bladder as well The second aim was to identify the cells responsible for this expression in
the organs under study
It was possible to verify that only the stomach showed values of normalized expression
different from zero This expression was lower than in the liver as expected
By immunohistochemistry immunofluorescence it was determined that cells expressing
TTR are present in the glandular region of the stomach of mice and also in human
stomach It was also determined that D cells responsible for secretion of somatostatin
synthesize TTR
Key-words Transthyretin Familial Amyloid Polyneuropathy Gastrointestinal Tract
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vi
Iacutendice
Abreviaturas vii
Introduccedilatildeo 1
Capiacutetulo I ndash Revisatildeo Bibliograacutefica 3
1 ndash Transtirretina 4
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo 4
ii - Expressatildeo de TTR 5
iii - Estrutura da TTR 5
iv - Funccedilotildees fisioloacutegicas da TTR 7
v - Metabolismo da TTR 10
vi - Variantes da TTR 11
2 - Sistema Gastrointestinal do murganho 14
Capiacutetulo II ndash Meacutetodos 17
1- Animais 18
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA 18
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA 19
4 - PCR em Tempo Real 20
5 - Imunohistoquiacutemica 20
6 - Hibridizaccedilatildeo in situ 22
Capiacutetulo III ndash Resultados 24
1 - PCR de cDNA 25
2 - PCR em Tempo Real 25
3 - Imunohistoquiacutemica 28
4 - Imunofluorescecircncia 29
5 - Hibridaccedilatildeo in situ 31
Capiacutetulo IV ndash Discussatildeo 32
Conclusatildeo 37
Bibliografia 39
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
vii
Abreviaturas
Aszlig ndash Peacuteptido szlig-amiloacuteide
Apo A1 ndash Apoliproteina A1
BSA ndash Albumina Seacuterica Bovina
cDNA ndash Aacutecido dexossiribonucleico complementar
CT ndash Treshold cycle
DEPC ndash Dietilpirocarbonato
FBS ndash Soro fetal bovino
HDL - Lipoproteina de elevada densidade
hTTR ndash Transtirretina humana
LCR ndash Liquido cefaloraquidiano
LDL ndash Lipoproteina de baixa densidade
mRNA ndash Aacutecido ribonucleico mensageiro
ORF ndash Quadro de leitura independente
mTTR ndash Transtirretina de murganho
PAF ndash Polineuropatia amiloidoacutetica familiar
PBS ndash Tampatildeo fosfato pH 75
PCR ndash Reacccedilatildeo em Cadeia da Polimerase
RBP ndash Proteina de ligaccedilatildeo ao retinol
RNA ndash Aacutecido ribonucleico
SSC ndash Tampatildeo citrato de soacutedio
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
viii
T3 ndash Triiodotironina
T4 ndash Tiroxina
TBG ndash Globulina de ligaccedilatildeo agrave tiroxina
TTR ndash Transtirretina
TTR KO ndash Murganho deficiente em TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
1
Introduccedilatildeo
A descoberta da transtirretina (TTR) no plasma (Seibert and Nelson 1942) e no liacutequido
cefaloraquidiano (LCR) (Kabat et al 1942) humano ocorreu em 1942 Esta eacute uma proteiacutena
constituiacuteda por quatro subunidades idecircnticas de 14 KDa cada e com uma massa molecular
de 55 KDa (Soprano et al 1985) Eacute principalmente sintetizada pelo fiacutegado e secretada
para o sangue (Murakami et al 1987) mas tambeacutem ao niacutevel do sistema nervoso pelas
ceacutelulas epiteliais do plexo coroacuteide e secretada para o LCR (Aleshire et al 1983) Em
menor quantidade verifica-se siacutentese noutros locais meninges cerebrais (Herbert et al
1986) ceacutelulas epiteliais da retina do olho (Martone et al 1988) pacircncreas (Kato et al
1985) saco vitelino visceral (Soprano et al 1986) placenta (McKinnon et al 2005)
intestino (Loughna et al 1995) e em pequena escala no estocircmago coraccedilatildeo muacutesculo e
baccedilo (Soprano et al 1985)
Eacute uma proteiacutena que possui duas funccedilotildees principais transporta a tiroxina (Andrea et al
1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al
1968) Os niacuteveis elevados de conservaccedilatildeo desta proteiacutena entre os mamiacuteferos e outros
vertebrados satildeo indicativos da sua importacircncia a niacutevel bioloacutegico (Richardson 2009)
Satildeo conhecidas mais de 100 mutaccedilotildees associadas a TTR com propriedades
amiloidogeacutenicas A primeira mutaccedilatildeo descrita foi associada agrave polineuropatia amiloidoacutetica
familiar (PAF) e foi identificada em 1952 por Corino de Andrade (Andrade 1952) eacute uma
patologia neurodegenerativa fatal e caracterizada pela deposiccedilatildeo de fibras de TTR mutada
principalmente no sistema nervoso perifeacuterico e autoacutenomo (Coimbra e Andrade 1971) A
sintomatologia inclui disfunccedilatildeo do sistema nervoso sensorial e autoacutenomo bem como dos
sistemas urogenital gastrointestinal e cardiacuteaco
Apesar de existirem referecircncias agrave expressatildeo de TTR no sistema gastrointestinal
desconhece-se a contribuiccedilatildeo da produccedilatildeo desta proteiacutena para a etiopatologia de PAF em
particular no que se refere agraves perturbaccedilotildees gastrointestinais observadas nesta doenccedila
Neste acircmbito os principais objectivos do presente trabalho foram (i) avaliar a expressatildeo de
transtirretina ao longo do tracto gastrointestinal ndash esoacutefago estocircmago duodeno coacutelon e
tambeacutem da bexiga de murganhos com cerca de 3 trecircs meses de idade e (ii) identificar
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
2
quais os locais principalmente no estocircmago e intestino onde ocorre a expressatildeo da
proteiacutena
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
3
Capiacutetulo I ndash Revisatildeo Bibliograacutefica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
4
1 ndash Transtirretina
A proteiacutena plasmaacutetica transtirretina (TTR) foi inicialmente designada de prealbumina
devido ao facto de migrar electroforeticamente antes da albumina Foi descoberta em 1942
no plasma (Seibert e Nelson 1942) e liquido cefaloraquidiano (LCR) (Kabat et al 1942)
humano O nome desta proteiacutena resulta da sua dupla funccedilatildeo fisioloacutegica como
transportadora de tiroxina (Andrea et al 1980) e de retinol (Vitamina A) (Kanai et al
1968) atraveacutes da proteiacutena de ligaccedilatildeo ao Retinol (RBP) (Raz et al 1970)
i - Estrutura do gene da transtirretina e a sua regulaccedilatildeo
O gene da TTR estaacute localizado no cromossoma 18 mais especificamente no 18q112-q121
(Whitehead et al 1984) e a proteiacutena eacute codificada por uma simples coacutepia geneacutetica (Tsuzuki
et al 1985) O gene eacute composto por 69 kilobases (kb) e consiste em 4 exotildees (E1 a E4)
cada um com cerca de 200 pares de bases (pb) 3 introtildees (I1 a I3) uma sequecircncia TATA
box na posiccedilatildeo de -30 a -24 pb seguida de uma regiatildeo rica em GC de aproximadamente 20
pb uma sequecircncia CAAT box na posiccedilatildeo -101 a -96 pb O sinal de poliadenilaccedilatildeo
(AATAAA) encontra-se a 123 pb a jusante da regiatildeo codificante (Sasaki et al 1985) Os
introtildees I1 e I3 contecircm dois quadros de leitura independentes (ORFs) contudo a sua funccedilatildeo
eacute ainda desconhecida (Tsuzuki et al 1985) Foi sugerido que estes ORFs podiam ser
responsaacuteveis pela expressatildeo de uma sequecircncia reguladora Mais tarde foi demonstrado que
estes ORFs correspondem a transcritos de TTR que natildeo foram sujeitos a splicing ou foram
sujeitos a um splicing incompleto e que por isso natildeo satildeo expressos nem como parte de um
polipeacuteptido ou independentemente (Soares et al 2003)
Estatildeo identificados dois locais principais de regulaccedilatildeo no gene da TTR uma sequecircncia
promotora proximal de -50 a -150 pb e um enhancer distal localizado entre 16 e 25 kb a
montante da regiatildeo codificante (Costa et al 1986) As mesmas sequecircncias reguladoras
estatildeo presentes no gene humano
O cDNA de TTR no murganho apresenta 82 de homologia com o cDNA humano e 90
com o de rato A sequecircncia de aminoaacutecidos apresenta ainda maior homologia 91 com o
humano e 96 com a sequecircncia de rato (Costa et al 1986)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
5
De um ponto de vista filogeneacutetico verifica-se que esta proteiacutena eacute extremamente
conservada O gene da TTR estaacute expresso nos peixes (Santos et al 1999) anfiacutebios reacutepteis
paacutessaros e ancestrais dos actuais mamiacuteferos (Richardson et al 1994)
ii - Expressatildeo de TTR
A TTR eacute expressa principalmente pelos hepatoacutecitos do fiacutegado (Murakami et al 1987) e
secretada para o sangue atingindo niacuteveis plasmaticos entre os 170 e 420 mgL (Vatassery
et al 1991) A concentraccedilatildeo plasmaacutetica de TTR eacute dependente da idade os receacutem nascidos
saudaacuteveis apresentam metade da concentraccedilatildeo encontrada nos adultos (Vahlquist et al
1975) e apoacutes os 50 anos verifica-se uma diminuiccedilatildeo dos niacuteveis desta proteiacutena (Ingenbleek
et al 1979) O fiacutegado eacute responsaacutevel pela produccedilatildeo de cerca de 90 da TTR presente no
sangue Adicionalmente a TTR eacute tambeacutem activamente sintetizada pelas ceacutelulas epiteliais
do plexo coroacuteide e secretada de forma unidireccional para o liacutequido cefaloraquidiano
(Aleshire et al 1983) Esta siacutentese representa cerca de 20 do total da proteiacutena no LCR
(Dickson et al 1986) O plexo coroacuteide apresenta os niacuteveis mais elevados de RNA
mensageiro cerca de 44 microg de mRNA por grama de peso corporal comparativamente aos
niacuteveis expressos pelo fiacutegado 030 microgg (Schreiber et al 1990)
Nos mamiacuteferos o gene da TTR eacute tambeacutem expresso nas meninges cerebrais (Herbert et al
1986) nos ilheacuteus de Langerhans do pacircncreas (Kato et al 1985) nas ceacutelulas do epiteacutelio da
retina do olho (Martone et al 1988) no saco vitelino visceral (Soprano et al 1986) na
placenta (McKinnon et al 2005) glacircndula pineal (Martone et al 1993) e em menores
quantidades no estocircmago coraccedilatildeo muacutesculo esqueleacutetico e baccedilo (Soprano et al 1985)
iii - Estrutura da TTR
A estrutura da TTR foi determinada por cristalografia de raios X em 1974 (Blake et al
1974) Eacute uma proteiacutena tetramerica constituiacuteda por quatro monoacutemeros idecircnticos distribuiacutedos
em torno de um canal central A massa molecular da proteiacutena eacute 55 KDa sendo que cada
monoacutemero tem de massa molecular 14KDa (Soprano et al 1985) e eacute constituiacutedo por 127
aminoaacutecidos (Kanda et al 1974)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
6
Em 1978 foi determinada por difracccedilatildeo de raio X a estrutura secundaacuteria e terciaacuteria de
cada monoacutemero (Blake et al 1978) Verifica-se que 55 dos aminoaacutecidos de cada
monoacutemero estatildeo associados agrave formaccedilatildeo de estruturas do tipo de folha szlig e 5 estatildeo
localizados no segmento de heacutelice α Cada monoacutemero eacute constituiacutedo por 8 cadeias szlig que
datildeo origem a duas folhas de quatro cadeias cada (Blake et al 1978)
Da associaccedilatildeo de dois monoacutemeros resulta um diacutemero formado por ligaccedilotildees de hidrogeacutenio
entre as cadeias F e H de duas folhas szlig de cada monoacutemero Os dois diacutemeros associam-se
originando um tetracircmero por interacccedilotildees do tipo diacutemero-diacutemero entre os dois loops no
topo de cada cadeia (Figura 1)
Figura 1 Representaccedilatildeo da estrutura quaternaacuteria da moleacutecula de TTR (subunidade A ndash cor
azul subunidade B - cor verde subunidade C ndash cor amarela subunidade D ndash cor
vermelha) Adaptado de (Foss et al 2005)
A formaccedilatildeo especiacutefica deste tetracircmero conduz agrave existecircncia de um canal central
hidrofoacutebico que atravessa a moleacutecula (Blake et al 1978 Nilsson e Peterson 1971) onde
ocorre a ligaccedilatildeo das hormonas da tiroacuteide
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
7
iv - Funccedilotildees fisioloacutegicas da TTR
A TTR tem duas funccedilotildees principais transporta a tiroxina ndash T4 (353rsquo5rsquo ndash tetraiodo-L-
tiroxina) (Andrea et al 1980) e a vitamina A (retinol) atraveacutes da proteiacutena de ligaccedilatildeo ao
Retinol (RBP) (Kanai et al 1968)
Transporte das hormonas da tiroacuteide
Eacute hoje reconhecida a enorme importacircncia que as hormonas da tiroacuteide tecircm sobre o
crescimento desenvolvimento metabolismo fertilidade sistema cardiovascular e
homeostasia do sistema nervoso central (Hulbert 2000)
A glacircndula da tiroacuteide sintetiza tiroxina (T4) que sofre deionizaccedilatildeo nos tecidos alvo dando
origem agrave forma biologicamente activa desta hormona a triiodotironina (T3) (Koumlhrle 1994)
No homem apoacutes ser secretada a hormona circula associada a trecircs proteiacutenas plasmaacuteticas
transtirretina (TTR) globulina de ligaccedilatildeo agrave tiroxina e albumina (TBG) (Robbins 1991)
Embora a TBG seja a proteiacutena plasmaacutetica que existe em maior quantidade nos humanos a
TTR eacute a maior transportadora de tiroxina no plasma de roedores e no LCR quer de
roedores quer de humanos Em humanos a TTR eacute responsaacutevel pelo transporte de 15 de
T4 no plasma e 80 no LCR (Robbins 1991)
A TTR apresenta no canal central de caraacutecter hidrofoacutebico dois locais de ligaccedilatildeo para a T4
(Nilsson e Peterson 1971 Blake et al 1978) Contudo apoacutes ligaccedilatildeo de uma moleacutecula de
T4 a afinidade da TTR para a segunda moleacutecula reduz-se drasticamente fazendo com que
cada tetracircmero de TTR apenas transporte uma moleacutecula de T4 (Robbins 1991) Verifica-se
tambeacutem que devido agrave elevada afinidade para a T4 a TTR natildeo liga agrave T3 (Andrea et al
1980) Para aleacutem da funccedilatildeo de transporte esta proteiacutena parece mediar a transferecircncia da
tiroxina seacuterica para os tecidos alvo principalmente no ceacuterebro atraveacutes das barreiras
sangue-ceacuterebro e sangue-plexo coroacuteide (Schreiber et al 1990 Hagen e Solberg 1974)
(Figura 2)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
8
Figura 2 Representaccedilatildeo do complexo TTR ndash tiroxina Adaptado de (Hamilton e Benson
2001)
Contudo embora os murganhos deficientes para TTR (TTR KO) apresentem diminuiccedilatildeo
dos niacuteveis de retinol e tiroxina apresentam um fenoacutetipo normal a ausecircncia de TTR natildeo
impede o transporte de tiroxina ateacute ao ceacuterebro (Palha et al 1997) bem como natildeo provoca
sinais de hipotiroidismo nem mesmo no parecircnquima cerebral (Palha et al 2000)
sugerindo a existecircncia de um mecanismo compensatoacuterio no metabolismo e transporte de
tiroxina que ocorreraacute na ausecircncia de TTR
Transporte de Retinol
O retinol tambeacutem designado de vitamina A eacute um micronutriente essencial envolvido em
inuacutemeras funccedilotildees fisioloacutegicas nomeadamente diferenciaccedilatildeo celular reproduccedilatildeo visatildeo e
desenvolvimento A TTR eacute a proteiacutena transportadora de retinol no plasma atraveacutes da
proteiacutena de ligaccedilatildeo ao retinol (RBP) (Kanai et al 1968) O RBP eacute uma proteiacutena
monomeacuterica de massa molecular de 21 kDa e consiste numa sequecircncia de 182 aminoaacutecidos
(Rask et al 1979) A ligaccedilatildeo do RBP agrave TTR aumenta a estabilidade do complexo
(Goodman 1984) e reduz os niacuteveis de filtraccedilatildeo glomerular da proteiacutena de baixo peso
molecular nos rins (Noy et al 1992) (Figura 3) O RBP previne a oxidaccedilatildeo do retinol e a
sua insolubilidade no plasma
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
9
Figura 3 Representaccedilatildeo do complexo TTR ndash RBP O tetracircmero de TTR (cinzento) com o
resiacuteduo 84 a amarelo e as duas moleacuteculas de ligaccedilatildeo (vermelho) e o retinol (preto)
Adaptado de (Hamilton e Benson 2001)
A concentraccedilatildeo de RBP no plasma humano eacute de cerca de 2 microM (Goodman 1984) enquanto
que a de TTR eacute de cerca de 45 microM (Ingenbleek et al 1994) e o complexo
TTRRBPRetinol estaacute presente no plasma na razatildeo de 111 (Blaner 1989)
Outras funccedilotildees da TTR
Recentemente tecircm sido descritas outras funccedilotildees importantes da TTR ao niacutevel do sistema
nervoso central Verificou-se que em animais TTR KO ocorre um aumento da actividade
exploratoacuteria e reduccedilatildeo de sinais depressivos provavelmente explicados por uma modulaccedilatildeo
do sistema noradreneacutergico (Sousa et al 2004) A TTR estaacute tambeacutem envolvida na
manutenccedilatildeo dos processos cognitivos durante o envelhecimento por actuaccedilatildeo na via de
sinalizaccedilatildeo do retinol (Brouillette et al 2008) Foi tambeacutem reportado que estes animais
apresentam niacuteveis de neuropeacuteptido Y superiores aos animais normais sendo este um
neuropeacuteptido com propriedades antidepressivas (Nunes et al 2006) A TTR tem tambeacutem
um papel importante no processo de regeneraccedilatildeo do nervo perifeacuterico in vivo apoacutes lesatildeo
(Fleming et al 2007)
Outra funccedilatildeo descrita da TTR eacute a capacidade de modular a agregaccedilatildeo do peacuteptido szlig-
amiloacuteide (Aszlig) formando complexos estaacuteveis com o Aszlig o que previne a formaccedilatildeo de
amiloacuteide (Schwarzman et al 1994) Para aleacutem disso os niacuteveis plasmaacuteticos de TTR estatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
10
significativamente diminuiacutedos em doentes com Alzheimer (Riisoslashen 1988 Serot et al
1997)
A TTR associa-se a lipoproteinas nomeadamente lipoproteinas de baixa densidade (LDL)
e de elevada densidade (HDL) (Tanaka et al 1994) e encontra-se tambeacutem associada agrave
apoliproteiacutena AI (apo AI) Estas observaccedilotildees sugerem uma relaccedilatildeo entre a TTR e a
biologia das lipoproteinas (Sousa et al 2000)
Para aleacutem das funccedilotildees acima descritas a TTR estaacute directamente envolvida nas fases de
resposta ao stress (Bernstein et al 1989) e em condiccedilotildees agudas de stress (Ingenbleek et
al 1994) Durante as fases de desnutriccedilatildeo ocorre uma diminuiccedilatildeo dos niacuteveis de TTR
(Ingenbleek et al 2002) O mesmo acontece durante os processos inflamatoacuterios onde se
verifica que os niacuteveis de TTR mRNA no fiacutegado decrescem (Qian et al 1995)
Outros estudos mostram a importacircncia do papel da TTR nos processos imunoloacutegicos
(Burton et al 1985) A TTR eacute capaz de inibir a secreccedilatildeo de interleucina I (Borish et al
1992) bem como estabelecer ligaccedilotildees com produtos de oxidaccedilatildeo da norepinefrina
(Boomsma et al 1991) com o aacutecido retinoacuteico (Smith et al 1994) flavonoides presentes
nas plantas (Lueprasitsakul et al 1990) contudo a importacircncia destas ligaccedilotildees natildeo eacute ainda
conhecida
v - Metabolismo da TTR
Em humanos a renovaccedilatildeo total de TTR eacute de cerca de 250-300 mgm2dia (Vahlquist et al
1973) O tempo meacutedio de vida desta proteiacutena eacute de 2 a 3 dias no ser humano (Vahlquist et
al 1973 Socolow et al 1965) e cerca de 29 horas em ratos (Dickson et al 1982) No
rato os principais locais de degradaccedilatildeo de TTR (proveniente do fiacutegado e do LCR) satildeo o
fiacutegado (36-38) o muacutesculo (12-15) e a pele (8-10) Os rins tecido adiposo testiacuteculos
e tracto gastrointestinal satildeo responsaacuteveis pela degradaccedilatildeo de cerca de 1-8 de TTR
Menos de 1 desta proteiacutena eacute degradada noutros tecidos (Makover et al 1988)
O mecanismo que conduz agrave internalizaccedilatildeo e degradaccedilatildeo de TTR ainda natildeo eacute conhecido
Alguns estudos indicam a existecircncia de um receptor responsaacutevel por mediar o transporte de
TTR para o meio intracelular (Divino e Schussler 1990 Vieira et al 1995) O receptor
designado de megalina pertencente agrave famiacutelia de receptores das lipoproteinas de baixa
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
11
densidade estaacute associado agrave internalizaccedilatildeo de TTR no rim prevenindo a sua filtraccedilatildeo a niacutevel
glomerular (Sousa et al 2000) A internalizaccedilatildeo de TTR ao niacutevel das ceacutelulas do fiacutegado
estaacute provavelmente associada ao metabolismo das lipoproteinas e parece ser mediada por
um receptor ainda natildeo identificado sensiacutevel para a proteiacutena RAP (Sousa et al 2001)
vi - Variantes da TTR
Ateacute agora satildeo conhecidas mais de 100 variantes de TTR com propriedades
amiloidogeacutenicas (httpamyloidosismutationscom) Haacute excepccedilatildeo de uma variante que
resulta de uma deleccedilatildeo de um aminoaacutecido na posiccedilatildeo 122 todas a outras resultam de
mutaccedilotildees pontuais na cadeia polipeptiacutedica
Satildeo conhecidas inuacutemeras mutaccedilotildees no gene de TTR sem qualquer manifestaccedilatildeo cliacutenica
um exemplo desta situaccedilatildeo eacute a mutaccedilatildeo Gly6Ser presente em 12 da populaccedilatildeo
caucasiana podendo por isso ser considerada um polimorfismo (Jacobson et al 1995)
Outras tecircm efeitos protectores como eacute o caso da mutaccedilatildeo Thr119Met esta forma de TTR
apresenta um aumento da afinidade de ligaccedilatildeo agrave T4 in vitro (Almeida et al 1997) e uma
maior capacidade de resistir agrave formaccedilatildeo de amiloacuteide (McCutchen et al 1995)
A maioria destas mutaccedilotildees conduz a situaccedilotildees de neuropatias perifeacutericas mas tambeacutem a
cardiomiopatias siacutendrome do tuacutenel caacuterpico e viacutetreopatia As mutaccedilotildees Leu55Pro e
Leu12Pro provocam quadros mais severos de doenccedila e aparecem mais precocemente As
diferenccedilas resultam muito provavelmente da instabilidade conformacional da proteiacutena
mutada (Brett et al 1999 Jacobson et al 1992) Algumas mutaccedilotildees no gene da TTR
resultam num grupo heterogeacuteneo de doenccedilas designadas por amiloidoses hereditaacuterias Em
1952 Corino de Andrade identificou a primeira forma destas doenccedilas e designou-a de
polineurapatia amiloidoacutetica familiar (PAF) (Andrade 1952) PAF eacute uma doenccedila
neurodegenerativa fatal caracterizada pela deposiccedilatildeo extracelular de fibras de TTR mutada
principalmente nos nervos e gacircnglios do sistema nervoso perifeacuterico e autoacutenomo (Coimbra
e Andrade 1971) A forma mais comum de PAF (nos doentes portugueses) resulta da
substituiccedilatildeo de uma valina por uma metionina na posiccedilatildeo 30 do gene da TTR (Val30Met)
(Saraiva et al 1984) Na regiatildeo de Portugal onde a doenccedila eacute mais comum a frequecircncia
desta mutaccedilatildeo estaacute estimada em 1 para 625 (Alves et al 1997) Para aleacutem do norte de
Portugal existem outros locais do mundo onde surgem doentes com PAF com a mutaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
12
Val30Met nomeadamente no Norte da Sueacutecia Japatildeo e Maiorca (Andersson 1970 Ando
et al 2000 Munar-Queacutes et al 1988)
De salientar tambeacutem que em alguns casos quando surge uma mutaccedilatildeo patogeacutenica e outra
natildeo patogeacutenica esta uacuteltima parece ter um efeito protector contra o desenvolvimento de
PAF (Coelho et al 1996) Eacute de notar tambeacutem que a TTR normal tem pouca tendecircncia a
depositar-se sobre a forma de fibras de amiloacuteide e a acontecer ocorre preferencialmente
no coraccedilatildeo numa patologia designada de amiloidose sisteacutemica senil em doentes muito
idosos (Westermark et al 1990)
Alguns estudos apontam no sentido do potencial amiloidogeacutenico da TTR resultar das
fracas interacccedilotildees entre as subunidades desta proteiacutena O tetracircmero de TTR na forma
mutada eacute pouco estaacutevel ocorrendo a dissociaccedilatildeo do mesmo em monoacutemeros com elevada
tendecircncia para se agregarem originando assim fibras de amiloacuteide insoluacuteveis (Quintas et
al 2001)
In vivo a formaccedilatildeo de amiloacuteide parece depender de outros factores para aleacutem do potencial
amiloidogeacutenico da proteiacutena (Cardoso et al 2008) Os depoacutesitos de amiloacuteide natildeo satildeo
constituiacutedos apenas por monoacutemeros de TTR mas tambeacutem por outros componentes
nomeadamente (i) componente amiloacuteide P seacuterico (Pepys 1988) (ii) glicosaminoglicanos
sulfonados (Snow et al 1987) (iii) apolipoproteina E (Gallo et al 1994) e J (Choi-Miura
et al 1992) (iv) α1-antiquimiotripsina (Abraham et al 1988) (v) componentes da
membrana ndash fibronectina laminina e colageacuteneo tipo IV (Scott et al 1983) (vi) proteiacutenas
complementares (Rogers et al 1992) e (vii) iotildees metaacutelicos (Bush et al 1994)
Os depoacutesitos de amiloacuteide podem ser facilmente identificados por coloraccedilatildeo de vermelho de
Congo observados com luz polarizada A existecircncia de depoacutesitos provoca birrrefringecircncia
de cor verde (figura 4) A acumulaccedilatildeo de fibras de amiloacuteide nos tecidos provoca danos
estruturais e funcionais nos mesmos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
13
Figura 4 Deposiccedilatildeo de TTR no nervo perifeacuterico de um doente PAF (A)
Imunohistoquiacutemica para TTR (B) Coloraccedilatildeo de Congo red Adaptado de (Sousa et al
2001)
Sintomatologia de PAF
Os sintomas cliacutenicos de PAF surgem na terceiraquarta deacutecada de vida do indiviacuteduo Os
sintomas iniciais estatildeo associados a uma disfunccedilatildeo do sistema nervoso sensorial e
autoacutenomo e incluem a perda de sensaccedilatildeo de dor e de temperatura nas extremidades
corporais Eacute uma doenccedila que afecta sobretudo o sistema nervoso perifeacuterico (Sousa e
Saraiva 2003) mas tambeacutem os sistemas urogenital gastrointestinal e coraccedilatildeo
No sistema urogenital surgem sintomas de disfunccedilatildeo ereacutectil e do sistema excretor que se
traduzem por uma urgecircncia em urinar e ao mesmo tempo em dificuldade na micccedilatildeo O
mecanismo responsaacutevel por esta disfunccedilatildeo do sistema excretor parece envolver os nervos
aferentes somaacuteticos e poacutes-ganglionares colineacutergicos (Ito et al 2006) Com o evoluir da
doenccedila surgem tambeacutem sintomas de disfunccedilatildeo renal (Ando et al 1993)
O sistema gastrointestinal eacute extremamente afectado em PAF As fibras de amiloacuteide surgem
em torno dos plexos mienteacutericos em muito maior quantidade do que em outro tipo de
amiloidose (Yoshimatsu et al 1998) No esoacutefago ocorre uma diminuiccedilatildeo dos movimentos
peristaacutelticos e um relaxamento anormal do esfiacutencter esofaacuteg ico inferior (Bjerle et al 1993)
Foi tambeacutem reportado que doentes com PAF apresentam um nuacutemero reduzido de ceacutelulas
endoacutecrinas que secretam serotonina colecistoquinina gastrina e secretina no duodeno o
que pode provocar uma desregulaccedilatildeo neuro-endoacutecrina responsaacutevel pela diminuiccedilatildeo da
motilidade intestinal (El-Salhy et al 1994) As perturbaccedilotildees no tracircnsito intestinal satildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
14
muito comuns tanto a diarreia como a obstipaccedilatildeo podem surgir em alternacircncia no mesmo
paciente A diarreia parece resultar de um crescimento bacteriano excessivo associado agrave
reduccedilatildeo da motilidade (Feurle 1987) e agrave reduccedilatildeo da absorccedilatildeo dos aacutecidos biliares (Suhr et
al 1992) A obstipaccedilatildeo parece estar associada ao fenoacutemeno de neuropatia enteacuterica (Ito et
al 2006) bem como a uma diminuiccedilatildeo do nuacutemero de ceacutelulas endoacutecrinas no intestino
grosso tal como jaacute tinha sido proposto para o intestino delgado (El-Salhy e Suhr 1996)
Ateacute ao momento o uacutenico tratamento para a PAF passa pelo transplante de fiacutegado que
elimina a produccedilatildeo de TTR mutada no fiacutegado e reduz drasticamente os seus niacuteveis no
plasma dos doentes (Holmgren et al 1991) Apoacutes o transplante ocorre uma melhoria
cliacutenica muito significativa na funccedilatildeo autoacutenoma bem como ao niacutevel dos sintomas
gastrointestinais O transplante deve ser realizado o mais cedo possiacutevel apoacutes o iniacutecio dos
primeiros sintomas cliacutenicos se possiacutevel logo no primeiro ano (Holmgren et al 1991)
2 - Sistema Gastrointestinal do murganho
O sistema gastrointestinal do murganho eacute similar ao dos restantes mamiacuteferos agrave excepccedilatildeo
dos ruminantes e consiste no esoacutefago estocircmago duodeno jejuno iacuteleo ceco coacutelon e
recto
O estocircmago encontra-se dividido em duas aacutereas distintas uma zona glandular e outra natildeo
glandular (Figura 5)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
15
Figura 5 Histologia do estocircmago A ndash Zona glandular B ndash Zona natildeo glandular
Todo o aparelho gastrointestinal desde o estocircmago ao coacutelon possui inuacutemeras ceacutelulas
endoacutecrinas As ceacutelulas enteroendoacutecrinas constituem cerca de 1 das ceacutelulas epiteliais
presentes neste sistema e encontram-se rodeadas pelas ceacutelulas parietais ceacutelulas principais e
ceacutelulas produtoras de muco no estocircmago Ao niacutevel do intestino para aleacutem das ceacutelulas
enteroendoacutecrinas existem tambeacutem ceacutelulas de goblet enteroacutecitos e ceacutelulas de Paneth (May e
Kaestner 2010) (Figura 6)
Figura 6 Ilustraccedilatildeo dos principais tipos de ceacutelulas presentes no epiteacutelio gastrointestinal
Adaptado de (May e Kaestner 2010)
Submucosa
A
A
B
Epiteacutelio secretor
Epiteacutelio secretor
Muscular
Muscular
Junccedilatildeo
esoacutefago
gaacutestrica
Enteroacutecitos
Ceacutelulas de Goblet
Ceacutelulas Endoacutecrinas
Ceacutelulas Estaminais
Ceacutelulas de Paneth
IntestinoEstocircmago
Ceacutelulas produtoras de muco
Ceacutelulas Principais
Ceacutelulas endoacutecrinas
Ceacutelulas Parietais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46
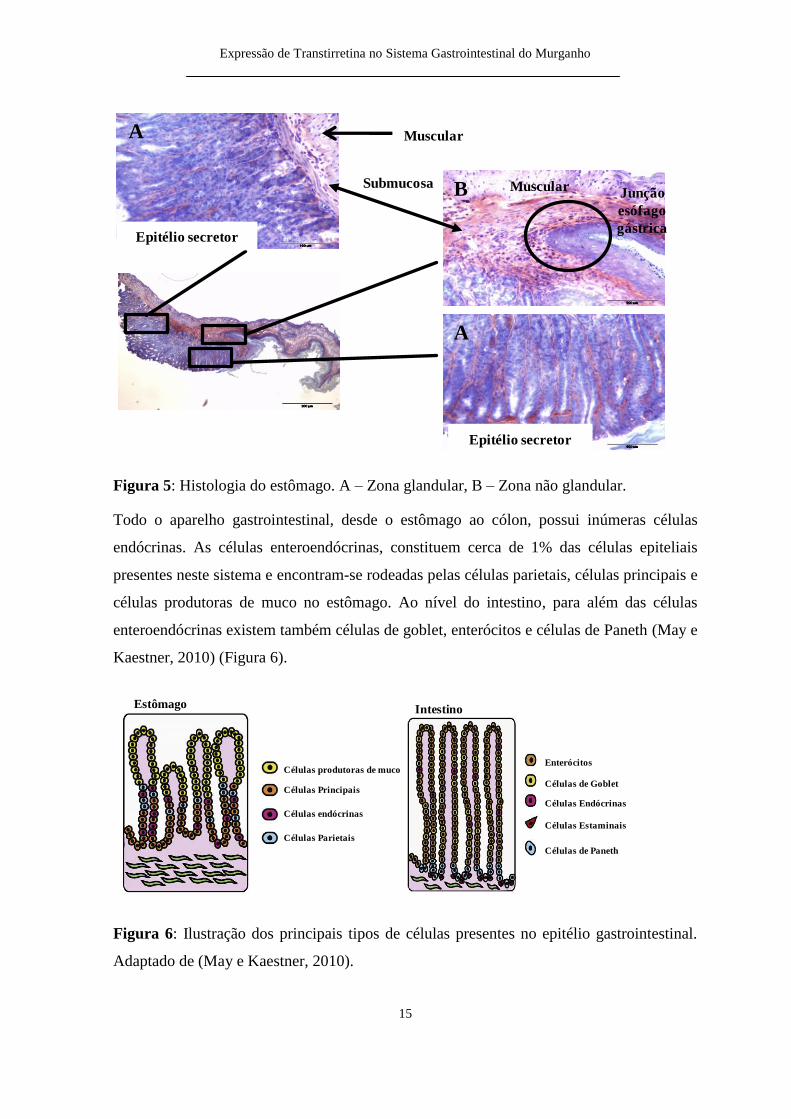
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
16
Existem pelo menos 15 tipos diferentes de ceacutelulas enteroendoacutecrinas classificadas de
acordo com o tipo de produtos hormonais e da ultraestrutura dos gracircnulos secretores (Rindi
et al 2004) Cada ceacutelula enteroendoacutecrina pode secretar uma ou mais hormonas bem como
substacircncias semelhantes a hormonas directamente para a lacircmina proacutepria do orgatildeo em
questatildeo e ser difundida ateacute aos capilares Essas hormonas incluem a gastrina histamina
serotonina colecistoquina somatostatina e peacuteptidos semelhantes ao glucagon (Rindi et al
2004) Embora estas ceacutelulas estejam presentes em pequena quantidade regulam a digestatildeo
a motilidade intestinal o apetite e o metabolismo (May e Kaestner 2010)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
17
Capiacutetulo II ndash Meacutetodos
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
18
1- Animais
Foram utilizados murganhos de background geneacutetico Sv129 e murganhos knockout para a
TTR (Episkopou et al 1993) Toda a manipulaccedilatildeo animal decorreu de acordo com as
directrizes e regras estipuladas pela Uniatildeo Nacional e Europeia para os cuidados e
manipulaccedilatildeo de animais de laboratoacuterio (Directiva 201063EU) Os animais foram
instalados em gaiolas apropriadas com uma temperatura ambiente constante (22ordmC) e um
fotoperiacuteodo de 12h de luz 12h de ausecircncia de luz A aacutegua e raccedilatildeo foram disponibilizadas
ad libitium Toda a manipulaccedilatildeo animal decorreu de forma a minimizar a dor e stress dos
animais envolvidos na experiecircncia bem como o nuacutemero de animais utilizados
A recolha de tecidos ocorreu apoacutes os animais serem anestesiados com uma mistura de
1mgKg de medetomidina (Sededorm) e 75 mgKg de ketamina (Clorketam 100)
Foram recolhidos os seguintes oacutergatildeos esoacutefago estocircmago duodeno coacutelon fiacutegado ceacuterebro
e bexiga Metade de cada oacutergatildeo foi imediatamente fixado em formalina neutra tamponada
e posteriormente os tecidos foram processados e incluiacutedos em parafina Apoacutes inclusatildeo os
tecidos foram cortados em secccedilotildees de 5 microm A outra metade foi imediatamente congelada
em gelo seco e mantida a -80ordmC ateacute ser usado
2 - Extracccedilatildeo de RNA total e siacutentese de cDNA
O RNA (aacutecido ribonucleico) total foi extraiacutedo de cada oacutergatildeo apoacutes homogeneizaccedilatildeo com
Reagente TRIzol (Invitrogen) de acordo com as instruccedilotildees do fabricante Foram usados
dois sistemas de homogeneizaccedilatildeo manual e com ultra turax (modelo VDI12 VWR) A
avaliaccedilatildeo quantitativa e qualitativa do RNA extraiacutedo foi realizada por espectrometria
(Nanodrop) e por electroforese em gel de agarose com brometo de etiacutedeo (Sigma)
A siacutentese de cDNA foi feita a partir de 1 microg de RNA total de cada uma das amostras A
esta quantidade de RNA foram adicionados 50 ng microl de Random hexacircmeros (Invitrogen)
10 mM de dNTP (dATP dCTP dGTP dTTP) e aacutegua ateacute perfazer um volume de reacccedilatildeo
de 10 microl As amostras foram incubadas a 65ordmC durante 5 minutos para ocorrer
desnaturaccedilatildeo De seguida foram colocadas sobre o gelo durante 1 minuto para manter a
desnaturaccedilatildeo das cadeias de RNA Apoacutes arrefecimento foi adicionado a cada volume de
reacccedilatildeo 10X RT buffer (Invitrogen) 25 mM de MgCl2 (Fermentas) 01M DTT
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
19
(invitrogen) e 40 U microl de RnaseOUT (invitrogen) Procedeu-se a uma incubaccedilatildeo a 25ordmC
durante 2 minutos e adicionou-se a cada tubo 50U de Superscripst II (Invitrogen)
Procedeu-se a trecircs ciclos de incubaccedilatildeo no primeiro as amostras foram mantidas a 25ordmC
durante 10 minutos no segundo foram mantidas a 42ordmC durante 50 minutos durante estes
dois ciclos ocorreu siacutentese de cDNA no terceiro ciclo as amostras foram mantidas a 70ordmC
durante 15 minutos ocorrendo assim a reacccedilatildeo de terminaccedilatildeo Apoacutes o uacuteltimo ciclo
procedeu-se ao arrefecimento em gelo Adicionou-se ainda a cada tubo 2U de RNase H
(New England Biolabs) e fez-se um uacuteltimo ciclo de aquecimento a 37ordmC durante 20
minutos para remover todo o RNA natildeo convertido em cDNA Apoacutes arrefecimento as
amostras foram conservadas a -80ordmC
3 - Reacccedilatildeo em cadeia da Polimerase (PCR) de cDNA
As sequecircncias de primers utilizadas foram desenhadas pelo Laboratoacuterio e satildeo as seguintes
gene de estudo ndash mTTR 5rsquoCCTTTGCCTCGCTGGACTGGTAT 3rsquo (sequecircncia sense) e 5rsquo
AATTCTGGGGGTTGCTGACGACA 3rsquo (sequecircncia antisense) gene de referecircncia ndash
HPRT (Hipoxantina-guanina fosforibosiltransferase) 5rsquo
GTAATGATCAGTCAACGGGGGAC 3rsquo (sequecircncia sense) e 5rsquo
CCAGCAAGCTTGCAACCTTAACCA 3 rsquo(sequecircncia antisense) O gene de estudo
conduz a uma banda de 419 e o de referecircncia a uma banda de 177 pares de bases Para um
volume de reacccedilatildeo de 50 microl por amostra foram utilizados 3 microl de cDNA 5 microl de 10X Taq
Buffer (Fermentas) 3 microl de Mgcl2 a 25 mM (Fermentas) 1 microl de dNTPs a 10mM
(Fermentas) 1 microl de cada um dos primers agrave concentraccedilatildeo de 10 OD 025 microl de Taq
Polimerase (Fermentas) O volume foi completado com aacutegua esteacuteril
A reacccedilatildeo de amplificaccedilatildeo ocorreu num termociclador (STRATAGENE) desnaturaccedilatildeo
inicial a 95ordmC durante 5 min 40 ciclos onde ocorreu uma desnaturaccedilatildeo a 95ordmC durante 30 s
um emparelhamento a 55ordmC durante 1 min e uma extensatildeo a 72ordmC durante 1min e 30 s (por
ciclo) e finalmente uma extensatildeo final a 72ordmC durante 6 min
Apoacutes adiccedilatildeo de loading buffer a cada amostra foi realizada uma electroforese em gel de
agarose a 2 com brometo de etiacutedeo (Sigma)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
20
4 - PCR em Tempo Real
Os primers utilizados foram desenhados usando o software Beacon gene de estudo -
mTTR 5rsquo-AGCCCTTTGCCTCTGGGAAGAC-3rsquo (sequecircncia sense) e 5rsquo-
TGCGATGGTGTAGTGGCGATGG-3rsquo (sequecircncia antisense) gene de referecircncia ndash 18S
5rsquo- AAATCAGTTATGGTTCCTTTGGTC-3rsquo (sequecircncia sense) e 5rsquo-
GCTCTAGAATTACCACAGTTATCCAA-3rsquo (sequecircncia antisense)
Os primers sense e antisense emparelham em diferentes exotildees O PCR em tempo real foi
executado num termociclador IQ5 (Bio-Rad Laboratories Hercules CA USA) Para um
volume final de reacccedilatildeo de 20 microl adicionou-se 10 microl de SYBR Green master mix (Bio-
Rad) 025 microl de cada primer de mTTR a 20 mM e 030 microl de cada primer de 18S a 10mM
1 microl de cada amostra de cDNA diluiacuteda num factor de 110 e aacutegua esteacuteril As condiccedilotildees
estabelecidas para o PCR foram desnaturaccedilatildeo inicial a 95ordmC durante 10 segundos seguido
de 40 ciclos a 95ordmC durante 15 segundos e de 20 segundos a 60ordmC (temperatura de
hibridaccedilatildeo) A mediccedilatildeo do produto fluorescente ocorreu num uacutenico modo de aquisiccedilatildeo a
72ordmC apoacutes cada ciclo
As curvas de calibraccedilatildeo do gene de estudo e do gene de referecircncia foram efectuadas
usando um conjunto de diluiccedilotildees seriadas de factor 14 em triplicado Foram aceites os
valores de eficiecircncia compreendidos entre 90 e 105 Os controlos negativos (onde se
substituiu cDNA por aacutegua esteacuteril) foram feitos para cada conjunto de reacccedilotildees Procedeu-
se agrave quantificaccedilatildeo da expressatildeo do gene de mTTR pelo meacutetodo de ΔCT (threshold cycle)
usando um gene de referecircncia Os resultados foram apresentados em unidades arbitraacuterias e
correspondem agrave meacutedia plusmn desvio padratildeo Os grupos de orgatildeos em estudo foram comparados
atraveacutes da utilizaccedilatildeo do teste t Studentrsquos Os resultados foram considerados
estatisticamente significantes quando plt 005 plt 001 e plt 0001
5 - Imunohistoquiacutemica
As secccedilotildees dos diferentes tecidos recolhidos foram analisados quanto agrave expressatildeo de TTR
utilizando para isso um anticorpo produzido em coelho contra TTR recombinante de
ratinho ndash mTTR - (11500 Q-Biogen llkirch Cedex France) e tambeacutem quanto agrave expressatildeo
de cromogranina A (1400 Abcam Cambridge UK) As secccedilotildees dos tecidos dos animais
TTR KO (TTR --) funcionaram como controlos negativos de toda a anaacutelise histoquiacutemica
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
21
Os procedimentos para a imunohistoquiacutemica envolveram a desparafinizaccedilatildeo de cerca de 30
min em Histoclear (National Diagnostics) hidrataccedilatildeo dos tecidos a partir de uma seacuterie
decrescente de aacutelcoois (100 - 90 - 80 -70 as lacircminas permaneceram 10 min em
cada aacutelcool) apoacutes hidrataccedilatildeo os tecidos foram imersos em tampatildeo fosfato a pH 75 (PBS)
A actividade da peroxidase endoacutegena foi bloqueada com uma soluccedilatildeo de 3 de H2O2 em
PBS De seguida procedeu-se agrave incubaccedilatildeo dos tecidos com um tampatildeo de bloqueio
contendo 1 de albumina seacuterica bovina (BSA) e 4 de soro fetal bovino (FBS) em PBS
durante 1h agrave temperatura ambiente O anticorpo primaacuterio foi diluiacutedo no mesmo tampatildeo de
bloqueio e a incubaccedilatildeo ocorreu durante toda a noite a 4ordmC Apoacutes lavagens em PBS
procedeu-se agrave incubaccedilatildeo das lacircminas com o anticorpo secundaacuterio ndash anti-coelho IgG
(Sigma St Louis MO USA) 1h agrave temperatura ambiente e posteriormente com
Extravidina peroxidade (Sigma) nas mesmas condiccedilotildees Como substrato de cor foi
utilizada a diaminobenzidina (DAB ndash Sigma) e como contrastante nuclear uma soluccedilatildeo de
hematoxilina modificada segundo Gill II (Merck) Apoacutes lavagem em aacutegua corrente
procedeu-se agrave desidrataccedilatildeo dos tecidos a partir de uma seacuterie crescente de aacutelcoois (70 -
80 - 90 - 100) De seguida as lacircminas foram colocadas novamente em histoclear
durante cerca de 30 min e finalmente foram montadas utilizando para isso meio de
montagem natildeo aquoso ndash Entellan (Merck)
Para a teacutecnica de imunofluorescecircncia foram utilizadas secccedilotildees dos tecidos dos animais em
estudo e estocircmago humano (cedido pelo Laboratoacuterio de Histologia do Centro de
Investigaccedilatildeo em Ciecircncias da Sauacutede da Universidade da Beira Interior) Procedeu-se a uma
dupla marcaccedilatildeo destas secccedilotildees utilizando os seguintes conjuntos de anticorpos anti-
mTTR (11500) com anti-Somatostatina (150 Santa Cruz Biotechnology Santa Cruz CA
USA) ou com anti-Beta ATPase de hidrogeacutenio e potaacutessio (150 Abcam Cambridge UK)
No tecido humano apenas se realizou uma dupla marcaccedilatildeo com um anticorpo anti
prealbumina humana (1 200 Dako Denmark) e o anticorpo anti-Somatostanina jaacute
referido Os anticorpos secundaacuterios incluem (i) um anticorpo anti-coelho 488 (ii) um
anticorpo anti-murganho 594 e (iii) um anticorpo anti-cabra 568 e foram utilizados numa
diluiccedilatildeo de 1500 O procedimento experimental envolveu a desparafinizaccedilatildeo hidrataccedilatildeo
dos tecidos incubaccedilatildeo com o tampatildeo de bloqueio e posterior incubaccedilatildeo com os anticorpos
primaacuterios durante toda a noite a 4ordmC (procedimentos idecircnticos aos realizados para
imunohistoquiacutemica) Os anticorpos secundaacuterios foram incubados durante 2h agrave temperatura
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
22
ambiente De seguida procedeu-se agrave montagem das lacircminas em meio de montagem para
fluorescecircncia ndash Vectashield (Vector laboratories Inc Burlingame) As lacircminas de tecido
de ratinho foram observadas com um microscoacutepio de fluorescecircncia AxioImager Z1 (Carl
Zeiss Germany) enquanto as lacircminas de tecido humano foram observadas com um
microscoacutepio confocal Leica SP2 AOBS SE (Leica Microsystems Germany) as imagens
forma analisados com os programas associados a cada microscoacutepio
6 - Hibridizaccedilatildeo in situ
Para a hibridizaccedilatildeo in situ foram utilizadas lacircminas contendo tecido incluiacutedo em parafina
Todo o processamento manuseamento e corte dos tecidos decorreu de forma a minimizar
possiacuteveis contaminaccedilotildees com RNA Foram utilizados secccedilotildees de ceacuterebro contendo plexo
coroacuteide que serviram de controlo positivo e secccedilotildees de estocircmago todos os tecidos eram
provenientes de animais com TTR endoacutegena (TTR++)
A detecccedilatildeo do RNA mensageiro (mRNA) de TTR foi realizada pelo meacutetodo da
digoxigenina (Roche Molecular Biochemicals Indianapolis IN USA) As sondas de
mTTR foram transcritas a partir de um plasmiacutedeo PGEM-4 (Promega) contendo o cDNA
completo de mTTR A sonda antisense foi transcrita a partir do promotor SP6 apoacutes
linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo Hind III (New England) e a sonda
sense a partir do promotor T3 apoacutes linearizaccedilatildeo do plasmiacutedeo com a enzima de restriccedilatildeo
Sma I (New England)
Apoacutes desparafinizaccedilatildeo e hidrataccedilatildeo dos tecidos (processos previamente descritos) os slides
foram tratados com uma soluccedilatildeo de HCl 02 M durante 10 min agrave temperatura ambiente 50
microgmL de proteinase K durante 10 min a 37ordmC 02 de glicina durante 5 min 4 de
paraformaldeiacutedo pH 7 durante 1 min e 133 de trietanolamina 025 de anidrido aceacutetico
durante 10 min Entre cada soluccedilatildeo os slides foram imersos duas vezes em PBS tratado
com Dietilpirocarbonato (DEPC) A preacute-hibridaccedilatildeo ocorreu durante 4h agrave temperatura de
58ordmC no tampatildeo de hibridizaccedilatildeo [10X cloreto de soacutedio tampatildeo citrato de soacutedio (SSC)
50 formamida] e a hibridaccedilatildeo ocorreu a 55ordmC durante toda a noite foi utilizada uma
concentraccedilatildeo de 25 ng microl de ribosondas No dia seguintes as lacircminas foram imersas em
4X 2X e 1X SSC durante 10 min e tratados com 20 microgmL de RNAse A durante 30 min a
37ordmC De seguida foram colocados numa soluccedilatildeo contendo 2xSSC e 50 de formamida
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
23
durante 1h a 52ordmC A detecccedilatildeo da hibridaccedilatildeo ocorreu utilizando um anticorpo conjugado
com fosfatase alcalina anti digoxigenina (Roche) de acordo com as instruccedilotildees do
fabricante Foram realizados 6 ensaios para optimizar as condiccedilotildees de hibridaccedilatildeo fazendo
variar a temperatura de hibridaccedilatildeo entre os 55ordmC e os 60ordmC e tambeacutem o tempo de exposiccedilatildeo
agrave proteinase K que variou entre os 10 min e os 30 min O ensaio descrito foi o uacutenico onde
ocorreu detecccedilatildeo de hibridaccedilatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
24
Capiacutetulo III ndash Resultados
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
25
1 - PCR de cDNA
Numa primeira fase para determinar se a TTR eacute expressa no tracto gastrointestinal de
murganho foi realizado um PCR para detecccedilatildeo de cDNA dos seguintes tecidos coacutelon
estocircmago pacircncreas bexiga esoacutefago duodeno e fiacutegado Os tecidos foram provenientes de
um murganho TTR ++ de 3 meses de idade (figura 7)
Figura 7 PCR de cDNA de um aninal TTR++ Legenda Cl ndash Coacutelon Et ndash Estocircmago Pc
ndash Pacircncreas Bx Bexiga Es ndash Esoacutefago Do ndash Duodeno Fg ndash Fiacutegado RT neg ndash Controlo
negativo do RT-PCR
Verificou-se que a banda correspondente a mTTR estava presente no coacutelon estocircmago e
bexiga com uma intensidade muito semelhante Essa intensidade estava diminuiacuteda no
duodeno O fiacutegado funcionou neste PCR como um controlo positivo uma vez que se sabe
ser um local de expressatildeo abundante de TTR Natildeo se observou banda de TTR no esoacutefago
A banda de HPRT (gene de referecircncia) estava presente em todas as amostras As bandas de
mTTR e HPRT natildeo apareceram no pacircncreas sendo este resultado inesperado sabendo que
o pacircncreas eacute tambeacutem um local de siacutentese de TTR Este resultado pode ter ficado a dever-se
agrave fraca qualidade do mRNA de pacircncreas usado no RT-PCR e devido ao facto deste tecido
ser rico em nucleases
Este resultado permitiu seleccionar os oacutergatildeos a utilizar nos procedimentos experimentais
seguintes tendo sido escolhidos o estocircmago coacutelon duodeno e bexiga O pacircncreas deixou
de ser usado como controlo positivo passando a ser o fiacutegado o controlo seleccionado
2 - PCR em Tempo Real
Apoacutes anaacutelise qualitativa pretendeu-se avaliar de forma quantitativa e comparativa a
expressatildeo de TTR nos tecidos em estudo Para este ensaio foi utilizado um grupo de 5
animais (3 fecircmeas e 2 machos) de 3-4 meses de idade
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
26
A B
O primeiro ensaio realizado mostrou uma enorme variabilidade na expressatildeo de TTR
entre os animais pelo que se tentou perceber o que poderia estar na base destas
observaccedilotildees Os RNA extraiacutedos tinham sido quantificados por espectrometria e a sua
qualidade avaliada atraveacutes do paracircmetro 260280 Procedeu-se por isso a uma electroforese
em gel de agarose com brometo de etiacutedeo para determinar a qualidade do RNA que estava
a ser usado O gel de agarose mostrou que os RNA extraiacutedos estavam muito degradados
natildeo sendo possiacutevel distinguir as bandas correspondes agraves subunidades ribossomais (Figura
8) Voltou-se por isso a extrair RNA dos oacutergatildeos em estudo Esta nova extracccedilatildeo foi
realizada apoacutes homogeneizaccedilatildeo dos tecidos com um homogeneizador Ultra Turax que
permite uma homogeneizaccedilatildeo muito mais raacutepida do que o meacutetodo usado na extracccedilatildeo das
amostras anteriores Apoacutes extracccedilatildeo o RNA foi quantificado por espectrometria e a sua
integridade avaliada por electroforese em gel de agarose (Figura 8)
Figura 8 Integridade das amostras de RNA obtida a partir de dois sistemas diferentes de
homogeneizaccedilatildeo Legenda Et ndash Estocircmago Cl ndash Coacutelon Do ndash Duodeno Fg ndash Fiacutegado e Bx
ndash Bexiga
O PCR em tempo real foi repetido com as novas amostras de RNA Em cada ensaio foi
realizada uma curva de calibraccedilatildeo para o gene de estudo (mTTR) e gene de referecircncia
(18S) os ensaios foram validados quando os valores de eficiecircncia estavam compreendidos
entre 90 e 105 (Figura 9)
Figura 9 Curvas de calibraccedilatildeo para mTTR (A) e 18S (B)
250 bp
500 bp
750 bp
1000 bp
1500 bp
Fg FgEt EtCl ClDo Do Bx
Extraccedilatildeo Manual Extraccedilatildeo com Ultra turax
28 S
18 S
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46
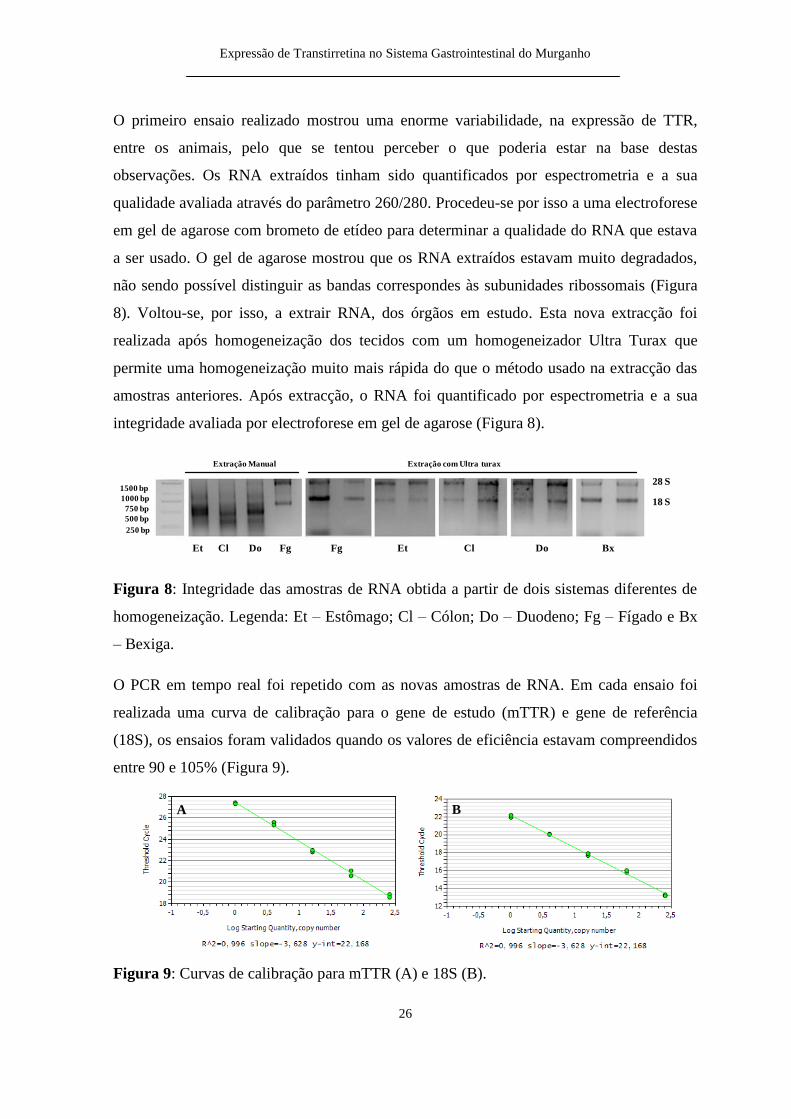
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
27
A especificidade da reacccedilatildeo foi confirmada atraveacutes da curva de fusatildeo que eacute um indicador
da temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias de mTTR e 18S
(Figura 10) Para cada amostra foram realizados triplicados e depois de analisados foram
descartados os valores de CT (treshold cycle) que diferissem entre si mais de 05 unidades
Figura 10 Exemplo de uma curva de fusatildeo para mTTR
A expressatildeo de TTR foi determinada por aplicaccedilatildeo do meacutetodo de ΔCT e conduziu ao
seguinte resultado (Figura 11)
Figura 11 Expressatildeo normalizada de TTR no sistema gastrointestinal Os dados
encontram-se em unidades arbitraacuterias e correspondem agrave meacutedia plusmn desvio padratildeo (plt
0001)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
28
O PCR em tempo real permitiu determinar a expressatildeo de TTR no estocircmago do murganho
embora esta expressatildeo tal como se esperava seja muito inferior aos niacuteveis encontrados no
fiacutegado Nos restantes oacutergatildeos em estudo ndash coacutelon duodeno e bexiga verificou-se que essa
expressatildeo ou natildeo existia ou era muito residual natildeo tendo por isso qualquer importacircncia
estatiacutestica Assim os ensaios seguintes incidiram apenas sobre estocircmago e fiacutegado
3 - Imunohistoquiacutemica
Apoacutes determinaccedilatildeo quantitativa da expressatildeo de mRNA de TTR no estocircmago do
murganho pretendeu-se determinar se esta expressatildeo de mRNA correspondia agrave presenccedila
de proteiacutena e no caso de ocorrer qual seria a localizaccedilatildeo celularhistoloacutegica dessa siacutentese
Para este efeito usaram-se teacutecnicas de imunohistoquiacutemicaimunofluorescecircncia
Na teacutecnica de imunohistoquiacutemica foi usado um anticorpo anti-TTR em lacircminas contendo
secccedilotildees transversais de estocircmago e pacircncreas O pacircncreas foi utilizado como controlo
positivo por ser mais faacutecil identificar neste oacutergatildeo os locais de siacutentese de TTR O controlo
negativo correspondeu aos mesmos oacutergatildeos (estocircmago e pacircncreas) de um animal TTR--
(Figura 12)
Figura 12 Imunohistoquimica para mTTR de (A) epiteacutelio secretor do estocircmago e (B)
ilheacuteus de Langerhans do pacircncreas As setas indicam o local de imunoreactividade positiva
para TTR
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46
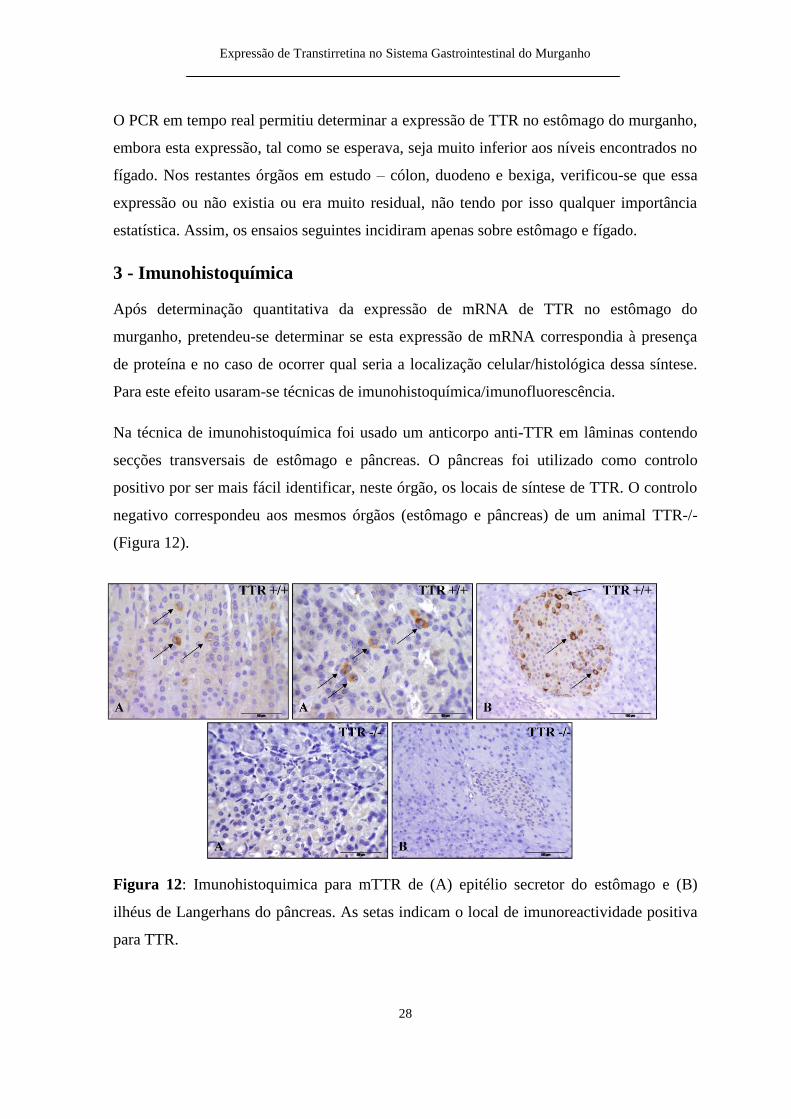
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
29
A marcaccedilatildeo de TTR no animal TTR++ surge na zona glandular do estocircmago do
murganho e parece coincidir com ceacutelulas muito especiacuteficas Natildeo eacute uma marcaccedilatildeo muito
abundante o que vem de encontro aos resultados obtidos com o PCR em tempo real O
animal TTR-- natildeo continha nenhuma marcaccedilatildeo para TTR como esperado
Com estes resultados surgiu a necessidade de identificar as ceacutelulas com reactividade para o
anticorpo anti-TTR A morfologia das ceacutelulas positivas para TTR parecia indicar tratar-se
de ceacutelulas endoacutecrinas Realizou-se por isso uma imunohistoquiacutemica para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e TTR em lacircminas contendo secccedilotildees sucessivas do
mesmo oacutergatildeo contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre as duas
proteiacutenas Para co-localizar dois antigeacutenios numa mesma lacircmina optou-se pela teacutecnica de
imunofluorescecircncia que o permite fazer de uma forma simples e eficaz
Natildeo foi possiacutevel proceder a ensaios de imunofluorescecircncia com o anticorpo de
cromogranina A e mTTR porque os anticorpos satildeo produzidos no mesmo animal
4 - Imunofluorescecircncia
O primeiro ensaio consistiu na localizaccedilatildeo de mTTR e de Beta ATPase de hidrogeacutenio e
potaacutessio Este uacuteltimo anticorpo permite identificar as ceacutelulas parietais do estocircmago
responsaacuteveis pela produccedilatildeo e secreccedilatildeo de aacutecido cloriacutedrico bem como de factor intriacutenseco
Natildeo se verificou qualquer co-localizaccedilatildeo a partir dos anticorpos utilizados (dados natildeo
apresentados)
Num segundo ensaio procedeu-se agrave localizaccedilatildeo de mTTR em ceacutelulas secretoras de
somatostatina utilizando para isso o anticorpo anti-mTTR e o anticorpo anti-somatostatina
(Figura 13)
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
30
Figura 13 Imunofluorescecircncia para mTTR e somatostatina em (A) estocircmago e (B)
pacircncreas
Verificou-se a localizaccedilatildeo de TTR nas ceacutelulas que secretam somatostatina no estocircmago
Mais uma vez o pacircncreas funcionou como controlo positivo da teacutecnica e verifica-se que
neste oacutergatildeo as ceacutelulas que sintetizam TTR satildeo diferentes das que secretam somatostatina
Foi utilizado um animal TTR-- como controlo negativo tendo-se verificado que natildeo surgia
qualquer tipo de marcaccedilatildeo para TTR (dados natildeo apresentados)
Com estes resultados questionou-se a hipoacutetese da localizaccedilatildeo de TTR nas ceacutelulas
secretoras de somatostatina ocorrer tambeacutem no estocircmago humano Pelo que se repetiu a
imunofluorescecircncia em lacircminas contendo tecido de estocircmago humano usando agora um
anticorpo anti-hTTR e o mesmo anticorpo anti-somatostatina (Figura 14) Foi possiacutevel
verificar que tambeacutem neste tecido ocorria co-localizaccedilatildeo entre TTR e somatostatina
Figura 14 Imunofluorescecircncia para hTTR e somatostatina em estocircmago humano
Somatostatina TTRSomatostatina
TTR Somatostatina TTRSomatostatina
TTRA AA
B B B
TTR Somatostatina TTRSomatostatina
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46
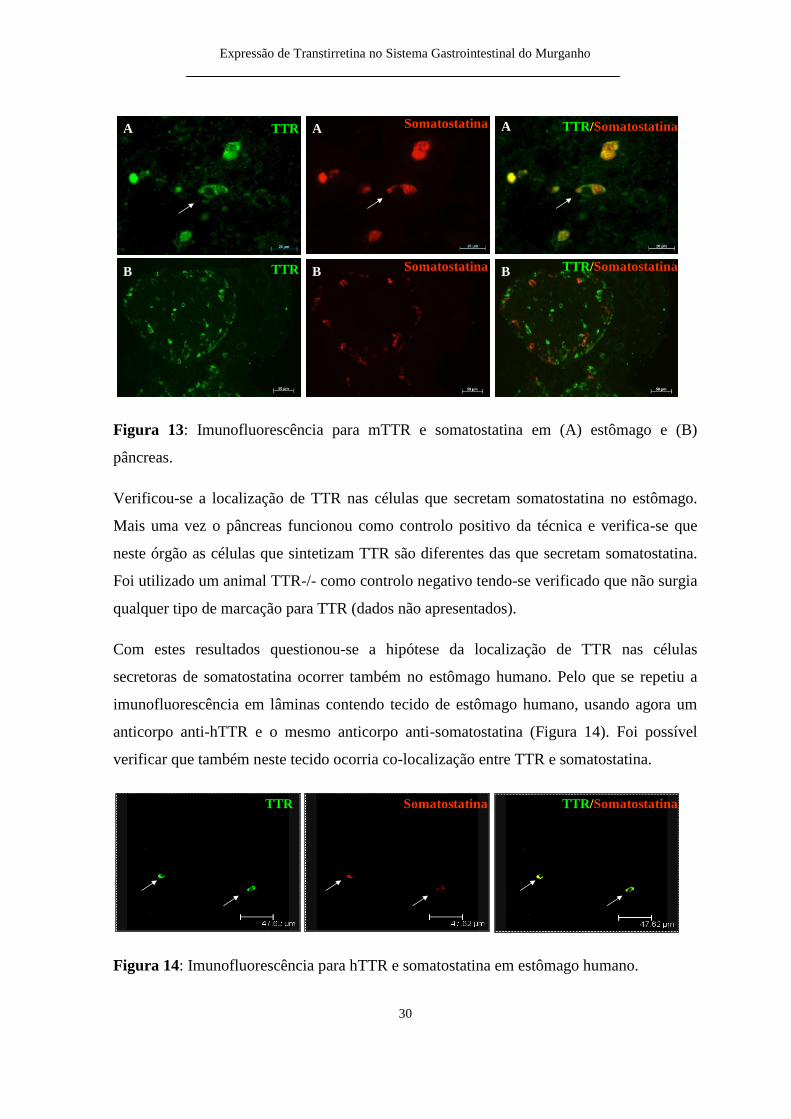
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
31
5 - Hibridaccedilatildeo in situ
O uacuteltimo procedimento experimental consistiu numa hibridaccedilatildeo in situ para confirmar a
siacutentese de TTR em ceacutelulas secretoras de somatostatina Esta teacutecnica foi aplicada a lacircminas
contendo estocircmago e ceacuterebro com plexo coroacuteide (como controlo positivo) incluiacutedos em
parafina Com esta teacutecnica eacute possiacutevel detectar a presenccedila de mRNA de TTR de murganho e
ao mesmo tempo identificar o local onde esse mRNA se encontra (Figura 15)
Figura15 Hibridaccedilatildeo in situ com uma sonda anti-mTTR (A) Plexo coroacuteide hibridado com
sonda antisense (AS) (B) Ampliaccedilatildeo da imagem (A) (C) Plexo coroacuteide hibridado com
sonda sense (SS) (D) Estocircmago hibridado com sonda antisense (AS) (E) Estocircmago
hibridado com sonda sense (SS)
A sonda antisense eacute complementar ao mRNA de TTR existente no murganho ocorrendo
emparelhamento das duas sequecircncias Esse emparelhamento eacute visiacutevel na imagem A onde
todo o plexo coroacuteide aparece marcado tal como era espectaacutevel A sonda sense tem a
mesma sequecircncia do mRNA pelo que natildeo ocorre emparelhamento funciona por isso como
um controlo negativo de todo o processo (imagens C e E) Na imagem D seria de esperar
observar marcaccedilatildeo das ceacutelulas identificadas anteriormente quando o slide foi incubado
com a sonda antisense mas tal natildeo se verificou
SAA SSCB
SAD SSE
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46
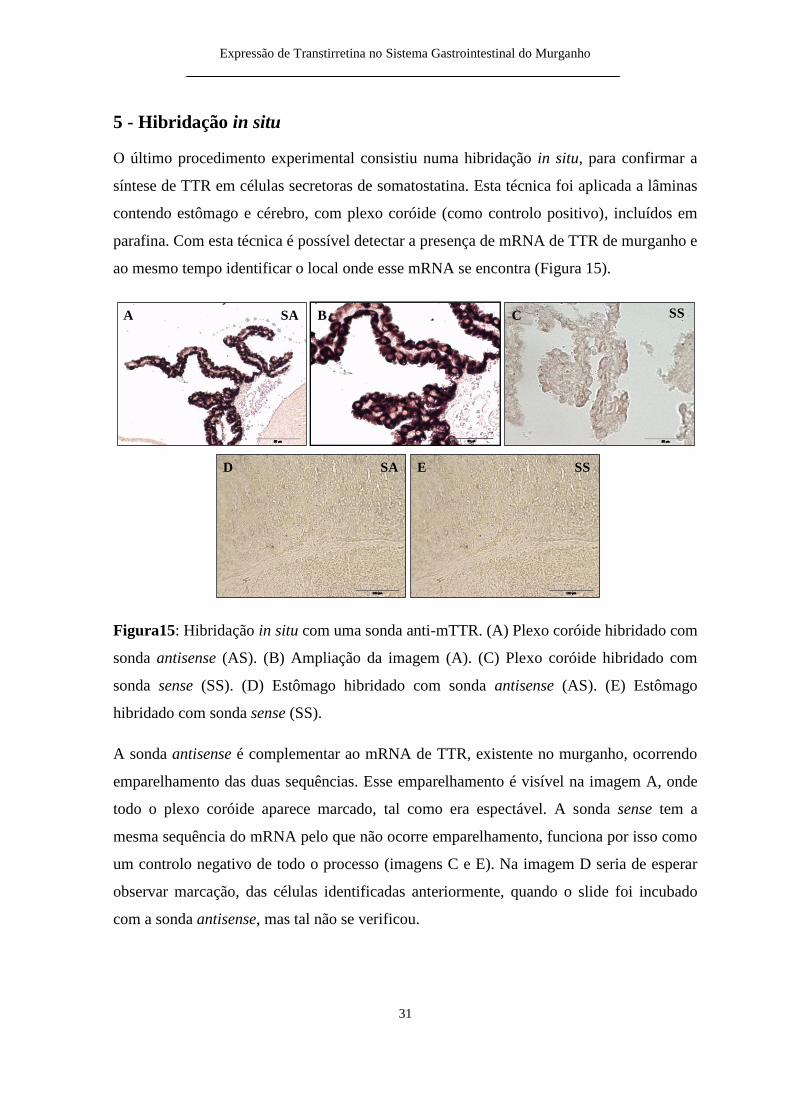
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
32
Capiacutetulo IV ndash Discussatildeo
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
33
A primeira parte deste trabalho visava sobretudo avaliar a expressatildeo de TTR ao longo do
tracto gastrointestinal de murganhos com cerca de 3 meses de idade Para isso foram
realizados dois tipos de procedimentos experimentais PCR de cDNA e PCR em tempo
real
A realizaccedilatildeo do PCR de cDNA de murganho permitiu de uma forma qualitativa
determinar que agrave excepccedilatildeo do esoacutefago todos os restantes oacutergatildeos do sistema
gastrointestinal investigado apresentavam a banda de mTTR bem como a do gene de
referecircncia Sabendo que o fiacutegado eacute um dos maiores sintetizadores de TTR do organismo
seria de esperar que a intensidade da banda de mTTR nos oacutergatildeos em estudo estivesse
diminuiacuteda em relaccedilatildeo ao fiacutegado mas tal natildeo se verificou Estes resultados permitem
concluir que este tipo de PCR tradicional apenas indica a existecircncia de cDNA de mTTR
sem estabelecer qualquer relaccedilatildeo entre a presenccedila e a quantidade de mRNA A expressatildeo
de TTR no estocircmago tinha jaacute sido referida (Soprano et al 1985) embora na mesma
publicaccedilatildeo seja referido o facto de o pacircncreas natildeo expressar TTR o que se sabe hoje de
facto acontecer
Apoacutes anaacutelise qualitativa foi necessaacuterio de uma forma quantitativa determinar a expressatildeo
de mRNA de TTR presente no estocircmago duodeno coacutelon e bexiga A quantificaccedilatildeo foi
realizada por PCR em tempo real Esta teacutecnica permite a amplificaccedilatildeo detecccedilatildeo e
quantificaccedilatildeo de DNA numa uacutenica etapa tornando mais raacutepido todo o processo de
obtenccedilatildeo de resultados e minimizando o risco de possiacuteveis contaminaccedilotildees A curva de
fusatildeo eacute uma medida da especificidade da reacccedilatildeo e indica o ponto correspondente agrave
temperatura de dissociaccedilatildeo dos primers especiacuteficos para as sequecircncias em estudo O
primeiro ensaio realizado conduziu a resultados pouco concordantes que se ficaram a dever
ao processo de homogeneizaccedilatildeo inicialmente utilizado Com a alteraccedilatildeo do meacutetodo de
extracccedilatildeo verificou-se que os resultados passaram a ser muito mais concordantes e com
uma menor variabilidade entre o grupo de animais uma vez que o RNA utilizado estava
integro Conclui-se portanto que eacute fundamental assegurar que o RNA a utilizar neste tipo
de teacutecnica seja de oacuteptima qualidade para garantir precisatildeo e exactidatildeo dos resultados Tal
como se esperava o fiacutegado foi o oacutergatildeo onde a expressatildeo normalizada de TTR foi superior
Nos restantes oacutergatildeos em estudo (estocircmago coacutelon duodeno e bexiga) apenas o estocircmago
apresentou valores de expressatildeo diferentes de zero
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
34
Os resultados obtidos atraveacutes dos dois tipos de PCR satildeo claramente distintos A ocorrecircncia
de bandas no PCR tradicional para coacutelon duodeno e bexiga pode ter ficado a dever-se a
uma contaminaccedilatildeo e por isso quando se passa para uma teacutecnica muito mais sensiacutevel (PCR
em tempo real) natildeo se verificam sinais de expressatildeo de TTR nestes oacutergatildeos O PCR em
tempo real eacute por isso uma teacutecnica muito mais precisa e reprodutiacutevel do que o PCR
convencional
A segunda parte deste trabalho tinha como objectivo identificar quais os locais
principalmente no estocircmago e intestino onde ocorre a expressatildeo da proteiacutena Para isso
procederam-se a teacutecnicas de imunohistoquiacutemica e imunofluorescecircncia
A imunohistoquimica forneceu dados importantes acerca da localizaccedilatildeo histoloacutegica da
TTR enquanto proteiacutena a imunoreactividade encontrada foi muito especiacutefica e pouco
abundante surgindo na mucosa do estocircmago do murganho tal como tinha sido
previamente descrito para o tecido humano (Liddle et al 1985) As ceacutelulas marcadas
poderiam por isso ser de quatro tipos diferentes ceacutelulas produtoras de muco ceacutelulas
principais ceacutelulas endoacutecrinas ou ceacutelulas parietais Foi necessaacuterio determinar que tipo de
ceacutelula presente na zona glandular do estocircmago expressava TTR A morfologia das ceacutelulas
marcadas para TTR era idecircntica agrave das ceacutelulas endoacutecrinas o que posteriormente se veio a
verificar Por imunohistoquimica foi realizada uma marcaccedilatildeo para cromogranina A
marcador de ceacutelulas enteroendoacutecrinas e para TTR em lacircminas contendo secccedilotildees
sucessivas de estocircmago contudo nada foi possiacutevel concluir acerca da co-localizaccedilatildeo entre
as duas proteiacutenas Natildeo se procederam a ensaios de imunofluorescecircncia devido ao facto de
ambos os anticorpos serem produzidos no mesmo animal
A partir de ensaios de co-localizaccedilatildeo por imunofluorescecircncia verificou-se que a TTR natildeo
se localiza em ceacutelulas parietais ocorrendo a sua localizaccedilatildeo nas ceacutelulas produtoras de
somatostatina ndash ceacutelulas D (ceacutelulas endoacutecrinas) tanto no murganho como no humano Estas
ceacutelulas possuem extensotildees citoplasmaacuteticas onde estatildeo armazenados os gracircnulos de
somatostatina (Lucey 1986) Um estudo realizado em 1992 (Kameko et al 1992) admitia
jaacute a existecircncia de ceacutelulas reactivas para TTR no estocircmago e intestino humano e mostrava a
ocorrecircncia de co-localizaccedilatildeo entre TTR e RBP sendo que as ceacutelulas que continham as
duas proteiacutenas apresentavam caracteriacutesticas morfoloacutegicas semelhantes agraves das ceacutelulas
endoacutecrinas Neste presente trabalho experimental natildeo se verificou a existecircncia de ceacutelulas
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
35
marcadas para TTR quer no intestino do murganho quer no intestino humano A
importacircncia bioloacutegica da expressatildeo da TTR neste tipo de ceacutelulas natildeo eacute ainda conhecida A
somatostatina funciona como um neurotransmissor inibitoacuterio do sistema nervoso e tem um
papel na regulaccedilatildeo neurohormonal (Sternini et al 1997) Da sua expressatildeo resulta a
inibiccedilatildeo de secreccedilotildees endoacutecrinas e exoacutecrinas de secreccedilotildees enzimaacuteticas e de electroacutelitos da
contracccedilatildeo muscular e proliferaccedilatildeo celular (Sternini et al 1997) No sistema digestivo a
somatostatina eacute expressa em neuroacutenios fibras e nas ceacutelulas D (Costa et al 1977 Costa et
al 1980 Ekblad et al 1988) e estaacute tambeacutem envolvida na regulaccedilatildeo da peristalsia e
motilidade interdigestiva confirmada pela presenccedila de neuroacutenios imunoreactivos a
somatostatina no plexo mienteacuterico e atraveacutes de estudos funcionais (McIntosh 1985) A
somatostatina eacute um regulador fisioloacutegico da secreccedilatildeo de aacutecido gaacutestrico por efeito directo
desta proteiacutena ao niacutevel das ceacutelulas parietais e de forma indirecta por inibiccedilatildeo da
libertaccedilatildeo de histamina e gastrina (McIntosh 1985) Seraacute por isso importante perceber se
existe alguma interacccedilatildeo entre a TTR e a somatostatina e a existir de que forma a TTR
poderaacute estar tambeacutem envolvida nas funccedilotildees acima mencionadas Tal como previamente
descrito (Liddle et al 1985) natildeo ocorreu co-localizaccedilatildeo entre TTR e somatostatina no
pacircncreas uma vez que neste oacutergatildeo satildeo as ceacutelulas α (sintetizadoras de glucagon) as
responsaacuteveis pela expressatildeo de TTR
No estocircmago natildeo eacute conhecida de que forma poderaacute a expressatildeo de TTR estar associada agrave
sintomatologia gastrointestinal caracteriacutestica de PAF nomeadamente retenccedilatildeo gaacutestrica
associada a naacuteuseas e voacutemitos bem como fenoacutemenos de obstipaccedilatildeo diarreia A niacutevel
histoloacutegico verifica-se que neste oacutergatildeo ocorre deposiccedilatildeo extracelular de fibras de TTR
mutada na camada submucosa e em torno dos plexos mienteacutericos (Yoshimatsu et al
1998) Ocorre tambeacutem a deposiccedilatildeo de amiloacuteide a niacutevel glandular originando depoacutesitos que
alteram a estrutura celular e morfoloacutegica das glacircndulas presentes no estocircmago A
deposiccedilatildeo de TTR provoca danos estruturais e tambeacutem funcionais nos tecidos A expressatildeo
da proteiacutena mutada no estocircmago poderaacute de alguma forma condicionar os processos
bioquiacutemicos que decorrem neste oacutergatildeo e assim influenciar os mecanismos bioloacutegicos
caracteriacutesticos deste sistema conduzindo aos sintomas referidos anteriormente Natildeo eacute
tambeacutem de descartar uma possiacutevel interacccedilatildeo entre a TTR na forma mutada e a
somatostatina podendo conduzir esta interacccedilatildeo agrave severidade dos sintomas
gastrointestinais caracteriacutesticos desta doenccedila
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
36
A uacuteltima metodologia experimental consistiu numa hibridaccedilatildeo in situ Com esta teacutecnica
apenas foi possiacutevel detectar a presenccedila de mRNA de TTR de murganho no plexo coroacuteide
utilizando a sonda antisense No estocircmago natildeo foi possiacutevel detectar qualquer tipo de
marcaccedilatildeo Estes resultados podem ter ficado a dever-se aos niacuteveis de expressatildeo de TTR
nos dois oacutergatildeos em questatildeo A sonda embora muito especiacutefica natildeo detectou a presenccedila de
mRNA na mucosa do estocircmago muito provavelmente devido ao reduzido nuacutemero de
ceacutelulas e agrave proacutepria estrutura histoloacutegica do tecido em questatildeo O plexo coroacuteide estaacute muito
mais acessiacutevel pelo que o processo de hibridaccedilatildeo ocorre naturalmente
O presente estudo permitiu detectar a expressatildeo de TTR no estocircmago bem como a
localizaccedilatildeo histoloacutegica da mesma Conduziu tambeacutem a novas questotildees que descrevemos
anteriormente e que teratildeo de ser respondidas em estudos futuros
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
37
Conclusatildeo
A transtirretina eacute uma proteiacutena extremamente conservada nos mamiacuteferos ao longo da
evoluccedilatildeo pelo que deveraacute ser uma proteiacutena importante para a funcionalidade bioloacutegica dos
organismos Satildeo conhecidas as duas principais funccedilotildees da TTR ndash transportar a tiroxina e a
vitamina A contudo sabe-se hoje que esta proteiacutena interveacutem noutros processos bioloacutegicos
importantes quer por associaccedilatildeo a outros peacuteptidos quer por ser um indicador de processos
inflamatoacuterios de stress etc Conhecer as interacccedilotildees que a TTR estabelece nos indiviacuteduos
portadoras de TTR natildeo mutada permite perceber que tipo de funccedilotildees poderatildeo estar
alteradas nos indiviacuteduos portadores de TTR mutada nomeadamente em PAF e se eacute a
ausecircncia ou disfunccedilatildeo dessas interacccedilotildees que conduz a muitos dos sintomas associados agrave e
doenccedila
Com este projecto foi possiacutevel investigar e caracterizar a expressatildeo de TTR ao longo do
tracto gastrointestinal do murganho nomeadamente no esoacutefago estocircmago duodeno coacutelon
e tambeacutem na bexiga Verificou-se que de todos os oacutergatildeos em estudo apenas o estocircmago
apresentou niacuteveis de expressatildeo normalizada de TTR com valores diferentes de zero
Conclui-se tambeacutem que a metodologia do PCR em tempo real eacute muito mais precisa e
especiacutefica do que o PCR convencional conduzindo este uacuteltimo a muitos falsos positivos
Foi tambeacutem possiacutevel determinar que tal como jaacute tinha sido descrito as ceacutelulas que
expressam TTR no estocircmago do murganho e humano existem em pequena quantidade e
estatildeo presentes na mucosa do oacutergatildeo em questatildeo Estas ceacutelulas co - localizam com as
ceacutelulas D responsaacuteveis pela secreccedilatildeo de somatostatina no estocircmago natildeo sendo ainda
conhecida nenhuma interacccedilatildeo funcional entre estes dois peacuteptidos Neste estudo natildeo se
observou quaisquer tipos de marcaccedilatildeo no intestino quer no murganho quer em tecidos
humanos
Os resultados obtidos entre as metodologias utilizadas (PCR em tempo real e
imunohistoquiacutemicaimunofluorescecircncia) satildeo concordantes entre si
A hibridaccedilatildeo in situ permitiu detectar a presenccedila de mRNA de TTR no plexo coroacuteide do
murganho natildeo sendo visiacutevel qualquer tipo de marcaccedilatildeo em ceacutelulas do estocircmago
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
38
Todos estes resultados colocam novas questotildees Ocorreraacute interacccedilatildeo entre TTR e
somatostatina De que tipo seraacute essa interacccedilatildeo Seraacute que na presenccedila de TTR mutada essa
interacccedilatildeo eacute alterada conduzindo agrave sintomatologia do sistema gastrointestinal caracteriacutestico
de PAF
Seria por isso interessante estudar futuramente a expressatildeo de TTR num modelo animal
de PAF embora o ideal seria poder utilizar o modelo humano para determinar se de
alguma forma esta expressatildeo pode estar aumentada e se ocorre nos mesmos locais
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
39
Bibliografia
Abraham CR Selkoe DJ Potter H ldquoImmunochemical identification of the serine protease inhibitor alpha
1-antichymotrypsin in the brain amyloid deposits of Alzheimers diseaserdquo Cell 1988 52(4)487-501
Aleshire SL Bradley CA Richardson LD Parl FF ldquoLocalization of human prealbumin in choroid
plexus epitheliumrdquo J Histochem Cytochem 1983 31(5)608-12
Almeida MR Damas AM Lans MC Brouwer A Saraiva MJ ldquoThyroxine binding to transthyretin
Met 119 Comparative studies of different heterozygotic carriers and structural analysisrdquo Endocrine 1997
6(3)309-15
Alves IL Altland K Almeida MR Winter P Saraiva MJ ldquoScreening and biochemical
characterization of transthyretin variants in the Portuguese populationrdquo Hum Mutat 1997 9(3)226-33
Andersson R ldquoHereditary amyloidosis with polyneuropathyrdquo Acta Med Scand 1970 1-2(1)85-94
Ando Y Araki S Ando M ldquoTransthyretin and familial amyloidotic polyneuropathyrdquo Intern Med 1993
32(12)920-2
Ando Y Ohtsu Y Terazaki H Kibayashi K Nakamura M Ando E Matsunaga N Obayashi K
Uchino M Ando M Tsunenari S ldquoJapanese monozygotic twins with familial amyloidotic polyneuropathy
(FAP) (ATTR Val30Met)rdquo Amyloid 2000 7(2)133-6
Andrade C ldquoA peculiar form of peripheral neuropathy familiar atypical generalized amyloidosis with
special involvement of the peripheral nervesrdquo Brain 1952 75(3)408-27
Andrea TA Cavalieri RR Goldfine ID Jorgensen EC ldquoBinding of thyroid hormones and analogues
to the human plasma protein prealbuminrdquo Biochemistry 1980 19(1)55-63
Bernstein LH Leukhardt-Fairfield CJ Pleban W Rudolph R ldquoUsefulness of data on albumin and
prealbumin concentrations in determining effectiveness of nutritional supportrdquo Clin Chem 1989 35(2)271-
4
Bjerle P Ek B Linderholm H Steen L ldquoOesophageal dysfunction in familial amyloidosis with
polyneuropathyrdquo Clin Physiol 1993 13(1)57-69
Blake CC Geisow MJ Oatley SJ Reacuterat B Reacuterat C ldquoStructure of prealbumin secondary tertiary and
quaternary interactions determined by Fourier refinement at 18 Ardquo J Mol Biol 1978 121(3)339-56
Blake CC Geisow MJ Swan ID Rerat C Rerat B ldquoStructure of human plasma prealbumin at 2-5 A
resolution A preliminary report on the polypeptide chain conformation quaternary structure and thyroxine
bindingrdquo J Mol Biol 1974 88(1)1-12
Blaner WS ldquoRetinol-binding protein the serum transport protein for vitamin Ardquo Endocr Rev 1989
10(3)308-16
Boomsma F Man in t Veld AJ Schalekamp MA ldquoNot norepinephrine but its oxidation products bind
specifically to plasma proteinsrdquo J Pharmacol Exp Ther 1991 259(2)551-7
Borish L King MS Mascali JJ Johnson S Coll B Rosenwasser LJ ldquoTransthyretin is an inhibitor of
monocyte and endothelial cell interleukin-1 productionrdquo Inflammation 1992 16(5)471-84
Brett M Persey MR Reilly MM Revesz T Booth DR Booth SE Hawkins PN Pepys MB
Morgan-Hughes JA ldquoTransthyretin Leu12Pro is associated with systemic neuropathic and leptomeningeal
amiloidosisrdquo Brain 1999 122 ( Pt 2)183-90
Brouillette J Quirion R ldquoTransthyretin a key gene involved in the maintenance of memory capacities
during agingrdquo Neurobiol Aging 2008 29(11)1721-32
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
40
Burton PM Hung P Lin T Lovelace C White A ldquoThe effects of homogeneous human prealbumin on
in vitro and in vivo immune responses in the mouserdquo Int J Immunopharmacol 1985 7(4)473-81
Bush AI Pettingell WH Multhaup G d Paradis M Vonsattel JP Gusella JF Beyreuther K
Masters CL Tanzi RE ldquoRapid induction of Alzheimer A beta amyloid formation by zincrdquo Science 1994
265(5177)1464-7
Cardoso I Brito M Saraiva MJ ldquoExtracellular matrix markers for disease progression and follow-up of
therapies in familial amyloid polyneuropathy V30M TTR-relatedrdquo Dis Markers 2008 25(1)37-47
Choi-Miura NH Ihara Y Fukuchi K Takeda M Nakano Y Tobe T Tomita M ldquoSP-4040 is a
constituent of Alzheimers amyloidrdquo Acta Neuropathol 1992 83(3)260-4
Coelho T Chorao R Sousa A Alves IL Torres MF Saraiva MJ ldquoCompound heterozygote of
transthyretin Met30 and transthyretin Met119 are protected from the devastating effects of familial
amyloidotic polyneuropathrdquo Neuromusc Disord 1996 6 20
Coimbra A Andrade C ldquoFamilial amyloid polyneuropathy an electron microscope study of the peripheral
nerve in five cases II Nerve fibre changesrdquo Brain 1971 94(2)207-12
Costa M JB Furness IJ Llewellyn-Smith B Davies and J Oliver ldquoAn immunohistochemical study of
the projections of somatostatin-containing neurons in the guinea-pig intestinerdquo Neuroscience 1980 5841-
852
Costa M Y Patel JB Furness and A Arimura ldquoEvidence that some intrinsic neurons of the intestine
contain somatostatinrdquo Neurosci Lett 1977 6215-222
Costa RH Lai E Darnell JE Jr ldquoTranscriptional control of the mouse prealbumin (transthyretin) gene
both promoter sequences and a distinct enhancer are cell specificrdquo Mol Cell Biol 1986 6(12)4697-708
Dickson PW Aldred AR Marley PD Bannister D Schreiber G ldquoRat choroid plexus specializes in
the synthesis and the secretion of transthyretin (prealbumin) Regulation of transthyretin synthesis in choroid
plexus is independent from that in liverrdquo J Biol Chem 1986 261(8)3475-8
Dickson PW Howlett GJ Schreiber G ldquoMetabolism of prealbumin in rats and changes induced by acute
inflammationrdquo Eur J Biochem 1982 129(2)289-93
Divino CM Schussler GC ldquoReceptor-mediated uptake and internalization of transthyretinrdquo J Biol Chem
1990 265(3)1425-9
Ekblad E Ekman R Hakanson R Sundler F ldquoProjections of peptide-containing neurons in rat colonrdquo
Neuroscience 1988 27655-674
El-Salhy M Suhr O ldquoEndocrine cells in rectal biopsy specimens from patients with familial amyloidotic
polyneuropathyrdquo Scand J Gastroenterol 1996 31(1)68-73
El-Salhy M Suhr O Stenling R Wilander E Grimelius L ldquoImpact of familial amyloid associated
polyneuropathy on duodenal endocrine cellsrdquo Gut 1994 35(10)1413-8
Feurle GE ldquoPathophysiology of diarrhea in patients with familial amyloid neuropathyrdquo Digestion 1987
36(1)13-7
Fleming CE Saraiva MJ Sousa MM ldquoTransthyretin enhances nerve regenerationrdquo J Neurochem 2007
103(2)831-9
Foss TR Wiseman RL Kelly JW ldquoThe pathway by which the tetrameric protein transthyretin
dissociatesrdquo Biochemistry 2005 44(47)15525-33
Gallo G Wisniewski T Choi-Miura NH Ghiso J Frangione B ldquoPotential role of apolipoprotein-E in
fibrillogenesisrdquo Am J Pathol 1994 145(3)526-30
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
41
Goodman DS ldquoVitamin A and retinoids in health and diseaserdquo N Engl J Med 1984 310(16)1023-31
Hagen GA Solberg LA Jr ldquoBrain and cerebrospinal fluid permeability to intravenous thyroid hormonesrdquo
Endocrinology 1974 95(5)1398-410
Hamilton JA Benson MD ldquoTransthyretin a review from a structural perspectiverdquo Cell Mol Life Sci
2001 58(10)1491-521
Herbert J Wilcox JN Pham KT Fremeau RT Jr Zeviani M Dwork A Soprano DR Makover
A Goodman DS Zimmerman EA et al ldquoTransthyretin a choroid plexus-specific transport protein in
human brain The 1986 S Weir Mitchell awardrdquo Neurology 1986 36(7)900-11
Holmgren G Steen L Ekstedt J Groth CG Ericzon BG Eriksson S Andersen O Karlberg I
Nordeacuten G Nakazato M et al ldquoBiochemical effect of liver transplantation in two Swedish patients with
familial amyloidotic polyneuropathy (FAP-met30)rdquo Clin Genet 1991 40(3)242-6
Hulbert AJ ldquoThyroid hormones and their effects a new perspectiverdquo Biol Rev Camb Philos Soc 2000
75(4)519-631
Ingenbleek Y De Visscher M ldquoHormonal and nutritional status critical conditions for endemic goiter
epidemiologyrdquo Metabolism 1979 28(1)9-19
Ingenbleek Y Young V ldquoTransthyretin (prealbumin) in health and disease nutritional implicationsrdquo Annu
Rev Nutr 1994 14495-533
Ingenbleek Y Young VR ldquoSignificance of transthyretin in protein metabolismrdquo Clin Chem Lab Med
2002 40(12)1281-91
Ito T Sakakibara R Ito S Uchiyama T Liu Z Yamamoto T Yamaguchi T Odaka T Higashi M
Hattori T ldquoMechanism of constipation in familial amyloid polyneuropathy a case reportrdquo Intern Med
2006 45(20)1173-5
Ito T Sakakibara R Yamamoto T Uchiyama T Liu Z Asahina M Higashi M Arai K Ito S
Awa Y Yamamoto K Kinou M Yamanishi T Hattori T ldquoUrinary dysfunction and autonomic control
in amyloid neuropathyrdquo Clin Auton Res 2006 16(1)66-71
Jacobson DR Alves IL Saraiva MJ Thibodeau SN Buxbaum JN ldquoTransthyretin Ser 6 gene
frequency in individuals without amyloidosisrdquo Hum Genet 1995 95(3)308-12
Jacobson DR McFarlin DE Kane I BuxbaumJN ldquoTransthyretin Pro55 a variant associated with
early-onset aggressive diffuse amyloidosis with cardiac and neurologic involvementrdquo Hum Genet 1992
89(3)353-6
Kabat EA Moore DH Landow H ldquoAn electrophoretic study of the protein components in cerebrospinal
fluid and their relationship to the serum proteinsrdquo J Clin Invest 1942 21(5)571-7
Kameko M Ota H Ishii K Nakayama J Katsuyama T Kanai M Tsutsumi Y ldquoDistribution of retinol-
binding protein in the human digestive tractrdquo Virchows Arch B Cell Pathol Incl Mol Pathol 1992
61(5)315-22
Kanai M Raz A Goodman DS ldquoRetinol-binding protein the transport protein for vitamin A in human
plasmardquo J Clin Invest 1968 47(9)2025-44
Kanda Y Goodman DS Canfield RE Morgan FJ ldquoThe amino acid sequence of human plasma
prealbuminrdquo J Biol Chem 1974 249(21)6796-805
Kato M Kato K Blaner WS Chertow BS Goodman DS ldquoPlasma and cellular retinoid-binding
proteins and transthyretin (prealbumin) are all localized in the islets of Langerhans in the ratrdquo Proc Natl
Acad Sci U S A 1985 82(8)2488-92
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
42
Koumlhrle J ldquoThyroid hormone deiodination in target tissues--a regulatory role for the trace element seleniumrdquo
Exp Clin Endocrinol 1994 102(2)63-89
Liddle CN Reid WA Kennedy JS Miller ID Horne CH ldquoImmunolocalization of prealbumin distribution in
normal human tissuerdquo J Pathol 1985 146(2)107-13
Loughna S Bennett P Moore G ldquoMolecular analysis of the expression of transthyretin in intestine and
liver from trisomy 18 fetusesrdquo Hum Genet 1995 95(1)89-95
Lucey M R ldquoEndogenous somatostatin and the gutrdquo Gut 1986 27(4)457-67
Lueprasitsakul W Alex S Fang SL Pino S Irmscher K Koumlhrle J Braverman LE ldquoFlavonoid
administration immediately displaces thyroxine (T4) from serum transthyretin increases serum free T4 and
decreases serum thyrotropin in the ratrdquo Endocrinology 1990 126(6)2890-5
Makover A Moriwaki H Ramakrishnan R Saraiva MJ Blaner WS Goodman DS ldquoPlasma
transthyretin Tissue sites of degradation and turnover in the ratrdquo J Biol Chem 1988 263(18)8598-603
Martone RL and Herbert J ldquoTransthyretin interacts with globin to form protein complexes with heme-
dependent solubilityrdquo Amyloid and Amyloidosis 1993 532-534
Martone RL Herbert J Dwork A Schon EA ldquoTransthyretin is synthesized in the mammalian eyerdquo
Biochem Biophys Res Commun 1988 151(2)905-12
May CL Kaestner KH ldquoGut endocrine cell developmentrdquo Mol Cell Endocrinol 2010 323(1)70-5
McCutchen SL Lai Z Miroy GJ Kelly JW Coloacuten W ldquoComparison of lethal and nonlethal
transthyretin variants and their relationship to amyloid diseaserdquo Biochemistry 1995 34(41)13527-36
McIntosh CHS ldquoMinireview Gastrointestinal somatostatin Distribution secretion and physiological
significance rdquo Life Sci 1985 37 2043-2058
McKinnon B Li H Richard K Mortimer R ldquoSynthesis of thyroid hormone binding proteins
transthyretin and albumin by human trophoblastrdquo J Clin Endocrinol Metab 2005 90(12)6714-20
Munar-Queacutes M Costa PP Saraiva MJ Viader-Farreacute C Munar-Bernat C ldquo[Familial type I
(Portuguese form) amyloidotic polyneuropathy in Majorca Study using the TTR (Met30) genetic markerrdquo
Med Clin (Barc) 1988 91(12)441-4
Murakami T Yasuda Y Mita S Maeda S Shimada K Fujimoto T Araki S ldquoPrealbumin gene
expression during mouse development studied by in situ hybridizationrdquo Cell Differ 1987 22(1)1-9
Nilsson SF Peterson PA ldquoEvidence for multiple thyroxine-binding sites in human prealbuminrdquo J Biol
Chem 1971 246(19)6098-105
Noy N Slosberg E Scarlata S ldquoInteractions of retinol with binding proteins studies with retinol-binding
protein and with transthyretinrdquo Biochemistry 1992 31(45)11118-24
Nunes AF Saraiva MJ Sousa MM ldquoTransthyretin knockouts are a new mouse model for increased
neuropeptide Yrdquo FASEB J 2006 20(1)166-8
Palha JA Fernandes R de Escobar GM Episkopou V Gottesman M Saraiva MJ ldquoTransthyretin
regulates thyroid hormone levels in the choroid plexus but not in the brain parenchyma study in a
transthyretin-null mouse modelrdquo Endocrinology 2000 141(9)3267-72
Palha JA Hays MT Morreale de Escobar G Episkopou V Gottesman ME Saraiva MJ
ldquoTransthyretin is not essential for thyroxine to reach the brain and other tissues in transthyretin-null micerdquo
Am J Physiol 1997 272(3 Pt 1)E485-93
Pepys MB ldquoAmyloidosis some recent developmentsrdquo Q J Med 1988 67(252)283-98
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
43
Qian X Samadani U Porcella A Costa RH ldquoDecreased expression of hepatocyte nuclear factor 3 alpha
during the acute-phase response influences transthyretin gene transcriptionrdquo Mol Cell Biol 1995
15(3)1364-76
Quintas A Vaz DC Cardoso I Saraiva MJ Brito RM ldquoTetramer dissociation and monomer partial
unfolding precedes protofibril formation in amyloidogenic transthyretin variantsrdquo J Biol Chem 2001
276(29)27207-13
Rask L Anundi H Peterson PA ldquoThe primary structure of the human retinol-binding proteinrdquo FEBS
Lett 1979 104(1)55-8
Raz A ShiratoriT Goodman DS ldquoStudies on the protein-protein and protein-ligand interactions involved
in retinol transport in plasmardquo J Biol Chem 1970 245(8)1903-12
Richardson SJ ldquoEvolutionary changes to transthyretin evolution of transthyretin biosynthesisrdquo FEBS J
2009 276(19)5342-56
Richardson SJ Bradley AJ Duan W Wettenhall RE Harms PJ Babon JJ Southwell BR Nicol
S Donnellan SC Schreiber G ldquoEvolution of marsupial and other vertebrate thyroxine-binding plasma
proteinsrdquo Am J Physiol 1994 266 1359-1370
Riisoslashen H ldquoReduced prealbumin (transthyretin) in CSF of severely demented patients with Alzheimers
diseaserdquo Acta Neurol Scand 1988 78(6)455-9
Rindi G Leiter AB Kopin AS Bordi C Solcia E ldquoThe normal endocrine cell of the gut changing
concepts and new evidencesrdquo Ann N Y Acad Sci 2004 10141-12
Robbins J ldquoTyroid hormone transport proteins and the physiology of hormone bindingrdquo In The Thyroid de
LE and Utiger RD Braverman pp 111-125 Philadelphia JB Lippincott Company 1991
Rogers J Cooper NR Webster S Schultz J McGeer PL Styren SD Civin WH Brachova L
Bradt B Ward P et al ldquoComplement activation by beta-amyloid in Alzheimer diseaserdquo Proc Natl Acad
Sci U S A 1992 89(21)10016-20
Santos CR Power DM ldquoIdentification of transthyretin in fish (Sparus aurata) cDNA cloning and
characterisationrdquo Endocrinology 1999 140(5)2430-3
Saraiva MJ Birken S Costa PP Goodman DS ldquoAmyloid fibril protein in familial amyloidotic
polyneuropathy Portuguese type Definition of molecular abnormality in transthyretin (prealbumin)rdquo J Clin
Invest 1984 74(1)104-19
Sasaki H Yoshioka N Takagi Y Sakaki Y ldquoStructure of the chromosomal gene for human serum
prealbuminrdquo Gene 1985 37(1-3)191-7
Schreiber G Aldred AR Jaworowski A Nilsson C Achen MG Segal MB ldquoThyroxine transport
from blood to brain via transthyretin synthesis in choroid plexusrdquo Am J Physiol 1990 258 338-345
Schwarzman AL Gregori L Vitek MP Lyubski SStrittmatter WJ Enghilde JJ Bhasin R
Silverman J Weisgraber KH Coyle PK et al ldquoTransthyretin sequesters amyloid beta protein and
prevents amyloid formationrdquo Proc Natl Acad Sci U S A 1994 91(18)8368-72
Scott DL Marhaug G Husby G ldquoComparative studies of the high molecular weight amyloid fibril
proteins and similar components from normal tissuesrdquo Clin Exp Immunol 1983 52(3)693-701
Seibert FB Nelson JW ldquoElectrophoretic study of the blood response in tuberculosisrdquo J Biol Chem 1942
143 29-38
Serot JM Christmann D Dubost T Couturier M ldquoCerebrospinal fluid transthyretin aging and late
onset Alzheimers diseaserdquo J Neurol Neurosurg Psychiatry 1997 63(4)506-8
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
44
Smith TJ Davis FB Deziel MR Davis PJ Ramsden DB Schoenl M ldquoRetinoic acid inhibition of
thyroxine binding to human transthyretinrdquo Biochim Biophys Acta 1994 1199(1)76-80
Snow AD Willmer J Kisilevsky R ldquoSulfated glycosaminoglycans a common constituent of all
amyloidsrdquo Lab Invest 1987 56(1)120-3
Soares ML Centola M Chae J Saraiva MJ Kastner DL ldquoHuman transthyretin intronic open reading
frames are not independently expressed in vivo or part of functional transcriptsrdquo Biochim Biophys Acta
2003 1626(1-3)65-74
Socolow EL Woeber KA Purdy RH Holloway MT Ingbar SH ldquoPreparation of I-131-labeled
human serum prealbumin and its metabolism in normal and sick patientsrdquo J Clin Invest 1965 44(10)1600-
9
Soprano DR Herbert J Soprano KJ Schon EA Goodman DS ldquoDemonstration of transthyretin
mRNA in the brain and other extrahepatic tissues in the ratrdquo J Biol Chem 1985 260(21)11793-8
Soprano DR Soprano KJ Goodman DS ldquoRetinol-binding protein and transthyretin mRNA levels in
visceral yolk sac and liver during fetal development in the ratrdquo Proc Natl Acad Sci U S A 1986
83(19)7330-4
Sousa JC Grandela C Fernaacutendez-Ruiz J de Miguel R de Sousa L Magalhatildees AI Saraiva MJ
Sousa N Palha JA ldquoTransthyretin is involved in depression-like behaviour and exploratory activityrdquo J
Neurochem 2004 88(5)1052-8
Sousa MM Berglund L Saraiva MJ ldquoTransthyretin in high density lipoproteins association with
apolipoprotein A-Irdquo J Lipid Res 2000 41(1)58-65
Sousa MM Cardoso I Fernandes R Guimaratildees A Saraiva MJ ldquoDeposition of transthyretin in early
stages of familial amyloidotic polyneuropathy evidence for toxicity of nonfibrillar aggregatesrdquo Am J Pathol
2001 159(6)1993-2000
Sousa MM Du Yan S Fernandes R Guimaraes A Stern D Saraiva MJ ldquoFamilial amyloid
polyneuropathy receptor for advanced glycation end products-dependent triggering of neuronal
inflammatory and apoptotic pathwaysrdquo J Neurosci 2001 21(19)7576-86
Sousa MM Saraiva MJ ldquoNeurodegeneration in familial amyloid polyneuropathy from pathology to
molecular signalingrdquo Prog Neurobiol 2003 71(5)385-400
Sternini C Wong H Wu SV de Giorgio R Yang M Reeve J Jr Brecha NC Walsh JH ldquoSomatostatin 2A
receptor is expressed by enteric neurons and by interstitial cells of Cajal and enterochromaffin-like cells of
the gastrointestinal tractrdquo J Comp Neurol 1997 386(3)396-408
Suhr O Danielsson A Steen L ldquoBile acid malabsorption caused by gastrointestinal motility dysfunction
An investigation of gastrointestinal disturbances in familial amyloidosis with polyneuropathyrdquo Scand J
Gastroenterol 1992 27(3)201-7
Tanaka Y Ando Y Kumamoto T Miyazaki A Nakamura M Nakayama M Araki S Ando M
ldquoChanged affinity of apolipoprotein AII to high density lipoprotein (HDL) in patients with familial
amyloidotic polyneuropathy (FAP) type Irdquo Biochim Biophys Acta 1994 1225(3)311-6
Tsuzuki T Mita S Maeda S Araki S Shimada K ldquoStructure of the human prealbumin generdquo J Biol
Chem 1985 260(22)12224-7
Vahlquist A Peterson PA Wibell L ldquoMetabolism of the viatmin A transporting protein complex I
Turnover studies in normal persons and in patients with chronic renal failurerdquo Eur J Clin Invest 1973
3(4)352-62
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
45
Vahlquist A Rask L Peterson PA Berg T ldquoThe concentrations of retinol-binding protein prealbumin
and transferrin in the sera of newly delivered mothers and children of various agesrdquo Scand J Clin Lab Invest
1975 35(6)569-75
Vatassery GT Quach HT Smith WE Benson BA Eckfeldt JH ldquoA sensitive assay of transthyretin
(prealbumin) in human cerebrospinal fluid in nanogram amounts by ELISArdquo Clin Chim Acta 1991
197(1)19-25
Vieira AV Sanders EJ Schneider WJ ldquoTransport of serum transthyretin into chicken oocytes A
receptor-mediated mechanismrdquo J Biol Chem 1995 270(7)2952-6
Westermark P Sletten K Johansson B Cornwell GG 3rd ldquoFibril in senile systemic amyloidosis is
derived from normal transthyretinrdquo Proc Natl Acad Sci U S A 1990 87(7)2843-5
Whitehead AS Skinner M Bruns GA Costello W Edge MD Cohen AS Sipe JD ldquoCloning of
human prealbumin complementary DNA Localization of the gene to chromosome 18 and detection of a
variant prealbumin allele in a family with familial amyloid polyneuropathyrdquo Mol Biol Med 1984 2(6)411-
23
Woeber KA Ingbar SH ldquoThe contribution of thyroxine-binding prealbumin to the binding of thyroxine in
human serum as assessed by immunoadsorptionrdquo J Clin Invest 1968 47(7)1710-21
Yoshimatsu S Ando Y Terazaki H Sakashita N Tada S Yamashita T Suga M Uchino M Ando
M ldquoEndoscopic and pathological manifestations of the gastrointestinal tract in familial amyloidotic
polyneuropathy type I (Met30)rdquo J Intern Med 1998 243(1)65-72
Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46

Expressatildeo de Transtirretina no Sistema Gastrointestinal do Murganho
46















![Motilidade gastrointestinal [Modo de Compatibilidade] · MOTILIDADE GASTROINTESTINAL Objetivo: Estudar os mecanismos fisiológicos responsáveis pela motilidade gastrointestinal Roteiro:](https://static.fdocumentos.tips/doc/165x107/5ba2c53109d3f208588c90c2/motilidade-gastrointestinal-modo-de-compatibilidade-motilidade-gastrointestinal.jpg)

