Estrada 2009 - Dissertaçãoarquivos.proderj.rj.gov.br/inea_imagens/downloads/pesquisas/RB... ·...
94
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CENTRO DE CIÊNCIAS DA SAÚDE INSTITUTO DE BIOLOGIA PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA ANÁLISE DA VARIABILIDADE ESTRUTURAL DE FLORESTAS DE MANGUE DE GUARATIBA, RIO DE JANEIRO - RJ GUSTAVO CALDERUCIO DUQUE ESTRADA Dissertação apresentada ao programa de pós- graduação em ecologia da universidade federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau de mestre em ciências biológicas (ecologia). RIO DE JANEIRO, RJ - BRASIL FEVEREIRO DE 2009
Transcript of Estrada 2009 - Dissertaçãoarquivos.proderj.rj.gov.br/inea_imagens/downloads/pesquisas/RB... ·...

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CENTRO DE CIÊNCIAS DA SAÚDE
INSTITUTO DE BIOLOGIA PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
ANÁLISE DA VARIABILIDADE ESTRUTURAL DE FLORESTAS DE MANGUE DE GUARATIBA, RIO DE JANEIRO - RJ
GUSTAVO CALDERUCIO DUQUE ESTRADA
Dissertação apresentada ao programa de pós-graduação em ecologia da universidade federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau de mestre em ciências biológicas (ecologia).
RIO DE JANEIRO, RJ - BRASIL FEVEREIRO DE 2009

ii
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO/UFRJ INSTITUTO DE BIOLOGIA PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA-PPGE CX.POSTAL 68.020 – ILHA DO FUNDÃO CEP: 21941-590 – RIO DE JANEIRO – RJ – BRASIL TEL./FAX: (21) 290-3308 TEL.: (21) 562-6320
Análise da variabilidade estrutural de florestas de mangue de Guaratiba, Rio de Janeiro - RJ
GUSTAVO CALDERUCIO DUQUE ESTRADA
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio de Janeiro, como parte dos requisitos
necessários à obtenção do grau de Mestre em Ciências Biológicas (Ecologia).
Defendida em 19 de fevereiro de 2009.
APROVADA POR:
________________________________________________Prof. Mário Luiz Gomes Soares, Dr. (Orientador)
________________________________________________Prof. Paulo da Cunha Lana, Dr.
________________________________________________Profa. Dorothy Sue Dunn De Araujo, Drª.

iii
ESTRADA, GUSTAVO CALDERUCIO DUQUE Análise da variabilidade estrutural de florestas de mangue de Guaratiba, Rio de Janeiro - RJ [Rio
de Janeiro] 2009 83 p. 29,7 cm (Instituto de Biologia/UFRJ, M.Sc., Ecologia, 1997)
Dissertação - Universidade Federal do Rio de Janeiro, PPGE
1. Florestas de Mangue 2. Estrutura Vegetal
I. IB/UFRJ II. Título ( série )

iv
Aos meu pais, Fernando e Ivone.

v
AGRADECIMENTOS
Aos meus pais, Fernando e Ivone, meus maiores incentivadores, por me conduzirem
até aqui, e aos meus irmãos, Eduardo, Bernardo e Guilherme, pelos papos e
brincadeiras durante os intervalos da dissertação.
À minha amada, Beatriz, pela dedicação incondicional na reta final deste trabalho e
pela compreensão nos momentos de ausência. Que o nosso cominho siga sempre
assim: SÓ AMOR!
Ao Mário, pela referência de ética, caráter e dedicação à profissão que tem sido para
mim há quase oito anos. Seu papel como orientador vai muito além do âmbito
acadêmico. É um grande amigo, quase um pai, sempre preocupado em dar o conselho
necessário.
À Dorothy, por aceitar a “roubada” de receber um aluno apenas para lhe dar a
oportunidade de ingressar no mestrado. Obrigado também pelos livros emprestados,
pela orientação no estágio em docência e pelos conselhos.
Aos grandes amigos e companheiros de NEMA e Instituto Marés, Filipe e Vivi, pela
compreensão na fase final da dissertação, fundamental para que eu conseguisse
defender a tempo. Espero que nossa parceria siga sempre evoluindo e prosperando.
À Paula e a todos os demais integrantes do NEMA, do presente e do passado: Fábio,
Júlio, Ana Margarida, Carlos Milton, Beatriz Barbosa e Daniel.
À todos os estagiários do NEMA, Jorge, Dan, Carol, Luiz, Patrícia, Nathalia, Lívia e,
Rita, que participaram das coletas, encarando, com muita raça, travessias
intermináveis de apicuns “chiclete” sob um sol de “rachar”, parcelas de estrutura

vi
debaixo de chuva, frio e maruins, e os atoleiros do rio Portinho. Faço também um
agradecimento especial à Rita, que colaborou de forma decisiva durante a fase de
digitação dos dados. A todos vocês serei eternamente grato.
Ao amigo Henrique Dias, por ter me ajudado a “convencer” a Dorothy a me receber
como aluno.
Ao Prof. José Roberto Verani pela ajuda durante as análises estatísticas e por ter me
recebido tão cordialmente na UFSCar.
Ao Prof. Alex Prast e a todos os editores executivos da Oecologia Brasiliensis pela
valiosa experiência de participar da editoração de um periódico científico. Espero,
assim, ter colaborado um pouco mais com o PPGE.
Ao Prof. Fernando Fernandez, pela colaboração fundamental para que eu pudesse
realizar o exame de qualificação a tempo e pelas valiosas contribuições na pré-banca.
À CAPES pela concessão da bolsa.
A Vale pelo apoio financeiro ao projeto.
Aos meus mentores espirituais e demais entidades que me acompanham, pela
proteção e pelos desafios impostos durante essa dissertação, todos necessários ao
meu amadurecimento.

vii
RESUMO
O presente estudo apresenta dois manuscritos (Capítulos I e II) que abordam o tema
“estrutura vegetal de florestas de mangue”. O primeiro Capítulo tem como objetivo
determinar a área amostral mínima necessária para caracterizar a estrutura de
florestas de mangue, utilizando como estudo de caso sete florestas típicas de
manguezais da região sudeste do Brasil. O Capítulo II teve como objetivo verificar a
existência de diferenças quanto ao desenvolvimento estrutural, à composição de
espécies, e à distribuição diamétrica entre as florestas de mangue de Guaratiba
agrupadas por tipos fisiográficos. No Capítulo I, os resultados demonstraram haver
uma tendência de redução da área amostral mínima entre as parcelas representativas
de florestas de franja até àquelas representativas de florestas de transição,
relacionada à redução do desenvolvimento estrutural. Com base nos testes realizados,
recomenda-se, para florestas de mangue da região sudeste do Brasil, a utilização de
parcelas com área de 100 a 450 m² para florestas de franja, 25 a 180 m² para florestas
de bacia e 15 a 50 m² para florestas arbustivas de transição. No Capítulo II, a
comparação através do teste de Kruskal-Wallis entre densidade, DAP médio, altura
média, área basal e área basal por espécie de 70 parcelas agrupadas por tipos
fisiográficos, demonstrou a existência de um gradiente de desenvolvimento estrutural
crescente, na ordem: transição - bacia - franja. A comparação da área basal por
espécie indicou que as florestas de franja e bacia são dominadas similarmente por A.
schaueriana e R. mangle, enquanto as de transição são dominadas por R. mangle.
Análises de regressão demonstraram haver uma relação inversa entre a densidade e o
DAP médio, que refletem o efeito da competição na estrutura da comunidade, para as
florestas de franja (R²=0,015; F(1,19)=162,5; p<0,0000001) e bacia (R²=0,83;
F(1,28)=139,6; p<0,0000001), mas não para as de transição (R²=0,02; F(1,16)=0,2;
p=0,63). O teste de Kolmogorov-Smirnov demonstrou que, quando são apresentadas
as distribuições de diâmetros utilizando-se todos os dados agrupados por tipo
fisiográfico, as distribuições referentes às florestas de franja, bacia e transição são

viii
diferentes entre si (p<0,05). A análise das distribuições aponta fortes indícios de que
as florestas de franja encontram-se em um estágio sucessional mais avançado do que
as florestas de bacia e transição, nessa ordem. Os parâmetros estruturais, portanto,
não refletem apenas um gradiente de rigor ambiental, mas também florestas em
estágios sucessionais gradativamente menos avançados (no sentido franja-transição).
A elevação do nível médio relativo do mar é o provável distúrbio em larga escala que
determina que as florestas de franja estejam em um estágio sucessional mais
avançado do que as florestas de bacia e de transição.

ix
ABSTRACT
This study presents two manuscripts (Chapters I and II) approaching the topic
“structure of mangrove forests”. The first chapter aims to determine the minimum area
necessary to characterize the structure of mangrove forests, using as case studies
seven typical mangrove physiognomies from southeast Brazil. Chapter II aimed to
verify the existence of differences in structural development, species composition and
diameter distribution between the mangrove forests of Guaratiba grouped into
physiographic types. In Chapter I, the results showed a tendency of reduction of the
minimum area, from the representative plots of fringe physiognomies to those of
transition physiognomies, which is related to the reduction of structural development.
Based on the performed tests, it is recommended, for mangrove forests from the
southeast of Brazil, 100 to 450 m² as plot areas for fringe forests, 25 a 180 m² for basin
forests and 15 a 50 m² for transition shrub forests. In Chapter II, the Kruskal-Wallis
comparison test between density, mean DBH, mean height, basal area and species-
specific basal area of 70 plots gathered into physiographic types demonstrated the
existence of a crescent structural development gradient in the order: transition - basin -
fringe. The comparison of species-specific basal area indicated that fringe and basin
forests are dominated similarly by A. schaueriana e R. mangle, while transition forests
are dominated by R. mangle. Regression analysis showed an inverse relation between
density and mean DBH, which reflects the effect of competition in the community
structure of fringe (R²=0.015; F(1,19)=162.5; p<0.0000001) and basin (R²=0.83;
F(1,28)=139.6; p<0.0000001) forests, but not for transition forests (R²=0,02;
F(1,16)=0,2; p=0,63). The Kolmogorov-Smirnov test showed that, when diameter
distributions are presented using all data grouped into physiographic type, the
distributions from fringe, basin and transition are different among them (p<0,05). The
analysis of distributions strongly indicated that fringe forests find themselves at a more
advanced successional stage than basin and transition forests, in this order. The
structural parameters, therefore, do not reflect only an environment gradient but also

x
forests at progressively less advanced successional stages (in the direction fringe-
transition). Relative mean sea level rise is the probable large scale disturbance
determining that fringe forests stay at a more advanced successional stage than basin
and transition forests.

xi
SUMÁRIO
Pág. 1-2
Introdução Geral
Pág. 3-4
Área de Estudo
Pág. 5-31
Capítulo I - Determinação da área amostral mínima para
caracterização estrutural de florestas de mangue
Pág. 32 - 64
Capítulo II - Análise da variabilidade estrutural das florestas de
mangue de Guaratiba - Rio de Janeiro, RJ
Pág. 65 - 67
Discussão Geral
Pág. 68 - 69
Conclusão Geral
Pág. 70 - 83
Referências Bibliográficas

Introdução Geral
1
INTRODUÇÃO GERAL
Manguezal é um ecossistema costeiro que ocorre em regiões tropicais e subtropicais
do mundo, ocupando as zonas entre-marés (Soares, 1997). Segundo Walsh (1974), a
ocorrência e o melhor desenvolvimento de florestas de mangue se dão onde a
temperatura média do mês mais frio é superior a 20o C e amplitude térmica anual
inferior a 5o C, em substrato lamoso, em ambientes abrigados, com presença de água
salgada e submetidas a grande amplitude de marés. Entretanto, embora tais fatores
controlem a ocorrência dos manguezais em escala global, os atributos estruturais e
funcionais de cada manguezal são regidos pela interação de fatores em escala
regional (“assinatura energética”), conforme citado por Schaeffer-Novelli et al. (1990) e
Twilley (1995) e fatores em escala local (associados à freqüência de inundação pelas
marés), que dão origem à classificação das florestas de mangue em tipos fisiográficos
(Lugo & Snedaker, 1974). Tais fatores, que incluem gradientes edáficos de salinidade
(Cintrón et al., 1978; Sam & Ridd, 1998; Ellison et al., 2000), concentração de H2S
(Boto & Wellington, 1984; Nickerson & Thibodeau, 1985; Ewel et al., 1998) e
concentração de nutrientes (Feller et al., 2003; Boyer, 2006; Lovelock et al., 2007) têm
sido descritos para manguezais mundialmente.
O presente estudo apresenta dois manuscritos (Capítulos I e II) que abordam o tema
“estrutura de florestas de mangue”. O primeiro capítulo, um teste metodológico, tem
como objetivo determinar a área amostral mínima necessária para caracterizar a
estrutura de florestas de mangue, utilizando como estudos de caso florestas de franja,
bacia e de porte arbustivo. Embora o tema “escalas e amostragem” seja bastante
discutido no âmbito da ecologia teórica (ex. Wiens, 1989; Levin, 1992) e, mais
especificamente, da ecologia de florestas terrestres (Dietvorst et al., 1982; Zenner,
2005), a determinação da área amostral mínima com base em testes estatísticos é
inédita para florestas de mangue.
No segundo capítulo, cujo título é o mesmo da dissertação, os resultados dos testes
realizados no Capítulo I foram utilizados como referência para a escolha da área das

Introdução Geral
2
parcelas. Esse capítulo teve como objetivo verificar a existência de diferenças quanto
ao desenvolvimento estrutural, à composição de espécies, e à distribuição diamétrica
entre as florestas de mangue de Guaratiba agrupadas por tipos fisiográficos. Para
florestas de mangue, a existência de diferenças estruturais entre tipos fisiográficos é
demonstrada por vários autores (Pool et al., 1977; Cintrón et al., 1978; Lugo et al.,
1988; López-Portillo & Ezcurra, 1989; Schaeffer-novelli et al., 1990; Jiménez & Sauter,
1991; Ewel et al., 1998; Ellison et al., 2000; Feller et al., 2003; Dahdough-Guebas,
2004; Lara-Domínguez et al., 2005; Krauss et al., 2006; Lara & Cohen, 2006; Suwa et
al, 2008). Entretanto, poucos autores testaram estatisticamente e em larga escala a
existência de tais diferenças, como se propõe o Capítulo II.
Os manuscritos aqui apresentados representam uma evolução na abordagem dos
estudos de estrutura vegetal desenvolvidos pelo Núcleo de Estudos em Manguezais
da UERJ (NEMA/UERJ) em Guaratiba desde 1992, não só pela elaboração de testes
estatísticos, conforme mencionado anteriormente, mas também pela ampliação da
escala de abordagem. Pellegrini (2000), Chaves (2001), Oliveira (2001), Portugal
(2002) demonstraram a existência de padrões de zonação do desenvolvimento
estrutural em escala local em Guaratiba, com estudos concentrados em poucas áreas.
No Capítulo II, os dados utilizados pelos citados autores foram unidos a dados obtidos
em novas amostragens realizadas em áreas ainda não caracterizadas pelo NEMA.
Assim, foi possível testar a existência de diferenças quanto à estrutura vegetal dos
tipos fisiográficos numa escala mais ampla.
Da mesma forma, a determinação estatística da área amostral mínima para
caracterização estrutural representa uma evolução na busca pelo aperfeiçoamento das
metodologias empregadas nos estudos de manguezais no Brasil.

Área de Estudo
3
ÁREA DE ESTUDO
A área de estudo localiza-se na Baixada de Sepetiba/ Guaratiba, distando cerca de 70
km do centro da cidade do Rio de Janeiro. O clima da região, segundo classificação de
Köppen pode ser dividido em Aw, tropical quente e úmido com estação seca no
inverno, típico da baixada e Af, tropical quente e úmido sem estação seca,
característico das encostas adjacentes (Ferreira e Oliveira, 1985). Segundo Estrada et
al. (2008), a temperatura média anual é de 23,5°C e a precipitação anual média de
1067 mm, sendo os meses de maior pluviosidade janeiro e março e, junho e agosto os
meses mais secos. O regime de marés é de micromarés, com amplitude inferior a dois
metros (Brasil, 2008). O manguezal de Guaratiba encontra-se na porção leste da Baía
de Sepetiba, entre o rio Piraquê e o morro de Guaratiba, integrando um sistema de
ilhas, rios e canais inserido na Reserva Biológica e Arqueológica de Guaratiba (Figura
1). As partes mais internas das zonas entre-marés são atingidas apenas pelas marés
de sizígia, dando origem a planícies hipersalinas (Pellegrini, 2000). Almeida (2007)
calculou em 28,3 Km2 a área total de manguezais em 2003, sendo 20,9 Km2 de
florestas de mangue e 7,4 Km2 de planícies hipersalinas. Schaeffer-Novelli et al.
(1990), com base no relevo, tipo de solo, cobertura vegetal, temperatura média anual,
evapotranspiração potencial, amplitude das marés médias e de sizígia, dividiram o
litoral brasileiro em oito unidades fisiográficas, estando as florestas de mangue
existentes entre Cabo Frio e Laguna inseridas na unidade VII, as quais, por estarem
submetidas a condições ambientais regionais similares, teriam um desenvolvimento
estrutural dentro de uma mesma faixa. Apenas três espécies típicas de mangue
ocorrem na unidade VII: Avicennia schaueriana Stapf & Leechm. ex Moldenke,
Laguncularia racemosa (L.) C.F. Gaertn. e Rhizophora mangle L.

Figur
quan
ra 1: Mapa
nto à localiz
a geral da
zação das e
área de es
stações de
studo. Nos
amostrage
Capítulos
em.
I e II o ma
Área de Es
apa é deta
studo
4
lhado

Capítulo I - Determinação da área amostral mínima
5
CAPÍTULO I
DETERMINAÇÃO DA ÁREA AMOSTRAL MÍNIMA PARA CARACTERIZAÇÃO
ESTRUTURAL DE FLORESTAS DE MANGUE
Introdução
O objetivo implícito em todas as tentativas de determinação da área amostral mínima
reside em estabelecer uma parcela cujo tamanho seja suficientemente representativo
da estrutura da comunidade em questão (Kenkel & Podani, 1991). Atualmente, é
comumente aceito que a vegetação pode ser estudada através de parcelas de
tamanhos variáveis, e a escolha de um tamanho específico determina a resolução do
estudo e afeta seus resultados (Wiens, 1989; Levin, 1992).
Økland (1990) considerou a determinação da área mínima de parcelas como um
compromisso entre homogeneidade, sugerindo pequenas parcelas, e
representatividade, sugerindo grandes parcelas. Em termos de homogeneidade, a
parcela deve ser pequena de forma que registre pequena variação ao longo do
gradiente de interesse. Em termos de representatividade, a parcela deve ser grande o
suficiente para garantir uma predição confiável das condições ambientais locais (ex:
zonas) a partir da estrutura da comunidade. Grandes parcelas filtram a variação
espacial que ocorre em escalas menores que o tamanho de suas unidades amostrais
(Bellehumeur et al., 1997).
Um dos métodos pioneiros, e ainda bastante utilizado, para determinação da área
mínima de parcelas é o da interpretação de curvas do tipo “espécie X área” (Cain,
1934, 1938, 1943; Hopkins, 1955; Poore Rice & Kelting, 1955). O método sugere um
desenho experimental chamado de “parcelas aninhadas”, no qual são posicionadas
parcelas cada vez maiores de tal forma que cada parcela maior englobe todas as
parcelas menores. A área mínima é então definida no ponto a partir do qual o número
de espécies tende a se estabilizar com o aumento da área. Esse ponto pode ser

Capítulo I - Determinação da área amostral mínima
6
determinado visualmente ou matematicamente, através do cálculo do ponto de
máxima curvatura ou encontrando-se o ponto no qual um aumento de 10% na área
provoca um aumento de 5 ou 10% no número de espécies (Cain, 1938).
Moravec (1973) desenvolveu um método baseado na similaridade florística. Para isso,
se baseou no pressuposto de que se uma parcela é representativa, sua composição
florística não deve apresentar grande diferença em relação a outra do mesmo
tamanho na mesma floresta. A área mínima é atingida quando a similaridade média
deixa de aumentar conforme a área da parcela aumenta. Dietvorst et al. (1982) aplicou
esse método, porém, considerando a parcela de tamanho ótimo aquela na qual a
similaridade média de 80% é atingida. Esse método é interessante do ponto de vista
teórico, mas é bastante trabalhoso, demandado um enorme gasto de tempo para
executá-lo. Nesse aspecto, Barkman (1989) chama atenção para a relação “custo X
benefício” na determinação do tamanho ótimo. Para o autor, o tamanho ótimo poderia
ser considerado como aquele no qual o ganho de informação, conforme a parcela é
aumentada, é muito pequeno em relação ao tempo investido, devendo-se considerar
também a dificuldade em penetrar em certos tipos de vegetação, como, por exemplo,
as florestas arbustivas densas. Na mesma linha, Kenkel & Podani (1991) sugerem que
o tamanho ótimo seja definido como o maior possível, dadas as limitações de tempo.
Os métodos anteriormente apresentados se baseiam apenas na composição de
espécies para determinar a área mínima das parcelas, desconsiderando a
variabilidade estrutural das florestas. Para florestas com alta diversidade de espécies,
como as Florestas Tropicais, tais métodos podem apresentar resultados satisfatórios.
Porém, para florestas de mangue, que possuem baixa diversidade de espécies típicas
(Tomlinson, 1986), torna-se fundamental a aplicação de métodos que utilizem
parâmetros estruturais.
Bormann (1953) desenvolveu um método pioneiro baseado na variância da área basal
entre tamanhos de parcela diferentes. O tamanho mínimo seria então definido com

Capítulo I - Determinação da área amostral mínima
7
base num valor mínimo de variância, definido a priori. Bellehumeur et al. (1997) propôs
um método semelhante, porém baseado na densidade de troncos por hectare.
Brower et al. (1998) sugere a confecção de curvas análogas às do tipo “espécies X
área”, porém, plotando parâmetros estruturais (“estrutura X área”). Zenner (2005)
também propõe a confecção de curvas “estrutura X área”, entretanto, com maior
embasamento estatístico. O autor se utiliza de dois índices estruturais, o Iog (Index of
Old-Growth), que mede a similaridade entre parâmetros estruturais de uma
determinada floresta com os valores tipicamente encontrados em florestas maduras
(Acker et al., 1998), e o SCI (Structural Complexity Index) que mede a complexidade
estrutural tri-dimensional de florestas (Zenner & Hibbs, 2000). A determinação da área
mínima se dá pela estabilização da variabilidade da média dos índices entre diferentes
tamanhos.
Não há registros na literatura sobre tentativas de determinação da área mínima de
parcelas para a caracterização de florestas de mangue, embora Jiménez (1990) tenha
demonstrado a relação direta entre o aumento da área da parcela e a tendência à
homogeneidade da distribuição de classes diamétricas através da aplicação do Índice
de Morisita.
Cintrón & Schaeffer-Novelli (1984) e Schaeffer-Novelli & Cintrón (1986) são as
principais referências metodológicas para a maioria dos trabalhos publicados sobre
estrutura de florestas de mangue mundialmente. Considerando que a acurácia de uma
amostragem fitossociológica é uma função da quantidade de enumerações e não da
área amostrada, Cintrón & Schaeffer-Novelli (1984) sugerem que a escolha da área da
parcela seja definida com base na densidade e na variabilidade da floresta em
questão. Dessa forma, em florestas maduras, cujos indivíduos são maiores e ocorrem
de forma esparsa, a área da parcela deve ser maior do que a utilizada em florestas
jovens, com alta densidade.
Com base no levantamento de 44 artigos científicos sobre estrutura de florestas de
mangue no mundo todo e constatamos que 37% usam tamanhos variáveis (Lugo et
Capítulo I - Determinação da área amostral mínima
8
al., 1988; McKee, 1993; Khoon & Eong, 1995; Fromard et al., 1998; Chen & Twilley,
1999; Soares, 1999; Fernandes, 2000; Cunha-Lignon, 2001; Berger et al., 2006;
Soares et al., 2003; Bernini & Rezende, 2004; Imai et al., 2006; Pellegrini et al., 2007;
Soares et al., 2007, 2008; Soares, Submetido), de acordo com a densidade e a
homogeneidade da floresta estudada, enquanto a maioria utiliza parcelas com
tamanho fixo (Golley et al., 1962; Pool et al., 1977; Ball, 1980; Roth, 1982; Jiménez,
1990; Jiménez & Sauter, 1991; Ukpong, 1991; Ball, 1998; Cox & Allen, 1999; Ellison et
al., 2000; Aubé & Caron, 2001; Baldwin et al., 2001; Blanco et al., 2001; Souza e
Sampaio, 2001; Kairo et al., 2002; Bosire et al., 2003; De Deus et al., 2003; Ellison &
Simmonds, 2003; Ong et al., 2005; Proffitt & Devlin, 2005; Rogers et al., 2005; Silva et
al., 2005; Bosire et al., 2006; Ferwerda et al., 2007; López-Hoffman et al., 2007;
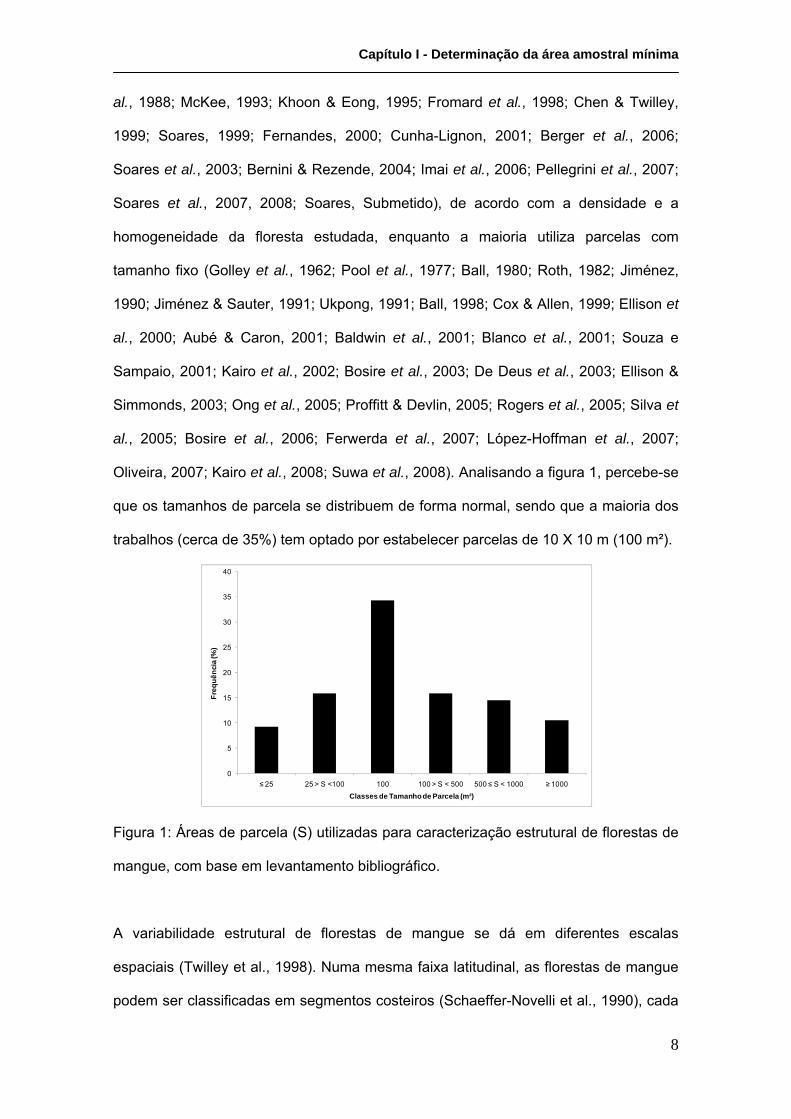
Oliveira, 2007; Kairo et al., 2008; Suwa et al., 2008). Analisando a figura 1, percebe-se
que os tamanhos de parcela se distribuem de forma normal, sendo que a maioria dos
trabalhos (cerca de 35%) tem optado por estabelecer parcelas de 10 X 10 m (100 m²).
Figura 1: Áreas de parcela (S) utilizadas para caracterização estrutural de florestas de
mangue, com base em levantamento bibliográfico.
A variabilidade estrutural de florestas de mangue se dá em diferentes escalas
espaciais (Twilley et al., 1998). Numa mesma faixa latitudinal, as florestas de mangue
podem ser classificadas em segmentos costeiros (Schaeffer-Novelli et al., 1990), cada
0
5
10
15
20
25
30
35
40
≤ 25 25 > S <100 100 100 > S < 500 500 ≤ S < 1000 ≥ 1000
Freq
uênc
ia (%
)
Classes de Tamanho de Parcela (m²)

Capítulo I - Determinação da área amostral mínima
9
um apresentando características climáticas, geológicas e oceanográficas distintas.
Cada segmento costeiro apresenta, numa escala menor, diferentes ambientes
geomorfológicos (Thom, 1984), nos quais a estrutura das florestas é definida pela
dominância relativa de ondas, marés ou rios no sistema em questão. Em cada
ambiente geomorfológico, as florestas de mangue podem ser analisados numa escala
local, na qual a hidrologia e a topografia determinam os regimes de inundação pelas
marés e de aporte fluvial, podendo-se classificá-las em tipos fisiográficos (Lugo &
Snedaker, 1974), que são resumidos por Schaeffer-Novelli et al. (2000) em: franja,
bacia e de porte arbustivo, os quais diferem basicamente quanto à freqüência de
inundação pelas marés. As florestas de franja são aquelas expostas a uma maior
freqüência de inundação pelas marés, o que favorece um maior desenvolvimento
estrutural. O oposto ocorre para as florestas de porte arbustivo, que têm seu
desenvolvimento limitado pela alta rigorosidade ambiental provocada pela baixa
freqüência de inundação pelas marés.
Dessa forma, o objetivo do presente estudo é determinar a área amostral mínima
necessária para caracterizar a estrutura de florestas de mangue, utilizando como
estudos de caso florestas de franja, bacia e de porte arbustivo.
Material e Métodos
Área de Estudo
A área de estudo localiza-se na Baixada de Sepetiba/ Guaratiba, distando cerca de 70
km do centro da cidade do Rio de Janeiro. O clima da região, segundo classificação de
Köppen pode ser dividido em Aw, tropical quente e úmido com estação seca no
inverno, típico da baixada e Af, tropical quente e úmido sem estação seca,
característicos das encostas adjacentes (Ferreira e Oliveira, 1985). Segundo Estrada
et al. (2008), a temperatura média anual é de 23,5°C e a precipitação anual média de
1067 mm, sendo os meses de maior pluviosidade janeiro e março e, junho e agosto os
meses mais secos. O regime de marés é de micromarés, com amplitude inferior a dois

metro
de S
ilhas
2). A
de s
calcu
flores
Scha
temp
de s
flores
quais
dese
de m
Mold
Figur
repre
os (Brasil, 2
Sepetiba, en
, rios e can
As partes m
sizígia, dand
ulou em 28
stas de man
aeffer-Nove
peratura mé
sizígia, divid
stas de ma
s, por estar
envolviment
mangue oc
denke, Lagu
ra 2: Map
esentativas
2008). O m
ntre o rio P
nais inserido
ais internas
do origem
8,3 Km2 a
ngue e 7,4
lli et al. (19
édia anual, e
diram o lito
ngue existe
rem submet
to estrutura
correm na
uncularia ra
pa da área
das floresta
anguezal d
Piraquê e o
o na Reserv
s das zonas
a planícies
área total
Km2 de pla
990), com
evapotrans
oral brasile
entes entre
tidas a con
l dentro de
unidade V
cemosa (L.
a de estud
as estudada
Capítulo I -
de Guaratiba
morro de
va Biológica
s entre-mar
s hipersalin
de mangu
nícies hiper
base no re
piração pot
eiro em oit
Cabo Frio
dições amb
uma mesm
II: Avicenn
) C.F. Gaer
do, com a
as.
Determinaç
a encontra-
Guaratiba,
a e Arqueo
rés são atin
as (Pellegr
ezais em 2
rsalinas.
elevo, tipo d
tencial, amp
to unidades
e Laguna in
bientais reg
ma faixa. Ap
ia schauer
rtn. e Rhizo
a posição
ção da área
-se na porçã
integrando
lógica de G
ngidas apen
rini, 2000).
2003, send
de solo, co
plitude das
s fisiográfic
nseridas na
gionais simi
penas três
riana Stapf
ophora mang
aproximada
amostral m
ão leste da
o um sistem
Guaratiba (F
nas pelas m
Almeida (2
do 20,9 Km
obertura veg
marés méd
cas, estand
a unidade V
lares, teriam
espécies tí
f & Leechm
gle L.
a das par
ínima
10
Baía
ma de
Figura
marés
2007)
m2 de
getal,
dias e
do as
VII, as
m um
ípicas
m. ex
rcelas

Capítulo I - Determinação da área amostral mínima
11
Desenho Experimental
Foram selecionadas sete florestas de mangue (Tabela I), sendo duas florestas de
franja, três florestas de bacia, e outras duas de florestas arbustivas, na zona de
transição com a planície hipersalina (Figura 2).
Em cada floresta foi estabelecida uma parcela com tamanho tal que caracterizasse
toda sua extensão de forma homogênea. O método de escolha da parcela com
tamanho máximo se baseia no método Relevé, descrito em Poore (1955) e Barbour et
al. (1980), que sugere a definição da área que melhor representa a fisionomia em
questão de forma subjetiva, porém com base na observação minuciosa das
características da mesma em termos de composição de espécies e arquitetura das
árvores. Ou seja, o método pressupõe um bom conhecimento da floresta estudada.
Tabela I - Descrição das florestas estudadas.
Floresta Tipo
Fisiográfico Descrição
F1 Franja Floresta bem desenvolvida, dominada por A.
schaueriana.
F2 Franja Floresta bem desenvolvida, com dominância mista
de A. schaueriana e R. mangle.
B1 Bacia Floresta com desenvolvimento intermediário,
dominada por A. schaueriana com arquitetura
ramificada, com ocorrência esparsa de indivíduos de
pequeno porte de L. racemosa e R. mangle.
B2 Bacia Floresta com desenvolvimento intermediário,
dominada por A. schaueriana, com presença
marcante de árvores de pequeno porte de R.
mangle.
B3 Bacia Floresta com desenvolvimento intermediário e
composição mista das três espécies.
T1 Arbustivo Florestas com baixo desenvolvimento, dominadas
por A. schaueriana.
T2 Arbustivo Florestas com baixo desenvolvimento, dominadas
por R. mangle.

Segu
deve
flores
expe
cada
meno
O tam
e 70
tanto
zona
pela
em s
Figur
sub-p
uindo o mé
e-se determ
sta esteja a
erimental ch
a vez maior
ores.
manho máx
0 m², tendo
o pela irreg
as associad
ocorrência
sub-parcela
ra 3: Desen
parcelas.
étodo Relev
inar a mínim
adequadam
hamado de
res de tal
ximo para a
o sido limita
gularidade
das à alta
de distúrbio
s (Figura 3,
nho experim
vé, após a
ma área na
ente repres
e “parcelas
forma que
a caracteriza
ado pela típ
longitudina
variabilidad
os (Soares
, Tabela II),
mental - exe
Capítulo I -
determinaç
a qual a com
sentada. Pa
aninhadas”
cada parc
ação das flo
pica disposi
al dos grad
de da micro
et al., 2003
seguindo o
emplo da flo
Determinaç
ção da par
mposição de
ara isso, o m
”, no qual
ela maior e
orestas con
ção em mo
dientes amb
otopografia
3). Cada pa
o método R
oresta B3. A
ção da área
rcela de ta
e espécies
método sug
são posicio
englobe tod
nsideradas
osaico das f
bientais, qu
(Pellegrini
arcela foi en
Relevé.
As letras A a
amostral m
manho má
e a estrutu
ere um des
onadas par
das as par
variou entre
floresta, de
ue resultam
i, 2000), qu
ntão segmen
a J referem
ínima
12
ximo,
ura da
senho
rcelas
rcelas
e 450
finida
m em
uanto
ntada
m-se a

Capítulo I - Determinação da área amostral mínima
13
Tabela II - Área das sub-parcelas (m²).
Sub-Parcela Florestas
F1 F2 B1 B2 B3 T1 T2
A 25 25 20 20 25 25 25
B 50 50 40 40 50 50 50
C 100 100 80 80 100 100 100
D 150 150 120 120 150 150 150
E 225 225 180 180 225 225 225
F 300 300 240 240 300 300 300
G 375 400 320 320 400 375 400
H 450 500 400 400 500 450 500
I 525 600 480 480 600 600
J 600 700 560 560
K 640 640
Caracterização da Estrutura Vegetal
O método seguido para a caracterização da estrutura vegetal é apresentado, de forma
geral, por Cintrón & Schaeffer-Novelli (1984) e Schaeffer-Novelli & Cintron (1986). Em
cada parcela, todos os indivíduos foram identificados em termos de espécie e medidos
quanto à altura dos mesmos e o diâmetro dos troncos, além de ter sido descrita a
condição (vivo ou morto) de cada tronco. O diâmetro dos troncos, por convenção, foi
medido a uma altura de 1,30 m do substrato, sendo denominado dap (diâmetro à
altura do peito). De posse dos dados levantados em campo, foram calculados, para
cada parcela, a altura média, o DAP (diâmetro à altura do peito, médio), a área basal,
a densidade de troncos, a dominância em área basal por espécie e condição (troncos
vivos ou mortos) e elaboradas as curvas diamétricas organizadas por espécie e
condição (troncos vivos ou mortos). A área basal representa a contribuição de madeira
em cada floresta, ou seja, é a área de troncos por área de terreno. Para o cálculo da
mesma, consideramos a área da seção transversal de cada tronco. A mesma é
calculada pela fórmula:
g = 0,00007854 (dap)²,

Capítulo I - Determinação da área amostral mínima
14
onde, g = área basal do tronco (em m²) e
dap = diâmetro à altura do peito (em cm).
O DAP é uma medida estrutural, que representa o diâmetro da árvore de área basal
média, portanto de grande importância para a caracterização das florestas. É
importante destacarmos que essa medida difere da média aritmética dos dap’s. O DAP
é obtido com os dados da área basal viva através da seguinte fórmula:
DAP = [(g). 12732,39/n]1/2,
onde n = número de troncos por hectare e
g = área basal por hectare.
Além dos parâmetros anteriormente descritos, foram calculadas também, para cada
parcela, as relações DAP/Altura Média e número de troncos/indivíduo, para que fosse
possível verificar possíveis diferenças de arquitetura das árvores entre as parcelas.
Análises Estatísticas
Para atingir o objetivo proposto, foram utilizadas duas abordagens metodológicas, que
serão detalhadas a seguir.
A primeira se refere à utilização do erro padrão percentual da média de dap’s. Os
valores de dap das sete florestas foram submetidos a uma transformação logarítmica,
para que pudessem se aproximar de uma distribuição normal e serem analisados de
forma paramétrica. A transformação logarítmica foi escolhida por ser a mais indicada
para distribuições deformadas à direita Sokal & Rolf (1980) e Zar (1996), o que foi
observado graficamente para os dados em questão. Para evitar a ocorrência de
valores negativos, em função da ocorrência de diâmetros inferiores a 1,0 cm, uma
transformação logarítmica adaptada foi utilizada:
dap’ = log (dap + 1),
onde dap = diâmetro à altura do peito

Capítulo I - Determinação da área amostral mínima
15
O erro padrão percentual da média de diâmetros foi então calculado através da
seguinte fórmula:
EPM% = (EPM / dap’)*100,
onde EPM% = erro padrão percentual da média,
EPM = erro padrão da média,
e dap’ = média aritmética dos diâmetros transformados.
Um erro máximo de 5% foi considerado para a determinação do tamanho ideal da
parcela. A área mínima foi determinada com base na regressão entre a área das sub-
parcelas e o erro padrão percentual, ajustando-se uma curva potencial negativa (y =
ax-b). Para a determinação da área mínima amostral, utilizou-se como referência o
ponto onde a curva ajustada cruza o horizonte de 5% de erro padrão percentual. A
opção pela curva potencial se deve ao fato de que a mesma tende ao infinito conforme
o tamanho da parcela se aproxima do zero, e tende a zero conforme a área da parcela
cresce infinitamente.
A segunda abordagem se baseia na comparação de distribuições de freqüência de
classes de dap. Para cada floresta, as distribuições diamétricas da menor sub-parcela
até a segunda maior sub-parcela foram comparadas com a parcela de tamanho
máximo. Essa comparação foi realizada através do teste de Kolmogorov-Smirnov
(Sokal & Rolf, 1980; Siegel, 2006). Esse teste se baseia na comparação entre as
freqüências acumuladas relativas de cada classe entre duas distribuições. A maior
diferença modular entre classes (Dmax) é comparada ao valor crítico do teste (Dα), que
é calculado da seguinte forma:
Dα = Kα * [(n1 + n2)/(n1 * n2)]1/2,
onde Kα = 1/2 * [-ln(α/2)]1/2.
Caso Dmax seja menor que Dα, a hipótese nula (de que as distribuições são similares)
é aceita, do contrário a hipótese alternativa (de que as distribuições são diferentes)
deve ser aceita.

Capítulo I - Determinação da área amostral mínima
16
Para a montagem do teste, as distribuições foram divididas em classes com amplitude
de 2 cm. No caso das florestas arbustivas (de transição), utilizou-se amplitude de 1
cm, a fim de proporcionar um número razoável de classes dado o baixo grau de
desenvolvimento típico desse tipo fisiográfico.
Para cada floresta, comparou-se cada sub-parcela à parcela de tamanho máximo,
considerando-se o tamanho mínimo aquele a partir do qual não se observa mais
diferença em relação à parcela de tamanho máximo.
Resultados e Discussão
Caracterização Estrutural das Parcelas de Tamanho Máximo
Os parâmetros estruturais das florestas consideradas para o teste são descritos na
Tabela III. De forma geral, observa-se uma clara distinção quanto ao desenvolvimento
estrutural das florestas em relação ao tipo fisiográfico correspondente. As florestas de
franja apresentam densidades mais baixas por serem as mais desenvolvidas,
apresentando os maiores valores de DAP, altura média e área basal. As florestas de
bacia apresentam desenvolvimento intermediário, enquanto as de transição
apresentam desenvolvimento bastante reduzido. Esse padrão de redução do
desenvolvimento estrutural entre florestas de franja e de transição é bem descrito na
literatura e está diretamente relacionado à freqüência de inundação pelas marés
(Cintron et al. 1978, 1980; Jimenez & Sauter, 1991; Cintron-Molero, 1993). As florestas
de franja, em níveis topográficos mais baixos, estão expostas a uma elevada
freqüência de inundação, o que determina uma maior homogeneidade temporal em
relação às condições edáficas (ex. salinidade, pH, nutrientes) e também um menor
rigor ambiental, favorecendo um maior desenvolvimento estrutural. Condições opostas
ocorrem nas florestas de transição, que em nível topográfico mais elevado estão
expostas a uma baixa freqüência de inundação pelas marés, o que determina maior
rigor ambiental, bem como maior heterogeneidade temporal das condições ambientais.

Capítulo I - Determinação da área amostral mínima
17
A influência do rigor ambiental na estrutura das florestas também pode ser percebida
pela arquitetura das árvores que compõem as florestas. Como pode-se perceber na
Tabela III, as florestas de transição apresentam as relações mais baixas entre DAP e
altura média e as mais elevadas entre o número de troncos e o número de indivíduos.
Tais características refletem a estratégia de maior investimento em crescimento
diamétrico (consolidação) em contrapartida ao crescimento vertical, que demandaria
maior disponibilidade hídrica, já que as folhas estariam numa posição ainda mais alta
em relação ao substrato.
Nas parcelas de franja estão representadas florestas com dominância em área basal
de A. schaueriana (F1) e mista de A. schaueriana e R. mangle (F2) (Tabela III). Em
Guaratiba não se observa florestas de franja dominadas por L. racemosa. As parcelas
de bacia representam florestas com dominância em área basal de A. schaueriana,
porém com contribuição expressiva de L. racemosa e R. mangle (Tabela III). Nas
parcelas T1 e T2 estão exemplificadas as florestas de transição dominadas por A.
schaueriana e R. mangle (Tabela III). Este padrão de distribuição de espécies é típico
de florestas de mangue das regiões sul e sudeste do Brasil (Schaeffer-Novelli 1989;
Schaeffer-Novelli et al. 1990), onde apenas estas três espécies ocorrem.
As distribuições diamétricas (Figuras 4A a 4G) demonstram o grau de
desenvolvimento das florestas, apresentando maior dispersão dos troncos entre todas
as classes de dap nas florestas de franja e concentração dos troncos apenas nas
classes inferiores de dap nas florestas de transição. As distribuições das florestas de
franja são bastante similares quanto à forma, aproximando-se de uma curva
exponencial suave, devido à abrangência de classes representadas. Diferem
basicamente quanto à composição de espécies, destacando-se na parcela F1 a
ocorrência de troncos de R. mangle apenas até a classe 17 cm e predominantemente
mortos. Esse padrão indica estar esta espécie ocupando principalmente o sub-bosque,
característica comum a R. mangle, descrita por McKee (1995) como mais tolerante ao
sombreamento quando comparada a L. racemosa e A. germinans, no sul da Flórida.

Capítulo I - Determinação da área amostral mínima
18
Nas florestas de bacia estão representados três níveis de desenvolvimento estrutural
ocorrentes nessa fisiografia, na ordem crescente: B1 – B2 – B3. Esse padrão pode ser
observado tanto pela variação dos parâmetros estruturais, especialmente DAP, altura
média, DAP/altura média e troncos/indivíduos (Tabela III), quanto pelas distribuições
diamétricas dessas florestas. Nas figuras 4C, 4D e 4E observa-se que o aumento do
desenvolvimento estrutural se traduz em um deslocamento da classe dominante para
a direita, na redução da freqüência associada à classe dominante e numa maior
abrangência de classes de dap.
Com relação à distribuição das espécies entre as classes, observa-se nas três
florestas a ocorrência exclusiva de troncos de A. schaueriana nas classes mais
superiores de dap. Na parcela B1, as classes inferiores e intermediárias são ocupadas
principalmente por A. schaueriana. Porém, em B2 e B3, observa-se uma contribuição
mais elevada de R. mangle em relação às demais espécies nessas classes. Nas
florestas de transição, o baixo desenvolvimento se traduz numa limitada abrangência
de classes de dap, não ultrapassando 5 cm.
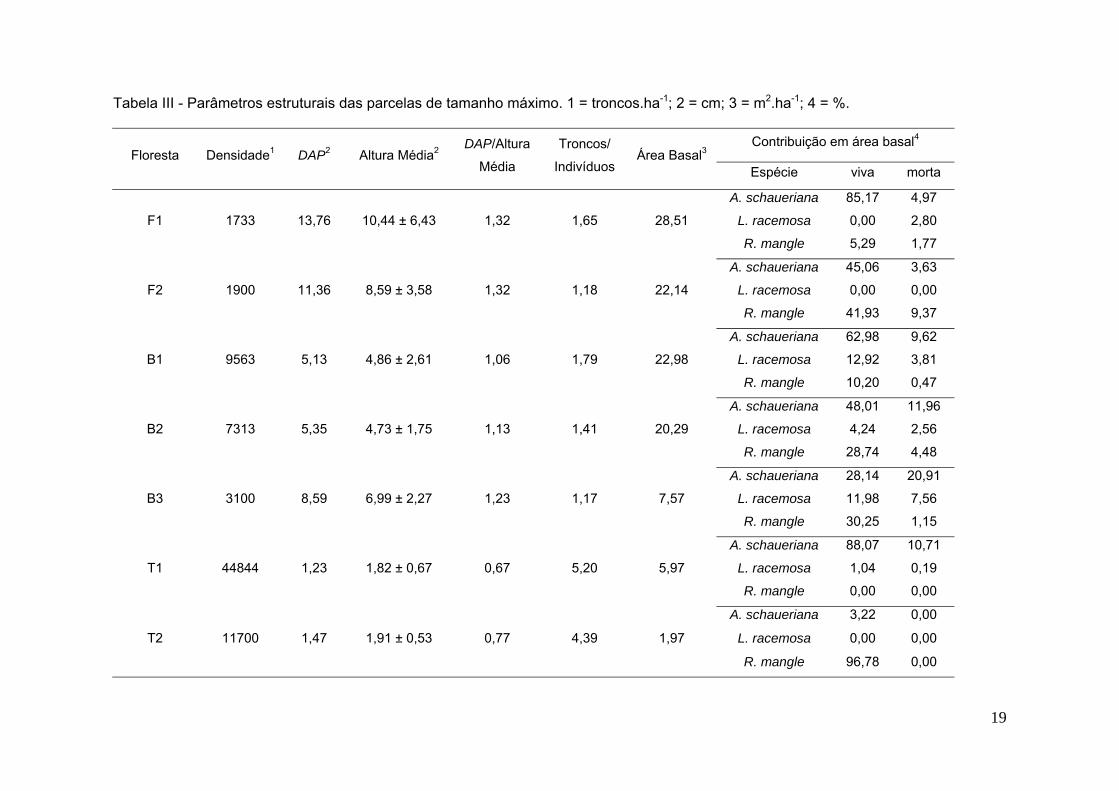
19
Tabela III - Parâmetros estruturais das parcelas de tamanho máximo. 1 = troncos.ha-1; 2 = cm; 3 = m2.ha-1; 4 = %.
Floresta Densidade1 DAP2 Altura Média2 DAP/Altura
Média
Troncos/
Indivíduos Área Basal3
Contribuição em área basal4
Espécie viva morta
F1 1733 13,76 10,44 ± 6,43 1,32 1,65 28,51
A. schaueriana 85,17 4,97
L. racemosa 0,00 2,80
R. mangle 5,29 1,77
F2 1900 11,36 8,59 ± 3,58 1,32 1,18 22,14
A. schaueriana 45,06 3,63
L. racemosa 0,00 0,00
R. mangle 41,93 9,37
B1 9563 5,13 4,86 ± 2,61 1,06 1,79 22,98
A. schaueriana 62,98 9,62
L. racemosa 12,92 3,81
R. mangle 10,20 0,47
B2 7313 5,35 4,73 ± 1,75 1,13 1,41 20,29
A. schaueriana 48,01 11,96
L. racemosa 4,24 2,56
R. mangle 28,74 4,48
B3 3100 8,59 6,99 ± 2,27 1,23 1,17 7,57
A. schaueriana 28,14 20,91
L. racemosa 11,98 7,56
R. mangle 30,25 1,15
T1 44844 1,23 1,82 ± 0,67 0,67 5,20 5,97
A. schaueriana 88,07 10,71
L. racemosa 1,04 0,19
R. mangle 0,00 0,00
T2 11700 1,47 1,91 ± 0,53 0,77 4,39 1,97
A. schaueriana 3,22 0,00
L. racemosa 0,00 0,00
R. mangle 96,78 0,00

Capítulo I - Determinação da área amostral mínima
20
Figura 4: Distribuições diamétricas das parcelas totais. A = F1; B = F2; C = B1; D = B2; E = B3; F
= T1; G = T2. Rh = Rhizophora mangle; Lg = Laguncularia racemosa; Av = Avicennia schaueriana.
0
10
20
30
40
50
≤2 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
0
10
20
30
40
50
≤2 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
0
10
20
30
40
50
≤2 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
0
10
20
30
40
50
≤2 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
0
10
20
30
40
50
≤2 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
0
10
20
30
40
50
60
70
≤1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
%
Centros de Classe (cm)
0
10
20
30
40
50
60
70
≤1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
%
Centros de Classe (cm)
A B
C D
E F
G

Capítulo I - Determinação da área amostral mínima
21
Erro Padrão Percentual da Média
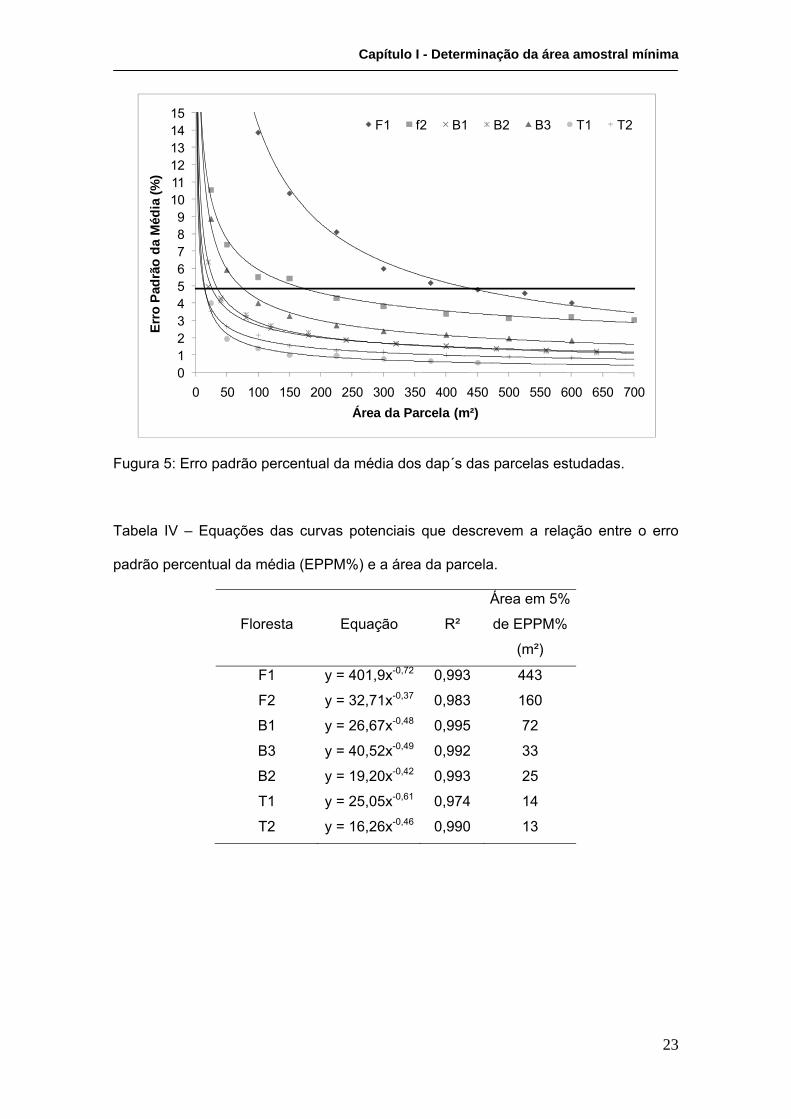
Os resultados referentes ao erro padrão percentual demonstraram uma forte relação
inversa entre o tamanho das sub-parcelas e o erro padrão percentual da média de
diâmetros (Figura 5), com R² variando entre 0,974 e 0,995. Os tamanhos mínimos que
caracterizam os tipos estruturais em questão variaram ente 13 e 443 m² (Tabela IV).
Pela análise da figura 4 e a tabela IV, pode-se observar que há uma relação entre o
tamanho mínimo da parcela e o tipo fisiográfico em questão. Para as parcelas
representativas de fisiografias de transição, a área mínima ficou definida entre 13 e 14
m²; para as representativas de bacia, 25 a 72 m²; e para as florestas de franja,
observou-se a maior variação, 160 a 443 m². A análise do erro padrão percentual da
média de diâmetros, além de demonstrar estatisticamente o proposto por Cintron &
Schaeffer-Novelli (1984), determina valores de referência para área amostral mínima
de florestas de mangue segundo o desenvolvimento. Dessa forma, as parcelas
mínimas devem ser maiores na franja e gradativamente se reduzir em direção às
florestas arbustivas, em zona de transição com a planície hipersalina.
Esse padrão pode ser explicado pelo grau de desenvolvimento das florestas,
associado às condições ambientais que prevalecem em cada tipo fisiográfico. Na
franja, a maior freqüência de inundação pelas marés permite a ocorrência de
condições ambientais menos rigorosas e mais estáveis, o que favorece um maior
desenvolvimento estrutural, que implica maior tamanho (altura e diâmetro) e menor
densidade (Tabela III). O oposto ocorre na transição com a planície hipersalina, onde
maior rigorosidade e instabilidade ambiental limitam o desenvolvimento da floresta.
Com árvores de menor porte e com maior número de ramificações, a densidade de
troncos se torna elevada nessas florestas. Esse padrão condiz com as tendências
gerais esperadas para a estrutura de ecossistemas estressados, descritas por Odum
(1985). Segundo o autor, a redução de tamanho é uma alteração esperada em termos
de estrutura das comunidades que compõe ecossistemas estressados, já que a
produtividade líquida se reduz.

Capítulo I - Determinação da área amostral mínima
22
Como o erro padrão da média é calculado dividindo-se o desvio padrão pela raiz
quadrada do número amostral, é de se esperar, portanto, que o aumento da densidade
de troncos provoque a redução do erro, mantendo-se o desvio padrão constante.
Conseqüentemente, com o aumento do erro padrão, mantendo-se a média constante,
o erro padrão percentual também tende a aumentar. Analisando-se a Tabela V,
observa-se que, embora não se mantenham constantes, a média e o desvio padrão
dos diâmetros não variam consideravelmente, ao contrário do número de troncos, que
aumenta progressivamente, determinando a diminuição do erro padrão percentual.
Biologicamente, esse padrão faz sentido, pois conforme a densidade de organismos
aumenta, é de esperar que a escala de variação espacial da estrutura da comunidade
se reduza (Peterson & Parker, 1998). Dessa forma, florestas com maior densidade de
troncos necessitariam de parcelas menores para representá-las quando comparadas a
florestas menos densas, padrão esse que foi encontrado no presente estudo.
Da Silva et al. (2003) encontrou padrão similar para uma floresta experimental de
Eucalyptus spp., porém, utilizando o coeficiente de variação dos parâmetros altura,
dap e volume em função do número de indivíduos. Os autores demonstraram que o
coeficiente de variação se reduzia exponencialmente conforme o número de indivíduos
aumentava. Além disso, para um mesmo número de plantas, o coeficiente de variação
aumentava conforme a idade da floresta aumentava, corroborando a relação entre
maturidade e número mínimo de indivíduos, ou tamanho mínimo de parcela,
encontrada no presente estudo.
Capítulo I - Determinação da área amostral mínima
23
Fugura 5: Erro padrão percentual da média dos dap´s das parcelas estudadas.
Tabela IV – Equações das curvas potenciais que descrevem a relação entre o erro
padrão percentual da média (EPPM%) e a área da parcela.
Floresta Equação R²
Área em 5%
de EPPM%
(m²)
F1 y = 401,9x-0,72 0,993 443
F2 y = 32,71x-0,37 0,983 160
B1 y = 26,67x-0,48 0,995 72
B3 y = 40,52x-0,49 0,992 33
B2 y = 19,20x-0,42 0,993 25
T1 y = 25,05x-0,61 0,974 14
T2 y = 16,26x-0,46 0,990 13
0123456789
101112131415
0 50 100 150 200 250 300 350 400 450 500 550 600 650 700
Erro
Pad
rão
da M
édia
(%)
Área da Parcela (m²)
F1 f2 B1 B2 B3 T1 T2

24
Tabela V – Número de troncos, média e desvio padrão dos diâmetros de cada sub-parcela.
Sub-parcela A B C D E F G H I J K
F1
N 2 5 20 29 42 61 91 107 136 158 -
Média 1,089 0,889 0,760 0,823 0,852 0,888 0,798 0,810 0,764 0,781 -
Desv. Pad. 0,471 0,492 0,471 0,459 0,448 0,416 0,394 0,401 0,408 0,394 -
F2
N 9 17 32 42 57 73 101 122 159 186 -
Média 0,945 0,957 0,915 0,895 0,921 0,919 0,879 0,894 0,831 0,824 -
Desv. Pad. 0,299 0,292 0,286 0,316 0,300 0,301 0,299 0,311 0,339 0,342 -
B1
N 36 70 136 188 268 414 526 615 741 845 959
Média 0,555 0,597 0,598 0,601 0,598 0,574 0,563 0,574 0,582 0,579 0,581
Desv. Pad. 0,165 0,213 0,221 0,210 0,209 0,216 0,213 0,216 0,218 0,218 0,221
B2
N 31 54 90 140 190 248 305 379 459 551 633
Média 0,572 0,581 0,621 0,607 0,636 0,660 0,681 0,684 0,686 0,684 0,682
Desv. Pad. 0,203 0,177 0,197 0,195 0,200 0,197 0,200 0,202 0,200 0,197 0,196
B3
N 14 23 43 68 101 129 160 199 232 - -
Média 0,767 0,815 0,835 0,806 0,832 0,850 0,850 0,842 0,855 - -
Desv. Pad. 0,255 0,232 0,219 0,216 0,227 0,230 0,234 0,234 0,239 - -
T1
N 82 252 496 797 1072 1503 1919 2386 - - -
Média 0,353 0,326 0,320 0,312 0,319 0,316 0,315 0,311 - - -
Desv. Pad. 0,128 0,100 0,099 0,089 0,101 0,096 0,092 0,087 - - -
T2
N 23 43 90 158 254 332 423 532 703 - -
Média 0,371 0,365 0,374 0,373 0,367 0,369 0,369 0,366 0,369 - -
Desv. Pad. 0,063 0,064 0,076 0,072 0,073 0,079 0,075 0,077 0,083 - -

Capítulo I - Determinação da área amostral mínima
25
Comparação de Distribuições de Freqüência de Classes de dap
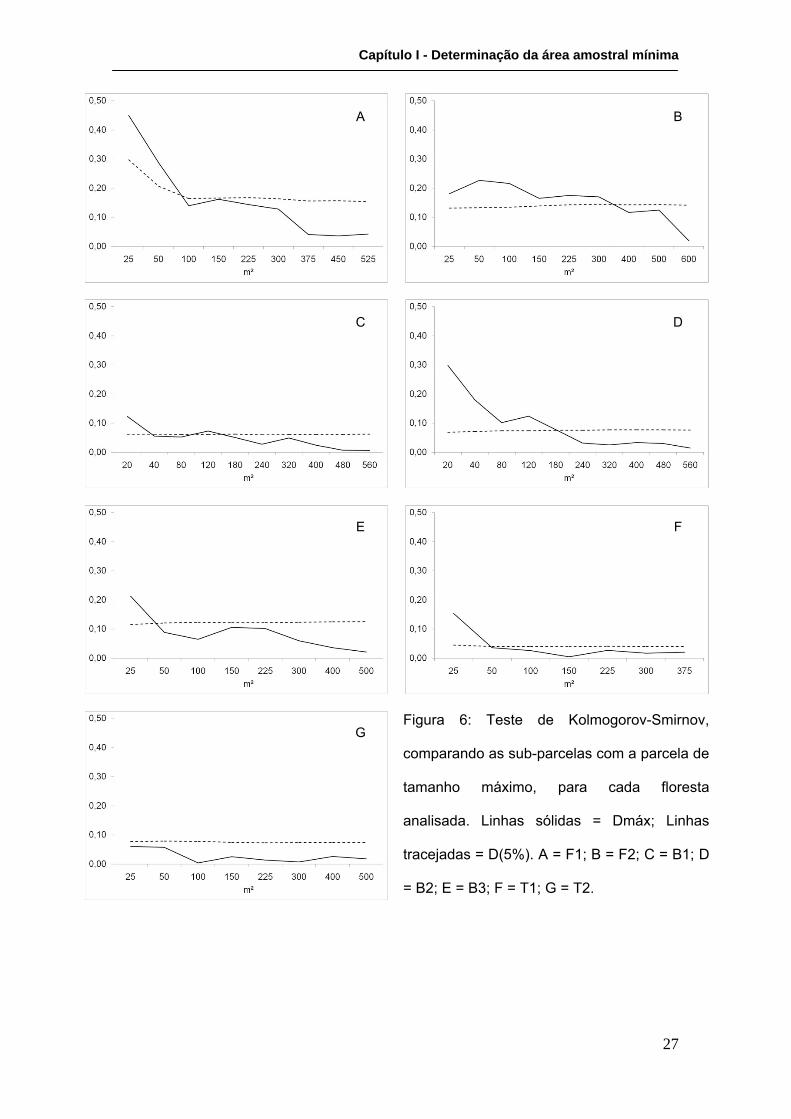
A aplicação do teste de Kolmogorov-Smirnov na comparação entre as distribuições
das sub-parcelas e a da parcela-total aponta para um padrão similar ao observado na
análise do erro padrão percentual da média de diâmetros no que se refere à relação
entre o tamanho ótimo e a fisiografia em questão. Ou seja, há uma tendência de que
as florestas de franja tenham um tamanho ótimo de parcela superior às de bacia e de
transição, nessa ordem (Figura 6; Tabela VI).
Nas parcelas representativas das florestas de franja, foi observada uma grande
diferença quanto ao tamanho ótimo. Na parcela F1, a distribuição de diâmetros da
sub-parcela de 100 m² foi a primeira a não mais apresentar diferença significativa em
relação à parcela-total. Já na parcela F2, apenas na sub-parcela de 400 m² não se
observou mais diferença significativa em reação à distribuição da parcela total. Tal fato
pode estar relacionado à dominância em área basal, já que em F1 a contribuição de A.
schaueriana chega a 90%, enquanto em F2 há um equilíbrio entre A. schaueriana
(49%) e R. mangle (51%), que poderia proporcionar uma maior heterogeneidade
estrutural. Porém, a possível influência da dominância relativa das espécies na área
amostral mínima precisa ainda ser analisada em estudos futuros para que se possa
confirmar esse padrão.
Para as parcelas representativas de florestas de bacia, o teste de Kolmogorov-
Smirnov indicou tamanhos ótimos de parcela inferiores aos encontrados para as
florestas de franja (B1 - 40 m²; B3 - 50 m²), com exceção para B2, na qual o tamanho
ótimo de 180 m² é superior ao de F1 (100 m²). Em B1, embora a sub-parcela de 120
m² tenha apresentado Dmáx ligeiramente superior a D(5%), foi considerado como
ótimo o tamanho 40 m², já que os tamanhos 80 m² e 180 m² em diante são inferiores a
D(5%).
Com relação às parcelas representativas de florestas de transição, T2 apresenta o
menor tamanho ótimo (25 m²) dentre as florestas analisadas, enquanto T1, com

Capítulo I - Determinação da área amostral mínima
26
tamanho ótimo de 50 m², apresenta tamanho similar ao encontrado para as parcelas
de bacia B3 e B1.
Gama et al. (2001), testando o tamanho ótimo de parcelas em florestas de várzea
amazônicas, concluiu que as parcelas devem ser maiores quanto maiores forem os
indivíduos em questão. Os autores encontraram um tamanho ótimo de 70 m² para os
indivíduos na classe de tamanho 0,3 m ≤ altura < 1,5 m, 80 m² para a classe de
tamanho altura ≥ 1,5 m a DAP < 5 cm, 90 m² para a classe de tamanho 5 cm ≤ DAP <
15 cm e 70 m² quando toda a população amostrada (h ≥ 0,3 m até DAP < 15 cm) foi
considerada.
Zhou et al. (2007), estudando florestas sub-tropicais da China, também considerou
classes de tamanho/tipos de vegetação para a definição tamanho ótimo e encontrou o
mesmo padrão. Os autores encontraram um tamanho ótimo de 4 m² para plântulas e
jovens (altura ≤ 1,3 m), 25 m² arbustos e árvores com altura > 1,3 m e DAP ≤ 10 cm,
100 m² para árvores com DAP ≤ 20 cm, 400 m² para árvores com DAP ≤ 40 cm, e 900
m² para árvores com DAP > 40 cm.
Capítulo I - Determinação da área amostral mínima
27
Figura 6: Teste de Kolmogorov-Smirnov,
comparando as sub-parcelas com a parcela de
tamanho máximo, para cada floresta
analisada. Linhas sólidas = Dmáx; Linhas
tracejadas = D(5%). A = F1; B = F2; C = B1; D
= B2; E = B3; F = T1; G = T2.
A B
C D
E F
G

Capítulo I - Determinação da área amostral mínima
28
Tabela VI – Comparação entre os tamanhos mínimos obtidos a partir das duas
análises empregadas.
Floresta Área Mínima
(K-S)
Área Mínima
(EPM%)
F1 100 443
F2 400 160
B3 50 72
B2 180 33
B1 40 25
T1 50 14
T2 25 13
Comparando as áreas mínimas definidas pelas duas análises, observa-se que ambas
apresentam, em linhas gerais, uma tendência de redução desde as parcelas
representativas de florestas de franja até àquelas representativas de florestas de
transição. Porém, observam-se também algumas diferenças bruscas entre os valores
(Tabela VI), principalmente nas parcelas F1, F2 e B2, que podem ser explicadas pela
diferença de objetivo entre as análises. Enquanto, a análise pelo EPM% é fortemente
influenciada pelo número amostral (troncos), a análise por K-S é sensível à
distribuição diamétrica da floresta, refletindo a amplitude de classes de diâmetro e a
distribuição das mesmas na floresta. No caso da parcela F2, a grande amplitude de
diâmetros (Figura 4B) torna necessária uma maior área de parcela, enquanto o oposto
ocorre em F1, na qual a grande amplitude de diâmetros compensou a influência da
baixa densidade na análise por EPM%, exigindo uma área mínima quatro vezes
menor. Tal fato demonstra que, provavelmente, as classes diamétricas em F1 se
distribuem de forma mais homogênea ao longo da floresta do que na parcela F2. No
caso de B2, embora não haja uma grande amplitude de classes de diâmetros, uma
possível heterogeneidade na distribuição das classes ao longo da floresta pode ser a
explicação para a disparidade dos valores de área mínima de parcela obtida pelas
duas análises (Tabela VI), já que para a análise por EPM% a área mínima é

Capítulo I - Determinação da área amostral mínima
29
condizente com a densidade da floresta, quando comparada às demais parcelas
representativas.
Os valores de área mínima de parcelas representativas obtidos neste estudo são
compatíveis com os utilizados por estudos previamente realizados em Guaratiba e em
regiões próximas, os quais se baseiam nas recomendações metodológicas descritas
por Cintrón & Schaeffer-Novelli (1984) e Schaeffer-Novelli & Cintrón (1986). Soares et
al. (Submetido), ao caracterizar florestas de mangue do rio Piracão, utilizou parcelas
com área variando entre 15 e 150 m² (altura média máxima = 7,63 ± 1,41 m), definidos
visualmente de acordo com aspectos de densidade e variabilidade estrutural da
floresta, de forma que as florestas de franja responderam pelas maiores áreas
enquanto as de transição tiveram as menores medidas. Portugal (2002) utilizou, para
florestas de mangue no rio Piraquê e na restinga da Marambaia, 35 a 225 m² (altura
média máxima = 7,58 ± 3,58 m) e 20 a 260 m² (altura média máxima = 9,57 ± 3,61 m),
respectivamente. Para a lagoa Tijuca, situada a cerca de 30 Km a norte de Guaratiba,
Soares (1999) utilizou parcelas cujas áreas variaram entre 50 e 200 m² (altura média
máxima = 7,66 m). Já para a baía de Guanabara, situada a cerca de 50 Km a norte de
Guaratiba, Soares et al. (2003) utilizaram parcelas de 25 a 400 m² (altura média
máxima = 7,27 ± 1,12 m). Para caracterizar as florestas de mangue do estuário do rio
Paraíba do Sul, a cerca de 300 Km a norte de Guaratiba, Bernini & Rezende (2004)
utilizaram parcelas com área entre 100 e 300 m² (altura média máxima = 9,90 ± 0,20
m), não havendo, entretanto, a ocorrência de florestas de transição dentre as
amostradas. Estudando florestas de franja localizadas no estuário do rio São Mateus
(700 Km a norte de Guaratiba), Silva et al. (2005) utilizaram parcelas de 400 m² (altura
média máxima = 12,00 ± 7,00 m).
Já em regiões mais afastadas (> 1.000 Km), ao norte de Guaratiba, onde o
desenvolvimento das florestas de mangue é substancialmente maior, observa-se a
utilização de áreas mínimas maiores do que as obtidas no presente estudo. Para o
estuário de Caravelas (17° 43’ S), Soares et al. (2008) utilizaram parcelas com área de

Capítulo I - Determinação da área amostral mínima
30
8 a 1.050 m², onde as florestas atingem altura média máxima de 13,94 ± 2,28 m. Para
caracterizar as florestas de mangue de Garapuá (13°29’ S), Pellegrini et al. (2007)
utilizaram parcelas com área variando entre 20 e 1.892 m². A altura média máxima
encontrada pelos autores nessas florestas foi de 18,17 ± 1,21 m. Fernandes (2000)
utilizou parcelas de 2.500 m² para caracterizar florestas de franja da Ilha de Maracá
(2°10’ N). Da mesma forma, considerando regiões mais afastadas, porém, ao sul de
Guaratiba, percebe-se que a área mínima é limitada pelo baixo desenvolvimento das
florestas. Soares et al. (2008) delimitaram parcelas de 20 e 25 m² para caracterizar a
estrutura das florestas de mangue de Laguna (28°28’ S), limite latitudinal de ocorrência
de manguezais na América do Sul, onde a altura média máxima atinge apenas 3,33 ±
0,54 m.
Pelo anteriormente exposto, percebe-se a existência de uma forte relação entre a área
mínima de parcelas para caracterização estrutural de florestas de mangue e o
desenvolvimento estrutural, considerando apenas os trabalhos que utilizam tamanhos
variáveis de parcela, de acordo com as recomendações metodológicas descritas por
Cintrón & Schaeffer-Novelli (1984) e Schaeffer-Novelli & Cintrón (1986). Além disso, os
valores de área de parcela mínima obtidos no presente estudo mostram-se
compatíveis com aqueles que vêm sendo utilizados nos manguezais da região sudeste
do Brasil.
Em outros manguezais do mundo, vários trabalhos têm utilizado tamanhos variáveis
de parcelas (Khoon & Eong, 1995; Fromard et al., 1998; Chen & Twilley, 1999; Cox &
Allen, 1999; Berger et al., 2006; Imai et al., 2006;), utilizando critérios muito similares
aos descritos anteriormente, porém, sem citar Cintrón & Schaeffer-Novelli (1984) e
Schaeffer-Novelli & Cintrón (1986) como referências metodológicas. Todos esses
trabalhos justificam esse método com base na relação entre o desenvolvimento
estrutural e a área necessária para representar a floresta. Embora não utilizem
tamanhos variáveis, Proffitt & Devlin (2005) e Rogers et al. (2005) delimitaram
parcelas de área reduzida (25 m²) e justificaram esse critério com base no baixo

Capítulo I - Determinação da área amostral mínima
31
desenvolvimento das florestas que estudavam (Flórida/EUA e Homebush
Bay/Austrália, respectivamente), já que se tratavam de florestas em estágio inicial de
regeneração, com elevados valores de densidade.
Conclusão
Os resultados das duas análises empregadas demonstraram haver uma tendência de
redução da área amostral mínima entre as parcelas representativas de florestas de
franja até àquelas representativas de florestas de transição, relacionada à redução do
desenvolvimento estrutural. Esse resultado enfatiza a importância de se utilizar
parcelas para caracterização da estrutura vegetal de florestas de mangue com
tamanho variável, confirmando de forma empírica a sugestão metodológica
recomendada por Cintrón & Schaeffer-Novelli (1984).
Para florestas de mangue da região sudeste do Brasil, ou para florestas de mangue de
outras regiões costeiras do mundo com características estruturais similares às
encontradas no segmento VII da costa brasileira (Schaeffer-Novelli et al., 1990),
recomenda-se, com base nos resultados do presente estudo, a utilização de parcelas
com área de 100 a 450 m² para florestas de franja, 25 a 180 m² para florestas de bacia
e 15 a 50 m² para florestas arbustivas de transição.
É importante ressaltar a necessidade de parcimônia ao se utilizar esses valores de
referência. Ao aplicar os valores de área mínima para uma determinada floresta de
franja, bacia ou transição, é preciso verificar se tal floresta apresenta grau de
desenvolvimento e arquitetura compatíveis com o apresentado pelas parcelas-teste do
presente estudo.

Capítulo II - Análise da Variabilidade Estrutural
32
CAPÍTULO II
ANÁLISE DA VARIABILIDADE ESTRUTURAL DAS FLORESTAS DE MANGUE DE
GUARATIBA - RIO DE JANEIRO, RJ
Introdução
A estrutura das florestas de mangue é definida em diferentes escalas espaciais.
Considerando uma escala global, as florestas de mangue apresentam gradientes
latitudinais de desenvolvimento (Cintrón et al., 1985; Schaeffer-Novelli et al., 1990;
Twilley, 1995), controlados basicamente pela radiação solar e temperatura. Em uma
mesma faixa latitudinal, os manguezais podem estar inseridos em diferentes
ambientes geomorfológicos, nos quais a estrutura das florestas é definida pela
dominância relativa de ondas, marés ou rios no sistema em questão (Thom, 1984).
Schaeffer-Novelli et al. (1990) descrevem ainda o conceito de segmentos costeiros
para a costa brasileira. Nesse conceito, cada um dos oito segmentos apresenta
características geomorfológicas, climáticas e oceanográficas similares.
Quando os manguezais são analisados numa escala ainda menor, constata-se que em
um mesmo ambiente geomorfológico a estrutura das florestas é influenciada pela
hidrologia e microtopografia locais, que determinam gradientes edáficos (ex.
salinidade, pH, nutrientes) associados ao gradiente de frequência de inundação pelas
marés e ao aporte de água doce, definindo tipos fisiográficos de florestas de mangue
(Lugo & Snedaker, 1974) que sumarizam processos de zonação. Sob esse aspecto,
Lugo & Snedaker (1974) classificam as florestas de mangue em 5 tipos fisiográficos:
fringe (franja), riverine (ribeirinho), overwash (ilhote), basin (bacia) e dwarf (anão ou de
transição), cada tipo apresentando características estruturais similares determinadas
por um conjunto de assinaturas energéticas similares. Schaeffer-Novelli et al. (2000)
resumem os tipos fisiográficos em: franja, bacia e anão. Segundo essa classificação,
as florestas de franja seriam aquelas localizadas em nível topográfico mais baixo,

Capítulo II - Análise da Variabilidade Estrutural
33
expostas a uma maior freqüência de inundação pelas marés e, consequentemente,
apresentando os mais altos níveis de desenvolvimento estrutural. Os tipos ribeirinho e
overwash seriam sub-tipos de franja, sendo o primeiro definido por um maior aporte de
água doce e nutrientes, o que aumentaria ainda mais o desenvolvimento estrutural das
florestas, e o segundo influenciado por uma maior carga de energia eólica e de ondas,
o que limitaria o desenvolvimento estrutural. As florestas de bacia seriam aquelas
localizadas em nível topográfico superior, submetidas a uma menor freqüência de
inundação pelas marés, reduzindo o desenvolvimento das árvores em relação às
florestas de franja. As florestas anãs (ou de transição) seriam aquelas expostas a
condições ambientais limítrofes do ponto de vista fisiológico para as espécies de
mangue, encontradas, por exemplo, na transição com planícies hipersalinas, ou em
regiões com limitação de nutrientes. Em função da alta rigorosidade ambiental, essas
florestas têm seu desenvolvimento estrutural limitado, raramente apresentando altura
média superior a 1,5 m.
Twilley et al. (1998) sugerem ainda escalas temporais que determinam a estrutura das
florestas de mangue. Essas escalas estão diretamente relacionadas com as escalas
espaciais e variam desde a fisiologia da planta (horas), passando pelos ciclos
sazonais de crescimento das árvores (1 ano), pela dinâmica de clareiras (décadas) e,
finalmente escalas temporais de centenas a milhares de anos, que englobam
mudanças ambientais de longo prazo e determinam a formação de tipos
geomorfológicos e fisiográficos distintos, além da própria evolução das populações
que compõem as florestas de mangue.
O objetivo do presente estudo é testar a existência de diferenças quanto ao
desenvolvimento estrutural e à composição de espécies entre as florestas de mangue
de Guaratiba agrupadas por tipos fisiográficos.

Capítulo II - Análise da Variabilidade Estrutural
34
Material e Métodos
Área de Estudo
A área de estudo localiza-se na Baixada de Sepetiba/ Guaratiba, distando cerca de 70
km do centro da cidade do Rio de Janeiro. O clima da região, segundo classificação de
Köppen pode ser dividido em Aw, tropical quente e úmido com estação seca no
inverno, típico da baixada e Af, tropical quente e úmido sem estação seca,
característicos das encostas adjacentes (Ferreira e Oliveira, 1985). Segundo Estrada
et al. (2008), a temperatura média anual é de 23,5°C e a precipitação anual média é
de 1067 mm, sendo os meses de maior pluviosidade janeiro e março e, junho e agosto
os meses mais secos. O regime de marés é de micromarés, com amplitude inferior a
dois metros (Brasil, 2008). O manguezal de Guaratiba encontra-se na porção leste da
Baía de Sepetiba, entre o rio Piraquê e o morro de Guaratiba, integrando um sistema
de ilhas, rios e canais inserido na Reserva Biológica e Arqueológica de Guaratiba
(RBAG) (Figura 1). As partes mais internas das zonas entre-marés são atingidas
apenas pelas marés de sizígia, dando origem a planícies hipersalinas ou apicuns
(Pellegrini, 2000). Almeida (2007) calculou em 28,3 Km2 a área total de manguezais
em 2003, sendo 20,9 Km2 de florestas de mangue e 7,4 Km2 de planícies hipersalinas.
Schaeffer-Novelli et al. (1990), com base no relevo, tipo de solo, cobertura vegetal,
temperatura média anual, evapotranspiração potencial, amplitude das marés médias e
de sizígia, dividiram o litoral brasileiro em oito unidades fisiográficas, estando as
florestas de mangue existentes entre Cabo Frio e Laguna inseridas na unidade VII, as
quais, por estarem submetidas a condições ambientais regionais similares, teriam um
desenvolvimento estrutural dentro de uma mesma faixa. Três espécies típicas de
mangue ocorrem na unidade VII: Avicennia schaueriana Stapf & Leechm. ex
Moldenke, Laguncularia racemosa (L.) C.F. Gaertn. e Rhizophora mangle L.

Figur
bran
Méto
Para
distri
âmbi
estud
estru
trans
planí
repre
trans
ra 1: Foto
cos e parce
odos
a atingir o o
ibuídas no
ito do prese
dos realiza
utural, as p
sição (Lugo
ície hipersa
esentativas
sição.
aérea da á
elas isolada
objetivo pro
interior da
ente estudo
ados por P
parcelas for
& Snedake
alina adjace
de floresta
área de es
as indicadas
posto, foi c
a RBAG. T
o, enquanto
ellegrini (20
ram classifi
er, 1974) -
entes. Send
as de franja
Capítulo
studo, com
s por pontos
caracterizad
Trinta e dua
os dados d
000) e Por
icadas em
de acordo
o assim, 21
a, 31 de flo
o II - Análise
as transve
s brancos (1
da a estrutu
as parcelas
das outras 3
rtugal (200
tipos fisiog
com a distâ
1 parcelas f
orestas de
e da Variabi
ersais indic
1: 3400000
ura vegetal
s foram ca
38 foram ob
02). Após a
gráficos –
ância do co
foram carac
bacia e 18
ilidade Estru
adas por t
0).
em 70 par
aracterizada
btidos a par
a caracteriz
franja, bac
orpo hídrico
cterizadas
de floresta
utural
35
raços
rcelas
as no
rtir de
zação
ia ou
o e da
como
as de

Capítulo II - Análise da Variabilidade Estrutural
36
Caracterização da estrutura vegetal
O método seguido para a caracterização da estrutura vegetal é apresentado, de forma
geral, por Cintron & Schaeffer-Novelli (1984) e Schaeffer-Novelli & Cintron (1986). Em
cada estação foi delimitada uma parcela, cuja área variou entre 20 e 1.224 m2 de
acordo com a densidade da floresta, de forma a ser amostrado um número
representativo de indivíduos (árvores), considerando-se ainda a homogeneidade da
mesma em termos de características estruturais (composição de espécies e
desenvolvimento estrutural dos indivíduos). A determinação da área das parcelas
delimitadas pelo presente estudo foi baseada no teste estatístico apresentado no
capítulo I. Em cada parcela, todos os indivíduos foram identificados em termos de
espécie e medidos quanto à altura dos mesmos e o diâmetro dos troncos, além de ter
sido descrita a condição (vivo ou morto) de cada tronco. O diâmetro dos troncos, por
convenção, foi medido a uma altura de 1,30 m do substrato, sendo denominado dap
(diâmetro à altura do peito do observador).
De posse dos dados levantados em campo, foram calculados, para cada estação de
amostragem, a altura média, o DAP (diâmetro à altura do peito, médio), a área basal, a
densidade de troncos, além da dominância em área basal. A área basal representa a
contribuição de madeira em cada floresta, ou seja, é a área de troncos por área de
terreno. Para o cálculo da mesma, consideramos a área da seção transversal de cada
tronco. A mesma é calculada pela fórmula:
0,00007854
Onde,
á
â à

Capítulo II - Análise da Variabilidade Estrutural
37
O DAP médio (DAP) é uma medida estrutural, que representa o diâmetro da árvore de
área basal média, portanto de grande importância para a caracterização das florestas.
É importante destacarmos que essa medida difere da média aritmética dos dap’s. O
DAP é obtido com os dados da área basal viva através da seguinte fórmula:
12.732,39
Onde,
ú
á
Comparação do desenvolvimento estrutural e da composição de espécies
Para a comparação do desenvolvimento estrutural entre os tipos fisiográficos, foram
considerados os parâmetros densidade, dap médio, altura média e área basal. Em
função da não-normalidade dos dados, foi aplicado o teste não-paramétrico de
Kruskal-Wallis (H), de acordo com recomendações de Zar (1996), através do software
Statistica 7.0 StatSoft®. Foi utilizado para os testes intervalo de confiança de 95% (p =
0,05). O mesmo teste foi aplicado para a comparação da composição de espécies
entre os tipos fisiográficos. Para tanto, foi utilizado o parâmetro área basal por espécie
de troncos vivos e mortos.
Foram realizadas ainda, no mesmo pacote estatístico, análises de regressão entre a
densidade de troncos e o DAP médio. Para facilitar a análise os dados foram
linearizados através de transformação logaritmica do tipo y = ln (x+1), com base em
recomendações de Zar (1996) e Sokal & Rolf (1980). A significância das regressões foi
testada utilizando-se intervalo de confiança de 95%.

Capítulo II - Análise da Variabilidade Estrutural
38
Comparação das distribuições diamétricas entre os tipos fisiográficos
Essa etapa consistiu inicialmente no ajuste das distribuições do número de troncos por
classe de diâmetro (classes de 2 cm) das 70 parcelas às seguintes funções:
- Exponencial (“J-Invertido”)
, 0 ; 0
Onde,
1
é
- Normal (distribuição unimodal simétrica)
1 √2
, 0
Onde,
é â
â é â
ã é â
- Log-Normal (distribuição unimodal assimétrica)
1 √2
, , 0
Onde,
é â ó çã í
â é â ó çã í
ã é â ó çã í

Capítulo II - Análise da Variabilidade Estrutural
39
A distribuição observada de cada parcela foi comparada com as distribuições
estimadas com base nas três funções apresentadas acima através do teste de
Kolmogorov-Smirnov (Sokal & Rolf, 1980; Siegel, 2006), utilizando como intervalo de
confiança 95% (α = 0,05). Esse teste se baseia na comparação entre as freqüências
acumuladas relativas de cada classe entre duas distribuições. A maior diferença
modular entre classes (Dmax) é comparada ao valor crítico do teste (Dα), que é
calculado da seguinte forma:
1 21 2
/
Sendo,
1/2 ln 2/
Quando Dmax é menor que Dα, a hipótese nula (de que as distribuições observada e
teórica são similares) é aceita, do contrário a hipótese alternativa (de que as
distribuições são diferentes) deve ser aceita. Caso a hipótese nula fosse aceita para
mais de uma função para uma mesma distribuição observada de uma determinada
parcelas, foi escolhida a função de melhor ajuste, com maior relação Dα/Dmax. A
partir desse teste, calculou-se o percentual de parcelas que melhor se ajustaram a
cada função, agrupando-as por tipo fisiográfico. Com essa análise, buscou-se verificar
se há diferença, entre os tipos fisiográficos, quanto ao tipo de distribuição dominante.
Por fim, os valores de diâmetro dos troncos de cada parcela foram agregados por tipo
fisiográfico e as distribuições diamétricas resultantes comparadas entre si e com as
distribuições teóricas calculadas com base nas funções descritas acima.

Capítulo II - Análise da Variabilidade Estrutural
40
Resultados e Discussão
Comparação do desenvolvimento estrutural e da composição de espécies
A análise das medianas dos parâmetros estruturais demonstra que há uma clara
distinção entre os tipos fisiográficos, sendo as florestas de franja as mais
desenvolvidas e as de transição as menos desenvolvidas. Esse padrão é
caracterizado pelo aumento da densidade de troncos e redução do DAP médio, da
altura média e da área basal entre as florestas de franja e as de transição (Figura 2A-
D). O teste de Kruskal-Wallis confirmou a existência de diferenças significativas entre
os 3 tipos fisiográficos para o DAP médio e para a altura média (Tabela 1). No caso da
densidade, não houve diferença significativa apenas entre as florestas de bacia e as
de transição. Já para a área basal, não foi observada diferença significativa apenas
entre franja e bacia.
Pellegrini (2000), Portugal (2002) e Soares et al. (submetido) encontraram esse
padrão de redução do desenvolvimento estrutural entre a franja e as florestas de
transição de Guaratiba, embora não tenham testado estatisticamente as diferenças
entre as zonas. Pellegrini (2000) analisou perfis microtopográficos e associou esse
padrão à ocorrência de elevações topográficas que determinam gradientes de
freqüência de inundação pelas marés e de salinidade da água intersticial, além de
outros possíveis gradientes físico-químicos (ex. H2S e nutrientes). De fato, gradientes
edáficos de salinidade (Cintrón et al., 1978; Sam & Ridd, 1998; Ellison et al., 2000),
concentração de H2S (Boto & Wellington, 1984; Nickerson & Thibodeau, 1985; Ewel et
al., 1998) e concentração de nutrientes (Feller et al., 2003; Boyer, 2006; Lovelock et
al., 2007) têm sido descritos para manguezais mundialmente.
De forma geral, Odum (1985) afirma que a redução do tamanho organismos é uma
tendência em ecossistemas estressados. Para florestas de mangue, a redução do
desenvolvimento estrutural entre florestas de franja e bacia/transição é demonstrada
por vários autores (Pool et al., 1977; Cintrón et al., 1978; Lugo et al., 1988; López-
Portillo & Ezcurra, 1989; Schaeffer-novelli et al., 1990; Jiménez & Sauter, 1991; Ewel
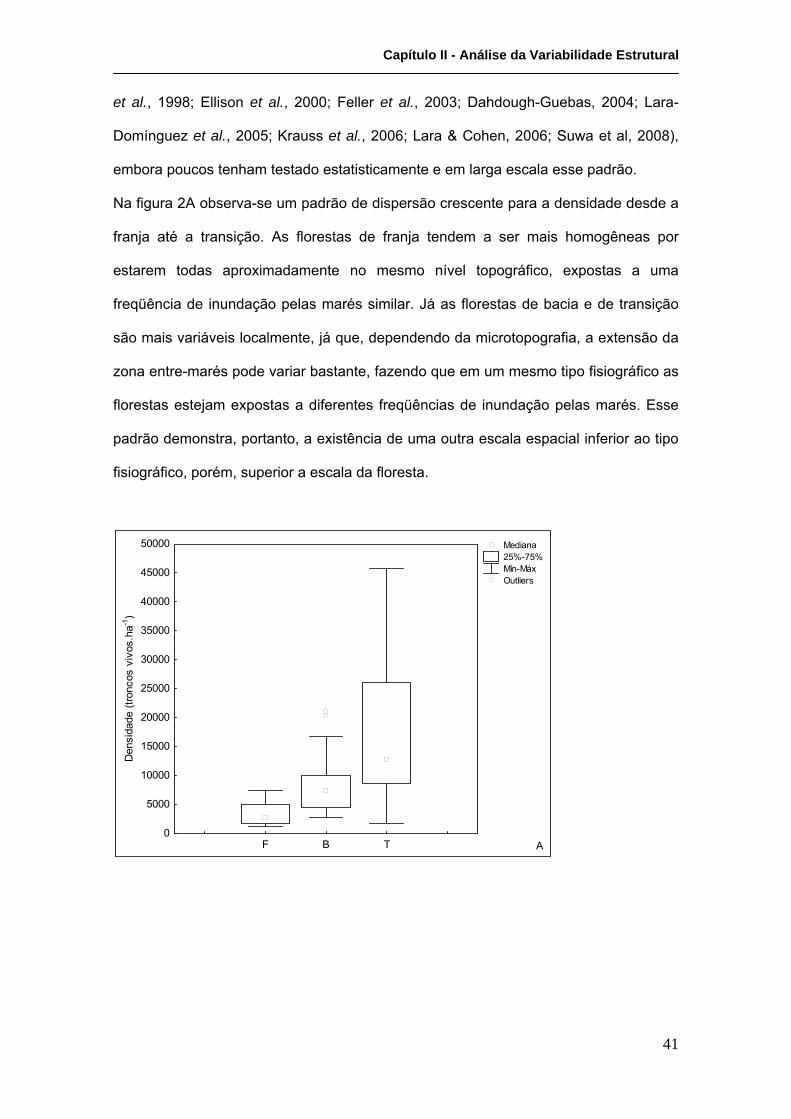
Capítulo II - Análise da Variabilidade Estrutural
41
et al., 1998; Ellison et al., 2000; Feller et al., 2003; Dahdough-Guebas, 2004; Lara-
Domínguez et al., 2005; Krauss et al., 2006; Lara & Cohen, 2006; Suwa et al, 2008),
embora poucos tenham testado estatisticamente e em larga escala esse padrão.
Na figura 2A observa-se um padrão de dispersão crescente para a densidade desde a
franja até a transição. As florestas de franja tendem a ser mais homogêneas por
estarem todas aproximadamente no mesmo nível topográfico, expostas a uma
freqüência de inundação pelas marés similar. Já as florestas de bacia e de transição
são mais variáveis localmente, já que, dependendo da microtopografia, a extensão da
zona entre-marés pode variar bastante, fazendo que em um mesmo tipo fisiográfico as
florestas estejam expostas a diferentes freqüências de inundação pelas marés. Esse
padrão demonstra, portanto, a existência de uma outra escala espacial inferior ao tipo
fisiográfico, porém, superior a escala da floresta.
Mediana 25%-75% Mín-Máx Outliers
F B T0
5000
10000
15000
20000
25000
30000
35000
40000
45000
50000
Den
sida
de (t
ronc
os v
ivos
.ha-1
)
A

Capítulo II - Análise da Variabilidade Estrutural
42
Mediana 25%-75% Mín-Máx
F B T0
2
4
6
8
10
12
14
16
DA
P M
édio
(cm
)
B
Mediana 25%-75% Mín-Máx
F B T0
2
4
6
8
10
12
Altu
ra M
édia
(m)
C
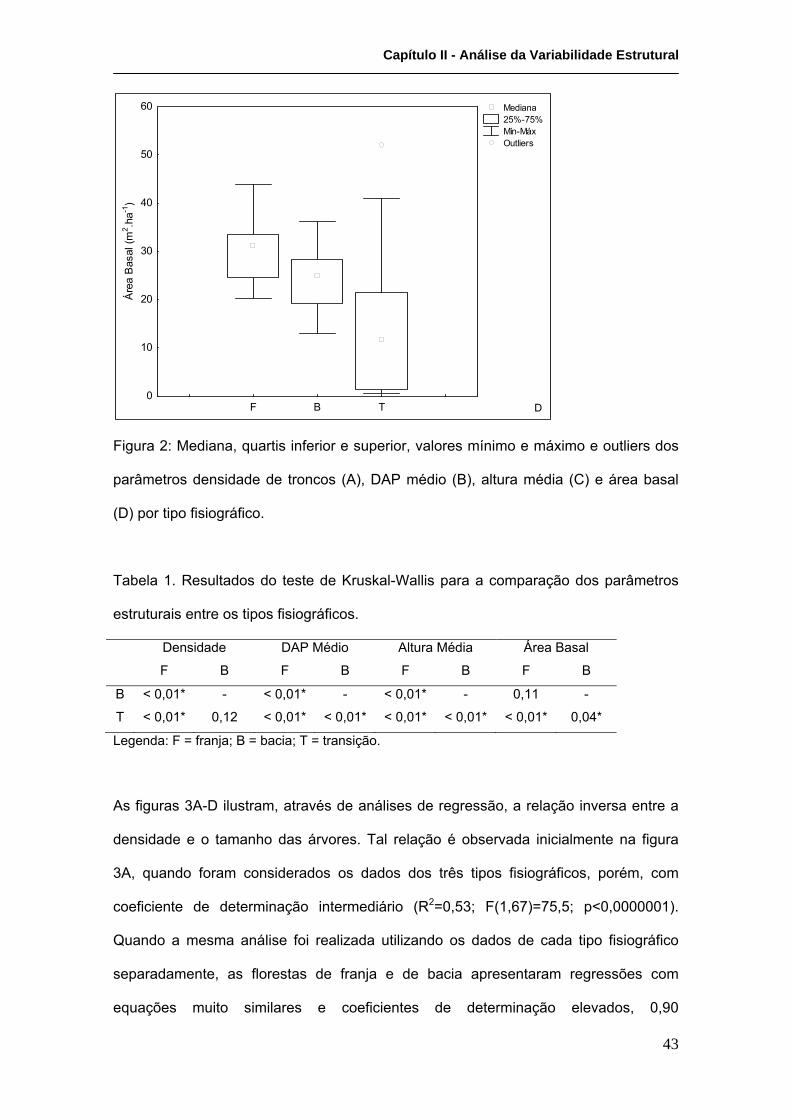
Capítulo II - Análise da Variabilidade Estrutural
43
Figura 2: Mediana, quartis inferior e superior, valores mínimo e máximo e outliers dos
parâmetros densidade de troncos (A), DAP médio (B), altura média (C) e área basal
(D) por tipo fisiográfico.
Tabela 1. Resultados do teste de Kruskal-Wallis para a comparação dos parâmetros
estruturais entre os tipos fisiográficos.
Densidade DAP Médio Altura Média Área Basal
F B F B F B F B
B < 0,01* - < 0,01* - < 0,01* - 0,11 -
T < 0,01* 0,12 < 0,01* < 0,01* < 0,01* < 0,01* < 0,01* 0,04*
Legenda: F = franja; B = bacia; T = transição.
As figuras 3A-D ilustram, através de análises de regressão, a relação inversa entre a
densidade e o tamanho das árvores. Tal relação é observada inicialmente na figura
3A, quando foram considerados os dados dos três tipos fisiográficos, porém, com
coeficiente de determinação intermediário (R2=0,53; F(1,67)=75,5; p<0,0000001).
Quando a mesma análise foi realizada utilizando os dados de cada tipo fisiográfico
separadamente, as florestas de franja e de bacia apresentaram regressões com
equações muito similares e coeficientes de determinação elevados, 0,90
Mediana 25%-75% Mín-Máx Outliers
F B T0
10
20
30
40
50
60
Áre
a B
asal
(m2 .h
a-1)
D

Capítulo II - Análise da Variabilidade Estrutural
44
(F(1,19)=162,5; p<0,0000001) e 0,83 (F(1,28)=139,6; p<0,0000001), respectivamente
(figuras 3B e 3C). Já as florestas de transição apresentaram alta variabilidade na
dispersão dos dados, com R² muito baixo e regressão não-significativa (R2=0,02;
F(1,16)=0,2; p=0,63 - figura 3D).
A relação inversa entre o DAP médio e a densidade observada para as florestas de
franja e bacia demonstra o efeito da competição na estrutura da comunidade. Quanto
mais desenvolvidas são as árvores que compõe a floresta, mais espaço ocupam e
mais recursos demandam. Dessa forma, a ocorrência de condições ambientais mais
rigorosas na bacia limita o desenvolvimento das árvores, permitindo que tais florestas
tenham uma maior densidade do que as florestas de franja, o que é corroborado na
figura 2A e na tabela 1, que mostram que de fato a densidade das florestas de franja é
menor. A análise da figura 3A mostra que, embora haja uma interseção na faixa
intermediária de desenvolvimento, na faixa inferior de desenvolvimento há um
predomínio de florestas de bacia, ao passo que na faixa superior de desenvolvimento,
acima de 10 cm de DAP médio, só ocorrem florestas de franja. Por outro lado, a
inexistência de uma relação entre a densidade e o DAP médio para as florestas de
transição reflete o elevado rigor ambiental que ocorre nessa zona, fazendo com que
algumas florestas de transição, apesar de terem baixo DAP médio, possuam baixa
densidade. Ou seja, o alto rigor ambiental, provavelmente, limita a ocupação ótima do
espaço disponível.
Os coeficientes angulares que descrevem a relação entre a densidade e o DAP médio
para as florestas de franja e bacia (-1,59 e -1,62, respectivamente) são próximos ao
coeficiente de -1,5 (ou -3/2) descrito por Niklas (1994) como característico da relação
densidade X biomassa (ou tamanho) para várias espécies de plantas, a partir de
observações de Yoda et al. (1963, apud. Niklas, 1994), chamado de regra do desbaste
natural (self thinning rule). Ward et al. (2006) também encontrou, para florestas de
mangue da Flórida (EUA), coeficiente próximo a -1,5 (-1,52; R² = 0,92). Weller (1987)
corroborou essa regra no âmbito interespecífico. Porém, o autor demonstrou que
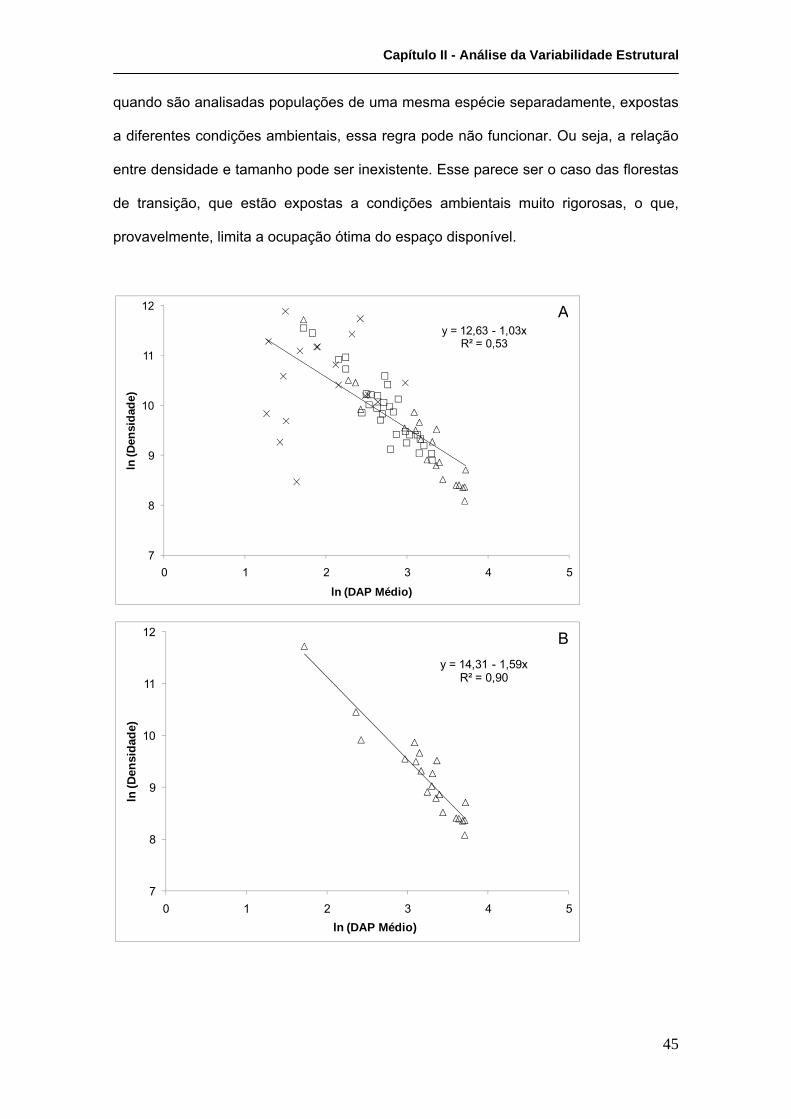
Capítulo II - Análise da Variabilidade Estrutural
45
quando são analisadas populações de uma mesma espécie separadamente, expostas
a diferentes condições ambientais, essa regra pode não funcionar. Ou seja, a relação
entre densidade e tamanho pode ser inexistente. Esse parece ser o caso das florestas
de transição, que estão expostas a condições ambientais muito rigorosas, o que,
provavelmente, limita a ocupação ótima do espaço disponível.
y = 12,63 - 1,03x R² = 0,53
7
8
9
10
11
12
0 1 2 3 4 5
ln (D
ensi
dade
)
ln (DAP Médio)
y = 14,31 - 1,59x R² = 0,90
7
8
9
10
11
12
0 1 2 3 4 5
ln (D
ensi
dade
)
ln (DAP Médio)
A
B
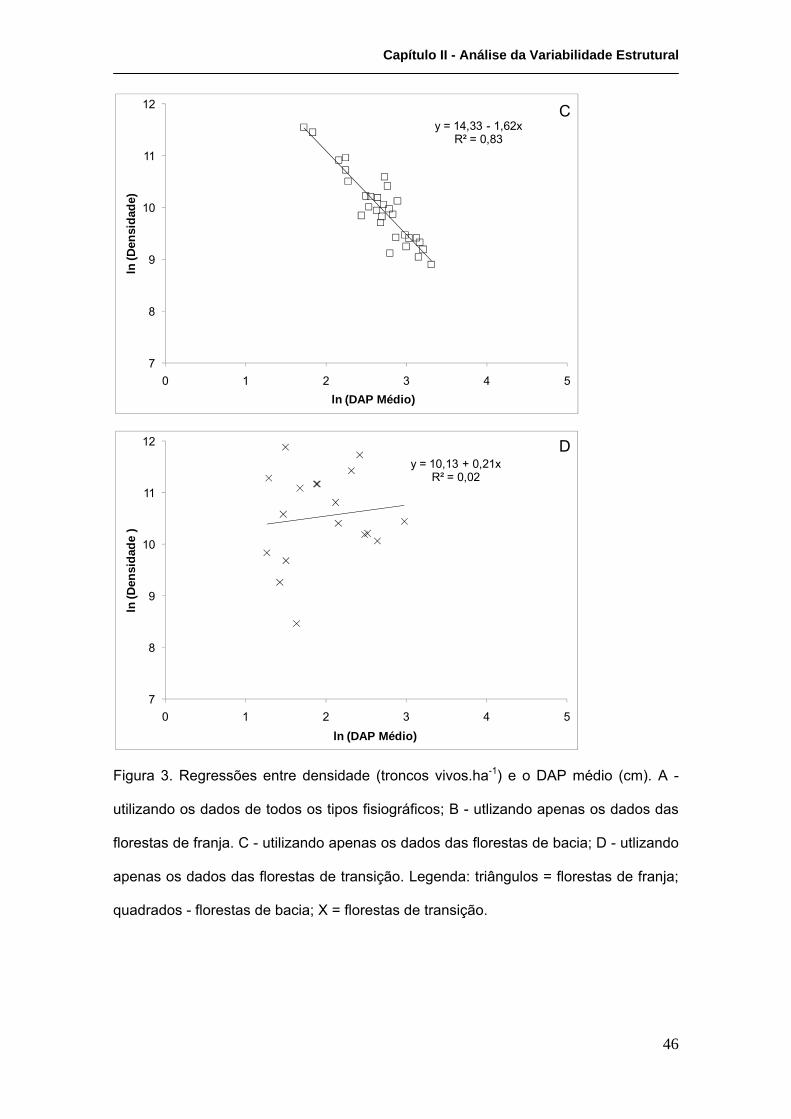
Capítulo II - Análise da Variabilidade Estrutural
46
Figura 3. Regressões entre densidade (troncos vivos.ha-1) e o DAP médio (cm). A -
utilizando os dados de todos os tipos fisiográficos; B - utlizando apenas os dados das
florestas de franja. C - utilizando apenas os dados das florestas de bacia; D - utlizando
apenas os dados das florestas de transição. Legenda: triângulos = florestas de franja;
quadrados - florestas de bacia; X = florestas de transição.
y = 14,33 - 1,62x R² = 0,83
7
8
9
10
11
12
0 1 2 3 4 5
ln (D
ensi
dade
)
ln (DAP Médio)
y = 10,13 + 0,21x R² = 0,02
7
8
9
10
11
12
0 1 2 3 4 5
ln (D
ensi
dade
)
ln (DAP Médio)
C
D

Capítulo II - Análise da Variabilidade Estrutural
47
Na tabela 2 são apresentados os valores de mediana, quartis inferior e superior e
valores máximo e mínimo de área basal por espécie em cada tipo fisiográfico,
enquanto na tabela 3 são apresentados os resultados do teste de Kruskal-Wallis,
comparando a área basal das três espécies em cada tipo fisiográfico. Não se observa
diferença significativa na área basal entre A. schaueriana e R. mangle nos três tipos
fisiográficos. A área basal de L. racemosa apresentou área basal significativamente
menor do que as demais espécies (tabelas 2 e 3), com exceção de A. schaueriana na
transição.
Tais resultados indicam que as florestas de franja e bacia são dominadas similarmente
por A. schaueriana e R. mangle, o que está de acordo com o descrito por Schaeffer-
Novelli et al. (1990) para florestas de mangue do segmento VII da costa brasileira, ao
passo que as florestas de transição possuem uma tendência a serem dominadas por
R. mangle. Lagucularia racemosa se mostrou a menos representativa nas florestas de
mangue de Guaratiba. Entretanto, ao se analisar a tabela 4, percebe-se que, embora a
área basal de A. schaueriana e R. mangle sejam similares nas florestas de franja e
bacia, A. schaueriana é a espécie dominante (área basal > 50%) na maioria das
parcelas (66,7% na franja e 61,3% na bacia). Já para as florestas de transição,
embora a área basal de R. mangle seja significativamente maior do que a de A.
schaueriana, há uma equivalência em termos de florestas dominadas por essas
espécies, ambas com 44,4%. Essa análise corroborou a baixa representatividade de L.
racemosa nas florestas de mangue de Guaratiba, já que a espécie apresentou baixos
percentuais de dominância nos três tipos fisiográficos. As florestas mistas também se
mostraram raras e sem nenhuma ocorrência entre as florestas de transição.
A ocorrência marcante de R. mangle nas florestas de transição é intrigante, já que esta
espécie é descrita por McKee (1995) como a menos tolerante à salinidade quando
comparada a A. germinans e a L. racemosa. Chaves (2001) e Portugal (2002)
demonstraram que em algumas florestas de Guaratiba localizadas às margens da baía
de Sepetiba ocorrem canais de maré que se prolongam até a transição com a planície
Capítulo II - Análise da Variabilidade Estrutural
48
hipersalina. A ocorrência de tais canais favoreceria então a chegada de propágulos de
R. mangle na transição com a planície hipersalina. Isso seria explicado pelo maior
tamanho dos propágulos desta espécie, quando comparados aos de A. schaueriana e
L. racemosa, corroborando a hipótese de “barreira topográfica à dispersão de
propágulos” postulada por Rabinowitz (1978). Além disso, a ocorrência desses canais
aumenta a frequência de inundação pelas marés transição, atenuando as
características físico-químicas do substrato, o que favoreceria competitivamente os
propágulos de R. mangle em relação aos das outras espécies.
A baixa representatividade de L. racemosa nas florestas de mangue de Guaratiba
pode ser um indicador de que tais florestas encontram-se bem preservadas. Soares et
al. (2003) citam que essa espécie é típica de florestas em processo de regeneração, e
que a mesma domina nas florestas de mangue sob forte pressão antrópica da baía de
Guanabara (a 50 Km de Guaratiba). Cavalcanti et al. (2009) demonstraram que nas
florestas de mangue localizadas no interior dos limites da APA Guapimirim, também na
baía de Guanabara, há um domínio de florestas dominadas por R. mangle, ao
contrário das florestas impactadas localizadas fora da APA e descritas por Soares et
al. (2003).
Tabela 2. Mediana, quartis (Q) inferior e superior, e valores máximo e mínimo de área
basal (m².ha-1) por espécie e tipo fisiográfico.
Franja Bacia Transição
Av Lg Rh Av Lg Rh Av Lg Rh
Mediana 20,7 0,4 9,9 12,1 0,0 8,3 0,5 0,0 1,5
Q inferior 12,2 0,0 4,1 6,2 0,0 3,8 0,0 0,0 0,1
Q superior 25,5 2,5 11,9 16,3 2,6 14,6 10,3 0,0 4,1
Máximo 38,8 20,7 32,5 122,1 34,2 23,7 37,2 53,7 13,4
Mínimo 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0

Capítulo II - Análise da Variabilidade Estrutural
49
Tabela 3. Resultados do teste de Kruskal-Wallis para a comparação da área basal
entre as espécies de cada tipo fisiográfico.
Franja Bacia Transição
Av Lg Av Lg Av Lg
Lg < 0,01* - < 0,01* - 0,10 -
Rh 0,07 < 0,01* 1,00 < 0,01* 1,00 0,04*
Legenda: Av = A. schaueriana; Lg = L. racemosa; Rh = R. mangle.
Tabela 4. Percentuais de florestas dominadas em área basal (> 50%) por (1) A.
schaueriana, (2) L. racemosa, (3) R. mangle e (4) florestas mistas das 3 espécies (sem
espécie dominante).
1 2 3 4
Franja 66,7 4,8 23,8 4,8
Bacia 61,3 9,7 22,6 6,5
Transição 44,4 11,1 44,4 0,0
Comparação das distribuições diamétricas entre os tipos fisiográficos
A análise da tabela 5 a 9 revela que quando as distribuições diamétricas das florestas
são analisadas apenas por tipo fisiográfico, sem separação por espécie, há um
predomínio da função log-normal como a de melhor ajuste, seguida da função
exponencial e da normal. Quando as distribuições são separadas por espécie, essa
tendência se mantém para A. schaueriana e R. mangle. Porém, para L. racemosa
observa-se que a função normal é a que melhor se ajusta às distribuições diamétricas
dessa espécie, com exceção àquelas provenientes de florestas de franja, nas quais
37,5% das distribuições se ajustam à função normal, enquanto 50,0% seguem a
tendência geral de ajuste à função log-normal.
Uma possível explicação para a função normal ser a predominante para L. racemosa
deva ser uma composição entre a sua grande habilidade como espécie pioneira de
processos de regeneração e sua baixa capacidade em tolerar sombreamento. McKee
(1995) demonstrou que L. racemosa apresenta maior taxa de crescimento em

Capítulo II - Análise da Variabilidade Estrutural
50
condições de grande disponibilidade luminosa quando comparada a R. mangle e A.
germinans. Além disso, essa espécie produz propágulos menores e em quantidade
superior a R. mangle e A. schaueriana (Schaeffer-Novelli & Cintrón, 1986). Essas duas
características favorecem a função dessa espécie como pioneira na regeneração. Por
outro lado, McKee (1995) demonstrou que L. racemosa é pouco tolerante ao
sombreamento, principalmente quando comparada a R. mangle e A. germinans, o que
dificulta a sobreposição de coortes dessa espécie após a fase inicial de colonização da
clareira. Por isso, é de se esperar que essa espécie apresente apenas uma coorte, a
que iniciou o processo de regeneração, o que explica a formação predominante de
distribuições unimodais simétricas. Kairo et al. (2008) encontrou uma distribuição
normal para uma floresta replantada de 12 anos monoespecífica de Rhizophora
mucronata no Quênia. Ou seja, a ocorrência de coorte única favorece a formação de
distribuições normais. Soares et al. (2003) também encontraram, para florestas de
mangue do Brasil (baía de Guanabara, Rio de Janeiro) dominadas por L. racemosa,
em processo de regeneração após derramamento de óleo, distribuição normal. Os
autores também associam a facilidade em se estabelecer no estágio inicial da
regeneração e a incapacidade da espécie em sobrepor coortes como provável
explicação para o desenvolvimento de distribuições normais para esta espécie.
A tendência geral de melhor ajuste à função log-normal revela que a maioria das
florestas de mangue de Guaratiba apresentam distribuições unimodais com assimetria
positiva. Harper & White (1974) e Lorimer & Kruger (1983) demonstram que florestas
temperadas de coorte única tipicamente apresentam distribuições de unimodais que
podem apresentar graus variados de assimetria quando jovens, mas gradualmente se
aproximam de uma distribuição simétrica (normal) conforme amadurecem.
No caso de A. schaueriana, embora a mesma siga o padrão geral de ter a função log-
normal como a de melhor ajuste às distribuições diamétricas, observa-se um elevado
percentual de florestas cujas distribuições melhor se ajustam a uma função
exponencial, com exceção daquelas classificadas como de transição. Oliver & Larson

Capítulo II - Análise da Variabilidade Estrutural
51
(1990) e Frelich (2002) citam que florestas que possuem múltiplas coortes tendem a
formar distribuições do tipo “J-invertido”, análogas a distribuições exponenciais. Frelich
(2002) descreve que esse tipo de distribuição resulta de uma alta taxa de mortalidade
para as classes menores de dap, que estão sofrendo desbaste natural no processo de
regeneração de pequenas clareiras ou não conseguem competir com as árvores
maduras que dominam o dossel.
Na figura 4A são apresentadas as distribuições de diâmetros utilizando-se todos os
dados agrupados por tipo fisiográfico. O teste de Kolmogorov-Smirnov demonstrou
que a distribuição referente às florestas de franja, bacia e transição são diferentes
entre si (p < 0,01). A análise das distribuições aponta fortes indícios de que as
florestas de franja encontram-se em um estágio sucessional mais avançado do que as
florestas de bacia e transição, nessa ordem. As florestas de transição apresentam uma
concentração de mais 60% dos troncos na classe de 1 cm e depois uma queda
abrupta até a classe de 5 cm, se extinguindo a seguir na classe de 11 cm. As florestas
de bacia também apresentam a maioria dos troncos na classe de 1 cm, porém com
menor concentração (38%) e maior amplitude de diâmetros (até a classe de 17 cm). Já
as florestas de franja apresentam uma distribuição com deslocamento da moda para a
classe de 3 cm e grande amplitude de diâmetros, englobando todas as classe
consideradas. Khoon & Eong (1995) estudaram florestas de mangue na Malásia em
diferentes estágios sucessionais (5 a 28 anos) expostas a manejo para silvicultura. Os
autores demonstraram que conforme a sucessão avança, a amplitude de diâmetros
aumenta e a distribuição diamétrica tende a passar de “J-invertido” para unimodal
assimétrica positiva e finalmente unimodal simétrica. Souza & Sampaio (2001)
compara florestas de mangue do Brasil (Suape, Pernambuco) impactadas pela
construção de um porto, e demonstra que as florestas mais preservadas e,
conseqüentemente, em estágio sucessional mais avançado, apresentam maior
amplitude de classes diamétricas do que as florestas mais impactadas. Oliver &
Larson (1990) descrevem esse padrão para florestas temperadas da América do

Capítulo II - Análise da Variabilidade Estrutural
52
Norte. Os autores demonstram que conforme a floresta avança ao longo da sucessão,
após a ocorrência de um distúrbio, a distribuição de diâmetros inicialmente é
caracterizada por uma alta densidade de troncos, concentrada nas classes iniciais de
diâmetro. Conforme a floresta amadurece, a classe dominante se desloca para a
direita e a distribuição se suaviza, englobando cada vez mais classes de diâmetro.
Harper & White (1974) confirma esse padrão demonstrando que as florestas
inicialmente apresentam distribuições exponenciais e depois passam a assumir
distribuições log-normais com maior amplitude de classes diamétricas conforme a
sucessão avança. Nogueira et al. (2006) apresenta o mesmo padrão para uma floresta
plantada de Tectona grandis no Mato Grosso. Frelich (2002) apresenta padrão similar,
para florestas temperadas da América do Norte, com aumento da amplitude de
classes diamétricas com o avanço da sucessão, porém, sem deslocamento da classe
dominante. O autor afirma que esse padrão ocorre em florestas nas quais ocorrem
pequenos, porém, freqüentes distúrbios ao longo do amadurecimento da floresta.
Esses distúrbios permitiriam, então, a constante formação e regeneração de pequenas
clareiras, favorecendo a manutenção da concentração dos troncos na menor classe
diamétrica.
Na figura 4B são apresentadas as distribuições teóricas que melhor se ajustaram às
distribuições observadas apresentadas na figura 4A: as distribuições das florestas de
transição e bacia melhor se ajustaram à função exponencial, enquanto a distribuição
das florestas de franja melhor se ajustou à função log-normal. Esse padrão condiz com
o deslocamento da classe dominante para a direita em função do amadurecimento, já
que a transformação de uma curva exponencial em log-normal implica na transferência
da classe dominante da menor classe para uma das seguintes. Frelich (2002)
demonstra que quando a re-colonização após um distúrbio é lenta, a distribuição de
diâmetros tende a assumir a forma de uma distribuição exponencial. Por outro lado,
quando a re-colonização é rápida, a distribuição de diâmetros tende a assumir uma
forma unimodal, que inicialmente pode apresentar assimetria positiva, assemelhando-

Capítulo II - Análise da Variabilidade Estrutural
53
se a uma distribuição log-normal. Oliver & Larson (1990) cita que o período de
colonização de uma clareira pode variar entre poucos anos a muitas décadas,
dependendo em quão rápido o espaço disponível é reocupado pelas espécies
colonizadoras. Os autores sugerem ainda que os fatores que levam a uma rápida re-
colonização são: condições ambientais favoráveis para o rápido crescimento; um
rápido aparecimento de novas plantas na clareira; a presença de espécies que
naturalmente crescem rapidamente. Portanto, a ocorrência de condições ambientais
mais favoráveis na franja, conforme explicado anteriormente, pode facilitar a rápida
regeneração de uma clareira em comparação às florestas que ocorrem nos tipos
fisiográficos de bacia e transição.
A elevação do nível médio relativo do mar (N.M.R.M.) é, possivelmente, distúrbio em
larga escala que determina que as florestas de franja estejam em um estágio
sucessional mais avançado do que as florestas de bacia e de transição, nessa ordem.
Oliveira (2001), Portugal (2002), Estrada et al. (2003) e Soares et al. (2005)
demonstram que de fato as florestas de mangue estão avançando em direção às
planícies hipersalinas adjacentes. Os autores apresentam como evidência o
monitoramento da colonização da planície hipersalina por espécies de mangue,
caracterizado pelo avanço, em pulsos, da vegetação e pela consolidação desses
pulsos. Portugal (2002) mostra ainda que é comum ocorrerem processos erosivos na
franja das florestas de mangue de Guaratiba, em função do aumento da energia
hidrológica provocado pela elavação do N.M.R.M. Dessa forma, os autores
supracitados concluiem que as florestas de mangue de Guaratiba estão avançando
em direção às planícies hipersalinas, sendo que em algumas áreas o manguezal está
se expandindo e em outras o avanço está sendo compensado pela erosão da franja.
Ellison & Stoddart (1991) e Twilley (1995) demonstram que esse tipo de resposta das
florestas de mangue à elevação do N.M.R.M é comum para os manguezais
globalmente.

Capítulo II - Análise da Variabilidade Estrutural
54
Baldwin et al. (2001) compararam as distribuições diamétricas antes e depois da
passagem de um furacão na Flórida e constatou que antes da passagem do furacão
as distribuições assumiam uma forma unimodal assimétrica positiva com grande
amplitude de classes diamétricas (até 45 cm). Após a passagem do furacão, no início
do processo de regeneração, as florestas assumiram distribuições diamétricas
similares a uma distribuição exponencial com amplitudes de classes diamétricas
reduzida (até 20 cm). Cole et al. (1999), estudando florestas de mangue da
Micronésia, compararam uma floresta que não sofreu efeito da última passagem de
um furacão (Kosrae) com outra que foi atingida pelo furacão (Pohnpei). Os autores
não encontraram diferença na forma das distribuições, porém demonstraram que a
classe diamétrica dominante era deslocada para a esquerda em Pohnpei em relação a
Kosrae. Cox & Allen (1999) encontraram, para florestas de mangue monoespecíficas
de R. mangle introduzidas no Havaí no início do século XX, uma distribuição unimodal
assimétrica positiva, similar a uma distribuição log-normal. Hauff et al. (2006)
compararam uma floresta preservada com outras sob influência de algum distúrbio. Os
autores encontraram, para a floresta preservada, uma distribuição unimodal
assimétrica positiva, enquanto que para as florestas impactadas foram observadas
florestas com distribuição aproximadamente exponencial. Iftekhar & Saenger (2008)
também encontraram para florestas preservadas do Sanderbuns (Bangladesh)
distribuições unimodais assimétricas positivas, sendo que as espécies dominantes
(Avicennia officinalis e Heritiera fomes) apresentaram maior amplitude de classes
diamétricas do que as espécies características de estágios iniciais de sucessão.
Jiménez (1990) e Jiménez & Sauter (1991), por outro lado, observaram, para florestas
de mangue preservadas da Costa Rica, mono-específicas e mistas de Avicennia
bicolor e Rhizophora racemosa, distribuições diamétricas com forma “J-invertido”,
similar a uma distribuição exponencial. Os autores, entretanto, utilizaram parcelas de 1
ha, o que provavelmente favoreceu a inclusão de muitas clareiras naturais de pequeno
porte em processo de regeneração e, conseqüentemente, alta densidade de árvores

Capítulo II - Análise da Variabilidade Estrutural
55
jovens de pequeno porte. Nesse sentido, Rubin et al. (2006) sugere que toda floresta
temperada apresenta distribuição exponencial se for caracterizada em larga escala,
com parcelas de 1 ha ou mairoes, dependendo do dap máximo, da taxa de
mortalidade base e da amplitude das classes diamétricas. Esse fato ressalta a
importância em se estabelecer parcelas de menor tamanho, porém adequadas às
características estruturais locais/regionais, conforme demonstrado no Capítulo I, para
demonstrar
Re-analisando as figuras 2A-D observa-se também que os parâmetros estruturais não
refletem apenas um gradiente de rigor ambiental, mas também florestas em estágios
sucessionais gradativamente menos avançados (no sentido franja-transição). Esse
padrão fica claro quando observamos que, conforme a altura e o DAP médio aumenta
entre as florestas de transição e as de franja, a densidade diminui, refletindo o
amadurecimento das florestas. Nesse sentido, Jiménez et al. (1985), Peet &
Christensen (1987) e Schaeffer-Novelli & Cintrón (1986) afirmam que a densidade de
uma floresta é função de sua idade e amadurecimento. As análises de regressão
apresentadas nas figuras 3A-D demonstram que essa relação é válida para as
florestas de franja e bacia, mas não para as florestas de transição, em função do alto
rigor ambiental que ocorre nesse tipo fisiográfico, conforme descrito anteriormente.
Porém, a análise da figura 3A sob enfoque de dinâmica, considerando as florestas de
transição em estágios sucessionais menos avançados, sugere uma interpretação
adicional. As florestas que encontram-se bem acima da linha de tendência da
regressão, ou seja, com densidade bem acima do esperado para um determinado
DAP médio, seriam aquelas que estariam em fase final de ocupação da planície
hipersalina, uma fase anterior a de desbaste natural. Já aquelas que se encontram
muito abaixo da linha de tendência, com baixa densidade, seriam aquelas em fase
inicial de ocupação da planície hipersalina. Essa alta variabilidade estrutural das
florestas de transição fica ainda mais clara ao observarmos os gráficos 2A-D. Embora
o DAP médio e a altura média não apresentem variabilidade superior a das florestas

Capítulo II - Análise da Variabilidade Estrutural
56
de franja e bacia, a densidade e, por conseqüência, a área basal apresentam altíssima
variabilidade. Já as florestas de franja e bacia estariam em estágios sucessionais mais
avançados, o que justifica a menor variabilidade estrutural apresentada por essas
florestas, acarretando em uma forte relação entre a densidade e o DAP médio.
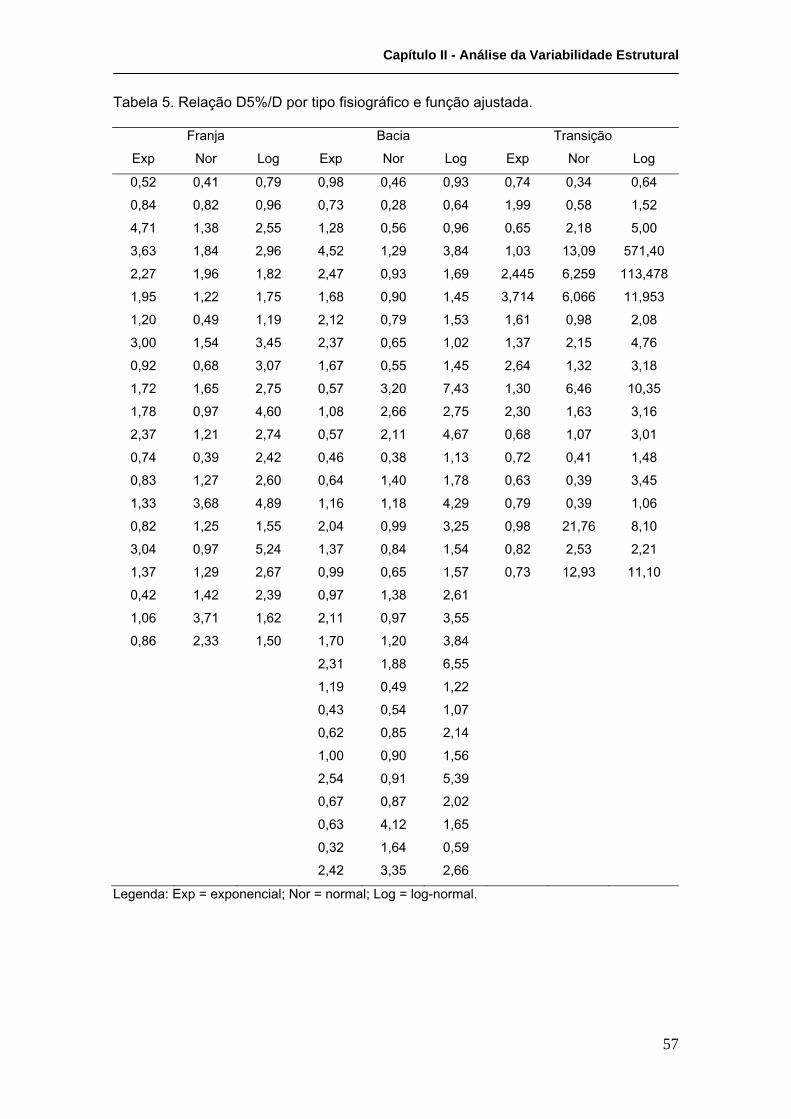
Capítulo II - Análise da Variabilidade Estrutural
57
Tabela 5. Relação D5%/D por tipo fisiográfico e função ajustada.
Franja Bacia Transição
Exp Nor Log Exp Nor Log Exp Nor Log
0,52 0,41 0,79 0,98 0,46 0,93 0,74 0,34 0,64
0,84 0,82 0,96 0,73 0,28 0,64 1,99 0,58 1,52
4,71 1,38 2,55 1,28 0,56 0,96 0,65 2,18 5,00
3,63 1,84 2,96 4,52 1,29 3,84 1,03 13,09 571,40
2,27 1,96 1,82 2,47 0,93 1,69 2,445 6,259 113,478
1,95 1,22 1,75 1,68 0,90 1,45 3,714 6,066 11,953
1,20 0,49 1,19 2,12 0,79 1,53 1,61 0,98 2,08
3,00 1,54 3,45 2,37 0,65 1,02 1,37 2,15 4,76
0,92 0,68 3,07 1,67 0,55 1,45 2,64 1,32 3,18
1,72 1,65 2,75 0,57 3,20 7,43 1,30 6,46 10,35
1,78 0,97 4,60 1,08 2,66 2,75 2,30 1,63 3,16
2,37 1,21 2,74 0,57 2,11 4,67 0,68 1,07 3,01
0,74 0,39 2,42 0,46 0,38 1,13 0,72 0,41 1,48
0,83 1,27 2,60 0,64 1,40 1,78 0,63 0,39 3,45
1,33 3,68 4,89 1,16 1,18 4,29 0,79 0,39 1,06
0,82 1,25 1,55 2,04 0,99 3,25 0,98 21,76 8,10
3,04 0,97 5,24 1,37 0,84 1,54 0,82 2,53 2,21
1,37 1,29 2,67 0,99 0,65 1,57 0,73 12,93 11,10
0,42 1,42 2,39 0,97 1,38 2,61
1,06 3,71 1,62 2,11 0,97 3,55
0,86 2,33 1,50 1,70 1,20 3,84
2,31 1,88 6,55
1,19 0,49 1,22
0,43 0,54 1,07
0,62 0,85 2,14
1,00 0,90 1,56
2,54 0,91 5,39
0,67 0,87 2,02
0,63 4,12 1,65
0,32 1,64 0,59
2,42 3,35 2,66
Legenda: Exp = exponencial; Nor = normal; Log = log-normal.
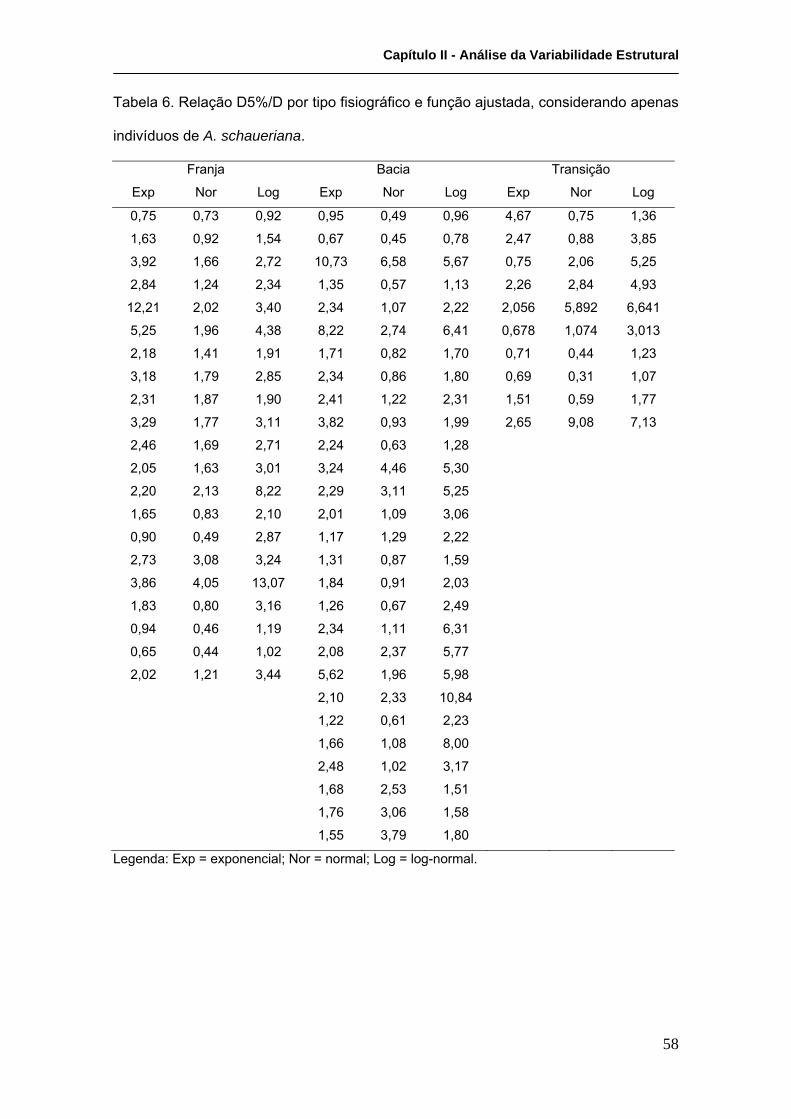
Capítulo II - Análise da Variabilidade Estrutural
58
Tabela 6. Relação D5%/D por tipo fisiográfico e função ajustada, considerando apenas
indivíduos de A. schaueriana.
Franja Bacia Transição
Exp Nor Log Exp Nor Log Exp Nor Log
0,75 0,73 0,92 0,95 0,49 0,96 4,67 0,75 1,36
1,63 0,92 1,54 0,67 0,45 0,78 2,47 0,88 3,85
3,92 1,66 2,72 10,73 6,58 5,67 0,75 2,06 5,25
2,84 1,24 2,34 1,35 0,57 1,13 2,26 2,84 4,93
12,21 2,02 3,40 2,34 1,07 2,22 2,056 5,892 6,641
5,25 1,96 4,38 8,22 2,74 6,41 0,678 1,074 3,013
2,18 1,41 1,91 1,71 0,82 1,70 0,71 0,44 1,23
3,18 1,79 2,85 2,34 0,86 1,80 0,69 0,31 1,07
2,31 1,87 1,90 2,41 1,22 2,31 1,51 0,59 1,77
3,29 1,77 3,11 3,82 0,93 1,99 2,65 9,08 7,13
2,46 1,69 2,71 2,24 0,63 1,28
2,05 1,63 3,01 3,24 4,46 5,30
2,20 2,13 8,22 2,29 3,11 5,25
1,65 0,83 2,10 2,01 1,09 3,06
0,90 0,49 2,87 1,17 1,29 2,22
2,73 3,08 3,24 1,31 0,87 1,59
3,86 4,05 13,07 1,84 0,91 2,03
1,83 0,80 3,16 1,26 0,67 2,49
0,94 0,46 1,19 2,34 1,11 6,31
0,65 0,44 1,02 2,08 2,37 5,77
2,02 1,21 3,44 5,62 1,96 5,98
2,10 2,33 10,84
1,22 0,61 2,23
1,66 1,08 8,00
2,48 1,02 3,17
1,68 2,53 1,51
1,76 3,06 1,58
1,55 3,79 1,80
Legenda: Exp = exponencial; Nor = normal; Log = log-normal.
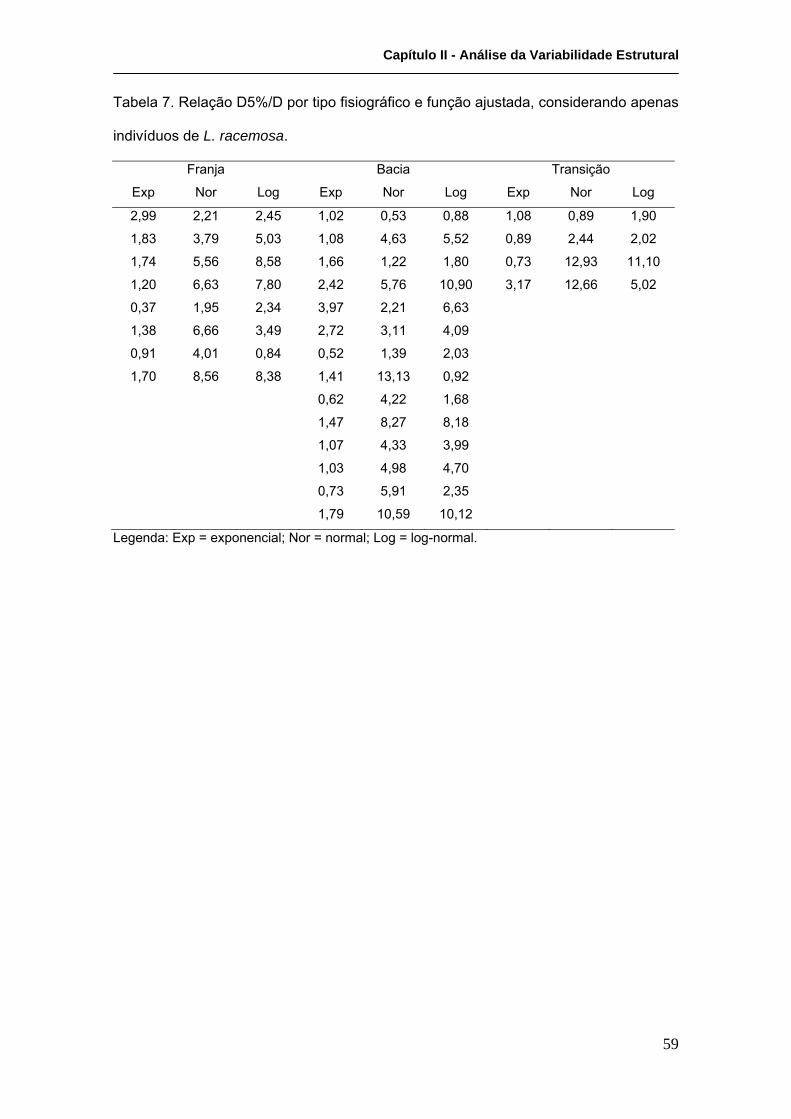
Capítulo II - Análise da Variabilidade Estrutural
59
Tabela 7. Relação D5%/D por tipo fisiográfico e função ajustada, considerando apenas
indivíduos de L. racemosa.
Franja Bacia Transição
Exp Nor Log Exp Nor Log Exp Nor Log
2,99 2,21 2,45 1,02 0,53 0,88 1,08 0,89 1,90
1,83 3,79 5,03 1,08 4,63 5,52 0,89 2,44 2,02
1,74 5,56 8,58 1,66 1,22 1,80 0,73 12,93 11,10
1,20 6,63 7,80 2,42 5,76 10,90 3,17 12,66 5,02
0,37 1,95 2,34 3,97 2,21 6,63
1,38 6,66 3,49 2,72 3,11 4,09
0,91 4,01 0,84 0,52 1,39 2,03
1,70 8,56 8,38 1,41 13,13 0,92
0,62 4,22 1,68
1,47 8,27 8,18
1,07 4,33 3,99
1,03 4,98 4,70
0,73 5,91 2,35
1,79 10,59 10,12
Legenda: Exp = exponencial; Nor = normal; Log = log-normal.
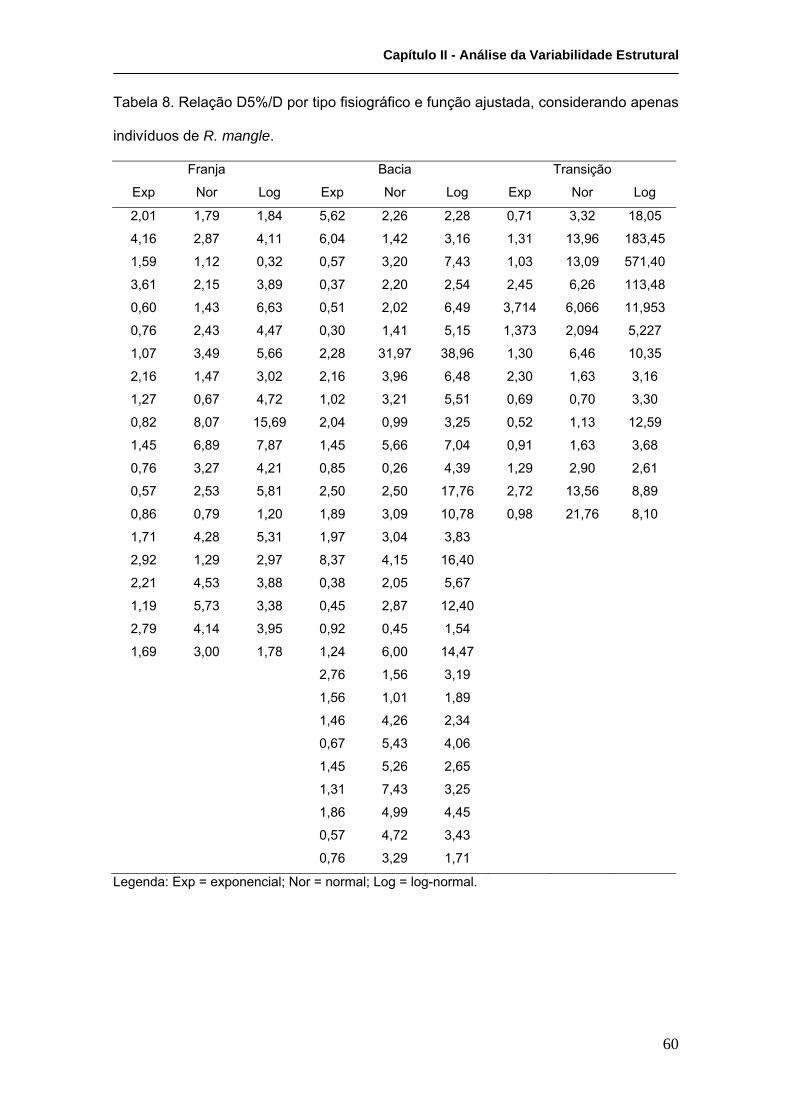
Capítulo II - Análise da Variabilidade Estrutural
60
Tabela 8. Relação D5%/D por tipo fisiográfico e função ajustada, considerando apenas
indivíduos de R. mangle.
Franja Bacia Transição
Exp Nor Log Exp Nor Log Exp Nor Log
2,01 1,79 1,84 5,62 2,26 2,28 0,71 3,32 18,05
4,16 2,87 4,11 6,04 1,42 3,16 1,31 13,96 183,45
1,59 1,12 0,32 0,57 3,20 7,43 1,03 13,09 571,40
3,61 2,15 3,89 0,37 2,20 2,54 2,45 6,26 113,48
0,60 1,43 6,63 0,51 2,02 6,49 3,714 6,066 11,953
0,76 2,43 4,47 0,30 1,41 5,15 1,373 2,094 5,227
1,07 3,49 5,66 2,28 31,97 38,96 1,30 6,46 10,35
2,16 1,47 3,02 2,16 3,96 6,48 2,30 1,63 3,16
1,27 0,67 4,72 1,02 3,21 5,51 0,69 0,70 3,30
0,82 8,07 15,69 2,04 0,99 3,25 0,52 1,13 12,59
1,45 6,89 7,87 1,45 5,66 7,04 0,91 1,63 3,68
0,76 3,27 4,21 0,85 0,26 4,39 1,29 2,90 2,61
0,57 2,53 5,81 2,50 2,50 17,76 2,72 13,56 8,89
0,86 0,79 1,20 1,89 3,09 10,78 0,98 21,76 8,10
1,71 4,28 5,31 1,97 3,04 3,83
2,92 1,29 2,97 8,37 4,15 16,40
2,21 4,53 3,88 0,38 2,05 5,67
1,19 5,73 3,38 0,45 2,87 12,40
2,79 4,14 3,95 0,92 0,45 1,54
1,69 3,00 1,78 1,24 6,00 14,47
2,76 1,56 3,19
1,56 1,01 1,89
1,46 4,26 2,34
0,67 5,43 4,06
1,45 5,26 2,65
1,31 7,43 3,25
1,86 4,99 4,45
0,57 4,72 3,43
0,76 3,29 1,71
Legenda: Exp = exponencial; Nor = normal; Log = log-normal.
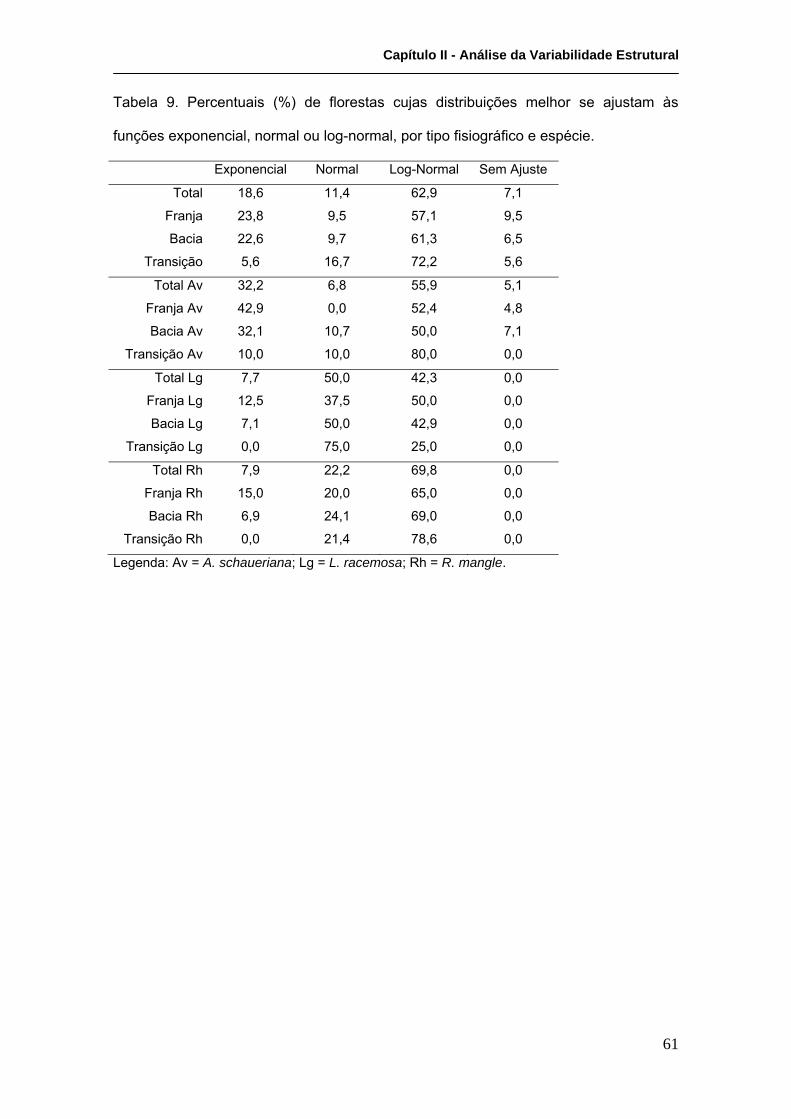
Capítulo II - Análise da Variabilidade Estrutural
61
Tabela 9. Percentuais (%) de florestas cujas distribuições melhor se ajustam às
funções exponencial, normal ou log-normal, por tipo fisiográfico e espécie.
Exponencial Normal Log-Normal Sem Ajuste
Total 18,6 11,4 62,9 7,1
Franja 23,8 9,5 57,1 9,5
Bacia 22,6 9,7 61,3 6,5
Transição 5,6 16,7 72,2 5,6
Total Av 32,2 6,8 55,9 5,1
Franja Av 42,9 0,0 52,4 4,8
Bacia Av 32,1 10,7 50,0 7,1
Transição Av 10,0 10,0 80,0 0,0
Total Lg 7,7 50,0 42,3 0,0
Franja Lg 12,5 37,5 50,0 0,0
Bacia Lg 7,1 50,0 42,9 0,0
Transição Lg 0,0 75,0 25,0 0,0
Total Rh 7,9 22,2 69,8 0,0
Franja Rh 15,0 20,0 65,0 0,0
Bacia Rh 6,9 24,1 69,0 0,0
Transição Rh 0,0 21,4 78,6 0,0
Legenda: Av = A. schaueriana; Lg = L. racemosa; Rh = R. mangle.

Capítulo II - Análise da Variabilidade Estrutural
62
Figura 4. Distribuições de troncos por classe de dap, utilizando-se os dados agrupados
por tipo fisiográfico. A = Distribuições a partir dos dados observados; B = Distribuições
a partir dos dados estimados com base nas funções exponencial (florestas de bacia e
transição) e log-normal (florestas de franja).
0
10
20
30
40
50
60
70
-1 1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
Franja
Bacia
Transição
A
0
10
20
30
40
50
60
-1 1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
%
Centros de Classe (cm)
Franja
Bacia
Transição
B

Capítulo II - Análise da Variabilidade Estrutural
63
Conclusão
A comparação através do teste de Kruskal-Wallis entre densidade, DAP médio, altura
média e área basal de 70 parcelas agrupadas por tipos fisiográficos, demonstrou a
existência de um gradiente estrutural crescente, na ordem: transição - bacia - franja. A
comparação da área basal por espécie indicou que as florestas de franja e bacia são
dominadas similarmente por A. schaueriana e R. mangle, enquanto as de transição
são dominadas por R. mangle. Análises de regressão demonstraram haver uma
relação inversa entre a densidade e o DAP médio, que refletem o efeito da competição
na estrutura da comunidade, para as florestas de franja (R²=0,895; p<0,001) e bacia
(R²=0,833; p<0,001), mas não para as de transição (R²=0,015).
A análise das distribuições diamétricas demonstrou que há uma tendência geral de
predomínio da função log-normal como a de melhor ajuste, revelando que a maioria
das florestas de mangue de Guaratiba apresentam distribuições unimodais com
assimetria positiva. Porém, quando são apresentadas as distribuições de diâmetros
utilizando-se todos os dados agrupados por tipo fisiográfico, o teste de Kolmogorov-
Smirnov demonstrou que a distribuição referente às florestas de franja, bacia e
transição são diferentes entre si (p<0,05).
A análise das distribuições aponta fortes indícios de que as florestas de franja
encontram-se em um estágio sucessional mais avançado do que as florestas de bacia
e transição, nessa ordem. Esse padrão é demonstrado principalmente pela maior
amplitude de classes diamétricas, observada nas florestas de franja em relação às de
bacia e transição e nas florestas de bacia em relação às de transição.
Dessa forma, os parâmetros estruturais não refletem apenas um gradiente de rigor
ambiental, mas também florestas em estágios sucessionais gradativamente menos
avançados (no sentido franja-transição). Esse padrão fica claro quando observamos
que, conforme a altura e o DAP médio aumenta entre as florestas de transição e as de
franja, a densidade diminui, refletindo o amadurecimento das florestas.

Capítulo II - Análise da Variabilidade Estrutural
64
A elevação do nível médio relativo do mar (N.M.R.M.) é o provável distúrbio em larga
escala que determina que as florestas de franja estejam em um estágio sucessional
mais avançado do que as florestas de bacia e de transição, nessa ordem.

Discussão Geral
65
DISCUSSÃO GERAL
No Capítulo I, o padrão de redução da área amostral mínima na ordem franja-bacia-
transição demonstra a necessidade de se utilizar parcelas com tamanho variável,
dependendo das características estruturais de cada área, para estudos de
caracterização estrutural de florestas de mangue. Os valores encontrados para área
amostral mínima no presente estudo são compatíveis com os valores que foram
utilizados por outros estudos na região sudeste e que utilizaram parcelas com
tamanho variável, embora tenham se baseado apenas na experiência e conhecimento
da área (Soares et al., Submetido; Portugal, 2002; Soares, 1999; Soares et al., 2003;
Bernini & Rezende, 2004; Silva et al., 2005). Foi demonstrado ainda que o
desenvolvimento estrutural de tais florestas encontra-se na mesma faixa do observado
para as florestas consideradas no presente estudo. Quando a comparação é feita com
estudos realizados nas regiões norte e nordeste, observa-se que o maior
desenvolvimento estrutural induz os autores a estabelecerem parcelas maiores do que
as utilizadas no sudeste. O oposto ocorre quando a comparação é feita com estudos
realizados na região sul, onde o desenvolvimento estrutural é menor.
Faz-se necessário, portanto, a realização de mais estudos em outras regiões do Brasil
e do mundo, que busquem encontrar, com embasamento estatístico, valores de
referência para a área amostral mínima necessária para a caracterização da estrutura
de florestas de mangue.
No Capítulo II, o padrão encontrado de redução do desenvolvimento estrutural desde a
franja até a transição é descrito por diversos autores para florestas de mangue
mundialmente, conforme o citado na discussão do capítulo. Esse gradiente estrutural é
determinado basicamente pela interação da microtopografia com o regime de marés,
embora em algumas regiões a precipitação e o escoamento superficial exerçam um
papel determinante também (Semeniuk, 1983; López-Portillo & Ezcurra, 1989), que
determina gradientes edáficos de salinidade (Cintrón et al., 1978; Sam & Ridd, 1998;
Ellison et al., 2000), concentração de H2S (Boto & Wellington, 1984; Nickerson &

Discussão Geral
66
Thibodeau, 1985; Ewel et al., 1998) e concentração de nutrientes (Feller et al., 2003;
Boyer, 2006; Lovelock et al., 2007).
A composição de espécies demonstrou um padrão típico para florestas de mangue de
franja e bacia que ocorrem no sudeste do Brasil, com dominância de A. schaueriana e
R. mangle. A dominância de R. mangle na florestas de transição é , provavelmente,
um reflexo da existência de canais que ligam a franja até a transição, conforme
descrito por Chaves (2001) e Portugal (2002). Esses canais permitem a chegada de
propágulos de R. mangle na transição e, provavelmente, atenuam as condições
edáficas favorecendo competitivamente esta espécie. A baixa representatividade de L.
racemosa é um indicador de que as florestas de mangue de Guaratiba encontram-se
bem preservadas, já que Soares et al. (2003) demonstram que esta espécie é típica de
estágios iniciais de sucessão e freqüentemente dominam florestas sob influência de
distúrbios antrópicos.
Embora a análise das distribuições diamétricas tenha demonstrado que há uma
tendência geral de predomínio da função log-normal como a de melhor ajuste, foram
observados desvios desse padrão quando a análise foi realizada por espécie. No caso
de L. racemosa, uma possível explicação para a função normal ser a predominante é a
composição entre a sua grande habilidade como espécie pioneira de processos de
regeneração e sua baixa capacidade em tolerar sombreamento, o que favorece
formação de coortes únicas para essa espécie, determinando distribuições unimodais
simétricas. No caso de A. schaueriana, o elevado percentual de florestas cujas
distribuições melhor se ajustam a uma função exponencial (J-invertido), revela uma
maior tendência em formar florestas com múltiplas coortes, conforme demonstrado por
Oliver & Larson (1990) e Frelich (2002).
A análise das distribuições de diâmetros utilizando-se todos os dados agrupados por
tipo fisiográfico, demonstrou que as florestas de mangue de Guaratiba encontram-se
em estágios sucessionais gradativamente mais avançados, na ordem: franja (mais
avançado) - bacia - transição. Esse resultado corrobora os estudos que têm sido

Discussão Geral
67
realizados pelo NEMA/UERJ na região sobre os impactos da elevação do nível médio
relativo do mar, porém, utilizando uma nova abordagem, e através de testes
estatísticos.

Conclusão Geral
68
CONCLUSÃO GERAL
No primeiro capítulo, os resultados das duas análises empregadas demonstraram
haver uma tendência de redução da área amostral mínima entre as parcelas
representativas de florestas de franja até àquelas representativas de florestas de
transição, relacionada à redução do desenvolvimento estrutural. Esse resultado
enfatiza a importância de se utilizar parcelas para caracterização da estrutura vegetal
de florestas de mangue com tamanho variável, confirmando de forma empírica a
sugestão metodológica recomendada por Cintrón & Schaeffer-Novelli (1984).
Para florestas de mangue da região sudeste do Brasil, ou para florestas de mangue de
outras regiões costeiras do mundo com características estruturais similares às
encontradas no segmento VII da costa brasileira (Schaeffer-Novelli et al., 1990),
recomenda-se, com base nos resultados do presente estudo, a utilização de parcelas
com área de 100 a 450 m² para florestas de franja, 25 a 180 m² para florestas de bacia
e 15 a 50 m² para florestas arbustivas de transição.
Cabe ressaltar que a determinação da área amostral mínima com base em testes
estatísticos, realizada pelo presente estudo, é inédita para florestas de mangue,
representando uma evolução na busca pelo aperfeiçoamento das metodologias
empregadas pelo NEMA/UERJ para o estudo da estrutura vegetal de manguezais.
No Capítulo II, a comparação através do teste de Kruskal-Wallis entre densidade, DAP
médio, altura média e área basal de 70 parcelas agrupadas por tipos fisiográficos,
demonstrou a existência de um gradiente estrutural crescente, na ordem: transição -
bacia - franja. A comparação da área basal por espécie indicou que as florestas de
franja e bacia são dominadas similarmente por A. schaueriana e R. mangle, enquanto
as de transição são dominadas por R. mangle. Análises de regressão demonstraram
haver uma relação inversa entre a densidade e o DAP médio, que refletem o efeito da
competição na estrutura da comunidade, para as florestas de franja (R²=0,895;
p<0,001) e bacia (R²=0,833; p<0,001), mas não para as de transição (R²=0,015).

Conclusão Geral
69
A demonstração estatística da existência de diferenças estruturais entre tipos
fisiográficos insere o Capítulo II na vanguarda dos estudos sobre a estrutura vegetal
de florestas de mangue mundialmente. Essa inserção é ainda reforçada pela utilização
de um escala de abordagem mais ampla, permitindo a demonstração da existência de
tais diferenças em escala sistêmica, o sistema estuarino de Guaratiba, que engloba a
Reserva Biológica e Arqueológica de Guaratiba.
A análise das distribuições diamétricas demonstrou que há uma tendência geral de
predomínio da função log-normal como a de melhor ajuste, revelando que a maioria
das florestas de mangue de Guaratiba apresentam distribuições unimodais com
assimetria positiva. Porém, quando são apresentadas as distribuições de diâmetros
utilizando-se todos os dados agrupados por tipo fisiográfico, o teste de Kolmogorov-
Smirnov demonstrou que a distribuição referente às florestas de franja, bacia e
transição são diferentes entre si (p<0,05). A análise das distribuições aponta fortes
indícios de que as florestas de franja encontram-se em um estágio sucessional mais
avançado do que as florestas de bacia e transição, nessa ordem. Esse padrão é
demonstrado principalmente pela maior amplitude de classes diamétricas, observada
nas florestas de franja em relação às de bacia e transição e nas florestas de bacia em
relação às de transição.
Dessa forma, os parâmetros estruturais não refletem apenas um gradiente de rigor
ambiental, mas também florestas em estágios sucessionais gradativamente menos
avançados (no sentido franja-transição). Esse padrão fica claro quando observamos
que, conforme a altura e o DAP médio aumenta entre as florestas de transição e as de
franja, a densidade diminui, refletindo o amadurecimento das florestas.
A elevação do nível médio relativo do mar (N.M.R.M.) é o provável distúrbio em larga
escala que determina que as florestas de franja estejam em um estágio sucessional
mais avançado do que as florestas de bacia e de transição, nessa ordem.

Referências Bibliográficas
70
REFERÊNCIAS BIBLIOGRÁFICAS
ACKER, S.A., SABIN, T.E., GANIO, L.M, MCKEE, W.A.,1998, “Development of old-
growth structure and timber volume growth trends in maturing Douglas-fir stands”.
Forest Ecology and Management, v.104, pp. 265-280.
ALMEIDA, P.M.M., 2007. Utilização de imagens de satélite para análise multi-temporal
do manguezal de Guaratiba-RJ. Monografia de Bacharelado, Universidade do
Estado do Rio de Janeiro, Rio de Janeiro, RJ, Brasil.
AUBÉ, M., CARON, L., 2001, “The mangroves of the north coast of Haiti. A preliminary
assessment”. Wetlands Ecology and Management, v.9, pp. 271-278.
BALDWIN, A., EGNOTOVICH, M., FORD, M., PLATT, W., 2001, “Regeneration in
fringe mangrove forests damaged by Hurricane Andrew”. Plant Ecology, v.157, pp.
149-162.
BALL, M.C., 1980, “Patterns of secondary succession in a mangrove forest of southern
Florida”. Oecologia, v.44, pp. 226-235.
BARBOUR, M. G., BURK, J.H., PITTS, W.D., 1980. Terrestrial Plant Ecology.
California, The Benjamin Cummings Publishing Company Inc. Menlo Park.
BARKMAN, J.J., 1989, “A critical evaluation of minimum area concepts”. Vegetatio, v.
85, pp. 89-104.
BELLEHUMEUR, C., LEGENDRE, P., MARCOTTE, D., 1997, “Variance and spatial
scales in a tropical rain forest: changing the size of sampling units”. Plant Ecology,
v.130, pp. 89-98.
BERGER, U., ADAMS, M., GRIMMC,V., HILDENBRANDT, H., 2006, “Modelling
secondary succession of neotropical mangroves: Causes and consequences of
growth reduction in pioneer species”. Perspectives in Plant Ecology, Evolution and
Systematics, v.7, pp. 243-252.
BERNINI, E., REZENDE, C.E., 2004, “Estrutura da vegetação em florestas de mangue
do estuário do rio Paraíba do Sul, Estado do Rio de Janeiro, Brasil”. Acta Botanica
Brasilica, v.18, n.3, pp. 491-502.

Referências Bibliográficas
71
BLANCO, J.F., BEJARANO, A.C., LASSO, J., CANTERA, J.R., 2001, “A new look at
computation of the complexity index in mangroves: do disturbed forests have clues
to analyze canopy height patchiness?”. Wetlands Ecology and Management, v.9, pp.
91-101.
BORMANN, F.H., 1953, “The Statistical Efficiency of Sample Plot Size and Shape in
Forest Ecology”. Ecology, v.34, n.3 (Jul.), pp. 474-487.
BOSIRE, J., DAHDOUH-GUEBAS, F., KAIRO J.G., KOEDAM, N., 2003, “Colonisation
of non-planted mangrove species into restored mangrove stands in Gazi Bay,
Kenya”. Aquatic Botany, v.76, pp. 267-279.
BOTO, K.G., WELLINGTON, J.T., 1984. “Soil Characteristics and Nutrient Status in a
Northern Australian Mangrove Forest”. Estuaries, v.7, n.1(Mar.), pp. 61-69.
BOYER, J.N., 2006, “Shifting N and P limitation along a north-south gradient of
mangrove estuaries in South Florida”. Hydrobiologia, v.569, pp.167-177.
BRASIL, 2008. Tábua de marés para o ano de 2008, costa do Brasil e portos
estrangeiros. Diretoria de Hidrografia e Navegação. Ministério da Marinha.
BROWER, J.E., ZAR, J.H., ENDE, C.N., 1997, “Field and Laboratory methods for
general ecology. WCB/McGraw-Hill, New York.
CAIN, S.A., 1934, “Studies on Virgin hardwood forest: II, a comparison of quadrat sizes
in a quantitative phytosociological study of nash’s wood, Posey County, Indiana”.
The American Midland Naturalist, v.15, n.5 (Sep.), pp. 529-566.
CAIN, S.A., 1938, “The species-area curve”. The American Midland Naturalist, v.19,
n.3 (May), pp. 573-581.
CAIN, S.A.,1943, “Sample-Plot Technique Applied to Alpine Vegetation in Wyoming”.
American Journal of Botany, v.30, n.3 (Mar.), pp. 240-247.
CHAVES, F.O., 2001, Análise dos padrões de zonação e sucessão de espécies de
mangue na região de Guaratiba, Baía de Sepetiba, RJ. Monografia de Bacharelado,
Universidade do Estado do Rio de Janeiro, Rio de Janeiro, RJ, Brasil.

Referências Bibliográficas
72
CHEN, R., TWILLEY, R.R., 1999, “Patterns of Mangrove Forest Structure and Soil
Nutrient Dynamics along the Shark River Estuary, Florida”. Estuaries, v. 22, n.4.
(Dec.), pp. 955-970.
CINTRON, G., SCHAEFFER-NOVELLI, Y., 1984, Methods for studying mangrove
structure. In: The mangrove ecosystem: Research Methods. UNESCO, Bungay,
United Kingdom.
CINTRON, G., LUGO, A.E., POOL, D.J., MORRIS, G., 1978, “Mangroves of Arid
Environments in Puerto Rico and Adjacent Islands”. Biotropica, v.10, n.2, pp. 110-
121.
CINTRON, G., GOENAGA, C., GONZALES-LIBOY, J., 1980a, “Ecologia del manglar
en una zona arida: exposición al oleaje y estructura del manglar”. Boletim do
Instituto Oceanográfico de São Paulo, v.29, n.2, pp. 113-127.
CINTRON, G., LUGO, A.E., MARTINEZ, R., 1985, Structural and Functional Properties
of Mangrove Forests. In: The Botany and Natural History of Panama. Missouri
Botanical Garden. Saint Louis, Missouri.
CINTRON-MOLERO, G., 1993, Mangroves of arid regions of Puerto Rico and the
Caribbean. In: Towards the rational use of high salinity tolerant plants. Zurich:
Kluwer Academic Publishers, Zurich.
COLE, T.G., EWEL, K.C., DEVOE, N.N., 1999, “Structure of mangrove trees and
forests in Micronesia”. Forest Ecology and Management , v.117, pp. 95-109.
COX, E.F., ALLEN, J.A., 1999, “Stand Structure and Productivity of the Introduced
Rhizophora mangle in Hawaii”. Estuaries, V.22, n.2 part A. (Jun), pp. 276-284.
CUNHA-LIGNON, M., 2001, Dinâmica do manguezal no Sistema de Cananéia-Iguape,
Estado de São Paulo, Brasil. Dissertação de M.Sc. Instituto Oceanográfico, USP,
São Paulo, SP, Brasil.
DAHDOUH-GUEBAS, F., DE BONDT, R., ABEYSINGHE, P. D., KAIRO, J. G.,
CANNICCI, S., TRIEST, L., KOEDAM, N., 2004, “Comparative study of the disjunct

Referências Bibliográficas
73
zonation pattern of the grey mangrove Avicennia marina (Forsk.) Vierh. in Gazi Bay
(Kenya)”. Bulletin of Marine Science, v.74, n.2, pp. 237–252.
DE DEUS, M.S.M., SAMPAIO, E.V.S.B., RODRIGUES, S.M.C.B, ANDRADE, V.C.,
2003, “Estrutura da vegetação lenhosa de três áreas de manguezal do Piauí com
diferentes históricos de antropização”. Brasil Florestal, v.78 (Dez), pp. 53-60.
DIETVORST, P., MAAREL, E., PUTTEN H., 1982, “A new approach to the minimal
area of a plant community”. Vegetatio, v.50, pp. 77-91.
ELLISON, J.C., STTODDART, D.R., 1991. “Mangrove ecosystem collapse during
predicted sea-level rise: Holocene analogues and implications”. Journal of Coastal
Research, v.7, n.1, pp. 151-165.
ELLISON, J.C., SIMMONDS, S., 2003, “Structure and productivity of inland mangrove
stands at Lake MacLeod, Western Australia”. Journal of the Royal Society of
Western Australia, v.86, pp. 25-30.
ESTRADA, G.C.D., OLIVEIRA, V.F., SOARES, M.L.G, CHAVES, F.O., SILVA JR.,
C.M.G, PELLEGRINI, J.A.C., CORRÊA, F.M., BARBOSA, B., PASSOS, E.N.S.,
2003, “Processo de colonização de uma planície hipersalina (apicum) por Avicennia
schaueriana e Laguncularia racemosa: 4 anos de monitoramento do manguezal de
Guaratiba – RJ”. Salvador, Bahia, Brazil, 20-24 May.
ESTRADA, G.C.D., CALLADO, C.H., SOARES, M.L.G., LISI, C.S., 2008, “Annual
growth rings in the mangrove Laguncularia racemosa (Combretaceae)’’. Trees, v.22,
n.5, pp. 663-670.
EWEL, K.C., ZHENG, S., PINZÓN, Z.S., BOURGEOIS, J.A., 1998, “Environmental
effects of canopy gap formation in high - rainfall mangrove forests”. Biotropica, v.34,
n.4, pp. 510-518.
FELLER, I.C., Mckee, K.L., WHIGHAM, D.F., O'NEILL, J.P., 2003, “Nitrogen vs.
phosphorus limitation across an ecotonal gradient in a mangrove forest”.
Biogeochemistry, v.62, n.2. (Feb.), pp. 145-175.

Referências Bibliográficas
74
FERNANDES, M.E.B., 2000, “A Structural analysis of Rhizophora, Avicennia, and
Laguncularia forests on Maracá Island, Amapá, Brazil”. In: German-Brazilian
Workshop on Neotropical Ecosystems – Achievements and Prospects of
Cooperative Research, pp. 565-572, Hamburg, Sep.
FERREIRA, A.A.M., OLIVEIRA, M.V., 1985, “Contribuição ao estudo arqueo-geológico
do Quaternário Superior da Baixada de Guaratiba-Sepetiba”. In: Coletores e
Pescadores Pré-Históricos de Guaratiba - Rio de Janeiro. Editora UFRJ/EDUFF.
Niterói, RJ, pp. 29-46.
FERWERDA, J. G., KETNER, P., MCGUINNESS, K. A., 2007, “Differences in
regeneration between hurricane damaged and clear-cut mangrove stands 25 years
after clearing”. Hydrobiologia, v.591, pp. 35-45.
BROWER, J.E., ZAR, J.H., ENDE, C.N., 1997, “Field and Laboratory methods for
general ecology”. 4ª ed. New York, WCB/McGraw-Hill.
FROMARD, F., PUIG, H., MOUGIN, E., MARTY, G., BETOULLE, J.L., CADAMURO,
L., 1998. “Structure, above-ground biomass and dynamics of mangrove ecosystems:
new data from French Guiana”. Oecologia, v.115, pp. 39-53.
GAMA, J.R.V., BOTELHO, S.A., BENTES-GAMA, M.M., SCOLFORO, J.R.S., 2001,
“Tamanho de parcela e suficiência amostral para estudo da regeneração natural em
floresta de várzea na Amazônia”. Cerne, v.7, n.2, pp.1-11
GOLLEY, F.B., ODUM, H.T.; WILSON, R.F., 1962. The structure and metabolism of a
Puerto Rican red mangrove forest in may. Ecology, 43 (1): 9-19.
HARPER, J.L., WHITE, J., 1974, “The Demography of Plants”. Annual Review of
Ecology and Systematics, v.5, pp. 419-463.
HAUFF, R.D., EWEL, K.C., JACK, J., 2006, “Tracking human disturbance in
mangroves: estimating harvest rates on a Micronesian Island”. Wetlands Ecology
and Management, v.14, pp. 95-105.
HOPKINS, B., 1955, “The Species-Area Relations of Plant Communities”. The Journal
of Ecology, v.43, n. 2 (Jul.), pp. 409-426.

Referências Bibliográficas
75
IFTEKHAR, M.S., SAENGER, P., 2008, “Vegetation dynamics in the Bangladesh
Sundarbans mangroves: a review of forest inventories”. Wetlands Ecology
Management, v.16, pp. 291-312.
IMAI, N., TAKYU, M., NAKAMURA, Y., NAKAMURA T., 2006, “Gap formation and
regeneration of tropical mangrove forests in Ranong,Thailand”. Plant Ecology, v.186,
pp. 37-46.
JIMENEZ, J.A., 1990, “The structure and function of dry weather mangroves on the
Pacific coast of Central America, with emphasis on Avicennia bicolor forests.
Estuaries, v.13, n.2, pp. 182-192.
JIMENEZ, J.A., LUGO, A.E., CINTRON, G, 1985, “Tree mortality in mangrove forests”.
Biotropica, v.17, n.3, pp. 177-185.
JIMENEZ, J.A. & SAUTER, K., 1991. “Structure and dynamics of mangrove forests
along a flooding gradient”. Estuaries, v.14, n.1, pp. 49-56.
KAIRO, J.G., LANG’AT, J.K.S., DAHDOUH-GUEBAS, F., BOSIRE, J., KARACHI, M.,
2008, “Structural development and productivity of replanted mangrove plantations in
Kenya”. Forest Ecology and Management, v.255, pp. 2670–2677.
KAIRO, J.G., DAHDOUH-GUEBAS, F., GWADA, P.O., OCHIENG, C., KOEDAM, N.,
2002, “Regeneration Status of Mangrove Forests in Mida Creek, Kenya: A
Compromised or Secured Future?”. Ambio v. 31, n.7, pp. 562-568.
KENKEL, N.C., PODANI, J., 1991, “Plot Size and Estimation Efficiency in Plant
Community Studies”. Journal of Vegetation Science, v.2, n.4, (Sep.), pp. 539-544.
KHOON, G.W., EONG, O.J., 1995, “The use of demographic studies in mangrove
silviculture”. Hidrobiologia, v.295, pp. 255-261.
KRAUSS, K.W., KEELAND, B.D., ALLEN, J.A., EWEL, K.C., JOHNSON, D.J., 2006,
“Effects of Season, Rainfall, and Hydrogeomorphic Setting on Mangrove Tree
Growth in Micronesia”. Biotropica, v.39, n.2, pp. 161-170.

Referências Bibliográficas
76
LARA, R.J., COHEN, M.C.L., 2006, “Sediment porewater salinity, inundation frequency
and mangrove vegetation height in Bragança, North Brazil: an ecohydrology-based
empirical model”. Wetlands Ecology and Management, v.14, pp. 349-358.
LARA-DOMÍNGUEZ, A.L., DAY, J.W., ZAPATA, G.V., TWILLEY, R.R., GUILLEN, H.A.,
YÁÑEZ-ARANCIBIA, A., 2005, “Structure of a unique inland mangrove forest
assemblage in fossil lagoons on the Caribbean Coast of Mexico”. Wetlands Ecology
and Management, v.13, pp. 111-122.
LEVIN, S.A., 1992, “The Problem of Pattern and Scale in Ecology: The Robert H.
MacArthur Award Lecture”. Ecology, v.73, n.6 (Dec.), pp. 1943-1967.
LÓPEZ-HOFFMAN, L., ACKERLY, D.D., ANTEN, N.P.R., DENOYER, J.L.,
MARTINEZ-RAMOS, M., 2007, “Gap-dependence in mangrove life-history
strategies: a consideration of the entire life cycle and patch dynamics”. Journal of
Ecology, v.95, pp.1222–1233.
LÓPEZ-PORTILLO, J., EZCURRA, E., 1989, “Zonation in Mangrove and salt marsh
vegetetion at Laguna de Mecoacan, Mexico”. Biotrópica, v.21, n.2 (Jun), pp. 107-
114.
LORIMER, C.G., KRUG, A.G., 1983, “Diameter distributions in even-aged stands of
shade-tolerant and midtolerant tree species”. American Midland Naturalist, v.109,
n.2 (Apr.), pp. 331-345.
LOVELOCK, C.E., FELLER, I.C., ELLIS, J., SCHWARZ, A.M., HANCOCK, N.,
NICHOLS, P., SORRELL, B., 2007, “Mangrove growth in New Zealand estuaries:
the role of nutrient enrichment at sites with contrasting rates of sedimentation”.
Oecologia, v.153, pp. 633-641.
LUGO, A.E., SNEDAKER, S.C., 1974. “The ecology of mangroves.” Annual Review of
Ecology and Systematics, v.5, pp. 39-64.
LUGO, A.E., BROWN, S., BRINSON, M.M.,1988, “Forested wetlands in freshwater and
salt-water environments”. Limnology and Oceanography, v.33, n.4 part 2, pp. 894-
909.

Referências Bibliográficas
77
MCKEE, K.L., 1993, “Soil physicochemical patterns and mangrove species distribution:
Reciprocal effects?”. Journal of Ecology, v.81, n.3. pp. 477-487.
MCKEE, K.L., 1995, “Interspecific variationin growth, biomass partitioning, and
defensive characteristics of neotropical mangrove seedlings: response to light and
nutrient availability”. American Journal of Botany, v.82, n.3, pp. 299-307.
MORAVEC, J., 1973, “The determination of the minimal area of phytocoenoses”. Folia
Geobotanica & Phytotaxonomica, v.8, pp. 23-47.
NICKERSON, N.H., THIBODEAU, F.R., 1985, “Association between Pore Water
Sulfide Concentrations and the Distribution of Mangroves”. Biogeochemistry, v.1,
n.2, pp. 183-192.
NIKLAS, K.J. 1994. Plant Allometry – the scaling of form and process. Chicago, The
University of Chicago Press.
NOGUEIRA, G.S., LEITE, H.G., CAMPOS, J.C.C., TAKIZAWA, F.H., COUTO, L.,
2006, “Avaliação de um modelo de distribuição diamétrica ajustado para
povoamentos de Tectona grandis submetidos a desbaste”. Árvore, v.30, n.3, pp.
377-387.
ODUM, EP., 1985, “Trends Expected in Stressed Ecosystems”. BioScience, v. 35, n.7,
pp. 419-422.
ØKLAND, R.H., EILERTSEN, O., ØKLAND, T., 1990, “On the relationship between
sample plot size and beta diversity in boreal coniferous forest”. Vegetatio, v.87, pp.
187-192.
OLIVEIRA, V.F., 2001, Monitoramento do ecossistema manguezal visando sua
utilização como indicador de variações do nível médio relativo do mar (Guaratiba –
RJ). Monografia de Bacharelado. Faculdade de Oceanografia. Universidade do
Estado do Rio de Janeiro. Rio de Janeiro, RJ, Brasil.
OLIVEIRA, A.E.S., 2007, “Caracterização do Mangue da Praia Gorda, Armação dos
Búzios, Estado do Rio de Janeiro – RJ”. Revista científica do Centro Universitário de
Barra Mansa, v. 9, n.17 (Jul.), pp. 51-56.

Referências Bibliográficas
78
OLIVER, C.D., LARSON, B.C., 1990, Forest Stand Dynamics. New York, McGraw-Hill
Inc..
JIN-EONG, O., KHOON, G.W., CLOUGH, B.F., 2005, “Structure and Productivity of a
20-Year-Old Stand of Rhizophora apiculata Bl. Mangrove Forest”. Journal of
Biogeography, v.22, n.2/3, Terrestrial Ecosystem Interactions with Global Change, v.
1, pp. 417-424.
PEET, R.K., CHRISTENSEN, N.L., 1987, “Competition and tree death”. Bioscience,
v.37, n.8, pp. 586-595.
PELLEGRINI, J.A.C. 2000. Caracterização da planície hipersalina (apicum) associada
a um bosque de mangue em Guaratiba, Baía de Sepetiba, Rio de Janeiro, RJ.
Dissertação de Mestrado. Instituto Oceanográfico, Universidade de São Paulo. São
Paulo, SP.
PELLEGRINI, J.A.C., SOARES, M.L.G., CHAVES, F.O., ESTRADA, G.C.D.,
BARBOSA, B., CAVALCANTI, V.F., 2007, “Caracterização estrutural das florestas
de mangue da porção oriental da Ilha de Tinharé – Bahia (Brasil)”. XII Congresso
Latino-Americano de Ciências do Mar, Florianópolis, Santa Catarina, Brasil, 15-19
Abril.
PETERSON, D.L., PARKER, V.T., 1998, Ecological Scale – theory and applications.
New York. Columbia University Press.
POOL, D.J., SNEDAKER, S.C., LUGO, A.E., 1977, “Structure of Mangrove Forests in
Florida, Puerto Rico, Mexico, and Costa Rica”. Biotropica, v.9, n.3. (Sep.), pp. 195-
212.
POORE, M.E.D., 1955, “The Use of Phytosociological Methods in Ecological
Investigations: I. The Braun-Blanquet System”. The Journal of Ecology, v.43, n.1
(Jan), pp. 226-244.
RICE, E.L., KELTING, R.W., 1955, “The Species--Area Curve”. Ecology, v.36, n.1,
(Jan), pp. 7-11.

Referências Bibliográficas
79
PORTUGAL, A.M.M., 2002, Manguezais de Guaratiba frente à perspectiva de
elevação do nível médio relativo do mar, Baía de Sepetiba, Estado do Rio de
Janeiro – Brasil. Dissertação de M.Sc. Instituto Oceanográfico, Universidade de São
Paulo. São Paulo, SP, Brasil.
PROFFITT, C. E., DEVLIN, D. J., 2005. “Long-term growth and succession in restored
and natural mangrove forests in southwestern Florida”. Wetlands Ecology and
Management , v.13, pp. 531-551.
RABINOWITZ, D., 1978, “Early growth of mangrove seedlings in Panama and
Hypothesis concerning the relationship of dispersal and zonation”. Biogeography,
v.5, pp.13-33.
ROGERS, K., SAINTILAN, N., CAHOON, D., 2005. “Surface elevation dynamics in a
regenerating mangrove forest at Homebush Bay, Australia”. Wetlands Ecology and
Management , v.13, pp. 587-598.
ROTH, L.C. 1982, “Hurricanes and Mangrove Regeneration: Effects of Hurricane Joan,
October 1988, on the Vegetation of Isla del Venado, Bluefields, Nicaragua”.
Biotropica, v. 24, n.3. (Sep), pp. 375-384.
RUBIN, B.D., MANION, P.D., FABER-LANGENDOEN, D., 2006, “Diameter
distributions and structural sustainability in forests”. Forest Ecology and
Management, v.222, v.1-3, pp. 427-438.
SAM, R., RIDD, P., 1998, “Spatial variations of groundwater salinity in a mangrove-salt
flat system, Cocoa Creek, Australia”. Mangroves and Salt Marshes, v.2, pp. 121-
132.
SCHAEFFER-NOVELLI, Y., 1989, “Perfil dos ecossistemas litorâneos brasileiros, com
especial ênfase sobre o ecossistema manguezal”. Publicação Especial, Instituto
Oceanográfico, v. 7, pp.1-16.
SCHAEFFER-NOVELLI, Y., CINTRON, G., 1986, Guia para estudo de áreas de
manguezal - estrutura, função e flora. São Paulo. Caribbean Ecological Research.
150 p.

Referências Bibliográficas
80
SCHAEFFER-NOVELLI, Y., CINTRON-MOLERO, G., ADAIME, R.R., CAMARGO,
T.M., 1990, “Variability of mangrove ecosystems along the Brazilian coast”.
Estuaries, v.13, n.2, pp. 204-218.
SCHAEFFER-NOVELLI, Y., CINTRÓN-MOLERO, G., SOARES, M.L.G., DE-ROSA T.,
2000, “Brazilian mangroves”. Aquatic Ecosystem Health and Management, v.3, pp.
561-570.
SEMENIUK, V., 1983, “Mangrove distribution in Northwestern Australia in relationship
to regional and local freshwater seepage”. Vegetatio, v.53, pp. 11-31.
SIEGEL, S., 2006, Estatística Não-Paramétrica para Ciências, Bookman Conhanhia
Ed.
SILVA, R.L., XAVIER, A., LEITE, H.G., PIRES, I.E., 2003, “Determinação do tamanho
ótimo da parcela experimental pelos métodos da máxima curvatura modificado, do
coeficiente de correlação intraclasse e da análise visual em testes clonais de
eucalipto”. Árvore, v.27, n.5, pp. 669-676.
SILVA, M.A.B., BERNINI, E., CARMO, T.M.S., 2005, “Características estruturais de
bosques de mangue do estuário do rio São Mateus, ES, Brasil”. Acta Botanica
Brasilica. v.19, n.3, pp. 465-471.
SOARES, M.L.G. 1997. Estudo da biomassa aérea de manguezais do sudeste do
Brasil – análise de modelos. 2 vol. Tese de D.Sc., Instituto Oceanográfico,
Universidade de São Paulo. São Paulo, SP, Brasil.
SOARES, M.L.G. 1999. “Estrutura vegetal e grau de perturbação dos manguezais da
Lagoa da Tijuca, Rio de Janeiro, RJ, Brasil”. Revista Brasileira de Biologia, v.59, n.3,
pp. 503-515.
SOARES, M.L.G., CHAVES, F.O., CORRÊA, F.M., SILVA JR., C.M.G., 2003,
“Diversidade estrutural de bosques de mangue e sua relação com distúrbios de
origem antrópica: o caso da Baía de Guanabara (Rio de Janeiro). Anuário do
Instituto de Geociências da UFRJ, v. 26, pp. 101-116.

Referências Bibliográficas
81
SOARES, M.L.G., TOGNELLA-DE-ROSA, M.M.P., OLIVEIRA, V.F., CHAVES, F.O.,
SILVA JR., C.M.G., PORTUGAL, A.M.M., ESTRADA, G.C.D., BARBOSA, B.,
ALMEIDA, P.M.M., 2005. Environmental Changes in South America in the Last 10k
Years: Atlantic and Pacific Controls and Biogeophysical Effects: Ecological Impacts
of Climatic Change and Variability: Coastal Environments - Mangroves and Salt
Flats. Inter-American Institute on Global Change - IAI.
SOARES, M.L.G., ESTRADA, G.C.D., CHAVES, F.O., CAVALCANTI, V.F.,
BARBOSA, B., PORTUGAL, A.M.M., 2007, “Densidade estrutural de manguezais
em um estuário adjacente ao banco de Abrolhos (Brasil). XII Congresso Latino-
Americano de Ciências do Mar, Florianópolis, Santa Catarina, Brasil, 15-19 Abril.
SOARES, M.L.G., ESTRADA, G.C.D., CAVALCANTI, V.F., SILVA Jr., C.M.G.,
TOGNELLA-De-ROSA, M.M.P., 2008, “Limite sul de ocorrência de manguezais no
Atlântico oeste”. III Congresso Brasileiro de Oceanografia, I Congresso Ibero-
Americano de Oceanografia, Fortaleza, Ceará, Brasil, 20-24 maio.
SOARES, M.L.G., CHAVES, F.O., ESTRADA, G.C.D., PELLEGRINI, J.A.C.,
CAVALCANTI, V.F., SILVA-JÚNIOR, C.M.G., “Mangrove Forests Associated with
Salt Flats: A Case Study from Rio de Janeiro (Brazil)”. Submetido ao periódico
Wetlands Ecology and Management em outubro de 2008.
SOKAL, R.R., ROHLF, F.J., 1979. Biometria: Principios y metodos estadisticos en la
investigacion biologica. Madrid, H.Blume.
SOUZA, M.M.A., SAMPAIO, E.V.S.B., 2001, “Variação temporal da estrutura dos
bosques de mangue de Suape - PE após a construção do porto”. Acta Botanica
Brasilica, v.15, n.1, pp.1-12.
SUWA, R., ANALUDDIN, K., 2008, “Structure and productivity along a tree height
gradient in a Kandelia obovata mangrove forest in the Manko Wetland, Okinawa
Island, Japan”. Wetlands Ecology Management, v.16, pp. 331-343.

Referências Bibliográficas
82
THOM, B.G., 1984, Coastal landforms and geomorphic processes. In: The mangrove
ecosystem: research methods. Monographs on oceanographic methodology 8,
UNESCO, Paris.
TOMLINSON, P.B., 1986, The botany of mangroves. Cambridge, Cambridge University
Press.
TWILLEY, R.R., 1995. Properties of mangrove ecosystems related to the energy
signature of coastal environments. In: Maximum Power. Colorado, University of
Colorado Press. Boulder.
TWILLEY, R.R., RIVERA-MONROY, V.H., CHEN, R., BOTERO, L., 1998, “Adapting an
ecological mangrove model to simulate trajectories in restoration ecology”. Marine
Pollution Bulletin, v.37, n.8-12, pp. 404-419.
UKPONG, I.E., 1991, “The performance and distribution of species along soil salinity
gradients of mangrove swamps in southeastern Nigeria”. Vegetatio, v.95, pp. 63-70.
WALSH, G.E., 1974, Mangroves: a review. In: Ecology of halophytes, New York,
Academic Press.
WARD, G.A., SMITH III, T.J., WHELAN, K.R.T., DOYLE, T.W., 2006, “Regional
processes in mangrove ecosystems: spatial scaling relationships, biomass, and
turnover rates following catastrophic disturbance”. Hydrobiologia, v.569, pp. 517–
527.
WELLER, D.E., 1987, “A reevaluation of the -3/2 power rule of plant self-thinning”.
Ecological Monographs v.57, pp. 23-43.
WIENS, J.A., 1989, “Spatial Scaling in Ecology”. Functional Ecology, v.3, n.4., pp. 385-
397.
Zar, J.H., 1996. Biostatistical Analysis. Upper Saddle River, NJ, Prentice Hall.
ZENNER, E.K., HIBBS, D.E., 2000, “A new method for modeling the heterogeneity of
forest structure”. Forest Ecology and Management, v.129, pp. 75-87.
ZENNER, E.K., 2005, “Investigating scale-dependent stand heterogeneity with
structure-area-curves”. Forest Ecology and Management, v.209, pp. 87-100.

Referências Bibliográficas
83
ZHOU, P., LUUKKANEN, O., TOKOLA, T., NIEMINEN, J., 2007, “Vegetation dynamics
and forest landscape restoration in the upper Min river watershed, Sichuan, China”.
Restoration Ecology, v.16, v.2, pp. 348-358.