
Estágio Curricular na Aquacria Piscícolas S.A ... · Palavras-chaves: FWAS, Bem-estar,...
109
Estágio Curricular na Aquacria Piscícolas S.A. Implementação de um plano de garantia de bem-estar na Aquacria Piscícolas S.A. Avaliação do impacto em Solea senegalensis expostos a luz de diferentes comprimentos de onda: crescimento, indicadores de stress e na pigmentação Pedro de Jesus Pires 2015
Transcript of Estágio Curricular na Aquacria Piscícolas S.A ... · Palavras-chaves: FWAS, Bem-estar,...

Estágio Curricular na Aquacria Piscícolas S.A.
Implementação de um plano de garantia de bem-estar na
Aquacria Piscícolas S.A.
Avaliação do impacto em Solea senegalensis expostos a luz
de diferentes comprimentos de onda: crescimento, indicadores de
stress e na pigmentação
Pedro de Jesus Pires
2015


Estágio Curricular na Aquacria Piscícolas S.A.
Implementação de um plano de garantia de bem-estar na
Aquacria Piscícolas S.A.
Avaliação do impacto em Solea senegalensis expostos a luz
de diferentes comprimentos de onda: crescimento, indicadores de
stress e na pigmentação
Pedro de Jesus Pires
Relatório de Estágio para obtenção do Grau de Mestre em Aquacultura
Projeto de Mestrado realizado sob a orientação de:
Licenciada (Pré-Bolonha) em Ciências do Meio Aquático Renata Serradeiro
Especialista em Pescas-Aquacultura Teresa Baptista
2015

II
Direitos de Cópia
Autorizo os direitos de copyright do presente relatório de estágio de mestrado.
A Escola Superior de Turismo e Tecnologia do Mar e o Instituto Politécnico de
Leiria têm o direito, perpétuo e sem limites geográficos, de arquivar e publicar este
relatório de estágio através de exemplares impressos reproduzidos em papel ou de forma
digital, ou por qualquer outro meio conhecido ou que venha a ser inventado, e de a
divulgar através de repositórios científicos e de admitir a sua cópia e distribuição com
objetivos educacionais ou de investigação, não comerciais, desde que seja dado crédito
ao autor e editor.

III
Agradecimentos
Para terminar este trabalho, contei com a disponibilidade, apoio e colaboração de
um grande grupo de pessoas, que se tornaram minhas amigas. Dentro de muitas, a toda a
equipa da SEA8, em especial ao pessoal da Aquacria Piscícolas S.A. por me receberem
tão bem e que me integraram melhor do que poderia imaginar. Para elas o meu
reconhecimento, sobretudo para me incentivarem a ultrapassar grandes dificuldades,
nesta fase da minha vida.
Manifesto a minha gratidão à Professora Teresa Baptista, orientadora desta tese,
pela sua disponibilidade desde o primeiro dia até o último, pelas críticas e conselhos na
escrita deste relatório de estágio, mas sobretudo pelo estímulo e ajuda na concretização
do mesmo.
Agradeço também à Renata Serradeiro, diretora de I+D do grupo SEA8, pela sua
simpatia desde o nosso primeiro encontro e por todas as oportunidades gratificantes que
me permitiu alcançar, pautadas por uma convivência saudável.
Agradeço ao Doutor Hans van de Vis pela revisão de todos os documentos e pela
orientação fundamental na elaboração deste projeto.
Por fim, agradeço aos meus pais, namorada e amigos, pela ajuda e pelo incentivo
dado durante todo este longo trajeto, por vezes difícil, por vezes sem vontade.
Muito Obrigado a todos.

IV

V
Resumo
A sobre pesca, o aumento da procura de proteína e o crescimento da população
levam a que o aumento na produção animal seja crucial, para combater estes problemas.
Hoje em dia, muitas autoridades apoiam que a aquacultura é uma maneira de ultrapassar
estes problemas, esta atividade apresenta a maior taxa de expansão a nível global, no
entanto com este crescimento novos obstáculos emergem, quer pela pressão para
corresponder aos padrões da sociedade, quer pela pressão de autoridades reguladoras.
Atualmente e cada vez mais, a sociedade preocupa-se com as condições de bem-estar
animal em produção, isto foi primeiramente direcionado para a produção de animais
terrestres, mas, com o aumento da aquacultura esta preocupação é agora aumentada para
este sector. Por isso, existem agora autoridades especializadas em bem-estar animal em
produção, inclusive no sector da aquacultura, que pode certificar que a produção preenche
os padrões de bem-estar animal. Contudo, esta certificação acontece, principalmente, nos
países nórdicos da Europa e é esperado que esta preocupação crie pressão em produtores
de outros países desenvolvidos. Devido a isso a direção da Aquacria Piscícolas S.A.
estava interessada na implementação de um plano Fish Welfare Assurance System
(FWAS), com o intuito de melhorar as condições de produção para salvaguardar o bem-
estar animal, este plano irá ajudar a instalação na obtenção de uma futura certificação para
o bem-estar e na competição em mercados internacionais. O linguado tem um grande
potencial para a aquacultura, pois apresenta um valor económico elevado contudo
apresenta diversas dificuldades entre as quais, a suscetibilidade a doenças, requisitos
nutricionais, grande dispersão no tamanho, entre outros. Para esclarecer alguns destes
parâmetros este trabalho teve como objetivo avaliar o efeito de luz de diferentes
comprimentos de onda no crescimento, bem como parâmetros hematológicos e
imunológicos e ainda alterações na coloração da pele em Solea senegalensis. Este ensaio
realizou-se nas instalações da Aquacria Piscícolas S.A. Vinte e sete mil juvenis foram
divididos em seis tanques com um sistema de recirculação de água e mantidos sobre um
regime intensivo de produção. Foram utilizadas lâmpadas azuis com um comprimento de
onda de 400nm, lâmpadas brancas com um comprimento de onda de 450nm e lâmpadas
revestidas com uma pelicula vermelha como controlo. Para analisar o crescimento foram
realizadas amostragens de duzentos peixes por tanque. Já para os parâmetros

VI
hematológicos e imunológicos retirou-se sangue de seis peixes por tanque e quanto à
coloração da pele a amostragem foi de cinquenta peixes por tratamento.
Em todos os aspetos calculados relacionados com o crescimento, a luz branca
parece ser a que promove um maior desenvolvimento, alcançando uma percentagem de
112 % comparando com a curva modelo da SEA8. Quantos aos parâmetros
hematológicos, os resultados mostram que existe uma diminuição da percentagem de
trombócitos e linfócitos, sendo que isto significa que os peixes se encontram em stress,
quanto aos dados imunológicos, a glucose elevada pode ser indício para agentes
stressantes. A análise à coloração da pele mostra que os peixes mantidos no controlo
apresentam a cor com mais brilho, tornando-a mais clara.
Concluindo, a luz azul parece não ter um efeito benéfico tanto no crescimento
como na coloração da cor da pele, sendo que a luz branca é aquela que influencia
positivamente no crescimento. O controlo é aquele que tem os resultados mais
satisfatórios para a coloração da pele.
Ao longo destes trabalhos, havia também o objetivo de realizar um estágio nesta
instalação, com a finalidade de perceber como é o trabalho rotineiro numa aquacultura.
Palavras-chaves: FWAS, Bem-estar, comprimento de onda, Pigmentação, stress, Solea
senegalensis

VII
Abstract
The overfishing, the increased protein demand and growth of the population, mean
that the increase in animal production is crucial to combat these problems. Nowadays,
many authorities support that aquaculture is a way to overcome these problems and this
activity has the highest global expansion rate. However new obstacles arrive with this
expansion, either the pressure to correspond to society's standards and regulatory
authorities.
Currently, society is concerned about the animal welfare conditions in production.
It was first directed to the production of land animals, but with increasing aquaculture this
concern is now extended to this sector. So there are now specialized authorities in animal
welfare at aquaculture sector which can make sure that the production meets the animal
welfare standards. However, this certification takes place mainly in the Nordic countries
of Europe and it is expected that this concern create pressure on producers in other
developed countries. Because of this, the direction of Aquacria Piscícolas SA was
interested in implementing a Fish Welfare Assurance System Plan (FWAs), in order to
improve production conditions to ensure animal welfare. This plan will help the facility
in obtaining future certification for the well-being and be able to compete in international
markets. Additionally, the Senegalese sole has a great potential for aquaculture, since it
has a high economic value. However presents several problems including, susceptibility
to disease, nutritional requirements and great dispersion in the size among others. To
clarify some of these parameters this study took place to evaluate the effect of light of
different wavelengths in growth as well as haematological and immunological parameters
and further changes in skin colouring at Solea senegalensis. This test was carried out at
Aquacria Piscícolas S.A. facilities using twenty-seven thousand young fishes which were
divided into six tanks with a water recirculation system and maintained on an intensive
system of production. Blue light was used with a wavelength of 400nm, white light with
a wavelength of 450nm and a red light coated film as a control. To analyse the growth,
samples of two hundred fish per tank were made. For haematological and immunological
parameters withdrew blood from six fish per tank. With regard to skin coloration fifty
fishes per treatment were used.
In all aspects related to the calculated growth in white light appears to be
promoting a higher development, reaching a percentage of 112% compared with the
model SEA8 curve. In haematological and immunological parameters, the results show

VIII
that there is a decrease in the percentage of lymphocytes and thrombocytes and high
glucose levels means that the fish are under stress. The analysis of the staining of the skin
shows that the fish held in the control exhibit the colour brighter, making it clearer.
In conclusion, the blue light appears to have no beneficial effect on both fish
growth and coloration of the skin. The white light seems to influence positively the fish
growth and the control is one that has the most satisfactory results for the coloration of
the skin.
Throughout these studies, there was also the aim of performing one stage in this
installation, in order to realize it as a routine work in aquaculture.
Key words: FWAS, welfare, wavelength, pigmentation, stress, Solea senegalensis

IX
Índice
I – Relatório de estágio na Aquacria Piscícolas S.A. ................................................... 1
A empresa ..................................................................................................................... 3
Pré-engorda .............................................................................................................. 5
Crescimento ............................................................................................................... 6
Embalagem e acondicionamento do pescado ......................................................... 11
II - Introdução ............................................................................................................... 13
Estado da aquacultura mundial ................................................................................... 15
Aquacultura em Portugal ............................................................................................ 16
A produção aquícola ............................................................................................... 18
Bem-Estar Animal ...................................................................................................... 19
Saúde: Fundamental para o bem-estar ................................................................... 21
Procedimentos que comprometem o bem-estar ...................................................... 21
Biologia da Espécie: Solea senegalensis .................................................................... 28
III – Implementação de um plano de garantia de bem-estar na Aquacria Piscícolas
S.A. ................................................................................................................................. 31
Implementação do FWAS ........................................................................................... 35
1. Reunir uma equipa ........................................................................................... 35
2. Descrição da espécie ....................................................................................... 35
3. Identificação do uso do produto ...................................................................... 36
4. Construção de um fluxograma ......................................................................... 36
5. Verificação do fluxograma no local ................................................................ 38
1. Condução de uma análise de perigos e avaliação de riscos ........................... 38
2. Determinação de pontos críticos de controlo (PCC) ...................................... 39
3. Estabelecer níveis-alvo (TLS) e limites críticos (CLS) para cada PCC. ......... 39
4. Estabelecer procedimentos de monitorização ................................................. 46
5. Estabelecer medidas corretivas ....................................................................... 47

X
6. Estabelecer procedimentos de verificação ...................................................... 48
7. Estabelecer um sistema de registo ................................................................... 49
IV - Avaliação do impacto em Solea senegalensis expostos a luz de diferentes
comprimentos de onda: crescimento, indicadores de stress e na pigmentação ...... 51
Introdução ................................................................................................................... 53
Alterações na Pigmentação ..................................................................................... 53
Stress: Indicador de bem-estar ............................................................................... 56
Imunologia .............................................................................................................. 58
Materiais e métodos .................................................................................................... 59
Desenho Experimental ............................................................................................ 59
Amostragens ............................................................................................................ 61
Parâmetros estudados ............................................................................................. 61
Resultados ................................................................................................................... 63
Discussão .................................................................................................................... 72
Conclusão .................................................................................................................... 77
Referências .................................................................................................................. 79
Anexos ........................................................................................................................ 87

XI
Índice de Figuras
Figura 1 - Localização da empresa Aquacria Piscícolas S.A. (Fonte: Google maps) ................. 3
Figura 2 - Ala da pré-engorda (Fonte: própria). ......................................................................... 5
Figura 3- Módulos cinco e seis da área da engorda (Fonte: própria)......................................... 6
Figura 4 - Esquema da vista lateral do crescimento, adaptado do panfleto Aquacria ................ 6
Figura 5- O alimentador automático (Esquerda) e o programa que o controla (Direita). (Fonte:
Própria) ......................................................................................................................................... 7
Figura 6 - Constituintes do sistema de tratamento de água. A) Tanques circulares da receção da
água proveniente do furo; B) Filtro de areia; C) Escumadores de proteínas; D) Torre de
oxigénio; E) Sedimentadores; F) Tanques de produção. (Fonte: Própria). ................................. 8
Figura 7- Determinação da concentração de amónia e nitritos (Esquerda) e computador
responsável pelos alarmes de oxigénio (Direita). (Fonte: própria) ............................................. 9
Figura 8 - Carro para distribuição de ração ao longo dos corredores (Fonte: própria). ......... 10
Figura 9 - Mesa de calibragem (Fonte: própria) ....................................................................... 11
Figura 10- Máquina calibradora na sala de abate e embalamento (Fonte: própria)................ 12
Figura 11 - Capturas mundiais na pesca e produção mundial em aquacultura (Fonte: FAO 2014).
..................................................................................................................................................... 16
Figura 12 - Tipos de estabelecimentos em águas marinhas e salobras em Portugal (Fonte:
INE/DGRM 2014). ...................................................................................................................... 17
Figura 13 - O plano FWAS é implementado quando os pré-requisitos são cumpridos, adaptado
de Van de Vis et al., 2012 ............................................................................................................ 34
Figura 14 - Grupo de símbolos que podem ser usados na elaboração de um fluxograma, adaptado
de van de Vis et al. (2012). .......................................................................................................... 36
Figura 15 - Fluxograma da Aquacria Piscícolas S.A., adaptado de van de Vis et al. (2012). ... 37
Figura 16 - Identificação dos pontos críticos de controlo (PCC), adaptado de Van de Vis et al.
(2012). ......................................................................................................................................... 39
Figura 17 - Esquema para explicar o limite crítico (M) e o limite operacional (m), adaptado de
van de Vis et al., (2012)............................................................................................................... 41
Figura 18 - Trajetos realizados na triagem no dia 5 de Maio. .................................................. 43
Figura 19- A esfera CIELAB (Fonte: Van der Salm, 2004) ....................................................... 54
Figura 20- Esquema dos tanques ............................................................................................... 60
Figura 21 - Locais na pele do peixe onde o colorímetro fez as medições .................................. 63
Figura 22- Diagrama da análise de componentes principais (ACP) para parâmetros
hematológicos e imunológicos para os diferentes tratamentos luminosos. Cada vetor representa
um parâmetro hematológico (NEUT – Neutrófilos; MON – Monócitos; THROM – trombócitos e
LYM – Linfócitos) e imunológico (Prot – Proteínas; Trigl – triglicéridos; Lisoz – Lisozima; Lact

XII
– Lactato; ActB – Atividade Bacteriana; Perox – Peroxidase e Gluc – Glucose), e cada ponto
representa os tanques (W1 – Tratamento com Luz branca no tempo 1, W2 – tratamento com luz
branca no tempo 2; B1 – Tratamento com luz azul no tempo 1, B2 – tratamento com luz azul no
tempo 2; C1 – Controlo no tempo 1, C2 – Controlo no tempo 2) com diferentes tratamentos
luminosos..................................................................................................................................... 71

XIII
Índice de Tabelas
Tabela I - Composição analítica da ração SKRETTING ELITE, adaptado do folheto
SKRETTING. ............................................................................................................................... 10
Tabela II- Evolução da produção aquícola (toneladas) por meios de cultura (Fonte INE/DGRM
2014). .......................................................................................................................................... 18
Tabela III - Exemplos de estudos com diferentes densidades e espécies. .................................. 23
Tabela IV - Dados obtidos durante o abate ............................................................................... 42
Tabela V - Média de tempos de exposição ao ar ........................................................................ 44
Tabela VI - Relação entre o desperdício e o aumento de ração disponibilizada vs stress. ........ 44
Tabela VII - Percentagem de tanques adjacentes afetados. ....................................................... 45
Tabela VIII - Relação entre diferentes tempos de exposição ao ar e desperdício de ração. ..... 46
Tabela IX - Sistema de monitorização numa instalação aquícola, adaptado de van de Vis et al.,
(2012). ......................................................................................................................................... 47
Tabela X - Ações corretivas que podem ser aplicadas numa aquacultura, adaptado de van de Vis
et al., (2012). ............................................................................................................................... 48
Tabela XI - Procedimentos de verificação numa aquacultura, adaptado de van de Vis et al.,
(2012). ......................................................................................................................................... 49
Tabela XII - Constituição de um plano FWAS, adaptado de van de Vis et al., (2012). ............. 50
Tabela XIII- Comparação do peso médio final (g) entre tanques com o mesmo regime luminoso.
..................................................................................................................................................... 64
Tabela XIV- Pesos médios finais comparando tratamentos, individualizando os tanques A e B de
cada regime luminoso (Azul, Branco e Controlo). ...................................................................... 65
Tabela XV- Comparação da soma dos tanques com a curva modelo da SEA 8, agrupando por
tratamento (Azul, Branco e Controlo). ........................................................................................ 65
Tabela XVI- Comparação entre tanques com a curva modela da SEA 8. .................................. 66
Tabela XVII- Condições do ensaio experimental - Número de peixes, peso médio (PM),
biomassa, densidade e comprimento standard, no dia inicial (8 Maio) e no dia final (22 Junho).
..................................................................................................................................................... 66
Tabela XVIII- Parâmetros calculados com base nas amostragens efetuadas – Peso, RGR and
SGR. ............................................................................................................................................ 67
Tabela XIX- Valores da leitura do colorímetro à superfície da pele de Solea senegalensis ..... 67
Tabela XX- Proporção relativa de células no sangue (trombócitos, leucócitos, linfócitos,
monócitos e neutrófilos) de Solea senegalensis de diferentes tratamentos durante 46 dias. ...... 68
Tabela XXI- Parâmetros da resposta imune inata de Solea senegalensis com diferentes
tratamentos luminosos durante 46 dias. ..................................................................................... 69

XIV

XV
Lista de Abreviaturas
FAO – Organização das Nações Unidas para Alimentação e Agricultura;
INE/DGRM – Instituto Nacional de Estatística/Direção Geral dos Recursos Marinhos
RSPCA – Sociedade Real para a Prevenção da Crueldade contra os Animais;
HACCP – Análise de perigos e controlo de pontos críticos;
FWAS – Sistema de garantia de bem-estar;
QA – Controlo de qualidade
CQ – Garantia de qualidade
GMP – Boas práticas de produção;
BCP – Plano de Biossegurança;
PCC – Pontos críticos de controlo;
RA – Avaliação de riscos;
OWI – Indicadores operacionais de bem-estar;
RAS – Sistema de Recirculação de água;
RGR – Taxa de crescimento relativo;
SGR – Taxa de crescimento específico;
HPA – Complexo Hipotálamo-Hipófise-Células interrenais;
αMSH – Hormona estimulante α melanócito;
MCH – Hormona concentradora de melanina;
ACTH – Hormona adrenocorticotropica.

XVI

1
I – Relatório de estágio na Aquacria Piscícolas S.A.

2

3
A empresa
A “SEA8” é uma empresa de capital Espanhol que desde o início de 2012 se dedica
à produção sustentável de linguado em Aquacultura. Tem como convicção que a aquacultura
pode ser o 8º mar, aquele que pode dar resposta às necessidades alimentares de uma
população em crescimento confrontada com a realidade que é a depleção dos stocks naturais
nos sete mares.
Presentemente a “SEA 8” dispõe de instalações produtivas no norte e no centro de
Portugal: uma maternidade, Safiestela - Sustainable Aqua Farming Investments, Lda.,
localizada no Lugar do Rio Alto, Póvoa do Varzim, com capacidade para produzir
anualmente mais de 5 milhões de juvenis e uma unidade de crescimento, Aquacria Piscícolas
S.A. sediada nas Quintas do Norte, Torreira (Figura 1) dimensionada para produzir 200
toneladas por ano de linguado para consumo.
O objetivo do grupo “SEA 8” é a produção em massa de juvenis na sua unidade da
Safiestela, seguida da fase de crescimento nas instalações da Aquacria. A produção é iniciada
na Safiestela, onde é feita a reprodução e a manutenção das larvas até à fase de pré-engorda.
Posteriormente os juvenis são transportados para a Aquacria, onde se efetua o crescimento
dos indivíduos.
Aquando do início desta empresa em 1997, esta empresa dedicava-se ao crescimento
de pregado (Psetta maxima), sendo os juvenis adquiridos a fornecedores de Espanha. No
entanto, a instalação da Acuinova, empresa pertencente ao grupo PESCANOVA com cerca
de 206 hectares, produzindo também pregado, inviabilizou do ponto de vista financeiro, a
produção de pregado. Por este motivo, a estratégia de expansão da empresa passou pela
requalificação das instalações, passando a produzir adicionalmente o linguado senegalês
Figura 1 - Localização da empresa Aquacria Piscícolas S.A. (Fonte: Google maps)

4
(Solea senegalensis). O enorme investimento na modernização e requalificação da unidade
da Safiestela – Sustainable Aqua Farming Investments, Lda. e da Aquacria Piscícolas S.A
colocam-nas na vanguarda da aquacultura em Portugal. É de prever, que num curto espaço
de tempo esta empresa passe a ser a número um, na produção de linguado da Península
Ibérica. O domínio completo de todo o ciclo produtivo do linguado dará uma enorme
vantagem económica e competitiva a esta empresa.
A empresa optou por apostar na inovação, na tecnologia de ponta e métodos
inovadores para proporcionar aos peixes as melhores condições de estabulação e a
estabilidade ambiental de que necessitam para fazer uma utilização ótima dos recursos
disponíveis. Ambas as instalações operam em regime de recirculação, ou seja a maioria da
água utilizada é continuamente tratada através de uma combinação de processos físicos e
biológicos. Isto permite por um lado tratar o pequeno volume de água adicionada a um nível
que inviabiliza a entrada na instalação de agentes patogénicos e por outro lado permite
também tratar o efluente gerado e reaproveitar nutrientes que de outra forma seriam
desperdiçados. Em paralelo esta tecnologia permite ainda manter os peixes num ambiente
de qualidade adequada e muito estável, maximizando o seu bem-estar e otimizando o seu
crescimento e a utilização dos recursos necessários, como sejam o alimento, a energia e o
oxigénio (Bravo, 2015).
Na unidade de crescimento, os juvenis produzidos na maternidade são recebidos com
o maior cuidado, para facilitar a sua ambientação às novas instalações. Durante mais de um
ano os linguados vão ser mantidos num ambiente estável e confortável e são alimentados
com dietas especialmente formuladas para a espécie, as quais asseguram as excelentes
características nutricionais e sensoriais do produto acabado. Periodicamente, são calibrados
por tamanho (triagem), de forma a garantir que fazem o melhor uso possível do espaço e do
alimento, otimizando o seu bem-estar e a utilização dos recursos consumidos. Cerca de um
ano e meio depois de nasceram, estão finalmente prontos para o mercado. Na “SEA 8” pesca-
se exclusivamente para o fornecimento de servir encomendas confirmadas, de forma a
garantir a máxima frescura do produto e a total satisfação do cliente. A embalagem é feita
na unidade de embalamento e acondicionamento do pescado, sendo que as caixas com o
produto final são expedidas para os clientes até 12h após a captura. Não havendo como tal,
stock de produto acabado: o peixe capturado é enviado no mesmo dia para o cliente que o
encomendou. Esta combinação de processos garante a máxima frescura, higiene e qualidade

5
do produto, bom como a sua conservação por um período de tempo adequado para que o
consumidor final possa disfrutar de todas as qualidades (Bravo, 2015).
Pré-engorda
A ala da pré-engorda é assim denominada porque no início da empresa quando ainda
produzia pregado era nesta ala que que se realizava a quarentena dos animais recebidos de
maternidades externas, só depois eram movidas para a unidade de crescimento.
A pré-engorda é constituída por três sistemas em que cada um possui dois tanques
(racks). Os tanques são retangulares de fibra de vidro e estão distribuídos verticalmente
(Figura 2). Os tanques são designados por “shallow raceways” e têm particularidades
positivas tais como: nível de água entre 1 e 20 cm dependendo do tamanho do peixe; elevada
densidade de cultivo, entre 100 e 300% onde os peixes usam toda a área disponível no fundo
e funciona com um regime de recirculação de água (RAS). No decorrer do estágio, esta ala
foi utilizada para a normal produção de linguado e ainda nestas mesmas instalações, decorreu
um ensaio para testar luzes de diferentes comprimentos de onda.
Figura 2 - Ala da pré-engorda (Fonte: própria).

6
Crescimento
Durante o período de estágio, os seis módulos da empresa estavam a funcionar. Cada
módulo é constituído por um conjunto de cinco tanques, dispostos verticalmente (Figura 3).
Entre módulos existe um corredor que os divide e que ainda divide os tanques Norte dos
tanques Sul. No total existem sessenta tanques a funcionar (Figura 4). Cada tanque possui
3,20 metros de largura, 30 cm de altura e 30 metros de comprimento. A água contida no
interior dos tanques tem por volta de 20 cm de altura.
Na tentativa de otimizar o processo de alimentação a empresa optou pela aquisição
de um alimentador automático controlado por um programa adaptado à realidade da empresa
(Figura 5)
Figura 3- Módulos cinco e seis da área da engorda (Fonte: própria).
Figura 4 - Esquema da vista lateral do crescimento, adaptado do panfleto Aquacria

7
A captação de água é feita através de dois furos que se localizam nas imediações da
empresa, mais precisamente nas dunas localizadas a sul do pavilhão do crescimento. Os furos
encontram-se a uma profundidade de cerca de 40 metros. Entretanto a água é encaminhada
para vários tanques circulares exteriores com capacidade de 9m3 (Figura 6) por onde passa
por um processo de arejamento para oxidação do ferro dissolvido. Depois deste sistema de
arejamento existe um filtro de areia que retém o ferro precipitado e outros sólidos em
suspensão (Figura 6). A existência deste passo no tratamento de água é de extrema
importância, uma vez que aqui ocorre a oxidação de Fe2+ para Fe3+ provocando um
precipitado. A água captada no local possui uma grande concentração de ferro, que tem de
ser removido porque se não o fosse, iria provocar mortalidade na produção, uma vez que o
ferro acumula-se nas brânquias dos peixes.
Em seguida a água segue para o bio filtro, depois deste, ela segue para os
escumadores de proteínas (Figura 6) em que a água vai ser misturada com a água proveniente
da coluna de ozonação e segue para os tanques de produção (Figura 6). A utilização dos
escumadores de proteínas deve-se à necessidade de retirar da água partículas finas que estão
dissolvidas na água e que por métodos mecânicos estas não seriam removidas.
Para maximizar a produção e evitar níveis baixos de oxigénio na água a empresa
possui uma torre de oxigénio (Figura 6). A entrada de água depois de tratada e efetuada pelas
entradas dos tanques, que se situam opostamente, percorre toda a superfície do tanque
levando consigo todos os desperdícios e produtos de excreção, saindo na zona de saída do
tanque, estas que se situam no meio do pavilhão do crescimento. De seguida a água segue
para os sedimentadores (Figura 6) onde as partículas provenientes do tanque são filtradas,
Figura 5- O alimentador automático (Esquerda) e o programa que o controla (Direita). (Fonte:
Própria)

8
sendo a água depois direcionada para os filtros mecânicos recomeçando assim todo o
processo de tratamento de água (RAS).
Outra etapa bastante importante é a limpeza do circuito. Esta realiza-se diariamente
e consiste na sifonagem das áreas de sedimentação criadas em cada tanque e na limpeza dos
sedimentadores. Estas duas zonas são aquelas que no circuito mais necessitam de proteção
em termos de limpeza. A análise da mortalidade é feita uma vez ao dia com a ajuda de um
camaroeiro e de uma lanterna, em que os operadores correm todos os tanques recolhendo os
mortos.
Para um bom desenvolvimento dos organismos aquáticos é fulcral ter o controlo do
meio ambiente onde estão inseridos, neste caso, a água dos tanques onde se vão desenvolver.
De modo a ter um maior controlo da qualidade da água, são realizadas análises diárias aos
parâmetros mais importantes como amónia, nitritos e bromos, tanto na zona de crescimento
como na pré-engorda (Figura 7). Procede-se ainda ao controlo da temperatura e do potencial
redox através de sondas que se encontram no tanque. O controlo do oxigénio é feito usando
sondas, caso as concentrações de oxigénio varie para valores críticos é acionado um alarme
sonoro (Figura 7).
F
Figura 6 - Constituintes do sistema de tratamento de água. A) Tanques circulares da receção da
água proveniente do furo; B) Filtro de areia; C) Escumadores de proteínas; D) Torre de
oxigénio; E) Sedimentadores; F) Tanques de produção. (Fonte: Própria).

9
O principal produto do metabolismo dos peixes, a amónia, é removida dos tanques
por nitrificação, este processo consiste na conversão da amónia em nitritos e posteriormente
em nitratos. A amónia proveniente da decomposição da matéria orgânica, ração,
excrementos é tóxica e deve ser medida com frequência. A medição de amónia baseia-se na
reação dos compostos de amónia com o cloro. Os nitritos como a amónia, é tóxica para os
peixes, daí ser necessário monitorizar a sua concentração. As amostras são retiradas
individualmente de cada módulo. Os valores referentes às amostras obtêm-se pela leitura de
um colorímetro.
Uma vez que é usado ozono para o tratamento da água a medição do bromo é fulcral
uma vez que este em concentrações altas pode ser prejudicial para a saúde dos peixes. Dessa
forma o controlo diário dos níveis de bromo previne riscos para a saúde dos peixes e para as
instalações e contribuiu para melhorar a sua função na água e a sua medição é feita com a
utilização do reagente DPD total chlorine.
Na Aquacria Piscícolas S.A., como acima referido, todo o alimento é dado aos peixes
com o auxílio de alimentadores automáticos. Os peixes na empresa Aquacria Piscícolas S.A.
são alimentados com ração seca SKRETTING ELITE. (Figura 8).
Figura 7- Determinação da concentração de amónia e nitritos (Esquerda) e computador
responsável pelos alarmes de oxigénio (Direita). (Fonte: própria)

10
Esta ração tem como principais ingredientes a farinha de peixe, óleo de peixe, óleo
vegetal, produtos e subprodutos de cereais, produtos e subprodutos de leguminosas,
vitaminas e minerais (Tabela I). Sendo que o tamanho da grão da ração depende do peso do
peixe. À medida que o peixe cresce, torna-se necessário aumentar a granulometria da ração.
No entanto esta transição não é imediata, uma vez que durante um período de tempo
(aproximadamente duas semanas) misturam-se dois tamanhos. A ração é armazenada numa
divisão separada do pavilhão do crescimento.
Tabela I - Composição analítica da ração SKRETTING ELITE, adaptado do folheto SKRETTING.
Para otimizar o rendimento da empresa é fundamental realizar amostragens e
calibragens para agrupar os peixes por classes de peso. As amostragens têm como objetivo
obter o peso e comprimento médios por tanque, sendo que estes valores vão ser uteis para
atualizar o programa que controla as biomassas e por sua vez ajustar a quantidade de ração
administrada. Os índices de crescimento são calculados pelo programa tendo em conta os
valores obtidos nas amostragens. Esta técnica consiste em capturar aleatoriamente um
Produto Proteína
Bruta Lípidos Cinzas Fibra Total P
Energia
Digestível
ELITE 57% 16% 10% 0,5% 1,6% 19,5 MJ/kg.
Figura 8 - Carro para distribuição de ração ao longo dos corredores (Fonte: própria).

11
número conhecido de peixes de uma divisão com o auxílio de um camaroeiro e depois
proceder a pesagem e medição dos peixes.
A calibragem é usada para agrupar os peixes por classes de tamanhos o que vai
maximizar o consumo de ração. Para isso são criadas várias categorias de pesos que são
introduzidas no sistema de balança que se encontra na mesa de calibragem. Na prática os
peixes são capturados com o camaroeiro, são levados para a mesa de calibragem (Figura 9)
e mediante um sistema de portas que abrem de acordo com o peso do peixe, estes são
separados para diferentes unidades.
Embalagem e acondicionamento do pescado
Por último, a empresa possui uma própria sala de embalagem e acondicionamento do
pescado (Figura 10). Os linguados que atingiram os pesos mínimos de comercialização são
capturados dos tanques e são colocados nos tanques de jejum, onde irão permanecer pelo
menos durante 48 horas. Após estar cumprido o tempo de jejum os peixes são capturados e
abatidos. O método de abate realizado é por choque térmico e consiste em introduzir os
peixes numa mistura de água e gelo.
Figura 9 - Mesa de calibragem (Fonte: própria)

12
Depois de mortos os peixes são introduzidos numa máquina calibradora (Figura 10),
que os vai separar, por tamanhos e para as caixas. Os peixes são acondicionados em caixas
de poliestireno e são envoltos numa folha de filme plástico e coberto com uma camada de
gelo. O uso de plástico é importante para que o gelo não esteja em contato direto com o peixe
e dessa forma evite causar queimaduras que possam afetar a qualidade do produto.
Seguidamente a serem embalados as caixas são dispostas em paletes e seguem para a câmara
de frio até serem transportadas para o seu destino.
O objetivo principal do estágio na empresa Aquacria Piscícolas S.A. baseou-se na
implementação de um plano de garantia de bem-estar, uma vez que os valores éticos deste
tema têm vindo a ganhar uma extrema importância por todo o mundo, proporcionando uma
busca intensa dos produtos certificados por parte de alguns países desenvolvidos. Deste
modo, a Aquacria Piscícolas S.A. está a conduzir os seus esforços para a implementação de
um plano de garantia de bem-estar para futuramente certificar o seu produto, Solea
senegalensis.
E ainda, foi proposto pela Aquacria Piscícolas S.A. um segundo projeto com o
objetivo de determinar o efeito da luz, com comprimentos de onda diferentes, em vários
parâmetros referentes ao desenvolvimento do peixe.
Figura 10- Máquina calibradora na sala de abate e embalamento (Fonte: própria)

13
II - Introdução

14

15
Estado da aquacultura mundial
A aquacultura consiste na produção de organismos aquáticos (peixes, crustáceos,
molúsculos, anfíbios e plantas aquáticas), tanto em água doce como em água salgada. Esta
atividade remonta há mais de 3500-4000 anos. Nas últimas duas décadas a aquacultura foi
ocupando uma parte cada vez mais importante no suprimento das necessidades alimentares
da população mundial devido ao estado de sobre-exploração da maior parte dos recursos
pesqueiros tradicionais. A aquacultura é uma atividade muito diversificada, que abrange uma
vasta gama de espécies, sistemas e práticas. A expansão económica do sector nos últimos
vinte anos contribuiu para a criação de emprego, para uma utilização mais eficiente dos
recursos locais e para o aumento de oportunidades de investimento no sector produtivo. Esta
expansão refletiu-se também num aumento da contribuição da aquacultura para o comércio
regional e mundial (FAO, 2014). A produção mundial de peixes de aquicultura tem crescido
rapidamente ao longo das últimas quatro décadas, contribuindo significativamente para
abastecer o mundo de peixe para consumo humano (FAO, 2014). Em 2012, a produção total
da aquacultura rondou os 66 milhões de toneladas (excluindo a produção de plantas aquáticas
e de produtos não alimentares) (FAO, 2014). Estima-se que esta participação da aquacultura,
no fornecimento de peixe para o consumo humano, atinja 65%, até 2030 (Fisheries -
European Commission, 2012).
Independentemente da variedade de espécies produzidas em todo o mundo, cerca de
567, apenas 20 espécies representam 74% de toda a produção. Da mesma forma do que
aconteceu com a produção animal terrestre, espera-se que a produção de aquacultura fique
restringida a um número limitado de espécies (Bostock et al., 2010; Diana et al., 2013; FAO,
2014).
Atualmente, a aquacultura é uma das mais importantes indústrias de produção de
alimento de elevado teor nutritivo. Em todo o mundo, a aquacultura tem contribuído para o
bem-estar das populações, representando uma importante fonte de riqueza e um meio de
subsistência (FAO, 2014). Paralelamente constitui uma alternativa às formas tradicionais de
abastecimento de peixe para consumo, sendo que atualmente existe uma preocupação face à
atual estabilização das capturas na pesca e no aumento da população mundial (FAO, 2014).

16
O desenvolvimento mundial da aquacultura foi bastante importante para o
suprimento de necessidades da população mundial sendo que o peixe de aquacultura
contribuiu com 42,2% de um total de 158 milhões de toneladas (Figura 11). Em comparação,
13,4 % em 1990 e 25,7% em 2000.
O crescimento mundial da Aquacultura permanece relativamente forte, existindo
uma crescente procura dos produtos aquícolas por todos os países. Contudo, as regiões
industrializadas que mais produziam como os Estados Unidos da América, Espanha, França,
Itália, Japão e Coreia estão a sofrer quebras na sua produção. A disponibilidade de peixe
produzido noutros países, com custos relativamente mais baixos, foi a causa da rutura na
produção dos países acima mencionados. O facto de ter havido essa quebra, permitiu que
outros países produzissem, focando-se em espécies para exportação (FAO, 2014). A
produção de proteína derivada da aquacultura expandiu 6,2% anualmente no período de
2000-2012, mais lentamente do que no período de 1980-1990 (10,8%) e no período de 1990-
2000 (9,5%). Entre 1980 e 2012, a produção da aquacultura mundial aumentou 8,6% por
ano. A produção de proteína proveniente da aquacultura aumentou mais do que o dobro, de
32,4 milhões de toneladas em 2000 para 66,6 milhões de toneladas em 2012 (FAO, 2014).
Aquacultura em Portugal
Os estabelecimentos aquícolas de culturas marinhas estão localizados nas zonas entre
marés e em terra, fora das zonas de influência direta das marés mas suficientemente próximas
Figura 11 - Capturas mundiais na pesca e produção mundial em aquacultura (Fonte: FAO 2014).

17
do mar para efeitos de captação de água (Plano estratégico para a aquicultura portuguesa,
2014). A aquicultura em Portugal carateriza-se pela existência de um elevado número de
estabelecimentos de pequena dimensão, nomeadamente viveiros de bivalves em zonas
estuarinas ou de ria. Em 2012 existiam 1432 estabelecimentos ativos, ou seja, 96% dos 1492
estabelecimentos licenciados. Daqueles, 94,2% são viveiros para produção de moluscos
bivalves; 4,7% são viveiros para a produção de peixe e 1 % são estruturas flutuantes, também
destinadas à produção de moluscos bivalves (Figura 12).
O facto de as salinas estarem desativadas, proporcionou aos produtores de peixe uma
forma alternativa de as usar, sendo que estas surgem como um processo barato e fácil para
as converter em pisciculturas. Contudo apresenta uma produtividade limitada devido ao
tamanho, tipo de fundos e os baixos fluxos de água, sendo por isso classificados como “semi-
extensiva”, nestes sistemas é usado um regime alimentar misto, ou seja, alimento natural
complementado com alimento inerte, dando por isso, um sabor e textura peculiar que os
diferencia de outros produtos de aquacultura. Em regime extensivo ainda podemos encontrar
nas zonas entre marés viveiros de bivalves.
Ultimamente, em Portugal foram criados algumas instalações em regime intensivo
na orla costeira, em que estas captam água do mar tendo como objetivo a produção de
pregado e linguado. Em questão à instalação de estabelecimentos em mar aberto, estes
apenas se localizam na costa algarvia, porque é onde as condições são mais favoráveis à
prática de aquacultura, tem vindo a ser promovida para a produção de bivalves (Plano
estratégico para a aquicultura portuguesa, 2014).
Figura 12 - Tipos de estabelecimentos em águas marinhas e salobras em Portugal (Fonte:
INE/DGRM 2014).
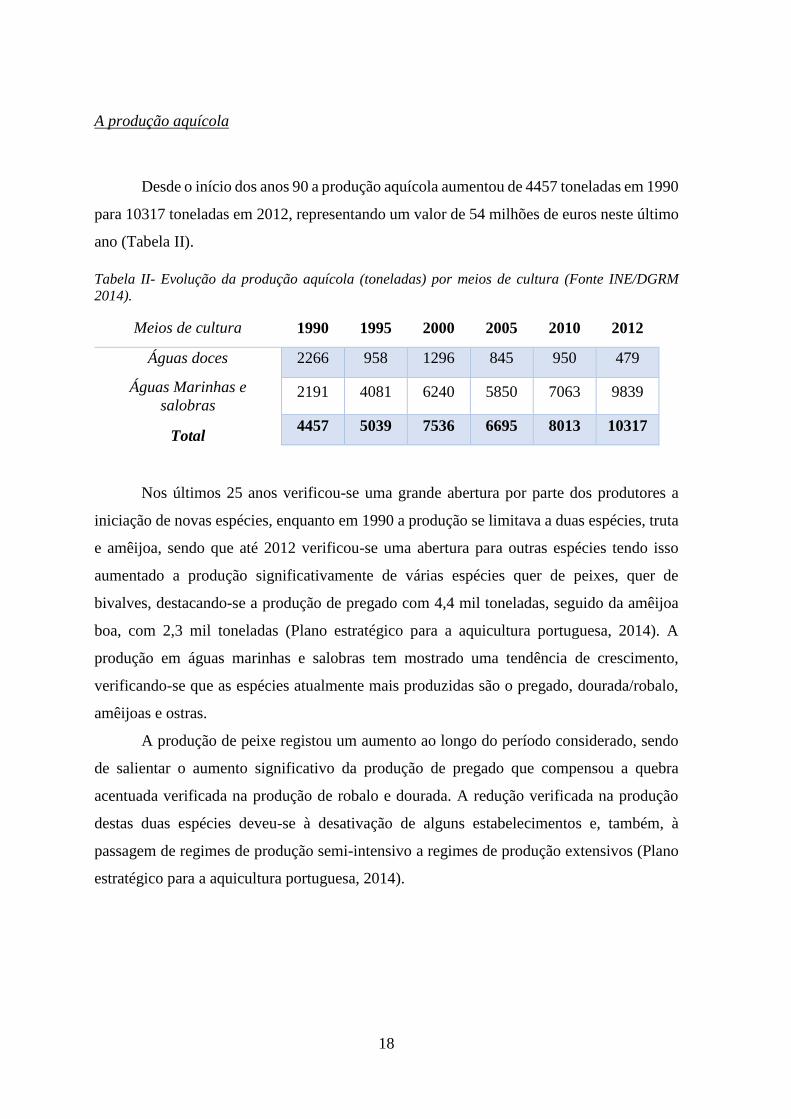
18
A produção aquícola
Desde o início dos anos 90 a produção aquícola aumentou de 4457 toneladas em 1990
para 10317 toneladas em 2012, representando um valor de 54 milhões de euros neste último
ano (Tabela II).
Tabela II- Evolução da produção aquícola (toneladas) por meios de cultura (Fonte INE/DGRM
2014).
Meios de cultura 1990 1995 2000 2005 2010 2012
Águas doces 2266 958 1296 845 950 479
Águas Marinhas e
salobras 2191 4081 6240 5850 7063 9839
Total 4457 5039 7536 6695 8013 10317
Nos últimos 25 anos verificou-se uma grande abertura por parte dos produtores a
iniciação de novas espécies, enquanto em 1990 a produção se limitava a duas espécies, truta
e amêijoa, sendo que até 2012 verificou-se uma abertura para outras espécies tendo isso
aumentado a produção significativamente de várias espécies quer de peixes, quer de
bivalves, destacando-se a produção de pregado com 4,4 mil toneladas, seguido da amêijoa
boa, com 2,3 mil toneladas (Plano estratégico para a aquicultura portuguesa, 2014). A
produção em águas marinhas e salobras tem mostrado uma tendência de crescimento,
verificando-se que as espécies atualmente mais produzidas são o pregado, dourada/robalo,
amêijoas e ostras.
A produção de peixe registou um aumento ao longo do período considerado, sendo
de salientar o aumento significativo da produção de pregado que compensou a quebra
acentuada verificada na produção de robalo e dourada. A redução verificada na produção
destas duas espécies deveu-se à desativação de alguns estabelecimentos e, também, à
passagem de regimes de produção semi-intensivo a regimes de produção extensivos (Plano
estratégico para a aquicultura portuguesa, 2014).

19
Bem-Estar Animal
Atualmente, uma crescente preocupação social, que ocorre predominantemente na
Europa, Canadá, Austrália e Nova Zelândia, resulta num crescente interesse no bem-estar
dos peixes em aquacultura (Kiessling et al., 2012). Os seres humanos afetam de muitas
formas o bem-estar dos peixes, através das pescas, aquacultura e uma série de outras
atividades contudo não existe uma medida para quantificar o bem-estar dos peixes
(Huntingford et al., 2006). Um dos problemas é que o bem-estar animal por vezes vai contra
os interesses humanos, mas recentemente, estruturas éticas estão a tentar inverter essa
tendência valorizando a importância do bem-estar (Huntingford et al., 2006).
O termo bem-estar foi possivelmente usado pela primeira vez em 1975 em
Shelbourne quando se discutia como aclimatar os reprodutores selvagens de Peuronectes
platessa. O conceito de bem-estar espalhou-se para outros países e por outras áreas em que
exista interação entre homem-peixe, como por exemplo, aquaculturas, oceanários, animais
de estimação, pesca para alimento ou pesca desportiva, indústria, consumidores,
organizações não-governamentais, distribuidores, cientistas e a classe governamental (Ellis
et al., 2012). Esta nova tendência tem vindo a ser alvo de pesquisa e investigação por parte
da comunidade científica para ver se os peixes têm a capacidade fisiológica para sentir o
sofrimento .
Uma questão surge quando se fala em bem-estar em peixes, esta passa por saber se
os animais expostos a condições adversas, tais como danos físicos ou outros, sofrem tal como
os humanos (Huntingford et al., 2006). O neocórtex, que nos seres humanos é uma parte
importante do mecanismo que gera a experiência subjetiva do sofrimento, nos peixes e em
animais não-mamíferos não existe, o que leva a ser pensado que estes grupos de animais
podem não sofrer. Uma forte visão alternativa, no entanto, afirma que os peixes são animais
complexos e possuem comportamentos sofisticados, logo, provavelmente têm a capacidade
de sofrimento, embora isso possa ser diferente em intensidade e tipo do que é vivido na
experiência humana (Huntingford et al., 2006). Em comum com todos os vertebrados, os
peixes respondem aos desafios ambientais, ou aos desafios impostos pelo homem, com uma
série de ajustes neuro-endócrinos adaptativos que são vulgarmente denominados de, resposta
ao stress. Estes por sua vez induzem alterações reversíveis, tanto comportamentais como
metabólicas que ajudam o peixe a superar ou evitar os desafios. Em contraste, a permanência

20
prolongada em estados adversos para o peixe faz com que a resposta ao stress seja
prejudicial, uma vez que as respostas são mais prolongadas levando à imunossupressão,
redução no crescimento e disfunção reprodutiva. Os indicadores associados à resposta ao
stress crónico fornece uma fonte de informação sobre o estado de bem-estar de um peixe.
Não existe uma definição clara para o bem-estar animal contudo a saúde física é a
medida de bem-estar mais universalmente aceite e é sem dúvida, um requisito necessário e
essencial para o bem-estar. É importante lembrar que os problemas de saúde podem ser
simultaneamente uma causa e uma consequência de mal-estar (Arlinghaus et al., 2007;
Ashley, 2007; Barber, 2007). Bem-estar pode ser definido como a qualidade de vida que um
animal pode vivenciar (Torgersen et al., 2011). Huntingford et al (2006) afirma que não é
fácil criar uma definição precisa, porque o conceito é complexo e o termo é usado com vários
sentidos contudo, a maioria das definições enquadra-se numa das três categorias e que
nenhuma delas é errada nem certa do ponto de vista científico.
1. Definições baseadas em sentimentos – Estas são definidas em estados mentais
subjetivos. Nestas, a exigência de um bom bem-estar é que o animal deve sentir-se
livre, livre de experiências negativas, como dor ou medo e ter acesso a experiências
positivas, tais como o companheirismo, no caso das espécies sociais (Dawkins,
1998).
2. Definições baseadas em funções – Estas são definidas sobre a capacidade de um
animal em se adaptar ao ambiente presente. Para um bom bem-estar, exige que o
animal esteja saudável, com o seu sistema biológico esteja a funcionar
adequadamente e não ser forçado a responder além da sua capacidade (Dawkins,
1998)
3. Definições baseadas na natureza – Estas surgem a partir da visão em que cada
espécie de animal tem uma natureza biológica inerente que deve expressar. Para um
bom bem-estar, este exige que o animal seja capaz de levar uma vida natural e
expressar o seu comportamento natural (Dawkins, 1998)
Com o crescente interesse no bem-estar nos peixes dentro da comunidade científica,
produtores e Governo Europeu, surgiu a European cost action 867 wellfish. Durante a
reunião científica inaugural em 2006, reconheceu-se que o stress é uma questão-chave nas
discussões sobre o bem-estar dos peixes e que o cortisol é o indicador mais utilizado (Ellis

21
et al., 2012). Contudo sendo uma área crescente de pesquisa, não significa que exista uma
avaliação abrangente, cobrindo todas as espécies de peixes de aquacultura e todas as
interações que possam por em causa do bem-estar (Ashley, 2007). Um dos principais temas
deste assunto é que diferentes espécies têm necessidades biológicas e ambientais diferentes
e apresentam respostas diferentes às condições em aquacultura(Arlinghaus et al., 2007;
Ashley, 2007; Barber, 2007; Ellis et al., 2012; Segner et al., 2012)
Saúde: Fundamental para o bem-estar
A saúde é uma medida fundamental do bem-estar (Ashley, 2007). Se existem
problemas de saúde, este estado pode levar a mais complicações que interferem com um
bom bem-estar, incluindo respostas agudas (adaptativas) a agentes de stress, a supressão do
sistema imunitário, menor taxa de alimentação e por fim as interações sociais também podem
ser afetadas (Ashley, 2007; Damsgard et al., 2004)
O stress é, portanto, um fator importante para a saúde dos peixes (Ashley, 2007).
Embora a saúde não esteja sempre relacionado com más condições ambientais, o objetivo
da aquacultura é minimizar a exposição dos seus produtos a agentes patogénicos e parasitas
e minimizar as fontes de dano físico, contudo existem muitas práticas em aquacultura que
potenciam o stress e a lesão física, o que aumenta a suscetibilidade a patógenos. A prevenção
é a forma ideal para evitar cenários de doença, e para isso ser alcançado é necessário ter uma
boa gestão de protocolos e ter um sistema otimizado (Ashley, 2007; Conte, 2004).
Procedimentos que comprometem o bem-estar
Para a aquacultura é fundamental reduzir os efeitos de certos procedimentos que
aumentam o stress nos peixes, apesar de alguns serem inevitáveis. Os efeitos de uma grande
variedade de práticas de aquacultura sobre os peixes estão muito bem documentados
(Andrade et al., 2015; Conte, 2004; Creţu et al., 2014; Iguchi et al., 2003; Menezes et al.,
2015; Sánchez et al., 2013; van de Nieuwegiessen et al., 2009; Villanueva et al., 2013;
Wunderink et al., 2011). Diferentes espécies apresentam uma grande variação nas respostas

22
fisiológicas ao stress associados a práticas de aquacultura, por exemplo, elevações dos níveis
de cortisol no plasma podem diferir até duas ordens de magnitude em espécies diferentes e
tendo o mesmo agente stressantes a atuar (Ashley, 2007; Barton, 2000, 2002; Congleton et
al., 2000; Conte, 2004).
Densidade
A aquacultura intensiva, por definição, é um tipo de produção que utiliza elevada
densidade sendo que esta é apenas viável porque utilizam RAS o que lhes permite manipular
alguns parâmetros da qualidade de água, tais como a temperatura, oxigénio, alimentação,
amónia e nitritos. (Colson et al., 2015; Kolarevic et al., 2014; Zhang et al., 2011).
Não existe legislação que determine a área mínima para as necessidades de cada
animal o que poderia favorecer as condições de produção. No entanto a rentabilidade de uma
aquacultura, em particular naquelas com regime intensivo, é estritamente ligado às boas
condições de produção. Assim o produtor tem o maior interesse em saber as melhores
condições de produção, sendo que isso é imperativo para uma maior rentabilidade e
qualidade (Andrade et al., 2015; Iguchi et al., 2003; Richard and Flemming, 2007; Saillant
et al., 2003).
De acordo com a espécie e com o estádio de vida, deve haver sempre um plano de
produção com base em estudos, a fim de, implementar as melhores densidades de produção.
Na literatura é descrito que existem espécies que não são afetadas em densidade elevada
como o contrário, e ainda de certas espécies que preferem baixas densidades, no entanto, de
acordo com vários estudos, densidades demasiado elevadas ou demasiado baixas afetam
negativamente o bem-estar e consequentemente a produção (Andrade et al., 2015; Daniels
et al., 1996; Iguchi et al., 2003; Luo et al., 2013; Menezes et al., 2015; Saillant et al., 2003;
Salas-Leiton et al., 2011; Turnbull et al., 2005). Comumente uma elevada densidade de
produção está associada ao bem-estar prejudicial, no entanto, existem estudos que provam
que, dependendo do estádio de vida, a densidade elevada de produção melhora o bem-estar
animal, por exemplo, nos juvenis de peixe-gato africano, refletindo-se no aumento da taxa
de crescimento e diminuição da agressão entre espécimes (van de Nieuwegiessen et al.,
2009). Outra espécie que aparentemente não é afetada pela densidade elevada é o linguado
senegalês (Andrade et al., 2015; Sánchez et al., 2010, 2013; Wunderink et al., 2011).

23
Na Tabela III, existem alguns exemplos de espécies e efeitos que diferentes níveis de
densidade causam.
Tabela III - Exemplos de estudos com diferentes densidades e espécies.
Espécie Densidade
inicial Parâmetro Conclusão
Solea
senegalensis
7, 17, 24 kg
m-2
Crescimento
e parâmetros
imunológicos
Densidade elevada não afetou a
taxa de crescimento e os
parâmetros imunológicos (Andrade
et al., 2015).
Scortum
barcoo
120, 180, 270
peixe m-3 Fisiológicos
A densidade de 180 peixes m-3
apresentou os melhores resultados
(Luo et al., 2013).
Plecoglossus
altivelis
100, 400,
1250 fish m-3
Parâmetros
Imunológicos
Densidades moderadas
apresentaram resultados positivos
para esta espécie (Iguchi et al.,
2003).
Rhamdia
quelen
8, 16, 32 kg
m-3 Fisiológicos
As densidades moderadas e
elevadas são mais adequadas para
esta espécie (Menezes et al., 2015).
Triagem
A triagem é uma operação que pode comprometer a produção e o bem-estar animal.
No entanto, esta operação é fundamental na aquacultura intensiva, já que em densidades de
produção elevadas, comumente encontradas nestes sistemas, há uma concorrência crescente
por alimento entre os animais, existindo o risco de propagação de doenças, danos acidentais
devido ao contato com outros indivíduos, o que pode levar a um aumento da agressividade
e competição pelo espaço (Barron et al., 2013; Daly et al., 2012; González et al., 2011;
Overton et al., 2010; Petrović et al., 2011; Qu et al., 2009; Saillant et al., 2003; Slavík et al.,
2011). Todas estas problemáticas podem ser melhoradas tendo implementado um sistema de

24
auto controlo que controlem por exemplo, o tamanho dos espécimes nas jaulas, tanques e/ou
lagoas.
Contudo, a triagem é benéfica para a produtividade uma vez que a dispersão de
tamanhos é evitada contribuindo para um melhor controlo da alimentação e existe o total
controlo sobre os tamanhos dos peixes em cada tanque. Este controlo é bastante útil também
na monitorização da biomassa na produção, podendo assim recalcular o nível de carga do
tanque (Barron et al., 2013; Lekang, 2013; Petrović et al., 2011; Saillant et al., 2003). Como
já referido acima, a triagem pode comprometer o bem-estar, uma vez que esta prática coloca
os peixes em contacto com utensílios abrasivos, como por exemplo, camaroeiros e também
os coloca por determinados períodos de tempo, fora de água. Isto pode causar danos físicos
e juntamente com as questões de stress, pode criar um foco para a entrada de doenças e
consequentemente diminuir a produtividade (Mustapha, 2014; Qu et al., 2009; Slavík et al.,
2011). O facto de a triagem separar indivíduos por classes de tamanhos compromete as
interações sociais entre peixes, como Slavik et al. (2011) comprovou. Este, utilizando peixe-
gato europeu concluiu que os peixes-gato desviavam mais energia do crescimento para
respostas ao stress por essas mesmas interações sociais se terem desfeito ao invés daqueles
que não eram sujeitos a quebra da interação social. Barro et al. (2013) mostrou também que
a triagem, aumenta em cerca de 15% a sobrevivência de larvas de Lota Lota maculosa. Por
fim, Salas-Leiton et al. (2010) mostrou que indivíduos da espécie Solea senegalensis têm
uma taxa de crescimento maior quando houve uma heterogeneidade nos tamanhos dos
indivíduos. Isto explica-se devido a relações sociais e hierarquia presentes nesta espécie, isto
é, a distribuição de tamanhos é causada pela hierarquia estabelecida.
Estes estudos comprovam a ambiguidade desta operação no processo de produção,
uma vez que é essencial para aumentar a produtividade mas no entanto se não for bem
executada pode comprometer o bem-estar e também a produtividade.
Alimentação
Um ponto que dita o sucesso e/ou o insucesso em todas as aquaculturas é a
alimentação. Esta operação, não só exige o conhecimento de quais são os requisitos
nutricionais da espécie em produção mas também é importante conhecer os processos
fisiológicos da mesma (Boglino et al., 2012; Luo et al., 2013; López-Olmeda et al., 2012;

25
Marinho et al., 2014; Mesquita, 2011; Trejchel et al., 2014; Turnbull and Huntingford,
2012). Alimentação, geralmente, está fortemente relacionada com o bem-estar, uma vez que
em condições de produção intensiva, um dos primeiros sinais de quando o bem-estar é
comprometido é a falta de consumo de ração pelos animais ou o aumento da quantidade de
resíduos. Assim, a alimentação, como as outras operações de produção, estão relacionadas e
é importante saber como variam para as diferentes espécies de produção (Liu et al., 2014;
Mesquita, 2011; Mustapha, 2014; Turnbull and Huntingford, 2012).
O consumo de ração é regulado pelo hipotálamo, esta parte do cérebro é responsável
por receber e interpretar estímulos internos e externos e, em seguida, transmitir a resposta
adequada. Estas respostas são, nomeadamente, regulação da secreção hormonal e
temperatura entre outros (Kulczykowska and Sánchez Vázquez, 2010). Portanto, o sistema
endócrino é fortemente regulado e afetado por estímulos externos. De uma forma geral, todas
as espécies, incluindo aos de aquacultura, são controladas pelo seu ciclo circadiano. Assim,
este ciclo controla um conjunto de comportamentos tais como ritmos de alimentação. Em
praticamente todos os peixes este comportamento ocorre apenas de dia ou de noite,
dependendo da espécie, classificando assim como espécies noturnas e/ou diurnas. No
entanto, em muitas espécies, o ciclo circadiano é flexível e pode ser alterado em condições
de produção, através de manipulação das condições ambientais (Ganzon-Naret, 2013;
Kulczykowska and Sánchez Vázquez, 2010; Liu et al., 2014; López-Olmeda et al., 2012).
Em aquacultura, nomeadamente em sistemas intensivos, a ingestão de alimentos está
condicionada por uma série de operações que ocorrem numa produção, uma delas é a
habituação ao alimento inerte. Estas operações podem afetar o bem-estar e
consequentemente a ingestão de alimento levando ao aumento da quantidade de resíduos o
que pode potencialmente alterar a qualidade da água para níveis não aceitáveis para a
biologia da espécie (Bonaldo et al., 2015; Ganzon-Naret, 2013; Kulczykowska and Sánchez
Vázquez, 2010; Liu et al., 2014; López-Olmeda et al., 2012; Marinho et al., 2014; Mesquita,
2011; Nilsson and Torgersen, 2010; Summerfelt et al., 2015; Turnbull and Huntingford,
2012). Na maioria das instalações de aquacultura, a distribuição do alimento é feito de forma
manual uma vez que este, a curto prazo, é mais barato do que os sistemas automáticos,
embora exija mais trabalho. O sistema manual de distribuição do alimento pode ser menos
eficaz, uma vez que depende da regularidade das distribuições, da quantidade de alimento e
do conhecimento por parte do operador (Mesquita, 2011).

26
Por todas as razões acima mencionadas, existem cada vez mais aquaculturas que
estão a investir em sistemas automáticos. Existem pelo menos dois tipos de sistemas, aqueles
que fornecem alimento quando o peixe vai acionar o mecanismo e aquele sistema que liberta
o alimento quando o produtor assim o deseja, podendo ser cronometrado. Este sistema requer
que a quantidade e o tipo de alimento sejam adequados para a fase de vida em causa, requer
ainda, um plano de gestão excelente, uma vez que o alimento dado é dependente da biomassa
presente em cada tanque. Se o plano de gestão não for adequado para a exploração, há o
risco de disponibilizar alimento a mais, o que pode causar a diminuição da qualidade da água
e um aumento da quantidade de partículas em suspensão na água, conduzindo a um risco
para a saúde dos peixes. Apesar destas desvantagens dos alimentadores automáticos, estes
requerem menos mão-de-obra, (Atoum et al., 2015; Bostock et al., 2010; Mesquita, 2011;
Nilsson and Torgersen, 2010).
Transporte de animais vivos
Este tipo de operação é indispensável numa aquacultura, no entanto, esta é a
prática que potencialmente induz mais stress nos organismos. Esta operação permite que os
peixes possam ser transferidos entre estabelecimentos aquícolas (transporte externo) e dentro
(transporte interno) de uma aquacultura. O transporte é visto como uma ameaça para o bem-
estar durante e após a operação, e é provável que exista um aumento da mortalidade, lesões
e deterioração da qualidade dos animais e da água (Barton and Peter, 1982; Dobsikova et
al., 2009; Iversen et al., 2009; King, 2009; Lekang, 2013; Mustapha, 2014; Tacchi et al.,
2015). Hoje em dia nalguns países existem veículos específicos para estas operações que
melhoram as condições de transporte e que cumprem certas regras para proteger o bem-estar
animal, tais como: densidade por cuba de transporte, design dos equipamentos envolvidos e
parâmetros da qualidade de água (FAO, 2014; Håstein, 2007; Ignacio Martín and Rasines,
2014; King, 2009; Lekang, 2013).
Os animais devem estar em jejum durante pelo menos 24 horas, antes do transporte
de forma a reduzir o metabolismo evitando a deterioração da qualidade da água durante o
transporte por aumento de resíduos metabólicos assim como diminuição do consumo de
oxigénio (Dobsikova et al., 2009; King, 2009; Lekang, 2013; Mustapha, 2014).

27
Para proceder ao transporte, os animais devem estar triados, esta operação, tal como
descrito acima, induz stress, sendo como tal necessário que os animais recuperem antes do
transporte (Lekang, 2013). Embora seja possível o transporte sem água (Ignacio Martín and
Rasines, 2014), na maior parte dos casos, o transporte dos animais aquáticos, requer a
utilização de água, assim a água utilizada é outro ponto crítico que deve ser tomado em conta
para o bem-estar animal. Esta deve ter as propriedades físico-químicas semelhantes da água
onde os peixes estavam, evitando choques térmicos, osmóticos entre outros. Contudo se
alterarmos a temperatura para valores mais baixos, desde que a variação não seja muita
brusca, pode beneficiar o bem-estar animal (Daniel et al., 2014; Farrell et al., 2010; Lekang,
2013; Tang et al., 2009). As temperaturas mais baixas diminuem a taxa metabólica dos
peixes, e consequentemente, diminui os resíduos metabólicos e de consumo de oxigénio.
Para Solea senegalensis foram feitos estudos no âmbito da realização de transportes sem
água, sendo este viável até 28 horas apenas com uma esponja que humedece a pele do animal
(Ignacio Martín and Rasines, 2014).
Alguns transportes utilizam alguns aditivos na água, como por exemplo, sal para
evitar problemas osmorregulatórios, cloreto de sódio que evita a formação de espuma em
água doce (Lekang, 2013; Tacchi et al., 2015). Existem outros compostos, nomeadamente
os sedativos, que podem ser usados de forma a melhorar o bem-estar durante o transporte,
como por exemplo o óleo de cravinho ou óleo de Aloysia triphylla (Daniel et al., 2014;
Iversen et al., 2009). O fornecimento de ar/oxigénio durante o transporte deve ser
garantido,a fim de garantir o bem-estar, neste caso, o ar pode ser fornecido por garrafas
presas ao veículo, desde que o veículo esteja equipado para tal (FAO, 2014; Lekang, 2013).
Quando os transportes são de longo curso, para garantir a qualidade há que
implementar rotinas para minimizar o stress nos peixes. Pode ser necessário realizar uma
troca parcial da água do tanque de transporte sendo que a nova água tem de ser em tudo
semelhante à antiga para evitar vários choques, ou fazer a reciclagem da água, sendo que
isto só é possível se o camião que realiza o transporte tiver equipamentos para esse efeito.
A biomassa e a densidade por tanque de transporte são em função da espécie. Para
maximizar a eficiência do transporte, a qualidade da água deve ser monitorizada durante o
transporte, com a ajuda de uma sonda paramétrica. Normalmente a primeira paragem é
realizada após 15 minutos e depois realizam-se a cada 2 horas. Isto acontece apenas quando
o camião não possui um sistema de alarmes para parâmetros da água (Lekang, 2013).

28
Jejum
Os peixes muitas vezes são privados de alimento antes da realização de certos
procedimentos numa aquacultura, com vista a reduzir o stress fisiológico durante esse
mesmo procedimento. Antes de um transporte, triagem, tratamento para determinada doença
e abate, os peixes são mantidos em jejum para esvaziar o trato intestinal, para reduzir o seu
metabolismo, e a produção de resíduos (Ashley, 2007; Lopez-Olmeda et al., 2012). A
relação entre jejum e stress ou comportamento é desconhecida (Ashley, 2007) e a maioria
os trabalhos nesta área diz respeito ao efeito do jejum prolongado sobre o crescimento, a
proteína muscular e composição da gordura (Lemieux et al., 2004; Pirhonen et al., 2003;
Rios et al., 2002). O impacto sobre o bem-estar em privar um peixe de se alimentar, quando
ele foi previamente alimentado regularmente não é conhecido. Portanto, a privação alimentar
em peixes de aquacultura por curtos períodos, em condições adequadas pode não causar
problemas de bem-estar (Ashley, 2007).
Biologia da Espécie: Solea senegalensis
O linguado da espécie Solea senegalensis é da ordem dos Pleuronectiformes e é
vulgarmente conhecido como peixe chato e não apresenta simetria bilateral, sendo
assimétrico. Possui boca redonda em posição inferior e arqueada (Castelo Branco et al.,
2010). Alguns aspetos relativamente a este peixe é o fato de os adultos possuírem ambos os
olhos na face direita do corpo, a face dorsal ou ocular. A fase ocular apresenta uma coloração
cinzenta – acastanhada e a face cega é totalmente branca. A metamorfose ocorre durante a
fase larvar. Esta metamorfose é caracterizada por grandes transformações anatómicas
durante o desenvolvimento que envolve a rotação de 90 graus na posição do corpo e a
migração de um olho para junto do outro olho. Estas transformações estão associadas á
fixação das larvas no substrato e a mudança do habitat pelágico para bentónico, como
consequência, há também a alteração da alimentação e fisiologia digestiva (Arjona et al.,
2010; Castelo Branco et al., 2010).
Esta espécie em Portugal é comum nos estuários do Tejo, Sado e na costa algarvia
(Castelo Branco et al., 2010). Adapta-se facilmente a ambientes de salinidade elevada, como

29
acontece nos reservatórios das salinas durante a época estival e faz parte das espécies
exploradas na aquacultura tradicional (Arjona et al., 2010; Castelo Branco et al., 2010)
O linguado, Solea senegalensis, distribui-se desde o Senegal – costa Atlântica de
África – até ao Golfo da Biscaia (Arjona et al., 2010). Os linguados adultos vivem em águas
marinhas próximo da costa a 80 metros de profundidade e preferem substrato de areia ou
vasa. É um peixe gonocórico, ovíparo, sendo os ovos fertilizados externamente (Castelo
Branco et al., 2010). O ciclo biológico do linguado começa com a desova dos reprodutores,
quando estes atingem a maturidade sexual, que ocorre aos três anos de idade, tendo as fêmeas
cerca de 32cm e 0.95kg de peso. As fêmeas apresentam uma fecundidade entre 1150000 a
1650000 ovos/kg/ano. Os períodos de reprodução são anuais e a desova escalonada. A época
de postura estende-se por um período de 4 meses, Abril a Julho com um máximo de emissão
de ovos em Maio. A reprodução ocorre no mar e a desova em águas costeiras. As larvas
entram para os estuários e lagoas costeiras por processos de transporte, quer passivos quer
ativos (Castelo Branco et al., 2010). No meio natural, alimenta-se sobretudo de poliquetas
(principalmente a espécie Hediste diversicolor), podendo ainda alimentar-se de anfípodes,
copépodes ou isópodes (Dinis et al., 1999).
A reprodução desta espécie em cativeiro tem sido tema de investigação em Espanha
e Portugal, desde o início da década de oitenta. A desova natural em unidades de reprodução
é atualmente uma rotina e a técnica da cultura larvar é relativamente simples comparada com
as outras espécies de peixes marinhos produzidos em maternidade, registando
sobrevivências elevadas, entre 60 – 80% sendo que em alguns casos chega a ser de 100%
(Castelo Branco et al., 2010).
Esta espécie pode ser facilmente produzida em regime extensivo, alimentando-se da
produção natural do meio (Castelo Branco et al., 2010), em tanques de terra ou em antigas
salinas (Castelo Branco et al., 2010)aguentando altos níveis de salinidade (Arjona et al.,
2010). Esta espécie pode ser produzida em regime de policultura conjuntamente com outras
espécies como a dourada (Sparus aurata) (Dinis et al., 1999)
Apesar da facilidade de produção em regime extensivo, em relação ao regime
intensivo, a produção torna-se mais complicada, uma vez que é necessário capturar os
reprodutores em meio natural. Estes têm de ser capturados fora da época da desova,
nomeadamente entre julho e dezembro, uma vez durante o período de desova as fêmeas têm
poucas probabilidades de sobrevivência ou podem, eventualmente, reabsorver as gónadas
(Cabrita et al., 2006; Dinis et al., 1999; Imsland et al., 2003)

30
À semelhança de outras espécies de água salgada, um dos constrangimentos à
produção de linguado centra-se na fase larvar, especialmente aquando a metamorfose ou a
passagem do alimento vivo para alimento inerte, que provoca grandes mortalidades (Blanco-
Vives et al., 2012; Canavate et al., 2006). Contudo, a produção de linguado está, também,
fortemente condicionada pela sua elevada sensibilidade a diferentes agentes de stress e a
doenças infeciosas, que resultam em perdas económicas para a piscicultura. No que se refere
às infeções, as principais que ocorrem no linguado são a pasteurelose, provocada pela
Photobacterium damselae ssp piscicida, e a vibriose, provocada por várias espécies do
género Vibrio, principalmente a Listonella anguillarum e a flexibacteriose, provocada por
Tenacibaculum maritimum (Imsland et al., 2003).

31
III – Implementação de um plano de garantia de bem-estar na
Aquacria Piscícolas S.A.

32

33
Atualmente, a maioria das aquaculturas monitorizam diariamente ou pelo menos de
forma rotineira vários parâmetros de qualidade da água, comportamento do peixe,
crescimento e estado de saúde (lesões na pele, parasitas etc.). Usando esta abordagem os
sintomas de mau-estar são observados, contudo a origem ou causa dos problemas associados
ao bem-estar podem não ser detetados.
Existem várias questões éticas na Europa que englobam o bem-estar animal, contudo
estas questões são manifestadas nos esquemas de certificação que visam esquemas de
produção sustentáveis (Kiessling et al., 2012; van de Vis et al., 2012; Veissier et al., 2008),
sendo que o bem-estar animal está incluído como parte dos princípios fundamentais,
juntamente com a saúde ambiental e humana, segurança e qualidade alimentar (FAO, 2014).
Os sistemas de certificação que incluem o bem-estar foram criados por organizações não-
governamentais (ONGs), retalhistas e pela indústria da aquacultura (Kiessling et al., 2012;
van de Vis et al., 2012; Veissier et al., 2008). No Reino Unido a entidade certificadora, a
Freedom Food é excecional a este respeito, uma vez que incide unicamente sobre o bem-
estar baseando-se nas Cinco Liberdades (van de Vis et al., 2012).
Estas preocupações criaram pressão no mercado de produtos de aquacultura, assim,
os produtos certificados com bem-estar estão a ser alvo de uma grande procura por parte dos
consumidores. Este aumento verificado já tinha sido evidenciado anteriormente na produção
terrestre, contudo com a grande procura de produtos vindos da aquacultura fez surgir
também um aumento da preocupação sobre o bem-estar dos peixes. Neste sentido foi criado
para peixes de aquacultura, um plano de garantia de bem-estar, FWAS – Fish welfare
assurance system (van de Vis et al., 2012).
A fim de criar um forte mas simples sistema de garantia de bem-estar (FWAS), este
deve ser apoiado por uma base composta por programas de pré-requisito ou seja é preciso
ter implementadas boas práticas de produção (GMP) e um plano que biossegurança (BCP),
o que significa que a aquacultura possua uma boa higiene, boa manutenção e um controlo
sobre todas as práticas que são realizadas nas instalações, uma vez que não é um sistema
autónomo (Figura 13). Se estes pré-requisitos já estiverem implementados serão necessários
apenas pequenos ajustes para alcançar o objetivo principal.

34
Figura 13 - O plano FWAS é implementado quando os pré-requisitos são cumpridos, adaptado de
Van de Vis et al., 2012
Nos mercados Europeus, há um aumento da preocupação com as condições de bem-
estar dos peixes, esta pressão fez com que os piscicultores a fim de cumprir essa exigência
tenham alterado certas rotinas. Para além disso, os piscicultores estão a investir cada vez
mais na certificação de bem-estar, para se tornarem mais competitivos em mercados mais
exigentes. Atualmente, a população está disposta a pagar mais por um produto que é
certificado com garantia de bem-estar. O Reino Unido foi um dos primeiros países a
implementar esta certificação e criar associações como a Sociedade Real para a Prevenção
da Crueldade contra os Animais (RSPCA) na tentativa de criar medidas preventivas para
garantir o bem-estar em aquacultura (Bovenkerk and Meijboom, 2013a).
É conhecido a partir da indústria alimentar e outras indústrias de fabrico que a
garantia de qualidade (QA) é focada na identificação de perigos e estabelecimentos das fases
críticas que necessitam de ser controladas de um determinado processo para evitar, eliminar
ou minimizar um perigo para um nível aceitável (van de Vis et al., 2012). Antes da
introdução do conceito da garantia de qualidade (QA) foram utilizados sistemas de controlo
de qualidade (CQ), contudo estes mostraram ser ineficazes na identificação de perigos com
uma baixa incidência. Posto isto, o CQ foi substituído por um sistema de garantia de
qualidade, conhecida como análise de perigos e pontos críticos de controlo (HACCP). O
sistema HACCP é a abordagem internacionalmente aceite para o controlo da segurança
alimentar (Comité do Códex sobre Higiene Alimentar, 2009; FAO, 2014).
O FWAS diverge do HACCP uma vez que o FWAS não só comprova a qualidade do
produto como também otimiza a produção (Fernández-Segovia et al., 2014; Psomas and
Kafetzopoulos, 2015).
O objetivo deste trabalho é implementar na Aquacria Piscícolas S.A. um plano de
garantia de bem-estar (FWAS) para potenciar o seu produto em mercados mais exigente,

35
isto apenas é conseguido devido a um preocupação por parte da direção da empresa em se
expandir para mercados onde empresas mais pequenas jamais conseguiriam competir.
Implementação do FWAS
Para iniciar a implementação de um sistema FWAS é necessário responder a cinco
estágios preliminares que antecedem os sete princípios principais. Estas cinco etapas
preliminares são consideradas relevantes para a implementação do FWAS.
1. Reunir uma equipa
Dependendo do tamanho da empresa esta equipa pode ser constituída por vários
membros, normalmente nunca mais de seis membros, ou por apenas uma pessoa (Dillon et
al., 1996). A seleção da equipa pode ser feita não só tendo em conta a experiência de cada
um, mas também no conhecimento sobre o controlo de qualidade, trabalhos de rotina, sobre
o plano FWAS e na identificação dos riscos, sendo estes pontos essenciais para a escolha da
equipa (Dillon et al., 1996; FAO, 2014; Miget, 2004; United et al., 2002; van de Vis et al.,
2012).
2. Descrição da espécie
No âmbito da garantia de qualidade, a descrição da espécie implica uma descrição
sobre as necessidades do animal no que diz respeito à qualidade da água, regime alimentar
e maneio, informações detalhadas sobre a tecnologia usada para efetuar o crescimento e
ainda do processo da abate (van de Vis et al., 2012).

36
3. Identificação do uso do produto
O uso pretendido do peixe é relevante isto é, é necessário que exista a identificação
da população alvo e se há algum perigo associado ao produto.
4. Construção de um fluxograma
A construção de um fluxograma é de extrema importância uma vez que vai descrever
todo o processo de produção e analisar todos os possíveis riscos ao longo da produção,
permitindo assim uma visão clara do ciclo de produção. Existe um conjunto de normas para
elaborar um diagrama de fluxo em que cada símbolo simboliza uma etapa diferente no ciclo
de produção (Figura 14)(van de Vis et al., 2012).
Nome de todo o processo.
Abre ou fecha o
fluxograma
Símbolo para um processo,
ou um passo no processo.
Símbolo para um produto
(um produto que constitui
a entrada ou saída de um
processo
Símbolo para um controlo
(exemplo: controlo da
temperatura
Símbolo para um
documento (exemplo:
instrução de trabalho)
Símbolo para uma decisão
no processo
Tarefa que resulta de uma
decisão (exemplo: ação
corretiva)
Símbolo que conecta duas
partes de um fluxograma
Figura 14 - Grupo de símbolos que podem ser usados na elaboração de um fluxograma, adaptado
de van de Vis et al. (2012).
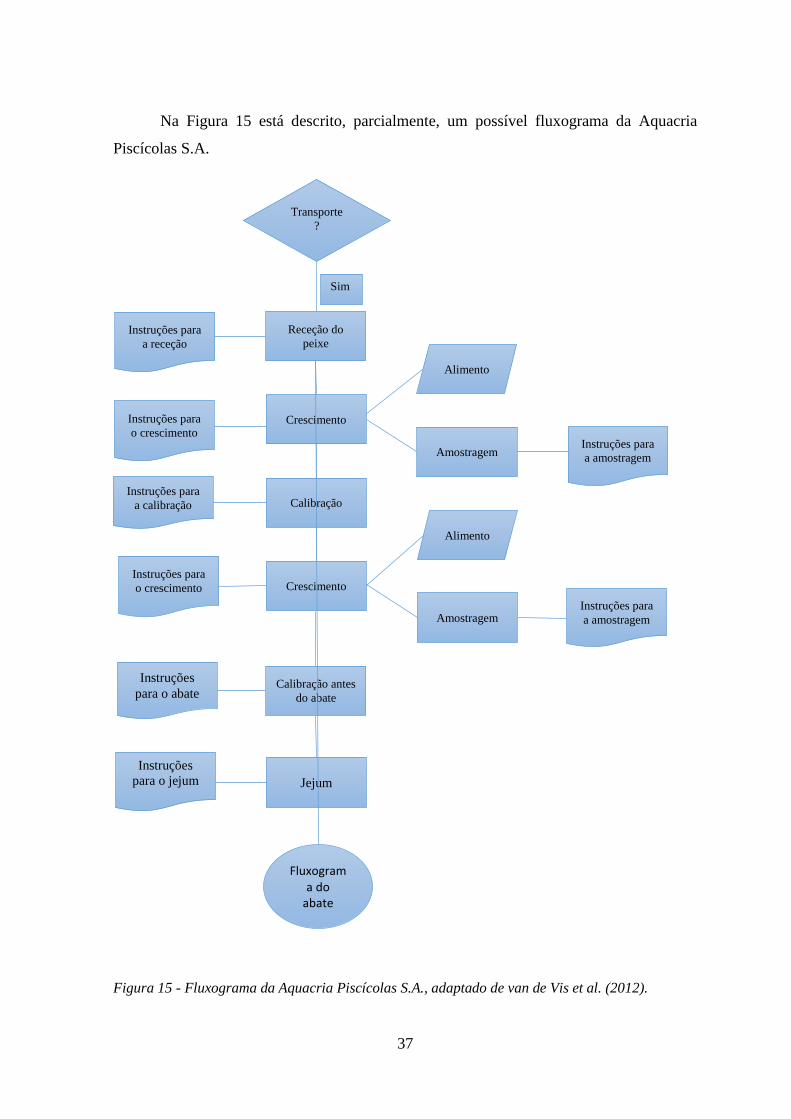
37
Na Figura 15 está descrito, parcialmente, um possível fluxograma da Aquacria
Piscícolas S.A.
Figura 15 - Fluxograma da Aquacria Piscícolas S.A., adaptado de van de Vis et al. (2012).
Receção do
peixe
Transporte
?
Sim
Instruções para
a receção
Crescimento
Calibração
Instruções para
o crescimento
Crescimento
Alimento
Instruções para
a calibração
Instruções para
o crescimento
Alimento
Amostragem
Amostragem Instruções para
a amostragem
Instruções para
a amostragem
Calibração antes
do abate
Instruções
para o abate
Jejum
Fluxograma do
abate
Instruções
para o jejum

38
5. Verificação do fluxograma no local
Uma vez que os fluxogramas foram produzidos precisam de ser verificados quanto à
sua precisão. A fim de conseguir isto é necessário que cada membro da equipa analise e
comprove a veracidade do fluxograma para tornar o plano mais preciso (van de Vis et al.,
2012).
Uma vez que as cinco etapas preliminares em que se baseia o FWAS estão completas,
é feito de seguida a análise aos sete princípios fundamentais do FWAS. Os sete princípios
são:
1. Condução de uma análise de perigos e avaliação de riscos
Num plano de FWAS o objetivo é melhorar o bem-estar animal na produção, no
entanto, este objetivo interfere também na qualidade alimentar (van de Vis et al., 2012).
Uma vez que o plano de FWAS deriva do plano HACCP existem muitas semelhanças entre
ambos, como os princípios que os regem, e em ambos, todos os passos acima referidos tem
de ser cumpridos.
O primeiro princípio é a análise de possíveis perigos para o bem-estar animal, estes
perigos, em aquacultura, podem assumir várias origens e podem ser classificados em:
abióticos, bióticos, operacional e ambientais (van de Vis et al., 2012). Não obstante esta
categorização é usado para facilitar a avaliação dos riscos, uma vez que estes fatores que
comprometem o bem-estar animal muitas das vezes estão relacionados. Por exemplo, a
concentração de oxigénio dissolvido na água depende se os peixes estão em repouso ou se
estão a alimentar. Cada espécie tem diferentes requisitos e é essencial conhecer os
parâmetros que vão atuar como limitantes para a produção e o bem-estar animal. Por
exemplo, o intervalo de temperaturas é apropriado para determinada espécie mas para outra
espécie é ótima. Gamas ótimas vão oferecer aos animais melhores condições de bem-estar o
que é o principal objetivo deste plano, no entanto quantificar o bem-estar é um desafio devido
à sua subjetividade (Bermejo-Poza et al., 2015; Bovenkerk and Meijboom, 2013b;
Grigorakis and Rigos, 2011; Huntingford and Kadri, 2014)

39
2. Determinação de pontos críticos de controlo (PCC)
Uma vez que todos os riscos de produção de uma aquacultura são identificados, estes
tem de ser alvo de uma análise para ver os que são pontos críticos de controlo, ou seja,
aqueles em que o controlo é essencial a fim de garantir o objetivo do plano, seja ele a
segurança alimentar ou o bem-estar animal (Dillon et al., 1996; van de Vis et al., 2012). Para
a identificação destes pontos críticos de controlo existe um esquema de decisão (Figura 16).
Figura 16 - Identificação dos pontos críticos de controlo (PCC), adaptado de Van de Vis et al.
(2012).
3. Estabelecer níveis-alvo (TLS) e limites críticos (CLS) para cada PCC.
A próxima etapa é estabelecer níveis-alvo e limites críticos para cada PCC. Apesar
de nalgumas produções este passo ser relativamente simples, num plano de FWAS, este
passo pode ser um desafio, devido à subjetividade do conceito de bem-estar animal. No
entanto este passo deve ser aplicado. Este estabelecimento de limites deve ser planeado de
acordo com o tipo de estabelecimento. Por exemplo, numa aquacultura pode-se estabelecer
um limite para a concentração de amónia nos tanques de produção, a fim de proteger o bem-

40
estar animal, sendo que estes limites são consoante a espécie, a fase de vida e o sistema de
produção. Nesta fase é essencial recolher dados suficientes durante um espaço de tempo
significativo para reconhecer qualquer padrão sobre a variação destes níveis, porque a
existência do mesmo pode significar se existe um problema ou não, no processo de produção.
Como mencionado acima, para implementar um plano FWAS existem quatro
categorias para agrupar os perigos. Dependendo da espécie produzida, estágio de vida e tipo
de produção, estes riscos têm diferentes limites, no entanto, em função dos riscos, por vezes
pode ser mais apropriado aplicar apenas um tipo de limite. Para os perigos operacionais a
aplicação de níveis-avo é mais apropriado do que limites críticos, como explicado por van
de Vis et al., (2012). Um exemplo disso é a necessidade de remover o salmão do Atlântico
da água quando é vacinado, nesse caso, o limite seria – remover o peixe quando for
estritamente necessário. Para implementar estes limites é necessário ter uma visão sobre
vários fatores e depois sim estabelecer limite, neste exemplo dado, se em vez de salmão do
atlântico fosse outra espécie que consegue resistir a baixos níveis de oxigénio, aí sim seria
possível determinar níveis-alvo (van de Vis et al., 2012).
Um dos principais focos para determinar o bem-estar animal em aquacultura é feito
pela avaliação dos parâmetros da água, uma vez que há uma série de relações entre esses
parâmetros e patologias. Há níveis de tolerância para os parâmetros da água de acordo com
cada espécie e é essencial para uma aquacultura conhecer e manter esses níveis para garantir
o ciclo de produção e do bem-estar animal. Essa faixa de tolerância pode ser mais estreita
ou larga, dependendo do grau de resistência da espécie, no entanto, todas as espécies
possuem um intervalo de tolerância, a fim de manter a estabilidade aquando de alterações
ambientais (McEwen and Wingfield, 2003; Schreck, 2010). Sob condições de produção o
intervalo de tolerância deve ser estreito para evitar qualquer problema para a produção.
Dessa forma são criados dois limites, (Figura 17) um limite com um intervalo de tolerância
maior – Limite operacional, em que dá ao operador algum tempo para ajustar os níveis para
os parâmetros normais – e um com um intervalo de tolerância menor – Limite Crítico – em
que certos casos é igual ao limite operacional, assim, nestes casos não há espaço para erros
(van de Vis et al., 2012).

41
Figura 17 - Esquema para explicar o limite crítico (M) e o limite operacional (m), adaptado de van
de Vis et al., (2012).
Para determinar os limites críticos e os limites operacionais, tendo em conta a
ausência de informação disponível sobre o linguado senegalês para certos parâmetros, foi
necessário realizar ensaios preliminares. Nestes, foram abordados aspetos como o abate e o
efeito de tempo de exposição ao ar.
Abate
A questão-chave na avaliação do bem-estar no momento do abate é que se existe
sofrimento, sendo que igualmente como o bem-estar este também é difícil de quantificar
(Poli et al., 2005). O peixe é pescado e colocado nas tinas que contêm gelo e água, sendo
que a relação entre gelo/água/kg peixe varia entre espécies. Este procedimento é fácil e
rápido e é vulgarmente usado no Mediterrâneo e Reino Unido. A temperatura corporal, taxa
metabólica e movimentos do peixe diminuem rapidamente, sendo que os peixes ficam num
estado de dormência, dando-se a morte por asfixia uma vez que os níveis de oxigénio baixam
drasticamente (Poli et al., 2005). Sendo o abate uma etapa de produção essencial onde o
bem-estar pode ser comprometido, foi desenvolvido um ensaio experimental para cálculo da
relação entre a quantidade de gelo, água e quantidade de peixe em kg.
Na empresa, o abate é realizado na madrugada de segunda-feira e eventualmente na
quarta-feira. O abate dá-se início quando os peixes são pescados com o auxílio de um
camaroeiro e colocados em caixas de transporte, sendo posteriormente pesados.
Paralelamente a tina, onde ocorre o abate, é cheia de gelo e água sendo que as quantidades
de cada elemento estavam estabelecidas por medidas pré-marcadas.

42
Um dos trabalhos desenvolvidos no âmbito deste estágio foi precisamente saber as
medidas exatas de cada elemento. Para tal, calculou-se o volume total da tina (V=600L),
antes e após a adição de gelo, fazendo a subtração de volume obteve-se a quantidade de gelo.
Como a adição da água é feita com o recurso de uma mangueira, foi necessário determinar
o volume, dessa forma, um recipiente com volume conhecido (80 litros) foi cheio e o tempo
desta operação foi cronometrado. Por sua vez, a quantidade de peixe era calculada pela soma
do peso das caixas que continham o peixe. Todos os dados obtidos estão descritos na tabela
IV.
Tabela IV - Dados obtidos durante o abate
Dados
Abate
19-
Fev
16-
Mar
23-
Mar
30-Mar 01-Abr
MÉDIA Tina
1
Tina
2
Tina
3
Tina
4
Tina
5
Tina
1
Tina
2
Tina
3
Tina
4
Kg
gelo 277 266 266 277 260 260 277 260 255 245 245 235 260
Kg
peixe 322 128 130 338 326 329 363 322 321 330 392 280 324
Litros
água 70 80 112 76 72 156 109 80 110 76 80 76 80
A temperatura interna do peixe também foi observada e registada, mas como em
todos os casos a temperatura interna era inferior a 4°C, foi desprezada da Tabela IV. Outra
preocupação neste método de abate é que a temperatura da tina vá aumentando, pois o peixe
fica nesta mistura de água e gelo durante 1h/1:30h. Dessa forma a temperatura da tina foi
medida no início e de meia em meia hora, sendo que os resultados obtidos também não foram
significativos pois a temperatura permaneceu na primeira hora nos -1° sendo que passado
1:30h, no momento do embalamento, a temperatura estava nos 0,8°.
Exposição prolongada do peixe ao ar
Ao longo do processo de produção, os peixes estão obrigatoriamente sujeitos a
procedimentos que implicam períodos fora de água (exposição ao ar). Neste parâmetro foram

43
avaliados duas vertentes, uma que relaciona a ração administrada com o desperdício
verificado, e a segunda vertente, é tempo que o peixe não se alimenta depois de ser exposto
ao ar.
O processo de triagem é aquele em que os peixes permanecem durante mais tempo
expostos ao ar. O objetivo do trabalho experimental neste parâmetro foi cronometrar o tempo
desde que a primeira caixa de peixe era pescada até o último peixe passar pela calibradora.
Foi igualmente feito o registo dos tanques adjacentes ao trajeto efetuado pela pessoa que
movimentava o carrinho com as caixas de peixe (Figura 18). Contudo, esta medição é
influenciada por algumas variáveis, entre as quais: eficácia da equipa, distância do tanque
onde o peixe está a ser pescado, a densidade do tanque (eficiência da pesca) e diferentes
calibres do peixe.
Na tabela V estão registados os tempos que uma caixa de peixe demora a ser
calibrada.
MOD1
MOD2
MOD3
MOD4
MOD5
MOD6
MOD1
MOD2
MOD3
MOD4
MOD5
MOD6
Norte
Sul
C
1
2
5
3
4
N24
Piso 0
Piso -1
Figura 18 - Trajetos realizados na triagem no dia 5 de Maio.

44
Tabela V - Média de tempos de exposição ao ar
8m08s 6m53s 7m01s 7m10s 8m15s
11m16s 7m22s 7m06s 8m57s 7m30s
Média: 8.02 min
Depois destas observações foi feita a comparação com os registos de desperdício de
ração (RD). Estes são realizados pelos trabalhadores, tendo sido adotada a seguinte chave: 0
– não existe desperdício, I – existe pouco desperdício, II e III – em que a quantidade de
desperdício vai aumentando. Existem diversas razões para existir um aumento de
desperdício de ração, entre os quais stress e a necessidade de ajustar a quantidade de ração
diária, sendo que essa análise está demonstrada na tabela VI.
Tabela VI - Relação entre o desperdício e o aumento de ração disponibilizada vs stress.
O parâmetro RD é o desperdício de ração e o parâmetro activity é a percentagem de
ração disponibilizada. Como se pode analisar, o único tanque onde se visualizou um aumento
de ração desperdiçada foi o tanque N14, tanque adjacente ao tanque calibrado (tanque N24).
Pode-se concluir que existe um aumento de RD no dia seguinte ao evento de calibragem
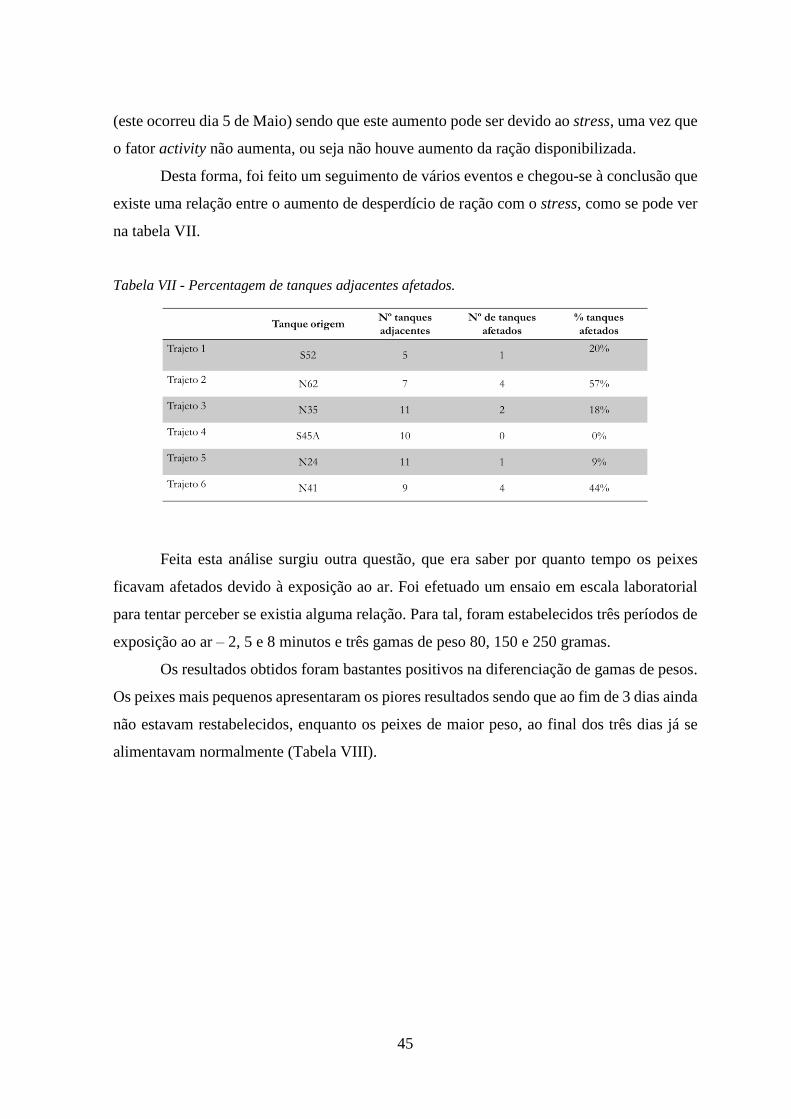
45
(este ocorreu dia 5 de Maio) sendo que este aumento pode ser devido ao stress, uma vez que
o fator activity não aumenta, ou seja não houve aumento da ração disponibilizada.
Desta forma, foi feito um seguimento de vários eventos e chegou-se à conclusão que
existe uma relação entre o aumento de desperdício de ração com o stress, como se pode ver
na tabela VII.
Tabela VII - Percentagem de tanques adjacentes afetados.
Feita esta análise surgiu outra questão, que era saber por quanto tempo os peixes
ficavam afetados devido à exposição ao ar. Foi efetuado um ensaio em escala laboratorial
para tentar perceber se existia alguma relação. Para tal, foram estabelecidos três períodos de
exposição ao ar – 2, 5 e 8 minutos e três gamas de peso 80, 150 e 250 gramas.
Os resultados obtidos foram bastantes positivos na diferenciação de gamas de pesos.
Os peixes mais pequenos apresentaram os piores resultados sendo que ao fim de 3 dias ainda
não estavam restabelecidos, enquanto os peixes de maior peso, ao final dos três dias já se
alimentavam normalmente (Tabela VIII).
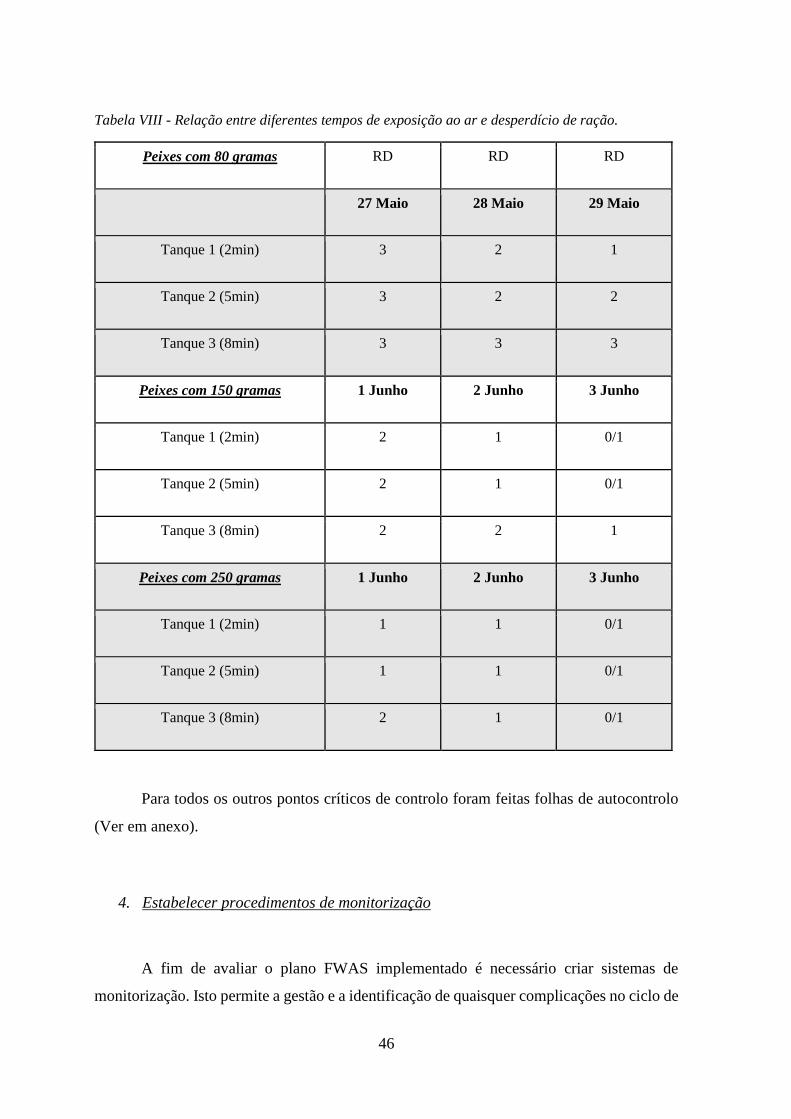
46
Tabela VIII - Relação entre diferentes tempos de exposição ao ar e desperdício de ração.
Peixes com 80 gramas RD RD RD
27 Maio 28 Maio 29 Maio
Tanque 1 (2min) 3 2 1
Tanque 2 (5min) 3 2 2
Tanque 3 (8min) 3 3 3
Peixes com 150 gramas 1 Junho 2 Junho 3 Junho
Tanque 1 (2min) 2 1 0/1
Tanque 2 (5min) 2 1 0/1
Tanque 3 (8min) 2 2 1
Peixes com 250 gramas 1 Junho 2 Junho 3 Junho
Tanque 1 (2min) 1 1 0/1
Tanque 2 (5min) 1 1 0/1
Tanque 3 (8min) 2 1 0/1
Para todos os outros pontos críticos de controlo foram feitas folhas de autocontrolo
(Ver em anexo).
4. Estabelecer procedimentos de monitorização
A fim de avaliar o plano FWAS implementado é necessário criar sistemas de
monitorização. Isto permite a gestão e a identificação de quaisquer complicações no ciclo de

47
produção e reconhecer algum tipo de padrão a qualquer momento, avaliando os limites
críticos estabelecidos previamente, e consequente identificação da causa (van de Vis et al.,
2012). Para avaliar o bem-estar animal, existem diversos indicadores que podem ser
aplicados num ciclo de produção, como por exemplo os indicadores de bem-estar
operacionais (OWI), sendo que o registo da mortalidade é um exemplo de um indicador
(Huntingford and Kadri, 2014). Esses indicadores devem ser práticos para não complicar os
trabalhos dos operadores a um ponto de ser contraproducente. No plano FWAS, é exigido
que existam indicadores que permitam a deteção de qualquer coisa (exemplo – observação
de lesões) que comprometa o bem-estar animal (Tabela IX).
Tabela IX - Sistema de monitorização numa instalação aquícola, adaptado de van de Vis et al.,
(2012).
5. Estabelecer medidas corretivas
A fim de controlar os PCC’s é necessário implementar ações corretivas (Tabela X).
Estas permitem recuperar o controlo sempre que os PCC’s estão além do limite crítico e/ou
operacional. Para aplicar estas ações, é necessário entender a causa do problema, determinar
o que fazer e registrar a ação corretiva tomada. Nos planos FWAS planear as ações corretivas
é muito importante e decisivo para os operadores quando as implementam (Tabela X) (Dillon
et al., 1996, van de Vis et al., 2012).

48
Tabela X - Ações corretivas que podem ser aplicadas numa aquacultura, adaptado de van de Vis et
al., (2012).
6. Estabelecer procedimentos de verificação
Como mencionado acima, os procedimentos de verificação são processos que
asseguram que os produtos se encontram dentro dos limites críticos. No caso do bem-estar
animal estes procedimentos podem ser mais difíceis de determinar porque o bem-estar
animal é subjetivo. No entanto, há um número de formas de analisar o bem-estar na
produção, como por exemplo, análise patológica, comportamentos alimentares, etc. (Tabela
XI). A frequência destes procedimentos deve ser regular e inesperada, para recolher a maior
diversidade possível, tornando assim a amostragem o mais realista possível (Dillon et al.,
1996, van de Vis et al., 2012).

49
Tabela XI - Procedimentos de verificação numa aquacultura, adaptado de van de Vis et al., (2012).
7. Estabelecer um sistema de registo
Um sistema de registos é essencial para garantir que o plano FWAS está a funcionar
corretamente, outra vantagem deste sistema é a criação de um plano FWAS dinâmico, uma
vez que ao longo do tempo a remoção ou adição de PCC é possível. De acordo com van de
Vis et al., (2012), há uma lista de pontos que ajudam a compreender se o sistema de registo
é eficaz ou não, estes são: (1) a responsabilidade da pessoa que efetua os registos; (2) Se as
normas são cumpridas; (3) Pré-requisitos se estão controlados e monitorizados; (4)
Documentação dos procedimentos; (5) Os registos dos procedimentos sanitários para
controlar os PCC; (6) As ações corretivas documentadas; (7) Documento que determina,
quando, por quem e como o plano FWAS é revisto.
Na tabela XII está descrito a constituição de um plano FWAS.
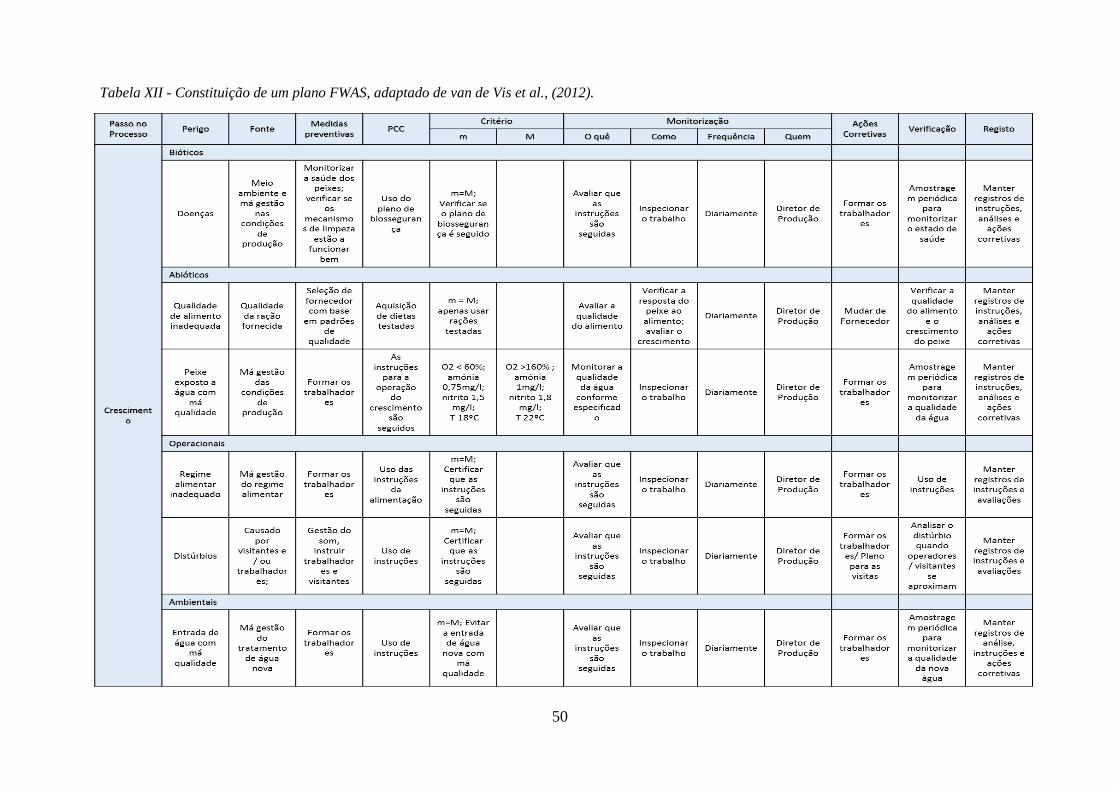
50
Tabela XII - Constituição de um plano FWAS, adaptado de van de Vis et al., (2012).

51
IV - Avaliação do impacto em Solea senegalensis expostos a luz
de diferentes comprimentos de onda: crescimento, indicadores de
stress e na pigmentação

52

53
Introdução
Na natureza, as características do espectro da luz são determinadas por uma
combinação entre a claridade do ambiente e as propriedades óticas da água. A luz com o
aumento da profundidade torna-se monocromática porque o perfil espectral é
seletivamente atenuado enquanto a luz passa pela água (Blanco-Vives et al., 2012). Dessa
forma, águas oceânicas transmitem em comprimentos de onda azul (470nm), enquanto as
águas costeiras transmitem melhor em comprimentos de onda azul-verde (500nm) e por
fim as águas estuarinas transmitem em comprimentos de onda verde (580nm) (Cohen &
Forward, 2002).
Nos peixes, os sistemas extrarretinais e os sistemas fotosensores são bastante
complexos e longe de ser totalmente compreendido, tal como a resposta à luz, que varia
entre espécies, dependendo de fatores filogenéticos e ecológicos (Marchesan et al., 2005).
Está provado que é uma tarefa difícil encontrar as melhores condições de luz para cada
espécie de peixe (Villamizar et al., 2011). Dependendo da ecologia das espécies,
comprimentos de onda específicos podem maximizar a sensibilidade visual quer para
peixes de água profundas como para peixes que vivem em ambientes foto-complexos,
como em águas rasas ou costeiras (Villamizar et al., 2011). No linguado senegalês a
melatonina plasmática é significativamente reduzida quando exposto a comprimentos de
onda curtos (violeta) durante a noite (Oliveira et al., 2007).
Alterações na Pigmentação
A cor é definida como os comprimentos de onda de luz no espectro visual (de 380
até 770 nm) que são detetados pela retina humana. Todas as cores detetadas pelo olho
humano estão associadas com a radiação da luz; a cor azul-violeta corresponde a valores
de absorvância entre 380 e 480nm; verde corresponde a 480-560nm; a amarela, a 560-
590 nm; laranja, a 590-630nm e vermelho, a 630-750nm (Sant'Anna et al., 2013).
Portanto, espectrofotómetros e colorímetros têm sido instrumentos úteis para a medição
da cor em diferentes produtos. Nestes instrumentos, os dados gerados são numericamente
categorizados de acordo com a absorvância, a distribuição de energia espectral relativa, a
geometria do objeto, entre outros fatores (Sant'Anna et al., 2013).

54
Entre os diferentes espaços de cores existentes (por exemplo, CIEXYZ, CIEUVW
e CIELUV), o CIELAB é atualmente o recomendado para ser utilizado para a maioria das
aplicações industriais, porque abrange uniformemente o espectro visível total do olho
humano. Os parâmetros relacionados com o CIELAB encontram-se resumidos na esfera
CIELAB na Figura 19. No espaço CIELAB, são definidos por um índice psicométrico de
luminosidade (L) e duas coordenadas de cor (a e b). O índice de L está relacionado com
a luminosidade, de acordo com esta propriedade, cada cor pode ser considerado como
equivalente a um membro da escala de cinza, ou seja, entre o preto (L = 0) e branco (L =
100). A coordenada a tem valores negativos para as cores esverdeadas e valores positivos
para cores avermelhadas. A coordenada b tem valores positivos para cores amareladas e
valores negativos para cores azuladas.
Figura 19- A esfera CIELAB (Fonte: Van der Salm, 2004)
Os atributos quantitativos e qualitativos da cor (chroma e hue, respetivamente)
são calculados através das coordenadas a e b. O chroma fornece informações sobre a
nitidez da cor, ao passo que o hue é o atributo de acordo com as cores que têm sido
tradicionalmente definidos como vermelho, laranja, amarelo, etc (Figura 19). O chroma
o hue são considerados os principais atributos da cromaticidade. O hue é o valor em grau
que corresponde ao esquema da cor tridimensional (0° para vermelho, 90° para amarelo,
180° para verde e 270° para o azul) (Sant'Anna et al., 2013).

55
Considerando que para algumas espécies de peixe, como por exemplo os
salmonídeos, a cor do filete constitui um importante parâmetro de qualidade, por outro
lado é a pigmentação da pele que melhora muito a aparência do peixe, sendo associado
por parte do consumidor, a um parâmetro de diferenciação e de frescura (Kalinowski et
al., 2007). A coloração da pele em teleósteos está sobre influência de vários parâmetros
e de uma serie de fatores externos e internos (físicos, nutricionais, genéticos, neuro-
hormonal) sendo que estes são fatores que influenciam o estado cromático da pele do
peixe (Szisch et al., 2002). Os peixes apresentam uma variedade de padrões de coloração
da pele que são de grande importância ecológica, fisiológica e comportamental.
De acordo com Pavlidis et al (2006), existem oito padrões gerais de cores que
podem ser observados em peixes: (1) a coloração; (2) poster colours; (3) disruptive
coloration; (4) countershading; (5) ornamentação ocular; (6) manchas oculares; (7) faixas
laterais; (8) policromia. As propriedades cromáticas sofisticadas observadas nesta classe
de vertebrados fornecem proteção contra predadores, impor a sua dominância na
população e ajuda na comunicação (Pavlidis et al., 2006). Além disso, o peixe pode alterar
a sua coloração em resposta às condições ambientais, desafios fisiológicos e estímulos de
stress (Szisch et al., 2002; Van der Salm et al., 2004), daí o padrão da pigmentação da
pele do peixe pode ser considerado como um índice de proteção animal em espécies de
aquacultura e um fator de consideração económica (Szisch et al., 2002; Van der Salm et
al., 2004).
O sistema nervoso simpático e/ou o sistema endócrino são os controladores da
motilidade dos cromatóforos, ou seja, controlam as mudanças de cor devido à ativação
intracelular do pigmento, esta mudança ajuda na adaptação cromática dos peixes quando
estes estão sujeitos a mudanças circadianas, a adaptabilidade às mudanças do fundo,
comunicação interespecífica e stress agudo. As mudanças morfológicas da cor, como a
alteração do número ou densidade dos cromatóforos e/ou a quantidade de material
pigmentado na pele, procedem de forma relativamente lente levando a mudanças
morfogenéticas (Szisch et al., 2002).
A hormona estimulante dos melanócitos (MSH) tem um papel chave na dispersão
dos pigmentos de melanina nos melanóforos na pele de muitos vertebrados exotérmicos,
ou seja tem um papel importante na mudança de cor (Szisch et al., 2002). As principais
hormonas que controlam a mudança na pigmentação são a hormona estimulante α-
melanócito (αMSH) e a hormona concentradora de melanina (MCH). Ambas são
pleiotrópicas e não apenas regulam a pigmentação mas também tem um papel regulador

56
na resposta ao stress (Van der Salm et al., 2004), e também atuam sobre a atividade das
células interrenais que produzem cortisol (Szisch et al., 2002). Durante uma resposta ao
stress, o eixo hipotálamo-hipófise-células interrenais é ativado. Para além da produção da
hormona adrenocorticotrópica (ACTH) a glândula pituitária liberta αMSH que vai induzir
a libertação de cortisol do tecido interrenal (Van der Salm et al., 2004). O cortisol é
libertado na corrente sanguínea e vai ligar-se aos recetores adrenérgicos dos
cromatóforos. Se os cromatóforos possuem α-adrenorecetores, vai haver um mudança na
tonalidade tornando-os mais claros, enquanto que, se os adrenorecetores forem do tipo β,
a mudança vai ser no sentido de os tornar mais escurso. Portanto a MSH pode ser um elo
essencial entre o stress e a pigmentação (Szisch et al., 2002).
Stress: Indicador de bem-estar
Existe uma extensa literatura sobre a biologia, sobre as condições fisiológicas e
respostas comportamentais de peixes para uma ampla variedade de caraterísticas físicas,
químicas, biológicas e fatores de stress (Ashley, 2007; Barton, 2000, 2002; Conte, 2004).
Este interesse considerável no stress dos peixes reflete o ganho de consciência que os
seus impactos na produção podem ser questões-chave para o sucesso ou insucesso de uma
aquacultura, de igual importância como a suscetibilidade à doença, o crescimento, a
eficiência de conversão alimentar, qualidade da carne e a reprodução (Ellis et al., 2012).
Stress é frequentemente discutido com diferentes definições (Ashley, 2007; Ellis et al.,
2012), mas muitas vezes se assemelha a “uma alteração da homeostasia, que é
reestabelecida por um complexo repertório de respostas adaptativas fisiológicas e
comportamentais por parte do organismo” (Ashley, 2007; Chrousos, 1998; Conte, 2004).
A resposta ao stress pode ser observado em diferentes níveis (Ellis et al., 2012):
1. A resposta neuroendócrina – Um estímulo recebido por um organismo vai levar
a uma libertação de hormonas (catecolaminas) e a ativação do complexo
hipotálamo-hipófise-células interrenais (HPA). A libertação de corticotropina a
partir do hipotálamo age sobre a glândula pituitária para sintetizar e libertar a
hormona corticotrópica que por sua vez vai estimular a síntese de hormonas
glucocorticoides (cortisol em teleósteos) a partir das células interrenais (Ashley,
2007; Ellis et al., 2012).

57
2. A alteração fisiológica – São alterações a nível de comportamento ou mesmo
alterações internas (aumento da concentração de glicose no sangue) que vão
permitir ao animal responder ao estímulo. Se estas alterações foram mantidas ao
longo de um período de tempo vai desencadear a terceira alteração (Ellis et al.,
2012; Schreck et al., 2001).
3. Alterações internas e gerais – Estas acontecem quando o animal é mantido em
condições de stress durante muito tempo. Uma vez que a ativação do complexo
hipotálamo-hipófise- células interrenais mobiliza energia, levando a um
esgotamento das reservas de glicogénio a um aumento das concentrações
plasmáticas de glicose, juntamente com a atividade muscular elevada, vai ativar a
glicólise anaeróbica que por sua vez vai aumentar as concentrações de ácido
láctico no plasma (Ashley, 2007; Schreck et al., 2001). Isto vai influenciar as taxas
de crescimento e no investimento reprodutivo podendo ganhar uma maior
suscetibilidade a doenças e mortalidade (Ellis et al., 2012).
Os níveis de glicose e de ácido láctico são utilizados frequentemente em paralelo
com o cortisol para avaliar os níveis de stress (Acerete et al., 2004; Ashley, 2007). A
expressão de genes relacionados com o stress pode proporcionar biomarcadores úteis e
sensíveis para diagnosticar o stress e melhorar o bem-estar futuro (Gornati et al., 2004;
Montero et al., 2001; Ribas et al., 2004). O stress crónico exerce um forte efeito sobre o
metabolismo, função neuroendócrina e osmorregulação (DiBattista et al., 2005).
Portanto, evitar a prolongada e repetida exposição ao stress por parte dos peixes é
claramente um objetivo central do bem-estar animal. Para tal, a avaliação dos métodos
possíveis para reduzir as respostas ao stress é uma área ativa de pesquisa (Ashley, 2007).
Em salmão, estes são criados seletivamente para minimizar a sua capacidade de resposta
ao stress (Ashley, 2007; Pottinger, 2010). Embora o processo de domesticação esteja num
estágio inicial, na produção de salmão, estes sofrem menos comparativamente a salmões
selvagens (Huntingford, 2004; Overli et al., 2005). A produção de cortisol em resposta
ao stress varia entre indivíduos sendo que esta característica apresenta uma moderada a
elevada hereditariedade (Fevolden et al., 1999; Overli et al., 2005; Pottinger et al., 1994;
Pottinger and Pickering, 1992; Pottinger et al., 1992).

58
Imunologia
Imunologia tem sido estudada há mais de 200 anos, a partir da descoberta da
varíola bovina em 1796 (Zhu et al., 2013). A maioria do que sabemos hoje sobre a
composição, funções e modulação do sistema imunológico são derivados principalmente
de investigações em mamíferos. De ponto de vista evolutivo, a pesquisa sobre sistemas
imunológicos em vertebrados inferiores vai ser inevitável pois só assim existirá uma
melhor compreensão da história evolutiva dos sistemas imunitários em todos vertebrados
como um todo (Zhu et al., 2013)
Os peixes, o primeiro grupo de vertebrados depois do período devónico, é aquele
que apresenta uma maior diversidade e sucesso. Este grupo heterogéneo de organismos
ocupa na biologia evolutiva uma encruzilhada entre a resposta imune inata e a resposta
imune adaptativa (Rauta et al., 2012). Dessa forma, órgãos imunes homólogos aos do
sistema imunitário dos mamíferos são encontrados nos peixes. O sistema imune do peixe
não só é condicionado pelo ambiente específico, mas também pela sua natureza
poiquilotérmica (Rauta et al., 2012). É habitual dividir o sistema imunitário em inato (não
específica) e adquirido (específico). No entanto, tanto em peixes como nos mamíferos,
estes dois sistemas dependem de combinações entre eles (Magnadottir, 2006). A resposta
inata geralmente precede a resposta adaptativa, ou seja a resposta inata ativa e determina
a natureza da resposta adaptativa e coopera na manutenção da homeostasia (Magnadottir,
2006).O sistema imunológico é um sistema de estruturas e processos biológicos dentro
de um organismo que deteta uma grande variedade de agentes, desde vírus até vermes
parasitas, protege contra as doenças. Nos animais, o sistema imunitário é um sistema
rudimentar ao contrário do que se verifica no Homem, este mais complexo (Magnadottir,
2006; Rauta et al., 2012).
O sistema imunológico possui mecanismos de defesa que aumenta a
especificidade no combate contra infeções. Em termos simples, possuem barreiras físicas
que evitam a entrada de agentes patogénicos, tais como bactérias e vírus. Se um agente
patogénico penetra no organismo, o sistema imune inato fornece uma resposta imediata,
mas não específica (Rauta et al., 2012). Se os patógenos resistem à resposta inata é
acionada a resposta adaptativa. Esta durante uma infeção é mais específica na
identificação do agente patogénico e possui ainda a capacidade de resposta sob a forma
de uma memória imunológica (Rauta et al., 2012). No nível celular, o sistema imunitário

59
é constituído por glóbulos brancos tais como macrófagos, neutrófilos e linfócitos. Uma
vez que aconteça uma lesão ou infeção eles são responsáveis pela iniciação de processos
como da fagocitose e a inflamação (Machado et al., 2015). O grupo de células fagocíticas
conhecidas como monócitos/macrófagos e neutrófilos é responsável pela produção de
uma variedade de mediadores celulares durante a inflamação, como por exemplo,
péptidos antibacterianos, fatores de complemento, citoquinas, proteínas de fase aguda
entre outros. Pele, brânquias e intestinos são os tecidos da mucosa com associação direta
com o sistema imunitário. As células fagocíticas produzem componente humorais,
capazes de inibir a adesão e liquidação dos microrganismos realizando um amplo espectro
de agentes antimicrobianos, tais como a lisozima, anti-proteases, espécies reativas de
oxigénio (ROS) e óxido nitroso (NO). A lisozima é uma enzima solúvel, originada a partir
de várias mucosas e células circundantes (Neutrófilos) e é capaz de atacar a camada de
peptidoglicano da parede celular bacteriana, levando-a a lise celular. Os fatores de
complemento também provocam a lise celular. Como medida para evitar a colonização
por parte dos agentes patogénicos o sistema imune inato também apresenta componente
solúveis denominados anti-proteases que são capazes de inibir as protéases libertadas
pelas bactérias limitando a digestão de proteínas (Machado et al., 2015).
O objetivo deste ensaio foi determinar o efeito da luz, com comprimentos de onda
diferentes, em vários parâmetros referentes ao desenvolvimento em juvenis de Solea
senagalensis, nomeadamente, o processo de crescimento, na pigmentação e indicadores
de stress, usando um fotoperíodo de 12 horas de luz e 12 horas de escuro.
Materiais e métodos
Desenho Experimental
Este ensaio realizou-se nas instalações da empresa Aquacria Piscícolas, SA,
sediada na Torreira, Aveiro, mais concretamente na antiga seção da pré-engorda da
empresa. O início do ensaio foi dia 8 de Maio e terminou dia 22 Junho, tendo um total de
46 dias. Os peixes foram cedidos pela empresa num total de 27 015 peixes com um
comprimento standard médio de (±SD) de 14,6±0,64 cm e um peso médio de 66±0,7g.
Os peixes foram mantidos durante algum tempo, antes do ensaio se iniciar, para estes se

60
aclimatarem às condições de produção. Durante a aclimatização e o ensaio, os peixes
foram mantidos em tanques com sistema de recirculação de água. Diariamente foram
feitas medições de vários parâmetros da água: salinidade de 20±0,4, temperatura de
20,1±1,2 °C, pH de 7,48±0,10, amónia 0,34±0,16 mg L−1, nitritos 0,118±0,042 mg L −1 e
por fim oxigénio dissolvido 12,3±1,63 mg L−1, estes parâmetros foram medidos com o
auxílio de sondas paramétricas e testes químicos. Utilizaram-se seis tanques, divididos
em três grupos, o RAQ1, RAQ2 e o RAQ3, cada um com dois tanques (Figura 20). Cada
tanque tem 15 metros de comprimento e 2,30 metros de largura, tendo uma área de
34,5m2.
Inicialmente foi feita a instalação das luzes e dos controladores (Aquaray
MultiControl 8), fornecido pela empresa TMC iberia. Cada tanque possuía três lâmpadas,
igualmente distribuídas, quer em distância quer em altura em todos os tanques que iam
ter tratamento luminoso. Com ajuda de um luxímetro (TES 1335 Light Meter) foi definida
a intensidade luminosa das luzes, fazendo os ajustes nos controladores. Depois de
realizados todos os ajustes a intensidade foi definida da seguinte forma: RAQ1 – A 209;
210; 210 lux, RAQ1 – B 209; 202; 193 lux, RAQ2 – A 189; 191; 186 lux, RAQ2 – B
193; 180; 204 lux e por fim o RAQ3 – A 279 lux e RAQ3 – B 0lux. O RAQ 1 teve o
tratamento com as luzes com comprimento de onda correspondente ao azul (450nm), o
RAQ 2 teve um comprimento de onda correspondente a um branco de baixa intensidade
(400nm) e por fim o RAQ 3, o controlo, que se assemelha às condições de produção, em
que este possuía lâmpadas por cima do tanque A sendo que estas estavam revestidas com
Figura 20- Esquema dos tanques

61
uma pelicula vermelha. Quanto ao fotoperíodo, as lâmpadas foram programadas para se
ligarem às 8h e se desligarem às 20h, fazendo por isso um ciclo de 12h de luz e um ciclo
de 12h de escuridão.
Inicialmente, em cada tanque experimental, o peixe foi mantido a diferentes
densidades, desde 10 a 6 kg m-2, em que os dois tanques do RAQ1 e o tanque A do RAQ2
apresentava a densidade máxima e o tanque B do RAQ3 tinha a densidade mais baixa. A
renovação de água foi mantida em 15,60 ± 4,98 L min-1 ou 0,9 ± 0,3 m3h-1, tendo um
volume total de 6555 litros de água salgada por cada tanque. Os peixes foram alimentados
utilizando alimentadores automáticos.
Amostragens
Durante o ensaio experimental, foram feitas duas amostragens. Antes do início,
uma amostragem inicial, dia 8 de Maio, em que foram recolhidos aleatoriamente 200
peixes de cada tanque e de cada tratamento. O procedimento consistia na pesca dos
mesmos para caixas, para serem posteriormente pesados e medidos com o auxílio de uma
balança e de uma caixa de esferovite graduada. Depois eram colocados noutra caixa para
não haver o risco de repetição da amostragem do mesmo peixe. A segunda amostragem
coincidiu com a final, dia 22 de Junho. Na medição apenas foi usado o tamanho standard.
Parâmetros estudados
Os parâmetros estudados foram agrupados em 5 categorias: avaliação do
crescimento – peso molhado, taxa de crescimento relativo, taxa de crescimento específica,
pigmentação, parâmetros hematológicos e resposta imune inata.

62
Avaliação do Crescimento
Este parâmetro, peso molhado (WW) é expresso em gramas. O WW é calculado
segundo esta fórmula:
WW (g) = (Wt – W0)
Em que W0 é o peso inicial do peixe em gramas, Wt é o peso final do peixe expresso
também em gramas.
O parâmetro, taxa de crescimento relativo (RGR) e a taxa de crescimento
específico (SGR) são calculados segundo a fórmula de Turker et al., 2005:
RGR (%) = % aumento em peso = [(Wt – W0)/W0] × 100
SGR (%/dia) = % aumento em peso por dia = [(ln Wt – ln W0)/ t] × 100
Em que t é os dias que se alimentou os peixes, W0 é o peso inicial expresso em gramas,
Wt peso final em gramas, ln(Wt) é o logaritmo natural do peso final do peixe e o ln(W0)
é o logaritmo natural do peso inicial do peixe.
Pigmentação
O sistema CIELAB proposto pela Comissão Internacional de Iluminação foi o
sistema aplicado para medir a cor da pigmentação da pele através do uso de um
colorímetro portátil (Konica Ninolta CR-400). Foram efetuadas nove medidas por peixe
de acordo com a Figura 21. Para realizar este teste, os animais foram mortos por
desconexão da coluna vertebral e rapidamente transferidos para gelo. Esta medição foi
feita na Escola Superior de Tecnologia do Mar em Peniche consistiu no disparo de um
feixe luminoso, que quando regressa ao aparelho fornece as características
tridimensionais da cor – brightness, hue e chroma.

63
Figura 21 - Locais na pele do peixe onde o colorímetro fez as medições
Parâmetros hematológicos e imunológicos
Todo o protocolo foi conduzido pelo CIIMAR - Centro Interdisciplinar de
Investigação Marinha e Ambiental em colaboração com a Aquacria Piscícolas S.A.
Análise Estatística
Todos os resultados são expressos como média±desvio padrão e foram analisados
com o programa SPSS. Os dados foram analisados quanto à normalidade e
homogeneidade da variância e, quando necessário, foram transformados antes de ser
tratados estatisticamente. Nível de significância estatística foi definido como P <0,05. Foi
ainda realizada uma análise multivariada para averiguar a relação entre os parâmetros
hematológicos e imunológicos.
Resultados
Crescimento
Para avaliar este parâmetro foi necessário recorrer a uma comparação com a curva
padrão para esta espécie, propriedade da SEA 8. Inicialmente, com a ajuda de análises
estatísticas foram realizados alguns testes para ver se existiam diferenças entre os tanques
com o mesmo tratamento (Tabela XIII). Existem diferenças estatisticamente

64
significativas entre tanques A e B com o mesmo tratamento de luz. O tanque A com o
espectro azul mostra uma média final de 113,8±34,9g, enquanto o tanque B tem uma
média final de 104,5±37,3g. A mesma situação acontece no espectro branco. No grupo
controlo não existem diferenças estatisticamente significativas entre os tanques.
Tabela XIII- Comparação do peso médio final (g) entre tanques com o mesmo regime luminoso.
Tanque
A B
Azul 113,8±34,9 a 104,5±37,3 b
Branco 100,8±43,3 a 78,7±34,2 b
Controlo 90,7±36,4 a 97,6±36,6 a
Valores são expressos como média±SD (n=200). Letras diferentes significam diferenças estatisticamente
significativas entre os tratamentos de luz (one-way ANOVA, p ≤ 0,05).
De seguida, foi feita a análise individualizada por tanques, separando o tanque A
do B. Foi feita a análise estatística, para cada tanque A por tratamento e o mesmo para
cada tanque B. Existem diferenças estatisticamente significativas entre todos os
tratamentos luminosos para o tanque A, sendo que o tanque A da luz azul tem um peso
médio de 113,80±34,94g, sendo este o tanque que possui um peso médio superior, por
outro lado o Tanque A do Controlo é aquele que possui o peso médio inferior
90,72±36,40g. Na análise aos tanques B existem diferenças estatisticamente significativas
entre tratamentos luminosos contudo, entre azul e controlo, não existem diferenças
estatisticamente significativas sendo que o tanque com a luz azul tem um peso médio de
104,49±37,30g e o tanque do Controlo tem um peso médio de 97,59±36,61g (Tabela
XIV).

65
Tabela XIV- Pesos médios finais comparando tratamentos, individualizando os tanques A e B de
cada regime luminoso (Azul, Branco e Controlo).
Tanque
A
Azul Branco
Tanque
B
Azul Branco
113,80±34,94a 100,85±43,29b 104,49±37,30a 78,71±34,21b
Azul Controlo Azul Controlo
113,80±34,94a 90,72±36,40b 104,49±37,30a 97,59±36,61a
Branco Controlo Branco Controlo
100,85±43,29a 90,72±36,40b 78,71±34,31a 97,59±36,61b
Valores são expressos como média±SD (n=200). Letras diferentes significam diferenças estatisticamente
significativas entre os tratamentos de luz (one-way ANOVA, p ≤ 0.05).
Com esta análise dos resultados obtidos, foi realizada a comparação com a curva
modelo da SEA 8, analisando a soma dos tanques (Tabela XV) e mais detalhadamente
tanque por tanque (Tabela XVI). É possível concluir que quando se comparam as taxas
de crescimento da soma dos tanques com tratamento azul, branco e o controlo, com a
curva modelo os valores obtidos são inferiores, no caso do azul é de 99%, no branco de
98% e no controlo de 97%.
Tabela XV- Comparação da soma dos tanques com a curva modelo da SEA 8, agrupando por
tratamento (Azul, Branco e Controlo).
TRATAMENTO NÚMERO DE
PEIXES
PESO MÉDIO
(G)
BIOMASSA
(KG)
CRESCIMENTO
(%)
AZUL 9 442 108,98 1 029 99%
BRANCO 10 726 90,02 966 98%
CONTROLO 6 768 94,16 637 97%
Na Tabela XVI, estão descritos os tanques individualizados, em que as melhores
taxas de crescimento apresentam percentagens superiores a 100%, significando
crescimentos superiores aos da curva modelo, isto verificou-se em todos os tanques A,
por sua vez, os tanques B obtiveram resultados inferiores do que a curva modelo, sendo
que o tanque B do tratamento com a luz branca e do controlo foram aqueles onde se
observaram taxas inferiores, 83% e 87%, respetivamente.
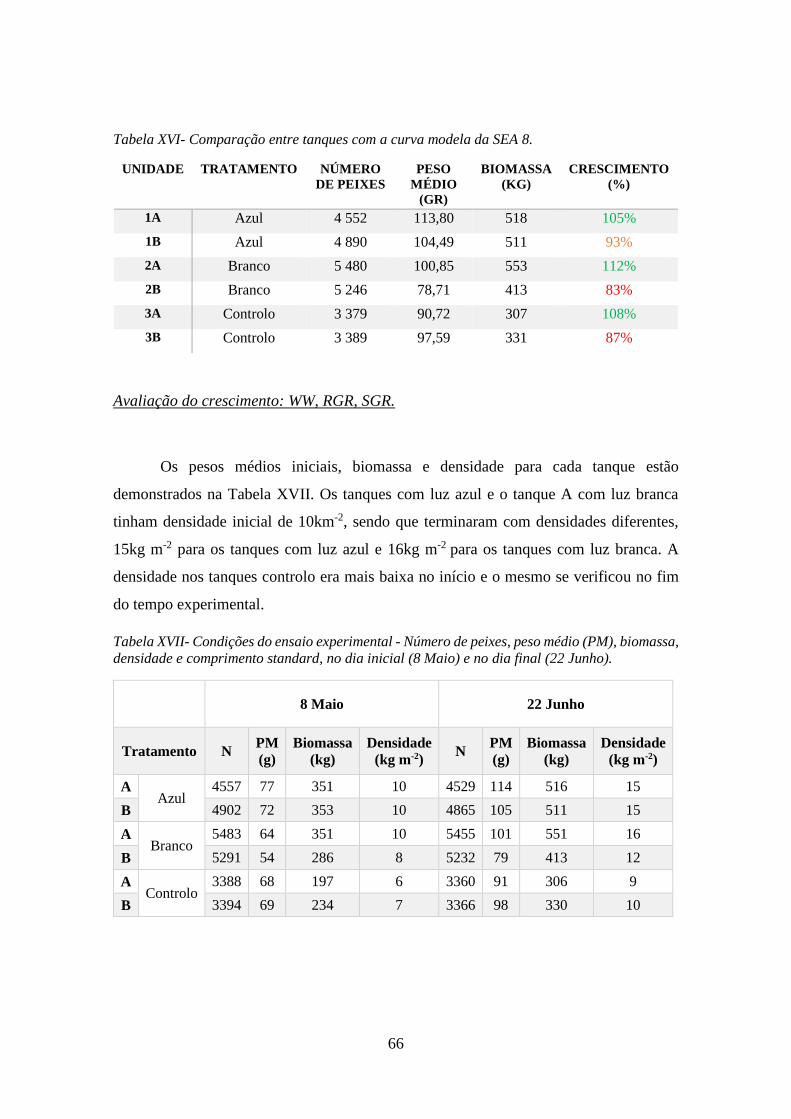
66
Tabela XVI- Comparação entre tanques com a curva modela da SEA 8.
UNIDADE TRATAMENTO NÚMERO
DE PEIXES
PESO
MÉDIO
(GR)
BIOMASSA
(KG)
CRESCIMENTO
(%)
1A Azul 4 552 113,80 518 105%
1B Azul 4 890 104,49 511 93%
2A Branco 5 480 100,85 553 112%
2B Branco 5 246 78,71 413 83%
3A Controlo 3 379 90,72 307 108%
3B Controlo 3 389 97,59 331 87%
Avaliação do crescimento: WW, RGR, SGR.
Os pesos médios iniciais, biomassa e densidade para cada tanque estão
demonstrados na Tabela XVII. Os tanques com luz azul e o tanque A com luz branca
tinham densidade inicial de 10km-2, sendo que terminaram com densidades diferentes,
15kg m-2 para os tanques com luz azul e 16kg m-2 para os tanques com luz branca. A
densidade nos tanques controlo era mais baixa no início e o mesmo se verificou no fim
do tempo experimental.
Tabela XVII- Condições do ensaio experimental - Número de peixes, peso médio (PM), biomassa,
densidade e comprimento standard, no dia inicial (8 Maio) e no dia final (22 Junho).
8 Maio 22 Junho
Tratamento N PM
(g)
Biomassa
(kg)
Densidade
(kg m-2) N
PM
(g)
Biomassa
(kg)
Densidade
(kg m-2)
A Azul
4557 77 351 10 4529 114 516 15
B 4902 72 353 10 4865 105 511 15
A Branco
5483 64 351 10 5455 101 551 16
B 5291 54 286 8 5232 79 413 12
A Controlo
3388 68 197 6 3360 91 306 9
B 3394 69 234 7 3366 98 330 10
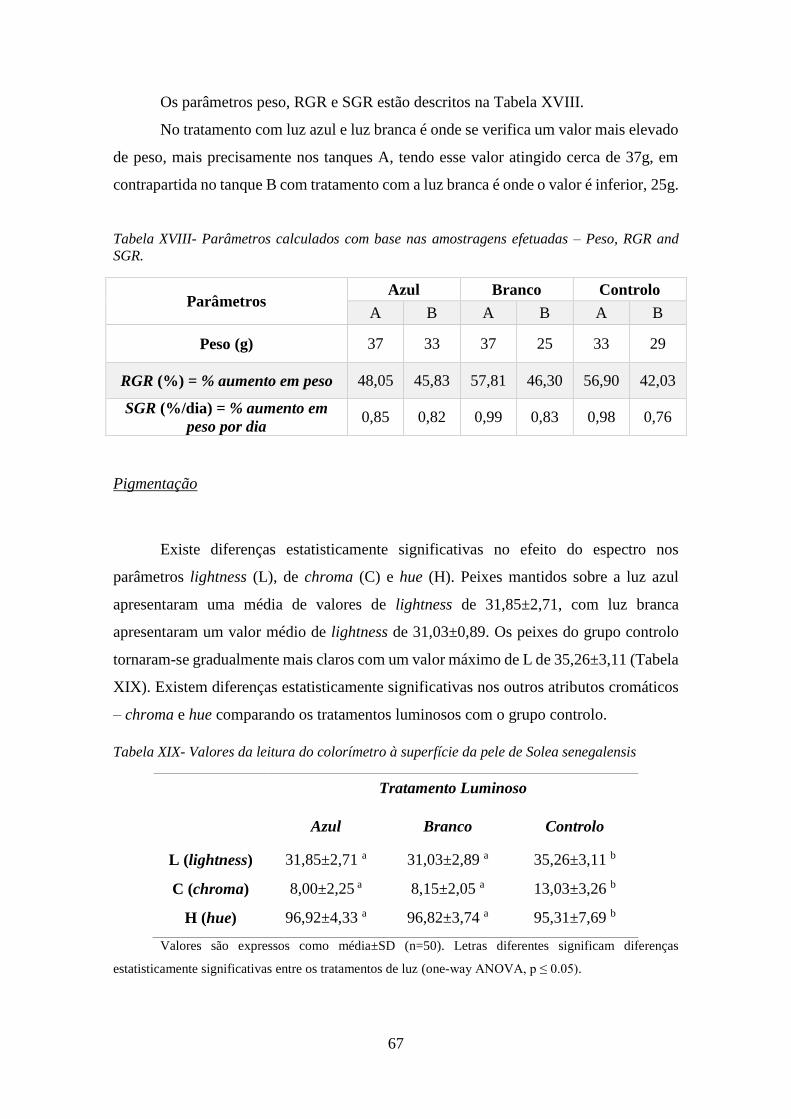
67
Os parâmetros peso, RGR e SGR estão descritos na Tabela XVIII.
No tratamento com luz azul e luz branca é onde se verifica um valor mais elevado
de peso, mais precisamente nos tanques A, tendo esse valor atingido cerca de 37g, em
contrapartida no tanque B com tratamento com a luz branca é onde o valor é inferior, 25g.
Tabela XVIII- Parâmetros calculados com base nas amostragens efetuadas – Peso, RGR and
SGR.
Parâmetros Azul Branco Controlo
A B A B A B
Peso (g) 37 33 37 25 33 29
RGR (%) = % aumento em peso 48,05 45,83 57,81 46,30 56,90 42,03
SGR (%/dia) = % aumento em
peso por dia 0,85 0,82 0,99 0,83 0,98 0,76
Pigmentação
Existe diferenças estatisticamente significativas no efeito do espectro nos
parâmetros lightness (L), de chroma (C) e hue (H). Peixes mantidos sobre a luz azul
apresentaram uma média de valores de lightness de 31,85±2,71, com luz branca
apresentaram um valor médio de lightness de 31,03±0,89. Os peixes do grupo controlo
tornaram-se gradualmente mais claros com um valor máximo de L de 35,26±3,11 (Tabela
XIX). Existem diferenças estatisticamente significativas nos outros atributos cromáticos
– chroma e hue comparando os tratamentos luminosos com o grupo controlo.
Tabela XIX- Valores da leitura do colorímetro à superfície da pele de Solea senegalensis
Tratamento Luminoso
Azul Branco Controlo
L (lightness) 31,85±2,71 a 31,03±2,89 a 35,26±3,11 b
C (chroma) 8,00±2,25 a 8,15±2,05 a 13,03±3,26 b
H (hue) 96,92±4,33 a 96,82±3,74 a 95,31±7,69 b
Valores são expressos como média±SD (n=50). Letras diferentes significam diferenças
estatisticamente significativas entre os tratamentos de luz (one-way ANOVA, p ≤ 0.05).
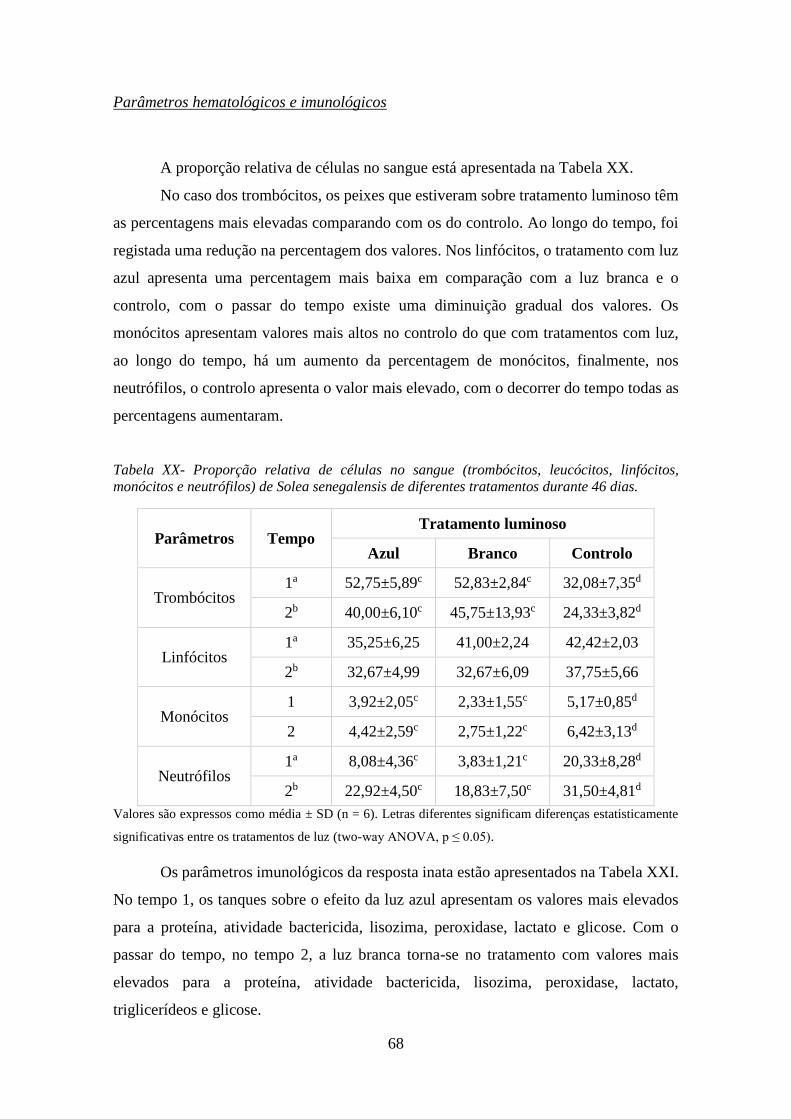
68
Parâmetros hematológicos e imunológicos
A proporção relativa de células no sangue está apresentada na Tabela XX.
No caso dos trombócitos, os peixes que estiveram sobre tratamento luminoso têm
as percentagens mais elevadas comparando com os do controlo. Ao longo do tempo, foi
registada uma redução na percentagem dos valores. Nos linfócitos, o tratamento com luz
azul apresenta uma percentagem mais baixa em comparação com a luz branca e o
controlo, com o passar do tempo existe uma diminuição gradual dos valores. Os
monócitos apresentam valores mais altos no controlo do que com tratamentos com luz,
ao longo do tempo, há um aumento da percentagem de monócitos, finalmente, nos
neutrófilos, o controlo apresenta o valor mais elevado, com o decorrer do tempo todas as
percentagens aumentaram.
Tabela XX- Proporção relativa de células no sangue (trombócitos, leucócitos, linfócitos,
monócitos e neutrófilos) de Solea senegalensis de diferentes tratamentos durante 46 dias.
Parâmetros Tempo Tratamento luminoso
Azul Branco Controlo
Trombócitos 1a 52,75±5,89c 52,83±2,84c 32,08±7,35d
2b 40,00±6,10c 45,75±13,93c 24,33±3,82d
Linfócitos 1a 35,25±6,25 41,00±2,24 42,42±2,03
2b 32,67±4,99 32,67±6,09 37,75±5,66
Monócitos 1 3,92±2,05c 2,33±1,55c 5,17±0,85d
2 4,42±2,59c 2,75±1,22c 6,42±3,13d
Neutrófilos 1a 8,08±4,36c 3,83±1,21c 20,33±8,28d
2b 22,92±4,50c 18,83±7,50c 31,50±4,81d
Valores são expressos como média ± SD (n = 6). Letras diferentes significam diferenças estatisticamente
significativas entre os tratamentos de luz (two-way ANOVA, p ≤ 0.05).
Os parâmetros imunológicos da resposta inata estão apresentados na Tabela XXI.
No tempo 1, os tanques sobre o efeito da luz azul apresentam os valores mais elevados
para a proteína, atividade bactericida, lisozima, peroxidase, lactato e glicose. Com o
passar do tempo, no tempo 2, a luz branca torna-se no tratamento com valores mais
elevados para a proteína, atividade bactericida, lisozima, peroxidase, lactato,
triglicerídeos e glicose.
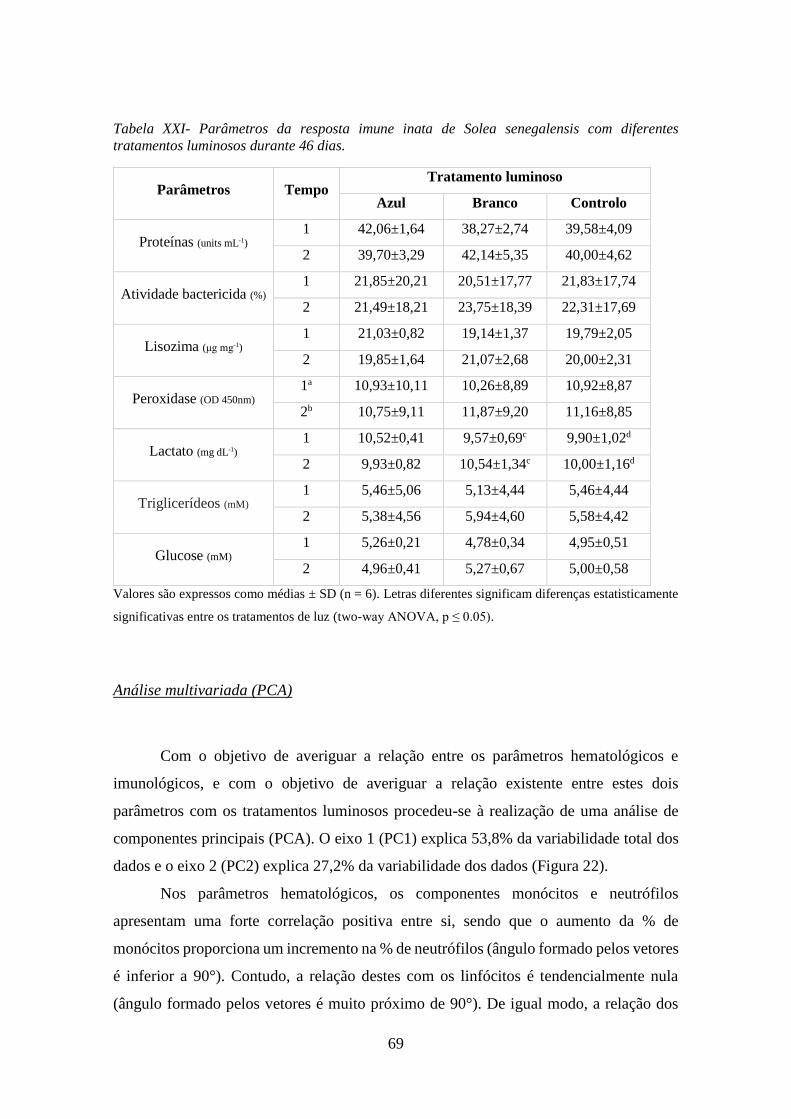
69
Tabela XXI- Parâmetros da resposta imune inata de Solea senegalensis com diferentes
tratamentos luminosos durante 46 dias.
Parâmetros Tempo Tratamento luminoso
Azul Branco Controlo
Proteínas (units mL-1) 1 42,06±1,64 38,27±2,74 39,58±4,09
2 39,70±3,29 42,14±5,35 40,00±4,62
Atividade bactericida (%) 1 21,85±20,21 20,51±17,77 21,83±17,74
2 21,49±18,21 23,75±18,39 22,31±17,69
Lisozima (μg mg-1) 1 21,03±0,82 19,14±1,37 19,79±2,05
2 19,85±1,64 21,07±2,68 20,00±2,31
Peroxidase (OD 450nm) 1a 10,93±10,11 10,26±8,89 10,92±8,87
2b 10,75±9,11 11,87±9,20 11,16±8,85
Lactato (mg dL-1) 1 10,52±0,41 9,57±0,69c 9,90±1,02d
2 9,93±0,82 10,54±1,34c 10,00±1,16d
Triglicerídeos (mM) 1 5,46±5,06 5,13±4,44 5,46±4,44
2 5,38±4,56 5,94±4,60 5,58±4,42
Glucose (mM) 1 5,26±0,21 4,78±0,34 4,95±0,51
2 4,96±0,41 5,27±0,67 5,00±0,58
Valores são expressos como médias ± SD (n = 6). Letras diferentes significam diferenças estatisticamente
significativas entre os tratamentos de luz (two-way ANOVA, p ≤ 0.05).
Análise multivariada (PCA)
Com o objetivo de averiguar a relação entre os parâmetros hematológicos e
imunológicos, e com o objetivo de averiguar a relação existente entre estes dois
parâmetros com os tratamentos luminosos procedeu-se à realização de uma análise de
componentes principais (PCA). O eixo 1 (PC1) explica 53,8% da variabilidade total dos
dados e o eixo 2 (PC2) explica 27,2% da variabilidade dos dados (Figura 22).
Nos parâmetros hematológicos, os componentes monócitos e neutrófilos
apresentam uma forte correlação positiva entre si, sendo que o aumento da % de
monócitos proporciona um incremento na % de neutrófilos (ângulo formado pelos vetores
é inferior a 90°). Contudo, a relação destes com os linfócitos é tendencialmente nula
(ângulo formado pelos vetores é muito próximo de 90°). De igual modo, a relação dos

70
trombócitos com os linfócitos é também praticamente nula. Ao analisar a relação dos
trombócitos com monócitos e neutrófilos, esta é inversa, sendo que o aumento na % dos
trombócitos implica uma diminuição na % do par monócitos e neutrófilos (ângulo
formado pelos vetores é superior a 90°).
Os indivíduos associados às amostras espécie-tratamento dos tanques dos
tratamentos com a luz azul e branca (B1; B2 e W1; W2) são caracterizados por apresentar
altos valores de trombócitos enquanto os tanques do controlo (C1 e C2) apresentam altos
valores para linfócitos, monócitos e neutrófilos.
Nos parâmetros imunológicos os componentes lisozima e triglicéridos apresentam
uma correlação forte positiva, enquanto com a peroxidase e a glucose estes têm uma
correlação praticamente nula. O lactato apresenta uma correlação positiva com a
peroxidase e com a lisozima, contudo o lactato apresenta uma correlação inversa com a
atividade bacteriana, sendo que esta última também apresenta correlação inversa com a
lisozima e com os triglicéridos. No que concerne à proteína esta apresenta uma correlação
nula com o lactato, sendo que possui uma correlação inversa com a glucose.
Os indivíduos associados às amostras espécie-tratamento dos tanques dos
tratamentos com a luz Azul e Branca (B1; B2 e W1; W2) são caracterizados por
apresentar altos valores de proteínas, triglicéridos, lisozima, lactato e peroxidase. Para os
tanques do controlo (C1; C2) estes são caracterizados por apresentar altos valores de
atividade bacteriana.

71
Figura 22- Diagrama da análise de componentes principais (ACP) para parâmetros
hematológicos e imunológicos para os diferentes tratamentos luminosos. Cada vetor representa
um parâmetro hematológico (NEUT – Neutrófilos; MON – Monócitos; THROM – trombócitos e
LYM – Linfócitos) e imunológico (Prot – Proteínas; Trigl – triglicéridos; Lisoz – Lisozima; Lact
– Lactato; ActB – Atividade Bacteriana; Perox – Peroxidase e Gluc – Glucose), e cada ponto
representa os tanques (W1 – Tratamento com Luz branca no tempo 1, W2 – tratamento com luz
branca no tempo 2; B1 – Tratamento com luz azul no tempo 1, B2 – tratamento com luz azul no
tempo 2; C1 – Controlo no tempo 1, C2 – Controlo no tempo 2) com diferentes tratamentos
luminosos

72
Discussão
Crescimento
Ao longo do tempo, todas as aquaculturas tentam maximizar a sua rentabilidade,
testando para tal vários parâmetros que possam controlar para que isso tenha um impacto
positivo na produção. A escolha deste parâmetro deveu-se à escassez de artigos científicos
específicos para esta espécie Solea senegalensis (Villamizar et al., 2011). Uma vez que a
única publicação com este parâmetro se centrava nas larvas desenvolveu-se um protocolo
para averiguar o efeito em juvenis de Solea senegalensis.
Sabendo que a produção de linguado apresenta vários desafios, entre as quais a
dispersão de tamanho (Dinis et al., 1999). Neste ensaio experimental foi evidenciado esse
mesmo desafio, uma vez que os pesos médios e as densidades iniciais verificadas em cada
tanque eram diferentes (Tabela XVII) sendo que estes pontos podem ter influenciado as
diferentes taxas de crescimento.
Dentro de cada tratamento (Azul, Branco) existem diferenças estatisticamente
significativas entre os tanques A e B. No controlo, não existem diferenças
estatisticamente significativas (Tabela XIII). Sendo que as lâmpadas estão igualmente
distribuídas por todos os tanques, tanto horizontalmente como verticalmente as diferenças
podem ser devido ao próprio design dos tanques e da canalização, uma vez que o
movimento da água é feito pelo início do tanque A, percorre todo o comprimento do
tanque, levando consigo todos os produtos de excreção presente na água, e por meio de
tubagens essa mesma água entra diretamente no tanque B, percorrendo todo o
comprimento do tanque B arrastando consigo todos os produtos de excreção do tanque A
e do tanque B. A zona de tratamento de água situa-se no final do tanque B. Isto pode
explicar a diferença (estatisticamente significativa) entre tanques A e B com o mesmo
tratamento luminoso. Já para o controlo, existe o mesmo design de tanque e tubagens do
que nos outros tratamentos. A única diferença, é o facto de os peixes sobre efeito das
luzes azul e brancas apresentarem muito mais atividade com um padrão natatório anormal
fazendo com que os produtos de excreção sejam muito superiores do que os peixes
presentes no controlo, que apresentam o comportamento normal para esta espécie,
excretando por isso menos, sendo que estes produtos vão ter menor efeito dos peixes dos
tanques B. Analisando os tanques individualmente, como podemos ver na Tabela XIV,

73
existem diferenças estatisticamente significativas, praticamente entre todos os tanques,
menos entre tanques B da luz azul e do controlo. Nos tanques A existem diferenças
estatisticamente significativas entre todos, contudo os peixes que possuem um peso médio
superior são os peixes do tanque azul e os peixes do controlo são aqueles que apresentam
um peso médio inferior, em relação ao tanque B, novamente os peixes com a luz azul
apresentam um peso médio superior enquanto os peixes do tanque com a luz branca
apresentam um peso médio inferior (Tabela XIV). Como referi, esta análise não é muito
fiável, uma vez que os pesos iniciais em todos os tanques diferia bastante (Tabela XVII),
levando a diferenças no metabolismo dos peixes, causando impacto nas taxas de
crescimento. Para que este erro fosse atenuado, foi realizada uma comparação de
tratamentos com a curva modelo de Solea senegalensis e os resultados estão expressos
nas Tabelas XV e XVI. Como se verifica na Tabela XV as taxas de crescimentos são
inferiores a 100%, isto significa que os peixes tiveram um crescimento inferior ao valor
dado pela curva modelo. Isto porque as taxas de crescimento referenciadas na Tabela XV
advém das somas dos dois tanques por tratamento. Como anteriormente, obtém-se
diferenças estatisticamente significativas entre tanques com o mesmo tratamento, foi feita
a comparação individualizando os tanques com a curva modelo (Tabela XVI). Nesta
Tabela, podemos concluir que todos os tanques A possuem uma taxa de crescimento
superior a 100%, sendo que isso significa que os peixes cresceram mais do que o valor
dado pela curva modelo, contudo, os tanques B, mais uma vez, tiveram uma performance
muito inferior (chegando a valores de 83% no tanque B com luz branca), isto vai de
acordo com a análise feita anteriormente (Tabela XIII). Tendo sido verificada esta
diferença entre tanques A e B, a Tabela XV que se baseia na soma dos dois tanques por
tratamento, é normal que tenha uma taxa de crescimento inferior ao valor dado pela curva
modela, pois todos os tanques B em todos os tratamentos ficam muito abaixo do valor
teórico, o que se esperava devido ao problema identificado do design dos tanques e das
tubagens.
Avaliação do crescimento WW, RGR, SGR
O parâmetro WW dos juvenis de Solea senegalensis é calculado através da
diferença entre peso inicial e final (Tabela XVIII). Como já foi referido acima, um
problema associado à produção de linguado é a dispersão dos tamanhos (Dinis et al.,1999)

74
e aqui mais uma vez, foi visível a influência desse facto. Da mesma forma acima referido,
a diferença no metabolismo associado a diferentes tamanhos faz com que este parâmetro
não seja semelhante entre tratamentos. Contudo, as mesmas tendências são verificadas,
os tanques A possuem um ganho de peso médio superior em comparação com os tanques
B. Contudo pode-se ver que com tratamento luminoso (Azul e Branco) existe um aumento
de peso médio superior, isto pode significar que talvez exista uma tendência para que
estes sobre tratamento luminoso possam crescer mais do que aqueles que estão no
controlo. Para retirar qualquer dúvida sobre o efeito, deveria ser realizada uma melhoria
do protocolo, sendo que certas medidas como, peso inicial e duração do ensaio deveria
ser alterado.
A taxa de crescimento relativo (RGR) e a tava de crescimento específico (SGR)
nos juvenis de Solea senegalensis, entre tratamentos, não difere de uma forma
significativa. O mesmo padrão verificado em cima existe também nestes dois parâmetros,
ou seja os tanques A tem valores superiores comparando com os tanques B dentro de cada
tratamento luminoso.
Estes parâmetros foram calculados na tentativa de perceber qual tratamento
luminoso potenciaria o crescimento dos juvenis de linguado, sendo que a Luz Branca, é
aquela que tem um SGR superior, ou seja tem o valor percentual de incremento de peso
por dia (0,99%) superior, mas mesmo este parâmetro pode ser influenciado pela diferença
de pesos iniciais, uma vez que essa diferença possa colocar os peixes em diferentes
estágios de crescimento.
Pigmentação
Os peixes criados em aquacultura são alvo de muitos preconceitos por parte do
consumidor, desde o teor em gordura até à cor da pele tendo por isso diferenças externas
comparativamente com os peixes do estado selvagem. Nesse sentido, surgiu a ideia de
testar luzes de diferentes comprimentos de onda, na tentativa de tornar o peixe de
aquacultura mais claro, idêntico ao selvagem, trazendo vantagens económicas para a
empresa que os produz. O mesmo se verifica noutra espécie, Pagrus pagrus, em que foi
testado o efeito da luz azul na pigmentação da pele do pargo (Szisch et al., 2002). A
realização deste ensaio iniciou-se pelas 6 horas da manhã com o abate de 150 peixes. O
abate foi feito pela desconexão da coluna vertebral. Esta técnica foi usada para sacrificar

75
os peixes por se pensar ser a maneira mais rápida minimizando assim o stress vivenciado
pelos peixes. A forma de abate foi bastante importante porque os peixes têm a capacidade
de alterar a sua coloração em resposta às condições ambientais ou condições stressantess
(Szisch et al., 2002; Van der Salm et al., 2004) uma vez que estas são controladas pelo
sistema endócrino e/ou sistema nervoso. Os resultados demonstram que existem
diferenças estatisticamente significativas entre os tratamentos luminosos e o controlo em
todos os parâmetros calculados (L, H e C). O parâmetro, hue é uma das principais
propriedades da cor, sendo que este pode assumir valores entre 0° a 360°, assumindo
cores como vermelho, verde, azul e amarelo. O parâmetro chroma é a saturação das
distintas tonalidades, sendo que assume valores de 0% a 100%. Por fim o parâmetro
lightness é a dimensão da cor de um objeto, através da qual o objeto parece refletir ou
transmitir mais ou menos a luz incidente, sendo que os valores variam de 0% a 100%,
sendo que valores superiores são mais claros. Para averiguar se a pele do Solea
senegalensis se torna mais clara, tornando-o mais vendável, o parâmetro mais importante
será o L (lightness), pois é este que avalia o brilho da cor.
Analisando a Tabela XIX podemos ver que existem diferenças estatisticamente
significativas entre tratamentos e o controlo para o parâmetro L, sendo que isso significa
que os tratamentos luminosos (azul e branco), como possuem valores inferiores
comparativamente ao controlo, são mais escuros do que os peixes do controlo.
Comparativamente, os nossos dados são contraditórios aos obtidos no ensaio do
pargo (Szisch et al., 2002), em que foi observado uma alteração na coloração do peixe
tornando-o mais claro usando a luz azul, com o linguado, essa tendência não foi
observada.
Como os dados não são concordantes com o trabalho de Szisch et al., 2002, podem
ser efetuados alguns ajustes ao protocolo desenvolvido nomeadamente a forma de
sacrificar o peixe, desde as horas a que foi feito, uma vez que a essa hora não existe
trabalho na zona, podendo ser por isso um motivo de stress que possa alterar a coloração
da pele. Como a pele dos linguados apresenta manchas de tonalidades diferente a medição
com o colorímetro pode ser falível uma vez que o feixe de luz pode medir diferentes
tonalidades, para evitar esse erro, uma medição de melanina deveria ter sido feita pois
esta é mais fiável e possui menos erros associados.

76
Parâmetros hematológicos e imunológicos
Com este ensaio, foi estudado o efeito da luz de diferentes comprimentos de onda,
no crescimento, contudo não se sabe o efeito no bem-estar nos linguados. Na tentativa de
eliminar estas dúvidas foi feita uma colheita de sangue para analisar parâmetros
biológicos tais como, parâmetros hematológicos e imunológicos. Neste estudo, e
analisando os parâmetros hematológicos, a percentagem de trombócitos, linfócitos e
neutrófilos varia estatisticamente significativamente durante o tempo do ensaio, ou seja
entre o tempo 1 e o tempo 2, sendo que nos trombócitos e linfócitos existe um diminuir
da percentagem relativa, enquanto nos neutrófilos existe um aumento. Entre tratamentos
luminosos existem diferenças estatisticamente significativas nos trombócitos, monócitos
e neutrófilos.
A composição dos glóbulos brancos é caracterizada pela predominância dos
trombócitos e linfócitos (Tabela XX). Contudo os valores de trombócitos dos peixes
presentes nos tanques com tratamento luminoso azul e branco, no tempo 1, tem valores
percentuais superiores aos valores de referência (Peres et al., 2015) para juvenis de Solea
senegalensis, o mesmo não se passa para os linfócitos. Já para os monócitos e neutrófilos
os peixes presentes no tanque de controlo e no tanque com luz azul apresentam valores
percentuais superiores aos valores de referência (Peres et al., 2015), contudo os tanques
com luz branca apresentam valores inferiores.
Os elevados valores percentuais de linfócitos e monócitos no tempo 1 sugere que
os peixes estariam sobre ação de agentes stressantes. A diminuição verificada ao longo
do tempo pode ser devida ao extravasamento destas células e a sua penetração no epitélio
das brânquias, da pele e do intestino (Wendelaar-Bonga, 1997). Diminuição da contagem
de linfócitos também foi observada em peixes famintos, sugerindo que a redução foi
causada por má nutrição (Rios et al., 2005).
Analisando os parâmetros imunológicos na tabela XXI, existem diferenças
estatisticamente significativas apenas na peroxidase ao longo do tempo e no lactato entre
o tanque com a luz branca e o controlo.
Em Solea senegalensis foi relatado que consegue manter a homeostasia da glucose
plasmática durante períodos de fome prolongada (até 21 dias) (Costas et al., 2011), devido
a esse facto, este parâmetro ganha um interesse extra como indicador fisiológico da
condição nutricional e stress. No presente estudo, os valores de glucose são superiores

77
aos valores de referência para esta espécie (Peres et al., 2015). Isto pode sugerir que os
peixes estão sobre condições stressantes. Como não existem diferenças estatisticamente
significativas entre tratamentos luminosos, o fator “luz” pode não ser o principal agente
stressante, levando para problemas nutricionais, como o jejum.
Em suma, verificou-se que a luz branca (400nm) é aquela que promove uma
melhor taxa de crescimento comparativamente ao controlo, no entanto, quanto à
coloração este tratamento não foi o indicado para obter uma coloração mais clara, tendo
sido verificado que mesmo assim o controlo é o melhor tratamento no caso da
pigmentação. Na análise feita aos parâmetros hematológicos e imunológicos conclui-se
que a diminuição dos valores de trombócitos e linfócitos, verificada em todos tratamentos,
ao longo do tempo deve-se á migração dessas mesmas células para outros tecidos o que
pressupõe um indício de stress. Contudo os valores de monócitos e neutrófilos parecem
mais concordantes com os valores de referência para esta espécie na luz branca.
Contudo neste trabalho não se conseguiu obter respostas concretas no que diz
respeito à influência da luz no crescimento, e uma das razões para tal, foi a discrepância
de médias iniciais no peso dos peixes o que levou a articular de forma diferente a
interpretação dos dados. Sendo assim, este estudo deveria realizar-se novamente com
pesos médios homogeneizados entre tanques e por um período de tempo mais extenso.
Quanto à coloração da pele presume-se que a forma de abate possa ter alguma influência
no resultado assim como a análise poderia ser mais precisa com a quantificação de
melanina.
Conclusão
O período de estágio que decorreu durante 9 meses (1 de Setembro de 2014 até
dia 30 de Junho de 2015), apesar de ter sido muito trabalhoso foi uma experiência muito
enriquecedora quer profissionalmente como pessoalmente. Foram-me transmitidas todas
as técnicas usadas para a produção de linguado em regime intensivo, como quer todas as
funções de cada equipamento instalado na produção.
Este estágio ainda me possibilitou muitas outras experiências enriquecedoras, quer
a presença em palestras em vários pontos do país, vários workshops e a presença e
participação em vários projetos inovadores em que a empresa participa, sendo que no

78
final deste período de estágio trabalhei não só com linguado mas também com ostras,
microalgas, douradas, robalos e plantas halófitas.
Diariamente, realizei todas as rotinas existentes da empresa, e fiquei a conhecer
melhor o sistema de produção utilizado pela empresa e aprendi mais sobre a espécie
produzida.
Tudo isto vivido num ambiente amigável e divertido, sempre acompanhado de
boa disposição e de muita solidariedade. Posso por isso afirmar que o objetivo do meu
estágio foi cumprido.

79
Referências
Acerete, L., J. C. Balasch, E. Espinosa, A. Josa, and L. Tort, 2004, Physiological
responses in Eurasian perch (Perca fluviatilis, L.) subjected to stress by transport
and handling: Aquaculture, v. 237, p. 167-178.
Andrade, T., A. Afonso, A. Pérez-Jiménez, A. Oliva-Teles, V. de las Heras, J. M.
Mancera, R. Serradeiro, and B. Costas, 2015, Evaluation of different stocking
densities in a Senegalese sole (Solea senegalensis) farm: Implications for growth,
humoral immune parameters and oxidative status: Aquaculture, v. 438, p. 6-11.
Anguis, V., and J. P. Canavate, 2005, Spawning of captive Senegal sole (Solea
senegalensis) under a naturally fluctuating temperature regime: Aquaculture, v.
243, p. 133-145.
Arjona, F. J., I. Ruiz-Jarabo, L. Vargas-Chacoff, M. P. Martin del Rio, G. Flik, J. M.
Mancera, and P. H. M. Klaren, 2010, Acclimation of Solea senegalensis to
different ambient temperatures: implications for thyroidal status and
osmoregulation: Marine Biology, v. 157, p. 1325-1335.
Arjona, F. J., L. Vargas-Chacoff, I. Ruiz-Jarabo, O. Goncalves, I. Pascoa, M. P. Martin
del Rio, and J. M. Mancera, 2009, Tertiary stress responses in Senegalese sole
(Solea senegalensis Kaup, 1858) to osmotic challenge: Implications for
osmoregulation, energy metabolism and growth: Aquaculture, v. 287, p. 419-426.
Arjona, F. J., L. Vargas-Chacoff, I. Ruiz-Jarabo, M. P. Martin del Rio, and J. M. Mancera,
2007, Osmoregulatory response of Senegalese sole (Solea senegalensis) to
changes in environmental salinity: Comparative Biochemistry and Physiology a-
Molecular & Integrative Physiology, v. 148, p. 413-421.
Arlinghaus, R., S. J. Cooke, A. Schwab, and I. G. Cowx, 2007, Fish welfare: a challenge
to the feelings-based approach, with implications for recreational fishing: Fish and
Fisheries, v. 8, p. 57-71.
Ashley, P. J., 2007, Fish welfare: Current issues in aquaculture: Applied Animal
Behaviour Science, v. 104, p. 199-235.
Atoum, Y., S. Srivastava, and X. Liu, 2015, Automatic feeding control for dense
aquaculture fish tanks: IEEE Signal Processing Letters, v. 22, p. 1089-1093.
Barber, I., 2007, Parasites, behaviour and welfare in fish: Applied Animal Behaviour
Science, v. 104, p. 251-264.
Barron, J. M., N. R. Jensen, P. J. Anders, J. P. Egan, and K. D. Cain, 2013, Suppression
of Cannibalism during Larviculture of Burbot through Size Grading: North
American Journal of Aquaculture, v. 75, p. 556.
Barton, B. A., 2000, Salmonid fishes differ in their cortisol and glucose responses to
handling and transport stress: North American Journal of Aquaculture, v. 62, p.
12-18.
Barton, B. A., 2002, Stress in fishes: A diversity of responses with particular reference to
changes in circulating corticosteroids: Integrative and Comparative Biology, v.
42, p. 517-525.
Barton, B. A., and R. E. Peter, 1982, Plasma cortisol stress response in fingerling rainbow
trout, Salmo gairdneri Richardson, to various transport conditions, anaesthesia,
and cold shock: Journal of Fish Biology, v. 20, p. 39-51.
Bermejo-Poza, R., J. De la Fuente, C. Perez, S. Lauzurica, E. Gonzalez, M. T. Diaz, and
M. Villarroel, 2015, The effect of intermittent feeding on the pre-slaughter fasting
response in rainbow trout: Aquaculture, v. 443, p. 24-30.

80
Blanco-Vives, B., M. Aliaga-Guerrero, J. P. Canavate, G. Garcia-Mateos, A. J. Martin-
Robles, P. Herrera-Perez, J. A. Munoz-Cueto, and F. J. Sanchez-Vazquez, 2012,
Metamorphosis Induces a Light-Dependent Switch in Senegalese Sole (Solea
senegalensis) from Diurnal to Nocturnal Behavior: Journal of Biological
Rhythms, v. 27, p. 135-144.
Bravo, André. Revista de Marinha, edição nº 984 Março/Abril, 2015
Boglino, A., M. J. Darias, A. Estévez, K. B. Andree, and E. Gisbert, 2012, The effect of
dietary arachidonic acid during the Artemia feeding period on larval growth and
skeletogenesis in Senegalese sole, Solea senegalensis: Journal of Applied
Ichthyology, v. 28, p. 411-418.
Bonaldo, A., L. Parma, F. Mongile, P. P. Gatta, P. Di Marco, T. Petochi, G. Marino, M.
G. Finoia, R. Fontanillas, and W. Koppe, 2015, Feeding turbot juveniles Psetta
maxima L. with increasing dietary plant protein levels affects growth performance
and fish welfare: Aquaculture Nutrition.
Bostock, J., B. McAndrew, R. Richards, K. Jauncey, T. Telfer, K. Lorenzen, D. Little, L.
Ross, N. Handisyde, I. Gatward, and R. Corner, 2010, Aquaculture: global status
and trends, v. 365, 2897-2912 p.
Bovenkerk, B., and F. B. Meijboom, 2013a, Fish Welfare in Aquaculture: Explicating the
Chain of Interactions Between Science and Ethics: Journal of Agricultural and
Environmental Ethics, v. 26, p. 41-61.
Bovenkerk, B., and F. L. B. Meijboom, 2013b, Fish Welfare in Aquaculture: Explicating
the Chain of Interactions Between Science and Ethics: Journal of Agricultural &
Environmental Ethics, v. 26, p. 41-61.
Cabrita, E., F. Soares, and M. T. Dinis, 2006, Characterization of Senegalese sole, Solea
senegalensis, male broodstock in terms of sperm production and quality:
Aquaculture, v. 261, p. 967-975.
Canavate, J. P., R. Zerolo, and C. Fernandez-Diaz, 2006, Feeding and development of
Senegal sole (Solea senegalensis) larvae reared in different photoperiods:
Aquaculture, v. 258, p. 368-377.
Castelo Branco, M. A., M. A. Arruda, and S. Gamito, 2010, Feeding habits of Solea
senegalensis in earthen ponds in Sado estuary: Journal of Sea Research, v. 64, p.
446-450.
Chrousos, G. P., 1998, Stressors, stress, and neuroendocrine integration of the adaptive
response - The 1997 Hans Selye Memorial Lecture: Stress of Life: from
Molecules to Man, v. 851, p. 311-335.
Colson, V., B. Sadoul, C. Valotaire, P. Prunet, M. Gaumé, and L. Labbé, 2015, Welfare
assessment of rainbow trout reared in a Recirculating Aquaculture System:
Comparison with a Flow-Through System: Aquaculture, v. 436, p. 151-159.
Comité do Códex sobre Higiene Alimentar, 2009
Congleton, J. L., W. J. LaVoie, C. B. Schreck, and L. E. Davis, 2000, Stress indices in
migrating juvenile Chinook salmon and steelhead of wild and hatchery origin
before and after barge transportation: Transactions of the American Fisheries
Society, v. 129, p. 946-961.
Conte, F. S., 2004, Stress and the welfare of cultured fish: Applied Animal Behaviour
Science, v. 86, p. 205-223.
Creţu, M., V. Cristea, L. Dediu, and S. M. Petrea, 2014, The Influence of Different
Stocking Densities on Biochemical Composition of Rainbow Trout Meat Reared
in a Recirculating Aquaculture System: Scientific Papers: Animal Science &
Biotechnologies / Lucrari Stiintifice: Zootehnie si Biotehnologii, v. 47, p. 200-
204.

81
Daly, B., J. S. Swingle, and G. L. Eckert, 2012, Increasing hatchery production of juvenile
red king crabs (Paralithodes camtschaticus) through size grading: Aquaculture,
v. 364-365, p. 206-211.
Damsgard, B., U. Sorum, I. Ugelstad, R. A. Eliassen, and A. Mortensen, 2004, Effects of
feeding regime on susceptibility of Atlantic salmon (Salmo salar) to cold water
vibriosis: Aquaculture, v. 239, p. 37-46.
Daniel, A. P., A. P. L. Veeck, B. Klein, L. F. Ferreira, M. A. da Cunha, T. V. Parodi, C.
C. Zeppenfeld, D. Schmidt, B. O. Caron, B. M. Heinzmann, B. Baldisserotto, and
T. Emanuelli, 2014, Using the essential oil of Aloysia triphylla (L'Her.) Britton to
sedate silver catfish (Rhamdia quelen) during transport improved the chemical
and sensory qualities of the fish during storage in ice: Journal Of Food Science,
v. 79, p. S1205-S1211.
Daniels, H. V., D. L. Berlinsky, R. G. Hodson, and C. V. Sullivan, 1996, Effects of
Stocking Density, Salinity, and Light Intensity on Growth and Survival of
Southern Flounder Paralichthys lethostigma Larvae: Journal of the World
Aquaculture Society, v. 27, p. 153-159.
Dawkins, M. S., 1998, Evolution and animal welfare: Quarterly Review of Biology, v.
73, p. 305-328.
Diana, J. S., H. S. Egna, T. Chopin, M. S. Peterson, L. Cao, R. Pomeroy, M. Verdegem,
W. T. Slack, M. G. Bondad-Reantaso, and F. Cabello, 2013, Responsible
Aquaculture in 2050: Valuing Local Conditions and Human Innovations Will Be
Key to Success: BioScience, v. 63, p. 255-262.
DiBattista, J. D., H. Anisman, M. Whitehead, and K. M. Gilmour, 2005, The effects of
cortisol administration on social status and brain monoaminergic activity in
rainbow trout Oncorhynchus mykiss: Journal of Experimental Biology, v. 208, p.
2707-2718.
Dillon, M. C., M. Dillon, and C. Griffith, 1996, How to HACCP: An Illustrated Guide,
M.D. Associates.
Dinis, M. T., L. Ribeiro, F. Soares, and C. Sarasquete, 1999, A review on the cultivation
potential of Solea senegalensis in Spain and in Portugal: Aquaculture, v. 176, p.
27-38.
Dobsikova, R. V. a. F. U. B., Z. V. a. F. U. B. Svobodova, J. V. a. F. U. B. Blahova, H.
V. a. F. U. B. Modra, and J. J. U. V. V. U. R. a. H. Velisek, 2009, The effect of
transport on biochemical and haematological indices of common carp (Cyprinus
carpio L.): Czech Journal of Animal Science - UZEI (Czech Republic).
Ellis, T., H. Y. Yildiz, J. Lopez-Olmeda, M. T. Spedicato, L. Tort, O. Overli, and C. I. M.
Martins, 2012, Cortisol and finfish welfare: Fish Physiology and Biochemistry, v.
38, p. 163-188.
FAO, 2014, The State of World Fisheries and Aquaculture.
Farrell, A., S. Tang, M. Nomura, and C. Brauner, 2010, Toward Improved Public
Confidence in Farmed Fish: A Canadian Perspective on Fish Welfare during
Marine Transport: Journal of the World Aquaculture Society.
Fernández-Segovia, I., A. Pérez-Llácer, B. Peidro, and A. Fuentes, 2014, Implementation
of a food safety management system according to ISO 22000 in the food
supplement industry: A case study: Food Control, v. 43, p. 28-34.
Fevolden, S. E., K. H. Roed, K. T. Fjalestad, and J. Stien, 1999, Poststress levels of
lysozyme and cortisol in adult rainbow trout: heritabilities and genetic
correlations: Journal of Fish Biology, v. 54, p. 900-910.
Fisheries policy – European commission 2012.

82
Ganzon-Naret, E. S., 2013, Influence of different feeding rates using commercial dry
pellets on growth, feed efficiency and survival for hatchery produced sea bass
Lates calcarifer: Animal Biology & Animal Husbandry, v. 5, p. 241-248.
González, R., J. D. Celada, J. M. Carral, V. García, M. Sáez-Royuela, and Á. González,
2011, Intensive rearing of juvenile crayfish ( Pacifastacus leniusculus, Astacidae)
during the first 6 months: effects of size grading: Aquaculture Research, v. 42, p.
1385-1392.
Gornati, R., E. Papis, S. Rimoldi, G. Terova, M. Saroglia, and G. Bernardini, 2004,
Rearing density influences the expression of stress-related genes in sea bass
(Dicentrarchus labrax, L.): Gene, v. 341, p. 111-118.
Grigorakis, K., and G. Rigos, 2011, Aquaculture effects on environmental and public
welfare - The case of Mediterranean mariculture: Chemosphere, v. 85, p. 899-919.
Huntingford, F. A., 2004, Implications of domestication and rearing conditions for the
behaviour of cultivated fishes: Journal of Fish Biology, v. 65, p. 122-142.
Huntingford, F. A., C. Adams, V. A. Braithwaite, S. Kadri, T. G. Pottinger, P. Sandoe,
and J. F. Turnbull, 2006, Current issues in fish welfare: Journal of Fish Biology,
v. 68, p. 332-372.
Huntingford, F. A., and S. Kadri, 2014, Defining, assessing and promoting the welfare of
farmed fish: Revue Scientifique Et Technique-Office International Des
Epizooties, v. 33, p. 233-244.
Håstein, T., 2007, OIE involvement in aquatic animal welfare: the need for development
of guidelines based on welfare for farming, transport and slaughter purposes in
aquatic animals: Developments In Biologicals, v. 129, p. 149-161.
Ignacio Martín, J. R. G., Juan Antonio Martos-Sitcha, Inmaculada, and C. R. Rasines,
Juan Miguel Mancera, Olvido Chereguini, 2014, is it possible to transport solea
senegalensis specimens without water?, Aquaculture Europe, Donostia - San
Sebastian, Spain.
Iguchi, K. i., K. Ogawa, M. Nagae, and F. Ito, 2003, The influence of rearing density on
stress response and disease susceptibility of ayu (Plecoglossus altivelis):
Aquaculture, v. 220, p. 515-523.
Imsland, A. K., A. Foss, L. E. C. Conceicao, M. T. Dinis, D. Delbare, E. Schram, A.
Kamstra, P. Rema, and P. White, 2003, A review of the culture potential of Solea
solea and S-senegalensis: Reviews in Fish Biology and Fisheries, v. 13, p. 379-
407.
Iversen, M., R. A. Eliassen, and B. Finstad, 2009, Potential benefit of clove oil sedation
on animal welfare during salmon smolt, Salmo salar L. transport and transfer to
sea: Aquaculture Research, v. 40, p. 233-241.
Kalinowski, C. T., M. S. Izquierdo, D. Schuchardt, and L. E. Robaina, 2007, Dietary
supplementation time with shrimp shell meal on red porgy (Pagrus pagrus) skin
colour and carotenoid concentration: Aquaculture, v. 272, p. 451-457.
Kiessling, A., H. van de Vis, G. Flik, and S. Mackenzie, 2012, Welfare of farmed fish in
present and future production systems: Fish Physiology and Biochemistry, v. 38,
p. 1-3.
King, H. R., 2009, 2008 Australian RSPCA Welfare Symposium: Fish transport in the
aquaculture sector: An overview of the road transport of Atlantic salmon in
Tasmania: Journal of Veterinary Behavior: Clinical Applications and Research,
v. 4, p. 163-168.
Kolarevic, J., G. Baeverfjord, H. Takle, E. Ytteborg, B. K. M. Reiten, S. Nergård, and B.
F. Terjesen, 2014, Performance and welfare of Atlantic salmon smolt reared in
recirculating or flow through aquaculture systems: Aquaculture, v. 432, p. 15-25.

83
Kulczykowska, E., and F. J. Sánchez Vázquez, 2010, Neurohormonal regulation of feed
intake and response to nutrients in fish: aspects of feeding rhythm and stress:
Aquaculture Research, v. 41, p. 654-667.
Lekang, O.-I., 2013, Transport of Live Fish, Aquaculture Engineering, John Wiley &
Sons, p. 328-338.
Lemieux, H., J. D. Dutil, H. Guderley, and R. Larocque, 2004, Growth, starvation and
enzyme activity in white muscle of Atlantic cod: At what point do muscle
metabolic capacities change?: Marine and Freshwater Behaviour and Physiology,
v. 37, p. 287-294.
Liu, Z., X. Li, L. Fan, H. Lu, L. Liu, and Y. Liu, 2014, Measuring feeding activity of fish
in RAS using computer vision: Aquacultural Engineering, v. 60, p. 20-27.
Lopez-Olmeda, J. F., C. Noble, and F. J. Sanchez-Vazquez, 2012, Does feeding time
affect fish welfare?: Fish Physiology and Biochemistry, v. 38, p. 143-152.
Luo, G., G. Liu, and H.-x. Tan, 2013, Effects of stocking density and food deprivation-
related stress on the physiology and growth in adult Scortum barcoo ( Mc Culloch
& Waite): Aquaculture Research, v. 44, p. 885-894.
López-Olmeda, J., C. Noble, and F. Sánchez-Vázquez, 2012, Does feeding time affect
fish welfare?: Fish Physiology & Biochemistry, v. 38, p. 143.
Machado, M., R. Azeredo, P. Diaz-Rosales, A. Afonso, H. Peres, A. Oliva-Teles, and B.
Costas, 2015, Dietary tryptophan and methionine as modulators of European
seabass (Dicentrarchus labrax) immune status and inflammatory response: Fish
& Shellfish Immunology, v. 42, p. 353-362.
Magnadottir, B., 2006, Innate immunity of fish (overview): Fish & Shellfish
Immunology, v. 20, p. 137-151.
Marinho, G., H. Peres, and A. P. Carvalho, 2014, Effect of feeding time on dietary protein
utilization and growth of juvenile Senegalese sole ( Solea senegalensis):
Aquaculture Research, v. 45, p. 828-833.
McEwen, B. S., and J. C. Wingfield, 2003, The concept of allostasis in biology and
biomedicine: Hormones and Behavior, v. 43, p. 2-15.
Menezes, C., I. Ruiz-Jarabo, J. A. Martos-Sitcha, C. Toni, J. Salbego, A. Becker, V. L.
Loro, G. Martínez-Rodríguez, J. M. Mancera, and B. Baldisserotto, 2015, The
influence of stocking density and food deprivation in silver catfish (Rhamdia
quelen): A metabolic and endocrine approach: Aquaculture, v. 435, p. 257-264.
Mesquita, F. O., 2011, Coping styles and learning in fish: developing behavioural tools
for welfare-friendly aquaculture.
Miget, R. J., 2004, HACCP seafood program and aquaculture.
Montero, D., L. Tort, L. Robaina, J. M. Vergara, and M. S. Izquierdo, 2001, Low vitamin
E in diet reduces stress resistance of gilthead seabream (Sparus aurata) juveniles:
Fish & Shellfish Immunology, v. 11, p. 473-490.
Mustapha, M. K., 2014, Aquaculture and Fish Welfare: Are the Rights of Fish
Compromised?/ Akwakultura I Dobrostan Ryb: Czy Prawa Ryb Są Szanowane?:
Zoologica Poloniae, v. 59, p. 49.
Nilsson, J., and T. Torgersen, 2010, Exploration and learning of demand-feeding in
Atlantic cod (Gadus morhua): Aquaculture, v. 306, p. 384-387.
Overli, O., S. Winberg, and T. G. Pottinger, 2005, Behavioral and neuroendocrine
correlates of selection for stress responsiveness in rainbow trout - a review:
Integrative and Comparative Biology, v. 45, p. 463-474.
Overton, J. L., S. J. Steenfeldt, and P. B. Pedersen, 2010, The effects of grading on the
growth and survival of juvenile Dover sole ( Solea solea L.): Aquaculture
Research, v. 42, p. 31-39.

84
Pavlidis, M., N. Papandroulakis, and P. Divanach, 2006, A method for the comparison of
chromaticity parameters in fish skin: Preliminary results for coloration pattern of
red skin Sparidae: Aquaculture, v. 258, p. 211-219.
Petrović, S., L. Bavčević, Z. Pasarić, and A. Mihovilović, 2011, Effects of successive size
grading on the growth of juvenile European sea bass ( Dicentrarchus labrax L.)
at three temperatures: Aquaculture Research, v. 42, p. 277-287.
Pirhonen, J., C. B. Schreck, P. W. Reno, and H. Ogut, 2003, Effect of fasting on feed
intake, growth and mortality of chinook salmon, Oncorhynchus tshawytscha,
during an induced Aeromonas salmonicida epizootic: Aquaculture, v. 216, p. 31-
38.
Poli, B. M., G. Parisi, F. Scappini, and G. Zampacavallo, 2005, Fish welfare and quality
as affected by pre-slaughter and slaughter management: Aquaculture
International, v. 13, p. 29-49.
Pottinger, T. G., 2010, A multivariate comparison of the stress response in three salmonid
and three cyprinid species: evidence for inter-family differences: Journal of Fish
Biology, v. 76, p. 601-621.
Pottinger, T. G., T. A. Moran, and J. A. W. Morgan, 1994, Primary And Secondary
Indexes Of Stress In The Progeny Of Rainbow-Trout (Oncorhynchus-Mykiss)
Selected For High And Low Responsiveness To Stress: Journal of Fish Biology,
v. 44, p. 149-163.
Pottinger, T. G., and A. D. Pickering, 1992, the influence of social-interaction on the
acclimation of rainbow-trout, oncorhynchus-mykiss (walbaum) to chronic stress:
Journal of Fish Biology, v. 41, p. 435-447.
Pottinger, T. G., A. D. Pickering, and M. A. Hurley, 1992, consistency in the stress
response of individuals of 2 strains of rainbow-trout, oncorhynchus-mykiss:
Aquaculture, v. 103, p. 275-289.
Psomas, E. L., and D. P. Kafetzopoulos, 2015, HACCP effectiveness between ISO 22000
certified and non-certified dairy companies: Food Control, v. 53, p. 134-139.
Qu, Y., X. Li, Y. Yu, M. Vandepeer, P. Babidge, S. Clarke, K. Bott, and H. Li, 2009,
effect of different grading equipment on stress levels assessed by catecholamine
measurements in Pacific oysters, Crassostrea gigas (Thunberg): Aquacultural
engineering.
Rauta, P. R., B. Nayak, and S. Das, 2012, Immune system and immune responses in fish
and their role in comparative immunity study: A model for higher organisms:
Immunology Letters, v. 148, p. 23-33.
Ribas, L., J. V. Planas, B. Barton, C. Monetti, G. Bernadini, A. Saroglia, L. Tort, and S.
MacKenzie, 2004, A differentially expressed enolase gene isolated from the
gilthead sea bream (Sparus aurata) under high-density conditions is up-regulated
in brain after in vivo lipopolysaccharide challenge: Aquaculture, v. 241, p. 195-
206.
Richard, R., and L. Flemming, 2007, Fin condition and growth among rainbow trout
reared at different sizes, densities and feeding frequencies in high-temperature re-
circulated water: Aquaculture International, v. 15, p. 97-107.
Rios, F. S., A. L. Kalinin, and F. T. Rantin, 2002, The effects of long-term food
deprivation on respiration and haematology of the neotropical fish Hoplias
malabaricus: Journal of Fish Biology, v. 61, p. 85-95.
Saillant, E., A. Fostier, P. Haffray, B. Menu, S. Laureau, J. Thimonier, and B. Chatain,
2003, Effects of rearing density, size grading and parental factors on sex ratios of
the sea bass (Dicentrarchus labrax L.) in intensive aquaculture: Aquaculture, v.
221, p. 183-206.

85
Salas-Leiton, E., V. Anguis, M. Manchado, and J. P. Canavate, 2008, Growth, feeding
and oxygen consumption of Senegalese sole (Solea senegalensis) juveniles
stocked at different densities: Aquaculture, v. 285, p. 84-89.
Salas-Leiton, E., V. Anguis, A. Rodriguez-Rua, and J. P. Canavate, 2010, Stocking
homogeneous size groups does not improve growth performance of Senegalese
sole (Solea senegalensis, Kaup 1858) juveniles: Individual growth related to fish
size: Aquacultural Engineering, v. 43, p. 108-113.
Salas-Leiton, E., V. Anguís, A. Rodríguez-Rua, and J. P. Cañavate, 2011, High stocking
density and food restriction have minimum impact on size dispersal of cultured
Senegalese sole (Solea senegalensis, Kaup 1858) juveniles. Evidence for
individual growth being regulated by population structure: Aquacultural
Engineering, v. 45, p. 43-50.
Sant'Anna, V., P. D. Gurak, L. D. Ferreira Marczak, and I. C. Tessaro, 2013, Tracking
bioactive compounds with colour changes in foods - A review: Dyes and
Pigments, v. 98, p. 601-608.
Schreck, C. B., 2010, Stress and fish reproduction: The roles of allostasis and hormesis:
General and Comparative Endocrinology, v. 165, p. 549-556.
Schreck, C. B., W. Contreras-Sanchez, and M. S. Fitzpatrick, 2001, Effects of stress on
fish reproduction, gamete quality, and progeny: Aquaculture, v. 197, p. 3-24.
Segner, H., H. Sundh, K. Buchmann, J. Douxfils, K. S. Sundell, C. Mathieu, N. Ruane,
F. Jutfelt, H. Toften, and L. Vaughan, 2012, Health of farmed fish: its relation to
fish welfare and its utility as welfare indicator: Fish Physiology and Biochemistry,
v. 38, p. 85-105.
Slavík, O., M. Pešta, and P. Horký, 2011, Effect of grading on energy consumption in
European catfish Silurus glanis: Aquaculture, v. 313, p. 73-78.
Summerfelt, S. T., A. Zühlke, J. Kolarevic, B. K. M. Reiten, R. Selset, X. Gutierrez, and
B. F. Terjesen, 2015, Effects of alkalinity on ammonia removal, carbon dioxide
stripping, and system pH in semi-commercial scale water recirculating
aquaculture systems operated with moving bed bioreactors: Aquacultural
Engineering, v. 65, p. 46-54.
Szisch, V., A. L. van der Salm, S. E. W. Bonga, and M. Pavlidis, 2002, Physiological
colour changes in the red porgy, Pagrus pagrus, following adaptation to blue
lighting spectrum: Fish Physiology and Biochemistry, v. 27, p. 1-8.
Sánchez, P., P. P. Ambrosio, and R. Flos, 2010, Stocking density and sex influence
individual growth of Senegalese sole (Solea senegalensis): Aquaculture, v. 300,
p. 93-101.
Sánchez, P., P. P. Ambrosio, and R. Flos, 2013, Stocking density affects Senegalese sole
( Solea senegalensis, Kaup) growth independently of size dispersion, evaluated
using an individual photo-identification technique: Aquaculture Research, v. 44,
p. 231-241.
Tacchi, L., L. Lowrey, R. Musharrafieh, K. Crossey, E. T. Larragoite, and I. Salinas,
2015, Effects of transportation stress and addition of salt to transport water on the
skin mucosal homeostasis of rainbow trout (Oncorhynchus mykiss): Aquaculture,
v. 435, p. 120-127.
Tang, S., C. J. Brauner, and A. P. Farrell, 2009, Using bulk oxygen uptake to assess the
welfare of adult Atlantic salmon, Salmo salar, during commercial live-haul
transport: Aquaculture, v. 286, p. 318-323.
Torgersen, T., M. B. M. Bracke, and T. S. Kristiansen, 2011, Reply to Diggles et al.
(2011): Ecology and welfare of aquatic animals in wild capture fisheries: Reviews
in Fish Biology and Fisheries, v. 21, p. 767-769.

86
Trejchel, K., D. Żarski, K. Palińska-Żarska, S. Krejszeff, B. Dryl, K. Dakowski, and D.
Kucharczyk, 2014, Determination of the optimal feeding rate and light regime
conditions in juvenile burbot, Lota lota (L.), under intensive aquaculture:
Aquaculture international.
Turnbull, J., A. Bell, C. Adams, J. Bron, and F. Huntingford, 2005, Stocking density and
welfare of cage farmed Atlantic salmon: application of a multivariate analysis:
Aquaculture.
Turnbull, J. F., and F. A. Huntingford, 2012, welfare and aquaculture: where benefish fits
in: Aquaculture Economics & Management (Taylor & Francis), v. 16, p. 433.
United, S., O. National, A. Atmospheric, P. Virginia Sea Grant College, and M. L.
Jahncke, 2002, Application of Hazard Analysis Critical Control Point (HACCP)
principles as a risk management tool to control viral pathogens at shrimp
aquaculture facilities / HACCP in shrimp facilities / Principles as a risk
management tool to control viral pathogens at shrimp aquaculture facilities.
van de Nieuwegiessen, P. G., J. Olwo, S. Khong, J. A. J. Verreth, and J. W. Schrama,
2009, Effects of age and stocking density on the welfare of African catfish,
Clarias gariepinus Burchell: Aquaculture, v. 288, p. 69-75.
van de Vis, J. W., M. Poelman, E. Lambooij, M. L. Begout, and M. Pilarczyk, 2012, Fish
welfare assurance system: initial steps to set up an effective tool to safeguard and
monitor farmed fish welfare at a company level: Fish Physiology and
Biochemistry, v. 38, p. 243-257.
Van der Salm, A. L., M. Martinez, G. Flik, and S. E. W. Bonga, 2004, Effects of
husbandry conditions on the skin colour and stress response of red porgy, Pagrus
pagrus: Aquaculture, v. 241, p. 371-386.
Veissier, I., A. Butterworth, B. Bock, and E. Roe, 2008, European approaches to ensure
good animal welfare: Applied Animal Behaviour Science, v. 113, p. 279-297.
Villanueva, R. R., M. E. Araneda, M. Vela, and J. C. Seijo, 2013, Selecting stocking
density in different climatic seasons: A decision theory approach to intensive
aquaculture: Aquaculture, v. 384-387, p. 25-34.
Wunderink, Y. S., S. Engels, S. Halm, M. Yúfera, G. Martínez-Rodríguez, G. Flik, P. H.
M. Klaren, and J. M. Mancera, 2011, Chronic and acute stress responses in
Senegalese sole (Solea senegalensis): The involvement of cortisol, CRH and
CRH-BP: General and Comparative Endocrinology, v. 171, p. 203-210.
Zhang, S.-Y., G. Li, H.-B. Wu, X.-G. Liu, Y.-H. Yao, L. Tao, and H. Liu, 2011, An
integrated recirculating aquaculture system (RAS) for land-based fish farming:
The effects on water quality and fish production: Aquacultural Engineering, v. 45,
p. 93-102.
Zhu, L. Y., L. Nie, G. Zhu, L. X. Xiang, and J. Z. Shao, 2013, Advances in research of
fish immune-relevant genes: A comparative overview of innate and adaptive
immunity in teleosts: Developmental and Comparative Immunology, v. 39, p. 39-
62.
87
Anexos
Tabela I – Ficha de autocontrolo do abate
FWAS - Fish Welfare Assurance System
Data Lote Tanque Limites OK / NOK Operador
Gelo 260kg
Peixe 350kg
Água 80 l
Temp. Int. <4˚
Tabela II – Ficha de autocontrolo do tempo de exposição ao ar
FWAS - Fish Welfare Assurance System
Dia Tipo de
Operação
Tempo Peixe fora de água
Operador
Limite OK / NOK
<150g – 5 min
>150g – 8 min

88
Tabela III – Ficha de autocontrolo das operações realizadas na empresa
FWAS - Fish Welfare Assurance System
Data
Tipo de Operação
Duração
Tanques afetados
Operador
Tabela IV – Folha de autocontrolo da qualidade da água (parte 1)
Data MOD
Temperatura
Data MOD
Amónia NH3
Limites OK/NOK Limites OK/NOK
18˚ <
Tem
p. <
22
˚
0,7
5 m
g/l

89
Tabela V – Folha de autocontrolo da qualidade da água (parte 2)
Data MOD
Nitritos NO2-
Data MOD
Bromo
Operador
Limites OK/NOK Limites OK/NOK
1,5
mg/l
0,0
8 m
g/l
Tabela VI – Folha de autocontrolo para as visitas
FWAS - Fish Welfare Assurance System
Dia Hora Nº de Visitantes Tanques afetados Operador
Tabela VII – Folha de autocontrolo para a necropsia
FWAS - Fish Welfare Assurance System
Dia Lote MOD
Sinais Externos Sinais Internos Operador
OK/NOK OK/NOK OK/NOK OK/NOK OK/NOK OK/NOK
34 39 40 1 21 26
34 39 40 1 21 26
34 39 40 1 21 26
90
Tabela VIII – Folha de autocontrolo para a qualidade da ração.
FWAS - Fish Welfare Assurance System
Data: Lote:
Resultados das análises externas à composição química
OK / NOK
Avaliar a qualidade física da ração
Flutuabilidade Presença de Finos
OK / NOK OK / NOK
Operador
Tabela IX – Folha de autocontrolo do fator de atividade geral
FWAS - Fish Welfare Assurance System
Data Tanque
Fator de Atividade Geral
Operador Limite OK / NOK
40%

91
Tabela X – Folha de autocontrolo do transporte
FWAS - Fish Welfare Assurance System
Data:
Transporte: Receção:
Cuba Lote
Oxigénio Temperatura Amónia
Limites OK/NOK Limites OK/NOK Limites OK/NOK
1
>100
Δ
3
< 3
mg/l
2
3
4
5
6
7
8
9
10
11
12


















