DISSERTAÇÃO ALINY LOBO LADD - USP · Barbosa, pois sempre disseram: “você vai conseguir; nós...
75
Aliny Antunes Barbosa Lobo Ladd Mecanismos de proliferação neuronal pós-natal no gânglio cervical cranial de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia. Dissertação apresentada ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Ciências Departamento: Cirurgia Área de concentração: Anatomia dos Animais Domésticos e Silvestres Orientador: Prof. Dr. Romeu Rodrigues De Souza São Paulo 2009
Transcript of DISSERTAÇÃO ALINY LOBO LADD - USP · Barbosa, pois sempre disseram: “você vai conseguir; nós...

Aliny Antunes Barbosa Lobo Ladd Mecanismos de proliferação neuronal pós-natal no gânglio cervical cranial
de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia.
Dissertação apresentada ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Ciências
Departamento: Cirurgia Área de concentração: Anatomia dos Animais Domésticos e Silvestres Orientador: Prof. Dr. Romeu Rodrigues De Souza
São Paulo 2009

Autorizo a reprodução parcial ou total desta obra, para fins acadêmicos, desde que citada a fonte.
DADOS INTERNACIONAIS DE CATALOGAÇÃO-NA-PUBLICAÇÃO
(Biblioteca Virginie Buff D’Ápice da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo)
T.2157 Ladd, Aliny Antunes Barbosa Lobo FMVZ Mecanismos de proliferação neuronal pós-natal no gânglio cervical cranial
de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia / Aliny Antunes Barbosa Lobo Ladd. – São Paulo : A. A. B. L. Ladd, 2009.
75 f. : il.
Dissertação (mestrado) - Universidade de São Paulo. Faculdade de Medicina Veterinária e Zootecnia. Departamento de Cirurgia, 2009.
Programa de Pós-Graduação: Anatomia dos Animais Domésticos e Silvestres.
Área de concentração: Anatomia dos Animais Domésticos e Silvestres.
Orientador: Prof. Dr. Romeu Rodrigues De Souza. 1. Gânglio cervical cranial. 2. Preás. 3. Estereologia. 4. Maturação.
5. Envelhecimento. I. Título.

Mecanismos de proliferação neuronal pós-natal no gânglio cervical cranial
de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia.
Esta Dissertação foi desenvolvida no Laboratório de Estereologia Estocástica
e Anatomia Química (L.S.S.C.A.) do Departamento de Cirurgia da Faculdade de
Medicina Veterinária e Zootecnia da Universidade de São Paulo e financiada com
Bolsa Mestrado pela Fundação de Apoio a Pesquisa do Estado de São Paulo
(FAPESP).
São Paulo 2009


FOLHA DE AVALIAÇÃO
Nome: LADD, Aliny Antunes Barbosa Lobo Ladd Título: Mecanismos de proliferação neuronal pós-natal no gânglio cervical
cranial de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia.
Dissertação apresentada ao Programa de Pós-Graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Ciências
Data: ______ /______/______
Banca Examinadora
Prof. Dr. _____________________________Instituição: ________________
Assinatura: __________________________Julgamento: ________________
Prof. Dr. ____________________________ Instituição: _________________
Assinatura: __________________________Julgamento: _________________
Prof. Dr. ____________________________ Instituição:__________________
Assinatura: _________________________ Julgamento:_________________

DEDICATÓRIAS

DEDICATÓRIAS
Dedico esta etapa da minha vida, representada por este trabalho, aos meus pais, Adelson Antunes Barbosa e Ilma Gomes J. Barbosa, pois sempre disseram: “você vai conseguir; nós confiamos em você”. Vocês dois são o meu grande orgulho; são exemplos de perseverança, honestidade e amor. Tudo que fazem por mim é muito mais do que qualquer filho no mundo pode querer. Aos meus irmãos, Alym Antunes Barbosa e Alisson Antunes Barbosa, por serem meus amigos e por dedicarem a mim os mais fraternos sentimentos. Nós somos uma família e exemplo da nossa união é que fazemos com que a nossa distância seja somente física. Vocês me apoiam incondicionalmente e vão estar eternamente em tudo o que eu fizer.

Dedico igualmente este trabalho ao Fernando Vagner Lobo Ladd, meu cúmplice, marido e companheiro em tudo que faço. Seu apoio é essencial. Sem você, as páginas da minha história não teriam aventura; com você, eu nunca me sinto sozinha. Por isto, saiba que... “Eu gosto de você, e gosto de ficar com você. Meu riso é tão feliz contigo. O meu melhor amigo é o meu amor.” Marisa Monte

AGRADECIMENTOS

AGRADECIMENTOS
Durante a minha vida, lutei diariamente e, durante este
processo, percorri caminhos tortuosos, com espinhos e cheios de
pedras; sofri derrotas, decepções e, em vários momentos, até
acreditei que não seria possível. Entre tantas lutas e lições,
aprendi que às vezes conquistamos o que não estava nos planos
iniciais. Aprendi que é preciso mudar de ideia, mudar de
objetivo, que é preciso simplesmente mudar... E que em toda
conquista existe um fator que se chama TEMPO... Ele é um
professor... Nos ensina a curar as feridas, a superar desafios, a
ter saudade de quem está longe e a valorizá-los. Ele nos faz
esperar para colher o fruto com a maturidade certa, nos
proporciona conquistar o que com a nossa falta de experiência
acreditamos não ser importante, mas que é imprescindível em
qualquer conquista, a SERENIDADE... E faz com que cada luta
dure o tempo certo para conquistarmos humildade suficiente
para não acreditarmos que realizamos algo sozinho...
Por isto, neste momento, é TEMPO de agradecer...

“Para os crentes, Deus está no princípio das coisas. Para os cientistas, no final de toda reflexão.”
Max Planck
Agradeço a DEUS por me iluminar e proteger; por minha família e
por ter colocado na minha vida as oportunidades que me trouxeram até
aqui...
“Se eu vi mais longe, foi por estar de pé sobre ombros de
gigantes.” Isaac Newton
Ao Prof. Dr. Antonio Augusto Coppi Ribeiro, agradeço de forma
especial pelo constante incentivo, por acreditar na minha capacidade, por
lapidar as minhas qualidades, por corrigir os meus erros. Pela coragem de
ousar trabalhar com novas ideias e conceitos, correndo os riscos inerentes
a esta atitude. Por ser um interlocutor interessado em me ajudar com as
minhas inquietações. A você, não só agradeço, dedico admiração por todo
seu brilhantismo acadêmico... É sob sua tutela que conquistei este título.
“Nenhum de nós é tão inteligente quanto todos nós juntos.”
Warren Bennis
Agradeço aos meus colegas de Laboratório, Fernando Ladd,
Andrea Almeida, Eliane Muniz, Silvio Gomes, Luciana Abraão,
Demilto Yamaguchi, Felipe Oliveira, Tais Sasahara, Fernanda Vieira
e Ana Paula Moura, por me aturarem nos meus momentos de stress,
por aprendermos a trabalhar em grupo, nos respeitando com as nossas
excentricidades. Pela convivência mínima de 40h semanais, que tanto
contribuiu para o meu desenvolvimento profissional e pessoal pelo
exercício diário da paciência.
Em especial, agradeço ao Silvio Gomes, Luciana Abrahão por
toda a animação que a presença de vocês causa no ambiente. Estes
momentos foram e são importantes para descontrairmos.

Deixo um abraço em forma de agradecimento a Eliane Muniz, com
seu jeito simples você conquistou seu espaço, com a sua determinação
conquistou o meu respeito. Você é muito batalhadora, continue assim.
Obrigada por toda ajuda e estímulo.
Agradeço ao Prof. Moacir Franco de Oliveira pela doação dos
animais que foram objeto desta pesquisa.
Agradeço a Profa. Dra. Julia Maria Matera por todo apoio
institucional e pessoal, sem os quais eu não teria concluído este mestrado.
Ao Prof. Dr. Romeu Rodrigues de Souza, que me forneceu,
institucionalmente, a legalidade de ser matriculada. Muito obrigada!
Aos funcionários, aos quais não pretendo nomear para não ser
indelicada ao deixar de citar algum. O meu profundo agradecimento a
todos pela contribuição, seja ela de que forma for.
Agradeço a FAPESP pelo apoio financeiro que ajudou na realização
desta pesquisa.
“Quem luta contra nós reforça os nossos nervos e aguça as nossas
habilidades. O nosso antagonista é quem mais nos ajuda.” Edmund Burke
Apesar de todas as dificuldades, agradeço também à coordenação
do Curso de Pós-graduação, por estimular minha formação politizada,
que me despertou o interesse por leis, direitos e deveres de um pós-
graduando.

“Eu aprendi que para se crescer como pessoa é preciso me cercar de gente mais inteligente do que eu”.
William Shakespeare
Agradeço, de forma carinhosa, a Andréia Almeida. Orgulho-me
muito de ter sido merecedora de tão grande confiança, apoio, amizade e
de compartilhar com você interesses comuns que nos uniram ainda mais.
Agradeço e deixo um abraço às minhas amigas, Elizete Lacerda
(Ely), Maria de Lurdes (Malu) e Berenice Raquel (Berê). Vocês três foram
uma das minhas fontes de força aqui em São Paulo... São exemplos de
mulheres heroínas, que dedicaram a mim o carinho e companheirismo de
forma gratuita. Conquistei um bem precioso com vocês, a amizade.
À minha amiga e primeira professora de anatomia, Rosimeire
Alves da Silva. Você me despertou para o mundo da ciência, me ensinou
o valor da ética, me educou para ser uma mulher e uma profissional de
respeito.
“A verdadeira família é aquela unida pelo espírito e não pelo sangue.”
Luiz Gasparetto
Às meninas da minha vida, Franciely (Fran), Geovana (BB), Ana
Carolina (Aninha), Emanuely (Manu), Emily (Porpeta), que despertam
sentimentos que até o nascimento de vocês, eram por mim
desconhecidos.
À minha família-amiga de Anápolis, Goiás, Adão Ladd, Maria
Celina, Frederico Ladd. Obrigada por todo amor, apoio e incentivo.

Às minhas “irmãs” e amigas, Fabrícia e Emiliane, por cuidarem da
“nossa” família na minha ausência.
Aos meus avós, Jason e Natalina; Elza e José (in memória), que
acreditaram em mim...
À madrinha Eliane, a Deucelina (Morena) e Edésia por todo
incentivo e apoio.
De forma geral, agradeço a todos de Goiânia, Goiás, que choram,
até hoje, com a minha partida e que se alegram todas as vezes com a
minha chegada.

"...não há coisa mais difícil de se
fazer, mais duvidosa de se alcançar,
ou mais perigosa de se manejar do
que ser o introdutor de uma nova
ordem, porque quem o é, tem por
inimigos todos aqueles que se
beneficiam com a antiga ordem..."
Maquiavel, O príncipe. .

RESUMO
LADD, A. A. B. L. Mecanismos de proliferação neuronal pós-natal do gânglio cervical cranial de preás (Galea spixii spixii) Wagler, 1831. Neurogênese x diferenciação neuronal tardia. [Mechanisms of post-natal neuronal proliferation in the cranial cervical ganglion of preás (Galea spixii spixii) Wagler, 1831. Neurogenesis vs. late neuronal differentiation ]. 2009. 75f Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2009
Neste estudo, foram abordados os aspectos quantitativos do gânglio cervical cranial
(GCC) esquerdo de preás (Galea spixii spixii) machos, oriundos do criatório da
Universidade Federal Rural do Semi-Árido Nordestino de Mossoró-RN. Os seguintes
parâmetros foram estimados: volume do GCC, número total e tamanho dos
neurônios (uni e binucleados) durante dois períodos do desenvolvimento pós-natal
(maturação e envelhecimento). Para tanto, utilizamos quatro grupos etários distintos
de animais: neonatos, jovens, adultos e senis. Após a eutanásia dos animais, os
GCCs foram fixados (perfusão sistêmica) com solução de formoldeído (4%) em PBS,
embebidos em solução de ágar (10%) e seccionados sistemática, uniforme e
aleatoriamente, aplicando-se o princípio de amostragem e contagem do fractionator.
Os principais resultados desta pesquisa foram: houve diferença significativa entre os
grupos etários para os parâmetros: Peso Neonato: 0,00056 ; Jovem: 0,00084;
Adulto: 0,00116; Senil: 0,00118. Comprimento Neonato: 1,294 ; Jovem: 2,076;
Adulto: 2,304; Senil: 2,082. Largura Neonato: 0,842; Jovem: 1,028; Adulto: 1,062;
Senil: 1,174. Volume ganglionar Neonato: 0,34 mm3 ; Jovem: 0,30 mm3; Adulto: 0,39
mm3; Senil: 0,63 mm3 e Volume neuronal médio de neurônios uninucleados Neonato:
2916,90 mm3 ; Jovem: 3550,00 mm3; Adulto: 7409,00 mm3; Senil: 6701,00 mm3. As
principais conclusões deste estudo foram: (i) peso, comprimento e largura do GCC
apresentaram aumento significativo de 185%, 60% e 79% respectivamente, durante
o período de maturação, enquanto durante o período de envelhecimento, o peso do
GCC apresentou queda de 33%, o comprimento um pequeno decréscimo de 10%,
entretanto a largura, durante este mesmo período, apresentou um aumento
progressivo de mais de 10%, tais alterações relacionadas com a idade refletiram no
volume ganglionar, causando hipertrofia; (ii) Essa hipertrofia do GCC de preás é
secundária ao desenvolvimento pós-natal (maturação e envelhecimento),

aumentando em 85%, acompanhado por um aumento alométrico, da ordem de 6
vezes em relação à massa corporal, aproximadamente 622%; (iii) Há hipertrofia dos
neurônios uninucleados que é secundária ao desenvolvimento pós-natal do GCC de
preás, aumentando em 154% durante o período de maturação com uma pequena
queda de 10% durante o envelhecimento. Finalmente, sugere-se que estudos
futuros possam investigar se o tamanho dos territórios de inervação destes gânglios
pode desencadear mudanças na micro e macroestrutura dos mesmos, incluindo a
possibilidade de ocorrência de neurogênese, que seria estudada por meio de
imunomarcadores específicos.
Palavras-chave: Gânglio cervical cranial, preás, estereologia, maturação,
envelhecimento, roedores.

ABSTRACT
LADD, A.A.B.L. Mechanisms of the post-natal neuronal proliferation in the cranial cervical ganglion of preás (Galea spixii spixii) Wagler, 1831. Neurogenesis vs late neuronal differentiation. [Mecanismos de proliferação neuronal pós-natal do gânglio cervical cranial de preás (Galea spixii spixii) Wagler, 1831. Neurogênese vs diferenciação neuronal. 2009. 75f Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2009
This study aimed at detecting whether post-natal development would exert any effect
on the size and number of cranial cervical ganglion (CCG) neurons of male preas
(Galea spixii spixii). Twenty left CCGs from twenty preas were harvested from the
Animal Facility of the Universidade Federal Rural do Semi-Árido Nordestino,
Mossoró- RN and were divided into four different age groups: newborn, young, adult
and aged subjects. The following parameters were estimated using Cavalieri's
principle and optical fractionator and planar rotator , respectively: the volume of CCG,
total number and mean volume of uni and binucleate neurons. After euthanasia, the
CCGs were perfusion-fixed with a 4% formaldehyde solution in PBS, agar-
embedded and, SUR vibrosectioned. The ganglion volume was 0.34 mm3 (newborn),
0.30 mm3 (young), 0.39 mm3 (adult) and 0.63 mm3 (aged animals) (p= 0,012). The
mean volume of uninucleate neurons was 2,917 mm3 (newborn), 3,550 mm3 (young),
7,409 mm3 (adult) and 6,701 mm3 (aged animals) (p=0,0001). The main conclusions
of this study were: (i) The CCG hypertrophy - 85% - is the result of post-natal
development (maturation and ageing), (ii) a 154% increase (hypertrophy) in the
volume of uninucleate neurons during the maturation is followed by a 10% atrophy of
them during ageing. Future studies may investigate whether the size of the CCG's
target-organs can affect the structural foundation of CCG and, therefore, add further
fuel to the notion that a post-natal neurogenesis may indeed exist in sympathetic
ganglia.
Keywords: Cranial cervical ganglion, preas, design-based stereology, maturation,
ageing, rodents.

LISTA DE FIGURAS
Figura 1- Fotografia de um Preá (Galea spixii spixii) adulto.-----------------------------38
Figura 2- Fotomacrografia evidenciando a cápsula conjuntiva do GCC esquerdo de preá jovem. Gânglio (Cabeça de seta) Asterisco (cápsula conjuntiva)--40
Figura 3- Fotomicrografia da secção de um GCC de um preá adulto onde se
exemplifica a aplicação do Princípio de Cavalieri para a estimativa do volume ganglionar. Giemsa. Escala de Barra 500µ.--------------------------43
Figura 4- Fotomicrografias de secções histológicas sequenciais do GCC de um preá
adulto, exemplificando a aplicação do método do disector óptico em que neurônios uni (M) e binucleados (B) foram amostrados em planos focais ópticos sucessivos (B,C). É possível identificar a posição de cada secção no eixo z na lateral (barra) esquerda da figura. Azul de Toluidina. Escala de barra: 20 µm ------------------------------------------------------------------------44
Figura 5- Fotomacrografia onde se evidencia a relação topográfica do gânglio
cervical cranial de um preá adulto na região cervical ventrolateral. Podem ser observadas as seguintes estruturas: medialmente o GCC (cabeça de seta preta), a bifurcação da artéria carótida comum e, dorsalmente, o gânglio distal do vago (cabeça de seta branca). Escala de barra 0,5 mm. ----------------------------------------------------------------------49
Figura 6- Microestrutura do gânglio cervical cranial de preá neonato (A) e senil (B)
em secções ópticas (40µm) no mesmo aumento. Onde se observam neurônios uninucleados (cabeças de seta cinzas) e binucleados (cabeças de seta pretas), os neurônios estão densamente distribuídos em neonatos, mas, em animais senis, estes neurônios estão mais espaçados e separados por tecido não neuronal (não marcado) e vasos sanguineos (v). Azul de Toluidina. Escala de barra: 20 µm.----------------51

Figura 7- Fotomicrografia de secção histológica de GCC de preá neonato (A) e senil (B), onde se evidenciam núcleos de neurônios Brd-U positivos, uninucleados (cabeça de seta preta) e binucleados (cabeça de seta branca). Pode-se observar também vasos sanguíneos (v). Reação de imunohistoquímica de campo claro com revelação por diaminobenzidina. Escala de barra 15 µm.---------------------------------------------------------------52
Figura 8- Volume do GCC em diferentes grupos etários. As diferenças entre os
grupos foram significantes (*p=0,012). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.-----53
Figura 9- Número total de neurônios do GCC em diferentes grupos etários. As diferenças entre os grupos não foram significantes (p=0,493). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.--------------------------------------------------------------------54
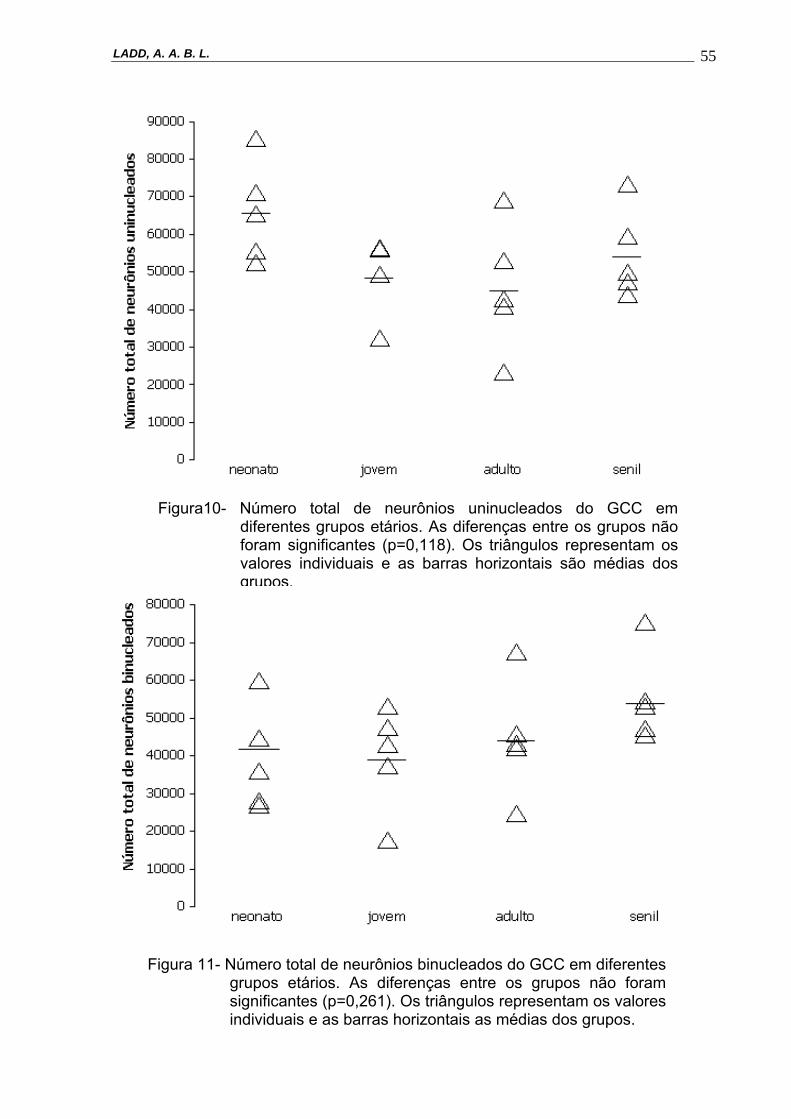
Figura 10- Número total de neurônios uninucleados do GCC em diferentes grupos
etários. As diferenças entre os grupos não foram significantes (p=0,118). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.--------------------------------------------------------------55
Figura 11- Número total de neurônios binucleados do GCC em diferentes grupos
etários. As diferenças entre os grupos não foram significantes (p=0,261). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.----------------------------------------------------------------55
Figura 12- Volume médio dos neurônios uninucleados do GCC em diferentes grupos
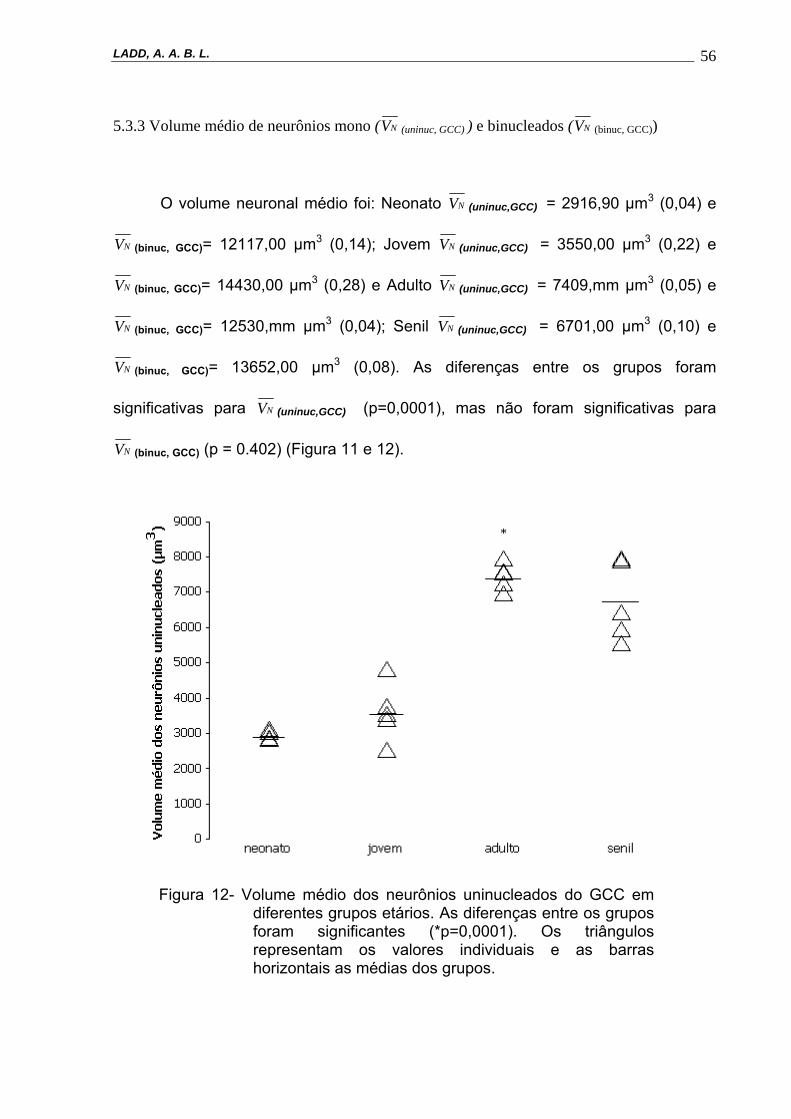
etários. As diferenças entre os grupos foram significantes (*p=0,0001). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.----------------------------------------------------------------56
Figura 13- Volume médio dos neurônios binucleados do GCC em diferentes grupos
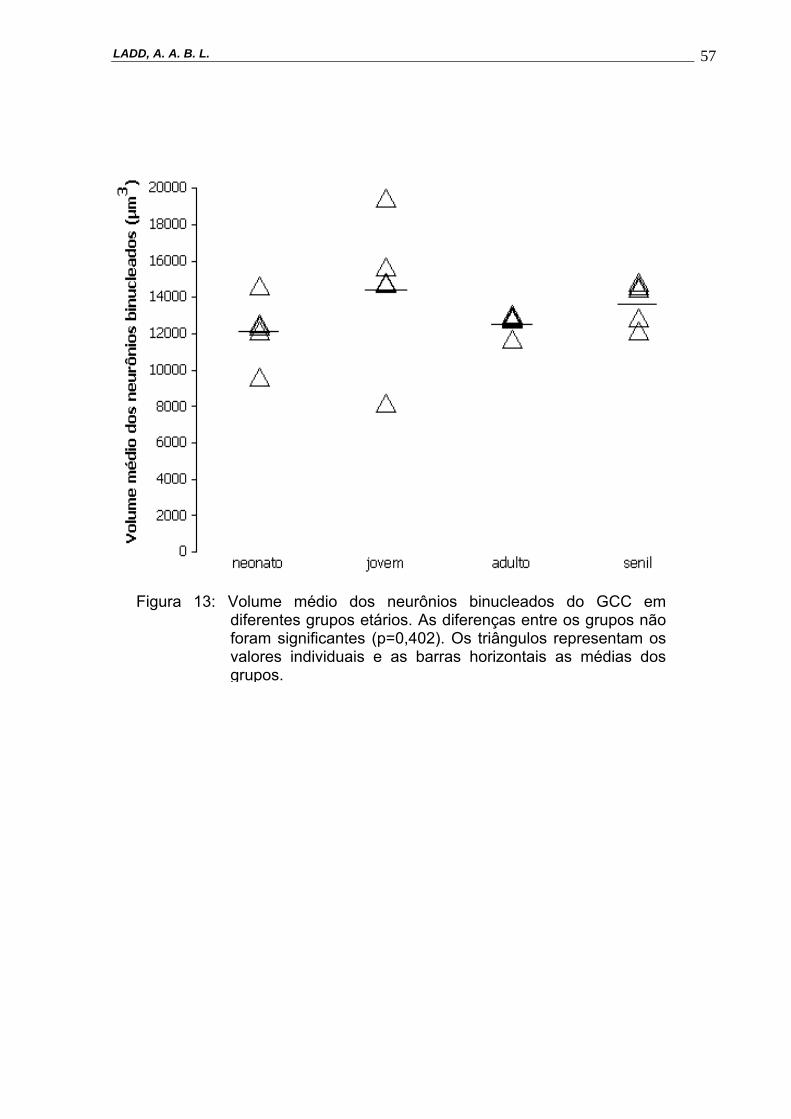
etários. As diferenças entre os grupos não foram significantes (p=0,402). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.----------------------------------------------------------------57
LISTA DE TABELAS
Tabela 1 - Peso corpóreo e peso do GCC esquerdo de preás. Os valores de cada
parâmetro são expressos pela média do grupo, seguida do seu coeficiente
de variação (CV) - São Paulo – 2009. ------------------------------------------------48

1 INTRODUÇÃO ......................................................................................................23 2 OBJETIVOS ..........................................................................................................27 Gerais:......................................................................................................................27 Específicos: ..............................................................................................................27 3 REVISÃO DE LITERATURA .................................................................................29 3.1 Características da macroestrutura e função.......................................................29 3.2 Aspectos microestruturais e quantitativos ..........................................................32 4 - MATERIAIS E MÉTODOS ..................................................................................37 4.1 Materiais.............................................................................................................37 4.2 Métodos..............................................................................................................38 4.2.1- Administração de bromodeoxyuridina (Brd-U) ...............................................38 4.2.2 Eutanásia ........................................................................................................39 4.2.3 Dissecação e individualização do GCC...........................................................39 4.2.4 – Reação de imunohistoquímica (Estudo qualitativo)......................................40 4.2.5 - Estudo estereológico.....................................................................................41 4.2.6 Análise Estatística ...........................................................................................46 5 RESULTADOS......................................................................................................48 5.1 Anatomia Macroscópica .....................................................................................48 5.2 Microestrutura ....................................................................................................49 5.3 ESTUDOS ESTEREOLÓGICO ..........................................................................53 5.3.1 Volume ganglionar (VGCC) ...............................................................................53 5.3.2 Número Total de neurônios (N(Total)), número total de neurônios uninucleados (N(uninuc, GCC)) e binucleados (N(binuc,GCC))...................................................................54 5.3.3 Volume médio de neurônios mono NV (uninuc, GCC) e binucleados NV (binuc, GCC) 56 6 DISCUSSÃO .........................................................................................................59 6.1 Anatomia ............................................................................................................59 6.2 Microestrutura ....................................................................................................60 6.3 Estereologia .......................................................................................................60 6.3.1 Volume Ganglionar (VGCC)...............................................................................61 6.3.2 Número Total de Neurônios (N) ......................................................................62 6.3.3 Volume médio dos neurônios ( NV ) ..................................................................63 7 CONCLUSÕES .....................................................................................................66 8 SUGESTÕES PARA ESTUDOS FUTUROS.........................................................67 REFERÊNCIAS........................................................................................................69

INTRODUÇÃO

LADD, A. A. B. L.
23
1 INTRODUÇÃO
O gânglio cervical cranial (GCC) do preá foi o objeto deste estudo
experimental, que serviu como modelo para analisarmos quantitativamente a
dinâmica neuronal pós-natal. Futuramente, estes resultados preliminares servirão de
base para um estudo específico sobre a neurogênese.
O preá (Galea Spixii spixii) (WANGLER, 1831) é um pequeno roedor da classe
dos mamíferos, com comprimento de 15 a 25 cm nos adultos. Seu peso ao nascer
varia de 30 a 70g, podendo atingir entre 300g e 800g, quando adultos. Pertencente à
Ordem Rodentia; à Família Cavidae e ao Gênero Gálea; são animais desprovidos de
calda, com superfície dorsal escura, acinzentada, e ventral branca, com manchas
infraoculares e pós-auriculares também brancas (MINISTÉRIO DA SAÚDE, 2002).
São encontrados em todos os estados da região nordeste do Brasil, também em
Minas Gerais e Mato Grosso (MOOJEN, 1952). Possui hábitos crepusculares
basicamente noturnos. Vive em capinzais, entre moitas e arbustos, às vezes, perto
de áreas alagadas, onde consegue mergulhar e nadar. Alimentam-se de brotos,
folhas, sementes, tubérculos e raízes. Quando seu ambiente natural sofre
alterações, invade arrozais e hortas e é considerado como praga. (CORRADELLO,
1987).
O sistema nervoso autônomo (SNA) supre o coração, vasos sanguíneos,
órgãos intestinais, genitais, urinários e as vias respiratórias. Os nervos regulam e
coordenam funções corpóreas como a atividade secretora de glândulas e a
contração e relaxamento dos músculos. O sistema nervoso autônomo é organizado
em grupos de gânglios, os quais podem ser divididos esquematicamente em quatro
grupos: paravertebrais, pré-vertebrais, paraviscerais e intramurais. Estes gânglios
não são apenas simples estações de retransmissão, mas locais de convergência,
divergência, facilitação e inibição de impulsos nervosos (MATTHEWS, 1983;
KRENLEN, 1984; BAKEWELL, 1995; GABELLA, 2004). Nos mamíferos, o SNA
possui uma estrutura comum, sendo compreendido pelos neurônios eferentes que
inervam toda a musculatura lisa do corpo e glândulas e pelos neurônios aferentes
que recebem e transmitem reflexos e sensações dos órgãos viscerais (GABELLA,
2004).

LADD, A. A. B. L.
24
O gânglio cervical cranial (GCC) é classificado como um gânglio paravertebral
e tem sido consideravelmente estudado durante as últimas quatro décadas. Estudos
sobre estrutura e função dos gânglios autônomos utilizam o GCC devido ao seu fácil
acesso e tamanho, o que permite a sua precisa localização. O GCC situa-se
dorsalmente ao nervo vago e próximo à bifurcação da artéria carótida comum.
Responsável pela inervação de toda a área da cabeça, inervando a túnica adventícia
de todas as artérias desta região (KÖNIG e LIEBICH, 2004), emite fibras pós-
ganglionares para os nervos cranianos óculomotor (III), glossofaríngeo (IX) e vago
(X) e dá origem a fibras simpáticas pós-ganglionares que se dirigem para a glândula
pineal.
Portanto, esta pesquisa experimental pode contribuir fornecendo base de
dados para outros estudos das áreas de cirurgia e neurocirurgia, quanto à
localização, topografia e aspectos microestruturais e quantitativos do GCC em
roedores silvestres (preás), evitando assim intervenções cirúrgicas inacuradas que
podem causar injúrias na região cervical, desencadeando disfunções simpáticas,
sendo a principal delas a Síndrome de Horner, representada por um grupo de sinais
clínicos que incluem anisocoria, enoftalmia, protrusão da terceira pálpebra, ptose e
aumento da temperatura da face como resultado da vasodilitação periférica. Esta
síndrome pode acometer tanto animais (principalmente gatos, cães e equinos)
quanto humanos (BOYDELL, 1995; BELL et al. 2001; OZEL, 2004)
Os neurônios autonômicos têm sido examinados sob microscopia de luz e
eletrônica (BARTON; CAUSEY, 1958; PICK et al., 1964).
Comumente há correlação alométrica entre o tamanho neuronal e o tamanho
do corpo do animal (GABELLA, 2004). Farel (2003) cita que, em ratos, o número de
neurônios no gânglio de raiz dorsal (DRGs) lombar é quase 35% maior em adultos
do que em neonatos, sem proliferação neuronal, resultado este que sugere que os
DRGs de ratos jovens contém uma população de neurônios que não sofrem
diferenciação completa, vindo esta ocorrer tardiamente. Neste estudo, os critérios de
contagem foram baseados em resultados de experimentos com marcação
retrógrada e complementada por métodos imunohistoquímicos, usando o marcador
de mitose Brd-U (Bromodeoxyuridina) e utilizando, a seguir, métodos estereológicos
para a quantificação das estruturas imunorreativas.
Através de estudos autorradiográficos em coelhos, Filichkina (1981) observou
que gânglios paravertebrais apresentaram neurônios mono e binucleados com

LADD, A. A. B. L.
25
quase a mesma intensidade no total de síntese protéica. Entretanto, Forsman et al.,
(1989) não detectou diferenças ultraestruturais nos núcleos de ambas as células,
entretanto, análises microdensitométricas mostraram que células binucleadas do
gânglio celíaco-mesentérico de cobaias contêm o dobro de DNA em relação às
células uninucleadas.
Em relação à frequência de neurônios binucleados, Eranko (1972) cita que
em ratos ocorrem algumas mitoses nos neurônios após o nascimento, sabendo-se
que o número final de neurônios é estabelecido durante a vida fetal, e que da
primeira semana após o nascimento, em ratos, o número de neurônios principais
permanece inalterado ou mostra um pequeno decréscimo. Com isso, Forsman et al.
(1989) sugere que a ocorrência de células binucleadas pode ser devido a uma
interrupção de mitoses durante o processo de diferenciação. A determinação dos
tipos neuronais pode ser verificada também quanto a sua funcionalidade (GUADIX et
al., 1979). Células binucleadas têm sido reportadas em roedores silvestres
brasileiros 36% e 13% em capivaras jovens e adultas (RIBEIRO; DAVIS; GABELLA,
2004 e RIBEIRO, 2006), respectivamente, porém seu significado funcional ainda é
obscuro.
Desta forma, a primeira fase desta pesquisa teve como objetivo analisar
morfoquantitativamente os GCCs de preás durante quatro períodos do
desenvolvimento pós-natal (neonatos, jovens, adultos e animais de meia-idade).
Concomitantemente à análise das fases etárias com a aplicação dos métodos
estereológicos, foi possível estabelecer uma relação entre idade, número total de
neurônios (uninucleados e binucleados), volume neuronal, volume ganglionar e
massa corporal.Preliminarmente, com o uso de marcadores mitóticos específicos,
como o Brd-U (Bromodeoxyuridina) Farel (2003) e Valero (2005), investigamos no
GCC de preás a resposta destes neurônios ao Brd-U. Em um estudo mais futuro,
pretendemos investigar, quimicamente e quantitativamente, a possível ocorrência de
divisão celular durante a maturação e o envelhecimento, investigando assim a
dinâmica neuronal pós-natal bem como os fenômenos que a regem: neurogênese x
diferenciação tardia.
Contudo, os dados desta pesquisa poderão ser comparados posteriormente
com capivaras e outros roedores, podendo assim fornecer bases citoarquiteturais
para o estudo, “in vivo”, do tecido nervoso destes roedores selvagens pertencentes a
fauna brasileira.

LADD, A. A. B. L.
26
OBJETIVOS

LADD, A. A. B. L.
27
2 OBJETIVOS Este capitulo destina-se a descrever os objetivos investigados neste estudo.
Gerais:
Analisar morfoquantitativamente (por meio de métodos estereológicos) o
desenvolvimento pós-natal no gânglio cervical cranial do preá (neonatos, jovens,
adultos e animais de meia-idade)
Específicos:
Estimar os seguintes parâmetros do gânglio cervical cranial em preás durante o
desenvolvimento pós-natal:
- Volume Ganglionar (VGCC)
- Número total de neurônios uninucleados (Nuninuc)
- Número total de neurônios binucleados (Nbinuc)
- Número total de neurônios (Ntotal)
- Volume médio de neurônios mononucleados ( Nv uninuc)
- Volume médio de neurônios binucleados ( Nv binuc)

LADD, A. A. B. L.
28
REVISÃO DE LITERATURA

LADD, A. A. B. L.
29
3 REVISÃO DE LITERATURA
Nesta seção faremos, à luz da literatura científica, uma abordagem descritiva
dos conceitos relacionados à forma e função do gânglio cervical cranial, que
subsidiaram a realização deste trabalho.
3.1 Características da macroestrutura e função
O sistema nervoso autônomo, em um contexto amplo e simplificado, é
considerado como o elo entre o sistema nervoso central e os órgãos, criando vias
aferentes e eferentes de ligação. Este sistema é organizado em feixes nervosos e
grupamentos ganglionares, que podem ser esquematicamente subdivididos em
quatro grupos: paravertebrais; pré-vertebrais; paraviscerais e intramurais. Estes
gânglios não são apenas simples estações de retransmissão, mas locais de
convergência, divergência, facilitação e inibição de impulsos nervosos (NICKEL et
al., 1975; MATTHEWS, 1983; KRENLEN, 1984; BAKEWELL, 1995; GABELLA,
2004).
O gânglio cervical cranial (GCC) é um gânglio paravertebral do sistema
nervoso autônomo simpático, podendo ser reconhecido macroscopicamente como
uma protusão ao longo dos troncos nervosos cervicais, direito e esquerdo,
denominado de tronco simpático (GABELLA, 2004).
O tronco simpático é uma estrutura bilateral simétrica, estendendo-se da base
do crânio até o sacro. No pescoço, o tronco simpático cervical encontra-se dorsal ao
nervo vago e à artéria carótida comum e ventral ao processo transverso das
vértebras e músculos epiaxiais. Cada tronco simpático cervical tem no mínimo dois
gânglios proeminentes, o gânglio cervical cranial e o gânglio cervical inferior (ou
caudal) que está fundido ao primeiro ou segundo gânglio torácico, sendo
denominado de cervicotorácico ou estrelado. Esporadicamente pode ocorrer o

LADD, A. A. B. L.
30
gânglio cervical médio (HEDGER; WEBBER, 1976; BALJET; DRUKKER, 1979;
EVANS; SURPRENANT, 1996).
Ao GCC têm sido direcionadas várias pesquisas científicas, pois este possui
um extenso território de inervação, inervando estruturas cefálicas e cervicais, tais
como: íris, glândula pineal, membrana nictitante, glândula submandibular, tireóide,
língua, mucosa oral, região cutânea da face, orelha e do pescoço (FLEET ; BELL,
1991; LUEBKE ; WRIGTH, 1992; HUBBARD et al 1999; MOLLER ; LIU, 1999;
HAYAKAWA et al 2000; LADIZESKY, et al 2000) e também devido a sua fácil
localização e acesso, tamanho e vascularização.
Vários estudos macroscópicos têm evidenciado a relativa semelhança quanto
à localização e posição do GCC em diferentes espécies animais de pequeno, médio
e grande porte, podendo estar sujeitos a pequenas alterações promovidas pela
espécie e idade. (CUI-SHENG et al. 1998; KABAK et al. 2005; BAO-PING et al.
2005; KABAK 2007; FIORETTO et al., 2007).
Em ratos, Ribeiro et al (2004) descrevem a localização do GCC na região
mais cranial do pescoço, dorsalmente em contato com o nervo vago e ventralmente
com a carótida interna, apresentando formato fusiforme. Kabak (2007) confirmou a
localização do GCC em cobaias (Cavia porcellus) caudoventralmente ao gânglio
distal do nervo vago, medialmente ao músculo digástrico e hioglosso, ventrolateral
ao músculo longo da cabeça, ventral à bula timpânica, apresentando formato oval.
Em cães, o GCC está coberto pelo músculo digástrico e situado cranialmente
ao gânglio distal do vago, em formato fusiforme, apresentando 4-6 mm de
comprimento no eixo longo e 2,5 mm no eixo curto; este apresenta ligação com o
nervo hipoglosso, e próximo à artéria carótida interna. (FIORETTO et al., 2007).
Neste mesmo trabalho foi analisado o GCC de gatos que, em termos topográficos,
apresentou-se semelhante, com exceção à sua proximidade com a artéria occipital e
não a carótida interna como nos cães.
Kabak et al. (2005) observaram em suínos o GCC posicionado crânio-
ventralmente ao gânglio distal do vago, medialmente à extremidade de processo
jugular, dorsal à base da cartilagem epiglote, ventral ao Atlas e medial à raiz comum,
formada pelas artérias carótida interna e artéria occipital. As dimensões do GCC
para suínos adultos foram 17,74 mm de comprimento, 3mm de largura e 2,58mm de
espessura.

LADD, A. A. B. L.
31
Os objetivos de Bao-Ping et al. (2005) foram descrever a forma, localização e
arranjo dos ramos do GCC da espécie Bos grunniens, bovino comumente
encontrado no Tibet. Nesta espécie, o GCC tem formato fusiforme, apresentando as
seguintes medidas 19,72 mm de comprimento, 7,65mm de largura e 4,55 mm de
espessura.
Cui-Sheng et al. (1998), estudando a anatomia deste órgão em camelos
(Camelus bactrianus), encontram uma estrutura bem desenvolvida com médias de
comprimento, largura e espessura variando respectivamente entre 15-20 mm, 4-6
mm, e 3 mm. Sua localização inclui a superfície rostrolateral do músculo largo da
cabeça, ventral ao músculo esternocefálico e coberto pela glândula mandibular; e o
eixo longo paralelo ao nervo vago.
A importância clinica no estudo do GCC pode ser também caracterizada por
sua relação com algumas disfunções simpáticas, ocasionadas pela ocorrência de
injúrias na região cervical e que as desencadeiam, sendo que a principal delas
associada a este gânglio é a Síndrome de Horner, representada por um grupo de
sinais clínicos (que incluem anisocoria, enoftalmia, protrusão da terceira pálpebra,
ptose e aumento da temperatura da face como resultado da vasodilitação periférica)
relacionados com a perda momentânea ou permanente da inervação autonômica
simpática, sendo secundária a processos inflamatórios e/ou compressivos do GCC
(devido a neoplasias benignas ou malignas). Ainda, destacam-se na
morfopatogênese da Síndrome de Horner os procedimentos de causa hiatrogênica,
ou seja, intervenção cirúrgica inacurada na região cervical, ocasionando lesão no
GCC e/ou em suas fibras. Esta síndrome pode acometer tanto animais
(principalmente gatos, cães e equinos) quanto humanos (BOYDELL, 1995; BELL et
al., 2001; OZEL, 2004).
Ozel (2004) relata um caso incomum, em que uma criança, após
toracostomia, um procedimento comum aplicado em várias patologias pulmonares,
apresentou sintomas compatíveis com a Síndrome de Horner, miose, ptose e
enoftalmia devido a uma interrupção do fluxo de inervação dos neurônios pré-
gaglionares de segunda ordem. Além disso, bloqueios anestésicos locorregionais na
região cefálica e cervical podem ocasionar sintomas clínicos relacionados à esta
síndrome (ELIAS, 2000).
A relação do gânglio cervical cranial com a Síndrome de Horner está bem
descrita em um estudo de 2002, no qual uma fêmea canina Airedale Terrier de 9

LADD, A. A. B. L.
32
anos de idade foi eutanasiada e sofreu necropsia após um progressivo caso clínico
que incluía a Síndrome de Horner (caracterizada clinicamente como uma doença
inflamatória restrita a nervos periféricos, raízes nervosas e gânglios específicos),
ptose palpebral unilateral e atrofia unilateral dos músculos mastigatórios. Esta
síndrome é comumente diagnosticada em cães e gatos, sendo a sua classificação, e
sua morfopatogênese ainda desconhecidas (PANCIERA et al. 2002).
3.2 Aspectos microestruturais e quantitativos
O gânglio cervical cranial de pequenos, médios e grandes mamíferos
apresenta composição interna variável em sua arquitetura geral, dependendo da
espécie, idade e antimeria (GABELLA et al., 1988; RIBEIRO et al., 2002; RIBEIRO et
al., 2004; RIBEIRO, 2006; ABRAHÃO et al., 2009; MELO et al., 2009; TOSCANO et
al., 2009).
No GCC encontram-se vários tipos celulares SIF cells (small intensely
fluorescent cells), células endoteliais, fibroblastos, mastócitos, (GABELLA., 2004) em
acréscimo aos neurônios ganglionares principais que podem ser uni ou binucleados.
Estes últimos têm sido descritos em gânglios simpáticos de pequenos a grandes
roedores: paca (Agouti paca), capivara (Hydrochaeris hydrochaeris), cobaia (Cavia
porcelus) e lagomorfos: coelhos (Oryctolagus cuniculus); (FORSMAN et al, 1989;
SASAHARA et al., 2003; RIBEIRO et al 2004; RIBEIRO, 2006; ABRAHÃO et al.,
2009; MELO et al., 2009; TOSCANO et al., 2009).
Dos métodos morfométricos aplicáveis para amostras biológicas, o método
estereológico é no momento o mais correto (STERIO, 1984; PAKKENBERG;
GUNDERSEN, 1988; MAYHEW; GUNDERSEN, 1996; HOWARD; REED, 2005).
Muitos dados quantitativos na literatura foram obtidos com métodos que não
poderiam ser aceitos hoje (ABERCROMBIE, 1946; EBBESSON, 1968; DAVIES,
1978; GORIN; JOHNSON, 1980; PURVES et al., 1986). Naturalmente, estes dados
não podem ser desconsiderados, mas seus resultados de tal forma não são
acurados e confiáveis.

LADD, A. A. B. L.
33
Um estudo comparativo do GCC, usando métodos estereológicos confiáveis,
entre espécies adultas com tamanhos distintos variando em quase 1000 vezes (rato,
capivara e cavalo), buscando associação entre volume ganglionar, tamanho e
número total de neurônios e analisando secções semifinas embebidas em resina
plástica, observaram uma arquitetura menos complexa em GCC de ratos do que em
GCC de capivaras e cavalos (RIBEIRO et al 2006).
Como aspecto geral, o GCC apresenta-se envolto por uma cápsula de tecido
conjuntivo, composta de colágeno e fibroblastos de espessura variável, sendo que
os neurônios desses gânglios apresentaram-se separados por fibras nervosas e
capilares intraganglionares, sendo que em capivaras e cavalos os neurônios estão
compartimentalizados por invaginações da cápsula externa formando septos
profundos nos gânglios dessas duas espécies (RIBEIRO et al., 2006). O mesmo
aspecto compartimentalizado com penetrações da cápsula conjuntiva no GCC foi
observado também em outros roedores silvestres como a paca (ABRAHÃO et al.,
2008). A idade também pode afetar a dinâmica tecidual do GCC. Em cobaias (Cavia
porcellus) foi observado um aumento de tecido não neuronal com o decorrer da
idade, alterando a densidade de volume dos neurônios e também o volume do
gânglio (TOSCANO et al., 2009). Sendo a hipertrofia um mecanismo adaptativo
amplamente conhecido durante o desenvolvimento pós-natal em muitos gânglios
autonômicos de mamíferos de porte médio e grande (MIOLAN e NIEL, 1996;
GAGLIARDO et al., 2005; FIORETTO et al., 2007).
O número e volume neuronal, dois importantes parâmetros de comparação,
estão sujeitos a variações interespecíficas, relacionados à idade ou ainda a fatores
individuais. Ribeiro (2006), relata um decréscimo no número total de neurônios de
16% durante a maturação e 23% para os neurônios binucleados com aumento do
peso dos animais em 98,5%.
O número total de neurônios do GCC de rato, cavalo e capivara, obtido por
Ribeiro et al., (2004) através de quantificação estereológica, baseada na utilização
do método do disector físico para determinar a densidade neuronal e no Princípio de
Cavalieri para estimar o volume ganglionar, em que a combinação destes dois
métodos possibilita estimar-se o número total de neurônios do gânglio cervical
superior de forma acurada. Os autores observaram uma correlação alométrica para
o número de neurônios dessas diferentes espécies, sendo que o número total de
neurônios para ratos foi 18840, para capivaras foi 1520000 e para cavalos foi

LADD, A. A. B. L.
34
3390000. O gânglio da capivara mostrou certas características distintas das outras
espécies. Como fato interessante, observou-se a ocorrência em média de 13% de
neurônios binucleados no gânglio cervical de capivaras adultas. A principal
conclusão deste estudo foi de que número total de neurônios é dependente da
massa corporal do animal, entretanto, isto não é necessariamente verdadeiro em
relação ao tamanho neuronal - 9000µm3 para ratos; 32 µm3 para capivaras e 20.200
µm3 para cavalos – medidos por métodos estereológicos indiretos.
Melo et al., (2009) reportaram um aumento de 95% no número de neurônios
mononucleados de pacas (Agouti paca), roedor silvestre de porte intermediário,
durante o desenvolvimento pós-natal, acompanhado do aumento do volume do
gânglio GCC em 154% e de 50% para o volume total de neurônios do GCC. Já
Abrahão et al (2009), estudando efeitos da antimeria no GCC, observaram um efeito
interessante da interação entre o peso e idade de pacas, em que o GCC direito de
pacas senis apresentaram mais neurônios mono e binucleados que o GCC esquerdo
de todas as idades, enquanto os neurônios do GCC esquerdo foram maiores que os
neurônios do GCC direito de qualquer idade.
Devor (1985) e colaboradores, usando métodos estereológicos para
contagem, concluíram que ocorreu um aumento de 50% no número de neurônios no
do gânglio da raiz dorsal lombar (DRG) em ratos que aumentaram o peso de 250
para 400g.
De outra parte, Coggeshall et al., (1994) e Pover et al., (1994), usando o
disector físico, não encontraram evidências de aumento do número neuronal em
dois estudos feitos separadamente em ratos; se os ratos adultos tivessem mais
neurônios, este número de neurônios poderia ter sido devido à neurogênese,
diferenciação tardia ou transdiferenciação. Em um ou outro evento, existiria a
possibilidade de que os mecanismos responsáveis para a adição do neurônio
poderiam ser recrutados para compensar os danos resultantes de ferimento ou de
doença, dizem os autores. Estes resultados discrepantes podem ter ocorrido devido
a inacurácias metodológicas, ainda que usando métodos estereológicos.
Popken e Farel (1997), utilizando métodos estereológicos e não
estereológicos de quantificação neuronal, verificaram um aumento no número de
neurônios em ratos adultos em relação a neonatos. O método de contagem não
estereológico (perfis de contagem núcleo/nucléolo) indicou um aumento de 19%
(8.805 neurônios) e o disector físico mostrou um aumento de 28% (10.720

LADD, A. A. B. L.
35
neurônios), contudo a diferença no número de neurônios entre ratos adultos e
neonatos pode ser devido à atividade mitótica ou à ocorrência de neurônios
incompletamente diferenciados em neonatos.
Farel (2003) também identificou um aumento de 35% no número total de
neurônios de ratos adultos quando comparados aos neonatos. A possibilidade da
neurogênese foi examinada por este autor, injetando subcutaneamente em ratos
neonatos um marcador específico para ciclo celular: bromodeoxyuridina (Brd-U). A
quantificação foi conduzida por meio de métodos estereológicos de contagem, cuja
validade e confiança têm sido confirmadas por reconstruções tridimensionais. Os
resultados revelaram que este aumento no número total de neurônios ocorreu pela
de diferenciação neuronal tardia e não devido à neurogênese (FAREL, 2003)
Este marcador celular (Brd-U) é utilizado na identificação de atividade
mitótica, pois o exame de protocolos da utilização deste marcador por MOMMA et.
al.(2002), sob microscopia eletrônica, mostrou que a marcação é sempre confinada
ao núcleo, indicando que a incorporação de Brd-u foi devido à síntese de DNA.
NGWENYA et.al (2005) descreveram as vantagens da utilização de Brd-u e os
motivos pelos quais ele está sendo amplamente utilizado para marcar células em
mitose, dentre os quais, o baixo risco de intoxicação celular e rapidez na análise
imunohistoquímica em contraste com outros métodos mais demorados, como os
autorradiográficos,e ainda permite a utilização de imunofluorescência.
Valero et al., (2005) utilizaram também como marcador de proliferação celular
o BrdU (5-bromo-2’-deoxyuridine) para estudar a proliferação celular nuclear
antigênica. Dois grupos de marcadores de proliferação celular podem ser
distinguidos: análogos de base nitrogenada e marcador de proliferação celular
endogênica. Atualmente, o análogo de base nitrogenada (BrdU) é o mais utilizado
para demonstrar a proliferação celular.
Em verdade, efeitos dissonantes quanto a parâmetros morfométricos durante
o envelhecimento, como, por exemplo, aumento ou diminuição do número total de
neurônios é assunto para discussões interessantes, e com o advento de técnicas
modernas de estereologia e imunohistoquímica, poderemos contribuir com a
elucidação dos mecanismos envolvidos nesse processo.

LADD, A. A. B. L.
36
Materiais e Métodos

LADD, A. A. B. L.
37
4 - MATERIAIS E MÉTODOS
Todos os procedimentos descritos neste estudo foram aprovados em
29/10/2008 pela Comissão de Bioética da Faculdade de Medicina Veterinária e
Zootecnia da Universidade de São Paulo, sob o número de protocolo 935/2006.
4.1 Materiais
Para este estudo, foram utilizados vinte gânglios (GCC) esquerdos de 20
preás (Galea spixii spixii), (Figura.1), machos, saudáveis (ausência clínica de
neuropatias), obtidos do Centro de Multiplicação de Animais Silvestres (CEMAS) da
Universidade Federal Rural do Semi-árido Nordestino (UFERSA), antiga ESAM
(Escola Superior de Agricultura), Mossoró – RN, entidade credenciada e registrada
pelo Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis
(IBAMA) como criadouro científico sob registro n° 12.492-000404.
Os animais foram separados em quatro grupos etários, de acordo com a fase
do desenvolvimento pós-natal, sendo assim:
Grupo I (neonatos): Cinco GCC de animais com 01 a 03 dias de idade (peso
corpóreo 0,37-0,63g)
Grupo II (jovem): Cinco GCC de animais com 01 mês de idade (peso corpóreo
148-215g)
Grupo III (adultos): Cinco GCC de animais com 12 meses de idade (peso
corpóreo entre 272-427 g)
Grupo IV (Meia idade): Cinco GCC de animais de 36 meses de idade (peso
corpóreo 284-403 g)

LADD, A. A. B. L.
38
4.2 Métodos
4.2.1- Administração de bromodeoxyuridina (Brd-U)
Tendo como base o protocolo de VALERO et al. (2005) para a administração
de bromodeoxyuridina (Brd-U), um marcador de ciclo celular, síntese de DNA e,
portanto sugestivo de atividade mitótica (FAREL, 2003; TAUPIN, 2007), os animais
foram injetados intraperitonealmente com doses de 50 µm/g de Brd-U numa
concentração de 25mg/ml, diluídas em tampão fosfato salino (PBS) (0,1 M e pH de
7,4), nos períodos de 24 e de 2h antes de serem eutanasiados.
Figura 1- Fotografia de um Preá (Galea spixii spixii) adulto.

LADD, A. A. B. L.
39
4.2.2 Eutanásia
Os animais foram previamente tranquilizados com aplicação de azaperone
(4mg/kg) intraperitonealmente (IP), seguida da aplicação de sulfato de atropina
(0,06mg/kg) via intraperitoneal (IP). Para a eutanásia ativa, o cloridrato de quetamina
(20mg/kg) foi associado ao cloridrato de xilazina (1,5mg/kg), ambos na mesma
seringa, para indução da anestesia na mesma via citada anteriormente. Logo em
seguida, os animais receberam uma overdose de anestésico, Tiopental sódico,
(80mg/100g de peso corpóreo) (IP).
4.2.3 Dissecação e individualização do GCC
Subsequentemente às injeções de Brd-U e à eutanásia, a cavidade torácica
foi aberta e uma cânula foi inserida dentro do ventrículo esquerdo do coração até
atingir a aorta ascendente. Primeiramente foram perfundidos 100ml de solução de
lavagem composta de tampão fosfato salino (PBS) (0,1M; pH 7,4) (Synth®) contendo
heparina (1%). A aurícula direita foi seccionada para extravasamento do sangue. A
seguir, os espécimes foram perfundidos com solução fixadora composta por
formoldeído (4%) [SigmaTM] em tampão fosfato salino PBS (0,1M; pH 7,4) (Synth®) à
temperatura de 10-15 °C, com volume total perfazendo 10% do peso corpóreo.
Os GCCs esquerdos foram dissecados e removidos do animal e suas
cápsulas conjuntivas foram retiradas por microdissecação, utilizando um microscópio
cirúrgico Leica® DM com aumento de 40x. (Figura. 2). Posteriormente, os GCCs
foram pesados e medidos nos seus três eixos: comprimento, largura e espessura,
(Tabela 1) usando o paquímetro digital Digimess®. Após a mensuração dos

LADD, A. A. B. L.
40
gânglios, os mesmos foram lavados com água destilada, verticalizados (BADDELEY
et al., 1986), embebidos em uma solução aquosa de Ágar (10%) e seccionados
seriada, uniforme e sistematicamente (SURS) em micrótomo de lâmina vibratória
(vibrótomo) Leica® (VT 1000S) com 40µm de espessura. As secções foram
montadas em lâminas com aderência eletrostática e coradas com giemsa (Merck®)
para o estudo quantitativo (estereológico). Contudo, as secções foram desidratadas
em séries crescentes de etanóis, montadas sob lamínulas e observadas ao
microscópio epi-fluorescência Leica® DMR e analisadas com o auxílio de um
software estereológico (CAST®) Versão 3.0.8.0.
4.2.4 – Reação de imunohistoquímica (Estudo qualitativo)
Bromodeoxiuridina (Brd-U) é atualmente um dos métodos de eleição para
estudos de proliferação e neurogênese in vivo (DEL RIO; SORIANO, 1989). O Brd-U
é sistematicamente injetado e incorporado como um análogo da Timidina dentro do
Figura 2- Fotomacrografia evidenciando a cápsula conjuntiva do GCC esquerdo de preá jovem. Gânglio (Cabeça de seta) Asterisco (cápsula conjuntiva).100µm.
*

LADD, A. A. B. L.
41
DNA de todas as células durante a síntese de DNA (DOLBEARE, 1995, 1996). Uma
vez incorporado ao DNA da célula, o Brd-U pode ser detectado
imunohistoquimicamente em células pós-mitóticas até o final de suas vidas, exceto
naquelas células que proliferam consecutivamente, as quais diluirão
exponencialmente o sinal do Brd-U no núcleo (WARD et al., 1991).
No entanto, mais recentemente tem sido apregoado que o Brd-U não é
primariamente um marcador de células em proliferação e, sim, um marcador de
síntese de DNA. Isto traz profundas consequências no estudo e análise da
neurogênese, visto que pode conduzir a interpretações errôneas se os controles
apropriados não forem conduzidos (TAUPIN, 2007).
Brevemente, o protocolo da reação consistiu do bloqueio da peroxidase
endógena, utilizando-se peróxido de hidrogênio (0,5% em etanol 50%) durante 30
minutos, seguido de bloqueio com soro normal de cabra (10% em PBS, pH 7.4,
0,1M) durante 30 minutos, outra lavagem em PBS (15 minutos) e incubação em
anticorpo primário monoclonal “anti-mouse” bromodeoxyuridina (Brd-U) (1:100)
durante 48 horas. A seguir, procedeu-se à lavagem em PBS (15 minutos), seguido
de incubação em anticorpo secundário “goat anti-mouse” conjugado com
“horseradish peroxidase” (HRP) (1:200) durante uma hora. Os cortes foram então
novamente lavados e a reação foi desenvolvida, utilizando-se a “3,3’-
diaminobenzidine tetrahydrochloride” (DAB, Sigma®) por 45 minutos.
Posteriormente, os cortes foram lavados em PBS (15 minutos), desidratados em
séries crescentes de etanóis e xilóis e montados sob lamínula com DPX (FLUKA®) e
a seguir, visualisados por meio da estação de microscopia de luz e epifluorescência
Leica DM 6000®.(VALERO et. al. 2005).
4.2.5 - Estudo estereológico
Os parâmetros estereológicos estimados foram: volume do gânglio, número
total de neurônios e volume neuronal médio (STERIO, 1984; PAKKENBERG ;
GUNDERSEN, 1988, 1989; POVER; COGGESHAL, 1991; MAYHEW;
GUNDERSEN, 1996; HOWARD ; REED, 2005)

LADD, A. A. B. L.
42
4.2.5.1- Volume do Gânglio (VGCC)
Para estimar o volume do gânglio, foi utilizado o Princípio de Cavalieri aplicado
aos cortes histológicos do GCC (Figura.03) (MAYHEW e OLSEN, 1991; HOWARD e
REED, 2005).
Inicialmente, cada GCC foi verticalizado, embebido em solução de Ágar (10%) e
seccionado exaustivamente (40µm de espesssura) por meio de um micrótomo de
lâmina vibratória Leica® VT100S. Este procedimento gerou um conjunto de secções
com espessura média T.
Assim como Ribeiro, Davis e Gabella. (2004) para o GCC de cavalos e capivaras
e por Mayhew e Olsen (1991) para o encéfalo humano, a seguinte fórmula foi usada
para estimar o volume do gânglio:
Vg= T x K x ∑A, onde:
T = espessura média das secções ganglionares
K= distância entre as secções
A = área das seções ganglionares
O coeficiente de erro (CE) para a estimativa de volume pelo Princípio de Cavalieri
CE(VGCC) foi estimado de acordo com a fórmula modificada por Gundersen et al.
(1999).
A fórmula utilizada foi:
Variância “noise” da contagem de pontos:
Var [noise] = c. (b/√a).(√n. ΣP) Onde c é uma constante (c = 0,0724)
b/√a expressa a forma ou complexidade dos perfis analisados
n é o número de secções utilizadas ΣP é o número total de pontos utilizados para estimar a área das secções.
Variância devido a amostragem sistemática e uniformemente aleatória (surs)
Var [srs] = (3(A-var[noise])-4B+C)/240 CE = √var[total]
Os valores médios para CE (VGCC) nos grupos etários estudados: neonatos,
jovens, adultos e senil, foram, respectivamente 2,97%, 2.88%, 3,18%, 2,19%.

LADD, A. A. B. L.
43
Para fins de ilustração, está representado na figura 4 o método do “optical
disector”, utilizado não somente para a quantificação do número total dos neurônios
mono e binucleados, mas, também para a amostragem das partículas, visando a
estimativa de seus volumes.
Figura 3- Fotomicrografia da secção de um GCC de um preá adulto onde se exemplifica a aplicação do Princípio de Cavalieri para a estimativa do volume ganglionar. Giemsa. Escala de Barra 500µ.

LADD, A. A. B. L.
44
Figura 4- Fotomicrografias de secções histológicas sequenciais do GCC de um preá adulto, exemplificando a aplicação do método do disector óptico em que neurônios uni (M) e binucleados (B) foram amostrados em planos focais ópticos sucessivos (B,C). É possível identificar a posição de cada secção no eixo z na lateral (barra) esquerda da figura. Azul de Toluidina. Escala de

LADD, A. A. B. L.
45
4.2.5.2 Número total de neurônios do GCC (N)
O fractionator óptico é uma combinação do fractionator, um princípio de
amostragem, com disector óptico. De acordo com o princípio do fractionator, o
número total de neurônios do GCC (N) foi estimado multiplicando-se o número total
de partículas contadas pelo inverso das frações amostradas (GUNDERSEN (1988),
GUNDERSEN (2002) E ANDERSEN (2004), utilizando-se a seguinte fórmula:
hsfasfssfQN 1.1.1.∑ −=
Onde:
ssf é a fração da amostragem das secções
asf é a fração das áreas amostradas para contagem
hsf é a fração da espessura do corte efetivamente utilizada para contagem
Q- é o número de partículas contadas pelo método do disector óptico que, neste
caso, são neurônios
Com o método descrito acima, foram estimados o número total de neurônios
uninucleados ( Nuninuc,GCC) e o número total de neurônios binucleados (Nbinuc, GCC).
Contudo, estimamos o número total de neurônios do GCC (Ntotal) obtido pelo
somatório de Nuninuc,GCC e Nbinuc, GCC.
4.2.5.3 Volume neuronal médio ( Nv ) O volume neuronal foi estimado pelo método do rotator planar. Este método
permite estimar o volume médio e a distribuição volumétrica das partículas
independentemente da sua forma, distribuição ou orientação (VEDEL, JENSEN e
GUNDERSEN, 1993).
A seguinte fórmula foi utilizada para a estimativa do volume neuronal:
∑=i
2l. t . iNv π onde, l2
i é uma distância medida a partir de um ponto fixo da
célula ou fora dela, até uma borda arbitrariamente escolhida na mesma. As
estimativas para Nv foram feitas separadamente: volume médio de neurônios
uninucleados ( Nv UNI) e volume médio de neurônios binucleados ( Nv bi).

LADD, A. A. B. L.
46
4.2.6 Análise Estatística A análise estatística foi feita com o auxílio do software estatístico Minitab 15®
(2007) e levando em conta os princípios de amostragem sistemática e
uniformemente aleatória (SURS) (GUNDERSEN et al. 1999) onde, para as variáveis
que apresentaram distribuição simétrica e igualdade de variância
(homocedasticidade), utilizamos o teste one-way ANOVA seguido pelo teste de
TUKEY e FISHER para comparações múltiplas ou “post-hoc test”. No entanto,
quando as variáveis em estudo apresentaram uma distribuição não simétrica e/ou
não apresentaram igualdade de variância, utilizamos os testes de mediana de
MOOD ou os testes de KRUSKAL–WALLIS e MANN-WHITNEY.
Diferenças entre grupos foram consideradas significativas quando (p<0,05).
No capítulo de resultados, as variáveis serão apresentadas como média (CV);
onde CV refere-se ao coeficiente de variação, sendo este o quociente entre o desvio
padrão e a média.

LADD, A. A. B. L.
47
RESULTADOS
LADD, A. A. B. L.
48
5 RESULTADOS
5.1 Anatomia Macroscópica
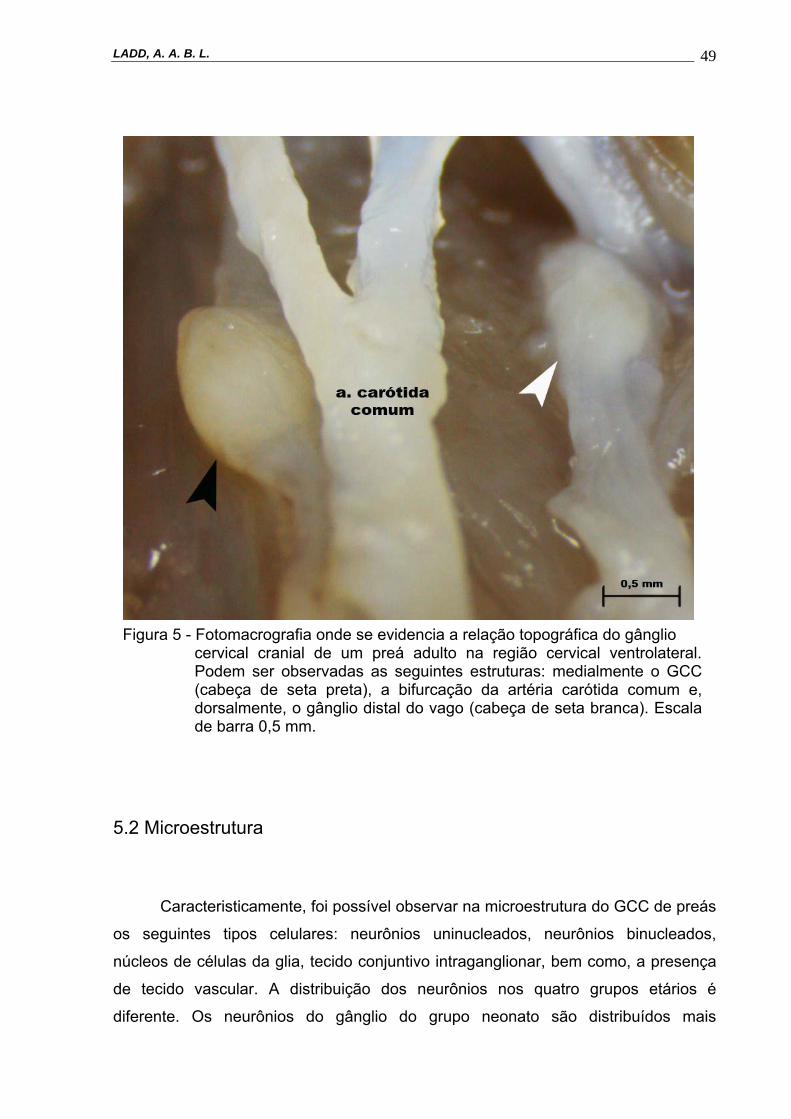
O gânglio cervical cranial GCC é localizado na região ventromedial da
carótida interna e também ventralmente ao gânglio distal do nervo vago. Nos preás
(Galea spixii spixii) foi facilmente identificável, macroscopicamente e
topograficamente, independentemente da idade, seguindo-se o tronco
vagosimpático até a bifurcação da artéria carótida comum na região proximal do
pescoço (Figura 5).
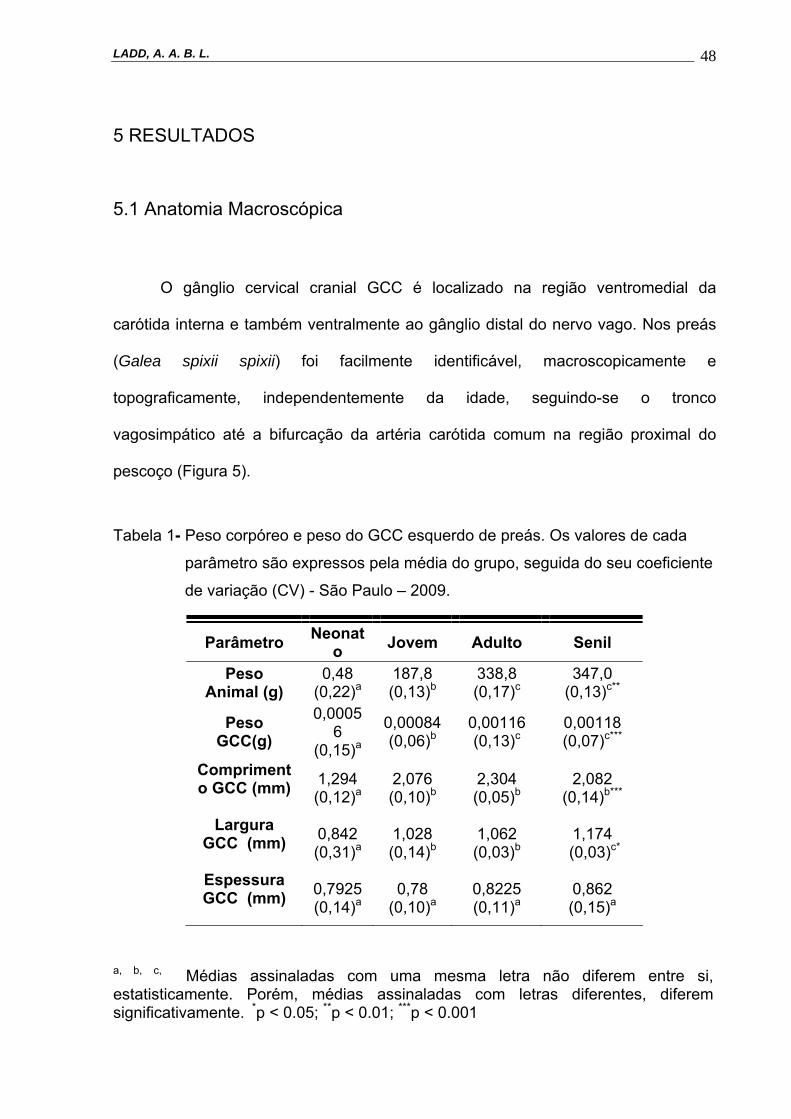
Tabela 1- Peso corpóreo e peso do GCC esquerdo de preás. Os valores de cada
parâmetro são expressos pela média do grupo, seguida do seu coeficiente
de variação (CV) - São Paulo – 2009.
a, b, c, Médias assinaladas com uma mesma letra não diferem entre si, estatisticamente. Porém, médias assinaladas com letras diferentes, diferem significativamente. *p < 0.05; **p < 0.01; ***p < 0.001
Parâmetro Neonato Jovem Adulto Senil
Peso Animal (g)
0,48 (0,22)a
187,8 (0,13)b
338,8 (0,17)c
347,0 (0,13)c**
Peso GCC(g)
0,00056
(0,15)a
0,00084 (0,06)b
0,00116 (0,13)c
0,00118 (0,07)c***
Comprimento GCC (mm)
1,294 (0,12)a
2,076 (0,10)b
2,304 (0,05)b
2,082 (0,14)b***
Largura GCC (mm)
0,842 (0,31)a
1,028 (0,14)b
1,062 (0,03)b
1,174 (0,03)c*
Espessura GCC (mm)
0,7925 (0,14)a
0,78 (0,10)a
0,8225 (0,11)a
0,862 (0,15)a
LADD, A. A. B. L.
49
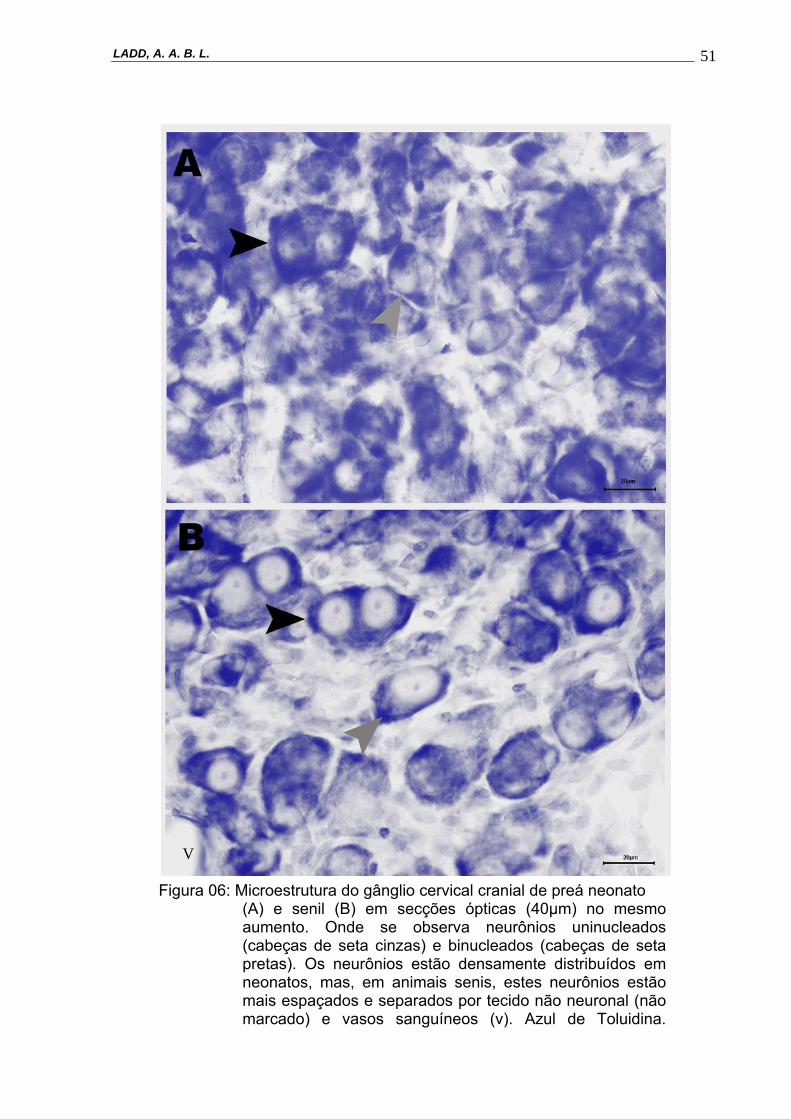
5.2 Microestrutura Caracteristicamente, foi possível observar na microestrutura do GCC de preás
os seguintes tipos celulares: neurônios uninucleados, neurônios binucleados,
núcleos de células da glia, tecido conjuntivo intraganglionar, bem como, a presença
de tecido vascular. A distribuição dos neurônios nos quatro grupos etários é
diferente. Os neurônios do gânglio do grupo neonato são distribuídos mais
Figura 5 - Fotomacrografia onde se evidencia a relação topográfica do gânglio cervical cranial de um preá adulto na região cervical ventrolateral. Podem ser observadas as seguintes estruturas: medialmente o GCC (cabeça de seta preta), a bifurcação da artéria carótida comum e, dorsalmente, o gânglio distal do vago (cabeça de seta branca). Escala de barra 0,5 mm.

LADD, A. A. B. L.
50
densamente. Com o desenvolvimento pós-natal, os neurônios estão difusamente
distribuídos, entre o tecido conjuntivo intraganglionar que se torna mais visível
(Figura 6). O núcleo dos neurônios uninucleados está predominantemente no centro
da célula, podendo estar deslocado do centro em alguns casos. Nos neurônios
binucleados, os núcleos apresentam-se em pólos distintos da célula, sendo que,
para cada núcleo, houve a presença de um a dois nucléolos, tanto para neurônios
uni, quanto para neurônios binucleados.
Foi possível identificar a presença de neurônios Brd-U-positivos em GCC de
preás neonatos e senis (Figura 7).
LADD, A. A. B. L.
51
Figura 06: Microestrutura do gânglio cervical cranial de preá neonato (A) e senil (B) em secções ópticas (40µm) no mesmo aumento. Onde se observa neurônios uninucleados (cabeças de seta cinzas) e binucleados (cabeças de seta pretas). Os neurônios estão densamente distribuídos em neonatos, mas, em animais senis, estes neurônios estão mais espaçados e separados por tecido não neuronal (não marcado) e vasos sanguíneos (v). Azul de Toluidina.
V

LADD, A. A. B. L.
52
Figura 07: Fotomicrografia de secção histológica de GCC de preá neonato (A) e senil (B), onde se evidenciam núcleos de neurônios Brd-U positivos, uninucleados (cabeça de seta preta) e binucleados (cabeça de seta branca). Pode-se observar também vasos sanguíneos (v). Reação de imunohistoquímica de campo claro com revelação por diaminobenzidina. Escala de barra 15 µm.

LADD, A. A. B. L.
53
5.3 ESTUDOS ESTEREOLÓGICO Os resultados dos parâmetros estereológicos investigados serão descritos nesta
seção.
5.3.1 Volume ganglionar (VGCC)
O volume do GCC (Figura 08) para os diferentes grupos etários foi: Neonato
VGCC= 0,34 mm3 (0,45); jovem VGCC= 0,30 mm3 (0,49); Adulto VGCC= 0,39 mm3
(0,42); Senil VGCC= 0,63 mm3 (0,19). As diferenças entre os grupos foram
significantes (p= 0,012).
Figura 8- Volume do GCC em diferentes grupos etários. As diferenças entre os
grupos foram significantes (*p=0,012). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.

LADD, A. A. B. L.
54
5.3.2 Número Total de neurônios (N(Total)), número total de neurônios uninucleados (N(uninuc, GCC)) e binucleados (N(binuc,GCC))
O número total de neurônios (N(Total)) (que é obtido pelo somatório de uni e
binucleados), o número total de neurônios mononucleados (N(uninuc,GCC)) e
binucleados (N(binuc,GCC)) foram: Neonato N(Total)=103680 (0,25), N(uninuc,GCC)=
65324,00(0.20) e N(binuc,GCC)= 38356,00 (0,35); Jovem N(Total)= 87102 (0,26),
N(uninuc,GCC)= 40083,00 (0,20) e N(binuc,GCC)= 39019,00 (0.35); Adulto N(Total)=89206
(0,35), N(uninuc,GCC)= 45226,00 (0,37) e N(binuc,GCC)= 43981,00 (0,34); Senil: N(Total)=
108540 (0,18), N(uninuc,GCC)= 54136,00 (0,21) e N(binuc,GCC)= 54404,00 (0,21) (Figuras
5, 6 e 7). As diferenças entre os grupos não foram estatisticamente significantes
N(Total)(p=0,493), N(uninuc,GCC) (p = 0,118) e N(binuc,GCC) (p=0,261). Figuras 9 a 11.
Figura 9- Número total de neurônios do GCC em diferentes grupos etários. As diferenças entre os grupos não foram significantes (p=0,493). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.
LADD, A. A. B. L.
55
Figura 11- Número total de neurônios binucleados do GCC em diferentes grupos etários. As diferenças entre os grupos não foram significantes (p=0,261). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.
Figura10- Número total de neurônios uninucleados do GCC em diferentes grupos etários. As diferenças entre os grupos não foram significantes (p=0,118). Os triângulos representam os valores individuais e as barras horizontais são médias dos grupos.
LADD, A. A. B. L.
56
5.3.3 Volume médio de neurônios mono ( NV (uninuc, GCC) ) e binucleados ( NV (binuc, GCC))
O volume neuronal médio foi: Neonato NV (uninuc,GCC) = 2916,90 µm3 (0,04) e
NV (binuc, GCC)= 12117,00 µm3 (0,14); Jovem NV (uninuc,GCC) = 3550,00 µm3 (0,22) e
NV (binuc, GCC)= 14430,00 µm3 (0,28) e Adulto NV (uninuc,GCC) = 7409,mm µm3 (0,05) e
NV (binuc, GCC)= 12530,mm µm3 (0,04); Senil NV (uninuc,GCC) = 6701,00 µm3 (0,10) e
NV (binuc, GCC)= 13652,00 µm3 (0,08). As diferenças entre os grupos foram
significativas para NV (uninuc,GCC) (p=0,0001), mas não foram significativas para
NV (binuc, GCC) (p = 0.402) (Figura 11 e 12).
Figura 12- Volume médio dos neurônios uninucleados do GCC em diferentes grupos etários. As diferenças entre os grupos foram significantes (*p=0,0001). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.
LADD, A. A. B. L.
57
Figura 13: Volume médio dos neurônios binucleados do GCC em diferentes grupos etários. As diferenças entre os grupos não foram significantes (p=0,402). Os triângulos representam os valores individuais e as barras horizontais as médias dos grupos.

LADD, A. A. B. L.
58
DISCUSSÃO

LADD, A. A. B. L.
59
6 DISCUSSÃO Neste capítulo, discutiremos os parâmetros divididos nos seguintes sub-
tópicos: anatomia, microestrutura e aspectos estereológicos.
6.1 Anatomia Neste estudo foi observado que o gânglio cervical cranial (GCC) de preás
(Galea spixii spixii) não apresentou alterações relevantes quanto à sua localização e
posicionamento durante a maturação e envelhecimento, estando sua topografia em
concordância com as descrições feitas em outros roedores e mamíferos como, por
exemplo: capivaras, ratos, equinos, cães, felinos, suínos, bovinos e camelos (CUI-
SHENG et al. 1998; RIBEIRO, DAVIS e GABELLA, 2004; BAO-PING, 2005; KABAK
et al.,2005; RIBEIRO, 2006; FIORETTO et al. 2007; KABAK, 2007).
Quanto aos dados macromorfométricos, observou-se o efeito da idade,
havendo um aumento significativo de 60% e 79% referentes ao comprimento e à
largura, respectivamente, durante o período de maturação (neonatos para adultos);
para o período de envelhecimento (adulto para senil), o comprimento apresentou um
pequeno decréscimo de 10% e para a largura, durante este mesmo período,
apresentou um aumento progressivo de mais de 10%. Quanto ao peso do gânglio,
observou-se um aumento de 185% durante a maturação, havendo uma queda de
33% durante o envelhecimento. Estas alterações morfométricas (similares) também
foram observadas em outros roedores (ABRAHÃO et al 2009; TOSCANO et al,

LADD, A. A. B. L.
60
2009; MELO et al, 2009) e se refletiram no volume do gânglio, que é discutido no
item volume ganglionar, a seguir.
6.2 Microestrutura A análise microscópica qualitativa do GCC de preás durante dois períodos
distintos do desenvolvimento pós-natal (maturação e envelhecimento) revelou
diferenças quanto a sua arquitetura, onde, principalmente, pôde ser observado
qualitativamente que, durante a maturação (neonato e jovem), a distribuição dos
neurônios neste período era mais aglutinada e, durante o período de envelhecimento
(jovem e adulto), estes neurônios apresentaram-se esparçamente distribuídos.
Quanto aos tipos celulares encontrados no GCC, estes corroboram com os
estudos publicados de GCC de várias espécies, tais como: capivaras, cães, eqüinos
e gatos (RIBEIRO et al, 2004, RIBEIRO, 2006; FIORETTO et al. 2007), assim como
aqueles publicados em outros gânglios simpáticos como o celíaco-mesentérico de
cães (RIBEIRO et al. 2002) e mesentérico inferior de cães (GAGLIARDO et al.
2005). Os neurônios binucleados observados no GCC do preá foram também
documentados no GCC de outros roedores silvestres, assim como, em outros
gânglios simpáticos (FORSMAN et al, 1989; SASAHARA et al., 2003; RIBEIRO et al,
2004; RIBEIRO, 2006; ABRAHÃO et al 2009; MELO et al 2009).
6.3 Estereologia
Nesta seção iremos discutir aspectos estereológicos abordados nesta pesquisa

LADD, A. A. B. L.
61
6.3.1 Volume Ganglionar (VGCC)
O volume do GCC durante o desenvolvimento pós-natal apresentou-se maior
em preás senis do que em neonatos, jovens e adultos, com um aumento progressivo
a partir do jovem (aumento significativo de 85% durante o envelhecimento)
acompanhado por um aumento alométrico, da ordem de 6 vezes em relação à
massa corporal, aproximadamente 622% do peso de massa corpórea entre os
animais neonatos e senis. No GCC de pacas Melo et al. (2009) identificaram um
aumento de 154% no seu volume durante a maturação e envelhecimento e Toscano
et al (2009) reportaram em GCC de cobaias um aumento volumétrico de 27% nesse
mesmo período. Estes e outros autores identificaram essa hipertrofia ganglionar em
outros mamíferos e roedores silvestres que não o preá, sendo este um mecanismo
adaptativo bem conhecido do desenvolvimento pós-natal de gânglios simpáticos de
pequenos, médios e grandes mamíferos (MIOLAN e NIEL 1996; GAGLIARDO et al
2005; RIBEIRO, 2006; FIORETTO et al, 2007; ABRAHÃO et al 2009; MELO et al
2009; TOSCANO et al, 2009).
Essas alterações volumétricas são decorrentes do aumento numérico e/ou
volumétrico de estruturas intraganglionares, em adição ao tecido neuronal
(neurônios uni e binucleados), e embora não tenhamos mensurado seu volume, o
tecido não neuronal torna-se mais evidenciado, por exemplo, de tecido vascular e
conjuntivo, representados principalmente por capilares sanguíneos e fibras
colágenas e fibroblastos em concordância com o observado em outros roedores
silvestres (RIBEIROet al, 2004; RIBEIRO, 2006; ABRAHÃO et al 2009; MELO et al
2009). Outro fator que contribui com o aumento do volume do GCC, em adição ao
aumento em número e tamanho de estruturas intraganglionares, é o volume total

LADD, A. A. B. L.
62
ocupado pelos neurônios, sendo este último o fator predominante na alteração
volumétrica do GCC (ABRAHÃO et al, 2009).
6.3.2 Número Total de Neurônios (N)
Usando o princípio do “fractionator”, associado ao método do “disector” óptico
para quantificar o número de células neuronais, garantimos acurácia e rapidez na
estimativa dos parâmetros investigados (WEST et al.1991; POVER e
COGGESHALL, 1991; MAYHEW; GUNDERSEN, 1996; GUNDERSEN, 2002;
HOWARD e REED, 2005). Sendo que o número total de neurônios uni e
binucleados, bem como, o número total de neurônios (aqui representados pelo
somatório entre uni e binucleados), embora apresentem um aparente decréscimo
durante a maturação, não apresentaram diferenças significativas relacionadas à
idade, permanecendo o número dessas células constante durante a maturação e o
envelhecimento.
Acerca das mudanças neuronais decorrentes da idade, algumas partes do
sistema nervoso reagem de forma diferente por razões ainda desconhecidas, por
exemplo, no GCC de capivaras, a maturação ocasionou redução de 16% no número
total de neurônios (uni e binucleados), acompanhado de uma redução de 23% no
número total de neurônios binucleados (RIBEIRO, 2006); Toscano et al. (2009)
encontraram no GCC de cobaias uma redução de 24% e 43% para neurônios uni e
binucleados, respectivamente, e também no sistema nervoso central as alterações
mais frequentes relacionadas à idade são atrofia e perda neuronal (ESIRI, 2007) e
perda de células de purkinje e células granulares no cerebelo (ANDERSEN, 2003).

LADD, A. A. B. L.
63
Em contraste a estes dados, Popken e Farel (1997) observaram um aumento
de 30% no número total de neurônios dos gânglios da raiz dorsal (DRGs) L3-L6 de
ratos Sprague-Dawley adultos em relação aos neonatos, usando o disector físico, e
Cecchini et. al. (1993) reportaram um aumento de 40% na população neuronal de
DRGs em ratos Wistar entre 1 e 5 meses, usando métodos quantitativos
morfométricos, portanto discutíveis. Farel (2003) reportou que o aumento no número
de neurônios nos gânglios da raiz dorsal de ratos não está associado à neurogênese
e, sim, à maturação tardia ou diferenciação incompleta.
Contudo, Melo et al. (2009) documentaram em GCC de pacas um aumento
de 95% no número de neurônios uninucleados durante o desenvolvimento pós-natal,
enquanto o número de neurônios binucleados permaneceu constante durante o
mesmo período, e Gagliardo et al. (2005) reportaram um aumento de 1700% no
número total de neurônios do gânglio mesentérico caudal de cães. Esses dados,
associados à presença de neurônios binucleados, sobretudo em gânglios simpáticos
de roedores silvestres (RIBEIRO et al 2004; RIBEIRO et al 2006; ABRAHÃO et al
2009), oferecem oportunidades concretas para futuras pesquisas sobre este tema.
6.3.3 Volume médio dos neurônios ( NV )
O desenvolvimento pós-natal afetou o volume dos neurônios uninucleados,
contudo, não alterou o volume de neurônios binucleados. Foi possível observar nos
neurônios uninucleados um aumento de 154% durante o período de maturação com
uma pequena queda de 10% durante o envelhecimento.
Os dados descritos na literatura referentes às alterações volumétricas em
neurônios de gânglios simpáticos e relacionadas à idade são contraditórios entre si.

LADD, A. A. B. L.
64
Por exemplo, Ribeiro (2006) e Melo et al. (2009) não reportaram alterações etárias
no volume dos neurônios do GCC em pacas e capivaras, respectivamente. No
entanto, Gagliardo et al. (2005) encontram um aumento de 217% no tamanho dos
neurônios do gânglio mesentérico caudal de cães, e embora a hipertrofia neuronal
seja bem conhecida em algumas partes do sistema nervoso central como um
mecanismo compensatório para a perda neuronal (CABELLO et al., 2002), isto pode
ocorrer também no sistema nervoso autônomo, além de outros mecanismos que
possam garantir a homeostase funcional dos gânglios simpáticos.

LADD, A. A. B. L.
65
CONCLUSÕES

LADD, A. A. B. L.
66
7 CONCLUSÕES Com base nos métodos empregados e resultados obtidos, é possível concluir
o que se segue:
- Os seguintes dados macromorfométricos: peso, comprimento e largura do gânglio
cervical cranial sofreram alterações relacionadas com a idade e que refletiram no
volume ganglionar.
- A hipertrofia do GCC de preás é secundária ao desenvolvimento pós-natal
(maturação e envelhecimento).
- A hipertrofia dos neurônios uninucleados é secundária ao desenvolvimento pós-
natal do GCC de preás.
- Detectou-se a presença de neurônios em ciclo celular no GCC de preás neonatos e
senil, o que sugere que estas células estão mitoticamente ativas durante o período
pós-natal.

LADD, A. A. B. L.
67
8 SUGESTÕES PARA ESTUDOS FUTUROS Em face da não existência de explicações conclusivas para as alterações
quantitativas nos neurônios e gânglios simpáticos durante a maturação e o
envelhecimento, sugere-se que estudos futuros possam investigar se o tamanho dos
territórios de inervação destes gânglios pode desencadear mudanças na micro e
macroestrutura dos mesmos, incluindo a possibilidade de ocorrência de
neurogênese, que seria estudada por meio de imunomarcadores específicos.

LADD, A. A. B. L.
68
REFERÊNCIAS

LADD, A. A. B. L.
69
REFERÊNCIAS
ABERCROMBIE, M., Estimation of nuclear population from microtome sections. The Anatomical Record. v. 94, p.239 – 247,1946. ABRAHÃO, L. M. B., NYENGAARD, J. R. , SASAHARA, T. H. C., GOMES S. P. , OLIVEIRA F. R. , LADD, F. V. L., LADD, A. A. B. L., MELO M. P. , MACHADO M. R. F., MELO S. R. , RIBEIRO A. A. C. M, Asymmetric post-natal development of superior cervical ganglion of paca (Agouti paca). Int. J. Devl Neuroscience. v. 27, p. 37–45, 2009. ANDERSEN, B.B; GUNDERSEN, H.J.G.; PAKKENBERG, B. Aging of the human cerebellum: A stereological study. J. Comp. Neurol. V. 466 p. 356-365 2003 ANDERSEN, B. B., FABRICIUS, K., GUNDERSEN, H. J., JELSING, J., STARK, A. K., No change in neuron numbers in the dentate nucleus of patients with schizophrenia estimated with a new stereological method--the smooth fractionator. J Anat. v. 205, p.313–321, 2004. BAKEWELL, S. The antonamic nervons system. Update in anesthesia, n. 5, art. 6, 2 p. 1995. BALJET, B., DRUKKER J., The extrinsic innervation of the abdominal organs in the female rat. Acta Anatomica. v.104, p.243-267, 1979. BAO-PING S.; YAN-PING.; D; ZHAO-HUI, X.; HONG-XIAN, Y.; BRAND-SABERI; JIAN-LIN, W. The cranial cervical ganglion and its branches in the yak (Bos-grunniens). The veterinary Jornal, v. 173, n. 1 , p. 174-177 , 2007. BARTON, A. A., CAUSEY, G., Electron microscopic study of the superior cervical ganglion. Journal Anatomy. v.92, p. 399–407, 1958.
BEDDELEY, A. J.; GUNDERSEN, H. J. G.; CRUZ-ORIVE, H. J. G. Estimation of surface area from vertical sections. Journal of Microscopy, v. 142, n. 3, p. 259-276, 1986.
BELL, R. L.; ATWEH, N.; IVY, M.; POSSENTI, P. Traumatic and iatrogenic horner syndrome: case reports and review of the literature. The Jornal of Trauma Injury, Infection and Critical Care, v. 51, n. 2, p. 400-404, 2001. BOYDELL, P. Idiopathic horner’s syndrome in the gold retriver. Journal of small animal practice n.36 p.382-384 1995. CABELLO, C. R., THUNE, J. J., PAKKENBERG, H., PAKKENBERG, B., Ageing of substantia nigra in humans: cell loss may be compensated by hypertrophy. Neuropathology & Applied Neurobiology. v. 28, p.283-291, 2002.

LADD, A. A. B. L.
70
CECCHINI, T; CUPPINIR, R; CIARONI, S.;DEL GRANDE, P. increased number of dorsal roots ganglion neurons in vitamin-E- deficient rats. Somatosens Mot. Res. V.10.433-443 1993. COGGESHALL, R. E., CHUNGA K., GREENWOODA D., HULSEBOSCHA, C. E., An empirical method for converting nucleolar counts to neuronal numbers. Journal of Neuroscience Methods. v. 12, p. 125-132, 1984. CORRADELLO, E., Criação de preá. p.83, 1987.
CUI-SHENG; JIAN-LING, W.; ZHEN-MINE, X. The gross anatoy of the cranial cervical ganglion and its branches in the Bactrian camel (camelus bactrianus). Veterinary Research Conmunications, v. 22, n. 1, p 1-5, 1998.
DAVIES, D. C. Neuronal numbers in the superior cervical ganglion of the neonatal rat. Journal Anatomy. v. 127, p. 43-51, 1978. DEL RIO, J. A., SORIANO, E., Immunocytochemical detection of 5'-bromodeoxyuridine incorporation in the central nervous system of the mouse. Brain research. v. 49, p. 311-317, 1989. DEVOR, M.; GOVRIN-LIPPMANN, R. Neurogenesis in adult rat DRGs. Neurosci. Lett, v. 61, p. 189-194, 1985. DOLBEARE, F., Bromodeoxyuridine: a diagnostic tool in biology and medicine, Part I: Historical perspectives, histochemical methods and cell kinetics. Histochemical Journal. v. 27, p.339-369, 1995. DOLBEARE, F., Bromodeoxyuridine: a diagnostic tool in biology and medicine, Part III. Proliferation in normal, injured and diseased tissue, growth factors, differentiation, DNA replication sites and in-situ hybridization. Histochemical Journal. v. 28, p. 531-575,1996.
EBESSON, S. O. E. Quantitative studies of the superior cervical sympathetic ganglia in a variety of primates including man. J. Morph., n. 124, p. 117-132 1968. ELIAS, M. D. Cervical sympathetic and stellate ganglion blocks. Pai Physician, v. 3, n. 3, p. 294-304, 2000. ERANKO, L., Postnatal development of histochemically demonstrable Catecholamines in the superior cervical Ganglion of the rat. Histochem.v.3, p. 225- 236, 1972. ESIRI, M. M., Ageing and the brain. The Journal of Pathology. v. 211, p.181 – 187, 2007. EVANS, R., J. F., SURPRENANT, A. P2X receptors in autonomic and sensory neurons. Seminars in Neuroscience. v. 8, p. 217-223, 1996

LADD, A. A. B. L.
71
FAREL, P. B. Late differentiation contributes to the apparent increase in sensory neuron number in juvenile rat. Developmental Brain Research. v. 144, p. 91-98, 2003. FILICHKINA, N. M., Comparative radioautographic analysis of synthesis and slow transport of protein axons of uni- anddouble nuclear neurocytes in the rabbit. Acta Anat HistolEmbryol. v. 11, p.49–52, 1981.
FIORETO, E. M.; GUIDI, W. L.; OLIVEIRA, P. C.; RIBEIRO, A. A. C. M. Macrostructure of the cranial cervical ganglionar complex and distal vagal ganglion during post natal development in dogs. Cells Tissues Organs, v.186 p. 129-40 2007. FLEET, D. L.; BELL, C. Topography of functional subpropulation of neurons in the superior cervical ganglion of the rat. Jornal of neurolytology. n. 21, p. 55-66, 1991.
FORSMAN, C. A.; LINDH, B.; ELFVIN, L.; HALLAMN, H. Measurements of the DNA amount in mono and binucleate cells in the celiac superior mesenteric ganglion of the guinea pig. Anat. Embryol., n. 179, p. 587-590, 1989. GABELLA, G., TRIGG, P., McPHAIL, H., Quantitative cytology of ganglion neurons and satellite glial cells in the superior cervical ganglion of the sheep. Relationship with ganglion neuron size. Journal of Neurocytology. v. 17, p.753-769,1988. GABELLA, G. The rat nervous system (Autonomic Nervous System). Elsevier, 2004. GAGLIARDO, K.M; BALIEIRO, J.C.C; SOUZA, R.R; RIBEIRO, A.A.C.M. Post natal related changes in the size and total number of neurons in the caudal mesenteric ganglion of dogs: total number of neurons can be predicted from body weight and ganglion volume Anatomical records part v.286 A p.917-929 2005. GUADIX, AR.; HERREROS, M.A.P.; GARRIDO, M.V.O.; ABADIA-FENOLL F. Quantitative Analysis of binucleate neurons in the lateral geniculate nucleus of the mouse. Trab. Inst. Cajal. Invest. Biol., n. 70, v. 3, p. 247-60, 1979. GUNDERSEN, H .J. G., BAGGER, P., BENDTSEN, T. F. The new stereoloical tools: disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. Acta Microb Imm Scand. v. 96, p. 857-881,1988. GUNDERSEN, H.J.G JENSEN, F.B KIEU,K. NIELSEN, J. The efficiency of systematic sampling in stereology-reconsidered. J Microsc. V.3 n.193 p.199-211 1999. GUNDERSEN, H. J. G .The smooth fractionator. Journal of Microscopy. v. 207, p.191-210, 2002

LADD, A. A. B. L.
72
HAYAKAWA, T., ITOH, M., MIKI T., KANETO T., TOMIYAMA H., TAKEUCHI Y. Sympathetic fibres innervating the extraocular muscles: cells of origin in the cat superior cervical ganglion. Okajimas Folia Anat Jpn. v. 77, p.119-124, 2000.
HEDGER, J. H., WEBBER, R. H., Anatomical study of the cervical sympathetic trunk and ganglia in the albino rat (Mus norvegicus albinus) Acta Anatomica. v. 96, p.206-217, 1976.
HOWARD, C. V.; REED, M. G.; Unbiased stereology. Three-dimensional measurement in microscopy. Oxford: Bios Scientific Publishers, 2005. HUBBARD, W.C., ROBINSON J.C., SCHMIDT, K., ROHEN, J.W., TAMM, E.R., KAUFMAN, P.L.. Superior cervical ganglionectomy in monkeys: effects on refraction and intraocular pressure. Exp Eye Res. v. 68, p.637-639, 1999 JOHNSON, J. R., GORIN, P. D., BRANDEIS, L. D., PEARSON, J., Dorsal root ganglion neurons are destroyed by exposure in utero to maternal antibody to nerve growth factor. Science. v. 210, p. 916-918, 1980.
KABAK, M. The Gross anatomy of the cranial cervical ganglion in the guinea pig (Cavia Porcellus) Veterinary Research communications, v. 31, p. 1-7, 2007.
KABAK, M.; ORHAN, I. O.; HAZIROLU, R. M.; Macro anatomical investigations of the cranial cervical ganglion in domestic pig. (sus serofa domesticus). Anat. Histol. Embryol. v. 34, p. 199-202, 2005.
KÖNIG, H .E.; LIEBICH, H. G. Anatomia dos animais domésticos. São Paulo: Artmed, 1999. v. 2. KONIG, H., LIEBICH, H., Veterinary anatomy of domestic animals. Textbook and color atlas. p. 495–7, 2004
KRENLEN, D. L. Integration in antonomic ganglia. Physiologist, v. 27. n. 1, p. 49-55, 1984. LADIZESKY M. G. , CUTRERA R. A., BOGGIO V. , MAUTALEN C. ,CARDINALI D. P., Effect of unilateral superior cervical ganglionectomy on bone mineral content and density of rat’s mandible Journal of the Autonomic Nervous System. v. 78, p. 113–116, 2000. LUEBKE, J. I., WRIGHT, L. L., Characterization of superior cervical ganglion neurons that project to the submandibular glands, the eyes, and the pineal gland in rats Brain Research. v. 589, p. 1-14, 1992
MATHEWS, M .R. The ultrastructure of the functions in sympathetic ganglia of mammls in. ELFVIN, G. (Ed.). Autonomic ganglia. Chichester: John Wiley,. p. 27-66 1983

LADD, A. A. B. L.
73
MAYHEW, T. M.; GUNDERSEN, H. J. G. ‘If you assume, you can make an ass out of u and me’: a decade of the dissector for stereological counting of particles in 3D space. J. Anat., v. 188, p. 1-15, 1996.
MAYHEW, T. M.; OLSEN, D. R. Magnetic resonance imaging (MRI) and model-free estimates of brain volume determined using the Cavalieri principle. J. Anat., v. 178, p. 133-144, 1991. MELO, S. R., NYENGAARD, J. R., DA ROZA, O. F., LADD, F. V., ABRAHÃO, L. M., MACHADO, M. R., SASAHARA, T. H., DE MELO, M. P., RIBEIRO, A. A., The developing left superior cervical ganglion of Pacas (Agouti paca). Anatomical Records. v. 292, p. 966-975, 2009. MINITAB (v.15) 2007. Minitab reference manual. Florence, Wadsworth MINISTÉRIO DA SAÚDE., Fundação Nacional de Saúde. Manual de controle de roedores Brasil, p.132, 2002. MIOLAN, J. P.; NIEL, J. P. The mammalian sympathetic pre-vertebral ganglia: integrative properties and role in the nervours control of digestive trac motility. J. Auton. Nerv. Syst. 58, p. 125-138. 1996. MOLLER, M., LIU, W. Innervation of the rat pineal gland by nerve fibres originating in the sphenopalatine, otic and trigeminal ganglia: a retrograde in vivo neuronal tracing study. Reprod Nutr Dev. v. 39, p.345-353, 1999. MOMMAA, S., VASILYEVAC, E., TOMILINC, N., SHUPLIAKOV,O. Ultrastructural identification of dividing cells in the adult mammalian central nervous system. Journal of Neuroscience Methods. v. 119, p. 59-63, 2002. MOOJEN, J., Os roedores do Brasil: Instituto Nacional do Livro Rio de Janeiro, Brazil. p. 214, 1952. NGWENYA, L. B., PETERS, A., ROSENE, D. L., Light and Electron Microscopic Immunohistochemical Detection of Bromodeoxyuridine-labeled Cells in the Brain : Different Fixation and Processing Protocols. Journal of Histochemistry and Cytochemistry. v 53 (7), p. 821-832, 2005. OZEL, S.K.; KAZEZ, A. Horner’s syndrome secondary to tube toracostomy. Turk J. Pediatr. V.46 p. 189-90 2004.
PANCIERA, R.J; RITCHEY,J.W; BAKER, J.E; DIGREGORIO, M.Trigeminal and polyradiculo neuritis in a dog presenting with masticatory muscle atrophy and horner’s syndrome. Vet. Pathol. v. 1 n. 39 p. 146-9 2002. PAKKENBERG, B., GUNDERSEN. H. J. G.,Total number of neurons and glial cells in human brain nuclei estimated by the disector and the fractionator. Journal of microscopy. v. 150, p. 1-20, 1988.

LADD, A. A. B. L.
74
PAKKENBERG, B., GUNDERSEN, H. J. G., New stereological method for obtaining unbiased and efficient estimates of total nerve cell number in human brain areas: exemplified by the mediodorsal thalamic nucleus in schizophrenics. APMIS. Acta pathologica, microbiologica et immunologica scandinavica. v. 97, p. 677-681, 1989. PICK, J., The fine structure of sympathetic neurons in x-irradiated frogs. J Cell Biol.v.26, p.335–351,1965. POVER, C. M.; COGGESHALL, R. E. Verification of the dissector method for counting neurons, whit comments on the empirical method. Anat. Rec., v 22, n. 231, p. 573-578, 1991.
POVER, C. M.; COGGESHALL, R. E.; FITZGERALD, M.; Dorsal root ganglion cell death and surviving cell numbers in relation to the development of sensory innervation in the rat hindlimb. Res Dev Brain, v. 82, n. 1-2, p. 193-212, 1994
POPKEN, G. J.; FAREL, P. B. Sensory neuron number in neonatal and Adult rats estimated by means of stereologic and profile-based methods. J. Comp. Neurol. v.386, p. 8-15, 1997.
PURVES, D.; RUBIN, E.; SNIDER, W. D.; LICHTMAN, J. Relation of animal size to convergence, divergence, and neuronal number in peripheral sympathetic pathways. The Journal of Neuroscience, v. 1 n. 6, p. 158-163, 1986.
RIBEIRO, A.A.C.M. Size and number of binucleate and mononucleate superior cervical ganglion neurons in Young capibaras. Anat embryol v.211 p. 207-217 2006
RIBEIRO, A. A. C. M.; DAVIS, C.; GABELLA, G. Estimate of size and total number of neurons in superior cervical ganglion of rat, capybara and horse. Anat. Embryol., v.208, n. 208, p. 367-380, 2004.
RIBEIRO, A.A.C.M; ELIAS, C.F.; LIBERTI, E.A.; GUIDI, W.L.; SOUZA,R.R.; Structure and ultrastructure os the celiac mesenteric ganglion complex in the domestic dog. Anat. Hitol. Embriol. V.31, p.344-349 2002.
SASAHARA, T. H. C.; DE SOUZA R. R.; MACHADO, M. R .F.; DA SILVA, R. A.; GUIDI, W. L.; RIBEIRO, A. A. C. M. Macro and microstructural organization of the rabbit’s celiac-mesenteric ganglion complex. Ann. Anat., v.23, n. 185, p. 441-448, 2003.
STERIO, D. C. The unbiased estimation of number and sizes of arbitrary particles using the dissector. J. Microsc., n. 134, p. 127-136, 1984.
TAUPIN, P.; GAGE, P. H. Adult neurogenesis and neural stem cells of the central nervous system in mammals. J.Neurosci.Res, v. 69, p. 745-749, 2007.

LADD, A. A. B. L.
75
TOSCANO, C. P., MELO. M. P., MATERA, J. M., LOESCH, A.; RIBEIRO, A. A. C. M.,The developing and restructuring superior cervical ganglion of guinea pigs (Cavia porcellus var. albina.) Int. Journal of Developmental Neuroscience. v. 27, p. 329-336, 2009. VALERO,J; WERUAGA,E; MURIAS,A.R; RECIO,J.S.; ALONSO,J.R.- Proliferation markers in the adult rodent brain: bromodeoxyuridine and proliferating cell nuclear antigen Brain research protocol, 2005.
VEDEL JENSEN, E.B.; GUNDERSEN, H.J.G. The rotator. Journal of Microscopy v.170, p.35-44, 1993 WARD, J. M., WEGHORST, C. M., DIWAN, B. A., KONISHI, N., LUBET, R. A., HENNEMAN, J. R, DEVOR, D. E. Evaluation of cell proliferation in the kidneys of rodents with bromodeoxyuridine immunohistochemistry or tritiated thymidine autoradiography after exposure to renal toxins, tumor promoters, and carcinogens. In: Chemically Induced Cell Proliferation: Implications for Risk Assessment. Butterworth BE, Slaga TJ, Farland W, McClain M (eds), p. 369—388, 1991.
WEST, M. J.; SLOMIANKA, L.; GUNDERSEN, H. J. Umbiased stereological estimation of the total number of neurons in the subdivisions of the rat hippocampus using the optical fractionator. The anatomical record, v. 231, p. 482-497,1991.