
CAPÍTULO 5. DISCUSSÃO DOS RESULTADOS · Quanto às espécies de Penicillium isoladas das uvas,...
30
CAPÍTULO 5. DISCUSSÃO DOS RESULTADOS Sumário Neste capítulo discutem-se os resultados obtidos na dissertação. 1. Micoflora das uvas ................................................................................................................. 269 2. Espécies de Aspergillus da secção Nigri produtoras de OTA ................................................ 285 3. Aspergillus ibericus................................................................................................................ 291 4. Concentração de OTA nas uvas portuguesas ......................................................................... 293 5. Avaliação do potencial ocratoxigénico das estirpes em meio de uva ..................................... 294 6. Outros perigos micotoxigénicos nas uvas .............................................................................. 295 268
Transcript of CAPÍTULO 5. DISCUSSÃO DOS RESULTADOS · Quanto às espécies de Penicillium isoladas das uvas,...

CAPÍTULO 5. DISCUSSÃO DOS RESULTADOS
Sumário
Neste capítulo discutem-se os resultados obtidos na dissertação.
1. Micoflora das uvas .................................................................................................................269
2. Espécies de Aspergillus da secção Nigri produtoras de OTA ................................................285
3. Aspergillus ibericus................................................................................................................291
4. Concentração de OTA nas uvas portuguesas .........................................................................293
5. Avaliação do potencial ocratoxigénico das estirpes em meio de uva.....................................294
6. Outros perigos micotoxigénicos nas uvas ..............................................................................295
268

CAPÍTULO 5. DISCUSSÃO
1. Micoflora das uvas
Nesta secção discutem-se os resultados obtidos quanto à composição da micoflora
das: i) uvas “normais”, sem podridão visível; ii) uvas “normais” após desinfecção
superficial; iii) fungos causadores de podridão visível nos bagos; iv) mosto de uvas.
1.1. Micoflora de uvas normais
Os géneros isolados nas uvas refletem as fontes de contaminação a que estas se
encontram sujeitas. As referências quanto aos habitats comuns dos géneros detectados
nas uvas está indicada na Tabela 5.1, bem como a indicação se foram detectados
noutros rastreios ao micobiota de folhas Portuguesas e de uvas noutros países.
A maior parte dos géneros detectados são de distribuição ubíqua, comuns no ar,
superfície de plantas, materiais em decomposição e solo. Os organismos que estão
associados à parte aérea das plantas (filoplano), como caules, folhas pétalas e frutos,
chegam a estas superfícies arrastadas pelo vento, chuva e/ou animais, e conseguem-se
fixar de alguma forma ao substrato e sobreviver às condições adversas, como de
radiação solar, variações de temperatura e variações de humidade, durante o dia e
durante o ano, bem como à exposição a compostos tóxicos, como pesticidas e
fungicidas. Alguns dos fungos detectados nas uvas compreendem espécies saprófitas
e patogénicas de plantas comummente detectadas em alimentos, como Aspergillus,
Botrytis, Penicillium e Rhizopus, que têm capacidade demonstrada de produzir
podridão em bagos.
269
CAPÍTULO 5. DISCUSSÃO
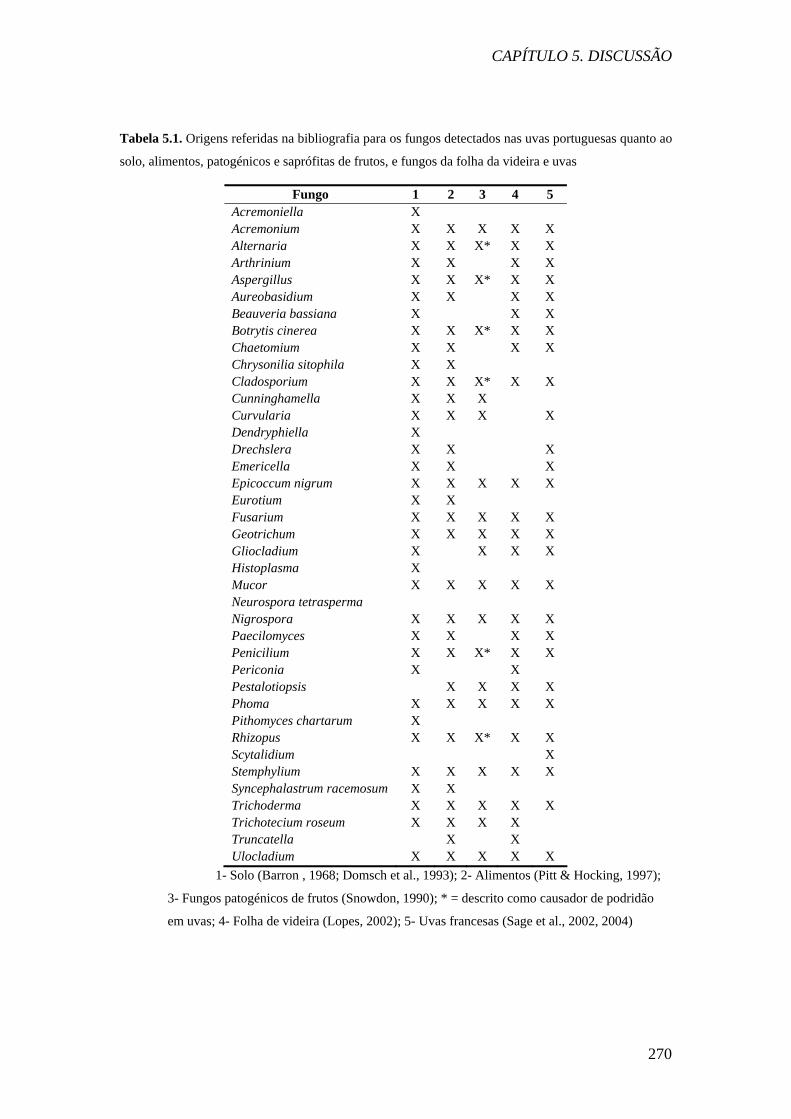
Tabela 5.1. Origens referidas na bibliografia para os fungos detectados nas uvas portuguesas quanto ao
solo, alimentos, patogénicos e saprófitas de frutos, e fungos da folha da videira e uvas
Fungo 1 2 3 4 5 Acremoniella X Acremonium X X X X X Alternaria X X X* X X Arthrinium X X X X Aspergillus X X X* X X Aureobasidium X X X X Beauveria bassiana X X X Botrytis cinerea X X X* X X Chaetomium X X X X Chrysonilia sitophila X X Cladosporium X X X* X X Cunninghamella X X X Curvularia X X X X Dendryphiella X Drechslera X X X Emericella X X X Epicoccum nigrum X X X X X Eurotium X X Fusarium X X X X X Geotrichum X X X X X Gliocladium X X X X Histoplasma X Mucor X X X X X Neurospora tetrasperma Nigrospora X X X X X Paecilomyces X X X X Penicilium X X X* X X Periconia X X Pestalotiopsis X X X X Phoma X X X X X Pithomyces chartarum X Rhizopus X X X* X X Scytalidium X Stemphylium X X X X X Syncephalastrum racemosum X X Trichoderma X X X X X Trichotecium roseum X X X X Truncatella X X Ulocladium X X X X X
1- Solo (Barron , 1968; Domsch et al., 1993); 2- Alimentos (Pitt & Hocking, 1997);
3- Fungos patogénicos de frutos (Snowdon, 1990); * = descrito como causador de podridão
em uvas; 4- Folha de videira (Lopes, 2002); 5- Uvas francesas (Sage et al., 2002, 2004)
270

CAPÍTULO 5. DISCUSSÃO
Todos os géneros detectados nas uvas com a excepção de 4 (35 géneros) já foram
isolados do solo. O solo é o reservatório dos fungos por excelência, por isso este facto
não é surpreendente. A riqueza de géneros encontrados nas uvas portuguesas é
semelhante à referida nas folhas de videira por Lopes (2002) e uvas francesas por
Sage e colaboradores (2002, 2004), em que também foram isolados estirpes de 39 e
40 géneros, respectivamente. Dos fungos detectados nas uvas portuguesas, 26 e 27
são comuns aos mencionados por estes investigadores, respectivamente.
Os 5 géneros mais frequentes detectados nas uvas portuguesas foram por ordem
decrescente Cladosporium, Alternaria, Botrytis, Penicillium e Aspergillus.
Cladosporium e Alternaria são géneros muito frequentes na natureza, frequentemente
dominantes em estudos de micoflora em condições de campo. Compreendem espécies
saprófitas muito comuns, como C. cladosporioides e C. herbarum, e A. alternata. As
espécies de Cladosporium e Alternaria podem viver como saprófitas, epífitas na
superfície de folhas vivas ou comportar-se como parasitas fracos. Alternaria e
Cladosporium são capazes de completar o seu ciclo de vida nas folhas mortas e
detritos vegetais e possuem características que os adaptam à vida na superfície das
plantas. Estes géneros podem alimentar-se de reservas endógenas e conteúdos de hifas
velhas, que são autolizadas e recicladas para as extremidades das hifas em
crescimento. Alternativamente, os esporos podem germinar para produzir esporos
secundários ou sofrer conidiação microcíclica (figura 4.1, capítulo 4). A presença de
melanina nas paredes das hifas e esporos é uma vantagem para a protecção da
radiação solar (Peberdy, 1990), e pensa-se que é por isso que os fungos do filoplano
são na sua maioria dematiáceos (Ellis & Ellis, 1997).
Como foi mencionado na introdução, Botrytis, Aspergillus e Penicillium
compreendem várias espécies saprófitas e parasitas oportunistas e patogénicas de
plantas. B. cinerea é por excelência o principal fungo patogénico da videira e
principal causa de podridão das uvas. Por esse motivo, a presença de propágulos
destes fungos nas vinhas é esperada. Na Tabela 5.2 e Tabela 5.3 estão indicadas as
espécies detectadas de Aspergillus e Penicillium nas uvas portuguesas bem como
informações quanto à sua presença noutros habitats e noutros rastreios à micoflora
normal das uvas.
271

CAPÍTULO 5. DISCUSSÃO
Tabela 5.2. Espécies de Aspergillus e seus teleomorfos detectadas nas uvas portuguesas e informações
quanto à sua presença noutros habitats
Espécie 1 2 3 4 5 6
A. auricomus X
A. candidus X X X
A. carbonarius X X X
A. carneus X
A. clavatus X X
A. flavipes X X X
A. flavus X X X X A. fumigatus X X X X A. japonicus X X X* X X A. niger (agregado) X X X* X X X A. ochraceus X X X A. ostianus
A. terreus X X X X
A. terreus var. africanus
A. ustus X X X X X A. versicolor X X X X X A. wentii X X
Emericella nidulans X X X
Eurotium amstelodami X X
Eurotium chevalieri X X
Pet. alliaceus X X
1- Solo (Klich et al., 1992); 2- Alimentos (Pitt & Hocking, 1997); 3- Fungos
patogénicos de frutos (Snowdon, 1990) * = causador de podridão em uvas; 4- Folha de videira
(Lopes, 2002); 5- Uvas francesas (Sage et al., 2002, 2004); 6- Uvas espanholas (Bau et al.,
2005)
272
CAPÍTULO 5. DISCUSSÃO
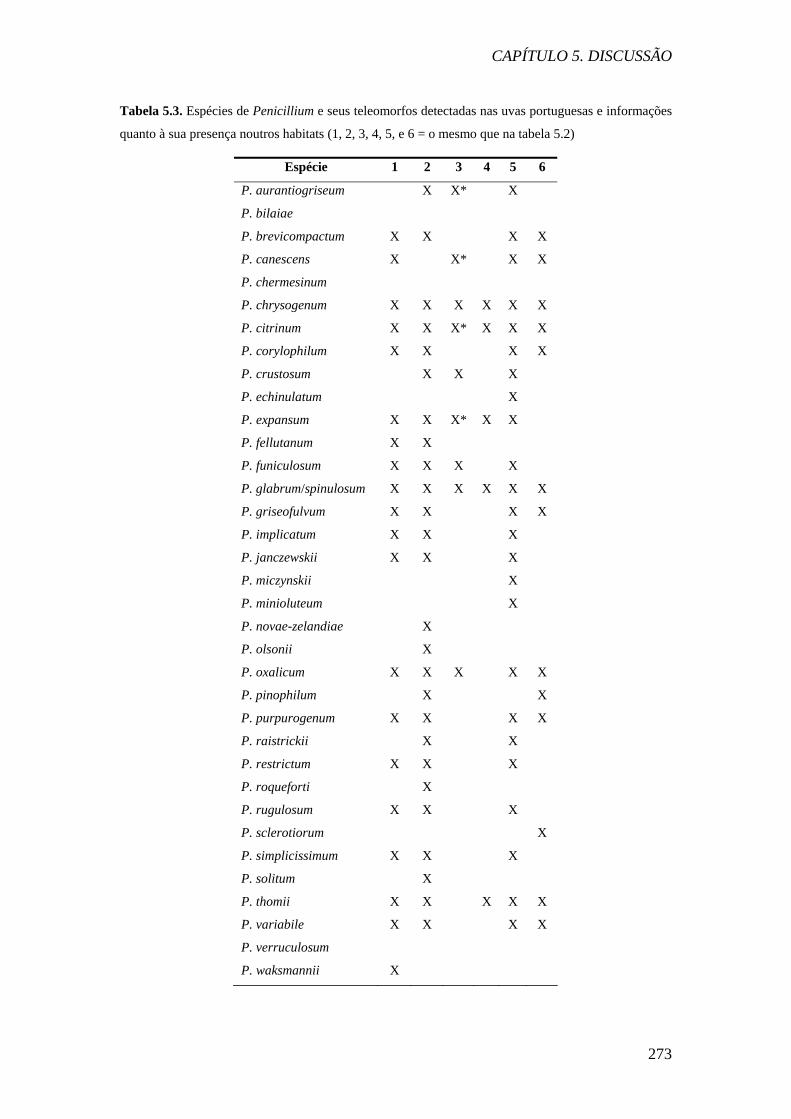
Tabela 5.3. Espécies de Penicillium e seus teleomorfos detectadas nas uvas portuguesas e informações
quanto à sua presença noutros habitats (1, 2, 3, 4, 5, e 6 = o mesmo que na tabela 5.2)
Espécie 1 2 3 4 5 6
P. aurantiogriseum X X* X
P. bilaiae
P. brevicompactum X X X X P. canescens X X* X X P. chermesinum
P. chrysogenum X X X X X X
P. citrinum X X X* X X X P. corylophilum X X X X P. crustosum X X X
P. echinulatum X
P. expansum X X X* X X
P. fellutanum X X
P. funiculosum X X X X
P. glabrum/spinulosum X X X X X X P. griseofulvum X X X X P. implicatum X X X
P. janczewskii X X X
P. miczynskii X
P. minioluteum X
P. novae-zelandiae X
P. olsonii X
P. oxalicum X X X X X P. pinophilum X X P. purpurogenum X X X X P. raistrickii X X
P. restrictum X X X
P. roqueforti X
P. rugulosum X X X
P. sclerotiorum X
P. simplicissimum X X X
P. solitum X
P. thomii X X X X X P. variabile X X X X P. verruculosum
P. waksmannii X
273

CAPÍTULO 5. DISCUSSÃO
Quanto às espécies de Aspergillus detectadas, todas à excepção de A. ostianus
foram descritas previamente em estudos do solo. Das 20 espécies isoladas nas uvas,
14 foram referidas em uvas de Espanha (Bau et al., 2005) e/ou França (Sage et al.,
2002, 2004), e muitas das espécies detectadas são de ocorrência comum em
alimentos.
A espécie mais frequente nas uvas portuguesas foi de longe o agregado A.
niger, que representou 76% das estirpes de Aspergillus isoladas. Esta espécie é vista
como um dos fungos mais comuns em alimentos, e apesar de estar presente no solo,
considera-se que não é habitante do solo, mas sim uma espécie invasora (Domsch et
al., 1993). A. niger é considerada a principal espécie de Aspergillus responsável pela
podridão de frutos, incluindo uvas (Pitt & Hocking, 1997; Snowdon, 1990; Zahavi et
al., 2000).
A segunda espécie de Aspergillus isolada com maior frequência foi A.
carbonarius (9%), outra espécie da secção Nigri. Esta espécie não é comummente
referida de outras fontes, tendo sido isolada de lama, solos pantanosos, solo e água
poluída na Índia e Iraque (Klich & Pitt, 1988). Recorda-se ainda que, tal como
mencionado na introdução, foi também na Índia que esta espécie foi referida como
causadora de podridão nas uvas (Raper & Fennell, 1965). As restantes espécies de
Aspergillus foram isoladas das uvas com pouca frequência. A terceira espécie mais
frequentemente isolada das uvas portuguesas, A. flavus, compreendeu 3% das estirpes
de Aspergillus isoladas. A. flavus é uma espécie produtora de aflatoxinas, e dado que
possui uma fisiologia semelhante a A. niger, pensa-se que compete com A. niger no
ambiente natural em muitas situações (Pitt & Hocking, 1997). No caso concreto das
uvas, dada a frequência destas espécies, A. niger venceu esta competição. Os
Aspergillus da secção Nigri tem esporos muito melanizados, o que lhes confere uma
vantagem adaptativa em ambientes expostos a radiação solar, e são xerófilos, capazes
de crescer com baixas actividades de água e temperaturas elevadas.
Quanto às espécies de Penicillium isoladas das uvas, das 35 espécies
detectadas, 26 foram isoladas de uvas na Espanha e/ou França. As espécies mais
frequentes foram P. brevicompactum, P. glabrum/spinulosum e P. thomii, que são
espécies xerófilas, comuns em materiais em decomposição e solo. P. brevicompactum
é das espécies mais xerófilas do género. Estas espécies já foram isoladas de uvas
podres, e a sua capacidade de causar podridão nas uvas é reconhecida (Sage et al.,
2002, 2004). A micoflora das uvas portuguesas de Aspergillus e Penicillium é pois
274

CAPÍTULO 5. DISCUSSÃO
dominada por espécies xerófilas, capazes de crescer até 45 ºC e 30 ºC,
respectivamente, e com capacidade para causar podridão nas uvas caso hajam
condições propícias à infecção dos bagos.
1.1.1. Variação da micoflora com a maturação
As uvas portuguesas estiveram expostas sensivelmente ao mesmo número de
propágulos fúngicos ao longo do período de maturação estudado, entre o bago ervilha
e a vindima: a média de propágulos fúngicos por bago foi sempre igual a 2.
No entanto, em termos de composição da micoflora, observaram-se diferenças.
No início da maturação, a micoflora do bago ervilha é dominada por fungos típicos do
filoplano de angiospérmicas, como Alternaria, Aureobasidium, Botrytis,
Cladosporium, Epiccocum e Stemphylium, e foi neste estado que se verificou a
incidência mais elevada de Arthrynium, Drechslera, e Fusarium. Com o avanço da
maturação, verificou-se um decréscimo nos fungos do filoplano e um aumento
significativo de agentes saprófitas e parasitas oportunistas causadoras de podridão nas
uvas, nomeadamente Aspergillus, Botrytis, Penicillium e Rhizopus. Na passagem do
bago ervilha para o pintor, observou-se um decréscimo significativo de Stemphylium e
um aumento significativo de Botrytis e Penicillium. Mas o maior número de
alterações significativas na incidência dos fungos deu-se na passagem do pintor para a
vindima, quando decrescem significativamente várias espécies comuns do filoplano,
entre as quais Alternaria, e aumenta a incidência de Aspergillus, Penicillium e
Rhizopus. No final da maturação, os bagos estão mais ricos em nutrientes e açúcares,
a película é fina e sujeita a fissuras. Do ponto de vista morfológico, o bago encontra-
se mais vulnerável ao ataque de agentes patogénicos com o avanço da maturação.
Além disso, a partir do momento em que o cacho fecha, proporciona-se no interior do
cacho um ambiente pouco arejado, abrigado dos agentes atmosféricos, que pode
facilitar a permanência de esporos de agentes saprófitas secundários, com dificuldade
em colonizar a superfície dos bagos. Se houver danos nos bagos, podem-se criar
condições favoráveis à podridão. É precisamente no momento mais vulnerável para os
bagos que os fungos descritos como causadores de podridão nas uvas estão presentes
com maior incidência.
275

CAPÍTULO 5. DISCUSSÃO
As únicas espécies de Aspergillus que aumentaram significativamente com a
maturação foram o agregado A. niger e A. carbonarius. Estirpes do agregado A. niger
são detectadas desde o início, mas nenhuma de A. carbonarius foi detectada neste
estado. No entanto, foram detectadas no bago ervilha 9 das 15 espécies de Aspergillus
detectadas na vindima. A. carbonarius só aumenta significativamente do pintor para a
vindima, quando se dá o aumento significativo na incidência de Aspergillus e dos
restantes fungos capazes de causar podridão nas uvas. O facto de praticamente não ser
detectado nos estados antes da vindima, quando os bagos não estão tão susceptíveis ao
ataque de fungos, juntamente com a informação de que a frequência desta espécie é
rara noutros materiais, parece indicar que A. carbonarius está dependente da infecção
das uvas para completar o seu ciclo de vida.
Relativamente a Penicillium, as únicas espécies que aumentaram signifi-
cativamente a sua incidência nos bagos com o avanço da maturação foram P.
brevicompactum e P. glabrum/spinulosum. P. thomii por sua vez aumentou
significativamente a incidência nas uvas exclusivamente na região do Douro. Todas as
espécies de Penicillium que estão presentes em mais de 1% dos bagos na vindima já
foram isoladas no bago ervilha. O facto de apenas 3 espécies de Penicillium
aumentarem significativamente com a maturação leva-nos a postular que estas
espécies poderão estar ligadas directamente à vinha e às uvas, enquanto as restantes
espécies de Penicillium detectadas parecem não depender exclusivamente das uvas
para variar os seus efectivos. No entanto, se existirem condições favoráveis à infecção
dos bagos, especialmente em fases de sobrematuração, é possível que espécies como
P. expansum, possam causar podridão nas uvas (V. secção 6.3 deste capítulo).
Rastreios a uvas em diferentes estados de maturação permitiram retirar
conclusões semelhantes noutros países. Bau e colaboradores (2005) verificaram um
aumento significativo de Aspergillus e Penicillium com o avanço da maturação e um
decréscimo significativo na incidência de Alternaria e Cladosporium nos bagos. Bellí
e colaboradores (2004b) reportaram um aumento de Aspergillus da secção Nigri nas
uvas com a maturação. Sage e colaboradores (2004) reportaram um decréscimo de
Cladosporium e um aumento de A. niger e de P. expansum do final do pintor para a
vindima. Nas uvas portuguesas, não se verificou um decréscimo de Cladosporium
com o avanço da maturação. As diferenças no método de detecção usado podem estar
na base desta observação: Bau e colaboradores (ob. cit.) referem que a inspecção dos
bagos foi feita apenas no final do tempo de incubação de 7 dias. As colónias de
276

CAPÍTULO 5. DISCUSSÃO
Cladosporium têm um crescimento mais lento que outros fungos, nomeadamente
Aspergillus, que aumentam com a maturação e que mascaram com frequência a
presença de Cladosporium nas amostras, levando a subestimar a incidência do género
nos bagos.
1.1.2. Variação da micoflora com a origem geográfica
Na vindima, observou-se que a população fúngica a que as uvas estão expostas
não difere significativamente entre regiões com a excepção de 3 géneros: Aspergillus,
Botrytis e Ulocladium. Observou-se que Aspergillus e Botrytis têm distribuições
inversas, isto é, as regiões onde se detectaram em média mais Botrytis foram aquelas
onde se detectaram em média menos Aspergillus nos bagos. As diferenças na
incidência de Aspergillus e Botrytis podem estar relacionadas com o clima das
regiões, em particular com o regime de precipitação e humidade. Por ordem
decrescente, as regiões em média com mais Botrytis e menos Aspergillus foram:
Vinhos Verdes, Ribatejo, Douro e Alentejo (as duas últimas regiões tiveram uma
média igual de Aspergillus nas amostras). A região mais húmida das estudadas é sem
dúvida a dos Vinhos Verdes, devido ao seu clima sofrer influências atlânticas e ser
Submediterrânico. Das regiões de clima Mediterrânico, a mais húmida é o Ribatejo,
seguida do Douro e Alentejo (tabela 3.4, capítulo 3, p. 135). Como foi mencionado na
introdução no capítulo 2, Botrytis é um patogénio da videira que requer humidade
para poder infectar as uvas, e é favorecido em ambientes húmidos e desfavorecido em
climas secos, como Douro e Alentejo. Por sua vez, os Aspergillus, em particular os
Aspergillus secção Nigri, as espécies dominantes na vinha, têm uma vantagem
adaptativa nestes climas, pois são capazes de crescer a temperaturas mais elevadas e
com menos humidade, e chegam a ser dominantes nas amostras.
Foi interessante verificar que apesar de não se verificarem diferenças
significativas quanto à incidência de Penicillium nas amostras, existiram diferenças
quanto à frequência das suas espécies. No caso dos Penicillium, as principais
diferenças não parecem estar relacionadas com o clima, mas sim com uma
distribuição geográfica Norte/Sul. Duas das espécies mais frequentemente detectadas
nas uvas, P. brevicompactum e P. thomii, têm uma distribuição inversa, isto é, P.
277

CAPÍTULO 5. DISCUSSÃO
brevicompactum é mais frequente nas uvas do Sul de Portugal e P. thomii mais
abundante no Norte de Portugal.
Os locais estudados dentro das regiões tiveram uma micoflora bastante
semelhante no que diz respeito aos géneros de fungos mais frequentemente detectados
na vindima. As principais excepções foram um local no Douro e outro no Ribatejo.
No Douro, o local Do3 diferiu dos restantes dois locais quanto à incidência de
Alternaria e Aspergillus. Neste local, a incidência de Aspergillus foi
significativamente menor que nos outros locais, enquanto a incidência de Alternaria
nas amostras foi significativamente maior. A vinha do local Do3 situa-se no Douro
Superior, num espaço bem ventilado. Foi-nos indicado que devido ao bom arejamento
do local, que a vinha analisada é pouco susceptível à podridão. O ambiente
circundante é dominado por 408 hectares de vinha, não sendo observada outra cultura
relevante. Parece-nos possível que devido à ventilação e condições excepcionais do
local, que existam piores condições para o estabelecimento de Aspergillus saprófitas
nos bagos e melhores condições para Alternaria, um fungo típico de campo que pode
completar o seu ciclo de vida nas folhas mortas.
No Ribatejo, no local R1, a presença mais elevada de Rhizopus na placa de
isolamento causa problemas na detecção de outros fungos de crescimento mais lento,
como Cladosporium e Alternaria, ou de identificação mais tardia, como Botrytis. Por
isso, é provável que as diferenças observadas digam respeito ao método de detecção
usado, e que a frequência destas espécies tenha sido subestimada.
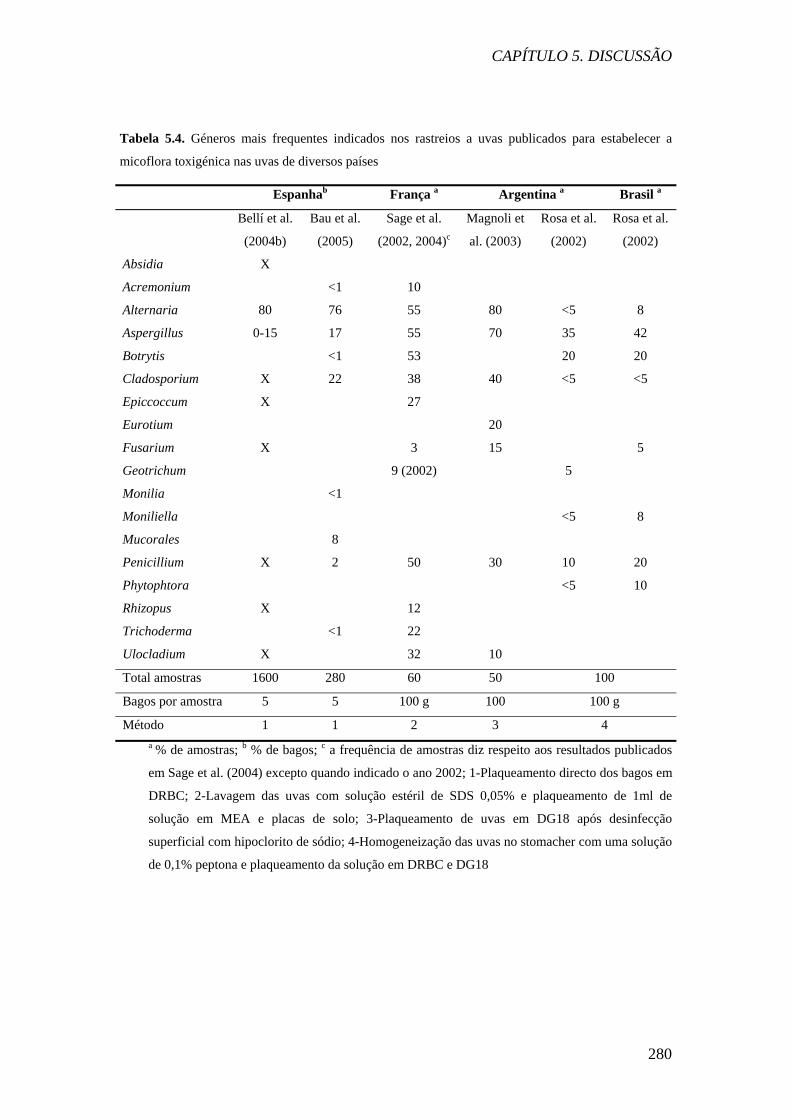
Na Tabela 5.4 estão indicados os fungos descritos como os mais frequentes nas
uvas nos rastreios feitos com o propósito de estabelecer a micoflora ocratoxigénica.
Sage e colaboradores (2004) obtiveram resultados semelhantes aos das uvas
portuguesas em uvas de diversas províncias francesas, tendo verificado que
Alternaria, Cladosporium, Botrytis, Aspergillus e Penicillium foram os géneros mais
frequentes nas amostras. Por outro lado, Magnoli et al. (2003), Bellí et al. (2004b) e
Bau et al. (2005) não reportaram Botrytis como um dos géneros mais frequentes nas
uvas. Aliás, na Argentina, Magnoli et al. (ob. cit.) enfatizam que não foi detectada a
presença de estirpes de B. cinerea durante a vindima de 2001 em 50 amostras de uvas
na província de Mendoza. No entanto, Rosa et al. (2002) verificaram que em uvas
originárias da Argentina da província de Mendonza e do Brasil (Rio Grande do Sul),
no ano de 1997-1998, os géneros mais frequentes foram Aspergillus, Penicillium e
Botrytis, e em menor abundância Alternaria e Cladosporium. Nos rastreios em que
278

CAPÍTULO 5. DISCUSSÃO
não se detectou Botrytis ou em que a sua frequência foi baixa, o género dominante foi
Alternaria, presente em cerca de 80% das amostras. Cladosporium e Penicillium
foram sempre descritos entre os fungos mais frequentemente isolados, mas Bau et al.
(2005) reportaram uma incidência de Penicillium mais baixa nas amostras que os
restantes investigadores.
A baixa incidência de Botrytis referida nas uvas da costa Mediterrânica
espanhola e na Argentina na província de Mendonza poderá estar relacionada com as
condições climáticas, visto que o clima é bastante seco. Nestes locais, o género
Alternaria foi dominante. No entanto, nas regiões portuguesas, não se verificou um
aumento significativo de Alternaria a par da baixa incidência de Botrytis.
Quanto às espécies de Aspergillus identificadas, os Aspergillus da secção
Nigri foram referidos por todos os investigadores como as espécies de Aspergillus
mais frequentes nas uvas, e são discutidos com mais detalhe no ponto 2.
Quanto às espécies de Penicillium, os investigadores referiram entre as
espécies mais frequentemente isoladas P. brevicompactum e P. expansum (Sage et al.,
2004), P. glabrum/spinulosum, P. funiculosum e P. thomii (Battilani et al., 2003), P.
chrysogenum (Magnoli et al., 2003) e P. glabrum, P. citrinum e P. decumbens (Rosa
et al., 2002), P. citrinum, P. glabrum e P. chrysogenum (Bau et al., 2005). Todas estas
espécies são agentes de deterioração comuns. Nenhum dos investigadores referidos
detectou a presença de P. verrucosum ou de P. nordicum.
279
CAPÍTULO 5. DISCUSSÃO
Tabela 5.4. Géneros mais frequentes indicados nos rastreios a uvas publicados para estabelecer a
micoflora toxigénica nas uvas de diversos países
Espanhab França a Argentina a Brasil a
Bellí et al.
(2004b)
Bau et al.
(2005)
Sage et al.
(2002, 2004)c
Magnoli et
al. (2003)
Rosa et al.
(2002)
Rosa et al.
(2002)
Absidia X
Acremonium <1 10
Alternaria 80 76 55 80 <5 8
Aspergillus 0-15 17 55 70 35 42
Botrytis <1 53 20 20
Cladosporium X 22 38 40 <5 <5
Epiccoccum X 27
Eurotium 20
Fusarium X 3 15 5
Geotrichum 9 (2002) 5
Monilia <1
Moniliella <5 8
Mucorales 8
Penicillium X 2 50 30 10 20
Phytophtora <5 10
Rhizopus X 12
Trichoderma <1 22
Ulocladium X 32 10
Total amostras 1600 280 60 50 100
Bagos por amostra 5 5 100 g 100 100 g
Método 1 1 2 3 4 a % de amostras; b % de bagos; c a frequência de amostras diz respeito aos resultados publicados
em Sage et al. (2004) excepto quando indicado o ano 2002; 1-Plaqueamento directo dos bagos em
DRBC; 2-Lavagem das uvas com solução estéril de SDS 0,05% e plaqueamento de 1ml de
solução em MEA e placas de solo; 3-Plaqueamento de uvas em DG18 após desinfecção
superficial com hipoclorito de sódio; 4-Homogeneização das uvas no stomacher com uma solução
de 0,1% peptona e plaqueamento da solução em DRBC e DG18
280

CAPÍTULO 5. DISCUSSÃO
1.1.3. Classificação das amostras de uvas portuguesas de acordo
com a origem geográfica com base na micoflora
Com as árvores de decisão, explorou-se a possibilidade de classificar a região
de origem das uvas com base em diferenças na composição da micoflora das
amostras. Caso essa tarefa fosse possível, significava que cada área geográfica podia
estar exposta a uma micoflora mais ou menos constante, dependente da região de
origem, o que poderia ter relevância para o estabelecimento de mapas de risco e de
identificação de perigos locais. Caso não fosse possível, seria interpretado como
reflexo de independência entre a micoflora das uvas e a região de origem.
Com base na micoflora total das uvas, criou-se um modelo de classificação em
que através das diferenças observadas na incidência de A. niger, Botrytis, P.
corylophilum e P.thomii nas amostras foi possível identificar a origem geográfica das
amostras no momento da vindima. A região dos Vinhos Verdes e do Douro surgem
definidas por apenas uma folha, o que significa que a micoflora dessas regiões pode
ser definida com base num critério. Mas para o Ribatejo e Alentejo temos duas folhas,
o que significa que não foi possível definir cada uma destas regiões apenas com um
critério. As amostras da região dos Vinhos Verdes foram identificadas com base na
sua baixa incidência de A. niger e incidência relativamente elevada de Botrytis. As
amostras do Douro foram distinguidas das dos Vinhos Verdes com base na sua
incidência mais elevada de A. niger, e as amostras do Norte de Portugal foram
distinguidas das do Sul com base na incidência de P. thomii nas amostras. Mas a
distinção entre as duas regiões do Sul, Alentejo e Ribatejo, foi principalmente feita
com base na incidência de Botrytis nas amostras destas regiões, mais elevada no
Ribatejo que no Alentejo. No entanto, a capacidade preditiva deste modelo foi baixa,
sendo o Alentejo a região com mais insucesso de classificação, onde 50% das
amostras foram classificadas erroneamente como sendo do Ribatejo, pelos critérios
deste modelo. Com a selecção dos melhores atributos, o modelo continuou a usar os
dois critérios que permitiam a distinção das amostras dos Vinhos Verdes e do Douro e
entre Norte/Sul (agregado A. niger e P. thomii, respectivamente). Por sua vez, as
regiões do Alentejo e Ribatejo foram distinguidas com base na frequência de
Ulocladium e de A. niger. Mas novamente se verificou que a região do Alentejo era
classificada com pouco sucesso com validação cruzada, sendo incorrectamente
281

CAPÍTULO 5. DISCUSSÃO
classificada como Ribatejo em 50% dos casos. Tornou-se claro que a distinção de
amostras do Ribatejo e Alentejo era complexa. Como tal, testou-se o modelo com
redefinição das classes: Douro, Sul e Vinhos Verdes.
O modelo com base apenas na frequência de A. niger e P. thomii nas amostras
revelou-se capaz de classificar correctamente 82% das amostras num estudo de
validação cruzada. Isto mostra que, aparentemente, as uvas das regiões dos Vinhos
Verdes, Douro e Sul estão expostas de forma diferente a estas espécies duma forma
bastante consistente, independentemente do local, casta ou ano estudados. As regiões
do Sul de Portugal- Ribatejo e Alentejo- surgem como uma região exposta à mesma
população fúngica e, consequentemente, aos mesmos riscos potenciais. Estas
diferenças na distribuição geográfica dos fungos, se provarem ser consistentes, podem
eventualmente ter aplicações na micologia forense, quanto à identificação da
proveniência da região das uvas pela sua composição micológica com base na
frequência de algumas espécies chave. Adicionalmente, a variação na micoflora que
se verificou nas regiões portuguesas reforça a ideia de que as diferenças na micoflora
das uvas referidas pelos investigadores de diferentes países podem não ser
exclusivamente devidas a diferenças no método usado ou ao clima, mas depender da
distribuição geográfica das espécies.
1.2. Micoflora das uvas após desinfecção superficial dos
bagos
Após desinfecção superficial, detectou-se uma menor diversidade de fungos
nas uvas, o que indica que muitos dos géneros estão apenas presentes na superfície
dos bagos. Foram detectados essencialmente fungos descritos como potenciais
causadores de podridão nos bagos, como Alternaria, Aspergillus, Botrytis,
Cladosporium, Penicillium, Stemphylium e Rhizopus (Lopes, 2002), incluindo fungos
saprófitas comuns em folhas verdes que em certas condições podem tornar-se
patogénicos para plantas, como Aureobasidium, Fusarium e Phoma (Lopes, ob. cit.).
Seis das 9 das espécies de Penicillium e estirpes do género Rhizopus só foram
detectadas na vindima, enquanto que outros fungos, como Alternaria, Aureobasidium,
Botrytis, Cladosporium, Epicoccum, P. glabrum/spinulosum, P. thomii e Stemphylium
282

CAPÍTULO 5. DISCUSSÃO
foram detectados em todos os estados de maturação, indicando que a colonização do
bago com alguns fungos se dá desde os estados de maturação iniciais. Bau et al.
(2005) estudaram a composição micológica de sementes de uva, e não detectaram
qualquer fungo, o que leva crer que a contaminação fúngica com as espécies
saprófitas e patogénicos oportunistas advém essencialmente da superfície do bago.
1.3. Podridão detectada nos bagos
Nas vinhas estudadas raras vezes se observou podridão. Em algumas das
ocasiões em que se observou podridão, associado a Botrytis, foram detectados A.
carbonarius (uma única vez) e T. roseum. T. roseum não é um fungo muito
frequentemente referido como causador de podridão nas uvas. A única referência que
conhecemos a este respeito foi em uvas provenientes da Alemanha (Schwenk et al.,
1989). A podridão de T. roseum associada a Botrytis pode passar despercebida à
primeira vista. Nas adegas, o aspecto róseo por cima da podridão cinzenta de Botrytis
foi considerado descoloração do fungo pelos engenheiros agrónomos e, como tal, é
possível que seja mais frequente que o referido. T. roseum foi descrito como um
micoparasita dos esclerócios de B. cinerea (Domsch et al., 1993). As nossas
observações não são suficientes para afirmar se T. roseum estava de facto a parasitar
as hifas de B. cinerea, visto que não foram detectadas ao microscópio estruturas de
penetração nas hifas de Botrytis por T. roseum. Aparentemente, T. roseum apenas
cresce sobre Botrytis. Na placa de isolamento, T. roseum foi detectado muitas vezes a
crescer sobre Botrytis, sendo detectado frequentemente no final do tempo de vida da
placa de isolamento.
Durante os anos do nosso estudo, não se observou podridão por P. expansum
nas vinhas estudadas ou de que tivéssemos conhecimento, mas Abrunhosa (2001)
detectou uvas podres com esta espécie na vinha Do4 no ano de 1999.
Apesar de a podridão por Aspergillus negros ser considerada em alguns países
(v.g., Israel) de igual importância económica que a por B. cinerea e de ser o fungo
predominante em uvas destinadas ao fabrico de vinho (Zahavi et al., 2000), durante o
nosso estudo a podridão com Aspergillus foi observada uma única vez num cacho (A.
carbonarius). A podridão por Aspergillus em Portugal é raramente referida, sendo a
Botrytis considerada a principal causa de podridão nas uvas (Amaro & Couto, 2001).
283

CAPÍTULO 5. DISCUSSÃO
1.4. Micoflora do mosto
A detecção e isolamento de fungos filamentosos no mosto de uvas antes e após a
primeira adição de sulfuroso foi dificultada pelo grande número de colónias de
leveduras presentes. Os principais fungos detectados no mosto foram Aspergillus,
Penicillium e Rhizopus. Foram detectadas duas espécies de Aspergillus da secção
Nigri, A. carbonarius e agregado A. niger. A presença de A. carbonarius só foi
detectada numa amostra, antes da adição de sulfuroso no mosto da casta alvarinho
proveniente da sub-região de Monção, onde nunca foi detectado este fungo nas uvas
da vinha estudada da mesma sub-região. Em todas as amostras foram detectadas
estirpes do agregado A. niger e do género Penicillium. Uma das espécies mais
frequentemente detectadas foi P. crustosum. Esta espécie é capaz de crescer em
condições bastante ácidas, a pH inferior a 2,2. Após a adição de sulfuroso, ainda
encontrámos propágulos viáveis de agregado A. niger, Mucor, Penicillium e Rhizopus.
O sulfuroso é um composto químico que tem acção fungistática, não fungicida. O
seu propósito é desinfectar o mosto e prevenir a proliferação de microrganismos
indesejáveis até ao arranque da fermentação, quando em condições industriais se
adicionam leveduras seleccionadas. Nas duas amostras de mosto em início de
fermentação da região dos Vinhos Verdes, não se detectaram colónias de fungos
filamentosos, apenas leveduras. Dada a dificuldade de detectar propágulos viáveis de
fungos filamentosos no mosto no início da fermentação devido ao grande número de
colónias de leveduras presentes nas placas, não se pode excluir que não possam
existir. No entanto, considera-se que mesmo que estivessem presentes propágulos de
fungos filamentosos viáveis no mosto nesta fase, estes não teriam condições para
crescer, dada a competição feroz com as leveduras.
284

CAPÍTULO 5. DISCUSSÃO
2. Espécies de Aspergillus da secção Nigri produtoras de
OTA
A. carbonarius foi sem dúvida a principal espécie responsável pela produção
de OTA detectada nas uvas portuguesas, compreendendo 73% das estirpes produtoras
de OTA isoladas. Apesar das estirpes do agregado A. niger serem muito mais
frequentes, a produção pelas estirpes foi muito mais rara que por A. carbonarius. No
agregado A. niger, foi detectada produção de OTA por 4% das estirpes, enquanto que
100% das estirpes de A. carbonarius foram capazes de produzir a micotoxina.
Nenhuma das 3 estirpes de A. japonicus foi capaz de produzir OTA em níveis
detectáveis pelo método usado.
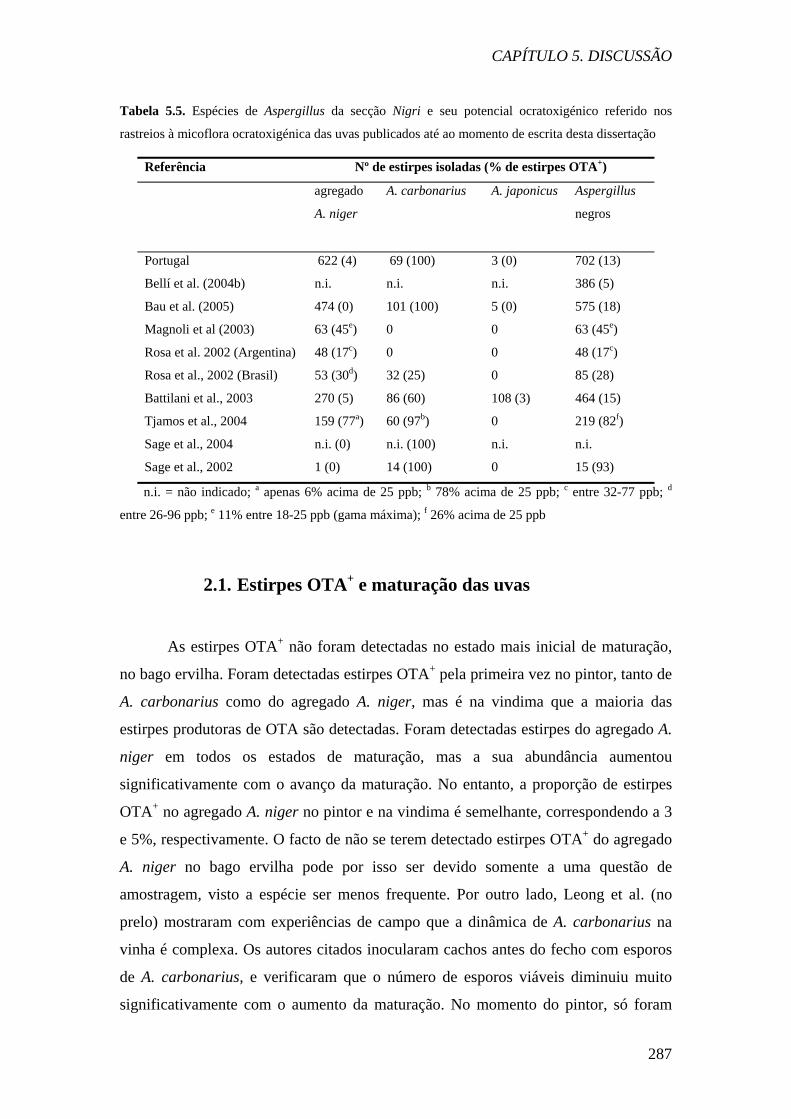
Os dados europeus são concordantes quanto ao facto de que a principal
espécie produtora de OTA isolada das uvas é A. carbonarius. No entanto, a proporção
de estirpes OTA+ referida variou consideravelmente. Nem todos os investigadores
referem 100% de produção de OTA pelas estirpes de A. carbonarius (Tabela 5.6).
Estes resultados são contrários aos obtidos no nosso estudo, bem como nos estudos
espanhóis e franceses, em que não se detectou uma única estirpe não produtora de
OTA desta espécie. Na literatura, outros estudos também não mencionaram 100% de
capacidade das estirpes de A. carbonarius produzirem a micotoxina (Abarca et al.,
2003; Joosten et al., 2001; Taniwaki et al., 2003; Téren et al., 1996). Este facto pode
ser devido a diferentes causas: 1) diferenças nos métodos e limites de detecção usados
em cada laboratório; 2) identificação errada dos metabolitos, visto que não é prática
corrente a confirmação dos metabolitos nas análises de ELISA e HPLC; 3)
identificação errada da estirpe; 4) diferenças locais quanto à proporção de estirpes
OTA+; 5) Algumas das estirpes verdadeiramente negativas poderão tratar-se de A.
ibericus, o taxon proposto que se pode confundir com A. carbonarius. Esta última
hipótese está na base da não detecção de OTA nas estirpes identificadas previamente
como A. carbonarius por Abarca et al. (2003).
Quanto às estirpes do agregado A. niger, a proporção de estirpes produtoras
nas uvas descrita na literatura também varia consideravelmente, entre 0 a 77%. As
causas para esta variação podem ser semelhantes às indicadas acima para A.
285

CAPÍTULO 5. DISCUSSÃO
carbonarius. Tjamos et al. (2004) reportaram que 77% de estirpes do agregado A.
niger produziram OTA em níveis detectáveis, mas mencionam que apenas 6% das
estirpes são fortemente positivas. Rosa et al. (2002) e Magnoli et al. (2003) referiram
uma proporção elevada de estirpes OTA+ do agregado A. niger nas uvas, tornando
esta espécie a principal produtora de OTA nas uvas da Argentina. Dalcero e
colaboradores (2002) obtiveram resultados semelhantes em rações animais, onde A.
carbonarius não foi detectado e 46% das estirpes do agregado A. niger produziram a
micotoxina em níveis detectáveis por HPLC. No entanto, A. carbonarius existe na
Argentina, tendo sido detectado em uvas passas recentemente (Magnoli et al., 2004).
A proporção de estirpes OTA+ no agregado A. niger nas uvas portuguesas está de
acordo com os estudos de Battilani et al. (2003), Téren et al. (1996) e Taniwaki et al.
(2003), que detectaram estirpes OTA+ entre 3 a 5% das estirpes do agregado.
Os únicos investigadores que reportaram produção de OTA por estirpes de A.
japonicus foram Battilani et al. (2003) e Dalcero et al. (2002). No entanto, outros
investigadores, como Bau et al. (no prelo), defendem que as únicas espécies da secção
Nigri capazes de produzir a micotoxina são A. carbonarius e A. niger.
Considerando a percentagem de estirpes produtoras de OTA face ao número
total de estirpes de Aspergillus secção Nigri isoladas, as uvas portuguesas apresentam
uma proporção de estirpes semelhante à referida por Bau et al. (2005) e Battilani et al.
(2003) na Europa. Os investigadores que referiram a incidência mais baixa de estirpes
OTA+ na secção Nigri foram Bellí et al. (2004b). Na Europa, os investigadores que
registaram a proporção de estirpes OTA+ mais elevada na secção Nigri foram Tjamos
et al. (2004), na Grécia, e Sage et al. (2002), na França.
286
CAPÍTULO 5. DISCUSSÃO
Tabela 5.5. Espécies de Aspergillus da secção Nigri e seu potencial ocratoxigénico referido nos
rastreios à micoflora ocratoxigénica das uvas publicados até ao momento de escrita desta dissertação
Referência Nº de estirpes isoladas (% de estirpes OTA+)
agregado
A. niger
A. carbonarius A. japonicus Aspergillus
negros
Portugal 622 (4) 69 (100) 3 (0) 702 (13)
Bellí et al. (2004b) n.i. n.i. n.i. 386 (5)
Bau et al. (2005) 474 (0) 101 (100) 5 (0) 575 (18)
Magnoli et al (2003) 63 (45e) 0 0 63 (45e)
Rosa et al. 2002 (Argentina) 48 (17c) 0 0 48 (17c)
Rosa et al., 2002 (Brasil) 53 (30d) 32 (25) 0 85 (28)
Battilani et al., 2003 270 (5) 86 (60) 108 (3) 464 (15)
Tjamos et al., 2004 159 (77a) 60 (97b) 0 219 (82f)
Sage et al., 2004 n.i. (0) n.i. (100) n.i. n.i.
Sage et al., 2002 1 (0) 14 (100) 0 15 (93)
n.i. = não indicado; a apenas 6% acima de 25 ppb; b 78% acima de 25 ppb; c entre 32-77 ppb; d
entre 26-96 ppb; e 11% entre 18-25 ppb (gama máxima); f 26% acima de 25 ppb
2.1. Estirpes OTA+ e maturação das uvas
As estirpes OTA+ não foram detectadas no estado mais inicial de maturação,
no bago ervilha. Foram detectadas estirpes OTA+ pela primeira vez no pintor, tanto de
A. carbonarius como do agregado A. niger, mas é na vindima que a maioria das
estirpes produtoras de OTA são detectadas. Foram detectadas estirpes do agregado A.
niger em todos os estados de maturação, mas a sua abundância aumentou
significativamente com o avanço da maturação. No entanto, a proporção de estirpes
OTA+ no agregado A. niger no pintor e na vindima é semelhante, correspondendo a 3
e 5%, respectivamente. O facto de não se terem detectado estirpes OTA+ do agregado
A. niger no bago ervilha pode por isso ser devido somente a uma questão de
amostragem, visto a espécie ser menos frequente. Por outro lado, Leong et al. (no
prelo) mostraram com experiências de campo que a dinâmica de A. carbonarius na
vinha é complexa. Os autores citados inocularam cachos antes do fecho com esporos
de A. carbonarius, e verificaram que o número de esporos viáveis diminuiu muito
significativamente com o aumento da maturação. No momento do pintor, só foram
287

CAPÍTULO 5. DISCUSSÃO
detectados 0,001% dos esporos inoculados. A partir do pintor, observou-se um
aumento do número de esporos e podridão. Os autores atribuiram a diminuição do
número de colónias viáveis à morte dos esporos de A. carbonarius por exposição à
radiação UV. No entanto, não exploraram até à data outra possibilidade que nos
parece viável: que os esporos de A. carbonarius tenham problemas em aderir e
manter-se na superfície da uva em estádios iniciais de desenvolvimento do bago. Esta
hipótese é suportada pela observação ao SEM de esporos de A. carbonarius, em que
se vê que as equinulações dos esporos se quebram facilmente, podendo prejudicar a
adesão. Quando o cacho fecha, é possível que se este comporte como uma armadilha
de esporos, aprisionando-os no seu interior, de forma que estes possam infectar os
bagos caso haja danos nas películas.
A contaminação dos bagos de uvas portuguesas com estirpes OTA+ é
essencialmente superficial em uvas aparentemente saudáveis. Após desinfecção
superficial de uvas normais, detectou-se A. carbonarius no momento da vindima em
apenas 3 dos 1200 bagos analisados (0,25%).
2.2. Variação da incidência de estirpes OTA+ com a região
de origem
Foram detectadas estirpes OTA+ em todas as regiões estudadas à excepção do
Dão, e em todas as vinhas das regiões do Douro, Ribatejo e Alentejo, mostrando que
todas as uvas de locais de clima Mediterrânico estão expostas a estes contaminantes.
Na região dos Vinhos Verdes foi detectada a presença de A. carbonarius numa
amostra de uvas e em mosto proveniente da sub-região de Monção, mostrando que
estes contaminantes também estão presentes na região dos Vinhos Verdes, mas com
menor frequência.
Verificou-se que A. carbonarius foi mais frequente nas vinhas do Sul do país
que no Douro, apesar de também ser detectado nesta região. No entanto, A.
carbonarius só foi detectado após desinfecção superficial no Alentejo e Ribatejo. Na
vinha 5 registou-se a incidência mais elevada de estirpes OTA+ do agregado A. niger,
e este foi o único local onde estas estirpes foram isoladas consistentemente na
vindima todos os anos. Este facto parece indicar que a população de estirpes OTA+ do
288

CAPÍTULO 5. DISCUSSÃO
agregado A. niger é residente nesta vinha. Por outro lado, a presença de estirpes OTA+
do agregado A. niger noutras vinhas foi ocasional. O mesmo se pode dizer da
incidência de A. carbonarius nas uvas, que variou consideravelmente, e só foi
detectado nos três anos na vinha 8.
A. carbonarius foi detectado a causar podridão no interior dum cacho podre da
vinha 5, e nesse ano, na vindima, não foi detectado em nenhum dos 50 bagos da
amostra. Este facto sugere que a capacidade de permanência de A. carbonarius na
superfície dos bagos é baixa, e só consegue manter-se no interior dos cachos mais
compactos, onde causa podridão quando as condições lhe são favoráveis. Sendo
assim, independentemente da sua incidência nos bagos, a presença de A. carbonarius,
principalmente nas vinhas de clima Mediterrânico, revela potencial para a ocorrência
de podridão mediante condições propícias, especialmente no interior dos cachos.
Battilani et al. (2003) não reportaram diferenças conspícuas na micoflora
ocratoxigénica do Norte e Sul da Itália, sendo a percentagem média de estirpes de
Aspergillus produtoras de OTA de 6 e 8% no Norte e Sul da Itália, respectivamente.
Por sua vez, Bau et al. (2005) e Bellí et al. (2004b) não reportaram diferenças na
incidência de Aspergillus negros na costa Mediterrânica de Espanha entre as regiões
de Barcelona, Tarragona, Valência, Múrcia e Cádiz, e entre Costers del Segre,
Penedés, Conca de Barberà e Utiel-Requena, respectivamente. No entanto, Sage et al.
(2002, 2004), no seu estudo de 6 regiões de França, só detectaram a presença de A.
carbonarius nas amostras nas regiões mais a sul (nomeadamente Aude e Languedoc).
Tjamos et al. (2004) reportaram diferenças entre a incidência de espécies de
Aspergillus negros nas castas Cabernet Sauvignon e Grenache Rouge na ilha de
Rhodes. A casta Grenache Rouge teve uma incidência muito maior de A. carbonarius
nas uvas que a casta Cabernet Sauvignon. Na Grenache Rouge, mais de 80% dos
bagos estavam colonizados com A. carbonarius, e menos de 10% dos bagos com
estirpes do agregado A. niger, enquanto na casta Cabernet Sauvignon, a contaminação
com A. carbonarius foi de 10 a 20%, e a contaminação com estirpes do agregado A.
niger mais de 80%. Desta forma, é na Grécia que foi referida a incidência mais
elevada de A. carbonarius numa vinha.
Em termos do conhecimento que se tem da distribuição geográfica das principais
espécies produtoras de OTA, do clima e da OTA detectada em vinhos, verifica-se que
foi referida a presença de A. carbonarius em todas as áreas onde foi detectada OTA
em vinhos (V. Capítulo 2).
289

CAPÍTULO 5. DISCUSSÃO
2.3. Relação entre estirpes OTA+ e a restante micoflora das
uvas
Considera-se que a presença de microrganismos epífitos na superfície dos
diferentes órgãos das plantas pode influenciar o crescimento de fungos parasitas
(Lopes, 2002). A possibilidade de se usarem alguns microrganismos como
antagonistas de patogénicos fúngicos nas uvas tem sido investigada (Zahavi et al.,
2000). Os microrganismos seleccionados para testar as potencialidades em termos de
luta biológica foram principalmente leveduras e o fungo dimórfico Aureobasidium
pullulans, apesar de alguns fungos filamentosos exibirem capacidade como
antagonistas, como Acremonium cephalosporium. Estirpes destas espécies, em
particular de Aureobasidium pullulans, têm efeito antagonista quanto à infecção por
Aspergillus (Zahavi et al., 2000), e fazem parte da micoflora das uvas portuguesas.
Por outro lado, como foi referido na introdução, o uso de estirpes não
toxigénicas no controlo de micotoxinas em culturas tem sido proposto como luta
biológica, visto que estas estirpes deverão ocupar o mesmo nicho ecológico em
campo. Observou-se que a proporção de estirpes toxigénicas para não toxigénicas de
Aspergillus negros variou nas vinhas estudadas na vindima variou entre 1:1 até 0, mas
os baixos níveis de OTA nas uvas e vinhos portugueses em geral não permitem tirar
conclusões quanto à influência da micoflora no teor de OTA das uvas.
Em termos de frequência nas amostras de uvas, as únicas correlações que
parecem relevantes quanto à relação entre espécies OTA+ e a restante micoflora são
entre A. carbonarius e o agregado A. niger e entre A. carbonarius e P.
brevicompactum. As estirpes do agregado A. niger estão significativamente
correlacionadas de forma positiva com a incidência de estirpes OTA+ nas uvas, pois
reflectem as preferências climáticas dos Aspergillus negros no geral. A correlação
positiva entre A. carbonarius e P. brevicompactum pode ser devida ao facto de que
ambas as espécies foram mais frequentes nas amostras do Sul de Portugal.
Quanto à situação nacional estudada, visto que as uvas estiveram em bom
estado fitossanitário e a concentração de OTA detectada nas uvas foi sempre baixa,
não se podem tirar conclusões quanto à influência da micoflora OTA- nestes
290

CAPÍTULO 5. DISCUSSÃO
parâmetros. Salienta-se apenas que de acordo com os nossos dados e com os estudos
previamente publicados sobre a micoflora ocratoxigénica das uvas, a presença de
Aspergillus negros nas uvas parece ser boa indicadora da possibilidade da presença de
estirpes OTA+, independentemente das espécies em questão.
3. Aspergillus ibericus
Visto que as diferentes espécies da secção Nigri possuem diferentes capacidades
de produzir OTA, e dada a proximidade morfológica das espécies, a necessidade de
identificar correctamente as estirpes desta secção é imperativa a fim de evitar a
confusão entre as espécies que produzem a micotoxina nesta secção.
As estirpes inicialmente identificadas como A. carbonarius que não produziram
OTA possuem um perfil molecular distinto das outras espécies da secção, que se
revelou por análise RAPD, análise da sequência do rDNA ITS-5.8S e por análise de
sequências do gene da calmodulina (Cabañes et al. (2004); Bau e colaboradores (no
prelo)). Considerou-se que estas estirpes poderiam representar uma nova espécie da
secção Nigri, designado provisoriamente de A. ibericus. A. ibericus foi isolado
raramente das uvas de climas Mediterrânicos, mas em estados de maturação e regiões
geográficas distintas, o que indica que está bem estabelecida nas vinhas portuguesas.
O nome A. ibericus foi por nós proposto por dois motivos: até à data, estas estirpes só
foram isoladas em Portugal e Espanha; por outro lado, não há nenhum aspecto
qualitativo a nível morfológico que lhe confira individualidade face às outras espécies
descritas na secção Nigri. A ausência de capacidade de produção de OTA pelas
estirpes de A. ibericus distingue-as das estirpes de A. carbonarius, mas não das
estirpes de A. niger. Por isso, apesar de diferenças quanto à produção da micotoxina, a
distinção destas estirpes tem de ser feita numa base molecular e/ou morfológica.
O estudo da morfologia dos esporos ao SEM permitiu verificar que não há
uma relação directa entre a ornamentação dos esporos, a taxinomia e a produção de
OTA quanto às duas classes definidas por Kozakiewicz (1989): verrucosos e
equinulados. Estes dois tipos de ornamentação foram detectados em estirpes do
agregado A. niger e A. carbonarius, por vezes na mesma estirpe. Por outro lado,
291

CAPÍTULO 5. DISCUSSÃO
confirmou-se que o tamanho dos esporos é uma característica valiosa na distinção dos
3 taxa bisseriados isolados das uvas: agregado A. niger, A. carbonarius e A. ibericus.
Detectaram-se diferenças significativas entre o tamanho médio dos esporos
dos 3 taxa mencionados. Não há sobreposição quanto ao tamanho dos esporos das
estirpes do agregado A. niger e de A. carbonarius, mas verificou-se sobreposição do
tamanho de esporos de A. ibericus com o limite superior do tamanho dos esporos do
agregado A. niger e com o limite inferior do tamanho dos esporos de A. carbonarius.
Apesar disso, foi possível classificar as estirpes negras bisseriadas isoladas das uvas
com 100% de exactidão nestas 3 espécies com base no modelo de classificação
produzido com o tamanho médio dos esporos: < 5 µm, agregado A. niger; entre 5 e
6µm, A. ibericus; > 6µm: A. carbonarius. Segundo este critério, as estirpes da espécie
A. ibericus foram correctamente classificadas, mas com base num intervalo de apenas
1 micrómetro. Este critério força a uma capacidade de medição exacta dos esporos, e
juntamente com a sobreposição do tamanho dos esporos individuais com as outras
espécies, enfatiza a necessidade de se classificar as estirpes com base no tamanho
médio dos esporos. A produção de OTA é uma característica que facilita a distinção
entre as estirpes de A. ibericus e A. carbonarius, mas a distinção entre as estirpes de
A. ibericus e agregado A. niger reside no tamanho dos esporos.
As estirpes de A. ibericus revelaram-se muito uniformes quanto ao tamanho
médio dos esporos, ao contrário das estirpes do agregado A. niger e A. carbonarius,
em que o tamanho médio dos esporos varia significativamente com a estirpe.
As estirpes de A. niger e A. tubingensis mostraram diferenças significativas
quanto ao tamanho médio dos esporos, com A. tubingensis a exibir esporos mais
pequenos que A. niger. Adicionalmente, verificou-se que os esporos de A. tubingensis
eram menos ornamentados. Estas observações vão de encontro às verificadas por
Accensi (2000) num estudo de 92 estirpes do agregado A. niger. O investigador
verificou que os esporos de A. tubingensis tem em média 3,6 µm de diâmetro
enquanto os de A. niger têm 4,0 µm de diâmetro. Tal como Accensi (ob. cit.),
consideramos que as diferenças morfológicas observadas são muito subtis e não
permitem uma distinção fidedigna entre as duas espécies.
Por outro lado, considera-se que a identificação morfológica do taxa A.
ibericus é possível, e que essa tarefa é facilitada quando aliada à avaliação da
capacidade de produção de OTA, visto que até agora não foi detectada a capacidade
292

CAPÍTULO 5. DISCUSSÃO
de produção de OTA por estas estirpes. Considera-se adicionalmente que este taxa
não complica a taxinomia das espécies bisseriadas da secção Nigri, mas ao invés
disso, estreita a relação entre a taxinomia e a produção de OTA das espécies, em
particular para A. carbonarius.
4. Concentração de OTA nas uvas portuguesas
As análises às uvas produziram resultados surpreendentes, na medida em que
se detectou OTA no bago ervilha, quando não foram isoladas estirpes OTA nesta fase
da maturação das uvas. Investigadores italianos também verificaram este fenómeno,
se bem que raramente (Battilani, comunicação pessoal). Adicionalmente, os níveis de
OTA detectados no bago ervilha e no pintor foram superiores aos detectados em uvas
normais na vindima. Apesar de a OTA ser detectada nas uvas desde os estados iniciais
de maturação, as implicações deste facto quanto aos níveis de OTA detectados na
vindima não são conhecidas. Visto que se detectaram níveis inferiores de OTA no
bagos maduro comparativamente aos estádios anteriores de maturação, é impossível
especular se este facto se deve a uma questão de amostragem ou se está relacionado
com outros fenómenos, como diluição com o aumento de volume do bago ou
degradação por fungos (Abrunhosa et al., 2002) ou pela própria planta. No entanto,
nenhum dos valores de OTA detectados em uvas normais excedeu o limite máximo de
2 µg/l de OTA proposto no vinho.
Em uvas aparentemente saudáveis, detectou-se OTA na vindima em 3 regiões
e uma sub-região: Dão, Madeira, Ribatejo e sub-região de Monção (região dos Vinhos
Verdes), respectivamente. Só foram detectadas diferenças significativas entre a
concentração de OTA nas uvas das regiões no momento da vindima. As amostras
oriundas do Ribatejo estiveram em média mais contaminadas que as provenientes das
restantes regiões. No entanto, a concentração média de OTA nas amostras do Ribatejo
não excedeu os 0,1 µg/kg.
Foi interessante verificar que na vindima, a OTA foi detectada nas regiões
mais húmidas de bioclima Mediterrânico Pluvio-estacional Oceânico, e de menor
continentalidade (mais oceânicas) (V. capítulo 3, Tabela 3.7, p. 138). Estes resultados
estão de acordo com a distribuição de OTA nos vinhos europeus, em que os vinhos
293

CAPÍTULO 5. DISCUSSÃO
mais contaminados são originários da bacia Mediterrânica, em particular, do Sul da
Itália e ilhas Gregas (V. capítulo 2). A presença de A. carbonarius foi detectada em
todas as regiões (e sub-região) mencionadas em amostras de uvas ou de mosto.
No entanto, a única situação em que se excedeu o limite de 2 µg/l nas uvas foi
num cacho em que foi detectada podridão com A. carbonarius numa vinha do Douro.
Estes resultados enfatizam a necessidade de se usarem uvas em bom estado
fitossanitário para vinificação em todas as regiões onde as uvas estão expostas a A.
carbonarius, visto que nenhuma das amostras de uvas sem sintomas de infecção
fúngica excedeu a concentração de 2 µg/l de OTA nos bagos.
5. Avaliação do potencial ocratoxigénico das estirpes em
meio de uva
A produção de OTA por estirpes da secção Nigri entre CYA e GJ50 foi
consistente, significando que o CYA é um bom meio para avaliar a capacidade de
produção de OTA pelas estirpes em meio de uva. Confirma também que todas as
estirpes ocratoxigénicas têm um comportamento semelhante quanto à capacidade de
produzir a micotoxina em meio de uva. Para estudar a influência das diferenças na
composição nutricional das uvas quanto à maturação e à casta, seleccionou-se uma
estirpe de A. carbonarius. Os resultados revelaram que a composição nutricional das
uvas permite a produção da micotoxina em todos os estados de maturação e castas,
sendo favorecida por concentrações de ácidos orgânicos elevadas e baixos teores de
açúcares redutores que se verificaram em estados de maturação mais precoces e na
vindima em castas mais ácidas, como Vinhão. Em uvas no estado de bago ervilha, a
estirpe seleccionada produziu cerca de 100 vezes mais OTA que nas uvas maduras.
Esta observação suporta os dados obtidos respeitantes à concentração de OTA das
amostras, no sentido em que confirma que o bago ervilha tem condições
nutricionalmente adequadas para que haja produção da micotoxina, e em quantidades
superiores que no bago maduro. Parece-nos possível que casos pontuais de infecção
por A. carbonarius em bagos danificados nesta fase (v.g., bagos rachados por oídio)
possam levar à contaminação dos bagos com a micotoxina nos níveis observados em
campo.
294

CAPÍTULO 5. DISCUSSÃO
6. Outros perigos micotoxigénicos nas uvas
Nesta secção discutem-se os principais perigos micotoxigénicos que
apresentam outros fungos filamentosos presentes nas uvas além dos Aspergillus.
6.1. T. roseum e seus metabolitos
O facto de se ter detectado podridão com T. roseum em uvas portuguesas
destinadas à vinificação, conjuntamente com o facto de se terem detectado
previamente metabolitos tóxicos produzidos por este fungo em vinhos, suscita
preocupação. A preocupação é particularmente acrescida na região dos Vinhos
Verdes, onde se observou podridão por este fungo num grande número dos cachos
que chegaram a adegas para serem vinificados, apesar de também ter sido detectada a
presença do fungo em cachos podres da região do Douro durante este estudo e em
1999 por Abrunhosa (2001). Apesar de não existir legislação sobre o assunto, e de se
considerar que o risco para a saúde dos metabolitos de T. roseum deva ser mínimo,
visto que a podridão por este fungo em alimentos é rara (Pitt & Hocking, 1997), trata-
se de compostos da família dos tricotecenos, compostos de reconhecida toxicidade
para animais. Adicionalmente, verificou-se que um dos metabolitos, a tricotecina,
apresenta toxicidade para leveduras, e que confere um gosto amargo ao vinho quando
em grandes quantidades, afectando as características organolépticas do vinho.
A podridão por este fungo só foi detectada associada a B. cinerea. A podridão
cinzenta de B. cinerea nos cachos causa um decréscimo enorme na qualidade das
uvas, dificultando o processo de vinificação. Visto ser a podridão cinzenta ser
fenómeno frequente na região dos Vinhos Verdes, algumas adegas possuem uma
avaliação da qualidade nas uvas pela quantificação da actividade de lacase, enzima
produzida por Botrytis e outros fungos, de forma a melhor avaliar a qualidade das
uvas. Por isso, especula-se que os metabolitos de T. roseum serão particularmente
importantes em vinhos de menor qualidade, feitos a partir de uvas com alguma
podridão cinzenta, e que é possível que a determinação da lacase esteja relacionada
com a eventual presença de metabolitos tóxicos na região, bem como outras regiões
295

CAPÍTULO 5. DISCUSSÃO
susceptíveis à podridão. A presença de T. roseum nos cachos podres leva-nos a
chamar a atenção de que vinificar uvas podres, além de ser uma má prática agrícola e
levar a vinhos de pior qualidade, pode conduzir à presença de compostos
potencialmente nocivos para a saúde.
6.2. Alternaria e alternariol
A presença de metabolitos produzidos por alternaria é particularmente relevante
em alimentos em que se detecta podridão visível por este fungo. Durante este estudo
nunca foi observada podridão nas uvas por Alternaria, mas existem registos de
algumas ocorrências associadas a Botrytis (Lopes, 2002). A autora considera que só é
provável que ocorram metabolitos de Alternaria, em particular o alternariol, caso se
vinifiquem cachos podres, e por isso, subscreve-se o que foi dito para T. roseum.
Apesar de a presença destes compostos não estar legislada em nenhum produto, a sua
presença é indicadora de má qualidade das matérias-primas (Scott, 2001).
6.3. Penicillium e patulina
As espécies do género Penicillium são capazes da produção de várias
micotoxinas, sendo as mais relevantes a OTA e a patulina. Como foi indicado na
secção dos resultados, não foram detectadas espécies OTA+ de Penicillium nas uvas
portuguesas. As espécies mais frequentemente detectadas nas uvas portuguesas, P.
brevicompactum, P. glabrum/spinulosum e P. thomii não produzem micotoxinas
consideradas relevantes para a saúde humana. No entanto, foram isoladas nas uvas
várias espécies capazes de produzir patulina, das quais a mais frequente foi P.
expansum.
Em 1999, Abrunhosa (2001) fez um rastreio a uvas das regiões demarcadas
dos Vinhos Verdes e Douro. As uvas da região dos Vinhos Verdes foram vindimadas
em meados de Setembro, cerca de mês e meio antes das da região do Douro, onde a
vindima se deu no início de Novembro. Apesar de Abrunhosa ter verificado que
Botrytis e Cladosporium foram dominantes nas uvas de ambas as regiões estudadas, a
incidência de Penicillium nas amostras do Douro foi elevada, sobretudo de P.
296

CAPÍTULO 5. DISCUSSÃO
expansum, particularmente nas amostras de colheita mais tardia, no início de Outubro
e Novembro. Foi postulado que a maior incidência de Penicillium nas amostras do
Douro se deveu ao estado de sobrematuração das uvas nesta região, mas a ausência de
P. expansum na região dos Vinhos Verdes requeria investigações futuras para se
determinar se a maturação foi de facto o factor justificativo das diferenças observadas
entre as regiões ou se poderiam estar envolvidos factores ambientais na distribuição
da espécie (Abrunhosa et al., 2001).
Durante este estudo verificou-se que a espécie P. expansum existe tanto na
região dos Vinhos Verdes como na região do Douro, e que nos anos e locais
estudados, foi inclusive mais frequente nas uvas da região dos Vinhos Verdes. Em
2003 foram analisadas uvas do mesmo local do Douro que foi estudado em 1999
(Do4) numa vindima corrente, e apesar de P. expansum ter sido detectado na amostra,
não foi detectada podridão e a sua frequência foi muito baixa. Com base nestes dados,
conclui-se que P. expansum só causa risco de podridão das uvas em estados de
sobrematuração ou em situações pós-colheita, e que, como tal, à semelhança do que se
passa com as maçãs e com as situações referidas atrás, a sua presença em uvas e
sumos é indicadora de má qualidade da matéria-prima. No entanto, este contaminante
não deverá estar presente no vinho, visto que é destruído com a fermentação, como foi
mencionado no capítulo 1.
297


















