
AVALIAÇÃO DE MODELOS DE DISTRIBUIÇÃO GEOGRÁFICA...
16
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 1 AVALIAÇÃO DE MODELOS DE DISTRIBUIÇÃO GEOGRÁFICA E SUA APLICAÇÃO PARA PREVER A OCORRÊNCIA DE ESPÉCIES DE MAMÍFEROS NO CORREDOR CENTRAL DA MATA ATLÂNTICA Paulo Inácio Prado 1 , Luiz Paulo Pinto 2 , Raquel Teixeira de Moura 3,4 , Elena Charlotte Landau 4 1. Núcleo de Estudos e Pesquisa Ambientais - Universidade Estadual de Campinas, Caixa Postal 6166, Cidade Universitária Zeferino Vaz, Campinas - SP, Brasil, CEP 13084-971. [email protected] 2. Instituto de Estudos Sócio-Ambientais do Sul da Bahia, Caixa Postal 84, Ilhéus - BA, Brasil, CEP 45653-970, [email protected] 3. Conservation International do Brasil, Av. Getúlio Vargas, 1300 7o Andar, Belo Horizonte - MG, CEP 30112- 021, [email protected] 4. Laboratório de Mastozoologia, Instituto de Ciências Biológica, Universidade Federal de Minas Gerais, A. Antônio Carlos 6627, Belo Horizonte - MG, Brasil, CEP 31270-901. [email protected] INTRODUÇÃO Em biogeografia, a área de ocorrência de um táxon é inferida a partir de informações pontuais, as localidades nas quais o táxon foi registrado (Cerqueira 1995). Assim, as distribuições geográficas são hipóteses, resultantes de algum modelo que extrapola uma área a partir de um conjunto de pontos. Como há vários modelos possíveis, temos um conjunto de hipótese alternativas, cuja eficácia pode ser comparada. Este trabalho realiza esse tipo de avaliação, usando registros de ocorrência de três espécies de mamíferos na área do Corredor Central da Mata Atlântica, na Bahia. O modelo que mostrou o melhor desempenho, foi, em seguida, usado para criar hipóteses de distribuição geográfica potencial para as três espécies usadas na avaliação, e outras oito espécies, todas de mamíferos endêmicos e/ou ameaçados de extinção. O modelo clássico de extrapolação de área de ocorrência é traçar um polígono que inclua todas as localidades de registro (Udvardy 1969, Brown & Lomolino 1998). Uma variante desta abordagem empírica é dividir a área em quadrículas e considerar a espécie presente em todas as quadrículas que contenham localidades de ocorrência. A distribuição geográfica é, então, definida pelo conjunto de quadrículas em que está registrada a presença da espécie. A única premissa desta classe de modelos é de que a ocorrência de uma espécie pode ser estendida para as proximidades das localidades em que foi registrada, embora os limites para isto (tamanho das quadrículas ou do polígono) sejam subjetivos. Quaisquer que sejam estes limites, todavia, a premissa não considera a heterogeneidade ambiental, que pode fazer com que áreas vizinhas aos pontos de ocorrência tenham características ambientais muito diferentes, que talvez impeçam a ocorrência da espécie. Em contraste, há muitos modelos que extrapolam a ocorrência da espécie apenas para áreas com condições ambientais equivalentes. Como estas áreas são similares àquelas onde a espécie foi registrada, presume-se que ela possa ocorrer aí também. Por essa razão, estes modelos estimam a distribuição potencial da espécie, que pode ser maior que a verdadeira, devido a extinções locais, interações bióticas, ou limitações por outras variáveis ambientais não consideradas no modelo (Cerqueira 1995). O modelo mais simples deste tipo é considerar a espécie presente em todos os locais que estejam dentro do intervalo dos valores ambientais das localidades de registro. As premissas do modelo são que as localidades de registro representam todo o limite de tolerância da espécie, e que os únicos fatores limitantes são as variáveis ambientais consideradas. Tratam-se dos modelos de envelopes ambientais, que têm muitas variações mais complexas, a partir desta premissa simples (Guisan & Zimmerman 2000).
Transcript of AVALIAÇÃO DE MODELOS DE DISTRIBUIÇÃO GEOGRÁFICA...

CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 1
AVALIAÇÃO DE MODELOS DE DISTRIBUIÇÃO GEOGRÁFICA E SUA APLICAÇÃO PARA PREVER A OCORRÊNCIA DE ESPÉCIES
DE MAMÍFEROS NO CORREDOR CENTRAL DA MATA ATLÂNTICA
Paulo Inácio Prado1, Luiz Paulo Pinto2, Raquel Teixeira de Moura3,4, Elena Charlotte Landau4
1. Núcleo de Estudos e Pesquisa Ambientais - Universidade Estadual de Campinas, Caixa Postal 6166, Cidade Universitária Zeferino Vaz, Campinas - SP, Brasil, CEP 13084-971. [email protected] 2. Instituto de Estudos Sócio-Ambientais do Sul da Bahia, Caixa Postal 84, Ilhéus - BA, Brasil, CEP 45653-970, [email protected] 3. Conservation International do Brasil, Av. Getúlio Vargas, 1300 7o Andar, Belo Horizonte - MG, CEP 30112-021, [email protected] 4. Laboratório de Mastozoologia, Instituto de Ciências Biológica, Universidade Federal de Minas Gerais, A. Antônio Carlos 6627, Belo Horizonte - MG, Brasil, CEP 31270-901. [email protected]
INTRODUÇÃO Em biogeografia, a área de ocorrência de
um táxon é inferida a partir de informações pontuais, as localidades nas quais o táxon foi registrado (Cerqueira 1995). Assim, as distribuições geográficas são hipóteses, resultantes de algum modelo que extrapola uma área a partir de um conjunto de pontos. Como há vários modelos possíveis, temos um conjunto de hipótese alternativas, cuja eficácia pode ser comparada. Este trabalho realiza esse tipo de avaliação, usando registros de ocorrência de três espécies de mamíferos na área do Corredor Central da Mata Atlântica, na Bahia. O modelo que mostrou o melhor desempenho, foi, em seguida, usado para criar hipóteses de distribuição geográfica potencial para as três espécies usadas na avaliação, e outras oito espécies, todas de mamíferos endêmicos e/ou ameaçados de extinção.
O modelo clássico de extrapolação de área de ocorrência é traçar um polígono que inclua todas as localidades de registro (Udvardy 1969, Brown & Lomolino 1998). Uma variante desta abordagem empírica é dividir a área em quadrículas e considerar a espécie presente em todas as quadrículas que contenham localidades de ocorrência. A distribuição geográfica é, então, definida pelo conjunto de quadrículas em que está registrada a presença da espécie. A única premissa desta classe de modelos é de que a ocorrência de uma espécie pode ser
estendida para as proximidades das localidades em que foi registrada, embora os limites para isto (tamanho das quadrículas ou do polígono) sejam subjetivos. Quaisquer que sejam estes limites, todavia, a premissa não considera a heterogeneidade ambiental, que pode fazer com que áreas vizinhas aos pontos de ocorrência tenham características ambientais muito diferentes, que talvez impeçam a ocorrência da espécie.
Em contraste, há muitos modelos que extrapolam a ocorrência da espécie apenas para áreas com condições ambientais equivalentes. Como estas áreas são similares àquelas onde a espécie foi registrada, presume-se que ela possa ocorrer aí também. Por essa razão, estes modelos estimam a distribuição potencial da espécie, que pode ser maior que a verdadeira, devido a extinções locais, interações bióticas, ou limitações por outras variáveis ambientais não consideradas no modelo (Cerqueira 1995).
O modelo mais simples deste tipo é considerar a espécie presente em todos os locais que estejam dentro do intervalo dos valores ambientais das localidades de registro. As premissas do modelo são que as localidades de registro representam todo o limite de tolerância da espécie, e que os únicos fatores limitantes são as variáveis ambientais consideradas. Tratam-se dos modelos de envelopes ambientais, que têm muitas variações mais complexas, a partir desta premissa simples (Guisan & Zimmerman 2000).

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 2
Modelos de distribuição potencial mais elaborados estimam a probabilidade de ocorrência em função de variáveis ambientais, e, em seguida, estabelecem a distribuição potencial como a área na qual esta probabilidade seja superior a um certo limite (normalmente entre 50% e 75%).
No momento, as regressões logísticas são modelo mais aceito e utilizado para tais estimativas de probabilidade de ocorrência (Manel et al. 1999). Regressões logísticas estimam a probabilidade de uma variável binária assumir um certo valor, em função de variáveis explanatórias contínuas (Hosmer e Lemeshow, 1989; Steinberg e Colla, 1991). No caso de modelos de ocorrência de espécies, a variável binária é a presença ou ausência da espécie nas localidades em que foi buscada, e as variáveis explanatórias são medidas tomadas em cada um destas localidades. A regressão, ajustada com os pontos de registro e de ausência, é então usada para estimar a probabilidade de ocorrência da espécie em toda a área.
A premissa deste modelo de distribuição potencial é que a probabilidade de ocorrência aumenta assintóticamente em função das variáveis ambientais, seguindo uma função logística (Figura 1). Em outras palavras, as chances de se registrar a espécie em um dado local aumentariam monotonicamente com as variáveis ambientais, até atingir 100% (p = 1,0). A partir deste ponto, as chances permaneceriam em 100% de ocorrência, indefinidamente (Figura 1), o que é uma previsão pouco plausível.
Um padrão reconhecido há bastante tempo pela teoria ecológica é que, para cada espécie, há um ponto de máxima abundância ao longo de um gradiente ambiental, que corresponderia às condições ótimas de sobrevivência e reprodução da espécie (Hutchinson 1957, Brown 1995). À medida que se afasta deste ponto ótimo, aumentando-se ou diminuindo-se as variáveis ambientais, a abundância se reduz gradativamente. Supondo-se que esta redução seja simétrica , a curva de abundância em função de variáveis ambientais teria a forma de um sino (Figura 1), o que, de fato, ocorre para várias espécies (e.g. Whittaker 1967, Hengeveld & Haeck 1982, Brown & Lomolino 1998). Na maioria
destes casos, há um bom ajuste à curva normal (Brown 1995).
Assim, como a probabilidade de se encontrar uma espécie em um local é proporcional à sua abundância, um modelo alternativo ao logístico é que esta probabilidade seja uma função normal de variáveis ambientais. Os parâmetros da curva normal (média e desvio-padrão) são estimados pelas média e desvio-padrão das variáveis nas localidades de registro. Com isto, o valor das variáveis ambientais em qualquer local pode ser transformado em variáveis normais-padrão (z), cuja probabilidade prevista pela curva normal estima a probabilidade de ocorrência da espécie. A premissa deste modelo é que a média das variáveis ambientais nas localidades de registro estimam o ótimo ambiental da espécie, e que a distribuição das probabilidades de ocorrência siga uma distribuição normal.
As hipóteses de distribuição geográficas podem ser avaliadas por sua taxa de acertos. Um conjunto de dados, independente do usado para gerar a hipótese, é usado para estimar a freqüência com que localidades de ausência e de ocorrência da espécie são corretamente classificadas como tal (Fielding & Bell 1997). Outro critério de avaliação das hipóteses de distribuição é a extensão da área em que ela prevê a ocorrência da espécie. Pelo princípio da parcimônia, é preferível a hipótese que
Figura 1 - Esquema dos modelos logístico (esq.) e de ótimo ambiental (dir.) de probabilidade de ocorrência de uma espécie em função de um gradiente ambiental. No primeiro, a probabilidade de ocorrência aumenta assintoticamente em função das variáveis ambientais, seguindo uma função logística. No segundo, a probabilidade de ocorrência é máxima em um dado ponto do gradiente, e decresce a medida que se distancia deste ponto, seja pela redução ou pelo aumento das variáveis ambientais.

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 3
obtenha a maior taxa de acerto com a menor área possível.
Este trabalho compara as hipóteses de distribuição geográfica geradas pelos modelos do polígono empírico, envelopes ambientais, de regressão logística e de ótimo ambiental, para três espécies de mamíferos, na área do Corredor Central na Bahia. Os objetivos são: 1. Identificar o modelo que gere as hipóteses mais acuradas e parcimoniosas, analisando as eventuais diferenças dos modelos, quanto a estes critérios. 2. Uma vez identificado o modelo mais adequado, gerar hipóteses de distribuição geográfica, na área do Corredor Central na Bahia, para espécies de mamíferos endêmicas e ameaçadas que tenham registros em densidade suficiente. 3. Identificar, pela sobreposição das áreas hipotetizadas de ocorrência, zonas de maior riqueza potencial de espécies na área. PROCEDIMENTOS Área de Estudo
O Corredor Central da Mata Atlântica compreende cerca de 80% da bio-região “Bahia” da Mata Atlântica (Silva & Casteleti 2003), que possui 120.954 km2 e estende-se desde Sergipe até o Espírito Santo. O Corredor Central da Mata Atlântica compõe a porção centro-sul dessa região biogeográfica, limitando-se ao norte pelo rio Jiquiriçá, onde se inicia o agrossistema do baixo-sul da Bahia, e estendendo-se pela região cacaueira tradicional, extremo sul da Bahia e centro norte do Estado do Espírito Santo (Fonseca et al. 2003).
O sul da Bahia é uma das áreas mais importantes para o efetivo estabelecimento do Corredor Central, devido aos seus elevados graus de riqueza de espécies, endemismo, e ameaça (CI & IESB 2000, Fonseca et al. 2003) e compreende a área de estudo deste trabalho. Está situada entre os paralelos 13o 05´S - 18o 05´S e meridianos 38o 50´W - 41o 33´W, e é delimitada ao Norte pelo Rio Jiquiriçá, ao Sul pelo Rio Mucuri, ao Leste pelo Oceano Atlântico, e a Oeste pela rodovia BR-116 (detalhes em Landau & Resende 2003a).
A região era originalmente ocupada por sete classes de fisionomias florestais da Mata Atlântica (Thomas & Carvalho 2003), que hoje cobrem 20% da área, entre florestas em estágio inicial (13% da área) e avançado de regeneração (7%). O restante da área é ocupado por agricultura, cultivos agro-florestais, pastos, solo exposto, e aglomerados urbanos, formando uma paisagem fragmentada e complexa (Landau 2003a).
O sul da Bahia é uma das áreas mais importantes para a conservação de mamíferos da Mata Atlântica. Inclui as áreas de distribuição de oito espécies de primatas: muriqui-do-norte (Brachyteles hypoxanthus), macaco-guigó (Callicebus melanochir), macaco-prego-do-peito-amarelo (Cebus xanthosternos, endêmico da região), macaco-prego-robusto (Cebus nigritus robustus, endêmico da região), mico-leão-da-cara-dourada (Leontopithecus chrysomelas, endêmico da região), bugio (Alouattta guariba), sagüi-da-cara-branca (Callithrix geoffroyi, endêmico da região), e sagüi-de-Wied (Callithrix kuhli). Destas, a primeira está provavelmente extinta, e as demais estão fortemente ameaçadas, a exceção, talvez, de C. kuhlii (Santos et al. 1987, Moura 2003). A área ainda abriga as principais populações da preguiça-de-coleira (Bradypus torquatus, Xenarthra: Bradypodidae), e do ouriço preto, Chaetomys subspinosus (Rodentia: Echimyidae), ambos ameaçados de extinção (Oliver & Santos 1991). Na região ocorrem, também, a preguiça comum (B. variegatus) e o ouriço-caixeiro (Sphiggurus insidiosus, Rodentia: Erethizontidae), de distribuição mais ampla (Oliver & Santos 1991). Registros de espécies usados na geração e avaliação das hipóteses
Os registros de espécies usados neste
trabalho foram compilados da literatura por Prado et al. (2003a), que caracterizam em detalhe este conjunto de dados. Estes registros estão disponíveis na Base Corredor Central, uma base de registros de espécies na região do Corredor Central da Mata Atlântica, distribuída em Prado et al. (2003b).
Foram geradas e avaliadas hipóteses de distribuição potencial para Bradypus

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 4
torquatus, Callicebus melanochir e Cebus xanthosternos. Estas três espécies foram escolhidas por terem os maiores números de registros de ausência, um dado raro e necessário para a avaliação das hipóteses, e para o modelo logístico.
Para construir as distribuições hipotéticas foram usados os registros de ocorrência na área do Corredor Central na Bahia que constam em Oliver & Santos (1991). Para a avaliação das hipóteses, foi usado um outro conjunto de dados, formado pelos registros de ocorrência de todas as outras fontes (Ver Prado et al. 2003), e os registros de ausências extraídos de Oliver & Santos (1991). Localidades onde uma espécie não ocorre raramente são informadas na literatura, embora sejam essenciais para a avaliação de hipóteses de distribuição. Para a área do Corredor Central, o melhor conjunto destes "pontos de ausência" é o que pode ser deduzido de Oliver & Santos (1991), que visitaram várias localidades em busca das mesmas espécies, e informam as encontradas em cada uma.
Assim, a avaliação das hipóteses foi feita com um conjunto de registros diferente do usado para gerá-las (Tabela 1). A única exceção foi o modelo logístico, que necessita de registros de ocorrência e também de ausências (ver a descrição dos modelos). Neste caso, os mesmos registros de ausência foram usados para gerar e avaliar a hipótese, embora os registros de ocorrência tenham sido diferentes.
Variáveis ambientais As variáveis ambientais usadas para
construir as hipóteses de distribuição potencial foram altitude em relação ao nível do mar, temperatura média anual, e pluviosidade média anual, extraídas, respectivamente, dos seguintes temas cartográficos digitais: • Zonas de altitude em classes de 100 m digitalizadas como polígonos, publicadas em arquivo para para o programa ArcView ( ESRI) (Landau & Resende 2003b). Tema preparado a partir da Carta de Relevo do Estado da Bahia (CEI, 1989), escala 1:1.500.000, altitudes em classes de 100 m. • Zonas de temperatura média compensada em classes de 2o C digitalizadas como polígonos, publicadas
em arquivo para ArcView (Landau 2003b). Tema preparado a partir da Carta de Temperatura Média Compensada Anual da Bahia (1961-1990), Escala: 1:6.000.000 (SEI et al. 1998), temperaturas em classes de 1o C. • Zonas de pluviosidade média compensada em classes de 200 mm digitalizadas como polígonos, publicadas em arquivo para ArcView (Landau 2003c). Tema preparado a partir da Carta de Pluviometria. Média Compensada Anual da Bahia (1943-1983), Escala: 1:6.000.000 (SEI et al. 1998), pluviosidade em classes de 100 mm.
Os valores das variáveis ambientais nas
localidades de registro de ocorrência e ausência das espécies foram obtidos em sistema de informação geográfica (ArcView), plotando-se os pontos das localidades sobre os temas, e extraindo-se uma tabela com os valores de altitude, temperatura, e pluviosidade em cada ponto (função "table joining").
Para a construção das hipóteses de distribuição, os temas de polígonos das variáveis ambientais foram transformados em um "grid" de quadrículas de 5 km2, sendo atribuído a cada quadrícula o valor de temperatura, pluviosidade e altitude lido em seu centro (função "convert to grid", módulo "Spatial Analyst" ESRI). Modelos avaliados
Hipóteses de distribuição potencial para cada espécie foram geradas pelos seguintes modelos: 1. Distribuição empírica: polígono mínimo que contém todos as localidades de registro da espécie. O polígono foi traçado manualmente, sobre as localidades plotadas em sistema de informação geográfica, a uma escala de 1:50.000. 2. Envelopes ambientais: a amplitude ambiental de ocorrência da espécie foi estimada pelos valores máximos e mínimos de altitude, pluviosidade e temperatura nas localidades em que a espécie foi registrada. a distribuição potencial foi definida como todas as quadrículas em que as três variáveis ambientais estivessem dentro deste intervalo. 3. Envelopes ambientais sem extremos: para as três espécies

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 5
analisadas, pelo menos 93% das localidades de registro tinham altitude igual ou inferior a 200 m, embora uma ou duas estivessem entre 400 e 700 m. O procedimento de limites ambientais foi repetido, excluindo-se este valores extremos. 4. Ótimo ambiental: a probabilidade de ocorrência de cada espécie em função de variáveis ambientais foi ajustada a distribuições normais, cujas médias representam o ótimo ambiental para a espécie. Como as três variáveis ambientais usadas eram muito correlacionadas, elas foram reduzidas a uma variável sintética, através de uma análise de componentes principais (PCA). A hipótese de distribuição potencial foi gerada da seguinte maneira: 4.1. Foi feito um PCA dos valores de temperatura, pluviosidade e altitude de 1000 pontos distribuídos aleatoriamente pela área de estudo. O primeiro eixo deste PCA, que representa a maior parte da variação (ver resultados), foi usado como variável ambiental sintética. As médias e desvios-padrão das variáveis ambientais nestes 1000 pontos foram usados para padronizar os escores, neste eixo do PCA, das localidades e quadrículas (próximos itens). 4.2. Os escores padronizados no primeiro eixo do PCA foram calculados para as localidades de ocorrência das três espécies. Para isto, as variáveis foram primeiramente padronizadas, subtraindo-se de cada um de seus valores a média da variável na área de estudo, e dividindo-a por seu desvio-padrão (estimados com pontos aleatórios, item 4.1.). Em seguida cada variável foi multiplicada pelo seu respectivo coeficiente no eixo do PCA. A soma destes produtos fornece os escores no eixo do PCA (Manly 1994). 4.3. Os escores no primeiro eixo do PCA foram calculados para cada quadrícula, da mesma maneira (ver Figura 2). 4.4. Para as três espécies, escores das localidades de ocorrência seguiram distribuições normais (ver resultados), cujas médias e desvios-padrão foram estimados. 4.5. Com as distribuições de probabilidade normais assim parametrizadas, foi calculada a probabilidade esperada de ocorrência de cada espécie em cada quadrícula.
4.6. A distribuição potencial da espécie foi definida como as quadrículas com pelo menos 75% de probabilidade de ocorrência. 5. Logístico: a probabilidade de ocorrência de cada espécie em função de variáveis ambientais foi ajustada a funções logísticas, da seguinte maneira: 5.1. Foram usadas todas as localidades de Oliver & Santos (1991) na área de estudo, sendo a variável dependente a presença ou ausência da espécie. As variáveis explanatórias testadas foram os valores de pluviosidade, temperatura e altitude nas localidades, bem como o escore padronizado da localidade no primeiro eixo do PCA (item 4.1.). 5.2. Para cada espécie, regressões logísticas (Hosmer e Lemeshow, 1989; Steinberg & Colla, 1991) foram usadas para testar o efeito das variáveis explanatórias sobre a variável de presença/ausência. Foi ajustada uma regressão para cada variável explanatória. 5.3. As regressões com efeitos significativos foram usadas para estimar a probabilidade de ocorrência da espécie em cada quadrícula, a partir dos valores das variáveis ambientais na quadrícula. 5.4. A distribuição potencial da espécie foi definida como as quadrículas com pelo menos 50% de probabilidade de ocorrência.
Para Cebus xanthosternos, as hipóteses de distribuição potencial foram restritas às quadrículas ao norte do Rio Jequitinhonha, considerado o limite sul de distribuição da espécie (Santos et al. 1987, Coimbra-Filho et al. 1991). Para B. torquatus e C. melanochir, espécies para as quais não há barreiras biogeográficas evidentes na área de estudo, não houve restrição de quadrículas. Avaliação das hipóteses
Os pontos de avaliação foram sobrepostos às áreas de distribuição geográfica previstas por cada hipótese, e classificados em: a. Ocorrências previstas corretamente: pontos de ocorrência abrangidos pela área de ocorrência prevista pela hipótese; b. Ocorrências não previstas: pontos de ocorrência fora da área prevista;

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 6
c. Ausências não previstas: pontos de registro de ausência dentro da área de ocorrência prevista; d. Ausências previstas corretamente: pontos de registro de ausência fora da área de ocorrência prevista O número de pontos em cada uma destas classes forma a matriz de acertos da hipótese (Cumming 2000, Manel et al. 2001):
Observado
Presente Ausente TOTAL
Presente a b
Ausente c d
Pre
vis
to
TOTAL N
Com esta matriz, foram calculados os seguintes índices, para cada hipótese (Cumming 2000, Manel et al. 2001): • Sensibilidade (proporção das ocorrências que foram corretamente previstas): Sn = a / (a + c) • Especificidade (proporção das ausências que foram corretamente previstas): Sp = d / (b + d) • Acerto (proporção de ocorrências e ausências corretamente previstas) : S = (a + d) / N • Kappa (Proporção de acertos não-casuais):
Os três primeiros índices são tradicionalmente usados na avaliação de modelos de ocorrência de espécies, e medem o total bruto de acertos (Manel et al. 2001). Já o índice kappa, ainda pouco conhecido dos biólogos, expressa apenas os acertos que não ocorreram pelo mero acaso, e parece ser um estimador bem mais robusto da eficácia de um modelo (Manel et al. 2001).
Para cada hipótese de distribuição, foi calculada também a extensão da área de ocorrência prevista das espécies. Para isto,
o total de quadrículas para as quais a ocorrência era prevista foi multiplicado pela área de cada quadrícula (5 km2).
Hipóteses de distribuição para as demais espécies
O modelo de ótimo ambiental (p = 0,75), que gerou as hipóteses mais acuradas e parcimoniosas (ver resultados), foi usado para estimar a distribuição para as mesmas três espécies usadas na avaliação dos modelos, e outras oito espécies de mamíferos. Assim, foram gerados temas cartográficos de distribuição potencial para onze espécies: Bradypus torquatus, Bradypus variegatus), Chaetomys subspinosus, Sphiggurus insidiosus Brachyteles hypoxanthus, Callicebus melanochir, Cebus xanthosternos, Cebus nigritus robustus, Leontopithecus chrysomelas, Callithrix kuhli, e Callithrix geoffroyi. Estas foram espécies registradas em pelo menos quinze localidades (Apêndice II de Prado et al. 2003), distribuídos por uma extensão representativa da área de estudo.
Antes do ajuste do modelo, foi testada a normalidade da distribuição de freqüência dos escores no primeiro do PCA de todas as localidades de ocorrência de cada espécie. Para algumas espécies, o ajuste à curva normal só foi satisfatório após a transformação dos escores para seus logaritmos naturais, ou raízes quadradas. Apenas para L. chrysomelas os escores das localidades não se ajustaram de maneira nenhuma a uma curva normal, mesmo transformados. Para esta espécie, o mapa de distribuição foi feito pelo modelo de polígono empírico.
Note que mesmo para as três espécies usadas na avaliação dos modelos foram geradas novas hipóteses de distribuição, agora com todas as localidades de ocorrência (incluindo as antes usadas para avaliação).
Para espécies que têm barreiras geográficas na área de estudo, estas foram usadas para delimitar os mapas de distribuição. Assim, a distribuição potencial de Callithrix geoffroyi e de Cebus nigritus robustus foram estimadas apenas ao sul do Rio Jequitinhonha; e a distribuição de Cebus xanthosternos e de Callithrix kuhli ao norte deste limite (conforme Santos et al. 1987).
[N – (((a+c)(a+b) + (b+d)(c+d))/N)]
[(a+d) – (((a+c)(a+b) + (b+d)(c+d))/N)] κ =

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 7
Mapas de riqueza potencial foram criados sobrepondo-se os mapas de distribuição potencial, e contando-se as espécies com ocorrência prevista em cada quadrícula. Para estes cálculos não foi considerada a hipótese de distribuição de Brachyteles hypoxanthus, provavelmente extinto na área de estudo.
RESULTADOS Variáveis ambientais
Segundo os temas cartográficos usados, há uma considerável variação de altitude (1000 m) e de pluviosidade (2200 mm) na área de estudo (Tabela 2). A temperatura é bem menos variável, indo de 21 a 24 oC (Tabela 2). As médias das altitude, temperatura e pluviosidade em toda área, estimadas por mil pontos distribuídos ao acaso, indicam a prevalência de regiões mais baixas, quentes e chuvosas (Tabela 2).
A análise de componentes principais indicou uma forte correlação entre pluviosidade, temperatura e altitude na área. Os pesos das três variáveis ambientais no primeiro eixo do PCA foram equivalentes, sendo que a altitude teve sinal negativo, e as demais sinal positivo (Tabela 3). Este eixo explica 75% da variação total dos dados. Assim, o principal vetor de variação ambiental na área é o aumento da temperatura e pluviosidade, com a redução da altitude. Os escores das quadrículas no primeiro eixo do PCA
(Figura 2) indicam que este vetor forma um gradiente leste-oeste, com as zonas costeiras mais baixas, chuvosas e quentes, e uma redução gradativa da pluviosidade e temperatura em direção ao interior, que tem áreas mais altas.
Ajuste e avaliação dos modelos
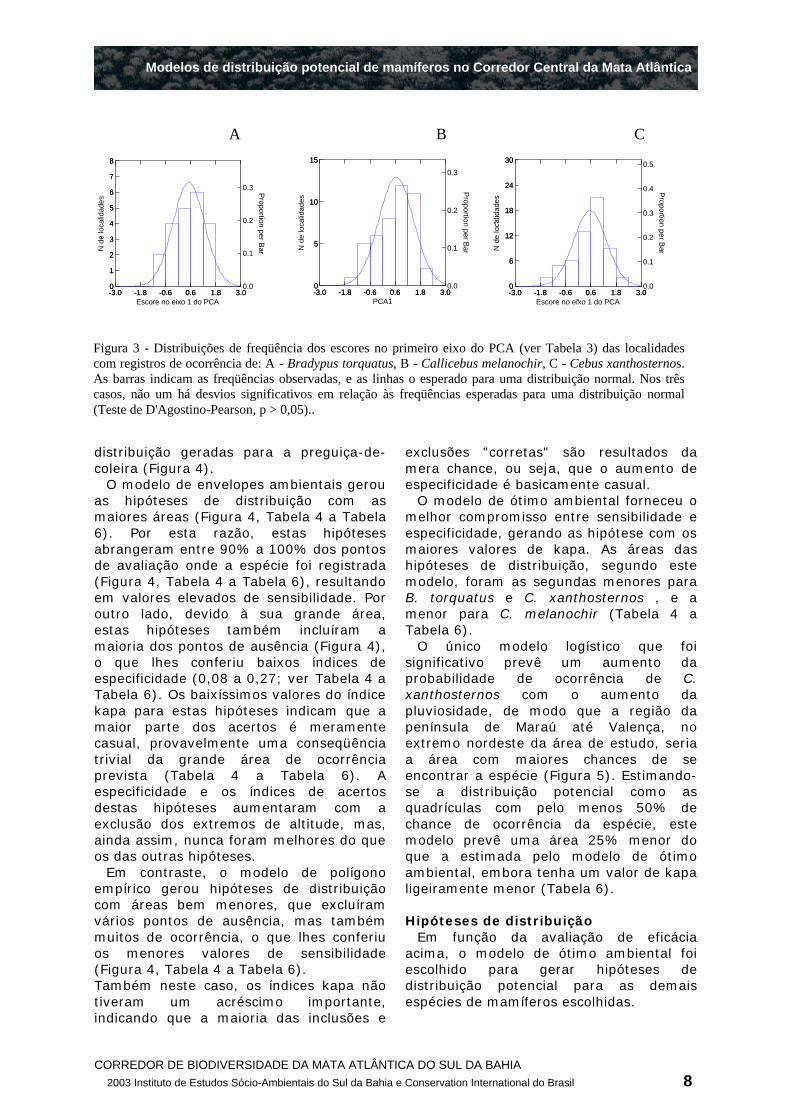
Para as três espécies usadas na avaliação, os escores no primeiro eixo do PCA das localidades de registro tiveram distribuições de freqüência que se ajustaram bem à uma curva normal (Figura 3). Por outro lado, a única regressão logística que teve um ajuste significativo foi a de probabilidade de ocorrência de C. xanthosternos em função da pluviosidade.
Os modelos tiveram resultados similares com as três espécies avaliadas, que são bem ilustrados pelas hipóteses de
Tabela 2 - Estatísticas das variáveis ambientais nos 1000 pontos sorteados na área de estudo, para realizar o PCA.
Altitude (m) Temperatura média anual (oC)
Pluviosidade média anual (mm)
Mínimo 0 21 500
Máximo 1000 24 2700
Média 226 23 1086
Desvio-padrão 181,3 0,95 424,0
Tabela 3 - Pesos das variáveis ambientais nos dois primeiros
eixos do PCA.
Variável Eixo 1 (λ = 2,327)
Eixo 2 (λ = 0,409)
Altitude - 0,3739 1,0748
Temperatura média anual 0,3915 - 0,0508
Pluviosidade média anual 0,3708 1,1349
Figura 2 - Escores no primeiro eixo do PCA das variáveis ambientais (ver Tabela 3 e texto) de quadrículas de 5 km2 da área de estudo. Escores altos (quadrículas mais escuras) indicam maior pluviosidade e temperaturas médias anuais, e menor altitude.
Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 8
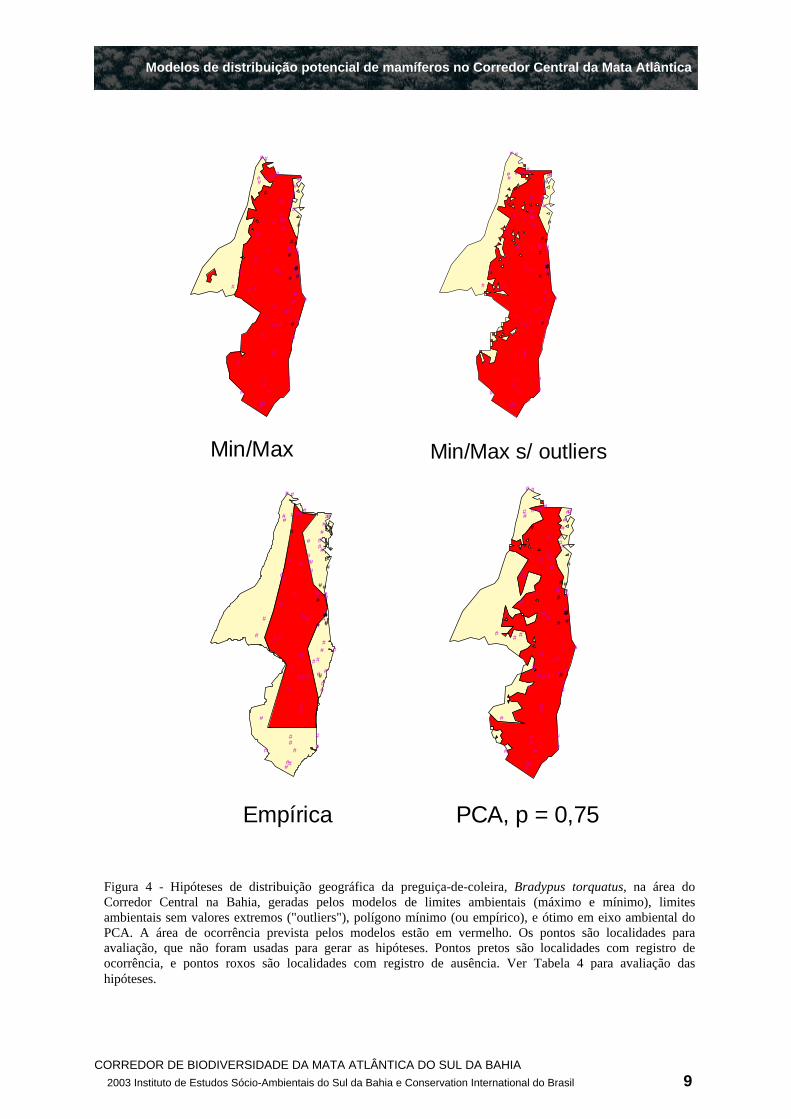
distribuição geradas para a preguiça-de-coleira (Figura 4).
O modelo de envelopes ambientais gerou as hipóteses de distribuição com as maiores áreas (Figura 4, Tabela 4 a Tabela 6). Por esta razão, estas hipóteses abrangeram entre 90% a 100% dos pontos de avaliação onde a espécie foi registrada (Figura 4, Tabela 4 a Tabela 6), resultando em valores elevados de sensibilidade. Por outro lado, devido à sua grande área, estas hipóteses também incluíram a maioria dos pontos de ausência (Figura 4), o que lhes conferiu baixos índices de especificidade (0,08 a 0,27; ver Tabela 4 a Tabela 6). Os baixíssimos valores do índice kapa para estas hipóteses indicam que a maior parte dos acertos é meramente casual, provavelmente uma conseqüência trivial da grande área de ocorrência prevista (Tabela 4 a Tabela 6). A especificidade e os índices de acertos destas hipóteses aumentaram com a exclusão dos extremos de altitude, mas, ainda assim, nunca foram melhores do que os das outras hipóteses.
Em contraste, o modelo de polígono empírico gerou hipóteses de distribuição com áreas bem menores, que excluíram vários pontos de ausência, mas também muitos de ocorrência, o que lhes conferiu os menores valores de sensibilidade (Figura 4, Tabela 4 a Tabela 6). Também neste caso, os índices kapa não tiveram um acréscimo importante, indicando que a maioria das inclusões e
exclusões "corretas" são resultados da mera chance, ou seja, que o aumento de especificidade é basicamente casual.
O modelo de ótimo ambiental forneceu o melhor compromisso entre sensibilidade e especificidade, gerando as hipótese com os maiores valores de kapa. As áreas das hipóteses de distribuição, segundo este modelo, foram as segundas menores para B. torquatus e C. xanthosternos , e a menor para C. melanochir (Tabela 4 a Tabela 6).
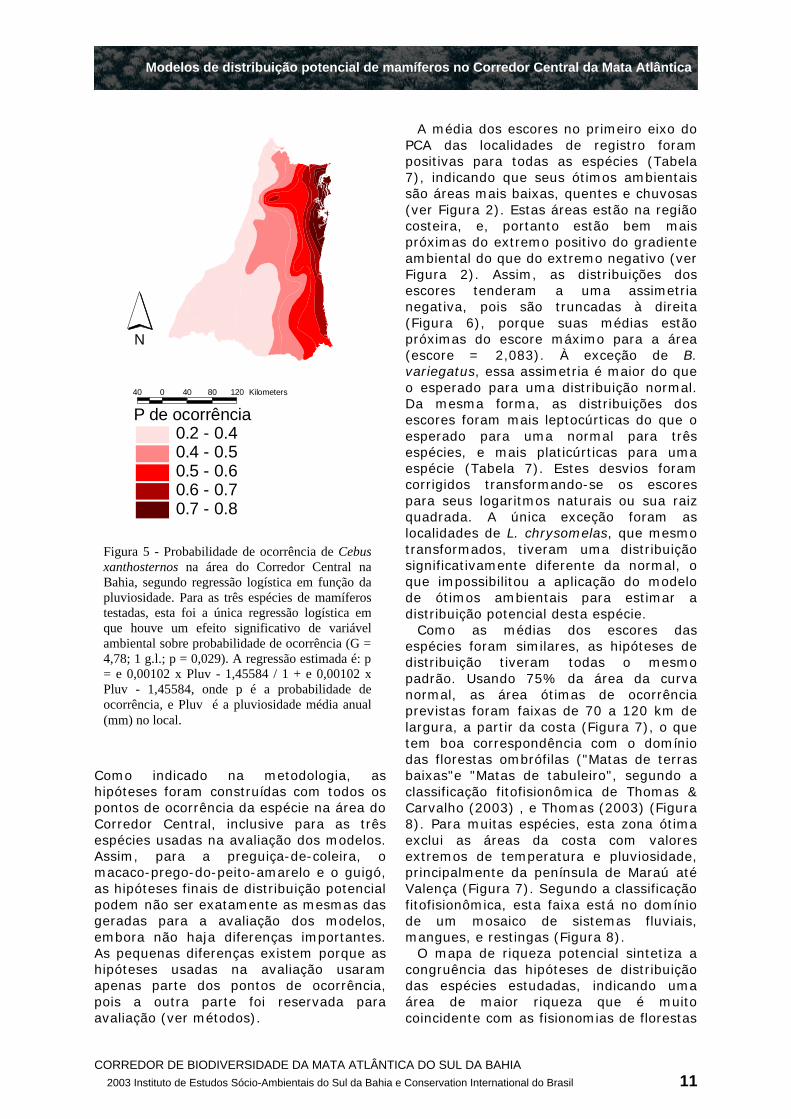
O único modelo logístico que foi significativo prevê um aumento da probabilidade de ocorrência de C. xanthosternos com o aumento da pluviosidade, de modo que a região da península de Maraú até Valença, no extremo nordeste da área de estudo, seria a área com maiores chances de se encontrar a espécie (Figura 5). Estimando-se a distribuição potencial como as quadrículas com pelo menos 50% de chance de ocorrência da espécie, este modelo prevê uma área 25% menor do que a estimada pelo modelo de ótimo ambiental, embora tenha um valor de kapa ligeiramente menor (Tabela 6).
Hipóteses de distribuição
Em função da avaliação de eficácia acima, o modelo de ótimo ambiental foi escolhido para gerar hipóteses de distribuição potencial para as demais espécies de mamíferos escolhidas.
Figura 3 - Distribuições de freqüência dos escores no primeiro eixo do PCA (ver Tabela 3) das localidades com registros de ocorrência de: A - Bradypus torquatus, B - Callicebus melanochir, C - Cebus xanthosternos. As barras indicam as freqüências observadas, e as linhas o esperado para uma distribuição normal. Nos três casos, não um há desvios significativos em relação às freqüências esperadas para uma distribuição normal (Teste de D'Agostino-Pearson, p > 0,05)..
A B C
-3.0 -1.8 -0.6 0.6 1.8 3.0Escore no eixo 1 do PCA
0
1
2
3
4
5
6
7
8
N d
e lo
calid
ades
0.0
0.1
0.2
0.3 Proportion per B
ar
-3.0 -1.8 -0.6 0.6 1.8 3.00
1
2
3
4
5
6
7
8
-3.0 -1.8 -0.6 0.6 1.8 3.0PCA1
0
5
10
15
N d
e lo
calid
ades
0.0
0.1
0.2
0.3
Proportion per B
ar
-3.0 -1.8 -0.6 0.6 1.8 3.00
5
10
15
-3.0 -1.8 -0.6 0.6 1.8 3.0Escore no eixo 1 do PCA
0
6
12
18
24
30
N d
e lo
calid
ades
0.0
0.1
0.2
0.3
0.4
0.5
Proportion per B
ar
-3.0 -1.8 -0.6 0.6 1.8 3.0''
0
6
12
18
24
30
''
Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 9
Figura 4 - Hipóteses de distribuição geográfica da preguiça-de-coleira, Bradypus torquatus, na área do Corredor Central na Bahia, geradas pelos modelos de limites ambientais (máximo e mínimo), limites ambientais sem valores extremos ("outliers"), polígono mínimo (ou empírico), e ótimo em eixo ambiental do PCA. A área de ocorrência prevista pelos modelos estão em vermelho. Os pontos são localidades para avaliação, que não foram usadas para gerar as hipóteses. Pontos pretos são localidades com registro de ocorrência, e pontos roxos são localidades com registro de ausência. Ver Tabela 4 para avaliação das hipóteses.
# #
#
#
##
######
####
# ###
####
##
##
# #
#
####
# ## ## #
###
#### # #
###
##
#
###
## #
###
#
##
#
#
#
#
#
#
#
#
#
#
#
Min/Max
##
#
#
##
######
####
# ###
####
##
##
# #
#
####
# ## ## #
###
#### # #
###
##
#
###
## #
###
#
##
#
#
#
#
#
#
#
#
#
#
#
Min/Max s/ outliers
##
#
#
# #
######
####
# ###
####
##
##
# #
#
####
# ## ## #
###
#### # #
###
##
#
###
## #
###
#
##
#
#
#
#
#
#
#
#
#
#
#
Empírica
##
#
#
##
######
####
# ###
####
##
##
# #
#
####
# ## ## #
###
#### # #
###
##
#
###
## #
###
#
##
#
#
#
#
#
#
#
#
#
#
#
PCA, p = 0,75

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 10
Tabela 4 - Avaliação de modelos de distribuição geográfica para Bradypus torquatus na região do Corredor Central na Bahia.. Para as hipóteses de distribuição geradas por cada modelo são indicados a área ocupada pela distribuição, e os índices de sensibilidade, especificidade, acertos totais, e de proporção de acertos específicos (kappa de Cohen). Os índices variam de zero a um (baixo a alto desempenho, respectivamente), e foram calculados com um conjunto de pontos independentes dos usados para propor a hipótese de distribuição. Número de pontos usados na avaliação: 10 registros de ocorrência e 63 registros de ausência.
Modelo Área (km2) Sensib. Especif. Acerto κ
Limites ambientais 62.525 0,90 0,14 0,25 0,013
Limites sem outliers 56.600 0,90 0,19 0,29 0,029
Empírica 34.250 0,40 0,62 0,38 0,011
Ótimo ambiental 48.850 0,80 0,29 0,36 0,031
Tabela 5 - Avaliação de modelos de distribuição geográfica para Callicebus melanochir na região do Corredor Central na Bahia.. Para as hipóteses de distribuição geradas por cada modelo são indicados a área ocupada pela distribuição, e os índices de sensibilidade, especificidade, acertos totais, e de proporção de acertos específicos (kappa de Cohen). Os índices variam de zero a um (baixo a alto desempenho, respectivamente), e foram calculados com um conjunto de pontos independentes dos usados para propor a hipótese de distribuição. Número de pontos usados na avaliação: 10 registros de ocorrência e 25 registros de ausência.
Modelo Área (km2) Sensib. Especif. Acerto κ
Limites ambientais 75.175 1,00 0,08 0,34 0,047
Limites sem outliers 60.775 1,00 0,16 0,40 0,098
Empírica 49.715 0,70 0,28 0,35 0,000
Ótimo ambiental 48.775 0,90 0,40 0,54 0,190
Tabela 6 - Avaliação de modelos de distribuição geográfica para Cebus xanthosternos na região do Corredor Central na Bahia.. Para as hipóteses de distribuição geradas por cada modelo são indicados a área ocupada pela distribuição, e os índices de sensibilidade, especificidade, acertos totais, e de proporção de acertos específicos (kappa de Cohen). Os índices variam de zero a um (baixo a alto desempenho, respectivamente), e foram calculados com um conjunto de pontos independentes dos usados para propor a hipótese de distribuição. Número de pontos usados na avaliação: 24 registros de ocorrência e 16 registros de ausência.
Modelo Área (km2) Sensib. Especif. Acerto κ
Limites ambientais 38.590 1,00 0,27 0,67 0,28
Limites sem outliers 31.240 0.94 0,33 0,67 0,29
Empírica 23.225 0,61 0,53 0,58 0,16
Logística* 16.900 0,71 0,44 0,65 0,27
Ótimo ambiental 22.390 0,89 0,40 0,64 0,30
* O modelo logístico usa os registros de ausências, que foram os mesmos usados para calcular os valores de especificidade, acerto e kapa. Neste caso, portanto, estes índices podem estar super-estimados, pois baseiam-se nos mesmos registros de ausência que foram usados para criar a hipótese.
Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 11
Como indicado na metodologia, as hipóteses foram construídas com todos os pontos de ocorrência da espécie na área do Corredor Central, inclusive para as três espécies usadas na avaliação dos modelos. Assim, para a preguiça-de-coleira, o macaco-prego-do-peito-amarelo e o guigó, as hipóteses finais de distribuição potencial podem não ser exatamente as mesmas das geradas para a avaliação dos modelos, embora não haja diferenças importantes. As pequenas diferenças existem porque as hipóteses usadas na avaliação usaram apenas parte dos pontos de ocorrência, pois a outra parte foi reservada para avaliação (ver métodos).
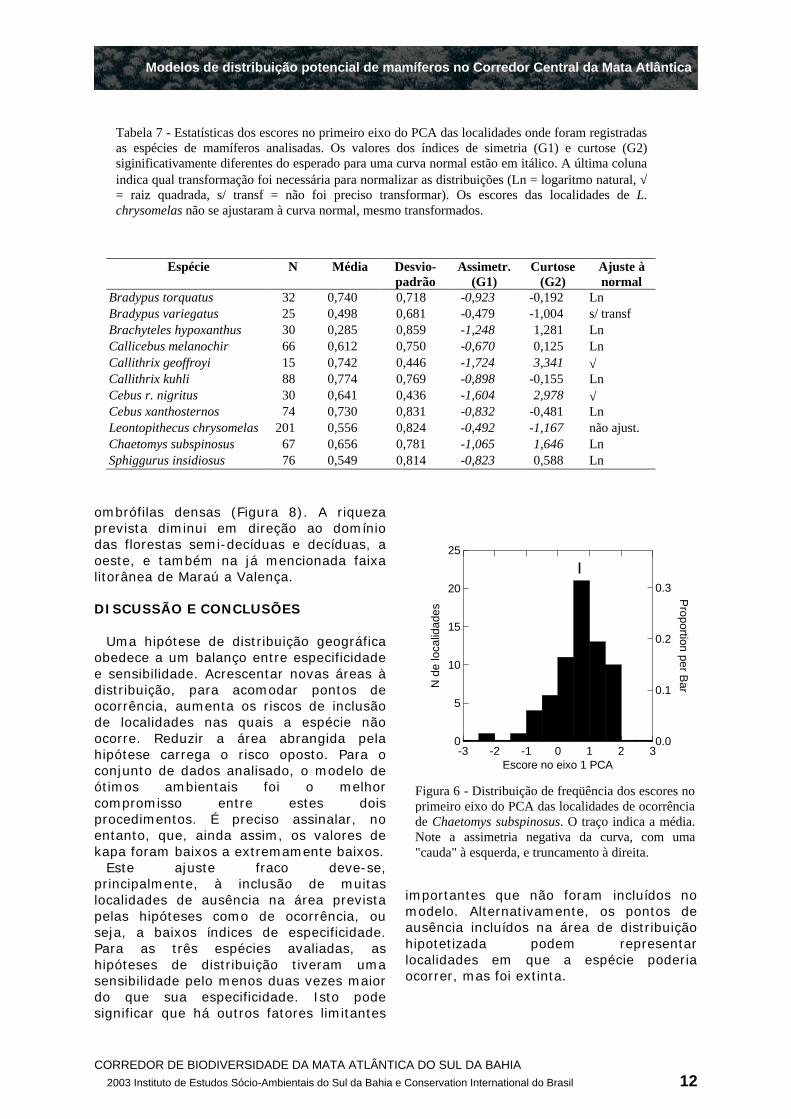
A média dos escores no primeiro eixo do PCA das localidades de registro foram positivas para todas as espécies (Tabela 7), indicando que seus ótimos ambientais são áreas mais baixas, quentes e chuvosas (ver Figura 2). Estas áreas estão na região costeira, e, portanto estão bem mais próximas do extremo positivo do gradiente ambiental do que do extremo negativo (ver Figura 2). Assim, as distribuições dos escores tenderam a uma assimetria negativa, pois são truncadas à direita (Figura 6), porque suas médias estão próximas do escore máximo para a área (escore = 2,083). À exceção de B. variegatus, essa assimetria é maior do que o esperado para uma distribuição normal. Da mesma forma, as distribuições dos escores foram mais leptocúrticas do que o esperado para uma normal para três espécies, e mais platicúrticas para uma espécie (Tabela 7). Estes desvios foram corrigidos transformando-se os escores para seus logaritmos naturais ou sua raiz quadrada. A única exceção foram as localidades de L. chrysomelas, que mesmo transformados, tiveram uma distribuição significativamente diferente da normal, o que impossibilitou a aplicação do modelo de ótimos ambientais para estimar a distribuição potencial desta espécie.
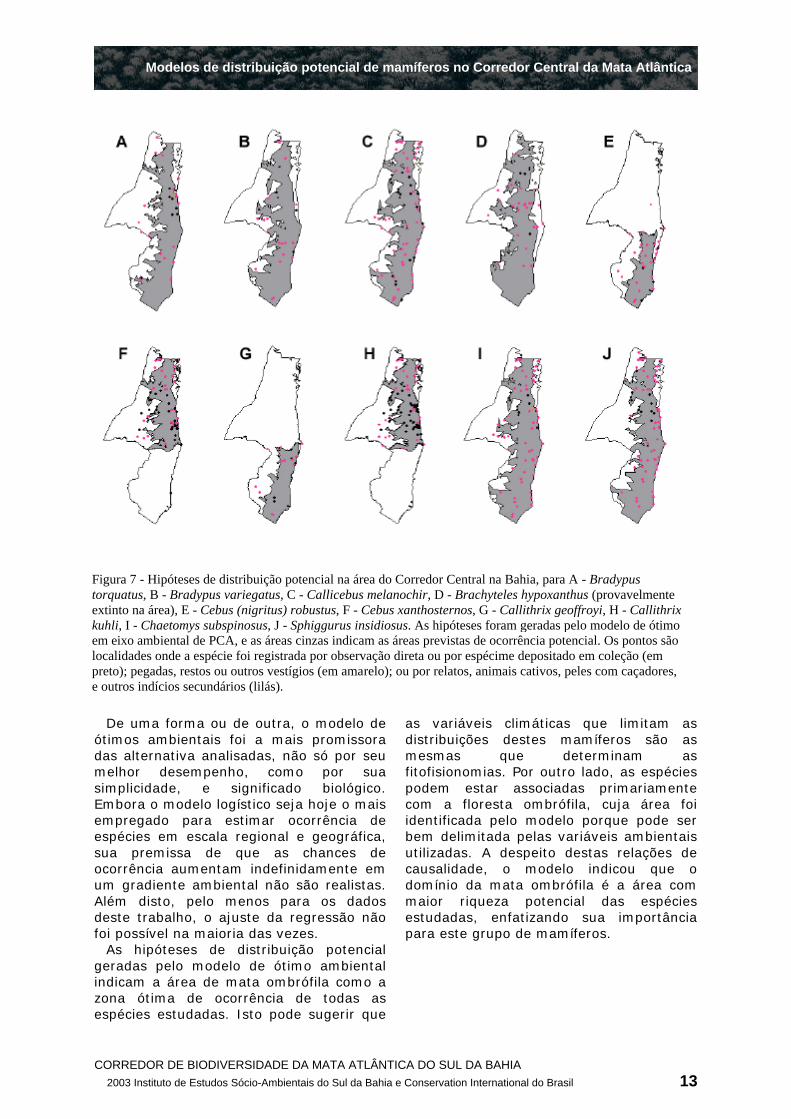
Como as médias dos escores das espécies foram similares, as hipóteses de distribuição tiveram todas o mesmo padrão. Usando 75% da área da curva normal, as área ótimas de ocorrência previstas foram faixas de 70 a 120 km de largura, a partir da costa (Figura 7), o que tem boa correspondência com o domínio das florestas ombrófilas ("Matas de terras baixas"e "Matas de tabuleiro", segundo a classificação fitofisionômica de Thomas & Carvalho (2003) , e Thomas (2003) (Figura 8). Para muitas espécies, esta zona ótima exclui as áreas da costa com valores extremos de temperatura e pluviosidade, principalmente da península de Maraú até Valença (Figura 7). Segundo a classificação fitofisionômica, esta faixa está no domínio de um mosaico de sistemas fluviais, mangues, e restingas (Figura 8).
O mapa de riqueza potencial sintetiza a congruência das hipóteses de distribuição das espécies estudadas, indicando uma área de maior riqueza que é muito coincidente com as fisionomias de florestas
Figura 5 - Probabilidade de ocorrência de Cebus xanthosternos na área do Corredor Central na Bahia, segundo regressão logística em função da pluviosidade. Para as três espécies de mamíferos testadas, esta foi a única regressão logística em que houve um efeito significativo de variável ambiental sobre probabilidade de ocorrência (G = 4,78; 1 g.l.; p = 0,029). A regressão estimada é: p = e 0,00102 x Pluv - 1,45584 / 1 + e 0,00102 x Pluv - 1,45584, onde p é a probabilidade de ocorrência, e Pluv é a pluviosidade média anual (mm) no local.
40 0 40 80 120 Kilometers
P de ocorrência0.2 - 0.40.4 - 0.50.5 - 0.60.6 - 0.70.7 - 0.8
N
Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 12
ombrófilas densas (Figura 8). A riqueza prevista diminui em direção ao domínio das florestas semi-decíduas e decíduas, a oeste, e também na já mencionada faixa litorânea de Maraú a Valença.
DISCUSSÃO E CONCLUSÕES
Uma hipótese de distribuição geográfica
obedece a um balanço entre especificidade e sensibilidade. Acrescentar novas áreas à distribuição, para acomodar pontos de ocorrência, aumenta os riscos de inclusão de localidades nas quais a espécie não ocorre. Reduzir a área abrangida pela hipótese carrega o risco oposto. Para o conjunto de dados analisado, o modelo de ótimos ambientais foi o melhor compromisso entre estes dois procedimentos. É preciso assinalar, no entanto, que, ainda assim, os valores de kapa foram baixos a extremamente baixos.
Este ajuste fraco deve-se, principalmente, à inclusão de muitas localidades de ausência na área prevista pelas hipóteses como de ocorrência, ou seja, a baixos índices de especificidade. Para as três espécies avaliadas, as hipóteses de distribuição tiveram uma sensibilidade pelo menos duas vezes maior do que sua especificidade. Isto pode significar que há outros fatores limitantes
importantes que não foram incluídos no modelo. Alternativamente, os pontos de ausência incluídos na área de distribuição hipotetizada podem representar localidades em que a espécie poderia ocorrer, mas foi extinta.
Tabela 7 - Estatísticas dos escores no primeiro eixo do PCA das localidades onde foram registradas as espécies de mamíferos analisadas. Os valores dos índices de simetria (G1) e curtose (G2) siginificativamente diferentes do esperado para uma curva normal estão em itálico. A última coluna indica qual transformação foi necessária para normalizar as distribuições (Ln = logaritmo natural, √= raiz quadrada, s/ transf = não foi preciso transformar). Os escores das localidades de L. chrysomelas não se ajustaram à curva normal, mesmo transformados.
Espécie N Média Desvio-
padrão Assimetr.
(G1) Curtose
(G2) Ajuste à normal
Bradypus torquatus 32 0,740 0,718 -0,923 -0,192 Ln Bradypus variegatus 25 0,498 0,681 -0,479 -1,004 s/ transf Brachyteles hypoxanthus 30 0,285 0,859 -1,248 1,281 Ln Callicebus melanochir 66 0,612 0,750 -0,670 0,125 Ln Callithrix geoffroyi 15 0,742 0,446 -1,724 3,341 √ Callithrix kuhli 88 0,774 0,769 -0,898 -0,155 Ln Cebus r. nigritus 30 0,641 0,436 -1,604 2,978 √ Cebus xanthosternos 74 0,730 0,831 -0,832 -0,481 Ln Leontopithecus chrysomelas 201 0,556 0,824 -0,492 -1,167 não ajust. Chaetomys subspinosus 67 0,656 0,781 -1,065 1,646 Ln Sphiggurus insidiosus 76 0,549 0,814 -0,823 0,588 Ln
Figura 6 - Distribuição de freqüência dos escores no primeiro eixo do PCA das localidades de ocorrência de Chaetomys subspinosus. O traço indica a média. Note a assimetria negativa da curva, com uma "cauda" à esquerda, e truncamento à direita.
-3 -2 -1 0 1 2 3Escore no eixo 1 PCA
0
5
10
15
20
25
N d
e lo
calid
ades
0.0
0.1
0.2
0.3
Proportion per B
ar
Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 13
De uma forma ou de outra, o modelo de ótimos ambientais foi a mais promissora das alternativa analisadas, não só por seu melhor desempenho, como por sua simplicidade, e significado biológico. Embora o modelo logístico seja hoje o mais empregado para estimar ocorrência de espécies em escala regional e geográfica, sua premissa de que as chances de ocorrência aumentam indefinidamente em um gradiente ambiental não são realistas. Além disto, pelo menos para os dados deste trabalho, o ajuste da regressão não foi possível na maioria das vezes.
As hipóteses de distribuição potencial geradas pelo modelo de ótimo ambiental indicam a área de mata ombrófila como a zona ótima de ocorrência de todas as espécies estudadas. Isto pode sugerir que
as variáveis climáticas que limitam as distribuições destes mamíferos são as mesmas que determinam as fitofisionomias. Por outro lado, as espécies podem estar associadas primariamente com a floresta ombrófila, cuja área foi identificada pelo modelo porque pode ser bem delimitada pelas variáveis ambientais utilizadas. A despeito destas relações de causalidade, o modelo indicou que o domínio da mata ombrófila é a área com maior riqueza potencial das espécies estudadas, enfatizando sua importância para este grupo de mamíferos.
Figura 7 - Hipóteses de distribuição potencial na área do Corredor Central na Bahia, para A - Bradypus torquatus, B - Bradypus variegatus, C - Callicebus melanochir, D - Brachyteles hypoxanthus (provavelmente extinto na área), E - Cebus (nigritus) robustus, F - Cebus xanthosternos, G - Callithrix geoffroyi, H - Callithrix kuhli, I - Chaetomys subspinosus, J - Sphiggurus insidiosus. As hipóteses foram geradas pelo modelo de ótimo em eixo ambiental de PCA, e as áreas cinzas indicam as áreas previstas de ocorrência potencial. Os pontos são localidades onde a espécie foi registrada por observação direta ou por espécime depositado em coleção (em preto); pegadas, restos ou outros vestígios (em amarelo); ou por relatos, animais cativos, peles com caçadores, e outros indícios secundários (lilás).

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 14
AGRADECIMENTOS
Este trabalho é resultado do Projeto de pesquisa "Abordagens ecológicas e instrumentos econômicos para o estabelecimento do Corredor do Descobrimento:uma estratégia para reverter a fragmentação florestal na Mata Atlântica do sul da Bahia", financiado pelo Ministério do Meio Ambiente (MMA), através do Projeto de Conservação e Utilização Sustentável da Diversidade Biológica Brasileira (PROBIO); e pelo Centro para Pesquisa Aplicada à Biodiversidade (CABS) da Conservation International. Nossos agradecimentos à toda equipe do projeto, pela constante colaboração, e pelo estimulante debate de idéias. Agradecemos ao IESB e CI do Brasil, executoras do projeto, e a João Carlos Pádua, gerente de finanças do projeto, pelo importante apoio logístico a este trabalho. Paulo Guedes, Júlia Nüschler e Camila Calheira prestaram valioso auxílio na digitação dos dados.
REFERÊNCIAS BIBLIOGRÁFICAS
Brown J. H. 1995. Macroecology. University of Chicago Press, Chicago.
Brown, J. H. & Lomolino, M. V. 1998. Biogeography. 2nd ed. Sinauer, Sunderland.
Cerqueira R. 1995. Determinação de distribuições potenciais de espécies, pp. 141-161. In: P. Peres-Neto, J. Valentin & F. Fernandez (eds.), Tópicos em Tratamentos de Dados Biológicos, Programa de Pós-Graduação em Ecologia - UFRJ, Rio de Janeiro.
CEI - Centro de Estatística e Informações 1989. Hipsometria: Estado da Bahia. Mapa impresso, Governo do Estado da Bahia, Salvador.
Coimbra-Filho A., Rylands A.B., Pissinatti A. & Santos I.B. 1991. The distribution and status of the Buff-Headed Capuchin Monkey, Cebus xanthosternos Wied 1820, in the Atlantic Forest Region of Eastern Brazil. Primate Conservation, [12-13]: 24-30.
Conservation International & Instituto de Estudos Sócio Ambientais do Sul da Bahia 2000. Designing Sustainable Landscapes. CABS/IESB, Washington, DC
Cumming G. S. 2000. Using between-model comparisons to fine-tune linear models of species ranges. J. Biogeog., 27: 441-455.
Fielding A. H. & Bell J. F. 1997, A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Cons., 24: 38-49.
Fonseca G.A.B., Alger K., Pinto L.P., Araújo M. & Cavalcanti R. 2003. Corredores de Biodiversidade: O Corredor Central da Mata Atlântica. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Guisan A. & Zimmermann N. E. 2000. Predictive habitat distribution models in ecology. Ecol. Mod. 135: 147-186.
Hengelved R. & Haeck J. 1982, The distribuition of abundance. I. Measurements. , 9: 303-316.
Hosmer D.W.J. & Lemeshow S. 1989. Applied logistic regression. John Wiley & Sons, New York.
Figura 8 – À esquerda: hipótese de zonas de riqueza potencial das espécies de mamíferos estudadas, exceto B. hypoxanthus, para a área do Corredor Central na Bahia. Foram contadas as espécies que potencialmente ocorreriam em cada quadrícula de 5 km2, segundo o modelo de ótimo ambiental (ver Figura 7). À direita: Classificação fitofisionômica da área do Corredor Central na Bahia, segundo Thomas & Carvalho (2003). Note a coincidência das zonas de maior riqueza potencial com o domínio de floresta ombrófila ("Lowland forest" e "tabuleiro forest", tons de verde), e a coincidência parcial da menor riqueza potencial em algumas áreas da costa com o domínio de restingas e mangues (amarelo e vinho).

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 15
Hutchinson G. E. 1957, Concluding remarks. Cold Spring Harb. Symp. Quant. Biol., 22: 415-427.
Landau E. C. 2003a. Padrões de ocupação espacial da paisagem na Mata Atlântica do sudeste da Bahia, Brasil. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Landau, E. C. 2003b. Normais de Precipitação no Sudeste da Bahia, Brasil.(mapa em formato digital, adaptado a partir de SEI, 1998), escala 1:6.000.000. Data dos Dados: 1943-1983. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Landau, E. C. 2003c. Normais de Temperatura no Sudeste da Bahia, Brasil.(mapa em formato digital, adaptado a partir de SEI, 1998), escala 1:6.000.000. Data dos Dados: 1961-1990. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Landau, E. C. & Resende, N. A. T. 2003b. Hipsometria do Sudeste da Bahia, Brasil.(mapa em formato digital, adaptado a partir de CEI,1989), escala 1:1.500.000. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Landau E. C. & Resende N. A. T. 2003a. Área de estudo do sub-projeto "Abordagens Ecologicas e Instumentos Economicos para o Estabelecimento do Corredor Central da Mata Atlântica: Uma Estratégia para Reverter a Fragmentação Florestal na Mata Atlântica do Sul da Bahia - PROBIO/MMA".(mapa em formato digital, adaptado a partir de IBGE, 1967 e SUDENE 1976; 1977), escala 1:100.000. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B.,
Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Manel, S., Williams, H. C. & Ormerod, S. J., 2001, Evaluating presence-absence models in ecology: the need to account for prevalence. J. Appl. Ecol., 38: 921-931.
Moura, R. T. 2003. Distribuição e ocorrência de mamíferos na Mata Atlântica do sul da Bahia . In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Oliver W. L. R. & Santos I. B. 1991. Threatened endemic mammals of the Atlantic Forest region of south-east Brazil Jersey Wildlife Preservation Trust, Jersey.
Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) 2003b. Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Prado P.I., Pinto L.P.S., Moura R.T., Carvalho A.M., Thomas W. W., Amorin A. 2003. Caracterização dos registros de ocorrência de espécies de plantas e mamíferos no sul da Bahia. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Santos I. B., Mittermeier R. A., Rylands A. B.& Valle C. M. 1987. The distribution and conservation status of primates in southern Bahia, Brazil. Primate Conservation 8: 126-142.
SEI - Superintendência de Estudos Econômicos e Sociais da Bahia et al. 1998. Pluviometria. Média Compensada Anual (1943-1983), Escala: 1:6.000.000 In: Atributos Climáticos do estado da Bahia, no.38, Salvador.
SEI Superintendência de Estudos Econômicos e Sociais da Bahia et al. 1998. Temperatura Média Compensada Anual (1961-1990), Escala: 1:6.000.000 In: Atributos Climáticos do estado da Bahia, no.38, Salvador.

Modelos de distribuição potencial de mamíferos no Corredor Central da Mata Atlântica
CORREDOR DE BIODIVERSIDADE DA MATA ATLÂNTICA DO SUL DA BAHIA 2003 Instituto de Estudos Sócio-Ambientais do Sul da Bahia e Conservation International do Brasil 16
Silva J.M.C. & Casteleti C.H.M. 2003. Status of the Biodiversity of the Atlantic Forest of Brazil. In: Galindo-Leal, C. & Câmara, I.G. (orgs.). State of the Hotspots: The Atlantic Forest of South America. pp. 43-59. Island Press, Washington, DC.
Steinberg D. & Colla P. 1991. LOGIT: a supplementary module for SYSTAT. SYSTAT Inc., Evanston.
Thomas W. W. 2003. Natural vegetation types in southern Bahia. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Thomas W. W. & Carvalho A. M. 2003. Zoneamento Ecológico do Sudeste da Bahia, Brasil.(mapa em formato digital). In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade na Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, IESB / CI / CABS / UFMG / UNICAMP, Ilhéus.
Udvardy M. D. F. 1969. Dynamic Zoogeography. With special reference to land animals. Van Nostrand, New York.
Whittaker R. H. 1967. Gradient analysis of vegetation. Biol. Rev., 42: 207-269.
Prado P.I., Pinto L.P., Moura R.T. e Landau E.C. 2003. Avaliação de modelos de distribuição geográfica e sua aplicação para prever a ocorrência de espécies de mamíferos no Corredor Central da Mata Atlântica. In: Prado P.I., Landau E.C., Moura R.T., Pinto L.P.S., Fonseca G.A.B., Alger K. (orgs.) Corredor de Biodiversidade da Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, Ilhéus, IESB / CI / CABS / UFMG / UNICAMP.


















