
Aula 1 - iq.usp.br · esquema abaixo . E + S ES E + P . Sabendo que a velocidade inicial é medida...
15
Aula 1 Leituras sugeridas Biochemistry Voet & Voet 3ª edição Capítulo 14 – parte 2 Enzyme kinetics Daniel Purich Capítulo 5 Gunawardena J. (2012). Mol. Biol of the Cell 23, 517- 519 Bar-Even et al. (2011). Biochemistry 50, 4402-4410 Wolfenden & Snider (2001). Acc. Chem. Res. 34, 938 - 945
Transcript of Aula 1 - iq.usp.br · esquema abaixo . E + S ES E + P . Sabendo que a velocidade inicial é medida...

Aula 1 Leituras sugeridas Biochemistry Voet & Voet 3ª edição Capítulo 14 – parte 2 Enzyme kinetics Daniel Purich Capítulo 5 Gunawardena J. (2012). Mol. Biol of the Cell 23, 517- 519 Bar-Even et al. (2011). Biochemistry 50, 4402-4410 Wolfenden & Snider (2001). Acc. Chem. Res. 34, 938 - 945

De modo geral E + S E + P Logo v = kobs1 [E] [S] Se o “consumo” de S é desprezível (< 5%), então podemos aproximar para v = kobs2 [E], sendo que a [S], que é constante, está incorporada em kobs2. Considerando que [E] é constante no experimento, então v0 é constante. Assim: Algumas lições práticas: 1 – A medida de velocidade deve ser feita ao longo de uma faixa de tempo a fim de avaliar experimentalmente que v seja inicial (v0), ou seja, que o produto acumula-se linearmente e portanto v0 será corretamente correlacionada com [E]. Uma medida de tempo único não é garantia de medida precisa de v0 2 – Se v é inicial, então inclinação é proporcional a [E] 3 – Se [P] versus t é uma reta, então a enzima é estável nas condições de ensaio
[P]
t [E]
v0
[P]
t
E
2E 4E
[P]
t
Triplicata; v0 subestimada
De modo simples uma reação catalisada por E poderia ser assim representada.

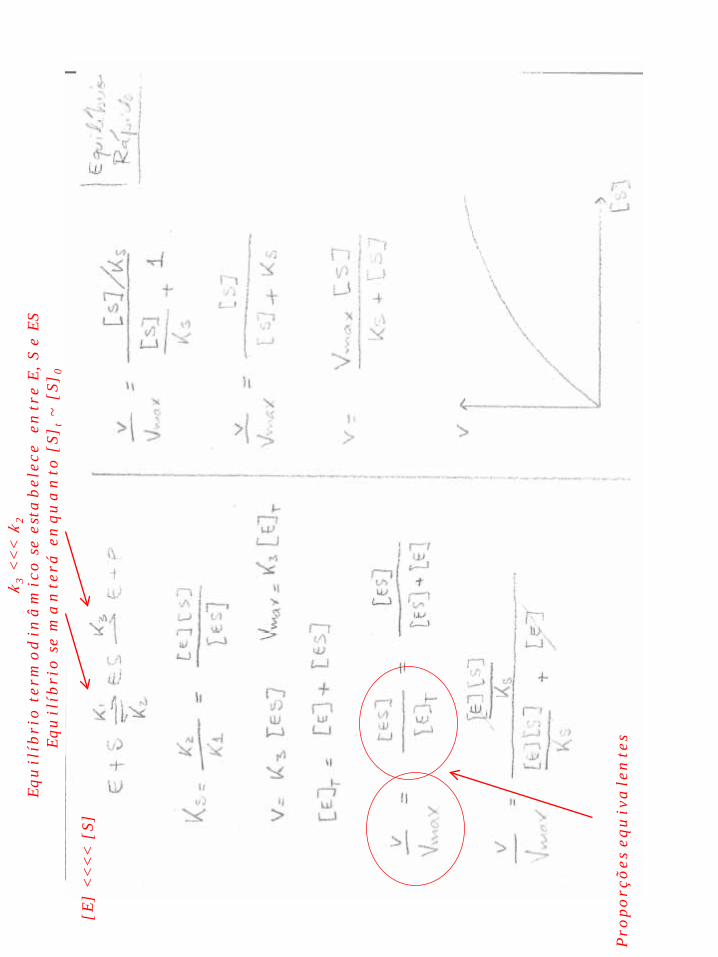
Por outro lado, observou-se experimentalmente que v0 correlaciona-se com [S] segundo esquematizado abaixo: Isto levou à hipótese da existência de um complexo ES. A velocidade estaria limitada a um máximo pela concentração de enzima. Este complexo poderia originar o produto ou dissociar-se gerando as formas livres do substrato e enzima. Assim a reação catalisada por uma enzima passa a ser descrita pelo esquema abaixo E + S ES E + P Sabendo que a velocidade inicial é medida em situação de consumo de no máximo cerca de 5 % do substrato inicial, então a etapa reversa de E + P para ES pode ser desprezada por ter velocidade muito pequena. Então o esquema simplifica-se para E + S ES E + P
v0
[S]
Partindo deste esquema há dois modelos que permitem a dedução de uma equação que correlacione v0 com [S]
k 3 <
<< k
2
Equ
ilíb
rio t
erm
od
inâ
mic
o s
e es
tabel
ece
en
tre
E, S
e E
S Eq
uil
íbri
o s
e m
an
terá
en
qua
nto
[S]
t ~ [
S]0
Pro
porç
ões
equ
iva
len
tes
[E]
<<<< [
S]
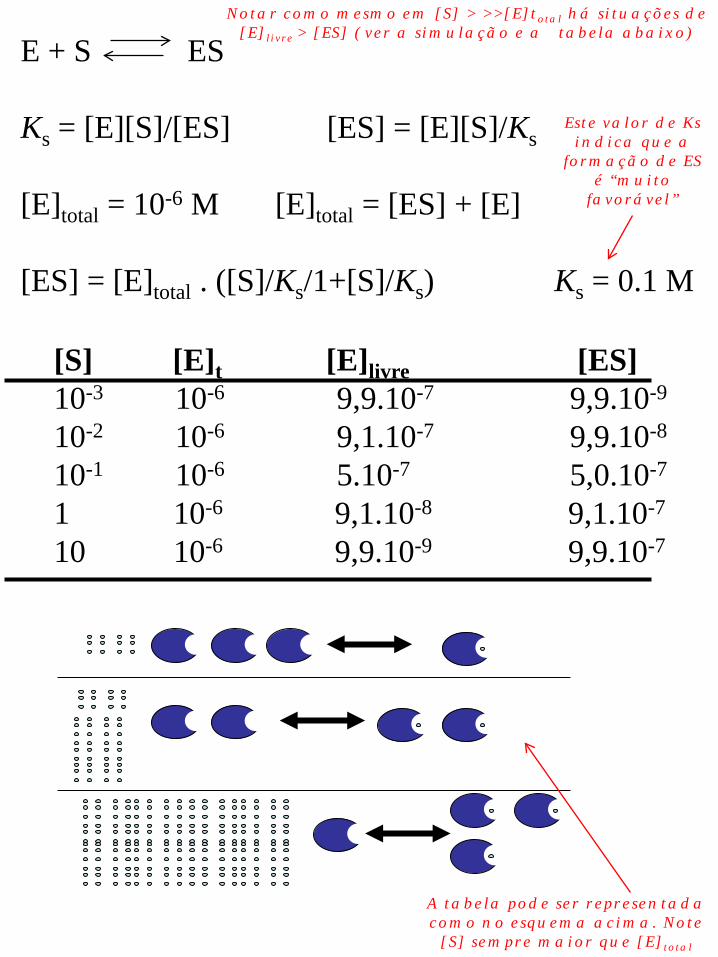
E + S ES Ks = [E][S]/[ES] [ES] = [E][S]/Ks [E]total = 10-6 M [E]total = [ES] + [E] [ES] = [E]total . ([S]/Ks/1+[S]/Ks) Ks = 0.1 M
[S] [E]t [E]livre [ES] 10-3 10-6 9,9.10-7 9,9.10-9
10-2 10-6 9,1.10-7 9,9.10-8
10-1 10-6 5.10-7 5,0.10-7
1 10-6 9,1.10-8 9,1.10-7
10 10-6 9,9.10-9 9,9.10-7
Este valor de Ks indica que a
formação de ES é “muito
favorável”
Notar como mesmo em [S] > >>[E]total há situações de [E]livre > [ES] (ver a simulação e a tabela abaixo)
A tabela pode ser representada como no esquema acima. Note [S] sempre maior que [E]total

Velo
cida
des d
e form
açã
o e “d
esapa
recimen
to” d
e ES se equ
ilibra
m.
k3 n
ão é o
brig
ato
riam
ente m
enor qu
e k2

Uma analogia é útil para comparar o equilíbrio rápido e o estado estacionário.
Em vasos comunicantes preenchidos a altura das colunas de água se manterá equilibrada durante certo tempo , mesmo com um pequeno furo em um dos vasos. Este vazamento, pequeno em relação à troca entre os dois compartimentos, ilustra k3 << k2. As alturas equilibradas ilustram o equilíbrio termodinâmico entre [ES] e [E] [S]
Já em vasos onde o furo é maior, ilustrando k3 ~ k2, a altura das colunas também poderá se manter estável no tempo enquanto o vazamento for semelhante a troca entre vasos (ilustrando v2 + v3 = v1). Neste caso o “equilíbrio” alcançado não é o mesmo que acima (termodinâmico), ilustrado pela menor altura das colunas de água.
[E] [S] [ES] E + P
[E] [S] [ES] E + P

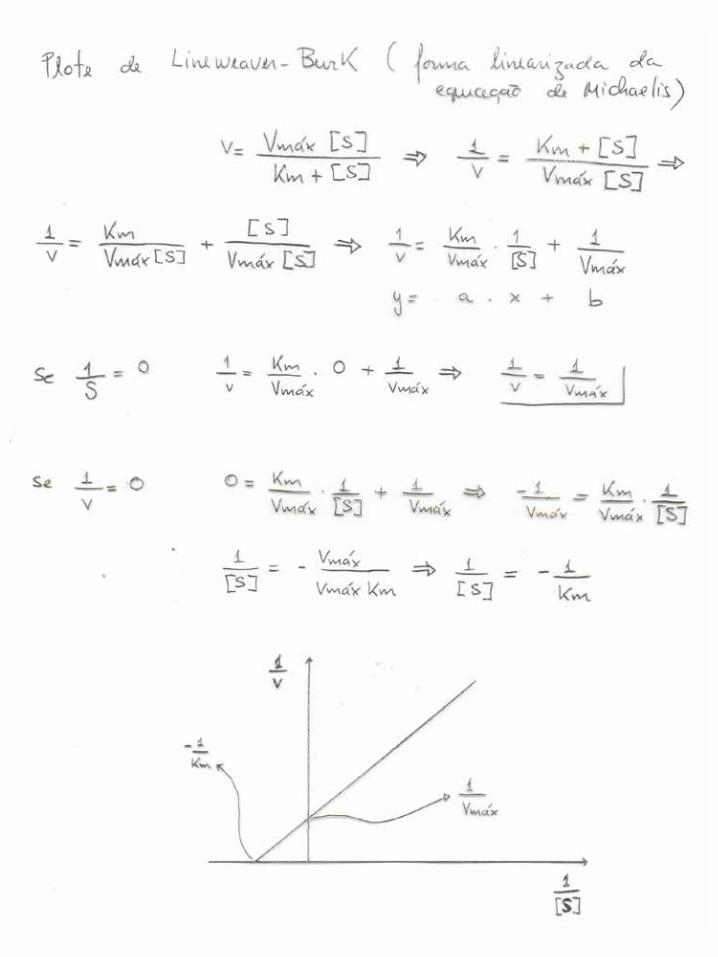
Ks = k2/k1 Km = k2+k3/k1 Ks é uma constante de equilíbrio “verdadeira” Km é uma constante cinética composta Note que Km = Ks quando k3 << k2 (Equilíbrio rápido) Ambos refletem a “abundância” de ES em relação ao produto E.S Quanto menor Ks, maior a afinidade entre E e S Dimensão de Ks e Km é “Molar”
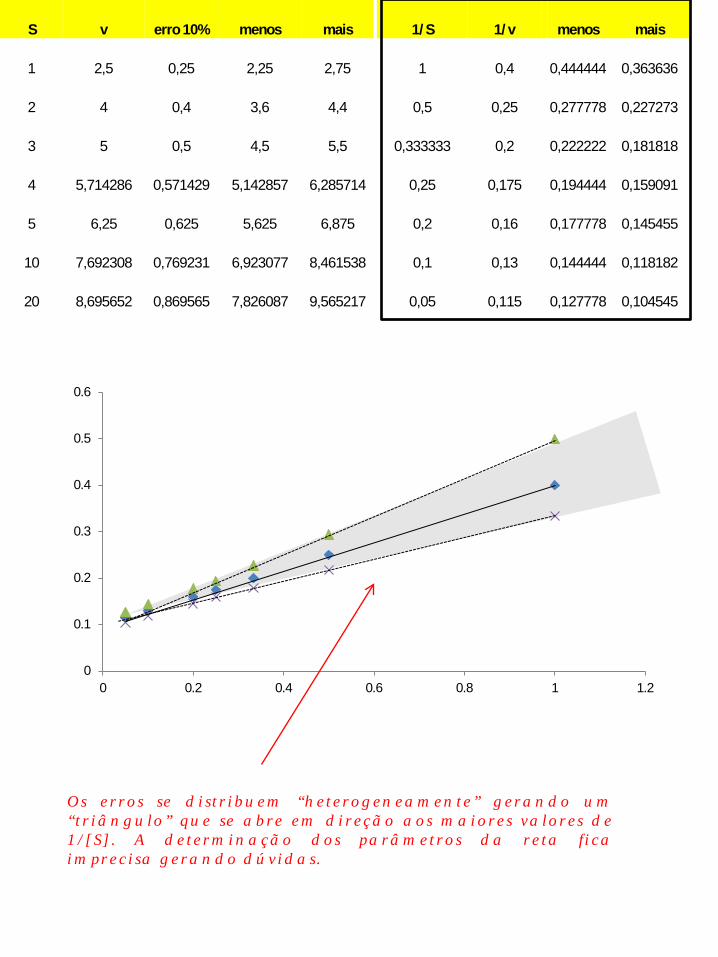
S v erro 10% menos mais 1/S 1/v menos mais
1 2,5 0,25 2,25 2,75 1 0,4 0,444444 0,363636
2 4 0,4 3,6 4,4 0,5 0,25 0,277778 0,227273
3 5 0,5 4,5 5,5 0,333333 0,2 0,222222 0,181818
4 5,714286 0,571429 5,142857 6,285714 0,25 0,175 0,194444 0,159091
5 6,25 0,625 5,625 6,875 0,2 0,16 0,177778 0,145455
10 7,692308 0,769231 6,923077 8,461538 0,1 0,13 0,144444 0,118182
20 8,695652 0,869565 7,826087 9,565217 0,05 0,115 0,127778 0,104545
0
0.1
0.2
0.3
0.4
0.5
0.6
0 0.2 0.4 0.6 0.8 1 1.2
Os erros se distribuem “heterogeneamente” gerando um “triângulo” que se abre em direção aos maiores valores de 1/[S]. A determinação dos parâmetros da reta fica imprecisa gerando dúvidas.
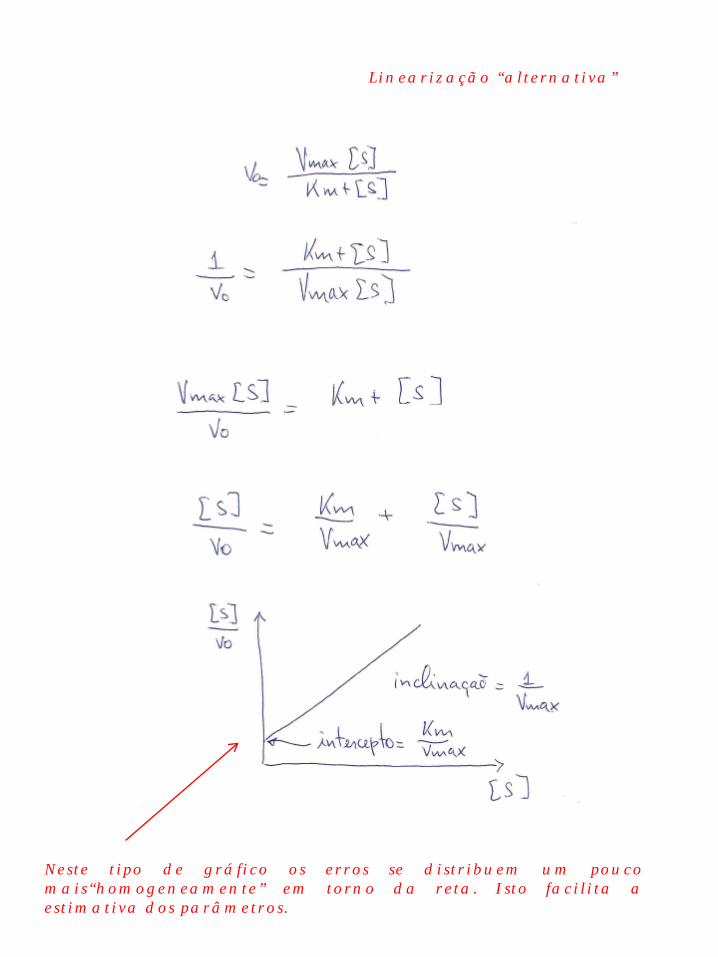
Linearização “alternativa”
Neste tipo de gráfico os erros se distribuem um pouco mais“homogeneamente” em torno da reta. Isto facilita a estimativa dos parâmetros.

menos mais[S] v erro 10% menos mais [S] [S]/v [S]/v [S]/v1 2.5 0.25 2.25 2.75 1 0.4 0.444444 0.3636362 4 0.4 3.6 4.4 2 0.5 0.555556 0.4545453 5 0.5 4.5 5.5 3 0.6 0.666667 0.5454554 5.714286 0.571429 5.142857 6.285715 4 0.7 0.777778 0.6363645 6.25 0.625 5.625 6.875 5 0.8 0.888889 0.72727310 7.6923 0.76923 6.92307 8.46153 10 1.300001 1.444446 1.18181920 8.6956 0.86956 7.82604 9.56516 20 2.300014 2.555571 2.090922
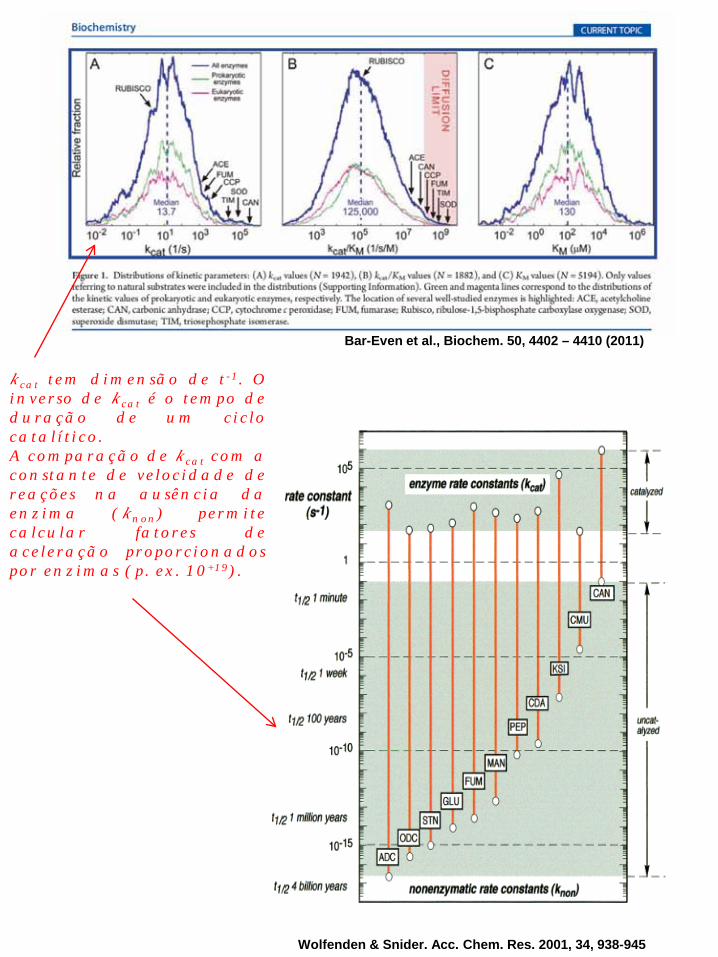
kcat tem dimensão de t-1. O inverso de kcat é o tempo de duração de um ciclo catalítico. A comparação de kcat com a constante de velocidade de reações na ausência da enzima (knon) permite calcular fatores de aceleração proporcionados por enzimas (p. ex. 10+19).
Bar-Even et al., Biochem. 50, 4402 – 4410 (2011)
Wolfenden & Snider. Acc. Chem. Res. 2001, 34, 938-945

kcat é usualmente maior em enzimas que catalisam reações de vias metabólicas centrais
Bar-Even et al., Biochem. 50, 4402 – 4410 (2011)
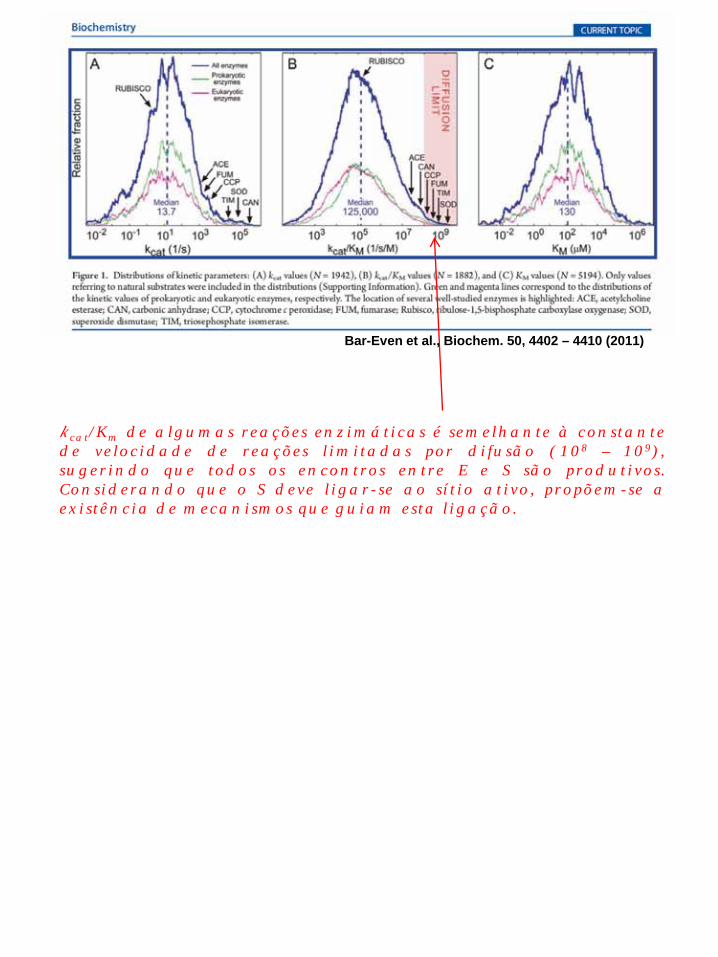
kcat/Km de algumas reações enzimáticas é semelhante à constante de velocidade de reações limitadas por difusão (108 – 109), sugerindo que todos os encontros entre E e S são produtivos. Considerando que o S deve ligar-se ao sítio ativo, propõem-se a existência de mecanismos que guiam esta ligação.
Bar-Even et al., Biochem. 50, 4402 – 4410 (2011)


















