
ANA RITA DE LIMA - USP€¦ · Ana Rita de Lima 4 DEDICATÓRIA À minha mãe Maria de Lourdes Lima...
109
ANA RITA DE LIMA Estudo quantitativo das células granulares adenohipofisárias associadas à produção do hormônio de crescimento e avaliação do perfil bioquímico do IGF-I em cães Golden Retriever com Distrofia Muscular (GRMD) São Paulo 2005
Transcript of ANA RITA DE LIMA - USP€¦ · Ana Rita de Lima 4 DEDICATÓRIA À minha mãe Maria de Lourdes Lima...

Ana Rita de Lima
0
ANA RITA DE LIMA
Estudo quantitativo das células granulares adenohipofisárias associadas à
produção do hormônio de crescimento e avaliação do perfil bioquímico do
IGF-I em cães Golden Retriever com Distrofia Muscular (GRMD)
São Paulo 2005

Ana Rita de Lima
1
ANA RITA DE LIMA
Estudo quantitativo das células granulares adenohipofisárias associadas à produção
do hormônio de crescimento e avaliação do perfil bioquímico do IGF-I em cães
Golden Retriever com Distrofia Muscular (GRMD)
Dissertação apresentada ao Programa de Pós-graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Ciências
Departamento: Cirurgia Área de Concentração: Anatomia dos Animais Domésticos e Silvestres
Orientador: Prof. Dr. Antonio Augusto Coppi Maciel Ribeiro
São Paulo
2005




Ana Rita de Lima
3
FOLHA DE AVALIAÇÃO
Nome: DE LIMA, Ana Rita.
Título: Estudo quantitativo das células granulares adenohipofisárias associadas à produção do hormônio de crescimento e avaliação do perfil bioquímico do IGF-I em cães Golden Retriever com Distrofia Muscular (GRMD)
Dissertação apresentada ao Programa de Pós-graduação em Anatomia dos Animais Domésticos e Silvestres da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para obtenção do título de Mestre em Ciências
Data: ____/____/____
Banca Examinadora
Prof. Dr. __________________________________ Instituição: ______________________
Assinatura: ________________________________ Julgamento: _____________________
Prof. Dr. __________________________________ Instituição: ______________________
Assinatura: ________________________________ Julgamento: _____________________
Prof. Dr. __________________________________ Instituição: ______________________
Assinatura: ________________________________ Julgamento: _____________________

Ana Rita de Lima
4
DEDICATÓRIA
À minha mãe Maria de Lourdes Lima e
meu pai José de Paula Lima,
Com vocês aprendi a andar, cair, levantar e a perseguir meus
objetivos. Vocês sempre me apoiaram nas minhas escolhas e
sempre estiveram por perto quando precisei. Muito obrigada por
existirem e por fazerem parte da minha vida.
Ao meu irmão Danilo, à minha irmã Eliani, meu cunhado Sinizeu
e às minhas sobrinhas Cíntia e Sara,
pelo apoio em todos os momentos. Com as pequenas eu aprendi
que ter uma criança adormecida em seus braços é um dos
momentos mais pacíficos do mundo e que algumas vezes tudo o
que precisamos é de uma mão para segurar e um coração para
nos entender. Muito obrigada por tudo.

Ana Rita de Lima
5
À minha grande amiga e irmã de coração Lillian de Jesus Oliveira,
Nestes anos que passamos juntas aprendi que verdadeiras amizades
continuam a crescer mesmo a longas distâncias, que aquilo que
realmente importa não é o que você tem na vida, mas quem você tem
na vida e que verdadeiros amigos são a família que nos permitiram
escolher... Você foi a primeira pessoa com quem fiz amizade aqui na
pós-graduação, sempre me ajudou em tudo o que precisei, sorrimos
juntas mas também sofremos juntas, você me adotou como irmã e
assim ficou até hoje e espero que isso não acabe... Pois tenha certeza
que você sempre terá um lugar muito especial em meu coração!!!
Ao meu grande amigo Emerson Ticona Fioretto,
Você é, e sempre será uma pessoa muito especial para mim... Sempre
disposto a ajudar a todos, está todo dia sorridente e sabe o que dizer
quando precisamos de consolo. Com você aprendi que falar pode
aliviar dores emocionais, descobri que as pessoas com quem você
mais se importa na vida são tomadas de você muito depressa e por
isso sempre devemos deixar as pessoas que amamos com palavras
amorosas, pois pode ser a última vez que as vejamos. Aprendi que
quando você está com raiva tem o direito de estar com raiva, mas
isso não te dá o direito de ser cruel. E que realmente a vida tem
valor e que você tem valor diante da vida!

Ana Rita de Lima
6
AGRADECIMENTOS
À Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo
pela oportunidade de realização deste curso.
À FAPESP
pelo apoio financeiro para o desenvolvimento desta pesquisa.
Ao Prof. Dr. Antonio Augusto Coppi Maciel Ribeiro
pela convivência, ensinamentos, dedicação e orientação no decorrer do mestrado
que nos possibilitou a finalização do mesmo.
A Profa. Dra. Maria Angélica Miglino
pela oportunidade de realização deste curso.
A Dra Mayana Zatz
pela oportunidade de realização deste trabalho com os animais do canil GRMD.
Ao Prof. Dr. Júlio César de Carvalho Balieiro
pelas explicações, orientação e realização das análises estatísticas.
A Profa. Dra. Ana Carolina Brandão de Campos Fonseca Pinto
pela dedicação e realização das radiografias e tomografias.

Ana Rita de Lima
7
Ao Dr. Alexander Augusto de Lima Jorge do Laboratório de Hormônios e Genética
Molecular do Hospital das Clínicas da Universidade de São Paulo
pela dedicação e ensinamentos.
A Profa. Dra. Berenice Bilharinho de Mendonça do Laboratório de Hormônios e
Genética Molecular do Hospital das Clínicas da Universidade de São Paulo
por autorizar o uso do laboratório para as dosagens hormonais.
A Profa. Sheila Canevese Rahal do Departamento de cirurgia da Faculdade de Medicina
Veterinária da Universidade Estadual Paulista (UNESP- Botucatu)
pela ajuda na realização do trabalho.
Ao Prof. Dr. Marco Antonio Zanini do Departamento de Neurologia e Psiquiatria da
Faculdade de Medicina da Universidade Estadual Paulista (UNESP- Botucatu)
pelas explicações quanto ao procedimento cirúrgico.
À equipe de técnicos e funcionários do Laboratório de Hormônios e Genética
Molecular do Hospital das Clínicas da Universidade de São Paulo.
À equipe de técnicos, funcionários e professores do setor de Anatomia Animal da
Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo
pela ajuda nos procedimentos da pós-graduação.

Ana Rita de Lima
8
Aos amigos Lílian de Jesus Oliveira, Emerson Ticona Fioretto, Janaína Munuera
Monteiro, Patrícia Orlandini Gonçalez, Robson Fortes Giglio, Tais Sasahara, Vivian
Samoto, Érica Branco, Adriana Morini, Daniele dos Santos Martins, Carlos Eduardo
Ambrósio, Karina Martinez Gagliardo, Renata de Britto Mari, Elizângela dos Anjos
Silva, Carolina do Prado, Rosemary Viola Bosch, Mônica Lotito, Priscilla Moraes,
Guilherme Ferreira e Kátia Viegas e a todos colegas da pós-graduação
pela amizade e companheirismo. Pois o valor de uma amizade é superior a qualquer
preço.
Aos amigos e colegas de trabalho no canil Adriana Morini (Drika), Angélica Grando
(Franguinho), Carla Casagrande, Carlos Eduardo Ambrósio (Caju), Daniele dos
Santos Martins (Dani), Guilherme Ferreira (Mamão) e Robson Giglio.
pelo aprendizado, trabalho em conjunto e principalmente pela amizade.
Aos cães do canil GRMD
pela possibilidade de realização desta pesquisa.

Ana Rita de Lima
9
“Você pode sonhar, criar e construir a idéia mais maravilhosa do mundo,
mas são necessárias pessoas para fazer o sonho virar realidade.”
WALT DISNEY

Ana Rita de Lima
10
RESUMO
LIMA, A.R. Estudo quantitativo das células granulares adenohipofisárias associadas à produção do hormônio de crescimento e avaliação do perfil bioquímico do IGF-I em cães golden retriever com distrofia muscular (GRMD) [Morphometric and stereological study of the adenohypophysis granular cells associated to GH producing and evaluation of the biochemical profile of IGF-I in golden retriever muscular dystrophy (GRMD)]. 2005. 107 f. Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2005. A Distrofia Muscular de Duchenne (DMD) é uma doença recessiva ligada ao cromossomo
“X”, causada pela ausência da proteína distrofina que ocorre em vários tecidos, sendo
caracterizada por uma severa disfunção da musculatura esquelética ocasionando morte
prematura do paciente. Embora controverso, alguns autores reportaram que o GH
(hormônio do crescimento) estaria implicado no desenvolvimento da doença e poderia ser
utilizado no tratamento da mesma. Desta forma, neste estudo a dosagem sérica de IGF-I
(Fator I de crescimento Similar à insulina) um peptídeo GH-dependente que regula as ações
do hormônio do crescimento, foi realizada no intuito de verificar se existe correlação ou
não entre o desenvolvimento da doença e a concentração sérica de IGF-I. As variações nos
níveis deste hormônio foram demonstradas com o decorrer da idade sendo que, nos três
primeiros meses todos os animais apresentaram comportamento semelhante com aumento
dos níveis de IGF-I, porém no quarto mês os animais Distróficos benignos apresentaram
redução média de 34% deste hormônio, enquanto ocorreu aumento de 1% no animal não
Distrófico. Ainda, as células granulares adenohipofisárias relacionadas à produção do
hormônio de crescimento em cães Golden Retriever com Distrofia Muscular apresentam-se
maiores do que nos cães Golden Retriever não distróficos quanto aos seguintes parâmetros
(eixo longo, área seccional e volume celular). À microscopia eletrônica de transmissão
observamos que as células estudadas apresentam grânulos elétron-densos de parede dupla e

Ana Rita de Lima
11
distribuídos por todo o citosol. Estes grânulos apresentaram-se maiores nos animais
Distróficos quando comparados aos animais não Distróficos.
Palavras-chave: Distrofia Muscular de Duchenne. hormônio do crescimento . IGF-I .
Golden Retriever . estereologia.

Ana Rita de Lima
12
ABSTRACT
LIMA, A.R. Morphometric and stereological study of the adenohypophysis granular cells associated to GH producing and evaluation of the biochemical profile of IGF-I in golden retriever muscular dystrophy (GRMD) [Estudo quantitativo das células granulares adenohipofisárias associadas à produção do hormônio de crescimento e avaliação do perfil bioquímico do IGF-I em cães golden retriever com distrofia muscular (GRMD)]. 2005. 107 f. Dissertação (Mestrado em Ciências) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2005. The Duchenne Muscular Dystrophy (DMD) is a X linked recessive disease, caused by the
absence of dystrophin which is found in a variety of tissues and characterized by a severe
disfunction of the skeletal musculature that results in a premature death of the patient.
Theoretically, the growth hormone (GH) is considered to be associated to the development
of Muscular Dystrophy and that could be used in its treatment. Hence in this study, a IGF-
I (Insulin like growth factor-I) seric dosage was performed to verify whether or not there
might be a between the evolution of the disease and IGF´s seric concentration. IGF-I is a
GH-peptide dependent that regulates the GH actions during the growth. Changes in IGF-I
levels were recorded during the dog’s post-natal development. On the first trimester, all
animals presented similar IGF-I levels, although in the fourth month, a stark 34% decrease
was observed in the dystrophic animals whereas a 1% increase was seen in the healthy dog.
Furthermore, the GRMD´s granule-containing cells were larger when compared to the
healthy animals. The following parameters in this comparison: long axis, cross-sectional
area and cell volume. The ultrastructural study showed electron-dense granules composed
by a double membrane and homogeneously distributed through the cell. These granules
were larger in the dystrophic animals then in healthy dogs.
Key words: Duchenne Muscular Dystrophy . growth hormone . IGF-I . Golden Retriever .
stereology.

Ana Rita de Lima
13
ÍNDICE DE FIGURAS
Figura 1- Esquema do ensaio Imunorradiométrico (IRMA) realizado para a dosagem de
IGF-I..................................................................................................................... 62 Figura 2- Esquema do imunoensaio utilizado para a dosagem de testosterona ................... 62 Figura 3 – Fotomacrografia detalhada mostrando situação da hipófise de Golden Retriever
não distrófico. Rostralmente observa-se o quiasma óptico (*), lateralmente os Tractos ópticos (cabeças de seta) e caudalmente os pedúnculos cerebrais (p)..67
Figura 4 – Fotografia de imagem radiográfica de crânio de um Golden Retriever não
distrófico. A seta indica a localização da hipófise que foi marcada com uma esfera de chumbo ............................................................................................... 68
Figura 5 - 5 A - Imagem tomográfica de cão Golden Retriever Distrófico onde podemos
observar a Hipófise (H), parte do Osso Basisfenóide (X) que foi removido na dissecação, Cavidade Timpânica (*), Bula Timpânica (B) e o Processo zigomático do osso temporal (T). 5 B - Imagem tomográfica de cão Golden Retriever Não Distrófico, podemos observar a Hipófise (H), o Osso Basisfenóide (X), o Processo zigomático do osso temporal (T) e o Processo condilar da mandíbula (M) ................................................................................ 69
Figura 6 – Fotomicrografias demonstrando as células granulares (g) adenohipofisárias e
vasos sanguíneos (seta) em um cão Golden Retriever distrófico (A e C) e não distrófico (B e D). Azul de toluidina. Escala de barra: 20µm (em A e B) e Escala de barra: 5µm (em C e D) ...................................................................... 71
Figura 7 - 7A -Eletromicrografia de células adenohipofisárias de cão Golden Retriever
Distrófico (Pars distalis) mostrando a presença de grânulos elétron-densos (g) distribuídos homogeneamente pelo citosol e ao redor do núcleo (N). Escala de Barra: 1µm. 7B - Eletromicrografia de duas células adenohipofisárias de cão Golden Retriever não Distrófico (Pars distalis), delimitadas pelo plasmalema (cabeças de seta cheia). Observa-se ainda o núcleo (N) e grânulos elétron-densos (g) distribuídos pelo citosol. Escala de Barra: 500nm........................... 77
Figura 8 - Eletromicrografias mostrando células adenohipofisárias (Pars proximalis) de cão
Golden Retriever Distrófico (A) e Golden Retriever não Distrófico (B) evidenciando grânulos elétron-densos (g) e presença do núcleo (N). Eletromicrografias de células adenohipofisárias (Pars distalis) de cão Golden Retriever Distrófico (C) e Golden Retriever não Distrófico (D) mostrando grânulos elétron-densos (g) ao redor do núcleo (N). Nota-se em B e D grânulos elétron-densos formados por parede dupla (cabeças de seta brancas). As cabeças de setas pretas indicam os limites do plasmalema externo. Escala de Barra: 200nm .................................................................................................................. 78

Ana Rita de Lima
14
Figura 8 - Seqüência de aminoácidos do IGF-I das espécies canina com a humana.
Podemos observar que as seqüências apresentam alta homologia (95,16%) diferindo apenas em seis aminoácidos ............................................................ 81

Ana Rita de Lima
15
ÍNDICE DE TABELAS
Tabela 1 - Número de células granulares adenohipofisárias (Pars proximalis e Pars distalis) de cães Golden Retriever distróficos e não distróficos utilizados no estudo estereológico, São Paulo – 2005 .......................................................................... 57
Tabela 2 - Dados de peso (Kg) e idade expressa em meses entre os animais Distróficos (1-
4) e não Distrófico (5) utilizados neste estudo, São Paulo – 2005....................... 59 Tabela 3 - Dados sumarizados de comprimento, largura e espessura das adenohipófises dos
cães Golden Retriever Distróficos e não Distróficos. São Paulo – 2005............. 66 Tabela 4 - Estatísticas descritivas para as variáveis avaliadas (comprimento, largura e
espessura) das adenohipófises estudadas, São Paulo – 2005............................ 66 Tabela 5 - Valores médios dos parâmetros quantitativos: eixo longo, área seccional e
volume para as células granulares da adenohipófise (Pars proximalis e Pars distalis) de cães Golden Retriever distróficos e não distróficos. São Paulo – 2005 ................................................................................................................... 73
Tabela 6 - Médias de quadrados mínimos para Área e Volume celular, na Pars proximalis e
distalis comparando animais distróficos e não-distróficos, São Paulo – 2005 .... 73 Tabela 7- Resumo das análises de variância das células granulares adenohipofisárias (Área
e volume celular na Pars proximalis e distalis), segundo os grupos comparativos. São Paulo, 2005. .................................................................................................. 74
Tabela 8- Dados sumarizados da densidade de perfis celulares da Pars proximalis e na Pars
distalis nos animais Distróficos e não Distróficos. São Paulo – 2005................. 75 Tabela 9 - Resumo das análises de variância das células granulares adenohipofisárias na
Pars proximalis, Pars distalis para a densidade de perfis celulares. São Paulo – 2005 ................................................................................................................... 75
Tabela 10 - Médias de quadrados mínimos para a Pars proximalis e Pars distalis para a
densidade de perfis celulares. São Paulo – 2005 ............................................... 75 Tabela 11 - Valores médios para eixo longo e área dos grânulos das células
adenohipofisárias (Pars proximalis e Pars distalis) em cães Golden Retriever distróficos e não distróficos. São Paulo – 2005 ............................ 76
Tabela 12 - Estatísticas descritivas para as variáveis avaliadas (eixo longo e área) dos
grânulos elétron–densos de células adenohipofisárias de cães Golden Retriever Distróficos e Golden Retriever não Distrófico, São Paulo – 2005..79

Ana Rita de Lima
16
Tabela 13 - Resumo das análises de variância para Eixo e Área dos grânulos elétron-densos das células adenohipofisárias na Pars proximalis e Pars distalis, segundo os grupos comparativos, São Paulo – 2005............................................................ 80
Tabela 14 - Médias de quadrados mínimos para Eixos e Áreas dos grânulos das células
adenohipofisárias (Pars proximalis e distalis), São Paulo – 2005 .................... 80 Tabela 15 - Dados da dosagem sérica do IGF-I nos cães Golden Retriever distróficos e não
distróficos. São Paulo – 2005 ............................................................................ 82 Tabela 16 - Resultado do teste t Student de comparações múltiplas para valores de IGF-I,
segundo os meses e grupos comparativos avaliados. Mostra-se que existem diferenças estatísticas significantes entre animais distróficos graves e não distrófico e animais distróficos benignos e não distrófico, São Paulo – 2005 ..84

Ana Rita de Lima
17
ÍNDICE DE GRÁFICOS
Gráfico 1 – Gráfico linear mostrando dados referentes à dosagem de IGF-I nos animais Distróficos graves (animais 01 e 02), animais distróficos benignos (animais 03 e 04) e não Distrófico (animal 05), durante os quatro primeiros meses e sétimo mês de vida. Nota -se em todos os animais um aumento nas concentrações séricas de IGF-I nos três primeiros meses, seguido de queda nos animais distróficos, ocorrendo um leve aumento no animal não distrófico no quarto mês, ocorrendo nova queda em todos os animais no sétimo mês, São Paulo – 2005 83
Gráfico 2 – Histograma demonstrando a distribuição dos valores médios das concentrações
séricas de IGF-I nos animais Distróficos graves, Distróficos benignos e não Distrófico, durante os quatro primeiros e sétimo mês de idade, São Paulo – 2005 ................................................................................................................... 84

Ana Rita de Lima
18
INDICE
1 INTRODUÇÃO................................................................................................................. 20 2 OBJETIVOS...................................................................................................................... 23 3 REVISÃO DE LITERATURA ......................................................................................... 25
3.1 ASPECTOS CLÍNICOS DA DISTROFIA MUSCULAR EM HUMANOS E EM CÃES GOLDEN RETRIEVER .................................................................................. 25
3.2 ASPECTOS MACROESTRUTURAIS, CITOLÓGICOS E HISTOLÓGICOS DA
ADENOHIPÓFISE ..................................................................................................... 29
3.3 ESTUDO MORFOMÉTRICO E ESTEREOLÓGICO............................................... 40
3.4 ANATOMIA DA IMAGEM (IMAGOLOGIA)......................................................... 42
3.5 ASPECTOS NEUROENDÓCRINOS ........................................................................ 43
3.6 ESTUDO PILOTO (ANATOMIA CIRÚRGICA) ..................................................... 46 4 MATERIAL E MÉTODOS............................................................................................... 49
4.1 MATERIAL ................................................................................................................ 49
4.2 MÉTODOS ................................................................................................................. 50
4.2.1 Estudo Macroscópico e Mesoscópico................................................................... 50
4.2.2 Anatomia da Imagem (Imagologia) ...................................................................... 51
4.2.2.1 Equipamento Tomográfico ............................................................................. 51
4.2.2.2 Técnica Tomográfica...................................................................................... 52
4.2.2.3 Equipamento Radiográfico............................................................................. 53
4.2.2.4 Técnica Radiográfica ..................................................................................... 53
4.2.3 Estudo Histológico................................................................................................ 54
4.2.3.1 Aspectos Microestruturais.............................................................................. 54
4.2.3.2 Aspectos Morfoquantitativos .......................................................................... 56
4.2.4 Aspectos Neuroendócrinos ................................................................................... 58

Ana Rita de Lima
19
4.2.5 Análise Estatística................................................................................................. 63
4.3 ESTUDO PILOTO (ANATOMIA CIRÚRGICA) ..................................................... 64 5 RESULTADOS ................................................................................................................. 65
5.1 ESTUDO MACROSCÓPICO E MESOSCÓPICO.................................................... 65
5.2 ANATOMIA DA IMAGEM (IMAGOLOGIA)......................................................... 67
5.3 ESTUDO HISTOLÓGICO ......................................................................................... 70
5.3.1 Aspectos Microestruturais .................................................................................... 70
5.3.2 Aspectos Morfoquantitativos................................................................................ 72
5.4 ASPECTOS NEUROENDÓCRINOS ........................................................................ 80
5.5 ESTUDO PILOTO (ANATOMIA CIRÚRGICA) ..................................................... 85 6 DISCUSSÃO..................................................................................................................... 86
6.1 ESTUDO MACRO E MESOSCÓPICO..................................................................... 86
6.2 ANATOMIA DA IMAGEM (IMAGOLOGIA)......................................................... 87
6.3 ESTUDO HISTOLÓGICO ......................................................................................... 88
6.3.1 Aspectos Microestruturais .................................................................................... 88
6.3.2 Aspectos Morfoquantitativos................................................................................ 89
6.4 ASPECTOS NEUROENDÓCRINOS ........................................................................ 91
5.5 ESTUDO PILOTO (ANATOMIA CIRÚRGICA) ..................................................... 94 7 CONCLUSÃO................................................................................................................... 95 REFERÊNCIAS ................................................................................................................... 97

Ana Rita de Lima
20
1 INTRODUÇÃO
A Distrofia Muscular de Duchenne (DMD) é uma desordem neuromuscular recessiva
ligada ao cromossomo “X” que acomete um menino a cada 3300 nascimentos. A
etiopatogenia está relacionada à ausência da proteína distrofina que ocorre em vários
tecidos, sendo mais evidente na musculatura estriada esquelética e em neurônios de regiões
específicas do Sistema Nervoso Central. Clinicamente, a DMD é caracterizada por uma
severa disfunção da musculatura esquelética que leva a morte prematura
(BOGDANOVICH et al., 2004; EMERY, 1991).
Na literatura médica há relatos de casos de pacientes apresentando um curso clínico
benigno da doença associado à comprovada deficiência de GH nestes pacientes (FRANK e
SMITH, 2001; GHAFOOR et al., 2003; KURTENBACH et al., 1989; ZATZ et al., 1981 a).
Desta forma, algumas hipóteses foram elaboradas de que este hormônio pode estar
relacionado à Distrofia muscular e atuar na melhora do quadro clínico dos pacientes.
Dentre os inibidores do GH destaca-se o Mazindol® (droga utilizada para inibir o
apetite que interfere no metabolismo do GH) que inicialmente foi benéfico em pacientes
com Distrofia Muscular de Duchenne (DMD). Entretanto, sua eficácia na redução dos
níveis de GH foi transitória e, devido aos seus efeitos colaterais, o uso prolongado no
tratamento de DMD não é recomendado (KURTENBACH et al., 1989; ZATZ et al., 1981
a; ZATZ, FROTA-PESSOA 1981 b).

Ana Rita de Lima
21
Segundo alguns estudos o GH pode ser usado como um possível tratamento para a
Distrofia Muscular de Duchenne já que o uso de GH humano em pacientes com distrofia
muscular e concomitante deficiência de GH ocasiona melhora na velocidade de
crescimento sem efeitos prejudiciais na resistência muscular (CHYATTE et al., 1973;
CHYATTE et al., 1974; FRANK e SMITH 2001; ZATZ, FROTA-PESSOA 1981b).
Devido aos fatos reportados anteriormente, neste estudo, a dosagem sérica do IGF-I
(Fator I de crescimento Símilar à insulina), que é um peptídeo GH-dependente e regula as
ações do hormônio do crescimento atuando como mediador de ações anabólicas e
mitogênicas do GH (BOQUETE et al., 2003; DAUGHADAY e ROTWEIN, 1989;
LISSETT et al., 2003; SHALET et al., 1998) foi utilizada para padronização na espécie
canina. Ainda, este procedimento foi utilizado com o intuito de verificar se existem
diferenças na concentração de hormônio do crescimento entre animais saudáveis e animais
distróficos portadores da forma benigna da doença.
Futuramente, se for comprovada a atuação do GH na melhora do quadro clínico de
pacientes com Distrofia muscular será útil conhecer suas alterações bioquímicas, bem como
aquelas relacionadas ao número e tamanho das células granulares adenohipofisárias
associadas à sua produção, nas diferentes formas clínicas da distrofia com o intuito de se
propor novas estratégias e esquemas terapêuticos visando, de igual forma, o paciente
humano e animal.
O presente estudo está inserido em um projeto mais amplo que é resultado da
colaboração científica entre o Instituto de Biociências da Universidade de São Paulo
(Centro de Estudos Genômicos Humanos), a Faculdade de Medicina Veterinária e
Zootecnia (FMVZ - USP) e a AADM (Associação de Amigos dos Portadores de Distrofia

Ana Rita de Lima
22
Muscular), concretizada pela construção de dois canis sendo um no setor de Anatomia do
Departamento de Cirurgia da Faculdade de Medicina Veterinária e Zootecnia da
Universidade de São Paulo e outro na cidade de Ribeirão Preto (SP) visando o estudo
circunstancial da distrofia muscular em cães da raça Golden Retriever.

Ana Rita de Lima
23
2 OBJETIVOS
No presente estudo os seguintes aspectos foram investigados comparativamente em
cães Golden Retriever não distróficos e em cães Golden Retriever acometidos pela
Distrofia muscular:
Avaliação do perfil bioquímico de GH por meio da dosagem sérica do IGF-I para
verificar se existem diferenças na concentração do hormônio de crescimento entre
cães saudáveis e cães distróficos portando as formas clínicas benigna e grave da
doença;
Caracterização macro, mesoscópica e anatomia da imagem (Imagologia) da
hipófise;
Caracterização microestrutural (morfométrica e estereológica) das células
granulares adenohipofisárias potencialmente associadas à produção do hormônio do
crescimento (GH).
Os parâmetros morfométricos e estereológicos analisados nesta pesquisa foram: área
seccional, eixo longo e estimativa global do volume das células granulares assim como
entender o comportamento destes parâmetros nos animais distróficos comparados aos
animais saudáveis. A Imagologia nos possibilitou determinar a localização da hipófise em
relação ao crânio para verificar se existiam modificações entre animais saudáveis e
distróficos.

Ana Rita de Lima
24
Finalmente, os parâmetros quantitativos destas células granulares adenohipofisárias
foram correlacionados aos dados referentes à variação do perfil sorológico de IGF-I nos
animais saudáveis e nos portadores benignos. Embora os animais utilizados para estudo
sorológico não foram os mesmos do estudo estereológico, pretendíamos verificar se em
uma mesma forma da doença as células associadas à produção de GH apresentariam ou não
alterações morfológicas qualitativas e quantitativas que pudessem ser correlacionadas às
possíveis variações séricas do próprio GH, estimadas pela dosagem de IGF-I.
Nossos objetivos iniciais incluíam viabilizar o acesso cirúrgico à adenohipófise, este
estudo do acesso cirúrgico pretendia embasar a aplicação de possíveis técnicas cirúrgicas
terapêuticas em relação à hipófise visando a ablação da mesma no intuito de avaliar o
progresso da doença visto que, conforme reportado anteriormente (CHYATTE et al., 1973;
CHYATTE et al., 1974; FRANK e SMITH 2001; ZATZ, FROTA-PESSOA 1981b) em
alguns pacientes com deficiência de GH os sintomas clínicos evoluem brandamente.

Ana Rita de Lima
25
3 REVISÃO DE LITERATURA
A revisão da literatura consultada segue abaixo.
3.1 ASPECTOS CLÍNICOS DA DISTROFIA MUSCULAR EM HUMANOS E EM CÃES
GOLDEN RETRIEVER
A Distrofia Muscular de Duchenne (DMD) é a mais comum desordem neuromuscular
recessiva associada ao cromossomo “X”, e caracterizada pela ausência da proteína
distrofina que causa degeneração progressiva do músculo esquelético e cardíaco. As
crianças afetadas locomovem-se por meio de cadeiras de rodas e geralmente morrem antes
dos trinta anos. A descoberta da proteína distrofina defeituosa na DMD permitiu a
avaliação crítica e a utilização de modelos animais. As formas de distrofias musculares
associadas ao “X” onde há ausência de distrofina foi descrita em ratos, cães e gatos. Nestes
modelos, a maioria dos sinais clínicos do Golden Retriever com Distrofia Muscular
(GRMD) é semelhante aos sinais da DMD. Os cães afetados possuem fraqueza progressiva
e desenvolvimento de contraturas debilitantes.Os sinais clínicos progridem rapidamente
entre 3 e 6 meses de idade. DMD e GRMD são caracterizadas por uma progressiva
evolução clínica que leva à morte prematura. Uma alta mortalidade é observada em GRMD

Ana Rita de Lima
26
durante as duas primeiras semanas de vida, quando uma alta concentração de creatinino-
quinase no soro permite um diagnóstico precoce da doença. Esta anormalidade bioquímica
sugere que todos os filhotes GRMD sofrem de mionecrose neonatal. Em neonatos GRMD,
o diafragma, músculos intercostais, a língua e a musculatura esquelética do tronco, do
pescoço e da porção proximal dos membros, são particularmente afetados (BERGMAN et
al 2002; BOGDANOVICH et al., 2004; KORNEGAY et al., 1994; NGUYEN et al 2002;
SHELTON et al., 2001).
A distrofina é uma macro proteína do citoesqueleto sendo importante para a
manutenção da integridade estrutural dos músculos durante a contração. O citoesqueleto é
uma rede complexa constituída de microtúbulos, microfilamentos e filamentos
intermediários. Essas proteínas estruturais condicionam a forma das células e, junto com as
proteínas motoras, possibilitam os movimentos de organelas e vesículas citoplasmáticas,
sendo também responsável pela contração celular e movimentação da célula inteira. Na
fibra muscular observa-se ao microscópio eletrônico filamentos finos de actina e filamentos
grossos de miosina dispostos longitudinalmente nas miofibrilas e organizados numa
distribuição simétrica e paralela. Essa organização é mantida por diversas proteínas, como
por exemplo os filamentos intermediários de desmina que ligam as miofibrilas umas às
outras. O conjunto de miofibrilas (actina e miosina) é preso à membrana plasmática da
célula muscular por meio de diversas proteínas que têm afinidade pelos miofilamentos e
por proteínas da membrana plasmática. Uma dessas proteínas, chamada distrofina liga os
filamentos de actina a proteínas integrais da membrana plasmática. A função precisa da
distrofina ainda não é conhecida, a falta desta proteína causa a desestabilização da
membrana e a ativação de múltiplos processos patofisiológicos. No músculo esquelético e

Ana Rita de Lima
27
cardíaco, a distrofina associada com várias proteínas forma complexos responsáveis pela
ligação do citoesqueleto de actina com a matriz extracelular, estabilizando o sarcolema
durante repetidos ciclos de contração e relaxamento (BERGMAN et al 2002; BLAKE et al.,
2002; BOGDANOVICH et al., 2004; EHMSEN, POON, DAVIES, 2002; JUNQUEIRA e
CARNEIRO, 1999; KORNEGAY et al., 1994; NGUYEN et al 2002; ROSS, KAYE,
PAWLINA, 2003; SHELTON et al., 2001).
A proteína distrofina também pode ser encontrada em neurônios em regiões
específicas do Sistema Nervoso Central. Estudos comportamentais mostram que crianças
com DMD possuem deterioração na cognição e baixo QI (média de 85). Histologicamente
ocorrem desordens na arquitetura do Sistema Nervoso Central, anormalidades nos dendritos
e perda de neurônios, todos associados com neurônios que normalmente expressam
distrofina. Camundongos mdx exibem memória curta e o número de neurônios reduz-se em
50% em regiões do córtex cerebral e tronco encefálico. Funcionalmente, crianças com
DMD apresentam anormalidades quando avaliadas com o uso de Eletroencefalograma e
apresentam evidências de que a função sináptica também é afetada pela ausência da
distrofina. Em camundongos mdx, estudos eletrofisiológicos mostraram que os neurônios
do hipocampo possuem um aumento na susceptibilidade à hipóxia (ANDERSON et al.,
2002).
Rudman et al. (1972) descreveram que o hGH exógeno causa efeitos catabólicos em
garotos com distrofia muscular de Duchenne. A resposta catabólica do hGH sugere não
somente que o hormônio exógeno não deve ser usado no tratamento de Distrofia Muscular,
mas também suscita a possibilidade de que o hGH endógeno pode estar envolvido na
patogênese da doença. Muitos estudos em ratos e no homem sugerem que o estrógeno inibe

Ana Rita de Lima
28
as ações periféricas do hGH. Isso demonstra que os efeitos adversos do hGH endógeno nas
células musculares de garotos com Duchenne podem ser inibidos pelo estrógeno.
ZATZ e FROTA-PESSOA (1981b) sugeriram que o GH pode ser usado como um
possível tratamento para a Distrofia Muscular de Duchenne, esta hipótese também foi
mantida por Chyatte et al. (1973,1974), quando testou o efeito do GH humano exógeno em
pacientes com distrofia miotônica (MyD), distrofia muscular de cintura e membros
(LGMD) e Distrofia Muscular de Duchenne (DMD). Em pacientes com DMD, foi obtida
uma resposta catabólica ao invés de produzir os efeitos anabólicos usuais, isso leva a
concluir que o GH endógeno pode estar envolvido na patogênese da doença. Em
contrapartida, CANAL et al., (1982) afirmaram que baixa ou nenhuma elevação no GH foi
descrita na distrofia miotônica (MD) durante a onda de sono.
Zatz et al. (1981a) descreveram o caso de um paciente afetado por Distrofia Muscular
de Duchenne (DMD) com um curso benigno incomum e apresentando deficiência de GH.
Os níveis séricos de creatinina fosfoquinase (CPK) e piruvato quinase (PK) estavam
grosseiramente elevados.
Evidências experimentais entre a ausência de hormônio do crescimento (GH) e a
progressão da distrofia foram descritas em pacientes com Distrofia Muscular de Duchenne
(DMD) e nanismo, e para um modelo animal com distrofia. Inversamente, a administração
de GH à pacientes com DMD acelera o declínio da performance muscular. O Mazindol,
uma droga que interfere no metabolismo do GH, foi benéfico em pacientes com DMD.
Contudo, seu efeito na redução dos níveis de GH foi transitório e devido aos seus efeitos
colaterais não é recomendado para uso prolongado no tratamento de DMD (GHAFOOR et
al., 2003; KURTENBACH et al., 1989).

Ana Rita de Lima
29
3.2 ASPECTOS MACROESTRUTURAIS, CITOLÓGICOS E HISTOLÓGICOS DA
ADENOHIPÓFISE
A hipófise é um corpo elipsoidal medindo cerca de 1 x 0,75 x 0,5cm em um cão de
tamanho médio. A glândula está suspensa na linha mediana do hipotálamo por uma haste
cilíndrica, o Tuber cinereum e está contida dentro de uma depressão (fossa hipofisária),
localizada no osso basisfenóide. Um revestimento de dura-máter envolve a glândula e
também o teto da depressão, estendendo-se das margens para logo em seguida abraçar e
limitar a haste hipofisária em todos os lados. A adenohipófise é composta por parênquima
glandular e possui um extensivo suprimento sangüíneo, aparecendo avermelhada e friável
em comparação à textura fibrosa da neurohipófise (DYCE et al., 2002; EVANS, 1993;
KÖNIG; LIEBICH, 2002).
A glândula está composta por duas partes que são funcionalmente distintas durante o
desenvolvimento: a adenohipófise, originária de uma evaginação dorsal do estomodeu, e a
neurohipófise, resultante do assoalho do diencéfalo. A porção da adenohipófise que se
funde com a neurohipófise é chamada de Pars intermedia. Em cães, um vestígio da fissura
neurohipofisária persiste separando a Pars intermedia da Pars nervosa. A porção
adenohipofisária é composta por três regiões (Pars distalis, Pars intermedia e Pars
proximalis). Os tipos celulares encontrados na Pars distalis são acidófilos, basófilos e
cromófobos. Os grânulos das células acidófilas são eosinofílicos e podem ser vistos
individualmente à microscopia de luz. Os somatotrofos predominam na porção lateral da
Pars distalis; possuem abundantes grânulos que possuem afinidade pelo corante Orange G.

Ana Rita de Lima
30
Os grânulos medem de 300 a 400nm de diâmetro e são corados imunohistoquimicamente
para GH (ADAM et al., 1970; DELLMANN e BROWN, 1987; MIKAMI, 1987).
As células presentes na Pars distalis são divididas dentro de dois grupos, células
cromófobas e células cromofílicas. As células cromófobas são pequenas, circulares com
citoplasma escasso e sem grânulos evidentes à microscopia de luz, sendo possível observar
pequenos grânulos à microscopia eletrônica. As células cromofílicas são divididas em dois
grupos: acidófilas e basófilas.
As células acidófilas são grandes, possuem citoplasma acidófilo granular
distinguindo-se assim, as células produtoras do hormônio do crescimento e da prolactina.
As basófilas são grandes e possuem grânulos PAS-positivos, estas células produzem
tirotropina, hormônio folículo estimulante, hormônio luteinizante e produzem moléculas
precursoras para a produção de hormônio adrenocorticotrófico (ACTH) e hormônio
estimulante de melanócitos (MSH) (BANKS, 1993).
A hipófise está localizada ventralmente ao hipotálamo. Rostralmente à mesma está o
quiasma óptico, lateralmente os tratos ópticos, e caudalmente observam-se os pedúnculos
cerebrais e o corpo mamilar. A protrusão ventral do hipotálamo que acompanha o terceiro
ventrículo é denominada de infundíbulo. A porção mais distal da protrusão infundibular é
chamada de processo infundibular ou mais comumente de lobo neural da hipófise. O
suprimento sanguíneo e a microcirculação das estruturas desta região são realizados pelas
artérias derivadas da artéria carótida interna e artéria basilar. A Pars distalis recebe um
pequeno número de fibras perivasculares do plexo carotídeo, mas supõe-se que estas fibras
não produzem efeitos diretos nos elementos secretórios da Pars distalis. Não ocorrem

Ana Rita de Lima
31
fibras nervosas originadas diretamente do hipotálamo para a Pars distalis (PORTER et al.,
1974).
A diferenciação dos tipos celulares hipofisários é feita usando vários corantes como o
ácido periódico de Schiff (PAS) e orange-G. Com hematoxilina e eosina podemos
diferenciar, em secções horizontais da glândula hipófise, a Pars distalis, pars intermédia e
pars nervosa. Com a coloração de PAS e orange-G aplicada a cortes de parafina é possível
distinguir células basófilas (PAS-positivas) de células acidófilas (orange-G positivas).
Quando observadas à microscopia eletrônica, as células-GH são arredondadas ou ovais e
contém muitos grânulos de secreção densamente corados e arredondados, com diâmetro
entre 300-350nm, distribuídos pelo citoplasma. Escasso retículo endoplasmático rugoso
está presente ao lado do núcleo (MORIARTY, 1973; OSAMURA,1996).
Em hipófises humanas foi descrita a existência de três tipos de células acidófilas, tipo
I, II e VIII, que correspondem às células alfa, eta e epsilon. O tipo I (células acidófilas ou
células alfa) contém grânulos citoplasmáticos caracteristicamente pequenos, os grânulos
são especificamente identificados por serem carminofílicos após uma seqüência de
coloração com azocarmine - orange-G. São mais numerosas na região posterior da
glândula. Outros tipos de células são pouco observados nas zonas acidofílicas e as células
aparecem em grandes e irregulares feixes associados aos sinusóides. O tipo I de célula varia
em tamanho, pode ser desde pequeno e arredondado com citoplasma escasso e um núcleo
central até células ovais grandes (20-25µm) que contém citoplasma abundante e núcleos
excêntricos. As células grandes freqüentemente são binucleadas e apresentam escassa
granulação na zona justanuclear e contém 1-2 vacúolos intracitoplasmáticos. O núcleo das
células grandes apresenta muitas variações morfológicas. Embora a maior parte dos núcleos

Ana Rita de Lima
32
seja vesicular com uma fina cromatina e um nucléolo basofílico, muitas células contêm um
núcleo picnótico escuro. O nucléolo nos demais tipos de células normalmente é acidófilo.
Os grânulos citoplasmáticos são carminofílicos. Azocarmine aplicada junto com Orange G
coram intensamente o tipo I de células (CONKLIN, 1966).
O tetracromo de Herlant’s (eritrosina, anilina azul, alizarina azul e orange G) cora e
diferencia os dois tipos de células acidófilas da hipófise, corando de laranja as células
produtoras de GH e vermelhas as produtoras de prolactina, o que foi comprovado com o
uso de anticorpo fluorescente (BAKER, 1970; STOKES; BODA, 1968).
A aplicação de anticorpo anti coelho para hormônio do crescimento humano em
hipófises de ratos machos adultos, revela numerosas células produtoras de hormônio do
crescimento, que se mostram grandes, ovóides, com citoplasma apresentando grânulos que
são intensamente corados (BAKER, 1970). Em ovinos o uso de anticorpo produzido com
GH de coelho anti-ovino associado com IgG biotinilado de suíno anti-coelho mostrou que
as células produtoras de GH aparecem distribuídas homogeneamente na Pars distalis da
adenohipófise em todos os animais estudados (BERNABÉet al., 2000).
Junqueira e Carneiro (1999) observaram que as células somatotróficas são células
acidófilas cujos grânulos de secreção são facilmente visíveis à microscopia de luz. Sua
identificação também é facilitada à microscopia eletrônica porque contêm numerosos
grânulos de secreção elétron-densos esféricos ou ovais. Apresentam núcleo centralizado e o
aparelho de Golgi é bem desenvolvido. A imunocitoquímica confirmou que estas células
produzem hormônio do crescimento ou somatotrofina, que estimula o metabolismo das
células em geral, tendo, porém um efeito muito mais pronunciado na cartilagem epifisária
dos ossos longos.

Ana Rita de Lima
33
A imunoreatividade da prolactina canina e das células produtoras de GH podem ser
distinguidas em duas diferentes populações de células acidófilas de acordo com sua
afinidade cromatográfica. Os tipos celulares diferem-se uns dos outros de acordo com sua
proporção relativa, morfologia e topografia, com variações pronunciadas em relação ao
sexo. As células produtoras de GH são pequenas, arredondadas ou ovais, com o citoplasma
mais denso e cheio de grânulos tênues. Seu núcleo possui elevado conteúdo de cromatina, o
que leva à afinidade específica pelo corante orange G. As células produtoras de GH estão
distribuídas isoladamente, ou tendem a se arranjar ao longo dos capilares. Isso ocorre mais
freqüentemente na região dorsal (porção rostral da Pars distalis) (EL ETREBY ; FATH EL
BAB, 1977).
As primeiras células GH imunoreativas aparecem entre 40 e 45 dias de gestação em
suínos, principalmente espalhadas individualmente, mas algumas vezes formam pequenos
grupos em justaposição com os vasos sanguíneos. Durante a vida fetal são caracterizadas
principalmente pela presença de grânulos secretórios. Entre os dias 45-90 ocorre um
aumento no número e tamanho das células imunoreativas com acúmulo de numerosos
grânulos secretórios levemente grandes em diâmetro, com aumento na concentração de GH
plasmático entre 70 e 105 dias de gestação (DACHEUX, 1984 a, b).
Com base nos estudos citofisiológicos e ultraestruturais da hipófise de ratos normais,
geralmente admite-se que as células mamotróficas contêm e secretam somente PRL, células
somatotróficas somente GH e que células mamosomatotróficas liberam ambos hormônios.
As células mamosomatotrópicas podem ter progenitoras comuns com diferenciação entre
mamotróficas e somatotróficas. Estas células agem como células-tronco e se transformam
em mamotróficas sobre estimulação apropriada. A proporção de mamotróficas,

Ana Rita de Lima
34
somatotróficas e mamosomatotróficas varia entre machos e fêmeas de ratos em diferentes
estágios fisiológicos. A grande heterogeneidade destas células indica que ocorre uma
interconversão de um tipo celular em outro, refletindo uma grande reserva funcional, isto é,
somente algumas células são estimuladas ao máximo, enquanto que outras são parcialmente
funcionais e outras são não funcionais (na categoria de célula tronco) (NIKITOVITCH-
WINER et al., 1987).
Células-GH e PRL se diferenciam de uma célula tronco comum, mas na
adenohipófise de galinhas adultas, as células-GH estão localizadas no lobo caudal e as
células-PRL no lobo rostral. No rato, as células-GH e PRL estão presentes em todas as
regiões da Pars distalis, mas com diferenças espaciais na concentração de ambos os tipos
celulares. Somente poucas células mamosomatotróficas podem ser vistas na Pars distalis
do rato, mas não são encontradas na Pars distalis da galinha (SASAKI et al., 2003).
Uma clara diferença sexual é observada na população de lactotrofos e somatotrofos
na hipófise anterior de ratos adultos; os lactotrofos são mais abundantes em fêmeas que em
machos e os somatotrofos são mais numerosos em machos (SASAKI; SANO, 1983).
As células produtoras de GH estão normalmente localizadas junto aos sinusóides
(BERNABÉ et al., 2000; NAKANE, 1970) e variam em sua forma de oval a piramidal com
um diâmetro aproximado de 10-13µm em ratos (NAKANE, 1970) e de 11-16µm em
ovelhas (BERNABÉ et al., 2000). O núcleo está localizado no centro da célula.
Ultraestruturalmente, as células são ovais e os grânulos de secreção têm diâmetro entre
300-350nm em todo o citoplasma. Um pouco de retículo endoplasmático pode ser
encontrado próximo ao núcleo. São encontradas mitocôndrias em grupos próximas ao
núcleo e ocasionalmente entre os grânulos de secreção (NAKANE, 1970).

Ana Rita de Lima
35
As células somatotróficas são caracterizadas por conterem grandes e arredondados
grânulos secretórios (por volta de 350 nm de diâmetro). O número de células
somatotróficas aumenta gradualmente com a idade. Os resultados obtidos com o estudo
estereológico à microscopia eletrônica sugerem que importantes mudanças ocorrem na
hipófise anterior de ratos no início da puberdade. O aumento no volume da hipófise anterior
com a idade é devido principalmente ao aumento no número de células e tamanho dos
somatotrofos e lactotrofos antes da puberdade, um aumento no tamanho das células
somatotróficas e no número de lactotrofos ao redor da puberdade, e o aumento no número
de ambos tipos celulares após a puberdade (SASAKI, 1988).
Na hipófise anterior de camundongos adultos e ratos, as células produtoras de PRL
são mais abundantes em fêmeas que em machos, a as células produtoras de GH são mais
abundantes em machos, de acordo com resultados de estudos morfométricos e
estereológicos com o uso de microscópio eletrônico. As considerações necessárias para a
amostragem visando estudo morfométrico e estereológico foram adotadas para a obtenção
do número total das células (produtoras de GH e PRL) neste estudo e, para assegurar que as
micrografias eletrônicas foram representativas de todas as partes da glândula. Metade foi
descartada, e a outra hemi pituitária foi cortada em cubos em seis partes. Três secções
seriadas, consistindo de uma ultra fina e duas semi-finas (0,6µm), foram cortadas de cada
bloco com um ultramicrótomo equipado com navalha de vidro. As secções semi-finas
foram montadas em lâminas de vidro, o Epon foi removido com sodium ethoxide
(C2H5ONa) em etanol, e as secções foram imersas em uma solução a 4% de H2O2 para
remover o tetróxido de ósmio. As secções foram lavadas em PBS e posteriormente tratadas
com anticorpos anti GH e anti PRL (SASAKI; IWAMA, 1988 a).

Ana Rita de Lima
36
Os hormônios da Pars distalis são polipeptídeos ou glicoproteínas. Os polipeptídeos
incluem a prolactina, hormônio do crescimento (GH), corticotropina (ACTH) e
melanotropina (MSH). Os hormônios glicoprotéicos são tireotropina, hormônio luteinizante
(LH) e hormônio folículo estimulante (FSH). A identificação microscópica das células
secretórias na Pars distalis depende primeiramente da capacidade destas células para
estocar os hormônios que são sintetizados. Os hormônios são acumulados na forma de
grânulos citoplasmáticos. As células parenquimais contêm muitos grânulos intensamente
corados e são chamados de cromófilas; células com pouco ou nenhum grânulo e pouco
coradas são chamadas de cromófobas. As células cromófilas são classificadas em acidófilas
e basófilas. A coloração histológica diferencial da hipófise está baseada na combinação de
três ou mais corantes ácidos, por exemplo, o Masson (fucsina ácida, ponceau R, anilina
azul) e Mallory (fucsina ácida, anilina azul, orange-G). Estas técnicas permitem uma clara
diferenciação entre acidófilos, que aparecem em vermelho (fucsina ácida) ou laranja
(orange-G), e basófilos que se coram em azul (anilina azul). As células produtoras de GH
normalmente são ovóides e estão dispostas ao longo dos capilares, seus grânulos reagem
intensamente com corantes ácidos incluindo eosina, eritrosina, orange-G e fucsina ácida. A
ultraestrutura das células produtoras de GH no rato é similar a outras células que secretam
proteínas. O retículo endoplasmático rugoso não é uma característica proeminente, os
grânulos secretórios maduros são arredondados, homogêneos, densos, envoltos por
membrana, e possuem diâmetro máximo de 350nm. O citoplasma possui lisossomos junto
com as mitocôndrias. O hipotálamo estimula a secreção de GH através do fator liberador de
hormônio do crescimento (GHRF). O GHRF acelera a extrusão dos grânulos secretórios
das GH células para dentro do espaço perivascular. O nível de GH circulante exerce um

Ana Rita de Lima
37
feedback negativo nas GH células. Algumas horas depois da administração de grandes
doses de GH em ratos, o conteúdo de GH na hipófise estará elevado. Se as injeções forem
continuadas por mais de cinco dias, o conteúdo de GH na hipófise reduz; o prolongamento
do tratamento causa redução no tamanho e número de acidófilos e sua granulação. Os
esteróides sexuais possuem uma significante influência sobre as “GH cells”. As GH células
são mais numerosas e maiores em machos adultos que em fêmeas de ratos (BAKER, 1974).
Arana et al. (1997) relataram que os métodos mais usados para a tipificação celular da
adenohipófise são os citoquímicos. As amostras obtidas foram processadas para exame
histológico coradas com Hematoxilina - eosina (HE) e para estudo citoquímico foram
utilizados os seguintes corantes Ácido Periódico de Shiff (PAS), Alcian Blue pH 2,5 e
Alcian Blue pH 0,5.
As células produtoras de GH possuem forma oval ou arredondada, tamanho médio e
distribuídas em toda a glândula. O número e tamanho das células produtoras de GH
aumenta com a idade em ambos os sexos. As células produtoras de GH são divididas em
três tipos baseado na ultraestrutura das células, em particular o tamanho dos grânulos
secretórios. O tipo I de células contém somente grânulos secretórios grandes (variando
entre 200 e 700nm), o tipo II contém tanto grânulos secretórios grandes quanto pequenos
(de 1 a 700nm) e estão distribuídos em todo o citoplasma das células. Estes dois tipos são
os somatotrofos clássicos, com forma oval e núcleo normalmente oval. O tipo III de célula
é angular ou oval e contêm somente grânulos secretórios pequenos (de 1 a 200nm) e
normalmente são menores que os tipos I e II (MANABE et al., 2000).
Sasaki e Iwama (1988 b) afirmaram que o número de células PRL e GH na
adenohipófise do rato aumenta durante o desenvolvimento pós-natal. Em animais adultos

Ana Rita de Lima
38
muitas células parenquimais são células-PRL ou GH, ocorrendo marcante diferença das
populações celulares com a diferenciação sexual. O estrógeno pode atuar na adenohipófise
durante este período para aumentar o número de células PRL e inibir as células GH. As
células PRL e GH estão em todas as áreas das secções horizontais da Pars distalis. As
células-PRL são predominantes nas áreas rostral e caudal, mas não nas porções central e
lateral. As células-GH são predominantes na região rostro-lateral.
Sasaki et al. (1992) descreveram que em suínos com 60 dias de gestação as células
produtoras de GH estão densamente distribuídas em todas as áreas exceto nas áreas das
secções medianas e paramedianas da Pars distalis, assim sendo, são distribuídas em todas
as áreas nas secções laterais. O padrão de distribuição das células pode ser visto na glândula
no final da gestação. Contrastando com as células produtoras de GH, as células LH estão
dispersas somente em pequenas regiões da área rostral da Pars distalis e Pars tuberalis e
visíveis aos 60 a 80 dias de gestação. No final da gestação, estão dispersas completamente
pela Pars distalis, mas uma população remanescente fica restrita à área rostral. A
contribuição do GH para o crescimento fetal é relativamente pequena, pois fetos de ratos
decapitados no útero continuam crescendo em ritmo normal até o nascimento.
Lee et al. (2004) realizaram estudo imunohistoquímico e descreveram que não
existem diferenças no número total de somatotrofos na hipófise de suínos recém nascidos e
pré-púberes. Contudo, ocorrem mudanças na distribuição espacial dos somatotrofos com o
decorrer da idade. O lobo anterior das hipófises foi dividido em cinco regiões e três
diferentes posições foram fotografadas em cada uma das cinco regiões. Os somatotrofos
foram contados em quinze posições (5 regiões x 3 posições por região) em três secções
(proximal, média e distal) em cada hipófise.

Ana Rita de Lima
39
Em embriões de galinha com 10 dias as células somatotróficas marcadas podem ser
vistas à microscopia de luz, mas a quantificação não é realizável. Durante o
desenvolvimento embrionário ocorre o aparecimento de grânulos de GH em células
marcadas. Embriões com 12 dias possuem grânulos pleomórficos sendo
predominantemente ovais, alongadas e poucos grânulos são esféricos. Com 15 dias a
maioria dos grânulos é esférica. A população de células somatotróficas passa abruptamente
de 3,6% para 20,7% entre o vigésimo dia de desenvolvimento embrionário e o primeiro dia
após o nascimento (MALAMED et al., 1993).
Os grânulos secretórios das adenohipofisárias estão ligados uns aos outros e
circundados pelo citoesqueleto. Muitos grânulos estão próximos à membrana e outros se
localizam abaixo dos microfilamentos periféricos ou estão presos nesta rede. A secreção de
hormônios rende um baixo ritmo de secreção de neurotransmissores e muitos grânulos
secretores são excluídos da região subplasmalemal. A dissolução da rede de actina cortical
pode ser necessária para a exclusão dos grânulos para próximo da membrana para que
ocorra a exocitose (CARBAJAL, VITALE, 1997).

Ana Rita de Lima
40
3.3 ESTUDO MORFOMÉTRICO E ESTEREOLÓGICO
A estereologia moderna promove eficiente e segura ferramenta para a descrição de
parâmetros quantitativos de estruturas em secções de tecidos. Muitos princípios que
permitem a estimativa do coeficiente de volume, área de superfície, coeficiente superfície-
volume e o número total de partículas são revistos e apresentados de forma geral
(HOWARD; REED, 2004).
Malamed et al. (1997) descreve que a área seccional em hipófises de frangos pode ser
determinada por micrografias de baixa capacidade. Estes valores foram multiplicados pela
distância entre as secções amostradas para calcular o volume de cada segmento cilíndrico
constituinte da hipófise anterior. A soma dos volumes de cada segmento das glândulas foi
coletado para estimar o volume total de cada glândula. A distância (K) entre as secções
amostradas foi de 150µm (30 secções com 5µm de espessura). Para cada micrografia de
alta capacidade três determinações foram feitas: área do campo exibido na micrografia, área
total de somatotrofos, e número de somatotrofos nucleados. A área seccional média dos
somatotrofos foi obtida pela divisão da área total ocupada pelos somatotrofos pelo número
total de somatotrofos por área de secção. A fração de área dos somatotrofos (porcentagem
de àrea ocupada pelos somatotrofos na àrea total da adeno hipófise) foi estimada pelos
coeficientes totais das áreas de somatotrofos nos campos amostrados e o total de área destes

Ana Rita de Lima
41
campos. A densidade numérica (número de somatotrofos / volume da adenohipófise) foi
estimado de acordo com a equação padrão (WEIBEL; BOLENDER, 1973):
Nv = K . NA
β VV
O valor de β foi determinado pelas secções dos somatotrofos sendo estimado através
do comprimento e diâmetro. N é o número somatotrofos contidos por unidade de área
seccional. V é a densidade de volume (volume dos somatotrofos / volume da glândula).
Para estimar o volume total dos somatotrofos a densidade de volume dos mesmos foi
multiplicada pelo volume da hipófise anterior. O número de somatotrofos em cada
segmento foi obtido através da densidade numérica dos somatotrofos multiplicado pelo
volume do segmento. Para o volume de um somatotrofo isolado, o volume total dos
somatotrofos na glândula foi dividido pelo número total de somatotrofos na mesma
(MALAMED et al., 1997).
O volume celular pode ser estimado pelo Princípio de Cavalieri multiplicando-se a
distância entre as secções histológicas pela somatória das áreas seccionais das células,
utilizando-se a seguinte fórmula (HOWARD; REED; 2004; RIBEIRO et al., 2004):
V= T x ∑A
Onde:
V= volume;
T= distância entre secções;
∑A= somatória das áreas seccionais.
Células produtoras de GH foram estudadas morfometricamente à microscopia de luz
e eletrônica em ovelhas lactantes e logo após o desmame. O estudo morfométrico revelou

Ana Rita de Lima
42
que a área celular variou de 66,2 a 114µm², com um diâmetro entre 11,1 e 15,9µm. Neste
estudo, não se observou diferença significante na densidade celular entre os grupos
estudados. As células produtoras de GH tiveram variações na forma mas em geral eram
esféricas ou ovais (BERNABÉ et al., 2000).
3.4 ANATOMIA DA IMAGEM (IMAGOLOGIA)
A tomografia computadorizada é uma modalidade diagnóstica que por meio de raios
X e do computador consegue formar imagens seccionais do paciente em diferentes planos
(transversal, dorsal e mediano), com a vantagem sobre a radiografia convencional, de
apresentar uma grande sensibilidade a pequenas diferenças de atenuação dos raios X e ser
isenta de sobreposição de estruturas, sendo utilizada para avaliação da hipófise em casos de
adenoma de hipófise e hiperadrenocorticismo hipófise dependente. Para a realização destes
exames é necessário o uso de contrastes radiopacos intravenosos para demarcar a região a
ser estudada (FEENEY et al., 1991; HATHCOCK; STICKLE, 1993; LOVE et al, 2000;
VLUGT-MEIJER et al., 2002; VLUGT-MEIJER et al., 2004 ).
O aparelho de tomografia é constituído basicamente por um “gantry” (abertura do
aparelho), onde estão colocados um tubo de raios X com um colimador e em sentido oposto
os detectores de raios X; um computador; um console de comando com um monitor e uma
câmara multiformato (HATHCOCK; STICKLE, 1993).

Ana Rita de Lima
43
A imagem tomográfica é adquirida através da emissão de raios X, colimados à
espessura desejada (1 a 10 mm), que sofrerão distintas atenuações ao atravessarem o
paciente em cada um dos 360° de rotação do tubo ao redor dele e serão captados pelos
detectores. Estes raios X captados serão transformados em um sinal elétrico e
posteriormente em números, que o computador utilizará para calcular, através de um
algoritmo matemático, o coeficiente linear de atenuação dos tecidos, para então produzir a
imagem dentro de uma escala de cinzas (BERRY, 2002; FEENEY et al., 1991;
HATHCOCK; STICKLE, 1993).
3.5 ASPECTOS NEUROENDÓCRINOS
O GH é um polipeptídeo de cadeia simples produzido pelos somatotrofos na
adenohipófise, com peso molecular de aproximadamente 22.000 daltons. A integridade do
eixo GH/IGF-I é essencial e um dos principais determinantes para o crescimento pós-natal
normal (EIGENMANN; PATTERSON, 1984; SIZONENKO et al., 2001). A liberação do
hormônio de crescimento é regulada por dois hormônios hipotalâmicos, o hormônio
liberador de GH (GHRH), que estimula a secreção pulsátil do GH (usualmente 6 pulsos
durante as 24 horas) e a somatostatina (SST) que atua inibindo sua liberação (BANKS,
1993).
Os esteróides sexuais (andrógenos e estrógenos) modulam a síntese e secreção de GH
pela hipófise anterior através de ações diretas nos somatotrofos ou via modulação dos

Ana Rita de Lima
44
neuropeptídeos hipotalâmicos que controlam a secreção de GH (Somatostatina e GHRH),
sendo seus efeitos observados com o início da puberdade (CHOWEN; FRAGO;
ARGENTE, 2004). No cão o início da puberdade varia de acordo com a raça ocorrendo
entre 5 e 12 meses de idade sendo os animais de pequeno porte mais precoces (LAING;
MORGAN; WAGNER, 1988).
Parte dos efeitos do GH é mediado por um polipeptídeo chamado de Fator I de
crescimento Símile à Insulina (IGF-I) também conhecido como somatomedina C, este
polipeptídeo é produzido ubiquamente, principalmente pelo fígado, rins e músculos sendo
regulado pelo GH. O IGF-I funciona como mediador de ações anabólicas e mitogênicas do
GH (DAUGHADAY; ROTWEIN, 1989). É encontrado na circulação periférica circulando
primariamente na forma de complexo ternário de alto peso molecular com a proteína 3 de
ligação ao IGF (IGFBP3) e a subunidade ácido lábil (ALS), ambos fatores GH dependentes
(BAXTER et al., 1989; RECHIER, 1993, YAKAR et al., 2002). Uma pequena proporção
de IGF-I pode circular em associação com outras proteínas ligadoras do IGF (RECHIER,
1993) e estima-se que menos de 5% do IGF-I plasmático circule na forma livre (ZAPF et
al., 1986). Os níveis plasmáticos de IGF-I apresentam uma variação diurna desprezível
(LEE; ROSENFIEL, 1987).
Dosagens isoladas do GH não representam sua secreção diária já que o mesmo é
secretado de maneira pulsátil com níveis extremamente baixos entre os pulsos. A
investigação laboratorial dos níveis séricos de GH baseia-se na sua análise direta através da
sua dosagem seriada (a cada 20 minutos) em ritmos de 24 horas, através de testes de
estímulo para sua secreção ou indiretamente com a dosagem das proteínas IGF-I e IGFBP-3
(ALBERTSSON-WIKLAND; ROSBERG, 1988).

Ana Rita de Lima
45
Devido a alta freqüência de falsos negativos e a complexidade em relação à análise
dos testes de liberação de GH e do ritmo de GH, na década de 90 as dosagens de IGF-I e
IGFBP-3 ganharam destaque como exames de avaliação de integridade do eixo GH/IGF-I
em estudos em seres humanos (ROSENFELD et al., 1995). As concentrações séricas de
IGF-I e IGFBP-3 estão diretamente relacionadas à ação do GH e por apresentarem níveis
sanguíneos relativamente estáveis ao longo de 24 horas, as suas determinações servem
como um espelho da secreção de GH. A avaliação do IGF-I e IGFBP-3 deve levar em conta
a idade e o sexo, pois tendem a se elevar com a idade e, em humanos, as dosagens são
maiores no sexo feminino que no masculino (JUUL et al.,1997; MAURAS et al., 2000).
A mensuração do IGF-I em seres humanos pode ser utilizada para avaliar os níveis de
GH (WACHARASINDHU et al., 2000) para diagnóstico de deficiência deste hormônio,
(CLEMMONS, 2001), para monitorar o tratamento com GH a curto e longo prazo (LEE;
ROSENFIEL, 1987) e para avaliar os riscos de doenças crônicas incluindo o câncer,
doenças cardiovasculares, diabetes mellitus e osteoporose (DELAFONTAINE, 1995;
JEHLE et al., 2003; PEACEY; SHALET, 2001; RENEHAN et al., 2003; SANDHU et al.,
2002; SHALET et al., 1998). Em animais de experimentação as dosagens de IGF-I e
IGFBP3 também são utilizadas para avaliar o eixo GH/IGF-I (HIGAKI et al., 1997; KIM et
al., 2004; SCHLEIM et al., 1999).
As concentrações séricas de IGF-I são determinadas por radioimunoensaio - RIA ou
ensaios imunorradiométricos – IRMA (BOQUETE et al., 2003; LISSETT et al., 2003;
SHALET et al., 1998). Os níveis plasmáticos de IGF-I são baixos na deficiência de GH
(LEE et al., 1990; LEE; ROSENFIEL, 1987), na desnutrição (LEE; ROSENFIEL, 1987;
SOLIMON et al., 1986) no hipotireoidismo, doenças hepáticas, diabetes mellitus

Ana Rita de Lima
46
descompensado (SHALET et al., 1998) e na síndrome de deficiência de receptores de GH
(GUEVARA-AGUIRRE et al., 1993).
A idade, níveis séricos de insulina, hormônios tireoidianos, esteróides sexuais e
estado nutricional regulam diretamente os níveis circulantes dos IGFs (GÓMEZ et al.,
2004; CLEMMONS, 2001). Em um estudo realizado para avaliar o efeito da idade e da
obesidade nas concentrações séricas de IGF-I, foi observado um contínuo e significante
declínio no total de IGF-I, IGF-I livre e IGFBP-3 entre 15 e 70 anos de idade em ambos
sexos; o avanço da idade foi o maior determinante das concentrações de IGF-I. Este
declínio no IGF-I pode ser resultante de alguns processos, tais como: a redução de secreção
de GH em função da idade ou a redução no número de receptores de GH no fígado
(GÓMEZ et al., 2004).
3.6 ESTUDO PILOTO (ANATOMIA CIRÚRGICA)
Nas hipofisectomias, as técnicas cirúrgicas são denominadas segundo a via de
acesso. Faz-se uma incisão de 3 a 4 cm no palato mole, no nível dos hâmulos do osso
pterigóide, que servem de referência. Posteriormente, em igual sentido e no mesmo nível do
palato mole, incisa-se o mucoperiósteo do canal craneofaríngeo, deixando descoberta parte
do corpo do basisfenóide. Neste momento inicia-se a trepanação da camada cortical externa
do basisfenóide, fazendo-se quatro perfurações que posteriormente se unem permitindo a
extração de um quadrilátero de 1cm2 de área. Na camada cortical interna extrai-se um
quadrilátero pequeno deixando-se a hipófise à mostra. É necessário ter muito cuidado ao

Ana Rita de Lima
47
extrair a cortical interna para não lesionar o seio cavernoso, pois se isso ocorrer será difícil
conter a hemorragia. O passo seguinte será incisar a lâmina ventral da dura-máter e aplicar
uma sucção moderada para manter o campo limpo de sangue e suspender levemente a
glândula; nestas condições, com um gancho de estrabismo pequeno se libera a hipófise de
suas envolturas meníngeas, sendo possível remover a glândula integralmente, deixando in
situ o túber cinerium. A trepanação foi tamponada com cera de Horsley e o palato mole foi
suturado com pontos simples separados. Durante o período pós-operatório administrou-se
antibióticos e analgésicos por via parenteral (COPPA et al.,1978) .
Dois acessos à hipófise canina podem ser realizados: intracranial e transbucal. A
menos invasiva é a trans esfenoidal (trans palatina, trans bucal) resultando estes acessos em
aumento na probabilidade de sobrevivência. Uma modificação do acesso trans esfenoidal,
usando a via retrofaríngea para alcançar o basisfenóide assepticamente foi utilizada neste
estudo. Apesar das vantagens do acesso transesfenoidal, é difícil identificar a localização da
hipófise precisamente, a osteotomia inexata pode resultar em hemorragia per-operatória
fatal. A hipofisectomia é considerada como uma técnica impraticável em raças
braquicefálicas. A hipófise canina é de difícil localização, a distância da hipófise com
pontos de referência palpáveis e visíveis tais como os hâmulos do pterigóide e a sutura
interesfenoidal varia com o tamanho e forma do crânio (NIEBAUER; EVANS, 1988).
Enemar (2003) efetuou a hipofisectomia fetal seletiva em ratos, que levou a baixa
taxa de sobrevivência. Como resultado, o desenvolvimento das glândulas tireóide, adrenal e
timo dos fetos foi retardado. Não obstante, o crescimento dos ossos longos não sofreu
alterações após o procedimento cirúrgico. No entanto, a seletividade de tal técnica cirúrgica

Ana Rita de Lima
48
foi questionada pelo próprio autor, uma vez que em alguns casos a adenohipófise não foi
excisada na sua totalidade.
Niebauer et al (1990) descreveram hipofisectomia experimental realizada em sete
cães clinicamente normais. Antes da realização da técnica foi utilizado radioimunoensaio
homólogo específico e as concentrações plasmáticas de hormônio do crescimento foram
mensuradas antes e em intervalos específicos depois da estimulação com clonidina HCl.
Meij et al (1997) observaram que devido à alta taxa de mortalidade associada com o
acesso intracraniano transtemporal, o acesso transesfenoidal para remoção da hipófise
tornou-se a técnica preferida em estudos experimentais em cães. Nenhum dos cães mostrou
resposta aos hormônios da hipófise posterior em dez semanas após a hipofisectomia. Não
ocorreram anormalidades no baço, fígado, coração, rins, ou pâncreas. A próstata e os
testículos estavam atrofiados, a tireóide e paratireóide estavam inativas.

Ana Rita de Lima
49
4 MATERIAL E MÉTODOS
O material e os métodos utilizados nesta pesquisa estão descritos abaixo.
4.1 MATERIAL
Para a realização deste trabalho foram utilizados quatro cães Golden Retriever não
distróficos e oito cães Golden Retriever acometidos pela distrofia muscular, todos do sexo
masculino e com idade variando entre um mês e um ano. No estudo histológico utilizamos
dois animais não distróficos (com um ano de idade) e três animais acometidos por Distrofia
muscular (entre sete meses e um ano de idade) apresentando a forma grave da doença. Estes
mesmos animais foram utilizados para o estudo macro e mesoscópico. Para o estudo
radiológico e tomográfico foram utilizados dois animais sendo um distrófico apresentando
a forma grave da doença e um não distrófico, ambos com um ano de idade. Na dosagem
sérica de IGF-I utilizamos cinco animais, sendo quatro animais Distróficos, inicialmente os
quatro animais apresentavam a forma clínica benigna da doença e posteriormente dois
destes animais evoluíram para a forma grave, e um animal não distrófico. Os animais
distróficos e não distróficos utilizados neste estudo foram obtidos no canil existente na
Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo (FMVZ-
USP), construído em colaboração do Instituto de Biociências da Universidade de São Paulo
para o estudo da Distrofia muscular em cães da raça Golden Retriever.

Ana Rita de Lima
50
4.2 MÉTODOS
Os métodos utilizados estão descritos nos itens que se seguem.
4.2.1 Estudo Macroscópico e Mesoscópico
Logo após o óbito dos animais, ocorrido como conseqüência da progressão da
doença, a hipófise foi fixada, utilizando-se os mesmos fixadores dos itens 4.2.3.1 e 4.2.3.2
sendo perfundida pela artéria carótida comum esquerda. Em seguida foi realizada a
dissecação do osso esfenóide e o acesso trans-esfenoidal para remoção da glândula hipófise
na sua totalidade, após a completa remoção esta foi seccionada em duas partes iguais por
meio de uma incisão mediana. A seguir, a neuro-hipófise foi separada da adenohipófise.
Espessura, comprimento e largura foram mensurados nas duas hemi-adenohipófises com o
uso de um paquímetro digital da marca Digimess®.

Ana Rita de Lima
51
4.2.2 Anatomia da Imagem (Imagologia)
Neste item utilizamos dois animais sendo um distrófico apresentando quadro grave e
um não distrófico que foram fixados em solução aquosa de formol a 10% logo após o óbito,
pelas artérias carótidas comuns direita e esquerda. Os dois animais tiveram a hipófise
dissecada pelo acesso trans-esfenoidal para exposição da mesma e colocação do marcador
(esfera de chumbo com 0,5mm de diâmetro) que foi utilizada para permitir a localização e
diferenciação da hipófise em relação às outras estruturas.
Os exames radiográficos e tomográficos foram realizados no Serviço de Diagnóstico
por Imagem do Hospital Veterinário do Departamento de Cirurgia da Faculdade de
Medicina Veterinária e Zootecnia da Universidade de São Paulo (FMVZ-USP).
4.2.2.1 Equipamento Tomográfico
O exame tomográfico foi realizado em equipamento CT-MAX 640 (General Eletric)
de terceira geração. As imagens foram fotografadas em câmera multiformato MFC640
(General Eletric®), em filmes de marcas MN NIF Agfa IBF Medix (IBF - Indústria
Brasileira de Filmes) e Kodak Ektanscan M (Kodak Brasileira Com. Ind. LTDA) tamanho

Ana Rita de Lima
52
35x43cm os quais foram revelados e fixados em Processadora Automática RPX-OMAT
Processor (Eastman Kodak Company).
4.2.2.2 Técnica Tomográfica
Os animais foram posicionados em decúbito ventral com os membros torácicos
tracionados cranialmente. Realizou-se radiografia digital (“scout”) em incidência
dorsoventral. Os cortes tranversais iniciaram-se a 8mm rostralmente em direção ao
marcador e extenderam-se por 7mm caudalmente.
O ajuste de técnica foi de 120kV e 22mA, com 3 segundos de tempo de exposição. A
espessura dos cortes foi de 5mm com incremento de 5mm entre os cortes. As imagens
adquiridas foram fotografadas em câmera multiformato MFC640 (General Eletric®), com
seleções de janela e nível que permitissem adequada avaliação de arcabouço ósseo para
localização da região hipofisária, onde a mesma encontrava-se marcada com uma esfera de
chumbo (0,5mm de diâmetro). Foi realizado um corte de 2mm de espessura exatamente em
cima do marcador. As imagens tomográficas foram realizadas com janelas e nível que
permitissem adequada avaliação de estruturas ósseas e partes moles.

Ana Rita de Lima
53
4.2.2.3 Equipamento Radiográfico
O exame radiográfico foi realizado em aparelho de raios-X da marca Ray-tec modelo
RT de 500mA e 125Kv, foram utilizados chassis de 18 x 24cm, com écrans intensificadores
Kodak Lanex X-Omatic Regular Screens (Kodak Eastman Company) e filmes radiológicos
no tamanho 18 x 24cm, RP-X-OMAT (Kodak Brás. Com. E Ind. Ltda). As radiografias
foram reveladas em Processadora Automática RPX-OMAT Processor (Eastman Kodak
Company).
4.2.2.4 Técnica Radiográfica
Os animais foram posicionados em decúbito ventral com os membros torácicos
tracionados cranialmente. Realizou-se radiografias convencionais com incidência dorso-
ventral o ajuste de técnica utilizado foi de 40 Kv e 100 mA.

Ana Rita de Lima
54
4.2.3 Estudo Histológico
Este capítulo foi dividido em vários tópicos para facilitar o entendimento do leitor.
4.2.3.1 Aspectos Microestruturais
A fixação neste caso consistiu de perfusão pela artéria carótida comum esquerda com
uma solução de Karnovsky modificada (5% glutaraldeído + 1% formoldeído em tampão
cacodilado de sódio - 0,125 M; pH 7,4) e imersão no mesmo fixador por 72 horas, antes de
realizar a fixação fizemos a lavagem do sistema com o uso de uma solução composta por
tampão fosfato, nitrito de sódio a 0,1% e 2% de heparina. O material foi lavado em solução
de tampão cacodilato de sódio, pós-fixado em solução de tetróxido de Ósmio a 2%
(Sigma®) em tampão cacodilato de sódio 0,1M por 90 minutos à temperatura ambiente. O
material sofreu outra lavagem em tampão cacodilato de sódio e água destilada por três
vezes e foi contrastado com solução saturada de acetato de uranila (Reagen®) por 90
minutos à temperatura ambiente e abrigado da luz.
As amostras foram desidratadas em concentrações crescentes de etanol (50% 70%
80% 90% 100%), logo após a desidratação o material foi submetido a dois banhos em
solução de Óxido de propileno (Aldrich®) puro e então incluído em uma solução de óxido
de propileno e Araldite 502 (Polyscience Inc.®), na proporção 9:1, e mantido sobre agitação

Ana Rita de Lima
55
constante por 10 minutos. Decorrido este período o material foi colocado em óxido de
propileno e resina Araldite (1:1) por 1 hora sob agitação constante, em seguida foi realizada
a troca de concentração de óxido propileno e Araldite na proporção (1:9) mantendo o
material sob mesma condição por 12 horas e posteriormente colocado em Araldite pura por
8 horas. O material foi colocado em nova solução pura de Araldite por 12 horas e
finalmente em resina pura por 2 a 3 dias em estufa (60º a 70ºC), sendo acondicionado em
moldes de silicone específicos para este procedimento.
Após a embebição, procedeu-se a trimagem do bloco e à seguir o material foi cortado
em um ultramicrótomo da marca RMC modelo MT- RL com 2 µm de espessura (secções
semi-finas) e secções ultra-finas (70-90 nm de espessura). Os cortes foram coletados em
lâminas de vidro, secos em placa térmica (Leica®), corados com solução de azul de
toluidina alcoólica (1%) e montados sob lamínula.
Para a microscopia eletrônica de transmissão seguimos o mesmo protocolo acima,
efetuando-se porém cortes ultra-finos com 70nm - 90 nm de espessura que foram colocados
em grids de cobre de 200 mesh, corados com acetato de uranila 3% e citrato de chumbo e
observados ao microscópio eletrônico de transmissão (JEOL Electron Microscopic® JEM-
1010) no Instituto de Ciências Biomédicas na Universidade de São Paulo (ICB-USP).

Ana Rita de Lima
56
4.2.3.2 Aspectos Morfoquantitativos
A adenohipófise (Pars proximalis e Pars distalis) foi cortada seriadamente e
exaustivamente (2 µm de espessura), de sorte que cada qüinquagésima secção (K=50) foi
amostrada juntamente com vinte outras secções vizinhas. As secções foram observadas e
analisadas em microscópio Leica® DMR com uso de software de Análise de Imagem Leica
QWin (Leica Microsystem Imaging Solution Ltd, Cambridge,UK). Após aplicação dos
planos de inclusão e exclusão tridimensionalmente, dezessete secções foram consideradas
para cada região. Portanto, a partir de dezessete secções seriadas com 2µm de espessura,
uma área (por região) foi escolhida sistematicamente e aleatoriamente (SURS) para
promover a análise morfométrica e estereológica. Um sistema teste composto por seis áreas
testes com área total de 56428µm² foi colocado sobre a imagem da secção histológica
captada e projetada na tela de um computador por meio da câmera digital Leica® DC 300 F
e todas as células granulares que estavam dentro das áreas testes (e que não tocavam as
linhas de exclusão) foram consideradas para o estudo quantitativo.
Os seguintes parâmetros foram estudados nas células granulares adenohipofisárias:
Eixo longo;
Área seccional;
Volume celular estimado globalmente pelo Princípio de Cavalieri;

Ana Rita de Lima
57
“Profile or Packing density” (densidade de perfis celulares – DP) estimado pela
seguinte fórmula:
DP = ∑ profiles / área total amostrada (HOWARD e REED, 2004).
Neste caso foi utilizado um sistema teste com seis frames, sendo a área total destas
frames de 56428µm².
Para a realização desta etapa foram mensurados o seguinte número de células para
cada região de cada animal (Tabela 1).
Tabela 1 - Número de células granulares adenohipofisárias (Pars proximalis e Pars distalis) de cães Golden Retriever distróficos e não distróficos utilizados no estudo estereológico, São Paulo – 2005
Número de células por região
Animal Estado Pars proximalis Pars distalis
01 Distrófico 22 89
02 Não Distrófico 31 254
03 Distrófico 34 440
04 Não Distrófico 8 369
05 Distrófico 34 230

Ana Rita de Lima
58
Tamanho médio dos grânulos das células adenohipofisárias
As mensurações dos grânulos foram realizados à microscopia eletrônica de
transmissão, com o uso de imagens capturadas no microscópio eletrônico da marca JEOL
uma área (por região) foi escolhida sistematicamente e aleatoriamente (SURS) com auxílio
de sistema teste para microscopia eletrônica de transmissão para promover a análise
quantitativa. Um sistema teste composto por seis áreas testes com área total de 6981µm² foi
colocado sobre a imagem projetada na tela de um computador e todos os grânulos elétron-
densos vistos em secção transversal e que estavam dentro das áreas testes (não tocavando as
linhas de exclusão) foram considerados. Desta forma, com o uso do software de Análise de
Imagem Leica QWin (Leica Microsystem Imaging Solution Ltd, Cambridge,UK)
realizamos a mensuração dos grânulos em seu eixo longo e área seccional.
4.2.4 Aspectos Neuroendócrinos
Amostras mensais (durante quatro meses consecutivos) foram coletadas de animais
distróficos (apresentando a forma benigna da doença) e de um animal não distrófico
(animal controle), filhotes machos e de mesma idade (a partir de um mês de idade). No
sétimo mês foi coletada outra amostra dos mesmos animais, sendo que dois animais
distróficos evoluíram para a forma grave da doença e dois permaneceram com a forma
benígna. Devido à interferência nutricional na dosagem sérica de IGF-I (SHALET et al;
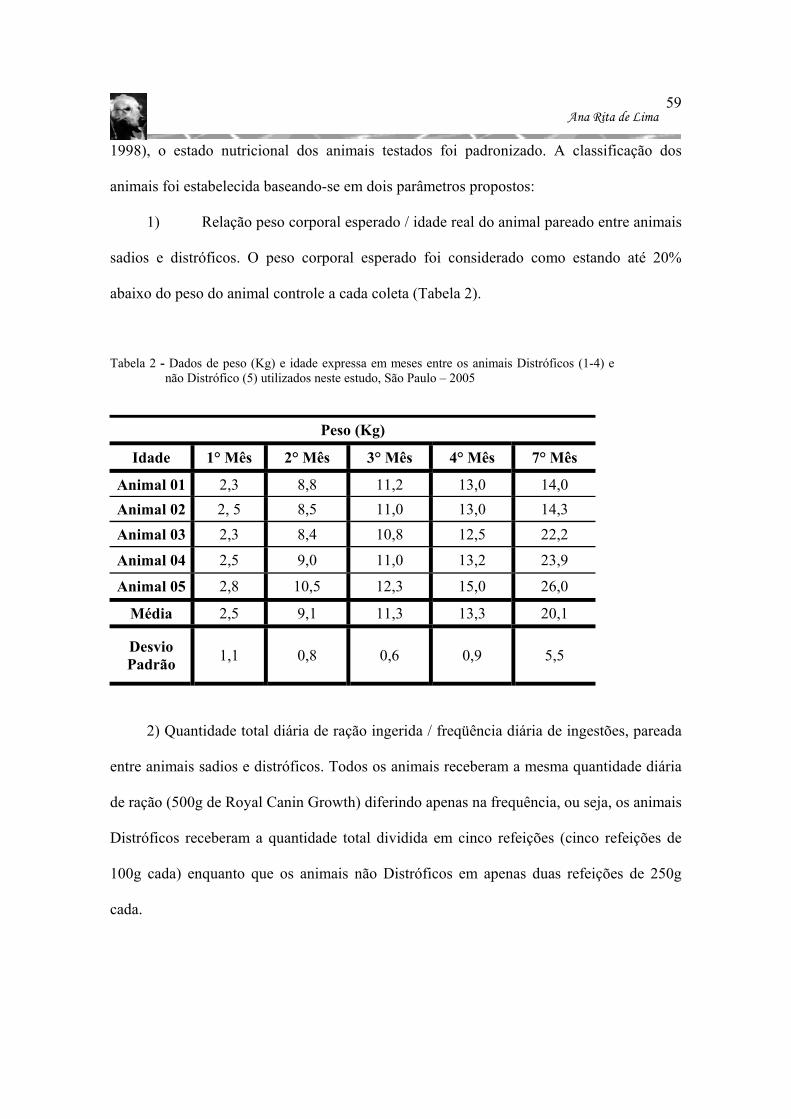
Ana Rita de Lima
59
1998), o estado nutricional dos animais testados foi padronizado. A classificação dos
animais foi estabelecida baseando-se em dois parâmetros propostos:
1) Relação peso corporal esperado / idade real do animal pareado entre animais
sadios e distróficos. O peso corporal esperado foi considerado como estando até 20%
abaixo do peso do animal controle a cada coleta (Tabela 2).
Tabela 2 - Dados de peso (Kg) e idade expressa em meses entre os animais Distróficos (1-4) e não Distrófico (5) utilizados neste estudo, São Paulo – 2005
Peso (Kg)
Idade 1° Mês 2° Mês 3° Mês 4° Mês 7° Mês
Animal 01 2,3 8,8 11,2 13,0 14,0 Animal 02 2, 5 8,5 11,0 13,0 14,3 Animal 03 2,3 8,4 10,8 12,5 22,2
Animal 04 2,5 9,0 11,0 13,2 23,9
Animal 05 2,8 10,5 12,3 15,0 26,0
Média 2,5 9,1 11,3 13,3 20,1
Desvio Padrão
1,1
0,8
0,6
0,9
5,5
2) Quantidade total diária de ração ingerida / freqüência diária de ingestões, pareada
entre animais sadios e distróficos. Todos os animais receberam a mesma quantidade diária
de ração (500g de Royal Canin Growth) diferindo apenas na frequência, ou seja, os animais
Distróficos receberam a quantidade total dividida em cinco refeições (cinco refeições de
100g cada) enquanto que os animais não Distróficos em apenas duas refeições de 250g
cada.

Ana Rita de Lima
60
Desta forma, a classificação do estado nutricional adotada foi:
Adequado - foram classificados como tendo estado nutricional adequado aqueles
animais que possuíam até 80% do peso do animal controle (não Distrófico).
Inadequado - foram classificados como tendo estado nutricional inadequado aqueles
animais que possuíam menos de 80% do peso do animal controle (não Distrófico).
As amostras de sangue foram coletadas em tubos sem anti-coagulante, e
centrifugados a 4.000 rpm durante 15 minutos sendo separado o soro e criopreservadas à –
20°C até sua dosagem (AGUSTSSON et al. 2000; BOQUETE et al., 2003; CORDIDO et
al., 2003; LISSETT et al., 2003; SHALET et al., 1999).
As dosagens foram efetuadas no Laboratório de Hormônios e Genética Molecular
Lim/42 da Disciplina de Endocrinologia e Metabologia do Hospital das Clínicas de São
Paulo da Faculdade de Medicina da Universidade de São Paulo, utilizando ensaio
Imunorradiométrico (IRMA) (Diagnostic Systems Laboratories – DSL-5600®, Webster,
USA). Este ensaio inclui uma etapa de extração simples na qual o IGF-I é separado das
suas proteínas de ligação no soro (IGFBP3 e ALS) através de acidificação seguida de
precipitação por etanol da fração de IGFBPs sendo o sobrenadante submetido a dosagem
hormonal. O IRMA é um ensaio não competitivo no qual a substância a ser determinada é
mantida ligada em dois anticorpos; o primeiro anticorpo está imobilizado nas paredes
internas dos tubos, o outro anticorpo é radiomarcado para a sua detecção. A substância a ser
determinada presente nas amostras (IGF-I) liga-se em ambos os anticorpos e os materiais

Ana Rita de Lima
61
não ligados são removidos por decantação e lavagem dos tubos, seguida de leitura dos
resultados obtidos (Figura 1).
As dosagens de testosterona foram realizadas utilizando-se um sistema automático
de imunoensaio com o kit AutoDELFIA®, este ensaio possui uma fase sólida de
fluoroimunoensaio baseado na competição entre europium marcado com testosterona e
amostras de testosterona para anticorpos policlonais anti-testosterona (originários de
coelhos). O agente bloqueador no ensaio de testosterona facilita a liberação da testosterona
das proteínas ligadoras, um segundo anticorpo, diretamente ligado a IgG de coelho, está
revestido na fase sólida, e a ligação do complexo IgG-testosterona promove uma
conveniente separação entre o anticorpo e o antígeno livre. A dissociação dos íons de
europium da testosterona ligada dentro da solução forma quelatos altamente fluorescentes
com os componentes da solução realçada. A fluorescência em cada um é mensurada, sendo
inversamente proporcional a concentração de testosterona da amostra (Figura 2).

Ana Rita de Lima
62
Figura 1- Esquema do ensaio Imunorradiométrico (IRMA) realizado para a dosagem de IGF-I. FONTE: www.portfolio.mvm.ed.ac.uk/.../ radidia.htm
Figura 2- Esquema do imunoensaio utilizado para a dosagem de testosterona

Ana Rita de Lima
63
4.2.5 Análise Estatística
Nas análises estatísticas considerou-se diferentes modelos de acordo com a natureza
das variáveis mensuradas. Para análises de dados paramétricos e de distribuição contínua
(área, volume e eixo para microscopia de luz de secções semi-finas e eixo e área à
microscopia eletrônica de transmissão), o modelo contemplou os efeitos dos grupos
comparativos (animais distróficos versus não-distróficos), segundo o Método de Quadrados
Mínimos por meio do procedimento PROC GLM do programa Statistical Analysis System,
versão 8.02 (SAS, 1995) . Para as variáveis de contagens celulares, o modelo contemplou
os mesmos efeitos dos grupos comparativos (animais ditróficos versus não-distróficos),
porém utilizando-se a metodologia de Modelos Lineares Generalizados, pressupondo
distribuição de Poisson com função de ligação logarítmica, por meio do procedimento
PROC GENMOD do programa Statistical Analysis System, versão 8.02 (SAS, 1995).
Para as dosagens de IGF-I, o modelo estatístico considerou os efeitos principais de
meses de idade (de um até quatro meses e sete meses), dos grupos comparativos (animais
distróficos versus não distróficos). Estas análises foram realizadas utilizando-se o teste t de
Student como procedimento de comparações múltiplas, onde utilizou-se a média amostral
dos animais distróficos comparando-se com o valor real obtido a cada mês do animal
controle. No sétimo mês comparou-se a média dos animais distróficos graves com o valor
real do animal não distrófico e a média dos animais distróficos benignos com o valor real
do animal não distrófico.

Ana Rita de Lima
64
4.3 ESTUDO PILOTO (ANATOMIA CIRÚRGICA)
Realizou-se estudos pilotos em dois cães S.R.D. com peso de 15Kg para a remoção
total da hipófise. Para a técnica anestésica utilizada empregou-se a aplicação de
Acepromazina (0,1mg/kg - IV) como medicação pré-anestésica, Midazolam (0,5mg/kg)
associado a Ketamina (5mg/kg) - IV como agentes de indução. Os animais foram então
entubados e mantidos à ventilação assistida em plano anestésico empregando-se Halotano.
O acesso cirúrgico realizado foi o transesfenoidal, o animal estava posicionado em
decúbito ventral com a cavidade oral aberta para o acesso ao palato mole que foi incisado
em sua linha média, após a incisão no palato mole localizamos os hâmulos do osso
pterigóide e traçando uma linha imaginária entre estas estruturas localizamos a posição da
hipófise no centro desta linha. Com o uso de uma broca o basisfenóide foi aberto até a
localização da hipófise que foi removida com o uso de pinça e tesoura oftálmica. Logo após
o procedimento cirúrgico ambos animais foram ortoeutanaziados mediante overdose do
mesmo anestésico usado na manutenção cirúrgica.

Ana Rita de Lima
65
5 RESULTADOS
Os resultados obtidos estão descritos nos itens deste capítulo.
5.1 ESTUDO MACROSCÓPICO E MESOSCÓPICO
No Golden Retriever a hipófise está localizada na Sella turcica do osso basisfenóide,
aderida ao túber cinerium por meio do infundíbulo hipofisário. Rostralmente à mesma está
o quiasma óptico, lateralmente os Tractos ópticos, e caudalmente observam-se os
pedúnculos cerebrais e os corpos mamilares (Figura 3). Um revestimento de dura-máter
envolve a glândula e também o teto da depressão. Macroscopicamente está composta pela
Adenohipófise que se divide em Pars distalis, Pars intermédia e Pars tuberalis e pela
Neuro-hipófise e foi possível isolarmos a Adeno da Neuro-hipófise. No material à fresco, a
Adenohipófise apresenta coloração amarelo-rosada e envolve a Neuro-hipófise látero-
rostro-caudalmente. A Neuro-hipófise possui coloração branco - amarelada. Os aspectos
macromorfométricos das adenohipófises estudadas estão sumarizados na Tabela 3. Estes
resultados foram não significativos quando analisados pelo teste t (p > 0,05) (Tabela 4).
Macroscopicamente não observamos diferenças entre os animais distróficos e não
distróficos quanto à coloração e localização da mesma.

Ana Rita de Lima
66
Tabela 3 - Dados sumarizados de comprimento, largura e espessura das adenohipófises dos cães Golden Retriever Distróficos e não Distróficos. São Paulo – 2005
Tabela 4 - Estatísticas descritivas para as variáveis avaliadas (comprimento, largura e espessura) das adenohipófises estudadas, São Paulo – 2005
Mínimo Máximo Média Desvio Padrão
Coeficiente de Variação
Comprimento Pars
proximalis
0,69 1,77 1,01 0,44 43,41
Comprimento Pars distalis
4,79 6,23 5,74 0,58 10,15
Largura 5,92 6,85 6,33 0,38 6,10
Espessura 3,77 4,54 4,12 0,37 8,97
Comprimento (mm)
Pars proximalis Pars distalis Largura (mm) Espessura (mm)
Animal 01 Distrófico
Grave 1,77 4,79 6,85 4,54
Animal 02 Não Distrófico 1,05 5,97 6,30 4,49
Animal 03 Distrófico
Grave 0,75 6,13 6,02 3,77
Animal 04 Não Distrófico 0,69 6,23 5,92 3,80
Animal 05 Distrófico
Grave 0,83 5,62 6,58 4,02
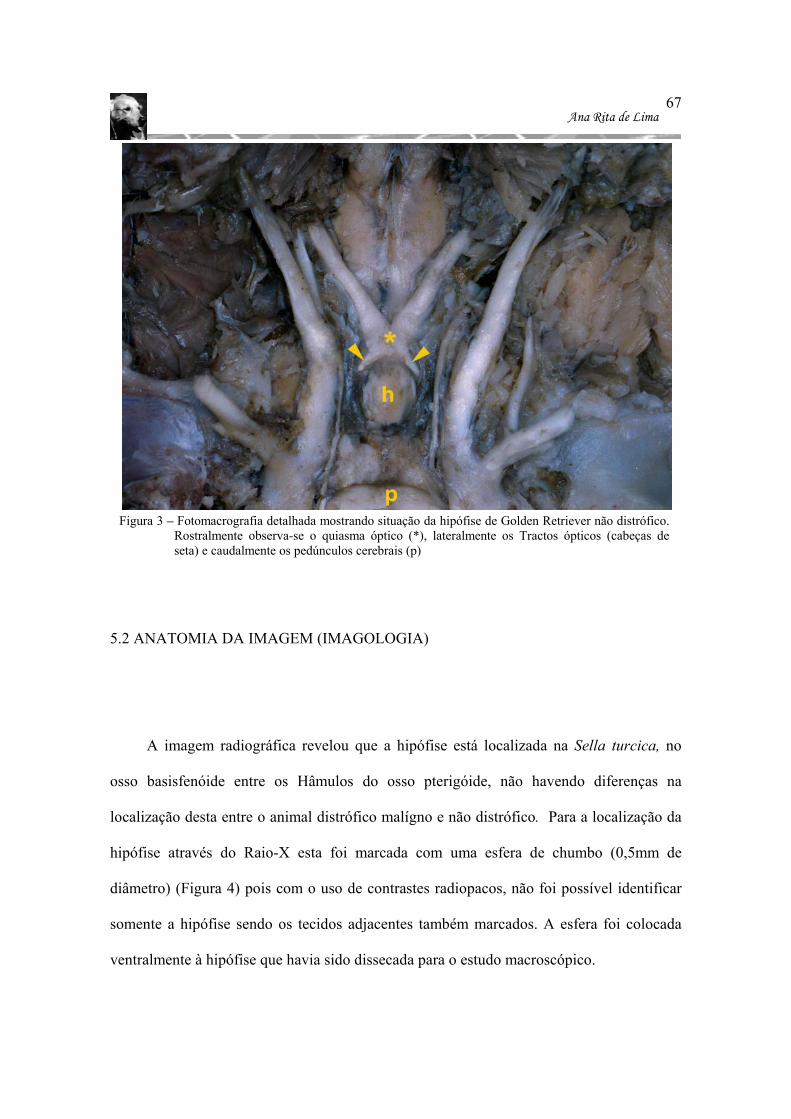
Ana Rita de Lima
67
Figura 3 – Fotomacrografia detalhada mostrando situação da hipófise de Golden Retriever não distrófico. Rostralmente observa-se o quiasma óptico (*), lateralmente os Tractos ópticos (cabeças de seta) e caudalmente os pedúnculos cerebrais (p)
5.2 ANATOMIA DA IMAGEM (IMAGOLOGIA)
A imagem radiográfica revelou que a hipófise está localizada na Sella turcica, no
osso basisfenóide entre os Hâmulos do osso pterigóide, não havendo diferenças na
localização desta entre o animal distrófico malígno e não distrófico. Para a localização da
hipófise através do Raio-X esta foi marcada com uma esfera de chumbo (0,5mm de
diâmetro) (Figura 4) pois com o uso de contrastes radiopacos, não foi possível identificar
somente a hipófise sendo os tecidos adjacentes também marcados. A esfera foi colocada
ventralmente à hipófise que havia sido dissecada para o estudo macroscópico.

Ana Rita de Lima
68
Figura 4 – Fotografia de imagem radiográfica de crânio de um Golden Retriever não distrófico. A seta indica a localização da hipófise que foi marcada com uma esfera de chumbo
A imagem tomográfica mostrou que a hipófise está localizada dentro da Sella
turcica (no osso basisfenóide), entre as bulas timpânicas e rostral ao conduto auditivo, não
ocorrendo diferenças entre o animal distrófico grave e o não distrófico (Figura 5 A e B). A
hipófise foi marcada com uma esfera de chumbo que foi colocada após a dissecação da
mesma.

Ana Rita de Lima
69
Figura 5 - 5 A - Imagem tomográfica de cão Golden Retriever Distrófico onde podemos observar a Hipófise (H), parte do Osso Basisfenóide (X) que foi removido na dissecação, Cavidade Timpânica (*), Bula Timpânica (B) e o Processo zigomático do osso temporal (T). 5 B - Imagem tomográfica de cão Golden Retriever Não Distrófico, podemos observar a Hipófise (H), o Osso Basisfenóide (X), o Processo zigomático do osso temporal (T) e o Processo condilar da mandíbula (M)
A
B

Ana Rita de Lima
70
5.3 ESTUDO HISTOLÓGICO
5.3.1 Aspectos Microestruturais
As células granulares, estão distribuídas homogeneamente na Pars proximalis e
distalis na adenohipófise de todos os animais, formando grupos (média de 7 células)
próximos aos vasos sanguíneos e às vezes aparecendo isoladas (Figura 6 A, B, C e D).
Estas células variaram na forma, sendo normalmente esféricas ou ovaladas.

Ana Rita de Lima
71
Figura 6 – Fotomicrografias demonstrando as células granulares (g) adenohipofisárias e vasos sanguíneos
(seta) em um cão Golden Retriever distrófico (A e C) e não distrófico (B e D). Azul de toluidina. Escala de barra: 20µm (em A e B) e Escala de barra: 5µm (em C e D)
A B
C D
g
g
g
g
g g

Ana Rita de Lima
72
5.3.2 Aspectos Morfoquantitativos
Nos animais distróficos graves, as células granulares da Pars proximalis apresentam
um eixo longo entre 4,6 e 14,4µm (n=90, média de 7,7µm e desvio padrão de 2,2) enquanto
que nos animais controle (não distróficos) o eixo longo variou entre 3,1 e 11,1µm (n=39,
média de 6,5µm e desvio padrão de 1,9). Em relação à área seccional destas células os
animais distróficos apresentaram uma área entre 34,5 e 121,7µm² (n=90, média de 66,3µm²
e desvio padrão de 18,7) e os animais controle de 15,1 e 116,1µm² (n=39, média de
50,4µm² e desvio padrão de 21,9). O volume das células granulares variou entre 337,5 e
1977,1µm³ (n=90, média 747,6µm³ e desvio padrão de 300) nos animais distróficos e
variou de 178,4 a 986,9µm³ (n=39, média 526,3µm³ e desvio padrão de 208) nos animais
não distróficos (Tabela 5).
O eixo longo das células granulares da Pars distalis nos animais distróficos variou de
3,5 a 14,7µm (n=759, média de 7,1µm e desvio padrão de 1,5), a área seccional variou de
18,6 a 135,2µm² (n=759, média de 64,1µm² e desvio padrão de 18,8). Nos animais controle
o eixo longo das células granulares da Pars distalis variou de 2,1 a 9,8µm (n=623, média
de 5,8µm com desvio padrão de 1,2) e a área seccional variou de 17,5 a 119,5µm² (n=623,
média de 47,1µm² e desvio padrão de 14,4) (Tabela 5). O volume das células granulares na
Pars distalis variou de 79,7 e 1574,8µm³ (n=759, média 702,1µm³ e desvio padrão de 239)
para os animais distróficos e de 141,2 a 1359,6µm³ (n=623, média 559,9µm³ e desvio
padrão de 203) para os animais controle (Tabela 5).
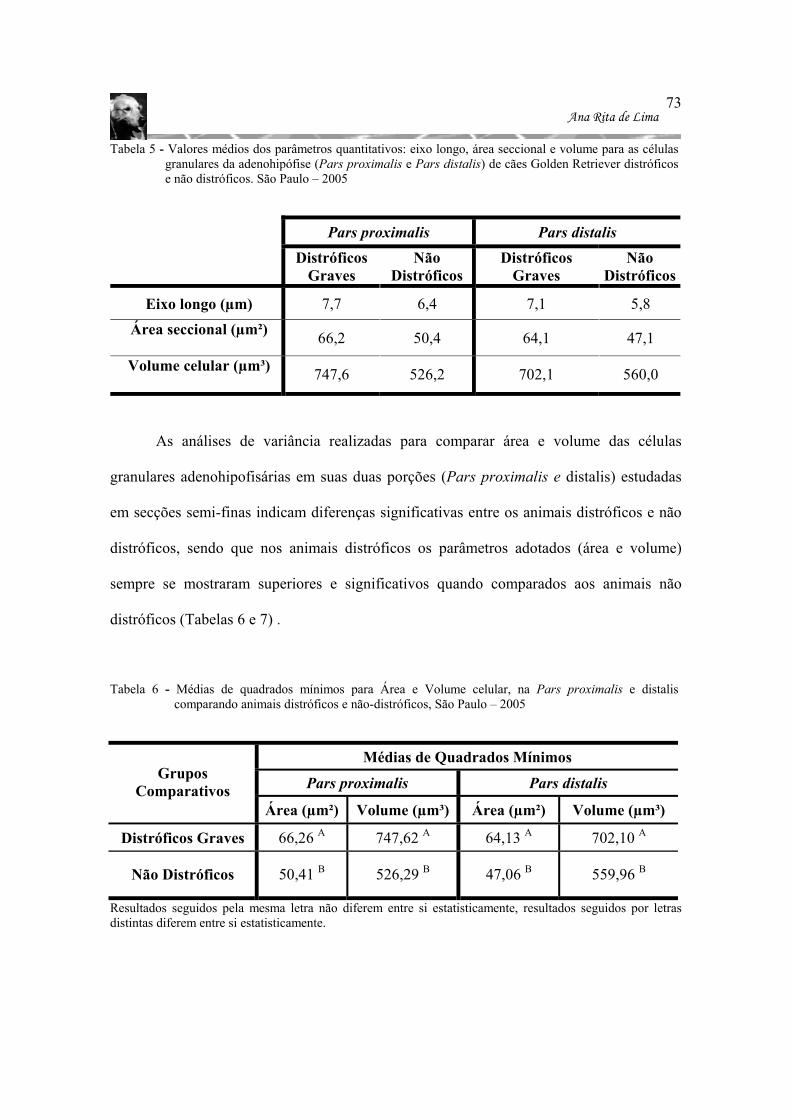
Ana Rita de Lima
73
Tabela 5 - Valores médios dos parâmetros quantitativos: eixo longo, área seccional e volume para as células granulares da adenohipófise (Pars proximalis e Pars distalis) de cães Golden Retriever distróficos e não distróficos. São Paulo – 2005
Pars proximalis Pars distalis
Distróficos Graves
Não Distróficos
Distróficos Graves
Não Distróficos
Eixo longo (µm) 7,7 6,4 7,1 5,8
Área seccional (µm²) 66,2 50,4 64,1 47,1
Volume celular (µm³) 747,6 526,2 702,1 560,0
As análises de variância realizadas para comparar área e volume das células
granulares adenohipofisárias em suas duas porções (Pars proximalis e distalis) estudadas
em secções semi-finas indicam diferenças significativas entre os animais distróficos e não
distróficos, sendo que nos animais distróficos os parâmetros adotados (área e volume)
sempre se mostraram superiores e significativos quando comparados aos animais não
distróficos (Tabelas 6 e 7) .
Tabela 6 - Médias de quadrados mínimos para Área e Volume celular, na Pars proximalis e distalis comparando animais distróficos e não-distróficos, São Paulo – 2005
Médias de Quadrados Mínimos
Pars proximalis Pars distalis Grupos Comparativos
Área (µm²) Volume (µm³) Área (µm²) Volume (µm³)
Distróficos Graves 66,26 A 747,62 A 64,13 A 702,10 A
Não Distróficos 50,41 B 526,29 B 47,06 B 559,96 B
Resultados seguidos pela mesma letra não diferem entre si estatisticamente, resultados seguidos por letras distintas diferem entre si estatisticamente.

Ana Rita de Lima
74
Tabela 7- Resumo das análises de variância das células granulares adenohipofisárias (Área e volume celular na Pars proximalis e distalis), segundo os grupos comparativos. São Paulo, 2005
Quadrados Médios Pars proximalis Pars distalis Fontes de
variação GL Área (µm²) Volume (µm³) Área (µm²) Volume (µm³)Grupo
Comparativo 1 6831,68** 1333001,11** 100061,13** 6941354,00** Resíduo 127 388,47 76172,54 287,34 50067,33
Resultados seguidos pelo mesmo número de asteriscos não diferem entre si estatisticamente, resultados seguidos por números distintos de asteriscos diferem entre si estatisticamente.
O número de células por área (profile density) variou de 3,5x10-5 a 3,2x10-4 na Pars
proximalis dos animais Distróficos (média 2,0x10-4, desvio padrão 9,6x10-5, coeficiente de
variação 48%) e de 3,5x10-5 a 3,7x10-4 nos animais não Distróficos (média 1,7x10-4, desvio
padrão 1,5x10-4, coeficiente de variação 88%). Na Pars distalis variou de 8,8x10-5 a
4,4x10-4 (média 4,4x10-4, desvio padrão 7,2x10-5, coeficiente de variação 16,3%) nos
animais Distróficos e de 3,5x10-5 a 3,7x10-4 (média 1,7x10-4, desvio padrão 1,5x10-4,
coeficiente de variação 88%) nos animais não distróficos (Tabela 8). Em relação ao número
de células estudadas a análise de variância revelou resultados não significativos (p > 0,01)
indicando que não existem diferenças entre os animais distróficos e não distróficos (Tabelas
9 e 10).

Ana Rita de Lima
75
Tabela 8- Dados sumarizados da densidade de perfis celulares da Pars proximalis e na Pars distalis nos animais Distróficos e não Distróficos. São Paulo – 2005
Pars proximalis Pars distalis
Distróficos Não Distróficos Distróficos Não
Distróficos Mínimo 8,8x10-5 3,5x10-5 3,5x10-5 3,5x10-5 Máximo 4,4x10-4 3,7x10-4 3,2x10-4 3,7x10-4 Média 2,5x10-4 1,7x10-4 2,0x10-4 1,7x10-4
Desvio padrão 7,1x10-5 1,4x10-4 9,6x10-5 1,5x10-4 Coeficiente de
variação 5,1x10-9 9,8x10-9 9,3x10-9 2,1x10-8
Tabela 9 - Resumo das análises de variância das células granulares adenohipofisárias na Pars proximalis, Pars distalis para a densidade de perfis celulares. São Paulo – 2005
Fontes de Variação QUI-QUADRADO
GL
Pars proximalis Pars distalis
Grupo Comparativo 1 0,57 NS 1,18 NS
GL – Grau de Liberdade NS – Resultado não significativo
Tabela 10 - Médias de quadrados mínimos para a Pars proximalis e Pars distalis para a densidade de perfis celulares. São Paulo – 2005
Médias de Quadrados Mínimos Grupos Comparativos
Pars proximalis Pars distalis
DISTRÓFICOS 11,25 A 14,06 A
NÃO DISTRÓFICOS 9,75 A 13,26 A
Resultados seguidos pela mesma letra não diferem entre si estatisticamente, resultados seguidos por letras distintas diferem entre si estatisticamente.

Ana Rita de Lima
76
À microscopia eletrônica de transmissão as células apresentaram grânulos elétron-
densos distribuídos por todo o citosol (Figura 7 A e B; Figura 8 A, B, C e D) apresentando
parede dupla e eixo longo entre 67 e 363nm (n=187, média de 175nm e desvio padrão de
69) na Pars proximalis da adenohipófise e eixo longo entre 102 e 448 (n= 145, média de
218nm e desvio padrão de 74) na Pars distalis nos animais distróficos. Nos animais não
distróficos eixo longo entre 47 e 293nm (n=78, média de 130nm e desvio padrão de 46) na
Pars proximalis da adenohipófise e eixo longo entre 64 e 329nm (n=198, média de 163nm
e desvio padrão de 47) na Pars distalis (Tabela 11).
Em relação à área dos grânulos os animais não distróficos apresentaram área entre 51
e 5684nm2 (n=78, média de 1664nm2 e desvio padrão de 1003) na Pars proximalis da
adenohipófise, na Pars distalis a área variou entre 833 e 6998nm2 (n=198, média de
2730nm2 e desvio padrão de 1363). Nos animais distróficos a área dos grânulos na Pars
proximalis variou entre 69 e 10392 nm2 (n=187, média de 2997 nm2 e desvio padrão de
2042) e na Pars distalis entre 1241 e 12448 nm2 (n= 145, média de 4344 nm2 e desvio
padrão de 2164) (Tabela 11).
Tabela 11 - Valores médios para eixo longo e área dos grânulos das células adenohipofisárias (Pars proximalis e Pars distalis) em cães Golden Retriever distróficos e não distróficos. São Paulo – 2005
Pars proximalis Pars distalis
Distróficos Não Distróficos Distróficos Não
Distróficos
Eixo longo (nm) 175 130 218 163
Área seccional (nm²) 2997 1664 4344 2730
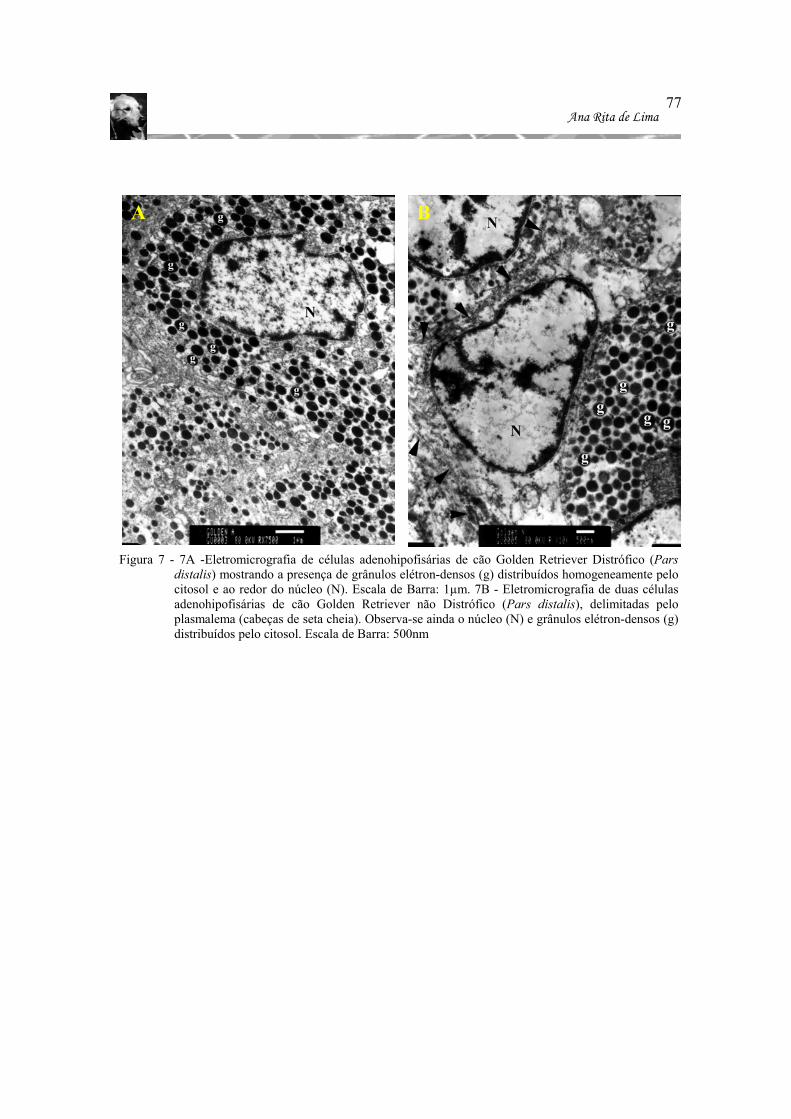
Ana Rita de Lima
77
A A B
Figura 7 - 7A -Eletromicrografia de células adenohipofisárias de cão Golden Retriever Distrófico (Pars distalis) mostrando a presença de grânulos elétron-densos (g) distribuídos homogeneamente pelo citosol e ao redor do núcleo (N). Escala de Barra: 1µm. 7B - Eletromicrografia de duas células adenohipofisárias de cão Golden Retriever não Distrófico (Pars distalis), delimitadas pelo plasmalema (cabeças de seta cheia). Observa-se ainda o núcleo (N) e grânulos elétron-densos (g) distribuídos pelo citosol. Escala de Barra: 500nm

Ana Rita de Lima
78
Figura 8 - Eletromicrografias mostrando células adenohipofisárias (Pars proximalis) de cão Golden Retriever
Distrófico (A) e Golden Retriever não Distrófico (B) evidenciando grânulos elétron-densos (g) e presença do núcleo (N). Eletromicrografias de células adenohipofisárias (Pars distalis) de cão Golden Retriever Distrófico (C) e Golden Retriever não Distrófico (D) mostrando grânulos elétron-densos (g) ao redor do núcleo (N). Nota-se em B e D grânulos elétron-densos formados por parede dupla (cabeças de seta brancas). As cabeças de setas pretas indicam os limites do plasmalema externo. Escala de Barra: 200nm
A B
C D

Ana Rita de Lima
79
Á microscopia eletrônica de transmissão, as análises de variâncias realizadas com o
uso das variáveis: número de grânulos mensurados - N, média, desvio padrão - DP,
coeficiente de variação - CV, valores mínimo e máximo dos grânulos para eixo longo e área
(Tabela 12) revelaram que as dimensões dos grânulos elétron-densos nas células
adenohipofisárias em suas duas porções (Pars proximalis e distalis) mostraram diferenças
significativas (p< 0,01) no teste de quadrados mínimos entre os animais distróficos e não
distróficos, sendo que nos animais distróficos os parâmetros (eixo longo e área seccional)
apresentaram valores maiores quando comparados com os animais não distróficos (Tabelas
13 e 14).
Tabela 12 - Estatísticas descritivas para as variáveis avaliadas (eixo longo e área) dos grânulos elétron–densos de células adenohipofisárias de cães Golden Retriever Distróficos e Golden Retriever não Distrófico, São Paulo – 2005
Variável N Mínimo Máximo Média DP CV (%)
Eixo(nm) 269 0,48 3,63 1,6 0,7 41,2 Pars proximalis Área (nm²) 269 5,12 103,92 26,0 18,9 72,9
Eixo(nm) 347 0,65 4,48 1,8 0,6 35,4 Pars distalis Área (nm²) 347 8,33 124,49 34,1 19,2 56,2
N – número total de grânulos mensurados DP - Desvio Padrão CV - Coeficiente de Variação

Ana Rita de Lima
80
Tabela 13 - Resumo das análises de variância para Eixo e Área dos grânulos elétron-densos das células
adenohipofisárias na Pars proximalis e Pars distalis, segundo os grupos comparativos, São Paulo – 2005
Pars proximalis Pars distalis Quadrados Médios Quadrados Médios
Fontes de Variação GL
Eixo(nm) Área(nm²)GL
Eixo(nm) Área(nm²) Grupo
Comparativo 1 11,24 ** 9981,48 ** 1 25,72 ** 22097,69 **
Resíduo 267 0,41 323,56 345 0,36 305,54 Resultados seguidos pelo mesmo número de asteriscos não diferem entre si estatisticamente, resultados seguidos por números distintos de asteriscos diferem entre si estatisticamente.
Tabela 14 - Médias de quadrados mínimos para Eixos e Áreas dos grânulos das células adenohipofisárias (Pars proximalis e distalis), São Paulo – 2005
Pars proximalis Pars distalis Grupos Comparativos Eixo(nm) Área(nm²) Eixo(nm) Área(nm²)
Distróficos Graves 1,76 A 29,98 A 2,19 A 43,45 A
Não distróficos 1,31 B 16,65 B 1,64 B 27,30 B
Resultados seguidos pela mesma letra não diferem entre si estatisticamente, resultados seguidos por letras distintas diferem entre si estatisticamente.
5.4 ASPECTOS NEUROENDÓCRINOS
As coletas foram realizadas em quatro cães Golden Retriever com distrofia muscular
na fase benígna e em um cão Golden Retriever não distrófico, todos jovens, machos e com
estado nutricional adequado. As amostras foram obtidas durante os quatro primeiros meses
de vida sendo realizada uma coleta por mês. No sétimo mês foi coletada outra amostra dos

Ana Rita de Lima
81
mesmos animais, sendo que dois animais distróficos evoluíram para a forma grave da
doença e os outros permaneceram com a forma benígna. A relação dos animais e resultados
dos testes de IGF-I e testosterona estão sumarizados na Tabela 15.
Os testes foram realizados com kit para dosagem de IGF-I humano (DSL-5600®),
pois verificamos por pareamento que as seqüências de IGF-I humano (NP 000609 -
COUTANT et al., 2004) e canino (P 33712 – DELAFONTAINE et al., 1993) apresentavam
alta homologia (95,16%), diferindo entre elas em apenas 6 aminoácidos (humano apresenta
124 aa e cão 122 aa) (Figura 8). Desta forma concluímos que é adequado o uso do kit de
IGF-I humano para a dosagem do IGF-I canino. Ainda, não se esperam reações cruzadas
para o IGF-II, insulina, pró-insulina e hormônio do crescimento. Todos os testes foram
feitos em duplicata em um único ensaio para não existirem variações inter-ensaios. O
coeficiente de variação intra-ensaio foi de 3,2%.
Figura 8 - Seqüência de aminoácidos do IGF-I das espécies canina com a humana. Podemos observar que as seqüências apresentam alta homologia (95,16%) diferindo apenas em seis aminoácidos

Ana Rita de Lima
82
Tabela 15 - Dados da dosagem sérica do IGF-I nos cães Golden Retriever distróficos e não distróficos. São Paulo – 2005
Identificação Idade Forma clínica Peso (Kg) IGF-I (ng/ml) Testosterona
(ng/dl) 1mês 2,3 267 7
2meses 8,8 386 --- 3meses 11,2 455 --- 4meses
Benigna
13,0 381 115 01 Distrófico
7meses Grave 14,0 7 --- 1mês 2,5 273 47
2meses 8,5 322 --- 3meses 11,0 474 --- 4meses
Benigna
13,0 249 150 02 Distrófico
7meses Grave 14,3 72 --- 1mês 2,3 191 7
2meses 8,3 324 --- 3meses 10,8 359 --- 4meses
Benigna
12,5 195 646 03 Distrófico
7meses Benigna 22,2 137 --- 1mês 2,5 259 7
2meses 9,0 305 --- 3meses 11,0 371 --- 4meses
Benigna
13,2 273 1092 04 Distrófico
7meses Benigna 23,9 187 XXX 1mês 2,8 338 7
2meses 10,5 445 --- 3meses 12,3 497 --- 4meses
XXX
15,0 502 258 05 Não Distrófico
7meses XXX 26,0 408 ---
Todos os animais distróficos apresentaram um aumento na concentração sérica de
IGF-I entre o primeiro e terceiro meses de idade, ocorrendo redução no quarto mês,
enquanto o animal não distrófico mostrou um aumento contínuo do primeiro ao quarto mês.
Estes resultados quando analisados estatisticamente pelo teste t mostraram-se

Ana Rita de Lima
83
0,00
100,0
020
0,00
300,0
040
0,00
500,0
060
0,00
Mês 1 Mês 2 Mês 3 Mês 4 Mês 7
Idade (meses)
IGF-
I (n
g/m
l) animal 01animal 02animal 03animal 04animal 05
estatisticamente significativos (p< 0,05) quando se comparou a média amostral dos animais
distróficos com o valor real mensal do animal não distrófico (Gráficos 1 e 2 e Tabela 16).
No sétimo mês ocorreu redução nos níveis séricos de IGF-I em todos os animais
sendo que nos distróficos graves ocorreu uma grande redução, estes resultados foram
estatisticamente significativos quando analisados pelo teste t (p< 0,05) onde se comparou a
média dos animais distróficos graves com o valor real do animal não distrófico e a média
dos animais distróficos benignos com o valor real do animal não distrófico (Gráficos 1 e 2,
Tabela 16).
Gráfico 1 – Gráfico linear mostrando dados referentes à dosagem de IGF-I nos animais Distróficos graves (animais 01 e 02), animais distróficos benignos (animais 03 e 04) e não Distrófico (animal 05), durante os quatro primeiros meses e sétimo mês de vida. Nota -se em todos os animais um aumento nas concentrações séricas de IGF-I nos três primeiros meses, seguido de queda nos animais distróficos, ocorrendo um leve aumento no animal não distrófico no quarto mês, ocorrendo nova queda em todos os animais no sétimo mês, São Paulo – 2005
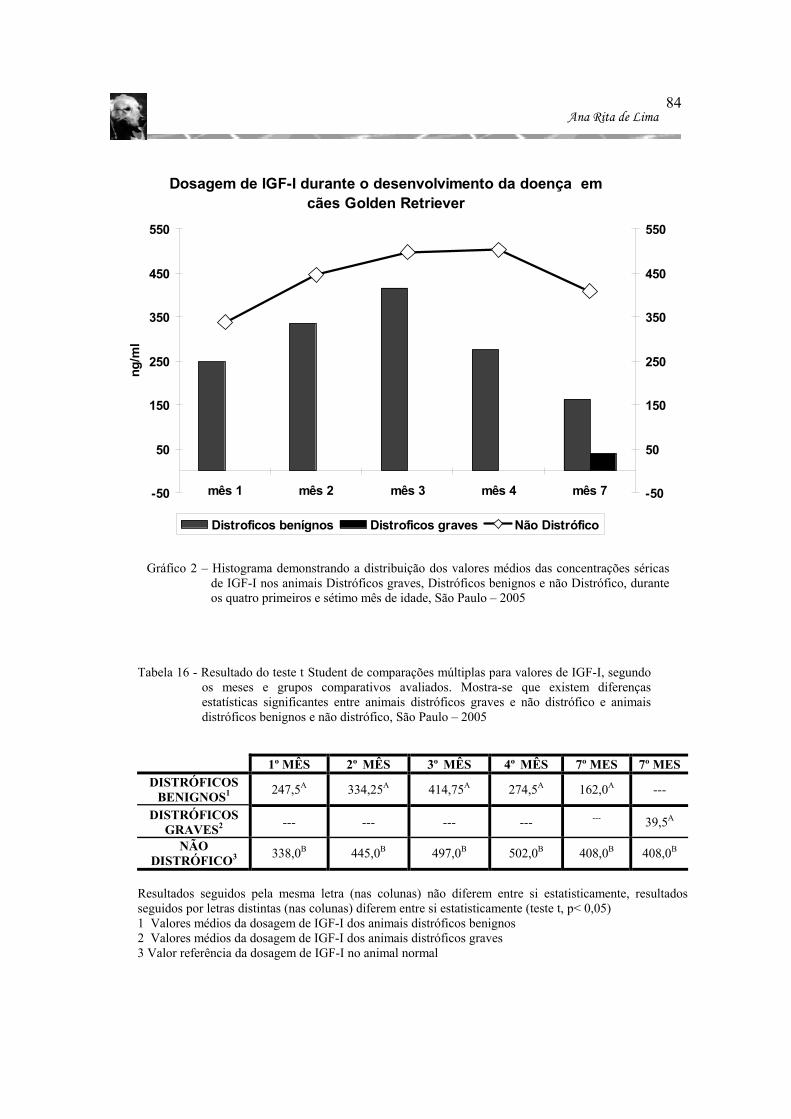
Ana Rita de Lima
84
Dosagem de IGF-I durante o desenvolvimento da doença em cães Golden Retriever
-50
50
150
250
350
450
550
mês 1 mês 2 mês 3 mês 4 mês 7
ng/m
l
-50
50
150
250
350
450
550
Distroficos benígnos Distroficos graves Não Distrófico
Gráfico 2 – Histograma demonstrando a distribuição dos valores médios das concentrações séricas de IGF-I nos animais Distróficos graves, Distróficos benignos e não Distrófico, durante os quatro primeiros e sétimo mês de idade, São Paulo – 2005
Tabela 16 - Resultado do teste t Student de comparações múltiplas para valores de IGF-I, segundo os meses e grupos comparativos avaliados. Mostra-se que existem diferenças estatísticas significantes entre animais distróficos graves e não distrófico e animais distróficos benignos e não distrófico, São Paulo – 2005
1º MÊS 2º MÊS 3º MÊS 4º MÊS 7º MES 7º MES DISTRÓFICOS
BENIGNOS1 247,5A 334,25A 414,75A 274,5A 162,0A ---
DISTRÓFICOS GRAVES2 --- --- --- --- --- 39,5A
NÃO DISTRÓFICO3 338,0B 445,0B 497,0B 502,0B 408,0B 408,0B
Resultados seguidos pela mesma letra (nas colunas) não diferem entre si estatisticamente, resultados seguidos por letras distintas (nas colunas) diferem entre si estatisticamente (teste t, p< 0,05) 1 Valores médios da dosagem de IGF-I dos animais distróficos benignos 2 Valores médios da dosagem de IGF-I dos animais distróficos graves 3 Valor referência da dosagem de IGF-I no animal normal

Ana Rita de Lima
85
Os resultados de testosterona mostraram que com o decorrer da idade ocorre
aumento nos níveis séricos deste hormônio, este aumento mostrou-se estatisticamente
significativo quando analisado pelo teste t de Student (p<0,05).
5.5 ESTUDO PILOTO (ANATOMIA CIRÚRGICA)
De acordo com a técnica cirúrgica empregada e discorrida no capítulo referente à
metodologia, observamos que o acesso cirúrgico da hipófise em cães caracteriza-se por ser
uma cirurgia de extrema delicadeza e de alto grau de dificuldade. A dificuldade de sucesso
em se individualizar a hipófise relaciona-se desde o posicionamento do animal em decúbito
dorsal e abertura da cavidade oral para o acesso do palato mole. Após a conseqüente
abertura da ferida cirúrgica observou-se que neste decúbito o animal apresenta intermitente
sangramento dificultando ainda mais a visualização das estruturas no foco cirúrgico. A
distinção entre as porções da neurohipófise e adenohipófise e, por conseguinte a
individualização das Pars proximalis e distalis desta, foi dificultada quando da aplicação da
abordagem cirúrgica sendo possível evidenciar a hipófise somente como um todo.

Ana Rita de Lima
86
6 DISCUSSÃO
Os resultados previamente descritos acima em cada um dos referentes capítulos serão
confrontados de acordo com a literatura consultada neste capítulo. Observa-se a
manutenção da divisão dos assuntos em sub itens para melhor abordagem do leitor.
6.1 ESTUDO MACRO E MESOSCÓPICO
A hipófise encontra-se situada ventralmente ao hipotálamo estando suspensa pelo
Tuber cinereum dentro da fossa hipofisária do osso basisfenóide coincidindo com os relatos
de Dyce et al, 2002 e Evans, 1993.
A análise macroscópica da adenohipófise nos animais estudados não revelou
diferenças entre os animais distróficos graves e os animais não distróficos, quanto à
coloração (amarelo-rosada), localização (fossa hipofisária) e tamanho. A avalição dos
dados macromorfométricos da adenohipófise (comprimento, largura e espessura) não
mostrou diferenças estatisticamente significativas ao Teste t de “Student” (p>0,05) entre os
animais distróficos graves e os animais não distróficos.

Ana Rita de Lima
87
6.2 ANATOMIA DA IMAGEM (IMAGOLOGIA)
As imagens radiográficas e tomográficas em relação à localização da hipófise
revelaram resultados congruentes com a descrição anatômica macroscópica e, estes
resultados estão de acordo com a literatura consultada demonstrando a localização da
hipófise dentro da Sella turcica no osso basisfenóide (FEENEY et al., 1991; KÖNIG;
LIEBICH, 2002). No paciente “in vivo” faz-se necessário o uso de contrastes intravenosos
para a realização da tomografia computadorizada (LOVE et al, 2000; VLUGT-MEIJER et
al., 2002; VLUGT-MEIJER et al., 2004). Desta forma, como não foi possível o uso de
contrastes neste estudo, pois os animais estavam mortos, foi utilizada uma bolinha de
chumbo antes do exame tomográfico para referenciar a localização da hipófise.

Ana Rita de Lima
88
6.3 ESTUDO HISTOLÓGICO
Os aspectos histológicos são discutidos em capítulos devido a diversas abordagens
aplicadas conforme descrito no capítulo referente ao método.
6.3.1 Aspectos Microestruturais
Com o uso da microscopia de luz de cortes semi-finos foram observadas células
granulares distribuídas homogeneamente na Pars proximalis e Pars distalis da
adenohipófise de todos os animais, estando estas células dispostas em grupos ao redor dos
capilares conforme descrito por Baker (1974); Bernabé et al. (2000); Conklin (1966);
Dacheux (1984 a, b); El Etreby; Fath El Bab (1977) e Nakane (1970). Nestes trabalhos
estão descritas as células acidófilas granulares produtoras de GH em diversas espécies de
animais (ratos, suínos, cães, homem e ovinos) que foram identificadas com o uso de
reações imunohistoquímicas com anticorpos específicos para cada espécie. As células
produtoras de GH variaram na forma, sendo normalmente esféricas ou ovaladas.
As células granulares distribuem-se ao longo da Pars proximalis e distalis nos
animais Distróficos graves e não Distróficos, sendo encontrado um menor número destas
células na Pars proximalis em relação a Pars distalis, porém esta diferença não se mostrou
estatisticamente significativa (p>0,05). Nos animais distróficos as freqüências de células

Ana Rita de Lima
89
granulares na Pars proximalis e na Pars distalis foram respectivamente 1,2 e 2,6% maiores
do que nos animais não distróficos.
6.3.2 Aspectos Morfoquantitativos
As células granulares adenohipofisárias (Pars distalis) relacionadas à produção do
hormônio de crescimento em cães Golden Retriever com Distrofia Muscular (forma grave)
apresentam-se maiores que nos cães Golden Retriever não distróficos quanto aos aspectos
mensurados. Nos animais distróficos, as células granulares da Pars proximalis
apresentaram um eixo longo de 7,7µm enquanto que nos animais não distróficos o eixo
longo foi de 6,5µm, sendo o eixo longo 1,2% maior nos animais distróficos que nos
animais não distróficos. A área e o volume foram 1,3% maiores nos animais distróficos
quando comparados com os animais não distróficos. O eixo longo das células granulares da
Pars distalis nos animais distróficos foi de 7,1µm e, nos animais não distróficos de 5,8µm,
sendo o eixo longo, área e volume 1,2% maiores nos animais distróficos que nos animais
não distróficos, estes resultados foram significativos quando comparados por meio do teste
de Quadrados Mínimos (Qui Quadrado - p< 0,01). Foi encontrado na literatura dados
referentes às células produtoras de GH em ratos e em ovelhas, onde estas apresentavam
diâmetro entre 10 a 13 µm em ratos (NAKANE, 1970) e de 11 a 15 µm em ovelhas
(BERNABÉ, 2000). Os resultados obtidos nesta pesquisa revelam que as células granulares
adenohipofisárias em cães, especificamente na raça Golden Retriever, são menores que as

Ana Rita de Lima
90
encontradas em ratos e em ovelhas, sendo o tamanho das células uma característica de cada
espécie.
À microscopia eletrônica de transmissão observamos que as células associadas à
produção de GH apresentaram grânulos elétron-densos distribuídos homogeneamente por
todo o citosol apresentando membrana de parede dupla e tamanho médio de 175nm na Pars
proximalis e 218nm na Pars distalis em cães Golden Retriever Distróficos e de 130nm na
Pars proximalis e 163nm na Pars distalis, em cães Golden Retriever não Distróficos,
diferindo do que encontramos na literatura onde os autores relatam que os grânulos
possuem um diâmetro médio de 300 a 400nm (ADAM et al., 1970; BAKER, 1974;
DELLMANN e BROWN, 1987; MIKAMI, 1987; MORIARTY, 1973; NAKANE, 1970;
OSAMURA, 1996 e SASAKI, 1988).
A presença de células e grânulos maiores nos animais Distróficos pode indicar um
acúmulo de grânulos e de seus produtos de extrusão no interior destas células devido à
dificuldade de exocitose, alterações na regulação da secreção dos peptídeos que controlam
a secreção de GH ou ainda, a falta de receptores periféricos para o GH como ocorre na
síndrome de Laron em humanos. A dificuldade na exocitose pode estar relacionada à
ausência da proteína distrofina, visto que esta faz parte do citoesqueleto das células e a
ausência desta ocasiona a desestabilização da membrana plasmática e alterações no
movimento de organelas e vesículas citoplasmáticas além de alterações na manutenção do
cálcio intracelular (BERGMAN et al 2002; BLAKE et al., 2002; BOGDANOVICH et al.,
2004; EHMSEN, POON, DAVIES, 2002; JUNQUEIRA e CARNEIRO, 1999;
KORNEGAY et al., 1994; NGUYEN et al 2002; ROSS, KAYE, PAWLINA, 2003;
SHELTON et al., 2001), promovendo desta forma o acúmulo de grânulos secretórios na

Ana Rita de Lima
91
interior das células que, conseqüentemente leva ao aumento no tamanho das células
associadas à produção de GH nos animais distróficos.
6.4 ASPECTOS NEUROENDÓCRINOS
A dosagem do IGF-I foi eleita para analisar as características do eixo GH/IGF-I, pois
este é mais estável durante o dia e não sofre picos de secreção como ocorre com o GH
(BOQUETE et al., 2003; LISSETT et al., 2003; SHALET et al., 1998). As dosagens foram
realizadas para avaliar se existem diferenças na secreção de GH entre animais não
Distróficos e acometidos pela Distrofia muscular nas diferentes formas clínicas da doença:
benigna e grave.
Os testes foram realizados com kit para dosagem de IGF-I humano (DSL-5600®),
pois verificamos por pareamento que as seqüências de IGF-I humano (NP 000609 -
COUTANT et al., 2004) e canino (P 33712 – DELAFONTAINE et al., 1993) apresentavam
alta homologia (95,16%), diferindo entre elas em apenas 6 aminoácidos. Desta forma
concluímos que o kit de IGF-I humano foi adequado para dosar o IGF-I canino, não sendo
esperadas reações cruzadas para o IGF-II, insulina, pró-insulina e hormônio do
crescimento. Todos os testes foram feitos em duplicata em um único ensaio para não
existirem variações inter-ensaios, sendo o coeficiente de variação intra-ensaio de 3,2%.
Os animais apresentaram aumento e posteriormente decréscimo nas concentrações
de IGF-I com o decorrer da idade o que está de acordo com os dados observados em
estudos com outros animais, bem como em humanos (GÓMEZ et al., 2004). O decréscimo

Ana Rita de Lima
92
nas concentrações de IGF-I pode estar relacionado à redução de secreção de GH ou a
redução no número de receptores de GH no fígado ou ainda, no caso dos animais
distróficos graves (no sétimo mês) pode ter ocorrido devido ao estado nutricional
inadequado, uma vez que a desnutrição leva a redução nos níveis de IGF-I.
Todos os animais seguiram um padrão, ocorrendo aumento seguido de queda da
concentração sérica de IGF-I. No entanto, o fato que diferencia os animais distróficos do
animal não distrófico é o mês em que esta queda ocorre, sendo mais tardia no animal não
distrófico (sétimo mês). Quanto aos animais distróficos graves e benignos, no sétimo mês
ocorreu uma queda bastante acentuada e abrupta nas concentrações de IGF-I dos animais
distróficos graves, sendo que esta queda pode estar relacionada ao estado nutricional visto
que nesta fase os animais apresentavam-se severamente desnutridos. No entanto, nos
animais distróficos benignos também foi constatada uma redução nos níveis de IGF-I,
porém não tão abrupta quanto nos animais com a forma grave da doença.
A puberdade também é um fator que interfere no IGF-I como descrito por Chowen;
Frago; Argente (2004), pois os esteróides sexuais modulam inibitoriamente a síntese, e
secreção de GH pela hipófise anterior. Em nossos resultados o IGF-I sofreu um aumento
progressivo independente da ação da testosterona, até o quarto mês. Tanto as concentrações
de IGF-I quanto de testosterona sofreram um aumento no decorrer dos meses sugerindo que
a testosterona não interferiu na concentração de IGF-I.
Além do músculo esquelético, a Distrofina encontra-se presente também em várias
regiões do sistema nervoso central (SNC) e a sua ausência (Distrofia muscular de
Duchenne) está relacionada com alterações na arquitetura do SNC, anormalidades
dendríticas e diminuição no número de neurônios no tronco encefálico e córtex cerebral

Ana Rita de Lima
93
tanto em seres humanos como em animais de laboratório (ANDERSON et al., 2002). Não
existe informação sobre a sua expressão na hipófise nem da sua função nos tecidos não
musculares. Podemos conjecturar que a sua ausência na hipófise e/ou no tronco encefálico e
córtex cerebral, possa levar a alterações na regulação da secreção dos
peptídeos que controlam a secreção de GH (GH-RH e SST), ocasionando uma
diminuição na secreção de GH o que resultaria em um acúmulo de grânulos de
secreção no interior dos somatotrofos e conseqüente diminuição nos níveis de
IGF-I circulante. Ou ainda, que devido à ausência da distrofina no citoesqueleto ocorra uma
redução na exocitose dos grânulos secretórios, pois sem a distrofina ocorre desestabilização
de suas membranas impedindo a exocitose dos mesmos e alterações na manutenção do
cálcio intracelular (CARBAJAL, VITALE, 1997).
Futuras investigações devem avaliar o papel da distrofina nos grânulos secretores
adenohipofisários de animais sadios para então inferir sobre o significado funcional de sua
ausência em animais distróficos. Finalmente, nossos resultados comprovam que através das
dosagens de IGF-I nos animais Golden Retriever é possível avaliar o eixo GH/IGF-I,
investigando-se possíveis alterações no mesmo que possam influenciar a evolução clínica
da distrofia muscular e analisar o efeito da aplicação do Hormônio de
Crescimento no tratamento clínico da doença. Até o presente momento, os dados obtidos
neste estudo sugerem que não há nenhuma relação entre a redução dos níveis séricos de GH
e o quadro clínico benigno da doença como foi proposto por alguns autores onde pacientes
com deficiência de GH apresentavam um quadro clínico benigno da doença (CHYATTE et
al., 1973; CHYATTE et al., 1974; FRANK e SMITH 2001; ZATZ, FROTA-PESSOA
1981b).

Ana Rita de Lima
94
Contrariamente, os dados obtidos nesta investigação, referentes à dosagem sérica de
IGF-I, revelaram uma maior concentração do mesmo nos animais portadores da forma
clínica benigna em relação aos animais com quadro clínico grave. Isto parece sugerir que o
GH poderá ser utilizado no futuro como uma alternativa terapêutica para a Distrofia de
Duchenne. Não obstante, estudos futuros devem investigar esta questão em um maior
número de cães.
5.5 ESTUDO PILOTO (ANATOMIA CIRÚRGICA)
A proposta inicial da hipofisectomia era a remoção seletiva da Pars distalis onde se
localizam as células produtoras do hormônio de crescimento (GH). Como descrito na
literatura é possível realizar a remoção total da hipófise em cães, entretanto, os resultados
deste estudo piloto demonstraram que o acesso específico à Pars distalis ou mesmo a
adenohipófise é inviável impossibilitando a sua remoção seletiva e pelo processo cirúrgico
ser excessivamente cruento. Ainda, a ablação total e não seletiva da hipófise ou da
adenohipófise privaria o animal não apenas do GH, mas também de outros hormônios
produzidos pela hipófise e mesmo que ocorresse uma melhora no quadro clínico, isto
poderia ser atribuído à deprivação do GH e/ou de outros hormônios hipofisários ou
adenohipofisários, o que não possibilitaria qualquer inferência acerca do papel
neuroendócrino do GH na patogênese da Distrofia Muscular de Duchenne.

Ana Rita de Lima
95
7 CONCLUSÃO
Com base nos métodos empregados e nos resultados obtidos podemos concluir que:
1. A localização da hipófise nos cães Golden Retriever Distróficos e Não Distróficos
não mostrou diferenças entre si e em relação ao que está descrito na literatura para
outras raças de cães.
2. As células granulares adenohipofisárias (Pars proximalis e distalis) associadas à
produção do GH e seus respectivos grânulos elétron-densos apresentaram-se 1,3%
maiores nos cães Golden Retriever com Distrofia Muscular do que nos cães Golden
Retriever não distróficos, tanto na Pars proximalis quanto na Pars distalis.
3. A dosagem de IGF-I em cães Golden Retriever foi padronizada e pode ser realizada
com o emprego de kit desenvolvido para a dosagem do IGF-I humano devido a alta
homologia (95,16%) entre estas duas espécies.
4. As concentrações séricas de IGF-I apresentaram um aumento progressivo até o
terceiro mês de idade tanto em animais Distróficos e quanto no não distrófico. O
decréscimo das concentrações séricas de IGF-I iniciou-se a partir do sétimo mês de
vida para o animal não Distrófico e a partir do terceiro mês em animais Distróficos,
o mesmo ocorrendo de modo abrupto em animais Distróficos (forma clínica Grave).

Ana Rita de Lima
96
5. Contrariamente ao sugerido por alguns autores, os dados referentes à concentração
sérica de IGF-I sugerem que não há nenhuma relação entre a diminuição dos níveis
séricos de GH e o estabelecimento de uma forma clínica benígna da doença.
Paradoxalmente, níveis mais altos de GH sugerem um quadro mais atenuado ou a
ausência da doença.
6. Este estudo sugere que o GH poderá ser utilizado no futuro como uma das
alternativas terapêuticas para a Distrofia de Duchenne
7. A ablação seletiva da Pars distalis da adenohipófise mostrou-se inviável do ponto de
vista cirúrgico tanto para se avaliar a evolução clínica quanto como possível
alternativa terapêutica para a Distrofia Muscular de Duchenne.

Ana Rita de Lima
97
REFERÊNCIAS
ADAM, W. S.; CALHOUN, M. L.; SMITH, E. M.; STINSON, A. W. Microscopic Anatomy of the dog. Springfield: Charles C. Thomas, 1970, p. 219-220. AGUSTSSON, T.; EBBESSON, L. O.; BJORNSSON, B. T. Dopaminergic innervation of the rainbow trout pituitary and stimulatory effect of dopamine on growth hormone secretion in vitro. Comparative Biochemistry and Physiology- Party A: Molecular & Integrative Physiology, v. 127, n. 3, 2000, p. 355-364. ALBERTSSON-WIKLAND, K.; ROSBERG, S. Analyses of 24-hour growth hormone profiles in children: relation to growth. The Journal of Clinal Endocrinology and Metabolism, v. 67, 1988, p. 493-500. ANDERSON, J. L.; HEAD, S. I.; RAE, C.; MORLEY, J. W. Brain function in Duchenne muscular dystrophy. Brain, v. 125, 2002, p. 4-13. ARANA, S.; FARIAS, E. C.; BORELLA, M. I.; PEREZ A. C. A.; GIAMAS, M. T. D. Morphological and cytochemical characterization of cell types of the adenohypophysis of Manjuba, Anchoviella lepidentostole (Fowler, 1911) (Osteichthyes, Engraulidae). Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 34, n. 2, 1997, p. 63-69. BAKER, B. L. Functional cytology of the hypophysial Pars distalis and pars intermedia. American Physiological Society. Handbook of Physiology: Endocrinology. Baltimore: Williams e Wilkins, 1974, p. 45-80. BAKER, B. L. Studies on Hormone Localization with Emphasis on the Hypophysis. The Journal of Histochemistry and Cytochemistry. v. 18, n. 1, 1970, p. 1-7. BANKS, W. J. Applied Veterinary Histology. 3. ed. Baltimore: Mosby Year Book, 1993, p. 408-413. BAXTER, R. C.; MARTIN, J. L.; BENLOC, V. A. High molecular weigth insulin-like factor binding protein complex. The Journal of Biological Chemistry, v. 264, 1989, p. 11843-11848. BERGMAN, R. L.; INZANA, K. D.; MONROE, W. E.; SHELL, L. G.; LIU, L. A.; ENGVALL, E.; SHELTON, G. D. Dystrophin-deficient muscular dystrophy in a Labrador Retriever. Journal of American Hospital Association, v. 38, n. 3, may-jun, 2002, p. 255-261.

Ana Rita de Lima
98
BERNABÉ, A.; GÓMEZ, M. A.; SEVA, J.; VÁSQUEZ, F.; PALLARES, F. J.; SERRANO, J. Light and Ultrastructural Immunocytochemical Study of Somatotropic Cell (GH Cells) in Ovine Adenohypophysis: Lactation and Weaning Influences. Anatomy Histology and Embryology, v. 29, 2000, p.13-18. BERRY, C. R. Physical principles of computed tomography and magnetic resonance imaging. In: THRALL, D. E. Textbook of veterinary diagnostic radiology. 4. ed. Philadelphia: W. B. Saunders, 2002, cap.3, p. 28-34. BLAKE, D. J.; WEIR, A.;NEWEY, S. E.; DAVIES, K. E. Function and Genetics of Dystrophin and Dystrophin-Related Proteins in Muscle. Physiological Reviews, v. 82, 2002, p. 291-329.
BOGDANOVICH, S.; PERKINS, K. J.; KRAG, T. O. B.; KHURANA, T. S. Therapeutics for Duchenne muscular dystrophy: current approaches and future directions. Journal of Molecular Medicine, v. 82, n. 2, 2004, p. 102-115. BOQUETE, H. R.; SOBRADO, P. G. V.; FIDELEFF, H. L.; SEQUERA, A. M.; GIACCIO, A. V.; SUÁREZ, M. G.; RUIBAL, G. F.; MIRAS, M. Evaluation of Diagnostic Accuracy of Insulin-Like Growth Factor (IGF)-I and IGF-Binding Protein-3 in Growth Hormone-Deficient Children and Adults Using ROC Plot Analysis. The Journal of Clinical Endocrinology e Metabolism, v. 88, n. 10, 2003, p. 4702-4708. CANAL, N.; SMIRNE, S.; COMI, G.; GUIDOBONO, F.; PECILE, A; CAVIEZEL, F. Study on Growth Hormone and Prolactin secretions in Myotonic Dystrophy. Acta Neurologica Belgica, v. 82, 1982, p. 178-184. CARBAJAL, M. E.; VITALE, M. L. The Cortical Actin Cytoskeleton of Lactotropes as an Intracellular Target for the Control of Prolactin Secretion. Endocrinology, v.138, n. 12, 1997, p. 5374-5384. CHOWEN, J. A.; FRAGO, L. M.; ARGENTE, J. The Regulation of GH Secretion by Sex Steroids. European Journal of Endocrinology, v. 151, 2004, p. 95-100. CHYATTE, S. B.; RUDMAN, D.; PATTERSON, J. H.; GERRON, G. G.; O’BEIRNE, I.; BARLOW, J; JORDAN A.; SHAVIN J. S. Human growth hormone and estrogens in boys with Duchenne muscular dystrophy. Archives of Physical Medicine and Rehabilitation, v. 54, 1973, p. 248-253. CHYATTE, S. B.; RUDMAN, D.; PATTERSON, J. H.; AHMANN, P.; JORDAN A. Human growth hormone in a myopathy: Myotonic dystrophy, Duchenne muscular dystrophy and limb-girdle muscular dystrophy. The Southern Medical Journal, v. 67, 1974, p. 170-172.

Ana Rita de Lima
99
CLEMMONS, D. R. Commercial Assays Available for Insulin-Like Growth Factor I and their Use in Diagnosing Growth Hormone Deficiency. Hormone Research, v. 55, 2001, p. 73-79. CONKLIN, J. L. The Identification of Acidophilic Cells in the Human Pars distalis. The Anatomical Records , v. 156, 1966, p. 347-360. COPPA, M.; FLORES, E.; PÉREZ, L. Tecnica de Hipofisectomia en Caninos Modificada. Archivos of Medicine Veterinary, v. 10, 1978, p. 155-159. CORDIDO, F.; ALVAREZ-CASTRO, P.; ISIDRO, M. L.; CASANUEVA, F. F.; DIEGUEZ, C.Comparison between insulin tolerance test, Growth hormone (GH)- releasing hormone (GHRH), GHRH plus acipimox and GHRH plus GH-releasing peptide-6 for the diagnosis of adult GH deficiency in normal subjects, obese and hipopituitary patients. European Journal of Endocrinology, v. 149, 2003, p. 117-121. COUTANT, R.; DE CASSON, F. B.; ROULEAU, S.; DOUAY, O.; MATHIEU, E.; GATELAIS, F., BOUHOURS-NOUET, N. ; VOINOT, C. ; AUDRAN, M. ; LIMAL, J. M. Divergent effect of endogenous and exogenous sex steroids on the insulin-like growth factor I response to growth hormone in short normal adolescents. The Journal of Clinical Endocrinology and Metabolism, v.89, n.12, 2004, p. 6185-6192. DACHEUX, F. Differentiation of cells producing polypeptide hormones (ACTH, MSH, LPH, α- e β-endorfina, GH e PRL) in fetal porcine anterior pituitary. Cell Tissue Research, v. 235, 1984 a, p. 615-621. DACHEUX, F. Functional differentiation of the anterior pituitary cells in the fetal pig. Cell and Tissue Research, v. 235,1984 b, p. 623-633. DAUGHADAY, E.; ROTWEIN, R. Insulin like growth factors I and II. Peptide, messenger ribonucleic acid and gene structures, serum and tissue concentrations. Endocrinology Review, v. 10, 1989, p. 68-91. DELAFONTAINE, P. Insulin-like growth factor I and its binding proteins in the cardiovascular system. Cardiovascular Research, v. 30, 1995, p. 825-834. DELAFONTAINE, P.; LOU, H.; HARRISON, D. G.; BERNSTEIN, K. E. Sequence of a cDNA encoding dog insulin-like growth factor I .Gene, v. 130, n. 2, 1993, p. 305-306. DELLMANN, H. D.; BROWN, E. M. Textbook of Veterinary Histology. 3. ed. Philadelphia: Lea e Febiger, 1987, p. 361-371. DYCE, K. M.; SACK, W. O.; WENSING, C. J. G. Textbook of Veterinary Anatomy. 3. ed. Philadelphia: Saunders, 2002, p. 210-21.

Ana Rita de Lima
100
EHMSEN J.; POON, E.; DAVIES, K. The dystrophin associated protein complex. Journal of Cell Science, v.115, 2002, p. 2801-2803. EINGENMANN, J. E.; PATTERSON, D. F. Growth hormone deficiency in the mature dog. Journal of American Animal Hospital Association, v. 20, 1984, p. 741-746. EL ETREBY, M. F.; FATH EL BAB, M. R. The utility of antisera to canine growth hormone and canine prolactin for immunocytochemical staining of the dog pituitary gland. Histochemistry, v. 53, 1977, p. 1-15. EMERY, A. H. Population frequencies of inherited neuromuscular diseases-a world survey. Neuromuscular Disorders, v. 1, 1991, p. 19-29. ENEMAR, A. Selective fetal hypophysectomy in utero: Microsurgical technique and comparisons with hypophysectomy by decapitation. Cells Tissues and Organs, v. 173, 2003, p. 172-183. EVANS, H. E. Miller’s Anatomy of the dog. 3. ed. Philadelphia: Saunders, 1993. FEENEY, D. A.; FLETCHER, T. F.; HAARDY, R. M. Atlas of correlative imaging anatomy of the dog ultrasound and computed tomography. Philadelphia: W.B. Saunders, 1991, p. 209-210, 335-352. FRANK, G. R.; SMITH, R. E. Effective growth hormone theraphy in a growth hormone deficient patient with Duchenne muscular dystrophy without evidence of acceleration of the dystrophic process. Journal of Pediatric Endocrinology & Metabolism, v. 14, n. 2, 2001, p. 211-214. GHAFOOR, T.; MAHMOOD, A.; SHAMS S. Duchenne muscular dystrophy with associated growth hormone deficiency. Journal of College Physicians and Surgeons Pakistan, v. 13, n. 12, 2003, p. 722-723. GOMEZ, J. M.; MARAVALL, F. J.; GOMEZ, N.; NAVARRO, M. A.; CASAMITJANA, R.; SOLER, R..The IGF-I system component concentrations that decrease with ageing are lower in obesity in relationship to body mass index and body fat. Growth Hormone and IGF Research, v. 14, 2004, p. 91-96. GUEVARA-AGUIRRE, J.; ROSENBLOOM, A. L.; FIELDER, P. J.; DIAMOND, F. B.; ROSENFELD, R. G. Growth hormone receptor deficiency in Ecuador: Clinical and biochemical phenotype in two populations. The Journal of Clinical Endocrinology and Metabolism, v. 76, 1993, p. 417-423. HATHCOCK, J. T.; STICKLE, R. L. Principles and concepts of computed tomography. Veterinary Clinics of North America. Small Animal Practice, v. 23, n. 2, 1993, p. 399-415.

Ana Rita de Lima
101
HIGAKI, K.; MATSUMOTO, Y.; FUJIMOTO, R.; KUROSAKI, Y.; KIMURA, T. Pharmacokinetics of recombinant human Insulin - Like Growth Factor -I in diabetic Rats. Drug Metabolism and Disposition, v. 25, n. 11, 1997, p. 1324-1327. HOWARD, C. V.; REED, M. G. Unbiased stereology. Three-dimensional measurement in microscopy. Oxford: Bios Scientific Publishers, 2004. JEHLE, P. M.; SCHULTEN, K.; SCHULZ,W.; JEHLE, D. R.; STRACKE, S.; MANFRAS, B. Serum levels of insulin-like growth factor (IGF)-I and IGF binding protein (IGFBP)-1 to -6 and their relationship to bone metabolism in osteoporosis patients. European Journal of Internal Medicine, v. 14, 2003, p. 32-38. JUNQUEIRA, L. C.; CARNEIRO, J. Histologia Básica. 9. ed. Rio de Janeiro: Guanabara Koogan, 1999, p. 162, 332-354. JUUL, A.; KASTRUP, K. W.; PEDERSEN, S. A.; SKAKKEBAEK, .N. E. Growth hormone (GH) provocative retesting of 108 young adults with childhood-onset GH deficiency and the diagnostic value of insulin-like growth factor I (IGF-I) and IGF-binding protein-3. The Journal of Clinical Endocrinology and Metabolism, v. 82,1997, p. 1195-1201. KIM, H.; BARTON, E.; MUJA, N.; YAKAR, S.; PENNISI, P.; Le ROITH, D. Intact Insulin and Insulin-Like Growth Factor -I Receptor Signaling is Required for Growth Hormone Effects on Skeletal Muscle Growth and Function In vivo. Endocrinology, 2004. KÖNIG, H. E.; LIEBICH, H.G. Anatomia dos Animais Domésticos. São Paulo: Artmed, v. 1, 2002, p. 35. KORNEGAY, J. N.; SHARP, N. J. L. H; BARTLETT, R. J.; VAN CAMP, S.; BURT, C. T.; HUNG, WU YEN; KWOCK, L.; ROSES, A. D. Golden Retriever Muscular Dystrophy: Monitoring for Success. Advances in Experimental Medicine and Biology, v. 280, 1990, p. 267-272. KORNEGAY, J. N.; SHARP, N. J.H.; BARTLETT, R. J.; VAN CAMP, S. D.; TYLER B., C.; HUNG, WU YEN; KWOCK, L.; ROSES, A. D. Tarsal Joint Contracture in Dogs with Golden Retriever Muscular Dystrophy. Laboratory Animal Science, v. 44, 1994, p. 331-333. KURTENBACH, E.; MORAES, S. S.; TROCADO, M. T.; LÔBO, G. F.; NASCIMENTO, P. S.; VERJOVSKI-ALMEIDA, S. Beneficial effects of anti-growth hormone antiserum in avian muscular dystrophy. The FASEB Journal, v. 3, 1989, p. 2189-2193. LAING, J. A.; MORGAN, W. J. B.; WAGNER, W. C. Fertility & Infertility in Veterinary Practice, 5. ed. London: Baillière Tindall, 1988, p.1-38.

Ana Rita de Lima
102
LEE, P. D. K.; ROSENFELD, R. G. Clinical utility of insulin-like growth factor assays. Pediatrician, v. 14, 1987, p. 154-161. LEE, P. D. K.; WILSON, D. M.; ROUNTREE, L.; ROSENFELD, R. G. Efficacy of insulin-like growth factor I levels in predicting the response to provocative growth hormone testing. Pediatric Research, v. 27, 1990, p. 45-51. LEE, JIN-SOOK ; JEFTINIJA, K; JEFTINIJA, S; STROMER, M. H.; SCANES, C. G.; ANDERSON, L. L. Immunocytochemical distribution of somatotrophs in porcine anterior pituitary. Histochemistry and Cell Biology, 2004. LISSETT, C. A.; JÖNSSON, P.; MONSON, J. P.; SHALET, S. M. Determinants of IGF-I status in a large cohort of growth hormone-deficient (GHD) subjects: the role of timing of onset of GHD. Clinical Endocrinology, v. 59, 2003, p. 773-778. LOVE, N.E.; FISHER, P.; HUDSON, L. The computed tomographic enhnancement pattern of the normal canine pituitary gland. Veterinary Radiolology & Ultrasound, v.41, n.6, 2000, p.507-510. MALAMED, S.; DEAVER, D.; PEREZ, F.; RADECKI, S.; GIBNEY, J.; SCANES, C. G. Quantitative studies of chicken somatotrophs during growth and development by morphometry, immunocytochemistry, and flow cytometry. General and Comparative Endocrinology, n. 108, 1997, p. 25-34. MALAMED, S.; GIBNEY, J. A.; CAIN, L. D.; PEREZ, F. M.; SCANES, C. G. Immucytochemical studies of the chicken somatotrophs and somatotroph granules before and after hatching. Cell Tissue Research, v. 272, 1993, p. 369-374. MALAMED, S.; GIBNEY, J. A.; LOESSER, K. E.; SCANES, C.G. Age-related changes of the somatotrophs of the domestic fowl Gallus gallus. Cell Tissue Research, n.239, 1985, p. 87-91. MAURAS, N.; WALTON, P.; NICAR, M.; WELCH, S.; ROGOL, A. D. Growth hormone stimulation testing in both short and normal statured children: use of an immunofunctional assay. Pediatric Research, v. 48, n. 5, 2000, p. 614-618. MANABE, Y.; NAKATOMI, K.; CHIKARAISHI, M.; TAKEUCHI, S.; KOBAYASHI, Y.; TAKAHASHI, S. Immunocytochemical and immunoelectron-microscopic study of somatotrophs in ICR and nonobese diabetic mice. Cells Tissues Organs, v. 166, 2000, p. 31-39. MEIJ, B. P; VOORHOUT, G.; VAN DEN INGH, TED S. G. A. M.; HAZEWINKEL, H. A. W.; VAN’T VERLAAT, JOHANA W. Transsphenoidal hypophysectomy in beagle dogs: evaluation of a microsurgical technique. Veterinary Surgery, v.26, 1997, p. 295-309.

Ana Rita de Lima
103
MIKAMI, S. I. Hypophysis. In: MATSUMOTO, A.; ISHII, S. Atlas of Endocrine Organs Vertebrates and Invertebrates. Tokyo: Springer-Verlag, 1987, p. 39-63. MORIARTY, G. C. Adenohypophysis: ultrastructural cytochemistry. The Journal of Histochemistry and Cytochemistry, v. 21, n. 10, 1973, p. 855-894. NAKANE, P. K. Classifications of anterior pituitary cell types with immunoenzyme histochemistry. The Journal of Histochemistry and Citochemistry, v. 18, n. 1, 1970. NGUYEN, F.; CHEREL, Y.; GUIGAND, L.; GOUBAULT-LEROUX, I.; WYERS, M. Muscle lesions associated with dystrophin deficiency in neonatal golden retriever Puppies. Journal of Comparative Pathology, v. 126, feb-apr, 2002, p. 100-108. NIEBAUER, G. W.; EIGENMANN, J. E.; VAN WINKLE, T. J. Study of long-term survival after transsphenoidal hypophysectomy in clinically normal dogs. American Journal of Veterinary Research, v. 51, 1990, p. 677-681. NIEBAUER, G. W.; EVANS, S. M. Transsphenoidal hypophysectomy in the dog. A new technique. Veterinary Surgery, v. 17, 1988, p. 296-303. NIKITOVITCH-WINER, M. B.; ATKIN, J.; MALEY, B. E. Colocalization of prolactin and growth hormone within specific adenohypophyseal cells in male, female and lactating female rats. Endocrinology, v. 121, n. 2, 1987, p. 625-630. OSAMURA, R. Y. Histology, ultrastructure, and immunochemistry, pituitary gland, rat. In: JONES, T. C.; CAPEN, C. C.; MOHR, U. Endocrine System. 2. ed. New York: Springer,1996, p. 33-41. PEACEY, S.R.; SHALET, S.M. Insulin-like growth factor I measurement in diagnosis and management of acromegaly. Annals of Clinical Biochemistry, v. 38, 2001, p. 297-303. PORTER, J. C.; ONDO, J.G.; CRAMER, O.M. Nervous and vascular supply of the pituitary gland. In: American Physiological Society. Handbook of Physiology: Endocrinology. Baltimore: Williams e Wilkins, 1974, p. 33-43. RECHLER, M. Insulin-like growth factor binding proteins. Vitamins and Hormones, v. 47, 1993, p.1-114. RENEHAN, A. G.; JONES, J.; O’DWYER, S. T.; SHALET, S. M. Determination of IGF-I, IGF-II, IGFBP-3 levels in serum and plasma: comparisons using the Bland-Altman method. Growth Hormone and IGF Research, v. 13, 2003, p. 341-346. REECE, W. O. Physiology of Domestic Animals. 2. ed. Baltimore: Williams e Wilkins, 1997, p. 426-431.

Ana Rita de Lima
104
RIBEIRO, A. A. C. M.; DAVIS, C.; GABELLA, G. Estimate of size and total number of neurons in superior cervical ganglion of rat, capybara and horse. Anatomy and Embryology, v. 208, 2004, p. 367-380. ROSENFELD, R.G.; ALBERTSSON-WIKLAND, K.; CASSORLA, F. Diagnostic controversy: the diagnosis of childhood growth hormone deficiency revisited. The Journal of Clinical Endocrinology and Metabolism, v. 80,1995, p.1532-40. ROSS, M.H.; KAYE, G.I.; PAWLINA, W. Histology a Text and Atlas with Cell and Mollecular Biology. Baltimore: Lippincott Williams &Wilkins, 2003, p. 254. RUDMAN, D.; CHYATTE, S. B.; PATTERSON, J. H.; GERRON, G. G.; O’BEIRNE, I.; BARLOW, J.; JORDAN, A.; SHAVIN, J. S. Metabolics effects of human growth hormone and of estrogens in boys with duchenne muscular dystrophy. The Journal of Clinical Investigation, v. 51, 1972, p.1118-1124. SANDHU, M. S.; HEALD, A. H.; GIBSON, J. M.; CRUICKSHANK, J. K.; DUNGER, D. B.; WAREHAM, N. J.Circulating concentrations of insulin-like growth factor-I and development of glucose intolerance: a prospective observational study. Lancet, v. 359, 2002, p.1740-1745. STATISTICAL ANALISYS SYSTEM. SAS user’s guide: basic and statistic. Cary, NC: SAS, 1995, p.1686. SASAKI, F.; SANO, M. Role of the ovary in sexual differentiation of lactotrophs and somatotrophs in the mouse adenohypophysis: a stereological morphometric study by electron microscopy. The Journal of Endocrinology, v. 99, 1983, p. 355-360. SASAKI, F.; IWAMA, Y. Sex difference in prolactin and growth hormone cells in mouse adenohyphophysis: stereological, morphometric, and immunohistochemical studies by light and electron microscopy. Endocrinology, v.123, n. 2, 1988 a, p. 905-912. SASAKI, F.; IWAMA, Y. Correlation of spatial differences in concentrations of prolactin and growth hormone cells with vascular patten in the female mouse adenohypophysis. Endocrinology, v. 122, n. 4, 1988 b, p. 1622-1630. SASAKI, F. Changes with age in the number and size of anterior pituitary cells in female mice from suckling to adulthood. Journal of Endocrinology, v.117, 1988, p. 5-10. SASAKI, F.; DOSHITA, A.; MATSUMOTO, Y.; KUWAHARA, S.; TSUKAMOTO, Y.; OGAWA, K. Embryonic development of the pituitary gland in the chick. Cells Tissues Organs, v.173, 2003, p. 65-74.

Ana Rita de Lima
105
SASAKI, F.; ICHIKAWA Y.; YAMAUCHI S. Imunohistological analysis in the distribuition of cells in the fetal porcine adenohypophysis. The Anatomical Record, v. 233, 1992, p.135-142. SCHLEIM, K. D.; JACKS, T.; CUNNINGHAM, P.; FEENEY, W.; FRAZIER, E. G.; NIEBAUER, G. W.; ZHANG, D.; CHEN, H.; SMITH, R. G.; HICKEY, G. Increases in circulating Insulin - Like Growth Factor - I levels by the oral growth hormone secretagogue MK- 0677 in the beagle are dependent upon pituitary mediation. Endocrinology, v. 140, n. 4, 1999, p. 1552-1558. SHALET, S. M.; TOOGOOD, A.; RAHIM, A.; BRENNAN, B. M. D. The diagnosis of growth hormone deficiency in children and adults. Endocrine Reviews, v. 19, n. 2, 1998, p. 203-223. SHELTON, G. D.; LIU, L. A.; GUO, L. T.; SMITH, G.; CHRISTIANSEN, J. S.; THOMAS, W. B.; SMITH, M. O.; KLINE, K. L.; MARCH, P. A.; FLEGEL, T.; ENGVALL, E. Muscular dystrophy in female dogs. Journal of Veterinary Internal Medicine, v.15, n. 3, may-jun, 2001, p. 240-244. SIZONENKO P. C., CLAYTON P. E., COHEN P., HINTZ R., TANAKA T., LARON Z. Diagnosis and management of growth hormone deficiency in childhood and adolescence. Part 1: Diagnosis of growth hormone deficiency. Growth Hormone and IGF Research, v. 11, 2001, p.137-165. SOLIMON, A. T.; HASSON, A. E. H. J.; AREL, M. K.; HINTZ,R. L.; ROSENFELD, R. G.; ROGOL, A. D. Serum insulin-like growth factors I and II concentrations and growth hormone and insulin responses to arginin infusion in children with protein-calorie malnutrition before and after nutritional rehabilitation. Pediatric Research, v.20, 1986, p. 1122-1130. STOKES, H.; BODA, J. M. Immunofluorescent localization of growth hormone and prolactin in the adenohypophysis of fetal sheep. Endocrinology, v. 83, 1968, p.1362-1366. VLUGT-MEIJER, R.H.; VOORHOUT, G.; MEIJ, B.P. Imaging of the pituitary gland in dogs with pituitary-dependent hyperadrenocorticism. Molecular and Cellular Endocrinology, v.197, n. 1-2, 2002, p. 81-87. VLUGT-MEIJER, R.H.; MEIJ, B.P.; VOORHOUT, G. Dynamic computed tomographic evaluation of the pituitary gland in healthy dogs. American Journal Veterinary Research, v.65, n. 11, 2004, p. 1518-1524. WACHARASINDHU, S.; CHAICHANWATANAKUL, K.; LIKITMASKUL, S.; ANGSUSINGHA, K. Measurement of serum free IGF-I in diagnosis of growth hormone deficiency. Journal of the Medical Association of Thailand, v.83, n. 5, 2000, p. 494-499.

Ana Rita de Lima
106
WEIBEL, E. R.; BOLENDER, R. P. Stereological techniques for electron microscopic morphometry. In: Principles and Techniques of Electron Microscopy. New York: Van Nostrand Reinhold, v. 3, 1973, p. 237-295. YAKAR S., ROSEN C. J., BEAMER W.G., ACKERT-BICKNELL C. L., WU Y., LIU J. L., OOI G.T., SETSER J., FRYSTYK J., BOISCLAIR Y. R., LEROITH D. Circulating levels of IGF-1 directly regulate bone growth and density. The Journal of Clinical Investigation, v. 110, 2002, p. 771-781. ZAPF, J.; HAURI, C.; WALDVOGEL, M.; FROESH, E. R. Acute metabolic effects and half-lives of intravenously administered insulin-like growth factors I and II normal and hypophysectomized rats. The Journal of Clinical Investigation, v. 77, 1986, p. 1768-1775. ZATZ M.; BETTI R. T. B.; LEVY J. A. Benign duchenne muscular dystrophy in a Patient with Growth Hormone Deficiency. American Journal of Medical Genetics, v.10, 1981 a, p. 301-304. ZATZ M.; FROTA-PESSOA O. Suggestion for a possible mitigating treatment of duchenne muscular dystrophy. American Journal of Medical Genetics, v. 10, 1981 b, p. 305-307.

















![Ana Rita Dias Efetividade da intervenção Patrocínio Sousa … Ana TM... · 2015. 10. 3. · Ana Rita Sousa – Abril 2012 II [DECLARAÇÕES] Declaro que este Relatório de Projeto](https://static.fdocumentos.tips/doc/165x107/5ff4bc060b1bdb42ff2a4c69/ana-rita-dias-efetividade-da-interveno-patrocnio-sousa-ana-tm-2015-10.jpg)
