







Línguas
Páginas
Legal


UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA
THIAGO BARROS GALVÃO
REGULAÇÃO COORDENADA DA MOBILIZAÇÃO DE LIPÍDIOS E PROTEÍNAS POR HORMÔNIOS, FONTES DE CARBONO E
NITROGÊNIO DURANTE O CRESCIMENTO PÓS-GERMINATIVO EM GIRASSOL
NATAL 2012

THIAGO BARROS GALVÃO
REGULAÇÃO COORDENADA DA MOBILIZAÇÃO DE LIPÍDIOS E PROTEÍNAS POR HORMÔNIOS, FONTES DE CARBONO E
NITROGÊNIO DURANTE O CRESCIMENTO PÓS-GERMINATIVO EM GIRASSOL
Dissertação apresentada ao Programa de Pós-Graduação em Bioquímica da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Mestre em Bioquímica. Orientador: Eduardo Luiz Voigt.
NATAL 2012

Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Galvão, Thiago Barros. Regulação coordenada da mobilização de lipídios e proteínas por hormônios, fontes de carbono e nitrogênio durante o crescimento pós-germinativo em girassol / Thiago Barros Galvão. – Natal, RN, 2012. 72 f.: il.
Orientador: Prof. Dr. Eduardo Luiz Voigt.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Departamento de Bioquímica.
1. Girassol (Helianthus annuus) – Dissertação. 2. Mobilização de reservas – Dissertação. 3.
Efeito cruzado – Dissertação. I. Voigt, Eduardo Luiz. II. Universidade Federal do Rio Grande do Norte. III. Título.
RN/UF/BSE-CB CDU 663.542

THIAGO BARROS GALVÃO
REGULAÇÃO COORDENADA DA MOBILIZAÇÃO DE LIPÍDIOS E PROTEÍNAS POR HORMÔNIOS, FONTES DE CARBONO E NITROGÊNIO DURANTE O
CRESCIMENTO PÓS-GERMINATIVO EM GIRASSOL Dissertação apresentada ao Programa de Pós-Graduação em Bioquímica da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Mestre em Bioquímica.
Aprovado em: / /
BANCA EXAMINADORA
___________________________________________________ Prof. Dr. Eduardo Luiz Voigt
Departamento de Biologia Celular e Genética - UFRN Orientador
____________________________________________________ Profa. Dra Katia Castanho Scortecci
Departamento de Biologia Celular e Genética – UFRN 1º Examinador
____________________________________________________ Prof. Dr Joaquim Albenísio Gomes da Silveira
Departamento de Bioquímica - UFC 2º Examinador

Dedico este trabalho
À minha família.

AGRADECIMENTOS
Ao professor Eduardo Luiz Voigt pelas ideias, paciência e amizade durante minha graduação e mestrado.
Aos amigos do laboratório por todo o apoio e aprendizado.
Aos professores do DBQ por todos os conhecimentos transmitidos durante o curso.
Aos amigos do mestrado pela amizade e carinho.
À CAPES pela bolsa concedida.

Embora eu não acredite que uma planta vá brotar onde nenhuma semente esteve, eu tenho muita fé
em uma semente. Convença-me que você tem uma semente lá, e eu estarei preparado para
esperar maravilhas. Henry D. Thoreau

RESUMO
A mobilização das reservas nutritivas nos tecidos de armazenamento e a alocação de seus produtos de hidrólise no eixo em crescimento são processos críticos para o estabelecimento das plântulas após a germinação das sementes. Assim sendo, é necessário que a mobilização das reservas seja sincronizada com o crescimento do eixo de forma que a atividade fotossintética tenha iniciado antes que as reservas sejam exauridas. Para isso, abordagens integrativas envolvendo as diferentes reservas, os diferentes produtos e o intercâmbio entre os tecidos de armazenamento e o eixo em crescimento, seja por intermédio de hormônios ou metabólitos com papel de sinalização, podem contribuir sobremaneira para o esclarecimento dos mecanismos que regulam a mobilização de reservas. Neste trabalho, foram levantadas as hipóteses de que o efeito de hormônios e de metabólitos sobre a mobilização das reservas é diferente e de que deve existir um efeito cruzado de açúcares sobre a mobilização de proteínas e de aminoácidos sobre a mobilização de lipídios e amido em plântulas de girassol. O presente trabalho foi desenvolvido com sementes de girassol (Helianthus annuus L.) híbrido Helio 253, utilizando sistema in vitro de cultura. As sementes foram germinadas em papel Germitest® e crescidas em meio ágar-água 4g/L sem adição de nutrientes durante 9 dias após a embebição (DAE) para a curva de crescimento. Para verificar o efeito de metabólitos e hormônios, as plântulas foram transferidas ao 2o DAE para meio ágar-água 4 g/L suplementado com concentrações crescentes de sacarose ou L-glutamina, ácido abscísico, ácido giberélico ou ácido indolbutírico. Os resultados deste trabalho confirmam que a mobilização dos lipídios e das proteínas de reserva ocorre de forma coordenada durante o crescimento pós-germinativo inicial em girassol, corroborando a hipótese de que a aplicação externa de fontes de carbono (sacarose) e nitrogênio (L-glutamina) é capaz de atrasar a mobilização dessas reservas nutritivas de forma cruzada. Além disso, considerando as mudanças nos padrões de mobilização das reservas e a partição dos seus produtos, proporcionadas pela aplicação externa de diferentes reguladores do crescimento, é evidente que os efeitos dos metabólitos e dos hormônios devem envolver, pelo menos em parte, mecanismos de ação distintos. Palavras-chave: Mobilização de reservas. Efeito cruzado. Helianthus annuus.

ABSTRACT
The mobilization of food reserves in storage tissues and allocation of their hydrolysis products in the growing axis are critical processes for the establishment of seedlings after germination. Therefore, it is crucial for mobilization of reserves to be synchronized with the growing axis, so that photosynthetic activity can be started before depletion of reserves. For this, integrative approaches involving different reserves, different hydrolysis products and interaction between storage and growing axis tissues, either through hormones or metabolites with signaling role, can contribute greatly to the elucidation of the regulation mechanisms for reserve mobilization. In this study, was hypothesized that hormones and metabolites have different actions on reserve mobilization, and there must be a crossed effect of sugars on the mobilization of proteins and amino acids on lipids and starch mobilization in sunflower seedlings. This study was conducted with seeds of sunflower (Helianthus annuus L.) hybrid Helio 253 using in vitro culture system. Seeds were germinated on Germitest® paper and grown on agar-water 4 g/L without addition of nutrients during 9 days after imbibition (DAI) for growth curve. To verify the effect of metabolites and hormones, seedlings were transferred in the 2nd DAI to agar-water 4 g/L supplemented with increasing concentrations of sucrose or L-glutamine, abscisic acid, gibberellic acid or indolebutyric acid. The results of this study confirm that the mobilization of lipids and storage proteins occurs in a coordinated manner during post-germination growth in sunflower, corroborating the hypothesis that the application of external carbon (sucrose) and nitrogen (L-glutamine) sources can delay the mobilization of these reserves in a crossed way. Moreover, considering the changes in the patterns of reserve mobilization and partition of their products in seedlings treated with different growth regulators, it is evident that the effects of metabolites and hormones must involve, at least in part, distinct mechanisms of action. Key-words: Reserve mobilization. Crossed effect. Helianthus annuus.

LISTA DE FIGURAS
FIGURA 1 Modelo que representa o sistema de cultivo e a coleta do
material vegetal.............................................................................
21
FIGURA 2 Aspecto morfológico de plântulas de girassol durante o
crescimento pós-germinativo.........................................................
28
FIGURA 3 Conteúdo de massa seca (MS) em plântulas de girassol durante
o crescimento pós-germinativo......................................................
29
FIGURA 4 Conteúdo de lipídios neutros (LN) e de proteínas solúveis (PS)
em plântulas de girassol durante o crescimento pós-germinativo.
30
FIGURA 5 Perfil eletroforético das proteínas solúveis (PS)............................ 31
FIGURA 6 Conteúdo de amido, de clorofilas totais (CT) e de massa seca
(MS) total em plântulas de girassol durante o crescimento pós-
germinativo....................................................................................
32
FIGURA 7 Conteúdo de açúcares solúveis totais (AST), de açúcares não-
redutores (ANR) e de aminoácidos livres totais (AALT) em
plântulas de girassol durante o crescimento pós-germinativo.......
34
FIGURA 8 Aspecto morfológico de plântulas de girassol tratadas com
concentrações crescentes de sacarose........................................
35
FIGURA 9 Conteúdo de massa seca (MS) em plântulas de girassol
tratadas com concentrações crescentes de sacarose..................
36
FIGURA 10 Conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e
de amido em plântulas de girassol tratadas com concentrações

crescentes de sacarose................................................................. 37
FIGURA 11 Conteúdo de açúcares solúveis totais (AST), de açúcares não-
redutores (ANR) e de aminoácidos livres totais (AALT) em
plântulas de girassol tratadas com concentrações crescentes de
sacarose........................................................................................
39
FIGURA 12 Aspecto morfológico de plântulas de girassol tratadas com
concentrações crescentes de L-glutamina....................................
40
FIGURA 13 Conteúdo de massa seca (MS) em plântulas de girassol
tratadas com concentrações crescentes de L-glutamina..............
41
FIGURA 14 Conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e
de amido em plântulas de girassol tratadas com concentrações
crescentes de L-glutamina............................................................
42
FIGURA 15 Conteúdo de açúcares solúveis totais (AST), de açúcares não-
redutores (ANR) e de aminoácidos livres totais (AALT) em
plântulas de girassol tratadas com concentrações crescentes de
L-glutamina....................................................................................
44
FIGURA 16 Aspecto morfológico de plântulas de girassol tratadas com ácido
giberélico (GA3), ácido abscísico (ABA) ou ácido indolbutírico
(IBA)..............................................................................................
45
FIGURA 17 Conteúdo de massa seca (MS) em plântulas de girassol
tratadas com ácido giberélico (GA3), ácido abscísico (ABA) ou
ácido indolbutírico (IBA)................................................................
46

FIGURA 18 Conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e
de amido em plântulas de girassol tratadas com ácido giberélico
(GA3), ácido abscísico (ABA) ou ácido indolbutírico (IBA).............
48
FIGURA 19 Conteúdo de açúcares solúveis totais (AST), de açúcares não-
redutores (ANR) e de aminoácidos livres totais (AALT) em
plântulas de girassol tratadas com ácido giberélico (GA3), ácido
abscísico (ABA) ou ácido indolbutírico (IBA).................................
50
FIGURA 20 Atividade de isocitrato liase (ICL) e de glutamina sintetase (GS)
em plântulas de girassol................................................................
52
FIGURA 21 Modelo integrativo proposto para as diferentes vias metabólicas
que utilizam os produtos de mobilização dos lipídios e das
proteínas........................................................................................
56

LISTA DE ABREVIATURAS / SIGLAS
α-KG α-cetoglutarato, do inglês α-ketoglutarate
β-O β-oxidação
AA Aminoácidos
AALT Aminoácidos livres totais
ABA Ácido abscísico, do inglês abscisic acid
AG Ácido graxo
ANR Açúcares não redutores
ASN Asparagina
ASP Aspartato
AST Açúcares solúveis totais
ATP Trifosfato de adenosina, do inglês adenosine triphosphate
CG Ciclo do glioxilato
CK Ciclo de Krebs
CL Corpos lipídicos
CT Clorofilas totais
DAE Dias após a embebição
EDTA Ácido etilenodiaminotetracético, do inglês ethylenediaminetetraacetic acid
GA3 Ácido giberélico, do inglês gibberellic acid
GA Giberelinas
GLN Glutamina

GLU Glutamato
GOX Glioxissomo
GS Glutamina sintetase, do inglês glutamine synthetase
IBA Ácido indolbutírico, do inglês indolebutyric acid
ICL Isocitrato liase, do inglês isocitrate lyase
LN Lipídios neutros
MF Massa fresca
MIT Mitocôndria
MS Massa seca
OAA Oxaloacetato, do inglês oxaloacetic acid
PRS Proteínas de reserva de sementes
PS Proteínas solúveis
SDS Dodecil sulfato de sódio, do inglês sodium dodecyl sulfate
TAG Triacilgliceróis
VEP Vacúolos de estocagem de proteínas

SUMÁRIO
1 INTRODUÇÃO.................................................................................................. 16
2 MATERIAIS E MÉTODOS................................................................................ 20
2.1 MATERIAL VEGETAL E CONDIÇÕES DE CULTIVO...................................... 20
2.2 TRATAMENTOS E COLETAS.......................................................................... 21
2.2.1 Curva de crescimento.................................................................................... 21
2.2.2 Efeito da sacarose.......................................................................................... 21
2.2.3 Efeito da L-glutamina..................................................................................... 22
2.2.4 Efeito dos hormônios..................................................................................... 22
2.3 DETERMINAÇÕES BIOQUÍMICAS.................................................................. 22
2.3.1 Lipídios neutros.............................................................................................. 22
2.3.2 Proteínas solúveis.......................................................................................... 23
2.3.3 Clorofilas totais............................................................................................... 23
2.3.4 Aminoácidos livres totais, açúcares solúveis totais e açúcares não
redutores.........................................................................................................
24
2.3.5 Amido............................................................................................................... 25
2.4 ELETROFORESE DE PROTEÍNAS EM GEL DE POLIACRILAMIDA SOB
CONDIÇÕES DESNATURANTES...................................................................
26
2.5 ATIVIDADES ENZIMÁTICAS........................................................................... 26
2.6 DELINEAMENTO EXPERIMENTAL E ANÁLISE ESTATÍSTICA 27
3 RESULTADOS 28

3.1 CARACTERIZAÇÃO DO PADRÃO DE MOBILIZAÇÃO DE RESERVAS E
PARTIÇÃO DE PRODUTOS EM GIRASSOL DURANTE O CRESCIMENTO
PÓS-GERMINATIVO........................................................................................
28
3.2 EFEITO DA CONCENTRAÇÃO EXTERNA DE SACAROSE NO
CONTEÚDO DAS RESERVAS NUTRITIVAS E PRODUTOS DE
HIDRÓLISE EM PLÂNTULAS DE GIRASSOL.................................................
35
3.3 EFEITO DA CONCENTRAÇÃO EXTERNA DE L-GLUTAMINA NO
CONTEÚDO DAS RESERVAS NUTRITIVAS E PRODUTOS DE
HIDRÓLISE EM PLÂNTULAS DE GIRASSOL.................................................
40
3.4 EFEITO DA APLICAÇÃO EXTERNA DE HORMÔNIOS NO CONTEÚDO
DAS RESERVAS NUTRITIVAS E PRODUTOS DE HIDRÓLISE EM
PLÂNTULAS DE GIRASSOL............................................................................
45
3.5 ATIVIDADE DE ISOCITRATO LIASE E GLUTAMINA SINTETASE
REGULADA POR HORMÔNIOS, FONTES DE CARBONO E NITROGÊNIO
DURANTE O CRESCIMENTO PÓS-GERMINATIVO EM GIRASSOL............
51
4 DISCUSSÃO..................................................................................................... 53
5 CONCLUSÃO................................................................................................... 63
REFERÊNCIAS................................................................................................ 64

16
1 INTRODUÇÃO
Na fase inicial de vida das plantas superiores, as necessidades energéticas e
os diversos eventos biossintéticos que ocorrem durante o estabelecimento da
plântula no ambiente são provenientes de grupos específicos de carboidratos,
lipídios e proteínas, os quais são acumulados durante o período de maturação das
sementes. Assim, as reservas nutritivas podem estar localizadas no próprio embrião
e/ou nos tecidos de armazenamento. Em dicotiledôneas não-endospérmicas, as
reservas são depositadas principalmente no mesofilo dos cotilédones, os quais
representam as primeiras folhas do embrião, e uma menor parcela é depositada no
próprio embrião (BEWLEY et al., 2013).
Dessa forma, a mobilização das reservas nutritivas nos tecidos de
armazenamento e a alocação dos seus produtos de hidrólise no eixo em
crescimento são processos críticos para o estabelecimento das plântulas após a
germinação das sementes (BUCKERIDGE et al., 2004). Para isso, é necessário que
o processo de mobilização das reservas seja sincronizado com o crescimento do
eixo, de forma que a atividade fotossintética tenha iniciado antes que as reservas
sejam exauridas (MELO et al., 2004).
Em cereais, muitos esforços já foram realizados e considerável conhecimento
já foi assimilado sobre o controle da mobilização de reservas, principalmente, no que
diz respeito à influência de hormônios sobre a biossíntese das enzimas hidrolíticas
na camada de aleurona. Como exemplo disso, já está estabelecido o papel
antagônico entre as giberelinas (GA) e o ácido abscísico (ABA) sobre a mobilização
do amido durante o crescimento pós-germinativo em cereais (BEWLEY et al., 2013).
Assim, após a germinação, as GA sintetizadas no eixo embrionário são

17
transportadas para a camada de aleurona, onde induzem a expressão dos genes, os
quais codificam as α-amilases. As α-amilases secretadas no endosperma amiláceo
hidrolisam o amido, e os produtos de mobilização são transportados para o eixo em
crescimento, atuando como substratos da respiração ou como precursores de
biomoléculas (THOMAS e RODRIGUEZ, 1994). A aplicação externa de ABA
contrabalança a ação das GA endógenas, inibindo a transcrição dos genes que
codificam as α-amilases e afetando a estabilidade dos respectivos RNAm
(BUCKERIDGE et al., 2004). Além disso, também já se sabe que a mobilização das
proteínas de reserva em cereais pode ser controlada pelo sistema tiorredoxina, que
é responsável por aumentar a solubilidade das proteínas de reserva e subsequente
proteólise (BEWLEY et al., 2013).
No entanto, muito menos se conhece a respeito do controle da mobilização de
reservas em dicotiledôneas. Nessas espécies, experimentos que demonstram o
papel de hormônios estimulando a mobilização de reservas ainda se mostram pouco
conclusivos e mal compreendidos, porque os hormônios geralmente são
consequência de um incremento na germinação e/ou crescimento da plântula e não
do controle direto sobre as enzimas hidrolíticas (BEWLEY et al., 2013).
A mobilização das reservas pode ser afetada pela biossíntese ou pela
atividade das enzimas hidrolíticas, contudo a regulação da mobilização pode ser
decorrente do controle de cada um desses eventos ou de ambos. Entretanto, não é
sempre possível determinar qual evento é responsável pela mobilização
simplesmente mensurando as taxas de redução das reservas nutritivas no decorrer
do tempo (BEWLEY et al., 2013). Além disso, muitos trabalhos sobre a mobilização
de reservas geralmente abordam a degradação de um tipo específico de reserva e
não correlacionam esse processo com as alterações bioquímicas concomitantes que

18
ocorrem no eixo em crescimento (RAMAKRISHNA e RAO, 2005; PENFIELD et al.,
2005; ELARBI et al., 2009).
Com esse panorama, em comparação com a hipótese hormonal, uma
segunda estratégia para avaliar a mobilização das reservas é através da hipótese
fonte-dreno. Conforme essa hipótese, o eixo em crescimento funciona como um
dreno, consumindo os produtos de mobilização transportados a partir das fontes, os
tecidos de armazenamento. Assim sendo, o eixo em crescimento pode limitar a
atividade das enzimas hidrolíticas nesses tecidos, por mecanismos de
retroalimentação negativa. Essa hipótese é confirmada por experimentos com
cotilédones destacados durante o crescimento pós-germinativo em leguminosas
(BEWLEY et al., 2013). A acumulação de açúcares solúveis em cotilédones
destacados pode reduzir a atividade das α-amilases, indicando o papel dos açúcares
como sinais que regulam a mobilização do amido (KARUNAGARAN e RAO, 1991).
De forma análoga, o aumento da concentração de aminoácidos livres em
cotilédones destacados pode estar relacionado com a diminuição da atividade das
proteinases, sugerindo que os aminoácidos podem mediar a regulação da
mobilização das proteínas (RAMAKRISHNA e RAO, 2005). Corroborando essas
evidências, já foi sugerido que a L-glutamina pode atuar como possível sinal para a
regulação da mobilização de proteínas (VOIGT et al., 2009) e que a aplicação
externa de açúcares reduz a expressão e a atividade de enzimas envolvidas na
degradação dos TAG, indicando que os açúcares podem também atuar como sinais
que regulam a mobilização dos lipídios (RYLOTT et al., 2001; BOREK et al., 2006).
A maioria dos trabalhos procura corroborar ou a hipótese hormonal ou a
hipótese do mecanismo fonte-dreno (KARUNAGARAN e RAO, 1991; MÜNTZ et al.,
2001; PRITCHARD et al., 2002). Contudo, alguns estudos indicam que deve haver a

19
interação entre hormônios e açúcares atuando como sinais para a regulação da
mobilização de amido (THOMAS e RODRIGUEZ, 1994) e de lipídios (TO et al.,
2002; BOREK et al., 2006) em mecanismos à longa distância.
O presente trabalho utilizou o girassol (Helianthus annuus L.) como novo
modelo experimental para dicotiledôneas, pois apresenta elevado teor de óleo e
proteínas (LIRA et al., 2009), fácil germinação, rápido crescimento, além de ser a
quarta oleaginosa mais consumida no mundo. De forma complementar, a
composição química do óleo é recomendável para os processos utilizados na
produção do biodiesel (DABDOUB e BRONZEL, 2009). Do ponto de vista
agronômico, o girassol é uma espécie produtiva sob diferentes condições climáticas,
podendo ser cultivada no Semiárido Nordestino, possibilitando a inclusão social por
meio do agronegócio (VARGAS, 2007).
A proposta desta dissertação foi de aplicar uma abordagem integrativa e de
varredura que envolvesse a mobilização das diferentes reservas, a alocação dos
diferentes produtos e o intercâmbio entre os tecidos de armazenamento e o eixo em
crescimento, seja por intermédio de hormônios ou metabólitos com papel de
sinalização. Para isso, levantamos as hipóteses de que o efeito de hormônios e de
metabólitos sobre a mobilização das reservas é diferente e de que deve existir um
efeito cruzado de açúcares sobre a mobilização de proteínas e de aminoácidos
sobre a mobilização de lipídios e amido em plântulas de girassol.

20
2 MATERIAIS E MÉTODOS
2.1 MATERIAL VEGETAL E CONDIÇÕES DE CULTIVO
O presente trabalho foi desenvolvido com sementes de girassol (Helianthus
annuus L.) híbrido Helio 253, oriundas da Empresa Heliagro Agricultura e Pecuária
Ltda. (Morumbi, SP). Para germinação das sementes, estas foram lavadas com
detergente comercial diluído, desinfetadas em câmara de fluxo laminar com etanol
70% (v/v) por 30s, seguido de NaClO 0,5% (m/v) por 5 min, sob agitação eventual.
Logo após, as sementes foram lavadas três vezes e embebidas durante 3h em água
destilada estéril. Em seguida, o semeio foi realizado em placas de Petri, entre folhas
de papel Germitest® (280 x 380 mm) umedecidas com água destilada estéril na
proporção de duas vezes e meia a massa do papel, distribuindo-se vinte sementes
por placa. As sementes foram incubadas em câmara de crescimento sob condições
controladas (radiação fotossinteticamente ativa de 80 μmol/m2/s, fotoperíodo de 12h
e temperatura de 28±2°C) durante 48h. Para o desenvolvimento das plântulas em
sistema in vitro de cultura, as sementes germinadas foram transferidas para frascos
de vidro com tampa de plástico, contendo meio ágar-água 4 g/L para o tratamento
controle e ágar-água 4 g/L suplementado para os demais tratamentos. As plântulas
foram mantidas sob as mesmas condições de crescimento. O material coletado foi
armazenado a -20°C até a realização das determinações.

21
2.2 TRATAMENTOS E COLETAS
Figura 1 - Modelo que representa o sistema de cultivo e a coleta do material vegetal. Esquema com os principais processos e pontos de coleta dos experimentos (A). Figura ilustrativa do modo de seção da plântula em cotilédones e eixo em crescimento (B).
2.2.1 Curva de crescimento
Com o intuito de acompanhar a mobilização das reservas e a partição de seus
produtos de hidrólise no decorrer do tempo, as sementes de girassol germinadas
foram cultivadas em meio ágar-água 4 g/L. As coletas foram realizadas aos 0, 1, 2,
3, 4, 5, 7 e 9 dias após a embebição (DAE), dividindo as plântulas em cotilédones e
eixo em crescimento, o que foi possível somente a partir do 2o DAE. A partir da
curva de crescimento, foi possível verificar que o 5o DAE representa um momento
crítico, no qual a maior parte das reservas nutritivas já foi mobilizada, sendo os
seguintes experimentos coletados nesse momento.
2.2.2 Efeito da sacarose
Para investigar o efeito da sacarose sobre a mobilização das reservas e a
partição de seus produtos de hidrólise, as sementes de girassol germinadas foram
cultivadas em meio ágar-água 4 g/L suplementado com 0 (controle), 50, 100 ou 200
mM de sacarose durante 3 dias. Assim sendo, as plântulas foram coletadas no 5o
DAE, separando os cotilédones do eixo em crescimento.
A B

22
2.2.3 Efeito da L-glutamina
Para testar o efeito da L-glutamina sobre a mobilização das reservas e a
partição de seus produtos de hidrólise, as sementes de girassol germinadas foram
cultivadas em meio ágar-água 4 g/L suplementado com 0 (controle), 5, 10 ou 20 mM
de L-glutamina durante 3 dias. Desse modo, no 5o DAE, as plântulas foram
coletadas e divididas em cotilédones e eixo em crescimento.
2.2.4 Efeito dos hormônios
No intuito de verificar o efeito dos hormônios sobre a mobilização das
reservas e a partição de seus produtos de hidrólise, as sementes de girassol
germinadas foram cultivadas em meio ágar-água 4 g/L sem adições (controle) ou em
meio ágar-água 4 g/L suplementado com ácido giberélico (GA3) 50 µM, ácido
abscísico (ABA) 50 µM ou ácido indolbutírico (IBA) 50 µM, durante 3 dias. Assim, as
plântulas foram coletadas no 5o DAE, separando os cotilédones do eixo em
crescimento.
2.3 DETERMINAÇÕES BIOQUÍMICAS
2.3.1 Lipídios neutros (LN)
A quantificação dos LN foi realizada pelo método gravimétrico. Para tanto,
amostras de cotilédones com aproximadamente 100 mg de massa seca foram
pulverizadas em gral e pistilo, e os LN foram extraídos com 8 mL de n-hexano a
60°C, durante 5 h, sob agitação eventual. Em seguida, o sobrenadante foi
transferido para tubos de plástico de massa conhecida. Após a evaporação do n-

23
hexano a 80°C, a massa de lipídios foi calculada a partir da diferença entre a massa
inicial e a final dos tubos e expressa em mg de LN/cotilédone.
2.3.2 Proteínas solúveis (PS)
Para a extração de PS a partir dos cotilédones, amostras com cerca de 200
mg de massa fresca foram maceradas durante 5 min com 1,5 mL de tampão Tris-
HCl 100 mM pH 7,0 com adição de NaCl 500 mM e 2-mercaptoetanol 2 mM. Após
centrifugação a 10.000 xg por 10 min, os sobrenadantes foram coletados, e os
precipitados foram reextraídos com 1 mL do tampão de extração por mais duas
vezes. Ao final, os sobrenadantes foram reunidos, perfazendo 3,5 mL de extrato total
por amostra. As PS do eixo em crescimento, por sua vez, foram extraídas com
tampão Tris-HCl 100 mM pH 7,0, seguindo os procedimentos supracitados.
A determinação das PS foi realizada de acordo com o método de Bradford
(1976), utilizando albumina sérica bovina (BSA) como padrão. O conteúdo de PS
dos cotilédones foi expresso em mg de PS/cotilédone e o do eixo em crescimento,
em mg de PS/g de MS.
2.3.3 Clorofilas totais (CT)
A extração de CT foi realizada a partir de amostras com aproximadamente
200 mg de massa fresca. Para isso, 5 mL de acetona 80% (v/v) com adição de
MgCO3 0,5% (m/v) foram adicionados em tubos com tampas, os quais continham as
amostras picotadas. Os tubos foram fechados e incubados à temperatura ambiente,
durante 30 min sob agitação eventual. Após a coleta dos sobrenadantes, os
resíduos foram reextraídos com mais 5 mL de acetona 80% (v/v) com adição de

24
MgCO3 0,5% (m/v) sob as mesmas condições. Em seguida, os sobrenadantes foram
reunidos, perfazendo 10 mL de extrato total por amostra.
A dosagem de CT foi realizada pelo método de Arnon (1949). Para cada
determinação, foram adicionados 2,5 mL da amostra, e as leituras foram realizadas
a 645 e 663 nm. O aparelho foi zerado com o meio extrator. A quantidade de CT foi
calculada de acordo com a fórmula: (CT (mg/L) = 0,0127 × A663 - 0,00269 × A645). O
conteúdo de CT foi expresso em mg de CT/g de MS.
2.3.4 Aminoácidos livres totais (AALT), açúcares solúveis totais (AST) e
açúcares não redutores (ANR)
A extração de AALT, AST e ANR foi realizada a partir de amostras com
aproximadamente 200 mg de massa fresca. Para isso, 5 mL de etanol 80% (v/v)
foram adicionados em tubos hermeticamente fechados, contendo as amostras
picotadas, e incubados a 60°C, durante 30 min. Após a coleta dos sobrenadantes,
os resíduos foram reextraídos com 5 mL de etanol 80% (v/v) sob as mesmas
condições. Em seguida, os sobrenadantes foram reunidos, perfazendo 10 mL de
extrato total por amostra, enquanto os resíduos foram utilizados para a extração e a
determinação de amido. A dosagem de AALT foi realizada pelo método de Peoples
(1989), com a utilização do reagente de ninidrina. Para cada determinação, foram
adicionados 100 μL da amostra;; 400 μL de água destilada;; 250 μL de tampão citrato
200 mM pH 5,0;; e 250 μL do reagente de ninidrina (1 mL de cianeto de potássio 10
mM; 59 mL de metoxietanol P.A.; 0,5 g de ninidrina). Os tubos foram vedados e
incubados a 100°C, durante 15 min. Após resfriamento, foram adicionados 4 mL de
etanol 50% (v/v) em cada amostra. As leituras foram realizadas a 570 nm, e a
quantidade de AALT foi calculada de acordo com uma curva padrão de L-glutamina,

25
sendo expressa em µmol de AALT/g de MS. Para a dosagem de AST, foi utilizado o
método de Dubois (1956) com algumas modificações. Em cada determinação, foram
adicionados 100 μL da amostra;; 400 μL de água destilada;; 500 μL de fenol 5%
(m/v); e 2,5 mL de ácido sulfúrico (H2SO4) 90% (v/v). A leitura foi realizada a 490 nm,
e o cálculo da quantidade de AST foi baseado em uma curva padrão de D-glicose,
sendo expressa em mmol de AST/g de MS. A dosagem de ANR foi realizada pelo
método de Morris (1948) com algumas modificações, utilizando o reagente de
antrona (MORRIS 1948;; YEMM e WILLIS 1954). Em cada determinação, 100 μL da
amostra;; 800 μL de água destilada;; e 100 μL de KOH 30% (m/v) foram pré-
incubados a 100°C, durante 10 min. Após resfriamento, 2,5 mL do reagente de
antrona foram adicionados em cada amostra, e foi realizada incubação a 40°C,
durante 15 min. A leitura foi realizada a 620 nm, e a quantidade de ANR das
amostras foi calculada a partir de uma curva padrão de sacarose, sendo expressa
em mmol de ANR/g de MS.
2.3.5 Amido
A extração de amido foi realizada utilizando os resíduos da extração dos
compostos solúveis de baixa massa molecular (AALT, AST e ANR). Para tanto, os
resíduos foram macerados durante 5 min com 1,5 mL de ácido perclórico 30% (v/v).
Após centrifugação a 10.000 xg, durante 10 min, os sobrenadantes foram coletados,
e os precipitados foram reextraídos com 1 mL de ácido perclórico 30% (v/v) por mais
duas vezes. Ao final, os sobrenadantes foram reunidos, perfazendo 3,5 mL de
extrato total por amostra.
A dosagem de amido foi realizada com a utilização do reagente de antrona
(MORRIS 1948; YEMM e WILLIS 1954). Para cada determinação, foram utilizados

26
100 μL da amostra;; 900 μL de água destilada;; e 2,5 mL do reagente de antrona. A
leitura foi realizada a 620 nm, utilizando uma curva padrão de D-glicose. Os valores
obtidos foram multiplicados pelo fator 0,9 para conversão em amido, segundo
McCready et al. (1950), sendo expressos em mg de amido/parte.
2.4 ELETROFORESE DE PROTEÍNAS EM GEL DE POLIACRILAMIDA SOB
CONDIÇÕES DESNATURANTES (SDS-PAGE)
Os experimentos de SDS-PAGE, em presença de 2-mercaptoetanol, foram
conduzidos seguindo-se a técnica descrita por Laemmli (1970). As PS dos
cotilédones de girassol foram separadas em géis de poliacrilamida 12% (m/v) sob
condições de corrida de 200 Volts, 50 mA, durante 1h20. A revelação dos géis foi
feita utilizando-se o corante Azul Brilhante de Coomassie R-250 0,1% (m/v) em
metanol 40% (v/v) e em ácido acético 10% (v/v), durante 30 min. A descoloração foi
realizada por meio de lavagens sucessivas em solução de metanol 40% (v/v) e ácido
acético 10% (v/v). Os géis foram digitalizados.
2.5 ATIVIDADES ENZIMÁTICAS
Para obter extratos enzimáticos, amostras de cotilédones com cerca de 200
mg de massa fresca foram pulverizadas em nitrogênio líquido, e as enzimas foram
extraídas com 1,5 mL de tampão de extração (isocitrato liase (ICL) - tampão fosfato
de potássio 100 mM pH 7,6 suplementado com MgCl2 1 mM e 2-mercaptoetanol 10
mM; glutamina sintetase (GS) - tampão Tris-HCl 100 mM pH 7,6 suplementado com
EDTA 1 mM, MgCl2 1 mM, PMSF 1 mM e PVP 2,5% [m/v]) por maceração, durante

27
5 min. Após centrifugação a 10.000 xg a 4°C, durante 10 min, os sobrenadantes
foram coletados e utilizados como extratos enzimáticos.
A atividade da ICL foi estimada pelo método de Chell et al. (1978). Para cada
determinação, foram adicionados 1 mL de tampão imidazol 50 mM pH 6,8; 200 µL
de MgCl2 50 mM; 200 µL de EDTA 10 mM; 200 µL de fenilhidrazina-HCl 40 mM; 200
µL de DL-isocitrato 10 mM; e 200 µL do extrato enzimático. A atividade enzimática
foi calculada a partir da absorbância do complexo glioxilato-fenilhidrazina a 324 nm,
a cada minuto, durante 10 min, e expressa em mmol de glioxilato-
fenilhidrazina/min/g de MS. A atividade da GS foi determinada pelo método descrito
por Berteli et al. (1995). Em cada determinação, foram utilizados 400 µL de tampão
Tris-HCl 250 mM pH 7,0; 100 µL de ATP 30 mM pH 7,0; 100 µL de MgSO4 500 mM;
100 µL de cloridrato de hidroxilamina 500 mM; 100 µL de glutamato de sódio 500
mM e 200 µL de extrato enzimático. As misturas foram incubadas a 30°C durante 30
min, e a atividade enzimática foi determinada pela absorbância do produto de reação
entre γ-glutamil hidroxamato e o FeCl3 a 540 nm e expresso em nmol de γ-glutamil
hidroxamato/min/g de MS.
2.6 DELINEAMENTO EXPERIMENTAL E ANÁLISE ESTATÍSTICA
O experimento foi realizado em delineamento inteiramente casualizado com
os tratamentos dispostos em fatorial 1 x 4, incluindo o híbrido Helio 253 de girassol e
quatro tratamentos (sacarose 0, 50, 100 ou 200 mM; ou L-glutamina 0, 5, 10 ou 20
mM; ou controle, ABA 50 µM, GA3 50 µM ou IBA 50 µM), com cinco repetições por
tratamento. Os dados foram submetidos ao teste de Tukey ao nível de significância
de 5%.

28
3 RESULTADOS
3.1 CARACTERIZAÇÃO DO PADRÃO DE MOBILIZAÇÃO DE RESERVAS E
PARTIÇÃO DE PRODUTOS EM GIRASSOL DURANTE O CRESCIMENTO PÓS-
GERMINATIVO
O aspecto morfológico foi acompanhado, e os padrões de mobilização das
reservas nutritivas e de partição dos seus produtos de hidrólise foram caracterizados
durante o crescimento pós-germinativo de plântulas de girassol híbrido Helio 253,
até o 9o dia após a embebição (DAE) em meio de cultura sem adição de nutrientes.
A partir da caracterização morfológica, foi possível verificar o crescimento das
plântulas no decorrer do tempo, o desenvolvimento da porção radicular a partir do 4o
DAE e da porção aérea pela mudança de coloração nos cotilédones, do amarelado
para o verde, entre o 3o e 4o DAE (Fig. 2).
Figura 2 - Aspecto morfológico de plântulas de girassol durante o crescimento pós-germinativo. Plântulas de girassol híbrido Helio 253 durante o crescimento pós-germinativo até o 9o dia após a embebição (DAE) crescidas em meio ágar-água 4 g/L sem adição de nutrientes. A barra vertical representa a escala de 1 cm. O conteúdo de massa seca (MS) foi diminuído nos cotilédones e
paralelamente aumentado no eixo em crescimento até o 9o DAE (Fig. 3). De fato,
1o DAE 2o DAE 3o DAE 4o DAE 5o DAE 7o DAE 9o DAE

29
houve diminuição de, aproximadamente, 1,4 vezes na MS dos cotilédones ao 9o
DAE em relação às sementes não germinadas (Fig. 3A), e ocorreu incremento de
cerca de 6 vezes na MS do eixo em crescimento ao 9o DAE em comparação com o
2o DAE (Fig. 3B).
Figura 3 - Conteúdo de massa seca (MS) em plântulas de girassol durante o crescimento pós-germinativo. Conteúdos de MS nos cotilédones (A) e no eixo em crescimento (B) de plântulas de girassol híbrido Helio 253 durante o crescimento pós-germinativo até o 9o dia após a embebição (DAE) em meio ágar-água 4 g/L sem adição de nutrientes. Os pontos representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. O conteúdo de lipídios neutros (LN) e de proteínas solúveis (PS) nos
cotilédones das plântulas de girassol, expresso em mg/parte, diminuiu de forma
coordenada durante o crescimento pós-germinativo inicial (Fig. 4A, B e D). Até o 3o
DAE, o conteúdo de LN e de PS foi reduzido de forma mais branda, apresentando
cerca de 25 e 9% de redução, respectivamente, em relação ao início do
experimento. No entanto, o conteúdo de LN e de PS foi diminuído de modo mais
drástico até o 5o DAE, mostrando aproximadamente 67 e 72% de diminuição, nessa
ordem, em comparação com as sementes não germinadas. Ao final do experimento,

30
restaram somente 15 e 2% do conteúdo inicial de LN e de PS, respectivamente. No
eixo em crescimento, o conteúdo de PS, expresso em mg/g MS, diminuiu
drasticamente do 2o ao 4o DAE, apresentando cerca de 73% de redução. A partir do
5o DAE até o final do experimento, o conteúdo de PS diminuiu de forma mais
branda, restando aproximadamente 13% do conteúdo inicial (Fig. 4C).
Figura 4 - Conteúdo de lipídios neutros (LN) e de proteínas solúveis (PS) em plântulas de girassol durante o crescimento pós-germinativo. Conteúdo de LN nos cotilédones (A) e de PS (PS) nos cotilédones (B) e no eixo em crescimento (C) de plântulas de girassol híbrido Helio 253 durante o crescimento pós-germinativo até o 9o dia após a embebição (DAE) em meio ágar-água 4 g/L sem adição de nutrientes. Os pontos representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. A sobreposição das curvas obtidas para LN e PS está representada em (D). Para acompanhar de forma qualitativa as alterações observadas no conteúdo
de PS cotiledonares, foi obtido o perfil eletroforético sob condições desnaturantes e
redutoras (Fig. 5). De acordo com esse ensaio, foi possível reconhecer os
polipeptídeos acídicos α e α’ (α, peso molecular aparente = 36,8 - 42,9 kDa;; α’, peso
molecular aparente = 31,0 - 35,3 kDa) e o polipeptídeo básico β (β, peso molecular
aparente = 21,0 - 29,6 kDa) da heliantinina, globulina 11S característica de girassol

31
(ŽILIĆ et al., 2010). Além disso, foi possível constatar que as subunidades α e α’ são
mobilizadas precocemente, a partir do 2o DAE, enquanto que a subunidade β é
mobilizada tardiamente, a partir do 4o DAE. Além da heliantinina, foi verificada a
presença da fração albumina 2S (peso molecular aparente = 11,5 - 21,1 kDa) (ŽILIĆ
et al., 2010), a qual parece ser mobilizada de forma tardia, a partir do 3o DAE (Fig.
5). O perfil de PS se manteve inalterado a partir do 7o DAE (dados não
apresentados).
Figura 5 - Perfil eletroforético das proteínas solúveis (PS). PS provenientes de cotilédones de girassol híbrido Helio 253 durante o desenvolvimento pós-germinativo, no decorrer de 7 dias após a embebição (DAE), em meio ágar-água 4 g/L sem adição de nutrientes. As PS foram separadas por eletroforese em gel de poliacrilamida 12% sob condições desnaturantes e redutoras (SDS-PAGE com 2-mercaptoetanol). α e α’ indicam os polipeptídeos acídicos e β indica o polipeptídeo básico da heliantinina, globulina 11S característica do girassol. M indica o marcador de massas moleculares em kDa. (indicação para volume)
O conteúdo de amido nos cotilédones diminuiu cerca de 61% até o 2o DAE
em comparação com as sementes não germinadas (Fig. 6A). Do 3o ao 4o DAE,
houve aumento de aproximadamente 39% no conteúdo de amido, o qual se manteve
inalterado até o final do experimento. De forma inversa, o conteúdo de amido
M

32
aumentou gradativamente no eixo em crescimento, após a germinação. Com efeito,
o conteúdo de amido apresentou incremento cerca de 11 vezes ao 9o DAE em
referência ao início do experimento (Fig. 6B).
Nos cotilédones, o conteúdo de clorofilas totais (CT) aumentou a partir do 4o
DAE, atingindo 5,5 vezes mais CT ao 9o DAE em comparação com o tempo inicial
(Fig. 3.1.5C). É importante destacar que o aumento do teor de CT (Fig. 6C) coincide
com o aumento do conteúdo de amido em ambas as partes (Fig. 6A e B) e o
aumento do conteúdo da MS total na plântula (Fig. 6D).
Figura 6 - Conteúdo de amido, de clorofilas totais (CT) e de massa seca (MS) total em plântulas de girassol durante o crescimento pós-germinativo. Conteúdo de amido nos cotilédones (A) e no eixo em crescimento (B), conteúdo de CT nos cotilédones (C) e conteúdo de MS total (D) de plântulas de girassol híbrido Helio 253 durante o crescimento pós-germinativo até o 9o dia após a embebição (DAE) em meio ágar-água 4 g/L sem adição de nutrientes. Os pontos representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições.
O conteúdo de açúcares solúveis totais (AST) foi incrementado nos
cotilédones e paralelamente diminuído no eixo em crescimento, durante o período
experimental (Fig. 7A e B). De fato, o conteúdo de AST nos cotilédones aumentou

33
cerca de 3,8 vezes até 9o DAE em comparação com as sementes não germinadas,
enquanto que o conteúdo de AST no eixo em crescimento diminuiu
aproximadamente 30% ao 9o DAE em relação ao 2o DAE.
Nos cotilédones e no eixo em crescimento, foram verificadas alterações
concomitantes no conteúdo de açúcares não-redutores (ANR) ao longo do
experimento. Embora o conteúdo de ANR tenha diminuído até o 2o DAE apenas nos
cotilédones, ocorreu acumulação de ANR no 4o DAE, seguida de diminuição do
conteúdo desses compostos ao 5o DAE, em ambas as partes. Além disso, é
importante destacar que a acumulação de AST e ANR nos cotilédones, a partir do 4o
DAE (Fig. 7A e C), coincide com a degradação dos LN (Fig. 4A), o aumento do
conteúdo de clorofilas totais (Fig. 6C) e a MS total (Fig. 6D) nos cotilédones no
mesmo período.
O conteúdo de aminoácidos livres totais (AALT) nos cotilédones aumentou de
forma coordenada com a mobilização das proteínas de reserva, verificada pelo perfil
eletroforético, durante o crescimento pós-germinativo (Fig. 7E; Fig. 5). Nos
cotilédones, o conteúdo de AALT aumentou cerca de 168 vezes no 2o DAE em
relação ao tempo inicial, manteve-se estável até o 5o DAE e aumentou novamente
ao 9o DAE, atingindo aproximadamente 302 vezes mais AALT que as sementes não
germinadas. No eixo em crescimento, o conteúdo de AALT sofreu diminuição de
cerca de 94% até o 4o DAE, restando aproximadamente 2% do conteúdo inicial no 9o
DAE (Fig. 7F).

34
Figura 7 - Conteúdo de açúcares solúveis totais (AST), de açúcares não-redutores (ANR) e de aminoácidos livres totais (AALT) em plântulas de girassol durante o crescimento pós-germinativo. Conteúdo de AST nos cotilédones (A) e no eixo em crescimento (B), de ANR nos cotilédones (C) e no eixo em crescimento (D) e de AALT nos cotilédones (E) e no eixo em crescimento (F) de plântulas de girassol híbrido Helio 253 durante o crescimento pós-germinativo até o 9o dia após a embebição (DAE) em meio ágar-água 4 g/L sem adição de nutrientes. Os pontos representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições.

35
3.2 EFEITO DA CONCENTRAÇÃO EXTERNA DE SACAROSE NO CONTEÚDO
DAS RESERVAS NUTRITIVAS E PRODUTOS DE HIDRÓLISE EM PLÂNTULAS DE
GIRASSOL
O aspecto morfológico foi acompanhado, e o conteúdo das reservas nutritivas
e de seus produtos de hidrólise foi caracterizado no 5o dia após a embebição (DAE)
em plântulas de girassol híbrido Helio 253, crescidas durante três dias em meio de
cultura sem adições (controle) ou suplementado com sacarose 50, 100 ou 200 mM.
As plântulas tratadas com diferentes concentrações de sacarose reduziram de
tamanho e apresentaram cotilédones com coloração mais amarelada, em relação ao
controle (Fig. 8).
Figura 8 - Aspecto morfológico de plântulas de girassol tratadas com concentrações crescentes de sacarose. Plântulas de girassol híbrido Helio 253 foram cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 50, 100 ou 200 mM de sacarose. A barra vertical representa a escala de 1 cm. O aumento da concentração externa de sacarose acarretou o incremento do
conteúdo de massa seca (MS) nos cotilédones e no eixo em crescimento em relação
ao tratamento controle (Fig. 9). Com efeito, houve aumento significativo de 32 e 13%
na MS dos cotilédones e do eixo em crescimento, respectivamente, em plântulas
tratadas com 200 mM de sacarose em relação ao controle (Fig. 9A).
Controle 50 mM 100 mM 200 mM
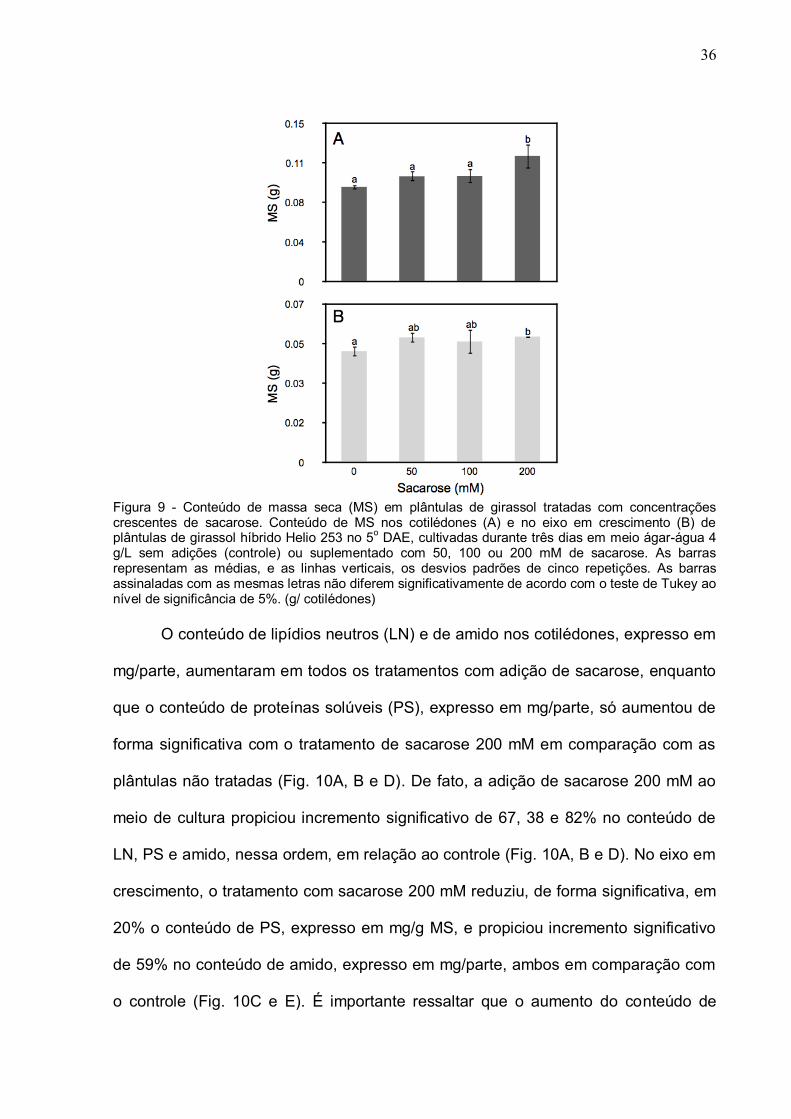
36
Figura 9 - Conteúdo de massa seca (MS) em plântulas de girassol tratadas com concentrações crescentes de sacarose. Conteúdo de MS nos cotilédones (A) e no eixo em crescimento (B) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 50, 100 ou 200 mM de sacarose. As barras representam as médias, e as linhas verticais, os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%. (g/ cotilédones) O conteúdo de lipídios neutros (LN) e de amido nos cotilédones, expresso em
mg/parte, aumentaram em todos os tratamentos com adição de sacarose, enquanto
que o conteúdo de proteínas solúveis (PS), expresso em mg/parte, só aumentou de
forma significativa com o tratamento de sacarose 200 mM em comparação com as
plântulas não tratadas (Fig. 10A, B e D). De fato, a adição de sacarose 200 mM ao
meio de cultura propiciou incremento significativo de 67, 38 e 82% no conteúdo de
LN, PS e amido, nessa ordem, em relação ao controle (Fig. 10A, B e D). No eixo em
crescimento, o tratamento com sacarose 200 mM reduziu, de forma significativa, em
20% o conteúdo de PS, expresso em mg/g MS, e propiciou incremento significativo
de 59% no conteúdo de amido, expresso em mg/parte, ambos em comparação com
o controle (Fig. 10C e E). É importante ressaltar que o aumento do conteúdo de

37
amido nos cotilédones e no eixo em crescimento foi dependente da concentração
externa de sacarose (Fig. 10D e E). Além disso, pode-se destacar que a diminuição
do conteúdo de PS no eixo em crescimento coincidiu com o aumento do conteúdo
de PS nos cotilédones para as plântulas tratadas com diferentes concentrações de
sacarose (Fig. 10B e C).
Figura 10 - Conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e de amido em plântulas de girassol tratadas com concentrações crescentes de sacarose. Conteúdo de LN nos cotilédones (A), de PS nos cotilédones (B) e no eixo em crescimento (C) e de amido nos cotilédones (D) e no eixo em crescimento (E) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 50, 100 ou 200 mM de sacarose. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

38
Apenas sob o tratamento com sacarose 200 mM, o conteúdo de açúcares
solúveis totais (AST) foi incrementado nos cotilédones de forma significativa,
atingindo 38% mais AST em relação às plântulas não tratadas (Fig. 11A). No eixo
em crescimento, o conteúdo de AST aumentou significativamente em 23 e 37% para
os tratamentos com sacarose 100 e 200 mM, nessa ordem, em comparação ao
controle (Fig. 11B). O conteúdo de açúcares não-redutores (ANR) nos cotilédones e
no eixo em crescimento aumentou de forma dependente da concentração externa de
sacarose (Fig. 11C e D). Com efeito, o conteúdo de ANR nos cotilédones e no eixo
em crescimento aumentou significativamente em 15 e 6 vezes, respectivamente, no
tratamento com sacarose 200 mM em referência ao controle. É notável que o
aumento no conteúdo de AST nos cotilédones e no eixo em crescimento (Fig. 11A e
B), resultante dos tratamentos com sacarose, foi representado principalmente pelo
incremento de ANR nos mesmos órgãos (Fig. 11C e D).
O conteúdo de aminoácidos livres totais (AALT) nos cotilédones aumentou,
de forma significativa, em 48% para todos os tratamentos com adição de sacarose
em comparação com as plântulas não tratadas (Fig. 11E). No eixo em crescimento,
o aumento da concentração externa de sacarose não ocasionou alterações
significativas no conteúdo de AALT (Fig. 11F). É importante evidenciar que o
aumento do conteúdo de AALT nos cotilédones de plântulas tratadas com sacarose
coincide com o atraso na mobilização de PS e com a diminuição do conteúdo de PS
no eixo em crescimento em referência ao controle (Fig. 10B e C; Fig. 11F).

39
Figura 11 - Conteúdo de açúcares solúveis totais (AST), de açúcares não-redutores (ANR) e de aminoácidos livres totais (AALT) em plântulas de girassol tratadas com concentrações crescentes de sacarose. Conteúdo de AST nos cotilédones (A) e no eixo em crescimento (B), de ANR nos cotilédones (C) e no eixo em crescimento (D) e de AALT nos cotilédones (E) e no eixo em crescimento (F) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 50, 100 ou 200 mM de sacarose. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

40
3.3 EFEITO DA CONCENTRAÇÃO EXTERNA DE L-GLUTAMINA NO CONTEÚDO
DAS RESERVAS NUTRITIVAS E PRODUTOS DE HIDRÓLISE EM PLÂNTULAS DE
GIRASSOL
Após três dias de incubação em meio de cultura sem adições (controle) ou
suplementado com L-glutamina 5, 10 e 20 mM, foi caracterizado o conteúdo das
reservas nutritivas e de seus produtos de hidrólise e acompanhado o aspecto
morfológico de plântulas de girassol híbrido Helio 253 no 5o dia após a embebição
(DAE). As plântulas tratadas com diferentes concentrações de L-glutamina
reduziram de tamanho, apresentaram uma menor quantidade de raízes secundárias
nos tratamentos de 10 e 20 mM de L-glutamina e, no tratamento com 20 mM de L-
glutamina, os cotilédones apresentaram coloração mais amarelada, isso quando
comparadas com o controle (Fig. 12).
Figura 12 - Aspecto morfológico de plântulas de girassol tratadas com concentrações crescentes de L-glutamina. Plântulas de girassol híbrido Helio 253 foram cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 5, 10 ou 20 mM de L-glutamina. A barra vertical representa a escala de 1 cm. Nos cotilédones, somente a aplicação externa de L-glutamina 20 mM
aumentou significativamente o conteúdo de massa seca (MS) em 20% em
comparação aos demais tratamentos (Fig. 13A). Já no eixo em crescimento, o
Controle 5 mM 10 mM 20 mM

41
conteúdo de MS foi significativamente reduzido em 34%, para o tratamento com L-
glutamina 20 mM em relação ao controle (Fig. 13B).
Figura 13 - Conteúdo de massa seca (MS) em plântulas de girassol tratadas com concentrações crescentes de L-glutamina. Conteúdo de MS nos cotilédones (A) e no eixo em crescimento (B) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 5, 10 ou 20 mM de L-glutamina. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%. O conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e de amido
nos cotilédones, expressos em mg/parte, aumentaram significativamente em quase
todos os tratamentos com adição de L-glutamina quando comparados com o
controle (Fig. 14A, B e D). A adição de L-glutamina 20 mM ao meio de cultura
propiciou o incremento de 67, 55 e 67% no conteúdo de LN, de PS e de amido,
nessa ordem, em relação às plântulas não tratadas (Fig. 14A, B e D). No eixo em
crescimento, o tratamento com L-glutamina 20 mM ocasionou o incremento de 14 e
53% no conteúdo de PS (mg/g MS) e de amido (mg/parte), respectivamente, em
referência ao controle (Fig. 14C e E). É importante ressaltar que os tratamentos com

42
aplicação externa de L-glutamina causaram incremento significativo no conteúdo das
reservas de carbono (LN e amido) nos cotilédones de plântulas de girassol (Fig. 14A
e D).
Figura 14 - Conteúdo de lipídios neutros (LN), de proteínas solúveis (PS) e de amido em plântulas de girassol tratadas com concentrações crescentes de L-glutamina. Conteúdo de LN nos cotilédones (A), de PS nos cotilédones (B) e no eixo em crescimento (C) e de amido nos cotilédones (D) e no eixo em crescimento (E) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 5, 10 ou 20 mM de L-glutamina. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

43
Em todos os tratamentos com L-glutamina, o conteúdo de açúcares solúveis
totais (AST) foi incrementado, de forma significativa, nos cotilédones atingindo, em
média, 19% mais AST em comparação com as plântulas não tratadas (Fig. 15A). Em
contraponto, concentrações externas crescentes de L-glutamina não acarretaram
mudanças significativas no conteúdo de AST no eixo em crescimento (Fig. 15B). De
forma adicional, o conteúdo de açúcares não-redutores (ANR) nos cotilédones e no
eixo em crescimento aumentou significativamente conforme o incremento da
concentração externa de L-glutamina (Fig. 15C e D). De fato, o conteúdo de ANR
nos cotilédones e no eixo em crescimento aumentou 4 vezes e 68%,
respectivamente, no tratamento com L-glutamina 20 mM em referência ao controle.
O conteúdo de aminoácidos livres totais (AALT) nos cotilédones não
apresentou alterações significativas entre os diferentes tratamentos (Fig. 15E). No
eixo em crescimento, o conteúdo de AALT aumentou de forma dependente da
concentração externa de L-glutamina (Fig. 15F). Com efeito, o conteúdo de AALT
aumentou 7 vezes no tratamento com L-glutamina 20 mM em relação ao controle. É
importante evidenciar que, sob L-glutamina 20 mM, a acumulação de AALT no eixo
em crescimento (Fig. 15F) coincide com o acúmulo de LN nos cotilédones, e com o
de PS e com o de amido nos cotilédones e no eixo em crescimento de plântulas de
girassol (Fig. 14).

44
Figura 15 - Conteúdo de açúcares solúveis totais (AST), de açúcares não-redutores (ANR) e de aminoácidos livres totais (AALT) em plântulas de girassol tratadas com concentrações crescentes de L-glutamina. Conteúdo de AST nos cotilédones (A) e no eixo em crescimento (B), de ANR nos cotilédones (C) e no eixo em crescimento (D) e de AALT nos cotilédones (E) e no eixo em crescimento (F) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com 5, 10 ou 20 mM de L-glutamina. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

45
3.4 EFEITO DA APLICAÇÃO EXTERNA DE HORMÔNIOS NO CONTEÚDO DAS
RESERVAS NUTRITIVAS E PRODUTOS DE HIDRÓLISE EM PLÂNTULAS DE
GIRASSOL
O aspecto morfológico foi acompanhado, e o conteúdo das reservas nutritivas
e de seus produtos de hidrólise foi caracterizado em plântulas de girassol híbrido
Helio 253, no 5o dia após a embebição (DAE), após três dias de tratamento em meio
de cultura ágar-água 4 g/L sem adições (controle) ou suplementado com ácido
giberélico (GA3) 50 µM, ácido abscísico (ABA) 50 µM ou ácido indolbutírico (IBA) 50
µM. O tratamento com GA3 proporcionou indução do crescimento em relação ao
controle (Fig. 16). As plântulas tratadas com ABA apresentaram retardo no
crescimento, ausência de raízes secundárias e cotilédones amarelados em
comparação com o controle (Fig. 16). Aquelas tratadas com IBA apresentaram
retardo no crescimento, desenvolvimento anormal da porção radicular e cotilédones
amarelados quando comparadas com o controle (Fig. 16).
Figura 16 - Aspecto morfológico de plântulas de girassol tratadas com ácido giberélico (GA3), ácido abscísico (ABA) ou ácido indolbutírico (IBA). Plântulas de girassol híbrido Helio 253 foram cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com GA3 50 µM, ABA 50 µM ou IBA 50 µM. A barra vertical representa a escala de 1 cm.
Controle GA3 ABA IBA

46
Somente a aplicação externa de ABA e IBA ocasionou o incremento
significativo do conteúdo de massa seca (MS) nos cotilédones em comparação com
o controle. De fato, o conteúdo de MS dos cotilédones aumentou 44 e 33% nos
tratamentos com ABA e IBA, nessa ordem, em relação ao controle (Fig. 17A). Já no
eixo em crescimento, o conteúdo de MS foi reduzido de forma significativa em 63 e
54%, para os tratamentos com ABA e IBA, respectivamente, em comparação com as
plântulas não tratadas (Fig. 17B).
Figura 17 - Conteúdo de massa seca (MS) em plântulas de girassol tratadas com ácido giberélico (GA3), ácido abscísico (ABA) ou ácido indolbutírico (IBA). Conteúdo de MS nos cotilédones (A) e no eixo em crescimento (B) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com GA3 50 µM, ABA 50 µM ou IBA 50 µM. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

47
O conteúdo de lipídios neutros (LN) nos cotilédones, expresso em mg/parte,
aumentou significativamente em 2,2 e 1,8 vezes nos tratamentos com ABA e IBA,
nessa ordem, em referência ao controle (Fig. 18A). Do mesmo modo, o conteúdo de
proteínas solúveis (PS), expresso em mg/parte, foi incrementado de forma
significativa em 2,0 e 1,4 vezes nos tratamentos com ABA e IBA, respectivamente,
em comparação com as plântulas não tratadas (Fig. 18B). O conteúdo de PS no eixo
em crescimento, expresso em mg/g MS, aumentou significativamente em 23 e 74%
nos tratamentos com ABA e IBA, nessa ordem, enquanto que houve diminuição
significativa de 29% no tratamento com GA3 em relação ao controle (Fig. 18C). O
conteúdo de amido nos cotilédones e no eixo em crescimento, expresso em
mg/parte, diminuiu significativamente em 38 e 45%, respectivamente, no tratamento
com ABA em comparação com as plântulas não tratadas (Fig. 18D e E). É
importante destacar que os tratamentos com ABA e IBA causaram aumento nos
conteúdos das reservas de LN e PS nos cotilédones, ao passo que o conteúdo de
amido foi reduzido, em comparação com o controle (Fig. 18A, B e D).

48
Figura 18 - Conteúdo de lipídios neutros (LN), proteínas solúveis (PS) e amido em plântulas de girassol tratadas com ácido giberélico (GA3), ácido abscísico (ABA) ou ácido indolbutírico (IBA). Conteúdo de LN nos cotilédones (A), de PS nos cotilédones (B) e no eixo em crescimento (C) e de amido nos cotilédones (D) e no eixo em crescimento (E) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com GA3 50 µM, ABA 50 µM ou IBA 50 µM. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.
O conteúdo de açúcares solúveis totais (AST) nos cotilédones diminuiu
significativamente em 90 e 39% nos tratamentos com ABA e IBA, respectivamente,
em comparação com o controle (Fig. 19A). No eixo em crescimento, o conteúdo de
AST foi reduzido, de forma significativa, em 56 e 79% nos tratamentos com ABA e
IBA, nessa ordem, em relação ao controle (Fig. 19B). Nos cotilédones de plântulas

49
tratadas com IBA, o conteúdo de açúcares não-redutores (ANR) aumentou
significativamente em 4 vezes em referência às plântulas não tratadas (Fig. 19C). Os
tratamentos com ABA e IBA ocasionaram incremento significativo de 2 e 3,1 vezes,
nessa ordem, no conteúdo de ANR do eixo em crescimento em relação ao controle
(Fig. 19D).
O conteúdo de aminoácidos livres totais (AALT) nos cotilédones aumentou
significativamente em 30% devido ao tratamento com ABA em comparação ao
controle (Fig. 19E). No eixo em crescimento, ocorreu aumento significativo do
conteúdo de AALT nos tratamentos com ABA e IBA, atingindo 9,6 e 2,9 vezes mais
AALT em relação às plântulas não tratadas (Fig. 19F). É importante evidenciar que,
nos cotilédones e no eixo em crescimento, o tratamento com ABA diminuiu o
conteúdo de AST e aumentou o conteúdo de AALT em referência ao controle (Fig.
19A e E).

50
Figura 19 - Conteúdo de açúcares solúveis totais (AST), de açúcares não-redutores (ANR) e de aminoácidos livres totais (AALT) em plântulas de girassol tratadas com ácido giberélico (GA3), ácido abscísico (ABA) ou ácido indolbutírico (IBA). Conteúdo de AST nos cotilédones (A) e no eixo em crescimento (B), de ANR nos cotilédones (C) e no eixo em crescimento (D) e de AALT nos cotilédones (E) e no eixo em crescimento (F) de plântulas de girassol híbrido Helio 253 no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com GA3 50 µM, ABA 50 µM ou IBA 50 µM. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%.

51
3.5 ATIVIDADE DE ISOCITRATO LIASE E GLUTAMINA SINTETASE REGULADA
POR HORMÔNIOS, FONTES DE CARBONO E NITROGÊNIO DURANTE O
CRESCIMENTO PÓS-GERMINATIVO EM GIRASSOL
As atividades das enzimas isocitrato liase (ICL) e glutamina sintetase (GS)
foram verificadas em cotilédones de plântulas de girassol híbrido Helio 253, no 5o dia
após a embebição (DAE), após três dias de tratamento em meio de cultura ágar-
água 4 g/L sem adições (controle) ou suplementado com sacarose 200 mM, L-
glutamina 20 mM, ácido giberélico (GA3) 50 µM ou ácido abscísico (ABA) 50 µM
(Fig. 3.5.1). A atividade de ICL nos cotilédones aumentou significativamente 94, 189,
265 e 130% nos tratamentos com sacarose, L-glutamina, GA3 e ABA, nessa ordem,
em relação ao controle (Fig. 20A). Entretanto, a atividade de GS nos cotilédones
tratados com sacarose e glutamina diminuiu de forma significativa 50 e 90%,
respectivamente, em comparação com o controle. No tratamento com ABA, a
atividade de GS aumentou significativamente 41% e permaneceu inalterada no
tratamento com GA3, ambas em comparação com o controle (Fig. 20B).

52
Figura 20 - Atividade de isocitrato liase (ICL) e de glutamina sintetase (GS) em plântulas de girassol. Atividade de ICL e de GS em cotilédones de plântulas de girassol híbrido Helio 253, no 5o DAE, cultivadas durante três dias em meio ágar-água 4 g/L sem adições (controle) ou suplementado com sacarose 200 mM, L-glutamina 20 mM, ácido giberélico (GA3) 50 µM ou ácido abscísico (ABA) 50 µM. As barras representam as médias, e as linhas verticais representam os desvios padrões de cinco repetições. As barras assinaladas com as mesmas letras não diferem significativamente de acordo com o teste de Tukey ao nível de significância de 5%. (micromol)

53
4 DISCUSSÃO
Os nossos resultados indicam que a transição do metabolismo heterotrófico
baseado nas reservas nutritivas para o metabolismo autotrófico fotossintetizante nos
cotilédones de girassol híbrido Hélio 253 depende da mobilização coordenada dos
lipídios e das proteínas de reserva durante o crescimento pós-germinativo inicial.
Nesse sentido, os padrões de degradação dessas reservas bem como de partição
dos seus produtos sustentam que o processo de mobilização é, pelo menos em
parte, regulado pelo mecanismo fonte-dreno. De forma complementar, o paralelismo
observado entre a mobilização dos lipídios e das proteínas de reserva pode ser
discutido com base em inter-relações recentemente demonstradas entre o
metabolismo do carbono e do nitrogênio (EASTMOND et al., 2000; BOREK et al.,
2003; LEHMANN e RATAJCZAK, 2008; BOREK e RATAJCZAK, 2010).
A transição para a atividade fotossintética em plântulas de girassol foi
evidenciada a partir do 4o dia após a embebição (DAE), com base na acumulação de
clorofilas e de carotenoides e na evolução de oxigênio (HEUPEL e KUTSCHERA,
1996). Nossos resultados confirmam que essa transição se dá no 4o DAE,
considerando o acúmulo de massa seca (MS) total nas plântulas (Fig. 6D) e o
acúmulo de clorofilas totais (CT) (Fig. 6C), de amido (Fig. 6A), de açúcares solúveis
totais (AST) (Fig. 7A) e de açúcares não-redutores (ANR) (Fig. 7C) nos cotilédones.
A mobilização coordenada das fontes de carbono e de nitrogênio, antes do
estabelecimento da autonomia fotossintética em plântulas de girassol, envolve o
mecanismo fonte-dreno. No tocante às fontes de carbono, a mobilização dos ANR
(Fig. 7C) e do amido (Fig. 6A) cumpre papel fundamental durante a germinação das
sementes nos momentos iniciais do crescimento das plântulas, isto é, até o 2o DAE.

54
Embora a mobilização dos LN (Fig. 4A) se inicie desde a germinação, esse processo
se mostra mais proeminente após a mobilização dos ANR e do amido. Nesse
sentido, o mecanismo fonte-dreno pode ser evidenciado ao longo do
estabelecimento das plântulas de girassol para as diferentes fontes de carbono, uma
vez que a mobilização sequencial dessas reservas nos cotilédones se dá em
paralelo com o consumo dos AST no eixo em crescimento (Fig. 7B).
É importante ressaltar que a mobilização dos lipídios de reserva ocorre
majoritariamente antes do estabelecimento do aparato fotossintético, pois nossos
resultados apontam que, a partir do 5o DAE, a degradação dos LN é diminuída (Fig.
4A). Esses resultados estão de acordo com um estudo prévio utilizando girassol, no
qual foi demonstrado que a acumulação de açúcares solúveis nos cotilédones no 5o
DAE modula negativamente a atividade da isocitrato liase (ICL), enzima marca-
passo da mobilização dos lipídios de reserva (PFEIFFER e KUTSCHERA, 1997).
De maneira semelhante, o mecanismo fonte-dreno parece atuar sobre as
fontes de nitrogênio durante o crescimento pós-germinativo em girassol, uma vez
que a mobilização das proteínas de reserva nos cotilédones (Fig. 4B), desde a
germinação, é acompanhada pelo consumo dos aminoácidos livres totais (AALT) no
eixo em crescimento (Fig. 7F). De forma complementar, verificamos que a
acumulação dos AALT nos cotilédones (Fig. 7E) ocorreu em um padrão bifásico,
corroborado pelo perfil eletroforético das proteínas solúveis (PS) (Fig. 5), o qual
mostra a degradação sequencial das cadeias acídicas e básica da heliantinina,
globulina 11S de girassol (ŽILIĆ et al., 2010). Além disso, a mobilização das
proteínas de reserva foi mais proeminente a partir do 3o DAE (Fig. 4B),
acompanhando de modo concomitante a degradação dos lipídios de reserva até o 5o
DAE (Fig. 4D). Contudo, após o estabelecimento do aparato fotossintético no 5o

55
DAE, o processo de mobilização das proteínas continuou até a quase exaustão (Fig.
4B; Fig. 5), provavelmente devido à carência de fontes externas de nitrogênio no
meio de cultura.
Baseado em estudos recentes, sugerimos que o paralelismo verificado entre a
mobilização dos lipídios e das proteínas de reserva nos cotilédones de girassol até o
estabelecimento da atividade fotossintética se deve à existência de vias alternativas
que utilizam os produtos de mobilização provenientes de ambas as reservas. Com
relação à mobilização dos lipídios de reserva, a assim chamada via clássica envolve
a utilização dos produtos de mobilização como precursores da gliconeogênese para
a síntese de sacarose a ser enviada para o eixo em crescimento (EASTMOND e
GRAHAM, 2001). De maneira complementar, a via clássica de mobilização das
proteínas de reserva gera aminoácidos a serem transportados para o eixo em
crescimento ou alocados na biossíntese de novo de proteínas nos tecidos de
armazenamento (MÜNTZ et al., 2001).
Estudos realizados com Lupinus luteus sugerem que as reservas lipídicas
podem ser convertidas não somente em sacarose, mas principalmente em
aminoácidos, a partir de duas rotas alternativas à via clássica. Assim, o isocitrato
gerado no citosol ou o malato proveniente da mitocôndria podem ser desviados para
a biossíntese das amidas de transporte, glutamina e asparagina (BOREK et al.,
2003; LEHMANN e RATAJCZAK, 2008; BOREK e RATAJCZAK, 2010). Além disso,
outro estudo, utilizando Arabidopisis thaliana como modelo, sugere que as rotas
alternativas podem atuar na ausência do ciclo do glioxilato, mantendo a mobilização
dos lipídios de reserva quando fontes externas de carbono são ofertadas
(EASTMOND et al., 2000).

56
Nesse sentido, propomos um modelo que integra as vias clássicas de
mobilização com as vias alternativas sugeridas na literatura, mostrando que essas
inter-relações metabólicas podem estar diretamente relacionadas com a mobilização
concomitante dos lipídios e das proteínas de reserva em girassol (Fig. 21).
Sugerimos que as vias metabólicas alternativas esquematizadas no modelo podem
ser responsáveis pela transformação parcial dos diferentes aminoácidos
provenientes da mobilização de proteínas em amidas para envio ao eixo em
crescimento, uma vez que as amidas são consideradas as principais formas de
transporte de nitrogênio em plantas (HELDT e PIECHULLA, 2011).
Figura 21 - Modelo integrativo proposto para as diferentes vias metabólicas que utilizam os produtos de mobilização dos lipídios e das proteínas. Corpo lipídico (CL), vacúolo de estocagem de proteínas (VEP), glioxissomo (GOX), mitocôndria (MIT), β-oxidação (β-O), ciclo do glioxilato (CG), ciclo de Krebs (CK), proteínas de reserva de sementes (PRS), triacilglicerol (TAG), ácido graxo (AG), aminoácido (AA), trifosfato de adenosina (ATP), α-cetoglutarato (α-KG), glutamato (GLU), glutamina (GLN), oxaloacetato (OAA), aspartato (ASP) e asparagina (ASN).

57
Além das mobilizações dos lipídios e das proteínas de reserva se mostrarem
coordenadas e paralelas nos cotilédones de plântulas de girassol híbrido Hélio 253,
durante o crescimento pós-germinativo inicial, os nossos resultados evidenciam que
a sacarose fornecida no meio externo modula negativamente a mobilização das
proteínas de reserva, assim como a L-glutamina adicionada no meio externo modula
negativamente a mobilização das reservas de carbono (LN e amido). De forma
complementar, a modulação negativa da mobilização das reservas de carbono e de
nitrogênio pelo tratamento com ácido abscísico (ABA) parece envolver um
mecanismo diferente daqueles proporcionados por fontes externas de sacarose ou
L-glutamina.
Nossos resultados apontam que a adição de sacarose ao meio de cultura
modula negativamente a mobilização do amido (Fig. 10D), dos lipídios (Fig. 10A) e
das proteínas (Fig. 10B) de reserva nos cotilédones de girassol, pois, já está
estabelecido na literatura que a sacarose pode possuir papel tanto como metabólito,
quanto como molécula sinalizadora (WIND et al., 2010). Em concordância com os
nossos resultados, a sacarose exógena também ocasiona o retardo da mobilização
dos lipídios (BOREK et al., 2006; BOREK e NUC, 2011) e das proteínas de reserva
(BOREK e RATAJCZAK, 2002) em plântulas de L. luteus, durante o crescimento
pós-germinativo.
Curiosamente, os nossos resultados apontam que o efeito modulador da
sacarose exógena sobre a mobilização dos lipídios durante o estabelecimento da
atividade fotossintética em plântulas de girassol não pode ser atribuído à diminuição
da atividade de ICL (Fig. 20A). De fato, a atividade de ICL é aumentada sob
tratamentos com sacarose externa em plântulas de L. luteus, e o retardo da
mobilização dos lipídios é atribuído à redução da transcrição e da atividade de

58
lipases (BOREK et al., 2006; BOREK e NUC, 2011). No entanto, durante o
crescimento pós-germinativo de plântulas de A. thaliana, o efeito modulador sobre a
mobilização dos lipídios parece não ser decorrente da ação direta da sacarose, mas
de seus produtos de hidrólise fosforilados (TO et al., 2002).
O retardo da mobilização das proteínas de reserva pela sacarose externa
durante o crescimento pós-germinativo inicial em girassol é acompanhado pela
acumulação de AALT (Fig. 11E). De fato, a acumulação dos produtos de hidrólise
diminui a atividade in vitro de proteases envolvidas na mobilização das proteínas de
reserva em Fagopyrum esculentum (DUNAEVSKY e BELOZERSKY, 1989). Assim
sendo, nossos resultados indicam que a mobilização das proteínas pode ser
retardada por um mecanismo de retroalimentação negativa mediado por
aminoácidos nos cotilédones de plântulas de girassol tratadas com sacarose
externa.
É possível que o efeito da sacarose externa seja indireto, envolvendo um
descompasso entre a mobilização dos lipídios e a das proteínas de reserva.
Levando em conta que a sacarose externa pode restringir a degradação dos lipídios
devido à repressão das lipases (BOREK et al., 2006; BOREK e NUC, 2011), a
acumulação de aminoácidos nos cotilédones de plântulas de girassol tratadas com
sacarose pode ser decorrente da desaceleração das vias alternativas que fornecem
esqueletos de carbono para a biossíntese de amidas a partir de ácidos graxos (Fig.
21) (BOREK et al., 2003; LEHMANN e RATAJCZAK, 2008; GAUFICHON et al.,
2010). Essa hipótese é corroborada pela diminuição da atividade de GS nos
cotilédones (Fig. 20B), uma vez que a atividade dessa enzima está relacionada com
a biossíntese das amidas de transporte (LEHMANN e RATAJCZAK, 2008;
GAUFICHON et al., 2010).

59
A adição de L-glutamina ao meio de cultura desencadeia efeitos mais
drásticos sobre o crescimento do eixo e a mobilização das reservas nutritivas, em
comparação com a sacarose exógena, durante o crescimento pós-germinativo inicial
em girassol. De fato, o conteúdo de MS do eixo em crescimento é diminuído em
34% na presença de L-glutamina 20 mM (Fig. 13B) e aumentado em 13% com a
adição de sacarose 200 mM ao meio externo (Fig. 9B). Além disso, a L-glutamina
externa modula negativamente a mobilização do amido (Fig. 14D), dos lipídios (Fig.
14A) e das proteínas (Fig. 14B) de reserva em concentração dez vezes mais baixa
que a da sacarose exógena.
Poucos trabalhos abordam o efeito de fontes externas de aminoácidos sobre
a mobilização das reservas nutritivas após a germinação das sementes. Em A.
thaliana, a adição de peptona ao meio de cultura ocasiona o retardo na mobilização
das globulinas (GARCIARRUBIO et al., 1997). De forma adicional, em Dolichos
lablab, o retardo da mobilização das proteínas de reserva pelo fornecimento de
caseína hidrolisada pode ser devido ao efeito inibidor de aminoácidos sobre a
atividade de proteases ácidas (RAMAKRISHNA e RAO, 2005). No entanto, os
mecanismos que modulam a mobilização das fontes de carbono na presença de
fontes externas de aminoácidos ainda continuam desconhecidos.
Curiosamente, a exposição das plântulas de girassol à L-glutamina externa
não proporciona a acumulação de AALT (Fig. 15E), mas ocasiona o aumento do
conteúdo de AST (Fig. 15A) e de ANR (Fig. 15C) e a diminuição da atividade de GS
(Fig. 20B) nos cotilédones. Neste sentido, é possível que a modulação negativa da
mobilização das reservas nutritivas pela L-glutamina exógena pode ser decorrente
de um efeito indireto envolvendo a alteração do balanço C:N. De maneira alternativa,

60
esse mecanismo de modulação pode estar relacionado com a alteração da
qualidade, e não da quantidade, do pool de aminoácidos presente nos cotilédones.
Já está claramente estabelecido que as giberelinas (GA) estimulam a
mobilização do amido e das proteínas de reserva em cereais após a germinação,
induzindo a expressão de α-amilases e proteinases cisteínicas, respectivamente.
Além disso, o papel do ácido abscísico (ABA) como antagonista das GA nesse
processo também está consolidado na literatura (BEWLEY et al., 2013). Neste
trabalho, a adição de GA3 ao meio de cultura não ocasiona alterações nos padrões
de mobilização das reservas nutritivas nos cotilédones (Fig. 18) ou de partição dos
seus produtos de hidrólise para o eixo em crescimento (Fig. 19), durante o
estabelecimento de plântulas de girassol. Tendo em vista que as GA são produzidas
pelo eixo em crescimento e transportadas para os tecidos de armazenamento após
a germinação (BEWLEY et al., 2013), é possível que o conteúdo endógeno das GA
nos cotilédones de girassol seja suficientemente elevado para mascarar o efeito do
GA3 adicionado ao meio externo em baixa concentração (50 µM).
Em contraste, a adição de ABA ao meio de cultura modula negativamente a
mobilização dos lipídios (Fig. 18A) e das proteínas de reserva (Fig. 18B), mas causa
a diminuição do conteúdo de amido (Fig. 18D) nos cotilédones de plântulas de
girassol durante o crescimento pós-germinativo. Embora alguns trabalhos indiquem
que o ABA afeta a mobilização dos lipídios restringindo a expressão de enzimas da
beta-oxidação e da malato sintase (PRITCHARD et al., 2002) ou a atividade da ICL
(MARRIOTT e NORTHCOTE, 1977; FINKELSTEIN e LYNCH, 2000), o retardo da
mobilização dos lipídios nas plântulas de girassol tratadas com ABA exógeno não
pode ser atribuído à alteração da atividade de ICL (Fig. 20A).

61
Algumas evidências indicam que o ABA é capaz de restringir a mobilização
das proteínas de reserva após a germinação (GARCIARRUBIO et al., 1997; TONINI
et al., 2010), modulando negativamente a atividade de proteases (BARDUCHE et al.,
1999). Em plântulas de girassol, sugerimos que o retardo da mobilização das
proteínas nos cotilédones (Fig. 18B) pode ser decorrente do efeito do ABA exógeno
sobre a mobilização dos lipídios. De fato, o atraso da degradação dos lipídios (Fig.
18A) induzido por ABA é corroborado pela diminuição do conteúdo de AST nos
cotilédones (Fig. 19A) e no eixo em crescimento (Fig. 19B). Em contraponto, a
acumulação de PS no eixo (Fig. 18C) e de AALT, em ambas as partes (Fig. 19E e
F), não favorecem a hipótese de que o ABA exógeno modula diretamente a
mobilização das proteínas nos cotilédones de plântulas girassol.
Já está estabelecido na literatura que as auxinas apresentam um papel
indutor no crescimento das plântulas, relacionado com todos os aspectos do
desenvolvimento vegetal (SIMON e PETRÁŠEK, 2011). Contudo, nossos dados
indicam que a aplicação externa de ácido indolbutírico (IBA) ocasiona retardo da
mobilização das reservas em plântulas de girassol, demonstrado pelo aumento dos
conteúdos de MS (Fig. 17A), de LN e de PS (Fig. 18A e B) nos cotilédones em
paralelo com a diminuição do conteúdo de MS no eixo em crescimento (Fig. 17B).
De forma complementar, verificamos uma alteração morfológica da parte radicular
das plântulas tratadas com IBA (Fig. 16) e um acúmulo de PS no eixo em
crescimento (Fig. 18C).
Em relação ao retardo do crescimento, um trabalho utilizando Aechmea
blanchetiana demonstrou que a aplicação externa de IBA em baixas e altas
concentrações ocasiona indução e retardo, nessa ordem, do desenvolvimento (CHU
et al., 2010). Quanto ao efeito morfológico, já é sabido que as auxinas auxiliam na

62
indução da formação de raízes laterais e de calos (INGENSIEP, 1982; CHENG et
al., 1992; WOODWARD e BARTEL, 2005). Nesse sentido, o acúmulo de proteínas
solúveis no eixo em crescimento está associado ao fato de as auxinas induzirem a
formação de raízes laterais e tem sido relacionado com um aumento na síntese de
enzimas durante o processo de regeneração de raízes (HUSEN, 2008). Assim, é
possível que a concentração de IBA utilizada no presente trabalho seja muito
elevada para promover o desenvolvimento das plântulas de girassol.
A utilização do girassol como espécie modelo para estudar os mecanismos de
mobilização de reservas em dicotiledôneas oleaginosas mostrou-se uma ótima
opção na medida em que propiciou um sistema experimental fácil e rápido, gerando
resultados reprodutíveis e evidentes.

63
5 CONCLUSÃO
Os resultados deste trabalho confirmam que a mobilização dos lipídios e das
proteínas de reserva ocorre de forma coordenada durante o crescimento pós-
germinativo inicial em girassol, corroborando a hipótese de que a aplicação externa
de fontes de carbono (sacarose) e nitrogênio (L-glutamina) é capaz de atrasar a
mobilização dessas reservas nutritivas de forma cruzada. Além disso, considerando
as mudanças nos padrões de mobilização das reservas e a partição dos seus
produtos, proporcionadas pela aplicação externa de diferentes reguladores do
crescimento, é evidente que os efeitos dos metabólitos e dos hormônios devem
envolver, pelo menos em parte, mecanismos de ação distintos.

64
REFERÊNCIAS
BARDUCHE, D.; PAIVA, R.; LOPES, M.A.; PAIVA, E. Effect of ABA and GA3 on
protein mobilization in embryos and cotyledons of angico [Anadenanthera peregrina
(L.) speg] seeds during germination. Brazilian Archives of Biology and
Technology, v.42, 1999.
BERTELI, F.; CORRALES, E.; GUERRERO, C.; ARIZA, M.J.; PILEGO, F.;
VALPUESTA, F.W. Salt stress increases ferredoxin-dependent glutamate synthase
activity and protein level in the leaves of tomato. Physiologia Plantarum, v.93,
p.259-264, 1995.
BEWLEY, J.D.; BRADFORD, K.J.; HILHORST, H.W.M.; NONOGAKI, H. SEEDS:
Physiology of Development, Germination and Dormancy. 3. ed. New York: Springer,
2013.
BOREK, S.; NUC, K. Sucrose controls storage lipid breakdown on gene expression
level in germinating yellow lupine (Lupinus luteus L.) seeds. Journal of Plant
Physiology, v.168, p.1795-1803, 2011.
BOREK, S.; RATAJCZAK, L. Storage lipids as a source of carbon skeletons for
asparagine synthesis in germinating seeds of yellow lupine (Lupinus luteus L.).
Journal of Plant Physiology, v.167, p.717-724, 2010.

65
BOREK, S.; RATAJCZAK, W.; RATAJCZAK, L. A transfer of carbon atoms from fatty
acids to sugars and amino acids in yellow lupine (Lupinus luteus L.) seedlings.
Journal of Plant Physiology, v.160, p.539-545, 2003.
BOREK, S.; RATAJCZAK, W.; RATAJCZAK, L. Ultrastructural and enzymatic
research on the role of sucrose in mobilization of storage lipids in germinating yellow
lupine seeds. Plant Science, v.170, p.441-452, 2006.
BOREK, S.; RATAJCZAK, W. Sugars as a metabolic regulator of storage protein
mobilization in germinating seeds of yellow lupine (Lupinus luteus L.). Acta
Physiologiae Plantarum, v.24, p.425-434, 2002.
BRADFORD, M.M. A rapid and sensitive for the quantitation of microgram quantitites
of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, v.
72, p.248-254, 1976.
BUCKERIDGE, M.S.; SANTOS, H.P.; TINÉ, M.A.S.; AIDAR, M.P.M. Mobilização de
Reservas. In: FERREIRA, A.G.; BORGHETTI, F. Germinação: do básico ao
aplicado, Porto Alegre: Artmed, 2004, p.163-185.
CHELL, R.M.; SUNDARAM, T.K.; WILKINSON, A.E. Isolation and characterization of
isocitrate lyase from a thermophilic Bacillus sp. Biochemical Journal, v.173, p.165-
177, 1978.

66
CHENG, B.; PETERSON, C.M.; MITCHELL, R.J. The role of sucrose, auxin and
explant source on in vitro rooting of seedling explants of Eucalyptus sideroxylon.
Plant Science, v.87, p.207-214, 1992.
CHU, E.P.; TAVARES, A.R.; KANASHIRO, S.; GIAMPAOLI, P.; YOKOTA, E.S.
Effects of auxins on soluble carbohydrates, starch and soluble protein content in
Aechmea blanchetiana (Bromeliaceae) cultured in vitro. Scientia Horticulturae,
v.125, p.451-455, 2010.
DABDOUB, M.J.; BRONZEL, J.L.; RAMPIN, M.A. Biodiesel: visão crítica do status
atual e perspectivas na academia e na indústria. Química Nova, v.32, p.776-792,
2009.
DUBOIS, M.; GILLES, K.A.; HAMILTON, J.K.; REBERS, P.A.; SMITH, F.
Colorimetric method for determination of sugars and related substances. Analytical
Chemistry, v.28, p.248-254, 1956.
DUNAEVSKY, Y.E.; Belozersky, M.A. The role of cysteine proteinase and
carboxypeptidasein the breakdown of storage proteins in buckwheat seeds. Planta,
v.179, p.316-322, 1989.
EASTMOND, P.J.; GERMAIN, V.; LANGE, P.R.; BRYCE, J.H.; SMITH, S.M.;
GRAHAM, I.A. Postgerminative growth and lipid catabolism in oilseeds lacking the
glyoxylate cycle. Proceedings of the National Academy of Sciences, v.97, p.5669-
5674, 2000.

67
EASTMOND, P.J.; GRAHAM, I.A. Re-examining the role of the glyoxylate cycle in
oilseeds. Plant Science, v.6, p.72-77, 2001.
ELARBI, M.B.; KHEMIRI, H.; JRIDI, T.; Hamida, J.B. Purification and characterization
of α-amylase from safflower (Carthamus tinctorius L.) germinating seeds. Comptes
Rendus Biologies, v.332, p.426-432, 2009.
FINKELSTEIN, R.; LYNCH, T. The Arabidopsis abscisic acid response gene ABI5
encodes a basic leucine zipper transcription factor. Plant Cell, v.12, p.599-609,
2000.
GARCIARRUBIO, A.; LEGARIA, J.P.; COVARRUBIAS, A.A. Abscisic acid inhibits
germination of mature Arabidopisis seeds by limiting the availability of energy and
nutrients. Planta, v.203, p.182-187, 1997.
HELDT, H.W.; PIECHULLA, B. Plant Biochemistry. 4. ed. London: Academic Press,
2011.
HEUPEL, T.; KUTSCHERA, U. Pigment accumulation, dark respiration and
photosynthesis during the greening of sunflower cotyledons. Journal of Plant
Physiology, v.147, p.567-572, 1996.

68
HUSEN, A. Clonal propagation of Dalbergia sissoo Roxb. and associated metabolic
changes during adventitious root primordium development. New Forests, v.36, p.13-
27, 2008.
INGENSIEP, H.W. The morphogenetic response of intact pea seedlings with respect
to translocation and metabolism of root-applied auxin. Zeitschrift für
Pflanzenphysiologie, v.105, p.149-164, 1982.
KARUNAGARAN, D.; RAO, P.R. Mode and control of starch mobilization during
germination of seeds of horse gram. Plant Science, v.73, p.155-159, 1991.
LAEMMLI, U.K. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, v.227, p.680-685, 1970.
LEHMANN, T.; RATAJCZAK, L. The pivotal role of glutamate dehydrogenase (GDH)
in the mobilization of N and C from storage material to asparagine in germinating
seeds of yellow lupine. Journal of Plant Physiology, v.165, p.149-158, 2008.
LIRA, M.A.; CHAGAS, M.C.M.; BRISTOT, G.; DANTAS, J.A.; HOLANDA, J.S.; LIMA,
J.M.P. Recomendações técnicas para o cultivo do girassol. In: LIRA, M.A.;
BARRETO, M.F.P. Oleaginosas como fonte de matéria-prima para a produção
de biodiesel. Natal: EMPARN, 2009.

69
MARRIOT, K.M.; NORTHCOTE, D.H. Influence of abscisic acid, cyclic AMP, and
gibberellic acid on induction of isocitrate lyase activity in endosperm of germinating
castor bean seeds. Journal of Experimental Botany, v.28, p.219-225, 1977.
MCCREADY, R.M., GUGGOLZ, J., SILVIERA, V., and OWENS H. S. Determination
of starch and amylose in vegetables. Analytical Chemistry, v.22, p.1156-1158,
1950.
MELO, F.P.L.; NETO, A.V.A.; SIMABUKURO, E.A.; TABARELLI, M. Recrutamento e
estabelecimento de plântulas. In: FERREIRA, A.G.; BORGHETTI, F. Germinação:
do básico ao aplicado, Porto Alegre: Artmed, 2004, p.237-250.
MORRIS, D.L. Quantitative determination of carbohydrates with Dreywood’s
anthrone reagent. Science, v.107, p.111-114, 1948.
MÜNTZ, K.; BELOZERSKY, M.A.; DUNAEVSKY, Y.E.; SCHLERETH, A.;
TIEDEMANN, J. Stored proteinases and the initiation of storage protein mobilization
in seeds during germination and seedling growth. Journal of Experimental Botany,
v.52, p.1741-1752, 2001.
PENFIELD, S.; GRAHAM, S.; GRAHAM, I.A. Storage reserve mobilization in
germinating oilseeds: Arabidopsis as a model system. Biochemical Society
Transactions, v.33, p.380-383, 2005.

70
PEOPLES, M.B.; FAIZAH, A.W.; REAKASEM, B.E.; HERRIDGE, D.F. Methods for
evaluating nitrogen fixation by nodulated legumes in the field. Canberra: Australian
Centre for International Agricultural Research, 1989.
PFEIFFER, I.; KUTSCHERA, U. Effect of white light on cell expansion and lipid
metabolism in sunflower cotyledons. Journal of Plant Physiology, v.151, p.590-
594, 1997.
PRITCHARD, S.L.; CHARLTON, W.L.; BAKER, A.; GRAHAM, I.A. Germination and
storage reserve mobilization are regulated independently in Arabidopsis. The Plant
Journal, v.31, p.639-647, 2002.
RAMAKRISHNA, V.; RAO, P.R. Axial control of protein reserve mobilization during
germination of indian bean (Dolichos lablab L.) seeds. Acta Biologica
Szegediensis, v. 49, p.23-27, 2005.
RYLOTT, E.L.; HOOKS, M.A.; GRAHAM, I.A. Co-ordinate regulation of genes
involved in storage lipid mobilization in Arabidopsis thaliana. Biochemical Society
Transactions, v.29, p.283-287, 2001.
SIMON, S.;; PETRÁŠEK, J. Why plants need more than one type of auxin. Plant
Science, v.180, p.454-460, 2011.

71
THOMAS, B.R.; RODRIGUEZ, R.L. Metabolite signals regulate gene expression and
source/sink relations in cereal seedlings. Plant Physiology, v. 106, p. 1235-1239,
1994.
TO, J.P.C.; REITER W.D.; GIBSON, S.I. Mobilization of seed storage lipid by
Arabdopsis seedling is retarded in the presence of exogenous sugars. BMC Plant
Biology, v.2, p.1-11, 2002.
TONINI, P.P.; PURGATTO, E.; BUCKERIDGE, M.S. Effects of abscisic acid,
ethylene and sugars on the mobilization of storage proteins and carbohydrates in
seeds of the tropical tree Sesbania virgata (Leguminosae). Annals of Botany, v.106,
p.607-616, 2010.
VARGAS, J.I. Vegetable oils as substitutes for diesel oil. Política Agrícola, n.1, p.
17-30, 2007.
VOIGT, E.L.; ALMEIDA, T.D.; CHAGASA, R.M.; PONTE, L.F.A.; VIÉGAS, R.A.;
SILVEIRA, J.A.G. Source-sink regulation of cotyledonary reserve mobilization during
cashew (Anacardium occidentale) seedling establishment under NaCl salinity.
Journal of Plant Physiology, v.166, p.80-89, 2009.
WIND, J.; SMEEKENS, S.; HANSON, J. Sucrose: metabolite and signaling molecule.
Phytochemistry, v.71, p.1610-1614, 2010.

72
WOODWARD, A.W., BARTEL, B. Auxin: regulation, action, and interaction. Annals
of Botany, v.95, p.707-735, 2005.
YEMM, E.W.; WILLIS, A.J. The estimation of carbohydrates in plant extracts by
anthrone. Biochemical Journal, v.57, p.508-514, 1954.
ŽILIĆ, S.;; BARAĆ, M.;; PEŠIĆ, M.;; CREVAR, M.;; STANOJEVIĆ, S.;; NIŠAVIĆ, A.;;
SARATLIĆ, G.;; TOLIMIR, M. Characterization of sunflower seed and kernel proteins.
Helia, v.33, p.103-114, (2010).
Top Related