







Línguas
Páginas
Legal


FRANCISCO FÁBIO BEZERRA DE OLIVEIRA
MECANISMOS ENVOLVIDOS NA SÍNDROME DA DOR AGUDA INDUZIDA POR
PACLITAXEL EM CAMUNDONGOS: PARTICIPAÇÃO DE MASTÓCITOS E
CÉLULAS SATÉLITES GLIAIS
FORTALEZA
2017
UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
DEPARTAMENTO DE FISIOLOGIA E FARMACOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM FARMACOLOGIA

1
FRANCISCO FÁBIO BEZERRA DE OLIVEIRA
MECANISMOS ENVOLVIDOS NA SÍNDROME DA DOR AGUDA INDUZIDA POR
PACLITAXEL EM CAMUNDONGOS: PARTICIPAÇÃO DE MASTÓCITOS E
CÉLULAS SATÉLITES GLIAIS
Tese apresentada ao Programa de Pós-
Graduação em Farmacologia do Departamento
de Fisiologia e Farmacologia da Faculdade de
Medicina da Universidade Federal do Ceará,
como parte dos requisitos para obtenção do
título de Doutor em Farmacologia. Área de
concentração: Farmacologia da Inflamação e
Dor.
Orientadora: Profa. Dra. Mariana Lima Vale
Coorientador: Prof. Dr. Thiago Mattar Cunha
FORTALEZA
2017

2

3
FRANCISCO FÁBIO BEZERRA DE OLIVEIRA
MECANISMOS ENVOLVIDOS NA SÍNDROME DA DOR AGUDA INDUZIDA POR
PACLITAXEL EM CAMUNDONGOS: PARTICIPAÇÃO DE MASTÓCITOS E
CÉLULAS SATÉLITES GLIAIS
Tese apresentada ao Programa de Pós-
Graduação em Farmacologia do Departamento
de Fisiologia e Farmacologia da Faculdade de
Medicina da Universidade Federal do Ceará,
como parte dos requisitos para obtenção do
título de Doutor em Farmacologia. Área de
concentração: Farmacologia da Inflamação e
Dor.
Aprovada em: 08/08/2017
BANCA EXAMINADORA
_________________________________________________
Profa. Dra. Mariana Lima Vale (Orientadora)
Universidade Federal do Ceará – UFC
_________________________________________________
Profa. Dra. Antoniella Souza Gomes
Universidade Federal do Ceará – UFC
_________________________________________________
Prof. Dr. Diego Veras Wilke
Universidade Federal do Ceará – UFC
_________________________________________________
Prof. Dr. José Carlos Farias Alves Filho
Faculdade de Medicina de Ribeirão Preto – FMRP/USP
_________________________________________________
Profa. Dra. Josenília Maria Alves Gomes
Universidade de Fortaleza – UNIFOR

4
Aos meus pais, Francisco e Maria, pelo amor
incondicional e aos meus irmãos Flávio,
Flaviano, Fabiano e Suzana pelo carinho e apoio
em todos os momentos.

5
AGRADECIMENTOS
Em primeiro lugar a Deus e a Nossa Senhora, por estarem sempre presentes em
todos os momentos de minha vida. Por sempre iluminarem meu caminho, por não terem me
deixarem fraquejar diante das dificulades ao longo dessa caminhada.
A Professora Dra. Mariana Lima Vale, minha orientadora, pela oportunidade em
ingressar na pós-graduação, por acreditar em mim e aceitar me orientar. Agradeço pelo tempo
dedicado à orientação e a correção do trabalho. Agradeço a você por me ensinar o caminho da
pesquisa, pela paciência, atenção, dedicação e pelos conhecimentos compartilhados. Obrigado
por todos esses anos de aprendizado.
Ao Professor Dr. Thiago Mattar Cunha, meu coorientador, por me receber em seu
laboratório, pela oportunidade, confiança e disponibilidade no desenvolvimento da minha
pesquisa de doutorado e por todo o aprendizado compartilhado. Obrigado por tudo.
Ao prof. Dr. Ronaldo Ribeiro (in memoriam), pelas contribuições e
ensinamentos, obrigado pela coorientação no inicio desse trabalho.
Aos professores da Pós-graduação em Farmacologia da UFC, pelos ensinamentos
compartilhados. Agradeço de forma especial a Profa. Dra. Geanne Matos de Andrade pelo
auxilio concedido para que eu pudesse ir a Ribeirão Preto para desenvolver parte da pesquisa.
Aos Professores Dr. José Carlos Alves Filho, Dra. Antoniella Souza Gomes, Dr.
Diego Veras Wilke e Dra. Josenília Maria Alves Gomes, por terem aceitado participar da
correção desse trabalho e pelas valiosas contribuições.
Aos amigos e colaboradores do Laboratório de Farmacologia da Inflamação e do
Câncer (LAFICA), Anamaria Falcão e Mario Lisboa. Obrigado pela colaboração, discussões,
ajuda na realização dos experimentos e pela amizade.
Aos amigos e colaboradores do Laboratório de Inflamação e Dor (LID) da
Faculdade de Medicina de Ribeirão Preto (FMRP/USP), Miriam Fonseca, Alexandre Lopes e
Rangel Leal. Obrigado pela presteza, disponibilidade, conhecimento, discussão e pela
amizade.

6
A Ieda Regina dos Santos, pelo apoio, pela ajuda constante desde que cheguei a
Ribeirão Preto. Muito obrigado pelos ensinamentos, pela realização dos experimentos e pela
amizade.
Aos meus pais, Francisco Costa de Oliveira e Maria Bezerra de Oliveira, pelos
ensinamentos, dedicação e amor. Obrigado por sempre acreditarem em mim e por permitirem
a realização de mais esse objetivo.
Aos meus irmãos, Flavio, Flaviano e Fabiano, e a minha irmã, Suzana, pela
dedicação e carinho em todos os momentos e por acreditarem sempre em mim. Vocês são
muito importantes em minha vida.
As minhas sobrinhas, Lurya, Livia e Eduarda que mesmo distante fisicamente
sempre estiveram presentes em minha vida. Obrigado por tornarem minha vida melhor.
A minha sobrinha e afilhada, Maria, que em toda a sua inocência consegue tornar
os dias mais fáceis, simples e alegres. Amo você.
Aos amigos do LAFICA, Carlos Wagner, Camila Meireles, Livia Talita, Kalina
Kelma e Rafael Santana. Obrigado pelo apoio, convivência e pela amizade.
Aos amigos do LID, Vanessa Borges, Marcela Davoli, Taty Cecilio, Rafaela
Guimarães, Nathália Quadros, Andreza Urba, Flávia Santa Cecília, Mateus Rossato, Cássia R.
Silva, André Saraiva, Maria Cláudia, David Colon, Bruno Marcel, Paulo H. Melo e a todos
que fazem parte deste laboratório. Obrigado pela receptividade.
Aos amigos da pós-graduação em Farmacologia da UFC, Cecília Mendes, Deysen
Girão, Kaira Emanuella, Samara Rodrigues e Renan Oliveira. Obrigado pelo apoio, ajuda,
conhecimentos, convivência e pela amizade.
Aos alunos de iniciação científica, Bruno Freitas e Jéssica Jamile Nogueira, pela
ajuda na realização dos experimentos, a participação de vocês foi essencial para realização da
pesquisa.
Aos meus familiares, em especial a minha tia Maria Irismar (in Memoriam), que
partiu muito cedo, mas nos deixou vários ensinamentos de vida. Tenho certeza de onde estiver
estará sempre torcendo por mim.

7
Aos amigos/irmãos, Thiago Lima e José Rodrigo, pela força, incentivo e pelo
encorajamento na busca da realização dos meus objetivos. Obrigado pela amizade,
cumplicidade e companheirismo de todas as horas.
Aos amigos da “Pensão da Jô”, Rodrigo Calderari, Lucas Faria, Luan Pereira,
Ivan Lorena, Joelma Cruz, Juliana Oliveira e Bruno Toschi. Vocês tornaram a caminhada
mais fácil durante o período em que estive em Ribeirão Preto. Obrigado pelos laços de
amizade construídos.
A Jô Santos, pela acolhida em sua casa e em sua vida. Muito obrigado pelo apoio,
incentivo, carinho e convivência. Você é muito especial, obrigado por tudo.
Aos amigos, Guilherme Almeida e Diangeles Chagas. Obrigado pela parceria,
pelas conversas, companheirismo, cumplicidade, apoio e amizade que levarei para sempre
comigo.
Aos funcionários do Departamento de Fisiologia e Farmacologia da UFC,
especialmente Laura Alves e Célia Gonzaga, pela disponibilidade constante em ajudar.
Aos técnicos do LID, Giuliana Bertozi, Sergio Rosa e Kátia Santos. Obrigado
pelo apoio e disponibilidade. Agradeço também aos demais funcionários do departamento de
Farmacologia da FMRP/USP por toda ajuda e apoio. Muito obrigado.
A CAPES, CNPq, FUNCAP e CRID pelo suporte financeiro.
Enfim, a todos que contribuíram direta ou indiretamente na realização desse
trabalho, muito OBRIGADO!

8
“Se você encontrar um caminho sem
obstáculos, ele provavelmente não leva a lugar
nenhum.”
Frank A. Clark

9
RESUMO
MECANISMOS ENVOLVIDOS NA SÍNDROME DA DOR AGUDA INDUZIDA POR
PACLITAXEL EM CAMUNDONGOS: PARTICIPAÇÃO DE MASTÓCITOS E
CÉLULAS SATÉLITES GLIAIS
Paclitaxel é um antineoplásico utilizado como primeira linha no tratamento de diversos
tumores sólidos, particularmente em carcinomas de mama, ovário, pulmão, cabeça e pescoço.
Entretanto, os pacientes que recebem tratamento com paclitaxel frequentemente desenvolvem
dor que ocorre imediatamente após o tratamento com este fármaco, conhecida como síndrome
da dor aguda associada ao paclitaxel. No entanto, os mecanismos pelos quais o paclitaxel
induz essa condição dolorosa, ainda não são conhecidos. Neste estudo objetivou-se investigar
o envolvimento de mastócitos e de células satélites gliais na síndrome de dor aguda induzida
pelo paclitaxel. Para isso foram utilizados camundongos C57BL/6 (selvagens), camundongos
que não expressam mastócitos (SH) e camundongos knockout TNFR1/R2; TLR4; IL-1R; IL-6
e CCR2 machos (20-25g). O modelo consiste na administração de paclitaxel por via
intravenosa (4 mg/kg, dose única). Após a administração de Paclitaxel a sensibilidade
mecânica foi avaliada utilizando filamentos de von Frey, mensurado em gramas de pressão. A
sensibilidade ao frio foi avaliada com um estímulo 10 ºC (acetona) aplicado na pata traseira
direita, observando-se comportamentos de agitação e elevação da pata, além de lambidas,
mensurados em segundos. E a sensibilidade ao calor foi avaliada através do teste de
hargreaves, onde uma fonte de luz infravermelha é posicionada sob a pata traseira do animal
durante 20s ou até que o animal exiba resposta positiva (flinch ou retirada da pata), então a
fonte de luz e para automaticamente. Além disso, foi realizada cultura de linhagem de
mastócitos e de células satélites gliais (cultura primária de gânglio da raiz dorsal) que foram
estimuladas com paclitaxel. Foram coletadas amostras (plasma, nervo isquiático, gânglios da
raiz doral e medula espinal) para determinação da expressão gênica e dosagem dos níveis de
citocinas/qumiocinas (IL-1β, TNF-α e IL-6, MCP-1 e KC/CXCL1). Também foi realizado
imunofluorescência (c-Fos, IL-6 e triptase). A concentração de citocinas/quimiocinas (IL-1β,
TNF-α e IL-6, MCP-1 e KC/CXCL1) também foi avaliada no sobrenadante das culturas de
mástocitos e de células satélites gliais. Os resultados demonstraram que a injeção intravenosa
de paclitaxel reduziu significativamente (p<0,05) o limiar nociceptivo, induzindo respostas
hiperalgesicas mecânica e térmica ao frio após a administração de paclitaxel. No entanto,
quando se avaliou a resposta térmica ao calor, verificou-se que a latência de retirada da pata
não foi alterada com o tratamento com paclitaxel, quando comparadas ao grupo controle. A
imunofluorescência para c-Fos nos gânglios da raiz dorsal e medula espinal demostrou

10
ativação de celular evidenciado por aumento da imunoexpressão nos grupos tratado com
paclitaxel. A administração de paclitaxel aumentou significativamente (p<0,05) as citocinas
IL-1β, TNF-α e IL-6, MCP-1 e KC/CXCL1. A hiperalgesia mecânica e térmica ao frio foram
reduzidas significativamente (p<0,05) em animais SH (não possuem mastócitos), quando
comparado aos animais selvagens, bem como o tratamento com cromoglicato de sódio
(estabilizador de membrana de mastócitos) foi eficaz em inibir significativamente (p<0,05) a
hiperalgesia mecânica e térmica ao frio após o tratamento com o quimioterápico. O pré-
tratamento com cromoglicato de sódio preveniu significativamente (p<0,05) o aumento de
citocinas e quimiocinas em camundongos tratados com paclitaxel. Nos camundongos SH
tratados com paclitaxel não foi observado aumento significativo (p<0,05) de citocinas. A
estimulação com paclitaxel provocou aumento significativo (p<0,05) de citocinas e
quimiocinas (TNF-α, IL-6 e MCP-1) em cultura de mastócitos. Além disso, a
imunofluorescência, no gânglio da raiz dorsal, demonstrou um aumento da imunoexpressão
de triptase (marcador de mastócito) e IL-6 após a administração de paclitaxel. Os resultados
mostraram também que o receptor TLR4 está envolvido no desenvolvimento de hiperalgesia
induzida por paclitaxel, pois o tratamento de animais knockout para o receptor TLR4 com este
quimioterápico não foi capaz de desenvolver hiperalgesia mecânica e térmica nesses animais.
Os resultados demonstram ainda, que nos camundongos knockout TLR4 tratados com
paclitaxel não houve aumento significativo (p<0,05) de citocinas e quimiocinas. E quando
comparados ao grupo de camundongos selvagens, observou-se significativa redução (p<0,05)
dos níveis de citocina e quimiocina. Além disso, o paclitaxel causou aumento significativo
(p<0,05) de citocinas (TNF-α e IL-6) e quimiocinas (MCP-1 e KC/CXCL1) em cultura de
células satélites gliais e a deleção gênica para o receptor TLR4 foi capaz de impedir o
aumento dos níveis de citocinas/quimiocinas. O estudo mostra, pela primeira vez, que
mastócitos e células satélites gliais estão envolvidos no desenvolvimento de dor aguda
induzida pelo paclitaxel em camundongos. Adicionalmente, o estudo revelou que a ativação
de mastócitos e de células satélites gliais, possivelmente se dá pela ligação do paclitaxel ao
TLR4, induzindo a liberação de citocinas/quimiocinas que, contribuem para o
desenvolvimento de dor aguda induzida pelo paclitaxel.
Palavras-chave: Paclitaxel; dor aguda; síndrome; mastócitos; células satélites gliais.

11
ABSTRACT
MECHANISMS INVOLVED IN PACLITAXEL-INDUCED ACUTE PAIN
SYNDROME IN MICE: PARTICIPATION OF MAST CELLS AND SATELLITE
GLIAL CELLS IN
Paclitaxel is an antineoplastic drug used as a first line in the treatment of several solid tumors,
particularly in breast, ovarian, lung, head and neck carcinomas. However, patients receiving
paclitaxel treatment often develop a painful condition that occurs immediately after the
treatment with this drug, known as acute pain syndrome associated with paclitaxel.
Nevertheless, the mechanisms by which paclitaxel induces this painful condition is not yet
known. The aim of this study was to investigate the involvement of mast cells and satellite
glial cells in the acute pain syndrome induced by paclitaxel. Adult male wild-type mice
(C57BL/6), SH mutant mice and TNFR1/R2, TLR4, IL-1R, IL-6 and CCR2 knockout mice
were used. Acute pain syndrome was induced by intravenous paclitaxel (4 mg / kg, single
dose). After administration of paclitaxel, mechanical and thermal sensitivity (cold) were
assessed. The mechanical sensitivity was evaluated using von Frey filaments, measuring
pressure in grams. The sensitivity to cold was evaluated with a stimulus of 10 °C (acetone)
applied to the right hind paw, leading to agitation and paw elevation behaviors, as well as
licking, measured in seconds. The heat sensitivity was assessed by the Hargreaves test, where
an infrared light source is positioned under the animal's hind paw for 20s or until the animal
exhibits a positive response (flinch or paw withdrawal), then the light source stop
automatically. In addition, mast cell line culture and satellite glial cells (dorsal root ganglia
primary culture) were stimulated with paclitaxel. Samples (plasma, sciatic nerve, dorsal root
ganglia and spinal cord) were collected for determination of gene expression and cytokine
levels (IL-1β, TNF-α, IL-6, MCP-1 and KC / CXCL1). Immunofluorescence (c-Fos, IL-6 and
tryptase) was also performed. The concentration of cytokines/chemokines (IL-1β, TNF-α e
IL-6, MCP-1 e KC/CXCL1) was also evaluated in the supernatant of mast cells and satellite
glial cells. The results demonstrated that intravenous injection of paclitaxel significantly
reduced (p < 0.05) the nociceptive threshold, inducing mechanical and thermal hyperalgesic
responses following administration of paclitaxel. However, when assessing the thermal
response to heat, it was found that paw withdrawal latency was not altered with paclitaxel
treatment when compared to the control group. Immunofluorescence reaction for c-Fos in the
dorsal root and spinal cord ganglia, demonstrated cell activation evidenced by an increased
immunoexpression was observed in the groups treated with paclitaxel. Administration of
paclitaxel caused a significant (p<0.05) increase in IL-1β, TNF-α, IL-6, MCP-1 and KC /
CXCL1 levels. Mechanical and thermal cold hyperalgesia were significantly reduced (p

12
<0.05) in SH animals (no mast cells) when compared to wild animals, and treatment with
sodium cromoglycate (mast cell membrane stabilizer) was effective (p <0.05) in inhibiting
mechanical and thermal hyperalgesia after the chemotherapic treatment. Pretreatment with
sodium cromoglycate significantly (p<0.05) prevented the increase of cytokines and
chemokines in mice treated with paclitaxel. In the SH mice treated with paclitaxel, no
significant increase (p<0.05) of cytokines was observed. Paclitaxel stimulation resulted in a
significant (p<0.05) increase in cytokines (TNF-α, IL-6) and chemokines (MCP-1)
concentration in mast cell culture. In addition, immunofluorescence of the dorsal root
ganglion demonstrated increased immunoexpression of tryptase and IL-6 after administration
of paclitaxel. The results also showed that the TLR4 receptor was involved in the
development of paclitaxel-induced hyperalgesia, since knockout animals to TLR4 receptor
treated with this chemotherapic did not develop mechanical and thermal hyperalgesia. The
results also demonstrated that, in the TLR4 knockout mice treated with paclitaxel, there was
no significant increase (p <0.05) in cytokines and chemokines; and when compared to the
group of WT mice, a significant reduction (p <0.05) in cytokine and chemokine levels was
observed. In addition, paclitaxel caused a significant (p <0.05) increase of cytokines (TNF-α
and IL-6) and chemokines (MCP-1 and KC/CXCL) in satellite glial cell culture and the
deletion of the TLR4 receptor gene was able to prevent this increase in cytokine/chemokine
levels. The study showed for the first time that mast cells and satellite glial cells are involved
in the development of acute pain induced by paclitaxel in mice. In addition, the study revealed
that the activation of mast cells and satellite glial cells is possibly due to the binding of
paclitaxel to TLR4, inducing the release of cytokines/chemokines that contribute to the
development of acute pain induced by paclitaxel.
Keywords: Paclitaxel; acute pain; syndrome; Mast cells; Glial satellite cells.

13
LISTA DE ILUSTRAÇÕES
Figura 1 - Imagem representativa do Taxus brevifolia ............................................................. 34
Figura 2 - Mecanismo de ação antitumoral do Paclitaxel. ....................................................... 36
Figura 3 - Delineamento esquemático do protocolo experimental 1 ........................................ 56
Figura 4 - Delineamento esquemático do protocolo experimental 2 ........................................ 57
Figura 5 - Delineamento esquemático do protocolo experimental 3 ........................................ 58
Figura 6 - Delineamento esquemático do protocolo experimental 4 ........................................ 59
Figura 7 - Delineamento esquemático do protocolo experimental 5 ........................................ 60
Figura 8 - Delineamento esquemático do protocolo experimental 6 ........................................ 61
Figura 9 - Fotomicrografia da imunoexpressão de c-Fos no gânglio da raiz dorsal de
camundongos submetidos à administração de paclitaxel ........................................................ 74
Figura 10 - Fotomicrografia da imunoexpressão de c-Fos no corno dorsal da medula espinal
de camundongos submetidos à administração de paclitaxel .................................................... 76
Figura 11 - Fotomicrografia da imunoexpressão de Triptase e IL-6 no gânglio da raiz dorsal
de camundongos submetidos à administração de paclitaxel .................................................. 114
Figura 12 - Participação de mastócitos e células satélites gliais na síndrome de dor aguda
induzida por paclitaxel............................................................................................................ 128

14
LISTA DE GRÁFICOS
Gráfico 1 - Paclitaxel induz hiperalgesia mecânica e térmica em camundongos ..................... 72
Gráfico 2 - Análise quantitativa da imunoexpressão para c-Fos no gânglio da raiz dorsal de
camundongos submetidos à administração de paclitaxel ......................................................... 75
Gráfico 3 - Análise quantitativa da imunoexpressão para c-Fos no corno dorsal da medula
espinal de camundongos submetidos à administração de paclitaxel ........................................ 77
Gráfico 4 - Paclitaxel induz aumento de IL-1β, TNF-α, IL-6 e KC/CXCL1 no plasma. ......... 79
Gráfico 5 - Paclitaxel induz aumento de IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL no nervo
isquiático. .................................................................................................................................. 81
Gráfico 6 - Paclitaxel aumenta a expressão relativa de RNAm de IL-6 no nervo isquiático. .. 83
Gráfico 7 - Paclitaxel induz aumento de IL-6, TNF-α e KC/CXCL1 em gânglios da raiz dorsal85
Gráfico 8 - Paclitaxel aumenta a expressão relativa de RNAm para IL-1β, TNF-α e CCR2 nos
gânglios da raiz dorsal .............................................................................................................. 87
Gráfico 9 - Paclitaxel induz aumento de IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL1 na medula
espinal ....................................................................................................................................... 88
Gráfico 10 - Paclitaxel aumenta a expressão relativa de RNAm para IL-1β, TNF-α, IL-6,
MCP-1 e CCR2, na medula espinal .......................................................................................... 90
Gráfico 11 - Papel da IL-6 na hiperalgesia mecânica e térmica induzida pelo paclitaxel ........ 93
Gráfico 12 - Envolvimento do receptor IL-1R na hiperalgesia induzida pelo paclitaxel ......... 95
Gráfico 13 - Envolvimento dos receptores TNFR1 e TNFR2 na hiperalgesia induzida pelo
paclitaxel ................................................................................................................................... 97
Gráfico 14 - Envolvimento do receptor CCR2 na hiperalgesia induzida pelo paclitaxel ........ 99
Gráfico 15 - Envolvimento de mástocitos na hiperalgesia induzida por paclitaxel ............... 101
Gráfico 16 - Cromoglicato de sódio atenua a hiperalgesia induzida pelo paclitaxel ............. 102

15
Gráfico 17 - Pré-tratamento com cromoglicato de sódio ou a ausência mastócitos previnem o
aumento de IL-1β, TNF-α, IL-6, KC/CXCL1 e MCP-1 no nervo isquiático de camundongos
tratados com paclitaxel ........................................................................................................... 105
Gráfico 18 - Pré-tratamento com cromoglicato de sódio ou a ausência de mastócitos previnem
o aumento dos níveis de citocinas e quimiocina nos gânglios da raiz dorsal de camundongos
tratados com paclitaxel ........................................................................................................... 107
Gráfico 19 - Pré-tratamento com cromoglicato de sódio ou a ausência de mastócitos previne o
aumento de IL-1β, TNF-α IL-6 e KC/CXCL1 na medula espinal de camundongos tratados
com paclitaxel ......................................................................................................................... 109
Gráfico 20 - Paclitaxel aumenta TNF-α, IL-6 e MCP1 em cultura de mastócitos (RBL-2H3)
estimulada com paclitaxel ...................................................................................................... 112
Gráfico 21 - Análise quantitativa da imunoexpressão para Triptase e IL-6 no gânglio da raiz
dorsal de camundongos submetidos à administração de paclitaxel ........................................ 115
Gráfico 22 - Participação do receptor TLR4 na hiperalgesia induzida pelo paclitaxel .......... 117
Gráfico 23 - A deleção gênica do TLR4 previne o aumento de IL-6 e KC/CXCL1 no plasma
de camundongos injetados com paclitaxel ............................................................................. 119
Gráfico 24 - A deleção gênica do TLR4 previne o aumento de IL-1β, IL-6, MCP-1 e
KC/CXCL1 no nervo isquiático de camundongos tratados com paclitaxel ........................... 120
Gráfico 25 - A deleção gênica do TLR4 previne o aumento de TNF-α e KC/CXCL1 nos
gânglios da raiz dorsal de camundongos tratados com paclitaxel .......................................... 121
Gráfico 26 - A deleção gênica do TLR4 previne o aumento de TNF-α, MCP-1 e KC/CXCL1
na medula espinal de camundongos tratados com paclitaxel ................................................. 122
Gráfico 27 - Paclitaxel aumenta IL-6, TNF-α, MCP1 e KC/CXCL1 em cultura primária de
células satélites gliais.............................................................................................................. 124
Gráfico 28 - A deleção gênica do receptor TLR4 previne o aumeno de IL-6, TNF-α, MCP-1 e
KC/CXCL1 em cultura primária de células satélites gliais estimulada com paclitaxel ......... 125
Gráfico 28 - A deleção gênica do receptor TLR4 previne o aumento de IL-6, TNF-α, MCP-1 e
KC/CXCL1 em cultura primária de células satélites gliais estimulada com LPS ou paclitaxel126

16
LISTA DE TABELAS E QUADROS
Quadro 1 - Sequência de primers utilizados no RT-PCR ......................................................... 65
Quadro 2 - Paclitaxel aumenta a expressão citocinas e/ou quimiocinas no plasma, no nervo
isquiático, nos gânglios da raiz dorsal e na medula espinal de camundongos ......................... 91
Quadro 3 - Papel dos mastócitos na produção/liberação de citocinas e/ou quimiocinas, no
nervo isquiático, gânglios da raiz dorsal e na medula espinal de camundongos injetados com
paclitaxel ................................................................................................................................. 110
Quadro 4 – A ausência do receptor TLR4 diminui a produção/liberação de citocinas e/ou
quimiocinas, no plasma, nervo isquiático, gânglios da raiz dorsal e na medula espinal de
camundongos injetados com paclitaxel ................................................................................. 123

17
LISTA DE ABREVIATURAS E SIGLAS
AMPA Ácido Alfa-Amino-3-Hidroxi-5-Metil-4-Isoxazolpropiónico
AMPc Adenosina Monofosfato Cíclico
ANOVA Análise de Variância
ATP Adenosina Trifosfato
B1 Receptores de Bradicinina 1
B2 Receptores de Bradicinina 2
BMS Myers Squibb Company
BSA Albumina de Soro Bovino
CAPES Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior
CaV Canais de Cálcio dependentes de Voltagem
CB1 Receptor Canabinóide 1
CCCE Centro de Criação de Camundongos Especiais
CCL2 Quimiocina (padrão C-C) do ligando 2
CCR2 Receptor de Quimiocinas CC Tipo 2
CD14 Cluster of Differentiation 14
cDNA DNA complementar
CEPA Comissão de Ética em Pesquisa Animal
CEUA Comissão de Ética para o Uso de Animais
CGRP Peptídeo Relacionado ao Gene da Calcitonina
cm Centímetro
CNPq Conselho Nacional de Desenvolvimento Científico e Tecnológico
CO2 Dióxido de Carbono
CONCEA Conselho Nacional de Controle de Experimentação Animal
COX-2 Cicloxigenase 2
CRID Centro de Pesquisa em Doenças Inflamatórias
CSG Células Satélites Gliais
CXCL1 Quimiocina (padrão C-X-C) ligando 1
DMEM Eagle modificado por Dulbecco
DNA Ácido Desoxirribonucleico
dNTP Desoxirribonucleotídeos Fosfatados
DTT Ditiotreitol
EDTA Ácido Etilenodiamino Tetra-acético

18
ELISA Enzyme-linked Immunosorbent Assay
EPM Erro Padrão da Média
et al. E colaboradores
EUA Estados Unidos da América
FAMED Faculdade de Medicina
FDA Food and Drug Administration
FMRP Faculdade de Medicina de Ribeirão Preto
g Gramas
GABA Ácido Gama-Aminobutírico
GAPDH Glyceraldehyde 3-phosphate dehydrogenase
GDP Guanosina Difosfato
GFAP Proteína Ácida Fibrilar Glial
GLAST Transportador de Glutamato-Aspartato
GRD Gânglios da Raiz Dorsal
GTP Guanosina Trifosfato
h hora
H2SO4 Ácido Sulfúrico
i.p. Intra-Peritonial
i.v. via intravenosa
IARC Agência Internacional para a Pesquisa em Câncer
IASP Associação Internacional para o Estudo da Dor
Iba-1 Molécula de adaptador de ligação de cálcio ionizado 1
IL-10 Interleucina 10
IL-13 Interleucina 13
IL-1R Receptor de Interleucina-1
IL-1β Interleucina 1 beta
IL-4 Interleucina 4
IL-5 Interleucina 5
IL-6 Interleucina 6
INCA Instituto Nacional do Câncer
iNOS Óxido Nítrico Sintase Induzível
IRAK Cinase Associada ao Receptor de Interleucina
KC Quimiocina Derivada de queratinócito
KCl Cloreto de Potássio

19
Kg Quilograma
KO Knockout
L3 Segmento Lombar 3
L4 Segmento Lombar 4
L5 Segmento Lombar 5
L6 Segmento Lombar 6
LPS Lipopolissacarídeo
m2
Metros quadrados
MAPK Proteína-quinase ativada por mitógenos
MCP-1 Proteína Quimioatraente de Monócitos
mg Miligrama
MgCl Cloreto de Magnésio
min Minutos
mL Mililitro
mM Milimolar
MRV Medula Rostral Ventromedial
NaCl Cloreto de Sódio
NCI National Cancer Institute
NF-κB Nuclear Factor Kappa B
NGF Fator de Crescimento de Nervos
NIH National Cancer Institute
NK1R Receptor de Neurocinina 1
NMDA N-metil-D-aspartato
NMDAr Receptor N-metil-D-aspartato
NPIQ Neuropatia Periférica Induzida Pela Quimioterapia
OMS Organização Mundial da Saúde
p Nível de significância
P2X7 Receptore Purinérgico P2X7
P2Y Receptore Purinérgico P2Y
PAF Fator de Agregação Plaquetária
P-APS Paclitaxel-Associated Acute Pain Syndrome
PBS Tampão Fosfato Salina
PCX Paclitaxel
pg Picograma

20
pH Potencial Hidrogeniônico
PKA Proteína Quinase A
PKC Proteína Quinase C
PMA Phorbol 12-Myristate 13-Acetate
RBL-2H3 Rat Basophilic Leukemia cells
RNA Ácido Ribonucleico
RNAm RNA mensagueiro
RT-PCR Reação em Cadeia da Polimerase em Tempo Real
s Segundos
SDAAP Síndrome da Dor aguda Associada ao Paclitaxel
SH Sash
SNC Sistema Nervoso Central
SNP Sistema Nervoso Periferico
TCZ Tocilizumabe
TLR Receptores Toll-Like
TLR2 Receptor Toll Like 2
TLR4 Receptor Toll Like 4
TMB 3,3′,5,5′-Tetramethylbenzidine
TNF- Fator de Necrose Tumoral alpha
TNFR1/R2 Recetor 1 e 2 do Fator de Necrose Tumoral
TRAF-6 Receptor de TNF Associado ao Fator 6
TRP Receptor de Potencial Transiente
TRPV1 Receptor de potencial transiente vanilóide 1
U Unidades
UFC Universidade Federal do Ceará
USP Universidade de São Paulo
WT Wild-Type
μg Micrograma
μL Microlitro
µM Micrometro
5-HT Serotonina
°C Grau Celsius
< Menor que

21
SUMÁRIO
1 Introdução ............................................................................................................................ 25
2 Revisão Bibliográfica. .......................................................................................................... 29
2.1 Dor ...................................................................................................................................... 30
2.1.1 Tipos de dor ..................................................................................................................... 31
2.2 Dor induzida por quimioterápicos ...................................................................................... 32
2.3 Paclitaxel ............................................................................................................................ 33
2.3.1 Mecanismo de ação antitumoral do paclitaxel................................................................ 35
2.3.2 Toxicidade associada ao paclitaxel ................................................................................. 37
2.3.3 Síndrome da dor aguda associada ao paclitaxel ............................................................ 39
2.4 Mastócitos ........................................................................................................................... 40
2.5 Células satélites gliais ......................................................................................................... 42
3 Justificativa .......................................................................................................................... 46
4 Objetivos ............................................................................................................................... 48
4.1 Objetivo Geral .................................................................................................................... 49
4.2 Objetivos Específicos ......................................................................................................... 49
5 Materiais e Métodos ............................................................................................................ 51
5.1 Animais ............................................................................................................................... 52
5.2 Aspectos Éticos .................................................................................................................. 53
5.3 Indução da síndrome da dor aguda associada ao paclitaxel ............................................... 53
5.4 Experimentos comportamentais ......................................................................................... 54
5.4.1 Avaliação da hiperalgesia mecânica ............................................................................... 54
5.4.2 Avaliação da hiperalgesia térmica ao frio ...................................................................... 54
5.4.3 Avaliação da hiperalgesia térmica ao calor ................................................................... 55
5.5 Delineamento experimental ................................................................................................ 55
5.5.1 Avaliação da hiperalgesia mecânica e térmica no modelo de síndrome da dor aguda
induzida pelo paclitaxel ............................................................................................................ 55
5.5.2 Investigação da participação de mastócitos na síndrome dolorosa aguda induzida pelo
paclitaxel .................................................................................................................................. 56
5.5.3 Avaliação do efeito do cromoglicato de sódi na síndrome dolorosa aguda induzida pelo
paclitaxel .................................................................................................................................. 57
5.5.4 Investigação da participação de citocinas na síndrome da dor aguda induzida pelo
paclitaxel .................................................................................................................................. 58

22
5.5.5 Coleta de sangue venoso periférico, nervo isquiático, gânglios da raiz dorsal e medula
espinal ....................................................................................................................................... 60
5.6 Cultura de mastócitos ......................................................................................................... 62
5.7 Cultura primária de células satélites gliais de gânglios da raiz dorsal ............................... 62
5.8 Expressão gênica por reação em cadeia da polimerase em tempo real .............................. 63
5.8.1 Coleta de tecidos ............................................................................................................. 63
5.8.2 Extração do RNA total ..................................................................................................... 63
5.8.3 Confecção do DNA complementar .................................................................................. 64
5.8.4 Reação em cadeia da polimerase em tempo real ............................................................ 64
5.9 Dosagem de citocinas pró-inflamatórias e quimiocinas ..................................................... 65
5.9.1 Coleta das amostras ........................................................................................................ 65
5.9.2 Preparação de amostras para análise de citocinas/quimiocinas no plasma .................. 66
5.9.3 Homogenato de Tecidos .................................................................................................. 66
5.9.4 Preparação de amostras para análise de citocinas/quimiocinas de cultura de mastócitos
(RBL-2H3) ................................................................................................................................ 66
5.9.5 Preparação de amostras para análise de citocinas/quimiocinas de cultura de células
satélites gliais ........................................................................................................................... 67
5.9.6 Determinação de citocinas/quimiocinas por ELISA ....................................................... 67
5.10 Imunofluorescência .......................................................................................................... 68
5.10.1 Coleta e preparação das amostras ................................................................................ 68
5.10.2 Imunofluorescência para Triptase, IL-6 e c-Fos ........................................................... 68
5.11 Análise Estatística ............................................................................................................ 69
6 Resultados ............................................................................................................................ 70
6.1 Paclitaxel induz hiperalgesia mecânica e térmica em camundongos ................................. 71
6.2 Imunofluorescência para c-Fos em gânglio da raiz dorsal de camundongos submetidos à
injeção de paclitaxel ................................................................................................................. 73
6.3 Imunofluorescência para c-Fos em corno dorsal da medula espinal de camundongos
submetidos à injeção de paclitaxel ........................................................................................... 75
6.4 Quantificação de citocinas e quimiocinas em camundongos tratados com paclitaxel ....... 78
6.4.1 Quantificação de citocinas e quimiocinas no plasma ..................................................... 78
6.4.2 Quantificação de citocinas e quimiocinas no nervo isquiático ....................................... 79
6.4.3 Expressão relativa de RNAm para citocinas e quimiocinas em nervo isquiático ........... 81
6.4.4 Quantificação de citocinas e quimiocinas nos gânglios da raiz dorsal .......................... 84
6.4.5 Expressão relativa de RNAm de citocinas e quimiocinas em gânglios da raiz dorsal ... 85

23
6.4.6 Quantificação de citocinas e quimiocinas na medula espinal ........................................ 87
6.4.7 Expressão relativa de RNAm de citocinas e quimiocinas em medula espinal ................ 88
6.5 Investigação da participação de citocinas e quimiocinas na hiperalgesia mecânica e
térmica induzida pelo paclitaxel ............................................................................................... 91
6.5.1 Investigação da participação da IL-6 na hiperalgesia mecânica e térmica induzida pelo
paclitaxel .................................................................................................................................. 91
6.5.2 Investigação da participação de IL-1 na hiperalgesia mecânica e térmica induzida pelo
paclitaxel .................................................................................................................................. 94
6.5.3 Investigação da participação TNF-α na hiperalgesia mecânica e térmica induzida pelo
paclitaxel .................................................................................................................................. 93
6.5.4 Investigação da participação de MCP-1 na hiperalgesia mecânica e térmica induzida
pelo paclitaxel .......................................................................................................................... 94
6.6 Investigação da participação de mastócitos na hiperalgesia mecânica e térmica induzida
pelo paclitaxel ......................................................................................................................... 100
6.7 Quantificação de citocinas e quimiocinas em camundongos SH e em camundongos
selvagens pré-tratados com cromoglicato de sódio e injetados com paclitaxel ..................... 103
6.7.1 Quantificação de citocinas e quimiocinas em nervo isquiático .................................... 103
6.7.2 Quantificação de citocinas e quimiocinas em gânglios da raiz dorsal ......................... 106
6.7.3 Quantificação de citocinas e quimiocinas em medula espinal ...................................... 108
6.8 Quantificação de citocinas e quimiocinas em cultura de mastócitos estimulada com
paclitaxel. ................................................................................................................................ 110
6.9 Imunofluorescência para Triptase e IL-6 em gânglio da raiz dorsal de camundongos
submetidos à injeção de paclitaxel ......................................................................................... 113
6.10 Investigação da participação de TLR4 na hiperalgesia mecânica e térmica induzida pelo
paclitaxel ................................................................................................................................. 116
6.11 Quantificação de citocinas e quimiocinas em camundongos TLR4-/- injetados com
paclitaxel ................................................................................................................................. 118
6.11.1 Quantificação de citocinas e quimiocinas no plasma ................................................. 118
6.11.2 Quantificação de citocinas e quimiocinas no nervo isquiático ................................... 119
6.11.3 Quantificação de citocinas e quimiocinas nos gânglios da raiz dorsal ...................... 120
6.11.4 Quantificação de citocinas e quimiocinas na medula espinal .................................... 121
6.12 Quantificação de IL-6, TNF-α, MCP1 e KC em cultura primária de células satélites gliais
estimulada com paclitaxel ...................................................................................................... 123
6.13 Modelo Conceitual ......................................................................................................... 127

24
7 Discussão ............................................................................................................................ 129
8 Conclusão ........................................................................................................................... 143
Referências ........................................................................................................................... 145
Anexos .................................................................................................................................... 163

25
1. INTRODUÇÃO

26
Câncer é um termo utilizado para definir doenças nas quais células anormais
dividem-se sem controle podendo invadir tecidos circundantes. As células cancerosas também
podem se espalhar para outras partes do corpo através do sangue e do sistema linfático (NIH -
NATIONAL CANCER INSTITUTE, 2015).
De acordo com estimativas da Organização Mundial da Saúde (OMS), cerca de
14,9 milhões de novos casos de câncer acometeram a população mundial, mais de 60%
ocorreram em países em desenvolvimento, resultando em 8,2 milhões de mortes por câncer,
sendo que 70% ocorreram nesses mesmos países. Os tipos de câncer mais incidentes no
mundo foram o câncer de pulmão, mama, intestino e próstata. O câncer de pulmão foi o que
mais acometeu a população masculina. Já entre as mulheres, o mais frequente foi o de mama.
Em 2025, estima-se que no mundo o número total de novos casos de câncer seja de 20
milhões que acarretaram em 13,2 milhões de mortes (INTERNATIONAL AGENCY FOR
RESEARCH ON CANCER, 2012).
No Brasil, estima-se que para o biênio 2016-2017, ocorram aproximadamente 600
mil novos casos de câncer, excluindo-se os casos de câncer de pele não melanoma (180 mil
novos casos). Em homens, o tipo mais incidente será o câncer de próstata e nas mulheres, o
câncer de mama (INSTITUTO NACIONAL DO CÂNCER, 2015).
Ainda que as estimativas sobre a epidemiologia do câncer sejam alarmantes, a
taxa de sobrevivência a estas doenças vem aumentando, e isso se deve aos avanços no
tratamento do câncer obtido nas últimas décadas, como por exemplo, no campo da
quimioterapia, na qual drogas são utilizadas para destruir as células doentes que formam o
tumor. A quimioterapia do câncer conta com um arsenal de fármacos que abrange centenas de
quimioterápicos que apesar da eficácia em limitar ou inibir o crescimento tumoral,
frequentemente é acompanhada de efeitos adversos que dificultam a adesão do paciente ao
tratamento. Portanto, faz-se necessário buscar melhorias para o tratamento do câncer não
apenas no sentido de desenvolver novas abordagens terapêuticas específicas, mas também em
descobrir maneiras de amenizar ou prevenir os efeitos adversos associados à quimioterapia.
O paclitaxel (PCX) é um alcaloide originalmente derivado do Taxus brevifolia, e é
utilizado como agente quimioterápico de primeira linha para o tratamento de diversos tipos de
tumores. No entanto, os pacientes que recebem tratamento com paclitaxel frequentemente
desenvolvem dor patológica, o que reduz significativamente a qualidade de vida desses
pacientes e dificulta a utilização deste quimioterápico. A dor patológica induzida pelo
paclitaxel inclui dor que ocorre imediatamente após o primeiro ciclo do tratamento com este

27
quimioterápico (REEVES et al., 2012; LONPRINZI et al., 2011), e dor que persiste durante
semanas a anos após o fim do tratamento (CATA et al., 2006; DOUGHERTY et al., 2004).
A dor aguda induzida pelo paclitaxel é uma morbidade significativa em pacientes
e, atualmente, não existe um padrão comprovado de cuidados para a sua prevenção ou
tratamento (REEVES et al., 2012; LONPRINZI et al., 2011), além disso, os mecanismos
pelos quais o paclitaxel induz dor aguda não são totalmente conhecidos. Atualmente o
entendimento dos mecanismos subjacentes da dor patológica induzida por paclitaxel em
modelos in vivo é baseado principalmente em estudos que examinam as mudanças patológicas
nos dias após o final do tratamento com essa droga. Até o momento, pouco se sabe sobre a
ação direta e imediata do paclitaxel no sistema de sinalização da dor aguda em modelos in
vivo, e poucos são os estudos que buscam esclarecer esses mecanismos utilizando esses
modelos (YAN et al., 2015, JATOI et al., 2016, BRUSCO et al., 2016). Assim, a
compreensão de tais mecanismos poderia fornecer fundamentos para o desenvolvimento de
tratamentos eficazes para essa síndrome dolorosa (POLOMANO et al., 2001; RIGO et al.,
2013).
Várias células imunitárias tais como macrófagos e mastócitos, têm sido
implicadas na patogênese e no processamento nociceptivo alterado característico de processos
dolorosos. Os mastócitos são agentes cruciais nas desordens alérgicas, no entanto, também
são importantes iniciadores e efetores da imunidade inata. Eles residem em muitos tecidos,
incluindo os nervos (GALLI, NAKAE, TSAI, 2005; METCALFE, BARAM, MEKORI,
1997). Os mastócitos contêm grânulos ricos em fatores pró-inflamatórios e tem sido relatado
que eles desempenham um papel vital no desenvolvimento de hiperalgesia térmica e alodinia
mecânica em camundongos (ZUO et al., 2003). Seus grânulos contêm mediadores tais como
histamina, proteases e citocinas (METCALFE, BARAM, MEKORI, 1997; GALLI et al.,
2005), vários dos quais são capazes de sensibilizar ou ativar neurônios. Os mastócitos
ativados contribuem diretamente para estados de dor, incluindo dor neuropática (XANTHOS
et al., 2011). Foi demonstrado in vitro que a quercetina inibe a liberação de histamina em
cultura de mastócito (RBL-2H3) estimulada com paclitaxel por estabilizar a membrana de
mastócitos (GAO et al., 2016). Uma grande quantidade de evidências suporta a hipótese de
que os mastócitos contribuem para o desenvolvimento de dor patológica. Estes resultados são,
portanto, uma forte indicação de que os mastócitos podem contribuir para a dor aguda
associada ao paclitaxel. Os mastócitos possuem estreita relação anatômica com fibras
nervosas em uma grande variedade de órgãos e tecidos e isso fornece uma comunicação
bidirecional entre mastócitos e neurônios (ALVING et al., 2001; PURCELL; ATTERWILL,

28
1995; ROZNIECKI et al., 1999). Os mastócitos são capazes de ativar neurônios através da
síntese e liberação de uma infinidade de mediadores, incluindo histamina, serotonina (5-HT),
citocinas e metabolitos do ácido araquidônico.
Além do envolvimento de células imunes na dor induzida por quimioterápicos, as
células da glia também podem estar envolvidas. Dentre elas as células satélites gliais (CSG),
estas são células especializadas que envolvem e se comunicam com neurônios sensoriais nos
gânglios da raiz dorsal (GRD), são capazes de modular o microambiente neuronal e a
transmissão sensorial dentro dos gânglios (HANANI, 2005; ZHANG et al., 2007; HANANI,
2010). Tem sido sugerido que as CSG nos gânglios sensoriais desempenham um papel
fundamental na geração e manutenção da dor, (HANANI et al., 2002; HUANG et al., 2010).
Evidencias sugerem ainda que as CSG possuem um papel crítico na dor induzida pela
quimioterapia (CAPUANO et al., 2009; NEEB et al., 2011; LAURSEN et al., 2014;
TAKEDA et al., 2011; WARWICK; HANANI, 2013).
Embora evidências tenham demonstrado o envolvimento de mastócitos e de CSG
na neuropatia induzida por quimioterápicos, não existem estudos que evidênciam a
participação destas células na dor aguda associada ao paclitaxel. Dessa forma, o presente
estudo se propôs a investigar a participação de mastócitos e de CSG na síndrome da dor aguda
associada ao paclitaxel. Essa investigação pode fornecer evidencias para uma melhor
compreensão dos mecanismos pelos quais o paclitaxel causa dor, especialmente a dor aguda.
Portanto, é interessante o estudo do papel dos mastócitos e das CSG na dor aguda induzida
por paclitaxel, tendo em vista que pouco se sabe sobre os mecanismos envolvidos no
desenvolvimento desta síndrome dolorosa.

29
2. REVISÃO BIBLIOGRÁFICA

30
2.1 Dor
A dor é considerada um mecanismo de defesa contra agentes nocivos ou lesão
tecidual. A capacidade de detectar estímulos nocivos é essencial para a sobrevivência e o
bem-estar de um organismo, além disso, a percepção da dor desempenha uma função
adaptativa e evolutiva, tendo em vista que estímulos térmicos, mecânicos e/ou químicos
precisam ser reconhecidos como potencialmente perigosos (BASBAUM et al, 2009;
BINGHAM et al., 2009). Embora seja uma sensação desagradável, a dor tem função
fundamental, que é a de avisar que ocorreu dano corporal ou que este é iminente. A percepção
de dor é o resultado da integração neuronal e do processamento de informação sensorial que é
geralmente iniciado na periferia e transferido para o Sistema Nervoso Central (SNC)
(BINGHAM et al., 2009). A dor pode ser modulada por vários fatores, tais como fatores
comportamentais, emocionais, sociais, culturais, ambientais e cognitivos (BASBAUM et al,
2009).
Considerando a subjetividade da dor, é importante diferenciar os termos dor e
nocicepção. Nocicepção refere-se ao processo neural de codificação e processamento de
estímulos nocivos, desencadeada por dano ou ameaça ao tecido não neural e é devido à
ativação de nociceptores. A dor é uma experiência humana singular influenciada por diversos
elementos tais como emoção, cognição, memória e construções sociais (INTERNATIONAL
ASSOCIATION FOR THE STUDY OF PAIN, 2012; FARQUHAR-SMITH, 2007). A dor,
de acordo com a Associação Internacional para o Estudo da Dor (IASP, sigla em inglês), é
definida como “uma experiência sensorial e emocional desagradável associada a uma lesão
tecidual real ou potencial, ou descrita em termos de tal dano”. Essa definição atual de dor,
forneceu uma sólida base conceitual para os avanços científicos e dos profissionais de saúde
em relação à compreensão da natureza e tratamento da dor aguda e crônica. No entanto, o
texto principal encontra-se inalterado desde a primeira publicação em 1979. Desde então,
houve avanços significativos na compreensão, avaliação e tratamento da dor, gerando uma
revolução na compreensão dos mecanismos da dor. Recentemente uma revisão da definição
de dor foi proposta: “A dor é uma experiência angustiante associada a uma lesão real ou
potencial dos tecidos com componentes sensoriais, emocionais, cognitivas e sociais”
(WILLIAMS; CRAIG, 2016). No entanto, os autores afirmam que essa nova definição
também exige revisão e que ela atende apenas elementos problemáticos e que o objetivo deste
relato era gerar debate e discussão que levará a uma melhor definição sobre dor.
Comumente, alterações na via da dor levam a uma hipersensibilidade, de tal forma
que a dor ultrapassa a sua utilidade como um sistema de alerta agudo se tornando crônica e

31
debilitante. Isso pode ser visto, em algum nível, como uma extensão do processo de cura
normal, em que danos no tecido ou nervos provocam hiperatividade para promover a
vigilância da área lesada. Por exemplo, queimaduras solares produzem sensibilização
temporária da área afetada. Como resultado, estímulos normalmente inócuos, como a
incidencia da luz ou calor, são percebidos como sensação dolorosa (fenômeno conhecido
como alodinia), ou estímulos dolorosos que normalmente provocam dor são percebidos com
maior intensidade (referido como hiperalgesia) (INTERNATIONAL ASSOCIATION FOR
THE STUDY OF PAIN, 2012; BASBAUM et al., 2009).
2.1.1 Tipos de dor
Em relação à duração, a dor pode ser classificada em transitória, aguda ou crônica.
Tanto na dor transitória como na dor aguda, a causa é bem definida e o curso de ambas são
limitados, podendo desaparecer antes mesmo da remoção da causa ou do reparo do dano
tecidual. Quando um estímulo nocivo é aplicado ao tecido normal, dor nociceptiva aguda
fisiológica é provocada. Esta dor protege o organismo para que o tecido não seja mais
danificado, através da geração de reflexos de retirada. A dor nociceptiva fisiológica ocorre
quando o tecido está inflamado ou ferido. Essa dor pode aparecer como dor espontânea (dor
na ausência de qualquer estímulo intencional) e/ou como hiperalgesia e/ou alodinia.
Hiperalgesia é um aumento da dor, em resposta a uma estimulação nociva, e alodinia é a
ocorrência de dor provocada por estímulos que são normalmente abaixo do limiar de dor, ou
seja, dor devido a um estímulo que normalmente não provocaria dor. Enquanto a dor
nociceptiva é provocada por estímulo nocivo nas terminações sensoriais nos tecidos e é
devido à ativação de nociceptores (INTERNATIONAL ASSOCIATION FOR THE STUDY
OF PAIN, 2012; SCHAIBLE; RICHTER, 2004), a dor crônica é geralmente causada por
lesões ou doenças que superam a capacidade do organismo de reverter o quadro, podendo
persistir até mesmo após o desaparecimento do trauma inicial, estendendo-se por meses ou
anos e comprometendo a qualidade de vida do indivíduo (LOESER; MELZACK, 1999). A
dor neuropática se desenvolve como resultado de lesão ou doença dos neurônios no SNC ou
Sistema Nervoso Periférico (INTERNATIONAL ASSOCIATION FOR THE STUDY OF
PAIN, 2012; SCHAIBLE; RICHTER, 2004). Esta condição pode apresentar uma combinação
de sintomas, que incluem sintomas negativos, como perda completa ou parcial da
sensibilidade, e sintomas positivos como parestesia (sensação anormal, seja espontânea ou
evocada), disestesia (sensações cutâneas subjetivas como formigamento, dormência e picada)

32
e dor. A sensação permanente de dor em queimação é relatada, e parece decorrer da atividade
espontânea das fibras C nociceptivas, as quais também promovem sensibilização dos
neurônios do corno dorsal na medula espinal. Da mesma maneira, a atividade espontânea de
fibras A, está relacionada à parestesia e, após sensibilização do SNC, à disestesia e à dor
(WOOLF; MANNION, 1999). Numerosos processos patológicos podem causar dor
neuropática, por exemplo, axotomia ou dano do nervo ou plexo, doenças metabólicas como
diabetes mellitus, herpes zoster, ou pós-acidente vascular encefálico (SCHAIBLE; RICHTER,
2004). Clinicamente, a dor neuropática é caracterizada por dor contínua ou espontânea e
respostas amplificadas de dor a estímulos nocivos ou não nocivos (BARON; BINDER;
WASNER, 2010). A dor neuropática pode surgir ainda devido a dano ao tecido neuronal
periférico em consequência da administração de quimioterápicos no tratamento de pacientes
com câncer (MANTYH, 2006). A dor neuropática provocada por quimioterápicos reduz a
qualidade de vida dos pacientes e muitas vezes esse sintoma é dose-limitante,
impossibilitando o tratamento correto do tumor, pois para amenizar o aparecimento da dor a
dose do quimioterápico é diminuída o que leva muitas vezes a perda da ação antitumoral
(QUASTHOFF; HARTUNG, 2002).
2.2 Dor induzida por quimioterápicos
A maioria dos pacientes com câncer utiliza a terapia quimioterápica na tentativa
de combater o tumor e isso se torna um dos fatores mais importantes na determinação da
sobrevivência e qualidade de vida dos mesmos (QUASTHOFF; HARTUNG, 2002). No
entanto, os quimioterápicos causam vários efeitos adversos, dentre eles, a neuropatia
periférica que é um efeito secundário de muitos agentes quimioterápicos. A neuropatia tem
sido descrita principalmente em pacientes tratados com os derivados de platina (cisplatina,
oxaliplatina, carboplatina), alcaloides da vinca (vincristina) e taxanos (paclitaxel, docetaxel,
nab-paclitaxel). A neuropatia periférica induzida pela quimioterapia (NPIQ) é um sintoma
comum e angustiante que precisa de opções preventivas ou terapêuticas eficazes
(ARGYRIOU et al., 2012; PACHMAN et al., 2011; DE IULIIS et al., 2015).
Os taxanos são alguns dos fármacos mais importantes no tratamento de tumores
sólidos. Entretanto, indivíduos submetidos ao tratamento com taxanos frequentemente são
acometidos por neuropatia que pode atingir até 70% dos pacientes. Estas taxas podem ser
alteradas de acordo com a intensidade da dose, número de ciclos de tratamento
quimioterápico, idade do paciente, duração do tratamento ou coadministração de outras
drogas neurotóxicas (ARGYRIOU, 2009; ARGYRIOU et al., 2012). Os taxanos causam dano

33
nos axônios dos neurônios, na mielina e nos gânglios da raiz dorsal (GRD). Os taxanos
estabilizam os microtúbulos e consequentemente interferem na mitose e no transporte
intracelular de proteínas e substâncias entre axônios e corpo celular (ARGYRIOU, 2009).
Uma vez que o transporte nos microtúbulos está danificado, a célula neuronal não pode
sobreviver e sofre apoptose (CARLSON; OCEAN, 2011).
A neuropatia causada pelos taxanos pode manifestar-se como parestesia ou
disestesia, dor em queimação, bem como alodinia mecânica e térmica ao frio e normalmente
envolve mãos e pés (distribuição em meia e luva). Os sintomas começam simetricamente nos
dedos dos pés, porque as extremidades dos nervos mais longos são primeiramente afetadas.
Após as doses conseguintes, os sintomas progridem de forma ascendente dos pés até o
tornozelo e pernas e, em seguida, também para as mãos, pulsos e braços (SWAIN; AREZZO,
2008). A capacidade de manusear objetos pode está comprometida, e, além disso, os pacientes
podem causar por si mesmos ferimentos mecânicos ou induzidos pela temperatura (calor ou
frio). Todos os nervos periféricos (sensoriais, motores e autonômicos) podem ser danificados
pelos taxanos, mas as fibras nervosas sensitivas mielinizadas são mais susceptíveis
(ARGYRIOU, 2009; CARLSON; OCEAN, 2011).
Outra toxicidade comumente relatada aos taxanos, principalmente ao paclitaxel, é
a dor aguda. Esta toxicidade é conhecida como síndrome dor aguda associada ao paclitaxel
(LONPRENZI et al., 2011) ou síndrome da dor aguda induzida por taxanos (FERNANDES
et al., 2016). Esta síndrome é tipicamente caracterizada por mialgias e artralgias que surgem
24-48 horas após o paciente receber a primeira dose da quimioterapia à base de taxano e tem
duração de 5-7 dias (SAIBIL et al., 2010). A síndrome da dor aguda associada ao paclitaxel
pode dificultar a adesão dos pacientes ao tratamento e, frequentemente, leva a uma
diminuição na dose eficaz contra o tumor ou, até mesmo, interrupção da quimioterapia.
2.3 Paclitaxel
O taxano antimitótico, paclitaxel é um antineoplásico utilizado como primeira
linha no tratamento de diversos tumores sólidos, particularmente em carcinomas de mama,
ovário, pulmão, cabeça e pescoço (ROWINSKY, 1997; ROWINSKY et al., 1993; VON
HOFF, 1997; CROWN; O’LEARY, 2000; MILLER; SLEDGE JUNIOR, 1999).
O paclitaxel foi o primeiro taxano usado em ensaios clínicos com atividade contra
uma variedade de cânceres refratários a quimioterapia convencional. Em 1963, através de um
programa do National Cancer Institute (NCI) dos Estados Unidos da América, um extrato
bruto da casca de um raro teixo do Pacífico, o Taxus brevifolia (Figura 1), foi descoberto, e

34
em estudos pré-clínicos esse extrato demonstrou ter atividade citotóxica contra muitos
tumores. O paclitaxel foi isolado originalmente como sendo constituinte desse extrato (WANI
et al., 1971). Atualmente, ele é obtido através de uma semissíntese do produto originado das
folhas de Taxus baccata, o precursor de paclitaxel e docetaxel.
Figura 1 - Imagem representativa do Taxus brevifolia.
Fonte: Adaptado de Sociedade Brasileira de Química.
Em 1979 foi identificado o mecanismo de ação exclusivo do paclitaxel como
droga antitumoral, que difere dos alcalóides da vinca e derivados da colchicina, pois o
paclitaxel induz uma hiperpolarização dos microtúbulos ao invés de inibir a formação destes,
e este achado representou o estímulo necessário, para que se intensificassem os estudos com a
droga pelo NCI (SCHIFF; HORWITZ, 1980; ROWINSKY, 1990).
O paclitaxel foi o agente antitumoral mais promissor desenvolvido nas décadas de
1980 e 1990. Foi aprovado pelo Food and Drug Administration (FDA) para o tratamento de
câncer ovariano refratário em 1992, e foi introduzido no mercado em 1993 através de Bristol-
Myers Squibb Company (BMS) operando sob uma pesquisa cooperativa e acordo de
desenvolvimento (CRADA) juntamente com o NCI. Foi aprovado para o tratamento do câncer
de mama em 1994, e é a primeira linha de tratamento para este tipo de câncer. Vendas
mundiais de paclitaxel alcançaram $1.2 bilhões em 1998. O paclitaxel também é utilizado em
grande escala no tratamento de linfomas, como primeira linha na abordagem do câncer de
pulmão e ovário (ROWINSKY; DONEWOHER, 1995).

35
2.3.1 Mecanismo de ação antitumoral do paclitaxel
O paclitaxel apresenta um mecanismo de ação exclusivo que o difere dos demais
inibidores mitóticos. Os microtúbulos são essenciais para a manutenção da forma celular,
sendo um dos componentes do fuso mitótico e do transporte dentro das células. A ligação do
paclitaxel aos microtúbulos estimula a polimerização da tubulina, que estabiliza os
microtúbulos. Dessa forma, o ciclo celular acaba sendo bloqueado na sua última fase (G2) na
mitose, impedindo a divisão e proliferação das células neoplásicas, ocasionando morte
subsequentemente por um mecanismo de apoptose (JORDAN; WILSON, 2004; JORDAN et
al., 1993). A ligação do paclitaxel ao longo do lúmen do microtúbulo estabiliza a rede de
microtúbulos, suprimindo assim a despolimerização e a instabilidade dinâmica (Figura 2). Em
concentrações mais elevadas do que as necessárias para suprimir a dinâmica, o paclitaxel
aumenta a polimerização dos microtúbulos (DERRY; WILSON; JORDAN, 1995;
DUMONTET; JORDAN, 2010; JORDAN et al., 1993). O paclitaxel é capaz de promover a
polimerização dos microtúbulos, mesmo na ausência de GTP, e estes microtúbulos são
resistentes à despolimerização (SCHIFF et al., 1979; SCHIFF; HORWITZ, 1981). O
mecanismo molecular descrito subjacente aos efeitos do paclitaxel é através da estabilização
da conformação helicoidal do anel M da β-tubulina, que fortalece os contatos laterais entre as
subunidades tubulínicas (PROTA et al., 2013).
Portanto, o paclitaxel é altamente eficaz contra proliferação de células cancerosas.
É amplamente utilizado, tanto sozinho como em combinação com outros agentes
quimioterápicos, no tratamento de muitos tipos de tumores sólidos, incluindo tumores da
mama, ovário, pulmão e cabeça e pescoço.
36
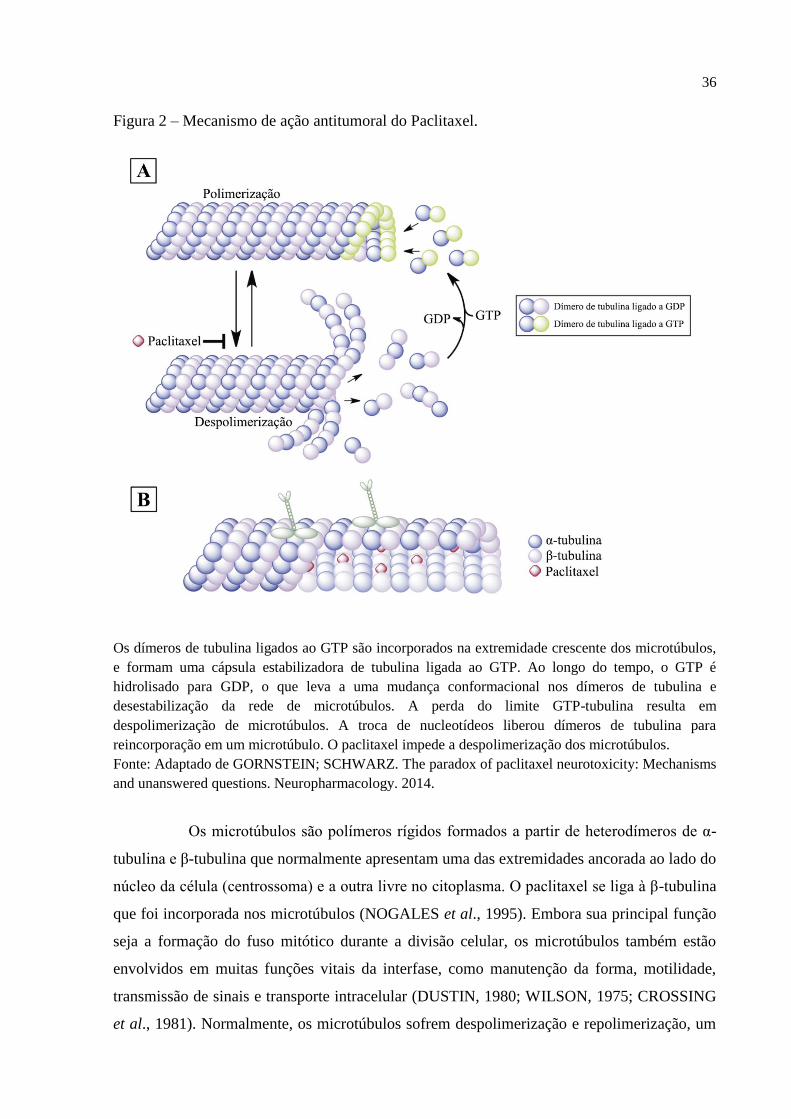
Figura 2 – Mecanismo de ação antitumoral do Paclitaxel.
Os dímeros de tubulina ligados ao GTP são incorporados na extremidade crescente dos microtúbulos,
e formam uma cápsula estabilizadora de tubulina ligada ao GTP. Ao longo do tempo, o GTP é
hidrolisado para GDP, o que leva a uma mudança conformacional nos dímeros de tubulina e
desestabilização da rede de microtúbulos. A perda do limite GTP-tubulina resulta em
despolimerização de microtúbulos. A troca de nucleotídeos liberou dímeros de tubulina para
reincorporação em um microtúbulo. O paclitaxel impede a despolimerização dos microtúbulos.
Fonte: Adaptado de GORNSTEIN; SCHWARZ. The paradox of paclitaxel neurotoxicity: Mechanisms
and unanswered questions. Neuropharmacology. 2014.
Os microtúbulos são polímeros rígidos formados a partir de heterodímeros de α-
tubulina e β-tubulina que normalmente apresentam uma das extremidades ancorada ao lado do
núcleo da célula (centrossoma) e a outra livre no citoplasma. O paclitaxel se liga à β-tubulina
que foi incorporada nos microtúbulos (NOGALES et al., 1995). Embora sua principal função
seja a formação do fuso mitótico durante a divisão celular, os microtúbulos também estão
envolvidos em muitas funções vitais da interfase, como manutenção da forma, motilidade,
transmissão de sinais e transporte intracelular (DUSTIN, 1980; WILSON, 1975; CROSSING
et al., 1981). Normalmente, os microtúbulos sofrem despolimerização e repolimerização, um

37
processo conhecido como instabilidade dinâmica (MITCHISON; KIRSCHNER, 1984). O
modelo atual para este processo é que as subunidades de tubulina ligadas a GTP são
incorporadas na extremidade do microtúbulo, formando uma tampa. Quando o GTP é
subsequentemente hidrolisado, a subunidade de tubulina muda de conformação, conduzindo à
desestabilização da rede de microtúbulos. Um tampão de tubulina GTP preservará o
microtúbulo, mas se ocorrer hidrólise de GTP das subunidades terminais, pode ocorrer uma
despolimerização extensa do microtúbulo durante o qual as subunidades GDP-tubulina se
dissociarão da extremidade dos microtúbulos (NOGALES; WANG, 2006).
Poucos agentes antineoplásicos exibem características farmacológicas tão
favoráveis e atrativas. Primeiro, porque o paclitaxel se liga a β-tubulina levando a
polimerização dos microtúbulos (SCHIFF, FANT; HORWITZ, 1979; CAPLOW, SHANKS;
RUHLEN, 1994). Essa estabilização dos microtúbulos causa parada da mitose e apoptose in
vitro, agindo independentemente da via p53 supressor, isso foi demostrado em ensaios
experimentais (JORDAN et al., 1996; WOODS et al., 1995; FAN et al.,1998;
VIKHANSKAYA et al., 1998) e clínicos (KING et al., 2000; GADDUCCI et al., 2000). O
paclitaxel induz ainda RNAm para TNF-α, interleucinas (DING et al., 1990; BURKHART et
al., 1994; WHITE et al., 1998; WATSON et al., 1998; COLLINS, LEE; TING, 2000), e
enzimas como iNOS (KIRIKAE et al., 1996; KIRIKAE et al., 2000; KIM; PAIK, 2005) e
COX-2 (MOOS et al., 1998; MOOS et al., 1999; SUBBARAMAIAH et al., 2000) que geram
mediadores da inflamação. A indução desses genes requer 10-30µM de paclitaxel.
2.3.2 Toxicidade associada ao paclitaxel
O paclitaxel exerce seus efeitos tóxicos principais na medula óssea. Neutropenia
geralmente ocorre em 8 a 11 dias após administração de uma dose, revertendo ou
recuperando-se rapidamente entre o 15º e o 21º dia. Reações de hipersensibilidade ocorrem
em pacientes que receberam infusões de paclitaxel de curta duração (1 a 6 horas), mas em
grande parte evitadas pelo pré-tratamento com difenidramina, cimetidina e dexametasona.
Bradicardia e taquicardia ventricular silenciosa também ocorrem, mas resolvem-se
espontaneamente durante as infusões de 3 a 24 horas. Mucosite é outro efeito tóxico comum
aos antineoplásicos e que na quimioterapia com paclitaxel é proeminente em esquemas de
infusões prolongadas.
Os pacientes que recebem tratamento com paclitaxel frequentemente desenvolvem
dor, o que reduz significativamente a sua qualidade de vida e dificulta o uso deste
quimioterápico. A dor induzida pelo paclitaxel inclui a dor que ocorre imediatamente após o

38
tratamento com este fármaco, conhecida como síndrome da dor aguda associada ao paclitaxel
(REEVES et al., 2012; LOPRINZI et al., 2011). Em adição a esta toxicidade aguda do
paclitaxel, neuropatia periférica pode ser uma consequência devastadora em longo prazo
(ARGYRIOU, 2009).
Os neurônios, apesar de não estarem em divisão constante, são suscetíveis ao
paclitaxel e isso provoca sérias complicações para seu uso como agente terapêutico. Como o
paclitaxel não atravessa a barreira hematoencefálica, esse fármaco afeta especificamente o
SNP, e leva a uma neuropatia axonal predominantemente sensorial (PARK et al., 2011). A
incidência e gravidade da neuropatia aumentam com doses únicas e cumulativas mais
elevadas. Embora a incidência relatada varie de acordo com o estudo, até 30% dos pacientes
apresentam sintomas graves em altas doses cumulativas (CARLSON; OCEAN, 2011). Os
sintomas neurológicos podem atingir tal gravidade que necessitam da cessação ou redução do
tratamento. Embora os sintomas geralmente se resolvam após a interrupção do tratamento,
eles podem persistir por meses ou anos, levando a uma menor qualidade de vida (LEE;
SWAIN, 2006).
Quando uma dose neurotóxica é atingida, os sintomas sensoriais começam nas
mãos e nos pés. Isto é reflexo de uma neuropatia, em que os axônios sensoriais distais se
degeneram (ARGYRIOU et al., 2008). Uma biópsia de um nervo periférico revelou uma
patologia de degeneração axonal, desmielinização secundária e perda de fibras nervosas em
casos de neuropatia grave (SAHENK et al., 1994). No entanto, em modelos animais foi
demonstrado que os sintomas da neuropatia podem estar presentes com a degeneração apenas
das fibras intraepidérmicas distais (SIAU et al., 2006). O paclitaxel pode afetar todas as
modalidades sensoriais, sendo as fibras maiores as mais afetadas (QUASTHOFF;
HARTUNG, 2002; DOUGHERTY et al., 2004). Os sintomas primários nos pacientes
incluem dormência, dor e formigamento (LEE; SWAIN, 2006). O sistema motor é
frequentemente menos afetado, com relatos de sintomas suaves com altas doses de paclitaxel,
e os efeitos no sistema autonômico são raros (FREILICH et al., 1996; WINER et al., 2004).
Embora estudos tenham identificado agentes que protegem contra a degeneração axonal
induzida por paclitaxel em cultura de neurônios, e degeneração axonal e sintomas sensoriais
em modelos animais (MELLI et al., 2006; ROVINI et al., 2010; VERSTAPPEN et al., 2004;
APFEL et al., 1991; BOYLE; WHEELER; SHENFIELD, 1999; PISANO et al., 2003;
WANG et al., 2004), os que foram testados em ensaios clínicos até agora produziram
resultados mistos e não existem atualmente intervenções eficazes ou aprovadas.

39
2.3.3 Síndrome da dor aguda associada ao paclitaxel
O paclitaxel está associado a uma síndrome peculiar de dores subagudas, que
inicialmente foi referida como artralgias e mialgias induzidas por paclitaxel (ROWINSKY,
1993; GARRISON et al., 2003). Atualmente essa síndrome está sendo designada como
"Síndrome da Dor Aguda Associada ao Paclitaxel" (Paclitaxel-Associated Acute Pain
Syndrome – P-APS) (REEVES et al., 2012; LOPRINZI et al., 2007; LOPRINZI et al., 2011).
Essa síndrome da dor aguda causa significativa morbidade com uma incidência de 70% dos
indivíduos, iniciando no 1º dia e culminando no 4º dia após o primeiro ciclo do paclitaxel. Os
sintomas desaparecem em grande parte dentro de uma semana (SAIBIL et al., 2010; REEVES
et al., 2012).
A síndrome da dor aguda associada ao paclitaxel pode resultar da sensibilização
do nociceptor, conforme observado em pacientes (LOPRINZI et al., 2007); além disso, em
modelos animais, ratos que receberam infusão de paclitaxel desenvolveram lesão do nervo
isquiático no prazo de 24 horas (JIMENEZ-ANDRADE et al., 2006; PETERS et al., 2007). A
natureza e o perfil temporal da P-APS distingue-se como uma entidade separada da neuropatia
periférica induzida por quimioterapia; no entanto, não se sabe se os pacientes que
desenvolvem P-APS são mais propensos a desenvolver neuropatia periférica (LOPRINZI et
al., 2011). Tal como acontece com P-APS, a neuropatia periférica foi descrito em até 70% dos
pacientes. (ARGYRIOU, 2009).
Um estudo randomizado comparou a eficácia de infusão curta ou prolongada de
paclitaxel no câncer de mama metastático e demonstrou uma incidência de 25% de dor aguda
no grupo de infusão de 3 horas, já no grupo que recebeu infusão de 96 horas a incidência de
foi de 2%. Isto pode sugerir que, os picos de concentração da droga em um tempo mais curto
são responsáveis pela P-APS (MOULDER et al., 2010).
Desde as duas últimas décadas, sabe-se que o tratamento com paclitaxel pode
induzir imediatamente sintoma de dor nos pacientes (ROWINSKY, 1993; GARRISON et al.,
2003). Nos últimos anos essa dor foi definida como P-APS (REEVES et al., 2012; LOPRINZI
et al., 2011). Até o presente, poucos estudos foram realizados a fim de investigar os
mecanismos relacionados com a dor aguda induzida por paclitaxel em modelos animais.
Os mecanismos pelos quais o paclitaxel induz P-APS, ainda hoje não são
conhecidos e nem existe um padrão comprovado de cuidados para a prevenção ou tratamento
da P-APS. Atualmente, o entendimento dos mecanismos subjacentes da dor aguda induzida
por pacliatxel em modelos animais é baseado principalmente em estudos que examinam as
mudanças patológicas nos dias após a interrupção do tratamento repetido com paclitaxel.

40
Pouco se sabe sobre a ação direta e imediata induzida pelo paclitaxel no sistema de
sinalização da dor em modelos in vivo. Uma maior compreensão da natureza e da etiologia da
P-APS pode fornecer esclarecimentos adicionais sobre maneiras de aliviar esta toxicidade
potencialmente incômoda.
Até o momento, poucos mecanismos foram descritos com o intuito de esclarecer o
desenvolvimento de dor aguda induzida por paclitaxel nos pacientes em tratamento com este
fármaco. Recentemente, um estudo demonstrou que o paclitaxel poderia provocar dor aguda
pela ativação do receptor toll like 4 (TLR4) na medula espinal e nos GRDs (YAN et al.,
2015). Outro estudo sugeriu que a ativação dos receptores de bradicinina (B1 e B2) no nervo
isquiático contribuem para o desenvolvimento da dor aguda induzida por paclitaxel
(BRUSCO et al., 2016). A compreensão de tais mecanismos poderia fornecer fundamentos
para o desenvolvimento de tratamentos eficazes para a síndrome dolorosa associada ao
paclitaxel (POLOMANO et al., 2001; RIGO et al., 2013).
2.4 Mastócitos
Os mastócitos são agentes cruciais nas desordens alérgicas, no entanto também
são importantes iniciadores e efetores da imunidade inata, residem em muitos tecidos,
inclusive nos nervos. (GALLI, NAKAE, TSAI, 2005; METCALFE, BARAM, MEKORI,
1997; GALLI et al, 2011). Derivam de células-tronco hematopoéticas e normalmente
circulam em sua forma imatura (CHEN et al., 2005; JAMUR, OLIVER, 2011; GALLI et al,
2011) e maturam em tecidos vascularizados ou cavidades serosas em que finalmente vão
residir sob a influência do fator de células-tronco e várias citocinas (METCALFE, BARAM,
MEKORI, 1997; KAWAKAMI et al., 2002; GALLI et al, 2011). Os mastócitos contêm
grânulos ricos em fatores pró-inflamatórios (ZUO et al., 2003). Além disso, são uma fonte
importante de várias citocinas e quimiocinas (THEOHARIDES, CONTI, 2004; MIDULLA et
al., 2006; THEOHARIDES, KALOGEROMITROS, 2006).
Até 97% dos mastócitos estão no lado abluminal dos vasos sanguíneos. Tal como
os linfoblastos, os mastócitos podem mover-se através do cérebro normalmente na ausência
de inflamação, com entrada rápida e com passagem livre através da barreira hematoencefálica
(LAMBRACHT-HALL, DIMITRIADOU, THEOHARIDES, 1990). Os mastócitos residentes
participam de reações inatas de defesa do hospedeiro e são encontrados em tecidos periféricos
inervados por fibras nervosas sensoriais de pequeno calibre e dentro do compartimento
endoneural, onde orquestram processos inflamatórios. Os mastócitos podem atravessar a
barreira sangue-medula espinal e barreira hematoencefálica quando a barreira está

41
comprometida como resultado de patologias do SNC. A inflamação sistêmica dá origem a
sinais que se comunicam com o cérebro e leva a alterações no metabolismo e comportamento,
incluindo a expressão de um fenótipo iniciado (pró-inflamatório) pela microglia
(SILVERMAN et al., 2000; ENGLER et al., 2011).
Vários tipos de células imunitárias têm sido implicados na patogênese e no
processamento nociceptivo alterado característico de estados dolorosos. Foi demonstrado por
Ribeiro e colaboradores (2000) que mastócitos e macrófagos residentes são cruciais para o
desenvolvimento da dor inflamatória e participam liberando citocinas e quimiocinas quando
ativados. Mastócitos desempenham um papel vital no desenvolvimento de hiperalgesia
térmica e alodinia mecânica em camundongos (ZUO et al., 2003). Os grânulos de mastócitos
contêm mediadores tais como histamina, proteases e citocinas (METCALFE, BARAM,
MEKORI, 1997; GALLI, NAKAE, TSAI, 2005), vários dos quais são capazes de sensibilizar
ou ativar neurônios. Os mastócitos ativados contribuem diretamente para estados de dor,
incluindo dor neuropática, liberando mediadores algogênicos após degranulação (XANTHOS
et al., 2011). Mastócitos residentes de nervos periféricos são as primeiras células ativadas no
local de lesão nervosa e contribuem para o recrutamento de neutrófilos e macrófagos (ZUO et
al., 2003). Os mastócitos também podem contribuir indiretamente, aumentando o
recrutamento de outros tipos de células imunitárias importantes que, por sua vez, liberam
mediadores pró nociceptivos, tais como IL-6 (VALLIE`RES, RIVEST, 1997; LEAL-
BERUMEN, CONLON, MARSHALL, 1994). Em modelos animais, a inibição da
degranulação de mastócitos reduziu a infiltração de neutrófilos e macrófagos e o
desenvolvimento de hiperalgesia após lesão do nervo periférico. Como células imunes
residentes nos nervos periféricos, os mastócitos têm um papel nos estágios iniciais da
inflamação e estabelecimento da hipersensibilidade da dor que acompanha a lesão do nervo
periférico (ZUO et al., 2003).
A estabilização dos mastócitos com cromoglicato de sódio atenua o
desenvolvimento de alodinia, hipersensibilidade mecânica e térmica. Além disso, reduz a
infiltração de neutrófilos e monócitos/macrófagos no local da lesão do nervo após ligadura
parcial do nervo isquiático (ZUO et al., 2003). No entanto, o efeito anti-alodínico com
antagonistas foi menos eficaz do que o tratamento com cromoglicato de sódio, sugerindo que
outros mediadores derivados de mastócitos (neurotrofinas, prostaglandinas, proteases e
citocinas) podem estar envolvidos na geração de dor neuropática (THACKER et al., 2007).
Estes estudos sugerem que os mastócitos ativados contribuem diretamente para a dor
neuropática. Os mastócitos também podem contribuir indiretamente aumentando o

42
recrutamento de outros tipos de células imunes importantes que, por sua vez, liberam
mediadores pró nociceptivos (THACKER et al., 2007).
Outro estudo demonstrou que o tratamento com estabilizador de mastócitos ou a
deleção destes reduziu a dor neuropática crônica em modelos animais (CHATTERJEA,
MARTINOV, 2015). Além disso, foi demosntrado in vitro que a estabilização de mastócitos
em cultura (Rat Basophilic Leukemia, RBL-2H3) com quercetina e posteriormente
estimulados com paclitaxel inibiu a liberação de histamina (GAO et al., 2016). Uma grande
quantidade de evidências suporta a hipótese de que os mastócitos contribuem para a dor
patológica. Estes resultados são uma forte indicação de que os mastócitos podem contribuir
para o desenvolvimento da síndrome da dor aguda induzida por paclitaxel, pois embora se
tenha demonstrado o seu envolvimento na dor neuropática, não há estudos sobre a sua
participação na síndrome dolorosa aguda induzida pelo paclitaxel.
2.5 Células satélites gliais
As células satélites gliais (CSG) são células especializadas que envolvem e se
comunicam com neurônios sensoriais nos gânglios da raiz dorsal. Acredita-se que as CSG são
capazes de modular o microambiente neuronal e a transmissão sensorial dentro dos gânglios
(HANANI, 2005; ZHANG et al., 2007; HANANI, 2010). Estas células são capazes de
expressar diferentes receptores e ainda respondem a substâncias pró-inflamatórias e liberam
transmissores que podem desempenhar um papel na patogênese de vários distúrbios
(HANANI, 2005; CAPUANO et al., 2009; HANANI, 2010; NEEB et al., 2011; HANANI
2012).
As CSG são derivadas a partir de células pluripotentes da crista neural, assim
como as células de Schwann (JESSEN; MIRSKY, 2005). Morfologicamente as CSG se
caracterizam por serem células de forma laminar, irregulares, geralmente mononucleares e
com expansões lamelares e microvilosidades que aumentam sua área de superfície (OHARA
et al., 2009; PANNESE, 2010). As CSG se dispõem em volta dos corpos dos neurônios
sensoriais e da porção proximal do seu axônio, geralmente formam uma bainha de uma única
camada em torno de cada corpo neuronal e se conectam entre si por junções
comunicantes. Cada CSG é separada do corpo do neurônio por um espaço de cerca de 20nm e
a face não neuronal das CSG possuem uma membrana basal de tecido conjuntivo formando
uma unidade estrutural (HANANI, 2005).
Muitas vezes as CSG são consideradas como o equivalente no SNP aos astrócito
do SNC, embora os astrócitos no SNC e as CSG no SNP compartilham funções comuns,

43
como isolamento, regulação da concentração iônica do espaço extracelular e reciclagem de
neurotransmissores, a morfologia dos dois tipos de células é bastante diferente, pois os
astrócitos são comumente multipolares com extensões citoplasmáticas muito atenuadas
(OHARA et al., 2009). CSG e astrócitos compartilham marcadores moleculares, como a
glutamina sintetase e a expressão da proteína ácida fibrilar glial (GFAP). No entanto, ambas,
CSG e astrócitos, parecem diferir quanto à expressão de GFAP que é detectável
constitutivamente em astrócitos, mas não em CSG. Porém, após uma lesão do nervo as CSG
passam a expressar GFAP, semelhante aos astrócitos, que aumentam a expressão de GFAP
após lesão do nervo (OHARA et al., 2009; HANANI, 2005). Além disso, CSG expressam
canais de cálcio e de potássio dependentes de voltagem e Kir 4.1 e componentes de
reciclagem de glutamato, tais como o transportador de glutamato-aspartato (GLAST)
(CHERKAS et al., 2004; ZHANG et al., 2009).
As células de Schwann que também são derivadas da crista neural mantém uma
estreita relação com as CSG (LE DOUARIN et al., 1991). Estudos tem mostrado que, em
vários tipos diferentes de cultura de gânglio, as CSG podem aumentar a expressão de
marcadores de células de Schwann, tais como a proteína básica de mielina de células de
Schwann e do fator de transcrição Krox-20 (MURPHY et al., 1996) e regulam negativamente
a expressão de marcadores específicos tais como Erm (HAGEDORN et al., 2000). Além
disso, as CSG expressam também inúmeros receptores de moléculas bioativas potencialmente
intervenientes em interações com outras células sendo que muitos deles foram, implicados na
gênese e manutenção da dor crônica, como os receptores purinérgicos P2Y e P2X7 (CERUTI
et al., 2008; VILLA et al., 2010; ZHANG et al., 2007), o receptor do peptídeo relacionado ao
gene de calcitonina (CGRP) (LI; VAUSE; DURHAM, 2008), da substância P (TAKEDA;
TAKAHASHI; MATSUMOTO, 2009), de citocinas e quimiocinas, como exemplo TNF-α e
IL-1β (DUBOVY et al., 2006; POMONIS et al., 2001) e do receptor N-metil-D-aspartato
(NMDAr) (CASTILLO et al., 2013).
Há fortes evidências de que a ativação de células gliais (astrócitos e microglia)
dentro do corno dorsal da medula espinal acarreta na subsequente produção de citocinas, tais
como IL-1β e TNF-α, sugerindo um papel fundamental dessas células na gênese e
manutenção da dor patológica (WATKINS; MILLIGAN; MAIER, 2001; WATKINS;
MAIER, 2002). Estudos tem demonstrado a importância das células gliais na modulação da
transmissão sensorial, incluindo a nocicepção (CHERKAS et al., 2004, HANANI, 2005;
TAKEDA et al., 2009). Na verdade, após a lesão local do nervo periférico, as células gliais
começam a apresentar aumento na expressão de GFAP (WOODHAM et al., 1989, LI e

44
ZHOU, 2001) e aumento da produção de citocinas (TAKEDA et al., 2007). Tem sido
sugerido ainda que não só as células gliais centrais estão envolvidas na geração e manutenção
da dor, mas também as CSG nos gânglios sensoriais desempenham esse papel (HANANI et
al., 2002; HUANG; BELZER; HANANI, 2010). Esta ideia tem sido apoiada firmemente por
estudos que demonstraram que as CSG possuem mecanismos para liberação de citocinas
(TAKEDA, TAKAHASHI, MATSUMOTO, 2009; ZHANG et al., 2007), ATP
(SUADICANI et al., 2010) e possivelmente outros mediadores químicos. As CSG sofrem
alterações importantes como resultado de lesão de nervos periféricos, e parecem contribuir
para a dor crônica como demonstrado em modelos animais (DUBLIN; HANANI, 2007;
HUANG; BELZER; HANANI, 2010; OHARA et al., 2009).
Evidências têm sugerido que as CSG possuem um papel crítico na dor induzida
pela quimioterapia e outras condições crônicas de dor (CAPUANO et al., 2009; NEEB et al.,
2011; LAURSEN et al., 2014; TAKEDA et al., 2011; WARWICK; HANANI, 2013). A
neuropatia periférica é um efeito adverso comum causado por diversos quimioterápicos,
incluindo os derivados de platina (cisplatina, oxaliplatina, carboplatina), alcaloides da vinca
(vincristina) e taxanos (paclitaxel, docetaxel). O paclitaxel é um agente quimioterápico que
tem sido conhecido por induzir distúrbios sensoriais periféricos incluindo dor neuropática que
é uma complicação comum do uso deste fármaco entre outros efeitos colaterais
(DOUGHERTY et al., 2004). Acredita-se que o paclitaxel e outros quimioterápicos se
acumulem nos GRD quando administrados para o tratamento de diferentes tipos de cânceres.
A dor induzida por quimioterápicos é acompanhada de distúrbios sensoriais dependentes da
dose cumulativa do agente utilizado no tratamento (WINDEBANK, GRISOLD, 2008;
CAVALETTI; MARMIROLI, 2010). Paclitaxel induz dor caracterizada por
hipersensibilidade a estímulos mecânicos e térmicos (especialmente a estímulos frios) que
começam logo após o início do tratamento e pode persistir por semanas ou anos após o
término da terapia (DOUGHERTY et al., 2004). Alguns mecanismos patológicos estão
associados à patogênese da neuropatia periférica induzida pela quimioterapia (CIPN),
incluindo neurotoxicidade, disfunção mitocondrial e desprendimento do transporte axonal
(FLATTERS; BENNETT, 2006; FIDANBOYLY; GRIFFITHS; FLATTERS, 2011; JAGGI;
SINGH, 2012). Estudos também sugerem o envolvimento de mediadores pró-inflamatórios
produzidos pela microglia na medula espinal no modelo da neuropatia induzida pelo
paclitaxel ou vincristina (JAGGI; SINGH, 2012). No entanto, novas evidências sugerem um
potencial papel das células gliais em associação com a CIPN (WARWICK; HANANI, 2013).

45
No entanto, até o momento, poucos estudos tem evidenciado a participação das
CSG na dor induzida por paclitaxel, especialmente na dor aguda. Estas podem fornecer
evidencias para uma melhor compreensão dos mecanismos pelos quais o paclitaxel causa dor,
principalmente dor aguda. Dessa forma, é interessante o estudo do papel das CSG na dor
aguda induzida por paclitaxel, tendo em vista que pouco se sabe sobre os mecanismos
envolvidos nessa síndrome dolorosa.

46
3. JUSTIFICATIVA

47
O paclitaxel é um antineoplásico derivado do Taxus brevifolia, possuindo potente
atividade citotóxica e utilizado como primeira linha no tratamento de diversos tumores
sólidos, particularmente em carcinomas de mama, ovário, pulmão, cabeça e pescoço. No
entanto, os pacientes que recebem tratamento com paclitaxel frequentemente desenvolvem
dor patológica, o que reduz significativamente a sua qualidade de vida e dificulta a utilização
deste quimioterápico. A dor patológica induzida pelo paclitaxel inclui a dor que ocorre
imediatamente após o primeiro ciclo do tratamento com este quimioterápico (REEVES et al.,
2012; LONPRINZI et al., 2011), e a dor que persiste durante semanas a anos após o fim do
tratamento (CATA et al., 2006; DOUGHERTY et al., 2004).
A dor aguda induzida pelo paclitaxel é uma morbidade significativa em pacientes
e atualmente, não existe um padrão comprovado de cuidados para a sua prevenção ou
tratamento (REEVES et al., 2012; LONPRINZI et al., 2011), além disso, os mecanismos
pelos quais o paclitaxel induz dor aguda não são totalmente conhecidos. Atualmente o
entendimento dos mecanismos subjacentes da dor patológica induzida por paclitaxel em
modelos animais é baseado principalmente em estudos que examinam as mudanças
patológicas nos dias após o final do tratamento com essa droga. Até o momento, pouco se
sabe sobre a ação direta e imediata do paclitaxel no sistema de sinalização da dor aguda em
modelos animais, e poucos são os estudos in vivo que têm investigado os mecanismos pelos
quais o paclitaxel induz dor aguda (YAN et al., 2015, JATOI et al., 2016, BRUSCO et al.,
2016). Assim, a compreensão de tais mecanismos poderia fornecer fundamentos para o
desenvolvimento de tratamentos eficazes para essa síndrome dolorosa (POLOMANO et al.,
2001; RIGO et al., 2013). Além disso, a investigação dos mecanismos envolvidos poderia
auxiliar na redução da intensidade, incidência ou ainda na prevenção de dor crônica.
Evidências clínicas de que a dor aguda pode está relacionada com o aparecimento e
severidade da dor neuropática foi observada após o tratamento com o paclitaxel em pacientes
(REEVES et al., 2012; LOPRINZI et al., 2011).
Diante disso, vê-se a importância de estudar os possíveis mecanismos envolvidos
na dor aguda induzida pelo paclitaxel, assim como uma maior compreensão da natureza e da
etiologia da dor aguda induzida por este quimioterápico pode fornecer esclarecimentos
adicionais sobre maneiras de aliviar esta toxicidade potencialmente incômoda.

48
4. OBJETIVOS

49
4.1 Geral
Investigar a possível participação de mastócitos e de células satélites gliais na
síndrome da dor aguda induzida pelo quimioterápico paclitaxel em camundongos.
4.2 Específicos
• Avaliar o decurso temporal da hiperalgesia mecânica e térmica no modelo de síndrome da
dor aguda induzida pelo paclitaxel em camundongos;
• Investigar o envolvimento dos mastócitos na hiperalgesia mecânica e térmica no modelo de
síndrome da dor aguda induzida pelo paclitaxel em camundongos;
• Investigar a participação da citocina pró-inflamatória IL-6 na hiperalgesia mecânica e
térmica no modelo de síndrome da dor aguda induzida pelo paclitaxel em camundongos, bem
como a participação dos receptores IL-1R, TNFR1/R2, CCR2 e do receptor TLR4;
• Avaliar a expressão gênica das citocinas pró-inflamatórias IL-6, IL-1β e TNF-α e da
quimiocina MCP-1 (CCL2) no nervo isquiático, gânglios da raiz dorsal (GRD) e medula
espinal de camundongos injetados com paclitaxel, assim como a participação do receptor
CCR2 nos GRD e medula espinal;
• Avaliar a concentração das citocinas pró-inflamatórias IL-6, IL-1β e TNF-α e quimiocinas
KC/CXCL1 e MCP-1 (CCL2) no plasma, nervo isquiático, gânglios da raiz dorsal (GRD) e
medula espinal de camundongos no modelo de síndrome da dor aguda induzida pelo
paclitaxel;
• Quantificar as citocinas pró-inflamatórias IL-6, IL-1β e TNF-α e quimiocinas KC/CXCL1 e
MCP-1 (CCL2) no plasma, nervo isquiático, gânglios da raiz dorsal (GRD) e medula espinal
de camundongos TLR4-/- no modelo de síndrome da dor aguda induzida pelo Paclitaxel;
• Quantificar as citocinas pró-inflamatórias IL-6, IL-1β e TNF-α e quimiocinas KC/CXCL1 e
MCP1 (CCL2) no nervo isquiático, gânglios da raiz dorsal (GRD) e medula espinal de
camundongos que não diferenciam mastócitos e em camundongos selvagens pré-tratados com
cromoglicato de sódio;

50
• Quantificar as citocinas pró-inflamatórias IL-6, IL-1β e TNF-α e das quimiocinas
KC/CXCL1 e MCP1 (CCL2) em cultura de linhagem de mastócitos (RBL-2H3) e em cultura
primária de células satélites gliais, ambas estimuladas com paclitaxel;
• Avaliar a expressão de c-Fos, IL-6 e triptase (marcador de mastócitos) e a localização por
imunofuorescência nos gânglios da raiz dorsal (GRD) e na medula espinal de camundongos
injetados com paclitaxel.

51
5. MATERIAIS E MÉTODOS

52
5.1 Animais
Foram utilizados camundongos C57BL/6 (20-25g), machos, provenientes do
biotério setorial do Departamento de Fisiologia e Farmacologia da Faculdade de Medicina da
Universidade Federal do Ceará – FAMED/UFC e do biotério central da Universidade de São
Paulo – USP, Campus de Ribeirão Preto.
Utilizou-se também camundongos knockouts IL-6, TNFR1/R2, CCR2, IL-1R,
TLR2 e TLR4. Além de camundongos que não diferenciam mastócitos, Kitw-sh
(SH),
provenientes do Centro de Criação de Camundongos Especiais (CCCE) da Faculdade de
Medicina de Ribeirão Preto da Universidade de São Paulo – FMRP/USP. Todos os
experimentos com animais knockouts foram realizados no Laboratório de Inflamação e Dor
(LID) do Departamento de Farmacologia da FMRP/USP, sob a orientação do Prof. Dr. Thiago
Mattar Cunha.
Os camundongos SH (ou “sash”) apresentam uma mutação que apareceu
espontaneamente em uma ninhada produzida de um cruzamento C3H/HEH x 101/H no
Radiobiology Unit at Harwell no Reino Unido (LYON; GLENISTER, 1982). A mutação
Kitw-sh
é uma inversão abrangendo um segmento próximo ao lócus c-kit causando a ruptura
das sequências reguladoras 5’, por conseguinte, a sinalização é reduzida através da tirosina
quinase c-kit (DUTTLINGER et al., 1993). Esta expressão reduzida de c-kit na medula óssea
provoca a incapacidade das células-tronco hematopoiéticas de se diferenciarem em
precursores de mastócitos, resultando numa abolição de mastócitos (GRIMBALDESTON et
al., 2005).
Os animais experimentais foram mantidos em microisoladores ventilados
apropriados a uma temperatura controlada (22 ± 2 ºC) no biotério do Departamento de
Farmacologia da FMRP/USP, num ciclo claro-escuro de 12h com ração padrão e água potável
ad libitum. Os animais foram aclimatizados no laboratório durante pelo menos 1h antes da
realização dos experimentos comportamentais. Todos os experimentos foram realizados entre
08:00 e 17:00h. Os animais geneticamente deficientes e seus respectivos controles selvagens
foram utilizados com idade entre 6-8 semanas, pesando entre 20-25g
5.2 Aspectos éticos
Todos os procedimentos experimentais foram conduzidos de acordo com as
diretrizes do Conselho Nacional de Controle de Experimentação Animal (CONCEA) e as
diretrizes éticas estabelecidas pela IASP sobre o uso de animais de laboratório em pesquisas
experimentais (ZIMMERMANN, 1983). Os protocolos experimentais foram aprovados pela

53
Comissão de Ética em Pesquisa Animal da UFC (CEPA, processo número 17/2014) e pela
Comissão de Ética para o Uso de Animais da FMRP/USP (CEUA, processo número
42/2015). Os estudos comportamentais seguiram as diretrizes segundo os Relatórios de
Experiências in vivo na Pesquisa Animal (ARRIVE) (MCGRATH et al., 2010). Além disso, o
número de animais e a intensidade dos estímulos usados foram os mínimos necessários para
demonstrar efeitos consistentes. Os animais foram distribuídos randomicamente entre os
grupos experimentais (número de 6-8 animais por grupo) e todos os experimentos
comportamentais foram conduzidos de maneira cega em relação à administração de drogas e
/ou linhagem dos animais a fim de reduzir viés experimental.
5.3 Indução da síndrome da dor aguda associada ao paclitaxel em camundongos
O modelo de síndrome de dor aguda associada ao paclitaxel foi baseado no
protocolo proposto por Polomano et al., (2001), adaptada para camundongos (SMITH et al.,
2004) e descrito por Yan et al., (2015) com algumas modificações. Resumidamente, a dor
aguda associada ao paclitaxel foi induzida pela administração do quimioterápico em uma
única injeção. O paclitaxel (Evotaxel®, Evolabis, 6mg/mL em Cremophor EL e etanol
desidratado) foi diluído em soro fisiológico (NaCl 0,9%) e administrado em camundongos por
via intravenosa (i.v.) na dose de 4mg/kg, através da veia caudal. A via intravenosa foi
escolhida para mimetizar a administração do paclitaxel utilizada na clínica. O veículo foi
composto de Cremophor EL e de etanol desidratado (Sigma Chemicals, St. Louis, MO, EUA)
diluído em solução de NaCl 0,9%. O volume de administração das soluções foi de 150µL.
Testes nociceptivos ou outros experimentos foram realizados antes da administração da droga
ou veículo (basal ou tempo zero) e 1, 2, 4, 6 e 24h após o tratamento com paclitaxel (4 mg/kg,
i.v.) ou veículo.

54
5.4 Experimentos comportamentais
5.4.1 Avaliação da hiperalgesia mecânica
O limiar de retirada da pata foi avaliado utilizando o método “up-and-down”, tal
como descrito por Chaplan e colaboradores (1994). Em uma sala silenciosa, os camundongos
foram aclimatizados (1 hora) em compartimentos de acrílico transparente individuais (9 x 7 x
11 cm) sobre uma plataforma elevada de malha de arame para permitir o acesso à superfície
plantar das patas traseiras. As medidas foram realizadas somente quando os animais estavam
quietos, não exibindo movimentos exploratórios ou defecação, e sem estarem descansando em
suas patas traseiras. Depois de aclimatizados, uma serie de filamentos de von Frey (North
Coast Medical) de intensidade crescente (0,008g – 1,0g) foram aplicados no centro da pata
posterior direita dos animais com uma pressão suficiente para dobrar o filamento. O estímulo
foi interrompido após a observação da resposta nociceptiva, caracterizada por flexão seguida
de retirada da pata, “flinch”. A ausência da retirada da pata depois de 3 segundos levou ao uso
do próximo filamento com o aumento de peso, ao passo que, quando houve resposta positiva
de retirada da pata, levou-se à utilização do próximo filamento mais fraco. Este método foi
continuado até um total de três medições. O filamento de menor força capaz de induzir
resposta foi determinado como sendo o limiar mecânico do animal. O valor do limiar de
resposta então foi anotado e a hiperalgesia mecânica foi considerada como uma diminuição do
limiar mecânico, quando comparado com valores basais (antes da administração das drogas).
O limiar mecânico foi avaliado antes da administração do paclitaxel (valores basais), e após
diferentes tempos da administração da droga (1, 2, 4, 6, e 24h) e os valores obtidos foram
expressos em miligramas (mg).
5.4.2 Avaliação da hiperalgesia térmica ao frio
A hiperalgesia ao frio foi avaliada conforme o método descrito por Flatters e
Bennett, (2004), Gauchan, et al., (2009) e Smith; Crager; Mogil (2004), com modificações.
Brevemente, os animais foram aclimatados em compartimentos de acrílico individuais (9 x 7
x 11 cm) em uma plataforma elevada de assoalho aramado, permitindo acesso à superfície
plantar das patas, e com auxilio de uma seringa de 1mL, 50μL de acetona foi aplicado de
modo que abrangesse tanto a superfície plantar, como o dorso da pata posterior direita do
animal. O estímulo frio causado pela acetona provoca comportamentos de agitação e elevação
da pata, além de lambidas (CASPANI et al., 2009). Esse tipo de resposta é caracterizada
como comportamento nocifensivo e o tempo gasto pelo animal com esse comportamento foi
mensurado em segundos, com o auxílio de um cronômetro – resposta ao frio. As respostas

55
podem ser intermitentes, por isso, o cronômetro foi parado e ativado conforme o
comportamento do animal. Cada animal foi observado por um total de 60 segundos. Um
espelho foi colocado em inclinação com a superfície, abaixo do compartimento de acrílico
para permitir a observação clara das patas. A hiperalgesia ao frio foi avaliada antes da
administração do paclitaxel (valores basais), e após diferentes tempos da administração da
droga (2, 4, 6, 24, 48 e 72 horas) e os valores obtidos foram expressos em segundos (s).
5.4.3 Avaliação da hiperalgesia térmica ao calor
Com o objetivo de avaliar o limiar de resposta ao estimulo térmico (calor) foi
realizado o teste plantar utilizando-se o aparelho Plantar Test (Ugo Basile, Varese, Itália),
como descrito previamente por Hargreaves et al., (1988). Para isso, os animais foram
alocados cuidadosamente em compartimentos individuais de acrílico e posicionados sobre
uma superfície de vidro especial, que permite a passagem homogênea da luz e do calor, os
animais foram deixados sobre o aparato durante 1h para adaptação ao ambiente. Após esse
período, uma fonte de luz infravermelha com potência previamente estabelecida de 50 watts
foi posicionada sob a pata traseira direita do animal. Já com o animal posicionado, a luz
infravermelha foi acionada simultaneamente com um cronômetro eletrônico, possibilitando a
contagem do tempo e do estímulo ao mesmo tempo. Quando o animal exibiu uma resposta
positiva (“flinch” ou retirada da pata) a fonte de luz e o relógio pararam automaticamente.
Um tempo máximo de 20 segundos de exposição da pata ao raio infravermelho foi
utilizado para evitar danos teciduais. Em cada animal foram realizadas duas medidas, sendo
estas obtidas no intervalo de 5 a 10 minutos e a média destas medidas representaram a
latência de retirada da pata. A hiperalgesia ao calor foi avaliada antes da administração do
paclitaxel (valores basais), e após diferentes tempos da administração da droga (2, 4, 6, 24, 48
e 72 horas) e os valores obtidos foram expressos em segundos (s).
5.5 Delineamento experimental
5.5.1 Avaliação da hiperalgesia mecânica e térmica no modelo de síndrome da dor aguda
induzida pelo paclitaxel
Para verificar se o paclitaxel era capaz de induzir hiperalgesia mecânica e térmica,
foram utilizados camundongos C57BL/6 selvagens. Os animais foram divididos em dois
grupos, cada grupo contendo 6 animais. O primeiro recebeu uma dose única de paclitaxel
(4mg/kg) por via intravenosa (i.v.) e o segundo grupo recebeu veículo (cremophor EL +
álcool absoluto). Em seguida os camundongos foram avaliados através da utilização dos testes

56
comportamentais nos diferentes tempos (1, 2, 4, 6, 24, 48 e 72 horas) após os tratamentos
(Figura 3).
Figura 3 – Delineamento esquemático do protocolo experimental 1
Os camundongos foram divididos em dois grupos, cada grupo contendo 6 animais. O primeiro recebeu
uma dose única de paclitaxel (4mg/kg) por via intravenosa (i.v.) e o segundo grupo recebeu veículo.
Em seguida os camundongos foram avaliados através da utilização dos testes comportamentais nos
diferentes tempos (1, 2, 4, 6, 24, 48 e 72 horas) após os tratamentos.
Fonte: Elaborado pelo próprio autor.
5.5.2 Investigação da participação de mastócitos na síndrome dolorosa aguda induzida por
paclitaxel
Para averiguar o possível envolvimento dos mastócitos na síndrome de dor aguda
induzida por paclitaxel, foram utilizados camundongos C57BL/6 selvagens divididos
aleatoriamente em 2 grupos: paclitaxel e veículo, cada grupo contendo 6 animais. O mesmo
ocorreu para camundongos SH (mutantes com deficiência em mastócitos). Após o tratamento
dos animais com paclitaxel ou veículo, foram avaliadas a hiperalgesia mecânica e térmica nos
tempo de 2, 4, 6 e 24h (Figura 4).
GRUPOS
Camundongos selvagens
C57BL/6
(n = 6)
Paclitaxel (4mg/kg, i.v.)
ou
Veículo (i.v.)
Von Frey
Teste da acetona
Hargreaves

57
Figura 4 – Delineamento esquemático do protocolo experimental 2
Camundongos selvagens (C57BL/6) foram divididos aleatoriamente em 2 grupos, cada grupo
contendo 6 animais. O primeiro foi tartado com paclitaxel e o segundo grupo foi tratado com o
veículo. O mesmo ocorreu para camundongos SH (mutantes, que sãodeficientes para mastócitos).
Após o tratamento, os animais de ambos os grupos foram submetidos a avaliação da hiperalgesia
mecânica e térmica nos tempo de 2, 4, 6 e 24h.
Fonte: Elaborado pelo próprio autor.
5.5.3 Avaliação do efeito do cromoglicato de sódio na síndrome dolorosa aguda induzida
pelo paclitaxel
Foram utilizados camundongos C57BL/6 selvagens divididos aleatoriamente em 2
grupos, cada grupo com 6 animais. Um grupo foi pré-tratado com cromoglicato de sódio
(30mg/kg) diluído em soro fisiológico e administrado via intra-peritonial (i.p.) diariamente,
durante 3 dias consecutivos (KIMURA et al., 2015). Outro grupo, denominado grupo controle
recebeu soro fisiológico ao invés da droga pela mesma via e período. Após o término do
tratamento, os animais de ambos os grupos foram injetados com paclitaxel ou veículo e a
hiperalgesia mecânica e térmica foram avaliadas nos tempo de 2, 4, 6 e 24h após a
administração dos estímulos (Figura 5).
GRUPOS
Camundongos Selvagens
C57BL/6
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona
Camundongos
SH
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona

58
Figura 5 – Delineamento esquemático do protocolo experimental 3
Camundongos selvagens (C57BL/6) foram divididos aleatoriamente em 2 grupos, cada grupo com 6
animais. Um grupo foi pré-tratado com cromoglicato de sódio (30mg/kg) administrado via i.p.
diariamente, durante 3 dias consecutivos. Outro grupo, denominado grupo controle recebeu soro
fisiológico nas mesmas condições que o grupo tratado com a droga. Após o término do tratamento, os
animais de ambos os grupos foram injetados com paclitaxel ou veículo e a hiperalgesia mecânica e
térmica (frio) foram avaliadas nos tempo de 2, 4, 6 e 24h após a administração dos estímulos. Fonte:
Elaborado pelo próprio autor.
5.5.4 Investigação da participação de citocinas na síndrome da dor aguda induzida pelo
paclitaxel
Camundongos C57BL/6 selvagens foram aleatoriamente divididos em 2 grupos,
cada grupo contendo 6 animais. Os grupos foram tratdos com paclitaxel ou veículo. O mesmo
ocorreu para camundongos deficientes (knockouts) para IL-6 (IL-6-/-) e para os camundongos
deficientes para os receptores IL-1β (IL-1R-/-), TNF-α (TNFR1/R2-/-) e MCP-1 (CCR2-/-).
Após o tratamento com paclitaxel ou veículo, foram avaliadas a hiperalgesia mecânica e
térmica (frio) nos tempo de 2, 4, 6 e 24h após a administração dos estímulos (Figura 6).
Camundongos Selvagens
C57BL/6
Cromoglicato de sódio
(10mg/kg, i.p.)
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona
Veículo (i.p.)
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona

59
Figura 6 – Delineamento esquemático do protocolo experimental 4
Camundongos selvagens (C57BL/6 ) foram aleatoriamente divididos em 2 grupos, cada grupo
contendo 6 animais. Os grupos foram tratados com paclitaxel ou veículo. O mesmo ocorreu para
camundongos deficientes para IL-6 (IL-6-/-) e para os camundongos deficientes para os receptores IL-
1β (IL-1R-/-), TNF-α (TNFR1/R2-/-) e MCP-1 (CCR2-/-). Após o tratamento com paclitaxel ou
veículo, foram avaliadas a hiperalgesia mecânica e térmica (frio) nos tempo de 2, 4, 6 e 24h após a
administração dos estímulos. Fonte: Elaborado pelo próprio autor.
Em outro delineamento experimental, camundongos C57BL/6 selvagens;
camundongos C57BL/6 selvagens foram pré-tratados com Tocilizumabe (8mg/kg) ou com o
seu veículo 30 minutos antes da administração de paclitaxel e em seguida foram
aleatoriamente divididos em grupos, cada grupo contendo 6 animais. O tocilizumabe (TCZ) é
um anticorpo monoclonal humanizado que se liga ao receptor de IL-6, inibindo o efeito
biológico da IL-6 (SINGH; BEG; LOPEZ-OLIVO, 2011). A dose recomendada de utilização
é de 8mg/kg de peso por dose (MOTA et al., 2012 ).
Após o tratamento, os grupos foram injetados com paclitaxel ou veículo e a
hiperalgesia mecânica e térmica (frio) foram avaliadas nos tempo de 1, 2, 4, 6 e 24h após a
administração dos estímulos (Figura 7).
GRUPOS
Camundongos
Selvagens
C57BL/6
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona
Camundongos
(IL-6-/-; IL-1R-/-; TNFR1/R2 -/- e CCR2-/-)
Paclitaxel
(4mg/kg, i.v.)
Von Frey
e
Teste da acetona
Veículo (i.v.)
Von Frey
e
Teste da acetona

60
Figura 7 – Delineamento esquemático do protocolo experimental 5
Camundongos selvagens (C57BL/6); camundongos selvagens (C57BL/6) foram pré-tratados com
Tocilizumabe (8mg/kg) ou com o seu veículo e em seguida foram aleatoriamente divididos em grupos,
cada grupo contendo 6 animais. Após o tratamento, os grupos foram injetados com paclitaxel ou
veículo e a hiperalgesia mecânica e térmica (frio) foram avaliadas nos tempo de 2, 4, 6 e 24h após a
administração dos estímulos. Fonte: Elaborado pelo próprio autor.
5.5.5 Coleta de sangue venoso periférico, nervo isquiático, gânglios da raiz dorsal e medula
espinal
Os animais foram anestesiados, via intraperitoneal (i.p), com uma mistura de
Ketamina (100 mg/kg) e xilazina (10 mg/kg); em seguida, foi realizada a coleta de sangue
através do plexo retro-orbital. O sangue então foi armazenado em tubos e acondicionado no
gelo. Após a coleta de sangue, procedeu-se a perfusão intracardíaca com solução de PBS. A
medula espinal lombar (L3 – L6), os gânglios da raiz dorsal (GRD; segmentos entre L3 – L5)
e o nervo isquiático foram removidos 2, 4, 6 e 24 horas após a administração de paclitaxel.
Em seguida, os tecidos foram acondicionados em gelo seco e posteriormente, todas as
GRUPOS
Camundongos
Selvagens
C57BL/6
_____
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Von Frey
e
Teste da acetona
Camundongos
Selvagens
C57BL/6
Tocilizumabe
(8mg/kg, i.p.)
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Von Frey
e
Teste da acetona
Veiculo (i.p.)
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Von Frey
e
Teste da acetona

61
amostras coletadas foram acondicionadas em freezer, à temperatura de -80°C para realização
dos ensaios de RT-PCR e/ou ELISA. Os mesmo experimentos realizados foram utilizados
camundongos knockout para o receptor TLR4, divididos em dois grupos, o primeiro grupo
recebeu PCX e segundo grupo recebeu veículo, após o tratamento seguiram-se os mesmo
procedimentos descritos anteriormente (Figura 8).
Figura 8 – Delineamento esquemático do protocolo experimental 6
Os animais foram tratados com as respectivas drogas, em seguida anestesiados para a coleta de
sangue. Após a coleta de sangue, procedeu-se a perfusão intracardíaca para remoção da medula espinal
lombar (L3 – L6), gânglios da raiz dorsal (GRD; segmentos entre L3 – L5) e nervo isquiático e
posteriormente, todas as amostras coletadas foram acondicionadas em freezer, à temperatura de -80°C
para realização dos ensaios de RT-PCR e/ou ELISA. Num outro momento, foram utilizados
camundongos knockout para o receptor TLR4, os animais foram divididos em 2 grupos, o primeiro
grupo recebeu PCX e segundo grupo recebeu veículo, após o tratamento seguiram-se os mesmo
procedimentos descritos anteriormente. Fonte: Elaborado pelo próprio autor.
GRUPOS
Camundongos
Selvagens
C57BL/6
____
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Coleta das amostras
(2, 4, 6 e 24h)
PCR
ou
ELISA
Camundongos
Selvagens
C57BL/6
Cromoglicato de sódio
(30mg/kg, i.p.)
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Coleta das amostras
(2, 4, 6 e 24h)
ELISA
Veículo( i.p.)
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Coleta das amostras
(2, 4, 6 e 24h)
ELISA
Camundongos
SH
____
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Coleta das amostras
(2, 4, 6 e 24h)
ELISA
Camundongos
TLR4-/-
____
Paclitaxel
(4mg/kg, i.v.) ou
Veículo
Coleta das amostras
(2, 4, 6 e 24h)
ELISA

62
5.6 Cultura de mastócitos
Foram utilizadas células da linhagem de mastócitos de rato RBL-2H3 (MORITA;
SIRAGANIAN, 1981). A linhagem celular foi cultivada em meio essencial de Eagle
modificado por Dulbecco – DMEM (Invitrogen, Gibco, Carlsbad, CA, EUA), suplementado
com 10% de soro fetal bovino (Sigma-Aldrich, St. Louis, Missouri, EUA), 0,434mg/mL de
glutamina, 100U/mL de penicilina e 100μg/mL de estreptomicina, como previamente descrito
(BASCIANO et al, 1986; PIERINI et al, 1996) e mantidas em atmosfera úmida contendo 5%
de CO2 a 37 ºC.
As células foram monitoradas com o auxílio de microscópio invertido DIAVERT -
Leitz (Leica Microsystemns, Wetzlar, Alemanha). Quando atingiram uma confluência de
90%, as células foram removidas dos frascos de cultivo utilizando-se Tripsina-EDTA
(Invitrogen) por 15 minutos a 37 ºC, em atmosfera úmida contendo 5% de CO2. Em seguida,
as células foram centrifugadas a 200 g por cinco minutos, o sobrenadante foi descartado e o
sedimento ressuspendido em meio de cultura para posterior quantificação e padronização do
número de células. As células foram contadas e 1x105
células foram semeadas em placa de 96
poços, após 24 horas do plaqueamento, as células foram tratadas com paclitaxel (10, 30 ou 90
μM), veículo, PMA (Phorbol 12-Myristate 13-Acetate) ou ionomicina (Ionóforo divalente de
cálcio que aumenta a permeabilidade das membranas celulares a íons cálcio). As culturas
foram mantidas a 37 ºC numa atmosfera saturada de água com 5% de CO2 e foram utilizadas
para o ensaio de Enzyme-Linked Immuno Assay (ELISA) para TNF-α, IL-6 e MCP-1, após 4
horas da incubação com paclitaxel, veículo, PMA ou ionomicina.
5.7 Cultura primária de células satélites gliais de gânglios da raiz dorsal
A cultura de células satélites gliais de GRD foi realizada conforme descrito por
Belzer, Shraer e Hanani (2010), com modificações. Para isso, foram utilizados camundongos
C57BL/6, os animais foram anestesiados e em seguida procedeu-se a eutanásia por
decaptação para remoção do sangue circulante. Foi então realizado a remoção dos músculos e
a parte óssea da coluna vertebral para expor a medula espinal e os GRD. Todos os gânglios
lombares e torácicos foram removidos e logo transferidos para um tubo de 2mL contendo
solução DMEM incompleto, pH 7,2, 2 °C. Após a coleta o tubo contendo os DRG foi
centrifugado a 400g por 10 min, 4° C, para realização da lavagem, a fim de remover possíveis
vestígios de sangue ou de outros tecidos. Em seguida o sobrenadante foi descartado e o pellet
de GRD foi incubado com colagenase tipo 1A (Sigma-Aldrich Corp, St. Louis, MO, USA) em
solução de DMEM incompleto, pH 7,2, por 1h, a 37 ºC, e em seguida essa solução foi

63
centrifugada a 400g por 10 min, 4 °C. A solução de colagenase foi substituída por DMEM
completo (10% de soro fetal bovino) e os gânglios foram então triturados por pipetagens
repetidas até a completa homogeneização do tecido, e em seguida esta solução foi filtrada em
peneira de 100 µm. O filtrado foi centrigudado a 400g por 10 min, 4° C, e o pellet foi
ressupendido em DMEM completo, adicionando 1 mL de meio para cada camundongo que
foi utilizado. Os GRD foram alocados em placas de 48 poços num volume de 250 uL por
poço. A cultura foi mantida em atmosfera úmida contendo 5% de CO2 a 37 ºC durante 7 dias
e o meio foi mudado pelo menos duas vezes por semana.
As culturas foram monitoradas utilizando um microscópio invertido DIAVERT -
Leitz (Leica Microsystemns, Wetzlar, Alemanha). Após o sétimo dia de cultivo as células
foram tratadas com paclitaxel (10, 30 ou 90 μM), veículo, PAm3cys, peptidioglicano ou LPS.
Em seguida, as culturas então foram mantidas a 37°C numa atmosfera saturada de água com
5% de CO2 e foram utilizadas para o ensaio de ELISA após 48 horas da incubação com
paclitaxel, veículo, PAm3cys, peptidioglicano ou LPS.
5.8 Expressão gênica por reação em cadeia da polimerase em tempo real (RT-PCR)
5.8.1 Coleta de tecidos
Camundongos tratados com o PCX ou veículo foram sacrificados (0, 2, 4, 6 e 24
horas) após a injeção de PCX. Os animais foram anestesiados, com uma mistura de Ketamina
(100 mg/kg) e xilazina (10 mg/kg); e, depois, foi realizada a perfusão intracardíaca com
solução de PBS para a coleta das seguintes amostras teciduais: medula espinal lombar,
gânglios da raiz dorsal (GRD; segmentos entre L3 – L5) e nervo isquiático. Após a coleta, as
amostras foram colocadas em gelo seco e em seguida armazenadas no freezer (-80 ºC) até a
extração do RNA total.
5.8.2 Extração do RNA total
A extração do RNA total foi realizada através da homogeneização dos tecidos em
1 mL de reagente de Trizol® (Invitrogen, SP, Brazil), conforme recomendações do fabricante.
Em seguida foram adicionados 200 μL de clorofórmio ao homogenato, sendo este
posteriormente submetido à agitação e centrifugação (14.000 rpm, durante 15 minutos a 4°C).
Após a centrifugação, a fase aquosa contendo o RNA foi transferida para um novo tubo, no
qual foram adicionados 500 μL de álcool isopropílico. O conteúdo do tubo foi misturado por
inversão e posteriormente mantido em repouso. Em seguida submetido à centrifugação

64
(14.000 rpm, 15 minutos, 4°C), o sobrenadante foi desprezado e acrescentado 1 mL de etanol
gelado. O RNA foi novamente centrifugado (7.000 rpm, 10 minutos, 4°C), o etanol foi
desprezado e o precipitado dissolvido em água ultrapura. A concentração e a pureza do RNA
foi determinadas por leitura espectrofotométrica a 260/280 nm. O RNA total foi estocado a -
80°C até o momento do uso.
5.8.3 Confecção do DNA complementar (cDNA)
Para determinar a expressão do RNAm codificador para os genes de IL-1β, TNF-
α, IL-6, GFAP, Iba-1, CCR2 e MCP-1 em diferentes tecidos após o tratamento com PCX, foi
realizado o ensaio de transcrição reversa seguido pela reação em cadeia da polimerase em
tempo real (RT-PCR). Para a reação da transcrição reversa, amostras contendo 1μg de RNA
total foram incubadas com 1μL de oligo dT, 1μL do mix de dNTP (10 mM) em água ultrapura
para um volume final de 12μL. Inicialmente, as amostras foram aquecidas por 5min a 65°C,
resfriadas por 5min a 4 ºC e acrescidas de 4μL de tampão de primeira fita (Tris-HCl 250 mM,
pH 8,3, KCl 375mM e MgCl2 15mM), 2μL de DTT 0,1mM e 1μL de inibidor de RNase
(2500 U). Posteriormente, essa mistura foi incubada a 37 ºC durante 2 minutos e a enzima M-
MLV (1μL, 200 U) foi adicionada. Após a adição da enzima, as amostras foram mantidas a 37
ºC durante 50 minutos e a inativação da reação foi realizada através da incubação das
amostras a 75 ºC durante 15 minutos. O cDNA foi estocado a -20 ºC até a realização da
reação de RT-PCR.
5.8.4 Reação em cadeia da polimerase em tempo real - PCR
O cDNA foi amplificado em duplicata utilizando o kit Master Mix Sibr Green®
Universal PCR, com sondas específicas para camundongos, de IL-1β, TNF-α, IL-6, GFAP,
Iba-1, CCR2 e MCP-1 e GAPDH utilizado como controle endógeno. A reação de PCR foi
realizada em placa de reação óptica de 96 poços. Cada reação continha: 1μL de cDNA,
3,1375μL de master mix, 0,5μL de primer e 1,6125μL de água ultrapura, em um volume final
de 6,25μL. As amplificações foram realizadas em um termociclador (StepOne Plus, Applied
Biosystems) para 60 ciclos. A fluorescência foram coletada a cada ciclo de amplificação e os
dados foram analisados utilizando o método 2-ΔΔCt
para a quantificação relativa. Os Primers
foram projetados com base em sequências de RNAm obtidas a partir do National Center for
Biotechnology Information (http://www.ncbi.nlm.nih.gov), como se segue no quadro 1.

65
Quadro 1 – Sequência de primers utilizados no RT-PCR
Alvos Primers
IL-1β
5’ - TGACAGTGATGAGAATGACCTGTTC - 3’ (F)
5’ – TTGGAAGCAGCCCTTCATCT - 3’ (R)
TNF-α
5’ – AGGGATAGAAGTTCCCAAATG – 3’ (F)
5’ – GGCTTGTCACTCGAATTTTGAGA – 3’ (R)
IL-6
5’ – TTCCTACCCCAATTTCCAAT – 3’ (F)
5’- CCTTCTGTGACTCCAGCTTATC – 3’ (R)
MCP-1
5’ – TAGAGATTGAGCTGTCTGCTCATTC – 3’ (F)
5’ – CCTAAAGTATGGGCTGGACTGTTT 3’ (R)
CCR2
5’- AGGGGGCCACCACACCGTATGA - 3’ (F)
5’- CATGTTGCCCACAAAACCAAAGATGAA - 3’(R)
GAPDH
5’- CATCTTCTTGTGCAGTGCCA - 3’(F)
5’- CGGCCAAATCCGTTCAC- 3’(R)
Sequência (Foward – “F”/ Reverse – “R”) dos primers utilizados no estudo na reação de RT-PCR.
5.9 Dosagem de TNF-α, IL-1β, IL-6, MCP-1 e KC/CXCL1
5.9.1 Coleta das amostras
Camundongos C57BL/6 selvagens; camundongos C57BL/6 selvagens pré-
tratados com cromoglicato de sódio ou com o seu veículo, camundongos SH e camundongos
TLR4-/- foram aleatoriamente divididos em 2 grupos. Os camundongos foram então tratados
com PCX ou veículo e foram eutanasiados (0, 2, 4 e 6 horas) após a injeção de PCX ou
veículo. Os animais foram anestesiados, via intraperitoneal (i.p), com uma mistura de
Ketamina (100 mg/kg) e xilazina (10 mg/kg); em seguida, foi realizada a coleta do sangue no
plexo retro-orbital. O sangue então foi armazenado em tubos e centrifugado para separação do
plasma que foi acondicionado no gelo. Após a coleta de sangue, procedeu-se a perfusão
intracardíaca com solução de PBS para a coleta das seguintes amostras teciduais: medula
espinal lombar, gânglios da raiz dorsal (GRD; segmentos entre L3 – L5) e nervo isquiático.

66
Após a coleta, todas as amostras foram congeladas (-80 ºC) até a realização do ensaio de
ELISA.
5.9.2 Preparação de amostras para análise de citocinas/quimiocinas no plasma
Os camundongos tratados de acordo como descrito anteriormente foram
eutanizados nos tempos conforme mencionados anteriormente. O sangue total foi colhido pelo
plexo retro-orbital, transferido para microtubos contendo 50µL de heparina e colocados no
gelo e em seguida as amostras foram centrifugadas a aproximadamente 6.000g durante 10
minutos. O plasma separado foi armazenado a -80 ºC até a análise.
5.9.3 Homogenato de Tecidos
Nos pontos de tempo indicados acima, os mesmos camundongos a partir dos quais
o plasma foi recolhido, foram eutanasiados e as amostras (nervo isquiático, gânglios da raiz
dorsal e medula espinal) foram removidas e armazenadas a -80 ºC até à análise. Os tecidos
foram homogeneizados como descrito em Famakin et al., (2012). Resumidamente, as
amostras foram homogeneizadas em soro fisiológico tamponado com fosfato (PBS) (pH 7,4),
contendo coquetel de inibidor de protease 1 X (Sigma-Aldrich, St Louis, MO, EUA) e 0,2%
de Triton X-100 (Sigma-Aldrich, St Louis, MO, EUA), durante 30 segundos, utilizando um
homogeneizador básico IKA T 10 (IKA Works, Wilmington, NC, EUA), todo processo de
hogeneização foi realizado no gelo. Os homogenatos resultantes foram centrifugados a
15.000g durante 30 minutos, a 4 ºC, e os sobrenadantes foram recolhidos e armazenados a -80
ºC até à análise de citocinas/quimiocinas por ELISA.
5.9.4 Preparação de amostras para análise de citocinas/quimiocinas de cultura de
mastócitos (RBL-2H3)
As células foram semeadas em placa de 96 poços em número de 1x105, após 24
horas do plaqueamento, as células foram tratadas com paclitaxel (10, 30 ou 90 μM), veículo,
PMA ou ionomicina. As culturas foram mantidas a 37ºC numa atmosfera saturada de água
com 5% de CO2. Após 4 horas de estimulação da cultura de mastócitos (RBL-2H3), com
paclitaxel, veículo, PMA ou ionomicina, o sobrenadante foi coletado e estocado a -20 ºC até a
realização do ensaio.

67
5.9.5 Preparação de amostras para análise de citocinas/quimiocinas de cultura primária de
células satélites gliais
As células foram semeadas em placa de 96 poços, após o sétimo dia de cultivo as
células foram tratadas com paclitaxel (10, 30 ou 90 μM), veículo, PAm3cys, peptidioglicano
ou LPS. As culturas foram mantidas a 37 ºC numa atmosfera saturada de água com 5% de
CO2 e após 48h de incubação foram utilizadas para o ensaio de ELISA após 48 horas da
incubação com paclitaxel, veículo, PAm3cys, peptidioglicano ou LPS. O sobrenadante foi
coletado e estocado a -20 ºC até a realização do ensaio.
5.9.6 Determinação de citocinas/quimiocinas por ELISA
O plasma, os extratos homogeneizados de nervo isquiático, gânglios da raiz dorsal
e medula espinal, os sobrenadantes de cultura de mastócitos (RBL-2H3) e da cultura primária
de células satélites gliais foram analisados para averiguar a produção/liberação de citocinas.
Foram avalidas as seguintes citocinas e quimiocinas: TNF-α, IL1-β, IL-6, KC/CXCL1 e
MCP-1, utilizado o método de ELISA.
Placas de 96 poços para ELISA foram incubadas over-nigth em temperatura
ambiente com 2μg/mL de anticorpo de captura diluído em PBS (pH 7.4) – 50 μL/poço. Em
seguida foram realizadas duas lavagens das placas com PBS – Tween 20, 0,05%. Após a
primeira lavagem é realizado o bloqueio com albumina bovina sérica (BSA) 1% diluída em
PBS, 100μl/poço por 2h à temperatura ambiente. Em seguida, duas lavagens das placas.
Procedeu-se então a incubação com a curva padrão das citocinas diluídas na solução de
PBS/BSA 1% e das amostras a serem dosadas (50μL/poço por over-nigth à 4 ºC). Mais duas
lavagens foram realizadas. Posteriormente houve a incubação com anticorpo biotinilado
diluído a em PBS/BSA 1% por 2h em temperatura ambiente. E duas lavagens das placas,
seguida de incubação com estreptoavidina diluída 1:40 em PBS/BSA 1% (100μL/poço) por
1h à temperatura ambiente. Em seguida procederam-se duas lavagens das placas e incubação
com o agente revelador 3,3′,5,5′-Tetramethylbenzidine (TMB), 100μl/poço. A placa é coberta
e deixada no escuro por 5-20 minutos em temperatura ambiente. A reação é parada com
50μL/poço de H2SO4 4M. A leitura foi feita em espectrofotômetro a 450 nm. E os resultados
são expressos em pg/mL/mg proteína.

68
5.10 Imunofluorescência
5.10.1 Coleta e preparação das amostras
Os animais foram anestesiados, via intraperitoneal, com uma mistura de ketamina
(100mg/kg) e xilazina (10 mg/kg); e, depois, foi realizada a perfusão intracardíaca com 40 mL
de solução tampão fosfato-salino (Phosphate Buffered Saline - PBS) e 40 mL de solução de
paraformaldeído (PFA) 4%. A medula espinal lombar e o gânglio da raiz dorsal (L5) foram
removidos e, em seguida, foi feita a crioproteção em sacarose 30% por 2 dias. Depois, os
tecidos foram acondicionados com tissue-tek®, em freezer, à temperatura de - 80°C.
5.10.2 Imunofluorescência para Triptase, IL-6 e c-Fos
Com os tecidos extraídos, foram feitos cortes com espessura de 10 μm (para gânglios)
ou 20 μm (para medula) no criostato (Leica CM1850), à temperatura de - 24°C. Os cortes foram
fixados por 2 minutos em metanol e ficaram imersos em PBS até o momento de iniciar a
imunofluorescência. Em seguida, foi feita a recuperação antigênica com tampão citrato 0,1M
(pH 6,0), sob aquecimento em forno micro-ondas, por 15 minutos, à temperatura de 95°C.
Após o resfriamento, obtido em temperatura ambiente, por 20 minutos, foi feita a
permeabilização da membrana nuclear (para Triptase, IL-6 e c-Fos) com triton X-100 (0,1%)
e foi feito o bloqueio com albumina sérica bovina (BSA) 5% acrescida de glicina 0,3M por 30
minutos para todas as lâminas. Os cortes foram lavados com PBS e incubados, durante a
noite, à temperatura de 4°C, com o anticorpo primário feito em coelho anti-Triptase
(Abcam®) ou anti-IL-6 (Santa Cruz Biotechnology®) ou anti-c-Fos (Santa Cruz
Biotechnology®) na diluição 1:200 em BSA 5%. As lâminas foram lavadas e incubadas,
durante 1 hora e 30 minutos, com o anticorpo secundário anti-igG de coelho Alexa flúor 568
(Invitrogen®) na diluição de 1:400 em BSA 5%. Para a marcação dos corpos neuronais, os
cortes foram incubados, durante 1 hora e 30 minutos, com o anticorpo NeuN conjugado com
Alexa Flúor 488 (Merck Millipore®) na diluição de 1:100 em BSA 5%. Em seguida, as
lâminas foram lavadas com PBS, montadas com “Prolong Gold” (Invitrogen®) e fotografadas
no microscópio confocal (LM 710, Zeiss).
A quantificação da área marcada nas fotos foi feita diferenciando as áreas
marcadas (pixels) pela maior saturação de cor associada à marcação (vermelha ou verde).
Para isso, foi utilizado o programa Fiji Image J. O procedimento foi baseado na saturação da
cor associada à marcação positiva para um determinado marcador. Os limites necessários para
definição de pixels marcados e não marcados foram definidos previamente. Para a
quantificação da área marcada foram utilizadas quatro fotos (de animais diferentes) por grupo

69
experimental. Os resultados dessa quantificação foram apresentados em porcentagem, a qual
foi calculada a marcação positiva de um determinado marcador em relação à área total da
foto, que foi considerada 100%. Os marcadores quantificados foram Triptase, IL-6 e c-Fos; o
NeuN foi utilizado para marcar os corpos neuronais, de modo que a co-marcação (“merge”)
foi utilizada para mostrar se os neurônios estariam expressando Triptase, IL-6 e c-Fos.
5.11 Análise estatística
Os resultados são apresentados como a média ± erro padrão da média (E.P.M.) de
3-6 animais para os experimentos moleculares e de 6-10 animais para os experimentos
comportamentais (conforme indicado em cada experimento). As porcentagens de inibição
estão apresentadas como a média ± erro padrão das inibições obtidas para cada experimento
individual. A análise estatística dos experimentos comportamentais com decurso temporal foi
realizada por meio de analise de variância (ANOVA) de duas vias com medidas repetidas,
seguida pelo teste de Bonferroni. Valores de P menores que 0,05 (p<0,05) foram considerados
como indicativos de significância. Todas as analises estatísticas foram realizadas através do
programa GraphPad Prism 5 (San Diego, CA, EUA).

70
6. RESULTADOS

71
6.1 Paclitaxel induz hiperalgesia mecânica e térmica (frio) em camundongos.
Para verificar se a administração intravenosa de paclitaxel era capaz de induzir
hiperalgesia aguda em camundongos, foram realizados testes nociceptivos comportamentais.
Observou-se que uma única injeção i.v. de paclitaxel foi capaz de reduzir (p<0,05) o limiar de
retirada da pata a estímulos mecânicos (filamento de von Frey, gráfico 1A), bem como
induziu um comportamento nocifencivo ao frio (teste da acetona, gráfico 1B). No entanto,
quando se avaliou a resposta térmica ao calor, verificou-se que a latência de retirada da pata
não foi alterada com o tratamento com paclitaxel, quando comparadas ao grupo controle
(Gráfico 1C). As respostas hiperalgesicas mecânica e térmica ao frio começaram dentro de 2
horas e atingiram o pico entre 4 e 6 horas, se resolvendo 24 horas após a administração do
paclitaxel (Gráfico 1A e 1B, respectivamente). Enquanto isso, o limiar de retirada da pata a
estímulos mecânicos do grupo controle permaneceu inalterado, assim como o comportamento
nocifensivo ao frio. Entretanto, em 24, 48 ou 72 horas após a administração única de
paclitaxel não foi observado respostas hiperalgesicas mecânica e térmica (Gráfico 1A e 1B,
respectivamente).
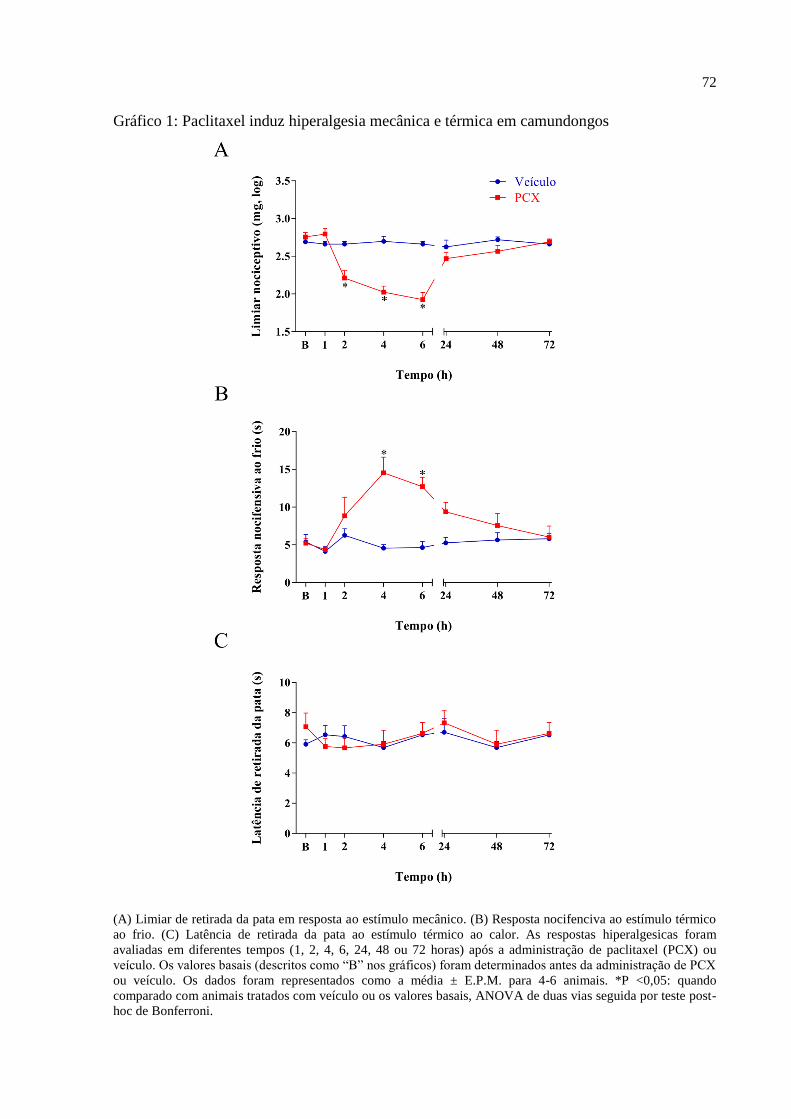
72
Gráfico 1: Paclitaxel induz hiperalgesia mecânica e térmica em camundongos
(A) Limiar de retirada da pata em resposta ao estímulo mecânico. (B) Resposta nocifenciva ao estímulo térmico
ao frio. (C) Latência de retirada da pata ao estímulo térmico ao calor. As respostas hiperalgesicas foram
avaliadas em diferentes tempos (1, 2, 4, 6, 24, 48 ou 72 horas) após a administração de paclitaxel (PCX) ou
veículo. Os valores basais (descritos como “B” nos gráficos) foram determinados antes da administração de PCX
ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando
comparado com animais tratados com veículo ou os valores basais, ANOVA de duas vias seguida por teste post-
hoc de Bonferroni.

73
6.2 Imunofluorescência para c-Fos em gânglio da raiz dorsal de camundongos
submetidos à injeção de paclitaxel
Na imunofluorescência para c-Fos, no gânglio da raiz dorsal, foi observado um
aumento da imunoexpressão nas em células que morfologicamnete assemelham-se a células
satélites. Esta marcação está aumentada nas células satélites de neurônios nos grupos tratado
com paclitaxel 4 e 6 horas, após o tratamento. Este aumento da imunoexpressão é evidenciado
quando comparado com o grupo controle (naive) (Figura 9). Na análise quantitativa da
imunoexpressão para c-Fos, a quantificação da área marcada, comparando com o grupo
controle, houve um aumentou significativo (p<0,05) no grupo tratados com paclitaxel, 4 e 6
horas, após o tratamento (Gráfico 2).
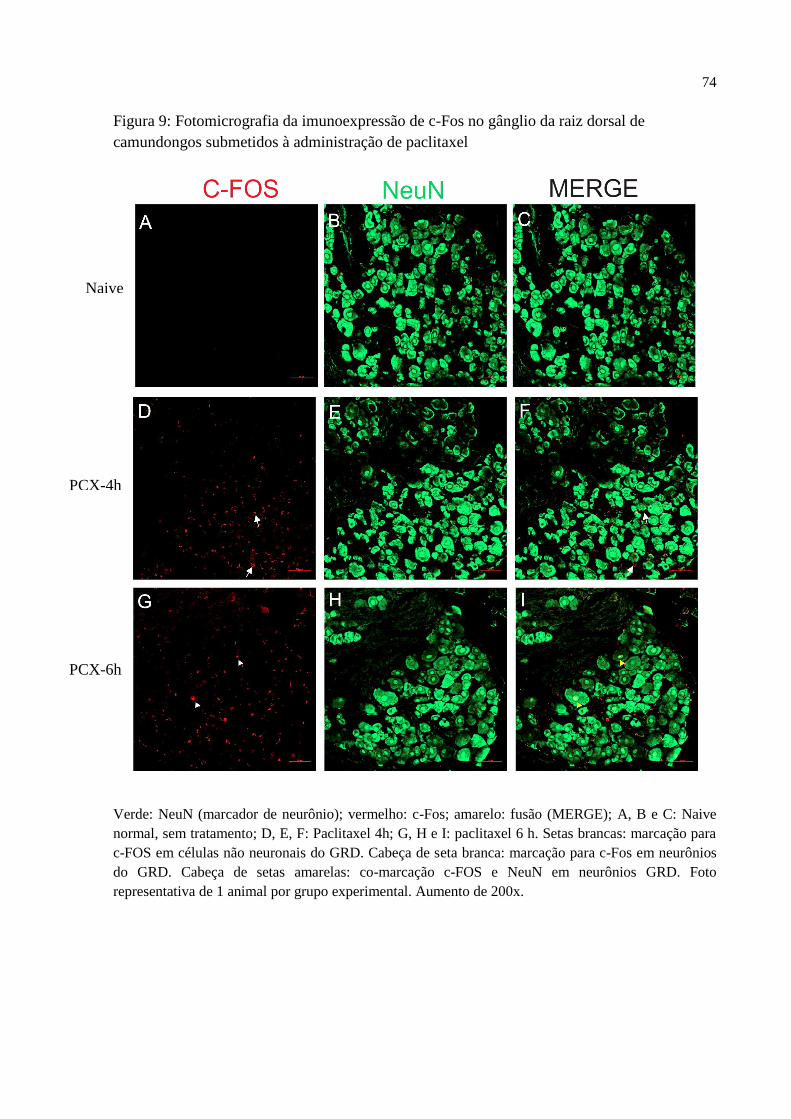
74
Figura 9: Fotomicrografia da imunoexpressão de c-Fos no gânglio da raiz dorsal de
camundongos submetidos à administração de paclitaxel
Verde: NeuN (marcador de neurônio); vermelho: c-Fos; amarelo: fusão (MERGE); A, B e C: Naive
normal, sem tratamento; D, E, F: Paclitaxel 4h; G, H e I: paclitaxel 6 h. Setas brancas: marcação para
c-FOS em células não neuronais do GRD. Cabeça de seta branca: marcação para c-Fos em neurônios
do GRD. Cabeça de setas amarelas: co-marcação c-FOS e NeuN em neurônios GRD. Foto
representativa de 1 animal por grupo experimental. Aumento de 200x.
Naive
PCX-4h
PCX-6h
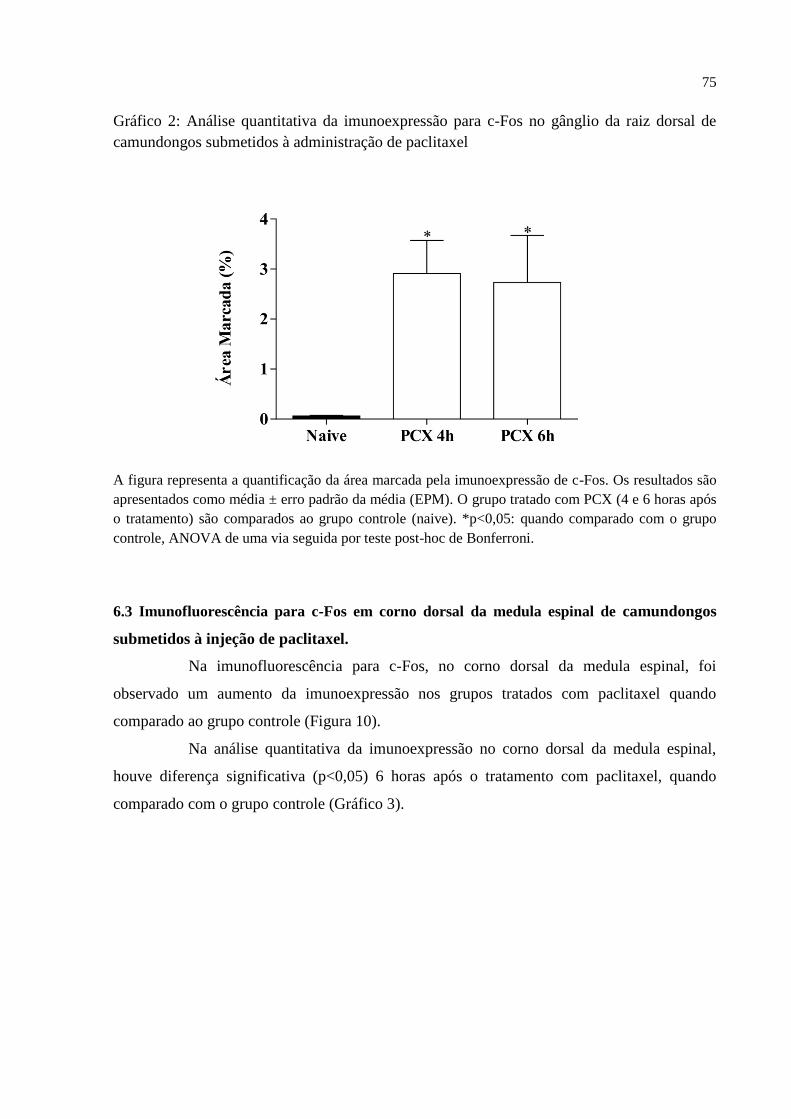
75
Gráfico 2: Análise quantitativa da imunoexpressão para c-Fos no gânglio da raiz dorsal de
camundongos submetidos à administração de paclitaxel
A figura representa a quantificação da área marcada pela imunoexpressão de c-Fos. Os resultados são
apresentados como média ± erro padrão da média (EPM). O grupo tratado com PCX (4 e 6 horas após
o tratamento) são comparados ao grupo controle (naive). *p<0,05: quando comparado com o grupo
controle, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.3 Imunofluorescência para c-Fos em corno dorsal da medula espinal de camundongos
submetidos à injeção de paclitaxel.
Na imunofluorescência para c-Fos, no corno dorsal da medula espinal, foi
observado um aumento da imunoexpressão nos grupos tratados com paclitaxel quando
comparado ao grupo controle (Figura 10).
Na análise quantitativa da imunoexpressão no corno dorsal da medula espinal,
houve diferença significativa (p<0,05) 6 horas após o tratamento com paclitaxel, quando
comparado com o grupo controle (Gráfico 3).

76
Figura 10: Fotomicrografia da imunoexpressão de c-Fos no corno dorsal da medula espinal de
camundongos submetidos à administração de paclitaxel
Verde: NeuN (marcador de neurônio); vermelho: c-Fos; laranja: fusão (MERGE); A, B e C: Naive
normal, sem tratamento; D, E, F: Paclitaxel 4h; G, H e I: paclitaxel 6 h. Setas brancas: marcação para
c-FOS em neurônios do corno dorsal da medula espinal. Setas amarelas: co-marcação c-FOS e NeuN
em neurônios do corno dorsal da medula espinal. Foto representativa de 1 animal por grupo
experimental. Aumento de 200x.
Naive
PCX-4h
PCX-6h

77
Gráfico 3: Análise quantitativa da imunoexpressão para c-Fos no corno dorsal da medula
espinal de camundongos submetidos à administração de paclitaxel
O gráfico representa a quantificação da área marcada pela imunoexpressão de c-Fos no corno dosal da
medula espinal. Os resultados são apresentados como média ± erro padrão da média (EPM). O grupo
tratado com PCX (4 e 6 horas) são comparados ao grupo controle (naive). *p<0,05: quando
comparado com o grupo controle; #p<0,05: quando comparado com o grupo PCX 6h, ANOVA de
uma via seguida por teste post-hoc de Bonferroni.

78
6.4 Quantificação de citocinas e quimiocinas em camundongos tratados com paclitaxel.
Mediante o resultado mostrando a indução de hiperalgesia aguda por paclitaxel,
procurou-se investigar a participação de mediadores da inflamação/hiperlagesia aguda nesse
fenômeno. Assim foram quantificadas as citocinas e quimiocinas no plasma, nervo isquiático,
gânglios da raiz dorsal e medula espinal de camundongos tratados com PCX pelo método de
ELISA. A quantificação dos mediadores da inflamação/hiperlagesia aguda é apresentada nos
gráficos 2, 3, 5 e 7, respectivamente.
Adicionalmente foi quantificada a expressão relativa de mediadores da
inflamação/hiperlagesia aguda no modelo de dor aguda induzida por PCX, através do método
de RT-PCR em amostras de nervo isquiático, gânglios da raiz dorsal e medula espinal. Os
RNAm das citocinas pró-inflamatórias IL-1β, TNF-α e IL-6, e da quimiocina MCP-1 e do seu
receptor CCR2 foram expressos (Gráficos 4, 6 e 8, respectivamente), e seus perfis de
expressão foram alteradas com o tratamento com PCX.
6.4.1 Quantificação de citocinas e quimiocinas no plasma.
A administração de paclitaxel provocou aumento significativo (p<0,05) das
citocinas pró-inflamatórias IL-1β, TNF-α e IL-6 no plasma de camundongos quando
comparados ao grupo naive. Os valores estão representados nos gráficos 4A, 4B e 4C,
respectivamente. Da mesma forma, também foi observado aumento significativo (p<0,05) da
quimiocina KC/CXCL1 (Gráfico 4D) quando comparados com o grupo naive. Foi observado
um aumento significativo (p<0,05) da IL-β (Gráfico 4A) a partir de 4 horas após
administração de paclitaxel, com pico 6 horas após o tratamento, retornando aos valores
basais 24 horas depois da administração. Já o TNF-α (Gráfico 4B) aumentou
significativamente (p<0,05), a partir de 2 horas e manteve-se até 24 horas após a
administração de paclitaxel. Quando se avaliou a IL-6, foi obervado um aumento
signignificativo (p<0,05) desta citocina 4 horas após a administração de PCX, e 6 horas após
a administração a concentração desta citocina começa a cair, retornando aos valores basais 24
horas após a administração. Os níveis de KC/CXCL1 aumentaram significativamente
(p<0,05) a partir de 2 horas após a administração de paclitaxel, foi observado uma pequena
redução da concentração, no entanto, os valores mantiveram-se significativamente altos
(p<0,05) até a 24ª hora após a administração da droga.
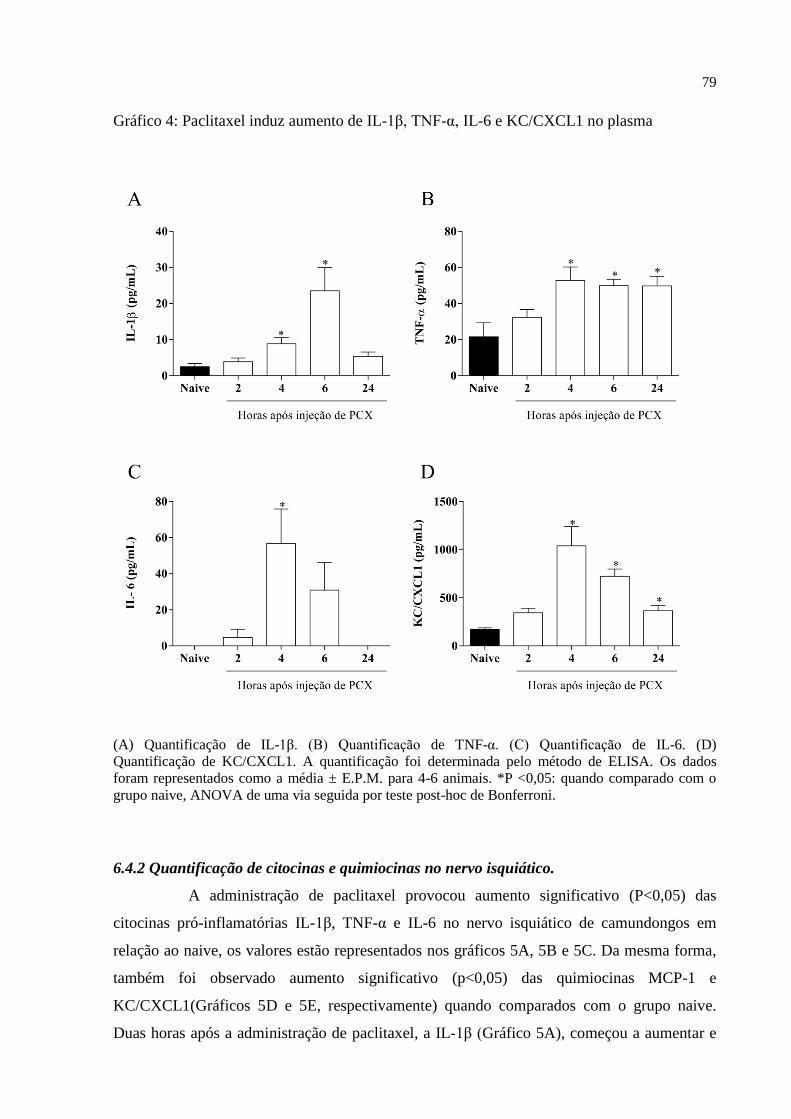
79
Gráfico 4: Paclitaxel induz aumento de IL-1β, TNF-α, IL-6 e KC/CXCL1 no plasma
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de KC/CXCL1. A quantificação foi determinada pelo método de ELISA. Os dados
foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando comparado com o
grupo naive, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.4.2 Quantificação de citocinas e quimiocinas no nervo isquiático.
A administração de paclitaxel provocou aumento significativo (P<0,05) das
citocinas pró-inflamatórias IL-1β, TNF-α e IL-6 no nervo isquiático de camundongos em
relação ao naive, os valores estão representados nos gráficos 5A, 5B e 5C. Da mesma forma,
também foi observado aumento significativo (p<0,05) das quimiocinas MCP-1 e
KC/CXCL1(Gráficos 5D e 5E, respectivamente) quando comparados com o grupo naive.
Duas horas após a administração de paclitaxel, a IL-1β (Gráfico 5A), começou a aumentar e

80
manteve-se alto até 6 horas depois da administração. Pode ser observado que esta citocina foi
reduzindo com o avanço do tempo, e com 24 horas após a administração a IL-1β retornou ao
valor basal em comparação ao naive. O TNF-α, mostrou-se alterado a partir de 2 horas
(Gráfico 5B) depois da administração de paclitaxel, no entanto um aumento significativo
(p<0,05) só foi observado 4 horas após a administração da droga. Daí em diante a
concentração de TNF-α começou a reduzir, não sendo obervado diferença signifivcativa com
6 e 24 horas após a administração do paclitaxel, apesar das concentrações terem se mantidas
elevadas. Após a administração de paclitaxel a IL-6 (Gráfico 5C) começou a aumentar no
nervo isquiático, entretanto, só foi detectado diferença significativa (p<0,05) 4 horas depois
da administração. A partir de 6 horas a concentração caiu e com 24 horas após o tratamento os
níveis retornaram aos valores basais. Quanto aos níveis das quimiocinas MCP-1 (Gráfico 5D)
e KC/CXCL1 (Gráfico 5E), ambas estavam alteradas a partir de 2 horas após o tratamento
com paclitaxel. MCP-1(Gráfico 5D) manteve um aumento significativo (p<0,05) até 6 horas
depois da administração de paclitaxel e houve uma leve redução 24 horas após o tratamento,
neste momento não houve diferença estatística quando comparado ao naive. Já a concentração
de KC/CXCL1(Gráfico 5E) foi reduzida a partir da 6 horas após a administração de
paclitaxel, o que também foi observado em 24 horas.

81
Gráfico 5: Paclitaxel induz aumento de IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL no nervo
isquiático
(A) Quantificação de TNF-α. (B) Quantificação de IL-1β. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com grupo naive, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.4.3 Expressão relativa de RNAm para citocinas e quimiocinas em nervo isquiático.
Foi avaliado se o tratamento com paclitaxel altera a expressão gênica das citocinas
pró-inflamatórias IL-1β, TNF-α e IL-6 no nervo isquiático de camundongos tratados com
paclitaxel. Para isso, detectou-se a expressão basal do RNAm para estas citocinas no nervo
isquiático, conforme observado no gráfico 6. O tratamento com paclitaxel induziu um
aumento significativo (p<0,05) da expressão relativa do RNAm para a IL-6 (Gráfico 6C)
quando comparados ao naive. No entanto, não houve aumento significativo na expressão das
citocinas IL-1β e TNF-α (Gráficos 6A e 6B).
Apesar de se observar uma tendência ao aumento na expressão relativa de RNAm
da IL-1β (Gráficos 6A) a partir de 2 horas após o tratamento com paclitaxel, esse aumento
não foi suficiente para que ocorresse uma expressão significativa dessa citocina, que
permaneceu nos níveis basais em todos os demais pontos temporais avaliados. A expressão
relativa de RNAm de TNF-α (Gráfico 6B), manteve-se em níveis basais até 4 horas após o
tratamento com paclitaxel, observando-se um aumento na expressão relativa de RNAm da

82
citocina 6 horas após a administração de paclitaxel, no entanto esse aumento na expressão
relativa de RNAm da citocina não foi significativamente diferente do naive. Observa-se ainda
que com 24 horas após o tratamento a expressão relativa do RNAm da citocina retorna ao
nível basal, quando comparado ao naive.
Com relação a IL-6 (Gráfico 6C) o tratamento com paclitaxel induziu aumento
significativo da expressão relativa do RNAm para esta citocina (p <0,05) quando comparados
ao naive. Observa-se um aumento na expressão relativa do RNAm de IL-6 a partir de 2 horas
após a administração de paclitaxel, porém um aumento significativo só é detectado 4 horas
após o tratamento com paclitaxel (p <0,05), mantendo-se até 6 horas e retornando a níveis
basais 24 horas após administração do paclitaxel.
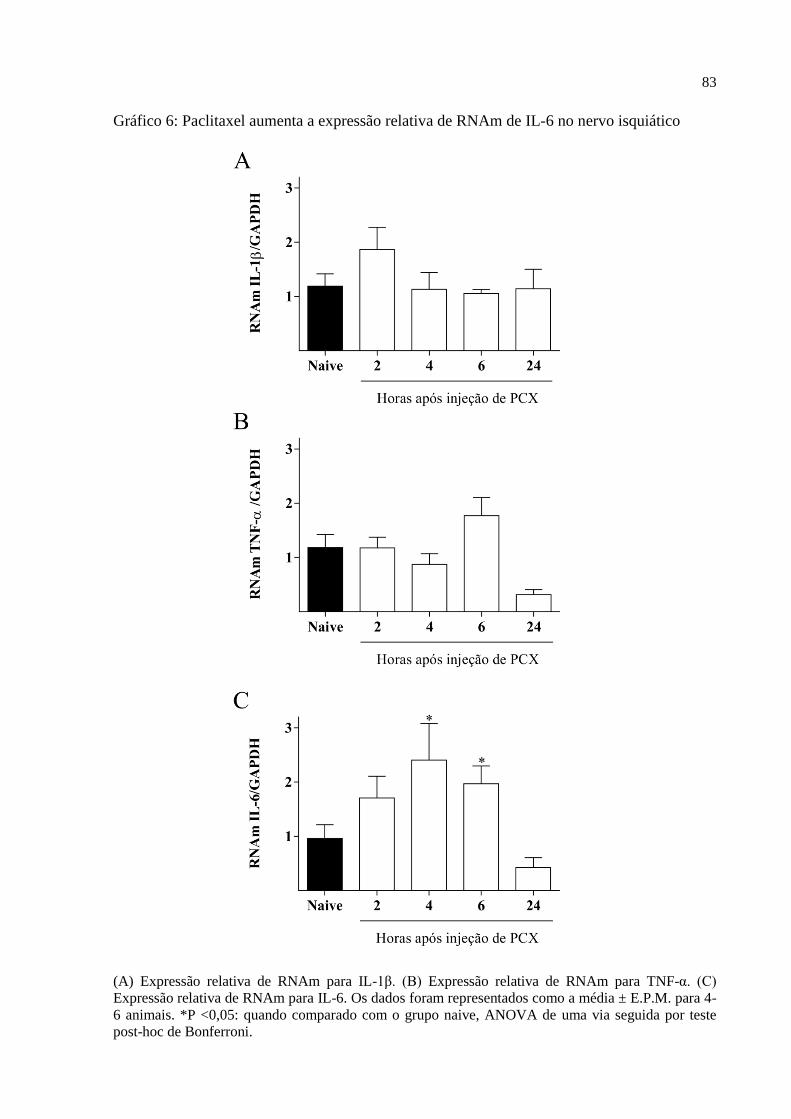
83
Gráfico 6: Paclitaxel aumenta a expressão relativa de RNAm de IL-6 no nervo isquiático
(A) Expressão relativa de RNAm para IL-1β. (B) Expressão relativa de RNAm para TNF-α. (C)
Expressão relativa de RNAm para IL-6. Os dados foram representados como a média ± E.P.M. para 4-
6 animais. *P <0,05: quando comparado com o grupo naive, ANOVA de uma via seguida por teste
post-hoc de Bonferroni.

84
6.4.4 Quantificação de citocinas e quimiocinas nos gânglios da raiz dorsal.
Observou-se que no grupo tratado com paclitaxel, houve um aumento
significativo (p<0,05) das citocinas pró-inflamatórias TNF-α e IL-6 nos gânglios da raiz
dorsal em comparação ao grupo naive (Gráfico 7B e 7C). Neste mesmo grupo, observou-se
ainda aumento significativo (p<0,05) da quimiocina KC/CXCL1 (Gráfico 7E) quando
comparados com o grupo naive. Porém a concentração da citocina IL-1β e da quimiocina
MCP-1 não foram aumentadas significativamente pela administração de paclitaxel (Gráficos
7A e 7D). A IL-1β (Gráfico 7A), aumentou moderadamente em relação ao naive, não sendo
observados aumentos significativos nos pontos de tempo avaliados. Já o TNF-α aumentou
significativamente (p<0,05) 2 horas após a administração de paclitaxel, esse aumento foi
observado até 6 horas, reduzindo significativamente 24 horas após a administração de
paclitaxel. A IL-6 teve aumento significativo (p<0,05) em 2 horas após a administração de
paclitaxel e esse aumento também foi observado com 4 horas depois do paclitaxel, com 6
horas não foi mais observada diferença significativa na concentração de IL-6, quando
comparado ao naive, e 24 horas depois da injeção de paclitaxel a concentração de IL-6 nos
GRD praticamente retornou aos valores basais.

85
Gráfico 7: Paclitaxel induz aumento de IL-6, TNF-α e KC/CXCL1 em gânglios da raiz dorsal
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com o grupo naive, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.4.5 Expressão relativa de RNAm de citocinas e quimiocinas em gânglios da raiz dorsal.
Foi avaliado se o tratamento com paclitaxel altera a expressão relativa de RNAm
das citocinas pró-inflamatórias IL-1β, TNF-α e IL-6, da quimiocina MCP-1 e do seu receptor
CCR2, nos gânglios da raiz dorsal (GRD). Os resultados da detecção da expressão basal do
RNAm em GRD para estes alvos estão representados na Gráfico 8. O tratamento com
paclitaxel foi capaz de aumentar significativamente (p<0,05) a expressão relativa de RNAm
para IL-1β e TNF-α (Gráficos 8A e 8B). O tratamento com paclitaxel ainda aumentou
significativamente (p<0,05) a expressão relativa de RNAm para CCR2 nos GRD (Gráfico
8E). Entretanto, não foi observada diferença estatística na expressão relativa do RNAm para
IL-6 (Gráfico 8C). Do mesmo modo que para as citocinas, detectou-se a expressão relativa do
RNAm para a quimiocina MCP-1 nos GRD (Gráfico 8D), porém não foi observado aumento
significativo da expressão relativa desta quimiocina após o tratamento com o paclitaxel.
Foi detectado aumento na expressão relativa de RNAm para IL-1β (Gráfico 8A) a
partir de 4 horas após a administração de paclitaxel, após 6 horas esse aumento se manteve,

86
entretanto, com 24 horas os níveis de expressão relativa de RNAm para IL-1β (Gráfico 8A)
retornou aos valores basais. Foi possível detectar aumento significativo (p<0,05) na expressão
relativa de RNAm para TNF-α (Gráfico 8B) a partir de 2 horas, mantendo-se até 6 horas e 24
horas após o tratamento com paclitaxel a expressão basal de RNAm para TNF-α havia
retornado ao valor basal. A expressão relativa de RNAm para IL-6 (Gráfico 8C) nos DRG não
foi alterada com o tratamento com paclitaxel em relação ao naive. A expressão relativa de
RNAm para a quimiocina MCP-1 (Gráfico 8D), também não foi alterada nos animais tratados
com paclitaxel, no entanto, a expressão relativa de RNAm do seu receptor (CCR2) (Gráfico
8E) foi aumenta significativamente (p<0,05) 6 horas após a administração de paclitaxel,
retornando ao nível basal em 24 horas.

87
Gráfico 8: Paclitaxel aumenta a expressão relativa de RNAm para IL-1β, TNF-α e CCR2 nos
gânglios da raiz dorsal
(A) Expressão relativa de RNAm para IL-1β. (B) Expressão do relativa de RNAm TNF-α. (C)
Expressão relativa de RNAm IL-6. (D) Expressão relativa de RNAm MCP-1. (E) Expressão relativa
de RNAm CCR2. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com o grupo naive, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.4.6 Quantificação de citocinas e quimiocinas na medula espinal.
O tratamento com paclitaxel provocou aumento significativo das citocinas pró-
inflamatórias IL-1β, TNF-α e IL-6 na medula espinal em comparação ao grupo naive, como
observado nos gráficos 9A, 9B e 9C, respectivamente. O tratamento com paclitaxel também
provocou aumento significativo dos níveis das quimiocinas MCP-1 e KC/CXCL1 (Gráficos
9D e 9E, respectivamente) na medula espinal, quando comparados ao grupo naive. A
concentração de IL-1β (Gráfico 9A) encontra-se aumentada significativamente (p<0,05) a
partir de 2 horas após o tratamento com paclitaxel, mantendo-se alterada até 24 horas. O TNF-
α (Gráfico 9B) aumenta moderadamente com 2 horas após o tratamento com paclitaxel, no
entanto, um aumento significativo (p<0,05) só é observado com 4 horas após a administração,
e é reduzido a partir de 6 horas, retornando aos valores basais 24 horas após a injeção de
paclitaxel. A IL-6 (Gráfico 9C) aumenta significativamente (p<0,05) na medula espinal de
camundongos tratados com paclitaxel após 2 horas, mantendo esse aumento até 6 horas, com
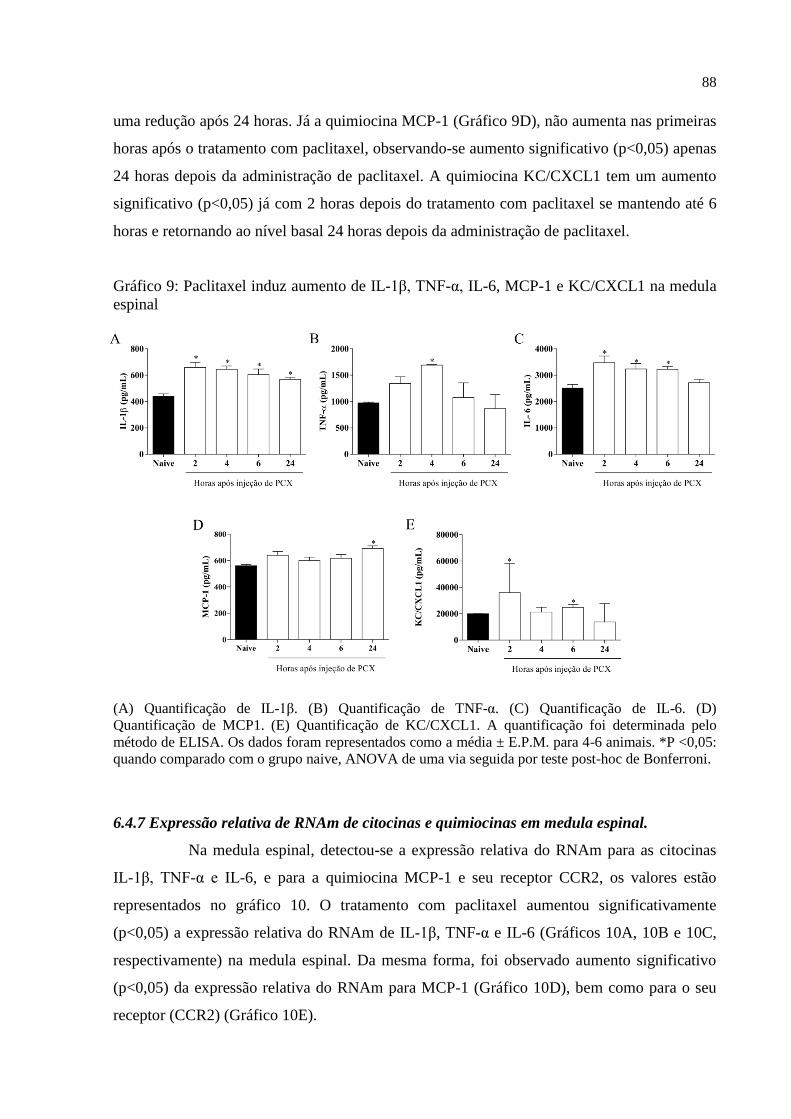
88
uma redução após 24 horas. Já a quimiocina MCP-1 (Gráfico 9D), não aumenta nas primeiras
horas após o tratamento com paclitaxel, observando-se aumento significativo (p<0,05) apenas
24 horas depois da administração de paclitaxel. A quimiocina KC/CXCL1 tem um aumento
significativo (p<0,05) já com 2 horas depois do tratamento com paclitaxel se mantendo até 6
horas e retornando ao nível basal 24 horas depois da administração de paclitaxel.
Gráfico 9: Paclitaxel induz aumento de IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL1 na medula
espinal
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de MCP1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com o grupo naive, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
6.4.7 Expressão relativa de RNAm de citocinas e quimiocinas em medula espinal.
Na medula espinal, detectou-se a expressão relativa do RNAm para as citocinas
IL-1β, TNF-α e IL-6, e para a quimiocina MCP-1 e seu receptor CCR2, os valores estão
representados no gráfico 10. O tratamento com paclitaxel aumentou significativamente
(p<0,05) a expressão relativa do RNAm de IL-1β, TNF-α e IL-6 (Gráficos 10A, 10B e 10C,
respectivamente) na medula espinal. Da mesma forma, foi observado aumento significativo
(p<0,05) da expressão relativa do RNAm para MCP-1 (Gráfico 10D), bem como para o seu
receptor (CCR2) (Gráfico 10E).

89
Um aumento significativo (p<0,05) na expressão relativa de RNAm para IL-1β
(Gráfico 10A) foi detectado a partir de 4 horas após a administração de paclitaxel, com 6
horas esse aumento se manteve, no entanto com 24 horas os níveis de expressão relativa de
RNAm para IL-1β (Gráfico 10A) retornou aos valores basais. Foi possível detectar aumento
significativo (p<0,05) na expressão relativa de RNAm para TNF-α (Gráfico 10B) com 4
horas, mas com 6 horas iniciou uma redução que com 24 horas após o tratamento com
paclitaxel o a expressão relativa de RNAm para TNF-α havia retornado ao valor basal. A
expressão relativa de RNAm para IL-6 (Gráfico 10C) apresentou aumento significativo
(p<0,05) em relação ao naive, iniciando 2 horas após a administração de paclitaxel, com um
pico de expressão relativa de RNAm para esta citocina com 4 horas e retonando ao basal com
24 horas. A expressão relativa de RNAm para a quimiocina MCP-1 (Gráfico 10D), também
aumentou significativamente em relação ao naive (p<0,05) com tratamento com o paclitaxel.
A expressão relativa de RNAm do seu receptor CCR2 (Gráfico 10E) aumentou
significativamente 4 horas após a administração de paclitaxel (p<0,05), retornando ao nível
basal em 24 horas.
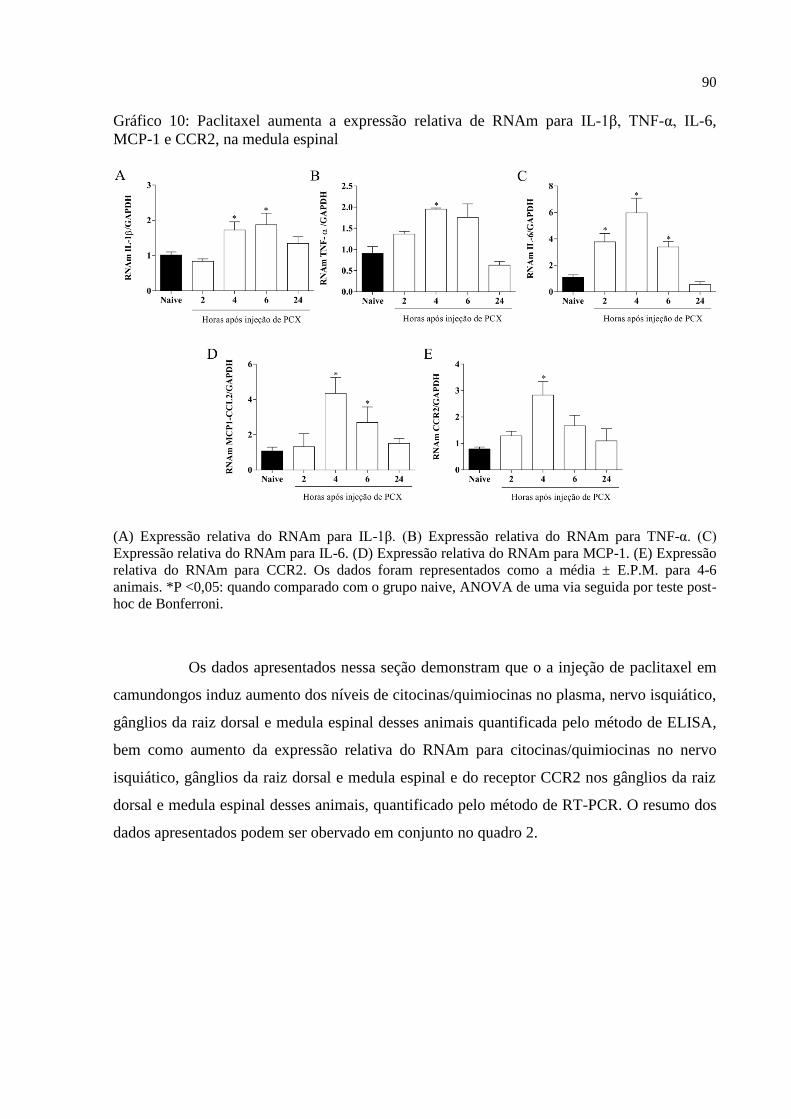
90
Gráfico 10: Paclitaxel aumenta a expressão relativa de RNAm para IL-1β, TNF-α, IL-6,
MCP-1 e CCR2, na medula espinal
(A) Expressão relativa do RNAm para IL-1β. (B) Expressão relativa do RNAm para TNF-α. (C)
Expressão relativa do RNAm para IL-6. (D) Expressão relativa do RNAm para MCP-1. (E) Expressão
relativa do RNAm para CCR2. Os dados foram representados como a média ± E.P.M. para 4-6
animais. *P <0,05: quando comparado com o grupo naive, ANOVA de uma via seguida por teste post-
hoc de Bonferroni.
Os dados apresentados nessa seção demonstram que o a injeção de paclitaxel em
camundongos induz aumento dos níveis de citocinas/quimiocinas no plasma, nervo isquiático,
gânglios da raiz dorsal e medula espinal desses animais quantificada pelo método de ELISA,
bem como aumento da expressão relativa do RNAm para citocinas/quimiocinas no nervo
isquiático, gânglios da raiz dorsal e medula espinal e do receptor CCR2 nos gânglios da raiz
dorsal e medula espinal desses animais, quantificado pelo método de RT-PCR. O resumo dos
dados apresentados podem ser obervado em conjunto no quadro 2.

91
Quadro 2: Paclitaxel aumenta a expressão citocinas e/ou quimiocinas no plasma, no nervo
isquiático, nos gânglios da raiz dorsal e na medula espinal de camundongos
Resultados dos níveis dos níveis de citocinas/quimiocinas no plasma, nervo isquiático, gânglios da raiz
dorsal e medula espinal de animais tratados com paclitaxel (PCX), quantificada pelo método de
ELISA. Expressão relativa do RNAm para citocinas/quimiocinas no nervo isquiático, gânglios da raiz
dorsal e medula espinal e do receptor CCR2 nos gânglios da raiz dorsal e medula espinal de animais
tratados com PCX, quantificado pelo método de RT-PCR. ( + ) Aumento significativo. ( - ) não
significativo. (na) Não Avaliado.
6.5 Investigação da participação de citocinas e quimiocinas na hiperalgesia mecânica e
térmica induzida pelo paclitaxel.
Como ficou demonstrado que citocinas e/ou quimiocinas estavam aumentadas
significativamente (p<0,05) após a administração de paclitaxel tanto no plasma, quanto no
nervo isquiático, GRD, assim como na medula, foi avaliado então o envolvimento das
citocinas IL-6, TNF-α, IL-1 e da quimiocina MCP-1 (os três últimos investigadas através da
avaliação de seus receptores) na hiperalgesia aguda induzida pelo paclitaxel, utilizando
animais knockout para esses alvos.
6.5.1. Investigação da participação da IL-6 na hiperalgesia mecânica e térmica induzida
pelo paclitaxel.
O gráfico 11 ilustra os efeitos do tratamento com paclitaxel em animais knockout
para a IL-6 (IL-6-/-). Como observado, a hiperalgesia mecânica e térmica ao frio foi reduzida
em animais com deleção da citocina IL-6 durante todo o período de avaliação, quando
comparado aos animais do tipo selvagem C57BL/6 (WT) (p<0,05). A ausência da IL-6

92
preveniu o aparecimento de respostas hiperalgesicas observadas nos animais IL-6-/- (Gráficos
11A e 11B, respectivamente). Além disso, camundongos WT foram previamente tratados com
um antagonista solúvel de IL-6 (tocilizumabe – TCZ) 30 minutos antes da injeção de
paclitaxel. Observou-se que o tratamento com TZC foi capaz de prevenir as respostas
hiperalgesicas induzidas pelo paclitaxel (Gráficos 11C e 11D, respectivamente).
O paclitaxel causou diminuição no limiar mecânico, quando comparado ao grupo
naive (p<0,05), caracterizado por uma maior sensibilidade ao estímulo mecânico de menor
intensidade aplicado (filamentos de von Frey). Também é observado que o tratamento com
paclitaxel induz hipernicepção térmica, caracterizada pelo aumento no tempo da resposta ao
frio 10 ºC (teste da acetona), quando comparado ao grupo controle (p<0,05). As respostas
hiperalgesicas se iniciaram a partir de 2 horas após o tratamento com paclitaxel, com o pico
de hiperalgesia mecânica e térmica 6 horas após, resolvendo-se completamente após 24 horas.
Foi observado que a ausência de IL-6 (Gráfico 11A) previne a hiperalgesia
mecânica desde o inicio em que esta se manifesta até 6 horas (pico da hiperalgesia) após o
tratamento com paclitaxel. Assim como na hiperalgesia mecânica, a ausência de IL-6 previne
o desenvolvimento de hiperalgesia térmica ao frio. Observou-se ainda que o pré-tratamento
com TZC apenas na maior dose utilizada foi capaz de prevenir as respostas hiperalgesicas
induzidas pelo paclitaxel (Gráficos 11C e 11D, respectivamente).

93
Gráfico 11: Papel da IL-6 na hiperalgesia mecânica e térmica induzida pelo paclitaxel
A ausência de IL-6, bem como o tratamento com um antagonista desta citocina inibe a hiperalgesia
induzida pelo PCX em camundongos. (A) Hiperalgesia mecânica causada pelo PCX em animais
selvagens (WT) e knockout para IL-6 (IL-6-/-). (B) Hiperalgesia térmica ao frio causada pelo PCX em
animais selvagens (WT) e knockout para IL-6 (IL-6-/-). (C) Efeito do tratamento com tocilizumabe na
hiperalgesia mecânica causada pelo PCX em animais selvagens (WT). (D) Efeito do tratamento com
tocilizumabe na hiperalgesia térmica ao frio causada pelo PCX em animais selvagens (WT). Os
valores basais (descritos como “B” nos gráficos) foram determinados antes da administração de PCX
ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando
comparado com animais tratados com veículo ou os valores basais, ANOVA de duas vias seguida por
teste post-hoc de Bonferroni.

94
6.5.2 Investigação da participação de IL-1 na hiperalgesia mecânica e térmica induzida
pelo paclitaxel.
A participação da IL-1 na hiperalgesia mecânica e térmica induzida pelo
paclitaxel foi investigada através de seu receptor (IL-1R). Utilizou-se então camundongos
knockout para o receptor de IL-1 (IL-1R-/-). Nos gráficos 12A e 12B, respectivamente, são
mostrados os efeitos do tratamento com paclitaxel em animais selvagens (WT) e em animais
IL-1R-/-. Como observado, a hiperalgesia mecânica iniciou 2 horas após a administração de
paclitaxel, culminando entre 4 e 6 horas e retornando aos valores basais após 24 horas da
administração de paclitaxel, quando comparado ao veículo. Observa-se nos gráficos 12A e
12B que a ausência do receptor de IL-1 foi capaz de prevenir o desenvolvimento de
hiperalgesia mecânica e térmica ao frio, quando comparados aos animais WT injetados com
paclitaxel (p<0,05).

95
Gráfico 12: Envolvimento do receptor IL-1R na hiperalgesia induzida pelo paclitaxel
A deleção do gene para o receptor IL-1R, inibe a hiperalgesia induzida pelo paclitaxel em
camundongos. (A) Hiperalgesia mecânica causada pelo PCX em animais selvagens (WT) e animais
IL-1R-/-. (B) Hiperalgesia térmica ao frio causada pelo PCX em animais selvagens (WT) e animais
IL-1R-/-. Os valores basais (descritos como “B” nos gráficos) foram determinados antes da
administração de PCX ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6
animais. *P <0,05: quando comparado com animais tratados com veículo ou os valores basais,
ANOVA de duas vias seguida por teste post-hoc de Bonferroni.

96
6.5.3 Investigação da participação TNF-α na hiperalgesia mecânica e térmica induzida
pelo paclitaxel.
A participação do TNF-α na hiperalgesia induzida pelo paclitaxel foi investigada
através da avaliação dos seus receptores (TNFR1 e TNFR2), para tanto foram utilizados
animais knockout para ambos os receptores. No gráfico 13, é mostrado os efeitos do
tratamento com paclitaxel em animais WT e em animais knockout para os receptores de TNF
(TNFR1/R2-/-). Como observado, a hiperalgesia mecânica e térmica ao frio foi
significativamente (p<0,05) reduzida nos animais com a dupla deleção gênica para os
receptores de TNF. Foi observado uma inibição significativa (p<0,05) das respostas
hiperalgesicas a partir de 2 horas após a administração de paclitaxel, com maior inibição
(p<0,05) entre 4 e 6 horas, onde é obervado o pico da hiperalgesia em animais WT. Após 24
horas da administração de paclitaxel observa-se que os valores retornam para aqueles
observados inicialmente, ou seja, antes da administração de paclitaxel. A ausência dos
receptores foi eficaz sobre as respostas de hiperalgesia mecânica e térmica, quando
comparados com os animais WT (p<0,05) (Gráficos 13A e 13B, respectivamente).
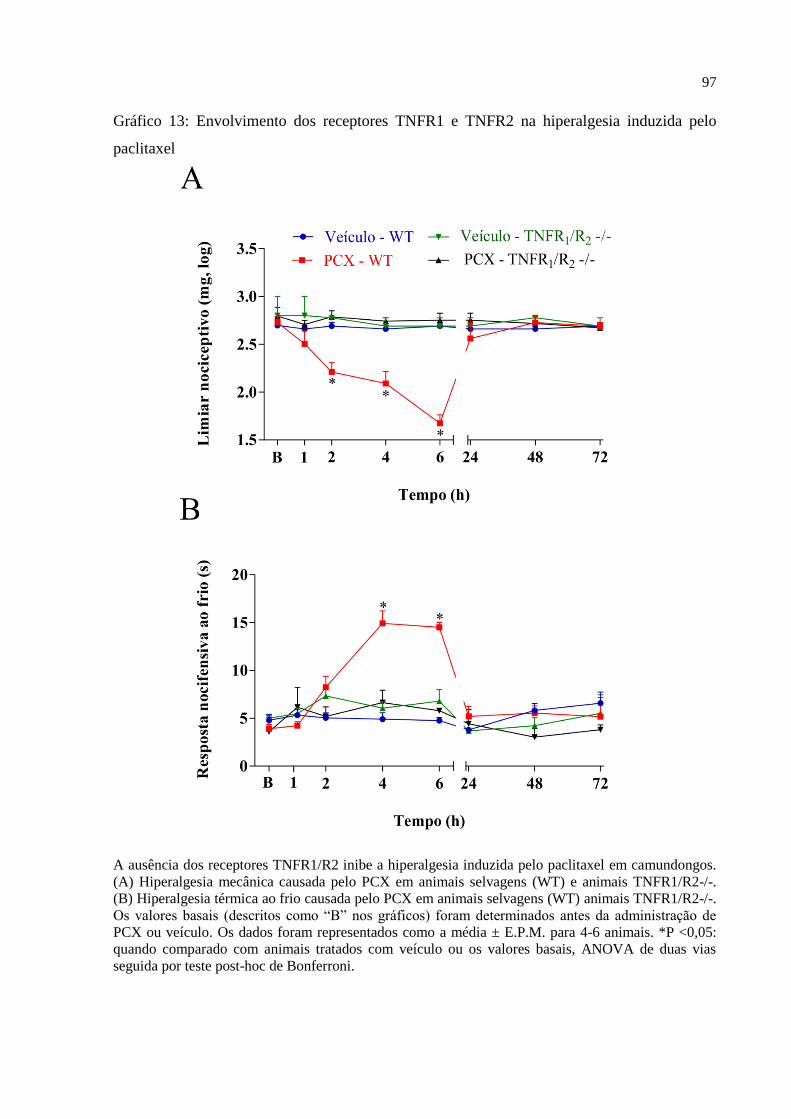
97
Gráfico 13: Envolvimento dos receptores TNFR1 e TNFR2 na hiperalgesia induzida pelo
paclitaxel
A ausência dos receptores TNFR1/R2 inibe a hiperalgesia induzida pelo paclitaxel em camundongos.
(A) Hiperalgesia mecânica causada pelo PCX em animais selvagens (WT) e animais TNFR1/R2-/-.
(B) Hiperalgesia térmica ao frio causada pelo PCX em animais selvagens (WT) animais TNFR1/R2-/-.
Os valores basais (descritos como “B” nos gráficos) foram determinados antes da administração de
PCX ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com animais tratados com veículo ou os valores basais, ANOVA de duas vias
seguida por teste post-hoc de Bonferroni.

98
6.5.4 Investigação da participação de MCP-1 na hiperalgesia mecânica e térmica induzida
pelo paclitaxel.
Foi avaliada a participação da quimiocina MCP-1 através da utilização do seu
receptor (CCR2) na hiperalgesia induzida pelo paclitaxel. Para tanto, foi utilizado animais
knockout para o receptor CCR2 (CCR2-/-). No gráfico 14 é mostrado os efeitos do tratamento
com paclitaxel em animais selvagens WT e em animais CCR2-/-. Como observado, a
hiperalgesia mecânica e térmica ao frio, foi alterada nos animais selvagens WT, a partir de 2
horas, com pico entre 4 e 6 horas e retornando aos valores basais 24 horas após a
administração do paclitaxel, quando comparados ao veículo (p<0,05). Já a ausência dos
receptores foi eficaz em prevenir as respostas hiperalgesicas mecânica e térmica ao frio
induzidas pelo paclitaxel quando comparado aos animais do tipo selvagem WT (p<0,05)
(Gráfico 14A e 14B, respectivamente).

99
Gráfico 14: Envolvimento do receptor CCR2 na hiperalgesia induzida pelo paclitaxel
A ausência do receptor CCR2 inibe a hiperalgesia induzida pelo paclitaxel em camundongos. (A)
Hiperalgesia mecânica causada pelo PCX em animais selvagens (WT) e animais CCR2-/-. (B)
Hiperalgesia térmica ao frio causada pelo PCX em animais selvagens (WT) e animais CCR2-/-. Os
valores basais (descritos como “B” nos gráficos) foram determinados antes da administração de PCX
ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando
comparado com animais tratados com veículo ou os valores basais, ANOVA de duas vias seguida por
teste post-hoc de Bonferroni.

100
6.6 Investigação da participação de mastócitos na hiperalgesia mecânica e térmica
induzida pelo paclitaxel.
Tendo em vista que citocinas pró-inflamatórias e quimiocinas estavam envolvidas
no processo de hiperalgesia aguda induzida por paclitaxel e sabendo-se que células imunes
residentes produzem esses mediadores, foi avaliado o possível envolvimento dessas células na
hiperalgesia aguda induzida pelo paclitaxel. Para isso, optou-se por investigar o envolvimento
de mastócitos na hiperalgesia induzida por paclitaxel. Para tanto, foram utilizados animais
mutantes que não diferenciam mastócitos (SH) (Gráfico 15), assim como também foram feitos
experimentos utilizando o estabilizador de membrana de mastócito, cromoglicato de sódio
(Gráfico 16).
O gráfico 15 ilustram os efeitos da administração do paclitaxel em animais SH e
selvagens (WT). Como pode ser observado, a hiperalgesia mecânica e térmica ao frio
(Gráficos 15A e 15B, respectivamente) foi reduzida significativamente (p<0,05) nos animais
SH, durante todo o período de avaliação, quando comparado aos animais WT.
Para confirmar que os mastócitos estavam envolvidos na hiperalgesia mecânica e
térmica, então animais WT foram pré-tratados com cromoglicato de sódio (CROMO) e
posteriormente injetados com PCX. O gráfico 16 mostra os efeitos do pré-tratamento com o
cromoglicato de sódio, sobre a hiperalgesia induzida pelo paclitaxel. Os resultados
demonstram que o cromoglicado de sódio, um estabilizador de membrana de mastócitos, foi
eficaz em inibir significativamente (p<0,05) a hiperalgesia mecânica (Gráfico 16A) e térmica
ao frio (Gráfico 16B) a partir de 2 horas e permanecendo durante todo o período de avaliação
após o tratamento com o quimioterápico. A inibição da resposta hipernociceptiva mecânica e
térmica dos animais tratados com cromoglicato de sódio sugere que a degranulação dos
mastócitos contribui para a gênese da hiperalgesia.
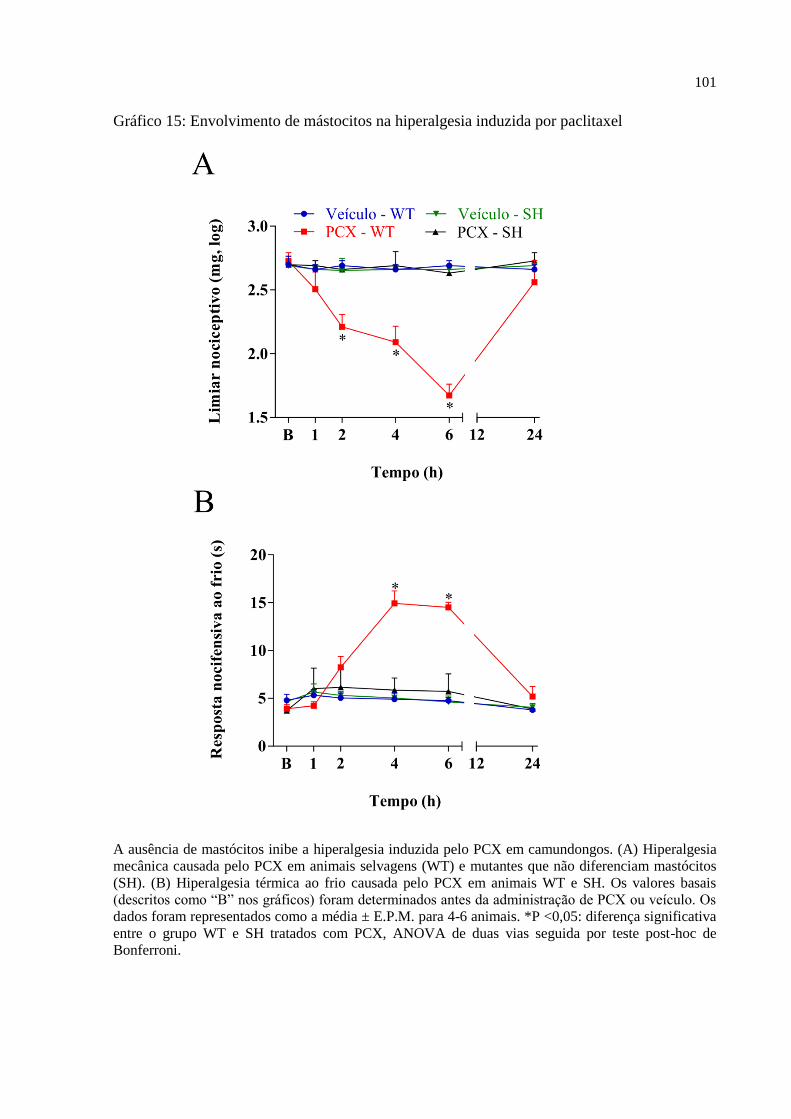
101
Gráfico 15: Envolvimento de mástocitos na hiperalgesia induzida por paclitaxel
A ausência de mastócitos inibe a hiperalgesia induzida pelo PCX em camundongos. (A) Hiperalgesia
mecânica causada pelo PCX em animais selvagens (WT) e mutantes que não diferenciam mastócitos
(SH). (B) Hiperalgesia térmica ao frio causada pelo PCX em animais WT e SH. Os valores basais
(descritos como “B” nos gráficos) foram determinados antes da administração de PCX ou veículo. Os
dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: diferença significativa
entre o grupo WT e SH tratados com PCX, ANOVA de duas vias seguida por teste post-hoc de
Bonferroni.

102
Gráfico 16: Cromoglicato de sódio atenua a hiperalgesia induzida pelo paclitaxel
O cromoglicato de sódio, um estabilizador de membrana de mastócito previne a hiperalgesia mecânica
e térmica induzida pelo PCX. (A) Efeito do pré-tratamento com cromoglicato de sódio (30mg/Kg, i.p.)
administrados durante 3 dias consecutivos antes do tratamento com PCX, sobre a hiperalgesia
mecânica. (B) Efeito do pré-tratamento com cromoglicato de sódio durante 3 dias consecutivos antes
do tratamento com PCX na hiperalgesia térmica ao frio. Os valores basais (descritos como “B” nos
gráficos) foram determinados antes da administração de PCX ou veículo. Os dados foram
representados como a média ± E.P.M. para 4-6 animais. *P <0,05: diferença significativa do grupo
tratado com PTX, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.

103
6.7 Quantificação de citocinas e quimiocinas em camundongos SH e camundongos pré-
tratados cromoglicato de sódio e injetados com paclitaxel.
Para confirmar que o aumento de citocinas e quimiocinas era devido à
estimulação dos mastócitos pelo paclitaxel, foi quantificado as citocinas pró-inflamatórias IL-
1β, TNF-α e IL-6 e quimiocinas MCP-1 e KC/CXCL1 no nervo isquiático, nos gânglios da
raiz dorsal e medula espinal de camundongos SH e camundongos selvagens (WT) tratados
com paclitaxel, ou ainda camundongos WT pré-tratados com cromoglicato de sódio e,
posteriormente, injetados compaclitaxel, como mostrado nos gráficos 17, 18 e 19.
6.7.1 Quantificação de citocinas e quimiocinas em nervo isquiático.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) das IL-1β,
TNF-α, IL-6, MCP-1e KC/CXCL1 no nervo isquiático de camundongos WT injetados com
paclitaxel quando comparados com o grupo naive (Gráficos 17A, 17B, 17C, 17D e 17E,
respectivamente). Entretanto, o pré-tratamento com cromoglicato de sódio foi capaz de
prevenir significativamente o aumento destas citocinas e quimiocinas (Gráficos 17A, 17B,
17C, 17D e 17E, respectivamente). Nos camundongos SH tratados com paclitaxel as citocinas
IL-6, IL-1β, TNF-α, MCP-1 e KC/CXCL1 tiveram um aumento moderado, não sendo
observado aumento significativo quando comparado ao grupo naive. Porém, quando
comparados ao grupo de camundongos selvagens tratados com paclitaxel, observa-se uma
redução significativa destas citocinas e quimiocinas (Gráficos 17A, 17B, 17C, 17D e 17E,
respectivamente). Duas horas após a administração de paclitaxel a IL-1β (Gráfico 17A),
começou a aumentar e sua concentração manteve-se alta até 6 horas depois da administração,
pode-se observar que a concentração desta citocina foi reduzinda com o avanço do tempo.
Como ficou demonstrado nos resultados anteriores, apenas com animais WT, que 24 horas
após a administração de paclitaxel a concentração da maioria das citocinas/quimiocinas
retornava aos valores basais, e que neste ponto os animais não mais apresentavam respostas
hiperalgesicas, a partir dessa seção optou-se por não mais avaliar o ponto de 24 horas após a
administração de paclitaxel.
Foi demonstrado que a ausência de mastócitos ou o tratamento com o
cromoglicato de sódio foram eficazes em prevenir o aumento significativo (p<0,05) de IL-1β
(Gráfico 17A) no nervo isquiático com inibição significativa (p<0,05) desta citocina em 2, 4 e
6 horas após o tratamento com paclitaxel. O TNF-α (Gráfico 17B), teve um aumento
significativo (p<0,05) a partir de 2 horas depois da administração de paclitaxel, mantendo-se
em 4 e 6 horas após a administração da droga. Em animais SH ou o tratamento com o

104
cromoglicato de sódio reduziram significativamente (p<0,05) as concentrações de TNF-α
(Gráfico 17B) após a administração de paclitaxel, em todos os pontos avaliados, quando
comparados ao grupo de animais WT. A IL-6 (Gráfico 17C) começou a aumentar no nervo
isquiático, 2 horas depois da administração, entretanto, só foi detectado diferença significativa
(p<0,05) 6 horas após administração do paclitaxel. Pode-se observar que a ausência de
mastócitos ou o tratamento com o cromoglicato de sódio foram eficazes em reduzir
significativamente (p<0,05) as concentrações de IL-6 em todos os pontos avaliados, quando
comparados ao grupo de animais WT. Quanto as quimiocinas MCP-1 (Gráfico 17D) e
KC/CXCL1 (Gráfico 17E), ambas estavam aumentadas significativamente (p<0,05) a partir
de 4 horas após o tratamento com paclitaxel e mantiveram-se altas até 6 horas depois do
tratamento. MCP-1(Gráfico 17D) foi reduzido significativamente (p<0,05), quando
comparado ao grupo de animais WT em 4 e 6 horas quando da ausência de mastócitos ou com
o tratamento com cromoglicato de sódio. A ausência de mastócitos ou o tratamento com o
cromoglicato de sódio foram eficazes em prevenir o aumento significativo (p<0,05) de
KC/CXCL1(Gráfico 17E) a partir de 4 horas após a administração de paclitaxel.
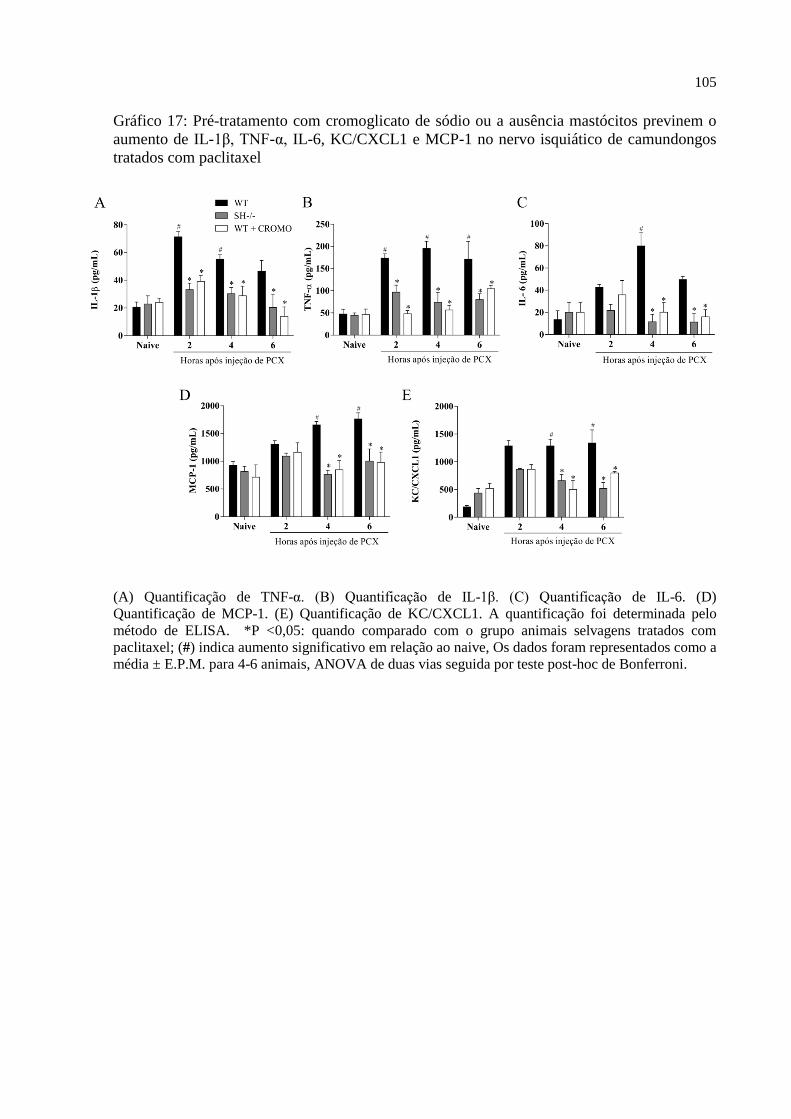
105
Gráfico 17: Pré-tratamento com cromoglicato de sódio ou a ausência mastócitos previnem o
aumento de IL-1β, TNF-α, IL-6, KC/CXCL1 e MCP-1 no nervo isquiático de camundongos
tratados com paclitaxel
(A) Quantificação de TNF-α. (B) Quantificação de IL-1β. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. *P <0,05: quando comparado com o grupo animais selvagens tratados com
paclitaxel; (#) indica aumento significativo em relação ao naive, Os dados foram representados como a
média ± E.P.M. para 4-6 animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.

106
6.7.2 Quantificação de citocinas e quimiocinas em gânglios da raiz dorsal.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) das
citocinas pró-inflamatórias TNF-α e IL-6 e da quimiocina KC/CXCL1, nos gânglios da raiz
dorsal de camundongos selvagens (WT) tratados com paclitaxel em comparação ao grupo
naive (Gráficos 18B, 18C e 18D, respectivamente). Porém, a citocina IL-1β (Gráfico 18A)
não aumentou significativamente. Todavia, o pré-tratamento com cromoglicato de sódio foi
capaz de prevenir de forma significativa (p<0,05) o aumento destas citocinas e da quimiocina
(Gráficos 18B, 18C e 18D, respectivamente). Nos camundongos SH tratados com paclitaxel,
as concentrações de TNF-α, IL-6 e KC/CXCL1 foram moderadas, não sendo observado
aumento significativo quando comparado ao grupo naive. Quando comparados ao grupo de
camundongos WT tratados com pacliitaxel, observa-se que não houve alteração nas
concentrações destas citocinas e quimiocina (Gráficos 18B, 18C e 18E, respectivamente).
Observou-se que no grupo tratado com paclitaxel, houve um aumento
significativo (p<0,05) das citocinas pró-inflamatórias TNF-α e IL-6, após o tratamento, nos
gânglios da raiz dorsal em comparação ao grupo naive (Gráfico 18B e 18C). Neste mesmo
grupo, observou-se ainda aumento significativo da quimiocina KC/CXCL1 (Gráfico 18D)
quando comparados com o grupo naive. Porém a citocina IL-1β não aumentou
significativamente pela administração de paclitaxel (Gráfico 18A). O TNF-α aumentou
significativamente (p<0,05) 6 horas após a administração de paclitaxel, no entanto, a ausência
de mastócitos ou o tratamento com o cromoglicato de sódio foram eficazes em prevenir o
aumento significativo (p<0,05) de TNF-α nos gânglios da raiz dorsal com inibição desta
citocina em 4 e 6 horas após o tratamento com paclitaxel . A IL-6 (Gráfico 18C) teve um
aumento significativo (p<0,05) 6 horas após a administração de paclitaxel, esse aumento foi
observado a partir de 2 horas depois do paclitaxel, quando comparado ao naive. Ficou
demonstrado que a ausência de mastócitos ou o tratamento com o cromoglicato de sódio
foram eficazes em prevenir o aumento significativo (p<0,05) de IL-6 (Gráfico 18C) em todos
os pontos avaliados, quando comparado aos animais WT. Quanto as quimiocina KC/CXCL1
(Gráfico 18D), esta estava aumentada significativamente (p<0,05) a partir de 4 horas após o
tratamento com paclitaxel e manteve-se alta em todos os pontos de tempo avaliados. No
entanto, a ausência de mastócitos ou o tratamento com o cromoglicato de sódio foram eficazes
em prevenir significativamente (p<0,05) aumento de KC/CXCL1(Gráfico 18D) em 4 e 6
horas após a administração de paclitaxel, quando comparados aos animais WT (p<0,05).

107
Gráfico 18: Pré-tratamento com cromoglicato de sódio ou a ausência de mastócitos previnem
o aumento de de IL-1β, TNF-α, IL-6, KC/CXCL1 e MCP-1nos gânglios da raiz de dorsal
camundongos tratados com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de KC/CXCL1. A quantificação foi determinada pelo método de ELISA.*P <0,05:
quando comparado com o grupo animais selvagens tratados com paclitaxel; (#) indica aumento
significativo em relação ao naive, Os dados foram representados como a média ± E.P.M. para 4-6
animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.

108
6.7.3 Quantificação de citocinas e quimiocinas em medula espinal.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) das
citocinas pró-inflamatórias IL-1β, TNF-α e IL-6 e da quimiocina KC/CXCL1, na medula
espinal de camundongos WT tratados com paclitaxel em comparação ao grupo naive (p<0,05)
(Gráficos 19A, 19B, 19C e 19E, respectivamente). Contudo, o pré-tratamento com
cromoglicato de sódio foi capaz de prevenir de forma significativa (p<0,05) o aumento de IL-
1β, TNF-α, IL-6 e KC/CXCL1 (Gráficos 19A, 19B, 19C e 19E, respectivamente). Porém
MCP-1 não foi alterado significativamente (Gráficos 19D). Nos camundongos SH tratados
com paclitaxel, IL-1β, TNF-α, IL-6 e KC/CXCL1 tiveram um aumento moderado, não sendo
observado aumento significativo quando comparado ao grupo naive. Já quando comparados
ao grupo de camundongos WT trattados com paclitaxel, observa-se que não houve alterações
nas concentrações destas citocinas e quimiocina (Gráficos 19A, 19B, 19C e 19E,
respectivamente). Já o MCP-1 não foi alterado.
IL-1β (Gráfico 19A) aumentou significativamente (p<0,05) a partir de 2 horas
após o tratamento com paclitaxel, e se mantive alterado até 6 horas. Observa-se que nos
animais que não possuíam mastócitos ou nos animais WT previamente tratados com
cromoglicato de sódio, a concentração de IL-1β foi reduzida significativamente (p<0,05) em
todos os pontos avaliados quando com parados aos animais WT sem tratamento prévio. TNF-
α (Gráfico 19B) aumentou moderadamente 2 horas após o tratamento com paclitaxel, no
entanto, um aumento significativo (p<0,05) só é observado 4 horas após a administração,
mantendo-se até 6 horas. No entanto a ausência de mastócito ou o pré-tratamento com
cromoglicato de sódio reduziu significativamente (p<0,05) a concentração de TNF-α nos
pontos analisados. IL-6 (Gráfico 19C) estava aumentada significativamente (p< 0,05) na
medula espinal de camundongos tratados com paclitaxel 4 horas após o tratamento, mantendo
esse aumento até 6 horas em camundongos WT, mas observa-se que na ausência de mastócito
esta citocina não se altera e quando os animais são pré-tratados com cromoglicato de sódio
observa-se uma redução significativamente (p<0,05) de IL-6 quando comparados ao grupo de
animais WT. Já a quimiocina MCP-1, não aumentado após o tratamento com paclitaxel.
Porém, KC/CXCL1 aumentou significativamente (p<0,05) 4 horas depois do tratamento com
paclitaxel em animais WT. Verificou-se que a ausência de mastócitos ou o pré-tratamento
com cromoglicato de sódio reduziu significativamente (p<0,05) KC/CXCL1 na medula
espinal quando comparados aos animais do grupo WT.

109
Gráfico 19: Pré-tratamento com cromoglicato de sódio ou a ausência de mastócitos previne o
aumento de IL-1β, TNF-α IL-6 e KC/CXCL1 na medula espinal de camundongos tratados
com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de KC/CXCL1. A quantificação foi determinada pelo método de ELISA. *P <0,05:
quando comparado com o grupo animais selvagens tratados com paclitaxel; (#) indica aumento
significativo em relação ao naive, Os dados foram representados como a média ± E.P.M. para 4-6
animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.
Os dados apresentados nessa seção demonstram que a ausência de mastócito ou o
pré-tratamento com cromoglicato de sódio, um estabilizador de membrana de mastócito,
previnem o aumento de citocinas/quimiocinas quantificados pelo método de ELISA no nervo
isquiático, gânglios da raiz dorsal e medula espinal de camundongos injetados com PCX. O
resumo dos dados apresentados podem ser obervado em conjunto no quadro 3.

110
Quadro 3: Papel dos mastócitos na produção/liberação de citocinas e/ou quimiocinas, no
nervo isquiático, gânglios da raiz dorsal e na medula espinal de camundongos injetados com
paclitaxel
Resultados da redução dos níveis de citocinas/quimiocinas no nervo isquiático, gânglios da raiz dorsal
e medula espinal de animais tratados com paclitaxel (PCX). Quantificação realizada pelo método de
ELISA. ( + ) Aumento significativo em relação ao naive. ( - ) não houve aumento significativo em
relação ao naive. ( < ) diminuição significativa em relação ao WT. ( na ) Não Avaliado.
6.8 Quantificação de citocinas e quimiocinas em cultura de mastócitos estimulada com
paclitaxel.
Com intuito de confirmar que a ativação dos mástócitos é responsável, pelo
menos em parte, pela produção/liberação de citocinas e quimiocinas na dor aguda induzida
pelo paclitaxel, foram quantificados as citocinas pró-inflamatórias IL-1β, TNF-α e IL-6 e
quimiocina MCP-1 em cultura de mastócitos (RBL-2H3) estimulada com paclitaxel, como
mostrado no gráfico 20.
A estimulação com paclitaxel provocou aumento significativo (p<0,05) de TNF-α,
IL-6 e MCP-1, em cultura de mastócitos (RBL-2H3), quando comparados com o grupo naive
(Gráficos 20A, 20B e 20C, respectivamente). Quando estimulados com diferentes doses de
paclitaxel, mastócitos em cultura liberam/produzem mediadores. Observa-se que TNF-α
(Gráfico 20A) é aumentado significativamente (P<0,05) com a dose de 30µM de paclitaxel,
apesar das outras doses utilizadas (10 e 90µM) também terem aumento significativamente
(p<0,05) esta citocina no sobrenadante da cultura em comparação com o meio sem o estímulo.
Cultura de mástocito também libera IL-6 (Gráfico 20B) quando estimulada com paclitaxel.

111
Observa-se que esta citocina alcançou seu pico com a utilização da dose de 30µM de
paclitaxel, apesar de se observar também diferença siginificativa (p<0,05) com a utilização de
outras doses (10 e 90µM), quando comparado ao meio sem estímulo. Em relação à
produção/liberação de MCP-1, a estimulação de mastócitos em cultura com paclitaxel,
aumentada significativamente (p<0,05) esta quimiocina nas doses de 10 e 30µM, quando
comparado ao meio sem estímulo. A dose de 90µM também foi capaz de aumentar
significativamente (p<0,05) MCP-1 no sobrenadante de mastócitos estimulados com
paclitaxel, no entanto, este valor foi menor quando comparado as outras doses utilizadas.
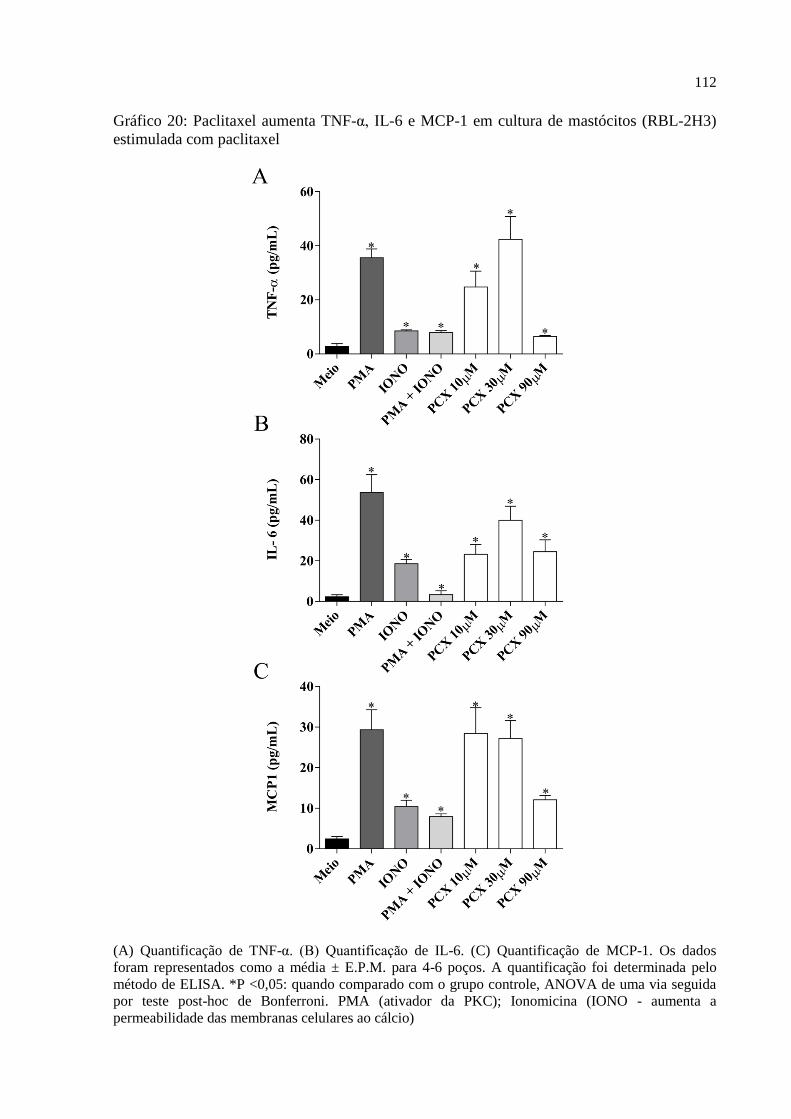
112
Gráfico 20: Paclitaxel aumenta TNF-α, IL-6 e MCP-1 em cultura de mastócitos (RBL-2H3)
estimulada com paclitaxel
(A) Quantificação de TNF-α. (B) Quantificação de IL-6. (C) Quantificação de MCP-1. Os dados
foram representados como a média ± E.P.M. para 4-6 poços. A quantificação foi determinada pelo
método de ELISA. *P <0,05: quando comparado com o grupo controle, ANOVA de uma via seguida
por teste post-hoc de Bonferroni. PMA (ativador da PKC); Ionomicina (IONO - aumenta a
permeabilidade das membranas celulares ao cálcio)

113
6.9 Imunofluorescência para Triptase e IL-6 em gânglio da raiz dorsal de camundongos
submetidos à injeção de paclitaxel
Na imunofluorescência para triptase e IL-6, no gânglio da raiz dorsal, foi
observado um aumento da imunoexpressão tanto de triptase quanto de IL-6, quando
comparado ao grupo naive 4 horas após a administração de paclitaxel (Figura 11). No entanto,
6 horas após a administração de paclitaxel houve uma redução da imunoexpressão para esses
marcadores quando comparado com o grupo tratado com paclitaxel 4 horas após o tratamento.
Na análise quantitativa da imunoexpressão para Triptase, a quantificação da área
marcada, comparando com o grupo naive, houve um aumento significativo (p<0,05) 4 horas
após a administração de paclitaxel (Gráfico 21A). Em relação a quantificação da área marcada
de IL-6, também observou-se um aumento significativo (p<0,05) 4 horas após a administração
de paclitaxel (Gráfico 21B).

114
Figura 11: Fotomicrografia da imunoexpressão de Triptase e IL-6 no gânglio da raiz dorsal de
camundongos submetidos à administração de paclitaxel
Verde: Triptase (marcador de mastócitos); vermelho: IL-6; amarelo: fusão (MERGE); A, B e C: Naive
normal, sem tratamento; D, E, F: Paclitaxel 4h; G, H e I: Paclitaxel 6 h. Setas brancas: marcação para
IL-6 em células não neuronais do DRG. Cabeça de seta amarela: marcação para IL-6 em células não
neuronais do GRD. Setas amarelas: co-marcação IL-6 e Triptase em células não neuronais do GRD.
Foto representativa de 1 animal por grupo experimental. Aumento de 200x
Naive
PCX-4h
PCX-6h
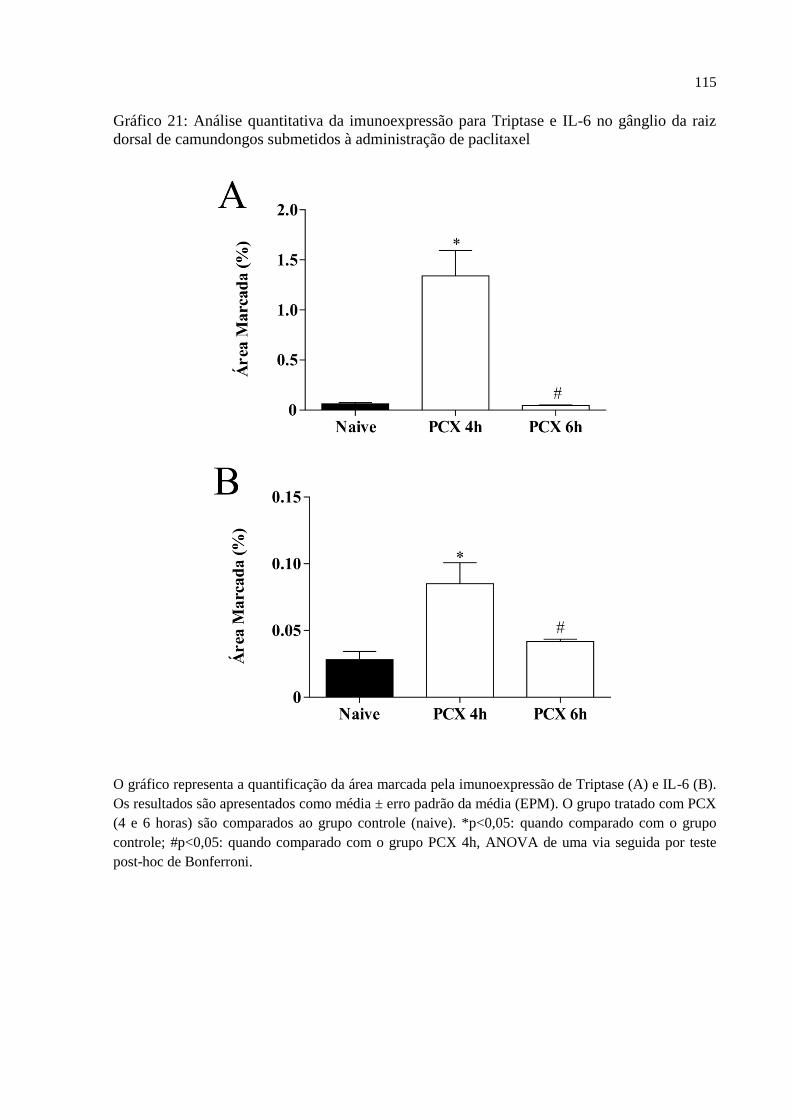
115
Gráfico 21: Análise quantitativa da imunoexpressão para Triptase e IL-6 no gânglio da raiz
dorsal de camundongos submetidos à administração de paclitaxel
O gráfico representa a quantificação da área marcada pela imunoexpressão de Triptase (A) e IL-6 (B).
Os resultados são apresentados como média ± erro padrão da média (EPM). O grupo tratado com PCX
(4 e 6 horas) são comparados ao grupo controle (naive). *p<0,05: quando comparado com o grupo
controle; #p<0,05: quando comparado com o grupo PCX 4h, ANOVA de uma via seguida por teste
post-hoc de Bonferroni.

116
6.10 Investigação da participação de TLR4 na hiperalgesia mecânica e térmica induzida
pelo paclitaxel.
Sabe-se que o paclitaxel provoca dor aguda através da ativação dos receptores
TLR4. De posse dessa evidencia, foi avaliada a participação desse receptor na hiperalgesia
induzida pelo paclitaxel, através do emprego de animais knockout para o receptor TLR4
(TLR4-/-). Verificou-se que as respostas hiperalgesicas mecânica e térmica ao frio
começaram dentro de 2 horas e atingiram o pico entre 4 e 6 horas, se resolvendo 24 horas
após a administração do paclitaxel (Gráficos 22A e 22B, respectivamente). Enquanto isso, o
limiar de retirada da pata a estímulos mecânicos do grupo controle permaneceu inalterado,
assim como o comportamento nocifensivo ao frio. Entretanto, em 24h após a administração
única de paclitaxel não foi observado o desenvolvimento de respostas hiperalgesicas mecânica
e térmica ao frio (Gráficos 22A e 22B, respectivamente) em animais TLR4-/-.
O tratamento de animais knockout para o receptor TLR4 (TLR4-/-) com paclitaxel
não foi capaz de desenvolver hiperalgesia mecânica e térmica nesses animais. Isto pode ser
observado pela redução significativa (p<0,05) das respostas hiperalgesicas nos animais com
deleção gênica para este receptor. Essa inibição das respostas hipernociceptiva mecânica e
térmica foi observado durante todo o período de avaliação, quando comparado aos animais
WT. A delação desse receptor, portanto foi eficaz na prevenção do desenvolvimento de
respostas hiperalgesicas ao paclitaxel em camundongos (Gráfico 22A e 22B).
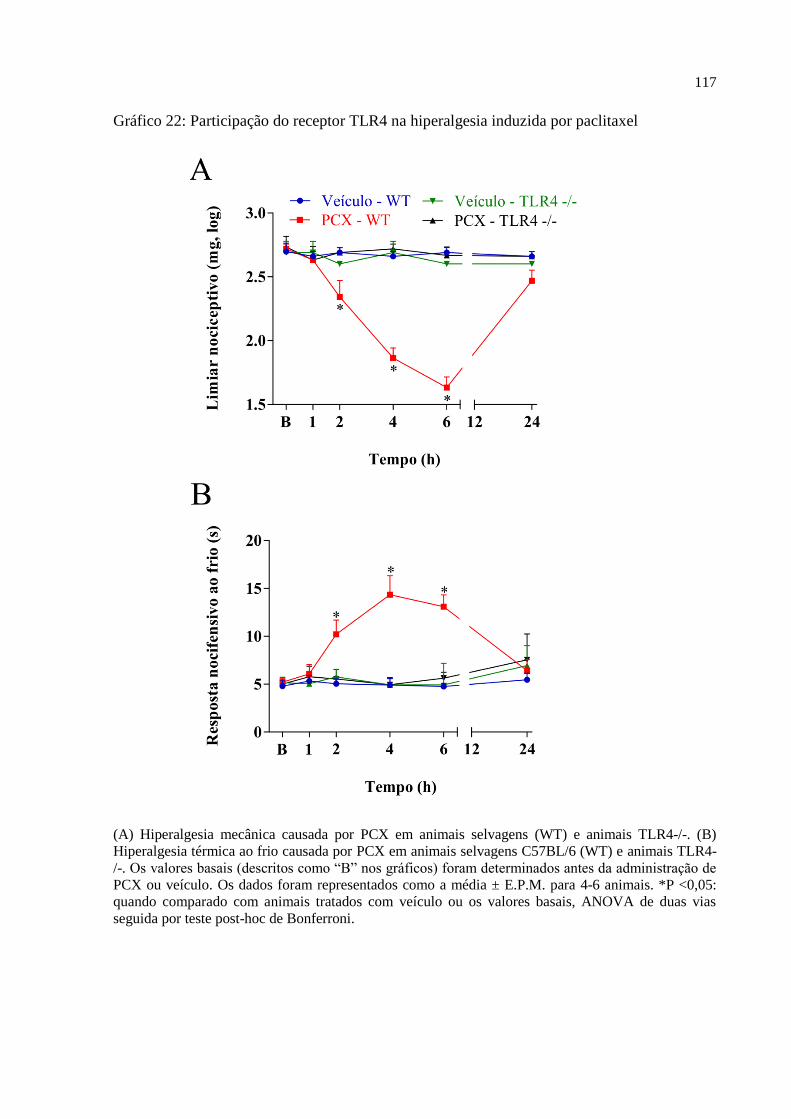
117
Gráfico 22: Participação do receptor TLR4 na hiperalgesia induzida por paclitaxel
(A) Hiperalgesia mecânica causada por PCX em animais selvagens (WT) e animais TLR4-/-. (B)
Hiperalgesia térmica ao frio causada por PCX em animais selvagens C57BL/6 (WT) e animais TLR4-
/-. Os valores basais (descritos como “B” nos gráficos) foram determinados antes da administração de
PCX ou veículo. Os dados foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05:
quando comparado com animais tratados com veículo ou os valores basais, ANOVA de duas vias
seguida por teste post-hoc de Bonferroni.

118
6.11 Quantificação de citocinas e quimiocinas em camundongos TLR4-/- tratados com
paclitaxel.
A ativação dos receptores do tipo toll 4 (TLR4), causa dor aguda induzida por
paclitaxel. Então foi avaliado se ativação desse receptor aumenta a expressão de citocinas ou
quimiocinas em camundongos tratados com paclitaxel. Para isso foi administrado em
camundongos TLR4-/- e em camundongos WT dose única de paclitaxel e foi verificado se à
droga provocava aumento de IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL1 no plasma, nervo
isquiático, gânglios da raiz dorsal e medula espinal nessa linhagem de camundongos, os
valores estão representados nos gráficos 23, 24, 25 e 26.
6.11.1 Quantificação de citocinas e quimiocinas no plasma.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) de IL-1β,
TNF-α e IL-6 em camundongos WT tratados com paclitaxel quando comparados com o grupo
naive (Gráficos 23A, 23B e 23C). Da mesma forma, foi observado aumento significativo
(p<0,05) da quimiocina KC/CXCL1 (Gráfico 23D) quando comparados com o grupo naive.
Entretanto, nos camundongos TLR4-/- tratados com paclitaxel as concentrações
de TNF-α, IL-6 e KC/CXCL1 encontraram-se moderadas, não sendo observado aumento
significativo quando comparado ao grupo naive. E quando comparados ao grupo de
camundongos WT tratados com paclitaxel, observa-se significativa redução (p<0,05) destas
citocinas e quimiocina, quatro horas após a administração de paclitaxel (Gráficos 23A e 23B).
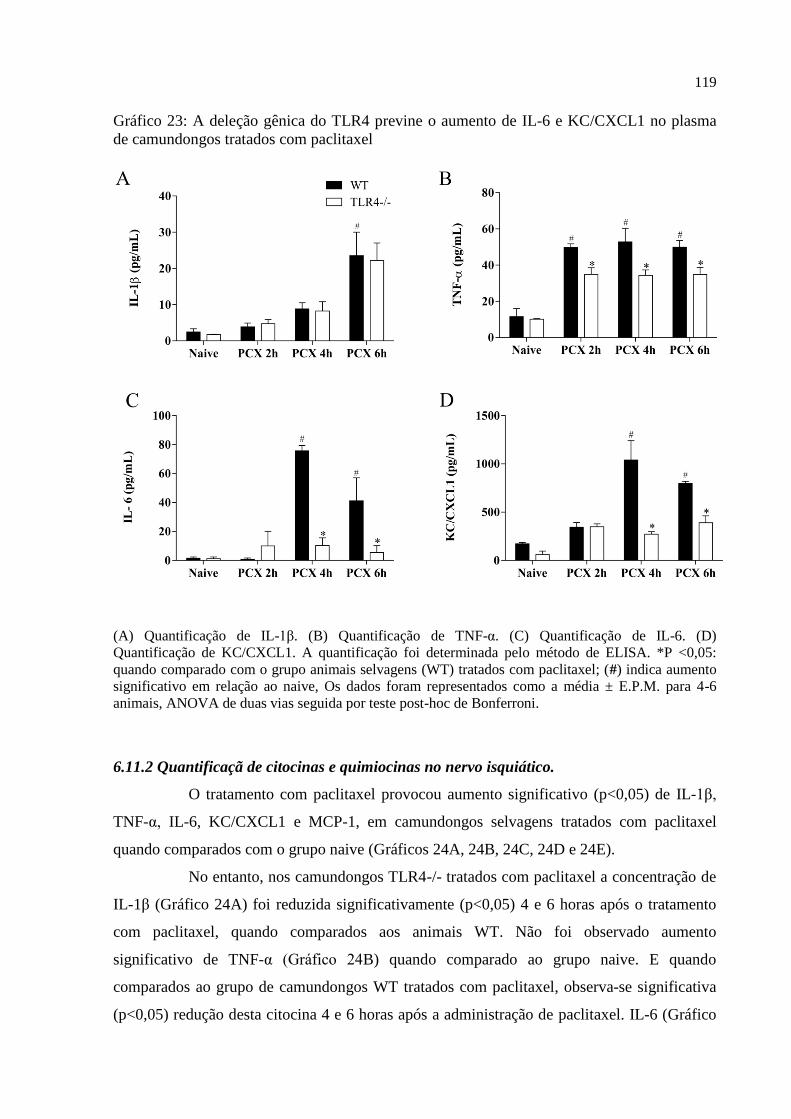
119
Gráfico 23: A deleção gênica do TLR4 previne o aumento de IL-6 e KC/CXCL1 no plasma
de camundongos tratados com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de KC/CXCL1. A quantificação foi determinada pelo método de ELISA. *P <0,05:
quando comparado com o grupo animais selvagens (WT) tratados com paclitaxel; (#) indica aumento
significativo em relação ao naive, Os dados foram representados como a média ± E.P.M. para 4-6
animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.
6.11.2 Quantificaçã de citocinas e quimiocinas no nervo isquiático.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) de IL-1β,
TNF-α, IL-6, KC/CXCL1 e MCP-1, em camundongos selvagens tratados com paclitaxel
quando comparados com o grupo naive (Gráficos 24A, 24B, 24C, 24D e 24E).
No entanto, nos camundongos TLR4-/- tratados com paclitaxel a concentração de
IL-1β (Gráfico 24A) foi reduzida significativamente (p<0,05) 4 e 6 horas após o tratamento
com paclitaxel, quando comparados aos animais WT. Não foi observado aumento
significativo de TNF-α (Gráfico 24B) quando comparado ao grupo naive. E quando
comparados ao grupo de camundongos WT tratados com paclitaxel, observa-se significativa
(p<0,05) redução desta citocina 4 e 6 horas após a administração de paclitaxel. IL-6 (Gráfico

120
24C) foi reduzido significativamente (p<0,05) em camundongos TLR4-/-, 2 e 6 horas após o
tratamento com paclitaxel, quando comparados aos animais WT. A quimiocina MCP-1
(Gráfico 24D) aumentou significativamente (p<0,05) no grupo de animais WT tratados com
paclitaxel. No entanto, em camundongos TLR4-/- foi observado uma redução
significativamente (p<0,05) desta quimiocina 4 e 6 horas após a administração de paclitaxel,
quando comparados aos animais WT. Já KC/CXCL1 (Gráfico 24E) foi significativamente
(p<0,05) reduzida apenas 6 horas após o tratamento com paclitaxel, quando comparados com
camundongos WT.
Gráfico 24: A deleção gênica do TLR4 previne o aumento de IL-1β, IL-6, MCP-1 e
KC/CXCL1 no nervo isquiático de camundongos tratados com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. Os dados foram representados como a
média ± E.P.M. para 4-6 animais. A quantificação foi determinada pelo método de ELISA. *P <0,05:
quando comparado com o grupo animais selvagens (WT) tratados com paclitaxel; (#) indica aumento
significativo em relação ao naive, Os dados foram representados como a média ± E.P.M. para 4-6
animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.
6.11.3 Quantificação de citocinas e quimiocinas nos gânglios da raiz dorsal.
O tratamento com paclitaxel provocou aumento significativo (p<0,05) de TNF-α,
IL-6 e KC/CXCL1, em camundongos WT tratados com paclitaxel quando comparados com o
grupo naive (Gráficos 25B, 25C e 25E, respectivamente).
No entanto, nos camundongos TLR4-/- tratados com paclitaxel não foi observado
aumento significativo de TNF-α (Gráfico 25B) quando comparado ao grupo naive. Quando
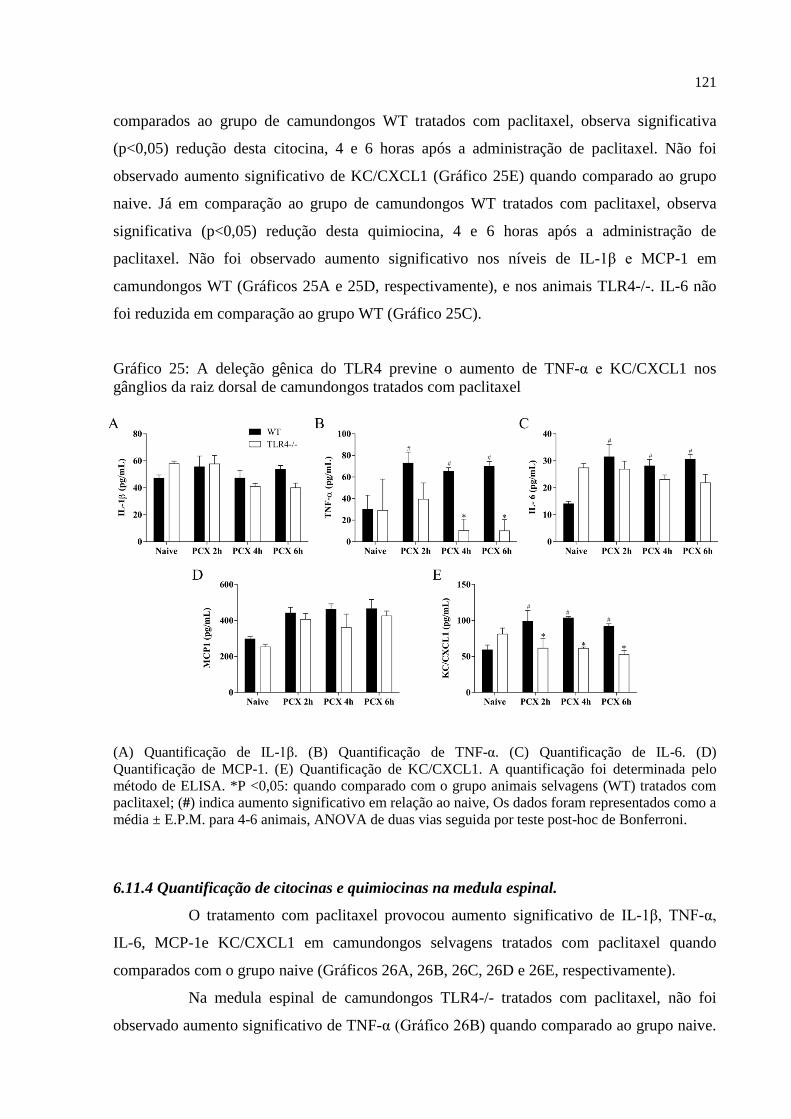
121
comparados ao grupo de camundongos WT tratados com paclitaxel, observa significativa
(p<0,05) redução desta citocina, 4 e 6 horas após a administração de paclitaxel. Não foi
observado aumento significativo de KC/CXCL1 (Gráfico 25E) quando comparado ao grupo
naive. Já em comparação ao grupo de camundongos WT tratados com paclitaxel, observa
significativa (p<0,05) redução desta quimiocina, 4 e 6 horas após a administração de
paclitaxel. Não foi observado aumento significativo nos níveis de IL-1β e MCP-1 em
camundongos WT (Gráficos 25A e 25D, respectivamente), e nos animais TLR4-/-. IL-6 não
foi reduzida em comparação ao grupo WT (Gráfico 25C).
Gráfico 25: A deleção gênica do TLR4 previne o aumento de TNF-α e KC/CXCL1 nos
gânglios da raiz dorsal de camundongos tratados com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. *P <0,05: quando comparado com o grupo animais selvagens (WT) tratados com
paclitaxel; (#) indica aumento significativo em relação ao naive, Os dados foram representados como a
média ± E.P.M. para 4-6 animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.
6.11.4 Quantificação de citocinas e quimiocinas na medula espinal.
O tratamento com paclitaxel provocou aumento significativo de IL-1β, TNF-α,
IL-6, MCP-1e KC/CXCL1 em camundongos selvagens tratados com paclitaxel quando
comparados com o grupo naive (Gráficos 26A, 26B, 26C, 26D e 26E, respectivamente).
Na medula espinal de camundongos TLR4-/- tratados com paclitaxel, não foi
observado aumento significativo de TNF-α (Gráfico 26B) quando comparado ao grupo naive.
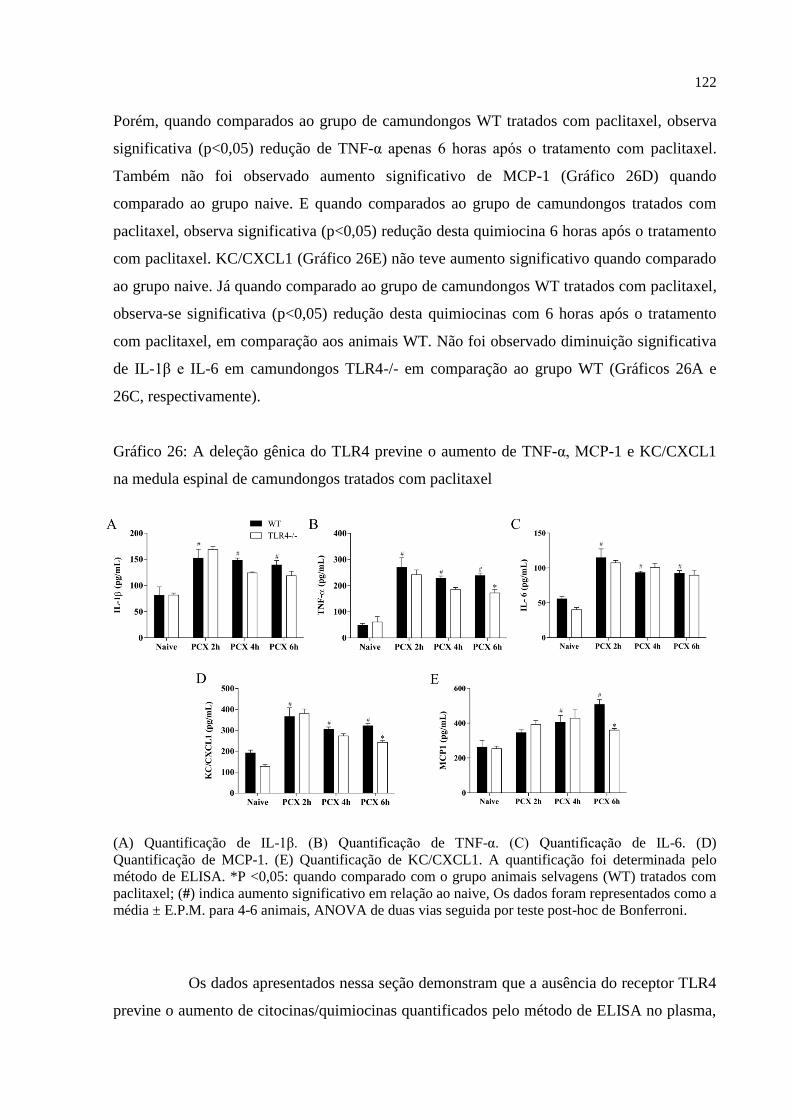
122
Porém, quando comparados ao grupo de camundongos WT tratados com paclitaxel, observa
significativa (p<0,05) redução de TNF-α apenas 6 horas após o tratamento com paclitaxel.
Também não foi observado aumento significativo de MCP-1 (Gráfico 26D) quando
comparado ao grupo naive. E quando comparados ao grupo de camundongos tratados com
paclitaxel, observa significativa (p<0,05) redução desta quimiocina 6 horas após o tratamento
com paclitaxel. KC/CXCL1 (Gráfico 26E) não teve aumento significativo quando comparado
ao grupo naive. Já quando comparado ao grupo de camundongos WT tratados com paclitaxel,
observa-se significativa (p<0,05) redução desta quimiocinas com 6 horas após o tratamento
com paclitaxel, em comparação aos animais WT. Não foi observado diminuição significativa
de IL-1β e IL-6 em camundongos TLR4-/- em comparação ao grupo WT (Gráficos 26A e
26C, respectivamente).
Gráfico 26: A deleção gênica do TLR4 previne o aumento de TNF-α, MCP-1 e KC/CXCL1
na medula espinal de camundongos tratados com paclitaxel
(A) Quantificação de IL-1β. (B) Quantificação de TNF-α. (C) Quantificação de IL-6. (D)
Quantificação de MCP-1. (E) Quantificação de KC/CXCL1. A quantificação foi determinada pelo
método de ELISA. *P <0,05: quando comparado com o grupo animais selvagens (WT) tratados com
paclitaxel; (#) indica aumento significativo em relação ao naive, Os dados foram representados como a
média ± E.P.M. para 4-6 animais, ANOVA de duas vias seguida por teste post-hoc de Bonferroni.
Os dados apresentados nessa seção demonstram que a ausência do receptor TLR4
previne o aumento de citocinas/quimiocinas quantificados pelo método de ELISA no plasma,
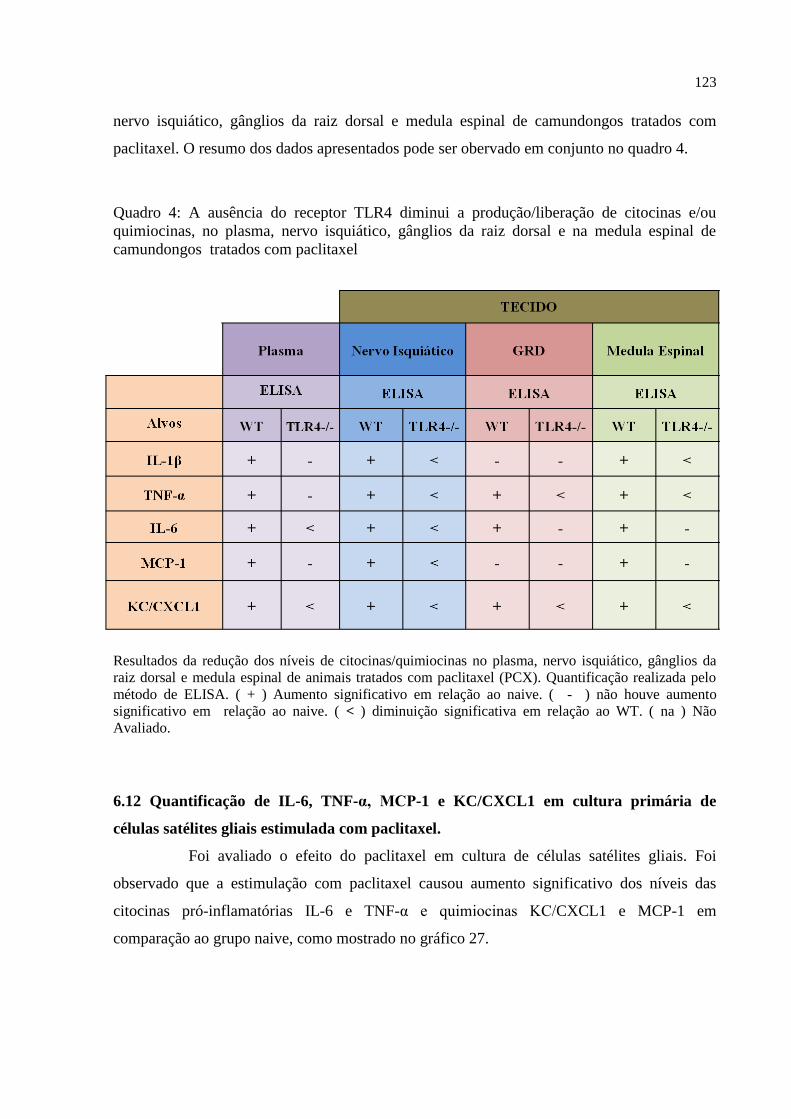
123
nervo isquiático, gânglios da raiz dorsal e medula espinal de camundongos tratados com
paclitaxel. O resumo dos dados apresentados pode ser obervado em conjunto no quadro 4.
Quadro 4: A ausência do receptor TLR4 diminui a produção/liberação de citocinas e/ou
quimiocinas, no plasma, nervo isquiático, gânglios da raiz dorsal e na medula espinal de
camundongos tratados com paclitaxel
Resultados da redução dos níveis de citocinas/quimiocinas no plasma, nervo isquiático, gânglios da
raiz dorsal e medula espinal de animais tratados com paclitaxel (PCX). Quantificação realizada pelo
método de ELISA. ( + ) Aumento significativo em relação ao naive. ( - ) não houve aumento
significativo em relação ao naive. ( < ) diminuição significativa em relação ao WT. ( na ) Não
Avaliado.
6.12 Quantificação de IL-6, TNF-α, MCP-1 e KC/CXCL1 em cultura primária de
células satélites gliais estimulada com paclitaxel.
Foi avaliado o efeito do paclitaxel em cultura de células satélites gliais. Foi
observado que a estimulação com paclitaxel causou aumento significativo dos níveis das
citocinas pró-inflamatórias IL-6 e TNF-α e quimiocinas KC/CXCL1 e MCP-1 em
comparação ao grupo naive, como mostrado no gráfico 27.

124
Gráfico 27: Paclitaxel aumenta IL-6, TNF-α, MCP-1 e KC/CXCL1 em cultura primária de
células satélites gliais
Paclitaxel aumenta os níveis de IL-6, TNF-α, MCP-1e KC/CXCL1 em cultura primária de células
satélites gliais. A quantificação foi determinada pelo método de ELISA. (A) Quantificação de TNF-α.
(B) Quantificação de IL-6. (C) Quantificação de MCP-1. (D) Quantificação de KC/CXCL1. Os dados
foram representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando comparado com o
grupo controle, ANOVA de uma via seguida por teste post-hoc de Bonferroni.
Adicionalmente foi realizada a dosagem de citocinas/quimiocinas em cultura de
células satélites gliais de camundongos knockout para os receptores do tipo toll (TLR2-/- e
TLR4-/-). Verificou-se que a estimulação com paclitaxel causou aumento significativo das
citocinas IL-6, IL-1β e TNF-α e quimiocinas KC/CXCL1 e MCP-1 em cultura de células
satélites gliais de camundongos selvagens (WT) em comparação ao grupo naive, como
mostrado no gráfico 28. No entanto a deleção gênica para o receptor TLR4 foi capaz de
impedir o aumento de citocinas/quimiocinas em cultura de células satélites gliais, essa
inibição não foi observada nos animais deficientes para TLR2.

125
Gráfico 28: A deleção gênica do receptor TLR4 previne o aumento de IL-6, MCP-1 e
KC/CXCL1 em cultura primária de células satélites gliais estimulada com paclitaxel
A deleção gênica do receptor TLR4 previne o aumento dos níveis de IL-6, TNF-α, MCP1 e
KC/CXCL1 em cultura primária de células satélites gliais. A quantificação foi determinada pelo
método de ELISA. (A) Quantificação de TNF-α. (B) Quantificação de IL-6. (C) Quantificação de
MCP-1. (D) Quantificação de KC/CXCL1. Os dados foram representados como a média ± E.P.M.
para 4-6 animais. *P <0,05: quando comparado com o grupo controle, ANOVA de uma via seguida
por teste post-hoc de Bonferroni.
Para confirmar se a deleção do gene para o receptor TLR4 era capaz de prevenir o
aumento dos níveis de citocinas/quimiocinas em cultura de células satélites gliais de GRD de
camundongos KO para os receptores TLR4, utilizou-se um agonista clássico desse receptor, o
LPS. Verificou-se que tanto a estimulação com LPS quanto com o paclitaxel causaram
aumento significativo das citocinas IL-6, IL-1β e TNF-α e quimiocinas KC/CXCL1 e MCP-1
em camundongos selvagens (WT) tratados com paclitaxel quando comparados ao grupo
naive, como mostrado no gráfico 29. Entretanto a deleção gênica para o receptor TLR4 foi
capaz de impedir o aumento de citocinas/quimiocinas nessa cultura. Em conjunto esses dados
sugerem que o paclitaxel pode sensibilizar os neurônios sensitivos através do aumento dos
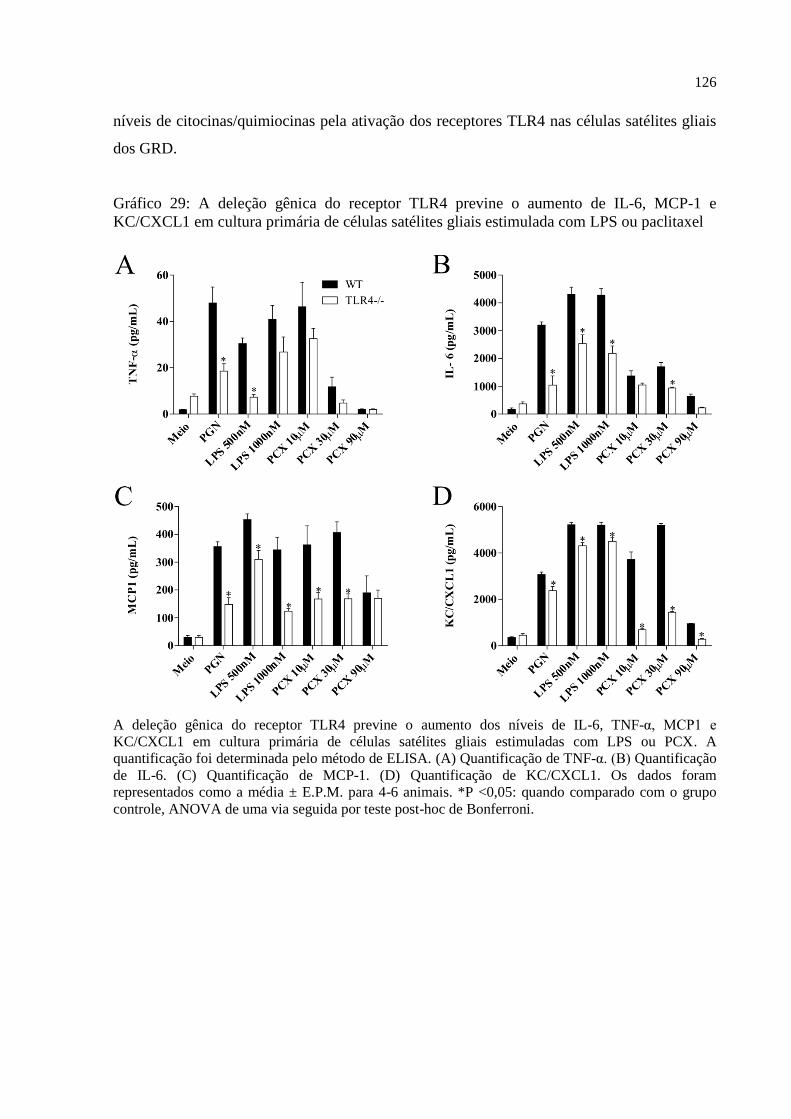
126
níveis de citocinas/quimiocinas pela ativação dos receptores TLR4 nas células satélites gliais
dos GRD.
Gráfico 29: A deleção gênica do receptor TLR4 previne o aumento de IL-6, MCP-1 e
KC/CXCL1 em cultura primária de células satélites gliais estimulada com LPS ou paclitaxel
A deleção gênica do receptor TLR4 previne o aumento dos níveis de IL-6, TNF-α, MCP1 e
KC/CXCL1 em cultura primária de células satélites gliais estimuladas com LPS ou PCX. A
quantificação foi determinada pelo método de ELISA. (A) Quantificação de TNF-α. (B) Quantificação
de IL-6. (C) Quantificação de MCP-1. (D) Quantificação de KC/CXCL1. Os dados foram
representados como a média ± E.P.M. para 4-6 animais. *P <0,05: quando comparado com o grupo
controle, ANOVA de uma via seguida por teste post-hoc de Bonferroni.

127
6.13 Modelo conceitual
Basedo nos resultados obtidos neste estudo, os seguintes mecanimos são
propostos: O paclitaxel se liga aos receptores TLR4 presentes em mastócitos na periferia,
estimulado essas células a produzirem/liberam mediadores pró-inflamatórios (TNF-α, IL-1β,
IL-6, MCP-1 e KC/CXCL1) que sensibilizam o nociceptor. Nos gânglios da raiz dorsal o
paclitaxel que se liga a receptores TLR4 em células satélites gliais, ativa essas células que
liberam mediadores (TNF-α, IL-6 e KC/CXCL1) sensibilizando neurônios sensoriais.
Adicionalmente mastócitos presentes nesses gânglios contribuem para a sensibilização dos
neuronios através da liberação de mediadores pró-inflamatórios (TNF-α, IL-6 E KC/CXCL1).
Além disso, no corno dorsal da medula espinal ocorre ainda um aumento da liberação de
mediadores pró-inflamatórios (TNF-α, IL-1Β, IL-6, MCP-1 e KC/CXCL1). Esses mediadores
produzindos pelos mastócitos e células satélites gliais contribuem então para o
desenvolvimento de dor aguda induzida por paclitaxel (Figura 12).
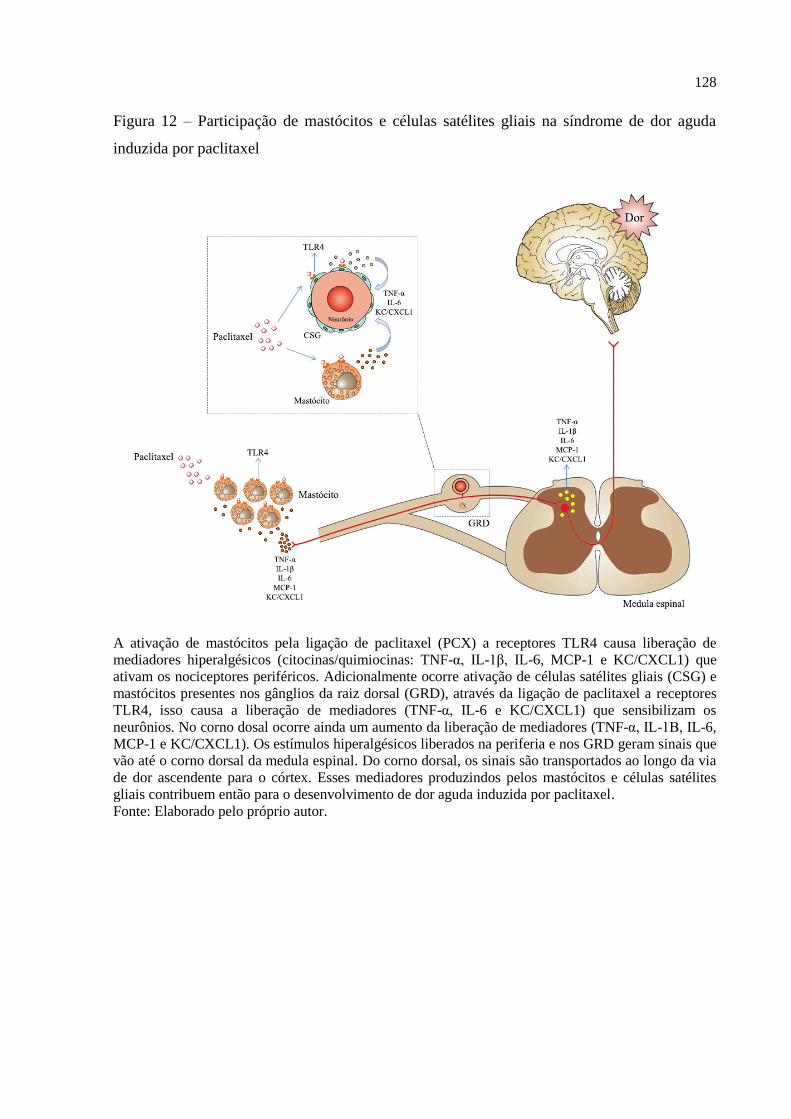
128
Figura 12 – Participação de mastócitos e células satélites gliais na síndrome de dor aguda
induzida por paclitaxel
A ativação de mastócitos pela ligação de paclitaxel (PCX) a receptores TLR4 causa liberação de
mediadores hiperalgésicos (citocinas/quimiocinas: TNF-α, IL-1β, IL-6, MCP-1 e KC/CXCL1) que
ativam os nociceptores periféricos. Adicionalmente ocorre ativação de células satélites gliais (CSG) e
mastócitos presentes nos gânglios da raiz dorsal (GRD), através da ligação de paclitaxel a receptores
TLR4, isso causa a liberação de mediadores (TNF-α, IL-6 e KC/CXCL1) que sensibilizam os
neurônios. No corno dosal ocorre ainda um aumento da liberação de mediadores (TNF-α, IL-1Β, IL-6,
MCP-1 e KC/CXCL1). Os estímulos hiperalgésicos liberados na periferia e nos GRD geram sinais que
vão até o corno dorsal da medula espinal. Do corno dorsal, os sinais são transportados ao longo da via
de dor ascendente para o córtex. Esses mediadores produzindos pelos mastócitos e células satélites
gliais contribuem então para o desenvolvimento de dor aguda induzida por paclitaxel.
Fonte: Elaborado pelo próprio autor.

129
7. DISCUSSÃO

130
Os resultados do presente estudo mostraram o envolvimento de citocinas como
TNF-α, IL-1β, IL-6, MCP-1 e KC/CXCL1, assim como do receptor TLR4 na hiperalgesia
induzida por paclitaxel, quando o mesmo é injetado de forma sistêmica em uma única dose,
mimetizando o que acontece na clínica. Mastócitos aparecem como células que participam
desse mecanismo, provavelmente liberando grande parte desses mediadores investigados. A
injeção intravenosa de paclitaxel induziu respostas hiperalgesicas mecânica e térmicas,
evidenciando que o paclitaxel diminui o limiar de resposta a dor quando esses animais são
testados com estímulos mecânicos e térmicos de frio. As respostas hiperalgésicas começaram
dentro de 2 horas e atingiram o pico entre 4 e 6 horas, se resolvendo 24 horas após a
administração do paclitaxel.
As evidências clínicas mostram que o tratamento com este quimioterápico
frequentemente leva o paciente a experimentar quadros de dor aguda, o que reduz
significativamente a sua qualidade de vida. A dor induzida pelo paclitaxel inclui dor que
ocorre imediatamente após o tratamento com este fármaco, conhecida como síndrome da dor
aguda associada ao paclitaxel (REEVES et al., 2012; LOPRINZI et al., 2011) e
posteriormente desenvolve-se a neuropatia periférica que pode ser uma consequência
devastadora em longo prazo (ARGYRIOU, 2009).
A síndrome da dor aguda associada ao paclitaxel é uma morbidade significativa
em pacientes que receberam tratamento com esse quimioterápico, atingindo cerca de 88% dos
pacientes tratados com este quimioterápico em doses altas (> 175mg/m2), desenvolvendo-se
logo após o primeiro ciclo de tratamento (REEVES et al., 2012; LOPRINZI et al., 2011). A
síndrome da dor aguda associada ao paclitaxel ocorre geralmente dentro de 1-3 dias após a
administração da droga e se resolve dentro de 7 dias. A importância dada a síndrome da dor
aguda associada ao paclitaxel aumentou quando foi demonstrado que a sua intensidade pode
estar ligada a severidade da neuropatia crônica por paclitaxel. (LOPRINZI et al., 2011;
REEVES et al., 2012). Em um estudo clinico que avaliou o aparecimento de dor em pacientes
após o primeiro ciclo do tratamento com o quimioterápico paclitaxel verificou-se que
pacientes com altos escores de dor aguda tinham uma baixa recuperação da neuropatia crônica
induzida por esse fármaco (REEVES et al., 2012).
Os dados do presente estudo encontram-se de acordo com os resultados obtidos
por Yan e colaboradores (2015). Os autores também observaram que o paclitaxel induzia dor
aguda em roedores com inicio e duração mais curtos do que aqueles descritos para seres
humanos. No entanto, outros modelos de síndrome da dor aguda induzida pelo paclitaxel são
propostos, no qual os autores observaram que o desenvolvimento da dor aguda ocorre apenas

131
24 a 72 horas após a administração do quimioterápico. Porém nestes casos foram utilizados
ratos Wistar e a injeção de paclitaxel foi realizada por via intraperitoneal e não intravenosa, e
também foi administrada uma dose menor do quimioterápico (1mg/kg) (RIGO et al., 2013,
BRUSCO et al., 2016). Isso pode refletir uma diferença entre as espécies e explicar a
diferença de tempo encontrado entre os pesquisadores.
Diversos estudos clínicos descrevem a síndrome da dor aguda associada ao
paclitaxel, no entanto, os mecanismos pelos quais esse quimioterápico induz dor aguda
continuam pouco esclarecidos (GARRISON et al., 2003; LOPRINZI et al., 2007; LOPRINZI
et al., 2011; PACHMAN et al., 2011; REEVES et al., 2012). Até o momento, poucos estudos
foram desenvolvidos com o intuito de investigar os mecanismos desta síndrome dolorosa
aguda em modelos animais. Recentemente, um estudo demonstrou que o paclitaxel poderia
provocar dor aguda devido à ativação do receptor TLR4 na medula espinal e nos gânglios da
raiz dorsal (GRD) de ratos (YAN et al., 2015). Outro estudo sugeriu que os receptores de
bradicinina (B1 e B2) estão envolvidos no desenvolvimento desta síndrome dolorosa aguda
(BRUSCO et al., 2016). A investigação dos mecanismos moleculares envolvidos na síndrome
da dor aguda associada ao paclitaxel pode fornecer fundamentos para o desenvolvimento de
tratamentos eficazes para essa síndrome dolorosa (POLOMANO et al., 2001; RIGO et al.,
2013). Além disso, a investigação dos mecanismos envolvidos pode auxiliar na redução da
intensidade, incidência ou ainda na prevenção de dor crônica, pois, há evidências que a dor
aguda pode está relacionada com o aparecimento e severidade da dor neuropática observada
após o tratamento com o paclitaxel (REEVES et al., 2012; LOPRINZI et al., 2011).
Os primeiros estudos clínicos que associaram o paclitaxel ao aparecimento de dor
aguda se referiam a essa condição como mialgias e artralgias causadas pelo paclitaxel,
indicando que este quimioterápico possivelmente estaria causando lesão nos músculos ou
articulações dos pacientes (GARRISON et al., 2003). Entretanto, foi demonstrado que o
paclitaxel não causava dor devido a lesões musculares ou articulares, mas sim porque,
possivelmente, o quimioterápico estaria induzindo hipersensibilidade sensorial, devido à
sensibilização dos nociceptores (LOPRINZI et al., 2007). Atualmente, essa morbidade
dolorosa, descrita antes como artralgias e/ou mialgias causadas pelo paclitaxel, é denominada
de síndrome da dor aguda associada ao paclitaxel (“Paclitaxel-associated Acute Pain
Syndrome, P-APS) (REEVES et al., 2012; LOPRINZI et al., 2011). Estudos realizados em
roedores indicaram que a administração de paclitaxel poderia causar dor neuropática, pois os
animais desenvolveram alodínia mecânica e hiperalgesia térmica (POLOMANO et al., 2001;
FLATTERS et al., 2006; CHEN, YANG, WANG, 2011), no entanto, a avaliação e descrição

132
de um modelo de síndrome de dor aguda em animais não era explorado. Então, para estudar
os mecanismos envolvidos nesta patologia foi desenvolvido um modelo de síndrome de dor
aguda induzida por paclitaxel em roedores (DINA et al., 2001; RIGO et al., 2013; YAN et al.,
2015).
Várias hipóteses são propostas para explicar o desenvolvimento de dor aguda
induzida pelo paclitaxel. A primeira possibilidade seria o envolvimento patológico dos
músculos, articulações, nervos ou outros tecidos. Outra hipótese seria a relação desta
síndrome com uma disfunção das fibras nociceptivas, ou do sistema espinotalâmico. No
entanto, a possibilidade de disfunção dos nociceptores seria o mecanismo primário para o
desenvolvimento da dor, ou ainda, o desenvolvimento da dor seria uma reposta secundária
como, por exemplo, a inflamação induzida pelo paclitaxel. As lesões nervosas provavelmente
se devem a um efeito do agente sobre os tecidos nervosos ou um efeito mediado por
inflamação induzida pelo paclitaxel (LONPRINZI et al., 2007).
A hipótese de que o paclitaxel causa dor aguda devido à indução de uma
inflamação é reforçada pelos dados obtidos no presente estudo. Os resultados mostraram que
a administração de paclitaxel aumenta significativamente a concentração de citocinas no
plasma, nervo isquiático, GRD e medula espinal de camundongos. Adicionalmente a
expressão relativa de RNAm da IL-6 estava alterada no nervo isquiático, GRD e medula
espinal, após o tratamento com paclitaxel. Além disso, ficou demostrado um aumento da
imunoexpressão de IL-6 no GRD de camandugos 4 horas após o tratamento com paclitaxel,
no entanto 6 horas após o tratamento essa marcação começa a desaparecer, passando então os
neurorios a expressá-la. O aumento desses mediadores coincide com o desenvolvimento da
hiperalgesia, apresentado o mesmo curso temporal. Esses mediadores encontram-se em
concentrações basais e aumentam consideravelmente entre 4 e 6 horas após a administração
de paclitaxel, desaparecendo quase que totalmente 24 horas após a administração da droga.
Os resultados mostraram que a administração de paclitaxel provocou aumento das
citocinas pró-inflamatórias IL-1β, TNF-α, IL-6 e KC/CXCL1 no plasma de camundongos que
receberam paclitaxel. No nervo isquiático o paclitaxel provocou aumento de IL-1β, TNF-α,
IL-6, MCP-1 e KC/CXCL1. A quantificação relativa do RNAm revelou que o tratamento com
paclitaxel induziu um aumento da expressão gênica para a IL-6. No entanto, não houve
aumento na expressão das citocinas IL-1β e TNF-α, o que pode evidenciar que a IL-6 liberada
é devido exclusivamente a estimulação direta dos mastócitos, já que esse mediador encontra-
se armazenado nos grânulos dessas células (MARSHALL, MCCURDY, OLYNYCH, 2003).

133
Nos gânglios da raiz dorsal de animais tratados com paclitaxel, TNF-α, IL-6 e
KC/CXCL1 estavam aumentados. Porém, IL-1β e MCP-1 não foram aumentados pela
administração de paclitaxel. Quanto a expressão relativa de RNAm, o tratamento com
paclitaxel foi capaz de aumentar a expressão relativa de RNAm para IL-1β, TNF-α e CCR2
nos GRD. Quando realizou-se immunofluorescencia para IL-6 nos GDR, foi observado um
aumento da imunoexpressão para essa citocina 4 horas após a administração de paclitaxel,
corroborando que a produção de citocinas está envolvida no processo de desenvolvimento de
dor induzida por paclitaxel. Entretanto, não foi observada diferença na expressão relativa do
RNAm para IL-6 e MCP-1.
Na medula espinal, o tratamento com paclitaxel provocou aumento de IL-1β,
TNF-α, IL-6, MCP-1 e KC/CXCL1. Detectou-se ainda aumento na expressão relativa do
RNAm para IL-1β, TNF-α e IL-6, MCP-1 e para o receptor CCR2. O paclitaxel pode causar
dor devido à produção desses mediadores que sensibilizaria o nociceptor, já que citocinas pró-
inflamatórias, incluindo TNF-α, IL-1β, e KC/CXCL1 são essenciais para o desenvolvimento
de doenças inflamatórias e hiperalgesia. Esta sensibilização deixa o neurônio mais suscetível a
estímulos que normalmente não causaria dor, processo conhecido como alodinia. O paclitaxel
poderia desenvolver dor aguda nos pacientes devido à indução de produção de citocinas e/ou
quimiocinas. Zhang e colaboradores (2013) mostraram que a neuropatia induzida por
paclitaxel está associada com uma indução da quimiocina MCP-1 e do seu receptor CCR2 em
neurônios sensoriais primários de GRD. Eles observaram que o MCP-1 estava expresso tanto
na medula espinal quanto nos GRD, mas o CCR2 só estava expresso nos GRD. E essa
expressão se iniciava 4 horas após a administração de paclitaxel. No entanto, no presente
estudo foi observado que o MCP-1 só aumentou significativamente 24 horas após a
administração de paclitaxel apenas na medula espinal. Já o CCR2 estava expresso em ambos,
GRD e medula espinal, isso pode indicar que principalmente o aumento da expressão do
receptor nas primeiras horas após a adiministração de paclitaxel contribui para o
desenvolvimento de dor aguda. E com o avanço do tratamento com paclitaxel essa quimiocina
e seu receptor passam a contribuir para o desenvolvimento da dor na neuropatia.
Separado dos seus efeitos sobre os microtúbulos, o paclitaxel, pode induzir genes
que codificam TNF- (DING et al., 1990; BODGAN et al., 1992; HWANG e DING, 1995;
MANTHEY et al., 1992, 1993; BURKHART et al., 1994; KIRIKAE et al., 1996, 1998),
interleucinas como a IL-1, IL-8 e IL-6 (BODGAN et al., 1992, MANTHEY et al., 1992,
LEE et al., 1996; WHITE et al., 1996; WATSON et l., 1998); e enzimas como a óxido nitrico
sintase (MANTHEY et al., 1992, KIRIKAE et al., 1996, 1998) e cicloxigenase-2 (MOOS e

134
FITZPATRICK, 1998, SUBBARAMAIAH et al., 2000) o que gera a produção de mediadores
inflamatórios e álgicos. O paclitaxel então estimula a produção e/ou a liberação desses
mediadores que sensibilizaram o nociceptor causando o aparecimento de dor aguda.
No presente estudo verificou-se que a deleção gênica para IL-6 ou o uso de um
antagonista dessa citocina, bem como a deleção gênica do receptor IL-1R, ou a deleção dupla
dos receptores de TNF (animais duplo-nocaute, TNFR1/R2), ou ainda a deleção gênica do
receptor da quimiocina MCP-1 (CCR2), inibiram a hiperalgesia aguda mecânica e térmica
induzida pelo tratamento com o paclitaxel, sugerindo o envolvimento destes mediadores na
síndrome aguda associada ao paclitaxel. Portanto, esses resultados são fortes evidencias que a
participação de citocinas/quimiocinas é primordial para o desenvolvimento da dor aguda
induzida por este quimioterápico, sugerindo que essa dor teria uma origem, pelo menos em
parte, inflamatória.
A dor de origem inflamatória resulta essencialmente da sensibilização dos
neurônios sensoriais primários (FERREIRA, 1972). Existe crescente evidência de que, além
da sensibilização periférica, a dor inflamatória pode ser dependente de um processo que
ocorre na medula espinal, assim como nos gânglios da raiz dorsal e gânglio trigeminal. Na
medula espinal, a plasticidade neuronal é modulada dinamicamente pela microglia ativada e
mediadores derivados de astrócitos, o que pode aumentar à capacidade de resposta a dor
(SOUZA et al., 2013). Os mecanismos envolvidos na sensibilização dos neurônios sensoriais
primários podem ser divididos em dois processos. O primeiro inclui eventos não neuronais
residentes que recrutam células imunes, as quais produzem uma sequência de mediadores da
hiperalgesia inflamatória iniciados pelo TNF-α. Isso provoca a liberação de IL-1β e
quimiocinas que por sua vez estimulam a liberação direta de mediadores que atuam no
nociceptor (VALÉRIO et al., 2009). O segundo processo inclui ativação das vias de
sinalização intracelular, tais como o monofosfato cíclico de adenosina (AMPc), proteína
quinase A (PKA) e proteína quinase C (PKC) (ALEY e LEVINE, 1999; KHASAR et
al.,1999).
Várias células imunes como, por exemplo, os mastócitos podem mediar ações
pró-inflamatórias ou imunorregulatórias através de sua degranulação quando ativados
(GALLI, NAKAE, TSAI, 2005). Além disso, os mastócitos são importantes reguladores
celulares das vias fisiológicas e patológicas da dor (WOOLF, 2010). A interação entre o
sistema nervoso e o sistema imunológico desempenha um papel central no processamento da
dor (REN e DUBNER, 2010). O mastócitos são particularmente importante neste aspecto,
uma vez que são frequentemente encontradas nas proximidades de neurônios nociceptivos

135
(FORSYTHE e BIENENSTOCK, 2012) e, portanto, podem participar na sinalização em
sinapses neuro-imunes (FORSYTHE e BIENENSTOCK, 2012). Os mastócitos induzem a
ativação do nociceptor através da liberação de mediadores químicos durante a degranulação e
podem ser ativados por mediadores liberados de nociceptores após lesão (REN e DUBNER,
2010; FORSYTHE e BIENENSTOCK, 2012).
Tem sido proposto que a degranulação de mastócitos pode desempenhar um papel
na dor neuropática induzida por paclitaxel (GAO et al., 2016), pois mediadores químicos
liberados de mastócitos pela sua degranulação ativam nociceptores (FORSYTHE e
BIENENSTOCK, 2012) e consequentemente ocorre geração de dor. Devido a essas
evidências de que os mastócitos participam da neuropatia induzida por paclitaxel, resolveu-se
investigar a participação dessas células imunes na dor aguda induzida por esse
quimioterápico, tendo em vista que os mastócitos estão envolvidos em respostas inflamatórias
e hiperalgesicas agudas. Os resultados do presente estudo demonstraram que em animais que
não que diferenciam mastócitos (SH), as respostas hiperalgesicas desencadeadas por
paclitaxel não se desenvolveram, quando comparados com camundongos selvagens (WT).
Além disso, animais WT foram pré-tratados com cromoglicato de sódio, um estabilizador de
membrana de mastócitos, e posteriormente injetados com paclitaxel. Observou-se então que o
tratamento com cromoglicato de sódio foi capaz de prevenir a hiperalgesia mecânica e
térmica, quando comparados com os animais que não receberam esse tratamento.
Também foi demostrado através de um aumento na imunoexpressão de triptase no
GRD de camundongos tratados com paclitaxel que os mástócitos podem estão envolvidos no
desenvolvimento da dor aguda induzida por esta droga. A triptase é uma protease de serina
tetramérica neutra que existe abundantemente nos mastócitos e, em menor concentração, em
outras linhagens celulares como os basófilos (SCHWARTZ, 2001). A triptase tem sido
vastamente utilizada como indicador do número e da ativação de mastócitos. Conjuntamente
esses resultados revelaram que os mastócitos estão envolvidos no desenvolvimento de
hiperalgesia mecânica e térmica no modelo de síndrome da dor aguda associada ao paclitaxel.
Provavelmente o paclitaxel estaria ativando os mastócitos que liberariam mediadores
químicos através da degranulação que ativariam nociceptores (REN e DUBNER, 2010;
FORSYTHE e BIENENSTOCK, 2012). Adicionalmente, o aumento da imunoexpressão de
triptase foi acompanhada pela maracação de IL-6, pricipalmente 4 horas após o tratamento
com o paclitaxel. Percebe-se que existem marcações coincidentes de triptase e IL-6, indicando
possivelmente que os mastócitos estão liberando esta citocina. Estudos relataram que a
resposta nociceptiva desencadeada pelos mastócitos está relacionada à liberação de

136
mediadores tais como a histamina, serotonina, IL-1β, TNF-α e IL-6. Estes mediadores têm a
capacidade de induzir hiperalgesia independentemente (CHATTERJEA et al., 2012; CUNHA
et al., 1992; VERRI et al., 2006) ou ainda induzir hiperalgesia evocada por alérgenos
(LAVICH et al., 2003, 2006), através de efeitos diretos em nociceptores, ou por estimulação
da produção de mediadores finais, tais como leucotrienos e prostanóides (CUNHA et al.,
1992). Além disso, os mastócitos que residem nas proximidades de fibras nervosas não
mielinizadas tais como as fibras C nociceptivas, podem sofrer alterações ultraestruturais que
permitem uma degranulação fragmentada diferencial ou seletiva (FORSYTHEAND e
BIENENSTOCK, 2012).
Foi demonstrado em um modelo de neuropatia que o tratamento com
cromoglicato de sódio, que impede que os mastócitos degranulem, eliminou total ou
parcialmente o desenvolvimento de hiperalgesia após a indução de neuropatia em ratos, o que
foi associado a uma redução do número de neutrófilos e macrófagos no local da lesão (ZUO
et al., 2003). Estes resultados contribuem para a hipótese de que os mastócitos estariam sendo
ativados pelo paclitaxel e consequentemente liberando mediadores que sensibilizariam os
nociceptores, isto é reforçado pelo fato que animais SH, não desenvolveram respostas
hiperalgesicas. Adicionalmente o tratamento de animais WT com cromoglicato de sódio, foi
capaz de inibir respostas nociceptivas mecânica e térmica em comparação a animais que não
receberam a droga, mas foram injetados com paclitaxel.
Para confirmar que os mastócitos estariam produzindo mediadores inflamatórios e
esses mediadores estariam contribuindo para o desenvolvimento da dor aguda induzida pelo
paclitaxel, foi realizada a quantificação de citocinas pró-inflamatórias e quimiocinas no nervo
isquiático, gânglios da raiz dorsal e medula espinal de camundongos SH tratados com
paclitaxel, ou ainda camundongos pré-tratados com cromoglicato de sódio e, posteriormente,
injetados com paclitaxel. Demonstrou-se então, que IL-1β, TNF-α, IL-6, MCP-1e KC/CXCL1
estavam diminuidas no nervo isquiático dos animais SH, bem como nos animais tratados com
cromoglicato de sódio. Nos gânglios da raiz dorsal de camundongos SH tratados com
paclitaxel, sa concentrações de TNF-α, IL-6 e KC/CXCL1 foram reduzidas. O pré-tratamento
com cromoglicato de sódio também foi capaz de prevenir o aumento destes mediadores. Na
medula espinal dos camundongos SH tratados com paclitaxel, IL-1β, TNF-α, IL-6 e
KC/CXCL1 foram reduzidos quando comparados ao grupo de camundongos WT tratados
com paclitaxel. Já a concentração MCP-1 não foi alterada. Estes dados revelaram que o
aumento da concetração de citocinas e quimiocinas que ocorreu nos camundongos

137
concomitantes ao desenvolvimento de hiperalgesia mecânica e térmica, evidencia que os
mastócitos podem contribuir para a dor aguda induzida por paclitaxel.
Gao e colaboradores (2016) demonstraram que o aumento de histamina no plasma
de ratos tratados com paclitaxel, ocorreu conjuntamente com a hiperalgesia mecânica e
térmica ao calor, os autores então sugerem que a histamina contribui para a dor neuropática
induzida pelo paclitaxel. No entanto, é sabido que os mastócitos produzem uma variedade de
mediadores, além de histamina, como outras aminas bioativas como a serotonina. Produzem
ainda enzimas (hidrolases ácidas, fosfolipases, quimase, triptase e outras proteases), citocinas
(IL-1, IL-6, TGF, GM-GCSF, TNF-α), metabolitos lipídicos (leucotrienos, prostaglandinas,
fator ativador de plaquetas - PAF), ATP (adenosina trifosfato), neuropeptídeos (peptídeo
intestinal vasoativo), fatores de crescimento (fator de crescimento de nervos - NGF) e óxido
nítrico (SCHWARTZ e AUSTEN, 1980; JOHNSON e KRENGER, 1992). Os resultados do
presente estudo demonstraram que além do plasma, o nervo isquiático, os gânglios da raiz
dorsal e a medula espinal apresentaram níveis elevados de citocinas e/ou quimiocinas, que
como demonstrado pode ser produzidas, pelo menos em parte, por mastócitos e liberados pela
quando ativados. Ativação esta induzida pelo paclitaxel. Esses mediadores liberados podem
estar induzindo a produção de novos mediadores pelos mastócitos. Fica claro que a ativação
de mastócitos pelo paclitaxel evidenciado pelo aumento da imunoexpressão para triptase nos
gânglios da raiz dorsal, especialmente 4 horas após o tratamento com esta droga causou um
aumento nos níveis de mediadores que possivelmente estariam sensibilizando o nociceptor.
Fato evidenciado pela comarcação de IL-6 e triptase, indicando possivelmente que são os
mastócitos que produzem/liberam esta citocina.
Com intuito de confirmar a que a ativação dos mastócitos é responsável, pelo
menos em parte, pela produção/liberação de citocinas e quimiocinas na dor aguda induzida
por paclitaxel, foram quantificados IL-1β, TNF-α, IL-6, MCP-1 em cultura de linhagem de
mastócitos (RBL-2H3) estimulada com paclitaxel. Os resultados mostraram que a estimulação
com paclitaxel provocou aumento de TNF-α, IL-6 e MCP-1. Quando estimulados com
diferentes doses de paclitaxel, mastócitos em cultura liberaram/produziram esses mediadores.
Mediadores esses, que também estavam aumentados no plasma, nervo isquiático, gânglios da
raiz dorsal e medula espinal.
Estudos anteriores demonstraram que essa linhagem de células produzia/liberava
várias citocinas e/ou quimiocinas tais como IL-1β, TNF-α e IL-6 (ZENG et al., 2016; TANG
et al., 2015), e MCP-1 (AHN et al., 2014). No entanto, não existem estudos publicados
mostrando que o paclitaxel ativa mastócito, e estes liberam mediadores hiperalgésicos. Um

138
estudo publicado recentemente demonstrou que a quercetina, melhora a dor neuropática
induzida por paclitaxel. O estudo relata que a quercetina estabiliza a membrana de mastócito
e, assim, a liberação excessiva de histamina é inibida. Sugerindo que a quercetina atenua a dor
neuropática induzida pelo paclitaxel. Esta ação da quercetina sobre os mastócitos impediria o
desenvolvimento de dor neuropática (GAO et al., 2016). Os autores testaram essa hipótese in
vivo e in vitro. Essa investigação pode confirmar a hipótese de que o paclitaxel estimula
mastócitos que por sua vez liberam mediadores hipernociceptivos que sensibilizam o
nociceptor gerando dor aguda. Esse mecanismo, portanto seria fundamental para o
desenvolvimento da síndrome da dor aguda associada ao paclitaxel.
No inicio da década de 1990, foi demonstrado que o paclitaxel mimetizava os efeitos
do lipopolissacarídeo (LPS) em macrófagos murinos, aumentando a liberação de TNF-α. Por
causa do comportamento comparável entre LPS e paclitaxel em células animais nas quais foi
determinado geneticamente o papel do LPS, os autores concluíram que o LPS e o paclitaxel
compartilhavam uma etapa inicial no processo de transdução de sinal (DING et al., 1990;
BODGAN et al., 1992).
Paclitaxel e outros taxanos não são quimicamente relacionados ao LPS, assim é
duvidoso que eles ocupem o mesmo sítio do LPS no receptor. No entanto, desde 1992, vários
autores vêm defendendo a hipótese de que o paclitaxel age através de um mecanismo receptor
dependente (MANTHEY et al., 1992; ZAKS-ZILBERMAN et al., 2001, 1998; PERERA et
al., 1997; MANTHEY et al., 1994; BHAT et al., 1999). Reforçando essa ideia, Manthey e
colaboradores (1993) foram os primeiros a referirem que antagonistas do receptor de LPS
inibem a expressão de genes e a fosforilação da proteína de tirosina pelo paclitaxel. Eles
também demonstraram que outro taxano, potente estabilizador de microtúbulos, o docetaxel,
não ativou as vias de sinalização dependentes de LPS ou induziu TNF-α em macrófagos
murinos. O contraste da potência e eficácia entre paclitaxel e docetaxel como indutores de
genes pró-inflamatórios em macrófagos murinos indicou que a estabilização de microtúbulos
per si era insuficiente para causar indução de gene ou mimetização da sinalização por LPS em
macrófagos murinos. Isto sugere que o mecanismo pelo qual o paclitaxel induz a estabilização
de microtúbulos não é o mesmo da indução de genes pró-inflamatórios. Embora o paclitaxel e
o LPS não sejam estruturalmente relacionados, eles compartilham sua ação no mesmo
receptor. Vários grupos de pesquisadores vêm estudando o envolvimento dos receptores Toll
like (Toll-like receptors - TLR) na via de transdução de sinal paclitaxel/LPS-símile em
camundongos (KAWAZAKI et al., 2000, 2001; BYRD-LEIFER et al., 2001).

139
A família de TLR apresenta um papel crucial nas respostas celulares ao LPS. No
reconhecimento de endotoxina das bactérias gram-negativas. O receptor Toll-like 4 (TLR4)
associa-se fisicamente ao complexo LPS-CD14, no qual o CD14 é o receptor específico para
LPS, ocorrendo ainda a ligação de uma proteína acessória extracelular, chamada MD2
(WRIGHT et al., 1900; KOPP; MEDZHITOV,1999). O TLR4 utiliza uma via de transdução
de sinal que consiste no recrutamento de várias proteínas intracelulares (MyD88, IRAK e
TRAF-6) que vão desencadear as vias JNK e ERK da cascata MAPK, envolvidas na ativação
dos fatores de transcrição AP-1 e NF-κB (nuclear factor kappa B), favorecendo a expressão
de genes envolvidos na resposta inflamatória (BOCHUD e CALANDRA, 2002).
Alguns alvos moleculares foram estudados como sendo mecanismos relevantes para
a dor aguda induzida por paclitaxel, dentre estes alvos pode citar o receptor TLR4.
Recentemente, foi demonstrando que o paclitaxel provoca a ativação deste receptor no corno
dorsal da medula e gânglios da raiz dorsal de ratos, aumentando as atividades sinápticas
glutamatérgicas, e reduzindo as atividades de transporte de glutamato gliais no corno dorsal
da medula e que esses eventos são críticos na gênese da dor aguda induzida por paclitaxel em
animais (YAN et al., 2015). No entanto, ainda não foi investigado se o paclitaxel ativa o
receptor TLR4 em mastócitos, ocasionando sua degranulação, o que contribuiria para o
desenvolvimento da dor aguda associada ao paclitaxel.
Para averiguar a hipótese de que o paclitaxel ativaria o receptor TLR4 em
mastócitos, foi avaliada a participação desse receptor na hiperalgesia induzida por este
quimioterápico. De acordo com o que é descrito na literatura, verificou-se que em animais
TLR4-/- as respostas hiperalgesicas induzidas por paclitaxel não se desenvolveram quando
comparado aos animais WT. Adicionalmente verificou-se que a ativação desse receptor
aumenta a expressão de citocinas ou quimiocinas em camundongos tratados com paclitaxel.
Foi verificado que esse quimioterápico não provocou aumento de IL-1β, TNF-α, IL-6, MCP-1
e KC/CXCL1 no plasma, nervo isquiático, gânglios da raiz dorsal e medula espinal de
camundongos TLR4-/-. No plasma desses camundongos tratados com paclitaxel, TNF-α, IL-6
e KC/CXCL1 tiveram uma significativa redução após a administração de paclitaxel. No nervo
isquiático IL-1β, TNF-α, IL-6, MCP-1 e KC/CXCL1 apresentam-se reduzidos, quando
comparados aos animais WT. Nos gânglios da raiz dorsal, observou-se que nos camundongos
TLR4-/- tratados com paclitaxel, TNF-α e KC/CXCL1 foram reduzidos quando comparados
ao grupo de camundongos WT. No entanto IL-6 não foi reduzido em comparação ao grupo
WT nos animais TLR4-/-. Na medula espinal de camundongos TLR4-/- tratados com

140
paclitaxel, TNF-α, MCP-1 e KC/CXCL1 apresentam-se reduzidas em comparação ao grupo
de camundongos WT, apenas 6 horas após o tratamento.
Os mastócitos expressam TLR funcionais (MARSHALL, 2004), o que pode explicar
a proteção conferida por mastócitos contra infecções bacterianas e parasitárias em modelos
animais (GALLI, NAKAE, TSAI, 2005; MARSHALL, MCCURDY, OLYNYCH, 2003). Os
mastócitos ativados liberam uma variedade de potentes mediadores inflamatórios por
descarga rápida de mediadores pré-formados presentes em seus grânulos, além da geração de
lipídios inflamatórios a partir do ácido araquidônico e a produção de numerosas citocinas e
quimiocinas de tipo Th2 (GALLI, NAKAE, TSAI, 2005). Todas estas respostas são evocadas
por alérgenos via FcεRI (receptor de alta afinidade para a IgE), que são membros da família
dos receptores multicadeia de reconhecimento imune (MIRRs) (SHIMIZU et al., 1988;
METZGER, 1992), enquanto que a estimulação de mastócitos através de TLR2 e TLR4
resulta essencialmente na produção de citocinas, tais como IL-4, IL-5, IL-6, IL-10, IL-13 e
TNF-α (MARSHALL, MCCURDY, OLYNYCH, 2003). Há relatos de que ligantes TLR2
também induzem a degranulação e geração de leucotrienos C4 (SUPAJATURA et al., 2002;
MCCURDY et al., 2003; VARADARADJALOU et al., 2003). No entanto, um estudo
realizado por Qiao e colaboradores (2006) descobriu que o antígeno interage sinergicamente
com ligandos de TLR2 e TLR4 para aumentar acentuadamente a produção de citocinas em
linhagens de mastócitos murinos.
Os dados obtidos demonstram que a ausência tando de mastócitos, quanto à do
receptor TLR4 previnem o aumento de citocinas/quimiocinas no plasma, nervo isquiático,
gânglios da raiz dorsal e medula espinal de camundongos tratados com paclitaxel. Reforçando
a hipótese de que o paclitaxel se liga ao TLR4 presente em mastócitos, ativando essas células
liberando mediadores, e isso levaria ao aumento de citocinas e quimiocinas que por sua vez
sensibilizariam o nociceptor, gerando a dor aguda associada ao paclitaxel. Essa hipótese
poderia se confimada por uma cultura de mastócitos KO para TLR4.
Os resultados do estudo sugerem ainda que o paclitaxel, estimula células satelites
gliais do gânglio da raiz dorsal que liberam citocinas/quimiocinas e a liberação dessas
citocinas/quimiocinas por essas células atuam em neurônios sensitivos do gânglio da raiz
dorsal resultando em uma mudança fenotípica que causa dor. Estudos anteriores sugerem o
envolvimento do TNF-α no desenvolvimento da dor neuropática induzida pelo paclitaxel
(LEDEBOER et al., 2007; CATA; WENG; DOUGHERTY, 2008). Para confirmar essa
hipótese, cultura primária de células satelites gliais de gânglios da raiz dorsal, foram
estimuladas com paclitaxel. Nossos resultados, demonstraram que o paclitaxel induziu a

141
liberação de TNF-α, IL-6, MCP-1 e KC/CXCL1 nessa cultura. Esses resultados sugerem que
a sinalização de citocinas/quimiocinas produzidas por células satelites gliais é necessária para
a facilitação da dor aguda induzida pelo paclitaxel.
Adicionalmente, os resultados sugerem que a liberação de citocinas/quimiocinas
pelas células satélites gliais é devido a ligação do paclitaxel ao receptores TLR4. Isto foi
evidenciado através da utilização de animais KO para esse receptor. Verificou-se então que a
deleção do gene para o receptor TLR4 foi capaz de prevenir o aumento de
citocinas/quimiocinas em cultura de células satélites gliais de gânglios da raiz dorsal de
camundongos KO para os receptores TLR4. Ficou demonstrado ainda que esse efeito é
seletivo para TLR4, tendo em vista que em animais KO para os receptores TLR2, não foi
observado diminuição dessas citocinas/quimiocinas quando estimulados com paclitaxel. Esses
resultados estão de acordo com o que observado in vivo, pois demostrado que o paclitaxel
aumenta a expressão e a liberação de citocinas/quimiocinas a partir de células satélites gliais,
um efeito que não foi observado em camundongos KO para os receptores TLR4. Estudos têm
demonstrado que o paclitaxel pode alterar o fenótipo dos neurônios e da glia através de
mecanismos de ação que diferem dependendo da concentração. In vitro, o paclitaxel (10 nM)
aumenta a liberação de CGRP atravéz de um agonista de TRPV1 (capsaicina) em neurônios
de gânglios da raiz dorsal. Enquanto o paclitaxel em doses mais altas (300 nM) inibe a
liberação de CGRP evocada por capsaicina (PITTMAN et al., 2014). Outro estudo, in vivo,
relatou que o paclitaxel em doses baixas (1-2 mg/kg) regula a expressão de TLR4 em
neurônios do gânglio da raiz dorsal e em astrócitos do corno dorsal da medula espinal (LI et
al., 2014). Ledeboer e colaboradores (2007) mostraram que citocinas inflamatórias estão
implicadas na patogênese da dor neuropática induzida por paclitaxel. Nós mostramos que isso
também ocorre na dor aguda.
No estudo, foi demonstrado um aumento da imunoexpressão de c-Fos, marcador
de ativação neuronal, nas células satélites glias de gânglio da raiz dorsal, 4 e 6 horas após a
administração de paclitaxel. Já no corno dorsal da medula espinal, houve aumento da
imunoexpressão de c-Fos apenas 6 horas após a administração da droga. A expressão de c-fos,
pode ser induzido por estímulos nocivos, tendo em vista que os níveis de Fos são baixos em
condições basais, no entanto, estímulos externos podem resultar em reprogramação fenotípica
da célula, aumentando os níveis de c-fos. Assim, Fos é um marcador útil para rastrear os
efeitos de estímulos farmacológicos, elétricos e fisiológicos no sistema nervoso e tem sido
utilizado para indicar dor e neuroplasticidade (AZEVEDO et al., 2013).

142
Diante disso, a realização da imunofluorescência para c-fos, no gânglio da raiz
dorsal e no corno dorsal da medula espinal, demonstram que o paclitaxel causa ativação
neuronal, observado pelo aumento da imunoexpressão de c-Fos em células satélites gliais e
em neurônios. Este aumento na expressão de c-Fos está relacionada à atividade neuronal e à
intensidade da dor (TEIXEIRA, 2009). Além de neurônios, células satélites gliais tembém
expressam c-Fos. A morfologia das células marcadas na imunofluorescência para c-fos
assemelham-se a células satélites gliais. Roivainen e Koistinaho (1990) demonstraram que
células satélites gliais expressam c-fos após a transecção do tronco simpático. Diante dessas
evidencias, sugere-se o paclitaxel possa estar ativando neurônios e células satélites gliais,
estas liberam mediadores inflamatórios que desencadeiam um processo doloroso logo após a
primeira administração de paclitaxel.
Os resultados obtidos no presente estudo indicaram que mastócitos e células
satélites gliais podem estar envolvidos na hipernociepção aguda causada pela administração
de paclitaxel em camundongos. Os dados indicam que o paclitaxel estimula os mastócitos
células satélites gliais que, por conseguinte produzem e/ou liberam mediadores que por sua
vez sensibilizaram os neurônios nociceptivos após a administração da droga, contribuindo
assim para o desenvolvimento da dor aguda. Estes mecanismos conjuntamente podem
contribuir para a nocicepção aguda que se manifesta após a administração de paclitaxel.
Portanto, esse estudo pode fornecer informações para se elucidar o mecanismo pelo qual o
PCX induz dor aguda e fornecer subsídios para a prevenção do aparecimento dessa condição.
Implica também na prevenção ou na atenuação da neuropatia, tendo em vista que a dor aguda
prediz o aparecimento e a severidade da neuropatia crônica induzida pelo paclitaxel.

143
8. CONCLUSÃO

144
Concluiu-se que a administração de paclitaxel induz hiperalgesia mecânica e
térmica ao frio em camundongos, iniciando 2h, com pico na 6ª hora e resolução 24h pós-
administração. O estudo, mostra pela primeira vez, que mastócitos e células satélites gliais
estão envolvidos na sídrome de dor aguda induzida por paclitaxel. Adicionalmente, o estudo
revelou que a ativação de mastócitos e de células satélites gliais, possivelmente pela ligação
do paclitaxel a TLR4 leva a liberação de mediadores inflamatórios, tais como IL-1β, TNF-α,
IL-6, MCP-1 e KC/CXCL1, que possivelmente sensibilizam o nociceptor, gerando dor aguda
após administração da droga.

145
REFERÊNCIAS

146
ALEY, K. O.; LEVINE, J. D. Role of Protein Kinase A in the Maintenance of Inflammatory
Pain. The Journal of Neuroscience, n. 6, v. 19, p. 2181-2186, mar. 1999.
ANH, K. B. et al. IgE in the absence of allergen induces the expression of monocyte
chemoattractant protein-1 in the rat basophilic cell-line RBL-2H3. Mol Immunol. n. 1, v. 62,
p. 114-121, nov. 2014.
APFEL, S. et al. Nerve growth factor prevents toxic neuropathy in mice. Ann Neurol., n. 1,
v. 29, p. 87-90, jan, 1991.
ARGYRIOU, A. A. et al. Chemotherapy-induced peripheral neurotoxicity (CIPN): an update.
Crit. Rev. Oncol. Hematol., n. 1, v. 82, p. 51-77, abril, 2012.
ARGYRIOU, A. A. Tracing the incidence of paclitaxel-induced peripheral neuropathy. Eur J
Cancer Care(Engl)., n. 5, v.18, p. 522–523, set. 2009.
ARGYRIOU, A. et al. Peripheral nerve damage associated with administration of taxanes in
patients with cancer. Crit Rev Oncol Hematol., n. 3, v. 66, p. 218-228, jun. 2008.
AZEVEDO, M. I. et al. The antioxidant effects of the flavonoids rutin and quercetin inhibit
oxaliplatin-induced chronic painful peripheral neuropathy. Molecular pain, n. 1, v. 9, p. 53,
2013.
BARON, R.; BINDER, A.; WASNER, G. Neuropathic pain: diagnosis, pathophysiological
mechanisms, and treatment. Lancet Neurol., n. 8, v. 9, p. 807 – 819, ago. 2010.
BASBAUM, A. I. et al. Cellular and Molecular Mechanisms of Pain. Cell., v. 139, n. 2, p.
267 – 284, out. 2009.
BASCIANO, L. K. et al. Monoclonal antibodies that inhibit IgE binding. J Biol Chem, n. 25,
v. 261, p. 11823-11831, set. 1986.
BINGHAM, B. et al. The molecular basis of pain and its clinical implications in
rheumatology. Nat Clin Pract Rheumatol. v. 5, n. 1, p. 28 – 37, jan. 2009.
BHAT, N. et al. Use of a photoactivatable taxol analogue to identify unique cellular targets in
murine macrophages: identification of murine CD18 as a major taxol-binding protein and a
role for Mac-1 in taxol-induced gene expression. J Immunol., n. 12, v. 162, p. 7335-7342,
jun, 1999.
BOCHUD, P. Y.; CALANDRA, T. Pathogenesis of sepsis: new concepts and implications for
future treatements. BMJ. n. 7383, v. 326, p. 262-266. fev. 2002.
BOGDAN, C.; DING, A. Taxol, a microtubule-stabilizing antineoplastic agent, induces
expression of tumor necrosis factor alpha and interleukin-1 in macrophages. J Leukoc Biol,
n. 1, v.52, p. 119– 21, jul. 1992.

147
BOYLE, F. M.; WHEELER, H. R.; SHENFIELD, G. M. Amelioration of experimental
cisplatin and paclitaxel neuropathy with glutamate. J Neurooncol., n. 2, v. 41, p. 107-116,
jan. 1999.
BRUSCO, I. et al. Potentiation of Paclitaxel-Induced Pain Syndrome in Mice by Angiotensin
I Converting Enzyme Inhibition and Involvement of Kinins. Mol Neurobiol., nov. 2016.
BURKHART, C. A. et al. Relationship between the structure of taxol and other taxanes on
induction of tumor necrosis factor-alpha gene expression and cytotoxicity. Cancer Res., n.
22, v. 54, p. 5779 – 5782, nov. 1994.
BYRD-LEIFER, C. A. et al. The role of MyD88 e TLR4 in the LPS-mimrtic activity of
Taxol. Eur Journal, n. 8, n. 31, p. 2448-2457, ago. 2001.
CAPLOW, M.; SHANKS, J.; RUHLEN, R. How Taxol modulates microtubule disassembly.
J. Biol. Chem., n. 38, v. 269, p. 23399-23402, set. 1994.
CAPUANO, A. et al. Proinflammatory activated trigeminal satellite cells promote neuronal
sensitization: relevance for migraine pathology. Molecular Pain. v. p. 43, 2009.
CARLSON, K.; OCEAN, A. J. Peripheral neuropathy with microtubule-targeting agents:
occurrence and management approach. Clin.Breast.Cancer.,n. 2, v. 11, p. 73-81, abril, 2011.
CASPANI, O. et al. The Contribution of TRPM8 and TRPA1 Channels to Cold Allodynia
and Neuropathic Pain. PLoS One, n. 10, v. 4, out. 2009.
CASTILLO, C. et al. Satellite gliacells in dorsal root ganglia express functional NMDA
receptors. Neuroscience. n. 240C, p. 135-146, 2013.
CATA, J. P. et al. Clinical and experimental findings in humans and animals with
chemotherapy-induced peripheral neuropathy. Minerva Anestesiol. v. 72, n. 3, p. 151 – 159,
mar., 2006.
CATA, J. P; WENG, H. R.; DOUGHERTY, P. M. The effects of thalidomide and
minocycline on taxol-induced hyperalgesia in rats. Brain Res., n. 10, v. 1229, p.100 – 110,
set, 2008.
CAVALETTI, G.; MARMIROLI, P. Chemotherapy-induced peripheral neurotoxicity. Nat
Rev Neurol, v. 6, p. 657-666, 2010.
CERUTI, S. et al. Purinoceptor-mediated calcium signaling in primary neuron–glia trigeminal
cultures. Cell Calcium , v. 43, p. 576–590, 2008.

148
CHAPLAN, S. R. et al. Quantitative assessment of tactile allodynia in the rat paw. J.
Neurosci Methods., n. 1, v. 53: 55-63, jul. 1994.
CHATTERJEA, D. et al. Mast cell degranulation mediates compound 48/80-induced
hyperalgesia in mice.Biochem. Biophys. Res. Commun. n. 2, v. 425, p. 237–243, ago. 2012.
CHATTERJEA, D.; MARTINOV, T. Mast cells: versatile gatekeepers of pain. Mol Immunol
n. 1, v. 63, p. 38–44, jan. 2015.
CHEN, C. C. et al. Identification of mast cell progenitors in adult mice. Proc Natl Acad Sci
USA., n. 32, v. 102, p. 11408-11413, ago. 2005.
CHEN, Y.; YANG, C.; WANG, Z. J. Proteinase-activated receptor 2 sensitizes transient
receptor potential vanilloid 1, transient receptor potential vanilloid 4, and transient receptor
potential ankyrin 1 in paclitaxel-induced neuropathic pain. Neuroscience., v. 193, p. 440-451,
out. 2011.
CHERKAS, P. S. et al. The effects of axotomy on neurons and satellite glial cells in mouse
trigeminal ganglion. Pain, v. 110, p. 290–298, 2004.
COLLINS, T. S.; LEE, L. F.; TING, J. P. Paclitaxel up-regulates interleukin-8 synthesis in
human lung carcinoma through an NF-kappaB- and AP-1-dependent mechanism. Cancer
Immunol Immunother, n. 2, v. 49, p.78–84, maio, 2000.
CROSSING, K. L.; CARNEY, D. H. Microtubule stabilization to taxol inhibits initiation of
DNA synthesis by thrombin and by epidermal growth factor. Cell, n. 2, v. 27, p. 341-350,
dec. 1981.
CROWN J., O’LEARY M. The taxanes: an update. Lancet, n. 9210, v. 355, p.1176-1178,
abril, 2000.
CUNHA, F. Q.; MONCADA, S.; LIEW, F. Y. Interleukin-10 (IL-10) inhibits the induction of
nitric oxide synthase by interferon-gamma in murine macrophages. Biochem Biophys Res
Commun. n. 182, v. 3, p. 1155-1159, fev. 1992.
DE IULIIS, F. et al. Taxane induced neuropathy in patients affected by breast cancer:
Literature review. Crit Rev Oncol Hematol., n. 1, v. 96, p. 34-45, out. 2015.
DERRY, W.; WILSON, L.; JORDAN, M. Substoichiometric binding of taxol suppresses
microtubule dynamics. Biochemistry, n. 7, v. 34, p. 2203-2211. fev. 1995.
DINA, O. A. et al. Role of protein kinase C epsilon and protein kinase A in a model of
paclitaxel-induced painful peripheral neuropathy in the rat. Neuroscience, n. 3, v. 108, p.
507-515, 2001

149
DING, A. H. et al. Shared actions of endotoxin and taxol on TNF receptors and TNF release.
Science, n. 4953, v. 248, p.370– 372, abril, 1990.
DOUGHERTY, P. M. et al. Taxol-induced sensory disturbance is characterized by
preferential impairment of myelinated fiber function in cancer patients. Pain. v. 109, n. 1-2, p.
132 – 142, maio, 2004.
DUBLIN, P.; HANANI, M. Satellite glial cells in sensory ganglia: their possible contribution
to inflammatory pain. Brain Behav. Immun., n. 21, p. 592–598, 2007.
DUBOVY, P. et al. Intra- and extraneuro-nial changes of immunofluorescence staining for
TNF-alpha andTNFR1 in the dorsal root ganglia of rat peripheral neuropathicpain models.
Cell Mol Neurobiol. n. 26, p. 1205-1217, 2006.
DUMONTET, C.; JORDAN, M. Microtubule-binding agents: a dynamic field of cancer
therapeutics. Nat Rev Drug Discov., n. 10, v. 9, p. 790-803, out. 2010.
DUSTIN, P. Microtubules. Sci Am, n. 2, v. 243, p. 66-76, ago. 1980.
DUTTLINGER, R. et al. W-sash affects positive and negative elements controlling c-kit
expression: ectopic c-kit expression at sites of kit-ligand expression affects melanogenesis.
Development., n. 3, v. 118, p. 705-717, jul. 1993.
ENGLER, H. et al. Acute amygdaloid response to systemic inflammation. Brain Behav.
Immun., n. 7, v. 25, p. 1384–1392, out. 2011.
FAMAKIN B et al., Downstream Toll-like receptor signaling mediates adaptor-specific
cytokine expression following focal cerebral ischemia. J Neuroinflammation., n. 9, v. 174,
jul. 2012.
FAN, S. Disruption of p53 function in immortalized human cells does not affect survival or
apoptosis after Taxol or vincristine treatment. Clin. Cancer Res., n. 4, v. 4, p. 1047-1054,
abril, 1998.
FARQUHAR-SMITH, W. P. Anatomy, physiology and pharmacology of pain.
ANAESTHESIA AND INTENSIVE CARE MEDICINE, n.1, v. 9, p. 3-7, ago. 2007.
FERNANDES, R. M. S. et al. Taxane acute pain syndrome (TAPS) in patients receiving
taxane based chemotherapy for breast cancer—a systematic review. Support Care Cancer,
n. 8, v. 24, p. 3633-50, ago. 2016.
FERREIRA, S. H. Prostaglandins, aspirin-like drugs and analgesia. Nat New Biol., n. 240, v.
102, p. 200-2003, dez. 1972.

150
FIDANBOYLU, M.; GRIFFITHS, L. A.; FLATTERS, S. J. Global inhibition of reactive
oxygen species (ROS) inhibits paclitaxel-induced painful peripheral neuropathy. PLoS One.
n. 6, v. 9, p. 25212, 2011.
FLATTERS, S. J. L.; BENNETT, G. J. Ethosuximide reverses paclitaxel and vincristine-
induced painful peripheral neuropathy. Pain., n. 1-2, v. 109, p.150-161, 2004.
FLATTERS, S. J.; BENNETT, G. J. Studies of peripheral sensory nerves in paclitaxel-
induced painful peripheral neuropathy: evidence for mitochondrial dysfunction. Pain. n. 3, v.
122, p. 245-57 jun. 2006.
FORSYTHE, P.; BIENENSTOCK, J. The mast cell-nerve functional unit: a key com-ponent
of physiologic and pathophysiologic responses. Chem. Imunol. Allergy, v. 98, p. 196–221.
2012.
FREILICH, R. Motor neuropathy due to docetaxel and paclitaxel. Neurology., n. 1, v. 47, p.
115-118, jul. 1996.
GADDUCCI, A. et al. p53 status is neither a predictive nor a prognostic variable in patients
with advanced ovarian cancer treated with a paclitaxel-based regimen. Anticancer Res., n.
6C, v. 20, p. 4793-4799, nov-dec. 2000.
GALLI, S. J.; BORREGAARD, N.; WYNN, T. A. Phenotypic and functional plasticity of
cells of innate immunity: macrophages, mast cells and neutrophils. Nat Immunol, n. 11, v.
12, p. 1035-1044, out. 2011.
GALLI, S. J.; NAKAE, S.; TSAI, M. Mast cells in the development of adaptive immune
responses. Nat. Immunol. n. 2, v. 6, p. 135–142, fev. 2005.
GAO, W. et al. Quercetin ameliorates paclitaxel-induced neuropathic pain by stabilizing mast
cells, and subsequently blocking PKCε-dependent activation of TRPV1. Acta Pharmacol
Sin., n. 9, v. 37, p. 1166–1177, set. 2016.
GARRISON, J. A. et al. Myalgias and arthralgias associated with paclitaxel. Oncology
(Williston Park). n. 2, v. 17, p. 271-277, fev. 2003.
GAUCHAN, P. et al. Involvement of increased expression of transient receptor potential
melastatin 8 in oxaliplatin-induced cold allodynia in mice. Neuroscience Letters, v. 458, n. 2,
p. 93–95, jul. 2009.
GLOBAL BURDEN OF DISEASE CANCER COLLABORATION. The Global Burden of
Cancer 2013. JAMA oncol., v. 1, n. 4, p. 505 – 527, jul. 2015.

151
GRIMBALDESTON, M. A. et al. Mast cell-deficient W-sash c-kit mutant Kit W-sh/W-sh
mice as a model for investigating mast cell biology in vivo. Am J Pathol, n. 3, v. 167, p. 835-
848, set. 2005.
HAGEDORN, L. et al. The Ets domain transcription factor Erm distinguishes rat satellite glia
from Schwann cells and is regulated in satellite cells by neuregulin signaling. Dev. Biol. n.
219, p. 44–58, 2000.
HAMEL, E. et al. Interactions of taxol,microtubule- associated proteins, and guanine
nucleotides in turbulin polymerization. J Biol Chem, n. 22, v. 256, p. 11887-11894, nov.
1981.
HANANI, M. et al. Glial cell plasticityinsensorygangliainducedbynervedamage.
Neuroscience, n. 114, p. 279–283, 2002.
HANANI, M. Intercellular communication in sensory ganglia by purinergic receptors and gap
junctions: implications for chronic pain. Brain Res, v. 1487, p. 183-191, 2012.
HANANI, M. Satellite glial cells in sensory ganglia: from form to function. Brain Research
Review v. 48, p. 457–476, 2005.
HANANI, M.Satelliteglialcellsinsympatheticandparasympatheticganglia: in searchoffunction.
Brain Res.Rev., n. 64, p. 304-327, 2010.
HUANG, T. Y., BELZER, V., HANANI, M.Gapjunctionsindorsalroot ganglia: possible
contribution to visceral pain. Eur. J. Pain, n. 14, p. 1–11, 2010.
HWANG, S.; DING, A. Activation of NF-kappa B in murine macrophages by taxol. Cancer
Biochem Biophys, n. 4, v.14, p. 265– 72, jan. 1995.
INSTITUTO NACIONAL DO CÂNCER. Estimativa 2016: incidência de câncer no
Brasil/Instituto Nacional de Câncer José Alencar Gomes da Silva. Rio de Janeiro, 2015.
INSTITUTO NACIONAL DO CÂNCER. O que é câncer. Disponível em:
<http://www1.inca.gov.br/conteudo_view.asp?id=322> Acesso em: 18 out. 2016.
INTERNATIONAL AGENCY FOR RESEARCH ON CANCER. Pharmaceuticals: a
review of human carcinogens. Lyon, 2012.
INTERNATIONAL ASSOCIATION FOR THE STUDY OF PAIN, 2012.
JAGGI, A. S.; SINGH, N. Mechanisms in cancer-chemotherapeutic drugs-induced peripheral
neuropathy. Toxicology. n. 291, v. 27, p. 1-9, jan. 2012.

152
JAMUR, M. C.; OLIVER, C. Origin, maturation and recruitment of mast cell precursors.
Front Biosci (Schol Ed), n. 3, v. 1, p. 1390-1406, jun. 2011.
JATOI, A. et al. A proof-of-concept trial of protein kinase C iota inhibition with auranofin for
the paclitaxel-induced acute pain syndrome. Support Care Cancer, n. 3, v. 25, p. 833-838,
out. 2016.
JESSEN, K. R.; MIRSKY, R. The origin and development of glial cells in peripheral nerves.
Nat. Rev. Neurosci. v. 6, p. 671–682, 2005.
JIMENEZ-ANDRADE, J. et al. Sensory neurons and their supporting cells located in the
trigeminal, thoracic and lumbar ganglia differentially express markers of injury following
intravenous administration of paclitaxel in the rat. Neuroscience Letters, v. 405, p. 62–67,
set. 2006.
JOHNSON, D.; KRENGER,W. Interactions of mast cells with the nervous system-recent
advances. Neurochem. Res. n. 9, v. 17, p. 939–951. set, 1992.
JORDAN, M. A. et al. Mitotic block induced in HeLa cells by low concentrations of
paclitaxel (Taxol) results in abnormal mitotic exit and apoptotic cell death. Cancer Res., n. 4,
v. 56, p. 816-825, fev. 1996.
JORDAN, M.; WILSON, L. Microtubules as a target for anticancer drugs. Nat Rev Cancer.,
n. 4, v. 4, p. 253-265, abril, 2004.
JORDAN, M. A. Mechanism of mitotic blook and inhibition of cell proliferation by taxol at
low concentrations. Proc Natl Acad Sci USA., n. 20, v. 90, p. 9552–9556, out. 1993.
KAWAKAMI, T.; GALLI, S. J. Regulation of mast-cell and basophil function and survival
by IgE. Nat. Rev. Immunol., n. 10, v. 2, p. 773–786, out. 2002.
KAWAZAKI, K. et al. Mouse toll-like receptor 4.MD-2 complex mediates lipopolysaccaride-
mimrtic signal transduction Taxol. J Biol Chem, n. 4, v. 275, p. 2251-2254, jan. 2000.
KAWAZAKI, K. et al., Involvement of TLR4/MD-2 complex in species-specific
lipopolysaccaride-mimetic signal transduction Taxol. J Endotoxin Res, n. 3, v. 7, p. 232-236,
jan. 2001.
KHASAR, S. G. et al. A novel nociceptor signaling pathway revealed in protein kinase C
epsilon mutant mice. Neuron., n. 24, v. 1, p. 253-260, set. 1999.
KIM, Y. M.; PAIK, S.G. Induction of expression of inducible nitric oxide synthase by taxol in
murine macrophage cell. Biochem Biophys Res Commur. n. 2, v. 326, p. 410-416, jan.
2005.

153
KIMURA, L. F. et al. Mast cells and histamine play an important role in edema and leukocyte
recruitment induced by Potamotrygon motoro stingray venom in mice. Toxicon., v. 103, p.
65-73, set. 2015.
KING, T. C. et al. p53 mutations do not predict response to paclitaxel in metastatic non-small
cell lung carcinoma. Cancer (Phila.), n. 4, v. 89, p. 769-773, ago. 2000.
KIRIKAE, T. et al. Structural requirements of taxoids for nitric oxide and tumor necrosis
factor production by murine macrophages. Biochem Biophys Res Commun, n. 1, v. 227, p.
227–235, out. 1996.
KIRIKAE, T. et al. Structural significance of the acyl group at the C-10 position and the A
ring of the taxane core of paclitaxel for inducing nitric oxide and tumor necrosis factor
production by murine macrophages. FEBS Lett, n. 3, v. 478, p. 221–226, ago, 2000.
KIRIKAE, T. et al. Structural significance of the benzoyl group at the C-3V-N position of
paclitaxel for nitric oxide and tumor necrosis factor production by murine macrophages.
Biochem Biophys Res Commun, n. 3, v. 245, p.698–704. abril, 1998.
KOPP, E. B.; Medzhitov, R. The Toll-receptor family and control of innate immunity. Curr.
Opin. Immunol. v. 1, n.11, p. 13 – 18, fev, 1999.
KUMAR, N. Taxol-induced polymerization of purified turbulin: mechanism of action. J Biol
Chem, n. 20, v. 256, p. 10435-10441, out. 1981.
LAMBRACHT-HALL, M.; DIMITRIADOU, V.; THEOHARIDES, T. C. Migration of mast
cells in the developing rat brain. Dev. Brain. Res., n. 2, v. 56, p. 151–159, nov. 1990.
LAURSEN, J. C. et al. Glutamate dysregulation in the trigeminal ganglion: A novel
mechanism for peripheral sensitization of the craniofacial region. Neuroscience, v. 256, p.
23-35, 2014.
LAVICH, T. R. et al. Combinedaction of vasoactive amines and bradykinin mediates
allergen-evoked thermalhyperalgesia in rats. Eur. J. Pharmacol. n. 1-3, v. 462, p. 185–192,
fev. 2003.
LAVICH, T. R. et al., Neutrophil infiltration is implicated in the sustained thermal hyper-
algesic response evoked by allergen provocation in actively sensitized rats. Pain, n. 1-2, v.
125, p. 180–187, nov. 2006.
LE DOUARIN, N. et al. Glial cell lineages in the neural crest. Glia 4, 175-184, 1991.

154
LEAL-BERUMEN, I.; CONLON, P.; MARSHALL, J. S. IL-6 production by rat peritoneal
mast cells is not necessarily preceded by histamine release and can be induced by bacterial
lipopolysaccharide. J. Immunol. n. 11, v. 152, p. 5468–5476, jun. 1994.
LEDEBOER, A. et al. Intrathecal interleukin-10 gene therapy attenuates paclitaxel-induced
mechanical allodynia and proinflammatory cytokine expression in dorsal root ganglia in rats.
Brain Behav Immun, n. 5, v. 21, p. 686–698, jul, 2007.
LEE, J.; SWAIN, S. Peripheral neuropathy induced by microtubule-stabilizing agents. J Clin
Oncol., n. 10, v. 24, p. 1633-1642, abril, 2006.
LEE, L. F. et al. Taxol-dependent transcriptional activation of IL-8 expression in a subset of
human ovarian cancer. Cancer Res, n. 6, v. 56, p. 1303–1308, mar. 1996.
LI, J.; VAUSE, C. V.; DURHAM, P. L. Calcitonin gene-related peptide sti-mulation of nitric
oxide synthesis and release from trigeminalganglion glial cells. Brain Res., v. 1196, p. 22-32,
2008.
LI, L; ZHOU, X. F. Pericellular Griffonia simplicifolia I isolectin B4- binding ring structures
in the dorsal root ganglia following peripheral nerve injury in rats. J Comp Neurol, n. 439, p.
259–274, 2001.
LI, Y. et al. Toll-like receptor 4 signaling contributes to paclitaxelinduced peripheral
neuropathy. J Pain., n. 7, v. 15, p. 712–725, jul, 2014.
LOESER, J. D.; MELZACK, R. Pain: an overview. Lancet., n. 9164, v. 353, p. 1607-1609,
maio, 1999.
LOPRINZI, C. L. et al. Natural history of paclitaxel-associated acute pain syndrome:
prospective cohort study NCCTG N08C1. J Clin Oncol. v. 29, n. 11, p. 1472 – 1478, mar.,
2011.
LOPRINZI, C. L. et al. The paclitaxel acute pain syndrome: sensitization of nociceptors as
the putative mechanism. Cancer J, n. 6, v. 13, p. 399–403. nov-dec. 2007.
LYON, M. F.; GLENISTER, P. H. A new allele sash (Wsh) at the W-locus and a spontaneous
recessive lethal in mice. Genet Res., n. 3, v. 39, p. 315-322, jun. 1982.
MANFREDI, J. J.; PARNESS, J.; HORWITZ. S. B. Taxol binds to cellular microtubules. J
Cell Biol, n. 3, v. 94, p. 688-696, set. 1982.
MANTHEY, C. L. et al. Lipopolysaccharide antagonists block taxol-induced signaling in
murine macrophages. J Exp Med, n. 2, v. 178, p. 695–702, ago. 1993.

155
MANTHEY, C. L. et al. Taxol increases steady-state levels of lipopolysaccharide-inducible
genes and protein-tyrosine phosphorylation in murine macrophages. J Immunol, n. 7, v. 149,
p. 2459– 65, out. 1992.
MANTHEY, C. L. et al. Taxol provides a second signal for murine macrophage tumoricidal
activity. J Immunol, n. 2, v. 152, p. 825–831, jan. 1994.
MANTYH, P. W. Cancer pain and its impact on diagnosis, survival and quality of life. Nat
Rev Neurosci., n. 10, v.7, p. 797-809, out. 2006.
MARSHALL, J. S. Mast-cell responses to pathogens. Nat Rev Immunol. n. 10, v. 4, p. 787-
799, out. 2004.
MARSHALL, J. S.; MCCURDY, J. D.; OLYNYCH, T. Toll-like receptor–mediated
activation of mast cells: implications for allergic disease?. Int Arch Allergy Immunol., n. 2,
v. 132, p. 87-97, out. 2003.
MCCURDY, J. D. et al. Distinct toll-like receptor 2 activators selectively induce different
classes of mediator production from human mast cells. J Immunol., n. 4, v. 170, p. 1625-
1629, fev. 2003.
MCGRATH, J. C. et al. Editorial: guidelines for reporting experiments involving animals:
the ARRIVE guidelines. Br J Pharmacol, n. 7, v. 160, p. 1573–1576, ago. 2010.
MELLI, G. Alpha-lipoic acid prevents mitochondrial damage and neurotoxicity in
experimental chemotherapy neuropathy. Exp Neurol., n. 2, v. 214, p. 276-84, dec. 2008.
METCALFE, D. D.; BARAM, D.; MEKORI, Y. A. Mast cells. Physiol. Rev., n. 4, v. 77, p.
1033–1079, out. 1997.
METZGER H. The receptor with high affinity for IgE. Immunol Rev, v. 125, p. 37-48, fev.
1992.
MIDULLA, F. et al. Cytokines in the nasal washes of children with respiratory syncytial virus
bronchiolitis. Int J Immunopathol Pharmacol, n. 1, v. 19, p. 231-235, jan-mar. 2006.
MILLER, K. D.; SLEDGE, G. W, JR. Taxanes in the treatment of breast cancer: a prodigy
comes of age. Cancer Investig., n. 2, v. 17, p. 121-136, 1999.
MITCHISON, T.; KIRSCHNER, M. Dynamic instability of microtubule growth. Nature, v.
312, p. 237 – 242, nov. 1984.
MOOS P. J.; FITZPATRICK F. A. Taxane-mediated gene induction is independent of
microtubule stabilization: induction of transcription regulators and enzymes that modulate

156
inflammation and apoptosis. Proc. Natl. Acad. Sci. USA, n. 7, v. 95, p. 3896-3901, mar.
1998.
MOOS P. J.; MUSKARDIN, D. T.; FITZPATRICK, F. A. Effect of Taxol and Taxotere on
gene expression in macrophages: induction of the prostaglandin H synthase-2 isoenzyme. J.
Immunol., n. 1, v. 162, p. 467-473, jan. 1999.
MORITA ,Y.; SIRAGANIAN, R. P. Inhibition of IgE-mediated histamine release from rat
basophilic leukemia cells and rat mast cells by inhibitors of transmethylation. J Immunol, n.
4, v. 127, p. 1339-1344, out. 1981.
MOTA, L. M. H. et al. Brazilian Society of Rheumatology Consensus for the treatment of
rheumatoid arthritis. Rev Bras Reumatol, v. 52, n. 2, p. 135-174, 2012.
MOULDER, S. L. et al. A randomized phase 2 trial comparing 3-h versus 96-h infusion
schedules of paclitaxel for the treatment of metastatic breast cancer. Cancer, n. 4, v. 116, p.
814–821, fev. 2010.
MURPHY, P. et al. The regulation of Krox-20 expression reveals important steps in the
control of peripheral glial cell development. Development. n. 9, v. 122, p. 2847 – 2857, set.
1996.
NEEB, L. et al. IL-1beta stimulates COX-2 dependent PGE(2) synthesis and CGRP release in
rat trigeminal ganglia cells. PLoS One, n. 6, p. 17360, 2011.
NIH - NATIONAL CANCER INSTITUTE. NCI Dictionary of Cancer Terms. Bethesda,
2015.
NOGALES, E. et al. Structure of tubulin at 6.5 A and location of the taxol-binding site.
Nature., n. 6530, v. 375, p. 424-427, jun. 1995.
NOGALES, E.; WANG, H. W. Structural mechanisms underlying nucleotidedependent self-
assembly of tubulin and its relatives. Curr Opin Struct Biol., n. 2, v. 16, p. 221-229, abril,
2006.
OHARA, P. T. et al. Gliopathic pain: when satellite glial cells go bad. Neuroscientist, v. 15,
p. 450–463. 2009.
PACHMAN, D. R. Chemotherapy-induced peripheral neuropathy: prevention and treatment.
Clin Pharmacol Ther, n. 11, v. 90, p.377–387, jul. 2011.
PANNESE, E. The structure of the perineuronal sheath of satellite glial cells (SGCs) in
sensory ganglia. Neuron Glia Biol., v. 6, p. 3–10, 2010.

157
PARK, S. et al. Early, progressive, and sustained dysfunction of sensory axons underlies
paclitaxel-induced neuropathy. Muscle Nerve, n. 3, v. 43, p. 367-374, mar. 2011.
PETERS, C. M. et al. Intravenous paclitaxel administration in the rat induces a peripheral
sensory neuropathy characterized by macrophage infiltration and injury to sensory neurons
and their supporting cells. Exp Neurol., n. 1, v. 203, p. 42–54, jan. 2007.
PIERINI, L.; HOLOWKA, D.; BAIRD, B. Fc epsilon RI-mediated association of 6-micron
beads with RBL-2H3 mast cells results in exclusion of signaling proteins from the forming
phagosome and abrogation of normal downstream signaling. J Cell Biol, n. 6, v. 134, p. 1427-
1439, set. 1996.
PISANO, C. et al. Paclitaxel and Cisplatin-induced neurotoxicity: a protective role of acetyl-
L-carnitine. Clin Cancer Res., n. 15, v. 9, p. 5756-5767, nov. 2003.
PITTMAN, S. K. et al. Paclitaxel alters the evoked release of calcitonin gene-related peptide
from rat sensory neurons in culture. Exp Neurol., v. 253, p. 146–153, mar, 2014.
POLOMANO, R. C. et al. A painful peripheral neuropathy in the rat produced by the
chemotherapeutic drug, paclitaxel. Pain.; n. 3, v. 94, p. 293-304. dez. 2001.
POMONIS, J. D. et al. Expression and localiza-tion of endothelin receptors: implications for
the involvementof peripheral glia in nociception. J Neurosci., n. 21, p. 999-1006, 2001.
PROTA, A. Molecular mechanism of action of microtubule-stabilizing anticancer agents.
Science., n. 6119, v. 339, p. 587-590, fev. 2013.
QIAO, H. et al. FcεR1 and toll-like receptors mediate synergistic signals to markedly
augment production of inflammatory cytokines in murine mast cells. Blood. n. 2, v. 107, p.
610–618, jan. 2006.
QUASTHOFF, S.; HARTUNG, H. Chemotherapy-induced peripheral neuropathy. J Neurol.,
n. 1, v. 249, p. 9-17, jan. 2002.
RAO, S. et al. 3’-(P-Azidobenzamido) taxol photolabels the N-terminal 1 amino ácids of
beta-turbulin. J Biol Chem, n. 5, v. 269, p. 3132-3134, fev. 1994.
RAO, S.; HORWITZ, S. B.; RINGRL, I. Direct photoaffinity labeling of turbulin with taxol.
J Natl Cancer Inst, n. 10, v. 84, p. 785-788, maio, 1992.
REEVES, B. N. et al. Further data supporting that paclitaxel-associated acute pain syndrome
is associated with development of peripheral neuropathy: North Central Cancer Treatment
Group trial N08C1. Cancer. v. 118, n. 20, p. 5171–51178, mar., 2012.

158
REN, K.; DUBNER, R. Interactions between the immune and nervous systems in pain. Nat.
Med., n. 11, v.16, p. 1267-76, nov. 2010.
RIBEIRO, R. A. et al. Involvement of resident macrophages and mast cells in the writhing
nociceptive response induced by zymosan and acetic acid in mice. Eur J Pharmacol. n. 1, v.
387, p. 111-118, jan 2000.
RIGO, F. K. et al. Effect of ω-conotoxin MVIIA and Phα1β on paclitaxel-induced acute and
chronic pain. Pharmacol Biochem Behav, v. 114-115, p. 16–22, dec. 2013.
ROIVAINEN, R.; KOISTINAHO, J. Decentralization induces long-term c-fos protein-like
immunoreactivity in non-neuronal cells in the rat superior cervical ganglion. Neuroscience
Letters, v. 119, n. 1, p. 105 – 108, out., 1990.
ROVINI, A. et al. Olesoxime prevents microtubule-targeting drug neurotoxicity: selective
preservation of EB comets in differentiated neuronal cells. Biochem Pharmacol., n. 6, v. 80,
p. 884-894, set. 2010.
ROWINSKY, E. K. et al. Clinical toxicities encountered with paclitaxel (Taxol). Semin
Oncol., v. 20, n. 3, p. 1 – 15, ago. 1993.
ROWINSKY, E. K. Paclitaxel pharmacology and other tumor types. Semin Oncol, n. 6, v.
24, 1– 12, dec. 1997.
ROWINSKY, E. K.; CAZENAVE, L. A.; DONEHOWER, R. C. Taxol: novel investigational
antimicrotubule agent. J Natl Cancer Inst, n. 15, v. 82, p. 247-1259, ago. 1990.
ROWINSKY, E. K.; DONEHOWER, R. C. Paclitaxel(taxol). New Engl J Med, n. 15, v. 332,
p. 1004-1014, abril, 1995.
SAHENK, Z. Taxol neuropathy. Electrodiagnostic and sural nerve biopsy findings. Arch
Neurol., n. 7, v. 51, p. 726-729, jul. 1994.
SAIBIL, S. et al. Incidence of taxane-induced pain and distress in patients receiving
chemotherapy for early-stage breast cancer: a retrospective, outcomes-based survey. Curr
Oncol, n. 4, v. 17, p. 42-47, ago. 2010.
SCHAIBLE, H. G. Spinal mechanisms contributing to joint pain. Novartis Found Symp, v.
260, p. 4–22, 2004.
SCHIFF, P. B.; HORWITZ, S. B. Taxol stabilizes microtubules in mouse fibroblast cells.
Proc Natl Acad Sci USA, n. 3, v. 77, p. 1561-1565, mar. 1980.

159
SCHIFF, P.; FANT, J.; HORWITZ, S. Promotion of microtubule assembly in vitro by taxol.
Nature., n. 5698, v. 277, p. 665-667, fev. 1979.
SCHIFF, P.; HORWITZ, S. Taxol assembles tubulin in the absence of exogenous guanosine
5’-triphosphate or microtubule-associated proteins. Biochemistry., n. 11, v. 20, p. 3247-3252,
maio, 1981.
SCHWARTZ, L. B. Clinical utility of tryptase levels in systemic mastocytosis and associated
hematologic disorders. Leuk Res, n. 7, v. 25, p. 553-562, 2001.
SCHWARTZ, L. B.; AUSTEN, K. F. Enzymes of the mast cell granule. J. Invest. Dermatol.
n. 5, v. 74, p. 349–353, maio, 1980.
SHIMIZU, A. et al. Human and rat mast cell high-affinity immunoglobulin E receptors:
characterization of putative alpha-chain gene products. Proc Natl Acad Sci USA, n. 6, v. 85,
p. 1907-1911, mar. 1988.
SIAU, C.; XIAO, W.; BENNETT, G. J. Paclitaxel- and vincristine-evoked painful peripheral
neuropathies: loss of epidermal innervation and activation of Langerhans cells. Exp Neurol.,
n. 2, v. 201, p. 507-14, out. 2006.
SILVERMAN, A. J. et al. Mast cells migrate from blood to brain. J. Neurosci., n. 1, v. 20, p.
401–408, jan. 2000.
SINGH, J. A.; BEG, S.; LOPEZ-OLIVO, M. A. Tocilizumab for rheumatoid arthritis: a
Cochrane systematic review. J Rheumatol, v. 8, n. 1, p. 10-20, 2011.
SMITH, S. B.; CRAGER, S. E.; MOGIL, J. S. Paclitaxel-induced neuropathic
hypersensitivity in mice: responses in 10 inbred mouse strains. Life Sci., n. 21, v. 74, p. 2593-
604, abril, 2004.
SOUZA, G. R. et al. Fractalkine mediates inflammatory pain through activation of satellite
glial cells. Proc Natl Acad Sci USA., n. 110, v. 27, p. 11193-11198, jul. 2013.
STEVENS, J.; LOUTIT, J. F. Mast cells in spotted mutant mice (W Ph mi). Proc R Soc
Lond B Biol Sci., n. 1200, v. 215, p. 405-409, jun. 1982.
SUADICANI, S. O. et al. Bidirectional calcium signaling between satellite glial cells and
neurons in cultured mouse trigeminal ganglia. Neuron Glia Biology, n. 6, p. 43–51, 2010.
SUBBARAMAIAH, K. et al. Microtubule-interfering agents stimulate the transcription of
cyclooxygenase: 2. Evidence for involvement of ERK1/2 and p38 mitogen-activated protein
kinase pathways. J Biol Chem, n. 20, v. 275, p. 14838–14845, maio, 2000.

160
SUPAJATURA, V. et al. Differential responses of mast cell Toll-like receptors 2 and 4 in
allergy and innate immunity. J Clin Invest, n. 10, v. 109, p. 1351-1359, maio, 2002.
SWAIN, S. M.; AREZZO, J. C. Neuropathy associated with microtubule inhibitors:
diagnosis, incidence, and management. Clin Adv Hematol Oncol, n. 6, n.6, p. 455-467, jun.
2008.
TAKEDA, M. et al. Enhanced excitability of nociceptive trigeminal ganglion neurons by
satellite glial cytokine following peripheral inflammation. Pain, n. 129, p. 155–166, 2007.
TAKEDA, M. et al. Potassium channels as a potential therapeutic target for tri geminal
neuropathic and inflammatory pain. Mol Pain, v. 7, p. 5. 2011.
TAKEDA, M.; TAKAHASHI, M.; MATSUMOTO, S. Contribution of the acti-vation of
satellite glia in sensory ganglia to pathological pain. Neurosci Biobehav Rev., n. 33, p. 784-
792, 2009.
TANG et al. Inhibitory effect of methyleugenol on IgE-mediated allergic inflammation in
RBL-2H3 cells. Mediators Inflamm., v. 2015, 2015.
TEIXEIRA, M. J. Dor: Princípios e Prática. São Paulo-SP: Artmed Editora, 2009. 165p.
THACKER, M. A. et al., Pathophysiology of Peripheral Neuropathic Pain: Immune Cells and
Molecules. Anesth Analg., n. 3, v. 105, set. 2007.
THEOHARIDES, T. C.; CONTI, P. Mast cells: the Jekyll and Hyde of tumor growth. Trends
Immunol., n. 5, v. 25, p. 235-241, maio, 2004.
THEOHARIDES, T. C.; KALOGEROMITROS, D. The critical role of mast cells in allergy
and inflammation. Ann N Y Acad Sci, v. 1088, p. 78-99, nov. 2006.
VALÉRIO, D. A. et al. Fructose-1,6-bisphosphate reduces inflammatory pain-like behaviour
in mice: role of adenosine acting on A1 receptors. Br J Pharmacol., n. 2, v. 158, p. 558-568,
set. 2009.
VALLIE`RES, L.; RIVEST, S. Regulation of the genes encoding interleukin-6, its receptor,
and gp130 in the rat brain in response to the immune activator lipopolysaccharide and the
proinflammatory cytokine interleukin-1b. J. Neurochem. n. 4, v. 69, p. 1668–1683. out.
1997.
VARADARADJALOU, S. et al. Toll-like receptor 2 (TLR2) and TLR4 differentially activate
human mast cells. Eur J Immunol., n. 4, v. 33, p. 899-906, abril, 2003.
VERRI, W. A. J. et al. Hypernociceptive role of cytokines and chemokines: targets for
analgesic drug development?. Pharmacol Ther., n. 112, v. 1, p. 116-138, out. 2006.
VERSTAPPEN, C. et al. Amifostine protects against chemotherapy-induced neurotoxicity: an
in vitro investigation. Anticancer Res., n. 4, v. 24, p. 2337-2341. jul-ago, 2004.

161
VIKHANSKAYA, F. et al. Inactivation of p53 in a human ovarian cancer cell line increases
the sensitivity to paclitaxel by inducing G2/M arrest and apoptosis. Exp. Cell Res., n. 1, v.
241, p. 96-101, maio, 1998.
VILLA. et al. Temporomandibular joint inflammation activates glial and immune cells in
both the trigeminal ganglia and in the spinal trigeminal nucleus. Molecular Pain, v. 6, p. 89,
2010.
VON HOFF, D. D. The taxoids: same roots, different drugs. Semin Oncol, n. 4, v. 24, p. 3–
10, ago. 1997.
WANG, M. et al. Calpain inhibition protects against Taxol-induced sensory neuropathy.
Brain. n. 3, v. 127, p. 671-679, mar. 2004.
WANI, M. C. et al. Plant antitumor agents. VI. The isolation and structure of taxol, a novel
antileukemic and antitumor agent fron Taxus brevifolia. JAM Chem Soc, n. 9, v. 93, p. 2325-
2327, maio, 1971.
WARWICK ,R. A.; HANANI, M. The contribution of satellite glial cells to chemotherapy-
induced neuropathic pain. Eur J Pain, v. 17, p. 571-580. 2013.
WATKINS, L. R.; MAIER, S. F. Beyond neurons: Evidence that immune and glial cells
contribute to pathological pain states. Physiol Rev., n. 82, p. 981–1011, 2002.
WATKINS, L. R.; MILLIGAN, E. D.; MAIER, S. F. Glial activation: A driving force for
pathological pain. Trends Neurosci, n. 24, p. 450–455, 2001.
WATSON, J. M. et al. Identification of the structural region of taxol that may be responsible
for cytokine gene induction and cytotoxicity in human ovarian cancer cells. Cancer
Chemother Pharmacol, n. 5, v. 41, p. 391–397, 1998.
WHITE, C. M. et al. Effects of paclitaxel on cytokine synthesis by unprimed human
monocytes, T lymphocytes, and breast cancer cells. Cancer Immunol Immunother, v. 46,
p.104–112, abril, 1998.
WILLIAMS, A. C. C.; CRAIG, K. D. Updating the definition of pain. Pain, v. 157, n. 11, p.
2420–2423, nov. 2016.
WILSON, L. et al. Taxol stabilization of microtubules in vitro: dynamics of turbulin addition
and loss at opposite microtubule ends. Biochemistry, n. 19, v. 24, p. 254-5262, set. 1985.
WILSON, L. Microtubules as drug receptors: pharmacological properties of microtubule
protein. Ann N Y Acad Sci.,v. 253, p. 213-251, jun. 1975.
WINDEBANK, A. J.; GRISOLD, W. Chemotherapy-induced neuropathy. J Peripher Nerv
Syst, v. 13, p. 27-46. 2008.
WINER, E. et al. Failure of higher-dose paclitaxel to improve outcome in patients with
metastatic breast cancer: cancer and leukemia group B trial 9342. J Clin Oncol., n. 11, v. 22,
p. 2061-2068, jun. 2004.

162
WOODHAM, P. et al. Satellite cells surrounding axotomised rat dorsal root ganglion cells
increase expression of a GFAP-like protein. Neurosci Lett., n. 98, p. 8–12, 1989.
WOODS C. M. Taxol-induced mitotic block triggers rapid onset of a p53-independent
apoptotic pathway. Mol. Med., n. 5, v. 1, p. 506-526, jul. 1995.
WOOLF C.J. Overcoming obstacles to developing new analgesics. Nat. Med., n. 11, v. 16, p.
1241-7, nov. 2010.
WOOLF, C. J.; MANNION, R. J. Neuropathic pain: aetiology, symptoms, mechanisms, and
management. Lancet., n. 9168, v. 353, p. 1959-1965, jun. 1999.
WRIGHT, S. D., R. A. et al. CD14, a receptor for complexes of lipopolysaccharide (LPS) and
LPS binding proteins. Science, v. 21, n. 249, p. 1431 - 1433, set. 1990.
XANTHOS, D. N. et al. Central nervous system mast cells in peripheral inflammatory
nociception. Mol. Pain, v. 7, jun. 2011.
YAN, X. et al. Paclitaxel induces acute pain via directly activating toll like receptor 4. Mol
Pain., n. 10, v. 11, mar. 2015.
ZAKS-ZILBERMAN, M. et al. Induction of adrenomedullin mRNA and protein by
lipopolysaccharide and paclitaxel (Taxol) in murine macrophages. Infect. Immun, n. 10, v.
66, p. 4669– 4675, out. 1998.
ZAKS-ZILBERMAN, M.; ZAKS, T. Z.; VOGEL, S. N. Induction of proinflammatory and
chemokine genes by lipopolysaccharide and paclitaxel (Taxol) in murine and human breast
cancer cell lines. Cytokine, n. 3, v. 15, p. 156–165, ago. 2001.
ZHANG, H. et al. Altered functional properties of satellite glial cells in compressed spinal
ganglia. Glia., v. 57, p. 1588–1599, 2009.
ZHANG, H. et al. Induction of Monocyte Chemoattractant Protein-1 (MCP-1) and Its
Receptor CCR2 in Primary Sensory Neurons Contributes to Paclitaxel-Induced Peripheral
Neuropathy. J Pain., n. 10, v. 14, p.1031-44, out. 2013.
ZHANG, X. et al. Neuronal somatic ATP release triggers neuron–satellite glial cell
communication in dorsal root ganglia. Proceedings of the National Academy of Sciences of
United States of America, n. 104, p.9864–9869, 2007.
ZIMMERMANN, M. Ethical guidelines for investigations of experimental pain in conscious
animals. Pain, n. 2, v. 16, p. 109–110, jun. 1983.
ZUO, Y. et al. Inflammation and hyperalgesia induced by nerve injury in the rat: a key role of
mast cells. Pain, n. 3, v. 105, p. 467–479, out. 2003.

163
ANEXOS

164

165
Top Related