







Línguas
Páginas
Legal


UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE ZOOLOGIA
CURSO DE MESTRADO DO PROGRAMA DE PÓS-GRADUAÇÃO
EM BIOLOGIA ANIMAL
COMPORTAMENTO DE FORRAGEIO DO BOTO-CINZA (Sotalia guianensis) EM OLINDA E
NO PORTO DO RECIFE, PERNAMBUCO.
MILENA SANTOS MONTEIRO
RECIFE, 2008.

2
MILENA SANTOS MONTEIRO
COMPORTAMENTO DE FORRAGEIO DO BOTO-CINZA (Sotalia guianensis) EM OLINDA E
NO PORTO DO RECIFE, PERNAMBUCO.
ORIENTADORA: Maria Adélia Borstelmann de Oliveira
CO-ORIENTADOR: Lídio França do Nascimento
RECIFE, 2008.
Dissertação apresentada ao Programa de Pós-Graduação em Biologia Animal da Universidade Federal de Pernambuco, como parte dos requisitos para obtenção do grau de Mestre em Ciências Biológicas na área de Biologia Animal.

3
Monteiro, Milena Santos Comportamento de forrageio do Boto-cinza (Sotalia
guianensis) em Olinda e no Porto do Recife, Pernambuco. / Milena Santos Monteiro. – Recife: O Autor, 2008.
92 folhas : il., fig. tab. Dissertação (mestrado) – Universidade Federal de
Pernambuco. CCB. Biologia Animal, 2008. Inclui bibliografia. 1. Boto-cinza - comportamento 2. Cetáceos – forrageio 3.
Sotalia guianensis I. Título. 599.53 CDU (2.ed.) UFPE 599.53 CDD (22.ed.) CCB – 2008- 093

4
MILENA SANTOS MONTEIRO
COMPORTAMENTO DE FORRAGEIO DO BOTO-CINZA (Sotalia guianensis) EM OLINDA E
NO PORTO DO RECIFE, PERNAMBUCO.
BANCA EXAMINADORA
Data: 25 de fevereiro de 2008
RECIFE, 2008

5
Dedico esta dissertação à minha mãe,
que guiou meus passos e esteve eternamente
presente em minha vida.
Ao meu namorado e à sua família, que
sempre vibraram pela minha felicidade e sucesso.

6
AGRADECIMENTOS
Meus sinceros agradecimentos:
A Deus, por ter iluminado meu caminho e concedido toda a força para que eu superasse todos as
dificuldades.
A toda minha família, especialmente a minha mãe, Carminha, por todo carinho, amizade,
companheirismo e ajuda incansável durante todas as fases desse trabalho. Sem ela sei que nada disso seria
possível.
A João Paulo, meu eterno companheiro, por todas às vezes, que me incentivou e me ajudou
incondicionalmente para que eu realizasse esse sonho. Também não poderia de esquecer de sua família,
principalmente do Sr. Severino, seu pai, que tão gentilmente me levava e trazia todos os dias para a Praia
de Bairro Novo, sempre na esperança de ver um golfinho.
A Maria Adélia, por haver me aceitado como sua orientanda e ajudado para que essa pesquisa
acontecesse. Obrigada também, pela amizade e carinho para comigo durante esses anos de convivência.
A Lídio França, por mais uma vez, co-orientar-me. Independente da distancia, sempre foi e será
para mim um exemplo de dedicação e sabedoria.
Ao Profº Manuel Flores e a toda sua equipe do Laboratório de Química, pelas análises de minhas
amostras e por todo o conhecimento que me trnsmitiram sobre monitoramento químico.
Ao Profº Gabriel Rivas de Melo, a Ricardo Emídio e ao amigo, Fernando Silva, pela ajuda com as
análises estatísticas tão importantes para elaboração deste trabalho.
A todos os meus queridos estagiários, Adriana, Andressa, Ariane, Bárbara, Elis Carla, Fábio,
Fernanda, Georgea, Hugo, Jefferson, Juliana, Julianne, Karla Monique, Maria Priscila, Marília, Raíssa,
Regina, Roberta, Sávio, Sérgio e Simone. Todos estes, que praticamente firmaram um pacto de
casamento, pois estiveram comigo na alegria de ver os golfinhos e na tristeza de sua ausência, na saúde e
na doença, sob o sol escaldante e as sob as chuvas que muitas vezes impediram até de nos alimentarmos.
Obrigada a todos. Vocês foram fundamentais na realização deste trabalho. Nunca me esquecerei de todos
os momentos em que passamos juntos.
À Janaína Araújo, pelos laços de amizade e pelas sugestões e “dicas” nos momentos de dúvidas e
por compartilhar na realização do I Simpósio Nordestino de Mamíferos Aquáticos (SINEMA), aqui em
Pernambuco.

7
A todos os meus amigos de turma, pelos momentos inesquecíveis de convivência e alegria, pela
paciência, incentivo e companheirismo, que sempre guardarei em um cantinho especial do meu coração.
Wel, Ana Karla e Vivi, obrigada por me ajudarem sempre nos meus momentos de desespero, vocês são
uns anjos da guarda para mim.
Ao Sr. Jarbas, por nos transportar todos os dias em seu barquinho “Paciência I”, com tanto
cuidado e competência e por muitas vezes ser o nosso anjo da guarda.
A essas criaturinhas tão especiais, os golfinhos, por me proporcionarem momentos tão
maravilhosos e mágicos que não me canso de viver.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), por financiar o meu
Projeto, tornando possível sua realização.

8
SUMÁRIO
Página
LISTA DE FIGURAS 10
LISTA DE TABELAS 12
RESUMO 13
ABSTRACT 14
1. INTRODUÇÃO
1.1. Aspectos Gerais 15
1.2. Sotalia guianensis 16
1.3. Comportamento de Forrageio 20
1.4. Influência de fatores Ambientais na Ocorrência e Comportamento de Forrageio 22
2. OBJETIVOS
2.1. Geral 24
2.2. Específicos 24
3. MATERIAIS E MÉTODOS
3.1. Caracterização das Áreas de Estudo 25
3.2. Metodologia em Campo
3.2.1. Local e Período de Observação 27
3.2.2. Coleta dos Dados 30
3.2.3. Definições Comportamentais 30
3.2.4. Parâmetros Ambientais 33
3.2.5. Composição de Grupo 36
3.2.6. Atividade de Forrageio e Produção de Pescado 36
3.3. Análise dos Dados 39
4. RESULTADOS
4.1. Presença dos Botos-cinza
4.1.1. Sazonalidade da Ocorrência dos Botos-cinza 40
4.1.2. Número de Botos Avistados Simultaneamente 41
4.1.3. Comparação entre os Parâmetros Ambientais do Porto do Recife e de
Olinda
42
4.2. Caracterização do Comportamento de Forrageio 49

9
4.3. Comportamento de Forrageio e Tamanho de Grupo 52
4.4. Influência das Variáveis Ambientais no Comportamento de Forrageio
4.4.1. Sazonalidade 54
4.4.2. Maré e Fases Lunares 56
4.4.3. Parâmetros Físicos e Químicos 57
4.5. Uso da Área do Porto do Recife para Forrageio 61
5. DISCUSSÃO
5.1. Ocorrência de Botos-cinza no Porto do Recife e em Olinda 62
5.2. Número de Botos Avistados Simultaneamente 64
5.3. Caracterização do Comportamento de Forrageio 65
5.4. Sazonalidade das Atividades de Forrageio 69
5.5. Uso da Área do Porto do Recife para Forrageio 73
6. CONCLUSÕES 75
7. REFERÊNCIAS BIBLIOGRÁFICAS 76
8. ANEXOS 86

10
LISTA DE FIGURAS Página Figura 1: Desenho esquemático do boto-cinza (Sotalia fluviatilis). 17
Figura 2: Mapa da distribuição geográfica da espécie Sotalia guianensis. 19
Figura 3. Áreas de estudo: Praia de Bairro Novo e Porto do Recife 26
Figura 4. Foto do Ponto de Observação no Porto do Recife. 28
Figura 5. Foto do Ponto de Observação em Olinda. 29
Figura 6. Estratégias de forrageio utilizadas pelos botos-cinza. 32
Figura 7. Divisão do Porto do Recife em áreas. 35
Figura 8: Freqüência absoluta de horas que os botos-cinza estiveram presentes
durante os meses de observação no Porto do Recife e em Olinda.
40
Figura 9: Freqüência absoluta de horas que os botos-cinza estiveram presentes
durante os turnos da manhã e tarde no Porto do Recife e em Olinda.
41
Figura 10. Freqüência absoluta do número de botos-cinza observados
simultaneamente no Porto do Recife e em Olinda.
42
Figura 11. Variações sazonais dos parâmetros ambientais. 43
Figura 12. Variação dos nutrientes inorgânicos dissolvidos. 45
Figura 13. Variação sazonal na quantidade total de presas do boto-cinza pescadas
no Recife e em Olinda nos meses de janeiro a dezembro de 2004.
47
Figura 14. Freqüências relativas das famílias de presas do boto-cinza pescadas no
Recife e em Olinda.
47
Figura 15. Freqüência relativa do número de comportamentos observados no
Porto do Recife e em Olinda.
50
Figura 16. Freqüência relativa dos tipos de comportamentos de forrageio
observados no Porto do Recife e em Olinda.
50
Figura 17. Comparação entre as freqüências de saltos na captura de presas pelos
botos-cinza no Porto do Recife e em Olinda.
51
Figura 18. Percentagens dos tipos de estratégias alimentares utilizadas pelos
botos-cinza no Porto do Recife em Olinda.
51
Figura 19. Freqüências relativas dos comportamentos de forrageio realizados na
presença das categorias de número de botos avistados no Porto do Recife e em
Olinda.
52

11
Figura 20.Variação das freqüências absolutas de atividades de pesca realizados
pelo número de golfinhos forrageando conjuntamente no Porto do Recife e em
Olinda.
53
Figura 21. Variação do número de forrageios cooperativos nos diferentes
tamanhos de grupos de boto-cinza presentes em Olinda e no Porto do Recife.
53
Figura 22. Variação do número de forrageios observados ao longo dos meses do
ano no Porto do Recife e em Olinda.
54
Figura 23. Comparação entre as freqüências de atividades de forrageio nas
estações seca e chuvosa no Porto do Recife.
54
Figura 24. Comparação entre as freqüências de forrageios nos turnos da manhã e
da tarde no Porto do Recife.
55
Figura 25. Variação no número de comportamentos de forrageio registrados ao
longo do dia no Porto do Recife e em Olinda.
55
Figura 26. Freqüências relativas do número de comportamentos de forrageio
observados durante os diferentes estados de maré no Porto do Recife.
56
Figura 27. Variação do número de forrageios registrados nos diferentes estados
de maré em função das marés de quadratura e sizígia no Porto do Recife.
57
Figura 28. Correlação entre salinidade e o número de comportamentos de
forrageio registrados no Porto do Recife.
58
Figura 29. Correlação entre os índices pluviométricos mensais e o número de
comportamentos de forrageio registrados no Porto do Recife.
58
Figura 30. Correlação entre as concentrações de nutrientes inorgânicos
dissolvidos e o número de forrageios registrados no Porto do Recife.
59
Figura 31. Correlação entre a quantidade total de presas potenciais do boto-cinza
pescadas e o número de forrageios observados no Porto do Recife.
60
Figura 32. Correlação entre o esforço pesqueiro direcionado a espécie Mugil
curema e a quantidade de forrageios registrados no Porto do Recife.
60
Figura 33. Número de forrageios realizados pelos botos-cinza em cada área do
Porto do Recife.
61
Figura 34. Freqüência absoluta de atividades de forrageio observadas para as
diferentes categorias de profundidades no Porto do Recife.
61

12
LISTA DE TABELAS
Página
Tabela 1. Repertório dos comportamentos de forrageio realizados pelo boto-cinza 31
Tabela 2. Espécies que compõe a dieta do boto-cinza presentes no Recife e
Olinda.
37
Tabela 3. Médias dos valores dos parâmetros ambientais no Porto do Recife e em
Olinda.
46
Tabela 4. Quantidade de presas do boto-cinza pescadas no Recife durante o
período de janeiro a dezembro de 2004.
48
Tabela 5. Quantidade de presas do boto-cinza pescadas em Olinda durante o
período de janeiro a dezembro de 2004.
48

13
RESUMO
O comportamento de forrageio compreende um conjunto de atividades realizadas pelos golfinhos
para obtenção de suas presas e pode variar de acordo com características específicas de cada região. O
presente trabalho objetivou realizar um estudo comparativo entre o comportamento de forrageio do boto-
cinza em Olinda e no Porto do Recife. A coleta dos dados foi realizada em pontos fixos. Os métodos de
amostragem utilizados foram “Todas as Ocorrências” e “Varredura”. O estudo foi conduzido entre
setembro de 2006 e agosto de 2007, com observações realizadas cinco dias consecutivos por mês no
período de 8h às 16 horas. Ao todo foram realizadas 399 horas de esforço amostral em cada área,
resultando em 177 horas de observação direta dos animais no Porto do Recife e 13 horas em Olinda. A
ocorrência de botos-cinza no Porto foi significativamente maior. O período do ano de maior freqüência de
eventos de forrageios do botos-cinza no Porto do Recife foi de setembro a dezembro. Em Olinda, os botos
foram observados apenas no período de novembro a fevereiro, sendo este último mês, o de maior
freqüência dos registros de forrageio. Em ambas as regiões, os golfinhos foram mais avistados e
forragearam mais durante o turno da manhã. Foram registrados ao todo 1264 comportamentos de
forrageio no Porto do Recife e 37 em Olinda. No Porto, foram registrados 11 comportamentos de
forrageio diferentes e 4 em Olinda. Em ambas as regiões, os eventos de forrageio ocorreram em maior
número na presença de 4 a 6 golfinhos. No Porto do Recife, a quantidade de forrageios parece ser
influenciada indiretamente por variáveis ambientais, como a estação do ano, os estados de maré, a
salinidade, o nível de precipitação pluviométrica e os teores de nutrientes inorgânicos dissolvidos e
diretamente pela sazonalidade de ocorrência das presas que compõem sua dieta.
Palavras-chave: comportamento; cetáceos; alimentação; ecologia; Sotalia.

14
ABSTRACT
Foraging behavior includes the assemblage of dolphin’s activities to obtain prey and it could vary
in dependence to specific characteristics of each region this cetacean lives. The main objective was to
realize a compartive foraging behavior study of the estuarine dolphin, Sotalia guianensis, in Olinda and
Recife Port. Data were obtained from fixed points. All Occurrence and Scan sampling observations were
carried out, five consecutive days per month, from 8:00 to 16:00 hours, totalizing 399 hours of sampling
effort for each point, resulting in 177 hours of dolphin direct observations at Recife Port and 13 hours at
Olinda (Bairro Novo Beach). These points were monitored between September, 2006 to August, 2007.
Estuarine dolphin occurrence at Recife Port was significantly bigger. A greater frequency of foraging
events of the estuarine dolphin at Recife Port was between September and December. At Olinda the
estuarine dolphins were observed only from November to February. This last month reach the greatest
frequency of foraging records. On both points the dolphins were visualized and spend more time on
foraging at day shift. We recorded 1264 foraging events at Recife Port and 37, at Olinda. Eleven different
patterns of foraging behavior were seen at Recife Port and four types at Olinda. For both regions the
majority of the foraging events involved 4 to 6 dolphins. At Recife Port the frequency of foraging events
seems to be indirectly influenced by environmental parameters as season, tide stage, salinity, rain level,
and the amount of dissolved inorganic nutrients; and it seems to be directly influenced by the seasonality
of occurrence on the preys that is part of its diet.
Key-words: behavior; cetaceans; diet; ecology; Sotalia.

15
1. INTRODUÇÃO:
1.1. Aspectos Gerais:
A ordem Cetacea é representada por mamíferos adaptados à vida aquática. Compreendem baleias,
golfinhos, botos e toninhas (PINEDO et al., 1992). No mundo existem cerca de 79 espécies de cetáceos,
(CASTELLO, 1996) e no Brasil chegam a 37 espécies descritas (IBAMA, 2001). São encontrados em
todos os oceanos do mundo, desde as águas geladas dos pólos até as águas quentes dos trópicos, bem
como em estuários e rios (PINEDO et al., 1992).
A ordem Cetacea está dividida em três sub-ordens: Archaeoceti, que compreende todos os
cetáceos extintos que viveram no Eoceno (55-38 m.a.) e no Oligoceno (38-28 m.a.). Os Archeoceti deram
origem aos cetáceos pertencentes às outras duas sub-ordens, os Misticetos e Odontocetos (FORDYCE &
BARNES, 1994).
A sub-ordem Mysticeti é composta por quatro famílias e distribuída em onze espécies de grandes
baleias, que na sua fase embrionária possuem um germe dentário, que é reabsorvido durante o
desenvolvimento embrionário e, na fase adulta, adquirem barbatanas córneas de queratina utilizadas para
filtrar seu alimento. Possuem dois orifícios respiratórios, os ossos da mandíbula são ligados frouxamente
e os ossos do crânio são simétricos (SIMÕES-LOPES, 2005). Alimentam-se de cardumes de pequenos
peixes, plâncton (microalgas e pequenos crustáceos como o krill e copépodos). Geralmente têm hábito
solitário, sendo apenas encontradas em grandes grupos durante migrações nos períodos de alimentação,
para altas latitudes, e de reprodução, para as baixas latitudes (HETZEL & LODI, 1993).
A sub-ordem Odontoceti compreende os cetáceos com dentes e apresentam apenas uma abertura
nasal, porém possuem duas passagens respiratórias cranianas. Os ossos da mandíbula são fusionados e o
crânio é assimétrico em alguns grupos e, em outros, não (SIMÕES-LOPES, 2005). Alimentam-se de
peixes, lulas, crustáceos e em alguns casos, de aves marinhas e de mamíferos marinhos (PINEDO et al.,
1992). Geralmente não apresentam diferenças entre machos e fêmeas. Apenas os Odontocetos possuem
um sistema de ecolocalização, que é utilizado para informar distância, tamanho, forma e textura
(SIMÕES-LOPES, 2005). Existem oito famílias e 66 espécies descritas para a sub-ordem Odontoceti
(PINEDO et al., 1992). A família Delphinidae é a maior família de odontocetos, possuindo 32 espécies,
dentre elas as mais conhecidas popularmente são: o golfinho-rotador (Stenella longirostris), o golfinho

16
nariz-de-garrafa (Tursiops truncatus), a Orca (Orcinus orca), e o boto-cinza (Sotalia guianensis)
(PINEDO et a.l, 1992; HETZEL & LODI, 1993).
1.2. Sotalia guianensis
No gênero Sotalia GRAY, 1866, foram reconhecidas inicialmente 58 formas nominais, das quais
22 foram transferidas para o gênero Souza GRAY, 1866 (HERSHKOVITZ, 1966 apud SIMÕES-LOPES,
2006). De acordo com a distribuição geográfica, foram consideradas posteriormente três espécies, Sotalia
fluviatilis (na Amazônia), Sotalia guianensis (na Guiana Francesa) e Sotalia brasiliensis (no Brasil), e
duas subespécies, S. fluviatilis guianensis (marinho) e S. fluviatilis fluviatilis (fluvial) (ALVES-JÚNIOR
& MONTEIRO-NETO, 1999). Contudo, recentes estudos morfométricos (MONTEIRO-FILHO et al.,
2002) e com DNA mitocrondial (CUNHA et al., 2005), evidenciaram a existência de duas espécies
distintas: uma fluvial denominada de tucuxi (Sotalia fluviatilis), endêmica à bacia dos rios Amazonas e
Orinoco e o boto-cinza (Sotalia guianensis), espécie marinha que ocorre na costa das Américas Central e
do Sul (BOROBIA et al., 1991; DA SILVA & BEST, 1996; ALVES-JÚNIOR & MONTEIRO-NETO,
1999).
O boto-cinza é uma das menores espécies de delfinídeos existentes, tendo em média 1,7 metros,
podendo atingir até no máximo 2,2 metros de comprimento (HETZEL & LODI, 1993; DA SILVA &
BEST, 1996). Apresenta como características morfológicas: nadadeira dorsal triangular, pequena, elevada
e situada no centro do dorso; melão arredondado e bem distinto do rostro; apresenta uma coloração que
varia do tom cinza escuro no dorso a tonalidades mais claras na região lateral do corpo, podendo a região
ventral apresentar-se na cor branca ou rosa, sendo a última, mais comum em animais imaturos (PINEDO
et al., 1992; HETZEL & LODI, 1993; DA SILVA & BEST, 1996) (Fig. 1).

17
Figura 1: Desenho esquemático do boto-cinza (Sotalia fluviatilis) (Projeto MAQUA).
O boto-cinza distribui-se geograficamente desde o litoral norte de Honduras, na América Central
(15058’S; 85042’W; DA SILVA & BEST, 1996) até o Estado de Santa Catarina, no Brasil (27o35’S;
48o34’W; SIMÕES-LOPES, 1988), apresentando uma distribuição extensa e possivelmente contínua ao
longo da costa brasileira (PINEDO et al., 1992) (Fig. 2). Habita regiões costeiras, principalmente baías,
canais entre baías e mar aberto, entradas de rios e áreas próximas a manguezais (HOELZEL, 1991;
TORRES & BEASLEY, 2003; FAVARO, 2004).
A dieta do boto-cinza é composta em grande parte por peixes das famílias Sciaenidae,
Haemulidae, Gerreidae, Mugilidae, Paralechtidae, Batrocoididae e Trichiuridae, sendo a maioria desses
demersais, associados a fundos arenosos ou barrentos, indicando a distribuição costeira dessa espécie. Em
menor proporção, o boto alimenta-se de cefalópodes (Lolligunla brevis – Loliginidae) e crustáceos das
espécies Penaeus paulensis, Penaeus schmitii. A presença de espécies pelágicas e demersais na dieta do
boto-cinza demostra que esses golfinhos podem se alimentar em diversas profundidades. (BOROBIA &
BARROS, 1989; DI BENEDITTO et al., 2001; ZANELATTO, 2001; SANTOS et al., 2002).
Geralmente, os botos-cinza são encontrados em grupos de dois a cinco animais, porém podem
formar grupos de dezenas de indivíduos, como foi reportado por GEISE et al. (1999) no sudeste do Brasil.
São extremamente sociais entre si, apresentando padrões de comportamento sociais bastante variados,
principalmente entre adultos, juvenis e filhotes, que podem constituir desde simples deslocamentos até
comportamentos de brincadeiras e forrageio (GEISE, 1991; SPINELLI et al., 2002).
Não se tem registro de predadores para o boto-cinza, no entanto o tubarão-cabeça-chata
(Carcharinus leucas), que utiliza sistemas estuarinos, além de tubarões costeiros de outras espécies e a
orca, que habitam a costa da América Central e do Sul, podem constituir predadores potenciais desses
golfinhos (DA SILVA & BEST, 1996).

18
Por ser uma espécie costeira, o boto-cinza vem sofrendo uma crescente ameaça decorrente do
aumento das atividades humanas na costa e conseqüente degradação do seu habitat. Dentre essas
ameaças, as principais são urbanização e industrialização das regiões costeiras; poluição do ambiente
marinho por produtos químicos e orgânicos; exploração inadequada de manguezais e estuários; aumento
do tráfego de barcos pesqueiros e turísticos; e as capturas acidentais por redes de pesca (DI BENEDITTO
et al., 2000; LODI, 2003).
A falta de informação levou o boto-cinza a ser classificado como espécie insuficientemente
conhecida na Lista Vermelha de Animais Ameaçados da União Internacional para Conservação da
Natureza e dos Recursos Naturais (IUCN, 2007); como espécie ameaçada de extinção no Apêndice I da
Convenção sobre o Comércio Internacional das Espécies da Flora e da Fauna Selvagens em Perigo de
Extinção (CITES, 2003); no Plano de Ação do Ibama para Mamíferos Aquáticos do Brasil (IBAMA,
2001); e espécie com estado de conservação desfavorável no Apêndice II da Convenção para
Conservação de Espécies Migratórias de animais Selvagens (CMS, 2002).

19
Figura 2: Mapa da distribuição geográfica da espécie Sotalia guianensis (Lídio França).

20
1.3. Comportamentos de Forrageio:
De maneira geral, os comportamentos estudados em Sotalia guianensis e em outros cetáceos
compreendem atividades aéreas (saltos, cambalhotas, spy hoping e batidas de partes do corpo),
deslocamentos (rápido, lento, mergulho em velocidade) e atividades de forrageio (perseguição, bote,
manipulação da presa, e estratégias de forrageio individuais ou em grupo). (ANDRADE et al., 1987;
GEISE, 1991; HAYES, 1999; ARAÚJO et al., 2001; MONTEIRO-FILHO, 1991; NASCIMENTO, 2002;
SPINELLI et al., 2002; POLETTO, 2003; SOUZA, 2004).
O termo “forrageio” tem sido utilizado para descrever um conjunto de atividades comportamentais
realizadas por cada golfinho ou por um grupo de golfinhos para obtenção de suas presas (HAYES, 1999).
As estratégias de obtenção de alimento podem ser divididas em duas grandes categorias: os padrões de
busca de alimento e os padrões de captura (SIMÕES-LOPES, 2005). Alguns autores consideram uma fase
intermediária, que é a de detecção de presas, porém esta categoria é bastante difícil de ser distinguida dos
padrões de captura (BEL’KOVICH et al., 1991).
Essas atividades de alimentação na superfície consistem em deslocamentos na superfície da água,
seguidos de mergulhos para captura de presas, perseguições a presas, arremesso da presa, mergulho em
áreas com possível abundância de alimento e consumação da presa (ARAÚJO et al., 2001;
BEL’KOVICH et al., 1991; GEISE, 1991; NASCIMENTO, 2002; NOWACEK, 2002).
Estratégias alimentares são reportadas para muitas espécies de cetáceos em todo o mundo.
Algumas são bastante peculiares, podendo até ocorrer variações entre populações diferentes de uma
mesma espécie. SAULITIS et al. (2000) observaram que a população residente de orcas (Orcinus orca)
captura salmões (Oncorhynchus tshanytscha) por meio de estratégias de agrupamentos e botes individuais
na superfície da água, enquanto mamíferos marinhos são obtidos por orcas transientes através de
movimentos de caça coordenados de vários membros de um mesmo grupo.
Golfinhos pintados do Atlântico (Stenella frontalis) formam sub-grupos associados entre si para
cercar e conduzir cardumes de peixes para a superfície da água produzindo bolhas de ar e utilizando saltos
e batidas de cauda (FERTL &WÜRSIG, 1995).
O golfinho-nariz-de-garrafa (Tursiops truncatus) apresenta um vasto repertório comportamental
de forrageio, utilizando várias estratégias de caça. BEL’KOVICH et al. (1991) reportaram em seu estudo,
vários tipos de estratégias individuais e em grupo. NOWACEK (2002) descreveu e analisou cinqüenta e
sete seqüências de comportamento de forrageio que antecediam o sucesso na captura de peixes por

21
golfinhos-nariz-de-garrafa na Baía de Sarasota. Técnicas de observações embaixo da água permitiram a
visualização de Golfinhos-nariz-de-garrafa procurando e escavando o sedimento com o rostro e parte do
corpo para capturar peixes bentônicos próximo à ilha de Bahamas (ROSSBACH & HERZINC, 1997).
As primeiras descrições de estratégias alimentares do boto cinza se referem a dois tipos básicos de
comportamento: a Pesca ao acaso ou aleatória e a Pesca cooperativa ou comunitária (ANDRADE et al.,
1987; GEISE, 1991; MONTEIRO-FILHO, 1992).
MONTEIRO-NETO et al. (1996), reportaram duas modalidades de comportamentos de pesca para
botos-cinza: a Pesca Individual ou Sincrônica, quando o indivíduo forrageia sem nenhuma organização
aparente entre os demais golfinhos presentes, e a Pesca em Grupo ou Sincrônica, quando vários golfinhos
são avistados trabalhando juntos para concentrar e cercar cardumes.
ROSSI-SANTOS (1997) registrou a ocorrência de seis estratégias de pesca para o boto-cinza em
Santa Catarina, as quais foram denominadas: Pesca aleatória individual, Pesca aleatória em grupo, Pesca
cooperativa em leque, Pesca cooperativa circular, Pesca cooperativa cruzada e Pesca cooperativa em
Zigue-Zague.
Na Praia de Pipa - RN (MONTEIRO et al., 2006), em Cananéia – SP e Ilha da Peças – PR
(DOMIT, 2006), os botos-cinza apresentaram um rico repertório comportamental de caça, no qual foram
observados 18, 20 e 19 comportamentos, respectivamente, relacionados à atividade de forrageio. Esses
comportamentos caracterizaram-se por um conjunto de diversas condutas, tais como saltos, batidas com
partes do corpo e periscópios realizadas individualmente ou em grupo, que também são comuns em
outros padrões comportamentais como deslocamentos e brincadeiras.

22
1.4. Influência de fatores Ambientais na Ocorrência e Comportamento de Forrageio
As estratégias alimentares adotadas pelos golfinhos podem variar de acordo com vários fatores,
tais como: espécie e tamanho do grupo de presas; seu habitat, comportamento e capacidade de fuga;
agilidade de perseguição do predador; experiência de vida adquirida; tamanho e interação do grupo de
predadores. (NASCIMENTO, 2002; NOWACEK, 2002; SOUTO, 2003).
A dispersão e densidade de recursos disponíveis em um ambiente variam espacial e
temporalmente. Estas flutuações quantitativas e qualitativas produzem mudanças em relação ao que é
consumido, influenciando o comportamento alimentar de vários mamíferos (OATES, 1987 apud
DAURA-JORGE et al., 2004). Assim, variáveis abióticas podem influenciar diretamente variáveis
bióticas, tais como distribuição e abundância de recursos alimentares, e, indiretamente, o comportamento
de forrageio dos predadores (WURSIG; WURSIG, 1980).
Alguns estudos relacionam a ocorrência, distribuição, uso do habitat e comportamento dos botos-
cinza com variáveis ambientais, como fotoperíodo (GEISE et al., 1999; HAYES, 1999; ARAÚJO, 2001;
LODI, 2002; SANTOS & LE PENDU, 2007), estado da maré (ARAÚJO, 2001; ROSSI-SANTOS, 2006),
fases da lua (ROSSI-SANTOS, 2006), profundidade (GEISE et al, 1999; ROSSI-SANTOS, 2006),
temperatura da água (DAURA-JORGE et al., 2004), salinidade (ROSSI-SANTOS, 2006), estações do ano
(HAYES, 1999; LINK, 2000; LODI, 2002), topografia e morfologia de fundo (DOMIT, 2006; ROSSI-
SANTOS, 2006), variações nas características físicas regionais (LODI, 2002) e tamanho de grupos
(POLETTO, 2003; LODI, 2003; DOMIT, 2006; ROSSI-SANTOS, 2006).
Dessa forma, o comportamento de forrageio pode apresentar variações de acordo com
características topográficas e hidrodinâmicas específicas de cada região. Em geral, áreas costeiras com
formação de baías, estuários e desembocaduras de rios são utilizadas pelo boto-cinza como áreas de
descanso, interação social, reprodução, cuidado parental e alimentação, pois representam áreas com águas
calmas, com risco mínimo ou ausente da ação de predadores e com grande quantidade de recursos
alimentares (BOROBIA et al., 1991; DA SILVA & BEST, 1996; GEISE et al., 1999; ARAÚJO, 2001;
NASCIMENTO, 2002; LODI, 2003; DAURA-JORGE et al., 2004; FLORES & BAZZALO, 2004;
ROSSI-SANTOS, 2006; CREMER, 2007).
O sucesso do gerenciamento de áreas ambientais depende da qualidade de informações disponíveis.
Para a preservação de regiões habitadas por cetáceos, é necessário o conhecimento sobre uso de área e de

23
como os fatores ambientais influenciam na sua distribuição e abundância, permitindo assim, a definição
de limites de áreas de preservação adequados (LINK, 2000).
Embora alguns trabalhos sobre o comportamento do boto-cinza tenham sido realizados, pouco ainda
se sabe sobre essa espécie, principalmente no que diz respeito ao comportamento de forrageio. No litoral
de Pernambuco, estudos recentes reportam o uso da região pelo boto-cinza (ARAÚJO et al., 2005), porém
ainda não existem dados comportamentais e ecológicos oriundos de um estudo sistemático sobre o boto-
cinza.
Dessa forma, o presente trabalho contribuirá para um melhor conhecimento dos aspectos
comportamentais e ecológicos do boto-cinza, fornecerá subsídios para elaboração de planos de
conservação para tal espécie, bem como fortalecerá a importância de um gerenciamento costeiro da
região, uma vez que desse habitat depende a sua sobrevivência.

24
2. OBJETIVOS:
2.1 Objetivo Geral:
O estudo visou realizar uma comparação do comportamento de forrageio do boto-cinza, Sotalia
guianensis entre duas regiões costeiras do estado de Pernambuco, Praia de Bairro Novo e Porto do Recife.
2.2. Objetivos Específicos:
2.2.1. Caracterizar o comportamento de forrageio do boto-cinza;
2.2.2. Identificar variações na freqüência do comportamento de forrageio do boto-cinza entre dois
diferentes habitas;
2.2.3. Verificar influências dos parâmetros ambientais (temperatura, salinidade, profundidade, estado
da maré, período do dia e nutrientes) sobre a freqüência de atividades de forrageio do boto-cinza;
2.2.4. Caracterizar variações na freqüência de forrageio considerando o tamanho de grupo e
estratégias utilizadas pelos botos;
2.2.5. Verificar a influência da disponibilidade de presas sobre a freqüência de atividade de forrageio
do boto-cinza.

25
3. MATERIAIS E MÉTODOS
3.1. Caracterização da área de estudo:
A Praia de Bairro Novo pertence à cidade de Olinda, enquanto o Porto do Recife à cidade de
Recife. As duas regiões estão distantes cerca de 2,4 km uma da outra. A praia de Bairro Novo caracteriza-
se por ser uma praia costeira e o Porto do Recife, uma região estuarina (Fig. 3).
O município de Olinda está compreendido entre os paralelos 7o57’30”e 8o02’30” de latitude sul e
os meridianos 39o49’41” e 39o55’00” de longitude oeste, perfazendo cerca de 40,83km2. Limita-se ao
norte com o município de Paulista, ao leste com o Oceano Atlântico, a oeste e ao sul com a cidade do
Recife (BELTRÃO et al., 1995). A Praia de Bairro Novo compreende o trecho da Rua do Farol com
imediação da R. Dr. Manoel Ramos Lima, apresentando uma extensão de aproximadamente 2 km.
Apresenta obras de proteção ao litoral com comprimento de cerca de 50m, eqüidistantes 50m uma das
outra e construídas perpendicular e paralelamente à praia (GOMES, 2002).
O Porto do Recife situa-se na parte interna da Bacia do Pina entre os paralelos 08o04’03” e
08o05’06” Sul e os meridianos 34o21’16” e 34o53’58” Oeste, compreendendo uma região estuarina em
plena área urbana, formada pela confluência dos rios Tejipió, Jiguá, Jordão, Pina e pelo braço sul do rio
Capibaribe. Limita-se ao norte com os bairros de Cabanga e São José, ao sul com os bairros do Pina e
Brasília Teimosa, a oeste com os rios acima mencionados, e a leste com um dique de arrecife de arenito
natural. Possui uma extensão de 3,6km aproximadamente, e larguras variáveis 0,26 a 0,86km, perfazendo
uma área total de 2,02km2 (CODEPE, 1980 apud FEITOSA, 1988). Constitui um ambiente
hidrodinâmico, cuja salinidade varia entre 0,42 a 37 ups, com profundidade máxima de 18m (FEITOSA,
1988).
O clima da região segundo a classificação de Köppen é do tipo As’, denominado quente e úmido,
caracterizado por dois períodos distintos no regime pluviométrico: uma estação seca, que se prolonga de
setembro a fevereiro (primavera-verão), e uma estação chuvosa, de março a agosto (outono-inverno). A
temperatura média anual é de 27oC (FEITOSA, 1988; GOMES, 2002).

26
Figura 3. Áreas de estudo 1. Praia de Bairro Novo; 2. Porto do Recife (Fonte: Carta Náutica Nº 902 –
DHN, 1989)
1
2

27
3.2. Metodologia em campo:
3.2.1. Local e período de observação:
As coletas dos dados foram realizadas em duas regiões: Praia de Bairro Novo, em Olinda, e porto
da cidade de Recife. O ponto fixo da Praia de Bairro Novo localiza-se na extremidade de um dique
artificial, denominado “dique Zé Pequeno” (7o59’74”S; 34o50’22”W) (Fig. 4) e o ponto fixo do Porto do
Recife, situa-se na extremidade de um arrecife artificial, em um farol no Molhe Sul do referido Porto, que
possui uma altura de 10 metros de altura (8o2’82”S; 34o51’51”W) (Fig. 5).
O estudo foi conduzido durante um ciclo anual, de setembro de 2006 a agosto de 2007, com
observações realizadas uma semana (cinco dias consecutivos) nos pontos de coleta. As coletas eram
sempre realizadas por dois ou três observadores, devidamente treinados. Em cada mês as semanas de
coleta ocorreram alternadamente em cada região. O período de observação foi de sete horas diárias no
horário de 8h às 16 horas.
Foi realizado um total de 798 horas de esforço amostral, do qual, 399 horas foram de observação
no Porto do Recife, totalizando 177 horas de observação efetiva dos animais e 399 horas de observação
em Olinda, sendo 13 horas de observação efetiva dos animais.

28
Figura 4. Foto do Ponto de Observação no Porto do Recife.
Foto: Flávio R. Berger

29
Figura 5. Foto do Ponto de Observação em Olinda.

30
3.2.2. Coleta dos dados:
Os dados foram coletados utilizando-se os métodos amostrais de “todas as ocorrências”
(ALTMAN, 1974), para registro das atividades relacionadas ao comportamento de forrageio, e,
“varredura” (ALTMAN, 1974), para verificar a presença e localização dos botos dentro das áreas em
estudo. Os registros de Varredura foram obtidos a cada dez minutos, e, após um período de trinta minutos
sem a presença de golfinhos foi considerado como ausência até que os botos fossem novamente avistados.
Para facilitar a visualização dos animais e dos comportamentos, foi utilizado um binóculo
SAKURA RD 10x–90x80. As observações dos tipos de comportamentos, número de indivíduos,
composição dos grupos, localização do animal no local de observação, hora e data foram registrados em
um gravador portátil. Posteriormente foram passadas para planilhas pré-estabelecidas (Anexo) e,
finalmente, analisadas estatisticamente.
3.2.3. Definições Comportamentais:
As estratégias foram denominadas de sincrônicas ou cooperativas, quando envolveram atividades
realizadas por grupos de golfinhos, e de pesca assincrônica, quando os botos forrageavam sem nenhuma
correlação aparente entre os demais botos presentes.
As descrições dos comportamentos de forrageio utilizadas nesta pesquisa estão listadas na tabela 1
e ilustradas na Figura 6.

31
Tabela 1. Repertório dos comportamentos de forrageio realizados pelo boto-cinza (adaptado de
MONTEIRO et al., 2006).
Tipos de Bote sem Salto
Lateral O Golfinho emerge o rostro movimentando-o lateralmente e captura a presa com
a boca.
Vertical* O golfinho emerge verticalmente parte do seu corpo (do rostro até nadadeiras
peitorais) e captura o peixe de baixo para cima.
Em baixo da Água O golfinho desloca-se com velocidade em direção à região mais rasa, mergulha,
captura o peixe em baixo da água e emerge com a presa na boca.
360º O golfinho emerge verticalmente, expondo parte do seu corpo e realizando um
giro de 360o captura a presa de baixo para cima.
Tipos de Botes com Salto
De baixo para cima* O golfinho executa um salto total ou parcial no momento em que a presa está no
ar e a captura de baixo para cima.
Adiantado* O golfinho persegue a presa de ventre para cima adianta-se em relação ao peixe,
então realiza um salto total ou parcial e de frente para a presa, a captura.
Com salto rasante
parcial lateral*
O golfinho realiza um salto parcial com o corpo posicionado lateralmente, e
captura a presa no ar ou na superfície da água.
Com salto de ventre
para cima*
O golfinho realiza um salto parcial de ventre para cima e captura a presa no ar.
Estratégias de Forrageio Cooperativas
Pente* Um grupo de golfinhos desloca-se paralelamente um em relação aos outros,
direcionando os peixes para barreiras.
Cerco* Um grupo de botos cerca um cardume, e logo após os golfinhos mergulham
independentes e emergem com um peixe em sua boca. Atividades aéreas como
saltos e batidas de cauda podem ser realizadas.
Controle de Presa
O golfinho joga o peixe para cima e antes que o animal caia na água, o golfinho o captura novamente,
prendendo-o em sua boca.
* Vertical = Log jump; Baixo para cima = Horinzontal leap; Adiantado = Wall; Salto rasante lateral parcial = 180º turn;
Ventre para cima = back jump; Pente = Front formation; Cerco = Horizontal carousel (BEL’KOVICH et al. , 1991).

32
Figura 6. Estratégias de forrageios realizadas pelo boto-cinza (MONTEIRO et al., 2006). I) Lateral; II)
Vertical; III) Em Baixo da Água; IV) 360º; V) Adiantado; VI) Ventre para Cima; VII) Baixo para Cima
com Salto; VIII) Rasante Lateral Parcial; IV) Controle de Presa; X) Cerco; XI) Pente.
III)
I)
II)
IV)
V)
VIII)
VI)
IV)
X)
XI)
VII)

33
3.2.4. Parâmetros Ambientais:
Para localizar os botos em cada área, foram mensuradas as distâncias entre o ponto
de observação e o boto, com um binóculo BUSNELL YARDAGE PRO 800 telêmetro laser
8x. Além disso, foi obtido o ângulo em que o boto encontra-se em relação ao observador,
com o auxílio de uma bússola portátil. Essas medidas foram plotadas na Carta Náutica
número 902 da Diretoria de Recursos Hídricos e Navegação da Marinha do Brasil (DHN,
1989), que contêm as profundidades de cada região em estudo.
Os estados da maré foram caracterizados como: alta (30 minutos antes e após o
maior pico da maré), vazante (30 minutos após a maior maré e 30 minutos antes da menor
altura), baixa (30 minutos antes e depois do menor pico da maré) e enchente (30 minutos
após a menor e 30 minutos antes da maior altura de maré) (PEREIRA, 1999 apud LODI,
2002). A altura das marés foi baseada na Tábua de Maré da Diretoria de Hidrografia e
Navegação da Marinha do Brasil, conforme a Tábua de Maré do Porto de Recife
(08º03',4S; 034º52',0W).
A temperatura foi medida com um termômetro de mercúrio, com escala de -10 a
110 ºC, e a salinidade com um refratômetro manual (RST-101ATC, com escala de 0,0 a
100,0) em todos os dias de observações nas duas localidades em estudo.
Os índices pluviométricos foram obtidos das Plataformas de Coleta de Dados
(PCD) de Olinda e de Recife. Esses dados foram provenientes do banco de dados do
Laboratório de Meteorologia de Pernambuco (LAMEPE) do Instituto de Tecnologia de
Pernambuco (ITEP, 2007).
Para a análise de sais nutrientes inorgânicos, foram coletadas amostras de 1 litro de
água no canal principal do Porto do Recife e na zona de arrebentação da Praia de Bairro
Novo, nos meses de setembro a outubro de 2006 e de junho a agosto de 2007, durante o
período do da baixa-mar. As amostras foram congeladas e posteriormente levadas ao
laboratório de Química do Departamento de oceanografia da UFPE, para serem
processadas. Os teores de nitrito (NO2), nitrato (NO3) e fosfato (PO4) dissolvidos na água
foram determinados de acordo com procedimentos descritos por STRICKLAND e
PARSONS (1972), e o Silicato (SiO2) através das técnicas utilizadas por GRASSHOFF et

34
al. (1983). O potencial hidrogeniônico (pH) foi determinado com o auxílio de um pH-metro
Beckman, tipo Zeromatic II.
Em decorrência da diversidade morfológica, o Porto do Recife foi dividido em 5
áreas, delimitadas por linhas imaginárias referentes a marcos já existentes no local e com
características ambientais peculiares a cada área. A área I compreendeu a parte mais interna
do Porto, onde está presente o canal do estuário dos rios Capibaribe e Beberibe; a área II
caracteriza-se por ser a região entre dois diques artificiais, o Molhe de Olinda, ao norte, e o
Molhe do Pina, ao sul, denominada popularmente de “Boca da Barra”; a área III refere-se a
uma região de grande influência marítima, não apresentando barreira física; a área IV
caracterizou-se pela presença de um dique artificial de contenção na área mais externa do
Porto, o qual possui dois faróis Náuticos em suas extremidades e a área V, localiza-se na
entrada principal do Porto do Recife em um canal entre o farol de observação e o dique de
contenção (Fig. 7).

35
Figura 7. Divisão do Porto do Recife em áreas.

36
3.2.5. Composição do Grupo:
Os animais foram considerados formando um grupo quando eram visualizados
juntos (cerca de 10 metros de distância), apresentando atividades coordenadas, podendo ou
não estar se deslocando para uma mesma direção (MANN et al., 2000). O número de botos
em cada grupo foi estimado através da contagem direta dos animais que os compunha.
O número de botos avistados simultaneamente nos locais de estudos foi agrupado
em cinco categorias: 1 a 3 indivíduos; 4 a 6; 7 a 10; 11 a 15 e ≥16, de acordo com os
critério adotados por ROSSI-SANTOS (2006):
3.2.6. Atividade de forrageio e produção de pescado:
As freqüências de comportamentos de forrageio foram analisadas e comparadas
mensalmente com a quantidade de pescado no litoral de Olinda e Recife. As espécies de
presas utilizadas para as análises foram determinadas de acordo com a composição
alimentar descrita na literatura para o boto-cinza em outras regiões do Brasil. Os dados de
produção de pescado em Recife e em Olinda do ano de 2004 foram obtidos através de uma
pesquisa no banco de dados do Centro de Pesquisa e Gestão de Recursos Pesqueiros do
Litoral do Nordeste – CEPENE/IBAMA e no Boletim Estatístico da Pesca Marítima e
Estuarina do Nordeste do Brasil - ESTATPESCA (Brasil, 2005). A utilização dos dados
referentes ao ano de 2004 ocorreu devido à ausência do registro dos dados de 2007 e a
semelhança no regime pluviométrico entre os dois anos ao longo dos meses. As espécies de
peixes que foram analisadas neste estudo estão listadas na tabela 2.

37
Tabela 2. Espécies que compõem a dieta do boto-cinza presentes no Recife e Olinda e que foram
analisadas no banco de dados do CEPENE/IBAMA e no ESTPESCA (BRASIL, 2005).
FAMÍLIA NOME COMUM
ESPÉCIE CLASSIFICAÇÃO ECOLOGICA*
FOTO
Centropomidae
Camurim ou Robalo
Centropomus undecimalis7
Marinha Dependente
Sardinella brasiliensis
2 Marinha Visitante
Clupeidae Sardinha
Harengula clupeola2 Marinha Visitante
Engraulidae Manjuba Anchoa filifera
2 Marinha Dependente
Biquara Haemulon steindachneri
3 Marinha Visitante
Haemulidae
Sapuruna Haemulon aurolineatum3 Marinha Visitante
Lutjanidae
Cioba- Vermelha
Lutjanus analis5 Marinha Visitante

38
FAMÍLIA NOME COMUM
ESPÉCIE CLASSIFICAÇÃO ECOLOGICA*
FOTO
Pargo -Verdadeiro
Lutjanus purpureus4 Marinha Visitante
Ariacó Lutjanus synagris
3 Marinha Visitante
Lutjanidae
Dentão Lutjanus jocu6 Marinha Visitante
Mugilidae Tainha ou
Saúna Mugil curema
1 Marinha Dependente
Cynoscion jamaicensis
4 Marinha Visitante
Scianidae Pescada
Macrodon ancylodon3 Marinha Visitante
Camarão Rosa
Farfantepenaeus subtilis Marinha Dependente Penaeidae
Camarão-branco
Litopenaeus schmitti Marinha Dependente
* Classificação Ecologia: Marinha Dependente: espécies de peixes de origem marinha que, obrigatoriamente, utilizam
águas estuarinas para alimentação e/ou para completar seu ciclo de reprodução; Marinha Visitante: espécies de origem
marinha que passam maior parte do ciclo de vida no mar, penetrando nas águas estuarinas em períodos regulares ou
ocasionais (VASCONCELOS-FILHO, 2001).
Fotos: www.fishbase.org (acesso: 29/12/2007) 1 - Calderón, S.; 2 – Carvalho Filho, A.; 3 – Flescher, D.; 4 – Matsuura, K.;
5 – Nirchio, M.; 6 – Nunes, J.L.S.; 7 – Patzner, R.; 8 – Ramjohn, D.D.

39
3.3. Análise Estatística:
Os comportamentos foram analisados a cada intervalo de uma hora, tanto no
período da manhã (8h às 12 horas) como no período da tarde (12h01min até 15 horas em
Olinda e 13h as 16 horas no Porto do Recife). Para minimizar os efeitos da diferença de
tempo entre o período do dia, bem como entre os estados de maré e, foram calculadas
proporções entre o tempo de observação efetiva dos botos pelo tempo total de observação
em cada período. Além disso, foram efetuados cálculos de densidade entre o tamanho de
cada área e a quantidade de comportamentos de forrageio registrados no Porto do Recife,
uma vez que as 5 áreas variam de dimensão.
Foi utilizado o teste U de Mann-Whitney para comparações entre a ocorrência de
botos nas áreas em estudo, realização de saltos durante a captura de presas, freqüência de
forrageio entre as estações do ano, período do dia e categorias de profundidade. Os dados
de parâmetros ambientais foram comparados as freqüências de comportamento de forrageio
utilizando o Teste de Correlação de Spearman. O Teste de Kruskal-Wallis foi usado para
verificar a relação entre estados de Marés e comportamento de forrageio e o de Qui-
quadrado para comparar os comportamentos de forrageio no diferentes categorias de maré
(enchente, cheia, vazante e seca), de acordo com as marés de quadratura e sizígia. Em todos
os testes foi adotando o índice de significância p ≤ 0,05 (bilateral),

40
4. RESULTADOS
4.1. Presença dos botos-cinza
4.1.1. Sazonalidade da Ocorrência de botos-cinza
No período de setembro de 2006 a agosto de 2007, os botos-cinza estiveram
presentes no Porto do Recife em 77,2 % dos dias (n = 44) e em 44,4% (177h) do tempo de
observação. Em Olinda, os botos estiveram presentes em apenas 19,3% (n = 11) dos dias e
em 3,26% (13h) do tempo total de observação na região. O teste de Mann-Whitney
demonstrou que a ocorrência de botos-cinza no Porto do Recife é significativamente maior
do que em Olinda (n1; n2 = 12; U = 18,0; p = 0,0018).
O Período do ano de maior ocorrência do botos-cinza no Porto do Recife foi de
setembro a dezembro (29,3h; 27,0h; 28,3h; 24,3h, respectivamente) e o de menor
ocorrência foi de março (6,8h) a abril (4,8h), seguido de maio e junho, onde não foi
observada a presença de botos. Em Olinda os botos foram observados apenas no período de
novembro de 2006 a fevereiro de 2007, sendo o último mês o de maior ocorrência (6,3h)
(Fig. 8). Em ambas as regiões os golfinhos foram mais avistados durante o turno da manhã
(Fig. 9).
0
5
10
15
20
25
30
Pre
sen
ça d
os
bo
tos
(h)
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul Ago
Meses
Porto do Recife Olinda
Figura 8: Freqüência absoluta de horas que os botos-cinza estiveram presentes durante os
meses de observação no Porto do Recife e em Olinda.

41
0
20
40
60
80
100
120F
req
üên
cia
Ab
solu
ta
(h)
Porto do Recife Olinda
Regiões
Manhã Tarde
Figura 9: Freqüência absoluta de horas em que os botos-cinza estiveram presentes durante
os turnos da manhã e tarde no Porto do Recife e em Olinda.
4.1.2. Número de botos avistados simultaneamente:
No Porto do Recife, o número de botos visualizados simultaneamente variou de 1 a
22 animais, sendo que os números de botos mais freqüentes foram 4 (n=165) e 5 (n=171) e
os números acima de 11 botos foram os menos freqüentes. Em Olinda, o número de botos-
cinza registrados simultaneamente variou de 1 a 6 animais, sendo 6 o número de botos mais
freqüente na região (n=24) (Fig. 10).
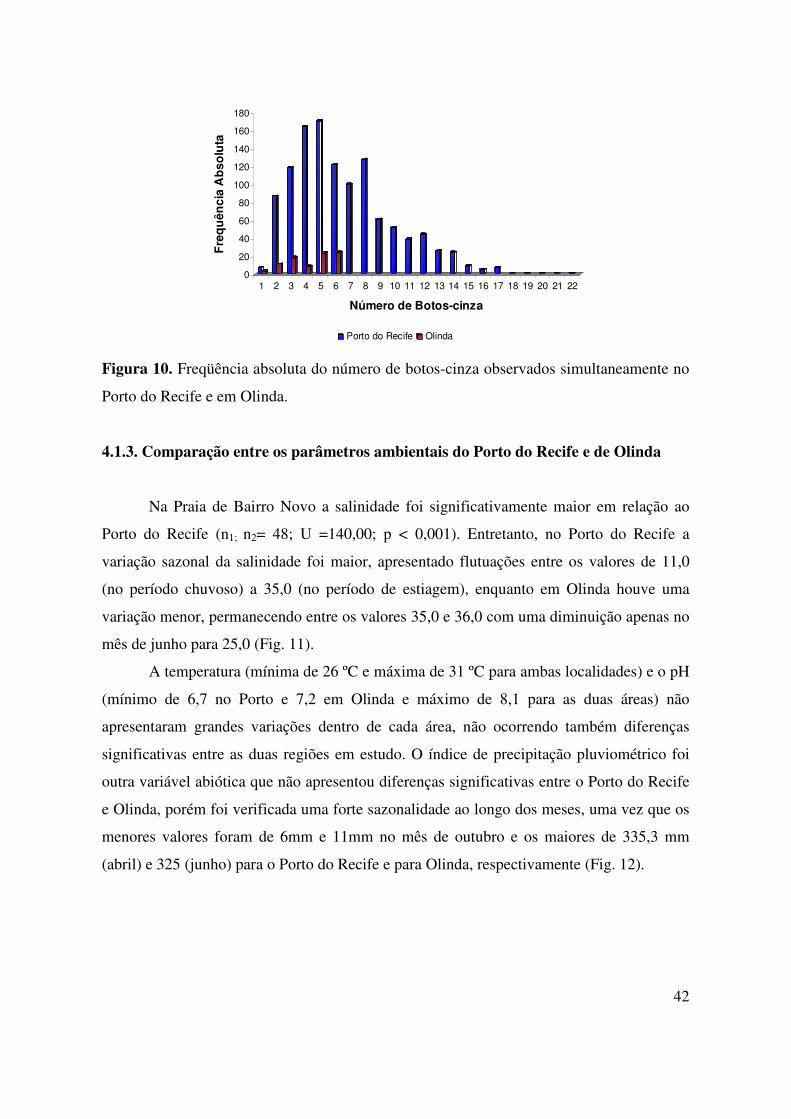
42
0
20
40
60
80
100
120
140
160
180
Fre
qu
ênci
a A
bso
luta
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Número de Botos-cinza
Porto do Recife Olinda
Figura 10. Freqüência absoluta do número de botos-cinza observados simultaneamente no
Porto do Recife e em Olinda.
4.1.3. Comparação entre os parâmetros ambientais do Porto do Recife e de Olinda
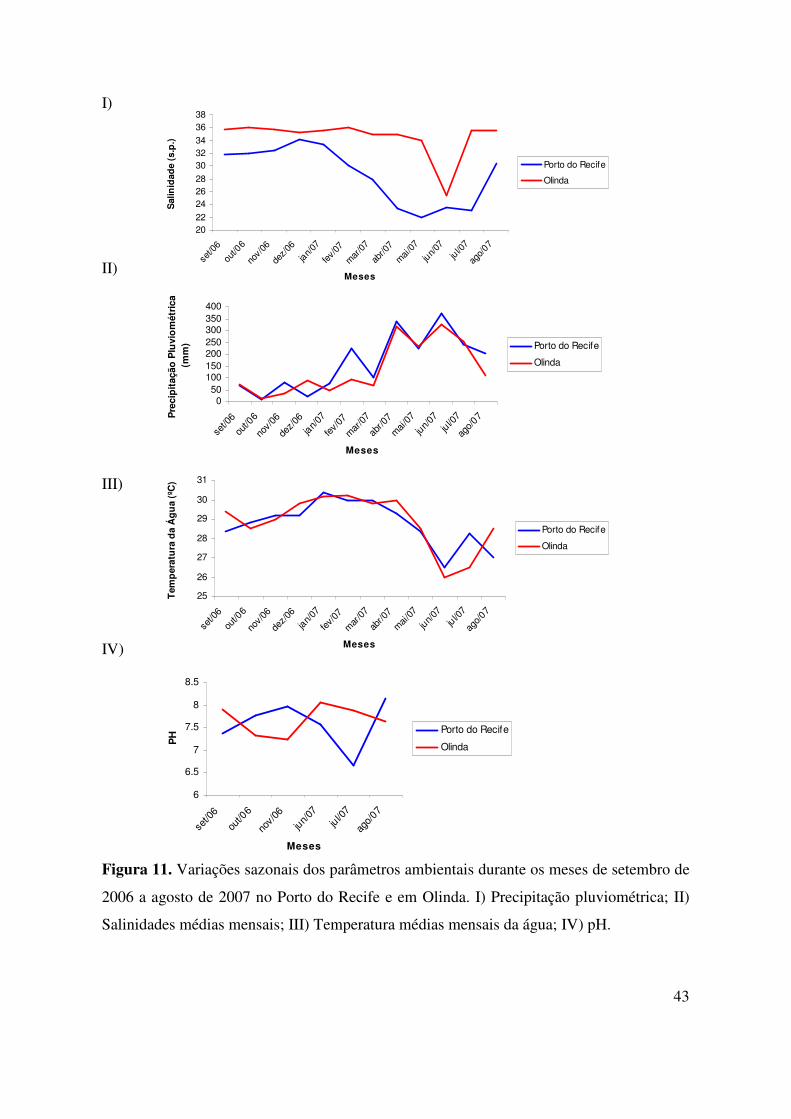
Na Praia de Bairro Novo a salinidade foi significativamente maior em relação ao
Porto do Recife (n1; n2= 48; U =140,00; p < 0,001). Entretanto, no Porto do Recife a
variação sazonal da salinidade foi maior, apresentado flutuações entre os valores de 11,0
(no período chuvoso) a 35,0 (no período de estiagem), enquanto em Olinda houve uma
variação menor, permanecendo entre os valores 35,0 e 36,0 com uma diminuição apenas no
mês de junho para 25,0 (Fig. 11).
A temperatura (mínima de 26 ºC e máxima de 31 ºC para ambas localidades) e o pH
(mínimo de 6,7 no Porto e 7,2 em Olinda e máximo de 8,1 para as duas áreas) não
apresentaram grandes variações dentro de cada área, não ocorrendo também diferenças
significativas entre as duas regiões em estudo. O índice de precipitação pluviométrico foi
outra variável abiótica que não apresentou diferenças significativas entre o Porto do Recife
e Olinda, porém foi verificada uma forte sazonalidade ao longo dos meses, uma vez que os
menores valores foram de 6mm e 11mm no mês de outubro e os maiores de 335,3 mm
(abril) e 325 (junho) para o Porto do Recife e para Olinda, respectivamente (Fig. 12).
43
050
100150
200250
300350
400
set/0
6
out/0
6
nov/
06
dez/
06
jan/
07
fev/
07
mar
/07
abr/0
7
mai
/07
jun/
07
jul/0
7
ago/
07
Meses
Pre
cip
itaç
ão P
luvi
om
étri
ca
(mm
) Porto do Recife
Olinda
20
22
24
26
28
30
32
34
36
38
set/0
6
out/0
6
nov/
06
dez/
06
jan/
07
fev/
07
mar
/07
abr/0
7
mai
/07
jun/
07
jul/0
7
ago/
07
Meses
Sal
inid
ade
(s.p
.)Porto do Recife
Olinda
25
26
27
28
29
30
31
set/0
6
out/0
6
nov/
06
dez/
06
jan/
07
fev/
07
mar
/07
abr/0
7
mai
/07
jun/
07
jul/0
7
ago/
07
Meses
Tem
per
atu
ra d
a Á
gu
a (º
C)
Porto do Recife
Olinda
6
6.5
7
7.5
8
8.5
set/0
6
out/0
6
nov/
06
jun/
07
jul/0
7
ago/
07
Meses
PH Porto do Recife
Olinda
I)
II)
III)
IV)
Figura 11. Variações sazonais dos parâmetros ambientais durante os meses de setembro de
2006 a agosto de 2007 no Porto do Recife e em Olinda. I) Precipitação pluviométrica; II)
Salinidades médias mensais; III) Temperatura médias mensais da água; IV) pH.

44
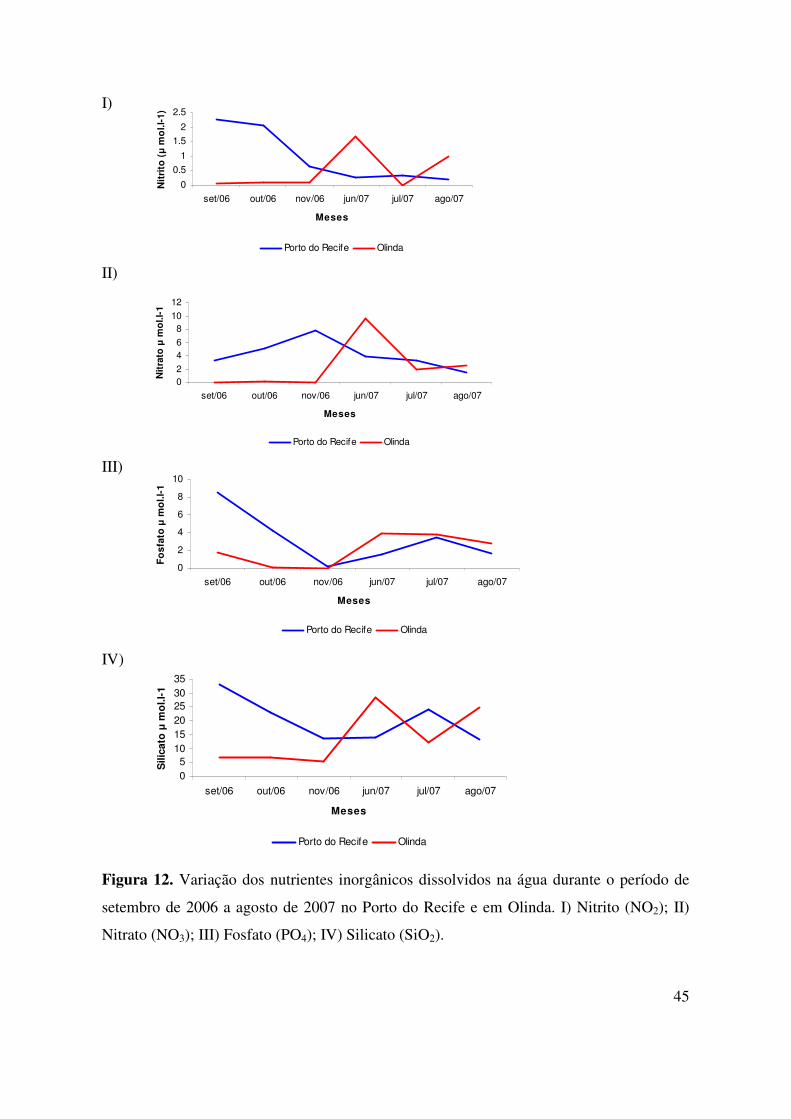
Em relação aos sais nutrientes dissolvidos, não ocorreram diferenças significativas
entre as duas regiões, porém considerando as médias dos meses de estudo, os teores obtidos
para todos os nutrientes dissolvidos foram mais elevados no Porto do Recife quando
comparados a Praia de Bairro Novo (Tab. 3). Além disso, as concentrações de sais tais
como o nitrito, nitrato e silicato sofreram variações sazonais inversas nas duas regiões. À
medida que no Porto houve uma tendência de redução desses nutrientes durante a estação
chuvosa, em Olinda esses teores tenderam a um aumento (Fig. 12)
45
0
0.5
1
1.5
2
2.5
set/06 out/06 nov/06 jun/07 jul/07 ago/07
Meses
Nit
rito
(µ
mo
l.l-1
)
Porto do Recife Olinda
0
2
4
6
8
10
12
set/06 out/06 nov/06 jun/07 jul/07 ago/07
Meses
Nit
rato
µ m
ol.l
-1
Porto do Recife Olinda
0
2
4
6
8
10
set/06 out/06 nov/06 jun/07 jul/07 ago/07
Meses
Fo
sfat
o µ
mo
l.l-1
Porto do Recife Olinda
0
5
10
15
20
25
30
35
set/06 out/06 nov/06 jun/07 jul/07 ago/07
Meses
Sili
cato
µ m
ol.l
-1
Porto do Recife Olinda
I)
II)
III)
IV)
Figura 12. Variação dos nutrientes inorgânicos dissolvidos na água durante o período de
setembro de 2006 a agosto de 2007 no Porto do Recife e em Olinda. I) Nitrito (NO2); II)
Nitrato (NO3); III) Fosfato (PO4); IV) Silicato (SiO2).

46
Tabela 3. Médias dos valores dos parâmetros ambientais no Porto do Recife e em Olinda.
NO2 NO3 PO4 SiO2 pH Pluviometria Temperatura Salinidade
MÉDIAS Porto Olinda Porto Olinda Porto Olinda Porto Olinda Porto Olinda Porto Olinda Porto Olinda Porto Olinda
Geral 0,95 0,48 4,13 2,39 3,23 2,09 20,29 14,10 7,58 7,67 163,58 137,42 28,90 29,15 29,27 35,02
Estação Seca 1,64 0,08 5,41 0,08 4,30 0,67 23,34 6,36 7,70 7,49 80,05 56,33 29,31 29,38 32,38 35,66
Estação Chuvosa 0,26 0,89 2,85 4,70 2,16 3,51 17,23 21,84 7,45 7,85 247,12 218,20 28,30 28,78 24,75 33,94

47
5000
10000
15000
20000
25000
30000
35000
jan/
04
fev/04
mar
/04
abr/0
4
mai/0
4
jun/
04
jul/0
4
ago/04
set/0
4
out/0
4
nov/04
dez/04
Meses
Qu
anti
dad
e d
e P
esca
do
(to
n)
Porto de Recife Olinda
As variações entre as quantidades de presas do boto-cinza nas áreas em estudo não foram
significantes quando comparadas entre si, porém as freqüências de pescados em Olinda
permaneceram mais estáveis ao longo do ano do que no Porto do Recife, onde o mês mais
representativo em relação à pesca foi o de dezembro (32.313,9 ton) e o de menor foi junho
(6.472,8 ton) (Fig. 13) (Tabs. 4 e 5).
As famílias mais freqüentes no Porto do Recife foram Lutjanidae (38%), Mugilidae
(19,6%) e Clupeidae (18,1%), e em Olinda foram Haemulidae (38,5%), Lutjanidae (36,6%) e
Mugilidae (14,5%) (Fig. 14).
Figura 13. Variação sazonal na quantidade total de presas do boto-cinza pescadas no Recife e em
Olinda durante o período de janeiro a dezembro de 2004.
05
10152025303540
Fre
qu
ên
cia
Re
lati
va
(%
)
Cen
tropo
mid
ae
Clupe
idae
Engra
ulid
ae
Hae
mulid
ae
Lutja
nida
e
Mug
ilidae
Scian
idae
Penae
idae
Famílias de Presas do Boto
Porto do Recife Olinda
Figura 14. Freqüências relativas das famílias de presas do boto-cinza pescadas no Recife e em
Olinda.

48
Tabela 4. Quantidade de presas do boto-cinza em toneladas pescadas em Recife durante o período de janeiro a dezembro de 2004.
Mês Ariacó Biquara Camurim Cioba Dentão Manjuba Pargo Pescada Sapuruna Sardinha Tainha Camarão Total/Mês
jan/04 131,4 407,2 437,9 3.261,3 138 0 91 0 2.455,5 4.480,1 821,6 817,5 13.041,5 fev/04 159,9 510,4 616,6 3.963,5 171,3 0 113,8 0 3201 5201 1305 995,1 16.237,6 mar/04 512,5 2.339,8 42,0 3.463,5 196,8 330,0 233,0 27,8 3.854,9 0 1.599,5 0 12.599,8 abr/04 63,0 164,4 667,0 3.251,5 65,5 1.180,6 0 0 2.251,0 4.959,2 1.075,0 1.720,0 15.397,2 mai/04 11,7 395,5 37,0 1.520,0 136,0 472,5 60,0 0 0 3.819,0 488,8 2.126,7 9.067,2 jun/04 3,0 289,0 34,0 1.490,0 0 0 0 0 0 0 2.488,0 2.168,8 6.472,8 jul/04 0 348,0 50,0 1.258,7 200,0 0 0 34,5 1.415,0 104,0 4.644,0 763,0 8817,2 ago/04 0 161,0 46,4 2.651,8 36,0 0 0 2,8 841,0 0 2.147,4 861,0 6.747,4 set/04 25,0 325,2 124,3 2.461,8 118,0 0 0 3,1 163,0 304,0 2.359,1 2.694,0 8.577,5 out/04 20,0 117,0 0 4.462,3 181,9 0 85,8 0 1.530,0 4.335,4 2.220,8 701,5 13.654,7 nov/04 30,0 170,3 55,8 6.724,4 243,3 1.413,6 313,1 0 2.577,3 3.475,0 10.695,0 2.868,0 28.565,8 dez/04 6,0 289,5 114,0 5.808,5 208,8 1.368,0 34,3 0 2.193,0 10.431,8 10.350,0 1.510,0 32.313,9
Total 962,5 5.517,3 2.225,0 40.317,3 1.695,6 4.764,7 931,0 68,2 20.481,7 37.109,5 40.194,2 17.225,6 171.492,6
Tabela 5. Quantidade de presas do boto-cinza em toneladas pescadas em Olinda durante o período de janeiro a dezembro de 2004.
Mês Ariacó Biquara Camurim Cioba Dentão Manjuba Pargo Pescada Sapuruna Sardinha Tainha Camarão Total/Mês
jan/04 1.038,0 2.464,6 72,0 2.135,1 227,7 420,0 95,0 20,9 2.338,8 280,0 700,6 0 9.792,7 fev/04 1.019,0 2.451,0 25,0 2135,1 227,7 845,0 95,0 10,8 2.338,8 311,9 704,8 1.014,0 11.178,1 mar/04 512,5 2.339,8 42,0 3.463,5 196,8 330,0 233,0 27,8 3.854,9 0 1.599,5 0 12.599,8 abr/04 377,9 1.521,5 47,6 1.878,0 246,0 1.409,0 134,7 3,0 4.735,6 50,6 603,3 858,0 11.865,2 mai/04 285,7 1.875,2 56,0 2.598,3 243,9 280,3 32,0 55,4 2.663,1 25,9 317,7 0 8.433,5 jun/04 146,5 1.862,5 64,2 3.116,8 101,5 0 0 9,7 2.748,8 2,7 1.044,0 0 9.096,3 jul/04 399,5 1.907,4 51,9 2.310,9 2.988,0 0 0 31,8 2.491,8 0 2.886,1 984,0 14.051,4 ago/04 800,5 1.957,1 43,1 3.794,9 159,4 1.406,1 49,7 2,3 2.724,2 4,0 1.311,7 1.194,5 13.447,5 set/04 262,6 1.594,4 44,2 2.772,4 140,6 1.189,8 5,2 7,0 1.762,1 7,9 1.394,0 1.010,7 10.190,9 out/04 142,0 1.445,4 31,5 4.463,7 155,6 0 104,1 5,9 1.471,0 75,6 1.008,0 666,0 9.568,8 nov/04 201,9 1.242,8 74,6 4.857,4 234,2 17,4 194,1 0 1.060,2 8,4 3.524,8 0 11.659,4 dez/04 510,4 1.865,5 51,4 4.269,2 169,0 592,8 12,9 29,6 1.374,8 73,0 3.768,4 320,0 13.841,6
Total 5.696,5 22.527,2 603,5 37.795,3 5.090,4 6.490,4 955,7 204,2 29.564,1 839,6 19.911,1 6.047,2 135.541,4

49
4.2. Caracterização do comportamento de forrageio
Foram registrados ao todo 1264 (97,16%) comportamentos de forrageio no Porto do
Recife e 37 (2,84%) em Olinda (Fig. 15). Destes, foi possível observar 337 sucessos na captura
de presas pelos botos no Porto e 18 em Olinda. No Porto do Recife, houve uma maior diversidade
de estratégias alimentares do que em Olinda. No Porto do Recife, foram observados oito tipos de
botes e duas estratégias de captura em grupo, enquanto em Olinda foram registrados três tipos de
bote e uma estratégia em grupo. Apenas no Porto do Recife foi avistado o comportamento de
arremesso de presa.
Os comportamentos registrados em Olinda foram: forrageio com captura em baixo da
água; lateral e de baixo para cima com salto, os quais foram, em ordem decrescente, os mais
freqüentes em ambas as localidades. Dos demais comportamentos registrados no Porto do Recife,
o forrageio com captura vertical foi o quarto mais freqüente (4,3%) e as capturas com bote
adiantado (0,1%) e 360º (0,1%) foram mais raras (Fig. 16).
Os comportamentos de forrageios sem utilização de saltos para captura de presas
apresentaram uma freqüência significativamente maior do que aqueles com uso de saltos tanto no
Porto do Recife (n1;n2= 36; U = 120,5; p< 0,001) quanto em Olinda (n1;n2 = 3; U = 0,00; p
=0,05). As investidas dos botos sem utilização de saltos foram significativamente mais bem
sucedidas no Porto (n1;n2= 31; U = 114,00; p< 0,001) e em Olinda (n1;n2= 3; U = 0,00; p= 0,05)
(Fig. 17).
Diferentes estratégias foram verificadas nas duas regiões: em Olinda, os botos foram
observados conduzindo as presas de regiões mais profundas (5m) para regiões mais rasas (menos
de 2m) próximas à zona de arrebentação, enquanto no Porto do Recife, os golfinhos eram
observados direcionando suas presas aos arrecifes artificiais. Além disso, grupos de botos eram
visualizados mergulhando e capturando peixes na zona de turbulência formada pela remoção de
substrato durante a passagem de alguns navios cargueiros no canal de entrada do Porto do Recife.
Botos-cinza também foram observados forrageando próximos a redes de espera e de embarcações
de pesca artesanal, no Porto do Recife.

50
0
20
40
60
80
100
Fre
qu
ênc
ia R
ela
tiva
(%
)
Porto do Recife Olinda
Ponto de Observação
Porto do Recife Olinda
Figura 15. Freqüência relativa do número de comportamentos observados no Porto do Recife e
em Olinda durante o período de setembro de 2006 a agosto de 2007.
0102030405060708090
100
Fre
quê
nci
a R
ela
tiva
(%
)
AD 360º BC EA LAT RLP VC VET
Tipos de Comportamento de Forrageio
Porto do Recife Olinda
Figura 16. Freqüência relativa dos tipos de comportamentos de forrageio observados no Porto do
Recife e em Olinda. AD= Bote adiantado; 360º= Bote com giro de 360º; B/C= Bote de baixo para
cima com salto; EA= Bote em baixo da água; LAT= Bote lateral; RLP= Bote rasante lateral
parcial; V/C= Bote de ventre para cima; VET= Bote vertical.

51
0
10
20
30
40
50
60
70
80
90
100
Fre
quê
ncia
Re
lativ
a (%
)
Porto do Recife Olinda
Regiões
Sem Salto
Com Salto
Figura 17. Comparação entre as freqüências de saltos na captura de presas no Porto do Recife e
em Olinda.
Ao todo, foram registrados 296 eventos de comportamentos de forrrageio cooperativos no
Porto do Recife e 22 em Olinda, sendo estes, de apenas um tipo, o cerco. A estratégia em grupo
no Porto do Recife mais representativa foi a de cerco (98,3%) (Fig. 18). Essa estratégia foi
realizada por grupos compostos de 2 a 10 indivíduos.
As estratégias alimentares assincrônicas foram mais freqüentes no Porto do Recife
(76,6%), enquanto que as estratégias realizadas cooperativamente pelos botos ocorreram em
maior proporção em Olinda (59,5%) do que no Porto do Recife (23,4%).
0
10
20
30
40
50
60
70
80
Fre
qu
ênci
a d
e F
orr
agei
os
(%)
Porto do Recife Olinda
Regiões
Assincrônicas Sincrônicas
Figura 18. Percentagens das estratégias alimentares utilizadas pelos botos-cinza no Porto do
Recife em Olinda.

52
4.3. Comportamento de forrageio e tamanho de grupo
As freqüências de comportamento de forrageio variaram de acordo com o número de
botos presentes no Porto do Recife (H = 40,77; gl = 5; p< 0,001), ocorrendo o maior número de
forrageios quando estavam presentes na região grupos contendo de 4 a 6 golfinhos quando
comparado as categorias 1 a 3 (Z = 3,91; p < 0,05), 10 a 12 (Z = 4,05; p < 0,05), 13 a 15 (Z =
4,69; p < 0,05) e ≥16 (Z = 5,58; p < 0,05). Em Olinda, o teste de Mann-Whitney demonstrou que
as freqüências de forrageio observadas foram significativamente maiores para o número de botos
entre 4 a 6 animais do que para a categoria de 1 a 3 indivíduos (n1; n2 = 3; U = 0,00; p = 0,05)
(Fig. 19).
O número de botos caçando cooperativamente variou de 1 a 10 no Porto do Recife, sendo
4 o número mais freqüente (n= 280), seguido de 2 (n= 255) e 3 (n= 215) e em Olinda o número
de botos forrageando juntos variou de 1 a 6 indivíduos, com maior freqüência do número máximo
de indivíduos (n= 12) para essa região (Figs. 20 e 21).
0
10
20
30
40
50
60
70
80
90
100
Fre
qu
ênci
a R
elat
iva
(%)
1 a 3 4 a 6 7 a 9 10 a 12 13 a 15 ≥16
Categorias de Grupos
Porto do Recife Olinda
Figura 19. Freqüências relativas dos comportamentos de forrageio realizados na presença das
categorias de número de botos avistados no Porto do Recife e em Olinda.

53
0
50
100
150
200
250
300
Fre
qu
ênci
a A
bso
luta
de
Fo
rrag
eio
s
1 2 3 4 5 6 7 8 9 10
Número de Botos Forrageando Juntos
Porto do Recife Olinda
Figura 20.Variação das freqüências absolutas de atividades de pesca realizadas pelo número de
golfinhos forrageando conjuntamente no Porto do Recife e em Olinda.
Total
0
20
40
60
80
100
120
140
160
1 3 1 2 3 4 5 6 1 2 3 1 2 3 4 5 6 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 1 2 3 4 5 10 1 2 3 4 6 10
1 a 3 4 a 6 1 a 3 4 a 6 7 a 9 10 a 12 13 a 15 ≥16
OLINDA P. RECIFE
Soma de N. Forrageio
Local Cat grupo Botos FOR juntos
Figura 21. Variação do número de forrageios cooperativos nos diferentes tamanhos de grupos de
boto-cinza presentes em Olinda e no Porto do Recife. (Barra interna do eixo das coordenadas:
número de botos forrageando cooperativamente; Barra externa: tamanho de grupo presente nas
regiões).

54
4.4. Influências das variáveis ambientais no Comportamento de Forrageio
4.4.1. Sazonalidade da ocorrência dos comportamentos de forrageio
Os meses com maior freqüência de comportamentos de forrageio foram os de dezembro
(n= 375) e setembro (n= 295) no Porto do Recife. Em Olinda, apenas nos meses de janeiro (n=
32) e fevereiro (n= 5) foram observados comportamentos de forrageio (Fig. 22).
A ocorrência de comportamentos de forrageio foi significativamente maior na estação
seca em relação à estação chuvosa no Porto (n1; n2= 29; U =225,5; p= 0,0024) (Fig. 23).
0
50
100
150
200
250
300
350
400
Fre
qu
ênci
a A
bso
luta
Set Out Nov Dez Jan Fev Mar Abr Maio Jun Jul Ago
Meses
Porto do Recife Olinda
Figura 22. Variação do número de forrageios observados ao longo dos meses do ano no Porto do
Recife e em Olinda.
0
100
200
300
400
500
600
700
800
900
1000
Nú
mer
o d
e F
orr
agei
os
Seca Chuvosa
Estações do Ano
Figura 23. Comparação entre as freqüências de atividades de forrageio nas estações seca e
chuvosa no Porto do Recife.
55
0
50
100
150
200
250
300
350
Fre
qu
ênci
a A
bso
luta
de
Fo
rrag
eio
s
8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00
Horários do Dia
0
5
10
15
20
25
30
freq
uên
cia
Ab
solu
ta d
e F
orr
agei
os
8:00 9:00 10:00 11:00 12:00 13:00
Horários do Dia
OlindaI)
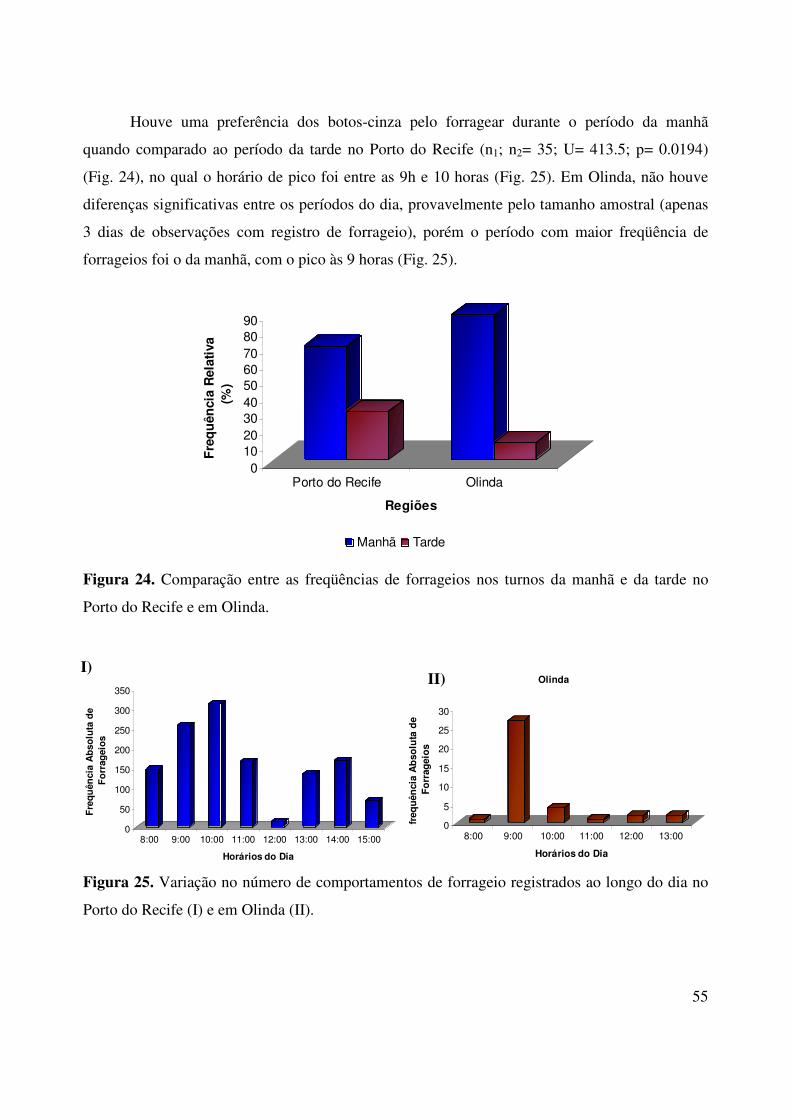
Houve uma preferência dos botos-cinza pelo forragear durante o período da manhã
quando comparado ao período da tarde no Porto do Recife (n1; n2= 35; U= 413.5; p= 0.0194)
(Fig. 24), no qual o horário de pico foi entre as 9h e 10 horas (Fig. 25). Em Olinda, não houve
diferenças significativas entre os períodos do dia, provavelmente pelo tamanho amostral (apenas
3 dias de observações com registro de forrageio), porém o período com maior freqüência de
forrageios foi o da manhã, com o pico às 9 horas (Fig. 25).
0
10
20
30
40
50
60
70
80
90
Fre
qu
ênci
a R
elat
iva
(%)
Porto do Recife Olinda
Regiões
Manhã Tarde
Figura 24. Comparação entre as freqüências de forrageios nos turnos da manhã e da tarde no
Porto do Recife e em Olinda.
Figura 25. Variação no número de comportamentos de forrageio registrados ao longo do dia no
Porto do Recife (I) e em Olinda (II).
II)

56
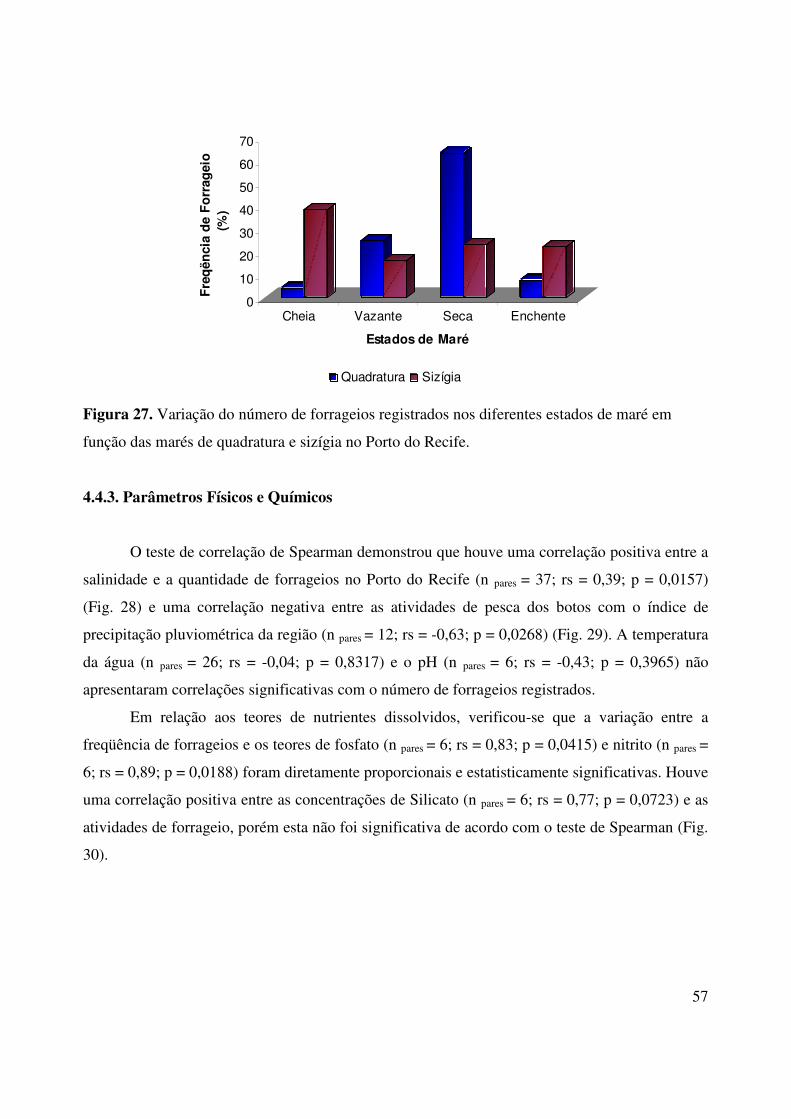
4.4.2. Maré e Fases Lunares
De acordo com o teste U de Kruskal-Wallis, a quantidade de forrageios observados variou
significativamente em relação aos estados de maré no Porto do Recife (H = 8,04; gl = 3; p = 0,05)
(Fig. 26). O teste de Dunn demonstrou que na maré vazante ocorre um aumento de atividades de
caça quando comparada à maré cheia (Z = 2,64; p < 0,05). Além disso, a maré seca foi a mais
representativa em termos de forrageio (22,1%) no Porto. Em Olinda não foi possível realizar
testes estatísticos nem cálculos de freqüências em decorrência do número reduzido de eventos de
forrageio.
No Porto do Recife, as atividades de pesca dos botos-cinza não sofreram influências das
fases lunares (Cheia, Minguante, Nova e Crescente) (H = 4,17; gl = 3; p = 0,2433). Contudo, os
comportamentos de forrageios variaram em relação aos diferentes estados de maré em função das
marés de quadratura e sizígia (241,57 = 2א; gl = 3; p< 0,001). Assim, as atividades de
alimentação foram mais intensas durante a baixa-mar (63,6%) na maré de quadratura e durante a
preamar (38,5%) na maré sizígia (Fig. 27).
0
10
20
30
40
50
60
70
80
90
100
Fre
qu
ênci
a d
e F
orr
agei
o
(%)
CHEIA VAZANTE SECA ENCHENTE
Estados de Maré
Figura 26. Freqüências relativas do número de comportamentos de forrageio observados durante
os diferentes estados de maré no Porto do Recife.
57
0
10
20
30
40
50
60
70
Fre
qën
cia
de
Fo
rrag
eio
(%
)
Cheia Vazante Seca Enchente
Estados de Maré
Quadratura Sizígia
Figura 27. Variação do número de forrageios registrados nos diferentes estados de maré em
função das marés de quadratura e sizígia no Porto do Recife.
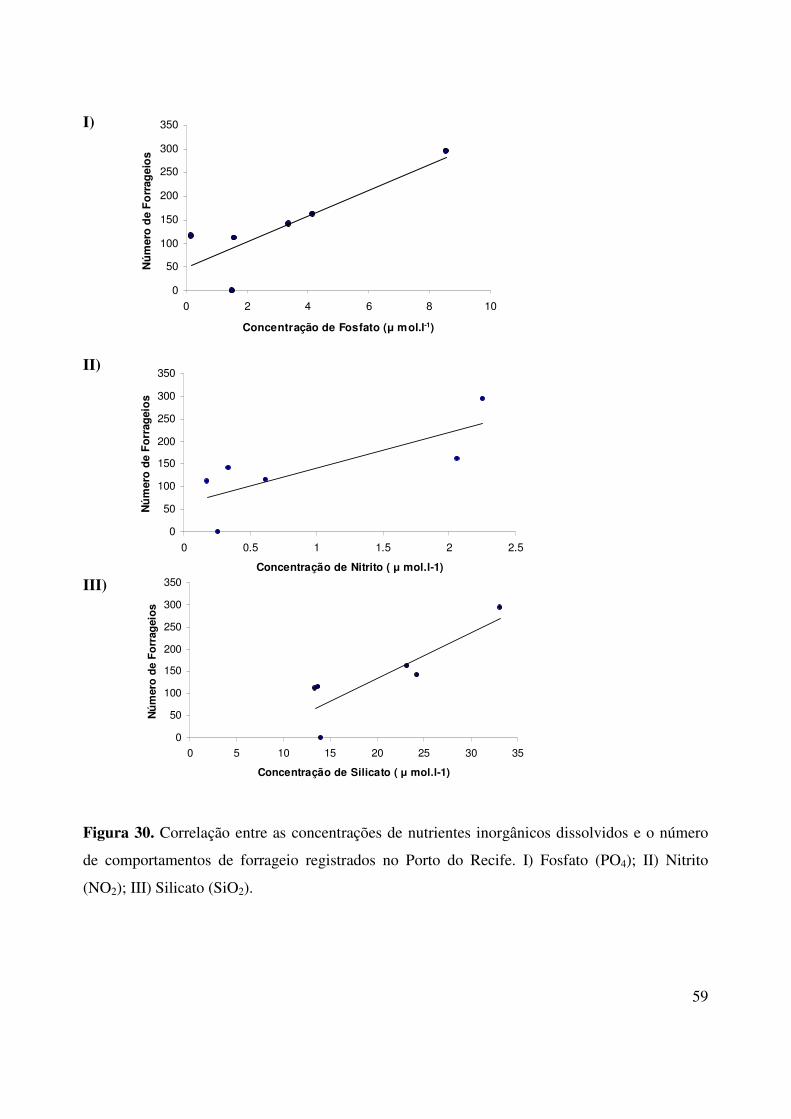
4.4.3. Parâmetros Físicos e Químicos
O teste de correlação de Spearman demonstrou que houve uma correlação positiva entre a
salinidade e a quantidade de forrageios no Porto do Recife (n pares = 37; rs = 0,39; p = 0,0157)
(Fig. 28) e uma correlação negativa entre as atividades de pesca dos botos com o índice de
precipitação pluviométrica da região (n pares = 12; rs = -0,63; p = 0,0268) (Fig. 29). A temperatura
da água (n pares = 26; rs = -0,04; p = 0,8317) e o pH (n pares = 6; rs = -0,43; p = 0,3965) não
apresentaram correlações significativas com o número de forrageios registrados.
Em relação aos teores de nutrientes dissolvidos, verificou-se que a variação entre a
freqüência de forrageios e os teores de fosfato (n pares = 6; rs = 0,83; p = 0,0415) e nitrito (n pares =
6; rs = 0,89; p = 0,0188) foram diretamente proporcionais e estatisticamente significativas. Houve
uma correlação positiva entre as concentrações de Silicato (n pares = 6; rs = 0,77; p = 0,0723) e as
atividades de forrageio, porém esta não foi significativa de acordo com o teste de Spearman (Fig.
30).

58
0
50
100
150
200
250
0 5 10 15 20 25 30 35 40
Salinidade
Nú
mer
o d
e F
orr
agei
os
Figura 28. Correlação entre salinidade e o número de comportamentos de forrageio registrados
no Porto do Recife.
0
50
100
150
200
250
300
350
400
0 50 100 150 200 250 300 350 400
Índice pluviométrico (mm)
Nú
mer
o d
e F
orr
agei
os
Figura 29. Correlação entre os índices pluviométricos mensais e o número de comportamentos
de forrageio registrados no Porto do Recife.
59
0
50
100
150
200
250
300
350
0 2 4 6 8 10
Concentração de Fosfato (µ mol.l-1)
Nú
mer
o d
e F
orr
agei
os
0
50
100
150
200
250
300
350
0 0.5 1 1.5 2 2.5
Concentração de Nitrito ( µ mol.l-1)
Nú
mer
o d
e F
orr
agei
os
0
50
100
150
200
250
300
350
0 5 10 15 20 25 30 35
Concentração de Silicato ( µ mol.l-1)
Nú
mer
o d
e F
orr
agei
os
I)
II)
III)
Figura 30. Correlação entre as concentrações de nutrientes inorgânicos dissolvidos e o número
de comportamentos de forrageio registrados no Porto do Recife. I) Fosfato (PO4); II) Nitrito
(NO2); III) Silicato (SiO2).

60
A quantidade de forrageios esteve correlacionada positivamente à freqüência mensal da
soma de todos os peixes que compõem a dieta do boto-cinza (n pares = 12; rs = 0,58; p = 0,05)
(Fig. 31), porém quando analisados separadamente, apenas para a espécie Mugil curema (tainha
ou saúna) o teste de Correlação de Spearman demonstrou uma correlação positiva e significante
(n pares = 12; rs = 0,64; p = 0,0257) (Fig. 32).
0
50
100
150
200
250
300
350
400
0 5000 10000 15000 20000 25000 30000 35000
Quantidade de Pescado (ton)
Nu
mer
o d
e F
orr
agei
os
Figura 31. Correlação entre a quantidade total de presas do boto-cinza pescadas e o número de
forrageios observados no Porto do Recife.
0
50
100
150
200
250
300
350
400
0 2000 4000 6000 8000 10000 12000
Mugil curema (ton)
Nú
mer
o d
e F
orr
agei
os
Figura 32. Correlação entre o esforço pesqueiro direcionado a espécie Mugil curema e a
quantidade de forrageios registrados no Porto do Recife.

61
4.5. Uso da área do Porto do Recife para Forrageio.
No Porto do Recife, a quantidade de forrageios variou significativamente com as áreas
(H= 12,59; gl= 4; p= 0,0134), Havendo uma preferência pela alimentação na área II (70,3%),
seguido da área V (10,8%) (Fig. 33).
Embora o teste de Mann-Whitney não tenha detectado diferenças significativas entre as
categorias de profundidades, houve um número maior de forrageios registrados em profundidades
de 7 a 10 metros (n=717) quando comparados a profundidades de 3 a 6 metros (Fig. 34). Em
Olinda, todos os comportamentos de forrageio foram observados em profundidades entre 3 e 5
metros.
0
10
20
30
40
50
60
70
80
Den
sid
ade
de
Fo
rrag
eio
(%
)
I II III IV V
Áreas do Porto do Recife
Figura 33. Número de forrageios realizados pelos botos-cinza em cada área do Porto do Recife.
0
100
200
300
400
500
600
700
800
Ati
vid
ades
de
Fo
rrag
eio
3 a 6 7 a 10
Profundidades (m)
Figura 34. Freqüência absoluta de atividades de forrageio observadas para as diferentes
categorias de profundidades no Porto do Recife.

62
5. DISCUSSÃO
5.1. Ocorrência de botos-cinza no Porto do Recife e em Olinda
Os oceanos formam um ecossistema aquático cujos processos químicos, físicos e
biológicos interagem, influenciando toda a comunidade marinha. Os parâmetros mais importantes
são a luz, a temperatura, as correntes e os elementos nutrientes, que exercem uma influência
direta ou indireta em toda a estrutura oceânica (CARTON, 1997 apud FLORES-MONTES,
2003).
A distribuição e abundância de alimento é um dos fatores ecológicos que determinam a
utilização de área, a organização social, a composição e o tamanho dos grupos de golfinhos em
uma determinada região (DI BENEDITTO et al., 2001). Águas costeiras são formadas por um
mosaico de habitats que diferem tanto nos aspectos físicos quanto biológicos. Isto fica ainda mais
evidente nos estuários, complexos lagunares, manguezais, pântanos alagados e desembocaduras
de rios, devido à grande concentração de nutrientes e produtividade nesses locais (NORIS &
DOHL, 1980; SIMÕES-LOPES, 2005).
As duas áreas apresentaram características físicas, químicas e biológicas bem distintas.
Segundo a classificação das águas adotada durante o Simpósio Internacional sobre Águas
Salobras 1958 (Veneza – Itália), Olinda se classificaria como uma área de águas euhalinas (teor
de salinidade de 30,0 a 40,0), com pouca variação sazonal, e o Porto do Recife como mixohalina
(0,5 a 30,0), (MACÊDO, 2004). Os estuários, diferente da plataforma continental, são
caracterizados por possuir marcadas variações sazonais mensais e diárias na salinidade, em
decorrência da oscilação das marés e pelo influxo de água doce dos rios e drenagem terrestre,
ocasionados pelas chuvas (PASSAVANTE & FEITOSA, 2004). Como resultado, os organismos
devem estar bem adaptados a estas variações de salinidade, que atuam indiretamente controlando
o metabolismo dos seres e, conseqüentemente, a produtividade do ecossistema (NASCIMENTO,
2001).
Em relação aos sais nutrientes dissolvidos, o Porto do Recife em média apresentou todos
os teores de nutrientes mais elevados em comparação com a Praia de Bairro Novo.
NASCIMENTO (2001) caracterizou o Porto como sendo um estuário supereutrofizado. Este fato

63
deve estar relacionado ao aporte de nutrientes carreados pelos rios que deságuam no estuário do
Porto e a grande entrada de matéria orgânica decorrente do lançamento diário de esgotos
domésticos provenientes dos bairros do Pina, Boa Viagem e parte do centro do Recife
(NASCIMENTO, 2001; COSTA et al., 2004).
Os grupos fosfatados, nitrogenados e, em menor proporção, o silicato, são elementos
estruturais e importantes no metabolismo de diversos organismos, principalmente daqueles da
base da cadeia alimentar. As concentrações dos nutrientes limitam a produção dos organismos
clorofilados e conseqüentemente os níveis de produção de toda teia trófica (FLORES-MONTES,
2003; MACÊDO, 2004).
Apesar de não serem constatadas diferenças entre as espécies de peixes presentes em
Olinda e no Porto do Recife, este último apresentou, em geral, maior concentração de presas
potenciais dos botos. Das espécies encontradas na plataforma continental, cerca de 60% penetram
nos estuários para a reprodução e criação, por ser uma área mais protegida, e para alimentação, e
por esta apresentar grande disponibilidade de recursos alimentares, principalmente zooplâncton,
fitoplâncton e integrantes da meiofauna (NEUMANN-LEITÃO et al., 2004; SANTOS et al.,
2004)
Assim, é possível sugerir que a maior ocorrência de botos no Porto do Recife possa estar
indiretamente associada a maior disponibilidade de nutrientes e diretamente relacionados à maior
concentração de presas nessa região, quando comparada à Praia de Bairro Novo. Provavelmente,
a Praia de Bairro Novo seja uma área de passagem dos botos do Porto para regiões mais ao norte
do Estado de Pernambuco com características mais favoráveis à alimentação e outras atividades.
Além disso, as observações de forrageios registradas em Olinda podem ter sido meramente
oportunistas, acontecendo durante esses deslocamentos.
Houve evidência de que os mesmos animais ocorrem em Olinda e no Porto do Recife,
pois foi possível videoidentificar um animal com marca natural visualizado no Recife e que foi
reconhecido, com o auxílio do binóculo, em Olinda. Estudos de fotoidentificação são de grande
importância para estimar o tamanho da população e determinar o uso e fidelidade de áreas, assim
como padrões de deslocamento do boto-cinza no litoral de Pernambuco.

64
MONTEIRO et al. (2006) realizaram um estudo etnobiológico, onde os pescadores das
Praias de Rio Doce e Casa Caiada relataram a ocorrência de botos próxima a uma área de
desembocadura de rio na Praia do Janga, porém estudos sobre a população de botos em outras
regiões costeiras do Estado são necessários para corroborar essa hipótese.
Além disso, a altura quase ao mesmo nível do mar do ponto de observação em Olinda,
dificultou a visualização dos botos em áreas mais afastadas da costa, onde os botos podem
realizar deslocamentos para regiões adjacentes.
5.2. Número de botos avistados
No Porto do Recife, o número máximo de botos visualizados simultaneamente foi de 22
animais e em Olinda, o número de botos-cinza variou de 1 a 6 animais. Em geral, os botos-cinza
são encontrados em pequenos grupos de 2 a 10 animais (ARAÚJO et al., 2001; REIS, 2002;
TORRES & BEASLEY, 2003; SANTOS & LE PENDU, 2007), porém podem formar grupos de
dezenas de indivíduos, como foi reportado por ANDRADE et al. (1987) e AZEVEDO et al.
(2005) na Baía de Guanabara – RJ (2 a 60 indivíduos). O maior tamanho grupo foi constituído de
450 botos, registrado na Baía de Paraty - RJ por LODI & HETZEL (1997).
O número de botos observados com maior freqüência no Porto do Recife variou entre 4 e
5 e, em Olinda alcançou 6 indivíduos. Em ambas as regiões, a quantidade de eventos de
forrageios foi maior na presença de 2 a 4 indivíduos. Grupos de 2 a 6 animais também foram os
mais visualizados em outros trabalhos no litoral do Brasil (ANDRADE et al., 1987; GEISE et al.,
1999; ARAÚJO et al., 2001; REIS, 2002). Possivelmente para a região nordeste, esta quantidade
de indivíduos em grupo seja suficiente para aumentar a eficiência na procura e captura de presas,
sem comprometer as necessidades alimentares dos indivíduos do grupo.

65
5.3. Caracterização do comportamento de forrageio
No Porto do Recife, os botos forragearam mais e apresentaram um repertório
comportamental mais rico quando comparado à Olinda. Talvez, a grande disponibilidade de
alimento somada à diversidade de microhabitats nessa região, proporcionem o desenvolvimento
de uma série de estratégias alimentares e benefícios energéticos que superem as desvantagens do
Porto do Recife ser uma área bastante degradada em decorrência da poluição marinha e do
intenso tráfego de embarcações.
Comportamentos semelhantes aos registrados no presente trabalho foram também
reportados para outras populações de boto-cinza e de outros cetáceos. GEISE (1989) relatou o
comportamento de Bote Lateral para o boto-cinza no Rio de Janeiro; DOMIT (2006) descreveu
praticamente todos os comportamentos reportados neste estudo para o boto-cinza em Cananéia e
Ilha de Perças, porém com diferentes denominações, tais como: “perseguição” (mistura de bote
lateral com controle de presas), “perseguição com captura na superfície” (Bote lateral);
“perseguição com giro do corpo” (bote com ventre para cima); “giro na superfície” (bote com
giro de 360º); “imersão” e “seqüência de mergulhos profundos” (bote em baixo da água).
BEL’KOVICH et al. (1991) observou tipos de forrageios semelhantes ao rasante lateral parcial
(180º “turn”), de baixo para cima (“log jump” e “horizontal leap”) e de ventre para cima (“back
jump”) para a população de golfinho-nariz-de-garrafa (Tursiops truncatus) no Mar Negro.
Os tipos de comportamentos mais freqüentes em ambas as localidades foram o forrageio
com captura em baixo da água, na lateral e de baixo para cima com salto. Dos comportamentos
registrados no Porto do Recife, as capturas com bote adiantado e com giro de 360º foram mais
raras. Resultados semelhantes a esses foram verificados na Praia de Pipa – RN por MONTEIRO
et al. (2006). Os autores sugeriram que as estratégias com bote lateral e de baixo para cima
seriam mais comuns por proporcionarem maior velocidade no momento da captura do peixe. O
bote adiantado, denominado de “wall” por BEL’KOVICH et al. (1991), foi observado sendo
realizado individualmente ou por grupos de golfinhos-naris-de-garrafa, de maneira que um grupo
conduzia os peixes para outro grupo que servia como barreira para conter o deslocamento das
presas.

66
Os botes sem a utilização de saltos foram mais freqüentes e apresentaram maiores índices
de sucesso do que os com saltos. Provavelmente os botes sem utilização de saltos proporcionem
maior velocidade e agilidade durante a perseguição, permitindo que o golfinho consiga alcançar o
peixe mais rapidamente sem muito gasto energético. Além disso, no caso dos Botes em Baixo da
Água, a utilização do biosonar pode ajudar na detecção e captura das presas, principalmente
quando se trata de águas turvas como as do Porto do Recife.
MONTEIRO et al. (2006), sugeriram que os saltos estariam associados às estratégias de
fuga dos peixes, porém neste estudo não foram verificadas diferenças significativas entre a
utilização de salto dos golfinhos e das suas presas. Provavelmente, a utilização de saltos esteja
muito mais associada à experiência do animal ou à escolha deste em relação a maior
probabilidade de sucesso na captura da espécie de peixe e das características do habitat onde se
encontra.
Na conduta controle de presa, observada apenas no Porto do Recife, os botos logo após
capturar suas presas realizavam sucessivos arremessos e depois consumiam as presas. Esse
comportamento, provavelmente, estaria relacionado ao posicionamento adequado durante a
deglutição e ingestão da presa, a fim de evitar que escamas e espinhos danifiquem o trato
digestivo dos golfinhos. Por esse motivo o peixe é arremessado algumas vezes para o ar e
recapturado em seguida. Outra explicação plausível é a de que os botos tentariam desnortear ou
até mesmo matar a presa, com o impacto na água, antes de consumi-la (BEL’KOVICH et al.,
1991; ARAÚJO, 2001; NASCIMENTO, 2002; SOUZA, 2004). DOMIT (2006) sugeriu que esta
conduta seria uma forma de dilacerar e de arrancar pedaços da presa, uma vez que muitas tainhas
com marcas de dentes e com pequenas partes do corpo suprimidas foram capturadas por
pescadores locais, porém no presente estudo não foi obtido nenhum indício que confirmasse essa
hipótese. BEL’KOVICH et al. (1991) e ARAÚJO et al. (2001), sugeriram que essa conduta
poderia ser utilizada como um comportamento de brincadeira pelos botos-cinza, principalmente
na presença de infantes.
Em relação aos comportamentos em grupo, a estratégia de cerco utilizada por grupos
maiores de golfinhos, foi também denominada de “horizontal carousel” por BEL’KOVICH et al.
(1991) de “caça em grandes grupos” por MONTEIRO-FILHO (1992) e de “arrebanhar” por
DOMIT (2006) e de “pesca cooperativa circular” por ROSSI-SANTOS (1997). Nesse

67
comportamento, esforços simultâneos de membros do grupo são alocados com a finalidade de
manter a estrutura do cardume, de modo que, enquanto alguns animais localizados na periferia do
cardume tentam manter o cardume coeso, outros golfinhos alimentam-se. Posteriormente, há um
revezamento dos golfinhos para que todos possam se alimentar. Além disso, os peixes são
conduzidos para a superfície da água, que pode ser utilizada como uma barreira (WÜRSIG,
1986). No presente trabalho a estratégia Em Dupla ocorreu basicamente com a mesmo função,
porém envolveu um número reduzido de animais.
Na estratégia de pente, a formação paralela dos animais parece estar associada à
diminuição do espaço de fuga dos peixes ao mesmo tempo em que as presas são direcionadas a
obstáculos ou áreas mais rasas, onde as chances de captura são maiores. Esse comportamento foi
reportado para o boto-cinza na Praia de Pipa – RN por NASCIMENTO (2002), que o denominou
de “arrastão” e por ROSSI-SANTOS (1997) em Santa Catarina com o nome de “pesca
cooperativa em leque”.
Comportamentos cooperativos entre grupos de boto-cinza também foram verificados em
Fortaleza (HAYES, 1999), Rio Grande do Norte (ARAÚJO et al., 2001; NASCIMENTO, 2002;
MONTEIRO et al., 2006), no Rio de Janeiro (ANDRADE et al., 1987; GEISE, 1989), São Paulo
(MONTEIRO-FILHO, 1992), em Santa Catarina (ROSSI-SANTOS, 1997). Outras espécies de
cetáceos também foram observadas realizando estratégias em grupo, como as descritas em
populações de golfinhos-pintados-do-Atlântico no Golfo do México (FERTL & WURSIG, 1995),
de orcas em Lofoten (NØTTESTAD et al., 2002), de golfinho-nariz-de-garrafa em águas
Oceânicas (WURSIG, 1979), de golfinhos-rotadores no Havaí (NORRIS & DOHL, 1980) e de
golfinho-nariz-de-garrafa e boto-de-burmeister (Phocoena phocoena) no Mar Negro
(BEL’KOVICH et al.,1991).
As estratégias alimentares assincrônicas foram mais freqüentes no Porto do Recife,
enquanto que as estratégias cooperativas ocorreram em maior proporção em Olinda. DOMIT
(2006) verificou que a pesca assincrônica foi a mais freqüente tanto para Cananéia quanto para a
Ilha das Peças, seguida pela de pesca em pequenos grupos e pela pesca em grandes grupos. Em
áreas abertas, a dificuldade em cercar os cardumes é maior e a formação de grupos pode ser uma
maneira de cooperação em busca de maior eficiência de captura e menor gasto energético
(SHANE et al., 1986).

68
De acordo com SHANE (1990), flexibilidades comportamentais em cetáceos podem
ocorrer de três formas distintas: indivíduos da mesma população com diferenças no
comportamento; diversos comportamentos com uma função em comum; e diferentes
comportamentos com a mesma função, porém sendo desenvolvidos por populações separadas em
habitats distintos.
Assim, os comportamentos individuais são limitados pela variabilidade genética,
características morfológicas e fisiológicas do organismo e pelo habitat, pois dentro da riqueza de
comportamentos que o organismo é capaz de realizar, ele seleciona um repertorio mais
apropriado quando se encontra em um determinado tipo de habitat ou diante de uma presa
(NOWACEK, 2002)
NORIS & DOHL (1980) sugeriram que a formação de grupos para forrageio pode indicar
mecanismos acústicos de comunicação, que possibilitam uma passagem mais rápida e eficiente
de informações de um indivíduo para o outro do grupo durante atividades de forrageio. Além
disso, a presença de uma grande quantidade de peixes pode exigir um maior número de golfinhos,
para que a pesca seja mais bem sucedida (WURSIG, 1986). Por outro lado, os golfinhos podem
efetuar perseguições individuais e obter sucesso, pois nem sempre é possível a presença de um
grande grupo de golfinhos no local.
O fato de os botos conduzirem as presas de regiões mais profundas para regiões mais
rasas próximas a zona de arrebentação em Olinda e aos arrecifes artificiais, ou, forragear
próximos a redes de espera e de embarcações de pesca artesanal no Porto do Recife, demonstra a
grande capacidade adaptativa que esses animais possuem diante das características ambientais.
Em Bairro Novo, por ser uma área de praia, os botos encontraram nos locais mais rasos e de
maior turbulência a possibilidade de diminuir o espaço de fuga dos peixes e interromper seus
deslocamentos (MONTEIRO et al., 2006). No Porto, ao invés de direcionar os peixes para a zona
de arrebentação, os golfinhos os direcionaram para os diques artificiais ou utilizaram peixes
presos próximos a redes de pesca na tentativa de diminuir a rota de fuga das presas e de quebrar a
estrutura dos cardumes facilitando a captura individual (MONTEIRO-FILHO, 1995).
Estratégias semelhantes a essa foram comumente observadas em grupos de orcas (Orcinus
orca) à procura de alimento em áreas rasas próximas à costa (HOELZEL, 1993). Para Tursiops
truncatus, também existem relatos do comportamento de encurralar cardumes em direção aos

69
bancos de areia e em direção à linha de costa (SHANE et al., 1986), assim como para redes de
pesca, beneficiando os pescadores artesanais de tainha (Mugil curema) (SIMÕES-LOPES et al.,
1998).
Grupos de botos foram visualizados mergulhando e capturando peixes na zona de
turbulência formada pela remoção de substrato durante a passagem de alguns navios cargueiros
no Porto do Recife. À medida que o substrato é removido ocorre a também a remoção de
materiais em suspensão, que podem ter proporcionado a atração de peixes para essa zona, uma
vez que são utilizados como fonte de alimento por algumas espécies de peixes. SIMÕES-LOPES
(2005) descreveu uma estratégia em que três botos-cinza forrageavam utilizando uma nuvem
subaquática de lodo. O autor explica que enquanto as presas se aproximavam dessas águas turvas,
tornavam-se mais suscetíveis à medida que não podiam enxergar as manobras de ataque dos
golfinhos, que possivelmente utilizavam seus sistemas de biosonar para detectá-las e escolher a
melhor estratégia de ataque. Além disso, a remoção do substrato pode ter também facilitado a
visualização de presas bentônicas que compõem a dieta do boto-cinza.
5.4. Sazonalidade das Atividades de Forrageio
Em ambas as regiões, os golfinhos foram mais observados e forragearam mais durante o
período da manhã. A preferência de uso do habitat durante o dia corrobora os resultados de
GEISE (1999) para Cananéia - SP; HAYES (1999) para Praia de Iracema - CE; ARAÚJO et al.
(2001), para Baía dos Golfinhos - RN; LODI (2002) para Baía de Paraty – RJ e SANTOS & LE
PENDU (2007) para Baía do Pontal - BA.
Esses padrões horários podem ser determinados pela utilização de uma área para
alimentação, descanso e socialização, pelo tamanho da área e variação diária na disponibilidade
de presas devido à influência de variáveis ambientais, tais como corrente, direção do vento e
influência das marés (GARCIÁ, 1998 apud REIS, 2002; DAURA-JORGE, et al., 2005).
A preferência dos botos em forragear durante as marés seca e vazante, provavelmente,
está relacionada à diminuição do volume de água, que proporciona uma maior facilidade em
capturar suas presas, uma vez que nessas condições a área de fuga do peixe encontra-se reduzida

70
(HAYES, 1999; ARAÚJO et al., 2001; SANTOS, et al., 2007). Além disso, a redução do volume
de água proporciona uma maior densidade de presas. Essa relação entre diminuição de volume de
água e o aumento na freqüência das atividades de alimentação é ainda mais evidenciada em
decorrência durante a baixa-mar, na maré de quadratura.
REIS (2002), ROSSI-SANTOS (2006) e DOMIT (2006) verificaram em suas respectivas
áreas de estudo, uma maior ocorrência de botos-cinza durante a maré enchente. As variações nas
freqüências no uso de uma área em diferentes regimes de marés demonstram que esta espécie
explora de maneira oportunista as direções e velocidades das correntes de maré, forrageando a
favor ou contra o sentido das correntes em estuários, uma vez que as presas podem utilizar o
sentido das correntes para entrar e sair do estuário atraindo os botos para as regiões (ROSSI-
SANTOS, 2006). BONIN (2001) observaram que a entrada de grupos de botos-cinza em águas
internas dos rios da Baía de Guaratuba (PR) coincidiu com os movimentos de cardumes de peixes
na Baía.
A dispersão e densidade de recursos disponíveis em um ambiente variam espacial e
temporalmente. Estas flutuações quantitativas e qualitativas produzem mudanças em relação ao
que é consumido, influenciando o comportamento alimentar de vários mamíferos (OATES, 1987
apud DAURA-JORGE et al., 2004). Assim, variáveis abióticas podem influenciar diretamente
variáveis bióticas, tais como distribuição e abundância de recursos alimentares, e, indiretamente,
o comportamento de forrageio dos predadores (WURSIG & WURSIG, 1980).
A produtividade nos ecossistemas aquáticos depende dos efeitos cinegéticos dos fatores
metereológicos, físicos, químicos e biológicos, que limitam ou estimulam a atividade dos
organismos de base de cadeia, principalmente os planctônicos autotróficos, e conseqüentemente
influenciam direta ou indiretamente toda a cadeia alimentar aquática (FLORES-MONTES, 2003;
PASSAVANTE & FEITOSA, 2004).
A teia trófica compreende uma série de relações entre os diversos níveis da cadeia
alimentar, podendo ser destacados o fluxo pelágico, que se inicia com o fitoplâncton e segue para
os copepodos, decapodos e micidaceos, depois para pequenos peixes e posteriormente para
grandes predadores; e o fluxo bentônico, que tem origem nos detritos e outras matérias orgânicas,
que são consumidas por pequenos peixes, e esses últimos são consumidos por predadores
maiores. Porém estes fluxos estão intimamente ligados, pois muitas espécies se alimentam tanto

71
de organismos pelágicos quanto de bentônicos, e o consumidor no topo da cadeia é
freqüentemente em ambos os fluxos o mesmo animal (VASCONCELOS-FILHO, 2001).
Na área estuarina da Bacia do Pina, FEITOSA (1988) verificou que as menores
produtividade e biomassa fitoplanctônicas ocorreram no período de maior precipitação (inverno),
proporcionando um maior carreamento de nutrientes continentais para o estuário. Como
conseqüência, os teores de sais nutrientes e a turbidez foram elevados culminando na redução da
camada fótica e, conseqüentemente, da produtividade fitoplanctônica. Assim, estima-se que a
abundância de peixes também tenha sofrido uma redução, pois segundo NEUMANN-LEITÃO et
al. (2004) é possível estabelecer uma relação estreita entre o índice pluviométrico e a produção de
peixes, uma vez que, muitos destes se alimentam de seres fito e zooplânctônicos.
Os resultados obtidos nesse estudo parecem corroborar essa hipótese, uma vez que
durante a estação chuvosa, as menores freqüências de forrageio no Porto do Recife estiveram
correlacionados ao aumento na pluviometria, diminuição na salinidade, redução dos nutrientes
dissolvidos e da quantidade de presas potenciais do boto-cinza. O inverso foi verificado durante a
estação seca, na qual as atividades de alimentação do boto-cinza apresentaram uma tendência a
aumentar conjuntamente com a diminuição dos índices pluviométricos, aumento da salinidade,
dos teores de nutrientes dissolvidos e da disponibilidade de presas na região.
Embora se esperasse que com o aumento da pluviometria, a quantidade de nutrientes
aumentasse devido à lixiviação continental, o ponto de coleta localizou-se na região mais externa
do canal, onde ocorre a mistura da água do canal principal do estuário com a água do mar. Desta
maneira, com o aumento do volume de água no canal principal, a concentração de sais minerais
provavelmente foi diluída. De acordo com NEUMANN-LEITÃO et al. (2004), nos meses
chuvosos, áreas costeiras pernambucanas próximas receberam influência da pluma estuarina dos
Rios Capibaribe e Beberibe, contendo grandes concentrações de nutrientes e de biomassa
fitoplantônica. Larvas de decápodes oriundas do estuário da Bacia do Pina foram predominantes
na plataforma durante a estação chuvosa, uma vez que essas larvas servem de alimento para
várias espécies de peixes (SCHWAMBORN et al., 1991; VASCONCELOS-FILHO, 2001).
A freqüência de botos-cinza foi relacionada à variação sazonal de abundância e
distribuição de presas no Rio Grande do Norte (ARAÚJO et al., 2001) em Fortaleza (HAYES,

72
1999), no Rio de Janeiro (LODI, 2002; AZEVEDO et al., 2005), em Santa Catarina (DAURA-
JORGE et al., 2004).
DAURA-JORGE et al. (2004) verificaram que durantes as estações frias (outono e
inverno) os botos-cinza percorreram uma área maior e se deslocaram mais intensamente. Os
autores sugeriram que o aumento no requerimento espacial e nos deslocamentos seria necessário
para compensar a ausência e/ou escassez de suas presas, principalmente aquelas dominantes.
A diminuição na ocorrência e na freqüência de forrageio durante o inverno e a ausência do
botos-cinza no Porto nos meses de maio e junho, poderiam estar associada ao deslocamento
desses animais do Porto para outras áreas costeiras com maior abundância de presas. Na Baía da
Babitonga, o inverno e a primavera foram os períodos mais críticos, em termos de recurso
alimentar, para as populações de botos-cinza e de toninhas (Pontoporia blaivillei). Assim, ambas
as espécies responderam a esta situação dispersando e ampliando sua área de forrageamento. Já
as estações de verão e outono apresentaram uma grande disponibilidade de recursos alimentares e
as populações das duas espécies se mantiveram mais agregadas em áreas de concentração de
presas (CREMER, 2007).
Das espécies de presas potenciais do boto-cinza analisadas, apenas a freqüência de pesca
da tainha ou sauna (Mugil curema) apresentou uma correlação com a quantidade de eventos de
forrageio ao longo dos meses. CREMER (2007) verificou que na Baía de Babitonga M. curema
foi a presa de maior relevância, seguida de Micropogonias furnieri (corvina) e Diapterus
rhombeus (carapeba). No Rio de Janeiro, as espécies mais representativas foram Trichiurus
lepturus (peixe-espada), Porichthys porosissimus (Piramangaba) e Mugil sp. ocupou a 19º
posição em relação ao índice de relevância (DI BENEDITTO et al., 2001).
Essa relação pode indicar que a tainha seja uma espécie de presa predominante na dieta do
boto e que a distribuição, comportamento e abundância do boto seja influenciada pela
disponibilidade desse recurso alimentar. Contudo, são necessários estudos sobre o conteúdo
estomacal e ecologia alimentar do boto-cinza no litoral de Pernambuco para corroborar essa
hipótese, pois muitas espécies que são importantes na dieta desse animal em outras regiões do
País, tais como o peixe-espada (Trichiurus lepturus) e a carapeba (Diapterus rhmobeus), habitam
a região costeira do Estado (LESSA et al., 2006) e, no entanto, não são registradas no banco de

73
dados do CEPENE. Essas espécies, e talvez outras que componham a dieta do boto, não sejam
alvos da pesca na região, porém representem grande relevância na alimentação dos botos.
Outro fator que pode ter influenciado na redução ou ausência de botos no Porto é a
presença de tubarões. HAZIN et al. (2000) registraram a ocorrência de oito espécies de tubarão
no litoral pernambucano, Carcharhinus acronotus (Flamengo); Carcharhinus plumbeus (Sucuri)
Carcharhinus leucas (Cabeça-chata); Carcharhinus limbatus (Galha-preta); Carcharhinus
porosus (Azeiteiro); Galeocerdo cuvier (Tigre); Ginglymostoma cirratum (Lixa) e Sphyrna
lewini (Martelo). Os golfinhos são itens alimentares, tanto para o tubarão Cabeça-chata, quanto
para o tubarão Tigre (COMPAGNO, 1984).
Várias espécies da família Carcharhinidae utilizam berçários, que são áreas
geograficamente discretas dentro da distribuição da espécie, para onde as fêmeas migram por
ocasião do parto, e onde os filhotes permanecem durante os estágios iniciais do ciclo de vida
(CASTRO, 1993 e SIMPFENDORFER & MILWARD, 1993 apud GARLA, 2003). Os berçários
normalmente são localizados em regiões costeiras com águas relativamente rasas e oferecem as
vantagens seletivas de uma baixa taxa de predação, abundância de presas e hábitat apropriado
para os jovens (MORRISSEY & GRUBER, 1993 e MERSON & PRATT, 2001 apud GARLA,
2003). Assim, o Porto do Recife constitui uma área com características atrativas para ser usada
como berçários. De acordo com COMPAGNO (1984) o tubarão Cabeça-chata utiliza áreas
estuarinas como berçários durante os meses de inverno. Além disso, HAZIN et al. (2000)
capturou esses animais em áreas um pouco ao Sul da entrada do Porto, durante os meses de
março (n = 1) e em junho (n = 3). Porém, estudos comparativos entre as freqüências de
ocorrências dos botos-cinza e de tubarões são necessários para corroborar essa hipótese.
5.5. Uso da área do Porto do Recife para Forrageio.
Em relação ao uso das áreas no Porto, houve uma preferência dos botos pelas áreas II e V
(vide Fig. 6), que se localiza em canais entre diques artificiais na entrada do Porto. De acordo
com LODI (2002), áreas de passagem proporcionam obstruções que agrupam as presas,
favorecendo emboscadas e produzem correntes favoráveis que reduzem o custo energético do
forrageamento. SHANE (1990), observou que o golfinho-nariz-de-garrafa após um período

74
nadando contra a corretenza, mergulhava e emergia com um peixe em sua boca. A autora
denominou esse comportamento de “alimentação contra corrente” e sugeriu que as presas eram
arrastadas para o predador devido à força das correntes. SIMÕES-LOPES (2005) relata a
ocorrência dessa estratégia pelo golfinho-nariz-de-garrafa na Baía de Tramandaí.
Embora o maior número de eventos de forrageio no Porto do Recife tenha sido registrado
em profundidades de 7 a 10 metros, não houve diferença significativa entre esta categoria e a de
profundidades de 3 a 6 metros. Provavelmente, a maior ocorrência de forrageios em áreas mais
profundas, seja devido a essas áreas se localizarem mais próximas à região de mar aberto,
favorecendo a concentração de presas marinhas. Em Olinda, os comportamentos foram
observados em profundidades de 3 a 5 metros. A preferência de utilização de botos variou com
diferentes profundidades em várias regiões do Brasil. ANDRADE et al. (1987), verificaram que
os botos ocorrem mais em áreas com profundidades maiores que 5 metros, REIS (2002) e
AZEVEDO et al. (2005) relataram a preferência dos botos em águas de 6 a 12m e 10m,
respectivamente. Contudo, SIMÃO & POLETTO (2002) constataram que os botos pescam mais
nas áreas de profundidade maior que 10 metros durante todo o ano. A presença de espécies de
presas tanto pelágicas quanto demersais na dieta do boto-cinza, demonstra que esse golfinho pode
se alimentar em diferentes profundidades (DI BENEDITTO et al., 2001).

75
6. CONCLUSÕES:
Por ser uma área estuarina, com grande disponibilidade de recursos alimentares, o Porto
do Recife constituiu um local muito utilizado pelos botos como área de forrageio, enquanto
Olinda caracterizou-se como uma área de deslocamento dos botos-cinza entre regiões adjacentes,
com atividades de forrageio ocorrendo de maneira oportunista.
Os botos-cinza apresentaram um repertório comportamental mais diversificado no Porto
do Recife quando comparado a Olinda.
Houve uma preferência dos botos por direcionar as presas para regiões mais rasas e com
presença de obstáculos, o que possivelmente proporcionou a diminuição da rota de fuga dos
peixes e o amento no sucesso da captura.
As freqüências de atividades de forrageio sofreram influência de vários fatores ambientais
em ambas as regiões, sendo mais freqüentes durante o turno da manhã e no verão. No Porto do
Recife, a quantidade de forrageios foi influenciada indiretamente por variáveis ambientais, como
a estação do ano, os estados de maré, a salinidade, o nível de precipitação pluviométrica e os
teores de nutrientes inorgânicos dissolvidos e, diretamente, pela sazonalidade de ocorrência das
presas que compõem a dieta dos botos.
Este trabalho constituiu um passo inicial para um melhor entendimento dos aspectos
comportamentais e ecológicos do boto-cinza. Porém faz-se necessária a realização de mais
estudos sobre a ecologia alimentar, o tamanho da população, o uso e fidelidade de áreas, bem
como sobre padrões de deslocamento do boto-cinza no litoral de Pernambuco. Locais de grande
importância para a alimentação, como o Porto do Recife, são consideradas como áreas-chave para
a conservação de cetáceos. Assim, espera-se que as informações contidas nesta pesquisa
forneçam subsídios para elaboração de planos de conservação e de gerenciamento costeiro da
região, uma vez que desse habitat depende a sobrevivência da espécie estudada.

76
7. REFERÊNCIAS BIBLIOGRÁFICAS:
ALTMANN, J. Observational study of behavior: sampling methods. Behaviour, 49(3/4): 227-
267. 1974.
ALVES-JUNIOR, T.T. & C. MONTEIRO-NETO. Caracterização morfológica e morfométrica
do boto-cinza, Sotalia fluviatilis (Gervais, 1853) da costa do Estado do Ceará, Brasil.
Arquivos de Ciências do Mar, Fortaleza, 32: 89-101. 1999.
ANDRADE, L.; SICILIANO, S. & CAPISTRANO, L. Movimentos e atividades do boto Sotalia
guianensis (Cetacea, Delphinidae) na Baía de Guanabara – Rio de Janeiro. In: Reunião de
Trabalhos de Especialista em Mamíferos Aquáticos da América do Sul, 2, 1987, Rio de
Janeiro. Anais... Rio de Janeiro: Fundação Brasileira para Conservação da Natureza, 1987.
p. 49-55.
ARAÚJO, J. P., PASSAVANTE, J. Z. O., & SOUTO, A. S. Behavior the estuarine dolphin,
Sotalia guianensis, at Dolphin Bay Pipa – Rio Grande do Norte – Brazil. Tropical
Oceanography, 29(2), 13-23. 2001.
ARAÚJO, P. J.; ARAÚJO, M. E. & FREITAS, D. A. Áreas de ocorrência de Sotalia fluviatilis
(Cetácea: Delphinidae) no litoral de Pernambuco. Resumos do VI Encontro Nacional
sobre Conservação e Pesquisa de Mamíferos Aquáticos, UNIVALI, Itajaí. p. 13. 2005.
AZEVEDO, A. F.; VIANA, S. C.; OLIVEIRA, A. M. & SLUYS, M. V. Group characteristics of
marine tucixis (Sotalia fluviatilis) in Guanabara Bay, south-eastern Brazil. Journal of the
Marine Biological Association of the Unitede Kingdom, 85: 209-212. 2005.
BEL´KOVIC, V. M.; IVOVA, E. E.; YEFREMENKOVA, O. V. ; KOZAROVITSKY, L. B. &
KHARITONOV, S. P.. Searching, behavior in bottlenose dolphin (Tursiops truncatus) in
the Black Sea. In: Prior, K & Norris, K. S. (Eds). Dolphin Societs: Discoveries and
Puzzles. University of California Press, Berkeley, C. A. 1991. p. 38-67.
BELTRÃO, A. DE L.; MAIA, J. T. A.& OLIVEIRA, M. L. Diagnóstico ambiental do
município de Olinda: uma contribuição ao Plano Diretor. Recife, CPRH. 1995. 160 p.
BONIN, C. A. Utilização de habitat pelo Boto-cinza, Sotalia fluviatilis guianensis (Cetacea,
Delphinidae), na porção norte do Complexo estuarino da baía de Paranaguá, PR.
Dissertação de Mestrado, Universidade Federal do Paraná, Curitiba. 2001.

77
BOROBIA, M. & BARROS, N. B. Notes on the diet of marine Sotalia fluviatilis. Marine
Mammal Science, 5: 395-399. 1989.
BOROBIA, M., S. SICILIANO, L. LODI & W. HOEK. Distribution of the South American
Dolphin Sotalia fluviatilis. Canadian Journal of Zoology, 69:1025-1038. 1991.
BRASIL. Boletim Estatístico da pesca marítima e estuarina do nordeste do Brasil.
CEPENE/IBAMA, Tamandaré. 2005.
CASTELLO, H. P. An introduction to the whales and dolphins. In: M. P. Simmonds and J. D.
Hutchinson Ed. The conservation of whales and dolphins. John Wiley & Sons. Lodon,
476p. 1996.
CITES. 2003. Convetion on Trade in Endagered Species of Wild Fauna and Flora, Appendix I.
Disponível em: <http://www.wcmc.org.uk/CITES/english/index.shtml>. Acesso em: 15 de
outubro de 2005.
CMS. 2002. Convention on Migratory Species. Appendix II of convention on the conservation
of migratory species of wild animals. Disponível em:
<http://www.cms.int/documents/appendix/cms_apphtm>. Acesso em: 15 de outubro de
2005.
COMPAGNO, L.J.V., FAO species catalogue. Vol. 4. Sharks of the world. An annotated and
illustrated catalogue of shark species known to date. Part 2. Carcharhiniformes. FAO Fish.
Synop., 125 (4), Pt.2: 251-655.1984.
COSTA, M. F.; ARAÚJO, M. C. B.; CHAGAS, A. C. O.; SANTI-ANNA Jr, N. & SOUZA, S.
T. Poluição Marinha, 287-318p. In: Oceanografia: um cenário tropical (EKINAZI-
LEÇA, E.; NEUMANN-LEITÃO, S.; COSTA, M. F. Orgs). Ed. Bagaço, Recife. 2004.
CUNHA, H. A; DA SILVA E. V. M. F.;. LAILSON-BRITO JR J.; SANTOS M.C.O.; FLORES
P.A.C.; MARTIN A.R.; AZEVEDO A.F.; FRAGOSO A.B.L.; ZANELATTO, R.C. &
SOLE´-CAVA A.M. Riverine and marine ecotypes of Sotalia dolphins are different species
Marine Biology, Berlin, 148: 449-457. 2005.
CREMER, M.J. Ecologia e Conservação do Golfinho Sotalia fluviatilis guianensis (Cetacea:
Delphinidae) na Baía de Babitonga, Litoral Norte de Santa Catarina. Dissertação de
Mestrado (Ecologia). Universidade Federal de São Carlos, São Carlos, SP. 2007.

78
DA SILVA, V. M. F. & BEST, R. C. Sotalia fluviatilis. Mammalian Species. American Society
of Mammalogists, 527: 1-17. 1996.
DAURA-JORGE, F. G.; WEDEKIN, L. L. & SIMÕES-LOPES, P. C. Variação sazonal na
intensidade dos deslocamentos do boto-cinza, Sotalia guianensis (Cetacea:Delphinidade) na
Baía da Ilha de Santa Catarina. Biotemas, 17(1): 203-216. 2004.
DAURA-JORGE, F. G., L. L. WEDEKIN, V. Q. PIACENTINI E P. C. SIMÕES-LOPES.
Seasonal and daily patterns of group size, cohesion and activity of the estuarine dolphin,
Sotalia guianensis (P.J. van Bénéden) (Cetacea, Delphinidae), in southern Brazil. Revista
Brasileira de Zoologia 22:1014-1021. 2005.
DHN. Carta Náutica nº 902: Porto do Recife. 5º Edição. Diretoria de Hidrografia e Navegação da
Marinha do Brasil. 1989.
DI BENEDITTO, A. M.; RAMOS, R. A.; SOUZA, C. M. M.; CARVALHO, C. E. V.de &
RESENDE, C. E. de. Heavy metal concentrations in two species of marine mammals and
their preys from the northern coast of Rio de Janeiro State, Brazil. International
Conference on Heavy Metals in the Enviromnent. Michigan, USA.6-10. 2000.
DI BENEDITTO, A. P. M.; RAMOS, R. M. A & LIMA, N. R. W. Os golfinhos:origem,
classificação, captura acidental, hábito alimentar. Editora Cinco Continentes LTDA,
Porto Alegre. 2001. 152p.
DOMIT, C. 2006. Comportamento de pesca do boto-cinza, Sotalia guianensis (van Bénéden,
1864). Dissertação de mestrado, Universidade Federal do Paraná, Curitiba.
FAVARO, E. G. P. Utilização da Baía dos Golfinhos (Pipa, RN) pelo boto Sotalia fluviatilis
(Cetacea: Delphinidae). Dissertação de mestrado, Universidade Federal de Pernambuco,
Recife, Brazil. 2004. 35 p.
FEITOSA, F. A. N.. Produtividade primária do fitoplâncton correlacionada com parâmetros
bióticos e abióticos na Bacia do Pina (Recife – Pernambuco - Brasil). Dissertação de
Mestrado, Universidade federal de Pernambuco, Recife. 1988. 270 p.
FERTL, D. & WÜRSIG, B. Coordinated feeding by atlantic spotted dolphins (Stenella frontalis)
in the Golfo of Mexico. Aquatic Mammals, v. 21, n. 1, p. 3-5. 1995.
FLORES-MONTES, M. J. P. Fatores que influenciam na produtividade dos oceanos: a
importância do fluxo de difusão dos nutrientes para a biomassa do fitoplâncton na

79
região oceânica do nordeste brasileiro. Tese de Doutorado, Universidade Federal de
Pernambuco, Recife. 2003. 179p.
FLORES, P. A. C. & BAZZALO, M. Home ranges and moviment patterns of the marine tucuxi
dolphin, Sotalia fluviatilis, in Baía Norte, Southern Brazil. Latin American Jornal of
Acuatic Mammals, 3 (1):.37-52. 2004.
FLORES, P. A. C. & FONTURA, N. F. Ecology of marine tucuxi, Sotalia guianensis, and
bottlenose dolphin, Tursiops truncatus, in Baía Norte, Santa Catarina State, southern Brazil.
LAJAN, 5(2): 105-115. 2006.
FORDYCE, R. E. & BARNES, L. G. The evolutionary history of whales and dolphins. Annual
review of earth and planetary science, 22: 419-455. 1994.
GARLA, R. C. Ecologia e conservação dos tubarões do arquipélago de Fernando de
Noronha, com ênfase no tubarão-cabeça-de-cesto Carcharhinus perezi (Poey, 1876)
(Carcharhiniformes, Carcharhinidae). Tese de Doutorado, Universidade Estadual
Paulista “Julio de Mesquita Filho”, Campus de Rio Claro São Paulo. 2003
GEISE, L. O Comportamento dos Golfinhos. Ciência Hoje. 13 (77): 27-33. 1991.
GEISE, L.; GOMES, N. & CERQUEIRA, R.. Behavior, habitat use and population size of
Sotalia fluviatilis (Gervais, 1853) (Cetacea: Delphinidae) in the Cananéia estuary region,
São Paulo, Brazil. Revista Brasileira de Biologia, 59(2): 183-194. 1999.
GOMES, D. N. F. Caracterização de fungos filamentosos isolados nas praias de Casa Caiada
e Bairro Novo, Olinda- PE, quanto aos fatores de patogenicidade. Dissertação de
mestrado, Universidade Federal de Pernambuco. 2002. 61 p.
GRASSHOF, K.; EHRHARDT, M.& KREMLING, K. Methods of seawater analysis. 2 ed.
New York: Verlag Chemie. 1983. 419p.
HAZIN, F. H. V.; WANDERLEY Jr, J. A. M. & MATTOS, S. M. G. Distribuição e abundância
relativa de tubarões no litoral do Estado de Pernambuco, Brasil. Arquivos de Ciência do
Mar, Fortaleza, 33: 33-42. 2000.
HAYES, A. J. S. Aspectos da atividade comportamental diurna da forma marinha do tucuxi
Sotalia fluviatilis Gervais, 1853 (Cetacea - Delphinidae), na Praia de Iracema
(Fortaleza - Ceará - Brasil). Relatório de Licenciatura, Universidade do Algarve, Faro.
1998. 52 p.

80
HETZEL, B. & LODI, L. Baleias, botos e golfinhos: guia de identificação para o Brasil. Rio
Janeiro: Nova Fronteira. 1993. 180 p.
HOELSEL, A R. Killer whale predation on marine mammals at Punta Norte, Argentina; food
sharing, provisioning and foraging strategy. Behavioral Ecology and Sociobiology, 29:
197-204. 1991.
IBAMA 2001. Plano de ação – Mamíferos Aquáticos do Brasil. Versão II, Brasília. 61 p.
IUNC 2007. Cetacean Specialist Group 1996. Sotalia fluviatilis. In: IUCN 2007. 2007 IUCN
Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 29 December
2007.
ITEP. 2007. Laboratório de Metereologia de Pernambuco (LAMEPE). Disponível em:
http://www.itep.br/LAMEPE.asp. Acesso em: 12 de janeiro de 2008.
LINK, L. O. & YANAMOTO, M. E. Fidelidade ao local de uso das baías por indivíduos de
Sotalia fluviatilis, na região de Pipa, Município de Tibau do Sul –RN. Resumos da 9 º
Reunión de Trabajo de Especialistas em Mamíferos Acuáticos da América Del Sul.
Bueno Aires, Argentina, p. 74. 2000.
LESSA, R.; VIEIRA, A. C. S.; MONTEIRO, A.; SANTOS, J. S.; LIMA, M. M.; CUNHA, E.
J.; SOUZA-JUNIOR, J. A. C.; BEZERRA, S.; TRAVASSOS, P. E. P. F. & OLIVEIRA, B.
A. B. R. Diagnóstico da pesca no litoral do Estado de Peranmbuco. In: A pesca marinha e
estuarina do Brasil no início do século XXI: recursos, tecnologias, aspectos
socioeconômicos e institucionais. Isaac, V. J.; Martins, A. M.; Haimovici, M.; Andriguetto-
Filho, J. M. (Orgs.). Belém: Universidade Federal do Pará – UFPA, 2006. 188 p.
LODI, F. L. Tamanho e composição de grupos dos botos-cinza Sotalia Guianensis (Van
Bénéden, 1864) (Cetacea, Delphinidae), na Baía de Paraty, RJ, Brasil. Atlântica, 25 (2):
135-146. 2003.
LODI, F. L. Uso do habitat e preferências do boto-cinza, Sotalia fluviatilis (Cetacea,
Delphinidae), na Baía de Paraty, Rio de Janeiro. Dissertação de Mestrado. Universidade
Federal Rural do Rio de Janeiro, Rio de Janeiro. 2002. 167 p.
LODI, L.; HETZEL, B. Uma grande agregação de botos-cinza (Sotalia fluviatilis) na baía da Ilha
Grande, Rio de Janeiro. In: Congresso Nordestino De Ecologia, 7., 1997, Ilhéus.
Resumos... Ilhéus: UFBA, UESC, SNE, CEPLAC, 1997, p. 337.

81
MACÊDO, S. J.; MUNIZ, K.& FLORES-MONTES, M. J. Hidrologia da região costeira e
plataforma continental do Estado de Pernambuco, 255-286p. In: Oceanografia: um
cenário tropical. EKINAZI-LEÇA, E.; NEUMANN-LEITÃO S.; COSTA, M. F. (Orgs).
Ed. Bagaço, Recife. 2004.
MANN, J. Unraveling the dynamics of social life, p. 45-64. In: Cetacean society - field studies
of dolphin and whales. J. MANN; R.C. CONNOR; P.L. TYACK & H. WHITEHEAD
(Eds). Chicago, University Chicago Press. 2000. 448p.
MONTEIRO, M. S.; SOUTO, A. & NASCIMENTO, L. F. Comparações entre os
comportamentos de forrageio nas diferentes faixas etárias do boto-cinza (Sotalia
guianensis) (Cetacea; Delphinidae) na Baia dos Golfinhos, Praia de Pipa, RN, Brasil.
Revista de Etologia, 8(1): 13-25. 2006.
MONTEIRO, M. S.; TARGINO, R. K. M. S. & CORREIA, R. C. F. Interação do boto-cinza
(sotalia guianensis) (Cetacea: Delphinidae) com atividades pesqueiras no litoral norte de
Pernambuco. In: Congresso Latino-Americano de Ciências do Mar, 12, 2007,
Florianópolis. Anais... Florianópolis: Associação Brasileira de Oceanografia, 2007. 3 p.
MONTEIRO-FILHO, E. L. A. Comportamento de caça e repertório sonoro do golfinho
Sotalia brasiliensis (Cetacea: Delphinidae) na região de Cananéia, Estado de São
Paulo. Tese de Doutorado, Universidade Estadual de Campinas, Campinas, SP. 99 p. 1991.
MONTEIRO-FILHO, E. L. A. Pesca associada entre golfinhos e aves marinhas. Revista
Brasileira de Zoologia, 9 (1/2): 29-37. 1992.
MONTEIRO-FILHO, E.L.A.; L.R. MONTEIRO & S.F. DOS REIS. Skull shape and size
divergence in dolphins of the genus Sotalia: a tridimensional morphometric analysis.
Journal of Mammalogy, Washington, 18 (1): 125-134. 2002.
MONTEIRO-NETO, C., T., ALVES-JÚNIOR, T. F.; ÁVILA, J. C., OLIVEIRA J. A. &
FURTADO-NETO, M. A. A. Registros de cetáceos para o litoral do Estado do Ceará,
Brasil. Arquivos de Ciência do Mar. Fortaleza, 30 (1-2): 79-92. 1996.
NASCIMENTO, F. A. F. Aspectos ecológicos da comunidade fitoplanctônica da Bacia do
Pina associados com alguns parâmetros abióticos (climatológicos e hidrológicos).
Dissertação de Mestrado, Universidade Federal de Pernambuco, Recife. 2001. 141p.

82
NASCIMENTO, L. F. Descrição comportamental do boto cinza (Sotalia
fluviatilis,GERVAIS,1853) no litoral sul do estado do Rio Grande do Norte.
Dissertação de Mestrado, Universidade Federal do Rio Grande do Norte, Natal. 2002. 60 p.
NEUMANN-LEITÃO, S.; SCHWAMBORN, R.; V. & VASCONCELOS-FILHO, A. L. Teia
Trófica Pelágica, 459-476p. In: Oceanografia: um cenário tropical. EKINAZI-LEÇA, E.;
NEUMANN-LEITÃO, S.; COSTA, M. F. (Orgs). Ed. Bagaço, Recife. 2004.
NORRIS, K. S. & DOHL, T. P. Behavior of the hawaihan spinner dolphin Stenella longirostris.
Fishery Bulletin, 77: 821-849. 1980.
NØTTESTAD, L., FERÖ, A. e BJØR, E. Digging in the deep: whales’s advanced huting tatic.
Polar Biology, 25: 939-941. 2002.
NOWACEK, D. P. Sequential foranging behaviour of bottlenose dolphins, Tursiops truncatus, in
Sarasota Bay, FL. Behaviour, 139: 1125-1145. 2002.
PASSAVANTE, J. Z. O. & FEITOSA, F. A. N. Dinâmica da produtividade fitoplanctônica na
zona costeira Marinha, 425-440p. In: Oceanografia: um cenário tropical. EKINAZI-
LEÇA, E.; NEUMANN-LEITÃO, S.; COSTA, M. F. (Orgs). Ed. Bagaço, Recife. 2004.
PINEDO, M. C., ROSAS, F. C. W. & MARMONTEL, M. Cetáceos e pinípedes do Brasil: uma
revisão dos registros e guia para identificação das espécies. Manaus, UNEP/FUA. 1992.
92 p.
POLETTO, F. R. Aspectos comportamentais da população de botos-cinza (Sotalia fluviatilis)
na Baía de Sepetiba (RJ). Dissertação de Mestrado. Universidade Federal do Rio de
Janeiro, Rio de Janeiro. 2003.
REIS, M. S. S. O boto sotalia fluviatilis (Gervais, 1853) (Cetácea: Delphinidae) no Litoral de
ilhéus, Bahia: Comportamento e interações com as atividades pesqueiras. Dissertação
de Mestrado, Universidade estadual de Santa Cruz, Ilhéus, Bahia. 2002.84 pp.
ROSSBACH, K. A. & HERZINC, D. L. Underwater observations of bethic-feeding bottlenose
(Tursiops truncatus) near Grand Bahamas Island, Bahamas. Marine Mammal Science,
13(3): 498-504. 1997.
ROSSI-SANTOS, M. R. Estudo quali-quantitativo do comportamento de alimentação do
golfinho ou boto-cinza Sotalia fluviatilis Gervais, 1853 (Cetacea, Delphinidae) na área

83
de proteção ambiental de Anhatomirim e Baía Norte de Santa Catarina. Monografia de
Bacharelado, Universidade Federal de Santa Catarina, Florianópolis. 1997.
ROSSI-SANTOS, M. R. Ecologia comportamental do boto-cinza, Sotalia guianensis (Van
Bénéden, 1874) (Cetacea: Delphinidae), na região extremo sul do Estado da Bahia,
Nordeste do Brasil. Dissertação de mestrado, Universidade Federal do Paraná, Curitiba.
2006.
SANTOS, M.C. DE O., ROSSO, S., SANTOS, R.A., LUCATO, S. H. B. & BASSOI, M.
Insights on small cetacean feeding habits in southeastern Brazil. Aquatic Mammals, 28:
38-45. 2002.
SANTOS, L. P. S.; SANTOS, P. J. P. & FONSECA-GENOVOIS, V. G. Meiofauna, 529-554. In:
Oceanografia: um cenário tropical. EKINAZI-LEÇA, E.; NEUMANN-LEITÃO, S.;
COSTA, M. F. (Orgs). Ed. Bagaço, Recife. 2004.
SANTOS, U.A. & LE PENDU, Y. Uso do habitat e características dos grupos de boto-cinza
(Cetacea, Delphinidae), na Baía do Pontal, Ilhéus, Bahia, Brasil. In: Congresso Latino-
Americano de Ciências do Mar, 12, 2007, Florianópolis. Anais... Florianópolis:
Associação Brasileira de Oceanografia, 2007. 3 p.
SAULITIS, E., MARTKIN, G., BARRETT-LENNARD, L., HEISE, K. e ELLIS, G. Foranging
Strategies of Sympatric Killer Whale (Orcinus orca) populations in Prince Willian Sound,
Alaska. Marine Mammal Science, v. 16, n. 1, p. 94-109. 2000.
SHANE, S.H.; WELLS, R.S. & WÜRSIG, B. Ecology, behavior and social organization of the
bottlenose dolphin: a review. Marine Mammal Science, 2(1): 34-63. 1986.
SHANE, S. H. Behavior and ecology of the bottlenose dolphin at Sinabel Island, Florida. In: The
bottlenose dolphin. LEATHERWOOD, S.; REEVES, R. R. (Eds.). San Francisco,
Academic Press, pp. 245-265. 1990.
SCHWAMBORNR, R.; ERAU, W.; SILVA, A. P.; SILVA,T. A. & SAINT-PAUL, U. The
contribuition of estuarine decapod larval to marine zooplankton communities in North-East
Brazil. Arch. Fish. Mar. Res., 47(2/3): 201-210. 1991.

84
SIMÃO, S. M., PIZZORNO, J. L. A., PERRY, V. N. & SICILIANO, S. Aplicação da técnica de
foto-identificação do boto-cinza (Sotalia fluviatilis) (Cetacea, Delphinidae) da Baía de
Sepetiba. Floresta e Ambiente, 7 (1), 31-39. 2000.
SIMÕES-LOPES, P. C. Ocorrência de uma População de Sotalia fluviatilis Cervais, 1853,
(CETACEAE, DELPHINIDAE) no Limite Sul da sua Distribuição, Santa Catarina, Brasil.
Biotemas, 1 (1): 57-62. 1988.
SIMÕES-LOPES, P.C., FÁBIAN, M.E. & MENEGHETI, J.O. Dolphin interactions with the
mullet artisanal fishing on Southern Brazil: a qualitative and quantitative approach. Revista
Brasileira de Zoologia, 15: 709-726. 1998.
SIMÕES-LOPES, P. C. O luar do delphin: a maravilhosa aventura da história natural. Ed.
Letradágua, Joinville. 2005. 304p.
SIMÕES-LOPES, P. C. Morfologia do sincrânio do boto-cinza, Sotalia guianensis (P.J. van
Bénéden) (Cetacea, Delphinidae). Revista Brasileira de Zoologia, 23(3): 652-660. 2006.
SOUZA, L. C. Quantificação e descrição de condutas comportamentais de infantes e adultos
do boto Sotalia guianensis na praia de baía Formosa (Rio Grande do Norte, Brasil).
Monografia de conclusão do curso de Ciências Biológicas Bacharelado, Universidade
Federal de Pernambuco, Recife, Brasil. 2004.30 p.
SOUTO, A. Etologia: princípios e reflexões. Editora Universitária UFPE, Recife. 343 p. 2003.
SPINELLI, L. H. P., NASCIMENTO, L. F. & YAMAMOTO, M. E. Interações entre adultos e
filhotes em grupos do boto cinza Sotalia fluviatilis na Praia de Pipa – RN. Estudos de
Psicologia, 7 (1): 165-171. 2002.
STRCKLAND, J. D. H. & PARSONS, T. R. A pratical handbook of seawater analisys. 2 ed.
Bulletin of Canada, Ottawa, 17: 207-211. 1972.
TORRES, D. & BEASLEY, C. R. Pattern of use of a small bay in northern Brazil by Sotalia
fluviatilis (Cetacea: Delphinidae). Amazoniana, 13 (3/4): 583-594. 2003.
VAN BÉNÉDEN, P.J. & P. GERVAIS. Osteographie des Cetaces, Vivants et Fossiles. Paris,
Arthus Bertrand, 634p. 1880.

85
VASCONCELOS-FILHO, A. L. Interações tróficas entre peixes do Canal de Santa Cruz
(Pernambuco – Brasil). Tese de Doutorado, Universidade Federal de Pernambuco, Recife.
2001.
ZANELATTO, R. C. Dieta do boto-cinza, Sotalia fluviatilis (CETACEA:DELPHINIDAE),
no complexo estuarino da Baía de Paranaguá e sua relação com a ictiofauna estuarina.
Dissertação de Mestrado, Universidade Federal do Paraná, Curitiba. 2001. 73p.
WÜRSIG, B. Dolphins. Scientific American, 240 (3): 108-119. 1979.
WÜRSIG, B. & WÜRSIG, M. Behavior and ecology of the dusky dolphins, Logenorhynchus
obscurus, in south Atlantic. Fishery Bulletin, 77(4): 817-890. 1980.
WÜRSIG, B. Delphinid foraging strategies. In: Dolphin cognitive and behavior: A
comparative approach. SCHUSTERMAN, R.J. (Ed.). Lawrence Erlbaum Associates,
Hillsdale, NJ. pp. 347-359. 1986.

86
A N E X O S

87
Planilha 1: Presença, localização e atividades dos botos-cinza, utilizando o método Varredura. Temp: _____ Salin: ______ Índice pluviométrico: _______ Insolação: _______ Tempo: ______________
Comportamentos Local Local Local LocalHora Nº boto
Nº grupo DE SU ST SP CA PE BC FOR B A D B A D B A D B A D
8:00 8:10 8:20 8:30 8:40 8:50 9:00 9:10 9:20 9:30 9:40 9:50
10:00 10:10 10:20 10:30 10:40 10:50 11:00 11:10 11:20 11:30 11:40 11:50 12:00 12:10 12:20 12:30 12:40 12:50 13:00 13:10 13:20 13:30 13:40 13:50 14:00 14:10 14:20 14:30 14:40 14:50 15:00 15:10 15:20 15:30 15:40 15:50 16:00
LEGENDA: Comportamentos: DE= deslocamento; SU =Surfe; SP= Salto parcial; ST= Salto total; CA= Cambalhota; PE= periscópio; BCD= batida corporais; FOR= Forrageio. Localização: B = Nº de botos; A = ângulo; D = distância do observador.

88
Planilha 2: Comportamentos de forrageio utilizando o método amostral Todas as Ocorrências.
Data
N. ind no local
N. de grupos
N. boto em Cada grupo
Ativ. Antes Do For.
N. boto For.
Tipo de Pers.
Tipo de Desloca
Tipos de Bote
Ang/dist/prof Tempo Pers
Sucesso CP Tipo CP
Ativ.depois do For
Tipo Fuga Peixe
Hora In/Fim D V LA R L P
LEGENDA In.= início; N.= número; Ativ.= atividade; For.= Forrageio; Pers.= perseguição; CP= controle da presa; Ang= Ângulo; Dist= Distancia; Prof.= Profundidade Faixa Etária: A= adulto; J= juvenil; F= filhote. Tipo de Perseguição: D= dorso para cima; V= ventre para cima; LA= lateral Tipo de Deslocamento: R= rápido; L= lento; P= porposing (deslocamento rápido com saltos) Tipos de Bote: VERT= Vertical; B/C S= baixo para cima com salto; LA= lateral; V/P= ventre para cima; EA= embaixo da água; AD=adiantado; AL= alicate; RPL= rasante lateral parcial Tipos de Controle da Presa: CA= para cima com recaptura na água; CAR= para cima com recaptura no ar; LA= Lateral; Co= Coletivo.

89
Tabela 6. Lista dos itens alimentares reportados na literatura que compõe a dieta de Sotalia guianensis, ao longo da costa brasileira (adaptada de LODI, 2002).
ESTADO FAMÍLIA GÊNERO e/ou ESPÉCIE NOME COMUM HÁBITAT
PEIXES
(25)
PR Antheridade Atherinella brasiliensis (23) Peixe-rei
PR, SP, RJ Ariidae Bagre bagre (7,21) Sciadeichthys luniscutis (21) Arius spixii (21)
Bagre Bagre-cabeçudo Bagre
Demersal /Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas
PR, SP, RJ, ES Batrachoididae (13,15)
Porichthys porosissimus
(2,4,6,7,13,20,21,24) Mamangá-liso,Bagre-sapo, Peixe-sapo
Demersal / Áreas neríticas e estuarinas
PR, SP, ES, BA, RN
Carangidae (13,16,19,22)
Oligoplites saliens (7) Oligoplites Spp. (23) Selene setapinnisI (23) Chloroscombrus chrysurus (23)
Guaivira Demersal-pelágico/Áreas neríticas e estuarinas
PR, RN
Centropomidae (22)
Centropomus spp. (17) Robalo
Demersal / Áreas neríticas e estuarinas
SC, PR, SP, RJ, ES, BA
Clupeidae (1,13,14,16,17,19)
Pellona harroweri (6,13,21,23) Harengula clupeola (7,11,24) Sardinella brasiliensis (11,21,23,24) Odontognatus mucronatus (21)
Olhuda, Apapá,Sardinha Sardinha-cascuda Sardinha-verdadeira Sardinha-dentuça,sardinha-de-dente
Demersal-pelágico / Áreas neríticas e estuarinas Demersal-pelágico / Áreas neríticas e estuarinas Pelágico / Áreas neríticas e estuarinas Demersal-pelágico / Áreas neríticas e estuarinas
SC, PR, SP, RJ, ES
Engraulidae (7,13,15,17)
Anchoa spp. (5,13,15,23,24) Anchoa filifera (20,21) Anchoviella epidentostole (3,5) Licengraulis grossidens (10,23) Cetengraulis edentulus (23)
Manjuba Manjuba-branca Manjuba Manjuba, manjubão Sardinha-xingó
Pelágico / Áreas neríticas e estuarinas Pelágico / Áreas neríticas e estuarinas Pelágico / Áreas neríticas e estuarinas Pelágico / Áreas neríticas e estuarinas
RN Exocoetidae (22) Peixe-agulha, Farnangaio Pelágico / Áreas neríticas PR Gadiidae Urophycis brasiliensis (23)
Bairdiella ronchus (23) Bacalhau Oveva
(CONTINUA)

90
ESTADO FAMÍLIA
GÊNERO e/ou ESPÉCIE
NOME COMUM HÁBITAT
SC, PR, SP, BA, AL, RN
Gerreidae (14,15,17,22)
Gerres sp. (18) Diapterus olisthostomus (9) Diapterus rhombeus (16,19,23) Eucinostomus gula (7) Eucinostomus argenteus (24) Eucinostomus Spp. (23)
Carapeba Carapeba Carapeba Parmitunga,Caratinga,Cara-picu
Demersal / Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas
SC, PR, SP, RJ, ES, BA
Haemulidae (13,14,15)
Orthopristis ruber (4,21,23,24) Bairdiella rhonchus (18) Haemulon steindachneri (7,23)
Conodon nobilis (21) Genyatremus luteus (23) Pomadasys corvinaeformis (23,24) Porichthys porossissimus (23)
Roncador-do-fundão Roncador Roncador-das-pedras, Corcoroca-boca-larga Roncador
Demersal / Áreas neríticas e estuarinas Demersal / Áreas neríticas e estuarinas Demersal-pelágico / Áreas neríticas Demersal / Áreas neríticas e estuarinas
RJ,ES Lutjanidae (13,21)
Lutjanus spp.
Lutjanus griséus (23)
Caranha, Pargo, Gariúba ,Baúna, Cioba-vermelho
Demersal, oceânico / Áreas neríticas e estuarinas
SC, PR, SP, RJ, BA, AL, RN
Mugilidae (12,15,17,22)
Mugil spp. (7,9, 21, 24) Mugil liza (9) Mugil curema (10,11)
Tainha Tainha Tainha, Parati ou Saúna
Demersal-pelágico / Áreas neríticas e estuarinas Demersal-pelágico / Áreas neríticas e estuarinas Demersal-pelágico / Áreas neríticas e estuarinas
RJ Muraenoscidae Ariosoma opisthophthalma (20,21)
Congro Demersal / Áreas neríticas e estuarinas
PR Ophichthiidae (17) Myrophis punctatus (23) Muçum,Iroró,Cobra-do-mar,Miriquitis,Muriongo
Demersal / Áreas neríticas e estuarinas
RJ Phycidae Urophicys brasiliensis (21) Abrotéia Demersal / Áreas neríticas SC Pleuronectidae (10) Linguado Demersal / Áreas neríticas PR, SP Paralichthyidae
(15) Paralichthys Spp. (23) Paralichthys orbgnyanus (24) Citharicthys Spp. (23)
Linguado Demersal / Áreas neríticas
RJ,ES Pomatomidae Pomatomus saltatrix (13,21) Enchova Pelágico / Áreas neríticas (CONTINUA)

91
ESTADO FAMÍLIA GÊNERO e/ou ESPÉCIE NOME COMUM HÁBITAT
SC, PR, SP, RJ, ES, AL
Scianidae (9,13,14,15,17)
Stellifer spp. (4,7,9,21,23,24) Stellifer brasiliensis (4,7,15,21,24) Stellifer rastrifer (4,7,15,21,23,24)
Cynoscion spp. (1,6,13)
Cynoscion leiarchus (1,24)
Cynoscion striatus (2,6,10) Cynoscion jamaicensis (2,6,10,15,21) Cynoscion virencens (21,23,24) Cynoscion guatucupa (21,23) Nebris microps (21) Macrodon ancylodon (21,24)
Umbrina coroides (13,21)
Ctenosciaena gracilicirrhus (4,13,21,23) Pogonias cromis (6,21) Micropogonias furnieri (4,7,10,15,21,23,24) Menticirrhus spp. (13) Menticirrhus americanus (10,21,23) Paralonchurrus brasiliensis (10,15,20,21,23,24) Larimus breviceps (7,24) Isopisthus parvipinnis (7,15,21,23,24)
Cabeça-dura, cangoá Pescada Pescada branca Pescada,Maria-mole Pescada,Pescada-goete Pescada Pescada Pescada-banana Pescada-foguete,Pescada-cambucu Curvininha,Castanha-baré, Castanhota Miraguaia Corvina Betara Papa-terra Maria-luíza,Roncador Oveva,Cabeça-torta Tortinha,Pescadinha
Demersal / Áreas neríticas e estuarinas Demersal, Áreas neríticas e estuarinas Demersal, Áreas neríticas e estuarinas Demersal pelágico /Áreas neríticas e estuarinas Demersal pelágico /Áreas neríticas e estuarinas Demersal pelágico /Áreas neríticas e estuarina Demersal pelágico/ Áreas neríticas e estuarinas Demersal pelágico/ Áreas neríticas e estuarinas Demersal pelágico/ Áreas neríticas e estuarinas Demersal/ áreas neríticas Demersal pelágico/Áreas neríticas e estuarinas Demersal / Áreas neríticas Demersal / Áreas neríticas Demersal / Áreas neríticas e estuarinas Demersal pelágico, Áreas neríticas e estuarinas Demersal, Áreas neríticas e estuarinas Demersal, Áreas neríticas e estuarinas Demersal, Áreas neríticas e estuarinas Demersal/ Áreas neríticas e estuarinas Demersal-pelágico/ Áreas neríticas e estuarinas
(CONTINUA)

92
ESTADO FAMÍLIA
GÊNERO e/ou ESPÉCIE
NOME COMUM HÁBITAT
SC, PR, SP Serranidae (14,17)
Dules auriga (7) Badejo-fita Demersal-pelágico / Áreas neríticas
ES, BA
Sparidae (13,16,19)
Pargo,sargo-de-dente, Peixe-pena,Marimbá
Demersal-pelágico /Áreas neríticas e estuarinas
RJ Stromateidae Peprilus paru (21) Gordinho Demersal-pelágico / Áreas neríticas PR Synodontidae Synodus foetens Peixe-lagarto SC, PR, SP, RJ, ES, AL
Trichiuridae (14,17)
Trichiurus lepturus
(2,6,7,9,10,13,15,16,18,20,21,23, 24)
Espada Demersal pelágico /Áreas neríticas e estuarinas
CEFALÓPODES
(24)
PR Argonautidae Argonauta nodosa (23) Polvo
SC, PR, SP, RJ, ES, AL
Loliginidae (2,14)
Loligo spp. (6,10,13) Loligo plei (4,17,20,23) Loligo sanpaulensis (7,17,20,23) Lolliguncula brevis
(6,7,9,15,17,23,24) Dorythenthis plei (7)
Lula Lula Lula Lula Lula
Demersal/Áreas neríticas e estuarinas Demersal-semipelágica/ Áreas neríticas Demersal / Áreas neríticas e estuarinas Semipelágica / Áreas neríticas
SP Ommastrephidae Illex argentinus (8) Lula Demersal / Áreas neríticas e profundidades acima de 200m
CRUSTÁCEOS
(23)
SC,PR,SP,RJ,ES,BA
Penaeidae (10,13,15,17,19)
Farfantepenaeus brasiliensis (7) Penaeus paulensis (24) Penaeus schmitii (24)
Camarão-rosa Áreas neríticas e estuarinas
SC = Santa Catarina , PR = Paraná , SP = São Paulo, RJ = Rio de Janeiro , ES = Espírito Santo, BA = Bahia, AL = Alagoas, RN = Rio Grande do Norte Fonte dos dados: 1 - Carvalho (1963), 2 - Borobia (1984), 3 - Andrade et al. (1987), 4 - Siciliano et al. (1988), 5 - Simões-Lopes (1988), 6 - Borobia & Barros (1989), 7 - Schmiegelow (1990), 8 - Geise & Gomes (1992), 9 - Barros & Teixeira (1994), 10 - Emerin (1994, apud. Rossi-Santos 1997), 11- Monteiro Filho (1995), 12 - Rautenberg et al. (1996), 13 - Barros et al. (1997), 14 - Barros et al. (1998), 15 - Bassoi et al. (1998), 16 - Moncôrvo et al. (1998), 17 - Oliveira et al. (1998), 18 - Reis et al. (1998) ,19 - Pereira et al. (1999), 20 - Di Beneditto et al. (2000a), 21 - Di Beneditto et al. (2001), 22 - Araújo (2001), L. Lodi (obs. pess.: tainha - RJ, BA e RN); 23 - Zenelatto (2001); 24 - Santos et al. (2002). Hábitat : 23 - Williams (1984) , 24 - Haimovici et al. (1989) , 25 - Carvalho Filho (1999).
Top Related