







Línguas
Páginas
Legal


UNIVERSIDADE DO ESTADO DO AMAZONAS FUNDAÇÃO DE MEDICINA TROPICAL DR. HEITOR VIEIRA DOURADO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL MESTRADO EM DOENÇAS TROPICAIS E INFECCIOSAS
XENOVIGILÂNCIA DA MALÁRIA: AVALIAÇÃO DE UM NOVO INDICADOR
MALARIOMÉTRICO BASEADO EM CULICÍDEOS NÃO-ANOFELINOS
JOABI ROCHA DO NASCIMENTO
MANAUS
2016

i
JOABI ROCHA DO NASCIMENTO
XENOVIGILÂNCIA DA MALÁRIA: AVALIAÇÃO DE UM NOVO INDICADOR MALARIOMÉTRICO BASEADO EM CULICÍDEOS NÃO-ANOFELINOS
Dissertação apresentada ao Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas em Convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, para obtenção grau de Mestre em Doenças Tropicais e Infecciosas.
Orientador: Prof. Dr. Wuelton Marcelo Monteiro
Co-orientador: Prof. Dr. Marcus Vinícius Guimarães de Lacerda
MANAUS
2016

ii
Ficha Catalográfica
N244 Nascimento, Joabi Rocha do.
Xenovigilância da malária: avaliação de um novo indicador
malariométrico baseado em culicídeos não-anofelinos ./Joabi
Rocha do Nascimento. -- Manaus : Universidade do Estado do
Amazonas, Fundação de Medicina Tropical, 2016.
xxi, 114f. : il.
Dissertação (Mestrado) apresentada ao Programa de Pós-
Graduação em Medicina Tropical da Universidade do Estado do
Amazonas – UEA/FMT, 2016.
Orientador: Prof. Dr. Wuelton Marcelo Monteiro
Co-orientador: Prof. Dr. Marcus Vinícius Guimarães de
Lacerda
1. Xenovigilância 2 Malária. 3. Plasmodium I.Título
CDU: 616.936(043.3)
Ficha Catalográfica elaborada pela Bibliotecária Maria Eliana N Silva, lotada na
Escola Superior de Ciências da Saúde - UEA

iii
FOLHA DE JULGAMENTO
XENOVIGILÂNCIA DA MALÁRIA: AVALIAÇÃO DE UM NOVO
INDICADOR MALARIOMÉTRICO BASEADO EM CULICÍDEOS
NÃO-ANOFELINOS
JOABI ROCHA DO NASCIMENTO
“Esta Dissertação foi julgada adequada para obtenção do Título de Mestre em
Doenças Tropicais e Infecciosas, aprovada em sua forma final pelo Programa de
Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas
em convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado”.
Banca Julgadora:
___________________________________________________
Presidente: Dr. Wuelton Marcelo Monteiro (UEA/FMT-HVD)
______________________________________________________
Membro: Dra. Maria das Graças Vale Barbosa (UEA/FMT-HVD)
________________________________________________
Membro: Dr. André Machado de Siqueira (INI-FIOCRUZ)

iv
DEDICATÓRIA
A minha Mãe Raimunda Rocha do Nascimento (in memoriam) que não
obstante as limitações e dificuldades vividas, nunca perdeu a Fé, sempre
acreditou e me fez acreditar num futuro melhor. Com seu sorriso lindo e meigo
me inspirava a ter esperança, e mesmo depois de descansar nos braços do
seu Deus, os seus sonhos continuam se concretizando. A esta Mulher Virtuosa
eu dedico!

v
AGRADECIMENTOS
Agradeço ao meu Deus, pois sem ele nada faz sentido.
Agradeço de forma muito especial a Jucy, a mulher de minha vida, por
seu amor, generosidade e compreensão para comigo.
Agradeço aos meus filhos, Talita, Tales e Taís, as “crianças” mais lindas
do mundo, por sempre alegrarem minha vida.
Agradeço ao meu Pai Paulo Marques do Nascimento, soldado da
borracha, empregado da construção civil, mas, sobretudo Pai, não um pai
qualquer, mas meu Pai exemplar. Pelo exemplo de Homem que é. E pelo
incentivo constante ao meu crescimento.
Agradeço aos meus irmãos e irmãs pelo amor e por sempre torcerem a
meu favor, independente do placar.
Agradeço ao meu Orientador Dr. Wuelton Marcelo Monteiro por ter me
aceito como orientando e pela sua atenção, paciência, objetividade e
sabedoria, que generosamente me concedeu.
Agradeço ao meu Co-Orientador e Líder do grupo de pesquisa da malária
na FMT-HVD, Dr. Marcus Vinícius Guimarães de Lacerda, por todo apoio que
dele recebi.
Agradeço ao meu amigo Vanderson Sampaio, grande incentivador deste
desafio. Na verdade bem mais que um incentivador, um apoiador em todos os
momentos. Obrigado amigo! Só Deus para retribuir sua generosidade.
Agradeço ao Sr. Nelson Fé por me receber de maneira tão amigável na
Gerência de Entomologia. Pude aprender muito com o senhor e em meio a
tantos ensinamentos, ganhei um novo amigo.
Agradeço aos colegas da turma 2014 do PPGMT pelo companheirismo e
amizade. Vocês são muito especiais!
Agradeço ao Raynner e ao Fabricio por me ajudarem no trabalho hercúleo
da coleta de campo.
Agradeço a toda equipe de laboratório da Gerência de Malária pela
acolhida e apoio que recebi.
Agradeço a toda equipe do projeto TransEpi pela parceria e apoio, desde
as coletas de campo até os trabalhos no laboratório. Em especial a Dra.
Andrea Kühn e a Mestre Anne Almeida pela imprescindível ajuda nos ensaios

vi
de biologia molecular. Além de excelentes profissionais, vocês são pessoas
muito generosas!
Agradeço também a Dra. Ana Paula Coelho, ao Biomédico Reinaldo Nery
e a Biomédica Teresa Sanchez pela ajuda na identificação das
espécies/gênero, na extração de DNA e no armazenamento das amostras,
respectivamente.
Agradeço a toda equipe da Gerência de Entomologia, em particular a
Doutoranda Keillen Monick Martins e a Bióloga Íria Cabral pela ajuda inicial no
estabelecimento da colônia e na infecção experimental dos mosquitos.
Agradeço a toda equipe envolvida neste projeto.
Agradeço ao Programa de Pós-Graduação em Medicina Topical –
UEA/FMT-HVD pela oportunidade, assim como pela atenção e orientação
adequada, prestada por toda equipe da secretaria e da coordenação.
Agradeço a Universidade do Estado do Amazonas pela segunda
oportunidade em minha formação, já que tive o privilégio de fazer minha
graduação também nesta Universidade.
Agradeço ao CNPq e a FAPEAM pelo financiamento do projeto. Assim
como à Fundação Bill e Melinda Gates, financiadora do projeto TransEpi, que
possibilitou os dados relativos a população humana.

vii
DECLARAÇÃO DAS AGÊNCIAS FINANCIADORAS
O projeto foi financiado com recursos do Conselho Nacional de
Desenvolvimento Científico e Tecnológico – CNPq (Edital Universal 2012) e
Fundação de Amparo à Pesquisa do Estado do Amazonas – FAPEAM
(CHAMADA N.001/2013 - Programa Pesquisa para o SUS - PPSUS).

viii
RESUMO
A malária é reconhecida como um grave problema de saúde pública no mundo.
No Brasil, a notificação dos casos de malária está concentrada predominantemente na Região Amazônica, conhecida por sua biodiversidade com diversas espécies de artrópodes, destacando-se a família Culicidae que tem em sua composição mosquitos com variados graus de antropofilia e endofagia. O desenvolvimento experimental de Plasmodium falciparum (parcial) em culicídeos não anofelinos, assim como a utilização de mosquitos refratários para identificar patógenos virais já foi testado anteriormente. O contato do homem com diversas espécies de culicídeos em áreas endêmicas para malária sugere que a identificação do material genético de Plasmodium sp. pode ser comum, dependendo provavelmente da prevalência de malária. Um estudo transversal foi realizado com coletas de mosquitos adultos no intradomicílio, utilizando-se aspirador entomológico elétrico e armadilha BG-Sentinel® para investigar a associação entre taxa de mosquitos carreadores de DNA de Plasmodium sp. em culicídeos não-anofelinos e a frequência de infecção por malária em uma área endêmica da doença na periferia do município de Manaus, Amazonas. Foram identificados mosquitos dos gêneros Culex sp., Culex (Anoedioporpa), Culex (Mel.), Anopheles sp., Uranotaenia sp., Psorophora sp., Limatus sp., Aedeomyia sp., Coquillettidia sp. e da subfamília Toxorhynchitinae. Suas frequências foram determinadas e Culex quinquefasciatus foi a espécie mais abundante. Uma colônia de Culex quinquefasciatus foi estabelecida e infecções experimentais foram realizadas chegando-se a padronização do uso somente de abdomes de mosquitos ingurgitados em Pools de 1 a 5, com extração de DNA usando Chelex®100. Os mosquitos da amostra de campo foram separados por espécie/gênero e seguindo a padronização tiveram seus abdomes seccionados para a extração do DNA total, sendo determinada a taxa de carreamento de Plasmodium sp.. Concomitantemente foi verificada taxa de infecção de malária na população humana da mesma área. Em ambos os casos, a amplificação da sequência alvo foi realizada em ensaios padronizados de q-PCR gênero-específico utilizando um sistema Applied Biosystems 7500 Fast System®. Os resultados evidenciaram uma taxa de carreamento de DNA de Plasmodium sp. de 3,4%.
No modelo de regressão logística multivariada hierarquizado, no bloco das
variáveis proximais, as variáveis “mosquitos positivo para Plasmodium”, “ter febre” e “uso de antimalárico”, tiveram associação positiva significativa com Odds Ratio de 3.09, 4.85 e 3.09 respectivamente, todos com p˂0.05. A concordância espacial entre a presença de mosquitos carreadores de DNA de Plasmodium foi semelhante à taxa de infecção em humanos especialmente nas áreas com maior prevalência de malária na população humana. Na estimativa da acurácia da xenovigilância, a sensibilidade considerando somente os assintomáticos foi de 17,50% com VPP 46,67%, enquanto no geral a sensibilidade foi de 14.58% com VPP de 46.67%. A ferramenta se mostrou operacional, técnica e economicamente viável. Palavras-chave: Xenovigilância. Malária. Plasmodium. Culicídeos. Padronização. Vigilância.

ix
ABSTRACT
Malaria is recognized as a serious public health problem worldwide. In Brazil, the notification of malaria cases is concentrated mainly in the Amazon region, known for its biodiversity, with many species of arthropods, especially the Culicidae family that has in its composition mosquitoes with varying degrees of anthropophily and endophagy. The experimental development of Plasmodium falciparum (partial) not to Anopheles mosquitoes, as well as the use of refractory mosquitoes to identify viral pathogens has been previously tested. The contact of the man with several species of mosquitoes in endemic areas for malaria, suggests that the identification of the genetic material of Plasmodium sp. it may be common, probably depending on the prevalence of malaria. A cross-sectional study was conducted with mosquito collections inside the home, using electric entomological aspirator and BG-Sentinel® trap to investigate the association between carrier mosquitoes rate of DNA Plasmodium sp. culicids in non-Anopheles and frequency of malaria infection in an endemic area of the disease on the outskirts of the city of Manaus, Amazonas. Mosquitoes were identified genera of Culex sp., Culex (Anoedioporpa), Culex (Mel.), Anopheles sp., Uranotaenia sp., Psorophora sp., Limatus sp., Aedeomyia sp., Coquillettidea sp. and Toxorhynchitinae subfamily. Their frequencies were determined and Cx. quinquefasciatus was the most abundant species. A colony of Cx. quinquefasciatus was established and experimental infections were performed reaching the standardization using abdomens of only engorged mosquitoes in pools 1 to 5, with DNA extraction using Chelex®100. Mosquitoes field sample were separated by species / genus and following the standardization their abdomens were sectioned for the extraction of total DNA and determined the rate of entrainment of Plasmodium sp.. Concurrently was verified malaria infection rates in the human population in the same area. In both cases, amplification of the target sequence was performed in standard assays q-PCR gender-specific using an Applied Biosystems 7500 Fast System®. The results showed a DNA entrainment rate of Plasmodium sp. 3.4%. In the regression model hierarchical multivariate logistic, the block of proximal variables, variables "positive mosquitoes to Plasmodium," "have a fever" and "use of antimalarial", they had a significant positive association with odds ratio 3.09, 4.85 and 03.09 respectively, all with P0.05. The spatial correlation between the presence of Plasmodium DNA carrier mosquitoes was similar to the rate of infection in humans particularly in the areas with the highest prevalence of malaria in the human population. In estimating the accuracy of xenosurveillance, sensitivity considering only asymptomatic was 17.50% with VPP 46.67%, while the overall sensitivity was 14:58% with PPV of 46.67%. The tool proved operational, technically and economically feasible.
Keywords: Xenosurveillance. Malaria. Falciparum. Culicids. Standardization.
Surveillance.

x
RESUMO LEIGO
Esse estudo investigou se os mosquitos não transmissores de malária, capturados em uma área periurbana do município de Manaus, onde há transmissão da doença, estavam infectados com o parasito causador da malária em uma proporção próxima da frequência de malária nesta área. Os resultados deste estudo foram satisfatórios e indicam que esta nova ferramenta de vigilância poderá ajudar no monitoramento e controle da transmissão da malária, apresentando ainda algumas vantagens operacionais, por dispensar coletas de sangue em humanos e a captura de mosquitos com pouso em humanos.

xi
LISTA DE FIGURAS
Figura 1. Distribuição espacial da malária causada por P. falciparum no mundo
em 2010. Fonte: Gething et al, 2011(2). .................................................................... 1
Figura 2. Distribuição espacial da malária causada por P. vivax no mundo em
2010. Fonte: Gething et al, 2012(3). ........................................................................... 2
Figura 3. Ilustração do ciclo biológico do plasmódio. Adaptada de Mueller et al.,
2009 (11). ........................................................................................................................ 4
Figura 4. Imagem de satélite da área do estudo (Google Earth®). ..................... 32
Figura 5. Número de casos de malária na área do projeto(linha azul) e o
percentual de participação (linha vermelha) destes casos no total de casos
notificados em Manaus no período de 2004 a 2014. Fonte de dados:
SIVEP/Malária. ............................................................................................................. 33
Figura 6. Número de casos de malária por espécie na área do projeto no
período de 2004 a 2014. Fonte de dados: SIVEP/Malária. .................................. 33
Figura 7. (A) Captura com armadilha BG-Sentinel®, (B) Captura com aspirador
entomológico, (C) Triagem dos mosquitos capturados (Acervo da Gerência de
Entomologia/FMT-HVD). ............................................................................................ 34
Figura 8. Fases imaturas de Cx. quinqufasciatus na manutenção da colônia. (A)
Jangada de ovos, (B) Bacia com larvas de I estádio (quase imperceptíveis) e
grânulos de ração, (C) Larvas de II e III estádio, e grânulos de ração (Acervo da
Gerência de Entomologia/FMT-HVD). ..................................................................... 37
Figura 9. Manutenção da colônia de Cx. quinqufasciatus. (A) Gaiola com copo
contendo pupas, das quais emergirão os mosquitos, e erlenmeyer com solução
açucarada para alimentação dos mosquitos; (B) Gaiola com mosquitos
alimentando de solução açucarada; (C) Mosquitos obtendo repasto sanguíneo
em camundongos (Acervo da Gerência de Entomologia/FMT-HVD). ................ 38
Figura 10. Infecção experimental. (A) Gaiolas com mosquitos e alimentadores
artificiais interligados em circuito com o banho-maria; (B) Mosquitos se
alimentando come sangue infectado (Acervo da Gerência de
Entomologia/FMT-HVD). ............................................................................................ 39

xii
Figura 11. Etapas do processo de maceração de mosquitos. (A) Mosquito
sendo colocado em tubo cônico para maceração; (B) Maceração de pool de
mosquitos em resina Chelex® 100 utilizando pistilo azul e Pellet pestles®; (C)
Suspensão obtida a partir do pool de mosquitos e da resina Chelex® 100
(Acervo da Gerência de Entomologia/FMT-HVD). ................................................. 40
Figura 12. Espacialização dos domicílios na área estudada, apresentando a
densidade populacional por domicílio nas comunidades do Ipiranga,
Puraquequara e Brasileirinho, e localização dos laboratórios de notificação e
diagnostico da malária. ............................................................................................... 49
Figura 13. Percentual de detecção de DNA de Plasmodium sp. em Culex
quinquefasciatus na linha do tempo do “Dia 0” ao “Dia 10” pós-infecção
experimental. ................................................................................................................ 53
Figura 14. Comparação espacial da taxa de infecção por Plasmodium sp. em
humanos por domicilio com o percentual de pools de mosquitos positivos para
Plasmodium sp. por domicilio. ................................................................................... 60

xiii
LISTA DE TABELAS
Tabela 1. Aspectos da Vigilância da malária quanto aos diferentes cenários
epidemiológicos e operacionais (Adaptado de WHO, 2012). ................................. 9
Tabela 2. Frequência de Culicídeos descrita em estudos de fauna de mosquitos
na Amazônia. ................................................................................................................ 20
Tabela 3. Índices de antropofilia de Anopheles spp. e de Culex
quinquefasciatus. ......................................................................................................... 22
Tabela 4. Frequências das variáveis estudadas. ................................................... 50
Tabela 5. Frequências das espécies/gênero coletadas segundo o método de
captura e o percentual de infecção de pools de mosquitos infectados
naturalmente. ................................................................................................................ 52
Tabela 7. Resultado do teste diagnóstico da xenovigilância utilizando
mosquitos, tendo como padrão ouro a q-PCR em humanos e em humanos
assintomáticos. ............................................................................................................. 55
Tabela 6. Análise de Regressão logística univariada e multivariada
hierarquizada das variáveis estudadas. Avaliação da associação com a
presença de humanos com q-PCR positiva para Plasmodium sp. nos imóveis
estudados (Pseudo R2=0.2775). .............................................................................. 57

xiv
LISTA DE ABREVIATURAS, SÍMBOLOS E UNIDADES DE MEDIDA
AM Amazonas
CNPq Conselho Nacional de Desenvolvimento Científico e Tecnológico.
CONEP Comissão Nacional de Ética em Pesquisa
DNA Ácido desoxiribonucleico
EIR Taxa de inoculação entomológica
FIOCRUZ Fundação Oswaldo Cruz
FMT-HVD Fundação de Medicina Tropical Dr. Heitor Vieira Dourado
FVS Fundação de Vigilância em Saúde
HBR Taxa de picadas em humanos
HLC Captura de mosquito com pouso em humanos
ICEMRs International Centers for Excellence in Malaria Research
IPA Incidência parasitária anual
IPHH Índice de picada homem hora
ISGlobal Barcelona Institute for Global Health
Km2 Quilômetro quadrado
MDA Administração de Drogas em Massa
MILD Mosquiteiro impregnado com inseticida de longa duração
MS Ministério da Saúde
MSAT Triagem em Massa e Tratamento
NIAID National Institutes for Allergies and Infectious Diseases
ng/µl Nanograma por microlitro
Odds Ratio Razão de chances
OMS Organização Mundial de Saúde
Parafilm® Membrana artificial
PCR Reação em cadeia de polimerase.
PGEM Programa Global de Erradicação da Malária
POP Procedimento operacional padrão.
q-PCR Reação em cadeia de polimerase em tempo real.
RJ Rio de Janeiro.
RT-PCR Transcrição reversa da reação em cadeia de polimerase

xv
rRNA Ácido ribonucleico ribossomal
® Marca Registrada.
SVS Secretaria de Vigilância em Saúde
SIVEP Sistema de Vigilância Epidemiológica da Malária do MS
SIG Sistemas de Informação Geográfica
SR Taxa de esporozoítos
TCLE Termo de Consentimento Livre e Esclarecido.
UEA Universidade do Estado do Amazonas.
UR% Umidade relativa do ar
VPP Valor Preditivo Positivo
VPN Valor Preditivo Negativo
ºC Graus Celsius
µl Microlitro

xvi
GLOSSÁRIO
Anautogenia – Estratégia reprodutiva de muitas espécies de artrópodes, que
consiste na exigência de ingestão de sangue de vertebrados para a produção
de ovos viáveis.
Antrópico - Relativo à ação do homem sobre o meio ambiente.
Apicomplexa - Grupo taxonômico (Filo) de protozoários, caracterizados pela
presença exclusiva de uma organela denominada apicoplasto.
Apicolplasto - O apicoplasto é uma organela exclusiva de protozoários do filo
Apicomplexa que tem a função de síntese de DNA e proteína.
Áreas Periurbanas - Espaços situados na periferia de uma cidade. Área de
transição do espaço urbano para o rural.
Autogenia - Estratégia reprodutiva de algumas espécies de artrópodes, que
consiste na dispensa da ingestão de sangue de vertebrados para a produção
de ovos viáveis, esta pode ser facultativa ou obrigatória.
Carreadores submicroscópicos – Indivíduos assintomáticos infectados com
Plasmodium vivax que desenvolvem níveis de parasitemia não detectáveis
pelos métodos convencionais de diagnóstico da malária (gota espessa e testes
rápidos), detectável somente pelas técnicas moleculares de diagnóstico.
Ciclo eritrocítico - Ciclo assexuado do Plasmodium spp. que ocorre nas
hemácias e determina o ciclo febril na malária.
Ciclo gonotrófico – Ciclo reprodutivo dos artrópodes que vai desde a procura
pelo repasto sanguíneo, passando pela maturação dos ovos até a oviposição.
Competência vetorial - A competência vetorial de um artrópode refere-se à
permissividade deste à infecção, replicação e transmissão de patógenos
específicos.
Diploide – Diz-se da célula cujos cromossomos se organizam em par de
cromossomos homólogos, e assim, para cada característica existem pelo
menos dois genes.
Distribuição espacial – Em epidemiologia, forma ou padrão que uma
determinada doença se encontra ordenada no espaço geográfico. Esse padrão

xvii
pode está relacionado aos fatores ambientais, sociais, econômicos de uma
população em determinada área geográfica.
Ecótopos – Corresponde ao ambiente ecológico de determinado ser vivo
(determinado por seus fatores físicos e químicos) ou a um tipo específico de
hábitat dentro de determinada área geográfica.
Ecótonos - Área de contato e de transição funcional entre duas comunidades
ecológicas adjacentes, como resultado da competição mútua entre organismos
comuns às duas.
Endotélio - camada celular que reveste interiormente os vasos sanguíneos e
linfáticos.
Epitélio - tecido animal, constituído por células justapostas, com pouca
substância intercelular, que reveste superfícies expostas, cavidades e dutos,
além de executar funções secretoras, sensoriais e de absorção; tecido epitelial.
Esporogonia – Formação sexuada de esporozoítos de Plasmodium spp. por
enquistamento e divisão subsequente de um zigoto (Oocisto).
Esporozoíto - Elemento fusiforme, alongado, nucleado, que resulta da divisão
subsequente de um zigoto de Plasmodium spp., sendo a forma infectante do
hospedeiro invertebrado para o hospedeiro vertebrado.
Esquizontes - Célula multinucleada do Plasmodium spp, formada pelo processo
esquizogônico, onde o trofozoíta ao parasitar a célula do hospedeiro se
segmenta diretamente em merozoítos.
Exflagelação – Processo de formação de flagelos dos gametas masculino de
Plasmodium spp., dando mobilidade a este para viabilizar a fecundação do
gameta feminino.
Gametócitos – Formas sexuadas no ciclo evolutivo do Plasmodium spp.
Haploide – Diz-se da célula que possui um único conjunto completo de
cromossomos, como é característico dos gametas.
Hepatócito - Principal célula do fígado com importantes funções metabólicas.
Hemóstase - É o conjunto de mecanismos que o organismo emprega para
coibir hemorragia.

xviii
Hemocele - Cavidade do corpo dos artrópodes e de alguns invertebrados, por
onde flui a hemolinfa.
Hemolinfa - Líquido existente no sistema circulatório dos animais invertebrados,
com funções semelhantes às do sangue e da linfa nos animais vertebrados
Hipnozoítos - Formas dormentes intra-hepáticas do ciclo evolutivo do
Plasmodium vivax.
Lâmina basal - Lâmina ou película de macromoléculas situada no espaço
extracelular, sobre a qual se apoiam células epiteliais.
Lúmen - O espaço interno entre as paredes de um vaso sanguíneo ou do
interior de um órgão.
Matriz peritrófica – membrana quitino proteica, presente no lúmen do intestino
médio dos insetos, que envolve o alimento ingerido, e que separa o conteúdo
luminal em dois compartimentos, o espaço endoperitrófico (dentro da
membrana) e o espaço ectoperitrófico (fora da membrana).
Merosomas - Grandes vesículas formadas pelo Plasmodium spp. nos
hepatócitos do hospedeiro vertebrado, que albergam os merozoítos.
Merozoítos - Esporo que se forma a partir da esquizogonia, no ciclo evolutivo
do Plasmodium spp..
Oocisto - Estádio evolutivo enquistado do ciclo sexuado do Plasmodium spp.
que ocorre no abdome do mosquito vetor.
Oocinetos – Fase móvel do zigoto do Plasmodium spp. que atravessa a parede
do intestino médio do mosquito vetor para a formação do Oocisto.
Trofozoítos - Corresponde ao estágio ativado de alimentação intracelular do
ciclo de vida do Plasmodium spp..
Xenovigilância – Vigilância entomo-epidemiológica através de mosquitos não
vetores de patógenos específicos.
Zoonótica - Diz-se de doença que pode ser transmitida ao ser humano por
animais.

xix
SUMÁRIO
1. INTRODUÇÃO .................................................................................................................... 1
1.1 A malária .............................................................................................................................. 1
1.2 O ciclo da malária ............................................................................................................... 3
1.3 A vigilância da malária ....................................................................................................... 5
1.4 O controle da malária ....................................................................................................... 11
1.5 Os culicídeos e a hematofagia ....................................................................................... 14
1.6 Xenovigilância ................................................................................................................... 21
1.7 Aspectos metodológicos da xenovigilância .................................................................. 25
1.8 A eliminação da malária .................................................................................................. 27
2. OBJETIVOS ...................................................................................................................... 31
2.1 Geral ................................................................................................................................... 31
2.2 Específicos ......................................................................................................................... 31
3. MATERIAL E MÉTODOS ............................................................................................... 32
3.1 Modelo de estudo ............................................................................................................. 32
3.2 Local de estudo ................................................................................................................. 32
3.3 Amostragem ...................................................................................................................... 34
3.4 Coleta e identificação de culicídeos no campo ............................................................ 34
3.5 Padronização dos testes moleculares ........................................................................... 36
3.5.1 Estabelecimento de colônia de Culex quinquefasciatus .......................................... 36
3.5.2 Infecção experimental de P. vivax em Cx. quinquefasciatus .................................. 38
3.5.3 Extração e purificação de DNA dos pools de culicídeos ......................................... 39
3.5.4 Realização dos ensaios de q-PCR .............................................................................. 40
3.5.5 Teste de tempo máximo de detecção ......................................................................... 41
3.5.6 Teste de limiar de detecção ......................................................................................... 42
3.6 Extração e purificação de DNA das amostras de campo ........................................... 43
3.7 Realização dos ensaios de q-PCR das amostras de campo ..................................... 44
3.8 Definição de positividade plasmodial em culicídeos e em humanos........................ 44
3.9 Plano analítico ................................................................................................................... 44
3.9.1 Registro e tratamento dos dados ................................................................................. 45
3.9.2 Frequência relativa das espécies de culicídeos ........................................................ 45
3.9.3 Unidade espacial de análise ......................................................................................... 45

xx
3.9.4 Determinação da taxa de encontro de DNA de Plasmodium spp. em culicídeos
por domicílio ...................................................................................................................... 45
3.9.5 Determinação da taxa de infecção de Plasmodium spp. na população por
domicílio ............................................................................................................................. 46
3.9.6 Avaliação da acurácia da xenovigilância .................................................................... 46
3.9.7 Análise espacial descritiva ............................................................................................ 46
3.9.8 Análise de regressão logística múltipla ...................................................................... 47
3.10 Questões éticas ................................................................................................................ 47
4. RESULTADOS ................................................................................................................. 49
4.1 Descrição das variáveis estudadas ............................................................................... 49
4.2 Frequência e diversidade dos Culicídeos capturados e análise da eficiência dos
métodos de captura utilizados. ............................................................................................... 51
4.3 Padronização das técnicas de extração de DNA e de q-PCR para a detecção de
Plasmodium sp. em culicídeos não anofelinos. ................................................................... 52
4.4 Concordância, sensibilidade, especificidade e valores preditivos da xenovilância. ..
............................................................................................................................................. 54
4.5 Avaliação da associação entre a positividade domiciliar de infecções em humanos
e a presença de parasitas em culicídeos não anofelinos. ................................................. 56
5. DISCUSSÃO ..................................................................................................................... 61
5.1 Características ambientais da amostra ......................................................................... 61
5.2 Técnicas de coleta ............................................................................................................ 61
5.3 Fauna de Culicídeos ........................................................................................................ 63
5.4 Infectividade ....................................................................................................................... 64
5.5 Padronização das técnicas ............................................................................................. 65
5.6 Associação ......................................................................................................................... 66
5.7 Análise espacial ................................................................................................................ 67
5.8 Acurácia ............................................................................................................................. 67
5.9 Viabilidade ......................................................................................................................... 68
6. CONCLUSÃO ................................................................................................................... 70
7. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................. 71
8. APÊNDICES ..................................................................................................................... 85
8.1 Ficha de controle de captura e identificação de culicídeos ....................................... 85
8.2 POP_MAL_LB_019_v01D_PT ....................................................................................... 86
8.3 POP_MAL_LB_020_v01D_PT ....................................................................................... 93

xxi
9. ANEXOS ........................................................................................................................... 97
9.1 POP_MAL_LB_013_v02D_PT ....................................................................................... 97
9.2 PARECER_CONSUBSTANCIADO_CONEP_349211 ............................................. 101
9.3 POP_MAL_LB_001_v02D_PT ..................................................................................... 107
9.4 POP_MAL_LB_021_v01D_PT ..................................................................................... 110

1
1. INTRODUÇÃO
1.1 A malária
A malária é reconhecida como um grave problema de saúde pública no mundo
com 3,2 bilhões de pessoas expostas ao risco de contrair a doença. A Organização
Mundial de Saúde–OMS estimou que em 2015 ocorreram cerca de 214 milhões de
casos, com incidência de 91 casos/1000 habitantes em risco, e 438 mil mortes, com
taxa de 19 mortes/100.000 habitantes em risco. Nesta estimativa, a Região Africana
da OMS concentra 90% das mortes, com 292 mil destas ocorrendo em crianças
abaixo de cinco anos de idade (1).
Figura 1. Distribuição espacial da malária causada por P. falciparum no mundo em 2010. Fonte: Gething et al, 2011(2).
As principais espécies de Plasmodium que causam malária em humanos
apresentam distribuição espacial distinta no mundo, destacando-se o Plasmodium

2
falciparum no Continente Africano (Figura 1) e o Plasmodium vivax nas Américas e
no Sudeste da Ásia (Figura 2) (2,3).
Figura 2. Distribuição espacial da malária causada por P. vivax no mundo em 2010. Fonte: Gething et al, 2012(3).
Estima-se que 121 milhões de pessoas em 21 países da Região das Américas
vivam em áreas com risco para malária, com as infecções por Plasmodium vivax
respondendo por 70% dos casos. Em 2014 esta região apresentou uma redução de
67,5% no número de casos confirmados em relação ao ano de 2000, reduzindo de
1,2 milhão para 390 mil casos. O Brasil foi responsável por 37% dos casos da região
em 2013 (1).
A transmissão da malária no Brasil concentra-se predominantemente na Região
Amazônica, que no período de 2000 a 2011 respondeu por 99,7% dos casos
registrados no país, com predominância das infecções causadas por Plasmodium
vivax. A zona rural é a mais afetada, ainda que áreas periurbanas de grandes

3
centros da região norte, como Manaus e Porto Velho, tenham participação
significativa no número de casos da doença. As áreas indígenas da Região
Amazônica tem apresentado crescente participação, principalmente nos Estados do
Pará, Roraima e Amazonas, neste último 49% dos registros de casos em 2.011
foram nas comunidades indígenas (4).
Uma visão geral da malária no Brasil ressalta seu aspecto histórico-
epidemiológico que se relaciona intimamente com os intensos processos sociais e
econômicos vivenciados na Amazônia, contribuindo com a manutenção e a
concentração da transmissão da doença nesta região. As complexidades ambientais
e socioeconômicas, e a escassez de centros de estudos e pesquisa na região,
representam um desafio atual para a eliminação da malária (5,6). A manutenção
desta endemia tem ainda implicações negativas no desempenho de escolares,
contribuindo para o subdesenvolvimento da região (7).
No município de Manaus, capital do Estado do Amazonas, em um estudo
epidemiológico do período 1986 a 2005, os autores observaram um forte
crescimento populacional, atingindo 105,2% no período estudado, e uma distribuição
dos casos de malária em área urbana, apresentando maior risco nas regiões leste e
oeste, ambas com Incidência Parasitária Anual (IPA) consideradas de alto risco (8).
1.2 O ciclo da malária
O parasito causador da malária é um protozoário Apicomplexa do gênero
Plasmodium e tem como vetor algumas espécies de mosquitos do gênero
Anopheles. O desenvolvimento do parasito no vetor começa quando este ao picar
um hospedeiro vertebrado infectado com Plasmodium sp., ingere formas sexuadas
do parasito, chamadas de gametócitos, que ao chegarem no lúmen do intestino
médio do mosquito, diferenciam-se em gametas feminino e masculino, este último
sofre exflagelação, replicação do DNA, formando oito microgametas haploides, os
quais procurarão ativamente os gametas femininos para a fertilização, originando
um zigoto diploide que subsequentemente passa por um ciclo de replicação de DNA
para se tornar tetraploide, diferenciando-se em oocinetos que apresentam

4
capacidade de mobilidade para invadir e atravessar, tanto a matriz peritrófica, quanto
o epitélio do intestino médio do mosquito. Chegando a lâmina basal do intestino,
ocorrerá sua transformação em oocisto e posteriormente sua maturação. Uma vez
que esta se complete, geralmente dentro de 10 a 24 dias, dependendo da espécie
de Plasmodium, ocorre a esporogonia liberando os esporozoítos na hemocele do
mosquito, onde circulam até invadirem a glândula salivar, se alojando em seu lúmen
(Figura 03) (9,10).
Figura 3. Ilustração do ciclo biológico do plasmódio. Adaptada de Mueller et al., 2009 (11).
Quando o mosquito infectado pica um novo hospedeiro vertebrado, os
esporozoítos deixam a glândula salivar, e a partir do tecido avascular da pele
chegam a um vaso sanguíneo, penetrando o endotélio desses vasos seguem até
chegarem ao fígado, onde invadem os hepatócitos e por divisão binária multiplicam-
se formando merosomas até o rompimento do hepatócito. No caso de P. vivax e P.
Ovale, estes apresentam um estágio de dormência que pode permanecer no fígado
por semanas ou anos antes do desenvolvimento dos merosomas. Os merozoítos,

5
liberados após este rompimento, migram para a corrente sanguínea, dando início ao
ciclo eritrocítico do parasito. Os merozoítos invadem as hemácias e por divisão
binária multiplicam-se formando esquizontes que ao romperem as hemácias liberam
os merozoítos na corrente sanguínea. Estes invadirão novas hemácias fazendo com
que esse ciclo eritrocítico se repita indeterminadamente, provocando o aparecimento
dos sintomas da doença que tem como característica a febre intermitente, sendo a
cada 48 horas para P. vivax e P. ovale, de 36 a 48 horas para P. falciparum, e de 72
horas para P. malarie. Depois de algum tempo de repetição deste ciclo eritrocítico
alguns trofozoítos diferenciam-se em gametócitos. No caso de P. vivax ocorre logo
nos primeiros ciclos, já em P. falciparum é mais tardio. Inicia-se assim a fase
sexuada do parasito que possibilitará a infecção do mosquito vetor ao picar este
hospedeiro infectado (Figura 3) (12,13).
1.3 A vigilância da malária
As diferenças biológicas no desenvolvimento de gametócitos nas duas principais
espécies de Plasmodium, pois em P. vivax a formação de gametas é mais precoce
que em P. falciparum, bem como o a particularidade do desenvolvimento de
hipnozoítos nas infecções por P. vivax, podem indicar a diferença no padrão de
transmissão entre estas espécies. A investigação comparativa da dinâmica de
transmissão co-endêmica de isolados destas espécies na Indonésia, com o uso de
abordagens moleculares, demonstrou contraste na diversidade e estrutura
populacional, indicativos de diferentes padrões de transmissão dentro e entre as
espécies (14).
P. vivax mostra-se mais amplamente distribuído do que P. falciparum, com 2,85
bilhões de pessoas vivendo em risco de infecção, a maioria no Sudeste Asiático. No
continente Africano a probabilidade de infecção é bastante reduzida devido à
frequência da característica Duffy negativo, mas a transmissão ocorre no continente.
(15).
A heterogeneidade na transmissão da malária no cenário mundial tem
demandado esforços para ampliar a compreensão dos aspectos regionais

6
relacionados ao vetor e consequentemente estabelecer vigilância adequada. Nesse
contexto o National Institutes for Allergies and Infectious Diseases (NIAID)
estabeleceu uma rede de centros de pesquisa da malária denominado International
Centers for Excellence in Malaria Research (ICEMRs) na busca por resultados nos
estudos de biologia do vetor e a padronização do monitoramento local. Foi
observado em quatro desses ICEMRs, evidências de novas espécies ou potenciais
vetores. Estudos epidemiológicos, nestes locais, são integrados com estudos
entomológicos, incluindo a medição da taxa de inoculação entomológica, índice de
picada humana, e resistência a inseticidas, assim como estudos de diversidade
genética do parasito. Para gerar dados relacionados a interação entre mosquito
vetor e hospedeiro humano, as avaliações entomológicas utilizam variadas
metodologias. Alguns métodos utilizados para a coleta de mosquitos são a captura
de atração/pouso em humanos, armadilhas luminosa do tipo CDC, aspiração na
casa, barreiras de tela e armadilha Shannon, esses métodos são baseados nos
hábitos de repouso e de picar das diferentes espécies de Anopheles. De forma
semelhante, vários métodos são utilizados para a identificação da espécie, das
fontes de refeições de sangue, e a determinação de infecção natural no mosquito,
nas várias fases do parasito. As diferenças ambientais locais e as características de
vetores predominantes, também contribuem para a variação dos métodos utilizados
(16,17).
Os levantamentos entomológicos podem fornecer uma rica fonte de dados e são
as únicas medidas diretas de transmissão, porém apresentam como desvantagens,
a falta de padronização nos métodos usados para coletar e interpretar dados, as
limitações dos métodos de captura de mosquitos em busca de repasto sanguíneo ou
ao ar livre durante o dia. A pouca precisão das pesquisas entomológicas podem
limitar as estimativas de indicadores, dela derivados, por não apresentarem a
capacidade de discriminar pequena escala de variabilidade espacial ou temporal,
principalmente em configurações de baixa transmissão, onde nem sempre é possível
a captura de um número suficiente de mosquitos. Esforços estão sendo envidados
para padronizar protocolos e métodos analíticos, com o intuito de preencher lacunas
de conhecimento e melhorar a qualidade dos dados entomológicos, visando uma
compreensão da dinâmica de transmissão da malária (17).

7
A definição conceitual categórica das abordagens e metas dos programas de
saúde relacionados aos níveis de transmissão da malária, torna-se necessária para
a compreensão e elaboração das estratégias adequadas. Controle trata da redução
deliberada de incidência da doença em determinado local para um nível aceitável e
controlável; Eliminação é a redução deliberada de incidência de infecção para zero
em uma área geográfica delimitada, sendo necessárias intervenções para
interromper possível reestabelecimento; já a Erradicação é a redução global e
permanente de incidência da infecção a zero, através de esforços deliberados, sem
necessidade de intervenções futuras. A vigilância no controle se limita a doença,
enquanto que na eliminação e a erradicação o foco deve estar na infecção, que se
torna mais difícil devido à ausência de sintomas e a difícil detecção pelos meios de
diagnósticos usuais (18).
Assim as métricas usadas para análise da transmissão da malária estão focadas
principalmente nas manifestações clínicas da doença na população humana, pois
aquelas baseadas em vetores raramente estão disponíveis, havendo escassez de
índices vetoriais graças à complexidade técnica de obtenção de métricas
entomológicas, às preocupações éticas relacionadas à exposição de seres humanos
à infecção da malária, e à possibilidade de erro de medição considerando o viés do
capturador. A medição da incidência da malária necessita que todos os casos
suspeitos sejam diagnosticados eficientemente de forma abrangente, tanto pela
detecção passiva de casos na rotina de todo e qualquer serviço de saúde, quanto
pela detecção ativa de casos através de visitas domiciliares em intervalos regulares.
O resultado é expresso como Incidência Parasitária Anual - IPA por 1.000 habitantes
de uma determinada área administrativa. O Programa Global de Erradicação da
Malária – PGEM só considera válida a IPA, se a taxa de proporção de examinados
na população-alvo ultrapassar os 10% (19).
Sistemas de vigilância baseados nos conhecimentos epidemiológicos da malária
buscam gerar informações sobre a ocorrência de casos e mortes para nortear o
planejamento, o monitoramento e a avaliação dos programas de controle da doença,
possibilitando a identificação de áreas mais afetadas, tendências nos casos e
mortes, assim como a avaliação do impacto das medidas de controle. A concepção

8
desses sistemas depende do nível de transmissão da malária e dos recursos
disponíveis para realizar a vigilância. A OMS tem adotado a classificação de alta
transmissão para indicar a malária hiper e holoendêmica, transmissão moderada
para indicar a malária mesoendêmica e baixa transmissão para indicar a malária
hipoendêmica, como um guia geral para os possíveis tipos de vigilância da malária
em diferentes níveis de endemicidade da doença (Tabela 1) (20).
A diversificação de instrumentos e tecnologias, bem como a integração na
vigilância da malária tornam-se necessárias para otimização e modernização do
processo de vigilância. Assim, a vigilância da malária integrada a programas de
agentes comunitários de saúde, lançando mão de novas tecnologias acessíveis, tais
como serviço de mensagens por telefonia móvel para envio de relatórios semanais
de casos, podem contribuir para a otimização do monitoramento de tendências da
transmissão da malária em determinada região (21). Também o uso de Sistemas de
Informação Geográfica (SIG) como ferramenta de vigilância integrada para os
programas de controle de doenças de transmissão vetorial, tais como malária e
filariose linfática (22), e a aplicação de algoritmos que integram sistemas de
vigilância, imagens de satélite, dados climáticos e ambientais, para produção rápida
de mapas preditivos de risco, são ferramentas cada vez mais utilizadas (23,24).
A prevalência parasitária da malária tem sido um importante indicador na
vigilância e monitoramento do controle da malária em populações endêmicas, sendo
tradicionalmente avaliada por microscopia. Porém, à medida que se caminha para
um cenário de eliminação da transmissão da malária, as técnicas convencionais de
vigilância tornam-se menos sensíveis. Nesta nova realidade, vem se utilizando de
forma mais frequente os métodos moleculares com maior capacidade de detecção
dos indivíduos infectados. Na análise dos fatores epidemiológicos associados à
baixa densidade, demonstrou-se que o carreamento submicroscópico de parasitos,
em ambientes de baixa endemicidade e nas infecções crônicas, é comum em
adultos. Em níveis muito baixos de transmissão, os carreadores submicroscópicos
podem responder por cerca de 20 a 50% de toda a transmissão homem-mosquito,
reforçando a importância do papel da triagem utilizando técnicas mais sensíveis no
cenário de eliminação da malária (25).
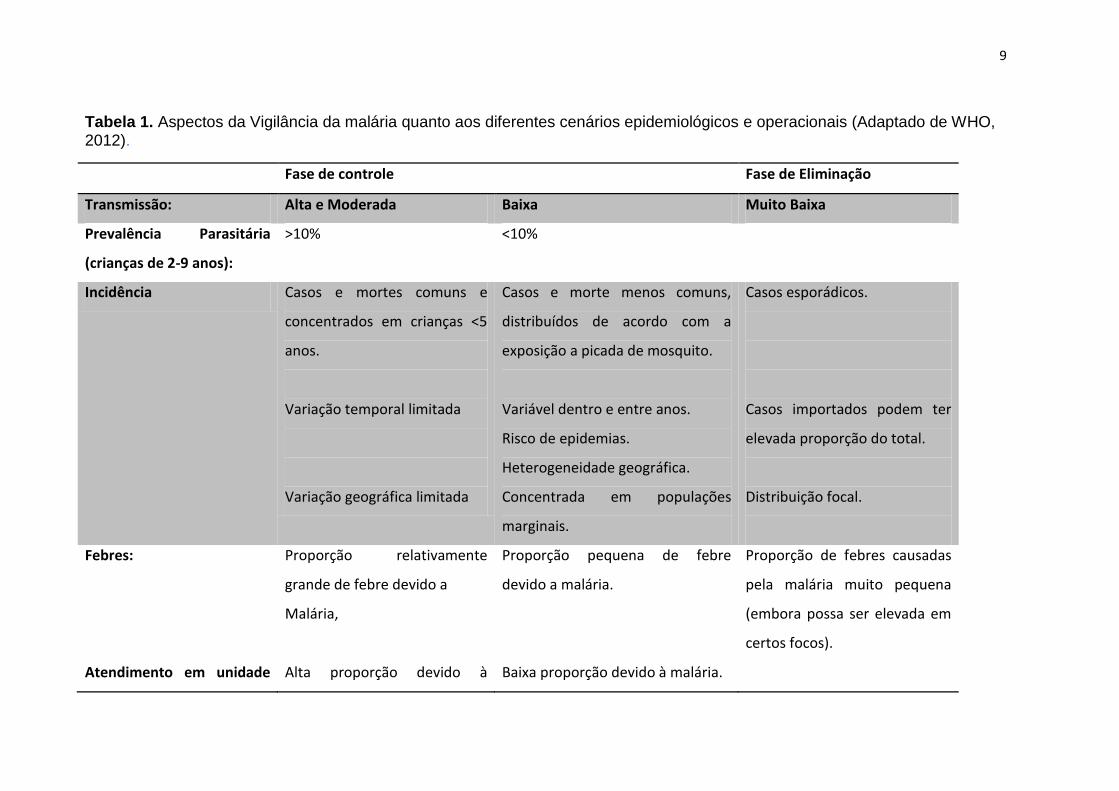
9
Tabela 1. Aspectos da Vigilância da malária quanto aos diferentes cenários epidemiológicos e operacionais (Adaptado de WHO, 2012).
Fase de controle Fase de Eliminação
Transmissão: Alta e Moderada Baixa Muito Baixa
Prevalência Parasitária
(crianças de 2-9 anos):
˃10% ˂10%
Incidência Casos e mortes comuns e
concentrados em crianças ˂5
anos.
Variação temporal limitada
Variação geográfica limitada
Casos e morte menos comuns,
distribuídos de acordo com a
exposição a picada de mosquito.
Variável dentro e entre anos.
Risco de epidemias.
Heterogeneidade geográfica.
Concentrada em populações
marginais.
Casos esporádicos.
Casos importados podem ter
elevada proporção do total.
Distribuição focal.
Febres:
Atendimento em unidade
Proporção relativamente
grande de febre devido a
Malária,
Alta proporção devido à
Proporção pequena de febre
devido a malária.
Baixa proporção devido à malária.
Proporção de febres causadas
pela malária muito pequena
(embora possa ser elevada em
certos focos).
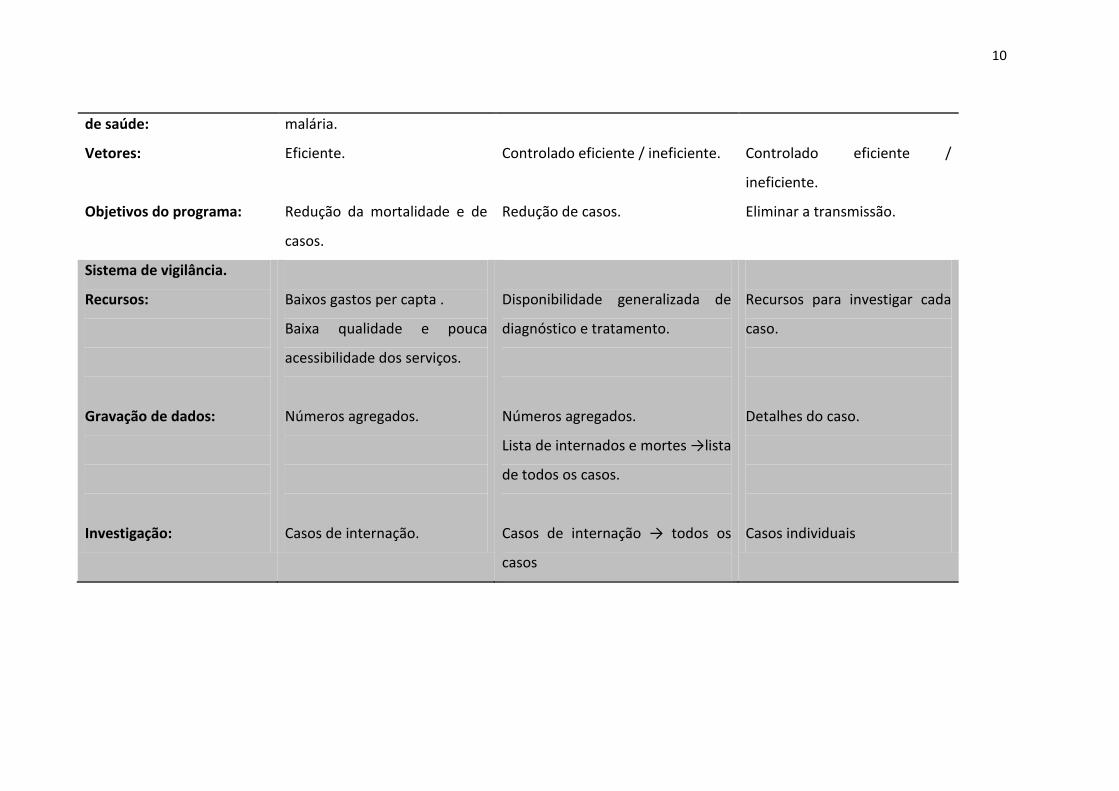
10
de saúde:
Vetores:
Objetivos do programa:
malária.
Eficiente.
Redução da mortalidade e de
casos.
Controlado eficiente / ineficiente.
Redução de casos.
Controlado eficiente /
ineficiente.
Eliminar a transmissão.
Sistema de vigilância.
Recursos:
Gravação de dados:
Investigação:
Baixos gastos per capta .
Baixa qualidade e pouca
acessibilidade dos serviços.
Números agregados.
Casos de internação.
Disponibilidade generalizada de
diagnóstico e tratamento.
Números agregados.
Lista de internados e mortes →lista
de todos os casos.
Casos de internação → todos os
casos
Recursos para investigar cada
caso.
Detalhes do caso.
Casos individuais

11
Em uma revisão sistemática sobre infecções submicroscópicas por P. vivax,
verificou-se que a prevalência de P. vivax e P. falciparum pela técnica de Reação em
Cadeia de Polimerase (PCR) foi estatisticamente mais elevada que a prevalência de
P. vivax pela técnica de microscopia ótica. A proporção relativa de infecções
submicroscópicas por P. vivax teve uma média de 69,5%, sendo significativamente
maior que a média observada em P. falciparum. Foi observada ainda uma relação
positiva entre a microscopia ótica e a PCR, sendo a prevalência de P. vivax pela
primeira, um fator significativo para a previsão da prevalência de P. vivax pela PCR,
ratificando a importância desses grupos de assintomáticos, principalmente em áreas
de baixa transmissão, como potenciais fontes de infecção para os vetores,
contribuindo para a manutenção do ciclo de transmissão (26).
Uma alternativa de vigilância no cenário de eliminação da malária seria a
aplicação de ferramentas e estratégias de sorovigilância, que apresenta um
potencial significativo para a aceleração deste processo e alguns modelos vem
sendo propostos (27). Porém, algumas lacunas de conhecimento precisam ser
preenchidas para o desenvolvimento e otimização destas ferramentas e sua
aplicação no controle da malária. O desenvolvimento de testes precisos e confiáveis,
que forneçam informações sobre exposições recentes e de longo prazo e que
tenham viabilidade em contextos de recursos limitados seria de grande valia (28).
1.4 O controle da malária
A redução da transmissão da malária a partir de níveis endêmicos, procurando
sua redução para níveis muito baixos, mas sem alcançar a interrupção da
transmissão endêmica ainda é uma realidade em grande parte do mundo. Este
controle implica em intervenções diretas, envolvendo diversos fatores operacionais
que precisam levar em conta as características biológicas e epidemiológicas de cada
espécie do parasito (29).
Considerando a instabilidade moderada das atuais dinâmicas de transmissão na
co-endemicidade de P. falciparum e P. vivax, onde surtos clonais do primeiro são

12
frequentes, abordagens direcionadas seriam viáveis para sua contenção utilizando
detecção reativa de casos e tratamento, borrifação intradomiciliar, no local de
ocorrência dos casos e na vizinhança, avaliando-se a compatibilidade da ação com
o comportamento das espécies predominantes do vetor. Mas para P. vivax que exibe
uma transmissão mais estável, sem evidência de surtos clonais, há indicativos de
que essas abordagens seriam menos eficazes. Os padrões observados em estudo
realizado na Indonésia sugerem que a necessidade de grandes intervenções com
cobertura em larga escala de mosquiteiros impregnados com inseticida de longa
duração – MILD, bem como o tratamento eficaz dos hipnozoítos, poderá ser crítica
para a eficiência das intervenções contra o P. vivax (14).
Controle e consolidação, eliminação, e prevenção da reintrodução, são as
classificações das fases do programa de malária que a OMS adotou. Para cada
fase, os focos são distintos. No controle as intervenções são diretas, diversificadas e
em larga escala visando redução acentuada de casos da doença, utilizando
diagnóstico rápido para malária e tratamento, assim como ações de controle vetorial.
Na eliminação as intervenções estão voltadas para a redução das infecções, com
possível uso de tratamento em massa, visando a eliminação da transmissão.
Enquanto que na fase de prevenção da reintrodução, as ações estão completamente
voltadas para a ampla vigilância de casos importados e possíveis focos de
reintrodução, com respostas rápidas e adequadas (30).
As medidas de intervenções adotadas para a redução de casos de malária
devem ser sustentadas por certo tempo, visando não somente a redução inicial, mas
também sua manutenção, para a mudança de linha de base epidemiológica,
possibilitando a transição de fase de controle e a implementação concomitante de
novas intervenções possíveis neste novo cenário. É certo que as linhas de base de
transmissão da malária também podem ser influenciadas por fatores sociais,
econômicos e ambientais, podendo contribuir, tanto para a redução quanto para a
elevação dessa linha de base, dependendo do tipo de alteração e consequente
impacto provocado na relação vetor hospedeiro, seja de bloqueio ou de
favorecimento da transmissão (31).

13
Nas modelagens matemáticas, o planejamento estratégico geral para atacar
agentes patógenos de transmissão vetorial deverá considerar a plena caracterização
epidemiológica e entomológica da linha de base de uma determinada configuração.
Identificando possíveis lacunas de cobertura eficazes, e também considerando a
interação entre os requisitos técnicos e capacidade operacional. Desta forma, a
política de controle adotada deverá levar em conta a linha de base como um fator
preponderante na escolha ideal das intervenções e seus possíveis resultados (32).
Neste sentido, a associação da transmissão da malária na Amazônia com os
fatores econômicos e sociais é consensual, sendo possível sua estratificação em
área rural, urbana, indígena, de assentamentos, de acampamentos, tais como
garimpos e expansão urbana desordenada, assim como grandes obras que
promovem alterações ambientais, como hidrelétricas, estradas, pontes e outras (33).
Essa associação de fatores ambientais e antrópicos, como condicionantes da
manutenção da transmissão da malária, é compatível com o resultado de um estudo
na cidade de Manaus sobre a incidência de malária e à influência desses fatores que
modificando o ecossistema, tornam possível a proliferação de mosquitos Anopheles
spp. em ambiente urbano, fomentando a alta incidência de malária na região (34).
Assim como os fatores determinantes da transmissão da malária são multi-
setoriais, as ações de controle também devem envolver as políticas de ocupação de
terras, reforma agrária, meio ambiente, atenção primária ao paciente com o
diagnóstico e o tratamento precoce, bem como o controle de vetores que envolvem
ações de manejo de criadouros, aplicação espacial e residual de inseticidas e
métodos de proteção como o uso de mosquiteiros e telas. Esses métodos de
controle atualmente utilizados apresentam benefícios e obstáculos, e ainda que,
quando usados de forma integrada e seletiva, demonstrem certa eficiência, é
consensual que são necessárias novas técnicas de controle e monitoramento
vetorial da malária (33,35).
Considerando que para algumas áreas a sobreposição e a concomitância de
doenças parasitárias de transmissão vetorial é uma realidade relativamente comum,

14
ocorrendo em alguns casos através de um mesmo vetor, alguns autores suscitam a
possibilidade/necessidade de controle e monitoramento integrado de vetores pelos
programas envolvidos, como forma de aperfeiçoar resultados e recursos (36–38).
1.5 Os culicídeos e a hematofagia
Os culicídeos são insetos dípteros, pertencentes à Família Culicidae. Seu ciclo
biológico compreende as seguintes fases: ovo, larva, pupa e adulto. As fases
imaturas são aquáticas, enquanto os adultos são alados com pernas e antenas
longas. Também conhecidos como mosquitos, usam açúcares vegetais e sangue de
vertebrados como recursos nutricionais, mas o sangue também tem função
essencial na reprodução destes, proporcionando proteínas necessárias para a
maturação de seus ovos que são postos em coleções de água, onde suas larvas se
desenvolvem em quatro estádios até passarem para o estágio de pupa, quando
ocorre a metamorfose, para então se transformarem em adultos. Essa característica
alimentar dos mosquitos na fase adulta os coloca como protagonistas na cadeia de
transmissão de várias doenças de importância para a saúde pública mundial (39,40).
As primeiras evidências científicas envolvendo mosquitos como transmissores
de agentes patógenos datam de 1877, com os trabalhos Patrick Manson mostrando
que o mosquito Culex fatigans era o hospedeiro intermediário da filariose. Em 1898,
Ronald Ross demonstrou que a fase infecciosa do parasita da malária era injetada
no hospedeiro, no momento da picada do mosquito, junto com a saliva. Com o
passar dos anos e os melhoramentos dos equipamentos microscópios e lentes de
imersão, possibilitaram novas evidências científicas para a relação entre insetos e
parasitos (41).
Atualmente com o desenvolvimento de novas tecnologias o conhecimento foi
bastante ampliado em várias áreas do conhecimento, permitindo inferências sobre a
história natural destes artrópodes, como recentes achados de fóssil de mosquito
com o abdome cheio de sangue, que data do Eoceno Médio. Fósseis de mosquitos
também são encontrados a partir de âmbar, mas em sua maioria foram preservados

15
em sedimentos de lagos, refletindo a sua relação com a água em seu ciclo biológico
(42,43).
No processo evolutivo dos mosquitos, o desenvolvimento adaptativo dos hábitos
alimentares guarda relação com a reprodução de sua prole. Neste processo algumas
espécies desenvolveram anautogenia e outras a autogenia, que é a capacidade das
fêmeas de desenvolverem a sua primeira oviposição sem a dependência de se
alimentarem de sangue de vertebrados, essa característica pode ser facultativa ou
obrigatória. Como exemplo temos o Culex molestus Forskal (Diptera: Culicidae) que
exibe autogenia obrigatória e apresenta um período relativamente curto entre o
surgimento das fêmeas e a oviposição de jangadas de ovos autógenos. Embora esta
espécie esteja perfeitamente adaptada para convivência com a habitação humana,
sua preferência alimentar por sangue humano é retardada em até 8 dias após sua
emergência, podendo reduzir sua importância relativa como um vetor de arbovírus.
(44,45).
A anautogenia é uma estratégia reprodutiva de sucesso utilizada por muitas
espécies de mosquitos e outros artrópodes, que consiste na exigência de ingestão
de substâncias proteicas para a produção de ovos viáveis, estabelecendo-se a
hematofagia, que é o hábito de alimentar-se de sangue, como fator determinante
para o ciclo gonotrófico das fêmeas de culicídeos hematófagos. Essa característica é
importante, pois a hematofagia, além de ser essencial para o ciclo gonotrófico das
fêmeas dos mosquitos, é fundamental para a transmissão de doenças vetoriais
(46,47).
O hábito hematofágico dos culicídeos, surgiu com a necessidade específica de
alimentar-se do sangue de vertebrados, o que levou a adaptações mecânicas
sofisticadas dos aparelhos bucais, bem como, a evolução de compostos anti-
hemostáticos salivares que neutralizassem a hemóstase no hospedeiro vertebrado
no momento do repasto sanguíneo do mosquito. Esse cenário de evolução
convergiu para hematofagia entre as mais de 14.000 espécies e 400 gêneros de
artrópodes com uma grande diversidade de anticoagulante salivar e substâncias
vasodilatadoras (48–52). O volume de sangue ingerido pelos culicídeos em cada

16
repasto sanguíneo é variável entre as espécies. Em experimentos com Aedes
aegypti alimentandos com sangue de camundongos foi estimada a ingestão de 1 a
8µl, com média de 5µl, de sangue em único repasto sanguíneo do mosquito (53). Em
Aedes albopictus (Skuse) e Culex tritaeniorhynchus Giles esse volume variou de 1 a
6µl de sangue, tendo como fontes alimentares humanos e cães (54). Os pesos
médios observados para as refeições de sangue por espécime, em Culex
quinquefasciatus e Aedes aegypti foram de 3,5 mg e 1,6 mg, respectivamente (55).
Além disso, o comportamento alimentar em múltiplas fontes num mesmo ciclo
gonotrófico, como demonstrado em Culex quinquefasciatus e Aedes aegypti, pode
favorecer a eficiência e rapidez na transmissão de patógenos (56,57).
Desta forma, a transmissão entre hospedeiros vertebrados é possível pelo hábito
hematofágico dos mosquitos, permitindo que agentes patogênicos tenham sucesso
no estabelecimento de sua transmissão pelos artrópodes, seus hospedeiros
intermediários. Assim, a seleção da fonte de alimentação de sangue é essencial
para o parasito completar o seu ciclo de vida. Esse hábito é parte do caráter
intrínseco do mosquito, pois as proteínas do sangue são nutrientes essenciais para
a produção de ovos e sua aptidão reprodutiva. Há uma plasticidade na preferência
alimentar destes artrópodes, sendo os mamíferos e as aves, os mais requeridos,
mas fatores extrínsecos como oferta, odor, gênero, massa corporal, parasito, clima,
humidade e qualidade do sangue do hospedeiro, podem influenciar essa seleção.
Esta plasticidade se torna uma característica chave para explicar o surgimento de
muitas doenças zoonótica transmissíveis por mosquito. (58,59).
Algumas espécies de culicídeos desenvolveram preferencia pelo sangue
humano, elevando a importância destes como vetores na transmissão de doenças.
Essa evolução comportamental antropofágica influenciou no processo de
domiciliação de determinadas espécies, culminando com o desenvolvimento do
hábito endofágico, de alimentar-se no interior dos domicílios. A evolução de
preferência pelo odor humano nesses mosquitos está firmemente ligada a um
aumento na expressão da sensibilidade do receptor que reconhece um composto
presente em níveis elevados no odor humano. Essas espécies são aquelas que

17
servem de vetores para importantes doenças infecciosas, sugerindo que este traço
comportamental pode ter evoluído em paralelo com a evolução parasito-hospedeiro.
(60,61).
Desta forma, a importância dos culicídeos para a saúde pública mundial, no que
tange as doenças transmissíveis, é reconhecidamente um tema relevante que tem
demandado estudos sobre as interações parasito-vetor que ocorrem na transmissão
de doenças infecto-parasitárias causadas por diferentes tipos de patógenos. Essas
relações são bastante complexas e envolvem interações coevolutivas de parasitos e
vetores que determinaram a especificidade vetor-patógeno na transmissão de
doenças, de tal forma que mesmo havendo sobreposição de espécies de mosquitos
hematófagos com graus semelhantes de antropofília, ou seja, com a mesma
possibilidade de se infectarem, somente os vetores específicos do patógeno
apresentam a capacidade de transmissão deste para um novo hospedeiro, a
exemplo do que acontece em doenças como malária e dengue, entre outras (62–
64).
Neste cenário mundial, destaca-se a ampla distribuição geográfica da subfamília
Anophelinae que tem relação direta com a transmissão da malária em humanos,
tanto no Continente Africano quanto nas Américas, sendo os principais vetores desta
doença membros dessa subfamília (65). Junto com Aedes aegypti transmissor dos
vírus da dengue e da febre amarela, e Cx. quinquefasciatus vetor da filariose
linfática, representam os principais culicídeos transmissores de doenças de
importância para a saúde pública mundial (66).
Em muitas regiões do mundo mosquitos do gênero Anopheles apresentam
índices elevados de antropofilia que lhe confere papel preponderante na transmissão
da malária. Resultados de estudo longitudinais no Iêmen confirmam um
comportamento antropofílico e endofágico do vetor naquela região(67).Resultados
semelhantes foram demonstrados na Zâmbia, onde a mesma espécie figura como
principal vetor (68), em Bangladesh (69), na Guiné Equatorial (70), no norte de
Brasil (71) no sul do Malawi (72), e em Burkina Faso (73),

18
Também grandes complexos de espécies, como o de Culex pipiens tem
relevante papel na transmissão de agentes patogênicos de importância clínica.
Farajollahi et al (2011), em seu trabalho de revisão sobre o papel da ecologia do
vetor na transmissão e sua influência sobre a evolução de patógenos, argumenta
que a adaptação destes mosquitos a ambientes antrópicos culminou com sua ampla
distribuição geográfica nas regiões tropicais e temperadas, especialmente Culex.
pipiens pipiens e Culex. quinquefasciatus, associada à dispersão humana nos
continentes, e somada ao padrão alimentar destes, que englobam aves e
mamíferos, contribuiu para a transmissão de patógenos de aves para a população
humana (74). Alguns trabalhos indicam um hábito preferencial, mas não exclusivo,
de mosquitos da espécie Cx. quinquefasciatus por sangue humano, com índices
elevedaos de antropofilia e um comportamento endofágico, apontando para sua
importância na transmissão de doenças (75,76).
Em um estudo realizado em aldeias no Paquistão Cx. tritaeniorhynchus foi a
espécie mais abundante compreendendo 51,8% das amostras totais, seguido por
Cx. quinquefasciatus (16,4%), Cx. pseudovishnui (6,8%), Anopheles subpictus
(4,8%) e An. culicifacies (4,7%) (77). Um estudo semelhante realizado na República
dos Camarões obteve 1.030 mosquitos, compreendendo 700 Cx. quinquefasciatus
(68%), 262 An. gambiae (25%) e outras espécies (7%) pertencentes ao gênero
Anopheles, Mansonia, Culex e Aedes (78).
No Brasil, a área de abrangência da Amazônia apresenta enorme potencial de
biodiversidade, incluindo diversas espécies de vetores artrópodes em seus
diferentes ecótopos. Este fato propicia o surgimento de novas doenças, como as
arboviroses, novos focos de endemias em locais onde não ocorriam e o
ressurgimento de focos de doenças como a malária a partir das alterações
ambientais provocadas pelo homem (79–81).
Além dos anofelinos, outras espécies ocupam nichos diversos, apresentando
variados graus de antropofilia e endofagia. Entre os estudos sobre a diversidade de
culicídeos na Região Amazônica estão os trabalhos de Natal et al. que evidenciaram
predomínio do gênero Anopheles, com espécimes infectados por P. vivax e P.

19
falciparum (82), e os de Fé et al que registraram a presença de Haemagogus
janthinomys, Hg. leucocelaenus, Sabethes belisarioi e Aedes fulvus, espécies
vetores do vírus da febre amarela silvestre (83).
Em um estudo sobre a frequência de culicídeos em ecótopos silvestres em uma
área periurbana do município de Manaus, a maioria dos exemplares coletados
pertencia à subfamília Culicinae (99%), seguida por Anophelinae (1%), havendo
predomínio da tribo Culicini (72,9%) entre os culicíneos. Cx. quinquefasciatus foi a
segunda espécie mais coletada nos diversos ambientes de coleta, apresentando
predominância no intradomicílio, mostrando tanto sua capacidade de dispersão,
quanto de domiciliação (84).
Outros trabalhos também apontam a ocorrência concomitante de diferentes
espécies de culicídeos em áreas de transmissão da malária na Amazônia Brasileira
(Tabela 2) (85–91).

20
Tabela 2. Frequência de Culicídeos descrita em estudos de fauna de mosquitos na Amazônia.
Localidade % de Anopheles % de Culex
% de outros gêneros de Culicídeos
Referência
Estação Ecológica do Rio Juami,
município de Japurá, Amazonas-Brasil 11 86 3 (91)
Município de Costa Marques,
Rondônia, Brasil - 61-peri/ 62-intra
39peri/ 38-intra
(89)
Município de Porto Velho, Rondônia,
Brasil 44,9 6,6 48,5 (88)
Hidrelétrica de Samuel, município de
Candeias do Jamari, Rondônia, Brasil - 24,5 75,5 (90)
Floresta Nacional de Caxiuanã,
município de Melgaço, Pará, Brasil 0,03 87,80 12,17 (86)
Município de Manaus, Amazonas,
Brasil - 26,5 73,5 (85)
Município de Porto Velho, Rondônia,
Brasil 27 8,4 67,6 (87)

21
1.6 Xenovigilância
Tradicionalmente índices entomológicos são usados como métricas para a
análise de transmissão da malária. Entre estes, estão a taxa de picadas em
humanos (HBR) que é a medida direta do número de mosquitos capturados por
pessoa por dia (ou tempo de exposição). Enquanto a taxa de esporozoítos (SR) é
calculada pelo número de mosquitos infectados com esporozoítos dividido pelo
número total de mosquitos examinados, para cada respectivo método de captura
utilizado, sendo expressa em porcentagem e denota a proporção de mosquitos
infectados. Destas duas resulta a taxa de inoculação entomológica – EIR, que
expressa à intensidade da transmissão do parasito da malária e também pode ser
utilizada para avaliar o envolvimento de Anopheles sp. na transmissão da malária
em determinada área. É o produto da HBR x SR. Para calcular EIR anual basta
multiplicar por 365 (92,93).
A intensidade de transmissão da malária na África é altamente variável com taxa
de inoculação entomológica anual que vão de <1 a >1.000, sendo demonstrada uma
relação linear entre a prevalência de P. falciparum e a EIR. Esta relação mostra que
reduções na prevalência da malária são possíveis de serem alcançadas apenas
quando houver redução para níveis inferiores a 1 picada infecciosa por pessoa por
ano. Destaca-se também que a EIR é uma medida mais direta da intensidade de
transmissão do que as medidas tradicionais de prevalência da malária (94).
O índice de antropofilia trata da proporção de alimentação de sangue humano
em relação ao total de fontes alimentares encontradas nos mosquitos capturados de
determinada espécie, em resumo mostra a preferência alimentar do mosquito pelo
homem. Os Anopheles apresentam altos índices de antropofilia (Figura 3) (72).
Diversos autores tem buscado desenvolver formas de monitoramento da
transmissão de doenças vetoriais através da vigilância entomológica, visando à
produção de informações, não somente sobre a presença do vetor e sua dispersão,
mas também sobre a circulação de patógenos, propondo indicadores para a

22
avaliação das medidas de controle e para a predição de risco da transmissão de
doenças em determinada área (95–106).
Tabela 3. Índices de antropofilia de Anopheles spp. e de Culex quinquefasciatus.
Espécies País IA (%) Referência
Anopheles gambiae s.s Anopheles
funestus
Anopheles arabiensis
Malawi
99,2 96,3
85,0
(72)
An. funestus s.s. Ghana 80 - 96 (107)
An. gambiae s.s. Guiné Equatorial 84,0 (70)
An. gambiae
An. funestus República dos Camarões
94.5
97,1 (108)
An. arabiensis República do Quénia 92,3 (68)
An. gambiae Senegal 83,0 (109)
Anopheles fluviatilis República da Índia 78,9 (110)
Anopheles culicifacies Cx. quinquefasciatus
República da Índia 24,7
26,3
(111)
A necessidade de avaliações entomológicas oportunas e baseadas em
evidências se torna imprescindível para a ponderação constante das ações de
controle e eliminação, assim como o redirecionamento de estratégias à medida que
se avança para um cenário de eliminação da malária, onde a manutenção de ações
evitam retrocessos e o balizamento entomológico pode garantir otimização dos
recursos e maior celeridade na tomada de decisão. Para que os países avancem
para a eliminação e prevenção de ressurgimento, é fundamental que a ligação entre
os dados de vigilância entomo-epidemiológica e controle de vetores como parte da
intervenção esteja bem estabelecida. As intervenções de controle são caras e
demoradas, mas um sistema de estratificação baseada em evidências, poderia

23
ajudar os programas de malária na decisão de onde e quando mantê-los, reduzindo
custos (112).
A existência de indivíduos assintomáticos carreadores de Plasmodium sp. em
proporções significativas, mesmo em área hipoendêmica, mostra que a prevalência
da infecção, quando baseada na gota espessa por microscopia, é subestimada. Isto
indica a importância destes reservatórios na dinâmica de transmissão da malária,
sugerindo que o reservatório do parasita submicroscópico pode ser importante para
a transmissão entre os períodos sazonais de alta e baixa transmissão, implicando
nas estratégias de controle e eliminação da malária, e alternativas para as medidas
convencionais de vigilância e controle devem ser perseguidas (113,114).
Há evidências experimentais que P. vivax, em infecções sub-microscópicas,
detectáveis apenas por técnicas de biologia molecular, são capazes de infectar
vetores. Ressaltando a importância destas infecções na transmissão da malária e
abrindo discussão para a necessidade de novas formas de vigilância da doença que
alcance estas infecções, com foco na eliminação da malária (115).
Neste sentido e considerando a ampla dispersão dos culicídeos, tem-se
estudado sua utilização como ferramenta importante no monitoramento de doenças
e até mesmo no cenário forense, devido à possibilidade de identificação e
quantificação de DNA humano mesmo depois de 48 horas após o repasto
sanguíneo. Assim, por técnicas moleculares, é possível identificar as múltiplas fontes
de refeição sanguínea desses mosquitos (57,116,117).
No contexto da diversidade e sobreposição de espécies em áreas geográficas
comuns, é natural esperar que os mosquitos tenham as mesmas possibilidades de
infectar-se com os mesmos patógenos durante o repasto sanguíneo em mamíferos
destas áreas. Embora a transmissão vetorial seja espécie-específica, alguns
culicíneos, como Aedes spp. e Culex spp. , vetores de patógenos de importância
médica, podem apresentar desenvolvimento parcial de patógeno não específico,
como por exemplo, Plasmodium spp., mas barreiras fisiológicas, por mecanismos
que ainda não são bem conhecidos, que determinam sua suscetibilidade ou

24
refratariedade ao agente patogênico, evitam o pleno desenvolvimento e a
transmissão destes patógenos. Isso sugere uma grande eficiência destas barreiras
nos culicíneos e/ou um elevado custo adaptativo para o parasito dispor de um novo
vetor, sendo provável que o sistema imunitário seja o responsável pela eliminação
destes patógenos nos culicíneos (118–121).
Com foco na pré-eliminação global da malária e reconhecendo a importância da
entomologia no balizamento, monitoramento e avaliação do controle da doença,
especialistas apontam, dentre outros aspectos, para a necessidade de
desenvolvimento de novas ferramentas para avaliar o contato homem-vetor e
proporção de mosquitos infecciosos, bem como novos tipos de armadilhas e novos
métodos de identificação de mosquitos (122).
Neste contexto, recente estudo experimental evidenciou o desenvolvimento das
formas sexuadas de P. falciparum no intestino médio de Cx. quinquefasciatus. O
seguimento pós-infecção foi realizado nos intervalos de tempo predeterminados
utilizando microscopia confocal com o registro das imagens de cortes do intestino
dessecado, comprovando que ocorreu a exflagelação dos gametas e a formação de
oocinetos no interior do intestino médio do mosquito, e os oocinetos conseguiram
atravessar a matriz peritrófica e a parede do intestino médio, porém sem a formação
de oocistos no lado basal do intestino médio do mosquito, muito provavelmente
devido a ação do sistema imune do mosquito (123).
Em uma aplicação prática desta linha de pensamento e utilizando o conceito de
mosquitos como “seringas biológicas”, apresentado por Kading et al. que em seu
estudo demonstrou que os mosquitos podem ser utilizados para quantificar viremias
em animais com precisão (124), Grubaugh et al apresentaram, em recente estudo,
resultados promissores baseados em experimentos laboratoriais e trabalhos de
campo na Libéria, utilizando An. gambiae como coletor indireto de material biológico
através do repasto sanguíneo destes, para verificar a presença de patógenos virais
para os quais o referido mosquito é refratário. O experimento em laboratório
demonstrou que 12 horas após o repasto do mosquito com sangue infectado foi
possível detectar espécies de vírus de famílias distintas, mesmo em condições de

25
coleta e armazenamento que simulavam as condições de campo. Posteriormente, os
autores realizaram coleta de mosquitos em vilas rurais da Libéria para avaliarem o
uso desta nova estratégia de vigilância, a qual eles denominaram de Xenovigilância,
obtendo resultados compatíveis com os achados nos experimentos laboratoriais
(125).
1.7 Aspectos metodológicos da xenovigilância
Neste sentido, protocolos de extração de DNA total utilizando pool de mosquitos
inteiros ou parte de suas peças anatômicas vem sendo aprimoradas para diminuir os
inibidores de amplificação na PCR, presentes na estrutura dos mosquitos,
promovendo o aumento da sensibilidade na detecção de DNA de parasitos em
ensaios de PCR, bem como maior praticidade em sua execução e desta forma,
viabilizar sua aplicação para estimar a prevalência de Plasmodium spp. em
populações de Anopheles spp. em áreas de baixa prevalência; para incriminar
espécies de anofelinos como vetores da malária; e para identificar os Plasmodium
spp. circulantes em vetores de uma determinada área (126–128).
O desenvolvimento da técnica de PCR em Tempo Real/q-PCR mostrou grande
praticidade e precisão em relação a outros métodos de PCR. Uma das técnicas é a
metodologia Taqman® que utiliza um termociclador capaz de medir a fluorescência
em tempo real, acoplado a um computador com software concebido para analisar a
intensidade de fluorescência dos corantes. A reação TaqMan® utiliza uma sonda de
hibridização marcada com dois corantes fluorescentes diferentes, um fluoróforo
(FAM) e um quencher (TAMRA), molécula que aceita energia do fluoróforo na forma
de luz e a dissipa em forma de luz e calor. Durante a fase de extensão do ciclo de
PCR, a sonda Taqman® hibridiza com a sequência da fita simples de DNA
complementar alvo para a amplificação; neste processo a sonda é degradada pela
atividade exonuclease 5'→3' da TaqDNA polimerase, separando o quencher
(TAMRA) da molécula fluorescente, o que resulta em um aumento do espectro de
emissão de fluorescência (129).

26
Novas técnicas de q-PCR para detecção múltipla de patógenos reforça a
viabilidade do seu uso além das fronteiras da pesquisa, constituindo-se em
importante ferramenta para uso em algumas situações de rotina e vigilância
(130,131). Assim como a busca por testes de PCR gênero-específico com maior
sensibilidade e praticidade que funcionam para triagem em grande escala de
amostras, quer seja para esta atividade em laboratórios de banco de sangue, ou
para trabalhos de campo em áreas rurais com pouca estrutura (132,133).
Na malária, com a evolução dessas técnicas moleculares, alguns trabalhos vêm
sendo realizados no sentido de utilizar e aprimorar essas técnicas para detecção de
infecção natural ou experimental em mosquitos do gênero Anopheles para
monitoramento e predição de risco de transmissão da malária. Martins-Campos et al
usou estas ferramentas para avaliar o impacto das ações integradas de controle em
um município da Região Amazônica (134). Pérez et al, propôs um desenho de
protocolo de extração de DNA de mosquitos que permitiria a amplificação em
ensaios de PCR de Plasmodium spp. visando a sua utilização como indicador
preditivo do risco de contrair a doença em áreas de atuação das forças militares da
Colômbia (135). Sorosjinda-Nunthawarasilp e Bhumiratana apresentam proposta de
xenomonitoramento de P. falciparum e P. vivax resistentes a multidrogas, utilizando
a captura de potenciais vetores em ecótopos e ecótonos situados na Tailândia,
aplicando técnicas moleculares para a identificação e mapeamento destas cepas
multirresistentes (136).
Como exemplo real de ferramentas importantes no avanço para a detecção de
infecções assintomáticas temos o desenvolvimento recente de uma RT-PCR
ultrassensível que em condições similares as de campo, simuladas em laboratório, é
capaz de aumentar substancialmente a sensibilidade do teste em comparação com
as q-PCR utilizadas atualmente, com quantidades inferiores de sangue coletado
(137). Outra técnica recém-desenvolvida é uma q-PCR que tem como alvo cópias do
elemento repetitivo telômero-associado – TARE-2 que contém aproximadamente
250 cópias por genoma. Nos ensaios, atingiu um limite de detecção de 0,03 a 0,15
parasitas/µl de sangue e foram 10 vezes mais sensível do que a qPCR padrão 18S
rRNA (138).

27
1.8 A eliminação da malária
Os recentes avanços na redução e controle da malária no mundo estimularam a
discussão de um cenário de eliminação mundial da doença. Por definição a
eliminação da malária é: “A redução a zero da incidência de infecção causada por
um parasita especifico da malária em uma área geográfica definida, como resultado
de esforços intencionais”; isso já é um cenário/meta possível de ser alcançado em
alguns países/regiões que tem apresentado o declínio anual da transmissão da
malária ou focos residuais de transmissão da doença. A OMS tem traçado
estratégias e metas para eliminação da malária nestes países com foco na
erradicação futura da malária no mundo, e para isto prevê vários cenários para o
cumprimento de metas no período de 2015 a 2030 (139–141).
Algumas estratégias experimentadas, como Triagem em Massa e Tratamento –
MSAT, demostraram que as limitações de sensibilidade dos métodos de
diagnósticos utilizados comprometem sua eficácia, demandando novas ferramentas
altamente sensíveis de teste rápido para a detecção de todas as espécies da
malária humana (142). Assim como o rastreio sistemático e tratamento de
portadores assintomáticos ao nível da comunidade não foi eficaz na redução da
incidência de casos de malária clínica no período sazonal subsequente de
transmissão, deduzindo-se também a necessidade de maiores níveis de
sensibilidade na detecção do parasito (143–145).
Modelos matemáticos ratificam essa necessidade de testes com sensibilidade
fora da nossa realidade atual, como ponto crucial para o alcance da eliminação da
malária utilizando as estratégias de rastreio de assintomáticos. Discute-se ainda a
possível eficácia da Administração de Drogas em Massa - MDA como melhor opção,
desde que tenha boa pela população, pois a MDA acrescenta o efeito profilático
adicional, mas pode estar relacionado ao aumento de resistência as drogas
utilizadas (146).
Nesse contexto de eliminação da malária algumas questões relevantes tem sido
foco de pesquisas cientificas para subsidiarem a discussão e viabilização deste

28
objetivo mundial (147–149); como exemplo de algumas destas questões tem-se: a
avaliação rigorosa dos programas de eliminação (150,151), o financiamento das
ações (152), sistemas eficientes de avaliação (153,154), produção de vacinas
(155,156), vigilância entomo-epidemiológica (157,158), produção de novos
medicamentos (159,160), produção de novas ferramentas operacionais para a
eliminação da malária (161).
Os casos importados de malária como fator de risco para a reintrodução da
transmissão em áreas antes interrompida, bem como o controle da malária
transfronteiriça (162,163), e a transmissão da malária em áreas hipoendêmicas, que
necessitam de abordagens diferenciadas para sua detecção e controle (164);
apresentam-se como importantes obstáculos a serem considerados e superados
para a eliminação da malária.
Esses obstáculos demandam a pesquisa de novas ferramentas operacionais
com a aplicação de novas tecnologias que atualmente são escassas, para preencher
as lacunas existentes e proporcionarem maior efetividade no controle e eliminação
da malária nestas áreas mais complexas e com maior custo operacional (161,165).
Também algumas estratégias de vigilância entomológica úteis para os
programas de malária esbarram nas questões éticas que envolvem a prática de
captura de mosquito com pouso em humanos (HLC). Essas questões são bastante
complexas, abrangendo ainda fatores econômicos e sociais, mas que precisam ser
superadas para garantir a viabilidade operacional desta metodologia, ainda que haja
indicativo de um menor risco de adoecimento dos capturadores que fazem uso de
quimioprofilaxia, em relação aos outros membros de sua comunidade (166,167).
Outros métodos que não envolvem problemas éticos têm sido testados
comparativamente para considerar sua eficácia como alternativas ao HLC (168–
170).
Além dessas questões técnicas e operacionais, a proposta de eliminação da
malária necessita ser integrada às politicas de desenvolvimento econômico e social
dos países para assegurar e garantir uma base sustentável dos ganhos obtidos no

29
controle da malária para viabilização dos passos seguintes rumo à eliminação da
malária (6,171).
Frente a essa nova realidade, da possível eliminação da malária no mundo, as
discussões junto a Organização Mundial de Saúde e as autoridades de saúde dos
países membros tem ganhado força, inclusive com o estabelecimento de metas de
médio prazo para alguns países onde a redução dos casos de malária tem sido
progressiva nos últimos anos e as condições epidemiológicas já permitem um plano
de eliminação.
O custeio para o desenvolvimento destes planos de eliminação, como não
poderia deixar de ser, é um dos pontos centrais dessa discussão, dado que esta
nova visão de eliminação exige novas e diferentes estratégias daquelas aplicadas
para os planos de controle, e que, por exigirem novas ferramentas com maior
acurácia, tanto para a detecção de casos autóctones em áreas hipoendêmicas,
quanto para a vigilância da reintrodução de casos em áreas livre de transmissão de
malária, tem consequentemente um custo mais elevado, exigindo maior
compromisso destes países signatários para o alcance das metas de
eliminação/erradicação da malária.
Um dos investimentos essenciais nesse contexto, é para a pesquisa aplicada ao
desenvolvimento de ferramentas de vigilância que utilizem novas técnicas para
preencher as lacunas existentes atualmente nas vigilâncias epidemiológica e
entomológica. Embora novas técnicas mais sensíveis, tais como a aplicação de
técnicas moleculares de q-PCR usadas na xenovigilância, que se apresenta como
uma ferramenta promissora na vigilância da transmissão de diversas doenças; há
uma completa ausência de trabalhos de pesquisa em xenovilância direcionados para
a malária.
O uso dessas novas ferramentas deverá ser explorado na eliminação da malária,
exigindo pesquisas na área que venham validar e balizar o uso da xenovigilância da
malária como um dos componentes do plano para a eliminação/erradicação da
malária.

30
Esse contexto aponta para a possibilidade de um novo indicador para a
transmissão da malária em áreas endêmicas. Considerando que os indicadores
malariométricos disponíveis, como a Incidência Parasitária Anual (IPA) são
influenciados pela qualidade da notificação, especialmente nas áreas hipoendêmicas
que apresentam uma grande porcentagem de assintomáticos, podem não refletir
adequadamente a vulnerabilidade da população à malária e a intensidade de
circulação do Plasmodium spp. na área.
Os inquéritos hemoscópicos, o uso de ferramentas moleculares como a PCR e
especialmente os estudos longitudinais, que poderiam corrigir os erros introduzidos
pelo sistema de notificação, são pouco viáveis nos seus aspectos logístico e
financeiro no contexto da Amazônia. Dessa forma, um novo indicador que tivesse
associação com a carga da malária numa área, e que fosse obtido de uma forma
aceitável pela população, seria um grande avanço para a implementação no sistema
de vigilância e controle da malária.
O contato de diversas espécies de culicídeos com as populações humanas que
vivem em áreas malarígenas sugere que a identificação do material genético do
parasito no repasto sanguíneo presente no intestino destes mosquitos pode ser
comum, dependendo provavelmente da incidência de malária. Diante disto, o
presente trabalho investigou a associação da taxa de culicídeos infectados com a
frequência de indivíduos infectados com Plasmodium sp. na população humana
relacionada, e a possível utilização desse parâmetro como um indicador
malariométrico.

31
2. OBJETIVOS
2.1 Geral
Avaliar uma ferramenta de xenovigilância para malária usando culicídeos não-
anofelinos em área endêmica da doença.
2.2 Específicos
Verificar a frequência e a diversidade das espécies de culicídeos capturados;
Padronizar as técnicas de extração de DNA e de q-PCR para a detecção de
Plasmodium sp. em culicídeos não anofelinos.
Estimar a concordância, sensibilidade, especificidade e valores preditivos da
xenovilância de infecção por malária baseada em culicídeos não anofelinos em
relação ao método q-PCR de detecção da infecção em humanos;
Avaliar a associação entre a positividade domiciliar de infecções em humanos e
a presença de parasitas em culicídeos não anofelinos.

32
3. MATERIAL E MÉTODOS
3.1 Modelo de estudo
Trata-se de um estudo transversal para a avaliação da acurácia de um método
de xenovigilância para malária baseada na detecção de DNA de Plasmodium em
culicídeos não anofelinos.
3.2 Local de estudo
O presente estudo foi realizado em uma área periurbana localizada na zona
Leste da cidade de Manaus - Amazonas (Brasil), abrangendo parte do bairro Distrito
Industrial II e ainda algumas áreas dos bairros Jorge Teixeira e Puraquequara. As
localidades comtempladas foram os ramais do Brasileirinho, do Puraquequara e do
Ipiranga; áreas contíguas que apresentam características ecológicas, sociais e
econômicas, bastante semelhantes, e transmissão ativa de malária. A área de
estudo tem uma população de 2.856 habitantes (1.805 no Brasileirinho, 621 no
Ipiranga e 430 no Puraquequara), segundo levantamento populacional realizado por
este grupo de pesquisa, distribuída em uma área aproximada de 72 km2 (Figura 4).
Figura 4. Imagem de satélite da área do estudo (Google Earth®).

33
A diversidade de coleções hídricas existentes nessa área, tais como igarapés,
nascentes, lagoas, lagos e mais recentemente tanques de aquicultura, bem como o
impacto das alterações ambientais e o aumento da exposição de humanos,
decorrente do processo de crescimento desordenado, configuram um quadro
bastante favorável para a transmissão de malária nesta área; fazendo sua
participação no registro de casos autóctones no município de Manaus aumentar nos
últimos quatro anos, chegando a 35,6% em 2014 (Figura 5). Historicamente, a
malária causada pelo P. vivax é a mais frequente na região, e nesta área, respondeu
por 95,3% dos casos autóctones de malária no mesmo ano (Figura 6).
Figura 5. Número de casos de malária na área do projeto(linha azul) e o percentual de participação (linha vermelha) destes casos no total de casos notificados em Manaus no período de 2004 a 2014. Fonte de dados: SIVEP/Malária.
Figura 6. Número de casos de malária por espécie na área do projeto no período de 2004 a 2014. Fonte de dados: SIVEP/Malária.
0
1000
2000
3000
4000
5000
6000
-
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
12
20
13
20
14
Cas
os
% d
e P
arti
cip
ação
% Participação em Manaus Casos na Área do Projeto
-
500
1.000
1.500
2.000
2.500
3.000
3.500
4.000
4.500
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
12
20
13
20
14
Cas
os
Falciparum
Vivax

34
3.3 Amostragem
Este estudo foi desenvolvido em paralelo ao Projeto Multicêntrico “Epidemiologia
comparativa da transmissão de P. falciparum e P. vivax no Brasil, Tailândia e Papua
Nova Guiné” (TransEpi) com vistas a utilização das informações ecológicas
referentes a população humana participante. Assim sendo, a unidade amostral
adotada foi a casa. Estas foram selecionadas pelo critério de ter um de seus
moradores participando do referido projeto. Desta forma, a amostra foi obtida com a
coleta simultânea de mosquitos e de sangue de 875 moradores, em 233 casas
distribuídas da seguinte forma: 103 no Ramal do Ipiranga, 81 no Ramal do
Brasileirinho e 49 no Ramal do Puraquequara.
3.4 Coleta e identificação de culicídeos no campo
A coleta de formas adultas de culicídeos nas casas foi simultânea à coleta de
sangue dos participantes do Projeto TransEpi. Foram utilizados dois métodos de
captura: o aspirador entomológico elétrico e a armadilha BG-Sentinel®, ambos no
intradomicílio (Figura 7AB). A identificação das amostras coletadas foi padronizada
por código numérico de 5 dígitos que indicavam o número da casa e o tipo de
captura. Com apenas dois técnicos foi possível realizar a coleta, enquanto um
preenchia o formulário de coleta e instalava a BG-Sentinel®, o outro realizava a
aspiração nos cômodos da casa sob a supervisão de um dos moradores.
Figura 7. (A) Captura com armadilha BG-Sentinel®, (B) Captura com aspirador entomológico, (C) Triagem dos mosquitos capturados (Acervo da Gerência de Entomologia/FMT-HVD).
A B C

35
Para a captura de mosquitos com a BG-Sentinel® foi utilizado acessório BG-
lure® e AgriSence® como atraentes. Estas foram instaladas em um dos cômodos
internos da casa, preferencialmente nos quartos. As coletas tiveram em média a
duração de 28 horas, pois eram instaladas na manhã do dia da coleta de sangue e
retiradas no inicio da tarde do dia seguinte, utilizou-se a energia elétrica das casas
através de adaptadores de voltagem para 12 volts, não havendo resistência ou
queixas da população, considerando o baixo nível de gasto energético do
equipamento e o uso de extensões que permitiam o compartilhamento da tomada
elétrica da casa.
As coletas com o aspirador entomológico foram realizadas no período da manhã,
durante a coleta de sangue, aspirando todos os recantos da casa e possíveis
esconderijos dos mosquitos, esse processo durava aproximadamente de 5 a 15
minutos, dependendo do número e tamanho dos cômodos. Para o funcionamento do
equipamento foram utilizadas 02 baterias (comuns para motocicleta), recarregáveis
de 12 volts. Considerando que a aspiração é bastante invasiva, pela necessidade de
adentrar em todos os ambientes da casa, havia uma expectativa de rejeição por
parte dos moradores, o que não se confirmou, havendo uma boa receptividade de
todos moradores, até mesmo quando algum membro da família ainda estava
dormindo.
Todos os procedimentos da coleta estão descritos de forma detalhada no
Procedimento Operacional Padrão-POP de código POP_MAL_LB_019_v01D_PT
(Apêndice 5.5).
Os culicídeos capturados foram levados para o Centro de Entomologia da FMT-
HVD e no mesmo dia era realizada a triagem e a identificação a nível de espécie ou
gênero (Figura 7C), separando-os em três grupos: Cx. quinquefasciatus, An. darlingi
e outros culicídeos, utilizando as chaves de Consoli e Lourenço-de-Oliveira (172) e
de Faran e Linthicum (173), em seguida foram inicialmente agrupados em pool com
10 mosquitos, acondicionados em microtubos contendo álcool etílico a 80ºGL e
armazenados em freezer a -20ºC. Quanto à identificação das espécies incluídas no
último grupo, dada à complexidade da identificação, estas só foram identificadas

36
posteriormente, devido a problemas de saúde do técnico responsável, o que
prejudicou em parte a identificação das mesmas, considerando que estavam
submersas em álcool e algumas partes anatômicas e cerdas se desprenderam.
Todos Estes procedimentos estão detalhados no POP_MAL_LB_020_v01D_PT
(Apêndice 5.6).
3.5 Padronização dos testes moleculares
Considerando que não havia nenhum protocolo de testes moleculares para
detecção de Plasmodium spp. em culicídeos não anofelinos, foi necessária a
padronização dos processos de extração de DNA e de q-PCR que assegurassem
um nível satisfatório de sensibilidade na detecção de Plasmodium spp. antes de
realizar os testes com as amostras coletadas.
Para tal, foi necessário o cultivo de uma colônia de Cx. quinquefasciatus, que é a
espécie mais abundante da amostra e de fácil manejo, viabilizando-se as infecções
experimentais e dando provimento ao material genético para as provas de
sensibilidade dos testes moleculares e do limiar de detecção referente à quantidade
máxima de mosquitos por pool e ao tempo máximo pós-infecção.
3.5.1 Estabelecimento de colônia de Culex quinquefasciatus
A colônia de Cx. quinquefasciatus foi estabelecida a partir de formas imaturas do
mosquito (larvas e pupas) coletadas em criadouros da zona centro sul da cidade de
Manaus. Esta escolha foi por conveniência, considerando que a equipe municipal de
controle de endemias trabalhava nesta área realizando a coleta de larvas e nos
doaram muitas destas para o projeto. Porém consideramos que não foi uma boa
escolha, pois a primeira geração de adultos apresentou um número muito maior de
machos, atrasando o início das infecções. Começar com a captura de mosquitos,
usando as jangadas de ovos destes para dar início a colônia seria uma escolha mais
apropriada.

37
A colônia de mosquito foi mantida em sala específica no Centro de Entomologia
da FMT-HVD, com o controle de temperatura entre 26 e 28ºC e umidade relativa
entre 60 e 80 UR%.
Eram separadas três jangadas de ovos, cerca de 350 a 500 ovos, por bacia de
dimensão18x27x5cm com água até um terço da capacidade máxima do recipiente.
Após a eclosão dos ovos, as larvas eram mantidas com ração granulada para peixes
ornamentais, inicialmente com 5 grânulos, aumentando o número destes conforme o
desenvolvimento larvário até o limite de 20. A limpeza das bacias era realizada
diariamente e a troca de água a cada dois dias (Figura 8).
À medida que passavam para o estágio de pupa, eram retiradas para um copo
médio de 180 ml com água e este era colocado em uma gaiola de dimensão
19x16x16cm até o limite de 300. A partir da primeira emersão de um mosquito era
disponibilizado dentro da gaiola recipiente com solução açucarada a 10%, os quais
eram trocados a cada três dias. O repasto sanguíneo das fêmeas era realizado uma
vez por semana em camundongos albinos (Mus musculus) anestesiados, garantindo
oviposições regulares para a manutenção e o crescimento da colônia (Figura 9)
(174–176).
Figura 8. Fases imaturas de Cx. quinqufasciatus na manutenção da colônia. (A) Jangada de ovos, (B) Bacia com larvas de I estádio (quase imperceptíveis) e grânulos de ração, (C) Larvas de II e III estádio, e grânulos de ração (Acervo da Gerência de Entomologia/FMT-HVD).
A B C

38
3.5.2 Infecção experimental de P. vivax em Cx. quinquefasciatus
Fêmeas com média de idade entre 5 a 7 dias foram separadas em gaiolas
teladas de dimensão 8x11x9,5cm para seu primeiro repasto sanguíneo, formando
grupos de 100 a 120 mosquitos; era retirada previamente a alimentação de solução
açucarada nas 36 horas que antecediam a infecção, quando era ofertado sangue
parasitado com P. vivax.
A amostra de sangue ficava disposta em alimentadores artificiais que utilizavam
membrana artificial Parafilm®, estes ficavam sobre as telas das gaiolas e estavam
ligados ao banho-maria por mangueiras, formando um sistema de circulação de
água, com controle de temperatura em 37ºC. O repasto era mantido por
aproximadamente 120 minutos, adaptado de Rios-Velásquez (177). Devido a
característica do ciclo circadiano do mosquito, as infecções foram realizadas a partir
das 20:00 horas. As infecções foram realizadas em triplicata com alíquotas da
mesma amostra de sangue parasitado (Figura 10). Estes procedimentos estão
detalhados no POP_MAL_LB_013_v02D_PT (Anexo 6.1).
Figura 9. Manutenção da colônia de Cx. quinqufasciatus. (A) Gaiola com copo contendo pupas, das quais emergirão os mosquitos, e erlenmeyer com solução açucarada para alimentação dos mosquitos; (B) Gaiola com mosquitos alimentando de solução açucarada; (C) Mosquitos obtendo repasto sanguíneo em camundongos (Acervo da Gerência de Entomologia/FMT-HVD).
A B C

39
3.5.3 Extração e purificação de DNA dos pools de culicídeos
Utilizando mosquitos infectados, conforme descrito no subitem anterior, foram
realizados testes para verificar qual protocolo de extração de DNA obtinha o melhor
rendimento e pureza. Os mosquitos eram macerados em pools com 10 mosquitos
utilizando pistilos de acrílico acoplado a um Pellet pestles® da SIGMA (Figura 11).
Foi utilizado o Kit comercial de extração por colunas Purelink™ Genomic DNA Mini
Kit da Invitrogen, seguindo as orientações do fabricante, e a resina Chelex® 100 da
BIO RAD, conforme padronização estabelecida no POP_MAL_LB_001_v02D_PT
(Anexo 6.3). Embora o produto genético obtido pelo kit comercial tenha apresentado
maior grau de pureza, seu rendimento foi bem abaixo do que foi obtido pelo Chelex®
100 que apresentou alto rendimento de DNA, com grau de pureza satisfatório.
Figura 10. Infecção experimental. (A) Gaiolas com mosquitos e alimentadores artificiais interligados em circuito com o banho-maria; (B) Mosquitos se alimentando come sangue infectado (Acervo da Gerência de Entomologia/FMT-HVD).
A B

40
Nesse processo de padronização, já definido que trabalharíamos com o
protocolo da resina Chelex®, foram realizados testes para determinar se usaríamos
os mosquitos inteiros ou certas peças anatômicas destes. Assim, foram realizadas
extrações de DNA a partir de mosquitos inteiros e de abdomes de mosquitos, estes
eram seccionados do corpo com pinças entomológicas, somente na hora da
extração e sem a necessidade de dissecação. O DNA extraído dos abdomes
apresentou maior grau de pureza. Desta forma, o POP_MAL_LB_001_v02D_PT
(Anexo 6.3) sofreu uma adaptação, padronizando-se o uso de abdomes, visando a
redução de inibidores da q-PCR, presentes nas peças anatômicas dos mosquitos,
principalmente na cabeça.
3.5.4 Realização dos ensaios de q-PCR
Nesta etapa, os trabalhos foram realizados no Laboratório de biologia molecular
da Gerência de Malária da FMT-HVD.
Inicialmente, utilizando DNA dos mosquitos infectados experimentalmente com
P. vivax, foi testado o protocolo utilizado por Martins-Campos et al (134), que
trabalhou com q-PCR Multiplex para a detecção simultânea de P. falciparum e P.
Figura 11. Etapas do processo de maceração de mosquitos. (A) Mosquito sendo colocado em tubo cônico para maceração; (B) Maceração de pool de mosquitos em resina Chelex® 100 utilizando pistilo azul e Pellet pestles®; (C) Suspensão obtida a partir do pool de mosquitos e da resina Chelex® 100 (Acervo da Gerência de Entomologia/FMT-HVD).
A B C

41
vivax em An. darlingi, vetor natural destes patógenos na região. Porém a
sensibilidade deste foi baixa para a detecção em Cx. quinquefasciatus, não se
obteve boas amplificações, considerando a comprovada refratariedade do deste
mosquito e consequente redução de material genético do patógeno. Outra
dificuldade neste teste foram os controles positivos, pois não eram padronizados e
para cada placa demandava amostra de DNA de sangue de voluntários com malária.
Em seguida foi testado o protocolo de um ensaio QMAL q-PCR gênero-
específico para a detecção dos principais parasitos da malária que infectam
humanos, utilizando uma reação já padronizada de PCR em Tempo Real baseada
no gene 18S rRNA, o mesmo utilizado na análise das amostras de sangue dos
participantes do projeto TransEpi, que utiliza plasmídeos como controle positivos
permitindo a quantificação das cópias de DNA em um sistema Applied Biosystems
7500 Fast System®, padronizado no Laboratório de biologia molecular da Gerência
de Malária da FMT-HVD conforme o POP_MAL_LB_021_v01D_PT (Anexo 6.4).
Com este último protocolo, obteve-se amplificações satisfatórias que permitiram
um próximo passo, a escolha da concentração mais adequada de DNA para melhor
amplificação. Como a quantidade de Material genético obtido com o Chelex® era
sempre alta, estes foram eluídos em água de injeção ultrapura nas seguintes
concentrações: 5ng/µl, 10ng/µl, 50ng/µl, 100ng/µl, 150ng/µl e 200ng/µl, todas em
duplicata. As melhores amplificações foram observadas nas concentrações 50ng/µl
e 100ng/µl. Ficou padronizada a concentração de 100ng/µl para a eluição do DNA
das amostras de campo para os ensaios de q-PCR.
3.5.5 Teste de tempo máximo de detecção
Considerando a refratariedade dos outros culicídeos, que não são vetores
naturais dos plasmódios que parasitam o homem, e a consequente redução
gradativa de material genético do parasito ao longo do tempo pós-ingestão de
sangue infectado, foi necessário definir com quais amostras da coleta de campo
trabalharíamos, se com todos os mosquitos fêmeas ou somente com os

42
ingurgitados. Para isso foram realizadas duas infecções experimentais de P. vivax
em Cx. quinquefasciatus, utilizando duas amostras de sangue, uma com “+” de
parasitemia e outra com “++”, todas em triplicata. O seguimento pós-infecção foi
realizado com a retirada de 10 mosquitos por gaiola, no D0, D1, D2, D3....D10 pós-
infecção.
Foi realizada a extração de DNA destas amostras e em seguida os ensaios de
QMAL q-PCR. O limite de tempo máximo de detecção de plasmódios humanos em
Cx. quinquefasciatus pós-infecção foi a última amostra na escala do tempo de
seguimento que apresentou amplificação satisfatória, que neste caso foi até o D2
para a amostra com “++” de parasitemia e até o D1 para aquela com “+”. Assim
sendo, ficou definido que os testes moleculares seriam realizados apenas das
amostras de campo com mosquitos ingurgitados.
3.5.6 Teste de limiar de detecção
Embora os mosquitos da amostragem, oriundos do trabalho de campo, tivessem
sido armazenados em pools de 1 a 10 indivíduos, houve a necessidade de se
determinar o número máximo de mosquitos por pool antes de realizar a extração de
DNA e a realização dos testes moleculares de detecção de Plasmodium sp.,
considerando as interferências que ocorrem nesses testes devido a certas
substâncias luminescentes presentes em peças anatômicas do mosquito,
principalmente na cabeça. Além disso, nesse caso particular em que os mosquitos
testados não são vetores naturais do parasito, a quantidade de DNA de Plasmodium
sp. que se esperava encontrar era relativamente baixa, já que este não completa
seu ciclo no mosquito e seu material genético vai degradando gradativamente.
Desta forma, foi realizada infecção experimental com sangue parasitado com P.
vivax, tal qual descrito anteriormente, acrescentando um grupo de mosquitos que
foram alimentadas com sangue não infectado. Na hora zero após o repasto

43
sanguíneo, foram retirados de ambos os grupos, aqueles com o abdome plenamente
distendido. Foram retirados os seus abdomes e colocados um do grupo de
infectados em cada tubo eppendorf, acrescentando “n” abdomes de mosquitos do
grupo controle, nas seguintes proporções: 1:1, 1:3, 1:5, 1:7 e 1:9, todas as amostras
em triplicata.
Foi realizada a extração de DNA desses pools com Chelex®100 e
posteriormente os ensaios de QMAL q-PCR gênero-específico. O limiar de detecção
foi considerado a amostra com a maior proporção de mosquitos alimentados com
sangue não parasitado que amplificou satisfatoriamente. A última amostra a
amplificar na escala de proporção foi a de 1:5. Com base nesse critério, ficou
padronizado que os pools da amostra de campo seriam compostos por 1 a 5
mosquitos ingurgitados.
3.6 Extração e purificação de DNA das amostras de campo
Com base na padronização já descrita, as amostras de campo foram
processadas formando pools de 1 a 5 abdomes de mosquitos ingurgitados que
foram macerados para a extração do DNA total seguindo o
POP_MAL_LB_001_v02D_PT (Anexo 6.3). Esse processo de reordenação dos
pools e retirada dos abdomes foi extremamente simples, pois era realizado somente
no momento da extração, quando os mosquitos tinham seus abdomes removidos
com pinças entomológicas, sem a necessidade de microscópios entomológicos e
aplicação de técnicas de dissecação, em seguida eram colocados em tubos
eppendorf com alíquotas da resina Chelex® previamente preparada, sendo por fim
macerados e extraído o material genético.

44
3.7 Realização dos ensaios de q-PCR das amostras de campo
Para a detecção dos principais parasitos da malária que infectam humanos, foi
utilizada um protocolo de uma reação já padronizada de PCR em Tempo Real
baseada no gene 18S rRNA, QMAL q-PCR gênero-específico. O mesmo utilizado na
análise das amostras de sangue dos participantes do Projeto TransEpi, que utiliza
um sistema Applied Biosystems 7500 Fast System® e foi padronizado no Laboratório
de biologia molecular da Gerência de Malária da FMT-HVD conforme o
POP_MAL_LB_021_v01D_PT (Anexo 6.4).
Os ensaios de QMAL q-PCR gênero-específico foram realizados em duplo-cego
para determinar a taxa de encontro de plasmódios em humanos e em culicídeos.
3.8 Definição de positividade plasmodial em culicídeos e em humanos
Para a população de culicídeos capturados, foi considerada amostra positiva
aquela que apresentou amplificação nos ensaios de QMAL q-PCR gênero-
específico.
Para a população humana participante do Projeto TransEpi, foi considerada
amostra positiva aquela que apresentou amplificação nos ensaios de QMAL q-PCR
gênero-específico, independente de sintomatologia destes participantes.
3.9 Plano analítico
A análise foi realizada a partir dos dados obtidos, que foram tratados e tabulados
para viabilizar os testes estatísticos visando responder aos objetivos desta pesquisa.
Para cumprir esta meta, o plano foi subdividido nas seguintes etapas:

45
3.9.1 Registro e tratamento dos dados
As fichas de coleta dos culicídeos foram processadas em um sistema de
reconhecimento inteligente de caracteres denominado Teleform® que realiza criação
de formulários, captura de imagem, processamento da imagem em dados e
exportação desses dados para planilhas do Excel®, reduzindo assim a possibilidade
de erros de digitação. Os resultados dos testes moleculares foram tabulados e
organizados e acrescentados em novas colunas na referida planilha. Após limpeza
do banco de dados foram procedidas análises estatísticas utilizando os softwares
Excel®, Stata/SE 13.1®, GeoDa® e ArcGis®.
3.9.2 Frequência relativa das espécies de culicídeos
A frequência relativa dos culicídeos foi calculada com base na proporção de
cada gênero encontrado em relação ao total de culicídeos coletados em toda a área
trabalhada.
3.9.3 Unidade espacial de análise
Para as análises espacial todo domicílio foi georreferenciado utilizando as
coordenadas geográficas dos locais de coleta, obtidas por ocasião do censo do
Projeto TransEpi. A esses pontos foram associadas informações populacionais e os
resultados dos testes moleculares, tanto em humanos como em culicídeos. Foram
utilizados os softwares GeoDa® e ArcGis®.
3.9.4 Determinação da taxa de encontro de DNA de Plasmodium spp. em
culicídeos por domicílio
A frequência relativa do encontro de DNA de Plasmodium spp. em culicídeos foi
calculada com base no número de pools que apresentaram amplificação nos
ensaios QMAL q-PCR gênero-específico por domicílio, em relação ao número total
de pools daquele domicílio.

46
3.9.5 Determinação da taxa de infecção de Plasmodium spp. na população por
domicílio
A taxa de infecção de Plasmodium spp. na população humana foi obtida a partir
dos resultados das coortes realizadas pelo Projeto TransEpi no mesmo período das
coletas de culicídeos. Foi calculada com base no número de amostras de humanos
que apresentarem amplificação nos ensaios de QMAL q-PCR gênero-específico no
domicílio em relação ao número total de amostras coletadas no domicílio.
3.9.6 Avaliação da acurácia da xenovigilância
Foi realizada uma avaliação da acurácia a partir dos resultados obtidos nos
ensaios de q-PCR das amostras de culicídeos, e para efeito desse teste usamos a
suposição de que a q-PCR da população humana do estudo era padrão ouro de
diagnóstico. Embora a técnica utilizada permitisse a quantificação do número de
cópias da sequência alvo, os resultados foram registrados na forma dicotômica,
presença ou ausência, tendo o domicílio como unidade de análise. Foram
determinados Sensibilidade, Especificidade, Valor Preditivo Positivo – VPP e o Valor
Preditivo Negativo - VPN.
3.9.7 Análise espacial descritiva
Para as análises espaciais, os domicílios conforme descrito anteriormente,
foram considerados como unidade de análise.
Foi realizada análise espacial descritiva das taxas de infecção nos domicílios
das comunidades do estudo, tanto de humanos quanto de mosquitos, em uma
mesma escala traduzida em círculos que expressão intensidade. Buscou-se
identificar semelhanças no padrão de distribuição espacial para verificar possível
associação entre taxas de infecção.

47
3.9.8 Análise de regressão logística múltipla
Para esta análise, o método stepwise de seleção foi aplicado de forma
hierarquizada em modelos separados em blocos: distal e proximal. A variável
dependente considerada na análise foi o domicílio com infecção detectada em
humanos pelo ensaio QMAL q-PCR.
Foram incluídas no modelo distal as variáveis ecológicas relacionadas à
características do domicílio e do ambiente, além de operacionais relacionadas ao
controle do vetor (material da casa, tipo de parede, localização, presença de frestas,
número de portas e janelas, presença de tela nas janelas, borrifação nos últimos 3
meses, comunidade de residência, uso de mosquiteiro na última noite e uso de
mosquiteiro na última noite). O modelo proximal incluiu variáveis relacionadas ao
inseto e a população humana, como sintomas e tratamento (número de mosquitos
coletados, número de Culex ingurgitados, número de pools positivos para o parasito,
febre e uso de antimalárico mês da coleta).
Foram selecionadas para compor os modelos proximal e distal, as variáveis com
nível de significância menor que 0.2 na análise univariada. O modelo final, incluindo
os dois níveis hierárquicos, foi composto pelas variáveis com nível de significância
menor que 0.05 em cada bloco (proximal e distal). O mesmo critério foi utilizado para
considerar estatisticamente significantes as variáveis do modelo final.
3.10 Questões éticas
Todas as amostras de sangue parasitado com P. vivax, que foram utilizadas na
infecção experimental para padronização dos testes moleculares, foram alíquotas
oriundas de amostras de sangue de participantes do Projeto multicêntrico
denominado “Epidemiologia comparativa da transmissão de P. falciparum e P. vivax
no Brasil, Tailândia e Papua Nova Guiné”, cujo centro no Brasil é a Fundação de

48
Medicina Tropical Dr. Heitor Vieira Dourado, Manaus-AM; o Termo de
Consentimento Livre e Esclarecido - TCLE desta pesquisa incluía a autorização do
uso da amostra de sangue em outros projetos, e foi aprovado pelo Comitê de Ética
em Pesquisa com Seres Humanos da FMT-HVD (parecer numero 51536/2012) e
pela Comissão Nacional de Ética em Pesquisa (CONEP) pelo parecer número
349.211/2013 (Anexo 6.2).
Desta forma foram comtempladas as exigências éticas para o uso de sangue
humano que foi utilizado sem a identificação do sujeito da pesquisa. Somente a
informação de parasitemia da amostra foi utilizada, e unicamente nesta fase de
padronização dos testes moleculares.

49
4. RESULTADOS
4.1 Descrição das variáveis estudadas
A distribuição espacial dos domicílios analisados no estudo apontou uma
variação, tanto na dispersão dos imóveis, quanto no número de habitantes por
domicílio entre as comunidades estudadas. A comunidade do Puraquequara
apresentou maior aglomeração e maior densidade populacional domiciliar, enquanto
o Brasileirinho, maior dispersão e menor densidade populacional domiciliar
(Figura 12).
Figura 12. Espacialização dos domicílios na área estudada, apresentando a densidade populacional por domicílio nas comunidades do Ipiranga, Puraquequara e Brasileirinho, e localização dos laboratórios de notificação e diagnostico da malária.
A amostra foi caracterizada por domicílios predominantemente de alvenaria
(133/57.08%), com paredes completas (221/94.85%), localizadas fora do eixo dos
ramais e que apresentavam portas e janelas (230/98.71%). Quanto às medidas
protetivas, observou-se que apenas 22.32% (52/233) apresentavam telas nas portas
e janelas e a cobertura de borrifação intradomiciliar era de 45.92% (107/233). O uso
de mosquiteiros foi identificado em 42.06% (98/233) dos domicílios, enquanto o uso

50
de mosquiteiros impregnados com inseticidas foi identificado na maioria dos imóveis
(138/59.23%). Embora a frequência de domicílios com número de mosquitos
capturados acima da mediana tenha sido alta, a frequência de domicílios com
número de Culex sp. ingurgitados acima da média foi de 24.03%. Já a frequência de
domicílios com mosquitos carreando DNA de Plasmodium sp. foi de 6.44%, valor
semelhante ao de domicílios com habitantes que apresentaram febre (Tabela 4).
Tabela 4. Frequências das variáveis estudadas.
Variáveis n %
Tipo de casa
Alvenaria 133 57.08
Madeira ou similar 100 42.92
Tipo de parede
Completa 221 94.85
Incompleta 12 5.15
Localização
No eixo do ramal 84 36.05
Fora do eixo do ramal 149 63.95
Frestas na parede
Não 175 75.11
Sim 58 24.89
Portas e janelas
Não 3 1.29
Sim 230 98.71
Uso de telas
Não 181 77.68
Sim 52 22.32
Borrifação intradomiciliar
Não 126 54.08
Sim 107 45.92
Comunidade
Ipiranga 103 44.21
Brasileirinho 81 34.76
Puraquequara 49 21.03

51
Uso de mosquiteiro na noite passada
Não 135 57.94
Sim 98 42.06
Uso de MILD na noite passada
Não 95 40.77
Sim 138 59.23
Número de mosquitos
< Mediana 112 48.07
> Mediana 121 51.93
Número de Culex ingurgitados
< Média 177 75.97
> Média 56 24.03
Mosquitos Plasmodium+
Não 218 93.56
Sim 15 6.44
Febre
Não 219 93.99
Sim 14 6.01
Uso de Antimaláricos
Não 203 87.12
Sim 30,0 12.88
4.2 Frequência e diversidade dos Culicídeos capturados e análise da eficiência
dos métodos de captura utilizados.
A frequência de Culex quinquefasciatus foi notavelmente maior que as outras
espécies e gêneros capturados em ambos os métodos de captura (94.2%),
demonstrando sua abundância no ambiente. O percentual de pools de mosquitos
positivos para Plasmodium sp. também foi maior nessa espécie, totalizando 3.4%.
Outros mosquitos do gênero Culex apresentaram frequência total de 2.4%, porém
concentrados no método de aspiração. De um modo geral, embora o número de
mosquitos capturados tenha sido maior com a captura por BG-Sentinel®, o
percentual de pools positivos foi maior com a aspiração. Entretanto, pode-se
considerar que os dois métodos se mostraram eficientes para a detecção de pools

52
positivos para Plasmodium sp.. A frequência de outros culicídeos, cuja identificação
em nível de gênero não foi possível, foi de 2.2% (Tabela 5).
Tabela 5. Frequências das espécies/gênero coletadas segundo o método de captura e o percentual de infecção de pools de mosquitos infectados naturalmente.
Espécies/gênero Aspiração BG-Sentinel® Total
n % % Pool
+ N %
% Pool +
N % % Pool
+
Cx. quinquefasciatus 1038 83.6 3.6 11561 95.3 3.3 12599 94.2 3.4
An. darlingi 12 1.0 0.0 122 1.0 1.9 134 1.0 1.5
Culex sp. 98 7.9 5.9 225 1.9 2.1 323 2.4 3.7
Culex (Anoedioporpa)
0 0.0 -- 1 0.0 -- 1 0.0 --
Culex (Mel.) spisseps 1 0.1 0.0 0 0.0 -- 1 0.0 0.0
Anopheles sp. 1 0.1 0.0 6 0,0 0.0 7 0.1 0.0
Uranotaenia sp. 0 0.0 -- 4 0.0 -- 4 0.0 --
Psorophora sp. 2 0.2 0.0 2 0.0 -- 4 0.0 0.0
Limatus sp. 0 0.0 -- 3 0.0 -- 3 0.0 --
Aedeomyia sp. 0 0.0 -- 2 0.0 -- 2 0.0 --
Coquillettidia sp. 0 0.0 -- 1 0.0 -- 1 0.0 --
Psorophora albigenu 1 0.1 0.0 0 0.0 -- 1 0.0 0.0
Toxorhynchitinae 0 0.0 -- 1 0.0 0.0 1 0.0 0.0
Outros culicídeos 89 7.2 0.0 204 1.7 0.0 293 2.2 0.0
Total 1242 -- 3.2 12132 -- 2.7 13374 -- 2.9
4.3 Padronização das técnicas de extração de DNA e de q-PCR para a detecção
de Plasmodium sp. em culicídeos não anofelinos.
Na busca pela definição de quais mosquitos da coleta de campo deveriam ser
usados para os ensaios de q-PCR gênero-específico, foram realizadas infecções
experimentais utilizando Cx. quinquefasciatus de uma colônia pré-estabelecida,
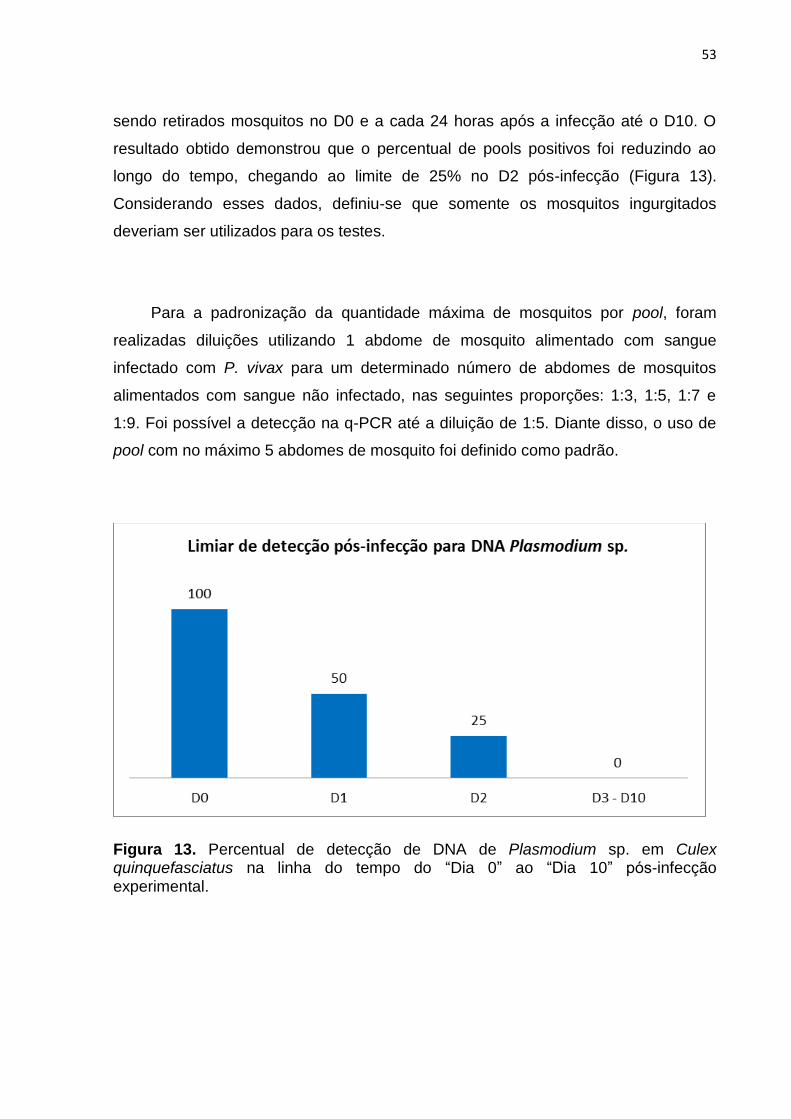
53
sendo retirados mosquitos no D0 e a cada 24 horas após a infecção até o D10. O
resultado obtido demonstrou que o percentual de pools positivos foi reduzindo ao
longo do tempo, chegando ao limite de 25% no D2 pós-infecção (Figura 13).
Considerando esses dados, definiu-se que somente os mosquitos ingurgitados
deveriam ser utilizados para os testes.
Para a padronização da quantidade máxima de mosquitos por pool, foram
realizadas diluições utilizando 1 abdome de mosquito alimentado com sangue
infectado com P. vivax para um determinado número de abdomes de mosquitos
alimentados com sangue não infectado, nas seguintes proporções: 1:3, 1:5, 1:7 e
1:9. Foi possível a detecção na q-PCR até a diluição de 1:5. Diante disso, o uso de
pool com no máximo 5 abdomes de mosquito foi definido como padrão.
Figura 13. Percentual de detecção de DNA de Plasmodium sp. em Culex quinquefasciatus na linha do tempo do “Dia 0” ao “Dia 10” pós-infecção experimental.

54
4.4 Concordância, sensibilidade, especificidade e valores preditivos da
xenovilância.
O teste diagnóstico englobando toda a área do estudo apresentou sensibilidade
de 14.58%, valor preditivo positivo-VPP de 46.67% e valor preditivo negativo-VPN de
81.19%. O teste mostrou desempenho semelhante quando se estratificou a análise
apenas para os assintomáticos, apresentando maior sensibilidade (17.50%), com
VPP de 46.67% e VPN 84.86%, com prevalência de 17.17% (Tabela 7).
Quando realizado de forma segmentada para cada comunidade estudada, o
teste mostrou que na comunidade com maior prevalência de infecção por
Plasmodium sp. em humanos (Puraquequara), o mesmo apresentou maior
sensibilidade (25.00%) com VPP de 60.00% e VPN de 53.85% (Tabela 7).

55
Tabela 6. Resultado do teste diagnóstico da xenovigilância utilizando mosquitos, tendo como padrão ouro a q-PCR em humanos e em humanos assintomáticos.
Comunidade Mês da coleta Prevalência (IC 95%)
Sensibilidade (IC 95%)
Especificidade (IC 95%)
VPP (IC 95%) VPN (IC 95%)
Puraquequara Janeiro 48.98
(34.98-62.98) 25.00
(12.88-37.12) 84.00
(73.74-94.26) 60.00
(46.28-73.72) 53.85
(39.89-67.80)
Ipiranga Fevereiro 16.50
(9.34-23.67) 5.88
(1.34-10.43) 98.84
(96.77-100.91) 50
(40.34-59.66) 84.16
(77.11-91.21)
Brasileirinho Março 8.64
(2.52-14.76) 0.00
(0.00-0.00) 95.95
(91.65-100.24) 0.00
(0.00-0.00) 91.03
(84.80-97.25)
Total Janeiro /Março 20.60
(15.41-25.79) 14.58
(10.05-19.12) 95.68
(93.06-98.29) 46.67
(40.26-53.07) 81.19
(76.18-86.21)
Total Assintomático
Janeiro /Março 17.17
(12.33-22.01) 17.50
(12.62-22.38) 95.85
(93.30-98.41) 46.67
(40.26-53.07) 84.86
(80.26-89.46)

56
4.5 Avaliação da associação entre a positividade domiciliar de infecções em
humanos e a presença de parasitas em Culicídeos não anofelinos.
Na análise de regressão logística univariada, a maioria das variáveis foi
selecionada para o modelo multivariado hierárquico (p<0.2), com exceção de “portas
e janelas”, “uso de telas”, “Comunidade Brasileirinho”, “uso de mosquiteiros
impregnados com inseticida de longa duração (MILD)”, “número de mosquitos” e
“número de mosquitos ingurgitados” (Tabela 6).
No modelo de regressão logística multivariada hierarquizado, no bloco de
variáveis distais, somente as variáveis “presença de frestas nas paredes” e “morar
na comunidade Puraquequara” apresentaram associação positiva significativa com
Odds Ratio de 4.41 e 5.29 respectivamente, ambos com p˂0.05. No bloco das
variáveis proximais, as variáveis “mosquitos positivo para Plasmodium”, “ter febre” e
“uso de antimalárico”, tiveram associação positiva significativa com Odds Ratio de
3.09, 4.85 e 3.09 respectivamente, todos com p˂0.05 (Tabela 6).
No modelo ajustado final, se mantiveram com forte associação positiva as
variáveis “frestas nas paredes”, “morar na comunidade Puraquequara” e “ter febre”,
independentemente dos demais fatores, todos com p˂0.05 (Tabela 6).

57
Tabela 7. Análise de Regressão logística univariada e multivariada hierarquizada das variáveis estudadas. Avaliação da associação com a presença de humanos com q-PCR positiva para Plasmodium sp. nos imóveis estudados (Pseudo R2=0.2775).
Positivo p/
Plasmodium %
Negativo p/ Plasmodium
% OR Bruto (IC 95%)
p OR Ajustado
(IC 95%) - Blocos p
OR Ajustado (CI 95%) -
Modelo Final p
Bloco Distal
Tipo de casa
Alvenaria 17 12.78 116 87.2 1 -- 1 -- 1 --
Madeira ou similar 31 31.0 69 69.0 3.07
(1.58-5.95) <0.001
1.58 (0.71-3.53)
0.263 1.60
(0.68-3.80) 0.284
Tipo de parede
Completa 42 19.0 179 81.0 1 -- 1 -- 1 --
Incompleta 6 50.0 6 50.0 4.26
(1.31-13.90) <0.05
2.18 (0.45-10.59)
0.337 2.14
(0.45-10.19) 0.338
Localização
No eixo do ramal 9 10.7 75 89.3 1 -- 1 -- 1 --
Fora do eixo 39 26.2 110 73.8 2.95
(1.35-6.46) <0.05
1.24 (0.48-3.17)
0.659 1.07
(0.40-2.88) 0.889
Frestas na parede
Não 25 14.3 150 85.7 1 -- 1 -- 1 --
Sim 23 39.7 35 60.3 3.94
(2.00-7.75) <0.001
4.41 (1.88-10.32)
<0.001 4.4
(1.77-10.91) <0.001
Portas e janelas
Não 2 66.7 1 33.3 1 -- 1 -- 1 --
Sim 46 20.0 184 80.0 8
(0.71-90.16) 0.092
1.89 (0.10-34.47)
0.669 2.5
(0.13-47.08) 0.541
Uso de telas

58
Não 35 19.3 146 80.7 1 -- 1 -- 1 --
Sim 13 25.0 39 75.0 1.39
(0.67-2.88) 0.375
1.31 (0.47-3.68)
0.603 1.33
(0.46-3.80) 0.597
Borrifação Residual
Não 20 15.9 106 84.1 1 -- 1 -- 1 --
Sim 28 26.2 79 73.8 1.88
(0.99-3.58) <0.05
0.76 (0.24-2.47)
0.648 0.58
(0.17-1.94) 0.373
Comunidade
Ipiranga 17 16.5 86 83.5 1 -- 1 -- 1 --
Brasileirinho 7 8.6 74 91.4 0.48
(0.19-1.22) 0.122
0.53 (0.15-1.84)
0.320 0.92
(0.24-3.46) 0.895
Puraquequara 24 49.0 25 51.0 4.86
(2.26-10.43) <0.001
5.29 (1.32-21.18)
<0.05 7.79
(1.65-36.77) <0.001
Uso de mosquiteiro na noite passada
Não 21 15.6 114 84.4 1 -- 1 -- 1 --
Sim 27 27.6 71 72.4 2.06
(1.09-3.93) <0.05
1.32 (0.46-3.78)
0.609 1.29
(0.42-3.98) 0.657
Uso de MILD na noite passada
Não 14 14.7 81 85.3 1 -- 1 -- 1 --
Sim 34 24.6 104 75.4 1.89
(0.95-3.76) 0.069
1.2 (0.50-2.87)
0.676 0.9
(0.35-2.30) 0.828
Bloco Proximal
Número de mosquitos
< Mediana 19 17.0 93 83.0 1 -- 1 -- 1 --
> Mediana 29 24.0 92 76.0 1.54
(0.81-2.94) 0.188
1.21 (0.37-2.16)
0.652 1.25
(0.46-3.34) 0.660
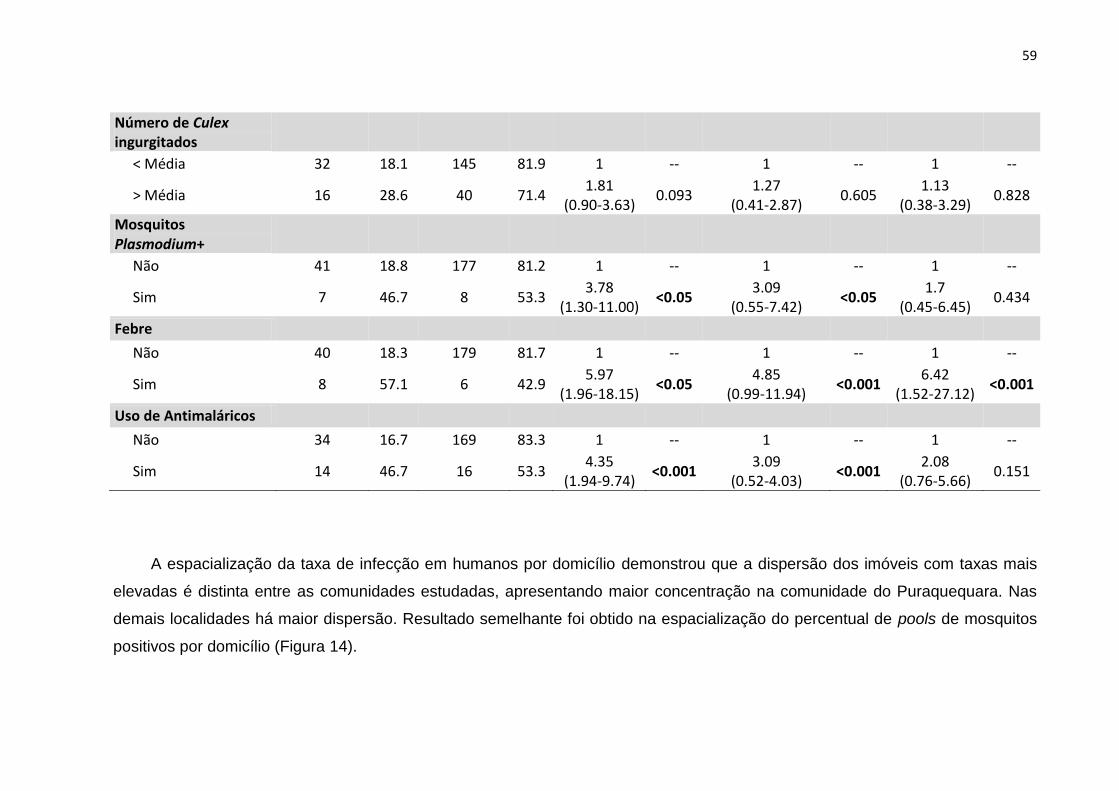
59
Número de Culex ingurgitados
< Média 32 18.1 145 81.9 1 -- 1 -- 1 --
> Média 16 28.6 40 71.4 1.81
(0.90-3.63) 0.093
1.27 (0.41-2.87)
0.605 1.13
(0.38-3.29) 0.828
Mosquitos Plasmodium+
Não 41 18.8 177 81.2 1 -- 1 -- 1 --
Sim 7 46.7 8 53.3 3.78
(1.30-11.00) <0.05
3.09 (0.55-7.42)
<0.05 1.7
(0.45-6.45) 0.434
Febre
Não 40 18.3 179 81.7 1 -- 1 -- 1 --
Sim 8 57.1 6 42.9 5.97
(1.96-18.15) <0.05
4.85 (0.99-11.94)
<0.001 6.42
(1.52-27.12) <0.001
Uso de Antimaláricos
Não 34 16.7 169 83.3 1 -- 1 -- 1 --
Sim 14 46.7 16 53.3 4.35
(1.94-9.74) <0.001
3.09 (0.52-4.03)
<0.001 2.08
(0.76-5.66) 0.151
A espacialização da taxa de infecção em humanos por domicílio demonstrou que a dispersão dos imóveis com taxas mais
elevadas é distinta entre as comunidades estudadas, apresentando maior concentração na comunidade do Puraquequara. Nas
demais localidades há maior dispersão. Resultado semelhante foi obtido na espacialização do percentual de pools de mosquitos
positivos por domicílio (Figura 14).
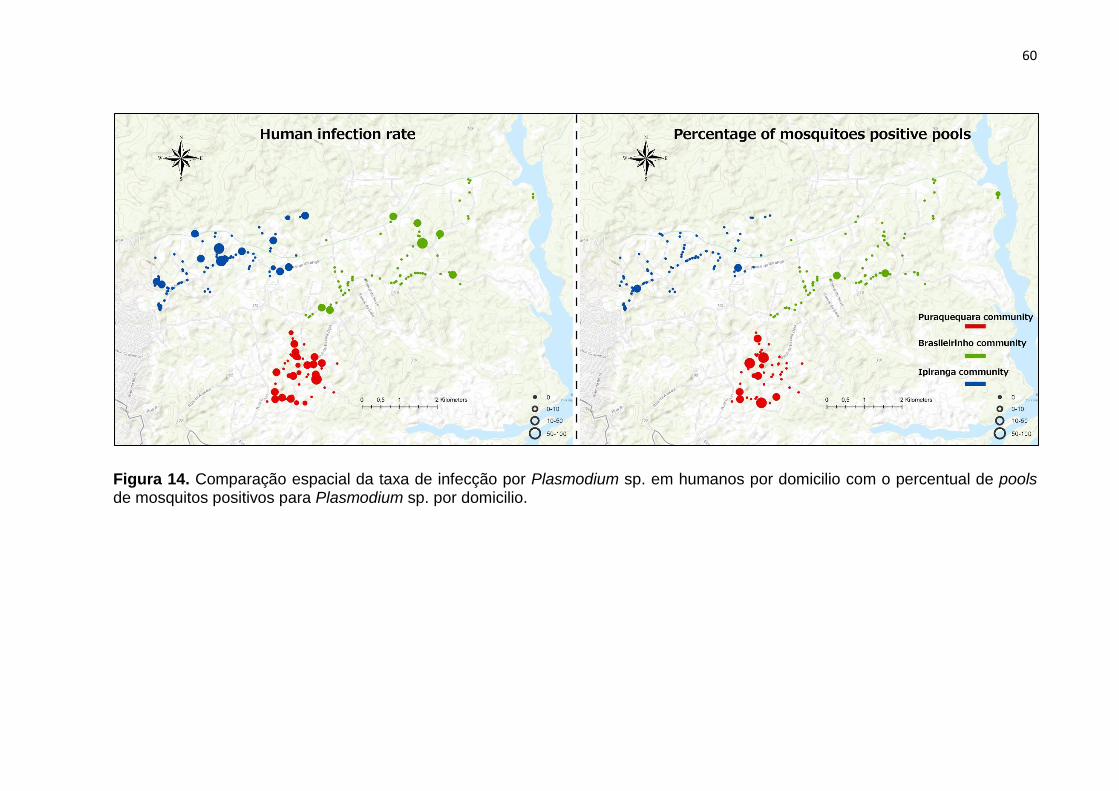
60
Figura 14. Comparação espacial da taxa de infecção por Plasmodium sp. em humanos por domicilio com o percentual de pools de mosquitos positivos para Plasmodium sp. por domicilio.

61
5. DISCUSSÃO
5.1 Características ambientais da amostra
Ao analisar as variáveis ambientais, denota-se que, embora a amostra
apresentasse condições relativamente boas de habitação, ainda estão presentes
condições que favorecem o contato homem-vetor, quer seja pela quase ausência de
algumas medidas protetivas, como no caso de vedação de frestas nas paredes e de
telas nas portas e janelas, ou pela baixa adesão a estas, como no caso do uso de
mosquiteiros e MILD’s, pois a área do estudo foi recentemente contemplada com a
implantação deste último, pelo Ministério da Saúde, no entanto este apresentou uma
frequência de uso de pouco mais de 50%, o que é incompatível para o impacto
pretendido com esta medida. De forma semelhante, a cobertura de borrifção
intradomiciliar observada não chega a 50% da amostra, não oferecendo a proteção
coletiva desejada, considerando que a maioria dos domicílios estão localizados fora
do eixo do ramal, onde o contato com o vetores provavelmente é maior, devido a
proximidade com a floresta. Essa realidade reflete as condições sociocultural e
econômica presentes em áreas periféricas de grandes centros na Amazônia,
reconhecidas como fatores importantes a serem considerados no controle da malária
(6,33,34).
5.2 Técnicas de coleta
O custo com as armadilhas BG Sentinel® foi somente com a aquisição dos
atraentes e de extensões elétricas, já que as mesmas haviam sido adquiridas por um
projeto anterior, sendo realizada somente sua manutenção e reparo por um membro
da equipe, sem custo adicional.
Desconsiderando o tempo de captura, pode-se creditar como vantagem
armadilhas BG Sentinel® a quantidade de mosquitos capturados, embora a grande
maioria destes não se encontre ingurgitados. Já as dificuldades dizem respeito à

62
exigência da logística, pois elas são volumosas, demandando veículos com maior
capacidade de carga/volume e uma segunda visita domiciliar para sua retirada, o
que ocasionou alguns desencontros com os moradores. Outra desvantagem é o
ressecamento dos mosquitos capturados logo no início da coleta.
Quanto aos aspiradores, também já haviam sido comprados anteriormente por
outro projeto, mas só agora foram usados, por isso o custo foi unicamente com as
baterias. As principais vantagens observadas foram:
A rapidez e praticidade da técnica permitindo que, com apenas um
técnico, se aspirasse até 15 casas numa manhã;
A portabilidade do equipamento com baixa exigência de logística,
possibilitando a realização das visitas até mesmo de motocicleta;
A necessidade de uma única visita para a captura;
Maior proporção de mosquitos ingurgitados entre os capturados.
Uma desvantagem do aspirador foi o fato que, nas casas sem mobília e com um
único cômodo, geralmente a captura era negativa, provavelmente pela ausência de
possíveis esconderijos para os mosquitos.
De modo geral, o número de mosquitos capturados por imóvel foi elevado, mas
com grande variação, estando a maioria acima da mediana. Em relação ao número
de mosquitos ingurgitados, houve menor variação e somente 24% dos imóveis
apresentaram números acima da média. Este fato reflete as diferenças das técnicas
de captura empregadas no estudo, já que a BG-Sentinel® de certa forma compete
com o homem na atração destes, coletando os mosquitos antes do repasto
sanguíneo (178). Os imóveis que apresentaram mosquitos carreando DNA de
Plasmodium sp. representou 6,4% da amostra, valor semelhante a frequência da
presença de pessoas com febre na casa (6%).
A diferença entre os métodos de captura, quanto ao número total de mosquitos e
o número de fêmeas ingurgitadas, mostrou-se acentuada, tendo a BG-Sentinel®

63
com medianas superiores a aspiração, mas a aspiração apresentou maior percentual
de ingurgitadas. Inicialmente poderíamos avaliar que a BG-Sentinel® seria a melhor
escolha, mas embora a aspiração tenha capturado menos mosquitos, apresentou
maior proporção de ingurgitadas. Considerando que se deseja um método
alternativo a HLC (168–170), sem os mesmos problemas éticos envolvidos
(166,167), e com boa capacidade de captura de culicídeos, possíveis carreadores de
Plasmodium sp.. O ideal seria, onde houver fornecimento regular de energia elétrica,
utilizar as duas metodologias em conjunto por seus aspectos econômicos e técnicos,
caso contrário a aspiração seria o método de primeira escolha.
É relevante considerar, além da viabilidade técnica e ética, a viabilidade
econômica de uma ferramenta, e neste caso tanto o custo de materiais, quanto os
rscursos humanos envolvidos são relativamente baixos comparados a HLC que
demanda um auxiliar de entomologia para cada ponto de captura por dia, enquanto
que neste método conjunto de captura utilizado em nosso estudo, com apenas dois
agentes é possível realizar a captura em até 15 pontos (casas) por dia,
proporcionando coberturas mais amplas no serviço de vigilância entomológica com
menor custo operacional.
5.3 Fauna de Culicídeos
A presença abundante de Cx. quinquefasciatus e outros mosquitos do gênero
Culex na amostra é compatível com um estudo realizado em outra área periurbana
da cidade de Manaus (84) e confirma sua dispersão demonstrada em diversos
estudos realizados na Amazônia Brasileira (85–91), revestindo de importância sua
participação na interação homem-mosquito-patógeno na região.
Ainda que os métodos utilizados no estudo não sejam os mais eficientes para a
captura de Anopheles darlingi, foram capturados 134 espécimes que correspondeu a
uma frequência de 1%, com taxa de infectividade de 1,5%.

64
5.4 Infectividade
A proporção de Cx. quinquefasciatus carreando DNA de Palsmodium sp. foi de
3,4% de positividade nos pools testados desta espécie. Embora, neste caso, não
podemos falar de infectividade, posto que a espécie não é vetor natural da doença.
Esse percentual de carreadores é compatível com as taxas de infectividade
encontradas em estudos com Anopheles spp..
Dentre estes estão os trabalhos de Martins-Campos et al, que realizou estudo
em uma área rural de um município do entorno de Manaus com elevada transmissão
de malária e obteve 7 pools positivos no universo de 3.189 mosquitos capturados
com HLC, formando pools com 10 mosquitos testados em ensaios de q-PCR-
Multiplex, resultando numa taxa de 2,2% de infectividade (134); de Sriwichai et al,
que coletou mosquitos com armadilhas luminosa CDC em diversas aldeias com
transmissão ativa de malária na Tailândia, esses mosquitos foram dissecados e
analisados individualmente por teste ELISA para detecção de infecções por
parasitos da malária, apresentando taxas de infectividade entre 0,37 e 1,74% nas
espécies de Anopheles spp. coletadas (179).
Outros trabalhos mais antigos, na região Amazônica, apresentaram taxa de
infectividade entre 0,4 a 3,41% para as diversas espécies de Anopheles spp.
estudadas (180–182). Encontramos percentuais superiores a estes, somente em um
estudo realizado no município de Anajás no Pará, que encontrou taxa de
infectividade de 6%, mas isso foi durante uma grande epidemia, onde se pode
identificar esporozoítos em de glândulas salivares dos mosquitos, o que é
relativamente raro de se encontrar em uma amostra relativamente pequena (183).
A vigilância entomológica utilizada para avaliar o impacto do uso de medidas de
controle em larga escala em Uganda apresentou taxas de infectividade em
mosquitos variando de 3,2%, antes da ação, e 1,8% no período pós-intervenção
(184). Em Angola, um estudo similar apresentou variação entre 0,9% e 3,41% (185).

65
Vale ressaltar que, diferentemente dos trabalhos supracitados, a amostra do
presente estudo foi realizada em um período sazonal de menor prevalência da
malária, em uma única rodada de coletas. Assim sendo, a informação gerada pode
ser importante para estimar a circulação do patógeno na área, mesmo em períodos
de baixa transmissão, corroborando para a possibilidade do uso desta ferramenta
como um novo indicador malariométrico.
5.5 Padronização das técnicas
O resultado obtido com o seguimento da infecção experimental de P. vivax em
Cx. quinquefasciatus, demonstrando o decaimento no percentual de detecção de
DNA desse Plasmodium, com detecção até 48 horas pós-infecção, são semelhantes
aos resultados apresentados por Knöckel et al, que observou por microscopia
confocal o desenvolvimento de P. falciparum em Cx. quinquefasciatus até 30 horas
depois da infecção experimental (123). Autoriza-nos a aplicar o conceito de “seringa
biológica” (124) não somente para os vetores naturais da malária, mas também para
espécies mais abundantes, com ampla distribuição e com hábito
antropofágico/endofágico, que é o caso de Cx. quinquefasciatus. Confirmando a
viabilidade metodológica para o uso da ferramenta proposta.
O limite de 5 (cinco) abdomes de mosquitos por pool demonstra a viabilidade
operacional da técnica, pois o simples corte do abdome do mosquito não exige
técnicas mais elaboradas de dissecação e com a formação destes pools há uma
aglutinação razoável da amostra, proporcionando a economia de materiais utilizados
na extração de DNA e nos ensaios de q-PCR, bem como a otimização do tempo
operacional (135), fundamentais para a viabilidade de qualquer ferramenta de
vigilância. A padronização dos ensaios de q-PCR utilizados já demonstrou boa
capacidade de amplificação mesmo com um baixo número de cópias, mas estudos
recentes apontam para uma possível otimização exponencial dos resultados
encontrados, pois as novas técnicas multiplicam em mais de 10 vezes essa
capacidade (137,138).

66
5.6 Associação
Na análise de regressão univariada, das variáveis observadas no estudo, a
maioria apresentou associação com a presença de infecções de Plasmodium spp.
em moradores da casa. Duas dessas associações que foram significativas parecem
estranhas, a “Borrifação Residual” e o “Uso de mosquiteiros”; no caso da primeira
existe uma explicação plausível, pois essa medida não é realizada de maneira
uniforme em toda a área, mas sim de forma seletiva nas micro-áreas com maior
prevalência da malária, daí a aparente associação.
No modelo multivariado, somente se mantiveram com associação significativa as
seguintes variáveis: “Frestas na parede”, “Morar na comunidade Puraquequara” e
“Febre” independentemente dos demais fatores.
Mas quando testamos as mesmas variáreis no modelo de regressão logística
multivariada hierarquizada, onde se separam as variáveis em blocos, pelo nível
racional-biológico de sua relação com a variável dependente, a fim de evitar uma
abordagem inteiramente baseada em associações estatísticas sem qualquer base
conceitual das inter-relações entre os fatores, já que nem todas as variáveis
explicativas pertencessem ao mesmo nível hierárquico (186). O resultado no bloco
proximal, aquele com possível relação no ciclo biológico do patógeno, apresentou
com associação significativa “Febre”, “Uso de Antimaláricos” e “Mosquitos
Plasmodium+”, este último com Odds Ratio de 3.09 e p˂0.05. Em relação ao bloco
das variáveis distais, aquelas que dizem respeito aos fatores ambientais que podem
estar indiretamente envolvidas na cadeia de transmissão da doença, se mantiveram
com forte associação somente “Frestas na parede” e “Morar na comunidade
Puraquequara”.

67
5.7 Análise espacial
A inspeção espacial comparativa da taxa de infecção por Plasmodium sp. em
humanos com o percentual de pools de mosquitos carreando Plasmodium sp.,
demonstra que onde a prevalência da infecção foi mais elevada, as taxas foram mais
semelhantes, embora nem sempre coincidentes. Importante registrar que o período
de coleta foi de janeiro a março de 2014, na seguinte sequência: Puraquequara,
Ipiranga e Brasileirinho, este período sazonal caracteriza-se pelo declínio natural da
transmissão da malária nesta área. Outro fator a ser considerado, é a distribuição
espacial da amostra que não foi da totalidade das casas das comunidades, deixando
lacunas espaciais, que somadas à capacidade de voo dos mosquitos que em geral
não é inferior a 100 metros (187–192). Podem ter limitado o potencial da ferramenta
no presente estudo.
5.8 Acurácia
Essa informação espacial é corroborada pelos resultados do teste diagnóstico da
xenovigilância utilizando mosquitos, tendo como padrão ouro a q-PCR em humanos
que indicou uma maior sensibilidade onde a prevalência foi mais elevada, chegando
a 25.00% de sensibilidade com um VPP de 60.00% na comunidade Puraquequara.
Considerando a importância dos casos assintomáticos submicroscópicos na
manutenção da transmissão da malária mesmo em períodos de baixa prevalência,
assim como sua detecção para a eliminação da malária (113–115). E que a amostra
da população humana era oriunda de uma coorte, que identificou com precisão os
casos assintomáticos submicroscópicos que não desenvolveram nenhuma
sintomatologia no mês da coleta. Comparamos a xenovigilância exclusivamente com
o diagnóstico desses assintomáticos observando uma sensibilidade de 17.50%, VPP
46.67% e VPN 84.86%. Essa sensibilidade é maior que a encontrada na
comparação com o diagnostico geral (sintomáticos+assintomáticos), mesmo frente a

68
uma menor prevalência dos assintomáticos. Este resultado, talvez seja a informação
mais importante deste trabalho, seguindo a ideia de xenovigilância através da
detecção da circulação de patógenos em mosquitos não vetores específicos (125).
5.9 Viabilidade
Embora neste estudo, tivemos o trabalho de identificar e separar os mosquitos
por espécie/gênero, para demostrar a real possibilidade de encontrar DNA de
Plasmodium sp. em outros culicídeos além de Anopheles spp., as vantagens
operacionais desta ferramenta de xenovigilância da malária está justamente em
poder prescindir destas etapas de identificação, que demandam trabalho árduo e
mão de obra especializada, e poder separar os mosquitos apenas por casa e por
pools de 1 a 5 espécimes.
Também outro aspecto operacional importante é a baixa exigência logística da
coleta com alta produtividade e sem as implicações éticas que envolvem atualmente
a HLC. Assim como a utilização de um método simples e barato de extração de DNA
utilizando Chelex® 100, que tem rendimento superior a outras técnicas de extração,
que somados a padronização dos ensaios de q-PCR com sensibilidade suficiente
para amplificar DNA do parasito mesmo com baixo número de cópias.
Além das vantagens operacionais, a xenovigilância da malária mostrou uma taxa
de carreamento de Plasmodium sp. compativel com as taxas de infectividade
encontradas nos estudos de campo com Anopheles spp., e no teste diagnóstico com
os assintomáticos apresentou sensibilidade razoável de 17% com VPP de 46.67%.
Por estes resultados podemos concluir que essa ferramenta é bastante promissora,
e pela simplicidade e baixo custo operacional podemos acreditar que sua
operacionalização em uma possível parceria dos programas de controle com as
entidades de pesquisa seria perfeitamente viável.

69
A que se considerar também, a perspectiva futura do uso desta ferramenta para
a vigilância integrada e simultânea de outros patógenos transmitidos por vetores,
quando houver sobreposição de áreas de transmissão com a malária, aproveitando
o mesmo material genético extraído e com o uso de outras técnicas moleculares que
sejam apropriadas para a detecção destes.

70
6. CONCLUSÃO
A frequência e a diversidade das espécies de culicídeos capturados foram
compatíveis com os resultados encontrados na literatura cientifica.
A padronização das técnicas de extração de DNA e do ensaio de q-PCR neste
trabalho se mostrou viável para a detecção de Plasmodium sp. em culicídeos não
anofelinos, desde de que estejam ingurgitados.
O teste diagnóstico realizado com os culicídeos não anofelinos apresentou
sensibilidade e VPP razoáveis em relação ao diagnóstico de infecção em humanos.
E a taxa de encontro de DNA de Plasmodium sp. nestes culicídeos foi compatível
com aquelas encontradas nos Anopheles sp., vetores naturais da malária.
A análise de regressão logística múltipla hierarquizada demonstrou associação
positiva entre a ocorrência de infecções domiciliar em humanos e a presença de
material genético de Plasmodium sp. em culicídeos não anofelinos no bloco
proximal.

71
7. REFERÊNCIAS BIBLIOGRÁFICAS
1. World Health Organization. WHO | World Malaria Report 2015. WHO Libr Cat Data. World Health Organization; 2015;243.
2. Gething PW, Patil AP, Smith DL, Guerra CA, Elyazar IRF, Johnston GL, et al. A new world malaria map: Plasmodium falciparum endemicity in 2010. Malar J . BioMed Central; 2011 Jan 20;10(1):378.
3. Gething PW, Elyazar IRF, Moyes CL, Smith DL, Battle KE, Guerra CA, et al. A long neglected world malaria map: Plasmodium vivax endemicity in 2010. PLoS Negl Trop Dis. Public Library of Science; 2012 Jan 9;6(9):e1814.
4. SVS-MS. Situação epidemiológica da malária no Brasil, 2000 a 2011 [Internet]. Boletim Epidemiológico. 2013. p. 16.
5. Oliveira-Ferreira J, Lacerda MVG, Brasil P, Ladislau JLB, Tauil PL, Daniel-Ribeiro CT. Malaria in Brazil: an overview. Malar J. 2010 Jan;9:115.
6. Sampaio VS, Siqueira AM, Alecrim M das GC, Mourão MPG, Marchesini PB, Albuquerque BC, et al. Malaria in the State of Amazonas: a typical Brazilian tropical disease influenced by waves of economic development. Rev Soc Bras Med Trop. SBMT; 2015 Jan;48 Suppl 1:4–11.
7. Vitor-Silva S, Reyes-Lecca RC, Pinheiro TRA, Lacerda MVG. Malaria is associated with poor school performance in an endemic area of the Brazilian Amazon. Malar J. 2009 Jan;8:230.
8. Saraiva M das GG, Amorim RDS, Moura MAS, Martinez-Espinosa FE, Barbosa M das GV. [Urban expansion and spatial distribution of malaria in the municipality of Manaus, State of Amazonas]. Rev Soc Bras Med Trop. 2009;42(5):515–22.
9. Pimenta PF, Orfano AS, Bahia AC, Duarte AP, Ríos-Velásquez CM, Melo FF, et al. An overview of malaria transmission from the perspective of Amazon Anopheles vectors. Mem Inst Oswaldo Cruz. 2015 Feb 13;110(1):23–47.
10. Aly ASI, Vaughan AM, Kappe SHI. Malaria parasite development in the mosquito and infection of the mammalian host. Annu Rev Microbiol. 2009 Jan;63:195–221.
11. Mueller I, Galinski MR, Baird JK, Carlton JM, Kochar DK, Alonso PL, et al. Key gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect Dis. Elsevier; 2009 Sep 1;9(9):555–66.
12. Cox FE. History of the discovery of the malaria parasites and their vectors. Parasit Vectors. 2010 Jan;3(1):5.
13. Wipasa J, Elliott S, Xu H, Good MF. Immunity to asexual blood stage malaria and vaccine approaches. Immunol Cell Biol. Nature Publishing Group; 2002 Oct 1;80(5):401–14.
14. Noviyanti R, Coutrier F, Utami RAS, Trimarsanto H, Tirta YK, Trianty L, et al. Contrasting Transmission Dynamics of Co-endemic Plasmodium vivax and P. falciparum: Implications for Malaria Control and Elimination. PLoS Negl Trop Dis. Public Library of Science; 2015 May 5;9(5):e0003739.

72
15. Guerra CA, Howes RE, Patil AP, Gething PW, Van Boeckel TP, Temperley WH, et al. The international limits and population at risk of Plasmodium vivax transmission in 2009. PLoS Negl Trop Dis. Public Library of Science; 2010 Jan 8;4(8):e774.
16. Conn JE, Norris DE, Donnelly MJ, Beebe NW, Burkot TR, Coulibaly MB, et al. Entomological Monitoring and Evaluation: Diverse Transmission Settings of ICEMR Projects Will Require Local and Regional Malaria Elimination Strategies. Am J Trop Med Hyg. 2015 Sep;93(3 Suppl):28–41.
17. Moss WJ, Dorsey G, Mueller I, Laufer MK, Krogstad DJ, Vinetz JM, et al. Malaria Epidemiology and Control Within the International Centers of Excellence for Malaria Research. Am J Trop Med Hyg. 2015 Sep;93(3 Suppl):5–15.
18. Dowdle WR. The principles of disease elimination and eradication. Bull World Health Organ. 1998 Jan;76 Suppl 2:22–5.
19. Hay SI, Smith DL, Snow RW. Measuring malaria endemicity from intense to interrupted transmission. Lancet Infect Dis. 2008 Jun;8(6):369–78.
20. World Health Organization. WHO | Disease surveillance for malaria elimination: operational manual. WHO Libr Cat Data. World Health Organization; 2012;48.
21. Hamainza B, Killeen GF, Kamuliwo M, Bennett A, Yukich JO. Comparison of a mobile phone-based malaria reporting system with source participant register data for capturing spatial and temporal trends in epidemiological indicators of malaria transmission collected by community health workers in rural Zambia. Malar J. 2014 Jan;13(1):489.
22. Okorie PN. Application of geographical information system for lymphatic filariasis and malaria control in Nigeria. Afr J Med Med Sci. 2014 Jun;43(2):161–6.
23. Tatem AJ, Huang Z, Narib C, Kumar U, Kandula D, Pindolia DK, et al. Integrating rapid risk mapping and mobile phone call record data for strategic malaria elimination planning. Malar J. 2014 Jan;13:52.
24. Mosha JF, Sturrock HJW, Greenwood B, Sutherland CJ, Gadalla NB, Atwal S, et al. Hot spot or not: a comparison of spatial statistical methods to predict prospective malaria infections. Malar J. 2014 Jan;13:53.
25. Okell LC, Bousema T, Griffin JT, Ouédraogo AL, Ghani AC, Drakeley CJ. Factors determining the occurrence of submicroscopic malaria infections and their relevance for control. Nat Commun. 2012 Jan;3:1237.
26. Cheng Q, Cunningham J, Gatton ML. Systematic Review of Sub-microscopic P. vivax Infections: Prevalence and Determining Factors. PLoS Negl Trop Dis. 2015 Jan;9(1):e3413.
27. Elliott SR, Fowkes FJI, Richards JS, Reiling L, Drew DR, Beeson JG. Research priorities for the development and implementation of serological tools for malaria surveillance. F1000Prime Rep. 2014 Jan;6:100.
28. Yman V, White MT, Rono J, Arcà B, Osier FH, Troye-Blomberg M, et al. Antibody acquisition models: A new tool for serological surveillance of malaria transmission intensity. Sci Rep. Nature Publishing Group; 2016 Jan 5;6:19472.

73
29. Cohen JM, Moonen B, Snow RW, Smith DL. How absolute is zero? An evaluation of historical and current definitions of malaria elimination. Malar J. 2010 Jan;9:213.
30. World Health Organization. WHO | Malaria elimination. A field manual for low and moderate endemic countries. WHO Libr Cat Data. World Health Organization; 2007 .
31. World Health Organization. WHO | From malaria control to malaria elimination: a manual for elimination scenario planning. WHO Libr Cat Data. World Health Organization; 2014.
32. Brady OJ, Godfray HCJ, Tatem AJ, Gething PW, Cohen JM, McKenzie FE, et al. Vectorial capacity and vector control: reconsidering sensitivity to parameters for malaria elimination. Trans R Soc Trop Med Hyg. 2016 Mar;110(2):107–17.
33. Ladislau JL de B, Leal M do C, Tauil PL. Evaluation of the plan for intensification of malaria control actions in the Brazilian Amazon region, based on a descentralization process. Epidemiol serv saúde. 2006;9–20.
34. Ferreira Gonçalves MJ, Alecrim WD. Non-planed urbanization as a contributing factor for malaria incidence in Manaus-Amazonas, Brazil. Rev Salud Publica (Bogota). 2004;6(2):156–66.
35. Tauil PL. Perspectivas de controle de doenças transmitidas por vetores no Brasil. Rev Soc Bras Med Trop. SBMT; 2006 Jun;39(3):275–7.
36. Ogoma SB, Lweitoijera DW, Ngonyani H, Furer B, Russell TL, Mukabana WR, et al. Screening mosquito house entry points as a potential method for integrated control of endophagic filariasis, arbovirus and malaria vectors. PLoS Negl Trop Dis. 2010 Jan;4(8):e773.
37. Muturi EJ, Mbogo CM, Ng’ang'a ZW, Kabiru EW, Mwandawiro C, Novak RJ, et al. Relationship between malaria and filariasis transmission indices in an endemic area along the Kenyan Coast. J Vector Borne Dis. 2006 Jun;43(2):77–83.
38. Manga L. Vector-control synergies, between “roll back malaria” and the Global Programme to Eliminate Lymphatic Filariasis, in the African region. Ann Trop Med Parasitol. 2002 Dec;96 Suppl 2:S129–32.
39. Harbach RE, Besansky NJ. Mosquitoes. Curr Biol. Elsevier; 2014 Jan 6;24(1):R14–5.
40. Consoli RAGB, Oliveira RL de. Principais mosquitos de importância sanitária no Brasil. Rio de Janeiro: EDITORA FIOCRUZ; 1994. 228 p.
41. Cirillo VJ. “Wonders unconceived”: reflections on the birth of medical entomology. Perspect Biol Med. 2011 Jan;54(3):381–98.
42. Briggs DEG. A mosquito’s last supper reminds us not to underestimate the fossil record. Proc Natl Acad Sci U S A. 2013 Nov 12;110(46):18353–4.
43. Greenwalt DE, Goreva YS, Siljeström SM, Rose T, Harbach RE. Hemoglobin-derived porphyrins preserved in a Middle Eocene blood-engorged mosquito. Proc Natl Acad Sci U S A. 2013 Nov 12;110(46):18496–500.
44. Kassim NFA, Webb CE, Russell RC. Culex molestus Forskal (Diptera: Culicidae) in Australia: colonisation, stenogamy, autogeny, oviposition and

74
larval development. Aust J Entomol. 2012 Feb 8;51(1):67–77.
45. Kassim NFA, Webb CE, Russell RC. Is the expression of autogeny by Culex molestus Forskal (Diptera: Culicidae) influenced by larval nutrition or by adult mating, sugar feeding, or blood feeding? J Vector Ecol. 2012 Jun;37(1):162–71.
46. Attardo GM, Hansen IA, Raikhel AS. Nutritional regulation of vitellogenesis in mosquitoes: implications for anautogeny. Insect Biochem Mol Biol. 2005 Jul;35(7):661–75.
47. Arcà B, Struchiner CJ, Pham VM, Sferra G, Lombardo F, Pombi M, et al. Positive selection drives accelerated evolution of mosquito salivary genes associated with blood-feeding. Insect Mol Biol. 2014 Feb;23(1):122–31.
48. Ciota AT, Ehrbar DJ, Matacchiero AC, Van Slyke GA, Kramer LD. The evolution of virulence of West Nile virus in a mosquito vector: implications for arbovirus adaptation and evolution. BMC Evol Biol. 2013 Jan;13:71.
49. Cohuet A, Harris C, Robert V, Fontenille D. Evolutionary forces on Anopheles: what makes a malaria vector? Trends Parasitol. 2010 Mar;26(3):130–6.
50. Galun R. The evolution of purinergic receptors involved in recognition of a blood meal by hematophagous insects. Mem Inst Oswaldo Cruz. Fundação Oswaldo Cruz; 1987;82:5–9.
51. Wu D-D, Wang G-D, Irwin DM, Zhang Y-P. A profound role for the expansion of trypsin-like serine protease family in the evolution of hematophagy in mosquito. Mol Biol Evol. 2009 Oct 1;26(10):2333–41.
52. Ribeiro JM. Blood-feeding arthropods: live syringes or invertebrate pharmacologists? Infect Agents Dis. 1995 Sep;4(3):143–52.
53. Maciel C, Fujita A, Gueroni DI, Ramos AD, Capurro ML, Sá-Nunes A. Evans blue as a simple method to discriminate mosquitoes’ feeding choice on small laboratory animals. PLoS One. Public Library of Science; 2014 Jan;9(10):e110551.
54. Konishi E. Size of blood meals of Aedes albopictus and Culex tritaeniorhynchus (Diptera: Culicidae) feeding on an unrestrained dog infected with Dirofilaria immitis (Spirurida: Filariidae). J Med Entomol. 1989 Nov;26(6):535–8.
55. Albuquerque CM, Cavalcanti VM, Melo MA, Vercosa P, Regis LN, Hurd H. Bloodmeal microfilariae density and the uptake and establishment of Wuchereria bancrofti infections in Culex quinquefasciatus and Aedes aegypti. Memórias do Inst Oswaldo Cruz. Jan;94(5):591–6.
56. Michael E, Ramaiah KD, Hoti SL, Barker G, Paul MR, Yuvaraj J, et al. Quantifying mosquito biting patterns on humans by DNA fingerprinting of bloodmeals. Am J Trop Med Hyg. 2001 Dec;65(6):722–8.
57. Harrington LC, Fleisher A, Ruiz-Moreno D, Vermeylen F, Wa C V, Poulson RL, et al. Heterogeneous feeding patterns of the dengue vector, Aedes aegypti, on individual human hosts in rural Thailand. PLoS Negl Trop Dis. Public Library of Science; 2014 Aug 8;8(8):e3048.
58. Takken W, Verhulst NO. Host Preferences of Blood-Feeding Mosquitoes. Annual Reviews; 2013 Jan 14.

75
59. Chaves LF, Harrington LC, Keogh CL, Nguyen AM, Kitron UD. Blood feeding patterns of mosquitoes: random or structured? Front Zool. 2010 Jan;7:3.
60. Forattini OP, Gomes A de C, Natal D, Kakitani I, Marucci D. [Food preferences and domiciliation of Culicidae mosquitoes in the Ribeira Valley, São Paulo, Brazil, with special reference to Aedes scapularis and Culex (Melanoconion)]. Rev Saude Publica. 1989 Feb;23(1):9–19.
61. McBride CS, Baier F, Omondi AB, Spitzer SA, Lutomiah J, Sang R, et al. Evolution of mosquito preference for humans linked to an odorant receptor. Nature. 2014 Nov 12;515(7526):222–7.
62. Bartholomay LC, Waterhouse RM, Mayhew GF, Campbell CL, Michel K, Zou Z, et al. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science. 2010 Oct 1;330(6000):88–90.
63. Zhou S-S, Zhang S-S, Wang J-J, Zheng X, Huang F, Li W-D, et al. Spatial correlation between malaria cases and water-bodies in Anopheles sinensis dominated areas of Huang-Huai plain, China. Parasit Vectors. 2012 Jan;5(1):106.
64. Perkins TA, Scott TW, Le Menach A, Smith DL. Heterogeneity, mixing, and the spatial scales of mosquito-borne pathogen transmission. Salathé M, editor. PLoS Comput Biol. Public Library of Science; 2013 Jan;9(12):e1003327.
65. Minár J. [Worldwide distribution of mosquitoes in the subfamily Anophelinae and its relation to the transmission of malaria]. Cesk Epidemiol Mikrobiol Imunol. 1987 Jul;36(4):244–51.
66. Severson DW, Behura SK. Mosquito genomics: progress and challenges. Annu Rev Entomol. 2012 Jan;57:143–66.
67. Al-Eryani SMA, Kelly-Hope L, Harbach RE, Briscoe AG, Barnish G, Azazy A, et al. Entomological aspects and the role of human behaviour in malaria transmission in a highland region of the Republic of Yemen. Malar J. 2016 Mar 1;15(1):130.
68. Kent RJ, Thuma PE, Mharakurwa S, Norris DE. Seasonality, blood feeding behavior, and transmission of Plasmodium falciparum by Anopheles arabiensis after an extended drought in southern Zambia. Am J Trop Med Hyg. 2007 Feb;76(2):267–74.
69. Bashar K, Tuno N, Ahmed TU, Howlader AJ. Blood-feeding patterns of Anopheles mosquitoes in a malaria-endemic area of Bangladesh. Parasit Vectors. 2012 Jan;5:39.
70. Pappa V, Reddy M, Overgaard HJ, Abaga S, Caccone A. Estimation of the Human Blood Index in malaria mosquito vectors in Equatorial Guinea after indoor antivector interventions. Am J Trop Med Hyg. 2011 Feb;84(2):298–301.
71. Barros FSM, Honório NA, Arruda ME. Mosquito anthropophily: implications on malaria transmission in the Northern Brazilian Amazon. Neotrop Entomol. Jan;39(6):1039–43.
72. Mzilahowa T, Hastings IM, Molyneux ME, McCall PJ. Entomological indices of malaria transmission in Chikhwawa district, Southern Malawi. Malar J. 2012 Jan;11:380.

76
73. Guelbeogo WM, Sagnon N, Liu F, Besansky NJ, Costantini C. Behavioural divergence of sympatric Anopheles funestus populations in Burkina Faso. Malar J. 2014 Jan;13:65.
74. Farajollahi A, Fonseca DM, Kramer LD, Marm Kilpatrick A. “Bird biting” mosquitoes and human disease: a review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect Genet Evol. 2011 Oct;11(7):1577–85.
75. Kanhekar LJ, Patnaik SK, Rao PK, Narayana M V, Raina VK, Kumar A. Some aspects of the bionomics of Culex quinquefasciatus (Diptera: Culicidae) in Rajahmundry town, Andhra Pradesh. J Commun Dis. 1994 Dec;26(4):226–30.
76. Dixit V, Gupta AK, Kataria OM, Prasad GB. Host preference of Culex quinquefasciatus in Raipur city of Chattisgarh state. J Commun Dis. 2001 Mar;33(1):17–22.
77. Reisen WK. A quantitative mosquito survey of 7 villages in Punjab Province, Pakistan with notes on bionomics, sampling methodology and the effects of insecticides. Southeast Asian J Trop Med Public Health. 1978 Dec;9(4):587–601.
78. Robert V, Le Goff G, Toto JC, Mulder L, Fondjo E, Manga L, et al. Anthropophilic mosquitoes and malaria transmission at Edea, Cameroon. Trop Med Parasitol. 1993 Mar;44(1):14–8.
79. Tadei WP, Dutary Thatcher B. Malaria vectors in the Brazilian amazon: Anopheles of the subgenus Nyssorhynchus. Rev Inst Med Trop Sao Paulo. 2000;42(2):87–94.
80. Tadei WP, Thatcher BD, Santos JM, Scarpassa VM, Rodrigues IB, Rafael MS. Ecologic observations on anopheline vectors of malaria in the Brazilian Amazon. Am J Trop Med Hyg. 1998 Aug;59(2):325–35.
81. Deane LM. Malaria vectors in Brazil. Mem Inst Oswaldo Cruz. Fundação Oswaldo Cruz; 1986;81:5–14.
82. Natal D, Barata JMS, Lagos CBT, Rocha RM. Nota sobre culicídeos (Diptera: Culicidae) da bacia do rio Purus, Acre, Amazônia (Brasil). Rev Saude Publica . Faculdade de Saúde Pública da Universidade de São Paulo; 1992 Apr;26(2):129–31.
83. Fé NF, Barbosa M das GV, Fé FAA, Guerra MV de F, Alecrim WD. Fauna de Culicidae em municípios da zona rural do estado do Amazonas, com incidência de febre amarela. Rev Soc Bras Med Trop. SBMT; 2003 Jun;36(3):343–8.
84. Barbosa M das GV, Fé NF, Marcião AHR, Silva APT da, Monteiro WM, Guerra MV de F, et al. [Record of epidemiologically important Culicidae in the rural area of Manaus, Amazonas]. Rev Soc Bras Med Trop. 2008;41(6):658–63.
85. Barbosa MGV, Fé NF, Jesus RDB de, Rodriguez IC, Monteiro WM, Mourão MPG, et al. Aedes aegypti e fauna associada em área rural de Manaus, na Amazônia brasileira. Rev Soc Bras Med Trop. SBMT; 2009 Apr;42(2):213–6.
86. Confalonieri UEC, Costa Neto C. Diversity of mosquito vectors (Diptera: culicidae) in caxiuanã, pará, Brazil. Interdiscip Perspect Infect Dis. 2012 Jan;2012:741273.

77
87. Cruz RMB, Gil LHS, de Almeida e Silva A, da Silva Araújo M, Katsuragawa TH. Mosquito abundance and behavior in the influence area of the hydroelectric complex on the Madeira River, Western Amazon, Brazil. Trans R Soc Trop Med Hyg. 2009 Nov;103(11):1174–6.
88. Gama RA, Silva IM da, Monteiro HA de O, Eiras ÁE. Fauna of Culicidae in rural areas of Porto Velho and the first record of Mansonia (Mansonia) flaveola (Coquillet, 1906), for the state of Rondônia, Brazil. Rev Soc Bras Med Trop. SBMT; 2012 Feb;45(1):125–7.
89. Klein TA, Lima JBP, Tang AT. Seasonal distribution and diel biting patterns of culicine mosquitoes in Costa Marques, Rondônia, Brazil. Mem Inst Oswaldo Cruz. Fundação Oswaldo Cruz; 1992 Mar;87(1):141–8.
90. Luz S, Lourenço-de-Oliveira R. Forest culicinae mosquitoes in the environs of samuel hydroeletric plant, state of Rondônia, Brazil. Mem Inst Oswaldo Cruz. Fundação Oswaldo Cruz; 1996 Aug;91(4):427–32.
91. Hutchings RSG, Hutchings RW, Sallum MAM. Culicidae (Diptera, Culicomorpha) from the western Brazilian Amazon: Juami-Japurá Ecological Station. Rev Bras Entomol. Sociedade Brasileira De Entomologia; 2010;54(4):687–91.
92. Kilama M, Smith DL, Hutchinson R, Kigozi R, Yeka A, Lavoy G, et al. Estimating the annual entomological inoculation rate for Plasmodium falciparum transmitted by Anopheles gambiae s.l. using three sampling methods in three sites in Uganda. Malar J. BioMed Central; 2014 Jan 21;13(1):111.
93. Galardo AKR, Arruda M, D’Almeida Couto AAR, Wirtz R, Lounibos LP, Zimmerman RH. Malaria vector incrimination in three rural riverine villages in the Brazilian Amazon. Am J Trop Med Hyg. 2007 Mar 1;76(3):461–9.
94. Beier J, Killeen G, Githure J. Short report: entomologic inoculation rates and Plasmodium falciparum malaria prevalence in Africa. Am J Trop Med Hyg. 1999 Jul 1;61(1):109–13.
95. Rutledge LC, Ward RA, Buckwalter RM. Plasmodium spp.: Dispersion of malarial oocyst populations in anopheline and culicine mosquitoes. Exp Parasitol. 1973 Aug;34(1):132–41.
96. Haji H, Smith T, Meuwissen JT, Sauerwein R, Charlwood JD. Estimation of the infectious reservoir of Plasmodium falciparum in natural vector populations based on oocyst size. Trans R Soc Trop Med Hyg. 1996 Sep;90(5):494–7.
97. Rao RU, Huang Y, Bockarie MJ, Susapu M, Laney SJ, Weil GJ. A qPCR-based multiplex assay for the detection of Wuchereria bancrofti, Plasmodium falciparum and Plasmodium vivax DNA. Trans R Soc Trop Med Hyg. 2009 Apr;103(4):365–70.
98. Intapan PM, Thanchomnang T, Lulitanond V, Maleewong W. Rapid detection of Wuchereria bancrofti and Brugia malayi in mosquito vectors (Diptera: Culicidae) using a real-time fluorescence resonance energy transfer multiplex PCR and melting curve analysis. J Med Entomol. 2009 Jan;46(1):158–64.

78
99. Farid HA, Morsy ZS, Helmy H, Ramzy RMR, El Setouhy M, Weil GJ. A critical appraisal of molecular xenomonitoring as a tool for assessing progress toward elimination of Lymphatic Filariasis. Am J Trop Med Hyg. 2007 Oct;77(4):593–600.
100. Khalifa RMA, El-Nadi NA, Ahmed AM, Hassan FA. Histological and PCR xenomonitoring of culicine mosquitoes for filarial infestation in Sohag Governorate, upper Egypt. J Egypt Soc Parasitol. 2013 Dec;43(3):591–600.
101. Korgaonkar NS, Kumar A, Yadav RS, Kabadi D, Dash AP. Mosquito biting activity on humans & detection of Plasmodium falciparum infection in Anopheles stephensi in Goa, India. Indian J Med Res. 2012 Jan;135:120–6.
102. Onyango SA, Kitron U, Mungai P, Muchiri EM, Kokwaro E, King CH, et al. Monitoring malaria vector control interventions: effectiveness of five different adult mosquito sampling methods. J Med Entomol. 2013 Sep;50(5):1140–51.
103. Schmaedick MA, Koppel AL, Pilotte N, Torres M, Williams SA, Dobson SL, et al. Molecular xenomonitoring using mosquitoes to map lymphatic filariasis after mass drug administration in american samoa. PLoS Negl Trop Dis. 2014 Aug;8(8):e3087.
104. Barbazan P, Thitithanyanont A, Missé D, Dubot A, Bosc P, Luangsri N, et al. Detection of H5N1 avian influenza virus from mosquitoes collected in an infected poultry farm in Thailand. Vector Borne Zoonotic Dis. 2008 Jan;8(1):105–9.
105. SCOTT TW, GITHEKO AK, FLEISHER A, HARRINGTON LC, YAN G. DNA PROFILING OF HUMAN BLOOD IN ANOPHELINES FROM LOWLAND AND HIGHLAND SITES IN WESTERN KENYA. Am J Trop Med Hyg. 2006 Aug 1;75(2):231–7.
106. Grubaugh ND, McMenamy SS, Turell MJ, Lee JS. Multi-gene detection and identification of mosquito-borne RNA viruses using an oligonucleotide microarray. PLoS Negl Trop Dis. 2013 Jan 15;7(8):e2349.
107. Dadzie SK, Brenyah R, Appawu MA. Role of species composition in malaria transmission by the Anopheles funestus group (Diptera: Culicidae) in Ghana. J Vector Ecol. 2013 Jun;38(1):105–10.
108. Tanga MC, Ngundu WI, Tchouassi PD. Daily survival and human blood index of major malaria vectors associated with oil palm cultivation in Cameroon and their role in malaria transmission. Trop Med Int Health 2011 Apr;16(4):447–57.
109. Robert V, Dieng H, Lochouran L, Traoré SF, Trape JF, Simondon F, et al. [Malaria transmission in the rural zone of Niakhar, Senegal]. Trop Med Int Health. 1998 Aug;3(8):667–77.
110. Parida SK, Hazra RK, Marai N, Tripathy HK, Mahapatra N. Host feeding patterns of malaria vectors of Orissa, India. J Am Mosq Control Assoc. 2006 Dec;22(4):629–34.
111. Kumar K, Katyal R, Gill KS. Feeding pattern of anopheline and culicine mosquitoes in relation to biotopes and seasons in Delhi and environs. J Commun Dis. 2002 Mar;34(1):59–64.
112. Smith Gueye C, Newby G, Gosling RD, Whittaker MA, Chandramohan D, Slutsker L, et al. Strategies and approaches to vector control in nine malaria-

79
eliminating countries: a cross-case study analysis. Malar J. 2016 Jan;15(1):2.
113. Imwong M, Nguyen TN, Tripura R, Peto TJ, Lee SJ, Lwin KM, et al. The epidemiology of subclinical malaria infections in South-East Asia: findings from cross-sectional surveys in Thailand-Myanmar border areas, Cambodia, and Vietnam. Malar J. 2015 Jan;14:381.
114. Vallejo AF, Chaparro PE, Benavides Y, Álvarez Á, Quintero JP, Padilla J, et al. High prevalence of sub-microscopic infections in Colombia. Malar J. 2015 Jan;14:201.
115. Vallejo AF, García J, Amado-Garavito AB, Arévalo-Herrera M, Herrera S. Plasmodium vivax gametocyte infectivity in sub-microscopic infections. Malar J. 2016 Jan;15(1):48.
116. Curic G, Hercog R, Vrselja Z, Wagner J. Identification of person and quantification of human DNA recovered from mosquitoes (Culicidae). Forensic Sci Int Genet. 2014 Jan;8(1):109–12.
117. Mukabana WR, Takken W, Seda P, Killeen GF, Hawley WA, Knols BGJ. Extent of digestion affects the success of amplifying human DNA from blood meals of Anopheles gambiae (Diptera: Culicidae). Bull Entomol Res. 2002 Jun;92(3):233–9.
118. Alavi Y, Arai M, Mendoza J, Tufet-Bayona M, Sinha R, Fowler K, et al. The dynamics of interactions between Plasmodium and the mosquito: a study of the infectivity of Plasmodium berghei and Plasmodium gallinaceum, and their transmission by Anopheles stephensi, Anopheles gambiae and Aedes aegypti. Int J Parasitol. 2003 Aug;33(9):933–43.
119. Yoeli M. Plasmodium berghei: Mechanisms and sites of resistance to sporogonic development in different mosquitoes. Exp Parasitol. 1973 Dec;34(3):448–58.
120. Molina-Cruz A, Lehmann T, Knöckel J. Could culicine mosquitoes transmit human malaria? Trends Parasitol. 2013 Nov;29(11):530–7.
121. Sinden RE, Alavi Y, Raine JD. Mosquito--malaria interactions: a reappraisal of the concepts of susceptibility and refractoriness. Insect Biochem Mol Biol. 2004 Jul;34(7):625–9.
122. James S, Takken W, Collins FH, Gottlieb M. Needs for monitoring mosquito transmission of malaria in a pre-elimination world. Am J Trop Med Hyg. 2014 Jan;90(1):6–10.
123. Knöckel J, Molina-Cruz A, Fischer E, Muratova O, Haile A, Barillas-Mury C, et al. An impossible journey? The development of Plasmodium falciparum NF54 in Culex quinquefasciatus. PLoS One. 2013 Jan;8(5):e63387.
124. Kading RC, Biggerstaff BJ, Young G, Komar N. Mosquitoes used to draw blood for arbovirus viremia determinations in small vertebrates. PLoS One. 2014 Jan 5;9(6):e99342.
125. Grubaugh ND, Sharma S, Krajacich BJ, Fakoli Iii LS, Bolay FK, Diclaro Ii JW, et al. Xenosurveillance: A Novel Mosquito-Based Approach for Examining the Human-Pathogen Landscape. PLoS Negl Trop Dis. 2015 Mar 16;9(3):e0003628.

80
126. Arez AP, Lopes D, Pinto J, Franco AS, Snounou G, do Rosário VE. Plasmodium sp.: optimal protocols for PCR detection of low parasite numbers from mosquito (Anopheles sp.) samples. Exp Parasitol. 2000 Apr;94(4):269–72.
127. Lardeux F, Tejerina R, Aliaga C, Ursic-Bedoya R, Lowenberger C, Chavez T. Optimization of a semi-nested multiplex PCR to identify Plasmodium parasites in wild-caught Anopheles in Bolivia, and its application to field epidemiological studies. Trans R Soc Trop Med Hyg. 2008 May;102(5):485–92.
128. Musapa M, Kumwenda T, Mkulama M, Chishimba S, Norris DE, Thuma PE, et al. A simple Chelex protocol for DNA extraction from Anopheles spp. J Vis Exp. 2013 Jan;(71).
129. Heid CA, Stevens J, Livak KJ, Williams PM. Real time quantitative PCR. Genome Res. 1996 Oct 1;6(10):986–94.
130. de Monbrison F, Angei C, Staal A, Kaiser K, Picot S. Simultaneous identification of the four human Plasmodium species and quantification of Plasmodium DNA load in human blood by real-time polymerase chain reaction. Trans R Soc Trop Med Hyg. 2003;97(4):387–90.
131. Perandin F, Manca N, Calderaro A, Piccolo G, Galati L, Ricci L, et al. Development of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for routine clinical diagnosis. J Clin Microbiol. 2004 Mar;42(3):1214–9.
132. Chua KH, Lee PC, Chai HC. Development of insulated isothermal PCR for rapid on-site malaria detection. Malar J. 2016 Jan;15(1):134.
133. Lima GFM de C, Lucchi NW, Silva-Flannery L, Macedo-de-Oliveira A, Hristov AD, Inoue J, et al. Still Searching for a Suitable Molecular Test to Detect Hidden Plasmodium Infection: A Proposal for Blood Donor Screening in Brazil. PLoS One. Public Library of Science; 2016 Jan 3;11(3):e0150391.
134. Martins-Campos KM, Pinheiro WD, Vítor-Silva S, Siqueira AM, Melo GC, Rodrigues IC, et al. Integrated vector management targeting Anopheles darlingi populations decreases malaria incidence in an unstable transmission area, in the rural Brazilian Amazon. Malar J. 2012 Jan;11:351.
135. Pérez Rico A, Lacasa Navarro J, Rubio Muñoz JM, Ruiz Contreras S, Vega Pla JL. Protocolo de extracción de ADN en lotes de 10 mosquitos para la identificación de Plasmodium spp. mediante qPCR. Sanid Mil. Ministerio de Defensa; 2013 Jun;69(2):78–81.
136. Sorosjinda-Nunthawarasilp P, Bhumiratana A. Ecotope-based entomological surveillance and molecular xenomonitoring of multidrug resistant malaria parasites in anopheles vectors. Interdiscip Perspect Infect Dis. 2014 Jan;2014:969531.
137. Adams M, Joshi SN, Mbambo G, Mu AZ, Roemmich SM, Shrestha B, et al. An ultrasensitive reverse transcription polymerase chain reaction assay to detect asymptomatic low-density Plasmodium falciparum and Plasmodium vivax infections in small volume blood samples. Malar J. BioMed Central; 2015 Jan 23;14(1):520.

81
138. Hofmann N, Mwingira F, Shekalaghe S, Robinson LJ, Mueller I, Felger I. Ultra-sensitive detection of Plasmodium falciparum by amplification of multi-copy subtelomeric targets. PLoS Med. Public Library of Science; 2015 Mar 3;12(3):e1001788.
139. WHO. Strategy for Malaria Elimination in the Greater Mekong Subregion (2015–2030). WHO Library Cataloguing-in-Publication Data. 2014. p. 64.
140. WHO. FROM MALARIA CONTROL TO MALARIA ELIMINATION. WHO Library Cataloguing-in-Publication Data. 2014. p. 53.
141. WHO. WHO | World Malaria Report 2014. WHO Library Cataloguing-in-Publication Data. World Health Organization; 2014. 243 p.
142. Cook J, Xu W, Msellem M, Vonk M, Bergström B, Gosling R, et al. Mass screening and treatment on the basis of results of a Plasmodium falciparum-specific rapid diagnostic test did not reduce malaria incidence in Zanzibar. J Infect Dis. 2015 May 1;211(9):1476–83.
143. Tiono AB, Ouédraogo A, Ogutu B, Diarra A, Coulibaly S, Gansané A, et al. A controlled, parallel, cluster-randomized trial of community-wide screening and treatment of asymptomatic carriers of Plasmodium falciparum in Burkina Faso. Malar J. 2013 Jan;12:79.
144. Tiono AB, Guelbeogo MW, Sagnon NF, Nébié I, Sirima SB, Mukhopadhyay A, et al. Dynamics of malaria transmission and susceptibility to clinical malaria episodes following treatment of Plasmodium falciparum asymptomatic carriers: results of a cluster-randomized study of community-wide screening and treatment, and a parallel entomology. BMC Infect Dis. 2013 Jan;13:535.
145. Tiono AB, Ouédraogo A, Diarra A, Coulibaly S, Soulama I, Konaté AT, et al. Lessons learned from the use of HRP-2 based rapid diagnostic test in community-wide screening and treatment of asymptomatic carriers of Plasmodium falciparum in Burkina Faso. Malar J. 2014 Jan;13:30.
146. Slater HC, Ross A, Ouédraogo AL, White LJ, Nguon C, Walker PGT, et al. Assessing the impact of next-generation rapid diagnostic tests on Plasmodium falciparum malaria elimination strategies. Nature. Nature Publishing Group, a division of Macmillan Publishers Limited. All Rights Reserved.; 2015 Dec 3;528(7580):S94–101.
147. González-Silva M, Bassat Q, Alonso PL. Getting ready for malaria elimination: a check list of critical issues to consider. Mem Inst Oswaldo Cruz. 2014 Aug;109(5):517–21.
148. Greenwood BM, Fidock DA, Kyle DE, Kappe SHI, Alonso PL, Collins FH, et al. Malaria: progress, perils, and prospects for eradication. J Clin Invest. 2008 Apr;118(4):1266–76.
149. Karunajeewa H, Waltmann A, Wini L, Mueller I. Missing malaria? Potential obstacles to diagnosis and hypnozoite eradication. Med J Aust. 2015 Apr 20;202(7):360.
150. Churcher TS, Cohen JM, Novotny J, Ntshalintshali N, Kunene S, Cauchemez S. Public health. Measuring the path toward malaria elimination. Science. 2014 Jun 13;344(6189):1230–2.

82
151. Gosling J, Case P, Tulloch J, Chandramohan D, Wegbreit J, Newby G, et al. Effective Program Management: A Cornerstone of Malaria Elimination. Am J Trop Med Hyg. 2015 May 26;ajtmh.14–0255.
152. Årdal C, Røttingen J-A. An open source business model for malaria. PLoS One. Public Library of Science; 2015 Jan 2;10(2):e0117150.
153. Ohrt C, Roberts KW, Sturrock HJW, Wegbreit J, Lee BY, Gosling RD. Information Systems to Support Surveillance for Malaria Elimination. Am J Trop Med Hyg. 2015 May 26;ajtmh.14–0257.
154. Murphy SC, Hermsen CC, Douglas AD, Edwards NJ, Petersen I, Fahle GA, et al. External quality assurance of malaria nucleic acid testing for clinical trials and eradication surveillance. PLoS One. Public Library of Science; 2014 Jan;9(5):e97398.
155. Crompton PD, Moebius J, Portugal S, Waisberg M, Hart G, Garver LS, et al. Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease. Annu Rev Immunol. 2014 Jan;32:157–87.
156. Halbroth BR, Draper SJ. Recent developments in malaria vaccinology. Adv Parasitol. 2015 Apr;88:1–49.
157. Mnzava AP, Macdonald MB, Knox TB, Temu EA, Shiff CJ. Malaria vector control at a crossroads: public health entomology and the drive to elimination. Trans R Soc Trop Med Hyg. 2014 Sep;108(9):550–4.
158. Bolscher JM, Koolen KMJ, van Gemert GJ, van de Vegte-Bolmer MG, Bousema T, Leroy D, et al. A combination of new screening assays for prioritization of transmission-blocking antimalarials reveals distinct dynamics of marketed and experimental drugs. J Antimicrob Chemother. 2015 May;70(5):1357–66.
159. Erath J, Gallego-Delgado J, Xu W, Andriani G, Tanghe S, Gurova K V, et al. Small-molecule xenomycins inhibit all stages of the Plasmodium life cycle. Antimicrob Agents Chemother. 2015 Mar;59(3):1427–34.
160. Wells TNC. Discovering and developing new medicines for malaria control and elimination. Infect Disord Drug Targets. 2013 Aug;13(4):292–302.
161. Zhou S, Rietveld AEC, Velarde-Rodriguez M, Ramsay AR, Zhang S, Zhou X, et al. Operational research on malaria control and elimination: a review of projects published between 2008 and 2013. Malar J. 2014 Jan;13:473.
162. Sturrock HJW, Roberts KW, Wegbreit J, Ohrt C, Gosling RD. Tackling Imported Malaria: An Elimination Endgame. Am J Trop Med Hyg. 2015 May 26;ajtmh.14–0256.
163. Wangdi K, Gatton ML, Kelly GC, Clements AC. Cross-border malaria: a major obstacle for malaria elimination. Adv Parasitol. 2015 Jun;89:79–107.
164. Sturrock HJW, Hsiang MS, Cohen JM, Smith DL, Greenhouse B, Bousema T, et al. Targeting asymptomatic malaria infections: active surveillance in control and elimination. PLoS Med. Public Library of Science; 2013 Jan;10(6):e1001467.
165. Tietje K, Hawkins K, Clerk C, Ebels K, McGray S, Crudder C, et al. The essential role of infection-detection technologies for malaria elimination and eradication. Trends Parasitol. 2014 May;30(5):259–66.

83
166. Gimnig JE, Walker ED, Otieno P, Kosgei J, Olang G, Ombok M, et al. Incidence of malaria among mosquito collectors conducting human landing catches in western Kenya. Am J Trop Med Hyg. 2013 Feb;88(2):301–8.
167. Kelly AH. Will He Be There?: Mediating malaria, immobilizing science. J Cult Econ. 2011 Feb;4(1):65–79.
168. Wong J, Bayoh N, Olang G, Killeen GF, Hamel MJ, Vulule JM, et al. Standardizing operational vector sampling techniques for measuring malaria transmission intensity: evaluation of six mosquito collection methods in western Kenya. Malar J. 2013 Jan;12:143.
169. Kilama M, Smith DL, Hutchinson R, Kigozi R, Yeka A, Lavoy G, et al. Estimating the annual entomological inoculation rate for Plasmodium falciparum transmitted by Anopheles gambiae s.l. using three sampling methods in three sites in Uganda. Malar J. 2014 Jan;13:111.
170. Tangena J-AA, Thammavong P, Hiscox A, Lindsay SW, Brey PT. The Human-Baited Double Net Trap: An Alternative to Human Landing Catches for Collecting Outdoor Biting Mosquitoes in Lao PDR. PLoS One. Public Library of Science; 2015 Jan;10(9):e0138735.
171. Kramer RA, Lesser A. Sustaining the gains made in malaria control and elimination. Infect Dis poverty. 2015 Jan;4:26.
172. Consoli R, Lourenço-de-Oliveira R. Principais mosquitos de importância sanitária no Brasil. Editora Fiocruz. Rio de Janeiro; 1994. p. 228.
173. Faran ME, Linthicum LK. A handbook of the Amazonian species of Anopheles Nyssorhynchus (Diptera: Culicidae). Mosquito Systematics. 13th ed. 1981. p. 81.
174. Klein TA, Lima JB, Toda-Tang A. Colonization and maintenance of Anopheles deaneorum in Brazil. J Am Mosq Control Assoc. 1990 Sep;6(3):510–3.
175. Knight JW, Nayar JK. Colonization of Culex nigripalpus theobald (Diptera: Culicidae) by stimulation of mating using males of other mosquito species. J Am Mosq Control Assoc. 1999 Mar;15(1):72–3.
176. KRISHNAN KS. A NOTE ON COLONIZATION OF CULEX. Bull World Health Organ. 1964 Jan;31:455–6.
177. Rios-Velásquez CM, Martins-Campos KM, Simões RC, Izzo T, dos Santos E V, Pessoa FAC, et al. Experimental Plasmodium vivax infection of key Anopheles species from the Brazilian Amazon. Malar J. 2013 Jan;12:460.
178. Arimoto H, Harwood JF, Nunn PJ, Richardson AG, Gordon S, Obenauer PJ. Comparison of Trapping Performance Between the Original BG-Sentinel® Trap and BG-Sentinel 2® Trap (1). J Am Mosq Control Assoc. 2015 Dec;31(4):384–7.
179. Sriwichai P, Samung Y, Sumruayphol S, Kiattibutr K, Kumpitak C, Payakkapol A, et al. Natural human Plasmodium infections in major Anopheles mosquitoes in western Thailand. Parasit Vectors. 2016 Jan;9(1):17.
180. Póvoa M, Wirtz R, Lacerda R, Miles M, Warhurst D. Malaria vectors in the municipality of erra do Navio, tate of Amapá, Amazon Region, Brazil. Memórias do Inst Oswaldo Cruz. 2001 Mar;96(2):179–84.

84
181. de Oliveira-Ferreira J, Lourenço-de-Oliveira R, Teva A, Deane LM, Daniel-Ribeiro CT. Natural malaria infections in anophelines in Rondonia State, Brazilian Amazon. Am J Trop Med Hyg. 1990 Jul;43(1):6–10.
182. Branquinho MS, Lagos CB, Rocha RM, Natal D, Barata JM, Cochrane AH, et al. Anophelines in the state of Acre, Brazil, infected with Plasmodium falciparum, P. vivax, the variant P. vivax VK247 and P. malariae. Trans R Soc Trop Med Hyg. 1993 Jan;87(4):391–4.
183. dos Santos RLC, Sucupira IMC, Lacerda RN da L, Fayal A da S, Póvoa MM. [Entomological survey and infectivity during malaria outbreak in the Anajás municipality, Pará State]. Rev Soc Bras Med Trop. 2005 Jan;38(2):202–4.
184. Helinski MEH, Nuwa A, Protopopoff N, Feldman M, Ojuka P, Oguttu DW, et al. Entomological surveillance following a long-lasting insecticidal net universal coverage campaign in Midwestern Uganda. Parasit Vectors. 2015 Jan;8:458.
185. Carnevale P, Toto J-C, Besnard P, Santos MA Dos, Fortes F, Allan R, et al. Spatio-temporal variations of Anopheles coluzzii and An. gambiae and their Plasmodium infectivity rates in Lobito, Angola. J Vector Ecol. 2015 Jul;40(1):172–9.
186. Victora CG, Huttly SR, Fuchs SC, Olinto MT. The role of conceptual frameworks in epidemiological analysis: a hierarchical approach. Int J Epidemiol. 1997 Feb;26(1):224–7.
187. Burton GJ. Observations on the habits and control of Culex pipiens fatigans in Guyana. Bull World Health Organ. 1967 Jan;37(2):317–22.
188. Cho S-H, Lee H-W, Shin E-H, Lee H-I, Lee W-G, Kim C-H, et al. A mark-release-recapture experiment with Anopheles sinensis in the northern part of Gyeonggi-do, Korea. Korean J Parasitol. 2002 Sep;40(3):139–48.
189. Fabian MM, Toma T, Tsuzuki A, Saita S, Miyagi I. Mark-release-recapture experiments with Anopheles saperoi (Diptera: Culicidae) in the Yona Forest, northern Okinawa, Japan. Southeast Asian J Trop Med Public Health. 2005 Jan;36(1):54–63.
190. Hamer GL, Anderson TK, Donovan DJ, Brawn JD, Krebs BL, Gardner AM, et al. Dispersal of adult culex mosquitoes in an urban west nile virus hotspot: a mark-capture study incorporating stable isotope enrichment of natural larval habitats. PLoS Negl Trop Dis. 2014 Mar;8(3):e2768.
191. HARRINGTON LC, SCOTT TW, LERDTHUSNEE K, COLEMAN RC, COSTERO A, CLARK GG, et al. DISPERSAL OF THE DENGUE VECTOR AEDES AEGYPTI WITHIN AND BETWEEN RURAL COMMUNITIES. Am J Trop Med Hyg. 2005 Feb 1;72(2):209–20.
192. Charlwood JD, Alecrim WA. Capture-recapture studies with the South American malaria vector Anopheles darlingi, Root. Ann Trop Med Parasitol. 1989 Dec;83(6):569–76.
85
8. APÊNDICES
8.1 Ficha de controle de captura e identificação de culicídeos

86
8.2 POP_MAL_LB_019_v01D_PT

87

88

89

90

91

92

93
8.3 POP_MAL_LB_020_v01D_PT

94

95

96

97
9. ANEXOS
9.1 POP_MAL_LB_013_v02D_PT

98

99

100

101
9.2 PARECER_CONSUBSTANCIADO_CONEP_349211

102

103

104

105

106

107
9.3 POP_MAL_LB_001_v02D_PT

108

109

110
9.4 POP_MAL_LB_021_v01D_PT
111

112

113

114
Top Related