







Línguas
Páginas
Legal


UNIVERSIDADE FEDERAL DE SANTA MARIA CENTRO DE CIÊNCIAS RURAIS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA DO SOLO
RELATÓRIO DE PÓS-DOUTORADO
PÓS-DOUTORADO JÚNIOR (PDJ)/CNPQ
PROJETO:
FUNGOS ECTOMICORRÍZICOS EM POVOAMENTOS
FLORESTAIS EXÓTICOS E FRAGMENTOS DE MATAS NATIVAS
DO BIOMA PAMPA
Dr. Ricardo Bemfica Steffen
Santa Maria, RS, novembro de 2012

2
SUMÁRIO 1 DADOS DO PROJETO ................................................................................................ 3
2 DADOS DO ESTÁGIO................................................................................................... 3
3 RESUMO......................................................................................................................... 5
4 INTRODUÇÃO............................................................................................................... 6
5 OBJETIVOS.................................................................................................................... 10
6 ATIVIDADES REALIZADAS...................................................................................... 11
7 ATIVIDADES PREVISTAS x ATIVIDADES DESENVOLVIDAS.......................... 20
8 PUBLICAÇÕES.............................................................................................................. 21
9 REFERÊNCIAS............................................................................................................ 22
10 ANEXOS ....................................................................................................................... 25

3
1 DADOS DO PROJETO
Projeto: Fungos ectomicorrízicos em povoamentos florestais exóticos e fragmentos de matas
nativas do bioma Pampa
Submetido ao: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), na
modalidade de bolsa de Pós-doutorado Júnior (PDJ)
Processo: 155132/2010-5
Coordenadora do Projeto:
Prof. Dra. Zaida Inês Antoniolli – Programa de Pós-Graduação em Ciência do Solo/UFSM
Instituição de Execução do Projeto:
Universidade Federal de Santa Maria - Centro de Ciências Rurais – Departamento de Solos
Av. Roraima, 1000 – Bairro Camobi – Santa Maria/RS
(55) 3220-8108 CEP 97.105-900
Vigência inicial:
Outubro de 2010 a setembro de 2011.
Prorrogação concedida:
Outubro de 2010 a setembro de 2011.
2. DADOS DO ESTÁGIO
Período de realização
De dezembro de 2010 a setembro de 2012.

4
Dados do pós-doutorando
Nome
Ricardo Bemfica Steffen
Formação profissional
Engenheiro Agrônomo Solo (Universidade Federal de Santa Maria, 2004), Mestre em Ciência
do Solo (Universidade Federal de Santa Maria, 2007) e Doutor em Ciência do Solo
(Universidade Federal de Santa Maria, 2010).
E-mail: [email protected]
Dados da orientadora
Nome
Zaida Inês Antoniolli
Formação profissional
Bióloga (Ciências Biológicas, Pontifícia Universidade Católica do Rio Grande do Sul, 1984),
Especialista em Biotecnologia (Universidade Federal do Rio Grande do Sul, 1988), Mestre
em Fitotecnia (Universidade Federal do Rio Grande do Sul, 1988), Doutora em Ecology of
Mycorrhizal Molecular Aspects (University of Adelaide, 1999).
Atuação profissional atual
Professora Associada do Departamento de Solos da Universidade Federal de Santa Maria.
Endereço profissional
Laboratório de Microbiologia e Biologia do Solo e Ambiente, Sala 3310, Departamento de
Solos, Centro de Ciências Rurais, Prédio 42. Avenida Roraima, número 1000, Cep: 97.105-
900. Telefone: (55) 3220 8108.
E-mail: [email protected]

5
3 RESUMO
O Bioma Pampa é o mais desconhecido dos biomas brasileiros. Sua fisionomia e
biodiversidade foram fortemente alteradas por pressões antrópicas e, frente à implantação de
grandes áreas de monoculturas florestais, o bioma permanece suscetível. As ectomicorrízas
encontram-se associadas às raízes de essências florestais e desempenham papel fundamental
na absorção de água e nutrientes por estas plantas. Contudo, dados referentes à diversidade
destes organismos no Pampa são extremamente escassos. Face ao exposto, torna-se urgente a
necessidade de pesquisas ligadas ao conhecimento da biodiversidade e prospecção destes
importantes recursos genéticos, os quais podem auxiliar na sua conservação e para o
desenvolvimento sustentável do país, além de servirem como indicadores da qualidade
ambiental. Este projeto de pós-doutoramento teve como objetivos: realizar uma revisão sobre
o atual estado do conhecimento de fungos ectomicorrizicos, com especial ênfase aos de hábito
hipógeo; avaliar a biodiversidade de fungos ectomicorrízicos ocorrentes em povoamentos de
monoculturas florestais e fragmentos de matas nativas do Bioma Pampa, utilizando-se
métodos morfológicos e moleculares; obter isolados que viabilizem a produção de
inoculantes; e avaliar a compatibilidade de ectomicorrizas em espécies silviculturais exóticas
e nativas por meio de técnicas de cultura de tecidos. Expedições foram realizadas em
povoamentos homogêneos de espécies exóticas e em fragmentos de matas nativas do Bioma
Pampa, onde foram coletados fungos (juntamente com o solo) os quais foram – e alguns estão
sendo - identificados morfologicamente. Além disso, realizou-se a extração do DNA
genômico destes fungos e a amplificação via reação em cadeia da polimerase da região do
Espaço Interno Transcrito (ITS) do rDNA destes fungos. Estes produtos de PCR foram
purificados e estão em fase de sequenciamento para a posterior comparação com dados do
GenBank e construção de análises filogenéticas. Da mesma forma, procurou-se isolar os
fungos coletados para posterior obtenção de inóculos. Os dados de pesquisa obtidos até o
presente momento estão em fase final de análise, visando posteriores publicações em
periódicos nacionais e internacionais.

6
4 INTRODUÇÃO
O Bioma Pampa, compartilhado pelo Brasil, Argentina e Uruguai, distribui-se de
forma predominante na maior parte do Rio Grande do Sul (64%), sendo sua ocorrência restrita
a este único Estado. O Pampa tem sido submetido à forte ação antrópica, tendo como
consequência a alteração significativa de sua fisionomia e biodiversidade. Poucas pesquisas
contemplam a avaliação dos organismos existentes neste ambiente, o qual pode ser
considerado como o menos conhecido dos biomas nacionais. Estudos nesse sentido podem
contribuir com novas descobertas científicas, que auxiliariam em estratégias de conservação e
na utilização de seres vivos de interesse no desenvolvimento científico e tecnológico.
Em diversos ambientes, observa-se a importância da presença de micorrizas no
desenvolvimento das plantas e na manutenção da dinâmica de ecossistemas. Dos vários tipos
de micorrizas existentes, as ectomicorrizas destacam-se em locais onde há presença de
espécies lenhosas. Contudo, a presença destes fungos nos fragmentos de mata nativa e em
povoamentos florestais exóticos ocorrentes no Bioma Pampa não é conhecida até o presente
momento. Dados dessa natureza podem representar informações fundamentais para a
sustentabilidade, para o conhecimento de organismos presentes e de novas espécies, como
indicadores da qualidade ambiental e, adicionalmente, no isolamento e utilização
biotecnológica destes fungos na indústria e na produção vegetal. No entanto, no tocante aos
campos sulinos, as áreas de florestas nativas, não frequentemente características destes
ecossistemas, encontram-se apenas como fragmentos esparsos, os quais necessitam de
medidas urgentes de conservação. Em contrapartida, o uso de monoculturas florestais tem
sido intensivo na região, como a descoberta de uma nova vocação econômica, porém, com
estudos ainda em discussão sobre os possíveis impactos desta atividade sobre a região.
Frente a esta realidade, destaca-se a necessidade de catalogação e conhecimento da
biodiversidade de fungos ectomicorrízicos do Pampa, o que pode ser realizado por meio de
técnicas clássicas – como estudos morfológicos baseados em microscopia estereoscópica e
ótica - e análises moleculares, as quais são, atualmente, essenciais frente ao grande
desenvolvimento nesta área, contribuindo de forma significativa para a sistemática. Desta
forma, pode-se também caracterizar espécies que estejam sobre forte pressão ambiental e em
risco de extinção. Adicionalmente, é possível a aquisição de informações de grande relevância
sobre a associação destes organismos com espécies florestais, exóticas ou nativas, que
ocorrem de forma natural ou cultivada no Pampa. Esta caracterização também permitirá traçar
comparativos entre a diversidade de ectomicorrizas em povoamentos florestais de

7
monoculturas (Eucalyptus spp., Pinus spp., e Acacia spp.) em relação a diversidade de
ectomicorrizas em fragmentos florestais nativos.
Outro fator de fundamental importância é a possibilidade de conhecimento destes
novos recursos genéticos, possibilitando a prospecção de ectomicorrizas para uso em plantios
florestais, por meio da produção de inóculos, contribuindo-se assim uma produção mais
sustentável. Sabendo-se que estes organismos são específicos quanto a sua atuação
mutualística, isolar e avaliar a sua especificidade e ação no desenvolvimento de mudas nativas
e exóticas por meio do cultivo de tecidos pode contribuir para a descoberta de novos isolados
e/ou espécies realmente compatíveis, que contribuirão para o desenvolvimento econômico de
base florestal no país, sabendo-se da importância desta atividade. Conforme a Sociedade
Brasileira de Silvicultura (SBS, 2007), o setor florestal possui participação em 3,5% do
Produto Interno Bruto do país, o que equivale a US$ 37,3 bilhões, gerando 4,33 milhões de
empregos em florestas plantadas e 2,58 milhões de empregos em florestas nativas.
4.1 O Bioma Pampa
De acordo com dados do Instituto Brasileiro de Geografia e Estatística (IBGE, 2004),
o Bioma Pampa, também conhecido como campos sulinos, abrange aproximadamente 176
km2, o que equivale a 64% do território gaúcho e a 2,07% do território do País. Este bioma
caracteriza-se por seu relevo ondulado contendo suaves elevações (SUERTEGARAY, 1998),
onde predomina a vegetação formada por campos - compostos principalmente por gramíneas
e plantas herbáceas – e, ocasionalmente, a presença de fragmentos florestais, arbustos e
árvores (CARVALHO; BATELLO, 2009). Atualmente, são criadas na região em torno de 13
milhões de cabeças de gado e cinco milhões de ovelhas, as quais utilizam a pastagem da
região (95%) como fonte de forragem.
Embora bastante antropizado, os poucos estudos realizados no Pampa mostram a
existência de uma biodiversidade bastante significativa. BOLDRINI (1997) estimou a
existência de aproximadamente três mil espécies de gramíneas, dados que, segundo
OVERBECH et al. (2007) devem estar subestimados. Ademais, o Bioma Pampa serve de
habitat para 385 espécies de pássaros e cerca de 90 espécies de mamíferos terrestres
(BILENCA; MIÑARRO, 2004). Apesar de haver preocupação com relação ao tema da
biodiversidade, o foco das atenções tem se voltado predominantemente para os recursos
animais e vegetais, sendo menor atenção dispensada à porção menos evidente, porém não
menos importante, que são os microrganismos do solo. Ao nosso conhecimento, são escassos

8
os trabalhos sobre a biodiversidade de fungos micorrízicos no Bioma Pampa. Estes dados
remetem a necessidade de urgentes discussões e pesquisas, buscando auxiliar na sua
conservação e no seu desenvolvimento econômico.
Face ao exposto, o estudo de ectomicorrizas associadas aos ambientes florestais
exóticos e nativos do Bioma Pampa, auxiliará significativamente para o melhor conhecimento
da biodiversidade destes organismos, corroborando com as necessidades de conservação
ambiental. Outrossim, conhecimentos importantes também podem ser gerados em termos de
avaliação da variabilidade genética destes fungos e da utilização destes recursos na
bioprospecção, tendo em vista seu importante papel na utilização biotecnológica.
4.2 Fungos ectomicorrízicos (fECM)
As micorrizas são associações mutualísticas entre fungos e as raízes das plantas
(BRUNDRETT et al., 1996), sendo que estas associações simbióticas são abundantes,
ocorrendo em 75 a 80% das plantas (SMITH; READ, 1997). Embora as micorrizas
arbusculares e as ectomicorrizas sejam de maior importância econômica e também as mais
conhecidas, Brundrett et al. (1996) relataram sete tipos de associações micorrízicas
conhecidas: a) micorrizas arbúsculo-vesiculares (MAV) – também chamadas de micorrizas
arbusculares (MA); b) ectomicorrizas (ECM); c) ectendomicorrizas; d) orquidóides; e)
ericóides; f) monotropóides; e g) arbutóides.
Os fungos ectomicorrízicos são muito importantes nos ecossistemas, pois sua
associação mutualística com plantas de diferentes espécies auxilia na absorção de nutrientes,
água e na proteção da planta hospedeira em relação aos patógenos e estresses abióticos
(SMITH; READ, 1997). Nos ecossistemas florestais, o papel das ectomicorrízas é ainda mais
pronunciado, atuando fortemente nos ciclos biogeoquímicos, na dinâmica das comunidades
vegetais e na manutenção da estrutura do solo (RILLIG; MUMMEY, 2006; MORRIS et al.,
2009).
Com aproximadamente 5000 espécies envolvidas em associações simbióticas com
plantas superiores, as ectomicorrizas, além de proporcionarem benefícios ao desenvolvimento
vegetal, atuam na estabilização de florestas naturais e cultivadas, servindo como fonte de
alimento e participando da ciclagem de nutrientes em florestas temperadas e tropicais
(TAYLOR, 1991; HOBBIE, 2006).
Os fungos ectomicorrízicos (fECM), embora se associem com um grupo restrito de
plantas (aproximadamente 5% das espécies conhecidas), são encontrados formando simbiose

9
com 90% das espécies arbóreas de clima temperado, principalmente com as pertencentes às
famílias Pinaceae (95%), Fagaceae (94%), Betulaceae (70%) e Salicaceae (83%)
(OLIVEIRA; GIACHINI, 1999; MOLINA et al., 2005).
Há indícios de que estes microrganismos contribuíram para a extensão de florestas
para áreas ambientalmente degradadas. Segundo Brunner (2001), o estabelecimento de mudas
florestais em solos deficientes em nutrientes ou contaminados pela deposição de certos
poluentes se torna possível devido à simbiose destas plantas com fECM.
Em solos que apresentam limitações quanto a sua fertilidade, a inoculação de fECM
promove aumento na sobrevivência das plantas após o transplante (SILVA et al., 2003;
MELLO et al., 2006).
A maioria, os fungos ectomicorrízicos pertencem à classe Basidiomycota, como os
gêneros: Amanita, Hebeloma, Hysterangium, Laccaria, Lactarius, Rhizopogon, Russula,
Scleroderma, Suillus, Tricholoma entre outros (VELLINGA et al., 2009). Têm sido
considerada extremamente favorável a associação destes fungos às espécies florestais
cultivadas como o Pinus spp., Eucalyptus spp. e a Acacia spp. Porém, não há relato no que se
refere às espécies ocorrentes no Bioma Pampa, o qual está inserido em um contexto de
produção silvícola, além da escassez de conhecimento sobre associações entre ectomicorrizas
e espécies florestais nativas, onde não são conhecidos, a priori, dados para o bioma em
questão. Conforme citam NARA e HOGETSU (2004), o papel das ectomicorrizas é
fundamental na recuperação de áreas impactadas ambientalmente, como é o caso do Pampa,
que possui a predominância de solos arenosos, de grande susceptibilidade à ação humana.
Ademais, sabendo-se da alta especificidade apresentada por ectomicorrizas, pode-se obter
isolados de interesse para a produção florestal brasileira por meio de testes neste sentido.
Nos campos sulinos, estudos morfológicos de ectomicorrizas caracterizam resultados
muito importantes no conhecimento da biodiversidade de espécies. Porém, nos dias atuais, o
uso da biologia molecular é indispensável, como pode ser observado nos trabalhos de Gomes
et al. (2002); Tedersoo et al. (2007); Morris et al. (2009), entre outros. Dentre as diversas
ferramentas moleculares disponíveis, tem sido consenso o uso daquelas baseadas no
sequenciamento do espaço interno transcrito (internal transcribed spacer – ITS) do rDNA de
fungos, auxiliando na caracterização da espécie e na construção de estudos filogenéticos.
A região ITS é composta por regiões não codificantes localizadas entre regiões
altamente conservadas do rDNA responsáveis pela codificação da subunidade menor e maior
dos ribossomos (BRIDGE; ARORA, 1998). São de fácil amplificação, possuem natureza

10
multicópia e permitem a diferenciação entre espécies. Além disso, em estudos de código de
barras (barcode) de fungos, a região ITS tem sido escolhida como prioritária.
5 OBJETIVOS
5.1 Objetivo geral
Caracterizar a biodiversidade de fungos ectomicorrízicos ocorrentes em povoamentos
de monoculturas florestais e fragmentos de matas nativas do Bioma Pampa por meio de
métodos morfológicos e moleculares, bem como obter isolados que viabilizem a produção de
inoculante e estudar a compatibilidade de ectomicorrizas em espécies silviculturais exóticas e
nativas por meio da cultura de tecidos, contribuindo para o conhecimento da biodiversidade e
utilização dos fungos micorrízicos do Bioma Pampa.
5.2 Objetivos específicos
• coletar fungos ectomicorrízicos associados a monoculturas florestais cultivadas e à
matas nativas do Bioma Pampa;
• identificar as espécies de fungos ectomicorrízicos coletadas;
• comparar a diversidade de ectomicorrizas presentes em solos com florestas nativas e
solos onde há presença de monocultivos florestais;
• determinar a compatibilidade entre isolados de fungos ectomicorrízicos e espécies
florestais exóticas e nativas, por meio do cultivo “in vitro”;
• capacitar recursos humanos para o desenvolvimento de pesquisas na área, abrangendo
o estudo de ectomicorrizas, biologia molecular e cultura de tecidos; treinando
estudantes de graduação, pós-graduação e um profissional em pós-doutoramento.

11
6 ATIVIDADES REALIZADAS
6.1 Coletas realizadas no Bioma Pampa e entorno
Com base na abrangência do bioma Pampa, realizaram-se incursões para os
municípios gaúchos de Alegrete, Barra do Quarai, Bororé, Rosário do Sul, Dilermano de
Aguiar, São Pedro do Sul, São Borja, Bororé, Cacequi, São Gabriel, Santiago, São Sepé,
Caçapava do Sul, Lavras do Sul, São Vicente do Sul, Itaqui, Maçambará, Unistalda, Santa
Margarida do Sul, Bagé, Pelotas, Santana da Boa Vista, Santana do Livramento, Dom Pedrito,
Candiota, Hulha Negra, Cerrito, Uruguaiana, Jaguari e Nova Esperança do Sul (Figura 1).
Estes dois últimos municípios estão em área de transição e, por esta razão, foram incluídos,
levando-se em consideração a importância e a grande biodiversidade, característica de regiões
ecotonais. Registros das coletas podem ser observados na Figura 2.
1- Santa Maria 2- Alegrete 3- Barra do Quaraí 4- Bororé 5- Rosário do Sul 6- Dilermando de Aguiar 7- São Pedro do Sul 8- São Borja 9- Cacequi 10- São Gabriel 11- Santiago 12- São Sepé 13- Caçapava do Sul 14- Lavras do Sul 15- São Vicente do Sul 16- Itaqui 17- Maçambará 18- Unistalda 19- Santa Margarida do Sul 20- Bagé 21- Pelotas 22- Santana da Boa Vista 23- Candiota 24- Hulha Negra 25- Uruguaiana 26- Cerrito 27- Jaguarí 28- Nova Esperança do Sul 29- Santana do Livramento
30- Dom Pedrito
Figura 1. Municípios amostrados no Bioma Pampa do Rio Grande do Sul.

12
Figura 2. Viagens prospectivas realizadas pelo grupo de pesquisas onde foram realizadas coletas nas áreas do Bioma Pampa (A), em diferentes ecossistemas (B), coletando-se diversos gêneros de fungos ectomicorrízicos (C e D). 6.2 Análises morfológicas
Para coletar os fECMs, realizaram-se incursões tanto em áreas de fragmentos florestais
nativos (de forma geral compostos por capões) como em áreas com povoamentos de
monoculturas florestais. Antecedendo a retirada dos basidiomas de seu local de origem, os
espécimes foram fotografados e realizou-se o registro de dados como: data, local de coleta,
coordenadas geográficas, coletor e característica das estruruturas reprodutivas (coloração,
consistência, inserção no solo, forma e mensurações). Os fungos foram removidos
cuidadosamente, sendo preservadas uma porção do solo onde estavam inseridos e a vegetação
circundante. O material foi numerado e acondicionado em sacos de papel para o transporte até
o Laboratório de Biologia do Solo e do Ambiente, do Departamento de Solos da Universidade
Federal de Santa Maria, onde, posteriormente, foi identificado (Tabela 1, Figura 3).

13
Figura 3. Fungos ectomicorrízicos Lactarius deliciosus (A), Russula sp. (B), Suillus luteus (C), Russula sp. (D), Suillus sp. (E), Scleroderma sp. (F) e gêneros de fungos não identificados (G e H) coletados em fragmentos de Pinus elliotti em municípios do Rio Grande do Sul localizados no Bioma Pampa.
Na maior parte das coletas realizadas em matas nativas não foram encontrados
exemplares de fungos ectomicorrízicos. Assim, optou-se por incluir a coleta de amostras de
solo contendo raízes de espécies arbóreas dos fragmentos florestais visitados. O objetivo
destas coletas foi observar a presença de associações micorrízicas nas raízes das árvores, a
qual se evidencia pela presença de morfotipos característicos.
Amostras do solo de aproximadamente 50 cm3 foram retiradas e armazenadas em
sacos plásticos com acréscimo de água destilada, objetivando-se manter a umidade por 24 h.
Posteriormente, o material foi peneirado deixando-se apenas as raízes presentes. Com o
auxílio de microscópio estereoscópico, procurou-se evidenciar a presença de morfotipos, bem
como de manto fúngico nas raízes.

14
No laboratório, as raízes presentes no solo coletado juntamente com os fungos foram
peneiradas e passaram por uma sequência de lavagens. Em seguida, receberam um número de
identificação e foram mantidas em solução de etanol a 70%. Posteriormente, procuraram-se
evidências da ocorrência de associações das raízes com hifas dos fungos coletados, utilizando-
se, para esta finalidade, microscópios estereoscópico e óptico (Figura 4).
Já para os basidiomas coletados, parte do material foi desidratada em estufa,
mantendo-se os mesmos a 50° C até atingirem massa constante (entre dois e sete dias,
dependendo do tamanho do fungo). Posteriormente, foram mantidos em sacos de papel e
caixas de papelão cuidadosamente armazenadas para manter o acervo herborizado. A
descrição dos basidiomas seguiu as metodologias usuais em micologia como, por exemplo, as
chaves de classificação e guias específicos para grupos de fungos.
As análises macroscópicas foram efetuadas a olho desarmado, bem como com o
auxílio de microscópio estereoscópico e equipamentos de mensuração como réguas e
paquímetros. Também foi utilizada a visualização de estruturas menores, como esporos e
hifas, em microscópio óptico, dispondo-se os cortes entre lâmina e lamínula, contendo
solução de azul de metileno a 1%.
Para o isolamento, seguiu-se a metodologia proposta por Brundrett et al. (1996) onde,
após desinfestação superficial dos basidiomas em etanol a 70%, estes foram partidos ao meio
retirando-se, com o auxílio de pinças e alça de platina, parte do interior do contexto, a qual foi
inoculada em meio de cultura MNM modificado (MARX, 1969). Todo este procedimento
ocorreu em câmara de fluxo laminar asséptico. Após, este material foi mantido para
crescimento em estufas com temperatura controlada de 25° C e com repicagens periódicas de
acordo com o crescimento micelial (Figura 5).
Durante as diversas incursões realizadas em povoamentos florestais do bioma Pampa
foi possível coletar 86 espécimes de fungos ectomicorrízicos. Apesar da dificuldade inicial de
encontrar as estruturas reprodutivas desses fungos devido à escassez hídrica na região do
Pampa entre novembro de 2010 e março de 2012, durante os meses de abril e julho de 2012
vários basidiomas foram coletados. Nas viagens realizadas nos meses mais secos foi
encontrado pouco ou nenhum material.
Dos 85 fungos coletados, 65 foram identificados ao menos em nível de gênero baseado
em características morfológicas macroscópicas, e dentre estes 15 já foram identificados em
nível de espécie com métodos de biologia molecular. No entanto ainda não foi possível
identificar 20 fungos, nem em nível de gênero, e estes aguardam para ser identificados por
métodos moleculares (Tabela1).
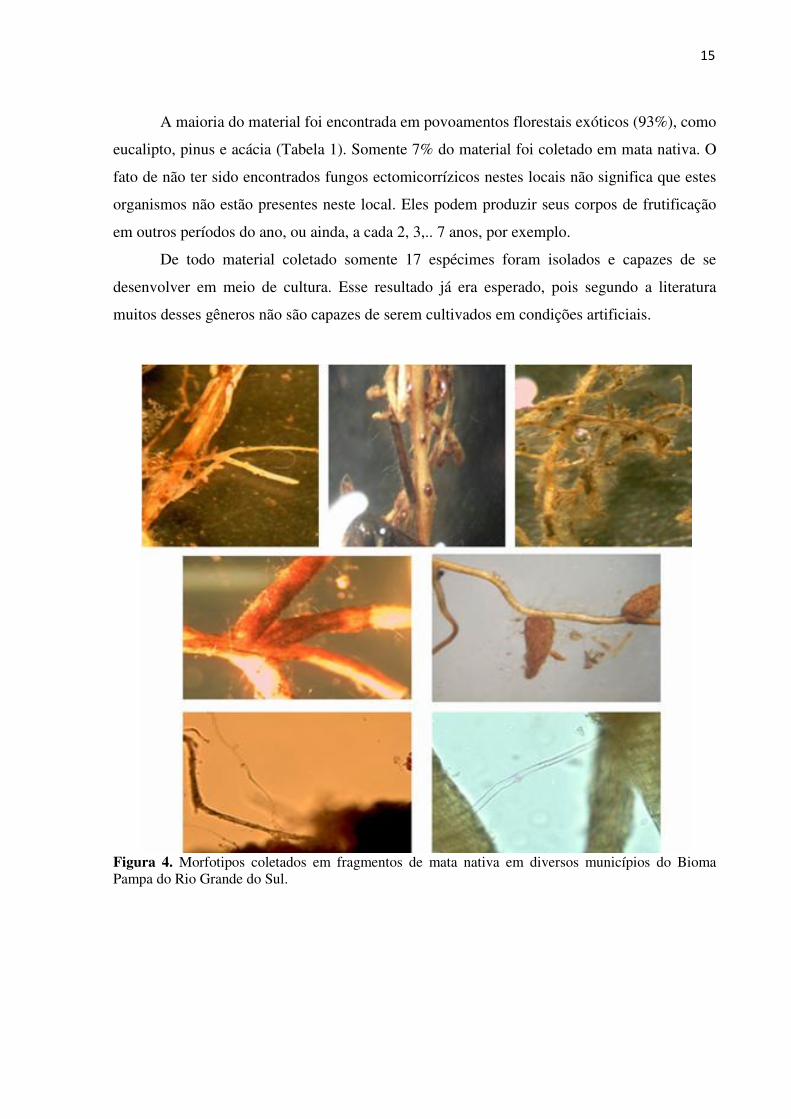
15
A maioria do material foi encontrada em povoamentos florestais exóticos (93%), como
eucalipto, pinus e acácia (Tabela 1). Somente 7% do material foi coletado em mata nativa. O
fato de não ter sido encontrados fungos ectomicorrízicos nestes locais não significa que estes
organismos não estão presentes neste local. Eles podem produzir seus corpos de frutificação
em outros períodos do ano, ou ainda, a cada 2, 3,.. 7 anos, por exemplo.
De todo material coletado somente 17 espécimes foram isolados e capazes de se
desenvolver em meio de cultura. Esse resultado já era esperado, pois segundo a literatura
muitos desses gêneros não são capazes de serem cultivados em condições artificiais.
Figura 4. Morfotipos coletados em fragmentos de mata nativa em diversos municípios do Bioma Pampa do Rio Grande do Sul.
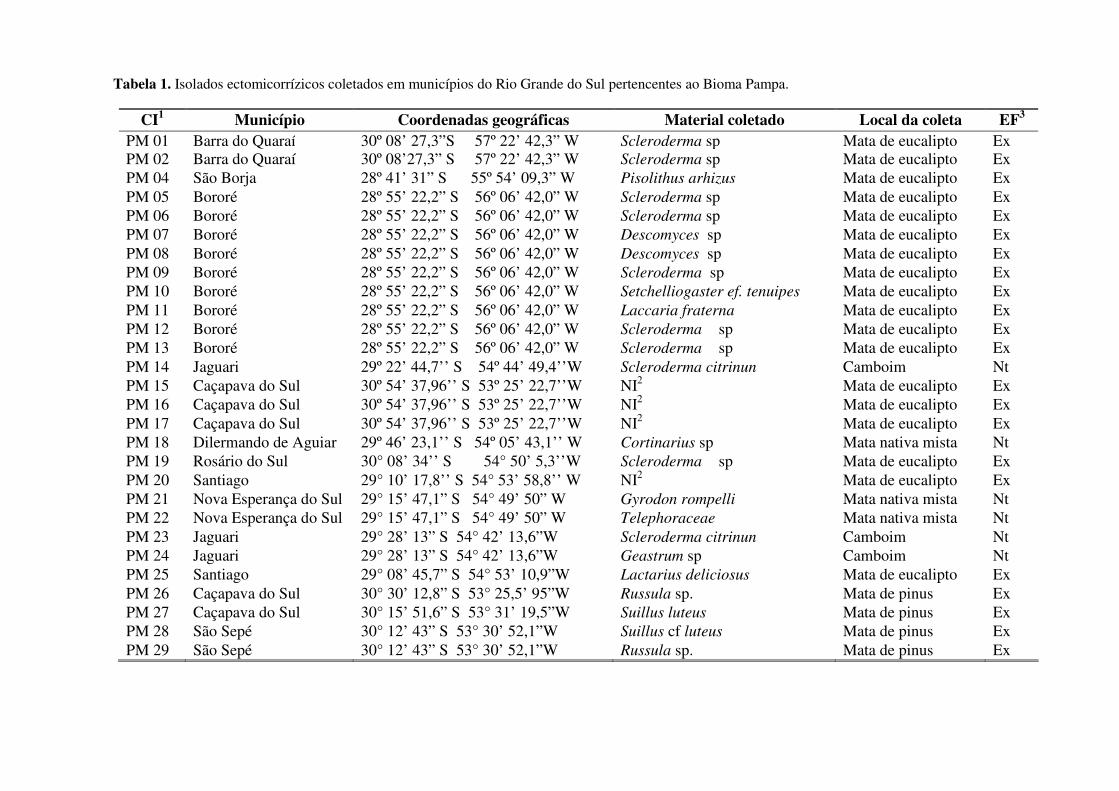
Tabela 1. Isolados ectomicorrízicos coletados em municípios do Rio Grande do Sul pertencentes ao Bioma Pampa.
CI1 Município Coordenadas geográficas Material coletado Local da coleta EF3 PM 01 Barra do Quaraí 30º 08’ 27,3”S 57º 22’ 42,3” W Scleroderma sp Mata de eucalipto Ex PM 02 Barra do Quaraí 30º 08’27,3” S 57º 22’ 42,3” W Scleroderma sp Mata de eucalipto Ex PM 04 São Borja 28º 41’ 31” S 55º 54’ 09,3” W Pisolithus arhizus Mata de eucalipto Ex PM 05 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Scleroderma sp Mata de eucalipto Ex PM 06 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Scleroderma sp Mata de eucalipto Ex PM 07 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Descomyces sp Mata de eucalipto Ex PM 08 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Descomyces sp Mata de eucalipto Ex PM 09 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Scleroderma sp Mata de eucalipto Ex PM 10 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Setchelliogaster ef. tenuipes Mata de eucalipto Ex PM 11 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Laccaria fraterna Mata de eucalipto Ex PM 12 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Scleroderma sp Mata de eucalipto Ex PM 13 Bororé 28º 55’ 22,2” S 56º 06’ 42,0” W Scleroderma sp Mata de eucalipto Ex PM 14 Jaguari 29º 22’ 44,7’’ S 54º 44’ 49,4’’W Scleroderma citrinun Camboim Nt PM 15 Caçapava do Sul 30º 54’ 37,96’’ S 53º 25’ 22,7’’W NI2 Mata de eucalipto Ex PM 16 Caçapava do Sul 30º 54’ 37,96’’ S 53º 25’ 22,7’’W NI2 Mata de eucalipto Ex PM 17 Caçapava do Sul 30º 54’ 37,96’’ S 53º 25’ 22,7’’W NI2 Mata de eucalipto Ex PM 18 Dilermando de Aguiar 29º 46’ 23,1’’ S 54º 05’ 43,1’’ W Cortinarius sp Mata nativa mista Nt PM 19 Rosário do Sul 30° 08’ 34’’ S 54° 50’ 5,3’’W Scleroderma sp Mata de eucalipto Ex PM 20 Santiago 29° 10’ 17,8’’ S 54° 53’ 58,8’’ W NI2 Mata de eucalipto Ex PM 21 Nova Esperança do Sul 29° 15’ 47,1” S 54° 49’ 50” W Gyrodon rompelli Mata nativa mista Nt PM 22 Nova Esperança do Sul 29° 15’ 47,1” S 54° 49’ 50” W Telephoraceae Mata nativa mista Nt PM 23 Jaguari 29° 28’ 13” S 54° 42’ 13,6”W Scleroderma citrinun Camboim Nt PM 24 Jaguari 29° 28’ 13” S 54° 42’ 13,6”W Geastrum sp Camboim Nt PM 25 Santiago 29° 08’ 45,7” S 54° 53’ 10,9”W Lactarius deliciosus Mata de eucalipto Ex PM 26 Caçapava do Sul 30° 30’ 12,8” S 53° 25,5’ 95”W Russula sp. Mata de pinus Ex PM 27 Caçapava do Sul 30° 15’ 51,6” S 53° 31’ 19,5”W Suillus luteus Mata de pinus Ex PM 28 São Sepé 30° 12’ 43” S 53° 30’ 52,1”W Suillus cf luteus Mata de pinus Ex PM 29 São Sepé 30° 12’ 43” S 53° 30’ 52,1”W Russula sp. Mata de pinus Ex
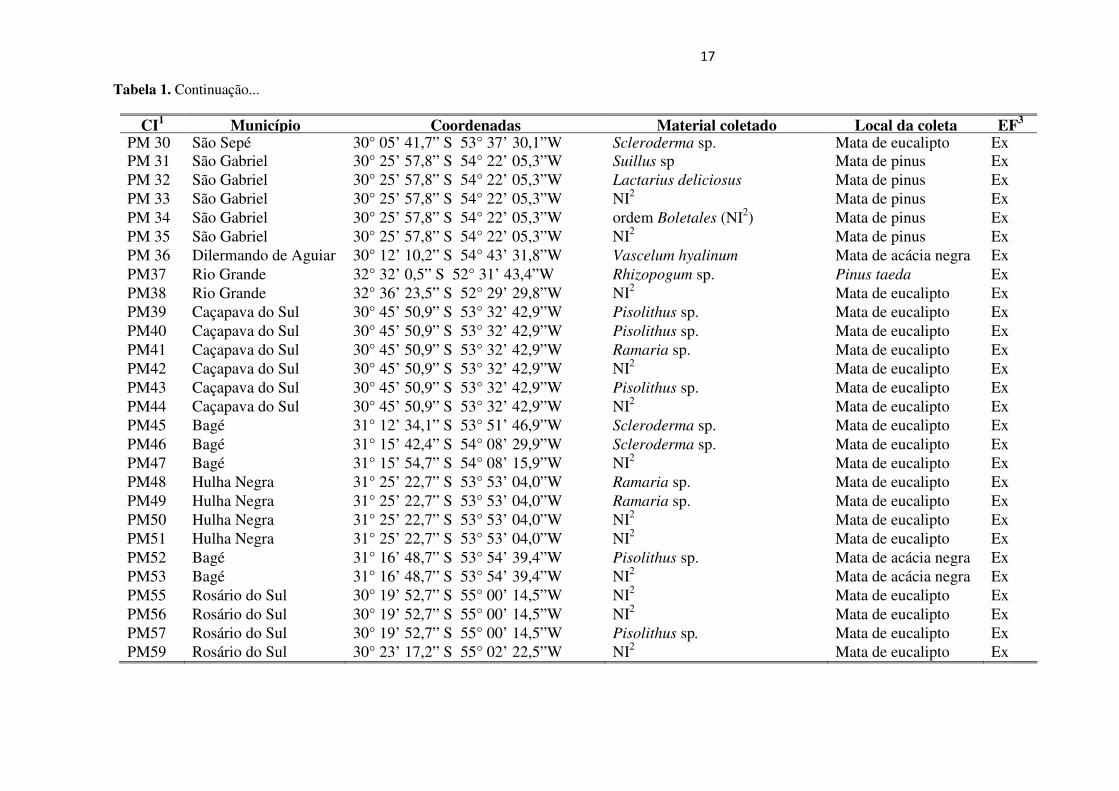
17
Tabela 1. Continuação...
CI1 Município Coordenadas Material coletado Local da coleta EF3 PM 30 São Sepé 30° 05’ 41,7” S 53° 37’ 30,1”W Scleroderma sp. Mata de eucalipto Ex PM 31 São Gabriel 30° 25’ 57,8” S 54° 22’ 05,3”W Suillus sp Mata de pinus Ex PM 32 São Gabriel 30° 25’ 57,8” S 54° 22’ 05,3”W Lactarius deliciosus Mata de pinus Ex PM 33 São Gabriel 30° 25’ 57,8” S 54° 22’ 05,3”W NI2 Mata de pinus Ex PM 34 São Gabriel 30° 25’ 57,8” S 54° 22’ 05,3”W ordem Boletales (NI2) Mata de pinus Ex PM 35 São Gabriel 30° 25’ 57,8” S 54° 22’ 05,3”W NI2
Mata de pinus Ex PM 36 Dilermando de Aguiar 30° 12’ 10,2” S 54° 43’ 31,8”W Vascelum hyalinum Mata de acácia negra Ex PM37 Rio Grande 32° 32’ 0,5” S 52° 31’ 43,4”W Rhizopogum sp. Pinus taeda Ex PM38 Rio Grande 32° 36’ 23,5” S 52° 29’ 29,8”W NI2
Mata de eucalipto Ex PM39 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W Pisolithus sp. Mata de eucalipto Ex PM40 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W Pisolithus sp. Mata de eucalipto Ex PM41 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W Ramaria sp. Mata de eucalipto Ex PM42 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W NI2
Mata de eucalipto Ex PM43 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W Pisolithus sp. Mata de eucalipto Ex PM44 Caçapava do Sul 30° 45’ 50,9” S 53° 32’ 42,9”W NI2
Mata de eucalipto Ex PM45 Bagé 31° 12’ 34,1” S 53° 51’ 46,9”W Scleroderma sp. Mata de eucalipto Ex PM46 Bagé 31° 15’ 42,4” S 54° 08’ 29,9”W Scleroderma sp. Mata de eucalipto Ex PM47 Bagé 31° 15’ 54,7” S 54° 08’ 15,9”W NI2
Mata de eucalipto Ex PM48 Hulha Negra 31° 25’ 22,7” S 53° 53’ 04,0”W Ramaria sp. Mata de eucalipto Ex PM49 Hulha Negra 31° 25’ 22,7” S 53° 53’ 04,0”W Ramaria sp. Mata de eucalipto Ex PM50 Hulha Negra 31° 25’ 22,7” S 53° 53’ 04,0”W NI2
Mata de eucalipto Ex PM51 Hulha Negra 31° 25’ 22,7” S 53° 53’ 04,0”W NI2
Mata de eucalipto Ex PM52 Bagé 31° 16’ 48,7” S 53° 54’ 39,4”W Pisolithus sp. Mata de acácia negra Ex PM53 Bagé 31° 16’ 48,7” S 53° 54’ 39,4”W NI2
Mata de acácia negra Ex PM55 Rosário do Sul 30° 19’ 52,7” S 55° 00’ 14,5”W NI2
Mata de eucalipto Ex PM56 Rosário do Sul 30° 19’ 52,7” S 55° 00’ 14,5”W NI2
Mata de eucalipto Ex PM57 Rosário do Sul 30° 19’ 52,7” S 55° 00’ 14,5”W Pisolithus sp. Mata de eucalipto Ex PM59 Rosário do Sul 30° 23’ 17,2” S 55° 02’ 22,5”W NI2
Mata de eucalipto Ex
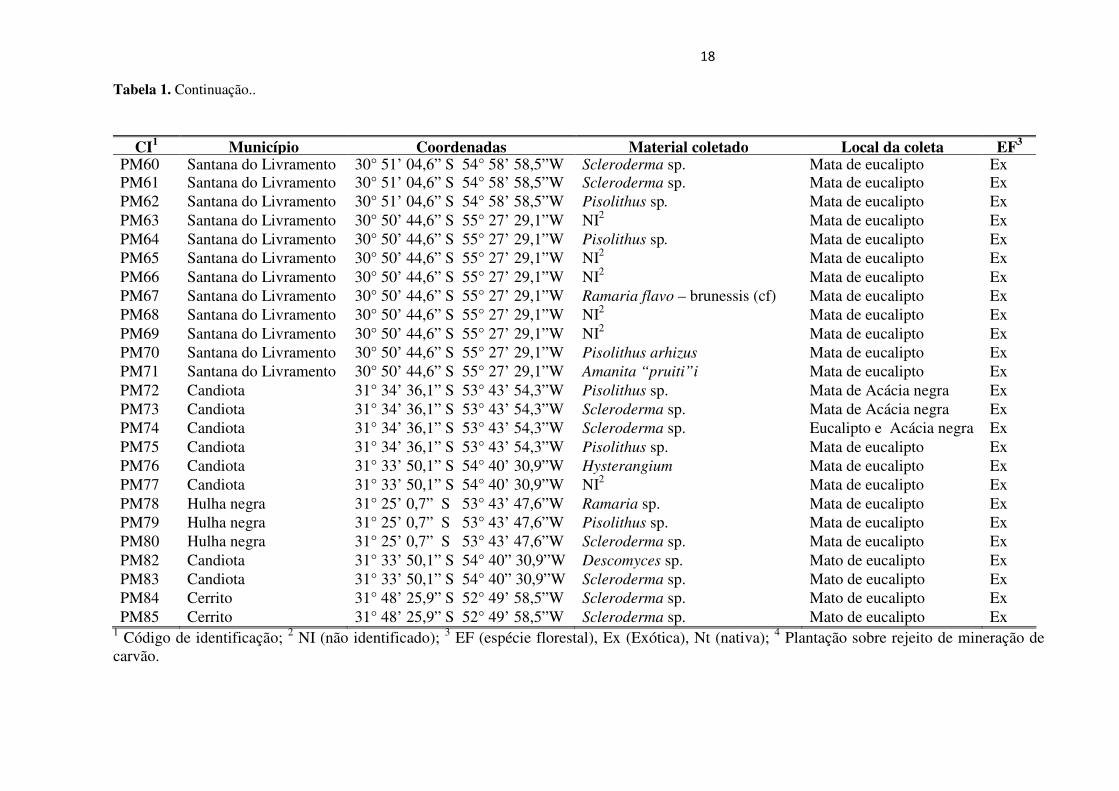
18
Tabela 1. Continuação..
1 Código de identificação; 2 NI (não identificado); 3 EF (espécie florestal), Ex (Exótica), Nt (nativa); 4 Plantação sobre rejeito de mineração de carvão.
CI1 Município Coordenadas Material coletado Local da coleta EF3 PM60 Santana do Livramento 30° 51’ 04,6” S 54° 58’ 58,5”W Scleroderma sp. Mata de eucalipto Ex PM61 Santana do Livramento 30° 51’ 04,6” S 54° 58’ 58,5”W Scleroderma sp. Mata de eucalipto Ex PM62 Santana do Livramento 30° 51’ 04,6” S 54° 58’ 58,5”W Pisolithus sp. Mata de eucalipto Ex PM63 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W NI2
Mata de eucalipto Ex PM64 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W Pisolithus sp. Mata de eucalipto Ex PM65 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W NI2
Mata de eucalipto Ex PM66 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W NI2
Mata de eucalipto Ex PM67 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W Ramaria flavo – brunessis (cf) Mata de eucalipto Ex PM68 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W NI2
Mata de eucalipto Ex PM69 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W NI2
Mata de eucalipto Ex PM70 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W Pisolithus arhizus Mata de eucalipto Ex PM71 Santana do Livramento 30° 50’ 44,6” S 55° 27’ 29,1”W Amanita “pruiti”i Mata de eucalipto Ex PM72 Candiota 31° 34’ 36,1” S 53° 43’ 54,3”W Pisolithus sp. Mata de Acácia negra Ex PM73 Candiota 31° 34’ 36,1” S 53° 43’ 54,3”W Scleroderma sp. Mata de Acácia negra Ex PM74 Candiota 31° 34’ 36,1” S 53° 43’ 54,3”W Scleroderma sp. Eucalipto e Acácia negra Ex PM75 Candiota 31° 34’ 36,1” S 53° 43’ 54,3”W Pisolithus sp. Mata de eucalipto Ex PM76 Candiota 31° 33’ 50,1” S 54° 40’ 30,9”W Hysterangium Mata de eucalipto Ex PM77 Candiota 31° 33’ 50,1” S 54° 40’ 30,9”W NI2 Mata de eucalipto Ex PM78 Hulha negra 31° 25’ 0,7” S 53° 43’ 47,6”W Ramaria sp. Mata de eucalipto Ex PM79 Hulha negra 31° 25’ 0,7” S 53° 43’ 47,6”W Pisolithus sp. Mata de eucalipto Ex PM80 Hulha negra 31° 25’ 0,7” S 53° 43’ 47,6”W Scleroderma sp. Mata de eucalipto Ex PM82 Candiota 31° 33’ 50,1” S 54° 40” 30,9”W Descomyces sp. Mato de eucalipto Ex PM83 Candiota 31° 33’ 50,1” S 54° 40” 30,9”W Scleroderma sp. Mato de eucalipto Ex PM84 Cerrito 31° 48’ 25,9” S 52° 49’ 58,5”W Scleroderma sp. Mato de eucalipto Ex PM85 Cerrito 31° 48’ 25,9” S 52° 49’ 58,5”W Scleroderma sp. Mato de eucalipto Ex
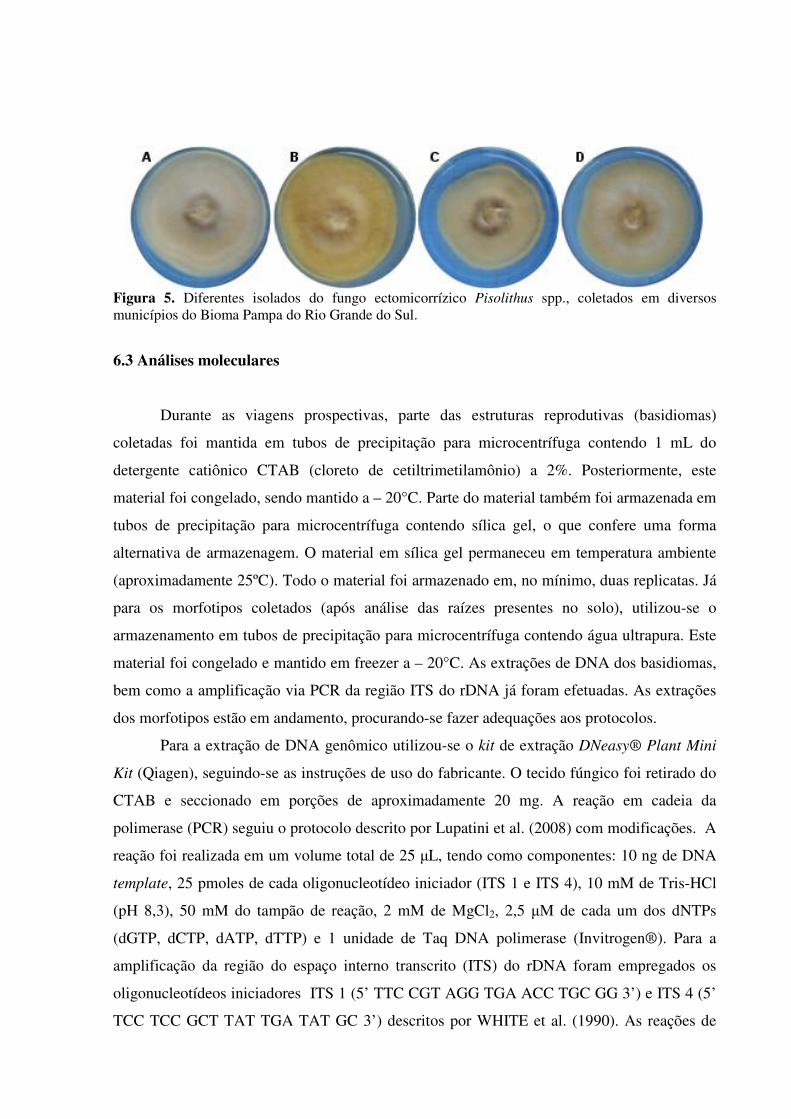
Figura 5. Diferentes isolados do fungo ectomicorrízico Pisolithus spp., coletados em diversos municípios do Bioma Pampa do Rio Grande do Sul.
6.3 Análises moleculares
Durante as viagens prospectivas, parte das estruturas reprodutivas (basidiomas)
coletadas foi mantida em tubos de precipitação para microcentrífuga contendo 1 mL do
detergente catiônico CTAB (cloreto de cetiltrimetilamônio) a 2%. Posteriormente, este
material foi congelado, sendo mantido a – 20°C. Parte do material também foi armazenada em
tubos de precipitação para microcentrífuga contendo sílica gel, o que confere uma forma
alternativa de armazenagem. O material em sílica gel permaneceu em temperatura ambiente
(aproximadamente 25ºC). Todo o material foi armazenado em, no mínimo, duas replicatas. Já
para os morfotipos coletados (após análise das raízes presentes no solo), utilizou-se o
armazenamento em tubos de precipitação para microcentrífuga contendo água ultrapura. Este
material foi congelado e mantido em freezer a – 20°C. As extrações de DNA dos basidiomas,
bem como a amplificação via PCR da região ITS do rDNA já foram efetuadas. As extrações
dos morfotipos estão em andamento, procurando-se fazer adequações aos protocolos.
Para a extração de DNA genômico utilizou-se o kit de extração DNeasy® Plant Mini
Kit (Qiagen), seguindo-se as instruções de uso do fabricante. O tecido fúngico foi retirado do
CTAB e seccionado em porções de aproximadamente 20 mg. A reação em cadeia da
polimerase (PCR) seguiu o protocolo descrito por Lupatini et al. (2008) com modificações. A
reação foi realizada em um volume total de 25 µL, tendo como componentes: 10 ng de DNA
template, 25 pmoles de cada oligonucleotídeo iniciador (ITS 1 e ITS 4), 10 mM de Tris-HCl
(pH 8,3), 50 mM do tampão de reação, 2 mM de MgCl2, 2,5 µM de cada um dos dNTPs
(dGTP, dCTP, dATP, dTTP) e 1 unidade de Taq DNA polimerase (Invitrogen®). Para a
amplificação da região do espaço interno transcrito (ITS) do rDNA foram empregados os
oligonucleotídeos iniciadores ITS 1 (5’ TTC CGT AGG TGA ACC TGC GG 3’) e ITS 4 (5’
TCC TCC GCT TAT TGA TAT GC 3’) descritos por WHITE et al. (1990). As reações de

20
amplificação foram efetuadas por meio de uma desnaturação inicial a 94° C por 2 minutos,
seguida por 35 ciclos que consistiram de: desnaturação a 94° C por 1 minuto, anelamento a
55° C por 1 minuto, e extensão a 72° C por 1 minuto e 30 segundos. Por fim, realizou-se uma
extensão final a 72° C por 10 minutos.
Posteriormente à amplificação, realizou-se a eletroforese dos produtos de PCR em gel
de agarose a 1,5%. A eletroforese foi conduzida em cuba horizontal contendo o gel submerso
em tampão TBE 1X (90 mM Tris-borato, 2 mM EDTA, pH 8,0). Para corar as bandas de
DNA e visualizá-las em transluminador, foi realizada a adição de 10 mg mL-1 de brometo de
etídeo, sendo o registro fotográfico efetuado com o uso do aparato Cânon Snot S2IS.
Para purificação dos produtos de PCR utilizou-se protocolo baseado no uso de PEG
8000 a 13% descrito por DUNN e BLATTNER (1987). Adicionou-se um volume de solução
de PEG 8000 (PEG a 13%, NaCl a 1,6M) homogeneizando-se cuidadosamente com o auxílio
de micropipeta. Este material permaneceu nesta condição overnight. Posteriormente, o
material foi centrifugado por 15 minutos a 13.000 rpm. Descartou-se a solução de PEG e o
material passou por uma etapa de limpeza com a adição de 200 µL de etanol a 70%, seguido
de uma centrifugação por 10 minutos a 13.000 rpm. O procedimento de limpeza foi repetido
por três vezes. Por fim, os pellets foram secos à temperatura ambiente com posterior eluição
em 8 µL de água ultrapura, conforme propõe SAMBROOK et al. (1989).
O material foi enviado para sequenciamento onde foram utlizados os
oligonucleotídeos iniciadores ITS 1 e ITS 4. O sequenciamento foi realizado no sequenciador
Mega BACE 5000 (Amersham Bioscience), seguindo-se o protocolo fornecido pelo
fabricante. Com as sequências analisadas obteve-se a sequência concenso, com auxílio do
programa STADEN (STADEN et al. 1992-2000), assim como o alinhamento de sequências
por meio do algoritmo ClustalW. As sequências do GenBank foram escolhidas utilizando-se o
Algoritmo Megablast, disponível na plataforma BLAST do NCBI
(http://www.ncbi.nlm.nih.gov) e as análises filogenéticas foram realizadas com auxílio do
programa MEGA (TAMURA et al., 2006). O andamento das análises moleculares dos
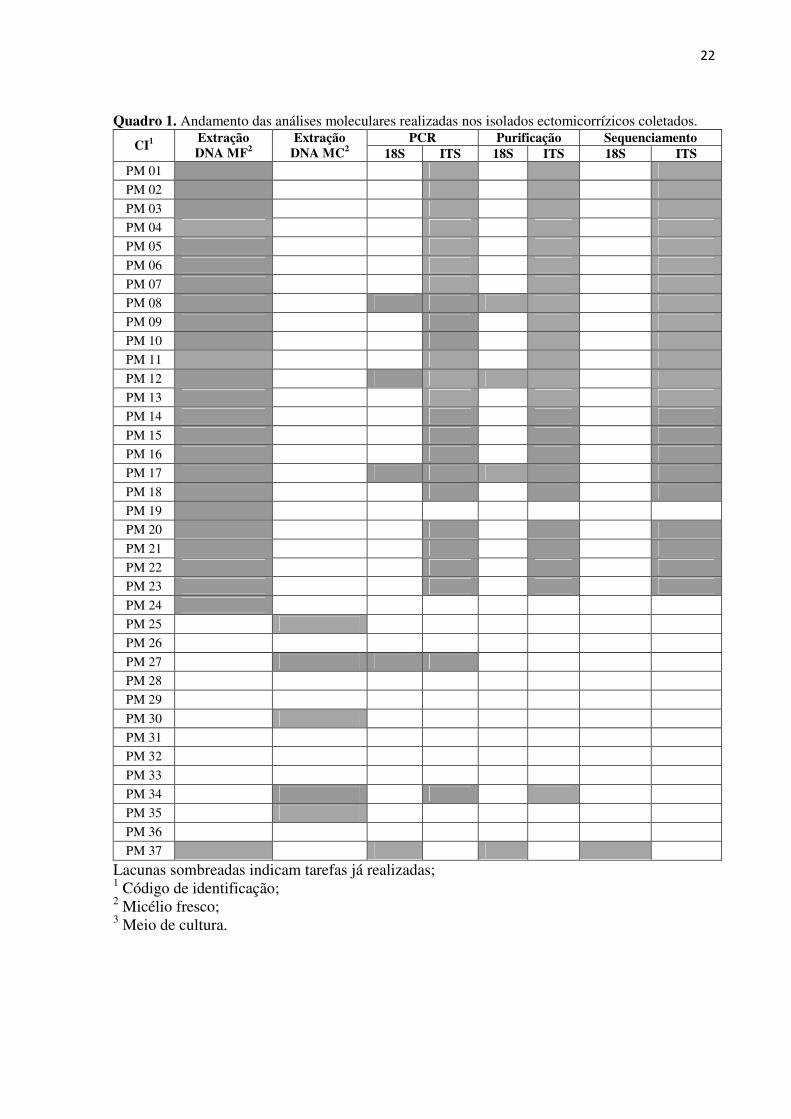
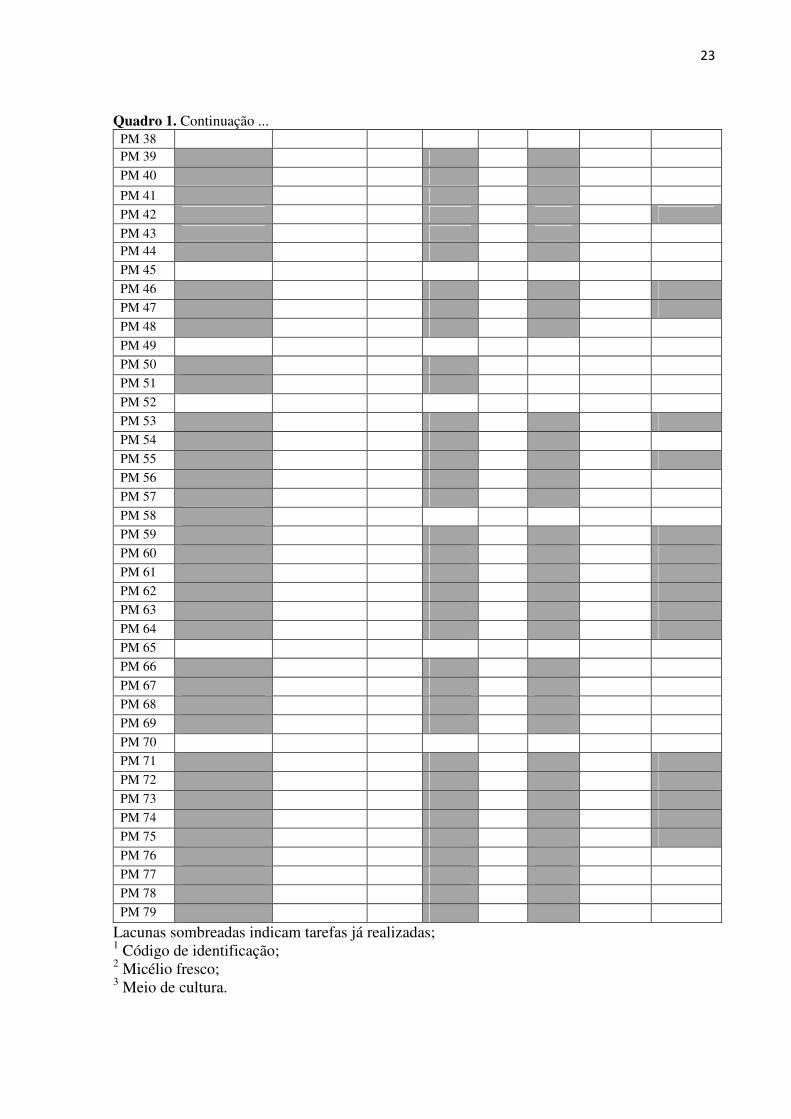
isolados está apresentado no quadro 1.
Na figura 6 é apresentado o gel obtido após a eletrofores do produto da PCR realizada
com os primers da região ITS. Durante a realização das análises foi decidido incluir também
um par de primers para amplificar a região 18S e posteriormente sequenciá-la, uma vez que
esse dado auxilia na identificação dos fungos.

21
Figura 6. Eletrofores dos produtos de PCR obtidos a partir do uso de primers que flanqueiam a região ITS do rDNA. A primeira coluna evidencia o padrão 100 pb, onde foram destacados os fragmentos de 850 e 650 pb. Observam-se as bandas formadas a partir da amplificação dos fungos onde as letras “PM”, seguida de diferentes números, representa o registro de coleta e “C-“ o controle negativo.
De maneira geral foi possível amplificar a região ITS da maioria dos fungos coletados.
Parte das sequencias já foi analisada e os resultados estão apresentados na tabela 2. É
importante salientar que apenas uma fração dos resultados estão apresentados neste
relalatório, já que muitas sequencias ainda estão sendo analisadas.
Tabela 2. Espécies de fungos ectomicorrízicos com a identificação baseada na sequencia ITS.
Amostra Espécie mais próxima N° acesso* Similaridade PM01 Scleroderma bovista HM237175 99% PM02 Scleroderma bovista HM237175 99% PM05 Scleroderma bovista HM237175 99% PM10 Setchelliogaster tenuipes AF325624 99% PM12 Scleroderma bovista HM237175 99% PM13 Scleroderma aurantium HM237174 99% PM14 Scleroderma bovista HM237175 99% PM42 Scleroderma aurantium HM237174 99% PM46 Scleroderma bovista HM237175 99% PM55 Scleroderma bovista HM237175 99% PM71 Amanita sp “prutitii” HQ625011 99% PM72 Pisolithus tinctorius AF374700 99% PM73 Scleroderma polyrhizum EU718123 100% PM74 Scleroderma polyrhizum EU718123 100% PM84 Scleroderma bovista HM237175 99% *N° de acesso no GenBank da espécie com maior similiaridade com a amostra.
22
Quadro 1. Andamento das análises moleculares realizadas nos isolados ectomicorrízicos coletados. PCR Purificação Sequenciamento
CI1 Extração DNA MF2
Extração DNA MC2 18S ITS 18S ITS 18S ITS
PM 01
PM 02
PM 03
PM 04
PM 05
PM 06
PM 07
PM 08
PM 09
PM 10
PM 11
PM 12
PM 13
PM 14
PM 15
PM 16
PM 17
PM 18
PM 19
PM 20
PM 21
PM 22
PM 23
PM 24
PM 25
PM 26
PM 27
PM 28
PM 29
PM 30
PM 31
PM 32
PM 33
PM 34
PM 35
PM 36
PM 37
Lacunas sombreadas indicam tarefas já realizadas; 1 Código de identificação; 2 Micélio fresco; 3 Meio de cultura.
23
Quadro 1. Continuação ... PM 38
PM 39 239
PM 40 30
PM 41
PM 42
PM 43
PM 44
PM 45
PM 46
PM 47
PM 48
PM 49 239
PM 50 30
PM 51
PM 52
PM 53
PM 54
PM 55
PM 56
PM 57
PM 58
PM 59 239
PM 60 30
PM 61
PM 62
PM 63
PM 64
PM 65
PM 66
PM 67
PM 68
PM 69 239
PM 70 30
PM 71
PM 72
PM 73
PM 74
PM 75
PM 76
PM 77
PM 78
PM 79 239
Lacunas sombreadas indicam tarefas já realizadas; 1 Código de identificação; 2 Micélio fresco; 3 Meio de cultura.
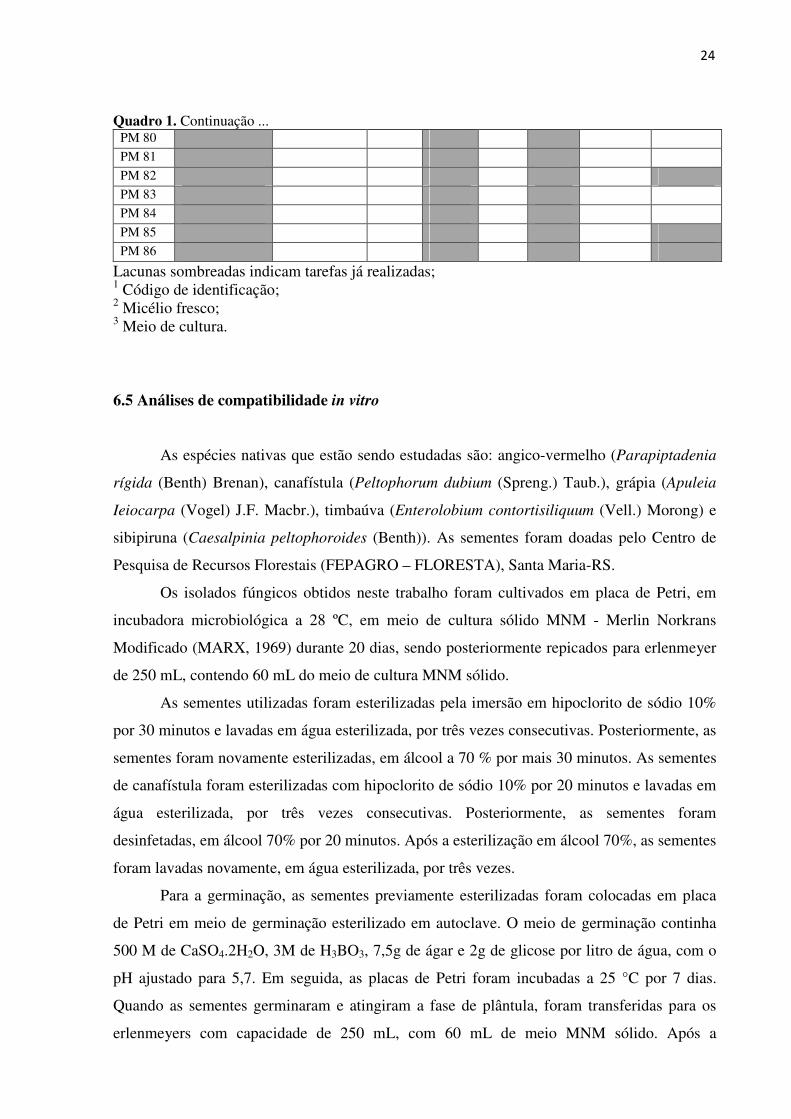
24
Quadro 1. Continuação ... PM 80 30
PM 81
PM 82
PM 83
PM 84
PM 85
PM 86
Lacunas sombreadas indicam tarefas já realizadas; 1 Código de identificação; 2 Micélio fresco; 3 Meio de cultura.
6.5 Análises de compatibilidade in vitro
As espécies nativas que estão sendo estudadas são: angico-vermelho (Parapiptadenia
rígida (Benth) Brenan), canafístula (Peltophorum dubium (Spreng.) Taub.), grápia (Apuleia
Ieiocarpa (Vogel) J.F. Macbr.), timbaúva (Enterolobium contortisiliquum (Vell.) Morong) e
sibipiruna (Caesalpinia peltophoroides (Benth)). As sementes foram doadas pelo Centro de
Pesquisa de Recursos Florestais (FEPAGRO – FLORESTA), Santa Maria-RS.
Os isolados fúngicos obtidos neste trabalho foram cultivados em placa de Petri, em
incubadora microbiológica a 28 ºC, em meio de cultura sólido MNM - Merlin Norkrans
Modificado (MARX, 1969) durante 20 dias, sendo posteriormente repicados para erlenmeyer
de 250 mL, contendo 60 mL do meio de cultura MNM sólido.
As sementes utilizadas foram esterilizadas pela imersão em hipoclorito de sódio 10%
por 30 minutos e lavadas em água esterilizada, por três vezes consecutivas. Posteriormente, as
sementes foram novamente esterilizadas, em álcool a 70 % por mais 30 minutos. As sementes
de canafístula foram esterilizadas com hipoclorito de sódio 10% por 20 minutos e lavadas em
água esterilizada, por três vezes consecutivas. Posteriormente, as sementes foram
desinfetadas, em álcool 70% por 20 minutos. Após a esterilização em álcool 70%, as sementes
foram lavadas novamente, em água esterilizada, por três vezes.
Para a germinação, as sementes previamente esterilizadas foram colocadas em placa
de Petri em meio de germinação esterilizado em autoclave. O meio de germinação continha
500 M de CaSO4.2H2O, 3M de H3BO3, 7,5g de ágar e 2g de glicose por litro de água, com o
pH ajustado para 5,7. Em seguida, as placas de Petri foram incubadas a 25 °C por 7 dias.
Quando as sementes germinaram e atingiram a fase de plântula, foram transferidas para os
erlenmeyers com capacidade de 250 mL, com 60 mL de meio MNM sólido. Após a

25
solidificação do meio, o frasco foi fechado com papel alumínio e plástico de PVC
transparente, para esterilização em autoclave a 1 atm., durante 20 minutos. Após a
autoclavagem, os erlenmeyers estavam prontos para a inoculação das plântulas com os fungos
ectomicorrízicos.
Três discos de 10 mm de diâmetro foram transferidos para os erlenmeyer de 250mL
contendo o meio MNM sólido. Esses erlenmeyer foram incubados em estufa a 28 ºC, durante
30 dias para o crescimento do micélio fúngico. Após a preparação das sementes, duas
sementes pré-germinadas foram adicionadas em cada erlenmeyer. Os erlenmeyers foram
mantidos em incubadora com fotoperíodo de 12 horas a 24±1 ºC, por período de 35 dias.
Durante a condução do experimento, realizou-se rodízio dos erlenmeyers, duas vezes
por semana, atendendo às exigências do delineamento experimental, cujo objetivo foi eliminar
possíveis diferenças quanto à incidência de luz, temperatura e sombreamento. O delineamento
experimental utilizado foi o inteiramente casualizado.
Ao término do experimento, as plântulas das cinco espécies foram avaliadas quanto à
formação de associação micorrízica com os isolados avaliados.
Nesta fase do trabalho, houve contratempos devido à constante adaptação de
metodologias para condução dos ensaios, em função de contaminações oriundas da incubação
das sementes florestais. De todas as espécies florestais avaliadas neste trabalho, observou-se
apenas associação ectomicorrízica “in vitro” da grápia (Apuleia Ieiocarpa) e da canafístula
(Peltophorum dubium) com o fungo Suillus luteus, e da espécie sibipiruna (Caesalpinia
peltophoroides) com o fungo Pisolithus sp. Nestas espécies, após o período de incubação,
observou-se a presença de rede de Hartig, o que evidencia a associação ectomicorrízica
(Figura 7).
Figura 7. Cortes transversais de raízes de mudas de sibipiruna (Caesalpinia peltophoroides) inoculadas com o fECM Pisolithus sp. Aumento de 200x.
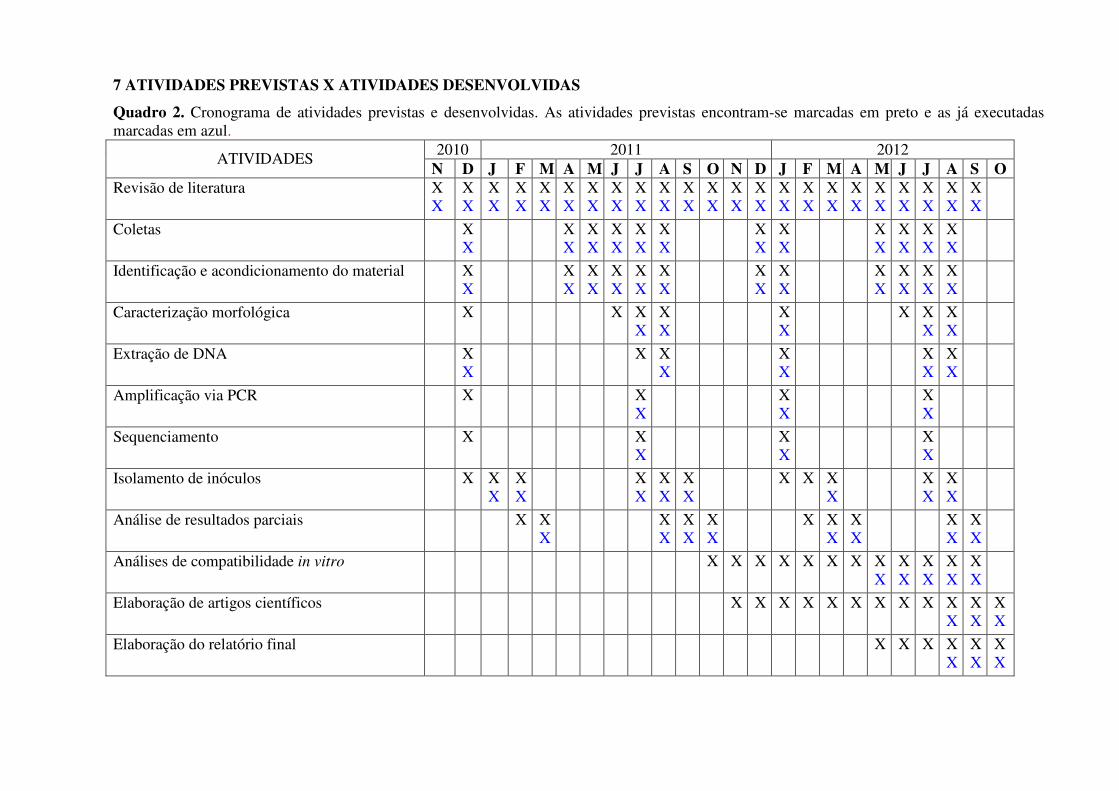
7 ATIVIDADES PREVISTAS X ATIVIDADES DESENVOLVIDAS
Quadro 2. Cronograma de atividades previstas e desenvolvidas. As atividades previstas encontram-se marcadas em preto e as já executadas marcadas em azul.
2010 2011 2012 ATIVIDADES
N D J F M A M J J A S O N D J F M A M J J A S O Revisão de literatura X
X X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
Coletas X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
Identificação e acondicionamento do material X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
X X
Caracterização morfológica X X X X
X X
X X
X X X
X X
Extração de DNA X X
X X X
XX
X X
X X
Amplificação via PCR X X X
X X
X X
Sequenciamento X X X
X X
X X
Isolamento de inóculos X X X
X X
X X
X X
X X
X X X X
X X
X X
Análise de resultados parciais X X X
X X
X X
X X
X X X
X X
X X
X X
Análises de compatibilidade in vitro X X X X X X X X X
X X
X X
X X
X X
Elaboração de artigos científicos X X X X X X X X X X X
X X
X X
Elaboração do relatório final X X X X X
X X
X X

8 PUBLICAÇÕES
Artigos publicados STEFFEN, G. P. K.; STEFFEN, R. B.; ANTONIOLLI, Z. I. Contaminação do solo e da água pelo uso de agrotóxicos. Tecno-lógica, Santa Cruz, v.15, n. 1, p. 15-21, 2011. STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; STEFFEN, R. B.; SCHIEDECK, G. Utilização de vermicomposto como substrato na produção de mudas de Eucalyptus grandis e Corymbia
citriodora. Pesquisa Florestal Brasileira, v. 31, n. 66, p. 75-82, 2011. STEFFEN, R. B.; STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; JACQUES, R. J. S. Ação do óleo essencial de eucalipto na micorrização e no estabelecimento de Eucalyptus grandis em solo contaminado por cobre. Pesquisa Florestal Brasileira, v. 31, n. 67, p. 245-255, 2011 STEFFEN, R. B.; STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; JACQUES, R. J. S.; SANTOS, M. L. dos. Efeito do óleo essencial de Eucalyptus grandis no crescimento de isolados de fungos ectomicorrízicos em diferentes concentrações de cobre, zinco e níquel. Pesquisa Florestal Brasileira, v. 31, n. 67, p. 227-234, 2011. STEFFEN, R. B.; STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; JACQUES, R. J. S. Influência do óleo essencial na micorrização e no crescimento de mudas de eucalipto. Pesquisa Florestal Brasileira, v. 31, n. 67, p. 235-243, 2011. STEFFEN, R. B.; STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; SILVA, R. F. da. Óleo essencial de Eucalyptus grandis Hill em Maiden no estímulo à micorrização de mudas de sibipiruna (Caesalpinia peltophoroides Benth.). Ciência Florestal, v. 22, n. 1, p. 69-78, 2012. SILVA, R. F.; SAIDELLES, F. L. F.; KEMERICH, P. D. C.; STEFFEN, R. B.; SWAROWSKY, A.; SILVA, A. S. Crescimento e qualidade de mudas de Timbó e Dedaleiro cultivadas em solo contaminado por cobre. Revista Brasileira de Engenharia Agrícola e Ambiental, v.16, n.8, p.881–886, 2012. Artigos aceitos para publicação SILVA, R. F.; BERTOLLO, G. M.; CORASSA, G. M.; COCCO, L. B.; STEFFEN, R. B. Influência de doses de dejetos de suínos na fauna do solo em sistema plantio direto e cultivo mínimo. Ciência Florestal, 2012. STEFFEN, R. B.; STEFFEN, G. P. K.; ANTONIOLLI, Z. I.; JACQUES, R. J. S.; SANTOS, M. L.; BOGUSZ JUNIOR, S.; GODOY, H. T. Óleo essencial de eucalipto como bioestimulador do crescimento de fungos ectomicorrízicos in vitro. Ciência Florestal, 2012.

28
9 REFERÊNCIAS
BILENCA, D.; MIÑARRO, F. Identificación de áreas valiosas de pastizal en las Pampas y Campos de Argentina, Uruguay y Sur de Brasil. Fundación Vida Silvestre, 2004. BOLDRINI, I. I. Campos no Rio Grande do Sul. Fisionomia e problemática ocupacional. Boletim do Instituto Central de Biociências, v. 56, 1997. p. 1-39. BRIDGE, P.D.; ARORA, D.K. Interpretation of PCR methods for species definition. In: BRIDGE, P.D.; ARORA, D.K.; REDDY, C.A.; ELANDER, R.P. Applications of PCR in mycology. CAB International: New York, 1998. p.63-84. BRUNDRETT, M. et al. Working with mycorrhizas in forestry and agriculture. Canberra, ACIAR, Monograph 32. 373p. BRUNNER, I. Ectomycorrhizas: their role in forest ecosystems under the impact of acidifying pollutants. Perspectives in Plant Ecology, Evolution and Systematics, Zürich, v. 4, n. 1, p. 13-27, 2001.
CARVALHO, P. C. F.; BATELLO, C. Access to land, livestock production and ecosystem conservation in the Brazilian Campos biome: The natural grasslands dilemma. Livestock Science, v. 120, p. 158-162, 2009. DUNN, I.S.; BLATTNER, F.R. Charon-36 to charon-40 multi enzyme, high-capacity, recombination deficient replacemet vectors with polylinkers and polystuffers. Nucleic Acids Research, n. 15, p. 2677-2698, 1987. GOMES, E.A. et al. Polymorphism in the internal transcribed spacer (ITS) of the ribosomal DNA of 26 isolates of ectomycorrhizal fungi. Genetics and molecular biology, v.25, n.4, 2002. p. 477-483. HOBBIE, E. A. Carbon allocation to ectomycorrhizal fungi correlates with belowground allocation in culture studies. Ecology, Davis, v. 87, n. 3, p. 563-569, 2006.
IBGE - Instituto brasileiro de geografia e estatística. Mapa dos biomas brasileiros. Acesso on line em 22 de março de 2010. Disponível em: <http://www.ibge.gov.br/home/presidencia/noticias/21052004biomas.shtm>. LUPATINI, M.; et al. Mycorrhizal morphotyping and molecular characterization of Chondrogaster angustisporus Giachini, Castellano, Trappe & Oliveira, na ectomycorrhizal fungus from Eucalyptus. Mycorrhiza, n. 18, p. 437 – 442, 2008. MARX, D. H. The influence of ectotrophic mycorrhizal fungi on the resistance of pine roots to pathogenic fungi and soil bacteria. I. Antagonism of mycorrhizal fungi to root pathogenic fungi and soil bacteria. Phytopathology, St. Paul, 59: 153-163, 1969. MELLO, A. H. de et al. Fungos arbusculares e ectomicorrízicos em áreas de eucalipto e de campo nativo em solo arenoso. Ciência Florestal, Santa Maria, v. 16, n. 3, p. 293-301, 2006.

29
MOLINA, M. L.; LEDESMA, L. M.; MEDINA, M. S. Importância del manejo de hongos micorrizógenos en el establecimiento de árboles en sistemas silvopastoriles. Revista Colombiana de Ciências Pecuárias, Medellín, v. 18, n. 2, p. 162-175, 2005.
MORRIS, M. H. et al. Influence of host species on ectomycorrhizal communities associated with two co-occuring oaks (Quercus spp.) in a tropical cloud forest. FEMS Microbiology ecology, v. 69, n.2, 2009. p. 274-287 NARA, K.; HOGETSU, T. Ectomycorrhizal fungi on established shrubs facilitate subsequent seedling establishment of successional plant species. Ecology, v. 85, n.6, 2004. p. 1700-1707. OLIVEIRA, V. L.; GIACHINI, A. J. Ecologia e aplicação de ectomicorrizas. In: SIQUEIRA, J. O. et al. Inter-relação fertilidade, biologia do solo e nutrição de plantas. UFLA: Lavras, 1999. 818p.
OVERBECK, G. E. et al. Brazil’s neglected biome: The south Brazilian Campos. Perspectives in Plant Ecology, Evolution and Systematics, v. 9, 2007. p. 101-116. RILLIG, M.C.; MUMMEY, D.L. Tansley review: mycorrhizas and soil structure. New Phytol, v.171, 2006. p. 41-53 SBS – Sociedade Brasileira de Silvicultura. Fatos e números do Brasil florestal. Sociedade Brasileira de Silviculta, 2007. 109 p. SILVA, R. F. da; ANTONIOLLI, Z. I.; ANDREAZZA, R. Efeito da inoculação com fungos ectomicorrízicos na produção de mudas de Eucalyptus grandis W. ex Maiden em solo arenoso. Ciência Florestal, Santa Maria, v. 13, n. 1, p. 33-42, 2003 SMITH, S. E.; READ, D.J. Mycorrhizal symbiosis. London: Academic press, 1997. 605p. SUERTEGARAY, D. M. A. Deserto Grande do Sul: Controvérsia. Porto Alegre: UFRGS, 1998. 130 p. TAMURA, K.; KUMAR, S.; NEI, M. Mega: integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics, Baltimore, v.5, p.150-163, 2006. TAYLOR, R. J. Plants, fungi and bettongs: A fire-dependent co-evolutionary relationship. Austral Ecology, Adelaide, v. 16, n. 3, p. 409-411, 1991. TEDERSOO, L. et al. Ectomycorrhizal fungi of the Seychelles: diversity patterns and host shifts from the native Vateriopsis seychellarum (Dipterocarpaceae) and Intsia bijuca (Caesalpiniaceae) to the introduced Eucalyptus robusta (Myrtaceae), but not Pinus caribea (Pinaceae). New Phytol, v. 175, 2007. p. 321-333. VELLINGA, E. C.; WOLFE, B. E.; PRINGLE, A. Global patterns of ectomycorrhizal introdictions. New Phytologist, 10: 1-14, 2009.

30
WHITE, T. J. et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A. et al. (Eds.) PCR protocols. A guide to methods and applications. Academic press, San Diego, 1990. p. 315-322.
Top Related