







Línguas
Páginas
Legal



PALOMA OLIVEIRA VIDAL
Estudo genético e morfológico de populações de Aedes aegypti (Culicidae) na área metropolitana de São Paulo – SP
Dissertação apresentada ao Departamento de Parasitologia do Instituto de Ciências Biomédicas da Universidade de São Paulo, para obtenção do Título de Mestre em Ciências. Área de concentração: Biologia da Relação Patógeno-Hospedeiro Orientador: Prof. Dr. Lincoln Suesdek
São Paulo 2010

DADOS DE CATALOGAÇÃO NA PUBLICAÇÃO (CIP) Serviço de Biblioteca e Informação Biomédica do
Instituto de Ciências Biomédicas da Universidade de São Paulo
© reprodução total
Vidal, Paloma Oliveira.
Estudo genético e morfológico de populações de Aedes aegypti (Culicidae) na área metropolitana de São Paulo (SP) / Paloma Oliveira Vidal. -- São Paulo, 2010.
Orientador: Lincoln Suesdek. Dissertação (Mestrado) – Universidade de São Paulo. Instituto de Ciências Biomédicas. Departamento de Parasitologia. Área de concentração: Biologia da Relação Patógeno-Hospedeiro. Linha de pesquisa: Culicídeos de interesse médico Versão do título para o inglês: Genetic and morphological study of Aedes aegypti (culicidae) population in São Paulo (SP) metropolitan area. Descritores: 1. Aedes aegypti 2. Morfometria 3. Evolução 4. Genética 5.Insetos vetores 6. Culicidae I. Suesdek, Lincoln II. Universidade de São Paulo. Instituto de Ciências Biomédicas. Programa de Pós Graduação em Biologia da Relação Patógeno-Hospedeiro III. Título.
ICB/SBIB0175/2010

UNIVERSIDADE DE SÃO PAULO INSTITUTO DE CIÊNCIAS BIOMÉDICAS
_________________________________________________________________________________________________________Candidato(a): Paloma Vidal Oliveira.
Título da Dissertação: Estudo genético e morfológico de populações de Aedes aegypti (Culicidae) na área metropolitana de São Paulo (SP).
Orientador(a): Lincoln Suesdek.
A Comissão Julgadora dos trabalhos de Defesa da Dissertação de Mestrado, em sessão pública realizada a .............../................./................., considerou
( ) Aprovado(a) ( ) Reprovado(a)
Examinador(a): Assinatura: ......................................................................................... Nome: ................................................................................................. Instituição: ..........................................................................................
Examinador(a): Assinatura: ......................................................................................... Nome: ................................................................................................. Instituição: ..........................................................................................
Presidente: Assinatura: ......................................................................................... Nome: ................................................................................................. Instituição: ..........................................................................................


Aos meus pais, Eudes e Nadja por todo apoio, dedicação e educação. Ao meu marido Jaime Henrique pelo amor, carinho e amizade. Ao meu irmão Ramon pela grande amizade.

AGRADECIMENTOS
Ao Professor Doutor Lincoln Suesdek, pela orientação, confiança, oportunidade de crescimento acadêmico/científico, amizade e conhecimento transmitidos durante esses anos de convivência, os quais foram fundamentais para a minha formação.
Ao programa de Pós-graduação Biologia da Relação Patógeno Hospedeiro, do
Instituto de Ciências Biomédicas da USP. Ao laboratório de Parasitologia do Instituto Butantan onde todo o trabalho foi
desenvolvido. Aos Centros de Controle de Zoonoses dos municípios de Guarulhos, Osasco e
Suzano por terem ajudado cedendo larvas coletadas nos pontos estratégicos de cada município.
Às funcionárias Ângela e Wilma do departamento de Parasitologia do ICB. Aos meus pais por serem meu exemplo de Vida, minha força, por estarem
sempre acreditando em mim e me apoiando. Ao meu amor Jaime Henrique, pela paciência, carinho, apoio, por ser um grande
companheiro, estando ao meu lado em todos os momentos. Sempre me incentivando e me encorajando a lutar pelos meus objetivos. Sem esquecer das nossas discussões científicas bastante enriquecedoras, muito obrigada!
Ao meu irmão pela amizade, momentos de diversão e por nunca me negar uma
ajuda quando eu precisava. Aos meus sogros que mesmo de longe sempre torceram pelo meu bom
desempenho. Às mestres Maria Cristina e Camila Moratore que me ajudaram a dar os
primeiros passos na área da morfometria geométrica. Aos “morfométricos” formado pelo Eduardo (Edu), Fábio (Fio), Mariana (Mari) e a
Vívian (Cidinha), por realmente ser uma equipe que trabalha unida, sempre dispostos a ajudar e proporcionando vários momentos de descontração.
Aos colegas, funcionários e pesquisadores do Laboratório de Parasitologia do
Instituto Butantan pela troca de experiências, ajuda, convívio e descontração. Ao CNPq pela concessão da bolsa. A Deus pelo dom da Vida e Saúde que possibilitaram a realização deste
trabalho.

O que vale na vida não é o ponto de partida e sim a caminhada. Caminhando e semeando, no fim terás o que colher.
Cora Coralina

RESUMO
Vidal PO. Estudo genético e morfológico de populações de Aedes aegypti (Culicidae) na área metropolitana de São Paulo (SP) [dissertação (Mestrado em Parasitologia)]. São Paulo: Instituto de Ciências Biomédicas da Universidade de São Paulo; 2010. Aedes aegypti é uma espécie de mosquito da família Culicidae (Diptera) proveniente da região africana. Atualmente está distribuída por quase todo o mundo, com ocorrência nas regiões tropicais e subtropicais. O controle das suas populações é considerado assunto de saúde pública por ser o vetor de agentes etiológicos de doenças graves como a dengue e a febre amarela. Devido à inexistência de uma vacina para combater o vírus dengue, a alternativa atual é controlar o vetor. Estudos populacionais são de grande importância para o desenvolvimento de estratégias de controle, no entanto, no Estado de São Paulo há uma carência de estudos populacionais referente ao Ae. aegypti. Devido a esse contexto, o objetivo do trabalho foi avaliar a variabilidade genética e fenotípica de quatro populações de Ae. aegypti na área metropolitana de São Paulo, Butantã, Guarulhos, Osasco e Suzano. As distâncias geográficas entre essas localidades são no máximo 50 km e no mínimo 13,5 km. A assimetria bilateral alar quanto à forma foi tênue e em relação ao tamanho não se mostrou mais acentuada em nenhuma das populações analisadas. As quatro populações revelaram alto dimorfismo sexual de forma e tamanho, sendo asas de fêmeas maiores que asas de machos. As análises interpopulacionais de forma em ambos os sexos mostraram variabilidade morfológica alar que, no entanto não indica estruturação populacional. Não foi observada correlação entre valores de distância fenética alar e distância geográfica para machos (r = -0,03) e fêmeas (r= 0,34). Nas análises genéticas foram utilizados cinco loci microssatélites. Foi encontrada uma média de cinco alelos por locus. A heterozigosidade média esperada variou de 0,512 a 0,609. Dos loci analisados 65 % não se encontram em equilíbrio de Hardy-Weinberg, devido ao déficit ou excesso de heterozigotos. Estruturação populacional foi detectada (Fst global = 0,062, p <0,05) e baixo fluxo gênico (Nm = 0,47) foi presumido nas quatro localidades. Esses resultados apontam para uma heterogeneidade genética entre essas populações, distanciadas em pequenas escalas geográficas. Foi observada correlação entre as distâncias genética e geográfica (r= 0,76). Os dois marcadores populacionais aqui empregados aparentemente têm graus de resolução distintos para eventos microevolutivos, tendo sido DNA microssatélite ligeiramente mais sensível para acusar estruturação populacional.
Palavras–chave: Morfometria geométrica alar. Microssatélites. Microevolução. Mosquitos. Culicídeos.

ABSTRACT
Vidal PO. Genetic and morphological study of Aedes aegypti (culicidae) population in São Paulo (SP) metropolitan area. [Master thesis (Parasitology)]. São Paulo: Instituto de Ciências Biomédicas da Universidade de São Paulo; 2010. Aedes aegypti is a mosquito species of Culicidae family (Diptera) originary from Africa. It currently occurs in most of the world, with more frequency in tropical and subtropical regions. The control of their populations is considered a public health subject because this mosquito is the vector of etiologic agents that causes serious diseases such as dengue and yellow fever. Since vaccines are still under development, dengue prevention depends primordially on vector control. Populational studies of Ae. aegypti are important to develop control strategies, but unfortunately the State of São Paulo lacks such studies. Despite this context, the objective of this work was to evaluate genetic and phenotypic variability of population samples of Ae. aegypti from four collecting sites in the metropolitan area of São Paulo city: Butantan, Guarulhos, Osasco and Suzano. Geographic distances between locations ranged from 13.5 to 50 km. The wing asymmetry is non-significant for size and tenuous for shape from four populations analyzed. The four populations revealed strong sexual dimorphism concerning shape and size, being the wings of females larger than those of males in all populational samples. The interpopulational analysis of shape for females and males showed morphological variability in wings, but did not indicate population structure. Pairwise fenetic distances among populations were not correlated to the geographic distances for males (r = -0.03) and females (r = 0.34). For genetic analysis we used five microsatellite loci. All five loci were polymorphic (mean alleles/locus= 5). The mean expected heterozygotes ranged from 0.512 to 0.609. Of the loci analized, 65 % were not in Hardy-Weinberg equilibrium due to either deficit or excess of heterozygote. Populational structure was detected (global Fst= 0.062; p<0.05) and low gene flow (Nm= 0.41) was presumed among four locations. These results indicate a genetic heterogeneity among these populations within a small geographic scale. Pairwise genetic distances among populations were correlated to the geographic distances (r = 0.76). The two populational markers used here apparently have different degrees of resolution for microevolutionary events, being that microsatellite DNA were slightly more sensitive in revealing populational structure.
Keywords: Wing geometric morphometrics. Microsatellites. Microevolution. Mosquitoes. Culicidae.

LISTA DE ABREVIATURAS E SIGLAS
SSRs BUT GUA OSA SUZ FD FE MD ME CP MG pb HW Nm
microssatélites população do bairro do Butantã, município de São Paulo população do município de Guarulhos população do município de Osasco população do município de Suzano asas direitas de fêmeas asas esquerdas de fêmeas asas direitas de machos asas esquerdas de machos componente principal morfometria geométrica pares de bases de DNA Hardy-Weinberg (modelo de genética populacional) número de migrantes

LISTA DE FIGURAS
Figura 1. Ciclo de Vida do Aedes aegypti ................................................................
19
Figura 2. Mapa da área metropolitana de São Paulo ..............................................
30
Figura 3. Pupas e larvas do mosquito Aedes aegypti .............................................
31
Figura 4. Fêmea de Aedes aegypti realizando o repasto sanguíneo ......................
32
Figura 5. Foto da asa de Aedes aegypti com os pontos anatômicos ......................
33
Figura 6. Assimetria bilateral do formato alar de fêmeas de Aedes aegypti ...........
41
Figura 7. Assimetria bilateral do formato alar de machos de Aedes aegypti ...........
42
Figura 8. Assimetria bilateral do tamanho alar de fêmeas de Aedes aegypti ..........
43
Figura 9. Assimetria bilateral do tamanho alar de machos de Aedes aegypti .........
44
Figura 10. Dimorfismo sexual alar na população do Butantã ..................................
46
Figura 11. Dimorfismo sexual alar na população de Guaruhos ...............................
47
Figura 12. Dimorfismo sexual alar na população de Osasco ..................................
48
Figura 13. Dimorfismo sexual alar na população de Suzano...................................
49
Figura 14. Dimorfismo sexual do tamanho alar .......................................................
51
Figura 15. Variação geográfica do formato alar de fêmeas .....................................
52

Figura 16. Variação geográfica do formato alar de machos ....................................
53
Figura 17. Fenograma de distância de Mahalanobis entre fêmeas .........................
54
Figura 18. Fenograma de distância de Mahalanobis entre machos ........................
54
Figura 19. Fenograma de distância geográfica .......................................................
55
Figura 20. Correlação entre distância geográfica e distância fenética ....................
56
Figura 21. Variação geográfica de tamanho alar de machos e fêmeas ..................
57
Figura 22. Locus AED19 – alelos e suas respectivas freqüências ..........................
61
Figura 23. Locus T3A7– alelos e suas respectivas freqüências ..............................
62
Figura 24. Locus 38/38 – alelos e suas respectivas freqüências ............................
62
Figura 25. Locus 34/72 – alelos e suas respectivas freqüências ............................
63
Figura 26. Locus C2A8 - alelos e suas respectivas freqüências .............................
63
Figura 27. Número de homozigotos esperado e observado para os cinco loci microssatélites na população do Butantã .................................................................
69
Figura 28. Número de homozigotos esperado e observado para os cinco loci microssatélites na população de Guarulhos .............................................................
69
Figura 29. Número de homozigotos esperado e observado para os cinco loci microssatélites na população do Osasco .................................................................
70

Figura 30. Número de homozigotos esperado e observado para os cinco loci microssatélites na população de Suzano .................................................................
70
Figura 31. Fenograma de distância genética ..........................................................
73
Figura 32. Fenograma de distância geográfica .......................................................
73
Figura 33. Correlação entre distância geográfica e distância genética para as quatro populações de Ae. aegypti.............................................................................
74

LISTA DE TABELAS
Tabela 1 - Dados de coleta das amostras populacionais de Aedes aegypti ............
31
Tabela 2 - Marcadores microssatélites utilizados para caracterizar população de Ae. aegypti.................................................................................................................
39
Tabela 3 - Resultados da aplicação do teste T estatístico: assimetria bilateral de tamanho alar de fêmeas ...........................................................................................
43
Tabela 4 - Resultados da aplicação do teste T estatístico: assimetria bilateral de tamanho alar de machos ..........................................................................................
44
Tabela 5 - Estatística descritiva com valores de média, mínimo, máximo e desvio padrão para tamanhos de centróide .........................................................................
45
Tabela 6 - Teste de reclassificação para as análises de dimorfismo sexual alar ....
50
Tabela 7 - Resultados da aplicação do teste T estatístico: dimorfismo sexual de tamanho alar .............................................................................................................
50
Tabela 8 - Teste de reclassificação para as análises de variação geográfica .........
53
Tabela 9 - Resultados da aplicação do teste T estatístico: Variação geográfica de tamanho alar .............................................................................................................
58
Tabela 10 - Tamanho dos fragmentos amplificados de microssatélites nas populações de Aedes aegypti ..................................................................................
59
Tabela 11 - Número de alelos encontrado em cada locus .......................................
59
Tabela 12 - Genótipos encontrados no locus AED19 .............................................. Tabela 13 - Genótipos encontrados no locus T3A7..................................................
64 65

Tabela 14 - Genótipos encontrados no locus 38/38 ................................................ Tabela 15 - Genótipos encontrados no locus 34/72 ................................................ Tabela 16 - Genótipos encontrados no locus C2A8 ................................................ Tabela 17 - Fis e desvios de Hardy – Weinberg observado em cinco locus microssatélites de Ae. aegypti .................................................................................. Tabela 18 - Estimativa de valores de freqüência de alelos nulos ............................ Tabela 19 - Estimativas de valores F-estatístico de Wright .....................................
65 66 66 68 68 71

SUMÁRIO
1 INTRODUÇÃO ....................................................................................................1.1 A espécie Aedes aegypti ...............................................................................1.2 O vetor Aedes aegypti ...................................................................................
1.3 Controle populacional versus microevolução ............................................1.4 Detectando variações populacionais com múltiplas abordagens ............
1.5 Morfometria geométrica alar .........................................................................1.6 DNA nuclear – Microssatélites ..................................................................... 1.7 Estudos de diferenciação populacional de Aedes aegypti no Brasil .......1.8 Aedes aegypti no estado de São Paulo .......................................................2 OBJETIVOS ....................................................................................................... 2.1 Objetivo geral .................................................................................................2.2 Objetivos específicos ....................................................................................3 MATERIAIS E MÉTODOS ..................................................................................3.1 Coleta e armazenamento dos exemplares .................................................. 3.2 Análise morfométrica alar .............................................................................3.2.1 Componentes principais ...............................................................................
3.2.2 Análises discriminantes ................................................................................
3.2.3 Distância de Mahalanobis .............................................................................
3.2.4 Tamanho de centróide ..................................................................................
3.3 Análises genéticas – Microssatélites ...........................................................3.3.1 Extração do DNA genômico ..........................................................................
3.3.2 Reação em cadeia da polimerase ................................................................
3.3.3 Análises microssatélites ................................................................................
3.4 Análises de correlação ..................................................................................
4 RESULTADOS ...................................................................................................4.1 Morfometria geométrica alar .........................................................................4.1.1 Assimetria bilateral ........................................................................................
4.1.1.1 Análise de forma ........................................................................................
4.1.1.2 Análise de tamanho....................................................................................
18
18
19
21
22
23
25
26
27
29
29
29
30
30
32
33
34
34
35
35
35
36
37
38
40
40
40
40
40

4.1.2 Dimorfismo sexual ........................................................................................
4.1.2.1 Análise de forma ........................................................................................
4.1.2.2 Análise de tamanho ...................................................................................
4.1.3 Variação geográfica ......................................................................................
4.1.3.1 Análise da variação interpopulacional de forma ........................................
4.1.3.1 Análise da variação interpopulacional de tamanho ...................................
4.1.4 Teste de repetibilidade ..................................................................................
4.2 Microssatélites ...............................................................................................4.2.1 Freqüências alélicas e genotípicas ...............................................................
4.2.2 Testes de equilíbrio de Hardy-Weinberg ......................................................
4.2.3 Desequilíbrio de ligação ................................................................................
4.2.4 Diferenciação populacional ..........................................................................
4.2.5 Distância genética versus distância geográfica ...........................................
5 DISCUSSÃO .......................................................................................................5.1 Morfometria geométrica alar .........................................................................5.2 Microssatélites ...............................................................................................6 CONCLUSÕES ...................................................................................................REFERÊNCIAS .....................................................................................................
45
45
50
51
51
57
58
58
59
67
71
71
72
75
75
76
81
82

18
1 INTRODUÇÃO 1.1 A espécie Aedes aegypti
A família Culicidae é representada por aproximadamente 3.500 espécies de
insetos. A grande importância dessa família é devida ao fato de seus representantes
estarem envolvidos na transmissão de patógenos ao homem. O Aedes (Stegomyia)
aegypti Linnaeus 1762 é natural da região afrotropical onde são encontrados os outros
membros do grupo (Belkin, 1962). Trata-se de uma espécie classicamente tida como
tropical e subtropical com sua distribuição que, beirando o cosmopolitismo, é em parte
resultante das atividades humanas. Originalmente descrito no Egito, o que lhe conferiu
seu epíteto específico (aegypti), a espécie tem acompanhado o homem em sua
permanente migração (Nelson, 1986; Christophers, 1960).
São insetos holometábolos, apresentando metamorfose completa durante o seu
desenvolvimento. Assim, como qualquer culicídeo, apresenta duas fases no seu ciclo
de vida, uma aquática (larva e pupa) e uma terrestre (ovo e o indivíduo adulto), como
ilustrado na Figura 1. As larvas se alimentam de partículas em suspensão na água e
tem duração de 4 a 8 dias, no estágio de pupa não ocorre alimentação, e tem duração
de 2 dias, o período que ocorre cada estágio depende principalmente da temperatura e
disponibilidade de alimento (Consoli e Lourenço-de-Oliveira, 1994; Forattini, 2002). Está
entre as espécies de mosquito que passa mais rapidamente pela fase imatura, já que
as fêmeas normalmente depositam os ovos em recipientes pequenos.
Machos e fêmeas adultas se alimentam de néctar de fluidos açucarados, mas
somente as fêmeas são hematófagas, picando durante todo o dia, apresentando hábito
diurno. O sangue é a fonte de proteínas que se fazem necessárias em sua alimentação
para a maturação dos ovos. Em geral a fêmea faz uma postura após cada repasto
sanguíneo.
Quanto à sua capacidade de dispersão, na sua forma ativa, apresenta-se
reduzida. Machos raramente atingem mais que 100 metros além do lugar de onde se
originou por isso a sua presença é um indicador seguro de criadouros próximos.
Havendo aglomeração humana e muitos criadouros disponíveis as fêmeas não

19
parecem necessitar de grandes deslocamentos, cerca de 100 a 500 metros, por outro
lado na falta desses recursos elas tendem a se deslocar por distâncias maiores (Maciel-
de-Freitas e Lourenço-de-Oliveira, 2006).
Figura 1. Ciclo de vida do Ae. aegypti. Fonte: Instituto Oswaldo Cruz (2010).
Em 2003, Braks et al. demonstraram que no Rio de Janeiro a distribuição,
densidade e freqüência de Ae. aegypti estavam associadas a presença humana, sendo
este um mosquito mais abundante no domicílio e peridomicilio. Esta espécie apresenta
hábitos sinantrópicos e antropofílicos, sendo considerado o culicídeo que tem maior
interação com os seres humanos.
1.2 O vetor Aedes aegypti
O Ae. aegypti foi reconhecido como transmissor da febre amarela em 1881
(Rodriguez e Finlay, 1971). Em 1906, Brancroft publicou as primeiras evidências de que
o mosquito também era o vetor de dengue, fato posteriormente confirmado por
Agramonte, em 1906, e por Simmons, em 1931 (Centers for Disease Control, 1979). O
Ae. aegypti é um eficiente vetor do vírus dengue especialmente devido ao

20
comportamento hematófago intermitente, podendo assim se alimentar em mais de um
hospedeiro durante um único ciclo gonotrófico, período que representa o tempo que
ocorre a ingestão de sangue até a postura dos ovos (Mackenzie et al., 2004). Estudo
mostrou que as fêmeas de Ae. aegypti tem preferência e freqüentemente se alimentam
de sangue humano devido à baixa concentração do aminoácido isoleucina (Harrington
et al., 2001)
Provavelmente esse vetor teve a introdução nas Américas através de barcos
vindos da Europa, que cruzavam o Atlântico durante as primeiras explorações e
colonizações européias ao Novo Mundo no século XV (Bisset, 2002). Os primeiros
registros de sua identificação em terras brasileiras foram em 1898, por Lutz, e em 1899,
por Ribas apud Franco, 1969.
A ocorrência de epidemias da febre amarela levou ao combate do vetor a partir
da década de 1940. A espécie foi erradicada em 1955, sendo encontrada mais tarde
em 1967 no estado do Pará. A segunda erradicação ocorreu em 1973. Em 1976 foi
detectado na cidade de Salvador, na Bahia e a partir dessa data nunca mais foi
eliminada totalmente do país (Tauil, 2002).
Atualmente, o Ae. aegypti é o principal vetor do vírus da dengue no Brasil,
presente nos 26 estados e no Distrito Federal (Secretaria de Vigilância em Saúde,
2010). Encontra-se em alta densidade próxima a áreas densamente povoadas por
humanos (Lourenço-de-Oliveira et al., 2004). A espécie exibe casos de resistência a
inseticidas (Luz et al., 2003; Braga et al., 2004; Da-Cunha et al., 2005; Macoris et al.,
2003) e apresenta uma alta capacidade vetorial para o vírus da dengue. Além disso, a
urbanização descontrolada e as migrações são fatores adicionais que favorecem tanto
o vetor quanto à disseminação do vírus (Gubler, 1998; Herrera et al., 2006). Esse fato
associado a péssimas condições de saneamento básico, moradia inadequada e a
fatores culturais/educacionais favorecem condições ecológicas à transmissão dos vírus
da dengue (Lines et al., 1994) pelo mosquito Ae. aegypti, seu principal vetor.
O vírus dengue e da febre amarela pertencem ao gênero Flavivirus da família
Flaviviridae, ambos transmitidos, em ciclos urbanos, pelo Ae. aegypti. O vírus da
dengue apresenta quatro sorotipos (DEN1, DEN2, DEN3 e DEN4) e é o único dentro da
família Flaviviridae, gênero Flavivirus, que não necessita manutenção de ciclos em

21
ambientes silvestres. Os sorotipos são antigenicamente diferentes, porém apresentam a
mesma epidemiologia (Gubler, 2002).
A dengue é uma arbovirose que se tornou um grave problema de saúde pública
no Brasil, assim como em outras regiões tropicais do mundo (World Health
Organization, 2010). Cerca de 100 milhões de casos ocorrem em cada ano no mundo
(Monath, 1994). Segundo Braga e Valle (2007) a doença tem se destacado entre as
enfermidades reemergentes e é considerada a mais importante das doenças virais
transmitidas por artrópodes.
1.3 Controle populacional versus microevolução
Tentativas de redução populacional de Ae. aegypti constituem a principal forma
de combate à dengue, já que as vacinas ainda estão em desenvolvimento. Os
mosquitos de modo geral, apresentam grande capacidade adaptativa, evidenciada pela
existência de mutantes resistentes a diversas condições ambientais e de complexos de
espécies filogeneticamente relacionadas ou crípticas. Estratégias para o controle desse
culicídeo têm sido desenvolvidas há décadas, entre elas, aplicação de inseticidas,
controle biológico e os recentes métodos alternativos de construção de mosquitos
transgênicos (Speranca e Capurro, 2007; Yakob et al., 2008) também utilizado para
outros mosquitos (Marrelli et al., 2007; Wilke et al., 2009). Os métodos utilizados até
hoje apresentaram eficácias diversas dependendo da situação em que são
empregados, e freqüentemente enfrentam como fator limitante a microevolução dos
mosquitos.
A “microevolução” é explicada como o conjunto de alterações evolutivas
ocorridas em níveis taxonômicos infra-específicos, como por exemplo, em populações
(Futuyma, 1997; Ridley, 2003). Devido à conjunção de fatores bióticos e abióticos que
ocorrem dinamicamente em populações, os processos microevolutivos surgem como
variações alélicas quantitativas ou qualitativas e mais raramente como amplificações
gênicas ou rearranjos cromossômicos. Com o tempo, e com a influência do fluxo
gênico, esses processos podem culminar em diferenciação ou convergência desses
padrões nas entidades biológicas envolvidas (Futuyma, 1997; Ridley, 2003).

22
Os Culicídeos em geral apresentam flutuações populacionais amplas e
freqüentes, e seu ciclo de vida é influenciado por condições ambientais instáveis como
temperatura, regime pluviométrico, disponibilidade de coleções hídricas e pela presença
humana (Hayes, 1975; Mouchet e Carnevale, 1997; Kovats, 2000; Forattini, 2002;
Ahumada et al., 2004). Devido ao conjunto desses fatos eles apresentam reprodução
rápida, podendo completar um ciclo de vida em poucos dias. Essas propriedades em
conjunto resultam freqüentemente em rápida evolução de suas populações. Um
exemplo é o culicídeo Culex quinquefasciatus e a evolução de sua resistência a
inseticida ao longo de poucos anos (Yebakima et al., 2004).
Outra questão importante na evolução de populações de mosquitos é a
diferenciação de populações geograficamente isoladas. É comum encontrar variações
geográficas entre populações de culicídeos cuja distribuição geográfica é ampla e
descontínua a ponto de constituir barreiras ao intenso fluxo gênico. Um exemplo de tal
situação é a variabilidade genética encontrada nas diversas populações de Ae. aegypti
ao redor do mundo (Powell et al., 1980).
Para estudo da microevolução, em geral, adota-se como passo inicial à
caracterização de populações, as quais são descritas quanto a padrões genéticos,
morfológicos, comportamentais e comparadas ao longo de transectos geográficos ou
temporais. Em um primeiro momento, o conjunto de dados permite o reconhecimento
de padrões microevolutivos, enquanto que com o acúmulo de informações ao longo dos
anos, existe a possibilidade de observar processos microevolutivos.
1.4 Detectando variação populacional com múltiplas abordagens
A utilização de múltiplos parâmetros comparativos pode fornecer maior robustez
aos testes de hipóteses em caracterização populacional que vise descrever padrões e
processos microevolutivos. Uma vez que os caracteres em uma espécie evoluem em
taxas e velocidades diversas, a utilização de múltiplos parâmetros permite encontrar o
marcador cujo nível de resolução é próximo do desejado, aumentando as chances de
serem detectadas variações fenotípicas e genotípicas (Sene et al., 1988; Reinert et al.,
1997; Rocha, 2005; Savard et al., 2006).

23
Um estilo de estudo populacional multiparamétrico que tem sido recentemente
valorizado e encorajado na entomologia médica é a combinação de genética
populacional e morfometria corporal (revisão em Dujardin, 2008). Tanto a morfometria
tradicional como a geométrica tem capacidade de detectar variação populacional
(Falconer, 1981; Dujardin e Slice et al., 2007). Com isso, informações necessárias
podem ser fornecidas para criar-se um mapa preliminar, apontando áreas em que se
devem aprofundar os estudos e investir na utilização de métodos moleculares. Além
disso, podem ser obtidas informações importantes para a entomologia médica.
Na Província de Barinas (Venezuela), a geometria alar de Rhodnius prolixus foi
utilizada recentemente para compreender a reinfestação das casas e do peridomicílio
após aplicação de inseticidas (Feliciangeli et al., 2007). As conclusões deste estudo
foram plenamente apoiadas por um posterior estudo genético (Fitzpatrick et al., 2008).
Estudo realizado em Niakhar, Senegal por Paupy et al. (2010) utilizou
marcadores microssatélites e análise do padrão abdominal para avaliar a variabilidade
genética e morfológica de populações de Ae. aegypti. Temu e Yan (2005) estudaram a
diferenciação de populações de Anopheles arabiensis, importante vetor da malária que
coexiste com outros vetores da malária em toda África, ao longo da costa do Kênia
através de marcadores microssatélites e da seqüência do gene mitocondrial ND5. Para
testar a variabilidade genética de populações de Anopheles darlingi em uma escala
microgeográfica (cerca de 100 km) no oeste da Colômbia, Gutiérrez et al. (2010)
utilizaram o gene mitocondrial COI e marcadores microssatélites para entender melhor
a contribuição genética potencial das populações locais aos programas de controle da
malária. A seguir serão descritos os marcadores utilizados no desenvolvimento do
presente trabalho.
1.5 Morfometria geométrica das asas
Espécies e populações podem eventualmente apresentar variações morfológicas
não notáveis à observação direta, como variações na razão entre múltiplas dimensões
de uma determinada estrutura corporal. Por isso torna-se necessário recorrer a
abordagens alternativas, como as análises morfométricas. A morfometria é a

24
formalização matemática das dissimilaridades entre formas geométricas de objetos,
uma ferramenta de alto poder de resolução que permite a comparação de padrões
corporais a partir de caracteres multivariados, ou seja, considerando-se
simultaneamente várias características de uma estrutura corporal complexa (Rohlf,
1993; Monteiro e Reis, 1999).
A partir dos dados gerados por morfometria podem-se efetuar estudos
estatísticos da variação morfométrica, comparar a magnitude das variações, entre
outras abordagens (Bookstein, 1997; Rohlf, 1993; Monteiro e Reis, 1999). As principais
propriedades da morfometria geométrica são as possibilidades de análises
multivariadas, com múltiplas variáveis simultaneamente e a possibilidade de recuperar
informações de forma e tamanho independentemente. Conseqüentemente seu poder
de resolução é alto e permite gerar diversos marcadores taxonômicos ou populacionais.
Em insetos, asas são preferenciais por se tratar de uma estrutura bidimensional,
evitando erros de posicionamento e deformação, além disso, as asas apresentam
características potencialmente herdáveis e assim sendo sujeita a evolução. Existem
trabalhos em que a morfometria geométrica demonstrou o caráter herdável e
presumivelmente poligênico do formato alar em Ae. aegypti mantidos em cativeiro
(Jirakanjanakit, 2008; Dujardin, 2008).
Estudo realizado por Morais et al. (2010) com populações de Culex
quinquefasciatus do Brasil e de La Plata, Argentina, utilizando morfometria geométrica,
observaram que as amostras brasileiras são morfologicamente diferentes da amostra
da Argentina quanto ao formato alar. Foi observada diferença em relação às
populações do Norte e do Sul do Brasil, sugerindo que a morfologia das asas pode ser
utilizada para uma avaliação preliminar da estrutura populacional de Cx.
quinquefasciatus no Brasil.
A forma da asa tem sido cada vez mais utilizada na identificação e
caracterização de espécies. Estudo realizado com vetores da dengue, Ae. aegypti e Ae.
albopictus coletados em várias parte do mundo, mostrou que a microevolução da forma
alar não produz sobreposição interespecífica significativa, independentemente da
origem geográfica da espécie (Henry et al., 2010). Naquele estudo, tanto a forma como
o tamanho das asas exibiram-se espécie-específicos. Exemplos como esse mostram a

25
utilidade da MG como complemento para diagnosticar exemplares desses mosquitos
quando não estão bem preservados.
1.6 DNA nuclear - Microssatélites
Os marcadores microssatélites também conhecidos como SSRs (sequências
simples repetidas) estão entre os marcadores mais polimórficos encontrados hoje nos
genomas de animais e plantas. São caracterizados por uma sequência de 1-6
nucleotídeos repetidos em tandem espalhados em todo o genoma (Weber e May, 1989;
Schlötterer, 2000). Por serem freqüentes, os loci microssatélites podem estar
intimamente associados a loci conservados que contêm regiões codificadoras. O
polimorfismo alélico ocorre em um locus SSR devido a mudanças no número de
repetições, que pode ser devido a mutações, permutação desigual e erros da
polimerase.
Os SSRs apresentam numerosas vantagens quando comparados com outros
marcadores moleculares, pois são altamente polimórficos e informativos, a herança é
codominante permitindo a discriminação entre homozigotos e heterozigotos. Além
disso, são multialélicos e possuem alta reprodutibilidade, características importantes
para análise baseada em marcadores moleculares. Esses marcadores são baseados
em PCR (amplificação pela reação da polimerase em cadeia) e, portanto, necessitam
de pequena quantidade de DNA, são abundantes e neutros.
Marcadores SSRs têm sido usados com sucesso para estudos genéticos
populacionais de insetos vetores de importância médica como Anopheles (Muturi et al.,
2010; Scarpassa e Conn, 2007), Glossina (Bouyer et al., 2007; Solano et al., 2000),
Culex (Venkatesan e Rasgon, 2010; Huang et al., 2009) e Ae. aegypti (Hlaing et al.,
2010; Paupy et al., 2008; Slotman et al., 2007; da Costa-Ribeiro et al., 2006; Herrera et
al., 2002; Ravel et al., 2001). A utilização desses marcadores em estudos genéticos
populacionais ajuda a investigar a base genética do vetor avaliando a variação
populacional e com isso desenvolver estratégias de controle genético.
Variações baseadas em análises SSRs têm fornecido evidências de que o
armazenamento de água e os hábitos humanos podem afetar na diferenciação genética

26
do Ae. aegypti (Huber et al., 2002). Além disso, marcadores SSRs fornecem uma
medida de divergência sensível e, portanto, podem distinguir populações que tenham
divergido recentemente (Lanzaro et al., 1995), já que essas regiões de microssatélites
evoluem rapidamente.
1.7 Estudos de diferenciação populacional de Aedes aegypti no Brasil
Devido ao fato de o Ae. aegypti ser vetor de doenças graves como a dengue e a
febre amarela vários estudos relacionados a genética populacional tem sido
desenvolvido no Brasil. Mais de 300 trabalhos já foram publicados endereçados ao
estudo populacional de Ae. aegypti (base de dados ISI Web of Science, 2010).
Citaremos alguns deles: Da Costa-Ribeiro et al. (2007) através de marcadores
isoenzimáticos observaram baixo fluxo gênico entre populações localizadas na região
sudeste e região sul não havendo evidência de dispersão passiva de Ae. aegypti por
veículos entre as diferentes rotas ligando as áreas metropolitanas analisadas. O
trabalho de Ayres et al. (2004) também utilizando isoenzimas, analisaram a estrutura
genética de populações naturais de Ae. aegypti em níveis micro e macrogeográfico no
Brasil, observando grande diferenciação genética populacional. No ano anterior, Ayres
et al. (2003) mostraram diferenciação genética micro e macrogeográfica no Brasil
utilizando marcadores RAPD (Random amplified polymorphic DNA).
Sousa-Polezzi e Bicudo (2005) analisaram a variação genética de duas
populações de Ae. aegypti em duas cidades brasileiras São José do Rio Preto (SP) e
Goiânia (GO) detectados através de alterações nos padrões de esterase ao longo do
tempo. Esses autores presumiram que, alterações nos padrões de esterase fossem
resultado de fatores como a seleção por condições ambientais e deriva genética. O
elevado uso de inseticidas causa alterações no padrão de esterase. Dados obtidos no
mesmo período pelas autoridades responsáveis para controlar o mosquito revelaram
uma crescente resistência dos mosquitos em São José do Rio Preto.
No Rio de Janeiro, Da Costa-Ribeiro et al. (2006), realizaram um trabalho
populacional comparando a variabilidade genética de populações separadas por uma
distância linear mínima de 8,2 km e máxima de 32,4 km utilizando marcadores

27
microssatélites e comparando com marcadores isoenzimáticos. Quando comparado
com polimorfismo, os microssatélites se mostraram mais eficientes que as isoenzimas,
mostrando uma grande variação genética.
1.8 Aedes aegypti no Estado de São Paulo
Considerado erradicado do Brasil em 1955, o Ae. aegypti foi reintroduzido
posteriormente. A reinfestação no estado de São Paulo foi identificada no período de
1980 e 1981, sendo os focos eliminados. Em 1985 através de um levantamento feito
em pontos estratégicos para instalação e desenvolvimento deste vetor (borracharias,
depósitos de pneus, ferros-velhos, etc.) foi detectada a presença do mosquito em 12
municípios do Estado. No final de 1985 já existiam 09 municípios com infestação
domiciliar pelo Ae. aegypti.
Entre 1986 e 1996, verificou-se contínua, rápida e ampla dispersão de Ae.
aegypti, ocupando principalmente as regiões Oeste, Norte e Central do Estado. Em
1995, o número de municípios que apresentavam infestação pela espécie era de 416,
distribuídos em áreas nas quais residiam aproximadamente 8 milhões de pessoas.
Desde então, a expansão geográfica desse vetor continua ocorrendo (Superintendência
do Controle de Endemias, 2009).
Nos últimos anos o Estado de São Paulo tem revelado grandes ocorrências de
dengue, só no primeiro semestre de 2010 ocorreram mais de 150.000 casos de dengue
autóctone (Centro de Vigilância Epidemiológica, 2010). Devido à falta de uma vacina
para combater a doença, o que precisa ser feito atualmente é controlar o vetor. Para
isso trabalhos referentes ao estudo populacional do Ae. aegypti são de grande
importância para entender o processo de evolução e dispersão desses mosquitos.
Apesar disso, as populações de Ae. aegypti no Estado de São Paulo ainda são pouco
estudadas.
Tendo em vista a questão do Ae. aegypti no Estado de São Paulo e as
problemáticas populacionais, propomos um estudo de caracterização populacional de
Ae. aegypti combinando marcadores moleculares do tipo microssatélites com
morfometria geométrica alar. A exemplo dos trabalhos já citados acreditamos que esses

28
métodos poderão ser úteis também aqui no estado de SP. Além disso, a correlação da
genética populacional com morfometria alar poderá trazer mais informações para os
métodos estimativos de estrutura populacional de Ae. aegypti.

29
2 OBJETIVOS 2.1 Objetivo geral Avaliar a estruturação populacional de Aedes aegypti na “Grande São Paulo”
amostrando-se quatro localidades: Bairro do Butantã (município de São Paulo) e
municípios vizinhos Guarulhos, Osasco e Suzano, mediante utilização de marcadores
genéticos e morfológicos reconhecidamente informativos do ponto de vista
microevolutivo.
2.2 Objetivos específicos
- Investigar o grau de polimorfismo dos loci microssatélites 38/38, 34/72, C2A8,
T3A7 e AED19 nas populações de Aedes aegypti;
- Analisar diferenciação morfológica alar entre as populações por meio da
morfometria geométrica;
- Estimar fluxo gênico e outros parâmetros de estrutura populacional,
verificando se há correlação entre os diferentes marcadores.

30
3 MATERIAIS E MÉTODOS 3.1 Coleta e armazenamento dos exemplares
Amostras de Ae. aegypti foram coletadas em quatro locais diferentes no estado
de São Paulo: Butantã (BUT) que é um distrito localizado na Zona Oeste do município
de São Paulo (23º 32’ 52’’ S 46º 38’ 09’’ W) e três municípios localizados na área
metropolitana da capital paulista, Guarulhos (GUA) (23º 27’ 46’’ S 46º 31’ 58’’ W),
Osasco (OSA) ( 23 º 31’ 58’’ S 46º 47’ 31’’ W) e Suzano (SUZ) (23º 32’ 33’’ S 46º 18’
39’’ W) (Figura 1). Os dados das coletas referentes a MG estão listados na Tabela 1.
Para as análises genéticas foram utilizados 30 indivíduos adultos de cada população,
exceto a população de SUZ, que foram analisados somente 26 indivíduos. A menor
distância em linha reta entre as populações analisadas é de 13,5 km entre BUT e OSA
e a maior distância de 50 km entre OSA e SUZ (Figura 2).
Figura 2. Mapa da área metropolitana de São Paulo. Os retângulos em vermelho representam
os locais em que foram feitas as coletas. O bairro do Butantã está assinalado “São Paulo”). Fonte modificada de Google maps (2010).
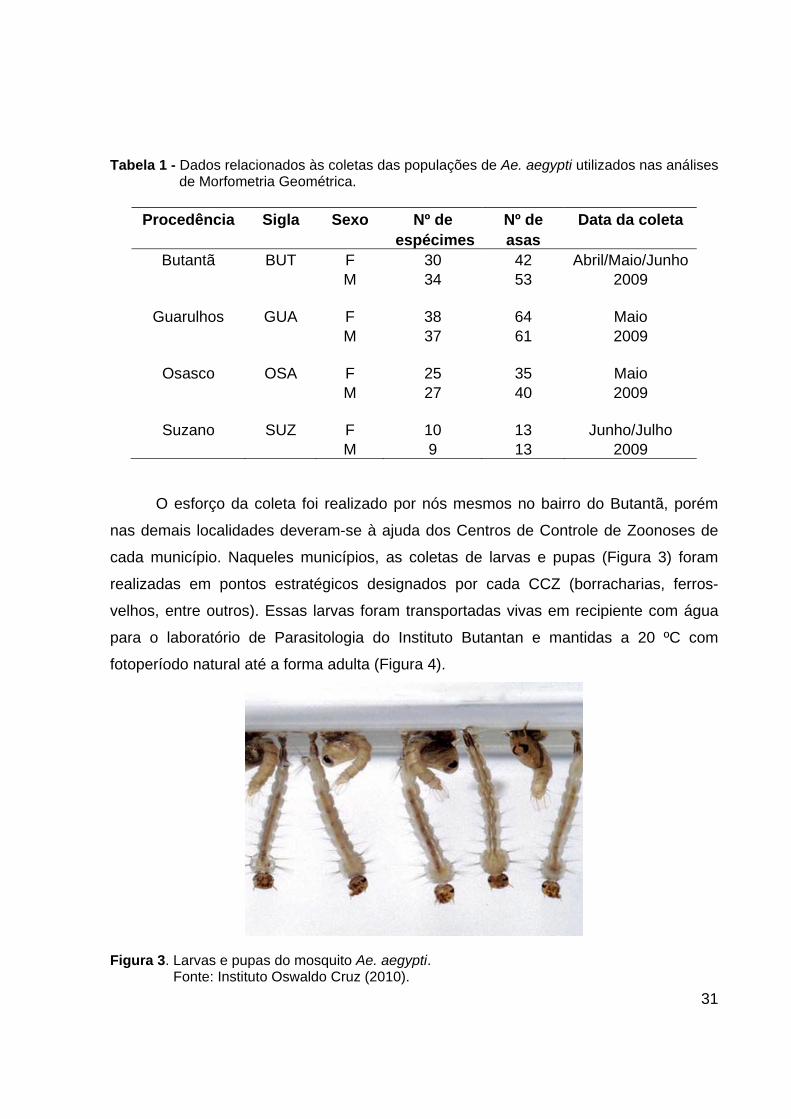
31
Tabela 1 - Dados relacionados às coletas das populações de Ae. aegypti utilizados nas análises de Morfometria Geométrica.
Procedência Sigla Sexo Nº de Nº de Data da coleta
espécimes asas Butantã BUT F 30 42 Abril/Maio/Junho
M 34 53 2009
Guarulhos GUA F 38 64 Maio M 37 61 2009
Osasco OSA F 25 35 Maio
M 27 40 2009
Suzano SUZ F 10 13 Junho/Julho M 9 13 2009
O esforço da coleta foi realizado por nós mesmos no bairro do Butantã, porém
nas demais localidades deveram-se à ajuda dos Centros de Controle de Zoonoses de
cada município. Naqueles municípios, as coletas de larvas e pupas (Figura 3) foram
realizadas em pontos estratégicos designados por cada CCZ (borracharias, ferros-
velhos, entre outros). Essas larvas foram transportadas vivas em recipiente com água
para o laboratório de Parasitologia do Instituto Butantan e mantidas a 20 ºC com
fotoperíodo natural até a forma adulta (Figura 4).
Figura 3. Larvas e pupas do mosquito Ae. aegypti. Fonte: Instituto Oswaldo Cruz (2010).

32
Figura 4. Fêmeas de Ae. aegypti realizando o repasto sanguíneo. Fonte: Encyclopedia Britannica Online ([2010]).
Assim que os adultos emergiam, eles eram colocados em tubos eppendorf e
armazenados em nitrogênio líquido a -196 ºC para preservação do material genético.
Antes do processo de extração do DNA de cada mosquito as asas eram retiradas para
as análises da morfometria alar.
3.2 Análise morfométrica alar Os métodos de MG seguiram em linhas gerais os empregados por Morais et al.
2010. As asas direitas e esquerdas de machos e fêmeas foram montadas com resina
sintética Entellan entre lâmina-lamínula. Imagens dessas asas foram capturadas
através de câmera fotográfica digital Leica 320 acoplada a um microscópio
estereoscópico Leica S6. Mais de 50 indivíduos de cada localidade foram utilizados
para as análises morfométricas, com exceção de SUZ que só foram utilizados 19
exemplares (Tabela 1). A população de SUZ só foi utilizada na análise de dimorfismo
sexual devido ao pequeno número amostral. Isso aconteceu porque alguns indivíduos
foram tentativamente acondicionados em nitrogênio líquido antes da remoção das asas.
Durante a manipulação dos mosquitos congelados, muitas asas dessa população
ficaram deformadas, impossibilitando de realizar as análises morfométricas.

33
Sobre as imagens das asas, com o auxílio do “software” TpsDig V.1.40 (QSC –
James Rohlf), foram tomadas as coordenadas posicionais de cada um dos 18 pontos
anatômicos sobre um plano cartesiano (Figura 5). Sobre esse dados foram analisados o
tamanho alar através do tamanho de centróide (TC) e o formato alar através das
variáveis canônicas (VC), com o auxílio dos programas de computador TpsUtil 1.26,
TpsRelw 1.36 (QSC - James Rohlf) e Statistica 7.0 (StatSoft) e componentes principais
(CP), com o auxílio dos programas BAC e PAD (JP Dujardin -
http://www.mpl.ird.fr/morphometrics/).
Figura 5. Asa de uma fêmea de Ae. aegypti. Acima, asa digitalizada com 18 pontos de encontro
das nervuras marcados em vermelho e numerada. Abaixo, conformação alar imaginária obtida com a ligação dos pontos.
3.2.1 Componentes Principais
Os Componentes Principais foram obtidos com os auxílios dos programas
tpsRelw 1.36 e BAC e empregados em comparações intra e interpopulacionais. Os
valores posicionais de cada ponto anatômico foram gerados no programa tps Relw
1.36, e através do programa BAC foram feitas às análises de CP. Dentre os
Componentes Principais gerados em cada comparação, apenas a primeira e a segunda

34
(CP1 e CP2) foram consideradas para interpretação, já que essas são as mais
informativas.
Consenso alar de cada população para ambos os sexos foram feitos através do
programa tps RelW. Agregados a esses consensos, para representar a distribuição dos
indivíduos em função dos CPs foram feitos gráficos de dispersão de pontos. Estes
gráficos ilustram as características biológicas avaliadas: assimetria alar bilateral,
dimorfismo sexual e diferenciação populacional.
3.2.2 Análises discriminantes
Testes de reclassificação foram realizados nas análises de dimorfismo sexual e
na diferenciação populacional, utilizando o programa PAD. O teste de reclassificação é
um teste com validação, pois se trata de um teste cego. Nesse tipo de teste cada
indivíduo é removido da análise, uma nova classificação é realizada com os outros
indivíduos e em seguida o indivíduo é reinserido para verificar em que grupo ele é
classificado. Nesse teste, a acurácia de reclassificação é representada em valor
percentual.
3.2.3 Distância de Mahalanobis
Comparações entre as amostras foram efetuadas mediante o cálculo das
distâncias de Mahalanobis seguindo o método de Ligação Completa, utilizando o
programa Statistica 7.0, conforme empregado por Moratore (2009) e Peruzin (2009). As
distâncias de Mahalanobis geradas entre as amostras populacionais foram organizadas
em tabelas e em seguida convertidas em fenogramas construídos pelo algoritmo de
neighbour-joining com o auxílio do software Phylip. Foi utilizada a distância de
Mahalanobis por ser adequada para comparações multivariadas, pois embora seja uma
distância linear, semelhante à distância Euclideana, é medida entre pontos através de
um espaço multidimensional.

35
3.2.5 Tamanho de Centróide
Os tamanhos de centróide e a construção dos gráficos de caixa (Box Plots)
foram obtidos com o auxílio dos programas de computador TpsRelw 1.36 e Statistica
7.0 e utilizados nas comparações intra e interpopulacionais das amostras. Com o auxílio
do programa de computador Graph Pad InsStat, análises estatísticas bicaudais ao nível
de rejeição 0,05 foram utilizadas nas comparações baseadas nos tamanhos dos
centróides.
Para testar a normalidade das distribuições amostrais, foi verificado se as
amostras analisadas apresentavam distribuição Gaussiana, empregando o teste de
Kolmogorov-Smirnov e, para as amostras de distribuição normal e variâncias não-
homogêneas, utilizou-se o Teste T paramétrico com correção de Welch, que é o
adequado para esse tipo de distribuição.
3.3 Análises genéticas – Microssatélites
3.3.1 Extração do DNA genômico
Trinta indivíduos das populações do BUT, GUA e OSA e vinte seis indivíduos da
população de SUZ foram utilizados para a extração do DNA genômico. A extração foi
feita isoladamente para cada indivíduo, segundo o método de Jowett (1986) com
algumas modificações que serão descritas a seguir.
Os indivíduos foram retirados do nitrogênio líquido e macerados em tubos
eppendorf de 1,5 mL contendo 100 µL de solução de homogeneização (Tris-HCl 10 mM
(pH= 7,5), NaCl 60 mM, EDTA 50 mM). Sobre o homogeneizado foi adicionado 100 µL
de solução de lise (SDS 1,25%, Tris-HCl 0,3 M (pH = 9,0), EDTA 0,1 M, sacarose 5%,
proteinase K a 100 µg/ml). A mistura foi incubada em banho-maria a 65 ºC por 1 hora
para “lise” das células e clivagem das proteínas em peptídeos menores pela proteinase
K.
Em seguida as amostras foram retiradas do banho maria e encubadas a 4 ºC por
45 minutos. Terminando a encubação os tubos foram centrifugados por 10 minutos a

36
12.000g na temperatura de 4 ºC. Após a centrifugação foram formadas duas fases na
mistura. O precipitado, que contém peptídeos, foi descartado e o sobrenadante
contendo ácidos nucléicos e sais foram transferidos para outro microtubo de 1,5 mL e
misturado com o dobro de seu volume de etanol 100% para a precipitação do DNA.
Após cinco minutos de incubação à temperatura ambiente, as amostras foram
novamente centrifugadas a 12.000 g à temperatura ambiente, por 5 minutos. O
sobrenadante foi descartado e ao precipitado foi adicionado 1 mL de etanol 70%, e
assim as amostras foram submetidas a uma última centrifugação à temperatura
ambiente e 12.000 g por 5 minutos.
O precipitado foi seco e cada amostra foi ressuspendida em 20 µL de H2O milli-
Q (Millipore). Para eliminar o RNA, as amostras de DNA recém-extraídas foram tratadas
com RNAse-free (Ribonuclease A, Sigma) a uma concentração final 100 µg/mL por 1
hora a 37 ºC em banho-maria. Em seguida as amostras foram armazenadas em freezer
a -20 ºC.
3.3.2 Reação em cadeia da polimerase (PCR)
Para cada população foram utilizados cinco loci microssatélites (C2A8, 34/72,
T3A7, AED19 e 38/38) os mesmos utilizados em trabalhos anteriores por Costa-Ribeiro
et al. (2006) e Huber et al. (2002) (Tabela 2). As regiões SSRs foram amplificadas por
PCR a partir das amostras de DNA genômico nas seguintes condições: O volume da
reação final foi de 20 µL. Os reagentes foram misturados na forma de coquetel,
separadamente do DNA genômico. Para cada reação foi utilizado 1x buffer (tampão de
reação) (Fermentas); 1,5 mM MgCl2; 0,2 mM de cada dNTP; 20 pmol de cada primer;
0,25 U/ µL Taq DNA polimerase (Fermentas) e água mili-Q totalizando 18 µL. Em
seguida foi adicionado 2 µL de DNA genômico em cada reação, totalizando 20 µL finais.
As condições para serem utilizadas no termociclador foram as seguintes: O
processo inicia com 5 ciclos consistindo em uma etapa de desnaturação (96 ºC por
2min), uma etapa de anelamento (T.A. por 30s) e uma etapa de extenção (72 ºC por
1min 15s ) em seguida a quantidade aumenta para 25 ciclos, com uma etapa de
desnaturação com o tempo menor (95 ºC por 30s), uma etapa de anelamento (T.A. por

37
30s) e uma etapa de extensão (72 ºC por 1min 15s ) no final foi adicionado uma etapa
de alongamento de 72 ºC por 5 minutos. Um primer de cada locus analisado foi
marcado no sentido 5’ com fluorescência (FAM, HEX ou TAMRA) apropriada para o
seqüenciador ABI 3730.
Após a amplificação, foram realizadas as diluições dos produtos de PCR. Como
eram 5 loci diferentes, uma diluição foi feita adicionando 3 µL de cada um dos produto
de PCR dos primers AED19, T3A7 e 38/38 adicionando 21 µL de água ultrapura para
um volume final de 30 µL. Outra diluição foi feita separadamente adicionando 2 µL dos
produtos de PCR referentes aos primers C2A8 e 34/72 e completando para um volume
final de 20 µL com água ultrapura. Depois de realizado as diluições, 2 µL da PCR
diluída foram ressuspendidas em 7,5 µL de Formamida HI-DI (Applied Biosystems). Em
seguida foi adicionado 0,5 µL do marcador de peso molecular GeneScan 500 ROX
(Applied Biosystems) para um volume final de 10 µL. As amostras eram encaminhadas
em microtubos de 0,2 mL para o Centro de Estudos do Genoma Humano da
Universidade de São Paulo onde era realizada a eletroforese das amostras no
seqüenciador automático ABI 3730.
3.3.3 Análises microssatélites
Para cada uma das quatro populações analisadas de Ae. aegypti foi calculado a
variação genética primeiramente através da proporção de loci polimórficos, número de
alelos por locus e heterozigosidade (Nei, 1973). Diferenças entre os valores estimados
para esses parâmetros foram analisadas estatisticamente pelo teste T de Student. Para
determinar se as populações analisadas estavam em equilíbrio de Hardy-Weinberg foi
feito o teste do Qui-quadrado para verificar se a distribuição das freqüências de
heterozigotos observadas desviava significativamente das freqüências de heterozigotos
esperadas. Para o processamento dessas estatísticas foi utilizado o software
GENEPOP v. 4.0 (Raymond e Rousset, 1995).
Diferenças entre as populações por F-statistics foram computadas de acordo
com Weir e Cockerham (1984). O desequilíbrio genotípico entre loci (comparados aos
pares) foi estimado utilizando um coeficiente de correlação comum (Weir, 1990) e

38
testado utilizando o teste exato de Fisher através de uma tabela de contigência
utilizando o software GENEPOP v. 4.0.
Estimativas de fluxo gênico foram obtidas a partir das estatísticas de Fst, onde
Nm é o número de migrantes por geração, assumindo um modelo de ilha infinita
(Wright, 1965) onde Nm = ((1 / Fst) - 1) / 4.
Isolamento genético por distância geográfica foi testado por estimar correlações
entre FST/ (1-FST) calculadas entre pares de amostras e as distâncias geográficas
logaritmizadas.
3.4 Análises de correlação Foi testada a existência de correlação entre distâncias geográficas e distâncias
fenéticas e entre as distâncias geográficas e distâncias genéticas. Usando-se o
programa Statistica 7.0. foi calculado o coeficiente de correlação de Pearson, o qual é
uma medida do grau de relação linear entre duas variáveis quantitativas. O intervalo de
confiança utilizado foi de 0,90.
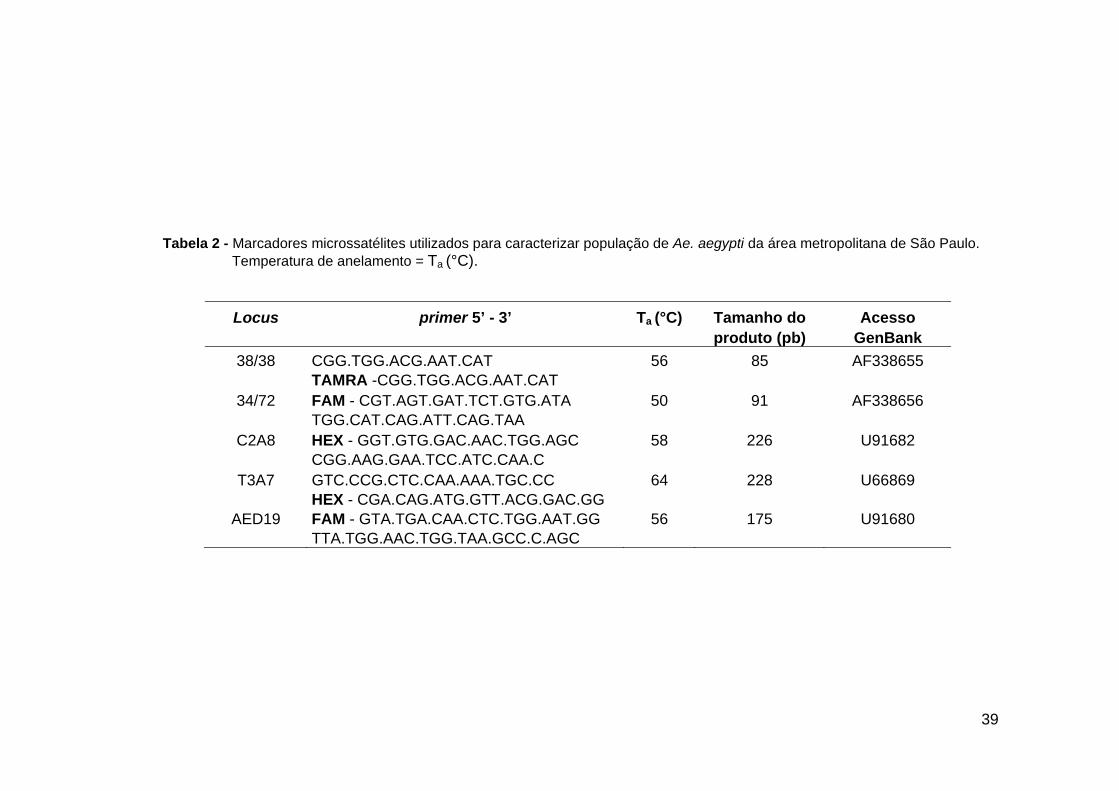
39
Tabela 2 - Marcadores microssatélites utilizados para caracterizar população de Ae. aegypti da área metropolitana de São Paulo.
Temperatura de anelamento = Ta (°C).
Locus primer 5’ - 3’ Ta (°C) Tamanho do Acesso produto (pb) GenBank
38/38 CGG.TGG.ACG.AAT.CAT 56 85 AF338655 TAMRA -CGG.TGG.ACG.AAT.CAT
34/72 FAM - CGT.AGT.GAT.TCT.GTG.ATA 50 91 AF338656 TGG.CAT.CAG.ATT.CAG.TAA
C2A8 HEX - GGT.GTG.GAC.AAC.TGG.AGC 58 226 U91682 CGG.AAG.GAA.TCC.ATC.CAA.C
T3A7 GTC.CCG.CTC.CAA.AAA.TGC.CC 64 228 U66869 HEX - CGA.CAG.ATG.GTT.ACG.GAC.GG
AED19 FAM - GTA.TGA.CAA.CTC.TGG.AAT.GG 56 175 U91680 TTA.TGG.AAC.TGG.TAA.GCC.C.AGC

40
4 RESULTADOS 4.1 Morfometria geométrica alar 4.1.1 Assimetria bilateral
4.1.1.1 Análise de forma
As comparações multivariadas de forma entre asas direitas e esquerdas de
machos e fêmeas das populações do BUT, GUA e OSA geraram componentes
principais, os quais estão representados em morfo-espaços (Bond et al., 2003)
ilustrados nas Figuras 6 e 7. A análise de assimetria bilateral alar de machos e fêmeas
em todas as populações analisadas mostrou uma elevada similaridade entre asas
esquerdas e direitas, não sendo possível diferenciá-las.
4.1.1.2 Análise de tamanho
Os estudos de assimetria bilateral de tamanho isométrico alar mostraram não
haver diferença significativa entre asas direitas e esquerdas de machos e fêmeas das
três populações amostradas (Tabelas 3 e 4). Os tamanhos de centróide das populações
do BUT, GUA e OSA encontram-se estatisticamente descritos nas Figuras 8 e 9 e
Tabela 5.
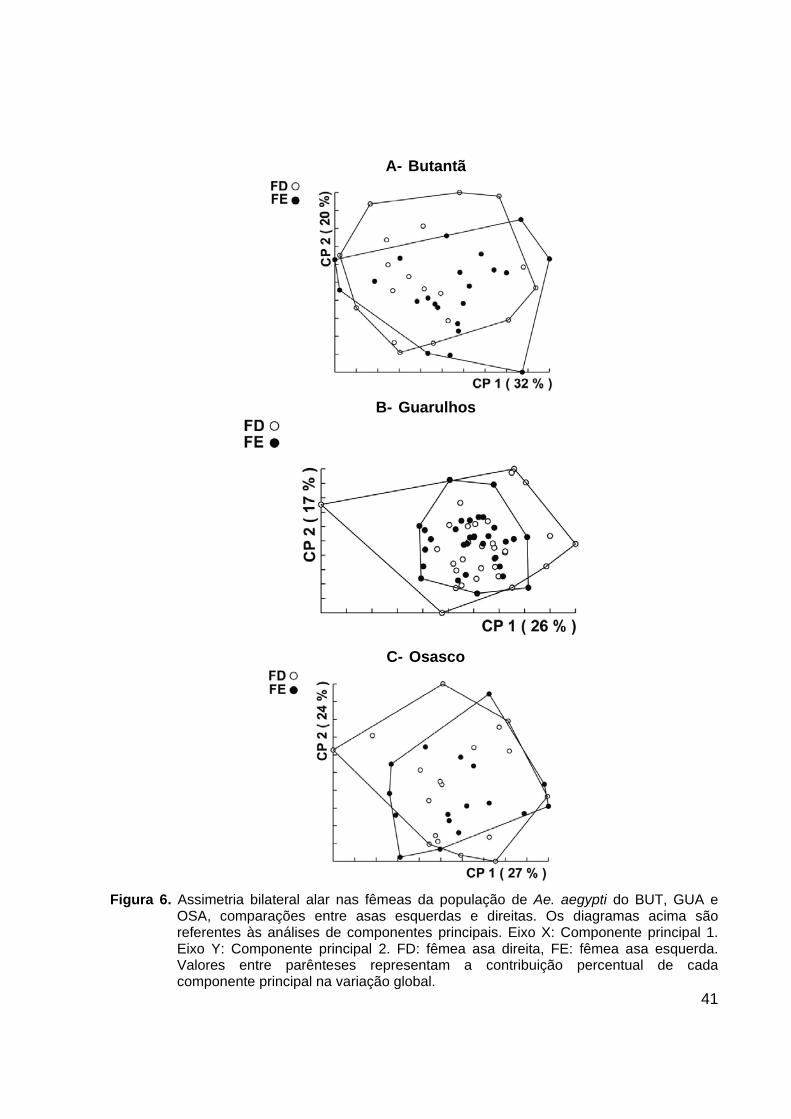
41
A- Butantã
B- Guarulhos
C- Osasco
Figura 6. Assimetria bilateral alar nas fêmeas da população de Ae. aegypti do BUT, GUA e
OSA, comparações entre asas esquerdas e direitas. Os diagramas acima são referentes às análises de componentes principais. Eixo X: Componente principal 1. Eixo Y: Componente principal 2. FD: fêmea asa direita, FE: fêmea asa esquerda. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global.
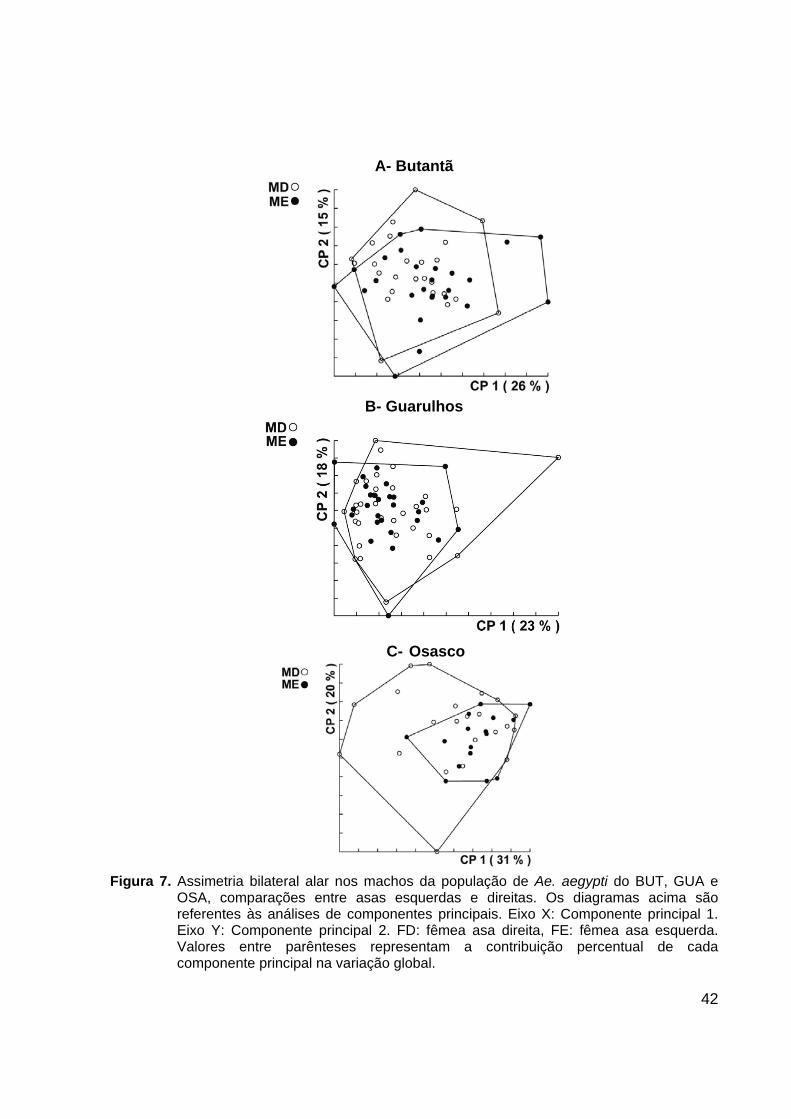
42
A- Butantã
B- Guarulhos
C- Osasco
Figura 7. Assimetria bilateral alar nos machos da população de Ae. aegypti do BUT, GUA e
OSA, comparações entre asas esquerdas e direitas. Os diagramas acima são referentes às análises de componentes principais. Eixo X: Componente principal 1. Eixo Y: Componente principal 2. FD: fêmea asa direita, FE: fêmea asa esquerda. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global.
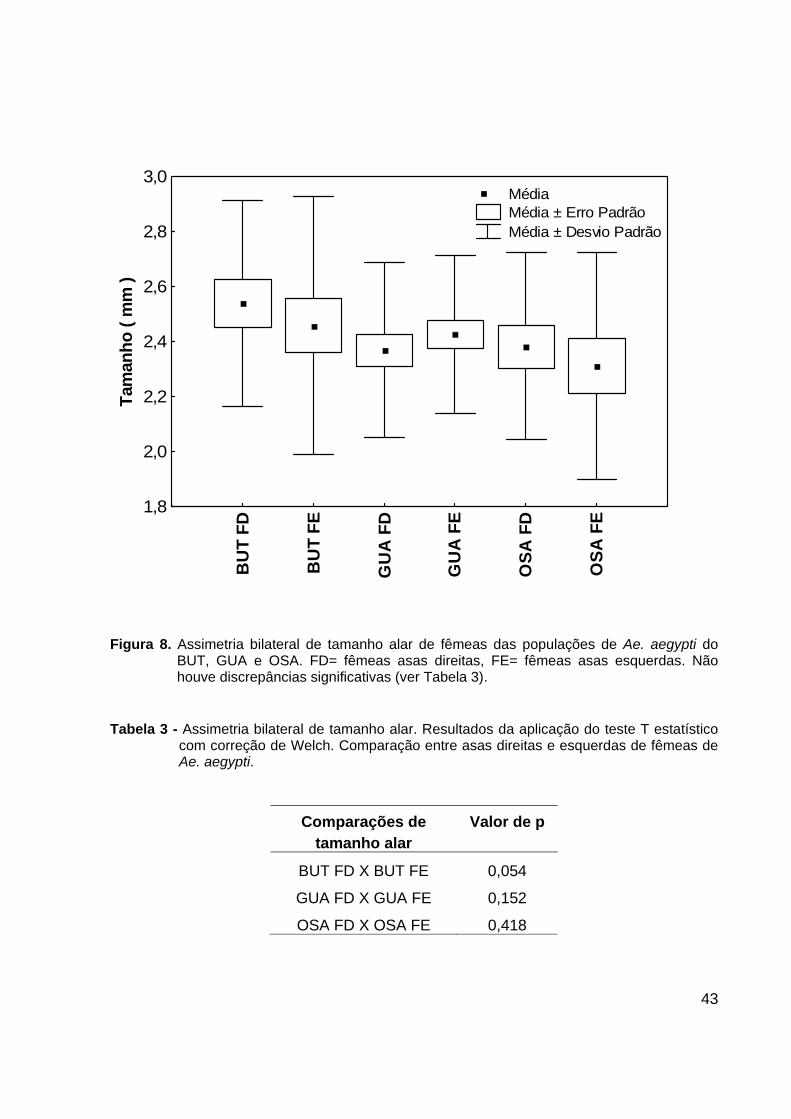
43
BU
T FD
BU
T FE
GU
A F
D
GU
A F
E
OSA
FD
OSA
FE1,8
2,0
2,2
2,4
2,6
2,8
3,0
Tam
anho
( m
m )
Média Média ± Erro Padrão Média ± Desvio Padrão
Figura 8. Assimetria bilateral de tamanho alar de fêmeas das populações de Ae. aegypti do
BUT, GUA e OSA. FD= fêmeas asas direitas, FE= fêmeas asas esquerdas. Não houve discrepâncias significativas (ver Tabela 3).
Tabela 3 - Assimetria bilateral de tamanho alar. Resultados da aplicação do teste T estatístico
com correção de Welch. Comparação entre asas direitas e esquerdas de fêmeas de Ae. aegypti.
Comparações de Valor de p tamanho alar
BUT FD X BUT FE 0,054
GUA FD X GUA FE 0,152
OSA FD X OSA FE 0,418
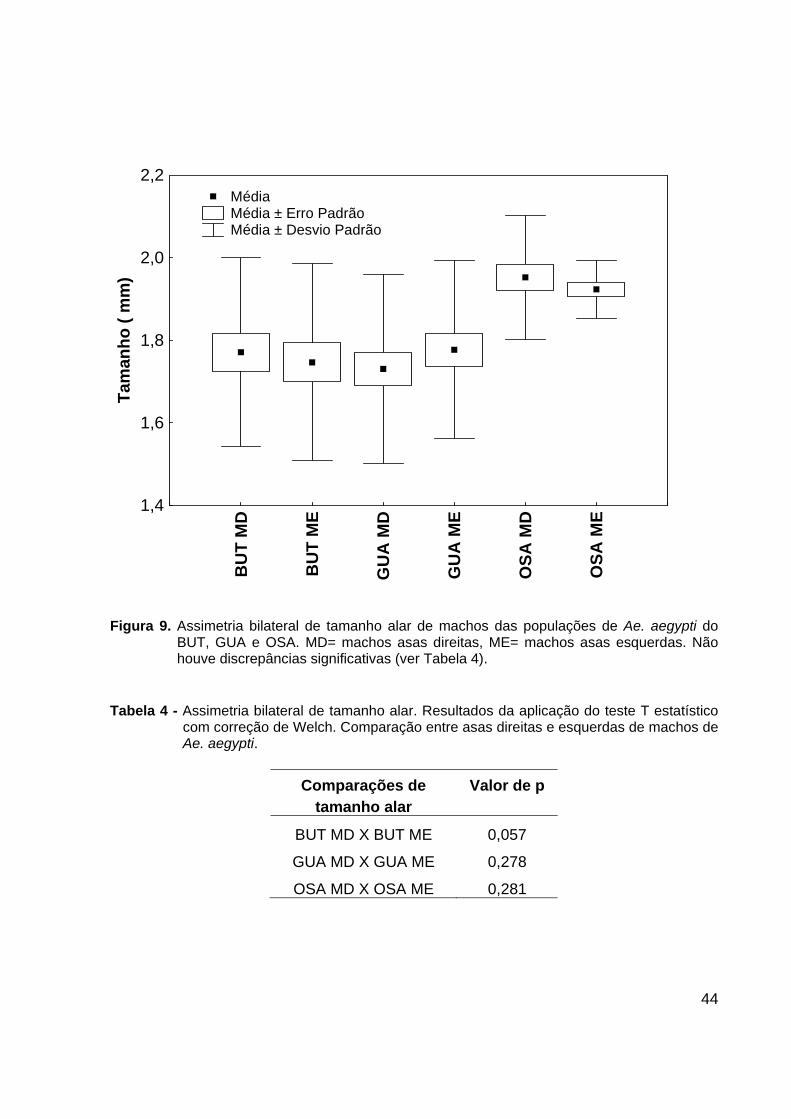
44
BU
T M
D
BU
T M
E
GU
A M
D
GU
A M
E
OSA
MD
OSA
ME1,4
1,6
1,8
2,0
2,2
Tam
anho
( m
m)
Média Média ± Erro Padrão Média ± Desvio Padrão
Figura 9. Assimetria bilateral de tamanho alar de machos das populações de Ae. aegypti do
BUT, GUA e OSA. MD= machos asas direitas, ME= machos asas esquerdas. Não houve discrepâncias significativas (ver Tabela 4).
Tabela 4 - Assimetria bilateral de tamanho alar. Resultados da aplicação do teste T estatístico
com correção de Welch. Comparação entre asas direitas e esquerdas de machos de Ae. aegypti.
Comparações de Valor de p
tamanho alar
BUT MD X BUT ME 0,057
GUA MD X GUA ME 0,278
OSA MD X OSA ME 0,281
45
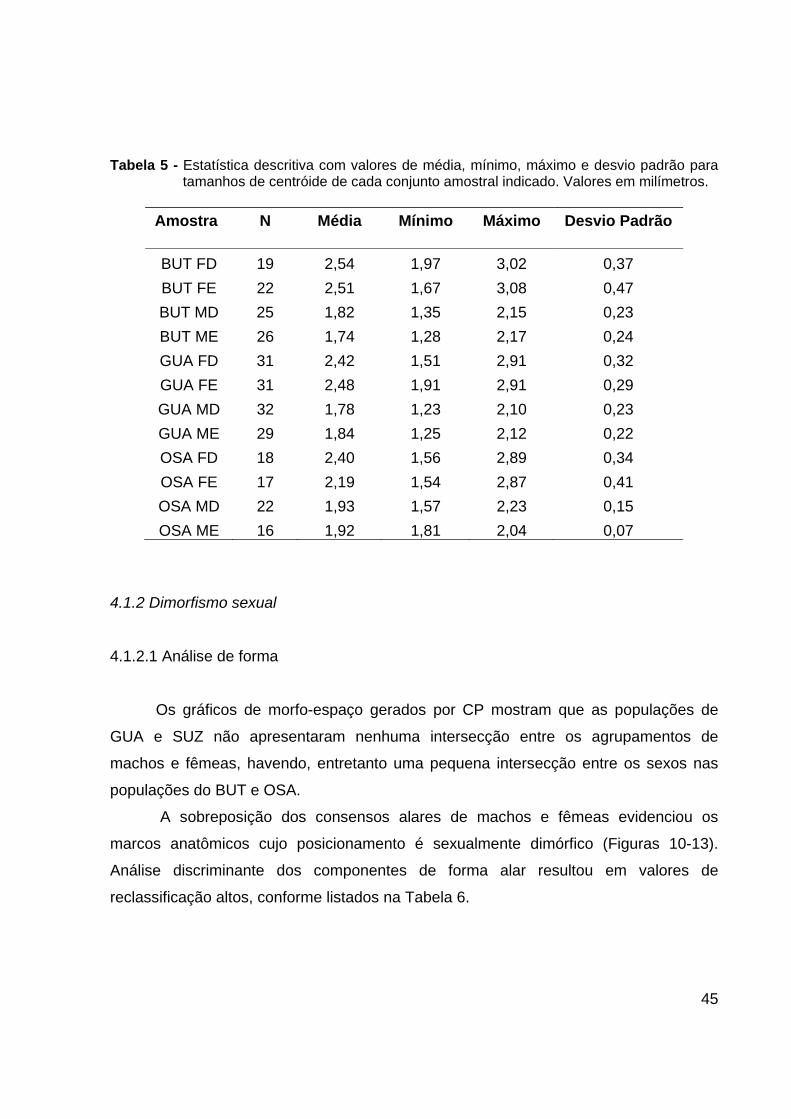
Tabela 5 - Estatística descritiva com valores de média, mínimo, máximo e desvio padrão para tamanhos de centróide de cada conjunto amostral indicado. Valores em milímetros.
Amostra N Média Mínimo Máximo Desvio Padrão
BUT FD 19 2,54 1,97 3,02 0,37 BUT FE 22 2,51 1,67 3,08 0,47 BUT MD 25 1,82 1,35 2,15 0,23 BUT ME 26 1,74 1,28 2,17 0,24 GUA FD 31 2,42 1,51 2,91 0,32 GUA FE 31 2,48 1,91 2,91 0,29 GUA MD 32 1,78 1,23 2,10 0,23 GUA ME 29 1,84 1,25 2,12 0,22 OSA FD 18 2,40 1,56 2,89 0,34 OSA FE 17 2,19 1,54 2,87 0,41 OSA MD 22 1,93 1,57 2,23 0,15 OSA ME 16 1,92 1,81 2,04 0,07
4.1.2 Dimorfismo sexual
4.1.2.1 Análise de forma
Os gráficos de morfo-espaço gerados por CP mostram que as populações de
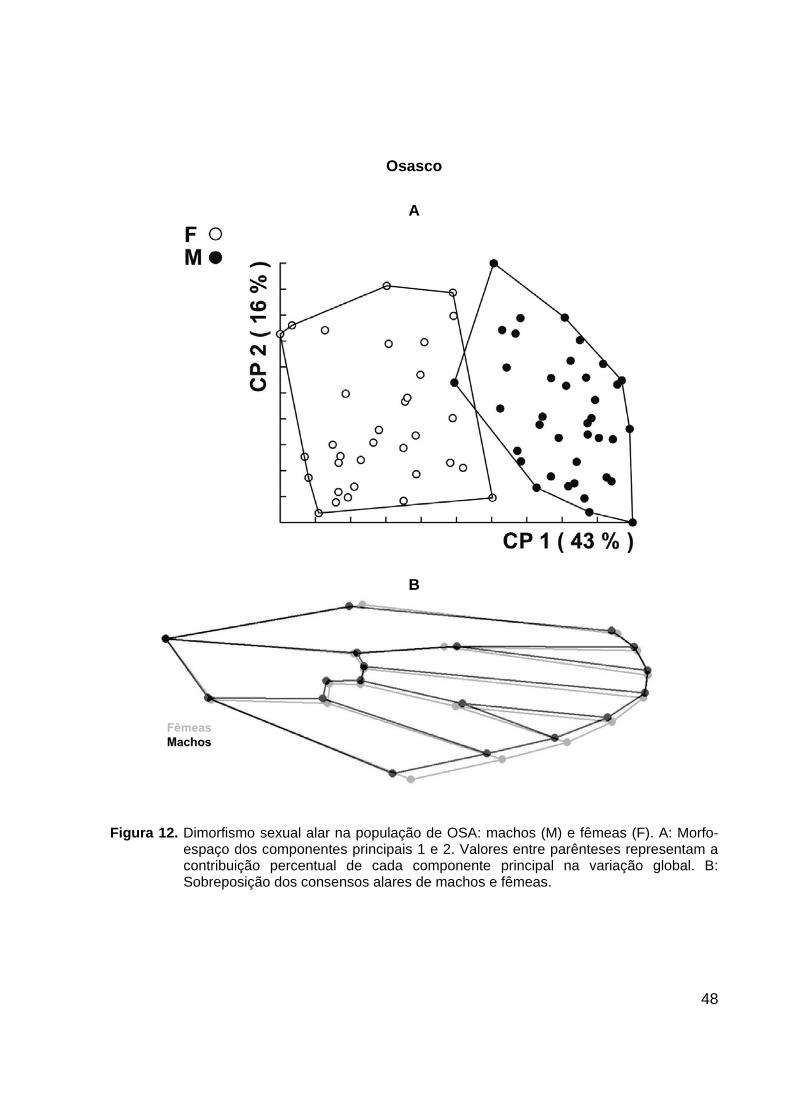
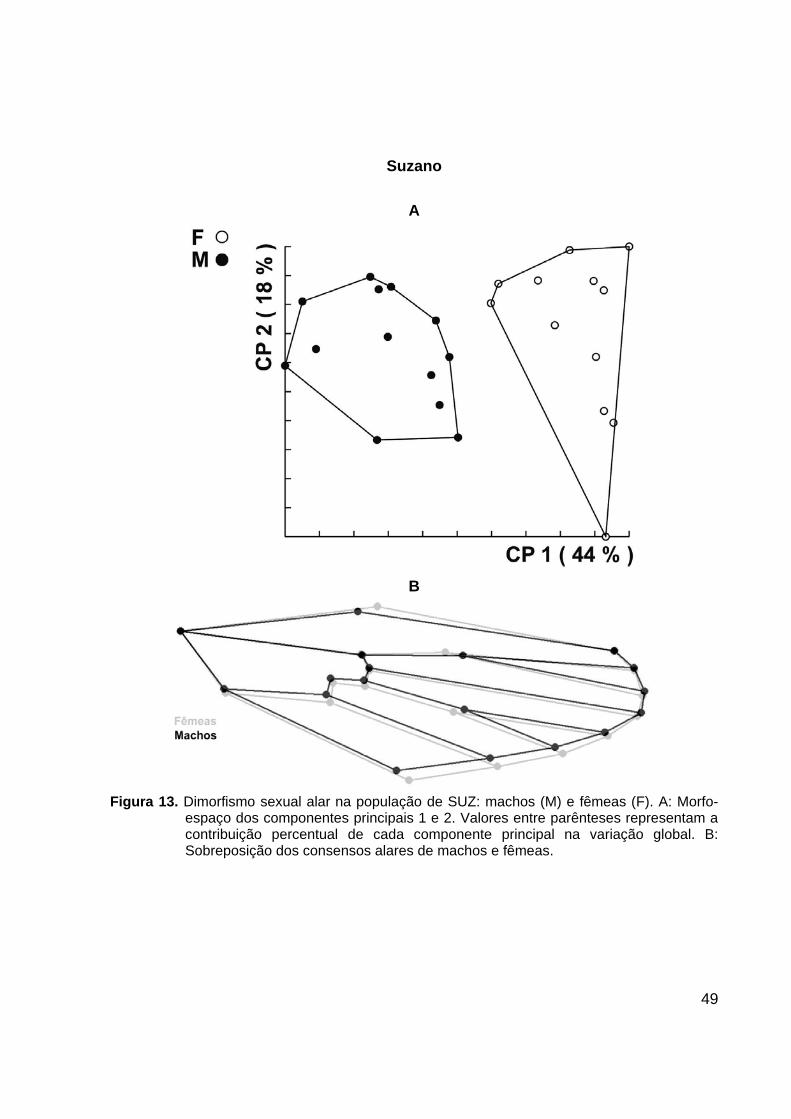
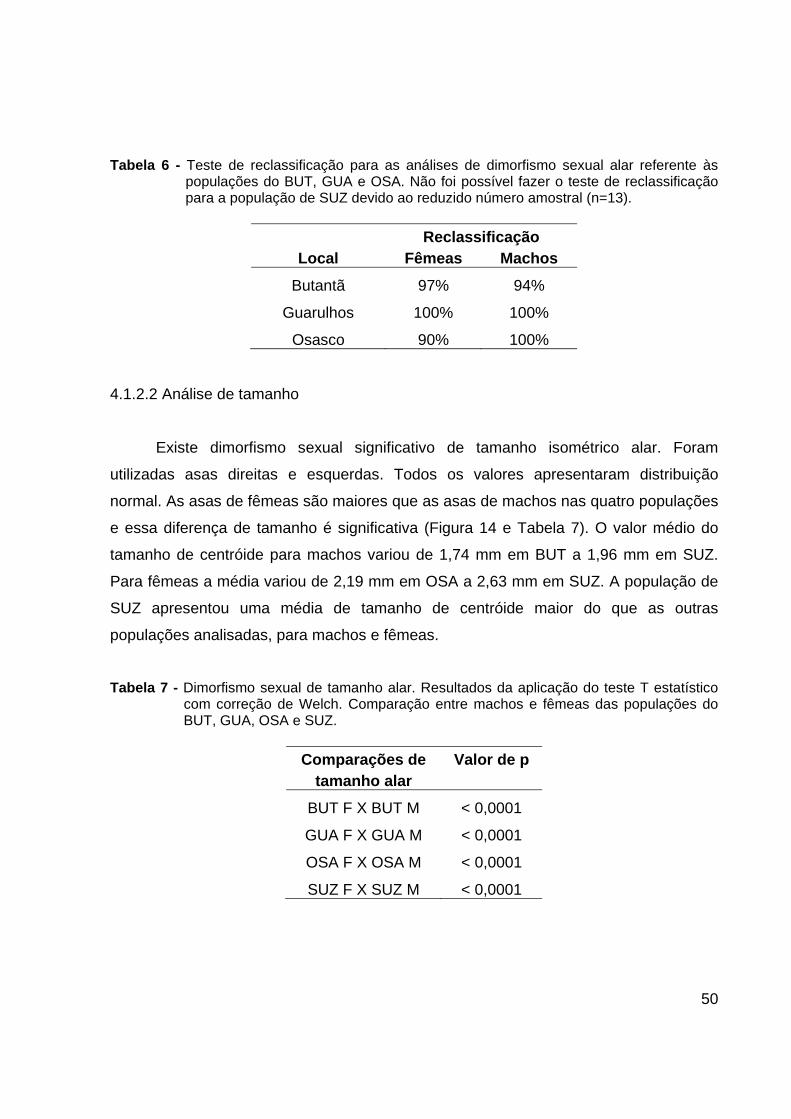
GUA e SUZ não apresentaram nenhuma intersecção entre os agrupamentos de
machos e fêmeas, havendo, entretanto uma pequena intersecção entre os sexos nas
populações do BUT e OSA.
A sobreposição dos consensos alares de machos e fêmeas evidenciou os
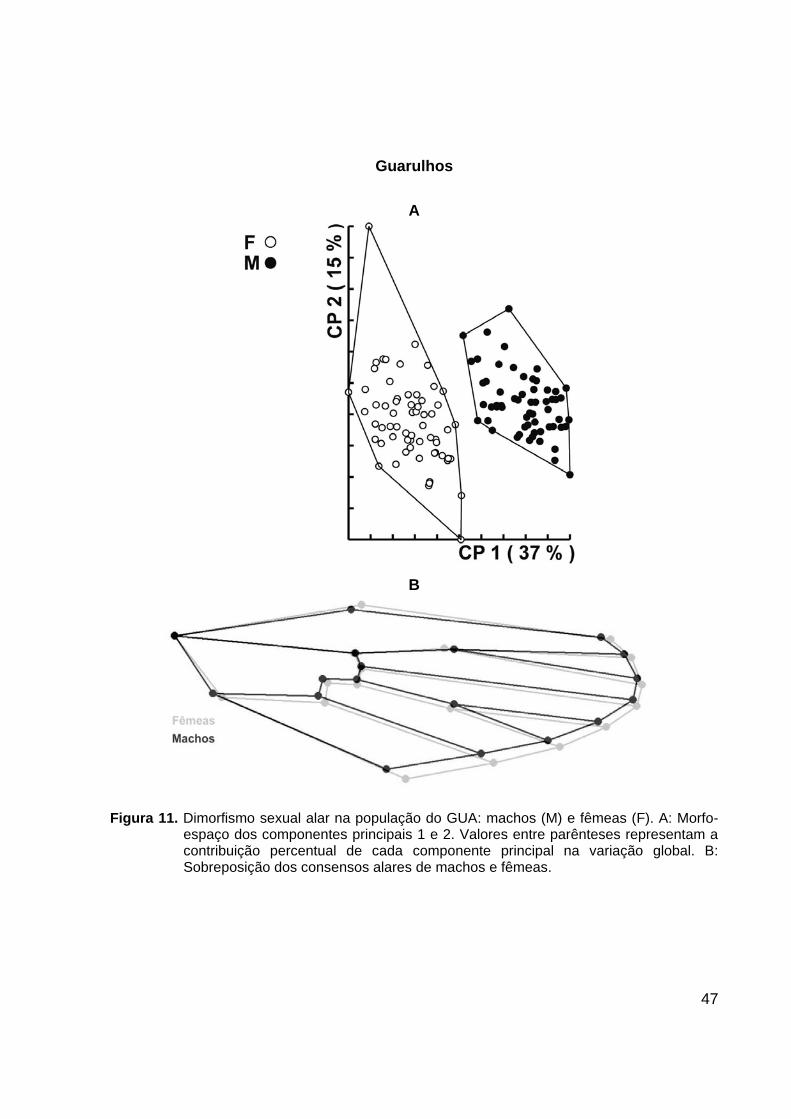
marcos anatômicos cujo posicionamento é sexualmente dimórfico (Figuras 10-13).
Análise discriminante dos componentes de forma alar resultou em valores de
reclassificação altos, conforme listados na Tabela 6.
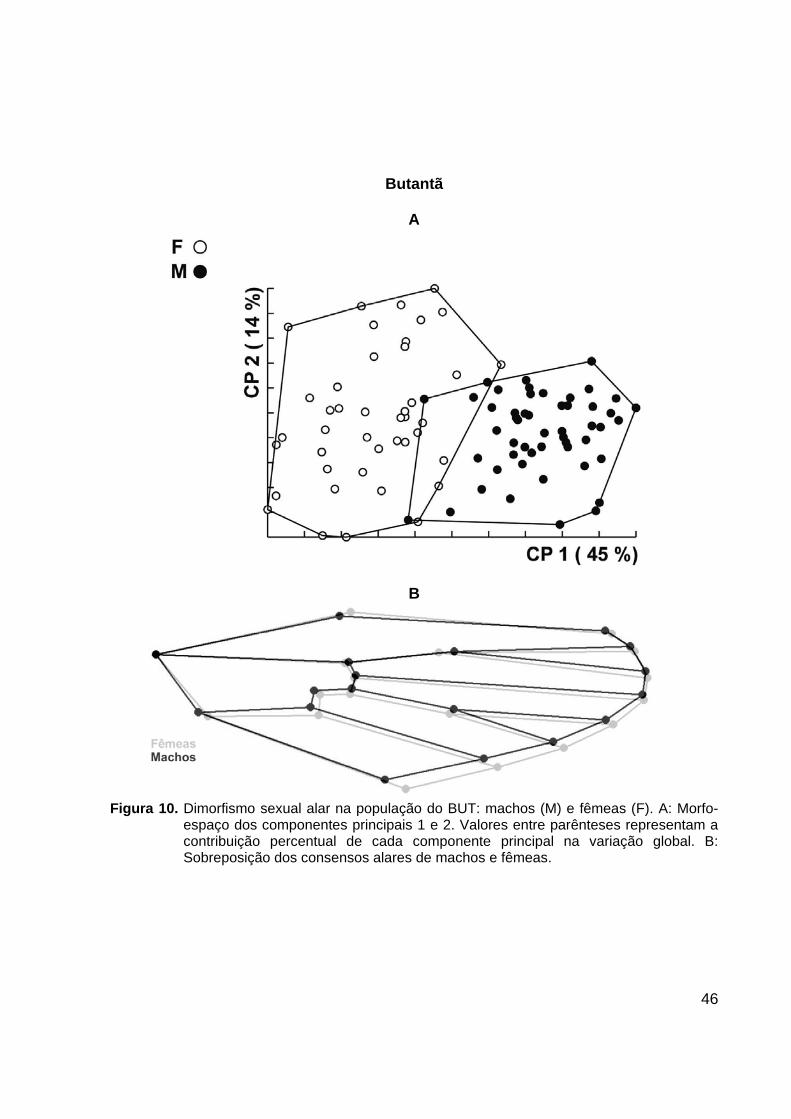
46
Butantã
A
B
Figura 10. Dimorfismo sexual alar na população do BUT: machos (M) e fêmeas (F). A: Morfo-
espaço dos componentes principais 1 e 2. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global. B: Sobreposição dos consensos alares de machos e fêmeas.
47
Guarulhos
A
B
Figura 11. Dimorfismo sexual alar na população do GUA: machos (M) e fêmeas (F). A: Morfo-
espaço dos componentes principais 1 e 2. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global. B: Sobreposição dos consensos alares de machos e fêmeas.
48
Osasco
A
B
Figura 12. Dimorfismo sexual alar na população de OSA: machos (M) e fêmeas (F). A: Morfo-
espaço dos componentes principais 1 e 2. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global. B: Sobreposição dos consensos alares de machos e fêmeas.
49
Suzano
A
B
Figura 13. Dimorfismo sexual alar na população de SUZ: machos (M) e fêmeas (F). A: Morfo-
espaço dos componentes principais 1 e 2. Valores entre parênteses representam a contribuição percentual de cada componente principal na variação global. B: Sobreposição dos consensos alares de machos e fêmeas.
50
Tabela 6 - Teste de reclassificação para as análises de dimorfismo sexual alar referente às populações do BUT, GUA e OSA. Não foi possível fazer o teste de reclassificação para a população de SUZ devido ao reduzido número amostral (n=13).
Reclassificação
Local Fêmeas Machos
Butantã 97% 94%
Guarulhos 100% 100%
Osasco 90% 100% 4.1.2.2 Análise de tamanho
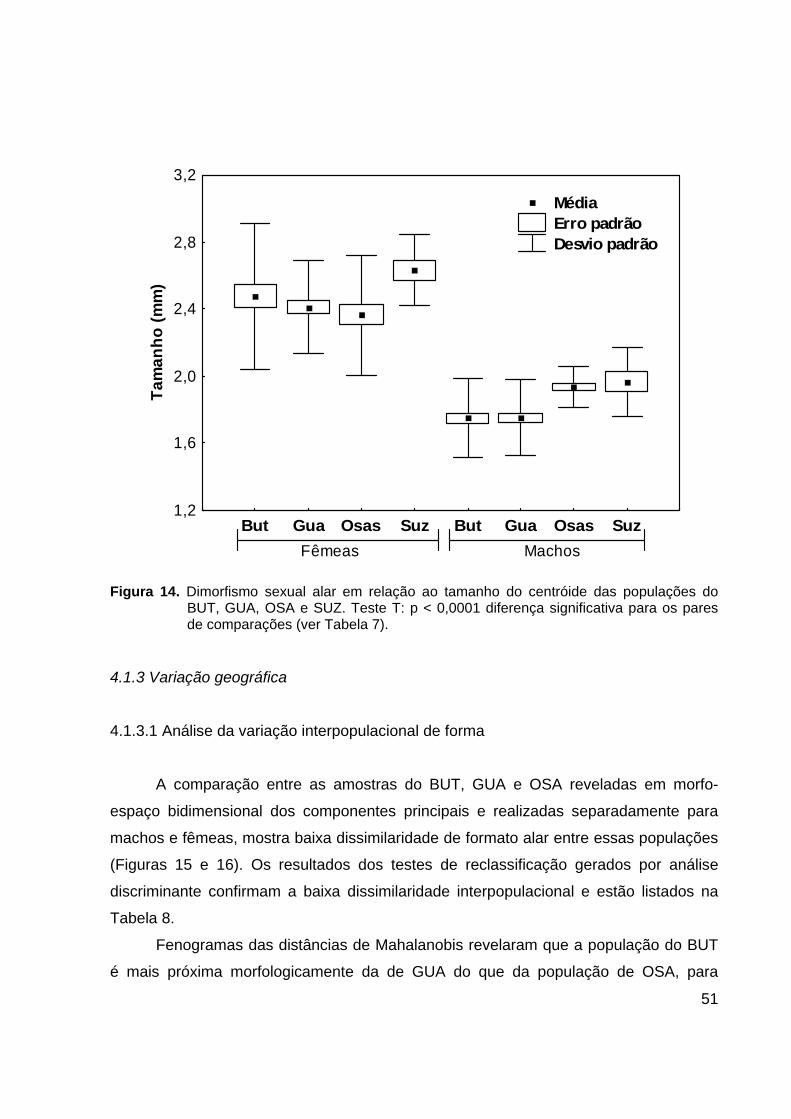
Existe dimorfismo sexual significativo de tamanho isométrico alar. Foram
utilizadas asas direitas e esquerdas. Todos os valores apresentaram distribuição
normal. As asas de fêmeas são maiores que as asas de machos nas quatro populações
e essa diferença de tamanho é significativa (Figura 14 e Tabela 7). O valor médio do
tamanho de centróide para machos variou de 1,74 mm em BUT a 1,96 mm em SUZ.
Para fêmeas a média variou de 2,19 mm em OSA a 2,63 mm em SUZ. A população de
SUZ apresentou uma média de tamanho de centróide maior do que as outras
populações analisadas, para machos e fêmeas.
Tabela 7 - Dimorfismo sexual de tamanho alar. Resultados da aplicação do teste T estatístico
com correção de Welch. Comparação entre machos e fêmeas das populações do BUT, GUA, OSA e SUZ.
Comparações de Valor de p
tamanho alar
BUT F X BUT M < 0,0001
GUA F X GUA M < 0,0001
OSA F X OSA M < 0,0001
SUZ F X SUZ M < 0,0001
51
But Gua Osas Suz But Gua Osas SuzFêmeas Machos
1,2
1,6
2,0
2,4
2,8
3,2Ta
man
ho (m
m)
Média Erro padrão Desvio padrão
Figura 14. Dimorfismo sexual alar em relação ao tamanho do centróide das populações do BUT, GUA, OSA e SUZ. Teste T: p < 0,0001 diferença significativa para os pares de comparações (ver Tabela 7).
4.1.3 Variação geográfica
4.1.3.1 Análise da variação interpopulacional de forma
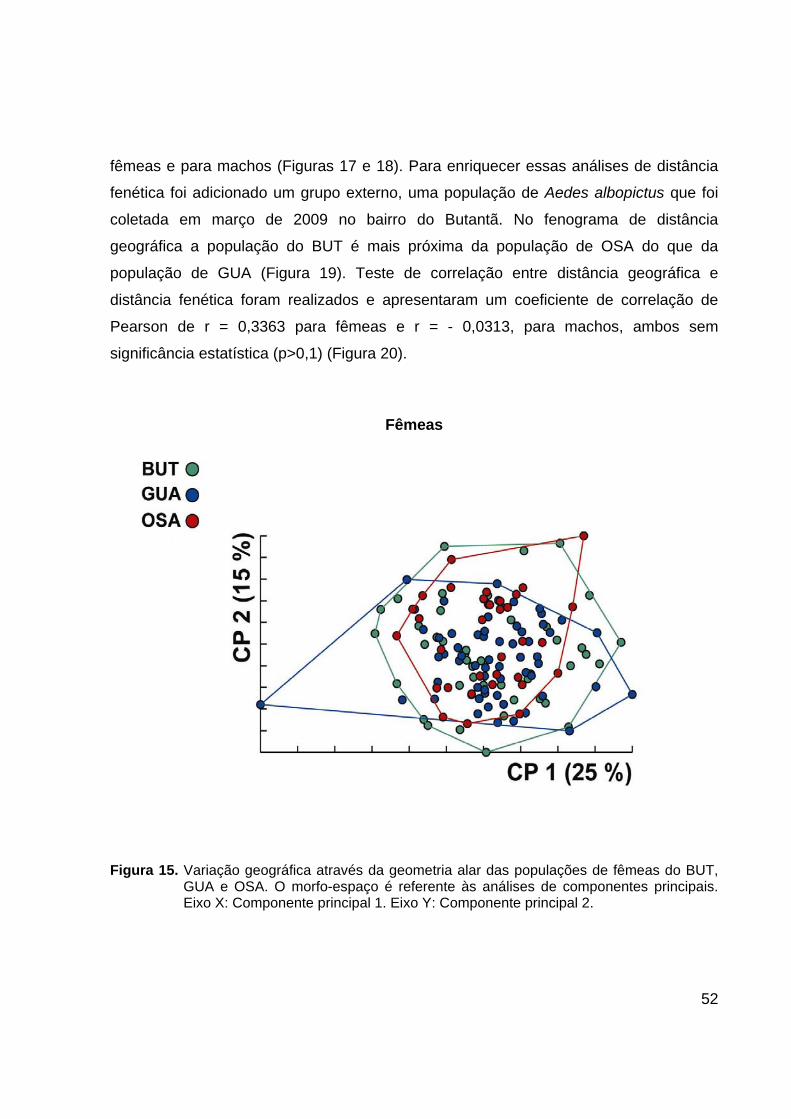
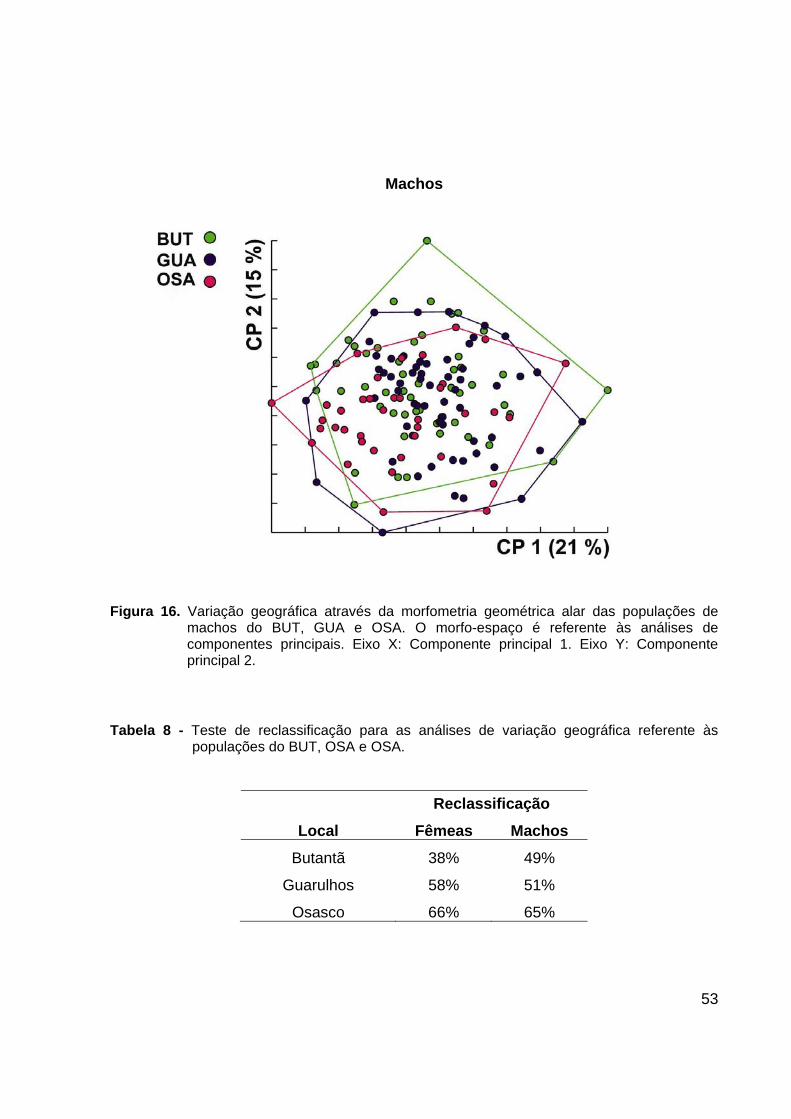
A comparação entre as amostras do BUT, GUA e OSA reveladas em morfo-
espaço bidimensional dos componentes principais e realizadas separadamente para
machos e fêmeas, mostra baixa dissimilaridade de formato alar entre essas populações
(Figuras 15 e 16). Os resultados dos testes de reclassificação gerados por análise
discriminante confirmam a baixa dissimilaridade interpopulacional e estão listados na
Tabela 8.
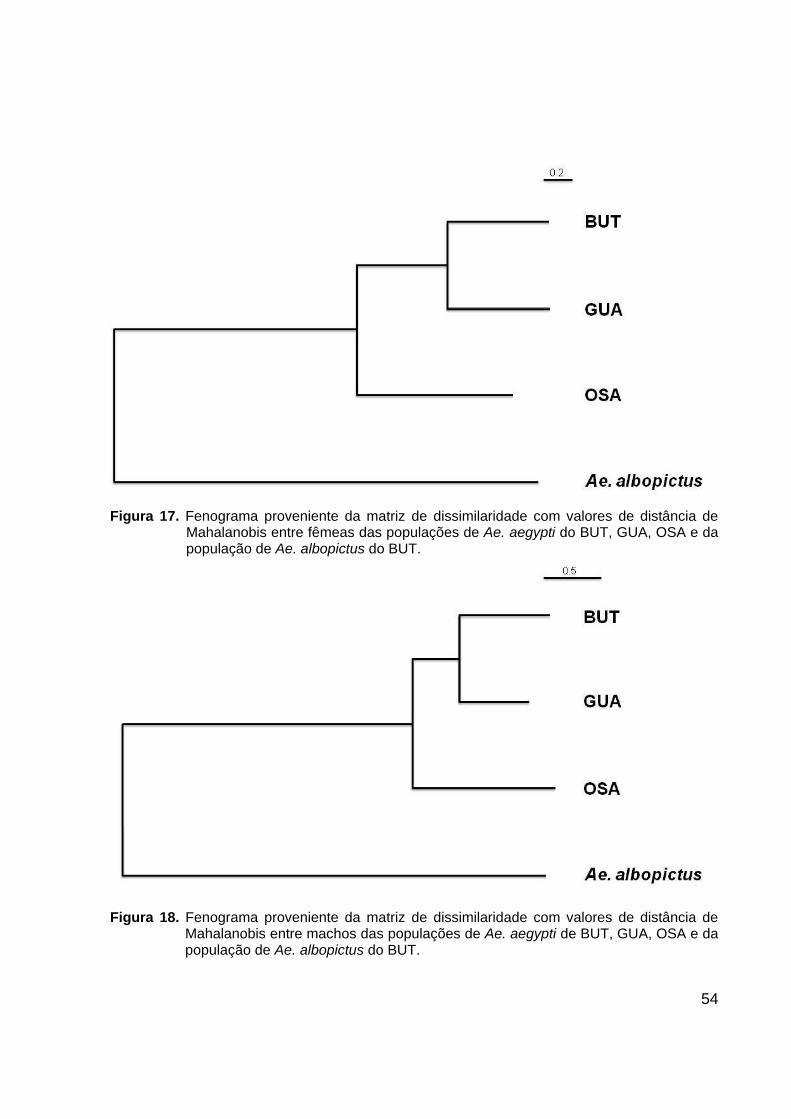
Fenogramas das distâncias de Mahalanobis revelaram que a população do BUT
é mais próxima morfologicamente da de GUA do que da população de OSA, para
52
fêmeas e para machos (Figuras 17 e 18). Para enriquecer essas análises de distância
fenética foi adicionado um grupo externo, uma população de Aedes albopictus que foi
coletada em março de 2009 no bairro do Butantã. No fenograma de distância
geográfica a população do BUT é mais próxima da população de OSA do que da
população de GUA (Figura 19). Teste de correlação entre distância geográfica e
distância fenética foram realizados e apresentaram um coeficiente de correlação de
Pearson de r = 0,3363 para fêmeas e r = - 0,0313, para machos, ambos sem
significância estatística (p>0,1) (Figura 20).
Fêmeas
Figura 15. Variação geográfica através da geometria alar das populações de fêmeas do BUT,
GUA e OSA. O morfo-espaço é referente às análises de componentes principais. Eixo X: Componente principal 1. Eixo Y: Componente principal 2.
53
Machos
Figura 16. Variação geográfica através da morfometria geométrica alar das populações de
machos do BUT, GUA e OSA. O morfo-espaço é referente às análises de componentes principais. Eixo X: Componente principal 1. Eixo Y: Componente principal 2.
Tabela 8 - Teste de reclassificação para as análises de variação geográfica referente às
populações do BUT, OSA e OSA.
Reclassificação
Local Fêmeas Machos
Butantã 38% 49%
Guarulhos 58% 51%
Osasco 66% 65%
54
Figura 17. Fenograma proveniente da matriz de dissimilaridade com valores de distância de
Mahalanobis entre fêmeas das populações de Ae. aegypti do BUT, GUA, OSA e da população de Ae. albopictus do BUT.
Figura 18. Fenograma proveniente da matriz de dissimilaridade com valores de distância de
Mahalanobis entre machos das populações de Ae. aegypti de BUT, GUA, OSA e da população de Ae. albopictus do BUT.
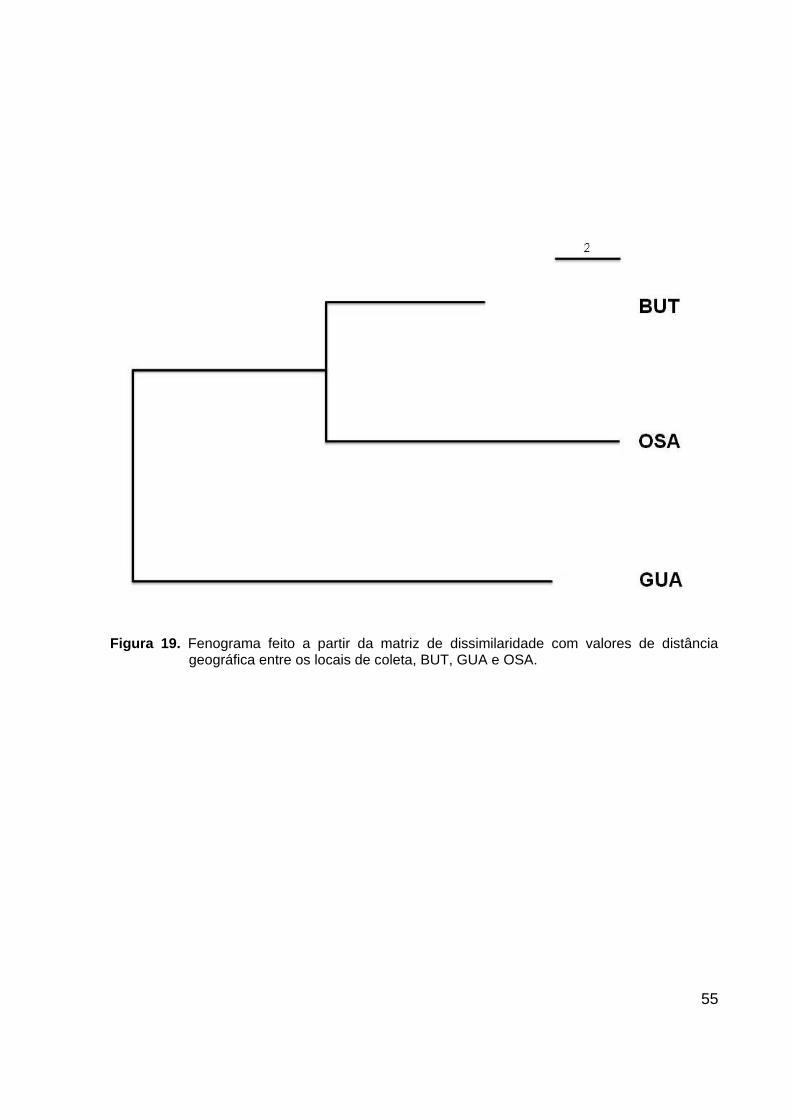
55
Figura 19. Fenograma feito a partir da matriz de dissimilaridade com valores de distância
geográfica entre os locais de coleta, BUT, GUA e OSA.
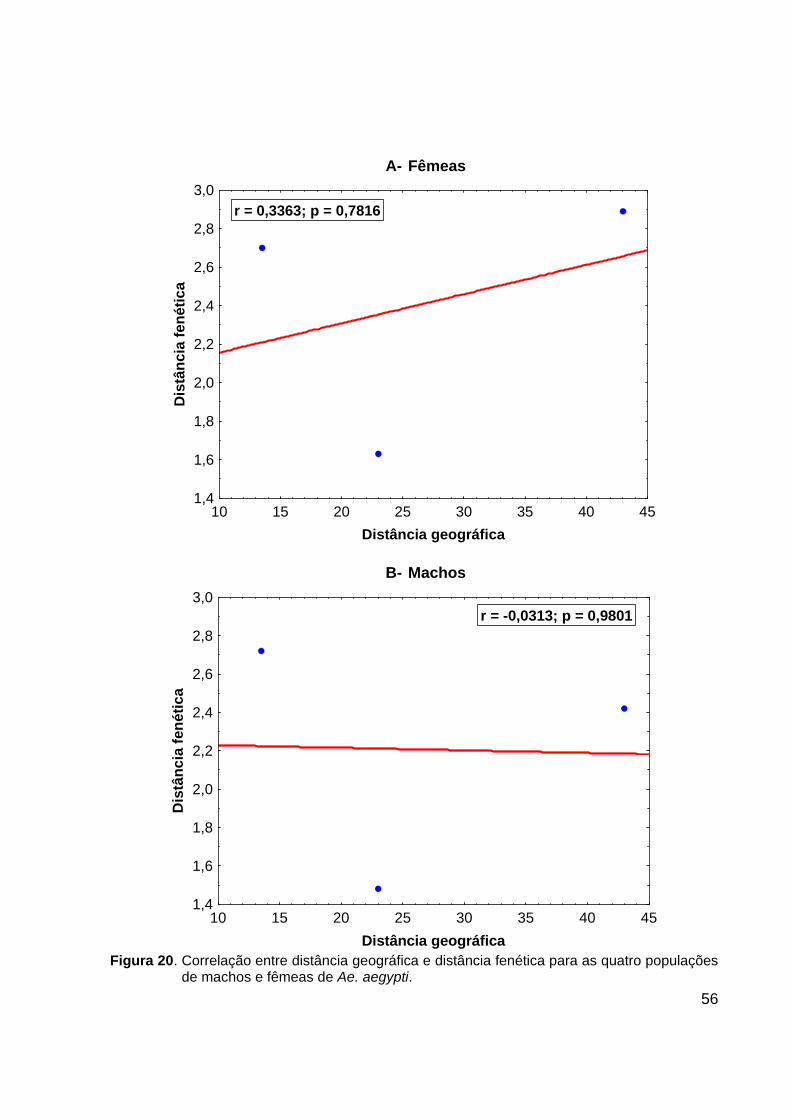
56
A- Fêmeas
10 15 20 25 30 35 40 45Distância geográfica
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0D
istâ
ncia
fené
tica
r = 0,3363; p = 0,7816
B- Machos
10 15 20 25 30 35 40 45Distância geográfica
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
Dis
tânc
ia fe
nétic
a
r = -0,0313; p = 0,9801
Figura 20. Correlação entre distância geográfica e distância fenética para as quatro populações
de machos e fêmeas de Ae. aegypti.
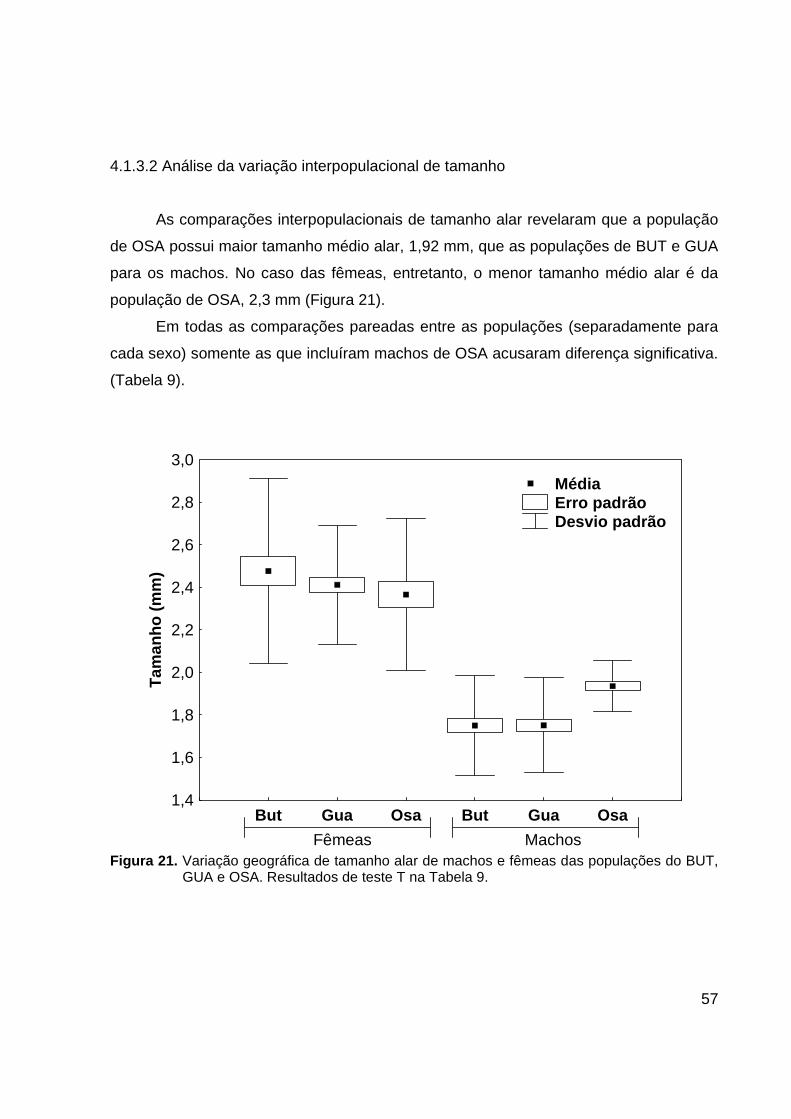
57
4.1.3.2 Análise da variação interpopulacional de tamanho
As comparações interpopulacionais de tamanho alar revelaram que a população
de OSA possui maior tamanho médio alar, 1,92 mm, que as populações de BUT e GUA
para os machos. No caso das fêmeas, entretanto, o menor tamanho médio alar é da
população de OSA, 2,3 mm (Figura 21).
Em todas as comparações pareadas entre as populações (separadamente para
cada sexo) somente as que incluíram machos de OSA acusaram diferença significativa.
(Tabela 9).
But Gua Osa But Gua Osa Fêmeas Machos
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
Tam
anho
(mm
)
Média Erro padrão Desvio padrão
Figura 21. Variação geográfica de tamanho alar de machos e fêmeas das populações do BUT,
GUA e OSA. Resultados de teste T na Tabela 9.

58
Tabela 9 - Variação geográfica de tamanho alar. Resultados da aplicação do teste T estatístico com correção de Welch. Comparação entre as populações de BUT, GUA e OSA em relação ao tamanho alar. Diferenças significativas indicadas por *.
Comparações de Valor de p
tamanho alar Fêmeas Machos
BUT X GUA p = 0,353 p = 0,985
BUT X OSA p = 0,241 p < 0,0001*
GUA X OSA p = 0,491 p < 0,0001* 4.1.4 Teste de repetibilidade
Foi realizado teste de repetibilidade das medidas utilizadas na coleta de dados
para análise morfométrica alar. Foram realizadas seis repetições de marcação de
pontos anatômicos. A única parte dessa análise que envolve interferência manual.
Uma única asa foi escolhida, de forma aleatória, e foram marcados dois pontos
anatômicos em cada repetição. Os pontos escolhidos foram o 2 e o 8, sendo o primeiro
de maior dificuldade de marcação correta devido às volumosas cerdas ao redor dele
(ver figura 5) . A análise de repetibilidade foi realizada no programa Statistica 7.0.
A análise de repetibilidade resultou em mais de 90 % de confiança nas medidas
realizadas, que de acordo com ASQC/AIAG (1991) p. 127, indica que o desempenho do
sistema de medidas aqui utilizado é aceitável. 4.2 Microssatélites Todos os cinco loci microssatélites (AED19, T3A7, 38/38, 34/72 e C2A8)
amplificaram em todas as populações de Ae. aegypti (BUT, GUA, OSA e SUZ) e foram
polimórficos. Os tamanhos dos fragmentos amplificados em cada locus estão
representados na Tabela 10.
O número de alelos observados para cada locus diferiu entre as populações
(Tabela 11). O locus que apresentou maior quantidade de alelos foi o 38/38, 6 alelos e
o locus que apresentou menor quantidade de alelos foi o AED19, 4 alelos.
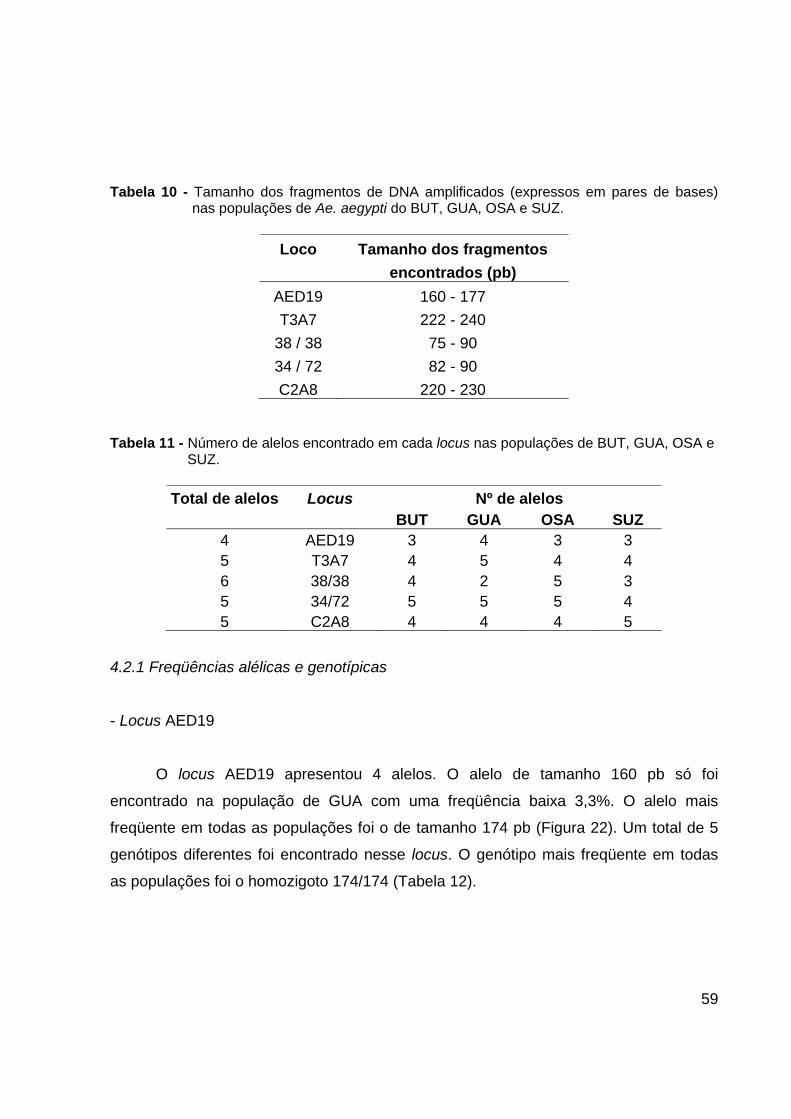
59
Tabela 10 - Tamanho dos fragmentos de DNA amplificados (expressos em pares de bases)
nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ.
Loco Tamanho dos fragmentos encontrados (pb)
AED19 160 - 177 T3A7 222 - 240
38 / 38 75 - 90 34 / 72 82 - 90 C2A8 220 - 230
Tabela 11 - Número de alelos encontrado em cada locus nas populações de BUT, GUA, OSA e
SUZ.
Total de alelos Locus Nº de alelos BUT GUA OSA SUZ 4 AED19 3 4 3 3 5 T3A7 4 5 4 4 6 38/38 4 2 5 3 5 34/72 5 5 5 4 5 C2A8 4 4 4 5
4.2.1 Freqüências alélicas e genotípicas
- Locus AED19
O locus AED19 apresentou 4 alelos. O alelo de tamanho 160 pb só foi
encontrado na população de GUA com uma freqüência baixa 3,3%. O alelo mais
freqüente em todas as populações foi o de tamanho 174 pb (Figura 22). Um total de 5
genótipos diferentes foi encontrado nesse locus. O genótipo mais freqüente em todas
as populações foi o homozigoto 174/174 (Tabela 12).

60
- Locus T3A7
O locus T3A7 apresentou 5 alelos. O alelo de tamanho 237 pb só foi encontrado
na população de GUA com uma freqüência baixa 1,7%. Os outros 4 alelos foram
encontrados em todas as quatro populações analisadas (Figura 23). Foi encontrado um
total de 10 genótipos diferentes para esse locus. O genótipo 222/237 foi observado em
um único indivíduo da população de GUA. O genótipo 222/225 foi encontrado nas
populações do BUT e GUA. Os genótipos 225/225 e o 222/240 foram encontrados em
BUT, GUA e SUZ. Os outros 6 genótipos foram encontrados em todas as quatro
populações analisadas (Tabela 13).
- Locus 38/38
O locus 38/38 apresentou 6 alelos. O alelo de tamanho 90 pb só foi encontrado
na população de SUZ com uma freqüência alta de 44,2%. O alelo de tamanho 88 pb só
foi encontrado na população de OSA com freqüência baixa de 6,7%. Os alelos de
tamanho 75 pb e 85 pb só foram encontrados nas populações de BUT e OSA. O alelo
mais freqüente em todas as populações foi o de tamanho 82 pb (Figura 24). Doze
genótipos foram encontrados nesse locus. O genótipo homozigoto 82/82 foi o único
encontrado em todas as populações com uma freqüência alta. O genótipo 82/90 só foi
observado na população de SUZ em 16 indivíduos (Tabela 14).
- Locus 34/72
O locus 34/72 apresentou 5 alelos. O alelo de tamanho 82 pb foi encontrado nas
populações do BUT, GUA e OSA. Os outros 4 alelos foram observados em todas as
quatro populações analisadas. O alelo de tamanho 88 pb foi o mais freqüente em todas
as populações (Figura 25). Foi observado um total de 11 genótipos nesse locus. Os
genótipos 86/88 e o 88/88 foram os mais comuns. Os genótipos 82/86 e o 86/86 só
foram encontrados em GUA. Os genótipos 82/90, 84/90 e o 90/90 só foram encontrados
no BUT (Tabela 15).
61
- Locus C2A8
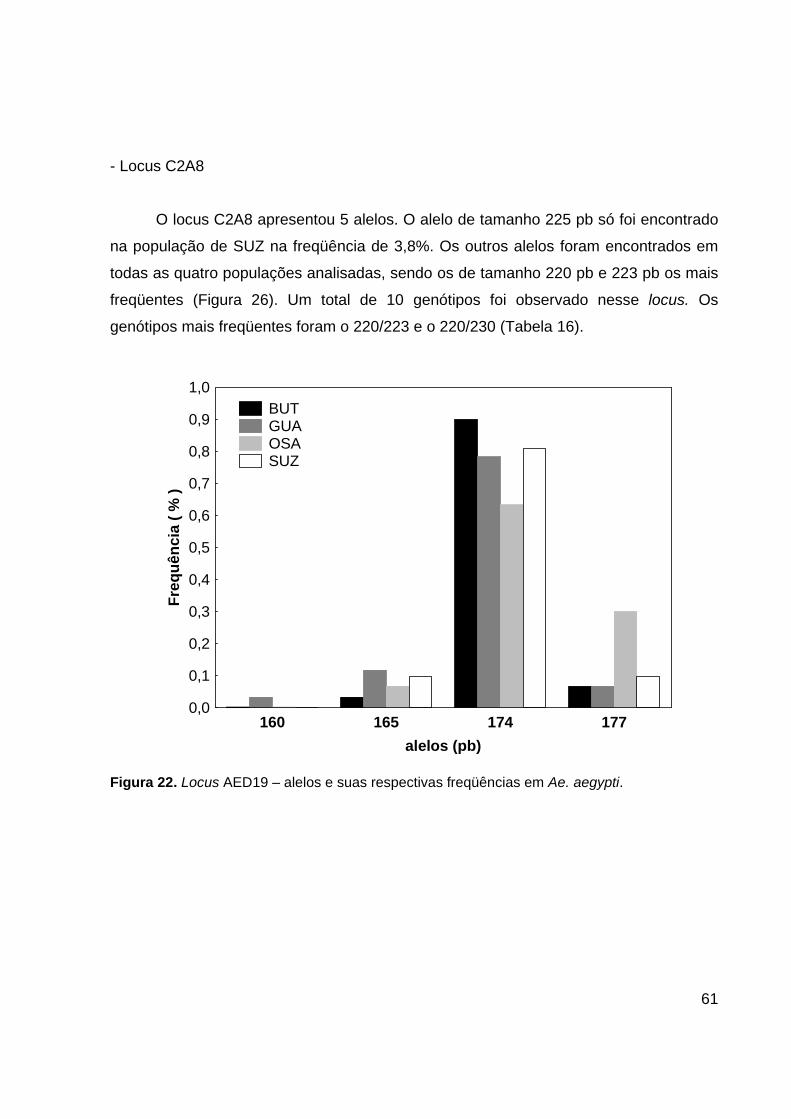
O locus C2A8 apresentou 5 alelos. O alelo de tamanho 225 pb só foi encontrado
na população de SUZ na freqüência de 3,8%. Os outros alelos foram encontrados em
todas as quatro populações analisadas, sendo os de tamanho 220 pb e 223 pb os mais
freqüentes (Figura 26). Um total de 10 genótipos foi observado nesse locus. Os
genótipos mais freqüentes foram o 220/223 e o 220/230 (Tabela 16).
160 165 174 177alelos (pb)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
Freq
uênc
ia (
% )
BUT GUA OSA SUZ
Figura 22. Locus AED19 – alelos e suas respectivas freqüências em Ae. aegypti.
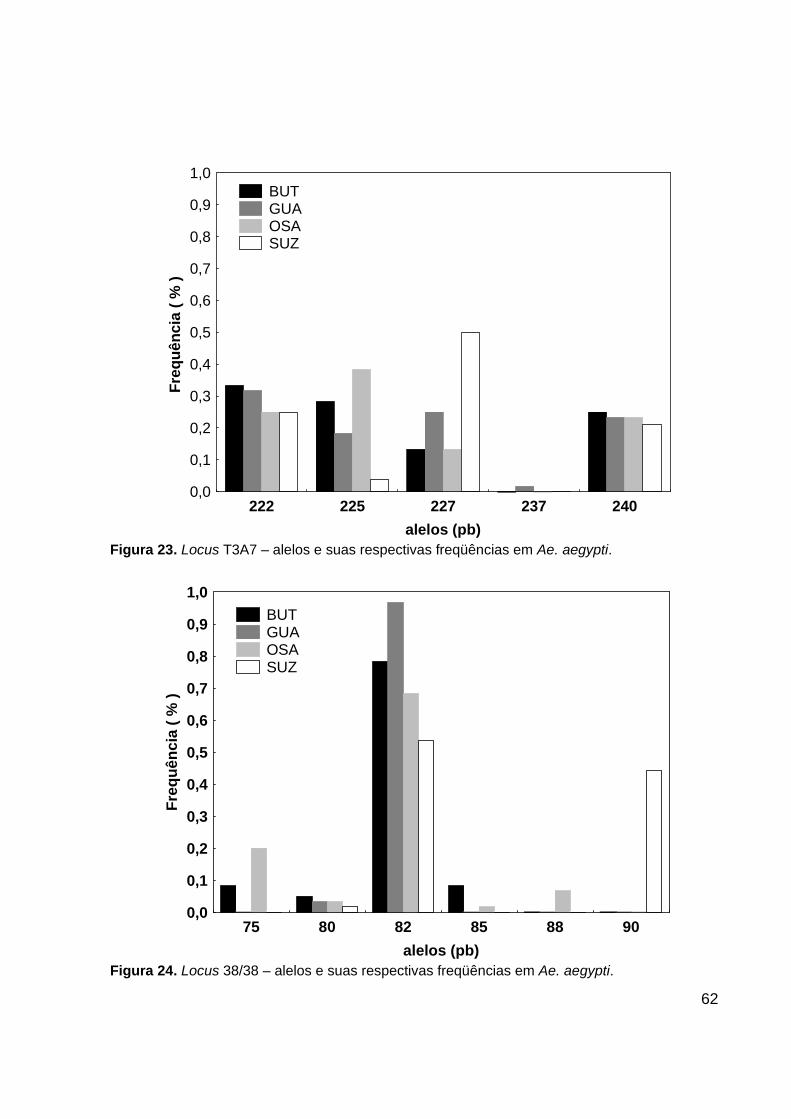
62
222 225 227 237 240alelos (pb)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0Fr
equê
ncia
( %
) BUT GUA OSA SUZ
Figura 23. Locus T3A7 – alelos e suas respectivas freqüências em Ae. aegypti.
75 80 82 85 88 90alelos (pb)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
Freq
uênc
ia (
% )
BUT GUA OSA SUZ
Figura 24. Locus 38/38 – alelos e suas respectivas freqüências em Ae. aegypti.
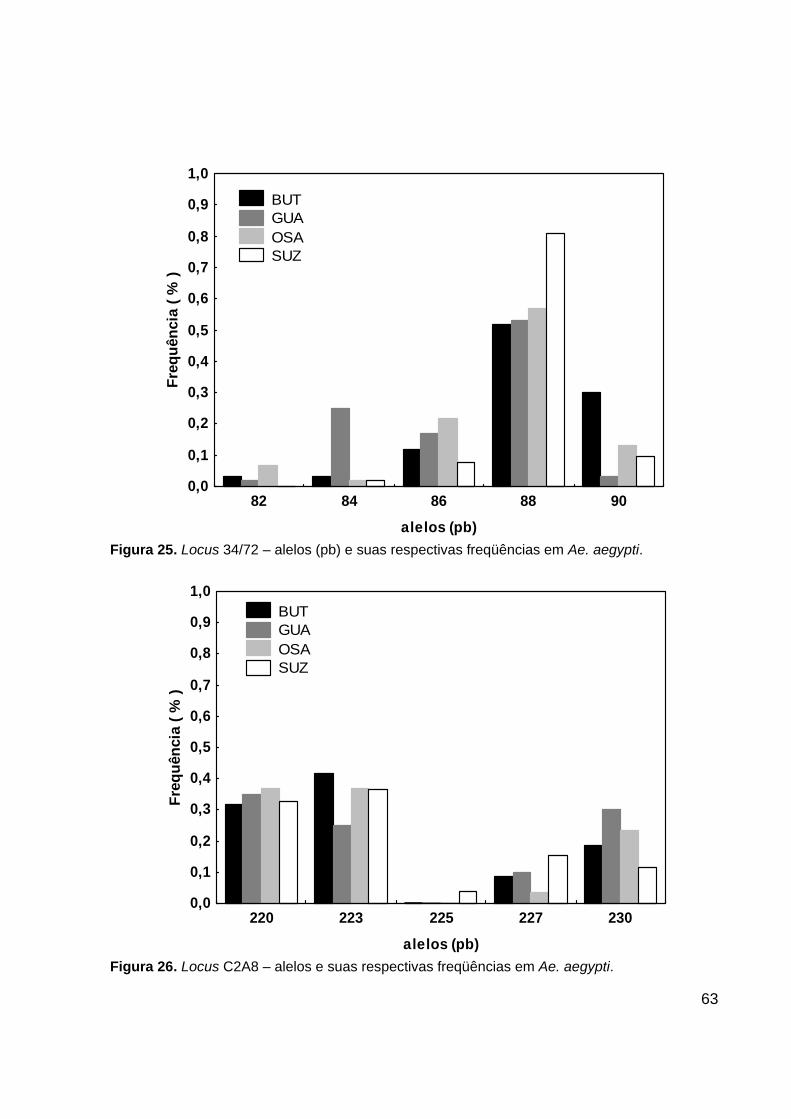
63
82 84 86 88 90
alelos (pb)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0Fr
equê
ncia
( %
)
BUT GUA OSA SUZ
Figura 25. Locus 34/72 – alelos (pb) e suas respectivas freqüências em Ae. aegypti.
220 223 225 227 230
alelos (pb)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
Freq
uênc
ia (
% )
BUT GUA OSA SUZ
Figura 26. Locus C2A8 – alelos e suas respectivas freqüências em Ae. aegypti.
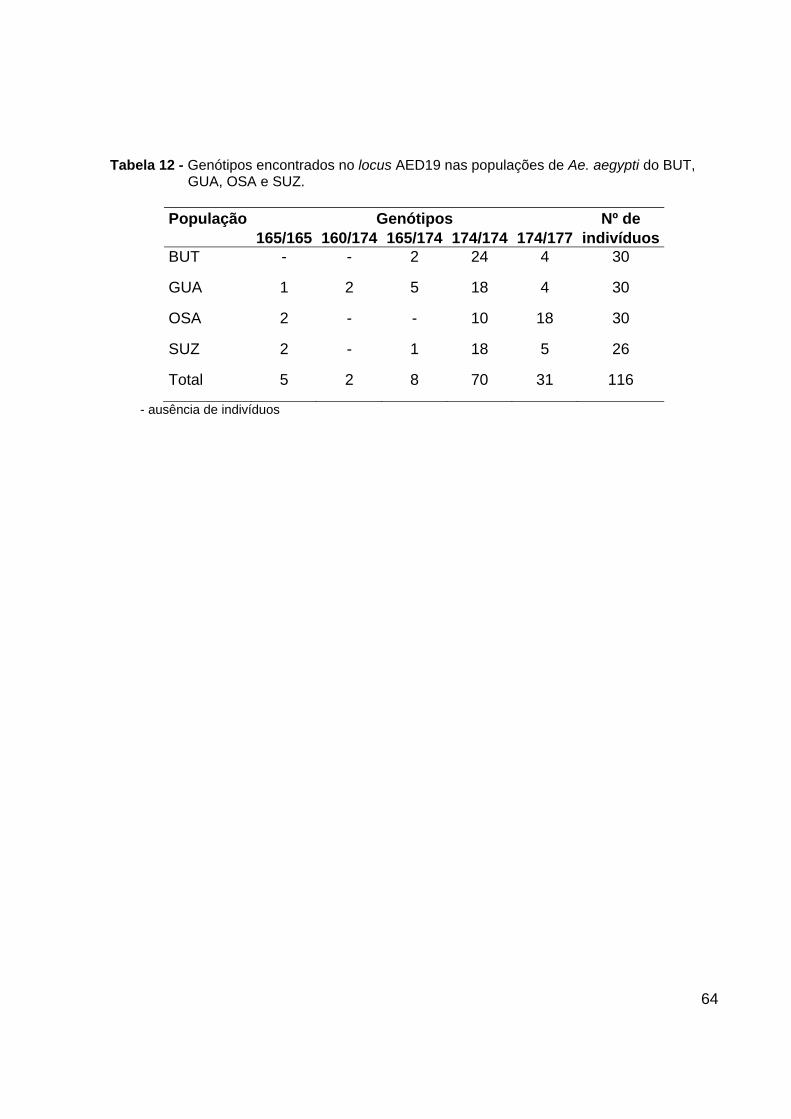
64
Tabela 12 - Genótipos encontrados no locus AED19 nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ.
População Genótipos Nº de
165/165 160/174 165/174 174/174 174/177 indivíduosBUT - - 2 24 4 30
GUA 1 2 5 18 4 30
OSA 2 - - 10 18 30
SUZ 2 - 1 18 5 26
Total 5 2 8 70 31 116
- ausência de indivíduos
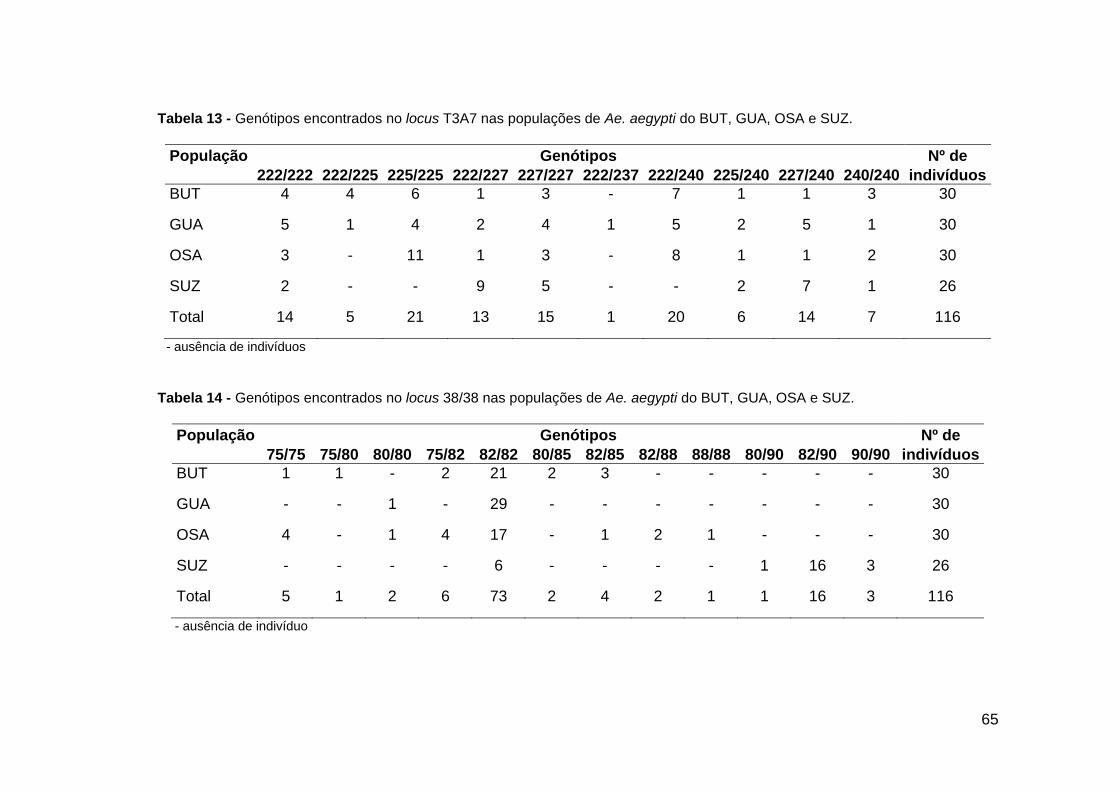
65
Tabela 13 - Genótipos encontrados no locus T3A7 nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ.
População Genótipos Nº de 222/222 222/225 225/225 222/227 227/227 222/237 222/240 225/240 227/240 240/240 indivíduos
BUT 4 4 6 1 3 - 7 1 1 3 30
GUA 5 1 4 2 4 1 5 2 5 1 30
OSA 3 - 11 1 3 - 8 1 1 2 30
SUZ 2 - - 9 5 - - 2 7 1 26
Total 14 5 21 13 15 1 20 6 14 7 116
- ausência de indivíduos Tabela 14 - Genótipos encontrados no locus 38/38 nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ.
População Genótipos Nº de 75/75 75/80 80/80 75/82 82/82 80/85 82/85 82/88 88/88 80/90 82/90 90/90 indivíduos
BUT 1 1 - 2 21 2 3 - - - - - 30
GUA - - 1 - 29 - - - - - - - 30
OSA 4 - 1 4 17 - 1 2 1 - - - 30
SUZ - - - - 6 - - - - 1 16 3 26
Total 5 1 2 6 73 2 4 2 1 1 16 3 116
- ausência de indivíduo
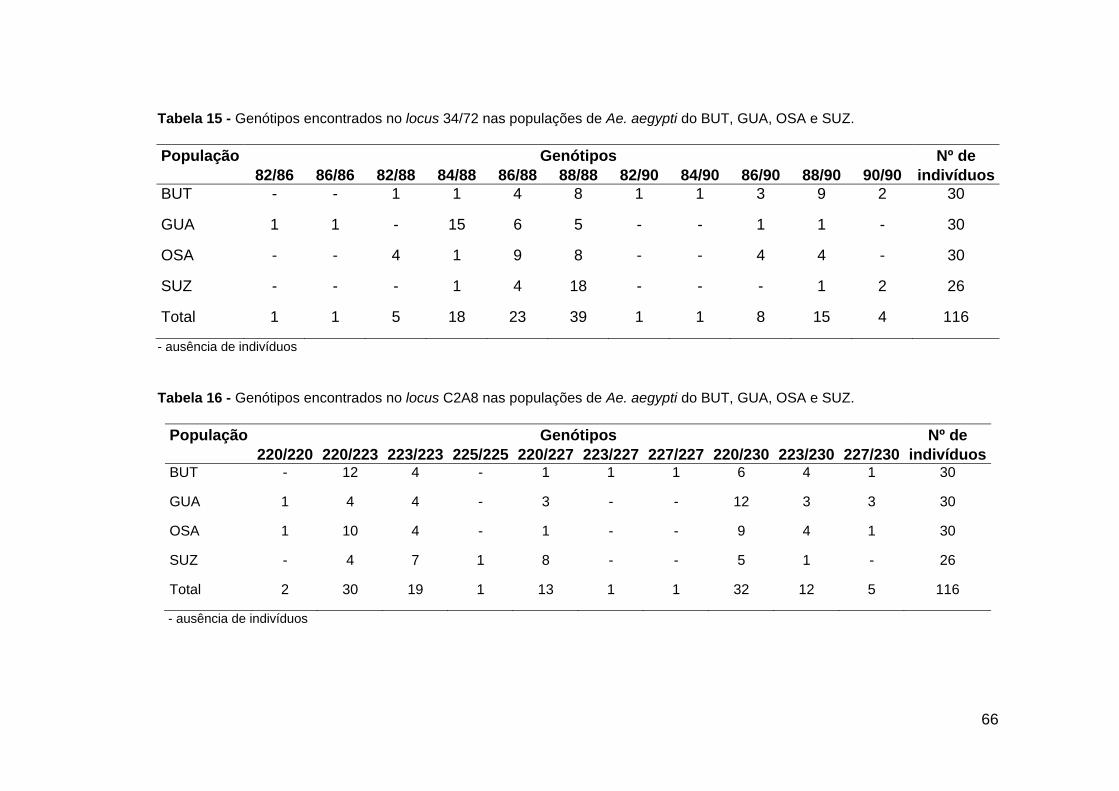
66
Tabela 15 - Genótipos encontrados no locus 34/72 nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ. População Genótipos Nº de
82/86 86/86 82/88 84/88 86/88 88/88 82/90 84/90 86/90 88/90 90/90 indivíduos BUT - - 1 1 4 8 1 1 3 9 2 30
GUA 1 1 - 15 6 5 - - 1 1 - 30
OSA - - 4 1 9 8 - - 4 4 - 30
SUZ - - - 1 4 18 - - - 1 2 26
Total 1 1 5 18 23 39 1 1 8 15 4 116
- ausência de indivíduos Tabela 16 - Genótipos encontrados no locus C2A8 nas populações de Ae. aegypti do BUT, GUA, OSA e SUZ.
População Genótipos Nº de 220/220 220/223 223/223 225/225 220/227 223/227 227/227 220/230 223/230 227/230 indivíduos
BUT - 12 4 - 1 1 1 6 4 1 30
GUA 1 4 4 - 3 - - 12 3 3 30
OSA 1 10 4 - 1 - - 9 4 1 30
SUZ - 4 7 1 8 - - 5 1 - 26
Total 2 30 19 1 13 1 1 32 12 5 116
- ausência de indivíduos

67
4.2.2 Testes de equilíbrio de Hardy- Weinberg
Equilíbrio de Hardy-Weinberg foi testado para cada locus em cada população
usando teste de probabilidade (Haldane, 1954). Dos loci analisados, 65%
apresentaram desvio significativo do equilíbrio de HW.
A população do BUT apresentou desvio do equilíbrio de HW em dois loci
(T3A7 e 38/38), ambos devido ao déficit de heterozigotos. A população de GUA não
apresentou equilíbrio de HW em quatro loci, dois loci devido ao déficit de
heterozigotos (T3A7 e 38/38), e dois loci devido ao excesso de heterozigotos (34/72
e C2A8). A população de OSA apresentou três loci com desvio do equilíbrio de HW,
um loci devido ao excesso de heterozigotos (AED19) e dois loci devido ao déficit de
heterozigotos (T3A7 e 38/38). A população de SUZ apresentou quatro loci com
desvio do equilíbrio de HW, três deles devido ao déficit de heterozigotos (AED19,
34/72 e C2A8) e um loci devido ao excesso de heterozigotos (T3A7) (Tabela 17 e
Figuras 27-30). O locus T3A7 apresentou desvio do equilíbrio de HW em todas as
populações analisadas. Do total de loci que não apresentou equilíbrio de HW,
69,23% foi devido ao déficit de heterozigotos e 30,77% devido ao excesso de
heterozigotos. Estimativas de freqüência de alelos nulos nos cinco loci
microssatélites estão mostrados na tabela 18.
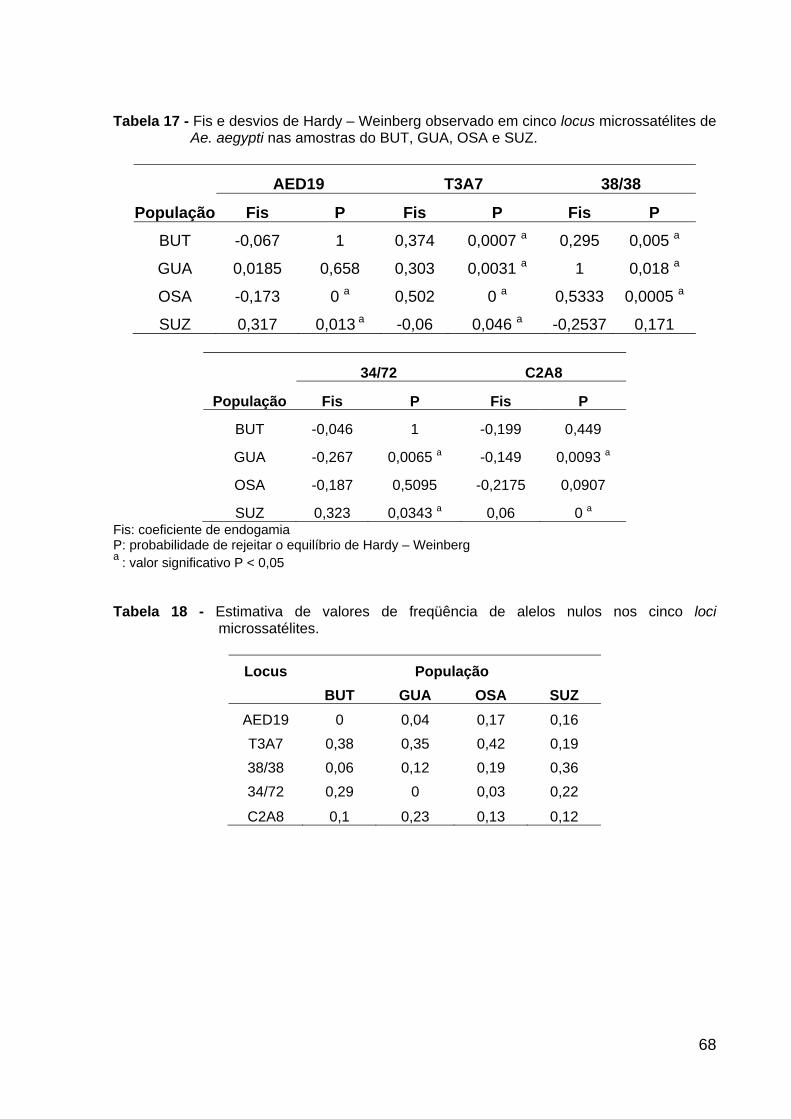
68
Tabela 17 - Fis e desvios de Hardy – Weinberg observado em cinco locus microssatélites de Ae. aegypti nas amostras do BUT, GUA, OSA e SUZ.
AED19 T3A7 38/38
População Fis P Fis P Fis P
BUT -0,067 1 0,374 0,0007 a 0,295 0,005 a
GUA 0,0185 0,658 0,303 0,0031 a 1 0,018 a
OSA -0,173 0 a 0,502 0 a 0,5333 0,0005 a
SUZ 0,317 0,013 a -0,06 0,046 a -0,2537 0,171
34/72 C2A8
População Fis P Fis P
BUT -0,046 1 -0,199 0,449
GUA -0,267 0,0065 a -0,149 0,0093 a
OSA -0,187 0,5095 -0,2175 0,0907
SUZ 0,323 0,0343 a 0,06 0 a Fis: coeficiente de endogamia P: probabilidade de rejeitar o equilíbrio de Hardy – Weinberg a : valor significativo P < 0,05
Tabela 18 - Estimativa de valores de freqüência de alelos nulos nos cinco loci microssatélites.
Locus População
BUT GUA OSA SUZ AED19 0 0,04 0,17 0,16 T3A7 0,38 0,35 0,42 0,19 38/38 0,06 0,12 0,19 0,36 34/72 0,29 0 0,03 0,22
C2A8 0,1 0,23 0,13 0,12
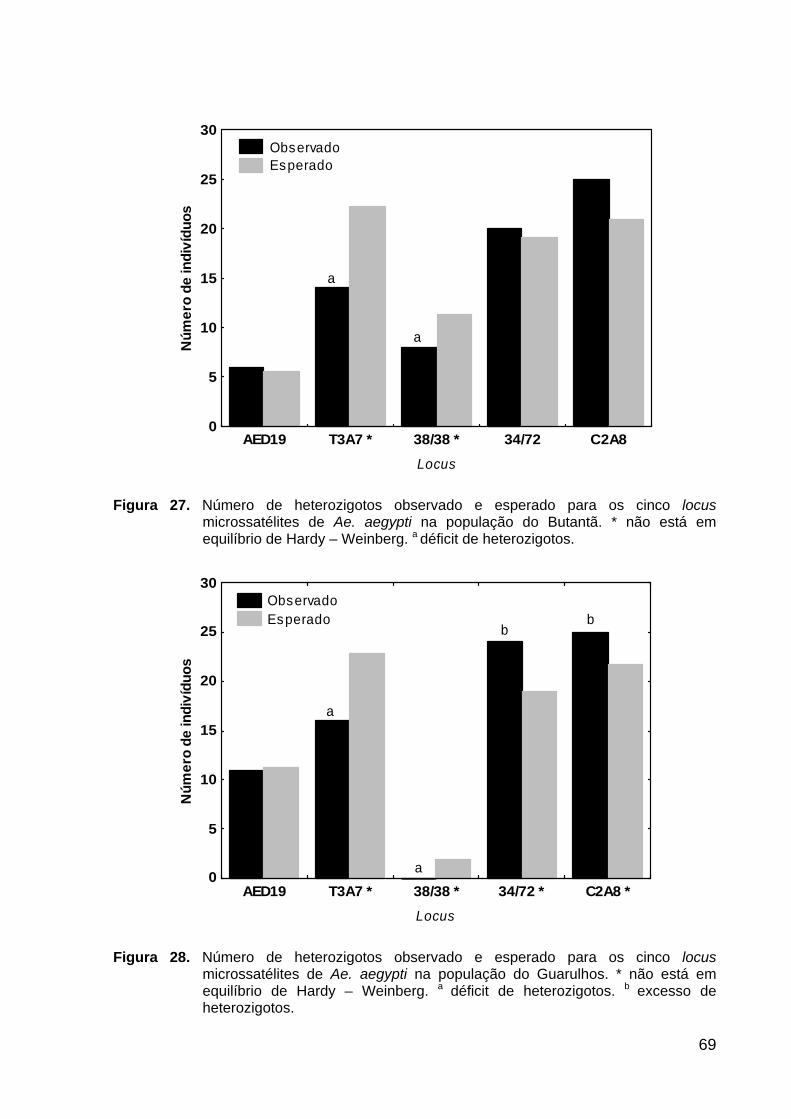
69
AED19 T3A7 * 38/38 * 34/72 C2A8
Locus
0
5
10
15
20
25
30
Núm
ero
de in
diví
duos
Observado Esperado
a
a
Figura 27. Número de heterozigotos observado e esperado para os cinco locus microssatélites de Ae. aegypti na população do Butantã. * não está em equilíbrio de Hardy – Weinberg. a déficit de heterozigotos.
AED19 T3A7 * 38/38 * 34/72 * C2A8 *
Locus
0
5
10
15
20
25
30
Núm
ero
de in
diví
duos
Observado Esperado
a
a
bb
Figura 28. Número de heterozigotos observado e esperado para os cinco locus microssatélites de Ae. aegypti na população do Guarulhos. * não está em equilíbrio de Hardy – Weinberg. a déficit de heterozigotos. b excesso de heterozigotos.
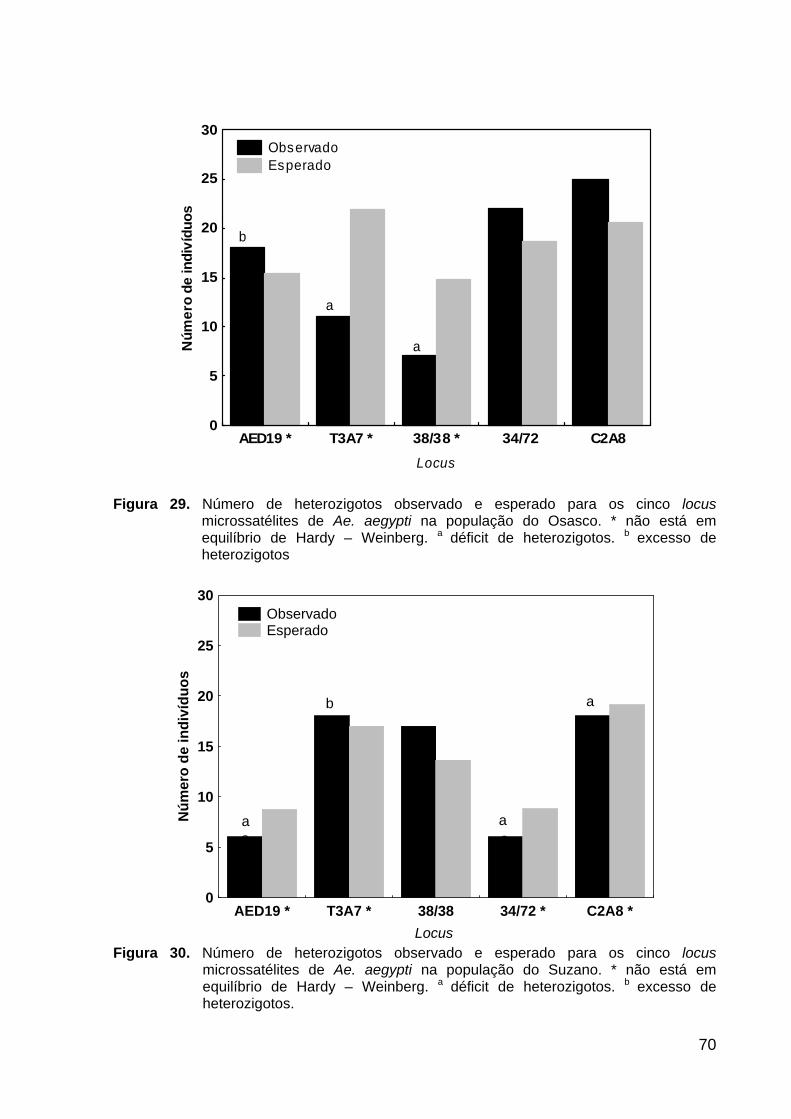
70
AED19 * T3A7 * 38/38 * 34/72 C2A8
Locus
0
5
10
15
20
25
30
Núm
ero
de in
diví
duos
Observado Esperado
a
b
a
Figura 29. Número de heterozigotos observado e esperado para os cinco locus microssatélites de Ae. aegypti na população do Osasco. * não está em equilíbrio de Hardy – Weinberg. a déficit de heterozigotos. b excesso de heterozigotos
AED19 * T3A7 * 38/38 34/72 * C2A8 *Locus
0
5
10
15
20
25
30
Núm
ero
de in
diví
duos
Observado Esperado
a
a
a aa
b
Figura 30. Número de heterozigotos observado e esperado para os cinco locus
microssatélites de Ae. aegypti na população do Suzano. * não está em equilíbrio de Hardy – Weinberg. a déficit de heterozigotos. b excesso de heterozigotos.

71
4.2.3 Desequilíbrio de ligação
Das 40 análises possíveis em relação ao desequilíbrio de ligação, apenas 4
foram significativos (AED19-C2A8, C2A8-T3A7, T3A7-34/72, AED19-T3A7) com p <
0,01 na população de Osasco.
4.2.4 Diferenciação populacional
Foram determinados valores F-estatístico de Wright (Fis, Fst e Fit) para os
cinco loci polimórficos. Heterogeneidade genética (Fst) refletiu altos níveis de
diferenciação genética entre as populações de Ae. aegypti da área metropolitana de
São Paulo, apresentando um valor médio de 0,0622, estatisticamente significativo p
< 0.05. O locus C2A8 foi o que apresentou menor valor de Fst. Desvio do equilíbrio
de HW nas subpopulações apresentou um valor médio de 0, 057 refletindo um déficit
de heterozigotos. O desvio de HW no total das populações também refletiu em
déficit de heterozigotos, com um valor médio de 0, 1159 (Tabela 19).
O fluxo gênico (Nm) estimado para as quatro populações estudadas foi de
0,47 indivíduos, valor relativamente baixo, o que é compatível com a hipótese de
estruturação populacional.
Tabela 19 - Estimativas de valores F-estatístico de Wright calculado em cada locus em
quatro populações de Ae. aegypti.
Locus Fis Fst Fit AED19 -0,0013 0,0568 0,0555
T3A7 0,3012 0,0456 0,3331
38/38 0,2364 0,1834 0,3765
34/72 -0,0998 0,0653 -0,028
C2A8 -0,1305 0,0051 -0,1247
Total 0,0572 0,0622 0,1159 Fis: Desvio de HW nas subpopulações; Fst: heterogeneidade genética; Fit: desvio de HW no total das populações.

72
4.2.4 Distância genética versus distância geográfica
Variações nos valores de Fst foram avaliadas de acordo com a distância
geográfica. Uma semi-matriz de Fst foi calculada para estimar a distância genética
entre as populações do BUT, GUA, OSA e SUZ. Fenogramas de distância
mostraram que as populações mais próximas geneticamente (Figura 31) também
são mais próximas geograficamente (Figura 32). Correlação entre distância genética
e geográfica mostrou um índice de correlação de r = 0,76, indicando uma alta
correlação positiva (Figura 33).
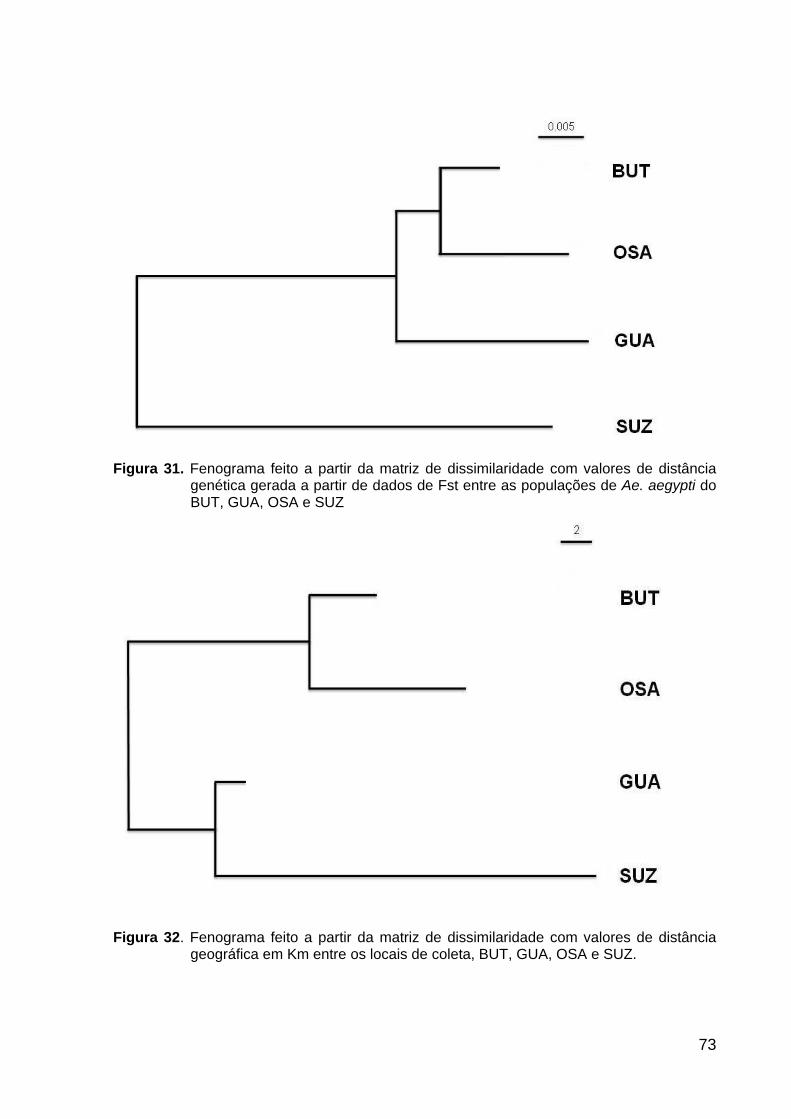
73
Figura 31. Fenograma feito a partir da matriz de dissimilaridade com valores de distância
genética gerada a partir de dados de Fst entre as populações de Ae. aegypti do BUT, GUA, OSA e SUZ
Figura 32. Fenograma feito a partir da matriz de dissimilaridade com valores de distância
geográfica em Km entre os locais de coleta, BUT, GUA, OSA e SUZ.
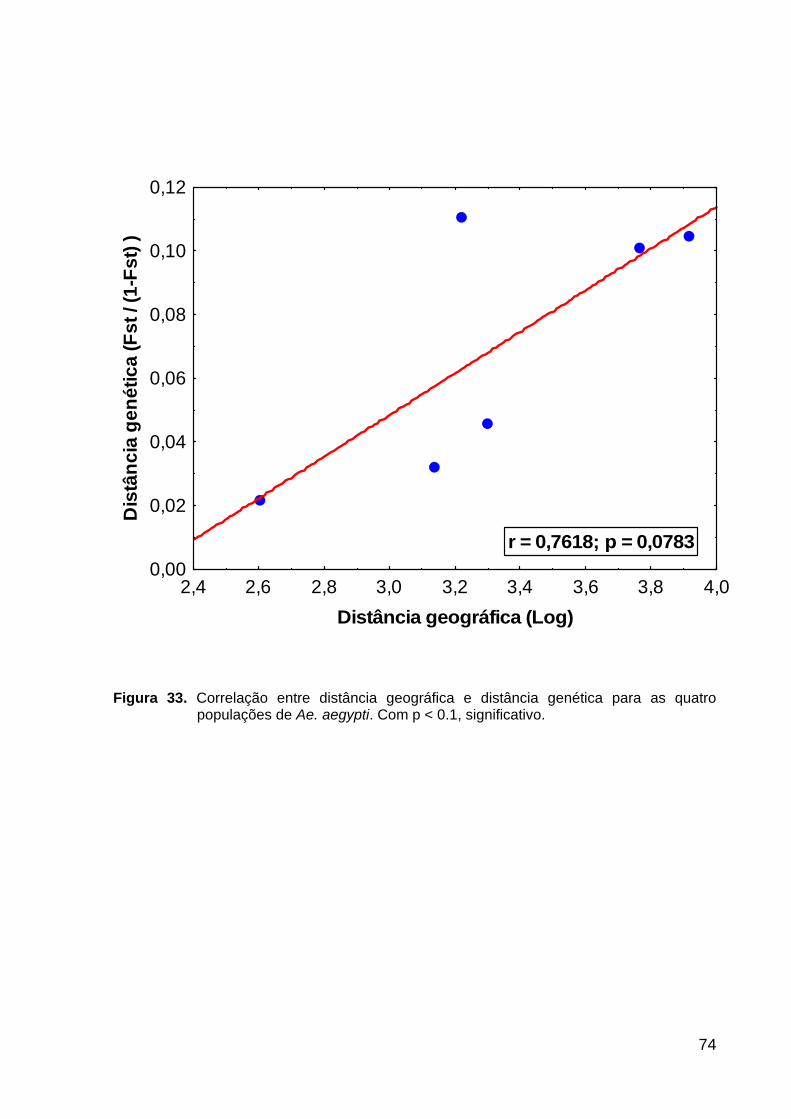
74
2,4 2,6 2,8 3,0 3,2 3,4 3,6 3,8 4,0Distância geográfica (Log)
0,00
0,02
0,04
0,06
0,08
0,10
0,12
Dis
tânc
ia g
enét
ica
(Fst
/ (1
-Fst
) )
r = 0,7618; p = 0,0783
Figura 33. Correlação entre distância geográfica e distância genética para as quatro
populações de Ae. aegypti. Com p < 0.1, significativo.

75
5 DISCUSSÃO 5.1 Morfometria geométrica alar
Análises intra e interpopulacionais foram realizadas com as quatro
populações de Ae. aegypti da área metropolitana de São Paulo. Dentro das
comparações intrapopulacionais, a assimetria bilateral de forma foi tênue e de
tamanho foi estatisticamente não-significativa. Há casos de insetos em que a
assimetria bilateral está associada a condições ambientais adversas, algumas delas
próprias de ambientes urbanos (Souza et al., 2007; Mpho et al., 2002; Mpho et al.,
2001). Isso poderia nos levar a subestimar as possíveis influências de eventual
estresse ambiental nessa espécie. Entretanto, antes de formular conclusões
prematuras, seria mais indicado avaliar a real exposição dos indivíduos a fatores de
stress, bem como a influência de pressão seletiva na manutenção da simetria alar.
O alto dimorfismo sexual alar detectado na análise de forma e tamanho de
todas as populações analisadas corrobora a suspeita levantada em estudos
taxonômicos tradicionais realizados em culicídeos (Lien, 1968) e com outras
observações de diversos autores sobre as diferentes funções que as asas podem
assumir para ambos os sexos. Os machos podem usar as asas na produção de
sons de corte sexual e as fêmeas para vôos precisos durante a busca de alimento,
dispersão e oviposição (revisão em Forattini, 2002). O tamanho das asas foi maior
em fêmeas do que em machos, o que é observado em alguns culicídeos (Agnew et
al., 2000; Jirakanjanakit et al., 2007).
Nas comparações interpopulacionais nota-se uma grande variabilidade
morfológica alar intrapopulacional. Foi possível reconhecer um ligeiro distanciamento
entre as populações que, no entanto não acusa diferenciação marcante e, portanto
não indica estruturação populacional evidente. Ausência de correlação foi observada
entre distâncias geográficas e fenéticas nas populações analisadas, indicando que a
distância geográfica não foi majoritariamente influente na evolução desses padrões
alares.
Foi observado nos fenogramas de distância de Mahalanobis o mesmo padrão
para ambos os sexos de Ae. aegypti. A população de Ae. albopictus utilizada como
grupo externo foi coletada no mesmo local e época que a população de Ae. aegypti

76
do BUT. Quando foi adicionada essa população como grupo externo, observou-se
que populações da mesma espécie se agruparam e a população de Ae. albopictus
ficou distanciada. Esses dados estão de acordo com o de Henry et al. (2010) em que
a forma da asa mostrou-se espécie-específica, independentemente da origem
geográfica da espécie.
Enquanto vários estudos mostram que a MG pode discriminar espécies
diferentes, estudos têm demonstrado que a morfometria geométrica alar tem
capacidade de discriminar populações de Culex quinquefasciatus em escalas
macrogeográficas (Peruzin, 2009; Morais et al., 2010). As populações de Ae. aegypti
de São Paulo utilizadas no estudo estão distanciadas no máximo de 50 km, por esse
motivo não era esperado uma diferenciação morfológica alar muito acentuada, além
disso são populações entre as quais o fluxo gênico ainda não foi totalmente
interrompido. Sendo assim, os resultados foram surpreendentes ao mostrar uma
diferenciação tênue. Isso indica que as asas são realmente marcadores sensíveis e
devem acusar evolução rápida como descrito anteriormente por Dujardin (2008).
5.2 Microssatélites
Vários tipos de marcadores moleculares têm sido utilizados para diversos
propósitos, tais como, estudos de estruturação populacional, identificação de
espécies, separação de espécies crípticas, reconstrução da história evolutiva de
organismos, etc. Os marcadores microssatélites utilizados no presente estudo estão
entre os marcadores mais polimórficos encontrado em genomas de animais.
As regiões SSRs selecionadas para serem utilizadas no estudo populacional
de Ae. aegypti na área metropolitana de São Paulo foram as mesmas utilizadas em
trabalhos anteriores com a mesma espécie (Da Costa-Ribeiro et al., 2006; Huber et
al., 2002). Os resultados mostraram que todos os loci amplificaram e foram
polimórficos, isso evidencia uma das vantagens de utilizar primers específicos. Bello
e Becerra, 2009 estudando a variabilidade genética de populações de Ochlerotatus
taeniorhynchus na Colômbia selecionaram oito primers que tinham sido
desenvolvidos para Ae. aegypti e Och. caspius, já que não tinha primers específicos
para a espécie que eles estavam estudando. Os resultados que eles obtiveram foi

77
que três primers que correspondiam ao Ae. aegypti não amplificaram, e os que
correspondiam ao mesmo gênero todos amplificaram.
O número de alelos por locus encontrados nas populações de Ae. aegypti na
área metropolitana de São Paulo foi menor (média de 5 alelos por locus), quando
comparados com populações de Ae. aegypti na Ásia (média de 10 alelos por locus)
(Huber et al., 2002; Paupy et al., 2004). Isso provavelmente acontece por essas
populações estarem mais próximas ao centro de origem do Ae. aegypti, a África.
Quando comparado com o trabalho realizado no Rio de Janeiro - Brasil, por Da
Costa-Ribeiro et al. (2006), utilizando os mesmos primers (média de 3 alelos por
locus), o número de alelos por locus foi maior. As populações de Ae. aegypti
estudadas em São Paulo foram mais polimórficas que populações de Ae. aegypti do
Rio de Janeiro.
Trabalho feito anteriormente com populações de Ae. aegypti de várias partes
do Mundo utilizando o gene mitocondrial NADH observaram que haplótipos
presentes na África central, Ásia, América do norte e central estão presentes em
cidades brasileiras com importantes portos comerciais como Santos, Belém, Manaus
e São Sebastião (Bracco et al., 2007). As análises feitas naquele estudo mostram a
existência de duas linhagens de DNA mitocondrial, nas populações de Ae. aegypti
do Brasil, uma relacionada com populações do Oeste Africano e a outra relacionada
com populações do leste Africano e Ásia. Segundo os autores a linhagem 1
apresentou menor polimorfismo quando comparado com a linhagem 2. No estado de
São Paulo foi encontrada as duas linhagens reconhecidas por Bracco et al. (2007),
enquanto que no estado do Rio de Janeiro foi encontrado somente a linhagem 1. A
presença das duas linhagens mitocondriais no estado de São Paulo condiz com o
maior polimorfismo de loci microssatélites nas populações de São Paulo quando
comparado com populações do Rio de Janeiro.
Os resultados obtidos entre pares de genótipos dos loci microssatélites
mostraram que só ocorreu desvio do equilíbrio de ligação em algumas comparações
dentro da população de Osasco, sugerindo a independência dos loci utilizados neste
estudo e portando a ausência de combinações alélicas entre os cromossomos do
inseto em subpopulações analisadas. É pouco provável que essa ligação foi
causada pelo enlace físico desses loci, já que só foi observado na população de
Osasco. Assim, as informações da variabilidade genética e heterozigosidade obtida

78
a partir de cada um dos marcadores polimórficos utilizados neste estudo são
relevantes.
As análises feitas para determinar se havia desvios em relação ao Equilíbrio
de Hardy-Weinberg nas populações analisadas mostraram que todas as populações
de Ae. aegypti não estavam em equilíbrio de HW, devido ao déficit ou excesso de
heterozigotos, embora a favor de genótipos homozigotos. Esta situação foi refletida
também pelo desvio do equilíbrio de HW em nível intrapopulacional que tinha um
valor positivo (Fis=0,0572). O mesmo foi observado ao nível da população total, um
valor positivo (Fit=0,1159) indicando desvio do equilíbrio de HW devido ao déficit de
heterozigotos. A situação geral indica que o desvio do equilíbrio de HW nas
populações de Ae. aegypti de São Paulo, não é igualmente afetado, por genótipos
heterozigoto ou homozigoto.
Desvio do equilíbrio de Hardy-Weinberg foi observado nas populações do
BUT, OSA e SUZ devido principalmente ao déficit de heterozigotos, mas não em
todos os loci, o que permite inferir a existência de eventos evolutivos naturais que
influenciam essa conclusão. Provavelmente, a subdivisão da população (efeito
Wahlund) seria uma das explicações mais coerente, desde outras causas como a
deriva genética, endogamia e a presença de alelos nulos. As hipóteses que
afetariam um marcador de forma igual foram descartadas, pois não foi observado
nessas populações, já que a presença de loci em desvio do equilíbrio de HW
também ocorreu devido ao excesso de heterozigotos. A existência simultânea de
déficit e excesso de heterozigotos na, com dominância de déficits nestas populações
são altamente compatíveis com o efeito de subdivisão genética presente em alguns
loci, mas não necessariamente em todos os loci, o mesmo foi observado em
trabalho anterior feito por Bello e Becerra (2009) utilizando marcadores
microssatélites em populações de Och. taeniorhynchus.
Os loci T3A7 e 38/38 apresentaram déficit de heterozigotos em três das
quatro populações analisadas. Isso pode estar relacionado devido a quatro fatores:
1- o locus está sob seleção; 2- presença de alelos nulos; 3- consangüinidade nas
populações; 4- a presença de populações subestruturadas que leva a um efeito
Wahlund. Da Costa Ribeiro et al. (2006) observaram em cinco populações de Ae.
aegypti do Rio de Janeiro déficit de heterozigotos no locus T3A7 e os dados do loci
38/38 não foram determinados. Esses loci provavelmente foram submetidos à

79
mutação nas regiões flanqueadoras resultando na inibição do anelamento do primer
com a seqüência complementar do DNA. As falhas na detecção dos produtos de
PCR de microssatélites geram alelos nulos e, portanto, déficit de heterozigotos
(Lanzaro et al., 1995). A endogamia não poderia ser a explicação do déficit de
heterozigotos já que não foi observado em todos os loci. O efeito Wahlund que
descreve os desvios devido à partilha de subpopulações em equilíbrio de HW
também poderia ser uma explicação.
A média do índice de Fst total correspondeu a um valor de 0,0622 indicando
uma diferenciação genética moderada entre as populações do BUT, GUA, OSA e
SUZ. Segundo Wright (1978), valores de Fst entre 0,05 e 0,15 indicam uma
moderada diferenciação genética entre as populações. Estudos populacionais
anteriores com Aedes aegypti utilizando marcadores microssatélites também
indicaram diferenciação genética populacional em escala microgeográfica. Da Costa
Ribeiro et al. (2006), analisando cinco populações no Rio de Janeiro com distâncias
variando de 8,2 a 32,4 km encontraram um Fst de 0,3304 significativo, mostrando
uma diferenciação genética alta. Ravel et al. (2002), estudando populações de Ae.
aegypti observaram a existência de populações geneticamente distintas ao longo de
um transecto norte-sul em Costa do Marfim (África) .
Fêmeas de Ae. aegypti tendem a colocar ovos em recipientes artificiais nas
proximidades de habitações humanas, assim a dispersão varia de acordo com a
distribuição dos locais de reprodução e disponibilidade de alimento (Tsuda et al.,
2001). O fluxo gênico contribui para homogeneizar as populações e, portanto reduz
a diferença genética entre as populações. O presente estudo indicou baixo fluxo
gênico (Nm = 0,47), ou seja, um número pequeno de migrantes por geração entre as
populações analisadas de Ae. aegypti na área metropolitana de São Paulo,
sugerindo uma diferenciação genética entre as populações desse mosquito,
apresentando alta estruturação de Ae. aegypti nas populações analisadas.
Os municípios onde foram realizadas as coletas (BUT, GUA, OSA e SUZ) são
intensamente urbanizados. Nesses centros urbanos existem vários habitats
importantes para o desenvolvimento do Ae. aegypti, como vaso de plantas, garrafas
plásticas, pneus, panelas e muitos outros. O número e a proximidade dos locais de
reprodução (para as fêmeas colocar os ovos) e densas populações humanas (para

80
alimentação sanguínea das fêmeas) tendem a limitar a dispersão dos mosquitos,
contribuindo assim para a diferenciação genética.
A distribuição da variabilidade genética de uma população subdividida
depende principalmente da distribuição espacial das populações, da sua dimensão,
do fluxo gênico e dos processos de extinção e recolonização (Slatkin, 1985; Wade e
Mccauley, 1988; Mccauley,1991).
Foi observada correlação entre distância genética e distância geográfica entre
as populações analisadas, indicando que os níveis de diferenciação genética são
dependentes da distância geográfica. Em trabalhos anteriores com populações de
Ae. aegypti utilizando microssatélites não foi observado correlação entre distância
genética e distância geográfica (Da Costa Ribeiro et al., 2006; Huber et al., 2002;
Ravel et al., 2002). A distância geográfica não é sempre um bom indicador de
distância genética entre as populações, embora o reduzido fluxo de alelos gênicos
contribua para reduzir as diferenças entre populações separadas por distâncias
compatíveis com a distância de migração da espécie.
A diferenciação de populações através do formato alar tem mostrado ser
possível até o momento para escalas macrogeográficas ou populações com
estruturação mais acentuada, já os microssatélites são capazes de diferenciar
populações em escalas micro e macrogeográficas. Os marcadores moleculares
utilizados no presente trabalho são de grande interesse para a caracterização
genética de várias populações de Ae. aegypti. A capacidade de discriminar
populações em escala microgeográficas é de grande importância em estudos que
envolvem taxonomia molecular e investigação epidemiológica de Ae. aegypti. Esse
estudo provavelmente será estendido a outras regiões do estado de São Paulo, e
possivelmente abrirá ainda mais os horizontes da aplicabilidade desses métodos.
Futuras investigações científicas ou trabalhos de vigilância epidemiológica poderão
fazer uso do método de MG em âmbito macrogeográfico e usar os microssatélites no
âmbito microgeográfico para entender a dispersão e a prevalência do vírus dengue.

81
6 CONCLUSÕES - Há acentuado dimorfismo sexual alar de forma e tamanho em Ae. aegypti;
- Fêmeas de Ae. aegypti apresentam tamanhos alares maiores que os machos;
- Assimetria bilateral alar foi tênue para ambos os sexos;
- Existe variabilidade morfológica alar que indica ligeira diferenciação mas não acusa
evidente estrutuação populacional;
- Todos os loci microssatélites foram polimórficos para as populações de Ae. aegypti
do BUT, GUA, OSA e SUZ;
- Há uma diferenciação genética moderada entre os grupos geográficos
considerados (Fst = 0,0622), indicando que essas populações estão estruturadas;
- Baixo fluxo gênico foi encontrado entre as populações analisadas de Ae. aegypti na
área metropolitana de São Paulo (Nm= 0, 47);
- Distância geográfica é um bom indicador de distância genética para as populações
de Ae. aegypti da área metropolitana de São Paulo, porém não se correlacionou
com distância fenética.
- Possivelmente as velocidades de evolução dos caracteres alares e das regiões
microssatélites são diferentes.
- A MG provavelmente poderá ser útil para estudos populacionais em níveis
macrogeográficos;
- Os resultados gerados no presente trabalho demonstraram que os marcadores
microssatélites são mais adequados para avaliar estrutura genética de amostras
coletadas em um pequeno espaço geográfico.

82
REFERÊNCIAS1 Agnew P, Haussy C, Michalakis Y. Effects of density and larval competition on selected life history traits of Culex pipiens quinquefasciatus (Diptera: Culicidae). J Med Entomol. 2000;37(5):732-5 Ahumada JA, Lapointe D, Samuel MD. Modeling the population dynamics of Culex quinquefasciatus (Diptera: Culicidae), along an elevational gradient in Hawaii. J Med Entomol. 2004;41(6):1157-70. Ayres CFJ, Melo-Santos MAV, Prota JRM, Sole-Cava AM, Regis L, Furtado AF. Genetic structure of natural populations of Aedes aegypti at the micro- and macrogeographic levels in Brazil. J Am Mosq Control Assoc. 2004;20(4):350-6. Ayres CFJ, Melo-Santos MAV, Sole-Cava AM, Furtado AF. Genetic differentiation of Aedes aegypti (Diptera : Culicidae), the major dengue vector in Brazil. J Med Entomol. 2003;40(4):430-5. Belkin JN. The mosquitoes of the South Pacific (Diptera: Culicidae). Berkeley: University of California Press; 1962. 576 p. Bello F, Becerra V. Genetic variability and heterogeneity of Venezuelan equine encephalitis virus vector Ochlerotatus taeniorhynchus (Diptera: Culicidae) populations of the Colombian Atlantic coast, based on microsatellite loci. Genet Mol Res. 2009;8(3):1179-90. Bisset JA. Uso correcto de insecticidas: control de la resistencia. Rev Cub Med Trop. 2002;54(3):202-19. Bond JE, Beamer DA, Hedin MC, Sierwald P. Gradual evolution of male genitalia in a sibling species complex of millipedes (Diplopoda: Spirobolida: Rhinocricidae: Anadenobolus). Invertebr Syst. 2003;17:711-17. Bookstein FL. Morphometric tools for landmark data: geometry and biology. Cambridge: Cambridge University press; 1997. 455 p.
1 De acordo com: International Committee of Medical Journal Editors. Uniform requirements for manuscripts submitted to Biomedical Journal: sample references. Available from: http://www.icmje.org [2010 May 22].

83
Bouyer J, Ravel S, Dujardin JP, Meeus T, Vial L, Thévenon S, Guerrini L, Sidibé I, Solano P. Population Structuring of Glossina palpalis gambiensis (Diptera: Glossinidae) According to Landscape Fragmentation in the Mouhoun River, Burkina Faso. J Med Entomol. 2007;44(5):788-95. Bracco JE, Capurro ML, Lourenço-de-Oliveira R, Sallum MAM. Genetic variability of Aedes aegypti in the Americas using a mitochondrial gene: evidence of multiple introductions. Mem Inst Oswaldo Cruz. 2007;102:573–80. Braga IA, Valle D. Aedes aegypti: histórico do controle no Brasil. Epidemiol Serv Saúde. 2007;16(2):113-18. Braga IA, Lima JBP, Soares SS, Valle D. Aedes aegypt iresistance to temephos during 2001 in several municipalitiesin the states of Rio de Janeiro, Sergipe and Alagoas, Brazil. Mem Inst Oswaldo Cruz. 2004;99(2):199-203. CDC - Centers for Disease Control. Control of Dengue. Vector Topics. 1979; 2:1-39. CVE - Centro de Vigilância Epidemiológica. Disponível em: http://www.cve.saude.sp.gov.br/htm/zoo/den10_import_autoc.htm. [2010 jul 27]. Christophers SR. Aedes aegypti (L.), the Yellow Fever Mosquito. Its life history, bionomics and structure. London: Cambridge University Press; 1960. 739 p. Da Costa-Ribeiro MCV, Lourenco-de-Oliveira R, Failloux AB. Low gene flow of Aedes aegypti between dengue-endemic and dengue-free areas in southeastern and southern Brazil. Am J Trop Med Hyg. 2007;77(2):303-9. Da Costa-Ribeiro MCV, Lourenço-de-Oliveira R, Failloux AB. Higher genetic variation estimated by microsatellites compared to isoenzyme markers in Aedes aegypti from Rio de Janeiro. Mem Inst Oswaldo Cruz. 2006;101(8):917-21. Da-Cunha MP, Lima JBP, Brogdon WG, Moya GE, Valle D. Monitoring of resistance to the pyrethroid cypermethrin in Brazilian Aedes aegypti (Diptera: Culicidae) populations collected between 2001 and 2003. Mem Inst Oswaldo Cruz. 2005;100(4):441-4. Dujardin JP. Morphometrics applied to medical entomology. Infection Genetics and Evolution. 2008;8(6):875-90.

84
Dujardin JP, Slice D. Geometric morphometrics. Contribuitions to medical entomology. In: Tibayrenc M. Encyclopedia of Infectious Diseases. Modern Methodologies. United States: Wiley & Sons; 2007. p. 435-47. Falconer DS. Introduction to quantitative genentics. Longman: London and New-York; 1981. 300 p. Feliciangeli M, Sanches-Martin M, Marrero R, Davies C, Dujardin J. Morphometric evidence for a possible role of Rhodnius prolixus from palm trees in house re-infestation in the State of Barinas (Venezuela). Acta Trop. 2007;101(2):169-77. Fitzpatrick S, Feliciangeli MD, Sanchez-Martin MJ, Monteiro FA, Miles MA Molecular genetics reveal that silvatic Rhodnius prolixus do colonise rural houses. PLoS Negl Trop Dis. 2008;2(4):e210 Forattini OP. Culicidologia Médica. São Paulo: EDUSP; 2002. 864 p. Franco O. Reinfestação do Pará por Aedes aegypti. Rev Bras Malariol Doencas Trop. 1969;21(4):729-31. Futuyma DJ. Evolutionary Biology. Sunderland, Mass.: Sinauer Associates; 1997. 763 p. Gubler DJ Dengue and dengue hemorrhagic fever. Clin Microbiol Rev. 1998;11(3):480-96. Gutiérrez LA, Gómez GF, González JJ, Castro MI, Luckhart S, Conn JE, CORREA MM. Microgeographic Genetic Variation of the Malaria Vector Anopheles darlingi Root (Diptera: Culicidae) from Córdoba and Antioquia, Colombia. Am J Trop Med Hyg. 2010;83(1):38-47. Hayes J. Seasonal changes in population structure of Culex pipiens quinquefasciatus Say (Diptera: Culicidae): study of an isolated population. J Med Entomol. 1975;12(2):167-78. Henry A, Thongsripong P, Fonseca-Gonzalez I, Jaramillo-Ocampo N, Dujardin JP. Wing shape of dengue vectors from around the world. Infect Genet Evol. 2010;10(2):207–14.

85
Herrera F, Urdaneta L, Rivero J, Zoghbi N, Ruiz J, Carrasquel G, Martinez JA, Pernalete M, Villegas P, Montoya A, Rubio-Palis Y, Rojas E. Population genetic structure of the dengue mosquito Aedes aegypti in Venezuela. Mem Inst Oswaldo Cruz. 2006;101(6):625-33. Herrera F, Urdaneta L, Rivero J, Zoghbi N, Ruiz J, Carrasquel G, Martinez JA, Pernalete M, Villegas P, Montoya A, Rubio-Huber K, Le Loan L, Hoang TH, Ravel S, Rodhain F, Failloux AB. Genetic differentiation of the dengue vector, Aedes aegypti (Ho Chi Minh City, Vietnam) using microsatellite markers. Mol Ecol. 2002;1(9):1629-35. Hlaing T, Tun-Lin W, Somboon P, Socheat D, Setha T, Min S, Thaung S, Anyaele O, Silva B, Chang MS, Prakash A, Linton Y, WALTON C. Spatial genetic structure of Aedes aegypti mosquitoes in mainland Southeast Ásia. Evolutionary Applications. 2010;3:319-39. Huang S, Hamer GL, Molai G, Walker ED, Goldberg TL, Kitron UD, Andreadis TG. Genetic variation associated with mammalian feeding in Culex pipiens from a West Nile virus epidemic region in Chicago, Illinois. Vector Borne Zoonotic Dis. 2009;9(6):637-42. Huber K, Luu Le L, Chanta N, Failloux AB. Human transportation influences Aedes aegypti gene flow in Southeast Asia. Acta Trop. 2004;90(1):23-29. Huber K, Le Loan L, Hoang TH, Ravel S, Rodhain F, Failloux AB. Genetic differentiation of the dengue vector, Aedes aegypti (Ho Chi Minh City, Vietnam) using microsatellite markers. Mol Ecol. 2002;11(9):1629-35.
Instituto Oswaldo Cruz. O mundo macro e micro do mosquito Ae. aegypti. Direção de Genilton José Vieira. Rio de Janeiro: Instituto Oswaldo Cruz; 2010. 1 DVD. Jirakanjanakit N, Leemingsawat S, Dujardin JP. The geometry of the wing of Aedes (Stegomyia) aegypti in isofemale lines through successive generations. Inf Gen Evol. 2008;8(4):414-21. Jirakanjanakit NS, Leemingsawat S, Thongrungkiat C, Apiwathnasorn S, Singhaniyom C, Bellec, Dujardin JP. Influence of larval density or food variation on the geometry of the wing of Aedes (Stegomyia) aegypti. Trop Med Int Health. 2007;12(11):1354–60. Jowett T. Preparation of Nucleic Acids. In: Roberts DB, editor. Drosophila, a practical approach. Oxford: Roberts DB; 1986. 389 p.

86
Kovats RS. El Nino and human health. Bull World Health Organ. 2000;78(9):1127-35. Lanzaro GC, Zheng L, Toure YT, Traore SF, Kafatos FC, Vernick KD. Microsatellite DNA and isozyme variability in a West African population of Anopheles gambiae. Insect Mol Biol. 1995;4(2):105-12. Lines J, Harpham T, Leake C, Schofield C. Trends, priorities and policy directions in the control of vector-borne diseases in urban environments. Health Policy Plann.1994;9(2):113-29. Lien JC. New Species of Mosquitoes from Taiwan (Diptera : Culicidae) Part V. Three New Subspecies of Aedes and Seven New Species of Culex. Trop Med. 1968;10(4):217-62. Lourenço-de-Oliveira R, Vazeille M, Filippis AMB, Failloux AB. Aedes aegypti in Brazil: genetically differentiated populations with high susceptibility to dengue and yellow fever viruses. Trans R Soc Trop Med Hyg. 2004;98(1):43-54. Luz PM, Codeco CT, Massad E, Struchiner CJ. Uncertainties regarding dengue modeling in Rio de Janeiro, Brazil. Mems Inst Oswaldo Cruz. 2003;98(7):871-8. Mackenzie JS, Gubler DJ, Petersen LR. Emerging flaviviruses: the spread and resurgence of Japanese encephalitis, West Nile and Dengue viruses. Nat Med. 2004;10(12):98-109. Macoris MLG, Andrighetti MTM, Takaku L, Glasser CM, Garbeloto VC, Bracco JE. Resistance of Aedes aegypti from the state of São Paulo, Brazil, to organophosphates insecticides. Mems Inst Oswaldo Cruz. 2003;98(5):703-08. Maciel-de-Freitas R, Lourenco-de-Oliveira R. Presumed unconstrained dispersal of Aedes aegypti in the city of Rio de Janeiro, Brazil. Rev Saude Publica. 2009;43(1):8-12. Marrelli MT, Rasgon J, Jacobs-Lorena M. Transgenic malaria-resistant mosquitoes have a fitness advantage when feeding on Plasmodium-infected blood. Proc Natl Acad Sci USA. 2007;104(13):5580-3. Monath TP. Dengue: the risk to developed and developing countries. Proc Natl Acad Sci USA. 1994;91(7):2395-400.

87
Monteiro LR, Reis SF. Princípios de morfometria geométrica. Ribeirão Preto: Holos; 1999. 189 p. Morais SA, Moratore C, Suesdek L, Marrelli MT. Genetic-morphometric variation in Culex quinquefasciatus from Brazil and La Plata, Argentina. Mems Inst Oswaldo Cruz. 2010;105(5):672-6. Mouchet J, Carnevale P. Impact des transformations de l'environnement sur les maladies à transmission vectorielle. Sante. 1997;7(4):263-9. Mpho M, Callaghan A, Holloway GJ. Temperature and genotypic effects on life history and fluctuating asymmetry in a field strain of Culex pipiens. Heredity. 2002;88:307–12 Mpho M, Holloway GJ, Callaghan A. A comparison of the effects of organophosphate insecticide exposure and temperature stress on fluctuating asymmetry and life history traits in Culex quinquefasciatus. Chemosphere. 2001;45(6-7):713-20. Muturi EJ, Kim CH, Baliraine FN, Musani S, Jacob B, Githure J, Novak RJ. Population Genetic Structure of Anopheles Arabiensis (Diptera: Culicidae) in a Rice Growing Area of Central Kenya. J Med Entomol. 2010;47(2):144-51. Nei M. Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci USA. 1973;70(12):3321-33. Nelson MJ. Aedes aegypti: biologia y ecologia. Washington, DC: Organización Panamericana de la Salud; 1986. Paupy C, Brengues C, Ndiath O, Toty C, Hervé JP, Simard F. Morphological and genetic variability within Aedes aegypti in Niakhar, Senegal. Infect Genet Evol. 2010;10(4):473-80. Paupy C, Brengues C, Kamgang B, Herve JP, Fontenille D, Simard F. Gene flow between domestic and sylvan populations of Aedes aegypti (Diptera : Culicidae) in North Cameroon. J Med Entomol. 2008;45(3)391-400. Paupy C, Chanta N, Huber K, Lecoz N, Reynes JM, Rodhain F, Failloux AB. Influence of breeding sites features on genetic differentiation of Aedes aegypti populations analyzed on a local scale in Phnom Penh municipality of Cambodia. Am J Trop Med Hyg. 2004;71(1):73-81.

88
Peruzin MCJ. Análises comparativas populacionais de Culex quinquefasciatus em dois locais do Estado de São Paulo. [dissertação (Mestrado em Biologia da Relação Patógeno-Hospedeiro)]. São Paulo: Universidade de São Paulo; 2009. 96 f. Powell JR, Tabachnick WJ, Arnold J. Genetics and the origin of a vector population: Aedes aegypti, a case study. Science. 1980;208(4450):1385-7. Ravel S, Monteny N, Velasco Olmos D, Escalante Verdugo J, Cuny G. A preliminary study of the population genetics of Aedes aegypti (Diptera: Culicidae) from Mexico using microsatellite and AFLP markers. Acta Trop. 2001;78(3):241-50. Raymond M, Rousset F. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J Heredity. 1995;86:248-9. Reinert Jf, Kaiser PE, Seawright JA. Analysis of the Anopheles (Anopheles) quadrimaculatus complex of sibling species (Diptera: Culicidae) using morphological, cytological, molecular, genetic, biochemical, and ecological techniques in an integrated approach. J Am Mosq Control Assoc. 1997;13:1-102. Ridley M. Evolution. Boston: Blackwell Scientific; 2003. 768 p. Rocha LS. Caracterização de populações de Anastrepha sororcula Zucchi 1979 (Diptera: Tephritidae) por análises cromossômicas, morfométricas e moleculares [tese (Doutorado em Genética e Biologia Evolutiva)]. São Paulo: Instituto de Biociências. Universidade de São Paulo; 2005. 127 f. Rohlf FJ. Relative warp analysis and example of its application to mosquito wings. In: Marcus, LF, Bello, E, Garcia-Valdcasas, editors. Contributions to morphometrics. Madrid: Museo Nacional de Ciencias Naturales; 1993. p. 135-59. Savard J, Tautz D, Lercher MJ. Genome-wide acceleration of protein evolution in flies (Diptera). BMC Evol Biol. 2006;6(7):1-6. Scarpassa VM, Conn JE. Population genetic structure of the major malaria vector Anopheles darlingi (Diptera: Culicidae) from the Brazilian Amazon, using microsatellite markers. Mem Inst Oswaldo Cruz. 2007;102(3):319-27. Schlötterer C. Evolutionary dynamics of microsatellite DNA. Chromosoma. 2000;109(6):365–71.

89
Sene FM, Pereira MAQR, Vilela CR. Contrasting Patterns of Differentiation Inferred from Traditional Genetic Markers in the Process of Speciation. Pacific Science. 1988;42:81-8. Slotman MA, Kelly NB, Harrington LC, Kitthawee S, Jones JW, Scott TW, Caccone A, Powell JR. (2007) Polymorphic microsatellite markers for studies of Aedes aegypti (Diptera: Culicidae), the vector of dengue and yellow fever. Mol Ecol Notes. 2007;7:168-71. Solano P, De La Rocque S, De Meeus T, Cuny G, Duvallet G, Cuisance D. Microsatellite DNA markers reveal genetic differentiation among populations of Glossina palpalis gambiensis collected in the agropastoral zone of Sideradougou, Burkina Faso. Insect Mol. Biol. 2000;9(4):433–39. Souza JMG, Gouveia M, Perondini ALP, Selivon D. Asymmetry of frontal bristles and postocular setae in species and hybrids of the Anastrepha fraterculus complex (Diptera: Tephritidae). Gen Mol Biol. 2007;30(1):145-51. Sousa-Polezzi RC, Bicudo HEMC. Genetic variation along time in a Brazilian population of Aedes aegypti (Diptera: Culicidae), detected by changes in the esterase patterns. Genetica. 2005;125(1):43–53. Speranca MA, Capurro ML. Perspectives in the control of infectious diseases by transgenic mosquitoes in the post-genomic era - a review. Mem Inst Oswaldo Cruz. 2007;102(4):425-33. StatSoft, Inc. Statistica for Windows [Computer program manual]. Tulsa, OK: StatSoft, Inc;. Available from: http://www.statsoft.com. 2004. [2010 ago 13]. SUCEN- Superintendência do Controle de Endemias. Dengue e Febre amarela. Disponível em: http://www.sucen.sp.gov.br/doencas/dengue_f_amarela/texto_dengue_pro3.htm. [25 mar 2009]. SVS- Secretaria de Vigilância em Saúde. Disponível em: http://portal.saude.gov.br/portal/arquivos/pdf/ano02_n03_dengue_brasil_si.pdf. [24 ago 2010]. Temu EA, Yan G. Microsatellite and mitochondrial genetic differentiation of Anopheles arabiensis (Diptera: Culicidae) from Western Kenya, the great rift Valley, and Coastal Kenya. Am J Trop Med Hyg. 2005;73(4):726-33.

90
Tsuda Y, Takagi M, Wang S, Wang Z, Tang L. Movement of Aedes aegypti (Diptera:Culicidae) released in a small isolated village on Hainan island, China. J Med Entomol. 2001;38:93-8. Venkatesan M, Rasgon JL. Population genetic data suggest a role for mosquito-mediated dispersal of West Nile virus across the western United States. Mol Ecol. 2010;19(8):1573–84. Weber JL, May PE. Abundant class of human DNA polymorphisms which can be typed using the polymerase chain reaction. Am J Hum Genet .1989;44(3):388–96. Weir BS. Genetic Data Analysis. Sunderland, Massachusetts: Sinauer Associates, Inc; 1990. 377 p. Weir BS, Cockerham CC. Estimating F-statistics for the analysis of population structure. Evolution. 1984;38(6):1358–70. WHO- World Health Organization. Dengue. Disponível em: http://www.who.int/csr/disease/dengue/en/ [2010 jul 27]. Wilke ABB, Nimmo DD, John OS, Kojin BB, Capurro ML, Marrelli MT. Genetic enhancements to the sterile insect technique to control mosquito populations. AsPac J Mol Biol Biotechn. 2009;17(3):65-74. Wright S. Evolution and genetics of populations. Chicago: University of Chicago Press; 1978. 511 p. Yakob L, Alphey L, Bonsall MB. Aedes aegypti control: the concomitant role of competition, space and transgenic technologies. Journal of Applied Ecology. 2008;45(4):1258-65. Yebakima A, Marquine M, Rosine J, Yp-Tcha MM, Pasteur N. Evolution of resistance under insecticide selection pressure in Culex pipiens quinquefasciatus (Diptera, Culicidae) from Martinique. J Med Entomol. 2004;41 (4):718-25.
Top Related