






Línguas
Páginas
Legal


i
NAICE ELEIDIANE SANTANA MONTEIRO
Efeito das proteínas do soro do leite bovino sobre alterações
metabólicas causadas por uma dieta hiperlipídica no
camundongo C57BL/6: disbiose intestinal, resposta inflamatória e
parâmetros associados
CAMPINAS
2015

ii

iii
UNIVERSIDADE ESTADUAL DE CAMPINAS
Faculdade de Engenharia de Alimentos
NAICE ELEIDIANE SANTANA MONTEIRO
Efeito das proteínas do soro do leite bovino sobre alterações
metabólicas causadas por uma dieta hiperlipídica no
camundongo C57BL/6: disbiose intestinal, resposta inflamatória e
parâmetros associados
Orientador: Profº. Dr. Jaime Amaya Farfan
Co-orientador: Profª. Dra. Fernanda de Pace
CAMPINAS
2015
Dissertação apresentada à Faculdade de
Engenharia de Alimentos da Universidade
Estadual de Campinas como parte dos
requisitos exigidos para obtenção do título
de Mestra em Alimentos e Nutrição na
Área de Nutrição Experimental e aplicada à
Tecnologia de Alimentos.
ESTE EXEMPLAR CORRESPONDE À
VERSÃO FINAL DA DISSERTAÇÃO
DEFENDIDA PELA ALUNA NAICE
ELEIDIANE SANTANA MONTEIRO, E
ORIENTADA PELO PROF. DR. JAIME
AMAYA FARFAN
ASSINATURA DO ORIENTADOR

iv

v
BANCA EXAMINADORA
__________________________________
Profº. Dr. Jaime Amaya Farfan
Orientador
__________________________________
Profº. Dr. Mário Roberto Maróstica Júnior
Titular
Universidade Estadual de Campinas (UNICAMP)
______________________________
Profª Dra. Julia Laura Delbue Bernardi
Titular
Pontifícia Universidade Católica de Campinas (PUC CAMPINAS)
_______________________________
Profª Dra. Elisa de Almeida Jackix
Suplente
Universidade Metodista de Piracicaba (UNIMEP)
______________________________
Profº Dr. Wilton Marlindo Santana Nunes
Suplente
Pontifícia Universidade Católica de Campinas (PUC CAMPINAS)

vi

vii
“Se procurar bem você acaba encontrando.
Não a explicação (duvidosa) da vida,
Mas a poesia (inexplicável) da vida”.
Carlos Drummond de Andrade

viii

ix
DEDICATÓRIA
A minha família, por ter permanecido ao
meu lado, me incentivando a percorrer
este caminho, por compartilhar todas as
angústias e dúvidas, me apoiando
incondicionalmente nos momentos mais
difíceis.

x

xi
AGRADECIMENTOS
A Deus, por todos os dias da minha vida e por me dirigir a cada passo, iluminando
constantemente meu caminho e me dando forças para seguir sempre em frente.
À minha mãe Maria Aparecida e a meus avós Leontina e Américo, que tanto se
esforçaram pela minha formação pessoal e profissional. Ao Nino, razão de muitos
momentos de alegria e descontração.
Ao meu esposo, Raildo Monteiro, por tanto amor e dedicação e por ser um ouvinte
atento de tantas dúvidas e inquietações, sempre me incentivando a prosseguir.
Ao Profº Dr. Jaime Amaya Farfan pela orientação e oportunidade de realização
deste trabalho.
À Profª. Dra. Fernanda de Pace pela co-orientação, sempre tão gentil e paciente.
À Aline Rissetti Roquetto, companheira de trabalho nestes últimos dois anos.
À Kelly Ferreira dos Santos pela amizade e acolhimento assim que cheguei ao
Laboratório de Fontes Proteicas/DEPAN.
Agradecimento especial ao Profº Dr. Mário Saad e sua equipe do Laboratório de
Investigação Clínica em Resistência à Insulina (Licri) da Faculdade de Ciências
Médicas (FCM–UNICAMP), pela fundamental participação no desenvolvimento
desta pesquisa.
Ao Profº Áureo Yamada e à Técnica Stephanie Federighi do Departamento de
Histologia e Embriologia/ IB – UNICAMP, por toda ajuda prestada na parte
histológica.
A Susana Correa, pelo auxílio no Laboratório de Ensaios Biológicos e a Carla
Greghi, Carolina Moura e Eder Muller pela colaboração nas análises
desenvolvidas no Laboratório de Fontes Proteicas DEPAN – FEA/UNICAMP.

xii
Ao CNPq pela concessão da bolsa de estudos, ao Departamento de Alimentos e
Nutrição e à Faculdade de Engenharia de Alimentos por viabilizarem a execução
do projeto.
À Hilmar Ingredients (EUA) pela doação dos produtos do soro de leite, através da
Doremus do Brasil, SP.
Agradeço a todos os membros da banca examinadora por aceitarem o convite e
pelas valiosas contribuições
Meu reconhecimento sincero a tantos outros que despretensiosamente
colaboraram na execução deste estudo.

i
SUMÁRIO
Resumo.....................................................................................................................v
Abstract...................................................................................................................vii
1. Introdução Geral.............................................................................................1
Referências...............................................................................................................4
CAPÍTULO 1: Revisão de Literatura.........................................................................6
1. Obesidade......................................................................................................7
1.1 Aspectos epidemiológicos da obesidade......................................................8
1.2 Inflamação crônica induzida pela obesidade..............................................11
1.3 A microbiota intestinal em indivíduos saudáveis.........................................17
1.4 A microbiota intestinal em indivíduos doentes............................................19
1.5 Tecido hepático, microbiota intestinal e NAFLD.........................................21
1.6 Relação entre obesidade, microbiota e inflamação....................................25
1.7 Tipificação da microbiota.............................................................................28
1.8 Proteínas do soro do leite...........................................................................29
Referências.............................................................................................................35
CAPÍTULO 2: Proteínas do soro do leite mitigam a deposição excessiva de
gordura epididimal, sem alterar a composição corporal de camundongos C57BL/6
alimentados com dieta hiperlipídica........................................................................47
Resumo...................................................................................................................48
Abstract...................................................................................................................49
1. Introdução....................................................................................................50
2. Objetivo........................................................................................................51
3. Material e métodos.......................................................................................51
3.1 Ensaio biológico.....................................................................................51
3.2 Composição das dietas experimentais...................................................52

ii
3.3 Ingestão dietética e ganho ponderal dos animais..................................53
3.4 Preparo das carcaças dos animais........................................................54
3.4.1 Determinação da composição centesimal das carcaças...............54
3.5 Análise estatística..................................................................................54
4. Resultados...................................................................................................55
4.1 Parâmetros antropométricos..................................................................55
4.2 Composição centesimal das carcaças...................................................57
5. Discussão.....................................................................................................57
6. Conclusão....................................................................................................60
Agradecimentos......................................................................................................60
Referências.............................................................................................................61
CHAPTER 3: Whey protein, but not casein, avert cecal dysbiosis,
lipopolysaccharide formation and inflammation marker expression in mice fat-rich
diet..........................................................................................................................63
Abstract...................................................................................................................64
1. Introduction...................................................................................................66
2. Materials and methods ................................................................................67
2.1 Animals and experimental protocol........................................................67
2.2 Experimental diets..................................................................................67
2.3 Blood for biochemical analysis and LPS determination.........................69
2.4 Biomarkers of the inflammatory process................................................69
2.4.1 Tissue extraction and protein quantification.................................69
2.4.2 Western Blotting...........................................................................70
2.5 Determination of serum concentrations of LPS......................................71
2.6 Sequencing the intestinal microbiota.....................................................72
2.7 Histological analysis of liver tissue.........................................................73

iii
2.8 Statistical analysis..................................................................................73
3. Results.........................................................................................................73
3.1 Biochemical parameters.........................................................................73
3.2 Inflammatory profile................................................................................74
3.3 Endotoxemia..........................................................................................77
3.4 Intestinal Microbiota...............................................................................77
3.5 Histopathologic profile............................................................................83
4. Discussion....................................................................................................84
5. Conclusion....................................................................................................92
Abbreviations…………………………………………………………………………..…92
Competing interests…………………………..……………………………………...…93
Author’s Contributions………………………..………………………………………….93
Acknowledgements.................................................................................................93
References..............................................................................................................94
Conclusão geral......................................................................................................97
ANEXO...................................................................................................................98

iv

v
RESUMO GERAL
Caracterizada como um dos mais importantes problemas que a saúde pública
enfrenta atualmente no Brasil e no mundo, a obesidade está associada a um
quadro de inflamação subclínica, que predispõe à resistência à insulina e ao
desenvolvimento de diabetes mellitus, além de representar fator etiológico para
diversas outras doenças crônicas não transmissíveis. Sabendo da importância que
os alimentos com propriedades bioativas podem ter no tratamento da obesidade é
que a utilização das proteínas do soro do leite na dieta vem sendo alvo de
diversos estudos, devido aos benefícios que podem trazer à saúde humana.
Dentre as propriedades da whey protein, citam-se a capacidade de regular a
função imune, atuar como agente antimicrobiano, estimular a síntese de proteína
muscular, suprimir o apetite e atuar na redução da gordura corporal, além das
propriedades de aumentar a expressão gênica de proteínas anti-estresse (HSPs)
e de ativar o transportador de glicose GLUT4, ambas descobertas pelo nosso
grupo da FEA. O presente estudo teve como objetivo investigar o impacto que as
proteínas do soro do leite, seja na sua forma íntegra ou na forma hidrolisada,
podem exercer sobre a homeostase e a proteção do organismo de camundongos,
contra os danos causados por uma dieta hiperlipídica. Para alcançar este objetivo,
34 camundongos C57BL/6 recém-desmamados foram divididos em quatro grupos
de forma aleatória e alimentados com as seguintes dietas: grupo controle
normolipídico (AIN93-G), grupo controle hiperlipídico (HFCAS), grupo hiperlipídico
com substituição da caseína pela proteína do soro de leite concentrada (HFWPC),
e grupo hiperlipídico, com adição da proteína do soro do leite hidrolisada
(HFWPH), por 9 semanas. Os camundongos foram acompanhados quanto ao
consumo alimentar e ganho de peso por meio de avaliação poderal em dias
alternados, parâmetros bioquímicos e de endotoxemia avaliados por técnicas
convencionais, biomarcadores inflamatórios analisados por western blot, além da
avaliação do perfil histopatológico do fígado e do estudo metagenômico da
microbiota intestinal. Os resultados demonstraram que alimentação com a
formulação contendo whey protein, em ambas as formas, por 9 semanas, não
diminuiu ganho de peso em comparação aos demais tratamentos, mas foi efetiva

vi
em reverter a disbiose causada pela dieta gordurosa, reduzir o processo
inflamatório para níveis indistinguíveis do controle, em atenuar a infiltração
gordurosa no tecido hepático, além de modular a microbiota intestinal. A caseína
não mostrou tais propriedades.
Palavras chave: obesidade, proteínas do soro do leite, inflamação, microbiota
intestinal, dieta hiperlipídica, resistência à insulina.

vii
ABSTRACT
Recognized as one of the most important issues currently facing public health in
Brazil and in the world, obesity is associated to a subclinical inflammation
framework, which predisposes to insulin resistance and the development of
diabetes mellitus, in addition to representing an etiological factor for several other
chronic diseases. Owing to the bioactive properties that the milk whey proteins
may have on human health, such as the ability to regulate immune function, act as
an antimicrobial agent, stimulate protein synthesis, suppress appetite thus helping
to reduce body fat, besides the two newly discovered functions from our laboratory
regarding the up-regulation of protective HSPs and activation of the glucose
transporter 4 (GLUT4), it is thought that these proteins may also have anti-
inflammatory action and, therefore, this study was designed to investigate the
impact that whey protein, either in its normal form or in the hydrolyzed form, may
exert on homeostasis protecting the mouse from the adverse effects of a high-fat
diet. To accomplish this, 34 C57BL/6 male mice were randomly divided into four
groups that received the following diets for 9 weeks: Normolipidic control (AIN93-
G), Hyperlipidic control (HFCAS), Hyperlipidic with whey protein concentrate
instead of casein (HFWPC) and a Hyperlipidic group with hydrolyzed whey protein
(HFWPH) as the only source of protein. Diet intake and weight gain were
monitored and recorded every other day. Biochemical parameters and
endotoxemia were evaluated using commercial kits, and inflammatory biomarkers
were analyzed by western blotting, besides the evaluation of histopathological liver
profile and metagenomic study of the intestinal microbiota. The results showed that
feeding the formulation contendowheyprotein, in both its forms, for 9 weeks, not
decreased weight gain compared to the other treatments, but was effective in
reversing dysbiosis caused by high-fat diet, reduce inflammation to levels
indistinguishable control and mitigate the fatty infiltration of the casein , in addition
to modulate the gut microbiota. Casein did not show such properties.
Keywords: obesity, whey protein, inflammation, gut microbiota, high fat diet,
diabetes, insulin resistance

viii

1
1. Introdução Geral
A prevalência da obesidade tem tomado proporções epidêmicas no
Brasil e no mundo. Um levantamento realizado pela Organização Mundial de
Saúde (OMS) estima que cerca de 500 milhões de adultos e 43 milhões de
crianças com até 5 anos de idade já possuam sobrepeso ou obesidade (WHO,
2012).
Atualmente a obesidade é entendida como um processo
inflamatório, onde há aumentada produção de citocinas pró-inflamatórias. Tais
substâncias são deletérias à ação da insulina no músculo, em demais tecidos
periféricos como fígado e tecido adiposo, além de ter importante atuação nas
funções centrais do hipotálamo, diminuindo a capacidade desta glândula de
controlar a ingestão alimentar e regular o gasto energético, tendendo sempre
ao balanço positivo (CINTRA, ROPELLE, MORAES, PAULI, 2011).
Em animais alimentados com dieta hiperlipídica foi demonstrado
que, nas primeiras 24 horas de dieta, ocorre produção exacerbada de
marcadores inflamatórios, a qual se mantém elevada durante os primeiros 3
dias, mas é seguida de uma normalização transitória, que pode durar de 2 a 3
semanas, para depois subir novamente aos níveis iniciais. Esse padrão
temporal equipara-se ao que é visto no consumo alimentar quando há
mudança de uma dieta normolipídica para uma hiperlipídica (THALER et al.,
2013). Estudos atuais também demonstram importante correlação entre
microbiota e o estado inflamatório na obesidade, devido à endotoxemia
metabólica que lipopolissacarídeos bacterianos causam ao ativar o sistema
imune (CANI; DELZENNE, 2010; CANI et al., 2012; KIM et al., 2012; LEE; LEE;
CHOUE, 2013; MCALLAN et al., 2013).
Visando à modulação do estresse oxidativo, da inflamação e da
disbiose intestinal propõe-se a hipótese de que a ingestão de alimentos
protéicos geradores de peptídeos bioativos, como as proteínas do soro do leite,
se traduza em potenciais benefícios para a saúde como: o controle glicêmico, o
controle do ganho de peso e a diminuição do processo inflamatório, que são

2
passos iniciais contra o desenvolvimento de doenças crônicas transmissíveis e
não transmissíveis.
Considerando as propriedades que as proteínas do soro lácteo
hidrolisadas têm demonstrado em proteger vários tecidos no rato contra os
efeitos danosos do estresse, aumentando a expressão de HSPs, e colocando
em funcionamento a via alternativa do transportador GLUT-4 para aumentar a
captação da glicose circulante, propõe-se as seguintes hipóteses:
Hipótese 1. A microbiota intestinal, de forma coerente com os
efeitos protetores apontados acima, possa também ser modulada de forma
positiva em função da dieta, resultando na possível remediação da inflamação
subclínica.
Hipótese 2. O consumo da proteína do soro do leite hidrolisada
possa reverter ou neutralizar o descontrole do peso corpóreo que resulta do
consumo de uma dieta obesogênica. De ser esse o caso, seria possível
observar um grau de saciedade normal, com aumento da sensibilidade à
leptina e a insulina, com a consequente melhora da inflamação.
A literatura fornece subsídios da existência de uma importante
correlação entre microbiota intestinal e o estado inflamatório que ocorre na
obesidade e, um estudo propondo a utilização das propriedades protetoras das
proteínas hidrolisadas do soro de leite estaria substanciado ainda no mesmo
princípio de que a dieta tem influência direta sobre a microbiota intestinal. Do
ponto de vista prático, a busca e o desenvolvimento de intervenções
alimentares que contribuam para a regularização da homeostase entre
hospedeiro e microbiota, seria suficiente justificativa para empreender um
estudo nesse sentido.
O trabalho teve como objetivo geral relacionar o consumo de
caseína e proteínas do soro do leite, nas formas concentrada e hidrolisada, à
produção de citocinas pró-inflamatórias e alterações na microbiota intestinal de
camundongos, verificando de forma mais específica a presença de
lipopolissacarídeos e parâmetros bioquímicos no soro (glicose, proteínas totais,
albumina, ácido úrico, triglicerídeos, colesterol total e colesterol-HDL), a

3
presença de esteatose hepática por meio de técnica histológica, a dosagem das
proteínas TNF (fator de necrose tumoral alfa), IL4 (interleucina quatro), IL6
(interleucina seis), IL8 (interleucina oito), IL10 (interleucina dez), IL1Ra
(antagonista do receptor da interleucina 1), IL1β (Interleucina 1 Beta), TLR4
(toll-like receptor 4), MyD88 (proteína adaptadora do fator de diferenciação
mielóide 88), todas em músculo gastrocnêmio pela técnica de western blot e a
caracterização da composição da microbiota intestinal por sequenciamento do
RNAr 16S.

4
Referências
CANI, P. D.; DELZENNE, N. M. Involvement of the gut microbiota in the
development of low grade inflammation associated with obesity: focus on this
neglected partner. Acta Gastroenterol Belg. v.73 n.2, p.267-9, 2010.
CANI, P. D.; OSTO, M.; GEURTS, L.; EVERARD, A. Involvement of gut
microbiota in the development of low-grade inflammation and type 2 diabetes
associated with obesity. Gut Microbes. v.3, n.4,p.279-88, 2012.
CINTRA, D. E.; ROPELLE, E. R.; MORAES, J. C.; PAULI, J. R. Obesidade e
Diabetes – Fisiopatologia e sinalização celular. São Paulo: Editora Sarvier,
2011, 424p.
KIM, K-A.; GU, W.; LEE, I-A; JOH, E-H.; KIM, D-H. High Fat Diet-Induced Gut
Microbiota Exacerbates Inflammation and Obesity in Mice via the TLR4
Signaling Pathway. PLoS One, v.7, n.10, 2012.
LEE,H.; LEE, I. S.; CHOUE, R. Obesity, Inflammation and Diet. Pediatr.
Gastroenterol Hepatol Nutr., v.16, n. 3, p.143-152, 2013.
MCALLAN, L.; KEANE, D.; SCHELLEKENS, H.; ROCHE, H. M.; KORPELA, R.;
CRYAN, J. F.; NILAWEERA, K. N. Whey protein isolate counteracts the effects
of a high-fat diet on energy intake and hypothalamic and adipose tissue
expression of energy balance-related genes. Br. J. Nutr. v.110, n.11,p. 2114-26,
2013.
THALER, J. P.; STEPHAN, J. G.; DORFMAN, M.; WISSE, B. E.; SCHWARTZ,
M. W. Hypothalamic inflammation: Marker or mechanism of obesity
pathogenesis? Diabetes v. 62, n. 8, p. 2629-34, 2013.

5
WORD HEALTH ORGANIZATION (WHO). Obesity and Overweight. Geneva,
Switzerland, 2012. Disponível em: < http://www.who.int/nutrition/challenges/en/
> acesso em 14 abr.2013.

6
CAPÍTULO 1
REVISÃO DE LITERATURA

7
1. Obesidade
Caracterizada pelo acúmulo excessivo de gordura corporal, de forma a
comprometer a saúde do indivíduo, a obesidade é consensualmente descrita
como doença de caráter multifatorial, onde fatores históricos, políticos,
socioeconômicos, culturais, psicossociais e biológicos estão agindo em conjunto,
devendo, portanto, serem alvos independentes dos esforços de prevenção e
intervenção. Em geral, os fatores mais estudados na etiologia da obesidade são
os biológicos relacionados ao estilo de vida, como dieta e prática de atividade
física (WANDERLEY; FERREIRA, 2010).
O Índice de massa corpórea (IMC) é uma das ferramentas mais
utilizadas para predição de obesidade. Seu cálculo é realizado por meio da divisão
do peso em quilos pelo quadrado da estatura em metros (kg/m2), sendo que a
classificação da Organização Mundial da Saúde (OMS) delimita IMC igual ou
superior a 25 kg/m2 como sobrepeso e IMC igual ou superior a 30 kg/m2 como
obesidade. O IMC apesar de simples é um importante referencial na avaliação
antropométrica do indivíduo, pois está positivamente correlacionado com a
porcentagem de gordura corporal, hiperinsulinemia e produção de citocinas pró-
inflamatórias (WHO, 2011).
A intensa transição demográfica, epidemiológica e nutricional que o
Brasil vem experimentando nos últimos 20 anos consolidou a obesidade como um
agravo nutricional associado a altas taxas de incidência de diabetes, doenças
cardiovasculares e câncer, o que modificou o perfil de morbi-mortalidade da
população. O comportamento alimentar inadequado, devido à ingestão de dieta
com alto teor de lipídios, em especial os de origem animal, carboidratos simples e
pobre em fibras, vitaminas e minerais, aliado à redução da atividade física, tem
promovido a obesidade principalmente entre as famílias de menor renda. Ao longo
do tempo, o perfil nutricional de crianças e adolescentes brasileiros também
mudou, a prevalência do baixo peso está em declínio, contrapondo-se aos altos
índices de sobrepeso e obesidade que predizem doenças de cunho nutricional na
vida adulta (PINHEIRO; FREITAS; CORSO, 2004; FLORES et al., 2013).

8
Estudos indicam que a exposição prolongada a dietas ricas em gordura
saturada promovem a apoptose de neurônios com função central no controle do
peso, além da resistência a sinais anorexígenos, que ocorre principalmente pela
ativação de TLR4, um indutor de estresse do retículo endoplasmático, que leva ao
aumento da expressão de citocinas pró-inflamatórias no hipotálamo. Tais
fenomenos explicam a dificuldade na manutenção do peso corporal em indivíduos
obesos quando conseguem emagrecer através de dietas de restrição calórica
(MORAES et al., 2009; MILANSKI et al., 2009). A presença na região hipotalâmica
de grandes quantidades de gordura saturada, proveniente da dieta ingerida, induz
a processos de apoptose. O hipotálamo, ao identificar longas cadeias lipídicas
saturadas, logo as associa àquelas presentes nas membranas de bactérias e o
organismo então as interpreta como ameaça respondendo de forma a ativar o
sistema imune com a liberação local de substancias pró inflamatórias, dentre as
quais se encontram as adipocinas, o TNFα, a IL6 e a leptina (VELLOSO, 2009).
Reconhecendo-se então a obesidade como um processo inflamatório complexo,
percebe-se que somente a prescrição de uma dieta reduzida em termos de
calorias é insuficiente para o efetivo controle do peso corporal, havendo a
necessidade da substituição dietética de gorduras saturadas por insaturadas, que
promovam a melhoria da inflamação (VELLOSO, 2009).
1.1 Aspectos Epidemiológicos da Obesidade
A prevalência de sobrepeso e de obesidade tem aumentando
progressivamente no Brasil e no mundo, sendo considerada como um importante
problema de saúde pública, tanto para países desenvolvidos como em
desenvolvimento. A obesidade é uma doença crônica, de caráter epidêmico e
multicausal associada ao surgimento de co-morbidades, como, por exemplo,
doenças coronarianas, diabetes, câncer e problemas músculo-esqueléticos, que
se não tratadas podem tornar-se incapacitantes, onerando a sociedade e o
sistema de saúde (PI-SUNYER, 1991; WHO, 2014). Além das implicações na

9
saúde, a obesidade e o sobrepeso trazem também conseqüências
socioeconômicas substanciais. Finkelstein et al. (2014) ao avaliarem os custos
médios durante a vida de uma criança obesa comparado-a a uma criança
eutrófica, observaram que o custo per capita de uma criança obesa corresponde a
cerca de 19 mil dólares a mais do que para uma criança com peso dentro da
normalidade.
Atribui-se à obesidade um custo direto de 2 a 8% do gasto total com
saúde, em países industrializados. Entretanto, os valores reais devem extrapolar
facilmente os estimados, visto que nesta porcentagem não são contabilizadas as
doenças associadas. Um estudo conduzido entre 2008 e 2010 sobre os custos
ligados a obesidade no Sistema Único de Saúde (SUS) revelou que o gasto
estimado com doenças relacionadas ao sobrepeso e obesidade atinge quase US$
2,1 bilhões por ano no Brasil, o equivalente a US$ 1,4 bilhões com hospitalizações
e US$ 679 milhões com procedimentos ambulatoriais (BAHIA et al., 2012).
Dados da Pesquisa de Orçamentos Familiares (POF) 2008/2009
também demonstraram importante aumento nos números da prevalência de
obesidade no país, principalmente entre crianças na faixa de 5 a 9 anos de idade.
O excesso de peso entre crianças foi de 33,5% e obesidade 14,3%, enquanto que
entre adolescente esses números são de 20,5% e 4,9%, respectivamente. Em
adultos observou-se aumento constante de excesso de peso e obesidade, onde
cerca de metade da população masculina (50,1%) e feminina (48%) possui
excesso de peso. Entre os indivíduos avaliados 14,8% estavam obesos, sendo
que as mulheres apresentaram-se mais obesas (16,9%) do que os homens
(12,5%) (IBGE, 2010).
Por conta da relevância das doenças crônicas não transmissíveis
(DCNT) e sua possibilidade de prevenção, o Ministério da Saúde tem realizado
anualmente desde 2006 o Vigitel – Vigilância de Fatores de Risco e Proteção para
Doenças Crônicas por Inquérito Telefônico, fruto de uma parceria entre Ministério
da Saúde e o Núcleo de Pesquisas Epidemiológicas em Nutrição e Saúde da
Universidade de São Paulo (Nupens/USP). O Vigitel 2011 trouxe dados
alarmantes, onde mais da metade da população brasileira maior de 18 anos

10
estava acima do peso, sendo 48,5% com sobrepeso/excesso de peso (IMC >
25kg/m2) e 15,8% com obesidade (IMC > 30kg/m2). Ao desmembrarmos este
resultado observa-se que o percentual masculino passou de 11%, em 2006, para
16% em 2011, mantendo os mesmos valores entre mulheres. De modo geral, o
estudo tem encontrado aumento crescente na prevalência de excesso de peso,
diabetes e hipertensão arterial sistêmica entre indivíduos adultos desde sua
concepção. Entretanto, a tendência de crescimento contínuo de excesso de peso
e obesidade pela primeira vez estabilizou entre 2012 e 2013, provavelmente
devido ao aumento no consumo recomendado de frutas e hortaliças (5 porções
diariamente) e também pelo possível aumento da prática de atividade física
atrelada ao lazer (VIGITEL, 2013).
Mesmo diante da estabilização dos índices de obesidade a estimativa
para o futuro não é animadora, pois estima-se que em 2025 cerca de 25% dos
brasileiros estarão obesos e esta projeção desfavorável não se restringe apenas
ao Brasil, pois a Organização Mundial da Saúde (OMS) projeta já para 2015 uma
população mundial de 2,3 bilhões de adultos com sobrepeso e 700 milhões de
obesos (WHO, 2011).
Segundo o Department of Health and Human Services (2001) indivíduos
obesos têm risco de morte prematura 50% a 100% maior que indivíduos
eutróficos, além de apresentarem três a quatro vezes mais sintomas de
ansiedade, depressão e serem estigmatizados e discriminados socialmente. A
prevalência de obesidade infantil também é algo preocupante, por ser comumente
vista em famílias onde pai e mãe também são obesos, compreende-se que não
somente as influências genéticas, mas também o comportamento obesogênico
adotado pela família é determinante. Um indivíduo obeso durante sua infância ou
adolescência possui 70% de chance de tornar-se um adulto com sobrepeso ou
obesidade. Por isso reduzir os índices desta doença é uma prioridade de saúde
pública que beneficia não somente a saúde, mas também a economia de um país,
sendo fundamental o investimento em campanhas contra a obesidade infantil, pois
este é o melhor modo de diminuir a obesidade na vida adulta.

11
1.2 Inflamação crônica induzida pela obesidade
A resposta inflamatória é classicamente definida por infecção e/ou
lesão tecidual que causa rubor, tumor, calor e dor, com tendência a evoluir para
resolução e homeostase (MEDZHITOV, 2008). Evidências atuais indicam que a
obesidade resulta de um quadro inflamatório crônico e subclínico de baixa
intensidade que se manifesta de forma sistêmica perpetuando-se e agravando-se
ao longo da vida do indivíduo de acordo com seu estilo de vida. Fatores como: o
descontrole da ingestão calórica e a diminuição do dispêndio energético, seja por
causas ambientais, emocionais ou genéticas determinam a obesidade em
indivíduos sedentários (EGGER; DIXON, 2011).
Do ponto de vista celular e molecular, a obesidade é uma doença
crônica, na qual os processos de infiltração de células inflamatórias e ativação da
rede de citocinas estão intimamente envolvidos, havendo exacerbada produção de
substâncias, em especial as pró-inflamatórias nos âmbitos central e periférico. O
tecido adiposo foi por muito tempo considerado como um tecido de função restrita
a de reservatório energético. Entretanto, diversos estudos vêm provando ao longo
dos anos que sua atividade fisiológica e importância é crucial para os processos
imunológicos e inflamatórios. Uma ampla variedade de hormônios, fatores de
crescimento, proteínas da via do complemento e citocinas de caráter pró e anti-
inflamatório são produzidas pelo tecido adiposo, como por exemplo, leptina,
adiponectina, resistina, visfatina, assim como as interleucinas, interferon gama
(IFN), fator de necrose tumoral (TNF), com distintos papéis fisiológicos o tornam
um importante órgão endócrino (LEE; LEE; CHOUE, 2013; KERSHAW; FLIER,
2004).
Em obesos, o tecido adiposo inflamado e hipertrofiado exibe sinais
como hipóxia, estresse do retículo endoplasmático, alterações da função
mitocondrial, produção elevada de espécies reativas de oxigênio (EROs), aumento
das concentrações intracelulares de ácidos graxos não esterificados (ácidos
graxos livres), produção descontrolada de citocinas pró-inflamatórias e ativação
das vias de sinalização envolvidas na apoptose celular. Entretanto, evidências
recentes têm colocado a inflamação hipotalâmica numa posição de destaque na

12
fisiopatologia da obesidade, sugerindo que a inflamação em tecidos periféricos
possa ser prevenida ou até mesmo revertida quando a inflamação central é
controlada (KAWASAKI et al., 2012).
A suposta relação entre inflamação em tecidos metabólicos atrelada
à obesidade e resistência à insulina originou-se em 1993, com a descoberta da
produção de TNF-α pelo tecido adiposo. Este mesmo grupo demonstrou em
trabalhos posteriores que a resistência à insulina induzida em animais obesos era
dependente dos níveis de TNF-α. Tais achados culminaram na descoberta de
outras citocinas inflamatórias, que corroboram a hipótese da inflamação como
base molecular da obesidade (HOTAMISLIGIL; SHARGILL; SPIEGELMAM, 1993).
Mecanismos distintos, porém sinérgicos, são responsáveis pela inicialização da
inflamação subclínica, induzindo a ativação das quinases IKK e JNK, que são as
mesmas ativadas na resposta inata pelo TLR4 em resposta a lipopolissacarídeos
e ácidos graxos. Esse processo mediado por moléculas inflamatórias também
pode interferir na sinalização de insulina, pela atuação da IKKβ na fosforilação
direta do substrato do receptor de insulina (IRS-1 e IRS-2) em resíduos de serina
e também pela ativação indireta do NF-кB, um fator de transcrição que estimula
produção de vários marcadores pró-inflamatórios. A sinalização de insulina ainda
pode ser prejudicada pela ativação da JNK, que pode ser ativada pela via do TLR4
ou também pela presença de TNF-α (PAULI et al, 2009).
A obesidade induz a uma importante infiltração de macrófagos tanto no
tecido adiposo quanto no tecido muscular (depósitos adiposos entre as fibras
musculares). Didaticamente os macrófagos podem ser classificados em dois
grupos distintos, M1 – que são macrófagos classicamente ativados, secretores de
citocinas pró-inflamatórias como IL1β, IL6 e TNF-α, recrutados e ativados durante
a obesidade; e o grupo M2 – macrófagos ativados de forma alternativa,
responsáveis por citocinas anti-inflamatórias, como IL1Ra e IL10. Entretanto,
evidências recentes revelam um cenário mais complexo na polarização de
macrófagos, que vão além da classificação binária M1/M2, expondo diferentes
perfis de macrófagos com diferentes atuações no contexto da obesidade (ESSER
et al, 2014).

13
Um estudo com animais alimentados com dieta hiperlipídica demonstra
que o início da produção exacerbada de marcadores inflamatórios se dá logo nas
primeiras 24 horas de ingestão da dieta, mantendo níveis altos nos primeiros 3
dias, seguido de uma faixa de normalização transitória, que pode durar de 2 a 3
semanas, mas retornando novamente aos elevados níveis iniciais, com tendência
a manter-se elevados. Esse padrão temporal equipara-se ao que é visto no
consumo alimentar quando há mudança de uma dieta normolipídica para uma
dieta hiperlipídica (THALER et al., 2013).
O estado inflamatório é caracterizado pela produção de uma ampla
variedade de mediadores bioquímicos que trabalham em uma complexa rede.
Evidencias demonstram que variações genéticas envolvidas na etiologia da
inflamação e distúrbios metabólicos podem interagir com fatores ambientais, como
por exemplo, a dieta, predispondo alguns indivíduos à maior vulnerabilidade que
outros. A citocina TNF-α envolvida na inflamação atrelada à obesidade é uma
citocina pleiotrópica, com papel central na inflamação, mediadora chave da
resistência à insulina devido a sua participação na fosforilação de IRS-1. O Fator
de necrose tumoral alfa (TNF-α) é primeiramente secretado por células mielóides
via ativação da MAPK e NFκB, resultando na liberação de outras citocinas
inflamatórias como IL1β e IL-6. Em humanos obesos os níveis de TNF- α
plasmáticos e no tecido adiposo são altos, mas com a redução de peso, os
indicadores tendem a diminuir. O TNF-α também induz indiretamente a resistência
à insulina, por inibir a diferenciação de pré-adipócitos a adipócitos maduros,
através de PPARy, além de atuar no metabolismo lipídico, promovendo lipólise e
secreção de ácidos graxos livres, os quais contribuem para um aumento na
produção de glicose hepática (CURTI et al, 2011; MAKKI;
FROGUEL;WOLOWCZUK, 2013).
IL-6 é secretada pelo tecido adiposo, músculo, fígado, além de outras
células como as pancreáticas, macrófagos, osteoblastos e miócitos. Citocina
pleiotrópica essencial no processo inflamatório, participa da síntese de outras
citocinas com potencial anti-inflamatório e regula negativamente alvos
inflamatórios. Sua expressão correlaciona-se com ganho de peso, aumento da

14
circunferência de cintura e dos níveis de ácidos graxos livres. Portanto, essa
proteína tem sido classificada como pró e anti-inflamatória, dependendo do
contexto em que está inserida e de seus níveis. Estudos demonstram que durante
o exercício, IL-6 atua de forma anti-inflamatória, pois aumenta a absorção de
glicose no músculo esquelético, levando à hipertrofia muscular, miogênese e
oxidação de ácidos graxos mediada por AMPK. Entretanto, no tecido adiposo e
fígado, a IL-6 age de forma pró-inflamatória, promovendo resistência à insulina
através da regulação da SOCS3 (supressor de sinalização citocina 3) que, por sua
vez, compromete o receptor de insulina IRS1 (MAUER et al., 2014; CURTI et al.,
2011; MAKKI; FROGUEL; WOLOWCZUK, 2013).
Normalmente produzida por monócitos em resposta à
lipopolissacarídeos, IL-1 ou TNF-α, a IL-8 são correlacionados a parâmetros como
IMC, circunferência dia cintura, Proteína C reativa (PCR), HDL e score HOMA, o
que faz dessas citocinas marcadores importantes em condições de aterosclerose,
obesidade e diabetes (KIM et al., 2006). A IL-4 é uma citocina de caráter anti-
inflamatório, que regula o metabolismo de lipídios e da glicose, aumentando a
sensibilidade à insulina, além promover lipólise em adipócitos maduros através da
fosforilação da perilipina e inibição da diferenciação de adipócitos através da
STAT6. A IL-4 ainda promove polarização de macrófagos, do estado M1 para o
M2, reduzindo a inflamação em tecidos adiposos e, portanto, impedindo a
incidência de resistência à insulina (TSAO et al, 2014).
Segundo Charles et al (2011) IL-10 é uma citocina de caráter anti-
inflamatória, produzida por macrófagos e células dendríticas que age atenuando
processos inflamatórios induzidos por TNF-α, IL-6 e IL-1, sendo um exemplo
clássico de feedback negativo, pois inibe a própria célula que a produz
(macrófagos), além de também inibir IL1-β, IL-6, IL-8, TNF-α e como mencionado,
se auto inibir.Negativamente correlacionada com o IMC e níveis de glicose em
jejum, baixos níveis de IL-10 estão associados com propensão a síndrome
metabólica e diabetes mellitus.
O IL1Ra é um inibidor de IL1β, sendo considerada uma citocina anti
inflamatória com função reguladora negativa no controle e atenuação da resposta

15
inflamatória. IL1β regula a produção de uma variedade de mediadores
inflamatórios, como IL-6, ICAM-1 e E selectin, sendo associada com distúrbios
metabólicos em obesos, como dislipidemia e resistência a insulina (CURTI et al,
2011).
Além dessas proteínas envolvidas na sinalização celular inflamatória,
alguns receptores e moléculas adaptadoras também possuem fundamental
importância dentro do processo. Exemplo disso são os Toll like receptors (TLR),
proteínas transmembranares responsáveis por detectar presença de patógenos
invasores no organismo. Expresso em várias células do sistema imune como
linfócitos, macrófagos, células dendríticas é possível encontrar esses receptores
em adipócitos, enterócitos e células endoteliais. O TLR4, éum subtipo responsável
pelo reconhecimento de lipopolissacarídeos de bactérias gram negativas e ácidos
graxos saturados. Os lipopolissacarídeos são um complexo glicolipídico composto
por um polissacarídeo hidrofílico e um domínio hidrofóbico (lipídio), a interação de
LPS com TLR4 presentes nas membranas de monócitos e macrófagos induz a
síntese das citocinas pró- inflamatórias já citadas (TNFα, IL1β,IL-6, IL-8), que
atuam mediando o processo inflamatório por meio de interações com receptores
em células alvo (AKIRA; TAKEDA, 2004; MILANSKI et al, 2009).
Quando um TLR associa-se a um de seus ligantes podem ser ativadas
duas vias sinalizatórias, dependente e não-dependente, do fator de diferenciação
mielóide 88 (MyD88) (AKIRA; TAKEDA, 2004). A MyD88 é uma molécula
adaptadora, que ao se unir à porção plasmática do receptor permite o
recrutamento de outras proteínas, como TRAF-6 (fator 6 associado ao receptor de
TNF-α) e IRAK, que proporcionaram a fosforilação do complexo IkB quinase (IKK).
No citoplasma da célula encontra-se o NFKB associado a uma proteína inibidora
IkB, a fosforilação desse complexo liberta o NFKB que migra para o núcleo e inicia
a transcrição de genes de citocinas inflamatórias, incluindo IL1β, IL-6 e TNF-α e
também para IkB, desse modo o ligante recém sintetizado suprime a atividade de
NFKB, o qual retorna a sua posição citosólica, cessando a produção de agentes
pró-inflamatórios, conforme esquematizado na Figura 1 (CINTRA; ROPPELE;
MORAES; PAULI, 2011).

16
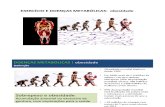
Figura 1. Ativação da via do NFKB por meio da ligação de
lipopolissacarídeos ao receptor do tipo Toll Like receptor 4 (TLR-4). DD=
domínio de morte, Tradd= TRAF associada ao domínio de morte, RIP=
proteína de interação com receptor, TRAF-2= fatores associados ao receptor
de TNF. Retirado de Cintra; Ropelle; Pauli, 2011.

17
1.3 A microbiota intestinal em indivíduos saudáveis
A microbiota intestinal humana é atualmente reconhecida como um
“órgão microbiótico” que abriga entre 10 e 100 trilhões de microorganismos,
das quais grande parte se aloja no cólon intestinal. A microbiota intestinal
compreende inúmeros microorganismos vivendo simbioticamente no trato
digestório do ser humano. Este complexo ecossistema é composto pela
microbiota residente ou endógena, que vive aderidas às membranas das
mucosas do trato gastrointestinal por um período prolongado e a microbiota
transitória ou exógena que atravessa a barreira gastrointestinal. Este
ecossistema complexo é composto por micro-organismos aeróbios, anaeróbios
e facultativos (THE HUMAN MICROBIOME PROJECT CONSORTIUM, 2012).
As diferentes regiões anatômicas intestinais muito diferem em
termos de pH, fluxo de digestão, disponibilidade de substrato e tensão de
oxigênio. O intestino grosso, caracterizado por um fluxo digestivo lento, pH de
neutro para levemente ácido, possui a maior quantidade de espécies
microbianas; comunidades estas dominadas por anaeróbios estritos. O
intestino delgado possui uma quantidade consideravelmente menor de micro-
organismos devido às condições ambientais não tão favoráveis, como o
trânsito digestivo relativamente curto (3-5 h) e a concentração alta de sais
biliares (FLINT et al., 2012).
Embora existam mais de 50 filos de bactérias descritos até o
momento, a microbiota intestinal humana é dominada pelos filos: Bacteroidetes
e Firmicutes, enquanto Proteobacteria, Verrucomicrobia, Actinobactérias,
Fusobactérias e Cianobactérias estão presentes em proporções menores. O
epitélio intestinal está recoberto por uma espessa camada de muco, e tanto o
lúmen, quanto o mucu, representam regiões distintas, contendo uma
microbiota específica. Por exemplo, Bacteroides, Bifidobacterium,
Streptococcus, membros da família Enterobacteriacea, Enterococcus,
Clostridium, Lactobacillus, e Ruminococcus são comumente encontrados no
conteúdo intestinal e nas fezes, enquanto que os Clostridium, Lactobacillus e

18
Enterococcus são detectados no muco e criptas epiteliais do intestino delgado
(SERIKOV et al, 2010).
O transplante de comunidades microbianas entre diferentes
hospedeiros tem demonstrado que as espécies da microbiota intestinal
apresentam um nível elevado de adaptação de um habitat para o outro,
exemplo disso é o estudo de Ridaura et al. (2013), que descreveu o transplante
de camundongos germ-free com a microbiota de humanos gêmeos, dos quais,
um apresentava fenótipo obeso e o outro magro, produzindo “camundongos
humanizados” e revelando que o fenótipo era transferido junto com a
microbiota e que a microbiota de indivíduos magros pode invadir outra
microbiota e reduzir tecido adiposo no camundongo obeso quando esta
recolonização é atrelada a uma dieta saudável.
O aumento do tecido adiposo de camundongos que receberam
microbiota de indivíduo obeso foi significantemente maior do que aquele
registrado nos que receberam microbiota de indivíduo magro, ambos os grupos
receberam dieta com 33% de gordura, inclusive notando-se maior peso da
gordura epididimal em animais colonizados com comunidade intestinal de
obesos. Outro achado foi sobre a microbiota de indivíduos obesos que pode
influenciar a produção de metabólitos que caracterizam o próprio estado de
obesidade, pois animais transplantados com microbiota de obeso exibiram alta
expressão de genes envolvidos na resposta ao estresse e nos processos de
desintoxicação, enquanto microbiota de magros induziu expressão de genes
envolvidos na digestão de polissacarídeos, bem como maior fermentação de
butirato e propionato. A maior capacidade para quebra e fermentação de
polissacarídeos (carboidratos não digeríveis) da microbiota de indivíduos
magros em comparação com a microbiota de obesos, está relacionada com
diminuição do peso corporal e menor adiposidade (RIDAURA et al., 2013).
Esta adaptação não surpreende, já que a densa comunidade
microbiana residente no intestino, referida como microbiota comensal,
relaciona-se intensamente com o hospedeiro sendo essencial para muitos
processos fisiológicos que incluem melhoria da barreira epitelial intestinal,

19
desenvolvimento do sistema imunológico, resposta ao estresse e aquisição de
nutrientes (KAMADA et al.; 2013).
O trato gastrointestinal durante a vida fetal é estéril, a partir do
nascimento as bactérias da mãe e do ambiente iniciam a colonização do
intestino do bebê. Essa colonização dependerá do tipo de parto (normal ou
cesárea) e da alimentação (aleitamento materno exclusivo, leite de vaca ou
fórmula industrial). Mesmo existindo filos bacterianos comuns entre humanos,
há uma clara diferença na colonização bacteriana entre indivíduos, o que
reforça a idéia de que cada pessoa, via de regra, tenha um subconjunto de
suas próprias bactérias colonizando diferentes partes do corpo. Essa
singularidade da comunidade microbiana de cada indivíduo parece ser estável
ao longo do tempo em relação à população total, sendo determinante do
estado de saúde (THE HUMAN MICROBIOME PROJECT CONSORTIUM,
2012)
No estado saudável os filos de bactérias dominantes em humanos
são os Firmicutes, Bacteroidetes e Actinobactérias, com Proteobacteria e
Verrucomicrobia presentes em menor número. Em adultos saudáveis o perfil
da microbiota fecal se mantém estável ao longo do tempo. Mudanças na
composição de macronutrientes da dieta claramente afetam a microbiota
intestinal, sendo que a intensidade da alteração irá depender da composição
original da microbiota, pois alguns grupos dominantes de bactérias são
nutricionalmente menos exigentes e serão, portanto, afetados em menor
proporção por mudanças dietéticas (FLINT et al., 2012).
1.4 A microbiota intestinal em indivíduos doentes
Evidências recentes demonstraram que existem diferentes
composições bacterianas para indivíduos magros e obesos (TSUKUMO et al.,
2009). Estes achados têm despertado grande interesse na comunidade
científica, ampliando o horizonte de hipóteses sobre a influência que a
microbiota intestinal exerçe na fisiopatologia da obesidade.

20
O descontrole na ingestão calórica, além de afetar o balanço
energético, promove mudanças na composição da microbiota intestinal, o que
pode gerar obesidade e doenças metabólicas associadas (GREINER;
BÄCKHED, 2011).
Indivíduos submetidos a uma mesma condição nutricional podem
reagir de maneiras distintas em relação ao ganho de peso, mesmo possuindo
características semelhantes. Essa maior susceptibilidade ao acúmulo de peso
e hiperglicemia pode ter maiores explicações do que simples inadequações
dietéticas e/ou sedentarismo. Estudos atuais demonstram uma importante
correlação entre microbiota intestinal e o estado inflamatório que ocorre na
obesidade (RODRIGUES, 2011).
Na obesidade humana, destaca-se a predominância de bactérias
dos Phyla Firmicutes e Bacteroidetes, composição considerada fundamental
para a homeostase energética do organismo, sistema imune e resposta
inflamatória (PISTELLI; COSTA, 2010).
Os micro-organismos do Phylum Bacteroidetes são gram-negativos,
possuem formato de bastonetes ou cocobacilos, são anaeróbios obrigatórios e
não esporulados, e são encontrados normalmente no trato intestinal e nas
mucosas. Os Bacteroidetes apresentam como produtos finais do metabolismo
da glicose os ácidos succínico e acético (TORTORA; FUNKE; CASE, 2006). As
bactérias do Phylum Firmicutes, maioria das quais possuem parede celular
gram-positiva, têm forma de cocos ou bacilos, muitas vezes produzindo
endósporos. Por ser um Phylum extenso costuma-se subdividi-lo em três
classes: 1) Bacilli - aeróbios facultativos ou obrigatórios; 2) Clostridia -
anaeróbios e 3) Mollicutes – não possuem parede celular. As classes Bacilli, e
em particular Lactobacillus e Enterococcus têm sido as de maior interesse
nutricional (TRABULSI; SAMPAIO, 2005).

21
1.5 Tecido hepático, microbiota intestinal e NAFLD
Fundamental no metabolismo de glicose, lipídios e aminoácidos, o
fígado participa não somente do metabolismo de macronutrientes, mas
também do armazenamento de substâncias (ferro, vitaminas A, D e B12), da
degradação e excreção de hormônios, além de realizar outras funções como
transformaçãoe excreção de drogas, produção de fatores hemostáticos e de
ser um componente essencial na regulação do sistema imune (SCHINONI,
2006).
O fígado é um dos maiores órgãos sólidos do corpo humano,
pesando cerca de 1,5 kg e consistindo de quatro lobos pouco definidos, que
recebem sangue parcialmente desoxigenado da veia porta. O sangue provém
do trato digestório, pâncreas e baço (75 a 80% do volume aferente) e da
artéria hepática, um ramo do tronco celíaco que transporta de 20 a 25% do
sangue oxigenado recebido pelo fígado. Nos capilares sinusóides, os sangues
da veia porta e da artéria hepática misturam-se, convergindo para a veia
central do lóbulo hepático. As veias centrais dão origem às veias sublobulares,
e o sangue retorna ao coração pela veia cava inferior. O fluxo sanguíneo
hepático total alcança entre 800 a 1200 mL/min, e embora a massa hepática
não ultrapasse 2,5% do peso corpóreo, o fígado recebe aproximadamente
25% do débito cardíaco (EIPEL; KERSTIN; VOLLMAR, 2010).
O fígado recebe suprimento sanguíneo da artéria hepática e da veia
porta, que por sua vez drenam o sangue das veias mesentéricas, este sangue
não contém apenas produtos da digestão de nutrientes, mas também produtos
oriundos de micro-organismos colonizadores do intestino. Portanto, o fígado é
o primeiro órgão de exposição aos produtos da microbiota, funcionando como
uma espécie de filtro (DUSEJA; CHAWLA, 2014).
Estruturalmente, o fígado é composto por hepatócitos, que atuam
na síntese e secreção de substâncias; células endoteliais que revestem os
sinusóides e ductos biliares revestidos por células epiteliais; células de
Kupffer, que possuem intensa atividade fagocitária, englobando eritrócitos
senescentes, materiais particulados e 99% dos micro-organismos presentes

22
no sangue portal; além das células de Ito (células estreladas), cuja principal
função é o armazenamento de vitamina A sob a forma de ésteres de retinol.
Em situação de inflamação hepática as células de Ito perdem sua capacidade
original e passam a secretar colágeno, exercendo importante função nos
mecanismos de injúria, regeneração e fibrose hepática (KMIEĆ, 2001;
CORMACK, 2003).
Estudos atuais demonstram forte associação entre alterações na
microbiota intestinal em indivíduos obesos e desenvolvimento da esteatose
hepática não-alcoólica (EHNA) ou NAFLD, do inglês "nonalcoholic fatty liver
disease", ou doença hepática gordurosa não-alcoólica (DHGNA). A DHGNA é
uma doença hepática crônica intimamente associada à obesidade e a
resistência à insulina, que pode ser exacerbada pela microbiota intestinal por
aumentar esteatose hepática, fibrose e inflamação (GANGARAPU; YILDIZ;
INCE; BAYSAL, 2014).
Muitos estudos conduzidos com humanos e animais têm
investigado a associação entre a EHNA e microbiota intestinal. A microbiota
intestinal pode iniciar esteatose hepática através do aumento na absorção de
monossacarídeos, produção de bioprodutos hepatotóxicos bacterianos, reação
inflamatória crônica de baixa intensidade ou pelo metabolismo de ácidos
biliares (PARK, SEO, YOUN; 2013).
Segundo Bäckhed et al. (2004) camundongos germ free possuem
menores níveis de triglicérides do que camundongos convencionalmente
colonizados, pois a alta absorção de monossacarídeos promove síntese de
novo de ácidos graxos e triglicerídeos, elevando atividade da acetil CoA
carboxilase (ACC) e ácido graxo sintase (FAS). Em camundongos obesos
encontrou-se aumento nas concentrações séricas de bioprodutos
hepatotóxicos como o etanol e dióis ativos, que estão diretamente
relacionados à esteatose hepática (AMAYA-FARFAN, 1999). A relação entre
inflamação, endotoxemia e esteatohepatite não alcoólica é associada a
mecanismos que envolvem lipopolissacarídeos bacterianos, células de
kuppfer, que são responsáveis por remover tais endotoxinas, CD14 e TLR4.

23
Além disso, a microbiota também pode promover esteatose e lipoperoxidação
através da estimulação de receptores farnesóides X (FXR) que provocam
mudanças na secreção biliar (PARK, SEO, YOUN; 2013).
Em indivíduos obesos, achados clínicos e bioquímicos de
alterações hepáticas são comuns. O principal produto bacteriano envolvido na
patogênese da esteatose não alcoólica são os lipopolissacarídeos (LPS),
componentes ativos das paredes celulares de bactérias gram negativas, mas
que também são produzidos continuadamente pelo intestino de forma
endógena através da morte destas mesmas bactérias. Essa translocação
ocorre através dos capilares presentes nas paredes intestinais e é um
mecanismo dependente de TLR4 (toll-like receptor 4). O transporte dos
lipopolissacarídeos do lúmen intestinal para órgãos alvos é facilitado por
lipoproteínas e quilomicrons, sintetizados pelos enterócitos em resposta ao
conteúdo de gordura da dieta, e a absorção dos lipopolissacarídeos acaba
ocorrendo durante a absorção de lipídios. Os LPS se ligam a
lipopolysaccharide binding protein (LBP), que se liga ao CD14, proteína que
pode estar na forma solúvel ou expressa em células inflamatórias. Juntos, o
complexo LPS-LBP-CD14 ativa o TLR4 presente em células como monócitos,
células de Kuppfer ou células estreladas, desencadeam uma cascata,
incluindo proteínas como a c-Jun N-terminal kinase (JNK), factor nuclear
kappa B (NFKB) e IFR3 (interferon regulatory factor-3). NFKB é translocado
para o núcleo induzindo a expressão de diversas proteínas inflamatórias,
como TNFα, IL1 e IL6. A sinalização do TLR4 pode promover resistência à
insulina, inflamação, fibrogenese e esteatose hepática (MACHADO; PINTO,
2012).
O epitélio intestinal é uma barreira especializada que com a ajuda
das proteínas de junção, seleciona a entrada de substâncias ao organismo,
com o objetivo maior de prevenir a entrada de toxinas e bactérias danosas.
Desordens na homeostase metabólica, entre microogranismos e hospedeiros,
como ocorre na SIBO (Small intestinal bacterial overgrowth) onde há um
crecimento anormal de bactérias no intestino delgado, podem causar ruptura

24
das proteínas de junção, o que aumenta permeabilidade intestinal induzindo a
translocação bacteriana, que é o transporte de micro-organismos e/ou de seus
produtos metabólicos do lúmen intestinal para a corrente sanguínea.
Obesidade é um importante fator de risco para desenvolvimento de NAFLD, e
uma microbiota desregulada pode tanto induzir obesidade, pelo aumento da
captação de energia da dieta, quanto causar SIBO, levando a maior
permeabilidade intestinal, também culminando em NAFLD (FEROLLA et al.,
2014).
Um exemplo da importância da microbiota intestinal no
desenvolvimento da esteatose hepática não alcoólica é o estudo de LE ROY
et al (2013) que avaliou dois grupos de ratos alimentados com uma mesma
dieta hiperlipídica, ambos os grupos (A e B) apresentaram ganho de peso
semelhante, porém um grupo (A) desenvolveu hiperglicemia e apresentou
altas concentrações séricas de citocinas pró-inflamatórias, enquanto que o
outro grupo (B) não teve alterações nas concentrações de glicose plasmática,
apresentando baixos níveis de marcadores inflamatórios sistêmicos. Os
autores transplantaram a microbiota do grupo A e B em ratos germ-free,
alimentando este novo grupo com a mesma dieta hiperlipídica. Os animais que
receberam a microbiota dos ratos do grupo A, apresentaram hiperglicemia em
jejum, hiperinsulinemia, esteatose hepática, concentrações elevadas de
triglicéridos no fígado e aumento da expressão de genes envolvidos na
lipogênese de novo. Os animais que receberam a microbiota dos ratos do
grupo B permaneceram normoglicêmicos com parâmetros metabólicos dentro
da normalidade. Os autores concluíram que a microbiota determina a
propensão ao desenvolvimento de NAFLD e ainda, enfatizaram que os filos
associados ao fenótipo propenso a desenvolver NAFLD e o não propenso, são
respectivamente, Firmicutes e Bacteroidetes.

25
1.6 Relação entre obesidade, microbiota e inflamação
Em pessoas obesas, há maior quantidade de microorganismos do
Phylum Firmicutes do que Bacteroidetes. Entretanto, com a perda de peso
ocorre uma inversão na estrutura da microbiota. As bactérias do Phylum
Bacteroidetes são benéficas, pois produzem ácidos graxos de cadeia curta,
que possuem propriedades antiinflamatórias (LEY et al., 2006). Há uma
importante relação entre as alterações da diversidade bacteriana intestinal e a
inflamação que ocorre na obesidade, esta relação fica ainda mais evidenciada
diante da introdução de dieta hiperlipídica, que acarreta no indivíduo obeso o
aumento da permeabilidade intestinal e consequente endotoxemia metabólica
(SNEDEKER; HAY, 2012).
A endotoxemia metabólica é causada pela entrada de
lipopolissacarídeos (LPS) na corrente sanguínea. Estas substâncias liberadas
durante a lise das células bacterianas ativam o sistema imune do indivíduo ao
serem reconhecidas como antígenos. Portanto, a microbiota intestinal influi na
imunidade inata e adaptativa, e na própria inflamação subclínica característica
da obesidade (CANI; DELZENNE, 2010).
A presença de determinadas bactérias ou nutrientes (proteínas e
lipídios) ativam os receptores Toll Like (TLRs), indicando às células imunes a
necessidade de resposta. Tanto os lipopolissacarídios dietéticos, quanto os
presentes nas paredes celulares das bactérias, possuem a capacidade de
ativar o TLR4 e desencadear a síntese de substâncias pró-inflamatórias
mediadoras da resistência à insulina e obesidade (CARICILLI; PICARDI;
ABREU, 2011).
A relação entre obesidade, microbiota e inflamação ainda tem sido
explicada por três possíveis mecanismos: o primeiro corresponde à excessiva
extração de energia de polissacarídeos não-digeríveis (fibras dietéticas) pelas
bactérias intestinais, o que resulta em maior glicose disponível para gerar
energia e, portanto, aumento da lipogênese. Excetuam-se algumas fibras que
possuem efeito prebiótico, como a inulina e o amido resistente, os quais são

26
capazes de auxiliarem no controle do peso (DELZENNE; CANI, 2011;
PEREIRA, 2007).
Dietas hipercalóricas favorecem bactérias intestinais produtoras de
enzimas capazes de quebrar polissacarídeos não digeríveis, introduzindo
calorias adicionais à dieta e diminuindo o valor energético eliminado nas fezes.
As interações das fibras dietéticas com a microbiota intestinal, o seu
metabolismo e o metabolismo do hospedeiro, dependerão da extensão em que
cada fibra seja fermentada. A incorporação de fibras à alimentação modifica o
ambiente intestinal, afetando a retenção de água, formação de gel e produção
de ácidos graxos de cadeia curta. A maioria das fibras dificulta a reabsorção de
ácidos biliares e gorduras, aumentando assim a sua excreção e diminuindo
consequentemente o colesterol circulante. Estas fibras também são capazes de
diminuir a população das bactérias eficientes em extrair energia, o que regula o
metabolismo de carboidratos que resulta da redução da glicemia pós-prandial e
dos níveis de insulina. Por sua vez, esse quadro diminui o estímulo da síntese
hepática de colesterol, promovendo também um efeito hipocolesterolemiante
(FAVA et al., 2006; CHAUD et al., 2007; COX et al., 2013).
O segundo mecanismo consiste no surgimento de uma inflamação
crônica subclínica, como resultado da maior quantidade de lipopolissacarídeos
para a microbiota intestinal, o que conduz à obesidade e diabetes (TSUKUMO,
2009). Em estudos pareados, onde se utilizaram animais germ-free e animais
cujos intestinos foram colonizados com bactérias típicas de indivíduos obesos,
percebeu-se maior quantidade de bactérias gram-negativas (possuidoras de
paredes celulares), as quais aproveitaram o excesso oportuno de
lipopolissacarídeos para o seu próprio metabolismo e proliferação. Foi
demonstrado, então, que o intestino destes animais possuía maior
permeabilidade aos lipopolissacarídeos que, ao adentrarem a mucosa e se
ligarem aos TLR4 e aos receptores CD14 das células, iniciaram reações em
cadeia, gerando citocinas pró-inflamatórias capazes de causar endotoxemia
metabólica (CANI et al, 2007).

27
O terceiro mecanismo propõe que a microbiota intestinal possa
induzir a regulação de genes do hospedeiro que modulam a forma em que a
energia é gasta e armazenada. A microbiota normalmente encontrada em
obesos se mostra capaz de suprimir a produção da enzima Fiaf (Fasting-
Induced Adipose Factor), responsável pelo controle da atividade da lípase
lipoproteica. A inibição da Fiaf faz com que a lipase lipoproteica aumente sua
atividade e inicie a estocagem de calorias na forma de gordura nos adipócitos
(CANI; DELZENNE, 2009).
A microbiota intestinal também promove maior absorção de
monossacarídeos e aumenta a lipogênese hepática do indivíduo (lipogênese
de-novo), em resposta ao aumento das enzimas acetil-CoA carboxilase (ACC)
e ácido graxo sintase (FAS), e de suas proteínas mediadoras: ChREBP
(carbohydrate-responsive element binding protein) e da SREBP-1 (sterol-
responsive element binding protein), fortemente associadas ao aumento de
tecido adiposo e de peso (BACKHED et al., 2004; RODRIGUES, 2011).
Camundongos germ-free, mesmo consumindo dieta rica em gordura
e açúcar, estão protegidos da obesidade por possuírem mecanismos que
aceleram o metabolismo de ácidos graxos. Este mecanismo baseia-se na
existência de maior concentração da enzima Fiaf nestes indivíduos,
promovendo a elevação da produção do co-ativador de PPAR-, responsável
por aumentar a expressão de genes envolvidos na oxidação mitocondrial de
ácidos graxos, além de aumentar a atividade da AMPK (adenosine
monophosphate–activated protein kinase), enzima reguladora mestre do
metabolismo energético (TURNBAUGH et al., 2006).
A microbiota adquirida pelo homem ou animal ao nascer pode ser
modificada ao longo da vida por fatores genéticos, imunológicos, dietéticos e
pelo uso de antibióticos. A dieta, em específico, influencia parâmetros como o
tempo de trânsito intestinal, pH e a própria população bacteriana, já que
mudanças na ingestão de algum macronutriente pode resultar em alterações
da microbiota (SCOTT et al., 2013).

28
Hábitos dietéticos são o maior fator de influência na diversidade da
microbiota humana. Contudo, a microbiota consegue por si só causar ganho de
peso, não sendo este ganho necessariamente mediado pelo aumento no
consumo de alimentos. Isso ocorre tanto pela maior eficiência da transferência
de calorias da dieta para o hospedeiro, quanto pela forma em que tais calorias
são metabolizadas (MILLION et al., 2013).
1.7 Tipificação da microbiota
A microbiota intestinal é um complexo ecossistema composto por
cerca de 500 a 1000 espécies diferentes, onde cada ser humano abriga entre
30 e 40 espécies predominantes, das quais constituídas em sua maior parte por
micro-organismos anaeróbios obrigatórios. A investigação por métodos
tradicionais de cultivo em muito restringe o conhecimento da microbiota
intestinal humana, pois além das técnicas convencionais estarem sujeita a
muitos erros de execução e interpretação, nem todos os micro-organismos que
habitam o intestino humano podem ser cultivados em condições de laboratório
(MUJICO et al., 2013; TANNOCK, 2002). A descoberta da existência de
características invariantes e específicas nas subunidades do RNA ribossomal
microbiano 16S, foi decisiva para a evolução das técnicas de análise das
comunidades bacterianas, representando um significativo avanço no
entendimento do microbioma humano. O RNAr 16S contém regiões ou
sequências de bases de nucleotídeos altamente conservadas que carregam a
assinatura do grupo filogenético e ainda da espécie, mesmo que intercaladas
com regiões hipervariáveis (regiões V). Para se identificar as sequências de
interesse, o DNA bacteriano é extraído da amostra e a região variável do RNAr
16S amplificada por PCR a partir do DNA. O uso de métodos moleculares tem
sido aplicado com sucesso para a identificação de bactérias, independentes de
serem ou não cultiváveis a partir de amostras como tecidos e fezes (MUJICO et
al., 2013, TANNOCK, 2002).

29
1.8 Proteínas do soro do leite
Alimentos funcionais com propriedades bioativas podem ter um
importante papel no combate à obesidade, dentre estes, destacam-se as
proteínas do soro do leite, que podem agir como agente antimicrobiano, anti-
hipertensivo e regulador da função imune, reduzindo gordura corporal, além de
atuar como supressor do apetite e estimulante da síntese proteica muscular
(MORENO et al., 2014).
O leite bovino possui cerca de 3,25% de proteínas, constituídas por
duas principais classes: caseína e proteínas do soro do leite (whey protein),
nas proporções de 80 e 20%, respectivamente. Essa relação caseína:proteínas
do soro é bastante variável entre as espécies de mamíferos. Por exemplo, no
leite humano a proporção de proteínas do soro se inverte em comparação ao
leite bovino, tendo 4 vezes mais caseina (20%) que proteinas do soro
(80%) (PAL, ELLIS, DHALIWAL, 2010; SGARBIERI, 2004). As proteínas do
soro do leite são proteínas globulares, estrututalmente estabilizadas por pontes
de dissulfeto, constituídas pela frações principais beta-lactoglobulina, alfa-
lactoalbumina, albumina do soro bovino (BSA), imunoglobulinas e
glicomacropeptideos (GMPs) (HARAGUCHI; ABREU; PAULA, 2006).
O soro do leite é obtido durante o processo de produção do queijo e
por muito tempo foi considerado um resíduo agroindustrial de baixo valor
comercial, entretanto a descoberta de importantes propriedades biológicas e
nutricionais permitiu a inclusão do soro de leite na fabricação de novos
produtos pela indústria, e coincidentemente, eliminando o impacto ambiental
negativo causado pelo descarte dos efluentes das fábricas de queijos.
O soro do leite bovino é um produto de alto valor nutritivo, rico em
vitaminas, minerais, α-lactoalbumina e altos teores de aminoácidos
indispensáveis, como os de cadeia ramificada. Além disso, as proteínas do
soro são melhores fontes de energia e de aminoácidos quando comparadas as
proteínas do ovo, carne, soja e até mesmo se comparadas à caseína
(SGARBIERI, 2004).

30
Proteínas do soro do leite (whey protein) são solúveis e apresentam
propriedades digestivas diferentes da caseína. A ingestão de proteína do soro
do leite leva a um aumento rápido nos níveis de aminoácidos do plasma
sanguíneo e consequentemente a um aumento na síntese proteica, isso
diminui o tempo de esvaziamento gástrico. Por sua vez, as caseínas são
proteinas que levam mais tempo para entrar no duodeno devido à sua
precipitação no estômago em condições ácidas. Tais características conferem
à caseína a denominação de proteína slow e às proteínas do soro, de
proteínas fast.
O consumo da proteína do soro do leite possui a capacidade de
síntese proteica, elevar absorção de minerais, otimizar a síntese hormonal,
além de contribuir para diminuição dos níveis de glicose e lipídios sanguíneos
(SOUSA et al., 2012). Em estudo recente executado em nossos laboratórios foi
demonstrado que o consumo das proteínas do soro do leite, especialmente na
forma hidrolisada, resulta na diminuição das gorduras epididimal e
mesentérica, embora sem haver alteração da gordura na carcaça de ratos
Wistar alimentados por 4 semanas (GASPARETTO et al., 2013).
Na ausência de carboidratos, os aminoácidos são os maiores
secretagogos de insulina. O consumo das proteínas do soro do leite promove
aumento mais rápido de seus aminoácidos no plasma, quando comparado a
outras proteínas. Diferenças na absorção e digestão de proteínas podem
impactar na resposta pós-prandial dos diversos aminoácidos, isso explica o
fato das proteínas do soro do leite, que são de rápida digestão, possuírem
efeito insulinotrópico maior em relação a outras fontes proteicas. O alto
conteúdo de aminoácidos de cadeia ramificada também colabora para essa
maior sensibilidade à insulina (PAL; RADAVELLI-BAGATINI, 2013).
Apesar de possuírem baixo índice glicêmico, produtos lácteos
produzem alta resposta à insulina. A concentração plasmática pós-prandial de
isoleucina, leucina, valina, lisina e treonina se elevam após ingestão de whey
protein, tais aminoácidos são conhecidos por serem mediadores do efeito
insulinogênico. Percebe-se também que uma refeição acrescida de whey

31
protein gera altos níveis pós-prandiais do hormônio Polipeptídeo inibitório
gástrico (GIP), também conhecido como polipeptídeo trópico insulínico
dependente de glicose e de peptídeo-1 tipo glucagon (GLP-1), quando
comparada a ingestão de pão branco (SALEHI et al., 2012).
O consumo de dieta hiperproteica demonstrou maior eficácia na
redução e controle do peso, através de maior secreção de hormônios
intestinais, efeito termogênico e aumento da saciedade. O aumento da síntese
proteica tem sido proposto como um dos possíveis mecanismos responsáveis
pelo aumento da termogênese (JAKUBOWICZ; FROY, 2013).
A síntese proteica após consumo de whey protein é duas vezes
maior que após o consumo de caseína e isso se deve ao alto conteúdo do
aminoácido leucina presente no soro lácteo, com níveis 50 a 75% maiores que
em outros tipos de proteínas. Substâncias como insulina, fatores de
crescimento e aminoácidos, em especial a leucina, um aminoácido de cadeia
ramificada, ativam a proteína mTOR (mammalian target of rapamycin), que por
sua vez, estimula a síntese proteica através de três proteínas regulatórias: a
proteína quinase ribossomal S6, de 70 kDa (p70S6k), a proteína 1 ligante do
fator de iniciação eucariótico 4E (4E-BP1) e o fator de iniciação eucariótico 4G
(eIF4G) (JAKUBOWICZ; FROY, 2013; LYNCH, 2001; ROGERO; TIRAPEGUI,
2008).
Os produtos de whey protein são frações do soro do leite ricas em
proteínas designados para funções específicas, com propriedades nutricionais e
nutracêuticas. Tais produtos podem possuir de 2,5 a 90% de proteína, além de
componentes não protéicos, como carboidratos, lipídios, sais minerais e outras
substâncias provenientes da fermentação bacteriana natural do leite
(FOEGEDING; LUCK, 2011).
Os produtos de whey protein são valorizados como excelentes
ingredientes alimentares por suas características funcionais únicas. Dentre os
produtos os que mais se destacam por suas alegações nutricionais são:
Concentrado do Whey protein (WPC) - Com teores de proteína variando entre
35 e 80%. Os passos básicos na produção destes produtos são o pré-

32
tratamento de soro de leite, a ultrafiltração/diafiltração, a concentração por
evaporação sob pressão reduzida e secagem por pulverização; Hidrolisado do
Whey protein (WPH) - A hidrólise enzimática do WPC com uma gama de
enzimas proteolíticas é usada para produzir WPHs com graus diferentes de
hidrólise, com teores de proteína na faixa de 75 a 95%, estes peptídeos podem
ser absorvidos de forma mais eficiente e rápida que aminoácidos livres ou
proteínas intactas, este tipo de produto possui maior valor nutricional e menor
potencial alergênico; Isolado do Whey protein (WPI) – Possui teor protéico
maior ou igual a 90% e 4-6% de água, os demais ingredientes (4-6%) são uma
combinação de gordura, lactose e minerais. WPIs são largamente usados em
suplementos nutricionais e bebidas enriquecidas devido ao elevado grau de
pureza de suas proteínas. O método geralmente utilizado na produção dos
WPIs é a cromatografia de troca iônica (ABD EL-SALAM et al., 2009).
As proteínas do soro do leite possuem efeito considerável sobre o
metabolismo de lipídios e na síntese de ácidos graxos no músculo, além de
diferentemente da caseína proporcionar uma importante diminuição na síntese
de ácidos graxos no fígado. Segundo Morato et al. (2013) as proteínas do soro
do leite possuem papel anti-hiperglicêmico, sendo capazes de quando inseridas
na dieta, aumentar as concentrações séricas de glicogênio e a translocação dos
transportadores de glicose, do tipo GLUT-4, para a membrana plasmática de
célula musculares de ratos sedentários ou exercitados, sendo que melhores
resultados foram visualizados neste último grupo. Considerando tais
propriedades das proteínas do soro do leite, encoraja-se seu uso na prevenção
ou tratamento da resistência a insulina e/ou diabetes tipo 2, patologias em que
o processo de translocação do GLUT-4 do citosol para a membrana plasmática
está prejudicado.
Moura et al. (2013) demonstraram que o consumo de proteínas do
soro do leite aumenta a expressão de proteínas de choque térmico 70 (HSP70),
capazes de modular o sistema enzimático antioxidante. Neste contexto, as
HSPs formam um sistema de defesa endógeno natural capaz de proteger
contra os danos celulares durante alterações na homeostase, meio com alta

33
concentração de espécies reativas de oxigênio (EROS), isquemia e hipóxia, que
são tipos de estresse fisiológico já bem documentados na obesidade. A partir
disso demonstram-se mais uma vez que o consumo de whey protein pode atuar
como coadjuvante no tratamento da obesidade.
A ingestão das proteínas do soro do leite possui importantes efeitos
no anabolismo muscular devido ao aumento das concentrações plasmáticas de
aminoácidos favorecer captação de insulina para o interior da célula muscular, o
que otimiza a síntese proteica. A redução de gordura corporal pode ser
alcançada pela ingestão do whey protein por meio de mecanismos relacionados
às altas concentrações de BCAAs e cálcio. O cálcio presente no soro do leite
reduz níveis de hormônios calcitrópicos, que quando estão em altas
concentrações incitam o influxo do cálcio para os adipócitos, estimulando
lipogênese (síntese de novo) e inibindo lipólise. A degradação dos BCAAs
aumenta concentrações plasmáticas dos aminoácidos alanina e glutamina,
participantes da gliconeogênese. O binômio alanina-glutamina contribui em até
70% com a glicose endógena em jejum noturno, tendo a função de estabilizar
glicemia durante jejuns prolongados e reduzir picos insulinêmicos pós-prandiais.
Em suma, as proteínas do soro do leite por serem ricas em BCAAs, reduzem
liberação de insulina pós-prandial, auxiliam o fígado no controle glicêmico, via
gliconeogênese hepática, além de favorecer manutenção de massa muscular
durante a perda de massa gorda (HARAGUCHI; ABREU; PAULA, 2006).
A composição de macronutrientes da dieta é uma variável importante
no balanço energético e alterações no tipo de proteína dietética têm sido pouco
investigadas no contexto da obesidade. Tramberg et al (2013) e McAllan et al
(2014) estudaram a função das proteinas do soro do leite na regulação
metabólica da obesidade envolvendo a microbiota intestinal. O primeiro estudo,
mais próximo aos objetivos deste trabalho, visou prioritariamente provar que
alterações metabólicas na obesidade estavam relacionadas à composição da
microbiota intestinal e que as proteinas do soro do leite poderiam de alguma
forma trazer benefícios, o segundo estudo buscou comprovar se o tipo de
proteína (caseína ou whey protein isolado) e a relação proteína/carboidrato

34
traria impacto ao balanço energético, parâmetros metabólicos e a microbiota
intestinal. Frente ao potencial das proteínas do soro do leite novos estudos são
necessários para esclarecer os mecanismos por trás de seus efeitos
metabólicos.

35
Referências
ABD EL-SALAM, M. H.; SAFINAZ EL-SHIBINY; AIDA SALEM.Factors affecting
the functional properties of whey protein products: A review. Food Reviews
International, v. 25, n.3, p.251-270, 2009.
AMAYA-FARFAN, J. The role of carbonyl compounds and active diols on
steatogenesis. The Lancet. v.353,n.9154,p.747-48,1999.
AKIRA, S.; TAKEDA, K. Functions of toll-like receptors: lessons from KO mice.
C R Biol. v.327, n.6, p.581-9, 2004.
BÄCKHED, F.; DING, H.; WANG, T.; HOOPER, L. V.; KOH, G. Y.; NAGY, A.;
SEMENKOVICH, C. F.; GORDON, J. I. The gut microbiota as an environmental
factor that regulates fat storage.Proc Natl Acad Sci USA; v.101, n. 44, p. 15718–
23, 2004.
BAHIA, L.; COUTINHO, E. S. F.; BARUFALDI, L. A.; ABREU, G. A.; MALHÃO,
T. A.; SOUZA, C. P. R.; ARAUJO, D. V. The costs of overweight and obesity-
related diseases in the Brazilian public health system: cross-sectional study.
BMC Public Health, v.12, n.440, 2012.
CANI, P. D.; AMAR, J.; IGLESIAS, M. A. et al. Metabolic endotoxemia initiates
obesity and insulin resistance.Diabetesv.56, n. 7, p.1761–1772, 2007.
CANI, P. D.; DELZENNE, N. M.The role of the Gut Microbiota in Energy
Metabolism and Metabolic Disease. Current Pharmaceutical Design, v.15, n.13,
p.1546-58, 2009.
CANI, P. D.; DELZENNE, N. M. Involvement of the gut microbiota in the
development of low grade inflammation associated with obesity: focus on this
neglected partner. Acta Gastroenterol Belg. v.73 n.2, p.267-9, 2010.

36
CARICILLI, A. M.; PICARDI, P.K.; ABREU, L. L. Gut microbiota is a key
modulator of insulin resistance in TLR 2 knockout mice. Plos Biology, volume
v.9,n.12, e1001212, 2011.
CHARLES, B. A.; DOUMATEY, A.; HUANG, H.; ZHOU, J.; CHEN,
G.; SHRINER, D.; ADEYEMO, A.; ROTIMI, C. N. The roles of IL-6, IL-10, and
IL-1RA in obesity and insulin resistance in African-Americans.J Clin Endocrinol
Metab.v.96,n.12, p. 2018-22, 2011.
CHAUD, S. G.; SGARBIERI, V. C.; VICENTE, E.; DA SILVA, N.; ALVES, A. B.;
DE MATTOS, J. A. R. Influência de frações da parede celular de levedura
(Saccharomyces cerevisiae) sobre os índices séricos de glicose e lipídios,
microbiota intestinal e produção de ácidos graxos voláteis (AGV) de cadeias
curtas de ratos em crescimento. Ciênc. Tecnol. Aliment., v..27, n.2, p. 338-348,
2007.
CINTRA, D. E.; ROPELLE, E. R.; MORAES, J. C.; PAULI, J. R.; MORARI,
J.; SOUZA C. T.; GRIMALDI, R.; STAHL, M.; CARVALHEIRA, J. B.; SAAD, M.
J.; VELLOSO L. A.Unsaturated fatty acids revert diet-induced hypothalamic
inflammation in obesity. PLoS One, v.7 n. 1, e30571, 2012.
CINTRA, D. E.; ROPELLE, E. R.; MORAES, J. C.; PAULI, J. R. Obesidade e
Diabetes – Fisiopatologia e sinalização celular. São Paulo: Editora Sarvier,
2011, 424p.
CORMACK, D. H. Fundamentos de Histologia. 2 ed. Rio de Janeiro: Ed.
Guanabara Koogan, p. 371, 2003.
COX, L. M.; CHO, I.; YOUNG, S. A.; ANDERSON W. H.; WATERS B. J.;
HUNG, S. C. et al. The nonfermentable dietary fiber hydroxypropyl

37
methylcellulose modulates intestinal microbiota. FASEB J. v. 27, n. 2, p. 692-
702, 2013.
CURTI, M.L.;JACOB, P.; BORGES, M. C.; ROGERO, M. M.; FERREIRA, S. R.
Studies of gene variants related to inflammation, oxidative stress, dyslipidemia,
and obesity: implications for a nutrigenetic approach.Journal of Obesity, doi:
10.1155/2011/497401, 2011.
DEPARTMENT OF HEALTH AND HUMAN SERVICES. The surgeon general’s
call to action to prevent and decrease overweight and obesity. [Rockville, MD]:
Department of Health and Human Services, Public Health Service, Office of the
Surgeon General, 2001. Disponível em
<http://www.ncbi.nlm.nih.gov/books/NBK44206/>. Acesso em 30 jun. 2014.
DELZENNE, N. M.; CANI, P. D. Interaction between obesity and the gut
microbiota: relevance in nutrition. Ann. Rev. Nutr.; v.31,n.15 p.15-31, 2011.
DUSEJA, A.; CHAWLA, Y. K.Obesity and NAFLD – The Role of Bacteria and
Microbiota.Clin Liver Dis., v.18, n. 1, p. 59-71, 2014.
EIPEL, C.; KERSTIN, A.; VOLLMAR, B. Regulation of hepatic blood flow: The
hepatic arterial buffer response revisited. World J. Gastroenterol., v. 16 n. 48, p.
6046-6057, 2010.
EGGER, G.; DIXON, J. Non-nutrient causes of low-grade, systemic
inflammation: support for a ‘canary in the mineshaft’ view of obesity in chronic
disease. Obesity Reviews., v.12, n.5, p.339-345, 2011.

38
ESSER, N.; LEGRAND-POELS, S.; PIETTE, J.; SCHEEN, A.J.; PAQUOT,
N.Inflammation as a link between obesity, metabolic syndrome and type 2
diabetes.Diabetes Res Clin Pract. v.105 n.2, p.141-50, 2014.
FAVA, F.; LOVEGROVE, J. A.; GITAU, R.; JACKSON, K. G.; TUOHY, K. M.
The gut microbiota and lipid metabolism: implications for human health and
coronary heart disease. Curr Med Chem. v.13,n,25, p.3005-21, 2006.
FEROLLA, S. M.; ARMILIATO, G. N.; COUTO, C. A.; FERRARI,
T.C.The Role of Intestinal Bacteria Overgrowth in Obesity-
Related Nonalcoholic Fatty Liver Disease.Nutrients. v.6, n.12, p.5583-5599,
2014.
FINKELSTEIN, E. A.; GRAHAM, W. C. K.; MALHOTRA, R. Lifetime Direct
Medical Costs of Childhood Obesity.Pediatrics, v.133, n.5, p.854-62, 2014.
FLINT, H. J.; SCOTT, K. P.; LOUIS, P.; DUNCAN, S. H.The role of the gut
microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol.v.9, n.10, p.
577–589, 2012.
FLORES, L. S.; GAYA, A. R.; PETERSEN, R. D.S.; GAYA, A. Trends of
underweight, overweight, and obesity in Brazilian children and adolescent. J.
Pediatr. v. 89, n.5, p. 456-461, 2013.
FOEGEDING, E. A. ; LUCK, P.; VARDHANABNUTI, B. Whey protein
products.Encyclopedia of Dairy Sciences (Second Edition), pages 873-878,
2011.
GANGARAPU, V.; YILDZ,K.; ÍNCE, A. T.; BAYSAL, B. Role of gut microbiota:
Obesity and NAFLD. Turk J. Gastroenterol; v. 25, n. 2, p.133-140, 2014.

39
GASPARETTO, D.; LOLLO, P.C.B.; MOURA, C. S.; PAULA, F. M.M.;
CARNEIRO, E. M.; AMAYA-FARFAN, J. Metabolic response of sedentary
normotrophic rats to dietary prehydrolyzed whey proteins. Food and Public
Health, v.3,n.1, p. 21-27, 2013.
GREINER, T.; BÄCKHED, F. Effects of the gut microbiota on obesity and
glucose homeostasis.Trends Endocrinol Metab.v.22, n.4, p.117-23, 2011.
HARAGUCHI, F. K.; ABREU, W. C.; PAULA, H. Whey protein: composition,
nutritional properties,applications in sports and benefits for human health. Rev.
Nutr., Campinas, v.19, n.4, p.479-488, 2006.
HOTAMISLIGIL, G. S.; SHARGILL, N. S., SPIEGELMAN, B. M.Adipose
expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin
resistance. Science, v.259, n.5091, p.87-91, 1993.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE). Pesquisa
de Orçamentos Familiares 2008-2009. Antropometria e estado nutricional de
crianças, adolescentes e adultos no Brasil, Rio de Janeiro, 2010. Disponível
em<
http://www.ibge.gov.br/home/estatistica/populacao/condicaodevida/pof/2008_20
09_encaa/pof_20082009_encaa.pdf>: Acesso em 20 abr. 2014.
JAKUBOWICZ, D.; FROY, O. Biochemical and metabolic mechanisms by wich
dietary whey protein may combat obesity and type 2 diabetes. Journal of
Nutritional Biochemistry v.24, n.1, p.1-5, 2013.
KAMADA, N.; CHEN, G. Y.; INOHARA, N.; NUNEZ, G. Control of pathogens
and pathobionts by the gut microbiota. Nature Immunology v.14,n.7,p.685–690,
2013.

40
KAWASAKI, N.; ASADA, R., SAITO, A.; KANEMOTO, S.; IMAIZUMI, K.
Obesity-induced endoplasmic reticulum stress causes chronic inflammation in
adipose tissue. Scientific Reports, v. 2, n. 799, 2012.
KIM, C. S.; PARK, H. S.;KAWADA, T.; KIM, J. H.;LIM, D.; HUBBARD,N.E.;
KWON,B.S;ERICKSON,K.L.; YU, R. Circulating levels of MCP-1 and IL-8 are
elevated in human obese subjects and associated with obesity-related
parameters. Int J Obes., v. 30, p.1347–1355, 2006.
KMIEĆ, Z. Cooperation of liver cells in health and disease.Adv Anat Embryol
Cell Biol.; v. 161, cap. III-XIII, p. 1-151, 2001.
LEE,H.; LEE, I. S.; CHOUE, R. Obesity, Inflammation and Diet.Pediatr.
Gastroenterol Hepatol Nutr., v.16, n. 3, p.143-152, 2013.
LE ROY, T.; LLOPIS, M.; LEPAGE, P.; BRUNEAU, A.; RABOT,
S.; BEVILACQUA, C.; MARTIN, P.; PHILIPPE, C.; WALKER, F.; BADO,
A.; PERLEMUTER, G.; CASSARD-DOULCIER, A. M.; GÉRARD P. Intestinal
microbiota determines development of non-alcoholic fatty liver disease in mice.
Gut. v.62, n.12, p.1787-94, 2013.
LEY, R.; HARRIS, J. K., WILCOX, J.; SPEAR, J.R.; MILLER, S.R, et al.
Unexpected diversity and complexity of the Guerrero Negro hypersaline
microbial mat.Applied and Environmental Microbiology,v.72, n.5, p.3685-95,
2006.
LYNCH, C. J. Role of leucine in the regulation of mTOR by amino acids:
revelations from structure–activity studies. J. Nutr. v.131, n.3, p. 861S–865S,
2001.

41
MACHADO, M.; PINTO, H. C. Gut microbiota and nonalcoholic fatty liver
disease.Annals of hepatology.v.11, n. 4, p.440-49, 2012.
MAKKI, K.; FROGUEL, P. ;WOLOWCZUK, I. Adipose Tissue in Obesity-Related
Inflammation and Insulin Resistance: Cells, Cytokines, and Chemokines. ISRN
Inflammation., 2013.
MCALLAN, L.; SKUSE, P.; COTTER, P. D.; O'CONNOR, P.; CRYAN, J.
F.; ROSS, R. P.; FITZGERALD, G.; ROCHE, H. M.; NILAWEERA, K. N.
Protein quality and the protein to carbohydrate ratio within a high fat diet
influences energy balance and the gut microbiota in C57BL/6J mice. Plos one,
v.9,n.2, e88904, 2014.
MAUER, J.; CHAURASIA, B.; GOLDAU, J.; VOGT, M. C.; RUUD, J.; NGUYEN,
K. D.; THEURICH, S.; HAUSEN, A. C. Signaling by IL-6 promotes alternative
activation of macrophages to limit endotoxemia and obesity-associated
resistance to insulin. NatImmunol. v.15, n.5 p.423-30, 2014.
MEDZHITOV, R. Origin and physiological roles of inflammation.Nature, v. 454 n.
7203, p. 428-35, 2008.
MILANSKI, M.; DEGASPERI, G.; COOPE, A.; MORARI, J.; DENIS, R.; CINTRA,
D. E.; TSUKUMO, D. M.; ANHE, G.; AMARAL, M. E.; TAKAHASHI, H. K.; CURI,
R.; OLIVEIRA, H. C.; CARVALHEIRA, J. B.;BORDIN, S.; SAAD, M.
J.; VELLOSO, L. A. Saturated fatty acids produce an inflammatory response
predominantly through the activation of TLR4 signaling in hypothalamus:
implications for the pathogenesis of obesity.J Neurosci., v.29, n.2, p.359-70,
2009.
MILLION, M.; LAGIER, J. C.; YAHAV, D.; PAUL, M. Gut Bacterial microbiota
and obesity.Clin Microbiol Infect.v.19, n.4, p.305-13, 2013.

42
MORAES, J. C.; COOPE, A; MORARI, J., CINTRA, D. E. ROMAN, E. A.;
PAULI, J. R.; ROMANATTO, T.; CARVALHEIRA, J. B.; OLIVEIRA A. L.; SAAD,
M. J.; VELLOSO, L. A. High-Fat Diet Induces Apoptosis of Hypothalamic
Neurons. PLoS ONE, v. 4, n.4 , 2009.
MOURA, C. S.; LOLLO, P. C. B.; MORATO, P. N.; CARNEIRO, E. M.; AMAYA-
FARFAN, J. Whey protein hydrolysate enhances the exercise-induced heat
shock protein (HSP70) response in rats. Food Chem v.136, n. 3-4, p.1350–
1357, 2013.
MORATO, P. N.; LOLLO, P. C. B; MOURA, C. S.; BATISTA, T. M.; CAMARGO,
R. L.; CARNEIRO, E. M.; AMAYA-FARFAN, J. Whey Protein Hydrolysate
Increases Translocation of GLUT-4 to the Plasma Membrane Independent of
Insulin in Wistar Rats. Plos one, v.8, n. 8, 2013.
MORENO, M . F.; SOUZA, G. I. M. H.; HACHUL, A. C. L.; SANTOS, B.;
OKUDA, M. H.; PINTO NETO, N. I.; BOLDARINE, V. T. et al. Coacervate whey
protein improves inflammatorymilieu in mice fed with high-fat diet. Nutrition &
Metabolism, v.11, n.15, 2014.
MUJICO, J. R.; BACCAN, G. C.; GHEORGHE, A., DÍAZ, L. E.; MARCOS, A.
Changes in gut microbiota due to supplemented fatty acids in diet-induced
obese mice. Br J Nutr., v. 110, n. 4, p. 711-20, 2013.
PAL, S.; ELLIS, V.; DHALIWAL, S. Effects of whey protein isolate on body
composition, lipids, insulin and glucose in overweight and obese individuals. Br J
Nutr, v.104,n.5, p.716–723, 2010.

43
PAL, S.; RADAVELLI-BAGATINI, S. The effects of whey
proteinoncardiometabolic risck factors.Obesity reviews, v.14,n.4, p.324-343,
2013.
PARK, J. S.; SEO, J. H.; YOUN, H. S. Gut Microbiota and Clinical Disease:
Obesity and Nonalcoholic Fatty Liver Disease. Pediatr Gastroenterol Hepatol
Nutr., v. 16, n. 1, p. 22-27, 2013.
PAULI, J. R.; CINTRA, D. E.; SOUZA, C. T.; ROPELLE, E. R. Novos
mecanismos pelos quais o exercício físico melhora a resistência à insulina no
músculo esquelético. Arq Bras Endocrinol Metab., v. 53, n.4, p. 399-408, 2009.
PEREIRA, K. D. Amido resistente, a última geração no controle de energia e
digestão saudável. Revista Cienc. Tecnol. ALiment. v. 27, n.1, p.88-
92.Campinas, 2007.
PINHEIRO, A. R. O. ; FREITAS, S. F. T.; CORSO, A. C. T. Uma abordagem
epidemiológica da obesidade. Rev. Nutr., vol.17, n.4, pp. 523-533,2004.
PISTELLI, G. C.; COSTA, C. E. M. Bactérias intestinais e obesidade. Revista
Saúde e Pesquisa. São Paulo. v. 3. n. 1. p. 115-116. 2010.
PI-SUNYER, F. X. Health implications of obesity. Am. J. Clin. Nutr., v.53, supl.
6, p.1595-1603, jun. 1991.
RIDAURA, V. K.; FAITH, J. J.; REY, F. E. et al. Gut microbiota from twins
discordant for obesity modulate metabolism in mice. Science. v.341, n. 6150,
2013.

44
RODRIGUES, A. Microbiota intestinal e sua possível relação com a obesidade.
ABESO v.53, 2011. Disponível em
<http://www.abeso.org.br/pdf/revista53/artigo_microbiota.pdf.> Acesso em: 03
mar. 2013.
ROGERO, M. M.; TIRAPEGUI, J. Aspectos atuais sobre aminoácidos de cadeia
ramificada e exercício físico. Rev. Bras. Cienc. Farm., vol.44, n.4, pp. 563-575,
2008.
SALEHI, A.; GUNNERUD, U.; MUHAMMED, S. J.; OSTMAN, E.; HOLST, J. J.;
BJORCK, I.; RORSMAN, P. The insulinogenic effect of whey protein is partially
mediated by a direct effect of aminoacids and GIP on β-cells. Nutrition &
Metabolism, v.9, n. 48, 2012.
SCHINONI, M. I. Fisiologia Hepática. Gaz. Med. Bahia, v.76, supl. 1, p .5-9,
2006.
SCOTT, K. P.; GRATZ, S. W.; SHERIDAN, P. O.; FLINT, H. J.; DUNCAN, S.
H.The influence of diet on the gut microbiota. Pharmacological Research, v.
v.69,n.1, p. 52– 60, 2013.
SGARBIERI, V. C. Propriedades fisiológicas funcionais das proteínas do soro
de leite. Revista de Nutrição, campinas, v. 17, n.4, p. 397-409, 2004.
SEKIROV, I.; RUSSEL, S. L.; ANTUNES, L. C.; FINLAY, B. B. Gut microbiota in
health and disease. Physiol Rev., v. 90; n. 3, p. 859-904, 2010.
SNEDEKER, S. M, HAY, A. G. Do Interactions Between Gut Ecology and
Environmental Chemicals Contribute to Obesity and Diabetes?.Environ Health
Perspect. v.120, n.3, p.332–339, 2012.

45
SOUSA, G. T. D.; LIRA, F. S.; ROSA, J. C.; OLIVEIRA, E. P.; OYAMA, L. M. et
al. Dietary whey protein lessens several risk factors for metabolic diseases: a
review. Lipids in Health and Disease, v.11, n.67, 2012.
TANNOCK, G. W. Analysis of the intestinal microbiota using molecular methods.
European Journal of Clinical Nutrition, v.56, Suppl. 4, p.S44–S49, 2002.
THALER, J. P.; STEPHAN, J. G.; DORFMAN, M.; WISSE, B. E.; SCHWARTZ,
M. W. Hypothalamic inflammation: Marker or mechanism of obesity
pathogenesis? Diabetes v. 62, n. 8, p. 2629-34, 2013.
THE HUMAN MICROBIOME PROJECT CONSORTIUM. Structure,function and
diversity of the healthy human microbiome.Nature., v. 486, n.7402, p.207–214,
2012.
TORTORA, G. J.; FUNKE, B. R.; CASE, C. L. Procariotos: Domínio Bactéria e
Archaea. In: MICROBIOLOGIA. 8. ed. São Paulo, SP: Artmed. p. 305-333.
2006.
TRABULSI, L. R.; SAMPAIO, M. C. Microbiota ou Microbiota Normal do Corpo
Humano. In: TRABULSI, L. R.; ALTERTHUM. Microbiologia, 4ª Ed. São Paulo:
Atheneu. 12, p. 101-109. 2005.
TRANBERG, B.; HELLGREN, L. I.; LYKKESFELDT, J.; SEJRSEN, K.; JEAMET,
A.; RUNE, I.; ELLEKILDE, M.; NIELSEN, D. S.; HANSEN, A. K.
Whey protein reduces early life weight gain in mice fed a high-fat diet. Plos one,
v.8, n.8, e71439, 2013.
TSAO, C. H.; SHIAU, M. Y.; CHUANG, P. H.; CHANG, Y. H.; HWANG, J.
Interleukin-4 regulates lipid metabolism by inhibiting adipogenesis and
promoting lipolysis. Journal of Lipid Research,v.55, n.3, p. 385-397, 2014

46
TSUKUMO, D. M.; CARVALHO, B. M.; MARCO A.; SAAD, M. J. A. Translational
research into gut microbiota: new horizons in obesity treatment. Arq Bras
Endocrinol Metab. v.53, n.2, p.139-44, 2009.
TURNBAUGH, P. J, LEY, R. E., MAHOWALD, M.A.; MAGRINI, V.; MARDIS
E.R. et al. An obesity-associated gut microbiome with increased capacity
forenergy harvest. Nature v.444, n.7122, p.1027–1031, 2006
VELLOSO, L. Obesidade: Uma doença inflamatória. Revista Ciência & Saúde,
Porto Alegre, v. 2, n. 2, p. 85-95, 2009.
VIGITEL - Vigilância de fatores de risco e proteção para doenças crônicas por
inquérito telefônico (2013).Disponível em
http://www.sbpt.org.br/downloads/arquivos/vigitel_2012.pdf Acesso em: 22 maio
2014.
WANDERLEY, E. N.; FERREIRA, V. A. Obesidade: uma perspectiva plural.
Ciênc. saúde coletiva. v.15, n.1, p. 185-194, 2010.
WORLD HEALTH ORGANIZATION.World Gastroenterology Organization
Global Guideline – Obesity, 2011. Disponível em
<http://www.worldgastroenterology.org/assets/export/userfiles/Obesity-
Master%20Document%20for%20Website.pdf> Acesso em 03 mai. 2014.
WORLD HEALTH ORGANIZATION (2014).Obesity and overweight.Disponível
em: <http://www.who.int/mediacentre/factsheets/fs311/en/> Acesso em: 03 mai
2014.

47
CAPÍTULO 2
PROTEINAS DO SORO DO LEITE MITIGAM A DEPOSIÇÃO EXCESSIVA DE
GORDURA EPIDIDIMAL, SEM ALTERAR A COMPOSIÇÃO CORPORAL DE
CAMUNDONGOS C57BL/6 ALIMENTADOS COM DIETA HIPERLIPÍDICA
Artigo submetido à Revista de Nutrição
Naice Eleidiane Santana Monteiro, Aline Rissetti Roquetto, Jaime Amaya-
Farfan
Faculdade de Engenharia de Alimentos, Universidade de Campinas
(UNICAMP), Caixa Postal 6121, 13083-862, Campinas-SP, Brasil.

48
Resumo
As proteínas do soro do leite possuem destacada posição entre atletas e
praticantes de atividade física devido a suas propriedades fisiológicas e conteúdo
de aminoácidos. Quando incorporadas à dieta, as proteínas do soro do leite
promovem síntese proteica muscular, auxiliam no controle do peso, melhoram
função imunológica do indivíduo, além de regular o metabolismo de glicose e
possuir capacidade antioxidante. Trinta e quatro camundongos C57BL/6 de 21
dias de vida foram divididos de forma aleatória em 4 grupos: Grupo controle
normolipídico (AIN 93-G), Grupo controle hiperlipídico (HFCAS), Grupo
hiperlipídico (38% de gordura suína) com proteína concentrada do soro do leite
(HFWPC), Grupo hiperlipídico com proteína hidrolisada do soro do leite (HFWPH),
por período de 9 semanas. Foram avaliados peso corporal, consumo alimentar,
energético e desperdício durante o protocolo experimental e, no sacrifício,
coletaram-se dados de gordura epididimal, comprimento naso-anal, além das
carcaças para avaliação da composição corporal. No curto período de 9 semanas,
o consumo de dieta hiperlipídica, independente do tipo de proteína, foi menor do
que para a dieta normolipídica, embora sem haver mudança no ganho ponderal
dos camundongos. Entretanto, com relação aos demais parâmetros, a intervenção
foi suficiente para sugerir que as proteínas do soro tendem a preservar a massa
magra do camundongo, melhorando o índice de Lee e controlando a deposição de
gordura epididimal, mesmo antes de se registrar qualquer alteração da massa
gorda global. Nessa ação, as diferenças entre as proteínas concentradas e
hidrolisadas foram mínimas.
Palavras chave: Proteínas do soro do leite, camundongos, ganho de peso,
composição corporal, obesidade

49
Milk-whey proteins control epididymal fat deposition, but without altering
global body composition of C57BL/6 mice consuming a high-fat diet
Abstract:
Owing to their nutritive and physiological properties, the milk whey proteins are
prime ingredients for sports supplements for athletes and physically active. When
incorporated to the diet whey proteins promote muscle protein synthesis, help
control body weight, improve the immune function and glucose metabolism of the
individual, besides exhibiting antioxidant power. Thirty-four 21 day-old male
C57BL/6 mice were randomly divided into 4 groups: Normolipidic control (AIN
93G), hyperlipidic control (HFCAS), hyperlipidic with whey protein concentrate
(HFWPC) instead of casein, and a hyperlipidic group with hydrolyzed whey protein
(HFWPH) instead of casein for 9 weeks. Body weight, net food and energy intakes
during the experimental period were recorded. At the time of sacrifice, epididymal
fat data, naso-anal length, in addition to the carcasses for body composition
determination. In the short period of 9 weeks, the intake of high fat diet, was lower
than for normolipidic diet, regardless of the type of protein, although there was no
change in weight gain. However, with respect to other parameters, the intervention
was suficient to suggest that whey proteins tend to preserve lean mass in the
mouse, improving the Lee index and controlling the deposition of epididymal fat,
even prior to registering any changes in overall fat mass. For this action,
differences between the concentrated and the hydrolyzed protein were minimal.
Keywords: whey protein, mice, weight gain, body composition, obesity

50
1. Introdução
A obesidade é uma doença crônica, decorrente de vários fatores
que podem agir isoladamente ou em conjunto, fatores como descontrole na
ingestão calórica, diminuição do dispêndio energético, questões emocionais e
genéticas acabam por determinara obesidade em indivíduos sedentários.
Determinados nutrientes e compostos bioativos presentes nos alimentos,
podem atuar na prevenção ou ser coadjuvante no tratamento de doenças,
principalmente as de cunho nutricional (BASTOS; ROGERO; AREAS, 2009).
O consumo de produtos lácteos tem sido associado a benefícios em
parâmetros relacionados à obesidade e síndrome metabólica. As proteínas do
leite bovino, particularmente as do soro tem demonstrado efeito positivo no
metabolismo de glicose, redução no ganho de peso e aumento da queima de
gordura durante restrição energética (TRANBERG, 2014).
Muitos estudos já demonstraram a tamanha eficiência com que o
whey protein e seus aminoácidos promovem a síntese proteica, o próprio
cálcio tem sido demonstrado como um nutriente chave no controle da
adiposidade corporal, sugerindo outra via pela qual a inclusão do soro do leite
à dieta possa otimizar a composição corporal, além de promover melhora
geral na saúde (HÁ;ZEMEL, 2003).
As proteínas do soro do leite além de modularem a lipogênese,
promovendo aumento de massa magra e diminuição da massa gorda, atuam
ainda como antioxidante, devido aos seus altos níveis de cisteína, aminoácido
precursor da glutationa, sendo um alimento cuja suplementação pode trazer
particulares benefícios para indivíduos em condições inflamatórias crônicas,
como por exemplo, a obesidade (LANDS; GREY;SMOUNTAS et al,1999).
A dieta ocidentalizada possui elevado teor de lipídios, o que pode
ocasionar vários prejuízos à saúde, tais como, obesidade, doenças
cardiovasculares e diabetes mellitus. As proteínas do soro de leite por
possuírem a capacidade de modular o apetite por meio de hormônios da
saciedade liberados após seu consumo podem atenuar parcialmente os
efeitos de uma dieta hiperlipídica em camundongos.

51
2. Objetivo
Avaliar o efeito de uma dieta hiperlipídica acrescida de proteínas
do soro do leite, nas formas concentrada e hidrolisada, parâmetros
metabólicos e antropométricos de camundongos C57BL/6, comparado a uma
dieta hiperlipídica e uma dieta normolipídica, contendo caseína como fonte
proteica.
3. Material e métodos
3.1 Ensaio biológico
Trinta e quatro camundongos da espécie C57BL/6, machos, saudáveis com 21
dias de idade, foram divididos em quatro grupos de forma aleatória, mantidos
em gaiolas individuais, com 2 dias de adaptação em dieta comercial seguido
das dietas experimentais pelo período de 9 semanas conforme o esquema a
seguir:
Os animais utilizados para o ensaio biológico foram cedidos pelo
Centro Multidisciplinar para Investigação Biológica (CEMIB) da Universidade
Estadual de Campinas, SP. O experimento foi aprovado pela Comissão de
Ética no uso de Animais (CEUA/UNICAMP) (Protocolo nº 3015-1), estando de
34 camundongos
C57BL/6
Grupo AIN 93G
Dieta Controle
Normolipídica
(n=10)
Grupo HFCAS
Dieta Hiperlipídica
de Caseína
(n=8)
Grupo HFWPC
Dieta Hiperlipídica
de Concentrado do
soro do leite (n=8)
Grupo HFWPH
Dieta Hiperlipídica
de Hidrolisado do
soro do leite (n=8)

52
acordo com os princípios éticos na experimentação animal adotados pela
Sociedade Brasileira de Ciência em Animais de Laboratório (SBCAL).
Os camundongos foram mantidos em livre acesso de água e
comida, em ciclo claro/escuro invertido de 12 horas (07h00 às 19h00
escuro/19h00 às 07h00 claro), a temperatura de 22±2ºC e umidade relativa de
55±10%.
3.2 Composição das dietas experimentais
Todas as dietas foram elaboradas de acordo com as
recomendações da American Institute of Nutrition (AIN-93G) (Reeves; Nielsen;
Fahey, 1993). Elaboraram-se quatro tipos de dietas: Grupo AIN 93G (Dieta
controle normolipídica); Grupo HFCAS (Dieta hiperlipídica com caseína como
fonte proteica); Grupo HFWPC (Dieta hiperlipídica com proteína do soro do
leite concentrada como fonte proteica); Grupo HFWPH (Dieta hiperlipídica com
proteína do soro do leite hidrolisada como fonte proteica). As dietas
hiperlipídicas foram formuladas de acordo com Cintra et al. (2012), sendo a
dieta hiperlipídica uma dieta AIN 93G modificada, com adição de
aproximadamente 38% de banha de suíno (referente ao valor calórico total)
em substituição aos carboidratos, e caseína como fonte proteica. As demais
dietas hiperlipídicas foram preparadas com: proteína concentrada do soro do
leite (WPC) ou a proteína concetrada hidrolisada (WPH) em substituição à
caseína. Todas as dietas foram elaboradas com teor protéico de 14%. As
dietas foram repostas em dias alternados aos animais, em quantidade
suficiente para garantir o consumo ad libitum.

53
Tabela 1. Formulação das dietas experimentais.
Ingredientes AIN 93G HFCAS HFWPC HFWPH
Amido 397,486 138,086 137,386 135,186
Amido dextrinizado 132 132 132 132
Caseina* 200 177,4 0 0
WPC* 0 0 178,1 0
WPH* 0 0 0 180,3
Fibra (celulose) 50 50 50 50
Sacarose 100 100 100 100
Óleo de soja 70 40 40 40
Banha 0 312 312 312
Mix vitamínico 10 10 10 10
Mix mineral 35 35 35 35
L-Cistina 3 3 3 3
Bitartarato de colina 2,5 2,5 2,5 2,5
t-butil hidroquinona 0,014 0,014 0,014 0,014
WPC, Whey protein concentrado; WPH, Whey protein hidrolisado. (*) 14% de proteína em todas as dietas
3.3 Ingestão dietética e ganho ponderal dos animais
Durante o período experimental os camundongos tiveram seu peso
corporal, consumo alimentar e energético e desperdício, avaliados em dias
alternados. As dietas eram repostas nesta mesma freqüência para minimizar
oxidação lipídica. No momento do sacrifício dados adicionais como gordura
epididimal, comprimento naso-anal e índice de Lee (peso corporal1/3
(g)/comprimento naso anal (cm)x100) também foram coletados.

54
3.4 Preparo das carcaças dos animais
Para avaliação da composição corporal as carcaças foram congeladas,
cortadas em postas, liofilizadas, trituradas em liquidificador até obtenção do
material em forma de pó homogêneo e armazenadas a -20ºC, até o momento
da determinação de umidade, cinzas, proteína e gordura.
3.4.1 Determinação da composição centesimal das carcaças
As análises de umidade e cinzas foram realizadas de acordo com
os métodos descritos pela AOAC. A análise lipídios pelo método de Bligh;
Dyer (1959) e a determinação de proteína pelo método de Dumas, que baseia-
se na conversão de nitrogênio em óxido de nitrogênio gasoso, redução a N2,
seguido da quantificação deste em cromatógrafo gasoso, através de um
detector de condutividade térmica (SIMONNE et al, 1997).
3.5 Análise Estatística
A análise dos resultados foi realizada utilizando-se o software
estatístico GraphPad Prism 5.0. Os resultados foram submetidos à análise de
variância (ANOVA) e as médias ± SEM dos tratamentos foram comparadas
pelo teste de Tukey (p<0,05).

55
4. Resultados
4.1 Parâmetros antropométricos
Figura 1. Características de consumo e evolução ponderal de camundongos
C57BL/6, grupos: AIN 93-G (controle normolipídico), HFCAS (dieta hiperlipídica
com caseína como fonte proteica), HFWPC (dieta hiperlipídica com proteína do
soro do leite concentrada), HFWPH (dieta hiperlipídica com proteína do soro do
leite hidrolisada). (A) Consumo de dieta, houve diferença estatistica significativa do
grupo AIN 93G em relação aos demais. (B) Ingestão Energética, diferença
estatistica significativa do grupo AIN 93-G, em relação aos demais. (C)
Desperdício de dieta, diferença estatistica significativa do grupo AIN 93-G em
relação aos demais grupos até a quarta semana. (D) Ganho de peso semanal, não
houve diferença estatística significativa entre os grupos. ANOVA seguido de teste
de tukey, * e # indicam diferença significativa com valor de (p<0,05).

56
Figura 2. Características antropométricas de camundongos C57BL/6, grupos:
AIN 93-G (controle normolipídico), HFCAS (dieta hiperlipídica com caseína como
fonte proteica), HFWPC (dieta hiperlipídica com proteína do soro do leite
concentrada), HFWPH (dieta hiperlipídica com proteína do soro do leite
hidrolisada). (A) Ganho total de peso, (B) Consumo dietético total, (C)
Comprimento naso-anal, (D) Indice de Lee, (E) Gordura epididimal. Letras

57
diferentes em cima das barras demonstram diferença estatisticamente
significativa com p<0,05 conforme ANOVA seguida de teste de Tukey.
4.2 Composição centesimal das carcaças
Tabela 1. Valores médios ± erro padrão da composição corporal
AIN 93G HFCAS HFWPC HFWPH
Proteína Bruta (%) 45,34±0,45a 45,57±0,15a 47,17±0,42a 45,25±1,44a
Lipídio (%) 47,10±1,16a 39,43±0,10ba 42,50±0,77a 46,02±0,93bc
Sólidos Totais (%) 96,50±0,29a 95,85±0,17a 96,35±0,17ab 97,84±0,15ba
Umidade (%) 3,50±0,29a 4,15±0,17a 3,65±0,17ab 2,16±0,15ba
Cinzas (%) 7,99±0,01a 7,63±0,31a 9,23±0,27ba 5,99±0,25bc
Valores com letras diferentes na mesma linha indicam diferença significativa (p < 0,05) ao testede
Tukey.
5. Discussão
É possível observar que em todas as semanas os grupos HFWPC e
HFWPH mantiveram-se com pesos abaixo dos grupos HFCAS e AIN 93G. O
ganho de peso total nos camundongos alimentados com dieta hiperlipídica e
proteínas do soro do leite por 9 semanas foi menor do que nos animais
alimentados com dieta hiperlipídica e caseína, embora superior ao controle
normolipídico (AIN 93G) (Figura 2 A). Ao término do protocolo experimental
observou-se que a evolução dos pesos dos animais não diferiu de forma
significativa entre os grupos em nenhum momento do protocolo experimental
(Figura 1 D).
Os animais do grupo controle (AIN 93G) apresentaram consumo
médio (em gramas e em kcal) superior aos demais grupos, isso se deve a
própria composição das dietas, uma vez que a oferta de maior quantidade de
lipídios implica em maior saciedade (BUETTNER et al., 2006).

58
Esperava-se que os animais do grupo HFCAS apresentassem o
maior ganho de peso, entretanto os grupos apresentaram ganho de peso
bastante similares, o que acabou sendo concordante com o relatado por Lin et
al. (2000), que observaram que o ganho significativo sobre o peso corporal e
coxins adiposos em camundongos C57BL/6J ocorre apenas após 19 semanas
de indução por dieta com 58,7% de lipídios. Também é importante ressaltar
que o peso dos animais em dieta hiperlipídica acrescida de caseína (HFCAS)
tenderia a aumentar com o passar do tempo, visto que mesmo nesse curto
período de 9 semanas já era possível observar níveis aumentados de gordura
epididimal no grupo HFCAS.
Esses autores mostraram que a ingestão energética do grupo
hiperlipídico é igual ao de um grupo em dieta normolipídica durante as
primeiras 3 semanas de tratamento, começando a diferir gradualmente após a
8ª semana, e atingindo diferença ponderal de 14% em relação aos controles
somente após 19 semanas. Em nosso estudo para uma melhor visualização
na diferença de peso entre os grupos o protocolo de indução de obesidade
poderia ser alterado com um período mais longo de indução e/ou com uma
dieta com teor lipídico maior do que a que foi utilizada (MORENO et al., 2014).
Tratamentos de indução superiores a 2 meses em camundongos, são por
demais irreais e estressantes, além de dificultar a pesquisa do possível efeito
protetor das fontes protéicas do soro do leite na progressão do processo de
indução.
Recentemente Moreno et al. (2014) também mostraram que o
tratamento de camundongos Swiss com dieta high fat por 9 semanas não foi
capaz de aumentar ganho de peso nestes animais. Porém, estes autores não
utilizaram fontes alternativas de proteína.
Em relação aos consumos médios de dieta, foi interessante notar
que os animais do grupo controle (AIN 93-G) ingeriram maior quantidade do
que os demais grupos, o qual pode ser explicado pela maior densidade
energética das dietas hiperlipídicas e a sua implicação em maior grau de
saciedade. Contudo, a evolução ponderal dos animais tratados com dieta

59
hiperlipídica com caseina (HFCAS) não diferiu da dos tratados com dietas
hiperlipídicas formuladas com proteína do soro do leite nas formas
concentrada (HFWPC) e hidrolisada (HFWPH) (Figura 1D).
A ingestão calórica do grupo AIN 93G foi superior à dos demais
grupos, devido à maior quantidade total de dieta consumida,
entretanto, independentemente do maior consumo energético sabe-se que foi
o tipo de gordura utilizada na composição da dieta o fator determinante nas
alterações metabólicas, já que gorduras saturadas e monoinsaturadas, como a
banha de porco utilizada neste estudo, são capazes de promover, de forma
mais pronunciada, obesidade, resistência à insulina e esteatose quando
comparadas às gorduras poli-insaturadas (BUETTNER et al, 2006).
Os dados coletados no sacrifício apresentaram diferença estatística
significativa para o peso final e gordura epididimal. Houve diferença para
comprimento naso anal e o índice de Lee, que é um preditor para obesidade e
tem forte correlação com massa gorda, podendo-se concluir através destes
parâmetros que o grupo HFCAS apresentou tendência a obesidade. Os
presentes achados indicam que no camundongo as proteínas do soro do leite
também tendem a controlar a deposição excessiva de gordura visceral e
epididimal, semelhante ao relatado por Gasparetto et al. (2013) sobre a
diminuição de gordura visceral e epididimal, que são gorduras fortemente
correlacionadas com resistência à insulina, em ratos Wistar normotróficos,
alimentados com WPH por um período de 6 semanas.
De forma geral, observou-se que, no final da nona semana, os
animais do grupo HF-caseína apresentavam já uma tendência a maior ganho
de peso e tecido adiposo, mesmo sem esta se traduzir em diferença
significativa de peso corporal, como pode ser apreciado na Figura 2D e 2E e
na Tabela 1. Os resultados apresentados na Figura 2 indicam que a
substituição da caseína pelas proteínas do soro do leite reverteu parcialmente
os efeitos da dieta rica em gordura, e os grupos HFWPC e HFWPH
equipararam-se aos animais do grupo controle (AIN 93G).

60
6. Conclusão
As proteínas do soro do leite, em ambas as formas: concentrada e
hidrolisada, diferem da proteína padrão da dieta, a caseína, no sentido de
mitigar os primeiros efeitos de uma dieta hiperlipídica. Quando as proteínas do
soro do leite são ofertadas ao camundongo C57BL/6, em substituição à
caseína durante 9 semanas, apresentam sinais benéficos na preservação de
massa magra, com menor acúmulo de gordura epididimal e índice de Lee,
porém não chegando a se traduzir em alteração ponderal global significativa.
Agradecimentos
Os autores NESM e ARR agradecem o suporte das bolsas de
estudo do CNPq e a Hilmar Ingredients pelo fonecimento gratuito do WPC e
WPH.

61
Referências
AOAC – Official Methods of Analysis of the Association of Official Analytical
Chemists. 17th ed. Gaithersburg, Maryland: Association of Official Analytical
Chemists, v.2, 2000, 1298p.
BASTOS, D. M.; ROGERO, M. M.; AREAS, J. A. G. Mecanismos de ação de
compostos bioativos dos alimentos no contexto de processos inflamatórios
relacionados à obesidade. Arq Bras Endocrinol Metab. v.53, n.5, p.646-656,
2009.
BLIGH, E. G.; DYER, W. J.A rapid method of total lipid extraction and
purification. Can J Biochem Physiol., v. 37, n. 8, p. 911-917, 1959. Disponível
em: <http://www.nrcresearchpress.com/doi/abs/10.1139/o59-099 >.
BUETTNER, R.; PARHOFER, K.G.; WOENCKHAUS, M.; WREDE, C. E.;
KUNZ-SCHUGHART, L. A.; SCHOÖLMERICH, J.; BOLLHEIMER, L. C.
Defining high-fat-diet rat models: metabolic and molecular effects of different
fat types. J. Mol Endocrinol. v. 36, n. 3, p. 485-501, 2006
CINTRA, D. E.; ROPELLE, E. R.; MORAES, J. C.; PAULI, J. R.; MORARI, J.; SOUZA,
C. T. et al. Unsaturated fatty acids revert diet-induced hypothalamic inflammation in
obesity. PLoS One.v.7,n.1. e30571, 2012
GASPARETTO, D.; LOLLO, P.C.B.; de MOURA CS, de PAULA, F.M.M.;
CARNEIRO, E.M.; AMAYA-FARFAN, J. Metabolic response of sedentary
normotrophic rats to dietary prehydrolyzed whey proteins. Food Pub Health.
v.3, n.1, p.21-27, 2013. e-ISSN2162-8440; p-ISSN2162-9412; doi:
10.5923/j.fph.20130301.03

62
HÁ, E.; ZEMEL, M. B. Functional properties of whey, whey components, and
essential amino acids: mechanisms underlying health benefits for active
people. J Nutr Biochem., v. 14, n.5, p. 251-258, 2003.
LANDS, L. C., GREY, V. L., SMOUNTAS, A. A. Effect of supplementation with
a cysteine donor on muscular performance. J Appl Physiol v.87, n.4,p.1381-85,
1999.
LIN, S.; THOMAS, T. C.; STORLIEN, L. H.; HUANG, X. F. Development of
high fat diet-induced obesity and leptin resistance in C57Bl/6J mice. Int J Obes
Relat Metab Disord. v. 24, n.5, p.639-46, 2000.
MORENO, M . F.; SOUZA, G. I. M. H.; HACHUL, A. C. L.; SANTOS, B.;
OKUDA, M. H.; PINTO NETO, N. I.; BOLDARINE, V. T. et al. Coacervate whey
protein improves inflammatory milieu in mice fed with high-fat diet. Nutrition &
Metabolism, v.11, n.15, 2014.
REEVES, P. G.; NIELSEN, F. H.; FAHEY, G. C. AIN-93 Purified Diets for Laboratory
Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee
on the Reformulation of the AIN-76A Rodent Diet. J Nutr., v.123,n.11,p.1939-51,1993.
SIMONNE, A.; SIMONNE, E.; EITENMILLER, R.; MILLS, H.; CRESMAN,
P. Could the Dumas method replace the Kjeldahl digestion for nitrogen and
crude determinations in foods?. J Sci Food Agric., v. 73, n.1, p.39-45, 1997.
TRANBERG, B.; MADSEN, A. N.; HANSEN, A. K. ;HELLGREN, L. Whey-
reduced weight gain is associated with a temporary growth reduction in young
mice fed a high-fat diet. J Nutr Biochem. v.26, n.1, p. 9-15, 2014.

63
CAPÍTULO 3
WHEY PROTEIN, BUT NOT CASEIN, AVERT CECAL DYSBIOSIS,
LIPOPOLYSACCHARIDE FORMATION AND INFLAMMATION MARKER
EXPRESSION IN MICE FED A FAT-RICH DIET
Artigo a ser submetido à Revista Nutrition
Naice Eleidiane Santana Monteiroa; Aline Rissetti Roquettoa; Fernanda de Paceb;
Carolina Soares Mouraa; Andrey dos Santosb; Aureo Tatsumi Yamadac; Jaime
Amaya Farfana
a Food & Nutrition Department, School of Food Engineering, University of
Campinas (UNICAMP), Campinas, SP, Brazil.
b Department of Clinical Medicine, Faculty of Medical Sciences, University of
Campinas (UNICAMP), Campinas, SP, Brazil.
c Department of Histology and Embriology, Institute of Biology, University of
Campinas (UNICAMP), Campinas, SP, Brazil.

64
ABSTRACT
The complexity of obesity and its comorbidities partly explains why
conventional approaches cannot by themselves control the advance of this
disease. Recent studies link obesity to subclinical inflammation and adverse
changes in the intestinal microbiota. In order to investigate the role of whey
proteins in this context, this work was designed to study their impact on the
modulation of these parameters.
We have investigated the ability of dietary milk-whey proteins, both intact and
hydrolyzed, to mitigate the subclinical inflammation linked to the onset of
obesity and to modulate the dominant phyla, genera and species of the
intestinal microbiota of mice fed a high-fat diet, and to compare the effects with
those produced by casein.
Male C57BL/6 mice were fed a control casein diet (AIN 93-G); a high-fat-
casein (HFCAS); a high-fat-whey protein concentrate (HFWPC) and a high-fat
whey-protein hydrolysate (HFWPH) for 9 weeks. Inflammatory parameters
were monitored by western blot, endotoxemic lipopolysaccharides (LPS) by
colorimetry and the intestinal microbiota analyzed by 16S rRNA of the invariant
(V1-V3) gene. In addition, liver histological analyses were determined.
The high-fat diet induced dysbiosis by inverting the predominance of the
phylum Firmicutes over Bacteroidetes, promoted inflammation, endotoxemia
and hepatic steatosis. The whey proteins, either intact or hydrolyzed, when
given as the only source of protein with the high-fat diet, resisted the
installation of dysbiosis, prevented the surge of circulating LPS and
overexpression of inflammatory biomarkers, and prevented fat infiltration in the
liver. The opposite was observed when the high-fat diet was formulated with
casein.
It is concluded that whey proteins exert metabolic effects that tend to preserve
the normal microbiota profile, control important inflammation biomarkers, while
mitigating fat deposition in hepatic tissue, in mice that consumed a high-fat diet
for nine weeks. Such responses were not seen when casein was the dietary

65
protein, and little differences between the intact and the hydrolyzed whey were
observed.
Keywords: insulin resistance, modern diet, steatosis, NAFLD, metagenome,
mucin

66
Introduction
Obesity is a clinical condition strongly associated with the risk of
insulin resistance. Its responsible for the production of lipopolysaccharides
(LPS), increased oxidative stress, overexpression of inflammatory cytokines as
it progresses on to insulin resistance and type-2 diabetes mellitus (DM2)[1].
The intestinal microbiota becomes the primary target of the amount and type of
fats delivered by the diet unleashing a series of consequences that include
inflammation and damage of the intestinal epithelium [2,3,4].
Since the inflammatory condition results from the presence of LPS
[1,3] derived mainly from the cell wall of gram-negative bacteria, the continued
ingestion of saturated fats tends to make inflammation a permanent condition.
Hepatic steatosis is also closely associated with obesity for promoting insulin
resistance and, furthermore, increasing the levels of VLDL, C-reactive protein
and coagulation factors that contribute to hyperlipidemia [5].
While the impact of dietary fats on the microbiota has been the
object of several studies, the role of proteins has been little studied, except for
the milk whey proteins. Tranberg et al. [6] compared the effect of casein to that
of a whey protein isolate in C57BL/6J mice receiving a high-fat diet and,
although there was an initial weight reduction due to the whey, no effect of the
different proteins on the microbiota was detected. Other recent findings on the
molecular mechanisms by which the complete set of milk whey proteins and
peptides contained there in can exert such specific benefits indicate that some
constituting peptides can promote translocation of GLUT4 to the plasma
membrane thus increasing glucose uptake and glycogen synthesis in Wistar
rats [7], while the hydrolyzate of the complete set of whey proteins can also
grant extra protection to sensitive body proteins mediated by HSP70 and
HSP90 [8].
Owing to the presence of bioactive peptides capable of acting in the
regulation of energy expenditure, and the several protective features attributed
to the whey proteins that help to reduce body fat, such as improving insulin
sensitivity, regulating liver steatosis and metabolic biomarkers [5], we wished

67
to investigate the milk whey proteins further in comparison to the hydrolyzed
whey protein and casein. Therefore, this study was designed to evaluate the
potential role of the bovine milk whey proteins, including the
glycomacropeptide, on the microbiota structure, expression of TLR-4, TNF-α,
IL-4, IL-6, IL-8, IL1Ra, MyD88, IL-10 and IL-1 proteins in gastrocnemius
muscle, as potentially damaging consequences of feeding a high-fat diet to
male C57BL/6 mice in comparison to casein and the peptide-bearing
hydrolysate.
Materials and Methods
Animals and experimental protocol
Animals used in the biological assay were purchased from CEMIB
(Multidisciplinary Center for Biological Research) of the University of
Campinas, SP. The experiments were approved by the Ethics Committee on
the use of Animals (CEUA/UNICAMP) (No 3015-1 Protocol), which is
consistent with the ethical principles for animal experimentation adopted by the
Brazilian Society of Laboratory Animal Science (SBCAL). Thirty-four C57BL/6
male mice were divided into 4 groups: Control (AIN 93G; n = 10), Hyperlipidic
control (HFCAS; n = 8), Group treated with the hiperlipidic diet containing only
whey protein (HFWPC; n = 8), Group treated with the hiperlipidic diet
containing only hydrolyzed whey protein (HFWPH; n = 8). The animals were
kept in individual cages with food and water "ad libitum", under an inverted 12-
hour light/dark cycle (07:00 to 19:00 dark / 19:00 to 7:00 a.m. light). The
temperature was maintained at 22 ± 2 °C and relative humidity at 55 ± 10 %.
Experimental diets
Diets were formulated according to the recommendations of the
American Institute of Nutrition (AIN 93-G) [9]. The compositions are shown in
Table 1. The experiment had two controls diets: a standard normolipidic diet

68
prepared with 14% protein, and a high-fat diet (AIN 93-G) modified by adding
lard to 38% lipids (11.4% of plant-derived lipids (soybean oil) and 88.6% of
animal fat, lard swine [10].
Table 1. Formulation of the experimental diets (g/kg)
Ingredients AIN 93G HFCAS HFWPC HFWPH
Corn starch 397.48 138.08 137.38 135.18
Dextrinized starch 132 132 132 132
Casein* 200 177.4 0 0
WPC* 0 0 178.1 0
WPH* 0 0 0 180.3
Fibre (cellulose) 50 50 50 50
Sacarose 100 100 100 100
Soybean Oil 70 40 40 40
Lard 0 312 312 312
Vitamin Mixture 10 10 10 10
Mineral Mixture 35 35 35 35
L-Cystine 3 3 3 3
Choline bitartrate 2.5 2.5 2.5 2.5
tert-
butylhydroquinone
0.014 0.014 0.014 0.014
WPC, Whey protein concentrated; WPH, Whey protein hydrolyzate.(*) 14% of proteins in all diet.

69
Table 2. Proximate composition of the diets (g/100g)
Diets
AIN 93G HFCAS HFWPC HFWPH
Proteins 13.99 ± 0.28a 14.23 ± 0.18a 14.70 ± 0.16a 14.50 ± 0.31a
Lipids 7.49 ± 0.01a 37.92 ± 0.37b 38.54 ± 0.06b 37.50 ± 0.26c
Carbohydrates 78.52±0.18a 47.85±0.28b 46.76±0.21cb 48.00±0.21b
Moisture 6.79 ± 0.14a 4.15 ± 0.34b 4.19 ± 0.25b 4.63 ± 0.17b
Ash 2.48 ± 0.01b 1.99 ± 0.01a 2.63 ± 0.16b 2.98 ± 0.00c
Energy 437.45 589.60 592.70 587.50
Tukey test at the error level of 5%. Means ± standard error followed by the same letter do not
differ statistically along the same line. * Carbohydrates calculated by difference. Energy
(kcal/100g) = (lipids x 9) + (proteins x 4) + (carbohydrates x 4).
Blood for biochemical analyses and LPS determination
After 9 weeks of the experimental protocol the animals were
anesthetized intraperitoneally using a mixture of ketamine and xylazine at a ratio
of 10 mg/kg and 150 mg/kg, respectively. After anesthesia, blood was collected
by vein puncture of the aorta. The blood samples were centrifuged at 3000 rpm
for 10 minutes at 4°C to obtain serum. Subsequently, serum glucose, albumin,
total protein, uric acid, triacylglycerols, total and HDL- cholesterol were
determined using commercial kits, following the manufacturer's instructions. Due
to the need for collection of stools animals were not fasted.
Biomarkers of the inflammatory process
Tissue extraction and protein quantification
The gastrocnemius muscles were removed and stored at -80 °C.
Extraction of tissue proteins for Western blotting was performed by
homogenization of organ (gastrocnemius) in an extraction buffer consisting of:
Trisma base, pH 7.5 (hydroxymethylaminomethane) 100 mM, EDTA (ethylene-
diaminetetraacetic acid) 10 mM, 100 mM sodium fluoride, 10 mMsodium

70
pyrophosphate, 2 mM PMSF (diluted in 1 mL ethanol), triton X-100, 1% sodium
orthovanadate 10 mMaprotinin and 0.1 mg/mL.
The samples remained in solution at 4 ºC until homogenized
(Polytron PT 1200 Kinematica®). A homogenate in the extraction buffer was
centrifuged at 11,000 rpm for 30 min at 4 ºC. Then, 400 μL of supernatant were
collected and solubilized in Laemmli buffer plus dithiothreitol (DTT; 200mM at a
ratio of 1:5). In the remaining supernatant the protein concentration was
determined by the biuret method, after determining the total extract for
application to the polyacrylamide gel. All samples solubilized in Laemmli were
stored at -80 °C, and before application to the gel, samples were boiled in
Laemmli at 100 °C for 5 minutes.
Western Blotting
The samples were applied on a polyacrylamide gel and separated
by electrophoresis (SDS-PAGE) and transferred to nitrocellulose membrane
transfer apparatus Bio-Rad®. After transfer the membranes were blocked for 2
hours with a 5% solution of skimmed milk (MOLICO®) in Tris-saline (base
solution), and then were washed with the basal solution in three sessions of 10
minutes. Then the membranes were incubated overnight with antibodies to the
proteins of interest under continuous stirring at 4 °C, and washed the next day
with the basal solution three times at 10 minute sessions. Subsequently, the
membrane was incubated with secondary antibody in blocking solution for 2
hours, washed with baseline solution for 2 hours, lastly placing a
chemiluminescent solution on the membrane. The method identified and
quantified the immunoreactive bands by densitometry, followed by image
capturing and analysis in a Bio-Rad® photodocumentator. The generated files
were analyzed using the Un-Scan-It Gel 6.1® program, and the graphs were
constructed using the Graph Pad Prism 5.0 and Corel draw 7.0. Antibodies
employed were: TNF-α (sc8301), IL-4 (sc1260), IL-6 (sc7920), IL-8 (sc7922),
TLR4 (sc30002), IL1Ra (sc25444), MyD88 (sc11356), GAPDH (sc32233)
(Santa Cruz Biotechnology, CA, USA), IL-10 (Ab9969) (AbCam Cambridge,

71
MA, USA),IL1-β (cell 8689), α-Tubulin (cell 2144) (Cell Signaling Technology,
Inc., Danvers, MA, USA).
Determination of serum concentrations of LPS
Blood samples were collected in sterile tubes and the serum was
separated by centrifugation, frozen at -80 ºC until analysis. Serum
concentrations of endotoxins were measured using the colorimetric quantitative
method LAL (LyzateAmebocyte from Limulus). Kinetic Chromogenic LAL
commercial kits were from Lonza (Lonza, Inc. Allendale, N.J., USA; cat nº 50-
647U). Briefly, the procedure involved the dilution of 20 μL at 1:5 in 80 μL
reagent water (Lonza), following inactivation at 70°C for 10 min. the diluted
samples were pipetted (25 μL) onto a 96-well sterile plate (LAL). The
commercial Kinetic Chromogenic kit LAL from Lonza (Lonza, Inc. Allendale,
N.J., USA; cat nº 50-647U).
Then, 50 μL of solution amebocyte lysate (LAL) was added to the
microplate was incubated for 10 min at 37 ± 1 °C. It is then pipetted 100 μL of
the solution substrate reaction was stopped after 6 min by adding 100 μl of
acetic acid 25%. Absorbance was read at 405 nm on a microplate reader.
The endotoxin concentrations (EU/mL) in the samples were
determined from a standard curve using standard endotoxin E. coli
concentration of 39 EU/mL. The endotoxin concentration curve was constructed
at 0.1 EU/mL, 0.25 EU/mL, 0.5 EU/mL and 1.0 EU/mL. The detection range of
this kit was linear in the range of 0.1-1.0 EU/mL.

72
Sequencing the intestinal microbiota
Total DNA was extracted from the gut with the QIAmp DNA Stool Kit.
The library construction and sequencing were performed using the same
procedure described for VLP, but without using the GenomiPhi.
For each sample, the gene for the 16S rRNA was amplified using the
direct amplifier corresponding to the following sequence: 5´-
TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGACTCCTACGGGAGGCA
GCAG -3´. In this sequence, the portion in italics corresponds to the adapter
Nextera® transposase sequences A, and the sequence in bold is the initiator
widely conserved 338F. The reverse initiator used was 5’-
GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGTTACCGCGGCTGCTG
GCAC -3’, the portion in italics corresponding to the adapter Nextera®
transposase sequences B and the sequence in bold is the initiator of ample
utilization 533R. For the preparation of the libraries, the kit “IlluminaTruseq DNA
Sample Preparation v2” was used, labeling each sample with a bar code.
Sequencing was performed on an equipment Illumina Hiseq 2000 at the
Laboratório Central de Tecnologias de Alto Desempenho em Ciências da Vida
(LaCTAD-Unicamp, Campinas, Brazil).
Phylotyping characterization and taxonomy - The identity of sequences was
calculated using the megablast technique. The taxonomy was filed seeking the
best megablast against Green Genes. To determine the best BLAST,
sequences derived from bacterial VLPs in custom databases were built with the
help of Green Genes database.
Clustering of the sample by UniFrac – A sequence representing each
phylotype was aligned using NAST in order to construct a phylogenetic tree by
means of a Fast Tree system with Kimura correction. The samples were
clustered using the UniFrac metric [11]. For phylogenetic reconstructions,
representative phylotype sequences were aligned using NAST [12].

73
Histological analysis of liver tissue
Fragments of the right hepatic lobe were collected, cut into slices
approximately 3 mm thick, fixed in buffered formalin, dehydrated in ascending
series of ethanol, diaphanized in xylene, embedded in paraffin, cut in the
microtome (5-μm thick) and stained with hematoxylin-eosin in order to
qualitatively assess hepatic steatosis using electron microscopy at 10x and 20x
objectives. A steatosis was graded according to the amount of vacuoles in
hepatocytes (fatty infiltration) along the cutting under evaluation.
Statistical analysis
The experimental results were expressed as means ± standard
errors, with all the variables initially assessed for normality of distribution by
ANOVA, followed by significance analysis using the Tukey test. In all analyses
we used the GraphPad PRISM ® software, version 5.01 (GraphPad Software
Inc., CA, USA), adopting a statistical significance of p < 0.05.
Results
Biochemical parameters
Means (± SEM) of the classical blood parameters showed that
replacing casein for the whole whey protein as source diet brought little
alteration to the general profile of the normal mice (Table 3). Total cholesterol
and uric acid were raised by the high-fat diets, but neither of the proteins were
capable of reverting this effect.

74
Table 3. Blood parameters
Parameter
AIN 93G HFCAS HFWPC HFWPH
Mean±SEM Mean±SEM Mean±SEM Mean±SEM
Glucose (mg.dL-1) 275.40±22.76 319.00±33.75 314.70±16.88 282.30±13.33
Albumin (g.dL-1) 1.67±0.06b 2.04±0.17a 1.68±0.06b 1.62±0.04b
Total protein (g.dL-1) 4.03±0.08 4.52±0.24 3.60±0.38 3.81±0.18
Uricacid (mg.dL-1) 2.49±0.37b 5.52±0.79a 4.01±0.40a 5.20±0.34a
Triglycerides (mg.dL-1) 42.18±7.06 59.74±9.73 35.81±5.67 34.95±3.13
Cholesterol(mg.dL-1) 109.60±6.46b 157.60±5.50a 158.30±5.51a 144.60±3.63a
HDL-c (mg.dL-1) 64.77±3.18b 94.09±5.59a 75.36±3.55b 77.15±3.54b
Different letters on the lines indicate statistical significant for Dunnett test (p< 0.05)
Inflammatory profile
In general, it was observed that the inflammatory cytokines were
increased in the group consuming the HF diet in a combination with casein,
while the groups consuming whey protein remained with cytokine levels close
to normal (Figure 3). These increased levels were matched by the also higher
levels of receptors TLR4 and IL1Ra.

75

76
Figure 1. Western blot analysis of TLR-4, TNF-alpha, IL-4, IL-6, IL-8, IL1Ra, MyD88, IL-10 and
IL-1 Beta proteins in gastrocnemius muscle in different groups. Data are means ± SEM,
different letters represent statistical differences.

77
Endotoxemia
Endotoxemia, measured by the concentration of lipopolysaccharide
(LPS), responded to the type of protein in the diet in a similar fashion to that of
the inflammatory cytokines and their receptors (Figure 1). Again, no difference
was noticed between the intact and the hydrolyzed whey protein, but the whey
dramatically impeded the rise in LPS.
Figure 2. Mean ± SEM values of lipopolysaccharide (EU/mL) in mouse serum,
different letters represent statistical differences. ANOVA and Tukey’s test (p<0,05).
Intestinal microbiota
As far as the cecal phyla, genera and species of microorganisms, it was
evident that the protein source brought considerable differences to the profiles.
Table 2 displays the internal percentages of classifiable organisms. Increases in
the Firmicutes phylum, coupled with decreasing Bacteroidetes, were evident as
the dietary protein was prepared with casein. The Verrucomicrobia phylum
exhibited significant increases with either casein or intact whey protein as dietary
proteins. Proteobacteria was a phylum that also decreased considerably in the HF
animals which consumed the diet with casein. For the remaining phyla, there were

78
no apparent tendencies. With respect to genera, a general observation was that
the-high fat content of the diet promoted the increase of some minor genera with
the high-fat diet, while others disappeared. Differences were observed between
the intact and the hydrolyzed whey proteins, but without any consistent tendency.
Species also responded to the type of protein in the diet in a similar fashion
highlighting the tendency to narrow the number of species
Table 4. Cecal microbiota composition as % of reads (Mean ± SEM) in mice fed a
control diet (AIN 93-G), a high-fat diet made with casein (HFCAS) and a high-fat
diet made with whey protein concentrate (HFWPC) or hydrolyzed whey protein
(HFWPH)
Phylum AIN 93G HFCAS HFWPC HFWPH
Firmicutes 12.96±1.10b 20.87±2.27
a 10.87±0.63
b 17.97±1.39
a
Bacteroidetes 15.58±0.85a 7.28±1.16
b 11.34±0.47
a 11.81±1.03
a
Verrucomicrobia 3.02±1.20b 7.92±1.85
a 11.13±0.84
a 2.06±0.98
b
Proteobacteria 12.56±0.52a 5.77±0.22
b 10.22±1.23
a 10.17±0.98
a
Thermotogae 0.52±0.09 1.91±0.60 0.92±0.56 1.14±0.29
Actinobacteria 1.52±0.21 2.11±0.18 1.56±0.13 2.46±0.19
Chloroflexi 0.25±0.14 0.08±0.08 0.21±0.07 0.13±0.13
Synergistetes 0.18±0.12 0.23±0.16 0.09±0.09 0.35±0.12
Tenericutes 0.00 0.7±0.7 0.00 0.00
Genus AIN 93G HFCAS HFWPC HFWPH
Akkermansia 2.14±1.33b 6.97±1.65
a 9.55±0.64
a 1.05±1.0
b
Blautia 0.00 3.74±0.51a 2.15±0.36
a 1.58±0.53
b

79
Butyricimonas 2.30±0.11a 2.33±0.39
a 0.00 0.51±0.51
b
Marinitoga 0.00 1.45±0.84 0.63±0.63 0.46±0.46
Oscillospira 3.44±0.94 1.74±0.64 2.27±0.36 3.56±0.69
Sedimentibacter 0.00 0.57±0.57 0.00 0.00
Clostridium 0.00 3.46±1.34 0.00 3.55±0.79
Ruminococcus 0.00 0.95±0.55 0.00 0.00
Alkaliphilus 0.00 0.62±0.36 0.00 0.00
Bacteroides 4.96±0.59a 0.70±0.70
b 3.32±0.44
a 3.54±0.41
a
Parabacteroides 1.47±0.52b 0.35±0.35
b 3.13±0.21
a 2.04±0.82
b
Helicobacter 7.85±0.62a 0.00 4.00±1.10
b 4.54±0.78
b
Desulfovibrio 0.00 0.00 0.24±0.24 0.00
Rikenella 0.00 0.00 0.29±0.29 0.00
Coraliomargarita 0.00 0.00 0.32±0.32 0.00
Natronincola 0.86±0.52 0.00 0.00 0.94±0.56
Actinocorallia 0.35±0.35 0.00 0.00 0.42±0.42
Flavobacterium 1.71±0.22 0.00 0.00 0.00
Species AIN 93G HFCAS HFWPC HFWPH
Akkermansia muciniphila 2.40±1.20b 6.90±1.64
a 9.46±0.64
a 1.38±0.98
b
Butyricimonas synergistica 1.76±0.26a 2.15±0.36
a 0.27±0.27
b 1.09±0.30
a
Marinitoga piezophila 0.00 1.75±0.60 0.60±0.60 0.71±0.42
Sedimentibacter hydroxybenzoicus 0.00 0.74±0.54 0.00 0.00
Alkaliphilus crotonatoxidans 0.00 0.77±0.28 0.00 0.00

80
Oribacterium sinus 0.00 0.51±0.17 0.00 0.00
Actinocorallia cavernae 0.59±0.35 0.64±0.26 0.18±0.18 1.35±0.16
Coraliomargarita akajimensis 0.00 0.31±0.18 0.52±0.31 0.00
Rikenella microfusus 0.60±0.35 0.11±0.11 0.29±0.29 0.32±0.19
Parabacteroides gordonii 0.00 0.00 0.65±0.38 0.00
Parabacteroides goldsteinii 0.72±0.42b 0.25±0.25
b 1.90±0.21
a 1.61±0.42
a
Ruminococcus gnavus 0.00 0.26±0.26 0.00 0.00
Blautia hydrogenotrophica 0.00 0.00 0.42±0.24 0.00
Bacteroides acidifaciens 0.29±0.29 0.00 0.36±0.21 0.00
Bacteroides dorei 0.00 0.00 0.29±0.29 0.00
Bacteroides rodentium 0.52±0.30 0.00 0.17±0.17 0.55±0.20
Helicobacter ganmani 0.56±0.33 0.00 0.19±0.19 0.36±0.21
Helicobacter rodentium 3.21±0.28a 0.00 1.44±0.61
b 1.62±0.22
b
Parapedobacter koreensis 0.22±0.22 0.00 0.25±0.25 0.00
Desulfovibrio fairfieldensis 0.00 0.00 0.15±0.15 0.00
Oscillospira eae 0.00 0.00 0.00 0.14±0.14
Anaerotruncus colihominis 0.74±0.26 0.00 0.00 0.00
Butyricimonas virosa 0.24±0.24 0.00 0.00 0.00
Johnsonella ignava 0.28±0.28 0.00 0.00 0.00

81
Figure 3. Percentage of predominant cecum bacterial phyla of the four diet groups.
Values are means ± SEM. Different letters on bars represent statistical differences for
each phylum among the four dietary groups (p<0,05).

82
Figure 4. Percentage of predominant cecum bacterial genus in the four diet groups. It
should be noticed that the presence of casein in the diet resulted in the suppression or
elimination of five genera. Values are means ± SEM. Different letters on bars represent
statistical differences for each genus among the four dietary groups (p<0,05).

83
Histopathologic profile

84
Figure 5. Representative photomicrographs of liver of mice obtained after nine weeks
of treatment showing that whey protein was conducive to less fat globules deposition
than casein: a), control group treated with AIN-93 diet, b) high-fat diet group treated with
casein (HFCAS) c) high-fat diet group treated with the whey concentrate (HFWPC);. d)
high-fat diet group treated with hydrolyzed whey (HFWPH). Slides were developed with
hematoxylin and eosin (HE) staining, 10x (I) and 20x (II).
Discussion
Subclinical inflammation, common place in obesity, is implicated in
the pathogenesis of such chronic diseases as diabetes, hypertension and
hepatic steatosis. In this study we investigated the potential benefits of dietary
whey protein in the development of obesity, the linked subclinical inflammation
and in the potential modulation of alterations in the structure of intestinal
microbiota in C57BL/6 mice fed a high-fat diet. Casein is the standard
reference protein in rat and mice diets owing to the property of promoting
maximal growth. Partially hydrolyzed whey protein was included in this
comparison because several studies have shown that released peptides can
raise the protein’s bioactivity [13,7,8].
The induction of obesity, insulin resistance and subclinical
inflammation by fatty diets has been reported in several studies. It is known
that even cytokine expression in adipose tissue already begins after the

85
second day of consuming a high-fat diet and, after long periods (6 to 20
weeks), the inflammation extends to other tissues such as muscle, liver and
hypothalamus [14]. Although treatment with whey proteins in either the intact or
hydrolyzed form is known not to produce changes in body weight, they were
effective in alleviating pro-inflammatory effects of the high-fat diet compared to
the normolipidic control diet with regard to the inflammatory markers.
According to Bleau et al [15], a wide variety of cytokines is produced by the
muscular fibers during obesity causing metabolic homeostasis damage. In our
study we investigated TNF-α, IL-4, IL-6, IL-8, IL-10, TLR4, ILR1α, MyD88 and
IL1 in the gastrocnemius muscle.
An important finding in this study was that by substituting the
standard casein of the diet for whey protein, as either WPC or WPH,
considerably decreased the inflammation induced by the high-fat diet in this
animal model, leaving biomarker levels indistinguishable from those of the
control diet (AIN 93-G). Although the HFWPH diet tended to reduce the
expression of inflammatory proteins more effectively, there was no significant
difference between WPC and WPH in their capacity to modulate inflammatory
parameters.
In the obese or pre-obese state, circulating lipopolysaccharides
and/or saturated fatty acids activate the toll-like receptors, in particular the
TLR4 subtype, thus stimulating the NFкB pathway and initiating the expression
of genes that encode pro-inflammatory proteins. Mediated by phosphorylation
and ubiquitination reactions of target proteins, the signal of the interaction
between receptor and agonist is passed on to the MyD88 protein, which
activates IRAK and TRAF-6 complexes, an essential step for uncoupling IкB,
the inhibitory protein of NFкB. Once released, the NFкB migrates to the
nucleus, binds to DNA, and initiates gene amplification proteins related to
inflammation [16].
To examine the mechanism of inhibition of cytokine expression after
treatment, TLR4 was measured in muscle, observing that those individuals
consuming whey proteins, regardless of hydrolysis, expressed lower levels of

86
TLR4, suggesting that something in whey suppressed the generation of factors
responsible for signaling the TLR-4 pathway thus avoiding the expression of
inflammatory cytokines. The anti-inflammatory effect and inhibitory whey
protein was also corroborated by the lower expression of MyD88 protein. Our
data suggest that the milk whey proteins may suppress the MyD88-dependent
TLR4 signaling in muscle, but this effect was not observed when the dietary
protein was casein.
Treatment with HFCAS diet also resulted in significant increase in
TNF-α expression, a cytokine related to insulin resistance, which stimulates
lipogenesis and promote hyperlipidemia. Mean while, TNF-α levels were
significantly decreased in groups HFWPC and HFWPH, becoming virtually
indistinguishable from normolipidic AIN 93-G.
While the whey proteins detained the development of inflammation,
cytokines IL4, IL6, IL8 and IL10 were more expressed in the animals of HFCAS
group. The joint evaluation of IL1 and IL6 can predict the risk for type-2
diabetes more reliably than when evaluated separately. A significant decrease
was found in the levels of both IL-6 and IL1 as a result of treating the sick
animals with either WPC or WPH suggesting a protective effect of the whey
proteins in the development of diabetes. IL-6 is a cytokine related to obesity
and insulin resistance, which will have its serum levels increased according to
the degree of obesity of the individual, but which has an ambiguous role,
because depending on its concentration it can be harmful or protective. Mauer
et al. [17] showed that mice genetically modified to not express receptors for
IL-6 were more likely to develop obesity with subclinical inflammation, besides
impaired glucose tolerance. However, IL-6 can also induce expression of the
IL4 receptor (IL4Ra) making cells more sensitive to IL-4, an anti-inflammatory
cytokine that induces activation of M2-macrophages responsible for the
production of the anti-inflammatory cytokines IL-10 and TGF-β.
Our results demonstrated that the IL-8 levels are in direct correlation
with IL-6 levels in individuals who received the high-fat diet with no whey
proteins (HFCAS), as was to be expected. IL-8 is strongly associated with

87
cardiovascular diseases and atherosclerosis, and correlates inversely with
HDL-cholesterol in obese [18,19]. Therefore, based on these findings, one can
infer that, in spite of the high-fat diet, the animals treated with the whey
proteins showed an attenuated inflammatory condition, almost
indistinguishable from that of the animals that consumed the normolipidic diet,
and very different from that seen in group HFCAS.
The lipopolysaccharides are essential factors for the systemic
inflammatory response. In this study, the use of whey proteins resulted in low
amounts of LPS, similar to the normolipidic group (AIN 93-G). Our findings are
consistent with reports that high-fat diets are associated with increased
intestinal permeability and increased levels of LPS [20].
Cani et al. [1] fed mice with a high-fat diet for a period of 4 weeks
and found that the diet increased the concentration of circulating LPS levels
1.4-fold causing metabolic endotoxemia. The enteral absorption mechanisms
of LPS and subsequent development of endotoxemia implicate the binding of
LPS to TLR4 receptors, event that triggers secretion of pro-inflammatory
cytokines and affects insulin action [21,22].
Lipopolysaccharides are strong inducers of hepatic steatosis, and in
our work, it was possible to directly relate the consumption of the fat-rich diet to
high serum LPS and the deposition of liver fat. In the histopathological
analyses, the AIN 93-G group had the smallest area occupied by the lipid,
while the HFCAS group showed a greater amount of lipids, indicating early
microgoticular steatosis. The HFWPC and HFWPH groups were quite similar
to each other, although treatment with HFWPC was the one that better
mitigated the effects of lipid droplet accumulation. The histological findings of
the livers of mice are in agreement with Hamad et al. [23], who also showed
that oral administration of whey protein decreases infiltration of lipids, causing
a protective effect against non-alcoholic steatosis attributed to the antioxidant
and anti-inflammatory effect of the whey proteins.
Regarding the blood parameters, glucose levels were lower in the
animals fed HFWPH, not differing from the normolipidic control (AIN 93-G).

88
Similar results were found in the literature that demonstrate the potent
insulinotropic activity of whey protein hydrolysate, Gaudel et al. [24] attribute
this to the better intestinal absorption of amino acids and peptides present in
the hydrolyzed protein, particularly of the insulinotropic leucine, arginine,
tyrosine and BCAAs.
De Moura et al. [13] worked with groups of animals treated with
nomolipidic diets containing casein and WPH and WPC as protein sources.
Their findings showed that while uric acid levels remained constant in
sedentary animals, regardless of the protein source, the levels increased in
exercised animals receiving whey protein. In our study, the increase in uric
acid was higher and more homogeneous among the high-fat diet groups. Our
results indicate that it was the high-fat diet, rather than the type of protein that
prompted extra antioxidant protection is crucial for uric acid.
With respect to total protein and albumin of groups where the
protein source was casein (AIN 93-G and HFCAS) the tendency was to favor
greater preservation of lean body mass when compared to the groups treated
with whey proteins (and HFWPC HFWPH) suggesting that when it comes to fat
diet, casein promoted greater muscle preservation.
In relation to total cholesterol, the control group (AIN 93-G) exhibited
the lowest values (109.6 mg/dL) in all groups treated only the HFWPH group
showed a decrease in cholesterol (144.6 mg/dL), but still well above the
control. Whey protein hydrolyzate potential to reduce blood cholesterol is
mainly due to high content of branched chain amino acids. HDL levels were
also lower in the groups receiving high-fat diet containing whey protein, being
closest to the figures provided by normolipidic control (64.77 mg/dL).
The triacylglycerol profiles of animals in groups HFWPC and
HFWPH were better than those of group AIN 93-G itself, which shows the
positive influence that the whey protein can have on carbohydrate metabolism.
The benefits of whey proteins for triglyceride was reported by Pal et al. [25]
showed that in adult humans that triacylglicerol levels were significantly lower
in the group receiving a protein isolate compared to the control group, which

89
received glucose solution. Fasting levels decreased 22% in whey protein group
compared to control group and these results milk have been observed in the
sixth week of treatment, remaining until the end of the study (twelfth week).
The data on the gut microbiota was carried out by sequencing the
16S RNA confirmed the changes in composition previously described in the
literature with regard to obesity in response to a high-fat diet. Nine phyla were
identified (Table 2) as predominant Firmicutes, Bacteroidetes,
Verrucomicrobia, Proteobacteria and Actinobacteria, and the less abundant
Thermotogae phyla Chloroflexi, Synergitetes and Tenericutes.
When the milk-whey protein concentrate, either intact or hydrolyzed,
(HFWPC or HFWPH) was used to prepare the high-fat diet, a significant
increase in the population of Bacteroidetes was found, while Firmicutes
decreased compared to the high-fat diet prepared with casein (HFCAS). The
HFWPC and HFWPH groups followed the same trend as the control group
normolipidic groups (AIN 93-G). These findings are supported by the literature,
with studies validated in humans and in experimental models, which propose
obesity as a crucial factor in changes in intestinal [26].
The first findings regarding the relationship microbiome-obesity were
the result of the work of Backhed et al. [26] and Ley et al. [27]. These workers
observed that the absence of intestinal microbiota in germ-free mice was a
protective factor against excessive weight gain, and that a higher proportion of
Firmicutes over Bacteroidetes was characteristic in obese mice when
compared to eutrophic mice. It can be seen that the Clostridium genus, phylum
Firmicutes, was present in groups HFCAS and HFWPH, but not in groups AIN
93-G and HFWPC (Figure 4).
Akkermansia muciniphila, a Verrucomicrobia phylum member, is a
bacterium with anti-inflammatory and Mucino-degrading properties, which
resides in the intestinal mucus layer and its presence is inversely correlated
with body weight in studies on humans and rodents. The abundance of A.
municiphila in obese and diabetic individuals is lower when compared to
normal individuals, and the presence of A. municiphila capable of preventing

90
metabolic disorders caused by high fat diet, including reversing weight gain,
metabolic endotoxemia, adipose tissue inflammation and insulin resistance.
The study of Everard et al. [28] demonstrated a significant decrease of A.
municiphila in obese mice (genetically or induced by a high-fat diet), and that
the inclusion of prebiotic (oligrofrutose) in the diet was able to restore the
population of A. municiphila improving barrier functions of the intestinal
epithelium by decreasing the levels of circulating lipopolysaccharides,
normalize metabolism of fatty acids, and promote greater glucose tolerance
and insulin sensitivity. In our study mice fed high-fat diet plus the concentrated
whey protein (HFWPC) had the highest levels of the bacteria in the cecum,
compared to HFCAS group, HFWPH and also to the normolipídic group, AIN
93-G. Three species of Bacteroides (phylum Bacteroidetes) were also
identified: B. acidifaciens, B. dorei and B. rodentium, found in groups AIN 93-
G, HFWPC and HFWPH, but not in the group receiving high fat diet formulated
with casein (HFCAS) (Table 2).
These findings are consistent with evidence reported here on the
inhibition of epididymal fat deposition in animals models [29] and the
conclusion is that the whey proteins act resisting the dysbiosis caused by the
high-fat diet or restructuring the microbiota, making it similar to that of the
normolipidic diet. Bacteroides acidifaciens, and Akkermansia, are described as
degrading important mucin in vivo, to form acetate, succinate and propionate
as end products of metabolism, altering the balance of short-chain fatty acids
(SCFAs) to acetate and propionate. The Bacteroides genus, in general, to
produce SCFAs acts as a protective factor against obesity induced by diet
because butyrate and propionate help regulate body weight through satiety
effects provided by anorexigenic gut hormones. In contrast, acetate inhibits
gain weight regardless of the suppression of food intake, showing no acute
effect on gut hormones [30,31]. Recent studies have also linked B. acidificiens
the production of immunoglobulin A (IgA) in the large intestine of rats, relating
to the existing intestinal tract inflammation in obesity. Ruminococcus gnavus
was found only in HFCAS group, finding consistent with studies that attribute to

91
this species a correlation with obesity and inflammation of the lower
gastrointestinal tract, such as diverticulitis [32].
In our findings, the Helicobacter genus was present in the
normolipídic control group, as well as in groups containing whey protein, but
was not identified in HFCAS group. Helicobacter genus is composed of several
species with gastric and intestinal habitat, despite being generally associated
with pathologies, recent studies have shown that Helicobacter genus is an
adjuvant in the balance of the gastrointestinal tract microbiota, and inversely
associated with obesity. Bassaganya-Riera et al. [33] showed that colonization
by specific strains of H. pylori decreased levels of blood glucose, increased
levels of leptin, and suppressed weight gain and improved inflammatory profile.
The incidence of obesity and diabetes coincides with the reduction of the
prevalence of H. pylori colonization, which suggests that gastric colonization
with this bacterium has its contribution portion for the balance of the metabolic
parameters.
Abundance of Parabacteroides spp. has been negatively correlated
with tight-junctions protein levels and inflammatory markers. Studies using diet
high in fat show induction of intestinal inflammation in part by the increased
expression of IL1β and, in part, by a decrease in the tight-junction proteins
occludin and ZO-1. Lee, Ham and Yim [34] showed that the addition of water-
soluble extract of soy and some fibers (psyllium, inulin and resistant starch) to
the diet exerted anti-inflammatory effect to recover intestinal microbiota
demonstrating low levels of Parabacteroides and increased levels for occludin,
ZO-1, Foxp3, adiponectin and IL-10. The whey proteins were efficient in
reducing levels of Blautia spp., which was elevated in HFCAS group,
decreased in HFWPC and HFWPH groups and not identified in the control
group AIN 93-G. Blautia spp. is positively correlated with intestinal
inflammation [34].

92
Conclusion
Detailed analysis of the cecal microbiota of C57BL/6 mice reveals that the
whey proteins differ substantially from the casein proteins in their ability to
resist the primary adverse effects of a diet rich in pork fat, namely, the disarray
of the intestinal microbiota and inflammatory cytokines, and the appearance of
circulating lipopolysaccharides. In contrast, the high-fat diet prepared with the
standard casein permitted the inversion of the dominant phyla Bacteroidetes
and Firmicutes and other significant alterations. Both the intact whey and the
hydrolyzate whey proteins granted the animal the ability to maintain its nearly
normal gut microbiota profile, allowed the presence of a greater diversity of
minor genera and species, repressed the appearance of LPS in serum thus
modulating the inflammatory response, as well as alleviating fat infiltration in
the liver, besides preserving other biochemical parameters.
Abbreviations
AIN-93G: American Institute of Nutrition 93 growth; WPC: Whey protein concentrated;
WPH: Whey protein hydrolyzate;HF: High fat; HFCAS: High-fat-casein; HFWPC: High-
fat-whey protein concentrate; HFWPH: High-fat-hydrolyzed whey protein; LPS:
Lipopolysaccharides; NAFLD:Non-alcoholic fatty liver disease; DM2: Type 2 diabetes
mellitus; VLDL: Very-low-density lipoprotein; HDL: High-density lipoprotein; GLUT4:
Type 4 Glucose transporter; HSP70: 70 kD heat shock protein; HSP90: 90 kD heat
shock protein; TLR4: Toll-like receptor-4; TNF-α: Tumor necrosis factor alpha;
IL4:Interleukin-4; IL6:Interleukin-6; IL8: Interleukin-8; IL10:Interleukin-10; IL1Ra:
Interleukin-1 receptor antagonist; IL1β: Interleukin-1 Beta; MyD88:Myeloid
differentiation factor 88; GAPDH: Glyceraldehyde 3-phosphate dehydrogenase; LAL:
Limulus amebocyte lysate; E. coli: Escherichia coli; BLAST: Basic Local Alignment
Search Tool; NAST: Nearest Alignment Space Termination; UniFRAC: Method to
calculate a distance measure between organismal communities;VLPs: Variable
lipoproteins; HE:Hematoxylin-eosin;IgA:Imunoglobulin A; H.pylori: Helicobacter pylori;
ZO-1: Zonula occludens-1; Foxp3:Forkhead box P3; NFKB: Factor nuclear kappa B;
IRAK: Interleukin-1 receptor-associated kinase; TRAF-6:Tumor necrosis factor

93
receptor-associated factor-6; IкB:Inhibitor kappa B; IL4Ra:Interleukin-4 receptor
antagonist; TGF-β: Transforming growth factor beta; BCAAs: Branch chain amino
acids.
Competing interests
The authors declare to have no competing interests
Author’s Contributions
All authors contributed significantly to the manuscript: N.E.S.M. and
A.R.R.participated in the project design, conducted the assay and most of the
experiments and wrote the first version of the manuscript;F.P.was in project
design co-advisor, supervised some westernblot analyses and revised the
manuscript; A.S. supervised the microbiota DNA analyses; C.S.M. supervised
complementary analyses and revised the manuscript, A.T.Y. supervised the
histological analysis, and J.A.F. designed the project, translated andcorrected
the manuscript.
Acknowledgments: the authors acknowledge the Brazilian CNPq for the
graduate fellowships of NESM and ARR, the support of CAPES PROAP to
PPGAN-FEA, the grants of FAPESP (Proc. 2012/05859-7 + JM Saad). Thanks
are also due to Hilmar Ingredients, Hilmar, CA, USA, for the gifts of the whey
products.

94
References
1. Cani P D, Amar J, Iglesias M A, Poggi M, Knauf C, Bastelica D et al.Metabolic
endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56:1761-72.
2. Kim K A, Gu W, Lee I A, Joh E H, Kim D H. High Fat Diet-Induced Gut Microbiota Exacerbates Inflammation and Obesity in Mice via the TLR4 Signaling Pathway. PLoS One. 2012; 7:10.
3. Anhê F F, Roy D, Pilon G, Dudonné S, Matamoros S, Varin T V et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut. 2014;doi: 10.1136/gutjnl-2014-307142.
4. Shen L, Black E D, Witkowski E D, Lencer W I, Guerriero V, Schneeberger E E et al. Myosin light chain phosphorylation regulates barrier function by remodeling tight junction structure. Journal of Cell Science.2006; 119: 2095-106.
5. Shi J, Tauriainen E, Martonen E, Finckenberg P, Ahlroos-Lehmus A, Tuomainen A et al. Whey protein isolate protects against diet-induced obesity and fatty liver formation. International.Dairy Journal.2011; 21: 513-22.
6. Tranberg B, Madsen A N, Hansen A K, Hellgren L. Whey-reduced weight gain is associated with a temporary growth reduction in young mice fed a high-fat diet.J NutrBiochem. 2014; 26:9-15.
7. Morato P N, Lollo P C B, Moura C S, Batista T M, Carneiro E M, Amaya-Farfan J. Whey protein hydrolysate increases translocation of GLUT-4 to the plasma membrane independent of insulin in Wistar rats. PLoS One.2013; 8:8.
8. Moura, C S, Lollo P C B, Morato P N, Nisishima l H, Carneiro E M et al.Whey Protein hydrolysate enhances HSP90 but does not alter HSP60 and HSP25 in skeletal muscle of rats. PLoS One. 2014; 9:1.
9. Reeves P G, Nielsen F H, Fahey G C. AIN-93 Purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. The Journal of Nutrition.1993; 123:1939-51.
10. Cintra D E, Ropelle E R, Moraes J C, Pauli J R, Morari J, Souza C Tet al.Unsaturated
fatty acids revert diet-induced hypothalamic inflammation in obesity. PLoS One.2012;7:1.
11. Lozupone C, Knight R.UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. ApplEnviron Microbiol.2005; 71:8228-35.
12. De Santis T Z Jr, Hugenholtz P, Keller K, BrodieE L, LarsenN, PicenoY M et al. NAST: a multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res.2006; 34:394-99.
13. De Moura C S, Lollo P C, Morato P N, Carneiro E M, Amaya-Farfan J. Whey
protein hydrolysate enhances the exercise-induced heat shock protein (HSP70) response in rats. Food Chem.2013;136:1350-7.
14. Li H, Lelliott C, Håkansson P, Ploj K, Tuneld A, Verolin-Johansson M et al.Intestinal, adipose, and liver inflammation in diet-induced obese mice. Metabolism.2008;57: 1704-
10.

95
15. Bleau C, Karelis A D, St-Pierre D H, Lamontagne L. Crosstalk between intestinal microbiota, adipose tissue and skeletal muscle as an early event in systemic low grade inflammation and the development of obesity and diabetes. Diabetes Metab Res Rev. [pub ahead of print] 2014; doi: 10.1002/dmrr.2617.15.
16. Takeda K, Akira S. TLR signaling pathways. Seminars in Immunology. 2004; 6:3-9.
17. Mauer J, Chaurasia B, Goldau J, Vogt M C, Ruud J, Nguyen K D et al. Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin. Nature Immunol. 2014; 15: 423-30.
18. Wermeling F, Anthony R M, Brombacher F, Ravetch J V. Acute inflammation primes myeloid effector cells for anti-inflammatory STAT-6 signaling. Proc. Nat. Acad. Sci USA. 2013; 110:13487-91.
19. Kim C S, Park H S, Kawada T, Kim J H, Lim D, Hubbard N E et al. Circulating levels of MCP-1 and IL-8 are elevated in human obese subjects and associated with obesity-related parameters. International Journal of Obesity.2006;30: 1347–55.
20. Cani P D, Bibiloni R, Knauf C, Waget A, Neyrinck A M, Delzenne N M et al. Changes in
gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes.2008;57:1470-81.
21. Moraes J C, Coope A, Morari J,Cintra D E, Roman E A, Pauli J R et al. High-Fat Diet Induces Apoptosis of Hypothalamic Neurons. PLoS One. 2009; 4:4.
22. Tsukumo D M, Carvalho B M, Marco A, Saad M J. Translational research into gut microbiota: new horizons in obesity treatment. Arq Bras EndocrinolMetab. 2009; 53: 139-44.
23. Hamad M E, Taha S H, Dawood A A, Sitohy M Z, Abdel-Hamid M. Protective effect of whey proteins against nonalcoholic fatty liver in rats. Lipids in health and disease.2011;10:57.
24. Gaudel C, Nongonierma A B,Maher S, Flynn S, Krause M, Murray B A et al. A whey
protein hydrolysate promotes insulinotropic activity in a clonal pancreatic β-cell line and enhances glycemic function in ob/ob mice. J. Nutr. 2013; 143: 1109-14.
25. Pal S, Ellis V, Dhaliwal S. Effects of whey protein isolate on body composition, lipids, insulin and glucose in overweight and obese individuals. British Journal of Nutrition. 2010; 104: 716–23.
26. Bäckhed F, Ding H, Wang T, Hooper L V, Koh G Y, Nagy A et al. The gut microbiota as an environmental factor that regulates fat storage. ProcNatlAcadSci USA. 2004;101:15718-23.
27. Ley R, Harris J K, Wilcox J, Spear J R, Miller S R,Bebout BMet al. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Applied and Environmental Microbiology. 2006;72:3685-95.
28. Everard A, Belzer C, Geurts L, Ouwerkerk J P, Druart C, Bindels L B et al.Cross-talk
between Akkermansiamuciniphila and intestinal epithelium controls diet-induced obesity. ProcNatlAcad Sci.2013.110:9066-71

96
29. Gasparetto D, Lollo P C B, Moura C S, Paula F M M, Carneiro E M, Amaya-Farfan J.
Metabolic Response of Sedentary Normotrophic Rats to Dietary Pre hydrolyzed Whey
Proteins. FoodandPublic Health. 2013; 3:21-27.
30. Schwab C, David B, Rauch I, Rennisch I, Ramesmayer J, Hainzl E et al. Longitudinal study of murine microbiota activity and interactions with the host during acute inflammation and recovery. The ISME Journal. 2014; 8: 1101-14.
31. Lin H V, Frasseto A, Kowalik E J, Nawrocki A R, Lu M M, Kosinski J R. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLoS One. 2012; 7:4.
32. Petriz B A, Castro A P, Almeida J, Gomes C P, Fernandes G R, Kruger R H et al. Exercise induction of gut microbiota modifications in obese, non-obese and hypertensive rats. BMC Genomics. 2014; 15:511.
33. Bassaganya-RieraJ, Dominguez-Bello M G, Kronsteiner B, Carbo A, Lu P, Viladomiu
Met al. Helicobacter pylori colonization ameliorates glucose homeostasis in mice through a PPAR γ-dependent mechanism. PLoS One. 2012;7:11.
34. Lee S M, Han H W, Yim S Y. Beneficial effects of soy milk and fiber on high cholesterol diet-induced alteration of gut microbiota and inflammatory gene expression in rats. FoodFunct. 2014; doi:10.1039/C4FO00731J.

97
Conclusão geral
As proteínas do soro do leite, tanto nas suas formas concentrada e
hidrolisada foram capazes de impedir a disbiose intestinal provocada por uma
dieta hiperlipídica, modulando a composição da microbiota intestinal e evitando a
elevação dos níveis de lipopolissacarídeos circulantes. Com isso, incorporando a
whey protein à dieta hiperlipídica inibiu-se significantemente a expressão de
proteínas envolvidas na resposta inflamatória subclínica registrada na obesidade.
As proteínas do soro do leite também foram capazes de suprimir ou evitar a
esteatose hepatica microgoticular e, embora não tenham contribuído para redução
no ganho de peso na fase inicial, as proteínas do soro do leite foram eficazes em
atenuar deposição de gordura epididimal e em normalizar parâmetros bioquímicos
como glicose e triacilgliceróis. Dessa forma, os achados sugerem que a
incorporação das proteínas do soro do leite à dieta habitual de indivíduos obesos
possa ter um impacto positivo na prevenção e manejo da obesidade e co-
morbidades associadas, como esteatose hepática e resistência à insulina.

98
ANEXO
Top Related