







Línguas
Páginas
Legal


TALITA TREVIZANI ROCCHETTI
Detecção Bacteriana e de Genes de Resistência a
Antimicrobianos pela Técnica de PCR em Tempo Real em
Infecções de Corrente Sanguínea de Pacientes Submetidos
a Transplante de Órgãos Sólidos
Tese apresentada à Universidade Federal de
São Paulo – Escola Paulista de Medicina para
obtenção do Título de Mestre em Ciências.
São Paulo
2011

TALITA TREVIZANI ROCCHETTI
Detecção Bacteriana e de Genes de Resistência a
Antimicrobianos pela Técnica de PCR em Tempo Real em
Infecções de Corrente Sanguínea de Pacientes Submetidos
a Transplante de Órgãos Sólidos
Tese apresentada à Universidade Federal de
São Paulo – Escola Paulista de Medicina para
obtenção do Título de Mestre em Ciências.
Orientador: Prof. Dr. Antonio Carlos Campos
Pignatari
Este trabalho foi realizado com auxílio
financeiro fornecido pela Fundação de Amparo
à Pesquisa do Estado de São Paulo (FAPESP)
Processo nº 2008/04761-8
São Paulo
2011

ROCCHETTI, Talita Trevizani
Detecção bacteriana e de genes de resistência a antimicrobianos pela técnica de
PCR em Tempo Real em infecções de corrente sanguínea de pacientes
submetidos a transplante de órgãos sólidos – Talita Trevizani Rocchetti - São
Paulo, 2011. xx, 156 p.
Tese (Mestrado) – Universidade Federal de São Paulo. Escola Paulista de
Medicina. Programa de Pós-graduação em Infectologia
Título em inglês: Real-time Polymerase Chain Reaction for Detection of Bacterial and Antimicrobial Resistance Genes in Blood Cultures of Solid Organ Transplanted Patients.
1. Diagnóstico molecular; 2. Bacteremia; 3.PCR em Tempo Real; 4. Transplante
de órgãos; 5. Genes de resistência.

iii
UNIVERSIDADE FEDERAL DE SÃO PAULO
ESCOLA PAULISTA DE MEDICINA
DISCIPLINA DE INFECTOLOGIA
CHEFE DO DEPARTAMENTO:
Prof. Dr. Angelo Amato Vicenzo de Paola
COORDENADOR DO CURSO DE PÓS-GRADUAÇÃO:
Prof. Dr. Ricardo Sobieh Diaz
São Paulo
2011

iv
TALITA TREVIZANI ROCCHETTI
Detecção Bacteriana e de Genes de Resistência a
Antimicrobianos pela Técnica de PCR em Tempo Real em
Infecções de Corrente Sanguínea de Pacientes
Submetidos a Transplante de Órgãos Sólidos
BANCA EXAMINADORA:
Titular: Prof. Dr Luis Fernando Aranha Camargo
Titular: Profa. Dra. Marinês Dalla Valle Martino
Titular: Prof. Dr Eliezer Silva
Suplente: Profa. Dra. Antonia Maria de Oliveira Machado

v
Não se deve ir atrás de objetivos fáceis.
É preciso buscar o que só pode ser alcançado
por meio dos maiores esforços.”
Albert Einstein

vi
Dedico este trabalho
ao meu amado pai Vanderlei (in memoriun),
a minha amada mãe, Maria Regina e
aos meus amados irmãos Thiago e Taisa...
Vocês são além da minha razão de vida,
minha inspiração profissional!!

vii
AGRADECIMENTOS
Ao meu orientador, Prof. Dr. Antonio Carlos Campos Pignatari, por todo
crescimento profissional que tive, pelas oportunidades de aprendizado, os
ensinamentos e pela grande orientação. Tenho no senhor uma imensa adimiração e
inspiração profissional.
À Liana Carbalo Menezes, por todos os ensinamentos, paciência e carinho
que recebi. Se eu aprendi um pouco do que você sabe de Biologia Molecular, saio
vitoriosa dessa primeira etapa no LEMC.
À Profª Dra. Ana Cristina Gales, pelos constantes ensinamentos, pelo
exemplo de caráter e profissionalismo.
Ao André Doi, por toda orientação, atenção, ajuda e amizade ao longo da
elaboração deste trabalho.
À equipe do Laboratório Central, Dra. Antonia, Eliete, Thomas, Flavia,
Fernando, Clarice, por todo aprendizado, paciência e dedicação na coleta e
separação das amostras deste projeto.
Ao grupo de Transplantes do Hospital São Paulo e Hospital do Rim, em
especial ao Prof. Dr. Luis Fernando Aranha Camargo e ao Dr. Moacyr Silva Jr, por
todo apoio e colaboração quanto ao rumo e perfil que este trabalho deveria tomar.
A equipe do SCIH do Hospital do Rim, principalmente à Dra Lucy Corrêa, por
toda ajuda e por me abrir as portas para acessar ao programa TASY onde pude
selecionar meu material de estudo.
Aos meninos do SAME do Hospital do Rim, por toda dedicação e ajuda na
coleta dos prontuários.
À Jussimara Monteiro, que proporcionou meu primeiro contato e meus
primeiros aprendizados dentro da Biologia Molecular. Você faz parte da inspiração que
tenho profissional.
Ao Paulo Bispo, um grande amigo que fiz e que me ensinou com todo seu
conhecimento, agradeço por toda confiança depositada em mim.
À Eloiza Helena Campana, minha amiga, vizinha, companheira de laboratório
e de baladas. Você me apoiou em cada momento que tive ao longo dessa minha

viii
jornada no mestrado. Conhecê-la foi uma das melhores coisas que me aconteceu
desde meu regresso a São Paulo.
À Karen Bauab, amiga querida, que tive prazer em conhecer. Deu-me todo
apoio que precisei, nos meus experimentos, na viajem ao Rio de Janeiro, com
conversas, você é uma grande amiga e companheira de laboratório.
Luiz Roberto Chirotto Filho, meu primeiro amigo do laboratório e que faço
questão de preservar. Fico feliz de podermos nos ajudar em nossos projetos e de
podermos manter essa nossa amizade. Você terá sempre meu apoio.
À Renata Picão, por todo ensinamento e ajuda valiosa que me proporcionou.
Tenho grande admiração por você e tive o prazer de ter convivio fora do laboratório.
Ao Danilo Elias, por toda amizade, momentos de conversa e descontração que tive
ao seu lado, fico triste de você não estar aqui para prestigiar esse momento comigo.
Às amigas Cynthea, Paula Peraro, Adryella, Thaís, Jéssica por toda
conversa e apoio que tive de vocês. Aos meus grandes amigos e colegas do
LEMC/ALERTA, Adriana Nicoletti, Lorena, Milene, Ana Carolina, Fernanda Inoue,
Raquel Girardello, Thiago, Rodrigo Cayô, Anderson, Loren, Kelly, Martha, Zonta,
Fernanda Marques, Vinicius, Thomas, Cecília Cergole, Cecília Godoy, Paula
Ignez, Paula Koga, Alinne, Katia, Mirian e Amilton e aos que recentemente
entraram para o nosso grupo, pelo convívio agradável e carinho demonstrado durante
a realização deste trabalho.
À Rosana Capecce, pela dedicação e valioso apoio em todos os momentos.
Ao Charles, por toda colaboração eu tive ao longo do meu mestrado.
Aos professores da DIPA por todo aprendizado.
Á minha prima Gabriela, e aos meus amigos Ademar e Monica, por
acreditarem em mim quando eu havia desacreditado.
A Rosalina, por todo companheirismo, carinho e apoio, durante toda essa
etapa da minha vida.
À minha família, Vanderlei, Maria Regina, Thiago e Taisa, obrigada por
sempre estarem ao meu lado.
A todos que participaram diretamente ou indiretamente do meu trabalho, o meu
muito obrigada!

ix
SUMÁRIO
ÍNDICE DE ABREVIATURAS E SIGLAS ........................................................ 13
ÍNDICE DE TABELAS ..................................................................................... 16
ÍNDICE DE FIGURAS ...................................................................................... 19
RESUMO.......................................................................................................... 21
1. INTRODUÇÃO ............................................................................................. 22
2. OBJETIVOS ................................................................................................. 24
3. REVISÃO BIBLIOGRÁFICA ........................................................................ 25
3.1. Transplante de Órgão Sólido ..................................................................... 25
3.1.1. Transplante no Brasil ...................................................................... 25
3.1.2. Epidemiologia em Transplantados .................................................. 26
3.2. Infecões de Corrente Sanguínea ............................................................... 28
3.2.1. Infecção de Corrente Sanguínea em Transplantados de Órgãos
Sólidos ...................................................................................................... 29
3.3. Genes de Resistência a Antimicrobianos .................................................. 32
3.3.1. Gene mecA ..................................................................................... 32
3.3.2. Genes vanA e vanB ........................................................................ 33
3.3.3. -Lactamases de Espectro Ampliado (ESLs) ............................... 36
3.3.5. Klebsiella pneumoniae Carbapenemase (KPC) .............................. 41
3.3.4. Metalo--Lactamases ...................................................................... 43
3.4. Sistemas Automatizados no Processamento de Hemoculturas ................ 47
3.5. Microbiologia Molecular Aplicada no Diagnóstico Clínico ......................... 50
3.5.1. Reação de Polimerização em Cadeia - PCR, PCR Multiplex e PCR
em Tempo Real ......................................................................................... 50
3.5.2. Validação de Novos Métodos de Diagnósticos ............................... 52

x
4 - MATERIAIS E MÉTODOS .......................................................................... 54
4.1. Casuística .................................................................................................. 54
4.1.1. Variáveis Clínicas ............................................................................ 54
4.2. Coleta das Hemoculturas de Acesso Periférico ........................................ 57
4.2.1. Coleta das Hemoculturas de Acesso Central – Cateteres .............. 57
4.3. Processamentos das Amostras pelo Método Fenotípico .......................... 57
4.3.1. Determinação da positividade e negatividade das hemoculturas pelo
aparelho Bactec® ...................................................................................... 57
4.3.2. Resultado Parcial da Hemocultura Positiva pela Coloração de Gram
.................................................................................................................. 58
4.3.3. Resultado Final da Hemocultura pelo Sistema Automatizado
Phoenix® ................................................................................................... 58
4.4. Pdronização da PCR em Tempo Real ...................................................... 59
4.4.1. Eficiência e Especificidade da PCR em Tempo Real ...................... 62
4.4.2. Determinação do Limite do Branco (LoB) ou “cutoff” ...................... 65
4.4.3. Determinação do Limite de Detecção - LoD (Sensibilidade) ........... 66
4.5. Aplicação nas Amostras de Hemocultura ................................................. 67
4.5.1. Extração do DNA Direto dos Frascos de Hemocultura ................... 68
4.5.2. Aplicação da PCR em Tempo Real para o Gene HFE de Controle
Interno de Extração ................................................................................... 69
4.5.3. Detecção do DNA Bacteriano PCR em Tempo Real pelo Sistema
TaqMan ..................................................................................................... 69
4.5.4. Detecção dos Genes que Conferem Resistências aos
Antimicrobianos por PCR em Tempo Real pelo Sistema SYBR Green .... 70
4.6. Testes Direto do Isolado Bacteriano Referente à Hemocultura ................. 71
4.6.1. Teste de Triagem com Disco-Difusão para Ertapenem ................... 72
4.6.2. Teste de Hidrólise para Detecção de Metalo-β-Lactamases ........... 73
4.6.3. Detecção Fenotípica de ESβL ......................................................... 74
4.6.4. Determinação da Suscetibilidade a Vancomicina ........................... 75
4.6.5. Determinação da Suscetibilidade a Oxacilina ................................. 76
4.6.6. PCR em Tempo Real Direto do Isolado Bacteriano ........................ 76
4.7. Análise Estatística dos Dados ................................................................... 77

xi
5. RESULTADOS ............................................................................................. 79
5.1. Amostras de Hemoculturas Incluídas no Estudo ....................................... 79
5.2. Validação Analítica da PCR em Tempo Real ............................................ 79
5.2.1. Eficiência e Especificidade da PCR em Tempo Real ...................... 80
5.2.2. Padronização da PCR em Tempo Real Sistema TaqMan para o
Gene Bacteriano 16S rDNA ...................................................................... 82
5.2.3. Determinação do Limite do Branco (LoB) ou “cutoff” ...................... 84
5.2.4. Determinação do Limite de Detecção – LoD ................................... 84
5.2.5. Determinação da Curva ROC para as Reações dos Genes de
Resistência a Antimicrobianos e para a Reação Multiplex Gram-positivo e
Gram-negativo por PCR em Tempo Real ................................................. 86
Na Figura 4 encontram-se as curvas Roc dos genes estudados. .................... 86
5.3. Aplicação da PCR em Tempo Real nas Amostras de Hemocultura .......... 89
5.3.1. Detecção do Gene HFE de Controle Interno da Extração ............... 89
5.3.2. Detecção do Gene Bacteriano 16S rDNA ....................................... 89
5.3.3. Detecção PCR dos Genes que Conferem Resistência aos
Antimicrobianos em Bactérias Gram Negativas ........................................ 93
5.3.4. Detecção PCR dos Genes que Conferem Resistência aos
Antimicrobianos em Bactérias Gram Positivas .......................................... 96
5.4. Testes Direto do Isolado Bacteriano ......................................................... 99
5.4.1. Confirmação de Susceptibilidade a Oxacilina e Gene de Resistência
mecA pelo Método da PCR em Tempo Real Direto do Isolado Bacteriano
Referente a Amostra de Hemocultura ....................................................... 99
5.4.2. Confirmação da Produção de Carbapenemase e Gene de
Resistência blaKPC pelo Método da PCR em Tempo Real Direto do Isolado
Bacteriano Referente a Amostra de Hemocultura ................................... 100
5.4.3. Confirmação da Produção de Metalo-β-Lactamases e Genes de
Resistência blaIMP, blaVIM e blaSPM pelo Método da PCR em Tempo Real
Direto do Isolado Bacteriano Referente a Amostra de Hemocultura ....... 101
5.4.4. Confirmação da Produção de ESβL e Genes de Resistência blaSHV,
blaTEM e blaCTX-M pelo Método da PCR em Tempo Real Direto do Isolado
Bacteriano Referente a Amostra de Hemocultura ................................... 102

xii
5.5. Variáveis Clínicas, Análise e Comparação do Tempo de Liberação dos
Resultados pelo Método Fenotipico e Tratamentos antes e Após os Resultados
em Relação ao Resultado Obtido pela PCR em Tempo Real ........................ 105
5.5.1. Hemoculturas Positivas para Isolados Gram Positivos ................. 106
5.5.2. Hemoculturas Positivas para Isolados Gram Negativos ................ 109
6. DISCUSSÃO .............................................................................................. 113
7. CONCLUSÕES .......................................................................................... 127
8. REFERÊNCIAS BIBLIOGRÁFICAS .......................................................... 128
9. ABSTRACT ................................................................................................ 149
10. ANEXOS .................................................................................................. 151
ANEXO 1. Dados referentes aos 117 pacientes e 185 hemoculturas incluídas
no estudo........................................................................................................ 151
ANEXO 2. Drogas Testadas para os respectivos Painéis do Sistema
automatizado Phoenix. ................................................................................... 154
ANEXO 3. Apresentação no 50 th ICAAC 2010. ............................................ 156

xiii
ÍNDICE DE ABREVIATURAS E SIGLAS
ABTO – Associação Brasileira de Transplantes de Órgãos
AIM – Australian Imipenemase
ATCC - American Type Cuture Collection
CCIH – Comissão de Controle de Infecção Hospitalar
CDC – Center for Disease Control
CIM - Concentração Inibitória Mínima
CLSI – Clinical Laboratory Standards Institute
Ct – Ciclo threshold
CTX-M – Cefotaximase
CVC – Cateter Venoso Central
DD – Disco Difusão
DM – Diabete Mellitus
DNA – Ácido Desoxirribonucleico
DIM – Dutch Imipenemase
EDTA – Ácido Etilenodiaminotetracético
ESL - Extended Spectrum -Lactamase
EUA – Estados Unidos da América
GIM – German Imipenemase
HAS – Hipertensão Arterial
HFE – Hemocromatose
ICS – Infecção de Corrente Sanguínea
IMP – Imipenemase
IS - Insertion Sequence
IUPAC – The International Union of Pure and Applied Chemistry

xiv
KHM – Kyorin Health Science metallo-β-lactamase
KPC - Klebsiella pneumoniae Carbapenemase
LB - Luria Bertani
LED – Diodos emissores de luz
LEMC – Laboratório Especial de Microbiologia Clínica
ML – Metallo--Lactamase
MRSA – Staphylococcus aureus resistente a meticilina
NDM – New Delhi metallo-β-lactamase
OMP – Outer Membrane Protein
PBP – Penicillin Binding Protein
PBP2a – Proteína Ligadora de Penicilina Adicional
PCR – Polymerase Chain ReaCtion
PFGE – Pulsed Field Gel EleCtrophoresis
SCOPE – Surveillance and Control of Pathogens of Epidemiological Importance
SCoN – Staphylococcus coagulase negativo
SDS-PAGE – Sodium Dodecyl Sulfate Polyacrylamide Gel EleCtrophoresis
SIM – Seul Imipenemase
SNT – Sistema Nacional de Transplantes
SPM – São Paulo metallo-β-lactamase
SUS – Sistema Único de Saúde
TBE – Tris-Borato-EDTA
TSB – Tryptone Soy Broth
TM – Temperatura de Melting
TMB – Tripoli metallo-β-lactamase
UFC – Unidade formadora de colônia

xv
UNIFESP – Universidade Federal de São Paulo
UTI – Unidade de Terapia Intensiva
UV – Ultravioleta
VIM – Verona Imipenemase
VRE – Enterococcus Resistente a Vancomicina
VRSA – Staphylococcus aureus Resistente a Vancomicina

xvi
ÍNDICE DE TABELAS
Tabela 1. Lista de iniciadores de amplificação da região codificadora dos
genes estudados.........................................................................................
60
Tabela 2. Relação dos microrganismos testados para validação das
reações de PCR em Tempo Real..........................................................
64
Tabela 3. Resultados dos controles dos genes de resistência e os Tm
dos produtos amplificados...........................................................................
80
Tabela 4. Desvio Padrão e media do Tm, Eficiência da reação de PCR,
Ct e Tm da ultima diluição............................................................................
82
Tabela 5. Resultados da padronização dos protocolos de PCR em
Tempo Real utilizando sondas TaqMan-MGB multiplex Gram
especificas..................................................................................................
83
Tabela 6. Resultados de cutoff e LoD para cada gene especifico dos
microrganismos avaliados...........................................................................
85
Tabela 7. Sensibilidade, especificidade e Ct delimitado pela curva ROC
para os genes que conferem resistência a antimicrobianos e para o gene
16S rDNA para Gram positivo e Gram negativo.........................................
88
Tabela 8. Sensibilidade, especificidade pela curva ROC de acordo com o
LoD para os genes que conferem resistência a antimicrobianos e para o
gene 16S rDNA para Gram positivo e Gram negativo.................................
88

xvii
Talela 9. Resultado da PCR em Tempo Real pelo sistema TaqMan para
o gene 16S rDNA nas 126 hemoculturas positivas pelo Bactec®................
91
Talela 10. Resultado da PCR em tempo real sistema SYBR Green para
os genes de resistência em Gram negativos e resultado do sistema
Phoenix®.....................................................................................................
95
Talela 11. Resultado da PCR em Tempo Real sistema SYBR Green para
os genes de resistência de Gram positivos e resultado do Phoenix®..........
97
Tabela 12. Resultado do Etest para Oxacilina e PCR em Tempo Real
sistema SYBR Green para o gene de resistência mecA direto da colônia
e comparação com a PCR direto do frasco de hemocultura.......................
99
Tabela 13. Resultado do método de disco difusão e PCR em Tempo
Real sistema SYBR Green para o gene de resistência blaKPC direto da
colônia e comparação com a PCR direto do frasco de hemocultura...........
101
Tabela 14. Resultado do teste de hidrólise e PCR em Tempo Real
sistema SYBR Green para os genes de resistência blaSPM, blaVIM e blaIMP
direto da colônia e comparação com a PCR direto do frasco de
hemocultura................................................................................................
102
Tabela 15. Resultado do disco aproximação para ESβL e PCR em
Tempo Real pelo sistema SYBR Green para os genes de resistência
blaSHV, blaCTX-M e blaTEM direto da colônia e comparação com a PCR
direto do frasco de hemocultura..................................................................
104

xviii
Tabela 16. Dados referentes aos resultados dos testes fenotípicos e da
PCR em Tempo Real liberados direto das hemoculturas e avaliação do
tratamento recebido pelos pacientes...........................................................
107
Tabela 17. Dados referentes aos resultados dos testes fenotípicos e da
PCR em Tempo Real liberados direto das hemoculturas e avaliação do
tratamento recebido pelos pacientes...........................................................
110

xix
ÍNDICE DE FIGURAS
Figura 1. Etapa da Transpeptidação: Formação das ligações cruzadas
nas cadeias peptídicas................................................................................
34
Figura 2. A - Amplificação do DNA de Gram positivo (curva em amarelo)
e Gram negativo (curva em vermelho); B- Sonda Gram Positivo (GP) e
Sonda Gram Negativo (GN).........................................................................
62
Figura 3. Fluxograma do processamento das amostras.............................
67
Figura 4. Curva ROC para genes de resistência a antimicrobianos e
sondas Gram Positivo e Sonda Gram Negativo: A – Gram Positivo; B-
Gram Negativo; C - blaTEM; D - vanA; E - vanB; F - blaCTX; G - blaSHV; H -
mecA; I - blaKPC............................................................................................
87
Figura 5. Prevalência de espécies Gram positivas identificadas pelo
sistema automatizado Phoenix®..................................................................
92
Figura 6. Prevalência de espécies Gram negativas identificadas pelo
sistema automatizado Phoenix®..................................................................
93
Figura 7. Total de amostras positivas pela metodologia da PCR em
Tempo Real pelo sistema SYBR Green para os respectivos genes de
resistência estudados..................................................................................
98

xx
Figura 8. Número e porcentagem de pacientes referentes a
hemoculturas em que foram isoladas bactérias Gram positivas,
recebendo tratamento inadequado no momento da coleta da
hemocultura, depois do resultado do Gram da hemocultura e depois do
resultado final da Hemocultura....................................................................
108
Figura 9. Número e porcentagem de pacientes referentes a
hemoculturas em que foram isoladas bactérias Gram negativas não
fermentador, recebendo tratamento inadequado no momento da coleta
da hemocultura, depois do resultado do Gram da hemocultura e depois
do resultado final da Hemocultura...............................................................
111
Figura 10. Número e porcentagem de pacientes referentes a
hemoculturas em que foram isoladas bactérias Gram negativas
enterobacteriaceas, recebendo tratamento inadequado no momento da
coleta da hemocultura, depois do resultado do Gram da hemocultura e
depois do resultado final da Hemocultura....................................................
112

RESUMO
21
RESUMO
Pacientes transplantados de órgãos sólidos apresentam alto risco para infecção da
corrente sanguínea. A instituição da terapia antimicrobiana apropriada guiada por um
diagnóstico rápido e preciso das infecções da corrente sangüínea está relacionada a
um resultado satisfatório. O objetivo deste estudo foi a detecção de bactérias Gram-
positivas e Gram-negativas em hemocultura automatizada (Bactec®, Becton
Dickinson), com a utilização do multiplex para reação da polimerase em cadeia (PCR)
em Tempo Real e detecção de genes de resistência. Métodos: Um total de 185
hemoculturas, 126 positivas e 59 negativas, foram obtidos de 117 pacientes
submetidos a transplante de órgãos sólidos, dois centros de transplante na cidade de
São Paulo, Brasil, Hospital São Paulo e Hospital do Rim e Hipertensão. DNA das
culturas de sangue foi extraído pelo método fenol clorofórmio (Brazol®, LGC, Brasil). A
detecção do DNA bacteriano foi realizada utilizando iniciadores universais do gene
16S rRNA. A diferenciação entre as bactérias Gram-positivas e Gram-negativas foi
feito por hibridização com sondas específicas por multiplex TaqMan em Tempo
Real. Os genes de resistência: blaSHV, blaTEM, blaCTX-M, blaKPC, blaSPM, blaVIM, blaIMP,
vanA, vanB e mecA foram detectadas utilizando iniciadores específicos, em Tempo
Real sitema SYBR Green. A adequação do tratamento antimicrobiano foi avaliada pela
revisão do prontuário dos pacientes. Resultados: Cinqüenta e nove amostras foram
positivas para bactérias Gram-positivas e sessenta e sete amostras foram positivas
para bactérias Gram-negativas. Todas as amostras (100%) foram concordantes entre
hemocultura e PCR. Trinta e duas amostras foram positivas para o gene mecA, cinco
para o gene blaCTX-M, uma para o gene blaKPC, uma para o gene blaSHV e uma para
blaTEM. Oitenta e oito foram negativas para todos os genes. A detecção de genes de
resistência a antimicrobianos favoreceria a adequação da antibioticoterapia,
particularmente no descalonamento no tratamento de bacteremias causadas por
bactérias Gram positivas e na adequação precoce no tratamento de bacteremias por
bacilos Gram negativas multiresistentes em pacienetes tranplantados de órgãos
sólidos. Conclusão: A PCR multiplex “in house” para bactérias Gram-positivas e
Gram-negativas e detecção de genes de resistência aos antimicrobianos por PCR em
Tempo Real poderia ser útil para o diagnóstico rápido da infecção da corrente
sangüínea em pacientes submetidos a transplante de órgão sólidos.

INTRODUÇÃO
22
1. INTRODUÇÃO
A introdução de técnicas moleculares na medicina diagnóstica e sua
aplicação no diagnóstico de doenças infecciosas estabelecem uma nova era na
detecção e caracterização de microrganismos na rotina de processamento de
amostras clínicas.
A presença de bacteremia agrava o quadro clínico do paciente e quando
se trata de pacientes transplantados há a necessidade do diagnóstico precoce
e rápido de Infecções de Corrente Sanguínea (ICS), uma vez que a taxa de
mortalidade relacionada a esta infecção nesta população é alta, principalmente
quando ocorre o agravamento do quadro clínico como o choque séptico
(Ferraresso & Berardineli, 2005).
Normalmente, as hemoculturas levam em média 24 a 36 h após a coleta
para positivar. A terapia antimicrobiana pode ser otimizada com base na
identificação presuntiva do agente bacteriano. Porém a completa identificação
do microrganismo e perfil de susceptibilidade normalmente não está disponível
antes de 24 a 72h. O uso da PCR tem sido sugerido em diagnóstico molecular,
pois além de ser uma técnica sensível também diminui o tempo de liberação
dos resultados quando comparado com o método fenotípico (Valones et al.,
2009).
A PCR convencional em pesquisa já é usada habitualmente e seu uso
no diagnóstico clínico cresce a cada dia. Porém no diagnóstico clínico a
sensibilidade e a rapidez na liberação dos resultados podem ser otimizados
pela técnica da PCR em Tempo Real, já que a amplificação e leitura são feitos
simultanemente.

INTRODUÇÃO
23
Em um recente trabalho, realizado por Lehmann e colaboradores (2010),
foi elaborado um método de detecção bacteriana, usando o gene 16s rDNA
diretamente da hemocultura. Os resultados apontaram que a técnica da PCR
foi duas vezes mais sensível que o método convencional, sugerindo assim a
importância e aplicabilidade da PCR nesses casos. Entretando, apesar da
disponibilidade de Kits comerciais para detecção de microrganismos e de
genes de resistência direto de amostras clínicas, até o momento, nenhum
trabalho relata a aplicação da PCR em Tempo Real, e a padronização “in
house” para o diagnóstico rápido de ICS em transplantados de órgãos sólidos.

OBJETIVOS
24
2. OBJETIVOS
2.1. Objetivo Principal
1. Padronizar a técnica de PCR em Tempo Real para a pesquisa de
bactérias e dos principais genes que codificam resistência aos
antimicrobianos;
2.2. Objetivos Secundários
1. Aplicar a técnica de PCR em Tempo Real padronizada para a pesquisa
de bactérias e dos principais genes que codificam resistência aos
antimicrobianos em amostras de frascos de hemocultura coletada de
pacientes submetidos a transplante de órgãos sólidos;
2. Correlacionar os resultados obtidos com a detecção molecular de
bactérias e de genes de resistência aos antimicrobianos, realizada
diretamente da amostras clínica com os resultados de identificação
bacteriana e teste de sensibilidade automatizado obtidos com o isolado
da respectiva hemocultura;
3. Avaliar o possível impacto dos testes moleculares no tratamento de
Infecções de Corrente Sanguínea nos pacientes submetidos a
transplantes de órgão sólidos;

REVISÃO BIBLIOGRÁFICA
25
3. REVISÃO BIBLIOGRÁFICA
3.1. Transplante de Órgão Sólido
3.1.1. Transplante no Brasil
O Brasil tem o maior programa público de transplantes de órgãos e
tecidos do mundo, sendo o Sistema Único de Saúde (SUS) responsável por
92% dos procedimentos. Este programa é composto por uma rede de 1.376
equipes médicas e 548 unidades credenciadas, porém, 63.866 pacientes
aguardam por um órgão no Brasil, de acordo com o Sistema Nacional de
Transplantes (SNT) em 2009 (Ministério da Saúde, 2009).
Em 2009, foram notificados 42.783 transplantes no Brasil, sendo 5.998
de órgãos sólidos. Destes, 4.259 foram transplantes de rim, ocupando o
primeiro lugar no número de transplantes nacionais, seguido por fígado (1.322),
coração (200), pâncreas (158) e pulmão (59) (Associação Brasileira de
Transplantes de Órgãos - ABTO, 2009).
Entre os centros de transplantes no Brasil, encontram-se o Hospital São
Paulo/Unifesp (HSP) e o Hospital do Rim e Hipertensão (HRH). No HSP são
realizados transplantes de rim, medula óssea, coração, osso, pulmão, córnea e
fígado. Sola e colaboradores (2007), em estudo retrospectivo, registraram 81
transplantes de órgãos sólidos no período de janeiro de 2004 a março de 2005.
Destes, 35 foram de rim-pâncreas, 17 apenas de rim, 17 de fígado, 20 de
coração.
O programa de transplante de rim do HSP teve início em 1976 como
uma modalidade de tratamento renal, pela divisão de Nefrologia da

REVISÃO BIBLIOGRÁFICA
26
Universidade Federal de São Paulo (UNIFESP). Na época, menos de 10
transplantes eram realizados. Este número aumentou para 178 em 1998,
mesmo ano em que foi inaugurado o HRH filiado a UNIFESP, e em 2004 foram
realizados 654 transplantes (Medina-Pestana, 2006), ocupando o primeiro lugar
no ranking mundial em transplantes de rim. Hoje o setor de transplantes do
Hospital do Rim e Hipertensão é referência em todo país.
O último levantamento de 2009 aponta que foram realizados 352
transplantes de rim, equivalente a 8,3% do total realizado em todo Brasil
(Associação Brasileira de Transplantes de Órgãos, 2009).
3.1.2. Epidemiologia em Transplantados
O transplante de órgãos sólidos representa hoje a melhor terapia
substitutiva para pacientes com doença crônica terminal. Apesar de o rim ser o
órgão mais transplantado no país, os transplantes de outros órgãos sólidos tem
crescido gradativamente nos últimos anos (Associação Brasileira de
Transplantes de Órgãos, 2009).
O transplante renal é uma alternativa terapêutica para pacientes com
doenças renais em estágio terminal, oferecendo assim melhor qualidade de
vida para aproximadamente 77.589 indivíduos registrados em programas de
diálise no Brasil (Sesso, 2010). No país, 6.500 pessoas apresentam doença
hepática avançada, risco de morte ou qualidade de vida comprometida e
aguardam na fila de transplante de fígado. No estado de São Paulo foram
realizados 200 transplantes de coração no ano de 2009. Entretanto, 305
pessoas se encontravam na lista de espera, sendo que aproximadamente 50%
não foram atendidos reservando-se o procedimento àqueles indivíduos com

REVISÃO BIBLIOGRÁFICA
27
alto risco de falecer de doença cardíaca em menos de um ou dois anos
(Ministério da Saúde, 2009).
O número de transplantes realizados por doadores falecidos se
sobrepõe ao numero realizado por doadores vivos. Este dado pode parecer
óbvio, porém há 10 anos, o número de doadores vivos para transplante de rim
era maior. Esse aumento se deve a inúmeras campanhas em todo país para
com intuito de incentivar a população à doadoção de órgãos (Associação
Brasileira de Transplantes de Órgãos, 2009). O transplante de rim, pâncreas,
medula óssea, fígado e pulmão intervivos pode ser realizado somente se não
trouxer prejuízo ou representar riscos à saúde do doador (Sistema Nacional de
Transplantes, 2011).
Os transplantes de órgãos sólidos apresentam alto risco de rejeição. As
rejeições agudas e crônicas são as principais causas de perda de órgãos
transplantados. A resposta imune pode ser tanto mediada por células T, sendo
a mais importante em rejeição aguda de órgãos transplantados, ou por
anticorpos produzidos por linfócitos B, que tem grande contribuição no
processo de rejeição (La Rosa et al., 2007).
Os avanços das técnicas cirúrgicas e dos tratamentos com terapia
imunossupressora levaram a diminuição nos índices de rejeição aguda, com a
sobrevivência dos enxertos após um ano em cerca de 90% dos transplantes.
(Danovitch, 2001; Mies, 1998). Os imunossupressores, apesar de reduzir
substancialmente as taxas de rejeição, apresentam efeitos colaterais
indesejáveis como imunodeficiência e toxicidade em outros órgãos (Halloran,
2004).

REVISÃO BIBLIOGRÁFICA
28
Apesar da excelente evolução na sobrevida dos pacientes e do enxerto,
o transplante de órgão sólido não é isento de complicações. A infecção é a
principal complicação no pós-transplante (Fishman & Rubin, 2005), sendo as
infecções de corrente sanguinea, uma das mais prevalentes (Nusair et
al.,2008). No primeiro mês pós-transplante, as infecções bacterianas são
responsáveis por 90% das complicações infecciosas, sendo que o risco destas
infecções ocorrerem varia com o tempo, principalmente em decorrência da
imunossupressão (Fishman, 2007).
3.2. Infecões de Corrente Sanguínea
Infecção de Corrente Sanguínea (ICS) é caracterizada pela presença de
bactéiras ou fungos em uma ou mais amostras de hemoculturas associada a
um quadro clínico compatível com sepse (Bone et al., 1992).
A microbiota humana normalmente não causa infecção e pode proteger
o organismo humano contra outros patógenos por competir pelos nutrientes e
locais de ligação nas superfícies celulares (Donnelly & De Paum, 2005). O
responsável pelo controle desta microbiota é o sistema imunológico. Quando a
imunidade celular e humoral encontra-se intacta, oferece uma proteção contra
a maioria dos microrganismos agressores devido a boas condições clínicas,
bom estado nutricional e função normal dos órgãos (Donnelly & De Paum,
2005). Entretanto um déficit específico de imunidade aumenta a suscetibilidade
a infecções, que seriam erradicadas em um organismo íntegro normalmente
pelo mecanismo de defesa (Halloran, 2004).
Como consequência da resposta imune celular deficitária, os pacientes
estão mais sujeitos a infecção por patógenos e apresentam também maior

REVISÃO BIBLIOGRÁFICA
29
gravidade no quadro clínico. Alguns dos fatores que agravam as condições de
imunossupressão são a dose e duração da terapia imunossupressora, doenças
auto-imunes, deficiências imunológicas funcionais, neutropenia e linfopenia,
condições metabólicas como diabetes, desnutrição e infecção por vírus
imunomoduladores como citomegalovírus (Fishman & Rubin, 1998; Halloran,
2004).
3.2.1. Infecção de Corrente Sanguínea em Transplantados de
Órgãos Sólidos
A incidência de infecção em transplantados é determinada pela
interação entre a exposição a agentes infecciosos e a terapia
imunossupressora. Esta varia devido a inúmeros fatores, tais como o tipo de
órgão transplantado, a quantidade de medicamento imunossupressor usado, a
necessidade do uso de terapia adicional contra rejeição e complicações pós-
cirurgicas (Patel & Paya, 1997).
ICS é considerada uma das causas mais importantes de mortalidade
em pacientes que realizaram transplante de órgãos sólidos (Singh et al, 2000;
Rodriguez et al., 2006; Husain et al., 2006). Estima-se que a incidência de
bacteremia seja de 28 a 30% em transplante hepático, 5 a 11% em transplante
renal e 10% em transplantados cardíacos (McClean et al., 1994; Moreno, et al,,
1994).
Moreno e colaboradores (2007), em um estudo realizado na Espanha,
no período de julho de 2003 a abril de 2005, incluíram 3.926 casos de
transplantes, sendo que destes, 2.935 eram de órgão sólido e 991 de células
hematopoiéticas. Dos 2.935 casos de transplantes, 321 (10%) apresentaram

REVISÃO BIBLIOGRÁFICA
30
ICS, sendo que 134 foram em transplantados hepáticos, 121 em
transplantados de rim, 32 em transplantados cardíacos, 17 em transplantados
de pulmão e 17 em transplantados de pâncreas. Com exceção do transplante
de rim, Staphylococcus coagulase negativo (SCoN) foi o microrganismo mais
isolado seguido de Escherichia coli. Outros patógenos como Acinetobacter
baumannii, Pseudomonas spp., Enterococcus spp., Staphylococcus aureus,
Klebsiella spp., Enterobacter spp., Streptococcus viridans, Streptococcus
pneumoniae, Corynebacterium spp. e Stenotrophomonas maltophilia também
foram identificados mas em menor freqüência. Entre os isolados de S. aureus,
16% eram resistentes a meticilina (MRSA), 14,5% das enterobactérias eram
resistentes a cefalosporinas e 10% dos bacilos gram negativos eram
multiresistentes.
Um estudo retrospectivo realizado em um hospital terciário da cidade de
São Paulo avaliou 81 transplantes de órgãos sólidos realizados no período de
janeiro de 2004 a março de 2005. Dos pacientes avaliados, 9% adquiriram ICS.
O transplante hepático apresentou a maior taxa de ICS (29%) e nenhum caso
foi relatado em transplantados renais. Os agentes infecciosos mais prevalentes
foram K. pneumoniae (33,3%), P. aeruginosa (33,3%) e A. baumannii (33,3%).
Porém, diferente do estudo de Moreno e colaboradores (2007), não foi
encontrado nenhum caso de infecção por bactéria Gram positiva (Sola et al.,
2007).
Em um recente estudo, realizado no Brasil, Silva Jr e colaboradores
(2010) avaliaram 3.308 tranpslantes realizados no HRH e HSP, entre os anos
de 2000 e 2006 e constataram uma incidência de 4,11% de ICS nestes
transplantes, sendo E. coli (30,3%) o principal agente isolado nessas infecções.

REVISÃO BIBLIOGRÁFICA
31
Cinco isolados de E. coli, 8 K.pneumoniae e 8 E. aerogenes foram
multiresistêntes aos antimicrobianos. Entre os Gram positivos, 5 SCoN e 6 S.
aureus foram resistentes a meticilina. Nenhum VRE foi relatado.
Alguns trabalhos demonstram um maior número de bactérias Gram
negativas isoladas (Candel et al, 2005; Rodriguez et al, 2006) enquanto outros
relatam um maior número de bactérias Gram positivas (Berger et al, 2006).
Entretanto há concordância entre as principais espécies encontradas e a
resistência aos antimicrobianos (Linares et al., 2007).
A ICS representa uma falência do hospedeiro em controlar a infecção de
um foco primário. Choque séptico é a complicação mais severa no contexto da
bacteremia, aumentando a chance de morte em até 50% (Candel et al, 2005).
Michalak e colaboradores (2005) estudaram 102 pacientes transplantados de
rim-pâncreas. Bacteremia ocorreu em 13 dos pacientes e destes, 5 (38%)
tiveram choque séptico, sendo que, 100% dos pacientes foram a óbito. Dentre
os sítios de infecção primária, o catéter venoso central aparece em maior
frequência em transplantados de órgão sólido (Rodriguez et al., 2006; Singh et
al., 2000). Outros trabalhos relatam que o órgão doado proveniente de um
doador apresentando infecção ou sendo um doador cadáver, aumenta a
possibilidade do paciente receptor apresentar infecção (Battaglia et al., 2004;
Dantas et al., 2006).
A presença de bacteremia em pacientes transplantados em vigência de
terapia imunossupressora requer atenção dos profissionais da área da saúde,
uma vez que é frequente a presença de microrganismos multirresistentes, o
que restringe a escolha da terapia antimicrobiana apropriada.

REVISÃO BIBLIOGRÁFICA
32
3.3. Genes de Resistência a Antimicrobianos
3.3.1. Gene mecA
Os antibióticos betalactâmicos produzem um efeito bactericida pela
inibição das enzimas responsáveis por catalisar um estágio vital da biossíntese
do peptideoglicano, principal componente da parede celular das bactérias
Gram positivas (Giesbrecht et al., 1998).
A resistência apresentada pelos estafilococos aos antibióticos
betalactâmicos deve-se principalmente a dois mecanismos distintos, porém não
dissociados, pois ambos podem interagir. O primeiro mecanismo desenvolvido
pela bactéria é produção da enzima betalactamase que hidrolisa o antibiótico.
O segundo mecanismo está associado à alteração do sítio de ação do
antibiótico betalactâmico pela produção de uma proteína ligadora de penicilina
adicional, a PBP2a (também denominada PBP2’), que está ausente nos
Staphylococcus spp. sensíveis à meticilina (Hackbarth et al., 1989).
A codificação desta nova PBP está relacionada à aquisição do gene
mecA por Staphylococcus spp. Este gene faz parte de um elemento genético
móvel encontrado em todos os isolados de S. aureus e Staphylococcus
Coagulase Negativo (SCoN) resistentes a meticilina. Katayama e
colaboradores, em 2000, demonstraram que o gene mecA faz parte de um
elemento genômico designado cassete estafilocócico do cromossomo mec
(“staphylococcal cassette chromosome mec” - SCCmec), integrado ao
cromossomo de S. aureus .
Atualmente, o gênero Staphylococcus compreende 40 espécies. (Cunha
et al., 2004). Entre seus representantes, S.aureus é considerado o patógeno

REVISÃO BIBLIOGRÁFICA
33
mais virulento e importante, mas a incidência de infecções causadas por SCoN
vem aumentando em todo o mundo (Sakai, 2004).
Em um recente estudo do Programa de Vigilância SENTRY ("SENTRY
“Antimicrobial Surveillance Program”) realizado com 3.907 amostras de
bactérias Gram positivas coletadas de centros médicos no Brasil, entre o
perído de janeiro de 2005 a setembro de 2008, constatou-se que S. aureus foi
o microrganismos mais prevalente em infecções de corrente sanguínea
seguido por SCoN. Neste mesmo estudo, 31% dos isolados de S. aureus e
80% dos SCoN apresentaram resistência a meticilina (Gales et al., 2009). Em
um estudo realizado em pacientes transplantados de órgãos sólidos, esta
prevalência também foi observada. Diversos autores relataram alta incidência
de infecção de corrente sanguínea por Staphylococcus spp. resistentes a
meticilina. (Linares et al., 2009; Bert et al., 2010).
3.3.2. Genes vanA e vanB
Da mesma maneira que os antimicrobianos β-lactâmicos, a classe dos
glicopepitideos (vancomicina) agem no metabolismo de construção da parede
celular bacteriana, através da ligação da porção terminal D-Ala-D-Ala de um
pentapeptídeo encontrado em precursores de peptidoglicano, interferindo na
etapa de transpeptidação (Figura 1) (Silveira et al., 2006; Eggert et al., 2001).

REVISÃO BIBLIOGRÁFICA
34
Figura 1. Etapa da Transpeptidação: Formação das ligações cruzadas nas
cadeias peptídicas (Eggert et al., 2001).
A resistência bacteriana à vancomicina em enterococos ocorre através
de uma modificação genética no gene bacteriano, sintetizando, como exemplo,
D-alanina-D-lactato (D-Ala-D-Lac) ao invés do dipeptídeo D-alanina-D-alanina
(D-Ala-D-Ala) (Silveira et al., 2006). A modificação do aminoácido terminal D-
alanina por D-lactato introduz uma interação eletrostática repulsiva no lugar da
ligação de hidrogênio. Em conseqüência, a afinidade da vancomicina com a
camada de peptidoglicano diminui em uma escala superior a 1000 vezes
(MacComas et al., 2003).
A aquisição do mecanismo de resistência a vancomicina é mediada por
vários genes (Werner et al., 2008). Embora a origem da sensibilidade de
enterococos frente à teicoplanina envolva mecanismos diferentes daquele da
vancomicina, essas características de sensibilidade e resistência frente aos
dois glicopeptídeos servem como base para uma classificação clínica (Silveira
et al., 2006).
Dos sete fenótipos descritos atualmente (A,B,C,D,E,G,L), os genes vanA
e vanB são, de longe, os mais prevalentes no mundo e são descritos

REVISÃO BIBLIOGRÁFICA
35
principalmente entre as espécies E. faecalis e E. faecium. Cepas de
enterococcus com fenótipo vanA apresentam alto grau de resistência a
vancomicina e teicoplanina e a resistência pode ser induzida pela presença de
glicopeptídeos (vancomicina, teicoplanina, avorpacina, ristocetina) e por
agentes não glicopeptídeos, como bacitracina e polimixina B (Cetinkaya et al.,
2000). O fenótipo vanB apresenta altos níveis de resistência a vancomicina
enquanto a sensibilidade a teicoplanina é mantida. Ambos os genes podem ser
encontrados em plasmídeos ou no cromossomo bacteriano e ser transferidos
entre espécies enterocócicas (Cetinkaya et al., 2000).
Uma situação mais preocupante foi o temor de que genes que conferem
resistência à vancomicina presentes nos VRE (Enterococcus Resistente a
Vancomicina) fossem transmitidos para os MRSA. Isto foi recentemente
confirmado em alguns casos isolados (n=7) surgidos na América do Norte,
sendo conhecidos como VRSA (S. aureus Resistente a Vancomicina) (Tenove,
2008; Zhu et al., 2008).
Em uma publicação de 2003, Chang e colaboradores reportaram em um
isolado de VRSA de uma ponta de cateter, a presença concomitante do gene
de resistência mecA e do gene vanA. Tal evidência foi confirmada através da
PCR localizado em um plasmídeo de 60 kb e a seqüência de DNA do gene
vanA VRSA foi idêntica à de uma amostra de E. faecalis resistente à
vancomicina isolada a partir da mesma cultura de ponta cateter (Chang et al.,
2003).
A consolidação da vancomicina como um antimicrobiano importante
frente a bactérias Gram positivas multiresistentes, trouxe um período de certa
tranqüilidade para a contenção destes agentes e tratamento nos centros

REVISÃO BIBLIOGRÁFICA
36
médicos. Entretanto, com o surgimento dos primeiros isolados de enterococos
resistentes a vancomicina e a emergência da sua disseminação no ambiente
hospitalar, levou a necessidade da busca de novos antimicrobianos para o
tratamento deste patógeno multirresistente (Silveira et al., 2008).
Este fato se agrava quando estes microrganismos são isolados em
infecções de corrente sanguínea. Dentre os Gram positivos, as espécies de
Enterococcus spp. aparecem entre os primeiros microrganismos mais isolados
nestas infecções (Cervera et al.,2009; Erdem et al.,2009). No Brasil Gales e
colaboradores, em um estudo multicêntrico em 2009, apontaram estes
microrganismos como oitavo entre todas as epécies encontradas e terceiro
entre os Gram positivos. De todos os isolados de enterococcus, 7,7% eram
VRE.
Enterococcos é frequentemente isolado em pacientes transplantados,
especialmente em transplantes de rim e fígado (Patel et al., 2001). Nestes
pacientes, infecções de corrente sanguínea é uma das mais comuns infecções
causadas por VRE. Esta infecção frequentemente é severa e associada a uma
bacteremia recorrente e persistente, e maior risco de morte (Nusair et al., 2008;
Asensio et al., 2008; Bhavinani et al., 2000; Lodise et al., 2002; Ghanen et al.,
2007).
3.3.3. -Lactamases de Espectro Ampliado (ESLs)
Os -lactâmicos são os agentes antimicrobianos utilizados com maior
frequência para o tratamento de infecções bacterianas. A ampla utilização
destes agentes principalmente das cefalosporinas de segunda e terceira
gerações, para o tratamento de infecções causadas por enterobactérias, foi

REVISÃO BIBLIOGRÁFICA
37
acompanhada pelo surgimento e disseminação de cepas bacterianas
resistentes produtoras de enzimas do tipo ESβL. A resistência aos -lactâmicos
dificulta o tratamento das infecções causadas por estes patógenos,
aumentando a morbi-mortalidade dos pacientes acometidos assim como os
custos diretos e indiretos relativos às internações (Tumbarello et al. 2006).
ESLs são enzimas capazes de hidrolisar todos os antimicrobianos β-
lactâmicos com exceção das cefamicinas (cefoxitina, cefotetan) e dos
carbapenenêmicos (imipenem, meropenem) e são inibidas pelo ácido
clavulânico, tazobactam e sulbactam. Pertencem à classe molecular A de
Ambler (1980) e estão distribuídas no grupo 2be de Bush e Jacoby (2010). As
ESL apresentam um resíduo serina no seu sítio ativo e são codificadas por
genes localizados em plasmídeos que podem também codificar resistência aos
aminoglicosídeos, tetraciclina, cloranfenicol, quinolonas e
trimetoprim/sulfametoxazol (Rossolini et al. 2008d;Winokur et al. 2001).
Outras ESLs tais como PER, VEB, BES, GES, BEL, TLA e SFO,
também foram descritas, porém SHV, TEM e CTX-M são as mais prevalentes
entre as enterobactérias (Harada et al.,2008).
3.3.3.1. TEM
As ESBLs do tipo TEM são derivadas das beta-lactamases de espectro
restrito do tipo TEM-1 e TEM-2. A descrição da primeira β-lactamase desta
classe, denominada TEM-1 ocorreu em 1960. Esta enzima foi encontrada em
uma amostra clínica de Escherichia coli isolada da corrente sanguínea de uma
jovem paciente grega chamada Temoniera, origem da sigla da β-lactamase
TEM (Turner 2005). A β-lactamase TEM-2 foi a primeira derivada da TEM-1

REVISÃO BIBLIOGRÁFICA
38
(Barthelemy et al. 1985). Já a β-lactamase TEM-3, reportada em 1988, foi a
primeira a apresentar o fenótipo ESβL (Sougakoff et al. 1988). Atualmente
existem mais de 160 derivados de TEM
(http://www.lahey.org/Studies/temtable.asp) sendo que algumas são resistentes
aos inibidores de β-lactamase e a grande maioria apresenta o fenótipo ESβL
(Bradford 2001b). Esta enzima é codificada por genes localizados, usualmente,
em elementos genéticos móveis, plasmídeos e transposons, que por meio de
uma série de eventos de transposição e rearranjos, migram para diferentes
plasmídeos que circulam entre microrganismos da mesma espécie ou de
espécies diferentes. Devido à grande facilidade de disseminação, esta enzima
tem sido descrita em diversas espécies de Enterobactérias, e em outras
espécies bacterianas (Paterson and Bonomo 2005). Embora essas β-
lactamases sejam mais frequentemente relatadas em E. coli, Klebsiella spp.,
EnterobaCter aerogenes, Morganella morganii, Proteus mirabilis, Proteus
rettgeri, e Salmonella spp. (Bonnet et al. 1999;Marchandin et al. 1999;Morosini
et al. 1995;Palzkill et al. 1995;Perilli et al. 2000;Tessier et al. 1998), têm sido,
também, reportadas entre bactérias Gram negativas não Enterobacteriaceae.
3.3.3.2. SHV
A denominação da sigla SHV vem de uma propriedade bioquímica da
enzima variável sulfidrila (“sulphydryl variable”) (Tzouvelekis and Bonomo
1999a). O fenótipo ESβL é caracterizado a partir de substituições de
aminoácidos que ocorrem no gene blaSHV-1, e estas alterações assemelham-se
as que acontecem com as ESβL do tipo TEM (Huletsky et al. 1993).

REVISÃO BIBLIOGRÁFICA
39
Em 1983, foi descrito o primeiro relato de uma SHV de espectro
ampliado, denominada SHV-2, encontrada em amostras clínicas de K.
pneumoniae, K. ozaenae e Serratia marcescens (Knothe et al. 1983). Em 1988,
SHV-3 e SHV-4 foram descritas por Jarlier e colaboradores (Jarlier et al. 1988)
e Buré e colaboradores (Bure et al. 1988), respectivamente. Ambas foram
encontradas em amostras clínicas de K. pneumoniae recuperadas de hospitais
franceses. SHV-5 foi descrita na Grécia e Tailândia (Chanawong et al., 2001; ,
Neonakis et al., 2003; Poirel et al., 2004a), enquanto SHV-12 foi identificada,
até o momento, somente na Tailândia (Chanawong et al., 2001). Desde 1982,
quando a ceftazidima foi disponibilizada para o uso clínico, até o presente
momento, foram descritas mais de 140 variantes da β-lactamase da classe
SHV (http://www.lahey.org/Studies/webt.asp#SHV).
A enzima SHV já foi relatada em vários membros da família
Enterobacteriaceae e em Acinetobacter spp. (Poirel et al., 2004a, Zavascki et
al., 2010). Em P. aeruginosa, poucos isolados foram descritos como produtores
dessa (Bush et al.,1995) enzima. Em K. pneumoniae, a enzima SHV-1 é
responsável por até 20% da resistência a ampicilina mediada por plasmídeos
nesta espécie (Tzouvelekis and Bonomo 1999b). Em muitas cepas de K.
pneumoniae o gene blaSHV-1, ou outro gene relacionado, apresenta-se integrado
ao cromossomo bacteriano (Livermore 1995). Inúmeros relatos de surtos
envolvendo K. pneumoniae produtoras de -lactamases de espectro ampliado
têm sido reportados (Coque et al. 2008).
Apesar de grande parte das -lactamases do tipo SHV apresentar
fenótipo ESβL, algumas, como por exemplo, SHV-10 e SHV-11, possuem
fenótipo de resistência aos inibidores das -lactamases e apesar de hidrolisar

REVISÃO BIBLIOGRÁFICA
40
as penicilinas, essa enzima apresenta uma hidrólise reduzida para as
cefalosporinas e, dessa forma, não é classificada como ESβL (Bradford 2001a).
3.3.3.3. CTX-M
As ESBL do tipo cefotaximase (CTX-M) são codificadas por genes
localizados em plasmídeos. Essas β-lactamases têm a propriedade de conferir
resistência a todas as cefalosporinas de espectro ampliado, porém,
apresentam como substratos preferenciais a cefotaxima e a ceftriaxona
(Bonnet 2004a;Cartelle et al. 2004;Rossolini et al. 2008c).
O primeiro relato da produção de CTX-M ocorreu em 1990, a partir de
um isolado clínico de E. coli recuperado na Alemanha (Bauernfeind et al.,
1990). Desde então, essas enzimas vêm se disseminando mundialmente em
diferentes gêneros de enterobactérias (Bonnet 2004b;Rossolini et al. 2008e).
Atualmente, encontram-se descritos na literatura mais de 80 tipos de CTX-M
diferentes (http://www.lahey.org/Studies/other.asp#table1).
No início da década de 1990, a β-lactamase CTX-M-2 disseminou-se
rapidamente na Argentina e em países vizinhos (Rossolini et al. 2008b). Outros
membros da família CTX-M estão distribuídos em diversos lugares do mundo;
CTX-M-9 e CTX-M-14 predominantemente na Ásia e na Espanha (Hernandez
et al. 2005;Munday et al. 2004), CTX-M-3 e CTX-M-15 na Europa. Apesar de
não ter sido isolada no Reino Unido antes de 2001, a enzima CTX-M-15 é
atualmente a ESβL mais prevalente em amostras bacterianas de E. coli e K.
pneumoniae nesta região (Livermore and Hawkey 2005).
Durante muitos anos, a identificação da enzima CTX-M esteve restrita
aos membros da família Enterobacteriaceae. Porém em isolados clínicos de P.

REVISÃO BIBLIOGRÁFICA
41
aeruginosa já foram descritas as enzimas CTX-M-1, CTX-M-43 (al Naiemi et
al., 2006; Celenza et al., 2006) e CTX-M-2 (Picão et al., 2009a; Picão et al.,
2009b). A disseminação das β-lactamases do tipo CTX-M está causando
importantes e imprevisíveis mudanças na epidemiologia da resistência aos
antibióticos β-lactâmicos (Rossolini et al. 2008a).
3.3.5. Klebsiella pneumoniae Carbapenemase (KPC)
Os carbapenêmicos são considerados os agentes mais potentes contra
infecções causadas por bactérias Gram negativas devido à sua elevada
afinidade pelas PBPs, estabilidade diante da maioria das β-lactamases e pela
excelente permeabilidade através da membrana externa bacteriana (Woodford
et al., 2004). No entanto, nos últimos anos, o isolamento de bactérias Gram
negativas resistentes também aos carbapenêmicos vêm se tornando mais
frequente em diversas partes do mundo (Nordmann and Poirel 2002).
Em enterobactérias, a resistência a esta classe de antimicrobianos pode
ser causada pela produção de carbapenemases. As carbapenemases do tipo
KPC (sigla para “Klebsiella pneumoniae carbapenemase”), incluídas na classe
A (Ambler 1980b) ou grupo 2f de Bush (Bush et al. 1995b) são enzimas que
são capazes de hidrolisar penicilinas, cefalosporinas, monobactâmicos e
carbapenêmicos (Queenan and Bush 2007b).
A primeira descrição da enzima KPC-1 ocorreu no ano de 2001, na
Carolina do Norte (EUA), a partir de um isolado de K. pneumoniae coletado em
1996, durante um estudo de vigilância denominado ICARE (“Intensive Care
Antimicrobial Resistance pidemiology”). No entanto, uma recente correção na
sequência de nucleotídeos do gene blaKPC-1 indicou que as enzimas KPC-1 e

REVISÃO BIBLIOGRÁFICA
42
KPC-2 são idênticas (Yigit et al., 2001). A partir de então, relatos de KPC-2
tornaram-se frequentes na Costa Leste dos Estados Unidos e rapidamente
expandiu-se, sendo descrito em diversas partes do mundo (Queenan and Bush
2007a). Até o momento, onze variantes de KPC foram descritas
(http://www.lahey.org/studies).
O gene que codifica a enzima KPC está frequentemente localizado em
um elemento genético móvel com grande potencial para disseminar-se. Esta
característica somada ao fato deste gene ser, frequentemente, encontrado em
cepas de K. pneumoniae, que é conhecida por sua capacidade de acumular e
transferir determinantes de resistência, é motivo de muita preocupação pelas
agências nacionais de saúde (Queenan and Bush 2007c). Embora as
carbapenemases do tipo KPC sejam predominantemente descritas em cepas
de K. pneumoniae, existem relatos dessa enzima em cepas de Enterobacter
spp. (Hossain et al. 2004); (Bratu et al. 2005b), Escherichia coli (Navon-
Venezia et al. 2006), Salmonella spp. (Miriagou et al. 2003), Citrobacter
freundii, Pseudomonas aeruginosa (Villegas et al. 2007), entre outras espécies
(Sacha et al., 2009). O tratamento de infecções causadas por microrganismos
que expressam esse determinante de resistência é extremamente difícil,
resultando muitas vezes em altas taxas de mortalidade, devido à resistência a
múltiplos agentes antimicrobianos (Bratu et al. 2005a).
A enzima KPC-2 foi primeiramente descrita no Brasil por Monteiro e
colaboradores (2009), em quatro amostras de K. pneumoniae isoladas entre
setembro e novembro de 2006 em um hospital de Recife. Desde então,
isolados de K. pneumoniae e E. cloacae produtores de KPC-2 também foram
descritas no Rio de Janeiro, São Paulo e Rio Grande do Sul, revelando que a

REVISÃO BIBLIOGRÁFICA
43
emergência deste mecanismo de resistência em hospitais brasileiros vem
ocorrendo desde o ano de 2005 (Pavez et al., 2009; Peirano et al., 2009;
Zavascki et al., 2009a).
Em um recente estudo do Programa de Vigilância SENTRY (SENTRY-
“Antimicrobial Surveillance Program”), realizado com 3.220 amostras de
bactérias Gram negativas, coletadas de centros médicos no Brasil, entre o
período de janeiro de 2003 a maio de 2008, constatou-se que Klebsiella spp. foi
o segundo microrganismo mais prevalente em infecções de corrente
sanguínea, porém nenhum isolado apresentou KPC (Andrade et al., 2008). Em
2010, Linares e colaboradores, avaliaram 1.057 transplantes de órgão sólidos
realizados em um centro Médico na Espanha, entre o período de julho de 2003
a dezembro de 2007,e observaram 116 episódios de infecções por K.
pneumoniae sendo que mais de 50% desses eram ICS. Dos isolados desse
estudo, nenhum apresentou produção de carbapenemase. O mesmo fato se
repetiu em outro estudo, agora realizado no Brasil, no qual K. pneumoniae foi a
segunda espécie de Gram negativo mais prevalente em ICS e nenhum isolado
de KPC foi relatado (Silva JR et al.,2010).
3.3.4. Metalo--Lactamases
As metalo-β-lactamases (MBL) são pertencentes ao grupo 3 de Bush
(Bush et al. 1995a) e à classe molecular B de Ambler (Ambler 1980a).
Apresentam potente atividade contra carbapenêmicos e hidrolisam todos os -
lactâmicos comercialmente disponíveis, exceto os monobactâmicos
(aztreonam). Este grupo requer íons Zn+2 ou outros cátions divalentes como
cofatores no sítio ativo. São resistentes à ação dos inibidores das serino-β-

REVISÃO BIBLIOGRÁFICA
44
lactamases como ácido clavulânico, sulbactam e tazobactam (Queenan and
Bush 2007d), embora sofram inibição por agentes quelantes (EDTA, derivados
de tiol e do ácido dipicolínico); (Payne et al., 1997a; Payne et al., 1997b; Laraki
et al., 1999; Bebrone, 2007).
As MBLs, como todas as β-lactamases, podem ser divididas em
cromossomais e codificadas por genes móveis. As MLs intrínsecas são
normalmente mediadas por cromossomo (Walsh et al. 1994) e são encontradas
em um número limitado de espécies, como: Bacilus cereus, Aeromonas
hydrophila, Flavobacterium odoratum, Legionella gormanii, Bacteroides fragilis,
Stenotrophomonas maltophilia (Hirakata et al. 1998). MLs adquiridas
apresentam-se em estruturas genéticas móveis que fornecem mobilidade ao
gene, facilitando a disseminação dessa enzima entre diferentes espécies
bacterianas. Atualmente, são conhecidas 10 sub-classes de MBLs adquiridas:
IMP (imipenemase), VIM (Verona imipenemase), SPM (São Paulo metallo-β-
laCtamase), GIM (German imipenemase), SIM (Seul imipenemase), AIM
(Australian imipenemase), KHM (Kyorin Health Science MBL), NDM (New Delhi
metallo-β-laCtamase), DIM (Dutch Imipenemase) e TMB (Tripoli metallo-β-
laCtamase) (Osano et al., 1994; Lauretti et al., 1999; Toleman et al., 2002;
Castanheira et al., 2004; Lee et al., 2005; Yong et al., 2007a, Sekiguchi et al.,
2008; Salabi et al., 2009; Poirel et al., 2009).
Embora essas enzimas sejam mais frequentemente isoladas em
espécies de bactérias Gram negativas não-fermentadoras, como Pseudomonas
aeruginosa e Acinetobacter baumannii, cada vez mais estas enzimas têm sido
reportadas em membros da família Enterobacteriaceae, incluindo K.
pneumoniae (Lincopan et al. 2005b;Morfin-Otero et al. 2009).

REVISÃO BIBLIOGRÁFICA
45
O primeiro relato de MβLs adquirida ocorreu em 1994 (Osano et al.
1994), sendo descrito como uma subclasse denominada IMP-1. Esta enzima foi
encontrada em uma cepa clínica de Serratia marcescens isolada no Japão, a
qual apresentava fenótipo de resistência a imipenem e cefalosporinas de amplo
espectro. Diversas variantes dessa enzima foram então descritas em P.
aeruginosa, Acinetobacter spp. e enterobactérias de praticamente todos os
continentes (Walsh et al., 2005; Walsh et al., 2008). As variantes IMP-1, IMP-16
e IMP-18 já foram reportadas em isolados clínicos brasileiros de P. aeruginosa
(Mendes et al., 2004; Mendes et al., 2007b; Xavier et al., 2007). Em 2005,
Lincopan e colaboradores relataram o primeiro caso de K. pneumoniae
produtora de IMP-1 isolada de uma única amostra, na América Latina
(Lincopan et al. 2005a).
Em 1999, a segunda subclasse descrita de MBL adquirida foi
denominada VIM foi observada em uma amostra de P. aeruginosa originária de
Verona, Itália (Lauretti et al., 1999). Muitas variantes de VIM já foram descritas,
tanto em microrganismos fermentadores da glicose, quanto em não
fermentadores. Embora a maioria das descrições tenha ocorrido na Europa,
algumas 25 variantes já foram identificadas na América e Ásia (Queenan &
Bush, 2007; Walsh et al., 2005; Walsh et al., 2008).
Em 2002, foi descrita uma nova subclasse de MBL, a SPM-1 (São Paulo
Metalo-β-lactamase). Esta enzima foi isolada de uma P. aeruginosa recuperada
do trato urinário de uma paciente de quatro anos hospitalizada no complexo
HSP/UNIFESP (Toleman et al., 2002). Esta enzima parece estar
especificamente relacionada à espécie de P. aeruginosa, uma vez que, até
então, não foi encontrada em demais patógenos. Amostras produtoras desta

REVISÃO BIBLIOGRÁFICA
46
enzima foram isoladas em diversas cidades brasileiras, tais como São Paulo,
Brasília, Salvador, Fortaleza, Santo André, Londrina, Maringá, Curitiba, Recife,
Rio de Janeiro, Porto Alegre e São Luiz (Gales et al., 2003; Pellegrino et al.,
2002; Poirel et al., 2004b; Nouér et al., 2005; Vieira et al., 2005; Zavascki et al.,
2005), sendo que na maioria destas descrições foi encontrado um perfil clonal
predominante, denominado clone SP. A única descrição da produção de SPM-
1 fora do Brasil ocorreu em 2010, na Suiça. Entretanto, a amostra de P.
aeruginosa, em que o gene blaSPM-1 foi detectado, foi isolada de um paciente
que havia sido atropelado e possuía internação prévia no Hospital Geral
Regional, em Recife (Salabi et al., 2010).
Em 2002, cinco isolados de P. aeruginosa foram recuperados de
diferentes pacientes de um centro médico em Dusseldorf, Alemanha. Estes
isolados codificavam uma nova β-lactamase da classe B, denominada GIM-1
(German imipenemase) (Castanheira et al., 2004). Em 2005, Lee e
colaboradores reportaram uma nova enzima do tipo MBL, SIM-1 que foi
identificada em sete isolados clínicos de Acinetobacter baumannii na Coréia, os
quais haviam sido isolados entre setembro de 2003 e novembro de 2004.
Recentemente, foram descritas novas subclasses de MBLs adquiridas:
AIM-1(Australian imipenemase), recuperada de um isolado clínico de P.
aeruginosa, em 2007, na Austrália (Yong et al., 2007b); KHM-1 (Kyorin Health
Science metalo-β-lactamase), descrita em 2008, no Japão, em um isolado
clínico de C.freundii (Seikiguchi et al., 2008); NDM-1 (MIM- 1) (New Delhi
metalo-β-lactamase), descrita em 2008, na Índia, em um isolado clínico de K.
pneumoniae, e posteriormente, sua ocorrência também foi relatada em
enterobactérias na Inglaterra (Doumith et al., 2009; Yong et al., 2009); TMB-1

REVISÃO BIBLIOGRÁFICA
47
(Tripoli MBL), recuperada de um isolado clínico de P. aeruginosa detectado da
Libia (África), em 2008 (El Salabi et al., 2009) e DIM-1 (Dutch Imipenemase),
descrita em 2009 a partir de um isolado de P. stutzeri originário da Holanda
(Poirel et al., 2010).
3.4. Sistemas Automatizados no Processamento de Hemoculturas
O sangue é um dos espécimes clínicos mais importantes dentro de um
laboratório de microbiologia. A hemocultura permite a identificação de
microrganismos na corrente sanguínea e é o método disponível mais sensível
na detecção da bacteremia.
A automação no laboratório de microbiologia ocorreu inicialmente em
1970 com a introdução do primeiro sistema semi-automatizado de hemocultura.
Posteriormente, em 1990 estes sistemas foram aperfeiçoados e criados os
equipamentos de hemocultura que realizam monitoramento contínuo do
crescimento microbiano.
Os métodos dos sistemas automatizados disponíveis comercialmente
baseiam-se na inoculação de amostra de sangue dentro de um frasco
hermeticamente fechado e estéril que contêm meio de cultura enriquecedor
que favorece o crescimento dos microrganismos, acrescido de substâncias que
inativam ou adsorvem os antibióticos.
Cada frasco possui meios específicos que, por suas características
químicas, permitem a seleção do microrganismo que deseja-se isolar: aeróbios,
anaeróbios, fungos ou micobactérias.
Os frascos de hemocultura são incubados a 37C dentro de
equipamentos automatizados que vão promover a agitação contínua do meio

REVISÃO BIBLIOGRÁFICA
48
de cultura. Na base do frasco de hemocultura existe um sensor de CO2 que
monitora a quantidade e CO2 no meio. Dentro do equipamento, próximo à base
onde o frasco de hemocultura é colocado, um diodo emissor e receptor de luz é
posicionado e vai detectar alterações de cor e/ou fluorescência.
Com o crescimento bacteriano e conseqüente produção de CO2, o
sensor localizado dentro do frasco promove, através de uma reação química,
alteração de cor ou emissão de fluorescência, com conseqüente alteração da
luz refletida detectada pelo diodo presente no equipamento. Este sinal é
transmitido a um computador acoplado ao equipamento. O computador possui
diversos algorítimos para reportar a detecção de um frasco positivo: (a) quando
a reflectância excede o ponto de corte estabelecido, (b) quando o instrumento
reconhece um aumento linear nos níveis de CO2 ou (c) quando ococrre
alteração nas taxas de CO2 produzido (Nolte et al.,1993).
Algumas metodologias, além de utilizar detecção colorimétrica ou
fluorimétrica podem também utilizar detecção manométrica da produção do
CO2.
Apesar do aperfeiçoamento das metodologias automatizadas para
aumentar a sensibilidade e reduzir o tempo de detecção do crescimento
microbiano a técnica é dependente de condições pré-analíticas ideais como:
volume da amostra, quantidade de amostras, momento e técnica de coleta.
Estudos revelam que 0,20 a 6% das amostras apresentam resultados
falso-negativos, onde o paciente apesar de apresentar episódio de bacteremia
o agente não é isolado (Shiguei et al., 1995; Smith et al., 1995; Ziegler et al.,
1998).

REVISÃO BIBLIOGRÁFICA
49
Após a detecção do crescimento do agente no equipamento de
hemocultura automatizado é realizado diretamente do frasco, pesquisa do
microrganismo com coloração pelo método de Gram e semeadura em meios
seletivos e enriquecedores para o isolamento do agente.
Após 24 horas de incubação é possível observar em placa o crescimento
dos microrganismos e proceder à etapas de identificação e teste de
sensibilidade.
A identificação dos agentes pode ser realizada por técnicas manuais,
onde características bioquímicas e físicas são observadas e interpretadas, ou
através de técnicas automatizadas, através de diversas provas bioquímicas em
que se estabelece um perfil bioquímico característico de cada.
Os métodos automatizados além de fornecer maior precisão nos
resultados proporciona rapidez na identificação, podendo esta ocorrer a partir
de 2 horas, a depender do microrganismo. No Brasil, os sistemas
automatizados mais utilizados são: Vitek®; Vitek-2® (bioMérieux, Hazelwood,
MO); Walk-Away® (DADE, West Sacramento, CA); BD Phoenix®, sendo que o
Phoenix apresenta no mesmo painel identificação e teste de susceptibilidade
(Anvisa, 2008).
O teste de sensibilidade também pode ser realizado através de
metodologia manual ou automatizada. A metodologia manual baseia-se na
técnica de Kirb-Bauer de disco difusão onde os halos de inibição são
interpretados de acordo com as recomendações do CLSI (Clinical Laboratory
Intitute). Outros métodos, tais como os Epsilométricos, como ETEST também
podem ser utilizados quando pretende-se realizar a determinação da
Concentração Inibitória Mínima (CIM) a determinados antimicrobianos.

REVISÃO BIBLIOGRÁFICA
50
Já as técnicas automatizadas baseiam-se na técnica de microdiluição
em caldo onde diversas concentrações de antibióticos são utilizadas para
chegar à determinação da CIM. Estas concentrações são interpretadas pelos
equipamentos de acordo com as recomendações vigentes do CLSI (2010).
Tanto a metodologia manual como a automatizada permitem a detecção
de alguns mecanismos de resistência específicos, seja pela técnica de disco
aproximação seja pela técnica de uso combinado de antibióticos. Os fenótipos
geralmente reportados são a presença de ESBL e MLSb (Macrolídeos,
Lincosaminas e Estreptograminas).
3.5. Microbiologia Molecular Aplicada no Diagnóstico Clínico
3.5.1. Reação de Polimerização em Cadeia - PCR, PCR Multiplex e
PCR em Tempo Real
A técnica da PCR, idealizada em 1983 por Kerry Mullis tornou-se
realidade após o isolamento e purificação da enzima Taq DNA polimerase a
partir da bactéria termofílica Thermus aquaticus (Saiki et al., 1988). Esta
técnica passou a ser a mais utilizada em laboratórios de pesquisa e consiste de
uma reação de amplificação do DNA extraído e adicionado com dNTPs
(desoxirribunucleotídeos trifosfatos), oligonucleotídeos (primers) iniciadores da
reação e Taqpolimerase. É facil de ser executada e é realizada em três fases:
desnaturação, anelamento e extensão. Na primeira etapa, desnaturação, a
dupla fita de DNA é separada, na segunda etapa, anelamento, os primers se
posicionam no DNA, na terceira etapa a Taq polimerase inicia a extensão do
DNA (Konenam et al. 2006b).

REVISÃO BIBLIOGRÁFICA
51
A partir do uso rotineiro da PCR, outros métodos e aplicações puderam
ser elaborados. Uma delas é a PCR Multiplex. Este consiste na amplificação de
mais de um alvo simultaneamente pela adição de diferentes conjuntos de
primers específicos para cada alvo no mesmo tubo de reação. Para isto,
devem-se tomar alguns cuidados com os conjuntos de primers escolhidos, tais
como a temperatura de anelamento dos primers usados deve ser semelhante,
os conjuntos de primers devem gerar produtos com tamanhos moleculares
distintos, e estes devem apresentar especificidade de anelamento apenas com
uma sequência alvo. Apesar de ser uma estratégia de amplificação de uso
conveniente para detecção de DNA de agentes infecciosos, apresenta maior
dificuldade no desenvolvimento da metodologia e menor sensibilidade quando
comparada com amplificação de uma única sequência alvo (Hayden, 2004).
A técnica de PCR em Tempo Real é um refinamento da técnica original
de PCR, pois combina a amplificação e a quantificação de uma sequência de
DNA alvo por meio da detecção de fluorescência utilizando sondas específicas
marcadas com fluoróforos, ou com base na determinação da temperatura de
desnaturação de uma seqüência de DNA dupla fita (“melting temperature” -
Tm) marcada com substância intercalante fluorescente (Klein, 2002; Kubista et
al., 2006). É considerado um método homogêneo de amplificação de DNA, no
qual amplificação e detecção são realizadas no mesmo tubo de reação,
podendo eliminar processamentos pós-PCR, diminuindo o manuseio de
produtos de amplificação e o risco de contaminação cruzada (Hayden, 2004).
Os princípios aplicados para detecção de uma sequência alvo por PCR em
Tempo Real são baseados na mensuração de fluorescência durante a reação.
A quantidade de produto formado é monitorada durante o decorrer da reação

REVISÃO BIBLIOGRÁFICA
52
por meio da detecção da fluorescência detectada por diferentes filtros de
captação em determinados comprimentos de onda. A fluorescência é
proporcional à quantidade de produto formado em cada ciclo e o número de
ciclos de amplificação necessários para obter uma determinada quantidade de
DNA é registrado (Cicldo Threshold- Ct). Com a alta eficiência dos reagentes,
sensibilidade dos instrumentos e otimização de técnicas, o número de
moléculas de DNA de uma determinada seqüência presente em uma amostra,
pode ser determinado com grande sensibilidade (Mackay, 2004; Kubista et al.,
2006).
Hoje se encotram disponíveis Kits que utilizam a metodologia da PCR
em Tempo Real para o diagnóstico rápido em infecções de corrente sanguínea,
sendo o SeptFast (LightCycler SeptiFast assay; Roche Diagnostics) o mais
conhecido. Porém se trata de um método comercial em que seu uso fica
restrito em laboratórios de analises clínicas, pois requer excelente habilidade
técnica para o manuseio e é muito caro quando comparado a cultura (Lehman
et al., 2009).
3.5.2. Validação de Novos Métodos de Diagnósticos
A introdução de um novo método de diagnóstico em laboratório clínico
necessita de uma validação analítica e clinica, principalmente com os métodos
in house, para garantir uma maior confiabilidade ao protocolo escolhido. A
validação analítica avalia a acurácia, precisão, sensibilidade e especificidade
do método, que são indicadores das características do teste e por isso, não
dependem da prevalência da doença ou de doenças correlacionadas com a
população em que o teste de diagnóstico vai ser utilizado. A sensibilidade

REVISÃO BIBLIOGRÁFICA
53
analitica é testada medindo o mais baixo limite de detecção do alvo ou do
microrganismo desejado. A especificidade analítica mede a capacidade do
teste em detectar outro DNA que não o do microrganismo desejado.
Cada protocolo de diagnóstico molecular, após ter seus parâmetros
analíticos testados, também deve ter sua validação clínica determinada na
população a que se destina. Essa validação clínica é realizada analisando-se o
desempenho do teste em um número adequado de pacientes. A utilidade
clínica de um teste de diagnóstico para a população em que será aplicado pode
ser avaliada pelos valores preditivos (positivo e negativo), que são calculados a
partir de análise dos parametros de validação clínica comparados com a
prevalência da doença (Rossetti et al., 2006).
De acordo como documento MM3-A2 do “Clinical Laboratory Standard
Institute” (CLSI, 2006), um roteiro de avaliação para introdução de novo método
de diagnóstico deve ser seguido, como a determinação da sensibilidade
analítica (limite de detecção – LoD), especificidade analítica, precisão, cutoff
(valor de corte), sensibilidade e especificidade diagnóstica, acurácia
diagnóstica, valores preditivos e diagnóstico.
A Agência Nacional de Vigilância Sanitária – ANVISA regulamenta
através da RDC 302/2005, que tanto laboratórios clínicos quanto de pesquisa
devem validar os métodos desenvolvidos no próprio laboratório fornecendo
evidências quanto ao desempenho (RDC 302/2005; Normas e Manuais
Técnicos, 2005).

MATERIAIS E MÉTODOS
54
4 - MATERIAIS E MÉTODOS
4.1. Casuística
Foram selecionadas para o estudo as hemoculturas positivas e as cinco
primeiras hemoculturas negativas de cada mês, independente do centro
referente, provenientes de pacientes submetidos a transplante de órgãos
sólidos internados no HSP e no HRH, coletadas entre fevereiro de 2009 a
fevereiro de 2010.
Foram incluídas no presente estudo 59 hemoculturas negativas e 126
hemoculturas positivas pelo sistema automatizado Bactec® (Becton Dickinson,
EUA). Destas, 4 hemoculturas negativas e 14 hemoculturas positivas foram
provenientes de 11 pacientes admitidos na unidade de transplantados de
órgãos sólidos (rim, rim e pâncreas, pulmão, fígado e coração) localizada no
10º andar do HSP e 55 hemoculturas negativas e 112 hemoculturas positivas
foram provenientes de 106 pacientes do serviço de transplante de órgãos
sólidos (rim e rim e pâncreas) do HRH.
4.1.1. Variáveis Clínicas
Os dados clínicos dos pacientes foram coletados no Same (Serviço de
Arquivo Médico) do HRH e no Same do HSP. O estudo foi retrospectivo
incluindo pacientes submetidos a transplantes de órgãos sólidos (rim,
rim/pâncreas, pulmão, coração e fígado), como também os pacientes
internados nos hospitais avaliados com intercorrências clínicas pós-transplante.
Os laudos com os resultados da hemocultura e antibiograma foram fornecidos
pelo Laboratório Central do HSP, UNIFESP.

MATERIAIS E MÉTODOS
55
Para descrever os episódios de ICS e analisar os fatores de risco
presentes no momento da coleta da hemocultura, foram coletadas as
informações relacionadas abaixo:
1- Dados do Paciente:
Nome;
Data de Nacimento;
Sexo;
2- Dados da Internação:
Hospital de Internação (HSP, HRH);
Data da Internação;
3- Dados do Transplante:
Data do Transplante;
Tipo de Doador: Vivo ou Falecido;
Órgão transplantado;
Tempo de Transplante até a bacteremia;
4- Dados da Infecção:
Data da coleta da bacteremia;
Tempo de Internação até a Bacteremia;
Resultado da coloração de Gram da hemocultura;
Data de validação do Gram;
Resultado da Hemocultura (microrganismos e antibiograma);

MATERIAIS E MÉTODOS
56
Data de validação da Hemocultura;
5- Dados do Tratamento:
Data de recebimento da coloração de Gram pelo médico;
Antibióticoterapia até quatro dias antes do resultado do Gram;
Antibióticoterapia até quatro dias após o resultado do Gram;
Avaliação da adequação da antibioticoterapia após o Gram;
Data de recebimento da Hemocultura pelo médico;
Antibióticoterapia até quatro dias antes do resultado da
Hemocultura;
Antibióticoterapia até quatro dias após o resultado da
Hemocultura;
Avaliação da adequação da antibioticoterapia após o resultado
do Gram;
Adequação da terapía empírica após o resultado da
Hemocultura;
A adequação da antibioticoterapia foi avaliada por 2 médicos e definida
como adequada quando o paciente utilizava pelo menos um dos
antimicrobianos ao qual a bactéria identificada era sensível “in vitro” e
inadequada quando o microrganismo isolado não era sensível “in vitro” a
nenhum dos antimicrobianos utilizado pelo paciente ou se este não estava em
uso de nenhum antimicrobiano, porém não foi avaliada quanto a abordagem
clínica da infecção (Pereira, 1997).

MATERIAIS E MÉTODOS
57
4.2. Coleta das Hemoculturas de Acesso Periférico
Frente a suspeita de infecção, as coletas de hemocultura foram
realizadas pela equipe de enfermagem seguindo normas preconizadas pelas
Comissões de Controle Infecção Hospitalar (CCIH) do HSP e do HRH.
Dos pacientes adultos, foram coletados 10 mL de sangue no frasco
BACTEC Plus Aerobic/F® e das crianças foram coletados 5 mL de sangue no
frasco BACTEC Plus Pediatric/F®.
Na persistência da febre uma coleta adicional de hemocultura foi
realizada a critério do médico assistente após 24 horas da primeira coleta.
4.2.1. Coleta das Hemoculturas de Acesso Central – Cateteres
A coleta de hemocultura de cateter foi realizada em pacientes portadores
de cateter venoso central (CVC), nos quais não é possível ou não está indicada
à punção periférica, para coleta de amostra sanguínea. O procedimento foi
realizado pela equipe de enfermagem seguindo normas preconizadas pelas
CCIH do HSP e do HRH.
4.3. Processamentos das Amostras pelo Método Fenotípico
As hemoculturas do HSP e do HRH foram processadas no setor de
microbiologia do Laboratório Central do Hospital São Paulo, UNIFESP.
4.3.1. Determinação da positividade e negatividade das
hemoculturas pelo aparelho Bactec®
Após a coleta, os frascos de hemocultura foram processados pelo
equipamento Bactec 9240® (Becton Dickinson, Maryland, EUA), um sistema

MATERIAIS E MÉTODOS
58
automatizado que realiza a monitorização contínua do crescimento bacteriano.
O meio de cultura do frasco possui nutrientes que são metabolizados frente à
presença de microrganismos viáveis na amostra. Esta metabolização resulta na
produção de CO2 e esta alteração pode ser detectada e interpretada pelo
sistema como amostra positiva.
4.3.2. Resultado Parcial da Hemocultura Positiva pela Coloração de
Gram
Frente à positivação da hemocultura pelo sistema Bactec®, foi realizada
coloração pelo método de Gram e semeadura em meios enriquecedores e
seletivos para cultura. Os meios utilizados para o crescimento bacteriano
foram: ágar chocolate, ágar sangue e ágar eosino-metileno-blue (EMB).
4.3.3. Resultado Final da Hemocultura pelo Sistema Automatizado
Phoenix®
A identificação dos microrganismos e o teste de susceptibilidade a
antimicrobianos foram realizadas pelo equipamento Phoenix® 100 (Becton
Dickinson Microbiology Systems, Coceysville, Md.) Este sistema utiliza painéis
(PMID-121 para isolados Gram negativo, PMID-123 para Gram negativo não
fermentador e PMID-104 para Gram positivo (Anexo 2)) nos quais há provas
para identificação bacteriana e drogas antimicrobianas desidratadas. O sistema
utiliza indicadores colorimétricos e fluorométricos para leitura dos testes de
identificação e antibiograma. Todas as amostras positivas e negativas foram
encaminhadas ao Laboratório Especial de Microbiologia Clínica - LEMC da

MATERIAIS E MÉTODOS
59
Disciplina de Infectologia da UNIFESP, onde as metodologias genotípicas
foram aplicadas.
4.4. Padronização da PCR em Tempo Real
Os iniciadores utilizados para detecção do controle interno da reação
foram desenhados para o gene da hemocromatose (HFE). A diferenciação
entre as bactérias Gram positivas e Gram negativas foi feita por multiplex
TaqMan (TaqMan-MGB) em Tempo Real utilizando iniciadores universais do
gene 16S rRNA (Bispo et al., 2011). Os iniciadores para Gram positivas, Gram
negativas, controle interno e os genes de resistência blaKPC, blaSHV, blaTEM,
blaCTX, blaIMP, blaSPM, blaVIM, vanA, vanB e mecA padronizados no presente
estudo, estão apresentados na Tabela 1.
Todos os iniciadores testados para a PCR em Tempo Real utilizando o
sistema TaqMan foram testado em concentrações diferentes.

MATERIAIS E MÉTODOS
60
Tabela 1. Lista de Iniciadores de amplificação da região codificadora dos genes
estudados.
Iniciador Sequencias (5’3’) Resistência Genes
Tamanha do Fragmento (PB)
HFE_ F GATGACCAGCTGTTCGTGTTCTAT -
HFE
HFE_R CCACATCTGGCTTGAAATTCTAC 93
16S NT-341Fw
16S 522Rv
GACTCCTACGGGAGGC
GCGGCTGCTGGCAC
-
16 S rDNA
192
Sonda GP CTGYSSAGCAACGCCGCG-MGB -
-
-
Sonda GN CCTGAYSCAGAMATGCCGCG-MGB -
-
blaSHV-F ATGCGTTATACGCCTGTG
Cefalosporina
blaSHV
blaSHV-R TGCTTTGTATTCGGGCCAA 1018
blaTEM-F TGCCGCATACACTATTCTCAGAATGA
Cefalosporina
blaTEM
blaTEM-R ACGCTCACCGGCTCCAGATTTAT 868
blaCTX-F ATGT GCAG(C/T))ACCAGTAA(A/G)GT(G/T)ATGGC
Cefalosporina
blaCTX
blaCTX-R TGG GT(A/G)AARTA(A/G)GTSACCAGAA(C/T)CAGCGG 600
blaKPC-F ATGTCACTGTATCGCCGTCTAGTTC
Carbapenêmicos
blaKPC
blaKPC-R CAATCCCTCGAGCGCGAGTC 800
blaIMP-F GAATAG(A\G) (A\G)TGGCTTAA(C\T)TCTC
Carbapenêmicos
blaIMP
blaIMP-R CAAAC(C\T)ACTA(G\C)GTTATC 188
blaVIM-F GTTTGGTCGCATATCGCAAC
Carbapenêmicos
blaVIM
blaVIM-R AATGCGCAGCACCAGGATAG 382
blaSPM-F CTAAATCGAGAGCCCTGCTTG
Carbapenêmicos
blaSPM
blaSPM-R CCTTTTCCGCGACCTTGATC 798
mecA-F AAAACTAGGTGTTGGTGAAGATATACC
Meticilina
mecA
mecA-R GAAAGGATCTGTACTGGGTTAATCAG 147
vanA-F CATGACGTAATCGGTAAAATC
Glicopeptidio
vanA
vanA-R ACCGGGCAGRGTATTGAC 732
vanB-F CATGATGTGTCGGTAAAATC
ACCGGGCAGRGTATTGAC
Glicopeptidio
vanB
vanB-R 635
PB= Pares de base
A amplificação da reação de PCR em Tempo Real foi realizada no
equipamento 7500 Real Time PCR System (Applied Biosystems, CA) utilizando
o Kit Platinum SYBR Green qPCR Super Mix (Invitrogen, California, EUA) e
TaqMan Universal PCR Master Mix (Applied Biosystems, Foster City, EUA). As
condições de termociclagem foram: 2 minutos a 50°C, seguidos de 10 minutos
a 95°C e 45 ciclos de 15 segundos a 95°C e 1 minuto a 60°C. Todos os

MATERIAIS E MÉTODOS
61
iniciadores testados para a PCR em Tempo Real utilizando o fluoroforo SYBR
Green foram submetidos à análise da curva de desnaturação (melting curve)
para garantir a correta amplificação de cada conjunto de iniciador e assim,
determinar a temperatura de desnaturação (Tm- Temperatura de melting) dos
produtos de PCR para cada gene. O ciclo de dissociação dos produtos para a
curva de desnaturação (melting curve) foi: 95°C por 10 minutos, seguidos por
40 ciclos de 40 segundos a 95°C, 1 minuto a 60°C, e uma etapa para
realização da curva de desnaturação que variou de 68°C até 95°C, com
aumento gradual na temperatura de 0,5°C/s. Dessa forma, foi determinada a
temperatura de desnaturação (Tm) dos produtos de PCR dos genes analisados.
Tanto para o sistema SYBR Green quanto para o sistema TaqMan foi
avaliado o Ct em que ocorreu a detecção da positividade.
Para a PCR pelo sistema TaqMan, a positividade foi analisada a partir
da sonda detectada. Na figura 2 está esquematizada a detecção de uma
bactéria Gram negativa e Gram positiva. Quando há amplificação e a sonda
VIC , trata-se de uma bactéria Gram negativa. Quando detectada a sonda FAM
na amplificação trata-se de uma bactéria Gram positiva.

MATERIAIS E MÉTODOS
62
Figura 2. A - Amplificação do DNA de Gram positivo (curva em amarelo) e
Gram negativo (curva em vermelho); B- Sonda Gram Positivo (GP) e Sonda
Gram Negativo (GN).
4.4.1. Eficiência e Especificidade da PCR em Tempo Real
A eficiência (E) das reações para os protocolos de PCR em Tempo Real
foi calculada após construção de uma curva padrão utilizando a fórmula: E =
10(-1/-slope) -1, onde E = especificidade, “slope” = inclinação da curva no 7500
Software v2.0.4 . O valor corresponde à eficiência da reação em porcentagem
quando multiplicado por 100. As curvas foram avaliadas por um ensaio com as
amostras em triplicata. A regressão linear foi construída com o mínimo de 5
pontos e foram avaliadas as diferenças intra ensaio, com intervalo de confiança
de 95%.
Para determinar a eficiência da PCR em Tempo Real foram utilizadas
cepas controles que apresentam genes de resistências que foram cedidas
gentilmente por outros grupos de pesquisas e que estão armazenadas no
banco de microrganismos do LEMC. Todas as amostras bacterianas foram
A
GP
B
GN

MATERIAIS E MÉTODOS
63
recuperadas em ágar sangue de carneiro (5%) após incubação por 24 horas
em atmosfera aeróbia ou microaerófila. Os respectivos controles utilizados para
os genes de resistência se encontram na Tabela 2. O limite mínimo de
detecção de DNA para o gene HFE foi avaliado utilizando DNA extraído dos
frascos Bactec®. Estes foram diluídos a partir de 10 ng/µl até 100 fg/µl. Em um
tubo contendo 4 mL de salina foi preparada uma suspensão bacteriana na
escala 0,5 de Mc Farland (1,5 x 108 UFC/mL). 1 mL da solução foi transferida
para tubo de 2mL estéril e assim foi extraído o DNA bacteriano por fervura, 10
minutos a 100°C e centrifugado a 16.000g por 5 minutos. O sobrenadante foi
transferido para um tubo de 2 mL estéril. Para a preparação das amostras para
a curva padrão, foi realizado uma diluição seriada de 10-1 a 10-7 a partir do tubo
da escala 0,5 de Mc Farland (1,5 x 108 UFC/mL) para cada gene testado. Para
a padronização as curvas foram realizadas em triplicata.
A determinação da especificidade analítica foi abordada em dois
aspectos: reação cruzada e falsos positivos, conforme o documento MM3 do
CLSI. Foram avaliados a capacidade de amplificação dos alvos amplificados e
a incapacidade de amplificação cruzada de microrganismos relacionados aos
microrganismos alvo. Uma reação contendo todos os reagentes, com exceção
do DNA da amostra, também foi utilizada como controle negativo.
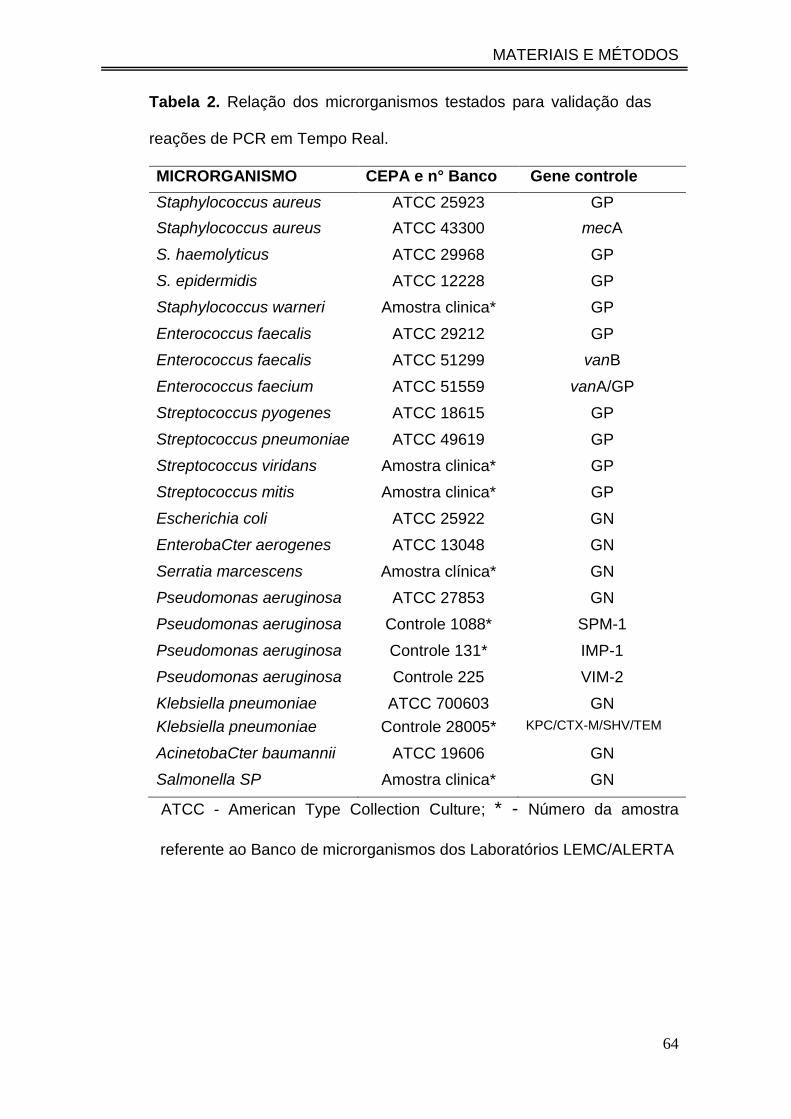
MATERIAIS E MÉTODOS
64
Tabela 2. Relação dos microrganismos testados para validação das
reações de PCR em Tempo Real.
MICRORGANISMO CEPA e n° Banco Gene controle
Staphylococcus aureus ATCC 25923 GP
Staphylococcus aureus ATCC 43300 mecA
S. haemolyticus ATCC 29968 GP
S. epidermidis ATCC 12228 GP
Staphylococcus warneri Amostra clinica* GP
Enterococcus faecalis ATCC 29212 GP
Enterococcus faecalis ATCC 51299 vanB
Enterococcus faecium ATCC 51559 vanA/GP
Streptococcus pyogenes ATCC 18615 GP
Streptococcus pneumoniae ATCC 49619 GP
Streptococcus viridans Amostra clinica* GP
Streptococcus mitis Amostra clinica* GP
Escherichia coli ATCC 25922 GN
EnterobaCter aerogenes ATCC 13048 GN
Serratia marcescens Amostra clínica* GN
Pseudomonas aeruginosa ATCC 27853 GN
Pseudomonas aeruginosa Controle 1088* SPM-1
Pseudomonas aeruginosa Controle 131* IMP-1
Pseudomonas aeruginosa Controle 225 VIM-2
Klebsiella pneumoniae ATCC 700603 GN
Klebsiella pneumoniae Controle 28005* KPC/CTX-M/SHV/TEM
AcinetobaCter baumannii ATCC 19606 GN
Salmonella SP Amostra clinica* GN
ATCC - American Type Collection Culture; * - Número da amostra
referente ao Banco de microrganismos dos Laboratórios LEMC/ALERTA

MATERIAIS E MÉTODOS
65
4.4.2. Determinação do Limite do Branco (LoB) ou “cutoff”
A determinação do Ct do Limite do Branco (LoB) ou “cutoff point” foi
realizada usando amostras negativas. O “cutoff”, limiar o qual um resultado
abaixo dele é relatado como positivo e acima como negativo, foi calculado de
acordo com o documento EP12 e EP17 do CLSI (Clinical and Laboratory
Standards Institute) e Chandelier et al.(2010).
Para dados paramétricos com distribuição dos Ct das amostras negativas
correspondentes a uma distribuição normal, com 95% de confiabilidade e valor
de α de 5%, o LoB foi calculado pela equação:
LoB = µCtN + 1.645 σCtN
CtN= Ct da amostras negativas.
Para dados não paramétricos com distribuição não correspondente a
distribuição normal e resultados quando truncado com zero, foi considerado p =
(100-α) = 95. Os valores mensurados foram ordenados de acordo com o
resultado e o percentil foi estimado pela observação dos valores classificados.
O LoB foi determinado pela posição do Ct correspondente ao 95 percentile
pela equação:
LoB = PctB(100-α)
LoB = NB (p/100) + 0,5
Quando o valor da posição não for um valor inteiro, para o cálculo da
posição do 95 percentile, foi realizado a interpolação linear. Se a posição foi
X1,5 a posição foi a média entre duas posições. Se a posição do 95 percentile
corresponder a uma posição X1,25 , a posição foi calculada como X1 + 0,25 (X2 –
X1).

MATERIAIS E MÉTODOS
66
O ciclo de “cutoff” foi determinado a partir do Ct de amostras negativas.
Os valores de Ct determinado pelas amostras negativas puderam ser
considerados quando determinados e quando foram “undetermined” os
resultados do Ct foram considerados como valores Ct correspondentes a 40,
ou seja, o último ciclo do PCR usando o método em tempo real (Chandelier et
al., 2010).
4.4.3. Determinação do Limite de Detecção - LoD (Sensibilidade)
Para métodos laboratoriais de análises clínicas foi recomendado pelo
CLSI (Clinical and Laboratory Standards Institute) no qual o limite inferior de
detecção pode ser determinado conforme o documento EP17. Foi sugerido
para determinar o LoD um número mínimo de 60 resultados de amostras
positivas com baixa concentração.
O limite inferior de detecção das reações de PCR foi obtido pela equação:
LoD = LoB + cβ SDS
LoD – limite de detecção
LoB – limite do branco
cβ – derivado do percentil de 95% da distribuição padrão Gaussiana e
fator de correção.
SDS – desvio padrão estimado para a população

MATERIAIS E MÉTODOS
67
4.5. Aplicação nas Amostras de Hemocultura
A figura 3 ilustra o fluxo do processamento das amostras.
Figura 3. Fluxograma do processamento das amostras.
LABORATÓRIO CENTRAL/ BACTEC
PCR EM TEMPO REAL
SYBER GREEN PARA O GENE HFE
EXTRAÇÃO DO DNA PELA TÉCNICA DO BRAZOL
LEMC
PCR EM TEMPO REAL TAQMAN PARA O GENE 16S rDNA
SISTEMA AUTOMATIZADO
PHOENIX
GENES DE RESISTÊNCIA A ANTIMICROBIANOS
CULTIVO DE BACTÉRIAS REFERENTES ÀS HEMOCULTURAS DISCORDANTES DO SISTEMA AUTOMATIZADO PHOENIX
TESTE FENOTIPICOS PARA OS RESPECTIVOS GENES DE RESISTÊNCIA
PCR EM TEMPO REAL PARA OS RESPECTIVOS GENES DE RESISTÊNCIA
HEMOCULTURA NEGATIVA BACTEC
HEMOCULTURA POSITIVA BACTEC
MICRORGANISMO ISOLADO ENCAMINHADO
PARA INCLUSÃO NO BANCO DE
MICRORGANISMO
COMPARAÇÃO DOS RESULTADOS PHOENIX/PCR EM TEMPO REAL
NEGATIVO POSITIVO
COLETA DE HEMOCULTURA EM TRASPLANTADOS DE ÓRGÃO SÓLIDO HSP/HRH
COLORAÇÃO DE GRAM

MATERIAIS E MÉTODOS
68
4.5.1. Extração do DNA Direto dos Frascos de Hemocultura
O DNA do microrganismo presente na corrente sanguínea de cada
paciente foi obtido a partir do sangue dos frascos de hemocultura positivos e
negativo detectados pelos sistemas automatizados. Antes de proceder à
extração do DNA foi realizada a anti-sepsia da tampa do frasco do sistema
Bactec®, com algodão ou gaze embebido em álcool a 70%. Uma agulha e
seringa foram acopladas à tampa do frasco e uma alíquota foi aspirada. Em um
microtubo foram colocados 500 µL de Brazol e a esse volume foram
adicionados 500 µL de sangue do frasco de hemocultura, e a mistura foi
homogeneizada de 3 a 5 segundos no vórtex. Em seguida foram adicionados
180 µL de clorofórmio gelado e novamente foi realizada homogeneização de 3
a 5 segundos no vórtex. O microtubo foi centrifugado a 20.000g por 12 min a
8oC. O sobrenadante foi retirado e transferido para outro microtubo contendo
500 µL de etanol gelado que foi agitado por 3-5 segundos no vórtex. O
microtubo foi centrifugado a 20.000g por 15min a 8oC. O sobrenadante foi
retirado com cuidado para não perder o pellet. O pellet foi lavado com 500 µL
de etanol gelado e em seguida foi centrifugado a 20.000g por 12 min a 8oC. O
sobrenadante foi retirado com cuidado e o pellet deixado a temperatura
ambiente para secagem e depois solubilizado em 50 µL de água dEPC e
incubado a 65º C por 30 minutos. Após o procedimento de extração, seguiu-se
diretamente para a realização das técnicas genotípicas ou as amostras foram
congeladas a -20ºC até o uso.

MATERIAIS E MÉTODOS
69
4.5.2. Aplicação da PCR em Tempo Real para o Gene HFE de
Controle Interno de Extração
Todas as hemoculturas positivas e negativas para o Bactec®, após a
extração do DNA, foram submetidas a PCR para o controle interno da reação.
A PCR em Tempo Real pelo sistema SYBR Green foi realizada em um volume
de reação de 25 μL contendo 12,5 μL Platinum kit SYBR Green qPCR Super
Mix (Platinum Taq DNA polimerase, SYBR Green I dye, Tris-HCl, KCl, 6 µM
MgCl2, 400 µM dGTP, 400 µM dATP, 400 µM dCTP, 800 µM dUTP, uracil DNA
glicosilase, e estabilizadores) (Invitrogen, California, EUA); 0,75 μL (10 µM) de
cada iniciador; 6 μL de água ultra pura e 5 μL de DNA. As reações foram
realizadas no aparelho 7500 Tempo Real PCR System (Applied Biosystems,
CA), utilizando o seguinte programa: 2 min a 50 º C, 10 min a 95 º C, seguida
de 40 ciclos de 15 segundos a 95 º C e 60 segundos a 60 º C. A dissociação
dos produtos para a curva de desnaturação (melting curve) foi a 95°C por 10
minutos, seguidos por 40 ciclos de 40 segundos a 95°C, 1 minuto a 60°C, e
uma etapa para realização da curva de desnaturação que variou de 68°C até
95°C, com aumento gradual na temperatura de 0,5°C/s. Dessa forma, foi
determinada a temperatura de desnaturação (Tm) dos produtos de PCR do
gene da hemocromatose.
4.5.3. Detecção do DNA Bacteriano PCR em Tempo Real pelo
Sistema TaqMan
As amostras de hemocultura com resultado positivo para o controle
interno de extração foram submetidas à detecção do DNA bacteriano. A PCR
em Tempo Real sistema TaqMan foi realizada em um volume de reação de

MATERIAIS E MÉTODOS
70
20μL contendo 10μL TaqMan Universal 2X Master Mix (Applied Biosystems,
CA); 0,5 μL (10 µM) de cada iniciador 0,6 μL (5 µM) da sonda para Gram
negativo 0,2 μL (5 µM) da sonda para Gram positivo, 5,2 μL de água ultra pura
e 3 μL de DNA. As reações foram realizadas no aparelho 7500 Tempo Real
PCR System (Applied Biosystems, CA), utilizando o seguinte programa: 2 min a
50 º C, 10 min a 95 º C, seguida de 40 ciclos de 15 segundos a 95 º C e 60
segundos a 60º C .
4.5.4. Detecção dos Genes que Conferem Resistências aos
Antimicrobianos por PCR em Tempo Real pelo Sistema SYBR
Green
Para as amostras identificadas pela sonda de Gram negativo pela PCR
em Tempo Real foi realizada a PCR dos genes que codificam as ESΒLs,
metalo-β-lactamases e KPC. Para as amostras identificadas pela sonda de
Gram positivoo pela PCR em Tempo Real foi realizada a PCR para os genes
vanA, vanB e mecA.
A amplificação da reação de PCR em Tempo Real para os genes blaSHV,
blaTEM, blaCTX, blaKPC, vanA, vanB e mecA foi realizada no equipamento 7500
Tempo Real PCR System, Applied Biosystems, CA, utilizando 12,5 µL Kit
Platinum SYBR Green qPCR Super Mix (Platinum Taq DNA polimerase, SYBR
Green I dye, Tris-HCl, KCl, 6 mM MgCl2, 400 µM dGTP, 400 µM dATP, 400 µM
dCTP, 800 µM dUTP, uracil DNA glicosilase, e estabilizadores) (Invitrogen,
California, EUA) e 0,75 μL (10 µM) de cada iniciador; 6 μL de água ultra pura
e 5 μL de DNA. As condições de termociclagem foram dois minutos a 50°C,
seguidos de 10 minutos a 95°C e 45 ciclos de 15 segundos a 95°C e um minuto

MATERIAIS E MÉTODOS
71
a 60°C. A dissociação dos produtos para a curva de desnaturação (melting
curve) foi a 95°C por 10 minutos, seguidos por 40 ciclos de 40 segundos a
95°C 1 minuto a 60°C, e uma etapa para realização da curva de desnaturação
que variou de 68°C até 95°C, com aumento gradual na temperatura de 0,5°C/s.
Dessa forma, foi determinada a temperatura de desnaturação (Tm) dos
produtos de PCR dos diferentes genes de resistências.
A detecção dos genes blaIMP, blaSPM, blaVIM por PCR multiplex em
Tempo Real pelo sistema SYBR Green foi realizada em um volume de reação
de 25 μL contendo 12,5 μL Platinum kit SYBR Green qPCR Super Mix
(Invitrogen, Califórnia, EUA), 0,125 μL (10 mM) de cada iniciador para os
genes blaSIM, blaSPM, blaVIM e blaGIM ; 1,25 μL (10 mM) de cada iniciador para
blaIMp; 6 μL de água ultra pura e 3 μL de DNA. As reações foram realizadas no
aparelho 7500 Tempo Real PCR System (Applied Biosystems, CA), utilizando o
seguinte programa: 2 min a 50 º C, 10 min a 95 º C, seguida de 40 ciclos de 20
segundos a 94 º C , 30 segundos a 53 º C e 60 segundos a 60 º C. Para o
SYBR Green PCR, a temperatura da amostra foi aumentada gradualmente
para 95°C para gerar curvas de dissociação (Mendes et al., 2007).
4.6. Testes Direto do Isolado Bacteriano Referente à Hemocultura
A metodologia da PCR Tempo Real foi comparada com os resultados
liberados pelo método automatizado Phoenix®. Para os resultados discordantes
entre as duas metodologias, foram realizadas testes fenotípicos e moleculares
a partir da cultura do isolado bacteriano referente à hemocultura testada. Os
exemplares bacterianos encontram-se armazenados no banco de

MATERIAIS E MÉTODOS
72
microrganismos do LEMC. Estas amostras foram retiradas do banco e
semeadas em meio ágar sangue e incubadas em estufa a 37o C por 24 horas.
Um segundo repique foi realizado, para as bactérias Gram negativas em meio
MacConkey e para as Gram positivas em meio ágar sangue e incubadas em
estufa a 37o C por mais 24 horas.
4.6.1. Teste de Triagem com Disco-Difusão para Ertapenem
Para amostras de K. pneumoniae sugestivas como produtoras de
carbapenemase, mas negativas para o gene blaKPC na metodologia de PCR em
Tempo Real, foi realizado o teste de sensibilidade por disco-difusão para
ertapenem. A preparação do inóculo bacteriano foi realizada após o
crescimento em placas de ágar MacConkey por 24 horas. Com o auxílio de
alça de semeadura, uma a três colônias isoladas de K. pneumoniae foram
transferidas para tubos contendo 4 mL de salina (NaCl - Sigma, St. Louis, MO).
A suspensão bacteriana foi homogeneizada e a turvação medida em
turbidímetro digital (Baxter®, Sacramento, EUA), para a obtenção de uma
concentração bacteriana correspondente a 0,5 da escala de McFarland, em
seguida as amostras foram semeadas em placa de ágar Müeller-Hinton (Oxoid,
Basingstoke, Inglaterra) com auxílio de swab estéril. O perfil de sensibilidade foi
realizado utilizando discos de ertapenem (10µg), (Oxoid, Basingstoke,
Inglaterra). As placas foram incubadas em aerobiose a 37ºC por 18 a 24 horas.
Após este período, a leitura foi realizada e o halo de inibição interpretado de
acordo com os critérios estabelecidos pelo CLSI para EnterobaCteriaceae.
Dessa forma, as amostras foram classificadas como sensíveis (S) ou
resistentes (R) (CLSI, 2010).

MATERIAIS E MÉTODOS
73
4.6.2. Teste de Hidrólise para Detecção de Metalo-β-Lactamases
O teste de hidrólise foi realizado nos isolados que foram sugestivos para
a produção de metalo-β-lactamases pelo sistema Phoenix®, porém com reação
de PCR em Tempo Real direto da amostra clínica para os genes blaSPM, blaIMP
e blaVIM negativa. O teste de hidrólise teve como objetivo principal a detecção
da atividade enzimática nas amostras avaliadas. Depois de isoladas, 10
colônias foram inoculadas em 10 mL caldo de TSB (Oxoid, Basingstoke,
Inglaterra) com 4 µg/mL de cefoxitina. Os tubos ficaram incubados na estufa a
37C por 18 horas sob agitação. A suspensão bacteriana foi transferida para
um tubo cônico e passou por uma centrifugação de 15 minutos à 3.000 rpm.
Em seguida, o sobrenadante foi desprezado e o sedimento ressuspenso em 1
mL de solução de hidrólise (Tris-HCl 1 mM e ZnSO4 1 mM). As amostras
suspensas foram ultrasonicadas com 4 ciclos de 30 segundos e o produto
sonicado foi transferido para tubo de microcentrífuga que então, foi
centrifugado por 3 minutos à 4ºC e 13.000 rpm (Centrifuge 5415R, Eppendorf,
Hamburg, Germany). O sobrenadante (extratos brutos de proteínas) foi
transferido para um novo tubo e as amostras mantidas no gelo até o momento
do ensaio.
Para verificar a atividade de hidrólise foram preparadas soluções de
imipenem em tampão fosfato 0,1 M, pH 7,0 a uma absorbância de
aproximadamente 1,5 a 2 unidades de absorbância a 260 nm. Em uma cubeta
de quartzo foram adicionados 900 µL da solução de antimicrobiano, juntamente
com 100 µL do extrato bruto de β-lactamase. O monitoramento da absorbância
da solução foi realizado por 1 minuto em espectrofotômetro. A diminuição da
absorbância da solução de antimicrobianos indica um resultado positivo para a

MATERIAIS E MÉTODOS
74
produção de metalo-β-lactamases. A variação da absorbância foi calculada
pela diferença entre o valor de absorbância final e inicial após um minuto de
leitura (Abs/min). Posteriormente, as amostras foram submetidas à inibição
por EDTA para confirmar se tratar de uma amostra produtora de metalo-β-
lactamases.
Como controle positivo do teste foi utilizada a amostra de P. aeruginosa
(Banco do LEMC número P1088) carreadora do gene blaSPM (Monteiro et al.
2009).
4.6.3. Detecção Fenotípica de ESβL
Em todos os isolados que foram identificados como produtores de ESβL
pelo método automatizado Phoenix® e que tiveram reações negativas para a
detecção dos genes blaSHV, blaCTX-M, blaTEM pela metodologia da PCR em
Tempo Real realizada diretamente da amostra clínica, foi realizada a detecção
fenotípica da produção de ESβL pela metodologia da disco-aproximação
(Jarlier et al., 1988). Foi preparada uma suspensão bacteriana com turbidez
correspondente a 0,5 da escala de McFarland. Após homogeneização, a
suspensão foi semeada em uma placa de ágar Müeller-Hinton utilizando swab
estéril. Foram dispensados os discos de aztreonam (30μg), ceftriaxona (30μg),
ceftazidima (30μg) e cefepima (30μg) a uma distância de 25 mm (centro a
centro) de um disco de amoxicilina/ácido clavulânico (30μg/10μg), que foi
colocado no centro da placa. Os discos utilizados foram os da Oxoid®
(Basingstoke, Inglaterra). As placas foram incubadas por 18 a 24 horas, em
temperatura de 35°C.

MATERIAIS E MÉTODOS
75
O teste da disco-aproximação foi considerado positivo para a produção
de ESβL quando houve o aparecimento de deformações do halo de inibição ou
o de uma zona fantasma entre o substrato e o inibidor. Como controle
positivodo teste foi incluída a cepa de K. pneumoniae ATCC 700603, produtora
de ESΒL, e a cepa de E. coli ATCC 25922 foi utilizada como controle negativo.
4.6.4. Determinação da Suscetibilidade a Vancomicina
Todos os isolados resistentes a vancomicina pelo método automatizado
Phoenix® e que tiveram resultado negativo para a pesquisa dos genes vanA e
vanB pelo método da PCR em Tempo Real realizado diretamente da amostra
clínica, foram submetidas a determinação da CIM, pela metodologia Etest®,
conforme recomendações do fabricante. Foi preparada uma suspensão
bacteriana ajustada a 0,5 da escala de McFarland foi uniformemente semeada
na placa de ágar Müeller-Hinton e, quinze minutos após, a fita de Etest®
contendo vancomicina foi dispensada sobre a placa. As placas foram
incubadas em aerobiose a 37ºC por 18 a 24 horas. A CIM foi determinada
como sendo a concentração indicada na intersecção entre a fita de Etest® e a
zona elíptica de inibição do crescimento bacteriano. As amostras foram
classificadas em sensíveis ou resistentes de acordo com os pontos de corte
preconizados pelo CLSI para métodos diluicionais. Como controle foi utilizada a
cepa Staphylococcus aureus ATCC® 29213.

MATERIAIS E MÉTODOS
76
4.6.5. Determinação da Suscetibilidade a Oxacilina
Todos os isolados sensíveis a oxacilina pelo método automatizado
Phoenix® e pesquisa positiva do gene mecA pela PCR em Tempo Real direto
da amostra clínica foram submetidos a determinação da CIM, pela metodologia
Etest®, conforme recomendações do fabricante. Foi preprarada uma suspensão
bacteriana ajustada a 0,5 da escala de McFarland foi uniformemente semeada
na placa de ágar Müeller-Hinton suplementado com 2% NaCl e, quinze minutos
após, a fita de Etest® contendo oxacilina foi dispensada sobre a placa. As
placas foram incubadas em aerobiose a 37ºC por 18 a 24 horas. A CIM foi
determinada como sendo a concentração indicada na intersecção entre a fita
de Etest® e a zona elíptica de inibição do crescimento bacteriano. As amostras
foram classificadas em sensíveis ou resistentes de acordo com os pontos de
corte preconizados pelo CLSI para métodos diluicionais. Como controle foi
utilizada a cepa Staphylococcus aureus ATCC® 29213.
4.6.6. PCR em Tempo Real Direto do Isolado Bacteriano
Os mesmos isolados submetidos ao teste fenotípico foram submetidos
ao PCR em Tempo Real sistema SYBR Green para detecção dos genes de
resistência. Após o isolamento das colônias, foi realizado em um tubo contendo
4 mL de salina uma suspensão bacteriana na escala 0,5 de Mc Farland (1,5 x
108 UFC/mL). 1 mL desta suspensão foi transferida para um tubo de 2mL
estéril e assim foi extraído o DNA bacteriano por fervura, 10 minutos a 100°C e
centrifugado a 20.000g por 5 minutos. O sobrenadante foi transferido para um
tubo de 2 mL estéril. Para a reação de PCR foi realizada um diluição a 10-1 do
DNA estraído. Para os isolados submetidos ao teste confirmatório para ESBL,

MATERIAIS E MÉTODOS
77
foi realizado a PCR para os genes blaCTX-M, blaSHV e blaTEM. Para os isolados
submetidos ao teste DD para ertapenem, foi realizado a PCR para blaKPC. Para
os isolados submetidos ai teste de hidrólise, foi realizado a PCR para os genes
blaIMP, blaSPM e blaVIM. Já para os isolados submetidos ao E-test para oxacilina
foi realizado a PCR para o gene mecA. As concetrações e condições de
ciclagem foram as mesmas dos testes realizados direto da hemocultura.
4.7. Análise Estatística dos Dados
A análise dos dados consistiu na descrição geral, a partir de Tabelas
utilizando o programa de planilha Microsoft Excel 2007 e SSPS 17.0 (SPSS
Inc., Chicago, IL).
Os valores preditivo positivo (VPP) e negativo (VPN) foram calculados
como medidas de eficácia diagnóstica da PCR em Tempo Real para o gene
16S rDNA em relação aos resultados do Phoenix®.
O grau de concordância entre os resultados da PCR em Tempo Real
para o gene 16S rDNA em relação aos resultados do Phoenix® foram
estabelecidos pelo cálculo do coeficiente kappa (K).
O Kappa é uma medida de concordância inter-observador e mede o grau
de concordância além do que seria esperado tão somente pelo acaso. Esta
medida de concordância tem como o valor máximo o “1”, valor que representa
o máximo de concordância e o “0” que indica ausência de concordância. Um
eventual valor de Kappa menor que zero sugere que a concordância
encontrada foi menor do que aquela esperada por acaso. Sugere, portanto,
discordância, mas seu valor não tem interpretação como intensidade de
discordância (Feinstein et al., 1990).

MATERIAIS E MÉTODOS
78
A acurácia analítica foi calculada pela curva ROC. Esse é o resultado
da plotagem dos resultados de sensibilidade (eixo y) e especificidade
subtraída de 1 (eixo x), para diferentes pontos de corte. A área abaixo da
curva (AAC) é comumente utilizada como uma medida do desempenho do
teste, podendo ser traduzida como a probabilidade do observador em
classificar amostras corretamente ou como a sensibilidade média entre toda a
faixa de valores possíveis de sensibilidade. Assim, para uma curva na qual a
AAC é igual a 1, o teste é 100% sensível e específico, e a chance de o
observador errar ao utilizar o teste para caracterizar uma determinada amostra
é nula. Se o teste não for discriminatório, a AAC aproxima-se a 0,5, o que
significa 50% de probabilidade de o observador acertar ao classificar uma
amostra por esse método, ou seja, a classificação é realizada ao acaso (Eng,
2005). Este foi elaborado tanto para os genes de resistência a
antimicrobianos pelo método da PCR em Tempo sistema SYBR Green quanto
para o gene 16S rDNA pelo método da PCR em Tempo sistema TaqMan,
utilizando apenas amostras controles bem caracterizadas como negativas e
positivas.

RESULTADOS
79
5. RESULTADOS
5.1. Amostras de Hemoculturas Incluídas no Estudo
Durante o período de fevereiro de 2009 a fevereiro de 2010, 185
hemoculturas de 117 pacientes transplantados de órgão sólidos foram incluídas
no estudo, das quais 18 hemoculturas (4 hemoculturas negativas e 14
hemoculturas positivas pelo sistema automatizado Bactec®) foram
provenientes de 11 pacientes admitidos na unidade de transplantados de
órgãos sólidos (rim, rim e pâncreas, pulmão, fígado e coração) localizada no
10º andar do HSP e 167 hemoculturas (55 hemoculturas negativas e 112
hemoculturas positivas) foram provenientes de 106 pacientes do serviço de
transplantados de órgãos sólidos (rim e rim/ pâncreas) do HRH.
5.2. Validação Analítica da PCR em Tempo Real
As temperaturas de desnaturação (Tm) obtidas na detecção do controle
interno (gene HFE) e dos genes que conferem resistências aos antimicobianos
para cada controle positivo ATCC testado foram observadas, permitindo o
estabelecimento de um intervalo experimental de Tm possíveis. O Tm médio e
o desvio padrão observados para os genes avaliados estão apresentados na
Tabela 3.
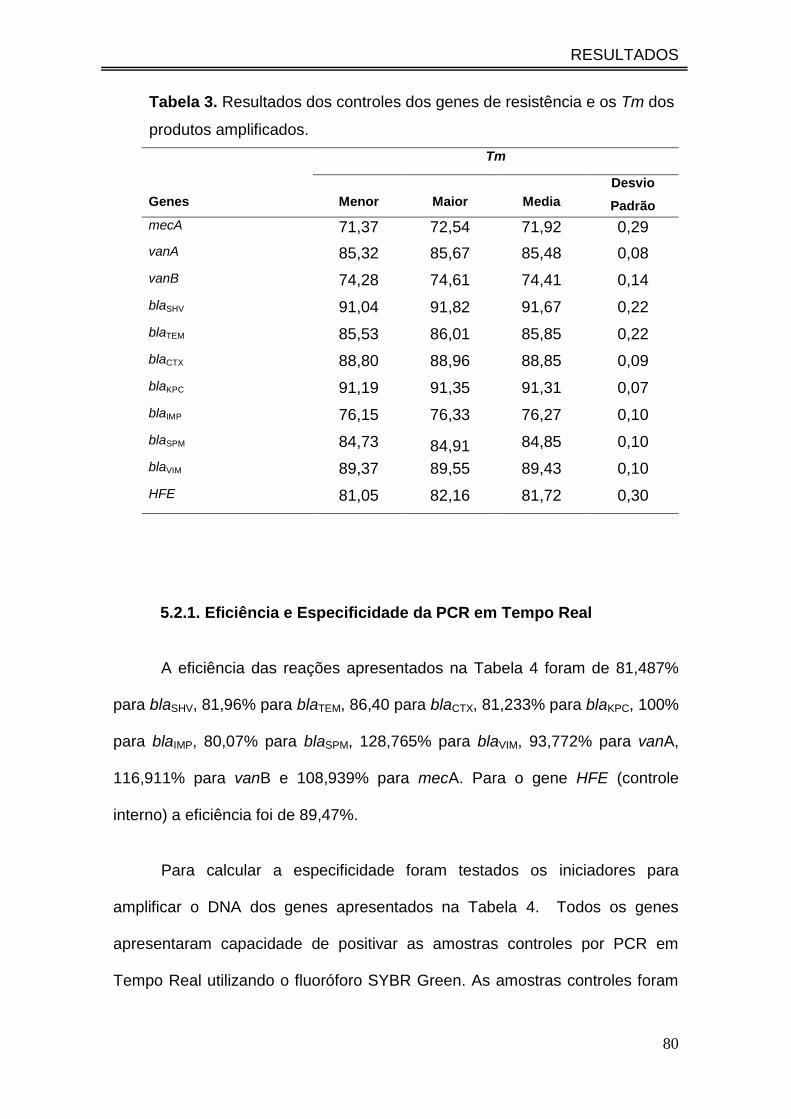
RESULTADOS
80
Tabela 3. Resultados dos controles dos genes de resistência e os Tm dos
produtos amplificados.
Genes
Tm
Menor Maior Media
Desvio
Padrão
mecA 71,37 72,54 71,92 0,29
vanA 85,32 85,67 85,48 0,08
vanB 74,28
74,61
74,41
0,14 blaSHV 91,04 91,82 91,67 0,22
blaTEM 85,53 86,01 85,85 0,22
blaCTX 88,80 88,96 88,85 0,09
blaKPC 91,19 91,35 91,31 0,07
blaIMP 76,15 76,33 76,27 0,10
blaSPM 84,73 84,91 84,85 0,10
blaVIM 89,37 89,55 89,43 0,10
HFE 81,05 82,16 81,72 0,30
5.2.1. Eficiência e Especificidade da PCR em Tempo Real
A eficiência das reações apresentados na Tabela 4 foram de 81,487%
para blaSHV, 81,96% para blaTEM, 86,40 para blaCTX, 81,233% para blaKPC, 100%
para blaIMP, 80,07% para blaSPM, 128,765% para blaVIM, 93,772% para vanA,
116,911% para vanB e 108,939% para mecA. Para o gene HFE (controle
interno) a eficiência foi de 89,47%.
Para calcular a especificidade foram testados os iniciadores para
amplificar o DNA dos genes apresentados na Tabela 4. Todos os genes
apresentaram capacidade de positivar as amostras controles por PCR em
Tempo Real utilizando o fluoróforo SYBR Green. As amostras controles foram

RESULTADOS
81
testadas e foram amplificadas para o gene HFE, controle interno. Todas as
amostras foram PCR positivas para os seus respectivos genes de resistência, e
os controles negativos apresentaram resultados negativos.
A reação para detecção do gene HFE, controle interno, não apresentou
reação cruzada com DNA de microrganismos. E uma reação contendo todos os
reagentes, com exceção do DNA da amostra, também foi utilizada como
controle negativo e apresentou PCR negativa
A detecção da ultima diluição foi calculada a partir da diluição seriada de
amostras controles para cada gene avaliado. Para o gene HFE, o controle
utilizado foi uma amostra de sangue periférico com concentração de DNA de
100 pg/uL. A diluição seriada variou de 10-1 a 10-7 e a menor diluição detectada
foi a 10-4 e a menor de DNA foi de 100 fg/uL. Para os genes blaSHV, blaTEM,
blaCTX, blaKPC, blaIMP, blaSPM, blaVIM, vanA, vanB, mecA e as menores diluições
foram de 10-6, 10-7, 10-6, 10-6, 10-7, 10-7, 10-7, 10-7, 10-6 e 10-7 respectivamente.
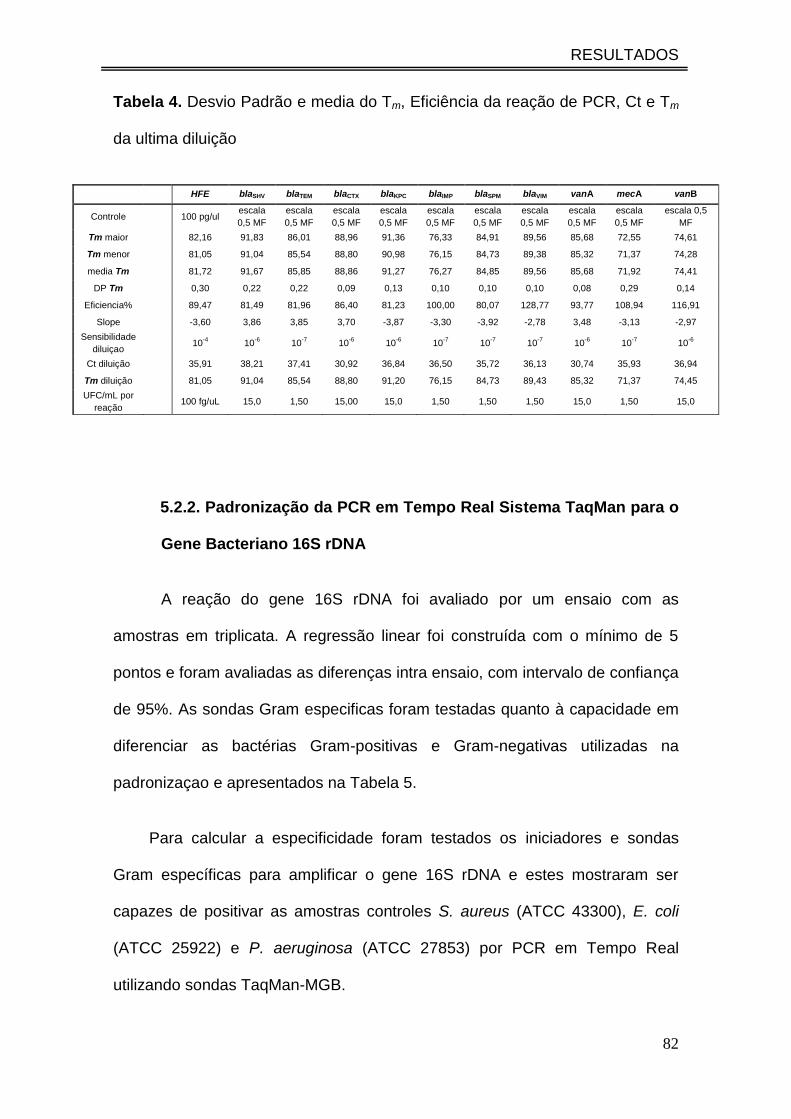
RESULTADOS
82
Tabela 4. Desvio Padrão e media do Tm, Eficiência da reação de PCR, Ct e Tm
da ultima diluição
5.2.2. Padronização da PCR em Tempo Real Sistema TaqMan para o
Gene Bacteriano 16S rDNA
A reação do gene 16S rDNA foi avaliado por um ensaio com as
amostras em triplicata. A regressão linear foi construída com o mínimo de 5
pontos e foram avaliadas as diferenças intra ensaio, com intervalo de confiança
de 95%. As sondas Gram especificas foram testadas quanto à capacidade em
diferenciar as bactérias Gram-positivas e Gram-negativas utilizadas na
padronizaçao e apresentados na Tabela 5.
Para calcular a especificidade foram testados os iniciadores e sondas
Gram específicas para amplificar o gene 16S rDNA e estes mostraram ser
capazes de positivar as amostras controles S. aureus (ATCC 43300), E. coli
(ATCC 25922) e P. aeruginosa (ATCC 27853) por PCR em Tempo Real
utilizando sondas TaqMan-MGB.
HFE blaSHV blaTEM blaCTX blaKPC blaIMP blaSPM blaVIM vanA mecA vanB
Controle 100 pg/ul escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala
0,5 MF
escala 0,5
MF
Tm maior 82,16 91,83 86,01 88,96 91,36 76,33 84,91 89,56 85,68 72,55 74,61
Tm menor 81,05 91,04 85,54 88,80 90,98 76,15 84,73 89,38 85,32 71,37 74,28
media Tm 81,72 91,67 85,85 88,86 91,27 76,27 84,85 89,56 85,68 71,92 74,41
DP Tm 0,30 0,22 0,22 0,09 0,13 0,10 0,10 0,10 0,08 0,29 0,14
Eficiencia% 89,47 81,49 81,96 86,40 81,23 100,00 80,07 128,77 93,77 108,94 116,91
Slope -3,60 3,86 3,85 3,70 -3,87 -3,30 -3,92 -2,78 3,48 -3,13 -2,97
Sensibilidade
diluiçao 10
-4 10
-6 10
-7 10
-6 10
-6 10
-7 10
-7 10
-7 10
-6 10
-7 10
-6
Ct diluição 35,91 38,21 37,41 30,92 36,84 36,50 35,72 36,13 30,74 35,93 36,94
Tm diluição 81,05 91,04 85,54 88,80 91,20 76,15 84,73 89,43 85,32 71,37 74,45
UFC/mL por
reação 100 fg/uL 15,0 1,50 15,00 15,0 1,50 1,50 1,50 15,0 1,50 15,0
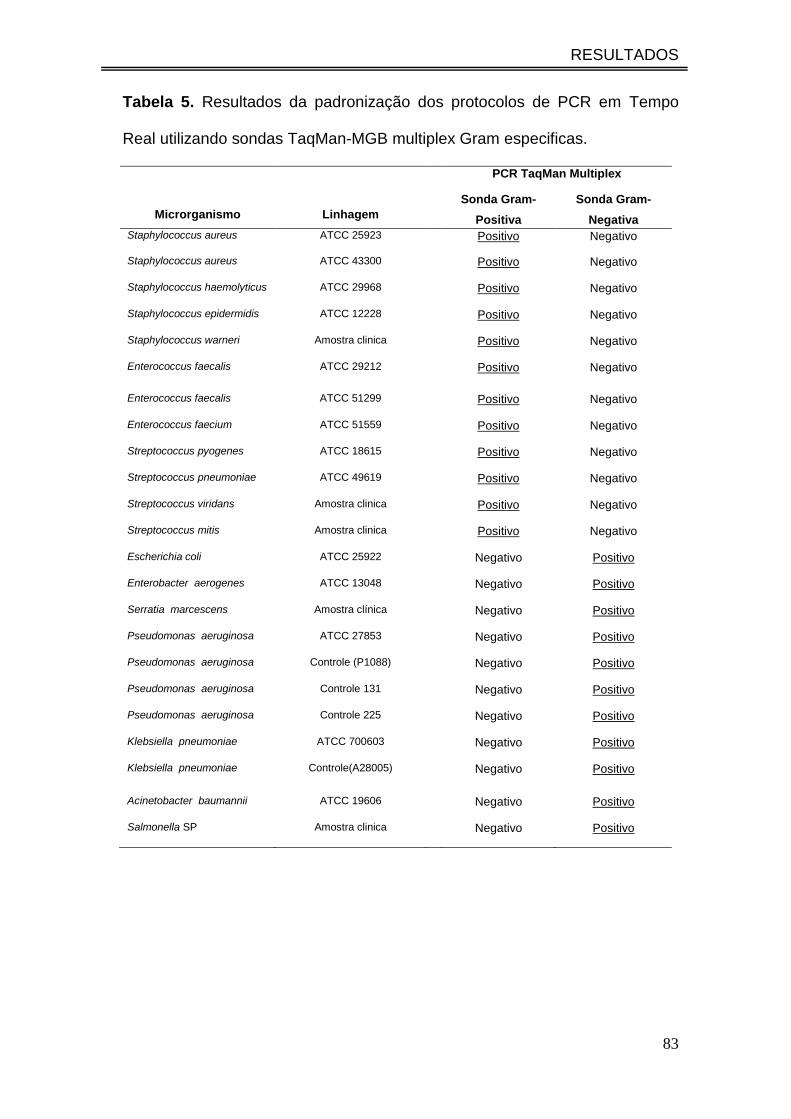
RESULTADOS
83
Tabela 5. Resultados da padronização dos protocolos de PCR em Tempo
Real utilizando sondas TaqMan-MGB multiplex Gram especificas.
Microrganismo Linhagem
PCR TaqMan Multiplex
Sonda Gram-
Positiva
Sonda Gram-
Negativa
Staphylococcus aureus ATCC 25923 Positivo Negativo
Staphylococcus aureus ATCC 43300 Positivo Negativo
Staphylococcus haemolyticus ATCC 29968 Positivo Negativo
Staphylococcus epidermidis ATCC 12228 Positivo Negativo
Staphylococcus warneri Amostra clinica Positivo Negativo
Enterococcus faecalis ATCC 29212 Positivo Negativo
Enterococcus faecalis ATCC 51299 Positivo Negativo
Enterococcus faecium ATCC 51559 Positivo Negativo
Streptococcus pyogenes ATCC 18615 Positivo Negativo
Streptococcus pneumoniae ATCC 49619 Positivo Negativo
Streptococcus viridans Amostra clinica Positivo Negativo
Streptococcus mitis Amostra clinica Positivo Negativo
Escherichia coli ATCC 25922 Negativo Positivo
Enterobacter aerogenes ATCC 13048 Negativo Positivo
Serratia marcescens Amostra clínica Negativo Positivo
Pseudomonas aeruginosa ATCC 27853 Negativo Positivo
Pseudomonas aeruginosa Controle (P1088) Negativo Positivo
Pseudomonas aeruginosa Controle 131 Negativo Positivo
Pseudomonas aeruginosa Controle 225 Negativo Positivo
Klebsiella pneumoniae ATCC 700603 Negativo Positivo
Klebsiella pneumoniae Controle(A28005) Negativo Positivo
Acinetobacter baumannii ATCC 19606 Negativo Positivo
Salmonella SP Amostra clinica Negativo Positivo

RESULTADOS
84
5.2.3. Determinação do Limite do Branco (LoB) ou “cutoff”
O Ct (ciclo threshold) de “cutoff” foi calculado para os genes resistência
a antimicrobianos e para as sondas Gram específicas. De acordo com o
número de amostras negativas, este número variou entre os genes e estão
apresentados na Tabela 6. A partir deste ciclo de “cutoff”, as amostras que
apresentarem resultados abaixo do Ct foram consideradas positivas e acima,
negativas.
5.2.4. Determinação do Limite de Detecção – LoD
O Ct (ciclo threshold) do limite inferior de detecção, o LoD foi calculado
para os genes resistência a antimicrobianos e para as sondas Gram
específicas a partir de amostras positivas mas com baixa concentração. O Ct
LoD está apresentado na Tabela 6.

RESULTADOS
85
Tabela 6. Resultados de cutoff e LoD para cada gene avaliados.
Nº Amostra
Negativa GP GN mecA bla SHV bla CTX-M vanA vanB bla KPC bla TEM
1 41,00 41,00 41 41 41 41 41 41 41
2 41,00 41,00 41 41 41 41 41 41 41
3 41,00 41,00 41 41 41 41 41 41 41
4 41,00 41,00 41 41 41 41 41 41 41
5 41,00 41,00 41 41 41 41 41 41 41
6 41,00 41,00 41 41 41 41 41 41 41
7 41,00 41,00 41 41 41 41 41 41 41
8 41,00 41,00 41 41 41 41 41 41 41
9 41,00 41,00 41 41 41 41 41 41 41
10 41,00 41,00 41 41 41 41 41 41 41
11 41,00 41,00 41 41 41 41 41 41 41
12 41,00 41,00 41 41 41 41 41 41 41
13 41,00 41,00 41 41 41 41 41 41 41
14 41,00 41,00 41 41 41 41 41 41 41
15 41,00 41,00 41 41 41 41 41 41 41
16 41,00 41,00 41 41 41 41 41 41 41
17 41,00 41,00 41 41 41 41 41 41 41
18 41,00 41,00 41 41 41 41 41 41 41
19 41,00 41,00 41 41 41 41 41 41 41
20 41,00 41,00 41 41 41 41 41 41 41
21 41,00 41,00 41 41 38,274689 41 41 41 41
22 41,00 41,00 41 41 37,055664 41 41 41 41
23 41,00 41,00 41 41 37,020847 41 41 41 41
24 41,00 41,00 41 41 37,003807 41 41 41 37,1618
25 41,00 41,00 41 41 36,766258 41 41 41 30,202
26 41,00 41,00 41 41 36,717575 41 41 41
27 41,00 41,00 41 41 36,521244 41 41 41
28 41,00 41,00 41 39,1427 36,477421 41 41 41
29 41,00 41,00 41 39,1427 36,187653 41 41 41
30 41,00 41,00 41 37,918 36,152454 41 41 41
31 41,00 41,00 41 37,4123 35,818134 41 41 41
32 41,00 41,00 36,93 36,5065 35,381996 41 41 41
33 41,00 41,00 36,93 35,6181 34,501266 39,25 41 41
34 41,00 41,00 35,94 35,3588 33,048428 39,2 41 41
35 41,00 41,00 35,06 35,0958 28,741648 37,29 41 41
36 41,00 41,00 33,6 34,2808 36,92 41 41
37 41,00 41,00 29,83 34,198 36,12 41 41
38 41,00 41,00 34,1476 35,62 41 41
39 41,00 41,00 33,8952 35,53 41 41
41 41,00 41,00 35,15 41 41
41 41,00 41,00 37,701 41
42 41,00 41,00 37,701 41
43 41,00 41,00 37,55 41
44 41,00 41,00 37,55 41
45 41,00 41,00 37,037 41
46 41,00 41,00 37,037 38,2736
47 41,00 41,00 36,615 38,1545
48 41,00 41,00 36,615 38,1545
49 41,00 39,44 36,26 37,0442
50 41,00 39,44 36,26 34,9422
51 41,00 39,29 34,982 34,9422
52 41,00 39,23 33,286
53 41,00 38,82 31,476
54 41,00 38,47 31,476
55 41,00 38,30
56 41,00 37,54
57 41,00 37,36
58 41,00 37,31
59 41,00 37,29
60 41,00 37,08
61 37,74 36,96
62 33,98 36,89
63 36,79
64 35,88
65 34,02
66 33,03
67
68
69
70
Nº Total 62 66 37 39 35 41 54 51 25
N B 59,40 63,20 35,65 37,55 33,75 38,50 51,80 48,95 24,25
Cutoff 40,00 36,61 34,97 34,19 33,77 35,57 33,32 37,98 36,04
LoD 38,68 34,47 33,91 33,26 32,62 33,67 32,29 36,67 34,53
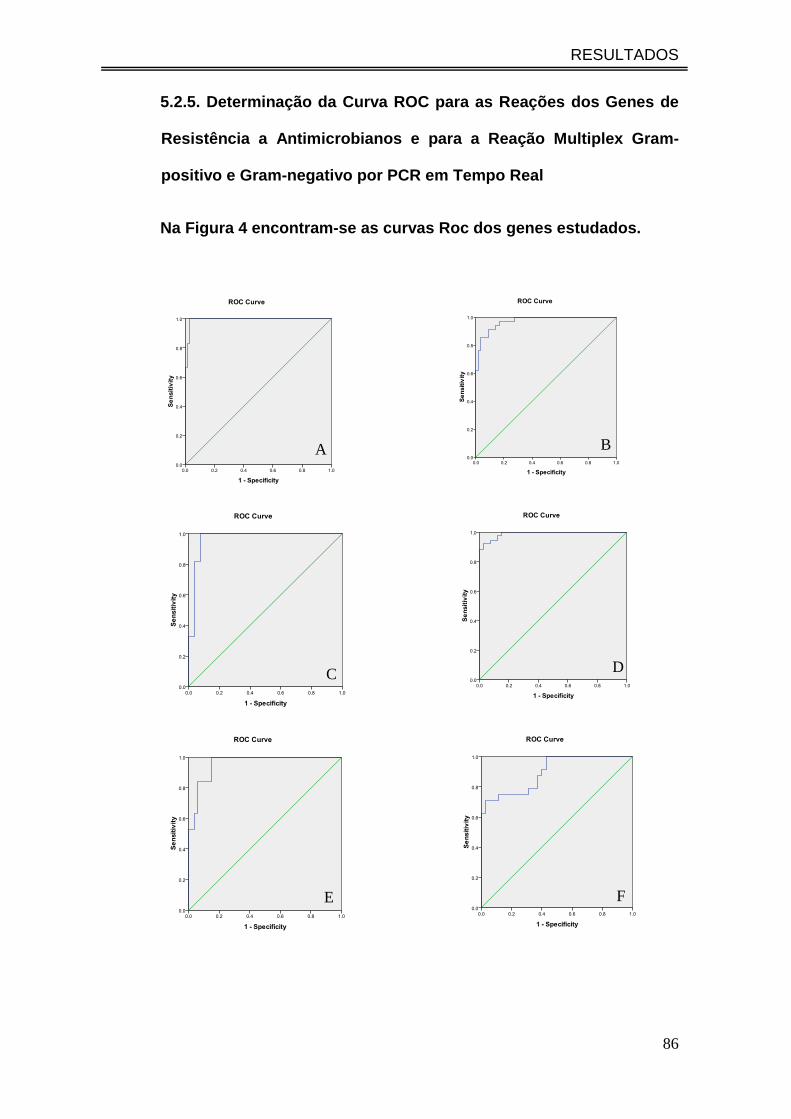
RESULTADOS
86
5.2.5. Determinação da Curva ROC para as Reações dos Genes de
Resistência a Antimicrobianos e para a Reação Multiplex Gram-
positivo e Gram-negativo por PCR em Tempo Real
Na Figura 4 encontram-se as curvas Roc dos genes estudados.
A B
D C
E F
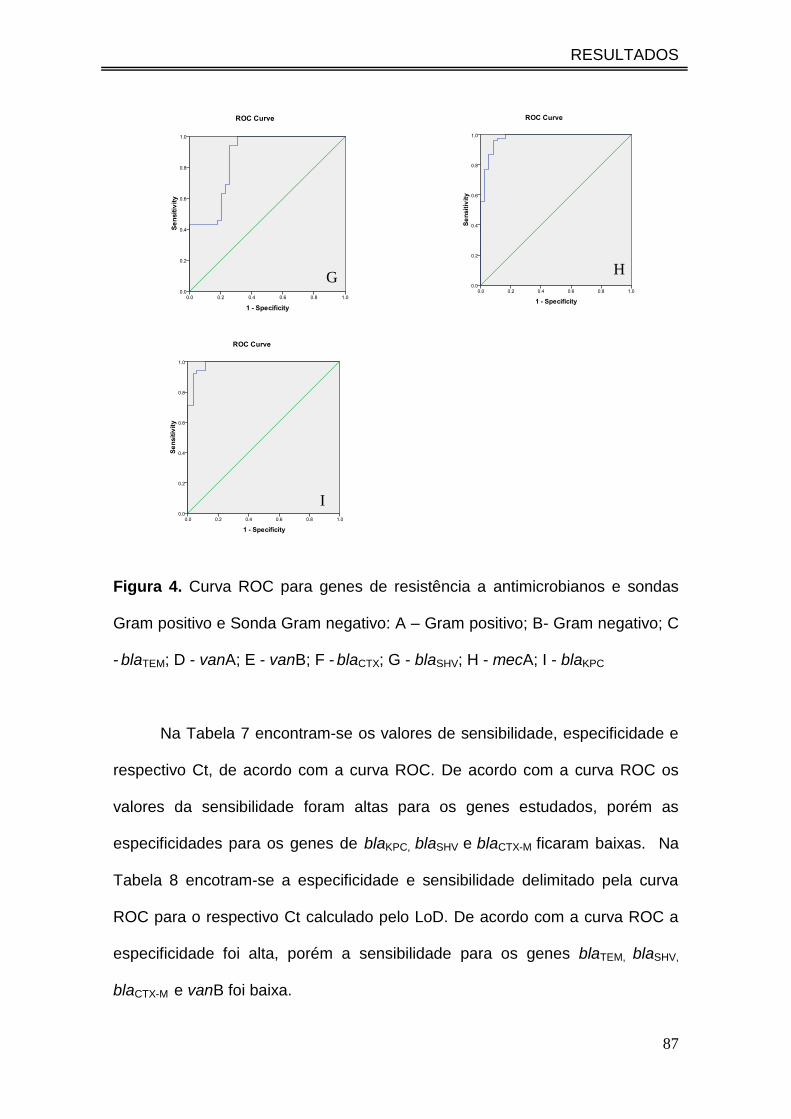
RESULTADOS
87
Figura 4. Curva ROC para genes de resistência a antimicrobianos e sondas
Gram positivo e Sonda Gram negativo: A – Gram positivo; B- Gram negativo; C
- blaTEM; D - vanA; E - vanB; F - blaCTX; G - blaSHV; H - mecA; I - blaKPC
Na Tabela 7 encontram-se os valores de sensibilidade, especificidade e
respectivo Ct, de acordo com a curva ROC. De acordo com a curva ROC os
valores da sensibilidade foram altas para os genes estudados, porém as
especificidades para os genes de blaKPC, blaSHV e blaCTX-M ficaram baixas. Na
Tabela 8 encotram-se a especificidade e sensibilidade delimitado pela curva
ROC para o respectivo Ct calculado pelo LoD. De acordo com a curva ROC a
especificidade foi alta, porém a sensibilidade para os genes blaTEM, blaSHV,
blaCTX-M e vanB foi baixa.
I
H G
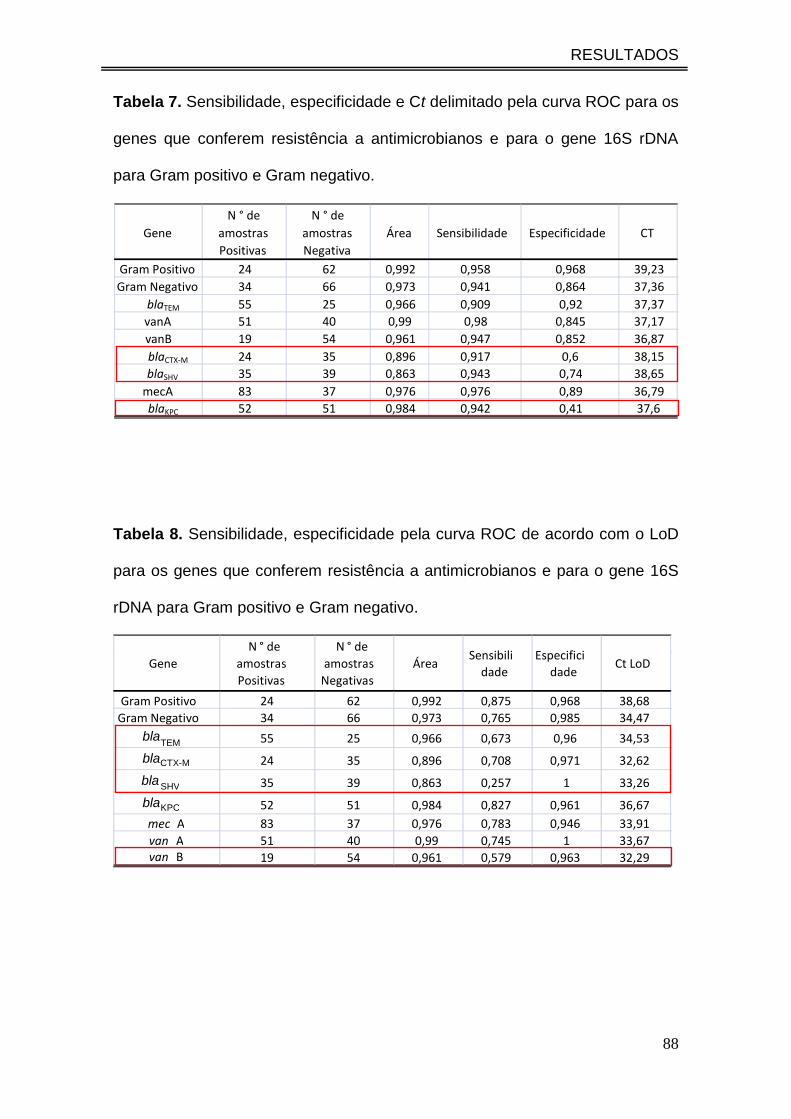
RESULTADOS
88
Tabela 7. Sensibilidade, especificidade e Ct delimitado pela curva ROC para os
genes que conferem resistência a antimicrobianos e para o gene 16S rDNA
para Gram positivo e Gram negativo.
Tabela 8. Sensibilidade, especificidade pela curva ROC de acordo com o LoD
para os genes que conferem resistência a antimicrobianos e para o gene 16S
rDNA para Gram positivo e Gram negativo.
Gene N ° de
amostras Positivas
N ° de amostras Negativa
Área Sensibilidade Especificidade CT
Gram Positivo 24 62 0,992 0,958 0,968 39,23 Gram Negativo 34 66 0,973 0,941 0,864 37,36
blaTEM 55 25 0,966 0,909 0,92 37,37 vanA 51 40 0,99 0,98 0,845 37,17 vanB 19 54 0,961 0,947 0,852 36,87 blaCTX-M 24 35 0,896 0,917 0,6 38,15 blaSHV 35 39 0,863 0,943 0,74 38,65
mecA 83 37 0,976 0,976 0,89 36,79 blaKPC 52 51 0,984 0,942 0,41 37,6
Gram Positivo 24 62 0,992 0,875 0,968 38,68 Gram Negativo 34 66 0,973 0,765 0,985 34,47
bla TEM 55 25 0,966 0,673 0,96 34,53
bla CTX-M 24 35 0,896 0,708 0,971 32,62
bla SHV 35 39 0,863 0,257 1 33,26
bla KPC 52 51 0,984 0,827 0,961 36,67
mec A 83 37 0,976 0,783 0,946 33,91 van A 51 40 0,99 0,745 1 33,67 van B 19 54 0,961 0,579 0,963 32,29
Ct LoD N ° de
amostras Positivas
N ° de amostras
Negativas Gene Área
Sensibili dade
Especifici dade

RESULTADOS
89
5.3. Aplicação da PCR em Tempo Real nas Amostras de
Hemocultura
5.3.1. Detecção do Gene HFE de Controle Interno da Extração
Todas as amostras de Hemoculturas, sendo elas negativas ou positivas
pelo aparelho Bactec®, foram submetidas à extração de DNA e a PCR em
Tempo Real para o gene HFE. O Tm dos controles positivos variou de 81,076 –
81,245. As amostras foram consideradas positivas quando a Temperatura de
melting (Tm) se encontrava neste intervalo. Somente quando o resultado de
PCR foi positivo para esta reação, é que as mesmas foram encaminhadas para
realizar a PCR em Tempo Real para os demais genes em estudo. Todas as
amostras incluídas foram positivas para o gene HFE indicando que houve
extração e evitando a possibilidade de falso negativo para os demais genes.
5.3.2. Detecção do Gene Bacteriano 16S rDNA
Todas as amostras que tiveram resultado positivo para o gene HFE
foram submetidas à detecção do gene 16S rDNA por PCR em Tempo Real.
Nenhuma das 59 hemoculturas negativas pelo sistema Bactec®
apresentou positividade por PCR para o gene 16S rDNA. Na Tabela 9 estão
apresentados os resultados da PCR para o gene 16S rDNA aplicada às 126
hemoculturas positivas. Das 66 amostras identificadas pelo sistema
automatizado Phoenix® como espécies de bactérias Gram negativas, em cinco
não foi possível a amplicação na PCR para o gene 16S rDNA, porém das 62
hemoculturas aplicadas houve concordância de 100% entre o resultado do
sistema automatizado Phoenix® e da PCR. Para as 59 amostras identificadas
pelo Phoenix® como espécies de bactérias Gram positivas, em 19 não foi

RESULTADOS
90
possível a aplicação da PCR para o gene 16S rDNA, porém, das 40
hemoculturas aplicadas houve concordância de 100% entre o resultado do
sistema automatizado Phoenix® e da PCR. Uma única amostra de hemocultura
que apresentou dois tipos de bactérias, uma Gram positiva e outra Gram
negativa, a PCR detectou apenas a bactéria Gram negativa. A média de Ciclo
threshold (CT) apresentada pelas amostras positivas para Gram positiva e para
Gram negativo foi 24.
Das 126 hemoculturas positivas pelo sistema automatizado Bactec®, 60
bactérias Gram positivas e 67 Gram negativas foram identificadas pelo sistema
automatizado Phoenix®. Cento e vinte e cinco hemoculturas apresentaram
apenas uma bactéria, sendo 59 Gram positivas e 66 Gram negativas e apenas
uma das hemoculturas teve crescimento de duas bactérias distintas, uma Gram
negativa e outra Gram positiva, identificadas como AcinetobaCter spp. e
Staphylococcus haemolyticus.
As 60 bactérias Gram positivas identificadas foram: 21 SCoN (9 SCoN, 5
Staphylococcus epidermidis, 4 Staphylococcus haemolyticus, 1 Staphylococcus
equorum, 1 Staphylococcus capitis, 1 Staphylococcus warneri; 14 S.aureus 5
Streptococcus pneumoniae; 3 Listeria monocytogenes; 3 Enterococcus
faecium; 1 Enterococcus faecalis; 1 Enterococcus durans e 1 Corynebaterium
jeikeium (figura 5).
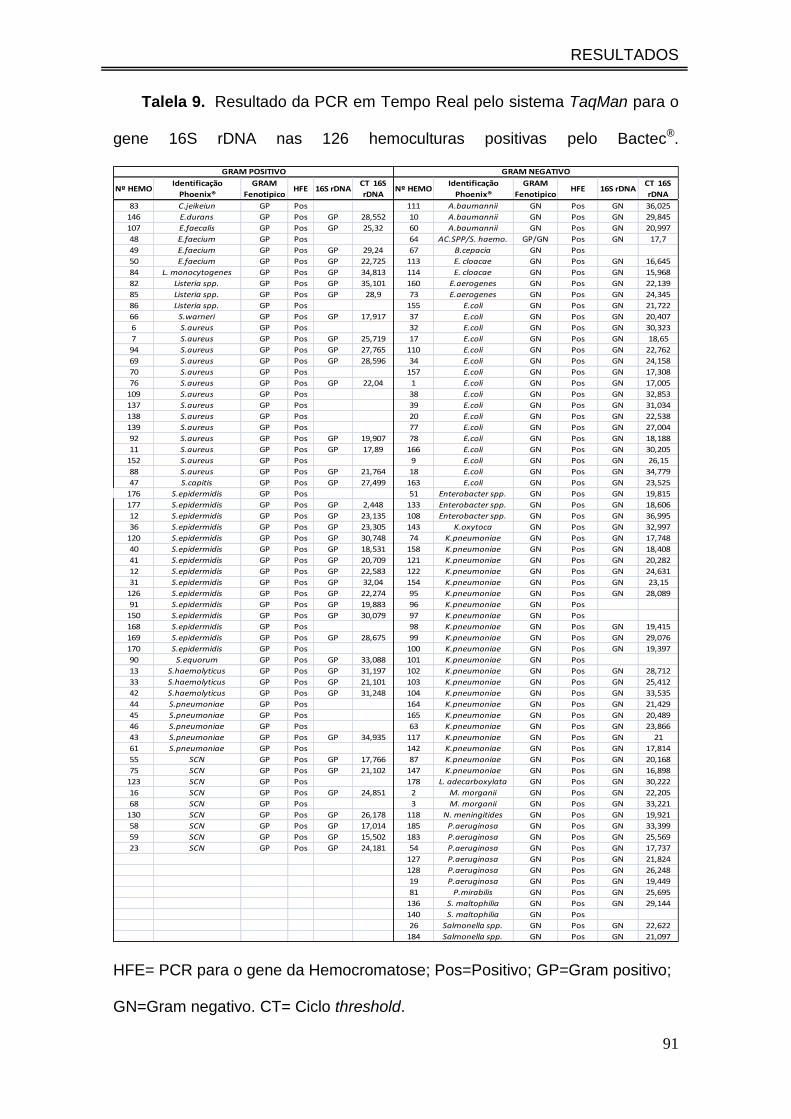
RESULTADOS
91
Talela 9. Resultado da PCR em Tempo Real pelo sistema TaqMan para o
gene 16S rDNA nas 126 hemoculturas positivas pelo Bactec®.
83 C.jeikeiun GP Pos 111 A.baumannii GN Pos GN 36,025
146 E.durans GP Pos GP 28,552 10 A.baumannii GN Pos GN 29,845
107 E.faecalis GP Pos GP 25,32 60 A.baumannii GN Pos GN 20,997
48 E.faecium GP Pos 64 AC.SPP/S. haemo. GP/GN Pos GN 17,7
49 E.faecium GP Pos GP 29,24 67 B.cepacia GN Pos
50 E.faecium GP Pos GP 22,725 113 E. cloacae GN Pos GN 16,645
84 L. monocytogenes GP Pos GP 34,813 114 E. cloacae GN Pos GN 15,968
82 Listeria spp. GP Pos GP 35,101 160 E.aerogenes GN Pos GN 22,139
85 Listeria spp. GP Pos GP 28,9 73 E.aerogenes GN Pos GN 24,345
86 Listeria spp. GP Pos 155 E.coli GN Pos GN 21,722
66 S.warneri GP Pos GP 17,917 37 E.coli GN Pos GN 20,407
6 S.aureus GP Pos 32 E.coli GN Pos GN 30,323
7 S.aureus GP Pos GP 25,719 17 E.coli GN Pos GN 18,65
94 S.aureus GP Pos GP 27,765 110 E.coli GN Pos GN 22,762
69 S.aureus GP Pos GP 28,596 34 E.coli GN Pos GN 24,158
70 S.aureus GP Pos 157 E.coli GN Pos GN 17,308
76 S.aureus GP Pos GP 22,04 1 E.coli GN Pos GN 17,005
109 S.aureus GP Pos 38 E.coli GN Pos GN 32,853
137 S.aureus GP Pos 39 E.coli GN Pos GN 31,034
138 S.aureus GP Pos 20 E.coli GN Pos GN 22,538
139 S.aureus GP Pos 77 E.coli GN Pos GN 27,004
92 S.aureus GP Pos GP 19,907 78 E.coli GN Pos GN 18,188
11 S.aureus GP Pos GP 17,89 166 E.coli GN Pos GN 30,205
152 S.aureus GP Pos 9 E.coli GN Pos GN 26,15
88 S.aureus GP Pos GP 21,764 18 E.coli GN Pos GN 34,779
47 S.capitis GP Pos GP 27,499 163 E.coli GN Pos GN 23,525
176 S.epidermidis GP Pos 51 Enterobacter spp. GN Pos GN 19,815
177 S.epidermidis GP Pos GP 2,448 133 Enterobacter spp. GN Pos GN 18,606
12 S.epidermidis GP Pos GP 23,135 108 Enterobacter spp. GN Pos GN 36,995
36 S.epidermidis GP Pos GP 23,305 143 K.oxytoca GN Pos GN 32,997
120 S.epidermidis GP Pos GP 30,748 74 K.pneumoniae GN Pos GN 17,748
40 S.epidermidis GP Pos GP 18,531 158 K.pneumoniae GN Pos GN 18,408
41 S.epidermidis GP Pos GP 20,709 121 K.pneumoniae GN Pos GN 20,282
12 S.epidermidis GP Pos GP 22,583 122 K.pneumoniae GN Pos GN 24,631
31 S.epidermidis GP Pos GP 32,04 154 K.pneumoniae GN Pos GN 23,15
126 S.epidermidis GP Pos GP 22,274 95 K.pneumoniae GN Pos GN 28,089
91 S.epidermidis GP Pos GP 19,883 96 K.pneumoniae GN Pos
150 S.epidermidis GP Pos GP 30,079 97 K.pneumoniae GN Pos
168 S.epidermidis GP Pos 98 K.pneumoniae GN Pos GN 19,415
169 S.epidermidis GP Pos GP 28,675 99 K.pneumoniae GN Pos GN 29,076
170 S.epidermidis GP Pos 100 K.pneumoniae GN Pos GN 19,397
90 S.equorum GP Pos GP 33,088 101 K.pneumoniae GN Pos
13 S.haemolyticus GP Pos GP 31,197 102 K.pneumoniae GN Pos GN 28,712
33 S.haemolyticus GP Pos GP 21,101 103 K.pneumoniae GN Pos GN 25,412
42 S.haemolyticus GP Pos GP 31,248 104 K.pneumoniae GN Pos GN 33,535
44 S.pneumoniae GP Pos 164 K.pneumoniae GN Pos GN 21,429
45 S.pneumoniae GP Pos 165 K.pneumoniae GN Pos GN 20,489
46 S.pneumoniae GP Pos 63 K.pneumoniae GN Pos GN 23,866
43 S.pneumoniae GP Pos GP 34,935 117 K.pneumoniae GN Pos GN 21
61 S.pneumoniae GP Pos 142 K.pneumoniae GN Pos GN 17,814
55 SCN GP Pos GP 17,766 87 K.pneumoniae GN Pos GN 20,168
75 SCN GP Pos GP 21,102 147 K.pneumoniae GN Pos GN 16,898
123 SCN GP Pos 178 L. adecarboxylata GN Pos GN 30,222
16 SCN GP Pos GP 24,851 2 M. morganii GN Pos GN 22,205
68 SCN GP Pos 3 M. morganii GN Pos GN 33,221
130 SCN GP Pos GP 26,178 118 N. meningitides GN Pos GN 19,921
58 SCN GP Pos GP 17,014 185 P.aeruginosa GN Pos GN 33,399
59 SCN GP Pos GP 15,502 183 P.aeruginosa GN Pos GN 25,569
23 SCN GP Pos GP 24,181 54 P.aeruginosa GN Pos GN 17,737
127 P.aeruginosa GN Pos GN 21,824
128 P.aeruginosa GN Pos GN 26,248
19 P.aeruginosa GN Pos GN 19,449
81 P.mirabilis GN Pos GN 25,695
136 S. maltophilia GN Pos GN 29,144
140 S. maltophilia GN Pos
26 Salmonella spp. GN Pos GN 22,622
184 Salmonella spp. GN Pos GN 21,097
GRAM POSITIVO GRAM NEGATIVO
Nº HEMOIdentificação
Phoenix®
GRAM
FenotipicoHFE 16S rDNA
CT 16S
rDNANº HEMO
Identificação
Phoenix®
GRAM
FenotipicoHFE 16S rDNA
CT 16S
rDNA
HFE= PCR para o gene da Hemocromatose; Pos=Positivo; GP=Gram positivo;
GN=Gram negativo. CT= Ciclo threshold.

RESULTADOS
92
Figura 5. Prevalência de espécies Gram positivas identificadas pelo sistema
automatizado Phoenix®.
As 67 bactérias identificadas pelo sistema automatizado Phoenix® como
Gram negativas foram: 22 Klebsiella pneumoniae; 17 Escherichia coli; 6
Pseudomonas aeruginosa; 3 Acinetobacter baumannii; 3 Enterobacter spp.; 2
Enterobacter cloacae; 2 Enterobacter aerogenes; 2 Morganella morgani; 2
Stenotrophomonas maltophilia; 2 Salmonella spp.; 1 Acinetobacter spp.; 1
Burkolderia cepacia; 1 Klebsiella oxytogenes; 1 Leclercia adecarboxylata; 1
Neisseria meningitidis e 1 Proteus mirabilis (Figura 6).
25%
24% 15%
8%
7%
5% 5%
1% 2% 2% 2% 2% 2%
Staphylococcus epidermidis Staphylococcus aureus Staphylococcus Coagulase Negativo Streptococcus pneumoniae Staphylococcus haemolyticus Listéria monocytogenes Enterococcus faecium Staphylocuccos equorum Staphylococcus capitis Staphylococcus warneri Enterococcus faecalis Enterococcus durans Corynebacterium jeikeium

RESULTADOS
93
Figura 6. Prevalência de espécies Gram negativas identificadas pelo sistema
automatizado Phoenix®.
A especificidade, a sensibilidade, o valor preditivo positivo, o valor
preditivo negativo e a concordância estimada pelo teste Kappa foi de 100%, já
que nenhuma amostra negativa pelo sistema automatizado foi considerada
positiva para a PCR em Tempo Real e as amostras consideradas Gram
positivo e Gram negativo pelo sistema automatizado concordaram com o
resultado da PCR.
5.3.3. Detecção PCR dos Genes que Conferem Resistência aos
Antimicrobianos em Bactérias Gram Negativas
Totas as 67 hemoculturas Gram negativas, foram submetidas a PCR em
Tempo Real pelo sistema SYBR Green para os genes de resistêcia de ESβL
(blaSHV, blaCTX-M, blaTEM), K. pneumoniae carbapenemase (blaKPC) e metalo-β-
lactamases (blaIMP, blaSPM e blaVIM), apesar de 5 não apresentarem resultado
33%
25%
9%
4%
4% 3%
3%
3% 3%
3%
1% 1% 1%
1% 1% 1%
Klebsiella pneumoniae Escherichia coli Pseudomonas aeruginosa Acinetobacter baumannii Enterobacter spp. Enterobacter cloacae Enterobacter aerogenes Morganella morgani Stenotrophomonas maltophilia Salmonella spp. Acinetobacter spp Burkolderia cepacia Klebsiella oxytogenes Leclercia adecarboxylata Neisseria meningitidis Proteus mirabilis

RESULTADOS
94
para a PCR de 16S rDNA. Das 67 hemoculturas Gram negativas, três amostras
apresentaram o gene blaCTX-M, uma blaKPC, uma blaCTX-M e blaTEM e uma blaCTX-
M e blaSHV. Os respectivos Tm e Ct estão na Tabela 10. Das cinco amostras
que apresentaram positividade na PCR para os genes de ESβL uma não foi
considerada produtora de ESβL pelo sistema automatizado. Dezenove
amostras consideradas produtoras de ESβL pelo sistema automatizado não
apresentaram positividade na PCR para os genes de ESβL. Apenas uma
amostra apresentando K.pneumoniae foi positiva na PCR para blaKPC, porém
no antibiograma liberado pelo sistema Phoenix® essa não foi resistente para os
carbapenêmicos meropenem e imipenem. Já duas amostras de K.pneumoniae
consideradas resistentes a estes carbapenêmicos não foram positivas na PCR
para blaKPC. Três amostras de P. aeruginosa e uma de A. baumannii
apresentaram resistênciaum a pelo menos um dos carbapenens (meropenem
e/ou imipenem), porém foram negativas na PCR para os genes de metalo-β-
lactamases.
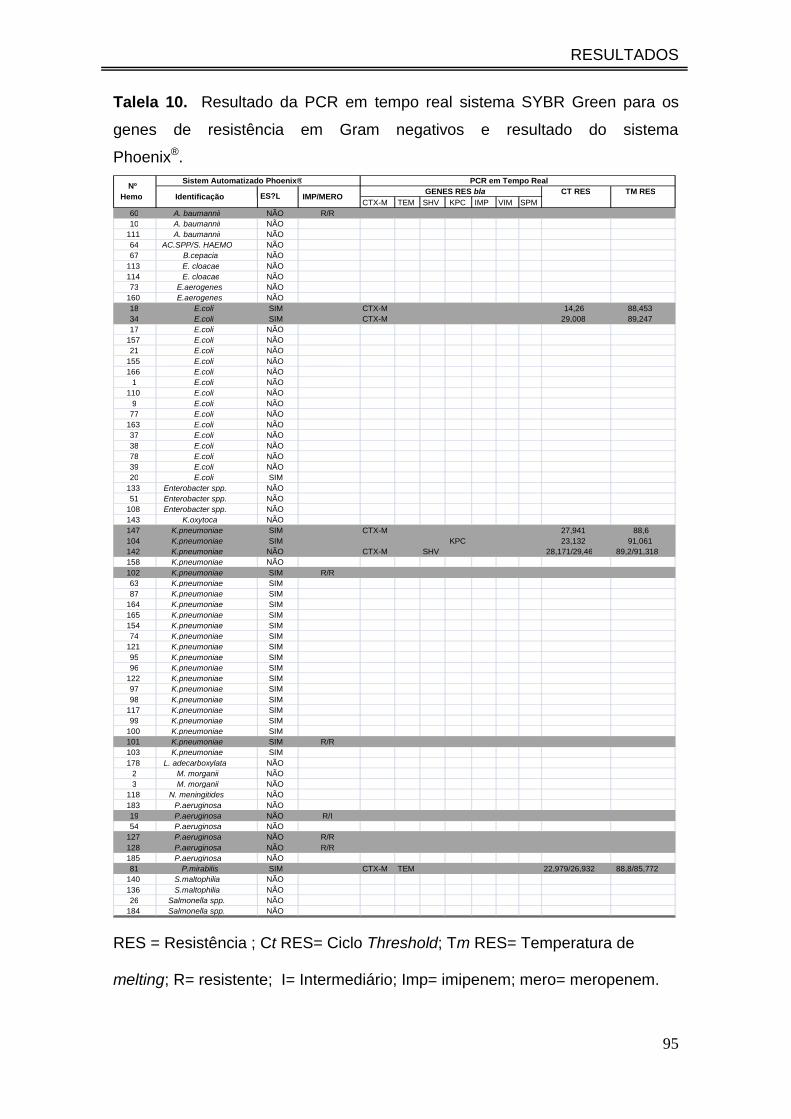
RESULTADOS
95
Talela 10. Resultado da PCR em tempo real sistema SYBR Green para os
genes de resistência em Gram negativos e resultado do sistema
Phoenix®.
RES = Resistência ; Ct RES= Ciclo Threshold; Tm RES= Temperatura de
melting; R= resistente; I= Intermediário; Imp= imipenem; mero= meropenem.
CTX-M TEM SHV KPC IMP VIM SPM 60 A. baumannii NÃO R/R 10 A. baumannii NÃO
111 A. baumannii NÃO 64 AC.SPP/S. HAEMO NÃO 67 B.cepacia NÃO
113 E. cloacae NÃO 114 E. cloacae NÃO 73 E.aerogenes NÃO
160 E.aerogenes NÃO 18 E.coli SIM CTX-M 14,26 88,453 34 E.coli SIM CTX-M 29,008 89,247 17 E.coli NÃO
157 E.coli NÃO 21 E.coli NÃO
155 E.coli NÃO 166 E.coli NÃO
1 E.coli NÃO 110 E.coli NÃO
9 E.coli NÃO 77 E.coli NÃO
163 E.coli NÃO 37 E.coli NÃO 38 E.coli NÃO 78 E.coli NÃO 39 E.coli NÃO 20 E.coli SIM
133 Enterobacter spp. NÃO 51 Enterobacter spp. NÃO
108 Enterobacter spp. NÃO 143 K.oxytoca NÃO 147 K.pneumoniae SIM CTX-M 27,941 88,6 104 K.pneumoniae SIM KPC 23,132 91,061 142 K.pneumoniae NÃO CTX-M SHV 28,171/29,46 89,2/91,318 158 K.pneumoniae NÃO 102 K.pneumoniae SIM R/R 63 K.pneumoniae SIM 87 K.pneumoniae SIM
164 K.pneumoniae SIM 165 K.pneumoniae SIM 154 K.pneumoniae SIM 74 K.pneumoniae SIM
121 K.pneumoniae SIM 95 K.pneumoniae SIM 96 K.pneumoniae SIM
122 K.pneumoniae SIM 97 K.pneumoniae SIM 98 K.pneumoniae SIM
117 K.pneumoniae SIM 99 K.pneumoniae SIM
100 K.pneumoniae SIM 101 K.pneumoniae SIM R/R 103 K.pneumoniae SIM 178 L. adecarboxylata NÃO
2 M. morganii NÃO 3 M. morganii NÃO
118 N. meningitides NÃO 183 P.aeruginosa NÃO 19 P.aeruginosa NÃO R/I 54 P.aeruginosa NÃO
127 P.aeruginosa NÃO R/R 128 P.aeruginosa NÃO R/R 185 P.aeruginosa NÃO 81 P.mirabilis SIM CTX-M TEM 22,979/26,932 88,8/85,772
140 S.maltophilia NÃO 136 S.maltophilia NÃO 26 Salmonella spp. NÃO
184 Salmonella spp. NÃO
Nº Hemo Identificação ES?L IMP/MERO
Sistem Automatizado Phoenix® PCR em Tempo Real GENES RES bla CT RES TM RES

RESULTADOS
96
5.3.4. Detecção PCR dos Genes que Conferem Resistência aos
Antimicrobianos em Bactérias Gram Positivas
Totas as 59 hemoculturas Gram positivas, foram submetidas a PCR em
Tempo Real pelo sistema SYBR Green para os genes de resistêcia mecA,
vanA e vanB, apesar de 19 não apresentarem resultado para a PCR de 16S
rDNA. Trinta e duas amostras foram positivas para o gene de resistência mecA.
No entanto, 5 amostras sendo 4 identificadas pelo sistema Phoenix® como S.
aureus e uma S. epidermidis foram sensíveis para oxacilina. Uma amostra de
E. durans foi postiva para a PCR do gene mecA e uma amostra de S.
pneumoniae foi resistente a oxacilina pelo sitema automatizado e negativa para
o gene mecA. Nenhuma amostra foi positiva para os genes de resistência vanA
e vanB nem resistente a vancomicina pelo sistema Phoenix®. Os respectivos
Tm e Ct estão apresentados na Tabela 11.
Com relação aos resultados de susceptibilidade a oxacilina liberados
pelo sistema automatizado Phoenix® e a detecção do gene mecA pelo método
da PCR em Tempo Real, houve 86,7% de concordância entre as duas
metodologias e 13,3% de discordância.

RESULTADOS
97
Talela 11. Resultado da PCR em Tempo Real sistema SYBR Green para os
genes de resistência de Gram positivos e resultado do Phoenix®.
mec A van A van B
83 C.jeikeiun NA S
146 E.durans NA S mec A 20,157 72,143
107 E.faecalis NA S
48 E.faecium NA S
49 E.faecium NA S
50 E.faecium NA S
84 L. monocytogenes NA S
85 Listeria spp. NA S
82 Listeria spp. NA S
86 Listeria spp. NA S
66 S. warneri R S mec A 11,545 72,143
137 S. aureus R S mec A 15,533 72,315
11 S.aureus S S mec A 29,331 71,799
76 S.aureus S S mec A 30,106 71,971
138 S.aureus S S mec A 30,186 72,315
139 S.aureus S S mec A 30,166 72,315
94 S. aureus R S mec A 19,763 72,83
88 S.aureus S S
109 S.aureus S S
92 S.aureus S S
69 S.aureus S S
70 S.aureus S S
152 S.aureus S S
6 S.aureus S S
7 S.aureus S S
47 S.capitis S S
36 S.epidermidis R S mec A 13,427 72,658
12 S.epidermidis R S mec A 13,581 72,658
40 S.epidermidis R S mec A 12,778 71,283
150 S.epidermidis R S mec A 13,322 72,315
176 S.epidermidis R S mec A 12,701 72,315
177 S.epidermidis R S mec A 17,97 72,315
91 S.epidermidis R S mec A 15,395 72,486
169 S.epidermidis S S mec A 30,047 72,143
41 S.epidermidis R S mec A 17,683 71,627
120 S.epidermidis R S mec A 28,299 72,143
126 S.epidermidis R S mec A 18,176 72,486
31 S.epidermidis R S mec A 19,396 72,43
168 S.epidermidis S S
125 S.epidermidis S S
170 S.epidermidis S S
90 S.equorum R S mec A 13,079 72,143
13 S.haemolyticus R S mec A 13,388 71,799
33 S.haemolyticus R S mec A 12,503 71,112
42 S.haemolyticus R S mec A 19,202 71,799
61 S.pneumoniae NA S
44 S.pneumoniae NA S
45 S.pneumoniae NA S
46 S.pneumoniae NA S
43 S.pneumoniae S S
58 SCN R S mec A 13,514 72,315
123 SCN R S mec A 16,351 72,315
59 SCN R S mec A 11,803 72,143
16 SCN R S mec A 12,589 71,283
55 SCN R S mec A 10,967 72,486
23 SCN R S mec A 14,197 72,658
130 SCN R S mec A 14,25 72,658
68 SCN R S mec A 18,48 72,19
75 SCN S S
TM RES
Sistema automatizado Phoenix® PCR em Tempo Real
GENES RESNº Hemo Identificação OXA VANCO CT RES
Ct RES= Ciclo Threshold; Tm RES= Temperatura de melting; R= resistente;
S= Sensível ; OXA= oxacilina; VANCO= Vancomicina; NA= Não aplicado.

RESULTADOS
98
No total, foram identificadas 5 amostras que apresentaram blaCTX-M, uma
blaTEM, uma blaSHV, uma blaKPC, 32 apresentaram mecA, e nenhuma
apresentou vanA, vanB, blaVIM, blaSPM e blaIMP pela PCR em Tempo Real
sistema SYBR Green (figura 7).
Figura 7. Total de amostras positivas pela metodologia da PCR em Tempo
Real pelo sistema SYBR Green para os respectivos genes de resistência
estudados

RESULTADOS
99
5.4. Testes Direto do Isolado Bacteriano
5.4.1. Confirmação de Susceptibilidade a Oxacilina e Gene de
Resistência mecA pelo Método da PCR em Tempo Real Direto do
Isolado Bacteriano Referente a Amostra de Hemocultura
Para as amostras que apresentaram positividade para o gene mecA pela
PCR em Tempo Real sistema SYBR Green, mas que foram consideradas
sensíveis a oxacilina pelo sistema Phoenix®, o exemplar bacteriano referente a
amostra de hemocultura, armazenada no Banco de Microrganismo do LEMC,
foi submetido aos testes confirmatórios de susceptibilidade a oxacilina e a PCR
em Tempo Real sistema SYBR Green para confirmação da presença do gene
mecA. Os resultados dos testes se encontram na Tabela 12.
Tabela 12. Resultado do Etest para oxacilina e PCR em Tempo Real sistema
SYBR Green para o gene de resistência mecA direto da colônia e comparação
com a PCR direto do frasco de hemocultura.
NT= Não Testado; S= sensível; R= resistente
Direto da Hemocultura
Nº HEMO Identificação do Microrganismo
Susceptibilidade a Oxacilina
Real Time para o gene mec A
Real Time para o gene
mec A
Etest Oxacilina
CIM E-test
76 S. aureus S mecA Negativo S 0,125 11 S. aureus S mecA Negativo S 0,125
138 S. aureus S mecA NT NT NT 139 S. aureus S mecA Positivo R >256 169 S. epidermidis S mecA Negativo S 0,125
Sistema automatizado Phoenix® Testes direto do Isolado Bacteriano

RESULTADOS
100
O exemplar referente à amostra 138 não foi localizado, portanto não
foram realizados os testes. Para as testadas, três amostras, duas de S. aureus
e uma de S. epidermidis confirmaram ser sensível a oxacilina, porém negativas
para o gene mecA. Em uma amostra observamos discordância do método
automatizado Phoenix® em que foi sensível a oxacilina, e no teste direto do
isolado apresentou resistência a oxacilina e a presença do gene mecA.
5.4.2. Confirmação da Produção de Carbapenemase e Gene de
Resistência blaKPC pelo Método da PCR em Tempo Real Direto do
Isolado Bacteriano Referente a Amostra de Hemocultura
Duas amostras de K. pneumoniae apresentaram resistência a imipenem
e meropenem pelo sistema automatizado Phoenix®. Porém, apresentaram
resultado negativo na PCR em Tempo Real pelo sistema SYBR Green para o
gene blaKPC. Outra amostra de K. pneumoniae apresentou sensibilidade a
imipenem e meropenem pelo sistema automatizado Phoenix®, mas apresentou
resultado positivo na PCR em Tempo Real pelo sistema SYBR Green para
blaKPC. As três amostras de K. pneumoniae foram isoladas de hemoculturas
provenientes de um mesmo paciente , porém os dois isolados resistentes aos
carbapenêmicos não se encontravam armazenados no banco de
microrganismo do LEMC. Assim, foi selecionado o isolado sensível a
carbapenêmicos com PCR em Tempo Real positivo para o gene blaKPC
adicionalmente a outros 4 isolados de K. pneumoniae provenientes do mesmo
paciente e que se encontravam no banco de microrganismo do LEMC, os quais
eram sensívies aos carbapenemicos e com resultado negativo para o gene
blaKPC pelo método da PCR em Tempo Real e todos foram submetidos a ao
A

RESULTADOS
101
teste de disco difusão para ertapenem e PCR em Tempo Real direto da
colônia para blaKPC. Os resultados estão apresentados na Tabela 13. Dos
cinco isolados retirados do banco de microrganismos do LEMC, todos
apresentaram resistência a ertapenem e positividade na PCR em Tempo Real
para blaKPC.
Tabela 13. Resultado do método de disco difusão e PCR em Tempo Real
sistema SYBR Green para o gene de resistência blaKPC direto da colônia e
comparação com a PCR direto do frasco de hemocultura.
Direto da
Hemocultura
Nº HEMO Nº PacienteData da
hemocultura
Identificaçao
do
Microrganismo
CIM/DD
para ERT
Susceptibilidad
e a IMP/MERO
CIM para
IMP/MERO
(µg/mL)
PCR em Tempo
Real para blaKPC
PCR em Tempo
Real para blaKPC
DD para
ERT
DD para ERT
(mm)
95 64 13/11/2009 K.pneumoniae 14mm S/S ≤1/≤1 Negativo KPC R 15mm
96 64 13/11/2009 K.pneumoniae 14mm S/S ≤1/≤1 Negativo KPC R 15mm
97 64 21/11/2009 K.pneumoniae S/S ≤1/2 Negativo KPC R 15mm
98 64 21/11/2009 K.pneumoniae S/S ≤1/2 Negativo NT NT NT
103 64 02/12/2009 K.pneumoniae Negativo NT NT NT
99 64 03/12/2009 K.pneumoniae S/S NL Negativo KPC R 15mm
100 64 03/12/2009 K.pneumoniae Negativo NT NT NT
104 64 10/12/2009 K.pneumoniae 22mm S/S NL KPC KPC R 15mm
101 64 15/12/2009 K.pneumoniae >4µg/mL R/R >8/>8 Negativo NT NT NT
102 64 15/12/2009 K. pneumoniae >4µg/mL R/R >8/>8 Negativo NT NT NT
Testes direto do Isolado BacterianoSistema automatizado Phoenix®Dados da Hemocultura e do Paciente
NT= Não Testado; S= sensível; I= Intermediário
5.4.3. Confirmação da Produção de Metalo-β-Lactamases e Genes
de Resistência blaIMP, blaVIM e blaSPM pelo Método da PCR em Tempo
Real Direto do Isolado Bacteriano Referente a Amostra de
Hemocultura
Quatro amostras, 3 de P. aeruginosa e 1 de A.baumannii foram
resistentes a pelo menos um carbapenem (imipenem e/ou meropenem),
sugerindo a produção de metalo-β-lactamases. Para esses exemplares foi
realizado o teste de hidrólise e a PCR em Tempo Real sistema SYBR Green
direto da colônia bacteriana referente à hemocultura para os genes blaIMP,
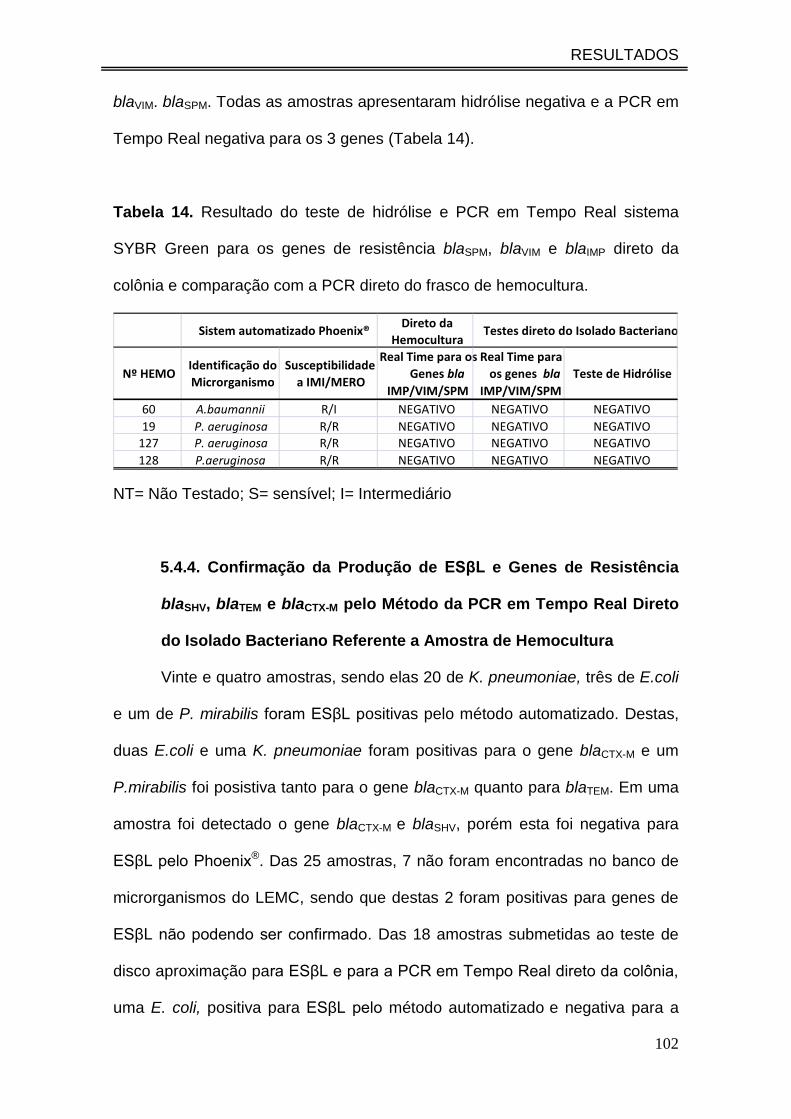
RESULTADOS
102
blaVIM. blaSPM. Todas as amostras apresentaram hidrólise negativa e a PCR em
Tempo Real negativa para os 3 genes (Tabela 14).
Tabela 14. Resultado do teste de hidrólise e PCR em Tempo Real sistema
SYBR Green para os genes de resistência blaSPM, blaVIM e blaIMP direto da
colônia e comparação com a PCR direto do frasco de hemocultura.
NT= Não Testado; S= sensível; I= Intermediário
5.4.4. Confirmação da Produção de ESβL e Genes de Resistência
blaSHV, blaTEM e blaCTX-M pelo Método da PCR em Tempo Real Direto
do Isolado Bacteriano Referente a Amostra de Hemocultura
Vinte e quatro amostras, sendo elas 20 de K. pneumoniae, três de E.coli
e um de P. mirabilis foram ESβL positivas pelo método automatizado. Destas,
duas E.coli e uma K. pneumoniae foram positivas para o gene blaCTX-M e um
P.mirabilis foi posistiva tanto para o gene blaCTX-M quanto para blaTEM. Em uma
amostra foi detectado o gene blaCTX-M e blaSHV, porém esta foi negativa para
ESβL pelo Phoenix®. Das 25 amostras, 7 não foram encontradas no banco de
microrganismos do LEMC, sendo que destas 2 foram positivas para genes de
ESβL não podendo ser confirmado. Das 18 amostras submetidas ao teste de
disco aproximação para ESβL e para a PCR em Tempo Real direto da colônia,
uma E. coli, positiva para ESβL pelo método automatizado e negativa para a
Direto da Hemocultura
60 A.baumannii R/I NEGATIVO NEGATIVO NEGATIVO 19 P. aeruginosa R/R NEGATIVO NEGATIVO NEGATIVO
127 P. aeruginosa R/R NEGATIVO NEGATIVO NEGATIVO 128 P.aeruginosa R/R NEGATIVO NEGATIVO NEGATIVO
Testes direto do Isolado Bacteriano
Nº HEMO Identificação do Microrganismo
Susceptibilidade a IMI/MERO
Real Time para os Genes bla
IMP/VIM/SPM
Real Time para os genes bla
IMP/VIM/SPM Teste de Hidrólise
Sistem automatizado Phoenix®

RESULTADOS
103
PCR em Tempo Real direto da hemocultura, não confirmou a positividade à
ESβL pelo método fenotípico e nem para a PCR direto da colônia. As outras
17 amostras confirmaram ser ESβL positiva pelo método do disco aproximação
e 9 apresentaram positividade para pelo menos um gene ESβL. Destas 9, 6
foram negativas para o gene ESβL direto da hemocultura. Em 3 amostras
houve detecção do gene ESβL tanto na amostra de hemocultura quanto direto
do isolado bacteriano, porém em uma destas amostras foi detectado direto da
hemocultura o gene blaCTX-M e na colônia apenas blaSHV e em outra havia sido
detectado os genes blaCTX-M e blaTEM e direto da colônia foi detectado apenas o
gene blaCTX-M (Tabela 15).
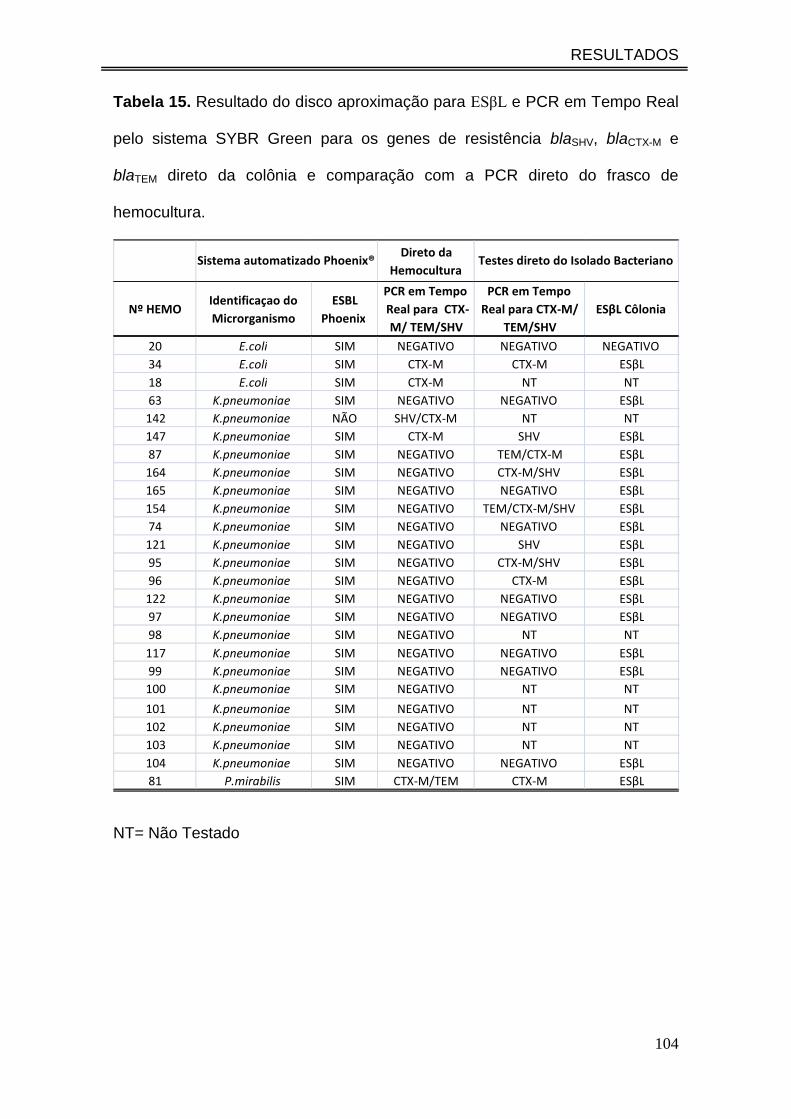
RESULTADOS
104
Tabela 15. Resultado do disco aproximação para ESβL e PCR em Tempo Real
pelo sistema SYBR Green para os genes de resistência blaSHV, blaCTX-M e
blaTEM direto da colônia e comparação com a PCR direto do frasco de
hemocultura.
NT= Não Testado
Direto da Hemocultura
Nº HEMO Identificaçao do Microrganismo
ESBL Phoenix
PCR em Tempo Real para CTX- M/ TEM/SHV
PCR em Tempo Real para CTX-M/
TEM/SHV ESβL Côlonia
20 E.coli SIM NEGATIVO NEGATIVO NEGATIVO 34 E.coli SIM CTX-M CTX-M ESβL
18 E.coli SIM CTX-M NT NT 63 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 142 K.pneumoniae NÃO SHV/CTX-M NT NT 147 K.pneumoniae SIM CTX-M SHV ESβL 87 K.pneumoniae SIM NEGATIVO TEM/CTX-M ESβL 164 K.pneumoniae SIM NEGATIVO CTX-M/SHV ESβL 165 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 154 K.pneumoniae SIM NEGATIVO TEM/CTX-M/SHV ESβL 74 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 121 K.pneumoniae SIM NEGATIVO SHV ESβL 95 K.pneumoniae SIM NEGATIVO CTX-M/SHV ESβL 96 K.pneumoniae SIM NEGATIVO CTX-M ESβL 122 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 97 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 98 K.pneumoniae SIM NEGATIVO NT NT
117 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 99 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 100 K.pneumoniae SIM NEGATIVO NT NT
101 K.pneumoniae SIM NEGATIVO NT NT 102 K.pneumoniae SIM NEGATIVO NT NT 103 K.pneumoniae SIM NEGATIVO NT NT
104 K.pneumoniae SIM NEGATIVO NEGATIVO ESβL 81 P.mirabilis SIM CTX-M/TEM CTX-M ESβL
Sistema automatizado Phoenix® Testes direto do Isolado Bacteriano

RESULTADOS
105
5.5. Variáveis Clínicas, Análise e Comparação do Tempo de
Liberação dos Resultados pelo Método Fenotipico e Tratamentos
antes e Após os Resultados em Relação ao Resultado Obtido pela
PCR em Tempo Real
Foram obtidas 185 amostras, 59 negativas e 126 positivas pelo sistema
automatizado Bactec®, coletadas de 117 pacientes submetidos a transplante de
órgão sólido. Dos 117 pacientes, 10,16% (11/117) foram admitidos na unidade
de transplantados de órgãos sólidos do HSP e 89,84% (106/117) no serviço de
transplantados de órgãos sólidos (rim e rim e pâncreas) do HRH. Em relação
ao sexo, 27,97% (32/117) dos pacientes eram do sexo feminino e 72,03%
(85/117) do sexo masculino. A idade média foi de 45,17 anos, sendo a idade
máxima de 74 anos e a mínima de 2 anos. Quanto ao tipo de transplante,
89,74% (105/117) pacientes foram submetidos a transplante de rim; 6,84%
(8/117) a transplante de rim e pâncreas; 1,70% (2/117) a transplante de fígado;
0,85% (1/117) a transplante de fígado e rim e 0,85% (1/117) a transplante de
coração. Em relação ao tipo de doador, 35,04% (41/117) pacientes receberam
o órgão transplantado de um doador vivo e 74,96% (76/117) pacientes de um
doador falecido.
Comparando as datas de coleta das 185 hemoculturas com a data de
transplante dos pacientes, o tempo médio em que os pacientes se
encontravam transplantados até a coleta da hemocultura foi de 331,15 dias
sendo que o menor tempo foi de 2 dias e o maior foi de 7300 dias (20 anos)
(Anexo 1).

RESULTADOS
106
5.5.1. Hemoculturas Positivas para Isolados Gram Positivos
Cinquenta e nove hemoculturas foram identificadas pelo sistema
automatizado Phoenix® com bactérias Gram positivas. A mediana em relação
ao tempo de liberação do resultado do Gram foi de 1 dia e para liberação final
de 4 dias.
A inadequação do tratamento, de acordo com o tempo de liberação dos
resultados parciais e finais é demonstrada na Figura 8. Durante o tratamento
empírico, isto é, antes da liberação do resultado do Gram, em 8 casos
(13,56%) os pacientes recebiam tratamento considerado inadequado; após a
liberação do resultado do Gram foi observada inadequação em 4 casos
(6,78%) e após a liberação final da hemocultura, 5 (8,48%) apresentavam
inadequação. Os critérios de inadequação adotados seguem padronização já
citada anteriormente.
A amostra 139 referente ao paciente 88, destacada na Tabela 15, foi
oxacilina sensível pelo método automatizado, porém o isolado retirado do
banco de microrganismos foi resistente a oxacilina pelo E-test. Neste caso o
paciente recebeu oxacilina no tratamento após o resultado final da hemocultura
e mesmo assim a antibioticoterapia foi adequada, pois foi considerado o
resultado do sistema automatizado para essa avaliação.
Dos cinco casos em que os pacientes receberam tratamento inadequado
mesmo após a liberação do resultado final, dois isolados eram SCoN
resistentes a oxacilina com gene mecA positivo tratados com levofloxacina e
bactrim respectivamente. Outros dois, um SCoN e um S. aureus, ambos
sensíveis a oxacilina e gene mecA negativos, não foram tratados. Um isolado
de Listeria spp. foi tratado com Bactrim (Tabela16).
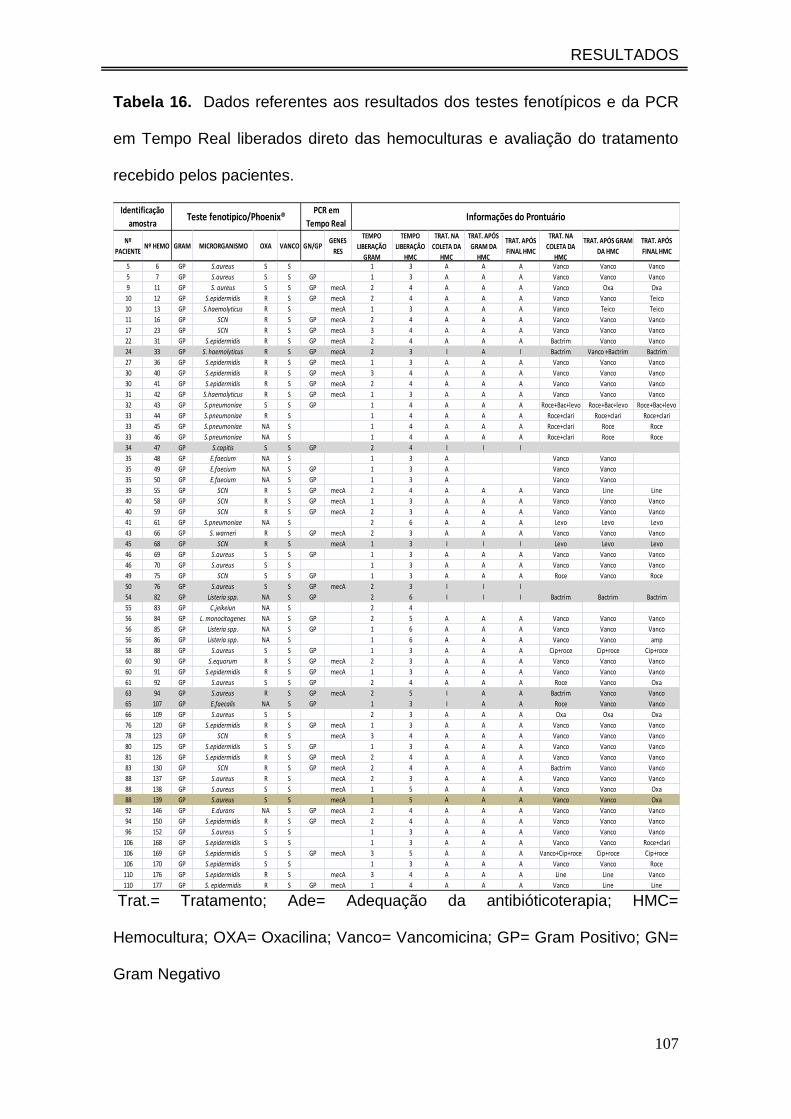
RESULTADOS
107
Tabela 16. Dados referentes aos resultados dos testes fenotípicos e da PCR
em Tempo Real liberados direto das hemoculturas e avaliação do tratamento
recebido pelos pacientes.
Nº
PACIENTENº HEMO GRAM MICRORGANISMO OXA VANCO GN/GP
GENES
RES
TEMPO
LIBERAÇÃO
GRAM
TEMPO
LIBERAÇÃO
HMC
TRAT. NA
COLETA DA
HMC
TRAT. APÓS
GRAM DA
HMC
TRAT. APÓS
FINAL HMC
TRAT. NA
COLETA DA
HMC
TRAT. APÓS GRAM
DA HMC
TRAT. APÓS
FINAL HMC
5 6 GP S.aureus S S 1 3 A A A Vanco Vanco Vanco
5 7 GP S.aureus S S GP 1 3 A A A Vanco Vanco Vanco
9 11 GP S. aureus S S GP mecA 2 4 A A A Vanco Oxa Oxa
10 12 GP S.epidermidis R S GP mecA 2 4 A A A Vanco Vanco Teico
10 13 GP S.haemolyticus R S mecA 1 3 A A A Vanco Teico Teico
11 16 GP SCN R S GP mecA 2 4 A A A Vanco Vanco Vanco
17 23 GP SCN R S GP mecA 3 4 A A A Vanco Vanco Vanco
22 31 GP S.epidermidis R S GP mecA 2 4 A A A Bactrim Vanco Vanco
24 33 GP S. haemolyticus R S GP mecA 2 3 I A I Bactrim Vanco +Bactrim Bactrim
27 36 GP S.epidermidis R S GP mecA 1 3 A A A Vanco Vanco Vanco
30 40 GP S.epidermidis R S GP mecA 3 4 A A A Vanco Vanco Vanco
30 41 GP S.epidermidis R S GP mecA 2 4 A A A Vanco Vanco Vanco
31 42 GP S.haemolyticus R S GP mecA 1 3 A A A Vanco Vanco Vanco
32 43 GP S.pneumoniae S S GP 1 4 A A A Roce+Bac+levo Roce+Bac+levo Roce+Bac+levo
33 44 GP S.pneumoniae R S 1 4 A A A Roce+clari Roce+clari Roce+clari
33 45 GP S.pneumoniae NA S 1 4 A A A Roce+clari Roce Roce
33 46 GP S.pneumoniae NA S 1 4 A A A Roce+clari Roce Roce
34 47 GP S.capitis S S GP 2 4 I I I
35 48 GP E.faecium NA S 1 3 A Vanco Vanco
35 49 GP E.faecium NA S GP 1 3 A Vanco Vanco
35 50 GP E.faecium NA S GP 1 3 A Vanco Vanco
39 55 GP SCN R S GP mecA 2 4 A A A Vanco Line Line
40 58 GP SCN R S GP mecA 1 3 A A A Vanco Vanco Vanco
40 59 GP SCN R S GP mecA 2 3 A A A Vanco Vanco Vanco
41 61 GP S.pneumoniae NA S 2 6 A A A Levo Levo Levo
43 66 GP S. warneri R S GP mecA 2 3 A A A Vanco Vanco Vanco
45 68 GP SCN R S mecA 1 3 I I I Levo Levo Levo
46 69 GP S.aureus S S GP 1 3 A A A Vanco Vanco Vanco
46 70 GP S.aureus S S 1 3 A A A Vanco Vanco Vanco
49 75 GP SCN S S GP 1 3 A A A Roce Vanco Roce
50 76 GP S.aureus S S GP mecA 2 3 I I I
54 82 GP Listeria spp. NA S GP 2 6 I I I Bactrim Bactrim Bactrim
55 83 GP C.jeikeiun NA S 2 4
56 84 GP L. monocitogenes NA S GP 2 5 A A A Vanco Vanco Vanco
56 85 GP Listeria spp. NA S GP 1 6 A A A Vanco Vanco Vanco
56 86 GP Listeria spp. NA S 1 6 A A A Vanco Vanco amp
58 88 GP S.aureus S S GP 1 3 A A A Cip+roce Cip+roce Cip+roce
60 90 GP S.equorum R S GP mecA 2 3 A A A Vanco Vanco Vanco
60 91 GP S.epidermidis R S GP mecA 1 3 A A A Vanco Vanco Vanco
61 92 GP S.aureus S S GP 2 4 A A A Roce Vanco Oxa
63 94 GP S.aureus R S GP mecA 2 5 I A A Bactrim Vanco Vanco
65 107 GP E.faecalis NA S GP 1 3 I A A Roce Vanco Vanco
66 109 GP S.aureus S S 2 3 A A A Oxa Oxa Oxa
76 120 GP S.epidermidis R S GP mecA 1 3 A A A Vanco Vanco Vanco
78 123 GP SCN R S mecA 3 4 A A A Vanco Vanco Vanco
80 125 GP S.epidermidis S S GP 1 3 A A A Vanco Vanco Vanco
81 126 GP S.epidermidis R S GP mecA 2 4 A A A Vanco Vanco Vanco
83 130 GP SCN R S GP mecA 2 4 A A A Bactrim Vanco Vanco
88 137 GP S.aureus R S mecA 2 3 A A A Vanco Vanco Vanco
88 138 GP S.aureus S S mecA 1 5 A A A Vanco Vanco Oxa
88 139 GP S.aureus S S mecA 1 5 A A A Vanco Vanco Oxa
92 146 GP E.durans NA S GP mecA 2 4 A A A Vanco Vanco Vanco
94 150 GP S.epidermidis R S GP mecA 2 4 A A A Vanco Vanco Vanco
96 152 GP S.aureus S S 1 3 A A A Vanco Vanco Vanco
106 168 GP S.epidermidis S S 1 3 A A A Vanco Vanco Roce+clari
106 169 GP S.epidermidis S S GP mecA 3 5 A A A Vanco+Cip+roce Cip+roce Cip+roce
106 170 GP S.epidermidis S S 1 3 A A A Vanco Vanco Roce
110 176 GP S.epidermidis R S mecA 3 4 A A A Line Line Vanco
110 177 GP S. epidermidis R S GP mecA 1 4 A A A Vanco Line Line
Identificação
amostraTeste fenotipico/Phoenix®
PCR em
Tempo Real Informações do Prontuário
Trat.= Tratamento; Ade= Adequação da antibióticoterapia; HMC=
Hemocultura; OXA= Oxacilina; Vanco= Vancomicina; GP= Gram Positivo; GN=
Gram Negativo
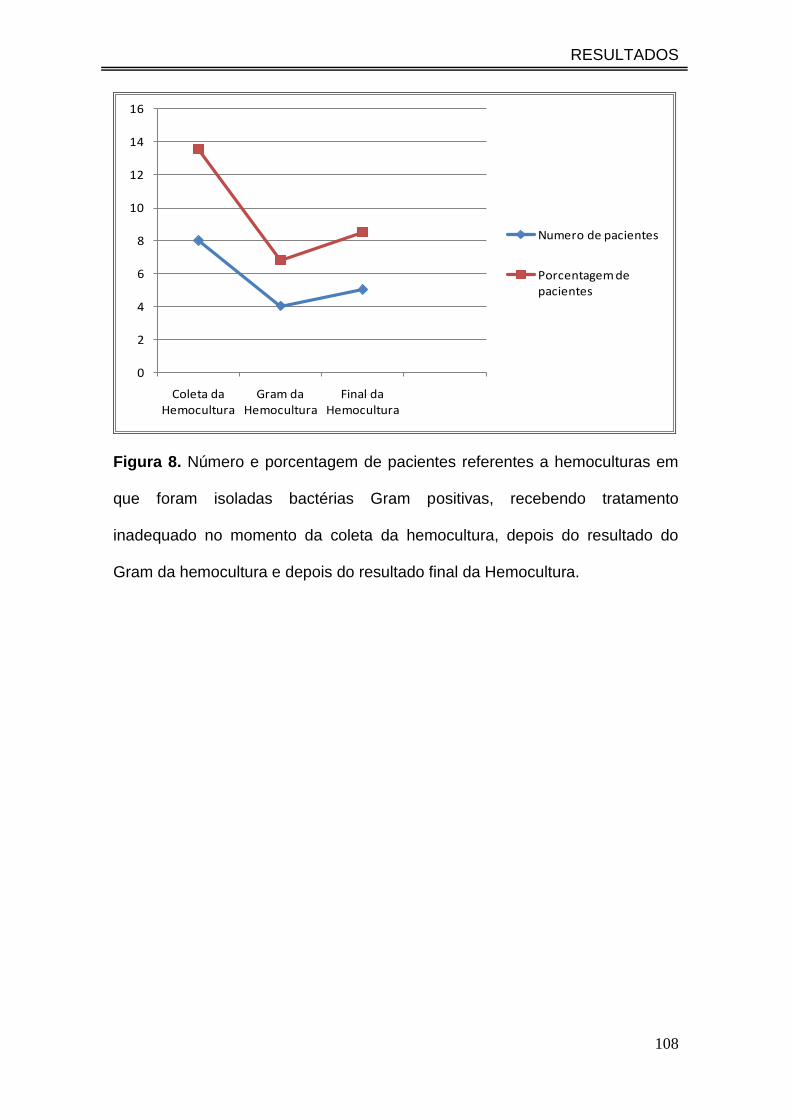
RESULTADOS
108
0
2
4
6
8
10
12
14
16
Coleta da Hemocultura
Gram da Hemocultura
Final da Hemocultura
Numero de pacientes
Porcentagem de pacientes
Figura 8. Número e porcentagem de pacientes referentes a hemoculturas em
que foram isoladas bactérias Gram positivas, recebendo tratamento
inadequado no momento da coleta da hemocultura, depois do resultado do
Gram da hemocultura e depois do resultado final da Hemocultura.

RESULTADOS
109
5.5.2. Hemoculturas Positivas para Isolados Gram Negativos
Sessenta e sete hemoculturas foram identificadas pelo sistema
automatizado Phoenix® apresentando bactérias Gram negativas. A mediana
em relação ao tempo de liberação do resultado do Gram foi de um dia. Com
relação ao tempo de liberação da espécie e susceptibilidade a antimicrobianos
do isolado foi de três dias. Durante o tratamento empírico, isto é, antes da
liberação do resultado do Gram, 19 (28,4%) dos casos de hemoculturas os
pacientes recebiam tratamento considerado inadequado. Destes, 14 (20,9%)
eram espécies de enterobacteriaceas e 4 (7,5%) espécies de não
fermentadores. Após a liberação do resultado do Gram, 14 (20,9%)
apresentavam-se em tratamento inadequado, sendo 11 (16,42%) de
enterobacteriaceas e 3 (4,48%) de não fermentadores. Após a liberação do
resultado final da hemocultura, 3 (4,5%) apresentavam-se em tratamento
inadequado sendo, 2 (2,99%) de enterobacteriaceas e 1 (1,51%) de não
fermentador (figuras 9 e 10). Destas 3 hemoculturas, 1 foi positiva para o gene
de resistência CTX-M e 2 negativas para qualquer gene de resistência
pesquisado, sendo que apenas a mostra CTX-M positiva foi ESβL positiva para
o sistema automatizado (Tabela 17).
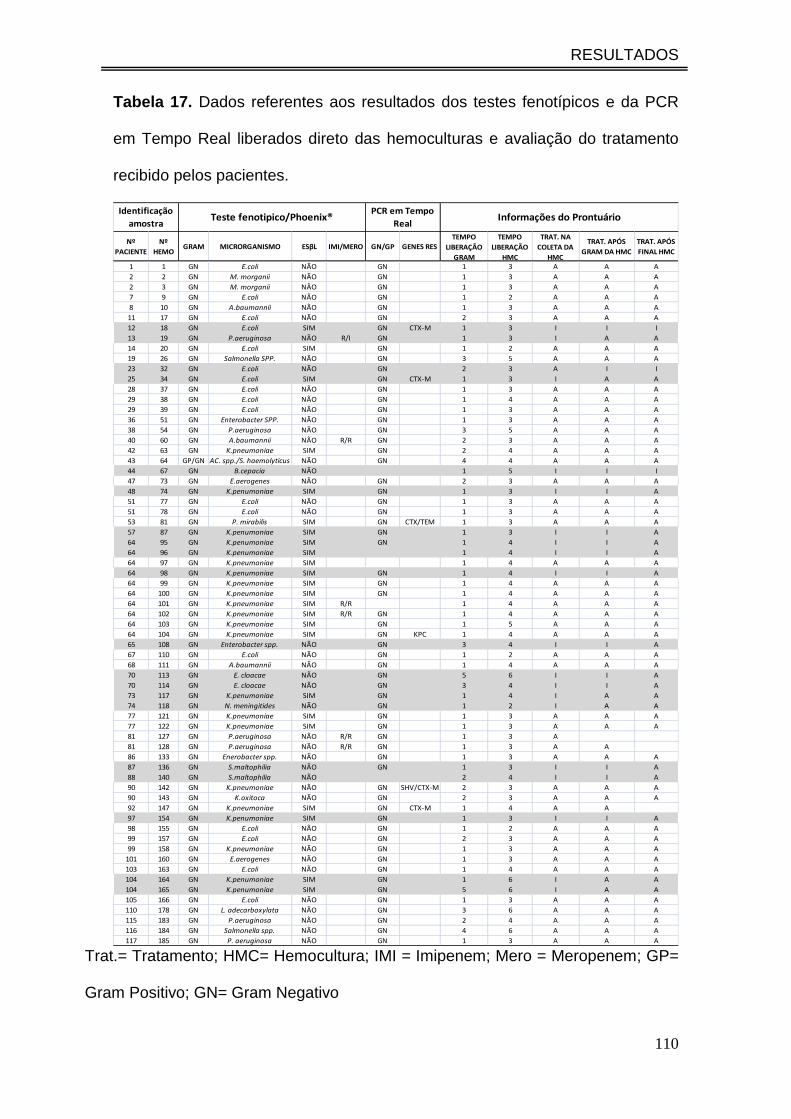
RESULTADOS
110
Tabela 17. Dados referentes aos resultados dos testes fenotípicos e da PCR
em Tempo Real liberados direto das hemoculturas e avaliação do tratamento
recibido pelos pacientes.
Nº
PACIENTE
Nº
HEMOGRAM MICRORGANISMO ESβL IMI/MERO GN/GP GENES RES
TEMPO
LIBERAÇÃO
GRAM
TEMPO
LIBERAÇÃO
HMC
TRAT. NA
COLETA DA
HMC
TRAT. APÓS
GRAM DA HMC
TRAT. APÓS
FINAL HMC
1 1 GN E.coli NÃO GN 1 3 A A A
2 2 GN M. morganii NÃO GN 1 3 A A A
2 3 GN M. morganii NÃO GN 1 3 A A A
7 9 GN E.coli NÃO GN 1 2 A A A
8 10 GN A.baumannii NÃO GN 1 3 A A A
11 17 GN E.coli NÃO GN 2 3 A A A
12 18 GN E.coli SIM GN CTX-M 1 3 I I I
13 19 GN P.aeruginosa NÃO R/I GN 1 3 I A A
14 20 GN E.coli SIM GN 1 2 A A A
19 26 GN Salmonella SPP. NÃO GN 3 5 A A A
23 32 GN E.coli NÃO GN 2 3 A I I
25 34 GN E.coli SIM GN CTX-M 1 3 I A A
28 37 GN E.coli NÃO GN 1 3 A A A
29 38 GN E.coli NÃO GN 1 4 A A A
29 39 GN E.coli NÃO GN 1 3 A A A
36 51 GN Enterobacter SPP. NÃO GN 1 3 A A A
38 54 GN P.aeruginosa NÃO GN 3 5 A A A
40 60 GN A.baumannii NÃO R/R GN 2 3 A A A
42 63 GN K.pneumoniae SIM GN 2 4 A A A
43 64 GP/GN AC. spp./S. haemolyticus NÃO GN 4 4 A A A
44 67 GN B.cepacia NÃO 1 5 I I I
47 73 GN E.aerogenes NÃO GN 2 3 A A A
48 74 GN K.penumoniae SIM GN 1 3 I I A
51 77 GN E.coli NÃO GN 1 3 A A A
51 78 GN E.coli NÃO GN 1 3 A A A
53 81 GN P. mirabilis SIM GN CTX/TEM 1 3 A A A
57 87 GN K.penumoniae SIM GN 1 3 I I A
64 95 GN K.penumoniae SIM GN 1 4 I I A
64 96 GN K.penumoniae SIM 1 4 I I A
64 97 GN K.pneumoniae SIM 1 4 A A A
64 98 GN K.penumoniae SIM GN 1 4 I I A
64 99 GN K.pneumoniae SIM GN 1 4 A A A
64 100 GN K.pneumoniae SIM GN 1 4 A A A
64 101 GN K.pneumoniae SIM R/R 1 4 A A A
64 102 GN K.pneumoniae SIM R/R GN 1 4 A A A
64 103 GN K.pneumoniae SIM GN 1 5 A A A
64 104 GN K.pneumoniae SIM GN KPC 1 4 A A A
65 108 GN Enterobacter spp. NÃO GN 3 4 I I A
67 110 GN E.coli NÃO GN 1 2 A A A
68 111 GN A.baumannii NÃO GN 1 4 A A A
70 113 GN E. cloacae NÃO GN 5 6 I I A
70 114 GN E. cloacae NÃO GN 3 4 I I A
73 117 GN K.penumoniae SIM GN 1 4 I A A
74 118 GN N. meningitides NÃO GN 1 2 I A A
77 121 GN K.pneumoniae SIM GN 1 3 A A A
77 122 GN K.pneumoniae SIM GN 1 3 A A A
81 127 GN P.aeruginosa NÃO R/R GN 1 3 A
81 128 GN P.aeruginosa NÃO R/R GN 1 3 A A
86 133 GN Enerobacter spp. NÃO GN 1 3 A A A
87 136 GN S.maltophilia NÃO GN 1 3 I I A
88 140 GN S.maltophilia NÃO 2 4 I I A
90 142 GN K.pneumoniae NÃO GN SHV/CTX-M 2 3 A A A
90 143 GN K.oxitoca NÃO GN 2 3 A A A
92 147 GN K.pneumoniae SIM GN CTX-M 1 4 A A
97 154 GN K.penumoniae SIM GN 1 3 I I A
98 155 GN E.coli NÃO GN 1 2 A A A
99 157 GN E.coli NÃO GN 2 3 A A A
99 158 GN K.pneumoniae NÃO GN 1 3 A A A
101 160 GN E.aerogenes NÃO GN 1 3 A A A
103 163 GN E.coli NÃO GN 1 4 A A A
104 164 GN K.penumoniae SIM GN 1 6 I A A
104 165 GN K.penumoniae SIM GN 5 6 I A A
105 166 GN E.coli NÃO GN 1 3 A A A
110 178 GN L. adecarboxylata NÃO GN 3 6 A A A
115 183 GN P.aeruginosa NÃO GN 2 4 A A A
116 184 GN Salmonella spp. NÃO GN 4 6 A A A
117 185 GN P. aeruginosa NÃO GN 1 3 A A A
Teste fenotipico/Phoenix® Informações do ProntuárioIdentificação
amostra
PCR em Tempo
Real
Trat.= Tratamento; HMC= Hemocultura; IMI = Imipenem; Mero = Meropenem; GP=
Gram Positivo; GN= Gram Negativo
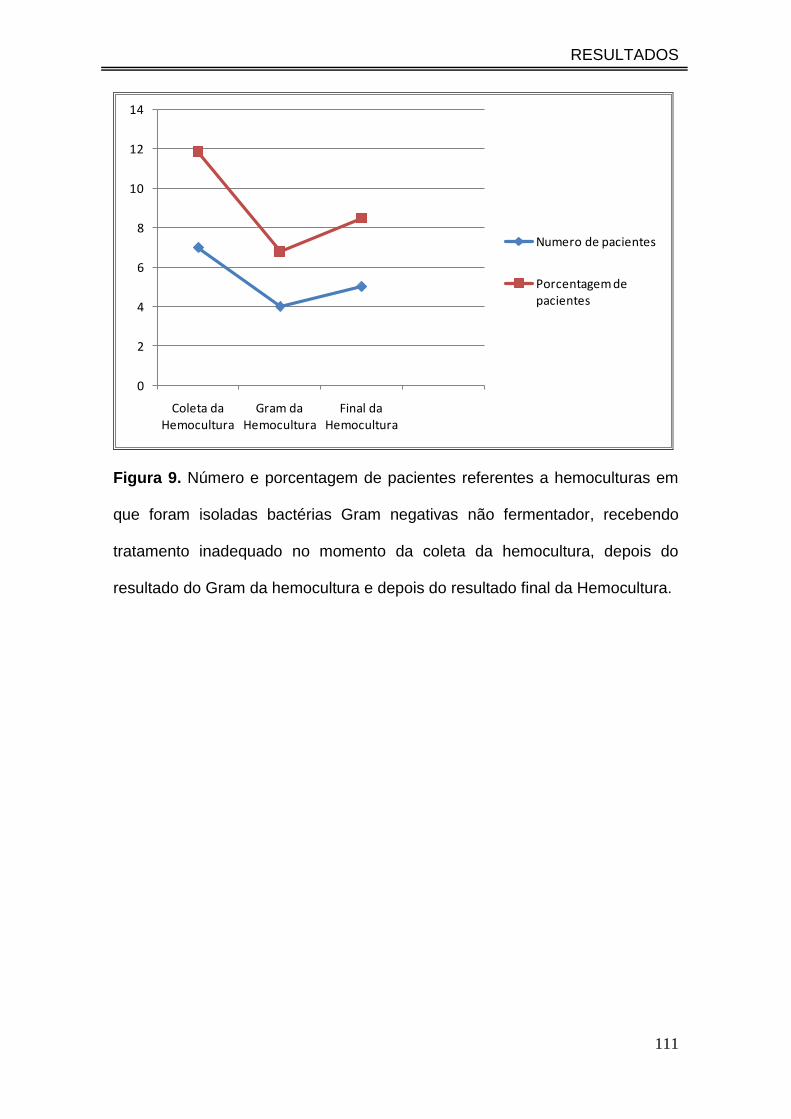
RESULTADOS
111
0
2
4
6
8
10
12
14
Coleta da Hemocultura
Gram da Hemocultura
Final da Hemocultura
Numero de pacientes
Porcentagem de pacientes
Figura 9. Número e porcentagem de pacientes referentes a hemoculturas em
que foram isoladas bactérias Gram negativas não fermentador, recebendo
tratamento inadequado no momento da coleta da hemocultura, depois do
resultado do Gram da hemocultura e depois do resultado final da Hemocultura.
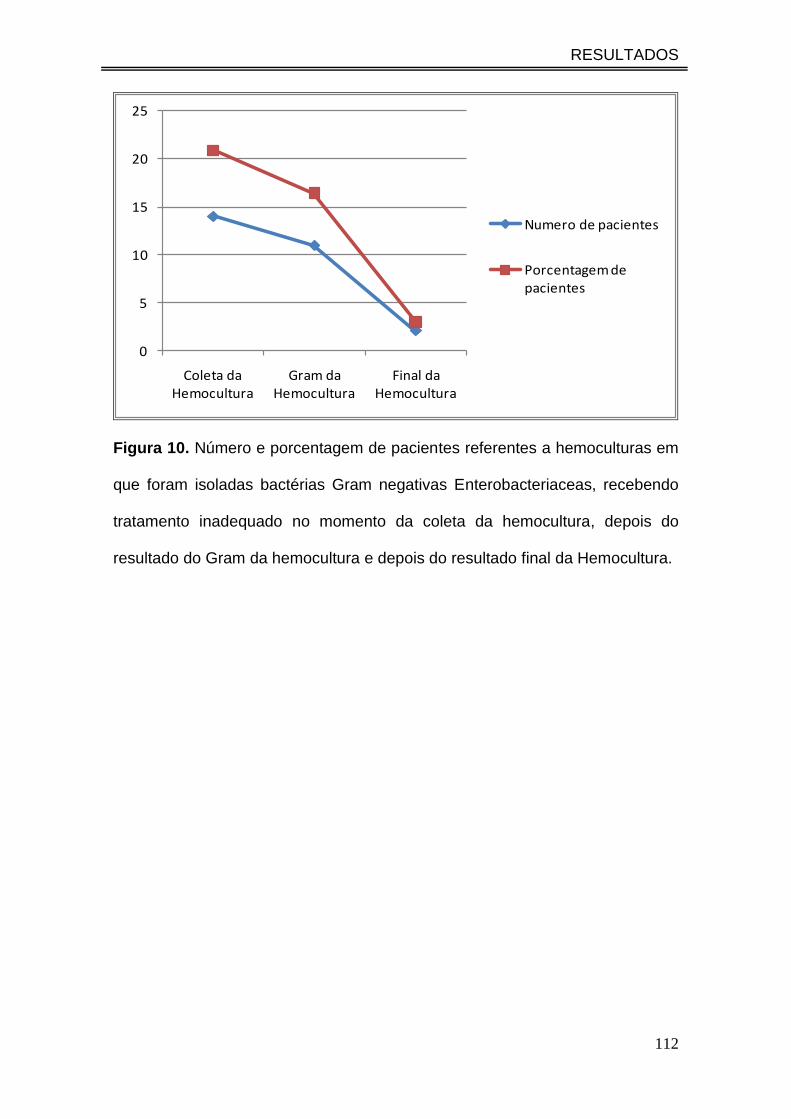
RESULTADOS
112
0
5
10
15
20
25
Coleta da Hemocultura
Gram da Hemocultura
Final da Hemocultura
Numero de pacientes
Porcentagem de pacientes
Figura 10. Número e porcentagem de pacientes referentes a hemoculturas em
que foram isoladas bactérias Gram negativas Enterobacteriaceas, recebendo
tratamento inadequado no momento da coleta da hemocultura, depois do
resultado do Gram da hemocultura e depois do resultado final da Hemocultura.

DISCUSSÃO
113
6. DISCUSSÃO
Infecções de Corrente Sanguínea é uma das infecções mais sérias que
um paciente hospitalizado pode apresentar, sendo o quadro agravado quando
se trata de pacientes em recuperação ou em condições debilitantes. Em
pacientes transplantados de órgão sólidos, bacteremia é uma das
complicações mais frequentes e está fortemente associada a altas taxas de
morbidade e mortalidade (Singh et al., 2000).
O diagnóstico precoce de ICS é essencial para a instituição de uma
terapia adequada em casos de pacientes transplantados favorecendo uma
maior chance de sobrevida e sucesso pós-transplante. Entretanto, a
terapêutica empírica pode ser dirigida a bactérias Gram positivas ou Gram
negativas, e mesmo para leveduras, dependendo das características clínicas
dos pacientes. Além disso, a probalilidade de bactérias carreando diversos
genes de resistência dificulta a adequação terapêutica que se confirma apenas
com o resultado final da identificação do microrganismo e antibiograma,
geralmente em não menos de 72 horas após a coleta da hemocultura mesmo
com a utiização de métodos fenotípicos automatizados. A utilização de testes
moleculares para a detecção precoce de microrganismo em corrente
sanguínea e a detecção de genes de resistência pode trazer contribuição para
a adequação do tratamento antimicrobiano com diminuição de
morbi/mortalidade (Ferraresso & Berardineli, 2005).
O uso da técnica da PCR para fins de diagnóstico estabelece a nova era
na detecção e caracterização de microrganismo direto de amostras clínicas. A
partir do uso rotineiro da PCR, outros métodos e aplicações puderam ser
elaborados entre elas a PCR Multiplex e a técnica da PCR em Tempo Real.

DISCUSSÃO
114
Esta ultima é um refinamento da técnica original de PCR, pois combinam a
amplificação e a quantificação de uma sequência de DNA alvo por meio da
detecção de fluorescência. É considerado um método homogêneo de
amplificação de DNA, no qual amplificação e detecção são realizadas no
mesmo tubo de reação, podendo eliminar processamentos pós-PCR (corrida
de eletroforese em agarose), diminuindo o manuseio de produtos de
amplificação e o risco de contaminação cruzada (Hayden, 2004).
A abordagem da PCR em Tempo Real para fins de diagnóstico tem
ganhado crescente aceitação como uma técnica de laboratório adequado,
porém, a interpretação dos resultados pode ser duvidosa. A amostra poderia
ser considerada negativa, enquanto que ela contém o patógeno (resultado falso
negativo). Tal situação pode ocorrer devido à baixa concentração de
patógenos, ou quando a amostra apresentar inibidores de PCR. Em contraste,
a amostra poderia ser considerada como positiva, enquanto ele não contém o
patógeno (resultado falso positivo). Esta situação pode ser encontrada quando
o desenho dos iniciadores e / ou sonda não são ideais, ou em caso de
contaminação do DNA, durante a etapa de extração, ou durante a PCR, como
mostrado por Stals e colaboradores (2009) para PCR em Tempo Real. Embora
estes problemas possam ser minimizados com o uso de controles adequados,
é de fundamental importância a determinação do cutoff (Limite de Detecção
Branco- LoB) e de um limite de detecção (LoD) do método para ajudar na
interpretação confiável.
No presente estudo foram realizados os cálculos do LoD e cutoff para o
gene 16S rDNA para detecção de bactérias Gram positivos e Gram negativos
além dos genes de resistência. Porém, para os genes blaIMP, blaSPM e blaVIM, a

DISCUSSÃO
115
interpretação dos resultados foi conforme Mendes e colaboradores (2007).
Além dos cálculos do LoD e Cutoff foi realizada, através da curva ROC, uma
análise direta de especificidade e sensibilidade dos iniciadores para os genes
estudados. Através do LoD e cutoff fica possível a interpretação confiável dos
resultados de forma qualitativa (presença e ausência).
O limite de detecção de Unidade Formadora de Colônia (UFC) por
reação para os respectivos genes em estudo mostrou o teste muito sensível,
sendo possível a detecção dos genes na presença de 15 á 1,5 UFC, porém
quando determinado o Cutoff e o LoD este limite aumenta a UFC por reação
para a maioria dos genes avaliados. Para o genes blaKPC, blaSHV e blaCTX-M a
especificidade mostrou-se mais baixa quando comparamos com os demais
genes, já os genes blaTEM, blaSHV, blaCTX-M e vanB mostrou uma sensibilidade
mais baixa quando comparados com os demais genes. Levando-se em conta
que se trata de um teste elaborado para ser aplicado diretamente em amostras
clínicas, há a prioridade de manter uma boa sensibilidade com uma boa
especificidade. Para PCR em Tempo Real quanto maior o tamanho do produto
de amplificação maior é a especificidade e menor a sensibilidade do teste. E a
baixa sensibilidade encontrada para os genes blaTEM, blaSHV, blaCTX-M e vanB
pode ser devido ao tamanho dos produtos amplificados (Dorak, 2006) pois, os
iniciadores utilizados para estes genes foram os mesmos utilizados em PCR
convencional, que formam produtos de amplificação entre 700 a 1000 pb, para
posteriormente serem direcionados para o seqüenciamento. Outro motivo para
esses genes não terem sido detectados é por haver muito DNA não do gene
procurado. Uma quantidade muito grande de DNA humano pode atrapalhar a
amplificação do gene procurado e também a presença de excessos de

DISCUSSÃO
116
inibidores que o método de extração não retirou. A solução dessas
interferências deve aumentar a sensibilidade do teste.
A diminuição da especificidade para detectar Gram negativos pode ser
devido ao fato de haver interferência quanto à Taq polimerase. As preparações
podem conter uma quantidade pequena de DNA contaminante proveniente de
Escherichia coli ou Thermus aquaticus (Okhravi et al., 2000b, Carroll et al.,
1999) e também de Pseudomonas spp., Sphingomonas spp., Moraxella spp. e
AcinetobaCter spp. (Mühl et al., 2008), que podem ser suficientes para
amplificação de sinal positivo após amplificação pela metodologia da PCR em
Tempo Real. Quando a técnica de PCR em Tempo Real é utilizada para a
detecção de patógenos em amostras clínicas com pequena quantidade de DNA
a presença de sinal positivo para os controles negativos pode dificultar a
interpretação dos resultados das amostras positivas (Klaschik et al., 2002).
Levando-se em conta que estes genes (blaCTX-M e blaSHV) podem ser
encontrados nessas bactérias, a interferência é ainda maior.
Em um recente estudo, Lehmann e colaboradores (2008) avaliaram a
aplicação da PCR em Tempo Real na detecção de patógenos no sangue pelo
Kit LightCycler SeptiFast® (Roche-Molecular Diagnosis) e comparou os
resultados com os obtidos pelo Bactec® 9240 (Becton Dickinson GmbH,
Germany). Kit LightCycler SeptiFast® é padronizado para utilização no aparelho
automatizado SeptiFast, no qual a partir da amplificação do DNA, realliza-se in
vitro a identificação das 25 espécies mais importantes de bactérias e fungos
diretamente do sangue. Das 114 amostras positivas para a PCR, 46 foram
negativas pelo Bactec®. Das 58 amostras positivas para ambos os testes, 18
foram discordantes, podendo ter ocorrido tanto por algumas espécies que

DISCUSSÃO
117
foram identificadas pelo Bactec® não se encontrarem no painel de identificação
do SeptiFast ou porque foram consideradas contaminações na hemocultura.
No presente estudo, não houve discordância quanto a detecção de
bactérias Gram positivas e Gram negativas pelo método fenotípico de
coloração de Gram e a PCR em Tempo Real para o gene 16S rDNA. Contudo,
para esta avaliação foram excluídas 5 amostras que apresentavam bactérias
Gram negativas e 19 amostras que apresentavam bactérias Gram positivas.
Esta conduta foi necessária porque durante o desenvolvimento do trabalho
houve necessidade de resintetizar as sondas de Gram positivo e Gram
negativo. Porém estas amostras não foram excluídas das outras etapas do
estudo e foram avaliadas quanto à presença de genes de resistência. Uma
amostra identificada com a presença de uma bactéria Gram positiva e outra
Gram negativa foi positiva apenas para a sonda de Gram negativo, sendo a
bactéria Gram positiva não detectada. A bactéria Gram positiva se tratava de
um S. haemolyticus. Em hemoculturas é comum a contaminação da amostra
por esses microrganismos no momento da coleta. Em estudos, a taxa de
amostras consideradas contaminadas por SCoN chega a ser de 26% (Ikegaya
et al,, 2010) já que se trata de bactérias colonizantes da pele. Apesar deste
dado demonstrar a dificuldade do método em detectar a presença de diferentes
microrganismos, houve 100% de concordância entre a detecção de bactérias
Gram positivas e Gram negativas quando comparado com o método fenotípico.
Trabalhos que avaliam a prevalência de bactérias Gram positivas e
Gram negativas presentes em hemoculturas referentes à pacientes
transplantados de órgão sólidos apontam para maiores taxas de
microrganismos Gram negativos (Bert et al., 2010; Silva Jr et al., 2010; Linares

DISCUSSÃO
118
et al., 2009; Al-Hasan et al., 2008). Porém, este dado pode variar de acordo
com o perfil epidemiológico da região estudada e do Hospital (Berger et al.,
2006). Neste estudo, a prevalência de bactérias Gram negativas foi de 53,17%,
e de Gram positivas de 46,83%, havendo pouca diferença entre as incidências.
Dentre as espécies Gram negativas identificadas no presente estudo
pelo sistema automatizado, K. pneumoniae foi a mais prevalente sendo que
destas, 90 % foram ESβL positivo e 2 resistentes a carbapenemicos. Na
literatura, esta espécie se apresenta entre os principais microrganismos isolado
em hemoculturas, inferior apenas a E. coli, sendo a taxa de exemplares ESβL
positivo bastante alta ( Linares et al., 2010; Andrade et al., 2008).
Dentre as espécies Gram positivas identificadas, o gênero
Staphylococcus foi a mais prevalente. Destas, 59% apresentaram resistência a
oxacilina pelo sistema automatizado Phoenix®. No presente estudo houve
predomínido de SCoN em relação a S. aureus, com altas taxas de resistência a
oxacilina, o mesmo encontrado na literatura (Gales et al., 2009). Em relação
ao gênero enterococcus, este é comumente isolado em pacientes
transplantados de órgão sólidos, especialmente em transplante de rim e fígado
(Patel et al.,2001).
Relatórios recentes mostram uma prevalência de colonização por VRE
entre 3,4% e 55% (Freitas et al., 2006; Hagen et al., 2003; Bakir et al.,
2001) e de infecção entre 4% e 11% (Patel et al., 2001; McNeil et al., 2005) em
transplantados de fígado e rim, respectivamente. Neste estudo houve 8,3% dos
casos de ICS por enterococos, porém nenhum isolado foi VRE.
Quando comparado os resultados dos testes de susceptibilidade em
isolados Gram negativos, liberados pelo sistema automatizado em relação a

DISCUSSÃO
119
PCR em Tempo Real para os genes de resistência, o método fenotípico
apontou a presença de 24 amostras ESβL positiva. Destas 24, apenas 4
apresentaram PCR positivo para ESβL direto da hemocultura. Uma amostra foi
ESβL positiva pelo método da PCR, mas não pelo método fenotípico. Porém,
quando avaliados os 15 isolados bacterianos referentes às hemoculturas em
que houve discordância entre a presença de ESβL pelo método automatizado e
a detecção molecular, uma amostra não confirmou ser ESβL pela técnica
fenotípica de disco aproximação.
Das 14 amostras em que foi confirmado a presença ESβL pelo método
da disco aproximação, 9 foram positivas para pelo menos um gene de ESβL,
sendo que destas, 6 que foram negativas para o gene ESβL direto da
hemocultura mas foram positivas direto da colônia. Em 3 amostras houve
detecção de ESβL tanto na amostra de hemocultura quanto direto da colônia,
porém em uma destas amostras foi detectado direto da hemocultura o gene
blaCTX-M e na colônia apenas blaSHV e em outra havia sido detectado os genes
blaCTX-M e blaTEM e direto da colônia foi detectado apenas o gene blaCTX-M. Para
os resultados que foram ESβL positivo pelo sistema, mas negativa para a PCR
direto da amostra clínica quanto do isolado, possivelmente se trata de outro
mecanismo de resistência ou uma ESβL diferente das pesquisadas no estudo.
Quanto aos resultados em que houve detecção do gene de ESβL direto do
isolado, mas não direto da amostra clínica, este fato pode ser explicado por se
tratar de concentrações diferentes de DNA, isto é, na hemocultura a quantidade
de DNA era inferior a necessária para ser detectada pelo teste. A sensibilidade
do teste pode ser otimizada diminuindo o tamanho do produto de amplificação
fazendo-se novos desenhos de iniciadores para os respectivos genes.

DISCUSSÃO
120
O mesmo ocorreu para o gene blaKPC. Duas amostras de K. pneumoniae
foram resistentes a carbapenemicos, porém negativas para a PCR do gene
blaKPC direto da amostra clínica. Outra amostra, sensível para os
carbapenemicos foi positiva para a PCR do mesmo gene. Ao avaliarmos estas
amostras, elas foram isoladas do mesmo paciente. Foram então estudados 5
isolados bacterianos pertencentes a este mesmo paciente e que se
encontravam armazenadas no banco de microrganismos do LEMC. Dos 5
isolados, todos foram resistentes a ertapenem pelo método da disco difusão e
positivo para blaKPC. Este fato pode ser explicado como para ESβL. A
sensibilidade do teste foi reduzida ou por não haver quantidade de DNA
suficiente para ser detectado direto da amostra clínica ou também pela redução
da sensibilidade direto da amostra clínica devido ao tamanho do produto de
amplificação para este gene.
Já para Gram positivos, no caso o gene mecA, a concordância foi maior.
O tamanho do produto de amplificação para este gene é menor interferindo
menos na sensibilidade do teste. Das 32 hemoculturas positivas para o gene
mecA pela técnica da PCR em Tempo Real, apenas 6 foram sensíveis a
oxacilina pelo sistema automatizado e uma se tratava de um Enterococcus.
Nesta amostra possivelmente houve interferência ou por alguma contaminação
na coleta ou por uma infecção prévia com algum microrganismo do gênero
Staphylococcus. Quanto as 5 hemoculturas citadas anteriormente, quatro eram
S. aureus e 1 S. epidermidis. Os isolados bacterianos referentes a essas
hemoculturas retirados do banco de microrganismos do LEMC foram testados
pelo método fenotípico E-test para avaliação da susceptibilidade a oxacilina e
PCR direto da colônia. Quatro destes isolados foram sensíveis a Oxacilina e

DISCUSSÃO
121
negativos para a PCR em Tempo Real para o gene mecA. Porém, uma
amostra de S.aureus, foi resistente ao antimicrobiano em questão e positivo
para o gene mecA, neste caso foi considerado erro “muito grave”. A
metodologia da PCR em Tempo Real para este gene se apresentou mais
sensível. É comum a presença de contaminação em hemoculturas de SCoN e
estes são avaliados como contaminantes ou causadores da bacteremia de
acordo com a análise do médico que acompanha ao paciente. Esta
interferência do gene mecA ou por contaminantes ou por infecções prévias
pode acarretar resultados falso positivos. Porém, nenhuma amostra
verdadeiramente resistente a oxacilina e positiva para o gene mecA deixou de
ser detectada, sendo este erro considerado erro “grave”. Em um recente
trabalho elaborado por Yanagihara e colaboradores, 2010, em que foi avaliada
a concordância entre a detecção de isolados de Staphylococcus resistentes a
oxacilina pelo método fenotípico e a detecção do gene mecA pelo Kit de
detecção SeptiFast®, o método molecular não foi capaz de detectar o gene
mecA em 2 amostras. Os autores sugerem que isto foi devido à diferença de
sensibilidade entre os dois testes (um foi realizado direto da amostra clinica e o
outro direto do isolado). De acordo com os resultados apresentados em nosso
estudo, a sensibilidade para o gene mecA foi capaz de detectar falso-positivo
mesmo direto de amostras clínicas, sendo alta a sensibilidade para o gene.
No Brasil, se encontram dois dos maiores centros que realizam
transplantes na América Latina. Estes são os HSP e HRH. Silva Jr e
colaboradores (2010) avaliaram todos os transplantes renais realizados nestes
dois centros entre os anos de 2000 a 2006. Um total de 3308 transplantes de
rim foram realizados. Mais de 50% dos casos foi referente a doador vivo e

DISCUSSÃO
122
receptor do sexo masculino. A incidência de ICS foi de 4,1% (185) sendo a
média de tempo pós-transplante para a ocorrência de infecção de 235 dias. No
presente estudo, só foram avaliados os transplantes em que ocorreu suspeita
de bacteremia. Dos 117 pacientes incluídos no estudo, a maioria (72%) foi do
sexo masculino e quase 90% foram submetidos a transplante de rim. O tempo
médio de pós-transplante até a coleta da hemocultura foi de 331 dias e quase
75% dos pacientes receberam transplante de doador falecido.
Não existe na literatura um consenso quanto ao maior risco de infecção
referente a transplante proveniente de doador vivo ou doador falecido. (Spees
et al., 1982; Dantas et al., 2006). Porém, alguns autores sugerem que o risco
infeccioso é maior em transplante de doador falecido. Isto se refere às
condições desfavoráveis do enxerto e a menor compatibilidade entre doador e
receptor, havendo a necessidade de tratamento imunossupressor mais
acentuado, facilitando os episódios de infecção (Dantas et al., 2006).
Normalmente, quando se refere a transplantes de rim, o número de
transplantes por doador vivo supera aos de doador falecido. Porém no presente
estudo, como referido anteriormente, só foram avaliados os episódios de
infecção, sendo assim a taxa de doador falecido foi superior.
O interesse da aplicação de um método molecular para o diagnóstico
direto de uma amostra clínica tem como principal objetivo a diminuição no
tempo do resultado, a escolha da terapia adequada em menor tempo, além de
maior sobrevida dos pacientes. A padronização aqui apresentada, leva em
média 8 horas para ser executada, desde o recebimento da hemocultura
positiva ou negativa pelo aparelho Bactec® até o resultado dos genes de
resistência. Levando-se em conta que quando o frasco de hemocultura é

DISCUSSÃO
123
aplicado ao método molecular já foi realizada a coloração de Gram pelo
laboratório de análises clínicas, o resultado de Gram positivo e Gram negativo
pelo método molecular pode não colaborar na rapidez do resultado.
Porém, no mesmo dia em que é realizada a PCR para a detecção da
presença de bactérias Gram negativas e Gram positivas pelo método
molecular, já é possível a avaliação da presença de genes que conferem
resistência. Considerando que a liberação do resultado final da hemocultura
levou de 3 a 4 dias para ser liberado pelo laboratório clínico após a positividade
pelo Bactec®, o método molecular poderia ser útil na antecipação do resultado
do gene de resistência.
Quanto ao tratamento dos pacientes apresentando hemoculturas
positivas para Gram positivos observamos uma inadequação da utilização
empírica em 11,86% na coleta da hemocultura, diminuindo para 6,78% quando
da liberação do resultado do Gram da hemocultura. O resultado final da
hemocultura incluindo o antibiograma não modificou a inadequação restante de
6,78% observada, sendo a maioria devido a hemoculturas positivas para
SCoN. Portanto, o nosso resultado molecular quando da realização do Gram
da hemocultura traria pouca contribuição para a adequação antimicrobiana
nessa população de pacientes e nessas instituições. Entretanto, poderia trazer
contribuição para racionalização do uso de antimicrobianos em hemoculturas
positivas para amostras sensíveis à oxacilina com gene mecA negativo. Em um
estudo realizado por Cattoir e colaboradores, 2011, houve 100% de
concordância entre o método fenotipico e molecular para detecção do gene
mecA em isolados do gênero Staphylococcus, porém muitos dos casos foram
definidos como uma possível contaminação da amostra pelo micorganismo

DISCUSSÃO
124
isolado. Neste mesmo estudo foi avaliada a possibilidade de interferência do
método molecular na adequação da antibióticoterapia e concluiu-se que a
rápida identificação de S. aureus e determinação de suscetibilidade à oxacilina
usando uma técnica de PCR não teve um impacto clínico importante devido ter
ocorrido 100% de concordância e pelo perfil de tratamento usado naquela
instituição.
No presente estudo avaliamos também o descalonamento da
antibioticoterapia. De acordo com os resultados apontados, a PCR poderia ser
útil em 16 dos casos em que pacientes se encontravam em uso de
vancomicina e eram sensíveis a oxacilina. Destes casos, 11 poderiam ter sido
beneficiados pelo resultado negativo para o gene mecA e a antibioticoterapia
poderia ter sido modificada para oxacilina.
Para bacilos Gram negativos não fermentadores observamos uma
inadequação de 7,5% no momento da coleta da hemocultura, diminuindo para
4,48% quando da liberação do resultado do Gram da hemocultura, e uma
queda importante da inadequação (1,51%) apenas quando do resultado final da
hemocultura. Não houve detecção de nenunha metalo-β-lactamase. Para
enterobacterias observamos uma inadequação de 20,9% no momento da
coleta da hemocultura, diminuído para 16,42% quando da liberação do
resultado do Gram da hemocultura, e uma queda ainda mais significante
quando do resultado final da hemocultura (2,99%). Destas 2,99% apenas uma
hemocultura foi positiva para ESβL pelo método da PCR porém, o tratamento
continuou inadequado. Uma amostra apresentou PCR positivo para blaKPC e
neste caso houve adequação na antibioticoterapia. Sendo assim, a PCR para
esta amostra poderia antecipar a escolha do tratamento.

DISCUSSÃO
125
Em algumas amostras, não houve a detecção do mecanismo de
resistência, ou porque como foi sugerido anteriormente o método da PCR para
alguns genes estudados não foi sensível o suficiente para detectá-los direto da
hemocultura ou porque se tratava de outro mecanismo. Em bactérias Gram
negativas há um extenso tipo de mecanismos que podem levar a resistência
nessas bactérias. Entre eles, outras cabapenemases sem ser as que foram
estudas como, por exemplo, as carbapenemases do tipo OXA, além de
alterações em porinas, outras ESβL além das estudadas (Bush, 2010) e
aumento da expressão de bombas de efluxo (Lister et al., 2009). Em Gram
negativos fica difícil a possibilidade do teste colaborar num descalonamento do
tratamento, pois quando a PCR for negativa para os genes estudados, poderá
haver outro mecanismo e sendo assim a bactéria poderá ser resistente ao
antibiótico de escolha.
Para Gram negativos, houve uma considerável redução da inadequação
do tratamento após o resultado do Gram na hemocultura. Os resultados
moleculares aqui apresentados concordaram em 100% com os fenotípicos
quando para detecção de bactérias Gram positivas e Gram negativas. Assim
sendo, se o teste fosse realizado de amostras extraídas do sangue, por EDTA,
seria possível contribuir para que fosse alterado o tratamento empírico. De
acordo com os dados apontados no presente estudo, o tratamento empírico foi
inadequado em 28, 4% dos casos em que houve ICS causada por bactérias
Gram negativas. Na literatura há relatos de taxas de até 23,5% de tratamentos
empíricos inadequados, sendo que 54% são em ICS por bactérias Gram
negativas (Zaragoza et al., 2003).

DISCUSSÃO
126
Em relação à racionalização do uso de antimicrobianos, 11 pacientes
com bactérias Gram positivas isoladas em hemocultura e sensíveis a oxacilina
foram tratados com vancomicina. Apesar de 2 isolados sensíveis terem
apresentado gene mecA (falsos positivos), em 9 a ausência na detecção
molecular deste gene poderia ter auxiliado o descalonamento do antibiótico
(troca da vancomicina pela oxacilina). Já em relação aos Gram negativos o
contrário foi observado, o escalonamento foi realizado na maioria dos casos de
inadequação.
O tempo mediano para liberação do resultado final foi de 4 dias. Com a
aplicação da detecção molecular dos genes de resistência pela técnica de PCR
multiplex é possível reduzir o tempo para reajuste da terapia empírica. Como já
demonstrado neste trabalho, poderíamos reduzir as taxas de inadequação aos
antimicrobianos de 13,56% para 8,48% para Gram positivos e de 28,4% para
4,5% em relação aos Gram negativos. Apesar de existir poucos dados de
literatura, acreditamos que o ajuste precoce da terapia empírica utilizada
contribui no curso do tratamento do paciente.
Pretendemos dar continuidade a este estudo, agora avaliando a
detecção bacteriana e de genes de resistência direto do sangue coletado para
hemoculturas, aproveitando a experiência adquirida com a padronização dos
testes moleculares.

CONCLUSÕES
127
7. CONCLUSÕES
Os ensaios moleculares foram capazes de detectar os genes de
interesse na diluição corresponde a 15 UFC até 1.5 UFC, porém com
baixa sensibilidade para os genes com produtos de amplificação entre
700 a 1.000 pares de bases e alta especificidade;
Observamos 100% de concordância entre a detecção bacteriana pelo
método de hemocultura automatizado com coloração de Gram e a PCR
em Tempo Real direto do frasco de hemocultura;
Para os genes de resistência a antimicrobianos em bactérias Gram
positivas observamos 87,6% de concordância na detecção do gene
mecA;
Para os genes de resistência a antimicrobianos em bactérias Gram
negativas observamos maior variabilidade entre a detecção fenotípica e
molecular;
A detecção de genes de resistência a antimicrobianos favoreceria a
adequação da antibioticoterapia, particularmente no descalonamento no
tratamento de bacteremias causadas por bactérias Gram positivas e na
adequação precoce no tratamento de bacteremias por bacilos Gram
negativas multiresistentes em pacientes tranplantados de órgãos
sólidos;

REFERÊNCIAS BIBLIOGRÁFICAS
128
8. REFERÊNCIAS BIBLIOGRÁFICAS
Al Hasan,M.N., Razonable,R.R., Eckel-Passow,J.E. and Baddour,L.M. Incidence rate
and outcome of Gram-negative bloodstream infection in solid organ transplant
recipients. Am J Transplant 2009; 9: 835-843.
al Naiemi N, B Duim, A Bart. A CTX-M extended-spectrum beta-lactamase in
Pseudomonas aeruginosa and Stenotrophomonas maltophilia. J Med Microbiol 2006;
55: 1607-1608.
Ambler RP. The structure of b-lactamases. Philos Trans R Soc Lond B Biol Sci 1980;
289: 321-331.
Andrade, S.S. ;Sader, H.S.;Barth, A.L.;Ribeiro, J.; Zoccoli, C.; Pignatari, A.C.C.; Gales,
A.C. Antimicrobial susceptibility of Gram negative bacilli isolate in Brazilian hospitals
participating in the SENTRY program (2003-2008). Braz J Infect Dis 2008; 12: 3-9.
ANVISA. Testes de sensibilidade aos antimicrobianos. Módulo 5. 2008.
http://www.anvisa.gov.br/servicosaude/controle/rede_rm/cursos/boas_praticas/
modulo5/introducao.htm
ANVISA. Resolução No 302/2005. Regulamento Técnico para funcionamento de
Laboratórios Clínicos. 13 de outubro de 2005.
http://lacen.saude.sc.gov.br/arquivos/RES_302B.pdf
Asensio A, Ramos A, Cuervas-Mons V et al. Effect of antibiotic prophylaxis on the risk
of surgical site infection in orthotopic liver transplant. Liver Transpl 2008; 14: 799–805.
Associação brasileira de transplante de órgãos. 2009. [texto da internet]. Disponível
em:http://www.abto.org.br/abtov02/portugues/populacao/rbt/2007/graficos.aspx?idCate
goria=2. Acesso em 10 jan 2011.
Bakir M, Bova JL, Newell KA, Millis JM, Buell JF, Arnow PM. Epidemiology and clinical
consequences of vancomycin-resistant enterococci in liver transplant patients
Transplantation 2001; 72: 1032–1037.

REFERÊNCIAS BIBLIOGRÁFICAS
129
Battaglia M, Ditonno P, Selvaggio O, et al. Kidney transplants from infected donors: our
experience. Transplant Proc 2004; 36:491-492.
Barthelemy,M., Peduzzi,J. and Labia,R. Distinction between the primary structures of
TEM-1 and TEM-2 beta-lactamases]. Ann Inst Pasteur Microbiol 1995; 136: 311-321.
Bauernfeind A, H Grimm, S Schweighart. A new plasmidic cefotaximase in a clinical
isolate of Escherichia coli. Infection 1990; 18(5): 294-298.
Bebrone C. Metallo-β-lactamases (classification, activity, genetic organization,
structure, zinc coordination) and their superfamily. Biochem Pharmacol 2007; 74(12):
1686-1701.
Berger N, Guggenbichler S, Steurer W, et al. Bloodstream infection following 217
consecutive systemic-enteric drained pancreas transplants. Infect Dis 2006; 6:1-9.
Bert F, Larroque B, Paugam-Burtz C, Janny S, Durand F, Dondero F et al. Microbial
epidemiology and outcome of bloodstream infections in liver transplant recipients: an
analysis of 259 episodes. Liver Transpl 2010; 16(3):393-401.
Bhavnani SM, Drake JA, Forrest A et al. A nationwide, multicenter, case-control study
comparing risk factors, treatment, and outcome for vancomycin-resistant and -
susceptible enterococcal bacteremia. Diagn Microbiol Infect Dis 2000; 36: 145–158.
Bispo,P.J., de Melo,G.B., Hofling-Lima,A.L. and Pignatari,A.C. Detection and gram
discrimination of bacterial pathogens from aqueous and vitreous humor using real-time
PCR assays. Invest Ophthalmol Vis Sci 2011; 52: 873-881.
Bone RC, Balk RA, Cerra FB, Dellinger RP, Fein AM, Knaus WA et al. Definitions for
sepsis and organ failure and guidelines for the use of innovative therapies in sepsis.
The ACCP/SCCM Consensus Conference Committee. American College of Chest
Physicians/Society of Critical Care Medicine Chest 1992; 101(6):1644-1655.

REFERÊNCIAS BIBLIOGRÁFICAS
130
Bonnet,R., De Champs,C., Sirot,D., Chanal,C., Labia,R. and Sirot,J. Diversity of TEM
mutants in Proteus mirabilis. Antimicrob Agents Chemother 1999; 43: 2671-2677.
Bonnet,R. Growing group of extended-spectrum beta-lactamases: the CTX-M
enzymes. Antimicrob Agents Chemother 2004; 48: 1-14.
Bradford, P.A. Extended-spectrum beta-lactamases in the 21st century:
characterization, epidemiology, and detection of this important resistance threat. Clin
Microbiol Rev 2001; 14: 933-951.
Bratu,S., Tolaney,P., Karumudi,U., Quale,J., Mooty,M., Nichani,S. and Landman,D.
Carbapenemase-producing Klebsiella pneumoniae in Brooklyn, NY: molecular
epidemiology and in vitro activity of polymyxin B and other agents. J Antimicrob
Chemother 2005; 56: 128-132.
Bure,A., Legrand,P., Arlet,G., Jarlier,V., Paul,G. and Philippon,A. Dissemination in five
French hospitals of Klebsiella pneumoniae serotype K25 harbouring a new transferable
enzymatic resistance to third generation cephalosporins and aztreonam. Eur J Clin
Microbiol Infect Dis 1988; 7: 780-782.
Bush K, GA Jacoby, AA Medeiros. A functional classification scheme for
betalactamases and its correlation with molecular structure. Antimicrob Agents
Chemother 1995; 39(6): 1211-1233.
Bush K, Jacoby GA. Updated functional classification of beta-lactamases. Antimicrob
Agents Chemother 2010; 54(3):969-976.
Candel FJ, Grima E, Matesanz M, et al. Bacteremia and septic shoch after solid-organ
transplantation. Transplant Proc 2005, 37:4097-4099.
Carroll NM, Adamson P, Okhravi N. Elimination of Bacterial DNA from Taq DNA
Polymerases by Restriction Endonuclease Digestion. J Clin Microbiol
1999;37(10):3402-4.
Cartelle,M., del Mar,T.M., Molina,F., Moure,R., Villanueva,R. and Bou,G. High-level
resistance to ceftazidime conferred by a novel enzyme, CTX-M-32, derived from CTX-

REFERÊNCIAS BIBLIOGRÁFICAS
131
M-1 through a single Asp240-Gly substitution. Antimicrob. Agents Chemother 2004; 48:
2308-2313.
Castanheira M, Toleman MA, Schmidt JF, Jones RN, Walsh TR. Molecular
characterization of a β-lactamase gene, blaGIM-1, encoding a new subclass of metallo-
β-lactamase. Antimicrob Agents Chemother 2004; 48(12): 4654-4661.
Cattoir,V., Merabet,L., Djibo,N., Rioux,C., Legrand,P., Girou,E. and Lesprit,P. Clinical
impact of a real-time PCR assay for rapid identification of Staphylococcus aureus and
determination of methicillin resistance from positive blood cultures. Clin Microbiol Infect
2011; 17: 425-431.
Celenza G, C Pellegrini, M Caccamo, B Segatore, G Amicosante, M Perilli.
2006.Spread of blaCTX-M-type and blaPER-2 beta-lactamase genes in clinical isolates
from Bolivian hospitals. J Antimicrob Chemother 57(5): 975-978.
Cetinkaya Y, Falk P, Mayhall CG. Vancomycin-Resistant Enterococci. Clin Microbiol
Rev 2000; 13 (4): 686–707.
Chanawong A, FH M’Zali, J Heritage, A Lulitanond, PM Hawkey. SHV-12, SHV-5,
SHV-2a and VEB-1 extended-spectrum B-lactamases in gram negative bacteria
isolated in a university hospital in Thailand. J Antimicrob Chemother 2001; 48(6): 924-
925.
Chandelier A, Planchon V and Oger R. Determination of cycle cut off in real-time PCR
for the detection of regulated plant pathogens. Bulletin OEPP/EPPO Bulletin 2010; 40:
52–58.
Chang S, Sievert DM, Hageman JC, Boulton ML, Tenover FC, Downes FP, Shah S,
Rudrik JT, Pupp GR, Brown WJ, Cardo D, Fridkin SK. Infection with vancomycin-
resistant Staphylococcus aureus containing the vanA resistance gene. N Engl J Med
2003; 348: 1342-1347.
CLSI. Clinical and Laboratory Standards Institute. Methods for dilution antimicrobial
susceptibility tests for bacteria that grow aerobically. Seventh Edition: Approved
Standard M100-S20. Clinical and Laboratory Standards Institute, Wayne, Approved
Standard M100-S20. (2010).

REFERÊNCIAS BIBLIOGRÁFICAS
132
Clinical and Laboratory Standards Institute EP17-A. Protocols for determination of
limits of detection and limits of quantitation. Approved guideline, Oct/2004. vol 24, No
34.
CLSI. Molecular Diagnostic Methods for Infectious Diseases; Approved Guideline -
Second Edition. CLSI document MM03-A2. Wayne PA: Clinical and Laboratory
Standard Institute; 2006.
Clinical and Laboratory Standards Institute. EP12-A2 User protocol for evaluation of
qualitative test performance. Approved guideline-second edition, Jan/2008. vol 28, No
3.
Coque,T.M., Baquero,F. and Canton,R. (2008) Increasing prevalence of ESBL-
producing Enterobacteriaceae in Europe. Euro Surveill 13.
Cunha Mde L, Sinzato YK, Silveira LV. Comparison of methods for the identification of
coagulase-negative staphylococci. Mem Inst Oswaldo Cruz 2004; 99(8):855-60.
Danovitch GM. Immunosuppressive medications for renal transplantation: A multiple
choice question. Kidney Int 2001; 59:388-402.
Dantas SRPE, Kuboyama RH, Mazzali M, Moretti ML. Nosocomial infections in renal
transplant patients: risk factors and treatment implications associated with urinary tract
and surgical site infections. J Hosp Infect 2006; 63:117-123.
Donnely JP, De Paum B. Infections in immunocompromised host: general principles.
In: Mandell L, Bennett J, Doli R, editors. Mandell, Bennett, & Doli: Principles and
practice of infectious diseases. 6th ed. Philadelphia: Churcill Livingstone. 2005; p.3421-
3432.
Dorak, MT. Real Time PCR. New Yorh: Taylor & Francis Group. 2006 Cap 5: 93-106.
Doumith M, M Warner, M Weinbren, DM Livermore, N Woodford. Dissemination of a
novel metallo-β-lactamase, NDM-1 (MIM-1) in diverse Enterobacteriaceae in the UK.
49th Interscience Conference in Antimicrobial Agents and Chemotherapy (ICAAC). San
Francisco - CA, USA. Setembro 12-15, 2009. Poster C1-058.

REFERÊNCIAS BIBLIOGRÁFICAS
133
Eggert US, Ruiz N, Falcone BV, Branstrom, AA.; Goldman, RC.; Silhavy, TJ.; Kahne,
D. Genetic basis for activity differences between vancomycin and glycolipid derivatives
of vancomycin. Science 2001; 294 (5541): 361-364.
El Salabi A, M Tolemam, TR Walsh. 2009. Novel subclass of a group B1 metallo-β-
lactamase, TMB-1, in clinical and non-clinical Gram-negative bacteria from Libya. 49th
Interscience Conference in Antimicrobial Agents and Chemotherapy (ICAAC). San
Francisco - CA, USA. Setembro 12-15, 2009. Poster C1-1365.
Erdem,I., Ozgultekin,A., Sengoz,I.A., Ozturk,E.D., Senbayrak,A.S., Turan,G.,
Dincer,E., Oguzoglu,N. and Goktas,P. Bloodstream infections in a medical-surgical
intensive care unit: incidence, aetiology, antimicrobial resistance patterns of Gram-
positive and Gram-negative bacteria. Clin Microbiol Infect. 2009; 15: 943-946.
Feinstein AR, Cicchetti DV. High agreement but low kappa: I. The problems of two
paradoxes. J Clin Epidemiol 1990; 43(6):543-9.
Ferraresso M, Berardinelli L. Nosocomial infection in kidney transplant recipients: a
retrospective analysis of a single-center experience. Transplant Proc 2005; 37:2495-
2496.
Fishman JA, Rubin RH. Infection in organ-transplant recipients. N Engl J Med 1998;
338(24):1741-1750.
FishmanJA, Rubin RH. Infection in Organ-Transplant Recipients. N Engl J Med 2005;
338:1741-1751.
Fishman JA. Infection in solid-organ transplant recipients. N Engl J Med 2007; 357(25):
2601-2614.
Freitas MC, Pacheco-Silva A, Barbosa D et al. Prevalence of vancomycin-resistant
Enterococcus fecal colonization among kidney transplant patients. BMC Infect Dis
2006; 6: 133.
Gales AC, Menezes LC, Silbert S, Sader HS. Dissemination in distinct Brazilian regions
of an epidemic carbapenem-resistant Pseudomonas aeruginosa producing SPM
metallo-β-lactamase. J Antimicrob Chemother 2003; 52(4): 699-702.

REFERÊNCIAS BIBLIOGRÁFICAS
134
Gales AC, Sader HS, Ribeiro J, Zoccoli C, Barth A, Pignatari AC. Antimicrobial
susceptibility of gram-positive bacteria isolated in Brazilian hospitals participating in the
SENTRY Program (2005-2008). Braz J Infect Dis 2009; 13(2):90-98.
Ghanem G, Hachem R, Jiang Y, Chemaly RF, Raad I. Outcomes for and risk factors
associated with vancomycin-resistant Enterococcus faecalis and vancomycin-resistant
Enterococcus faecium bacteremia in cancer patients. Infect Control Hosp Epidemiol
2007; 28: 1054–1059.
Giesbrecht P, Kersten T, Maidhof H, Wecke J. Staphylococcal cell wall: morphogenesis
and fatal variations in the presence of penicillin. Microbiol Mol Biol Rev 1998;
62(4):1371-1414.
Hackbarth CJ, Chambers HF. Methicillin-resistant staphylococci: detection methods
and treatment of infections. Antimicrob Agents Chemother 1989; 33(7): 995-999.
Hagen EA, Lautenbach E, Olthoff K, Blumberg EA. Low prevalence of colonization with
vancomycin-resistantEnterococcus in patients awaiting liver transplantation. Am J
Transplant 2003; 3: 902–905.
Halloran PF. Imunossupressive drugs for kidney transplantation. N Engl J Med. 2004;
351: 2715-2729.
Harada S, Ishii Y, Yamaguchi K. Extended-spectrum beta-lactamases: implications for
the clinical laboratory and therapy. Korean J Lab Med 2008; 28(6):401-412.
Hayden RT. In Vitro Nucleic Acid Amplification Techniques. In: Persing DH, Tenover
FC, Versalovic J, Tang Y-W, Unger ER, Relman DA, White TJ. Molecular Microbiology:
Diagnostic Principles and Practice. Washington DC: ASM Press; 2004.
Hernandez,J.R., Martinez-Martinez,L., Canton,R., Coque,T.M. and Pascual,A.
Nationwide study of Escherichia coli and Klebsiella pneumoniae producing extended-
spectrum beta-lactamases in Spain. Antimicrob. Agents Chemother 2005; 49: 2122-
2125.

REFERÊNCIAS BIBLIOGRÁFICAS
135
Hirakata,Y., Izumikawa,K., Yamaguchi,T., Takemura,H., Tanaka,H., Yoshida,R.,
Matsuda,J., Nakano,M., Tomono,K., Maesaki,S., Kaku,M., Yamada,Y., Kamihira,S. and
Kohno,S. Rapid detection and evaluation of clinical characteristics of emerging
multiple-drug-resistant gram-negative rods carrying the metallo-beta-lactamase gene
blaIMP. Antimicrob Agents Chemother 1998; 42: 2006-2011.
Hossain,A., Ferraro,M.J., Pino,R.M., Dew,R.B., III, Moland,E.S., Lockhart,T.J.,
Thomson,K.S., Goering,R.V. and Hanson,N.D. Plasmid-mediated carbapenem-
hydrolyzing enzyme KPC-2 in an Enterobacter sp. Antimicrob Agents Chemother 2004;
48: 4438-4440.
Huletsky,A., Knox,J.R. and Levesque,R.C. Role of Ser-238 and Lys-240 in the
hydrolysis of third-generation cephalosporins by SHV-type beta-lactamases probed by
site-directed mutagenesis and three-dimensional modeling. J Biol Chem 1993; 268:
3690-3697.
Husain S, Chan KM, Palmer SM et al. Bacteremia in lung transplant recipients in the
current era. Am J Transplant 2006; 6:3000-3007.
Ikegaya,S., Iwasaki,H. and Ueda,T. Clinical significance of coagulase-negative
Staphylococci isolated from blood culture samples of patients with hematological
disorders; true bacteremia or contamination. Rinsho Ketsueki 2010; 51: 398-401.
Jarlier,V., Nicolas,M.H., Fournier,G. and Philippon,A. Extended broad-spectrum beta-
lactamases conferring transferable resistance to newer beta-lactam agents in
Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev Infect Dis
1988; 10: 867-878.
Katayama Y, Ito T, Hiramatsu K. A new class of genetic element, staphylococcus
cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus.
Antimicrob Agents Chemother 2000; 44(6):1549-55.
Klaschik S, Lehmann LE, Raadts A, Hoeft A, Stuber F. Comparison of different
decontamination methods for reagents to detect low concentrations of bacterial 16S
DNA by real-time-PCR. Mol Biotechnol 2002b;22(3):231-42.

REFERÊNCIAS BIBLIOGRÁFICAS
136
Klein D. Quantification using real-time PCR technology: applications and limitations.
TRENDS in Molecular Medicine 2002; 8(6):257-60.
Knothe,H., Shah,P., Krcmery,V., Antal,M. and Mitsuhashi,S. Transferable resistance to
cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of Klebsiella
pneumoniae and Serratia marcescens. Infection 1983; 11: 315-317.
Konenan, E. Et al. Diagnóstico Microbiológico, Guanabara koogan SA, 6ª ediçao,
capitulo 12, parte 1, pag 618-651. 2006.
Kubista M, Andrade JM, Bengtsson M, Forootan A, Jonák J, Lind K et al. The real-time
polymerase chain reaction. Molecular aspects of Medicine 2006;27:95-125.
Laraki N, Franceschini N, Rossolini GM, Santucci P, Meunier C, Pauw E, Amicosante
G, Frere JM, Galleni M. Biochemical characterization of the Pseudomonas aeruginosa
101/1477 metallo-β-lactamase IMP-1 produced by Escherichia coli. Antimicrob Agents
Chemother 1999; 43(4): 902-906.
LaRosa DF, Rahman AH, Turka LA. The Innate Immune System in Allograft Rejection
and Tolerance. J Immunol 2007; 178: 7503–7509.
Lauretti L, ML Riccio, A Mazzariol, G Cornaglia, G Amicosante, R Fontana, GM
Rossolini. Cloning and characterization of blaVIM, a new integronborne metallo-
betalactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob
Agents Chemother 1999; 43(7): 1584-1590.
Lee K, Yum JH, Yong D, Lee HM, Kim HD, Docquier JD, Rossolini GM, Chong Y.
Novel acquired metallo-beta-lactamase gene, blaSIM-1, in a class 1 integron from
Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother
2005; 49(11): 4485-4491.
Lehmann,L.E., Hunfeld,K.P., Emrich,T., Haberhausen,G., Wissing,H., Hoeft,A. and
Stuber,F. A multiplex real-time PCR assay for rapid detection and differentiation of 25
bacterial and fungal pathogens from whole blood samples. Med Microbiol Immunol
2008; 197:313-324.

REFERÊNCIAS BIBLIOGRÁFICAS
137
Linares L, Cervera C, Cofán F, et al. Epidemiology and Outcomes of Multiple
Antibiotic–Resistant Bacterial Infection in Renal Transplantation. Transplant Proc 2007;
39:2222–2224.
Linares L, Garcia-Goez JF, Cervera C, Almela M, Sanclemente G, Cofan F et al. Early
bacteremia after solid organ transplantation. Transplant Proc 2009; 41(6):2262-2264.
Linares,L., Cervera,C., Hoyo,I., Sanclemente,G., Marco,F., Cofan,F., Ricart,M.J.,
Navasa,M. and Moreno,A. Klebsiella pneumoniae infection in solid organ transplant
recipients: epidemiology and antibiotic resistance. Transplant Proc 2010; 42: 2941-
2943.
Lincopan N, McCulloch JA, Reinert C, Cassettari VC, Gales AC, Mamizuka EM. First
isolation of metallo-β-lactamase-producing multiresistant Klebsiella pneumoniae from a
patient in Brazil. J Clin Microbiol 2005; 43(1): 516-519.
Lister PD, DJ Wolter, ND Hanson. Antibacterial-resistant Pseudomonas aeruginosa:
clinical impact and complex regulation of chromosomally encoded
resistancemechanisms. Clin Microbiol Rev 2009; 22(4): 582-610.
Livermore,D.M. Beta-Lactamases in laboratory and clinical resistance. Clin
Microbio.1995; 8: 557-584.
Livermore,D.M. & Hawkey,P.M. CTX-M: changing the face of ESBLs in the UK. J
Antimicrob Chemother 2005; 56: 451-454.
Lodise TP, McKinnon PS, Tam VH, Rybak MJ. Clinical outcomes for patients with
bacteremia caused by vancomycin-resistant Enterococcus in a level 1 trauma center.
Clin Infect Dis 2002; 34: 922–929.
Mackay IM. Real-time PCR in the microbiology laboratory. Clin Microbiol Infect
2004;10:190-212.
Marchandin,H., Carriere,C., Sirot,D., Pierre,H.J. and Darbas,H. TEM-24 produced by
four different species of Enterobacteriaceae, including Providencia rettgeri, in a single
patient. Antimicrob Agents Chemother 1999; 43: 2069-2073.

REFERÊNCIAS BIBLIOGRÁFICAS
138
McClean K, Kneteman N, Taylor G. Comparative risk of bloodstream infection in organ
transplant recipients. Infect Control Hosp Epidemiol 1994; 15(9):582-584.
McComas CC, Crowley BM, Boger DL. Partitioning the loss in vancomycin binding
affinity for D-Ala-D-Lac into lost H-bond and repulsive lone pair contributions. J Am
Chem Soc 2003; 125 (31): 9314 -9315.
McNeil SA, Malani PN, Chenoweth CE et al. Vancomycin-resistant enterococcal
colonization and infection in liver transplant candidates and recipients: A prospective
surveillance study. Clin Infect Dis 2006; 42: 195–203.
Medina-Pestana JO. Organization of a High-Volume Kidney Transplant Program-The
“Assembly Line” Approach. Transplantation 2006; 81: 1510-1520.
Mendes RE, MA Toleman, J Ribeiro, HS Sader, RN Jones, TR Walsh. Integron
carrying a novel metallo-beta-lactamase gene, blaIMP-16, and a fused form of
aminoglycosideresistant gene aac(6')-30/aac(6')-Ib': report from the SENTRY
Antimicrobial Surveillance Program. Antimicrob Agents Chemother 2004; 48(12): 4693-
4702.
Mendes ER, Kiyota AK, Monteiro J, Castanheira M, Andrade SS, Gales CA, Pignatari
CCA, Tufik S. Rapid Detection and Identification of Metallo-β-lactamase-Encoding
Genes by Multiplex Real-Time PCR Assay and Melt Curve Analysis. J Of Clin Microbiol
2007a; 45:544–547.
Mendes RE, M Castanheira, MA Toleman, HS Sader, RN Jones, TR Walsh.
Characterization of an integron carrying blaIMP-1 and a new aminoglycoside
resistance gene, aac(6')-31, and its dissemination among genetically unrelated clinical
isolates in a Brazilian hospital. Antimicrob Agents Chemother 2007b; 51(7): 2611-2614.
Michalak G, Kwiatkowski A, Bieniasz M, et al. Infectious Complications After
Simultaneous Pancreas–kidney transplantation. Transplant Proc 2005; 37: 3560-3563.
Mies S. Transplante de fígado. Rev Ass Med Brasil 1998; 44(2): 127-34.

REFERÊNCIAS BIBLIOGRÁFICAS
139
Ministério da Saúde. Brasil. [texto da internet]. 2009. Disponível em:
http://portal.saude.gov.br/saude/. Acesso em 10 jan 2011.
Miriagou,V., Tzouvelekis,L.S., Rossiter,S., Tzelepi,E., Angulo,F.J. and Whichard,J.M.
Imipenem resistance in a Salmonella clinical strain due to plasmid-mediated class A
carbapenemase KPC-2. Antimicrob Agents Chemother 2003; 47: 1297-1300.
Monteiro,J., Santos,A.F., Asensi,M.D., Peirano,G. and Gales,A.C. First report of KPC-
2-producing Klebsiella pneumoniae strains in Brazil. Antimicrob Agents Chemother
2009; 53: 333-334.
Moreno A, Mensa J, Amela M, et al. 138 episodes of bacteremia or fungemia in
patients with solid organ (renal or hepatic) transplantation. Med Clin (Barc) 1994;
103(5):161-4.
Moreno A, Cervera C, Gavald ´a J, et al. Bloodstream infections among transplant
recipients: results of nationwide surveillance in Spain. Am J Trans 2007; 7:2579-2586.
Morfin-Otero,R., Rodriguez-Noriega,E., Deshpande,L.M., Sader,H.S. and
Castanheira,M. Dissemination of a bla(VIM-2)-Carrying Integron Among
Enterobacteriaceae Species in Mexico: Report from the SENTRY Antimicrobial
Surveillance Program. Microb Drug Resist 2009; 15: 33-35.
Morosini,M.I., Canton,R., Martinez-Beltran,J., Negri,M.C., Perez-Diaz,J.C., Baquero,F.
and Blazquez,J. New extended-spectrum TEM-type beta-lactamase from Salmonella
enterica subsp. enterica isolated in a nosocomial outbreak. Antimicrob Agents
Chemother 1995; 39: 458-461.
Mühl H, Kochem AJ, Disqué C, Sakka SG. Activity and DNA contamination of
commercial polymerase chain reaction reagents for the universal 16S rDNA real-time
polymerase chain reaction detection of bacterial pathogens in blood. Diagn Microbiol
Infect Dis 2010; 66:41-49.
Munday,C.J., Xiong,J., Li,C., Shen,D. and Hawkey,P.M. Dissemination of CTX-M type
beta-lactamases in Enterobacteriaceae isolates in the People's Republic of China. Int.
J Antimicrob Agents 2004; 23: 175-180.

REFERÊNCIAS BIBLIOGRÁFICAS
140
Navon-Venezia,S., Chmelnitsky,I., Leavitt,A., Schwaber,M.J., Schwartz,D. and
Carmeli,Y. Plasmid-mediated imipenem-hydrolyzing enzyme KPC-2 among multiple
carbapenem-resistant Escherichia coli clones in Israel. Antimicrob Agents Chemother
2006; 50: 3098-3101.
Neonakis IK, EV Scoulica, SK Dimitriou, AI Gikas, YJ Tselentis. Molecular
epidemiology of extended-spectrum beta-lactamases produced by clinical isolates in a
university Hospital in Greece: detection of SHV-5 in Pseudomonas aeruginosa and
prevalence of SHV-12. Microbial Drug Resistance 2003; 9(2): 161-165.
Nolte, FS; Williams, JM; Jerris, RC; Morello, JA; Leitch; Matushek, S; Schwabe,
LD; Dorigan, F; kocka, FE. Multicenter Clinical Evaluation of a Continuous
Monitoring Blood Culture System Using Fluorescent-Sensor Technology (BACTEC
9240). J Clin Microbiol 1993; 31: 552-557.
Nordmann, P. and Poirel,L. Emerging carbapenemases in Gram-negative aerobes.
Clin Microbiol Infect 2002; 8: 321-331.
Nouér SA, Nucci M, Oliveira MP, Pellegrino FLPC, Moreira BM. Risk factors for
acquisition of multidrug-resistant Pseudomonas aeruginosa producing SPM metallo-β-
lactamase. Antimicrob Agents Chemother 2005; 49(9): 3663-3667.
Nusair A, Jourdan D, Medcalf S et al. Infection control experience in a cooperative care
center for transplant patients. Infect Control Hosp Epidemiol 2008; 29: 424–429.
Okhravi N, Adamson P, Carroll N, Dunlop A, Matheson MM, Towler HM, Lightman S.
PCR-based evidence of bacterial involvement in eyes with suspected intraocular
infection. Invest Ophthalmol Vis Sci 2000a; 41(11):3474-9.
Osano E, Arakawa Y, Wacharotayankun R, Ohta M, Horii T, Ito H, Yoshimura F, Kato
N. 1994. Molecular characterization of an enterobacterial metallo β- lactamase found in
a clinical isolate of Serratia marcescens that shows imipenem resistance. Antimicrob
Agents Chemother 1994; 38(1): 71-78.

REFERÊNCIAS BIBLIOGRÁFICAS
141
Palzkill,T., Thomson,K.S., Sanders,C.C., Moland,E.S., Huang,W. and Milligan,T.W.
(1995) New variant of TEM-10 beta-lactamase gene produced by a clinical isolate of
proteus mirabilis. Antimicrob Agents Chemother 1995; 39: 1199-1200.
Patel R, Paya CV. Infections in solid organ transplantation recipients. Cli Microbiol Ver
1997; 10:86-124.
Patel R, Allen SL, Manahan JMet al. Natural history of vancomycinresistant
enterococcal colonization in liver and kidney transplant recipients. Liver Transpl 2001;
7: 27–31.
Paterson,D.L. and Bonomo,R.A. Extended-spectrum beta-lactamases: a clinical
update. Clin Microbiol Rev, 2005; 18: 657-686.
Pavez M, Mamizuka EM, Lincopan N. Early dissemination of KPC-2-producing
Klebsiella pneumonia strains in Brazil. Antimicrob Agents Chemother 2009; 53(6):
2702.
Payne DJ, Bateson JH, Gasson BC, Khushi T, Proctor D, Pearson SC, Reid R.
Inhibition of metallo-β-lactamases by a series of thiol ester derivatives of
mercaptophenylacetic acid. FEMS Microbiol Lett 1997a; 157(1): 171-175.
Payne DJ, Bateson JH, Gasson BC, Proctor D, Khushi T, Farmer TH, Tolson DA, Bell
D, Skett PW, Marshall AC, Reid R, Ghosez L, Combret Y, Marchand- Brynaery J.
Inhibition of metallo-β-lactamases by a series of mercaptoacetic acid thiol ester
derivatives. Antimicrob Agents Chemother 1997b; 41(1): 135- 140.
Peirano,G., Seki,L.M., Val,P., V, Pinto,M.C., Guerra,L.R. and Asensi,M.D.
Carbapenem-hydrolysing beta-lactamase KPC-2 in Klebsiella pneumoniae isolated in
Rio de Janeiro, Brazil. J Antimicrob Chemother 2009; 63: 265-268.
Pellegrino FLPC, Teixeira LM, Carvalho MGS, Nouér AS, Oliveira MP, Sampaio JLM,
Freitas ADF, Ferreira ALP, Amorim ELT, Riley LW, Moreira BM. Ocurrence of a
multidrug-resistant Pseudomonas aeruginosa clone in different hospitals in Rio de
Janeiro, Brazil. J Clin Microbiol 2002; 40(7): 2420-2424.

REFERÊNCIAS BIBLIOGRÁFICAS
142
Pereira, CAP. Impacto da Implementação de métodos automatizados em
hemoculturas na evolução clínica de pacientes com infecções de corrente sanguínea.
1997. 109f. Tese ( Doutorado em Medicina) – Disciplina de Doença Infecciosas e
Parasitárias, Universidade Federal de São Paulo, São Paulo. 1997.
Perilli,M., Segatore,B., de Massis,M.R., Riccio,M.L., Bianchi,C., Zollo,A.,
Rossolini,G.M. and Amicosante,G. TEM-72, a new extended-spectrum beta-lactamase
detected in Proteus mirabilis and Morganella morganii in Italy. Antimicrob Agents
Chemother 2000; 44: 2537-2539.
Picão RC, L Poirel, AC Gales, P Nordmann. Further identification of CTX-M-2
extended-spectrum beta-lactamase in Pseudomonas aeruginosa. Antimicrob Agents
Chemother 2009a 53(5): 2225-2226.
Picão RC, LCC Fehlberg, PP Barbosa, R Girardello, KC Bauab, AC Gales.
Cefotaximases (CTX-M) em Pseudomonas aeruginosa: devemos nos preocupar? XXV
Congresso Brasileiro de Microbiologia. Porto de Galinhas, PE. Novembro, 8-12. Poster
A03- 090. 2009b.
Poirel L, E Lebessi, M Castro, C Fevre, M Foustoukou, P Nordmann. Nosocomial
outbreak of extended-spectrum beta-lactamase SHV-5 producing isolates of
Pseudomonas aeruginosa in Athens, Greece. Antimicrob Agents Chemother 2004a;
48(2): 2277- 2279.
Poirel L, Magalhaes M, Lopes M, Nordmann P. Molecular analysis of metallobeta-
lactamase gene blaSPM-1-surrounding sequences from disseminated Pseudomonas
aeruginosa isolates in Recife, Brazil. Antimicrob Agents Chemother 2004b; 48(4):
1406-1409.
Poirel L, Rodriguez-Martinez J, Naiemi NA, Debets-Ossenkopp Y, Nordmann P.
Characterization of blaDIM-1, a novel integron-located metallo-β-lactamase gene from
a Pseudomonas stutzeri clinical isolate in the Netherlands. Abstracts of the Nineteenth
European Congress of Clinical Microbiology and Infectious Diseases, 2009: Helsinki,
Finland. Basel, Switzerland: European Society of Clinical Microbiology and Infectious
Diseases. Abstract O-309, p. S69.

REFERÊNCIAS BIBLIOGRÁFICAS
143
Poirel L, J Rodriguez-Martinez, N al Naiemi, Y Debets-Ossenkopp, P Nodmann.
Characterization of DIM-1, an integrin-encoding metallo-beta-lactamase from a
Pseudomonas stutzeri clinical isolate in the Netherlands. Antimicrob Agents Chemother
2010; 54(6): 2420-2424.
Queenan AM & K Bush. Carbapenemases: the versatile beta-lactamases. Clin
Microbiol Rev 2007; 20(3): 440-458.
Rodriguez C, Munoz P, Rodriguez-Creixems M, Yanes JF, Palomo J, Bouza E.
Bloodstream infections among heart transplant recipients. Transplantation 2006;
81:384-391.
Rossetti ML; da Silva CMD; Rodrigues JJS. Doenças Infecciosas- Diagnóstico
Molecular. Rio de janiero: Ed Guanabara koogan. 2006 Cap:3: 41-46.
Rossolini,G.M., D'Andrea,M.M. and Mugnaioli,C. The spread of CTX-M-type extended-
spectrum beta-lactamases. Clin Microbiol Infect 2008; 14: 33-41.
Sacha, P., Ostas, A., Jaworowska, J., Wieczorek, P., Ojdana, D., Ratajczak, J. and
Tryniszewska,E. The KPC type beta-lactamases: new enzymes that confer resistance
to carbapenems in Gram-negative bacilli. Folia Histochem Cytobiol 2009; 47: 537-543.
Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT et al. Primer-directed
enzymatic amplification of DNA with a thermostable DNA polymerase. Science 1988;
239: 487-491.
Sakai H, Procop GW, Kobayashi N, Togawa D, Wilson DA, Borden L, Krebs V, Bauer
TW. Simultaneous detection of Staphylococcus aureus and coagulase negative
staphylococci in positive blood cultures by real-time PCR with two fluorescence
ressonance energy transfer probe sets. J Clin Microbiol 2004; 42(12):5739-5744.
Salabi AE, Toleman M, Walsh TR. Novel subclass of a group B1 metallo-β- lactamase,
TMB-1, in clinical and non-clinical Gram-negative bacteria from Libya. In: Annual
Intescience Conference on Antimicrobial Agents and Chemotherapy, 49., 2009, San
Francisco. Anais… Washington: American Society for Microbiology, 2009.

REFERÊNCIAS BIBLIOGRÁFICAS
144
Salabi AE, Toleman MA, Weeks J, Bruderer T, Frei R, Walsh TR. First report of the
metallo-β-lactamase SPM-1 in Europe. Antimicrob Agents Chemother 2010; 54(1):
582.
Sekiguchi J, Morita K, Kitao T, Watanabe N, Okazaki M, Miyoshi-Akiyama T, Kanamori
M, Kirikae T. KHM-1, a Novel Plasmid-Mediated Metallo-β-Lactamase from a
Citrobacter freundii Clinical Isolate. Antimicrob Agents Chemother 2008; 52(11): 4194-
4197.
Sesso RCC; Lopes AA; Thomé FS; Lugon JR; Burdmann EA. Censo Brasileiro de
Diálise, 2009. J Bras Nefro 2010; 32(4): 380-384.
Shigei, J. T., J. A. Shimabukuro, M. T. Pezzlo, L. M. de la Maza, and E. M., Peterson.
Value of terminal subcultures for blood cultures monitored by BACTEC 9240. J Clin
Microbiol 1995; 33:1385–1388.
Silva Jr M, Marra AR, Pereira CAP, Pestana JOM, Camargo LFA. Bloodstream
Infection after Kidney Transplantation: Epidemiology, Microbiology, Associated Risk
Factors, and Outcome. Transplantation 2010; 20(10): 1-7.
Silveira GP, Nome, F, Gesser JC, Sá, MM. Estratégias utilizadas no combate a
resistência bacteriana. Quim Nova 2006; 29 (4): 844-855.
Singh N, Paterson DL, Gayowski T, Wagner MM, Marino IR. Predicting bacteremia and
bacteremic mortality in liver transplantation recipients. Liver Transpl 2000; 6:54-61.
Sistema Nacional de Transplantes. [texto da internet]. Disponível em:
http://portal.saude.gov.br/portal/saude/area.cfm?id_area=1004. Acesso em 10 jan
2011.
Smith, J. A., E. A. Bryce, J. H. Ngui-Yen, and F. J. Roberts. Comparison of BACTEC
9240 and BacT/Alert blood culture systems in an adult hospital. J Clin Microbiol 1995;
33:1905–1908.
Sola AF, Bittencourt ARC, Guerra CM, Godoy HL, Medeiros EAS. Health Care-Related
Infections in Solid Organ Transplants. Brazilian J Infect Dis 2007; 11(6):567-570.

REFERÊNCIAS BIBLIOGRÁFICAS
145
Sougakoff,W., Goussard,S., Gerbaud,G. and Courvalin,P. Plasmid-mediated
resistance to third-generation cephalosporins caused by point mutations in TEM-type
penicillinase genes. Rev Infect Dis 1988; 10: 879-884.
Spees,E.K., Light,J.A., Oakes,D.D. and Reinmuth,B. Experiences with cadaver renal
allograft contamination before transplantation. Br J Surg 1982; 69: 482-485.
Stals,A., Werbrouck,H., Baert,L., Botteldoorn,N., Herman,L., Uyttendaele,M. and Van
Coillie,E. (2009) Laboratory efforts to eliminate contamination problems in the real-time
RT-PCR detection of noroviruses. J. Microbiol. Methods 2009; 77: 72-76.
Tenover FC. Vancomycin-resistant Staphylococcus aureus: a perfect but
geographically limited storm? Clin Infect Dis 200; 46(5): 675-7.
Tessier,F., Arpin,C., Allery,A. and Quentin,C. Molecular characterization of a TEM-21
beta-lactamase in a clinical isolate of Morganella morganii. Antimicrob Agents
Chemother 1998; 42: 2125-2127.
Toleman,M.A., Simm,A.M., Murphy,T.A., Gales,A.C., Biedenbach,D.J., Jones,R.N. and
Walsh,T.R. Molecular characterization of SPM-1, a novel metallo-beta-lactamase
isolated in Latin America: report from the SENTRY antimicrobial surveillance
programme. J. Antimicrob Chemother 2002; 50: 673-679.
Tumbarello,M., Spanu,T., Sanguinetti,M., Citton,R., Montuori,E., Leone,F., Fadda,G.
and Cauda,R. Bloodstream infections caused by extended-spectrum-beta-lactamase-
producing Klebsiella pneumoniae: risk factors, molecular epidemiology, and clinical
outcome. Antimicrob Agents Chemother 2006, 50: 498-504.
Turner,P.J. Extended-spectrum beta-lactamases. Clin Infect Dis 2005; 41(4): 273-275.
Tzouvelekis,L.S. and Bonomo,R.A. SHV-type beta-lactamases. Curr Pharm 1999; 5:
847-864.
Vieira VV, Fonseca EL, Vicente ACP. Metallo-β-lactamases produced by carbapenem-
resistant Pseudomonas aeruginosa in Brazil. Clin Microbiol Infect 2005; 11(11): 937.

REFERÊNCIAS BIBLIOGRÁFICAS
146
Villegas,M.V., Lolans,K., Correa,A., Kattan,J.N., Lopez,J.A. and Quinn,J.P. (2007) First
identification of Pseudomonas aeruginosa isolates producing a KPC-type carbapenem-
hydrolyzing beta-lactamase. Antimicrob Agents Chemother 2007; 51: 1553-1555.
Valones, MAA; Guimarães, RL; Brandão, LAC; Souza PRE; Carvalho AAT; Crovela, S.
Principles and applications of polymerase chain reaction in medical diagnostic fields: a
review. Brazilian Journal of Microbiology 2009; 40:1-11.
Walsh,T.R., Hall,L., Assinder,S.J., Nichols,W.W., Cartwright,S.J., MacGowan,A.P. and
Bennett,P.M. Sequence analysis of the L1 metallo-beta-lactamase from Xanthomonas
maltophilia. Biochim Biophys Acta 1994; 1218: 199-201.
Walsh TR, MA Toleman, L Poirel, P Nordmann. Metallo-betalactamases: the quiet
before the storm? Clin Microbiol Rev 2005; 18(2): 306-325.
Walsh TR. Clinically significant carbapenemases: an update. Curr Opin Infect Dis
2008; 21(4): 367-371.
Werner G, Coque TM, Hammerum AM, Hope R, Hryniewicz W, Johnson A, Klare I,
Kristinsson KG, Leclercq R, Lester CH, Lillie M, Novais C, Olsson-Liljequist B, Peixe
LV, Sadowy E, Simonsen GS, Top J, Vuopio-Varkila J, Willems RJ, Witte W, Woodford
N. Emergence and spread of vancomycin resistance among enterococci in Europe.
Euro Surveill 2008; 13 (47): pii 19046.
Winokur,P.L., Canton,R., Casellas,J.M. and Legakis,N. Variations in the prevalence of
strains expressing an extended-spectrum beta-lactamase phenotype and
characterization of isolates from Europe, the Americas, and the Western Pacific region.
Clin Infect Dis 2001; 3(2): 94-103.
Woodford N, Tiemo PM, Young K, Tysall L, Palepou MFI, Ward E, Painter RE, Suber
DF, Shungu D, Silver LL, Inglima K, Komblum J, Livermore DM. Outbreak of Klebsiella
pneumonia producing a new carbapenem-hydrolyzing class A β-lactamase, KPC-3, in
a New York Medical Center. Antimicrob Agents Chemother 2004; 48(12):4793-4799.
Xavier DE, AC Gales, RE Mendes, ACC Pignatari, LS Filho, LF Cirilo, M Castanheira.
2006. IMP-18-Producing Pseudomonas aeruginosa (PSA): increasing diversity of
mobile metallo-beta-lactamase (MBL) in Brazil. 46th Intescience Conference on

REFERÊNCIAS BIBLIOGRÁFICAS
147
Antimicrobial Agents and Chemotherapy (ICAAC). San Francisco - CA. Setembro, 27-
30, 2007. Poster C2-419.
Yanagihara,K., Kitagawa,Y., Tomonaga,M., Tsukasaki,K., Kohno,S., Seki,M.,
Sugimoto,H., Shimazu,T., Tasaki,O., Matsushima,A., Ikeda,Y., Okamoto,S., Aikawa,N.,
Hori,S., Obara,H., Ishizaka,A., Hasegawa,N., Takeda,J., Kamihira,S., Sugahara,K.,
Asari,S., Murata,M., Kobayashi,Y., Ginba,H., Sumiyama,Y. and Kitajima,M. Evaluation
of pathogen detection from clinical samples by real-time polymerase chain reaction
using a sepsis pathogen DNA detection kit. Crit Care 2010; 14, R159.
Yigit, H, AM Queenan, GJ Anderson, A Domenech-Sanchez, JW Biddle, CD Steward,
S Alberti, K Bush, FC Tenover. Novel carbapenem-hydrolyzing betalactamase, KPC-1,
from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents
Chemother 2001; 45(4): 1151–1161. (Erratum, 52(2): 809, 2008.).
Yong D, Bell JM, Ritchie B, Pratt R, Toleman MA, Walsh T R. A novel subgroup
metallo-β-lactamase (MBL), AIM-1 emerges in Pseudomonas aeruginosa (PSA) from
Australia. In: Annual Intescience Conference on Antimicrobial Agents and
Chemotherapy, 47., 2007, Chigaco. Anais… Washington: American Society for
Microbiology, 2007a.
Yong D, JM Bell, B Ritchie, R Pratt, MA Toleman, TR Walsh. 2007. A Novel Sub-
Group Metallo-β-Lactamase (MβL), AIM-1 Emerges in Pseudomonas aeruginosa
(PSA) from Australia. 47th Interscience Conference in Antimicrobial Agents and
Chemotherapy (ICAAC). Chicago, IL, USA. Setembro 17-20, 2007b. Poster C1-539.
Yong D, MA Toleman, CG Giske, HS Cho, K Sundman, K Lee, TR Walsh.
Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel
erythromycin esterase gene carried on a unique genetic structure in Klebsiella
pneumoniae Sequence Type 14 from India. Antimicrob Agents Chemother 2009;
53(12): 5046-5054.
Zaragoza,R., Artero,A., Camarena,J.J., Sancho,S., Gonzalez,R. and Nogueira,J.M.
The influence of inadequate empirical antimicrobial treatment on patients with
bloodstream infections in an intensive care unit. Clin Microbiol Infect 2003; 9: 412-418.

REFERÊNCIAS BIBLIOGRÁFICAS
148
Zavascki AP, Gaspareto PB, Martins AF, Gonçalves AL, Barth AF. Outbreak of
carbapenem-resistant Pseudomonas aeruginosa producing SPM-1 metallo-β-
lactamase in a Teaching Hospital in Southern Brazil. J Antimicrob Chemother 2005;
56(6): 1148-1151.
Zavascki AP, Machado AB, Oliveira KR, Superti SV, Pilger DA, Cantarelli W, Pereira
PR, Lieberkmecht AC, Barth AL. KPC-2-producing Enterobacter cloacae in two cities
from Southern Brazil. Int J Antimicrob Agents 2009; 34(3): 286-288.
Zavascki AP, CG Carvalhaes, RC Picão, AC Gales. Multi-drug resistant Pseudomonas
aeruginosa and Acinetobacter baumannii: resistance mechanisms and implications for
therapy. Expert Rev Anti Infect Ther 2010; 8(1): 71-93.
Zhu W, Clark NC, McDougal LK, Hageman J, McDonald LC, Patel JB. Vancomycin
resistant Staphylococcus aureus isolates associated with inc18-like vanA plasmids in
Michigan. Antimicrob Agents Chemother 2008; 52 (2): 452-457.
Ziegler, R., I. Johnscher, P. Martus, D. Lenhardt, and H.-M. Just. Controlled clinical
laboratory comparison of two supplemented aerobic and anaerobic media used in
automated blood culture systems to detect blood- stream infections. J Clin Microbiol
1998; 36: 657–661.

ABSTRACT
149
9. ABSTRACT
Solid organ transplanted patients are at high risk for blood stream infection. The
institution of appropriate antimicrobial therapy guided by a rapid and accurate
microbiological diagnosis of blood stream infections is related to a successful outcome.
The aim of this study was the detection of Gram-positive and Gram-negative bacteria
from automated blood cultures (Bactec®, Becton Dickinson) with the use of the
multiplex real-time polymerase chain reaction (PCR) and detection of antimicrobial
resistance genes. Methods: A total of 185 blood cultures, 126 positive and 59
negative, were obtained of 117 patients submitted to solid organ transplant at two
transplant centers in the city of São Paulo, Brazil, Hospital São Paulo and Hospital do
Rim e Hipertensão. DNA from the blood cultures was extracted by phenol chloroform
method (Brazol®, LGC, Brazil). The detection of bacterial DNA was performed using
universal primers of 16S rRNA gene. The differentiation between Gram-positive and
Gram-negative bacteria was done by hybridization with Gram-specific probes by
multiplex real-time TaqMan. The resistance genes: blaSHV, blaTEM, blaCTX-M, blaKPC,
blaSPM, blaVIM, blaIMP, vanA, vanB and mecA were detected using specific primers by
real-time SYBR Green. The adequacy of antimicrobial treatment was evaluated by
reviewing the records of patients. Results: Fifty nine samples were positive for Gram-
positive and sixty seven samples were positive for Gram-negative. All samples (100%)
were concordant between blood culture and PCR. Thirty two samples were positive for
mecA gene, five for blaCTX-M gene, one for blaKPC gene, one for blaSHV gene and one for
blaTEM. Eighty eight were negative for all genes. The detection of antimicrobial
resistance genes could enhance the appropriateness of antibiotic therapy,
particularly in descalation the treatment of bacteremia caused by Gram positive and
early adequacy in the treatment of Gram negative bacteremia of solid organ
transplanted patients. Conclusion: The in house multiplex PCR for Gram-
positive/Gram-negative bacteria and detection of antimicrobial resistance genes by
Real Time PCR could be useful for rapid diagnosis of bloodstream infection in patients
undergoing solid organ transplant.

ANEXOS
150
ANEXOS
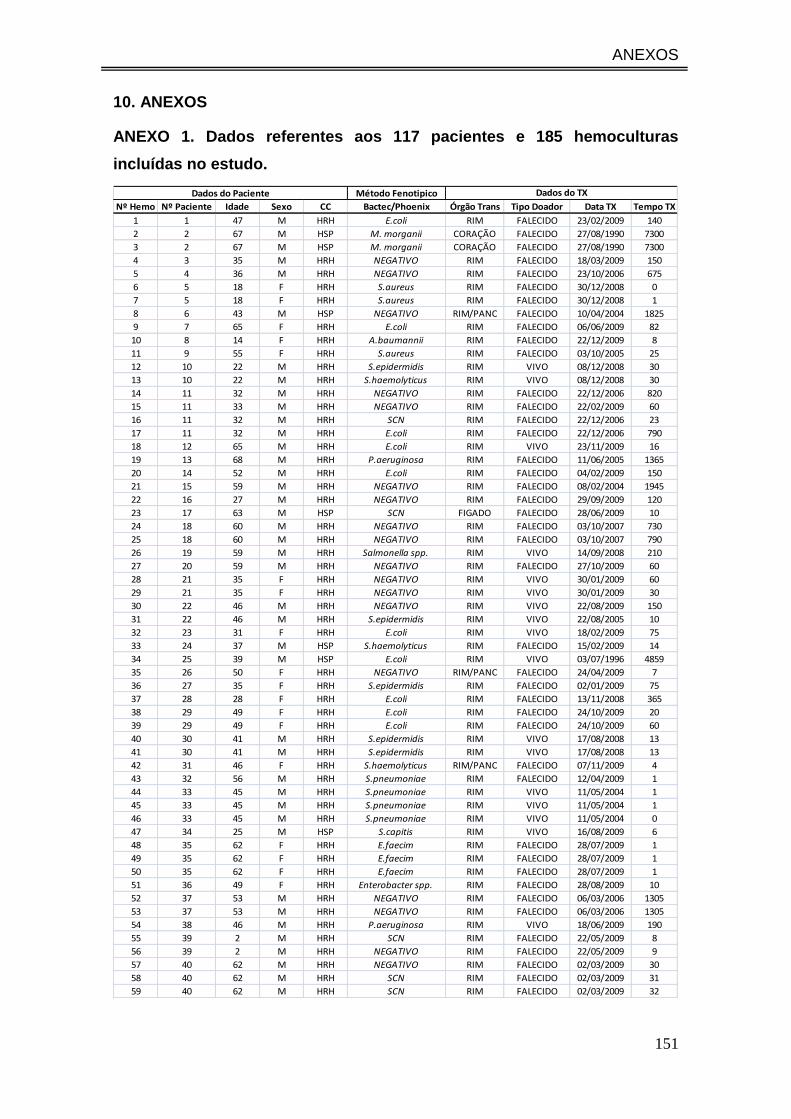
ANEXOS
151
10. ANEXOS
ANEXO 1. Dados referentes aos 117 pacientes e 185 hemoculturas
incluídas no estudo.
Método Fenotipico
Nº Hemo Nº Paciente Idade Sexo CC Bactec/Phoenix Órgão Trans Tipo Doador Data TX Tempo TX
1 1 47 M HRH E.coli RIM FALECIDO 23/02/2009 140
2 2 67 M HSP M. morganii CORAÇÃO FALECIDO 27/08/1990 7300
3 2 67 M HSP M. morganii CORAÇÃO FALECIDO 27/08/1990 7300
4 3 35 M HRH NEGATIVO RIM FALECIDO 18/03/2009 150
5 4 36 M HRH NEGATIVO RIM FALECIDO 23/10/2006 675
6 5 18 F HRH S.aureus RIM FALECIDO 30/12/2008 0
7 5 18 F HRH S.aureus RIM FALECIDO 30/12/2008 1
8 6 43 M HSP NEGATIVO RIM/PANC FALECIDO 10/04/2004 1825
9 7 65 F HRH E.coli RIM FALECIDO 06/06/2009 82
10 8 14 F HRH A.baumannii RIM FALECIDO 22/12/2009 8
11 9 55 F HRH S.aureus RIM FALECIDO 03/10/2005 25
12 10 22 M HRH S.epidermidis RIM VIVO 08/12/2008 30
13 10 22 M HRH S.haemolyticus RIM VIVO 08/12/2008 30
14 11 32 M HRH NEGATIVO RIM FALECIDO 22/12/2006 820
15 11 33 M HRH NEGATIVO RIM FALECIDO 22/02/2009 60
16 11 32 M HRH SCN RIM FALECIDO 22/12/2006 23
17 11 32 M HRH E.coli RIM FALECIDO 22/12/2006 790
18 12 65 M HRH E.coli RIM VIVO 23/11/2009 16
19 13 68 M HRH P.aeruginosa RIM FALECIDO 11/06/2005 1365
20 14 52 M HRH E.coli RIM FALECIDO 04/02/2009 150
21 15 59 M HRH NEGATIVO RIM FALECIDO 08/02/2004 1945
22 16 27 M HRH NEGATIVO RIM FALECIDO 29/09/2009 120
23 17 63 M HSP SCN FIGADO FALECIDO 28/06/2009 10
24 18 60 M HRH NEGATIVO RIM FALECIDO 03/10/2007 730
25 18 60 M HRH NEGATIVO RIM FALECIDO 03/10/2007 790
26 19 59 M HRH Salmonella spp. RIM VIVO 14/09/2008 210
27 20 59 M HRH NEGATIVO RIM FALECIDO 27/10/2009 60
28 21 35 F HRH NEGATIVO RIM VIVO 30/01/2009 60
29 21 35 F HRH NEGATIVO RIM VIVO 30/01/2009 30
30 22 46 M HRH NEGATIVO RIM VIVO 22/08/2009 150
31 22 46 M HRH S.epidermidis RIM VIVO 22/08/2005 10
32 23 31 F HRH E.coli RIM VIVO 18/02/2009 75
33 24 37 M HSP S.haemolyticus RIM FALECIDO 15/02/2009 14
34 25 39 M HSP E.coli RIM VIVO 03/07/1996 4859
35 26 50 F HRH NEGATIVO RIM/PANC FALECIDO 24/04/2009 7
36 27 35 F HRH S.epidermidis RIM FALECIDO 02/01/2009 75
37 28 28 F HRH E.coli RIM FALECIDO 13/11/2008 365
38 29 49 F HRH E.coli RIM FALECIDO 24/10/2009 20
39 29 49 F HRH E.coli RIM FALECIDO 24/10/2009 60
40 30 41 M HRH S.epidermidis RIM VIVO 17/08/2008 13
41 30 41 M HRH S.epidermidis RIM VIVO 17/08/2008 13
42 31 46 F HRH S.haemolyticus RIM/PANC FALECIDO 07/11/2009 4
43 32 56 M HRH S.pneumoniae RIM FALECIDO 12/04/2009 1
44 33 45 M HRH S.pneumoniae RIM VIVO 11/05/2004 1
45 33 45 M HRH S.pneumoniae RIM VIVO 11/05/2004 1
46 33 45 M HRH S.pneumoniae RIM VIVO 11/05/2004 0
47 34 25 M HSP S.capitis RIM VIVO 16/08/2009 6
48 35 62 F HRH E.faecim RIM FALECIDO 28/07/2009 1
49 35 62 F HRH E.faecim RIM FALECIDO 28/07/2009 1
50 35 62 F HRH E.faecim RIM FALECIDO 28/07/2009 1
51 36 49 F HRH Enterobacter spp. RIM FALECIDO 28/08/2009 10
52 37 53 M HRH NEGATIVO RIM FALECIDO 06/03/2006 1305
53 37 53 M HRH NEGATIVO RIM FALECIDO 06/03/2006 1305
54 38 46 M HRH P.aeruginosa RIM VIVO 18/06/2009 190
55 39 2 M HRH SCN RIM FALECIDO 22/05/2009 8
56 39 2 M HRH NEGATIVO RIM FALECIDO 22/05/2009 9
57 40 62 M HRH NEGATIVO RIM FALECIDO 02/03/2009 30
58 40 62 M HRH SCN RIM FALECIDO 02/03/2009 31
59 40 62 M HRH SCN RIM FALECIDO 02/03/2009 32
Dados do Paciente Dados do TX
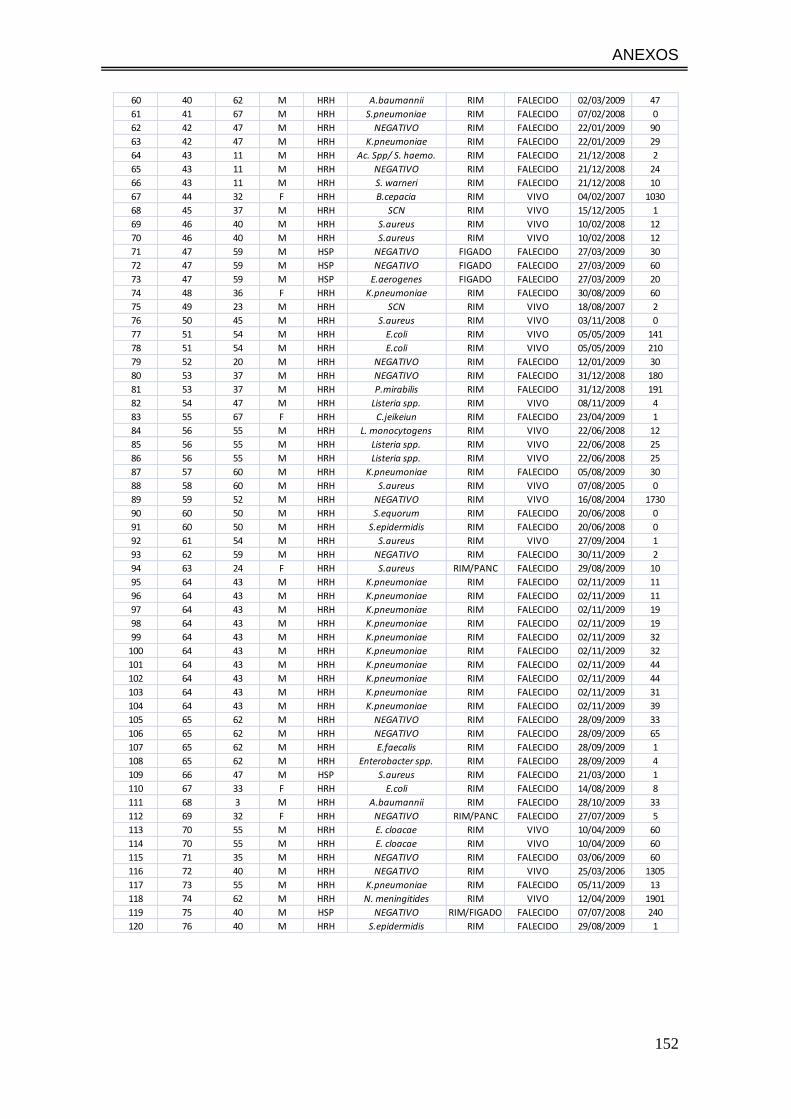
ANEXOS
152
60 40 62 M HRH A.baumannii RIM FALECIDO 02/03/2009 47
61 41 67 M HRH S.pneumoniae RIM FALECIDO 07/02/2008 0
62 42 47 M HRH NEGATIVO RIM FALECIDO 22/01/2009 90
63 42 47 M HRH K.pneumoniae RIM FALECIDO 22/01/2009 29
64 43 11 M HRH Ac. Spp/ S. haemo. RIM FALECIDO 21/12/2008 2
65 43 11 M HRH NEGATIVO RIM FALECIDO 21/12/2008 24
66 43 11 M HRH S. warneri RIM FALECIDO 21/12/2008 10
67 44 32 F HRH B.cepacia RIM VIVO 04/02/2007 1030
68 45 37 M HRH SCN RIM VIVO 15/12/2005 1
69 46 40 M HRH S.aureus RIM VIVO 10/02/2008 12
70 46 40 M HRH S.aureus RIM VIVO 10/02/2008 12
71 47 59 M HSP NEGATIVO FIGADO FALECIDO 27/03/2009 30
72 47 59 M HSP NEGATIVO FIGADO FALECIDO 27/03/2009 60
73 47 59 M HSP E.aerogenes FIGADO FALECIDO 27/03/2009 20
74 48 36 F HRH K.pneumoniae RIM FALECIDO 30/08/2009 60
75 49 23 M HRH SCN RIM VIVO 18/08/2007 2
76 50 45 M HRH S.aureus RIM VIVO 03/11/2008 0
77 51 54 M HRH E.coli RIM VIVO 05/05/2009 141
78 51 54 M HRH E.coli RIM VIVO 05/05/2009 210
79 52 20 M HRH NEGATIVO RIM FALECIDO 12/01/2009 30
80 53 37 M HRH NEGATIVO RIM FALECIDO 31/12/2008 180
81 53 37 M HRH P.mirabilis RIM FALECIDO 31/12/2008 191
82 54 47 M HRH Listeria spp. RIM VIVO 08/11/2009 4
83 55 67 F HRH C.jeikeiun RIM FALECIDO 23/04/2009 1
84 56 55 M HRH L. monocytogens RIM VIVO 22/06/2008 12
85 56 55 M HRH Listeria spp. RIM VIVO 22/06/2008 25
86 56 55 M HRH Listeria spp. RIM VIVO 22/06/2008 25
87 57 60 M HRH K.pneumoniae RIM FALECIDO 05/08/2009 30
88 58 60 M HRH S.aureus RIM VIVO 07/08/2005 0
89 59 52 M HRH NEGATIVO RIM VIVO 16/08/2004 1730
90 60 50 M HRH S.equorum RIM FALECIDO 20/06/2008 0
91 60 50 M HRH S.epidermidis RIM FALECIDO 20/06/2008 0
92 61 54 M HRH S.aureus RIM VIVO 27/09/2004 1
93 62 59 M HRH NEGATIVO RIM FALECIDO 30/11/2009 2
94 63 24 F HRH S.aureus RIM/PANC FALECIDO 29/08/2009 10
95 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 11
96 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 11
97 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 19
98 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 19
99 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 32
100 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 32
101 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 44
102 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 44
103 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 31
104 64 43 M HRH K.pneumoniae RIM FALECIDO 02/11/2009 39
105 65 62 M HRH NEGATIVO RIM FALECIDO 28/09/2009 33
106 65 62 M HRH NEGATIVO RIM FALECIDO 28/09/2009 65
107 65 62 M HRH E.faecalis RIM FALECIDO 28/09/2009 1
108 65 62 M HRH Enterobacter spp. RIM FALECIDO 28/09/2009 4
109 66 47 M HSP S.aureus RIM FALECIDO 21/03/2000 1
110 67 33 F HRH E.coli RIM FALECIDO 14/08/2009 8
111 68 3 M HRH A.baumannii RIM FALECIDO 28/10/2009 33
112 69 32 F HRH NEGATIVO RIM/PANC FALECIDO 27/07/2009 5
113 70 55 M HRH E. cloacae RIM VIVO 10/04/2009 60
114 70 55 M HRH E. cloacae RIM VIVO 10/04/2009 60
115 71 35 M HRH NEGATIVO RIM FALECIDO 03/06/2009 60
116 72 40 M HRH NEGATIVO RIM VIVO 25/03/2006 1305
117 73 55 M HRH K.pneumoniae RIM FALECIDO 05/11/2009 13
118 74 62 M HRH N. meningitides RIM VIVO 12/04/2009 1901
119 75 40 M HSP NEGATIVO RIM/FIGADO FALECIDO 07/07/2008 240
120 76 40 M HRH S.epidermidis RIM FALECIDO 29/08/2009 1

ANEXOS
153
121 77 41 M HSP K.pneumoniae RIM/PANC FALECIDO 31/11/2009 15
122 77 41 M HSP K.pneumoniae RIM/PANC FALECIDO 31/11/2009 15
123 78 30 M HRH SCN RIM FALECIDO 18/02/2009 0
124 79 18 M HRH NEGATIVO RIM FALECIDO 07/09/2008 120
125 80 44 M HRH S.epidermidis RIM FALECIDO 24/06/2009 0
126 81 49 M HRH S.epidermidis RIM VIVO 30/11/2006 20
127 81 49 M HRH P.aeruginosa RIM/PANC FALECIDO 08/11/2009 41
128 81 49 M HRH P.aeruginosa RIM/PANC FALECIDO 08/11/2009 41
129 82 38 M HRH NEGATIVO RIM VIVO 04/03/2005 1460
130 83 42 M HRH SCN RIM FALECIDO 05/12/2008 9
131 84 52 F HRH NEGATIVO RIM FALECIDO 04/04/2009 60
132 85 53 F HRH NEGATIVO RIM VIVO 03/08/2007 90
133 86 57 F HRH Enterobacter SPP. RIM FALECIDO 01/04/2009 137
134 87 46 F HRH NEGATIVO RIM VIVO 14/02/2008 665
135 87 46 F HRH NEGATIVO RIM VIVO 14/02/2008 730
136 87 46 F HRH S.maltophilia RIM VIVO 14/02/2008 730
137 88 51 M HSP S.aureus RIM FALECIDO 27/02/2009 15
138 88 51 M HSP S.aureus RIM FALECIDO 27/02/2007 0
139 88 51 M HSP S.aureus RIM FALECIDO 27/02/2007 0
140 88 51 M HSP S.maltophilia RIM FALECIDO 27/02/2007 730
141 89 58 M HRH NEGATIVO RIM FALECIDO 13/11/2008 60
142 90 58 M HRH K.pneumoniae RIM VIVO 28/12/2004 1640
143 90 58 M HRH K.oxytoca RIM VIVO 28/12/2004 1640
144 91 21 F HRH NEGATIVO RIM VIVO 27/07/2009 34
145 92 70 M HRH NEGATIVO RIM FALECIDO 07/04/2009 30
146 92 70 M HRH E.durans RIM FALECIDO 07/04/2009 28
147 92 70 M HRH K.pneumoniae RIM FALECIDO 07/04/2009 48
148 93 39 F HRH NEGATIVO RIM FALECIDO 10/05/2009 120
149 93 39 F HRH NEGATIVO RIM FALECIDO 10/05/2009 120
150 94 56 M HRH S.epidermidis RIM FALECIDO 19/04/2009 33
151 95 65 M HRH NEGATIVO RIM VIVO 25/01/2009 8
152 96 58 M HRH S.aureus RIM FALECIDO 27/07/2009 12
153 97 42 M HRH NEGATIVO RIM FALECIDO 29/09/2009 34
154 97 42 M HRH K.pneumoniae RIM FALECIDO 29/09/2009 32
155 98 20 M HRH E.coli RIM FALECIDO 12/02/2006 1215
156 99 40 F HRH NEGATIVO RIM VIVO 09/11/2008 280
157 99 40 F HRH E.coli RIM VIVO 09/11/2008 131
158 99 40 F HRH K.pneumoniae RIM VIVO 09/11/2008 216
159 100 52 M HRH NEGATIVO RIM VIVO 19/02/2009 14
160 101 36 M HRH E.aerogenes RIM FALECIDO 11/09/2009 11
161 102 38 M HRH NEGATIVO RIM VIVO 20/08/2009 11
162 102 38 M HRH NEGATIVO RIM VIVO 20/08/2009 15
163 103 67 F HRH E.coli RIM FALECIDO 16/03/2003 2430
164 104 46 M HRH K.pneumoniae RIM FALECIDO 29/07/2009 16
165 104 46 M HRH K.pneumoniae RIM FALECIDO 29/07/2009 15
166 105 57 M HRH E.coli RIM VIVO 15/08/2006 1030
167 106 64 M HRH NEGATIVO RIM VIVO 17/03/2004 1975
168 106 64 M HRH S.epidermidis RIM VIVO 17/03/2004 0
169 106 64 M HRH S.epidermidis RIM VIVO 17/03/2004 3
170 106 64 M HRH S.epidermidis RIM VIVO 17/03/2004 0
171 107 57 M HRH NEGATIVO RIM FALECIDO 16/11/2009 46
172 107 57 M HRH NEGATIVO RIM FALECIDO 16/11/2009 47
173 108 32 F HRH NEGATIVO RIM/PANC FALECIDO 18/09/2007 790
174 108 32 F HRH NEGATIVO RIM/PANC FALECIDO 18/09/2007 46
175 109 54 F HRH NEGATIVO RIM FALECIDO 31/10/2008 74
176 110 17 F HRH S.epidermidis RIM FALECIDO 19/07/2009 15
177 110 17 F HRH S.epidermidis RIM FALECIDO 19/07/2009 30
178 110 17 F HRH L. adecarboxylata RIM FALECIDO 19/07/2009 40
179 111 55 M HRH NEGATIVO RIM FALECIDO 14/06/2009 77
180 112 31 F HRH NEGATIVO RIM FALECIDO 06/12/2008 60
181 113 74 M HRH NEGATIVO RIM VIVO 11/04/2009 30
182 114 51 F HRH NEGATIVO RIM FALECIDO 15/12/2006 740
183 115 42 M HRH P.aeruginosa RIM VIVO 08/06/2009 36
184 116 72 M HRH Salmonella spp. RIM VIVO 20/01/2004 2125
185 117 17 M HRH P.aeruginosa RIM FALECIDO 29/03/2009 300
CC=Centro de Custo; HSP= Hospital São Paulo; HRH= Hospital do Rim e Hipertensão; TX= Transplante; Rim/Panc= Transplante de Rim e Pâncreas
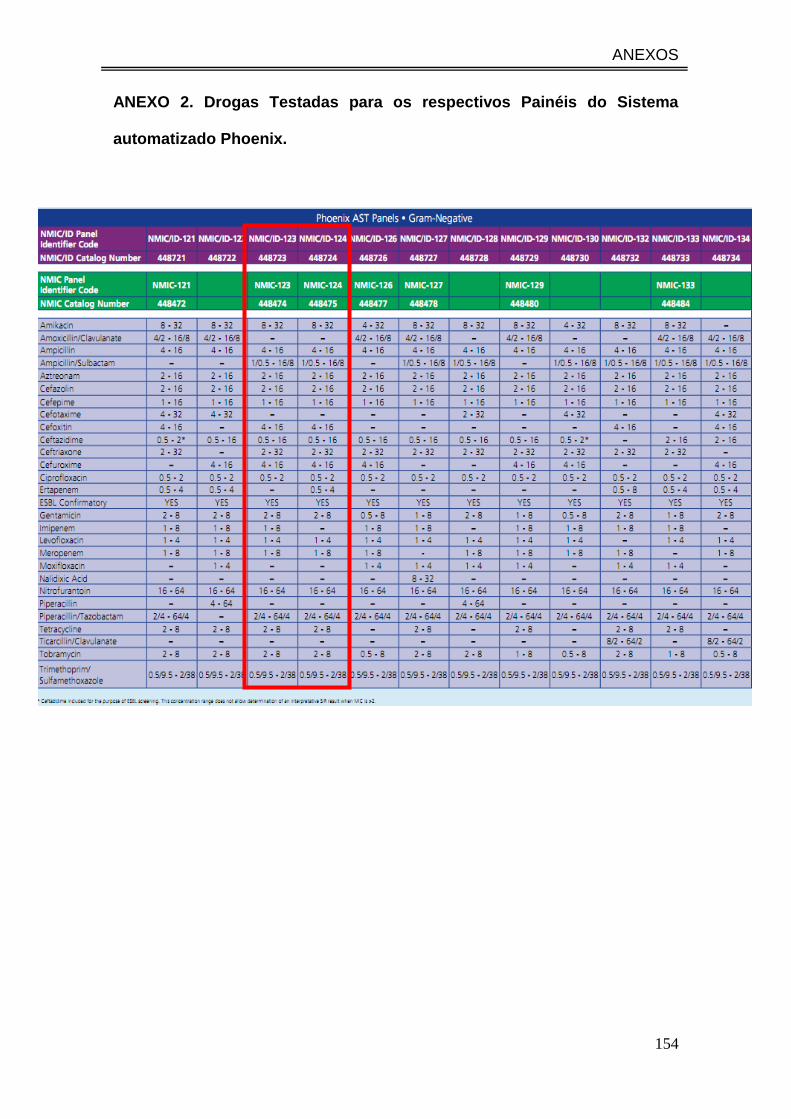
ANEXOS
154
ANEXO 2. Drogas Testadas para os respectivos Painéis do Sistema
automatizado Phoenix.

ANEXOS
155

ANEXOS
156
ANEXO 3. Apresentação no 50 th ICAAC 2010.
Real-time Polymerase Chain Reaction for Detection of Bacterial Pathogens and Antimicrobial Resistance Genes in Blood Cultures of Solid Organ Transplanted Patients. Talita Trevizani Rocchetti1,2, Liana Carballo Menezes1,2, Luiz Roberto Chirotto Filho1,2, Karen De
Castro Bauab1,2, Luis Fernando Aranha Camargo2, Antonio Carlos Campos Pignatari1,2.
1. Special Clinical Microbiology Laboratory (LEMC), Federal University of São Paulo/UNIFESP
2. Division of Infectious Diseases, Federal University of São Paulo/UNIFESP
Solid organ transplanted patients are at high risk for blood stream infection. The institution of
appropriate antimicrobial therapy guided by a rapid and accurate microbiological diagnosis of
blood stream infections is related to a successful outcome. The aim of this study was the
detection of Gram-positive and Gram-negative bacteria from automated blood cultures
(Bactec®, Becton Dickinson) with the use of the multiplex real-time polymerase chain reaction
(PCR) and detection of antimicrobial resistance genes. Methods: A total of 154 blood cultures,
113 positive and 41 negative, were obtained of 109 patients submitted to solid organ transplant
at two transplant centers in the city of São Paulo, Brazil, Hospital São Paulo and Hospital do
Rim e Hipertensão. DNA from the blood cultures was extracted by phenol chloroform method
(Brazol®, LGC, Brazil). The detection of bacterial DNA was performed using universal primers of
16S rRNA gene. The differentiation between Gram-positive and Gram-negative bacteria was
done by hybridization with Gram-specific probes by multiplex real-time TaqMan. The resistance
genes: blaSHV, blaTEM, blaCTX, blaKPC and mecA were detected using specific primers by real-time
SYBR Green. Results: Fifty four samples were positive for Gram-positive and fifty nine samples
were positive for Gram-negative. All samples (100%) were concordant between blood culture
and PCR. Forty six samples were positive for mecA gene, seven for blaCTX gene, eight for blaKPC
gene and fifty three were negative for all genes. Conclusion: The results of this study suggest
that multiplex PCR for Gram-positive/Gram-negative bacteria and detection of antimicrobial
resistance genes by Real Time PCR could be useful for rapid diagnosis of bloodstream infection
in patients undergoing solid organ transplant.
Top Related