


![HSM MANAGEMENT Peter Diamandis V.16, N.95, NOVEMBRO ... · 132 4 liçöes sobre plataformas Outro modo de entender —e de replicar— os modelos estratégicos do iPhone (Apple].](https://static.fdocumentos.tips/doc/165x107/601a4618e9df9230b70ae237/hsm-management-peter-diamandis-v16-n95-novembro-132-4-lies-sobre-plataformas.jpg)




Línguas
Páginas
Legal


1
Base Física da Evolução
• DNA pode replicar• DNA pode mutar e recombinar• DNA codifica informação que interage com
o ambiente para influenciar o fenótipo
DNA pode replicar
Por causa da replicação, umúnico tipo de gene podeexistir tanto no tempo quantono espaço em uma maneiraque transcende os indivíduosque temporariamente contémos genes.
Identidade por Descendência
Alguns alelos são idênticos porque sãodescendentes replicados de um únicoalelo ancestral
A existência de genes no espaço e no tempo
• Manifesta-se apenas no nível da populaçãoque está se reproduzindo
• Fornece a continuidade espacial e temporalque é necessária à evolução
Evolução
• É o fado de formas alternativas de genes oucombinações gênicas no espaço e no tempoem uma população que está sereproduzindo
• É um processo que se manifesta apenas nonível de uma população que está sereproduzindo
• Nunca pode ser entendida apenas emtermos individuais.
• Requer Variação Genética
“Aleatoriedade” significa que mutações tem umamplo espectro de ação em seus carreadores
Númerode
Linhagens
Habilidade competitiva relativa a linhagem original
Efeito de 50 linhagens com mutações expontâneasderivadas de uma linhagem de levedura crescendo em
laboratório.
0
2
4
6
8
10
0.6 0.8 1.0

2
Aleatoriedade refere-se às conseqüênciasfenotípicas. O ambiente pode e influencia ataxa e o tipo de mutação no nível molecular.
Mutação cria variação alélica,recombinação e diploidia a amplificam
• P.ex., MHC é um agrupamento autossômicode 20 Loci com um conjunto total de 698Alleles (até 1997);
• Recombinação pode produzir 1.21×1021
combinações gaméticas destes 698 Alelos• Tais gametas podem criar 1042 genótipos• Existem 6 ×109 humanos na terra
Single Nucleotide Polymorphisms (SNPs)“Polimorfismos de base única”
• Até Fev de 2001, cerca de 1.420.000 SNPsbialélicos haviam sido identificados emhumanos
• Tais SNPs definem 21.420.000 = 10427.463 tipospossíveis de gametas
• Tais SNPs definem 31.420.000 = 10677.512
genótipos potenciais• Existem 1080 elétrons no universo
Alta variação genética implica que:
• Cada indivíduo é único
• Evolução pode ocorrer
O que é uma população?
• Pode ser qualquer agrupamento de indivíduos– Todos os indivíduos de uma localidade– Os indivíduos de um estado– Os indivíduos de um ecossistema– Indivíduos de uma espécie
Deme
Um Deme é uma população local deindivíduos que reproduzem e têmcontinuidade física no tempo e no
espaço. Demes são o nível biológicomais baixo que podem evoluir.

3
Demes
• Têm em comum o sistema de acasalamento:
• regras que indivíduos usam para o acasalamento(reprodução sexual)
Demes são caracterizados por freqüências genotípicas.P.ex., considere uma população de índios Pueblo
tipados para o marcador sangüíneo MN
111/140=0.08
46/140=0.33
83/140=0.59
Freq.Genot.
140114683N
NNMNMMGenótipoTotal
NMNMTipoSangüíneo
Demes são caracterizados por freqüências genotípicas.P.ex., considere uma população de Aborígenes
australianos tipados para o marcador sangüíneo MN
1250/ 372= 0.672
113/ 372= 0.304
9/372 =0.024
Freq.Genot.
3722501139N
NNMNMMGenótipoTotal
NMNMTipo
Sangüíneo
Demes com os mesmos alelos podem terfreq. genotípicas muito diferentes:
NN.08
MN0.33
MM0.59
Índios Pueblo
NN0.672
MN0.304
Aborígines AustralianosMM
0.024
Pool Gênico
Um Pool Gênico é a população decópias gênicas que são
coletivamente mantidas pelosindivíduos de um deme.
Pools gênicos são caracterizados por Freqüências gaméticas (Freq.alélicas quando considerando apenas1 locus). P.ex., considere
aqueles índios Pueblo tipados para MN
N MAlelo (Tipogamético)
1(2*11+46)/280= 0.24
(2*83+46)/280= 0.76
Freq. Alélica
140114683N
NNMNMMGenótipoTot
NMNMTipo Sangüíneo

4
Pools gênicos são caracterizados por Freqüências gaméticas (Freq.alélicas quando considerando apenas1 locus). P.ex., considere
aqueles Aborígenes Australianos tipados para MN
N MAllele (GameteType)
1(2*250+113)/744
= 0.824(2*9+113)/744
= 0.176Allele Freq.
3722501139Number
NNMNMMGenotypeSum
NMNMBlood Type
Pool Gênico(definição alternativa)
Pool Gênico é a população de gametaspotenciais que podem ser produzidos
pelos indivíduos de um deme.
Pool gênico como uma população degametas potenciais
Deme
Pool gênico
Demes e Pools gênicos:
NN.08
MN0.33
MM0.59
Deme de índios Pueblo
N1(.08) + 1/2(.33) = .24
M1(0.59) + 1/2(0.33) = 0.76
Pool gênico de índios Pueblo
ProbabilidadesMendelianas
na meiose
diplóide
haplóide
Meiose 1 11/21/2
N1(0.672) + 1/2(0.304) = 0.824
M1(.024) + 1/2(.304)
= 0.176
Pool gênico de Aborígenes Australianos
1 11/21/2
NN0.672
MN0.304
Deme de Aborígenes AustralianosMM
0.024
Demes e Pools gênicos:
ProbabilidadesMendelianas
na meiose
diplóide
haplóide
Meiose
Demes com os mesmos alelos podem ter poolsgênicos com freqüências alélicas diferentes
Índios Pueblo
Aborígenes Australianos
N0.24
M0.76
N0.824
M 0.176

5
Demes e Pools gênicos:
• Meiose interconecta o deme ao pool gênico• Logo, dadas as leis de Mendel e uma meiose
normal, pode-se sempre calcular as freq.alélicas no pool gênico a partir das freq.genotípicas do deme
• Pode-se também prever o deme (freq.genotípicas) a partir do pool gênico ( freq.alélicas)?
aa1/4
Aa1/2
AA1/4
1 11/21/2
aa1/2
AA1/2
a1/2
A1/2
1 1
a1/2
A1/2
Demes
Pools Gênicos
Definição de Evolução
A definição operacional deevolução em nível de deme é
mudanças na freqüência alélicaou genotípica ao longo do tempo.
Evolução
• É uma propriedade emergente dos organismos quese reproduzem e não dos indivíduos.
• 3 mecanismos principais devem ser investigados– como produzir gametas– como unir gametas– como criar fenótipos
Arquitetura Genética
• número de loci e localização genômica• número de alelos por locus• taxa e modo de mutação• regras de herança dos elementos genéticos
Estrutura Populacional
• Sistema de acasalamento da população• Tamanho populacional• Presença, quantidade e padrão de troca genética
com outras populações• Estrutura etárea de indivíduos na população.

6
Contexto Ambiental
• fenótipos são interações entre genes e ambientes
• Desenvolvimento fenotípico - os mecanismos quedescrevem como zigotos adquirem fenótiposnaquele contexto ambiental
Punnett disse que braquidactilia era uma doença autossômicadominante com alta penetrância. Yule disse que o Mendelismo
estava errado, pois não existe uma razão de 1:1 entrebraquidáctilos e pessoas não afetadas.
1908: Punnett x Yule
I am reluctant tointrude…
Ao Editor de Science: I am reluctant to intrude in adiscussion concerning matters which I have no expertknowledge, and I should have expected the verysimple point which I wish to make to have beenfamiliar to biologists. However, some remarks of Mr.Udny Yule, to which Mr. R. C. Punnett has called myattention, suggest that it may be worth making...
De Hardy, G. H. Mendelian Proportions in a Mixed Population
Science 28:49-50. 1908.
G. H. Hardy
Hardy (e Weinberg) resolveram oproblema de Punnett
Para ir do Pool gênico (Gametas) ao Deme (Zigotos inicialmente),precisa-se explicitar as regras pelas quais os gametas se unem
FERTILIZAÇÃO
Pressupostos de Hardy Weinberg
• Produção de alelos: 1 locus autossômico2 alelossem mutação1ª Lei de Mendel
• União de alelos:– Sistema de acasalamento aleatório– Tamanho populacional infinito– Troca genética Ausente– Estrutura etária Nenhuma (gerações discretas)
• Criação de fenótipos: Genótipos têm fenótipos idênticos (sem Seleção Natural)
Sistemas de Acasalamento
Sistemas de Acasalamento são as regras em nívelde deme que regulam como gametas se unem nafertilização, portanto, definindo a transição dehaploidia a diploidia.

7
Acasalamento ao acaso
Acasalamento ao acaso ocorre quando ambosos gametas unidos em um gameta são retiradosao acaso, e independentemente do pool gênico.
Isto significa que a probabilidade do gameta terum alelo específico é igual à freq. daquele alelono pool gênico, e isto é verdade para todos os
gametas envolvidos na fertilização.
Acasalamento ao acaso
aaq×q=q2
aAqp
aq
Aapq
AAp×p=p2
Ap
aq
Ap
aq = 1-p
Ap
Gameta Materno
Gam
eta
Pate
rno
Pool Gênico
Freqüências Genotípicas de Hardy-Weinberg
aaq2
Aa2pq
AAp2
O ciclo de vida de uma população
aaPaa
AaPAa
AAPAA
1 11/21/2
aq=Paa+ 1/2PAa
Ap=PAA+ 1/2PAa
Deme de indivíduosdiplóides
Pool gênico de gametas Haplóides
Meiose
Fertilização
Deme de indivíduosdiplóides
ProbabilidadesMendelianas
aaq2
Aa2pq
AAp2
Acasalamentoao acaso p p p q q q
Teste de Hardy-Weinberg das freq genotípicas. P.ex., a populaçãode índios Pueblo tipados para MN
1400.06(140)= 8.4
0.36(140)= 50.4
0.58(140)= 81.2N Esp.
2(0.76)(0.24)= 0.36
1 (0.24)2 =0.06
(0.76)2 =0.58Freq. H.-W.
140114683NNNMNMMGenótipo
SomaNMNMTipo Sangüíneo
Acasalamento ao acaso
• Hardy assumiu que indivíduos são monoécios
• Weinberg assumiu que freqüências são iguais emambos os sexos

8
Hardy-Weinberg implica que NÃO se esperaque caracteres Mendelianos apresentem
razões Mendelianas nos demes
aaq2
Aa2pq
AAp2
Fenótipo Dominante Fenótipo Recessive
Razão Dominante:Recessivo é p2+2pq : q2
Evolução em populações em EHW
P’ = p2 + pq= p (p+q)= p
Ou seja, basta uma geração de acasalamento aoacaso para a população entrar em Equilíbrio deHardy-Weinberg.
Genótipos MM MN NN Experado 294 496 209
Observado 298 489 213
χ2 = Σ(O -E)2/E
As freqüências genotípicas se conformam aoesperado no EHW?
M M MN NN
p2 2pq q2
0.294 0.496 0.209
Teste de Hardy-Weinberg das freq genotípicas. P.ex., a populaçãode índios Pueblo tipados para MN
1.23(11-8.4)2
8.4(46-50.4)2
50.4(83-81.2)2
81.2(Obs.-Esp.)2
Esp.
1400.06(140)= 8.4
0.36(140)= 50.4
0.58(140)= 81.2N Esp.
2(0.76)(0.24)= 0.36
1 (0.24)2 =0.06
(0.76)2 =0.58Freq. H.-W.
140114683NNNMNMMGenótipo
SomaNMNMTipo Sangüíneo
Graus de Liberdade = 3 Categorias - 1 - 1 parâmetro estimado = 1
Testes de Equilíbrio de HW
• Qui Quadrado
• Teste exato de Fisher
• Cadeias de Markov de Monte Carlo
• etc...
9
Razão defenótipos
dominantes arecessivos
p = Freq do alelo dominante
Razão 3:1
Taxa de eliminação de alelo letal a = q2
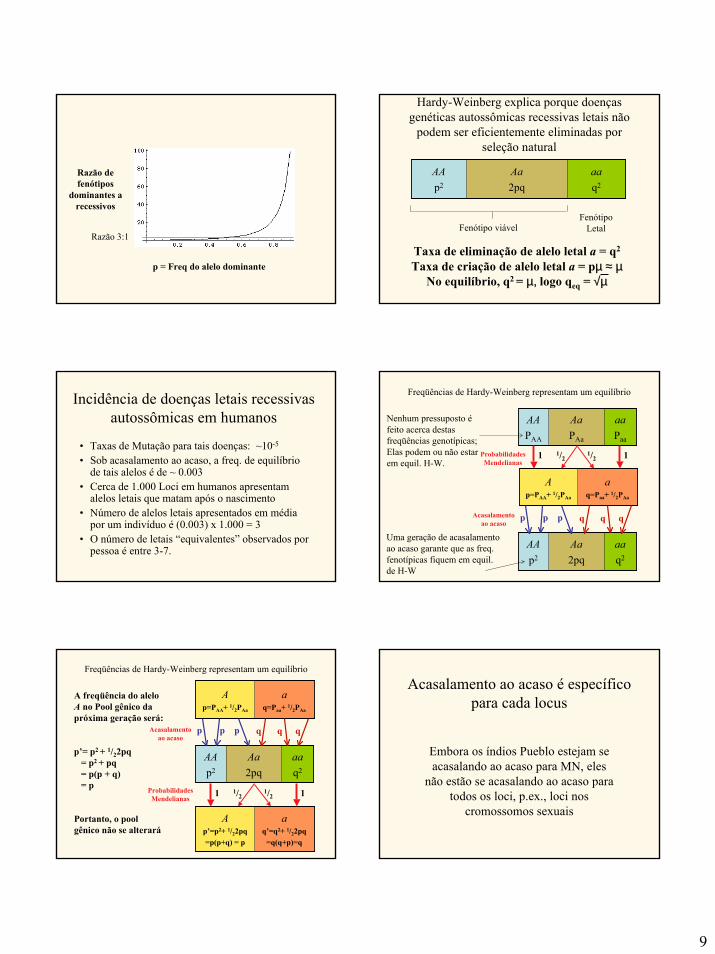
Taxa de criação de alelo letal a = pµ ≈ µNo equilíbrio, q2 = µ, logo qeq = √µ
Hardy-Weinberg explica porque doençasgenéticas autossômicas recessivas letais não
podem ser eficientemente eliminadas porseleção natural
aaq2
Aa2pq
AAp2
Fenótipo viávelFenótipo
Letal
Incidência de doenças letais recessivasautossômicas em humanos
• Taxas de Mutação para tais doenças: ~10-5
• Sob acasalamento ao acaso, a freq. de equilíbriode tais alelos é de ~ 0.003
• Cerca de 1.000 Loci em humanos apresentamalelos letais que matam após o nascimento
• Número de alelos letais apresentados em médiapor um indivíduo é (0.003) x 1.000 = 3
• O número de letais “equivalentes” observados porpessoa é entre 3-7.
aaPaa
AaPAa
AAPAA
1 11/21/2
aq=Paa+ 1/2PAa
Ap=PAA+ 1/2PAa
ProbabilidadesMendelianas
aaq2
Aa2pq
AAp2
Acasalamentoao acaso
p p p q q q
Nenhum pressuposto éfeito acerca destasfreqüências genotípicas;Elas podem ou não estarem equil. H-W.
Uma geração de acasalamentoao acaso garante que as freq.fenotípicas fiquem em equil.de H-W
Freqüências de Hardy-Weinberg representam um equilíbrio
aq=Paa+ 1/2PAa
Ap=PAA+ 1/2PAa
aaq2
Aa2pq
AAp2
p p p q q q
1 11/21/2
aq’=q2+ 1/22pq
=q(q+p)=q
Ap’=p2+ 1/22pq=p(p+q) = p
A freqüência do aleloA no Pool gênico dapróxima geração será:
p’= p2 + 1/22pq = p2 + pq = p(p + q) = p
Portanto, o poolgênico não se alterará
Freqüências de Hardy-Weinberg representam um equilíbrio
ProbabilidadesMendelianas
Acasalamentoao acaso
Acasalamento ao acaso é específicopara cada locus
Embora os índios Pueblo estejam seacasalando ao acaso para MN, eles
não estão se acasalando ao acaso paratodos os loci, p.ex., loci nos
cromossomos sexuais

10
Pode também ser usado para caracteresligados ao X
• Q: If 8% of men are colorblind, whatpercent of women are color blind?
Sexo HomogaméticoXA XA p2
XA Xa 2pqXa Xa q2
Sexo HeterogaméticoXA Y p
Xa Y q
Genotype Allele
frequency
Genotype
frequency
XA Y p p
Xa Y q q=0.08
If 8% of m en are color blind =>Xa allele frequency q = 0.08
W hat proportion of wom en are color blind?
Genotype Expected frequency
XA XA p2
XA Xa 2pq
Xa Xa q2
0.84 64
0.14 72
0.00 64
1.00 00
XA allele frequency p = 1 - q = 0.08
W hat proportion of wom en are heterozygous carriers?
Equilíbrio em genes ligados ao sexo.
Dois Loci
• Mantém-se mesmos pressupostos de umlocus, porém, permite-se recombinação.
• Pool Gênico pode ser definido pelasfreqüências dos 4 gametas: AB, Ab, aB, ab
• Da mesma forma que p e q, a soma destasfreqüências também é 1
Dois Loci

11
Dois Loci
• Prob de um genótipo particular é também oproduto das freqüências gaméticas
• Duas formas de se obter AaBb.• Regras iguais ao locus único, mas temos agora 10
combinações genotípicas• Tudo muda quando vamos de uma geração de
adultos a outra!
Dois Loci
• Duplo homozigoto - apenas um tipo de gameta• Heterozigoto - 2 tipos de gametas• Duplo heterozigoto - 4 tipos de gametas!
• Prob definida pela recombinação r (0 ≤ r ≤ 0.5)
• Não significa que não esteja ocorrendorecombinação em outras combinações
Dois Loci
• A diferença do modelo de locus único criaum fato importante:
• As duas definições de pool gênico não sãomais equivalentes!– Genes comuns aos adultos de uma população– Gametas potenciais produzidos pelos adultos
Conseqüências para a evolução
• Quando alcançará o equilíbrio?
g'AB = 1. g2AB + ½ (2gABgAb) + ½(2gABgaB ) + ½(1- r)(2gABgab) + ½r(2gAbgaB )
= gAB[ gAB + gAb + gaB + (1- r)gab] + r.gAbgaB
= gAB[ gAB + gAb + gaB + gab] + rgAbgaB - rgABgab
= gAB + r(gAbgaB - gABgab) = gAB - rD
Onde D é gAbgaB - gABgab sendo comumente conhecido comodesequilíbrio de ligação, ou desbalanço de fase gamética
Conseqüências para a evoluçãog'Ab = 1. g2
Ab + ½ (2gABgAb) + ½(2gAbgab ) + ½(1- r)(2gAbgaB) + ½r(2gABgab )= gAb + rD
g'aB = 1. g2AB + ½ (2gABgaB) + ½(2gaBgab ) + ½(1- r)(2gAbgaB) + ½r(2gABgab )
= gaB + rD
g'ab = 1. g2AB + ½ (2gABgab) + ½(2gaBgab ) + ½(1- r)(2gABgab) + ½r(2gAbgaB )
= gab - rD
E está ocorrendo evolução?
Conseqüências para a evolução
• Enquanto r >0 e D ≠0 --> g'ab ≠ gab
• Se r = 0, funciona como locus único
• Se D = 0 populações estão em equilíbrio (o quenão quer dizer que não esteja havendorecombinação!)

12
Conseqüências para a evolução
• E quanto a D na próxima geração?
D’ = gABgab - gAb gaB
= [(gAb + rD)(gaB + rD)-(gAB + rD)(gab + rD)= D(1-r)
D” = D(1-r)2
Dt = D0(1-r)t tende a 0
Conseqüências para a evolução
LD reduz a ritmo que depende de (1-r)Mesmo loci independentes podem possuir LD> 0 e
equilíbrio não é alcançado imediatamente.
Quando r é grande, queda é rápida (5 gerações para 3%)
Quando r é pequeno, queda é lenta (345 gerações para 3%)
Conseqüências para a evolução
CorolárioO estado do pool gênico de uma população e sua
evolução atual são influenciados por sua históriapassada.
O passado não pode ser ignorado para se entender opresente e predizer o futuro em sistemasbiológicos.
Conseqüências para a evolução
• Vantagem: sistemas multi-loci contém informaçãosobre origem e história da variação hoje existente
• Desvantagem: efeitos duradouros do desequilíbrioinicial podem limitar nossas inferências
Dt= D0(1-r)t
Fatores que aumentam D
• Deriva genética
• Seleção Natural
• Migração

13
Blondes 'to die out in 200 years'
Scientists believe the last blondes will be in Finland
Dyed rivalsThe researchers also believe that so-called bottle blondes may be toblame for the demise of their natural rivals.They suggest that dyed-blondes are more attractive to men whochoose them as partners over true blondes.
Bottle-blondes like AnnWiddecombe may be toblame
The last natural blondes will die out within 200years, scientists believe.
A study by experts in Germany suggests peoplewith blonde hair are an endangered species andwill become extinct by 2202.
Researchers predict the last truly natural blondewill be born in Finland - the country with thehighest proportion of blondes.
But they say too few people now carry the genefor blondes to last beyond the next two centuries.
The problem is that blonde hair is caused by arecessive gene.
Friday, 27 September, 2002, 11:51 GMT 12:51 UK
http://news.bbc.co.uk/1/hi/health/2284783.stm
Fairly specific conditions need to be met for a recombination event to bedetectable: genetic diversity of the region, age of the event, sample sizeand demographic history are all important factors. The issue of samplesize is particularly problematic — small samples are unlikely to containthe rarest gamete necessary for the detection of recombination events, butthe rate at which adding extra chromosomes improves the estimate isextremely low (of the order of the log of the log of the sample

14
• (i) Simulate recombinant genealogies by using the coalescent withrecombination.
• (ii) Evolve nucleotide sequences on the simulated genealogy to obtaina sequence alignment.
• (iii) Apply 14 different methods to the simulated data and record howmany times, of 100 replicates, a method infers the presence ofrecombination.
Most methods showed more power with increased rates of recombination,which is the expected behavior for efficient methods.However, some methods are more efficient than others. Mostmethods showed better performance at higher levels of divergence,probably because in such cases there is more information availableto recognize the footprint of recombination. The only method thatshowed decreased power with more sequence divergence was theHOMOPLASY RATIO. For most methods, a minimum sequencedivergence of 5% seems necessary to attain substantial power.When the number of recombination events in the history of thesequences is around 3 ( 1), the most powerful methods inferredthe presence of recombination only 50% of the time, whichindicates that several recombination events are needed in order for
There are two different contexts in which we may wish to detectrecombination: rare recombination or frequent repeated recombination.Simulated data over a wide range of recombination rate values. Notsurprisingly, most methods have trouble detecting rare recombinationalevents, especially when sequence divergence is low. Indeed, recent eventsshould be more easily identifiable than older events, as the later may beobscured by subsequent mutation.On the other hand, when recombination rates are very high (higher thanthose simulated here), leading to situations close to linkage equilibrium,substitution methods might have trouble identifying site patterns.
Linkage analysis is based upon associations in well-characterized pedigrees, LD refers to associations withinpopulations of ‘unrelated’ individuals.In general, chromosomes sampled from ‘unrelated’ individualsin a population will be much more distantly related than thosesampled from members of traditional pedigrees.More opportunities for recombination to take place - pedigreestudies work with recombination events that exchangemegabase chunks of chromosomes, LD studies deal withsegments measured in kilobases.

15
In linkage mapping the pedigree is known, making it possible to model thegenealogy of the chromosomes using basic genetics.When dealing with unrelated individuals, however, the pedigree is almostcompletely unknown, and different models are needed.Pairwise LD is expected to be extremely variable. This variation isattributable both to the history of recombination and to the history ofmutations.For mapping purposes, it is the recombination history that is important.The mutations are of interest only to the extent that they reveal somethingabout this unobservable history. However, the recombination history isitself variable, and the pattern of LD reflects this too.
The relationship between LD and distance is far from smooth, inagreement with what is typically observed. LD is expected to varybetween chromosomal regions as well as between pairs of loci. It is clearthat, although there is a statistical relationship between the LD measureand distance, the variability is too great for reliable estimates to be madefrom any particular pair of loci. It will rarely be possible in practice toestimate the genetic distance between two markers using LD .
Top Related