







Línguas
Páginas
Legal


Curso de Pós-graduação em Patologia Humana
DISSERTAÇÃO DE MESTRADO
AVALIAÇÃO DAS HISTONAS DE L. infantum COMO CANDIDATOS À VACINAS NA INFECÇÃO POR L.infantum
EM HAMSTERS
Laís da Silva Pereira
Salvador – Bahia – Brasil 2013
UFBA
UNIVERSIDADE FEDERAL DA BAHIA
FACULDADE DE MEDICINA FUNDAÇÃO OSWALDO CRUZ - FIOCRUZ
CENTRO DE PESQUISAS GONÇALO MONIZ
FIOCRUZ

Curso de Pós-graduação em Patologia Humana
AVALIAÇÃO DAS HISTONAS DE L. infantum COMO CANDIDATOS À VACINAS NA INFECÇÃO POR L.infantum
EM HAMSTERS
Laís da Silva Pereira
Orientador: Dra. Cláudia Ida Brodskyn
Dissertação apresentada ao Colegiado do Curso de Pós-graduação em Patologia Humana, como pré-requisito obrigatório para obtenção do grau de Mestre.
Salvador – Bahia – Brasil 2013
UFBA
UNIVERSIDADE FEDERAL DA BAHIA
FACULDADE DE MEDICINA FUNDAÇÃO OSWALDO CRUZ - FIOCRUZ
CENTRO DE PESQUISAS GONÇALO MONIZ
FIOCRUZ

Ficha Catalográfica elaborada pela Biblioteca do
Centro de Pesquisas Gonçalo Moniz / FIOCRUZ - Salvador - Bahia.
Pereira, Laís da Silva Pereira P436a Avaliação das histonas de L.infantum. Como candidatos à vacinas na infecção por L.
Infantum em hamsters[manuscrito] / Laís da Silva Pereira.- 2013. 86 f.; 30 cm
Dissertação (Mestrado) – Universidade Federal da Bahia. Centro de Pesquisa
Gonçalo
Moniz. Curso de Pós-Graduação em Patologia Humana, 2013.
Orientadorª: Drª. Claúdia Ida BrodsKyn .
1. Leishmania 2. Vacina 3. Histonas 4. Hamsters I. Título
CDU 616.993.166:615.371

“AVALIAÇÃO DAS HISTONAS DE L.INFANTUM COMO CANDIDATOS A VACINAS CONTRA A INFECÇÃO POR L.INFANTUM EM HAMSTERS”
LAÍS DA SILVA PEREIRA
FOLHA DE APROVAÇÃO
COMISSÃO EXAMINADORA
Dra. Deborah Bittencourt Mothé Fraga Professora Adjunta
UFBA
Dr. Daniel Abensur Athanázio Professor Adjunto
UFBA
Dra. Cláudia Ida Brodskyn Pesquisadora Titular CPqGM/FIOCRUZ

À Deus, o autor e mantenedor da vida.
Aos meus pais, Noelia e José, pelo amor incondicional dedicado a mim.

AGRADECIMENTOS
À professora Dra. Cláudia Brodskyn, pelos conhecimentos, os quais tanto domina com
maestria, e sua participação fundamental no meu processo de formação profissional durante
esses anos de pesquisa.
Aos coordenadores Dra. Valéria Borges, Dra. Aldina Barral e Dr. Barral-Neto, pelo
acolhimento e a convivência harmoniosa.
Ao Dr. Manoel Soto, por haver cedido a mim as proteínas e os plasmídeos das
histonas.
Ao Dr. Washington Conrado, por sua competência durante as análises histopatológicas
realizadas neste trabalho.
À Dra. Clarissa Teixeira, o quanto sua experiência profissional e serenidade foram
valiosas na transição do mestrado para a seleção do doutorado.
À minha colega de trabalho e amiga, Melissa Abbehusen, pelas vivências, risos e por
ter sido um braço direito diante das dificuldades que foram superadas.
À Dra. Natália, por sua solicitude e indispensáveis conhecimentos compartilhados.
À Equipe Brodskyn: Jurema,Virna, Martha, Suzane, Claire e Lilian , pela amizade e as
alegrias vividas nesta jornada.
A Kyioshi Ferreira, pelas colaborações nas instruções da técnica do Real Time PCR.
Aos amigos Eder e Valter, pelo companheirismo.
Aos técnicos Jorjão Clarêncio e Natalí Alexandrino, pela organização e manutenção
do laboratório LIMI, facilitando o trabalho de todos os estudantes.
Às secretárias LIMI-LIP, Andressa e Elze, pela administração hábil.
Aos meus professores e colegas da pós- graduação, pelos conhecimentos trocados
durante as disciplinas.
Aos funcionários pertencentes aos setores de pós-graduação, biotério, biblioteca,
histotecnologia e esterilização no CPqGM.
Aos hamsters, esse modelo experimental, esses seres, essas vidas utilizadas nesta
pesquisa.
À Fonte financiadora CNPq e Fiocruz, pela viabilização da pesquisa.

Respirar é prova de existir e não de viver. Você respira com o corpo e vive com a alma.
Alejandro Bullón, Janelas para a Vida

SUMÁRIO
LISTA DE ABREVIATURAS ..............................................................................................10 LISTA DE FIGURAS .............................................................................................................12 LISTA DE QUADROS ...........................................................................................................14 RESUMO.................................................................................................................................15 ABSTRACT ............................................................................................................................16 1 INTRODUÇÃO ................................................................................................................. 17 1.1 Aspectos gerais da leishmaniose......................................................................................17
1.2 Estratégias de Controle para Leishmaniose Visceral ....................................................18
1.3 Resposta imune na Leishmaniose Visceral ....................................................................19 1.4 O hamster como modelo experimental na Leishmaniose Visceral...............................21 1.5 As limitações para a elaboração de uma vacina para LV .............................................23
1.6 Proteínas recombinantes como candidatas para vacinas de LV..................................24
1.7 Candidatos a vacina de DNA para estratégia prime-booster.........................................26
1.8 Os antígenos das histonas como candidatos vacinais para leishmaniose.....................28
1.9 Oligonucleotídeos de CpG como adjuvante de antígenos vacinais...............................30
2 JUSTIFICATIVA .................................................................................................................32
3 HIPÓTESE...........................................................................................................................33
4 OBJETIVOS........................................................................................................................33
4.1 Objetivo Geral...................................................................................................................33
4.2 Objetivos Específicos........................................................................................................33
5 DESENHO EXPERIMENTAL ..........................................................................................34
6 MATERIAIS E MÉTODOS ...............................................................................................36 6.1 População de estudo.........................................................................................................36

6.2 Plasmídeos que codificam para as proteínas de histonas nucleossomais e proteínas recombinantes das histonas de L. infantum.........................................................................36 6.3 Imunizações dos animais/Grupos....................................................................................36 6.3.1 Imunização Homóloga.....................................................................................................37 6.3.2 Imunização Heteróloga....................................................................................................37 6.4 ELISA para detecção de IgG anti-histonas....................................................................37 6.5 Parasitas.............................................................................................................................38 6.6 Separação das formas metacíclicas de L.infantum usando Gradiente Descontínuo de Ficoll........................................................................................................................................38 6.7 Flebótomos e preparação do sonicado de glândula salivar (SGC) ..............................40 6.8 Desafio dos animais...........................................................................................................40 6.9 Eutanásia dos animais......................................................................................................40
6.10 Avaliações histopatológicas............................................................................................41
6.11 Determinação da carga parasitária por diluição limitante ........................................41 6.12 Extração de RNA e preparo do cDNA para detecção de citocinas.............................42 6.13 Quantificação da Produção de Citocinas por PCR em Tempo Real.........................43 6.14 Análise estatística............................................................................................................44
7 RESULTADOS.....................................................................................................................45
7.1 Detecção de anticorpos da classe IgG anti-histonas, em soros de hamsters, pelo método de ELISA...................................................................................................................45
7.2 Avaliação da expressão de citocinas por PCR, em tempo real, no linfonodo drenante do local de imunização e baço, após a terceira imunização................................................46
7.2.1 Expressão de citocinas (TGF-ß, IFN-γ e IL-10) no linfonodo.........................................46
7.2.2 Expressão de citocinas (IFN-γ, TGF-ß e IL-10) no baço.................................................47
7.3 Razão da produção de citocinas IFN-γ/ IL-10 e IFN-γ /TGF-ß....................................47
7.4 Correlação linear entre a produção de anticorpos e a expressão de citocinas no linfonodo..................................................................................................................................49

7.5 Correlação da produção de anticorpos e citocinas no baço..........................................50
7.6 Análise da carga parasitária............................................................................................52
7.6.1 Análise da carga parasitária no linfonodo........................................................................52
7.6.2 Análise da carga parasitária no baço................................................................................53
7.6.3 Análise da carga parasitária no fígado.............................................................................54
7.7 Expressão de citocinas (TGF-ß, IFN-γγγγ E IL-10) no baço, 5 meses após o desafio......55
7.8 Razão da produção de citocinas IFN-γ/ IL-10 e IFN-γ /TGF-ß no baço, 5 meses após o desafio....................................................................................................................................57
7.9 Correlação da carga parasitária e produção de citocinas (IFN-γγγγ, TGF-β, IL-10) no baço, 5 meses após o desafio...................................................................................................57
7.10 Análise histológica de 5 e 7 meses após o desafio.........................................................60
7.10.1 Análise histológica do linfonodo...................................................................................61
7.10.2 Análise histológica do baço dos animais infectados......................................................62 7.10.3- Quantificações de folículos e centro germinativo........................................................63
7.10.4 Análise histológica do fígado dos animais infectados...................................................63
8 DISCUSSÃO.........................................................................................................................66
9 CONCLUSÃO......................................................................................................................75
REFERÊNCIAS......................................................................................................................76

LISTA DE ABREVIATURAS
______________________________________________________________________
APC Células Apresentadoras de Antígenos
BSA Do inglês “Bovine Serum Albumin”
cDNA Ácido Desoxirribonucléico Complementar
CpG ODN Do inglês “Cytosine-phosphate-Guanosine Oligodeoxynucleotides”
DNA Ácido Desoxirribonucléico
DTH Do inglês “Delayed Type Hipersensitivity”
ELISA Ensaio Imunoenzimático
gp63 Glicoproteína 63
H Histona
H&E Hematoxilina & Eosina
HIS Proteínas de Histonas
HIV/AIDS Vírus da Imunodeficiência Humana/Síndrome da Imunodeficiência Adquirida.
ID Intradérmica
IFN-γ Interferon gama Ig Imunoglobulina
IL Interleucina
iNOS Do inglês “Inducible Nitric Oxide Synthase”
KMP11 Proteína de membrana de cinetoplastídeos de 11 kilodáltons
LACK Receptor para cinase C ativada homóloga da Leishmania
LeIF Do inglês “Leishmania elongation and Initiation Factor”
LiESAp Do inglês Leishmania infantum Excreted Secreted Antigens proteins
LiP0 Proteína ácida ribossomal
LmSTI1 Do inglês “Leishmania. major stress inducible protein”
LPS Lipopolissacaridio
LV Leishmaniose Visceral

NK “Natural killer”
NO Óxido Nítrico
OD Densidade Ótica
OMS Organização Mundial de Saúde
PBS Do inglês “Phosphate Buffered Saline”
PCR Do inglês “Polymerase Chain Reaction”
PSA-2 Antígeno de superfície parasitária
RNA Ácido Ribonucléico
RNAm Ácido Ribonucléico mensageiro
RPM Rotações por minuto
SBF Soro Bovino Fetal
SGS Sonicado de glândula salivar
TCD4+ Co-receptor de linfócito T auxiliar
TCD8+ Co-receptor de linfócito T citolítico
TGF-β Fator de crescimento tumoral beta
Th Linfócito T auxiliary
Th1 “T helper 1”
Th2 “T helper 2”
TLR Do inglês “Toll-Like Receptors”
TNF-α Fator de necrose tumoral alfa
TSA Do inglês “thiol-specific antioxidant”

LISTA DE FIGURAS
______________________________________________________________________
Figura 1 – Análise de promastigotas de L.infantum estacionária, por gradiente de Ficoll. --39 Figura 2 – Avaliação da resposta imune humoral anti-histonas, através da quantificação de
anticorpos no soro por ELISA.-----------------------------------------------------------45 Figura 3 – Avaliação da expressão de citocinas no linfonodo, após imunizações por PCR em
tempo real.----------------------------------------------------------------------------------47 Figura 4 – Avaliação da expressão de citocinas no baço, após imunização por PCR em tempo
real.-------------------------------------------------------------------------------------------48 Figura 5 – Avaliação da razão das citocinas IFN-γ/IL-10 e IFN-γ/TGF-ß no linfonodo e baço,
após imunização por PCR em tempo real.----------------------------------------------49 Figura 6 – Regressão linear simples, relativa à correlação da produção de IgG anti-
histonas totais e citocinas IFN-γ, TGF-ß e IL-10 no linfonodo.---------------------51
Figura 7 – Regressão linear simples, relativa à correlação da produção de IgG anti- histonas totais e citocinas IFN-γ, TGF-ß e IL-10 no baço.--------------------------------------52
Figura 8 – Carga parasitária no linfonodo, após imunização e desafio.--------------------------54
Figura 8 – Carga parasitária no baço, após imunização e desafio.--------------------------------55
Figura 10 – Carga parasitária no fígado, após imunização e desafio.-----------------------------56
Figura 11 – Avaliação da expressão de citocinas no baço por PCR, em tempo real, cinco meses após o desafio.----------------------------------------------------------------------57
Figura 12 – Avaliação da razão das citocinas IFN-γ/IL-10 e IFN-γ/TGF-ß no baço, por PCR em tempo real, 5 meses após o desafio. ------------------------------------------------58
Figura 13 – Regressão linear simples relativa à correlação da produção de citocinas IFN-γ, TGF-ß e IL-10 e carga parasitária no baço, 5 meses após o desafio.---------------60
Figura 14A – Linfonodo do grupo salina, 5 meses após o desafio. Seta: folículo primário. Aumento de 10x, coloração com H&E.------------------------------------------------61.
Figura 14B – Linfonodo do grupo salina cinco meses após o desafio com folículo atrofiados sem centro germinativo e macrófagos com citoplasma amplo. Aumento de 40x, coloração com H&E.----------------------------------------------------------------------61
Figura 14C – Linfonodo com arquitetura preservada, folículo grande com centro germinativo, hiperplasia da área paracortical, infiltrado subcapsular. Grupo heterólogo 5 meses após o desafio. Aumento de 20x, coloração com H&E.------------------------------62

Figura 14D – Cordões medulares com plasmócitos, macrófagos de citoplasma amplo e eosinofílico. Grupo heterólogo 5 meses após o desafio. Aumento de 100x, coloração com H&E.----------------------------------------------------------------------62
Figura 15A – Baço com folículo e centro germinativo desenvolvidos. Grupo homólogo 5 meses após o desafio. Aumento de 20x, coloração com H&E.----------------------63
Figura 15B – Baço com centro germinativo evidente. Grupo homólogo 5 meses após o desafio. Aumento de 40x, coloração com H&E.---------------------------------------63
Figura 15C – Baço com folículo desorganizado, sem centro germinativo. Grupo salina 5 meses após o desafio. Aumento de 40x, coloração com H&E.----------------------63
Figura 15D – Esplenócito com inclusões de amastigotas no citoplasma. Grupo heterólogo7 meses após o desafio. Aumento de 100x, coloração com H&E.---------------------63
Figura 16– Contagem de número de folículos com centro germinativo no baço.---------------64
Figura 17A– Fígado com leucocitose intrasinusoidal. Grupo homólogo 7 meses após o desafio. Aumento de 40x, coloração com H&E.---------------------------------------66
Figura 17B – Fígado com granuloma e com inclusões de materiais amorfos no hepatócitos. Aumento de 100x, coloração com H&E.-----------------------------------------------66
Figura 17C – Mensuração das áreas lesionada do fígado, 7 meses após o desafio.-------------66

LISTA DE QUADROS
_____________________________________________________________________
Tabela 1. Descrição e identificação dos genes estudados, sequência dos primers utilizados e
características do produto de amplificação.

RESUMO
A leishmaniose visceral é uma doença infecciosa grave, causada por protozoários intracelulares obrigatórios do gênero Leishmania. Vários antígenos de Leishmania têm sido avaliados como candidatos vacinais, destacando-se as proteínas de histonas (HIS), antígenos altamente conservados. A exposição de HIS pela Leishmania induz uma resposta imune potente no hospedeiro vertebrado. Desse modo, neste estudo, avaliamos, em hamsters, a capacidade imunoprotetora dos antígenos de histonas contra a infecção por Leishmania infantum. Os animais foram vacinados com estratégia homóloga, utilizando-se plasmídeos de DNA que codificam para HIS (pcDNA3LiH2A-H3, pcDNA3LiH2B-H4) ou heteróloga (DNA/proteínas HIS) mais 1nM de CpG. Quinze dias após a última imunização, os animais foram desafiados pela via intradérmica com 105 Leishmania infantum metacíclicas mais 0,5 par de glândula salivar de Lutzomya longipalpis. Após a última imunização e durante a infecção, realizaram-se dosagens de citocinas por PCR em tempo real (linfonodo e baço), sorologia por ELISA (soro), carga parasitária por diluição limitante e análise histopatológica de tecidos (linfonodo, baço e fígado). Detectou-se produção de anticorpos IgG anti HIS nos grupos imunizados com a estratégia homóloga e heteróloga, quando comparados aos hamsters não imunizados. As imunizações homóloga e heteróloga diferiram na razão IFN-γ/IL-10 no linfonodo em relação ao grupo controle. Não houve diferença significativa na expressão dessas citocinas no baço após a imunização, entretanto, cinco meses após o desafio o grupo homólogo apresentou um aumento na produção de IL-10 nesse órgão. Na análise histopatológica do baço, verificou-se formação de mais folículos com centro germinativo, evidentes nos animais imunizados independentemente do grupo analisado. Observou-se, também, leucocitose intrasinusoidal e periportal no fígado, e folículos reativos no linfonodo. Nenhuma das estratégias de imunizações com antígenos de histonas acarretou em diminuição da carga parasitária no linfonodo, baço e fígado. As estratégias de imunização homóloga e heteróloga, com antígenos de histonas nucleossomais, não foram capazes de proteger contra infecção por L. infantum no modelo do hamster.
Palavras-chave: Leishmania; vacina; histonas; hamsters.

ABSTRACT
Visceral leishmaniasis is a serious infectious disease caused by obligatory intracellular protozoan of the Leishmania genus. Many antigens from Leishmania have been evaluated as vaccine candidates mainly the histones (HIS) that are highly conserved. Exposure to HIS from Leishmania elicits a strong host immune response in the vertebrate host. Threfore, in this study, we evaluated the immunoprotective ability of HIS antigens against infection by Leishmania infantum in hamsters. The animals were immunized with homologous strategy using plasmid of the DNA coding for HIS (pcDNA3LiH2A-H3, pcDNA3LiH2B-H4) or heterologous strategy using DNA and recombinant protein plus CpG. Fifteen days after last immunization, the animals were challenged by the intradermal route with 105Leishmania infantum metacyclics plus 0,5 pair of Lutzomya longipalpis salivary gland. After the last immunization and in the course of the infection, we determined cytokine production by Real time PCR (lymph node and spleen), serology by ELISA (sera), parasite burden was estimated by limiting dilution assay and histopathological analysis (lymph node, spleen and liver). Production of antibody anti HIS IgG was detected in the immunized groups treated with the homologous and heterologous strategies when compared to non-immunized hamsters. Homologous and heterologous immunizations altered the ratio of IFN-γ/IL-10 in the lymph node compared to the control group. There is not a significant difference in the expression of this cytokine in the spleen after the immunization, however, five months after the challenge the homologous group presented a higher expression of IL-10 in this organ. The histopathological analysis of the spleen showed the formation of high number of follicles with a germinative center in the immunized animals independently of the immunization employed. Intrasinusoidal and periportal leukocytosis were observed in the liver and reactive follicles in the lymph node. Both immunization strategies with histone antigen did not result in reduction of parasite burden in the lymph node, spleen and liver. Immunization with homologous and heterologous strategies using histone antigen were not able to confer protection in the hamster model against L. infantum infection.
Key words: Leishmania; Vaccine; Histone; hamsters.

18
1 INTRODUÇÃO
1.1 Aspectos gerais da leishmaniose
A leishmaniose é uma doença infecto-parasitária, considerada um grave
problema de saúde pública. Aproximadamente 350 milhões de pessoas estão expostas
ao risco de contrair essa infecção, principalmente os moradores de áreas tropicais e
subtropicais, de regiões da Ásia, Oriente Médio, África e América (CUNNINGHAM,
2002).
Amplamente distribuída no mundo, a leishmaniose visceral está entre as três
mais importantes enfermidades transmitidas por vetores e entre as seis endemias
consideradas prioritárias pela Organização Mundial de Saúde. A incidência em
humanos é de aproximadamente 500.000 novos casos e 59.000 mortes por ano
(DESJEUX et al., 2004,WHO, 2002). A leishmaniose visceral ocorre em cerca de 65
países. A maioria dos casos ocorre em zonas pobres, suburbanas e rurais dos países de
Bangladesh, Índia, Sudão e Brasil (DESJEUX et al., 1996).
Os parasitos protozoários do gênero Leishmania possuem ciclo digenético
(heteróxeno). São encontrados na forma livre flagelada, denominada promastigota,
presente no lúmen do trato digestivo do vetor e na forma amastigota, sem flagelo, no
interior de células do sistema fagocítico mononuclear do hospedeiro vertebrado
(LAINSON; SHAW, 1987).
A Leishmania infantum é transmitida ao vertebrado no momento em que o
flebótomo Lutzomya infectado faz seu repasto sanguíneo, injetando na pele as formas
promastigotas metaciclícas. Essas últimas são fagocitadas por macrófagos e se instalam
no interior de fagolisossomos, onde se diferenciam na forma amastigota, proliferando
por divisão binária. A Leishmania desenvolveu uma variedade de mecanismos de
adaptação para sobreviver dentro do macrófago e evadir da sua resposta imune (revisto
em ALMEIDA et al. 2003).
As espécies que podem causar a Leishmaniose Visceral, doravante LV,
pertencem ao subgênero Leishmania (L.), sendo L. donovani e L. infantum presentes no
Velho Mundo e L. chagasi no Novo Mundo. Embora diferentes no nome e origem

19
geográfica, estudos moleculares indicam que L. infantum e L. chagasi são da mesma
espécie (MAURÍCIO et al., 2000).
A LV é uma doença grave e pode ser fatal se não tratada, consistindo em uma
infecção generalizada que acomete o sistema reticulo-endotelial envolvendo baço,
fígado, medula óssea e linfonodo, podendo culminar em anemia, leucopenia,
trombocitopenia, caquexia e hepatoesplenomegalia. A forma e a gravidade da doença
dependem do estado imunológico, nutricional e idade do hospedeiro, dentre outros
fatores intrínsecos da espécie da Leishmania (BITTENCOURT; BARRAL-NETTO,
1995).
A desnutrição é apontada como fator de risco para LV, influenciando na
gravidade da doença e na resposta imune do individuo. No Brasil, observou-se que
crianças desnutridas têm nove vezes mais riscos de desenvolver a LV com toda
sintomatologia clássica (CREF et al, 1987). Nas últimas décadas, é visto o aumento no
número de casos de pacientes com leishmaniose visceral co-infectados com HIV/AIDS,
principalmente nos grupos de riscos (DESJEUX, 2001).
A ausência de planejamento urbano, em bairros periféricos, onde se concentram
pessoas de baixa renda, oriundas do massivo êxodo rural, associadas as precárias
condições sociais e econômicas aumentam a taxa de incidência da LV em centros
urbanos (CREF et al., 1987). De fato, atualmente, verificou-se a presença do vetor em
ambientes urbanos, tanto na periferia como nos grandes centros. Observa-se também a
adaptação do vetor em ambiente rural, no peridomicílio, em galinheiros, chiqueiros,
canis, paióis e, também, no intradomicílio (MINISTÉRIO DA SAÚDE, 2006).
1.2 Estratégias de Controle para Leishmaniose Visceral
As estratégias atuais, preconizadas pela Organização Mundial de Saúde (OMS),
como medidas de controle para a LV consistem em detecção e tratamento de casos
humanos, diagnóstico e eliminação dos cães infectados e controle do vetor.

20
A OMS recomenda diagnosticar pacientes suspeitos de LV utilizando testes de
ELISA, imunofluorescência, parasitológico de aspirado citológico de punções de baço,
medula e linfonodo. O tratamento dos pacientes positivos consiste na terapêutica com
drogas de primeira eleição, Antimoniato de Meglubina, Estibogluconato de Sódio e os
casos que não respondem aos antimoniais são tratados com os fármacos de segunda
linha, como Anfotericina B ou Pentamidina, os quais são mais tóxicos (WHO, 2002).
Tendo em vista que os cães são reservatórios de LV, eliminam-se os cães com
diagnóstico positivo, na tentativa de se controlar a incidência da doença em seres
humanos. No Brasil, 850.000 cães são selecionados e suas amostras são coletadas,
20.000 são eutanasiados anualmente, após diagnosticados positivos para LV. São
principalmente nas áreas suburbanas de cidades brasileiras, como Fortaleza, Natal,
Teresina, São Luis, Santarém, Belo Horizonte e Rio de Janeiro, onde são registrados o
maior número de cães positivos. Contudo, o impacto desta medida interventiva sobre o
controle da LV é passível de questionamento, pois não há comprovada correlação de tal
intervenção com o decréscimo nos números de incidência da doença humana (NUNES
et al., 1991).
Outra medida de controle preconizada é o uso de inseticidas residuais nos
domicílios e peridomicílios, com a finalidade de reduzir a população de flebotómos e
consequentemente a transmissão do parasita. Contudo, o efeito dessa medida tem se
mostrado temporário, devido principalmete a ocorrência de resistência ao inseticida ou
quando a aplicação é descontinuada. Entretanto, o prolongamento de tal estratégia, não
é desejavel, porque implica em consideráveis questões de desequilíbrio ambiental e
elevado custo financeiro de insumos e mão de obra (ROUSH, 1993).
1.3 Resposta imune na Leishmaniose Visceral
A imunidade anti-Leishmania é mediada tanto pela via inata (macrófagos e
neutrófilos), quanto pela via adaptativa (linfócitos B e T e células dendríticas). A
resolução da infecção na LV está associada à apresentação de antígenos de Leishmania
por macrófagos e células dendríticas (CD), além da ação de linfócitos TCD4 e CD8

21
(revisto em PETERS; SACKS, 2006). A dicotomia da resposta Th1/Th2 é um dogma
controverso da resposta imune na leishmaniose visceral, pois, existe um padrão misto de
resposta, com a participação de células Th1 e Th2, em seres humanos e modelos
experimentais. Alguns autores evidenciaram que é necessária a persistência de um
número reduzido de parasitas para manter a resposta imune duradoura e protetora
(ENGWERDA; ATO; KAYE, 2004).
Um das primeiras células a chegarem no sitio da inoculação da Leishmania são
as células NK, que promovem a maturação das células dendríticas, com consequente
fagocitose, apresentação de antígenos e a diferenciação de células T. As células NK
atuam na interação entre os componentes da resposta imune inata e adaptativa
(NYÉLEN et al.,2007). Em camundongos infectados com L.infantum, as células
dendríticas mielóides expressam TLR9 e produzem IL-12, levando à ativação das
células NK. Estas células secretam IFN-γ no paracórtex do linfonodo, recrutando células
TCD4+, necessárias para o controle da infecção (HAEBERLEIN et al., 2010). A
inibição da produção de IL-12 é uma tentativa de escape dos parasitas nesse processo,
pois esta citocina é essencial para a indução de IFN-γ e fundamental para a ação
leishmanicida do macrófago (AHUJA et al., 1999). Estudos evidenciam que pacientes
com LV crônica apresentam baixa freqüência de células NK, agravando o seu
prognóstico (NYÉLEN et al.,2007).
Além disso, citocinas anti-inflamatórias como a IL-10, atuam na supressão das
células efetoras Th1 e na inibição da síntese das citocinas IL-1β, IL-6, IL-8 e TNF-α.
Assim, ocorre redução na ativação de macrófagos, ocasionando uma infecção latente
(BOGDAN et al., 1991; BELKAID et al., 2001). Pacientes com LV, que apresentam
agravamento progressivo da doença, manifestam um aumento na expressão de mRNA
para IL-10 na medula e nos linfonodos (SAHA et al.,2007).
Observou-se que, em macrófagos humanos infectados com L.infantum,
L.amazonensis e L.braziliensis, ocorre uma resposta anti-inflamatória com ativa
produção de TGF-β. Um estudo in vitro apontou que concentrações fisiológicas de
TGF-β não inibiram a explosão respiratória induzida pela fagocitose, mas suprimiram a
expressão de iNOS, que tem ação crítica sobre a destruição das amastigotas (GANTT et
al., 2003).

22
Estudos ressaltaram que citocinas como, IL-13, IL-15, IL-17 e IL-22 estão
associadas à proteção contra infecção por L.donovani (BABALOO et al., 2001,
MILANNO et al.,2002, MURUGAIYAN et al.,2009, PITTA et al.,2009). Por outro
lado, a IL-27 produzida por macrófagos e células dendríticas na LV, induziu a produção
de IL-21 e influenciou a expansão de células T secretoras de IL-10 no baço,
favorecendo a multiplicação do parasita (ANSARI et al., 2011).
Observou-se, em um estudo com clones de células T oriundos de pacientes
humanos, apresentando LV ativa ou assintomáticos (sorologia positiva e teste de DTH
positivo), uma maior população de células CD8+ e quantidades reduzidas na população
de células CD4+. Uma situação oposta foi constatada em indivíduos resistentes a LV
(SAHA et al., 2006). A persistência dos parasitas nestes pacientes com infecções
crônicas está associada à inibição da fusão do lisossomo com o fagossomo em
macrófagos, como também à redução na produção de NO e ROS, influenciando o
recrutamento de células T reguladoras (T-regs) CD4+CD25+FoxP3+ (WILSON;
JERONIMO; PEARSON, 2005).
1.4 O hamster como modelo experimental na Leishmaniose Visceral
O hamster golden syrian (Mesocricetus auratus) tem sido amplamente utilizado
como modelo experimental para uma série de patologias humanas como sífilis,
micobactérias, micoses, arboviroses e leishmaniose visceral (BARBOSA JUNIOR et
al., 1987). Esses roedores também são utilizados para estudar diversas formas de
leishmaniose causadas por diferentes espécies de parasitas (CHILDS et al., 1984).
Distintas rotas de infecção também têm sido utilizadas, sendo as mais comuns a
intradérmica, a intracardíaca e a intraperitoneal (MATHIAS et al., 2001; SHARMA et
al., 2004).
Hamsters infectados com Leishmania (L. donovani ou L. infantum) apresentam
infecção progressiva no fígado, baço e medula óssea, levando-os ao desenvolvimento de
manifestações clínicas similares aquelas desenvolvidas pelo homem. Seres humanos,
cães e hamsters frequentemente exibem sinais clínicos e sintomas durante a LV,

23
diferente do camundongo que praticamente não apresenta sinal clínico, dependendo do
número de parasitas utilizados no inóculo (CARRION et al., 2006).
Dados da literatura mostram que uma porcentagem dos hamters é capaz de
resistir à infecção, por desenvolverem uma resposta imune protetora espontânea. Em um
trabalho, monitorado por um ano, os hamsters infectados por via intracardíaca, com 103
e 105 parasitas de L.infantum, desenvolveram sintomatologia clínica. Em ambos os
grupos, houve animais sintomáticos e assintomáticos. Os animais sintomáticos
apresentaram emaciação, desidratação, astenia, hepatoesplenomegalia e alta carga
parasitária no fígado e baço. Já os animais oligossintomáticos desenvolveram
esplenomegalia acompanhada de moderada carga parasitária em baço e fígado, sem
sinais clínicos; ao passo que nos animais assintomáticos não foram detectados parasitas
(REQUENA et al., 2000).
As características patológicas observadas durante o curso da LV em hamsters
incluem, desestruturação, depleção linfóide e hipoplasia de polpa branca em baço,
granulomas hepáticos e depósito secundário de substância amilóide em fígado e baço
(RICA-CAPELA et al., 2003). Muitos hamsters desenvolvem ascite grave antes de
sucumbirem e estudos histológicos têm revelado a presença de glomerulonefrite
mediada por imunocomplexos e amiloidose disseminada, que conduzem ao
desenvolvimento de síndrome nefrótica (SARTORI et al., 1991, FERNÁNDEZ et al.,
2013).
Outro estudo mostrou que a infecção de hamsters com 107 promastigotas de
L.infantum, pela via intracardíaca, levou ao desenvolvimento de lesões mucocutâneas
localizadas, acompanhadas de úlceras nas áreas próximas ao focinho (NIETO et al.,
2011). Foram observados diferentes alterações como anemia, leucopenia,
trombocitopenia, caquexia associada com o acúmulo de parasitas nas vísceras,
hipergamaglobulinemia, hepatoesplenomegalia (PEARSON et al., 1990; MELBY et al.,
2001).
Um trabalho avaliando a resposta imune de hamsters contra a LV indicou que
esplenócitos infectados com L. donovani expressam quantidade significante de citocinas
do tipo Th1, como IFN-γ, IL-2 e TNF-α. Embora possuindo baixa ou nenhuma

24
expressão de IL-4, quantidades substanciais de mRNA para TGF-β e IL-10 estão
presentes. Associado a isto, estudos com hamsters, verificaram que alguns animais
apresentam uma resposta imune do perfil Th1, e mesmo assim ocorre progressão da
doença, pois a atividade de mRNA para a síntese de iNOS é reduzida no fígado e baço
nesses animais, levando a um agravamento do quadro da LV (BELKAID et al., 2000,
MELBY et al., 2001). Assim, a incapacidade em controlar a infecção pode estar
relacionada à falta das funções efetoras dos macrófagos infectados (PEREZ, 2006).
A investigação com hamsters constitui um modelo apropriado para o estudo das
características patológicas manifestas da LV. Por serem susceptíveis, os hamsters, têm
sido utilizados em estudos de fármacos, vacinas e mecanismos de imunossupressão na
LV. Além disso, são animais de fácil manipulação, de manutenção prática e de baixo
custo, com possibilidade de obtenção em grande escala em curtos períodos de tempo
(BASU et al., 2005; GARG et al., 2006). Entretanto, a carência de muitos reagentes e
anticorpos ainda constitui um obstáculo, os quais são superados utilizando-se novas
abordagens moleculares para a avaliação imunológica desses animais (MELBY et al.,
2001).
1.5 As Limitações para a elaboração de uma vacina para LV
A Leishmania é um parasita eucarioto intracelular complexo, que possui
mecanismos muito elaborados para sobreviver e multiplicar-se dentro de células do
sistema mononuclear fagocítico. A maioria dos antígenos de Leishmania é reconhecida
pelo hospedeiro, pois ocorre a produção de anticorpo e/ou resposta imune celular.
Entretanto, a resposta a alguns antígenos não induz proteção, podendo inclusive
contribuir para exacerbação da resposta patológica, devido à reação cruzada com
moléculas do próprio hospedeiro (HANDMAN, 2001).
De acordo com a literatura, existem indivíduos com capacidades diferentes de
resposta frente ao mesmo antígeno de Leishmania, como é observado em indivíduos
baixo ou altos respondedor. Uma vacina de Leishmania para ser utilizada em seres
humanos, requer diferentes antígenos (polivalentes), capazes de induzir uma resposta

25
protetora para a maior parte da população e que seja produzida em larga escala. O
desenvolvimento de uma vacina parece ser o mais efetivo e factível enfoque profilático
como método de controle para a LV (LARA et al., 1991).
Atualmente, os ensaios clínicos com candidatos a vacinas de Leishmania, são
classificados em vacinas de primeira geração (antígeno total do parasita morto), vacinas
de segunda geração (Leishmania vivas atenuadas, vírus ou bactéria com inserção de
genes expressando antígenos, frações nativas ou recombinantes de antígenos da
Leishmania) e, por fim, vacinas de terceira geração (vacinas de DNA/Gênicas).
1.6 Proteínas recombinantes como candidatas para vacinas de LV
Vacinas de segunda geração, que utilizam proteínas recombinantes, têm sido
testadas desde 1990. A administração dessas proteínas tem sido avaliada em
combinação, ou como poliproteínas ou quiméricas, sendo que a maioria deve ser
associada a adjuvantes para induzir maior proteção. Geralmente, a imunização com
proteínas induz apenas uma fraca resposta de células T. Por outro lado, a adição de
adjuvantes com propriedades agonistas para os receptores de reconhecimento de
padrões (PRRs), induz uma resposta mais intensa e duradoura contra patógenos
intracelulares como a Leishmania (GURUNATHAN et al., 1998).
Uma das primeiras proteínas testadas contra leishmaniose foi a gp63, um
antígeno conservado em todas as espécies de Leishmania. Um estudo recente,
imunizando camundongos BALB/c com antígeno rgp63 mais adjuvante CpG, conferiu
proteção quando desafiados com L.donovani (MAZUNDER et al.,2012). Tewary e
colaboradores (2005) identificaram um gene ORFF (do inglês, open reading frame),
localizado, no locus LD1 do cromossomo 35 de amastigotas de L.donovani, que
expressa uma proteína nuclear ORFF. A imunização apenas com a proteína rORFF no
modelo murino, protegeu parcialmente (45-60%) contra a infecção por L.infantum.
Um estudo com um antígeno de membrana da promastigota de L.donovani
(LAg) encapsulado em lipossomas catiônicos, induziu níveis significantes de proteção
contra LV ao imunizar camundongos e hamsters diminuído respectivamente em 83% e

26
54% a carga parasitária no baço (AFRIN et al., 2000). A proteína ácida ribossomal
purificada de L. infantum (LiP0) é um componente estrutural da subunidade maior do
ribossomo e é um antígeno imunodominante capaz de induzir a produção de anticorpos
em pacientes e cães infectados com L. infantum (IBORRA et al., 2005). Do mesmo
modo, foi demonstrado, em estudos utilizando-se a região C-terminal de LiP0
administrada com BCG como adjuvante, que houve proteção parcial dos cães,
diminuído o número de parasitos em 42% contra infecção por L. infantum (MOLANO
et al., 2003). Essa vacina é um potente imunógeno em coelhos e camundongos
(DANTAS-TORRES, 2006).
A utilização da vacina Leish-111f resultante da fusão de diferentes proteínas
como os antígenos TSA (proteína oxidante tiol específico), LmSTI1 (proteína de
estresse induzível da L. major) e LeIF (fator de iniciação de Leishmania), formulados
com o adjuvante MPL-SE (Lipídio A, do LPS atenuado) protegeu camundongos e
hamsters contra LV, levando a diminuição da carga parasitária em 99,6%. Essa mesma
formulação foi testada em cães expostos à infecção natural por L.infantum e observou-
se uma resposta imunogênica, entretanto esta não foi eficaz na prevenção da infecção,
contudo tal vacina embora tenha aumentado a taxa de sobrevivência dos cães (COLER
et al., 2002; SKEIKY et al., 2002; COLER et al., 2007).
Dentre outros antígenos candidatos vacinais para LV, destaca-se a proteína de
superfície rHASPB1, que conferiu proteção em camundongos contra um desafio
experimental com L. donovani, sem uso de adjuvante (STAGER et al., 2000). Outra
proteína candidata vacinal promissora, é o antígeno LeIF (fator de iniciação de
eucarioto) expresso em promastigotas e amastigotas das diferentes espécies de
Leishmania. Um estudo com a proteína recombinante LeIF de L.infantum, conferiu nos
hamsters proteção de 65% contra a LV, devido ao aumento da transcrição de mRNA
para síntese de NO e diminuição nas concentrações das citocinas IL-4, IL-10 e TFG-β
(KUSHAWAHA et al., 2011). Em outro trabalho esta mesma proteína, induziu uma
resposta de perfil Th1 mediada por IL-12, quando células mononucleares de sangue
periférico de pacientes com LV foram estimuladas pela LeiF. Além disso, também
observou-se que linfócitos de camundongos BALB/c imunizados quando estimulados

27
pela rLeIF apresentavam alta expressão de IL-12, e uma baixa produção de IL-4
(BARHOUMI et al., 2006).
1.7- Candidatos a vacinas de DNA para leishmaniose e estratégia prime-boost
Nos últimos anos, as vacinas gênicas ou vacinas de DNA têm representado uma
nova estratégia para o controle de doenças infecciosas, pois essas vacinas induzem
respostas imunes protetoras para o hospedeiro, principalmente contra microrganismos
intracelulares (PLOTKIN, 2005; PALATNIK-DE-SOUSA, 2008). São várias as
vantagens da utilização de vacinas de DNA, incluindo sua construção e a produção em
larga escala, além da possibilidade de manipulação, que permite a expressão de
múltiplos antígenos (GARMORY et al., 2005).
O gene que codifica a proteína é clonado em um vetor de expressão de bactérias
e o DNA é injetado diretamente na pele ou no músculo do hospedeiro (ULMER et al.,
1996). O DNA plasmidial é absorvido pela célula e translocado até o núcleo, onde é
transcrito e, posteriormente, traduzido no citoplasma. A eficiência da captação e da
expressão do DNA plasmidial é baixa, mas é suficiente para induzir uma resposta imune
celular e humoral (AZEVEDO et al., 1999).
Atualmente, muitos antígenos na forma de vacina gênica têm sido testados como
candidatos promissores para utilização no controle e prevenção da leishmaniose,
incluindo antígenos parasitários, de vetores e/ou adjuvantes (OLIVEIRA et al., 1999).
Entre os antígenos de Leishmania que exibiram atividade protetora quando
administrados como vacinas de DNA, encontram-se diferentes proteínas de membrana e
intracelulares tais como, histonas, proteínas de choque térmico e ribossomais
(KHAMESIPOUR et al., 2006; DUMONTEIL, 2007).
As vacinas de DNA codificam antígenos específicos com capacidade de
ativarem as células T essenciais para a proteção contra microrganismos intracelulares
(GURUNATHAN, et al., 2000). Contudo, quando administradas exclusivamente, as
proteínas possuem limitação para manutenção de uma resposta imune intensa e durável.
Um enfoque visando uma estratégia de imunização mais elaborada, com permanência

28
do antígeno no organismo por mais tempo, encontra-se no princípio do prime-booster.
Este princípio consiste na exposição de um mesmo antígeno de diferentes formas para o
sistema imune, durante a dose inicial e a de reforço. O objetivo deste protocolo é que a
dose inicial induza a resposta imune com o padrão requerido para conferir proteção e
que a dose de reforço mantenha e expanda tal resposta. Este tipo de imunização tem
sido usada experimentalmente contra doenças como malária, tuberculose, influenza e
HIV (WANG et al., 2001).
O uso das vacinas de DNA é a metodologia mais atual na busca por proteção
contra leishmaniose visceral. RAMIRO e colaboradores (2003) obtiveram proteção em
60% dos animais contra a infecção por L. infantum, utilizando um protocolo de
vacinação heteróloga de prime-booster, no qual cães foram vacinados inicialmente com
DNA codificando a proteína LACK de L. infantum e, posteriormente, por vírus
recombinante de Vaccínia, contendo o gene da proteína. Um regime de vacinação
semelhante também induziu proteção em cães com duas doses da vacina de DNA, que
codifica cisteína-proteases do tipo I e II do parasita, e um reforço comas proteínas
recombinantes. Observou-se reação positiva ao teste de DTH e ausência de parasitas na
medula óssea 12 meses após desafio (RAFATI et al., 2005).
Um outro estudo comparou o potencial do antígeno gp63 de Leishmania nas
estratégias de vacinação homóloga (DNA/DNA ou Proteína/ Proteína e adjuvante CpG)
ou heteróloga (DNA/proteína) em camundongos BALB/c contra a infecção por
L.infantum. Foi observado controle da carga parasitária nos animais imunizados com as
estratégias heterólogas, com elevada produção de IFN-γ, IL-12, óxido nítrico e redução
de IL-4 e IL-10 (MAZUMDER et al., 2011).
Outra pesquisa mostrou o potencial da imunização com a estratégia heteróloga
em hamsters. O conjunto dos antígenos KMP-11 (proteína de membrana do
cinetoplasto), TRYP (Peroxidase Triparadoxina), PAPLE22 (Proteína de Agravamento
Potencial de L.infantum) e LACK induziu produção significante de óxido nítrico em
macrófagos de hamsters. Além disso, foi observada redução na carga parasitária do baço
e no sangue, quando comparado ao grupo imunizado com estratégia homóloga
(DNA/DNA e Proteína/ Proteína) (TODOLÍ et al., 2012).

29
1.8 Os antígenos das histonas como candidatos vacinais para leishmaniose
As histonas nucleossomais constituem proteínas estruturais com papel
importante na organização e regulação dos genes. Localizam-se no núcleo das células
eucarióticas e existem quatro classes principais de histonas (H2A, H2B, H3 e H4) de
Leishmania que formam a unidade básica da cromatina, o nucleossomo (REQUENA et
al., 2000). As histonas de Leishmania destacam-se como antígenos protetores quando
administradas em vacinas de DNA, em camundongos Balb/c, induzindo uma resposta
imune do perfil Th1 (KHAMESIPOUR et al., 2006).
As histonas são antígenos altamente conservados, produzidos por várias espécies
de Leishmania, que não são secretados, entretanto, são capazes de induzir uma intensa
resposta imune (SANTAREM et al., 2007). Essas proteínas são liberadas durante a
infecção, após a destruição das amastigotas intracelulares pelo macrófago ativado ou
pela citólise espontânea de amastigotas dentro da célula infectada. Além disso, são
capazes de modular a resposta imune do hospedeiro, devido ao fato de não sofrerem
pressão seletiva pela resposta imune, ao contrário das proteínas de superfície e das
proteínas secretadas pela Leishmania (CHANG et al., 2003).
Um estudo recente mostrou que peptídeos das histonas de seres humanos podem
atuar como microbicidas contra bactérias, fungos e promastigotas de Leishmania. Em
um estudo, utilizando diferentes concentrações das histonas H2A e H2B no tratamento
in vitro de promastigotas de L.major, L.braziliensis, e L.mexicana, foi observada
diminuição da infectividade dos macrófagos de camundongo por estes parasitas tratados
(WANG et al., 2011).
As histonas de Leishmania também podem ser reconhecidas por outras espécies
de hospedeiros vertebrados, além do camundongo. Ao utilizar soros de cães infectados
com L.infantum, observou-se que as histonas H2A (72%) e H3 (68%) são mais
imunogênicas, pois induziram maior produção de anticorpos quando comparadas às
histonas H2B (60%) e H4 (44%). Além disso, investigações com proteínas de histonas
de Leishmania, demonstraram que as proteínas H2A e H2B são capazes de levar à
produção de IFN-γ, quando utilizadas na re-estimulação de células mononucleares

30
provenientes de pacientes com leishmaniose cutânea, citocina esta que é indispensável
para que ocorra a morte das amastigotas (DE CARVALHO et al., 2003).
Ainda vale mencionar que um estudo experimental, com uma vacina de DNA
polivalente composta por 10 antígenos diferentes (H1, H2A, H2B, H3, H4, p36, PSA-2,
peroxidoxin/TSA, STI1 e ARP-1) de isolados de L. donovani, resultou na indução de
uma resposta imune celular tipo Th1 em cães. Esses animais apresentaram evidente
linfo-proliferação antígeno-induzida e síntese de IFN-γ, entretanto, sem ocorrer
produção de IL-4. Além disso, houve resposta do tipo DTH contra amastigota de L.
donovani, redução da carga parasitária em monócitos in vitro e nos linfonodos drenantes
destes animais (SALDARIGA et al., 2006).
O efeito protetor das histonas também foi demonstrado quando camundongos
BALB/c foram imunizados com células dendríticas pulsadas e associadas aos antígenos
histonas, 50% dos animais imunizados quando desafiados com L.infantum, reduziu a
carga parasitária no baço para 55% quando comparados ao grupo controle
(BERBERICH et al., 2003). Iborra e colaboradores, em 2004, também mostraram no
modelo BALB/c que ao utilizar imunização com vacinas gênicica codificando para os
antígenos de histonas induziu redução no tamanho da lesão após desafio com L.major
(IBORRA et al., 2004).
Outra estratégia de imunização pela via endovenosa e intradémica em
camundongos BALB/c, utilizando células dendríticas pulsadas e proteínas de histonas e
CpG, reduziu em 50% a carga parasitária no baço e fígado quatro semanas após a
infecção dos animais por L.infantum (CARRIÓN et al., 2008). Esse mesmo grupo
demonstrou que a adição de anticorpo monoclonal anti-CD8 em cultura de esplenócitos,
diminui a produção de IFN-γ, apenas nos camundongos imunizados com DNA
codificantes para H2A e H3. Por outro lado não houve diminuição significativa na
produção desta citocina nos esplenócitos dos animais imunizados com H2B e H4,
sugerindo que CD8 também pode participar na proteção conferida por estas proteínas,
pela produção de IFN-γ (CARRIÓN et al., 2008).
Um estudo recente do nosso grupo imunizando camundongos BALB/c com os
antígenos de histonas nucleossomais (H2A, H2B, H3, H4) de L.infantum em estratégias

31
homóloga (DNA/DNA) e heteróloga (DNA/proteína) com CpG mostrou que ambas
abordagens levaram à inibição do desenvolvimento da lesão, controle da carga
parasitária na orelha e linfonodo dos animais infectados com L. braziliensis. Observou-
se também ausência de macrófagos infectados em cortes histológicos no local do
desafio (CARNEIRO et al., 2012).
1.9 Oligonucleotídeos de CpG como adjuvante de antígenos vacinais
O CpG ODN (do inglês Cytosine-phosphorothioate-
guanineoligodeoxynucleotides) é uma sequência padrão encontrada no DNA de
bactérias que possuem a capacidade de ativar células Natural Killer (NK)a secretar IFN-
γ (KLINMAN et al., 1994). O CpG é reconhecido pelas células do sistema imune via
Toll-like 9 (TLR9), presente no endossoma de células B e células dendríticas
plasmocitóides. A ativação desta via desencadeia a expressão de genes relacionados à
produção de IL-1, IL-6, IL-12, IL-18 e TNF-α, citocinas que caracterizam o perfil de
resposta Th1(KRIEG et al., 1995).
Estudos recentes evidenciam a importância da associação de adjuvantes aos
antígenos vacinais, devido a sua capacidade de atuar nos receptores do sistema imune
inato (SALEN et al., 2005). Dados da literatura mostram que CpG ODN têm sido
extensivamente utilizados como adjuvante e imunoterápico, pois acelera e estimula a
resposta vacinal. Foram observados bons resultados na ativação da resposta imune em
animais neonatos ou imunocomprometidos, após o protocolo terapêutico com CpG
ODN (KRISHNAMACHARI; SALEM, 2009).
Os experimentos de Sane e colaboradores (2010), com hamsters e camundongos,
demonstraram a capacidade imunomoduladora do CpG. Essa estratégia de tratamento
dos animais induziu a produção de óxido nítrico e citocinas do perfil Th1, quando
associado ao fármaco Miltefosina no tratamento da LV.
Do mesmo modo, diferentes abordagens experimentais têm sido utilizadas com
CpG ODN, co-administrado com antígenos vacinais contra antraz, brucelose, influenza
vírus, vírus da coriomeningite linfocítica, varíola, sarampo, antígenos de superfície da

32
hepatite C e toxina tetânica (OXENIUS et al.,1999). Essas diferentes abordagens
ressaltam que a imunização com CpG ODN induz uma intensa resposta de padrão Th1,
com maturação e ativação de células dendríticas, acompanhada de níveis elevados de
IgG2a (HARN, 2005).
Além disso, o CpG-ODN possui a capacidade de induzir uma resposta imune
celular e humoral de longa duração, quando adicionados à vacina de DNA ou a
estratégia prime-booster. De fato, uma vez que sequências de DNA podem ser
reconhecidas por receptores do tipo Toll, elas possuem um papel importante na ativação
de células da imunidade inata (BABUIK et al., 2004). Além disso, ocorre ativação das
células apresentadoras de antígeno, com produção de IL-12, uma citocina fundamental
na diferenciação de células Th1(BASTOS et al., 2004).
Em um estudo realizado por Mendez e colaboradores, em 2003, camundongos
receberam baixas doses de promastigotas de L.major associado a CpG-ODN. Foi
observada uma resposta intensa de células TCD4+ e TCD8+ com produção de IFN-γ,
levando à redução na carga parasitária. Adicionalmente, outro estudo com camundongos
C57BL/6 demonstrou que, após imunização com LiP0-DNA de L.infantum associado ao
CpG, não houve desenvolvimento de lesão e ocorreu redução da carga parasitária nos
animais desafiados com L.major na orelha. Nesse mesmo estudo, observou-se que a
imunização de camundongos BALB/c com rLiP0 e CpG também induziu proteção
parcial quando desafiados com L. major (IBORRA et al., 2005). Todos estes trabalhos
evidenciam a capacidade imunoduladora do adjuvante CpG, quando associado a
antígenos vacinais.

33
2 JUSTIFICATIVA
As espécies de Leishmania que infectam o homem estão presentes em quase
todos os continentes e são endêmicas em 65 países. A incidência anual estimada de LV
é de, aproximadamente, 500.000 casos (WHO, 2002). Atualmente, temos um quadro
limitado e oneroso de fármacos utilizados no tratamento dos pacientes com
leishmaniose, de efeitos colaterais adversos. A atual estratégia de controle da LV,
preconizada pela Organização Mundial de Saúde, está baseada na detecção e eliminação
de cães infectados, associado ao tratamento dos casos humanos e ao controle vetorial
(TESH,1995). Contudo, tais medidas não têm sido suficientes para diminuir a incidência
da doença. A viabilidade de uma vacina eficaz para animais e seres humanos seria um
método profilático de controle desta endemia. Por isso, a utilização de vacinas, em todo
o mundo, foi proposta como medida profilática em Saúde Pública para o controle de
várias doenças em distintas situações epidemiológicas.
É sabido da necessidade da elaboração de uma vacina para LV, tanto para seres
humanos quanto para animais. Com esse propósito, no presente estudo, foi avaliado se
as vacinas de DNA que codificam histonas nucleossomais de Leishmania infantum
(H2A, H2B, H3 e H4) ou seguido de reforço com as proteínas recombinantes (rH2A,
rH2B, rLH3 e rH4) em combinação com o adjuvante CpG-ODN protegem hamsters da
infecção por L. infantum. Esses animais após as imunizações foram infectados por via
intradérmica com Leishmania infantum na presença da saliva de Lutzomyia longipalpis;
um modelo experimental de LV descrito anteriormente pelo nosso grupo (GOMES et
al., 2008). Foram avaliadas a carga parasitária no baço, fígado e linfonodos, bem como
a resposta imune proveniente da imunização e infecção dos animais.
Sabe-se que ainda inexistem vacinas eficazes capazes de controlar a LV em cães
e humanos e o desenvolvimento das mesmas tem exigido um esforço por parte de
diversos grupos de pesquisa em diferentes partes do mundo. Neste contexto, uma das
possibilidades que se abrem estar relacionada com o desenvolvimento de uma vacina
com os antígenos nucleossomais de Leishmania, os quais indicam novas perspectivas na
profilaxia da LV, em animais e no homem.

34
3 HIPÓTESE
A imunização de hamsters com os antígenos de histonas nucleossomais protege
estes animais de uma subsequente infecção por Leishmania infantum.
4 OBJETIVOS
4.1 Objetivo Geral
Avaliar se a imunização com plasmídeos de DNA que codificam histonas
nucleossomais de Leishmania (H2A, H2B, H3 e H4) de forma isolada ou em
combinação com reforço das proteínas recombinantes (rH2A, rH2B, rH3 e rH4) com
CpG induz resposta protetora contra a infecção por Leishmania infantum em hamsters.
4.2 Objetivos Específicos
• Avaliar a resposta imune induzida em hamsters após imunização com
plasmídeos de DNA que codificam histonas nucleossomais de Leishmania
(H2A, H2B, H3 e H4) ou em combinação com reforço constituído pelas
proteínas recombinantes associadas ao CpG.
• Avaliar se os candidatos vacinais utilizados na estratégia homóloga e
heteróloga são capazes de reduzir a carga parasitária em pelo menos 80%, contra
a infecção por L.infantum em relação aos controles não imunizados.

35
5. Desenho experimental
Grupo Homólogo 3 doses com 50µg de cada
pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4
Grupo Heteróloga 2 doses com 50µg de cada
pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4 + 1 dose de 5 µg cada proteína H2A,H3,H2B, H4 + 1nM CpG)
Grupo Plasmídeo vazio 3 doses de 100µg de pcDNA3+1nM CpG
Grupo Salina
Avaliação da Imunogenicidade
Quantificação das citocinas por Real Time PCR : IFN-γ, IL-10, TGF-β
- Linfonodo e baço dos animais apenas imunizados -Baço dos animais 5 meses após o desafio
Elisa:
IgG total anti-histonas
15 hamsters Golden syrius/ Grupo
1º imunização
15 dias
2º imunização 3º imunização
15 dias 15 dias
Eutanásia
15 dias após á ultima imunização

36
Avaliação da Capacidade Protetora
Quantificação da carga parasitária, por
LDA no: linfonodo, baço e fígado
Grupo Homólogo 3 doses com 50µg de cada
pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4
Grupo Heteróloga 2 doses com 50µg de cada
pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4 + 1 dose de 5 µg cada proteína H2A,H3,H2B, H4 + 1nM CpG)
Grupo Plasmídeo vazio 3 doses de 100µg de pcDNA3+1nM
CpG
1º imunização
15 dias
2º imunização 3º imunização
15 dias
Desafio105 de L.infantum +
1/2 glândula de L.longipalpis pela via ID
15 hamsters Golden syrius/Grupo
Grupo Salina
Avaliação histopatológica:
linfonodo, baço e fígado
15 dias 2, 5 e 7 meses 15 dias
Eutanásia

37
6 MATERIAIS E MÉTODOS
6.1 População de estudo
Hamsters (Mesocricetus auratus) da raça Golden Syrius, machos, com idade
entre 2 e 4 meses foram obtidos do biotério do Centro de Pesquisas Gonçalo Moniz
(CPqGM– FIOCRUZ). Os procedimentos experimentais foram avaliados e aprovados
pela Comissão de Ética no Uso de Animais (CEUA) do CPqGM, protocolado com a
numeração L-IGM-005/2011, em 05-05-2011.
6.2 Plasmídeos que codificam para as proteínas de histonas nucleossomais e
proteínas recombinantes das histonas de L. infantum
As proteínas recombinantes das histonas de Leishmania infantum (rH2A, rH2B,
rH3 e rH4), bem como os plasmídeos contendo os insertos que codificam os genes das
histonas de Leishmania infantum (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) e o
plasmídeo vazio (pcDNA3) foram gentilmente cedidos pelo Dr. Manoel Soto, da
Universidad Autonoma de Madrid, Espanha.
As bactérias Escherichia coli competentes foram transformadas com os
plasmídeos e do sedimento do cultivo bacteriano a purificação destes foi realizada
utilizando-se um kit de isolamento de plasmídeo “Giga Prep Endofree” (Qiagen,
Alemanha), de acordo com as recomendações do fabricante.
6.3 Imunizações dos animais/Grupos
Os ensaios foram realizados em hamsters divididos em 4 grupos
experimentais(Homólogo, Heterólogo, Plasmídeo vazio e Salina). Vale ressaltar que
todos os ensaios foram repetidos 3 vezes. Realizou-se dos ensaios para as dosagens das
citocinas após imunização e um outro grupo de animais foram imunizados e desafiados
por L.infantum, totalizando 15 animais por grupo em cada experimento realizado.

38
6.3.1 Imunização Homóloga
Os animais foram imunizados três vezes com os plasmídeos de DNA com
intervalos de quinze dias entre as doses. A imunização com DNA foi realizada com
injeção intramuscular na pata traseira direita, nos músculos tibiais anteriores, contendo
100µg de um coquetel de plasmídeos recombinantes (50ug de cada tipo de plasmídeo
recombinante (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) diluídos em 50uL de salina.
6.3.2 Imunização heteróloga
Na estratégia heteróloga (DNA/proteína), os animais receberam duas
imunizações na perna direita nos músculos tibiais anteriores, com os plasmídeos
recombinantes seguidas de uma imunização contendo 20µg de proteínas recombinantes
de histonas (5µg de cada um dos tipos H2A, H2B, H3 e H4), associado com 1 nM de
CpG ODN 1826 (18-24 pb - 5´TCC ATG ACG TTC CTG ACG TT-3´ mol wt
6364,1G/mol) (SANE et al, 2010), a dose de reforço ( Prime-booster) feita por via
intradérmica na orelha direita, diluídos em 30uL. Cada dose foi administrada com um
intervalo de quinze dias. O grupo controle recebeu 30uL de salina nas mesmas
condições.
Para estas imunizações, foram utilizadas seringas de insulina BD ULTRAFINE
(BD Pharmingen, EUA) e agulha hipodérmica de 30G (271/2G). Cada grupo era
composto de 15 animais.
6.4 ELISA para detecção de IgG anti-histonas
Os títulos de anticorpos foram mensurados antes da primeira imunização e 15
dias após a terceira imunização, nos animais imunizados com plasmídeos recombinantes
(pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4 ou pcDNA3), tanto na estratégia homóloga
como na heteróloga.
Para a realização do ELISA, para detectarmos anticorpos anti-histonas, as placas
de 96 poços foram sensibilizadas com 1µg/mL de cada histona recombinante (rH2A;
rH2B, rH3 e rH4) em tampão carbonato-bicarbonato pH 9.6, 100µL/poço, incubadas
durante à noite a 4ºC. Foram realizadas três lavagens, com 250µl/poço, de PBS Tween

39
20 0,05% e os sítios inespecíficos foram bloqueados, utilizando-se 200µl/poço, de PBS
Tween BSA, contendo 1% de BSA por 2h a 37ºC. Foram realizadas mais quatro
lavagens com PBS Tween 0,05% (250µl/poço) e adicionou-se 100µL/poço dos soros
diluídos 1/100µL em PBS Tween 0,05% e incubou-se por 1h a 37ºC. Após 3 lavagens
com PBS Tween 0,05% (250µl/poço), adicionou-se 100µl/poço do anticorpo anti-
imunoglobulina G (IgG) total de hamster diluído a 1/100 em PBS Tween 0,05% e as
placas foram incubadas por 1h a 37ºC. Em seguida, os poços das placas foram lavados 3
vezes com PBS Tween0,05% e incubadas com 100µl/poço do substrato p-nitrofenil
fosfato na concentração de 1mg/mL dissolvido em tampão carbonato-bicarbonato mais
0.2mM de cloreto de magnésio pH 9.6 por 15 minutos à temperatura ambiente. A reação
de cor foi interrompida após adição de 50µl/poço de 3M NaOH. A leitura dos poços das
placas foi realizada em um espectrofotômetro leitor de ELISA em comprimento de onda
de 450nm. Os resultados foram expressos em Densidade Ótica (OD). O ponto de corte
foi calculado a partir de soros de hamster negativos, calculando-se a média da OD mais
duas vezes o desvio padrão.
6.5 Parasitas
Promastigostas de L.infantum (MCAN/BR/00/BA262) foram mantidas em meio
de cultura Schneider (LGC, São Paulo, BRASIL) suplementado com 10% de soro
bovino fetal inativado (SBF), 100 UI/ml de penicilina, 100 µg/ml de estreptomicina e
2mM de L-glutamina a 25ºC (todos da Gibco, Carlsbad, CA, USA), denominado
Schneider completo (SACKS & MELBY, 2001). Quando os parasitas atingiram a fase
estacionária, promastigotas foram coletadas e separadas por gradiente descontinuo de
Ficoll 400, para obtenção de uma população rica em promastigotas metacíclicas (YAO
et al., 2008).
6.6 Separação das formas metacíclicas de L.infantum usando Gradiente
Descontínuo de Ficoll
Promastigotas de L. infantum em fase estacionária foram lavadas em meio
DMEM 1% e contadas em câmara de Neubauer. Em um tubo de 15 ml foi preparado o
gradiente de Ficoll 400, constituído de 2mL de Ficoll a 40% diluído em PBS, e sobre
este adicionou-se 2 ml de Ficoll a 10% diluído em Meio 199, utilizando-se seringas de 5

40
ml e agulhas 40x12. Sobre o gradiente, foram colocadas 109promastigotas ressuspensas
em 2mL de DMEM 1% e submetidas à centrifugação a 365g, a temperatura ambiente
por 10 minutos. As formas metacíclicas que se acumularam na fase superior do Ficoll
10% foram lavadas a 1700g por 5 minutos a 4o C. A amostra foi analisada por
citometria de fluxo no aparelho de FACSCAN para confirmar que a população
encontrava-se enriquecida de formas metacíclicas. Após a separação, analisou-se os
parâmetros de tamanho (SSC) e granulosidade (FSC) (SARAIVA et al., 2005).
Observou-se que o purificado possuía uma maior quantidade de formas de
L.infantum metacíclicas que apresentavam um maior tamanho (SSC) e menor
granulosidade (FSC), parâmetros resultantes das características de um flagelo alongado,
corpo estreito, curto e delgado, morfologia presente na fase metacícilica das
Leishmanias (Figura 1). O dotplot representa a análise das subpopulações de parasitas,
correspondendo ao tamanho e granulosidade das células, cuja aquisição foi de 10.000
eventos.
Figura 1 – Análise de promastigotas de L.infantum na estacionária, por gradiente de
Ficoll.

41
6.7 Flebótomos e preparação dos sonicados de glândula salivar (SGS)
Os flebótomos da espécie Lutzomyia longipalpis (Diptera: Psychodidae) (local
de coleta: Corte de Pedra e Cavunge, Bahia) foram mantidos em nosso insetário em
condições controladas, a 26ºC e 80% de umidade relativa.
O sonicado de glândula salivar (SGS) foi obtido por dissecação de glândulas
salivares de flebótomos Lutzomyia longipalpis, 5 a 7 dias após eclosão dos ovos. Após a
dissecação, as glândulas foram re-suspensas em solução salina e estocadas a –70ºC até o
momento do uso. Imediatamente antes de sua utilização, as glândulas foram sonicadas
por 40 ciclos em aparelho ultrasonicador Branson Sonifier 450 (Connecticut, USA),
centrifugadas a 8.160g/ 4ºC por cinco minutos em centrifuga Eppendorf Centrifuge
5415C, (Brinkman Instruments Inc., New York, USA) e os sobrenadantes foram
utilizados nos experimentos.
6.8 Desafio dos animais
Duas semanas após a última imunização, os animais foram desafiados por via
intradérmica na orelha direita, com 1x105 promastigotas enriquecidas de metacíclicas de
L. infantum, separadas por gradiente descontinuo de Ficoll400 e ressuspensas em salina,
mais o equivalente a 0.5 par de sonicado de glândula salivar (SGS) de Lutzomyia
longipalpis mimetizando a infecção natural.
6.9 Eutanásia dos animais
Todos os animais foram eutanasiados na câmera de CO2, seguido de
deslocamento cervical. Para avaliação da resposta imune induzida pela imunização
homóloga ou heteróloga, mensurou-se a produção de anticorpos anti-proteína de
histonas nucleossomais, bem como a dosagem de citocinas, 15 dias após cada
imunização e posteriormente ao desafio, obtendo-se para este fim o soro, baço e
linfonodo drenante auricular do local da imunização. Com relação aos grupos que foram
imunizados e posteriormente desafiados, os tempos de sacrifícios aconteceram nos dias
60, 150 e 210 após o desafio.

42
6.10 Avaliações histopatológicas
Os fragmentos de tecidos (fígado, baço e linfonodo) foram coletados após 5 e 7
meses após o desafio dos animais. Os fragmentos foram fixados em formalina ácida a
10% por 24 horas e processados pela técnica histológica de rotina para inclusão em
parafina no setor de histotecnologia, do Centro de Pesquisa Gonçalo Moniz. Realizou-se
microtomia com obtenção de cortes com 4-5 micrômetros de espessura que foram
corados pela Hematoxilina-Eosina (H&E), montados com auxílio de lamínula e
Bálsamo do Canadá sintético. Os cortes histológicos foram posteriormente avaliados à
luz da microscopia ótica quanto à presença de alterações relevantes no quadro de LV.
6.11 Determinação da carga parasitária por diluição limitante
Após a eutanásia dos animais, foram retirados o baço, fígado e linfonodo
auricular. O baço e o fígado foram pesados e retirou-se um fragmento de 0,035 a 0,045g
de cada. As amostras foram posteriormente maceradas com 1mL de meio Schneider
(Sigma, St Louis, MO, USA) em peneira ultrafina e homogeneizadas, adicionando-se
mais 4ml de meio Schneider (Sigma, St Louis, MO, USA), suplementado com soro
bovino fetal a 10%, 2 mM de L-glutamina, 100 UI/mL de penicilina e 100U/mL de
estreptomicina (Gibco, Carlsbad, CA, USA). Ao macerado do linfonodo foi adicionado
2mL de solução salina para lavagem das células (300g por 10 minutos a 4ºC), que, em
seguida, foram ressuspensas em 1mL de meio completo para contagem de 107 células.
Com os homogeneizados foram realizadas diluições seriadas que variaram de 1:5 à
1:1000 em placas de 96 poços contendo meio sólido NNN ágar sangue (Novy-
MacNeall-Nicolle) (NICOLLE, 1909). Seis replicações foram feitas para cada diluição.
As placas foram analisadas em microscópio invertido para verificação do aparecimento
de formas promastigotas de L.infantum a partir do terceiro até o décimo dia de cultura.
Neste período, as placas foram incubadas numa estufa BOD a 23oC. A carga parasitária
do baço, fígado e linfonodos foi analisada, utilizando-se a técnica de diluição limitante
descrita previamente por TITUS e colaboradores (1991) e a determinação da carga
parasitária baseada na diluição limitante foi estimada utilizando-se o programa ELIDA
(LIMA et al., 1997).

43
6.12 Extração de RNA e preparo do cDNA para detecção de citocinas
O perfil das citocinas (TGF-ß, IFN-γ e IL-10), produzidas pelas células do
linfonodo e baço dos hamsters imunizados com plasmídeos de DNA que codificam para
diferentes antígenos de histonas nucleossomais de Leishmania (H2A, H2B, H3 e H4)
e/ou em combinação com um reforço constituído pelas proteínas recombinantes
associadas ao adjuvante CpG, foi avaliado por PCR em tempo real. Assim, 15 dias após
a última imunização, os animais foram eutanasiados, e o baço, o linfonodo drenante
auricular (imunização heteróloga) e poplíteo (imunização homóloga), e cinco meses
após o desafio o baço também foram coletados para extração de RNA e posterior síntese
do cDNA.
O RNA total foi extraído utilizando-se o reagente Trizol (Invitrogen, Carlsbad,
CA, USA), de acordo com as recomendações do fabricante. O linfonodo drenante foi
macerado e centrifugado a 300g por 10 minutos a 4ºC. Em seguida, as células foram
homogeneizadas em 250µL de Trizol e para isolar-se o RNA, adicionou-se 500µL de
clorofórmio e precipitou-se na presença de isopropanol. Após a precipitação foi
realizado lavagem com etanol a 70%, o RNA foi dissolvido em 10 µL de água tratada
com dietilpirocarbonato. A concentração e a razão de pureza da extração foram
determinadas após a leitura em espectofotômetro, utilizando-se comprimentos de ondas
de 260 e 280nm. Após quantificação, o material foi mantido a -70ºC. A síntese de
cDNA foi realizada acrescentando-se 500ng de RNA a 5µL de uma mistura contendo
oligonucleotídeos (dt) 12-18, 2,5 µM, dNTPs, 1mM (Invitrogen, Carlsbad, CA, USA),
tampão 1X (Tris-HCl 20mM, pH 8,4, KCl 50 mM, MgCl2 2mM), 20U de inibidor de
ribonuclease e 50U da transcriptase reversa Superscript II (Gibco, Carlsbad, CA, USA).
A reação foi incubada no termociclador a uma temperatura de 42ºC durante 50 minutos,
depois desta incubação a enzima foi desnaturada a temperatura de 95ºC durante 5
minutos. Em seguida as amostras de cDNA foram armazenadas a -20°C até o momento
do uso.

44
6.13 Quantificação da Produção de Citocinas por PCR em Tempo Real
Os primers (oligonucleotídeos ou iniciadores) utilizados nas reações de
amplificação foram desenhados pelo programa Primer Express (Applied Biosystems,
EUA). O tamanho dos primers variou de 18 a 23 bases, a temperatura de anelamento
(Tm, do inglês melting temperature) variou de 59º-61ºC, o conteúdo de GC foi de 40-
60% e foram utilizados na concentração de 250nM. O tamanho dos produtos de
amplificação gerados variou de 90 a 110pb. As reações de PCR em tempo real foram
realizadas em placas de 96 poços, usando o reagente SYBR-Green PCR Master MIX
(Applied Biosystems, Foster City, CA, USA) e o equipamento “Perkin-Elmer ABI
Prism 7500 Sequence Detection System”. A reação foi realizada em 40 ciclos com a
ciclagem de acordo com o manual do fabricante ABI PRISM 7500. A determinação da
intensidade de fluorescência na reação foi feita pelo cálculo do ∆Rn (∆Rn=Rn+ - Rn-),
onde Rn+ = intensidade de emissão do SYBR-Green / intensidade de emissão do ROX
em um dado momento da reação, e Rn- = intensidade de emissão do SYBR-Green/
intensidade de emissão do ROX, antes da amplificação. O composto ROX é utilizado
como controle interno passivo, pois a fluorescência que emite possui intensidade
constante durante toda a reação, enquanto que a fluorescência emitida pelo SYBR-
Green aumenta à medida que este se liga às duplas fitas de DNA. Durante os ciclos
iniciais da reação, não há acúmulo de produtos de amplificação e os valores de ∆Rn
permanecem na linha de base (fluorescência do ROX >SYBR-Green). O cálculo da
quantificação relativa foi feito pelo método de 2-∆∆Ct, onde ∆Ct = Ct gene alvo – Ct
gene referência, e ∆∆Ct = ∆Ct amostra – ∆Ct controle.
A especificidade dos primers foi avaliada pela curva de dissociação. A curva de
dissociação consiste na monitorização da fluorescência das amostras em relação ao
aumento de temperatura. A fluorescência das amostras decresce com o aumento da
temperatura, pois à medida que as pontes de hidrogênio, que mantém as duplas fitas
unidas se rompem ocorre a liberação do SYBR-Green. Assim, quando observamos
somente um pico de fluorescência em uma dada temperatura significa que houve
amplificação de um produto específico. Esta temperatura é a temperatura de anelamento
ou melting point (Tm) do produto de amplificação (amplicon). A Tabela 1 mostra a
sequência e as características dos primers utilizados.

45
Tabela 1 Descrição e identificação dos genes estudados, sequência dos primers
utilizados e características do produto de amplificação.
6.14 Análise estatística
A análise estatística foi realizada utilizando-se o programa GraphPad-Prism 5.0
(GraphPad Software, USA).
Os dados de correlação foram tratados, com o coeficiente de correlação para
postos de Spearman (rs). O cálculo (rs) baseia-se nas diferenças entre os postos (valores
das variáveis analisadas) de x e y, resultando em um coeficiente de variação entre -1
(correlação perfeita negativa) e +1 (correlação perfeita positiva) passando pelo valor
zero (ausência de correlação).
Todos os dados não paramétricos foram apresentados como média e desvio
padrão. As diferenças observadas entre os grupos foram analisados pelo teste de
Kruskall Wallis (one-way ANOVA), seguido pelo pós teste de Dunns. As diferenças
estatísticas foram consideradas significantes quando p<0.05 utilizando um intervalo de
confiança de 95%.

46
7 RESULTADOS
7.1 Detecção de anticorpos da classe IgG anti-histonas, em soros de hamsters, pelo
método de ELISA
Com a finalidade de avaliar a imunogenicidade das histonas nucleossomais de
L.infantum, coletou-se soros dos animais imunizados e controles, antes das imunizações
e, 15 dias após a terceira imunização, para a detecção de anticorpos específicos contra
tais proteínas.
Constatou-se que apenas alguns dos hamsters, pertencentes ao grupo que
recebeu a estratégia heteróloga (9/19) e homóloga (7/19), apresentaram respectivamente
maior produção de anticorpos IgG total anti-histonas (p<0,0079) e (p<0,0075),
diferença que foi estatisticamente significante quando comparada a produção do grupo
controle que recebeu apenas salina, evidenciando a capacidade imunogênica deste
veículo na indução da resposta imune (Fig. 2).
Figura 2 – Avaliação da resposta imune humoral anti-histonas pela quantificação de anticorpos no soro por ELISA. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeo de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular (n=19). Na estratégia heteróloga (■) receberam duas doses de 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG (n=19). Os grupos controles plasmídeo vazio (o) receberam três doses de 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG (n=12) ou 30µL
de salina (•) pela via intramuscular (n=12). Os soros dos animais imunizados, e seus respectivos controles, foram coletados quinze dias, após a terceira imunização, para a detecção de anticorpos específicos contra as histonas nucleossomais. A linha pontilhada refere-se ao ponto de corte (0.023365) obtido com soros de hamsters saudáveis não infectados. A análise estatística foi feita pelo teste Kruskall-Wallis com pós-teste de Dunns. (**, p<0,001).

47
7.2 Avaliação da expressão de citocinas por PCR em tempo real no linfonodo drenante do local de imunização e no baço, após a terceira imunização
7.2.1 Expressão de citocinas (TGF-ß, IFN-γ e IL-10) no linfonodo
Alguns hamsters pertencentes ao grupo imunizado com a estratégia homóloga
apresentaram uma resposta imune local, em relação ao grupo controle salina, com tais
porcentagens de animais respondendo para as citocinas IFN-γ (44%), TGF-ß (30,7%), e
IL-10 (60%). Detectou-se o grupo imunizado com estratégia heteróloga, pois
apresentaram uma leve diminuição com diferença estatística na quantificação relativa
para expressões dos genes de IFN-γ (p<0,0026) TGF-β (p<0,0417), e IL-10(p<0,0277),
em relação ao grupo homólogo (Fig. 3A, B, C).
No grupo controle, que recebeu o plasmídeo vazio (pcDNA3), observou-se que
os animais (63%) apresentaram alta expressão do gene para citocina IL-10, quando
comparado com o grupo heterólogo (Fig. 3C). Além disso não houve diferença
significativa na estratégia heteróloga, e o grupo controle (pcDNA3), em relação às
citocinas IFN-γ e TGF-β, pois poucos animais(23%) expressaram o gene desta citocina
(Fig. 3A,B).
Figura 3 – Avaliação da expressão de citocinas no linfonodo após imunizações por PCR em tempo real. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (o) receberam três doses de 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG e grupo salina recebeu 30µL de salina pela via intramuscular. Os animais foram eutanasiados 15 dias após a última imunização e o linfonodo drenante foi coletado para extração de RNA. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa do gene (QR), empregando-se como controle a expressão dos genes dos hamsters não imunizados do grupo salina (representado pela linha pontilhada). Os valores obtidos são referentes a quantidade de vezes que aumentou a expressão destes genes nos animais imunizados em relação a média do grupo salina. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=10-13. *p<0,05, **p<0,01.

48
7.2.2 Expressão de citocinas (IFN-γ, TGF-ß e IL-10) no baço
Após a última imunização, a produção das citocinas no baço foi avaliada e
observou-se que os hamsters do grupo homólogo (40%), expressaram concentrações
superiores de IFN-γ e IL-10, quando comparado ao grupo controle plasmídeo vazio e
heterólogo. Entretanto, não houve diferença estatística (Fig. 4 A,C). Ao analisar a
produção da citocina TGF-ß, observou-se que os animais do grupo homólogo (55%) e
grupo heterólogo (50%) apresentaram baixas concentrações desta citocina, em relação a
grupo controle plasmídeo vazio, embora, a diferença não tenha sido estatisticamente
significante (Fig. 4B).
Figura 4 – Avaliação da expressão de citocinas no baço após imunização por PCR em tempo real. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses de 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG. Os grupos controles plasmídeo vazio (o) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3)+1nM CpG e grupo salina recebeu 30µL de salina pela via intramuscular. Os animais foram eutanasiados 15 dias após a última imunização e o baço foi coletado para extração de RNA. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa do gene (QR), empregando-se como controle a expressão dos genes dos hamsters não imunizados do grupo salina (representado pela linha pontilhada). Os valores obtidos são referentes a quantidade de vezes que aumentou a expressão destes genes nos animais imunizados em relação a média do grupo salina. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=9.
7.3 Razão da produção de citocinas IFN-γ/ IL-10 e IFN-γ /TGF-ß.
A partir da análise dos resultados da quantificação das citocinas, após a 3ª
imunização, realizou-se o cálculo da razão da produção de citocina pró-inflamatória,
IFN-γ, com as citocinas anti-inflamatórias TGF-ß e IL-10 no baço e linfonodo, com os
valores individuais de cada hamster.
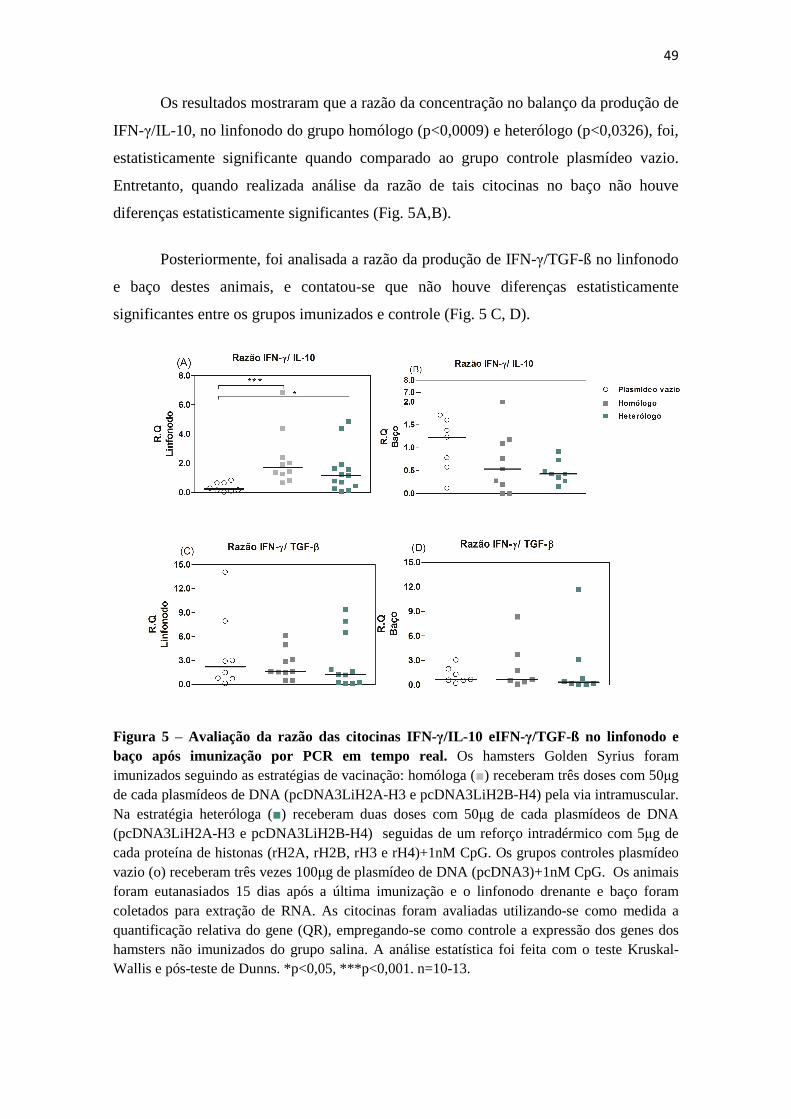
49
Os resultados mostraram que a razão da concentração no balanço da produção de
IFN-γ/IL-10, no linfonodo do grupo homólogo (p<0,0009) e heterólogo (p<0,0326), foi,
estatisticamente significante quando comparado ao grupo controle plasmídeo vazio.
Entretanto, quando realizada análise da razão de tais citocinas no baço não houve
diferenças estatisticamente significantes (Fig. 5A,B).
Posteriormente, foi analisada a razão da produção de IFN-γ/TGF-ß no linfonodo
e baço destes animais, e contatou-se que não houve diferenças estatisticamente
significantes entre os grupos imunizados e controle (Fig. 5 C, D).
Figura 5 – Avaliação da razão das citocinas IFN-γ/IL-10 eIFN-γ/TGF-ß no linfonodo e baço após imunização por PCR em tempo real. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (o) receberam três vezes 100µg de plasmídeo de DNA (pcDNA3)+1nM CpG. Os animais foram eutanasiados 15 dias após a última imunização e o linfonodo drenante e baço foram coletados para extração de RNA. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa do gene (QR), empregando-se como controle a expressão dos genes dos hamsters não imunizados do grupo salina. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. *p<0,05, ***p<0,001. n=10-13.

50
7.4 Correlação linear entre a produção de anticorpos e a expressão de citocinas no
linfonodo
Conforme os resultados anteriormente descritos, observou-se que os hamsters
imunizados com antígenos de histonas nucleossomais de Leishmania, na estratégia
homóloga e heteróloga, produziram títulos de anticorpos, IgG anti-histonas. Detectou-
se, também, maior produção de citocinas no linfonodo do grupo homólogo (IFN-γ, IL-
10, TGF-β), quando comparado ao heterólogo.
Assim, foi realizada a análise de relação causa-efeito, entre uma possível
dependência das variáveis quantitativas, produção de citocinas no linfonodo e títulos de
anticorpos dos animais, que encontravam-se acima do ponto de corte do ensaio de
ELISA. Construiu-se um diagrama de dispersão, em um modelo de regressão linear
simples buscando avaliar a intensidade da correlação, seguidos do teste do coeficiente
de correlação para postos de Spearman (rs).
O grupo homólogo mostrou uma correlação negativa, nas variáveis de produção
de anticorpos e citocinas, IFN-γ (rs=-0,1429)/(p=0,7825) (Fig 6A), porém, não
significativa. Neste mesmo grupo, observou-se a existência de uma correlação positiva
estatisticamente significante, quanto maior os títulos de anticorpos anti-histonas, maior
a concentração de TGF-β (rs=0,8929)/(*p=0.0123) (Fig. 6B). A correlação com a
citocina IL-10 mostrou que alguns dos animais que produziram IgG anti- histonas,
apresentaram maior expressão do gene IL-10 (rs=0,4643)/(p=0,3024), contudo, esta
correlação positiva não foi significante (Fig. 6C).
As correlações das variáveis para os postos, quando verificadas no grupo
heterólogo, evidenciaram uma correlação negativa que não foi estatisticamente
significante, sugerindo que quanto maior é produção de anticorpos menor é a produção
das citocinas IFN-γ (rs=-0,1916)/(p=0,6646) e TGF-β (rs=-0,1757)/(p=0,6436) no
linfonodo. Neste mesmo grupo, verificou-se correlação positiva para citocina IL-10
(rs=0,08368)/(p=0,8432) (Fig. 6D,E,F).Todavia não observou-se diferenças
estatísticamente significantes em nenhuma das correlações.
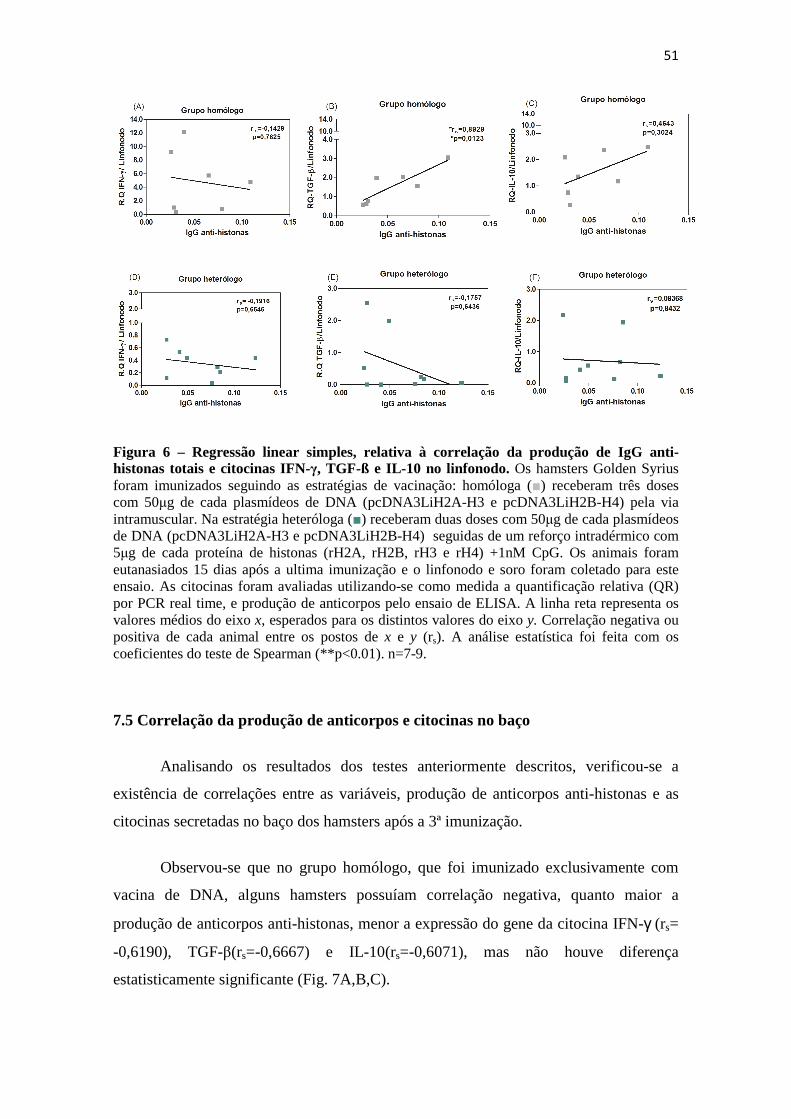
51
Figura 6 – Regressão linear simples, relativa à correlação da produção de IgG anti-histonas totais e citocinas IFN-γ, TGF-ß e IL-10 no linfonodo. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG. Os animais foram eutanasiados 15 dias após a ultima imunização e o linfonodo e soro foram coletado para este ensaio. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa (QR) por PCR real time, e produção de anticorpos pelo ensaio de ELISA. A linha reta representa os valores médios do eixo x, esperados para os distintos valores do eixo y. Correlação negativa ou positiva de cada animal entre os postos de x e y (rs). A análise estatística foi feita com os coeficientes do teste de Spearman (**p<0.01). n=7-9.
7.5 Correlação da produção de anticorpos e citocinas no baço
Analisando os resultados dos testes anteriormente descritos, verificou-se a
existência de correlações entre as variáveis, produção de anticorpos anti-histonas e as
citocinas secretadas no baço dos hamsters após a 3ª imunização.
Observou-se que no grupo homólogo, que foi imunizado exclusivamente com
vacina de DNA, alguns hamsters possuíam correlação negativa, quanto maior a
produção de anticorpos anti-histonas, menor a expressão do gene da citocina IFN-γ (rs=
-0,6190), TGF-β(rs=-0,6667) e IL-10(rs=-0,6071), mas não houve diferença
estatisticamente significante (Fig. 7A,B,C).

52
Assim, ao verificar-se essas mesmas variáveis no grupo heterólogo, constatou-se
que a secreção de todas as citocinas IFN-γ, TGF-β e IL-10 apresentaram o coeficiente
de Spearman correlacionado negativamente com a produção de anticorpos anti-histonas
(rs=-0,7186), (rs=-0,07186) e (rs=-0,2727) respectivamente, sem diferenças
estatísticamente significantes (Fig.7D,E, F).
Figura 7– Regressão linear simples, relativa à correlação da produção de IgG anti-
histonas totais e citocinas IFN-γ, TGF-ß e IL-10 no baço. Os hamsters Golden Syrius foram
imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg
de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular.
Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA
(pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de
cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os animais foram eutanasiados
15 dias após a ultima imunização, o baço e o soro foram coletados para este ensaio. As citocinas
foram avaliadas utilizando-se como medida a quantificação relativa (QR) por PCR real time, e
produção de anticorpos pelo ensaio de ELISA. A linha reta representa os valores médios do eixo
x, esperados para os distintos valores do eixo y. Correlação negativa ou positiva de cada animal
entre os postos de x e y (rs). A análise estatística foi feita com os coeficientes do teste e
Spearman. (**p<0.01). n=7-9.

53
7.6 Análise da carga parasitária
7.6.1 Análise da carga parasitária no linfonodo
Com o objetivo de verificar a capacidade protetora da imunização dos animais
com as histonas nucleossomais, tanto na abordagem homóloga como heteróloga, grupos
de hamsters foram imunizados e posteriormente desafiados com L. infantum mais SGS.
A carga parasitária presente nos linfonodos auriculares direitos (linfonodos drenantes),
foi verificada uma vez que, a via do desafio foi a injeção intradérmica na orelha direita.
A partir da análise dos resultados, foi possível observar que, 2 meses após o desafio, o
grupo dos animais imunizados com a estratégia heteróloga possuía uma carga
parasitária menor (104), quando comparado ao grupo homólogo (107), mas esta
diferença não foi estatisticamente significativa.
Após 5 meses de infecção, o número de parasitas no linfonodo no grupo de
animais imunizados com a estratégia homóloga igualou-se ao do grupo heterólogo. Aos
7 meses, todos os grupos apresentaram aumento progressivo e não diferiram entre os
animais imunizados e controles (Fig. 8A). Quando avaliamos a carga parasitária,
calculada pela comparação da área sob a curva resultante não observamos diferença
significativa (Fig. 8B).
Desse modo, temos que: as linhas do gráfico A representam a média mais o
desvio padrão da carga parasitárias; o gráfico B representa a área sob a curva da carga
parasitária. Vale ressaltar que os resultados foram obtidos de três experimentos
diferentes totalizando 15 hamsters por grupo a cada tempo de observação.

54
Figura 8– Carga parasitária no linfonodo após imunização e desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (□) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3) +1nM
CpG e grupo salina(▆) recebeu 30µL de salina pela via intramuscular. Os animais foram infectados com 105 de L. infantum metacíclicas pela via intradérmica, quinze dias após a terceira imunização. Nos tempos de 2, 5 e 7 meses, os animais foram eutanasiados e o linfonodo drenante coletado para quantificação da carga parasitária por diluição limitante. As linhas do gráfico A representam a média somada ao desvio padrão da carga parasitária. O gráfico B representa a área sob a curva da carga parasitária. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=15.
7.6.2 Análise da carga parasitária no baço
Na análise da carga parasitária, aos 2 meses, nos baços dos hamsters, não houve
redução de carga parasitária entre os grupos imunizados na estratégia homóloga e
heteróloga, quando comparado aos grupos controles. Surpreendentemente, aos 5 meses,
o grupo controle que recebeu a salina, apresentou a menor carga parasitária (106) em
relação aos demais grupos vacinais (1010). Porém, essa diferença não mostrou ser
significativa. Entretanto, no decorrer da infecção, no tempo de 7 meses, percebeu-se que
a carga parasitária dos grupos controles e vacinais ficou estabilizada, em torno de 1012
parasitas (Fig. 9A).
Quando avaliamos a carga parasitária, calculada pela comparação da área sob a
curva resultante, observamos que a imunização não resultou em diferenças

55
significativas (Fig. 9B). Com isso, constatamos que não houve diferenças estatísticas
entre os grupos imunizados e os grupos controles.
Figura 9 – Carga parasitária no baço após imunização e desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (o) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG e grupo salina (•) recebeu 30µL de salina pela via intramuscular. Os animais foram infectados com 105 de L. infantum metacíclicas pela via intradérmica, quinze dias após a terceira imunização. Nos tempos de 2, 5 e 7 meses, os animais foram eutanasiados e o baço coletado para quantificação da carga parasitária por diluição limitante. As linhas do gráfico A representam a média somada ao desvio padrão da carga parasitária. O gráfico B representa a área sob a curva da carga parasitária A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=15.
7.6.3 Análise da carga parasitária no fígado
Concomitantemente, avaliou-se após 2 meses do desafio, a carga parasitária no
fígado, observou-se que o grupo imunizado com a estratégia homóloga, não apresentava
parasitas, mas essa diferença não foi estatisticamente significativa. No entanto, aos 5
meses, o número de parasitas do grupo homólogo apresentou um aumento da carga,que
atingiu valores em torno de 106, semelhante aos apresentados pelos demais grupos
controle e imunizado (Fig. 10A). Aos 7 meses pós infecção, ocorreu aumento
progressivo, duplicando-se a carga parasitária no fígado em todos os grupos desafiados
(1015).

56
Quando foi realizado o cruzamento de dados e calculado a Área sob a Curva, foi
observado que o grupo heterólogo se destaca, apresentando uma carga parasitária de
1013 parasitas, equivalente a uma diferença de três logaritmos menor que os demais
grupos, mas essa diferença não foi estatisticamente significante (Fig. 10B).
Figura 10 – Carga parasitária no fígado após imunização e desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (□) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG e grupo salina (■) recebeu 30µL de salina pela via intramuscular. Os animais foram infectados com 105 de L. infantum metacíclicas pela via intradérmica, quinze dias após a terceira imunização. Nos tempos de 2, 5 e 7 meses, os animais foram eutanasiados e o fígado coletado para quantificação da carga parasitária por diluição limitante. As linhas do gráfico A representam a média somadas desvio padrão da carga parasitária. O gráfico B representa a área sob a curva da carga parasitária A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=15.
7.7 Expressão de citocinas (TGF-ß, IFN-γγγγ E IL-10) no baço 5 meses após o desafio
Foi verificada a quantificação das expressões das citocinas no baço dos animais
imunizados e infectados há 5 meses e observou-se que alguns animais imunizados, com
a estratégias homóloga (63%) e heteróloga (61%), apresentaram produção um pouco
mais elevada de IFN-γ, resultados semelhantes foram visto para citocina TGF-ß, no
grupo homólogo (50%) e heterólogo (61%), contudo não houve diferenças estatísticas
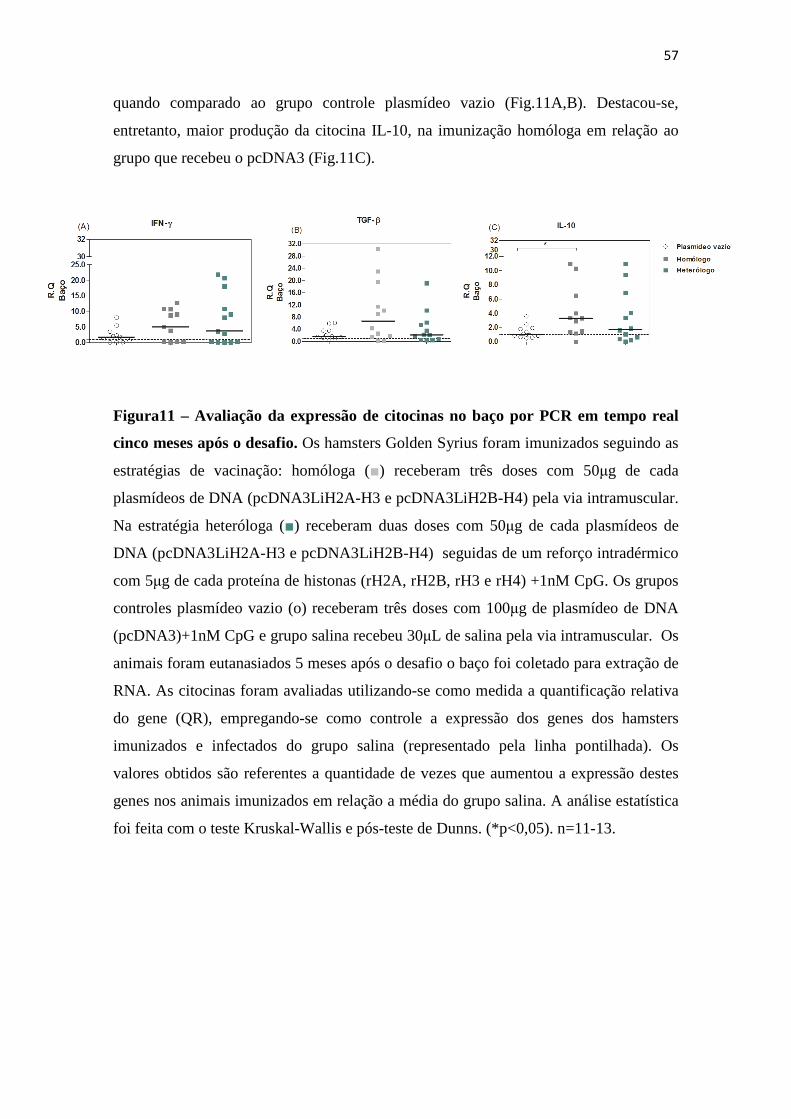
57
quando comparado ao grupo controle plasmídeo vazio (Fig.11A,B). Destacou-se,
entretanto, maior produção da citocina IL-10, na imunização homóloga em relação ao
grupo que recebeu o pcDNA3 (Fig.11C).
Figura11 – Avaliação da expressão de citocinas no baço por PCR em tempo real
cinco meses após o desafio. Os hamsters Golden Syrius foram imunizados seguindo as
estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada
plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular.
Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de
DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico
com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG. Os grupos
controles plasmídeo vazio (o) receberam três doses com 100µg de plasmídeo de DNA
(pcDNA3)+1nM CpG e grupo salina recebeu 30µL de salina pela via intramuscular. Os
animais foram eutanasiados 5 meses após o desafio o baço foi coletado para extração de
RNA. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa
do gene (QR), empregando-se como controle a expressão dos genes dos hamsters
imunizados e infectados do grupo salina (representado pela linha pontilhada). Os
valores obtidos são referentes a quantidade de vezes que aumentou a expressão destes
genes nos animais imunizados em relação a média do grupo salina. A análise estatística
foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. (*p<0,05). n=11-13.

58
7.8 Razão da produção de citocinas IFN-γ/ IL-10 e IFN-γ /TGF-ß no baço 5 meses
após o desafio
Com o objetivo de se avaliar a diferença entre a secreção das citocinas pró-
inflamatórias e anti-inflamatórias, no baço dos animais infectados após 5 meses, foi
realizada a razão de IFN-γ, com IL-10 e TGF- ß, e não foi constatada nos grupos
imunizados e controles nenhuma diferença entre as razões (Fig.12A, B).
Figura 12– Avaliação da razão das citocinas IFN-γ/IL-10 e IFN-γ/TGF-ß baço por PCR em tempo real cinco meses após o desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) receberam três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4)+1nM CpG. Os grupos controles plasmídeo vazio (o) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG. Os animais foram eutanasiados cinco meses após o desafio e coletou-se o baço para extração de RNA. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa do gene (QR), empregando-se como controle a expressão dos genes dos hamsters não imunizados do grupo salina. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=10-13.
7.9 Correlação da carga parasitária e produção de citocinas (IFN-γγγγ, TGF-β, IL-10)
no baço cinco meses após o desafio
Após o período de 5 meses de infecção, verificou-se a possibilidade de
correlação entre as diferentes variáveis analisadas, carga parasitária e produção de
citocinas secretadas no baço dos hamsters.

59
Observou-se que os animais do grupo controle que receberam o plasmídeo
vazio, juntamente com o grupo heterólogo apresentaram respectivamente uma
correlação positiva (rs=0,2392)/(p=0,5135), (rs=0,2802)/(p=0,3777) entre a carga
parasitária e a produção de IFN-γ (Fig.13A,B), porém sem significância estatística.
Quando analisado estes parâmetros no grupo que recebeu apenas a vacina gênica,
verificou-se um correlação positiva (rs=0,5397)/(p=0,0876), pois a maioria dos animais
apresentaram maior produção de IFN-γ e maior carga parasitária (Fig.13B), mas
novamente sem significância estatística.
Ao analisar a variável carga parasitária e a expressão do gene da citocina TGF-β,
o grupo que recebeu o plasmídeo vazio e os grupos vacinais, que receberam a estratégia
heteróloga e homóloga, apresentaram correlação positiva irrelevante para a carga
parasitária, não havendo diferença estatística (Fig.13.D,E,F).
Dando continuidade à análise dos experimentos, constatou-se que o grupo
controle teve a menor carga parasitária, com diferentes concentrações de produção de
IL-10, havendo uma correlação negativa sem diferença estatística (rs=-
0,1951)/(p=0,5456) (Fig.13G). Identificamos, no grupo homólogo, uma correlação
positiva estatisticamente significante (rs=0,7025)/(*p=0,0159), para as variáveis quanto
maior a carga parasitária, maior a produção da citocina IL-10 (Fig.13H). Todavia, não
identificamos no grupo heterólogo correlação entre as variáveis supramencionadas
(rs=0,2378)/(p=0,4568) (Fig.13I).

60
Figura 13–Regressão linear simples, relativa à correlação da produção de citocinas IFN-γ, TGF-ß e IL-10 e carga parasitária no baço, cinco meses após o desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG, Os grupos controles plasmídeo vazio (o) receberam três doses com 100µg de plasmídeo de DNA (pcDNA3) +1nM CpG. Os animais foram eutanasiados 15 dias após a ultima imunização, e o baço foi coletado para este ensaio. As citocinas foram avaliadas utilizando-se como medida a quantificação relativa (QR) por PCR real time, e a carga parasitária pelo ensaio de diluição limitante. A linha reta representa os valores médios do eixo x, esperados para os distintos valores do eixo y. Correlação negativa ou positiva de cada animal entre os postos de x e y (rs). A análise estatística foi feita com os coeficientes do teste e Spearman. (**p<0.01). n=7-9.
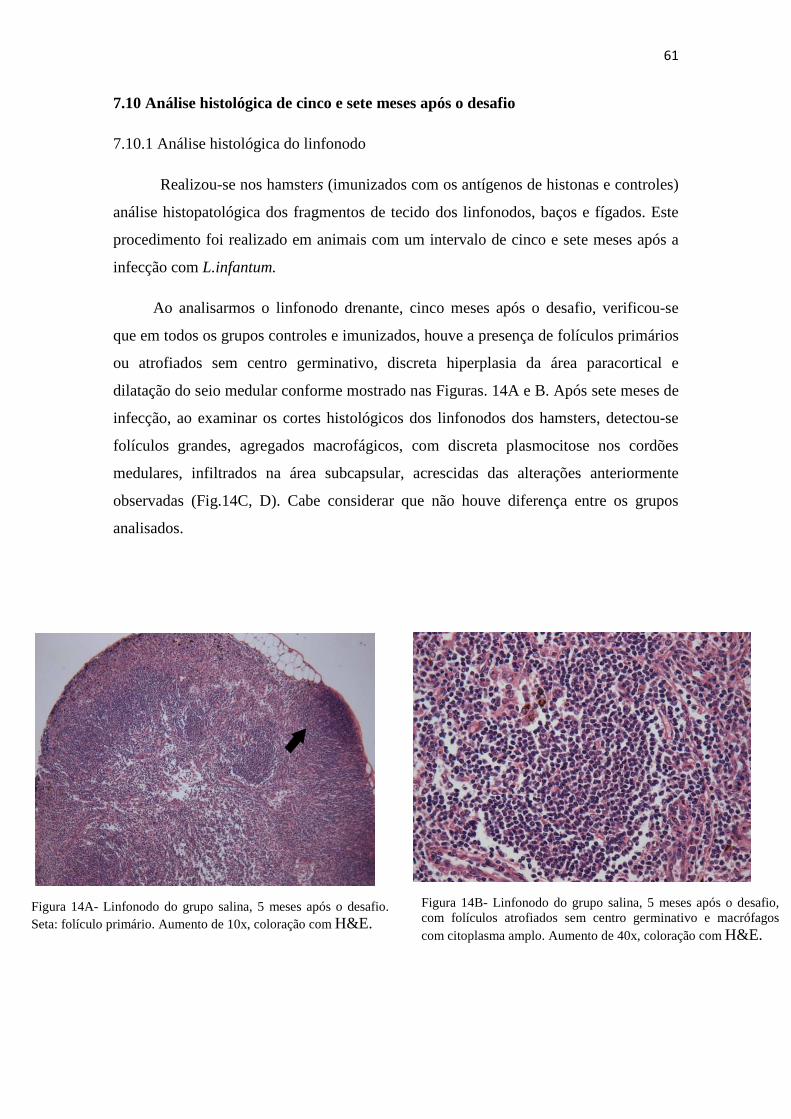
61
7.10 Análise histológica de cinco e sete meses após o desafio
7.10.1 Análise histológica do linfonodo
Realizou-se nos hamsters (imunizados com os antígenos de histonas e controles)
análise histopatológica dos fragmentos de tecido dos linfonodos, baços e fígados. Este
procedimento foi realizado em animais com um intervalo de cinco e sete meses após a
infecção com L.infantum.
Ao analisarmos o linfonodo drenante, cinco meses após o desafio, verificou-se
que em todos os grupos controles e imunizados, houve a presença de folículos primários
ou atrofiados sem centro germinativo, discreta hiperplasia da área paracortical e
dilatação do seio medular conforme mostrado nas Figuras. 14A e B. Após sete meses de
infecção, ao examinar os cortes histológicos dos linfonodos dos hamsters, detectou-se
folículos grandes, agregados macrofágicos, com discreta plasmocitose nos cordões
medulares, infiltrados na área subcapsular, acrescidas das alterações anteriormente
observadas (Fig.14C, D). Cabe considerar que não houve diferença entre os grupos
analisados.
Figura 14A- Linfonodo do grupo salina, 5 meses após o desafio. Seta: folículo primário. Aumento de 10x, coloração com H&E.
Figura 14B- Linfonodo do grupo salina, 5 meses após o desafio, com folículos atrofiados sem centro germinativo e macrófagos com citoplasma amplo. Aumento de 40x, coloração com H&E.

62
7.10.2 Análise histológica do baço dos animais infectados
Nos fragmentos de baço dos animais controles foram identificados folículos sem
centro germinativo que estavam atrofiados e desorganizados no quinto e sétimo mês
após o desafio. Em relação aos hamsters imunizados com as histonas nucleossomais
(estratégia homóloga e heteróloga), diferentemente do grupo controle, identificou-se a
presença de folículos organizados com grandes centros germinativos evidentes no baço
dos animais imunizados, cinco e sete meses após o desafio com L.infantum (Fig.15A,B).
Neste estudo, ficou evidente que em alguns animais infectados há 7 meses,
independente do grupo, verificou-se a formação de agregados macrofágicos contendo
inclusões de amastigotas de L.infantum, presença de células epiteliódes na polpa branca
e vermelha, assim como formação de granulomas (Fig.15D).
Figura 14C- Linfonodo com arquitetura preservada, folículo grande com centro germinativo, hiperplasia da área paracortical, infiltrado subcapsular. Grupo heterólogo 5 meses após o desafio. Aumento de 20x, coloração com H&E.
Figura 14D- Cordões medulares com plasmócitos, macrófagos de citoplasma amplo e eosinofílico. Grupo heterólogo 5 meses após o desafio. Aumento de 100x, coloração com H&E.
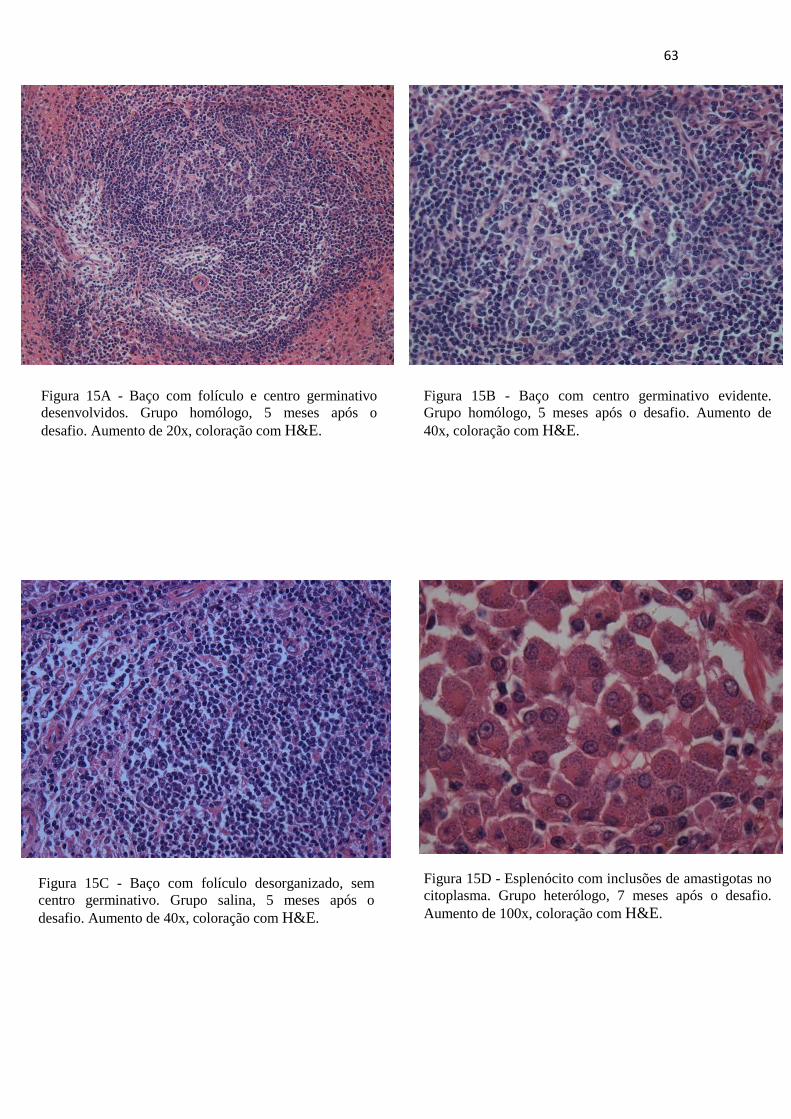
63
Figura 15A - Baço com folículo e centro germinativo desenvolvidos. Grupo homólogo, 5 meses após o desafio. Aumento de 20x, coloração com H&E.
Figura 15B - Baço com centro germinativo evidente. Grupo homólogo, 5 meses após o desafio. Aumento de 40x, coloração com H&E.
Figura 15C - Baço com folículo desorganizado, sem centro germinativo. Grupo salina, 5 meses após o desafio. Aumento de 40x, coloração com H&E.
Figura 15D - Esplenócito com inclusões de amastigotas no citoplasma. Grupo heterólogo, 7 meses após o desafio. Aumento de 100x, coloração com H&E.
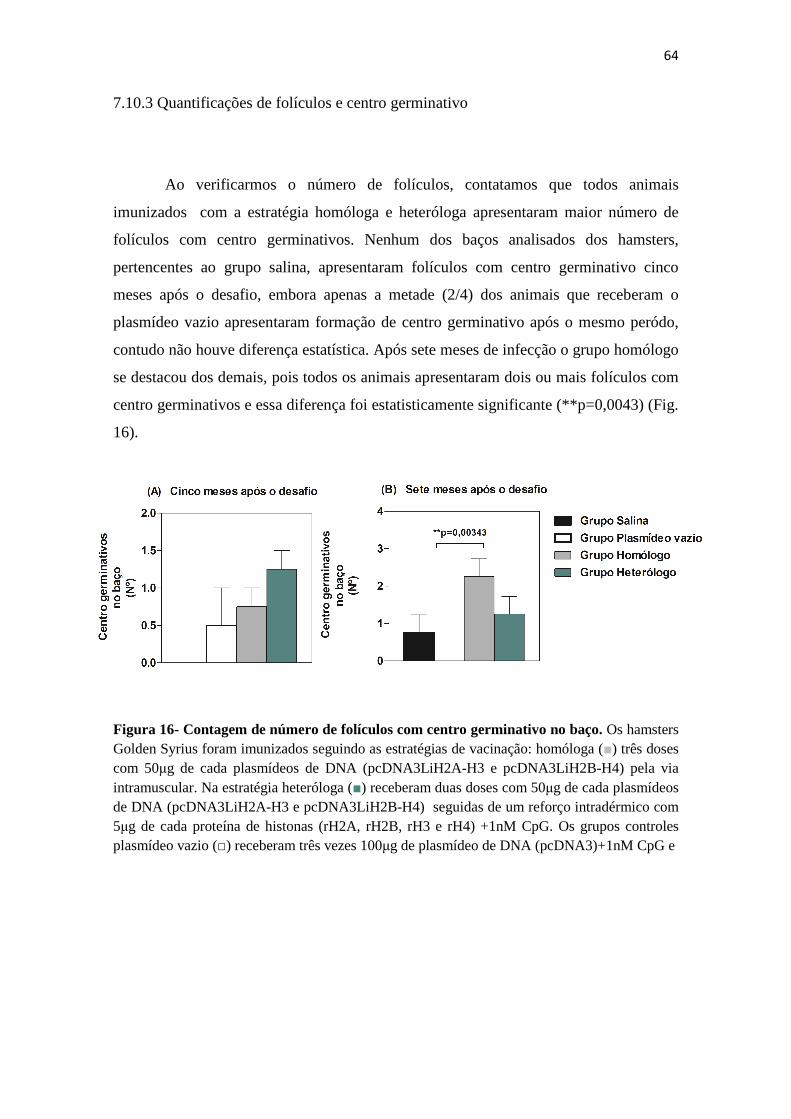
64
7.10.3 Quantificações de folículos e centro germinativo
Ao verificarmos o número de folículos, contatamos que todos animais
imunizados com a estratégia homóloga e heteróloga apresentaram maior número de
folículos com centro germinativos. Nenhum dos baços analisados dos hamsters,
pertencentes ao grupo salina, apresentaram folículos com centro germinativo cinco
meses após o desafio, embora apenas a metade (2/4) dos animais que receberam o
plasmídeo vazio apresentaram formação de centro germinativo após o mesmo peródo,
contudo não houve diferença estatística. Após sete meses de infecção o grupo homólogo
se destacou dos demais, pois todos os animais apresentaram dois ou mais folículos com
centro germinativos e essa diferença foi estatisticamente significante (**p=0,0043) (Fig.
16).
Figura 16- Contagem de número de folículos com centro germinativo no baço. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: homóloga (■) três doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG. Os grupos controles plasmídeo vazio (□) receberam três vezes 100µg de plasmídeo de DNA (pcDNA3)+1nM CpG e
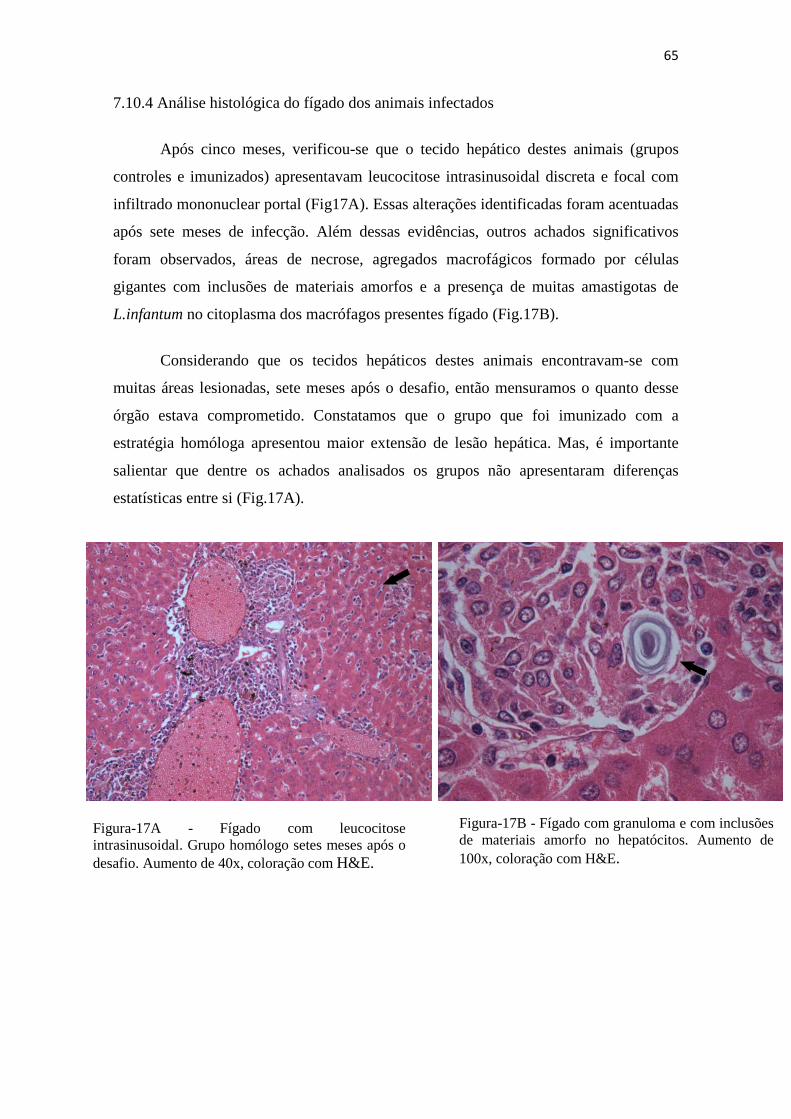
65
7.10.4 Análise histológica do fígado dos animais infectados
Após cinco meses, verificou-se que o tecido hepático destes animais (grupos
controles e imunizados) apresentavam leucocitose intrasinusoidal discreta e focal com
infiltrado mononuclear portal (Fig17A). Essas alterações identificadas foram acentuadas
após sete meses de infecção. Além dessas evidências, outros achados significativos
foram observados, áreas de necrose, agregados macrofágicos formado por células
gigantes com inclusões de materiais amorfos e a presença de muitas amastigotas de
L.infantum no citoplasma dos macrófagos presentes fígado (Fig.17B).
Considerando que os tecidos hepáticos destes animais encontravam-se com
muitas áreas lesionadas, sete meses após o desafio, então mensuramos o quanto desse
órgão estava comprometido. Constatamos que o grupo que foi imunizado com a
estratégia homóloga apresentou maior extensão de lesão hepática. Mas, é importante
salientar que dentre os achados analisados os grupos não apresentaram diferenças
estatísticas entre si (Fig.17A).
Figura-17A - Fígado com leucocitose intrasinusoidal. Grupo homólogo setes meses após o desafio. Aumento de 40x, coloração com H&E.
Figura-17B - Fígado com granuloma e com inclusões de materiais amorfo no hepatócitos. Aumento de 100x, coloração com H&E.

66
Figura 17C- Mensuração das áreas lesionada do fígado sete meses após o desafio. Os hamsters Golden Syrius foram imunizados seguindo as estratégias de vacinação: (■) receberam três vezes com 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) pela via intramuscular. Na estratégia heteróloga (■) receberam duas doses 50µg de cada plasmídeos de DNA (pcDNA3LiH2A-H3 e pcDNA3LiH2B-H4) seguidas de um reforço intradérmico com 5µg de cada proteína de histonas (rH2A, rH2B, rH3 e rH4) +1nM CpG. Os grupos controles plasmídeo vazio (□) receberam três doses de 100µg de plasmídeo de DNA (pcDNA3) +1nM
CpG e grupo salina (▆) recebeu 30µL de salina pela via intramuscular. Os hamsters foram
infectados com 105 de L. infantum metacíclicas pela via intradérmica, quinze dias após a terceira imunização. Sete meses após o desafio, os animais foram eutanasiados e o fígado foi coletado para analise histopatológica. A análise estatística foi feita com o teste Kruskal-Wallis e pós-teste de Dunns. n=4.

67
8 DISCUSSÃO
A leishmaniose visceral é uma doença grave que vem se expandido
geograficamente, com repercussões na saúde coletiva, atingindo populações em vários
países. Caso não ocorra intervenção, os sinais e sintomas podem progredir, evoluindo
para óbito (DESJEUX et al., 1996, WHO, 2002).
Pesquisas no campo das ciências biológicas buscam desenvolver uma vacina
contra Leishmania. Estudos vêem sendo direcionados, objetivando-se identificar
antígenos indutores de uma resposta imunológica, que sejam capazes de impedir a
transmissão da doença para o homem e animais, e consequentemente, a infecção do
vetor. Diferentes formulações de antígenos candidatos vacinais para LV tem sido
testados, principalmente em modelos experimentais, como camundongos, hamsters e
cães (LARA et al.,1991, HANDMAN, 2001, de MOURA et al., 2007).
Entre os antígenos de Leishmania que exibiram atividade protetora, quando
administrados como vacinas de DNA, destacam-se as diferentes proteínas de membrana
e várias proteínas intracelulares, tais como: histonas, proteínas de choque térmico e
diferentes proteínas ribossomais (revisto por KHAMESIPOUR et al., 2006; e por
DUMONTEIL, 2007). As proteínas de choque térmico, proteínas ribossomais e histonas
são antígenos altamente conservados, produzidos pela Leishmania, que não são
secretados, mas que, no entanto, são capazes de induzir uma potente resposta imune
(revisto por SANTAREM et al., 2007). Estes antígenos chamados pan-antígenos são
liberados durante a infecção após a destruição das amastigotas intracelulares pelo
macrófago ativado, ou, pela citólise espontânea de amastigotas dentro da célula
infectada. Ademais, eles são capazes de modular a resposta imune do hospedeiro devido
ao fato de não sofrerem pressão seletiva pela resposta imune, ao contrário das proteínas
de superfície e das proteínas secretadas (CHANG et al., 2003).
Existem cinco principais classes de histonas de Leishmania. Quatro delas
formam a unidade central do nucleossoma da cromatina (H2A, H2B, H3 e H4),
enquanto a H1, as mantêm ligadas ao DNA (REQUENA et al., 2000). As histonas
nucleossomais de Leishmania têm sido consideradas moléculas imunogênicas e que
estão envolvidas no mecanismo do parasita para estabelecimento da doença. Assim, os

68
pan-antígenos podem prover a capacidade imunomodulatória necessária ao
desenvolvimento de vacinas.
Assim, este estudo avaliou a capacidade protetora das histonas nucleossomais
em vacinas de DNA, utilizando estratégia homóloga (composta de plasmídeo de DNA)
e heteróloga (composta de plasmídeo de DNA mais proteína recombinante e CpG-
ODN), contra a infecção por L. infantum em hamsters.
Inicialmente, foi investigado se as proteínas de histonas, quando utilizadas na
estratégia homóloga e heteróloga, possuíam propriedades imunogênicas, capazes de
induzir a produção de anticorpos. Constatou-se que, alguns hamsters imunizados com
estratégia homóloga (36%) e heteróloga (50%) apresentaram resposta imune humoral,
com produção de IgG anti-histonas de Leishmania infantum. Logo, pode-se inferir que
as diferentes respostas imunogênicas detectadas pelas distintas concentrações de títulos
de anticorpos anti- histonas de Leishmania, dentro do mesmo grupo podem ser
justificadas pelo fato dos hamsters não serem isogênicos apresentando variabilidade de
resposta frente ao mesmo antígeno e/ou pela diferenças imunogênicas das proteínas
nucleossomais.
É importante mencionar que estudos anteriores, já descartaram a hipótese de
reação cruzada entre as proteínas de histonas de mamíferos e as de Leishmania.
Trabalhos com anticorpos específicos, obtidos de cães com LV, reconheceram as
histonas nucleossomais H2A, H2B, H3 e H4, que não apresentaram homologia com as
histonas das células de mamíferos (SOTO et al., 1999). Pesquisas com soro de pacientes
com leishmaniose cutânea e mucocutânea, foram capazes de reconhecer H1 de
Leishmania, mas sem reação cruzada com H1 de humanos (CARMELO et al., 2006).
Outro estudo mostrou a resposta humoral de cães através da detecção de anticorpos em
soros de animais infectados por Leishmania, onde as proteínas H2A (72%) e H3 (68%)
apresentaram maiores títulos do que H2B (60%) e H4 (44%).
Um estudo recente mostrou a variabilidade da resposta imune em 4 hamsters
frente ao mesmo antígeno, imunizando-os com o plasmídeo pVAX1, expressando os
genes dos antígenos de Leishmania, KMPII, TRYP, LACK, e PAPLE22. Constatou-se
que entre os 4 hamsters, apenas um foi capaz de produzir anticorpos anti-KMPII com

69
esta imunização com a vacina de DNA. Neste mesmo estudo, outro grupo de hamsters
foram imunizados com a mesma vacina de DNA, seguido de um reforço com as
proteínas dos antígenos (KMPII, TRYP, LACK, e PAPLE22), detectou-se anticorpos
específicos no soro destes animais contra os antígenos utilizados no prime-booster
(TODOLÍ et al., 2012). Os resultados destes trabalhos ratificam a variabilidade da
resposta humoral dos hamsters expostos a um mesmo antígeno. Em nosso estudo
imunizando-os com os antígenos de histonas nucleossomais de L.infantum, observamos
que somente alguns hamsters foram respondedores com produção de anticorpos IgG na
estratégia heteróloga (7/15) e homóloga (8/15).
Resultados semelhantes aos nossos, mostrando a capacidade indutora de vacinas
de DNA na produção de anticorpos, foram vistos por Mazumder e colaboradores (2011)
ao detectarem em soros de camundongos imunizados com gp63, utilizando a estratégia
homóloga composta de vacina de DNA e adjuvante CpG, um aumento significativo de
IgG2a; resultados equivalentes foram observados na estratégia prime-booster
(DNA/Proteína). Nesse mesmo trabalho, verificou-se que, três meses após o desafio, os
títulos de anticorpos (IgG) aumentaram no grupo que recebeu a imunização heteróloga e
diminuíram no grupo que recebeu a vacina de DNA/DNA.
Do mesmo modo, resultados foram obtidos em um trabalho utilizando apenas
vacinas de DNA codificante para os antígenos de histonas, no modelo experimental
BALB/c, mostrando que apenas após a terceira imunização, detectou-se anticorpos,
restringindo a resposta humoral apenas ao isotipo IgG2a, cujo perfil é indicador da
resposta Th1(CARRIÓN et al., 2008). Os resultados da nossa presente pesquisa, que
similarmente detectou anticorpos (IgG) nos animais imunizados com vacina gênica
codificando para antígenos de histonas nucleossomais.
As imunizações com a estratégia heteróloga, utilizando o reforço com proteínas
culminam na produção de anticorpos, a depender da imunogenicidade do antígeno
utilizado. Por sua vez, as imunizações com DNA favorecem mais o desenvolvimento de
uma resposta celular do que humoral (AGUILAR-BE et al., 2005). Alguns trabalhos
demonstraram a produção de anticorpos IgG anti-KMP11 em hamsters e camundongos,
imunizados com plasmídeo de DNA que codifica a KMP-11, algumas semanas
posteriores ao desafio, o que pode ter levado à participação do antígeno parasitário na

70
indução de resposta humoral (JARDIM et al., 1995, BASU et al., 2005). IBORRA e
colaboradores (2003) observaram a presença de anticorpos anti-LiP0 apenas em
camundongos imunizados com estratégia heteróloga composta de DNA e proteína. Em
nossos experimentos adicionamos no booster da imunização heteróloga com as
diferentes proteínas de histonas (rH2A, rH2B, rH3 e rH4) o adjuvante CpG, que ativar
as células da imunidade inata, como também estimula a produção de citocinas
essenciais para diferenciação de células do perfil Th1. Entretanto somente 36% dos
hamsters imunizados com a estratégia heteróloga responderam com produção de
anticorpos anti-histonas.
Em relação às citocinas, sabe-se que na leishmaniose cutânea experimental
murina existem duas respostas bem distintas, as quais determinam o curso da infecção
pela Leishmania. Uma das respostas mediadas por células Th1 é caracterizada pela
secreção das IFN-γ, TNF-α e IL-2 que levam à destruição do parasita e resolução da
infecção. Diferentemente, a resposta induzida por células Th2, secretoras de citocinas
como IL-4, IL-13 e TGF-β, estimula respostas humorais e que, juntamente com a
citocina regulatória IL-10, inibem respostas celulares do tipo Th1, promovendo a
sobrevivência do parasita, ocorrendo o progresso da doença. (MURRAY et al., 1983;
CHANNON et al., 1984; LIEW et al., 1990; PASSWELL et al., 1994).
No modelo de hamster empregado na leishmaniose visceral, detectam-se
concentrações significativa das citocinas Th1 (IFN-γ, IL-2 e TNF-α) no baço e fígado e
baixos níveis de IL-4. Um dos fatores que tornam o hamster um modelo susceptível
para LV é a expressão reduzida do gene codificante para expressão de iNOS, em
resposta ao IFN-γ, e isso acarreta uma baixa síntese de NO, essencial no macrófago
infectado, para eliminação do parasita (MELBY et al., 2001; DEA-AYUELA et al.,
2007). Entretanto, para que suceda a multiplicação do parasita, há ativação de citocinas
do perfil Th2 (TGF-β e IL-10), que podem atuar em macrófagos infectados, bem como
na indução de anticorpos anti-Leishmania. Esse último opsoniza amastigotas e induz a
produção de IL-10 em macrófagos (GOTO e PRIANT, 2009).
No decorrer do nosso estudo, ao se analisar o perfil das citocinas, constatou-se
que apenas os hamsters pertencentes ao grupo imunizado com antígenos de histonas,
com estratégia hetróloga, diminui a resposta imune local (linfonodo drenante) com

71
diferenças significativas na produção das citocinas IFN-γ, IL-10 e TGF-β, quando
comparados ao grupo imunizado com a estratégia homológo. Contudo, a razão da
expressão das citocinas IFN-γ/IL-10 no linfonodo dos hamsters imunizados, com
estratégia homóloga e heteróloga, foi significativamente mais alta, comparada com o
controle, imunizado com o plasmídeo vazio. Conclui-se que ambas as estratégias
induziram uma resposta imune celular local no linfonodo, com alta expressão de
citocina pró-inflamatória (IFN-γ) no linfonodo.
Entretanto, após a imunização, ao se analisar, no baço, a resposta imune
sistêmica desses mesmos grupos supracitados, observou-se que nenhuma das estratégias
foram capazes de desencadear aumento sistêmico das citocinas IFN-γ, IL-10 e TGF-β,
quando comparadas ao grupo controle, e, também não houve diferenças estatísticas na
razão das citocinas IFN-γ/IL-10 e IFN-γ/TGF-β entre os grupos imunizados e controle
no baço.
Um trabalho publicado recentemente pelo nosso grupo, mostrou que a
imunização com estratégia homóloga e heteróloga, com as mesmas concentrações dos
antígenos de histonas nucleossomais mais CpG, porém, utilizados no modelo BALB/c,
não foi capaz de induzir uma resposta imune intensa (IFN-γ,IL-4, TNF-α) no linfonodo,
em relação ao grupo controle. Todavia, ambas as estratégias de imunizações foram
capazes de inibir, significativamente, o desenvolvimento da lesão após o desafio com
inóculo de 105L. braziliensis na derme da orelha dos animais imunizados (CARNEIRO
et al., 2012). Provavelmente, no caso da leishmaniose cutânea causada pela L.
braziliensis, a resposta imune não tão intensa é capaz de modular a resposta inflamatória
causada pelo parasita, protegendo os camundongos.
Entretanto, Iborra e colaboradores (2004), testando a imunogenicidade das
histonas nucleossomais de Leishmania em camundongos BALB/c, mostraram
resultados opostos aos de Carneiro e colaboradores (2012), ao detectarem a produção in
vitro das citocinas de esplenócitos dos BALB/c imunizados com plasmídeos que
codificam para histonas nucleossomais. Os esplenócitos quando estimulados com as
histonas recombinantes e antígeno total do parasita, produziram alta concentração
específica de IFN-γ, mas não IL-4. Neste mesmo trabalho de Iborra, ao adicionar-se

72
anticorpo monoclonal anti-IL-12 na cultura de esplenócitos, observou-se inibição da
produção de IFN-γ, de aproximadamente 80%, com as quatro condições diferentes dos
estímulos de histonas (H2A, H2B, H3 e H4). Nesse trabalho, pode-se concluir que a
produção de IFN-γ específico de histonas é dependente de IL-12. Em nossos
experimentos, ao utilizarmos no modelo hamster, as mesmas estratégias das
imunizações, houve uma resposta imune celular local no linfonodo drenante, com
produção de citocina pró-inflamatória como IFN-γ, todavia a imunização não foi capaz
de induzir uma resposta imune sistêmica no baço, imprescindível para o controle da
carga parasitária que não foi observado posteriormente.
O sistema imune tem a capacidade de distinguir diferentes tipos de antígenos,
conhecidos com Padrões Moleculares Associados aos Patógenos (PAMPs), através das
células da imunidade inata. Os PAMPs, podem ser encontrados em RNA de alguns
vírus, na parede celular de bactérias e também em DNA rico em CpG, os quais podem
estar presentes em bactérias. O CpG ODN, têm sido utilizado amplamente em vários
estudos como adjuvante de candidatos vacinais, devido a sua capacidade de ativar a
imunidade inata, sinalizando para os Receptores de Reconhecimento Padrão (PRRs). O
CpG têm a capacidade de ligar-se às células, via adsorção da endocitose, e ativa os
receptores de Toll-like 9 (TLR9) presentes no endossoma de compartimentos de células
B e células dendríticas plasmocitóides.
A literatura mostra que a adição de adjuvantes, como o CpG nos plasmídeos de
DNA e proteínas de antígenos de Leishmania, pode propiciar o desenvolvimento da
ativação da resposta imune com a produção de citocinas pró-inflamatórias. Além disso,
o CpG tem a capacidade de ativar as células apresentadoras de antígenos, estimulando,
assim, a produção de IL-12, uma citocina fundamental na ligação da imunidade inata
com a adaptativa (BASTOS et al., 2004). A IL-12 pode estimular a produção de IFN-γ
por células NK, mecanismo importante para a geração de respostas protetoras contra
parasitas intracelulares (VIVIER et al., 2005). Diferentes trabalhos comprovaram a
capacidade imunomoduladora do CpG-ODN quando adicionados a antígenos vacinais
contra antraz, brucelose, influenza vírus, vírus da coriomeningitelinfocítica, varíola,
sarampo, antígenos de superfície da hepatite C e toxina tetânica, utilizando vacinas de

73
proteínas, gênicas ou uma estratégia prime-booster composta de DNA e proteína
(OXENIUS et al.,1999)
A literatura relata resultados satisfatórios do CpG para a formulação de vacinas
ou fármacos em modelos experimentais de leishmanioses. Um estudo avaliou de
tratamento para LV com Miltefosina e CpG ODN em hamster se camundongos
infectados com L. infantum, teve capacidade leishmanicida mais eficaz, quando
comparado ao uso isolado da droga (SANE et al., 2010). Um estudo com a proteína
LiP0 de Leishmania infantum mais CpG, induziu proteção contra leishmaniose cutânea
em camundongos C57Bl/6, no desafio empregando L.major. (IBORRA et al., 2005).
Outro trabalho mostrou a eficácia do antígeno gp63, utilizado como vacina de DNA,
seguido do reforço da proteína na presença de CpG, diminuindo a carga parasitária no
fígado e baço dos BALB/c desafiados com L.donavani (MAZUNDER et al., 2011).
Em nosso trabalho com hamsters, nenhuma das estratégias vacinais (DNA/DNA
ou DNA/Proteína mais CpG) com os antígenos de histonas nucleossomais foram
capazes de diminuir as cargas parasitárias nos linfonodos, baços e fígados, dos animais
desafiados com L.infantum. Resultados semelhantes foram observados por CARRIÓN e
colaboradores (2007), onde camundongos imunizados com vacina de DNA codificantes
para antígenos de histonas e posteriormente desafiados com L.infantum, não
apresentaram redução da carga parasitária. Neste mesmo trabalho, foram utilizadas
outras estratégias de imunização, pela via endovenosa e intradémica em camundongos
BALB/c, utilizando células dendríticas pulsadas, associadas às proteínas das histonas
mais CpG. Esta estratégia foi capaz de diminuir em 50% a carga parasitária no baço e
fígado, quatro semanas após infecção por L.infantum. Todavia, eles observaram que o
CpG não influenciou na eficácia da vacina.
Resultados opostos ao nosso trabalho foram vistos na leishmaniose cutânea,
mostrando a capacidade protetora das quatro proteínas de histonas nucleossomais,
(rH2A, rH2B, rH3 e rH4), associadas ao CpG em BALB/c, através de vacina DNA ou
DNA/ proteínas recombinantes. Nos animais imunizados e desafiados com L.
braziliensis, empregando-se ambas estratégias vacinais observou-se inibição no
desenvolvimento de lesão, diminuição da carga parasitária nos linfonodos e ausência de
macrófagos infectados na análise histológica (CARNEIRO et al., 2012). Outro estudo

74
com camundongos BALB/c observou que a utilização de vacina de DNA com genes
codificantes para histonas mais CpG, foi capaz de diminuir o número de parasitas
L.major, no linfonodo drenante e baço, quando comparados ao grupo controle
(IBORRA et al., 2004).
Diante dos resultados do nosso experimento, comparando-o a outros estudos
realizados com histonas, é fundamental atentar as possíveis discrepâncias genéticas
existentes entre os modelos de camundongos e hamsters, como também as diferenças
biológicas nos mecanismos de virulência e patogenia entre as espécies de L.major e/ou
L. braziliensis e L.infantum.
Ao analisarmos nestes animais o perfil de citocinas no baço, cinco meses após o
desafio, observamos que apenas o grupo homólogo destacou-se com concentrações
elevadas de IL-10, em relação ao grupo controle que recebeu o plasmídeo vazio.
A partir da constatação das pesquisas anteriormente realizadas, avaliou-se na
presente investigação, a possível correlação das respostas - local e sistêmica - das
citocinas produzidas no linfonodo e baço, com a produção de anticorpos dos animais
imunizados com estratégia homóloga e heteróloga, que apresentaram títulos de IgG no
ELISA acima do ponto de corte . No experimento realizado, destacou-se que os animais
do grupo homólogo, imunizados apenas com vacina de DNA codificante para os
antígenos de histonas, apresentaram significância na correlação positiva, quanto maior a
produção de anticorpos anti-histonas, maior a produção de TGF-β, no linfonodo. Após
cinco meses de infecção neste mesmo grupo observou-se significância estatística na
correlação, quanto maior carga parasitária, maior a produção das citocinas TGF-β no
baço. A alta produção das citocinas anti-inflamatórias, TGF-β, condiz com a resposta
imune do perfil Th2, e a indução da resposta imune humoral, com alta produção de
anticorpos, justificando-se os resultados obtidos na correlação.
Apesar das imunizações com os antígenos de histonas nucleossomais não diferir
nos resultados da carga parasitária no linfonodo, baço e fígado, observou-se nas análises
histopatológicas diferenças estatísticas no aumento do número de folículos com centros
germinativos no baço, sete meses após o desafio, quando comparado aos grupos
controles.

75
É sabido que as células dendríticas na bainha linfática periarteriolar capturam
antígenos e os apresentam aos linfócitos T. As células T e B interagem nas margens dos
folículos, da polpa branca, estimulando a geração de plasmócitos secretores de
anticorpos, encontrados no interior da polpa branca no baço. Ao ocorrer estimulação
antigênica estas bainhas linfáticas se expandem, geralmente em um dos lados das
artérias, e formam-se os nódulos linfóides compostos principalmente por linfócito B,
por fim, formando os centros germinativos (ROBBINS et al., 2005, JUNQUEIRA,
2010). Assim, podemos inferir que no nosso estudo, ambas imunizações homóloga e
heteróloga foram capazes de não somente aumentar o número de folículos, como
concomitantemente induzir a proliferação de, uma maior população de linfócitos B, nos
centros germinativos nos folículos esplênicos, após cinco e sete meses de infecção,
Ao analisarmos o linfonodo drenante, cinco meses após o desafio, verificou-se
em todos os grupos controles e imunizados a presença de folículos primários ou
atrofiados. Após sete meses este órgão apresentou mudanças nas características
morfológicas, dos folículos em todos os grupos experimentais, mostrando-se grandes,
com agregados macrofágicos e discreta plasmocitose nos cordões medulares.
Verificou-se, igualmente, nos tecidos hepáticos destes animais presença de muitas áreas
lesionadas, com leucocitose e formação de granulomas, contudo não apresentando
diferenças estatísticas entre os grupos. Nossos achados são compatíveis com a literatura,
que relata que as características histopatológicas observadas durante o curso da LV em
hamsters incluem, formação de granulomas em órgãos como baço e fígado (RICA-
CAPELA et al., 2003).
Assim, apesar dos resultados obtidos por outros grupos de pesquisa utilizando as
histonas nucleossomais nas diferentes estratégias de imunização, conferirem proteção
no modelo experimental de leishmaniose cutânea, nossos resultados mostram que tais
antígenos não foram capazes de proteger no modelo hamster, frente ao desafio por
L.infantum. Estes achados podem ser justificados, pelo fato da leishmaniose visceral, ser
uma doença sistêmica que afeta vários órgãos, e cuja imunopatogenia difere da
leishmaniose cutânea, que se manifesta de forma localizada ou os antígenos de histonas
nucleossomais não são imunogênicos o suficiente a tal ponto de proteger o hamster.

76
9 CONCLUSÃO
As estratégias de imunização homóloga e heteróloga, utilizando-se vacinas de
DNA e proteínas de histonas nucleossomais de L. infantum, com o adjuvante CpG, não
foram capazes de diminuir a carga parasitária e o agravamento do quadro
histopatológico em hamsters infectados com L. infantum.

77
REFERÊNCIAS
AGUILAR-BE, I.; DA SILVA, Z, R.; PARAGUAI, S.E.; BORJA-CABRERA, G. P.; ROSADO, V. M.; MUT, M.M.; GARCIA, M.M. R.; PALATNIK, S.C.B.; DUMONTEIL, E. Cross-protective efficacy of a prophylactic Leishmania donovani DNA vaccine against visceral and cutaneous murine Leishmaniasis. Infect Immun, v. 73, n. 2, p. 812-819, 2005.
AHUJA, S.S.; REDDICK, R.L.; SATO, N.; MONTALBO, E.; KOSTECKI, V.; ZHAO, W.; DOLAN, M.J.; MELBY, P.C.; AHUJA, S.K. Dendritic cell (DC)-based anti-infective strategies: DCs engineered to secrete IL-12 are a potent vaccine in a murine model of an intracellular infection. J Immunol , v.163, n.7, p. 3890-3897, 1999.
ANSARI, N.A.; KUMAR, R.; GAUTAM, S.; NYLÉN, S.; SINGH, O.P.; SUN-DAR, S.; ANDSACKS, D. IL- 27 and IL-21 are associated with T cell IL-10 responses in human leishmaniasis. J. Immunol, 186, p. 3977–3985, 2011.
AZEVEDO, V.; LEVITUS, G.; MIYOSHI, A.; CANDIDO, A.L.; GOES, A.M., OLIVEIRA, S.C. Main features of DNA-based immunization vectors. Braz. J. Med. Biol. Res, v.32, n.2, p. 147-153, 1999.
BABUIK, S.; MOOKHERJEE, N.; PONTAROLLO, R.; GRIEBEL, P.; VAN DEN, H, S.; HECKER, R.; BABIUK, L. TLR9-/- and TLR9+/+ mice display similar immune responses to a DNA vaccine. Immunology, n.113, p.114-120, 2004.
BARBOSA, J.; ANDRADE, Z. A; REED, S. G. The pathology of experimental visceral leishmaniasis in resistant and susceptible lines of inbred mice. Braz J Med Biol Res, v.20, p. 63-72, 1987.
BARHOUMI, M.; TANNER, N. K.; BANROQUES, J.; LINDER, P.; GUIZANI, I. Leishmania infantum LeIF protein is an ATP-dependent RNA helicase and an eIF4A-like factor that inhibits translation in yeast. FEBS J. 273(22), p.5086-6100, 2006.
BASTOS, K.; MARINHO C.R.; BARBOZA R.; RUSSO M.; ALVAREZ J.M.; D¦IMPERIO-LIMA M.R. What kind of message does Il-12/IL-23 bring to macrophages and dendritic cells? Microbes Infect, n. 6, p. 630-636, 2004.
BASU, R.; BHAUMIK, S.; BASU, J.M.; NASKAR, K, DE T.; ROY, S. Kinetoplastid membrane protein-11 DNA vaccination induces complete protection against both pentavalent antimonial-sensitive and-resistant strains of Leishmania donovani that correlates with inducible nitric oxide synthase activity and IL-4 generation: evidence for mixed th1- and th2-like responses in visceral leishmaniasis. J Immunol , v.174, n.11, p.7160-717, 2005.
BASU, R.; BHAUMIK, S.; HALDAR, A. K.; NASKAR, K.T.; DANA, S. K., WALDEN, P.; ROY, S. Hybrid cell vaccination resolves Leishmania donovani infection by eliciting a strong CD8+ cytotoxic T-lymphocyte response with concomitant suppression of interleukin-10 (IL-10) but not IL-4 or IL-13. Infect Immun , v.75, n.12, p. 5956-5966, 2007.

78
BELKAID, B. Y.; KAMHAWI, S.; MODI, G.; VALENZUELA, J.; TRAUTH, N. N.; ROWTON, E.; RIBEIRO, J.; SACKS, D. L. Development of a natural model of cutaneous leishmaniasis: powerful effects of vector saliva and saliva pre exposure on the long-term outcome of Leishmania major infection in the mouse ear dermis. Journal of Experimental Medicine, v. 88, n. 10, p.1941-1953, 1998.
BELKAID, Y.; HOFFMANN, K. F.; MENDEZ, S.; SACKS, D. L.; The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med, v.194, n.10, p. 1497-1506, 2001.
BELKAID, Y.; MENDEZ, S.; LIRA, R.; KADAMBI, N.; MILON, G.; SACKS, D. A natural model of Leishmania major infection reveals a prolonged "silent" phase of parasite amplification in the skin before the onset of lesion formation and immunity. J Immunol, v.165, n.2, p.969-977, 2000.
BERBERICH, C., J. R.; RAMIREZ-PINEDA, C.; HAMBRECHT, G.; ALBER, Y. A.; SKEIKY, MOLL, H. Dendritic cell (DC)-based protection against an intracellular pathogen is dependent upon DC-derived IL-12 and can be induced by molecularly defined antigens. J Immunol, n.170, p.3171-3179, 2003.
BITTENCOURT, A.; BARRAL-NETTO, M. Leishmaniasis. Tropical Pathology, v. 8, n. 6, p. 597-644, 1995.
BOGDAN, C.; DONHAUSER, N.; DORING, R.; ROLLINGHOFF, M.; DIEFENBACH, A.; RITTIG, M. G. Fibroblasts as host cells in latent leishmaniosis. J. Exp. Med., v.191, n.12, p.2121-2130, 2000.
BOGDAN, C.; VODOVOTZ, Y.; NATHAN, C. Macrophage deactivation by interleukin 10. J. Exp. Med., v.174, n.6, p. 1549-1555, 1991.
CARMELO, E.; ZURITA, A.I.; MARTINEZ, E.; VALLADARES, B. The sera from individuals suffering from cutaneous leishmaniasis due to Leishmania brazilensis present antibodies against parasitic conserved proteins, but not their human counterparts. Parasite, 13, p-231–236, 2006.
CARNEIRO, M.W.; SANTOS, D.M.; FUKUTANI, K.F.; CLARENCIO, J.; MIRANDA, J.C.; BRODSKYN, C.; BARRAL, A.; BARRAL-NETTO, M, SOTO M.; DE OLIVEIRA, C.I. Vaccination with L. infantum Nucleosomal Histones Confers Protection against New World Cutaneous Leishmaniasis Caused by Leishmania braziliensis. PLoS One, 52296, p.7-12. 2012.
CARRION, J.; NIETO, A.; IBORRA, S.; INESTIA, V.; SOTO,M.; FOLGUEIRA, C.; ALBANADES, D.R.; REQUENA, J.M.; ALONSO, C. Immunohistological feature of visceral leishmaniasis in BALB/c mice. Parasite Immunol, 28, p 173-183, 2006.
CARRIÓN, J.; NIETO, A.; SOTO, M.; ALONSO, C. Adoptive transfer of dendritic cells pulsed with Leishmania infantum nucleosomal histones confers protection against cutaneous leishmaniosis in BALB/c mice. Microbes Infect, v.9, n.6, p. 735-43, 2007.

79
CHANG, K.P.; REED, S.G.; MCGWIRE, B.S.; SOONG, L. Leishmania model for microbial virulence: the relevance of parasite multiplication and pathoantigenicity. Acta Trop , 85(3), p.375-390, 2003.
CHANNON, H. W. Interaction of Leishmania with a macrophage cell line. Correlation between intracellular killing and the generation of oxygen intermediates. J Exp Med, v.153, n.6, Jun 1, p.1690-1695, 1981.
CHANNON, J. Y.; ROBERTS, M. B.; BLACKWELL, J. M. A study of the differential respiratory burst activity elicited by promastigotes and amastigotes of Leishmania donovani in murine resident peritoneal macrophages. Immunology, v.53, n.2, p.345-355, 1984.
CHILDS, G. E.; LIGHTNER, L. K.; MCKINNEY, L.; GROVES, M. G.; PRICE, E. E; HENDRICKS, L. D. Inbred mice as model hosts for cutaneous leishmaniasis. I. Resistance and susceptibility to infection with Leishmania braziliensis, L. mexicana, and L. aethiopica. Ann Trop Med Parasitol, v.78, n.1, p.25-34, 1984.
COLER, R. N.; SKEIKY, Y. A.; BERNARDS, K.; GREESON, K.; CARTER, D.; CORNELLISON, C. D.; MODABBER, F.; CAMPOS-NETO, A.; REED, S. G. Immunization with a polyprotein vaccine consisting of the T-Cell antigens thiol-specific antioxidant, Leishmania major stress-inducible protein 1, and Leishmania elongation initiation factor protects against leishmaniasis. Infect Immun , v. 70, n. 8, p. 4215-4225, 2002.
COLER, R.N.; GOTO, Y.; BOGATZKI, L.; RAMAN, V.; REED, S.G. Leish-111f, a recombinant polyprotein vaccine that protects against visceral Leishmaniasis by elicitation of CD4+ T cells. Infect Immun , v.75, p.4648-54, 2007.
COURTNEY, S. C.; SUSTA, L.; GOMEZ, D.; HINES, L. N, PEDERSEN, J. C.; BROWN, C. C.; MILLER, P. J.; AFONSO, C.L. Highly Divergent Virulent Isolates of Newcastle Disease Virus from the Dominican Republic are Members of a New Genotype that May have Evolved Unnoticed for Over Two Decades. Journal of clinical microbiology, (51), 2, p.508-507, 2013
CREF, B.J T.; JONES, C.; BADAR, D.; SAMPAIO, R. T.; JOHNSON, W.D.J.; Malnutrition as a risk factor for severe visceral leishmaniasis. J Infect Dis, 156, p. 1030-1033, 1987.
CUNNINGHAM, A. C. Parasitic adaptive mechanisms in infection by leishmania. Exp Mol Pathol, v.72, n.2, p.132-141, 2002.
DA'DARA, A. A.; HARN, D. A. DNA vaccines against tropical parasitic diseases. Expert Rev Vaccines, v.4, n.4, p.575-589, 2005.
DANTAS-TORRES, F. Leishmune vaccine: the newest tool for prevention and control of canine visceral leishmaniosis and its potential as a transmission-blocking vaccine. Vet. Parasitol, v.141, n.2, p.1-8, 2006.

80
DE CARVALHO, L.P.; SOTO, M.; JERONIMO, S.; DONDJI, B.; BACELLAR, O.; LUZ, V.; ORGE, G.; ALONSO, C.; JESUS, A.R.; CARVALHO, E.M. Characterization of the immune response to Leishmania infantum recombinant antigens. Microbes Infect, 5(1), p.7-12, 2003.
DEA-AYUELA, M.A.; RAMA-INIGUEZ, S.; ALUNDA, J.M.; BOLAS-FERNANDEZ, F. Setting new immunobiological parameters in the hamster model of visceral leishmaniasis for in vivo testing of antileishmanial compounds. Vet Res Commun, 31, p-703-717, 2007.
DESJEUX, P. Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis , 305, p.18-27, 2004.
DESJEUX, P. Leishmaniosis: public health aspects and control. Clin. Dermatol, 14, p.417-423, 1996.
DESJEUX, P. The increase in risk factors for leishmaniosis world wide. Trans. R. Soc. Trop. Med. Hyg, 95, p.239-243, 2001.
DUMONTEIL, E. DNA Vaccines against Protozoan Parasites: Advances and Challenges. J Biomed Biotechnol, 9, p.520-528, 2007.
ENGWERDA,C.R.; ATO, M.; KAYE, P.M. Macrophages, pathology and parasite persistence in experimental visceral leishmaniasis. Trends Parasitol, v.20, n.11, p.524-530, 2004.
FERNÁNDEZ, C. J.; INIESTA, V.; BELINCHÓN, L, S.; MUÑOZ, M.R.; PAREJO, J.C.; GÓMEZ-GORDO, L.; SOTO, M.; ALONSO, C.; GÓMEZ, N. L.C. Experimental model for reproduction of canine visceral leishmaniosis by Leishmania infantum. Vet Parasitol. 18, p192, 2013.
GARG, R.; DUBE, A. Animal models for vaccine studies for visceral leishmaniasis. Indian J. Med. Res, 123 (3), p. 439-454, 2006.
GARMORY, H. S.; PERKINS, S. D.; PHILLPOTTS, R. J.; TITBALL, R.W. DNA vaccines for biodefence. Adv. Drug Deliv. Rev, v.57, n.9, p.1343-1361, 2005.
GARMORY, H. S.; PERKINS, S. D.; PHILLPOTTS, R.; J.TITBALL, R. W. DNA vaccines for biodefence. Adv Drug Deliv Rev, v.57, n.9, p.1343-1361, 2005.
GOMES, R.; TEIXEIRA, C.; TEIXEIRA, M. J.; OLIVEIRA, F.; MENEZES, M. J.; SILVA, C.; DE OLIVEIRA, C. I.; MIRANDA, J. C.; ELNAIEM, D. E. KAMHAWI, S., VALENZUELA, J. G; BRODSKYN, C. I. Immunity to a salivary protein of a sand fly vector protects against the fatal outcome of visceral leishmaniasis in a hamster model. Proc Natl Acad Sci U S A, v.105, n.22, p.7845-7850, 2008.
GOTO, H.; PRIANTI, M.G. Immunoactivation and immunopathogeny during active visceral leishmaniasis. Rev Inst Med Trop, 51, p.241-246, 2009.

81
GURUNATHAN, S.; PRUSSIN, C.; SACKS, D.L.; SEDER, R.A. Vaccine requirements for sustained cellular immunity to an intracellular parasitic infection. Nat med, v.4, n.1, p-1409- 1405, 1998.
GURUNATHAN, S.; WU, C.Y.; FREIDAG, B.L.; SEDER, R.A. DNA vaccines: a key for inducing long-term cellular immunity. Curr Opin Immunol , 12, p.442-447, 2000.
HAEBERLEIN, S.; SEBALD, H,; BOGDAN, C.; SCHLEICHER, U. IL-18, but not IL-15, contributes to the IL-12-dependent induction of NK-cell effector functions by Leishmania infantum in vivo. Eur J Immunol, 40, p.1708-1717, 2010.
HANDMAN, E. Leishmaniasis: current status of vaccine development. Clin Microbiol Rev, v.14, n. 2, p. 229-243, 2001.
IBORRA, S. J.; CARRION, C.; ANDERSON, C.; ALONSO, D.; SACKS, D.; M, SOTO. Vaccination with the Leishmania infantum acidic ribosomal P0 protein plus CpG oligodeoxynucleotides induces protection against cutaneous leishmaniasis in c57bl/6 mice but does not prevent progressive disease in BALB/c mice. Infect Immun 73, p-5842-5852, 2005.
IBORRA, S.; SOTO, M.; CARRION, J.; ALONSO, C.; REQUENA, J.M. Vaccination with a plasmid DNA cocktail encoding the nucleosomal histones of Leishmania confers protection against murine cutaneous leishmaniosis. Vaccine, 22 (29-30) p.3865-3876, 2004.
JARDIM, A.; FUNK, V.; CAPRIOLI, R.; M.OLAFSON, R. W. Isolation and structural characterization of the Leishmania donovani kinetoplastid membrane protein-11, a major immunoreactive membrane glycoprotein. Biochem J, v.305, n. 1, p.307-313, 1995.
JERONIMO, S. M. An urban outbreak of visceral leishmaniasis in Natal, Brazil. Trans Royal Society of Tropical Medicine and Hygiene, Natal, v. 88, n. 4, p. 386- 388, 1994.
JUNQUEIRA,J.; CARNEIRO, L.C.U. Histologia Básica, Rio de Janeiro; Guanabara Koogan, 10 ed. 2010.
KHAMESIPOUR, A.; RAFATI, S.; DAVOUDI, N.; MABOUDI, F.; MODABBER, F. Leishmaniasis vaccine candidates for development: a global overview. Indian J. Med. Res, v.123, n.3, p.423-438, 2006.
KLINMAN, D.M.; CURRIE,D.; GURSEL,I.; VERTHELYI, D. Use of CpG oligodeoxynucleotides as immune adjuvants. Immunological Reviews, 199(1), p.201-216, 2004.
KRIEG, A.M.; YAK, M.S.; WALDSCHMIDT, T.J.; BISHOP, G.A, TEASDALE, R.; KORETZKY, G.A, KLINMAN, D.M. CpG Motifs in Bacterial-DNA Trigger Direct B-Cell Activation. Nature, 374 (6522), p.546–549, 1995

82
KRISHNAMACHARI, Y.; SALEM, A.K. Innovative strategies for co-delivering antigens and CpG oligonucleotides. Advanced Drug Delivery Reviews, v.61, p. 205-217, 2009.
KUTZLER, M.A.; WEINER, D.B. Developing DNA vaccines that call to dendritic cells. J. Clin. Invest, v.114, n.9, p.1241-1244, 2004
LAINSON, R.; SHAW, J. J; SILVEIRA, F. T. Dermal and visceral leishmaniasis and their causative agents. Trans R Soc Trop Med Hyg, v.81, n.4, p.702-703, 1987.
LARA, M. L.; LAYRISSE, J. V.; SCOIRZA, E.; GARCIA, Z.; STOIKOW, J.; GRANADO, J.; BIA, W. Immunogenitics of human American cutanneous leishmaniasis. Study of HLA haplotypes in 24 families from Venezuela. Hum. Immuno, p.129-135. v.30, 1991.
LIEW, F.Y.; LI, Y.; MILLOTT, S. Tumor necrosis factor-alpha synergizes with IFN-gamma in mediating killing of Leishmania major through the induction of nitric oxide. J Immunol , v.145, n.12, p.4306-4310, 1990.
LINDOSO, J.A.L.; GOTO, H. Immunity and immunosuppression in experimental visceral leishmaniasis. Braz J Med Biol Res, v.37, p.615-623, 2004.
MASINA, S. M. G.; DEMOTZ, M, S. O.; FASEL, N. J. Protection against cutaneous leishmaniasis in outbred vervet monkeys using a recombinant histone h1 antigen. J Infect Dis, 188, p.1250-1257, 2003.
MATHIAS, R.; COSTA, F.A.; GOTO, H. Detection of immunoglobulin G in the lung and liver of hamsters with visceral leishmaniasis. Braz j Med Biol Res. v. 34, p. 539-43, 2001.
MAURICIO, I. L.; STOTHARD, J. R.; MILES, M. A. The strange case of Leishmania chagasi. Parasitol, v. 16, p. 188-189, 2000.
MAZUMDER, S.; MAJI, M.; DAS, A.; ALI, A. Potency, Efficacy and Durability of DNA/DNA, DNA/Protein and Protein/Protein Based Vaccination Using gp63 Against Leishmania donovani in BALB/c Mice. PLoS ONE, v.6, p.1-15, 2011.
MELBY, P. C.; YANG, J.; ZHAO, W.; PEREZ, L. E.; CHENG, J. Leishmania donovani p36 (LACK) DNA vaccine is highly immunogenic but not protective against experimental visceral Leishmaniasis. Infect Immun , v. 69, n. 8, p. 4719-4725, 2001.
MENDEZ, S.; TABBARA, K.; BELKAID, Y.; BERTHOLET, D.; VERTHELYI, D. Co-injection with CpG-contining immunostimulatory reduces the pathogenicity of a live vaccine against cutaneous Leishmaniasis but its potency and durability. Infect. Immune, (71), p. 5121-5129, 2003.
MILANO, S.; DIBELLA, G.; D’AGOSTINO, P.; BARBERA, C.;CARUSO, R.; LAROSA, M., FERLAZZO, V.; VITALE, G.; LARUSSA, C.; GAMBINO, G.; CHIFARI, N.; MANSUETO, S.; ANDCILLARI, E. IL-15 in human visceral leishmaniasis caused by Leishmania infantum. Clin. Exp. Immunol , 127, p.360-365, 2002.

83
MOLANO, I.; GARCIA ALONSO, M.; MIRON, C.; REDONDO, E.; REQUENA, J.M.; SOTO, M.; GOMEZ NIETO,C. & ALONSO, C. A. Leishmania infantum multi-component antigenic protein mixed with live BCG confers protection to dogs experimentally infected with L.infantum. Veterinary Immunology and Immunopathology, 92, p.1-13, 2003.
MURRAY, H. W.; CARTELLI, D. M. Killing of intra cell Leishmania donovani by human mononuclear phagocytes. Evidence for oxygen-dependent and -independent leishmanicidal activity. J Clin Invest, v. 72, n. 1, p. 32-44, 1983.
MURUGAIYAN, G.; MITTAL, A.; LOPEZ-DIEGO, R.; MAIER, L.M.; ANDERSON, D.E.; WEINER, H.L. IL-27 is a key regulator of IL-10 and IL-17 production by human CD4+T cells. J. Immunol, 183, p-2435-2443, 2009.
NIETO, A.; DOMÍNGUEZ BERNAL, G.; ORDEN, A,J.; FLUENTE, R.D.L.; MADRID-ELENA, N.; CARRIÓN, J. Mechanismo of resistence and susceptibility to experimental visceral leishmaniasis: BALB/c mouse versus Syrian hamsters model. Veterinary Research, 39-42, 2011.
NUNES, M. P.; JACKSON, J. M.; CARVALHO, R. W.; FURTADO, N. J.; COUTINHO S. G. Serological survey for canine cutaneous and visceral leishmaniasis in areas at risk for transmission in Rio de Janeiro where prophylactic measures had been adopted. Mem do Instituto Oswaldo Cruz, 86(4), p-411-417, 1991.
NYLEN, S.; MAURYA, R.; L EIDSMO,L.; KRISHNA, D.M.; SACKS,D. Splenic accumulation of IL-10 mRNA in T cells distinct from CD4+CD25+ (Foxp3) regulatory T cells in human visceral leishmaniasis. J Exp Med, 204 (4) p. 805–817, 2007.
OLIVEIRA, S.C.; ROSINHA, G.M.; DE-BRITO, C.F.; FONSECA, C.T.; AFONSO, R.R.; COSTA, M.C.; GOES, A.M.; RECH, E.L.; AZEVEDO, V. Immunological properties of gene vaccines delivered by different routes. Braz. J. Med. Biol. Res., v.32, n.2, p. 207-214, 1999.
OXENIUS, A.; MARTINIC, M.M.; HENGARTNER, H.; KLENERMAN, P. CpG containing oligonucleotides are efficient adjuvants for induction of protective antiviral immune responses with T-cell peptide vaccines. J. Virol , 73, p.4120-4126, 1999.
PALATNIK-DE-SOUSA, C. B. Vaccines for Leishmaniasis in the fore coming 25 years. Vaccine, v. 26, n. 14, p. 1709-1724, 2008.
PASSWELL, J. H.; SHOR, R.; SMOLEN, J; .JAFFE, C. L. Infection of human monocytes by Leishmania results in a defective oxidative burst. Int J Exp Pathol, v.75, n.4, p.277-284, 1994.
PEARSON R.D.; ROBERTS, D. Host immunoglobulin on spleen-derived Leishmania donovani amastigotes. Am Journal Tropical Medicine, v.4, n. 3, p.263-265, 1990.
PEARSON, R. D.; SOUSA, A. Q. Clinical spectrum of Leishmaniasis. Clin Infect Dis, v.22, n.1, p.1-13. 1996.

84
PEREZ, L. E.; CHANDRASEKAR, B.; SALDARRIAGA, O. A.; ZHAO, W.; ARTEAGA, L. T.; TRAVI. B. T.; MELBY, P.C. Reduced Nitric Oxide Synthase 2(NOS2) Promoter Activity in the Syrian Hamster Renders the Animal Functionally Deficient in NOS2 Activity and Unable to Control an Intracellular Pathogen. The Jornal of immunology, v.176, p.5519-5528, 2006.
PETERS, N.; SACKS, D. Immune privilege in sites of chronic infection: Leishmania and regulatory T cells. Immunol Rev, 213, p.159–179, 2006.
PITTA, M. G.; ROMANO, A.; CABANTOUS, S.; HENRI, S.; HAMMAD, A.; KOURIBA, B.,ARGIRO, L.; EL KHEIR, M.; BUCHETON, B.; MARY, C.; EL-SAFI, S. H.; Des, S.A. IL-17 and IL-22 are associated with protection against human kalaazar caused by Leishmania donovani. J. Clin.Invest, 119, p.2379-2387, 2009.
PLOTKIN, S.A. Vaccines: past, present and future. Nat. Med., v.11, n.4, p.5-11, 2005
RAFATI, S.; NAKHAEE, A.; TAHERI, T.; TASLIMI Y.; DARABI, H.; ERAVANI, D.; SANOS, S.; KAYE, P.; TAGHIKHANI, M.; JAMSHIDI, S.; RAD, M.A. Protective vaccination against experimental canine visceral leishmaniasis using a combination of DNA and protein immunization with cysteine proteinases type I and II of L. infantum. Vaccine, 23 (28), p-3716-3725, 2005.
RAMIRO, M.J.; ZARETE, J.J.; HANKE, T.; RODRIGUEZ, D.; RODRIGUEZ, J.R.; ESTEBAN, M.; LUCIENTES, J.; CASTILLO, J.A.; LARRAGA, V. Protection in dogs against visceral leishmaniasis caused by Leishmania infantum is achieved by immunization with a heterologous prime-boost regime using DNA and vaccinia recombinant vectors expressing LACK. Vaccine, v.21, n.19-20, p. 2474-2484, 2003.
REQUENA, J. M.; ALONSO, C.; SOTO, M. Evolutionarily conserved proteins as prominent immunogens during Leishmania infections. Parasitol Today, v. 16, n. 6, p. 246-250, 2000.
RICA-CAPELA, M. J.; CORTES, S.; LEANDRO,; PELETEIRO, M. C.; SANTOS-GOMES, G., CAMPINO. Immunological and histopathological studies in a roedent model infected with Leishmania infantum promastigotes or amastigostes. Res Parasitol, 89, p.163-169, 2003.
ROBBINS E COTRAN. Patologia: bases patológicas das doenças. 7ª ed. Rio de Janeiro: Elsevier; 2005.
ROGERS, K. A.; DEKREY, G. K.; MBOW, M. L.; GILLESPIE, R. D.; BRODSKYN C.I.; TITUS, R.G. Type 1 and type 2 responses to Leishmania major. FEMS Microbiol , v.19; n.209, p- 1-7, 2002.
ROUSH, R.T. Occurrence genetic and management of insecticide resistance. Parasitol Today, 9, p174-179, 1993.
SAHA, S.; MONDAL, S.; RAVINDRAN, R.; BHOWMICK, S.; MODAK, D.; MALLICK, S.; RAHMAN, M.; KAR, S.; GOSWAMI, R.; GUHA, S. K.; PRAMANIK, N.; SAHA, B.; ALI, N. IL-10- and TGF-beta-mediated susceptibility in

85
kala-azar andpost-kala-azarder- mal leishmaniasis: the significance of amphotericin Binthecontrol of Leishmania donovani infection in India. J. Immunol, 179, p-5592-5603, 2007.
SALDARRIAGA, O.A.; TRAVI, B.L., PARK, W.; PEREZ, L.E.; MELBY, P.C. Immunogenicity of a multicomponent DNA vaccine against visceral leishmaniasis in dogs. Vaccine. v.10, n.24, p.1928, 2006.
SALEN, A.K.; HUNG, T.W.; KIM, T.C.; SEARSON, K.W. Leong, multicomponent nanorods for vaccination applications. Nanatechnology, v.4, n.16, p. 484-487, 2005.
SANTAREM, N.; SILVESTRE, R.; TAVARES, J.; SILVA, M.; CABRAL, S.; MACIEL, J.; CORDEIRO-DA-SILVA, A. Immune response regulation by Leishmania secreted and nonsecreted antigens. J Biomed Biotechnol, 6, p-85154, 2007.
SARTORI, A.; ROQUE-BARREIRA, M.C.; COE, J.; CAMPOS-NETO, A. Immune complex glomerulonephritis in experimental kala-azar II: Detection and characterization of parasite antigens and antibodies eluted from kidneys of Leishmania donovani-infected hamsters. Clinical & Experimental Immunology , v.87, p.386-392, 1991.
SHARMA, P.; SINGH, N.; GARG, R.; HAQ, W.; DUBE, A. Efficacy of human beta-casein fragment (54-59) and its synthetic analogue compound 89/215 against Leishmania donovani in hamsters. Peptides, v. 25, n. 11, p.1873-1881, 2004.
SKEIKY, Y.A.W.; COLER, R.N.; BRANNON, M.; STROMBERG, E.; GREESON, K.; CRANE, T.; CAMPOS NETO, A.; REED,S. Protective efficacy of a tandemly linked, multi-subunit recombinant leishmanial vaccine (Leish-111f) formulated in MPL adjuvant. Vaccine, 20, p.3292-3303, 2002.
SOLIOZ, N.U.; BLUM-TIROUVNZIAM, R.; JACQUET, S.; RAFATI, G.; CORRADIN, J.; MAUEL, AND N. FASEL. The protective capacities of histone h1 against experimental murine cutaneous leishmaniasis. Vaccine 18, p.850-859, 1999.
SOTO. M.; REQUENA, J.M.; QUIJADA, L.; PEREZ, M.J.; NIETO, C.G. Antigenicity of the Leishmania infantum histones H2B and H4 during canine viscerocutaneous leishmaniasis. Clin Exp Immunol , 115, p.342-349, 1999.
STAGER, S.; SMITH, D.F.;KAYE, P.M. immunization with a recombinant stage-regulated surface protein from Leishmania donovani induces protection against visceral leishmaniasis. Journal of Immunology. v.165, p. 7064-7071, 2000.
TESH, R. Control of zoonotic visceral leishmaniasis: its time to change estrategies? Am. J. Trop. Med. and Hyg, 52(3), p.287-292, 1995.
TEWARY, P.; JAIN, M.; SAHANI, M.H.; SAXENA, S.; MADHUBALA, R. A heterologous prime-boost vaccination regimen using ORFF DNA and recombinant ORFF protein confers protective immunity against experimental visceral leishmaniasis. J Infect Dis. v.191, n.12, p.2130-2137, 2005.

86
TODOLÍ,F.; RODRÍGUEZ-CORTÉS,A.; NÚÑEZ,M.C.; LAURENTI,M.D.; GÓMEZ,S.S.; RODRÍGUEZ,F.; PÉREZ,M.E.; ESCRIBANO, .M.; ALBEROLA, J. Head-to-head comparison of three vaccination strategies based on DNA and raw insect-derived recombinant proteins against Leishmania. PLoS One. v.7, n.12, p.5118, 2012.
ULMER, J. B.; SADOFF, J C.; LIU, M. A. DNA and RNA-based vaccines: principles, progress and prospects. Gene vaccine Immunotecnology , v.8, n.4, p.531-536. 1996.
VIVIER, E.; MALISSEN, B. Innate and adaptive immunity: specificities and signaling hierarchies revisited. Nat Immun , v.7, p.17-21, 2005 .
WANG, R.; EPSTEIN, J.; BARACEROS, F.M.; GORAK, E.J.; CHAROENVIT, Y. Induction of CD4 (+) T cell-dependent CD8(+) type 1 responses in humans by a malaria DNA vaccine. Proc Natl Acad Sci U S A, 98, p.10817-10822, 2001.
WANG, Y.; CHEN, Y.; XIN, L.; BEVERLY, S.M.; CARLSEN, E.D.; POPOV, V.; CHANG, K.P.; WANG, M.; SOONG, L. Infection and immunity , v. 79, p. 1124-1133, 2011.
WHO, The World Health Report, Geneva. p. 192, v.7, 2002.
WILSON, M. E.; JERONIMO, S. M.; PEARSON, R. D. Immunopathogenesis of infection with the visceralizing Leishmania species. Microb Pathog, 38 (4), p.147-160. 2005.
YAO, C.; CHEN, Y.; SUDAN, B.; DONELSON, J. E.; WILSON, M. E. Leishmania chagasi: homogenous metacyclic promastigotes isolated by buoyant density are highly virulent in a mouse model. Exp Parasitol, 118(1), p.129-133, 2008.
Top Related