







Línguas
Páginas
Legal


UNIVERSIDADE FEDERAL FLUMINENSE FACULDADE DE FARMÁCIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS APLICADAS A PRODUTOS PARA SAÚDE
CAROLINA SERRA JOGAIB CABO
EXTRATO DICLOROMETANO DE Eugenia punicifolia: MODULAÇÃO DO
FENÓTIPO COLINÉRGICO NA RETINA DE RATOS NEONATOS IN VITRO
Orientadores: Prof. Dr. Wilson da Costa Santos Profa Dra. Carla Valéria Vieira Guilarducci Ferraz
Niterói 2014

1
CAROLINA SERRA JOGAIB CABO
EXTRATO DICLOROMETANO DE Eugenia punicifolia: MODULAÇÃO DO
FENÓTIPO COLINÉRGICO NA RETINA DE RATOS NEONATOS IN VITRO
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Aplicadas a Produtos para Saúde, da Faculdade de Farmácia, da Universidade Federal Fluminense, como requisito para obtenção do título de Mestre.
Dissertação desenvolvida no Laboratório de Farmacologia da Faculdade de Farmácia da Universidade Federal Fluminense
Orientadores: Prof. Dr. Wilson da Costa Santos Profa Dra. Carla Valéria Vieira Guilarducci Ferraz
Niterói 2014

2
CAROLINA SERRA JOGAIB CABO
EXTRATO DICLOROMETANO DE Eugenia punicifolia: MODULAÇÃO DO
FENÓTIPO COLINÉRGICO NA RETINA DE RATOS NEONATOS IN VITRO
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Aplicadas a Produtos para Saúde, da Faculdade de Farmácia, da Universidade Federal Fluminense, como requisito para obtenção do título de Mestre.
Aprovada em 30 de maio de 2014
BANCA EXAMINADORA
_________________________________________________ Dra. Karinne Cristinne da Silva Cunha
UNIRIO
_________________________________________________ Dra. Aline Araujo dos Santos Rabelo
UFF
_________________________________________________ Dr. Wilson da Costa Santos (Orientador)
UFF
Niterói 2014

3
Lâmpada para os meus pés é tua palavra, e luz para o meu caminho.
Salmos 119:105

4
Dedico este trabalho aos meus pais, Fátima e Ronald,
ao meu irmão, Ronald e àqueles que de forma direta ou indireta colaboraram para sua
realização.

5
AGRADECIMENTOS
À Deus pela oportunidade de concluir mais uma etapa de crescimento pessoal e profissional, e por ser minha fonte de forças, equilíbrio e paz nos momentos de dificuldade.
Aos meus pais, Fátima e Ronald, por proverem meus estudos e me
incentivarem sempre. A minha mãe por estar presente a cada instante em minha vida com seu apoio e amor incondicionais. Ao meu pai por sua ajuda e apoio para que eu me dedique aos estudos. A ambos pelo exemplo de vida e de trabalho. Amo vocês mais que o infinito!
Ao meu irmão, Roninho, por ser um exemplo de esforço e dedicação no
trabalho ao buscar sempre o melhor desempenho. Te admiro e amo demais, Ninho! A minha cunhada, Marta, por suas palavras de incentivo. Sei que vocês estão sempre mandando bons pensamentos para mim. Obrigada!
Aos meus orientadores, Wilson e Carla. Wilson por confiar a mim o trabalho
que possibilitou a realização do meu mestrado. Obrigada pela oportunidade e por seus conselhos! Carla por estar comigo durante o desenvolver do trabalho, me apoiando e passando seus conhecimentos em cada etapa da pesquisa. Obrigada por sua paciência, incentivo e confiança!
A professora Elizabeth Giestal por ter me acolhido em seu laboratório de
forma tão especial. Obrigada por sua ajuda durante o desenvolver do meu projeto e, além disso, obrigada pelo exemplo de profissional, que gosta do que faz, e pelo exemplo de pessoa por toda a sua generosidade! Jamais esquecerei tudo o que você fez por mim! A professora Aline por sua ajuda e atenção sempre que foi precisei!
Aos meus amigos pela compreensão nos momentos de ausência, pelas
risadas nos momentos de distração e por acreditarem em mim! Vocês tornam tudo mais fácil e leve: Rachel, Gabi Veiga, Munick, Ana Renata, Roberta, Fabiana, Raísa, Raquel, Camilly, Júlia Mattos e Júlia Hauaji.
Ao Anderson por seu incentivo e companheirismo nas horas de estudo e desenvolvimento dessa dissertação, e por todos os momentos de descontração.
As amigas que a faculdade me presenteou: Natália, Flaviane e Thais. Sei que
mesmo que cada uma esteja trilhando o seu caminho, uma torce pela outra da mesma forma. A Gabriela Deutsch por sua amizade e ajuda durante o mestrado.
Aos amigos do Laboratório de Cultura de Tecidos Hertha Meyer, que me
receberam de braços abertos, me ajudaram no aprendizado das técnicas desenvolvidas, transmitiram seus conhecimentos quando necessário e proporcionaram muitas risadas. Vocês tornam o ambiente de trabalho mais leve e agradável. Sentirei saudades! Muito obrigada Gérsica, Leandro, Babu, Luis, Patrícia, Marcelo, Thayana, Sheila, Lucienne, Karen, Thalita, Amanda, Érica, Adriano, Eliezer, Gustavo Mataruna e Alex. Aos amigos do Laboratório de Farmacologia, que apesar da pouca convivência também me acolheram de forma especial.

6
Aos técnicos do laboratório pela preparação do material do laboratório. Aos responsáveis pelo cuidado com os animais do biotério. Aos envolvidos nos serviços gerais prestados em nosso ambiente de pesquisa.

7
RESUMO
Estudos sobre os efeitos do extrato aquoso da Eugenia punicifolia (EP) demonstram sua ação na neurotransmissão da junção neuromuscular. Entretanto, apesar de saber que o aumento da neurotransmissão colinérgica pode apresentar atividade neuroprotetora e atuar na plasticidade neuronal, não existem estudos sobre o efeito do extrato de EP em células do Sistema Nervoso Central. O objetivo deste trabalho foi estudar o efeito do extrato diclorometano de EP sobre células da retina de ratos neonatos in vitro no que tange à proliferação celular, modulação do fenótipo colinérgico e aos níveis de fator de crescimento do nervo (NGF), fator neurotrófico derivado do cérebro (BDNF) e da interleucina IL-4. Foram realizadas culturas primárias de células da retina de ratos neonatos da linhagem Lister Hooded de ambos os sexos, dia pós-natal 0-2. As culturas foram plaqueadas em placas de Petri pré-tratadas com poli-L-ornitina, na densidade de 1,0x105 cel/cm2, receberam meio 199 ou 1µg/mL do extrato diclorometano de Eugenia punicifolia (EP 1µg/mL) e foram mantidas por 48 horas a 37°C, em atmosfera de 95% de ar e 5% de CO2. O método bioquímico utilizado para análise da proliferação celular foi a incorporação de [3H]-timidina. Os níveis dos receptores muscarínicos, de neurotrofinas e citocinas foram determinados por Western Blot. Todos os dados são apresentados em relação à porcentagem do controle (100%). Os procedimentos experimentais foram aprovados pelo Comitê de Ética no Uso de Animais da UFF (Projeto nº 186/2012). Os resultados mostram que o tratamento das culturas com diferentes concentrações do extrato diclorometano de EP por 48h induziu aumento dependente da concentração na proliferação celular, sendo o aumento mais significativo observado na concentração de 1µg/mL, concentração que foi utilizada em todos os experimentos. Foi observada a redução nos níveis dos receptores muscarínicos M1 e M4, e nos níveis de transportador de acetilcolina associado à vesícula, aumento na expressão do receptor M3 e nenhuma alteração nos níveis do receptor M5. Observou-se, também, que o extrato diclorometano de EP aumenta os níveis de NGF e da interleucina-4, e diminui dos níveis de BDNF. Os resultados sugerem que o extrato diclorometano de EP exerce efeito proliferativo e de diferenciação através da alteração do fenótipo colinérgico da retina e da participação de fatores neurotróficos. Palavras-chave: Eugenia punicifolia, receptores colinérgicos, neurotrofinas, interleucina-4.

8
ABSTRACT
Studies on the effects of the Eugenia punicifolia (EP) aqueous extract demonstrate its action on neurotransmission in the neuromuscular junction. However, despite knowing that increased cholinergic neurotransmission may have neuroprotective activity and act in neuronal plasticity, there are no studies on the effect of the extract of EP in cells of the Central Nervous System. The aim of this work was to study the effect of the dichloromethane extract of EP on retinal cells of neonatal rats in vitro with respect to cell proliferation, modulation of the cholinergic phenotype and levels of nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and interleukin IL-4. Primary cell cultures of neonatal rat retina from Hooded Lister strain of both sexes, postnatal day 0-2 were performed. Cultures were plated on pre-treated Petri plates with poly-L- ornithine at a density of 1.0 x105 cel/cm2 received medium 199 or 1μg/mL of dichloromethane extract of E. punicifolia (EP 1μg/mL) and kept for 48 hours at 37°C in an atmosphere of 95% air and 5% CO2. The method used for biochemical analysis of cellular proliferation was the incorporation of [3H]-thymidine. The levels of muscarinic receptors, neurotrophins and cytokines were determined by Western blot. All data are presented in relation to the percentage of control (100%). The experimental procedures were approved by the Ethics Committee on Animal Use of UFF (Project nº 186/2012). The results show that treatment of cultures with different concentrations of the dichloromethane extract of EP for 48h induces an increase in cell proliferation, with the most significant increase observed with the concentration of 1μg/mL, concentration, which was used in all experiments. A reduction in levels of muscarinic receptors M1 and M4, and VAChT was observed, an increase in expression of the M3 receptor and no change in the levels of the M5 receptor were observed. Also was observed that the dichloromethane extract of EP increased NGF levels and interleukin-4, and decreases levels of BDNF. The results suggest that the dichloromethane extract of EP exerts a proliferative effect and differentiation by altering the cholinergic phenotype of the retina and the involvement of neurotrophic factors. Keywords: Eugenia punicifolia, cholinergic receptors, neurotrophins, interleukin-4.
.

9
LISTA DE ILUSTRAÇÕES
Figura 1: Imagens da Eugenia punicifolia (Kunth) DC. A) árvore; B) frutos p.14 Figura 2: Sistema colinérgico (encéfalo humano) p.17 Figura 3: Estrutura química da acetilcolina p.18 Figura 4: Representação esquemática da sinapse colinérgica p.20 Figura 5: Estrutura dos receptores nicotínicos neurais p.23 Figura 6: Estrutura dos receptores muscarínicos p.25 Quadro 1: Subtipos de receptores muscarínicos centrais e periféricos, transdução de sinal, perfil de expressão e doenças relacionadas p.26 Figura 7: Diagrama esquemático das interações dos receptores com neurotrofinas maduras e pró-neurotrofinas p.30 Figura 8: Histogênese da retina p.33 Figura 9: Esquema simplificado da retina de vertebrados recém-nascidos (A) e adultos (B) p.34 Figura 10: Regulação do ciclo celular p.40 Figura 11: Curva concentração-resposta do extrato de E. punicifolia em células da
retina mantidas em cultura por 48h p.51 Figura 12: Fotomicrografia em contraste de fase da morfologia das culturas de
céuluas da retina após 48 horas in vitro p.52 Figura 13. Níveis de receptor M1 em culturas de células da retina tratadas com E.
punicifolia por 48h p.53 Figura 14. Níveis de receptor M3 em culturas de células da retina tratadas com E.
punicifolia por 48h p.54 Figura 15. Níveis de receptor M4 em culturas de células da retina tratadas com E.
punicifolia por 48h p.55 Figura 16. Níveis de receptor M5 em culturas de células da retina tratadas com E.
punicifolia por 48h p.56 Figura 17. Níveis da subunidade α7 do receptor nicotínico (Nα7) em culturas de
células da retina tratadas com E. punicifolia por 48h p.57 Figura 18. Níveis do transportador de acetilcolina associoado à vesícula
VAChT em culturas de células da retina tratadas com E. punicifolia p.58 Figura 19. Níveis do fator de crescimento do nervo (NGF) em culturas de células da
retina tratadas com E. punicifolia p.59 Figura 20. Níveis do fator neurotrófico derivado do cérebro (BDNF) em culturas de
células da retina tratadas com E. punicifolia p.60 Figura 21. Níveis de interleucina IL-4 em culturas de células da retina tratadas com
E. punicifolia p.61

10
LISTA DE ABREVIATURAS, SIGLAS E SÍMBOLOS
[3H] – Trítio 5–HT3 – 5-hidroxitriptamina 3 α-Bgtx – Alfa bungarotoxina Acetil CoA – Acetil coenzima A ACh – Acetilcolina AChE – Acetilcolinesterase AChRs – Receptores de acetilcolina AMPc – Adenosina monofosfato cíclico ATP – Adenosina trifosfato BDNF – Fator neurotrófico derivado do cérebro BSA – Albumina de soro bovino BuChE – Butirilcolinesterase CAK – Ativador da cinase CCG – Camada de células ganglionares CDKs – Cinases dependentes de ciclina CGR – Células ganglionares da retina ChAT – Colina acetiltransferase CHT / CHT1 – Transportador de colina de alta afinidade CKI – Inibidores de CDK Cl- – Íon cloro CMF – Solução salina sem cálcio e sem magnésio CNE – Camada nuclear externa CNI – Camada nuclear interna CNTF – Fator trófico para neurônios do gânglio ciliar CPE – Camada plexiforme externa CPI – Camada plexiforme interna CT – Controle DAG – Diacilglicerol DMD – Distrofia muscular de Duchenne DMSO – Dimetilsulfóxido DA – Doença de Alzheimer DNA – Ácido desoxirribonucleico E – Período embrionário ECL – Solução reagente de Luminol EGFRs – Fator de crescimento epidermal EP – Eugenia punicifolia EPR – Epitélio pigmentar da retina ERK – Cinase regulada por sinal extracelular Fase M – Mitose FGFb / FGF-2 – Fator de crescimento de fibroblasto básico GABA – Ácido gama aminobutírico GRKs – Cinase de receptores acoplados a proteína G HC-3 – Hemicolinium-3 IFN-γ – Interferon gama IL-1β – Interleucina 1β IL-2 – Interleucina 2 IL-4 – Interleucina 4 IL-4R – Receptor de IL-4

11
IL-4Rα – Subunidade alfa do receptor de IL-4 IL-13Rα1 – Subunidade alfa 1 do receptor de IL-13 IP3 – Inositol 1,4,5-trifosfato LIF – Fator inibitório de leucemia M1, M2, M3, M4, M5 – Subtipos de receptores muscarínicos mAChRs – Receptores muscarínicos MMP – Metaloproteinases de matriz ms – Septo medial Nα7 – Receptor nicotínico de subunidade alfa 7 Na+ – Íon sódio nAChRs – Receptores nicotínicos nb – Núcleos basais NF-κB – Fator nuclear kappa B NGF – Fator de crescimento do nervo NTs – Neurotrofinas NT-3 – Neurotrofina 3 NT-4/5 – Neurotrofina 4/5 NT-6 – Neurotrofina 6 NT-7 – Neurotrofina 7 P – Período pós-natal p75 NTR – Receptor de neurotrofina p75 p53 – Gene suppressor de tumor p53 pRb/Rb – Proteína de retinoblastoma PKC – Proteína cinase C PMA – 13-acetato de forbol 12-miristato Proteína G – Proteínas ligantes de nucleotídeo de guanina PVDF – Polímero de fluoreto de vinilideno Raf-1 cinase – Cinase que ativa a MEK RNAm – Ácido ribonucleico mensageiro SDS – Dodecil sulfato de sódio SFB – Soro fetal bovino SNC – Sistema Nervoso Central SNP – Sistema Nervoso Periférico Src – Protooncogene proteína cinase tirosina TBS – Solução salina tamponada por Tris TCA – Ácido tricloroacético TEMED – N,N,N’,N’-tetrametiletilenodiamina TGFα – Fator de transformação de crescimento alfa TNF – Fator de necrose tumoral TNFα – Fator de necrose tumoral alfa Trk – Receptor com atividade cinase relacionado à tropomiosina VAChT – Transportador de acetilcolina associoado à vesícula VAT – Transportador associado à vesícula

12
SUMÁRIO
1 INTRODUÇÃO p. 13 2 REVISÃO DE LITERATURA p. 14 2.1 Eugenia punicifolia p.14 2.2 Sistema colinérgico p. 17 2.2.1 Acetilcolina p. 18 2.2.2 Receptores colinérgicos p. 21 2.3 Sistema Nervoso Central p. 27 2.3.1 Neurotrofinas e citocinas p.28 2.3.2 Retina p.32 2.3.2.1 Atividade colinérgica na retina p.35 2.3.2.2 Ações das neurotrofinas na retina p.37 2.4 Ciclo celular p.39 3. OBJETIVOS p. 42 3.1 Objetivo geral p. 42 3.2 Objetivos específicos p. 42 4 MATERIAL E MÉTODOS p. 43 4.1 Material p. 43 4.2 Extrato diclorometano da Eugenia punicifolia p. 44 4.3 Animais experimentais p. 44 4.4 Cultura de células da retina p. 45 4.5 Incorporação de timidina p. 45 4.6 Western blot p. 46 4.6.1 Extração de proteínas p. 47 4.6.2 Dosagem de proteínas p. 47 4.6.3 Preparação do gel de poliacrilamida p. 47 4.6.4 Transferência das proteínas do gel para a membrana de PVDF p. 48 4.6.5 Bloqueio e imunodetecção p. 48 4.6.6 Revelação p. 48 4.7 Fotomicrografia em contraste de fase p. 49 4.8 Análise estatística dos resultados p.49 4.9 Questões éticas p. 50 5 RESULTADOS p. 51 6 DISCUSSÃO p. 62 7 CONCLUSÕES p. 74 8 REFERÊNCIAS BIBLIOGRÁFICAS p. 75 9 ANEXOS 9.1 Anexo 1 - Folha de aprovação do projeto pelo Comitê de Ética no Uso de Animais p. 95

13
1 INTRODUÇÃO
Historicamente, há uma longa tradição de pesquisas sobre o perfil
farmacológico de extratos de plantas, e consequente busca por novos compostos
bioativos com interesse terapêutico. Esta estratégia tem conduzido à descoberta de
diversos compostos que deram origem a várias moléculas para tratamento de uma
série de doenças (MARCAURELLE & JOHANNES, 2008). Neste aspecto, nosso
grupo de pesquisas tem interesse na determinação de extratos de produtos naturais
com atividade farmacológica. Nos últimos anos, interessamo-nos por estudar os
efeitos colinérgicos de Eugenia punicifolia, uma planta pertencente à família
Myrtaceae, de larga ocorrência na região amazônica do Brasil bem como no
Nordeste do Brasil. A planta é amplamente empregada, embora de forma empírica,
no tratamento de distúrbios estomacais (JORGE et al., 2000) e na diabetes mellitus
(BRITO et al., 2007). Recentemente foi demonstrada a importância de extratos da E.
punicifolia sobre a transmissão colinérgica periférica (GRANGEIRO et al., 2006;
LEITE et al., 2010).
Sabendo que o aumento da neurotransmissão colinérgica pode apresentar
atividade neuroprotetora e atuar na plasticidade neuronal (RAFII & AISEN, 2009), é
racional realizar experimentos com extratos de E. punicifolia em células do Sistema
Nervoso Central (SNC). Esta investigação é relevante uma vez que não há nenhum
estudo utilizando este extrato neste tecido, o qual apresenta grande dependência da
atividade colinérgica, tanto durante o desenvolvimento quanto na manutenção da
homeostasia em adultos (DESCARRIES et al., 1997; PERRY et al., 1999). Para o
desenvolvimento do estudo foi utilizado, como modelo experimental, retina de ratos
neonatos, pertencente ao SNC e de fácil obtenção, livre de outras populações
celulares.

14
2 REVISÃO DE LITERATURA
2.1 Eugenia punicifolia
Eugenia punicifolia (Humb., Bonpl. & Kunt) DC é uma planta arbustiva
pertencente à família Myrtaceae, constituída por mais de 3.000 espécies dos
gêneros Syzigium e Eugenia, largamente distribuída em toda a Amazônia (Figura 1).
A família Myrtaceae é uma das mais características da flora brasileira e entre suas
espécies são encontrados vegetais de uso medicinal, ornamentais, produtores de
madeiras e de frutos comestíveis. As folhas das mirtáceas são simples e
hipoestomáticas, com estômatos do tipo ranunculáceo (JORGE et al., 2000). Além
das caracteríscas gerais, a Eugenia punicifolia apresenta folhas com contorno
predominantemente oboval. Quanto às características microscópicas a espécie
apresenta células epidérmicas de contorno onduloso, recobertas por cutícula
estriada, em secção paradérmica; nervuras medianas plano-convexas com feixes
bicolaterais envoltos por bainha esclerenquimática; esporadicamente há presença de
cristais dos tipos drusas e prismáticos, e o parênquima paliçádico em duas camadas
(JORGE et al., 2000, OLIVEIRA et al., 2005).
A B Figura 1 – Imagens da Eugenia punicifolia (Kunth) DC. A) árvore; B) frutos. Fonte: A)
http://www.kew.org/science/tropamerica/imagedatabase/large1/cat_single1-1636.html B) http://www.kew.org/science/tropamerica/imagedatabase/index.html

15
A composição química do óleo essencial das folhas secas de E. punicifolia
varia conforme a região de coleta da planta, devido a diferenças nos parâmetros
climáticos e geográficos. Considerando plantas colhidas na região de Matas
Serranas próximas à Pernambuco, a composição química consiste principalmente
de monoterpenos oxigenados como o 1,8-cineol, o linalol e o α-terpineol;
sesquiterpenos oxigenados como o β-cariofileno, elemol e α-cadinol (OLIVEIRA et
al., 2005). Por outro lado, nas plantas colhidas próximas a Manaus, a composição
química é constituída majoritariamente de benzenóides e β-cariofileno (MAIA et al.,
1997).
Popularmente denominada “pedra-ume caá”, as folhas frescas são utilizadas
na forma de infusão ou decocção no tratamento de distúrbios estomacais (JORGE et
al., 2000) e da diabetes mellitus (BRITO et al., 2007; BOPP et al., 2009). Outras
espécies do gênero Eugenia são utilizadas na terapêutica popular e, estudos
comprovaram a eficácia da E. uniflora como agente anti-hipertensivo (CONSOLINI et
al., 1999), antidiarréico, diurético e antirreumático (PIO CORREA, 1984;
SCHAPOVAL et al., 1994). As propriedades farmacológicas descritas para os
membros do gênero Eugenia são atribuídas à ação de terpenóides, taninos,
antraquinonas e flavonóides (mircitina, quercitina) (CONSOLINI & SARUBBIO,
2002).
O uso da E. punicifolia na medicina tradicional foi comprovado em ratos com
diabetes induzida por estreptozotocina. Nesses animais, a administração crônica do
extrato metanólico de folhas de E. punicifolia apresentou efeito benéfico com a
redução dos sintomas da diabetes, melhorando o metabolismo de carboidratos e de
proteína. Por outro lado, o extrato aquoso induziu um efeito anoréxico, uma vez que
houve significativa redução da ingestão de alimentos em todos os animais do grupo
tratado. Cabe ressaltar que ambos os extratos não induziram nenhum tipo de lesão
hepatocelular, muscular, pancreática ou microvascular (BRUNETTI et al., 2006).
Em preparações de diafragma de ratos, as contrações induzidas por
estímulos elétricos são bloqueadas pelos antagonistas nicotínicos galamina e

16
pancurônio. A utilização de extrato aquoso de E. punicifolia nessa preparação
induziu reversão completa dos efeitos observados com os antagonistas nicotínicos,
galamina e pancurônio, diferente daquela observada quando foi utilizado a
neostigmina (inibidor reversível da enzima acetilcolinesterase). Também foi proposta
atividade inibitória do extrato aquoso na acetilcolinesterase (GRANGEIRO et al.,
2006). Estes resultados sugerem uma participação importante dos receptores
nicotínicos nas respostas induzidas pela E. punicifolia.
O tratamento de camundongos com distrofia muscular de Duchenne (DMD),
com a fração de diclorometano de E. punicifolia, exerce efeito antiinflamatório no
músculo gastrocnêmico, através da redução da produção de mediadores da
inflamação e redução de apoptose (LEITE et al., 2010). Outro efeito observado com
o extrato diclorometano de E. punicifolia foi a capacidade de aumentar a liberação
exocitótica de catecolaminas desencadeada pela acetilcolina ou com soluções
enriquecidas de potássio em células cromafins da glândula adrenal de bovinos
(PASCUAL et al., 2011). Outra atividade importante descrita para as folhas de
Eugenia punicifolia é a ação antioxidante e inibidora de enzimas como α-amilase, α-
glicosidase e xantina oxidase. Essa última pode levar à redução da taxa de absorção
de carboidratos e diminuir fatores de risco relacionados a doenças cardiovasculares,
proporcionando assim uma nova alternativa dietética para a prevenção da síndrome
metabólica (GALENO et al., 2013).
A partir dos resultados descritos, pode ser sugerido que a E. punicifolia é uma
importante ferramenta para estudos da neurotransmissão colinérgica. Em distúrbios
como doença de Alzheimer, Síndrome de Tourette, dentre outras, sabe-se que há
uma alteração importante da atividade do sistema colinérgico, porém as alterações
fisiopatológicas não estão totalmente elucidadas. A interferência no sistema
colinérgico é uma das bases clínicas para o tratamento de algumas destas doenças.
Sendo assim, torna-se importante ponderar a utilização de extratos da E. punicifolia
como importante ferramenta experimental para o desenvolvimento de novas
estratégias terapêuticas para tais doenças.

17
2.2 Sistema Colinérgico
O sistema colinérgico tem papel fundamental em várias funções vitais, tanto
centrais quanto periféricas, o que faz deste sistema, atualmente, alvo de inúmeras
pesquisas (MESULAM et al., 2002). Seu bloqueio agudo é geralmente letal;
enquanto sua perda gradual, como observada na Doença de Alzheimer (DA), é
associada com a deterioração progressiva de funções neurais (HOFFMAN &
TAYLOR, 2001). Sua importância é observada na manutenção e modulação da
homeostasia do Sistema Nervoso Central (SNC), além de desempenhar funções
importantes durante o desenvolvimento. No SNC regula a proliferação e
diferenciação de células progenitoras (ANGELIS et al., 2012), o desenvolvimento de
células gliais, a maturação e a plasticidade neuronal (BROIDE & LESLIE, 1999), o
crescimento axonal, a expressão gênica e a sobrevivência celular (PEZET &
MCMAHON, 2006), influenciando em processos cognitivos como atenção, memória,
aprendizado e vigília (PEPEU & GIOVANINNI, 2004). As diferentes regiões cerebrais
envolvidas nas transmissões colinérgicas estão descritas na Figura 2.
Figura 2: Sistema colinérgico (encéfalo humano). A localização dos principais grupos de corpos celulares e tratos de fibras colinérgicas, núcleos septo medial (ms) e núcleo basal (nb) (vermelho). O núcleo pedúnculo pontino (azul) e os interneurônios estriatais (laranja). (Adaptado de PERRYet al. (1999), modificado por MAZZANTI, 2007).

18
Os neurônios colinérgicos e suas projeções se encontram amplamente
distribuídos através do prosencéfalo, região na qual estão os hemisférios cerebrais,
núcleos da base, diencéfalo e porção frontal do mesencéfalo (BAGLEY, 2005). O
mais significante grupo de neurônios colinérgicos se encontra nos núcleos do septo
medial (ms) e nos núcleos basais (nb), e suas projeções vão para a região do
hipocampo, amígdala e córtex cerebral. Os neurônios colinérgicos que se encontram
no núcleo pedúnculo pontino se projetam para o tálamo e os interneurônios estriatais
apresentam um circuito colinérgico intrínseco (MESULAM, 1995; PERRY et al, 1999)
As funções do sistema colinérgico estão associadas à ação de seus diferentes
componentes como: acetilcolina (ACh), colina acetiltransferase (ChAT),
transportador associado à vesícula (VAT), transportadores de colina de alta
afinidade (CHT), acetilcolinesterase (AChE), receptores nicotínicos (nAChR) e
muscarínicos (mAChR) (ABREU-VILLAÇA et al., 2011 para revisão).
2.2.1 Acetilcolina
Em 1921, a acetilcolina foi a primeira molécula descrita como
neurotransmissor (BENNETT, 2000), e passou a ser amplamente estudada nas
sinapses centrais e periféricas (DESCARRIES et al., 1997; FERREIRA et al., 2008).
Figura 3: Estrutura química da acetilcolina. (FERREIRA et al., 2008)
Quimicamente, a acetilcolina, é um éster de ácido acético e colina (Figura 3),
cuja simplicidade estrutural subestima a complexidade das ações que exerce. É
sintetizada a partir da colina e acetil coenzima A (Acetil CoA) nos neurônios pré-
sinápticos, pela ação da colina acetiltransferase (ChAT) (ABREU-VILLAÇA et al.,
2011) (Figura 4). Os níveis de acetil-CoA são mantidos através do metabolismo da

19
glicose, sob ação da enzima ATP-citrato-liase, que apresenta sua expressão
aumentada em neurônios colinérgicos (TOMASZEWICZ et al., 2003; BEIGNEUX et
al., 2004). O suprimento de colina é provido principalmente da dieta, devido à
incapacidade de neurônios em sintetizar colina (FERNSTROM, 1981; ZEISEL,
1981). A ACh é armazenada em vesículas por meio do transportador associado à
vesícula (VAChT), sendo liberada de forma quântica, após influxo de cálcio no
terminal pré-sináptico que vai induzir alterações de diversas proteínas, favorecendo
a fusão da vesícula com o terminal pré-sináptico. Uma vez liberada na fenda
sináptica, a ACh interage com receptores nicotínicos (nAChRs), causando
despolarização e propagação do potencial de ação na célula pós-sináptica, e com
receptores muscarínicos (mAChRs) (ODA, 1999). O término da ação da ACh ocorre
por hidrólise pela enzima acetilcolinesterase (AChE) gerando colina e acetato
(FERREIRA et al., 2008). Grande parte da colina resultante é captada pelo terminal
colinérgico pré-sináptico pelo transportador de colina de alta afinidade (CHT1) e
reutilizada na síntese de nova ACh (ABREU-VILLAÇA et al., 2011 para revisão;
FERREIRA et al., 2008; MESULAM et al., 2002).

20
Figura 4: Representação esquemática da sinapse colinérgica. VAChT: transportador vesicular de acetilcolina; CHT: transportador de colina; AChE: acetilcolinesterase; ChAT: colina acetil transferase; Acetyl CoA: acetilcoenzima A; ACh: acetilcolina; nAChR: receptor nicotínico de acetilcolina; mAChR: receptor muscarínico de acetilcolina. Fonte: Abreu-Villaça et al., 2011.
O término da transmissão sináptica pela hidrólise do neurotransmissor é uma
característica substancial das sinapses colinérgicas. Este mecanismo único faz da
acetilcolinesterase (AChE), uma serina hidrolase encontrada predominantemente no
cérebro, junção neuromuscular e eritrócitos, encarregada de executar a hidrólise da
ACh, peça chave da sinalização colinérgica (ZIMMERMAN & SOREQ, 2006). A
hidrólise da ACh também pode ser feita por uma enzima menos específica chamada
butirilcolinesterase (BuChE) (ou pseudocolinesterase), que se encontra no plasma,
rins, fígado, intestino, coração, pulmão e tem distribuição neuronal mais restrita em
relação a AChE (MESULAM et al., 2002)
Como mencionado anteriormente, a colina resultante da hidrólise da
acetilcolina é recaptada pelo terminal pré-sináptico, e isso se dá através do
transporte ativo realizado pelo transportador de colina de alta afinidade, CHT1, que
depende dos íons Na+ e Cl- (YAMAMURA & SNYDER,1973; COLLIER & ILSON,

21
1977). Este transporte é específico de neurônios colinérgicos e pode ser inibido por
baixas concentrações de hemicolinium-3 (HC-3). Em outras células há outro tipo de
transporte de colina, que é inibido por doses maiores de HC-3 (YAMAMURA &
SNYDER,1973)
A respeito da caracterização do transporte de colina de alta afinidade,
trabalhos mostraram que o aumento da atividade de neurônios colinérgicos gera
aumento no transporte de colina e síntese de ACh (COLLIER & KATZ, 1974; SIMON
& KUHAR, 1975; SIMON et al., 1976; O’REGAN & COLLIER 1981; SALTARELLIi et
al., 1987). Segundo Fergunson e colaboradores, 2003 o aumento da atividade
neuronal acarreta em aumento da fusão de vesículas sinápticas e maior fluxo de
CHT1 em direção à membrana plasmática. Maior captação de colina ocorre com o
aumento da concentração do CHT1 na membrana pré-sináptica, e com isso a
síntese de ACh é favorecida. (RIBEIRO et al., 2003; FERGUNSON et al., 2003).
O VAChT é outro transportador colinérgico responsável por transportar a ACh
sintetizada no citoplasma para o interior das vesículas sinápticas através de
transporte ativo, que utiliza gradiente de prótons gerado pela ATPase do tipo V. Esse
transporte realizado pelo VAChT possui relação de dois prótons por molécula de
acetilcolina (PARSONS et al., 1993).
O isômero levógiro do vesamicol é capaz de inibir o transporte de ACh
impedindo o preenchimento de vesículas colinérgicas, caracterizando assim uma
inibição alostérica e não competitiva. (PARSONS et al., 1993).
2.2.2 Receptores colinérgicos
A ACh exerce suas diversas ações fisiológicas através da interação com duas
classes distintas de receptores: os nicotínicos (nAChRs) e os muscarínicos
(mAChRs). Tais receptores foram divididos nessas classes através da

22
caracterização da atividade farmacológica de alcaloides como a nicotina e a
muscarina (VENTURA et al., 2010).
Receptores nicotínicos
Os receptores nicotínicos fazem parte da superfamília de canais iônicos
ativados por ligante, que também inclui receptores de ácido gama aminobutírico
(GABAA e GABAB), glicina e receptores de serotonina (5–HT3) (FERREIRA et al.,
2008, CHANGEUX, 2003). Os receptores nicotínicos compreendem combinações
pentaméricas de subunidades individuais (NAI et al., 2003). Por sua distribuição
anatômica e propriedades farmacológicas, esses receptores podem ser divididos em
três grupos: receptores musculares, encontrados na junção neuromuscular
esquelética; receptores ganglionares, nos terminais pós-sinápticos dos gânglios
autônomos; e receptores neuronais, amplamente distribuídos no sistema nervoso
central (HOGG et al., 2003).
No SNC, a inervação colinérgica regula diferentes processos tais como
liberação de neurotransmissores, excitabilidade celular e integração neuronal. Tais
ações envolvem os receptores nicotínicos em funções e estados fisiológicos como
sono, alerta, fadiga, ansiedade, processamento central da dor e uma série de
funções cognitivas (GOTTI & CLEMENTI, 2004). Tem sido demonstrado que a
perturbação da neurotransmissão colinérgica nicotínica pode levar a doenças
envolvendo disfunção do sistema colinérgico durante o desenvolvimento, como o
vício, epilepsia, esquizofrenia, mal de Alzheimer, miastenia, entre outras
(CHANGEUX & EDELSTEIN, 2001; GOTTI et al, 1997; HOGG et al, 2003;
LINDSTROM, 1997).

23
Figura 5: Estrutura dos receptores nicotínicos neurais. A) Topologia das subunidades nAChR dos receptores neuronais. B) Formação do poro do canal. C) Receptores homoméricos. D) receptores heteroméricos. Os triângulos vermelhos em C e D indicam os locais de interação da acetilcolina nos receptores. Fonte: HENDRICKSON et al., 2013.
Os subtipos de nAChR neuronais são localizados nas membranas pré- e pós-
sinápticas dos neurônios e em locais extra-sinápticos no SNC (AMENTA &
TAYEBATI, 2008). Múltiplos subtipos de nAChRs podem ser formados a partir das
várias combinações de subunidades, e esses subtipos podem ser classificados
como heteropentaméricos ou homopentaméricos (ABREU-VILLAÇA et al., 2011 para
revisão; FERREIRA et al., 2008). Para os receptores nicotínicos (nAChR) neuronais
as subunidades contém 4 domínios transmembrana (M1-M4) carboxiterminal, uma
região extracelular aminoterminal e uma proeminente alça intracelular (M3-M4) de
tamanhos variados (Figura 5A). Cinco subunidades iguais (receptores
homopentaméricos) ou distintas (receptores heteropentaméricos) se organizam para
formar o poro do canal (Figura 5B), o qual é permeável aos íons Na+ e Ca+2. Os
receptores homopentaméricos consistem de subunidades α somente e apresentam
baixa afinidade pelo agonista (Figura 5C). Enquanto que a maioria dos receptores de
alta afinidade são heteropentaméricos, formados pela combinação de subunidades α
e β que se organizam entre si, formando, por exemplo, os receptores α4α6β2β3,
α4β2 e α3β4 (Figura 5D) (HENDRICKSON et al., 2013).
A família de subunidades de AChRs é agrupada em duas classes principais:
Sensíveis à toxina de serpente (α-bungarotoxina-AChRs), formando AChRs
homopentaméricos (α7-α9), com cinco subunidades idênticas formando um canal

24
iônico; e heteropentaméricos (α7, α8 ou α9 e α10) (ZHANG et al., 1994;
LINDSTROM, 2000; HENDRICKSON et al., 2013). O receptor homomérico é ativado
pela acetilcolina e bloqueado por concentrações nanomolares de α-bungarotoxina
com alta permeabilidade ao Ca2+ e rápida taxa de dessensibilização. A classe não-
sensível à α-bungarotoxina (nAChRs) possui ligação à nicotina com alta afinidade,
formando sempre AChRs heteropentaméricos: contém um par de subunidades α
idênticas (α2-α6) como ligantes e requer associação com três subunidades β
idênticas (β2-β4) para serem um AChR funcional (LINDSTROM, 2000;
HENDRICKSON et al., 2013).
A maior seletividade dos receptores nicotínicos (α7)5 do sistema nervoso
central ao cálcio permite que sejam desencadeadas, por esse íon, respostas a longo
e médio prazo, como regulação da liberação de neurotransmissores, mecanismos de
plasticidade neuronal e memória, dependência a drogas, neuroproteção e morte
celular (ROLE & BERG, 1996; HENDRICKSON et al., 2013).
Receptores muscarínicos
As ações muscarínicas da acetilcolina são mediadas por cinco subtipos
distintos de receptores (FERREIRA et al., 2008; NEUBIG & SIDEROVSKI, 2002;
CAULFIELD & BIRDSALL, 1998) (Figura 6). Baseado na sua habilidade em ativar
diferentes classes de proteína G heterotriméricas, os cinco subtipos podem ser
agrupados em duas principais classes funcionais. Os receptores M2 e M4 mostram
seletividade para proteína Gi, inibem a atividade da adenilil ciclase, enquanto os
receptores M1, M3 e M5 estão seletivamente acoplados à proteína Gq, que ativa
fosfolipase C, levando à formação de inositol 1,4,5-trifosfato (IP3) e diacilglicerol
(DAG) (NEUBIG & SIDEROVSKI, 2002; ABREU-VILLAÇA et al., 2011 para revisão).

25
Figura 6: Estrutura dos receptores muscarinicos. Adaptado de: Neubig & Siderovski, 2002.
Os receptores muscarínicos estão presentes em diferentes órgãos, tecidos ou
tipos celulares (WOLFE & YASUDA, 1995; LEVEY, 1983). Receptores muscarínicos
periféricos estão envolvidos nas ações clássicas da ACh em tecidos ou órgãos que
recebem inervação parassimpática (CAULFIELD, 1993), enquanto os mAChRs
centrais participam da regulação de um grande número de funções cognitivas,
comportamentais, sensoriais, motoras e autonômicas (FELDER et al. 2003).
Na periferia, os receptores do tipo M3 são responsáveis por respostas
funcionais como a contração dos músculos lisos e estimulação da secreção
glandular, enquanto que os receptores do tipo M2 medeiam a inibição de canais de
cálcio voltagem-dependentes no miocárdio, sendo este resultado da inibição da
adenilato ciclase (MATSUI el at, 2004) modulando a resposta crono e inotrópica dos
músculos cardíacos.
De modo geral, no Sistema Nervoso Central, os receptores do tipo M1 são
mais abundantes, enquanto os receptores M2 e M4 são expressos de forma
moderada e os receptores M3 e M5 apresentam baixos níveis de expressão. No
hipocampo os receptores M1 e M3 são mais expressos em neurônios, os receptores
M2 e M4 são localizados em interneurônios, e os receptores M5 são pouco
expressos. A diversidade dos efeitos muscarínicos somada à presença destes
subtipos moleculares no hipocampo sugere que estes receptores possuam
importante papel na memória e no processo de aprendizado (DREVER et al., 2011).
No Quadro 1 são descritos os subtipos de receptores muscarínicos, suas
respectivas transduções de sinal, regiões onde são expressos e doenças a eles
relacionadas.
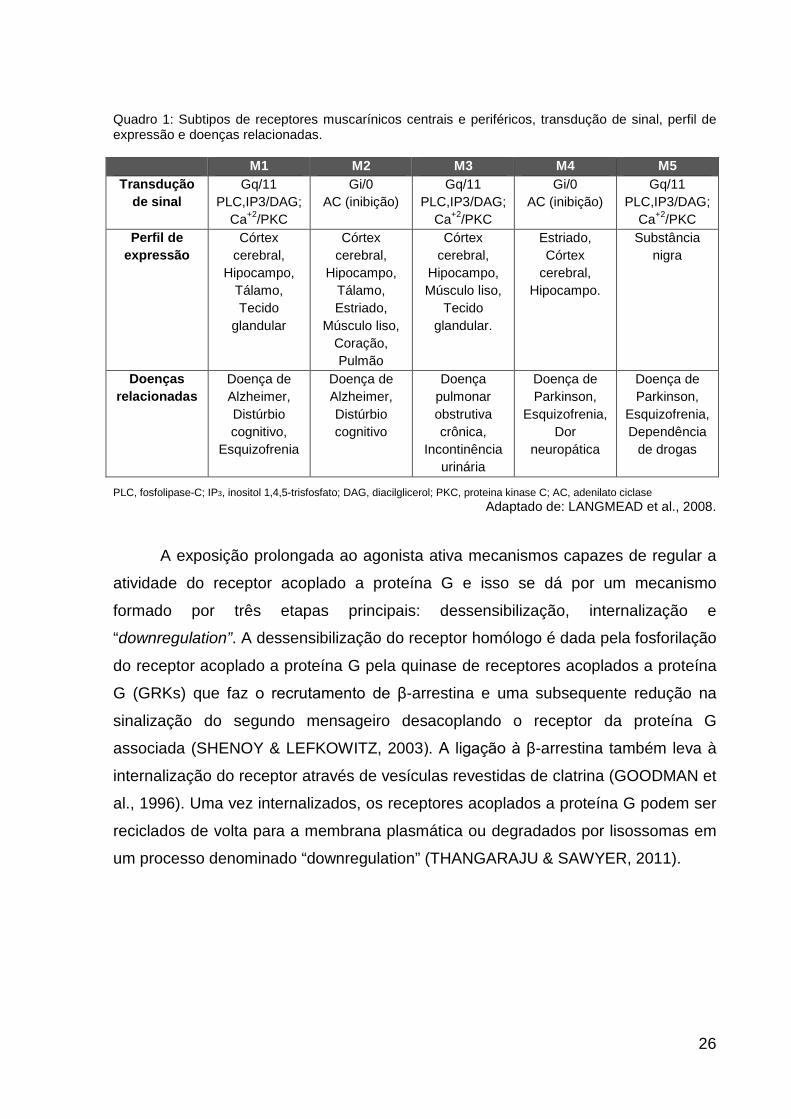
26
Quadro 1: Subtipos de receptores muscarínicos centrais e periféricos, transdução de sinal, perfil de expressão e doenças relacionadas.
M1 M2 M3 M4 M5 Transdução
de sinal Gq/11
PLC,IP3/DAG; Ca+2/PKC
Gi/0 AC (inibição)
Gq/11 PLC,IP3/DAG;
Ca+2/PKC
Gi/0 AC (inibição)
Gq/11 PLC,IP3/DAG;
Ca+2/PKC Perfil de
expressão Córtex
cerebral, Hipocampo,
Tálamo, Tecido
glandular
Córtex cerebral,
Hipocampo, Tálamo, Estriado,
Músculo liso, Coração, Pulmão
Córtex cerebral,
Hipocampo, Músculo liso,
Tecido glandular.
Estriado, Córtex
cerebral, Hipocampo.
Substância nigra
Doenças relacionadas
Doença de Alzheimer, Distúrbio cognitivo,
Esquizofrenia
Doença de Alzheimer, Distúrbio cognitivo
Doença pulmonar obstrutiva crônica,
Incontinência urinária
Doença de Parkinson,
Esquizofrenia, Dor
neuropática
Doença de Parkinson,
Esquizofrenia, Dependência
de drogas
PLC, fosfolipase-C; IP3, inositol 1,4,5-trisfosfato; DAG, diacilglicerol; PKC, proteina kinase C; AC, adenilato ciclase
Adaptado de: LANGMEAD et al., 2008.
A exposição prolongada ao agonista ativa mecanismos capazes de regular a
atividade do receptor acoplado a proteína G e isso se dá por um mecanismo
formado por três etapas principais: dessensibilização, internalização e
“downregulation”. A dessensibilização do receptor homólogo é dada pela fosforilação
do receptor acoplado a proteína G pela quinase de receptores acoplados a proteína
G (GRKs) que faz o recrutamento de β-arrestina e uma subsequente redução na
sinalização do segundo mensageiro desacoplando o receptor da proteína G
associada (SHENOY & LEFKOWITZ, 2003). A ligação à β-arrestina também leva à
internalização do receptor através de vesículas revestidas de clatrina (GOODMAN et
al., 1996). Uma vez internalizados, os receptores acoplados a proteína G podem ser
reciclados de volta para a membrana plasmática ou degradados por lisossomas em
um processo denominado “downregulation” (THANGARAJU & SAWYER, 2011).

27
2.3 Sistema Nervoso Central
A formação das estruturas do Sistema Nervoso Central (SNC) depende do
balanço controlado entre a neurogênese e a morte celular durante o
desenvolvimento. No SNC de mamíferos, a interação de vários tipos neuronais e
suas terminações sinápticas, bem como a plasticidade e a rede de sinalização que
envolve diferentes citocinas e neurotransmissores, são responsáveis pelos
processos de desenvolvimento e funcionamento deste complexo sistema (REUSS &
HALBACH, 2003).
Durante o desenvolvimento do SNC, ocorrem fenômenos progressivos de
proliferação celular e neuritogênese, de forma que as células geradas buscam
contato com as populações alvo, na formação da árvore dendrítica. O crescimento
dos prolongamentos até o alvo depende de moléculas tróficas liberadas no meio
pelo próprio neurônio, por neurônios vizinhos e por células gliais. Esta sinalização
autócrina e parácrina é necessária para a manutenção da sobrevida dos neurônios
ao longo da vida (OPPENHEIM, 1999).
Durante esse período de chegada ao alvo, acontece a morte celular natural
que é um processo morfológico e bioquímico inerente ao desenvolvimento, a qual
pode ser classificada em três tipos: necrose, apoptose (LO et al., 1995) e autofagia
(MAIURI et al., 2007). Neste período ocorre um fenômeno regressivo, onde
aproximadamente 50% dos neurônios inicialmente gerados morrem, uma vez que as
células alvo liberam quantidades limitadas de fatores tróficos, levando os neurônios
que não conseguem captar estes fatores a ativar o programa de morte celular.
(OPPENHEIM, 1991; HENDERSON, 1996).
Os três tipos de morte celular apresentam características distintas entre si. Na
necrose celular observa-se tumefação do citoplasma e das organelas, sem
mudanças nucleares facilmente detectáveis. Tal tumefação leva à destruição das
organelas e ruptura da membrana plasmática com o extravasamento do conteúdo
citoplasmático, levando a resposta inflamatória importante e acometendo todas as

28
células vizinhas (SCHWARTZMAN & CIDLOWSKI, 1993). A autofagia é
caracterizada pela eliminação de organelas danificadas ou em excesso, podendo
evoluir para a destruição completa da célula (LINDEN & GUIMARÃES, 2004;
MAIURI et al., 2007). Observa-se a formação de vacúolos que se fundem aos
lisossomas levando à degradação de seus conteúdos (MAIURI et al., 2007). A
apoptose é caracterizada por perda de volume celular, condensação da cromatina,
fragmentação organizada do DNA em oligonucleossomos provocada pela ativação
das endonucleases, manutenção da estrutura das organelas e da membrana
plasmática até seu rompimento. Por último ocorre fagocitose dos corpúsculos
apoptóticos por macrófagos e por células vizinhas (KERR, 2002), sem a ocorrência
de resposta inflamatória expressiva no tecido, bem como sem dano ao micro-
ambiente (AMARANTE-MENDES & GREEN, 1999). O programa apoptótico ocorre
através da ativação das caspases (AMARANTE-MENDES & GREEN, 1999; MAIURI
et al., 2007), que podem ser classificadas como iniciadoras da cascata de
sinalização e efetoras da resposta que agem nos substratos nucleares (SALVESEN,
1999).
Com o avanço dos estudos sobre mecanismos envolvidos no processo de
morte celular observou-se que algumas moléculas, denominadas moléculas tróficas,
são capazes de controlar essa degeneração. As neurotrofinas são moléculas com
funções importantes tanto durante o desenvolvimento do sistema nervoso quanto na
homeostasia deste tecido, desempenhando funções de controle de proliferação,
diferenciação, sobrevida, crescimento axonal e plasticidade sináptica, dentre outras
(ARÉVALO & WU, 2006).
2.3.1 Neurotrofinas e citocinas
Na década de 1950, Rita Levi-Montalcini e Victor Hamburger descreveram o
fator de crescimento do nervo (NGF), o primeiro membro de uma grande família,
denominada família de neurotrofinas (OPPENHEIM, 1999). Além do NGF, o fator
neurotrófico derivado do cérebro (BDNF) (BARDE et al., 1982), a neurotrofina 3 (NT-
3) (HOHN et al., 1990), a neurotrofina 4/5 (NT-4/5) (HALLBÖÖK et al., 1991;

29
BERKEMEIER et al., 1991; EBENDAL; 1992), a neurotrofina 6 (NT-6) (GÖTZ et al.,
1994) e a neurotrofina 7 NT-7 (NILSSON et al., 1998) também foram descritos em
mamíferos.
As neurotrofinas (NTs) constituem um grupo de pequenos polipeptídeos
solúveis relacionados a fenômenos de sobrevivência, diferenciação e sobrevida
neuronal, apoptose, crescimento axonal, plasticidade sináptica e regeneração axonal
de neurônios do sistema nervoso, podendo agir de forma parácrina ou autócrina
(MENDELL, 2001; TERENGHI, 1999; RICHARDSON et al., 1991). São sintetizadas
como precursores, as pró-neurotrofinas, que podem ser clivadas intracelularmente
pela furina e outras pró-convertases ou, alternativamente, podem ser secretadas na
sua forma não clivada como pró-neurotrofinas (FRIEDMAN, 2010).
As NTs exercem seus efeitos através da ligação com dois receptores
diferentes - receptor com atividade cinase relacionado à tropomiosina (Trk) e o
receptor de neurotrofina p75 (p75NTR) da superfamília de receptores para fator de
necrose tumoral (TNF) (ARÉVALO & WU, 2006) (Figura 7). Todas as neurotrofinas
descritas ligam-se ao receptor p75, porém, com diferentes cinéticas de ligação
(TERENGHI, 1999). A família dos receptores Trk compreende três membros, o TrkA
o TrkB e o TrkC, e as neurotrofinas ligam-se a eles com seletividades diferentes.
NGF liga-se com maior seletividade ao TrkA ;o BDNF e a NT-4/5 ao TrkB e o NT-3
ao TrkC (BOYD & GORDON, 2003; CHAO, 2003). A pró-neurotrofina desempenha
um papel de ligante para o receptor p75NTR (RATTENHOLL et al., 2001), sendo
capaz de desencadear ações celulares como apoptose (NYKJAER et al., 2005),
crescimento e sobrevida neuronal (YAMASHITA et al., 1999). As formas “maduras”
clivadas de neurotrofinas se ligam com alta afinidade aos receptores Trk, que podem
estar complexados com p75NTR, enquanto as pró-neurotrofinas se ligam
preferencialmente à p75NTR em complexo com o co-receptor sortilina (FRIEDMAN,
2010).
A sinalização mediada pelo receptor p75, em determinadas situações
experimentais, leva à morte neuronal, através da ação do pró-NGF (IBÃNÉZ, 2002)
(Figura 7). Enquanto que a sinalização mediada pelos receptores Trk desencadeia
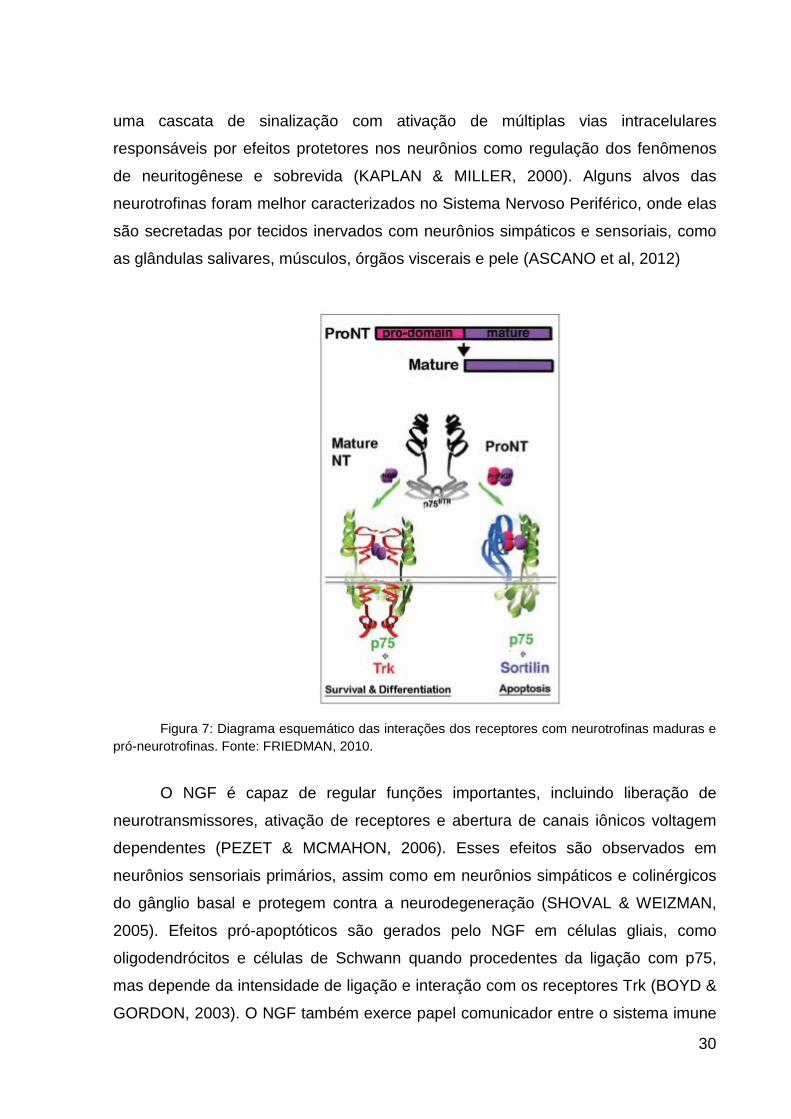
30
uma cascata de sinalização com ativação de múltiplas vias intracelulares
responsáveis por efeitos protetores nos neurônios como regulação dos fenômenos
de neuritogênese e sobrevida (KAPLAN & MILLER, 2000). Alguns alvos das
neurotrofinas foram melhor caracterizados no Sistema Nervoso Periférico, onde elas
são secretadas por tecidos inervados com neurônios simpáticos e sensoriais, como
as glândulas salivares, músculos, órgãos viscerais e pele (ASCANO et al, 2012)
Figura 7: Diagrama esquemático das interações dos receptores com neurotrofinas maduras e
pró-neurotrofinas. Fonte: FRIEDMAN, 2010.
O NGF é capaz de regular funções importantes, incluindo liberação de
neurotransmissores, ativação de receptores e abertura de canais iônicos voltagem
dependentes (PEZET & MCMAHON, 2006). Esses efeitos são observados em
neurônios sensoriais primários, assim como em neurônios simpáticos e colinérgicos
do gânglio basal e protegem contra a neurodegeneração (SHOVAL & WEIZMAN,
2005). Efeitos pró-apoptóticos são gerados pelo NGF em células gliais, como
oligodendrócitos e células de Schwann quando procedentes da ligação com p75,
mas depende da intensidade de ligação e interação com os receptores Trk (BOYD &
GORDON, 2003). O NGF também exerce papel comunicador entre o sistema imune

31
e nervoso, onde se observa aumento em seus níveis teciduais e séricos em
respostas inflamatórias, doenças auto-imunes e reações alérgicas podendo, dessa
maneira, agir como modulador (ALOE, 2004; BONINI et al., 1999).
O BDNF é importante para a formação, maturação e sobrevida neuronal na
fase inicial do desenvolvimento fetal e, na fase adulta, tem papel bastante conhecido
nos fenômenos cognitivos como no processo de consolidação da memória e
aprendizado (POST, 2007). Estudos mostram que há diminuição na sua expressão e
de seu receptor, TrkB, na Doença de Alzheimer (SCHINDOWSKI et al., 2008), além
de alteração nos níveis séricos do BDNF em pacientes com esquizofrenia (REIS et
al., 2008). A ativação de receptores TrkB e p75 promove ou suprime o crescimento
dendrítico e gera potenciação ou depressão sináptica. Sendo o balanço entre as
ligações da forma madura no receptor TrkB e a sua forma pró-BDNF no receptor p75
importante para as alterações das estruturas sinápticas (LU et al., 2005).
Assim como as interleucinas, as citocinas são pequenas proteínas liberadas
por diferentes tipos celulares e responsáveis por diferentes respostas como
sobrevivência, crescimento e diferenciação assim como respostas imunes
inflamatórias. São cruciais para a homeostasia, mas também desempenham papel
importante em condições patológicas (LUHESHI & ROTHWELL, 1996).
A interleucina 4 (IL-4) foi descoberta em 1982 sendo inicialmente chamada de
fator de crescimento da célula B. Com o conhecimento de seus efeitos pleiotrópicos
foi definitivamente denominada de IL-4 (PAUL, 1991). Essa citocina é uma proteína
glicosilada formada por 129 aminoácidos com peso molecular de aproximadamente
15 kDa (CHOI & REISER, 1998). A presença de receptores IL-4 (IL-4R) foi
observada em células hematopoiéticas, endoteliais, hepáticas e neuronais. Foram
identificados dois tipos de receptores de IL-4, I e II, sendo o tipo I formado pela
heterodimerização da subunidade alfa (IL-4Rα) com a subunidade γc (NELMS et al,
1999). O tipo II é um complexo envolvendo a subunidade alfa do receptor IL-4 e a
subunidade alfa I do receptor IL-13 (IL-13Rα1) (KEEGAN & PIERCE, 1994).

32
A IL-4 possui papel chave na resposta imune (PAUL, 1991), além de exercer
influência sobre o ciclo celular, induzindo as células a saírem da fase G1 (NELMS et
al., 1999). Além disso, ela atua como fator de crescimento autócrino e de
diferenciação, sendo promotora para proliferação de células T imaturas e
diferenciação destas mesmas células em efetoras (CHOI & REISER, 1998). Essa
interleucina tem capacidade tanto de suprimir a inflamação no sistema imune
(PONOMAREY et al., 2007) como de exercer função anti-inflamatória no cérebro.
Neste contexto a IL-4 age inibindo a produção de citocinas pró-inflamatórias como a
IL-1β e o TNF-α (LEE et al., 1995; MORI et al., 1995) e antagoniza os efeitos do IFN-
γ no sistema nervoso (FIORENTINO et al., 1991).
2.3.2 Retina
A retina pertence ao Sistema Nervoso Central e desenvolve-se a partir do
ectoderma neural. O tecido retiniano é translúcido e sua localização periférica
privilegiada permite fácil obtenção, livre do tecido conjuntivo adjacente e de outras
populações neuronais, o que facilita em muito sua obtenção para a realização de
diferentes tipos de experiências. A retina possui uma rica circuitaria de
neurotransmissores e de citocinas que a coloca na posição de um excelente modelo
experimental para o estudo do desenvolvimento, da diferenciação e da fisiologia de
células do sistema nervoso central, tanto in vivo quanto in vitro. Além disso, sua
organização morfo-funcional em camadas sinápticas e nucleares é muito semelhante
ao observado em outras estruturas do sistema nervoso (DOWLING, 1991). Dentre
todas as complexas estruturas do sistema nervoso central, nenhuma oferece tantas
vantagens a estudos do desenvolvimento como a retina (CEPKO, 1993).
A neurogênese das células da retina de ratos (Figura 8) se dá da seguinte
maneira: inicialmente são formadas as células ganglionares no período embrionário
(E) entre E14 e E20; em seguida as células amácrinas (REESE & COLELLO, 1992)
e horizontais entre E14 e E22; os cones são gerados entre E14 a P0 (dia do

33
desenvolvimento pós-natal 0) e, por fim as células bipolares, bastonetes e as células
de Müller no período entre E20 a P13 (CEPKO, 1993).
Figura 8: Histogênese da retina. Desenvolvimento das células progenitoras originam os sete tipos principais de células da retina em diferentes espécies. Período embrionário (E); Período pós-natal (P). Retirado de MARTINS & PEARSON, 2008.
No período pós-natal (P0 a P2) a retina é imatura, formada por uma
população de neuroblastos e apenas a camada de células ganglionares está
formada (Figura 9A). O processo de diferenciação retiniana depende de um
programa genético pré-determinado e de pistas, sinais ambientais intraretinianos,
como a expressão de moléculas de adesão e da matriz extracelular, bem como da
liberação de diversas citocinas (FARAH, 2006; DOWLING, 1991).
Após o desenvolvimento (Figura 9B), a retina de vertebrados organiza-se em
camadas: na camada nuclear externa (CNE) encontram-se os corpos celulares dos
fotorreceptores (os cones, que possuem baixa sensibilidade à luz e permitem a visão
cromática, e os bastonetes, responsáveis pela adaptação da visão ao claro-escuro);
na camada plexiforme externa (CPE) observam-se os prolongamentos de
fotorreceptores, células bipolares e horizontais, que se comunicam através de
conexões sinápticas; a camada nuclear interna (CNI) é composta por corpos
celulares das células horizontais, das células bipolares e das amácrinas; os corpos
celulares das células ganglionares e de células amácrinas deslocadas formam a

34
camada mais interna da retina, a camada de células ganglionares (CCG). As células
ganglionares da retina (CGR), cujos axônios formam o nervo óptico, fazem sinapses
com outras regiões centrais do sistema visual; na camada plexiforme interna (CPI),
os prolongamentos de células bipolares, amácrinas e ganglionares fazem sinapses
entre si (DOWLING, 1991; FARAH, 2006).
As células de Müller são o principal tipo de célula glial encontrado na retina de
todas as espécies de vertebrados (WON et al., 2000). O corpo celular da célula de
Müller está localizado na CNI e seus prolongamentos se estendem por todas as
camadas da retina, formando a membrana limitante interna, adjacente ao humor
vítreo, e a membrana limitante externa, adjacente à camada nuclear externa
(NEWMAN & REICHENBACH, 1996).
A B
Figura 9: Esquema simplificado da retina de vertebrados recém-nascidos (A) e adultos (B). Representação dos diferentes tipos celulares, bem como as camadas celulares e sinápticas. Modificados de: (A) Dowling, 1991 e (B) Yang, 2004.
Posterior à retina, no fundo do olho, encontra-se a camada de células
epiteliais pigmentadas, conhecida como epitélio pigmentar da retina (EPR), rico em
melanina e responsável, dentre outras funções, por absorver a luz não capturada
pelos fotorreceptores. Desta forma, a degradação e a distorção da imagem pela
retina neural são evitadas, uma vez que não permite a reflexão da luz não absorvida
por cones e bastonetes (DOWLING, 1991).

35
2.3.2.1 Atividade colinérgica na retina
A acetilcolina (ACh) é um importante neurotransmissor na retina dos
mamíferos, estando presente principalmente nas células amácrinas (VANEY, 1990).
O envolvimento dessa molécula na neurotransmissão na retina de vertebrados está
bem estabelecido, em particular com a participação dos receptores nicotínicos
(nAChRs) na sinalização nos estágios iniciais do desenvolvimento, bem como na
etapa de crescimento e maturação neuronal (FELLER, 2002).
Os nAChRs na retina de vertebrados desempenham papel na sinalização
durante os estágios iniciais do desenvolvimento e mais tarde durante o crescimento
neuronal e sinaptogênese (FELLER, 2002; ZHOU, 2001). Na retina de vertebrados
adultos, ACh liberada por células amácrinas ativa grande variedade de nAChRs.
Estudos de hibridização in situ e imunolocalização em conjunto com análises de
Northern blot mostraram que a retina expressa quase todas as subunidades
nicotínicas presentes em receptores homoméricos e heteroméricos (FELLER, 2002).
Estudos bioquímicos e farmacológicos usando ligantes nicotínicos e
anticorpos policlonais específicos para cada subunidade mostraram a presença de
três subtipos que se ligam à α-Bgtx na retina de galinha: os homoméricos α7 e α8 e
os heteroméricos com os subtipos α7 e α8 (KEYSER et al., 1993). Utilizando a retina
adulta de coelho, Keyser e colaboradores (2000) mostraram que muitos dos
receptores heteroméricos contém a subunidade β2, que é parcialmente associada à
subunidade α3. Em conjunto, estes resultados indicam que nAChRs contendo a
subunidade β2 são essenciais para o desenvolvimento anatômico e funcional do
sistema visual de roedores (MORETTI et al., 2004).
Os receptores muscarínicos (mAChRs) são expressos por células amácrinas,
ganglionares, bipolares em retinas de animais adultos. Marcadores de neurônios
colinérgicos (colina acetil transferase e colinesterase) são expressos na camada
neuroblástica da retina de embrião (E3) de pinto. Marcadores colinérgicos também
são expressos na retina de mamíferos mesmo antes do período de sinaptogênese,

36
sugerindo que células da retina neural imatura podem também liberar acetilcolina
(MARTINS et al., 2008).
Strang e colaboradores (2010) verificaram a presença de todos os sub-tipos
de receptores muscarínicos em células amácrinas em análise imunocitoquímica na
retina de coelhos. As células amácrinas, bipolares e ganglionares apresentaram
marcação para M1, enquanto que as células ganglionares, horizontais e algumas
bipolares, apresentaram marcação para M3, além das células amácrinas.
Imunorreatividade para receptores M2 e M4 foram observadas na retina de tritões
adultos. Sendo o M2 encontrado na somata em ambos os lados da camada
plexiforme interna (CPI), principalmente em células ganglionares, e também
distribuídas em duas bandas dentro da CPI. O M4 foi localizado próximo à somata
oposta aos lados da CPI, provavelmente somata de células amácrinas, e nenhuma
imunorreatividade foi detectada na CPI (CHEON et al., 2001)
Pereira e colaboradores (2001) avaliaram o papel do sistema colinérgico no
aumento da sobrevida das CGR. Eles demonstraram que a estimulação por
agonistas colinérgicos aumenta a sobrevida das CGR em cultura e que o bloqueio
dos receptores M1 é capaz de abolir totalmente esse efeito. O efeito da estimulação
colinérgica é abolido pela inibição dos receptores Trk e pela inibição da secreção
vesicular de polipeptídeos. Esses resultados sugerem que a estimulação colinérgica
pode mediar a liberação de fatores tróficos capazes de regular a sobrevida das
CGR. Esses dados estão de acordo com outros dados da literatura que demonstram
o envolvimento de neurotransmissores na liberação de fatores tróficos de células do
tecido neuronal (BERZAGHI et al., 1993; ELLIOT et al., 1994).
A via da proteína cinase C (PKC) também está envolvida no controle da
sobrevida das CGR. SANTOS e ARAUJO (2000) descreveram que a ativação da
PKC por PMA (13-acetato de forbol 12-miristato), tem papel importante na sobrevida
das CGR mantidas em culturas mistas por 48 horas. Por outro lado, a ativação da
PKC por PMA reduz a proliferação celular de células da retina, mediada por ativação
de receptor muscarínico (M1). O tratamento das culturas com carbamilcolina e BDNF
também reduz a proliferação dessas células de forma semelhante ao PMA,

37
sugerindo que a ativação da PKC induz redução da proliferação celular através da
liberação de acetilcolina e de BDNF na cultura, bem como ativação de receptores
Trk (SANTOS et al., 2000). Em 2009, Santos e colaboradores demonstraram que o
PMA induz redução na expressão de receptor M1 e de BDNF em culturas de células
da retina. Porém, observou-se que em culturas tratadas por 45min com PMA, houve
aumento transiente na expressão de BDNF e redução na expressão de receptores
M1 (SANTOS et al., 2009).
2.3.2.2 Ações de neurotrofinas e citocinas na retina
No sistema visual, dentre as neurotrofinas citadas, o BDNF tem papel de
destaque, coordenando eventos de sobrevivência e diferenciação em todas as
áreas, desde a retina até o córtex visual (von BARTHELD, 1998). Seus efeitos foram
caracterizados em estudos in vitro e in vivo, e podem ser observados durante o
desenvolvimento até a vida adulta (ISENMANN et al., 2003).
O desenvolvimento das CGR depende de diferentes fatores tróficos,
produzidos por diferentes populações celulares, na sobrevida das células
ganglionares. De fato, as células de Müller são capazes de liberar fatores tróficos
que aumentam a sobrevivência das células ganglionares de ratos neonatos em
cultura (RAJU & BENNETT, 1986; ARMSON et al., 1987). Esses fatores atuam
durante o desenvolvimento e a maturação da retina, bem como podem se constituir
em importantes agentes terapêuticos em doenças degenerativas da retina como
após lesão de nervo óptico (von BARTHELD, 1998).
Estudos mostram que o NGF, o BDNF e a NT-4/5 exercem efeito de
sobrevida nesses neurônios in vivo. O NGF e o BDNF têm efeito após lesão
isquêmica e o BDNF e a NT-4/5 promovem crescimento de neuritos dos neurônios
em regeneração. Em experimentos in vitro, o BDNF mantém a sobrevida desses
neurônios de diferentes espécies animais, enquanto a NT-4/5 tem esse efeito
somente em células ganglionares de ratos. A NT-3 parece não ter papel importante

38
na sobrevida, ramificação de neuritos ou crescimento axonal de células ganglionares
de mamíferos (von BARTHELD, 1998). Ugolini e colaboradores (1995) detectaram a
expressão de RNA mensageiro (RNAm) para os receptores (Trk e p75) para as
neurotrofinas na camada de células ganglionares da retina de ratos P0.
O BDNF e as Neurotrofinas 4/5 são capazes de proteger as células
ganglionares, impedindo sua degeneração (von BARTHELD, 1998). Essa
degeneração ocorre após lesão do nervo óptico (axotomia das células ganglionares)
de ratos recém-nascidos, quando aproximadamente 80% dessas células morrem
nas primeiras 24 horas (RABACCHI et al., 1994).
Em cultura de células ganglionares purificadas foi observado que os níveis de
3´5´-adenosina monofosfato cíclico (AMPc) regulam o efeito trófico de inúmeras
citocinas: BDNF, NT-4/5, FGFb (fator de crescimento de fibroblasto básico, também
chamado de FGF-2), CNTF (fator trófico para neurônios do gânglio ciliar), LIF (fator
inibidor de leucemia) e TGFα (fator de transformação de crescimento alfa) (MEYER-
FRANKE et al., 1995). Santos e Araujo (2000) demonstraram que o tratamento de
culturas mistas de células da retina com forscolina, agente que eleva os níveis de
AMPc, induz aumento na sobrevida das células ganglionares, sem a necessidade de
adicionar fatores tróficos às culturas.
Em relação às ações das citocinas em células da retina, Sholl-Franco e
colaboradores (2001) demonstraram que o tratamento de culturas mistas de células
da retina com IL-2 ou com IL-4 por 48h induz aumento da sobrevida das CGR de
maneira dependente da concentração. O efeito da IL-2 não depende da atividade
colinérgica e também não foi afetado pela inibição da proliferação celular, enquanto
o efeito da IL-4 depende tanto da atividade colinérgica quanto da proliferação celular
(SHOLL-FRANCO et al., 2001). Posteriormente foi observado também em células
da retina, que essa citocina é capaz de modular o fenótipo GABAérgico, sugerindo
importante papel desta citocina durante o desenvolvimento normal da circuitaria
retiniana ou durante a neuroproteção após lesões (SHOLL-FRANCO et al., 2002).

39
2.4 CICLO CELULAR
Nos eucariontes o ciclo celular é dividido em duas fases distintas: a intérfase e
a mitose. A mitose, ou fase M, é quando ocorre a divisão celular. A interfase é o
período em que a célula se prepara para a divisão, aumentando a sua massa e
duplicando seu material genético. A mitose estende-se desde o início da prófase até
o final da telófase e é o período no qual não há síntese de DNA, a síntese de
proteínas está reduzida ao mínimo e a de RNA está limitada. A interfase é o período
mais longo do ciclo celular, representando o intervalo entre duas mitoses, sendo
subdividido em três fases (G1, S, G2) (GALLI-MUHTASIB, 2002).
• G1: fase pós-mitótica durante a qual as células sintetizam ácido ribonucleico
(RNA) e proteínas;
• S: fase na qual o DNA é duplicado enquanto a síntese de proteínas e RNA
continua Nesta fase as células podem ser marcadas com [3H]-timidina, um
nucleotídeo derivado da timina, utilizado exclusivamente para a síntese de DNA;
• G2: fase pós-sintética, período entre o fim da fase S e o início da mitose,
caracterizado por não apresentar síntese de DNA e a síntese de RNA e proteínas
continua assim como na fase pós-mitótica.
As células em fase G1 podem começar o processo de divisão celular ou
mudar para G0 (fase de repouso), onde podem permanecer por dias, semanas ou
anos. O ciclo celular é estreitamente controlado por vários mecanismos que podem
tanto permitir ou restringir a sua progressão (GALLI-MUHTASIB, 2002).

40
Figura 10: Regulação do ciclo celular. A progressão pela fase G1 do ciclo celular ocorre pela e fosforilação de ciclinas e cinases dependentes de ciclina (CDKs). Os inibidores de CDK agem parando as ciclinas e o ciclo. (Retirado de BOEHM & NABEL, 2001).
O ciclo celular é altamente controlado contendo vários pontos de checagem,
sendo o ponto de restrição encontrado na etapa de transição entre as fases G1 e S,
e é nesse período que os progenitores saem do ciclo celular e se diferenciam. Esta
decisão é determinada por fatores extrínsecos e intrínsecos e suas interações
(Figura 10). As principais famílias de proteínas reguladoras da progressão do ciclo
celular são as ciclinas, cinases dependentes de ciclina (CDKs), os inibidores de CDK
(CKI) e o produto dos genes supressores de tumor, p53 e pRb (GALLI-MUHTASIB,
2002). Sinais provenientes do meio extracelular podem regular a expressão destas
proteínas fazendo com que as células entrem ou saiam do ciclo mitótico (GOLIAS et
al., 2004).
Primeiramente identificadas em invertebrados marinhos, as ciclinas
receberam este nome por oscilarem durante o ciclo celular (HUNT, 1989). As CDKs
são cinases serina-treonina, e sua atividade cinase acontece quando estão
complexadas com as ciclinas. A formação deste complexo induz a passagem da
fase G1 para S pela fosforilação das proteínas retinoblastoma (Rb), p107 e p130

41
(Figura 10). A hiperfosforilação dessas proteínas libera o fator de transcrição E2F
permitindo a transcrição de genes pró-mitóticos (DYER & CEPKO, 2001).
Algumas CDKs, tais como as CDK1-CDK4, CDK6 e talvez CDK11, estão
envolvidas na progressão do ciclo celular, ao passo que a CDK7 possui papel duplo
como ativador de cinase CDK (CAK) e regulador da maquinaria transcricional. CDK8
e CDK9 parecem ter papel fundamental no controle da transcrição pela RNA
polimerase II. Outras CDKs e proteínas que agem como as CDK atuam em
processos mais específicos em tipos particulares de células. A atividade da CDK
está aumentada em doenças proliferativas tais como câncer, devido à frequente
superexpressão de reguladores positivos (ciclinas) e a frequente inativação de
inibidores de CDK (MALUMBRES et al., 2007).
A proliferação celular que é induzida por fatores de crescimento ou citocinas
pode ocorrer apenas na presença de diferentes sinais de sobrevida. As células que
recebem o sinal proliferativo na ausência de sinal de sobrevida não proliferam, ao
contrário, morrem por apoptose. Assim, a apoptose age como importante
mecanismo para eliminação de células que sofreram mutações durante o ciclo
celular e que podem resultar em proliferação celular descontrolada (VERMEULEN et
al., 2003).

42
3 OBJETIVOS
3.1 Objetivo geral
Analisar o efeito do extrato diclorometano de Eugenia punicifolia em células
da retina de ratos neonatos.
3.2 Objetivos específicos
• Pesquisar o efeito do extrato na proliferação celular;
• Analisar o perfil de expressão de receptores colinérgicos muscarínicos (M1, M3,
M4 e M5);
• Investigar o perfil de expressão da subunidade α7 do receptor colinérgico
nicotínico;
• Analisar a expressão do transportador vesicular de acetilcolina (VAChT);
• Investigar os níveis de NGF;
• Averiguar nos níveis de BDNF;
• Analisar os níveis de IL-4.

43
4 MATERIAL E MÉTODOS
4.1. Material
Materiais utilizados nos procedimentos experimentais desta dissertação com a
respectiva origem.
PRODUTO EMPRESA LOCALIZAÇAO SDS (Sodium Dodecyl Sulfate) Tris(hidroximetil)aminometano
USB USB
NJ, USA NJ, USA
Penicilina G Sigma MO, USA Sulfato de estreptomicina Sigma MO, USA L-Glutamina Sigma MO, USA poli-L-Ornitina Sigma MO, USA Meio 199 Gibco MD, USA Soro Fetal Bovino Gibco MD, USA Tripsina Worthington NJ, USA Placas de Petri TPP Switzerland Hiperfilme Amersham Bucknghamshre, UK Kit ECL Amersham Bucknghamshre, UK 2-Mercaptoetanol Amersham Bucknghamshre, UK Anticorpo Secundário Anti-rabbit ligado a HRP
Amersham Bucknghamshre, UK
Glicina Queel SP, Brazil Tween 20 Ácido tricloroacético (TCA)
Vetec Vetec
RJ, Brazil RJ, Brazil
Azul de Cromassie Vetec RJ, Brazil Glicerol Vetec RJ, Brazil Anti-Actina Sta. Cruz Biotecnology Santa Cruz, USA Anti-M1 Sta. Cruz Biotecnology Santa Cruz, USA Anti-M3 Anti-M4
Sta. Cruz Biotecnology Sta. Cruz Biotecnology
Santa Cruz, USA Santa Cruz, USA
Anti-M5 Anti-α7
Sta. Cruz Biotecnology Sta. Cruz Biotecnology
Santa Cruz, USA Santa Cruz, USA
Anti-VAChT Albumina sérica bovina (BSA) Anti-NGF Anti-BDNF Anti-IL-4
Sta. Cruz Biotecnology Sta. Cruz Biotecnology Peprotech Peprotech Peprotech
Santa Cruz, USA Santa Cruz, USA NJ, USA NJ, USA NJ, USA
Temed Invitrogem Carlsbad, USA Padrão de Peso Molecular Invitrogem Carlsbad, USA Luminata Millipore MA, USA Metanol [3H-metil]-timidina Filtros de fibra de vidro DMSO
J.T.Baker GE Sigma Sigma
NJ, USA NJ, USA MO, USA MO, USA

44
Estudos sobre a atividade colinérgica induzida por EP (GRANGEIRO et al.,
2006, PASCUAL et al., 2011) e sobre a importância da transmissão colinérgica na
retina de mamíferos (VANEY, 1990; FELLER, 2002; SANTOS et al., 2009) indicam a
necessidade de se estudar o efeito do extrato diclorometano de Eugenia punicifolia
na modulação do fenótipo colinérgico de células da retina de ratos neonatos,
mantidas em cultura. Foram realizadas culturas de células da retina para se avaliar
as possíveis alterações na expressão de receptores muscarínicos (M1, M3, M4, M5)
e da subunidade α7 do receptor nicotínico (Nα7), além da expressão do
transportador vesicular de acetilcolina (VAChT), induzidas pelo extrato.
4.2 Extrato diclorometano de Eugenia punicifolia
O extrato diclorometano de Eugenia punicifolia (EP) foi obtido na Faculdade
de Farmácia, segundo metodologia descrita (YOO et al., 2005) e foi estocado em
quantidade suficiente para a realização do presente trabalho. O extrato foi diluído em
10% de dimetilsulfóxido (DMSO). A solução obtida foi filtrada em filtro 0,22µm e
armazenada em alíquotas de 1mg/mL (50µL), a uma temperatura de -20oC.
4.3 Animais experimentais
Foram utilizados ratos neonatos pigmentados da linhagem Lister Hooded nas
primeiras 72 horas após o nascimento (P0-P2), provenientes do Biotério da
Faculdade de Farmácia da UFF ou do Biotério do Departamento de Neurobiologia
(Instituto e Biologia – UFF), os quais receberam ração e água ad libidum e foram
mantidos a um ciclo claro/escuro de 12h. Os procedimentos experimentais foram
aprovados pelo Comitê de Ética no Uso de Animais da UFF (Projeto nº 186/2012).

45
4.4 Cultura de células da retina
Em capela de fluxo laminar, ratos neonatos (dia pós-natal 0-2) foram
sacrificados por decapitação, seus olhos removidos por enucleação. As retinas
foram dissecadas e colocadas em um tubo contendo solução salina sem cálcio e
sem magnésio (CMF: NaCl 131mM, KCl 4,09mM, Na2HPO4.7H2O 0,92mM,
KH2PO4 0,45mM, glicose.H2O 12,2mM, NaHCO3 9,4mM, vermelho de fenol
10mg/mL.) acrescida de penicilina 100U/mL e estreptomicina 100μg/mL, e a
dissociação química foi realizada por ação da tripsina 0,1% por aproximadamente
20min a 37°C. Posteriormente, para inativação da tripsina, as células foram lavadas
duas vezes com meio de cultura 199 acrescido de 5% de soro fetal bovino (SFB),
glutamina 2,0mM, estreptomicina 100µM e penicilina 100U/mL (M199 completo). As
células foram dissociadas mecanicamente utilizando pipeta Pasteur e plaqueadas,
na densidade de 1,25x106 células/placa, em placas de Petri de 35 mm, previamente
tratadas com poli-L-ornitina, em um volume inicial de 1 mL de M199 completo. A
seguir, as placas receberam mais 1 mL de M199 completo (culturas controle) ou
mais 1 mL de M199 completo contendo extrato diclorometano de E. punicifolia (EP).
Todas as culturas foram mantidas a 37°C, em atmosfera de 95% de ar e 5% de CO2.
As células eram mantidas in vitro por 48h quando então eram processadas para
incorporação de [3H]-timidina ou para Western blot.
4.5 Incorporação de [3H]-timidina
Para a determinação da concentração do extrato que seria utilizado nos
experimentos, optou-se pelo estudo da proliferação celular através da utilização do
método de incorporação de [3H]-timidina ao DNA das células. Esta é uma técnica
útil, uma vez que a [3H]-timidina é capaz de marcar todas as células que se
encontram na fase S do ciclo celular, indiscriminadamente. Uma vez que a célula
tem o marcador incorporado ao DNA, suas células filhas terão a metade desse
marcador em seu DNA após sofrerem mitose, e assim a divisão vai acontecendo, o
marcador vai se diluindo e as células geradas serão igualmente marcadas (FARAH,
2006). A utilização dessa técnica permite a quantificação da timidina marcada

46
radioativamente incorporada ao DNA, porém não é possível identificar as células em
divisão em um dado momento.
A incorporação de [3H]-timidina nas culturas mantidas por 48 horas in vitro foi
realizada segundo o seguinte protocolo:
Após o período de incubação, as culturas foram lavadas duas vezes com 2
mL de M199 sem SFB, a temperatura ambiente, e foram incubadas com 0,5µCi/mL
de [3H]-timidina em 1mL de M199 sem SFB por 60 minutos a 37°C em atmosfera de
5% de CO2 e 95% de ar. Neste período as células que estavam se dividindo tiveram
a [3H]-timidina incorporada ao seu DNA. As culturas foram então lavadas quatro
vezes com M199 sem SFB, a fim de remover a [3H]-timidina que não foi incorporada
ao DNA. O M199 foi removido e 200µL de NaOH 0,4N foram adicionados às culturas
e mantido por quinze minutos para lisar as células. Posteriormente, foi feita a
raspagem das células aderidas a essas placas e o conteúdo de cada placa foi
transferido para tubos de vidro contendo 3mL de água MilliQ gelada, aos quais
foram adicionados 600µL de ácido tricloroacético (TCA) 50%. Após 30 minutos, cada
amostra foi filtrada sob pressão negativa em membrana de filtro de vidro (Whatman
GF/B). Os filtros foram colocados para secar overnight e a radioatividade foi
determinada em cintilador de fase líquida (Packard, USA) em vials contendo 3mL de
tolueno + 2,4-difeniloxazol (PPO). As culturas controle apresentaram contagens por
minuto (CPM) em torno de 1100. Os resultados foram expressos em porcentagem
do controle.
4.6 Western blot
A técnica foi utilizada para a detecção dos níveis dos receptores colinérgicos
(M1, M3, M4, M5, Nα7), VAChT, das neurotofinas (NGF e BDNF), da interleucina-4 e
actina.

47
4.6.1 Extração de proteínas
As culturas foram lavadas com solução Hank’s e as proteínas totais extraídas
com 100 µL de tampão de amostra (glicerol; 2-mercaptoetanol; SDS; Tris 0,5M pH
6,8 e água destilada). As amostras foram fervidas a 100°C por 10 min, para
desnaturar as proteínas.
4.6.2 Dosagem de proteínas
A dosagem de proteínas foi realizada pelo método de Bradford (BRADFORD,
1976), no qual a curva padrão é feita com 0, 5, 10, 15 e 20µg de uma solução de
albumina (BSA) na concentração de 1mg/mL. A leitura das amostras foi feita em
espectrofotômetro em 595nm.
4.6.3 Gel de poliacrilamida
O gel de corrida foi preparado com Tris base 1,5M (pH 8,8), acrilamida 30% +
0,9% de metileno bisacrilamida, SDS 10%, TEMED, persulfato de amônia 10%. O
gel foi aplicado e, após polimerização por 30 min de polimerização, adicionou-se o
gel de compactação (Tris base 0,5M pH 6,8; acrilamida 30% + 0,9% de metileno
bisacrilamida, SDS 10%, TEMED, persulfato de amônia 10%). Após aplicar o gel, o
pente foi colocado e, mantido 30 min para que ocorresse a polimerização. O pente
foi retirado e o suporte colocado na cuba contendo tampão de corrida (glicina, tris
base e SDS). As amostras foram aplicadas nos poços em volumes variados das
amostras das experiências representando 60 µg de proteínas por poço. Foi utilizado
10µl do padrão de peso molecular no primeiro poço. A corrida foi realizada com
corrente constante de 10mA por aproximadamente 15min seguido por 15mA por
60min. Após a corrida o gel foi removido e transferido para uma cuba contendo
tampão de transferência onde foi mantido por 30 minutos.

48
4.6.4 Transferência das proteínas do gel para a membrana de polímero de fluoreto
de vinilideno (PVDF)
A membrana de PVDF foi incubada por 30 segundos em metanol e transferida
para uma cuba contendo tampão de transferência (glicina, Tris base, SDS 10% e
20% de metanol), onde ficou equilibrando por aproximadamente 30 min. O
“sandwich” com o gel e a membrana foi montado e a transferência realizada em
sistema Semi-Dry utilizando voltagem constante de 10V por 30 min.
4.6.5 Bloqueio e imunodetecção
O bloqueio foi realizado com 5% de leite desnatado em solução salina
tamponada de Tris (TBS) pH 7,6 com 0,1% tween 20, por 2h, em temperatura
ambiente. Após este período, a solução para o bloqueio era retirada e a membrana
lavada três vezes em TBS com 1% de leite e 0,1% de tween. A membrana era,
então, incubada com o anticorpo primário específico em TBS (1% de leite e 0,1%
tween), overnight. Foram utilizados vários anticorpos primários separadamente (anti-
M1; anti-M3, anti-M4, anti-M5, anti-α7N, anti-NGF, anti-BDNF, anti-IL4, anti-actina) e,
após a incubação com o primário, a membrana era lavada três vezes em TBS (1%
de leite e 0,1% tween) por 10 min cada lavagem e posteriormente incubada com o
anticorpo secundário específico, ligado a peroxidase por 1 hora. As membranas
eram lavadas duas vezes, por 10 min cada, em TBS + 0,1% tween, e a última
lavagem era realizada com TBS por 10 min.
4.6.6 Revelação
A revelação foi realizada com exposição da membrana à solução reagente de
Luminol (ECL) por 5 minutos. Esta reação é baseada na quimioluminescência que
resulta da oxidação do luminol, catalisada pela peroxidase presente no anticorpo
secundário. Posteriormente a membrana ficou exposta ao filme (Hiperfilme) por 5

49
minutos, pois esta luz resultante foi detectada pelo filme em segundos ou minutos. O
filme foi retirado e a revelação realizada com revelador GBX Kodak 1:10 por 15
segundos, seguido de fixação (Fixador Kodak 1:5) por 1 minuto. A quantificação das
bandas obtidas foi feita através do ImageJ.
4.7 Fotomicrografia em contraste de fase
Após 48h, as culturas controle e tratadas com EP foram fixadas em Karnovski
(paraformaldeído 1% e glutaraldeído 2%) por um período de 5 a 10 minutos. Em
seguida, as células foram lavadas três vezes em tampão fosfato 0,1M pH 7,2-7,4.
Cada placa foi armazenada por um período de aproximadamente uma semana em
geladeira, com 1mL de tampão fosfato. As culturas foram observadas em
microscópio óptico (LEICA DFC 310FX) em contraste de fase.
4.8 Análise estatística dos resultados
Os resultados foram expressos na forma de histogramas e, cada experimento
foi repetido no mínimo 3 vezes, sendo utilizadas no mínimo 3 amostras por ponto
experimental. O resultado da [3H]-timidina foi apresentado em porcentagem do
controle, como média ± erro padrão da média e a análise estatística foi feita através
da análise de variância (ANOVA) seguida do pós-teste, Newman-Keuls, que faz a
comparação entre as diferentes condições experimentais. Os testes e gráficos foram
feitos através do programa GraphPad Prism®.
Os resultados do Western blot representam os níveis de expressão das
proteínas estudadas em unidades arbitrárias (em porcentagem do controle) como
média ± erro padrão da média. A análise estatística foi feita através da comparação

50
entre dois grupos através do teste t-student. Os testes e gráficos foram feitos através
do programa GraphPad Prism®.
4.9 Questões éticas
O projeto foi submetido à apreciação do Comitê de Ética no Uso de Animais
da UFF sob o número 186, tendo sido aprovado em 17/05/2012 (Anexo 1). Todos os
experimentos foram realizados reduzindo ao máximo qualquer sofrimento e tendo
sido utilizado apenas o número suficiente de animais.
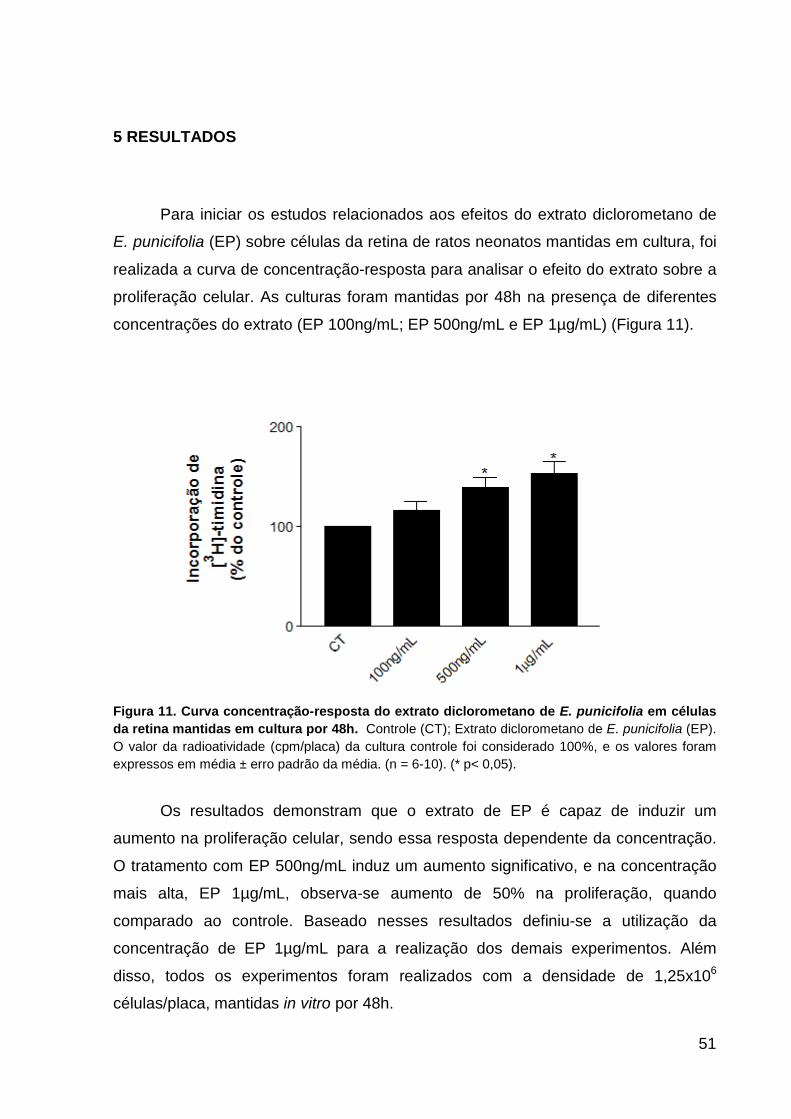
51
5 RESULTADOS
Para iniciar os estudos relacionados aos efeitos do extrato diclorometano de
E. punicifolia (EP) sobre células da retina de ratos neonatos mantidas em cultura, foi
realizada a curva de concentração-resposta para analisar o efeito do extrato sobre a
proliferação celular. As culturas foram mantidas por 48h na presença de diferentes
concentrações do extrato (EP 100ng/mL; EP 500ng/mL e EP 1µg/mL) (Figura 11).
Figura 11. Curva concentração-resposta do extrato diclorometano de E. punicifolia em células da retina mantidas em cultura por 48h. Controle (CT); Extrato diclorometano de E. punicifolia (EP). O valor da radioatividade (cpm/placa) da cultura controle foi considerado 100%, e os valores foram expressos em média ± erro padrão da média. (n = 6-10). (* p< 0,05).
Os resultados demonstram que o extrato de EP é capaz de induzir um
aumento na proliferação celular, sendo essa resposta dependente da concentração.
O tratamento com EP 500ng/mL induz um aumento significativo, e na concentração
mais alta, EP 1µg/mL, observa-se aumento de 50% na proliferação, quando
comparado ao controle. Baseado nesses resultados definiu-se a utilização da
concentração de EP 1µg/mL para a realização dos demais experimentos. Além
disso, todos os experimentos foram realizados com a densidade de 1,25x106
células/placa, mantidas in vitro por 48h.

52
Com o objetivo de analisar a morfologia das culturas, estas foram tratadas
com EP 1µg/mL, mantidas por 48h e posteriormente fixadas e observadas em
microscopia de contraste de fase (Figura 12).
Figura 12: Fotomicrografia em contraste de fase mostrando a morfologia das culturas após 48 horas in vitro. A: Cultura controle; B: Cultura tratada com Ep1µg/mL. Barra de calibração: 20μm. Setas: Processos neuronais; cabeça de seta: células gliais.
Analisando as fotomicrografias das culturas de células pode-se constatar na
cultura controle (Figura 12A) a presença de poucos grumos com a formação de
pequenos processos neuronais. Por outro lado, a cultura tratada com EP1µg/mL
(Figura 12B) apresentou em sua morfologia a presença de grumos espraiados
(cabeça de seta), maior número de células, além da presença de muitos processos
(seta). Nota-se presença de células gliais (cabeça de seta) sobre as quais as células
neuronais se localizam e podem ser individualizadas, sendo este padrão morfológico
observado em toda a extensão da placa.
A partir deste resultado, pode-se sugerir que o extrato de EP 1µg/mL é capaz
de aumentar o número e tamanho de processos e, possivelmente, a sobrevida das
células da retina de ratos neonatos in vitro.

53
MODULAÇÃO DO FENÓTIPO COLINÉRGICO
Os níveis de receptor M1 reduzem ao longo dos primeiros 14 dias de
desenvolvimento pós-natal em células da retina de ratos neonatos mantidas em
cultura (SANTOS et al., 2009). Para avaliar se o extrato diclorometano é capaz de
induzir alterações na expressão desse receptor, culturas foram tratadas com extrato
EP 1µg/mL por 48h (Figura 13).
CT
EP 1µg/m
L0
50
100
*
Nív
eis
de r
ecep
tor
M1
Uni
dade
s A
rbitr
ária
s(%
do
cont
role
)
50 kDa M1
49 kDa actina Figura 13. Níveis de receptor M1 em culturas tratadas com extrato diclorometano de E. punicifolia por 48h. Controle (CT); Extrato diclorometano de E. punicifolia (EP) 1µg/mL. A: Níveis da expressão de receptores M1, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 4). B: Imagem do filme radiográfico representativo da imunodetecção obtidos nessas experiências. (*p< 0,001)
Os resultados demonstram que culturas tratadas com o extrato apresentam
redução significativa (32%) nos níveis dos receptores M1, quando comparadas com
cultura controle.
A
B
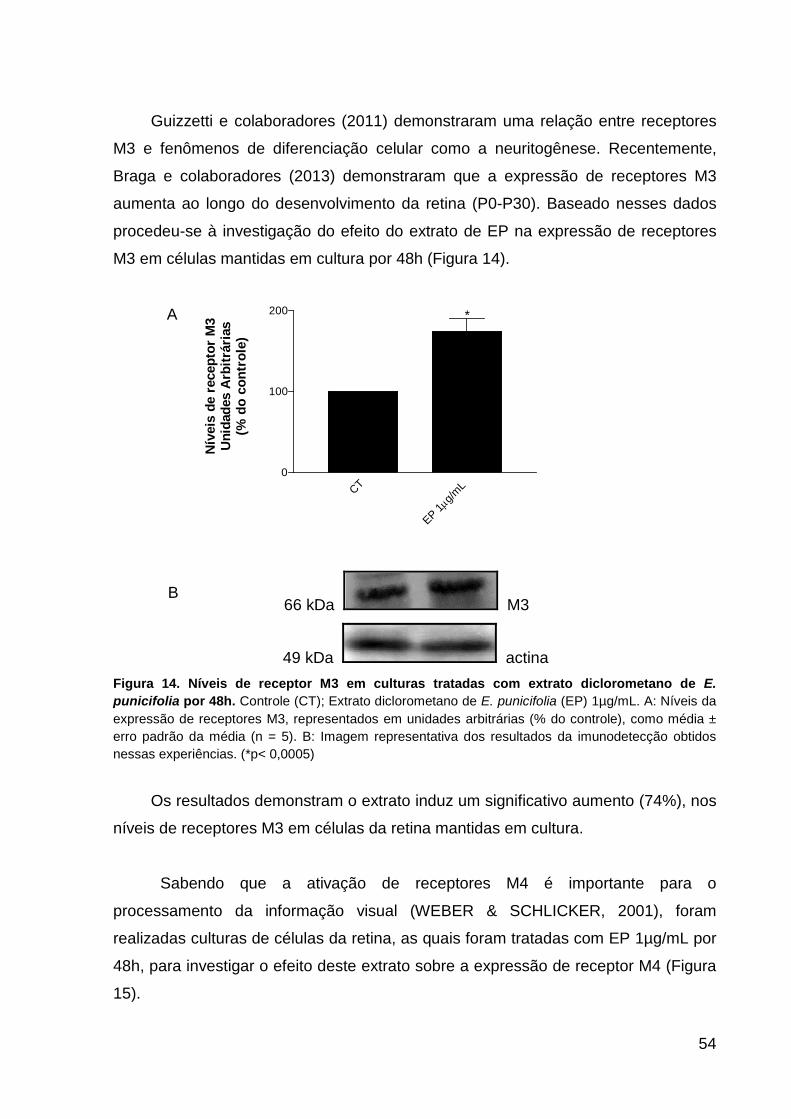
54
Guizzetti e colaboradores (2011) demonstraram uma relação entre receptores
M3 e fenômenos de diferenciação celular como a neuritogênese. Recentemente,
Braga e colaboradores (2013) demonstraram que a expressão de receptores M3
aumenta ao longo do desenvolvimento da retina (P0-P30). Baseado nesses dados
procedeu-se à investigação do efeito do extrato de EP na expressão de receptores
M3 em células mantidas em cultura por 48h (Figura 14).
CT
EP 1µg/m
L0
100
200
Nív
eis
de r
ecep
tor
M3
Uni
dade
s A
rbitr
ária
s(%
do
cont
role
)
*
66 kDa M3
49 kDa actina Figura 14. Níveis de receptor M3 em culturas tratadas com extrato diclorometano de E. punicifolia por 48h. Controle (CT); Extrato diclorometano de E. punicifolia (EP) 1µg/mL. A: Níveis da expressão de receptores M3, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 5). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,0005)
Os resultados demonstram o extrato induz um significativo aumento (74%), nos
níveis de receptores M3 em células da retina mantidas em cultura.
Sabendo que a ativação de receptores M4 é importante para o
processamento da informação visual (WEBER & SCHLICKER, 2001), foram
realizadas culturas de células da retina, as quais foram tratadas com EP 1µg/mL por
48h, para investigar o efeito deste extrato sobre a expressão de receptor M4 (Figura
15).
A
B
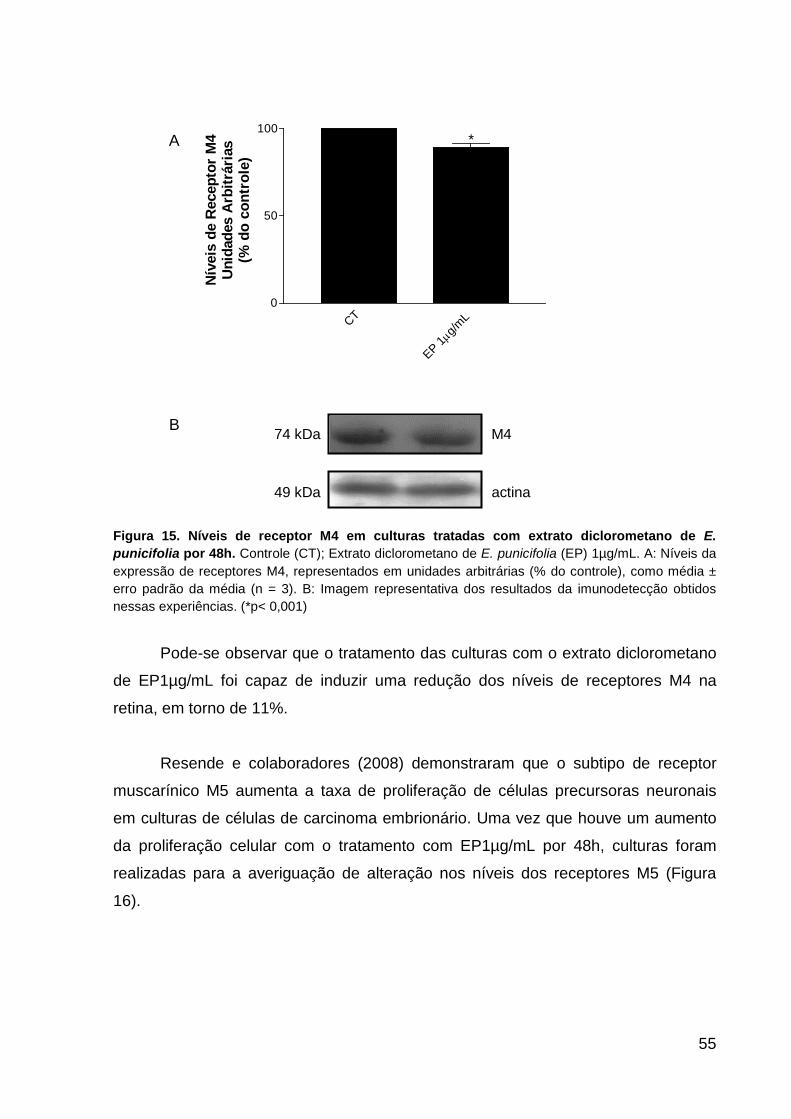
55
CT
EP 1µg/m
L0
50
100
Nív
eis
de R
ecep
tor
M4
Uni
dade
s Ar
bitr
ária
s(%
do
cont
role
)
*
74 kDa M4
49 kDa actina
Figura 15. Níveis de receptor M4 em culturas tratadas com extrato diclorometano de E. punicifolia por 48h. Controle (CT); Extrato diclorometano de E. punicifolia (EP) 1µg/mL. A: Níveis da expressão de receptores M4, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 3). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,001)
Pode-se observar que o tratamento das culturas com o extrato diclorometano
de EP1µg/mL foi capaz de induzir uma redução dos níveis de receptores M4 na
retina, em torno de 11%.
Resende e colaboradores (2008) demonstraram que o subtipo de receptor
muscarínico M5 aumenta a taxa de proliferação de células precursoras neuronais
em culturas de células de carcinoma embrionário. Uma vez que houve um aumento
da proliferação celular com o tratamento com EP1µg/mL por 48h, culturas foram
realizadas para a averiguação de alteração nos níveis dos receptores M5 (Figura
16).
A
B
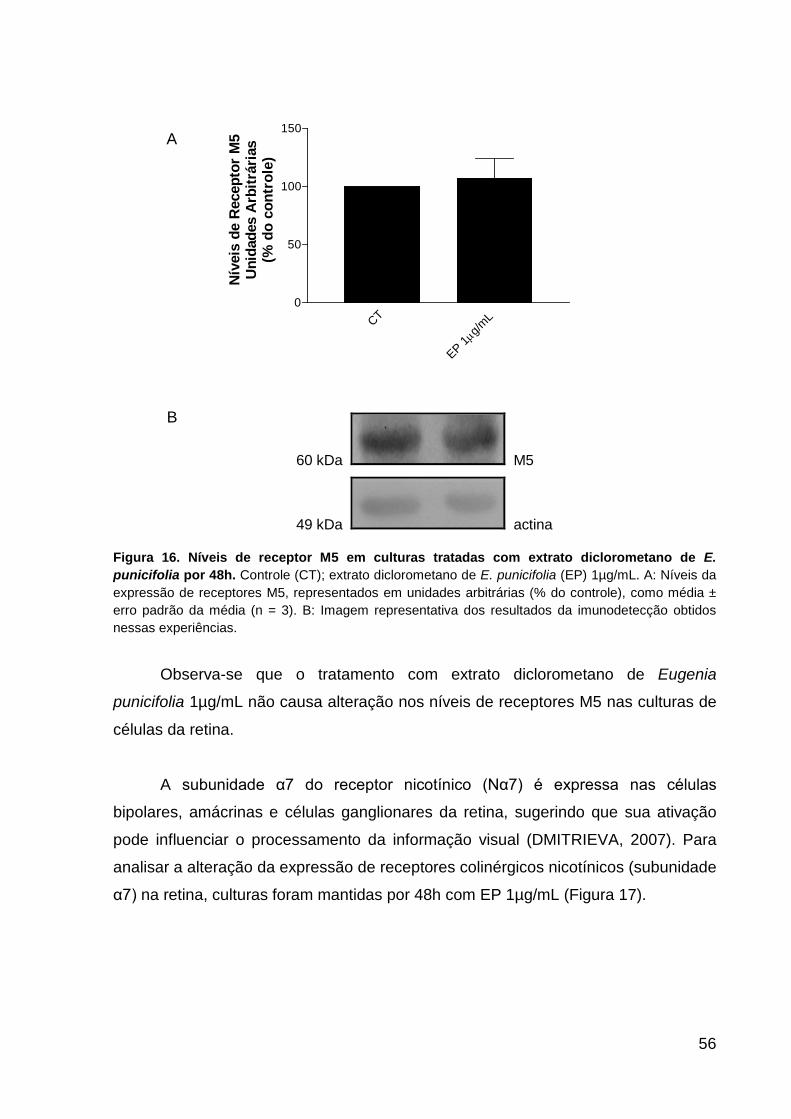
56
CT
EP 1µg/m
L0
50
100
150
Nív
eis
de R
ecep
tor
M5
Uni
dade
s Ar
bitr
ária
s(%
do
cont
role
)
60 kDa M5
49 kDa actina Figura 16. Níveis de receptor M5 em culturas tratadas com extrato diclorometano de E. punicifolia por 48h. Controle (CT); extrato diclorometano de E. punicifolia (EP) 1µg/mL. A: Níveis da expressão de receptores M5, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 3). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências.
Observa-se que o tratamento com extrato diclorometano de Eugenia
punicifolia 1µg/mL não causa alteração nos níveis de receptores M5 nas culturas de
células da retina.
A subunidade α7 do receptor nicotínico (Nα7) é expressa nas células
bipolares, amácrinas e células ganglionares da retina, sugerindo que sua ativação
pode influenciar o processamento da informação visual (DMITRIEVA, 2007). Para
analisar a alteração da expressão de receptores colinérgicos nicotínicos (subunidade
α7) na retina, culturas foram mantidas por 48h com EP 1µg/mL (Figura 17).
A
B
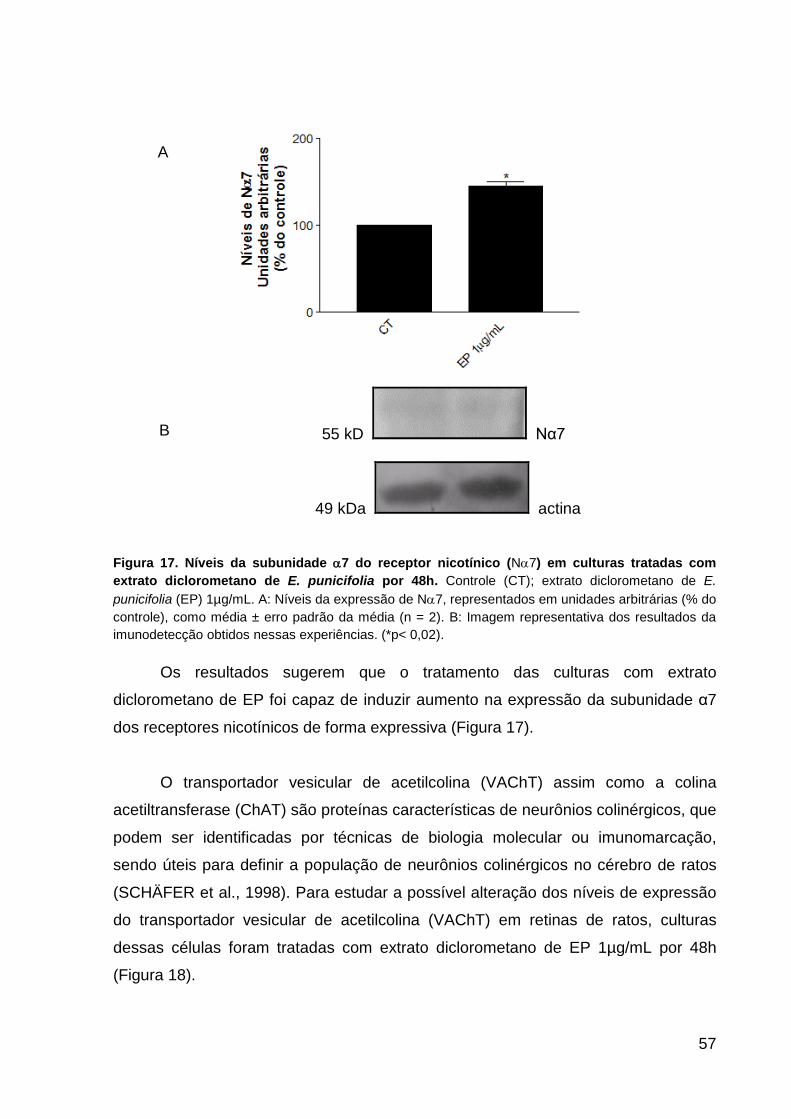
57
55 kD Nα7
49 kDa actina
Figura 17. Níveis da subunidade α7 do receptor nicotínico (Nα7) em culturas tratadas com extrato diclorometano de E. punicifolia por 48h. Controle (CT); extrato diclorometano de E. punicifolia (EP) 1µg/mL. A: Níveis da expressão de Nα7, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 2). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,02). Os resultados sugerem que o tratamento das culturas com extrato
diclorometano de EP foi capaz de induzir aumento na expressão da subunidade α7
dos receptores nicotínicos de forma expressiva (Figura 17).
O transportador vesicular de acetilcolina (VAChT) assim como a colina
acetiltransferase (ChAT) são proteínas características de neurônios colinérgicos, que
podem ser identificadas por técnicas de biologia molecular ou imunomarcação,
sendo úteis para definir a população de neurônios colinérgicos no cérebro de ratos
(SCHÄFER et al., 1998). Para estudar a possível alteração dos níveis de expressão
do transportador vesicular de acetilcolina (VAChT) em retinas de ratos, culturas
dessas células foram tratadas com extrato diclorometano de EP 1µg/mL por 48h
(Figura 18).
A
B

58
55 kDa VAChT
49 kDa actina Figura 18. Níveis de VAChT em culturas tratadas com extrato diclorometano de E. punicifolia. Controle (CT); Extrato diclorometano E. punicifolia (EP). A: Níveis de VAChT, representados em unidades arbitrárias (% do contr'ole), como média ± erro padrão da média (n = 4). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (* < 0,001).
Os resultados demonstram que o tratamento com extrato diclorometano de
EP induziu uma diminuição de 11% nos níveis do transportador vesicular de
acetilcolina (VAChT) em culturas de células da retina.
NÍVEIS DA EXPRESSÃO DE NEUROTROFINAS
Durante o desenvolvimento retiniano, os níveis de NGF aumentam
gradualmente, atingindo um máximo em 14 dias pós-natal (aumento de 100% em
relação ao dia do nascimento – P0), e reduzindo para os níveis do controle em 30
dias. Esses resultados sugerem o envolvimento do NGF na maturação do tecido
retiniano (BRAGA, 2013). Para avaliar o possível envolvimento do extrato na
A
B
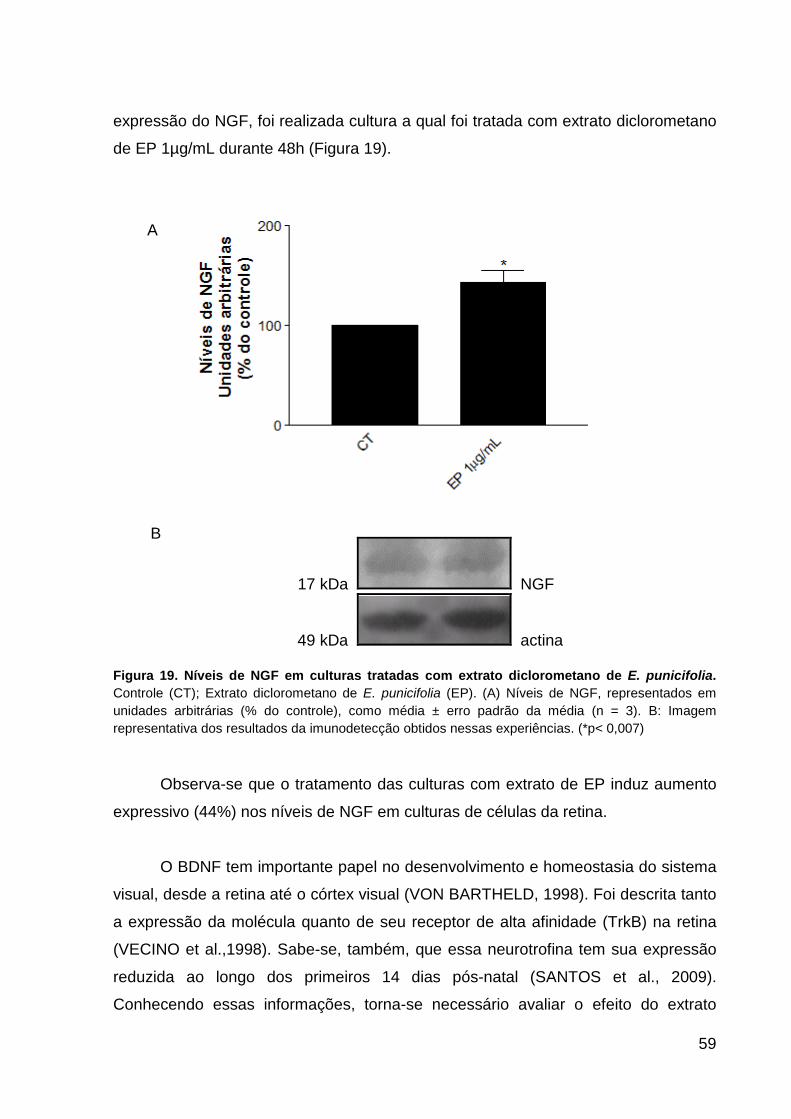
59
expressão do NGF, foi realizada cultura a qual foi tratada com extrato diclorometano
de EP 1µg/mL durante 48h (Figura 19).
17 kDa NGF
49 kDa actina
Figura 19. Níveis de NGF em culturas tratadas com extrato diclorometano de E. punicifolia. Controle (CT); Extrato diclorometano de E. punicifolia (EP). (A) Níveis de NGF, representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 3). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,007)
Observa-se que o tratamento das culturas com extrato de EP induz aumento
expressivo (44%) nos níveis de NGF em culturas de células da retina.
O BDNF tem importante papel no desenvolvimento e homeostasia do sistema
visual, desde a retina até o córtex visual (VON BARTHELD, 1998). Foi descrita tanto
a expressão da molécula quanto de seu receptor de alta afinidade (TrkB) na retina
(VECINO et al.,1998). Sabe-se, também, que essa neurotrofina tem sua expressão
reduzida ao longo dos primeiros 14 dias pós-natal (SANTOS et al., 2009).
Conhecendo essas informações, torna-se necessário avaliar o efeito do extrato
A
B
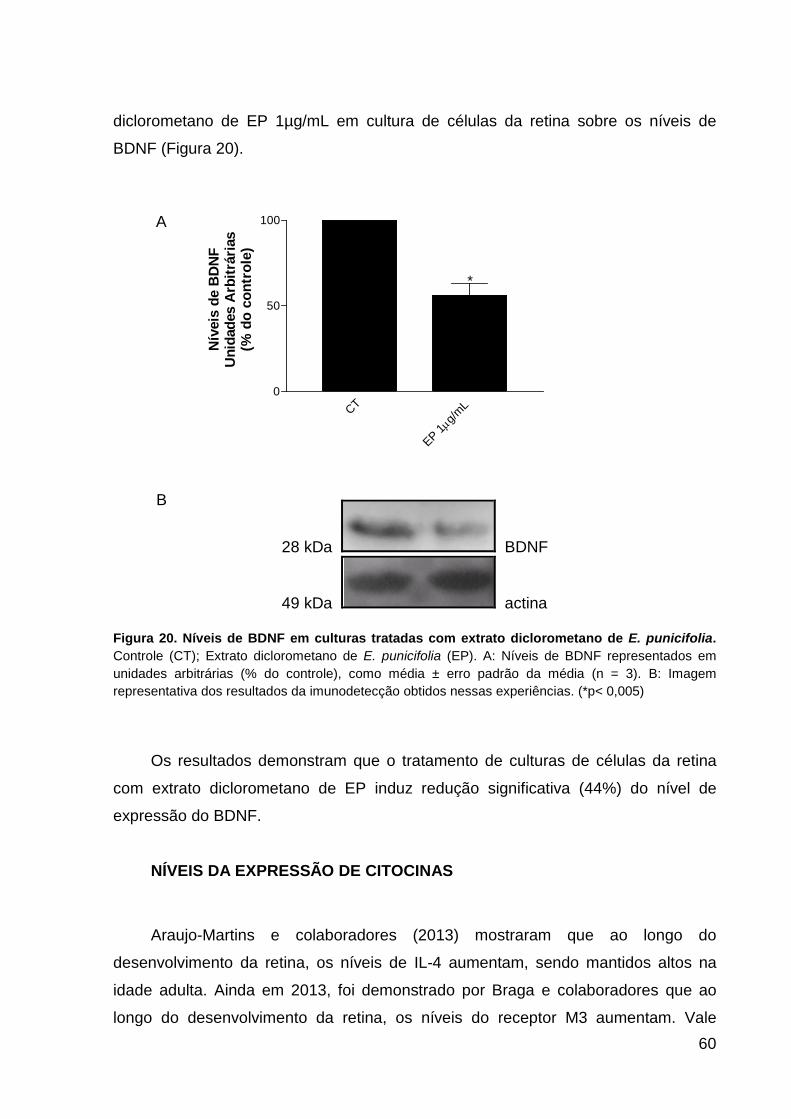
60
diclorometano de EP 1µg/mL em cultura de células da retina sobre os níveis de
BDNF (Figura 20).
CT
EP 1µg/m
L0
50
100
*
Nív
eis
de B
DN
FU
nida
des
Arb
itrár
ias
(% d
o co
ntro
le)
28 kDa BDNF
49 kDa actina Figura 20. Níveis de BDNF em culturas tratadas com extrato diclorometano de E. punicifolia. Controle (CT); Extrato diclorometano de E. punicifolia (EP). A: Níveis de BDNF representados em unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 3). B: Imagem representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,005)
Os resultados demonstram que o tratamento de culturas de células da retina
com extrato diclorometano de EP induz redução significativa (44%) do nível de
expressão do BDNF.
NÍVEIS DA EXPRESSÃO DE CITOCINAS
Araujo-Martins e colaboradores (2013) mostraram que ao longo do
desenvolvimento da retina, os níveis de IL-4 aumentam, sendo mantidos altos na
idade adulta. Ainda em 2013, foi demonstrado por Braga e colaboradores que ao
longo do desenvolvimento da retina, os níveis do receptor M3 aumentam. Vale
A
B
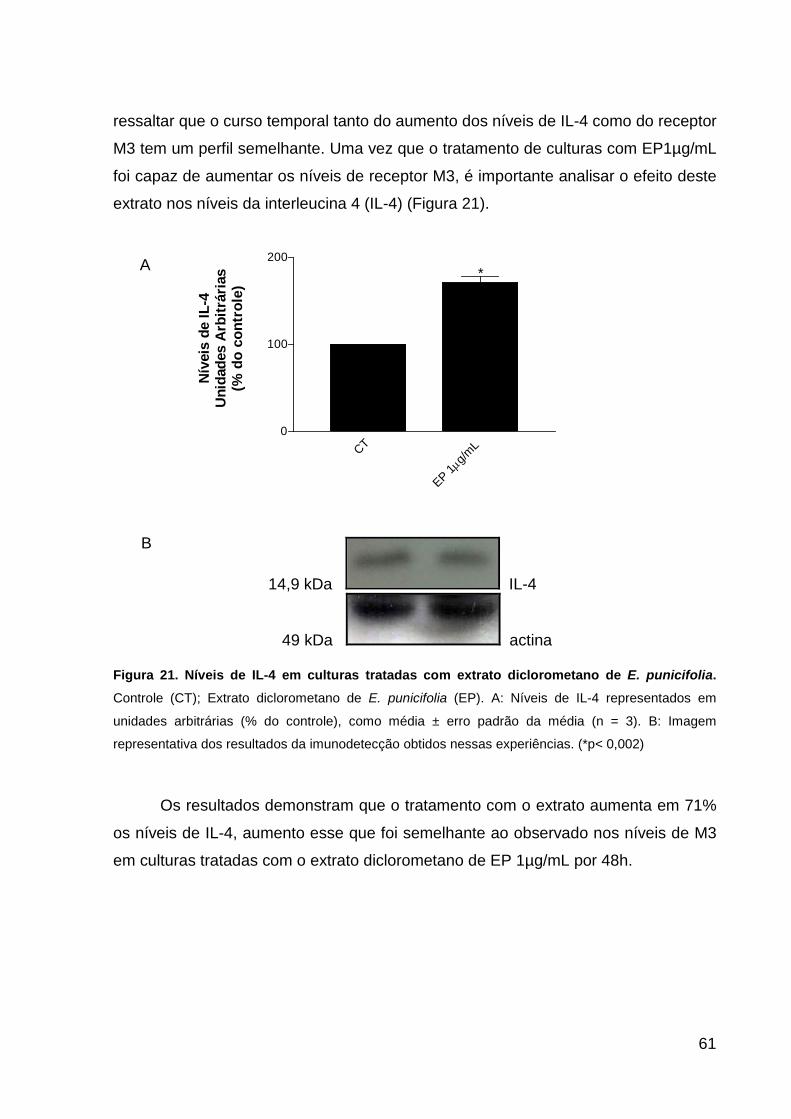
61
ressaltar que o curso temporal tanto do aumento dos níveis de IL-4 como do receptor
M3 tem um perfil semelhante. Uma vez que o tratamento de culturas com EP1µg/mL
foi capaz de aumentar os níveis de receptor M3, é importante analisar o efeito deste
extrato nos níveis da interleucina 4 (IL-4) (Figura 21).
CT
EP 1µg/m
L0
100
200
Nív
eis
de IL
-4U
nida
des
Arb
itrár
ias
(% d
o co
ntro
le)
*
14,9 kDa IL-4
49 kDa actina Figura 21. Níveis de IL-4 em culturas tratadas com extrato diclorometano de E. punicifolia. Controle (CT); Extrato diclorometano de E. punicifolia (EP). A: Níveis de IL-4 representados em
unidades arbitrárias (% do controle), como média ± erro padrão da média (n = 3). B: Imagem
representativa dos resultados da imunodetecção obtidos nessas experiências. (*p< 0,002)
Os resultados demonstram que o tratamento com o extrato aumenta em 71%
os níveis de IL-4, aumento esse que foi semelhante ao observado nos níveis de M3
em culturas tratadas com o extrato diclorometano de EP 1µg/mL por 48h.
A
B

62
6. DISCUSSÃO
Neste trabalho foi demonstrado que o tratamento de culturas de células da
retina com o extrato diclorometano de EP interfere na modulação do fenótipo
colinérgico e na expressão de fatores tróficos como neurotrofinas e a citocina IL-4.
O Extrato da Eugenia punicifolia induz efeitos pronunciados sobre a
transmissão colinérgica periférica (GRANGEIRO et al., 2006; LEITE et al., 2010) e,
sabe-se que compostos que aumentam a neurotransmissão colinérgica central
podem apresentar atividade neuroprotetora e atuar na plasticidade neuronal (RAFII
& AISEN, 2009). No presente trabalho foram realizados experimentos a fim de
observar o efeito do tratamento com extrato diclorometano de Eugenia punicifolia em
células do Sistema Nervoso Central, utilizando como modelo experimental células da
retina de ratos neonatos. Essas células apresentam grande dependência da
atividade colinérgica, tanto durante o desenvolvimento quanto na manutenção da
homeostasia em adultos (ZHOU, 2001; FELLER, 2002).
Dados da literatura mostram que receptores de acetilcolina muscarínicos
(mAChRs) são expressos, em retina de animais adultos, por células amácrinas,
ganglionares, bipolares (STRANG et al., 2010), e que durante o desenvolvimento
tecidual há uma alteração na expressão dos subtipos de receptores (SANTOS et al.,
2009; BRAGA et al., 2013). Inicialmente foi analisado se o tratamento com o extrato
diclorometano de E. punicifolia seria capaz de interferir nos níveis de expressão
desses receptores.
Inicialmente fez-se uma avaliação da proliferação celular induzida pelo
tratamento de culturas com diferentes concentrações do extrato diclorometano de E.
punicifolia. Observou-se aumento da proliferação dessas células de forma
dependente da concentração, sendo o melhor efeito (50% de aumento em relação
ao controle) obtido com o tratamento com EP 1µg/mL. Posteriormente foi
investigado o efeito do extrato na expressão dos marcadores colinérgicos. Os
resultados demonstraram que o tratamento induz redução dos níveis de receptores
M1 e M4; aumento expressivo nos níveis de receptores M3, sem induzir alterações

63
significativas nos níveis de receptores M5. Também foi demonstrada redução dos
níveis do transportador vesicular de acetilcolina, o VAChT, além de aumento nos
níveis dos receptores Nα7. Por fim, foi observado aumento nos níveis do fator de
crescimento do nervo (NGF) e da interleucina (IL-4), e diminuição da expressão do
fator neurotrófico derivado do cérebro (BDNF).
A utilização da técnica de incorporação de [3H]-timidina nesse trabalho
permitiu que fosse determinada a concentração do extrato que seria utilizada na
análise da atividade colinérgica, na expressão de neurotrofinas e de citocinas
induzida pelo extrato diclorometano de E. punicifolia.
Os resultados mostraram aumento da proliferação celular com a utilização de
100ng/mL-1µg/mL, sendo o melhor efeito observado com a utilização de EP 1µg/mL.
Essa concentração foi compatível com a utilizada por Pascual e colaboradores
(2011) que utilizaram o extrato diclorometano de E. punicifolia para estudar a
influência na secreção de catecolaminas por células cromafins da adrenal bovina e
trataram essa células com concentrações que variavam de 0,1ng/mL a 10µg/mL.
Um dos mediadores mais importantes da organização do sistema nervoso e
da plasticidade são as células gliais, cujo tipo principal da retina é representado
pelas células de Müller, que desempenham papéis dos astrócitos, oligodendrócitos e
células ependimais em outras regiões do SNC. As células de Müller derivam do
neuroectoderma da retina e pertencem à linhagem astrocítica e são geradas do
período E20 a P13 (CEPKO, 1993). Cabe lembrar que neste período (E20-P13)
também são gerados os fotorreceptores. As células de Müller respondem ao
tratamento com diferentes fatores de crescimento e tem grande importância na
manutenção e suporte de vários tipos celulares da retina, através da regulação da
atividade metabólica, concentração iônica, dentre outros (WAHLIN et al, 2000).
Ao analisar a morfologia das culturas, observa-se um padrão morfológico
semelhante em toda a placa, com a presença de um grande tapete glial sobre o qual
se encontram neurônios capazes de emitir processos longos. Ao contrário, as
culturas controle, mantidas também por este período, evidenciam uma morfologia

64
menos exuberante, com presença de poucos processos. Considerando o período do
desenvolvimento em que a retina é estudada (P0-P2), pode-se sugerir que as
células que estão proliferando nessas culturas sejam as células gliais ou
fotorreceptores, porém, o estudo da proliferação celular não foi o foco deste
trabalho.
Estudos demonstraram que o extrato aquoso da E. punicifolia tem efeito
pronunciado sobre a atividade colinérgica de diafragma de ratos (GRANGEIRO et
al., 2006) e que o extrato diclorometano da EP aumenta a secreção de
catecolaminas induzida pela acetilcolina ou por K+ sem causar bloqueio da
acetilcolinesterase ou butirilcolinesterase, além de não ativar diretamente o sistema
colinérgico em células cromafins da adrenal de bovinos em nenhuma concentração
utilizada (PASCUAL et al., 2011).
Na retina de mamíferos, a neurotransmissão colinérgica é importante tanto no
desenvolvimento quanto na manutenção da homeostasia (VANEY, 1990)
envolvendo diretamente a participação dos nAChRs (FELLER, 2002) e mAChR
(SANTOS et al., 2009). Todos os subtipos de mAChR são expressos em
subpopulações de células amácrinas, bipolares e ganglionares de retina de coelho,
incluindo células dos circuitos glicinérgico e colinérgico (SHANG et al., 2010). Além
disso, foi observado que o receptor muscarínico do subtipo M2 foi localizado na
camada nuclear interna (CNI) e na camada plexiforme interna (CPI) em retinas de
rato, mais especificamente, nas células amácrinas glicinérgicas (STRANG et al.,
2010). Os subtipos de receptores muscarínicos M2, M3, M4 foram localizados em
células amácrinas de retina (GLEASON, 2012). Também foi demonstrada a
presença da subunidade α7 do receptor nicotínico em células bipolares, amácrinas e
ganglionares na retina de coelhos (DMITRIEVA, 2007). Sabendo da importância da
atividade colinérgica na retina, foram realizadas culturas de células da retina de
ratos neonatos a fim de analisar a ação do extrato diclorometano de E. punicifolia na
atividade colinérgica dessas células.
O tratamento de culturas de células da retina de ratos neonatos com forbol
12-miristato 13-acetato (PMA), um agente mitogênico, promotor de tumor (GRINER

65
& KAZANIETZ, 2007) induz redução da proliferação celular e este efeito que é
mediado pela ativação da PKC, depende da atividade colinérgica (receptores M1) e
da ativação de receptores Trk (SANTOS et al., 2003). Em 2009, Santos e
colaboradores, analisando o nível de expressão de receptores M1, demonstraram
que há redução dos níveis ao longo do desenvolvimento, desde o nascimento até o
dia pós-natal P30 (P0 a P30). No presente trabalho foi demonstrado que o
tratamento das culturas de células da retina com o extrato diclorometano de E.
punicifolia 1µg/mL por 48h, induz diminuição na expressão de receptores M1,
semelhante ao que foi observado por Santos e colaboradores (2009). Apesar desta
semelhança, estes autores observaram redução da proliferação celular pelo
tratamento das células da retina com PMA, enquanto o tratamento dessas células
com extrato de E. punicifolia induziu aumento da proliferação.
A ativação de receptores colinérgicos M1 e M3 está diretamente envolvida no
aumento da proliferação em vários tipos de tumor como, por exemplo, tumor de
próstata (RAYFORD et al., 1997), de cólon (CHENG e RAUFMAN, 2005), de mama
(ESPANOL et al., 2007). Em linhagem de células de astrocitoma, a ativação de
receptor M3 por agonista colinérgico é capaz de aumentar a proliferação celular
(GUIZZETTI et al, 1996), sinalizando via ERK e NF-kB (GUIZZETTI et al., 2003). Em
experimentos usando células humanas de câncer de cólon, Peng e colaboradores
(2013) mostraram aumento da proliferação celular e altos níveis de expressão de
receptor M3, induzido pelo agonista muscarínico betanecol. Esse efeito é mediado
ativação de receptores M3 e de receptores do fator de crescimento epidermal
(EGFRs), sinalizando via proteína cinase ativada por mitógenos p44/42. Além disso,
os receptores M3 estimulam uma robusta expressão de genes de metaloproteinases
de matriz (MMP), que desempenham papéis chave na mediação da proliferação
celular, migração celular e invasão in vitro (PENG et al, 2013). O tratamento de
células da retina com o extrato diclorometano de E. punicifolia induz aumento da
proliferação celular com respectivo aumento dos níveis dos receptores M3. E apesar
de não se tratar de células tumorais, sugerimos que esse aumento do receptor M3
em células da retina, esteja envolvido no efeito de proliferação através de
mecanismos semelhantes.

66
A ativação de receptores M3 está envolvida com a diferenciação celular de
astrócitos de ratos tratados com carbacol, um análogo da acetilcolina (GUIZZETTI et
al., 2011), e a acetilcolina liberada pelas células amácrinas induz liberação de cálcio,
estabilizando o desenvolvimento dos neuritos de células ganglionares da retina
(LOHMANN et al., 2002). Ao analisar as culturas tratadas com extrato diclorometano
de EP, observa-se aumento no número de processos celulares, o que pode sugerir
que o aumento nos níveis de M3 nessas culturas também possa estar relacionado
com as alterações morfológicas observadas nas culturas tratadas em comparação
com as culturas controle.
A ativação dos receptores M4 é capaz de induzir a diferenciação de células
progenitoras neurais em culturas de células de carcinoma embrionário (P19) em
ratos (RESENDE, et al., 2008). ANGELIS e colaboradores (2012) mostraram que em
células progenitoras de oligodendrócitos, o receptor M4 medeia o aumento da
proliferação celular na presença de muscarina. Na retina de pintos, os receptores
M4 foram identificados nas células da camada nuclear interna e camada de células
ganglionares, havendo uma co-localização com células amácrinas quando utilizado
marcadores específicos tais como somatostatina, tirosina hidroxilase (FISCHER, et
al, 1998). Braga e colaboradores (2013) demonstraram que o tratamento de células
da retina de ratos neonatos com PMA, não foi capaz de induzir alterações nos níveis
dos receptores M4. Ao contrário do tratamento com PMA, o tratamento dessas
células com extrato diclorometano da EP foi capaz de induzir pequena redução nos
níveis dos receptores M4, sugerindo envolvimento deste receptor no efeito
proliferativo observado.
Ashkenazi e colaboradores (1989) descreveram que a ativação de receptores
muscarínicos como M1, M3 e M5 aumenta a proliferação em astrócitos corticais de
ratos. Os subtipos de receptores muscarínicos M1, M3 e M5, acoplados a proteína
Gαq/11, aumentam a taxa proliferação de diversos tipos celulares como, por exemplo,
células precursoras neurais (RESENDE et al., 2008). Apesar de ser verificada
redução da expressão de M1 e aumento da expressão de M3, não foram
observadas alterações dos níveis dos receptores M5 com o tratamento com extrato

67
diclorometano de E. punicifolia em células da retina de ratos neonatos, sugerindo
que não há participação deste subtipo de receptor nos efeitos observados.
As respostas celulares verificadas frente à ativação de receptores
muscarínicos dependem do fenótipo celular em questão (van KOPPEN & KAISER,
2003). Muitos destes tipos celulares bem como as respostas frente à ativação de
receptores colinérgicos foram citados. O tratamento dessas células com o extrato
diclorometano de E. punicifolia induz aumento nos níveis dos receptores M3 e
redução dos níveis de M1 mimetizando, tanto o que acontece durante o
desenvolvimento do tecido quanto o que é observado com o tratamento com PMA
(SANTOS et al., 2009; BRAGA et al., 2013). Porém, apesar da semelhança
observada frente às alterações morfológicas de ambos os tratamentos, sugerindo
indução da diferenciação celular, houve um efeito distinto sobre a proliferação
celular: o EP aumenta enquanto o PMA reduz a proliferação. É possível especular
que os constituintes do extrato da EP utilizem vias de sinalização distintas daquelas
observadas pelo tratamento com PMA e que isso seja capaz de induzir aumento da
proliferação celular, enquanto o PMA reduz. Outro fator a ser considerado é que o
PMA é uma droga purificada, enquanto o extrato de EP utilizado é composto por
grande número de constituintes ainda não identificados (somente o ácido
barbinérvico e o daucosterol foram isolados). Pode ser sugerido que os constituintes
do extrato estejam atuando de forma coordenada, ativando e/ou inibindo vias de
sinalização distintas sendo elas responsáveis pelo aumento da proliferação celular.
Receptores acoplados a proteína G, quando expostos a estímulos
prolongados de agonistas podem passar por um processo composto por etapas tais
como a dessensibilização, “down-regulation” e degradação (MOLINA-HOLGADO et
al., 2003). Santos e colaboradores (2009) observaram diminuição na expressão de
receptores M1 após 45 minutos de tratamento com PMA, um período considerado
suficiente para induzir a degradação desses receptores. Estudos anteriores haviam
descrito que a internalização e/ou degradação de receptores muscarínicos ocorre a
partir dos 30 minutos após a adição do agonista (LILES et al., 1986). Nota-se então
que esse “down-regulation” ocorre até que seja atingida estabilidade, que em alguns
estudos ocorre após 30 minutos de tratamento (FEIGENBAUM & EL-FAKAHANY,

68
1984; FEIGENBAUM & EL-FAKAHANY, 1985), mostrando que esse tempo é
suficiente para induzir a internalização e “down-regulation” do receptor (SANTOS et
al., 2009). No presente estudo foram observadas redução do nível de expressão de
receptores M1 e M4, o que pode sugerir que haja dessensibilização dos mesmos
após tratamento por 48h com extrato de EP. Porém, não foram realizados
tratamentos de curta duração com o extrato diclorometano da EP, não sendo
possível inferir se o mesmo é capaz de causar dessensibilização dos receptores por
tratamento de curtos períodos.
Receptores nicotínicos neuronais são expressos nos gânglios do Sistema
Nervoso Autônomo e no Sistema Nervoso Central, em localização pós-, pré- e extra
sinápticas. Dados da literatura mostram que a nicotina é capaz de estimular a
proliferação em células HT-29 de adenocarcinoma de cólon, e que a metil-
licaconitina, um antagonista de nAChRα7 é capaz de reverter este efeito (WONG et
al., 2007). A indução da proliferação se dá através do aumento na síntese de
catecolaminas, via ativação da subunidade α7 do receptor nicotínico (WONG et al.,
2007). Pascual e colaboradores (2011) mostraram que o aumento da secreção de
catecolaminas evocada pela acetilcolina em células cromafins de bovino pode ser
devido ao efeito direto do extrato diclorometano de EP em receptores nicotínicos,
comportando como um modulador alostérico da subunidade α7 do receptor
nicotínico.
O subtipo de receptor nicotínico, especialmente aquele contendo a
subunidade α7, é amplamente expresso em regiões do cérebro responsáveis pelo
aprendizado e memória, como o hipocampo e o neocórtex (SEGUELA et al., 1993).
Esse subtipo possui alta permeabilidade aos íons Ca+2 e Na+, aumenta a
neurotransmissão glutamatérgica e modula a plasticidade neuronal influenciando o
crescimento de axônios (BROIDE & LESLIE, 1999). Resende e colaboradores
(2008) ainda relacionam a expressão genes da subunidade α7 em células P19
indiferenciadas, sugerindo o envolvimento desse subtipo de receptor em processos
iniciais da proliferação e da diferenciação neuronal. Na retina, a subunidade α7 do
receptor nicotínico (Nα7) é expressa nas células bipolares, amácrinas e células

69
ganglionares, sugerindo que sua ativação pode influenciar o processamento da
informação visual (DMITRIEVA, 2007).
O tratamento de células da retina de ratos neonatos com o extrato
diclorometano de E. punicifolia é capaz de induzir aumento da expressão da
subunidade α7 do receptor nicotínico de forma significativa, sugerindo uma possível
ação desse receptor no processo de proliferação celular observado e possivelmente
na diferenciação. Como os nAChRs não possuem atividade tirosina cinase intrínseca
(GOTTI & CLEMENTI, 2004), os mecanismos moleculares envolvidos em seus
efeitos na proliferação ainda não estão muito claros. Foi demonstrado que a indução
da proliferação celular mediada por nicotina envolve recrutamento de β-arrestina,
que facilita a ativação de Src. Isto por sua vez leva à ligação da Raf-1 cinase ao Rb,
levando à entrada no ciclo celular (DASGUPTA et al., 2006).
Estudos em neurônios do hipocampo mostraram uma relação entre a
presença do VAChT e a liberação de acetilcolina, na qual o aumento do VAChT está
acompanhado do aumento na liberação do neurotransmissor (NAGY & AUBERT,
2012), o que é importante na consolidação da memória e do aprendizado (JAEGER
et al., 2013). O tratamento das culturas de células da retina de ratos neonatos com o
extrato diclorometano de E. punicifolia por 48h, induz redução nos níveis de VAChT,
o que pode indicar que o efeito da acetilcolina independe da liberação da acetilcolina
mediada por VAChT nessas células e que estes efeitos podem ser diretos via
ativação de receptores muscarínicos e/ou nicotínicos. Também pode ser sugerido
que a redução da expressão de VAChT pode estar diminuindo a liberação de
acetilcolina, corroborando a ideia de que o efeito seja direto em receptores
colinérgicos.
As atividades da ChAT e do VAChT podem ser reguladas por vários fatores
extracelulares que são capazes de influenciar o fenótipo colinérgico e exercer efeitos
tróficos sobre o desenvolvimento de neurônios colinérgicos. Um dos fatores que
influencia a sobrevida e diferenciação de neurônios colinérgicos é o fator de
crescimento do nervo (NGF) (TIAN et al., 1996). O aumento nos níveis de NGF no
desenvolvimento cerebral de ratos precede o aumento da atividade de ChAT por

70
aumentar a expressão dos genes da ChAT e VAChT em neurônios colinérgicos
(TIAN et al., 1996). Além do NGF, o fator neurotrófico derivado do cérebro (BDNF)
também modula o fenótipo colinérgico do cérebro de ratos (LI et al., 1995). Outros
fatores tróficos como a NT-3 e o CNTF podem agir modulando os receptores de
acetilcolina e promovendo a diferenciação colinérgica de neurônios derivados de
células tronco embrionário humano (NILBRATT et al., 2009). Diversos fatores
neurotróficos e receptores incluindo as neurotrofinas e receptores Trk (LEWIN &
BARDE, 1996) são expressos em padrões temporais e celulares específicos na
retina de aves durante o período de desenvolvimento (HALLBÖÖKet al., 1996; von
BARTHELD, 1998). O NGF, receptores TrkA e p75 são expressos por células
horizontais e amácrinas na retina em desenvolvimento (KARLSSON et al., 1998),
enquanto p75 tem sua expressão em diferentes tipos celulares na retina de recém-
nascidos e adultos (LARGE et al., 1989; HALLBÖÖK et al., 1990; von BARTHELD et
al., 1991; KARLSSON et al., 1998).
Dentre as ações de regulação de proliferação, diferenciação e manutenção da
função de diferentes populações neuronais descritas para o NGF, sabe-se que ele é
capaz de regular o tamanho do corpo celular, sua arborização dendrítica e sua
conectividade (MENDELL, 2001). Além disso, esta neurotrofina induz diferenciação
de neurônios colinérgicos no SNC (SHOVAL & WEIZMAN, 2005) e é importante nos
processos de plasticidade e sobrevivência neuronal (PEZET & MCMAHON, 2006).
Braga e colaboradores (2013) demonstraram que os níveis do NGF, ao longo do
desenvolvimento, tem aumento gradual a partir de P0, sendo o máximo em P14,
sugerindo envolvimento do NGF na maturação do tecido retiniano. Neste trabalho foi
observado aumento nos níveis de NGF em culturas de células tratadas com extrato
diclorometano de E. punicifolia, sugerindo envolvimento do NGF na indução da
diferenciação celular. Porém, diferente do que foi observado por TIAN e
colaboradores (1996) com relação à expressão de VAChT, em cérebro de ratos
durante o desenvolvimento, o extrato induz aumento na expressão de NGF sem que
haja aumento subsequente da expressão de VAChT, sugerindo uma sinalização
distinta nas células da retina.

71
O fator neurotrófico derivado do cérebro (BDNF) é importante para a
formação, maturação e sobrevida neuronal na fase inicial do desenvolvimento fetal
e, na fase adulta, tem papel bastante conhecido nos fenômenos cognitivos como na
consolidação da memória e aprendizado (POST, 2007). No sistema visual, dentre as
neurotrofinas citadas, o BDNF tem papel de destaque, coordenando eventos de
sobrevivência e diferenciação em todas as áreas, desde a retina até o córtex visual
(von BARTHELD, 1998). Seus efeitos foram caracterizados em estudos in vivo e in
vitro, e podem ser observados durante o desenvolvimento até a vida adulta
(ISENMANN et al., 2003).
Estudos in vitro demonstraram que o BDNF é uma neurotrofina presente nas
células ganglionares da retina de várias espécies incluindo ratos (VON BARTHELD,
1998). Em culturas de células da retina de ratos neonatos o tratamento com BDNF
reduz a proliferação celular após 48h, sendo o mesmo efeito observado no
tratamento com PMA, (SANTOS et al., 2003). O tratamento de células da retina de
ratos com extrato diclorometano da E. punicifolia foi capaz de induzir redução nos
níveis de BDNF, semelhante ao que ocorre durante o desenvolvimento. Porém, o
BDNF induz redução da proliferação celular (SANTOS et al., 2003), enquanto o
extrato diclorometano da EP induziu aumento da proliferação celular. Esses efeitos
contraditórios em relação à proliferação celular podem ser explicados pela
diversidade de constituintes do extrato que poderia estar modulando vias de
sinalização distintas na geração desses efeitos, ao contrário do que acontece com o
BDNF que é uma molécula purificada e com ativação de receptor e vias de
sinalização bem caracterizadas. Pode-se sugerir que essa redução da expressão de
BDNF induzida por extrato diclorometano de EP possa estar envolvida na indução
da diferenciação celular observada com tratamento das culturas com o extrato.
No Sistema Nervoso Central, a IL-4 tem ação importante sobre as células
gliais, inibindo a liberação de moléculas neurotóxicas como o TNF alfa por microglia
ativada (DE ARAUJO et al., 2009) além de aumentar a expressão de BDNF em
células do hipocampo (HALL et al., 2000). Também foi observado que a IL-4 reduz a
deposição de proteína AB bem como a formação de oligômeros (KIYOTA et al.,
2010), importante para o desenvolvimento e progressão da Doença de Alzheimer.

72
O interesse nos efeitos da IL-4 no desenvolvimento do sistema nervoso vem
aumentando e a literatura mostra a ação desta interleucina em eventos como a
proliferação celular (ESTES et al., 1993; BARNA et al., 1995; BRODIE et al., 1998),
diferenciação celular (BRODIE & GOLDREICH, 1994), controle da sobrevida
neuronal de diferentes tipos celulares (ARAUJO & COTMAN, 1993; SHOLL-
FRANCO et al., 2001), arranjo do citoesqueleto e fenótipo das células gliais
(WIRJATIJASA, 2002). Esta citocina induz diferenciação de bastonetes em roedores
através da ativação de tirosina cinase, PKC e MAP cinase, independente da
liberação de fatores tróficos (DA SILVA et al., 2008). Em relação à proliferação de
células da retina de ratos neonatos, não foi observado aumento considerável neste
parâmetro quando do tratamento com IL-4 (SILVA, 2011). Porém, a IL-4 aumenta a
sobrevida das células ganglionares da retina, desempenhando importante papel
neuroprotetor nessas células após axotomia, e sua expressão na retina aumenta
durante o período pós-natal (ARAUJO-MARTINS et al., 2013).
A IL-4 é capaz de controlar a síntese e secreção de NGF in vitro bem como
regular a proliferação de células gliais (DE ARAUJO et al., 2009), e induzir a
secreção de NGF em células astrogliais de ratos (AWATSUJI et al., 1993). Araujo-
Martins e colaboradores (2013) mostraram que a IL-4 pode estar induzindo a
liberação de neurotrofinas e/ou aumentando a expressão de receptores Trk nas
membranas das células da retina de ratos neonatos, uma vez que a utilização do
anticorpo anti-BDNF inibiu o efeito da IL-4 de resgate da morte de células
ganglionares da retina induzida por axotomia. O mesmo estudo ainda observou
aumento na expressão de BDNF em culturas tratadas com IL-4, bem como um
aumento da expressão de IL-4 ao longo do desenvolvimento pós-natal (P0 até 1ano)
O tratamento de células da retina de ratos neonatos com extrato diclorometano da
EP induz um aumento expressivo dos níveis da IL-4, sugerindo que a IL-4 esteja
induzindo a síntese e secreção de NGF e que ambos estão atuando em conjunto
para a indução da proliferação e diferenciação celular observadas.
É possível sugerir que a diferenciação esteja sendo induzida através da ação
direta do extrato em receptores muscarínicos do tipo M1 e M3, além da liberação de

73
fatores tróficos como o NGF, BDNF e a IL-4. E por outro lado, a proliferação seja
mediada por uma ação do extrato na subunidade α7 do receptor nicotínico. No
entanto, por não haver nenhum outro tipo de trabalho utilizando o extrato
diclorometano de EP em células do Sistema Nervoso Central, esses resultados são
referentes a etapa inicial. É necessário que seja realizada uma investigação das vias
de sinalização que estão sendo ativadas ou inibidas pelo extrato e ainda um estudo
com os compostos isolados. Conhecendo mais detalhadamente as ações do extrato
ou seus componentes no desenvolvimento retiniano, pode-se sugerir o
desenvolvimento de estratégias terapêuticas para o tratamento de diferentes
doenças que acometem esse tecido.

74
7. CONCLUSÕES Baseados nos resultados do tratamento de culturas mistas de células da
retina de ratos neonatos com o extrato diclorometano de E. punicifolia, pode-se
concluir que este é capaz de aumentar a proliferação de células da retina de ratos
neonatos de forma dependente da concentração.
No que diz respeito ao perfil fenotípico colinérgico o extrato induz aumento a
expressão de receptores colinérgicos muscarínicos M3, reduz a expressão de
receptores colinérgicos muscarínicos M1 e M4, não altera a expressão de receptor
colinérgico M5 e aumenta a expressão da subunidade α7 do receptor nicotínico.
O tratamento com o extrato ainda induz aumento na expressão de NGF,
reduz a expressão de BDNF e aumenta a expressão de interleucina-4.
Por se tratar de um extrato, cuja composição ainda não se encontra
totalmente caracterizada, pode-se sugerir que o extrato diclorometano de EP esteja
atuando de modo a induzir uma diferenciação e proliferação das células da retina, e
tais processos estão sendo desencadeados por vias de sinalização distintas.

75
8 REFERÊNCIAS BIBLIOGRÁFICAS
ABREU-VILLAÇA Y., FILGUEIRAS C.C., MANHÃES A.C. Developmental aspects of
the cholinergic system. Behavioural Brain Research, v.221, p.367–378, 2011.
ALOE L. Rita Levi-Montalcini: the discovery of nerve growth factor and modern
neurobiology. Trends Cell Biol., v.14, p.395-399, 2004.
AMARANTE-MENDES G.P., GREEN D.R. The regulation of apoptotic cell death.
Braz. J. Med. Biol. Res., v.32, p.1035-1061, 1999.
AMENTA F., TAYEBATI S.K. Pathways of acetylcholine synthesis, transport and
release as targets for treatment of adult-onset cognitive dysfunction. Curr Med
Chem., v.15, p.488–98, 2008.
ANGELIS F., BERBARDO A., MAGNAGHI V., MINGHETTI L., TATA A.D. Muscarinic
receptor subtypes as potential targets to modulate oligodendrocyte progenitor
survival, proliferation and differentiation. Developmental Neurobiology, v.72,
p.713-728, 2012.
ARAUJO D.M., COTMAN C.W. Trophic effects of interleukin-4, -7 and -8 on
hippocampal neuronal cultures: potential involvement of glial-derived factors.
Brain Res., v.600, p.49–55, 1993.
ARAUJO-MARTINS, L., OLIVEIRA, R.M., SANTOS, G.V.G., SANTOS, R.C.C.,
SANTOS, A.A., ARAUJO, E.G. Treatment In vitro of Retinal Cells with IL-4
Increases the Survival of Retinal Ganglion Cells: The Involvement of BDNF.
Neurochemistry Research, v.38, (1), p.162-173, 2013.
ARÉVALO J.C., WU S.H. Neurotrophin signaling: many exciting surprises! Cell. Mol.
Life Sci., v.63, p.1523-1537, 2006.
ARMSON P.F., BENNET M.R., RAJU T.R. Retinal ganglion cell survival and neurite
regeneration requirements: the changes from Müller cell dependence to
superior colliculli dependence during development. Dev. Brain Res., v.32,
p.207-216, 1987.
ASCANO M.,BODMER D.,KURUVILLA R. Endocytic trafficking of neurotrophins in
neural development. Trends Cell Biol., v.22, p.266–273, 2012.

76
ASHKENAZI, A., RAMACHANDRAN J., CAPON D.J. Acetylcholine analogue
stimulates DNA synthesis in brain-derived cells via specific muscarinic receptor
subtypes. Nature, v.340, p.146, 1989
BAGLEY R.S. Fundamentals of Veterinary Clinical Neurology, Oxford, p.570, 2005.
BARDE Y.A., EDGAR D., THOENEN H. Purification of a new neurotrofic factor from
mammalian brain. EMBO J., v.1, p.549-553, 1982.
BARNA B.P., ESTES M.L., PETTAY J., IWASAKI K., ZHOU P., BARNETT G.H.
Human astrocyte growth regulation: interleukin-4 sensitivity and receptor
expression. J. Neuroimmunol., v.60, p.75–81, 1995.
BEIGNEUX A.P., KOSINSKI C., GAVINO B., HORTON J.D., SKARNES W.C.,
YOUNG S.G. ATP-citrate lyase deficiency in the mouse. Journal of Biological
Chemistry, v.279, p.9557-9564, 2004.
BENNETT M. R. The concept of transmitter receptors: 100 years on.
Neuropharmacology, v.39, p.523-46, 2000.
BERKEMEIER L.R., WINSLOW J.W., KAPLAN D.R., NIKOLICS K., GOEDDEL D.V.,
ROSENTHAL A. Neurotrophin-5: a novel neurotrophic factor that activates Trk
and TrkB. Neuron, v.7, p.857-866, 1991.
BERZAGHI, M.P., COOPER, J., CASTRÉN, E., ZAFRA, F., SOFRONIEW, M.,
THOENEN, H. and LINDHOLM, D. Cholinergic regulation of brainderived
neurotrophic factor (BDNF) and nerve growth factor (NGF) but not neurotrophin-
3 (NT-3) mRNA levels in the developing rat hippocampus. J. Neurosci. 13:
3818-3826. (1993).
BOEHM M., NABEL E.G. Cell Cycle and Cell Migration: New Pieces to the Puzzle.
Circulation, v.103, p.2879-2881, 2001.
BONINI S., LAMBIASE A., BONINI S., LEVI-SCHAFFER F., ALOE L. Nerve growth:
an important molecule in allergic inflammation and tissue remodeling. Arch.
Allergy Immunol., v.118, p.159-162, 1999.
BOPP A., De BONA K.S., BELLE L.P., MORESCO R.N., MORETTO M.B.BONA KS,
BELLE L.P., MORESCO R.N., MORETTO M.B.. Syzygium cumini inhibits
adenosine deaminase activity and reduces glucose levels in hyperglycemic
patients. Fundam Clin Pharmacol., v.23, p.501–507, 2009.

77
BOYD J.G.; GORDON, T. Neurotrophic factors and their receptors in axonal
regeneration and functional recovery after peripheral nerve injury, Mol.
Neurobiol., v.27, p.277-323, 2003.
BRADFORD M.M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem.,
v.72, p.248-254, 1976.
BRAGA L.E.G., GRANJA M.G., DA SILVA G.M., GIESTAL DE ARAÚJO E.,
SANTOS A.A. PMA increases M3 muscarinic receptor levels and decreases
retinal cells proliferation through a change in the levels of cell-cycle regulatory
proteins. Neuroscience Letters, v.550, p.29–34, 2013.
BRAGA L.E.G. Papel da PKC na diferenciação e modulação do fenótipo colinérgico
de células da retina de ratos neonatos: Envolvimento de neurotrofinas. Niterói,
95p. Dissertação de Mestrado, Programa de Pós-graduação em Neurociências,
Universidade Federal Fluminense, 2013.
BRITO F.A., LIMA L.A., RAMOS M.F., NAKAMURA M.J., CAVALHER-MACHADO
S.C., SIANI A.C., HENRIQUES M.G., SAMPAIO A.L. Pharmacological study of
antiallergic activity of Syzygium cumini (L.). Skeels. Braz J Med Biol Res., v.40,
p.105–115, 2007.
BROID R.S., LESLIE F.M. The alpha7 nicotinic acetylcholine receptor in neuronal
plasticity. Mol. Neurobiol., v.20, p.1-16, 1999.
BROIDE C., GOLDREICH N. Interleukin-4 modulates the proliferation and
differentiation of glial cells. J. Neuroimmunol., v.55, p.91–97, 1994.
BROIDE C., GOLDREICH N., HAIMAN T., KAZIMIRSKY G. Functional IL-4 receptors
on mouse astrocytes: IL-4 inhibits astrocytes activation and induces NGF
secretion. J. Neuroimmunol., v.81, p.20-30, 1998.
BRUNETTI I.L. VENDRAMINI, R.C., JANUÁRIO A.H., FRANÇA S.C., PEPATO, M.T.
Effects and toxicity of Eugenia punicifolia extracts in streptozotocin-diabetic rat.
Pharmac Biol. , v. 44 (1), p. 35-43, 2006.
CAULFIELD M.P. Muscarinic Receptors—Characterization, coupling and function.
Pharmacology & Therapeutics, v.58, p.319–379, 1993.

78
CAULFIELD M.P., BIRDSALL N.J.M. Classification of muscarinic acetylcholine
receptors. International Union of Pharmacology XVII. Pharmacological Reviews,
v.50, p.279-290, 1998.
CEPKO C.L. Retinal cell fate determination. Prog. Retinal Res., v.12, p.1-12, 1993.
CHANGEUX J.P. Molecular mechanisms of nicotine dependence. Biofutur, v.234,
p.22-25, 2003.
CHANGEUX J.P., EDELSTEIN S.J. Allosteric mechanisms in normal and
pathological nicotinic acetylcholine receptors. Opin. Neurobiol., v.11, p.369-377,
2001.
CHAO M.V. Neurotrophins and their receptors: a converge point for many signaling
pathways. Nat. Rev. Neurosci., v.4, p.199-309, 2003.
CHAO M.V. The p75 neurotrophin factor. J. Neurobiol., v.25, p.1373-1385, 1994.
CHENG K, RAUFMAN JP. Bile acid-induced proliferation of a human colon cancer
cell line is mediated by transactivation of epidermal growth factor receptors.
Biochem Pharmacol 70: 1035–1047, 2005.
CHEON E.W., KUWATA O.,SAITO T. Muscarinic acetylcholine receptors in the
normal, developing and regenerating newt retinas. Developmental Brain
Research, v.127, p.9-21, 2001.
CHOI P., REISER H. IL-4: role in disease and regulation of production. Clin Exp
Immunol v.113, p.317–319, 1998.
COLLIER B., ILSON D. The effect of preganglionic nerve stimulation on the
accumulation of certain analogues of choline by a sympathetic ganglion. J.
Physiol., v.264, n.2, p.489-509. 1977.
COLLIER B., KATZ H.S. Acetylcholine synthesis from recaptured choline by a
sympathetic ganglion. J. Physiol., v.238, n.3, p.639-55, 1974.
CONSOLINI A.E., SARUBBIO M.G. Pharmacological effects of Eugenia uniflora
(Myrtaceae) aqueous crude extract on rat's heart. J Ethnopharmacol., v.81,
p.57-63, 2002.
CONSOLINI, A.E., BALDINI, O.A., AMAT, A.G., Pharmacological basis for the
empirical use of Eugenia uniflora L. (Myrtaceae) as antihypertensive. J
Ethnopharmacol 66, 33-39. 1999.
DA SILVA A.G., CAMPELLO-COSTA P., LINDEN R, SHOLL-FRANCO A.
Interleukin-4 blocks proliferation of retinal progenitor cells and increases rod

79
photoreceptor differentiation through distinct signaling pathways. J.
Neuroimmunol. v.196, p.82–93, 2008.
DALE H.H.: The action of certain esters and ethers of choline, and their relation to
muscarine. J. Pharmacol. Exp Ther. 6, 147-190, (1914).]
DASGUPTA P., RASTOGI S., PILLAI S., ORDONEZ-ERCAN D., MORRIS M.,
HAURA E., CHELLAPPAN S. Nicotine induces cell proliferation by beta-
arrestin-mediated activation of Src and Rb-Raf-1 pathways. J. Clin. Invest.,
v.116, p.2208-2217, 2006.
DE ARAUJO EG, DA SILVA GM, DOS SANTOS AA. Neuronal cell survival: the role
of interleukins. Ann N Y Acad Sci, v.11. n. 53, p.57–64, 2009.
DESCARRIES L., GISIGER V., STERIADE, M. Diffuse transmission by acetylcholine
in the CNS. Prog Neurobiol, v.53, p.603-325, 1997.
DMITRIEVA N.A., STRANG C.E., KEYSER K.T. Expression of Alpha 7 Nicotinic
Acetylcholine Receptors by Bipolar, Amacrine, and Ganglion Cells of the Rabbit
Retina. Journal of Histochemistry & Cytochemistry. v.55, p. 461-476, 2007.
DOWLING J E. Retina. Enc. Human Biol., v.6 p.615-31, 1991.
DREVER B.D., RIEDEL G., PLATT B. The cholinergic system and hippocampal
plasticity. Behav. Brain Res., v.221, p.505-514, 2011.
DYER M.A., CEPKO C.L. Regulating proliferation during retinal development. Nat.
Rev. Neurosci., v.2, p.333-42, 2001.
EBENDAL T. Function and evolution in the NGF family and its receptors. J. Neurosci.
Res., v.32, p.461-470, 1992.
ELLIOT, K., NINAMI, N., KOLESNIKOV, Y.A., PASTERNAK, G.W. AND INTURRISI,
C.E. The NMDA receptor antagonists, LY 274614 and MK- 801, and the nitric
oxide synthase inhibitor, NG-nitro- -arginine attenuate analgesic tolerance to the
mu-opioid morphine but not to the kappa opioids. Pain. v.56, p.69-75, 1994.
ESPANOL AJ, DE LA TORRE E, FISZMAN GL, SALES ME. Role of nonneuronal
cholinergic system in breast cancer progression. Life Sci 80: 2281–2285, 2007.
ESTES M.L., IWASAKI K., JACOBS B.S., BARNA B.P. Interleukin-4 down-regulates
adult human astrocyte DNA synthesis and proliferation. Am. J. Pathol. V.143,
p.337–34, 1993.
FARAH M.H. Neurogenesis and cell death in the ganglion cell layer of vertebrate
retina. Brain Res. Rev., v.52, p.264-274, 2006.

80
FEIGENBAUM .P, EL-FAKAHANY E.E. Regulation of muscarinic cholinergic receptor
density in neuroblastoma cells by brief exposure to agonist: possible
involvement in desensitization of receptor function. J. Pharmacol. Exp. Ther.
v.233, p.134–140, 1985.
FEIGENBAUM P., EL-FAKAHANY E.E. Short-term regulation of muscarinic
acetylcholine receptor binding cultured nerve cells.Res. Commun. Chem.
Pathol. Pharmacol. V.43, p.519–522, 1984.
FELDER C.C., BYMASTER F.P., MCKINZIE D.L., WESS J. Use of M1–M5
Muscarinic Receptor Knockout Mice as Novel Tools to Delineate the
Physiological Roles of the Muscarinic Cholinergic System. Neurochemical
Research, v.28, p.437-442, 2003.
FELLER M.B. The role of nAChR-mediated spontaneous retinal activity in visual
system development. J Neurobiol., v.53, p.556–567,2002.
FERGUNSON S.M., SAVCHENKO V., APPARSUNDARAM S., ZWICK M., WRIGHT
J., HEILMAN C.J., YI H., LEVEY A.I., BLAKELY R.D. Vesicular localization and
activitydependent trafficking of presynaptic choline transporters. J. Neurosci.,
v.23, n.30, p.9697-709, 2003.
FERNSTROM J.D. Effects of Precursors on Brain Neurotransmitter Synthesis and
Brain Functions. Diabetologia, v.20, p.281-289, 1981.
FERREIRA V.F., ROCHA D.R., ARAÚJO K.G.L., Santos W.C. Advances in Drug
Discovery to Assess Cholinergic Neurotransmission: A Systematic Review. Current Drug Discovery Technologies, v.5, p.236-249. 2008,
FIORENTINO D.F., ZLOTNIK A., MOSMANN T.R., HOWANR M., O’HARA A. IL-10
inhibits cytokine production by activated macrophages. J. Immunol., v.147, p.
3815-3822, 1991.
FISCHER A.J., MCKINNON L.A., NATHANSON N.M., STELL W.K. Identification and
localization of muscarinic acetylcholine receptors in the ocular tissue of the
chick. J. Comp. Neurol., v.392, p.273–284, 1998.
FRIEDMAN W.J. Proneurotrophins, Seizures, and Neuronal Apoptosis.
Neuroscientist. v.16, p.244–252, 2010.

81
FRUCHT H., JENSEN R.T., DEXTER D., YANG W.L., XIAO Y. Human Colon Cancer
Cell Proliferation Mediated by the M3 Muscarinic Cholinergic Receptor. Clin.
Cancer Res., v.5, p.2532-2539, 1999
GALENO D.M.L., CARVALHO R.P., BOLETI A.P.A., LIMA A.S., ALMEIDA P.D.O.,
PACHECO C.C., SOUZA T.P., LIMA E.S. Extract from Eugenia punicifolia is an
Antioxidant and Inhibits Enzymes Related to Metabolic Syndrome. Appl.
Biochem. Biotechnol. 2013. (PUBLICADO ONLINE)
GALI-MUHTASIB H., BAKKAR N. Modulating Cell Cycle: Current Applications and
Prospects for Future Drug Development. Current Cancer Drug Targets, v.2,
p.309-336, 2002.
GLEASON, E. The influences of metabotropic receptor activation on cellular
signaling and synaptic function in amacrine cells. Vis. Neurosci., v.29, p.31-39,
2012.
GOLD P.E. Acetylcholine modulation of neural systems involved in learning and
memory. Neurobiology of Learning and Memory, v.80, p.194-210, 2003.
GOLIAS C. H., CHARALABOPOULOS A., CHARALABOPOLOS K. Cell proliferation
and cell cycle control: a mini review. Int. J. Clin. Practice, v.58, p.1134-1141,
2004.
GOODMAN O.B.J., KRUPNICK J.G., SANTINI F., GUREVICH V.V., PENN R.B.,
GAGNON A.W., KEEN J.H., BENOVIC J.L. Beta-arrestin acts as a clathrin
adaptor in endocytosis of the beta2-adrenergic receptor. Nature, v.383, p.447–
450, 1996.
GOTTI C., CLEMENTI F. Neuronal nicotinic receptors: from structure to pathology.
Progress in Neurobiology, v.74, p.363–396, 2004.
GOTTI C., FORNASARI D., CLEMENTI F. Human neuronal nicotinic receptors. Prog.
Neurobiol., v.53, p.199-237, 1999.
GÖTZ R., KOSTER R., WINKLER C., FRIEDICH R., LOTTSPEICH F., SCHARTL
M., THOENEN H. Neurotrophin-6 is a new member of the nerve growth factor
family. Nature, v.372, p.266-269, 1994.
GRANGEIRO M.S., LIMA A.P.C., MARTINS M.F., ARRUDA L.F., do CARMO L.G.,
SANTOS W.C. Pharmacological effects of Eugenia Punicifolia (Myrtaceae) in
cholinergic nicotinic neurotransmission. J. Ethnopharmacol., v.108, p.26-30,
2006.

82
GRANJA M.G. Efeito da Interleucina-4 in vitro na modulação do fenótipo colinérgico
em células da retina de ratos. Niterói, 80p. Dissertação de Mestrado, Programa
de Pós-graduação em Neurociências, Universidade Federal Fluminense, 2013.
GUIZZETTI M, BORDI F, DIEGUEZ-ACUNA FJ, VITALONE A, MADIA F, WOODS
JS, COSTA LG. Nuclear factor kappaB activation by muscarinic receptors in
astroglial cells: effect of ethanol. Neuroscience 120: 941–950, 2003.
GUIZZETTI M, COSTA P, PETERS J, COSTA LG. Acetylcholine as a mitogen:
muscarinic receptor-mediated proliferation of rat astrocytes and human
astrocytoma cells. Eur J Pharmacol., 297: 265–273, 1996.
GUIZZETTI M.; MOORE N.H.; VANDEMARK K. L.; GIORDANO G.; COSTA L.G.
Muscarinic receptor-activated signal transduction pathways involved in the
neuritogenic effect of astrocytes in hippocampal neurons. Eur. J. Pharmacol.,
v.659, p.102-107, 2011
HALL J., THOMAS K.L., EVERITT B.J. Rapid and selective induction of BDNF
expression in the hippocampus during contextual learning. Nat Neurosci
3(6):533–535, 2000.
HALLBÖÖK F., AYER-LELIÉVRE C., EBENDAL T., PERSSON H., Expression of
nerve growth factor receptor mRNA during early development of the chicken
embryo: emphasis on cranial ganglia. Development, v.108, p.693-704, 1990.
HALLBÖÖK F., BÄCKSTRÖM A., KULLANDER K., EBENDAL T., CARRI N. G.
Expression of neurotrophins and Trk receptors in the avian retina.J. Comp.
Neurol. v.364, p.664-676, 1996.
HALLBÖÖK F., IBANÉZ C.F., PERSON H. Evolutionary studies of the nerve growth
factor family reveal a novel member abundantly expressed in xenopus ovary.
Neuron., v.6, p.845-858, 1991.
HENDERSON, C.E. Role of neurotrophic factors in neuronal development. Curr.
Opin. Neurobiol. v.6, p. 64-70, 1996.
HENDRICKSON L.M., LIU L., GUILDFORD M., ZHAO-SHEA R., GARDNER P.D.,
TAPPER A.R. Nicotinic Acetylcholine Receptors Containing the α4 Subunit
Modulate Alcohol Reward.Biological Psychiatry, v.73(8), p.738–746, 2013.
HOFFMAN, B.B. & TAYLOR, P. Goodman and Gilman´s: The Pharmachological
Basis of Therapeutics. McGraw-Hill, New York, 2001.

83
HOGG R.C., RAGGENBASS M., BERTRAND D. Nicotinic acetylcholine receptors:
from structure to brain function. Reviews of Physiology, Biochemistry and
Pharmacology, v.147, p.1-46, 2003.
HOHN A., LEIBROCK J., BAILEY K., BARDE Y.A. Identification and characterization
of a novel member of the nerve growth factor / brain derived neurotrophic factor
family. Nature, v.344, p.339-341, 1990.
HUNT T. Maturation promoting factor, cyclin and the control of M-phase. Curr. Opin.
Cell Biol., v.1, p.268-74, 1989.
IBÃNÉZ C.F. Jekyll-hyde neurotrophins: the story of pro-NGF. Trends Neurosci.,
v.25, p.284-286, 2002.
ISENMANN S., KRETZ A., CELLERINO A. Molecular determinants of retinal
ganglion cell development survival, and regeneration. Ret. Eye Res., v.22,
p.483-543, 2003.
JAEGER, X., CAMMAROTA, M., PRADO, M.A.M., IZQUIERDO, I., PRADO, V.F.,
PEREIRA, G.S. Decreased acetylcholine release delays the consolidation of
object recognition memory. Behav Brain Res. v. 238 p. 62-68, 2013.
JORGE L.I.F., AGUIAR J.P.L., SILVA M.L.P. Anatomia foliar de Pedra-hume-caá
(Myrcia sphaerocarpa, Myrcia guianensis, Eugenia punicifolia – MYRTACEAE).
Acta Amazonica, v.30, p. 49-57, 2000.
KAPLAN D.R., MILLER F.D. Neurotrophin signal transduction in the nervous system.
Curr. Opin. Neurobiol., v.10, p.381-391, 2000.
KARLSSON M., CLARY D.O., LEFCORT F.B., REICHARDT L.F., KARTEN H.J.,
HALLBÖÖK F. Nerve growth factor receptor TrkA is expressed by horizontal
and amacrine cells during chicken retinal development. J. Comp. Neurol., v.400,
p.408-416, 1998.
KEEGAN A.D., PIERCE J.H. The interleukin-4 receptor: signal transduction by a
hematopoietin receptor. J Leukoc Biol v.55, p.272–279, 1994.
KERR J.F.R. History of the events leading to the formulation of the apoptosis
concept. Toxicol., v.181-182, p.471-474, 2002.
KEYSER K.T., MACNEIL M.A., DMITRIEVA N., WANG F., MASLAND R.H.,
LINDSTROM J.M. Amacrine, ganglion and displaced amacrine cells in the
rabbit retina express nicotinic acetylcholine receptors. Vis Neurosci., v.17,
p.743–752, 2000.

84
KIYOTA T, OKUYAMA S, SWAN RJ, JACOBSEN MT, GENDELMAN HE, IBKEZU T.
CNS expression of anti-inflammatory cytokine interleukin-4 attenuates
Alzheimer’s disease-like pathogenesis in APP-PS1 bigenic mice. FASEB J.,
v.24(8), p.3093–3102, 2010.
LANGMEAD C.J., WATSON J., REAVILL C. Muscarinic acetylcholine receptors as
CNS drug targets. Pharmacology & Therapeutics, v.117(2), p.232-243, 2008.
LARGE T.H., WESKAMP G., HELDER J.C., RADEKE M.J., MISKO T.P., SHOOTER
E.M., REICHARDT L.F. Structure and developmental expression of the nerve
growth factor receptor in the chicken central nervous system. Neuron., v.2,
p.1123-1134, 1989.
LEE J.D., RHOADES K., ECONOMOU J.S. Interleukin-4 inhibits the expression of
tumor necrosis factor alpha and beta, interleukins-1 beta and -6 and interferon-
gama. Immunol. Cell Biol., v.73, p. 57-61, 1995.
LEITE P.E.C. et al. DE ALMEIDA, K.B., LAGROTA-CANDIDO, J., TRINDADE, P.,
DA SILVA, R.F., RIBEIRO, M.G., LIMA-ARAUJO, K.G., SANTOS, W.C.,
QUIRICO-SANTOS, T.,Anti-Inflammatory Activity of Eugenia punicifolia Extract
on Muscular Lesion of mdx Dystrophic Mice. J. Cel Biochem, v.111, p.1652–
1660, 2010.
LEVEY A.I., WAINER B.H., MUFSON E.J., MESULAM M.M. Co-Localization of
Acetylcholinesterase and Choline-Acetyltransferase in the Rat Cerebrum.
Neuroscience, v.9, p.9-22, 1983.
LEWIN G.R., BARDE Y.A. Physiology of the neurotrophins. Annu. Rev. Neurosci.,
v.19, p.289-317, 1996.
LILES W.C., HUNTER D.D., MEIER K.E. NATHANSON N.M. Activation of protein
kinase C induces rapid internalization and subsequent degradation of
muscarinic acetylcholine receptors in neuroblastoma cells. J. Biol. Chem.
V.261, p.5307–5313, 1986.
LINDEN R., GUIMARÃES C.A. Programmed cell death Apoptosis and alternative
death styles. Eur. J. Biochem., v. 271, p.1638–1650, 2004.
LINDEN R., PERRY V.H. Ganglion cell death within the developing retina: a
regulatory role for retina dendrites? Neurosci., v.11, p.2813-2827, 1982.

85
LINDEN, R., MARTINS, R.A.P., SILVEIRA, M.S. Control of programmed cell death
by neurotransmitters and neuropeptides in developing mammalian retina.
Progress in Retinal and Eyes Research. v. 24, p. 457-491, 2005.
LINDSTROM J. Nicotinic acetylcholine receptors in health an disease. Mol.
Neurobiol., v.15, p.193-222, 1997.
LINDSTROM J. The structures of nicotinic receptors. Handbook of Experim
Pharmacol v.Neuronal Nicotinic Receptors, p.101-62, 2000.
LO A.C., HOUENOU L.J., OPPENHEIM R.W. Apoptosis in the nervous system:
morphological features, methods, pathology and prevention. Arch. Histol. Cytol.,
v.58, p.139-149, 1995.
LOHMANN C., MYHR K.L., WONG R.O. Transmitter-evoked local calcium release
stabilizes developing dendrites. Nature, v.418, p.177-181, 2002.
LU B., PANG P.T., WOO N.H. The yin and yang of neurotrophin action. Nat.
Rev.Neurosci., v.6, p.603–614, 2005.
LUHESHI G, ROTHWELL N Cytokines and fever. Int. Arch. Allergy Immunol. v.109,
p.301–307, 1996.
MAIA, J. G. S., ZOGHBI, M. G. B, LUZ, A. I. R. Essential oil of Eugenia punicifolia
(HBK) DC. J Essent Oil Res, v. 9, p. 337-338, 1997.
MAIURI M.C., ZALCKVAR E., KIMCHI A., KROEMER G. Self-eating and self-killing:
crosstalk between autophagy and apoptosis. Molecular Cell Biol., v.8, p.741-
752, 2007.
MALUMBRES M., PEVARELLO P., BARBACID M., BISCHOFF J. CDK inhibitors in
cancer therapy: what is next? Trends in Pharmacological Sciences, v.29, p.16-
21, 2007.
MARCAURELLE L.A., JOHANNES C.W. Application of natural product-inspired
diversity-oriented synthesis to drug discovery. Prog Drug Res., v.66, p.187-216,
2008.
MARTINS R.A.P, PEARSON R.A. Control of cell proliferation by neurotransmitters in
the developing vertebrate retina. Brain Res., v.1192, p.37-60, 2008.
MASSOULIÉ, J. et al. Molecular and cellular biology of cholinesterase. Progress in
Neurobiology, v.41, p.31-91, 1993.

86
MATSUI M., YAMADA S., OKI T., MANABE T., TAKETO M.M., EHLERT F.J.
Functional analysis of muscarinic acetylcholine receptors using knockout mice,
Life Sci., v.75, p.2971-2981, 2004.
MAZZANTI, C.M.A. Efeito do interferon beta, da ciclosporina A, do ebselen e da
vitamina E no sistema colinérgico e purinérgico de ratos normais e submetidos
à desmielinização pelo brometo de etídio, 2007. 160 f. Tese (Doutorado em
Bioquímica) - Universidade Federal do Rio Grande do Sul, Porto Alegre.
MENDELL, L.M. Neurotrophins and synaptic plasticity in the mammalian spinal cord.
Journ. Phisiol., v.533, p.91-97, 2001.
MESULAM M.M. The cholinergic contribution to neuromodulation in the cerebral
cortex. Neuroscience, v.7, p.297-307,1995.
MESULAM, M.M., GUILLOZET A., SHAW P., LEVEY A., DUYSENC E.G.,
LOCKRIDGE O. Acetylcholinesterase knockouts establish central cholinergic
pathways and can use butyrylcholinesterase to hydrolyse acetylcholine.
Neuroscience, v.110, p.627-639, 2002.
MEYER-FRANKE A., KAPLAN M.R., PFRIEGER F.W., BARRES B.A.
Characterization of the signaling interactions that promote the survival and
growth of developing retinal ganglion cells in culture. Neuron, v.15, p.805-819,
1995.
MILLER F.D., KAPLAN D.R. Life and death decisions: a biological role for the p75
neurotrophin receptor. Cell Death Differ., v.5, p.343-345, 1998.
MOLINA-HOLGADO E., KHORCHID A., LIU H.N., ALMAZAN G. Regulation of
muscarinic receptor function in developing oligodendrocytes by agonist
exposure. Br. J. Pharmacol., v.138, p.47–56, 2003.
MORETTI M., VAILATI S., ZOLI M., LIPPI G., RIGANTI L., LONGHI R., VIEGI A.,
CLEMENTI F., GOTTI C. Nicotinic Acetylcholine Receptor Subtypes Expression
during Rat Retina Development and Their Regulation by Visual Experience. Mol
Pharmacol., v.66, p.85–96, 2004.
MORI N., SHIRAKAWA F., MURAKAMI S., ODA S., ETO S. Interleukin-4 inhibits the
production of interleukin-1 by adult T-cell leukemia cells. European Journal of
Hematology, v.55, p. 121-125, 1995.

87
NAGY, P.M., AUBERT, I. Overexpression of the vesicular acetylcoline transporter
increased acetylcoline release in the hippocampus. Neurosci. v.218, p.1-11,
2012.
NAI Q., MCLNTOSH J.M., MARGIOTTA J.F. Relating neuronal nicotinicacetylcholine
receptor subtypes defined by subunit composition and channel function.
Molecular Pharmacology, v.63, p.311-324, 2003.
NELMS K., KEEGAN A.D., ZAMORANO J., RYAN J.J., PAUL W.E. The IL-4
receptor: signaling mechanisms and biologic functions. Annu. Rev. Immunol.
v.17, p.701–738, 1999.
NEUBIG R.R., SIDEROVSKI D.P. Regulators of G-Protein signalling as new central
nervous system drug targets. Nature Reviews Drug Discovery, v.1, p.187-197,
2002.
NEWMAN E., REICHENBACH A. The Müller cell: a funcional element of the retina.
Trends Neurosci., v.19, p.307-312, 1996.
NILBRATT M., PORRAS O., MARUTLE A., HOVATTA O., NORDBERG A.
Neurotrophic factors promote cholinergic differentiation in human embryonic
stem cell-derived neurons J. Cell. Mol. Med. v.14, p.1476-1484, 2010.
NILSSON A.S., FAINZILBER M., FALCK P., IBÁÑEZ C.F. Neurotrophin-7: a novel
member of the neurotrophin family from the zebrafish. FEBS Lett., v.13, p.285-
290, 1998.
NYKJAER A., WILLNOW T.E, PETERSEN C. M. p75NTR-live or let die. Curr. Opin.
Neurobiol., v.15, p.49-57, 2005.
ODA Y. Choline acetyltransferase: the structure, distribution and pathologic changes
in the central nervous system. Pathol Int, v.49, p.921-937, 1999.
OLIVEIRA R.N., DIAS I.J.M., CÂMARA C.A.G. Estudo comparativo do óleo essencial
de Eugenia punicifolia (HBK) DC. de diferentes localidades de Pernambuco.
Revista Brasileira de Farmacognosia. v.15, p.39-43, 2005.
OPPENHEIM R.W. Cell death during development of the nervous system. Annu.
Rev. Neurosci., v.14, p.453-501, 1991.
OPPENHEIM R.W. Programmed Cell Death. In Fundamental Neuroscience.
Academic Press, p.581-610, 1999.

88
O'REGAN S., COLLIER B. Effect of increasing choline, in vivo and in vitro, on the
synthesis of acetylcholine in a sympathetic ganglion. J. Neurochem., v.36, n.2,
p.420-30, 1981.
PARSONS S.M., BAHR B.A., ROGERS G.A., CLARKSON E.D., NOREMBERG K.,
HICKS B.W. Acetylcholine Transporter Vesamicol Receptor Pharmacology and
Structure. Progress in Brain Research, v.98, p.175-181, 1993.
PASCUAL R. COLMENA I., de los RIOS C., ROSA J.M., LEITE P.E.C., ARAUJO
K.G.L., FERREIRA V.F., ROCHA D.R., GONZAGA D.T.G., GARCÍAL A.G.,
SANTOS W.C., GANDÍAL L. Augmentation of catecholamine release elicited by
an Eugenia punicifolia extract in chromaffin cells. Braz J. Pharmacog, v.22(1),
p.1-12, 2011.
PAUL W.E. Interleukin-4: a prototypic immunoregularegulatory lymphokine. Blood
v.77, p.1859–1870, 1991.
PENG Z., HEATH J., DRACHENBERG C., RAUFMAN J.P., XIE G. Cholinergic
muscarinic receptor activation augments murine intestinal epithelial cell
proliferation and tumorigenesis.BMC Cancer, v.13, p. 1-8, 2013.
PEPEU G., GIOVANNINI M. G. Changes in acetylcholine extracellular levels during
cognitive processes. Learn Mem., v.11, p.21, 2004.
PEREIRA S.P.F., MEDINA S.V., ARAUJO E.G. Cholinergic activity modulates the
survival of retinal ganglion cells in culture: the role of M1 muscarinic receptors.
Int. J. Devl. Neurosci., v.19, p.559-567, 2001.
PERRY E., WALKER M., GRACE J., PERRY R. Acetylcholine in mind: a
neurotransmitter correlate of consciousness? Trends in Neuroscience, v.22,
n.6, p.273-280, 1999.
PEZET, S. & MCMAHON, S.B. Neurotrophin: mediators and modulators of pain.
Annu. Rev. Neurosci., v.29, p.507-538, 2006.
PIO CORREA, M., Dicionário de plantas úteis do Brasil e das exóticas cultivadas.
Ministério da agricultura. Instituto Brasileiro de desenvolvimento Florestal, Rio
de Janeiro, pp. 129. 1984.
PONOMAREV E.D., MARESZ K., TAN Y., DITTEL B.N. CNS derived interleukin-4 is
essential for the regulation of autoimmune inflammation and induces a state of
alternative activation in microglial cells. The J. of Neurosci. v.27, p.10714-
10721, 2007.

89
POST R.M. Role of BDNF in bipolar and unipolar disorder: clinical and theoretical
implications. J Psychiatr Res., v.41, p.979-990, 2007.
RABACCHI S.A., ENSINI M., BONFANTI L., GRAVINA A., MAFFEI L. Nerve growth
factor reduces apoptosis of axotomized retinal ganglion cells in the neonatal rat.
Neurosci., v.63, p.969-973, 1994.
RAFII M.S., AISEN P.S. Recent developments in Alzheimer's disease therapeutics.
BMC Med., v.7, p.7, 2009.
RAJU T.R., BENNETT M.R. Retinal ganglion cell survival requirements: a major but
transient dependence on Muller glia during development. Brain Res., v.24,
p.165-176, 1986.
RATTENHOLL A., RUOPPOLO M., FLAGIELLO A., MONTI M., VINCI F., MARINO
G., LILIE H., SCHWARZ E., RUDOLPH R. Pro-sequence assisted folding and
disulfide bond formation of human nerve growth factor. J. Mol. Biol., v.305,
p.523–533, 2001.
RAYFORD W, NOBLE MJ, AUSTENFELD MA, WEIGEL J, MEBUST WK, SHAH GV. Muscarinic cholinergic receptors promote growth of human prostate cancer
cells. Prostate 30: 160–166, 1997.
REESE B.E., COLELLO R.J. Neurogenesis in the retinal ganglion cell layer of the rat.
Neurosci., v.45, p.419-429, 1992.
REIS H.J., NICOLATO R., BARBOSA I.G., TEIXEIRA DO PRADO P.H.,
ROMANOSILVA M.A.. TEIXEIRA A.L. Increased serum levels of brain-derived
neurotrophic factor in chronic institutionalized patients with schizophrenia.
Neurosci Lett., v.439, p.157-159, 2008.
RENDE M., BRIZI E., CONNER J., TREVES S., CENSIER K., PROVENZANO C.,
TAGLIALATELA G., SANNA P.P., DONATO R. Nerve growth factor (NGF)
influences differentiation and proliferation of myogenic cells in vitro via TrKA.
Int. J. Devl. Neuroscience, v.18, p869–885, 2000.
RESENDE R.R., ADHIKARI A. Cholinergic receptor pathways involved in apoptosis,
cell proliferation and neuronal differentiation. Cell Communication and Signaling
v.7, p.1-20, 2009.
RESENDE R.R., ALVES A.S., BRITTO L.R., ULRICH H. Role of acetylcholine
receptors in proliferation and differentiation of P19 embryonal carcinoma cells.
Exp Cell Res., v.314, p.1429–1443, 2008.

90
RESENDE R.R., GOMES K.N., ADHIKARI A., BRITTO L.R.G., ULRICH H.
Mechanism of acetylcholine-induced calcium signaling during neuronal
differentiation of P19 embryonal carcinoma cells in vitro. Cell Calcium, v.43, p.
107-121, 2008.
REUSS B. & HALBACH O.B. Fibroblast growth factor and their receptors in the
central nervous system. Cell Tissue Res., v.313, p.139-157, 2003.
RIBEIRO F.M., ALVES-SILVA J., VOLKNANDT W, MARTINS-SILVA C., MAHMUD
H., WILHELM A., GOMEZ M.V., RYLETT R.J., FERGUSON S.S., PRADO V.F.,
PRADO M.A. The hemicholinium-3 sensitive high affinity choline transporter is
internalized by clathrin-mediated endocytosis and is present in endosomes and
synaptic vesicles. J. Neurochem., v.87, n.1, p.136-46, 2003.
RICHARDSON, P. M. Neurotrophic factors in regeneration. Curr. Op. Neurob., v.1,
p.401-406, 1991.
ROLE L.W., BERG D.K. Nicotinic receptors in the development and modulation of
CNS synapses. Neuron, v.16, p.1077-1085, 1996.
SALTARELLI M.D., LOWENSTEIN P.R., COYLE J.T. Coyle. Rapid in vitro
modulation of [3H]hemicholinium-3 binding sites in rat striatal slices. Eur. J.
Pharmacol., v.135, n.1, p.35-40, 1987.
SALVESEN G.S. Programmed cell death and caspases. APMIS, v.107, p.73-79,
1999.
SANTOS A.A., ARAUJO E.G. The effect of PKC activation on survival of rat retinal
ganglion cells in culture. Brain Res., v.853, p.338-343, 2000.
SANTOS A.A., MEDINA S.V., MARTINS L.A., ARAUJO A.G. Protein kinase C
regulates the expression of M1 receptors and BDNF in rat retinal cells.
Neurochem Res., v.34, p.884-890, 2009.
SANTOS A.A., MEDINA S.V., SHOLL-FRANCO A., ARAÚJO E.G. PMA decreases
the proliferation of retinal cells in vitro: the involvement of acetylcholine and
BDNF. Neurochem. Int., v.42, p.73–80, 2003.
SARTER M., HASSELMO M.E., Bruno J.P., GIVENS B. Unraveling the attentional
functions of cortical cholinergic inputs: interactions between signal-driven and
cognitive modulation of signal detection. Brain Research Reviews, v.48, p.98-
111, 2005.

91
SCHAFER, M.K., EIDEN L.E. & WEIHE, E. Cholinergic neurons and terminal fields
revealed by immunohistochemistry for the vesicular acetylcholine transporter. II.
The peripheral nervous system. Neuroscience, v.84, p.361-376, 1998.
SCHAPOVAL, E.E., SILVEIRA, S.M., MIRANDA, M.L., ALICE, C.B., HENRIQUES,
A.T. Evaluation of some pharmacological activities of Eugenia uniflora L. J
Ethnopharmacol 44, 137-142. 1994
SCHINDOWSKI K., BELARBI K., BUE´E L. Neurotrophic factors in Alzheimer’s
disease: role of axonal transport. Genes Brain Behav., v.7, p.43–56, 2008.
SCHWARTZMAN R.A., CIDLOWSKI J.A. Apoptosis: The biochemistry and molecular
biology of programmed cell death. Endocr. Rev., v.4, p.133-151, 2003.
SEGUELA P., WADICHE J., DINELEY-MILLER K., DANI J.A., PATRICK J.W.
Molecular cloning, functional properties, and distribution of rat brain alpha 7: a
nicotinic cation channel highly permeable to calcium. J. Neurosci., v.13, p.596-
604, 1993.
SHENOY S.K., LEFKOWITZ R.J. Multifaceted roles of beta-arrestins in the regulation
of seven-membrane-spanning receptor trafficking and signaling. Biochemical
Journal, v.375, p.503–515, 2003.
SHOLL-FRANCO A., FIGUEIREDO K.G.A., ARAUJO E.G. Interleukin-2 and
interleukin-4 increase the survival of retinal ganglion cells in culture.
Neuroreport, v.12, p.109-112, 2001.
SHOLL-FRANCO A., MARQUES P.M., FERREIRA C.M., ARAUJO E.G. IL-4
increases GABAergic phenotype in rat retinal cell cultures: involvement of
muscarinic receptors and protein kinase C. J. Neuroimmunol. v.133, p.20-29,
2002.
SHOVAL G, WEIZMAN A. The possible role of neurotrophins in pathogenesis and
therapy of schizophrenia. Eur Neuropsychopharmacol., v.15, p.319-329, 2005.
SILVA, G. M. IL4 modula o efeito proliferativo do FGFb e EGF em células da retina
de ratos em cultura. Tese de Doutorado apresenta ao Programa de Pós-
Graduação em Neuroimunologia do Departamento de Neurobiologia,
Universidade Federal Fluminense, 2011.
SIMON J.R., ATWEH S., KUHAR M.J. Sodium-dependent high affinity choline
uptake: a regulatory step in the synthesis of acetylcholine. J. Neurochem., v.26,
n.5, p.909-22, 1976.

92
SIMON J.R., KUHAR M.G. Impulse-flow regulation of high affinity choline uptake in
brain cholinergic nerve terminals. Nature, v.255, n.5504, p.162-3, 1975.
STRANG, C.E., RENNA, J.M., AMTHOR, F.R. & KEYSER, K.T. Muscarinic
acetylcholine receptor localization and activation effects on ganglion response
properties. Investigative Ophthalmology & Visual Science., v.5, p.2778–2789,
2010.
TERENGHI, G. Peripheral nerve regeneration and neurotrophic factors. J. Anat.,
v.194, p.1-14, 1999.
THANGARAJU A., SAWYER G.W. Comparison of the kinetics and extent of
muscarinic M1–M5 receptor internalization, recycling and downregulation in
Chinese hamster ovary cells. Eur. J. of Pharmacology, v.650, p.534-543, 2011.
TIAN X., SUN X., SUSZKIW J.B. Developmental age-dependent upregulation of
choline acetyltransferase and vesicular acetylcholine transporter mRNA
expression in neonatal rat septum by nerve growth factor. Neuroscience
Letters, v.209(2), p.134–136, 1996.
TOMASZEWICZ M., ROSSNER S., SCHLIEBS R., CWIKOWSKA J., SZUTOWICZ
A. Changes in cortical acetyl-CoA metabolism after selective basal forebrain
cholinergic degeneration by 192IgG-saporin. Journal of Neurochemistry, v.87,
p.318-324, 2003.
TORRES P.M.M., ARAUJO E.G. Interleukin-6 increases the survival of retinal
ganglion cells in vitro. J. Neuroimmunol., v.117, p.43-50, 2001.
UGOLINI G., CREMISI F., MAFFEI L. TrkA, TrkB and p75 mRNA expression is
developmentally regulated in the rat retina. Brain Res., v.704, p.121-124, 1995.
van Koppen C.J., Kaiser B: Regulation of muscarinic acetylcholine receptor signaling.
Pharmacol. Ther., v.98, p.197-220, 2003.
VANEY D.I. The mosaic of amacrine cells in the mammalian retina. In: Progress in
retinal research, v.9, p. 49–100, 1990.
VECINO E., CAMINOS E., UGARTE M., MARTIN-ZANCA D., OSBORNE N.N.
Immunohistochemical distribution of neurotrophins and their receptors in the rat
retina and the effects of ischemia and reperfusion General Pharmacology: The
Vascular System, v.30, p.305-314, 1998.
VENTURA A.L.M., ABREU P.A., FREITAS R.C.C., SATHLER P.C., LOUREIRO N.,
CASTRO H.C. Sistema colinérgico: revisitando receptores, regulação e a

93
relação com a doença de Alzheimer, esquizofrenia, epilepsia e tabagismo. Rev.
psiquiatr. clín., v..37(2), p.74-80, 2010.
VERMEULEN, K., DIRK, R., BOCKSTAELE, V., BERNEMAN, Z. N. The cell cycle: a
review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif.,
v.36, p.131–149, 2003.
von BARTHELD C.S. Neurotrophins in the development and regenerating visual
system. Histol. Histopathol., v.13, p.437-459, 1998.
von BARTHELD C.S., HEUER J.G., BOTHWELL M. Expression of nerve growth
factor (NGF) receptors in the brain and retina of chick embryos: comparison
with cholinergic development. J. Comp. Neurol., v.310, p.103-129, 1991.
WAHLIN K.J., CAMPOCHIARO P.A., ZACK D.J., ADLER R. Neurotrophic Factors
Cause Activation of Intracellular Signaling Pathways in Müller Cells and Other
Cells of the Inner Retina, but not Photoreceptors. Invest. Ophthalmol. Vis. Sci.
v.41(3), p.927-936, 2000.
WEBER B., SCHLICKER E. Modulation of dopamine release in the guinea-pig retina
by G(i)- but not by G(s)- or G(q)-protein-coupled receptors. Fundam Clin
Pharmacol., v.15, p.393– 400,2001.
WIRJATIJASA F., DEHGHAMI F., BLAHETA R.A., KORF H.W., HAILER N.P.
Interleukin-4, interleukin-10, and interleukin-1-receptor antagonist but not
transforming growth factor-beta induce ramification and reduce adhesion
molecule expression of rat microglial cells. J. Neurosci. Res. v.68, p.579–587,
2002.
WOLFE B.B., YASUDA R.P. Development of Selective Antisera for Muscarinic
Cholinergic Receptor Subtypes. Diversity of Interacting Receptors, v.757,p.186-
193, 1995.
WON M.H., KANG T.C., CHO S.S. Glial cells in the bird retina: immunochemical
detection. Microsc Res Tech., v.50, p.151-160, 2000.
WONG H.P.S., YU L., LAM E.K.Y., TAI E.K.K., WU Y.K.K., CHO C.H. Nicotine
promotes cell proliferation via α7-nicotinic acetylcholine receptor and
catecholamine-synthesizing enzymes-mediated pathway in human colon
adenocarcinoma HT-29 cells. Toxicology and Applied Pharmacology, v.221, p.
261–267, 2007.

94
YAMAMURA H.I., SNYDER S.H. High affinity transport of choline into synaptosomes
of rat brain. J Neurochem., v.21, n.6, p.1355-74, 1973.
YAMASHITA T., TUCKER K.L., BARDE Y.A. Neurotrophin binding to the p75
receptor modulates Rho activity and axonal outgrowth. Neuron., v.24, p.585-
593, 1999.
YANG X.L. Characterization of receptors for glutamate and GABA in retinal neurons.
Prog Neurobiol., v.73, p.127-150, 2004.
YOO C.B. HAN K.T., CHO K.S., HA J., PARK H.J., NAM J.H., KIL U.H., LEE K.T.
Eugenol isolated from the essential oil of Eugenia caryophyllata induces a
reactive oxygen species-mediated apoptosis in HL-60 human promyelocytic
leukemia cells. Cancer Letters, v.225, p.41–52, 2005.
ZEISEL S.H. Dietary Choline - Biochemistry, Physiology, and Pharmacology. Annual
Review of Nutrition, v.1, p.95-121, 1981.
ZHANG Z-W., VIJAYARAGHAVAN S., BERG D. K. Neuronal acetylcholine receptors
that bind α-bungarotoxin with high affinity function as ligandgated ion channels.
Neuron, v.12, p.167-77, 1994.
ZHIVOTOVSKY, B. Apoptosis, necrosis and between. Cell Cycle, v. 3, n. 1, p. 64-66.
2004.
ZHOU Z.J. The function of the cholinergic system in the developing mammalian
Retina. Prog Brian Res., v.131, p.599–613, 2001.
ZIMMERMAN, G.; SOREQ, H. Termination and beyond: acetylcholinesterase as a
modulator of synaptic transmission. Cell Tissue Research, v.326, p.655-669,
2006.

95
9 ANEXOS
9.1 Anexo 1 – Folha de aprovação do projeto pelo Comitê de Ética no Uso de
Animais