Lectinas: caracterização quimiotaxinomica de microalgas de água ...

UM
inho |
2016
Universidade do Minho
Escola de Engenharia
Vânia Isabel Novais Cunha Pôjo
Seleção de Microalgas com Potencial
de Produção de Biocombustíveis
Janeiro de 2016
Vânia
Isa
bel N
ova
is C
unha P
ôjo
Se
leçã
o d
e M
icro
alg
as c
om
Po
ten
cia
l d
e P
rod
uçã
o d
e B
ioco
mb
ustí
ve
is


Dissertação de Mestrado
Mestrado em Bioengenharia
Trabalho efetuado sob a orientação do
Professor Doutor Giuliano Marcelo Dragone Melnikov
e do
Professor Doutor José António Teixeira
Vânia Isabel Novais Cunha Pôjo
Seleção de Microalgas com Potencial
de Produção de Biocombustíveis
Janeiro de 2016
Universidade do Minho
Escola de Engenharia

ii
DECLARAÇÃO
Nome: Vânia Isabel Novais Cunha Pôjo
Endereço eletrónico: [email protected]
Número do Bilhete de Identidade: 134359895
Título da Dissertação: Seleção de Microalgas com Potencial de Produção de Biocombustíveis
Orientadores:
Professor Doutor Giuliano Marcelo Dragone Melnikov
Professor Doutor José António Teixeira
Ano de conclusão: 2016
Designação do Mestrado: Mestrado em Bioengenharia
É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA DISSERTAÇÃO APENAS PARA EFEITOS DE
INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE
COMPROMETE;
Universidade do Minho, 11 de Janeiro de 2016
Assinatura: ______________________________________________________________

iii
AGRADECIMENTOS
Gostaria de expressar o meu profundo agradecimento ao meu orientador, Dr. Giuliano
Dragone, pela simpatia, disponibilidade, conhecimento, dedicação, conselhos e sugestões.
Às minhas amigas, Marisa, Ana, Diana e Joana que sempre me apoiaram, encorajaram e
me proporcionaram muitos momentos felizes. Muito obrigada, sem vocês teria sido mais difícil!
Para finalizar, gostaria de agradecer aos meus pais, Isabel e Cipriano, e ao meu irmão
Miguel, que ao longo da minha formação académica sempre me apoiaram e incentivaram, nunca
deixando de acreditar em mim. Pedro, obrigada pelo conforto que sempre encontrei junto de ti e
pelo teu apoio incondicional.
Obrigada por tudo!


Seleção de microalgas com potencial de produção de biocombustíveis v
RESUMO
A dependência contínua dos combustíveis fósseis é insustentável, devido ao esgotamento das
reservas mundiais e às emissões de gases de efeito de estufa associadas à sua utilização. Portanto,
existem iniciativas de investigação vigorosas destinadas a desenvolver alternativas renováveis. A biomassa é
a fonte de energia renovável mais promissora porque está naturalmente disponível, pode ser produzida em
grandes quantidades e permite a produção de combustíveis líquidos, gasosos ou sólidos com potencial em
diferentes setores. Além disso, é um recurso capaz de ser totalmente renovável e ambientalmente,
socialmente e economicamente sustentável. A biomassa microalgal tem sido reconhecida recentemente.
Microalgas são microrganismos fotossintéticos, com requisitos mínimos de crescimento, que
podem produzir lípidos, proteínas e carbohidratos em grandes quantidades, durante um curto período de
tempo. Estes podem ser processados em biocombustíveis e em coprodutos de valor acrescentado.
Esta dissertação teve como principal objetivo avaliar o potencial de produção de biocombustíveis
de algumas espécies de microalgas. As microalgas selecionadas, Chlorella emersonii ACOI 516, Chlorella
vulgaris ACOI 879 e Chlorella saccharophila ACOI 96, foram cultivadas em cinco meios de cultura
diferentes, MP12, TAP, BG-11, BBM e Chu_13 mod., em fotobiorreatores, com 6 % (v/v) de CO2, uma taxa
de arejamento de 0,2 vvm e 100 µmol de fotões m-2.s-1. Após o cultivo das microalgas nestas condições,
estas foram transferidas para os mesmos meios, no entanto, desprovidos de azoto, durante cerca de 4
dias, a fim de aumentar a concentração de lípidos e/ou amido nas células. O crescimento das microalgas
selecionadas foi monitorizado através da determinação da concentração celular e quantificação da
biomassa produzida. No final dos cultivos nas condições com e sem azoto, as quantidades de lípidos,
amido e proteínas obtidas foram registadas.
Os resultados obtidos demonstraram que o meio MP12 foi um dos melhores para o crescimento
das microalgas. C. vulgaris foi a microalga que obteve valores de concentração celular e peso da biomassa
seca superiores. Para além disso, esta microalga apresentou um conteúdo lipídico de 39,2 % na condição
sem azoto do meio TAP, tendo também atingido o maior valor de produtividade lipídica, nas mesmas
condições (109,57 mg L-1 d-1). Em relação ao conteúdo de amido, C. emersonii foi a microalga que obteve
percentagens superiores – 42,7 % na condição sem azoto do meio BG-11. No entanto, C. vulgaris
demonstrou ser mais adequada para a produção de amido, por ter obtido valores de produtividade
superiores. Estes resultados fazem valer a pena a concretização de mais estudos relativos à otimização das
condições de cultura destas microalgas, visando ainda maiores produções de lípidos e amido e são
passíveis de levar a futuros estudos de ampliação de escala com possibilidade de diminuição dos custos de
produção de biomassa e seus produtos sem necessidade de investimentos adicionais.


Screening of microalgae with potential for biofuels production vii
ABSTRACT
The continuous reliance on fossil fuels is unsustainable, due to the depletion of global
reserves and the greenhouse gas emissions associated with their use. Therefore, there are
vigorous research initiatives intended to develop renewable alternatives. Biomass is the most
promising source of renewable energy because it is naturally available, can be produced in large
quantities and allows the production of liquid, gaseous or solid fuels with potential in different
sectors. Besides, it is a resource capable of being completely renewable and environmentally,
socially and economically sustainable. Microalgal biomass has been recently recognized.
Microalgae are photosynthetic microorganisms with minimum growth requirements that
can produce lipids, proteins and carbohydrates in large quantities in a short period of time. These
products can then be processed into biofuels and in co-products of added value.
The present dissertation aimed to evaluate the potential for biofuels production of some
microalgae species. The selected microalgae, Chlorella emersonii ACOI 516, Chlorella vulgaris
ACOI 879 e Chlorella saccharophila ACOI 96, were grown in five different culture mediums MP12,
TAP, BG-11, BBM and Chu_13 mod., in photobioreactors, with 6 % (v/v) of CO2 at an aeration rate
of 0.2 vvm and 100 µmol photons m-2 s-1. After cultivation of microalgae in these conditions, they
were transferred to the same media, however, with nitrogen deficiency, for about 4 days, in order
to increase the concentration of lipids and/or starch in cells. The microalgae growth was monitored
by determining the cell concentration and quantification of biomass. At the end of cultivation with
nitrogen supply and nitrogen deficiency, the amounts of lipids, starch and proteins obtained were
registered.
The results obtained showed that MP12 is one of the best mediums for microalgae growth.
The highest cellular concentration and biomass dry weight obtained was found to be in C. vulgaris.
In addition, this microalga produced a lipid content of 39.2 % in TAP nitrogen deficiency, having
also reached the highest lipid productivity value, under the same conditions (109.57 mg L-1 d-1). In
relation to starch content, C. emersonii was the one who got higher values – 42.7 % in BG-11
nitrogen deficiency. However, C. vulgaris proved to be the most suitable for starch production by
having higher productivity values. These results makes worthwhile the realization of more studies
on optimization of culture conditions of these microalgae aiming higher yields of lipid and starch
and are likely to lead to future studies of expanding scale with the possibility of reducing production
costs of biomass and its products without need for additional investment.


ix
ÍNDICE
AGRADECIMENTOS...................................................................................................................... iii
RESUMO ......................................................................................................................................v
ABSTRACT .................................................................................................................................. vii
ÍNDICE ........................................................................................................................................ ix
LISTA DE FIGURAS .................................................................................................................... xiii
LISTA DE TABELAS ..................................................................................................................... xv
ABREVIATURAS......................................................................................................................... xvii
PREFÁCIO ................................................................................................................................. xix
1. CAPÍTULO I – INTRODUÇÃO.. .............................................................................................. 1
1.1. Questões ambientais salientes .................................................................................. 3
1.2. Desenvolvimento de recursos de biocombustíveis ...................................................... 5
1.3. Características das microalgas .................................................................................. 6
1.4. Vantagens de usar microalgas para a produção de biocombustíveis ........................... 9
1.5. Biocombustíveis derivados de microalgas ................................................................ 10
1.5.1. Biodiesel ......................................................................................................... 11
1.5.2. Bio-óleo e Gás de Síntese ................................................................................ 14
1.5.3. Bioetanol ........................................................................................................ 15
1.5.4. Bio-hidrogénio ................................................................................................. 19
1.5.5. Biometano ...................................................................................................... 20
1.6. Impacto da seleção da estirpe ................................................................................. 21
1.7. Tecnologias de produção de microalgas .................................................................. 21
1.7.1. Sistemas de produção fotoautotróficos ............................................................ 21
1.7.1.1. Sistemas de cultura abertos ........................................................................ 22
1.7.1.2. Sistemas de cultura fechados ..................................................................... 23
1.7.1.3. Sistemas híbridos ....................................................................................... 24
1.8. Tecnologias de recuperação da biomassa microalgal ............................................... 25

x
1.9. Biocombustíveis de microalgas: que futuro? ............................................................. 26
2. CAPÍTULO II – MATERIAIS E MÉTODOS .............................................................................. 29
2.1 Microrganismos ............................................................................................................ 31
2.2 Meios de crescimento ................................................................................................... 31
2.3 Cultivo fotoautotrófico ................................................................................................... 37
2.3.1 Crescimento em meio com azoto ........................................................................... 37
2.3.2 Crescimento em meio sem azoto ........................................................................... 38
2.4 Monitorização do crescimento celular ........................................................................... 38
2.5 Caracterização da biomassa ......................................................................................... 38
2.5.1 Análise de amido ................................................................................................... 39
2.5.2 Análise de Proteínas .............................................................................................. 40
2.5.3 Análise de Lípidos ................................................................................................. 40
2.6 Determinação das Produtividades ................................................................................. 41
3. CAPÍTULO III – RESULTADOS ............................................................................................ 43
3.1 Crescimento das microalgas em meio com azoto .......................................................... 45
3.1.1 Chlorella emersonii ................................................................................................ 45
3.1.2 Chlorella vulgaris ................................................................................................... 46
3.1.3 Chlorella saccharophila .......................................................................................... 47
3.2 Crescimento das microalgas em meio sem azoto .......................................................... 48
3.3 Análise da Biomassa .................................................................................................... 51
3.3.1 Proteínas ............................................................................................................... 51
3.3.2 Lípidos .................................................................................................................. 52
3.3.3 Amido ................................................................................................................... 53
3.4 Determinação das Produtividades ................................................................................. 54

xi
4. CAPÍTULO IV – DISCUSSÃO DOS RESULTADOS .................................................................. 57
5. CAPÍTULO V – CONSIDERAÇÕES FINAIS E PERSPETIVAS FUTURAS ................................... 71
6. BIBLIOGRAFIA ................................................................................................................... 75


xiii
LISTA DE FIGURAS
CAPÍTULO I - INTRODUÇÃO
Figura 1 - Vias de produção de biocombustíveis a partir de microalgas. ...................................... 11
Figura 2 - Transesterificação do óleo para biodiesel. R1-3 são grupos hidrocarboneto. ................... 14
Figura 3 - Procedimento para a produção de bioetanol a partir de microalgas. ............................ 18
Figura 4 - Vista aérea de um raceway pond. ............................................................................... 22
CAPÍTULO III - RESULTADOS
Figura 5 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C.
emersonii ACOI 516, ao longo do tempo, em diferentes meios com azoto. .................................. 45
Figura 6 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. vulgaris
ACOI 879, ao longo do tempo, em diferentes meios com azoto. ................................................. 47
Figura 7 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C.
saccharophila ACOI 89, ao longo do tempo, em diferentes meios com azoto. ............................. 48
Figura 8 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C.
emersonii ACOI 516, ao longo do tempo, em diferentes meios sem azoto. .................................. 49
Figura 9 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. vulgaris
ACOI 879, ao longo do tempo, em diferentes meios sem azoto. .................................................. 50
Figura 10 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C.
saccharophila ACOI 96, ao longo do tempo, em diferentes meios sem azoto. .............................. 51
Figura 11 - Percentagem de proteínas em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e
C. saccharophila ACOI 96 (C), nos diferentes meios em estudo, nas condições com e sem azoto e
com e sem pré-tratamento com NaOH. N – Não determinado. ................................................... 52
Figura 12 - Percentagem de lípidos em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e C.
saccharophila ACOI 96 (C), nos diferentes meios em estudo, no inóculo e nas condições com e
sem azoto. N – Não determinado. .............................................................................................. 53
Figura 13 - Percentagem de amido em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e C.
saccharophila ACOI 96 (C), nos diferentes meios em estudo, no inóculo e nas condições com e
sem azoto. N – Não determinado. .............................................................................................. 54


xv
LISTA DE TABELAS
CAPÍTULO II – MATERIAIS E MÉTODOS
Tabela 1 - Concentração dos reagentes usados no meio MP12. .................................................. 31
Tabela 2 - Concentração dos reagentes usados na preparação das soluções stock do meio TAP. 32
Tabela 3 - Composição do meio BG-11. ...................................................................................... 33
Tabela 4 - Composição da solução de micronutrientes do meio BG-11. ....................................... 33
Tabela 5 - Composição do meio Chu_13 modificado. ................................................................. 34
Tabela 6 - Soluções stock constituintes do meio BBM. ................................................................ 34
Tabela 7 - Soluções stock constituintes do meio BBM. ................................................................ 35
Tabela 8 - Concentração dos reagentes utilizados nos cinco diferentes meios de cultura. ............ 35
CAPÍTULO III - RESULTADOS
Tabela 9 - Produtividades obtidas (biomassa, lípidos, amido e proteínas) para as microalgas C.
emersonii ACOI 516, C. vulgaris ACOI 879 e C. saccharophila ACOI 96 nos meios MP12, TAP, BG-
11, BBM e Chu_13 mod. nas condições com e sem azoto. ........................................................ 56


xvii
ABREVIATURAS
MMbbl Million barrels
EISA Energy Independence and Security Act
ATP Adenosina trifosfato
AGL Ácidos gordos livres
TAG Triacilglicerídeo
FBR Fotobiorreator
vvm Volume de gás por volume de suspensão de cultura por minuto
rpm Rotações por minuto


xix
PREFÁCIO
Tendo em conta a crise energética em que se vive atualmente, causada pelo aumento da
população mundial e pelas tendências atuais de consumo (nomeadamente de combustíveis
fósseis), surgiu a necessidade de investigar energias renováveis que possam substituir
parcialmente os recursos petroquímicos. As microalgas têm sido reconhecidas recentemente
como fontes promissoras de biocombustíveis. Estas caracterizam um grupo diversificado de
microrganismos aquáticos, capazes de produzir O2, aumentando a sua concentração na atmosfera.
O crescimento de uma população de microalgas está dependente de vários fatores,
nomeadamente, o tempo de exposição à luz, a intensidade e o comprimento de onda da fonte
luminosa, a presença de nutrientes essenciais no meio, tais como, azoto (N) e fósforo (P); o local
onde as microalgas são cultivadas, o arejamento das culturas e as variações de pH. Sob todas as
condições favoráveis, estes microrganismos são capazes de elevadas velocidades de crescimento.
Para além disso, quando comparadas com outras fontes de biocombustíveis, como é o caso dos
biocombustíveis de 1ª e 2ª geração, algumas das suas principais vantagens são o facto de
permitirem a melhoria da qualidade do ar; possuírem alta capacidade de adsorção de carbono;
alto teor de lípidos; exigirem um meio de crescimento simples; apresentarem taxas de crescimento
rápidas e tempos de geração curtos; não necessitarem de terrenos aráveis e poderem ser
cultivadas em águas salobres; a sua produtividade não ser suscetível a variações sazonais e
geográficas; não necessitarem de herbicidas; terem capacidade de remover fosfatos e nitratos de
águas residuais e produzirem subprodutos valiosos para fins comerciais ou com propósito
farmacêutico.
O conceito de bioconversão em microalgas traduz-se na utilização do processo
fotossintético para a produção de biomassa para ser usada como uma fonte de energia e de
produtos químicos. Algumas espécies de microalgas são conhecidas como produtores naturais de
ácidos gordos e hidratos de carbono, o que faz destas, candidatas ideais para a produção de
biodiesel e bioetanol. O teor de óleo nalgumas espécies do género Chlorella pode variar de cerca
de 14 a 63 % do peso seco. Estima-se que aproximadamente entre 46,760 - 140,290 L ha-1 de
etanol podem ser produzidos a partir de microalgas. Adicionalmente, a biomassa microalgal pode
ser submetida a um processo de digestão anaeróbio, fazendo, desta forma, com que possa ser
usada para a produção de biometano. A produção de hidrogénio também é possível graças à
produtividade fotossintética reconhecida nestes microrganismos.

xx
O conteúdo em lípidos e hidratos de carbono pode aumentar nalgumas espécies de
microalgas, se algumas condições do cultivo forem alteradas, como por exemplo, reduzindo a
composição de elementos químicos necessários ao crescimento e divisão das células. Para além
disso é possível aumentar o conteúdo destas macromoléculas no interior das células, otimizando
alguns fatores determinantes como é o caso da intensidade da luz, da temperatura, salinidade e
da concentração de CO2 no meio de crescimento.
Desta forma, o principal objetivo da presente dissertação foi avaliar o potencial de
produção de biocombustíveis de três espécies diferentes de Chlorella – C. emersonii, C. vulgaris e
C. saccharophila. Para isso, estas espécies foram colocadas a crescer em diferentes meios de
cultura, em condições específicas. A concentração celular e a quantidade de biomassa produzida
foram analisadas ao longo do tempo. No final de cada crescimento, procedeu-se à extração de
lípidos, amido e proteínas presentes nas células. Para além disso, metade da biomassa obtida no
final do crescimento em condições normais foi transferida para meios desprovidos de azoto, com o
objetivo de aumentar a concentração de lípidos e/ou amido nas células.
A presente dissertação está organizada em cinco capítulos. O capítulo 1 apresenta uma
breve revisão das bases teóricas associadas a este trabalho, incluindo as questões ambientais
salientes; o desenvolvimento de recursos de biocombustíveis; características das microalgas e as
vantagens do seu uso para a produção de biocombustíveis. O capítulo 2 inclui os materiais e
métodos utilizados no trabalho experimental. No capítulo 3, todos os resultados obtidos são
apresentados. O capítulo 4 apresenta a discussão dos resultados obtidos. Finalmente, o capítulo 5
destaca as principais conclusões obtidas a partir do trabalho realizado e apresenta algumas
sugestões para trabalho futuro.

1. CAPÍTULO I
INTRODUÇÃO


INTRODUÇÃO | 3
1.1. Questões ambientais salientes
O rápido crescimento da população mundial e a ascensão de países em desenvolvimento
tem levado ao aumento das necessidades energéticas (Harun, Danquah, et al. 2010). Por isso,
hoje em dia, a escassez dos recursos da petroquímica e o problema de poluição ambiental são
dois desafios críticos, que precisam de ser abordados pela nossa sociedade.
Segundo a revisão estatística da energia mundial referente ao ano de 2013, da British
Petroleum (BP), o petróleo continua líder do combustível no mundo, com 32,9 % do consumo
mundial de energia. Globalmente, o gás natural representa 23,7 % do consumo de energia
primária. A participação do carvão no consumo global de energia primária atingiu 30,1 %,
enquanto a energia nuclear e a hidroeletricidade contam com 4,4 % e 6,7 % do consumo total de
energia primária, respetivamente (BP 2014).
No Reference case do International Energy Outlook 2014 (IEO2014), prevê-se que o
consumo de combustíveis líquidos no mundo aumente por mais de um terço (33 milhões de barris
de petróleo por dia (MMbbl d-1), de 87 MMbbl d-1 registados em 2010 para 119 MMbbl d-1 em 2040
(EIA-DOE 2014).
Todas as fontes de energia não-renováveis que existem para além do petróleo, como é o
caso do gás natural, da energia nuclear (proveniente do urânio, por exemplo) e do carvão mineral
são limitadas e inevitavelmente a sua disponibilidade também diminuirá. O carvão é o candidato
imediato provável para substituir o petróleo como fonte de energia quando este se esgotar de
forma definitiva. É rico em energia, pode ser convertido em combustível líquido, e ainda é muito
abundante. No entanto, como o uso de carvão, em relação ao petróleo produz ainda maior
quantidade de emissões de CO2, o esgotamento do petróleo não irá ajudar na diminuição dos
níveis de CO2 na atmosfera (Campbell 2008).
De acordo com os resultados da literatura científica, a poluição ambiental é causada
principalmente pela utilização de enormes quantidades de combustíveis fósseis, e o impacto dos
seus co - produtos causa a poluição do ar, o aquecimento global, poluição da água e dos solos,
derramamentos de óleo e efeitos na saúde adversos (Arora 2012).
Os principais produtos provenientes da combustão dos combustíveis fósseis são o dióxido
de carbono (CO2), óxidos de azoto e hidrocarbonetos, incluindo uma mistura de gases comumente
denominados por gases do efeito de estufa, que provocam desequilíbrio ambiental e alterações
climáticas. Entre todos os gases emitidos pelos combustíveis fósseis, o CO2 é um dos mais

4 | INTRODUÇÃO
significantes, tendo aumentado a sua percentagem na atmosfera 25 % desde que se iniciou a
combustão dos combustíveis fósseis e prevê-se que aumente 1,9 % cada ano (Arora 2012).
Desta forma, a emissão de CO2 deve ser consideravelmente alta num futuro próximo.
Assim há uma necessidade urgente de neutralizar o efeito do CO2 na atmosfera para continuar o
crescimento económico e manter os padrões de vida. Nesta perspetiva, várias técnicas podem ser
aplicadas. Uma técnica óbvia é reduzir o consumo de energia, no entanto, o aumento da
população e o estilo de vida humana são os principais obstáculos para implementar isso. O
armazenamento do CO2 libertado pode ser outra solução (Chiu et al. 2008; Abbas et al. 2014). No
entanto, tanto barreiras técnicas como económicas limitam esta aplicação.
Devido à preocupação com o aquecimento global, que pode ser atribuído, principalmente,
ao elevado nível de CO2 na atmosfera (Kondili & Kaldellis 2007; Román-Leshkov et al. 2007), a
Organização das Nações Unidas promoveu o Protocolo de Quioto (1997), com o objetivo de
reduzir os gases de efeito estufa 5,2 % com base na emissão de 1990, e mais de 170 países
ratificaram o protocolo (Gutiérrez et al. 2008).
Vários investigadores começaram a procurar alternativas de energias renováveis que
podem substituir parcialmente os recursos dos combustíveis fósseis para o estabelecimento de
uma sociedade mais sustentável e promover a recuperação económica no mundo.
A biomassa é uma das mais importantes fontes de energia renováveis. Esta faz uso do ar,
água, solo e da fotossíntese para produzir bioenergia. Pode ser extraída a partir de cultivos
alimentares, resíduos de cultivos, madeira, lixo, excremento de animais e algas marinhas, etc. As
principais vantagens da utilização de biomassa são o facto de ser renovável e menos poluidora.
Biocombustíveis e biogases são produzidos a partir de biomassa como uma alternativa aos
combustíveis provenientes do petróleo. Atualmente, a nível mundial, o desenvolvimento da
biomassa tornou-se um caminho importante para ajustar a estrutura de energia e reduzir as
emissões dos gases com efeito de estufa a fim de encontrar a sustentabilidade ambiental e
económica (Berndes et al. 2003).
Consequentemente, muitos países estabeleceram metas nacionais de produção de
biocombustíveis e proporcionam incentivos e apoios para acelerar o crescimento da indústria de
bioenergia. Por exemplo, nos EUA a Renewable Fuels Standard (RFS), parte da Energy
Independence and Security Act (EISA) de 2007, estabeleceu a meta de produção anual de ≈ 136
mil milhões de litros de biocombustíveis até 2022 (U.S. Congress 2007).

INTRODUÇÃO | 5
Para evitar impactos adversos sobre a oferta de alimentos para consumo do Homem, a
EISA especifica ainda que 60 dos 136 mil milhões de litros de combustíveis renováveis produzidos
em 2022 devem ser biocombustíveis avançados produzidos a partir de biomassa algal, por
exemplo. No entanto, a capacidade de produção atual de biocombustíveis avançados é inferior a
37 mil milhões de litros em todo o mundo (Yue et al. 2014).
1.2. Desenvolvimento de recursos de biocombustíveis
A biomassa é a fonte de energia renovável mais promissora porque está naturalmente
disponível, pode ser produzida em grandes quantidades e permite a produção de combustíveis
líquidos, gasosos ou sólidos com potencial em diferentes sectores. Além disso, é um recurso
capaz de ser totalmente renovável e ambientalmente, socialmente e economicamente sustentável
(Demirbas 2008).
Os cultivos alimentares têm sido amplamente utilizados na produção industrial de
biocombustíveis, como os principais representantes da primeira geração de biocombustíveis. As
matérias-primas podem ser divididas em três categorias, culturas de açúcar (cana-de-açúcar e
beterraba), culturas de amido (milho, trigo e sorgo) e oleaginosas (colza, soja e girassol). Todas
estas matérias-primas podem ser convertidas para biocombustíveis por meio de técnicas
diferentes. Mas o desenvolvimento da primeira geração de biocombustíveis apresenta alguns
problemas. As culturas alimentícias não só têm um ciclo mais longo de crescimento, como
também exigem grandes áreas de terra arável. À medida que a primeira geração de
biocombustíveis foi alargando a escala de produção, um grande lote de terra arável foi ocupada, o
que conduziu, por um lado, à desflorestação, degradação dos solos e erosão (impactos causados
pela aplicação de fertilizantes, pesticidas e herbicidas) e à devastação da biodiversidade (Puppán
2002; Escobar et al. 2009); por outro lado, ao agravamento da fome e da pobreza nos países em
desenvolvimento, uma vez que, os terrenos aráveis antes dedicados ao cultivo de bens alimentares
estão destinados à produção de matéria-prima para a produção de biocombustíveis para alimentar
os mercados dos países desenvolvidos (C. Ford & Senauer 2007).
Portanto, utilizar culturas alimentares para resolver a crise energética aumenta a crise
alimentar.
A produção de biocombustíveis a partir de Jatropha, mandioca, Miscanthus, materiais
lignocelulósicos, resíduos de exploração florestal, resíduos agrícolas (folhas, palha ou aparas de

6 | INTRODUÇÃO
madeira), bem como os componentes não comestíveis do milho ou da cana-de-açúcar são
exemplos de biocombustíveis de segunda geração (Dragone et al. 2010; de Vries et al. 2010).
Uma vez testada a viabilidade de produção dos biocombustíveis de segunda geração,
espera-se que estas matérias-primas possam desacelerar a escassez de alimentos e os problemas
de recuperação de terras. Mas, mesmo que o preço das matérias-primas dos biocombustíveis de
segunda geração sejam inferiores e que essas matérias-primas apresentem grandes quantidades
de celulose, o custo de produção é muito mais elevado em relação à primeira geração de
biocombustíveis, porque a conversão da biomassa em açúcares fermentáveis envolve um pré-
tratamento com enzimas especiais (Sims & Taylor 2008; Brennan & Owende 2010).
As matérias-primas para os biocombustíveis de terceira geração são os microrganismos
aquáticos. As microalgas estão a liderar o desenvolvimento da terceira geração de
biocombustíveis. As principais vantagens da sua utilização são a alta capacidade de adsorção de
carbono; alto teor de lípidos; meio de crescimento simples e tempo de crescimento curto. O seu
ciclo de colheita varia aproximadamente de entre 1 a 10 dias, dependendo do processo,
permitindo colheitas múltiplas ou contínuas com aumento significativo no rendimento (Schenk et
al. 2008). E, em comparação com a soja, por exemplo, o teor de lípidos de microalgas pode ser
25 a 200 vezes superior. O óleo extraído das microalgas pode ser transformado em biodiesel; os
seus hidratos de carbono podem ser fermentados para álcool, e o azoto e o fósforo podem ser
reciclados e usados como fertilizantes (Maity et al. 2014).
Desta forma, para que os recursos de biocombustíveis sejam tecnicamente e
economicamente viáveis devem cumprir alguns requisitos, tais como (Himmel et al. 2007): ser
competitivos ou custar menos do que os combustíveis de petróleo; exigir baixo ou nenhum uso
adicional de terras; permitir a melhoria da qualidade do ar e exigir o uso mínimo de água. Utilizar
microalgas como fonte de biocombustíveis pode atender a estas condições e, portanto, fazer uma
contribuição significativa para responder à demanda de energia primária, proporcionando
simultaneamente benefícios ambientais (Bai et al. 2008).
1.3. Características das microalgas
Ao contrário das plantas, as microalgas não possuem raízes, caules, folhas e vasos
condutores (xilema/floema) (Croft et al. 2006). O termo microalga é normalmente uma referência

INTRODUÇÃO | 7
a formas eucariotas mas muitas vezes é estendido na literatura para incluir cianobactérias
(Mutanda et al. 2011).
As microalgas caracterizam um incrível e diversificado grupo polifilético de organismos
com representantes procariotas e eucariotas (Croft et al. 2006). Acredita-se que existam entre 200
a 800 mil espécies (Ratha & Prasanna 2012), no entanto, apenas alguns milhares são mantidos
em coleções e apenas algumas são cultivadas em escala industrial (Parmar et al. 2011).
De acordo com Tomaselli (2004), as microalgas têm sido tradicionalmente classificadas
por vários critérios, tais como o tipo de pigmentos, a natureza química dos produtos de reserva e
pelos constituintes da parede celular. Também tem sido tomado em conta aspetos morfológicos e
citológicos, tais como a ocorrência de células flageladas, a estrutura dos flagelos, os processos de
formação do núcleo e a divisão celular, a presença de um envelope no retículo endoplasmático em
torno do cloroplasto e a conexão entre o retículo endoplasmático e a membrana nuclear (Tomaselli
2004; Dragone et al. 2010). Muitas espécies são altamente adaptáveis na sua morfologia, facto
que pode tornar duvidosa a sua classificação (Surek 2008).
As três classes mais importantes em termos de abundância são as diatomáceas
(Bacillariophyceae), as algas castanhas (Chrysophyceae), e as algas verdes (Chlorophyceae). O
grupo maior de Chlorophyceae é constituído por várias formas marinhas e de água doce,
unicelulares e filamentosas, com uma área geográfica ampla e taxas de crescimento rápido. Este
grupo inclui as espécies mais estudadas, tais como Chlorella spp., Chlamydomonas spp.,
Dunaliella spp. e Haematococcus spp. (Pulz & Gross 2004).
As cianobactérias também são referidas como microalgas, que no fitoplâncton,
representam a forma dominante de vida e provavelmente o maior grupo produtor de biomassa na
Terra.
Todas elas são capazes de produzir O2, aumentando a sua concentração na atmosfera,
através do uso de pigmentos fotossintéticos; têm gamas de tamanhos celulares que podem variar
entre 2 a 200 μm e várias estratégias de alimentação, das autotróficas às heterotróficas (Barsanti
& Gualtieri 2006). A grande maioria das microalgas apresenta um metabolismo autotrófico
fotossintético que se caracteriza pela sua capacidade de sintetizar matéria orgânica de reserva a
partir de carbono inorgânico (nomeadamente CO2 atmosférico), através do processo de
fotossíntese (Richmond 2004; Sukahara & Awayama 2005; Eshaq et al. 2010). Dentro das
autotróficas, podem ser fotoautotróficas, usando a luz como fonte de energia, ou
quimioautotróficas, oxidando compostos inorgânicos para obtenção de energia (Dragone et al.

8 | INTRODUÇÃO
2010). Para as algas autotróficas, a fotossíntese é um componente chave na sua sobrevivência,
através do qual convertem radiação solar e CO2, absorvido pelos cloroplastos, em adenosina
trifosfato (ATP) e O2, a energia utilizável a nível celular, que é depois usada na respiração para
produzir energia e apoiar o crescimento celular (Brennan & Owende 2010; Dragone et al. 2010).
Muitas outras espécies apresentam outros tipos de metabolismo, nomeadamente as microalgas
heterotróficas que utilizam somente compostos orgânicos como fonte de carbono e de energia.
Dentro desta classe, podem ser fotoheterotróficas, usando a luz como fonte de energia, ou
quimioheterotróficas, oxidando compostos orgânicos para receber energia. O meio de cultura
heterotrófico é bastante similar ao meio de cultura autotrófico com a exceção de que é adicionado
um composto de carbono orgânico, normalmente glucose, acetato ou glicerol e crescem na
ausência de luz. Por norma, estas culturas heterotróficas apresentam algumas desvantagens,
nomeadamente na limitação de espécies de microalgas que conseguem crescer desta forma; no
aumento dos custos devido à adição de mais um composto no meio, o carbono orgânico; na
contaminação e competição por outros microrganismos e inibição por excesso de substrato
orgânico (Lee Y.K. 2004; Perez-Garcia et al. 2011). Algumas microalgas fotossintéticas são
mixotróficas, ou seja, são capazes de viver de modo autotrófico ou heterotrófico, dependendo da
concentração de compostos orgânicos e da intensidade de luz disponível, podendo também ser
designadas por autotróficas facultativas (Ferreira & Sousa 1998; Lee 2008).
As respostas aos estímulos ou às mudanças ambientais são inerentes a todos os
organismos vivos. Nas microalgas, as condições ambientais afetam o crescimento celular,
podendo reduzir a sua taxa de crescimento ou alterar alguma reação bioquímica. Pode também
provocar um desequilíbrio metabólico, desencadeando ajustes bioquímicos, induzindo as células a
estabelecer um novo estado de crescimento e biossíntese (Vonshak & Torzillo 2004).
O crescimento de uma população de microalgas é resultado da interação entre fatores
biológicos, físicos e químicos. Os fatores biológicos estão relacionados com as taxas metabólicas
da espécie cultivada. Os fatores físico-químicos são principalmente a luz, a temperatura, a
salinidade e a disponibilidade de nutrientes (Lourenço 2006).
O tempo de exposição à luz, conhecido como fotoperíodo, a intensidade e o comprimento
de onda da fonte luminosa é de importância fundamental para as microalgas, porque o aumento
da luz proporcional ao nível de saturação ou de máxima produção celular pode causar inibição da
atividade fotossintética (Boney 1989)

INTRODUÇÃO | 9
Em relação à nutrição, para um crescimento ótimo, as microalgas têm necessidade de
uma série de nutrientes (Lourenço 2006). O meio de crescimento deve fornecer os elementos
inorgânicos que constituem a célula das algas. Elementos essenciais incluem azoto (N) e fósforo
(P). Requisitos nutricionais mínimos podem ser estimados usando a fórmula molecular
aproximada da biomassa microalgal, que é CO0.48H1.83N0.11P0.01 (Chisti 2007; Dragone et al. 2010;
Grobbelaar 2007). O azoto é muitas vezes fornecido na forma de nitrato (NO3-), mas também
frequentemente através da amónia (NH4+) e da ureia. A ureia é mais favorável como fonte de azoto
porque, para uma concentração equivalente de azoto, proporciona rendimentos mais elevados e
provoca variações de pH inferiores no meio, durante o crescimento celular (Shi et al. 2000;
Dragone et al. 2010). Por outro lado, nutrientes como o fósforo (P) devem ser fornecidos em
excesso significativo porque os fosfatos adicionados ligam-se aos iões metálicos, por isso nem
todos os P ficam disponíveis para consumo (Chisti 2007). Além disso, o crescimento das
microalgas depende não só de um fornecimento adequado de macronutrientes essenciais como
carbono, azoto, fósforo e silício e iões maiores como Mg2+, Ca2+, Cl-, e SO42- mas também de um
número de micronutrientes como ferro, manganésio, zinco, cobalto, cobre e molibdénio (Sunda et
al. 2005; Dragone et al. 2010). Algumas microalgas também necessitam de baixas concentrações
de vitaminas no meio de cultura (Robert R. L. Guillard 1975).
Além dos fatores anteriormente descritos, outros podem influenciar o desenvolvimento das
culturas, como o local onde as microalgas são cultivadas, o arejamento das culturas e as variações
de pH (Richmond 1990).
Sob todas as condições favoráveis, as microalgas podem apresentar elevadas velocidades
de crescimento. Em geral, a maioria das microalgas tem um tempo de duplicação da biomassa de
cerca de 24 h mas, em casos muito favoráveis, poderá atingir 3,5 h durante a fase exponencial de
crescimento (Chisti 2007).
1.4. Vantagens de usar microalgas para a produção de biocombustíveis
As microalgas têm rendimentos elevados de energia solar; taxas rápidas de crescimento e
tempos de geração curtos e são vulgarmente conhecidas por duplicarem a sua biomassa no
período de 24 h, o que permite colheitas múltiplas e contínuas de biomassa durante todo o ano
(Chisti 2007; Schenk et al. 2008). Algumas espécies têm tempos de duplicação tão curtos quanto
3,5 h, o que resulta em múltiplas divisões da sua biomassa por dia; necessitam de menos água

10 | INTRODUÇÃO
doce para cultivo, do que as plantas terrestres (Ahmad et al. 2011; Li, Horsman, Wu, Lan &
Dubois-calero 2008); o cultivo de microalgas pode ocorrer em terrenos não aráveis e em águas
salobres (Li, Horsman, Wu, Lan & Dubois-calero 2008), reduzindo a pressão sobre os recursos
necessários para a produção de culturas alimentares, assim como, prevenindo efeitos ambientais
adversos. Não têm necessidade de utilização de produtos químicos, como herbicidas ou
pesticidas, diminuindo, por isso, os custos e os impactos ambientais (Rawat et al. 2011);
adicionalmente, estes microrganismos requerem menor espaço terrestre para serem cultivados,
cerca de 2 % da terra necessária para produzir a mesma quantidade de biodiesel a partir de
culturas oleaginosas (Ahmad et al. 2011); durante o crescimento das microalgas, estas têm
capacidade de remover fosfatos e nitratos de águas residuais, que usam como substrato para o
seu crescimento, o que consequentemente favorece a produção de biocombustíveis e, ao mesmo
tempo, faz com que estas contribuam para o tratamento terciário de águas residuais. Algumas
microalgas produzem subprodutos valiosos na forma de proteínas, pigmentos, biopolímeros,
incluindo substâncias antioxidantes para fins comerciais ou com propósito farmacêutico (Brennan
& Owende 2010; Ahmad et al. 2011; Rawat et al. 2011; Kumar et al. 2014); o custo associado à
colheita e ao transporte da biomassa é relativamente baixo quando comparado com o de culturas
oleaginosas. A biomassa residual pode ser usada como fertilizante ou na produção de outros
produtos de alta energia, podendo desta forma contribuir para melhoras na economia (Ahmad et
al. 2011).
1.5. Biocombustíveis derivados de microalgas
A biomassa pode ser convertida em formas utilizáveis usando uma variedade de maneiras
diferentes (Figura 1). As reações de conversão da biomassa em energia podem ser bioquímicas e
termoquímicas. A conversão termoquímica é a decomposição térmica dos componentes orgânicos
da biomassa para produzir produtos de combustão. Exemplos deste tipo de conversão são a
gaseificação, a pirólise e a liquefação. Exemplos de conversões bioquímicas são a fermentação, a
digestão anaeróbia, as células de combustível, e outros processos de produção de combustível
utilizando o metabolismo dos organismos. As vias biológicas para conversão da biomassa são
geralmente mais eficientes em termos de reciclagem de nutrientes e de matéria orgânica.
A produção fotobiológica de combustíveis e produtos químicos a partir de microalgas é,
provavelmente, uma das iniciativas mais importantes no sentido de estabelecer uma importante

INTRODUÇÃO | 11
fonte de fornecimento de energia renovável. O conceito de bioconversão por microalgas é a
utilização do processo fotossintético para a produção de biomassa para ser usada como uma fonte
de energia e de produtos químicos. Um rendimento de produção de microalgas de 15 - 25 tpeso-seco
ha-1 ano-1 foi alcançado ao longo de um período relativamente longo e algumas microalgas são
conhecidas como produtores naturais de ácidos gordos e hidratos de carbono (Du et al. 2008).
Figura 1 – Vias de produção de biocombustíveis a partir de microalgas (Mussatto et al. 2010).
1.5.1. Biodiesel
O conceito de utilização de combustível à base de óleos vegetais remonta a 1895, quando
o Dr. Rudolf Diesel desenvolveu o primeiro motor de ignição por compressão especificamente para
funcionar com óleos vegetais. Uma vez que o biodiesel tem propriedades semelhantes ao diesel
convencional, pode ser misturado em qualquer proporção com o diesel proveniente do petróleo e
pode ser utilizado em motores diesel sem grandes modificações (Puppán 2002).
Trata-se de um combustível de queima limpa, oxigenado e à base de ésteres
monoalquílicos, que pode ser feito a partir dos lípidos extraídos das microalgas. É renovável,
simples de usar, apresenta características de alta combustão, biodegradável, não tóxico, e
essencialmente livre de enxofre e compostos aromáticos (Puppán 2002).

12 | INTRODUÇÃO
A produção de biodiesel a partir de microalgas é um processo de quatro etapas: cultivo
das microalgas, colheita, extração dos lípidos, e conversão em biodiesel. A extração dos lípidos
exige rompimento celular por métodos que podem ser mecânicos, químicos, ou biológicos ou
podem também ser extraídos com a utilização de solventes.
Para maior rendimento deste processo, já foram avaliadas técnicas que envolvem a
utilização de enzimas, radiação micro-ondas, sonicação, água quente, e organoclorados
(compostos orgânicos que contêm, pelo menos, um átomo de cloro ligado covalentemente) (Halim
et al. 2012).
Pelo facto da parede celular das microalgas ser tipicamente espessa e forte, foram
sugeridos métodos de rutura celular severos, que usam peróxido de hidrogénio como agente
oxidante ou água quente contendo ácido sulfúrico, para obtenção de rendimentos de extração de
lípidos superiores (Lee et al. 2013; Park et al. 2014). Os lípidos extraídos das microalgas podem
ser de diversos tipos, tais como, lípidos neutros (triacilglicerídeos), glicolípidos, fosfolípidos e
ácidos gordos livres (AGL) (Prabhakara Rao et al. 2013). Entre todos, os triacilglicerídeos e os AGL
são os principais recursos para a conversão em biodiesel.
Muito do trabalho de investigação em curso foca-se num pequeno número de espécies de
microalgas de rápido crescimento que tenham sido descritas como capazes de acumular
quantidades substanciais de lípidos, embora sob condições específicas. Dentro das algas verdes,
espécies típicas incluem Chlamydomonas reinhardtii, Dunaliella salina, e várias espécies de
Chlorella, bem como Botryococcus braunii, que apesar de crescimento lento podem acumular
grandes quantidades de lípidos (Dragone et al. 2010).
O rendimento de óleo das microalgas e o perfil dos ácidos gordos pode variar de acordo
com as espécies e com as condições do cultivo. O teor de óleo nalgumas espécies do género
Chlorella (Chlorophyta) pode variar de cerca de 14 a 63 % do peso seco, e a composição de ácidos
gordos foi relatada como podendo variar de C-14:0 a C-20:0 (Tang et al. 2011).
O conteúdo lipídico pode aumentar nalgumas espécies de microalgas, se algumas
condições do cultivo forem alteradas, como por exemplo, reduzindo a composição de elementos
químicos necessários ao crescimento e divisão das células (Sheehan et al. 1998). Nesta situação,
as células param de proliferar, mas é mantida a fixação de CO2, que é então convertido em
glucose, em seguida em ácidos gordos e, finalmente, em triacilglicerídeos (TAG) (Hu et al. 2008).
Isto pode ser explicado pelo facto de, por exemplo, o azoto ser essencial para a síntese de
proteínas e ácidos nucleicos e a sua privação faz com que a célula deixe de ter substrato para se

INTRODUÇÃO | 13
multiplicar, continuando a contar, no entanto, com o mecanismo desenvolvido para a fixação de
CO2 (Xin et al. 2010). Desta forma, o conteúdo lipídico pode aumentar até 2 vezes (Illman et al.
2000; Fidalgo et al. 1998).
De forma a aumentar a concentração de lípidos no interior das células de microalgas, é
possível otimizar alguns fatores de crescimento determinantes, como a intensidade da luz, os
níveis de azoto, a temperatura, a salinidade, a concentração de CO2 e os procedimentos de
colheita (Dragone et al. 2010).
No entanto, aumentar a acumulação de lípidos não resultará no aumento da produtividade
de lípidos, uma vez que a produtividade da biomassa e a acumulação de lípidos não estão
necessariamente correlacionados (Rodolfi et al. 2009; Sheehan et al. 1998). A acumulação de
lípidos refere-se ao aumento da concentração de lípidos no interior das células de microalgas sem
ter em conta a produção total de biomassa. A produtividade de lípidos leva em conta tanto a
concentração de lípidos no interior das células, como a biomassa produzida por estas células e é,
portanto, um indicador mais útil dos potenciais custos de produção de biocombustíveis líquidos
(Brennan & Owende 2010; Dragone et al. 2010).
A adaptação das microalgas a mudanças ambientais é normalmente resultado de uma
mudança no padrão dos lípidos e na síntese de vários compostos incomuns (Guschina & Harwood
2006). Esta é provavelmente a razão pela qual as microalgas têm uma superprodução de ácidos
gordos quando sujeitas a condições de stresse.
A caracterização dos lípidos é um requisito, uma vez que, espécies diferentes de
microalgas produzem diferentes tipos de ácidos gordos. Alguns ácidos gordos são mais adequados
para a transesterificação em biodiesel do que outros. A maior parte dos óleos das microalgas são
ricos em ácidos gordos polinsaturados com quatro ou mais ligações duplas. O problema associado
a este grau de polinsaturação é, estes ácidos gordos e os ésteres metílicos de ácidos gordos são
suscetíveis à oxidação durante o armazenamento, reduzindo assim a sua aceitabilidade para
utilização no biodiesel (Chisti 2007).
Para a produção de biodiesel, três moléculas de ácidos gordos são esterificadas com uma
molécula de glicerol (Chisti 2007). Os TAG são colocados a reagir com metanol numa reação
conhecida como transesterificação ou alcoólise. A transesterificação produz ésteres metílicos de
ácidos gordos, que são biodiesel, e glicerol (Figura 2). A reação ocorre por etapas: inicialmente, os
TAG são convertidos para diglicerídeos, depois para monoglicerídeos e, em seguida, para gliceróis.
A transesterificação requer 3 mol de álcool por cada mol de TAG, para produzir 1 mol de glicerol e

14 | INTRODUÇÃO
3 mol de ésteres metílicos (Figura 2). A reação é um equilíbrio. Os processos industriais usam 6
mol de metanol por cada mol de TAG (Fukuda et al. 2001). Este excesso de metanol garante que a
reação é conduzida na direção dos ésteres metílicos, ou seja, no sentido do biodiesel. A
transesterificação é catalisada por ácidos, bases (Fukuda et al. 2001; Meher et al. 2006) ou por
lipases (Sharma et al. 2001). Quando catalisada por uma base é cerca de 4000 vezes mais rápida
do que quando catalisada por um ácido (Fukuda et al. 2001). O uso de lipases oferece vantagens
importantes, mas não é atualmente possível por causa do custo relativamente alto deste
catalisador (Fukuda et al. 2001).
Figura 2 - Transesterificação do óleo para biodiesel. R1-3 são grupos hidrocarboneto (Chisti 2007).
Outros álcoois, em vez do metanol, podem ser usados, mas o metanol é o menos caro.
Para evitar a perda de rendimento devido a reações de saponificação (ou seja, formação de
sabão), o óleo e o álcool devem estar secos e o óleo deve ter um mínimo de AGL. O biodiesel é
recuperado por lavagens repetidas com água para remover o glicerol e o metanol (Chisti 2007).
A colheita das microalgas é vista como um dos principais desafios da sua utilização para a
produção de biodiesel. As microalgas que armazenam lípidos são geralmente unicelulares, têm
baixas densidades e são encontradas em suspensão, tornando a separação difícil. Os
procedimentos de extração em grande escala para os lípidos de microalgas são complexos e ainda
estão em fase de desenvolvimento (Rawat et al. 2011). Outros desafios no que diz respeito à
utilização do biodiesel como combustível são a sua suscetibilidade a oxidação bacteriana, podendo
causar corrosão interna dos tanques de armazenamento (Antoni et al. 2007). No entanto, o
biodiesel tem potencial para ser um combustível 100 % biológico no futuro (Antoni et al. 2007).
1.5.2. Bio-óleo e Gás de Síntese
Quando a biomassa é processada sob altas temperaturas (entre ≈ 400 °C e 800 °C) na
ausência de oxigénio, os produtos são produzidos em três fases: a fase de vapor (≈ 800 °C), a
fase líquida (≈ 500 °C) e a fase sólida (≈ 400 °C) (Li, Horsman, Wu, Lan & Dubois-Calero 2008;

INTRODUÇÃO | 15
Bridgwater 2003). A fase líquida é uma mistura complexa chamada bio-óleo, também conhecido
como óleo de pirólise (Ringer et al. 2006). O gás de síntese, derivado da biomassa microalgal, a
partir do processo de gaseificação, é um gás de mistura composto principalmente por CO e H2.
O bio-óleo tem várias características indesejáveis, tais como, conteúdo de oxigénio (35 % –
40 % p/p) e água (15 % - 30 %) muito elevado, é altamente corrosivo e quimicamente instável
devido ao elevado teor de sólidos (Czernik & Bridgwater 2004; Bridgwater et al. 1999; Chiaramonti
et al. 2007; Bridgwater 2003). Além disso, o seu teor de metais alcalinos pode conduzir a
incrustações e contribuir para o envenenamento do catalisador. A elevada viscosidade do óleo de
pirólise em comparação com o petróleo bruto requer maior trabalho de bombeamento. A sua
polaridade devido ao alto teor de oxigénio faz com que o óleo de pirólise seja mais propenso a
agarrar-se às paredes dos tanques. Devido ao seu promissor potencial para substituir o petróleo
bruto, há vários trabalhos de pesquisa sobre a estabilização e modernização dos óleos de pirólise
(Bunting et al. 2010).
Recentemente, alguns estudos têm sido realizados sobre o potencial da biomassa de
microalgas para a produção de bio-óleo (Demirbaş 2006; Miao & Wu 2004; Miao et al. 2004). Foi
demonstrado que, em geral, os bio-óleos de microalgas são de melhor qualidade do que o bio-óleo
da madeira, por exemplo (Demirbaş 2006).
1.5.3. Bioetanol
O bioetanol, sendo um combustível limpo e renovável, é considerado uma boa alternativa
para substituir o petróleo (Mussatto et al. 2010; Bai et al. 2008). Embora a energia equivalente de
etanol seja 68 % inferior ao combustível de petróleo, a combustão do etanol é mais limpa (porque
contém oxigénio). Por conseguinte, a emissão de substâncias tóxicas é inferior (Krylova et al.
2008). O uso de bioetanol como combustível de transporte também pode ajudar a reduzir a
formação de CO2, por substituir a utilização dos combustíveis fósseis, e pela reciclagem do CO2
lançado. Ao usar o bioetanol em vez de combustíveis fósseis, as emissões resultantes da utilização
de combustíveis fósseis são evitadas.
A queima de etanol em vez da queima de gasolina reduz as emissões de carbono em mais
de 80 %, enquanto elimina inteiramente a libertação de chuva ácida causada pelo dióxido de
enxofre (Lashinky & Schwartz 2006).

16 | INTRODUÇÃO
O uso de etanol como combustível de automóveis tem uma longa história. Os primeiros
protótipos de motores de combustão interna construídos no século XIX por Samuel Morey em
1826 e por Nicholas Otto em 1876 foram construídos de forma a ser possível usar etanol como
combustível (Demirbas et al. 2009). O primeiro carro produzido por Henry Ford em 1896 podia
usar etanol puro como combustível e em 1908 o Ford Modelo – T, primeiro carro manufaturado
em série, era um veículo flexível, disponível para a utilização de etanol como combustível, da
mesma forma que também podia usar gasolina ou qualquer mistura de ambos.
O uso de bioetanol para combustível foi utilizado de forma generalizada na Europa e nos
Estados Unidos até ao início do ano 1900. Após a Primeira Guerra Mundial, houve uma
diminuição na demanda por etanol, porque se tornou mais caro para produzir do que o
combustível à base de petróleo, no entanto, houve um interesse (por exemplo, da General Motors
Corporation e DuPont) em etanol como agente antidetonante (ou seja, aumento de octanagem) e
como possível substituto para combustíveis derivados do petróleo (Demirbas et al. 2009; Balat &
Balat 2009; Solomon et al. 2007).
O desejo de promover a produção e o uso de bioetanol recomeçou no início de 1980, em
grande parte para revitalizar o setor agrícola num momento de excesso de oferta de produtos
agrícolas (Johnson & Rosillo-Calle 2007). Os Estados Unidos refizeram a sua indústria de
combustível de etanol, e são hoje em dia líderes mundiais na sua produção e uso (Mussatto et al.
2010).
Atualmente o etanol é o principal biocombustível utilizado no mundo e seu uso está cada
vez mais generalizado, as perspetivas mundiais são a expansão da produção e do consumo de
etanol (Mussatto et al. 2010).
A produção de etanol tem aumentado em todo o mundo desde a crise do petróleo em
1970. O seu mercado cresceu desde menos de mil milhões de litros em 1975 para mais de 39
mil milhões de litros em 2006, e espera-se chegar a 100 mil milhões de litros no presente ano
(2015) (Licht 2006). O continente americano é o maior produtor mundial de etanol, com os
Estados Unidos e o Brasil representando um papel importante neste setor.
Certas espécies de microalgas têm capacidade de produzir altos níveis de carbohidratos
em vez de lípidos como polímeros de reserva. Estas espécies são candidatas ideais para a
produção de bioetanol, uma vez que os hidratos de carbono das microalgas podem ser extraídos
para produzir açúcares fermentáveis. Estimou-se que aproximadamente entre 46,760 - 140,290 L

INTRODUÇÃO | 17
ha-1 de etanol podem ser produzidos a partir de microalgas (Cheryl 2008). Este rendimento é
bastante maior do que o que se obtém para produções com outras matérias-primas.
Mesmo Panicum virgatum (“switchgrass”), considerado o biocombustível celulósico da
segunda geração de biocombustíveis, atinge rendimentos de produção de etanol que são apenas
uma fração dos rendimentos atingidos por microalgas.
Chlorella vulgaris, particularmente, tem sido considerada como uma matéria-prima
promissora para a produção de bioetanol, porque consegue acumular até 37 % (peso seco) de
amido (Hirano et al. 1997). Em condições climáticas favoráveis, podem ser alcançados
rendimentos entre 80 - 100 t de biomassa seca de Chlorella por ha, para um tempo de cultura de
300 dias (Doucha & Lívanský 2009). Segundo esses autores a estirpe Chlorella sp. é capaz de
acumular amido até 70 % de peso seco de algas sob condições de supressão de síntese de
proteínas.
A produção de etanol é geralmente realizada em três passos: (1) obtenção de uma
solução de açúcares fermentáveis, (2) fermentação dos açúcares em etanol, (3) separação do
etanol e purificação, geralmente por destilação - retificação – desidratação (Demirbas 2005). O
passo anterior à fermentação, para obtenção de açúcares fermentáveis, constitui a principal
diferença entre os processos de produção de etanol a partir de açúcares simples, amido ou
material lignocelulósico.
A produção de etanol a partir de microalgas pode ser realizada de acordo com o seguinte
procedimento (Figura 3). Na primeira etapa, o cultivo de microalgas usando a energia da luz solar
é realizado em tanques abertos ou cobertos (open or covered ponds) ou em fotobiorreatores
fechados, com base tubular, placa plana ou outros designs. Na segunda etapa, a biomassa tem de
ser concentrada por um fator inicial de pelo menos cerca de trinta vezes, requerendo, desta forma,
processos de colheita de baixo custo. Após a colheita, o amido microalgal é extraído das células
com o auxílio de um dispositivo mecânico ou através da utilização de enzimas. Após a extração do
amido, as enzimas amilolíticas são utilizadas para promover a formação de açúcares fermentáveis.
S. cerevisiae é então adicionada para iniciar a fermentação alcoólica. No final da fermentação, o
caldo fermentado que contém etanol é drenado do tanque e bombeado para um tanque de
retenção, para ser alimentado para uma unidade de destilação (Amin 2009).
À parte disto, há também espécies de algas capazes de realizar autofermentação. A
Seambiotic, em colaboração com a Inventure Chemicals, demonstrou com sucesso a produção de
etanol por fermentação de polissacarídeos de microalgas.

18 | INTRODUÇÃO
Seambiotic é a primeira empresa no mundo que está a utilizar gases de combustão de
centrais elétricas, provenientes da queima de carvão, como fonte de CO2, para o cultivo de
microalgas (Goh & Lee 2010).
Figura 3 - Procedimento para a produção de bioetanol a partir de microalgas (Mussatto et al. 2010).
Nas espécies de microalgas capazes de produzir amido, a sua acumulação pode ser
estimulada pela indução de estados de stresse metabólico. O estado de tensão resulta tipicamente
da privação de nutrientes essenciais. Alguns autores (Douskova et al. 2008) induziram uma
acumulação de amido na microalga Chlorella vulgaris pela privação de fósforo, azoto e enxofre e o
teor em amido nas células aumentou de 83 %, 50 % e 33 %, respetivamente. Também estudaram
o efeito de inibir a proteossíntese, o que se refletiu no dobro da quantidade de amido na biomassa.
A adição de ferro às culturas de microalgas induz a produção de hidratos de carbono,
pelas microalgas, devido ao aumento da eficiência fotossintética. No entanto, esta acumulação
depende da luz incidente e é resultado de um efeito sinérgico entre a suplementação da cultura e
da luz incidente (Van Oijen et al. 2005). Fatores como o aumento da temperatura ou da salinidade
também têm sido descritos como tendo efeito estimulador sobre a acumulação de amido nas
microalgas (Warr et al. 1985).
Todos estes passos concomitantemente aumentam os custos de energia de produção de
bioetanol (González-Fernández et al. 2012).
Hoje em dia, este biocombustível é amplamente distribuído e utilizado como um
componente de mistura de combustível pelos EUA. O seu ponto baixo de congelação torna-o
adequado para utilização em climas frios. Pode ser misturado com gasolina, em qualquer
combinação, e atualmente está aprovado como integrante dos combustíveis (10 %) de todos os
veículos e, como uma mistura de 85 % nos veículos de combustível duplo (Yue et al. 2014).

INTRODUÇÃO | 19
A Agência de Proteção Ambiental (EPA) dos Estados Unidos também aprovou uma mistura
de 15 % em todos os veículos a partir do ano 2001, no entanto, a disponibilidade de bioetanol
ainda é muito limitada e há vários desafios relacionados com a sua distribuição (Yue et al. 2014).
Adicionalmente, vários estudos têm citado o papel do etanol na corrosão das paredes das
tubagens. Devido a todos estes desafios, muitas das principais operadoras de tubos dos EUA
proíbem expressamente etanol e misturas de etanol-gasolina nas tubagens (Bunting et al. 2010).
1.5.4. Bio-hidrogénio
Relacionadas com a sua produtividade fotossintética e eficiência de utilização da luz, as
microalgas podem realizar reações bioquímicas e fotoquímicas especiais com requisitos mínimos
que tornam a produção de hidrogénio possível em ambientes aeróbios e anaeróbios (Das &
Veziroǧlu 2001; Benemann 2000).
Elas usam nitrogenases e hidrogenases, que estão intimamente relacionadas com o
processo final de geração de hidrogénio na fotossíntese (Masukawa et al. 2002; Kosourov et al.
2005). A nitrogenase é uma enzima que participa na fixação biológica do azoto e a hidrogenase é
capaz de catalisar as reações de consumo de hidrogénio. A produção de hidrogénio com base
nestas enzimas difere no consumo de energia. Reações mediadas pela hidrogenase são cerca de
três vezes mais eficientes do que as catalisadas pela nitrogenase, com base na energia
despendida sob a forma de ATP. Por outro lado, as nitrogenases são relativamente menos
sensíveis ao oxigénio em comparação com as hidrogenases (Miyake et al. 2004; Dasgupta et al.
2010), no entanto, ambas são sensíveis ao oxigénio. É importante controlar as condições de
cultura para a produção ótima de hidrogénio.
As vias metabólicas para a produção de hidrogénio diferem de acordo com o
microrganismo (Murugesan et al. 2009; Levin et al. 2004). No caso das microalgas, a produção de
hidrogénio ocorre através da fotólise (neste processo, a energia luminosa é utilizada para separar
os átomos que compõem a molécula de água, havendo libertação de eletrões, que são depois
usados para repor os perdidos pela clorofila na sua excitação. A molécula de oxigénio (O2)
resultante é libertada para a atmosfera, como resíduo da reação química) direta e indireta da
água. A fotólise direta envolve a energia da luz e os sistemas fotossintéticos das microalgas para
converter água em energia química. No entanto, durante a fotossíntese, o oxigénio produzido pode
inibir a atividade da enzima hidrogenase. A fotólise indireta supera a limitação da inibição da

20 | INTRODUÇÃO
hidrogenase pelo oxigénio através da produção de oxigénio fotossintético e hidrogénio em duas
fases separadas. O hidrogénio é produzido na segunda fase onde a anaerobiose é forçada pela
privação de enxofre (González-Fernández et al. 2012):
(a) Fotólise direta: 2H2O Luz 2H2 + O2
(b) Fotólise indireta:
(1) 12H2O + 6CO2 Luz C6H12O6 + 6O2
(2) C6H12O6 + 12H2O 12H2 + 6CO2
Obviamente, a estratégia indireta confere uma complexidade adicional ao sistema. Neste
ponto, deve-se ressaltar que de acordo com Benemann (2000) para a biofotólise ser uma fonte de
energia competitiva, a eficiência da conversão de energia solar deve ser em torno dos 10 %. Os
valores mais elevados encontrados na literatura foram relatados por Kruse et al. (2005) que
alcançou 2 % de eficiência de conversão em escala laboratorial através da fotólise direta. Assim, a
produção de hidrogénio por microalgas ainda tem de ser desenvolvida para que esta tecnologia
possa desempenhar um papel importante no desenvolvimento da produção de biocombustíveis
sustentáveis.
1.5.5. Biometano
O biometano é produzido a partir da digestão anaeróbia da matéria orgânica (neste caso,
microalgas), por bactérias anaeróbias, ou seja, que se desenvolvem em ambientes sem oxigénio.
Os principais produtos deste processo são os gases metano (55 - 75 %) e dióxido de carbono (25 –
45 %) (biogás), uma parte sólida que decanta no fundo do tanque (biofertilizante), e uma parte
líquida que corresponde ao efluente mineralizado (tratado) (Pienkos & Darzins 2009; GOLUEKE et
al. 1957; Mussgnug et al. 2010).
O metano é uma forma de energia renovável, enquanto os outros dois produtos (efluente
mineralizado e o dióxido de carbono) podem ser reciclados para o sistema de cultivo de
microalgas (GOLUEKE et al. 1957; Schenk et al. 2008).
A biomassa microalgal, tendo nas suas células quantidades altas de lípidos, amido e
proteínas, baixa quantidade de celulose e, por outro lado, a ausência de lenhina, possibilita uma
digestão anaeróbia fiável e eficaz. Estas especificações fazem das microalgas uma boa alternativa

INTRODUÇÃO | 21
para a produção eficaz de biometano em comparação com as outras (Pienkos & Darzins 2009;
Mussgnug et al. 2010; Schenk et al. 2008). A biomassa de microalgas que resta após a digestão
anaeróbia pode ainda ser processada para fabricar fertilizantes que podem acrescentar um valor
adicional ao processo global (Mussgnug et al. 2010; Doušková et al. 2010).
1.6. Impacto da seleção da estirpe
Na produção de biocombustíveis, para se obter sucesso, é essencial escolher a estirpe de
microalga apropriada. Geralmente, a microalga deve ser capaz de sobreviver a uma grande
variedade de temperaturas de forma a sobreviver às variações sazonais; possuir alta eficiência
fotossintética; não necessitar de muitos nutrientes; ser capaz de dominar estirpes selvagens em
sistemas de produção open pond; sobreviver às tensões de cisalhamento comuns nos
fotobiorreatores; elevada produção de lípidos, hidratos de carbono (especialmente amido); alta
capacidade de captação de CO2; características de autofloculação e produção de coprodutos
valiosos. No entanto, atualmente, nenhuma estirpe de microalgas possui todas estas
características ao mesmo tempo (Brennan & Owende 2010).
1.7. Tecnologias de produção de microalgas
O princípio por trás do desenvolvimento e produção de microalgas é relativamente
simples. Para responder aos requisitos biológicos destas culturas, foram desenvolvidos vários tipos
de tecnologias. A tecnologia mais favorável à produção de microalgas depende do produto alvo,
por isso, é necessário ter atenção cuidada na seleção da espécie e no conhecimento da sua
fisiologia (Dimitrov 2007).
1.7.1. Sistemas de produção fotoautotróficos
Neste tipo de sistemas de produção, as microalgas usam luz como fonte de energia única
e CO2 como fonte de carbono. Foram os primeiros sistemas a ser criados, sendo atualmente os
mais difundidos. Os sistemas de produção fotoautotróficos podem ser abertos, fechados ou
híbridos.

22 | INTRODUÇÃO
1.7.1.1. Sistemas de cultura abertos
Os sistemas de cultura abertos são os mais baratos para produção de biomassa
microalgal em grande escala porque, como podem ser implementados em áreas com potencial
produtivo de cultura marginal, não competem forçosamente por terra com culturas agrícolas.
Quando comparados com os sistemas de cultura fechados, os sistemas de cultura abertos
são menos dispendiosos para construir e operar, mais duráveis e com grande capacidade de
produção. No entanto, geralmente são mais suscetíveis às condições meteorológicas, devido a
ciclos diurnos e variações sazonais, não permitindo o controlo da temperatura da água, nem o
controlo dos níveis de CO2, devido à sua difusão na atmosfera. Para além disso, normalmente
possuem mecanismos de agitação ineficientes; apresentam riscos de contaminação elevados,
nomeadamente por outras espécies de algas, protozoários, bactérias e outros contaminantes
biológicos. Para as microalgas receberem energia solar suficiente para crescer, o nível de água
não pode ser mantido muito mais alto do que 15 cm (Benemann & Oswald 1996; Carlsson et al.
2007). Todos estes fatores tornam estes sistemas menos eficientes, quando comparados com os
sistemas fechados (Richmond 2004).
Normalmente, os sistemas de cultura abertos são projetos "raceway", nos quais as
microalgas são cultivadas (Figura 4). Estes representam o design mais eficiente para a produção
em larga escala (Spolaore et al. 2006). A água é normalmente mantida em movimento por rodas
de pás ou estruturas rotativas (Mata et al. 2010). Os nutrientes podem ser fornecidos através de
águas de escoamento de terras próximas, ou através da canalização da água das estações de
tratamento de resíduos (Spolaore et al. 2006).
Figura 4 - Vista aérea de um raceway pond (Brennan & Owende 2010).

INTRODUÇÃO | 23
As lagoas circulares representam os sistemas mais antigos de cultivo de algas, em grande
escala. Estes sistemas possuem um agitador rotativo articulado central e a sua profundidade é de
cerca de 0,3 m. A conceção destes sistemas limita o tamanho da lagoa a cerca de 10,000 m2
porque misturar usando o braço rotativo não é possível em tanques maiores. Estes sistemas têm a
limitação de requerer uma construção em cimento caro e entradas de alta energia para satisfazer
as necessidades de mistura (Borowitzka 2005).
No que diz respeito aos sistemas inclinados, a cultura é continuamente bombeada a partir
do topo para o fundo de uma superfície inclinada, obtendo um fluxo altamente turbulento, que
conduz à existência de camadas muito finas de cultura (menos de 1 cm). Estas características
conduzem a concentrações de células mais elevadas (até 10 g L-1) (Šetlík et al. 1970).
1.7.1.2. Sistemas de cultura fechados
Devido às dificuldades encontradas na operação dos sistemas abertos, foi proposta uma
outra abordagem para a produção de biomassa de microalgas. Esta nova abordagem baseia-se em
sistemas fechados, mais conhecidos como fotobiorreatores (FBRs), onde a maioria dos problemas
associados aos sistemas abertos são resolvidos. No entanto, isso não significa que os sistemas
fechados não têm obstáculos técnicos e operacionais a serem superados. Os FBRs são
caracterizados pela regularização e controlo de parâmetros quase todos bióticos e abióticos
importantes para o cultivo de microalgas e têm uma série de benefícios, quando comparados com
os sistemas abertos (Pulz 1992). A maioria da investigação atual e experiências relativas ao cultivo
de microalgas acontecem em FBRs (Eriksen 2008). Os mais utilizados são os FBRs tubulares,
planos e em coluna (Eriksen 2008).
Os FBRs tubulares podem ser dispostos na horizontal, inclinados, na vertical, ou em forma
de espiral. Em relação ao modo de operação, existem dois tipos de FBRs tubulares: os reatores
em que a mistura é levada a cabo através de bombeamento mecânico, ou tecnologia airlift, o meio
líquido existe apenas no reator e as trocas de gás ocorrem num compartimento separado; e os
reatores nos quais o líquido e a fase de gás estão presentes no mesmo compartimento, ocorrendo
por isso a transferência contínua de gás no reator. Hoje em dia, os FBRs maiores usam a
configuração tubular (Pulz 2001). Este FBR permite a troca de gases com o meio ambiente onde
ocorre a saída de O2 e entrada de CO2 no meio de cultura. Além disso, este sistema proporciona
meios para a recuperação da biomassa produzida. A série de tubos tem o objetivo de expor a

24 | INTRODUÇÃO
cultura de microalgas à luz solar. Tipicamente, os tubos têm diâmetros limitados a 0,1 m ou
menos, porque com diâmetros maiores, a luz torna-se um fator limitante, uma vez que não pode
penetrar completamente na cultura (Chisti 2007). O tamanho dos tubos também é limitado devido
à potencial acumulação de O2 que limita a fotossíntese e, por conseguinte, provoca uma
diminuição na produtividade, esgotamento do CO2 e mudanças de pH no meio de cultura (Eriksen
2008).
No que diz respeito aos FBRs planos, estes ganharam atenção especial devido a dois
fatores muito importantes que influenciam a produção de microalgas: maior superfície de captação
solar e elevada densidade de biomassa obtida. Este facto é extremamente importante, uma vez
que reduz muito o custo do processo. Além disso, os FBRs planos têm baixa acumulação de
oxigénio dissolvido e alta eficiência fotossintética (Brennan & Owende 2010). As desvantagens
associadas a FBRs planos incluem dificuldade em controlar a temperatura da cultura, algum grau
de crescimento na parede, possibilidade de stresse hidrodinâmico para algumas estirpes de
microalgas e o aumento de escala requer muitos compartimentos e materiais de apoio.
Em relação aos FBRs estes são constituídos por uma unidade de geração de bolhas de ar,
que tem duas funções distintas: misturar o meio de cultura para aumentar a eficiência
fotossintética e realizar a transferência de gases. Em comparação com os outros tipos de FBRs,
estes proporcionam uma mistura mais eficiente da cultura, maior transferência de gás volumétrico
e um maior controlo sobre as condições do meio de cultura (Eriksen 2008; Mata et al. 2010;
Suzuki et al. 1995).
1.7.1.3. Sistemas híbridos
Sistemas híbridos combinam os FBRs fechados e os sistemas abertos para a produção de
biomassa de microalgas. Estes sistemas foram concebidos a fim de maximizar as vantagens e
minimizar as desvantagens inerentes a ambos, sem aumentar muito o custo de produção (Huntley
& Redalje 2007). A operação dos sistemas híbridos ocorre em duas fases distintas. A primeira fase
consiste na produção de biomassa em FBRs num meio de cultura rico em nutrientes e com as
condições de cultura controladas. A segunda fase opera em sistemas de lagoas abertas, onde a
biomassa produzida no passo 1 é sujeita a pressões ambientais, tais como o défice de azoto e de
fósforo e de elevada luminosidade, que aumenta a síntese do produto lipídico e dos carbohidratos.

INTRODUÇÃO | 25
Os stresses ambientais podem ocorrer naturalmente por meio da transferência da cultura do FBR
para o sistema aberto (Huntley & Redalje 2007; Rodolfi et al. 2009).
1.8. Tecnologias de recuperação da biomassa microalgal
Recolher a biomassa do meio de cultura é um dos passos mais caros da produção de
microalgas e envolve um custo que pode rondar 20 a 30 % do custo total de produção (Molina
Grima et al. 2003). Isto acontece devido às baixas concentrações de biomassa encontradas nos
sistemas de produção e ao tamanho das células das microalgas que tipicamente varia desde 3 a
30 µm. A concentração ideal para a conversão industrial da biomassa rondará entre 300 g L-1 –
400 g L-1.
No entanto, o custo de colheita nos FBRs é uma fração do valor nos sistemas abertos,
porque a concentração de biomassa é cerca de 30 vezes superior nos FBRs (Packer 2009). Vários
métodos de recolha de biomassa são praticados, mas a centrifugação, filtração e floculação são os
mais adequados à larga escala, embora também apresentem limitações e possam não ser sempre
os menos dispendiosos. Não existe um método de recolha de biomassa que sirva para todos os
sistemas de produção, por isso, selecionar a tecnologia mais apropriada é crucial, de um ponto de
vista económico, técnico e operacional.
A floculação é a primeira etapa do processo de recuperação de biomassa e destina-se a
agregar as microalgas para facilitar e aumentar a eficiência dos processos que se seguem
(Elmaleh et al. 1991). Como as microalgas têm carga negativa, existem forças de repulsão que
não permitem a agregação das células. No entanto, esta carga pode ser neutralizada ou reduzida
através da adição de produtos químicos, melhor conhecidos como floculantes. Os floculantes
normalmente utilizados são os sais de sulfato de alumínio (Al2(SO4)3), cloreto de ferro (FeCl3), e
sulfato de ferro (Fe2(SO4)3) (Molina Grima et al. 2003). A autofloculação é um processo que
também já começou a ser investigado e verificou-se que algumas espécies de microalgas podem
flocular naturalmente, em resposta a estímulos ambientais, tais como alterações no pH do meio
de cultura (Benemann & Oswald 1996).
A flotação corresponde a um método simples através do qual as microalgas podem ser
forçadas a flutuar na superfície do meio para de seguida serem removidas. São usadas
microbolhas de ar, dispersas no meio aquoso, que vão adsorver as células e arrastá-las para a
superfície do meio. Ao contrário da floculação, não exige qualquer adição de produtos químicos

26 | INTRODUÇÃO
(Brennan & Owende 2010). Há poucas provas da sua viabilidade técnica ou económica (Brennan
& Owende 2010).
A centrifugação envolve a aplicação de forças centrífugas para separar a biomassa
microalgal do meio de crescimento. Uma vez separadas, as microalgas podem ser removidas por
simples drenagem do excesso de meio (Harun, Singh, et al. 2010). A recuperação centrífuga é um
método rápido de recuperação das células das algas. No entanto, as forças gravitacionais e de
cisalhamento durante o processo de centrifugação podem danificar a estrutura da célula. Outra
desvantagem deste método é o elevado consumo de energia, especialmente quando se considera
grandes volumes (Harun, Singh, et al. 2010).
No que diz respeito aos processos de filtração, estes consistem na separação da fase
sólida (algas) da fase líquida (meio de cultura), passando a mistura através de um filtro, onde as
algas são retidas. Os processos de filtração convencionais incluem filtração dead end,
microfiltração, ultrafiltração, filtração sob pressão, filtração sob vácuo e filtração de fluxo tangencial
(TFF) (Dragone et al. 2010). Para recuperar a biomassa em culturas de microalgas com
dimensões mais pequenas, são comummente utilizadas as técnicas de microfiltração e
ultrafiltração que fazem uso de membranas com poros de menores dimensões (Petrusevski et al.
1995). Os principais custos desta técnica estão associados à substituição das membranas e ao
sistema de bombeamento. Em comparação com outras opções de colheita, a filtração é o método
mais competitivo (Dragone et al. 2010).
1.9. Biocombustíveis de microalgas: que futuro?
Numa série de artigos publicados em várias edições da Biofuels International Magazine,
Thurmond destacou vários passos estratégicos necessários para a comercialização bem-sucedida
de biocombustíveis de algas (Thurmond 2009a; Thurmond 2009b; Thurmond 2010). Segundo o
autor, a principal necessidade dos produtores é identificar espécies que possuam alto teor de óleo,
e que cresçam rapidamente, para produzir biodiesel.
Se os produtores de algas usarem espécies mais “gordas”, com 60 % de teor de óleo,
podem reduzir o tamanho e a pegada ecológica dos sistemas de produção de biocombustíveis até
metade, reduzindo os custos operacionais e facilitando a entrada no mercado dos biocombustíveis.
Uma vez que os sistemas de produção de algas são uma combinação complexa de vários
subconjuntos de sistemas (ou seja, produção, colheita, extração, sistemas de secagem), reduzir o

INTRODUÇÃO | 27
número de etapas na produção de biocombustíveis de algas é essencial para fornecer sistemas de
produção com custo inferior.
Um desafio económico crucial é descobrir métodos de colheita de algas e extração de óleo
de baixo custo. Por exemplo, a Origin Oil desenvolveu uma tecnologia que combina os sistemas de
colheita e extração num processo único, que é desenhado para reduzir a complexidade do sistema
global e os custos para os produtores. Outro exemplo é a utilização de um método que utilize as
células de algas como miniprocessadores que irão consumir CO2 e excretar combustíveis de
hidrocarbonetos diretamente.
De acordo com Singh & Gu 2010, o futuro dos biocombustíveis de algas prevê-se brilhante
de várias maneiras. Alguns analistas do mercado sentem que apesar de todos os fatores positivos
associados à utilização de microalgas para os biocombustíveis, o sucesso da implementação
comercial dos biocombustíveis de algas deverá depender do desenvolvimento de coprodutos de
alto valor, por exemplo, polímeros renováveis ou pigmentos. As algas contêm produtos químicos
valiosos ou compostos moleculares que podem ser utilizados na produção de plásticos verdes,
detergentes verdes, produtos de limpeza, e polímeros biodegradáveis, não-tóxicos, e que podem
ser vendidos a um preço premium em relação aos produtos tradicionais.


2. CAPÍTULO II
MATERIAIS E MÉTODOS


MATERIAIS E MÉTODOS | 31
2.1 Microrganismos
As microalgas utilizadas para a elaboração do trabalho experimental foram a Chlorella
emersonii ACOI 516, Chlorella vulgaris ACOI 879 e Chlorella saccharophila ACOI 96. As estirpes
foram adquiridas à Algoteca do Departamento de Botânica da Universidade de Coimbra (ACOI -
Coimbra Collection of Algae), tendo sido repicadas periodicamente e mantidas à temperatura
ambiente.
2.2 Meios de crescimento
O cultivo autotrófico das microalgas referidas decorreu em cinco meios de cultura
diferentes (MP12, TAP, BG-11, Chu_13 mod. e BBM).
O meio baseado na composição elementar de C. vulgaris P12, e denominado no presente
trabalho como MP12 foi preparado de acordo com Douskova et al. (2008). Para a preparação
deste meio, a 900 mL de água destilada adicionaram-se 2 mL de uma solução de micronutrientes,
assim como os reagentes KH2PO4, MgSO4.7H2O, C10H12O8N2NaFe e CaCl2.2H2O, nas concentrações
apresentadas na Tabela 1. À parte, a 100 mL de água destilada juntaram-se 1,10 g de (NH2)2CO
(ureia). Depois de autoclavar as soluções a 121ºC durante 15 min, aos 900 mL de solução
adicionaram-se os 100 mL da solução de ureia. A solução de ureia foi autoclavada separadamente
para evitar a precipitação de sais.
Tabela 1 - Concentração dos reagentes usados no meio MP12.
Reagentes Concentração (g L-1)
KH2PO4 0,237
MgSO4.7H2O 0,204
C10H12O8N2NaFe 0,040
CaCl2.2H2O 0,116
(NH2)2CO 1,100
Solução de micronutrientes
H3BO3 0,416
CuSO4.5H2O 0,473
MnCl2.4H2O 1,647
(NH4)6Mo7O24.4H2O 0,086
ZnSO4.7H2O 1,339
CoSO4.7H2O 0,308
(NH4)VO3 0,007

32 | MATERIAIS E MÉTODOS
O meio Tris - Acetate - Phosphate (TAP) elaborou-se segundo Starr & Zeikus (1993). Para
isso, fizeram-se quatro soluções stock, cujos reagentes e correspondentes concentrações podem
ser consultados na Tabela 2.
Tabela 2 - Concentração dos reagentes usados na preparação das soluções stock do meio TAP.
Reagentes Concentração (g L-1)
Solução Beijerinck’s
NH4Cl 8
CaCl2·2H2O 1
MgSO4·7H2O 2
Solução tampão fosfato
Na2HPO4 11,62
KH2PO4 7,26
Solução Hunter's Trace
Na2EDTA·2H2O 50
ZnSO4·7H2O 22
H3BO3 11,4
MnCl2·4H2O 5,1
FeSO4·7H2O 5
CoCl2·6H2O 1,6
CuSO4·5H2O 1,16
(NH4)6Mo7024·4H2O 1,1
Solução Tris Acetato
Trisma Base 242 Ácido acético
glacial 100 1
1 Unidade em mL L-1.
Na elaboração deste meio, a um volume de 400 mL de água destilada, adicionaram-se 50
mL L-1 da solução Beijerinck’s; 8,5 mL L-1 da solução tampão fosfato; 1 mL L-1 da solução Hunter's
Trace e 10 mL L-1 da solução Tris Acetato, com agitação contínua. No final perfez-se o volume até 1
L com água destilada.
O meio BG-11 também foi preparado segundo Starr & Zeikus (1993), conforme o que vem
descrito nas tabelas 3 e 4.

MATERIAIS E MÉTODOS | 33
Tabela 3 - Composição do meio BG-11.
Reagentes Quantidade (mL L-1) Concentração da
solução stock
NaNO3 10 30 g 200 mL-1
dH2O
K2HPO4 10 0,8 g 200 mL-1
dH2O
MgSO4·7H2O 10 1,5 g 200 mL-1
dH2O
CaCl2·2H2O 10 0,72 g 200 mL-1
dH2O
Citric Acid·H2O 10 0,12 g 200 mL-1
dH2O C6H8O7.xFe3+.yNH3 (citrato de ferro
amoniacal) 10
0,12 g 200 mL-1 dH2O
C10H14N2Na2O8· 2H2O 10 0,02 g 200 mL-1
dH2O
Na2CO3 10 0,4 g 200 mL-1
dH2O Solução de
micronutrientes1 1
Na2S2O3·5H2O (usado só em meio
com agar)
1
49,8 g 200 mL-1 dH2O
1 Ver Tabela 4.
Tabela 4 - Composição da solução de micronutrientes do meio BG-11.
Reagentes Concentração (g L-1)
H3BO3 2,86
MnCl2·4H2O 1,81
ZnSO4·7H2O 0,22
Na2MoO4·2H2O 0,39
CuSO4·5H2O 0,079
Co(NO3)2·6H2O 0,049
No que diz respeito ao meio Chu_13 modificado, este foi preparado de acordo com
Kaewkannetra et al. (2012). Os reagentes utilizados e respetivas concentrações apresentam-se na
Tabela 5.

34 | MATERIAIS E MÉTODOS
Tabela 5 - Composição do meio Chu_13 modificado.
Reagentes Concentração (g L-1)
KNO3 0,2
K2HPO4 0,04
MgSO4·7H2O 0,1
CaCl2·2H2O 0,054
C6H5FeO7 0,01
C6H8O7 0,1
Solução de micronutrientes
H3BO3 2,85
MnCl2 1,80
ZnSO4·7H2O 0,02
CuSO4·5H2O 0,08
CoCl2·6H2O 0,08
Na2MoO4·2H2O 0,05
Em relação ao meio Bold’s Basal Medium (BBM), este foi preparado de acordo com Starr
& Zeikus (1993). Para a elaboração deste meio, fizeram-se 6 soluções stock com os reagentes e
respetivas concentrações mencionados na Tabela 6, sendo que se retiraram 10 mL de cada uma
para o volume final (1 L). Adicionalmente e de acordo com o que vem descrito na Tabela 7,
também se prepararam 4 soluções stock diferentes, sendo que destas, o volume retirado para a
preparação do meio foi 1 mL.
Tabela 6 - Soluções stock constituintes do meio BBM.
Reagentes Concentração da
solução stock
NaNO3 2,5 g 100 mL-1
dH2O
MgSO4.7H2O 0,75 g 100 mL-1
dH2O
NaCl 0,25 g 100 mL-1
dH2O
K2HPO4 0,75 g 100 mL-1
dH2O
KH2PO4 1,75 g 100 mL-1
dH2O
CaCl2.2H2O 0,25 g 100 mL-1
dH2O

MATERIAIS E MÉTODOS | 35
Tabela 7 - Soluções stock constituintes do meio BBM.
Reagentes Concentração (g L-1)
Solução Stock 1
ZnSO4.7H2O 8,82
MnCl2.4H2O 1,44
MoO3 0,71
CuSO4.5H2O 1,57
Co(NO3)2.6H2O 0,49
Solução Stock 2
H3BO3 11,42
Solução Stock 3
EDTA 50,0
KOH 31,0
Solução Stock 4
FeSO4.7H2O 4,98 H2SO4
(concentrado) 1,0 1
1 Unidade em mL L-1.
Também foi preparado meio sólido de todos os meios de crescimento acima
mencionados. Para isso, bastou adicionar 1,5 % de agar a cada meio.
Na Tabela 8 apresentam-se todos os reagentes usados na elaboração dos cinco meios de
crescimento, assim como a sua concentração, de forma a ser possível comparar as diferentes
concentrações utilizadas para o mesmo reagente.
Tabela 8 - Concentração dos reagentes utilizados nos cinco diferentes meios de cultura.
Reagentes1 Meios de Cultura
Nomenclatura Símbolo químico
Meio M (µM)
TAP (µM)
BBM (µM)
Chu_13 mod. (µM)
BG-11 (µM)
Ácido acético glacial CH3CO2H 1700
Ácido bórico H3BO3 0,027 184 185 46 46
Ácido cítrico C6H8O7 520
Ácido cítrico monohidratado C6H8O7.H2O 240
Ácido sulfúrico H2SO4 18,76
Carbonato de sódio Na2CO3 190
Citrato de ferro C6H5FeO7 41
Citrato de ferro amoniacal C6H8O7.xFe3+.yN
H3 21
Cloreto de amónio NH4Cl 7500 1 Todos os reagentes foram adquiridos à empresa Sigma-aldrich®.

36 | MATERIAIS E MÉTODOS
Tabela 8 – Continuação.
Reagentes1 Meios de Cultura
Nomenclatura Símbolo químico
Meio M (µM)
TAP (µM)
BBM (µM)
Chu_13 mod. (µM)
BG-11 (µM)
Cloreto de cálcio dihidratado CaCl2·2H2O 790 350 170 340 240
Cloreto de cobalto hexahidratado
CoCl2·6H2O 7 0,34
Cloreto de manganês (II) MnCl2 14
Cloreto de manganês (II) tetrahidratado
MnCl2·4H2O 0,033 26 7,3 9
Cloreto de sódio NaCl 430
EDTA (ácido etilenodiaminotetracético)
C10H16N2O8 170
EDTA dissódico dihidratado C10H14N2Na2O8·
2H2O 134 2,7
EDTA férrico sódico C10H12O8N2NaFe 110
Fosfato de potássio dibásico K2HPO4 430 230 230
Fosfato de potássio monobásico
KH2PO4 1740 450 1290
Fosfato de sódio dibásico Na2HPO4 690
Hidróxido de potássio KOH 550
Molibdato de amónio tetrahidratado
(NH4)6Mo7024·4H2O
0,000275 0,8
Molibdato de sódio dihidratado
Na2MoO4·2H2O 0,21 1,6
Nitrato de cobalto hexahidratado
Co(NO3)2·6H2O 1,7 0,17
Nitrato de potássio KNO3 1980
Nitrato de sódio NaNO3 2940 17600
Sulfato de cobre heptahidratado
CoSO4·7H2O 0,0043
Sulfato de cobre pentahidratado
CuSO4·5H2O 0,0076 5 6,3 0,32 0,3
Sulfato de ferro heptahidratado
FeSO4·7H2O 18 18
Sulfato de magnésio heptahidratado
MgSO4·7H2O 830 400 300 410 300
Sulfato de zinco heptahidratado
ZnSO4·7H2O 0,019 77 30,60 0,070 0,77
Tiossulfato de sódio pentahidratado2
Na2S2O3·5H2O 1000
Trióxido de molibdénio MoO3 4,90
Tris base NH2C(CH20H)3 20000
Ureia (NH2)2CO 18310
Vanadato de amónio (NH4)VO3 0,000239 1 Todos os reagentes foram adquiridos à empresa Sigma-aldrich®. 2 Usado somente em meio sólido.

MATERIAIS E MÉTODOS | 37
2.3 Cultivo fotoautotrófico
2.3.1 Crescimento em meio com azoto
Para a preparação dos inóculos, retirou-se uma ansada de microalgas a crescer nos meios
de cultura em estado sólido e inocularam-se em 35 mL de cada meio, em tubos de ensaio com
capacidade de 50 mL. Os inóculos foram preparados em duplicado para cada um dos meios. O
seu tempo de crescimento foi de cerca de 7 dias e ocorreu à temperatura ambiente
(aproximadamente 25 °C). Durante esse tempo, estiveram sujeitos a uma irradiação de 100 µmol
de fotões m-2 s-1 fornecidos por duas lâmpadas fluorescentes (Sylvania Standard F18W). Todos os
inóculos estiveram sob arejamento usando ar enriquecido com 6 % (v/v) de CO2 a uma taxa de 0,2
vvm (volume de gás por volume de suspensão de cultura por minuto). Assim, foi fornecido um
caudal de ar de 7 mL min-1 (0,42 mL min-1 CO2 e 6,58 mL min-1 ar) a cada tubo de ensaio.
No final do tempo de crescimento das microalgas nos inóculos, procedeu-se à passagem
das células para os fotobiorreatores. Assim, após contagem celular das amostras retiradas dos
inóculos, no microscópio, com o auxílio da câmara de Neubauer (Carl Zeiss, West Germany), e
determinação do volume a retirar dos mesmos, as células foram recolhidas por centrifugação
(4000 rpm, 10 min) e ressuspendidas em 350 mL de meio de cultura, em provetas com
capacidade de 500 mL. A concentração celular inicial para cada cultivo foi 1 x 106 cél. mL-1 e todos
os cultivos foram realizados em duplicado. O cultivo das microalgas ocorreu nas mesmas
condições referidas para os inóculos (100 µmol de fotões m-2 s-1 e 6 % (v/v) de CO2 a uma taxa de
0,2 vvm). Desta forma, foi fornecido um caudal de ar de 70 mL min-1 (4,2 mL min-1 CO2 e 65,8 mL
min-1 ar), para cada proveta.
Durante o tempo em que o ensaio decorreu (≈ 15 dias) retirou-se um volume diário de 3
mL de amostra, que foram usados para a medição do pH, determinação do peso da biomassa
seca e determinação da concentração celular, tal como descrito no ponto 2.4.
Tendo em conta que o cultivo em cada meio foi realizado em duplicado, no final do
mesmo uma das réplicas foi utilizada para o crescimento em meio sem azoto e a outra para
caracterização da biomassa.

38 | MATERIAIS E MÉTODOS
2.3.2 Crescimento em meio sem azoto
No final de cada ensaio em meio com azoto, as células foram centrifugadas a 9000 rpm
durante 2 min em frascos de 250 mL. Depois, foram ressuspendidas no volume de 350 mL de
cada um dos meios descritos no ponto 2.2, sendo que a cada um destes foi retirada a sua
principal fonte de azoto. Foram retirados os reagentes (NH2)2CO, NH4Cl, NaNO3, KNO3 e NaNO3 aos
meios MP12, TAP, BG-11, Chu_13 modificado e BBM, respetivamente.
O ensaio em meio sem azoto também foi realizado em duplicado e decorreu durante cerca
de 4 dias nas mesmas condições de arejamento, temperatura e irradiação de luz referidos para o
ensaio em meio com azoto. A monitorização do crescimento celular em meio sem azoto também
foi feita de acordo com o que está descrito no ponto 2.4.
2.4 Monitorização do crescimento celular
Para a monitorização do crescimento celular, retiraram-se, diariamente, 3 mL de amostra
dos fotobiorreatores. Após centrifugação (4000 rpm, 5 min) procedeu-se à medição do pH do
sobrenadante. No final, o pellet foi ressuspendido com o auxílio do vortex.
A determinação do peso da biomassa seca efetuou-se usando filtros de nitrocelulose de
0,45 µm de poro. Estes filtros foram colocados no micro-ondas durante 15 min à potência de 90
W. Em seguida, foram colocados a arrefecer no exsicador durante 5 a 10 min e foram pesados.
Depois filtraram-se 2 mL de amostra e colocaram-se os filtros novamente no micro-ondas, nas
mesmas condições. O peso da biomassa seca foi obtido subtraindo ao peso final o peso do filtro.
Para a determinação da concentração celular, através da contagem na câmara de
Neubauer, procedeu-se a uma diluição inicial das amostras em 1,5 M de EDTA (pH = 8,2) de
forma a facilitar a contagem, uma vez que as espécies estudadas demonstraram ser
extremamente floculantes, à exceção de C. vulgaris, embora também tenha sido diluída na mesma
concentração de EDTA. Este esteve 2 h em contacto com as células, que foram agitadas
periodicamente durante 1 min.
2.5 Caracterização da biomassa
No final de cada ensaio, tanto na condição com azoto como na condição sem azoto, as
células foram centrifugadas a 9000 rpm durante 2 min em frascos de 250 mL. Depois, foram

MATERIAIS E MÉTODOS | 39
liofilizadas e moídas com o auxílio do almofariz, para se proceder à sua caracterização no que diz
respeito ao conteúdo de lípidos, amido e proteínas.
2.5.1 Análise de amido
O conteúdo de amido nas células foi determinado por um método de hidrólise enzimática,
tendo sido utilizado um kit produzido pela Megazyme (Wicklow, Ireland) e aceite pela AOAC
(Método Oficial 996.11) e AACC (Método 76.13), baseado no uso das enzimas termoestáveis α-
amilase e amiloglucosidade.
O protocolo disponibilizado pela Megazyme foi seguido e iniciado na alínea (e). Pesaram-se
10 mg de biomassa liofilizada e moída (em vez de 100 mg, como indica o protocolo). Adicionaram-
se 5 mL de etanol aquoso (80 % v/v) e incubou-se a 80 - 85 °C durante 5 min. Misturou-se depois
a amostra no vortex e adicionaram-se mais 5 mL de etanol aquoso (80 % v/v). Centrifugou-se a
amostra durante 10 min a 1800 x g e descartou-se o sobrenadante. Ressuspendeu-se o pellet em
10 mL de etanol aquoso (80 % v/v) e misturou-se no vortex. Centrifugou-se novamente e
descartou-se o sobrenadante. Retomou-se o procedimento no passo 4 da alínea (b) do protocolo,
tendo-se, por isso, adicionado 3 mL da enzima termoestável α-amilase diluída 1:30 no reagente
MOPS 50 mM a pH 7,0. Incubou-se num banho de água a ferver durante 12 min. De seguida,
colocou-se a amostra num banho a 50 °C, adicionou-se 4 mL de tampão acetato de sódio 200
mM a pH 4,5, seguido de 0,1 mL da enzima amiloglucosidase 20 U. Misturou-se tudo no vortex e
manteve-se no banho a 50 °C durante 30 min. Passou-se, depois, para a alternativa do passo 6,
alínea (a). Ajustou-se o volume da amostra para 10 mL com água destilada e depois centrifugou-se
a 3000 rpm durante 10 min. Transferiram-se 0,1 mL da solução diluída para tubos de vidro e
adicionaram-se 3 mL do reagente GOPOD a cada tubo. Também se adicionaram 3 mL de GOPOD
a 0,1 mL de D-glucose padrão (controlo) e a 0,1 mL de água (branco) e incubaram-se todos os
tubos a 50 °C durante 20 min. No final, leu-se a absorvância a 510 nm.
O cálculo do conteúdo de amido presente nas amostras das microalgas foi obtido a partir
da utilização do programa de cálculo disponibilizado pela Megazyme “Mega-calc™ Total Starch
Determination (Solids)”.

40 | MATERIAIS E MÉTODOS
2.5.2 Análise de Proteínas
Para a análise do conteúdo proteico das células, utilizou-se o método de Bradford. Para C.
emersonii, pesou-se 0,1 g de biomassa liofilizada e moída e, para otimizar a extração das
proteínas, juntaram-se 2 mL de H3PO4 0,15 M e levou-se a mistura a um banho a 100 °C durante
10 min. Parou-se a reação com gelo e centrifugou-se a 3000 rpm durante 10 min. Separou-se o
sobrenadante e diluiu-se o mesmo numa proporção de 1:20. Adicionaram-se 300 µL do reagente
de Coomassie a 10 µL da amostra diluída. Incubou-se durante 5 min à temperatura ambiente e,
no final, leu-se a absorvância a 595 nm.
Para além disso, também se testou outro químico a fim de otimizar a extração de
proteínas, tendo-se adicionado às células moídas 2 mL de NaOH 1 M e levado a mistura a um
banho a 80 °C durante 10 min. As restantes etapas do procedimento seguiram como descrito
anteriormente. Para C. vulgaris e C. saccharophila, apenas foi aplicado este procedimento.
As concentrações de proteína foram estimadas por referência às absorvâncias obtidas por
uma série de diluições da proteína padrão – Bovine Serum Albumin (BSA) – analisadas
juntamente com as amostras desconhecidas.
2.5.3 Análise de Lípidos
Para a análise de lípidos, seguiu-se o método descrito por Folch et al. (1957), tendo-se
pesado 0,08 g de biomassa liofilizada e moída. Homogeneizou-se a amostra com uma mistura de
clorofórmio e metanol, numa proporção de 2:1, até se perfazer um volume final 20 vezes superior
ao volume da amostra. Centrifugou-se a solução para recuperar a fase líquida. Lavou-se a fase
líquida com um volume 0,2 x volume final, de uma solução de KCl 0,88 % de forma a facilitar a
extração de contaminantes não lipídicos para a fase aquosa (metanol), diminuindo também a
tendência de dissociação dos lípidos com carácter ácido, que se mantiveram deste modo, na fase
orgânica. Centrifugou-se a 2000 rpm para separar as duas fases. Descartou-se a fase superior
(aquosa). Lavou-se a interface 3 vezes, com uma mistura de clorofórmio, metanol e água nas
proporções (3:48:47 v:v) para remover alguns resíduos ainda presentes. No final, removeu-se o
solvente, deixando-se a evaporar na hotte de um dia para o outro. Depois, colocou-se 15 min na
estufa a 80 °C, retirou-se e deixou-se arrefecer no exsicador. O conteúdo lipídico das amostras
obteve-se através da subtração do peso final ao peso inicial. Estes valores foram posteriormente
convertidos em percentagem.

MATERIAIS E MÉTODOS | 41
De notar que nas Figuras apresentadas no Capítulo 3 referentes à análise da biomassa, a
ausência de alguns resultados é explicada pela falta de biomassa suficiente.
2.6 Determinação das Produtividades
A determinação das produtividades volumétricas de biomassa foi efetuada de acordo com
a Equação 1.
Pbiomassa (g L-1 d-1) = (X2 – X1) · (t2 – t1)-1 (Equação 1)
Onde X1 e X2 correspondem ao peso da biomassa seca (g L-1) nos dias t1 (início do cultivo) e
t2 (fim do cultivo), respetivamente.
As produtividades de lípidos, amido e proteínas foram calculadas de acordo com a
Equação 2.
Plípidos; amido; proteínas (mg L-1 d-1) = Pbiomassa · Cf (Equação 2)
Onde Pbiomassa é a produtividade da biomassa e Cf é o conteúdo final de lípidos, amido ou
proteínas (valores percentuais).


3. CAPÍTULO III
RESULTADOS

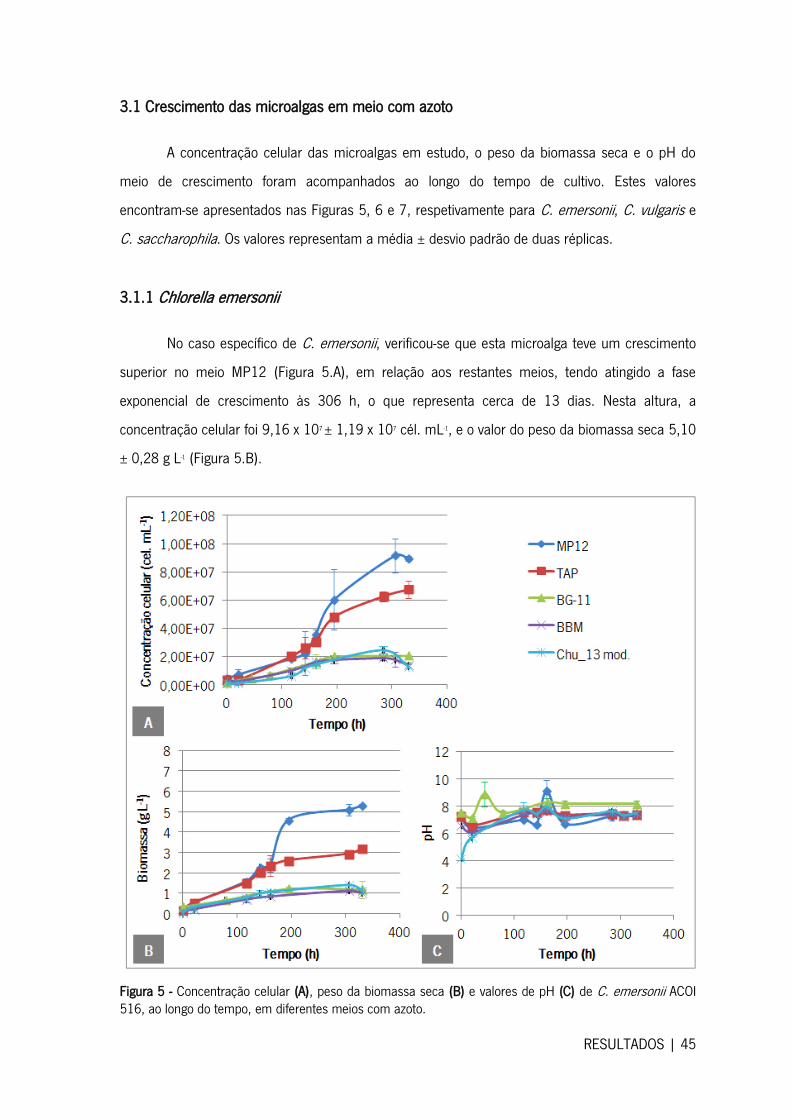
RESULTADOS | 45
3.1 Crescimento das microalgas em meio com azoto
A concentração celular das microalgas em estudo, o peso da biomassa seca e o pH do
meio de crescimento foram acompanhados ao longo do tempo de cultivo. Estes valores
encontram-se apresentados nas Figuras 5, 6 e 7, respetivamente para C. emersonii, C. vulgaris e
C. saccharophila. Os valores representam a média ± desvio padrão de duas réplicas.
3.1.1 Chlorella emersonii
No caso específico de C. emersonii, verificou-se que esta microalga teve um crescimento
superior no meio MP12 (Figura 5.A), em relação aos restantes meios, tendo atingido a fase
exponencial de crescimento às 306 h, o que representa cerca de 13 dias. Nesta altura, a
concentração celular foi 9,16 x 107 ± 1,19 x 107 cél. mL-1, e o valor do peso da biomassa seca 5,10
± 0,28 g L-1 (Figura 5.B).
Figura 5 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. emersonii ACOI 516, ao longo do tempo, em diferentes meios com azoto.

46 | RESULTADOS
Mais próximo deste crescimento esteve o registado no meio TAP, com o valor 6,75 x 107 ± 6,19 x
106 cél. mL-1 (330 h), sendo que, nesta fase, o valor do peso seco foi 3,20 ± 0,07 g L -1. Nos
restantes meios, o crescimento de C. emersonii foi bastante semelhante apesar de ligeiramente
inferior no meio BBM, com 1,89 x 107 ± 1,94 x 106 cél. mL-1. O valor do peso da biomassa seca
mais baixo também se verificou no meio BBM, com 0,80 ± 0,07 g L -1. No que diz respeito às
variações de pH registadas em cada meio, foi possível observar que, inicialmente, o pH do meio
Chu_13 mod. encontrava-se mais ácido do que o pH dos restantes meios (Figura 5.C). Durante o
tempo de cultivo, observaram-se variações nos valores de pH de cada meio e, no final, os seus
valores estiveram compreendidos entre 7,31 ± 0,01 (no meio BBM) e 8,16 ± 0,20 (no meio BG-
11).
3.1.2 Chlorella vulgaris
No que diz respeito ao crescimento de C. vulgaris, constatou-se que esta microalga atingiu
um valor de concentração celular superior no meio BG-11 (Figura 6.A). Na fase exponencial, o seu
valor foi 3,42 x 108 ± 7,64 x 107 cél. mL-1 e o valor do peso seco 3,80 ± 0,49 g L-1 (Figura 6.B).
Verificou-se também que nos meios Chu_13 mod. e BBM, C. vulgaris atingiu a fase exponencial
primeiro, em comparação com os restantes, com concentrações celulares inferiores.
Quanto aos valores do peso da biomassa seca, representados na Figura 6.B, estes foram
superiores nos meios MP12, BG-11 e TAP, correspondendo a 6,33 ± 1,03 g L-1, 4,30 ± 0,35 g L-1 e
4,05 ± 0,14 g L-1, respetivamente. À semelhança do que aconteceu com a concentração celular, o
peso da biomassa seca nos meios BBM e Chu_13 mod. foi inferior ao observado nos outros meios
de crescimento. Durante o tempo de cultivo, observaram-se variações nos valores de pH de cada
meio, sendo que, no final, estes valores encontraram-se entre 6,68 ± 0,12 no meio MP12 e 7,89
± 0,03 no meio BG-11 (Figura 6.C).
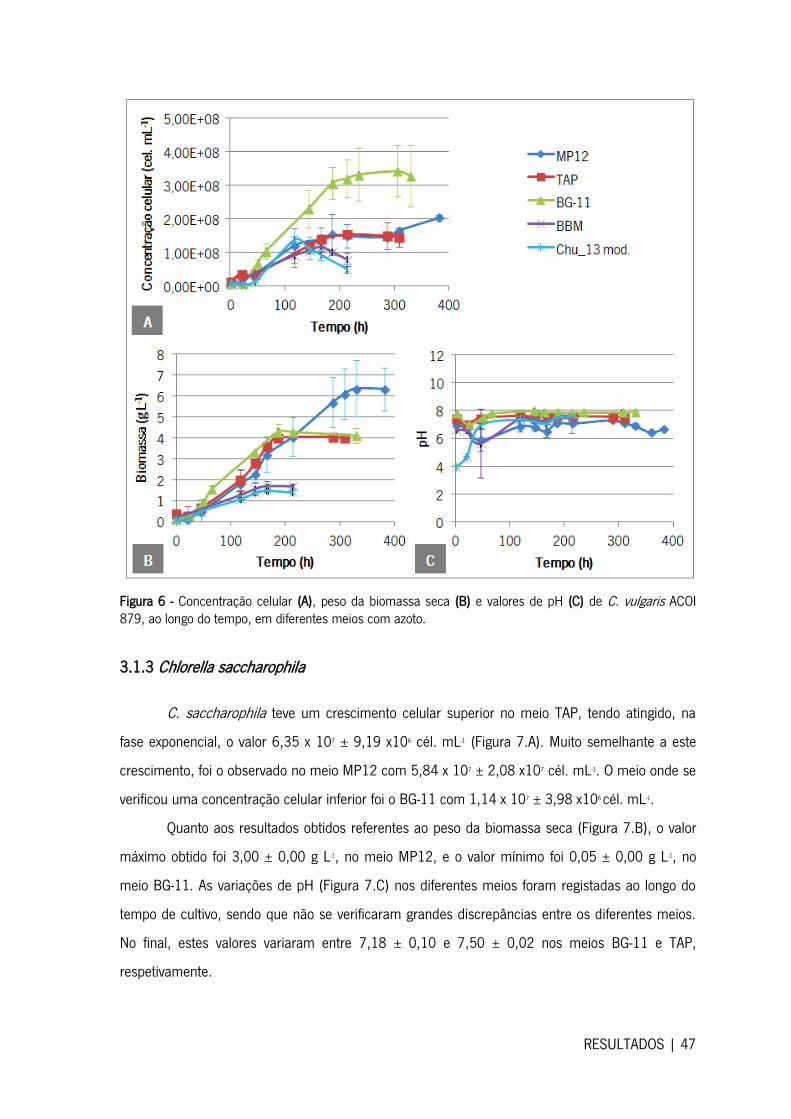
RESULTADOS | 47
Figura 6 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. vulgaris ACOI 879, ao longo do tempo, em diferentes meios com azoto.
3.1.3 Chlorella saccharophila
C. saccharophila teve um crescimento celular superior no meio TAP, tendo atingido, na
fase exponencial, o valor 6,35 x 107 ± 9,19 x106 cél. mL-1 (Figura 7.A). Muito semelhante a este
crescimento, foi o observado no meio MP12 com 5,84 x 107 ± 2,08 x107 cél. mL-1. O meio onde se
verificou uma concentração celular inferior foi o BG-11 com 1,14 x 107 ± 3,98 x106 cél. mL-1.
Quanto aos resultados obtidos referentes ao peso da biomassa seca (Figura 7.B), o valor
máximo obtido foi 3,00 ± 0,00 g L-1, no meio MP12, e o valor mínimo foi 0,05 ± 0,00 g L-1, no
meio BG-11. As variações de pH (Figura 7.C) nos diferentes meios foram registadas ao longo do
tempo de cultivo, sendo que não se verificaram grandes discrepâncias entre os diferentes meios.
No final, estes valores variaram entre 7,18 ± 0,10 e 7,50 ± 0,02 nos meios BG-11 e TAP,
respetivamente.
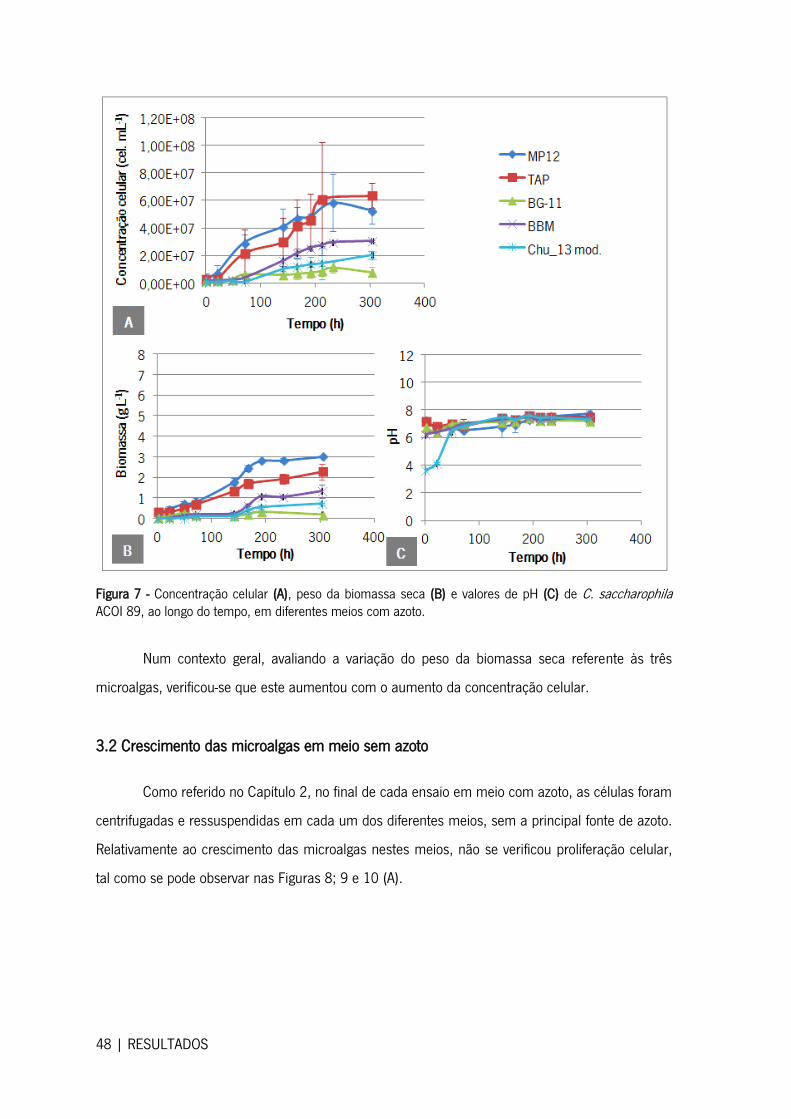
48 | RESULTADOS
Figura 7 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. saccharophila ACOI 89, ao longo do tempo, em diferentes meios com azoto.
Num contexto geral, avaliando a variação do peso da biomassa seca referente às três
microalgas, verificou-se que este aumentou com o aumento da concentração celular.
3.2 Crescimento das microalgas em meio sem azoto
Como referido no Capítulo 2, no final de cada ensaio em meio com azoto, as células foram
centrifugadas e ressuspendidas em cada um dos diferentes meios, sem a principal fonte de azoto.
Relativamente ao crescimento das microalgas nestes meios, não se verificou proliferação celular,
tal como se pode observar nas Figuras 8; 9 e 10 (A).

RESULTADOS | 49
Figura 8 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. emersonii ACOI 516, ao longo do tempo, em diferentes meios sem azoto.
Em C. emersonii (Figura 8.B), apenas se verificaram ligeiras variações do peso da
biomassa seca, em todos os meios de crescimento. Em C. vulgaris (Figura 9.B), somente no meio
MP12 se observou um aumento de cerca de 4 g L-1 e, nos restantes meios, o peso da biomassa
seca manteve-se constante. Em C. saccharophila (Figura 10.B), verificou-se um aumento de cerca
de 1 g L-1 para todos os meios, exceto para o meio BG-11, que se manteve constante e muito
próximo de 0 g L-1.
Seria expectável que a concentração celular inicial na cultura das microalgas na condição
sem azoto fosse aproximadamente igual à concentração celular final observada na condição com
azoto. No entanto, isto não se verificou, o que pode dever-se a perda de material biológico
aquando da transferência das células para os meios sem azoto.
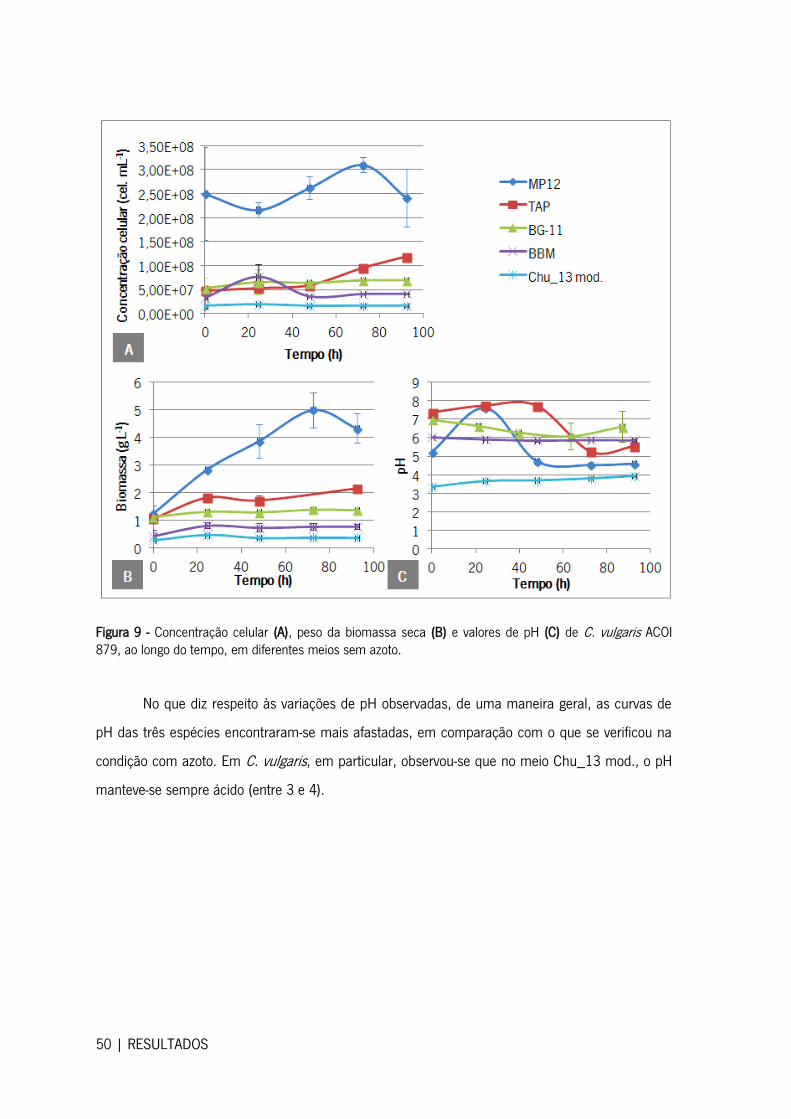
50 | RESULTADOS
Figura 9 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. vulgaris ACOI 879, ao longo do tempo, em diferentes meios sem azoto.
No que diz respeito às variações de pH observadas, de uma maneira geral, as curvas de
pH das três espécies encontraram-se mais afastadas, em comparação com o que se verificou na
condição com azoto. Em C. vulgaris, em particular, observou-se que no meio Chu_13 mod., o pH
manteve-se sempre ácido (entre 3 e 4).

RESULTADOS | 51
Figura 10 - Concentração celular (A), peso da biomassa seca (B) e valores de pH (C) de C. saccharophila ACOI 96, ao longo do tempo, em diferentes meios sem azoto.
3.3 Análise da Biomassa
Os resultados obtidos após extração de proteínas, lípidos e amido estão representados nas
Figuras 11, 12 e 13, respetivamente. Os valores correspondem à média ± desvio padrão de, pelo
menos, duas réplicas de medições. Os resultados que não apresentam desvio padrão
correspondem a apenas uma medição, sendo que não foi possível fazer mais medições por falta
de biomassa suficiente.
3.3.1 Proteínas
A análise da Figura 11.A permitiu verificar que nas condições em que foi efetuado um pré-
tratamento com NaOH, C. emersonii apresentou valores percentuais superiores, em comparação
com os valores obtidos na extração inicial, com H3PO4. Observou-se também que em meio sem

52 | RESULTADOS
azoto a percentagem de proteínas diminuiu em todos os meios estudados. Em relação a C.
vulgaris, observou-se maior percentagem de proteínas em meio sem azoto, contrariamente ao
observado em C. emersonii. Na microalga C. saccharophila, foi possível observar que a
percentagem de proteínas nos meios TAP e BBM foi superior na condição sem azoto.
Figura 11 - Percentagem de proteínas em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e C. saccharophila ACOI 96 (C), nos diferentes meios em estudo, nas condições com e sem azoto e com e sem pré-tratamento com NaOH. N – Não determinado.
3.3.2 Lípidos
O conteúdo lipídico de C. emersonii, no meio MP12, não apresentou variações relevantes
nas três condições. Nos meios TAP, BG-11 e Chu_13 mod., a percentagem de lípidos foi superior
na condição sem azoto, tendo-se verificado o caso contrário no meio BBM. Em C. vulgaris
verificou-se um aumento desta percentagem nos meios MP12, TAP e BBM, na ausência de azoto.
No entanto, no meio BG-11 a percentagem de lípidos foi superior na condição com azoto. A

RESULTADOS | 53
percentagem de lípidos nas amostras de C. saccharophila permitiu aferir que, na ausência de
azoto, este valor foi superior nos meios BBM e Chu_13 mod..
Figura 12 - Percentagem de lípidos em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e C. saccharophila ACOI 96 (C), nos diferentes meios em estudo, no inóculo e nas condições com e sem azoto. N – Não determinado.
3.3.3 Amido
O conteúdo de amido de C. emersonii aumentou na condição sem azoto, em todos os
meios de cultura. Em C. vulgaris, a percentagem de amido foi superior na presença de azoto nos
meios MP12, TAP, BG-11 e BBM. Para C. saccharophila verificaram-se percentagens superiores
na ausência de azoto nos meios MP12, TAP, BBM e Chu_13 mod..

54 | RESULTADOS
Figura 13 - Percentagem de amido em C. emersonii ACOI 516 (A), C. vulgaris ACOI 879 (B) e C. saccharophila ACOI 96 (C), nos diferentes meios em estudo, no inóculo e nas condições com e sem azoto. N – Não determinado.
3.4 Determinação das Produtividades
A Tabela 9 apresenta as produtividades de biomassa, lípidos, amido e proteínas obtidas
para as microalgas C. emersonii, C. vulgaris e C. saccharophila nos meios MP12, TAP, BG-11,
BBM e Chu_13 mod., nas condições com e sem azoto. De um modo geral, na condição com
azoto, C. vulgaris obteve produtividades de biomassa superiores, em relação às restantes
microalgas, tendo atingido uma produtividade de 0,41 ± 0,34 g L-1 d-1 no meio MP12. C. emersonii
e C. saccharophila também apresentaram produtividades de biomassa superiores neste meio de
cultura. Na condição sem azoto, o maior valor de produtividade de biomassa obtido registou-se em
C. vulgaris, 0,80 ± 0,06 g L-1 d-1, no meio MP12. Em C. emersonii, os valores da produtividade da
biomassa diminuíram da condição com azoto para a condição sem azoto em todos os meios,
exceto no BG-11. O mesmo se verificou em C. vulgaris, exceto nos meios MP12 e TAP. Em C.

RESULTADOS | 55
saccharophila, os valores da produtividade da biomassa aumentaram da condição com azoto para
a condição sem azoto, em todos os meios de cultura. Em relação aos valores de produtividade
lipídica e de amido obtidos, na condição com azoto, estes foram superiores em C. vulgaris,
comparativamente com as restantes microalgas, tendo sido 77,20 ± 0,02 mg L-1 d-1 e 106,82 ±
0,01 mg L-1 d-1 os maiores valores de produtividade lipídica e de amido obtidos, respetivamente.
Num contexto geral, a produtividade de lípidos aumentou da condição com azoto para a condição
sem azoto em C. vulgaris (exceto nos meios BG-11 e BBM) e em C. saccharophila, contrariamente
ao que se verificou para C. emersonii, cuja produtividade de lípidos diminuiu da condição com
azoto para sem azoto (exceto no meio BG-11). Em relação à produtividade de amido, verificou-se a
sua diminuição em C. vulgaris, após cultivo em meio sem azoto, ao contrário do que aconteceu
em C. saccharophila. Em C. emersonii, a produtividade de amido aumentou na condição sem
azoto, nos meios MP12 e BG-11. No que diz respeito à produtividade de proteínas, o maior valor
obtido na condição com azoto foi 56,36 ± 0,01 mg L-1 d-1 em C. emersonii, no meio MP12 e na
condição sem azoto foi 160,50 ± 0,00 mg L-1 d-1 em C. vulgaris, no mesmo meio de cultura. Em C.
vulgaris e C. saccharophila, a produtividade de proteínas aumentou da condição com azoto para a
condição sem azoto, no entanto, em C. emersonii, isso não se verificou.

56 | RESULTADOS
Tabela 9 - Produtividades obtidas (biomassa, lípidos, amido e proteínas) para as microalgas C. emersonii ACOI 516, C. vulgaris ACOI 879 e C. saccharophila ACOI 96 nos meios MP12, TAP, BG-11, BBM e Chu_13 mod. nas condições com e sem azoto.
Meio de crescimento
Produtividade Biomassa (g L-1 d-1)
Produtividade Lipídica (mg L-1 d-1)
Produtividade Amido (mg L-1 d-1)
Produtividade Proteínas (mg L-1 d-1)
Com Azoto Sem Azoto Com Azoto Sem Azoto Com Azoto Sem Azoto Com Azoto Sem Azoto
C. e
mer
soni
i
MP12 0,37 ± 0,00 0,13 ± 0,20 53,66 ± 0,01 18,24 ± 0,00 36,36 ± 0,00 46,19 ± 0,00 27,34 ± 0,00 1,81 ± 0,00
MP12 * 0,37 ± 0,00 0,13 ± 0,20 56,36 ± 0,01 15,08 ± 0,00
TAP 0,22 ± 0,01 0,06 ± 0,01 30,34 ± 0,00 11,39 ± 0,00 48,44 ± 0,01 17,83 ± 0,00 4,01 ± 0,00 0,62 ± 0,00
TAP * 0,22 ± 0,01 0,06 ± 0,01 15,69 ± 0,00 ND
BG-11 0,06 ± 0,03 0,23 ± 0,05 7,75 ± 0,00 33,76 ± 0,00 9,95 ± 0,00 97,95 ± 0,01 5,57 ± 0,00 3,97 ± 0,00
BBM 0,07 ± 0,01 0 10,76 0 15,19 ± 0,00 0 0,41 0
Chu_13 mod. 0,08 ± 0,01 0,01 ± 0,07 12,29 ± 0,00 1,80 23,60 ± 0,00 4,99 ± 0,00 0,86 0,09
C. v
ulga
ris
MP12 0,41 ± 0,34 0,80 ± 0,06 50,22 ± 0,00 99,60 ± 0,01 106,82 ± 0,01 65,49 ± 0,00 3,65 ± 0,00 160,50 ± 0,00
TAP 0,28 ± 0,00 0,28 ± 0,03 77,20 ± 0,02 109,57 ± 0,01 54,96 ± 0,01 34,48 ± 0,00 6,74 ± 0,00 72,02 ± 0,01
BG-11 0,29 ± 0,02 0,07 ± 0,02 74,09 ± 0,00 13,97 ± 0,00 60,32 ± 0,01 10,03 ± 0,00 26,24 ± 0,01 ND
BBM 0,17 ± 0,00 0,09 ± 0,08 66,77 ± 0,00 36,44 ± 0,00 26,09 ± 0,00 13,31 ± 0,00 ND 10,13
Chu_13 mod. 0,15 ± 0,00 0,02 ± 0,03 57,62 ± 0,00 ND 23,78 ± 0,00 ND ND ND
C. s
acch
arop
hila
MP12 0,22 ± 0,00 0,34 ± 0,08 28,62 ± 0,00 30,93 ± 0,00 7,51 ± 0,00 19,82 ± 0,00 47,26 ± 0,01 68,07 ± 0,00
TAP 0,15 ± 0,04 0,28 ± 0,11 29,70 ± 0,00 34,10 ± 0,00 3,00 ± 0,00 17,22 ± 0,00 29,07 ± 0,01 71,65 ± 0,01
BG-11 0,01 ± 0,00 0,04 ± 0,02 ND 5,17 0,70 ± 0,00 ND ND ND
BBM 0,10 ± 0,03 0,22 ± 0,02 23,75 ± 0,00 66,17 3,40 ± 0,00 9,97 5,81 24,51
Chu_13 mod. 0,06 ± 0,00 0,14 ± 0,02 11,37 35,07 2,09 ± 0,00 7,32 ± 0,00 ND ND
* Ensaios em que a extração de proteínas foi efetuada com pré-tratamento com NaOH.

4. CAPÍTULO IV
DISCUSSÃO DOS RESULTADOS


DISCUSSÃO DOS RESULTADOS | 59
A presente dissertação teve como objetivo monitorizar o crescimento celular das
microalgas C. emersonii, C. vulgaris e C. saccharophila e avaliar o potencial da biomassa
microalgal como matéria-prima para a produção de biocombustíveis. Para isso, as microalgas
foram cultivadas em cinco meios de cultura diferentes, e depois transferidas para os mesmos
meios, sem a sua principal fonte de azoto.
Relativamente à concentração celular das microalgas nos diferentes meios, na presença
de azoto, observou-se que C. emersonii, C. vulgaris e C. saccharophila atingiram valores
superiores nos meios MP12 e TAP; BG-11 e MP12; e TAP e MP12, respetivamente. Desta forma, é
notório que o MP12 foi um dos melhores meios de crescimento para o cultivo das três microalgas.
O azoto e o fósforo são os principais nutrientes essenciais para o crescimento das microalgas
(Dragone et al. 2010). Em cada um dos meios, existe uma fonte de azoto diferente, o que pode
ser um dos motivos diferenciadores da concentração celular obtida para cada microalga. No meio
MP12, a principal fonte de azoto é a ureia, numa concentração de 18310 µM. No meio TAP, é o
NH4Cl (7500 µM), enquanto que nos meios BBM e BG-11 a principal fonte de azoto encontra-se no
composto NaNO3, nas concentrações 2940 µM e 17600 µM, respetivamente. No meio Chu_13
mod., as microalgas consomem o azoto a partir do composto KNO3 (1980 µM). No entanto, a
ureia está descrita, na literatura, como sendo a fonte mais favorável de azoto porque, para uma
concentração de azoto equivalente, permite atingir rendimentos superiores e causa variações de
pH inferiores durante o crescimento das microalgas (Shi et al. 2000). No que diz respeito à
presença de fósforo nos diferentes meios, é também no MP12 que este se encontra em maior
concentração, estando associado ao composto KH2PO4 numa concentração de 1740 µM. Como se
pode verificar na Tabela 8, nenhum outro meio tem uma concentração em fósforo tão elevada
como a que aqui se verifica. É importante que o fósforo esteja presente numa quantidade
significativa visto que, por vezes, encontra-se ligado a iões metálicos, o que dificulta o seu
consumo na totalidade (Chisti 2007).
Ainda sobre a evolução do crescimento celular ao longo do tempo, observou-se uma
característica comum às três microalgas, no meio Chu_13 mod.. Estas mantiveram-se em fase lag
durante mais tempo, o que pode ser justificado pelo valor de pH deste meio no tempo inicial, que
se verificou inferior (≈ 4), em relação aos outros meios (> 6).
Para além disto, verificou-se o fenómeno de auto-floculação com as microalgas C.
emersonii e C. saccharophila, o que poderá ser a causa destas não terem atingido uma
concentração celular e peso da biomassa seca superiores, uma vez que, não estando em

60 | DISCUSSÃO DOS RESULTADOS
circulação permanente por todo o meio de cultura, estão privadas do acesso apropriado aos
nutrientes. A auto-floculação pode ter sido provocada por vários fatores, tais como: valores de pH
moderadamente básicos (< 8) ou básicos (≈ 10 – 11) (Dupré et al. 1995; Spilling et al. 2011);
ligações entre as células das microalgas através de matéria orgânica extracelular excretada pela
própria microalga (Tenney & Verhoff 1973); precipitação de sais contidos num meio de cultura
com pH superior (≈ 8,5 – 10,5) causado pela fotossíntese e remoção do CO2 dissolvido nas bolhas
de ar (Sukenik & Shelef 1984).
Como referido acima, no final de cada cultivo, as células foram transferidas para meios
desprovidos de azoto. Sabe-se que o crescimento celular em meio sem azoto é importante para
induzir estados de stresse metabólico nas microalgas (Sheehan et al. 1998). Nesta situação, as
células param de proliferar, mas é mantida a fixação de CO2. Desta forma, a concentração celular
das microalgas em estudo, nesta condição, evoluiu como previsto (Herrera-Valencia et al. 2011;
Widjaja et al. 2009) uma vez que não se verificou crescimento celular. Em relação ao peso da
biomassa seca, num contexto geral, verificou-se um decréscimo destes valores para as três
microalgas em estudo, o que está de acordo com o observado em estudos anteriores (Chu et al.
2013), que registaram uma diminuição destes valores, devido à interrupção do crescimento
celular. Contudo, os autores Illman et al. (2000); Fidalgo et al. (1998); Guschina & Harwood
(2006); Douskova et al. (2008) e Herrera-Valencia et al. (2011) reportaram que, nesta condição, é
expectável o aumento do peso da biomassa seca, uma vez que o estado de stresse induz a
produção de lípidos e / ou amido.
A eficiência de produção das microalgas é estimada com base nos valores de
produtividade (g L-1 d-1) do material gerado, incluindo biomassa, lípidos e hidratos de carbono (Ho
et al. 2010; Fu et al. 2012; Converti et al. 2009; Rodolfi et al. 2009; S.-H. Ho et al. 2012). Desta
forma, o cálculo das produtividades de biomassa, lípidos, amido e proteínas foi efetuado e os
resultados foram analisados. Em relação aos valores de produtividade de biomassa obtidos em C.
emersonii, estes variaram entre 0,06 g L-1 d-1 (no meio BG-11) e 0,37 g L-1 d-1 (no meio MP12), na
condição com azoto. No entanto, na condição sem azoto os valores atingidos foram inferiores e
variaram entre 0,01 ± 0,07 g L-1 d-1 (no meio Chu_13 mod.) e 0,23 ± 0,05 g L-1 d-1 (no meio BG-11).
Produtividades inferiores na condição sem azoto devem-se à interrupção do crescimento celular
nesta condição. Em condições normais de crescimento, as produtividades obtidas foram
superiores às publicadas por Illman et al. (2000), que registaram 0,03 g L-1 d-1 em C. emersonii
cultivada em condições normais de crescimento (no meio Watanabe; biorreator agitado a 200

DISCUSSÃO DOS RESULTADOS | 61
rpm; intensidade de luz de 25 µmol m-2 s-1 na superfície do biorreator, arejamento de 1 L min-1 com
5 % (v/v) CO2 e Tª 25 °C).
De notar que estes autores forneceram ao cultivo apenas ¼ da intensidade de luz utilizada
no presente trabalho. Valores de produtividade de biomassa bastante superiores foram publicados
pelos autores Takeshita et al. (2014), que obtiveram 1,00 g L-1 d-1 em C. emersonii (células
cultivadas no meio TAP, em condições normais de crescimento, com irradiação de 600 µmol m-2 s-
1 incidente na superfície do fotobiorreator; Tª 23 ºC; 20 mL min-1 de ar contendo entre 2 - 3 % de
concentração de CO2; 0,25 vvm; agitação a 2 rpm, utilizando um agitador rotativo), ou seja, um
valor de produtividade de biomassa elevado, associado a uma intensidade de luz igualmente
elevada e a uma concentração de CO2 inferior à utilizada neste trabalho.
No que diz respeito aos valores de produtividade de biomassa obtidos em C. vulgaris, na
condição com azoto, estes variaram entre 0,15 ± 0,00 g L-1 d-1 (no meio Chu_13 mod.) e 0,41 ±
0,34 g L-1 d-1 (no meio MP12). As produtividades obtidas nesta condição mostraram-se superiores a
alguns valores publicados na literatura, nomeadamente pelos autores Illman et al. (2000), que
obtiveram uma produtividade máxima de 0,04 g L-1 d-1 (condições de crescimento supracitadas). O
mesmo se verificou no trabalho realizado por Scragg et al. (2002), que também registaram 0,04 g
L-1 d-1, quando cultivaram C. vulgaris no meio Watanabe, com 6 % (v/v) CO2 (16 h de luz com
intensidade de 76 µmol m−2 s−1; Tª 25 °C; células agitadas a 60 rpm num agitador rotativo), assim,
apesar destes autores Scragg et al. (2002) terem testado a mesma concentração de CO2 que se
usou neste trabalho, a intensidade de luz utilizada foi inferior e foi aplicada durante menos tempo.
A variação da produtividade de biomassa da condição com azoto para a condição sem azoto
também foi analisada, tendo-se verificado uma diminuição destes valores, exceto nos meios MP12
e TAP. O mesmo se verificou nos estudos realizados por Illman et al. (2000), que obtiveram uma
diminuição da produtividade de 0,041 g L-1 d-1 (condição com azoto) para 0,037 g L-1 d-1 (condição
sem azoto) (condições do crescimento mencionadas anteriormente) e por Scragg et al. (2002) que
observaram uma diminuição de 0,040 g L-1 d-1 para 0,024 g L-1 d-1 (condições do cultivo
supracitadas). Esta diminuição da produtividade de biomassa na condição sem azoto é uma
consequência da diminuição do peso da biomassa seca, causada pela interrupção da proliferação
celular. Uma possível explicação para o facto da produtividade de biomassa ter aumentado para o
dobro, no meio MP12 sem azoto, e de se ter mantido no meio TAP sem azoto, foi a constância da
concentração celular e o aumento do conteúdo de lípidos e proteínas, nestes meios, embora o
peso da biomassa seca tenha diminuído cerca de 2 g L-1. Produtividades de biomassa superiores

62 | DISCUSSÃO DOS RESULTADOS
às observadas no presente trabalho, em condições normais de crescimento foram publicadas por
vários autores. No entanto, em todos os casos analisados em que isso se verificou, os autores
forneceram intensidades de luz superiores, como foi o caso de Ho et al. (2013), que obtiveram
1,36 g L-1 d-1 para C. vulgaris, cultivada sob uma intensidade de luz de 450 µmol m -2 s-1 (no meio
Basal Modificado; 2 % CO2; 0,2 vvm; Tª 28 °C; células agitadas a 300 rpm num agitador rotativo).
Os autores Přibyl et al. (2012) registaram 0,91 g L-1 d-1 de produtividade de biomassa com as
células cultivadas no meio SŠ, com 2 % CO2 (v/v), irradiação de 500 μmol m−2 s−1 e Tª 28 °C. Os
autores Takeshita et al. (2014) observaram em C. vulgaris uma produtividade de biomassa de
0,55 g L-1 d-1 (condições do cultivo supracitadas).
Em relação às produtividades de biomassa obtidas para C. saccharophila, na condição
com azoto, os valores variaram entre 0,06 ± 0,00 g L-1 d-1 (no meio Chu_13 mod.) e 0,22 ± 0,00 g
L-1 d-1 (no meio MP12). No meio TAP com azoto, obteve-se 0,15 g L-1 d-1. O mesmo valor foi
publicado pelos autores Herrera-Valencia et al. (2011) (células cultivadas no meio TAP, a 25 ± 2 º
C, sob um ciclo de luz de 16:8 h; intensidade de luz de 20 µmol m -2 s-1 cultivo agitado a 140 rpm),
no entanto, estes autores referiram que, na condição sem azoto, a produtividade aumentou
ligeiramente para 0,16 g L-1 d-1. No presente trabalho, o aumento registado foi superior, para 0,28
± 0,11 g L-1 d-1. Chinnasamy et al. (2010) obtiveram uma produtividade de 0,02 g L-1 d-1, com as
células cultivadas no meio BG-11 (meio suplementado com 5 mL de água residual; crescimento a
25 º C, sob uma intensidade de luz de 75 – 80 µmol m-2 s-1 e um fotoperíodo de 12:12 h), ou seja,
semelhante ao obtido neste trabalho, no mesmo meio (0,01 ± 0,00 g L-1 d-1).
De forma a avaliar o potencial de C. emersonii, C. vulgaris e C. saccharophila para a
produção de biocombustíveis, após crescimento celular em meio com e sem azoto, procedeu-se à
extração de lípidos, amido e proteínas.
Uma variedade de fatores ambientais pode influenciar a acumulação de lípidos nas
microalgas, nomeadamente a disponibilidade de nutrientes. Como referido no Capítulo 1, há
nutrientes presentes no meio, cuja carência pode induzir maior acumulação de lípidos, que
funcionam como energia de reserva para ser utilizada após terminar a fase de stresse (Guschina &
Harwood 2006).
Neste trabalho, verificou-se que o conteúdo de lípidos obtido em C. emersonii foi
semelhante em todos os meios, no entanto, observou-se um ligeiro aumento na condição sem
azoto, nos meios TAP, BG-11 e Chu_13 mod.. Os valores variaram de 13,9 % para 19,3 %; 12,5 %
para 14,7 % e 15,4 % para 18,0 %, respetivamente. O aumento do conteúdo de lípidos é

DISCUSSÃO DOS RESULTADOS | 63
expectável em meio sem azoto, no entanto, os valores obtidos ainda ficam aquém dos publicados
pelos autores Illman et al. (2000), que verificaram o aumento de 29 % para 63 % do conteúdo
lipídico, em C. emersonii. Para além disso, os mesmos autores obtiveram, em meio com azoto,
um conteúdo lipídico superior (29 %), em comparação com o obtido no presente trabalho, ainda
que em diferentes condições de crescimento (supracitadas). De lembrar que em cima referiu-se
que estes autores (Illman et al. 2000) obtiveram valores de produtividade de biomassa, em C.
emersonii, inferiores aos obtidos no presente trabalho, no entanto, o conteúdo lipídico foi superior,
o que poderá significar que as condições de cultivo usadas por estes poderão ser mais favoráveis
à acumulação de lípidos nas células.
A limitação de azoto pode induzir a produção de um elevado conteúdo lipídico, no entanto,
também pode diminuir a produtividade da biomassa. Como consequência, a produtividade de
lípidos também é afetada, ficando inferior ao obtido sob condições normais de crescimento
(Griffiths & Harrison 2009; Widjaja et al. 2009; Wijffels & Barbosa 2010), o que se verificou em C.
emersonii, exceto no meio BG-11. Na condição com azoto, os valores de produtividade lipídica
variaram entre 12,29 ± 0,00 mg L-1 d-1 (no meio Chu_13 mod.) e 53,66 ± 0,01 mg L-1 d-1 (no meio
MP12). No entanto, na condição sem azoto, a variação esteve entre 1,80 mg L-1 d-1 (no meio
Chu_13 mod.) e 33,76 ± 0,00 mg L-1 d-1 (no meio BG-11). Os valores obtidos em condições
normais de crescimento, em comparação com o que foi publicado pelos autores Takeshita et al.
(2014), são bastante baixos, uma vez que estes obtiveram em C. emersonii, uma produtividade
lipídica de 230 mg L-1 d-1, com crescimento no meio TAP (irradiação de 600 µmol m-2 s-1, incidente
na superfície do fotobiorreator), contudo, a intensidade de luz utilizada por estes foi 6 vezes
superior à fornecida no presente trabalho.
Em relação ao conteúdo lipídico obtido para C. vulgaris, é de notar que, para a produção
de lípidos, em condições normais de crescimento, os meios BBM e Chu_13 mod. destacaram-se
em relação aos restantes, embora o crescimento celular desta microalga tenha sido inferior
precisamente nestes dois meios, o que pode ser justificado pelo facto da concentração da principal
fonte de azoto ser mais baixa nestes do que nos restantes, indicando que concentrações inferiores
de azoto no meio de crescimento são mais favoráveis à acumulação de lípidos (Yeh & Chang
2011; Hsieh & Wu 2009; Rodolfi et al. 2009; Takagi et al. 2000). O aumento do conteúdo lipídico
da condição com azoto para a condição sem azoto, foi mais visível no meio TAP, tendo-se
verificado um aumento de 27,8 % para 39,2 %. Em estudos anteriores, verificou-se que, após
diminuir a concentração de azoto até 75 %, a percentagem de lípidos aumentou de 5,9 % para

64 | DISCUSSÃO DOS RESULTADOS
16,4 %, no meio BBM (usando como fonte de carbono o CO2 presente no ar – 300 ppm) (Converti
et al. 2009), ou seja, em comparação com o que foi obtido, neste trabalho, no meio TAP, o
aumento do conteúdo lipídico foi semelhante, ainda que, para os autores mencionados, os valores
percentuais atingidos tenham sido inferiores. No entanto, no presente trabalho, no meio BBM, o
aumento do conteúdo lipídico, na condição sem azoto foi de apenas 1 %. Illman et al. (2000)
verificaram, em C. vulgaris, que a percentagem de lípidos aumentou de 18 % (condição com azoto)
para 40 % no meio Watanabe sem azoto (condições de crescimento acima citadas), pelo que este
aumento foi superior ao maior aumento observado neste trabalho, em C. vulgaris. Conteúdos
lipídicos superiores aos obtidos no presente trabalho, em condições normais de crescimento,
foram publicados pelos autores Přibyl et al. (2012), que obtiveram uma média de 50,80 %
(condições de crescimento supracitadas) e pelos autores Chu et al. (2013), que registaram um
conteúdo lipídico de 37,60 % no meio BG-11 (células cultivadas com fotoperiodicidade 16:8 h; 0,5
vvm com 4 % CO2; Tª 25 ± 2 °C), sendo que estes últimos verificaram o aumento do conteúdo
lipídico para 50,27 %, na condição sem azoto. No meio BG-11 do presente trabalho verificou-se o
oposto, ou seja, diminuição do conteúdo lipídico na condição sem azoto. Regra geral, tem-se
verificado que os autores que utilizam uma concentração de CO2 inferior à utilizada no presente
trabalho, assim como intensidades de luz superiores, obtêm conteúdos lipídicos mais elevados.
Para além disso, o facto de não se estar a comparar com as mesmas estirpes e adicionalmente as
condições de crescimento serem diferentes, em comparação com as usadas neste trabalho, torna
expectável diferenças nos resultados obtidos.
Em relação aos valores de produtividade lipídica obtidos em C. vulgaris, em condições
normais de crescimento, estes variaram entre 50,22 ± 0,00 mg L-1 d-1 (meio MP12) e 77,20 ± 0,02
mg L-1 d-1 (meio TAP). Produtividades lipídicas superiores às obtidas neste trabalho foram
publicadas por Přibyl et al. (2012), que obtiveram em C. vulgaris uma média de 460 mg L-1 d-1
(condições do cultivo acima mencionadas). Os autores Chu et al. (2013) obtiveram em C. vulgaris
uma produtividade lipídica de 43,17 mg L-1 d-1, no meio BG-11, em condições normais de
crescimento (condições de crescimento supracitadas), tendo este valor aumentado para 58,39 mg
L-1 d-1, na condição sem azoto. Neste trabalho, a produtividade lipídica de C. vulgaris no meio BG-11
variou de 74,09 ± 0,00 mg L-1 d-1 para 13,97 ± 0,00 mg L-1 d-1, ou seja, foi superior na condição
com azoto, em relação aos mesmos autores, no entanto, diminuiu bastante na condição sem
azoto. Esta queda da produtividade lipídica na condição sem azoto foi uma consequência da
diminuição da produtividade de biomassa, no mesmo meio, assim como do conteúdo lipídico.

DISCUSSÃO DOS RESULTADOS | 65
Estudos anteriores (Widjaja 2010) demonstraram que expondo C. vulgaris a condições de
privação de azoto, são obtidas produtividades lipídicas inferiores, causadas pela diminuição do
crescimento celular, nesta situação. No entanto, estes também verificaram que após exposição a
períodos superiores de limitação de azoto (17 dias), é possível obter não só um conteúdo lipídico
superior como também o aumento da produtividade lipídica para valores semelhantes e até
superiores aos obtidos no final do cultivo sob condições normais. Pelo que, produtividades lipídicas
superiores poderão ser obtidas variando não só a duração do período de privação de azoto, assim
como, do período de cultivo normal (Widjaja 2010; Yeh & Chang 2011).
No que diz respeito ao conteúdo lipídico obtido em C. saccharophila, os valores
percentuais aumentaram da condição com azoto para a condição sem azoto nos meios BBM e
Chu_13 mod., de 22,7 % para 30,5 % e de 19,9 % para 25,9 %, respetivamente, sendo que
também foi nestes meios que se observou o maior conteúdo lipídico. Griffiths & Harrison (2009)
reportaram uma percentagem lipídica média de 23,0 % para algas verdes (Chlorophyta). Assim, as
percentagens obtidas para esta microalga encontram-se próximas desta média. Contudo, o mesmo
não se verificou em meio sem azoto, uma vez que a média referida pelos mesmos autores é 41,0
%. Herrera-Valencia et al. (2011) verificaram na condição com azoto, do meio TAP, uma
percentagem lipídica de 50 %. Na ausência de NH4Cl, observaram o aumento desta percentagem
para 63,6 % (condições de crescimento mencionadas anteriormente). Outros estudos também
obtiveram valores percentuais de lípidos superiores em C. saccharophila, em relação ao que foi
obtido no presente trabalho, como é o caso dos autores Hempel et al. (2012), que obtiveram um
conteúdo lipídico de 27,6 % (células cultivadas no meio Tamiya modificado, com 2 % (v/v) CO2;
irradiação de 200 µmol fotões m-2 s-1). Em condições heterotróficas de crescimento, os autores Tan
& Johns (1991), também registaram um conteúdo lipídico superior – 46,7 %. No presente
trabalho, não foi possível fazer extração de lípidos do meio BG-11 na condição com azoto, por falta
de biomassa suficiente, no entanto, na literatura, vem descrito que, neste meio (suplementado
com solo), C. saccharophila apresenta um conteúdo lipídico aproximado de 12,9 % (células
cultivadas a 25 °C sob uma intensidade de luz a variar entre 75 – 80 µmol fotões m-2 s-1 e com
fotoperiodicidade 12:12 h) (Chinnasamy et al. 2010).
No que diz respeito aos valores de produtividade lipídica obtidos com esta microalga,
verificou-se que, na condição com azoto, variaram entre 11,37 mg L-1 d-1 (no meio Chu_13 mod.) e
29,70 ± 0,00 mg L-1 d-1 (no meio TAP). Estes valores encontram-se abaixo da média reportada para
microalgas cultivadas sob condições normais de crescimento – 50 mg L-1 d-1 (Griffiths & Harrison

66 | DISCUSSÃO DOS RESULTADOS
2009). Na condição sem azoto, do presente trabalho, as produtividades variaram entre 5,17 mg L-1
d-1 (no meio BG-11) e 66,17 mg L-1 d-1 (no meio BBM). Estes valores também se encontram abaixo
do que publicaram os autores Herrera-Valencia et al. (2011), uma vez que estes registaram em C.
saccharophila uma produtividade lipídica de 99,33 mg L-1 d-1 quando as células foram cultivadas
em meio sem azoto (condições do cultivo acima mencionadas). No entanto, neste trabalho, em C.
saccharophila, verificou-se em todos os meios de crescimento, o aumento da produtividade
lipídica, na condição sem azoto, o que pode ser o resultado da síntese de novo lipid synthesis,
uma característica de algumas microalgas oleaginosas, que consiste em direcionar o excesso de
carbono e energia em lípidos de armazenamento (principalmente TAG), quando cultivadas em
condições de carência de azoto ou sob uma tensão que limita o crescimento (Rodolfi et al. 2009).
Várias microalgas do género Chlorella estão descritas como sendo capazes de produzir
níveis mais elevados de amido do que de lípidos, como polímeros de reserva (Fernandes et al.
2012). Sabe-se que a acumulação de amido pelas microalgas é dependente da estirpe e varia de
acordo com o meio e com as condições da luz (Takeshita et al. 2014; Liu et al. 2008; Dragone et
al. 2011). Várias espécies foram descritas como sendo ideais para a produção de bioetanol, por
produzirem grandes quantidades de amido sob privação de azoto (Mussatto et al. 2010).
No presente trabalho, extraiu-se o amido presente nas microalgas, nas condições com e
sem azoto. Os resultados obtidos em C. emersonii mostraram-se bastante satisfatórios, uma vez
que se verificou em todos os meios o aumento da percentagem de amido na condição sem azoto,
o que de acordo com a literatura é expectável (Douskova et al. 2008; Converti et al. 2009; Lv et al.
2010; Přibyl et al. 2012; Widjaja et al. 2009; Behrens et al. 1989). Para além disso, nos meios
MP12, BG-11 e Chu_13 mod., observou-se um aumento de 9,9 % (com azoto) para 35,2 % (sem
azoto); de 16,1 % para 42,7 % e de 29,5 % para 38,0 %, respetivamente. Assim, o maior conteúdo
de amido obtido é semelhante, ainda que ligeiramente superior, à percentagem máxima registada
pelos autores Dragone et al. (2011), que obtiveram um conteúdo de amido de 41,0 % (na
condição sem azoto) (C. vulgaris cultivada com ar enriquecido com 2 % (v/v) CO2, taxa de
arejamento de 0,83 vvm; irradiação de 70 µmol m-2 s-1 de luz).
Em relação aos valores das produtividades de amido obtidas em C. emersonii, na
condição com azoto, estes variaram entre 9,95 ± 0,00 mg L-1 d-1 (no meio BG-11) e 48,44 ± 0,01
mg L-1 d-1 (no meio TAP). Estes valores encontraram-se bastante abaixo do publicado na literatura
pelos autores Takeshita et al. (2014), que obtiveram uma produtividade de amido de 180 mg L -1 d-
1, em C. emersonii (condições do crescimento referidas anteriormente). No entanto, de notar que

DISCUSSÃO DOS RESULTADOS | 67
os autores citados forneceram uma intensidade de luz 6 vezes superior à utilizada neste trabalho e
intensidades de luz superiores induzem a produção de polissacáridos em microalgas (Friedman et
al. 1991; Tredici et al. 1991).
Em relação ao conteúdo de amido obtido em C. vulgaris, verificou-se maior percentagem
deste na condição com azoto, em todos os meios. Nesta condição, os valores obtidos são
semelhantes aos reportados pelos autores Cheng et al. (2015), que verificaram nesta microalga
uma percentagem de amido de 21,0 %, numa cultura com 2 % (v/v) CO2. Um conteúdo de amido
superior foi publicado pelos autores Ho et al. (2013), que obtiveram 31,25 % (C. vulgaris cultivada
sob uma intensidade de luz elevada, 450 µmol m -2 s-1). No que diz respeito à variação do conteúdo
de amido da condição com azoto para a condição sem azoto, os estudos encontrados na literatura
apresentam resultados que contrariam os obtidos no presente trabalho. Douskova et al. (2008)
induziram uma acumulação de amido na microalga C. vulgaris (cultivada no meio MP12, numa
concentração de CO2 > 10 %) pela privação de azoto e o teor em amido nas células aumentou 50
%. Os autores Dragone et al. (2009) verificaram que o conteúdo de amido em C. vulgaris (cultivada
no meio MP12, nas mesmas condições utilizadas no presente trabalho) sob condições de privação
de azoto, constituiu 36 % da biomassa algal, enquanto que, sob condições normais, esta
percentagem foi apenas 8 %, o que enfatiza o facto da privação de azoto ser um fator chave para a
acumulação de amido pelas microalgas. De notar que estes autores, não determinaram o
conteúdo de amido pelo mesmo método utilizado no presente trabalho. Ainda assim, tendo em
conta que os resultados obtidos, em C. vulgaris, não foram de encontro à literatura, tudo indica
que alguns dos fatores que podem influenciar a produção de amido pelas células, se possam ter
alterado, nomeadamente a intensidade da luz, a concentração de CO2, a temperatura e / ou a
concentração de ferro (aumenta a eficiência fotossintética) (Van Oijen et al. 2005; Warr et al.
1985) ou até a possibilidade de existência de erros no procedimento de extração de amido das
células.
Em relação aos valores de produtividade de amido obtidos com esta microalga, na
condição com azoto, estes variaram entre 23,78 ± 0,00 mg L-1 d-1 (no meio Chu_13 mod.) e
106,82 ± 0,01 mg L-1 d-1 (no meio MP12). Uma produtividade superior foi obtida pelos autores
Dragone et al. (2011), que registaram em C. vulgaris, 199 mg L-1 d-1 (condições de crescimento
referidas anteriormente), que pode estar relacionado com o facto destes autores terem usado uma
taxa de arejamento superior e uma concentração de CO2 inferior, em relação ao que se utilizou
neste trabalho. No entanto, um valor próximo dos que foram obtidos com os meios TAP e BG-11

68 | DISCUSSÃO DOS RESULTADOS
foi reportado pelos autores Takeshita et al. (2014), que obtiveram 69 mg L-1 d-1 (condições do
cultivo supracitadas).
No que diz respeito ao conteúdo de amido obtido em C. saccharophila, verificou-se que os
valores obtidos foram inferiores aos observados nas outras microalgas. Nos meios em que foi
possível analisar o conteúdo de amido, na condição sem azoto, verificou-se que estes valores
foram ligeiramente maiores, em comparação com os observados na condição com azoto. Como
demonstram os estudos acima citados, é possível obter conteúdos de amido superiores (para as
microalgas, na generalidade) às observadas em C. saccharophila.
Uma troca entre a acumulação de lípidos e de amido tem sido observada em algumas
espécies de Chlorella (Mizuno et al. 2013). Os autores Ho et al. (2013) verificaram o aumento da
concentração de carbohidratos em condições de privação de azoto. No entanto, o conteúdo lipídico
não variou significativamente durante este período. Alguns estudos demonstraram que o fluxo de
carbono nas microalgas está alocado em compostos ricos em energia, e há uma competição entre
a síntese de lípidos e de amido (Rismani-Yazdi et al. 2011; Siaut et al. 2011). No entanto, o
aumento da síntese de lípidos ou de carbohidratos sob condições de stresse difere entre espécies
(S. H. Ho et al. 2012). No presente trabalho, em C. emersonii foi predominante a produção de
amido, contudo, em C. vulgaris e C. saccharophila, predominou a produção de lípidos.
O elevado conteúdo de proteínas presente em várias espécies de microalgas é uma das
principais razões para considerar estes microrganismos fontes convencionais de proteínas (Soletto
et al. 2005). Como as células são capazes de sintetizar todos os aminoácidos, podem fornecer os
essenciais para os seres humanos e os animais (Guil-Guerrero et al. 2004), desta forma Chlorella
é reconhecida como ingrediente alimentar seguro a nível mundial (Guccione et al. 2014; Spolaore
et al. 2006; Cornet 1998; Soletto et al. 2005). Vários estudos já demonstraram que a alteração na
concentração de nutrientes pode modificar o crescimento e o metabolismo secundário das
microalgas. Como referenciado acima, condições de limitação de azoto induzem a acumulação de
lípidos e / ou amido, o que não se verifica no caso das proteínas, sendo que, nestas condições,
ocorre uma diminuição da sua concentração nas células (Biller et al. 2011; Lv et al. 2010; da Silva
et al. 2009; Pruvost et al. 2011).
No que diz respeito ao procedimento de extração de proteínas efetuado no presente
trabalho, é importante lembrar que foram testadas duas condições diferentes em C. emersonii. Na
primeira condição, as proteínas foram extraídas com o auxílio de uma solução de H3PO4 (0,15 M),
no entanto, após observação dos resultados obtidos, verificou-se que este procedimento não

DISCUSSÃO DOS RESULTADOS | 69
deveria ser adequado, uma vez que estavam a obter-se percentagens de proteínas muito baixas,
comparativamente com o que vem descrito na literatura (Dragone et al. 2010). Desta forma,
testou-se uma segunda condição que implicou a extração das proteínas com NaOH (1 M). Após
testar a segunda condição, os resultados mostraram-se mais apropriados (ainda assim, aquém do
que se esperaria), pelo que, para a extração das proteínas das microalgas C. vulgaris e C.
saccharophila, apenas se utilizou a segunda condição. No entanto, do ponto de vista prático, o
segundo procedimento apresentou uma desvantagem em relação ao primeiro. Após adicionar a
solução de NaOH à biomassa liofilizada, levou-se a mistura a um banho de 80 °C e, no final,
terminou-se a reação com gelo. Este último passo provocou a solidificação da solução de NaOH
com as amostras de microalgas, pelo que foi difícil retirar volume de sobrenadante suficiente para
a determinação das proteínas. Esta desvantagem pode estar na origem dos resultados obtidos
para as três microalgas em estudo. C. vulgaris apresentou um conteúdo de proteínas superior na
condição sem azoto, o que contradiz os resultados publicados por Ho et al. (2012) e Siaut et al.
(2011) que demonstraram que a privação de azoto diminui o conteúdo de proteínas das células,
uma vez que força estes microrganismos a transformar as proteínas em lípidos ou carbohidratos.
O conteúdo de proteínas médio publicado na literatura para C. vulgaris, em condições normais de
crescimento, encontra-se entre 41 % e 58 % (Dragone et al. 2010), desta forma verificou-se que os
resultados obtidos com esta microalga ficaram abaixo do expectável.
No que diz respeito aos valores de produtividade de proteínas obtidos em C. vulgaris,
variaram entre 3,65 ± 0,00 mg L-1 d-1 (no meio MP12) e 26,24 ± 0,01 mg L-1 d-1 (no meio BG-11).
Na literatura encontraram-se valores bastante superiores a estes, nomeadamente os que foram
publicados pelos autores Chen et al. (2015), que obtiveram em C. vulgaris uma produtividade de
proteínas de 146 mg L-1 d-1 (cultivada no meio Basal Modificado, com uma intensidade de luz de
150 µmol m-2 s-1; concentração inicial de ureia de 9,3 mM e de Fe2+ de 180 µM) e por Mahboob et
al. (2012), que registaram uma produtividade de proteínas de 2260 mg L -1 d-1 com uma espécie de
C. vulgaris termotolerante cultivada no meio N-8 otimizado. Uma produtividade inferior e
semelhante à que foi obtida, neste trabalho, na condição com azoto do meio BG-11, foi publicada
pelos autores Wang et al. (2014), que obtiveram com outra espécie de Chlorella (ellipsoidea) uma
produtividade de proteínas de 21,67 ± 2,24 mg L-1 d-1 (com células cultivadas no meio BG-11, 16 h
luz por dia e irradiação de 35 µmol m_2 s_1).
Em relação ao conteúdo de proteínas obtido em C. saccharophila, verificou-se que, nos
meios MP12 e TAP, na condição com azoto, os valores foram superiores aos obtidos em C.

70 | DISCUSSÃO DOS RESULTADOS
emersonii e C. vulgaris. No entanto, também ficaram aquém dos resultados publicados na
literatura, nomeadamente pelos autores Hempel et al. (2012) (condições de crescimento acima
citadas), que reportaram para a microalga C. saccharophila um conteúdo de proteínas de 42,4 % e
para Chlorella sp. 44,3 %. Guccione et al. (2014) demonstraram que o conteúdo de proteínas em
nove estirpes de Chlorella em meio com azoto foi, em média, 39,7 %. Após um período de
limitação de azoto, a percentagem de proteínas registada foi 24,6 % (valor médio). Illman et al.
(2000) obtiveram em C. vulgaris uma percentagem de proteínas de 29,0 %, na condição com
azoto, tendo este valor diminuído até 7,0 % na condição sem azoto. Em C. emersonii, os mesmos
autores obtiveram 32,0 % e 28,0 % nas condições com e sem azoto, respetivamente (condições do
cultivo supracitadas). Ho et al. (2013) verificaram uma diminuição no conteúdo de proteínas de
58,8 % para 20,1 %, quando colocaram C. vulgaris em meio com carência de azoto. Uma razão
possível para justificar que o conteúdo de proteínas diminua na condição sem azoto é o facto de
todas as estruturas de carbono produzidas durante o processo metabólico serem direcionadas
para a produção de carbohidratos e lípidos, enquanto que, na presença de azoto, a maior parte
das estruturas de carbono são incorporadas em compostos azotados (como aminoácidos,
proteínas e ácidos nucleicos) (Dragone et al. 2011; El-Moneim M. R. Afify et al. 2010; Widjaja
2010).

5. CAPÍTULO V
CONSIDERAÇÕES FINAIS E PERSPETIVAS FUTURAS


CONSIDERAÇÕES FINAIS E PERSPETIVAS FUTURAS | 73
A capacidade de produção atual de biocombustíveis provenientes de microalgas ainda é
inferior ao necessário para o estabelecimento de uma sociedade mais sustentável. Desta forma,
este trabalho surge na tentativa de acrescentar informação nesta área, com a seleção de
microalgas com potencial de produção de biocombustíveis.
A elaboração da presente dissertação permitiu concluir que para as condições avaliadas, o
meio MP12 demonstrou ser um dos melhores para o crescimento das microalgas estudadas, pelo
facto de apresentar concentrações de azoto e fósforo superiores, em relação aos outros meios de
crescimento. Concluiu-se também que C. vulgaris foi a microalga que obteve valores de
concentração celular e peso da biomassa seca superiores, tendo, desta forma atingido o maior
valor de produtividade de biomassa, no meio MP12.
Analisando os conteúdos lipídicos obtidos, concluiu-se que, de uma maneira geral, C.
vulgaris apresentou percentagens superiores (inclusive 39,2 % na condição sem azoto do meio
TAP). O facto de ter sido a microalga que atingiu maior peso da biomassa seca, juntamente com o
facto de ter produzido o maior conteúdo lipídico, refletiu-se na maior produtividade lipídica obtida,
pelo que revelou ter o maior potencial de produção de lípidos. No entanto, para poder afirmar que
tem potencial de produção de biodiesel, por exemplo, seria necessário dar continuidade ao
presente trabalho no sentido de obter a caracterização dos lípidos, uma vez que espécies
diferentes de microalgas produzem diferentes tipos de ácidos gordos e alguns ácidos gordos são
mais adequados para a transesterificação em biodiesel do que outros, nomeadamente os TAG e os
AGL.
O conteúdo de amido obtido em C. emersonii faria crer que esta seria a melhor microalga
para a produção de amido, no entanto, a análise dos valores de produtividade de amido obtidos
demonstraram que C. vulgaris é mais adequada porque, embora tenha produzido, em média,
menos amido, o facto de ter atingido uma concentração de células superior, tornou esta espécie
mais rentável.
Os resultados obtidos referentes ao conteúdo de proteínas não permitiram tirar conclusões
em relação à capacidade de produção das microalgas em estudo.
Estudos experimentais adicionais serão necessários para determinar com melhor precisão
as condições de crescimento ideais, por exemplo, através da alteração de condições ambientais
(pH, temperatura, nível de nutrientes, percentagem de CO2, intensidade da luz e arejamento).
Idealmente, as microalgas deveriam ser colhidas num ponto de elevada concentração de
biomassa e elevada produtividade de lípidos e/ou amido, no entanto, sabe-se que as microalgas

74 | CONSIDERAÇÕES FINAIS E PERSPETIVAS FUTURAS
aumentam o conteúdo de lípidos e/ou amido sob privação de nutrientes e, nestas condições,
estes microrganismos produzem menos biomassa. Desta forma, é importante encontrar um
equilíbrio entre este dilema, através da otimização da concentração de azoto no meio de
crescimento.
De forma a compreender melhor a relação entre o crescimento celular e a acumulação de
lípidos seria útil medir a clorofila presente nas células durante o processo de cultivo, uma vez que
esta é um componente essencial das algas verdes, responsável por capturar CO2 e energia solar
para gerar um fluxo metabólico tanto para o crescimento celular como para a acumulação de
lípidos. Para além disso, e tendo em conta que o cultivo de microalgas em condições mixotróficas
tem sido considerado um processo muito eficiente para a produção de biomassa microalgal,
comparativamente com as condições fotoautotróficas, seria também interessante cultivar as
microalgas estudadas nessas condições.
Um rápido progresso no campo de produção de biocombustíveis a partir de microalgas
tem sido feito a todos os níveis, desde a identificação de espécies de alto rendimento à melhoria
das técnicas de colheita e extração. No entanto, a utilização industrial de microalgas para a
produção de biocombustíveis poderá ser rentabilizada através da utilização de co-produtos de alto
valor produzidos pelas mesmas, para outros fins. Adicionalmente, utilizar espécies de microalgas
de rápido crescimento para a fixação de CO2 é uma alternativa muito promissora para a mitigação
de CO2. O mérito principal desta estratégia reside no facto de que, através do cultivo de
microalgas, a mitigação de CO2 e a produção de biocombustíveis poderiam ser combinados de
uma forma economicamente viável e ambientalmente sustentável. A viabilidade desta estratégia
pode ser reforçada através da fixação de CO2 dos gases industriais e pela integração do cultivo de
microalgas no tratamento de águas residuais. Seria também importante identificar espécies
capazes de manter o cultivo estável, sob uma intensidade de luz elevada, utilizando
fotobiorreatores ao ar livre, uma vez que a luz solar exterior pode variar entre 100 µmol fotões m_2
s_1 (em tempo nublado) e mais de 1000 µmol fotões m_2 s_1 em clima moderado. Pode ainda atingir
um máximo de mais de 2000 µmol fotões m_2 s_1 na luz direta do sol do solstício de verão.

6. BIBLIOGRAFIA


BIBLIOGRAFIA | 77
Abbas, T. et al., 2014. Greener energy: Issues and challenges for Pakistan-geothermal energy prospective. Renewable and Sustainable Energy Reviews, 31, pp.258–269.
Ahmad, A.L. et al., 2011. Microalgae as a sustainable energy source for biodiesel production: A review. Renewable and Sustainable Energy Reviews, 15, pp.584–593.
Amin, S., 2009. Review on biofuel oil and gas production processes from microalgae. Energy Conversion and Management, 50(7), pp.1834–1840.
Antoni, D., Zverlov, V. V. & Schwarz, W.H., 2007. Biofuels from microbes. Applied Microbiology and Biotechnology, 77(1), pp.23–35.
Arora, R., 2012. Microbial Biotechnology: Energy and Environment, CABI.
Bai, F.W., Anderson, W.A. & Moo-Young, M., 2008. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnology Advances, 26, pp.89–105.
Balat, M. & Balat, H., 2009. Recent trends in global production and utilization of bio-ethanol fuel. Applied Energy, 86(11), pp.2273–2282.
Barsanti, L. & Gualtieri, P., 2006. Algae anatomy biochemistry and biotechnology, London UK: Taylor & Francis Group.
Behrens, P.W. et al., 1989. Studies on the incorporation of CO2 into starch by Chlorella vulgaris. Journal of Applied Phycology, 1(2), pp.123–130.
Benemann, J.R., 2000. Hydrogen production by microalgae. Journal of Applied Phycology, 12, pp.291–300.
Benemann, J.R. & Oswald, W.J., 1996. Systems and Economic Analysis of Microalgae Ponds for Conversion of CO2 to Biomass. Final report to the Department of Energy, Pittsburgh Energy Technology Center, pp.DOE/PC/93204–T5.
Berndes, G., Hoogwijk, M. & Van Den Broek, R., 2003. The contribution of biomass in the future global energy supply: A review of 17 studies. Biomass and Bioenergy, 25, pp.1–28.
Biller, P., Riley, R. & Ross, A.B., 2011. Catalytic hydrothermal processing of microalgae: Decomposition and upgrading of lipids. Bioresource Technology, 102(7), pp.4841–4848.
Boney, A.D., 1989. Phytoplankton: new studies in biology 2nd ed., London: Edward Arnold.
Borowitzka, M., 2005. Culturing microalgae in outdoor ponds. In A. Richmond & Q. Hu, eds. Algal Culturing Techniques. London: Elsevier Academic Press, pp. 205–218.
BP, 2014. BP statistical review of world energy.

78 | BIBLIOGRAFIA
Brennan, L. & Owende, P., 2010. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renewable and Sustainable Energy Reviews, 14(2), pp.557–577.
Bridgwater, A. V., Meier, D. & Radlein, D., 1999. An overview of fast pyrolysis of biomass. Organic Geochemistry, 30(12), pp.1479–1493.
Bridgwater, A., 2003. Renewable fuels and chemicals by thermal processing of biomass. Chemical Engineering Journal, 91(2-3), pp.87–102.
Bunting, B. et al., 2010. Fungible and compatible biofuels: literature search, summary, and recommendations, pp.112.
C. Ford, R. & Senauer, B., 2007. How Biofuels Could Starve the Poor. Foreign Affairs.
Campbell, 2008. Biodiesel: Algae as a Renewable Source for Liquid Fuel. Guelph Engineering Journal, (1).
Carlsson, A. et al., 2007. Micro- and Macro-Algae: Utility for Industrial Applications A. S. Carlsson, ed., CPL Press.
Chen, C.-Y. et al., 2015. Improving protein production of indigenous microalga Chlorella vulgaris FSP-E by photobioreactor design and cultivation strategies. Biotechnology Journal, 10(6), pp.905–914.
Cheng, Y.-S., Labavitch, J.M. & VanderGheynst, J.S., 2015. Elevated CO2 concentration impacts cell wall polysaccharide composition of green microalgae of the genus Chlorella. Letters in Applied Microbiology, 60(1), pp.1–7.
Cheryl, 2008. Algae becoming the new biofuel of choice.
Chiaramonti, D., Oasmaa, A. & Solantausta, Y., 2007. Power generation using fast pyrolysis liquids from biomass. Renewable and Sustainable Energy Reviews, 11(6), pp.1056–1086.
Chinnasamy, S. et al., 2010. Microalgae cultivation in a wastewater dominated by carpet mill effluents for biofuel applications. Bioresource Technology, 101(9), pp.3097–3105.
Chisti, Y., 2007. Biodiesel from microalgae. Biotechnology Advances, 25(3), pp.294–306.
Chiu, S.Y. et al., 2008. Reduction of CO2 by a high-density culture of Chlorella sp. in a semicontinuous photobioreactor. Bioresource Technology, 99, pp.3389–3396.
Chu, F.F. et al., 2013. Phosphorus plays an important role in enhancing biodiesel productivity of Chlorella vulgaris under nitrogen deficiency. Bioresource Technology, 134, pp.341–346.
Converti, A. et al., 2009. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chemical Engineering and Processing: Process Intensification, 48(6), pp.1146–1151.

BIBLIOGRAFIA | 79
Cornet, J.F., 1998. Le technoscope: les photobioréacteurs. Biofutur, 176, pp.1–10.
Croft, M.T., Warren, M.J. & Smith, A.G., 2006. Algae need their vitamins. Eukaryotic Cell, 5, pp.1175–1183.
Czernik, S. & Bridgwater, A. V., 2004. Overview of applications of biomass fast pyrolysis oil. Energy and Fuels, 18(2), pp.590–598.
Das, D. & Veziroǧlu, T.N., 2001. Hydrogen production by biological processes: A survey of literature. International Journal of Hydrogen Energy, 26, pp.13–28.
Dasgupta, C.N. et al., 2010. Recent trends on the development of photobiological processes and photobioreactors for the improvement of hydrogen production. International Journal of Hydrogen Energy, 35, pp.10218–10238.
Demirbas, A., 2005. Bio-ethanol from cellulosic materials: a renewable motor fuel from biomass. Energy Source, 27, pp.327–37.
Demirbas, A., 2008. Biofuels sources, biofuel policy, biofuel economy and global biofuel projections. Energy Conversion and Management, 49, pp.2106–2116.
Demirbaş, A., 2006. Oily Products from Mosses and Algae via Pyrolysis. Energy Sources, Part A: Recovery, Utilization, and Environmental Effects, 28(10), pp.933–940.
Demirbas, M.F., Balat, M. & Balat, H., 2009. Potential contribution of biomass to the sustainable energy development. Energy Conversion and Management, 50(7), pp.1746–1760.
Dimitrov, K., 2007. GreenFuel Technologies: A Case Study for Industrial Photosynthetic Energy Capture.
Doucha, J. & Lívanský, K., 2009. Outdoor open thin-layer microalgal photobioreactor: Potential productivity. Journal of Applied Phycology, 21(1), pp.111–117.
Douskova, I. et al., 2008. Microalgae As a Means for Converting Flue Gas CO2 Into Biomass With a High Content of Starch. International Conference and Exhibition on Bioenergy, Universidade do Minho, Guimarães, Portugal.
Doušková, I. et al., 2010. Utilization of distillery stillage for energy generation and concurrent production of valuable microalgal biomass in the sequence: Biogas-cogeneration-microalgae-products. Energy Conversion and Management, 51, pp.606–611.
Dragone, G. et al., 2009. Effect of nitrogen limitation on starch accumulation in Chlorella vulgaris. Book of abstracts of MicroBiotec09 Meeting, 3, Vilamoura. Universidade do Minho Departamento de Engenharia Biológica, pp.157
Dragone, G. et al., 2011. Nutrient limitation as a strategy for increasing starch accumulation in microalgae. Applied Energy, 88(10), pp.3331–3335.

80 | BIBLIOGRAFIA
Dragone, G. et al., 2010. Third generation biofuels from microalgae. Applied Microbiology and Microbial Biotechnology, pp.1355–1366.
Du, W. et al., 2008. Perspectives for biotechnological production of biodiesel and impacts. Applied Microbiology and Biotechnology, 79(3), pp.331–337.
Dupré, C., Guary, J.C. & Grizeau, D., 1995. Culture of an autoflocculent microalga in a vertical tubular photobioreactor for phycoerythrin production. Biotechnology Techniques, 9(3), pp.185–190.
EIA-DOE, 2014. Energy Information Administration (EIA) – US Department of Energy (DOE), Washington DC, USA.
Elmaleh, S. et al., 1991. Magnesium Induced Algal Flocculation in a Fluidized Bed. Water Science & Technology, 23(7-9), pp.1695–1702.
El-Moneim M. R. Afify, A., Shalaby, E.A. & Shanab, S.M.M., 2010. Enhancement of biodiesel production from different species of algae. Grasas y Aceites, 61(4), pp.416–422.
Eriksen, N.T., 2008. The technology of microalgal culturing. Biotechnology Letters, 30(9), pp.1525–1536.
Escobar, J.C. et al., 2009. Biofuels: Environment, technology and food security. Renewable and Sustainable Energy Reviews, 13, pp.1275–1287.
Eshaq, F.S., Ali, M.N. & Mohd, M.K., 2010. Spirogyra biomass a renewable source for biofuel (bioethanol) production. International Journal of Engineering Science and Technology, 2(12), pp.7045–7054.
Fernandes, B. et al., 2012. Starch determination in Chlorella vulgaris - a comparison between acid and enzymatic methods. Journal of Applied Phycology, 24(5), pp.1203–1208.
Ferreira, W.F.C. & Sousa, J.C.F., 1998. Microbiologia – Volume 1, Lisboa: Lidel.
Fidalgo, J.P. et al., 1998. Effects of nitrogen source and growth phase on proximate biochemical composition, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture, 166(1-2), pp.105–116.
Folch, J., Lees, M. & Sloane Stanley, G.H., 1957. A simple method for the isolation and purification of total lipides from animal tissues. The Journal of biological chemistry, 226(1), pp.497–509.
Friedman, O., Dubinsky, Z. & Arad, S. (Malis), 1991. Effect of light intensity on growth and polysaccharide production in red and blue-green Rhodophyta unicells. Bioresource Technology, 38(2-3), pp.105–110.
Fu, W. et al., 2012. Maximizing biomass productivity and cell density of Chlorella vulgaris by using light - emitting diode - based photobioreactor. Journal of Biotechnology, 161(3), pp.242–249.

BIBLIOGRAFIA | 81
Fukuda, H., Kondo, A. & Noda, H., 2001. Biodiesel fuel production by transesterification of oils. Journal of Bioscience and Bioengineering, 92, pp.405–416.
Goh, C.S. & Lee, K.T., 2010. A visionary and conceptual macroalgae-based third-generation bioethanol (TGB) biorefinery in Sabah, Malaysia as an underlay for renewable and sustainable development. Renewable and Sustainable Energy Reviews, 14(2), pp.842–848.
Golueke, C.G., Oswald, W.J. & Gotaas, H.B., 1957. Anaerobic digestion of Algae. Applied microbiology, 5, pp.47–55.
González-Fernández, C. et al., 2012. Impact of microalgae characteristics on their conversion to biofuel. Part II: Focus on biomethane production. Biofuels, Bioproducts and Biorefining, 6, pp.205–218.
Griffiths, M.J. & Harrison, S.T.L., 2009. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology, 21, pp.493–507.
Grobbelaar, J.U., 2007. Algal Nutrition ‒ Mineral Nutrition. In Handbook of Microalgal Culture. pp.
95–115.
Guccione, A. et al., 2014. Chlorella for protein and biofuels: from strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnology for biofuels, 7(1), p.84.
Guil-Guerrero, J.L. et al., 2004. Functional properties of the biomass of three microalgal species. Journal of Food Engineering, 65(4), pp.511–517.
Guschina, I.A. & Harwood, J.L., 2006. Lipids and lipid metabolism in eukaryotic algae. Progress in Lipid Research, 45(2), pp.160–186.
Gutiérrez, R., Gutiérrez-Sánchez, R. & Nafidi, A., 2008. Trend analysis using nonhomogeneous stochastic diffusion processes. Emission of CO2; Kyoto protocol in Spain. Stochastic Environmental Research and Risk Assessment, 22, pp.57–66.
Halim, R., Danquah, M.K. & Webley, P.A., 2012. Extraction of oil from microalgae for biodiesel production: A review. Biotechnology Advances, 30, pp.709–732.
Harun, R., Singh, M., et al., 2010. Bioprocess engineering of microalgae to produce a variety of consumer products. Renewable and Sustainable Energy Reviews, 14(3), pp.1037–1047.
Harun, R., Danquah, M.K. & Forde, G.M., 2010. Microalgal biomass as a fermentation feedstock for bioethanol production. Journal of Chemical Technology and Biotechnology, 85, pp.199–203.
Hempel, N., Petrick, I. & Behrendt, F., 2012. Biomass productivity and productivity of fatty acids and amino acids of microalgae strains as key characteristics of suitability for biodiesel production. Journal of Applied Phycology, 24(6), pp.1407–1418.

82 | BIBLIOGRAFIA
Herrera-Valencia, V. A. et al., 2011. The Green Microalga Chlorella saccharophila as a Suitable Source of Oil for Biodiesel Production. Current Microbiology, 63(2), pp.151–157.
Himmel, M.E. et al., 2007. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science (New York, N.Y.), 315, pp.804–807.
Hirano, A. et al., 1997. CO2 fixation and ethanol production with microalgal photosynthesis and intracellular anaerobic fermentation. In Energy. pp. 137–142.
Ho, S.H. et al., 2013. Bioethanol production using carbohydrate-rich microalgae biomass as feedstock. Bioresource Technology, 135, pp.191–198.
Ho, S.H., Chen, C.Y. & Chang, J.S., 2012. Effect of light intensity and nitrogen starvation on CO2 fixation and lipid/carbohydrate production of an indigenous microalga Scenedesmus obliquus CNW-N. Bioresource Technology, 113, pp.244–252.
Ho, S.H., Chen, W.M. & Chang, J.S., 2010. Scenedesmus obliquus CNW-N as a potential candidate for CO2 mitigation and biodiesel production. Bioresource Technology, 101(22), pp.8725–8730.
Hsieh, C.H. & Wu, W.T., 2009. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresource Technology, 100(17), pp.3921–3926.
Hu, Q. et al., 2008. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant Journal, 54, pp.621–639.
Huntley, M.E. & Redalje, D.G., 2007. CO2 mitigation and renewable oil from photosynthetic microbes: A new appraisal. Mitigation and Adaptation Strategies for Global Change, 12(4), pp.573–608.
Illman, A.M., Scragg, A.H. & Shales, S.W., 2000. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme and Microbial Technology, 27(8), pp.631–635.
Johnson, F.X. & Rosillo-Calle, F., 2007. Biomass, Livelihoods and International Trade.
Kaewkannetra, P., Enmak, P. & Chiu, T., 2012. The effect of CO2 and salinity on the cultivation of Scenedesmus obliquus for biodiesel production. Biotechnology and Bioprocess Engineering, 17(3), pp.591–597.
Kondili, E.M. & Kaldellis, J.K., 2007. Biofuel implementation in East Europe: Current status and future prospects. Renewable and Sustainable Energy Reviews, 11, pp.2137–2151.
Kosourov, S. et al., 2005. The effect of sulfur re-addition on H2 photoproduction by sulfur-deprived green algae. Photosynthesis Research, 85, pp.295–305.
Kruse, O. et al., 2005. Photosynthesis: a blueprint for solar energy capture and biohydrogen production technologies. Photochemical & photobiological sciences : Official journal of the

BIBLIOGRAFIA | 83
European Photochemistry Association and the European Society for Photobiology, 4, pp.957–970.
Krylova, A.Y., Kozyukov, E.A. & Lapidus, A.L., 2008. Ethanol and diesel fuel from plant raw materials: A review. Solid Fuel Chemistry, 42(6), pp.358–364.
Kumar, R.R. et al., 2014. Enzymatic and non-enzymatic antioxidant potentials of Chlorella vulgaris grown in effluent of a confectionery industry. Journal of Food Science and Technology, 51, pp.322–328.
Lashinky, A. & Schwartz, N., 2006. How to beat the high cost of gasoline. Forever!
Lee, R., 2008. Phycology 4th ed. Cambridge University Press, ed.
Lee, Y.C. et al., 2013. Oil extraction by aminoparticle-based H2O2 activation via wet microalgae harvesting. RSC Adv., 3, pp.12802–12809.
Lee Y.K., 2004. Algal Nutrition: Heterotrophic Carbon Nutrition. In A. Richmond, ed. Handbook of Microalgal Culture: Biotechnology and Applied Phycology. London: Blackwell, pp. 116–121.
Levin, D.B., Pitt, L. & Love, M., 2004. Biohydrogen production: Prospects and limitations to practical application. International Journal of Hydrogen Energy, 29, pp.173–185.
Li, Y., Horsman, M., Wu, N., Lan, C.Q. & Dubois-calero, N., 2008. Articles : Biocatalysts and
Bioreactor Design. Biotechnology Prog, 24, pp.815–820.
Li, Y., Horsman, M., Wu, N., Lan, C.Q. & Dubois-Calero, N., 2008. Biofuels from Microalgae. Biotechnology Progress, 24, pp.815–820.
Licht, F., 2006. World ethanol markets: the outlook to 2015.
Liu, Z.Y., Wang, G.C. & Zhou, B.C., 2008. Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresource technology, 99(11), pp.4717–4722.
Lourenço, S.O., 2006. Cultivo de microalgas marinhas: princípios e aplicações Rima, ed.
Lv, J.M. et al., 2010. Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresource Technology, 101(17), pp.6797–6804.
Mahboob, S. et al., 2012. High-density growth and crude protein productivity of a thermotolerant Chlorella vulgaris: Production kinetics and thermodynamics. Aquaculture International, 20(3), pp.455–466.
Maity, J.P. et al., 2014. Microalgae for third generation biofuel production, mitigation of greenhouse gas emissions and wastewater treatment: Present and future perspectives - A mini review. Energy.

84 | BIBLIOGRAFIA
Masukawa, H., Mochimaru, M. & Sakurai, H., 2002. Hydrogenases and photobiological hydrogen production utilizing nitrogenase system in cyanobacteria. In International Journal of Hydrogen Energy. pp. 1471–1474.
Mata, T.M., Martins, A. A. & Caetano, N.S., 2010. Microalgae for biodiesel production and other applications: A review. Renewable and Sustainable Energy Reviews, 14(1), pp.217–232.
Meher, L.C., Vidya Sagar, D. & Naik, S.N., 2006. Technical aspects of biodiesel production by transesterification - A review. Renewable and Sustainable Energy Reviews, 10, pp.248–268.
Miao, X. & Wu, Q., 2004. High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. Journal of Biotechnology, 110(1), pp.85–93.
Miao, X., Wu, Q. & Yang, C., 2004. Fast pyrolysis of microalgae to produce renewable fuels. Journal of Analytical and Applied Pyrolysis, 71(2), pp.855–863.
Miyake, J., Igarashi, Y. & Ro gner, M., 2004. The Potential of Using Cyanobacteria as Producers of
Molecular Hydrogen. In Elsevier B.V., ed. Biohydrogen III. Renewable Energy System by Biological Solar Energy Conversion.
Mizuno, Y. et al., 2013. Sequential accumulation of starch and lipid induced by sulfur deficiency in Chlorella and Parachlorella species. Bioresource Technology, 129, pp.150–155.
Molina Grima, E. et al., 2003. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnology Advances, 20(7-8), pp.491–515.
Murugesan, A. et al., 2009. Production and analysis of bio-diesel from non-edible oils - A review. Renewable and Sustainable Energy Reviews, 13, pp.825–834.
Mussatto, S.I. et al., 2010. Technological trends, global market, and challenges of bio-ethanol production. Biotechnology Advances, 28(6), pp.817–830.
Mussgnug, J.H. et al., 2010. Microalgae as substrates for fermentative biogas production in a combined biorefinery concept. Journal of Biotechnology, 150, pp.51–56.
Mutanda, T. et al., 2011. Bioprospecting for hyper-lipid producing microalgal strains for sustainable biofuel production. Bioresource Technology, 102, pp.57–70.
Van Oijen, T. et al., 2005. Enhanced carbohydrate production by Southern Ocean phytoplankton in response to in situ iron fertilization. Marine Chemistry, 93(1), pp.33–52.
Packer, M., 2009. Algal capture of carbon dioxide; biomass generation as a tool for greenhouse gas mitigation with reference to New Zealand energy strategy and policy. Energy Policy, 37(9), pp.3428–3437.
Park, J.Y. et al., 2014. Acid-catalyzed hot-water extraction of lipids from Chlorella vulgaris. Bioresource Technology, 153, pp.408–412.

BIBLIOGRAFIA | 85
Parmar, A. et al., 2011. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresource Technology, 102, pp.10163–10172.
Perez-Garcia, O. et al., 2011. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Research, 45, pp.11–36.
Petrusevski, B. et al., 1995. Tangential flow filtration: A method to concentrate freshwater algae. Water Research, 29(5), pp.1419–1424.
Pienkos, P.T. & Darzins, A., 2009. The promise and challenges of microalgal-derived biofuels. Biofuels, Bioproducts and Biorefining, 3, pp.431–440.
Prabhakara Rao, P.G. et al., 2013. Lipid classes, fatty acid and phospholipid composition of roe lipids from Catla catla and Cirrhinus mrigala. International Food Research Journal, 20(1), pp.275–279.
Přibyl, P., Cepák, V. & Zachleder, V., 2012. Production of lipids in 10 strains of Chlorella and
Parachlorella, and enhanced lipid productivity in Chlorella vulgaris. Applied Microbiology and Biotechnology, 94(2), pp.549–561.
Pruvost, J. et al., 2011. Systematic investigation of biomass and lipid productivity by microalgae in photobioreactors for biodiesel application. Bioresource Technology, 102(1), pp.150–158.
Pulz, O., 1992. Cultivation techniques for microalgae in open and closed ponds. In Proceedings of the 1st European workshop on microalgal biotechnology, pp. 61.
Pulz, O., 2001. Photobioreactors: Production systems for phototrophic microorganisms. Applied Microbiology and Biotechnology, 57(3), pp.287–293.
Pulz, O. & Gross, W., 2004. Valuable products from biotechnology of microalgae. Applied Microbiology and Biotechnology, 65, pp.635–648.
Puppán, D., 2002. Environmental evaluation of biofuels. Periodica Polytechnica, Social and Management Sciences, 10(1), pp.95–116.
Ratha, S.K. & Prasanna, R., 2012. Bioprospecting microalgae as potential sources of “Green Energy”— challenges and perspectives (Review). Applied Biochemistry and Microbiology, 48, pp.109–125.
Rawat, I. et al., 2011. Dual role of microalgae: Phycoremediation of domestic wastewater and biomass production for sustainable biofuels production. Applied Energy, 88(10), pp.3411–3424.
Richmond, A., 1990. Cell response to environmental factores. In A. Richmond, ed. CRC Handbook of Microalgal Mass Culture. Boca Raton: CRC, pp. 69–100.
Richmond, A., 2004. Handbook of microalgae culture: Biotechnology and applied phycology. Analytical biochemistry, 296.

86 | BIBLIOGRAFIA
Ringer, M. et al., 2006. Large-Scale Pyrolysis Oil Production: A Technology Assessment and Economic Analysis.
Rismani-Yazdi, H. et al., 2011. Transcriptome sequencing and annotation of the microalgae Dunaliella tertiolecta: pathway description and gene discovery for production of next-generation biofuels. BMC genomics, 12, pp.148.
Robert R. L. Guillard, 1975. Culture of Phytoplankton for Feeding Marine Invertebrates. In W. L. Smith & Matoira H. Chanley, eds. Culture of Marine Invertebrate Animals. Springer US, pp. 29–60.
Rodolfi, L. et al., 2009. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnology and Bioengineering, 102(1), pp.100–112.
Román-Leshkov, Y. et al., 2007. Production of dimethylfuran for liquid fuels from biomass-derived carbohydrates. Nature, 447, pp.982–985.
Schenk, P.M. et al., 2008. Second Generation Biofuels: High-Efficiency Microalgae for Biodiesel Production. BioEnergy Research, 1, pp.20–43.
Scragg, A.. et al., 2002. Growth of microalgae with increased calorific values in a tubular bioreactor. Biomass and Bioenergy, 23(1), pp.67–73.
Šetlík, I., Šust, V. & Málek, I., 1970. Dual Purpose Open Circulation Units for Large Scale Culture of Algae in Temperate Zones. I. Basic Design Considerations and Scheme of a Pilot Plant. Algological Studies 1, pp.111–164.
Sharma, R., Chisti, Y. & Banerjee, U.C., 2001. Production, purification, characterization, and applications of lipases. Biotechnology Advances, 19, pp.627–662.
Sheehan, J. et al., 1998. A look back at the U. S. Department of Energy’s aquatic species program: biodiesel from algae. U.S. Department of Energy, 328, pp.291.
Shi, X.M., Zhang, X.W. & Chen, F., 2000. Heterotrophic production of biomass and lutein by Chlorella protothecoides on various nitrogen sources. Enzyme and Microbial Technology, 27, pp.312–318.
Siaut, M. et al., 2011. Oil accumulation in the model green alga Chlamydomonas reinhardtii: characterization, variability between common laboratory strains and relationship with starch reserves. BMC biotechnology, 11, pp.7.
Da Silva, A.F., Lourenço, S.O. & Chaloub, R.M., 2009. Effects of nitrogen starvation on the photosynthetic physiology of a tropical marine microalga Rhodomonas sp. (Cryptophyceae). Aquatic Botany, 91(4), pp.291–297.
Sims, R. & Taylor, M., 2008. From 1st to 2nd Generation Biofuel Technologies: An overview of current industry and RD&D activities, November, pp.16–38.

BIBLIOGRAFIA | 87
Singh, J. & Gu, S., 2010. Commercialization potential of microalgae for biofuels production. Renewable and Sustainable Energy Reviews, 14(9), pp.2596–2610.
Soletto, D. et al., 2005. Batch and fed-batch cultivations of Spirulina platensis using ammonium sulphate and urea as nitrogen sources. Aquaculture, 243(1-4), pp.217–224.
Solomon, B.D., Barnes, J.R. & Halvorsen, K.E., 2007. Grain and cellulosic ethanol: History, economics, and energy policy. Biomass and Bioenergy, 31(6), pp.416–425.
Spilling, K., Seppälä, J. & Tamminen, T., 2011. Inducing autoflocculation in the diatom Phaeodactylum tricornutum through CO2 regulation. Journal of Applied Phycology, 23(6), pp.959–966.
Spolaore, P. et al., 2006. Commercial applications of microalgae. Journal of bioscience and bioengineering, 101(2), pp.87–96.
Starr, R.C. & Zeikus, J.A., 1993. Utex - the culture collection of algae at the University of Texas at Austin 1993 list of cultures. Journal of Phycology, 29, pp.1–106.
Sukahara, K.T. & Awayama, S.S., 2005. Liquid Fuel Production Using Microalgae. Journal of the Japan Petroleum Institute, 48(5), pp.251–259.
Sukenik, A. & Shelef, G., 1984. Algal autoflocculation--verification and proposed mechanism. Biotechnology and bioengineering, 26(2), pp.142–147.
Sunda, W. et al., 2005. Trace metal ion buffers and their use in culture studies. In Andersen RA, ed. Algal Culturing Techniques. Amsterdam: Elsevier, pp. 35–63.
Surek, B., 2008. Meeting Report: Algal Culture Collections 2008. An International Meeting at the Culture Collection of Algae and Protozoa (CCAP), Dunstaffnage Marine Laboratory, Dunbeg, Oban, United Kingdom; June 8-11, 2008. Protist, 159, pp.509–517.
Suzuki, T. et al., 1995. Gas-sparged bioreactors for CO2 fixation of Dunaliella tertiolecta. Journal of Chemical Technolgoy & Biotechnology, 62, pp.351–358.
Takagi, M. et al., 2000. Limited feeding of potassium nitrate for intracellular lipid and triglyceride accumulation of Nannochloris sp. UTEX LB1999. Applied Microbiology and Biotechnology, 54(1), pp.112–117.
Takeshita, T. et al., 2014. Starch and lipid accumulation in eight strains of six Chlorella species under comparatively high light intensity and aeration culture conditions. Bioresource Technology, 158, pp.127–134.
Tan, C.K. & Johns, M.R., 1991. Fatty acid production by heterotrophic Chlorella saccharophila. Hydrobiologia, 215(1), pp.13–19.
Tang, H. et al., 2011. Culture of microalgae Chlorella minutissima for biodiesel feedstock production. Biotechnology and Bioengineering, 108(10), pp.2280–2287.

88 | BIBLIOGRAFIA
Tenney, M.W. & Verhoff, F.H., 1973. Chemical and autoflocculation of microorganisms in biological wastewater treatment. Biotechnology and bioengineering, 15(6), pp.1045–1073.
Thurmond, W., 2010. Algae. Advanced biofuel insight. Biofuels International.
Thurmond, W., 2009a. Five drop in fuels: the next generation. Biofuels International.
Thurmond, W., 2009b. Five key strategies for algae biofuels commercialization. Biofuels International. Emerging Markets Online.
Tomaselli, L., 2004. The microalgal cell. In A. Richmond, ed. Handbook of Microalgal Culture: Biotechnology and Applied Phycology. Oxford: Blackwell Publishing Ltd, pp. 3–19.
Tredici, M.R. et al., 1991. A vertical alveolar panel (VAP) for outdoor mass cultivation of microalgae and cyanobacteria. In Bioresource Technology. pp. 153–159.
U.S. Congress, 2007. Energy independence and security act of 2007. Public Law, pp.110–140.
Vonshak, A. & Torzillo, G., 2004. Environmental stress physiology. In A. Richmond, ed. Handbook of Microalgal Culture: Biotechnology and Applied Phycology. Oxford: Blackwell Science, pp. 57–82.
De Vries, S.C. et al., 2010. Resource use efficiency and environmental performance of nine major biofuel crops, processed by first-generation conversion techniques. Biomass and Bioenergy, 34, pp.588–601.
Wang, S.K. et al., 2014. Scale-up cultivation of Chlorella ellipsoidea from indoor to outdoor in bubble column bioreactors. Bioresource Technology, 156, pp.117–122.
Warr, S., Reed, R. & Stewart, W., 1985. Carbohydrate accumulation in osmotically stressed cyanobacteria (blue - green algae): interactions of temperature and salinity. New phytologist, 100(3), pp.285–292.
Widjaja, A., 2010. Lipid production from microalgae as a promising candidate for biodiesel production. MAKARA Journal of Technology Series, 13(1), pp.47–51.
Widjaja, A., Chien, C.C. & Ju, Y.H., 2009. Study of increasing lipid production from fresh water microalgae Chlorella vulgaris. Journal of the Taiwan Institute of Chemical Engineers, 40(1), pp.13–20.
Wijffels, R.H. & Barbosa, M.J., 2010. An Outlook on Microalgal Biofuels. Science, 329(5993), pp.796–799.
Xin, L., Hong-ying, H. & Jia, Y., 2010. Lipid accumulation and nutrient removal properties of a newly isolated freshwater microalga, Scenedesmus sp. LX1, growing in secondary effluent. New Biotechnology, 27(1), pp.59–63.

BIBLIOGRAFIA | 89
Yeh, K.L. & Chang, J.S., 2011. Nitrogen starvation strategies and photobioreactor design for enhancing lipid content and lipid production of a newly isolated microalga Chlorella vulgaris ESP-31: Implications for biofuels. Biotechnology Journal, 6(11), pp.1358–1366.
Yue, D., You, F. & Snyder, S.W., 2014. Biomass - to - bioenergy and biofuel supply chain optimization: Overview, key issues and challenges. Computers and Chemical Engineering, 66, pp.36–56.