
UNIVERSIDADE REGIONAL DE BLUMENAU CENTRO DE … · manustrito da qualificação com tanto carinho...
155
UNIVERSIDADE REGIONAL DE BLUMENAU CENTRO DE CIÊNCIAS TECNOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA FLORESTAL O CONCEITO DE NATURALIDADE APLICADO AOS REMANESCENTES DE FLORESTA MISTA LATI-ACICULIFOLIADA DE SANTA CATARINA DISSERTAÇÃO DE MESTRADO LAIO ZIMERMANN OLIVEIRA BLUMENAU 2015
-
Upload
hoangthien -
Category
Documents
-
view
216 -
download
0
Transcript of UNIVERSIDADE REGIONAL DE BLUMENAU CENTRO DE … · manustrito da qualificação com tanto carinho...

UNIVERSIDADE REGIONAL DE BLUMENAU
CENTRO DE CIÊNCIAS TECNOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA FLORESTAL
O CONCEITO DE NATURALIDADE APLICADO AOS REMANESCENTES DE
FLORESTA MISTA LATI-ACICULIFOLIADA DE SANTA CATARINA
DISSERTAÇÃO DE MESTRADO
LAIO ZIMERMANN OLIVEIRA
BLUMENAU
2015

LAIO ZIMERMANN OLIVEIRA
O CONCEITO DE NATURALIDADE APLICADO AOS REMANESCENTES DE
FLORESTA MISTA LATI-ACICULIFOLIADA DE SANTA CATARINA
Dissertação apresentada ao Programa de Pós-
Graduação em Engenharia Florestal do Centro de
Ciências Tecnológicas da Universidade Regional
de Blumenau - FURB, como requisito parcial
para a obtenção do grau de Mestre em Engenharia
Florestal, área de concentração Conservação e
Restauração de Ecossistemas Florestais.
Orientador: Alexander Christian Vibrans
Coorientadora: Lúcia Sevegnani
BLUMENAU
2015

Ficha Catalográfica elaborada pela
Biblioteca Universitária da FURB
______________________________________________________________________
Oliveira, Laio Zimermann, 1989-
O49c O conceito de naturalidade aplicado aos remanescentes de Floresta Mista lati-
aciculifoliada de Santa Catarina / Laio Zimermann Oliveira. - 2015.
154 f.: il.
Orientador: Alexander Christian Vibrans.
Coorientador: Lúcia Sevegnani.
Dissertação (mestrado) - Universidade Regional de Blumenau,
Centro de Ciências Tecnológicas, Programa de Pós-Graduação em
Engenharia Florestal.
Inclui bibliografia.
1. Floresta - Conservação. 2. Inventários Florestais Nacionais.
3. Biomassa. 4. Vegetação - Mapeamento. I. Vibrans, Alexander Christian. II.
Sevegnani, Lúcia. II. Universidade Regional de Blumenau. Programa de Pós-
Graduação em Engenharia Florestal. III. Título.
CDD 634.9
______________________________________________________________________


“Louvado seja Ele, que é tal que, quando nossas mentes tentam
vislumbrar Sua essência, nossa inteligência se converte em
imbecilidade; quando estudamos a conexão entre Sua ação e Sua
vontade, nosso conhecimento se converte em ignorância, e quando
nossas línguas desejam declarar Sua grandeza por meio de termos
descritivos, toda a eloquência torna-se impotência e imbecibilidade.”
Rabi Moshe ben Maimon (de abençoada memória)

AGRADECIMENTOS
A Hashem, O Criador do Universo, por toda a benevolência para com a minha alma.
À minha querida família, Vilson, Estela e Caio e Vó Maria, por todo o amor, suporte
e incentivo.
Aos demais membros da família, tios e primos, os quais também me suportaram
nessa caminhada.
Ao amigo e grande incentivador desse meu início de carreira acadêmica: Alexander
Christian Vibrans. Chefe, obrigado pelas oportunidades, confiança e por todas as discussões.
À Lúcia Sevegnani; sempre admirei-a, desde a graduação. O ato de ter corrigido o
manustrito da qualificação com tanto carinho me fez convidá-la para ser minha coorientadora.
Ao amigo Paolo Moser (Paulus) pela grande participação no meu crescimento
intelectual (extensions) nesses últimos dois anos. Mais importante que o conhecimento
acadêmico é a amizade verdadeira.
Ao amigo João Paulo de Maçaneiro por toda ajuda, amizade e companheirismo no
infinito caminho do conhecimento sobre (nossas) florestas.
À amiga Débora Lingner pela amizade e prestatividade no que fosse preciso.
Ao amigo Gustavo Piazza pelas ajudas e pelas parcerias acadêmicas.
Ao amigo Lucas Vedana pelo incentivo que sempre me dá na vida acadêmica e na
vida musical.
Ao amigo Airton Sutil, por toda a constante caminhada.
Aos amigos Kanda Kuppa, Guilherme Treis, Ana Treis, Nashschon Kalala Kuppa,
Plamedi Lusembo, por todo amor e incentivo nesse período.
Aos amigos Átila Costa e Ricardo de Pauli e aos amigos Rafael da Costa, Gabriel
Ugarte e Raphael Fogagnoli, pelo incentivo e irmandade.
Ao amigo André Luis de Gasper por todo o conhecimento compartilhado.
Ao amigo Ronald McRoberts por toda ajuda e aprendizado.
Ao amigo Pedro V. Eisenlohr, por todo conhecimento compartilhado nesse período.
Aos colegas da turma de mestrado.
Aos professores Ary T. Oliveira-Filho, Joberto V. de Freitas e Eduardo Adeneski
Filho pela participação na banca avaliadora.

SUMÁRIO
APRESENTAÇÃO ................................................................................................................... 8
OBJETIVOS ........................................................................................................................... 10
ESTRUTURA DA DISSERTAÇÃO ..................................................................................... 10
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 11
CAPÍTULO 1: O CONCEITO DE NATURALIDADE, SUA APLICAÇÃO E
QUANTIFICAÇÃO EM REMANESCENTES FLORESTAIS: É POSSÍVEL APLICÁ-
LO À REALIDADE DA FLORESTA MISTA COM ARAUCÁRIA DE SANTA
CATARINA? ........................................................................................................................... 14
RESUMO ................................................................................................................................. 14
ABSTRACT ............................................................................................................................ 15
1.1. INTRODUÇÃO ........................................................................................................... 16
1.2. REVISÃO BIBLIOGRÁFICA .................................................................................. 17
1.2.1. A NATURALIDADE DE ECOSSISTEMAS FLORESTAIS ................................ 17
1.2.1.1. A biodiversidade e a humanidade ............................................................................ 18
1.2.1.2. O conceito de naturalidade....................................................................................... 20
1.2.1.3. Estudos que adotaram o conceito de naturalidade ................................................... 25
1.2.1.4. Indicadores de naturalidade ..................................................................................... 28
1.2.1.5. Florestas referência de naturalidade......................................................................... 30
1.2.2. FITOFISIONOMIA ALVO: FLORESTA MISTA COM ARAUCÁRIA .............. 33
1.2.2.1 Caracterização geral ................................................................................................. 33
1.2.2.2 Histórico de degradação da Floresta Mista com Araucária ..................................... 38
1.2.2.3 Atual estado de conservação da Floresta Mista com Araucária .............................. 40
1.3. A PROPOSTA: A APLICAÇÃO DO CONCEITO DE NATURALIDADE
COMO MEIO DE ACESSAR O ESTADO DE CONSERVAÇÃO DOS
REMANESCENTES DE FLORESTA MISTA COM ARAUCÁRIA DE SANTA
CATARINA ............................................................................................................................. 43
1.3.1. APRESENTAÇÃO .................................................................................................. 43
1.3.2. INTRODUÇÃO ....................................................................................................... 43
1.3.3. A DEFINIÇÃO DE NATURALIDADE ................................................................. 47
1.3.4. METODOLOGIA DE ACESSO À NATURALDADE .......................................... 53
1.3.4.1 Área de estudo ......................................................................................................... 53
1.3.4.2 Dados do Inventário Florístico-Florestal de Santa Catarina .................................... 55
1.3.4.3 Indicadores potenciais de naturalidade .................................................................... 57

1.3.4.4 Escolha dos indicadores de naturalidade ................................................................. 67
1.3.4.5 Considerações finais acerca dos indicadores de naturalidade.................................. 90
1.3.4.6 Quantificação da naturalidade ................................................................................. 92
1.3.4.7 Definição e escolha das florestas de referência ....................................................... 94
1.4. CONCLUSÕES ........................................................................................................... 99
1.5. REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................... 99
CAPÍTULO 2: QUANTIFICAÇÃO DA NATURALIDADE DOS REMANESCENTES
DE FLORESTA MISTA COM ARAUCÁRIA DE SANTA CATARINA ...................... 122
RESUMO ............................................................................................................................... 122
ABSTRACT .......................................................................................................................... 123
2.1. INTRODUÇÃO ......................................................................................................... 124
2.2. MATERIAL E MÉTODOS ..................................................................................... 125
2.2.1. ESCALAMENTO DA MEDIDA DE NATURALIDADE ................................... 125
2.2.1.1 Índice de naturalidade I .......................................................................................... 125
2.2.1.2 Índice de naturalidade II ........................................................................................ 126
2.2.2. PADRÕES ESPACIAIS DE NATURALIDADE ................................................. 126
2.3. RESULTADOS ......................................................................................................... 128
2.3.1. INDICADORES DE NATURALIDADE ............................................................. 128
2.3.2. FLORESTAS DE REFERÊNCIA ......................................................................... 130
2.3.3. A NATURALIDADE QUANTIFICADA ............................................................. 132
2.3.3.1 Mapas temáticos de naturalidade ................................................................................ 135
2.4. DISCUSSÃO ............................................................................................................. 137
2.4.1 OS ÍNDICES DE NATURALIDADE ................................................................... 137
2.4.2 UMA NOVA VISÃO SOBRE AS FLORESTAS NATIVAS .............................. 138
2.4.2.1 Observações acerca da naturalidade da Floresta Mista com Araucária ................. 138
2.4.2.2 Alternativa para a análise de remanescentes florestais .......................................... 142
2.4.3 PERSPECTIVAS PARA O USO DO CONCEITO DE NATURALIDADE ....... 143
2.5. CONCLUSÕES ......................................................................................................... 144
2.6. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................... 144
APÊNDICE ........................................................................................................................... 146

8
APRESENTAÇÃO
A dependência do ser humano pelas florestas é conhecida há muito tempo, pois desde
os primórdios ele mantém uma estreita relação com elas. As florestas desempenham muitos
serviços e funções ambientais, além de abrigar grande porção da biodiversidade da Terra
(COSTANZA et al., 1997; HOLDRIDGE, 1967; REIS et al., 2014; RICKETTS et al., 2005).
Vivenciamos uma drástica mudança do uso do solo nos últimos 50 anos e, consequentemente,
os ecossistemas foram afetados (MILLENNIUM ECOSYSTEM ASSESSMENT, 2005;
WRIGHT, 2005). Tal mudança foi, e continua sendo, regida pelo crescimento demográfico e
econômico e pelo desenvolvimento da ciência e tecnologia, que são os principais fatores
atuantes nas alterações dos ecossistemas florestais e na perda de habitats, afetando
diretamente a biodiversidade (FOLEY et al., 2005; GIBSON et al., 2011; NELSON, 2005;
PFEIFER et al., 2014). O crescimento populacional é, muitas vezes, acompanhado por
atividades degradadoras, tais como a exploração madeireira indiscriminada e a transformação
de florestas em áreas agrícolas e centros urbanos. A conservação dos ecossistemas florestais
tornou-se uma preocupação no decorrer das últimas décadas, e esta preocupação induziu o
desenvolvimento de pesquisas que objetivam identificar os agentes causadores, os impactos e
as consequências da degradação, tanto na esfera biológica como na social (CHAZDON, 2008;
GARDNER et al., 2009; ASWANI e LAUER, 2014; HUNTER JR. et al., 2014).
Os Inventários Florestais Nacionais (IFNs) são uma fonte primária importante de
informações sobre as florestas, em uma escala geográfica mais ampla; eles apresentam uma
imagem do estado das florestas, constituindo as bases para a criação de políticas e programas
florestais, tanto em escala regional como nacional (TOMPPO et al., 2010). Em uma
perspectiva histórica, os IFNs evoluíram em direção ao levantamento e monitoramento da
biodiversidade, de modo que no passado eram restritamente conduzidos com base na
necessidade da obtenção de informações sobre a produtividade de madeira das florestas
(CHIRICI et al., 2012). Este passo dado em direção à biodiversidade é justificado pelo
incremento da exploração e degradação dos ecossistemas florestais, a ponto de medidas
conservacionistas terem sido tomadas em várias regiões do planeta, através de ações internas
ou acordos internacionais (CHIRICI et al., 2012; McROBERTS et al., 2008; VIBRANS et al.,
2013a). Desta maneira, os IFNs nas últimas décadas vêm desempenhando a função de prover
uma ampla gama de informações que possam retratar não só o estoque de madeira disponível,
como também, os aspectos estruturais, florísticos, dinâmicos, os estoques de biomassa

9
disponíveis, dentre outros, podendo melhor embasar a gestão das florestas (CHIRICI et al.,
2011; McROBERTS et al., 2010).
Perante esse contexto, surge uma questão importante: Como monitorar o estado de
conservação das florestas nativas em larga escala utilizando dados de IFNs?
O acesso e a coleta de dados que contemplem todos os componentes de um dado
ecossistema florestal não podem ser atingidos (RONDEUX, 2012), assim, todos esses
componentes dificilmente poderão ser precisamente medidos (BOUTIN et al., 2009). Uma
saída para este obstáculo é selecionar variáveis indicadoras que podem relacionar e sintetizar
tais componentes (NOSS, 1990; CHIRICI et al., 2011; CHIRICI et al., 2012; MARCHETTI,
2004).
Os dados gerados pelos IFNs da América do Norte, Europa e, recentemente, na
América do Sul e Ásia, possibilitam verificar o estado de conservação das florestas através da
quantificação e/ou monitoramento da naturalidade destes ecossistemas (CHIRICI et al., 2011;
EEA, 2014; McROBERTS et al., 2008; McROBERTS et al., 2012; WINTER, 2012). O
conceito de naturalidade refere-se ao quanto algo é livre de influência antropogênica,
especialmente a tecnológica. Assim, ela pode ser representada por um gradiente contínuo que
estende-se entre dois extremos, o totalmente artificial (ou alterado) e o totalmente natural
(ANGERMEIER, 2000; SJÖRS, 1986; WINTER, 2012). Poucos métodos objetivos de
quantificação da naturalidade foram desenvolvidos (MCROBERTS et al., 2012; WINTER,
2012).
Dentro do universo dos dados coletados pelos IFNs, é importante determinar quais
variáveis (ou indicadores) têm o potencial de expressar a naturalidade de ecossistemas
florestais de uma forma quantitativa? E ainda, quais destas variáveis são coletadas pela
maioria dos IFNs?
Por exemplo, segundo Winter et al. (2011), os IFNs europeus preferem coletar
variáveis relacionadas à estrutura da floresta como a composição de espécies, estrutura
horizontal e vertical e necromassa do que variáveis de outros componentes da diversidade
biológica, como espécies de pássaros, invertebrados, briófitas, fungos, dentre outras; estas
variáveis são ecologicamente relevantes, mas seu acesso torna-se trabalhoso, demorado e
dispendioso.
Onde se enquadra o estado de Santa Catarina nesta proposta de monitoramento do
estado de conservação através de um estimador quantitativo de naturalidade?
A conclusão do primeiro ciclo do Inventário Florístico-Florestal de Santa Catarina
(IFFSC) gerou um precioso banco de informações sobre as florestas nativas do estado e

10
possibilitou a execução de diversas análises sobre a atual situação das florestas (VIBRANS et
al. 2012a; VIBRANS et al., 2012b; VIBRANS et al., 2013b; VIBRANS et al., 2013c). A
conformidade metodológica do IFFSC com o IFN-Brasil constitui um incentivo a mais para o
início da aplicação de estimadores de naturalidade dos remanescentes florestais, pois outros
estados podem utilizar o mesmo estimador, possibilitando resultados comparáveis (EEA,
2014; KÖHL et al., 2000; McROBERTS et al., 2012; TOMPPO e SCHADAUER, 2012).
Em florestas tropicais ou subtropicais, o conceito de naturalidade ainda não foi
utilizado como um meio de determinar o estado de conservação de ecossistemas florestais
(WINTER, 2012). Tal fato motiva o presente trabalho a investigar a viabilidade da aplicação
de uma metodologia de determinação da naturalidade para a Floresta Mista Lati-
aciculifoliada (OLIVEIRA-FILHO, 2009) de Santa Catarina, a partir dos dados coletados pelo
IFFSC.
OBJETIVOS
Diante do contexto apresentado, os objetivos deste trabalho são: (i) apresentar e
discutir o conceito de naturalidade e conceitos associados; (ii) propor uma metodologia de
quantificação da naturalidade para os remanescentes de Floresta Mista Lati-aciculifoliada de
Santa Catarina, com base nos dados levantados pelo IFFSC; (iii) aplicar a metodologia de
quantificação com o intuito de criar um índice de naturalidade para os remanescentes de
Floresta Mista Lati-aciculifoliada amostrados pelo IFFSC.
ESTRUTURA DA DISSERTAÇÃO
Este manuscrito está divido em dois capítulos. O primeiro capítulo é dedicado ao
cumprimento dos dois primeiros objetivos propostos, e é constituído de:
uma revisão da literatura sobre o conceito de naturalidade, sua definição e sua
aplicação no acesso do estado conservação de ecossistemas florestais naturais, e sobre
a Floresta Mista Lati-aciculifoliada, sua descrição, histórico de exploração e atual
situação de conservação;
uma proposta para a quantificação da naturalidade. A proposta é constituída por uma
fundamentação teórica introdutória, pela estruturação de princípios fundamentais e
pela descrição geral da abordagem metodológica a ser adotada no processo de
quantificação da naturalidade dos remanescentes de Floresta Mista Lati-aciculifoliada
de Santa Catarina.

11
O segundo capítulo é dedicado à aplicação da metodologia desenvolvida no capítulo
anterior e a criação de um índice numérico de naturalidade, de fácil interpretação. São
apresentados os resultados obtidos e a discussão dos mesmos, seguidos de recomendações
para a conservação dos remanescentes de Floresta Mista Lati-aciculifoliada de Santa Catarina.
REFERÊNCIAS BIBLIOGRÁFICAS
ANGERMEIER, P.L. The natural imperative for biological conservation. Conservation
Biology, v.14: p.373-381, 2000.
ASWANI, S.; LAUER, M. Indigenous people‟s detection of rapid ecological change.
Conservation Biology, v.28, p.820-828, 2014.
BOUTIN, S.; HAUGHLAND, D.L.; SCHIECK, J.; HERBERS, J.; BAYNE, E. A new
approach to forest biodiversity monitoring in Canada. Forest Ecology and Management,
v.258, p.168-175, 2009.
CHAZDON, R.L. Beyond deforestation: Restoring forests and ecosystem services on
degradated lands. Science, v. 320, p.1458-1460, 2008.
CHIRICI, G.; MCROBERTS, R.E.; WINTER, S.; BERTINI, R.; BRÄNDLI, U.; ASENSIO,
I.A.; BASTRUP-BIRK, A.; RONDEUX, J.; BARSOUM, N.; MARCHETTI, M. National
Forest Inventory Contributions to Forest Biodiversity Monitoring. Forest Science, v.58, n.3,
2012.
CHIRICI, G.; WINTER, S.; McROBERTS, R.E. (Eds.). National forest inventories:
contributions to forest biodiversity assessments. Springer, 2011.
COSTANZA, R.; D‟ARGE, R.; GROOT, R.; FARBERK, S.; GRASSO, M.; HANNON, B.;
LIMBURG, K.; NAEEM, S.; O‟NEILL, R.V.; PARUELO, J.; RASKIN, R.G.; SUTTONKK,
P.; VAN DEN BELT, M. The value of the world‟s ecosystem services and natural capital.
Nature, v. 387, p. 253-260, 1997.
FOLEY, J.A.; DEFRIES, R.; ASNER, G.P.; BARFORD, C.; BONAN, G.; CARPENTER,
S.R. Global consequences of land use. Science, v.309, p.570–574, 2005.
EEA. European Environment Agency. Developing a forest naturalness indicator for Europe -
Concept and methodology for a high nature value (HNV) forest indicator. EEA Technical
report, n.13, 2014.
GARDNER, T.A.; BARLOW, J.; CHAZDON, R.L; EWERS, R.M.; HARVEY, C.A.;
PERES, C.A.; SODHI, N.S. Prospects for tropical forest biodiversity in a human-modified
world. Ecology Letters, v.12, p.561–582, 2009.
GIBSON, L., LEE, T.M., KOH, L.P., BROOK, B.W., GARDNER, T.A., BARLOW, J.,
PERES, C.A., BRADSHAW, C.J.A., LAURANCE, W.F., LOVEJOY, T.E.; SODHI, N.S.
Primary forests are irreplaceable for sustaining tropical biodiversity. Nature, v.478, p.378-
381, 2011.

12
HOLDRIDGE, L.R. Life zone ecology. San Jose: Tropical Science Center, 1967.
HUNTER JR., M.L.; REDFORD, K.H.; LINDENMAYER, D.B. The complementary niches
of anthropocentric and biocentric conservationists. Conservation Biology, v.28, n.3, p.641-
645, 2014.
KÖHL, M.; TRAUB, B.; PÄIVINEN, R. Harmonization and standardization in multi-national
environmental statistics - Mission impossible? Environmental Monitoring and Assessment,
v.63, p.361-380, 2000.
MARCHETTI, M. Monitoring and Indicators of Forest Biodiversity in Europe - From Ideas to
Operationality. EFI Proceedings, n.51, 2004.
McROBERTS, R.E.; TOMPPO, E.O; NAESSET, E. Advances and emerging issues in
national forest inventories. Scandinavian Journal of Forest Research, v.25, p.368-381,
2010.
McROBERTS, R.E.; WINTER, S.; CHIRICI, G.; LAPOINT, E. Assessing Forest
Naturalness. Forest Science, v.58, p.294-309, 2012.
McROBERTS, R.E.; WINTER, S.; CHIRICI, G.; HAUK, E.; PELZ, D.R; MOSER, K.;
HATFIELD, M.A. Large-scale spatial patterns of forest structural diversity. Canadian
Journal of Forest Research, v.38, p.429-438, 2008.
MILLENNIUM ECOSYSTEM ASSESSMENT. Ecosystems and Human Well-being:
Synthesis. Washington DC: Island Press, 2005.
NELSON, G.C. Drivers of Ecosystem Change: Summary Chapter. In: HASSAN, R.;
SCHOLES, R.; ASH, N. (Eds.) Ecosystems and Human Well-being: Current State and
Trends, Volume 1. Washington DC: Island Press, 2005.
NOSS, R.F. Assessing and monitoring forest biodiversity: A suggested framework and
indicators. Forest Ecology and Management, v.115, p.135–146, 1999.
OLIVEIRA-FILHO, A.T. Classificação das fitofisionomias da América Do Sul cisandina
tropical e subtropical: Proposta de um novo sistema – prático e flexível – ou uma injeção a
mais de caos? Rodriguésia, v.60, p. 237-258, 2009.
PFEIFER, M. et al. BIOFRAG – a new database for analyzing BIOdiversity responses to
forest FRAGmentation. Ecology and Evolution, v.4, n.9, p.1524-1537, 2014.
REIS, M.S; LADIO, A.; PERONI, N. Landscapes with Araucaria in South America: evidence
for a cultural dimension. Ecology and Society, v.19, n.2, p.43-56, 2014.
RICKETTS, T; BROOKS, T; HOFFMANN, M.; STUART, S.; BALMFORD, A.; PURVIS,
A.; REYERS, B.; WANG, J.; REVENGA, C.; KENNEDY, E.; NAEEM, S.; ALKEMADE,
R.; ALLNUTT, T.; BAKARR, M.; BOND, W.; CHANSON, J.; COX, N.; FONSECA, G.;
HILTON-TAYLOR, C.; LOUCKS, C.; RODRIGUES, A.; SECHREST, W.;
STATTERSFIELD, A.; VAN RENSBURG, J.B; WHITEMAN, C. Biodiversity. In:

13
HASSAN, R.; SCHOLES, R.; ASH, N. (Eds.) Ecosystems and Human Well-being: Current
State and Trends, Volume 1. Washington DC: Island Press, 2005.
RONDEUX, J.; BERTINI, R.; BASTRUP-BIRK, A.; CORONA, P.; LATTE, N.;
MCROBERTS, R.E.; STÅHL, G.; WINTER, S.; CHIRICI, G. Assessing Deadwood Using
Harmonized National Forest Inventory Data. Forest Science¸ v.58, n.3, 2012.
SJÖRS, H. On the gradient from near-natural to man-made. Transactions of the Botanical
Society of Edinburgh, v.45, p.77-84, 1986.
TOMPPO, E.; GSCHWANTNER, T.; LAWRENCE, M.; MCROBERTS, R.E. National
Forest Inventories. Pathway for Common Reporting. Heidelberg: Springer, 2010.
TOMPPO, E. O.; SCHADAUER, K. Harmonization of National Forest Inventories in Europe:
Advances under COST Action E43. Forest Science, v.58, n.3, p.191-200, 2012.
VIBRANS; A.C.; McROBERTS; R.E.; MOSER; P.; NICOLETTI; A. How much remains of
the Brazilian Atlantic forest in the state of Santa Catarina? Assessing the accuracy of forest
cover maps using ground data from the Santa Catarina Forest and Floristic Inventory. Remote
Sensing of Environment, v.130, p.87-95, 2013a.
VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V. (Eds.). Inventário
Florístico Florestal de Santa Catarina: Diversidade e conservação dos remanescentes
florestais. Blumenau: Edifurb, 2012a.
VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V. (Eds.). Inventário
Florístico Florestal de Santa Catarina: Floresta Estacional Decidual. Blumenau: Edifurb,
2012b.
VIBRANS, A. C.; SEVEGNANI, L.; GASPER, A. L.; LINGNER, D. V. (Eds.). Inventário
Florístico Florestal de Santa Catarina: Floresta Ombrófila Mista. Blumenau: Edifurb,
2013b.
VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V. (Eds.). Inventário
Florístico Florestal de Santa Catarina: Floresta Ombrófila Mista. Blumenau: Edifurb,
2013c.
WINTER, S.; MCROBERTS, R.E.; CHIRICI, G.; BASTRUP-BIRK, A.; RONDEUX, J.;
BRÄNDLI, U.; NILSEN, J.O.; MARCHETTI, M. The need for harmonized of forest
biodiversity indicators. In: CHIRICI, G.; WINTER, S.; MCROBERTS, R.E. National forest
inventories: Contributions to forest biodiversity assessments. Springer, 2011.
WINTER, S. Forest naturalness assesment as a component of biodiversity monitoring and
conservation management. Forestry, v.85, n.2, p.293-304, 2012.
WRIGHT, S.J. Tropical forests in a changing environment. Trends in Ecology and
Evolution, v.20, n.10, 2005.

14
CAPÍTULO 1: O CONCEITO DE NATURALIDADE, SUA APLICAÇÃO E
QUANTIFICAÇÃO EM REMANESCENTES FLORESTAIS: É POSSÍVEL APLICÁ-
LO À REALIDADE DA FLORESTA MISTA COM ARAUCÁRIA DE SANTA
CATARINA?
RESUMO
Nas últimas décadas os Inventários Florestais Nacionais (IFN) vêm desempenhando a função
de prover uma gama de informações que retratam não só o estoque de madeira disponível,
mas também os aspectos estruturais, florísticos, e os estoques de biomassa disponíveis nas
florestas nativas. Em resposta a esta demanda, o Inventário Florístico-Florestal de Santa
Catarina (IFFSC), inserido no programa IFN-Brasil, foi conduzido com o intuito de gerar
informações sobre as florestas nativas do estado. A partir do contexto social e histórico do
estado de Santa Catarina, e de dados qualitativos e quantitativos provenientes de 155
Unidades Amostrais levantadas na Floresta Mista com Araucária pelo IFFSC, adaptamos e
aplicamos o conceito de naturalidade, amplamente utilizado na Europa. Esse conceito está
relacionado ao grau em que algo é alterado do seu estado original. Assim, a naturalidade pode
ser quantificada através de um gradiente que estentede-se do menos ao mais natural.
Buscamos por indicadores quantitativos de naturalidade que têm o poder de sintetizar
processos e aspectos ecológicos importantes. Dos 16 indicadores potenciais elencados foram
escolhidos seis. De posse dos indicadores, definimos “florestas de referência” (Unidades
Amostrais), as quais foram consideradas como mais naturais e serviram de base para a criação
da medida quantitativa de naturalidade. A medida foi gerada a partir de duas abordagens: (i)
da distância euclidiana do vetor gerado pelos seis indicadores (das Unidades Amostrais
ordinárias) até o vetor gerado para dada floresta de referência (Unidades Amostrais de
referência); (ii) da distância euclidiana do vetor gerado pelos seis indicadores (das Unidades
Amostrais ordinárias) até o vetor gerado para a floresta de referência teórica, 100% natural.

15
ABSTRACT
The ongoing Brazilian National Forest Inventory (NFI) is being performed as a tool for
providing wide-range information, reporting not only the available timber stock, but also
information about forest ecosystem components, conservation status and biomass stock. In
response to this demand, the Forest and Floristic Inventory of Santa Catarina (IFFSC),
inserted in IFN-Brazil program, was conducted in order to gather information about native
forests. Based on the social and historical aspects of Santa Catarina state and also on
qualitative and quantitative data from 155 sample plots measured by the IFFSC in Mixed
Needle-broadleaved Forest, we adapted and applied the concept of naturalness which is
widely used in Europe. This concept is related to the degree to which something is changed
from its original state. Thus, naturalness can be quantified through a gradient that goes from
least to most natural. We searched for quantitative naturalness indicators which have power to
synthesize important ecological aspects and processes. We chosed six out of the 16 listed
indicators. After the calculation of the indicadors, we defined "reference forests" (sample
plots), which were considered to be closest to natural. These references were estabilished as
the basis for creation of a naturalness quantitative measure. The measure was generated based
on two approaches: (i) the Euclidean distance of the vector generated by the six indicators (of
an ordinary sample plot) to the vector generated for an observed reference forest (reference
sample plot); (ii) the Euclidean distance of the vector generated by the six indicators (of an
ordinary sample plot) to the vector generated for a teorethical 100% natural reference forest.

16
1.1. INTRODUÇÃO
Após um recente processo de intensa exploração e degradação dos ecossistemas
naturais (FOLEY et al., 2005; SALA et al., 2000), a humanidade começa a perceber a
magnitude e as consequências de seus atos destrutivos, muitas vezes motivados pela força de
mercado e falta de conhecimento. Dentre os muitos ecossistemas, os florestais vêm
despertando a atenção, talvez por sua biodiversidade e extensão (FAO, 2012; RICKETTS et
al., 2005), e por serem provedores diretos, ou indiretos, de tantos recursos necessários à vida
humana (GONZALEZ et al., 2005). Adicionamente, as florestas nos chamam atenção por sua
exuberância e complexidade, que nos intriga e leva a refletir sobre suas origens, sejam através
de princípios criacionistas ou evolucionistas (LOURENÇO, 2012).
A conservação dos remanescentes florestais e a restauração de áreas anteriormente
cobertas por vegetação florestal vêm sendo comentada pela mídia, e as instituições ligadas ao
meio ambiente, sejam acadêmicas ou não, têm se ocupado no desenvolvimento de técnicas de
levantamento e análises de dados ecológicos, de conservação, restauração e monitoramento
dos elementos bióticos e abióticos que integram um sistema ecológico. Apesar deste
movimento em prol da conservação, a atual política e as ações de conservação ainda são
insuficientes. Um exemplo desse fato é a biodiversa Floresta Atlântica sensu lato
(OLIVEIRA-FILHO e FONTES, 2000), a qual se encontra intensamente fragmentada, com
remanescentes de pequeno tamanho e afetada por diversos efeitos causados pela fragmentação
(RIBEIRO et al., 2009; TABARELLI et al., 2010).
Perante este contexto, os Inventários Florestais Nacionais (IFNs, ou IFN, no singular)
tornam-se os principais instrumentos de geração de informações em larga escala sobre as
florestas, tanto informações quantitativas como qualitativas (McROBERTS et al., 2010b).
Muitos países já possuem programas de IFN implantados, como, por exemplo, a Alemanha,
República da Coréia do Sul, Estados Unidos da América, Finlândia, França, Grã-Bretanha,
México, entre outros (TOMPPO et al., 2010).
As informações coletadas pelos IFNs possibilitam determinar parâmetros e
indicadores do estado de conservação das florestas, entretanto, por questões operacionais,
uma coleta de dados que contempla todos os componentes do ecossistema florestal não pode
ser realizada (RONDEUX, 2012). Logo, uma alternativa a esta limitação é identificar e
selecionar variáveis (ou indicadores) que podem relacionar e sintetizar os componentes do
ecossistema florestal (CHIRICI, et al., 2012; MARCHETTI, 2004; NOSS, 1990, 1999;
WINTER et al., 2011).

17
A determinação e o monitoramento da biodiversidade e do estado de conservação das
florestas nativas podem ser realizados através do emprego do conceito de naturalidade
(McROBERTS et al., 2012; WINTER, 2012). Angermeier (2000) e Hunter (1996) definem o
conceito de naturalidade baseado em quanto algo é livre de influência antrópica,
especialmente a tecnológica. A naturalidade pode ser vista como uma medida de
biodiversidade, mas também como um descritor do estado de conservação de um ecossistema
(MACHADO, 2004), e pode ser representada por um gradiente contínuo que estende-se entre
dois extremos, o totalmente artificial (ou alterado) e o totalmente natural (ANGERMEIER,
2000). Apesar dos muitos trabalhos que abordam o conceito (WINTER, 2012), existe a
carência de uma definição comum do conceito e de métodos de determinação/quantificação
da naturalidade (MACHADO, 2004; McROBERTS et al., 2012; WINTER, 2012). Poucos
métodos explícitos de quantificação foram propostos até o momento (McROBERTS et al.,
2012; WINTER, 2012), de forma que estes autores incentivam a realização de estudos que
tratem da naturalidade e de formas de quantificá-la.
McRoberts et al. (2012) justificam a importância de determinar a naturalidade de
florestas por três razões: (i) estimar adequadamente e reportar a sanidade ecológica das
florestas para fins de avaliação da conservação e manejo florestal, (ii) para desenvolver metas
e padrões para a conservação de florestas e (iii) identificar florestas primárias ou em estágio
avançado de regeneração que poderão ser designadas como áreas de conservação.
Diante do contexto apresentado, os objetivos deste capítulo são: (i) apresentar e
discutir o conceito de naturalidade e conceitos associados; ii) propor uma abordagem
metodológica para a utilização da naturalidade visando avaliar o estado de conservação dos
remanescentes florestais de Floresta Mista Lati-aciculifoliada de Santa Catarina.
1.2. REVISÃO BIBLIOGRÁFICA
1.2.1. A NATURALIDADE DE ECOSSISTEMAS FLORESTAIS
O processo envolvido na determinação da naturalidade de florestas requer um forte
embasamento teórico do próprio conceito de naturalidade, e de conceitos relacionados.
Diante desta premissa, serão abordadas definições importantes que fundamentarão a proposta
de determinação da naturalidade de remanescentes de Floresta Mista Lati-aciculifoliada de
Santa Catarina.
Este tópico está estruturado em subtópicos que contemplarão o conjunto de temas que
se fazem necessários para o desenvolvimento de um pensamento sistêmico em relação à

18
naturalidade, sua utilização e aplicação dentro do contexto apresentado. Ainda, os subtópicos
trazem um panorama da biodiversidade florestal no mundo, definições de conceitos
fundamentais, revisão de estudos que de alguma forma, sejam com uma abordagem
qualitativa ou quantitativa, adotaram a naturalidade como um princípio fundamental.
1.2.1.1. A biodiversidade e a humanidade
A Terra possui cerca de 3,8 bilhões de hectares de florestas, correspondendo a 30% da
superfície total (FAO, 2012). Seis dos 12 grandes biomas da Terra são caracterizados como
floresta e dois terços de todas as ecorregiões do planeta estão contidos nestes seis biomas
(MATTHEWS et al., 2000). Sabe-se que os ecossistemas florestais prestam grandes
contribuições para a biodiversidade existente no planeta, estando entre os mais ricos nos
aspectos biológicos e genéticos - 70% das espécies animais e de plantas conhecidas estão
contidas nas florestas (MATTHEWS et al., 2000). De acordo com The Plant List (2014)
existem cerca de 350.000 espécies de plantas no planeta.
O que vem ser a tão comentada biodiversidade? A definição de biodiversidade pode
variar, dependendo de cada área do conhecimento e, ou, de cada pessoa ou especialista
(NOSS, 1990). A definição de biodiversidade dada pela Convenção de Diversidade Biológica
(CBD, 1992) refere-se à:
"[...] variedade de organismos vivos de todas as origens,
compreendendo, dentre outros, os ecossistemas terrestres, marinhos e
outros ecossistemas aquáticos e os complexos ecológicos de que
fazem parte; compreendendo ainda a diversidade dentro de espécies,
entre espécies e de ecossistemas."
Tratando-se de florestas, o termo biodiversidade florestal, de acordo com Hunter
(1990), refere-se à “diversidade de vida em todas as suas formas e níveis de organização
dentro de áreas florestadas”. Tratando-se de definições mais específicas, algumas se
destacam. Whittaker (1972) propôs três subdivisões para a diversidade em escalas espaciais:
alpha (α) se refere à diversidade de um ecossistema ou remanescente; beta (β) se refere à
mudança na diversidade entre ecossistemas ou remanescentes; gamma (γ) se refere à
diversidade geral entre os diferentes ecossistemas ou remanescentes presentes em uma região.
A CBD (1992) aponta três níveis de biodiversidade: o nível de ecossistema, o de espécie e o
de diversidade genéticas das espécies. Noss (1990) define biodiversidade em três classes:
composição (relativo à identificação e à variedade de componentes), funcional (refere-se aos

19
processos ecológicos e evolutivos) e estrutural (refere-se à organização espacial dos
componentes). Gaston e Spier (2004) também caracterizam a biodiversidade em três níveis:
genética, espécies e ecossistêmica.
A mobilização em prol da conservação dos ecossistemas naturais tomou proporções
globais através de acordos, encontros e convenções, os quais passaram a exigir que os países
signatários apresentassem estimativas nacionais de parâmetros relacionados à área florestada
e o estoque de madeira, principalmente (McROBERTS et al., 2010a; VIBRANS et al.,
2012a). Em relação às medidas de conservação dos ecossistemas naturais, muitos acordos e
conferências passaram a exigir o monitoramento dos componentes da biodiversidade para fins
conservacionistas e de uso sustentável; entre estes acordos e conferências se destacam:
Conventionon Biological Diversity (CBD) em 1992 e 2009, Protocolo de Kioto em 1998,
Montreal Process em 2005, United Nations Framework Conventionon Climate Change
(UNFCCC) em 1994, Ministerial Conference on the Protection of Forests in Europe
(MCPFE) em 2009 e Global Forest Resources Assessment (FAO, 2010).
Os ecossistemas florestais são particularmente vulneráveis à perda de integridade
ecológica dada a complexidade intrínseca de sua estrutura e função (KARNOSKY et al.,
2001). Logo, as ações do homem nos ecossistemas florestais, tais como, o desflorestamento, a
exploração madeireira, a introdução de espécies exóticas (as quais podem tornar-se invasoras,
sensu Richardson et al. [2000]), assim como a poluição, representam ameaças à
biodiversidade florestal (GIBSON et al., 2011; McROBERTS et al., 2012). Com o
crescimento da dimensão do cenário ambiental, a biologia da conservação emergiu com o
enfoque de aplicar princípios científicos para solucionar problemas relativos às perturbações
feitas pelos homens nos ecossistemas naturais (SOULÉ, 1985).
Gaston e Spicer (2004) subdividiram as ameaças à biodiversidade em quarto causas
principais: (i) exploração direta, (ii) perda de habitats e degradação, (iii) espécies
introduzidas, e (iv) processos de extinção de espécies.
Sendo assim, a conservação baseia-se fortemente na observação e no estudo do estado
da integridade ecológica dos ecossistemas e diversas definições deste conceito são sugeridas
pela literatura (FISCHMAN, 2004). A primeira menção do termo integridade ecológica foi
feita por Aldo Leopold (1949 apud DORREN et al., 2004), o qual propôs que um ecossistema
está saudável quando este tende a preservar a integridade e estabilidade da comunidade
biótica (ODUM, 2004); quando esta tendência se mostra inversa, a integridade ecológica está
afetada.

20
Karr e Dudley (1981) definem integridade ecológica, de forma semelhante a Frey
(1975), como o poder de um ecossistema suportar e manter as comunidades de organismos
que têm composição de espécies, diversidade e organização (funcional) comparáveis à
habitats conservados presentes em uma região. Segundo Tierney et al. (2009), a integridade
ecológica avalia a composição, estrutura e função de um ecossistema, em comparação com o
seu natural potencial ou ao longo de uma série temporal de variação.
Noss (1990) apresenta sua definição de integridade ecológica de uma maneira
exemplificada, afirmando que uma comunidade tem integridade se esta é dominada por
espécies nativas e possui estabilidade (ODUM, 2004) e potencial de resiliência
(CARPENTER et al., 2001). Para Angermeier e Karr (1994) a integridade ecológica refere-se
à totalidade do sistema, onde todos os elementos interagem em um nível espacial e temporal.
Segundo Steedman (1995), a integridade biológica é definida, especificamente e
operacionalmente, como o estado de um ecossistema exposto ao mínimo de influência
antrópica possível.
Estes conceitos de biodiversidade e integridade ecólogica estão relacionados ao
conceito de naturalidade (McROBERTS et al., 2012) que será apresentado e discutido no
tópico seguinte.
1.2.1.2. O conceito de naturalidade
A definição de natural pode diferir entre autores, embora sempre esteja intimamente
ligada a princípios filosóficos e semânticos (MACHADO, 2004); Williams (2002) acrescenta
que a definição pode variar também conforme o contexto social considerado. Winter (2012)
aponta a inconsistência observada na literatura em relação à definição do termo. A definição
de natural dada por Angermeier (2000) e Hunter (1996) reside na premissa de que pouca
influência foi aplicada em alguma coisa para que esta seja considerada natural, especialmente
a tecnologia desenvolvida pelos humanos. Angermeier (2000) cita wild como sinônimo e
artificial, ou antropogênico, como antônimos de natural. O autor fundamenta seu argumento
na ideia de que o ser humano não somente vive em ecossistemas, mas altera sua estrutura
física, seus componentes bióticos e seus processos de organização. Diegues (2001) discorda
da visão do movimento preservacionista brasileiro, de que a natureza em seu estado "puro"
(ou natural) seja intocada e intocável, e não se possa admitir que uma área de conservação, ou
seja, uma unidade de conservação proteja também a diversidade cultural, além da diversidade
biológica.

21
Do termo natural, deriva-se um conceito denominado naturalidade, o qual Winter
(2012), em uma ampla revisão sobre o tema, aponta duas definições principais observadas na
literatura.
A primeira definição está relacionada ao grau em que algo é alterado do seu estado
original, logo, pode ser representada por um gradiente contínuo que vai entre extremos, do
totalmente natural ao artificial (ANGERMEIER, 2000; HUNTER, 1996; MACHADO, 2004;
SJÖRS, 1986; WINTER et al., 2010). Winter et al. (2010) define a naturalidade como a
distância entre o estado atual de um ecossistema e seu estado natural potencial. Para o Global
Forest Resources Assessment (FRA, 2000) a naturalidade é expressa pela ausência de
distúrbios antrópicos.
Para Angermeier (2000), a naturalidade não é uma qualidade que só existe em seus
extremos, ou seja, “tudo ou nada”; mesmo ecossistemas intensamente manejados podem
conter alguma naturalidade. O autor salienta que a complexidade dos ecossistemas não
permite que todas as ações do homem sejam precisamente mensuradas, de forma que a
classificação de um ecossistema num gradiente de naturalidade pode ser imperfeita; apesar
disso, o autor defende o uso do conceito para fins de conservação de ecossistemas.
A segunda definição, segundo Winter (2012), refere-se ao estado (de conservação, ou
similar) de um ecossistema sem a influência ou ações perturbadoras aplicadas pelos humanos.
Tratando-se de ecossistemas florestais, algumas definições para este estado são adotadas
(Tabela 1.1 e 1.2), como o termo "floresta primária" e “floresta virgem”, por exemplo.
Segundo a autora, esta definição não é tão adequada quanto a primeira definição (exposta
anteriormente), pois o conceito de naturalidade engloba toda a amplitude entre o artificial ou
alterado (antropizado) e o estado natural de um dado ecossistema.
De acordo com Winter (2012), a ideia de comparar o estado atual de um ecossistema
com um equivalente em um estado natural foi desenvolvido nos países ligados ao idioma
alemão da Europa Central durante o último século. Segundo a autora, a palavra alemã
Naturnähe refere-se à comparação entre o estado atual e um estado natural de referência e
Natürlichkeit refere-se ao estado atual de conservação de uma floresta.
Os níveis (ou categorias) de naturalidade variam conforme o autor. Por exemplo, para
Angermeier (2000) e Šaudytè et al. (2005) a naturalidade das florestas pode ser determinada
através dos seguintes termos/níveis: floresta primitiva, floresta natural, floresta semi-natural,
floresta cultural e floresta artificial. Já a Conferência Ministerial sobre a Proteção das
Florestas na Europa (MCPFE, 2002) definiu três níveis de naturalidade para as florestas
europeias: i) florestas sem distúrbios antrópicos - os processos naturais e a composição de
22
espécies se mantiveram naturais a um nível considerável ou se regeneraram; ii) florestas
seminaturais - podem conter algumas características ecológicas que possibilitam a
manutenção da dinâmica e da biodiversidade, semelhante ao esperado em um estado original;
iii) reflorestamentos - florestas artificiais, distintas do ecossistema original.
Na Tabela 1.1 sintetizamos as categorias observadas na literatura e na Tabela 1.2
constam as descrições delas. Segundo McRoberts et al. (2012) os limites que separam estas
categorias são difíceis de serem definidos, e dependem das caracteristicas locais e da
abordagem que se faz em relação à categorização. Tal fato torna-se crucial neste estudo –
vamos abordá-lo no tópico 1.3.3.
Tabela 1.1. Categorias de naturalidade encontradas na literatura e categorias equivalentes
adotadas no Brasil. Informações extraídas e adaptadas de McRoberts et al. (2011).
Referência Categoria de floresta Categoria equivalente
Lindenmayer e Franklin (1997),
Angermeier (2000),
Šaudytè et al. (2005),
Liira et al. (2007),
Reitz (1965)
Primitiva (Primeval),
Pristina (Pristine),
Virgem (Virgin),
Primária
Floresta primária,
Floresta virgem,
Floresta climácica
Montford (2002) Quase-natural (Near
natural)
Floresta primária alterada,
Floresta secundária
Anderson (1991) Intacta (Intact)
Floresta virgem,
Floresta primária,
Floresta climácica
Angermeier (2000),
Uotila et al. (2002),
Šaudytè et al. (2005),
MCPFE (2007)
Semi-natural (Semi-
natural)
Floresta alterada,
Floresta secundária,
Floresta secundária alterada
Liira et al. (2007)
Convencionalmente
manejada (Conventionally
managed)
Floresta alterada,
Floresta manejada
Angermeier (2000),
Šaudytè et al. (2005) Plantada (Plantation)
Reflorestamentos,
Floresta artificial

23
Tabela 1.2. Complementação da Tabela 1.1 Descrição das categorias de naturalidade
utilizadas por países europeus. Informações extraídas e adaptadas de McRoberts et al. (2011).
Categoria de floresta Descrição
Primitiva (Primeval),
Pristina (Pristine),
Virgem (Virgin)
Floresta nativa que evoluiu sem a intervenção de humanos e
que apresenta estruturas semelhantes a uma floresta climácica
Quase-natural (Near
natural)
Floresta nativa que regenerou naturalmente, que por um longo
período está exposta a um baixo nível de influencias antrópicas
Intacta (Intact) Floresta que apresenta todos os componentes, funções e
estruturas inerentes ao ecossistema
Semi-natural (Semi-natural)
Floresta que se desenvolveu influenciada por atividades
humanas fundamentadas em princípios ecológicos, resultando
em uma floresta com composição e estrutura semelhantes a
uma floresta natural
Convencionalmente
manejada (Conventionally
managed)
Floresta que apresenta certo nível de intensidade de manejo
Plantada (Plantation) Floresta plantada artificialmente e com espaçamento regular,
composta por espécies nativas ou exóticas
Qual a serventia do conceito de naturalidade? McRoberts et al. (2012), em sua revisão
sobre o conceito, sintetizam, baseados em muitos estudos, que o conceito é empregado por
três propósitos: (i) para descrever o valor ecológico, visando elaboração de propostas e ações
de manejo para manter ou aumentar este valor; (ii) para avaliar ações de manejo da
conservação da biodiversidade; (iii) para identificar áreas com florestas conservadas para fins
de estabelecimento de áreas destinadas à conservação.
Dentro dos propósitos citados, muitos estudos focaram em avaliar a naturalidade do
ecossistema florestal como um todo. Entretanto, Šaudytè et al. (2005) afirmam que existe a
possibilidade de avaliar a naturalidade de componentes específicos do ecossistema, como por
exemplo, da composição de espécies e da estrutura. Para determinar a naturalidade, existem
duas maneiras (MCROBERTS et al., 2012): a primeira tem como fundamento o acesso aos
processos ecossistêmicos (PETERKEN, 1996), e a segunda fundamenta-se no grau de
influência humana, a qual é a propulsora das perturbações em um dado ecossistema
(ANDERSON, 1991; ANGERMEIER, 2000; WINTER, 2012). Assim, a naturalidade tem

24
como hemerobia o seu conceito oposto (WINTER, 2012). A palavra hemerobia tem origem
no idioma grego e significa cultivado, domesticado, refinado.
Jalas (1955 apud SUKKOP, 1976; MACHADO, 2004; MCROBERTS et al., 2012;
REIF e WALENTOWSKY, 2008; TROPPMAAIR, 1995) foi o primeiro a propor o conceito.
Os graus de naturalidade e hemerobia podem ser expressos em níveis hierárquicos para
paisagens, espécies ou ecossistemas (JALAS, 1955 apud WINTER, 2012). Jalas propôs
quatro níveis de hemerobia:
i) ahemeorobio – refere-se às paisagens naturais ou que sofreram pequena interferência
antrópica, por exemplo, florestas primárias que sofreram influência de emissões de poluentes;
ii) oligohemeorobio – refere-se às paisagens mais naturais do que artificiais, por
exemplo, florestas manejadas por povos tradicionais;
iii) mesohemeorobio – refere-se às paisagens mais artificiais do que naturais, por
exemplo, pastagens plantadas;
iv) euhemeorobio – refere-se às paisagens artificiais, como plantios florestais
monoespecíficos, ou monoculturas agrícolas.
Para Sukkop (1972) a hemerobia consiste nos efeitos das intervenções antrópicas
diretas ou indiretas nas paisagens ou ecossistemas. O autor adota uma classificação baseada
nos graus de naturalidade e de hemerobia: (i) natural (ahemerobio), (ii) quase-natural
(oligohemerobio), (iii) semi-natural (mesohemerobio), (iv) agricultural (euhemerobio), (v)
quase-cultural (polihemerobio) e (vi) cultural (metahemerobio). O autor leva em consideração
as mudanças na cobertura do solo (uso do solo) e as mudanças na vegetação.
Winter (2012) conclui que o conceito de hemerobia possui menos inconsistências nas
definições adotadas em estudos do que o conceito de naturalidade, provavelmente porque
uma definição concreta foi proposta desde o início. Winter (2012) conclui ainda que existe
uma tendência dos estudos enfatizarem o lado natural do gradiente ou escala, sendo que a
hemerobia enfatiza o lado oposto do gradiente, ou seja, das influências antrópicas em um
dado ecossistema.
As Figuras 1.1 e 1.2 trazem uma representação gráfica de um gradiente teórico de
naturalidade/hemerobia:

25
Figura 1.1. Representação de um gradiente de naturalidade, considerando as influências
antrópicas e os processos ecológicos naturais. Adaptado de Winter et al. (2010).
Figura 1.2. Representação em três dimensões de um gradiente de naturalidade (representado
pela linha cinza). Os componentes e processos ecológicos de um ecossistema florestal são
representados pelos “Elementos e processos naturais”, os quais são expostos às ações
antrópicas perturbadoras (“Elementos antrópicos e “Ações antrópicas”). Adaptado de
Machado (2004).
1.2.1.3. Estudos que adotaram o conceito de naturalidade
De acordo com Winter et al. (2010), a maioria dos estudos que contemplaram o
conceito de naturalidade focaram na detecção de indicadores confiáveis e em descrever
florestas em seu estado (mais) natural, as quais tornaram-se florestas de referência; os autores
afirmam que ambos os procedimentos são necessários antes de desenvolver uma abordagem
aplicável de avaliação da naturalidade de florestas.
Hemerobia
Naturalidade
0% 100%
100% Influência antrópica
Natureza
0%

26
Anderson (1991) desenvolveu uma classificação categórica de naturalidade para o
Parque Nacional Yellowstone. Ele propôs três critérios para avaliar a naturalidade: (i) o grau
de alteração esperado se os seres humanos fossem removidos; (ii) o custo de manutenção da
paisagem para sustentar o sistema em seu estado atual; e (iii) a proporção de espécies nativas
que ainda estão presentes.
Para Loidi (1994) a naturalidade pode ser expressa por uma distância conceitual de
uma floresta alterada até uma climácica, de máxima expressão ecológica. Fisher e Young
(2007) afirmaram, categoricamente, que no caso da Europa, as florestas intocadas não
existem, de modo que a naturalidade só pode ser construída de forma hipotética. Esta questão
levantada sobre ecossistemas que serviriam de referência, ou seja, apresentariam maior
naturalidade, é discutida por muitos autores que julgam que não é possível definir a
naturalidade, pois, segundo eles, não existem mais ecossistemas intocados pelo ser humano
(LANDRES et al., 1998). Segundo Landres et al. (1998) é possível definir a naturalidade sem
ter como base ecossistemas intocados; os ambientes naturais de referência, nos dias atuais, são
a base para o entendimento do sistema ecológico que se deseja estudar ou avaliar o estado de
conservação.
Edarra (1997) propôs uma escala (0 – 10) para estimar a naturalidade de formações
vegetacionais, levando em consideração o grau de influência antrópica. Áreas muito alteradas
pelo meio urbano, onde se observava a ausência de espécies de plantas, receberam o valor 0.
As florestas primárias não exploradas ou outras manifestações de comunidades de plantas em
seu estado natural ganharam o valor 10. As classes intermediárias não foram contempladas
pelo autor.
Para Angermeier (2000) e Šaudytè et al. (2005) a naturalidade das florestas pode ser
determinada através dos seguintes termos/níveis: floresta primitiva; floresta natural; floresta
semi-natural; floresta cultural e floresta artificial. Uma classificação de naturalidade de
florestas reconhecida internacionalmente foi proposta pela UM-ECE/FAO (2000), a qual foi
constituída por três classes categóricas de naturalidade; Šaudytè et al. (2005) fazem uma
observação em relação a esta classificação, argumentando que esta não se adéqua à todas as
situações observadas nos vários países, pois cada um possui suas particularidades.
Uotila et al. (2002) estudaram o grau de naturalidade de florestas boreais protegidas e
não manejadas na Finlândia, concentrando nos efeitos das práticas de manejo na estrutura da
floresta; os autores definiram as florestas como naturais ou seminaturais, de acordo com a
ausência ou presença de sinais históricos de manejo.

27
Machado (2004) elaborou uma escala qualitativa de naturalidade com o intuito de ser
um método de aplicação prático. A escala partiu do valor mínimo de “0” (totalmente artificial)
até “10” (máxima naturalidade), definindo os parâmetros que devem ser levados em
consideração para cada um dos níveis da escala. O autor afirma que seu método pode ser
aplicado de forma consideravelmente rápida se informações como mapas de vegetação, mapas
cartográficos e fotografias aéreas estiverem disponíveis.
Liira e Sepp (2009) investigaram 50 variáveis (qualitativas e quantitativas)
relacionadas à floresta, sub-bosque e chão da floresta. Após uma rotina de modelagens
matemáticas, os autores chegaram a uma lista de indicadores estatisticamente significativos
que podem estimar a naturalidade de Florestas Boreais da Finlândia.
Moravčík et al. (2010) adotaram sete classes de naturalidade para avaliação de
florestas de coníferas na Eslováquia a partir de 25 indicadores, sendo 15 deles ligados à
estrutura e dez ligados à composição de espécies. Os autores ajustaram uma função
discriminante para classificar remanescentes em uma das sete classes de naturalidade,
utilizando os indicadores quantitativos como variáveis preditoras.
Winter et al. (2010) apresentaram um método de acesso a naturalidade baseado no que
denominaram de Relative Quantitative Reference. Este método foi criado para superar a falta
de florestas virgens para comparação; os autores definiram florestas de referência e diversos
indicadores importantes como quantidade de necromassa, conectividade dos remanescentes,
variedade de microhabitats, dentre outros.
Merganič et al. (2012) avaliaram a naturalidade na Reserva Natural Babia Hora na
Eslováquia utilizando uma classificação objetiva baseada em um modelo preditivo gerado
através de uma análise discriminante. Os indicadores utilizados estavam relacionados à
estrutura da floresta, cobertura de gramíneas e líquens, agregação espacial das espécies
arbóreas e necromassa.
McRoberts et al. (2012) criaram um estimador de naturalidade baseado em três
indicadores (diversidade de espécies arbóreas, desvio-padrão dos diâmetros e assimetria da
distribuição de diâmetros) os quais foram eleitos de um conjunto de indicadores considerados
ecologicamente relevantes e aplicáveis pelos IFNs europeus. Como a naturalidade é expressa
por um gradiente que estende-se entre dois extremos (o alterado e o natural), os autores
basearam-se na proposta de Loidi (1994), de que o grau de naturalidade pode ser expresso
pela distância conceitual entre condições observadas em uma dada área (ou unidade amostral),
até uma área referência, considerada como mais próxima do natural. Assim, o estimador
baseou-se na distância euclidiana entre os indicadores de uma floresta que se deseja acessar a

28
naturalidade até os de uma floresta de referência que expressa as características mais
próximas das originais.
Bartha et al. (2006) investigaram se a riqueza de espécies, a estrutura (vertical, etária e
dossel) e a heterogeneidade dos estratos da floresta estão relacionados com critérios de
naturalidade individual (para remanescentes), e naturalidade geral (para o conjunto de
remanescentes).
Para as florestas da Lituânia, Šaudytè et al. (2005) criaram seis classes de naturalidade
que vão desde as florestas primárias até as florestas artificiais. Os indicadores adotados
possuem um caráter qualitativo e estão ligados a componentes estruturais da floresta e a
perturbações antrópicas.
Smelko e Fabrika (2007) desenvolveram uma abordagem sobre a conversão numérica
de indicadores categóricos dentro do programa computacional Natura 2000, o qual foi
desenvolvido para avaliar a naturalidade de florestas da Eslováquia, a princípio. Tierney et al.
(2009) avaliaram a integridade ecológica de área protegidas e criaram um sistema de
caracterização da naturalidade baseado em três classes. Grabherr et al. (1998) avaliaram a
naturalidade de florestas da Áustria baseados no conceito de hemerobia. Nesta abordagem os
autores não utilizaram florestas de referência, pois se dedicaram em avaliar o impacto
antrópico sobre as florestas. No entanto, os autores consideraram uma floresta natural
potencial como um referencial teórico.
1.2.1.4. Indicadores de naturalidade
Segundo McRoberts et al. (2012) muitos critérios são considerados para determinar a
utilidade e relevância de variáveis que podem ser consideradas indicadores de naturalidade,
embora muitas vezes o discernimento dos critérios naturais e antropogênicos não tenham sido
expressos de forma explicita pelos pesquisadores (MACHADO, 2004).
Uma única variável não é suficiente para expressar a naturalidade, assim como todas
as possíveis variáveis não são capazes de serem mensuradas, ainda mais dentro do contexto de
um IFN (McROBERTS et al., 2012). Então, resta selecionar variáveis que podem relacionar e
sintetizar a biodiversidade florestal e o estado de conservação do ecossistema, através da
utilização de indicadores (CHIRICI et al., 2011; CHIRICI et al., 2012; McROBERTS et al.,
2012; NOSS, 1990; WINTER et al., 2010). Seguindo as indicações feitas por Winter et al.
(2010), os indicadores candidatos precisam atender as seguintes premissas:

29
i) os indicadores têm de ser ecologicamente relevantes e indicadores importantes da
naturalidade;
ii) os indicadores têm de ser representativos para a área - por exemplo, a presença de
epífitos em uma floresta pluvial pode expressar maior naturalidade;
iii) os indicadores têm ser passíveis de serem acomodados no contexto de um
programa de monitoramento cíclico.
A abordagem ecossistêmica de acesso à naturalidade (McROBERTS et al., 2012;
PETERKEN, 1996) requer a determinação de poucos indicadores que descrevam os principais
aspectos e processos inerentes ao ecossistema. McRoberts et al. (2012) afirmam que tal
procedimento é difícil e tem de ser cuidadosamente realizado de modo a encontrar indicadores
realmente representativos. Já na abordagem ligada à hemerobia, os indicadores estão
geralmente associados a sinais de intervenção e exploração realizados pelos humanos (FRA,
2000; UOTILA et al., 2002).
Os estudos que trataram da naturalidade utilizaram uma gama de variáveis
indicadoras. Na Tabela 1.3 condensamos um pacote de indicadores adotados nos estudos mais
recentes.
Para exemplificar como os indicadores são determinados, citamos McRoberts et al.
(2012), os quais avaliaram dez variáveis (diversidade de espécies, número de estratos de
altura, número de árvores de grande porte, idade das árvores (dendrocronologia), número de
microhabitats, necromassa, volume, sinais de exploração de madeira e sinais de manejo
silvicultural) com o intuito de avaliar seus potenciais e praticabilidade para estimar a
naturalidade utilizando dados provenientes de IFNs participantes da Ação COST E431. Os
autores realizaram uma triagem dos indicadores baseados na aplicabilidade e relevância
ecológica destes.
Winter et al. (2010) selecionaram os indicadores candidatos baseados em informações
de várias fontes, incluindo a Convenção sobre Diversidade Biológica de 1992, os indicadores
de manejo florestal sustentável da Europa (MCPFE, 1997, 2003ab), dentre outros. Os autores
também consultaram a literatura especializada em ecologia florestal e o conhecimento
especializado de cientistas ligados à Bavarian Forest National Park.
1 European CoOperation in Science and Technology (COST). A Ação COST E43
(Harmonisation of National Forest Inventories in Europe: Techniques for Reporting) está
ligada a harmonização dos IFNs da Europa.

30
Tabela 1.3. Síntese dos indicadores utilizados em estudos de naturalidade. Qt: variável
quantitativa; Ql: variável qualitativa; E: ligada a processos ecossistêmicos; H: ligada a
processos hemeróbicos.
Referência Indicador adotado Natureza Fonte
McRoberts et al.
(2012)
Assimetria da distribuição diamétrica (gdap)
Desvio padrão dos diâmetros (Sdap)
Diversidade - Índice de Shannon (H')
Estrutura vertical
Qt
Qt
Qt
Qt
E
E
E
E
Winter et al.
(2010)
Necromassa
Conectividade dos remanscentes
Sanidade das árvores
Microhábitats
Árvores bifurcadas
Comparação com uma floresta potencial
Qt
Qt
Ql
Qt
Qt
Qt
Qt
E
E
E
E
E
E
E
Liira e Sepp
(2009)
Presença e ausência de distúrbios antrópicos
Índice de intensidade de manejo
Variáveis estruturais
Ql
Qt
Qt
H
H
E
Moravčík et al.
(2010)
Variáveis estruturais Qt E
Composição (riqueza e uniformidade) Qt E
Machado (2004) Aspectos e propriedades ecológicas
Influência antrópica
Ql
Ql
E
H
Šaudytè et al.
(2005)
Aspectos estruturais
Composição de espécies
Influência de elementos antrópicos
Qt
Ql
Ql
E
E
H
Uotila et al.
(2002)
Dados históricos sobre manejo
Dados históricos sobre incêndios
Sinais de manejo
Idade da floresta (dendrocronologia)
Estoque de volume
Distribuição diamétrica
Ql
Ql
Ql
Ql
Qt
Qt
H
H
H
E
E
E
1.2.1.5. Florestas referência de naturalidade
As florestas de referência são o marco para as comparações entre florestas mais
alteradas e florestas naturais (McROBERTS et al., 2012; LANDRES et al., 1998; LOIDI,
1994; WINTER et al., 2010). Porém, em toda a Terra, sabemos que poucas áreas florestadas
estão em seu estado natural; no caso da Floresta Atlântica sensu strictu, as florestas são, em
sua maioria, resultantes da sucessão secundária (RIBEIRO et al., 2011). Ellen (1989) afirma
que a natureza em estado puro já não existe, e Winter et al. (2010) são rigorosos e afirmam
que não existem mais habitats intocados pelas emissões de gases; consequentemente, uma

31
floresta 100% natural não pode mais ser encontrada para servir de referência em estudos de
naturalidade (WINTER et al., 2010). Além das emissões, outros estresses têm atingido os
processos naturais dos ecossistemas (FREEDMAN, 1989), como, por exemplo, a
contaminação por defensivos agrícolas (CDB, 2010).
Segundo Landres et al. (1998), é possível definir a naturalidade sem ter como base
ecossistemas intocados; os ambientes de referência, nos dias atuais, são a base para o
entendimento do sistema ecológico e que se deseja estudar ou avaliar o estado de
conservação. Entretanto, Winter (2012) afirma que, antes de conduzir um estudo de
determinação da naturalidade de um ecossistema florestal, é necessário definir concretamente
o conceito de floresta de referência. Winter et al. (2010) propôs que, mesmo não existindo
florestas de referência (100% naturais), é possível considerar remanescentes florestais mais
naturais (mais próximos do seu estado natural) como referências. Tal procedimento também
foi adotado por McRoberts et al. (2012) em florestas temperadas do norte do Estados Unidos.
O diagrama abaixo representa essa ideia:
Ecossistema não alterado:
Ecossistema alterado A:
Naturalidade
Hemerobia
100% 0%
0% 100%
Natureza
Influência antrópica
Naturalidade
Hemerobia
100% 0%
0% 100%
Natureza
Influência antrópica

32
Ecossistema alterado B:
Figura 1.3. Diagrama referente aos remanescentes de referência considerados como 100%
naturais no gradiente de naturalidade. Adaptado de Winter et al. (2010).
Gibbons et al. (2008) e McRoberts et al. (2012) apresentaram métodos para
identificação de parcelas com maior naturalidade (referências) sem o uso de classes pré-
estabelecidas. Winter et al. (2010) realizaram um estudo de avaliação da naturalidade das
florestas do Bavarian Florest National Park na Alemanha e determinaram áreas de florestas
primárias como sendo as referências para comparações com áreas manejadas, áreas de borda,
áreas core e áreas de recreação do parque.
Para Gibbons et al. (2008), o estado de referência está relacionado à variação na
vegetação nativa que demonstra pouca evidência de modificação por humanos desde a
colonização européia na Austrália. A partir de dados de 462 locais de vegetação nativa em
uma paisagem fragmentada no sudeste da Austrália, os autores propuseram um método
objetivo de determinar condições de referência de variáveis utilizadas como indicadoras de
biodiversidade (por exemplo, densidade de árvores por classe de diâmetro, árvores ocas,
regeneração de espécies arbóreas, necromassa, cobertura vegetal por estrato vertical,
cobertura de serapilheira, cobertura de criptógamas e riqueza de espécies nativas). Os autores
utilizaram modelos aditivos generalizados (GAM) calibrados com indicadores que
representam as medidas de modificação humana desde a colonização européia (cobertura de
plantas exóticas, número de tocos, evidências de extração de lenha, evidência da presença de
coelhos, evidências de pastoreio recente de gado e uso do solo no entorno) e dados de
variação ambiental (precipitação anual média, temperatura anual média, insolação e
inclinação). Para estes autores, a determinação das referências baseia-se nos valores mínimos
das variáveis explicativas e significativas previstas pelo modelo, as quais representam as
modificações desde a colonização.
Naturalidade
Hemerobia
100% 0%
0% 100%
Natureza
Influência antrópica

33
McRoberts et al. (2012) determinaram referências através da avaliação de Unidades
Amostrais do IFN dos Estados Unidos que apresentaram maior naturalidade. Foi utilizado um
processo de triagem das Unidades Amostrais, de acordo com o cumprimento de critérios
condicionais como ausência de espécies exóticas, ausência de sinais de atividade humana,
diversidade da estrutura vertical, dentre outros. Bartha et al. (2006) definiram florestas
referenciais hipotéticas e, baseados nelas, por meio de um processo hierárquico de três níveis,
determinaram indicadores quantitativos e qualitativos de naturalidade. Os autores não
deixaram explicita a definição destas florestas de referencia e como as determinaram.
1.2.2. FITOFISIONOMIA ALVO: FLORESTA MISTA COM ARAUCÁRIA
1.2.2.1 Caracterização geral
A Floresta Mista Lati-aciculifoliada (OLIVEIRA-FILHO, 2009), também classificada
como, Floresta com Araucária (KLEIN, 1978) e Floresta Ombrófila Mista (LEITE e KLEIN,
1990; IBGE, 2012), é uma fitofisionomia florestal que tem maior expressão territorial nos
planaltos do Paraná, Santa Catarina e Rio Grande do Sul. Ela também ocorre na forma de
pequenos relictos em pontos altos das serras da Mantiqueira, dos Órgaos, do Caparaó,
Paranapiacaba e Paranapanema nos estados de Minas Gerais, Rio de Janeiro, Espírito Santo,
São Paulo, e em países vizinhos como Argentina e Paraguai (BACKES, 2009). No estado de
Santa Catarina (Figura 1.4) ela é principalmente encontrada entre as altitudes de 500 e 1000 m
(BACKES, 2009), ocorrendo também em áreas com altitude superior a 1000 m (UHLMANN
et al., 2013). No oeste do estado, entre os paralelos 25°30‟ e 27°27‟ S, ela adentra o território
da Argentina, na província de Missiones e, em direção ao litoral brasileiro, chega até os
municípios de Sombrio e Lauro Muller, e em certos municípios situados na Serra do
Tabuleiro em Santa Catarina (BACKES, 2009).
No sistema de classificação de Oliveira-Filho (2009) a concepção do termo Floresta
Mista Lati-aciculifoliada, que daqui em diante será referida como Floresta Mista com
Araucária, está vinculada à mistura de angiospermas e gimnospemas. O sistema também
considera os aspectos topológicos das espécies arbóreas, neste caso das folhas - as acículas
são vistas em espécies características desta tipologia florestal, que coexistem com espécies
latifoliadas. Mais especificamente, o termo “mista” vem da fusão de elementos provenientes
da flora afro-brasileira, a qual exerce maior participação na composição florística, e de
elementos da flora austral-andino-antártica, que, em certas regiões, podem contribuir
significativamente para o aspecto fisionômico da vegetação (REITZ e KLEIN, 1966; LEITE,

34
2002; IBGE, 2012). Dentro do conjunto de espécies de origem austral-andino-antártica, a
conífera Araucaria angustifolia Bert. O.Kuntze imprime grande importância ao aspecto
fisionômico e estrutural da floresta, podendo constituir agrupamentos densos e quase puros.
Em formações florestais multiestratificadas esta espécie apresenta-se como elemento
emergente (KLEIN, 1960) ao dossel, composto, predominantemente, por espécies de
Lauraceae (KLEIN, 1966). Outras espécies de origem austral-andino-antártica frequentemente
encontradas são Dicksonia sellowiana Hook., Drimys brasiliensis Miers., Lamanonia ternata
Vell., Podocarpus lambertii Klotzsch ex Endl., Prunus sellowii Koehne, Weinmania humilis
Engl. e espécies do gênero Roupala (KLEIN, 1978; LEITE, 2002).
Figura 1.4. Mapa fitogeogrpafico de Santa Catarina (KLEIN, 1978). Fitofisionomias
classificadas segundo o sistema de Oliveira-Filho (2009), exceto “Vegetação litorânea”
(KLEIN, 1978).
De acordo com Leite (2002), as variações climáticas parecem determinar uma
tendência dos elementos florísticos temperados ocuparem as regiões mais altas do planalto
meridional; enquanto isso, os elementos da flora tropical tendem a expandir-se através dos
vales e das encostas das serras, sobre os flancos planaltinos. A Floresta Mista com Araucária,
segundo Klein (1960), divide-se em associações que retratam os diferentes estágios temporais
de desenvolvimento, do menos para o mais desenvolvido. Klein (1978) definiu dois grandes
grupos de formações para esta fitofisionomia no estado de Santa Catarina:

35
a) Floresta de Araucária, caracterizada por indivíduos de A. angustifolia emergentes e
de grande porte, dossel denso, composto por angiospermas bem desenvolvidas, com
predomínio da família Lauraceae;
b) Florestas dos Faxinais, caracterizada por indivíduos de A. angustifolia de menor
porte e dossel menos denso formado, predominantemente, por espécies de Myrtaceae e
Aquifoliaceae, entremeadas de taquarais e carazais (Merostachys spp. e Chusquea spp.).
Dentro destes grupos, Klein (1978) prossegue com a descrição das formações e subdivide os
grandes grupos de forma mais específica.
Na região norte de Santa Catarina (município de Canoinhas), na bacia dos Rios Iguaçu
e Negro, bem como na parte superior das bacias dos afluentes do Rio Uruguai, existiam
grandes formações contínuas de A. angustifolia, correspondendo ao estrado das
megafanerófitas, com o denso estrato das macrofanerófitas dominado por Ocotea porosa
(Nees) Barroso (canela-imbuia) e Sloanea lasiocoma K. Schum. (sapopema). No estrato das
mesofanerófitas ocorria (predominantemente) Ilex paraguariensis A. St.-Hil. (erva-mate).
Na bacia drenada pelo Rio Canoas, a floresta caracterizava-se pelos agrupamentos
densos e em manchas de A. angustifolia (pinhais), por vezes interrompidos por campos com
capões. Nos pinhais densos, o estrato das macrofanerófitas era dominado por Ocotea
pulchella (Nees) Mez (canela-lageana), com a presença representativa de outras Lauraceae
como Nectandra lanceolata Ness (canela-amarela), Ocotea puberula (Rich.) Nees (canela-
guaicá), Cryptocaria aschersoniana Mez (canela-fogo) e Nectandra grandifolia Nees (canela-
fedida). Espécies seletivas como Capsicodendron dinisii Schwanke (pimenteira), Matayba
elaeagnoides Radlk. (camboatá-branco), Myrcianthes gigantea (D. Legrand) D. Legrand
(araçazeiro), dentre outras, possuiam afinidade com o ambiente formado por O. pulchella.
A floresta no extremo oeste do estado apresentava agrupamentos de A. angustifolia
com o estrato das macrofanerófitas com abundância de espécies de Fabaceae, como Apuleia
leiocarpa (Vogel) J.F.Macbr. (grápia), Parapitadenia rigida (Benth.) Brenan (angico-
vermelho), Cordia americana (L.) Gottshling & J.E.Mill. (guajuvira), Diatenopteryx
sorbifolia Radik. (maria-preta) e Lonchocarpus campestris Mart. ex Benth. (rabo-de-mico).
Outras espécies como Cedrela fissilis Vell. (cedro), Cordia trichotoma (Vell.) Arrab. ex
Steud. (louro-pardo), Myrocarpus frondosus Allemão (cabreúna), dentre outras, eram
expressivas no estrato das macrofanerófitas.
Na zona da Floresta Latifoliada Pluvial (OLIVEIRA-FILHO, 2009), ocorriam os
núcleos de pinhais formados por indivíduos antigos e elementos da Floresta Mista com
Araucária e da Floresta Latifoliada Pluvial compondo o estrato das macrofanerófitas.

36
As Florestas de Faxinais são consideradas por Klein (1978) áreas de transição entre a
Floresta Latifoliada Pluvial e a Floresta Mista com Araucária, sendo esta formação composta
por espécies que ocorrem em solos pouco profundos e encostas declivosas. As Florestas de
Faxinais ocorrem nas encostas da Serra do Mar e Serra Geral e podem ser encontradas em
áreas de pinhais das outras formações citadas anteriormente. O autor divide esta formação em
quatro subgrupos de regiões, argumentando que existem semelhanças sensíveis na
composição florística, entretanto, são notadas diferenças na distribuição e abundância das
espécies. São estes subgrupos: Serra do Tabuleiro, Serra Geral, Campo Erê (Serra da Fartura e
Serra do Capanema) e Guedes (Serra do Tigre).
O Manual Técnico da Vegetação Brasileira (IBGE, 2012) reconhece quatro formações
distintas2 para a Floresta Mista com Araucária, as quais foram determinadas pela faixa
altitudinal:
Aluvial: ocorre em planícies aluviais, onde a espécie A. angustifolia associa-se com
diferentes espécies, conforme os aspectos geomorfológicos e topográficos. Em altitudes mais
elevadas são encontradas Podocarpus lambertii e Drimys brasiliensis. Em altitudes menos
elevadas a A. angustifolia associa-se com espécies de Lauraceae, Luehea divaricata Mart. ex
Zucc, Blepharocalyx salicifolius (Kunt) O. Berg e Sebastiania commersoniana (Baill.) L. B.
Sm. ex Dows;
Submontana: essa formação ocorre em altitudes inferiores a 400 m e consiste em
pequenas disjunções localizadas nas bordas da Serra Geral. Tais disjunções são vistas no
município de Lauro Muller (Santa Catarina), onde, na década de 1950, havia 12.000
indivíduos de A. Angustifolia. Atualmente, restam no local poucos indivíduos desta espécie
associados às culturas agrícolas e às florestas secundárias;
Montana: ocorre entre 400 e 1000 m de altitude e é caracterizada pela associação de A.
angustifolia e Ocotea pulchella, Cryptocarya aschersoniana, Ilex paraguariensis e Nectandra
megapotamica (Spreng.) Mez na bacia do Rio Canoas; no planalto norte de Santa Catarina,
observa-se A. angustifolia associada a Ocotea porosa;
Altomontana: ela ocorre em altitudes superiores a 1000 m nas encostas das colinas
diabásicas em mistura com arenitos que constituem a Formação Serra Geral. Espécies
características do estrato arbóreo são: Podocarpus lambertii, Drimys brasiliensis, Cedrela
2 Para uma representação gráfica das formações, ver IBGE (2012, p.83).

37
fissilis, e espécies de Lauraceae e Myrtaceae. Famílias características do estrato arbustivo:
Rubiaceae, Myrtaceaee, Winteraceae, Lauraceae e Meliaceae.
Saint-Hilare (1978, p.49) e Leite (2002) salientam que a Floresta Mista com Araucária
coexiste com a Campina (OLIVEIRA-FILHO, 2009), Campos Naturais (KLEIN, 1978) ou
Estepe Ombrófila (LEITE, 2002), de forma que a A. angustifolia, e outras espécies pioneiras,
invadem esta fitofisionomia campestre (DUARTE et al., 2009; KLEIN, 1960; REITZ e
KLEIN, 1966). Este fato dificulta o estabelecimento preciso dos limites florísticos entre as
formações florestais e campestres. Pesquisadores afirmam (baseados em métodos de datação3)
que a expansão da Floresta Mista com Araucária pela Campina, a qual (supostamente) ocorria
abundantemente no último glacial máximo, se deu através de formações florestais que
ocorriam ao longo de corpos d‟água lóticos, seguindo uma mudança climática, de um clima
mais seco para um clima mais úmido (BAUERMANN e BEHLING, 2009; BEHLING et al.,
2004; BEHLING et al., 2001). A ocorrência da Campina nos dias atuais, diante de um clima
quente e úmido, é alvo de pesquisas (citadas em PILLAR et al., 2009). Este fato está,
provavelmente, ligado à utilização histórica do fogo pelos povos pré-Colombianos e pela ação
de animais pastadores (BEHLING et al., 2004; GALVÃO e AUGUSTIN, 2011; PILLAR et
al., 2009).
Nas escarpas rochosas dos Aparados da Serra Geral, geralmente em altitudes acima de
1000 m (IBGE, 2012), ocorre uma fitofisionomia florestal peculiar denominada Nanofloresta
Latifoliada Nebular (OLIVEIRA-FILHO, 2009), a qual frequentemente encontra-se encoberta
por neblina. A Nanofloresta Latifoliada Nebular apresenta dossel de no máximo 8 m, formado
por árvores tortuosas devido às condições climáticas e edáficas adversas. São espécies
características: Drimys brasiliensis e D. angustifolia; Myrceugenia euosma (O. Berg) D.
Legrand, Ilex microdonta Reissek, Euplassa nebularis Rambo & Sleumer, Podocarpus
lambertii e Weinmannia humilis (MMA/IBAMA, 2004).
Falkenberg (2003) caracteriza a Nanofloresta Latifoliada Nebular como uma
vegetação florestal em posição intermediária entre a Floresta Latifoliada Pluvial
Superomontana e a Floresta Mista com Araucária, ou como um ecótono das vegetações
florestais desenvolvidas (Floresta Latifoliada Pluvial nas encostas altas e Floresta Mista com
Araucária no planalto) com formações de Campina. Oliveira-Filho et al. (2013) constataram
que a Nanofloresta Latifoliada Nebular compartilha espécies das formações superomontanas
da Floresta Latifoliada Pluvial e da Floresta Mista com Araucária. 3 Para uma exposição sobre métodos de datação e questionamentos sobre a acurácia dos
mesmos, veja o capítulo 6 em Lourenço (2012).

38
A segregação da Nanofloresta Latifoliada Nebular como uma fitofisionomia particular
é motivo de discussão (FALKENBERG e VOLTOLINI, 1995). Oliveira-Filho (2009) a
considera uma fitofisionomia distinta, assim como Klein (1978). Entretanto, Leite (2002) a
considera como uma formação da Floresta Mista com Araucária. Tais divergências de
opiniões não reduzem a importância desta vegetação - Falkenberg (2003) registrou uma
enorme riqueza de espécies e mostrou que pelo menos 10% das espécies vasculares
encontradas na Nanofloresta Latifoliada Nebular são endêmicas, o que reforça sua
peculiaridade.
1.2.2.2 Histórico de degradação da Floresta Mista com Araucária
O processo de degradação da Floresta Mista com Araucária iniciou-se com a
colonização européia e intensificou-se no decorrer da ocupação progressiva das terras, a qual
foi impulsionada pela atratividade do desenvolvimento da economia na região sul (MÄHLER
JÚNIOR e LAROCCA, 2009). Esta fitofisionomia, no inicio do processo de colonização do
Brasil, cobria, estimadamente, cerca de 200.000 km² (BAUERMANN e BEHLING, 2009);
desta área total, cerca de 60% ocorriam no Paraná (INOUE et al, 1984 apud BACKES, 2009),
30% em Santa Catarina e 10% no Rio Grande do Sul (REITZ et al., 1983 apud BACKES,
2009).
Certamente, os primeiros visitantes "estrangeiros" que chegaram aos planaltos do sul
do Brasil impressionaram-se com a exuberância das associações de A. angustifolia.
Observando as descrições feitas pelos primeiros botânicos e naturalistas que contemplaram o
planalto sul do Brasil, é possível imaginar a floresta antes de ser intensamente explorada. A
seguinte descrição, feita pelo botânico francês Saint-Hilaire4, nos permite ter uma visão da
beleza da vegetação (SAINT-HILARE, 1978):
“É a Araucaria brasiliensis que, por sua altura, pela imponência e
elegância de suas formas, por sua imobilidade e pelo verde-escuro de
suas folhas contribui, mais do que qualquer outra coisa, para dar uma
fisionomia característica aos Campos Gerais. Em alguns trechos essa
pitoresca árvore, elevando-se isolada ao meio das pastagens, expõe à
nossa admiração toda a beleza do seu talhe e faz ressaltar, pelos
matizes sombrios de suas folhas, o verde tenro de relva que cresce
sobre ela.”
4 Augustin François César Prouvençal de Saint-Hilaire (1779-1853).

39
Com um sentimento não diferente dos primeiros visitantes, diante das belas formações
florestais, estudiosos se dedicaram em analisar e compreender a composição e os aspectos
ecológicos da Floresta Mista com Araucária (KLEIN, 1960, 1978, 1984; LINDMAN, 1906;
MAACK, 1953; RAMBO, 1951, 1953, 1956; REITZ e KLEIN, 1966). Estes traçaram as
bases do conhecimento sobre a composição, estrutura e dinâmica do ecossistema florestal em
questão.
Infelizmente, apesar do conhecimento desenvolvido, Schaaf et al. (2006b) apontaram
um fato verdadeiro que pouco é comentado entre os autores que estudam esta floresta e
revisam a história de sua exploração. Os autores afirmam que a floresta não foi conhecida
adequadamente antes de ser explorada intensivamente. A maior parte dos remanescentes
existentes de hoje encontra-se descaracterizada e simplificada. Eduardo (1974) fez uma busca
ampla em relação aos aspectos históricos da exploração e da economia madeireira em Santa
Catarina no período de 1930 a 1972 e Cabral e Cesco (2008) relatam interessantes
informações históricas e recentes sobre a exploração madeireira do Sul e Sudeste do Brasil.
A Floresta Mista com Araucária foi a mais significante para a indústria madeireira do
Sul do país, pelo menos por 150 anos de exploração, quando a araucária e a canela-imbúia
foram as espécies de destaque durante este período exploratório (REIS et al., 2007). O setor
madeireiro de Santa Catarina foi um dos impulsionadores do crescimento econômico do
estado no início do século XX até a década de 1970 (EDUARDO, 1974; GOULARTI FILHO,
2002, 2008; MÄHLER JÚNIOR e LARROCA, 2009): entre os anos de 1915 e 1929 o valor
de madeira exportada aumentou 22 vezes, principalmente devido à colonização do oeste do
vale do Rio do Peixe; aumentaram as quantias exportadas para Argentina (via Rio Uruguai) e
para o Rio de Janeiro, no decorrer da construção de ferrovias integrando a Região Sul à
Sudeste e ligando o interior catarinense aos portos (EDUARDO, 1974; GOULARTI FILHO,
2002).
Uma preocupação com a intensa e frenética exploração foi expressa pelo Instituto
Nacional do Pinho (fundado em 1941), o qual já via a necessidade de promover e subsidiar o
reflorestamento (INSTITUTO NACIONAL DO PINHO, 1948). Os benefícios fiscais
oferecidos pelo governo entre as décadas de 1960 e 1980, regulamentados pelo Instituto
Brasileiro de Desenvolvimento Florestal (IBDF), criado pelo decreto-lei nº 289 de 1967,
foram, sem dúvida, outro fator que impulsionou a derrubada de florestas nativas para a
implantaçao de reflorestamentos monoespecíficos, que em sua maioria foram de espécies
exóticas, como as do gênero Pinus. Por exemplo, no estado do Paraná, entre os anos de 1966 e

40
1976 a área reflorestada com Pinus spp., fruto desses incentivos fiscais, passou de 221,30 ha
para 655.545,82 ha (BREPOHL, 1980).
Alguns dados históricos são assombrosos e nos mostram a intensidade da exploração
madeireira ocorrida no Sul do Brasil (EDUARDO, 1974; GOULARTI FILHO, 2008). Mähler
Júnior e Larroca (2009) citam, em uma revisão sobre o tema, dados estimados da exploração
de araucárias. Segundo os autores, entre 1930 e 1940 a Floresta Mista com Araucária de Santa
Catarina era responsável por mais da metade da exportação de madeira do país; no Estado
havia mais de 381 serrarias que produziam cerca de quatro milhões de dúzias de tábuas ao
ano. Mähler Júnior e Larroca (2009) citam ainda outro dado espantador: no ano de 1968 o
Brasil exportou mais de um bilhão de metros cúbicos de madeira, e 45% destes foram
retirados das florestas de Santa Catarina.
O fogo foi outro agente degradador. Após a exploração madeireira, o fogo era uma
prática comum aplicada com o intuito de “limpar” a área, uma espécie de preparo para outros
usos do solo, como cultivos agrícolas e pastagens (BORGES, 2005).
Após a exploração das araucárias no vale do rio do Peixe pela empresa Lumber5, os
pinhais do município de Lages (Santa Catarina) e região atraíram olhares na década de 1950,
quando ocorreu um surto migratório de colonos vindos do Rio Grande do Sul, os quais
alavancaram o processo de exploração madeireira na região (MARTENDAL, 1980). Este
mesmo autor relata que além de inúmeras serrarias, a região de Lages sediava grandes
depósitos que armazenavam as madeiras exploradas na região. Tais depósitos tinham a
finalidade de armazenar a madeira explorada que seria transportada e comercializada em
outras regiões, e até mesmo exportada via porto de Itajaí (Santa Catarina). Do ponto de vista
social, Moretto (2010) aponta a importância da exploração da Floresta Mista com Araucária
no desenvolvimento da região do planalto de Santa Catarina, devido ao surgimento de um
novo grupo social ascendente formado pelos donos das madeireiras. Segundo Martendal
(1980), entre 1950 e 1970, no pico da atividade madeireira, a população da região do
município de Lages quadruplicou, de 20.000 para 80.000 habitantes.
1.2.2.3 Atual estado de conservação da Floresta Mista com Araucária
Qual a atual situação dos remanescentes de Floresta Mista com Araucária de Santa
Catarina?
5 A Lumber era uma empresa (americana) madeireira e colonizadora que subsidiou a empresa
construtora da ferrovia do Sul do Brasil, a São Paulo – Rio Grande do Sul.

41
Esta questão torna-se fundamental para o desenvolvimento deste estudo, pois a história
do ciclo de exploração madeireira em Santa Catarina é conhecida e temos uma base de
conhecimento sobre a floresta, a começar pelos estudos pioneiros de Klein6 e Reitz
7, além dos
estudos de outros autores. Destacam-se os trabalhos de Jarenkow e Baptista (1987), Longhi et
al. (2006), Schaaf et al. (2006ab), Canalez et al. (2006), Ribeiro et al. (2007), Vibrans et al.
(2008), Bauermann e Behling (2009), Jarenkow e Budke (2009), Fonseca et al. (2009),
Klauberg et al. (2010) e Vibrans et al. (2011).
Apesar dos estudos de Klein e Reitz terem sido desenvolvidos no final do ciclo da
exploração madeireira, eles foram realizados em florestas conservadas, ou seja, com
composição e estrutura características do ecossistema florestal em questão. Não obstante, eles
servem como referência para os estudos mais recentes relacionados à fitofisionomia.
Existe um número considerável de trabalhos que estudaram a Floresta Mista com
Araucária, entretanto, estes estão restritos a níveis locais, e possuem metodologias de
amostragem diferentes entre si, o que dificulta a integração e comparação dos dados entre as
regiões. O IFFSC trouxe uma nova dimensão ao entendimento da atual situação das florestas
do estado, devido à sua amostragem sistemática que cobriu todo o estado e à sua dedicação
em amostrar e levantar muitos dados.
Sevegnani et al. (2013a) discorrem sobre a atual situação da Floresta Mista com
Araucária no estado de Santa Catarina com base nos dados obtidos pelo IFFSC.
Primeiramente, apontaram uma diminuição da cobertura desta tipologia florestal, de uma área
de aproximadamente 42.851 km² (KLEIN, 1978) para 13.741 km² (VIBRANS et al., 2013).
Esta cobertura florestal remanescente é, em sua quase totalidade, resultado da sucessão
vegetal secundária e encontra-se altamente fragmentada e degradada. Os levantamentos de
campo realizados pelo IFFSC registraram 1.107 espécies de plantas vasculares, o que aponta a
alta biodiversidade contida nestes remanescentes florestais (GASPER et al., 2013).
Entretanto, esta riqueza de espécies está exposta ao declínio perante as muitas pressões
antrópicas; muitas das espécies arbóreas são singletons ou doubletons (representadas por um e
dois indivíduos na amostra, respectivamente) (OLIVEIRA et al., submetido). Rezende et al.
(2014) encontraram que aproxidamente 72% das espécies arbóreas presentes em Santa
Catarina possuem distribuição geográfica restrita. Tal fato é preocupante, pois, mesmo que
tais espécies sejam naturalmente raras (sensu Rabinowitz et al. [1986]), a questão da
necessidade de medidas conservacionistas não se altera.
6 Roberto Miguel Klein (1923-1992)
7 Raulino Reitz (1919-1990)

42
Em toda amostragem da Floresta Mista com Araucária, o IFFSC encontrou 368
espécies (com DAP ≥ 10,0 cm) de 176 gêneros, distribuídos em 69 famílias. Cinco das 368
espécies são pteridófitas (Dicksonia sellowiana, Alsophila setosa Kaulf., Cyathea
corcovadensis (Raddi) Domin, Cyathea phalerata Mart. e Cyathea degaldii Sternb.), três são
gimnospermas (A. angustifolia, Podocarpus lambertii e P. sellowii Klotzsch ex Endl.) e 360
são angiospermas (MEYER et al., 2013).
Uhlmann et al. (2013) investigaram a existência das formações descritas por Klein
(1978) nos dias atuais; os autores aplicaram uma técnica multivariada de ordenação (DCA)
em dados florísticos do IFFSC e concluíram que as formações descritas por Klein (1978)
parecem emergir dos dados; entretanto, foi constatada uma grande homogeneização estrutural
da floresta, a qual encontra-se floristica e estruturalmente simplificada.
A diversidade genética é muitas vezes esquecida pelos ecólogos conservacionistas,
porém detém grande importância na conservação da biodiversidade em longo prazo. Reis et
al. (2012), dentro do projeto IFFSC, estudaram a diversidade genética de algumas espécies e
constataram que são claras as evidências de limitações de fluxo gênico entre os fragmentos.
Apesar de tais conclusões, os autores afirmam que a diversidade genética ainda é passível de
ser resgatada; eles sugerem a identificação de áreas com grande diversidade genética para
conservação e futuras coletas de sementes para fins de restauração da diversidade genética das
florestas.
A atual fragilidade deste ecossistema, causada pelas pressões antrópicas, reflete não
somente na redução da biodiversidade e dos outrora definidos padrões estruturais da
vegetação. Atributos como a produtividade primária e secundária, e a eficiência de ciclagem
de nutrientes também são afetados, e a dominância de espécies exóticas e oportunistas é
facilitada (RAPPORT e WHITFORD, 1999). Sabemos que ambos os conjuntos de processos
(perturbações e consequências) tornam-se cumulativos, aumentando a vulnerabilidade do
ecossistema e da sociedade, principalmente as partes que possuem relação mais estreita com a
floresta (KASPERSON et al., 1995)
O conjunto de informações apresentados neste tópico, e nos dois anteriores a este,
serve como um panorama do ecossistema alvo deste estudo sobre naturalidade. Apresentamos
as riquezas, as pressões contrárias à biodiversidade e o status de conservação. Por fim, este
parágrafo assume um carácter de desfecho e “prelúdio” para a proposta que nos
comprometemos a elaborar.

43
1.3. A PROPOSTA: A APLICAÇÃO DO CONCEITO DE NATURALIDADE COMO
MEIO DE ACESSAR O ESTADO DE CONSERVAÇÃO DOS REMANESCENTES
DE FLORESTA MISTA COM ARAUCÁRIA DE SANTA CATARINA
1.3.1. APRESENTAÇÃO
O presente capítulo tem o objetivo de responder a pergunta inicial: É possível adaptar
o conceito de naturalidade e a sua determinação aos remanescentes de Floresta Mista com
Araucária de Santa Catarina?
E mais: Qual é a importância desta pesquisa para nossa realidade?
Além de esclarecer estas questões, uma proposta para a quantificação da naturalidade
será redigida, seguindo uma disposição lógica e progressiva, conforme sugestões observadas
na literatura. Na introdução são abordados o contexto do estado de Santa Catarina e o IFFSC,
assim como proposições iniciais. No decorrer deste capítulo serão apresentados:
nos tópicos 1.3.2 e 1.3.3 - definições de princípios e conceitos básicos que
fundamentam a quantificação da naturalidade, adaptados à realidade histórica, social e
ecológica de Santa Catarina e da Floresta Mista com Araucária;
nos tópicos 1.3.4 a 1.3.7 - propostas metodológicas em relação à escolha dos
indicadores, das florestas de referência e desenvolvimento da medida de naturalidade.
1.3.2. INTRODUÇÃO
Informações qualitativas e quantitavias sobre as florestas, em um contexto geográfico
amplo, são geradas e fornecidas por programas de inventário florestal. O estado de Santa
Catarina saiu na frente e foi o primeiro estado do Brasil a finalizar seu inventário florestal, de
acordo com a metodologia do IFN-Brasil (FREITAS et al., 2010). Podemos afirmar que Santa
Catarina tem uma grade adensada de pontos amostrais medidos em campo, o que viabiliza a
visualização do estado de conservação das florestas nativas. Na região fitoecológica da
Floresta Mista com Araucária foram implantadas 155 Unidades Amostrais de área nominal de
4.000 m² (VIBRANS et al., 2010), de acordo com a seguinte definição de floresta: áreas
contínuas de vegetação arbórea com um dossel de no mínimo 10,0 m de altura e área basal de
no mínimo 10,0 m².ha-1
(VIBRANS et al., 2013).
Posto que os ecossistemas florestais do Domínio Atlântico do Brasil foram e
continuam sendo explorados, degradados e fragmentados (como o exemplo da Floresta Mista
com Araucária, descrito anteriormente), estudos mostram que a conservação e a restauração

44
de fragmentos são de grande importância (BRASIL, 2003; RIBEIRO et al., 2009 e PFEIFER
et al., 2014). O patrimônio biológico remanescente possui valor inestimável: 15.782 espécies
de plantas foram catalogadas por Stehmann et al. (2009) para o Domínio Atlântico, e cerca de
2.341 espécies de plantas vasculares e 800 espécies arbóreas foram identificadas no estado de
Santa Catarina (GASPER et al., 2012). Na Floresta Mista com Araucária o IFFSC catalogou
1.107 espécies vasculares (GASPER et al., 2013). A importância biológica vai além da flora;
Fonseca et al. (2009) realizaram estudos que contemplam a biodiversidade da fauna silvestre,
de fungos e líquens. As florestas, além de conter grande biodiversidade, nos propiciam
inúmeros benefícios (CHOMITZ e KUMARI, 1998; COSTANZA et al., 1997; CREPALDI e
PEIXOTO, 2010; HUNTER, 1999); num sentido mais amplo, elas estão fortemente aliadas à
manutenção da esfera social e de suas ramificações (ARCHER et al., 2005; DIEGUES, 2001).
A riqueza de espécies é a medida de biodiversidade mais difundida e comumente
utilizada (BROOK et al., 2003; BROSE et al., 2003; WILSEY et al., 2005), porém, ela não é
um componente isolado, de forma que a estrutura da comunidade, a estrutura da paisagem, a
diversidade funcional e a genética também possuem participação relevante e equivalente
(GASTON e SPIER, 2004; NOSS, 1990). Esses elementos que constituem um ecossistema
estão esquematizados na Figura 1.5, juntamente com elementos ativos que influenciam os
processos ecológicos de formação, retroalimentação e manutenção (elementos preditores), e,
os que, a um dado nível, perturbam algum componente do ecossistema (elementos
perturbadores).
Um método de avaliação do estado de conservação de um ecossistema florestal deve
considerar o complexo de interações entre seus componentes (PETERKEN, 1996; ŠAUDYTÈ
et al., 2005). Dentro do sistema que propomos e do contexto ecológico de uma floresta, temos
de focar em condensar informações sobre os elementos do ecossistema (“elementos do
ecossistema florestal”, Figura 1.5), ou seja, precisamos encontrar os componentes essenciais,
os quais podem ser expressos através de indicadores majoritários, altamente representativos
do todo. Desta forma, a identificação do grau de conservação de um dado remanescente
florestal, pode ser atingido através das respostas que os elementos do ecossistema
demonstram (BARTHA et al., 2006), assim como, através dos sinais de perturbação antrópica
(“elementos perturbadores” na Figura 1.5) presentes nesse remanescente (ANGERMEIER,
2000). Os conceitos de naturalidade e hemerobia (WINTER, 2012) podem ser utilizados
complementarmente como ferramentas de diagnóstico para a detecção desses sinais
supracitados. Os efeitos dos “elementos preditores” (Figura 1.5) influenciam os componentes
do ecossistema (BALDEK et al., 2013; DINIZ-FILHO et al., 2012; COTTENIE, 2005), logo,

45
precisam ser controlados na quantificação da naturalidade (LIIRA e SEPP, 2009; WINTER et
al., 2010), de forma que não venham a adicionar tendências aos resultados.
Figura 1.5. Esboço do (eco)sistema (objeto de estudo), seus componentes e interações.
Modificado de Tierney et al. (2009).
Em um âmbito global, a aplicação do conceito de naturalidade ainda é restrita às
florestas temperadas e boreais do hemisfério Norte. Sua utilização em florestas subtropicais
do hemisfério Sul, as quais possuem características ecológicas, históricas e sociais diferentes
será um feito pioneiro. Além deste fato, a falta de estudos que abordam a quantificação da
naturalidade nos incentiva a desenvolver um método de quantificação através de um
algoritmo específico (McROBERTS et al., 2012; WINTER, 2012).
O conceito de naturalidade incorporado ao pensamento e às ações conservacionistas
pode ser considerado uma ferramenta para planos de manutenção, monitoramento e
utilização/manejo de ecossistemas florestais (BARTHA et al., 2006), podendo adequar-se a
diferentes contextos ambientais, políticos e sociais. Winter (2012) afirma que poucos são os
estudos que utilizam esta ferramenta para fins de conservação e monitoramento da

46
biodiversidade e de processos ecológicos. Dentro da proposta do IFN-Brasil e do escopo do
IFFSC, tal ferramenta pode expandir as concepções sobre a análise de florestas nativas,
podendo envolver várias dimensões do ecossistema em si, da sociedade e das políticas
públicas acerca dos recursos naturais.
As justificativas apresentadas por McRoberts et al. (2012) acerca da relevância da
determinação da naturalidade de florestas nativas podem ser adaptadas à realidade do
contexto sócio-ambiental e econômico da Floresta Mista com Araucária de Santa Catarina: (i)
ela pode ser empregada para reportar o estado ecológico das florestas nativas visando a
avaliação da conservação e da exploração dos recursos florestais; (ii) ela pode servir para
identificar áreas de grande expressão ecológica e, consequentemente, para delinear metas de
conservação e utilização das mesmas.
Nossa perspectiva para este estudo tende a observar as alterações e descaracterizações
mais explícitas observadas nas florestas do planalto de Santa Catarina. Na revisão
anteriormente apresentada salientamos pontos preocupantes como a baixa riqueza de espécies,
baixa regeneração natural, pequeno tamanho e baixa conectividade dos fragmentos, reduzida
área basal, entre outros. Tais aspectos serão priorizados na metodologia de determinação da
naturalidade, diante da vital importância destes para a manutenção dos remanescentes
florestais.
Ao revisarmos a literatura, identificamos princípios e recomendações para a
elaboração de um estudo de naturalidade (McROBERTS et al., 2012; WINTER et al., 2011;
WINTER et al., 2010; WINTER, 2012). Seguindo o processo ilustrado na Figura 1.6,
partimos do propósito inicial que é ter conhecimento sobre o estado de conservação das
florestas, o que foi proporcionado pelo IFFSC, a partir de todo seu esforço amostral. Ao
escolhermos a naturalidade como o fundamento que expressa o estado de conservação de um
remanescente florestal, precisamos:
i) definir concretamente o conceito de naturalidade a ser adotado;
ii) detectar indicadores (variáveis) potenciais de naturalidade;
iii) definir o conceito de florestas referências e identificá-las;
iv) determinar um método de quantificação da naturalidade e sumarizar as diferenças entre os
remanescentes de referência e os demais.
Ao definirmos essa direção ao estudo, dedicaremos os próximos tópicos deste capítulo
ao desenvolvimento de cada um dos passos essenciais citados acima.

47
Figura 1.6. Processo envolvido na determinação da naturalidade de remanescentes florestais
de Floresta Mista com Araucária de Santa Catarina, a partir dos dados coletados pelo IFFSC.
1.3.3. A DEFINIÇÃO DE NATURALIDADE
A definição de naturalidade adotada neste estudo é a proposta por Winter (2012): Um
ecossistema possui maior naturalidade se o seu estado atual é mais próximo ao seu estado
conservado, ou seja, sem influências antrópicas perturbadoras e degradadoras. Após
uma intensa revisão sobre o assunto, a autora sugere esta definição por vias de padronização,
evitando inconsistências entre estudos.
Em nossa abordagem, damos ênfase na visão de Peterken (1996), de que, em um
ecossistema natural, existe o predomínio de processos naturais, juntamente com

48
características e recursos modelados por eles. Assim, a naturalidade não equivale à
originalidade, a qual exclui a interferência humana de qualquer remanescente; logo, só pode
ser considerada dentro de um contexto histórico e inatingível para as florestas brasileiras,
como para outras regiões do planeta, como a Europa Central (KOWARIK, 1999), por
exemplo.
De acordo com McRoberts et al. (2012) e Chirici et al. (2012), existem duas maneiras
de determinar a naturalidade: a primeira tem como fundamento o acesso aos processos
ecossistêmicos (PETERKEN, 1996) - essa abordagem requer a determinação de poucos
indicadores que descrevam os principais aspectos e processos ecológicos em um ecossistema
florestal. A segunda maneira fundamenta-se no grau de influência humana, a qual é a
propulsora das perturbações em um dado ecossistema (ANDERSON, 1991; ANGERMEIER,
2000; WINTER, 2012). Indicadores ligados à hemerobia estão geralmente associados a sinais
de intervenção e exploração humana (FRA, 2000; LIIRA e SEPP, 2009; MACHADO, 2004;
UOTILA et al., 2002); logo, podem ser utilizados como uma ferramenta para identificar
florestas mais impactadas pelos humanos (WINTER, 2012).
Utilizaremos as duas maneiras de determinação, ou seja, a determinação do grau de
hemerobia será utilizada em conjunto com a naturalidade. Assim, estes dois conceitos
antagônicos serão utilizados de forma complementar neste estudo.
Ao se trazer à luz o conceito de hemerobia ligado à naturalidade, surge uma questão
intrigante e delicada: Qual a influência dos humanos nas florestas? Eles fazem parte da
natureza ou não?
Os critérios que definem o que é um ecossistema natural não são fixos e podem variar
conforme o contexto social; a educação e a experiência ambiental de cada indivíduo, ou de
um grupo, influenciam na formação da concepção do que é uma floresta dita natural
(WILLIAMS, 2002). Para exemplificar, citamos Lamb e Purcell (1990), os quais realizaram
um estudo com o objetivo de coletar diversas percepções sobre a naturalidade de vegetações,
expondo fotografias para colaboradores. Os participantes entrevistados consideraram uma
vegetação alta e densa como mais natural do que uma vegetação mais aberta e baixa. Nota-se
que houve uma tendência de os aspectos estruturais serem captados em detrimento de outros
componentes do ecossistema que não foram considerados.
A inclusão ou não do ser humano aos ecossistemas é tema de discussão. No enfoque
da Antropologia Ecológica a sociedade é um subsistema (ou um componente) de uma
totalidade maior, onde os seres humanos, animais e plantas mantêm relações bioenergéticas
(DIEGUES, 2001). Logo, os humanos não diferem dos demais seres vivos em relação ao

49
equilíbrio homeostático com o ambiente. Para Vibrans (2003), a premissa epistemológica de
que as sociedades humanas e a natureza integram-se de forma recíproca formando um todo
supera a visão ocidental antropocêntrica, de que a sociedade e a natureza vivem à parte uma
da outra.
Diegues (2001) cita as sociedades tradicionais do Brasil, a saber, índios e camponeses
(definição conforme Queiroz [1973]), como um exemplo da relação entre o homem e a
natureza em oposição ao modelo de colonização predatório realizado pelos europeus. Gomez-
Pompa et al. (1972) lançaram a hipótese de que atividades do homem dito "primitivo", como,
por exemplo, a agricultura itinerante, tem favorecido a diversidade de espécies em florestas
tropicais do México. Entretanto, os mesmos autores afirmam que as atividades
contemporâneas ligadas ao uso do solo são extremamente perigosas no que diz respeito à
intensa perturbação dos ecossistemas florestais, causando então, a perda de habitats e
espécies, como aferido também por Mähler Júnior e Larocca (2009) para a Floresta Mista com
Araucária. Na visão de Luztemberger (1976, p.6) vemos a participação e até a contribuição
das sociedades tradicionais nas florestas:
"O índio, muitos milênios antes da chegada do homem branco, já
havia alcançado situações de equilíbrio estável em seu ambiente. No
mar da Natureza intata, as derrubadas do índio, pequenas e a grande
distância uma das outras, constituíam até vantagem ecológica, pois
acrescentavam diversidade ao sistema. Nestas clareiras vingavam os
organismos pioneiros, as plantas e os animais das comunidades de
recuperação, que são os tecidos cicatrizantes dos ecossistemas."
Reis et al. (2014) discorrem sobre a influência das sociedades pré-Colombianas na
formação da Floresta Mista com Araucária, as quais coexistiam há, pelo menos, 3.000 anos.
Os autores afirmam que são muitas as evidências (etno)botânicas e etnohistóricas que
apontam a participação desses povos (Kaigang e Xokleng) na ampliação da floresta,
envolvendo predominantemente uma estreita relação de dependência deles com os pinhões de
A. angustifolia. Nesse sentido, a Floresta Mista com Araucária poderia ser classificada como
uma paisagem cultural. Schmitz (2009) discute brevemente tais evidências; porém conclui
que estas não são confirmatórias da interferência dos povos citados na ampliação da Floresta
Mista com Araucária.
Os “faxinais” e as “caívas” são exemplos de paisagens culturais que foram criadas
pelos colonos europeus e caboclos em Santa Catarina e nos outros estados da Região Sul,

50
onde a vegetação florestal é mantida para o manejo de espécies de interesse como a erva-
mate, além da criação de animais pastejadores (CHANG, 1998; HANISCH et al., 2010;
MELLO, 2013).
Apesar das inegáveis evidências da participação do homem na formação de certas
fitofisionomias, de forma semelhante a Angermeier (2000), vemos Luztemberger (1976, p.3)
denunciando a posição tomada pelo homem moderno em suas atividades predatórias e
geograficamente abrangentes:
"A continuar a atual cegueira ambiental e exploração irresponsável de
nosso outrora pródigo meio natural, serão inevitáveis calamidades de
magnitude nunca vista. Somente uma transição rápida a atitudes
fundamentalmente novas, atitudes de respeito e integração ecológica
poderão ainda evitar o desastre. Encontramo-nos num divisor de
águas."
Hoehne (1930, p.5), numa visão biocêntrica, presenciando a devastação das florestas
com Araucária no Sul do Brasil, a qual denominou de “Araucarilândia”, expressa seus
pensamentos:
“As selvas naturais e virgens encerram vidas e riquezas, cujo
verdadeiro valor e importância real ainda se não podem avaliar
devidamente. Protege-las é defender um patrimônio, que, uma vez
perdido, jamais pode ser adquirido, quer pela força, quer pelo
dinheiro.”
“O homem precisa entender que é parte colaborante, elemento do
conjunto de vidas sobre a face da terra e saber que, se se excede,
deixando de respeitar o direito alheio, as consequências serão sentidas,
não somente pelos oprimidos e prejudicados, mas mesmo por ele e por
todo o conjunto. O direito de viver assiste a cada ser; mas, o de
abusar, a nenhum deles.”
Estas exposições nos incentivam a considerar que existiu, ou, ainda existe um
manejo/exploração dito "consciente" ou "equilibrado" feito por pessoas que mantêm contato
com a floresta. Ao lado deste modo de utilização dos recursos naturais, observamos a
exploração predatória a e degradação dos ecossistemas, visando benefícios econômicos, sem a
preocupação com a conservação e preservação. Embora essas duas modalidades pareçam ser

51
claramente distinguíveis, encontrar o limiar entre a exploração (e/ou manejo) ditos
"conscientes" ou "predatórios" é uma tarefa complexa, assim como é complicado avaliar os
efeitos de cada uma dentro de um contexto geográfico mais amplo, como um estado. Estamos
de acordo com Oliver e Larson (1990) de que existe um gradiente contínuo que vai desde
pequenas até grandes explorações de recursos florestais ou de distúrbios no ecossistema. Um
exemplo interessante da percepção entre o limiar do considerado natural e não natural foi
apresentado por Dunk (1994 apud WILLIAMS, 2002). O autor entrevistou pessoas da área
rural no Canadá acerca de suas opiniões sobre o corte de árvores:
"Eu acho a coisa mais natural do mundo cortar uma árvore."
Em contrapartida, o mesmo entrevistado declarou:
"O corte raso da floresta não me parece natural."
Trazendo o foco da discussão para a realidade do Sul do Brasil, analisamos o
comportamento cultural dos colonizadores do planalto de Santa Catarina, dentro de um
contexto histórico, e observamos a estreita relação das atividades de pecuária e as florestas.
Uma prática comum era a conversão de florestas em pastagens para criação de gado
(BORGES, 2005). Esta prática foi de certa forma perpetuada, incluindo, atualmente, outros
motivos para a derrubada de florestas, como culturas agrícolas extensivas e plantios de
florestas artificiais. O levantamento sócio-ambiental-histórico de Siminski (2004), que teve
como alvo os agricultores (idade média de 50 anos) residentes na região da Floresta Pluvial
Latifoliada de Santa Catarina (Figura 1.4), apontou diversas maneiras e intensidades de
utilização dos recursos naturais, desde a coleta de plantas ornamentais até a intensa extração
de lenha para engenhos de açúcar e extração de madeiras nobres para fins de construção civil.
Atualmente, de uma forma geral, a relação do pequeno agricultor, ou proprietário
rural, com a floresta em Santa Catarina, resume-se ao uso de recursos florestais para lenha e o
consumo esporádico de frutas e plantas medicinais. Na Floresta Mista com Araucária, a erva-
mate e o pinhão desempenham um papel importante, onde muitas vezes sua exploração e
comercialização são utilizadas como complemento para a renda de famílias que vivem no
meio rural (SIMINSKI, 2009). Siminski (2009), entrevistando proprietários de áreas rurais,
elencou alguns motivos pelos quais as florestas nativas de Santa Catarina estão sendo
convertidas em outros usos; dentre eles foram citados a baixa rentabilidade econômica e as
barreiras e complicações impostas pela legislação ambiental.
Ao analisarmos alguns relatos presenciados em nossa vivência pela região do planalto
de Santa Catarina, vemos o espírito deliberadamente explorador dos homens:

52
"... encurralei e matei a última paca da região."
"...até os pinheiros das grotas eram arrastados com cabos de aço."
"...na repartição das terras, os pinheiros com mais de 30 cm de
diâmetro foram cortados."
"...daqui de casa até a casa do pai só tinha um carreiro, era só pinheiro
e taquara."
Enfim, diante desta breve exposição, novamente mencionando e considerando a
dificuldade de decretar um limiar entre níveis "conscientes" e "displicentes" de exploração
dos recursos florestais e afins, tomamos uma posição conservadora e propomos que:
Os seres humanos são, sem dúvida, integrantes históricos dos ecossistemas florestais,
porém não estão adaptados aos mesmos. As populações tradicionais, que outrora
habitavam o Sul do Brasil, geravam baixo impacto sobre os ecossistemas. Atualmente, os
seres humanos (em um sentido global) convertem os ecossistemas florestais em outros
tipos, alterando sua composição, estrutura e funcionalidade.
(Axioma 1)
Desta forma, consideramos as ações (de uma maneira geral) dos humanos (pós-
colonização) nas florestas de Santa Catarina, como sendo opostas aos processos
ecossistêmicos naturais. Deixamos claro que a discussão filosófica sobre os humanos e a
natureza não é o foco principal deste estudo. Diante das divergências de opiniões sobre o
tema, direcionamos nosso enfoque ao Axioma 1, de forma que nossa abordagem de
naturalidade tem um caráter conservador, porém não biocêntrico em sua essência. Esperamos
contribuir no que diz respeito à abordagem metodológica de acesso à naturalidade, a qual
poderá ser adaptada para outras perspectivas relacionadas à interação humano-natureza.

53
1.3.4. METODOLOGIA DE ACESSO À NATURALDADE
1.3.4.1 Área de estudo
O Estado de Santa Catarina (Figura 1.7) encontra-se nas latitudes 26° e 29° S e entre
as longitudes 48° e 53°W e possui área total de 95.346 km². São descritos por Klein (1978)
três regiões fitoecológicas para o Estado (Figura 1.4): Floresta Mista com Araucária, Floresta
Latifoliada Pluvial e Floresta Latifoliada Estacional (classificação segundo Oliveira-Filho
[2009]), as quais possuem área de ocorrência de 56,39 km², 31,28 km² e 7,67 km² (VIBRANS
et al., 2013), respectivamente.
Figura 1.7. Estado de Santa Catarina, Brasil, com preto indicando a cobertura florestal (SAR,
2005).
Brasil

54
De acordo com a classificação climática de Köppen (KOTTEK et al., 2006), Santa
Catarina tem dois tipos climáticos: Cfa - clima temperado úmido, com verão quente e Cfb -
clima temperado úmido, com verão fresco. Esses tipos climáticos sao definidos
principalmente por diferenças de temperatura devido a gradientes de altitude. A temperatura
média anual varia de 10 a 22 °C, com precipitação média anual de 1.100 a 2.900 mm bem
distribuídos durante o ano e umidade relativa média anual entre 74 e 88% (EPAGRI, 2002).
No diagrama de Walter elaborado para o município de Curitibanos, localizado no planalto
central de Santa Catarina, não existem períodos de déficit hídrico (Figura 1.8).
Figura 1.8. Diagrama de Walter gerado a partir de dados climáticos (série histórica de 50
anos) do município de Curitibanos, situado na região de ocorrência da Floresta Mista com
Araucária em Santa Catarina.
Quanto aos aspectos geológicos, o Planalto Ocidental de Santa Catarina pertence às
formações geológicas Botucatu e Serra Geral, constituindo a área mais extensa do estado,
sendo caracterizado pela presença de altitudes superiores a 800 m na área limite com a serra
litorânea. Conforme avança em sentido sudoeste ao vale do rio Uruguai, nos limites com a
Argentina e Rio Grande do Sul, vai reduzindo gradativamente a altitude, até cotas de 200 m.

55
O planalto de Santa Catarina, devido à suas características geológicas e geomorfológicas, foi
subdividido em Zona Basáltica, Planalto de Canoinhas e Planalto de Lages (NETTO, 1984).
Na Zona Basáltica, a qual constitui a maior parte do planalto, é observada uma maior
variação da altitude. Entre as cotas de 200 a 800 m observa-se uma topografia movimentada
que forma vales em “V”; entre as cotas de 900 a 1000 m a topografia ondulada com longos
declives predomina. O Planalto de Lages é constituído por rochas sedimentares e ígneas
alcalinas localizadas na escarpa, adentrando de forma suave na Zona Basáltica. A altitude
varia, em média, na faixa dos 900 m, sendo o relevo ondulado com a presença de campos de
altitude. O Planalto de Canoinhas localiza-se ao norte do Planalto Ocidental e apresenta
relevo suave ondulado formado por rochas sedimentares; as altitudes médias oscilam entre
800 a 900 m (NETTO, 1984).
1.3.4.2 Dados do Inventário Florístico-Florestal de Santa Catarina
As equipes de campo do IFFSC coletaram os dados referentes à Floresta Mista com
Araucária entre o período de 2007 a 2010, adotando a metodologia do Inventário Florestal
Nacional do Brasil (FREITAS et al., 2010). A metodologia de amostragem utilizada baseou-
se na amostragem de múltiplas ocasiões com possibilidade de repetição total da amostragem.
As Unidades Amostrais foram distribuídas sistematicamente a partir de uma grade de pontos
com distância de aproximadamente 10 km x 10 km, cobrindo todo o território do estado
(VIBRANS et al., 2010; VIRABNS et al., 2012a).
Para a região de ocorrência da Floresta Mista com Araucária, a grade gerou 202 pontos
amostrais dos quais 143 foram efetivamente amostrados (Figura 1.10). Os pontos amostrais
foram previamente analisados através da interpretação de imagens orbitais multiespectrais de
média resolução advindos dos sensores LandSat-5, LandSat-7 e Spot-4 (SAR, 2005;
GEOAMBIENTE, 2008), e classificados como “floresta” e “não floresta”. A definição de
“floresta” adotada pelo IFFSC baseou-se nos seguintes critérios: áreas cobertas por vegetação
arbórea contínua, com altura do dossel maior que 10 m e área basal maior que 10m².ha-1
(VIBRANS et al., 2013). Serão incluídas neste estudo as 12 Unidades Amostrais
complementares alocadas em florestas consideradas mais conservadas, instaladas fora da
grade de pontos amostrais. Dessas 12, quatro delas se encontram no município de Três Barras
(FLONA de Três barras), uma no município de Caçador (Reserva Florestal
EMBRAPA/EPAGRI), uma no município de Anita Garibaldi (em uma propriedade

56
particular), quatro no município de Palma Sola (em uma propriedade particular) e duas no
município de São Domingos (Parque Estadual das Araucárias).
A Unidade Amostral do IFFSC é composta por um conglomerado com área nominal
total de 4.000 m², formado por quatro subunidades com área de 1.000 m², com as dimensões
de 20 m x 50 m (orientadas na direção dos pontos cardeais: norte, sul, leste e oeste), com área
de inclusão de 2,56 ha (Figura 1.9). Cada subunidade foi estabelecida a uma distância de 30 m
do centro do conglomerado e divida em dez subparcelas de 10 m x 10 m (100 m²), onde foram
levantados todos os indivíduos com DAP ≥ 10,0 cm.
As variáveis levantadas foram espécie, número de fustes, DAP, altura do fuste, altura
total, qualidade do fuste, sanidade da árvore e posição sociológica. Em cada subunidade foi
destinada uma subparcela de 5 m x 5 m (25 m²), localizada na extremidade da subunidade,
destinada ao levantamento da regeneração natural; foram registradas a espécie e a altura dos
indivíduos com DAP < 10 cm e altura ≥ 1,5 m.
Figura 1.9. Unidade Amostral em forma de conglomerado adotada pelo IFFSC para o
levantamento do componente arbustivo-arbóreo da Floresta Mista com Araucária.

57
Figura 1.10. Distribuição das 155 Unidades Amostrais do IFFSC nas regiões fitoecológicas
da Floresta Mista com Araucária e Campina.
1.3.4.3 Indicadores potenciais de naturalidade
Dentro da concepção de ecossistema de Noss (1990) os indicadores têm de
contemplar três componentes básicos: composição, estrutura e funcionalidade. Šaudytè et al.
(2005) propõe que ao avaliar a naturalidade temos que estar cientes dos principais
componentes de uma floresta.
Além destas premissas iniciais, segundo a revisão de McRoberts et al. (2012), os
indicadores devem produzir resultados consistentes, precisos e objetivos (SMITH e
THEBERGE 1987; LIIRA et al. 2007; UOTILA et al., 2012). Devem ser facilmente e
prontamente calculáveis a partir de dados disponíveis (BARTHA et al. 2006; LIIRA et al.
2007) e devem ser adequados para variadas escalas espaciais (BARTHA et al., 2006).
Devemos ainda considerar os diferentes fatores climáticos, geológicos e tradições regionais de
manejo nos quais as Unidades Amostrais do IFFSC estão inseridas; tais fatores podem ser um
empecilho na adoção ou criação de indicadores de naturalidade (LIIRA e SEPP, 2009).
Estes apontamentos remetem a um questionamento: O que se espera, em relação às
características e propriedades de uma floresta mais próxima do seu estado natural?
Elencando alguns pontos observados na literatura:

58
A diversidade de espécies em áreas conservadas pode ser superior quando comparada
com áreas menos conservadas (GIBSON et al., 2011; SEVEGNANI et al., 2013b).
Santos et al. (2008) observou que áreas bem conservadas da Floresta Atlântica
apresentaram o dobro da diversidade de espécies que fragmentos florestais com idade
de regeneração entre 5 e 65 anos;
Em relação à riqueza de espécies, Sevegnani et al. (2013a), baseados em
levantamentos históricos, afirmam que, para um remanescente de Floresta Mista com
Araucária conservado, o valor estaria em torno de 60 espécies arbóreas;
Leite (2002) aponta a A. angustifolia como uma espécie que imprime a fisionomia da
Floresta Mista com Araucária e Klein (1978) aponta espécies características como D.
sellowiana e indicadoras de formações mais específicas, como O. pulchella, O. porosa
e P. rigida.
Segundo Klein (1978), a estrutura vertical de uma Floresta Mista com Araucária
conservada apresenta um estrato emergente composto por A. angustifolia, um estrato
abaixo formado por angiospermas (geralmente lauráceas) e um estrato médio;
A distribuição de árvores em classes diamétricas de florestas conservadas apresenta
uma tendência em assumir uma distribuição exponencial negativa, ou J-invertido,
onde a frequência de árvores é alta para o lado esquerdo da média e baixa para o lado
direito da média (KUULUVAINEN et al., 1996; UOTILA et al., 2002);
A distribuição de frequências em classes diamétricas de A. angustifolia tende a possuir
assimetria à esquerda, segundo o modelo Losango de Ogden e Stewart (1995);
Florestas conservadas apresentam espécies de vários grupos ecológicos, com
predomínio de espécies mais exigentes em recursos ecológicos e tolerantes à sombra -
espécies caracterizadas como secundárias tardias e climácicas (FINEGAN, 1992);
A área basal e o estoque de biomassa em florestas mais conservadas tende a ser maior
do que em florestas perturbadas (D‟OLIVEIRA, et al., 2011; HUGHES et al., 1999);
A regeneração natural em florestas maduras e conservadas deve ocorrer
abundantemente, com a presença de espécies características de florestas desenvolvidas
(CHAZDON et al., 1998);
A necromassa é um componente importante da biodiversidade florestal, fornecendo
habitats para muitas espécies de fungos, briófitas e invertebrados (CHRISTENSEN et
al., 2005; ÓDOR et al., 2006), e também pode representar a história recente da
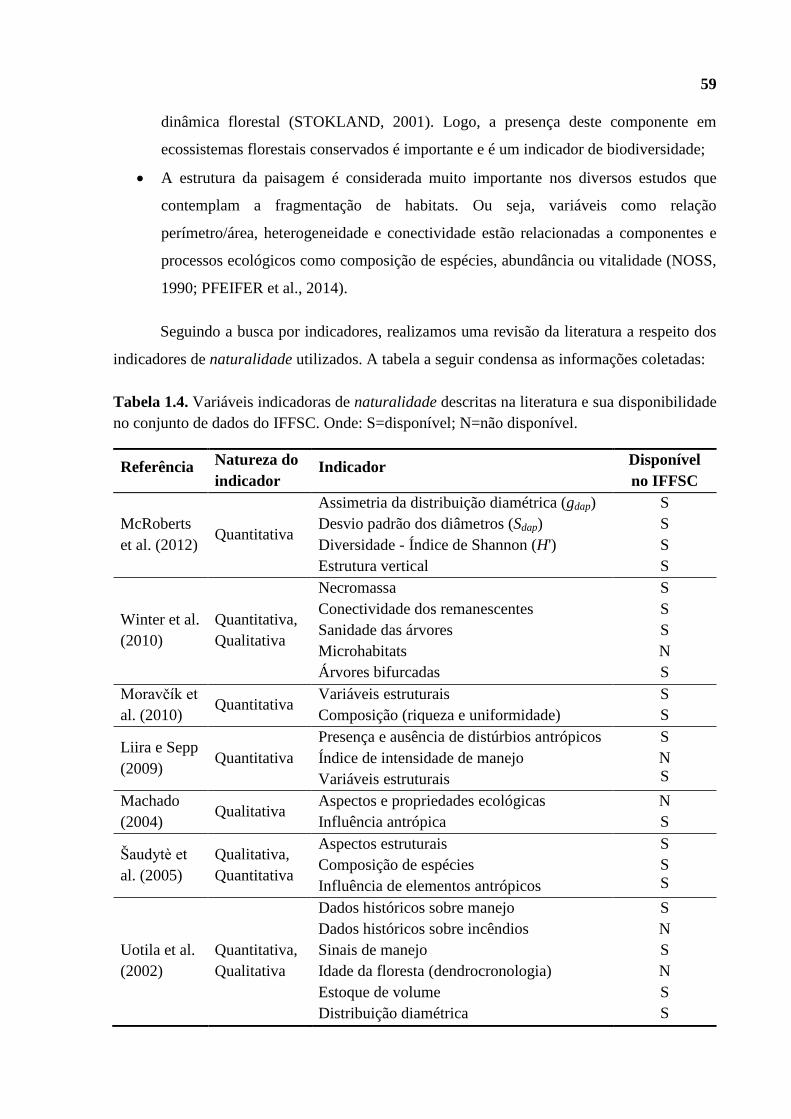
59
dinâmica florestal (STOKLAND, 2001). Logo, a presença deste componente em
ecossistemas florestais conservados é importante e é um indicador de biodiversidade;
A estrutura da paisagem é considerada muito importante nos diversos estudos que
contemplam a fragmentação de habitats. Ou seja, variáveis como relação
perímetro/área, heterogeneidade e conectividade estão relacionadas a componentes e
processos ecológicos como composição de espécies, abundância ou vitalidade (NOSS,
1990; PFEIFER et al., 2014).
Seguindo a busca por indicadores, realizamos uma revisão da literatura a respeito dos
indicadores de naturalidade utilizados. A tabela a seguir condensa as informações coletadas:
Tabela 1.4. Variáveis indicadoras de naturalidade descritas na literatura e sua disponibilidade
no conjunto de dados do IFFSC. Onde: S=disponível; N=não disponível.
Referência Natureza do
indicador Indicador
Disponível
no IFFSC
McRoberts
et al. (2012) Quantitativa
Assimetria da distribuição diamétrica (gdap)
Desvio padrão dos diâmetros (Sdap)
Diversidade - Índice de Shannon (H')
Estrutura vertical
S
S
S
S
Winter et al.
(2010)
Quantitativa,
Qualitativa
Necromassa
Conectividade dos remanescentes
Sanidade das árvores
Microhabitats
Árvores bifurcadas
S
S
S
N
S
Moravčík et
al. (2010) Quantitativa
Variáveis estruturais
Composição (riqueza e uniformidade)
S
S
Liira e Sepp
(2009) Quantitativa
Presença e ausência de distúrbios antrópicos
Índice de intensidade de manejo
Variáveis estruturais
S
N
S
Machado
(2004) Qualitativa
Aspectos e propriedades ecológicas
Influência antrópica
N
S
Šaudytè et
al. (2005)
Qualitativa,
Quantitativa
Aspectos estruturais
Composição de espécies
Influência de elementos antrópicos
S
S
S
Uotila et al.
(2002)
Quantitativa,
Qualitativa
Dados históricos sobre manejo
Dados históricos sobre incêndios
Sinais de manejo
Idade da floresta (dendrocronologia)
Estoque de volume
Distribuição diamétrica
S
N
S
N
S
S

60
A partir dos indicadores apresentados na tabela anterior realizamos uma triagem, de
acordo com a disponibilidade destes indicadores, ou seja, quais foram levantados ou podem
ser calculados a partir do banco de dados do IFFSC. Adicionamos a estes indicadores os que
julgamos como importantes e que não constam na literatura; os indicadores estão expressos na
Tabela 1.5.
Criamos duas categorias de indicadores: os descartados e os potenciais. Expomos e
discutimos as propriedades e características de cada um, e por fim, concluímos sobre a sua
relevância ou irrelevância ecológica, de acordo com os conhecimentos sobre ecologia florestal
e sobre a Floresta Mista com Araucária. Os indicadores descartados foram excluídos (a
priori) do repertório de alternativas. Nessa primeira apresentação dos indicadores potenciais
focamos apenas em descrever suas propriedades e potencialidades, e em sintetizar os que
estão ligados a elementos fundamentais do ecossistema.
Tabela 1.5. Indicadores de naturalidade potenciais acessados pelo IFFSC. * = Indicador
sugerido pelos autores do presente estudo.
Indicador de naturalidade Natureza Presente na literatura
Árvores bifurcadas Ecossistêmica Sim
Assimetria da distribuição diamétrica (gdap) Ecossistêmica Sim
Composição de espécies Ecossistêmica Sim
Desvio padrão dos diâmetros (Sdap) Ecossistêmica Sim
Diversidade de espécies Ecossistêmica Sim
Estoque de volume Ecossistêmica Sim
Estrutura de população de espécies* Ecossistêmica Não
Estrutura vertical (estratos) Ecossistêmica Sim
Necromassa Ecossistêmica Sim
Número de árvores velhas e (ou) grossas Ecossistêmica Sim
Regeneração natural Ecossistêmica Sim
Riqueza de espécies Ecossistêmica Sim
Sanidade das árvores Ecossistêmica Sim
Métricas de paisagem de remanescentes Hemerobia Sim
Presença e ausência de distúrbios antrópicos Hemerobia Sim
Histórico de manejo/exploração Hemerobia Sim
Indicadores de naturalidade descartados:
Estoque de volume:
Esta variável tem potencial para ser um bom indicador de naturalidade, porém, a
obtenção de dados do volume individual de árvores em um inventário em larga escala é

61
realizada através de modelos estatísticos, os quais adicionam um grau de incerteza nas
predições (McROBERTS e WESTFALL, 2014). No IFFSC as estimativas de volume foram
feitas através de modelos estatísticos (VIBRANS et al., em revisão), logo, a incerteza nos
valores estimados torna-se um obstáculo para a utilização desta variável como um indicador
(BREIDENBACH et al., 2014; McROBERTS et al., 2012).
Sanidade das árvores e árvores bifurcadas:
Esses dois indicadores estão estreitamente relacionados com o indicador microhabitat.
Segundo Winter e Möller (2008) e Michel e Winter (2009), as árvores vivas bifurcadas e a
perda de vitalidade das árvores são pré-requisitos para a formação de troncos mortos, os quais
podem formar importantes microhabitats. Apesar de o IFFSC ter coletado tais informações
sobre sanidade e bifurcação das árvores, não existem estudos relacionados a microhabitats
para a Floresta Mista com Araucária, dificultando a adoção destes indicadores em nosso
estudo.
Histórico de manejo/exploração:
Os indicadores relacionados ao histórico de manejo florestal, como os adotados nos
estudos citados, estão ligados à Europa, que há decênios pratica o manejo florestal (CHIRICI
et al., 2011; TOMPPO et al., 2010, LIIRA et al., 2007). No Brasil, a exploração teve um
caráter indiscriminado e descontrolado (DEAN, 1996); logo, possuímos poucas informações
precisas e fidedignas sobre o histórico de exploração de remanescentes florestais, o que torna
este indicador impossibilitado de ser utilizado.
Indicadores de naturalidade não descartados:
Assimetria da distribuição de diâmetros:
A assimetria (g) de uma distribuição de frequências é obtida pela raiz cúbica do
terceiro momento central da distribuição das observações (GOTELLI e ELLISON, 2011). Na
área florestal a assimetria pode descrever se a distribuição de frequências em classes
diâmétricas (gdap) de uma floresta é simétrica, ou não. Uma distribuição normal apresenta gdap
= 0. Já uma distribuição com gdap > 0 apresenta assimetria à direita; ou seja, possui uma cauda
de observações à direta da média. Um valor de gdap < 0 aponta assimetria à esquerda,

62
indicando uma cauda de observações à esquerda da média (BENDEL et al., 1989; SOUZA,
2009; WRIGHT et al., 2003).
Souza (2009) usou esse parâmetro com o intuito analisar a estrutura de populações de
A. angustifolia; McRoberts et al. (2008) utilizaram-o como uma medida de diversidade
estrutural e McRoberts et al. (2012) adotoram-o como um indicador de naturalidade para
florestas nativas dos Estados Unidos. Adaptando este indicador ao nosso contexto, as florestas
com maior naturalidade tendem a apresentar distribuições de diâmetros com assimetria à
direita (J-invertido, seguindo a distribuição convencional de uma floresta climácica), sendo
expressas por gdap > 0. A distribuição diamétrica da floresta pode apresentar um panorama do
processo dinâmico de sucessão ou de mortes, egressos e ingressos, visto que está relacionada
com o crescimento dos indivíduos em seus diversos estágios de vida, como observado por
Schaff et al. (2006a) em um período de 21 anos na Floresta Mista com Araucária.
Composição de espécies do estrato arbóreo:
A composição de espécies pode revelar o estágio de sucessão ecológica, o grau de
conservação, ou ainda o histórico de distúrbios ocorrentes em uma floresta, mediante a
avaliação dos hábitos e funções das espécies (CHAZDON et al., 1998; CHAZDON et al.,
2007; FINEGAN, 1996; IMAI et al., 2014; SIMINSKI, 2004).
Estrutura de população de espécies:
A distribuição de tamanho dos indivíduos de uma população é uma síntese dos
processos dinâmicos ao longo do tempo e, para a Floresta Mista com Araucária,
remanescentes que se encontram conservados podem ser caracterizados pela abundância de A.
angustifolia (BACKES, 2009). Pode-se considerar que a presença desta espécie é um
indicador do estado de conservação de remanescentes. As florestas mistas são caracterizadas
pela coexistência de angiospermas e gimnospermas e a estratégia de regeneração natural das
coníferas de grande porte é motivo de discussão, não estando a A. angustifolia à parte. Klein
(1960) considerou-a pioneira; outros autores (por exemplo, Soares [1979]) afirmaram que esta
depende de perturbações para se regenerar e outros afirmaram que ela regenera sem maiores
limitações em florestas fechadas (por exemplo, Seitz [1982]).
Apesar do comportamento de uma espécie poder variar conforme o processo
sucessional (DILLENBURG et al., 2009), nota-se que a A. angustifolia parece adequar-se ao
modelo Losango (OGDEN e STEWART, 1995), o qual foi proposto a partir de observações

63
da regeneração natural de coníferas de grande porte em florestas mistas da Nova Zelândia;
este modelo, é baseado em indícios que mostram que o recrutamento destas coníferas dentro
da floresta é reduzido, pois as angiospermas possuem vantagens competitivas. O modelo
sugere que as coníferas caracterizadas como pioneiras longevas dependem de perturbações na
floresta (por exemplo, clareiras) para se regenerarem, formando coortes que são seguidas por
coortes de angiospermas. Consequentemente, as coníferas acabam por imprimir a fisionomia
da vegetação, sendo grandes contribuintes nos componentes estruturais, porém, sua
regeneração é reduzida devido ao domínio das angiospermas nos estratos inferiores.
Souza (2009) estudou a estrutura de populações de A. angustifolia em remanescentes
de Floresta Mista com Araucária do estado do Rio Grande do Sul e confirmou as proposições
do modelo Losango. A assimetria da distribuição de diâmetros à esquerda, ou seja, gdap < 0
aponta, segundo o modelo Losango, que a população de A. angustifolia se encontra em um
estágio climácico, portanto, pode ser considerada um indicador de naturalidade para a
Floresta Mista com Araucária.
Desvio padrão dos diâmetros (Sdap):
As florestas do hemisfério Norte foram manejadas conscientemente por decênios, e as
práticas de manejo foram utilizadas para modelar a estrutura da floresta (BARTHA et al.,
2006). Como resultado, tais florestas tornaram-se estruturalmente homogêneas - maiores
valores de Sdap para estas florestas podem indicar uma maior naturalidade (McROBERTS et
al., 2012). Este indicador é utilizado pelos IFNs da Europa participantes da Ação COST E43 e
foi utilizado por McRoberts et al. (2008) e McRoberts et al. (2012) nos Estados Unidos. Este
indicador aplicado nas florestas subtropicais também pode gerar resultados ecologicamente
interpretáveis, porque a variabilidade de diâmetros remete à complexidade da estrutura da
vegação, a qual está intimamente relacionada com microhábitats e a biodiversidade
(BOUNGIORNO et al. 1994; McROBERTS et al. 2012; MICHEL e WINTER, 2009; NOSS,
1990; WINTER e MÖLLER, 2008).
Diversidade de espécies do estrato arbóreo:
A diversidade de espécies leva em consideração dois atributos: riqueza e equabilidade
(HILL, 1978; MELO, 2008). Por décadas, este aspecto vem sendo estudado (LANDE, 1996),
pois duas comunidades podem ter a mesma riqueza e o mesmo número de indivíduos, mas
podem possuir estruturas diferentes. Portanto, a combinação da riqueza e da distribuição

64
relativa das espécies (em um índice) torna-se mais interessante do que conhecer somente a
riqueza (HURLBERT, 1971).
Buckland et al. (2011) assumem que a diversidade de espécies é um atributo escencial
para o monitoramento da biodiversidade, e tal atributo vem sendo utilizado em estudos sobre
naturalidade (CHIRICI et al., 2011; McROBERTS et al., 2012). Gibson et al. (2011) e
Sevegnani et al. (2013b) afirmam que florestas mais conservadas possuem maior diversidade
de espécies, embora, Imai et al. (2014) e Maçaneiro et al. (manuscrito em preparação) não
concordam com os apontamentos dos autores citados anteriormente, diante da evidência que
florestas conservadas (que possuem espécies com alta abundância relativa) podem apresentar
menor diversidade que florestas alteradas que não possuem tais espécies, como, por exemplo,
Euterpe edulis Martius para Floresta Latifoliada Pluvial de Santa Catarina, ou Dicksonia
sellowiana para Floresta Mista com Araucária.
Estrutura vertical (estratos):
Florestas conservadas possuem uma organização estrutural vertical definida por
estratos de altura. Klein (1960, 1978) cita que o estrato superior da Floresta Mista com
Araucária (formado por A. angustifolia) atingia até 35 m de altura. A partir dos dados atuais
do IFFSC, Vibrans et al. (2013) afirmaram que a altura média do dossel é de 15 m e a altura
total média das árvores é de 10 m. Souza (2009) encontrou evidências estatísticas
significativas em relação aos apontamentos de Reitz e Klein (1966) e Klein (1978) em relação
ao comportamento emergente da A. angustifolia.
Métricas de paisagem de remanescentes:
A literatura aponta vários impactos causados pela área reduzida e a fragmentação de
florestas (BROADBENT et al., 2008; HADLEY e BETTS, 2012; PFEIFER et al., 2014).
Winter et al. (2010) segregaram áreas de florestas manejadas, áreas de borda e áreas core
como tendo diferentes graus de naturalidade. As bordas impõem condições ambientais e
biológicas diferentes das encontradas no interior de um fragmento; o efeito de borda pode
afetar a fenologia das espécies, interferindo nas interações planta-dispersor e reprodutivas
(REZNIK et al., 2012, LAURANCE et al., 2003; TABARELLI e PERES, 1999). A grande
variação em um gradiente pode alterar as propriedades microclimáticas do sub-bosque
próximas às bordas (POHLMAN et al., 2007), pode alterar a incidência de luz, temperatura,

65
umidade, taxa de decomposição da necromassa, riqueza de espécies e interações bióticas
borda-interior (MURCIA, 1995; SCHAADT, 2012).
Engel et al. (1998) e Jordão (2009) expõem o problema do desequilíbrio de populações
de lianas, as quais são muito beneficiadas pelas condições criadas por bordas. Jordão (2009)
apresenta o problema das lianas nas bordas das florestas do estado de São Paulo. Santos et al.
(2008) apresentam a erosão funcional em fragmentos de Floresta Pluvial Latifoliada na região
nordeste do Brasil.
Necromassa:
A necromassa é um importante componente da biodiversidade florestal (HARMON et
al., 1986; SAMUELSSON et al., 1994); diferentes quantidades de necromassa estão
relacionadas a diferentes graus de naturalidade (LAARMAN et al., 2009). Uotila et al. (2002)
adotou o critério presença/ausência para avaliar a naturalidade de florestas boreais na
Finlândia; Nilsson et al. (2002) se referem à proporções de árvores mortas e vivas. Outros
estudos que abordaram a necromassa podem ser encontrados em McRoberts et al. (2012).
Estudos sobre valores de referência de necromassa em florestas primárias ou com pouca
intervenção antrópica são numerosos na Europa (VANDEKERKHOVE et al., 2009), por
exemplo.
Entretanto, no Brasil, são escassos os estudos que determinaram quantidades de
necromassa em florestas mais e menos conservadas. As variáveis levantadas pelo IFFSC
referentes a este componente são: volume (m³.ha-1
), peso (Mg.ha-1
), carbono (Mg.ha-1
) e nível
de decomposição (três classes: material novo; decomposição inicial; decomposição
avançada). Mediante essa exposição, observamos que este componente apresenta relevância
ecológica e potencial de ser um indicador de naturalidade.
Número de árvores velhas e/ou grossas:
O número de árvores velhas e/ou grossas pode retratar o estado de conservação do
ecossistema; ele pode sugerir a magnitude de possíveis explorações feitas na floresta, além de
árvores de grande porte contribuirem para a diversidade estrutural e serem importantes fontes
de microhábits para diversos organismos (GUSTAFSSON, 2002; McCOMB et al., 2003;
MICHEL e WINTER, 2009; NILSSON et al., 2002; von OHEIMB et al., 2007).

66
Presença e ausência de distúrbios antrópicos:
Este indicador está relacionado à hemerobia e pode ser útil para indicar influências
antrópicas na floresta. O IFFSC levantou (em nível de Unidade Amostral) a ausência e a
presença de distúrbios antrópicos como roçada, exploração, presença de gado, entre outros
(Tabela 1.9). Apesar da discussão da utilização de tais variáveis como indicadores de
naturalidade em florestas do hemisfério Norte (CHIRICI et al., 2011; McROBERTS et al.,
2012), nós as consideramos potenciais para o contexto do Brasil, e mais especificamente para
Santa Catarina (de acordo com o Axioma 1 – tópico 1.3.3).
Regeneração natural:
A manutenção da composição de espécies de uma floresta, ou especificamente do
dossel, com o desenrolar da sucessão ecológica ou do tempo, depende dos indivíduos que
regeneram abaixo do dossel (RICHARDS, 1996; SWAINE e HALL, 1988). Meyer et al.
(2013), baseados nos dados coletados pelo IFFSC, concluíram que a regeneração natural da
Floresta Mista com Araucária abriga uma diversidade significativa e que metade das espécies
encontradas no estrato arbóreo foram encontradas na regeneração natural. Entretanto, a
regeneração natural encontra-se altamente perturbada em muitos remanescentes. O pastoreio,
certamente, é o maior perturbador da regeneração natural em Santa Catarina - Sampaio e
Guarino (2007) observaram que a densidade de plântulas de espécies arbóreas é menor em
remanescentes com maior intensidade de pastoreio quando comparado a remanescentes
expostos a uma menor intensidade de pastoreio.
Riqueza de espécies do estrato arbóreo:
É uma variável muito importante e utilizada dentro em investigações de biodiversidade
e conservação (BROOK et al., 2003; BROSE et al., 2003; MAY, 1988), porém sofre com
diferentes tamanhos amostrais, devido à não linearidade do aumento de indivíduos, além da
abundância relativa das espécies ser diferente (CHAO et al., 2014; COLWELL et al., 2012;
COLWELL e CODDINGTON, 1994). Por muitas vezes, a amostragem não reflete a real
riqueza, por conseguinte, a utilização de métodos de estimação da riqueza torna-se necessária
(CHAO et al., 2014; COLWELL e CODDINGTON, 1994; COLWELL et al., 2012;
MAGURRAM, 2004). Ainda, a riqueza de espécies não incorpora informações sobre a
abundância relativa das espécies, visto que considera todas as espécies da mesma forma (raras
e comuns). Apesar destas desvantagens é um indicador importante e passível de ser adotado.

67
1.3.4.4 Escolha dos indicadores de naturalidade
Este tópico tem uma característica peculiar, pois mesclamos aspectos metodológicos,
resultados e discussões acerca da escolha dos indicadores de naturalidade. Cada indicador é
tratado de maneira independente e, por fim, é apresentado um diagnóstico individual final
sobre a aceitação dos mesmos para a quantificação da naturalidade.
Julgamos que nem todas as Unidades Amostrais são passíveis de serem incluídas nas
análises apresentadas a seguir. Optamos por excluir as quatro Unidades Amostrais (UA165,
UA167, UA188 e UA214) implantadas em Nanofloresta Latitifoliada Nebular devido às
discussões em relação à sua independência das demais fitofisionomias florestais
(FALKENBERG e VOLTOLINI, 1995), além de ter sido pouco amostrada pelo IFFSC
(VIBRANS et al., 2012a).
Algumas das Unidades Amostrais complementares foram implantadas no mesmo
remanescente (fragmento) (Figura 1.11). Nesses casos, sorteamos uma Unidade Amostral para
representar o remanescente; foram excluídas seis delas (UA2001, UA3001, UA6002,
UA6003, UA7001 e UA7002).
Assim, das 155 Unidades Amostrais, serão consideradas 145 para as futuras análises.
A B
C
D

68
Figura 1.11. Unidades Amostarais complementares (UA) implantadas em um mesmo
remanescente. (A) UA – 7001, 7002 e 926; (B) UA – 3001 e 3002; (C) UA – 2001 e 2002;
(D) UA – 6003 e 6004; (E) UA – 6001 e 6002.
A partir dos apontamentos citados no primeiro e segundo parágrafos do tópico 1.3.4.3,
acerca das premissas para a escolha dos indicadores de naturalidade, definimos três critérios a
serem cumpridos na escolha:
critério A: relevância ecológica (estrutura, composição e função);
critério B: objetividade e consistência dos resultados;
critério C: passível de ser prontamente calculável.
Adicionamos, ainda, três níveis de cumprimento dos critérios:
i) totalmente satisfeito;
ii) parcialmente satisfeito;
iii) não satisfeito.
Consideramos que a premissa de Winter et al. (2010), de que os indicadores têm ser
passíveis de serem acomodados no contexto de um programa de monitoramento cíclico, já
está cumprida, visto que em futuros ciclos do IFFSC as informações coletadas serão as
mesmas, ou adicionais, conforme os requisitos do IFN-Brasil.
Para ser aceito, o indicador precisa satisfazer os três critérios na totalidade, ou um
deles apenas parcialmente.
Reforçamos que a naturalidade possui diferentes dimensões, e que indicadores
utilizados de forma combinada podem exprimir estas dimensões, gerando resultados
expressivos (McROBERTS et al., 2012; ŠAUDYTÈ et al., 2005; WINTER et al., 2010). Na
sequência, trataremos de cada um dos indicadores.
E

69
Composição de espécies do estrato arbóreo:
A composição de espécies pode ser um bom indicador de naturalidade; por exemplo,
Liira et al. (2007) associou a presença de determinadas espécies com a presença de ações
hemeróbicas na floresta. Do ponto de vista ecológico, cada espécie representa uma função
para a comunidade (por exemplo, Almeida-neto et al. [2008]; Munguía-Rosas et al. [2014]).
Um método de avaliar este componente é pelo grupo ecológico, ou tolerância à intensidade de
radiação solar (por exemplo, Imai et al. [2014]; Silva et al. [2003]). Porém, a falta de
informações quantitativas de referência para florestas conservadas, como, por exemplo, a
proporção de espécies pioneiras e tolerantes à sombra, torna este indicador limitado para
nossa aplicação. Outro fator limitante é a inconsistência das informações sobre a classificação
das espécies em grupos ecológicos, visto que esta classificação varia entre autores (por
exemplo, Reitz [1965]; Reitz et al. [1979]; Swaine e Withmore [1988]; Sevegnani [não
publicado]).
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: não satisfeito
Diversidade e riqueza de espécies do estrato arbóreo:
Trataremos desses dois indicadores de forma conjunta, porque são intimamente
relacionados.
Devido ao fato de que comparar a riqueza de espécies observada, ou seja, somente a
contagem das espécies encontradas em uma comunidade, pode gerar resultados ilusórios
(COLWELL et al., 2012; GOTELLI e COLWELL, 2001), geramos uma curva de acumulação
de espécies extrapolada baseada em amostras para cada Unidade Amostral, utilizando o
modelo-produto de Bernoulli (COLWELL et al., 2012), considerando as subparcelas (10 x 10
m) como amostras. Adotamos o método de extrapolação para contornar o fato de que algumas
Unidades Amostrais apresentaram um número baixo de árvores, o que reduz demasiadamente
o número comum de indivíduos no qual será extraída a riqueza estimada, para fins de
padronização de diferentes intensidades amostrais (COLWELL et al., 2004; GOTELLI e
COLWELL, 2001, 2011).

70
A riqueza estimada na amostra extrapolada, denotada como *)(~
tTSamostra onde T é a
amostra de referência (ou original) e t* é a porção extrapolada da amostra original (t* > 0), é
obtida pelo algoritmo:
01
10 ˆ
*exp1ˆ*)(
~
QtQStTS obsamostra
onde, Q1 é o número de espécies dectectadas em somente uma amostra (unique) e 0Q é o
número de espécies que não foram dectectadas na amostra T. Para Q2 (número de espécies
dectectadas em somente duas amostras - duplicates) > 0, 210 21²1ˆ QQTTQ .
Para Q2 = 0, 1211ˆ2110 QQQTTQ .
Adotamos um fator de extrapolação de dois, ou seja, o dobro da amostra de referência
(t*=T), segundo as recomendações de Chao et al. (2014).
O número mínimo de indivíduos encontrado foi de 58 (UA 301), logo, tornou-se o
número comum da extração dos valores de riqueza para as Unidades Amostrais. Nessa
perspectiva, correlacionamos a riqueza com a diversidade de espécies obtida pelos índices de
Simpson (1-D), calculado através do algoritmo não enviesado proposto por Lande (1996), e o
índice de Shannon (H’, MAGURRAM, 2004) para as 145 Unidades Amostrais. O primeiro
índice dá menor peso à riqueza de espécies e maior peso às espécies dominantes e o segundo
dá maior peso à riqueza e dá maior peso para as espécies comuns (BUCKLAND et al., 2005;
CHAO et al., 2014).
Encontramos uma correlação alta e significativa entre a riqueza e 1-D (r de
Spearman=-0,87; p <0,001) e H‟ (r de Pearson=-0,94; p <0,001); ambas as significâncias
foram obtidas pelo teste de Dutilleul (1993). Estas relações estão apresentadas na Figura 1.12.
A riqueza estimada para um número comum de indivíduos amostrados, se analisada
em outra pespectiva, pode ser considerada uma métrica de diversidade, pois, para dado
número de indivíduos amostrados obtem-se um número n de espécies; Imai et al. (2014)
adotaram essa ideia para determinar a diversidade de comunidades florestais. Esse fato é mais
um que explica a alta correlação entre diversidade e riqueza.

71
Figura 1.12. Relação entre os índices de diverside H‟ e 1-D com a riqueza de espécies,
calculados para as 145 Unidades Amostrais do IFFSC na Floresta Mista com Araucária.
A utilização da diversidade de espécies como um indicador traz à tona um
questionamento fundamental. Há um conflito entre naturalidade e diversidade que necessita
ser esclarecido. Tal conflito está relacionado a aspectos inerentes às comunidades florestais
que contêm uma espécie (ou mais de uma) que possuem alta abundância relativa. Ao se
aplicar um índice de diversidade tal como o 1-D ou o H’ para uma comunidade florestal
dominada por uma ou poucas espécies, se obteria um valor baixo para dado índice. Entretanto,
comunidades dominadas por poucas espécies podem ser resultado de processos naturais de
dispersão e ocupação (HUBBELL, 2001), de adaptações que essas espécies possuem
(PITMAN et al., 2001), de fatores ambientais (BAUERMANN e BEHLING, 2009; KLEIN,
1984), da sucessão ecológica (CHAZDON, 2009; KLEIN, 1960; VOLKOV et al., 2007) e da
competição interespecífica e por recursos (GOTELLI, 1999; McGILL et al., 2007).
Considerando tais comunidades, seria errôneo afirmar que por apresentarem baixa diversidade
são menos naturais que comunidades que apresentam valores altos em um índice de
diversidade.
Na Floresta Mista com Araucária, esse fato mostra-se presente nas diversas formações
descritas por Klein (1978). Klein (1960, 1966, 1978) descreve suas observações referentes aos
densos agrupamentos de A. angustifolia e Dicksonia sellowiana (GASPER et al., 2011;
y = 0.0954x + 0.9009 p < 0.05 R² = 0.87 r = 0.78 p < 0.001
y = -0.0009x2 + 0.0497x + 0.2862 p < 0.05 r = 0.60 p < 0.001
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
1.00
1.50
2.00
2.50
3.00
3.50
4.00
0.0 5.0 10.0 15.0 20.0 25.0 30.0 35.0
Índ
ice
de
Sim
pso
n (
1-D
)
Índ
ice
de
Sh
an
no
n (
H')
Riqueza de espécies
H'
1-D

72
MANTOVANI, 2004), e de espécies do gênero Ocotea, como O. porosa e O. pulchella
(KLEIN, 1966, 1978).
Este conflito entre naturalidade e diversidade foi observado nos dados coletados pelo
IFFSC. Para investigar essa hipótese, aplicamos dois procedimentos: i) calculamos a
abundância relativa (AR%) e a área basal relativa (AB%) das espécies A. angustifolia, D.
sellowiana, O. pulchella e O. porosa das 145 Unidade Amostrais; ii) correlacionamos a soma
da abundância e a área basal relativa (das quatro espécies) com os valores obtidos para os
índices de diversidade (1-D e H’).
A correlação entre AR% das quatro espécies e H’ foi de -0,74 (r de Pearson, p <
0,001), e para AB% foi de -0,74 (r de Pearson, p < 0,001). A correlação entre AR% das quatro
espécies e 1-D foi de -0,56 (r de Spearman, p < 0,001), e para AB% foi de -0,56 (r de
Spearman, p < 0,001).
Como densos agrupamentos das espécies citadas, principalmente de D. sellowiana,
podem estar relacionados a remanescentes conservados, descartamos a possibilidade da
utilização desses indicadores, corroborando as observações de Imai et al. (2014), os quais
consideraram que riqueza e diversidade não são bons indicadores do estado de conservação de
florestas mistas de dipterocarpáceas na Ásia.
Diagnóstico final: - critério A: parcialmente satisfeito
- critério B: parcialmente satisfeito
- critério C: satisfeito

73
y = -166.17x2 + 96.78x + 70.52 p < 0.01 R² = 0.55 r = -0.56 p < 0.001
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
0.30 0.40 0.50 0.60 0.70 0.80 0.90 1.00
Figura 1.13. Correlação negativa entre a soma da área basal relativa (A), e soma da
abundância relativa (B), de Araucaria angustifolia, Dicksonia sellowiana, Ocotea porosa e
Ocotea pulchella com os índices de diversidade Simpson e Shannon.
Estrutura de população de A. angustifolia:
Esse indicador mostra-se, teoricamente, muito interessante, entretanto, ao analisarmos
os dados do IFFSC, verificamos que sua aplicabilidade é limitada neste estudo. Das 145
Unidades Amostrais consideradas, 94 apresentaram indivíduos de A. angustifolia e 47,9%
apresentaram uma abundância maior que 10 indivíduos por Unidade Amostral. Julgamos que
10 indivíduos (na melhor hipótese) não são suficientes para a análise da estrutura da
população, de forma que Wright et al. (2003) e Souza (2009) sugerem que ao menos 25
indivíduos sejam considerados. Diante desses fatos, ficamos limitados para fazer inferências
y = -30.05x + 107.23 p < 0.01 R² = 0.52 r = - 0.74 p < 0.01
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
0.50 1.00 1.50 2.00 2.50 3.00 3.50 4.00
Áre
a b
asa
l R
ela
tiva
(%
)
y = -26.25x + 91.69 p < 0.01 R² = 0.54 r = - 0.74 p < 0.01
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
0.50 1.00 1.50 2.00 2.50 3.00 3.50 4.00
Ab
un
dâ
ncia
Rela
tiva
(%
)
Índice de Shannon (H')
B
A
y = -80.81x2 - 23.00x + 103.73 p < 0.01 R² = 0.63 r = -0.56 p < 0.001
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
0.30 0.40 0.50 0.60 0.70 0.80 0.90 1.00
Índice de Simpson (1-D)

74
sobre a estrutura de população da espécie para os remanescentes amostrados; por estas razões,
este indicador foi desconsiderado.
Diagnóstico final: - critério A: parcialmente satisfeito
- critério B: não satisfeito
- critério C: satisfeito
Estrutura vertical (estratos):
A estrutura vertical expressa uma dimensão da diversidade estrutural de uma
comunidade florestal, logo, torna-se um indicador importante (BUONGIORNO et al. 1994;
McROBERTS et al., 2008; POMMERENING, 2002; LÄHDE et al., 1999, VARGA e
KLINKA, 2001).
O primeiro obstáculo no uso deste indicador está no fato de que a altura total (ht) das
árvores amostradas pelo IFFSC foi estimada pelas equipes de campo, mediante medições
prévias com hipsômetro como referência. Não obstante esse fato, as médias de ht estimadas
mostraram-se estatisticamente equivalente às médias de ht medidas com hipsômetro (para
construção de modelos hipsométricos) pelo teste z (VIBRANS et al., no prelo). O primeiro
obstáculo pode ser considerado negligivel, porém, a dificuldade em adotar este estimador está
em como manipulá-lo numericamente, devido à falta de dados de florestas de referência.
Por ser frequentemente levantado por IFNs (CHIRICI et al., 2011), utilizamos a
Estrutura vertical como um critério na escolha das florestas de referência, adotando a proposta
de McRoberts et al. (2012). Estabelecemos três estratos de altura para cada Unidade
Amostral, seguindo os seguintes passos: i) calculamos a altura média de 5% dos indivíduos
mais altos e dividimos o valor obtido em terços; ii) cada indivíduo da Unidade Amostral é
atribuído a um dos três estratos.
Segundo a proposta de McRoberts et al. (2012), as Unidades Amostrais que
representam as florestas mais naturais apresentam os três estratos, e consequentemente,
indivíduos nos três estratos. No tópico 1.3.4.7 (Determinação e escolha das florestas de
referência) consta a aplicação deste indicador.
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito

75
Estrutura diamétrica:
Devido às várias formas de como abordar esse indicador (Estrutura diamétrica),
subdividimos-o em quatro indicadores, os quais serão tratados de forma independente:
a) Assimetria da distribuição de diâmetros
Nossas investigações acerca deste indicador mostraram resultados interessantes.
Calculamos o gdap para seis Unidades Amostrais com número de indivíduos amostrados
variando de 296 a 308, e seis variando de 177 a 189 indivíduos amostrados. Chegamos à
conclusão de que não é possível utilizá-lo tal como McRorterts et al. (2012), os quais
relacionaram florestas mais naturais com maiores valores positivos de gdap. Percebemos que
os maiores valores de gdap nem sempre significam distribuições que reportam florestas
(tropicais ou subtropicais) mais conservadas, as quais tendem a apresentar uma distribuição
exponencial negativa. Mesmo em valores mais elevados de gdap notamos descontinuidades na
função exponencial negativa (por exemplo, Figura 1.14: UA 843 e UA 1056 e Figura 1.15:
UA 551), o que significa que alguma perturbação está sendo aplicada à comunidade florestal,
ou esta se encontra em estágios de sucessão marcados por espécies heliófitas, as quais são
caracterizadas por conter menos indivíduos nas classes menores de DAP (FINGEGAN, 1996).
A comunidade também pode estar em processo de substituição de espécies heliófitas para
espécies tolerantes à sombra (CHAZDON, 2008), como observado por Klein (1980, 1984)
para a Floresta Latifoliada Pluvial de Santa Catarina.
Comparamos, para exemplicar nossos apontamentos, a Unidade Amostral 843 e a
7002 (Figura 1.14); notamos que a 7002 apresenta uma característica esperada em uma
floresta subtropical mais natural, embora, possua menor valor de gdap que a 843.
Diagnóstico final: - critério A: parcialmente satisfeito
- critério B: parcialmente satisfeito
- critério C: satisfeito
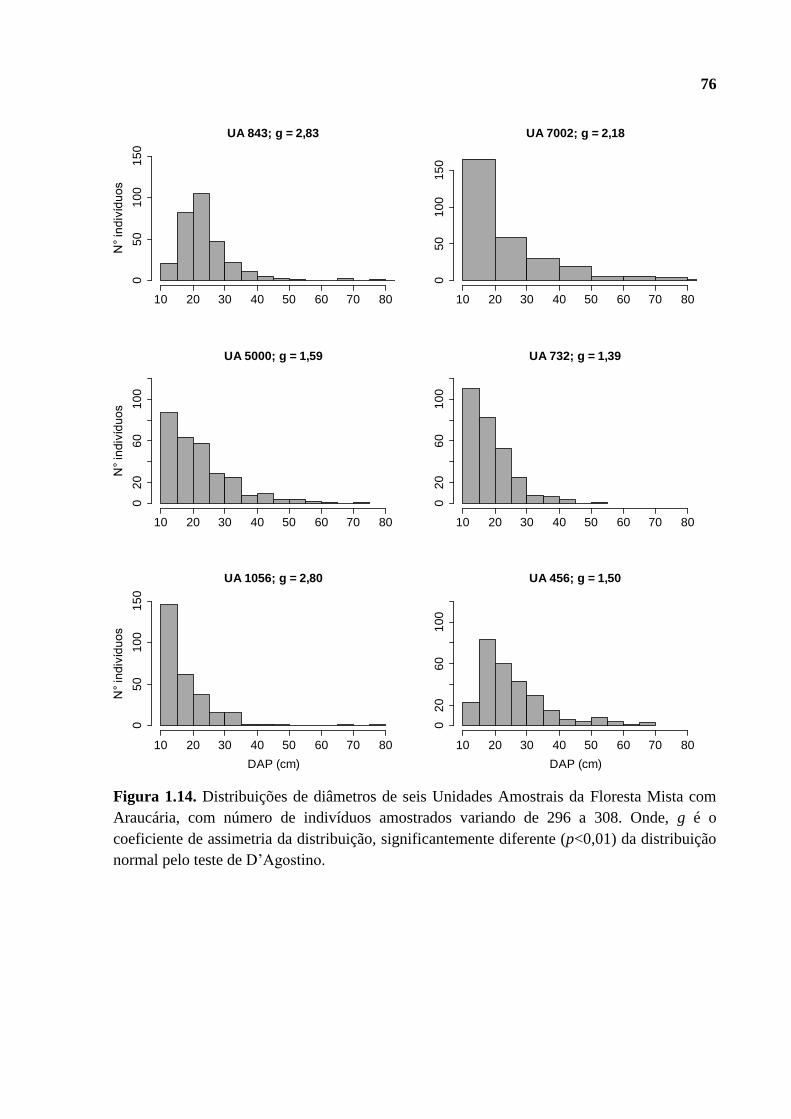
76
Figura 1.14. Distribuições de diâmetros de seis Unidades Amostrais da Floresta Mista com
Araucária, com número de indivíduos amostrados variando de 296 a 308. Onde, g é o
coeficiente de assimetria da distribuição, significantemente diferente (p<0,01) da distribuição
normal pelo teste de D‟Agostino.
UA 843; g = 2,83
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
05
01
00
15
0UA 7002; g = 2,18
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
05
01
00
15
0
UA 5000; g = 1,59
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
06
01
00
UA 732; g = 1,39
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
06
01
00
UA 1056; g = 2,80
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
05
01
00
15
0
UA 456; g = 1,50
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
06
01
00

77
Figura 1.15. Distribuições de diâmetros de seis Unidades Amostrais da Floresta Mista com
Araucária, com número de indivíduos amostrados variando de 177 a 189. Onde, g é o
coeficiente de assimetria da distribuição, significantemente diferente (p<0,01) da distribuição
normal pelo teste de D‟Agostino.
UA 673; g = 0,84
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0UA 894; g = 1,64
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0
UA 3002; g = 1,29
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0
UA 856; g = 1,45
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0
UA 551; g = 2,12
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0
UA 87; g = 0,91
DAP (cm)
N°
ind
ivíd
uo
s
10 20 30 40 50 60 70 80
02
04
06
08
0

78
b) Ajuste à função exponencial negativa
A partir das constatações não favoráveis sobre o gdap, propomos avaliar o ajuste da
distribuição de diâmetros observada e a estimada pela função exponencial negativa (ZAR,
2010), através do modelo de Meyer (SCHNEIDER, 1993). A modelo é expresso por:
i1.x
0.eiN
onde Ni é a frequência observada para a i-ésima classe de diâmetro, xi é o centro da i-ésima
classe de diâmetro (cm) e ε é o resíduo aleatório com média 0.
Para o ajuste do modelo pelo método de mínimos quadrados, aplicamos a
transformação logarítmica (ln) na variável resposta e na preditora, logo, o modelo foi
reformulado (ou linearizado) para:
.x)ln()ln( i10 iN
onde n são os parâmetros a serem estimados.
Voltando à escala original, temos:
2
ˆ.xˆˆexpˆ
i10res
iN
onde iN é número de indivíduos estimado para a i-ésima classe de diâmetro, res é o erro
padrão residual calculado na escala ln-ln, e o termo 2
ˆ res compensa o viés da transformação
ln-ln para a escala original (BASKERVILLE, 1972).
Para avaliar o ajuste do modelo às distribuições observadas (Figura, 1.16) calculamos
a raiz do erro padrão residual (RMSE) na escala original, que é calculado por:
n
i
ii YYn
RMSE1
2)ˆ(1
onde iY é o valor predito, Yi é o valor observado e n é o número de observações (ou de classes
de DAP).
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito
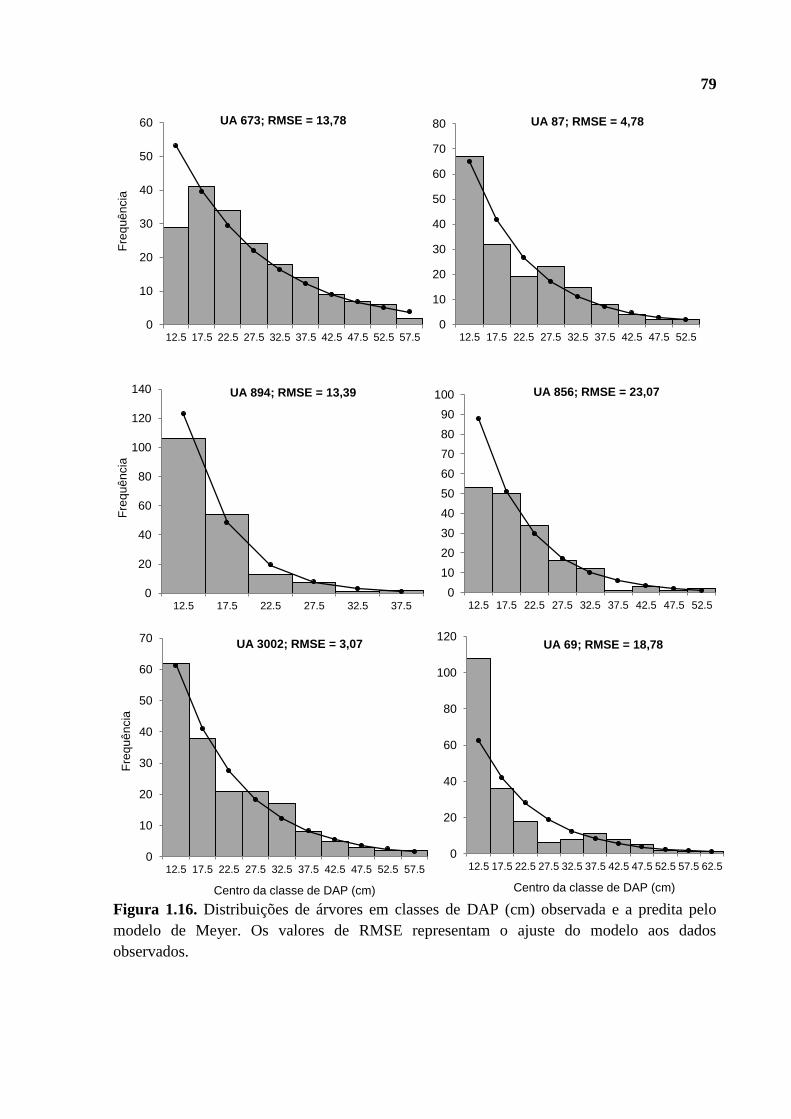
79
0
10
20
30
40
50
60
70
80
12.5 17.5 22.5 27.5 32.5 37.5 42.5 47.5 52.5
0
10
20
30
40
50
60
70
80
90
100
12.5 17.5 22.5 27.5 32.5 37.5 42.5 47.5 52.5
0
20
40
60
80
100
120
12.5 17.5 22.5 27.5 32.5 37.5 42.5 47.5 52.5 57.5 62.5
Centro da classe de DAP (cm)
Figura 1.16. Distribuições de árvores em classes de DAP (cm) observada e a predita pelo
modelo de Meyer. Os valores de RMSE representam o ajuste do modelo aos dados
observados.
0
10
20
30
40
50
60
12.5 17.5 22.5 27.5 32.5 37.5 42.5 47.5 52.5 57.5
Fre
qu
ên
cia
0
20
40
60
80
100
120
140
12.5 17.5 22.5 27.5 32.5 37.5
Fre
qu
ên
cia
0
10
20
30
40
50
60
70
12.5 17.5 22.5 27.5 32.5 37.5 42.5 47.5 52.5 57.5
Fre
qu
ên
cia
Centro da classe de DAP (cm)
UA 673; RMSE = 13,78 UA 87; RMSE = 4,78
UA 894; RMSE = 13,39 UA 856; RMSE = 23,07
UA 3002; RMSE = 3,07 UA 69; RMSE = 18,78

80
c) Desvio padrão dos diâmetros (Sdap)
O ajuste à função exponencial negativa por si só não é capaz de estimar toda a
naturalidade do componente estrutural relacionado à diversidade estrutural (neste caso a
estrutra horizontal - distribuição de diâmetros das árvores). O Desvio padrão dos diâmetros
(Sdap) leva em conta outra dimensão da diversidade estrutural, ou seja, da variabilidade de
tamanhos das árvores (estreita relação com a biodiversidade - Buongiorno et al. [1994];
COST [2006]; McRoberts et al. [2008]; Michel e Winer [2009]; Uotila et al. [2002]).
O Sdap é calculado por:
1
²)(1
_____
n
DAPDAPS
n
i i
dap
onde, DAPi é o diâmetro à altura do peito da i-ésima árvore, _____
DAP é a média da Unidade
Amostral e n é o número de árvores medidas na Unidade Amostral.
A Unidade Amostral 894 (Figura 1.16) apresenta uma situação interessante para
exemplificar o argumento recém lançado: possui um baixo valor para o RMSE, embora
apresente um valor baixo para o Sdap (4,86), indicando baixa variabilidade de DAP das árvores
e, consequentemente, menor diversidade estrutural, obviamente, quando comparada com
outras Unidades Amostrais.
Ao calcularmos o Sdap para as 145 Unidades Amostrais, observamos uma relação deste
com o DAP médio das mesmas (Figura 1.17), com correlação de Pearson forte e altamente
significativa (r=0,81; p<0,001 - obtido do teste t de Dutilleul [1993]), semelhante ao
encontrado por McRoberts et al. (2008). Empiricamente, sabemos que pode existir uma
tendência de florestas mais conservadas, com árvores mais velhas, apresentarem DAP médio
maior do que florestas menos conservadas. Sendo assim, existem evidências significativas de
que o Sdap possa expressar esta outra dimensão da naturalidade.
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito
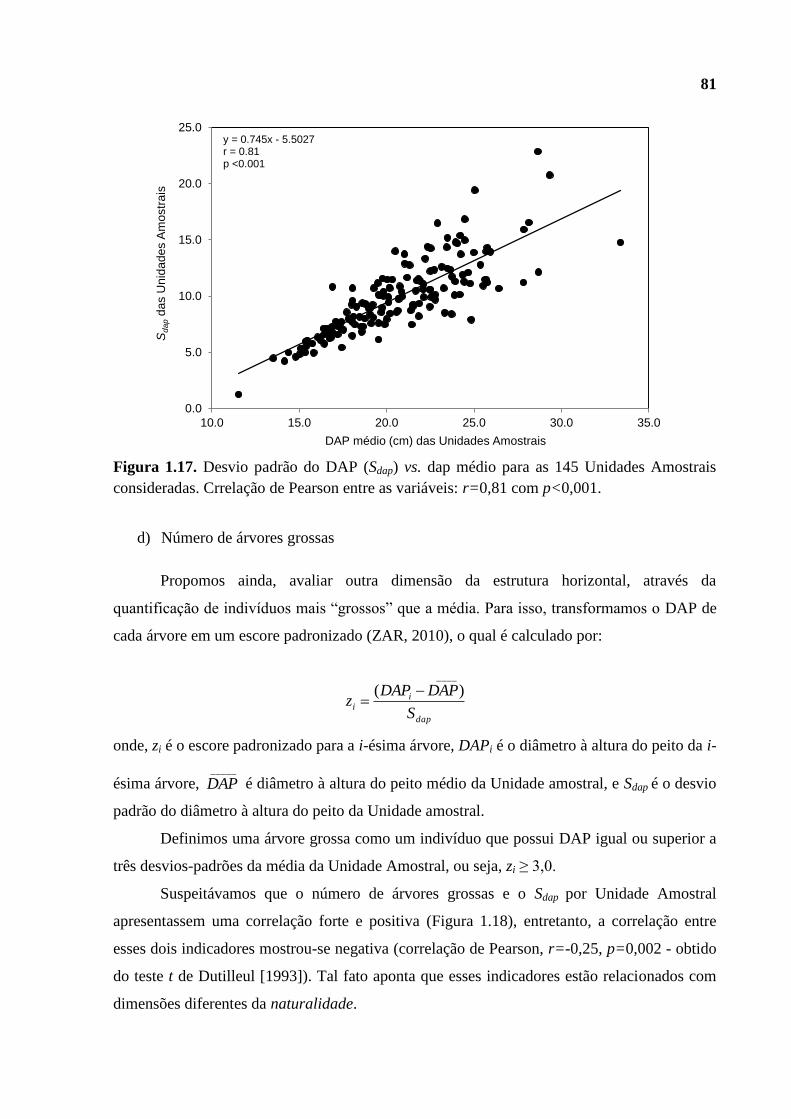
81
Figura 1.17. Desvio padrão do DAP (Sdap) vs. dap médio para as 145 Unidades Amostrais
consideradas. Crrelação de Pearson entre as variáveis: r=0,81 com p<0,001.
d) Número de árvores grossas
Propomos ainda, avaliar outra dimensão da estrutura horizontal, através da
quantificação de indivíduos mais “grossos” que a média. Para isso, transformamos o DAP de
cada árvore em um escore padronizado (ZAR, 2010), o qual é calculado por:
dap
ii
S
DAPDAPz
____
)(
onde, zi é o escore padronizado para a i-ésima árvore, DAPi é o diâmetro à altura do peito da i-
ésima árvore, _____
DAP é diâmetro à altura do peito médio da Unidade amostral, e Sdap é o desvio
padrão do diâmetro à altura do peito da Unidade amostral.
Definimos uma árvore grossa como um indivíduo que possui DAP igual ou superior a
três desvios-padrões da média da Unidade Amostral, ou seja, zi ≥ 3,0.
Suspeitávamos que o número de árvores grossas e o Sdap por Unidade Amostral
apresentassem uma correlação forte e positiva (Figura 1.18), entretanto, a correlação entre
esses dois indicadores mostrou-se negativa (correlação de Pearson, r=-0,25, p=0,002 - obtido
do teste t de Dutilleul [1993]). Tal fato aponta que esses indicadores estão relacionados com
dimensões diferentes da naturalidade.
y = 0.745x - 5.5027 r = 0.81 p <0.001
0.0
5.0
10.0
15.0
20.0
25.0
10.0 15.0 20.0 25.0 30.0 35.0
Sd
ap d
as U
nid
ad
es A
mo
str
ais
DAP médio (cm) das Unidades Amostrais
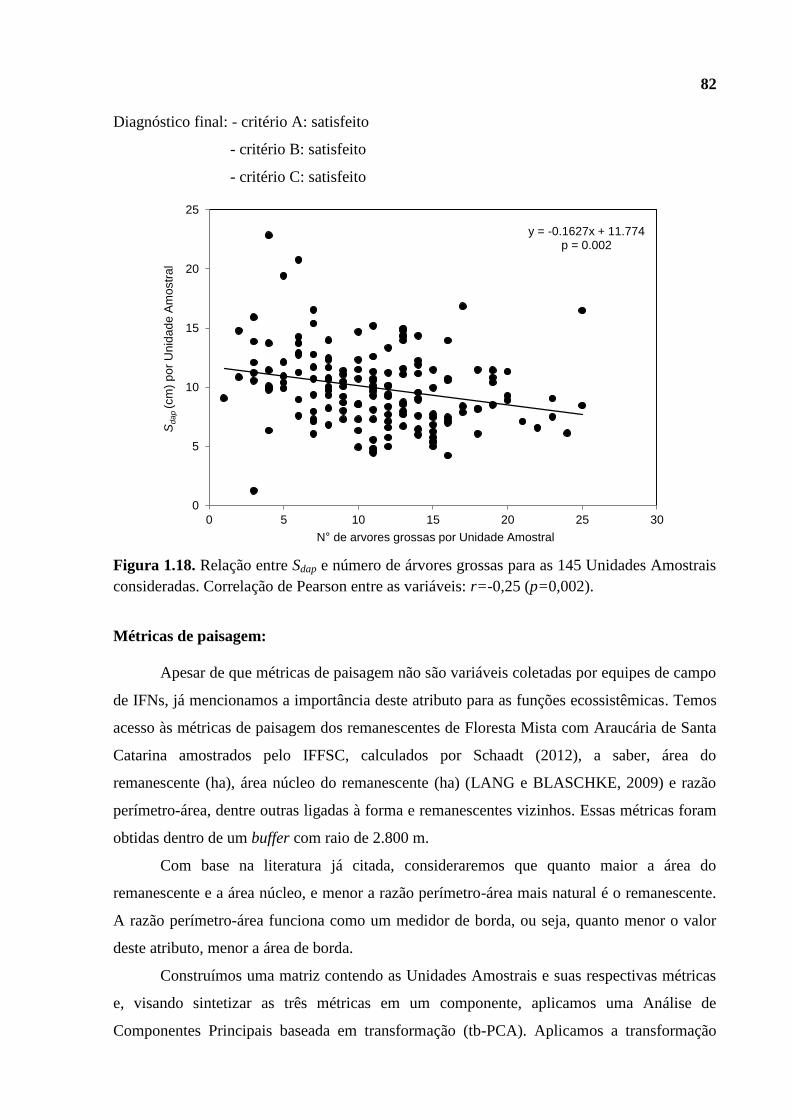
82
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito
Figura 1.18. Relação entre Sdap e número de árvores grossas para as 145 Unidades Amostrais
consideradas. Correlação de Pearson entre as variáveis: r=-0,25 (p=0,002).
Métricas de paisagem:
Apesar de que métricas de paisagem não são variáveis coletadas por equipes de campo
de IFNs, já mencionamos a importância deste atributo para as funções ecossistêmicas. Temos
acesso às métricas de paisagem dos remanescentes de Floresta Mista com Araucária de Santa
Catarina amostrados pelo IFFSC, calculados por Schaadt (2012), a saber, área do
remanescente (ha), área núcleo do remanescente (ha) (LANG e BLASCHKE, 2009) e razão
perímetro-área, dentre outras ligadas à forma e remanescentes vizinhos. Essas métricas foram
obtidas dentro de um buffer com raio de 2.800 m.
Com base na literatura já citada, consideraremos que quanto maior a área do
remanescente e a área núcleo, e menor a razão perímetro-área mais natural é o remanescente.
A razão perímetro-área funciona como um medidor de borda, ou seja, quanto menor o valor
deste atributo, menor a área de borda.
Construímos uma matriz contendo as Unidades Amostrais e suas respectivas métricas
e, visando sintetizar as três métricas em um componente, aplicamos uma Análise de
Componentes Principais baseada em transformação (tb-PCA). Aplicamos a transformação
y = -0.1627x + 11.774 p = 0.002
0
5
10
15
20
25
0 5 10 15 20 25 30
Sd
ap
(cm
) p
or
Un
ida
de
Am
ostr
al
N° de arvores grossas por Unidade Amostral

83
Box-Cox nos dados (OSBORNE, 2010), a fim de maximizar a relação linear entre as
variáveis. Testamos a significância dos eixos da tb-PCA por meio de 999 permutações de
Monte Carlo, sendo que o primeiro eixo se mostrou altamente significativo (p=0,001).
O processo de sintetização das três variáveis mostrou-se eficiente; o eixo 1 explicou
grande parte da variação dos dados (88,84%, Tabela 1.6), logo, temos uma componente
confiável. O indicador de naturalidade relacionado às métricas de paisagem é o escore
extraído do eixo 1 para cada Unidade Amostral.
Para exemplificar, consideramos as Unidades Amostrais 1013 e 336, as quais estão
posicionadas nos extremos do diagrama de ordenação da tb-PCA (Figura 1.19). Vemos que a
1013 possui maior área que a 336, em contrapartida, a 336 possui maior valor para a razão
perímetro-área (Figura 1.20); os escores destas são 1,84 e -5.31, respectivamente. Observando
a Tabela 1.7 e o diagrama de ordenação, vemos que as variáveis área do remanescente (ha) e
área núcleo do remanescente (ha) possuem peso positivo e a razão perímetro-área peso
negativo no componente criada (eixo 1). Logo, quanto menor o escore da Unidade amostral
menor sua área e maior a razão perímetro-área.
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito
Tabela 1.6. Autovalores e variância explicada (%) de cada eixo da tb-PCA.*valor-p
significativo (α=0,05) após 999 permutações de Monte Carlo.
Eixo Autovalor Variância
explicada (%) p
1 2,66 88,84 0,001*
2 0,25 8,50 1,000
3 0,08 2,65 1,000
Tabela 1.7. Autovetores correspondentes a cada variável, somente para o eixo 1 da tb-PCA.
Variável Autovetor
Área do remanescente (ha) 0,574
Área núcleo do remanescente (ha) 0,596
Razão perímetro-área -0,562

84
Figura 1.19. Análise de componentes principais (tb-PCA) das métricas de paisagem, visando
a sintetização das mesmas em um único componente (eixo 1 da tb-PCA). RPA = razão
perímetro-área.
Figura 1.20. Remanescentes correspondentes às Unidades Amostrais 1013 (A) e a 336 (B)
com escores de 1,85 e -5,32 no eixo 1 da tb-PCA. Representação das métricas de paisagem
adotadas.
Necromassa:
O IFFSC levantou a necromassa por meio de dois transectos de 10 m de comprimento,
localizados no ponto central do conglomerado e dispostos em forma de cruz. Foi medido o
material com diâmetro maior que 1 cm (CARDOSO et al., 2012; VIBRANS et al., 2010).
Com o intuito de avaliar este indicador, observamos a variável peso (Mg.ha-1
) de
necromassa levantada pelo IFFSC. As demais variáveis (volume [m³.ha-1
], carbono [Mg.ha-1
]
e nível de decomposição [três classes: material novo; decomposição inicial; decomposição
A B
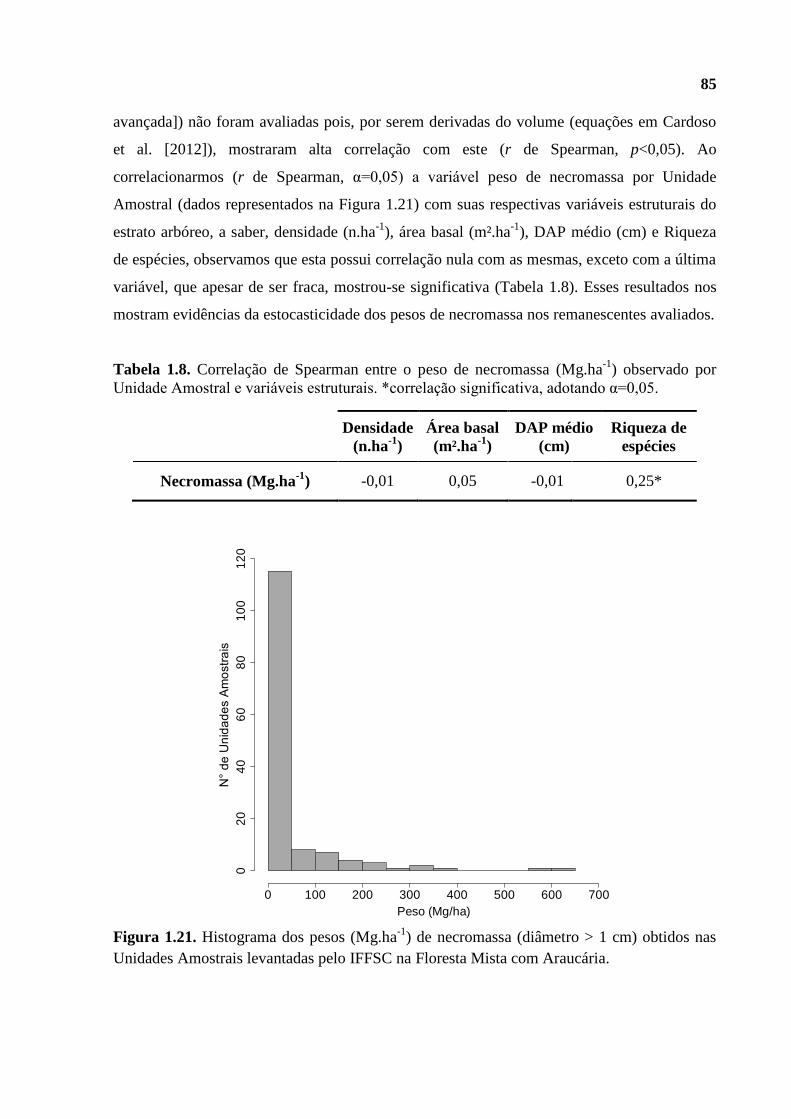
85
avançada]) não foram avaliadas pois, por serem derivadas do volume (equações em Cardoso
et al. [2012]), mostraram alta correlação com este (r de Spearman, p<0,05). Ao
correlacionarmos (r de Spearman, α=0,05) a variável peso de necromassa por Unidade
Amostral (dados representados na Figura 1.21) com suas respectivas variáveis estruturais do
estrato arbóreo, a saber, densidade (n.ha-1
), área basal (m².ha-1
), DAP médio (cm) e Riqueza
de espécies, observamos que esta possui correlação nula com as mesmas, exceto com a última
variável, que apesar de ser fraca, mostrou-se significativa (Tabela 1.8). Esses resultados nos
mostram evidências da estocasticidade dos pesos de necromassa nos remanescentes avaliados.
Tabela 1.8. Correlação de Spearman entre o peso de necromassa (Mg.ha-1
) observado por
Unidade Amostral e variáveis estruturais. *correlação significativa, adotando α=0,05.
Densidade
(n.ha-1
)
Área basal
(m².ha-1
)
DAP médio
(cm)
Riqueza de
espécies
Necromassa (Mg.ha-1
) -0,01 0,05 -0,01 0,25*
Figura 1.21. Histograma dos pesos (Mg.ha-1
) de necromassa (diâmetro > 1 cm) obtidos nas
Unidades Amostrais levantadas pelo IFFSC na Floresta Mista com Araucária.
Peso (Mg/ha)
N°
de
Un
ida
de
s A
mo
str
ais
0 100 200 300 400 500 600 700
02
04
06
08
01
00
12
0

86
Apesar da importância ecológica da necromassa e da relação desta com a naturalidade
(CHIRICI et al., 2011; LAARMANN et al., 2009; WINTER et al., 2010), McRoberts et al.
(2012), observaram que os critérios adotados em relação à quantidade ou forma de
necromassa (por exemplo, troncos caídos, galhos, etc.) diferem entre estudos, tornando
obscuros os valores de referência. Ainda, de acordo com von Oheimb et al. (2007), a
produção de necromassa é influenciada por questões climáticas e de sítio. No Brasil,
levantamentos de necromassa ainda são raros e pouco explorados. Consequentemente, são
escassos os estudos que determinaram quantidades de necromassa em florestas mais
conservadas e não conservadas do Brasil. Os resultados obtidos, descritos anteriormente, não
nos fornecem embasamento para inferir sobre quais fatores predizem a produção de
necromassa, ou se esta tem ligação direta com a naturalidade dos remanescentes avaliados.
Embora o IFFSC tenha sido o primeiro a conduzir este levantamento no IFN do Brasil,
Cardoso et al. (2012) afirmam que a metodologia utilizada pelo IFFSC ainda precisa ser
aprimorada. Descartamos, também, a possibilidade levantada anteriormente de utilizar este
indicador na forma de presença e ausência.
Diagnóstico final: - critério A: parcialmente satisfeito
- critério B: não satisfeito
- critério C: satisfeito
Regeneração natural:
Propomos avaliar a regeneração natural dos remanescentes (Unidades Amostrais)
através de sua diversidade (sensu Hurlbert [1971]), já que a riqueza de espécies é
correlacionada com a diversidade. Consideramos os mesmos índices de diversidade avaliados
anteriormente, 1-D e H‟.
Constatamos uma resposta interessante do H‟ aplicado à regeneração natural. Em
comparação ao 1-D, o H‟ respondeu de forma mais eficiente à variação da riqueza de
espécies, conforme apresentamos na Figura 1.22. Nesta mesma figura, nota-se o caráter do 1-
D, o qual dá mais peso à uniformidade das abundâncias das espécies dominantes (CHAO et
al., 2014). Esse comportamento apresentado pelo H‟ nos assegura que ambas as dimensões da
diversidade serão bem relatadas. Sendo a regeneração natural um componente fundamental e
bastante afetado na Floresta Mista com Araucária, principalmente pelo pastoreio do gado,
consideramos este um indicador de naturalidade escencial.
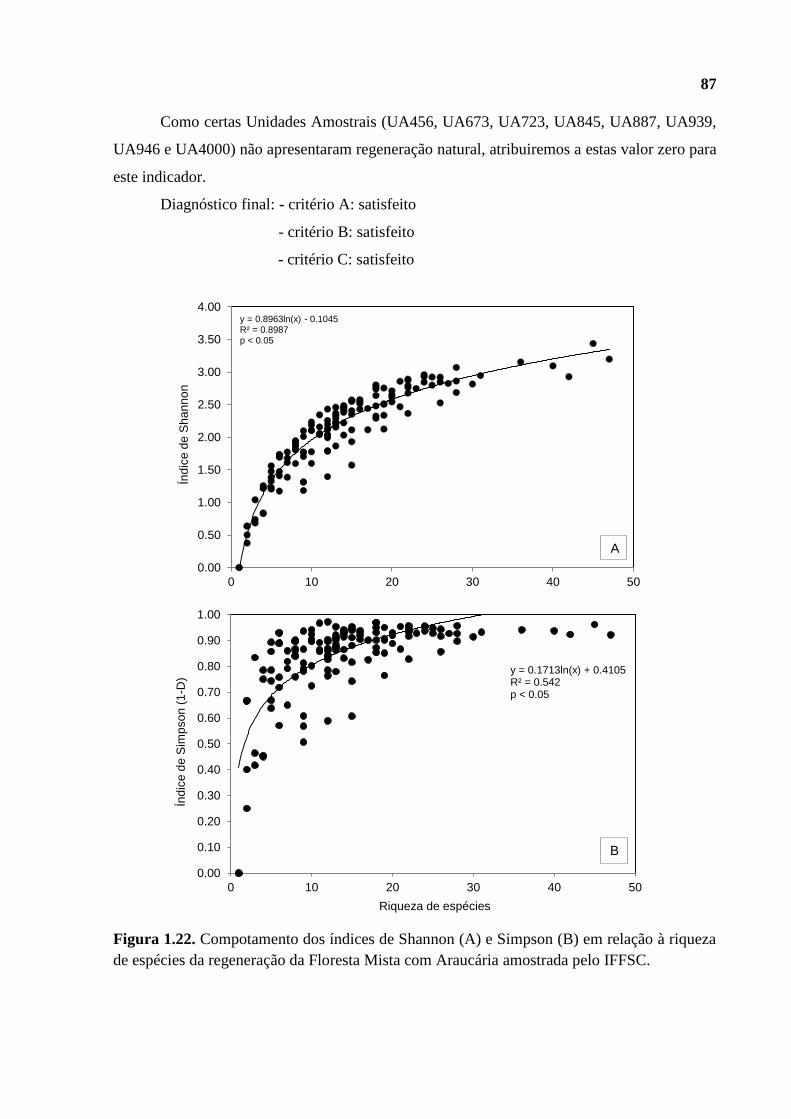
87
Como certas Unidades Amostrais (UA456, UA673, UA723, UA845, UA887, UA939,
UA946 e UA4000) não apresentaram regeneração natural, atribuiremos a estas valor zero para
este indicador.
Diagnóstico final: - critério A: satisfeito
- critério B: satisfeito
- critério C: satisfeito
Figura 1.22. Compotamento dos índices de Shannon (A) e Simpson (B) em relação à riqueza
de espécies da regeneração da Floresta Mista com Araucária amostrada pelo IFFSC.
y = 0.8963ln(x) - 0.1045 R² = 0.8987 p < 0.05
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
0 10 20 30 40 50
Índ
ice
de
Sh
an
no
n
y = 0.1713ln(x) + 0.4105 R² = 0.542 p < 0.05
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
0 10 20 30 40 50
Índ
ice
de
Sim
pso
n (
1-D
)
Riqueza de espécies
A
B

88
Presença e ausência de distúrbios antrópicos:
Chirici et al. (2011) e McRoberts et al. (2012) fazem um apontamento interessante.
Eles afirmam que variáveis ligadas à hemerobia podem ser indicadoras de florestas
manejadas/alteradas, porém, podem não ser indicadoras de florestas mais naturais. Como já
apresentamos nossas reflexões sobre as influências antrópicas nas florestas de Santa Catarina
(sintetizadas no Axioma 1, presente no tópico 1.3.3 deste capítulo), consideramos que
perturbações antrópicas podem ser incluídas no processo de acesso à naturalidade.
Propomos um Índice Quantitativo de Hemerobia, tratado daqui em diante como IQH,
baseado na avaliação de variáveis qualitativas (ausência e presença) ligadas à hemerobia
levantadas pelo IFFSC em nível de Unidade Amostral (VIBRANS et al., 2009). Em campo, as
variáveis foram avaliadas de acordo com sua ausência e presença dentro dos limites da
Unidade Amostral. Iniciando a explanação do índice, consideramos, de forma arbitrária,
baseados em considerações já feitas sobre as principais ações perturbadoras/degradadoras
aplicadas à Floresta Mista com Araucária, pesos que vão de um a cinco para cada perturbação
(Tabela 1.9). Apesar da exploração da erva-mate ser uma atividade cultural e econômica, e
que incentiva proprietários rurais a manter a floresta nativa em pé, consideramos que as
atividades ligadas à exploração, como priorização da espécie (I. paraguariensis) no sub-
bosque, as roçadas, aberturas de estradas para acessar os remanescentes, dentre outras ações,
podem prejudicar processos ecossistêmicos importantes.
Tabela 1.9. Variáveis ligadas à hemerobia coletadas em campo pelo IFFSC e seus respectivos
pesos arbitrários.
Perturbação Ausência ou presença Peso
Roçada de sub-bosque 0 ou 1 5
Corte seletivo 0 ou 1 5
Corte raso 0 ou 1 5
Exploração de erva-mate 0 ou 1 3
Pastejo 0 ou 1 5
Caça 0 ou 1 2
Espécie exótica 0 ou 1 4
Estradas 0 ou 1 2
Sinais de fogo 0 ou 1 3
Quando todas as perturbações mostram-se presentes, temos uma hemerobia máxima
(algoritimo i), e quando todas as perturbações mostraram-se ausentes, temos uma hemerobia
mínima (algoritmo ii) (Figura 1.23).

89
Matematicamente, podemos expressar isto como:
onde Hmáx é o valor assumido pela hemerobia máxima, Hmín é o valor assumido pela
hemerobia mínima, p é a perturbação, np é o número total de perturbações, vppres é o valor
obtido pela presença de todas as perturbações multiplicadas pelo seus respectivos pesos e
vpaus é o valor obtido pela ausência de todas as perturbações multiplicadas pelo seus
respectivos pesos.
Para chegar no IQH escalado para o intervalo [0,1], aonde 0 represanta uma hemerobia
máxima (algoritmo iii) e 1 uma hemerobia mínima (algoritmo iv), temos:
Figura 1.23. Representação do Índice de quantitativo de hemerobia (IQH) baseado em
variáveis qualitativas levantadas em campo pelo IFFSC.
Na Tabela 1.10 apresentamos o caso de uma Unidade Amostral hipotética, com seus
respectivos valores para presença ou ausência de perturbações, onde o IQH apresenta valor
mínimo (0). Na Tabela 1.11 apresentamos outro caso hipotético, onde a Unidade Amostral
possui um valor de IQH intermediário.
0 1
Índice Quantitativo de Hemerobia
Baixa hemerobia
Alta hemerobia
01
máx
máxmáx
H
HIQH
11
máx
mínmín
H
HIQH
(iii)
341
pn
p
presmáx vpH
01
pn
p
ausmín vpH
(i)
(ii)
(iv)

90
Tabela 1.10. Representação do valor mínimo obtido para o Índice Quantitativo de
Hemerobia. Onde: P é o resultado da multiplicação do peso atribuído à perturbação por sua
ausência ou presença (0 ou 1); IQH* é o Índice Quantitativo de Hemerobia escalado para o
intervalo [0,1].
Perturbação Peso Ausência ou
presença P
Roçada de sub-bosque 5 1 5
Corte seletivo 5 1 5
Corte raso 5 1 5
Exploração de erva-mate 3 1 3
Pastejo 5 1 5
Caça 2 1 2
Espécie exótica 4 1 4
Estradas 2 1 2
Sinais de fogo 3 1 3
∑ - - 34
IQH* 0,0
Tabela 1.11. Situação hipotética para representação do Índice Quantitativo de Hemerobia.
Onde: P é o resultado da multiplicação do peso atribuído à perturbação por sua ausência ou
presença (0 ou 1); IQH* é o Índice Quantitativo de Hemerobia escalado para o intervalo [0,1].
Perturbação Peso Ausência ou
presença P
Roçada de sub-bosque 5 0 0
Corte seletivo 5 1 5
Corte raso 5 1 5
Exploração de erva-mate 3 0 0
Pastejo 5 0 0
Caça 2 0 0
Espécie exótica 4 1 4
Estradas 2 0 0
Sinais de fogo 3 1 3
∑ 34 - 17
IQH* 0,48
1.3.4.5 Considerações finais acerca dos indicadores de naturalidade
Na Tabela 1.12 trazemos um resumo sobre os indicadores aceitos e não aceitos; dos 13
indicadores considerados após a primeira triagem, seis foram aceitos. Como os indicadores
lidam com diferentes dimensões da naturalidade, McRoberts et al. (2012) sugerem que a

91
independência dos mesmos deve ser assegurada, para que, quando combinados em um
estimador único, este venha gerar resultadados robustos. Calculamos a correlação entre os
indicadores (Tabela 1.13), com significância avaliada pelo teste t corrigido proposto por
Dutilleul (1993). As correlações entre os indicadores mostraram-se fracas, com valores que
variaram de -0,35 até 0,24. Esse resultado nos mostra evidências de que os indicadores são
independentes e retratam diferentes dimensões da naturalidade. Em futuras referências os
indicadores aceitos serão tratados pela sua abreviação (Tabela 1.12).
Tabela 1.12. Resumo final dos indicadores de naturalidade descartados e adotados; *adotado
somente na triagem das florestas de referência.
Indicador de
naturalidade Método
Diagnóstico
final Abreviação
Ajuste à função
exponencial negativa Modelo de Meyer Aceito J-invertido
Assimetria da distribuição
de diâmetros gdap Não aceito -
Composição
de espécies Grupo ecológico Não aceito -
Desvio-padrão
dos diâmetros Sdap Aceito Sdap
Estrutura de população A.
angustifolia gdap Não aceito -
Estrutura
vertical Estratificação Aceito*
Estrutura
vertical
Diversidade do estrato
arbóreo H‟ e 1-D Não aceito -
Métricas de paisagem Escores tb-PCA Aceito Métricas de
paisagem
Necromassa Peso (Mg) Não aceito -
Árvores grossas Escore-z (dap) Aceito Árvores
grossas
Pres./aus. de distúrbios
antrópicos IQH Aceito IQH
Regeneração
natural Índice de Shannon Aceito
H‟ reg.
natural
Riqueza de espécies do
estrato arbóreo
Rarefação -
Extrapolação Não aceito -

92
Tabela 1.13. Correlação de Spearman entre os indicadores de naturalidade. Valor da
correlação na diagonal inferior e significância corrigida pelo teste t de Dutilleul (1993) na
diagonal superior, com α=0,05. *correlação significativa; **correlação altamente
significativa; ns = correlação não significativa.
Indicador J-invertido Sdap Árvores
grossas
Métricas de
paisagem IQH
H‟ reg.
natural
J-invertido 1 < 0.001 0.01 0.01 0.77 0.9
Sdap -0.35** 1 0.14 0.14 0.06 0.32
Árvores
grossas 0.24** 0.07
ns 1 0.93 0.09 0.21
Métricas de
paisagem 0.20** 0.07
ns -0.04
ns 1 0.02 0.16
IQH -0.02ns
-0.17ns
-0.12ns
-0.13* 1 0.92
H‟ reg.
natural 0.01
ns -0.15
ns 0.24
ns -0.11
ns 0.05
ns 1
1.3.4.6 Quantificação da naturalidade
A quantificação da naturalidade será baseada na distância conceitual entre uma dada
floresta mais alterada e uma floresta mais conservada, de maior (ou “máxima”) expressão
ecológica (LOIDI, 1994). Utilizaremos a distância euclidiana, conforme McRoberts et al.
(2012) e Chirici et al. (2011). A medida de distância irá determinar o grau de naturalidade
dos remanescentes florestais de Floresta Mista com Araucária em comparação com florestas
referenciais consideradas mais naturais.
A construção da medida de distância consiste nos seguintes processos:
(I) transformação dos indicadores de naturalidade;
(II) determinação de Florestas de referência teórica e de Florestas de referência
observadas;
(III) aplicação da medida de distância a todos os remanescentes amostrados pelo
IFFSC.
A seguir serão descritos os três processos:
I) Transformação dos indicadores: os indicadores de naturalidade serão escalados para o
intervalo [0,1] com o intuito de assumirem o mesmo peso, possibilitando a utilização deles em
conjunto. O valor “1” ou próximo a este expressa alta naturalidade na dimensão considerada

93
(sintetizada pelo indicador). Os passos a seguir trazem o processo de escalamento dos
indicadores:
Serão agrupadas as Unidades Amostrais com seus respectivos valores já calculados
para cada um dos seis indicadores;
Será calculada a média da proporção p=0,05 dos maiores valores para dado indicador;
Para cada Unidade Amostral serão geradas seis razões Rk, onde k é o indicador (k = 1,
2, ..., 6), obtidas pelo valor do indicador k divido pela média da proporção pk = 0,05
dos maiores valores obtidos. Tal procedimento pode ser expresso porp
ik
x
xR , onde
Rk ≤ 1, xi é o valor do indicador obtido para a i-ésima Unidade Amostral e é a
média dos maiores valores da proporção pk da i-ésima Unidade Amostral.
II) Determinação das Florestas de referência: dois tipos de Florestas de referência serão
considerados:
Floresta de referência teórica: é uma referência teórica (100% natural), a qual
apresenta valores máximos em todos os indicadores de naturalidade; é expressa pelo
vetor de valor máximo N (1, 1, 1, 1, 1, 1);
Floresta de referência observada: é uma referência real de naturalidade, a qual não
apresenta (necessariamente) indicadores com valores máximos. O processo de
quantificação da naturalidade deste tipo de referência consiste em:
A distância euclidiana (Nref) em um espaço multidimensional será calculada, com
número de dimensões definido pelo número de indicadores; considerando os indicadores
eleitos, temos um espaço com seis dimensões (R6). A distância do vetor gerado pelas seis
razões (Rk) até o vetor N (1, 1, 1, 1, 1, 1), será calculada pelo algoritmo:
onde Nref é a medida adimensional de naturalidade, Rk (k = 1, 2, ..., 6) é o indicador escalado
para o intervalo [0,1].
A distância euclidiana (Nref) do vetor (das razões) até o vetor de valor máximo indica o
grau de naturalidade das Florestas de referência observadas. Quanto menor o valor de Nref,
maior a naturalidade.
2
6
2
3
2
2
2
1 )1(....)1()1()1( RRRRNref

94
III) Aplicação da medida de distância aos remanescentes ordinários: este procedimento
consiste na aplicação da medida de distância euclidiana aos demais remanescentes. A
naturalidade de cada um dos remanescentes florestais ordinários será quantificada de duas
maneiras:
Abordagem A: Através da distância euclidiana dos indicadores até os indicadores
obtidos para a Floresta de referência teórica:
Abordagem B: Através da distância euclidiana dos indicadores até os indicadores
obtidos para as Florestas de referência observadas:
onde Rk é o indicador escalado calculado para a i-ésima Unidade Amostral e Rok (k = 1, 2, ...,
6) é o indicador escalado, obtido para a Floresta de referência observada.
1.3.4.7 Definição e escolha das florestas de referência
Definimos o conceito de floresta de referência como: remanescente florestal com
alta expressão ecológica localizado numa região pré-determinada, o qual apresenta
características esperadas numa floresta conservada sem influências antrópicas
degradadoras.
Como temos poucas florestas que atendem a definição acima e, numa perspectiva mais
rigorosa, não temos mais florestas completamente naturais e inafetadas por elementos
perturbadores, temos que considerar os remanescentes florestais mais naturais existentes
como se estivessem posicionados no extremo do gradiente de naturalidade. Em outras
palavras, estas florestas serão, teoricamente, consideradas 100% naturais (mediante uma
criteriosa triagem), mesmo podendo ser originárias da sucessão vegetal secundária8
(GIBBONS et al., 2008; LANDRES et al., 1998; McROBERTS et al., 2012; WINTER et al.,
2010). Os esquemas das Figuras 1.24 e 1.25 exemplificam graficamente a ideia. Na Figura
1.24, na situação "A", temos uma floresta 100% natural, sem distúrbios antrópicos; na
8 sensu Chokkalingam e De Jong (2001)
2
6
2
3
2
2
2
1 )1(....)1()1()1( RRRRNA
2
66
2
33
2
22
2
11 )(....)()()( RoRRoRRoRRoRNB
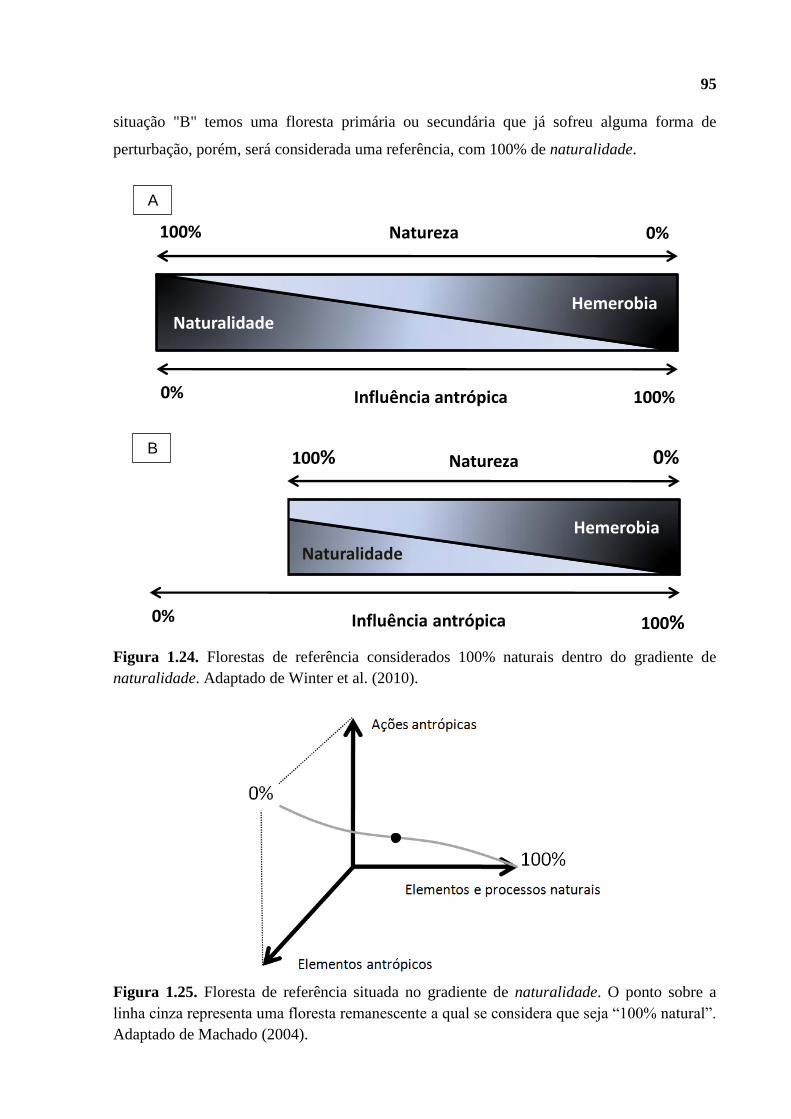
95
situação "B" temos uma floresta primária ou secundária que já sofreu alguma forma de
perturbação, porém, será considerada uma referência, com 100% de naturalidade.
Figura 1.24. Florestas de referência considerados 100% naturais dentro do gradiente de
naturalidade. Adaptado de Winter et al. (2010).
Figura 1.25. Floresta de referência situada no gradiente de naturalidade. O ponto sobre a
linha cinza representa uma floresta remanescente a qual se considera que seja “100% natural”.
Adaptado de Machado (2004).
Naturalidade
Hemerobia
100% 0%
0% 100%
Natureza
Influência antrópica
Naturalidade
Hemerobia
100% 0%
0% 100%
Natureza
Influência antrópica
A
B

96
Na literatura encontramos uma abordagem comum em relação às florestas de
referência – esta deve representar a floresta mais natural em relação às condições de
crescimento (elementos preditores), tais como clima, solo, elevação, aspectos florísticos,
dentre outros (WINTER, 2012).
Em florestas tropicais e subtropicais, um dos principais fatores que atuam nas
mudanças dos padrões de riqueza/composição de espécies é a heterogeneidade ambiental,
cujos efeitos podem estar relacionados às características climáticas, edáficas e espaciais
(BALDECK et al., 2013; EISENLOHR et al., 2013; JONES et al., 2011; OLIVEIRA-FILHO
et al., 2013; PENÃ-CLAROS et al., 2012; SANCHEZ et al., 2013; THUILLER, 2013).
Sabemos que o clima e o espaço influenciam na distribuição espécies em Santa
Catarina (GASPER et al., 2013; REZENDE, 2013; REZENDE et al., 2014), embora, o clima
do sul do Brasil, de uma maneira geral, é homogêneo (LEITE e KLEIN, 1990). Uhlmann et
al. (2013) levantaram uma hipótese de que os remanescentes de Floresta Mista com Araucária
de Santa Catarina são (atualmente) homogêneos floristicamente e estruturamente, em resposta
à exploração e degradação.
Para contornarmos o efeito das influências ambientais em nossas análises, seguimos
alguns procedimentos. Primeiramente, classificamos as Unidades Amostrais segundo o
sistema de classificação de Oliveira-Filho (2009), o qual permite a agregação de até cinco
atributos hierárquicos: regime climático (a), regime de renovação foliar (b), domínio térmico
(c), faixa altitudinal (d) e substrato (e). Conseguimos classificar os remanescentes nos
atributos b, c, e d. Os atributos b e c são os mesmos para os remanescentes - perenifólias e
subtropical, respectivamente; para o atributo d, 71 Unidades Amostrais foram classificadas
como inferomontanas e 74 como superomontanas. Não foi possível usar os atributos a e e
devido à falta de informações precisas.
Em busca de evidências estatísticas sobre a segregação dos grupos inferomontana e
superomontana, no que diz respeito a padrões estruturais da vegetação, aplicamos uma análise
de similaridade (ANOSIM; LEGENDRE e LEGENDRE, 1998) em uma matriz de abundância
de 354 espécies presentes nas Unidades Amostrais classificadas em um dos grupos. Adotamos
a medida de distância de Bray-Curtis, com 9999 permutações e α=0,05. O teste mostrou-se
significativo (R=0,099; p=0,0001), mostrando evidências para a segregação das faixas
altitudinais.

97
De forma complementar, visando ainda minimizar influências do ambiente,
agrupamos as Unidades Amostrais por bacias hidrográficas (Figura 1.26). Algumas bacias
hidrográficas apresentaram poucas Unidades Amostrais (de uma até duas): Rio Irani, Rio
Itapocu, Rio Peperi-Guaçú, Rio Tubarão e Rio Jacutinga. Devido a este fato, essas bacias
hidrográficas e, consequentemente, suas respectivas Unidades Amostrais serão agregadas às
bacias mais próximas (Tabela 1.14 e Figura 1.26 e 1.27). Assim, será eleita uma Unidade
Amostral referência para cada faixa altitudinal dentro de cada bacia hidrográfica
considerada.
Tabela 1.14. Bacias hidrográficas consideradas na quantificação de naturalidade e seus
respectivos números de Unidades Amostrais, classificadas conforme o atributo faixa
altitudinal de Oliveira-Filho (2009). As bacias com número pequeno de Unidades Amostrais
foram agregadas à bacia mais próxima.
Bacia
hidrográfica
Faixa altitudinal N° total
de UA
Agregada
Inferomontana Superomontana à bacia
Rio Canoas 15 22 37 -
Rio Canoinhas 5 3 8 -
Rio Chapecó 2 6 8 -
Rio das Antas 3 0 3 Rio Chapecó
Rio do Peixe 5 11 16 -
Rio Irani 1 1 2 Rio Chapecó
Rio Itajaí-açu 11 6 17 -
Rio Itapocú 0 1 1 Rio Negro
Rio Jacutinga 1 0 1 Rio Chapecó
Rio Negro 9 4 13 -
Rio Pelotas 9 10 19 -
Rio Peperi-Guaçú 1 0 1 Rio Chapecó
Rio Tijucas 3 1 4 Rio Itajaí-açu
Rio Timbó 5 8 13 -
Rio Tubarão 1 1 2 Rio Canoas
Σ 71 74 145 -
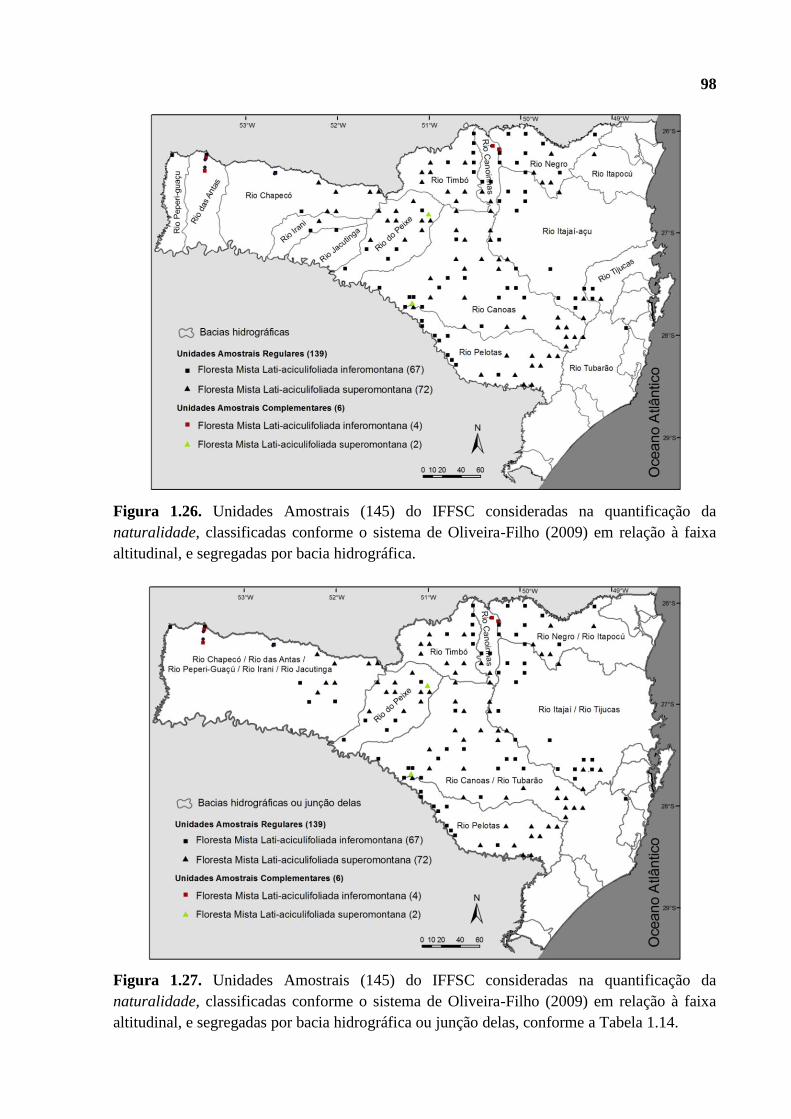
98
Figura 1.26. Unidades Amostrais (145) do IFFSC consideradas na quantificação da
naturalidade, classificadas conforme o sistema de Oliveira-Filho (2009) em relação à faixa
altitudinal, e segregadas por bacia hidrográfica.
Figura 1.27. Unidades Amostrais (145) do IFFSC consideradas na quantificação da
naturalidade, classificadas conforme o sistema de Oliveira-Filho (2009) em relação à faixa
altitudinal, e segregadas por bacia hidrográfica ou junção delas, conforme a Tabela 1.14.

99
A determinação final das florestas de referência observadas (Unidades Amostrais)
utilizará de parâmetros condicionais, os quais estão relacionados a aspectos inerentes à
naturalidade, e à hemerobia aplicada à floresta. O processo de determinação consistirá em
etapas de triagem das Unidades Amostrais, inspiradas em McRoberts et al. (2012):
Serão eliminadas as Unidades Amostrais com um ou mais indivíduos de espécie(s)
exótica(s);
Será avaliada a diversidade da estrutura vertical através do indicador Estrutura vertical
(descrito no tópico 1.3.4.4); a Unidade Amostral canditada deve conter pelo menos
10% do número total de indivíduos em cada um dos três estratos de altura;
Será feito um ranking das Unidades Amostrais com as menores distâncias Euclidianas
(Nref) ao vetor máximo (ou Floresta de referência teórica - tópico 1.3.4.6.);
Para ser selecionada, a Unidade Amostral terá de passar pelos dois primeiros critérios
de seleção; então, para a seleção final, será avaliada a Nref - as Unidades Amostrais
com menor valor serão selecionadas como referências;
Em casos de empate (Unidades Amostrais com mesmos valores para Nref) o valor final
da naturalidade (Nref*) será o resultado da soma de Nref com peso de 80% e do IQH
com peso de 20%: Nref* = Nref . (0,80) + IQH . (0,20).
1.4. CONCLUSÕES
Quatro conclusões podem ser tiradas desse capítulo: (i) o conceito de naturalidade
pôde ser coerentemente adaptado ao contexto histórico, social e ambiental da Floresta Mista
com Araucária do estado de Santa Catarina; (ii) os dados coletados pelo IFFSC são capazes de
fornecer informações para o desenvolvimento de vários indicadores de naturalidade; (iii) nem
todos os indicadores aparentemente relevantes estão relacionados com a naturalidade, ou por
questões “operacionais” não puderam ser adotados; (iv) a metodologia proposta para a
quantificação da naturalidade integrou várias dimensões desta em uma só medida.
1.5. REFERÊNCIAS BIBLIOGRÁFICAS
ALMEIDA-NETO, M.; CAMPASSI, F.; GALETTI, M.; JORDANO, P.; OLIVEIRA-FILHO,
A.T. Vertebrate dispersal syndromes along the Atlantic forest: broad-scale patterns and
macroecological correlates. Global Ecology and Biogeography, v.17, p.503–513, 2008.
ANDERSON, J.E. A conceptual framework for evaluating and quantifying naturalness.
Conservation Biology, v.5, p.347–352, 1991.

100
ANGERMEIER, P.L. The natural imperative for biological conservation. Conservation
Biology, v.14, p.373–381, 2000.
ANGERMEIER, P.L.; KARR, J.R. Biological integrity versus biological diversity as policy
directives. BioScience, v.44, p.690 – 697, 1994.
ARCHER, E.R.M.; CÁCERES, D.; DOWNING, T.E.; ELMQVIST, T.; ERIKSEN, S.;
FOLKE, C.; HAN, G.; IYENGAR, K.; VOGEL, C.; WILSON, K.A.; ZIERVOGEL, G.
Vulnerable Peoples and Places. In: HASSAN, R.; SCHOLES, R.; ASH, N. (Eds.) Ecosystems
and Human Well-being: Current State and Trends, Volume 1. Washington DC: Island Press,
2005, p. 145-164.
BACKES, A. Distribuição geográfica atual da Florestal com Araucária: condicionamento
climático. In: FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.;
BACKES, A; GRANADE G. (Eds.). Floresta com Araucária: Ecologia, Conservação e
Desenvolvimento Sustentável. Ribeirão Preto: Holos, 2009, p. 39-44.
BALDECK, C.A.; HARMS, K.E.; YAVITT, J.B.; JOHN, R.; TURNER, B.L.; VALENCIA,
R.; NAVARRETE, H.; DAVIES, S.J.; CHUYONG, G.B.; KENFACK, D.; THOMAS, D.W.;
MADAWALA, S.; GUNATILLEKE, N.; GUNATILLEKE, S.; BUNYAVEJCHEWIN, S.;
KIRATIPRAYOON, S.; YAACOB, A.; SUPARDI, M.N.N.; DALLING, J.W. Soil resources
and topography shape local tree community structure intropical forests. Proceedings of the
Royal Society B, v.280, p.25-32, 2013.
BARTHA, D.; ÓDOR, P.;HORVÁTH, T.; TÍMÁR, G.; KENDERES, K.; STANDOVÁR, T.;
BÖLÖNI, J.; SZMORAD, F.; BODONCZI, L.; ASZALÓS, R. Relationship of Tree Stand
Heterogeneity and Forest Naturalness. Acta Silvatica & Lingaria Hungarica, v.2, p.7-22,
2006.
BASKERVILLE, G.L. Use of logarithmic regression in the estimation of plant biomass.
Canadian Journal of Forest Research, v.2, n.1, p.49-53, 1972.
BAUERMANN, S.G.; BEHLING, H. Dinâmica paleovegetacional da Florestal com
Araucária a patir do final do Pleistoceno: oque mostra a palinologia. In: FONSECA, C.R.;
SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (eds.).
Floresta com Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão
Preto: Holos, 2009, p. 35-38.
BENDEL, R.B.; HIGGINS, S.S.; TEBERG, J.E.; PYKE, D.A. Comparison of skewness
coefficient, coefficient of variation and Gini coefficient as inequality measures within
populations. Oecologia, v.78, p.394-400, 1989.
BEHLING, H.; BAUERMANN, S. G.; NEVES, P. C. Holocene nvironmental changes from
the São Francisco de Paula region, southern Brazil. Journal of South American Earth
Sciences, v.14, p.631-639, 2001.
BEHLING, H.; PILLAR, V.; ORLÓCI, L.; BAUERMANN, S.G. Late Quaternary Araucaria
forest, grassland (Campos), fire and climate dynamics, studied by high resolution pollen,
charcoal and multivariate analysis of the Cambará do Sul core in southern Brazil.
Palaeogeography, Palaeoclimatology, Palaeoecology, v.203, p.277-297, 2004.

101
BORGES, N.C.O. Terra, gado e trabalho: sociedade e economia escravista em Lages, SC
(1840-1865). Dissertação (Mestrado em História). Universidade Federal de Santa Catarina,
2005.
BRASIL. Ministério do Meio Ambiente. Fragmentação de ecossistemas: causas, efeitos
sobre a biodiversidade e recomendações de políticas públicas. Brasília, 2003.
BREIDENBACH, J.; ANTÓN-FERNÁNDEZ, C.; PETERSSON, H.; ASTRUP, P.;
McROBERTS, R.E. Quantifying the contribution of biomass model errors to the uncertainty
of biomass stock and change estimates in Norway. Forest Science, v.60, p-25-33, 2014.
BREPOHL, D. O reflorestamento com incentivos fiscais no estado do Paraná. Floresta, v.11,
n.1, p.62-66, 1980.
BROADBENT, E.N.; ASNER, G.P.; KELLER, M.; KNAPP, D.E.; OLIVEIRA, P.J.C.;
SILVA, J.N. Forest fragmentation and edge effects from deforestation and selective logging
in the Brazilian Amazon. Biological Conservation, v.4, p.745-757, 2008.
BROOK, B.W.; SODHI, N.S.; NG, P.K.L. Catastrophic extinctions follow deforestation in
Singapore. Nature, v.424, p.420-426, 2003.
BROSE, U.; MARTINEZ, N. D.; WILLIAMS, R. J. Estimating species richness: sensitivity
to sample coverage and insensitivity to spatial patterns. Ecology, v.84, p.2364-2377, 2003.
BUCKLAND, S.T; MAGURRAN, A.E.; GREEN, R.E.; FEWSTER, R.M. Monitoring
change in biodiversity through composite indices. Philosophical Transactions of the Royal
Society, v.360, p.243-254, 2005.
BUCKLAND, S.T.; STUDENY, A.C.; MAGURRAM, A.; NEWSON, S.E. Biodiversity
monitoring: the relevance of detectability. In: MAGURRAN, A.E.; McGILL, B.J.
(Eds.) Biological diversity: frontiers in measuring biodiversity. New York: Oxford
University Press, 2011, p.25-33.
BUONGIORNO, J.; DAHIR, S.; LY, H-C.; LIN, C.-R. Tree size diversity and economic
returns in uneven-aged forest stands. Forest Science, v.40, n.1, p.83–103, 1994.
CABRAL, D.C; CESCO, S. Notas para uma história da exploração madeireira na Mata
Atlântica do Sul-Sudeste. Ambiente & Sociedade, v.11, n.1, p.33-48, 2008.
CANALEZ, G.G.; CORTE, A.P.D.; SANQUETTA, C.R. Dinâmica da estrutura da
comunidade de Lauráceas no período 1995-2004 em uma floresta de araucária no sul do
estado do Paraná, Brasil. Ciência Florestal, v. 4, p. 357-367, 2006.
CARDOSO, D. J.; VIBRANS, A.C.; LINGNER, D.V. Inventário da necromassa florestal
caída no chão nos remanescentes florestais em Santa Catarina. In: VIBRANS, A.C.;
SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V (Eds.). Diversidade e conservação dos
remanescentes florestais. Blumenau: Edifurb, 2012.

102
CARPENTER, S.R.; WALKER, B.; ANDERIES, J.M.; ABEL, N. From metaphor to
measurement: Resilience of what to what? Ecosystems, v.44, p.765–781, 2001.
CDB. Convenção de Diversidade Biológica. A convenção sobre Diversidade Biológica -
CDB. Brasilia: MMA, 1992.
CHANG, M.Y. Sistema faxinal: uma forma de organização camponesa em desagregação no
centro-sul do Paraná. Londrina: IAPAR, 1988.
CHAO, A.; GOTELLI, N.J.; HSIEH, T.C.; SANDER, E.L.; MA, K.H.; COLWELL, R.K.;
ELLISON, A.M. Rarefaction and extrapolation with Hill numbers: a framework for sampling
and estimation in species diversity studies. Ecological Society of America, pré-impressão,
2014.
CHAZDON, R.L. Chance and Determinismin Tropical Forest Succession. In: CARSON,
W.P.; SCHITZER, S.A. (Eds.). Tropical Forest Community Ecology. Blackwell Publishing
Ltd, 2008, p.384-408.
CHAZDON, R.L.; COLWELL, R.K.; DENSLOW, J.S.; GUARIGUATA, M.R. Statistical
methods for estimating species richness of woody regeneration in primary and secondary rain
forests of NE Costa Rica. In: DALLMEIERAND, F.; COMISKEY, J. (Eds.). Forest
Biodiversity Research, Monitoring and Modeling: Conceptual Background and Old World
Case Studies. Paris: Parthenon Publishing, 1998, p.285–309.
CHAZDON, R.L.; LETCHER, S.G.; BREUGEL, M.; RAMOS, M.M.; BONGERS, F;
FINEGAN, B. Rates of change in tree communities of secondary tropical forests following
major disturbances. Proceedings of the Royal Society of London, v.362, p.273–289, 2007.
CHAZDON, R.L.; PERES, C.A; SHEIL, D.; LUGO, A.E.; LAMB, D.; STORKN.E.;
MILLER,E. The potential for species conservation in tropical secondary forests.
Conservation Biology, v.23, p. 1406-1417, 2009.
CHIRICI, G.; MCROBERTS, R.E.; WINTER, S.; BERTINI, R.; BRÄNDLI, U.; ASENSIO,
I.A.; BASTRUP-BIRK, A.; RONDEUX, J.; BARSOUM, N.; MARCHETTI, M. National
Forest Inventory Contributions to Forest Biodiversity Monitoring. Forest Science, v.58, n.3,
2012.
CHIRICI, G.; WINTER, S.; McROBERTS, R.E. (Eds.). National forest inventories:
contributions to forest biodiversity assessments. Springer, 2011.
CHOKKALINGAM, U.; DE JONG, W. Secondary forest: a working definition and typology.
International Forestry Review, v.3, n.1, p.19-26, 2001.
CHOMITZ, K.M.; KUMARI, K. The Domestic Benefits of Tropical Forests: A Critical
Review. World Bank Research Observer, v.13, p.13-35, 1998.
CHRISTENSEN, M.; HAHN, K.; MOUNTFORD, E.; ÓDOR, P.; ROZENBERGER, D.;
DIACI, J.; STAN-DOVAR, T.; WIJDEVEN, S.; WINTER, S.; VRSKA, T.; MEYER, P.
Dead wood in European beech (Fagus sylvatica) forest reserves. Forest Ecology and
Management, v. 210, p.267–282, 2005.

103
COLWELL, R.K.; MAO, C.X.; CHANG, J. Interpolating, extrapolating, and comparing
incidence-based species accumulation curves. Ecology, v.85, p.2717-2727, 2004.
COLWELL, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species
from Samples. Version 9. User‟s Guide and application. 2011. Disponível em:
http://purl.oclc.org/estimates, acesso em: 23 de Maio de 2014.
COLWELL, R.K.; CHAO, A.; GOTELLI, N.J.; LIN, S.; MAO, C.X.; CHAZDON, R.L.,
LONGINO, J.T. Models and estimators linking individual-based and sample-based
rarefaction, extrapolation and comparison of assemblages. Journal of Plant Ecology, v.5,
n.1, p.3–21, 2012.
COLWELL, R.K.; CODDINGTON, J.A. Estimating terrestrial biodiversity through
extrapolation. Philosophical transactions of the Royal Society (Series B), v.345, p.101-118,
1994.
COST Action E43 - Cooperation in the Field of Science and Technology. Harmonisation of
national forest inventories in Europe: techniques for common reporting. COST Action E43,
2006.
COSTANZA, R. et al. The value of the world‟s ecosystem services and natural capital.
Nature, v.387, p.253-260, 1997.
COTTENIE, K. Integrating environmental and spatial processes in ecological community
dynamics. Ecology Letters, v.8, p.1175–1182, 2005.
CREPALDI, M.O.S.; PEIXOTO, A.L. Use and knowledge of plants by “Quilombolas” as
subsidies for conservation efforts in an area of Atlantic Forest in Espı´rito Santo State, Brazil.
Biodiversity Conservation, v.19, p.37–60, (2010).
DEAN, W. A ferro e fogo: A história e a devastação da Mata Atlântica brasileira. São Paulo:
Companhia das Letras, 1996.
DIEGUES, A.C.S. O mito moderno da natureza intocada. São Paulo: Editora Hucitec,
2001.
DILLENBRUG, L.R.; FRANCO, A.M.; COUTINHO, A.L.; KÖRNDORFER, C.L;
CLEBSCH, C.C.; DUARTE, L.S.; FERLA, L.; ROSA, L.M.G; DA SILVA, L.G.R.,
GARBIN, M.L.; MÓSENA, M.; ZANDAVALLI, R.B.; YAMASKI, S. Aspectos
ecofisiológicos de regeneração de Araucaria angustifolia. In: FONSECA, C.R.; SOUZA,
A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.). Floresta
com Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão Preto:
Holos, 2009. p. 57-66.
DINIZ-FILHO, J.A.F.; SIQUEIRA, T.; PADIAL, A.A.; RANGEL, T.F.; LANDEIRO, V.L.;
BINI, L.M. Spatial autocorrelation analysis allows disentangling the balance between neutral
and niche processes in metacommunities. Oikos, v.121, p.201-210, 2012.

104
DORREN, L.K.A.; BERGER, F.; IMESON, A.C.; MAIER, B.; REY, F. Integrity, stability
andmanagement of protection forests in the European Alps. Forest Ecology and
Management, v.195, p.165–176, 2004.
D‟OLIVEIRA, M.V.N.; ALVARADO, E.C.; SANTOS, J.C.; CARVALHO JR., J.A. Forest
natural regeneration and biomass production after slash and burn in a seasonally dry forest in
the Southern Brazilian Amazon. Forest Ecology and Management, v.261, p.1490–1498,
2011.
DUARTE, L.S.; HARTZ, S.M.; PILLAR, V.P. A dinâmica de nucleação dos pinhais sobre ps
campos no planalto sul-riograndense. In: FONSECA, C.R.; SOUZA, A.F.; LEAL-
ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.). Floresta com
Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão Preto: Holos,
2009. p.67-74.
DUTILLEUL, P. Modifying the t test for assessing the correlation between two spatial
processes. Biometrics, v.49, p.305-314, 1993.
EDARRA, T.D. Botánica ambiental aplicada. Pamplona: Eunsa, 1997.
EDUARDO, R.P. A madeira em Santa Catarina, 1930-1972. Dissertação (Mestrado em
História). Departamento de História, Universidade Federal do Paraná, 1974.
EISENLOHR, P.V.; ALVES, L.F.; BERNACCI, L.C.; PADGURSCHI, M.C.G.; TORRES,
R.B.; PRATA, E.M.B.; SANTOS, F.A.M.; ASSIS, M.A.; RAMOS, E.; ROCHELLE, A.L.C.;
MARTINS, F.R.; CAMPOS, M.C.R.; PEDRONI, F.; SANCHEZ, M.; PEREIRA, L.S.;
VIEIRA, S.A.; GOMES, J.A.M.A.; TAMASHIRO, J.Y.; SCARANELLO, M.A.S.; CARON,
C.J.; JOLY, A.C. Disturbances, elevation, topography and spatial proximity drive vegetation
patterns along an altitudinal gradient of a top biodiversity hotspot. Biodiversity and
Conservation, v.22, p.2767-2783, 2013.
ELLEN, R. Environment, Subsistence and System: the ecology of small scale Social
formations. New York: Cambridge University Press, 1989.
ENGEL, V.L.; FONSECA, R.C.B.; OLIVEIRA, R.E. Ecologia de lianas e o manejo de
fragmentos florestais. Série Técnica IPEF, v.12, n.32, p.43-64, 1998.
EPAGRI. Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina. Atlas
Climatológico do Estado de Santa Catarina. Florianópolis. CD-ROM, 2002.
FALKENBERG, D.B. Matinhas nebulares e vegetação ripícola dos Aparados da Serra Geral
(SC/RS), sul do Brasil. Tese (Doutorado em Biologia Vegetal), Instituto de Biologia,
UNICAMP, 2003.
FALKENBERG, D.B.; VOLTOLINI, J. The Montane Cloud Forest in Southern Brazil. In:
HAMILTON, L.; JUVIK, J.; SCATENA, F.N. (Eds.). Tropical Montane Cloud Forests.
Springer, 1995. p. 138–149.
FELFILI, J.M.; ROITMAN, I.; MEDEIROS, M.M.; SANCHEZ, M. Procedimentos e
métodos de amostragem de vegetação. In: FELFILI, J. M.; EISENLOHR, P. V.; MELO, M.

105
M. R. F.; ANDRADE, L. A.; MEIRA-NETO, J. A. A. (Eds.). Fitosssociologia no Brasil.
Métodos e estudos de caso. Viçosa: Editora UFV, 2011. p.86-121.
FINEGAN, B. Pattern and process neotropical secondary rain florests: the first 100 years of
succession. Tree, v.11, n.3, p.119-124, 1996.
FINEGAN, B. The management potential of neotropicalsecondary lowland rain forest. Forest
Ecology and Management, v.47, p. 295-321, 1992.
FISCHER, A.; YOUNG, J.C. Understanding mental constructs of biodiversity: Implications
for biodiversity management and conservation. Biological Conservation, v.136, p.271-286,
2007.
FISCHMAN, R.L. The meanings of Biological Integrity, Diversity, and Environmental
Health.Natural Resources Journal, v.44, p.989-1026, 2004.
FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A;
GRANADE, G. (Eds.). Floresta com Araucária: Ecologia, Conservação e Desenvolvimento
Sustentável. Ribeirão Preto: Holos, 2009.
FOOD AND AGRICULTURE ORGANIZATION – FAO. Global Forest Resources
Assessments 2010: Terms and Definitions. Italy, Rome, 2010.
FOOD AND AGRICULTURE ORGANIZATION – FAO. Global Forest Resources. Italy,
Rome, 2000. Disponível em: http://www.fao.org/forestry/fo/fra/index.jsp.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS - FAO.
State of the world's forests. Rome, 2012.
FOLEY, J.A.; DEFRIES, R.; ASNER, G.P.; BARFORD, C.; BONAN, G.; CARPENTER,
S.R. Global consequences of land use. Science, v.309, p.570–574, 2005.
FOREST RESOURCES ASSOCIATION - FRA. Assessing forest integrity and naturalness
in relation to biodiversity: Global forest resources assessment, 2000. Disponível em:
<www.fao.org/docrep/006/ad654e/ad654e05.htm>. Acessado em: Fevereiro de 2014.
FREEDMAN, B. Environmental ecology: The impacts of pollution and other stresses on
ecosystem structure and function. San Diego: Academic Press, 1989.
FREITAS, J.V.; OLIVEIRA, Y.M. ; ROSOT, M.A.D. ; GOMIDE, G. L. A. ; MATTOS, P. P.
National Forest Inventory Reports: Brazil. In: TOMPPO, E.; GSCHWANTNER, T.;
LAWRENCE, M.; MCROBERTS, R.E. (Org.). National Forest Inventories: Pathways for
Common Reporting. London: Springer, p. 89-96, 2010.
FREY, D. Biological integrity of water: An historical perspective. In: BALLENTINE, R.K.;
GUARRAIA, L.J. (Eds.). The integrity of water. Washington DC: Environmental Protection
Agency, 1975, p.127–139.
GALVÃO, F.; AUGUSTIN, C. A gênese dos campos sulinos. Floresta, v.41, n.1, p.191-200,
2011.

106
GASPER, A.L.; EISENLOHR, P.V.; SALINO, A. Climate-related variables and geographic
distance affect fern species composition across a vegetation gradient in a shrinking hotspot.
Plant Ecology & Diversity, p.1-11, 2013.
GASPER, A.L.; MEYER, L; SEVEGNANI, L.; SOBRAL, M.G; BONNET, A. Flora vascular
de Santa Catarina. In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V
(Eds.). Inventário Florístico Florestal de Santa Catarina: Diversidade e conservação dos
remanescentes florestais. Blumenau: Edifurb, 2012.
GASPER, A.L.; SEVEGNANI, L.; SOBRAL, M.G.; MEYER, L.; VERDI, M.; SANTOS,
A.S. DOS; DREVECK, S.; KORTE, A. Flora vascular da Floresta Floresta Ombrófila Mista.
In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L. DE; LINGNER, D.V. (Eds.).
Inventário Florístico Florestal de Santa Catarina: Floresta Ombrófila Mista. Blumenau:
Edifurb, 2013.
GASPER, A.L.; SEVEGNANI, L.; VIBRANS, A.C.; UHLMANN, A.; LIGNER, D.V.;
VERDI, M.; DREVECK, S.; STIVAL-SANTOS, A.; BROGNI, E.; SCHMITT, R.; KLEMZ,
G. Inventário de Dicksonia sellowiana Hook. em Santa Catarina. Acta Botanica Brasilica,
v.25, n.4, p.776-784, 2011.
GASTON, K.J.; SPICER, J.I. Biodiversity: An introduction, 2ed.Oxford: Blackwell Science
Ltd, 2004.
GEOAMBIENTE SENSORIAMENTO REMOTO LTDA. Projeto de Proteção da Mata
Atlântica em Santa Catarina (PPMA/SC) - Relatório Técnico do Mapeamento Temático Geral
do Estado de Santa Catarina. São José dos Campos, 2008.
GIBBONS, P.; BRIGGS, S.V.; AYERS, D.A.; DOYLE, S.; SEDDON, J.; MCELHINNY,
C.; JONES, N.; SIMS, R.; DOODY, J.S. Rapidly quantifying reference conditions in
modified landscapes. Biological Conservation, v.141, p.2483–2493, 2008.
GIBSON, L.; LEE, T.M.; KOH, L.P.; BROOK, B.W.; GARDNER, T.A.; BARLOW, J.;
PERES, C.A.; BRADSHAW, C.J.A.; LAURANCE, W.F.; LOVEJOY, T.E.; SODHI, N.S.
Primary forests are irreplaceable for sustaining tropical biodiversity. Nature, v.478, p.378-
381, 2011.
GOMEZ-POMPA, A.; VASQUEZ-YANES, C.; GUEVARA, S. The Tropical Rainforest: a
Nonrenewable Resource. Science, v. 177, p. 762-765, 1972.
GONZALEZ, P.; HASSAN, R.; LAKYDA, P.; MCCALLUM, I.; NILSSON, S.; PULHIN, J.;
VAN ROSENBURG, D.; SCHOLES, B. Forest and Woodland Systems. In: HASSAN, R.;
SCHOLES, R.; ASH, N. (Eds.) Ecosystems and Human Well-being: Current State and
Trends, Volume 1. Washington DC: Island Press, 2005.
GOTELLI, N.J. Ecology: How Do Communities Come Together? In: Weiher, E; Keddy, P.
(Eds.) Ecological Assembly Rules: perspectives, advances, retreats. Cambridge: Cambridge
University Press, 1999.

107
GOTELLI, N.J; COLWELL, R.K. Quantifying biodiversity: procedures and pitfalls in the
measurement and comparison of species richness. Ecology Letters, v.4, p.379-39, 2001.
GOTELLI, N.J; COLWELL, R.K Estimating species richness. In: MAGURRAN, A.E.;
McGILL, B.J. (Eds.) Biological diversity: frontiers in measuring biodiversity. New York:
Oxford University Press, 2011, p.39-54.
GOTELLI, N.J.; ELLINSON, A.M. Princípios de Estatística em Ecologia. Porto Alegre:
Artmed, 2011.
GOULARTI FILHO, A. Formação econômica de Santa Catarina. Ensaios FEE, v.23, n.2,
p.977-1007, 2002.
GOULARTI FILHO, A ferrovia no planalto catarinense: um território de passagem. Artigos
científicos do II Encontro de Economia Catarinense, p.238-251, 2008.
GRABHERR, G.; KOCH, G.; KIRCHMEIR, H.; REITER, K. Hemerobie österreichischer
Waldökosysteme. Austria: Publication of the Austrian MaB-Programme 17, 1998.
GUSTAFSSON, L. Presence and abundance of red-listed plant species in Swedish forests.
Conservation Biology, v.6, n.2, p.377–388, 2002.
HADLEY, A.S.; BETTS, M.G. The effects of landscape fragmentation on pollination
dynamics: absence of evidencenot evidence of absence. Biological Reviews, v. 87, p.526–
544, 2012.
HANISCH, A.L.; VOGT, G.A.; MARQUES, A.Q.; BONA, L.C.; BOSSE, D.D. Estrutura e
composição florística de cinco áreas de caíva no Planalto Norte de Santa Catarina. Pesquisa
Florestal Brasileira, v.10, n.64, p.303-310, 2010.
HARMON, M.E.; FRANKLIN, J.F.; SWANSON, F.J.; SOLLINS, P.; GREGORY, S.V.;
LATTIN, J.D.; ANDERSON, N.H.; CLINE, S.P.; AUMEN, N.G.; SEDELL, J.R.;
LIENKAEMPER, G.W.; CROMACK, K.; CUMMINS, K.W. Ecology of coarse woody
debris in temperate ecosystems. Advances in Ecological Research, v.15, p.133–276, 1986.
HILL, M. O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology,
v.54, n.2, p. 427-432, 1978.
HOEHNE. F.C. Araucarilândia. São Paulo: Companhia Melhoramentos, 1930.
HUBBELL, S.P. The Unified Theory of Biogeography and Biogeography. Princeton:
Princeton University Press, 2001.
HUGHES, R.F.; KAUFFMAN, J.B.; JARAMILLO, V.J. Biomass, carbon, and nutrient
dynamics of secondary forests in a humid tropical region of Mexico. Ecology, v.80, p.1892–
1907, 1999.
HUNTER, M.L. Wildlife, forests, and forestry: Principles of managing forests for biological
diversity. Englewood Cliffs: Prentice-Hall, 1990.

108
HUNTER, M.L.Jr. Benchmarks for managing ecosystems: are human activities natural?
Conservation Biolology, v.10, p.695–697, 1996.
HUNTER, M.L.Jr. (Ed.). Maintaining biodiversity in forest ecosystem. Cambridge:
Cambridge University Press, 1999.
HURLBERT, S.H. The Nonconcept of Species Diversity: A Critique and Alternative
Parameters. Ecology, v.52, n.4, p.577-586, 1971.
IMAI, N.; TANAKA, A.; SAMEJIMA, H.; SUGAU, J.B.; PEREIRA, J.T.; TITIN, J.;
KURNIAWAN, Y.; KITAYAMA, K. Tree community composition as an indicator in
biodiversity monitoring of REDD+. Forest Ecology and Management, v.313, p.169–179,
2014.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Manuais Técnicos
em Geociências: Manual Técnico da Vegetação Brasileira. 2ª ed. Rio de Janeiro: IBGE, 2012.
INSTITUTO NACIONAL DO PINHO. O Instituto Nacional do Pinho: origem da criação e
desenvolvimento das atividades da autarquia madeireira. Anuário Brasileiro de Economia
Florestal. Rio de Janeiro: Instituto Nacional do Pinho, 1948.
JARENKOW, J.A.; BAPTISTA, L.R.M. Composição florística e estrutura da mata com
Araucariaangustifolia na Estação Ecológica de Aracuri, Esmeralda, Rio Grande do Sul.
Napaea: Revista de Botânica, v.3, p. 9-18, 1987.
JARENKOW, J.A; BUDKE, J.C. Padrões florísticos e análise estrutural de remanescentes de
Florestas com Araucária no Brasil. In: FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET,
A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (eds.). Floresta com Araucária:
Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão Preto: Holos, 2009. p. 113-
126.
JONES, M.M.; SZYSKA, B.; KESSLER, M. Microhabitat partitioning promotes plant
diversity in a tropical montane forest. Global Ecology and Biogeography, v.20, p.558-569,
2011.
JORDÃO, S.M.S. Manejo de lianas em bordas de floresta estacional semidecidual e de
cerradão, Santa Rita do Passa Quatro, SP. 247p. Tese (Doutorado em Ciências) Escola
Superior de Agricultura Luiz de Queiroz - ESALQ, 2009.
KARNOSKY, D.F.; CEULEMANS, R.; SCARASCIA-MUGNOZZA, G.E.; INNES, J.L.
(Eds.). The Impact of Carbon dioxide and Other Greenhouse Gasses on Forest
Ecosystems. New York: CABI Publishing, 2001.
KARR, J.R.; DUDLEY, D.R. Ecological perspective on water quality goals. Environment
Management, v.5, p.55–68, 1981.
KASPERSON, J.X.; KASPERSON, R.E.; TURNER, B.L. Regions at Risk: Comparisons of
Threatened Environments. New York: United Nations University Press, 1995.

109
KLAUBERG, C.; PALUDO, G.F.; BORTOLUZZI, R.L.C.; MANTOVANI, A. Florística e
estrutura de um fragmento de Floresta Ombrófila Mista no Planalto Catarinense. Biotemas,
v.23, p.35-47, 2010.
KLEIN, R. M. Mapa fitogeográfico de Santa Catarina. Itajaí: Herbário Barbosa Rodrigues,
1978.
KLEIN, R.M. Importância e fidelidade das Lauráceas na "Formação de Araucária" do
estado de Santa Catarina. Trabalho apresentado no XVII Congresso de Botânica de Brasília,
1966.
KLEIN, R.M. O aspecto dinâmico do pinheiro-brasileiro. Sellowia, v.12, n.12, p.17-48, 1960.
KLEIN, R.M. Ecologia da flora e vegetação do Vale do Itajaí. Sellowia, v.32, p.165-389,
1980.
KLEIN, R.M. Aspectos dinâmicos da vegetação do sul do Brasil. Sellowia, v.36, p.5-54,
1984.
KOTTEK, M.; GRIESER, J.; BECK, C.; RUDOLF, B.; RUBEL, F. World Map of the
Köppen-Geiger climate classification updated. Meteorologische Zeitschrif, v.15, p.259-263,
2006.
KOWARIK, I. Natürlichkeit, Naturnähe und Hemerobie als Bewertungskriterien. In:
KONOLD, W.; BÖCKER, R.; HAMPICKE, U. (Eds.): Handbuch Naturschutz und
Landschaftspflege. Landsberg: Ecomed, 1999.
KUULUVAINEN, T.; PENTTINEN, A.; LEINONEN, K.; NYGREN, M. Statistical
opportunities for comparing stand structural heterogeneity in managed and primeval forests:
An example from boreal spruce forest in southern Finland. Silva Fennica, v.30, p.315–328,
1996.
LAARMANN, D.; KORJUS, H.; SIMS, A.; STANTFURF, J.A.; KVISTE, A.; ÖSTER, K.K.
2009. Analysis of forest naturalness and tree mortality patterns in Estonia. Forest Ecology
and Management, v.258, p.187–195, 2009.
LAMB, R.J.; PURCELL, A.T. Perception of naturalness in landscape and its relationship to
vegetation structure. Landscape and Urban Planning, v.19, p.333-352, 1990.
LANDE, R. Statistics and partitioning of species diversity, and similarity among multiple
communities. Oikos, v.76, p.5-13, 1996.
LANDRES, P.B.; WHITE, P.S.; APLET, G.; ZIMMERMANN, A. Naturalness and Natural
Variability: Definitions, Concepts, and Strategies for Wilderness Management. In:
KULHAVY, D.L.; LEGG, M.H. (Eds.). Wilderness & natural areas in eastern North
America: research, management and planning. Nacogdoches: Austin State University, Arthur
Temple College of Forestry, 1998, p.41-50.
LANG, S.; BLASCHKE, T. Análise da paisagem com SIG. Tradução Hermann Kux. São
Paulo: Oficina de Textos, 2009.

110
LAURANCE, W.F.; RANKIN-DE-MERONA, J.M.; ANDRADE, A.; LAURANCE, S.G.;
D‟ANGELO S.; LOVEJOY, T.E.; VASCONCELOS, H.L. Rain-forest fragmentation and the
phenology of Amazonian tree communities. Journal of Tropical Ecology, v.19, p.343-347,
2003.
LEGENDRE, P.; LEGENDRE, L. Numerical Ecology (2nd English Edition). Amsterdam:
Elsevier, 1998.
LEITE, P.F. Contribuição ao conhecimento fitoecológico do sul do Brasil. Ciência &
Ambiente, v.24, p.51–73, 2002.
LEITE, P.F.; KLEIN, R.M. Vegetação. In: IBGE. Instituto Brasileiro de Geografia e
Estatística. Geografia do Brasil: Região Sul. Rio de Janeiro, 1990. p.113-150.
LIIRA, J.; SEPP, T.; PARREST, O. The forest and ecosystem quality in conditions of
anthropogenic disturbance along productivity gradient. Forest Ecology and Management,
v.250, p. 34-46, 2007.
LIIRA, J.; SEPP, T. Indicators of structural and habitat natural quality in boreo-nemoral
forests along the management gradient. Annales Botanici Fennici, v.46, p.308–325, 2009.
LINDENMAYER, D.B.; FRANKLIN, J.F. Managing stand structure as part of ecologically
sustainable forest management in Australian mountain ash forests. Conservation Biology,
v.11, n.5, p.1053–1068, 1997.
LINDMAN, C.A.M. A vegetação no Rio Grande do Sul. Porto Alegre: Universal, 1906.
LOIDI, J. Phytosociology applied to nature conservation and land management. In: SONG,
U.; DIERSCHKE, H; WANT, X (Eds). Applied vegetation ecology. Shanghai: East China
Normal University Press, 1994.
LONGHI, S.J.; BRENA, D.A.; GOMES, J.F.; NARVAES, I.S.; BERGER, G.; SOLIGO, A. J.
Classificação e caracterização de estágios sucessionais em remanescentes de Floresta
Onbrófila Mista na FLONA de São Francisco de Paula, RS, Brasil. Ciência Florestal, v.16,
p.113-125, 2006.
LOURENÇO, A. Como tudo começou: Uma introdução ao criacionismo. São José dos
Campos: Editoral Fiel, 2012.
LUTZEMBERG, J. Manifesto Ecológico Brasileiro. Porto Alegre: Lançamento, 1976.
MAACK, R. O aspecto fitogeográfico atual do Paraná. Anuário Brasileiro de Economia
Florestal. Rio de Janeiro: Instituto Nacional do Pinho, 1953.
MACHADO, A. An index of naturalness. Journal for Nature Conservation, v.12, p.95-110,
2004.
MAGURRAN, A.M. Measuring Biological Diversity. Oxford: Blackwell Science Ltd.,
2004.

111
MÄHLER JUNIOR, J.K.F.; LAROCCA, J.F. Fitofisionomias, desmatamento e fragmentação
da Floresta com Araucária. In: FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET, A.M.;
DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.). Floresta com Araucária: Ecologia,
Conservação e Desenvolvimento Sustentável. Ribeirão Preto: Holos, 2009. p. 243-252.
MANTOVANI, M. Caracterização de populações naturais de xaxim (Dicksonia sellowiana
(Presl.) Hooker), em diferentes condições edafo-climáticas no estado de Santa Catarina.
Dissertação (Mestrado). Universidade Federal de Santa Catarina, 2004.
MARCHETTI, M. Monitoring and Indicators of Forest Biodiversity in Europe - From ideas to
operationality. EFI Proceedings, n.51, 2004.
MARTENDAL, J.A.C. Processos produtivos e trabalho-educação: A Incorporação do
Caboclo Catarinense na Indústria Madeireira. 92f. Dissertação (Mestrado em Educação) -
Departamento de Administração de Sistemas Educacionais, Fundação Getúlio Vargas
Instituto de Estudos Avançados em Educação, 1980.
MATTHEWS, E. PAYNE, R.; ROHWEE, M.; SIOBAHN, M. Pilot analysis of global
ecosystems: forests ecosystems. Washington DC.: World Resources Institute, 2000.
MAY, M. How many species are there on Earth? Science, v.241, p.1441-1448, 1988.
McCOMB, W.C.; SPIES, T.A.; EMMINGHAM, W.H. Douglas-fir forests: Managing for
timber and mature-forest habitat. Jornal of Forestry, v.91, n.12, p.31– 42, 1993.
McGILL, B. J.; ETIENNE, R. S.; GRAY, J. S.; ALONSO, D.; ANDERSON, M. J.;
BENECHA, H. K.; DORNELAS, M.; ENQUIST, B. J.; GREEN, J. L.; HE, F.; HURLBERT,
A. H.; MAGURRAN, A. E., MARQUET, P. A.; MAURER, B. A.; OSTLING, A.; SOYKAN,
C. U., UGLAND, K. I.; WHITE, E. P. Species abundance distributions: moving beyond
single prediction theories to integration within an ecological framework. Ecology Letters,
v.10, n.10, p. 995-1015, 2007.
MCPFE - Ministerial Conference on the Protection of Forests in Europe. Improved Pan-
European Indicators for Sustainable Forest Management. MCPFE Expert Level Meeting.
Vienna, 2002.
MCPFE - Ministerial Conference on the Protection of Forests in Europe. Work-programme
on the conservation and enhancement of biological and landscape biodiversity in forest
ecosystems. Adopted at Expert-Level by the Third Meeting of the Executive Bureau of the
Pan-European Biological and Landscape Diversity Strategy. Geneva, 1997.
MCPFE - Ministerial Conference on the Protection of Forests in Europe. Vienna declaration
and Vienna resolutions. Adopted at the Fourth Ministerial Conference on the Protection of
Forests in Europe. Vienna, 2003a.
MCPFE - Ministerial Conference on the Protection of Forests in Europe. Improved Pan-
European indicators for sustainable forest management. Adopted by the MCPFE Expert
Level Meeting, Vienna, 2003b.

112
McROBERTS, R.E.; CHIRICI, G.; WINTER, S.; BARBATI, A.; CORONA, P.;
MARCHETTI, M.; HAUK, E.; BRÄNDLI. U.; BERANOVA, J.; RONDEUX, J.;
SANCHEZ, C.; BERTINI, R.; BARSOUM, N.; ASENSIO, I.A., CONDÉS, S.; SAURA, S.;
NEAGU, S.; CLUZEAU, C.; HAMZA, N. Prospects for harmonized biodiversity assessments
using National Forest Inventory data. In: CHIRICI, G.; WINTER, S.; MCROBERTS, R.E.
National forest inventories: Contributions to forest biodiversity assessments. Springer, 2011.
McROBERTS, R.E.; TOMPPO, E.O; NAESSET, E. Advances and emerging issues in
National Forest Inventories. Scandinavian Journal of Forest Research, v.25, p.368-381,
2010a.
McROBERTS, R.E.; WINTER, S.; CHIRICI, G.; HAUK, E.; PELZ, D.R.; MOSER, W.K.;
HATFIELD, M.A. Large-scale patterns of forest structural diversity. Canadian Journal of
Forest Research, v.38, p.429-438, 2008.
McROBERTS, R.E; STAHL, G.; VIDAL, C.; LAWRENCE, M.; TOMPPO, E.;
SCHAUDER, K.; CHIRICI, G; BASTRUP-BIRK, A. 2010b. National Forest Inventories:
Prospects for Harmonized International Reporting. In: TOMPPO, E.; GSCHWANTNER, T.;
LAWRENCE, M.; MCROBERTS, R.E. National Forest Inventories. In: Pathway for
Common Reporting. Heidelberg: Springer, 2010b.
McROBERTS, R.E.; WINTER, S.; CHIRICI, G.; LAPOINT, E. Assessing Forest
Naturalness. Forest Science, v.58, p.294-309, 2012.
McROBERTS, R.E.; WESTFALL, J. Effects of Uncertainty in Model Predictions of
Individual Tree Volume on Large Area Volume Estimates. Forest Science, v.60, p.34–42,
2014.
MELLO, A.J.M. Etnoecologia e manejo local de paisagens antrópicas da Floresta Ombrófila
Mista, Santa Catarina, Brasil. Dissertação (Mestrado em Ecologia), Programa de Pós-
Graduação em Ecologia, Universidade Federal de Santa Catarina, 2013.
MELO, A.S. O que ganhamos „confundindo‟ riqueza de espécies e equabilidade em um índice
de diversidade? Biota Neotropical, v.8, n.3, 2008.
MERGANIČ, J.; MERGANIČOVÁ, K.; MORAVČÍK, M.; VORČÁK, J.; MARSUŠÁK, R.
Objetive evaluation of forest naturalness: case study in Slovak Nature Reserve. Polish
Journal of Environmental Studies, v.21, n.5, p.1327-1337, 2012.
MEYER, L.; GASPER, A.L.; SEVEGNANI, L.; SCHORN, L.A.; VIBRANS, A.C.;
LINGNER, D.V.; VERDI, M.; SANTOS, A.S.; DREVECK, S.; KORTE, A. Regeneração
natural da Floresta Ombrófila Mista em Santa Catarina. In: VIBRANS, A.C.; SEVEGNANI,
L.; GASPER, A.L.; LINGNER, D.V (Eds.). Inventário Florístico Florestal de Santa
Catarina: Floresta Ombrófila Mista. Blumenau: Edifurb, 2013. p.191-222.
MICHEL, A.; WINTER, S. Tree microhabitat structures as indicators of biodiversity in
Douglas-fir forests of different stand ages and management histories in the Pacific Northwest,
USA. Forest Ecology and Management, v.257, p.1453–1464, 2009.

113
MMA - MINISTÉRIO DO MEIO AMBIENTE/IBAMA – INSTITTO DO MEIO
AMBIENTE E DOS RECURSOS NATURAIS RENOVÁVEIS. Plano de Manejo dos
Parques Nacionais de aparados da Serra e Serra Geral. Brasília: MMA, 2004.
MONTFORD, E.P. Fallen deadwood levels in the near-natural beech forest at La Tillaie
reserve, Fontainbleau, France. Forestry, v.75, p.203-208, 2002.
MORAVČÍK, M.; SARVAŠOVÁ, Z.; MERGANIČ, J.; SCHWARZ, M. Forest Naturalness:
Criterion for Decision Support in Designation and Management of Protected Forest Areas.
Environmental Management, v.46, p.908-919, 2010.
MORETTO, S.P. Remontando a floresta: a implementação do pinus e as práticas de
reflorestamento na região de Lages (1960 – 1990). Dissertação (Mestrado em História),
Programa de Pós-Graduação em História, Universidade Federal de Santa Catarina, 2010.
MUNGUÍA-ROSAS, M.A; JURADO-DZIB, S.G.; MEZETA-COB, C.R.; MONTIEL, S.;
ROJAS, A.; PECH-CANCHÉ, L.M. Continuous forest has greater taxonomic, functional and
phylogenetic plant diversity than an adjacent naturally fragmented forest. Journal of
Tropical Ecology, first view article, p.1-11, 2014.
MURCIA, C. Edge effects in fragmentted forest: implications for conservation. Tree, v.10,
n.2, p.58-62, 1995.
NETTO, S.P. Instituto Brasileiro de Desenvolvimento Florestal. Inventario florestal
nacional: florestas nativas, Paraná, Santa Catarina. Brasília: IBDF, 1984.
NOSS, R.F. Can we maintain biological and ecological integrity? Conservation Biology, v.4,
p.241-243, 1990.
NOSS, R.F. Assessing and monitoring forest biodiversity: A suggested framework and
indicators. Forest Ecology and Management, v.115, p.135–146, 1999.
NILSSON, S.G.; NIKLASSON, M.; HEDIN, J.; ARONSSON, G. GUTOESKI, J.M.;
LINDER, P.; LJUNGBERG, H.; MIKUSINSKY, G.; RANIUS, T. Densities of large living
and dead trees in old-growth temperate and boreal forests. Forest Ecology and
Management, v.161, p.189-204, 2002.
ÓDOR, P.; HEILMANN-CLAUSEN, J.; CHRISTENSEN, M.; AUDE, E.; VAN DORT,
K.W.; PILTAVER, A.; SILLER, I.; VEERKAMP, M.T.; WALLEYN, R.; STANDOVÁR,
T.; van HEES, A.F.M.; KOSEC, J.; MATOČEC, N.; KRAIGHER, H.; GREBENC, T.
Diversity of dead wood inhabiting fungi and bryophytes in semi-natural beech forests in
Europe. Biological Conservation, v.131, p.58-71, 2006.
ODUM, E.P. Fundamentos em ecologia (7ª edição). Lisboa: Fundação Calouste Gulbenkian,
2004.
OGDEN, J.; STEWART, G.H. Community dynamics of the New Zeale conifers. In:
ENRIGHT, N.; HILL, R.S. (Eds.) Ecology of the southern conifers. Washington:
Smithsonian Institution Press, 1995. p.81-119.

114
OLIVEIRA-FILHO, A.T. Classificação das fitofisionomias da América Do Sul cisandina
tropical e subtropical: Proposta de um novo sistema – prático e flexível – ou uma injeção a
mais de caos? Rodriguésia, v.60, p.237-258, 2009.
OLIVEIRA-FILHO, A.T.; BUDKE, J.C.; JARENKOW, J.A.; EISENLOHR, P.V.; NEVES,
D.R.M. Delving into the variations in tree species composition and richness across South
American subtropical Atlantic and Pampean forests. Journal of Plant Ecology, 2013.
OLIVEIRA-FILHO, A.T.; FONTES, M.A. Patterns of floristic differentiation among Atlantic
Forests in Southeastern Brazil and the influence of climate. Biotropica, v 32, n.4b, p.793-810,
2000.
OLIVEIRA, L.Z.; MOSER, P.; VIBRANS, A.C.; GASPER, A.L.; PIAZZA, G.A. Measuring
the performance of nonparametric species richness estimators using a large scale forest
inventory dataset. Biodiversity and Conservation, manuscrito submetido.
OLIVER, C.D.; LARSON, B.C. Forest stand dynamics. New York: McGraw-Hill Inc, 1990.
OSBORNE, J.W. Improving your data transformations: Applying the Box-Cox
transformation. Practical Assessment, Research & Evaluation, v.15, n.12, p.1-9, 2010.
PENÃ-CLAROS, M.; POORTER, L.; ALARCÓN, A.; BLATE, G.; CHOQUE, U.;
FREDERICKSEN, T.S.; JUSTINIANO, M.J.; LEAÑO, C.; LICONA, J.C.; PARIONA, W.;
PUTZ, F.E.; QUEVEDO, L.; TOLEDO, M. Soil effects on forest structure and diversity in a
moist and a dry Tropical Forest. Biotropica, v.44, p.276-283, 2012.
PETERKEN, G.F. Natural woodland: ecology and conservation in temperate regions.
Cambridge: University Press, 1996.
PFEIFER, M. et al. BIOFRAG – a new database for analyzing BIOdiversity responses to
forest FRAGmentation. Ecology and Evolution, v.4, n.9, p.1524–1537, 2014.
PILLAR, V.P.; MÜLLER, S.C.; OLIVEIRA, J.M. MACHADO, R.E. Moisaicos de Cmapos e
Floresta com Araucária: dilemas para conservação. In: FONSECA, C.R.; SOUZA, A.F.;
LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.). Floresta com
Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão Preto: Holos,
2009, p.273-283.
PITMAN, N. C. A.; TERBORGH, J. W.; SILMAN, M. R.; NÚNEZ, P.; NEILL, D. A.;
CERÓN, C. E.; PALACIOS, W. A.; AULESTIA, M. Dominance and distribution of tree
species in upper Amazonian Terra Firme Forests. Ecology, v.82, n.8, p. 2101-2117, 2001.
POHLMAN, C.L.; TURTON, S.M.; GOOSEM, M. Edge effects of linear canopy openings on
tropical rain forest understory microclimate. Biotropica, v. 39, p.62-71, 2007.
POMMERENING, A. Approaches to quantifying forest structure. Forestry, v.75, p.305–324,
2002.

115
QUEIROZ, M.I.P. O campesinato brasileiro: ensaios sobre civilização e grupos rústicos no
Brasil. Petrópolis-São Paulo: Vozes-Edusp, 1973.
RABINOWITZ, D.; CAIRNS, S.; DILLON, T. Seven forms of rarity and their frequency in
the flora of the British Isles. In SOULÉ, M.E. (ed.) Conservation Biology: The Science of
Scarcity and Diversity. Sinauer, 1986, p.182-204.
RAMBO, B. O elemento andino no pinhal riograndense. Anais Botânicos do Herbário
Barbosa Rodrigues, v.3, p.7-39, 1951.
RAMBO, B. História da flora do planalto riograndense. Sellowia, v.5, p.185-232I, 1953.
RAMBO, B. A flora fanerógama dos Aparados riograndenses. Sellowia, v.7/8, p.98-235,
1956.
RAPPORT, D.J.; WHITFORD, W.G. How Ecosystems Respond to Stress. BioScience, v.49,
n.3, p.193–203, 1999.
REIF, A; WALENTOWSKI, H. The assessment of naturalness and its role for nature
conservation and forestry in Europe. Waldökologie, Land schafts forschungund
Naturschutz, v.6, p.63-76, 2008.
REIS, A.; TRES, D.R.; SCARIOT, E.C. Restauração na Floresta Ombrófila Mista através da
sucessão natural. Pesquisa Florestal Brasileira, v.55, p.67-73, 2007.
REIS, M.S; LADIO, A.; PERONI, N. Landscapes with Araucaria in South America: evidence
for a cultural dimension. Ecology and Society, v.19, n.2, p.43-56, 2014.
REIS, M.S.; MANTOVANI, A.; SILVA, J.Z.; MARIOT, J.; BITTENCOURT, R.;
NAZARENO, A.G.; FERREIRA, D.K.; STEINER, F.; MONTAGNA, T.; SILVA, F.A.L.S.;
FERNANDES, C.D.; ALTRAK, G.; FIGUEIREDO, G.U. Distribuição da diversidade
genética e conservação de espécies arboreas em remanescentes florestais de Santa Catarina.
In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V (Eds.). Inventário
Floristico Florestal de Santa Catarina: Diversidade e conservação dos remanescentes
florestais. Blumenau: Edifurb, 2012.
REITZ, R. Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues, 1965.
REITZ, R.; KLEIN, R.M.; REIS, A. Madeiras do Brasil. Editora Lunardelli, 1979.
REITZ, R.; KLEIN, R.M. Flora Ilustrada Catarinense: Araucariáceas. Itajaí: Herbário
Barbosa Rodrigues, 1966.
REZENDE, V.L. Padrões fitogeográficos da flora arbórea de Santa Catarina. Dissertação
(Mestrado em Biologia Vegetal). Belo Horizonte: UFMG, 2013.
REZENDE, V.L.; OLIVEIRA-FILHO, A.T.; EISENLOHR, P.V.; KAMINO, L.H.Y.;
VIBRANS, A.C. Restricted geographic distribution of tree species calls for urgent

116
conservation efforts in the Subtropical Atlantic Forest. Biodiversity Conservation,
publicação online, 2014.
REZNIK, G.; PIRES, J.P.A.; FREITAS, L. Efeito de bordas lineares na fenologia de espécies
arbóreas zoocóricas em um remanescente de Mata Atlântica. Acta Botanica Brasilica, v.26,
n.1, p. 65-73, 2012.
RIBEIRO, M.C.; METZGER, J.P.; MARTENSEN, A.C.; PONZONI, F.J.; HIROTA, M.M.
The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed?
Implications for conservation. Biological Conservation, v. 142, p. 1141–1153, 2009.
RIBEIRO, M.C.; MARTENSEN, A.C.; METZGER, J.P.; TABARELLI, M.; SCARANO, F.;
FORTIN, M.J. The Brazilian Atlantic forest: a shrinking biodiversity hotspot. In: ZACHOS,
F.E.; HABEL, J.C. (Eds.) Biodiversity hotspots. Heidelberg: Springer, 2011. p.405–434.
RIBEIRO, S.B.; LONGHI, S.J.; BRENA, D.A.; NASCIMENTO, A.R.T. Diversidade e
classificação da comunidade arbórea da Floresta Ombrófila Mista da FLONA de São
Francisco de Paula, RS. Ciência Florestal, v.17, n.2, 2007.
RICHARDS, P.W. The Tropical Rain Forest. Cambridge: Cambridge University Press,
1996.
RICHARDSON, D.M.; PYSEK, P.; REJMÁNEK, M.; BARBOUR, M.G.; PANETTA, F.D.;
WEST, C.J. Naturalization and invasion of alien plants: concepts and definitions. Diversity
and Distributions, v.6, p.93-107, 2000.
RICKETTS, T; BROOKS, T; HOFFMANN, M.; STUART, S.; BALMFORD, A.; PURVIS,
A.; REYERS, B.; WANG, J.; REVENGA, C.; KENNEDY, E.; NAEEM, S.; ALKEMADE,
R.; ALLNUTT, T.; BAKARR, M.; BOND, W.; CHANSON, J.; COX, N.; FONSECA, G.;
HILTON-TAYLOR, C.; LOUCKS, C.; RODRIGUES, A.; SECHREST, W.;
STATTERSFIELD, A.; VAN RENSBURG, J.B; WHITEMAN, C. Biodiversity. In:
HASSAN, R.; SCHOLES, R.; ASH, N. (Eds.) Ecosystems and Human Well-being: Current
State and Trends, Volume 1. Washington DC: Island Press, 2005.
SAINT-HILAIRE, A. Viagem a Curitiba e Santa Catarina. Belo Horizonte: Editora Itatiaia,
1978.
SALA, O.E.; CHAPIN, I.F.S.; ARMESTO, J.J.; BERLOW, E.; BLOOMfiELD, J.; DIRZO,
R.; HUBER SANWALD, E.; HUENNEKE, L.F.; JACKSON, R.B.; KINZIG, A.;
LEEMANS, R.; LODGE, D.H.; MOONEY, H.A.; OESTERHELD, M.; LEROY POFF, N.;
SYKES, M.T.; WALKER, B.H.; WALKER, M.; WALL, D.H. Global biodiversity scenarios
for the year 2100. Science, v.287, p.1770-1774, 2000.
SAMPAIO, M.B.; GUARINO, E.S.G. Efeitos do pastoreio de bovinos na estrutura
populacional de plantas em fragmentos de Floresta Ombrófila Mista. Revista Árvore, v.31,
n.6, p.1035-1046, 2007.
SAMUELSSON, J.; GUSTAFSSON, L.; INGELÖG, T. Dying and dead trees: a review of
their importance for biodiversity. Uppsala: Swedish Threatened Species Unit, 1994.

117
SANCHEZ, M.; PEDRONI, F.; EISENLOHR, P.V.; OLIVEIRA-FILHO, A.T. Changes in
tree community composition and structure of Atlantic rain forest on a slope of the Serra do
Mar range, Southeastern Brazil, from near sea level to 1000 m of altitude. Flora, v.208,
p.184-196, 2013.
SANTOS, B.A.; PERES, C.A.; OLIVEIRA, M.A.; GRILLO, A.; ALVES-COSTA, C.P.;
TABARELLI, M. Drastic erosion in functional attributes of tree assemblages in Atlantic
forest fragments of northeastern Brazil. Biological Conservation, v.141, n.1, p.249-260,
2008.
SAR. Secretaria de Agricultura e Abastecimento de Santa Catarina. Inventário Florístico-
Florestal de Santa Catarina: Relatório do Projeto Piloto. Florianópolis: Mimeo, 2005.
ŠAUDYTÈ, S.; KARAZIJA, S.; BELOVA, O. An Approach to Assessment of Naturalness
for forest Stands in Lithuania. Baltic Forestry, v.11, n.1, p.39-45, 2005.
SCHAAF, L.B.; FIGUEIREDO FILHO, A.; GALVÃO, F.; SANQUETTA, C. R.; LONGHI,
S. J. Modificações florístico-estruturais de um remanescente de Floresta Ombrófila Mista
montana no período entre 1979 e 2000. Ciência Florestal, v. 16, n.3, 2006b.
SCHAAF, L.B.; FIGUEIREDO-FILHO, A.; GALVÃO, F.; SANQUETTA, C.R. Alteração na
estrutura diamétrica de uma Floresta Ombrófila Mista no período entre 1979 e 2000. Revista
Árvore, v.30, n.2, p.283-295, 2006a.
SCHAADT, S.S. A fragmentação da Floresta Ombrófila Mista em Santa Catarina, analisada a
partir de dados do Inventário Florístico Florestal, fotografias aéreas e mapa temático.
Dissertação (Mestrado em Engenharia Ambiental). Universidade Regional de Blumenau,
2012.
SCHNEIDER, P.R. Introdução ao manejo florestal. Santa Maria: UFSM, 1993.
SCHIMITZ, P.I. Povos indígenas associados à Floresta com Araucária. In: FONSECA, C.R.;
SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.).
Floresta com Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão
Preto: Holos, 2009, p. 45-54.
SEITZ, R.A. A regeneração natural de Araucaria angustifolia. Silvicultura em São Paulo,
v.16, p.1412-420, 1982.
SEVEGNANI, L.; VIBRANS, A.C.; GASPER, A.L. Considerações finais sobre a Floresta
Ombrófila Mista em Santa Catarina. In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.;
LINGNER, D.V. (Eds.) Inventário Florístico Florestal de Santa Catarina: Floresta
Ombrófila Mista. Blumenau: Edifurb, 2013a, p.275-278.
SEVEGNANI, L.; GASPER, A. L.; BONNET, A.; SOBRAL, M. G.; VIBRANS, A. C.;
VERDI, M.; SANTOS, A. S.; DREVECK, S.; KORTE, A.; SCHMITT, J.; CADORIN, T.;
LOPES, C. P.; CAGLIONI, E.; TORRES, J. F.; MEYER, L. Flora vascular da Floresta
Ombrófila Densa em Santa Catarina. In: VIBRANS, A. C.; SEVEGNANI, L.; GASPER, A.
L.; LINGNER, D. V. (Ed.). Inventário Florístico Florestal de Santa Catarina: Floresta
Ombrófila Densa. Blumenau: Edifurb, 2013b, p. 127-139.

118
SILVA, A.F.; OLIVEIRA, R.V.; SANTOS, N.R.L.; PAULA, A. Composição florística e
grupos ecológicos das espécies de um trecho de Floresta Semidecídua Submontana da
Fazenda São Geraldo, Viçosa-MG. Revista Árvore, v.27, n.3, p.311-319, 2003.
SIMINSKI, A. A floresta do futuro: conhecimento, valorização e perspectivas de uso das
formações florestais secundárias no Estado de Santa Catarina. Tese (Doutorado em Recursos
Genéticos Vegetais). Universidade Federal de Santa Catarina, 2009.
SIMINSKI, A. Formações florestais secundárias como recurso para o desenvolvimento rural e
a conservação ambiental no litoral de Santa Catarina. 117p. Dissertação (Mestrado em
Recursos Genéticos Vegetais). Universidade Federal de Santa Catarina, 2004.
ŠMELKO, Š; FABRIKA, M. Evaluation of qualitative attributes of forest ecosystems by
means of numerical quantifiers. Journal of Forest Science, v.53, p.529–537, 2007.
SMITH, P.G.R.; THEBERGE, B. Evaluating natural áreas using multiple criteria: Theory and
practice. Environmental Management, v.11, n.4, p.447– 460, 1987.
SOARES, R.V. Considerações sobre a regeneração natural de Araucaria angustifolia.
Floresta, v.10, p.12-18, 1979.
SJÖRS, H. On the gradient from near-natural to man-made. Transactions of the Botanical
Society of Edinburgh, v.45, p.77–84, 1986.
SOULÉ, M.E. What is conservation biology? BioScience, v.35, n.11, p.727–734, 1985.
SOUZA, A.F. Estrutura de populacões de Araucaria angustifolia. In: FONSECA, C.R.;
SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.).
Floresta com Araucária: Ecologia, Conservação e Desenvolvimento Sustentável. Ribeirão
Preto: Holos, 2009, p. 67-74.
STEEDMAN, R.J. Ecosystem health as management goal. Journal of the North American
Benthological Society, v.13, p.605-610, 1995.
STEHMANN, J.R.; FORZZA, R.C; SALINO, A.; SOBRAL, M.; COSTA, D.P.; KAMIRO,
L.H.Y. Plantas da Floresta Atlântica. Rio de Janeiro: Jardim Botânico do Rio de Janeiro,
2009.
STOKLAND, J.N. The coarse woody debris profile: an archieve of recent forest history and
an important biodiversity indicator. Ecological Bulletins, v.49, p.71-84, 2001.
SUKOPP, H. Dynamik und Konstanz in der Flora der Bun-desrepublik Deutschland.
Schriftenreihe für Vegetationskunde, v.10, p. 9–27, 1972.
SWAINE, M.D.; HALL, J.B. The mosaic theory of forest regeneration and the determination
of forest composition in Ghana. Journal of Tropical Ecology, v.4, p.253-269, 1988.
SWAINE, M.D.; WHITMORE, T.C. On the definition of ecologiacal species groups in
tropical rain forests. Vegetation, v.75, n.1-12, p.81-86, 1988.

119
TABARELLI, M.; AGUIAR, A.V.; RIBEIRO, M.C.; METZGER, J.P.; PERES, C.A.
Prospects for biodiversity conservation in the Atlantic Forest: Lessons from aging human-
modified landscapes. Biological Conservation, v.143, p.2328-2340, 2010.
TABARELLI, M.; MANTOVANI, W.; PERES, C.A. Effects of habitat fragmentation on
plant guild structure in the montane Atlantic Forest of southeastern Brazil. Biological
Conservation, v.91, p.119-127, 1999.
THE PLANT LIST. The Angiosperms (Flowering plants). Disponível em:
<http://www.theplantlist.org>. Acesso em: 08/09/2014
THUILLER, W. On the importance of edaphic variables to predict plant species distributions
– limits and prospects. Journal of Vegetation Science, v.24, p.591-592, 2013.
TIERNEY, G., FABER-LANGENDOEN, D., MITCHELL, B.R., SHRIVER, W.G. AND
GIBBS, J.P. Monitoring and evaluating the ecological integrity of forest ecosystems.
Frontiers in Ecology and the Environment, v.7, p.308–316, 2009.
TOMPPO, E.; GSCHWANTNER, T.; LAWRENCE, M.; MCROBERTS, R.E. National
Forest Inventories: Pathway for Common Reporting. Heidelberg: Springer, 2010.
TROPPMAIR, H. Biogeografia e Meio Ambiente. Rio Claro: Edição do autor, 1995.
UHLMANN, A.; GASPER, A.L.; SEVEGNANI, L.; VIBRANS, A.C. Grupos florísticos
estruturais da Floresta Ombrófila Mista em Santa Catarina. In: VIBRANS, A.C.;
SEVEGNANI, L.; GASPER, A.L.; LINGNER, D.V. (Eds.) Inventário Florístico Florestal
de Santa Catarina: Floresta Ombrófila Mista. Blumenau: Edifurb, 2013, p.145-153.
UOTILA, A.; KOUKI, J.; KONTKANEN, H.; PULKKINEN, P. Assessing the naturalness of
boreal forests in eastern Fen-noscandia. Forest Ecology Management, v.161, p.257–277,
2002.
VANDEKERKHOVE, K.; KEERSMAEKER, L.DE; MENKE, N.; MEYER, P.;
VERSCHELDEA, P. When nature takes over from man: Dead wood accumulation in
previously managed oak and beech woodlands in North-western and Central Europe. Forest
Ecology and Management, v.258, p.425-425, 2009.
VARGA, P.; KLINKA, K. The structure of high-elevation, old-growth stands in west central
British Columbia. Canadian Journal of Forest Research, v.31, p.2098–2106, 2001.
VIBRANS, A.C. A cobertura florestal da bacia do Rio Itajaí – Elementos para uma análise
histórica. 231p. Tese (Doutorado em Geografia), Universidade Federal de Santa Catarina –
UFSC, Florianópolis, 2003.
VIBRANS, A. C.; UHLMANN, A.; SCHORN, L.A.; SEVEGNANI, L.; MARCOLIN, M.;
GASPER, A. L. de; LINGNER, D. V. Inventário Florístico Florestal de Santa Catarina
(IFFSC): Fase II, Meta 2 (2007/2008) – Relatório Final. Blumenau, 2009.

120
VIBRANS, A.C.; SEVEGNANI, L.; LINGNER, D.V.; GASPER, A.L; SABBAGH, S.
Inventário Florístico Florestal de Santa Catarina (IFFSC): aspectos metodológicos e
operacionais. Pesquisa Florestal Brasileira, v.30, n.64, p. 291-302, 2010.
VIBRANS, A.C.; GASPER, A.L.; MÜLLER, J.J.V. Para que inventariar florestas? Reflexões
sobre a finalidade do Inventário Florístico Florestal de Santa Catarina. REA – Revista de
estudos ambientais (Online), v.14, p.6-13, 2012a.
VIBRANS, A.C.; ULHMANN, A.; SEVEGNANI, L.; MARCOLIN, M.; NAKAJIMA, N.;
GRIPPA, C.R.; BROGNI, E.; GODOY, M.B. Ordenação dos dados estruturais da Floresta
Ombrófila Mista partindo de informações do Inventário Florístico-Florestal de Santa Catarina.
Ciência Florestal, v.18, n.4, p. 511-523, 2008.
VIBRANS, A.C; MOSER, P.; LINGNER, D.V.; GASPER, A.L. Metodologia do Inventário
Florístico Florestal de Santa Catarina. In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER,
A.L.; LINGNER, D.V. Inventário Florístico Florestal de Santa Catarina: Diversidade e
conservação dos remanescentes florestais. Blumenau: Edifurb, 2012a.
VIBRANS; A.C.; MCROBERTS; R.E.; MOSER; P.; NICOLETTI; A. How much remains of
the Brazilian Atlantic forest in the state of Santa Catarina? Assessing the accuracy of forest
cover maps using ground data from the Santa Catarina Forest and Floristic Inventory. Remote
Sensing of Environment, v.130, p.87-95, 2013.
VIBRANS, A.C.; MOSER, P.; OLIVEIRA, L.Z; MAÇANEIRO, J.P. Generic and specific
stem volume models for three subtropical forest types in southern Brazil. Annals of Forest
Science, em revisão.
VIBRANS, A.C.; MOSER, P.; OLIVEIRA, L.Z; MAÇANEIRO, J.P. Height-diameter models
for three subtropical forest types in southern Brazil. Ciência e Agrotecnologia, no prelo.
VIBRANS; A.C.; SEVEGNANI; L.; UHLMANN; A.; SCHORN; L.A.; SOBRAL; M.;
GASPER; A.L.; LINGNER; D.V.; BROGNI; E.; KLEMTZ; G.; GODOY; M.B.; VERDI; M.
Structure of Mixed Ombrophylous Forests with Araucaria angustifolia (Araucariaceae) under
external stress in southern Brazil. Revista de Biología Tropical, v.9, n.3, p.1371-1387, 2011.
VOLKOV, I.; BANAVAR, J. R.; HUBBELL, S. P.; MARITAN, A. Patterns of relative
species abundance in rainforests and coral reefs. Nature, v.450, n.7166, p.45-49, 2007.
von OHEIMB, G.; WESTPHAL, C.; HÄRDTLE, W. Diversity and spatio-temporal dynamics
of dead wood in a temperate near-natural beech forest (Fagus sylvatica). European Journal
of Forest Research, v.126, p.359-370, 2007.
WHITTAKER, R.H. Evolution and measurement of species diversity. Taxon, v.21, p.213-
251, 1972.
WILLIAMS, K. Beliefs about natural forest systems. Austraian Forestry, v.64, n.2, p.81-86,
2002.

121
WILSEY, B.J.; CHALCRAFT, D.R.; BOWLES, C.M.; WILLIG, M.R. Relationships among
indices suggest that richness is an incomplete surrogate for grassland biodiversity. Ecology,
v.86, n.5, p.1178-1184, 2005.
WINTER, S.; FISCHER, H.S.; FISCHER, A. Relative Quantitative Reference Approach for
Naturalness Assessment. Forest Ecology Management, v. 259, p. 1624–1632, 2010.
WINTER, S.; MCROBERTS, R.E.; BERTINI, R.; BASTRUP-BIRK, A.; SACNHEZ, C.;
CHIRICI, G. Essential features of forest biodiversity for assessment purposes. In: CHIRICI,
G.; WINTER, S.; MCROBERTS, R.E (Eds.). National forest inventories: Contributions to
forest biodiversity assessments. Springer, 2011.
WINTER, S.; MÖLLER, G. Microhabitats in lowland beech forests as monitoring tool for
nature conservation. Forest Ecology Management, v.255, p.1251–1261, 2008.
WINTER, S. Forest naturalness assesment as a component of biodiversity monitoring and
conservation management. Forestry, v.85, n.2, p.293-304, 2012.
WRIGHT, S.J.; MULLER-LANDAU, H.C.; CONDIT, R.; HUBBELL, S.P. Gap-dependent
recruitment, realized vital rates, and size distributions of tropical trees. Ecology, v.84, n.12,
p.3174 –3185, 2003.
ZAR, J.H. Biostatistical Analysis. 5th
Edition. New Jersey: Pearson Prentice Hall, 2010.

122
CAPÍTULO 2: QUANTIFICAÇÃO DA NATURALIDADE DOS REMANESCENTES
DE FLORESTA MISTA COM ARAUCÁRIA DE SANTA CATARINA
RESUMO
A naturalidade é um atributo que está relacionado ao grau em que algo é alterado do seu
estado original; ela pode ser quantificada através de um gradiente, que estende-se do menos
ao mais natural. A partir das duas abordagens de quantificação baseadas em distância
euclidiana (floresta de referência teórica e observada) propostas no primeiro capítulo deste
manuscrito, criamos dois Índices de naturalidade que vão do valor zero até um (naturalidade
mínima e máxima, respectivamente). Os Índices foram calculados para 145 remanescentes
(Unidade Amostrais) de Floresta Mista com Araucária amostrados pelo Inventário Florístico-
Florestal de Santa Catarina (IFFSC). A maioria dos remanescentes avaliados possui
naturalidade média (~0,50), levando em consideração a referência teórica (100% natural).
Quando comparados às florestas de referência observadas, os remanescentes apresentaram
valores em torno de 0,60. Ambos os Índices apresentaram resultados coerentes e que
corroboraram o conhecimento já existente sobre o estado de conservação (prejudicado) da
Floresta Mista com Araucária de Santa Catarina. Algumas das bacias hidrográficas avaliadas
mostraram-se menos naturais que a média e merecem atenção em futuras ações e políticas
ligadas à conservação, restauração e manejo. Diante dos resultados satisfatórios gerados pelos
Índices, a abordagem proposta tem potencial para ser adaptada aos diversos contextos
(ecológicos e sociais) das florestas nativas do Brasil. Os dados provenientes do Inventário
Florestal Nacional do Brasil (IFN-Brasil) são uma rica matéria prima para o desenvolvimento
de novos índices quantitativos de naturalidade. Assim, tais índices podem ser ferramentas
úteis para avaliação de remanescentes e, por conseguinte, podem ser utilizados em análises
multitemporais dentro do programa IFN-Brasil.

123
ABSTRACT
The naturalness is an atribute related to the degree to which something is changed from its
original state and it can be quantified through a gradient that range from the least to the most
natural. From the two quantification approaches based on Euclidean distance (theoretical and
observed reference forests) proposed in the first chapter of this manuscript, we created two
indexes of naturalness ranging from zero to one (minimal and maximum naturalness,
respectively). The indexes were calculated for 145 sample plots of the Forest and Floristic
Inventory of Santa Catarina in Mixed Needle-broadleaved Forest. Most forest remnants
presented an average naturalness (~0.50) when considering the theoretical reference (100%
natural). When forest remnants were compared to the observed reference forests they
presented naturalness values around 0.60. Both indexes showed consistent results that
corroborated the existing knowledge about the (degraded) condition of the Mixed Needle-
broadleaved Forest on Santa Catarina state. Some of the evaluated watersheds are less natural
than the average; therefore, they deserve attention in future actions and policies related to the
conservation, restoration and management. Given the satisfactory results generated by the
indexes, the proposed approach has potential for being adapted to different contexts
(ecological and social) Brazil‟s native forests. Data from Brazilian National Forest Inventory
(NFI-Brazil) are a rich resourse for the development of new quantitative naturalness indexes.
Thus, these indexes may be useful tools for assessing ecological integrity of native forests;
they can also be used for multi-temporal analysis on the NFI-Brazil program.

124
2.1. INTRODUÇÃO
A partir de dados coletados por Inventários Florestais Nacionais (IFNs, ou IFN, no
singular) pode-se quantificar a naturalidade de remanescentes florestais (CHIRICI et al. 2011,
2012; MARCHETTI, 2004; McROBERTS et al., 2012; EEA, 2014). Este atributo está
relacionado ao grau em que algo é alterado do seu estado original, e pode ser quantificado
através indicadores que sintetizam informações acerca de um eco(sistema) e das pressões
contrárias (antrópicas) aplicadas nele (LIRA e SEPP, 2009; WINTER et al., 2010; WINTER,
2012).
Diante desses fatos e da qualidade e abrangência dos dados coletados pelo Inventário
Florístico-Florestal de Santa Catarina (IFFSC) (VIBRANS et al., 2010), um estimador
quantitativo de naturalidade pode se tornar uma ferramenta inovadora, prática e efetiva na
identificação de remanescentes de alta relevância para a conservação (McROBERTS et al.,
2012). Não só áreas potenciais para conservação são relevantes - áreas com baixo grau de
conservação devem ser identificadas e tratadas (TABARELLI et al., 2010). Ainda, os
resultados obtidos em estudos de naturalidade podem auxiliar na formação de diretrizes para
políticas públicas envolvendo a conservação e o manejo das florestas nativas. Sabemos sobre
as peculiaridades ecológícas e sobre o grande potencial de uso das florestas nativas brasileiras
- a Floresta Mista com Araucária (OLIVEIRA-FILHO, 2009) de Santa Catarina, que é a
fitofisionomia considerada no presente estudo, possui espécies com grande potencial de uso e
contém alta biodiversidade (FONSECA et al., 2009).
Apesar dos muitos trabalhos que abordam esse atributo, existe a carência de uma
definição comum do conceito e de métodos de determinação/quantificação da naturalidade
(MACHADO, 2004; McROBERTS et al., 2012; WINTER, 2012). Poucos métodos explícitos
de quantificação foram propostos até o momento (McROBERTS et al., 2012; WINTER,
2012; EEA, 2014), de forma que estes autores incentivam a realização de estudos que tratem
da naturalidade e de formas de quantificá-la. A ausência de estudos que tratam do conceito na
América do Sul é outro fator que impulsiona a realização do presente estudo.
Diante deste contexto, os objetivos deste capítulo são: (i) aplicar a metodologia de
quantificação da naturalidade apresentada no primeiro capítulo desse manuscrito; (ii) escalar
a medida de naturalidade com o intuito de torná-la um índice de interpretação facilitada; (iii)
discutir sobre a metodologia e os resultados obtidos acerca dos Índices de naturalidade.

125
2.2. MATERIAL E MÉTODOS
A metodologia proposta no primeiro capítulo desse manuscrito foi utilizada para
quantificar a naturalidade dos remanescentes de Floresta Mista com Araucária de Santa
Catarina amostrados pelo IFFSC.
A partir da medida baseada em distância euclidiana criamos dois Índices de
naturalidade, que são basicamente a medida escalada para o intervalo [0,1]. Os próximos
tópicos tratam dos procedimentos adotados para o escalamento da medida.
2.2.1. ESCALAMENTO DA MEDIDA DE NATURALIDADE
O procedimento de escalamento da medida de naturalidade tem a finalidade de torná-
la um índice de fácil interpretação. Conforme as duas abordagens propostas (tópico 1.3.4.6 do
capítulo 1), temos dois índices: (i) Índice de naturalidade, baseado na floresta de referência
teórica e (ii) Índice de naturalidade, baseado na floresta de referência observada. Daqui em
diante, tais índices serão chamados de Índice de naturalidade I e Índice de naturalidade II,
respectivamente.
2.2.1.1 Índice de naturalidade I
Ao aplicarmos a medida de quantificação da naturalidade apresentada no capítulo
anterior, verificamos que a distância Euclidiana máxima ao vetor máximo (1, 1, 1, 1, 1, 1) é
de 2,45. Logo, este valor representa um remanescente florestal com naturalidade mínima,
quando comparado à floresta de referência teórica. O primeiro procedimento de escalamento
das medidas para um intervalo [0,1] consiste na aplicação do algoritmo:
(i)
onde N é a medida de naturalidade escalada para o intervalo [0,1], a é o limite inferior do
intervalo (0), b é o limite superior do intervalo (1), xi é o valor de naturalidade para a i-ésima
Unidade Amostral, mín é a distância Euclidiana mínima (0) até a referência e máx é a
distância Euclidiana máxima (2,45) até a referência.
Finalizando o processo, a medida N será submetida à transformação:
onde Nfinal é a medida escalada, conforme a Figura 2.1.
amínmáx
mínxabN i
))((
NN final 1 (ii)

126
Agora, a medida mínima de naturalidade torna-se “0” e a máxima “1”, facilitanto a
interpretação.
Figura 2.1. Representação do Índice de naturalidade (gradiente) em sua forma final.
2.2.1.2 Índice de naturalidade II
O escalamento do Índice de naturalidade II para o intervalo [0,1] segue os mesmos
procedimentos do Índice I, porém, a distância máxima até a floresta de referência observada
varia, conforme a Tabela 2.1.
Tabela 2.1. Distâncias máximas até as florestas de referência de cada bacia hidrográfica,
utilizadas para o escalamento do Ìndice de naturalidade II para o intervalo [0,1].
Bacia hidrográfica Dist. máx. até a referência
Inferomontana Superomontana
Rio Pelotas 1,77 1,63
Rio Canoas 1,64 1,77
Rio Itajaí 1,76 1,72
Rio do Peixe 1,49 1,72
Rio Chapecó 1,83 1,76
Rio Canoinhas 1,51 1,80
Rio Timbó 1,65 1,66
Rio Negro 1,70 1,78
2.2.2. PADRÕES ESPACIAIS DE NATURALIDADE
Buscamos por padrões espaciais de naturalidade através de correlogramas
expressando o coeficiente I de Moran em classes de distância; avaliamos esses correlogramas
com base no nível de significância corrigido pelo método sequencial de Bonferroni (FORTIN
e DALE, 2005).
O Índice de naturalidade I não apresentou autocorrelação espacial, ou seja, não
existem padrões espaciais significativos (correlograma não significativo [Figura 2.2 - A];
0 1
edida de naturalidade
Alta naturalidade
Baixa naturalidade
Índice de naturalidade
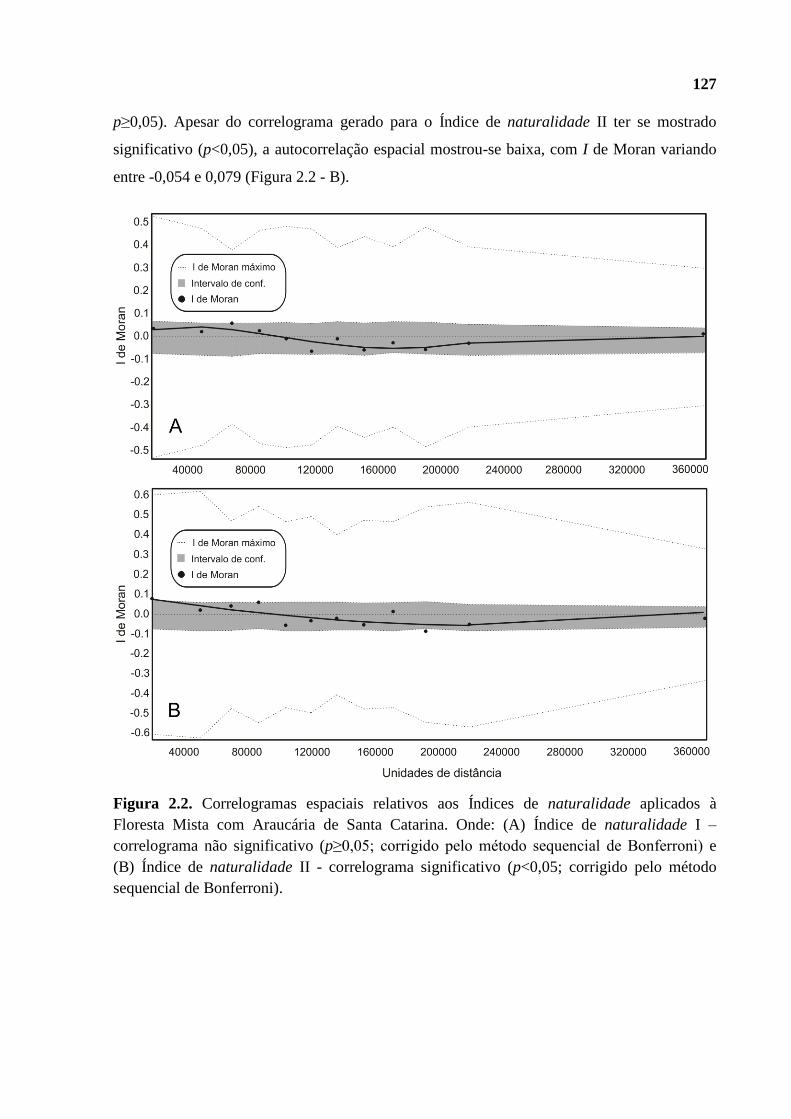
127
p≥0,05). Apesar do correlograma gerado para o Índice de naturalidade II ter se mostrado
significativo (p<0,05), a autocorrelação espacial mostrou-se baixa, com I de Moran variando
entre -0,054 e 0,079 (Figura 2.2 - B).
Figura 2.2. Correlogramas espaciais relativos aos Índices de naturalidade aplicados à
Floresta Mista com Araucária de Santa Catarina. Onde: (A) Índice de naturalidade I –
correlograma não significativo (p≥0,05; corrigido pelo método sequencial de Bonferroni) e
(B) Índice de naturalidade II - correlograma significativo (p<0,05; corrigido pelo método
sequencial de Bonferroni).
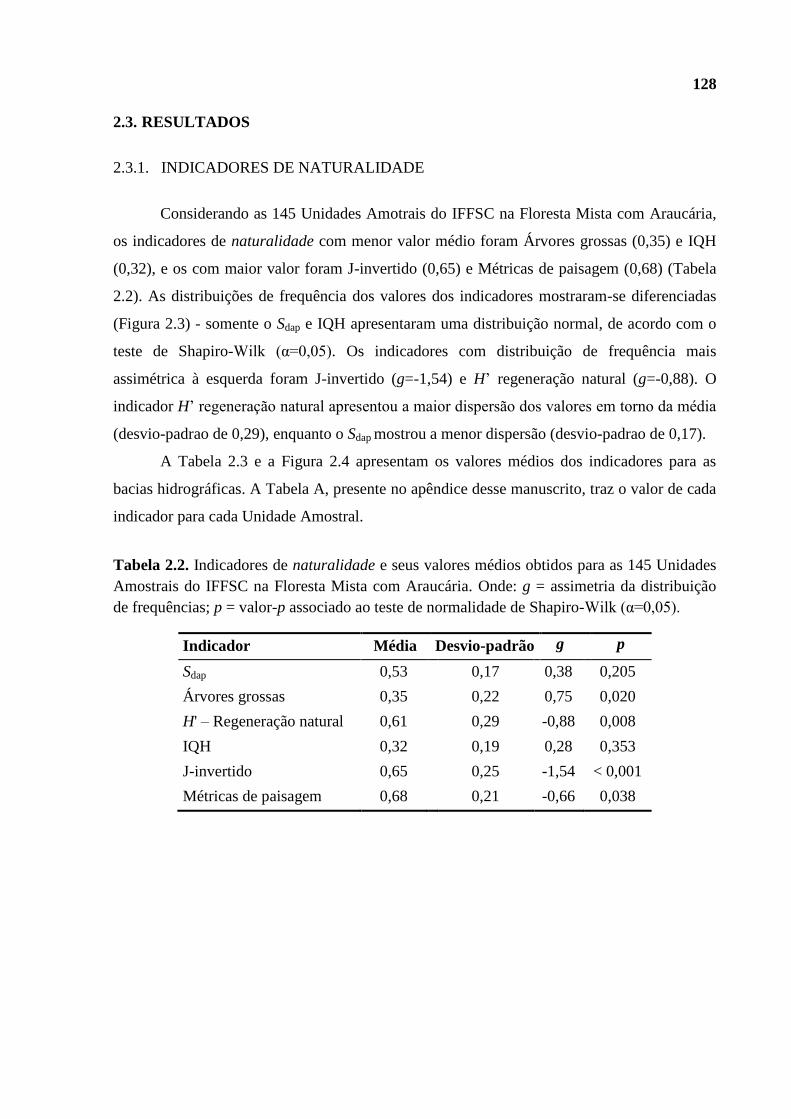
128
2.3. RESULTADOS
2.3.1. INDICADORES DE NATURALIDADE
Considerando as 145 Unidades Amotrais do IFFSC na Floresta Mista com Araucária,
os indicadores de naturalidade com menor valor médio foram Árvores grossas (0,35) e IQH
(0,32), e os com maior valor foram J-invertido (0,65) e Métricas de paisagem (0,68) (Tabela
2.2). As distribuições de frequência dos valores dos indicadores mostraram-se diferenciadas
(Figura 2.3) - somente o Sdap e IQH apresentaram uma distribuição normal, de acordo com o
teste de Shapiro-Wilk (α=0,05). Os indicadores com distribuição de frequência mais
assimétrica à esquerda foram J-invertido (g=-1,54) e H‟ regeneração natural (g=-0,88). O
indicador H‟ regeneração natural apresentou a maior dispersão dos valores em torno da média
(desvio-padrao de 0,29), enquanto o Sdap mostrou a menor dispersão (desvio-padrao de 0,17).
A Tabela 2.3 e a Figura 2.4 apresentam os valores médios dos indicadores para as
bacias hidrográficas. A Tabela A, presente no apêndice desse manuscrito, traz o valor de cada
indicador para cada Unidade Amostral.
Tabela 2.2. Indicadores de naturalidade e seus valores médios obtidos para as 145 Unidades
Amostrais do IFFSC na Floresta Mista com Araucária. Onde: g = assimetria da distribuição
de frequências; p = valor-p associado ao teste de normalidade de Shapiro-Wilk (α=0,05).
Indicador Média Desvio-padrão g p
Sdap 0,53 0,17 0,38 0,205
Árvores grossas 0,35 0,22 0,75 0,020
H' – Regeneração natural 0,61 0,29 -0,88 0,008
IQH 0,32 0,19 0,28 0,353
J-invertido 0,65 0,25 -1,54 < 0,001
Métricas de paisagem 0,68 0,21 -0,66 0,038
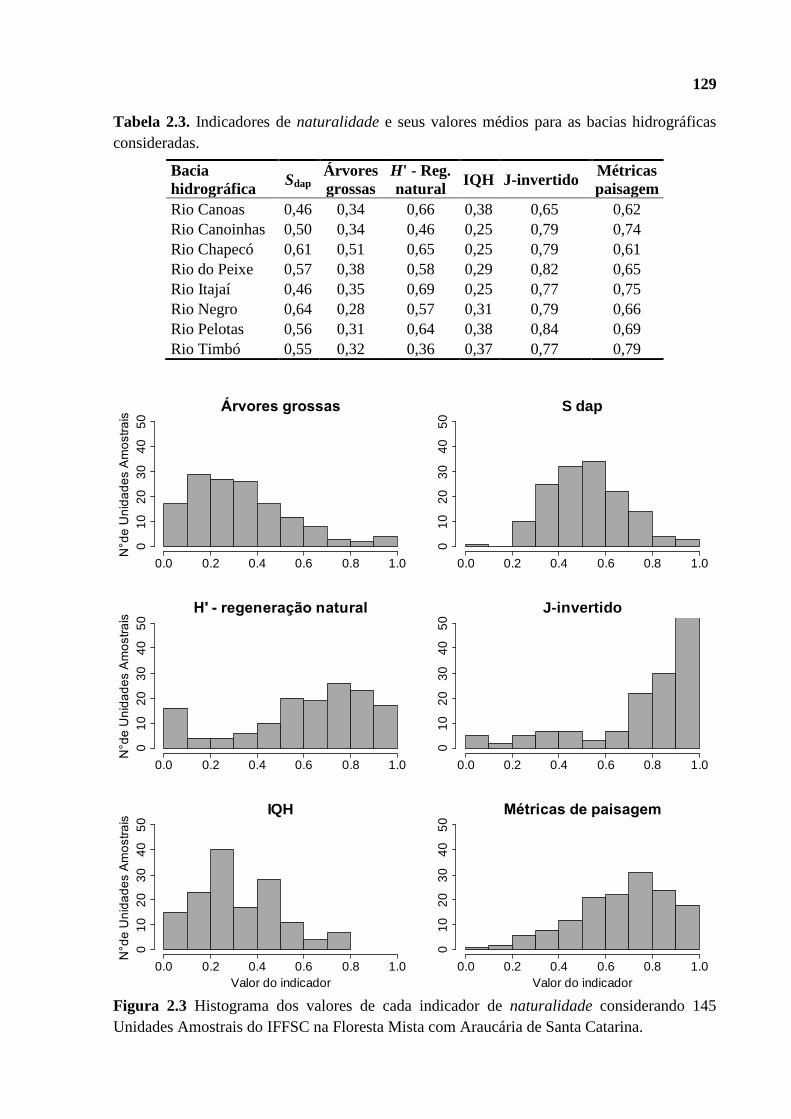
129
Tabela 2.3. Indicadores de naturalidade e seus valores médios para as bacias hidrográficas
consideradas.
Bacia
hidrográfica Sdap
Árvores
grossas
H' - Reg.
natural IQH J-invertido
Métricas
paisagem
Rio Canoas 0,46 0,34 0,66 0,38 0,65 0,62
Rio Canoinhas 0,50 0,34 0,46 0,25 0,79 0,74
Rio Chapecó 0,61 0,51 0,65 0,25 0,79 0,61
Rio do Peixe 0,57 0,38 0,58 0,29 0,82 0,65
Rio Itajaí 0,46 0,35 0,69 0,25 0,77 0,75
Rio Negro 0,64 0,28 0,57 0,31 0,79 0,66
Rio Pelotas 0,56 0,31 0,64 0,38 0,84 0,69
Rio Timbó 0,55 0,32 0,36 0,37 0,77 0,79
Figura 2.3 Histograma dos valores de cada indicador de naturalidade considerando 145
Unidades Amostrais do IFFSC na Floresta Mista com Araucária de Santa Catarina.
Árvores grossas
N°d
e U
nid
ad
es A
mo
str
ais
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0
S dap
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0
H' - regeneração natural
N°d
e U
nid
ad
es A
mo
str
ais
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0
J-invertido
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0
IQH
Valor do indicador
N°d
e U
nid
ad
es A
mo
str
ais
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0
Métricas de paisagem
Valor do indicador
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
05
0

130
Figura 2.4. Valores médios dos indicadores de naturalidade por bacia hidrográfica. As barras
de erro expressam o desvio-padrão.
2.3.2. FLORESTAS DE REFERÊNCIA
Os remanescentes florestais mais naturais, segundo os critérios de triagem
estabelecidos, estão apresentados na Tabela 2.4, e suas localizações geográficas na Figura 2.5.
Seguindo a triagem proposta, duas Unidades Amostrais foram escolhidas para cada uma das
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
Va
lor
mé
dio
do
in
dic
ad
or
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
Va
lor
mé
dio
do
in
dic
ad
or
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
Va
lor
do
mé
dio
do
in
dic
ad
or
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
Árvores grossas S dap
H’ regeneração natural J-invertido
IQH Métricas de paisagem
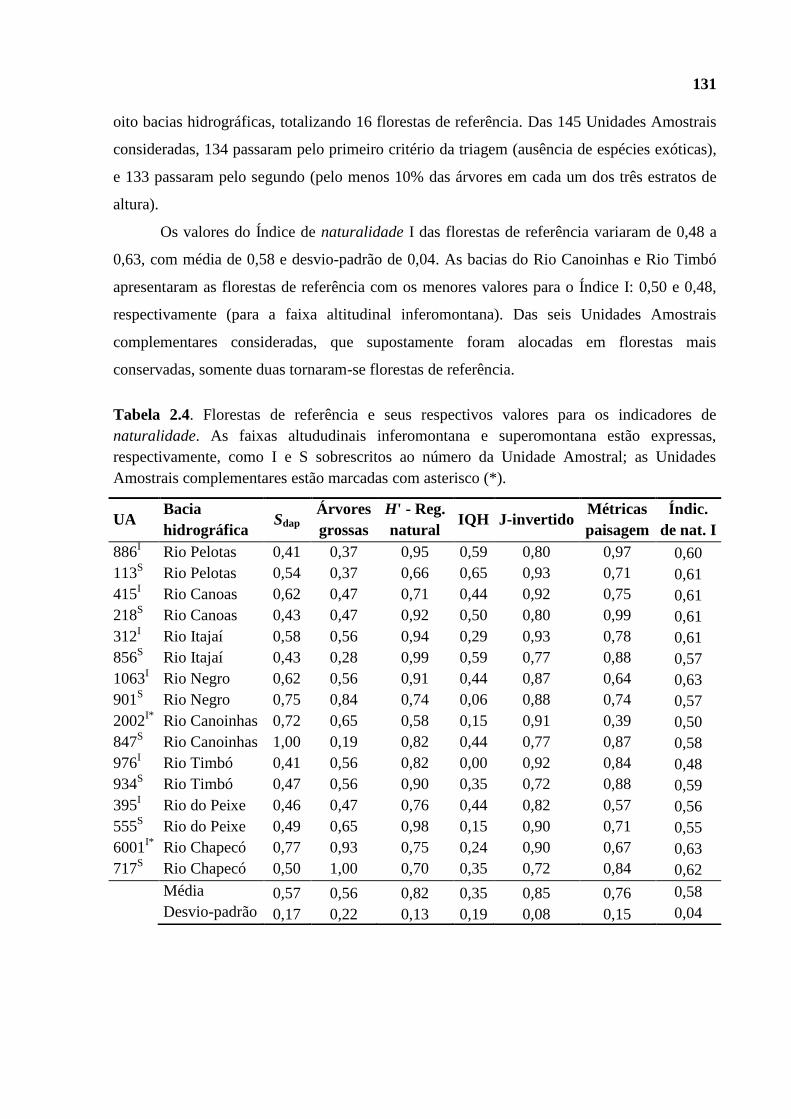
131
oito bacias hidrográficas, totalizando 16 florestas de referência. Das 145 Unidades Amostrais
consideradas, 134 passaram pelo primeiro critério da triagem (ausência de espécies exóticas),
e 133 passaram pelo segundo (pelo menos 10% das árvores em cada um dos três estratos de
altura).
Os valores do Índice de naturalidade I das florestas de referência variaram de 0,48 a
0,63, com média de 0,58 e desvio-padrão de 0,04. As bacias do Rio Canoinhas e Rio Timbó
apresentaram as florestas de referência com os menores valores para o Índice I: 0,50 e 0,48,
respectivamente (para a faixa altitudinal inferomontana). Das seis Unidades Amostrais
complementares consideradas, que supostamente foram alocadas em florestas mais
conservadas, somente duas tornaram-se florestas de referência.
Tabela 2.4. Florestas de referência e seus respectivos valores para os indicadores de
naturalidade. As faixas altududinais inferomontana e superomontana estão expressas,
respectivamente, como I e S sobrescritos ao número da Unidade Amostral; as Unidades
Amostrais complementares estão marcadas com asterisco (*).
UA Bacia
hidrográfica Sdap
Árvores
grossas
H' - Reg.
natural IQH J-invertido
Métricas
paisagem
Índic.
de nat. I
886I Rio Pelotas 0,41 0,37 0,95 0,59 0,80 0,97 0,60
113S Rio Pelotas 0,54 0,37 0,66 0,65 0,93 0,71 0,61
415I Rio Canoas 0,62 0,47 0,71 0,44 0,92 0,75 0,61
218S Rio Canoas 0,43 0,47 0,92 0,50 0,80 0,99 0,61
312I Rio Itajaí 0,58 0,56 0,94 0,29 0,93 0,78 0,61
856S Rio Itajaí 0,43 0,28 0,99 0,59 0,77 0,88 0,57
1063I Rio Negro 0,62 0,56 0,91 0,44 0,87 0,64 0,63
901S Rio Negro 0,75 0,84 0,74 0,06 0,88 0,74 0,57
2002I*
Rio Canoinhas 0,72 0,65 0,58 0,15 0,91 0,39 0,50
847S Rio Canoinhas 1,00 0,19 0,82 0,44 0,77 0,87 0,58
976I Rio Timbó 0,41 0,56 0,82 0,00 0,92 0,84 0,48
934S Rio Timbó 0,47 0,56 0,90 0,35 0,72 0,88 0,59
395I Rio do Peixe 0,46 0,47 0,76 0,44 0,82 0,57 0,56
555S Rio do Peixe 0,49 0,65 0,98 0,15 0,90 0,71 0,55
6001I*
Rio Chapecó 0,77 0,93 0,75 0,24 0,90 0,67 0,63
717S Rio Chapecó 0,50 1,00 0,70 0,35 0,72 0,84 0,62
Média 0,57 0,56 0,82 0,35 0,85 0,76 0,58
Desvio-padrão 0,17 0,22 0,13 0,19 0,08 0,15 0,04

132
Figura 2.5. Localização geográfica das 16 Unidades Amostrais consideradas como florestas
referência de naturalidade para a Floresta Mista com Araucária de Santa Catarina.
2.3.3. A NATURALIDADE QUANTIFICADA
Levando em consideração todos os remanescentes florestais considerados, o Índice de
naturalidade I (referência teórica) apresentou um valor médio de 0,47 com desvio-padrão de
0,09. De forma diferente da maioria dos indicadores, o Índice I apresentou uma distribuição
normal (p>0,05; Shapiro-Wilk), conforme a Figura 2.6 (A). O Índice de naturalidade II
(referência observada) apresentou um valor médio de 0,59 com desvio-padrão de 0,14;
diferentemente do Índice I, esse não apresentou uma distribuição normal (p<0,05; Shapiro-
Wilk), sendo esta assimétrica à esquerda com g=-0,30 (Figura 2.6, B). Apesar da aparente
diferença nas distribuições dos valores dos dois Índices, elas se mostraram equivalanentes de
acordo com o teste de Kolmogorov-Smirnov (D=0,30; p=0,67), adotando α=0,05.
Os valores dos Índices para cada uma das 145 Unidades Amostrais estão presentes na
Tabela A, que se encontra no apêndice desse manuscrito.
Ao analisarmos os valores de forma pontual, os do Índice II superaram os do Índice I,
pois as referências observadas são, obviamente, menos naturais que a referência teórica
(100% natural), diminuindo o valor da medida de distância. Essa tendência pôde ser
observada no gráfico de dispersão dos Índices (Figura 2.7), onde a maioria dos pontos
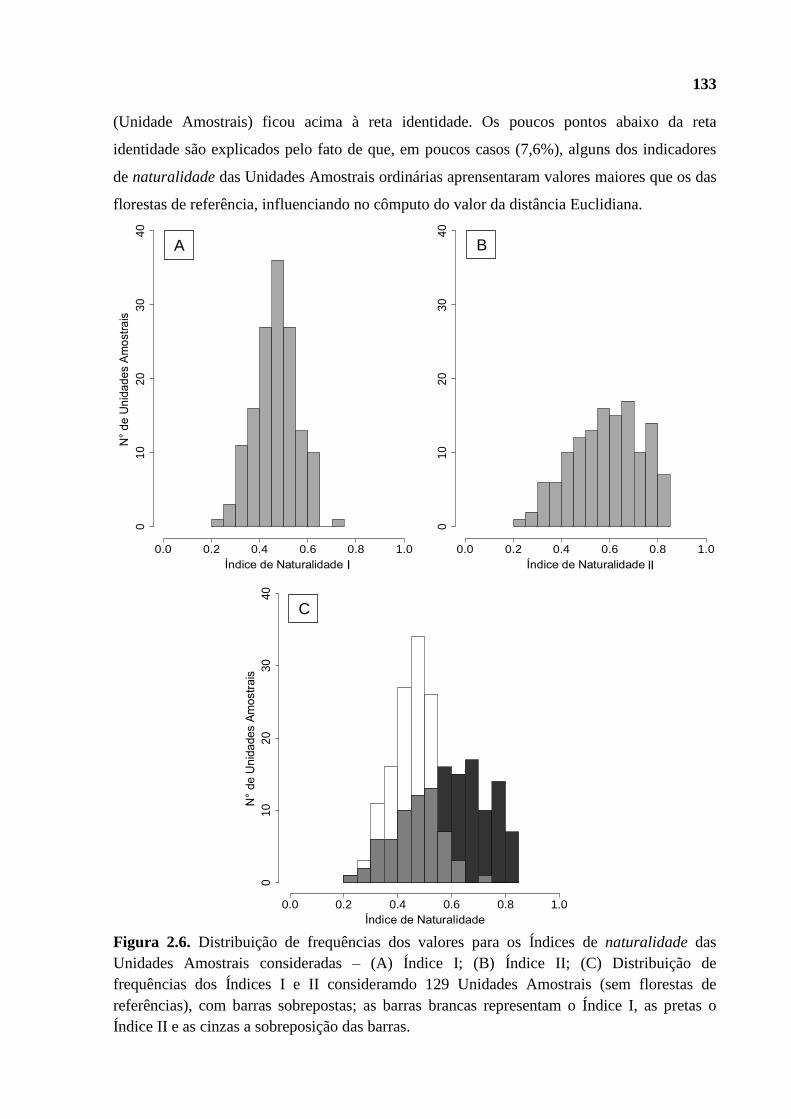
133
(Unidade Amostrais) ficou acima à reta identidade. Os poucos pontos abaixo da reta
identidade são explicados pelo fato de que, em poucos casos (7,6%), alguns dos indicadores
de naturalidade das Unidades Amostrais ordinárias aprensentaram valores maiores que os das
florestas de referência, influenciando no cômputo do valor da distância Euclidiana.
Figura 2.6. Distribuição de frequências dos valores para os Índices de naturalidade das
Unidades Amostrais consideradas – (A) Índice I; (B) Índice II; (C) Distribuição de
frequências dos Índices I e II consideramdo 129 Unidades Amostrais (sem florestas de
referências), com barras sobrepostas; as barras brancas representam o Índice I, as pretas o
Índice II e as cinzas a sobreposição das barras.
Índice de Naturalidade
N°
de
Un
ida
de
s A
mo
str
ais
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
0
Índice de Naturalidade
0.0 0.2 0.4 0.6 0.8 1.0
01
02
03
04
0
Índice de Naturalidade
N°
de U
nid
ades A
mostr
ais
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
A B
Índice de naturalidade I
N°
de U
nid
ades
Am
ost
rais
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
Índice de naturalidade II
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
Índice de naturalidade I
N°
de U
nid
ades
Am
ost
rais
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
Índice de naturalidade II
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
Índice de naturalidade I
N°
de U
nid
ades
Am
ost
rais
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
Índice de naturalidade II
0.0 0.2 0.4 0.6 0.8 1.0
010
20
30
40
C
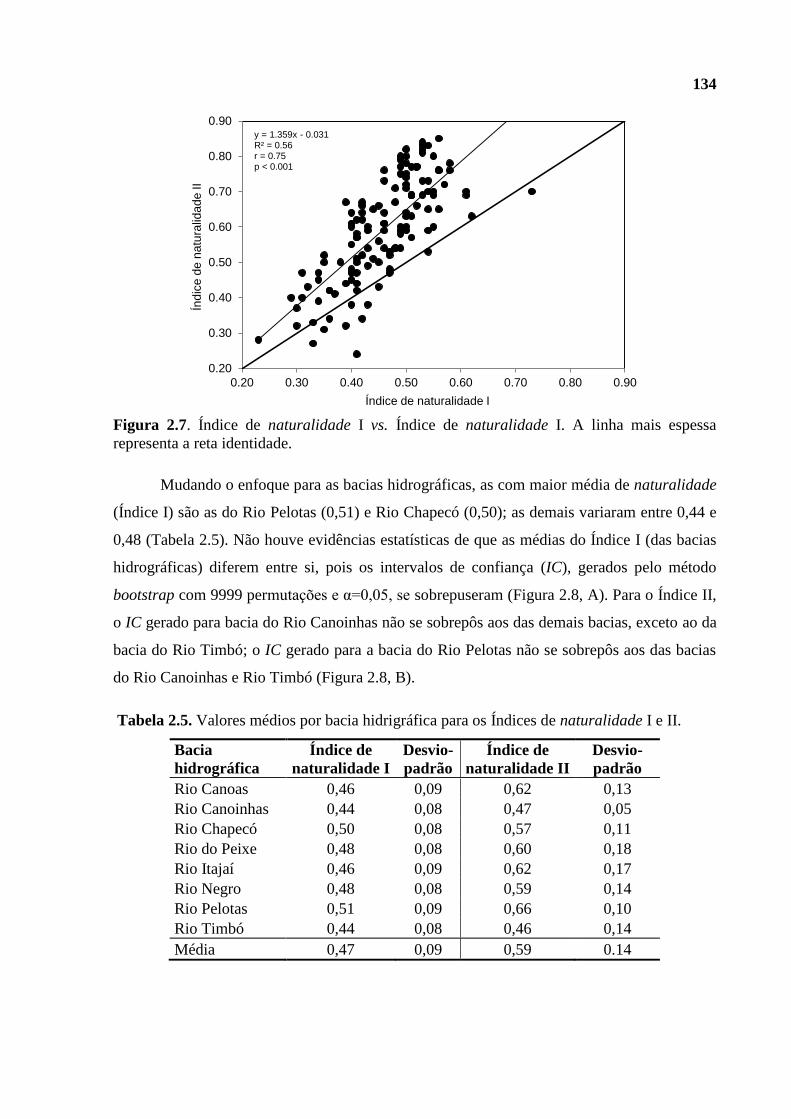
134
Figura 2.7. Índice de naturalidade I vs. Índice de naturalidade I. A linha mais espessa
representa a reta identidade.
Mudando o enfoque para as bacias hidrográficas, as com maior média de naturalidade
(Índice I) são as do Rio Pelotas (0,51) e Rio Chapecó (0,50); as demais variaram entre 0,44 e
0,48 (Tabela 2.5). Não houve evidências estatísticas de que as médias do Índice I (das bacias
hidrográficas) diferem entre si, pois os intervalos de confiança (IC), gerados pelo método
bootstrap com 9999 permutações e α=0,05, se sobrepuseram (Figura 2.8, A). Para o Índice II,
o IC gerado para bacia do Rio Canoinhas não se sobrepôs aos das demais bacias, exceto ao da
bacia do Rio Timbó; o IC gerado para a bacia do Rio Pelotas não se sobrepôs aos das bacias
do Rio Canoinhas e Rio Timbó (Figura 2.8, B).
Tabela 2.5. Valores médios por bacia hidrigráfica para os Índices de naturalidade I e II.
Bacia
hidrográfica
Índice de
naturalidade I
Desvio-
padrão
Índice de
naturalidade II
Desvio-
padrão
Rio Canoas 0,46 0,09 0,62 0,13
Rio Canoinhas 0,44 0,08 0,47 0,05
Rio Chapecó 0,50 0,08 0,57 0,11
Rio do Peixe 0,48 0,08 0,60 0,18
Rio Itajaí 0,46 0,09 0,62 0,17
Rio Negro 0,48 0,08 0,59 0,14
Rio Pelotas 0,51 0,09 0,66 0,10
Rio Timbó 0,44 0,08 0,46 0,14
Média 0,47 0,09 0,59 0.14
y = 1.359x - 0.031 R² = 0.56 r = 0.75 p < 0.001
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
0.20 0.30 0.40 0.50 0.60 0.70 0.80 0.90
Índ
ice
de
na
tura
lida
de
II
Índice de naturalidade I

135
Figura 2.8. Valores médios por bacia hidrográfica dos Índices de naturalidade I (A) e II (B).
As barras de erros expressam o intervalo de confiança gerado pelo método bootstrap com
9999 permutações e α=0,05.
2.3.3.1 Mapas temáticos de naturalidade
Dois mapas temáticos de naturalidade (Figuras 2.9 e 2.10) foram gerados, um para
cada Índice. Eles foram gerados pelo método de interpolação kriging, através do modelo
exponencial (semivariograma); a interpolação foi gerada utilizando 12 pontos amostrais mais
próximos.
Os mapas gerados para ambos os Índices concordam com os resultados já
aprensentados, onde as florestas menos naturais estão concentradas nas bacias do Rio
Canoinhas e Rio Timbó. Já as florestas mais naturais, considerando o Índice I estão
concentradas na bacia do Rio Pelotas e Chapecó. As florestais mais naturais de acordo com o
Índice II estão concentradas nas bacias do Rio Pelotas e Rio Canoas.
0.35
0.40
0.45
0.50
0.55
RioCanoas
RioCanoinhas
RioChapecó
Rio doPeixe
Rio Itajaí Rio Negro RioPelotas
Rio Timbó
Va
lor
mé
dio
do
s Ín
dic
es d
e
na
tura
lida
de
I ±
I.C
.
Índice de naturalidade I
A
0.30
0.35
0.40
0.45
0.50
0.55
0.60
0.65
0.70
0.75
RioCanoas
RioCanoinhas
RioChapecó
Rio doPeixe
Rio Itajaí Rio Negro RioPelotas
Rio Timbó
Va
lor
mé
dio
do
s Ín
dic
es d
e
na
tura
lida
de
II
± I
.C
Índice de naturalidade II
B
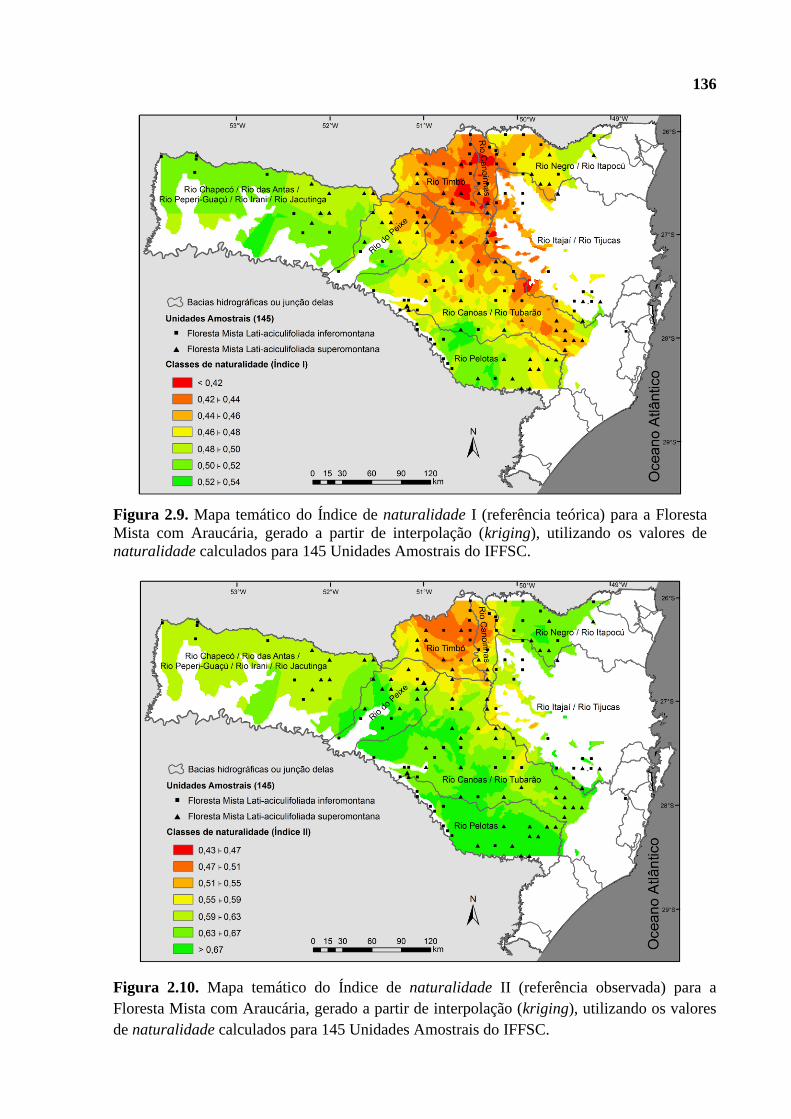
136
Figura 2.9. Mapa temático do Índice de naturalidade I (referência teórica) para a Floresta
Mista com Araucária, gerado a partir de interpolação (kriging), utilizando os valores de
naturalidade calculados para 145 Unidades Amostrais do IFFSC.
Figura 2.10. Mapa temático do Índice de naturalidade II (referência observada) para a
Floresta Mista com Araucária, gerado a partir de interpolação (kriging), utilizando os valores
de naturalidade calculados para 145 Unidades Amostrais do IFFSC.

137
2.4. DISCUSSÃO
2.4.1 OS ÍNDICES DE NATURALIDADE
A partir de dados coletados por IFNs, eleger indicadores quantitativos da naturalidade
de um sistema complexo como um ecossistema florestal constitui um grande desafio. Com
base na ilustração apresentada no presente estudo, alguns desses indicadores possuem
restrições no que diz respeito à metodologia de coleta (por exemplo, necromassa), outros em
relação à subjetividade das estimativas (por exemplo, estoque de volume). Além desses,
existem os que, à primeira vista, podem parecer indispensáveis (por exemplo, diversidade de
espécies), mas podem não estar relacionados à naturalidade. O fato de não possuírmos
florestas de referência (dados de referência) pré-estabelecidas dificulta a utilização de certos
indicadores (por exemplo, composição de espécies). Dos indicadores que restam, eles devem
satisfazer os pressupostos propostos pelos pesquisadores com experiência no tema
(McROBERTS et al., 2012; WINTER el al., 2010; WINTER, 2012).
Apesar da simplicidade da concepção dos Índices propostos, concretizamos uma
previsão de Chirici et al. (2011), os quais desenvoveram uma medida de naturalidade de
forma semelhante a nossa, adotando a distância Euclidiana, porém utilizando poucos
indicadores: “Even if the number and particular subindicators used change, the general
approach and the manner in which the data are aggregated to derive a complex naturalness
index have future potential for operational applications.”
Em termos operacionais, os procedimentos de cálculo dos indicadores e dos Índices
requeriram certo esforço computacional, entretanto, são totalmente passíveis de serem
calculados para dados de novos ciclos do IFFSC, com a finalidade de monitorar as florestas.
Os dados utilizados para a geração do indicador Métricas de paisagem não estão na lista
básica do IFN-Brasil (IFN-BRASIL, 2014), embora este possa ser facilmente calculado
mediante interpretação de imagens de sensoriamento remoto e das ferramentas de
geoprocessamento disponíveis atualmente.
Os resultados obtidos apontam que a medida de distância euclidiana sintetizou os
indicadores em um único valor de forma satisfatória, pois a distribuição normal dos valores
dos Índices I, com média em torno de 0,5, aponta um fato esperado – grande parte dos
remanescentes de Floresta Mista com Araucária de Santa Catarina apresenta naturalidade
mediana quando comparados a uma referência teórica (100% natural). Essa afirmação,
embora tenha traduzido o grau de conservação de uma maneira diferente dos métodos

138
habituais, concorda com conclusões já encontradas pelo IFFSC (SEVEGNANI et al., 2013;
VIBRANS et al., 2011). De fato, os resultados obtidos através dos Índices propostos remetem
à intensa exploração e alteração que outrora foram conduzidas (e continuam sendo
conduzidas) nessa fitofisionomia caracterizada pela Araucaria angustifolia (MÄHLER
JÚNIOR e LAROCCA, 2009; THOMÉ, 2010).
Segundo o Índice I, as florestas consideradas como referências mostraram-se com
naturalidade mediana (0,58), não muito acima da média geral (0,47). Esse resultado mostra
que os “melhores” remanescentes de hoje podem ser considerados descaracterizados e
simplificados quando comparados a uma referência 100% natural. Nessa perspectiva,
considerando agora o Índice II, observamos que em média os remanescentes ordinários são
mais próximos às florestas de referêcia observadas, como o esperado. Ambas as abordagens
de quantificação são úteis para o monitoramento e análises multitemporais, pois refletem dois
“padrões de qualidade” das florestas nativas.
2.4.2 UMA NOVA VISÃO SOBRE AS FLORESTAS NATIVAS
2.4.2.1 Observações acerca da naturalidade da Floresta Mista com Araucária
Das seis Unidades Amostrais complementares consideradas, três estão em unidades de
conservação (UC) - duas na FLONA de Três Barras e uma na Reserva Florestal
EMBRAPA/EPAGRI de Caçador. Somente uma destas Unidades Amostrais foi eleita como
floresta de referência, de acordo com nossa proposta de triagem. Nossa expectativa era de que
outras Unidades Amostrais em UC fossem eleitas como mais naturais. Algumas hipóteses são
levantadas: (i) que ocorreu exploração madeireira e de outros produtos nas UC, logo, essas
apresentam naturalidade não muito superior aos remanescentes fora de UC; (ii) as florestas
das UC estão em estágios de sucessão intermediários ou avançados; (iii) as UC ainda sofrem
exploração clandestina de madeira (e/ou outros produtos florestais) ou outras perturbações
antrópicas; (vi) os índices criados não representaram fielmente a naturalidade.
Chamamos a atenção para algumas bacias hidrográficas analisadas. As bacias dos Rios
Canoinhas e Timbó (planalto norte de Santa Catarina) apresentaram as menores médias para
os dois Índices de naturalidade. Isso mostra que estas bacias possuem florestas mais alteradas
e, assim, merecem atenção em políticas ligadas à conservação, restauração e manejo. Em
concordância com os resultados obtidos, o Projeto Microbacias 2, conduzido pela Fundação
do Meio Ambiente de Santa Catarina (FATMA), observou que essas bacias apresentam

139
florestas consideravelmente degradadas pela influência de atividades como a pecuária,
agricultura e silvicultura. Diante deste cenário, ações conservacionistas, fundamentadas em
um mecanismo de pagamento por serviços ambientais (PSA), foram formuladas e
implementadas. No ano de 2010 foram criados Corredores Ecológicos (CE) no estado: o CE
Chapecó e o CE Timbó (detalhes e histórico dos CE e do PSA em Alarcon et al. [2013]).
Levando em consideração certos fatos históricos, observamos que estes podem ter
contribuído para a atual situação concernente à menor naturalidade das florestas do planalto
norte e de partes da bacia do Rio Canoas. Os “tropeiros” conduziam as manadas de gado e
mulas dos campos do Rio Grande do Sul para o comércio em Sorocaba (THOMÉ, 2012). O
principal caminho das tropas passava pela região do planalto norte de Santa Catarina, assim
como nos campos da região de Lages (Figura 2.11). Em consequência da atividade, realizada
por pelo menos 200 anos, Thomé (2012) afirma:
“Nas invernadas, durante os descansos das tropas, a paisagem natural
do Espaço Livre do Contestado reteve muitos tropeiros paulistas e
paranaenses, que aqui se estabeleceram ao longo dos caminhos, os
pousos foram dando origem a currais, fazendas e povoados”.
“Fixando-se, promoviam o aparecimento de núcleos populacionais e,
assim, no decorrer do tempo, fizeram surgir as primeiras povoações,
mais tarde vilas, no Espaço Livre do Contestado. [...] Muitos dos ex-
tropeiros, agora novos fazendeiros, seguindo tradições familiares,
lançaram-se à criação de gado bovino, oportunizando emprego para
outros tipos de homens: o peão, especializado nas lidas campeiras com
o gado vacum; o mateiro, profundo conhecedor das matas e
explorador dos ervais; e, o roceiro, o plantador de cereais [...]”
A construção da ferrovia que liga a região Sudeste a Sul (do Brasil), de Itararé (SP) até
Santa Maria (RS) (Figura 2.11), entre os anos de 1907 e 1910 e.c., desempenhou um papel
fundamental na colonização e exploração das florestas nativas de Santa Catarina. A empresa
Southern Brazil Lumber and Colonization Company, conhecida popularmente como Lumber,
subsidiou a Brazil Railway Company (construtora da ferrovia) e recebeu uma concessão para
explorar a madeira (e a erva-mate) dos terrenos que margeavam a linha principal da estrada de
ferro. Esta concessão, juntamente com a inovadora tecnologia de exploração e processamento
da madeira, tornou a Lumber a maior madeireira da América do Sul (GOULARTI FIHLO,

140
2009; TOMPOROSKI, 2006). Goularti Filho (2009) acrescenta que não só a Lumber extraia a
madeira das florestas catarinenses:
“As primeiras pequenas serrarias, de propriedade dos colonos
imigrantes, que inicialmente se instalaram como comerciantes,
transformaram-se mais tarde em industriais”.
A construção da linha São Francisco que segue até Porto União (Ramal Porto União -
São Francisco do Sul), construída entre 1906 e 1917 e.c., e sua integração com Tronco Sul da
Rede Ferroviária Federal Sociedade Anônima (Figura 2.11), que segue de Mafra até Lages,
construída entre 1961 a 1968, também acelerou o processo de crescimento
econômico/populacional e a exploração dos recursos naturais (GOULARTI FIHLO, 2009).
Figura 2.11. Representação das ferrovias construídas no início e meados do século XX e do
caminho dos tropeiros. Onde, tracejado amarelo: ferrovia São Paulo-Rio Grande do Sul;
tracejado vermelho: Ramal Porto União-São Francisco e Tronco Sul; tracejado azul: caminho
dos tropeiros.
Outro fator pode estar ligado à naturalidade; existem evidências de que a quantidade
(e o tipo) de rodovias estão relacionados ao grau de alteração das florestas nativas, inclusive
Lages
Vacaria
Sorocaba
Lapa
Três Barras
Itararé
Santa Maria
S. Fco. do Sul

141
em UC (FREITAS et al., 2013). Duas importantes rodovias passam pelo planalto norte de
Santa Catarina: a BR 116 e a BR 280. Três rodovias consideravelmente trafegadas passam
pela bacia do Rio Canoas: a BR 470, a BR 282 e a BR 116. Segundo Goularti Filho (2009), a
partir da década de 1960 (já na “era rodoviária”), a pavimentação dessas rodovias auxiliou a
distribuição dos produtos provenientes da agricultura e da pecuária para diversas regiões (e
para os portos) do estado.
Atividades (em ascenção) na bacia do Rio Canoas devem ser consideradas e,
possivelmente, são inversamente correlacionadas à naturalidade dos remanescentes de
Floresta Mista com Araucária: a agricultura mecanizada de grãos na primavera/verão aliada à
pecuária no outono/inverno. Estas atividades estão tornando-se lucrativas e vem substituindo
os investimentos em plantios florestais de espécies do gênero Pinus e Eucalyptus, que outrora
foram incentivados pelo governo do país (BREPOHL, 1980). Não obstante a este fato
(esfriamento do setor florestal), os plantios florestais ainda cobrem grande porção da bacia,
devido à alta demanda das empresas de papel e celulose, aliado a alta taxa de incremento (em
volume de madeira) das espécies de Pinus e Eucalyptus na região.
Resultados que despertam atenção são, também, os referentes aos indicadores Árvores
grossas e IQH. Ambos apresentaram valores médios baixos (~0,30), mostrando que grande
parte dos remanescentes avaliados possui poucas árvores velhas, o que é, provavelmente,
consequência da exploração madeireira intensiva (décadas passadas) e seletiva (dias atuais).
Além dessa “marca” deixada pela exploração, observamos que muitas atividades antrópicas
estão atualmente presentes nas florestas. Apesar de não podermos quantificar precisamente as
influências das atividades antrópicas na vegetação, ao adotarmos uma posição conservadora
em relação a tais atividades, alertamos sobre esse fato.
Como procederemos diante destas situações? Sabemos da burocracia do sistema
político e legislativo do Brasil, sendo que informações preciosas sobre as florestas, as quais
são levantadas com recursos públicos (em sua maioria), retornam vagarosamente de forma
concreta à sociedade e à própria floresta. Entretanto, respostas a essa indagação foram
apresentadas nas Diretrizes para a Política Florestal Catarinense9, a qual propõe diversas
medidas para conservação e utilização dos recursos florestais nativos de Santa Catarina, como
o incentivo ao manejo de florestas secundárias e a implantação de um serviço de extensão
florestal. Embora tal documento tenha sido elaborado antes da publicação da abordagem do
9 Dispoível em:
<http://ciram.epagri.sc.gov.br/ciram_arquivos/arquivos/iff/pdf/diretrizes_gt_pefsc_dez_2013.
pdf>

142
conceito de naturalidade para a Floresta Mista com Araucária catarinense, acreditamos que
esta irá contribuir para a manutenção das propostas e na parte de monitoramento
multitemporal dos remanescentes florestais.
2.4.2.2 Alternativa para a análise de remanescentes florestais
É certo que o conceito de naturalidade traz uma nova maneira de lidar com o espectro
ecológico e ambiental concernente às florestas nativas do Brasil. Ela pode ser uma ferramenta
alternativa em situações que, por vezes, certos métodos ou abordagens podem não gerar
resultados satisfatórios. Inspirados pelos apontamentos de Magurram e Queiroz (2010),
apresentamos três situações onde o conceito pode contribuir ou vir a ser um método
alternativo:
(i) A classificação de remanescentes florestais em estágios de sucessão é um dos paradigmas
atuais acerca das florestas nativas (SIMINSKI et al., 2013). Existem estudos que trazem novas
abordagens para a classificação (ZANINI et al., 2014), porém, o problema está na aplicação
de resoluções (como a do CONAMA) para classificação de estágios de sucessão.
Empiricamente, sabemos que os parâmetros sugeridos por essas resoluções podem não ser
adequados para classificar coerentemente remanescentes florestais em estágios de sucessão
ecológica. Existe a possibilidade, por exemplo, de: (a) criar informações de referência de
naturalidade e relacioná-los com estágios de sucessão ecológica; (b) criar classes de
naturalidade compatíveis com os estágios de sucessão ecológica; (c) usar uma medida de
distância (como a utilizada nesse estudo) entre dada vegetação que se deseja classificar em
um estágio de sucessão até uma vegetação climácica – assim, criariam-se classes de distância,
as quais seriam relacionadas com estágios de sucessão ecológica.
(ii) Mostramos evidências de que a diversidade de espécies pode gerar resultados precipitados
quando é adotada como descritor do estado de conservação de remanescentes; Maçaneiro et
al. (manuscrito em preparação) e Imai et al. (2014) chegaram à conclusões semelhantes. A
utilização deste parâmetro não deveria ser aplicada sem ser justificada. Portanto, é prudente
utilizar métodos mais abrangentes quando o objetivo é retratar o estado de conservação de
florestas que possuem espécies dominantes.
(iii) A utilização do conceito de naturalidade tem serventia e potencial para auxiliar na
compreensão da interação homem-natureza, pelo fato de poder incluir o fator hemerobia

143
(WINTER, 2012). As visões acerca dessa interação estão ligadas a uma perspectiva
biocêntrica e antropocêntrica (HUNTER et al., 2014). Na ilustração do presente estudo
concatenaram-se as duas perspectivas. Como o conceito de hemerobia pode ser adotado em
diversas escalas espaciais (BARTHA et al., 2006), existe a possibilidade da criação de índices
(ou estimadores) específicos, respondendo a demanda atual de informações e respostas sobre
as florestas nativas.
2.4.3 PERSPECTIVAS PARA O USO DO CONCEITO DE NATURALIDADE
A naturalidade é um atributo considerado por muitos IFNs da Europa como descritor
do estado de conservação, resiliência e estabilidade de ecossistemas florestais (CHIRICI et al.
2011; EEA, 2014). Diante da novidade do conceito no Brasil, e também por Santa Catarina
ser o primeiro estado a realizar o primeiro ciclo de levantamento de dados dentro do programa
IFN-Brasil, esperamos que nossa abordagem ecológica e filosófica do conceito (adaptada para
a realidade histórica, social e ambiental do Brasil) abra novas perspectivas para a análise
integrada de dados dendrométricos e de perturbações antrópicas.
Tomando por base o exemplo europeu, o qual vem desenvolvendo metodologias de
acesso à naturalidade a partir de dados de IFNs e de sensores remotos (EEA, 2014), existe a
possibilidade de nosso estudo ser um piloto, para que futuramente, o conceito seja integrado
ao programa IFN-Brasil. Apostamos na utilização da arborgem proposta em variadas escalas
espaciais e para outras fitofisionomias do Brasil, utilizando dados do IFN-Brasil, com o
intuito de aprofundar as análises e diagnósticos sobre o estado de conservação das florestas.
Uma das vantagens dessa abordagem é a possibilidade da harmonização dos Índices
(ou estimadores) gerados, objetivando comparações entre estados, dentro dum programa de
IFN, por exemplo. A compatibilidade entre definições, conceitos e estimativas geradas por um
IFN pode ser atingida pela harmonização. O processo de harmonização busca por um acordo
sobre como os dados podem ser convertidos para atender a uma definição harmonizada,
tornando possível a geração de estimativas harmonizadas de variáveis dendrométricas, por
exemplo (EEA, 2014; KÖHL et al., 2000; McROBERTS et al., 2009, 2012).
Outro exemplo de abordagem muitidimensional quantitativa é o Índice de
Sustentabilidade Florestal (Forest Sustainability Index) criado pelo Serviço Florestal da
Coréia do Sul (KOREA FOREST SERVICE, 2009). Esse índice tem um caráter quantitativo e
funciona como um estimador (global) da sustentabilidade florestal, levando em conta aspectos
físicos, ecológicos, sociais e econômicos, em nível local e nacional. Dezenove indicadores são

144
utilizados em conjunto para o cálculo do valor final. Certas ações contidas na política florestal
da Coréia do Sul estão voltadas para recuperar florestas que apresentam valores baixos para o
Índice de Sustentabilidade Florestal. Quando ocorre um declínio nos valores dos índices, a
politica florestal também prevê o desenvolvimento de contraplanos que visam recuperar o
índice.
Na área científica, relacionada às florestas, esperamos contribuir com a vertente de
pesquisadores que se dedicam ao conceito (naturalidade), pois, em nosso conhecimento, não
existem estudos referentes à aplicação do conceito em florestas subtropicais com propriedades
ecológicas muito diferentes das florestas temperadas e boreais do hemisfério Norte.
2.5. CONCLUSÕES
A partir da ilustração apresentada neste capítulo, cinco conclusões podem ser tiradas:
(i) os Índices de naturalidade apresentaram resultados coerentes e que corroboraram o
conhecimento prévio sobre o estado de conservação da Floresta Mista com Araucária de Santa
Catarina; (ii) a maior parte dos remanescentes avaliados possui naturalidade média, levando
em consideração a referência teórica (100% natural); (iii) os dois Índices desenvolvidos
trazem duas abordagens de quantificação da naturalidade (referências teóricas e observadas),
as quais podem ser úteis para questões relativas ao monitoramento de remanescentes florestais
em larga escala; (iv) a abordagem proposta para a quantificação da naturalidade tem potencial
para ser aplicada no contexto do IFN-Brasil; (v) existem possibilidades da criação de Índices
de naturalidade harmonizados para fitofisinomias inteiras ou regiões específicas do Brasil.
2.6. REFERÊNCIAS BIBLIOGRÁFICAS
ALARCON, G.G.; DA-RÉ, M.A.; FUKAHORI, S.T.I. Análise de instrumentos de mercado
na gestão do corredor ecológico Chapecó, Santa Catarina, Brasil. Sustentabilidade em
Debate, v.4, n.1, p.117-138, 2013.
BARTHA, D.; ÓDOR, P.;HORVÁTH, T.; TÍMÁR, G.; KENDERES, K.; STANDOVÁR, T.;
BÖLÖNI, J.; SZMORAD, F.; BODONCZI, L.; ASZALÓS, R. Relationship of Tree Stand
Heterogeneity and Forest Naturalness. Acta Silvatica & Lingaria Hungarica, v.2, p.7-22,
2006.
BREPOHL, D. O reflorestamento com incentivos fiscais no estado do Paraná. Floresta, v.11,
n.1, p.62-66, 1980.
CHIRICI, G.; MCROBERTS, R.E.; WINTER, S.; BERTINI, R.; BRÄNDLI, U.; ASENSIO,
I.A.; BASTRUP-BIRK, A.; RONDEUX, J.; BARSOUM, N.; MARCHETTI, M. National

145
Forest Inventory Contributions to Forest Biodiversity Monitoring. Forest Science, v.58, n.3,
2012.
CHIRICI, G.; WINTER, S.; McROBERTS, R.E. (Eds.). National forest inventories:
contributions to forest biodiversity assessments. Springer, 2011.
EEA. European Environment Agency. Developing a forest naturalness indicator for Europe -
Concept and methodology for a high nature value (HNV) forest indicator. EEA Technical
report, n.13, 2014.
FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET, A.M.; DUTRA, T.L.; BACKES, A;
GRANADE, G. (Eds.). Floresta com Araucária: Ecologia, Conservação e Desenvolvimento
Sustentável. Ribeirão Preto: Holos, 2009.
FORTIN, M.J.; DALE, M.R.T. Spatial analysis: a guide for ecologists. Cambridge:
University Press, 2005.
FREITAS, S.R.; SOUSA, C.O.M.; BOSCOLO, D.; METZGER, J.P. How are native
vegetation and reserves affected by differente road types in a southeastern Brazilian state?
Oecologia Australis, v.17, n.4, p.447-458, 2013.
GOULARTI FILHO, A. A Estrada de Ferro São Paulo-Rio Grande na formação econômica
regional em Santa Catarina. Geosul, v.24, n.48, p.103-128, 2009.
HUNTER JR., M.L.; REDFORD, K.H.; LINDENMAYER, D.B. The complementary niches
of anthropocentric and biocentric conservationists. Conservation Biology, v.28, n.3, p.641–
645, 2014.
IFN-BRASIL. Manual de Campo - Procedimentos para coleta de dados biofísicos e
socioambientais. Disponível em: <https://www.fao.org.br/download/annex4.pdf>. Acessado
em: 16 de Setembro de 2014.
IMAI, N.; TANAKA, A.; SAMEJIMA, H.; SUGAU, J.B.; PEREIRA, J.T.; TITIN, J.;
KURNIAWAN, Y.; KITAYAMA, K. Tree community composition as an indicator in
biodiversity monitoring of REDD+. Forest Ecology and Management, v.313, p.169–179,
2014.
KÖHL, M.; TRAUB, B.; PÄIVINEN, R. Harmonization and standardization in multi-national
environmental statistics - Mission impossible? Environmental Monitoring and Assessment,
v.63, p.361–380, 2000.
KOREA FOREST SERVICE. National Report on Sustainable Forest Management in Korea
2009. Disponível em: <http://www.rinya.maff.go.jp/j/kaigai/pdf/2009p_4_k.pdf>. Acessado
em: 02 de Outubro de 2014.
LIIRA, J.; SEPP, T. Indicators of structural and habitat natural quality in boreo-nemoral
forests along the management gradient. Annales Botanici Fennici, v.46, p.308–325, 2009.
MACHADO, A. An index of naturalness. Journal for Nature Conservation, v.12, p.95-110,
2004.

146
MAGURRAN, A.E.; QUEIROZ, H. Evaluating tropical biodiversity: Do we need a more
refined approach? Biotropica, v.42, p.537-539, 2010.
MÄHLER JUNIOR, J.K.F.; LAROCCA, J.F. Fitofisionomias, desmatamento e fragmentação
da Floresta com Araucária. In: FONSECA, C.R.; SOUZA, A.F.; LEAL-ZANCHET, A.M.;
DUTRA, T.L.; BACKES, A; GRANADE G. (Eds.). Floresta com Araucária: Ecologia,
Conservação e Desenvolvimento Sustentável. Holos, Ribeirão Preto, 2009, p. 243-252.
MARCHETTI, M. Monitoring and Indicators of Forest Biodiversity in Europe - From ideas to
operationality. EFI Proceedings, n.51, 2004.
McROBERTS, R.E.; TOMPPO, E.; SCHADAUER, K.; VIDAL, C.; STÅHL, G.; CHIRICI,
G.; LANZ, A.; CIENCIALA, E.; WINTER, S.; SMITH, B. Harmonizing National Forest
Inventories Journal of Forestry, p.179–187, 2009.
McROBERTS, R.E.; WINTER, S.; CHIRICI, G.; LAPOINT, E. Assessing Forest
Naturalness. Forest Science, v.58, p.294-309, 2012.
OLIVEIRA-FILHO, A.T. Classificação das fitofisionomias da América Do Sul cisandina
tropical e subtropical: Proposta de um novo sistema – prático e flexível – ou uma injeção a
mais de caos? Rodriguésia, v.60, p. 237-258, 2009.
SEVEGNANI, L.; VIBRANS, A.C.; GASPER, A.L. Considerações finais sobre a Floresta
Ombrófila Mista em Santa Catarina. In: VIBRANS, A.C.; SEVEGNANI, L.; GASPER, A.L.;
LINGNER, D.V. (Eds.) Inventário Florístico Florestal de Santa Catarina: Floresta
Ombrófila Mista. Blumenau: Edifurb, 2013, p.275-278.
SIMINSKI, A.; FANTINI, A.C.; REIS, M.S. Classificação da vegetação secundária em
estágios de regeneração da Mata Atlântica em Santa Catarina. Ciência Florestal, v.23, p.369-
378, 2013.
TABARELLI, M.; AGUIAR, A.V.; RIBEIRO, M.C.; METZGER, J.P.; PERES, C.A.
Prospects for biodiversity conservation in the Atlantic Forest: lessons from aging human-
modified landscapes. Biological Conservation, v.143, p. 2328-2340, 2010.
THOMÉ, N. O ciclo da madeira no Contestado: Um retrato do século XX. Curitiba: Gestão
em comunicação, 2010.
THOMÉ, N. Caminhos de tropeiros nos séculos XVIII e XIX como fatores pioneiros de
desbravamento do Contestado. Desenvolvimento Regional em Debate, v.2, n. 1, 2012.
TOMPOROSKI, A. A. Um estudo acerca dos trabalhadores da Southern Brazil Lumber
and Colonization Company e sua atuação no planalto norte de Santa Catarina, 1910 –
1929. Dissertação (Mestrado em História) Universidade Federal de Santa Catarina, 2009.
VIBRANS, A. C.; SEVEGNANI, L.; LINGNER, D.V.; GASPER, A.L; SABBAGH, S.
Inventário Florístico Florestal de Santa Catarina (IFFSC): aspectos metodológicos e
operacionais. Pesquisa Florestal Brasileira, v.30, n.64, p.291-302, 2010.

147
VIBRANS; A.C.; SEVEGNANI; L.; UHLMANN; A.; SCHORN; L.A.; SOBRAL; M.;
GASPER; A.L.; LINGNER; D.V.; BROGNI; E.; KLEMTZ; G.; GODOY; M.B.; VERDI; M.
Structure of Mixed Ombrophylous Forests with Araucaria angustifolia (Araucariaceae) under
external stress in southern Brazil. Revista de Biología Tropical, v.9, n.3, p.1371-1387, 2011.
WINTER, S. Forest naturalness assesment as a component of biodiversity monitoring and
conservation management. Forestry, v.85, n.2, p.293-304, 2012.
WINTER, S.; FISCHER, H.S.; FISCHER, A. Relative Quantitative Reference Approach for
Naturalness Assessment. Forest Ecology Management, v. 259, p. 1624–1632, 2010.
ZANINI, K.J.; BERGAMIN, R.S.; MACHADO, R.E.; PILLAR, V.D.; MÜLLER, S.C.
Atlantic rain forest recovery: successional drivers of floristic and structural patterns of
secondary forest in Southern Brazil. Journal of Vegetation Science, v.25, n.4, p.1056–1068,
2014.
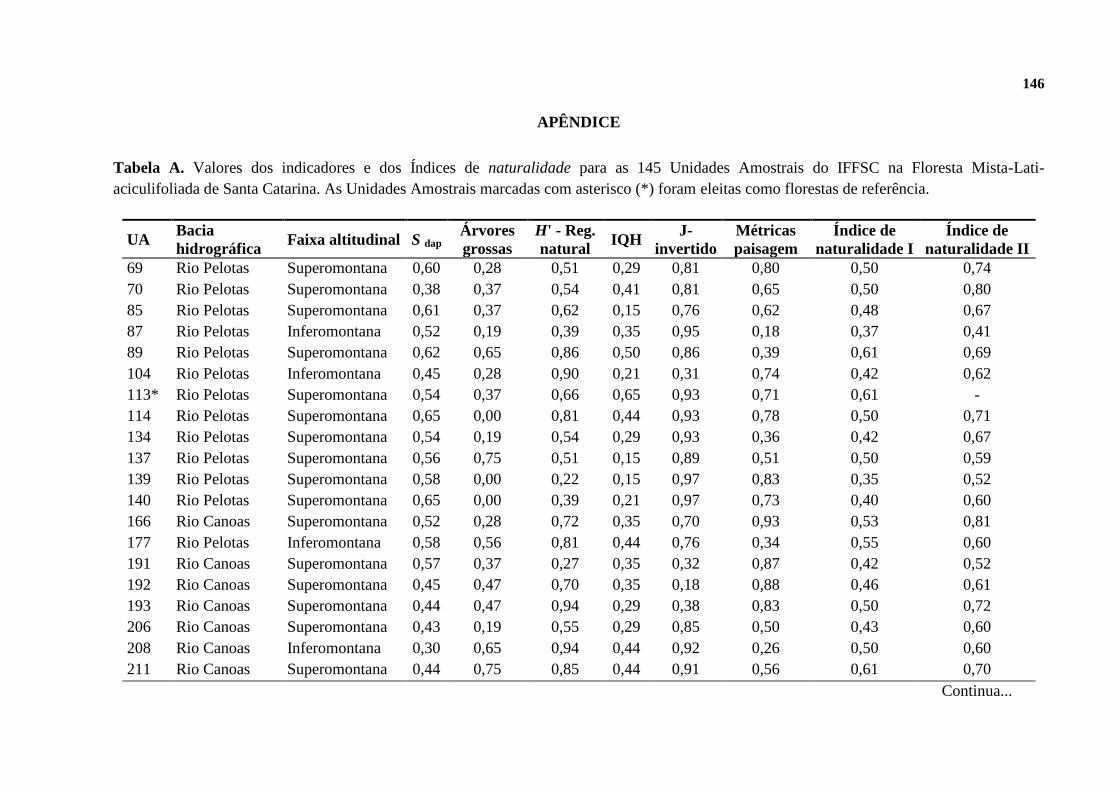
146
APÊNDICE
Tabela A. Valores dos indicadores e dos Índices de naturalidade para as 145 Unidades Amostrais do IFFSC na Floresta Mista-Lati-
aciculifoliada de Santa Catarina. As Unidades Amostrais marcadas com asterisco (*) foram eleitas como florestas de referência.
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
69 Rio Pelotas Superomontana 0,60 0,28 0,51 0,29 0,81 0,80 0,50 0,74
70 Rio Pelotas Superomontana 0,38 0,37 0,54 0,41 0,81 0,65 0,50 0,80
85 Rio Pelotas Superomontana 0,61 0,37 0,62 0,15 0,76 0,62 0,48 0,67
87 Rio Pelotas Inferomontana 0,52 0,19 0,39 0,35 0,95 0,18 0,37 0,41
89 Rio Pelotas Superomontana 0,62 0,65 0,86 0,50 0,86 0,39 0,61 0,69
104 Rio Pelotas Inferomontana 0,45 0,28 0,90 0,21 0,31 0,74 0,42 0,62
113* Rio Pelotas Superomontana 0,54 0,37 0,66 0,65 0,93 0,71 0,61 -
114 Rio Pelotas Superomontana 0,65 0,00 0,81 0,44 0,93 0,78 0,50 0,71
134 Rio Pelotas Superomontana 0,54 0,19 0,54 0,29 0,93 0,36 0,42 0,67
137 Rio Pelotas Superomontana 0,56 0,75 0,51 0,15 0,89 0,51 0,50 0,59
139 Rio Pelotas Superomontana 0,58 0,00 0,22 0,15 0,97 0,83 0,35 0,52
140 Rio Pelotas Superomontana 0,65 0,00 0,39 0,21 0,97 0,73 0,40 0,60
166 Rio Canoas Superomontana 0,52 0,28 0,72 0,35 0,70 0,93 0,53 0,81
177 Rio Pelotas Inferomontana 0,58 0,56 0,81 0,44 0,76 0,34 0,55 0,60
191 Rio Canoas Superomontana 0,57 0,37 0,27 0,35 0,32 0,87 0,42 0,52
192 Rio Canoas Superomontana 0,45 0,47 0,70 0,35 0,18 0,88 0,46 0,61
193 Rio Canoas Superomontana 0,44 0,47 0,94 0,29 0,38 0,83 0,50 0,72
206 Rio Canoas Superomontana 0,43 0,19 0,55 0,29 0,85 0,50 0,43 0,60
208 Rio Canoas Inferomontana 0,30 0,65 0,94 0,44 0,92 0,26 0,50 0,60
211 Rio Canoas Superomontana 0,44 0,75 0,85 0,44 0,91 0,56 0,61 0,70
Continua...
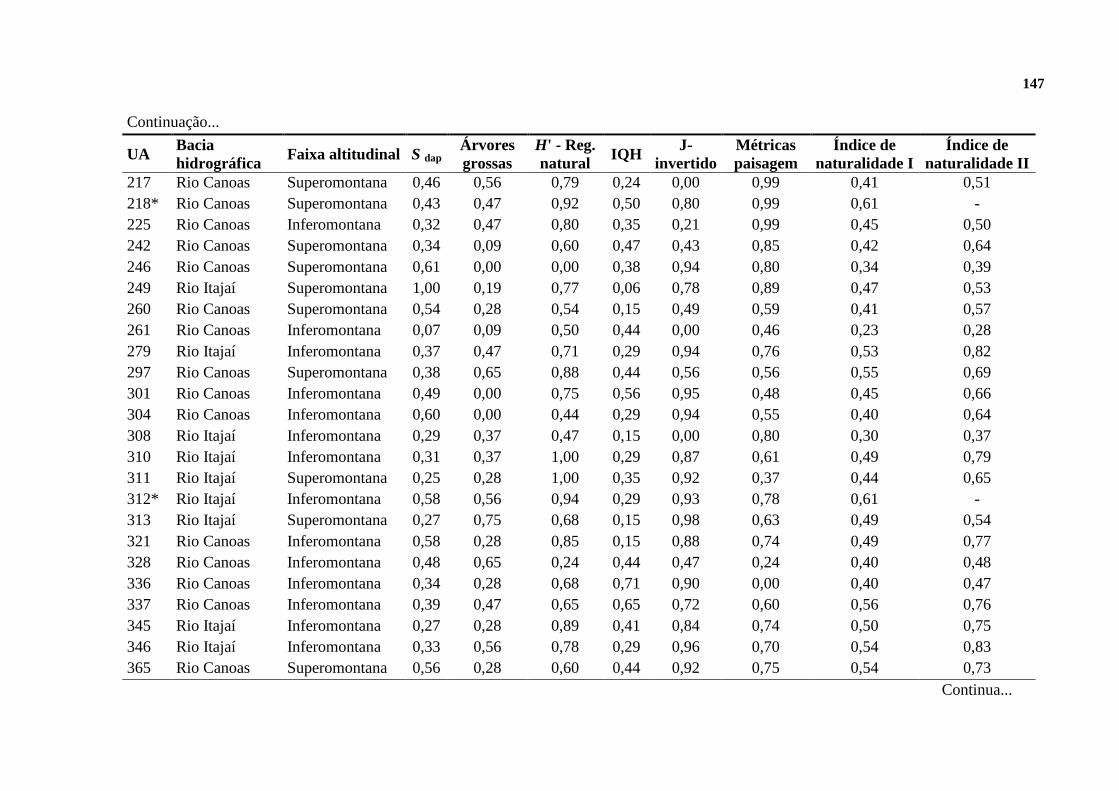
147
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
217 Rio Canoas Superomontana 0,46 0,56 0,79 0,24 0,00 0,99 0,41 0,51
218* Rio Canoas Superomontana 0,43 0,47 0,92 0,50 0,80 0,99 0,61 -
225 Rio Canoas Inferomontana 0,32 0,47 0,80 0,35 0,21 0,99 0,45 0,50
242 Rio Canoas Superomontana 0,34 0,09 0,60 0,47 0,43 0,85 0,42 0,64
246 Rio Canoas Superomontana 0,61 0,00 0,00 0,38 0,94 0,80 0,34 0,39
249 Rio Itajaí Superomontana 1,00 0,19 0,77 0,06 0,78 0,89 0,47 0,53
260 Rio Canoas Superomontana 0,54 0,28 0,54 0,15 0,49 0,59 0,41 0,57
261 Rio Canoas Inferomontana 0,07 0,09 0,50 0,44 0,00 0,46 0,23 0,28
279 Rio Itajaí Inferomontana 0,37 0,47 0,71 0,29 0,94 0,76 0,53 0,82
297 Rio Canoas Superomontana 0,38 0,65 0,88 0,44 0,56 0,56 0,55 0,69
301 Rio Canoas Inferomontana 0,49 0,00 0,75 0,56 0,95 0,48 0,45 0,66
304 Rio Canoas Inferomontana 0,60 0,00 0,44 0,29 0,94 0,55 0,40 0,64
308 Rio Itajaí Inferomontana 0,29 0,37 0,47 0,15 0,00 0,80 0,30 0,37
310 Rio Itajaí Inferomontana 0,31 0,37 1,00 0,29 0,87 0,61 0,49 0,79
311 Rio Itajaí Superomontana 0,25 0,28 1,00 0,35 0,92 0,37 0,44 0,65
312* Rio Itajaí Inferomontana 0,58 0,56 0,94 0,29 0,93 0,78 0,61 -
313 Rio Itajaí Superomontana 0,27 0,75 0,68 0,15 0,98 0,63 0,49 0,54
321 Rio Canoas Inferomontana 0,58 0,28 0,85 0,15 0,88 0,74 0,49 0,77
328 Rio Canoas Inferomontana 0,48 0,65 0,24 0,44 0,47 0,24 0,40 0,48
336 Rio Canoas Inferomontana 0,34 0,28 0,68 0,71 0,90 0,00 0,40 0,47
337 Rio Canoas Inferomontana 0,39 0,47 0,65 0,65 0,72 0,60 0,56 0,76
345 Rio Itajaí Inferomontana 0,27 0,28 0,89 0,41 0,84 0,74 0,50 0,75
346 Rio Itajaí Inferomontana 0,33 0,56 0,78 0,29 0,96 0,70 0,54 0,83
365 Rio Canoas Superomontana 0,56 0,28 0,60 0,44 0,92 0,75 0,54 0,73
Continua...
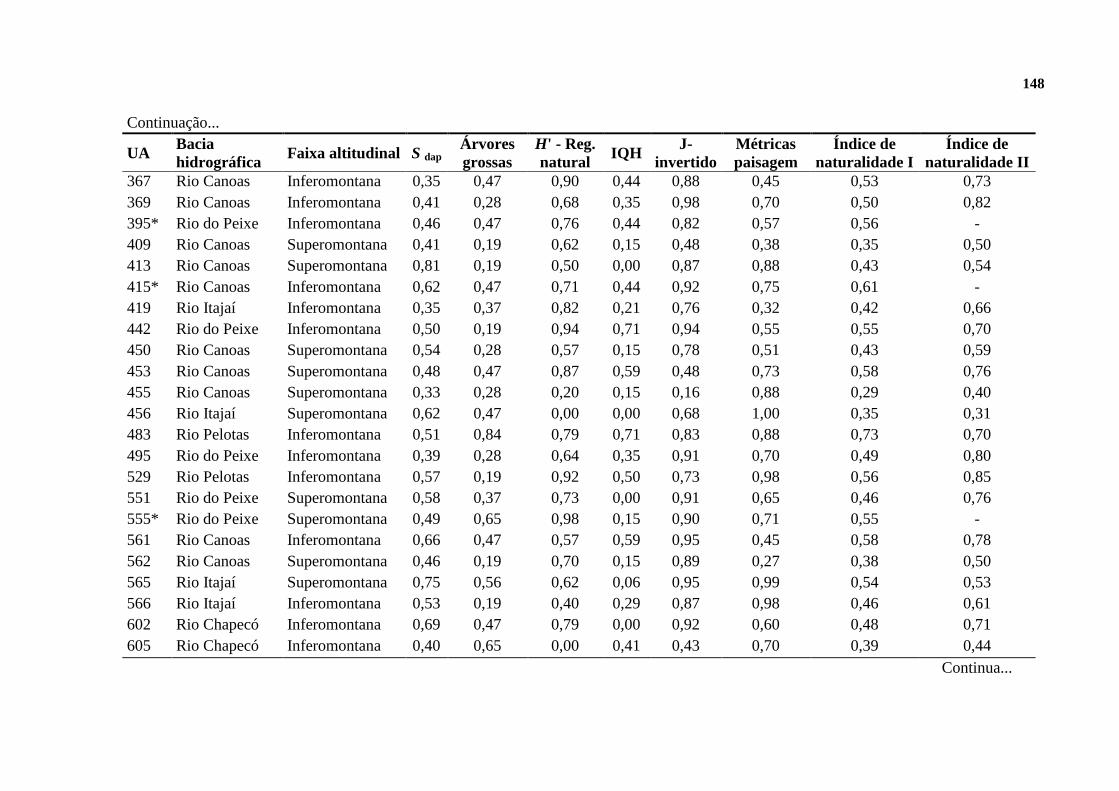
148
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
367 Rio Canoas Inferomontana 0,35 0,47 0,90 0,44 0,88 0,45 0,53 0,73
369 Rio Canoas Inferomontana 0,41 0,28 0,68 0,35 0,98 0,70 0,50 0,82
395* Rio do Peixe Inferomontana 0,46 0,47 0,76 0,44 0,82 0,57 0,56 -
409 Rio Canoas Superomontana 0,41 0,19 0,62 0,15 0,48 0,38 0,35 0,50
413 Rio Canoas Superomontana 0,81 0,19 0,50 0,00 0,87 0,88 0,43 0,54
415* Rio Canoas Inferomontana 0,62 0,47 0,71 0,44 0,92 0,75 0,61 -
419 Rio Itajaí Inferomontana 0,35 0,37 0,82 0,21 0,76 0,32 0,42 0,66
442 Rio do Peixe Inferomontana 0,50 0,19 0,94 0,71 0,94 0,55 0,55 0,70
450 Rio Canoas Superomontana 0,54 0,28 0,57 0,15 0,78 0,51 0,43 0,59
453 Rio Canoas Superomontana 0,48 0,47 0,87 0,59 0,48 0,73 0,58 0,76
455 Rio Canoas Superomontana 0,33 0,28 0,20 0,15 0,16 0,88 0,29 0,40
456 Rio Itajaí Superomontana 0,62 0,47 0,00 0,00 0,68 1,00 0,35 0,31
483 Rio Pelotas Inferomontana 0,51 0,84 0,79 0,71 0,83 0,88 0,73 0,70
495 Rio do Peixe Inferomontana 0,39 0,28 0,64 0,35 0,91 0,70 0,49 0,80
529 Rio Pelotas Inferomontana 0,57 0,19 0,92 0,50 0,73 0,98 0,56 0,85
551 Rio do Peixe Superomontana 0,58 0,37 0,73 0,00 0,91 0,65 0,46 0,76
555* Rio do Peixe Superomontana 0,49 0,65 0,98 0,15 0,90 0,71 0,55 -
561 Rio Canoas Inferomontana 0,66 0,47 0,57 0,59 0,95 0,45 0,58 0,78
562 Rio Canoas Superomontana 0,46 0,19 0,70 0,15 0,89 0,27 0,38 0,50
565 Rio Itajaí Superomontana 0,75 0,56 0,62 0,06 0,95 0,99 0,54 0,53
566 Rio Itajaí Inferomontana 0,53 0,19 0,40 0,29 0,87 0,98 0,46 0,61
602 Rio Chapecó Inferomontana 0,69 0,47 0,79 0,00 0,92 0,60 0,48 0,71
605 Rio Chapecó Inferomontana 0,40 0,65 0,00 0,41 0,43 0,70 0,39 0,44
Continua...
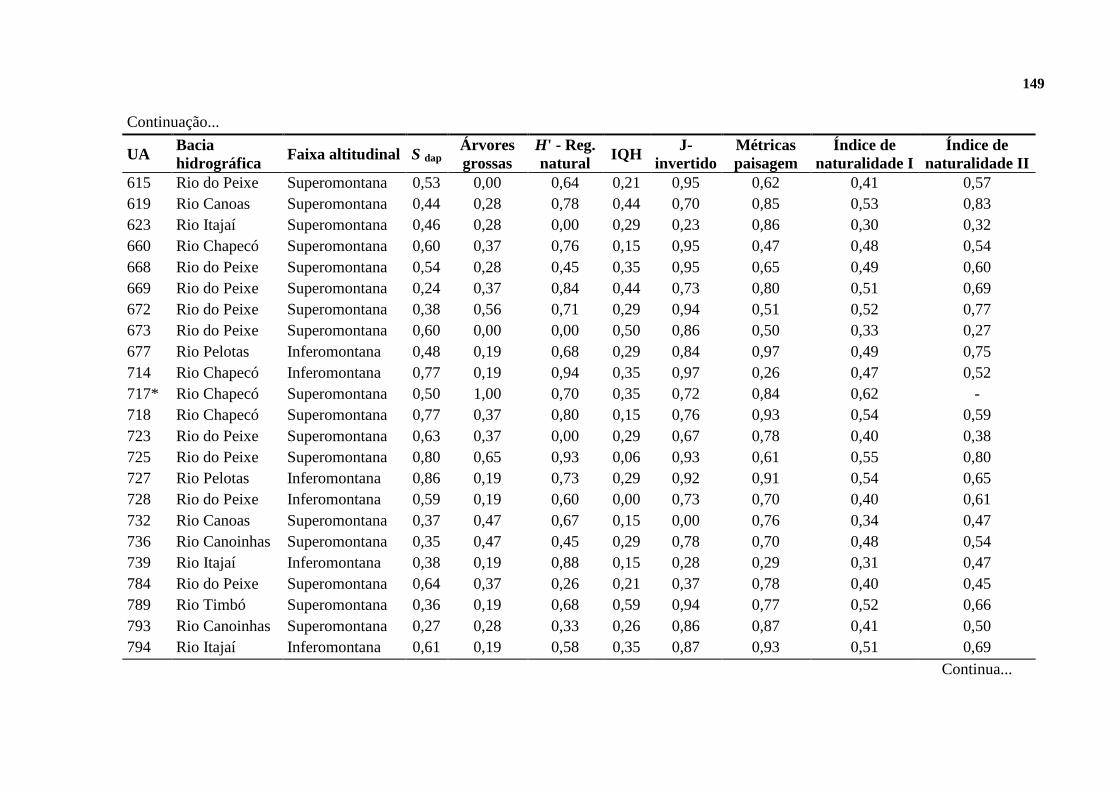
149
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
615 Rio do Peixe Superomontana 0,53 0,00 0,64 0,21 0,95 0,62 0,41 0,57
619 Rio Canoas Superomontana 0,44 0,28 0,78 0,44 0,70 0,85 0,53 0,83
623 Rio Itajaí Superomontana 0,46 0,28 0,00 0,29 0,23 0,86 0,30 0,32
660 Rio Chapecó Superomontana 0,60 0,37 0,76 0,15 0,95 0,47 0,48 0,54
668 Rio do Peixe Superomontana 0,54 0,28 0,45 0,35 0,95 0,65 0,49 0,60
669 Rio do Peixe Superomontana 0,24 0,37 0,84 0,44 0,73 0,80 0,51 0,69
672 Rio do Peixe Superomontana 0,38 0,56 0,71 0,29 0,94 0,51 0,52 0,77
673 Rio do Peixe Superomontana 0,60 0,00 0,00 0,50 0,86 0,50 0,33 0,27
677 Rio Pelotas Inferomontana 0,48 0,19 0,68 0,29 0,84 0,97 0,49 0,75
714 Rio Chapecó Inferomontana 0,77 0,19 0,94 0,35 0,97 0,26 0,47 0,52
717* Rio Chapecó Superomontana 0,50 1,00 0,70 0,35 0,72 0,84 0,62 -
718 Rio Chapecó Superomontana 0,77 0,37 0,80 0,15 0,76 0,93 0,54 0,59
723 Rio do Peixe Superomontana 0,63 0,37 0,00 0,29 0,67 0,78 0,40 0,38
725 Rio do Peixe Superomontana 0,80 0,65 0,93 0,06 0,93 0,61 0,55 0,80
727 Rio Pelotas Inferomontana 0,86 0,19 0,73 0,29 0,92 0,91 0,54 0,65
728 Rio do Peixe Inferomontana 0,59 0,19 0,60 0,00 0,73 0,70 0,40 0,61
732 Rio Canoas Superomontana 0,37 0,47 0,67 0,15 0,00 0,76 0,34 0,47
736 Rio Canoinhas Superomontana 0,35 0,47 0,45 0,29 0,78 0,70 0,48 0,54
739 Rio Itajaí Inferomontana 0,38 0,19 0,88 0,15 0,28 0,29 0,31 0,47
784 Rio do Peixe Superomontana 0,64 0,37 0,26 0,21 0,37 0,78 0,40 0,45
789 Rio Timbó Superomontana 0,36 0,19 0,68 0,59 0,94 0,77 0,52 0,66
793 Rio Canoinhas Superomontana 0,27 0,28 0,33 0,26 0,86 0,87 0,41 0,50
794 Rio Itajaí Inferomontana 0,61 0,19 0,58 0,35 0,87 0,93 0,51 0,69
Continua...
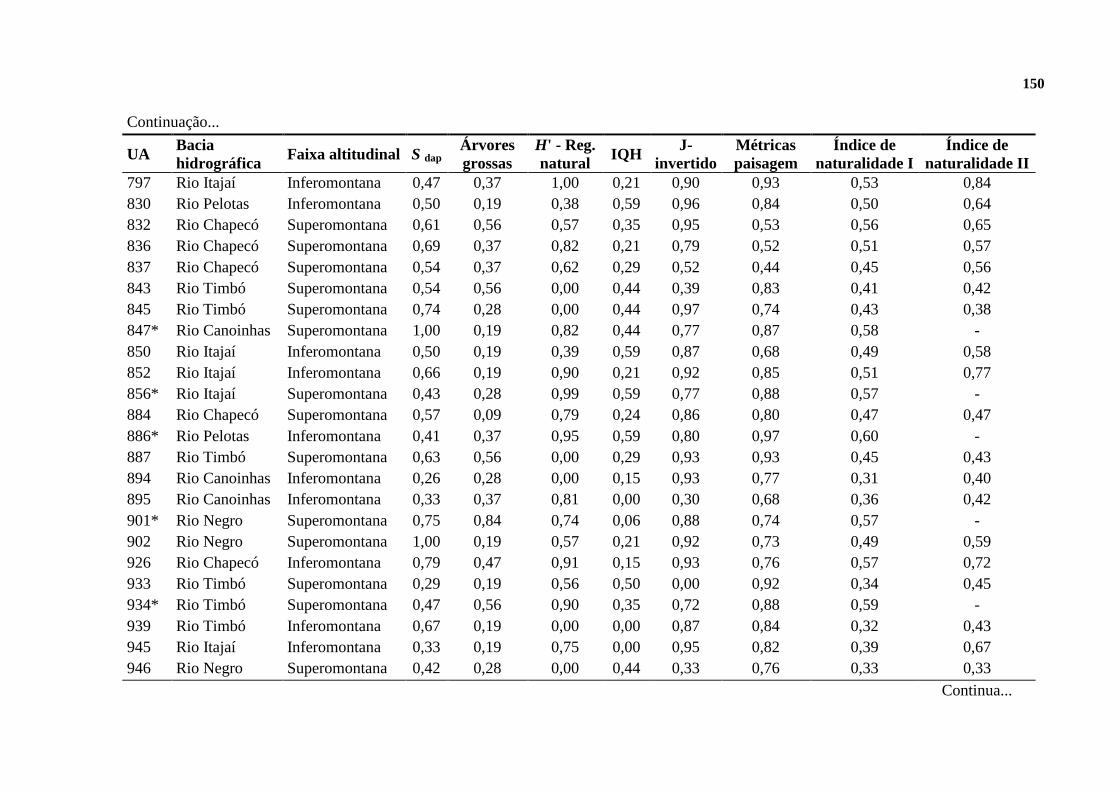
150
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
797 Rio Itajaí Inferomontana 0,47 0,37 1,00 0,21 0,90 0,93 0,53 0,84
830 Rio Pelotas Inferomontana 0,50 0,19 0,38 0,59 0,96 0,84 0,50 0,64
832 Rio Chapecó Superomontana 0,61 0,56 0,57 0,35 0,95 0,53 0,56 0,65
836 Rio Chapecó Superomontana 0,69 0,37 0,82 0,21 0,79 0,52 0,51 0,57
837 Rio Chapecó Superomontana 0,54 0,37 0,62 0,29 0,52 0,44 0,45 0,56
843 Rio Timbó Superomontana 0,54 0,56 0,00 0,44 0,39 0,83 0,41 0,42
845 Rio Timbó Superomontana 0,74 0,28 0,00 0,44 0,97 0,74 0,43 0,38
847* Rio Canoinhas Superomontana 1,00 0,19 0,82 0,44 0,77 0,87 0,58 -
850 Rio Itajaí Inferomontana 0,50 0,19 0,39 0,59 0,87 0,68 0,49 0,58
852 Rio Itajaí Inferomontana 0,66 0,19 0,90 0,21 0,92 0,85 0,51 0,77
856* Rio Itajaí Superomontana 0,43 0,28 0,99 0,59 0,77 0,88 0,57 -
884 Rio Chapecó Superomontana 0,57 0,09 0,79 0,24 0,86 0,80 0,47 0,47
886* Rio Pelotas Inferomontana 0,41 0,37 0,95 0,59 0,80 0,97 0,60 -
887 Rio Timbó Superomontana 0,63 0,56 0,00 0,29 0,93 0,93 0,45 0,43
894 Rio Canoinhas Inferomontana 0,26 0,28 0,00 0,15 0,93 0,77 0,31 0,40
895 Rio Canoinhas Inferomontana 0,33 0,37 0,81 0,00 0,30 0,68 0,36 0,42
901* Rio Negro Superomontana 0,75 0,84 0,74 0,06 0,88 0,74 0,57 -
902 Rio Negro Superomontana 1,00 0,19 0,57 0,21 0,92 0,73 0,49 0,59
926 Rio Chapecó Inferomontana 0,79 0,47 0,91 0,15 0,93 0,76 0,57 0,72
933 Rio Timbó Superomontana 0,29 0,19 0,56 0,50 0,00 0,92 0,34 0,45
934* Rio Timbó Superomontana 0,47 0,56 0,90 0,35 0,72 0,88 0,59 -
939 Rio Timbó Inferomontana 0,67 0,19 0,00 0,00 0,87 0,84 0,32 0,43
945 Rio Itajaí Inferomontana 0,33 0,19 0,75 0,00 0,95 0,82 0,39 0,67
946 Rio Negro Superomontana 0,42 0,28 0,00 0,44 0,33 0,76 0,33 0,33
Continua...
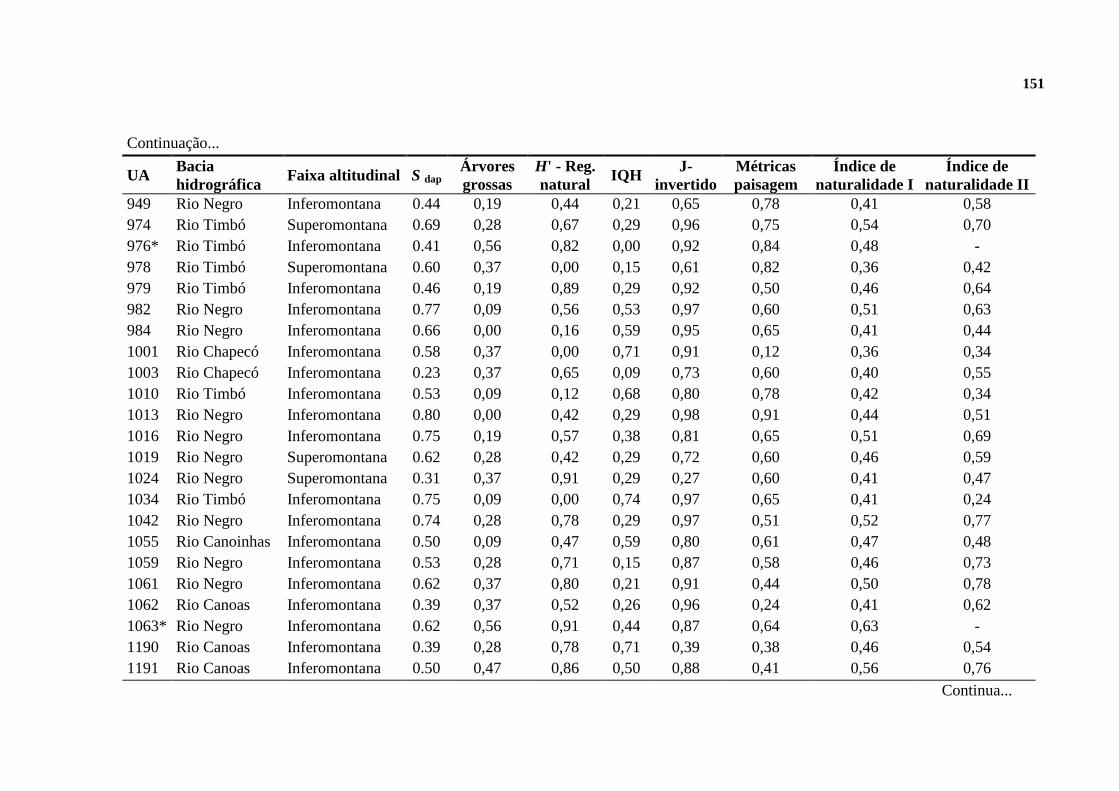
151
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
949 Rio Negro Inferomontana 0.44 0,19 0,44 0,21 0,65 0,78 0,41 0,58
974 Rio Timbó Superomontana 0.69 0,28 0,67 0,29 0,96 0,75 0,54 0,70
976* Rio Timbó Inferomontana 0.41 0,56 0,82 0,00 0,92 0,84 0,48 -
978 Rio Timbó Superomontana 0.60 0,37 0,00 0,15 0,61 0,82 0,36 0,42
979 Rio Timbó Inferomontana 0.46 0,19 0,89 0,29 0,92 0,50 0,46 0,64
982 Rio Negro Inferomontana 0.77 0,09 0,56 0,53 0,97 0,60 0,51 0,63
984 Rio Negro Inferomontana 0.66 0,00 0,16 0,59 0,95 0,65 0,41 0,44
1001 Rio Chapecó Inferomontana 0.58 0,37 0,00 0,71 0,91 0,12 0,36 0,34
1003 Rio Chapecó Inferomontana 0.23 0,37 0,65 0,09 0,73 0,60 0,40 0,55
1010 Rio Timbó Inferomontana 0.53 0,09 0,12 0,68 0,80 0,78 0,42 0,34
1013 Rio Negro Inferomontana 0.80 0,00 0,42 0,29 0,98 0,91 0,44 0,51
1016 Rio Negro Inferomontana 0.75 0,19 0,57 0,38 0,81 0,65 0,51 0,69
1019 Rio Negro Superomontana 0.62 0,28 0,42 0,29 0,72 0,60 0,46 0,59
1024 Rio Negro Superomontana 0.31 0,37 0,91 0,29 0,27 0,60 0,41 0,47
1034 Rio Timbó Inferomontana 0.75 0,09 0,00 0,74 0,97 0,65 0,41 0,24
1042 Rio Negro Inferomontana 0.74 0,28 0,78 0,29 0,97 0,51 0,52 0,77
1055 Rio Canoinhas Inferomontana 0.50 0,09 0,47 0,59 0,80 0,61 0,47 0,48
1059 Rio Negro Inferomontana 0.53 0,28 0,71 0,15 0,87 0,58 0,46 0,73
1061 Rio Negro Inferomontana 0.62 0,37 0,80 0,21 0,91 0,44 0,50 0,78
1062 Rio Canoas Inferomontana 0.39 0,37 0,52 0,26 0,96 0,24 0,41 0,62
1063* Rio Negro Inferomontana 0.62 0,56 0,91 0,44 0,87 0,64 0,63 -
1190 Rio Canoas Inferomontana 0.39 0,28 0,78 0,71 0,39 0,38 0,46 0,54
1191 Rio Canoas Inferomontana 0.50 0,47 0,86 0,50 0,88 0,41 0,56 0,76
Continua...

152
Continuação...
UA Bacia
hidrográfica Faixa altitudinal S dap
Árvores
grossas
H' - Reg.
natural IQH
J-
invertido
Métricas
paisagem
Índice de
naturalidade I
Índice de
naturalidade II
1195 Rio Canoas Superomontana 0,38 0,19 0,81 0,65 0,91 0,44 0,50 0,63
1980 Rio do Peixe Inferomontana 0,89 0,28 0,77 0,71 0,95 0,58 0,62 0,63
2002* Rio Canoinhas Inferomontana 0,72 0,65 0,58 0,15 0,91 0,39 0,50 -
3002 Rio Canoinhas Inferomontana 0,56 0,37 0,20 0,15 0,97 0,98 0,43 0,49
4000 Rio do Peixe Superomontana 0,89 1,00 0,00 0,00 0,59 0,77 0,39 0,32
5000 Rio Canoas Superomontana 0,57 0,37 0,89 0,35 0,69 0,51 0,53 0,69
6001* Rio Chapecó Inferomontana 0,77 0,93 0,75 0,24 0,90 0,67 0,63 -
6004 Rio Chapecó Inferomontana 0,58 1,00 0,57 0,15 0,44 0,82 0,51 0,69


















