
UNIVERSIDADE FEDERAL DO PARANÁ MARCOS FERNANDO …
157
UNIVERSIDADE FEDERAL DO PARANÁ MARCOS FERNANDO GLÜCK RACHWAL URAL E CURITIBA 2013 DRENADO - PARANÁ GASES DE EFEITO ESTUFA EM ORGANOSSOLO NAT
Transcript of UNIVERSIDADE FEDERAL DO PARANÁ MARCOS FERNANDO …

UNIVERSIDADE FEDERAL DO PARANÁ MARCOS FERNANDO GLÜCK RACHWAL
URAL E
CURITIBA 2013
DRENADO - PARANÁ GASES DE EFEITO ESTUFA EM ORGANOSSOLO NAT

MARCOS FERNANDO GLÜCK RACHWAL
URAL E
Tese apresentada ao Curso de Pós-Graduação em Engenharia Florestal, Setor de Ciências Agrárias, Universidade Federal do Paraná, como requisito parcial à obtenção do título de Doutor em Engenharia Florestal - Conservação da Natureza. Orientador: Dr. Gustavo Ribas Curcio Co-orientadores: Dra. Josiléia Acordi Zanatta Dr. Jeferson Dieckow
CURITIBA 2013
GASES DE EFEITO ESTUFA EM ORGANOSSOLO NATDRENADO – PARANÁ

ERRATA: na página 119, Figura 3, eixo Y, onde se lê µg N2O, leia-se mg CO2


Dedico
Ao ar, à água, ao solo, à flora, à fauna e aos seres humanos.

AGRADECIMENTOS
Elaborar este ítem me proporcionou muito prazer por perceber que quando
temos um objetivo nobre pela frente, aparecem inúmeros colaboradores, sem os
quais as metas propostas jamais seriam atingidas.
Mas o início se deu muito antes. Agradeço à Deus, aos antepassados, a
“todos” os professores, a “todos” os familiares e a “todos” os amigos. À maravilhosa
filosofia de vida proposta pela Seicho-No-Iê por ter contribuído muito para o meu
crescimento espiritual, familiar, social, pessoal e econômico. À Danielle Alpendre
Daher pelo treinamento psicológico que expandiu minhas habilidades para lidar com
desafios e direcionar o foco para as soluções.
Tive o privilégio de contar com a preciosa contribuição de pessoas ligadas a
várias instituições e empresas e a este vasto elenco passo a agradecer com muita
gratidão.
EMBRAPA FLORESTAS (Colombo - PR): Roberto Carletto, Arnaldo de
Oliveira Soares, João Altair Torques, Leonildo Tadeo Tanner, Ari Gonçalvez e Michele
Ribeiro Ramos pelo auxílio nos trabalhos de campo.
Onécimo Nunes da Veiga, Renato Antônio Taverna, Nadia Novadzki Venafro,
Daiane Rigoni Kestring, Fabrício Augusto Hansel, Anne Elise Previdi Dotaf, Tielidy
Angelina de Morais de Lima, Marcos Wigo, Cristiane Vieira Helm e Cláudia Mara
Pereira pela contribuição nos trabalhos dos laboratórios.
Renato Antônio Dedecek, Sergio Gaiad, Maria Izabel Radomski, Marcos
Silveira Wrege, Osmir José Lavoranti, Luziane Franciscon, Patrícia Povoa de Mattos,
Rosana Clara Victoria Higa, Cláudia Maria Branco de Freitas Maia, Luís Cláudio
Maranhão Froufe, Itamar Antônio Bognola, Guilherme Andrade, Marilice Cordeiro
Garrastazu e Sandra Bos Mikich pela colaboração técnico-científica.
Betania Busato, Ivar Wendling, Marcus Vinícius Sidoruk Vidal, Alison Luiz
Skroch, José Amauri Moreira Antunes, Waldemar Stival, José Elias Lunardon,
Mieceslau Siegesmundo Stefankowski, Aparicio Schileider, Eleusis Borba Antunes,
Antônio Carlos Wilke, Maria Paraguaçu de Souza Cardoso, Claudiana Eugênio
Rogério, Natália Lordello de Aguiar Vieira, Iclea de Brito, Arielly Francillene do
Nascimento, Rosana Marques e Jackline Paula de Godoi Degani pelo apoio
logístico.

Sérgio Ahrens pela elaboração do abstract geral da tese.
Francisca Rasche, Simone Aparecida Sopchaki e Elizabeth Denise Roskamp
Câmara pelas buscas bibliográficas.
EMBRAPA SPM - EPGA (Ponta Grossa - PR): Osmar Paulo Beckert por ter
gentilmente cedido a área experimental e juntamente a José Luciano Bail, Paulo
Alexandre Virgolino, Vitor Luiz Pereira, Inês Kieltika Maier, Angélica Terezinha Trentin,
Ana Flávia Mellem, Deraldo Vargenski, João Carlos Cardoso, Renato Carlos Wantroba,
Wilson Felipe da Silva, Anísio Pedroso da Silva, Diego Marcelo Ferreira e Josias, por
terem viabilizado os recursos humanos e logísticos para os trabalhos de campo.
EMBRAPA SOLOS (Rio de Janeiro - RJ): Daniel Vidal Perez por
disponibilizar o laboratório para as análises de solos. Ademir Emídio da Silva, André
Marcelo de Souza, Andréa Campos de Souza, Andrea Matos dos Santos, Damaris
de Araújo Alencar, Fátima Barbosa Paulino, Felicia Miranda de Jesus, Jean Marcel
Rodrigues Pinho, Jorge Luis Nunes Ramilo, Marcelo Teixeira de Andrade, Rogerio
Tavares de Campos, Rojane Chapeta Peixoto, Sérgio Gomes, Viviane Escaleira
pelas análises laboratoriais. Etelvino Henrique Novotny pelas inúmeras discussões
sobre matéria orgânica do solo. Aos pedólogos do antigo Serviço Nacional de
Levantamento e Conservação de Solos da EMBRAPA, Américo Pereira de Carvalho,
Alcides Cardoso, Delcio Perez Hochmüller, Moacyr de Jesus Rauen, Pedro Jorge
Fasolo, Reinaldo Oscar Pötter, Marcelo Nunes Camargo, Paulo Klinger Tito
Jacomine, Jorge Olmos Iturri Larach e João Carlos ker, por terem participado
efetivamente de minha formação em pedologia e contribuído com meu crescimento
pessoal.
UFPR (Curitiba - PR): Fernando Rodrigo Bortolozo pela coleta de amostras
de solos em anéis volumétricos. Edenilson Nascimento pelo auxílio na instalação
das câmaras coletoras de gás e poços hídricos. Carina Kozera e Bianca Ott Andrade
pela caracterização da vegetação. Letícia di Pierri pela participação na preparação
de várias amostras de solos para análise. Fabiana de Medeiros Silveira e Daniel
Carvalho pelas orientações técnicas e práticas sobre análises de amônio e de
nitrato. Iackson Oliveira Borges pelas orientações sobre a drenagem. Franklin
Galvão, Carlos Vellozo Roderjan e Yoshico Saito Kuniyoshi pela constante
motivação e pelo amor e dedicação aos alunos. Omar Fereira Lopes e Claudinei
Taborda da Silveira por contriburem para a compreensão do contexto geológico da

área em estudo. Vander de Freitas Melo e Valmiqui Costa Lima pelo estímulo
constante e profissionalismo. Letícia Gonçalvez pelas orientações sobre a
determinação do potencial redox do solo. Aldair Marty Munhoz, Reginaldo Bodziak,
Flori Roberto Margraf Barberi, Elda Nazaré Lubasinski, Cleusa Maria Correa Lopes,
pelo auxílio nos trabalhos de laboratório. Simone Amadeu pela revisão das
referências bibliográficas e formatação geral do texto. “Baiano das Agrárias”, pela
sua maravilhosa lição de vida.
UFRGS (Porto Alegre - RS): Cimélio Bayer pela troca de idéias e pela
disponibilização do cromatógrafo de fase gasosa; Genuir Luis Denega por ter
“encarado” o cromatógrafo durante várias madrugadas.
USP/ESALQ (Piracicaba - SP): Luiz Antonio Silva Junior pela determinação
dos teores de amônio e de nitrato no solo e Jorge de Castro Kiehl pelas discussões
sobre amônio e nitrato.
UEPG (Ponta Grossa - PR): Rafael Mazer Etto pelas explicações sobre os
organismos metanogênicos e metanotróficos do solo.
UFPEL (Pelotas - RS): Rogério Oliveira de Sousa pelas discussões sobre o
potencial redox do solo.
IAPAR: Leocádio Grodzki e Rui Carlos Maranhão Biscaia pelas dicas sobre a
estação meteorológica.
EMPRESAS: Raulindo Brusamolin da Metalúrgica Brusamolin por suas
idéias e pelo capricho na construção das câmaras coletoras de gases, dos poços
hídricos e da extensão do pHmetro de campo para determinação do potencial redox
do solo. Silvio José Barchick do Boa Vista Materiais de Construção pelas sugestões
de inúmeros materiais apropriados para confecção de diversos equipamentos.
Amilton Hamirisi da Drenos Hamirisi, Osmar Petersen e Alexandre Petersen da
Osmar Petersen e Cia Ltda, pela construção dos drenos.
Carlos Alberto de Oliveira Júnior, Alan Borges Vieira e Thifani Clasen da
Agrosystem que não mediram esforços para o perfeito funcionamento da estação
meteorológica.
Éder David Borges da Silva pelas análises estatísticas e as inúmeras
discussões para interpretação dos resultados.
Ao meu orientador Gustavo Ribas Curcio (EMBRAPA FLORESTAS) e aos
co-orientadores Jeferson Dieckow (UFPR) e Josiléia Acordi Zanatta (EMBRAPA
FLORESTAS) pelas constantes discussões e correções de rumo. Aos membros da

banca examinadora Carlos Eduardo Pellegrino Cerri (USP/ESALQ), Cimélio Bayer
(UFRGS), Josiléia Acordi Zanatta, Antônio Carlos Vargas Motta (UFPR), Jeferson
Dieckow (UFPR) e Rafael Mazzer Etto (UEPG) por suas preciosas sugestões e
questionamentos.
FAMILIARES e AMIGOS: A minha esposa Mirta Malczewski, filho Matheus
Malczewski Rachwal (“Firico”), enteada Carolina Izabel Malczewski Santos e genro
Marcos Vinicius Marschalk, por terem “aguentado” heroicamente as minhas neuras e
rabugices durante estes quatro anos. Aos dois primeiros ainda sou grato pela ajuda
que prestaram na coleta de dados no campo.
Aos meus pais Pedro Rachwal (“Pedrão”) e Neusa Glück Rachwal (“Bruxa”),
a minha sogra Elvira Santos Malczewski, ao irmão Luiz Augusto Glück Rachwal
(“Guggo”), a cunhadinha Ana Maria Schier Dória Rachwal, aos sobrinhos Gabriel
Dória Rachwal (“Gabirú”) e Anna Dória Rachwal (“Polaca”) por estarem sempre
muito presentes nos momentos mais importantes de minha vida.
Ao meu irmão “mais querido”, “Guggo” pela elaboração das figuras.
A Eva Gonçalves, nossa “governanta” por deixar em ordem a “bagunça” do
retorno das viagens de campo, pelos lanches das “infindáveis” tardes e por escutar
minhas explicações sobres os gases. A Berenice (a nossa gata) por dormir todas as
tardes enquanto eu estudava, calculava e escrevia.
Aos amigões Luiz Sergio Piovesan, Daniela Fatima Michelon Piovesan e
seus filhos, Luiza Michelon Piovesan e Arthur Michelon Piovesan pelos momentos
de total descontração e pelas sessões de terapia de grupo.
A minha amiga, colega de Embrapa, afiliada e parceira Letícia Penno Dereti
pela constante troca de idéias e por compartilhar dias de sol intenso, mas também
de muito frio e chuva, durante as campanhas de campo. Ao amigo, colega de
Embrapa e afiliado Rogério Morcelles Dereti, pela sua ajuda no campo e por ter
rapidamente simulado a banca examinadora, poucos dias antes da defesa.
A Gustavo Ribas Curcio, Annete Bonnet e Murilo Barddal pelo
profissionalismo e espírito científico acurado, nos quais me espelhei.
A Daniel Ghenov Filho pela ajuda na coleta de dados no campo.
A Ana Maria Banks pelo apoio logístico e Claudio Ferreira Banks Machado
pela elaboração dos abstracts dos capítulos.

Agradeço ainda a todos aqueles que em função de minha ingratidão ou “data
avançada de nascimento”, foram esquecidos. Me perdoem, mas não deixem de
aceitar meu sincero agradecimento.
E para finalizar, quero demonstrar minha gratidão aos que compartilharam
dos momentos produtivos, emocionantes e alegres da defesa da tese e da posterior
confraternização.
MUITO OBRIGADO!

“Montes, rios, árvores, campos e demais elementos da natureza não agem como
partes que disputam umas contra as outras, mas sim formam um conjunto em que
se ajudam e se complementam e, assim, mantêm a natureza rica e generosa.”
Masanobu Taniguchi.

RESUMO
Organossolos estocam quantidades elevadas de carbono orgânico, e podem ser fontes ou sumidouros de metano, óxido nitroso e dióxido de carbono. Características pedológicas intrínsecas, mudanças no uso do solo e formas de manejo, além dos fatores climáticos, interferem nos fluxos destes gases, sendo a drenagem artificial uma das práticas que mais alteram o comportamento dos mesmos. O objetivo desta pesquisa foi avaliar a influência da drenagem sobre os fluxos de metano (CH4), óxido nitroso (N2O) e dióxido de carbono (CO2), considerando as variações no nível freático e na umidade gravimétrica do solo, bem como a influência da temperatura do ar e da precipitação. Durante um período de dois anos foi realizado um experimento em área de Organossolo Háplico sáprico típico localizada na fazenda experimental da Embrapa SPM-EPGA (Ponta Grossa, PR), segundo planalto paranaense, na região dos Campos Gerais. No solo em condição natural, com hidromorfia plena, houve emissão de CH4, a qual variou entre 238 e 5365 µg de CH4 m-2 h-1. Na área drenada, a retirada da água determinou o consumo líquido de metano entre 39 a 242 µg de CH4 m-2 h-1. No solo sob condição natural ocorreu consumo líquido de N2O cujas taxas variaram de -0,13 a -31,2 µg de N2O m-2 h-1. Por sua vez, na área drenada, a retirada da água determinou a emissão líquida em taxas que variaram de 0,19 a 55,7 µg de N2O m-2 h-1. O aumento do número de dias após a drenagem determinou a elevação da emissão de N2O. Na condição natural houve consumo de óxido nitroso devido ao excesso de umidade e aos baixos valores de potencial redox. No solo natural a taxa de emissão de dióxido de carbono variou entre 244 e 870 mg de CO2 m
-2 h-1. Na área drenada o fluxo oscilou de 238 a 1018 mg de CO2 m
-2 h-1. Embora não tenha ocorrido diferença significativa nas taxas de emissão de dióxido de carbono, entre o Organossolo natural e o drenado, o efeito acumulado da drenagem proporcionou um aumento de 17,9 Mg C-CO2-eq ha-1 ano-1 no potencial de aquecimento global, em função da perda de 37,8 Mg C ha-1 do estoque de carbono do solo, até um metro de profundidade, em apenas dois anos. Somente no Organossolo natural o fluxo de metano foi afetado significativamente pela temperatura do ar ao longo do tempo. No Organossolo drenado o efeito da temperatura foi secundário, pois a diminuição da umidade foi o fator que determinou o consumo de CH4. A temperatura do ar teve efeito marcante sobre o fluxo de óxido nitroso apenas no Organossolo natural. Por outro lado, a precipitação só teve efeito pronunciado sobre o fluxo de N2O no Organossolo drenado. A precipitação praticamente não afetou os fluxos de CH4 nos solos natural e drenado. No primeiro, porque a chuva não aumentou significativamente a elevada umidade natural do solo e, no segundo, porque a água da chuva era rapidamente removida devido à alta permeabilidade. No caso do CO2, tanto a temperatura do ar quanto a precipitação influenciaram intensamente os fluxos em ambos os solos. Embora a drenagem tenha transformado o Organossolo drenado em um pequeno sumidouro de metano, o mesmo passou a emitir óxido nitroso e grande quantidade de dióxido de carbono. Como os Organossolos de campo hidrófilo possuem importantes funções no ecossistema, como acumular carbono orgânico, armazenar água e contribuir na diminuição da quantidade de nitrato, podendo reduzi-lo à N2, não devem ser drenados, mas, efetivamente preservados. A capacidade dos Organossolos sob condição natural de estocar 581 Mg ha-1 de carbono orgânico e de armazenar 8,6 milhões de litros de água por hectare, até a profundidade de um metro, justifica plenamente que os mesmos não devam ser drenados.

Palavras chave: Gases do efeito estufa. Perda de carbono. Nascentes. Temperatura e precipitação. Nível freático e potencial redox. Potencial de aquecimento global.

ABSTRACT
GREENHOUSE FLUXES IN HISTOSOL UNDER NATURAL CONDITIONS AND AFTER DRAINAGE - PARANA - BRAZIL
Large quantities of organic carbon are trapped in histosols which can be either a source or a sink of methane, nitrous oxide and carbon dioxide. Intrinsic soil characteristics, changes in soil use and management, in addition to climate factors, influence intensively in the flow of these gases, but artificial drainage is one of the practices that most affect their behavior. This research was carried out in order to evaluate the influence of drainage in the flow of methane (CH4), nitrous oxide (N2O) and carbon dioxide (CO2), considering variations in groundwater level, in the soil gravimetric moisture, as well as air temperature and rainfall. An experiment was carried out over a two year period in an Typic Haplosaprists within an experimental farm located in Ponta Grossa, PR, a town situated in the second plateau of that state. Under natural conditions, and with full hydromorphic, there was an emission of CH4, from 238 to 5365 µg of CH4 m
-2 h-1. In the drained area, the removal of water has determined the consumption of liquid methane between 39 and 242 µg of CH4 m
-2 h-
1. Net consumption of N2O in soil under natural conditions varied from -0,13 to -31,2 µg of N2O m-2 h-1. However, in the drained area, the removal of water has determined a net emission in rates that may vary between 0,19 to 55,7 µg of N2O m-2 h-1. In drained soil the increase in the number of days after drainage has determined the increase of the emission of N2O. Under natural conditions, consumption of nitrous oxide was observed due to the excessive moisture and low values for redox potential. Under natural conditions emission rates of carbon dioxide ranged from 244 to 870 mg of CO2 m
-2 h-1. In the drained area the same flow has varied from 238 to 1018 mg of CO2 m
-2 h-1. Although there was no significant difference in the rate of carbon dioxide emission in both soil, the effect of accumulated drainage resulted in an increase of 17,9 Mg C- CO2 eq ha-1 yr-1 in the global warming potential overall, due to the loss of 37,8 Mg C from the stock of soil carbon, up to a meter deep, in just two years. The flow of methane was significantly affected by air temperature over time in natural histosols only. In histosol drained the temperature effect was secondary, once the decreased moisture was the factor that determined the consumption of CH4. Air temperature had a major effect on the flow of nitrous oxide only in natural histosol. On the other hand, rainfall had a notorious effect over the flow of N2O in drained histosol. Rainfall had no major effect on CH4 flow for both, natural and drained soil. In the first, the rainfall did not increased the high natural moisture of the soil and in the second, the rainfall water was rapidly removed due to the high soil permeability. In the case of CO2, air temperature and the rainfall intensively has influenced the flows in both soils. Although the drainage has transformed drained histosol in a methane sink, this soil started releasing nitrous oxi de and large quantities of carbon dioxide. Given that histosols located in hydrophilous grassland perform important functions for the ecosystem, such as acting as a carbon sink, retaining water and contributes in the decrease of the quantities of nitrate, could possibly change it into N2, so they should not be drained but effectively preserved. The capacity of histosol, under natural conditions, of trapping 581 Mg ha-1 of organic carbon and 8,6 million liter of water per hectare, at one meter depth, justifies entirely that they must not be drained.

Keywords: Greenhouse Gases. Carbon loss. Springs. Air temperature. Rainfall. Waterbed level. Redox potential and global warming potential.

LISTA DE TABELAS
CAPÍTULO 1 TABELA 1 -
ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE......................
31
TABELA 2 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE VARIÁVEIS PEDOLÓGICAS E METEOROLÓGICAS E O EFLUXO DE CH4 EM ORGANOSSOLO NATURAL E O INFLUXO DE CH4 NO ORGANOSSOLO DRENADO...................
39
TABELA 3 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE OS FLUXOS DE CH4, PELO MODELO ADITIVO GENERALIZADO - GAM................................................................................................
47
TABELA 4 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE POTENCIAL REDOX, NAS PROFUNDIDADES DE 5, 10, 15 , 20, 30 E 40 cm E O EFLUXO DE CH4 NO ORGANOSSOLO NATURAL E O INFLUXO DE CH4 NO ORGANOSSOLO DRENADO........................................................
50
CAPÍTULO 2
TABELA 1 -
ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE......................
69
TABELA 2 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE OS FLUXOS DE N2O PELO MODELO ADITIVO GENERALIZADO - GAM................................................................................................
76
TABELA 3 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE FATORES PEDOLÓGICOS E METEOROLÓGICOS E O INFLUXO DE N2O, NO ORGANOSSOLO NATURAL E O EFLUXO DE N2O NO ORGANOSSOLO DRENADO.....................
77
TABELA 4 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE POTENCIAL REDOX, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm E O INFLUXO DE N2O NO ORGANOSSOLO NATURAL E O EFLUXO DE N2O NO ORGANOSSOLO DRENADO........................................................
85
TABELA 5 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE N-NH4
+ E N-NO3-, NA PROFUNDIDADE DE 0 A 10 cm,
EFLUXO DE CO2 E TEMPO, EM ORGANOSSOLO NATURAL E DRENADO, POR PERÍODO..........................................................
92
TABELA 6 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE N-NO3
- E N-NH4+ NA PROFUNDIDADE DE 0 A 10 cm E
O INFLUXO DE N2O EM ORGANOSSOLO NATURAL E EFLUXO DE N2O NO ORGANOSSOLO DRENADO....................
93

CAPÍTULO 3 TABELA 1 -
ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE......................
114
TABELA 2 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE FATORES PEDOLÓGICOS E METEOROLÓGICOS E O EFLUXO DE CO2............................................................................
120
TABELA 3 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE O EFLUXO DE CO2, PELO MODELO ADITIVO GENERALIZADO - GAM................................................................................................
127
TABELA 4 - BALANÇO DE CARBONO EM ORGANOSSOLO DRENADO E SUA RELAÇÃO COM O POTENCIAL DE AQUECIMENTO GLOBAL - PAG...............................................................................
131

LISTA DE FIGURAS
CAPÍTULO 1
FIGURA 1 - LOCALIZAÇÃO DA ÁREA DE ESTUDO........................................ 30 FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E
TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO......................
36
FIGURA 3 - FLUXO DE METANO (µg CH4 m-2
h-1) EM ORGANOSSOLO EM
CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR (a). CONSUMO DE METANO EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR (b)............................................................................
38
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR...................................................................................................
44
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR.........................
45
FIGURA 6 - POTENCIAL REDOX (mV) EM ORGANOSSOLO NATURAL, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR................................................................................
49
FIGURA 7 - POTENCIAL REDOX (mV) EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR...............................................
51
FIGURA 8 - FLUXO ACUMULADO DE METANO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR..................................
53
CAPÍTULO 2
FIGURA 1 -
LOCALIZAÇÃO DA ÁREA DE ESTUDO........................................
68
FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO......................
73
FIGURA 3 - FLUXO DE ÓXIDO NITROSO (µg N2O m-2 h-1) EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR.........................
75
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM, PONTA GROSSA, PR..............
78
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR.........................
79
FIGURA 6 - POTENCIAL REDOX (mV) EM ORGANOSSOLO NATURAL NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR.................................................................................
84
FIGURA 7 - POTENCIAL REDOX (mV) EM ORGANOSSOLO DRENADO ARTIFICIALMENTE NAS PROFUNDIDADES DE 5, 10, 15, 20,

30 E 40 cm, PONTA GROSSA, PR............................................... 86 FIGURA 8 - TEOR DE N-NH4
+ (mg kg-1) E DE N-NO3- (mg kg-1) NA
PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO NATURAL, PONTA GROSSA, PR..................................................
87
FIGURA 9 - TEOR DE N-NH4+ (mg kg-1) E DE N-NO3
- (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR...............
89
FIGURA 10 - TEOR DE N-NH4+ (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm
E CONSUMO DE N2O (µg m-2 h-1) EM ORGANOSSOLO NATURAL, PONTA GROSSA, PR.................................................
94
FIGURA 11 - TEOR DE N-NH4+ (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm
E EMISSÃO DE N2O (µg m-2 h-1) EM ORGANOSSOLO DRENADO, PONTA GROSSA, PR................................................
95
FIGURA 12 - FLUXO ACUMULADO DE ÓXIDO NITROSO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR..............
96
CAPÍTULO 3
FIGURA 1 -
LOCALIZAÇÃO DA ÁREA DE ESTUDO........................................
113
FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO......................
118
FIGURA 3 - FLUXO DE DIÓXIDO DE CARBONO (mg CO2 m-2 h-1) EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR.........................
119
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR...................................................................................................
122
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR.........................
123
FIGURA 6 - TEORES DE CARBONO (g/kg) NOS HORIZONTES H1 A H5 NO ORGANOSSOLO DRENADO ARTIFICIALMENTE EM FUNÇÃO DO TEMPO DE DRENAGEM........................................
126
FIGURA 7 - FLUXO ACUMULADO DE DIÓXIDO DE CARBONO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR...................................................................................................
128

SUMÁRIO
1 INTRODUÇÃO GERAL.................................................................................... 20 REFERÊNCIAS.................................................................................................... 22 2 CAPITULO I FLUXO DE METANO EM ORGANOSSOLO SOB ESTEPE HIDRÓFILA EM CONDIÇÃO NATURAL E APÓS DRENAGEM - PARANÁ - BR........................
25
RESUMO.............................................................................................................. 25 ABSTRACT.......................................................................................................... 26 2.1 INTRODUÇÃO............................................................................................... 27 2.2 MATERIAL E MÉTODOS............................................................................... 30 2.2.1 Experimento de campo............................................................................... 30 2.2.2 Coleta de amostras de ar............................................................................ 32 2.2.3 Parâmetros pedológicos............................................................................. 33 2.2.4 Variáveis meteorológicas........................................................................... 34 2.2.5 Análise estatística....................................................................................... 34 2.3 RESULTADOS E DISCUSSÃO...................................................................... 36 2.3.1 Efeito dos fatores meteorológicos sobre o fluxo de CH4............................. 36 2.3.2 Efeito da drenagem sobre o fluxo de CH4................................................... 42 2.3.3 Efeito do potencial redox sobre o fluxo de CH4........................................... 47 2.3.4 Fluxo acumulado de metano....................................................................... 51 2.3.5 Considerações adicionais........................................................................... 53 2.4 CONCLUSÕES.............................................................................................. 55 REFERÊNCIAS.................................................................................................... 56 3 CAPÍTULO II FLUXO DE ÓXIDO NITROSO EM ORGANOSSOLO SOB ESTEPE HIDRÓFILA EM CONDIÇÃO NATURAL E APÓS DRENAGEM - PARANÁ - BR........................................................................................................................
62
RESUMO.............................................................................................................. 62 ABSTRACT.......................................................................................................... 63 3.1 INTRODUÇÃO............................................................................................... 64 3.2 MATERIAL E MÉTODOS............................................................................... 68 3.2.1 Experimento de campo............................................................................... 68 3.2.2 Coleta de amostras de ar............................................................................ 70 3.2.3 Parâmetros pedológicos............................................................................. 71 3.2.4 Variáveis meteorológicas............................................................................ 72 3.2.5 Análise estatística....................................................................................... 72 3.3 RESULTADOS E DISCUSSÃO...................................................................... 73 3.3.1 Efeito dos fatores meteorológicos sobre o fluxo de N2O............................. 73 3.3.2 Efeito da drenagem sobre o fluxo de N2O................................................... 78 3.3.3 Efeito do potencial redox sobre o fluxo de N2O.......................................... 83 3.3.4 Teores de N-NH4
+ e N-NO3- no solo............................................................ 87
3.3.5 Influência dos teores de N-NH4+ e N-NO3
- no solo sobre o fluxo de N2O... 93 3.3.6 Fluxo acumulado de óxido nitroso.............................................................. 95 3.3.7 Considerações adicionais........................................................................... 97 3.4 CONCLUSÕES.............................................................................................. 98 REFERÊNCIAS.................................................................................................... 99 4 CAPÍTULO III

FLUXO DE CO2 E ESTOQUE DE CARBONO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM - PARANÁ - BR........................
107
RESUMO.............................................................................................................. 107 ABSTRACT.......................................................................................................... 108 4.1 INTRODUÇÃO............................................................................................... 109 4.2 MATERIAL E MÉTODOS............................................................................... 112 4.2.1 Experimento de campo............................................................................... 112 4.2.2 Coleta de amostras de ar............................................................................ 112 4.2.3 Parâmetros pedológicos............................................................................. 115 4.2.4 Variáveis meteorológicas............................................................................ 116 4.2.5 Análise estatística....................................................................................... 116 4.3 RESULTADOS E DISCUSSÃO...................................................................... 117 4.3.1 Efeito dos fatores meteorológicos sobre o fluxo de CO2............................. 117 4.3.2 Efeito da drenagem sobre o fluxo de CO2................................................... 121 4.3.3 Fluxo acumulado de dióxido de carbono no solo........................................ 127 4.3.4 Efeito da drenagem sobre o estoque de carbono no solo.......................... 129 4.3.5 Efeito da drenagem sobre o potencial de aquecimento global................... 130 4.3.6 Considerações adicionais........................................................................... 132 4.4 CONCLUSÕES.............................................................................................. 133 REFERÊNCIAS.................................................................................................... 134 5 CONSIDERAÇÕES FINAIS.............................................................................. 140 6 RECOMENDAÇÕES......................................................................................... 140 ANEXOS.............................................................................................................. 142

20
1 INTRODUÇÃO GERAL
A compreensão dos processos de produção de gases de efeito estufa a
partir de solos hidromórficos é importante para completar o entendimento do ciclo
global do carbono (PAGE; DALAL, 2011).
O dióxido de carbono, o metano e o óxido nitroso são os mais importantes
gases de efeito estufa, contribuindo em 60%, 20% e 6%, respectivamente, com o
aquecimento global (IPCC, 2007).
Durante o período de 1750 a 2005 a concentração de CO2 na atmosfera
aumentou de aproximadamente 280 para 380 ppm, a de CH4 se elevou de 715 ppb
para 1774 ppb, além de um acréscimo de 270 ppb para 319 ppb na quantidade de
N2O (FORSTER et al., 2007). Esta elevação ocorreu principalmente por ações
antrópicas como incremento do uso de combustíveis fósseis e da poluição industrial,
aumento das queimadas, agricultura praticada de forma insustentável, uso irracional
dos recursos naturais e mudanças no uso da terra (IPCC, 2001, 2007).
O excesso de água no solo limita a difusão de oxigênio e cria condições
anaeróbicas propícias para a produção de metano e de óxido nitroso (REDDY;
DELAUNE, 2008). Por outro lado, esta deficiência de oxigênio permite baixos índices
de decomposição microbiana, resultando no armazenamento de grandes
quantidades de carbono orgânico (ALONGI et al., 2001; WHITING; CHANTON,
2001).
A drenagem de solos saturados com água afeta a maioria dos fatores que
controlam a produção, o consumo e o fluxo líquido dos gases de efeito estufa
(PAAVILAINEN; PÄIVÄNEN, 1995; MINKKINEN et al., 2002). A oxidação do solo
provocada pela drenagem pode causar tanto o decréscimo na produção de metano
(WANG et al., 1996) quanto a perda de carbono orgânico (ARMENTANO; MENGES,
1986; FURUKAWA et al., 2005). Organossolos drenados e cultivados deixam de ser
sumidouros de CO2 e de nitrogênio e transformam-se em persistentes fontes dos
mesmos (LOHILA et al., 2003; MALJANEN et al., 2002, 2004).
No Estado do Paraná é comum a presença de nascentes em áreas situadas
em altitudes superiores a 800m, sobretudo, naquelas localizadas em reversos de
cuestas, sujeitas a baixa temperatura e a elevada precipitação, onde ocorrem os
Organossolos dispostos sob estepes hidrófilas, os quais constituem importantes
reservatórios de carbono e água, além de abrigarem elevada biodiversidade.

21
Estes solos podem armazenar uma quantidade de água equivalente a 4,5 a
30 vezes o seu peso seco (SOIL SURVEY STAFF, 1992). Scheer et al. (2011)
registraram que Organossolos sob campo altomontano estocam 315 Mg ha-1 de
carbono e 3,4 milhões de litros de água por hectare até a profundidade de 58 cm. No
que se refere a riqueza de espécies vegetais, Andrade (2011) e Kozera (2008)
registraram, respectivamente, 146 espécies em 0,65 ha e 87 espécies em
aproximadamente 4 ha, em Organossolos de altitude sob estepe.
Entretanto, a maioria dessas nascentes estão sujeitas a vários tipos de
intervenções antrópicas, entre elas, a exploração agropecuária, a qual tem alterado
as funções ecológicas destes ambientes. No Brasil, informações sobre fluxos de
gases de efeito estufa em nascentes com alto grau de hidromorfia onde ocorrem
Organossolos e os efeitos da drenagem sobre tais fluxos, são inexistentes.
Para este estudo supôs-se que a drenagem artificial modificaria os fluxos de
CO2, CH4 e N2O em diferentes intensidades e alteraria o potencial de aquecimento
global e que as variações sazonais na temperatura do ar e na precipitação também
influenciariam os fluxos destes gases.
Os objetivos desta pesquisa foram quantificar, durante um período de 26
meses, os fluxos de CO2, CH4 e N2O em Organossolo Háplico sáprico típico sob
vegetação estépica em condições naturais e após drenagem e indicar as alterações
pedológicas e as variáveis climáticas que mais influenciam estes fluxos.

22
REFERÊNCIAS
ALONGI, D. M.; WATTAYAKORN, G.; PFITZNER, J.; TIRENDI, F.; ZAGORSKIS, I.; BRUNSKILL, G. J.; DAVIDSON, A.; CLOUGH, B. F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in Southern Thailand. Marine Geology, Amsterdam, v. 179, p. 85-103, 2001. ANDRADE, B. O.; KOZERA, C.; CURCIO, G. R.; GALVÃO, F. Vascular gassland plants of Tibagi River Spring, Ponta Grossa, Brazil. Chek List, Jaboticabal, v. 7, n. 3, p. 257-262, 2011. ARMENTANO, T. V.; MENGES, E. S. Patterns of changes in the carbono balance of organic soils-wetlands of temperate zone. Journal of Ecology, Oxford, v. 74, p. 755-774, 1986. FORSTER, P.; RAMASWAMY, V.; ARTAXO, P.; BERNSTEN, T.; BETTS, R.; FAHEY, D. W.; HAYWOOD, J.; LEAN, J.; LOWE, D. C.; MYHRE, G.; NGANGA, J.; PRINN, R.; RAGA, G.; SCHUTZ, M.; van DORLAND, R. Changes in atmospheric constituents and in radiative forcing. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z.; MARQUIS, M.; AVERYT, K. B.; TIGNOR, M.; MILLER, H. L. (Ed.). Climate change 2007: the physical science basis - contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge, UK: University Press, 2007. p. 129-234. FURUKAWA, Y.; INUBUSHI, K.; ALI, M.; ITANG, A. M.; TSURUTA, H. Effect of changing groundwater levels caused by land-use changes on greenhouse gas fluxes from tropical peat lands. Nutrient Cycling Agroecosystems, Dordrecht, v. 71, p. 81-91, 2005. INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC). Climate Change 2001: synthesis report. In: WATSON, R. T. (Ed.). A Contribution of Working Groups I, II and III to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University, 2001. INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC). Climate Change 2007: the physical science basis. Cambridge: Cambridge University, 2007. KOZERA, C. Florística e fitossociologia de uma formação pioneira com influência fluvial e de uma estepe gramíneo-lenhosa em diferentes unidades geopedológicas, município de Balsa Nova, Paraná - Brasil. 267 f. Tese (Doutorado) - Universidade Federal do Paraná, Curitiba, 2008.

23
LOHILA, A.; AURELA, M.; REGINA, K.; LAURILA, T. Soil and total ecosystem respiration in agricultural fields: effects of soil and crop type. Plant Soil, The Hague, v. 251, p. 303-317, 2003. MALJANEN, M.; MARTIKAINEN, P. J.; AALTONEN, H.; SILVOLA, J. Short-term variation in fluxes of carbon dioxide, nitrous oxide and methane in cultivated and forested organic boreal soils. Soil Biology & Biochemistry, Oxford, v. 34, p. 577-584, 2002. MALJANEN, M.; KOMULAINEN, V. M.; HYTONEN, J.; MARTIKAINEN, P. J.; LAINE, J. Carbon dioxide, nitrous oxide and methane dynamics in boreal organic agricultural soils with different soil characteristics. Soil Biology & Biochemistry, Oxford, v. 36, p. 1801-1808, 2004. MINKKINEN, K.; KORHONEN, R.; SAVOLAINEN, I.; LAINE, J. Carbon balance and radiative forcing of Finnish peatlands 1900-2100 – the impact of forestry drainage. Global Change Biology, West Sussex, v. 8, p. 785-799, 2002. PAAVILAINEN, E.; PÄIVÄNEN, J. Peatlands forestry-ecology and principles. Berlim: Springer, 1995. (Ecological Studies, 111). PAGE, K. L.; DALAL, R. C. Contribution of natural and drained wetland systems to carbon stocks, CO2, N2O and CH4 fluxes: an Australian perspective. Soil Research, Rome, v. 49, p. 377-388, 2011. REDDY, K. R.; DELAUNE, R. D. Biogeochemistry of wetlands: science and applications. London: Taylor & Francis, 2008. SCHEER, M. B.; CURCIO, G. R.; RODERJAN, C. V. Funcionalidades ambientais de solos altomontanos na Serra da Igreja, Paraná. Revista Brasileira de Ciência do Solo, Campinas, v. 35, p. 1013-1026, 2011. SOIL SURVEY STAFF. Keys to soil taxonomy. 5. ed. Virginia: Pocahontas, 1992. WANG, Z. P.; ZENG, D.; PATRICK, W. H. Methane emissions from natural wetlands. Environmental Monitoring and Assessment, Dordrecht, v. 42, p. 143-161, 1996. WHITING, G. J.; CHANTON, J. P. Greenhouse carbon balance of wetlands: methane emission versus carbon sequestration. Tellus. Serie B: Chemical and Physical

24
Meteorology, Hoboken, v. 53, p. 521-528, 2001.

25
2 CAPÍTULO I
FLUXO DE METANO EM ORGANOSSOLO SOB ESTEPE HIDRÓFILA EM
CONDIÇÃO NATURAL E APÓS DRENAGEM - PARANÁ - BR
RESUMO
Organossolos estocam quantidades expressivas de carbono orgânico e podem atuar como fonte ou sumidouro de metano (CH4), dependendo do balanço entre metanogênese e metanotrofia. Atributos pedológicos e fatores meteorológicos, além das práticas de manejo, influenciam os fluxos de metano no solo. O objetivo deste estudo foi avaliar os fluxos de metano em Organossolo Háplico sáprico típico sob estepe hidrófila de altitude, em condição natural e após drenagem artificial. Variáveis pedológicas e meteorológicas foram avaliadas para auxiliar a interpretação dos fluxos. Uma pesquisa experimental foi conduzida em uma nascente do rio Tibagi (Ponta Grossa, PR), onde foram avaliados os fluxos de CH4. Seis câmaras estáticas foram instaladas em cada área, possibilitando a coleta de amostras de ar durante 26 meses, sendo que a concentração de CH4 foi determinada por cromatografia gasosa. No solo em condição natural ocorreu efluxo líquido de CH4 cujas taxas variaram de 238 a 5365 µg de CH4 m-2 h-1. Por sua vez, na área drenada artificialmente, a oxigenação do solo determinou o influxo líquido em taxas que variaram de 39 a 242 µg de CH4 m
-2 h-1. No solo drenado, a menor altura média do nível freático (-57,6 cm) e, consequentemente, a menor umidade gravimétrica média na camada de 0-10 cm (5,1 kg kg-1), foram os principais fatores determinantes para o consumo acumulado de 10 kg ha-1 ano-1 de CH4. Em condição natural, o nível freático médio ficou 6 cm abaixo da superfície e a umidade gravimétrica média foi 9,9 kg kg-1 na camada de 0-10 cm, sendo que a emissão de CH4 foi relacionada à temperatura média do ar, totalizando 167 kg ha-1 ano-1 de CH4. Apesar de Organossolos sob estepe hidrófila atuarem como fonte de metano, os mesmos possuem importantes funções no ecossistema, como acumular carbono orgânico e armazenar água, não devendo, portanto, ser drenados como alternativa para redução da emissão de metano, mas efetivamente preservados. Palavras-chave: Gás de efeito estufa. Nascentes. Nível freático. Umidade
gravimétrica. Temperatura do ar e precipitação. Potencial redox.

26
METHANE FLUXES IN A WETLAND HISTOSOL UNDER NATURAL AND
DRAINED CONDITIONS - PARANÁ - BRAZIL
ABSTRACT
Histosols traps significant amounts of organic carbon and may serve as source or sink of methane (CH4), depending upon the balance between methanogenesis and methanotrophy. Pedological attributes and climate factors, as well as management practices, influence the fluxes of CH4 in the soil. The purpose of this study was to assess CH4 flow in a Haplic Histosol of a high altitude wetland, either under natural or drained conditions. Soil and climate variables were also assessed. An experiment was conducted in the source of Tibagi river (Ponta Grossa, PR), where CH4 emissions were assessed in a natural and drained area. Six static chambers were installed in each area, that allowed air sampling during 26 months, and the CH4 concentration was determined by gas chromatography. The natural soil showed a net efflux of CH4, with rates varying from 238 to 5365 µg CH4 m
-2 h-1. In drained soil, the lowest water table level (-57,6 cm) and the lower gravimetric water content in 0-10 cm (5,1 kg kg-1) were the main causes of the cumulative consumption of 10 kg ha-1 yr-1 of CH4. In natural condition, the water table level was in average -6 cm and the gravimetric water content was 9.9 kg kg-1 in 0-10 cm, being the CH4 emission related to the average air temperature, totalizing 167 kg ha-1 yr-1 of CH4. Despite Histosol of wetlands are CH4 sources, those soils have important ecosystem functions, like storage of organic carbon and water, so that they must not be drained as an alternative to curb CH4 emission, but effectively preserved instead.
Keywords: Greenhouse gases. Springs. Water table. Water content. Air temperature. Precipitation. Redox potential.

27
2.1 INTRODUÇÃO
Depois do dióxido de carbono e do vapor de água o metano (CH4) é o gás
mais importante para o efeito estufa (BAIRD, 2002). Uma molécula de metano causa
um aquecimento 25 vezes maior do que o CO2 (IPCC, 2007).
O CH4, cuja emissão é afetada por mudanças no uso do solo, contribui com
15 a 20% das emissões globais de gases de efeito estufa (WUEBBLES et al., 2000;
IPCC, 2007), enquanto os ecossistemas naturais, como terras úmidas e lagos, são
responsáveis por aproximadamente 1/3 da emissão global de CH4 (FORSTER et al.,
2007). Solos hidromórficos, incluindo Organossolos, correspondem a 3% da
superfície terrestre e apenas 1% do território brasileiro (VALLADARES, 2003). Esses
solos têm grande capacidade de estocar carbono (C) em comparação aos solos que
apresentam nível freático baixo (RAICH; POTTER, 1995; ALONGI et al., 2001;
WHITING; CHANTON, 2001). Por outro lado, podem ser significativas fontes de CH4
(HAYES; CLAPP, 2001; WHALEN, 2005).
Os fluxos de CH4 do solo representam o balanço entre os processos de
metanogênese e metanotrofia. Estes processos são regulados pelo suprimento de
oxigênio no solo e disponibilidade de C lábil, sendo que em condições anaeróbicas
predomina a metanogênese. Atributos e processos pedológicos (ROULET; MOORE,
1995; WHALEN, 2005; JAUHIAINEN et al., 2008), microbiológicos (Le MER;
ROGER, 2001; KÖGEL-KNABNER et al., 2010; PAGE; DALAL, 2011) e fatores
climáticos (von ARNOLD et al., 2005; DALAL; ALLEN, 2008; JIANG et al., 2009),
além da vegetação (FURUKAWA et al., 2005; KOH et al., 2009) e do manejo das
áreas (ELDER; LAL, 2008; HUANG et al., 2010), controlam os fluxos de CH4 em
solos hidromórficos.
A oscilação do nível freático e a altura em que o mesmo permanece em
Organossolos altera a intensidade e a duração dos processos de produção e
oxidação de CH4 (FURUKAWA et al., 2005; von ARNOLD et al., 2005; JAUHIAINEN
et al., 2008). Quando a altura do nível freático apresenta pequena amplitude de
variação, o potencial máximo de produção de CH4 em Organossolo ocorre em torno
de 20 cm abaixo do nível de água, enquanto o potencial máximo de oxidação pode
ocorrer até 10 cm abaixo da lâmina de água (KETTUNEN et al., 1999), porque ainda

28
há oxigênio nesta profundidade.
Nestes solos, a emissão de CH4 diminui logaritmicamente com o
rebaixamento do nível freático (MOORE; DALVA, 1993; FURUKAWA et al., 2005), e
com o aumento nos valores de potencial redox (FURUKAWA et al., 2005; KOH et al.,
2009).
A drenagem artificial também afeta a capacidade destas áreas em fixar
carbono e armazenar água. Huang et al. (2010) estimaram que os solos dos
pântanos que foram convertidos em lavouras, entre 1950 e 2000, perderam
aproximadamente 50% do carbono orgânico, sendo que 70% desta perda se deu
entre 0 e 20 cm de profundidade. Andrade (2010) registrou redução de 6 para 2 kg
kg-1 na umidade gravimétrica na camada de 0 a 10 cm, comparando Organossolo
natural e drenado.
Entre os fatores climáticos que condicionam os fluxos de CH4, a temperatura
e a precipitação são muito citados na literatura, embora a influência dos mesmos
sobre os fluxos seja muito variável (Le MER; ROGER, 2001; WHALEN, 2005;
DALAL; ALLEN, 2008; KOH et al., 2009; HUANG et al., 2010).
No Estado do Paraná, áreas de nascentes são de extrema recorrência nas
paisagens situadas em altitudes superiores a 800m, principalmente nos reversos de
cuestas, onde ocorrem Organossolos sob estepe hidrófila, os quais constituem
importantes reservatórios de carbono e água, além de abrigarem, segundo Kozera
(2008) e Andrade (2010), elevada biodiversidade. Dependendo do grau de
decomposição da matéria orgânica, estes solos podem armazenar uma quantidade
de água equivalente a 4,5 a 30 vezes o seu peso seco (SOIL SURVEY STAFF,
1992). Scheer et al. (2011) indicam que Organossolos sob campo altomontano
estocam 314,7 Mg ha-1 de carbono e 3,4 milhões de litros de água por hectare até a
profundidade de 58 cm. No que se refere a riqueza de espécies vegetais, Andrade
(2011) e Kozera (2008) registraram, respectivamente, 146 espécies em 0,65 ha e 87
espécies em aproximadamente 4 ha, em Organossolos de altitude sob vegetação
campestre.
Entretanto, a maioria dessas nascentes encontram-se alteradas por
eventuais usos agropecuários, que promovem a alteração das suas funções
ecológicas. No Brasil, informações sobre fluxos de CH4 em ambientes úmidos
naturais de ocorrência de Organossolos e os efeitos da drenagem artificial sobre tais
fluxos, são inexistentes.

29
Elaborou-se a hipótese de que a drenagem alteraria intensamente os fluxos
de metano e que as oscilações na temperatura do ar e na precipitação também
afetariam estes fluxos, tanto no organossolo natural como no drenado.
Este estudo objetivou avaliar os fluxos de CH4 em Organossolo Háplico
sáprico típico, sob condição natural e drenado, considerando a influência de
variáveis pedológicas e climáticas ao longo do tempo.
30
2.2 MATERIAL E MÉTODOS
2.2.1 Experimento de campo
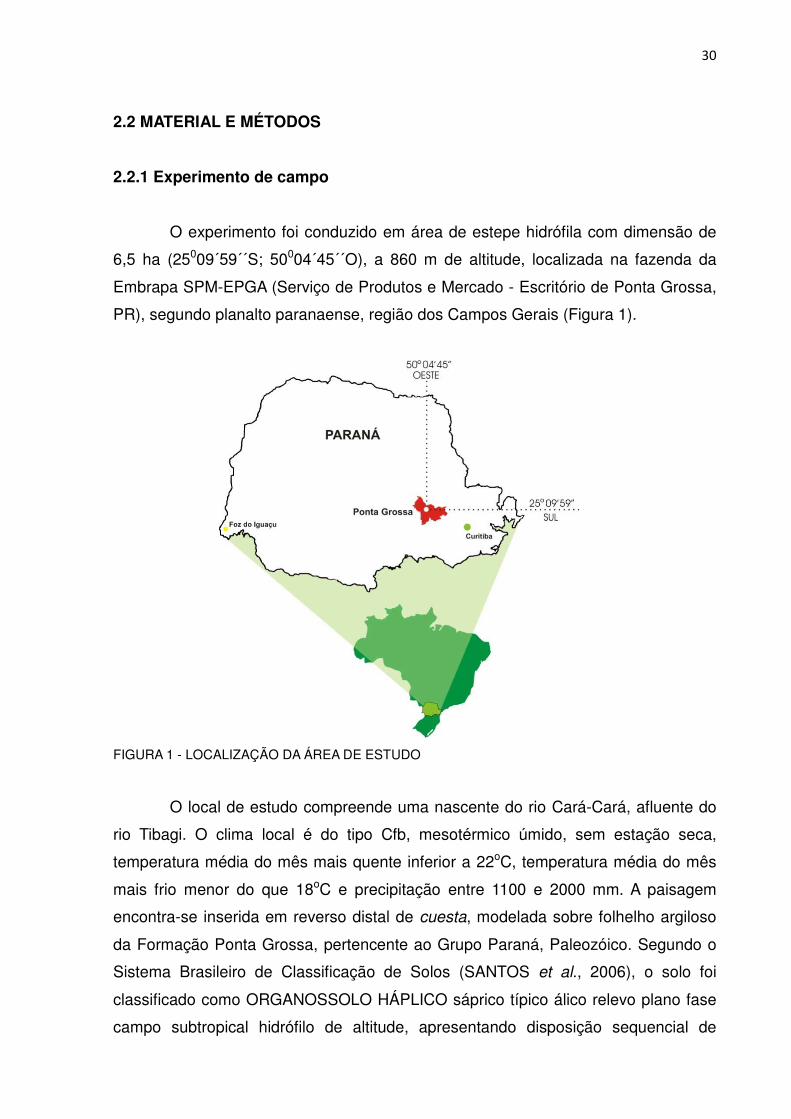
O experimento foi conduzido em área de estepe hidrófila com dimensão de
6,5 ha (25009´59´´S; 50004´45´´O), a 860 m de altitude, localizada na fazenda da
Embrapa SPM-EPGA (Serviço de Produtos e Mercado - Escritório de Ponta Grossa,
PR), segundo planalto paranaense, região dos Campos Gerais (Figura 1).
FIGURA 1 - LOCALIZAÇÃO DA ÁREA DE ESTUDO
O local de estudo compreende uma nascente do rio Cará-Cará, afluente do
rio Tibagi. O clima local é do tipo Cfb, mesotérmico úmido, sem estação seca,
temperatura média do mês mais quente inferior a 22oC, temperatura média do mês
mais frio menor do que 18oC e precipitação entre 1100 e 2000 mm. A paisagem
encontra-se inserida em reverso distal de cuesta, modelada sobre folhelho argiloso
da Formação Ponta Grossa, pertencente ao Grupo Paraná, Paleozóico. Segundo o
Sistema Brasileiro de Classificação de Solos (SANTOS et al., 2006), o solo foi
classificado como ORGANOSSOLO HÁPLICO sáprico típico álico relevo plano fase
campo subtropical hidrófilo de altitude, apresentando disposição sequencial de
31
horizontes hísticos, em elevado grau de decomposição, com 100 cm de espessura.
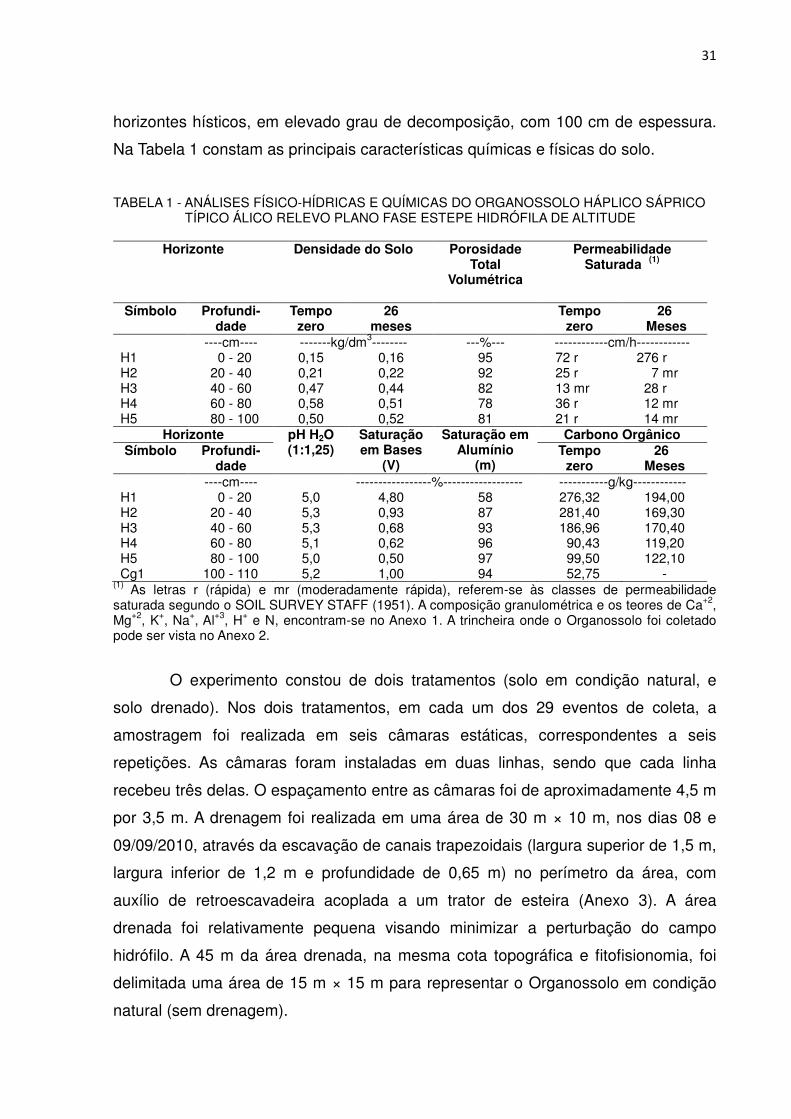
Na Tabela 1 constam as principais características químicas e físicas do solo.
TABELA 1 - ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE
Horizonte Densidade do Solo Porosidade
Total Volumétrica
Permeabilidade Saturada (1)
Símbolo Profundi-dade
Tempo zero
26 meses
Tempo zero
26 Meses
----cm---- -------kg/dm3-------- ---%--- ------------cm/h------------ H1 0 - 20 0,15 0,16 95 72 r 276 r H2 20 - 40 0,21 0,22 92 25 r 7 mr H3 40 - 60 0,47 0,44 82 13 mr 28 r H4 60 - 80 0,58 0,51 78 36 r 12 mr H5 80 - 100 0,50 0,52 81 21 r 14 mr
Horizonte pH H2O (1:1,25)
Saturação em Bases
(V)
Saturação em Alumínio
(m)
Carbono Orgânico Símbolo Profundi-
dade Tempo
zero 26
Meses ----cm---- -----------------%------------------ -----------g/kg------------
H1 0 - 20 5,0 4,80 58 276,32 194,00 H2 20 - 40 5,3 0,93 87 281,40 169,30 H3 40 - 60 5,3 0,68 93 186,96 170,40 H4 60 - 80 5,1 0,62 96 90,43 119,20 H5 80 - 100 5,0 0,50 97 99,50 122,10 Cg1 100 - 110 5,2 1,00 94 52,75 -
(1) As letras r (rápida) e mr (moderadamente rápida), referem-se às classes de permeabilidade saturada segundo o SOIL SURVEY STAFF (1951). A composição granulométrica e os teores de Ca+2, Mg+2, K+, Na+, Al+3, H+ e N, encontram-se no Anexo 1. A trincheira onde o Organossolo foi coletado pode ser vista no Anexo 2.
O experimento constou de dois tratamentos (solo em condição natural, e
solo drenado). Nos dois tratamentos, em cada um dos 29 eventos de coleta, a
amostragem foi realizada em seis câmaras estáticas, correspondentes a seis
repetições. As câmaras foram instaladas em duas linhas, sendo que cada linha
recebeu três delas. O espaçamento entre as câmaras foi de aproximadamente 4,5 m
por 3,5 m. A drenagem foi realizada em uma área de 30 m × 10 m, nos dias 08 e
09/09/2010, através da escavação de canais trapezoidais (largura superior de 1,5 m,
largura inferior de 1,2 m e profundidade de 0,65 m) no perímetro da área, com
auxílio de retroescavadeira acoplada a um trator de esteira (Anexo 3). A área
drenada foi relativamente pequena visando minimizar a perturbação do campo
hidrófilo. A 45 m da área drenada, na mesma cota topográfica e fitofisionomia, foi
delimitada uma área de 15 m × 15 m para representar o Organossolo em condição
natural (sem drenagem).

32
2.2.2 Coleta de amostras de ar
A avaliação das emissões de CH4 iniciaram um dia após a drenagem do solo
e no período de um pouco mais de dois anos (10/9/2010 a 16/10/2012) foram
realizadas 29 amostragens de ar. No primeiro mês, a frequência de amostragem foi
mais intensa, para não se perder o efeito inicial da drenagem. Com este objetivo
foram realizadas avaliações aos 1, 4, 8 e 14 dias, seguidas de 25 coletas mensais.
O primeiro ano de estudo refere-se ao período compreendido entre 10/9/2010 e
26/9/2011 e o segundo, entre 27/9/2011 e 16/10/2012 (Anexo 4).
Imediatamente após a drenagem do solo, seis bases circulares de metal
(Anexo 5) foram instaladas em cada tratamento. As bases utilizadas tinham diâmetro
interno de 38 cm e foram inseridas a 15 cm de profundidade no solo. Na parte
inferior da base, junto a superfície do solo, foram abertos quatro orifícios circulares
com 2 cm de diâmetro, para permitir a circulação de água dentro das bases. No
momento das coletas de amostras de ar estes orifícios foram fechados com rolha de
borracha.
Amostras de ar foram coletadas em câmaras estáticas baseando-se nos
modelos utilizados por Gomes et al. (2009) e Elder e Lal (2008). Em cada evento de
coleta, câmaras de PVC (Anexo 5) com 40 cm de altura foram assentadas sobre
uma canaleta situada na parte superior da base com 4 cm de altura e 6 cm de
largura. Essa canaleta foi preenchida com água para vedar o sistema câmara-base.
Em sua parte superior, as câmaras foram equipadas com termômetro, ventilador
para homogeneizar o ar e torneira de três vias acoplada a um septo de borracha
para a tomada de amostras, que foi feita por seringas de polipropileno de 20 ml. As
amostras de ar foram coletadas aos 0, 15, 30 e 45 minutos após o fechamento da
câmara, no período entre 9:00 e 11:00 horas da manhã.
As seringas contendo as amostras foram acondicionadas em caixa de isopor
com gelo reutilizável a base de polímero e enviadas imediatamente, por SEDEX-10,
para o Laboratório de Biogeoquímica Ambiental da UFRGS, em Porto Alegre, RS. A
partir da coleta 20 (04/01/2012) o conteúdo das seringas coletoras de gás passou a
ser transferido para vials de vidro da marca LABCO, com volume de 12 ml.
A concentração de CH4 foi determinada em cromatógrafo de fase gasosa
(Shimadzu GC-2014) equipado com colunas Porapak-Q com temperatura de 70 °C,

33
gás de arraste N2 a vazão de 30 mL min-1, temperatura do metanador de 380 °C e
detector de ionização de chama (FID) de 250 °C.
Os fluxos de CH4 foram calculados pela equação F(CH4) = d[CH4]/dt x 12 x P
x V/ R x T, onde d[CH4]/dt é a mudança na quantidade do gás dentro da câmara no
intervalo de tempo; V e T são o volume (L) e temperatura interna (0K) da câmara; P é
a pressão no interior da câmara, assumida como 1 atm e R é a constante universal
dos gases (0,0821 L.atm K-1 mol-1). A variação da concentração do gás no tempo foi
obtida pelo coeficiente angular da equação da reta ajustada. Considerando-se a
área de solo contido na base da câmara, calculou-se a massa do gás que foi emitida
por unidade de área e de tempo.
Os fluxos obtidos em µg CH4 m-2 h-1 foram considerados como
representativos do fluxo médio diário, conforme proposto por Jantália et al. (2008), e
o efluxo ou influxo líquido acumulado por estação e anual foram calculados pela
integração da área sob a curva, a qual foi estabelecida por interpolação dos valores
diários dos fluxos.
2.2.3 Parâmetros pedológicos
As análises químicas e físico-hídricas do solo foram realizadas conforme
metodologia proposta por Claessen (1997). O teor de carbono orgânico foi
determinado por oxidação sob atmosfera de gás oxigênio em analisador automático
Analytik Jena.
Em cada evento de amostragem de ar, três amostras de solo de 0 a 10 cm
foram coletadas por parcela para determinação da umidade gravimétrica.
Visando o monitoramento da altura do nível freático, foram instalados quatro
poços hídricos nas parcelas de cada tratamento. Os poços, feitos com tubos de PVC
de 75 mm de diâmetro e 1,20 m de altura, alcançavam o horizonte mineral Cg. Os
tubos possuíam quatro aberturas circulares de aproximadamente 1 cm de diâmetro,
dispostas a cada 10 cm ao longo do comprimento do tubo, para permitir o rápido
fluxo de água. Cada poço hídrico foi instalado entre duas câmaras. Leituras da altura
do nível freático foram feitas a cada coleta do ar, com uso de uma trena rígida.
A quantidade de carbono estocada nos horizontes do solo foi calculada pela
fórmula: estoque de C = teor de C (%) x densidade do solo (g/cm3) x espessura do

34
horizonte (cm). O estoque total de C até 1 metro de profundidade foi obtido pela
soma do C estocado em cada horizonte e expresso em Mg ha-1.
O volume de água armazenado por hectare em cada horizonte do solo foi
obtido pela fórmula: volume de água = [porosidade total (cm3/cm3)/1000] x espessura
do horizonte (cm) x 100.000.000 (cm3). O somatório dos cinco horizontes foi
convertido em milhões de litros/ha, representando o volume total de água
armazenado no solo até a profundidade de 1 metro.
Nas coletas 24, 25, 26, 28 e 29 foi determinado o potencial de oxi-redução
(ORP) dos solos nas profundidades de 5, 10, 15, 20, 30 e 40 cm utilizando um
pHmetro portátil modelo HI 8424 acoplado a um eletrodo combinado de ORP com
corpo de vidro e sensor de platina, ambos da Marca HANNA e faixa de leitura entre
699,9 a 1999 mV, aproximadamente. Utilizou-se as classes de estado de redução,
oxidado > 400 milivolts (mV), fracamente reduzido (400 a 200 mV), moderadamente
reduzido (200 a -100 mV) e fortemente reduzido (< -100 mV), adaptadas de Tian-Yen
(1985).
2.2.4 Variáveis meteorológicas
A temperatura do ar e a precipitação foram monitoradas diariamente, durante
dois anos, por meio de uma estação meteorológica portátil Vantage Vue marca Davis
instalada a aproximadamente 1 km da área experimental. O equipamento
armazenava 24 registros diários de temperatura e de precipitação gerados com a
média de cada hora.
2.2.5 Análise estatística
Os valores dos fluxos de CH4, umidade gravimétrica, altura do nível freático
e potencial redox foram submetidos à análise de variância. As diferenças entre as
médias do organossolo natural e drenado foram avaliadas pelo teste de Tukey (5%).
Para avaliar o efeito isolado das variáveis pedológicas e climáticas, foram
determinados os coeficientes de correlação linear de Pearson (r) entre os fluxos de
CH4 e o nível freático, a umidade gravimétrica na profundidade de 0 a 10 cm, a

35
temperatura média do ar e a precipitação acumulada, entre às 9 e 12 horas na
ocasião das coletas e durante os períodos de 1, 3, 5, 7, 15 e 30 dias que
antecederam as coletas.
Para avaliar conjuntamente os efeitos dos tratamentos (com e sem
drenagem), dos componentes principais, os quais foram compostos pelos fatores
pedológicos e meteorológicos e, do tempo (número de dias), sobre os fluxos de CH4,
utilizou-se o Modelo Aditivo Generalizado (GAM) o qual além de analisar todas as
variáveis simultaneamente, considera o fato de que as determinações dos fluxos ao
longo do tempo não são isoladas entre si.
Todas as análises estatísticas foram realizadas com o Software R (R
DEVELOPMENT CORE TEAM 2011).
36
2.3 RESULTADOS E DISCUSSÃO
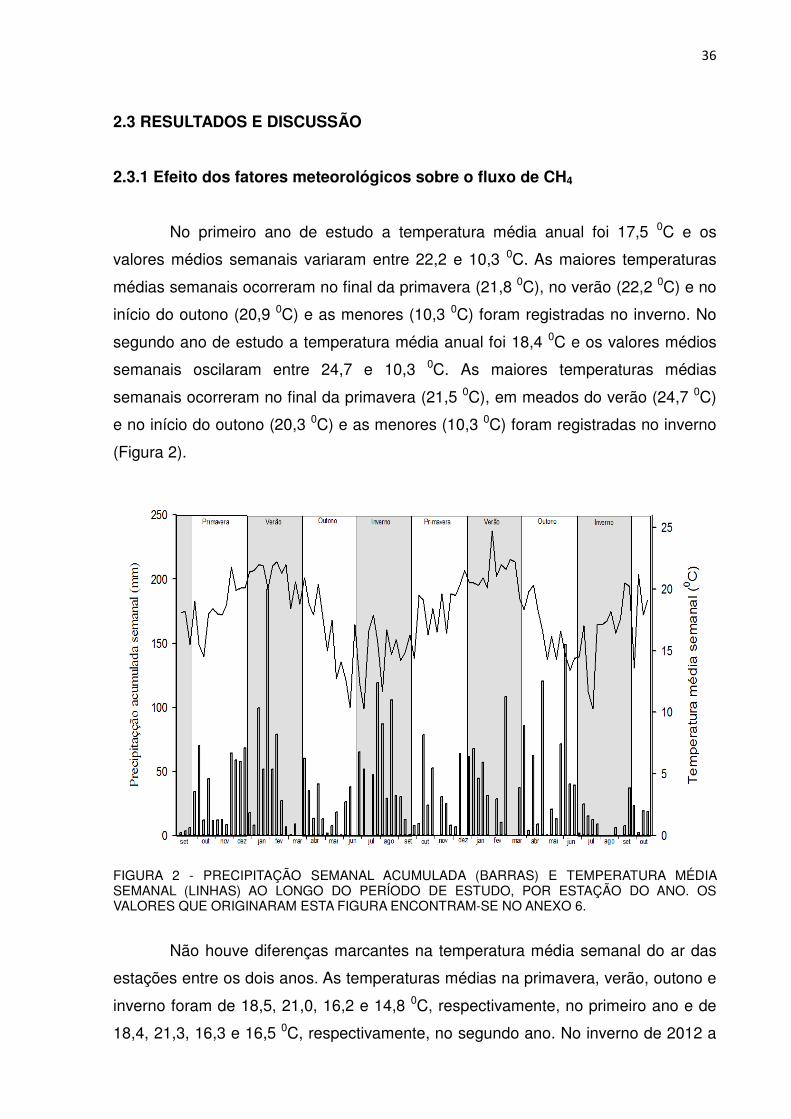
2.3.1 Efeito dos fatores meteorológicos sobre o fluxo de CH4
No primeiro ano de estudo a temperatura média anual foi 17,5 0C e os
valores médios semanais variaram entre 22,2 e 10,3 0C. As maiores temperaturas
médias semanais ocorreram no final da primavera (21,8 0C), no verão (22,2 0C) e no
início do outono (20,9 0C) e as menores (10,3 0C) foram registradas no inverno. No
segundo ano de estudo a temperatura média anual foi 18,4 0C e os valores médios
semanais oscilaram entre 24,7 e 10,3 0C. As maiores temperaturas médias
semanais ocorreram no final da primavera (21,5 0C), em meados do verão (24,7 0C)
e no início do outono (20,3 0C) e as menores (10,3 0C) foram registradas no inverno
(Figura 2).
FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO. OS VALORES QUE ORIGINARAM ESTA FIGURA ENCONTRAM-SE NO ANEXO 6.
Não houve diferenças marcantes na temperatura média semanal do ar das
estações entre os dois anos. As temperaturas médias na primavera, verão, outono e
inverno foram de 18,5, 21,0, 16,2 e 14,8 0C, respectivamente, no primeiro ano e de
18,4, 21,3, 16,3 e 16,5 0C, respectivamente, no segundo ano. No inverno de 2012 a

37
temperatura média semanal foi apenas 1,7 0C mais elevada do que no inverno de
2011.
A precipitação total acumulada ao longo do primeiro ano foi de 1837 mm. As
estações mais chuvosas foram a primavera (442 mm), o verão (543 mm) e,
atipicamente, o inverno (580 mm). No outono a precipitação acumulada foi 254 mm.
A precipitação total acumulada ao longo do segundo ano foi de 1518 mm. As
estações mais chuvosas foram o verão (446 mm) e atipicamente o outono (615 mm),
seguidos pela primavera (298 mm) e inverno (114 mm). A precipitação anual no
segundo ano de estudo foi 319 mm menor do que no primeiro, sendo que com
exceção do outono, as outras estações foram menos chuvosas.
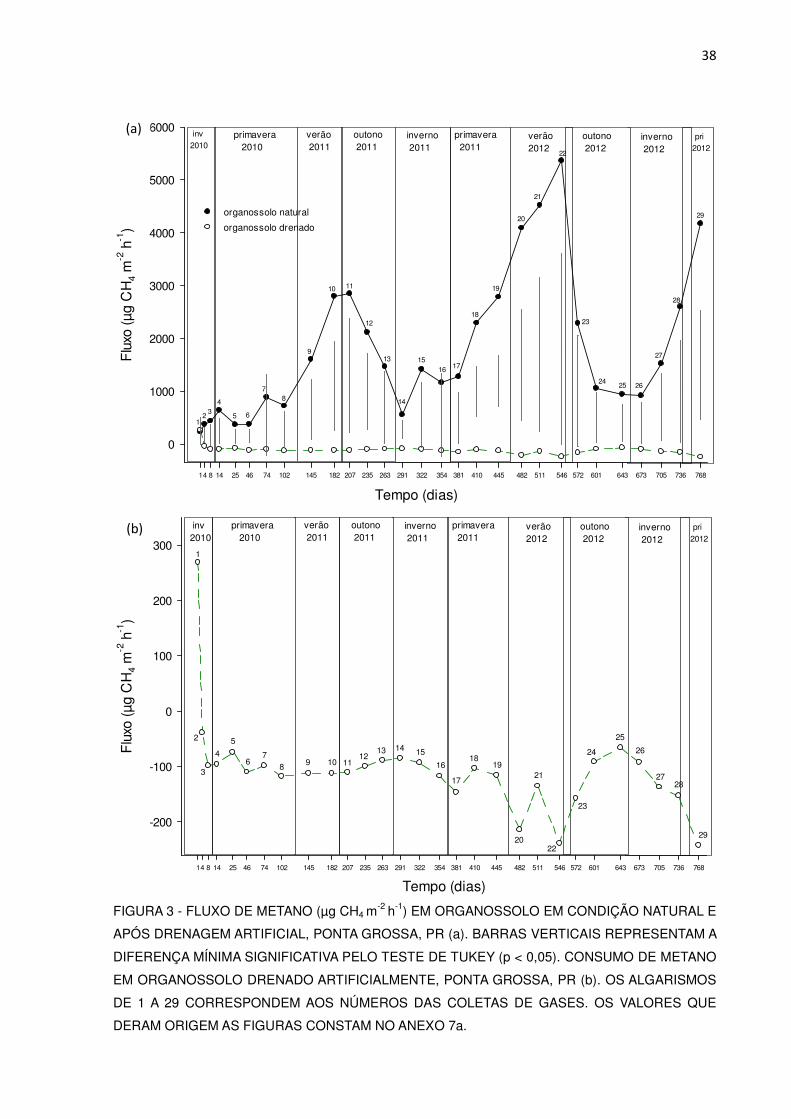
No Organossolo natural, no primeiro ano de estudo, a emissão de metano
oscilou entre 238 e 2850 µg m-2 h-1 de CH4 (Figura 3), com média de 1.137 µg m-2 h-1
e no segundo entre 918 e 5.365 µg m-2 h-1 de CH4, e média de 2.714 µg m-2 h-1, ou
seja, 2,4 vezes superior.
Os valores mais elevados de emissão de todo o estudo ocorreram entre os
meses de janeiro a março de 2012, oscilando entre 4.087 a 5.365 µg m-2 h-1.
Considerando os dois anos de estudo, os picos de emissão ocorreram nos meses
mais quentes (Figura 3a), tendo-se ocorrido correlação positiva entre a emissão de
metano e a temperatura média do ar (Tabela 2).
No primeiro ano os fluxos mais elevados (2.800 e 2850 µg m-2 h-1, coletas 10
e 11, respectivamente, Figura 3) coincidiram com os meses mais quentes,
correspondentes ao final do verão e início de outono (Figura 2). Do mesmo modo, no
segundo ano, os maiores fluxos (coletas 20 a 22 e 29), também ocorreram nos
meses mais quentes, o que neste caso, correspondeu aos meses do verão e da
primavera. Em ambos os anos as menores emissões ocorreram no inverno.
Huang et al. (2010) também encontraram maiores emissões durante os
meses de verão e primavera em lavouras de arroz no nordeste da China.
Houve uma redução drástica na emissão de CH4 na coleta 14, quando se
registrou taxa de 559 µg m-2 h-1 (Figura 3), provavelmente devido à ocorrência de
geada, a qual baixou a temperatura para 4,1 0C no momento da coleta, reforçando
novamente a interferência da temperatura. O aumento na emissão de CH4 na coleta
15 pode estar associado ao aumento na quantidade de carbono facilmente
decomponível proveniente da biomassa de comunidades de microrganismos mortos
com a geada que houve no mês anterior.
38
Tempo (dias)/Coletas
14 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Flu
xo (µ
g C
H4
m-2
h-1
)
0
1000
2000
3000
4000
5000
6000 inv2010
primavera 2010
verão 2011
outono 2011
inverno 2011
primavera 2011
verão 2012
outono 2012
inverno 2012
pri2012
organossolo natural
organossolo drenado
12
34
5 6
78
9
10 11
12
13
14
1516 17
18
19
20
21
22
23
2425 26
27
28
29
Tempo (dias)
14 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Flu
xo (µ
g C
H4
m-2
h-1
)
-200
-100
0
100
200
300
inv2010
primavera 2010
verão 2011
outono 2011
inverno 2011
primavera 2011
verão 2012
outono 2012
inverno 2012
pri2012
1
2
3
4
5
67
8 9 10 1112
13 14 15
16
17
1819
20
21
22
23
24
25
26
2728
29
FIGURA 3 - FLUXO DE METANO (µg CH4 m-2
h-1) EM ORGANOSSOLO EM CONDIÇÃO NATURAL E
APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR (a). BARRAS VERTICAIS REPRESENTAM A
DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). CONSUMO DE METANO
EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR (b). OS ALGARISMOS
DE 1 A 29 CORRESPONDEM AOS NÚMEROS DAS COLETAS DE GASES. OS VALORES QUE
DERAM ORIGEM AS FIGURAS CONSTAM NO ANEXO 7a.
(b)
(a)(a)
39
Na coleta 26 houve outra geada, porém menos intensa, com temperatura
média no momento da coleta de 8,9 oC. A elevação na emissão de metano na coleta
27 também pode estar relacionada ao aumento na quantidade de carbono
prontamente acessível, oriundo de organismos mortos pelo frio.
Panikov e Dedysh (2000) trabalhando com Organossolos, associaram uma
elevação na emissão de metano ao incremento na quantidade de carbono disponível
para os microrganismos, proveniente da matéria orgânica liberada pela quebra de
agregados ou de microrganismos mortos pelos ciclos de congelamento/degelo.
No segundo ano os fluxos se elevaram a partir de meados da primavera de
2011 (Figura 3a) atingindo o pico máximo no final do verão (5.365 µg m-2 h-1, coleta
22), retraindo-se no outono e inverno para ascender novamente no início da
primavera de 2012 (4.174 µg m-2 h-1, coleta 29). A emissão média durante o verão do
segundo ano (4.658 µg m-2 h-1) foi duas vezes maior do que no primeiro (2.203 µg m-
2 h-1).
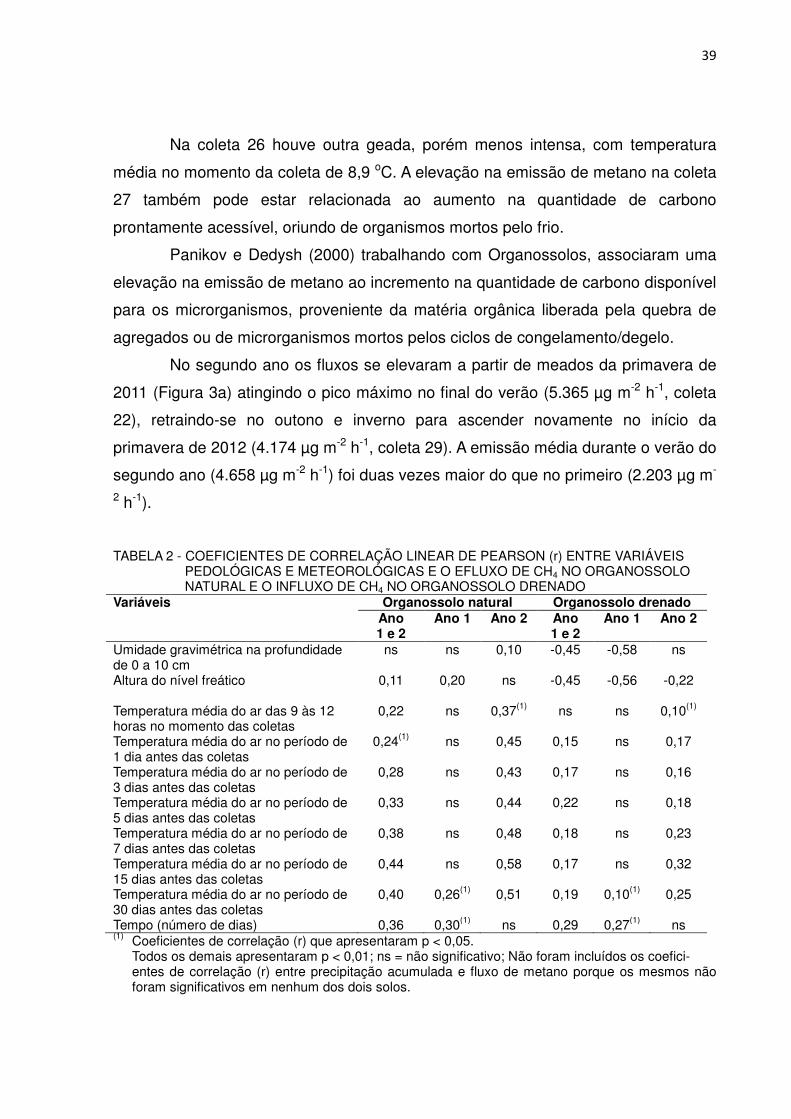
TABELA 2 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE VARIÁVEIS PEDOLÓGICAS E METEOROLÓGICAS E O EFLUXO DE CH4 NO ORGANOSSOLO NATURAL E O INFLUXO DE CH4 NO ORGANOSSOLO DRENADO
Variáveis Organossolo natural Organossolo drenado Ano 1 e 2
Ano 1 Ano 2 Ano 1 e 2
Ano 1 Ano 2
Umidade gravimétrica na profundidade de 0 a 10 cm
ns ns 0,10 -0,45 -0,58 ns
Altura do nível freático 0,11 0,20 ns -0,45 -0,56 -0,22 Temperatura média do ar das 9 às 12 horas no momento das coletas
0,22
ns
0,37(1)
ns
ns
0,10(1)
Temperatura média do ar no período de 1 dia antes das coletas
0,24(1) ns 0,45 0,15 ns 0,17
Temperatura média do ar no período de 3 dias antes das coletas
0,28 ns 0,43 0,17 ns 0,16
Temperatura média do ar no período de 5 dias antes das coletas
0,33 ns 0,44 0,22 ns 0,18
Temperatura média do ar no período de 7 dias antes das coletas
0,38 ns 0,48 0,18 ns 0,23
Temperatura média do ar no período de 15 dias antes das coletas
0,44 ns 0,58 0,17 ns 0,32
Temperatura média do ar no período de 30 dias antes das coletas
0,40 0,26(1) 0,51 0,19 0,10(1) 0,25
Tempo (número de dias) 0,36 0,30(1) ns 0,29 0,27(1) ns (1) Coeficientes de correlação (r) que apresentaram p < 0,05.
Todos os demais apresentaram p < 0,01; ns = não significativo; Não foram incluídos os coefici- entes de correlação (r) entre precipitação acumulada e fluxo de metano porque os mesmos não foram significativos em nenhum dos dois solos.

40
A influência do aumento da temperatura na elevação da emissão de metano
no solo natural, ficou fortemente evidenciada, uma vez que as temperaturas médias
semanais mais elevadas em 2012, no verão, no final do inverno e no início da
primavera (Figura 2), contribuíram para o aumento na emissão de metano, nas
coletas 21, 28 e 29, respectivamente (Figura 3).
Não houve correlação entre a emissão de metano e a precipitação
acumulada em ambos os anos. A precipitação praticamente não afetou os fluxos de
CH4, provavelmente porque no solo natural a elevada umidade natural se manteve
constante, não sofrendo acréscimo considerável quando chovia.
A relação entre o fluxo de CH4 e os fatores climáticos é muito variável (Le
MER; ROGER, 2001; JOABSSON; CHRISTENSEN, 2001; WHALEN, 2005; DALAL;
ALLEN, 2008; KOH et al., 2009). Em pesquisas com Organossolos naturais, Jiang et
al. (2009) demonstraram que nos meses mais quentes e chuvosos a emissão de
CH4 foi maior, enquanto Inubushi et al. (2003) encontraram correlação positiva
apenas com a precipitação. Por outro lado, também em Organossolos, von Arnold et
al. (2005) indicaram que em solo não drenado a temperatura do ar teve grande
influência na emissão de CH4. Por sua vez, Furukawa et al. (2005) não verificaram
relação entre a emissão de CH4 e a temperatura do ar, mas sim, com a precipitação,
em lavoura de arroz irrigado.
No Organossolo drenado, no primeiro ano de estudo, os fluxos oscilaram
entre -99 e -146 µg m-2 h-1 de CH4, com média de -104 µg m-2 h-1, desconsiderando-
se os valores das coletas 1 e 2, os quais ainda não representavam o efeito da
drenagem (Figura 3b). No segundo ano os fluxos variaram entre -66 e -242 µg m-2 h-
1 de CH4, com média de -145 µg m-2 h-1, ou seja, o consumo médio foi 1,4 vezes
maior do que no primeiro ano.
Em todas as coletas, com exceção da primeira, da sétima e da décima
sexta, os fluxos de metano no solo drenado foram estatisticamente diferentes dos
fluxos no solo natural, tendo ocorrido consumo no primeiro e emissão no segundo,
exceto na coleta 1, na qual em ambos os solos houve emissão (Figura 3a), porque o
efeito da drenagem ainda não havia se manifestado.
No primeiro ano, praticamente não houve picos de consumo com exceção
da coleta 17, no início da primavera de 2011 (-146 µg m-2 h-1), enquanto no segundo
ano ocorreram dois picos de consumo no verão (-214 e -239 µg m-2 h-1, coletas 20 e
22 respectivamente), o que em média, representa quase o dobro do consumo médio

41
(-112 µg m-2 h-1) ocorrido no verão do ano anterior (Figura 3b). Entretanto, não houve
diferença estatística no consumo de metano entre as coletas nos dois anos (Anexo
7a).
No primeiro ano houve uma influência pequena da variação da temperatura
durante os 30 dias que antecederam as coletas sobre o consumo de metano, mas
durante o segundo ano a temperatura passou a induzir um maior incremento no
consumo de metano (Tabela 2), o qual apresentou maior oscilação (Figura 3b). Jang
et al. (2006) afirmaram que o efeito da temperatura sobre a oxidação do metano não
é consistente, sendo muito maior sobre a produção do mesmo.
No Organossolo drenado, onde o ambiente era favorável para os
organismos metanotróficos, as variações nos fatores climáticos ao longo do tempo
alteraram pouco as taxas de consumo de CH4. Este fenômeno sugere que os
organismos metanotróficos sejam menos suscetíveis às oscilações de temperatura e
precipitação, do que os organismos metanogênicos predominantes no solo sob
condição natural.
As populações de organismos metanogênicos e metanotróficos são
sensíveis às variações de temperatura do ar (DUNFIELD et al., 1993; van HULZENA
et al., 1999; MEGONIGAL; SCHLESINGER, 2002), embora organismos
metanotróficos pareçam ser menos sensíveis à temperatura do solo em
Organossolos, bem como à temperatura do ar, do que os metanogênicos
(DUNFIELD et al., 1993; KOH et al., 2009). Isto ajuda a explicar que no Organossolo
em condições naturais, do presente estudo, o aumento na emissão de CH4 (Figura
3a) acompanhou as elevações de temperatura do ar (Figura 2), tendo as reduções
na mesma ocasionado retração na produção. Porém, como existem outros fatores
que afetam a metanogênese e a metanotrofia, como variações sazonais na estrutura
da comunidade microbiana e composição da matéria orgânica, é possível que haja
uma relação não linear entre o fluxo de CH4 e a temperatura (WILSON et al., 1989;
JOABSSON; CHRISTENSEN, 2001).
Durante os meses de janeiro a março do segundo ano (coletas 20 a 22),
houve uma tendência de incremento no consumo de metano no Organossolo
drenado (Figura 3b). A temperatura média semanal na primeira semana de fevereiro
foi de 24,7 0C e de 21,9 0C, respectivamente em 2012 e 2011. Isto induz a pensar
que houve um efeito conjunto do tempo de drenagem e da elevação da temperatura,
no segundo ano, sobre o consumo de metano. Além disto, as comunidades de

42
organismos metanotróficos possivelmente tiveram condição para se adaptarem às
mudanças ambientais que se estabeleceram no solo após este maior tempo de
drenagem. Ainda na Figura 3b, observa-se nitidamente a tendência descendente dos
fluxos, a partir do final do outono de 2012 (coleta 25), o que indica o incremento no
consumo de metano com o passar do tempo. Isto é comprovado pela correlação
positiva existente entre o consumo de metano e o número de dias decorridos a partir
da drenagem, quando considerados os dois anos conjuntamente (Tabela 2). Isto
possivelmente esteja associado à redução da umidade gravimétrica com o passar do
tempo.
A precipitação também não influenciou os fluxos de metano no solo drenado
pois a água da chuva foi rapidamente removida em função da rápida permeabilidade
no solo (Tabela 1).
2.3.2 Efeito da drenagem sobre o fluxo de CH4
No Organossolo em condições naturais ocorreu emissão líquida de CH4
variando de 238 e 5.365 µg CH4 m-2 h-1, durante todo o período de condução do
experimento (Figura 3). Por sua vez, a drenagem do solo determinou a ocorrência
de consumo de CH4 no Organossolo drenado, cujas taxas variaram de 39 e 242 µg
CH4 m-2 h-1 (Figuras 3a e 3b).
No Organossolo natural o nível freático variou ao longo do primeiro ano entre
0,3 e -16,3 cm, permanecendo em média a -7 cm da superfície do solo e entre 0,25
e -14,8 cm, com média de -5 cm, no segundo ano (Figura 4), indicando que a
oscilação foi muito similar nos dois anos e bem inferior à que ocorreu no
Organossolo drenado. Foi pequeno o coeficiente de correlação entre a oscilação do
nível freático e a emissão de metano analisando-se os dois anos em conjunto
(Tabela 2), pois a variação na altura do freático não foi grande.
A umidade gravimétrica média na profundidade de 0 a 10 cm, no
Organossolo natural, foi de 9,9 kg kg-1 e variou entre 7,6 e 11,7 kg kg-1, no primeiro
ano. No segundo ano a média ficou em 9,8 kg kg-1 tendo oscilado entre 8,1 e 11,9 kg
kg-1 (Figura 5), mostrando também muita similaridade entre os dois anos. Não houve
correlação entre a emissão de CH4 e a umidade gravimétrica na profundidade de 0 a

43
10 cm (Tabela 2), pois possivelmente a produção do mesmo ocorreu em
profundidades maiores.
Deve-se ressaltar que a capacidade de retenção de água do Organossolo
natural é realmente muito elevada, uma vez que na camada de 0 a 10 cm, cada
quilo de solo armazena 10 quilos de água. Comparativamente a um Latossolo
Vermelho Amarelo no qual a umidade gravimétrica na capacidade de campo é de
0,43 kg kg-1 (MILDE et al., 2008), a quantidade de água armazenada no Organossolo
natural é 23 vezes maior.
No Organossolo drenado o nível freático variou ao longo do primeiro ano
entre -25 e -64,8 cm, permanecendo em média a 57 cm abaixo da superfície do solo
e entre -54 e -64,5 cm, com média de -59 cm, no segundo ano (Figura 4).
A umidade gravimétrica média na profundidade de 0 a 10 cm foi de 5,7 kg
kg-1 e variou entre 4,5 e 6,8 kg kg-1, no primeiro ano. No segundo ano a média ficou
em 4,2 kg kg-1 tendo oscilado entre 3,6 e 4,7 kg kg-1 (Figura 5). A umidade
gravimétrica além de ter sido significativamente menor do que no Organossolo
natural, mostrou nítida tendência descendente.
O consumo de CH4 no solo drenado foi atribuído à retirada da água, sendo
que o suprimento de oxigênio provavelmente estimulou o incremento da população
de organismos consumidores de CH4. Um dia após a drenagem, na primeira
avaliação, o nível freático rebaixou para -25 cm (Figura 4), mas isso possivelmente
ainda não tenha sido suficiente para afetar os fluxos de CH4 (Figura 3).
Organismos anaeróbicos como os metanogênicos não se desenvolvem nas
camadas aeradas da turfa, mas parecem manter sua viabilidade durante períodos
nos quais o solo não se encontra saturado com água (KETUNNEN et al., 1999). Por
outo lado, organismos metanotróficos precisam de oxigênio, mas podem sobreviver
a condições anóxicas (ROSLEV; KING, 1994). Entretanto, deve-se levar em conta
que organismos metanogênicos e metanotróficos são sensíveis às oscilações do
nível freático (WHALEN, 2005), sofrendo o efeito imediato da retirada da água.
Quatro dias após a drenagem, o nível rebaixou para -49 cm (Figura 4),
quando então foi observado o início do consumo de CH4 (-39 µg CH4 m-2 h-1) (Figura
3). No oitavo dia, o nível freático foi rebaixado para -65 cm e o consumo aumentou
para -99 µg CH4 m-2 h-1.
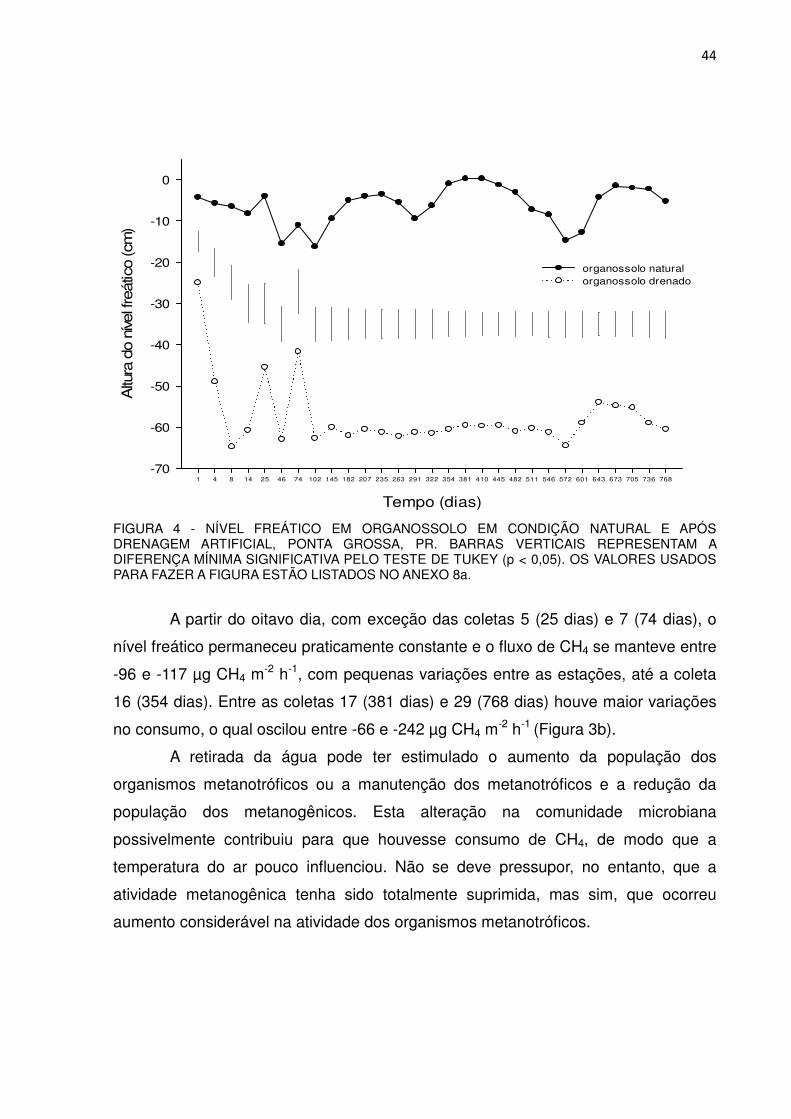
44
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Altu
ra d
o ní
vel f
reát
ico
(cm
)
-70
-60
-50
-40
-30
-20
-10
0
organossolo naturalorganossolo drenado
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA FAZER A FIGURA ESTÃO LISTADOS NO ANEXO 8a.
A partir do oitavo dia, com exceção das coletas 5 (25 dias) e 7 (74 dias), o
nível freático permaneceu praticamente constante e o fluxo de CH4 se manteve entre
-96 e -117 µg CH4 m-2 h-1, com pequenas variações entre as estações, até a coleta
16 (354 dias). Entre as coletas 17 (381 dias) e 29 (768 dias) houve maior variações
no consumo, o qual oscilou entre -66 e -242 µg CH4 m-2 h-1 (Figura 3b).
A retirada da água pode ter estimulado o aumento da população dos
organismos metanotróficos ou a manutenção dos metanotróficos e a redução da
população dos metanogênicos. Esta alteração na comunidade microbiana
possivelmente contribuiu para que houvesse consumo de CH4, de modo que a
temperatura do ar pouco influenciou. Não se deve pressupor, no entanto, que a
atividade metanogênica tenha sido totalmente suprimida, mas sim, que ocorreu
aumento considerável na atividade dos organismos metanotróficos.
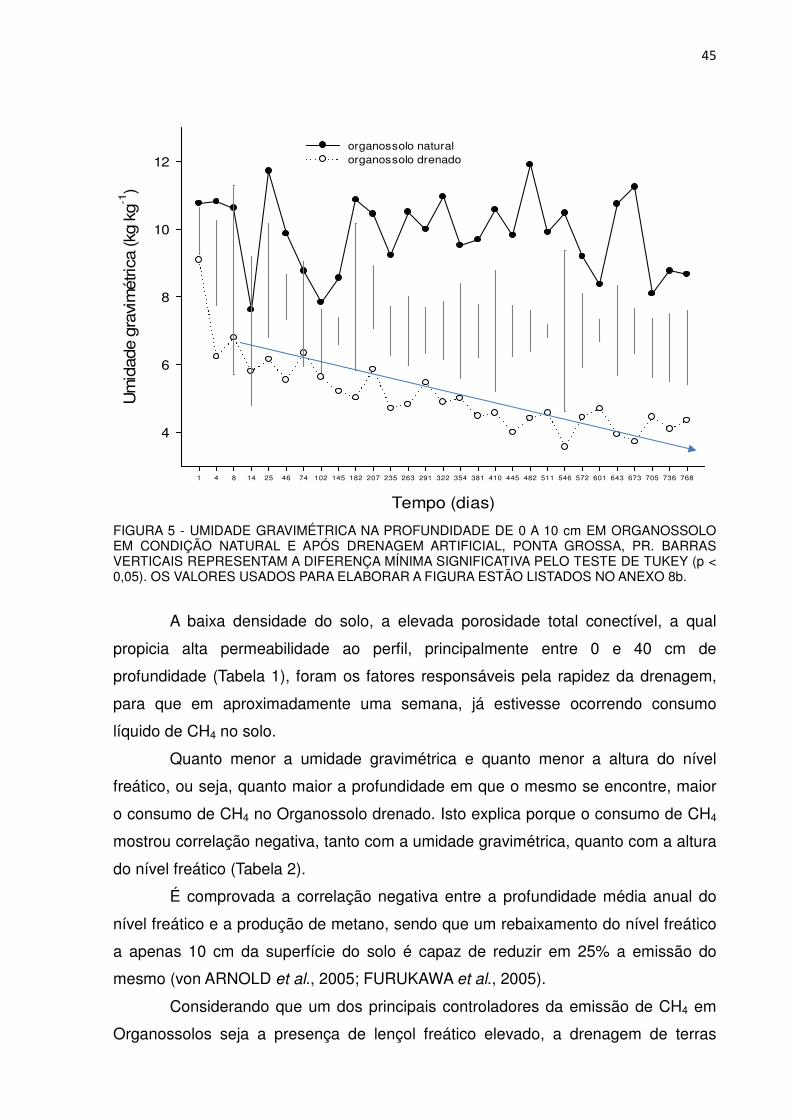
45
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Um
idad
e gr
avim
étric
a (k
g kg
-1)
4
6
8
10
12organossolo naturalorganossolo drenado
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA ELABORAR A FIGURA ESTÃO LISTADOS NO ANEXO 8b.
A baixa densidade do solo, a elevada porosidade total conectível, a qual
propicia alta permeabilidade ao perfil, principalmente entre 0 e 40 cm de
profundidade (Tabela 1), foram os fatores responsáveis pela rapidez da drenagem,
para que em aproximadamente uma semana, já estivesse ocorrendo consumo
líquido de CH4 no solo.
Quanto menor a umidade gravimétrica e quanto menor a altura do nível
freático, ou seja, quanto maior a profundidade em que o mesmo se encontre, maior
o consumo de CH4 no Organossolo drenado. Isto explica porque o consumo de CH4
mostrou correlação negativa, tanto com a umidade gravimétrica, quanto com a altura
do nível freático (Tabela 2).
É comprovada a correlação negativa entre a profundidade média anual do
nível freático e a produção de metano, sendo que um rebaixamento do nível freático
a apenas 10 cm da superfície do solo é capaz de reduzir em 25% a emissão do
mesmo (von ARNOLD et al., 2005; FURUKAWA et al., 2005).
Considerando que um dos principais controladores da emissão de CH4 em
Organossolos seja a presença de lençol freático elevado, a drenagem de terras

46
úmidas reduz o potencial de emissão de CH4 (BRIDGHAM et al., 2006; HUANG et
al., 2010), quando o lençol freático é rebaixado e uma camada aerada mais espessa
do solo passa a contribuir para a oxidação do CH4. Segundo Whalen (2005), 20 a
40% do CH4 produzido nos sítios anaeróbicos do solo é oxidado na rizosfera e nas
camadas superficiais oxigenadas, durante o transporte difuso do gás para a
superfície do solo.
Os resultados do presente estudo, de acordo com a queda das emissões
nos primeiros dias, sugerem que houve redução da população dos organismos
metanogênicos após a drenagem, o que resultou no consumo líquido de metano.
Vários estudos evidenciaram reduções de 50% a 100% na emissão de CH4
em Organossolos após a drenagem (MARTIKAINEN et al., 1995; NYKANEN et al.,
1995; JIANG et al., 2009; PAGE; DALAL, 2011), sendo que alguns destes solos
passam a ser sumidouros de CH4 (MALJANEN et al., 2004; ELDER; LAL, 2008).
Von Arnold et al. (2005), trabalhando com Organossolo sob floresta no Sul
da Suécia, observaram uma emissão média de 79 µg CH4 m-2 h-1 quando o nível
freático permaneceu entre -13 e -23 cm após a drenagem, enquanto no solo não
drenado, cujo nível freático permaneceu entre 2 e -4 cm, a emissão média foi de
1.118 µg de CH4 m-2 h-1. Na Indonésia, Furukawa et al. (2005) registraram redução
de 25% na emissão de CH4 em Organossolo quando o nível freático foi rebaixado a
apenas 10 cm. Organossolos convertidos em áreas de lavouras por meio de
drenagem, deixaram de emitir 1991 kg CH4 ha-1 ano-1 e passaram a consumir 1,4 kg
CH4 ha-1 ano-1 (JIANG et al., 2009). A drenagem fez com que Organossolos
deixassem de ser fontes de metano e passassem a atuar como sumidouros
(MALJANEN et al. 2002, 2004).
A drenagem compromete outras funcionalidades ecológicas como a
produção de biomassa, o armazenamento e a filtragem de água e a conservação da
biodiversidade. Sousa et al. (no prelo)1, trabalhando na mesma área onde foi
realizada esta pesquisa, determinaram que a quantidade média de biomassa foi de
42,4 Mg ha-1, incluindo a biomassa aérea e a subterrânea até 20 cm de
profundidade.
1 SOUSA, L. P.; RACHWAL, M. F. G.; MARQUES, R.; CURCIO, G. R. Above and belowground biomass of wet subtropical grassland in a river spring: relationship with geo-pedological factors, Ponta Grossa, Paraná State. Biota Neotropica. (no prelo)

47
Não se pode deixar de enfatizar que em cada hectare o Organossolo natural
armazena até a profundidade de um metro, 581 Mg C e 8,6 milhões de litros de
água.
A análise estatística efetuada pelo modelo GAM (Tabela 3), com todas as
variáveis em conjunto, permitiu concluir que a drenagem causou diferenças
marcantes nos fluxos de CH4 entre o Organossolo natural e o drenado.
O efeito da drenagem foi muito mais importante do que as variações que
ocorreram no nível freático, na umidade gravimétrica na profundidade de 0 a 10 cm,
na temperatura do ar e na precipitação. Apenas no Organossolo natural ocorreram
diferenças significativas no fluxo de metano ao longo do tempo (Tabela 3 e Figura 3).
TABELA 3 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE OS FLUXOS DE CH4, PELO MODELO ADITIVO GENERALIZADO - GAM
Efeitos paramétricos p valor Tratamento (solo natural e drenado) 0,0296 * Nível freático (Componente principal 1) 0,1979 ns Temperatura do ar + umidade relativa do ar (Componente principal 2)(1) 0,4753 ns Temperatura do ar + umidade relativa do ar + precipitação (Componente principal 3)(2)
0,4109 ns
Efeitos não paramétricos p valor Tempo(3) (solo drenado) 0,966 ns Tempo(4) (solo natural) 2,0 x 10-16 ***
*** - altamente significativo (p < 0,001); * - significativo (p < 0,05); ns - não significativo; (1) Temperatura máxima e mínima do ar das 9 às 12 horas no momento das coletas + temperatura máxima e média do ar nos períodos de 1 e 3 dias que antecederam as coletas + temperatura mínima do ar no período de 3 dias que antecederam as coletas + umidade relativa média do ar no período das 9 às 12 horas e nos períodos de 1, 3 e 5 dias que antecederam as coletas. (2) Temperatura máxima e mínima do ar das 9 às 12 horas no momento das coletas + temperatura máxima e média do ar nos períodos de 1, 3 e 5 dias que antecederam as coletas + temperatura mínima do ar nos períodos de 3 e 5 dias que antecederam as coletas + umidade relativa média do ar das 9 às 12 horas no momento das coletas e nos períodos de 3 e 5 dias que antecederam as coletas + precipitação acumulada no período de 1 dia que antecedeu as coletas. (3) Número de dias a partir da drenagem. (4) Número de dias a partir do início do experimento.
2.3.3 Efeito do potencial redox sobre o fluxo de CH4
Quando o solo é submetido à hidromorfia plena a concentração de oxigênio
dissolvido decresce rapidamente e organismos anaeróbicos facultativos e
obrigatórios se desenvolvem. Estes organismos usam sucessivamente vários
aceptores de elétrons para sua respiração: NO3- a valores de Eh < 350 mV, Mn4+ a
Eh < 200 mV, Fe3+ a Eh < 100 mV e SO4-2 a Eh em torno de -150 mV (PATRICK;

48
JUGSUJINDA, 1992). O sulfato é o último aceptor de elétrons antes da
metanogênese. Esta sucessão de reações de redução pode levar rapidamente a um
potencial redox (Eh) em torno de -200 mV, favorável para a redução do CO2 a CH4
(NEUE; ROGER, 1994).
São variáveis os valores de potencial redox para metanogênese e
metanotrofia indicados na literatura.
Le Mer (2001) afirma que a faixa ideal de potencial redox para a produção
de metano está entre -200 e -300 mV, enquanto para Koh et al (2009) os valores se
estendem entre 300 e -300 mV, sendo que acima de 300 mV começa a ocorrer
consumo. Em solos cultivados com arroz irrigado, no norte da China, ocorreu
emissão de CH4 quando o potencial redox era menor do que -100 mV (HOU et al.,
2000).
Inubushi et al. (2003) registraram em Organossolos, emissões de 0,6 ± 0,7 g
C-CH4 m-2 ano-1 em lavouras de sequeiro, para valores de potencial redox de 358, 7
e 157 mV, nas profundidades de 0-10, 10-20 e 20-30 cm, respectivamente. Em
campos de arroz irrigado a emissão foi de 1,9 ± 0,5 g C-CH4 m-2 ano-1 para
potenciais redox de 124, -10 e -81 mV, respectivamente, nas mesmas
profundidades. Em contra partida, Furukawa et al. (2005), indicaram emissão de
aproximadamente 2,25 mg C-CH4 m-2 ano-1 para potencial redox de
aproximadamente 300 mV e consumo de 0,2 mg C-CH4 m-2 ano-1 quando o potencial
redox aumentou para aproximadamente 650 mV.
Koh et al. (2009) registraram potencial redox (Eh) entre -230 e -99 mV em
solo permanentemente inundado, entre 68 e 188 mV em solo inundado
ocasionalmente e entre 273 a 343 mV em solo não inundado. Nestes solos os fluxos
de CH4 foram de aproximadamente 4 a 21 mg CH4 m-2 h-1, de 2,5 a 10 e de -4 a 3,0
mg CH4 m-2 h-1, respectivamente.
No Organossolo natural em todas as determinações efetuadas nas
profundidades de 5 a 40 cm os valores de potencial redox oscilaram entre 267 e -
178 mV. Foram detectados valores negativos nas coletas 25, 28 e 29 e conforme
esperado, o potencial de oxi-redução foi diminuindo em profundidade (Figura 6).
Na coleta 28 na qual o nível freático médio era de -2,3 cm, o segundo mais
elevado, não houve diferença significativa no potencial redox entre as
profundidades. Mesmo mostrando valores positivos em algumas coletas e
profundidades, o potencial redox do solo encontrava-se moderadamente e
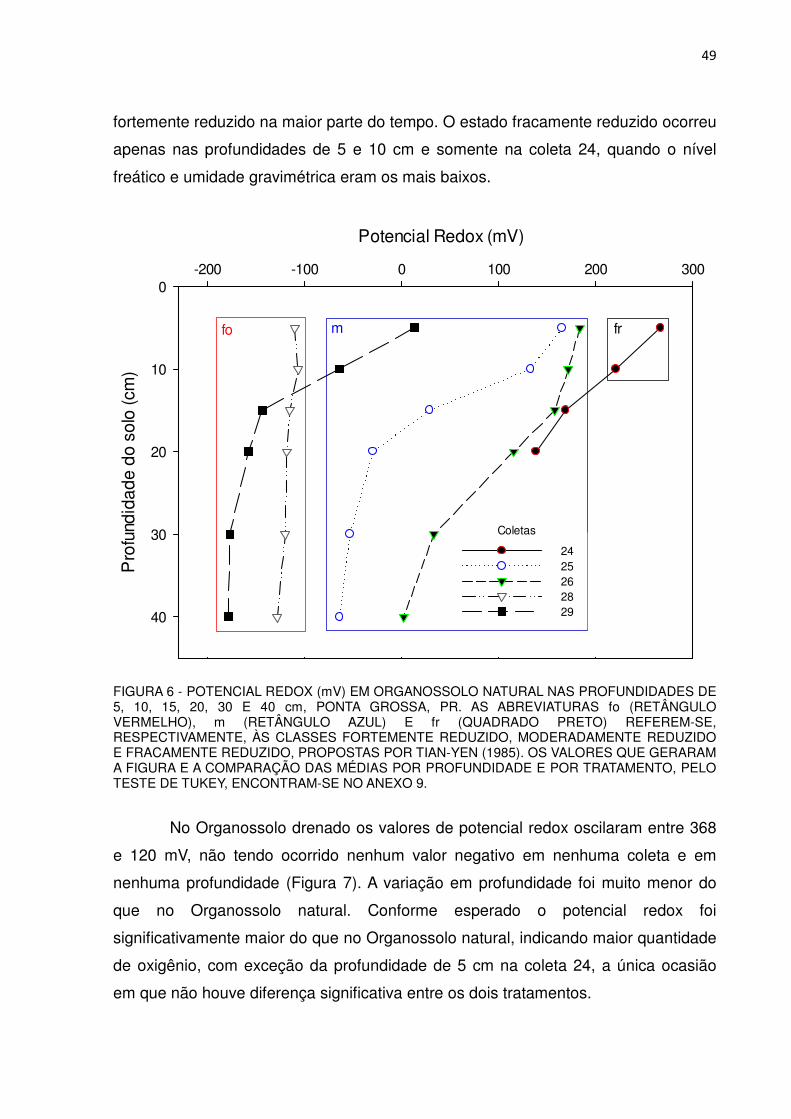
49
fortemente reduzido na maior parte do tempo. O estado fracamente reduzido ocorreu
apenas nas profundidades de 5 e 10 cm e somente na coleta 24, quando o nível
freático e umidade gravimétrica eram os mais baixos.
Potencial Redox (mV)
-200 -100 0 100 200 300
Pro
fund
idad
e do
sol
o (c
m)
0
10
20
30
40
242526 28 29
Coletas
frmfo
FIGURA 6 - POTENCIAL REDOX (mV) EM ORGANOSSOLO NATURAL NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR. AS ABREVIATURAS fo (RETÂNGULO VERMELHO), m (RETÂNGULO AZUL) E fr (QUADRADO PRETO) REFEREM-SE, RESPECTIVAMENTE, ÀS CLASSES FORTEMENTE REDUZIDO, MODERADAMENTE REDUZIDO E FRACAMENTE REDUZIDO, PROPOSTAS POR TIAN-YEN (1985). OS VALORES QUE GERARAM A FIGURA E A COMPARAÇÃO DAS MÉDIAS POR PROFUNDIDADE E POR TRATAMENTO, PELO TESTE DE TUKEY, ENCONTRAM-SE NO ANEXO 9.
No Organossolo drenado os valores de potencial redox oscilaram entre 368
e 120 mV, não tendo ocorrido nenhum valor negativo em nenhuma coleta e em
nenhuma profundidade (Figura 7). A variação em profundidade foi muito menor do
que no Organossolo natural. Conforme esperado o potencial redox foi
significativamente maior do que no Organossolo natural, indicando maior quantidade
de oxigênio, com exceção da profundidade de 5 cm na coleta 24, a única ocasião
em que não houve diferença significativa entre os dois tratamentos.

50
No Organossolo drenado não houve diferença significativa no potencial
redox entre as profundidades em nenhuma das determinações.
Em todas as coletas e profundidades os valores de potencial redox
indicaram que o Organossolo drenado se enquadrava nos estados de redução
fracamente reduzido, principalmente, e moderadamente reduzido, não ocorrendo a
classe fortemente reduzido, conforme observado no solo natural. Na grande maioria
das determinações, em todas as profundidades, o potencial redox no Organossolo
natural foi significativamente menor do que no Organossolo drenado, indicando
maior deficiência de oxigênio.
No Organosolo natural o fluxo de metano respondeu de modo inverso ao
potencial redox. A diminuição dos valores de potencial redox em todas as
profundidades ocasionaou aumento na emissão de CH4 (Tabela 4). Nas coletas 24 a
26 o solo estava moderadamente reduzido em praticamente todas as profundidades,
tornando-se fortemente reduzido nas duas coletas seguintes.
TABELA 4 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE POTENCIAL REDOX, NAS PROFUNDIDADE DE 5, 10, 15, 20, 30 E 40 cm E O EFLUXO DE CH4 NO ORGANOSSOLO NATURAL E O INFLUXO DE CH4 NO ORGANOSSOLO DRENADO
Solo Profundidades (cm)
5 10 15 20 30 40 Natural -0,23
(p<0,05) -0,37
(p<0,01) -0,49
(p<0,01) -0,48
(p<0,01) -0,53
(p<0,01) -0,53
(p<0,01)
Drenado 0,55 (p<0,01)
0,41 (p<0,01)
0,40 (p<0,01)
0,42 (p<0,01)
0,36 (p<0,01)
0,33 (p<0,01)
Valores de r obtidos om os dados dos dois anos de avaliação.
No Organossolo drenado o consumo de metano correlacionou-se
positivamente com o potencial redox (Tabela 4).
Os valores de potencial redox determinados no Organossolo natural e no
drenado estão compatíveis com os encontrados na literatura para ocorrência de
emissão de metano no primeiro e de consumo no segundo.

51
Potencial Redox (mV)
100 150 200 250 300 350 400
Pro
fund
idad
e do
sol
o (c
m)
0
10
20
30
40
242526 28 29
Coletas
frm
FIGURA 7 - POTENCIAL REDOX (mV) EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR. AS ABREVIATURAS m (RETÂNGULO AZUL) E fr (RETÂNGULO PRETO) REFEREM-SE, RESPECTIVAMENTE, ÀS CLASSES MODERADAMENTE REDUZIDO E FRACAMENTE REDUZIDO, PROPOSTAS POR TIAN-YEN (1985). OS VALORES QUE GERARAM A FIGURA E A COMPARAÇÃO DAS MÉDIAS POR PROFUNDIDADE E POR TRATAMENTO, PELO TESTE DE TUKEY, ENCONTRAM-SE NO ANEXO 9.
2.3.4 Fluxo acumulado de metano
Em todas as estações nos dois anos de pesquisa o fluxo acumulado de
metano no solo natural diferiu do fluxo acumulado no solo drenado, tendo ocorrido
emissão de metano no primeiro e consumo no segundo. No solo natural a emissão
acumulada de metano variou de 13 a 40 kg ha-1 no primeiro ano e de 30 a 100 kg ha-
1, no segundo (Figura 8).
No primeiro ano, as maiores emissões acumulados ocorreram no outono e
no verão de 2011, com temperaturas médias de 16,2 e 21 0C, respectivamente. No
segundo ano as maiores emissões acumuladas ocorreram no verão de 2012 e na
primavera de 2011 nos quais a temperatura média foi mais elevada (21,3 e 18,4 0C,

52
respectivamente). No inverno de 2011 e de 2012, a temperatura média semanal de
14,8 e 16,5 0C, respectivamente, contribuiu para que as emissões fossem menores.
No outono do primeiro ano de estudo, estação menos chuvosa (254 mm), a
emissão acumulada mostrou-se superior as demais estações. Por outro lado, no
outono do segundo ano, o qual foi atipicamente a estação mais chuvosa (615 mm) o
fluxo acumulado não foi o mais elevado (33 kg ha-1). O maior fluxo acumulado
ocorreu no verão do segundo ano de estudo (100 kg ha-1), no qual a precipitação
acumulada foi de 446 mm, ou seja, 169 mm menor do que no outono. A precipitação
não influenciou os fluxos acumulados (Tabela 2), nos dois anos, pois as maiores
precipitações acumuladas no inverno de 2011 (580 mm) e outono de 2012 (615 mm),
não estiveram associadas às maiores emissões acumuladas de metano. A emissão
acumulada anual no segundo ano (218 kg ha-1) foi praticamente o dobro da ocorrida
no ano anterior (116 kg ha-1), embora neste último, a precipitação anual tenha sido
superior em 319 mm.
No solo drenado o consumo acumulado de metano nas estações oscilou
entre -2,05 a -2,37 kg ha-1 no primeiro ano e de -2,34 a -4,17 kg ha-1, sem diferença
estatística entre as estações ao longo dos anos. Entretanto, no verão de 2012 o
consumo acumulado foi quase o dobro do verão do ano anterior embora a
temperatura média das duas estações tenha-se mantido praticamente a mesma e a
precipitação acumulada no segundo tenha sido 97 mm menor. A diferença no
consumo acumulado anual no segundo ano (-11,7 kg ha-1) foi 24% superior ao do
segundo (-8,92 kg ha-1).
A precipitação também não influenciou o consumo acumulado de metano
(Tabela 2) porque a água da chuva percolava rapidamente devido à pequena
densidade e porosidade elevada do solo, as quais permitiram uma alta taxa de
infiltração de água.
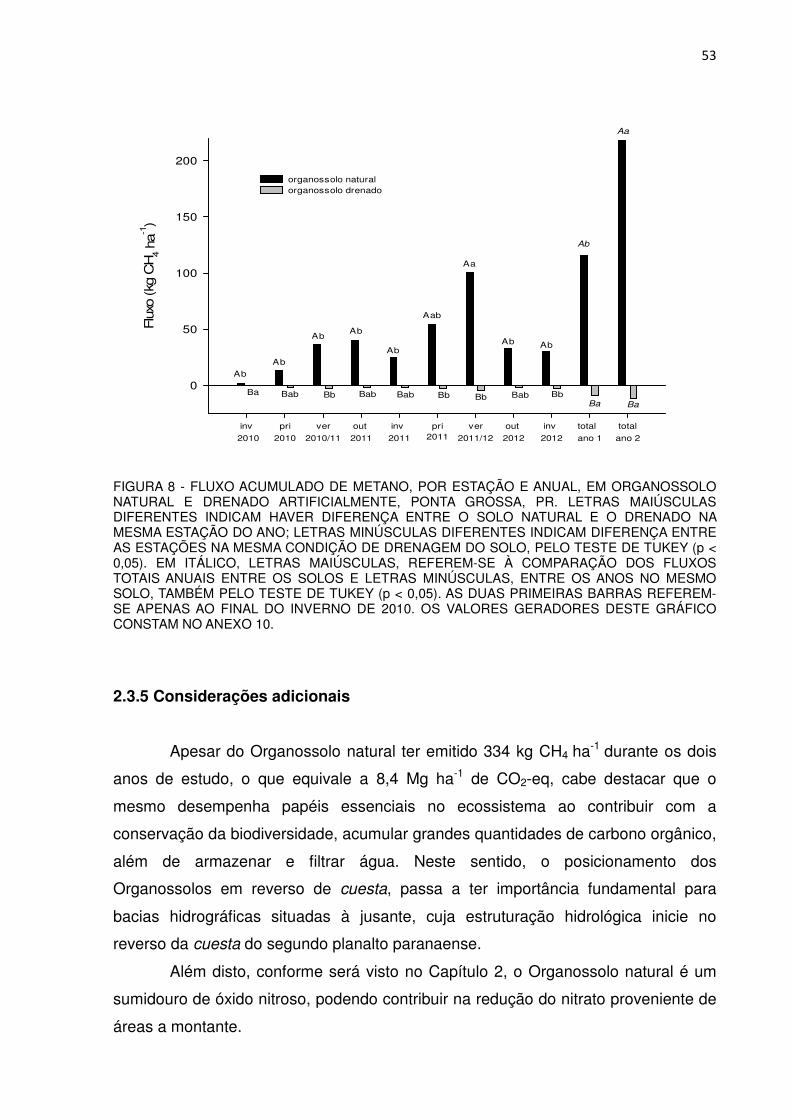
53
inv
2010
pri
2010
ver
2010/11
out
2011
inv
2011
pri2011
ver
2011/12
out
2012
inv
2012
total
ano 1
total
ano 2
Flu
xo (kg
CH
4 ha
-1)
0
50
100
150
200
organossolo naturalorganossolo drenado
Ab Ab
AbAb
Aa
Ab
Aab
Ab Ab
Ba Bab Bb Bab Bab BabBb Bb Bb
Ab
Aa
Ba Ba
FIGURA 8 - FLUXO ACUMULADO DE METANO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR. LETRAS MAIÚSCULAS DIFERENTES INDICAM HAVER DIFERENÇA ENTRE O SOLO NATURAL E O DRENADO NA MESMA ESTAÇÃO DO ANO; LETRAS MINÚSCULAS DIFERENTES INDICAM DIFERENÇA ENTRE AS ESTAÇÕES NA MESMA CONDIÇÃO DE DRENAGEM DO SOLO, PELO TESTE DE TUKEY (p < 0,05). EM ITÁLICO, LETRAS MAIÚSCULAS, REFEREM-SE À COMPARAÇÃO DOS FLUXOS TOTAIS ANUAIS ENTRE OS SOLOS E LETRAS MINÚSCULAS, ENTRE OS ANOS NO MESMO SOLO, TAMBÉM PELO TESTE DE TUKEY (p < 0,05). AS DUAS PRIMEIRAS BARRAS REFEREM-SE APENAS AO FINAL DO INVERNO DE 2010. OS VALORES GERADORES DESTE GRÁFICO CONSTAM NO ANEXO 10.
2.3.5 Considerações adicionais
Apesar do Organossolo natural ter emitido 334 kg CH4 ha-1 durante os dois
anos de estudo, o que equivale a 8,4 Mg ha-1 de CO2-eq, cabe destacar que o
mesmo desempenha papéis essenciais no ecossistema ao contribuir com a
conservação da biodiversidade, acumular grandes quantidades de carbono orgânico,
além de armazenar e filtrar água. Neste sentido, o posicionamento dos
Organossolos em reverso de cuesta, passa a ter importância fundamental para
bacias hidrográficas situadas à jusante, cuja estruturação hidrológica inicie no
reverso da cuesta do segundo planalto paranaense.
Além disto, conforme será visto no Capítulo 2, o Organossolo natural é um
sumidouro de óxido nitroso, podendo contribuir na redução do nitrato proveniente de
áreas a montante.

54
Por outro lado, embora o Organossolo drenado tenha consumido 0,52 Mg
ha-1 CO2-eq de metano em dois anos, o mesmo sofreu uma redução considerável no
estoque de carbono, de acordo com o que será mostrado no Capítulo 3.
Levando em conta as funções que os Organossolos desempenham no
ecossistema, os mesmos devem ser preservados e para isso não devem ser
drenados. Estratégias de mitigação das emissões de metano devem ser
concentradas nas inúmeras atividades antrópicas e em hipótese alguma, drenando
áreas onde ocorrem Organossolos.

55
2.4 CONCLUSÕES
a) Em condições naturais, o Organossolo sob estepe hidrófila, em função do
excesso de água, apresentou emissão líquida de metano em todas as estações do
ano.
b) A drenagem do Organossolo sob estepe hidrófila determinou o
rebaixamento do nível freático, elevando o potencial redox o que ocasionou o
consumo de metano, ao longo de todo o ano.
c) O aumento da temperatura do ar teve maior influência sobre a elevação
dos fluxos de metano no Organossolo natural do que no drenado.
d) A precipitação não teve influência sobre os fluxos de metano nos
Organossolos natural e drenado.

56
REFERÊNCIAS
ALONGI, D. M.; WATTAYAKORN, G.; PFITZNER, J.; TIRENDI, F.; ZAGORSKIS, I.; BRUNSKILL, G. J.; DAVIDSON, A.; CLOUGH, B. F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in Southern Thailand. Marine Geology, Amsterdam, v. 179, p. 85-103, 2001. ANDRADE, B. O.; KOZERA, C.; CURCIO, G. R.; GALVÃO, F. Vascular gassland plants of Tibagi River Spring, Ponta Grossa, Brazil. Chek List, Jaboticabal, v. 7, n. 3, p. 257-262, 2011. BAIRD, C. O efeito estufa e o aquecimento global. In: BAIRD, C. Química ambiental. 2. ed. Porto Alegre: Bookman, 2002. p. 195-243. BRIDGHAM, S. D.; MEGONIGAL, J. P.; KELLER, J. K.; BLISS, N. B.; TRETTIN, C. The carbon balance of North American wetlands. Wetlands, Dordrecht, v. 26, p. 889-916, 2006. CLAESSEN, M. E. C. (Org.). Manual de métodos de análises de solo. 2. ed. rev. atual. Rio de Janeiro: EMBRAPA-CNPS, 1997. (Documentos, 1). DALAL, R. C.; ALLEN, D. E. Greenhouse gas fluxes from natural ecosystems. Australian Journal of Botany, Collingwood, v. 56, p. 369-407, 2008. DUNFIELD, P.; KNOWLES, R.; DUMONT, R.; MOORE, T. R. Methane production and consumption in temperate and subarctic peat soils – response to temperature and pH. Soil Biology & Biochemistry, Oxford, v. 25, p. 321-326, 1993. ELDER, J. W.; LAL, R. Tillage effects on gaseous emissions from an intensively farmed organic soil in North Central Ohio. Soil and Tillage Research, Amsterdam, v. 98, p. 45-55, 2008. FORSTER, P.; RAMASWAMY, V.; ARTAXO, P.; BERNSTEN, T.; BETTS, R.; FAHEY, D. W.; HAYWOOD, J.; LEAN, J.; LOWE, D. C.; MYHRE, G.; NGANGA, J.; PRINN, R.; RAGA, G.; SCHUTZ, M.; VAN DORLAND, R. Changes in atmospheric constituents and in radiative forcing. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z; MARQUIS, M.; AVERYT, K. B.; TIGNOR, M.; MILLER, H. L. (Ed.). Climate change 2007: the physical science basis: contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge: Cambridge University, 2007. p. 129-234.

57
FURUKAWA, Y.; INUBUSHI, K.; ALI, M.; ITANG, A. M.; TSURUTA, H. Effect of changing groundwater levels caused by land-use changes on greenhouse gas fluxes from tropical peat lands. Nutrient Cycling in Agroecosystems, Dordrecht, v. 71, p. 81-91, 2005. GOMES, J.; BAYER, C.; COSTA, F. S.; PICCOLO, M. C.; VIEIRA, F. C. B.; SIX, J. Soil nitrous oxide emission as affected by long term tillage, crop rotations and fertilization in a subtropical environment. Soil and Tillage Research, Amsterdam, v. 106, p. 36-44, 2009. HAYES, M. H. B.; CLAPP, C. E. Humic substances: considerations of compositions, aspects of structure and environmental influences. Soil Science, Philadelphia, v. 166, n. 11, p. 723-727, 2001. HOU, A. X.; CHEN, G. X.; WANG, Z. P.; VAN CLEEMPUT, O.; PATRICK, W. H. Methane and nitroux oxide emissions from a rice field in relation to soil redox and microbiological processes. Soil Science Society of America Journal, Madison, v. 64, n. 6, p. 2180-2186, 2000. HUANG, Y.; SUN, W. J.; ZHANG, W.; YU, Y. Q.; SU, Y. H.; SONG, C. C. Marshland conversion to cropland in northeast China from 1950 to 2000 reduced the greenhouse effect. Global Change Biology, West Sussex, v. 16, p. 680-695, 2010. INUBUSHI, K.; FURUKAWA, Y.; HADI, A.; PURNOMO, E.; TSURUTA, H. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use changes in tropical peatlands located in coastal area of South Kalimantan. Chemosphere, Oxford, v. 52, p. 603-608, 2003. INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC). Climate Change 2007: the physical science basis. Cambridge: Cambridge University, 2007. JANG, I.; LEE, S.; HONG, J. H.; KANG, H. Methane oxidation rates in forest soils and their controlling variables: e review and a case study in Korea. Ecological Research, Tokyo, v. 21, p. 849-854, 2006. JANTALIA, C. P.; SANTOS, H. P.; URQUIAGA, S.; BODDEY, R. M.; ALVES, B. J. R. Fluxes of nitrous oxide from soil under different crop rotations and tillage systems in the South of Brazil. Nutrient Cycling in Agroecosystems, Dordrecht, v. 82, p. 161-173, 2008.

58
JAUHIAINEN, J.; LIMIN, S.; SILVENNOINEN, H.; VASANDER, H. Carbon dioxide and methane fluxes in drained tropical peat before and after hydrological restoration. Ecology, Ithaca, v. 89, n. 12, p. 3503-3514, 2008. JIANG, C.; WANG, Y.; HAO, Q.; SONG, C. Effect of land-use change on CH4 and N2O emissions from freshwater marsh in Northeast China. Atmospheric Environment, Oxford, v. 43, p. 3305-3309, 2009. JOABSSON, A.; CHRISTENSEN, T. R. Methane emissions from wetlands and their relationship with vascular plants: an Arctic example. Global Change Biology, West Sussex, v. 7, p. 919-932, 2001. KETTUNEN, A.; KAITALA, V.; LEHTINEN, A.; LOHILA, A.; ALM, J.; SILVOLA, J.; MARTIKAINEN, P. J. Methane production and oxidation potentials in relation to water table fluctuations in two boreal mires. Soil Biology & Biochemistry, Oxford, v. 31, p. 1741-1749, 1999. KOH, H. S.; OCHS, C. A.; YU, K. W. Hydrologic gradient and vegetation controls on CH4 and CO2 fluxes in a spring-fed forested wetland. Hydrobiologia, Dordrecht, v. 630, p. 271-286, 2009. KÖGEL-KNABNER, I.; AMELUNG, W.; CAO, Z. H.; FIEDLER, S.; FRENZEL, P.; JAHN, R.; KALBITZ, K.; KOLBL, A.; SCHLOTER, M. Biogeochemistry of paddy soils. Geoderma, Amsterdam, v. 157, p. 1-14, 2010. KOZERA, C. Florística e fitossociologia de uma formação pioneira com influência fluvial e de uma estepe gramíneo-lenhosa em diferentes unidades geopedológicas, município de Balsa Nova, Paraná - Brasil. 267 f. Tese (Doutorado) - Universidade Federal do Paraná, Curitiba, 2008. LE MER, J.; ROGER, P. Production, oxidation, emission and consumption of methane by soils: A review. European Journal of Soil Biology, Cedex, v. 37, p. 25-50, 2001. MALJANEN, M.; MARTIKAINEN, P. J.; AALTONEN, H.; SILVOLA, J. Short-term variation in fluxes of carbon dioxide, nitrous oxide and methane in cultivated and forested organic boreal soils. Soil Biology & Biochemistry, Oxford, v. 34, p. 577-584, 2002. MALJANEN, M.; KOMULAINEN, V. M.; HYTONEN, J.; MARTIKAINEN, P. J.; LAINE, J. Carbon dioxide, nitrous oxide and methane dynamics in boreal organic agricultural

59
soils with different soil characteristics. Soil Biology & Biochemistry, Oxford, v. 36, p. 1801-1808, 2004. MARTIKAINEN, P. J.; NYKANEN, H.; ALM, J.; SILVOLA, J. Changes in fluxes of carbon dioxide, methane and nitrous oxide due to forest drainage of mire sites of different trophy. Plant Soil, The Hague, v. 168, p. 571-577, 1995. MEGONIGAL, J. P.; SCHLESINGER, W. H. Methane-limited methanotrophy in tidal freshwater swamps. Global Biogeochemical Cycles, Hoboken, v. 16, n. 4, p. 35/1-35/10, 2002. MILDE, G. A.; DEDECEK, R. A.; GAVA, J. L. Diâmetro médio ponderado de partículas para estimativa de propriedades físico-hídricas dos solos. Pesquisa Florestal Brasileira, Colombo, v. 57, p. 21-27, 2008. MOORE, T. R.; DALVA, M. The influence of temperature and water-table position on carbon-dioxide and methane emission from laboratory columns of peat land soils. Journal of Soil Science, Oxford, v. 44, p. 641-664, 1993. NEUE, H. U.; ROGER, P. A. Potencial of methane emission in major rice ecologies. In: ZEPP, R. G. (Ed.). Climate Biosphere Interaction: biogenic emissions and environmental effects of climate change. New York: John Wiley and Sons, 1994. p. 65-93. NYKANEN, H.; ALM, J.; LANG, K.; SILVOLA, J.; MARTIKAINEN, P. J. Emissions of CH4, N2O and CO2 from a virgin fen and a fen drained for grassland in Finland. Journal of Biogeography, Oxford, v. 22, p. 351-357, 1995. PAGE, K. L.; DALAL, R. C. Contribution of natural and drained wetland systems to carbon stocks, CO2, N2O and CH4 fluxes: an Australian perspective. Soil Research, Rome, v. 49, p. 377-388, 2011. PANIKOV, N. S.; DEDYSH, S. N. Cold season CH4 and CO2 emissions from boreal peat bogs (West Siberia): winter fluxes and thaw activation dynamics. Global Biogeochemical Cycles, Hoboken, v. 14, p. 1071-1080, 2000. PATRICK, W. H.; JUGSUJINDA, A. Sequencial reduction and oxidation of inorganic nitrogen, manganese and iron in flooded soil, Soil Science Society of American Journal, Madison, v. 56, p. 1071-1073, 1992.

60
R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing, 2011. Disponível em: <http://www.R-project.org/>. Acesso em: 25/05/2012. RAICH, J. W.; POTTER, C. S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles, Hoboken, v. 9, p. 23-36, 1995. ROULET, N. T.; MOORE, T. R. The effect of forestry drainage practices on the emission of methane from northern peatlands. Canadian Journal of Forest Research, Ottawa, v. 25, p. 491-499, 1995. ROSLEV, P.; KING, G. M. Survival and recovery of methanotrophic bacteria starved under oxic and anoxic conditions. Applied and Environmental Microbiology, Washington, DC, v. 60, p. 2602-2608, 1994. SANTOS, H. G. dos; JACOMINE, P. K. T.; ANJOS, L. H. C. dos; OLIVEIRA, V. A. de; OLIVEIRA, J. B. de; COELHO, M. R.; LUMBRERAS, J. F.; CUNHA, T. J. F. (Ed.). Sistema Brasileiro de Classificação de Solos. 2. ed. Rio de Janeiro: Embrapa Solos, 2006. SCHEER, M. B.; CURCIO, G. R.; RODERJAN, C. V. Funcionalidades ambientais de solos altomontanos na Serra da Igreja, Paraná. Revista Brasileira de Ciência do Solo, Campinas, v. 35, p. 1013-1026, 2011. SOIL SURVEY STAFF. Soil survey manual. Washington, DC: U.S. Department of Agriculture, 1951. (Handbook, No. 18). SOIL SURVEY STAFF. Keys to soil taxonomy. 5. ed. Virginia: Pocahontas, 1992. TIAN-YEN, Y. Soil and plants. In: PHYSICAL chemistry of paddy soils. Berlin: Springer-Verlag, 1985. p. 197-214. VALLADARES, G. S. Caraterização de Organossolos, auxílio à sua classificação. 129 f. Tese (Doutorado) - Universidade Federal Rural do Rio de Janeiro, Seropédica, 2003. VAN HULZENA, J. B.; SEGERSA, R.; VAN BODEGOMA, P. M.; LEFFELAAR, P. A. Temperature effects on soil methane production: an explanation for observed variability. Soil Biology & Biochemistry, Oxford, v. 31, p. 1919-1929, 1999.

61
VON ARNOLD, K.; NILSSON, M.; HANELL, B.; WESLIEN, P.; KLEMEDTSSON, L. Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biology & Biochemistry, Oxford, v. 37, p. 1059-1071, 2005. WHALEN, S. C. Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environmental Engineering Science, New Rochelle, v. 22, p. 73-94, 2005. WHITING, G. J.; CHANTON, J. P. Greenhouse carbon balance of wetlands: methane emission versus carbon sequestration. Tellus. Series B: Chemical and Physical Meteorology, Hoboken, v. 53, p. 521-528, 2001. WILSON, J. O.; CRILL, P. M.; BARTLETT, K. B.; SEBACHER, D. I.; HARRISS, R. C.; SASS, R. L. Seasonal variation of methane emissions from a temperate swamp. Biogeochemistry, Dordrecht, v. 8, p. 55-71, 1989. WUEBBLES, D. J.; HAYHOE, K. A. S.; KOTAMARTHI, R. Methane in the global environment. In: KHALIL, M. A. K. (Ed.). Atmospheric Methane: its role in the global environment. Berlin: Springer-Verlag, 2000. p. 304-341.

62
3 CAPÍTULO II
FLUXO DE ÓXIDO NITROSO EM ORGANOSSOLO SOB ESTEPE HIDRÓFILA EM
CONDIÇÃO NATURAL E SOB DRENAGEM - PARANÁ - BR.
RESUMO
Atributos pedológicos e fatores climáticos, além das práticas de manejo, influenciam os fluxos de óxido nitroso no solo. Organossolos tropicais podem ser fonte de N2O em condições aeróbicas e sumidouro, em condições anaeróbicas. O fluxo de N2O apresenta grande variação porque os processos de produção do mesmo são muito complexos e determinados por inúmeros fatores. O objetivo deste estudo foi avaliar os fluxos de óxido nitroso em Organossolo Háplico sáprico típico sob estepe hidrófila de altitude, em condição natural e após drenagem artificial. Variáveis de solo e de clima foram avaliadas para auxiliar a interpretação dos fluxos. Um experimento foi conduzido em uma nascente nascente do rio Tibagi (Ponta Grossa, PR), onde foram avaliados os fluxos de N2O em área natural e em área drenada. Seis câmaras estáticas foram instaladas em cada área, e no decorrer de 26 meses foram realizadas 29 coletas de amostras de ar, sendo a concentração de N2O determinada por cromatografia gasosa. No solo em condição natural ocorreu consumo líquido de N2O cujas taxas variaram de 0,13 a 31,24 µg de N2O m-2 h-1. Por sua vez, na área drenada, a retirada da água determinou o efluxo líquido em taxas que variaram de 0,19 a 55,72 µg de N2O m-2 h-1. No solo drenado o aumento no tempo de drenagem foi o principal fator que determinou o aumento da emissão de N2O. Na condição natural, o consumo de óxido nitroso teve relação com o excesso de umidade e com os baixos valores de potencial redox. No Organossolo natural houve consumo acumulado de 0,92 kg N2O ha-1 ano-1, enquanto o Organossolo drenado emitiu 1,65 kg N2O ha-1 ano-1. Organossolos em estepe hidrófila possuem importantes funções no ecossistema, como acumular carbono orgânico, armazenar água e reduzir nitrato, não devendo portanto ser drenados, mas efetivamente preservados. Palavras Chave: Gás de efeito estufa. Nascentes. Nível freático. Umidade
gravimétrica. Temperatura do ar e precipitação. Potencial redox.

63
NITROUS OXIDE FLUXES IN A WETLAND HISTOSOL UNDER NATURAL AND DRAINED CONDITIONS - PARANA - BRAZIL
ABSTRACT
Pedological attributes and climate factors, as well as management practices, influenced the fluxes of nitrous oxide (N2O) in the soil. Tropical Histosols can be a source of N2O in aerobic and sink in anaerobic conditions. The N2O presents variations because its production process is very complex and determined by many factors. The purpose of this study was to evaluate the fluxes of nitrous oxide in a typical Haplic Histosol of a high altitude wetland in natural condition and under artificial drainage. Variables soil and climate were evaluated to assist in the interpretation of fluxes. An experiment was conducted in the spring of the river Tibagi (Ponta Grossa, PR) were evaluated in N2O flow in natural area and drained. Six static chambers were installed in each area, and in the course of 26 months were taken 29 samples of the air, and the N2O concentration in the samples determined by gas chromatography. In soil under natural condition occurred net consumption of N2O which rates range from -0,13 to -31,24 mg N2O m-2 h-1. In turn, in the drained area, the removal of the water has determined the net efflux in the rates that has ranged from 0,19 to 55,72 mg N2O m-2 h-1. In drained soil increased the drainage time was the main factor that determined the increase of N2O emissions. In natural conditions, the consumption of nitrous oxide was related to excess moisture and the low redox potential values. In natural histosol were accumulated consumption of 0,92 kg N2O ha-1 yr-1, while the drained histosol issued 1,65 kg N2O ha-1 yr-1. Histosols steppe hydrophilic play important roles in the ecosystem, such accumulate organic carbon, to store water and reduce nitrate, and therefore should not be drained, but effectively preserved.
Keywords: Greenhouse gas. Springs. Groundwater level. Gravimetric moisture. Air temperature. Precipitation and redox potential.

64
3.1 INTRODUÇÃO
O ciclo do nitrogênio no sistema solo-planta-atmosfera é muito complexo
sendo regido por fatores físicos, químicos e biológicos e influenciado por condições
climáticas difíceis de prever e controlar (CANTARELLA, 2007). O nitrogênio
apresenta vários estados de oxidação desde formas mais reduzidas como o NH4+
onde possui valência -3, até formas mais oxidadas como o NO3- no qual sua valência
é +5, assumindo importância especial nos ciclos biogeoquímicos (EPSTEIN;
BLOOM, 2005). O potencial de aquecimento global do N2O é 298 vezes maior em
comparação com o CO2 (IPCC, 2007).
O óxido nitroso é um gás estável na atmosfera mas instável no solo onde
pode ser reduzido a N2 durante a desnitrificação. Por isso os solos encharcados
podem também funcionar como dreno de N2O o qual é muito solúvel em água
(GROFMAN, 2000).
De um modo geral, os ecossistemas naturais respondem por 55% das
emissões de N2O (FORSTER et al., 2007). No entanto, Organossolos tropicais
podem funcionar como fonte de N2O, em condições aeróbicas e como sumidouro em
condições anaeróbicas (HADI et al., 2000; INUBUSHI et al., 2003). Page e Dalal
(2011) ressaltam que as emissões esperadas de N2O são baixas em sistemas
saturados com água.
Características do solo como estrutura, compactação, textura, presença de
superfície de selamento, impedimento de drenagem e nível freático, podem afetar o
fluxo de N2O (CHAPUIS-LARDY et al., 2007), além da umidade do solo (JIANG et
al., 2009; DAVIDSON, 1991). O teor de carbono (MATHIEU et al., 2006), de nitrato e
de amônio (INUBUSHI et al., 2003; ROSENKRANZ et al., 2005; CASTALDI et al.,
2006), além do potencial redox (HOU et al., 2000; FURUKAWA et al., 2005;
WLODARCZYK et al., 2005), também interferem no fluxo de óxido nitroso. Do
mesmo modo, os fatores climáticos, principalmente a temperatura e a precipitação,
podem influenciar os fluxos de N2O, mas com tendências diferentes em situações
específicas.
A presença simultânea de sítios anaeróbicos e aeróbicos favorece a
produção de N2O e gera grande variabilidade nos fluxos de óxido nitroso (DALAL;
ALLEN, 2008). Von Arnold et al. (2005) trabalhando com Organossolo registraram

65
grandes variações nos fluxos de óxido nitroso sendo que as câmaras individuais
mostraram em todos os sítios, captação e liberação.
São muito variáveis os valores de potencial redox citados na literatura para o
enquadramento dos solos como aeróbicos e anaeróbicos, para propiciar os
processos de nitrificação e desnitrificação e para haver emissão ou consumo de
N2O. Curi e Kampf (2012) enquadram como aeróbicos solos com potencial redox em
torno de 600 mV, enquanto em solos anaeróbicos estes valores oscilam entre 400 e
-300 mV. Para Tian-Yen (1985) solos com valores de potencial redox acima de 400
mV são considerados oxidados.
A desnitrificação pode ocorrer a potenciais redox abaixo de 300 mV (SMITH
et al., 1983; PATRICK; JUGSUJINDA, 1992) ou em torno de 400 mV (GLINSKI;
STEPNIEWSKA, 1986).
Furukawa et al. (2005), indicaram que para valores de potencial redox
oscilando entre 260 e 660 mV, aproximadamente, na profundidade de 10 cm, pode
ocorrer tanto emissão como consumo de óxido nitroso. Em potenciais redox acima
de 200 mV ocorre produção de N2O (SMITH et al., 1983; HOU et al., 2000;
WLODARCZYK et al., 2005).
Elder e Lal (2008) e Huang et al. (2010) encontraram correlação positiva
entre temperatura do ar e o fluxo de óxido nitroso. Porém, Von Arnold et al. (2005)
indicaram que em Organossolo não drenado com nível freático próximo à superfície,
os fluxos de N2O não se correlacionaram com a temperatura do ar e nem com a
precipitação. Entretanto, Inubushi et al. (2003) detectaram aumento no consumo de
N2O em solos cultivados com arroz irrigado quando ocorria aumento na precipitação.
A drenagem e o cultivo de Organossolos são práticas de manejo que
contribuem diretamente com o incremento de N2O na atmosfera (USEPA, 2007). Em
Organossolo drenado o fluxo médio de N2O foi de 131 µg m-2 h-1 e no Organossolo
não drenado os fluxos sofreram brusca redução, caindo para 13 µg m-2 h-1 em média
(VON ARNOLD et al., 2005). Para Furukawa et al. (2005), no entanto, o efeito da
drenagem sobre os fluxos de óxido nitroso não foi claro.
O nível freático também não interfere de maneira padronizada nos fluxos de
óxido nitroso. Embora possa afetar o ciclo do nitrogênio, a relação é mais complexa
do que para o ciclo do carbono (FURUKAWA et al., 2005), sendo que Hadi et al.
(2000 e 2001) não encontraram inter-relação clara entre o nível freático e os fluxos
de N2O.

66
Von Arnold et al. (2005) concluíram que em Organossolos são necessárias
outras variáveis além da temperatura e do nível freático para explicar os fluxos de
N2O, embora o rebaixamento do freático em Organossolo drenado tenha aumentado
a emissão. Em Organossolo não drenado onde o nível freático oscilou entre 12 e -30
cm, o fluxo variou entre 17 e 8 µg N2O m-2 h-1, enquanto em Organossolo drenado,
com nível freático oscilando entre -5 e -45 cm, o fluxo médio variou de 90 a 171 µg
N2O m-2 h-1.
Inúmeros autores explicam que o consumo de óxido nitroso em solos com
excesso de umidade ocorre devido à redução total do N2O à N2 (ROLSTON et al.,
1978; SMITH et al., 2003; INUBUSHI et al., 2003; DALAL et al., 2003; ROBERTSON;
GROFMAN, 2007; DALAL; ALLEN, 2008). A emissão de N2O e N2 via desnitrificação
tem sido amplamente estudada devido o potencial de remoção de nitrato das terras
úmidas (GUTKNECHT et al., 2006), embora pouca atenção tenha sido dada para a
quantificação de N2O em solos anóxicos típicos de terras úmidas os quais são
geralmente considerados desfavoráveis para a produção de óxido nitroso (BLAIS et
al., 2005).
A interação complexa entre os inúmeros fatores e processos apresentados
anteriormente é responsável pela grande variabilidade espacial e temporal que
comumente ocorre nos fluxos de N2O (GROFFMAN, 2000; HUANG et al., 2010;
VON ARNOLD et al., 2005; JIANG et al., 2009). Esta complexidade faz com que o
solo possa ser tanto uma fonte como um sumidouro de N2O (HADI et al., 2000;
CHAPUIS-LARDY et al., 2007) e é responsável pela grande variação encontrada nos
fluxos, a saber: -0,0014 µg N2O m-2 h-1 (FENN et al., 1996) a -484 µg N2O m-2 h-1
(LONGORIA et al., 2003); -0,04 a 0,03 mg N m-2 h-1 (INUBUSHI et al., 2003); -18 a
842 µg N m-2 h-1 (FURUKAWA et al., 2005); -10 a 584 mg N m-2 h-1 (HADI et al.,
2005); 8 a 171 µg N2O m-2 h-1 (VON ARNOLD et al., 2005) e 275 e 307 mg N2O m-2
h-1 (JIANG et al., 2009).
A atuação dos fatores que controlam as emissões de N2O não está bem
clara, embora o padrão estacional mostre algumas tendências (INUBUSHI et al.,
2003). Baseado no conhecimento atual não é ainda possível definir claramente as
condições que promovem o consumo de N2O (CHAPUIS-LARDY et al., 2007).
Na presente pesquisa, formulou-se a hipótese de que a drenagem e as
variações na temperatura do ar e na precipitação, ao longo do tempo, provocariam
mudanças nos fluxos de N2O.

67
O objetivo do estudo foi avaliar os fluxos de N2O em Organossolo Háplico
sáprico típico em campo hidrófilo de altitude, em condições naturais e após a
drenagem, considerando a influência do clima e das alterações provocadas no solo.
68
3.2 MATERIAL E MÉTODOS
3.2.1 Experimento de campo
Instalou-se um experimento em área de estepe hidrófila com dimensão de
6,5 ha (25o09´59´´S; 50o04´45´´O), a 860 m de altitude, na fazenda da Embrapa
SPM-EPGA (Serviço de Produtos e Mercado - Escritório de Ponta Grossa, PR),
segundo planalto paranaense, região dos Campos Gerais (Figura 1).
FIGURA 1 - LOCALIZAÇÃO DA ÁREA DE ESTUDO
O local de estudo situa-se em uma nascente do rio Cará-Cará, afluente do
rio Tibagi. O clima local é do tipo Cfb, mesotérmico úmido, sem estação seca,
temperatura média do mês mais quente inferior a 22oC, temperatura média do mês
mais frio menor do que 18oC e precipitação entre 1100 e 2000 mm. A área encontra-
se inserida em reverso distal de cuesta, sobre folhelho argiloso da Formação Ponta
Grossa, pertencente ao Grupo Paraná, Paleozóico. O solo foi classificado como
ORGANOSSOLO HÁPLICO sáprico típico álico relevo plano fase campo subtropical
hidrófilo de altitude, de acordo com o Sistema Brasileiro de Classificação de Solos
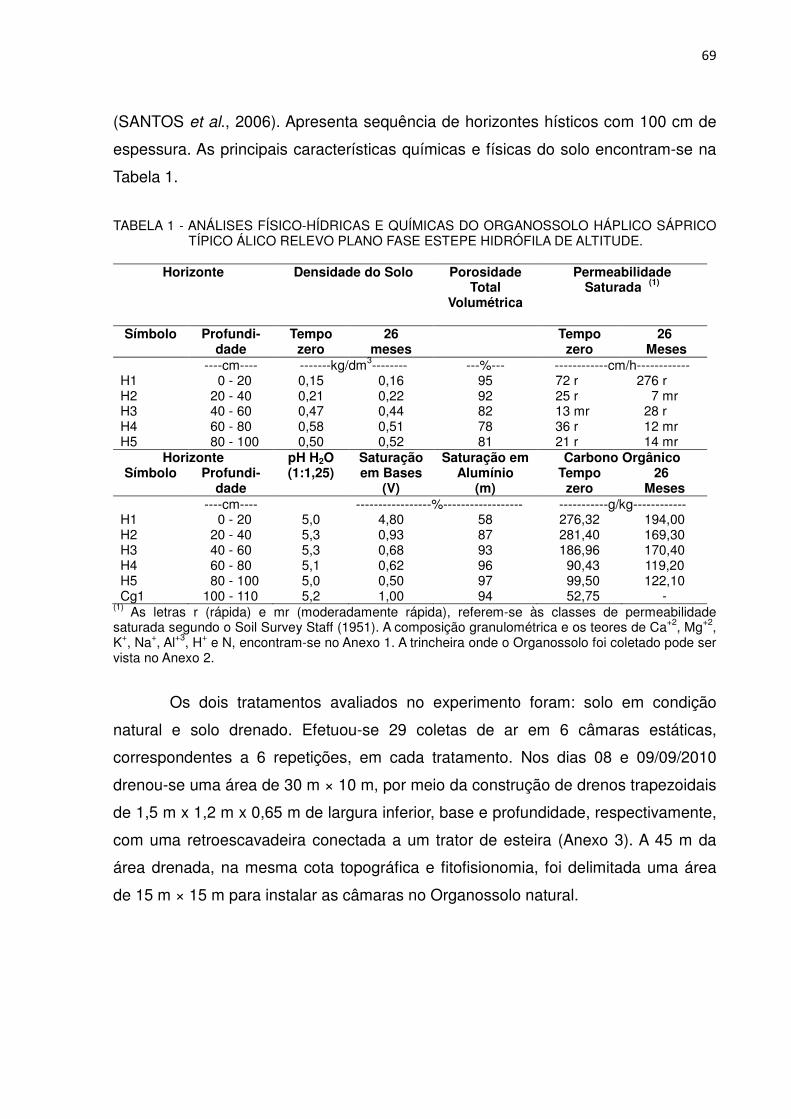
69
(SANTOS et al., 2006). Apresenta sequência de horizontes hísticos com 100 cm de
espessura. As principais características químicas e físicas do solo encontram-se na
Tabela 1.
TABELA 1 - ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE.
Horizonte Densidade do Solo Porosidade
Total Volumétrica
Permeabilidade Saturada (1)
Símbolo Profundi-dade
Tempo zero
26 meses
Tempo zero
26 Meses
----cm---- -------kg/dm3-------- ---%--- ------------cm/h------------ H1 0 - 20 0,15 0,16 95 72 r 276 r H2 20 - 40 0,21 0,22 92 25 r 7 mr H3 40 - 60 0,47 0,44 82 13 mr 28 r H4 60 - 80 0,58 0,51 78 36 r 12 mr H5 80 - 100 0,50 0,52 81 21 r 14 mr
Horizonte pH H2O (1:1,25)
Saturação em Bases
(V)
Saturação em Alumínio
(m)
Carbono Orgânico Símbolo Profundi-
dade Tempo
zero 26
Meses ----cm---- -----------------%------------------ -----------g/kg------------
H1 0 - 20 5,0 4,80 58 276,32 194,00 H2 20 - 40 5,3 0,93 87 281,40 169,30 H3 40 - 60 5,3 0,68 93 186,96 170,40 H4 60 - 80 5,1 0,62 96 90,43 119,20 H5 80 - 100 5,0 0,50 97 99,50 122,10 Cg1 100 - 110 5,2 1,00 94 52,75 -
(1) As letras r (rápida) e mr (moderadamente rápida), referem-se às classes de permeabilidade saturada segundo o Soil Survey Staff (1951). A composição granulométrica e os teores de Ca+2, Mg+2, K+, Na+, Al+3, H+ e N, encontram-se no Anexo 1. A trincheira onde o Organossolo foi coletado pode ser vista no Anexo 2.
Os dois tratamentos avaliados no experimento foram: solo em condição
natural e solo drenado. Efetuou-se 29 coletas de ar em 6 câmaras estáticas,
correspondentes a 6 repetições, em cada tratamento. Nos dias 08 e 09/09/2010
drenou-se uma área de 30 m × 10 m, por meio da construção de drenos trapezoidais
de 1,5 m x 1,2 m x 0,65 m de largura inferior, base e profundidade, respectivamente,
com uma retroescavadeira conectada a um trator de esteira (Anexo 3). A 45 m da
área drenada, na mesma cota topográfica e fitofisionomia, foi delimitada uma área
de 15 m × 15 m para instalar as câmaras no Organossolo natural.

70
3.2.2 Coleta de amostras de ar
Foram realizadas 17 coletas de amostras de ar durante o primeiro ano de
estudo, entre 10/09/2010 e 26/09/2011 e 12 coletas ao longo do segundo ano, entre
27/09/2011 e 16/10/2012 (Anexo 4).
Seis bases de metal com 38 cm de diâmetro foram inseridas no solo para
receberem as câmaras coletoras de gás, em cada tratamento. Na parte superior
destas bases havia uma canaleta circular de 6 cm de largura por 4 cm de altura, a
qual era preenchida com água, após a colocação da câmara, para evitar vazamento
dos gases, na ocasião das coletas.
As coletas de gases foram efetuadas em câmaras de PVC (Anexo 5) com 40
cm de altura equipadas com termômetro, ventilador para homogeneizar o ar e
torneira de três vias para coleta de amostras de ar. Seringas de polipropileno de 20
ml acopladas à torneira três vias eram usadas para conter as amostras de ar que
foram retiradas a 0, 15, 30 e 45 minutos, após a câmara ter sido fechada. As coletas
eram feitas entre às 9:00 e 12:00 horas.
Para determinar a concentração de N2O utilizou-se um cromatógrafo de fase
gasosa (Shimadzu GC-2014) com detector de captura de elétrons (ECD), colunas
empacotadas em temperatura de 70 ºC, N2 como gás de arraste com fluxo de 26 mL
min-1, temperatura do injetor de 250 ºC e temperatura do detector de 325 ºC. Estas
análises foram realizadas no Laboratório de Biogeoquímica Ambiental da UFRGS,
em Porto Alegre (RS).
Os fluxos de N2O foram calculados pela equação F(N2O) = d[N2O]/dt x 28 x
P x V/ R x T, onde d[N2O]/dt é a mudança na quantidade do gás dentro da câmara
no intervalo de tempo; V e T são o volume (L) e temperatura interna (0K) da câmara;
P é a pressão no interior da câmara, assumida como 1 atm e R é a constante
universal dos gases (0,0821 L.atm K-1 mol-1). A variação da concentração do gás no
tempo foi obtida pelo coeficiente angular da equação da reta ajustada.
Considerando-se a área de solo contido na base da câmara, calculou-se a massa do
gás que foi emitida por unidade de área e de tempo em µg N2O m-2 h-1.
Os fluxos determinados foram considerados como representativos do fluxo
médio diário, conforme proposto por Jantália et al. (2008). O fluxo acumulado por

71
estação e anual foram calculados no programa Sigmaplot, pela integração da área
sob a curva, a qual foi estabelecida por interpolação dos valores diários dos fluxos.
3.2.3 Parâmetros pedológicos
O teor de carbono orgânico foi determinado por oxidação sob atmosfera de
gás oxigênio em determinador automático Analytik Jena. As análises químicas e
físico-hídricas do solo foram realizadas conforme metodologia proposta por
Claessen (1997).
Para determinar a umidade gravimétrica e a quantidade de amônio e de
nitrato no solo em cada coleta de ar, três amostras de solo de 0 a 10 cm foram
coletadas por parcela. O teor de amônio e de nitrato foi determinado pelo método de
microdestilação a vapor proposto por Bremner (1965), utilizando-se óxido de
magnésio para alcalinizar o meio e Liga de Devarda para reduzir o nitrato a amônia.
Os valores de amônio e de nitrato foram expressos em mg kg-1 de N-NH4+ e de N-
NO3-, respectivamente, tendo sido calculados com base no peso úmido das
amostras, devido a natureza hidromórfica dos Organossolos estudados.
O monitoramento da altura do nível freático foi realizado em quatro poços
hídricos instalados em cada tratamento. Estes poços, construídos com tubos de
PVC de 75 mm de diâmetro e 1,20 m de altura, foram introduzidos até o horizonte
Cg. Visando acelerar a passagem da água dentro dos tubos, foram feitos quatro
furos de 1 cm de diâmetro a cada 10 cm, ao longo do comprimento dos mesmos. A
cada duas câmaras foi colocado um poço hídrico, no qual determinou-se a altura do
nível freático em todas as coletas de ar.
O potencial redox foi determinado no campo, por um pHmetro portátil
modelo HI 8424 equipado com um eletrodo de vidro com sensor de platina, ambos
da marca Hanna. As leituras foram efetuadas nas profundidades de 5, 10, 15, 20, 30
e 40 cm, nas coletas 24, 25, 26, 28 e 29. Para o enquadramento dos solos nas
classes de redução utilizou-se as classes de estado de redução propostas por Tian-
Yen (1985), a saber: oxidado > 400 milivolts (mV), fracamente reduzido (400 a 200
mV), moderadamente reduzido (200 a -100 mV) e fortemente reduzido (< -100 mV).

72
3.2.4 Variáveis meteorológicas
A temperatura do ar e a precipitação foram monitoradas diariamente, durante
os dois anos de estudo, por meio de uma estação meteorológica portátil Vantage
Vue marca Davis instalada a aproximadamente 1 km da área experimental.
3.2.5 Análise estatística
Utilizou-se o teste de Tukey (p < 0,05) para verificar as diferenças entre os
dois tratamentos nos fluxos de N2O, umidade gravimétrica do solo na profundidade
de 0 a 10 cm, altura do nível freático, potencial redox do solo e teor de N-NH4+ e N-
NO3-.
Determinou-se os coeficientes de correlação linear de Pearson (r) entre os
fluxos de N2O e a temperatura média do ar e precipitação acumulada, entre às 9 e
12 horas na ocasião das coletas e durante o período de 1, 3, 5, 7, 15 e 30 dias que
antederam as coletas, altura do nível freático, e umidade gravimétrica na
profundidade de 0 a 10 cm.
Aplicou-se o Modelo Aditivo Generalizado (GAM) para avaliar os efeitos dos
tratamentos, dos componentes principais (fatores climáticos e pedológicos) e do
tempo, sobre o fluxos de N2O. Este modelo possibilitou uma análise conjunta de
todas as variáveis envolvidas.
As análises estatísticas foram realizadas com o Software R (R
DEVELOPMENT CORE TEAM, 2011).
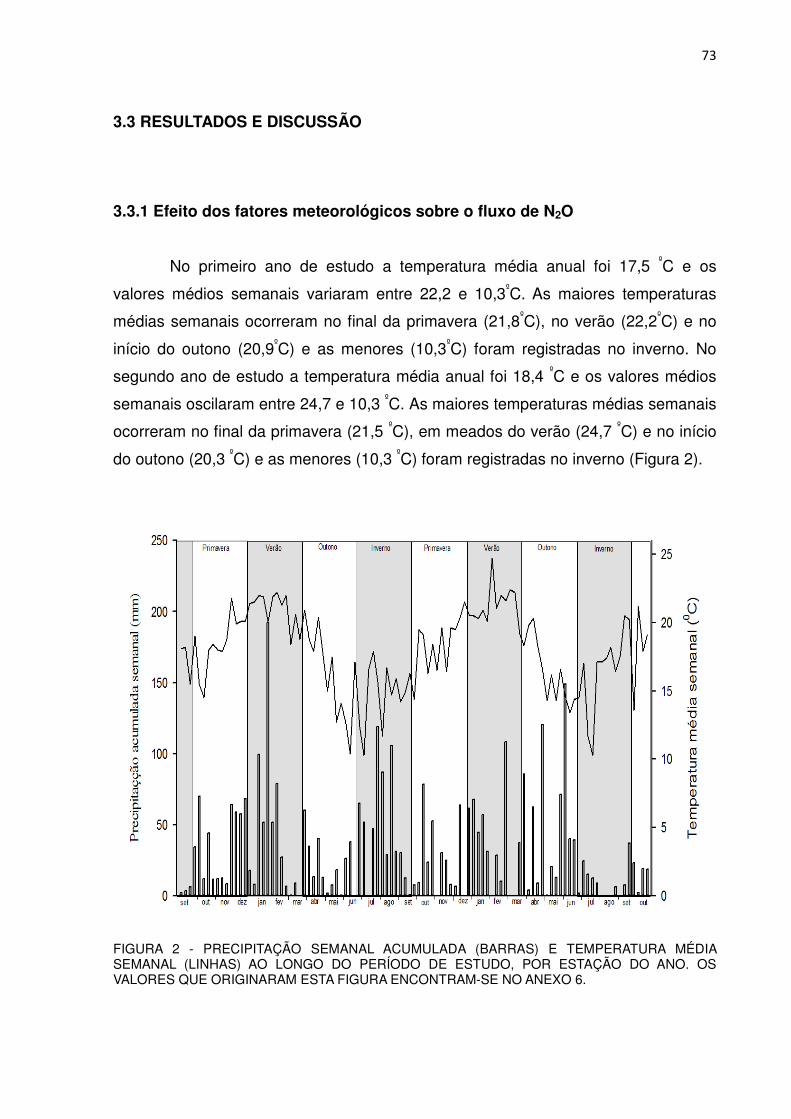
73
3.3 RESULTADOS E DISCUSSÃO
3.3.1 Efeito dos fatores meteorológicos sobre o fluxo de N2O
No primeiro ano de estudo a temperatura média anual foi 17,5 ºC e os
valores médios semanais variaram entre 22,2 e 10,3ºC. As maiores temperaturas
médias semanais ocorreram no final da primavera (21,8ºC), no verão (22,2ºC) e no
início do outono (20,9ºC) e as menores (10,3ºC) foram registradas no inverno. No
segundo ano de estudo a temperatura média anual foi 18,4 ºC e os valores médios
semanais oscilaram entre 24,7 e 10,3 ºC. As maiores temperaturas médias semanais
ocorreram no final da primavera (21,5 ºC), em meados do verão (24,7 ºC) e no início
do outono (20,3 ºC) e as menores (10,3 ºC) foram registradas no inverno (Figura 2).
FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO. OS VALORES QUE ORIGINARAM ESTA FIGURA ENCONTRAM-SE NO ANEXO 6.

74
Não houve diferenças marcantes na temperatura média semanal do ar nas
estações entre os dois anos. As temperaturas médias na primavera, verão, outono e
inverno foram de 18,5, 21,0, 16,2 e 14,8 0C, respectivamente, no primeiro ano e de
18,4, 21,3, 16,3 e 16,5 0C, respectivamente no segundo ano. No inverno de 2012 a
temperatura média semanal foi apenas 1,7 0C mais elevada do que no inverno de
2011.
A precipitação total acumulada ao longo do primeiro ano foi de 1837 mm
(Figura 2). As estações mais chuvosas foram a primavera (442 mm), o verão (543
mm) e, atipicamente, o inverno (580 mm). No outono a precipitação acumulada foi
254 mm. A precipitação total acumulada ao longo do segundo ano foi de 1518 mm.
As estações mais chuvosas foram o verão (446 mm) e atipicamente o outono (615
mm), seguidos pela primavera (298 mm) e inverno (114 mm). A precipitação anual no
segundo ano de estudo foi 319 mm menor do que no primeiro, sendo que com
exceção do outono, as outras estações foram menos chuvosas.
No solo natural os fluxos variaram entre 1,6 e -29,2 µg m-2 h-1 de N2O, com
média de -11,9 µg m-2 h-1 no primeiro ano e de -0,13 a -31,24 µg m-2 h-1 com média
de -10,27 µg m-2 h-1 no segundo (Figura 3). Não houve diferença estatística nos
fluxos entre as coletas em ambos os anos. Isto indica que houve homogeneidade
entre os fluxos de um ano para o outro o que é compatível com a condição natural
deste solo, o qual não sofreu nenhuma intervenção antrópica. Entretanto, deve-se
considerar que a variabilidade entre os valores das câmaras foi muito grande, sendo
que as médias foram geradas, algumas vezes, por valores negativos e positivos.
Os picos de consumo de óxido nitroso (Figura 3) se deram em novembro
(final de primavera - coleta 7) e fevereiro (verão - coleta 9), no primeiro ano, e em
janeiro e março (verão - coletas 20 e 22) do segundo ano, coincidindo com as
temperaturas mais elevadas (Figura 2).
Resultados similares foram encontrados por Inubushi et al. (2003) os quais
registraram os maiores valores de consumo de óxido nitroso entre fevereiro e junho,
correspondendo ao final do inverno, a primavera e ao início do verão.
No solo drenado os fluxos variaram entre -0,70 e 38,2 µg m-2 h-1 de N2O com
média de 8,7 µg m-2 h-1, no primeiro ano e de 13,0 a µg m-2 h-1, e 55,7 µg m-2 h-1 com
média de 27,9 µg m-2 h-1, no segundo (Figura 3). Os valores mínimos e máximos
foram superiores no segundo ano e a média foi 3,2 vezes maior do que no primeiro,
o que indica o forte efeito da drenagem sobre a emissão de óxido nitroso. Percebe-
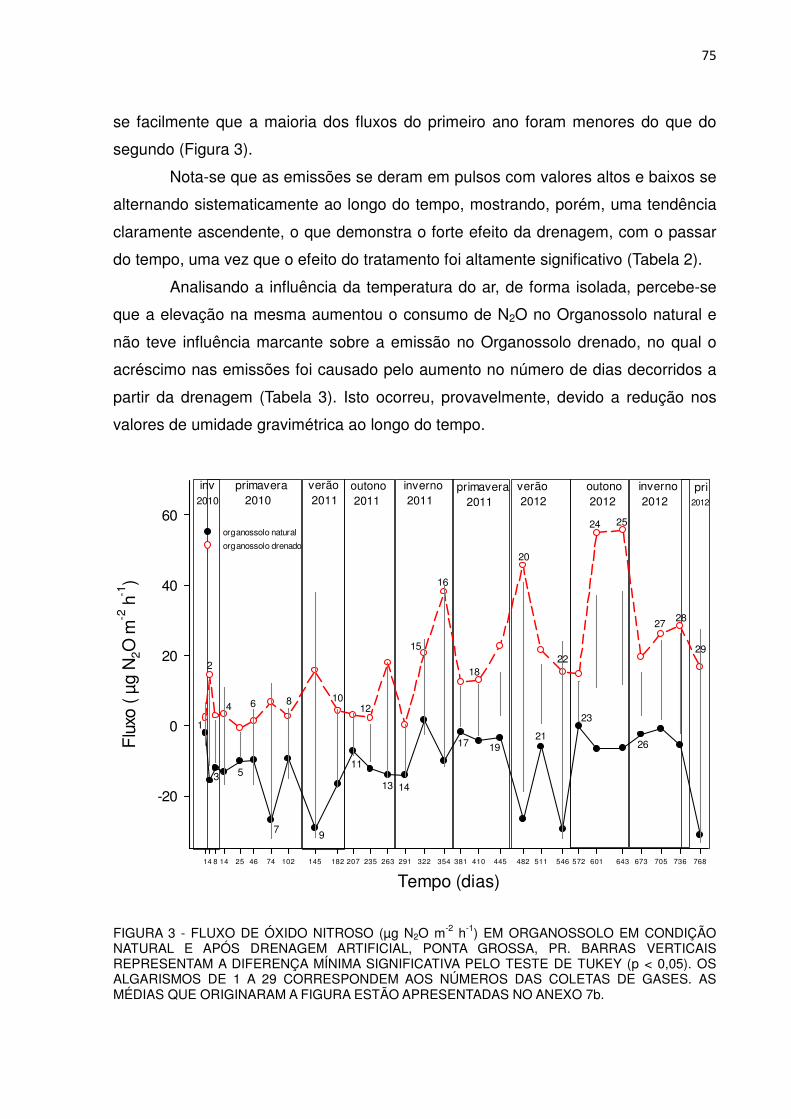
75
se facilmente que a maioria dos fluxos do primeiro ano foram menores do que do
segundo (Figura 3).
Nota-se que as emissões se deram em pulsos com valores altos e baixos se
alternando sistematicamente ao longo do tempo, mostrando, porém, uma tendência
claramente ascendente, o que demonstra o forte efeito da drenagem, com o passar
do tempo, uma vez que o efeito do tratamento foi altamente significativo (Tabela 2).
Analisando a influência da temperatura do ar, de forma isolada, percebe-se
que a elevação na mesma aumentou o consumo de N2O no Organossolo natural e
não teve influência marcante sobre a emissão no Organossolo drenado, no qual o
acréscimo nas emissões foi causado pelo aumento no número de dias decorridos a
partir da drenagem (Tabela 3). Isto ocorreu, provavelmente, devido a redução nos
valores de umidade gravimétrica ao longo do tempo.
organossolo natural
Tempo (dias)14 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Flu
xo (
µg N
2O m
-2 h
-1)
-20
0
20
40
60
inv 2010
primavera 2010
verão 2011
outono 2011
inverno 2011
primavera 2011
verão 2012
outono 2012
inverno 2012
pri 2012
organossolo drenado
1
2
3
4
5
6
7
8
9
10
11
12
13 14
15
16
17
18
19
20
21
22
23
24 25
26
2728
29
FIGURA 3 - FLUXO DE ÓXIDO NITROSO (µg N2O m-2 h-1) EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS ALGARISMOS DE 1 A 29 CORRESPONDEM AOS NÚMEROS DAS COLETAS DE GASES. AS MÉDIAS QUE ORIGINARAM A FIGURA ESTÃO APRESENTADAS NO ANEXO 7b.

76
A precipitação, por sua vez, não interferiu intensamente sobre o consumo de
N2O no solo natural, pois como o mesmo se manteve saturado com água durante
todo o tempo, o acréscimo de água pelas chuvas não aumentava significativamente
a quantidade de água no solo. Por outro lado, no Organossolo drenado, o aumento
na precipitação resultava em elevação na emissão de N2O (Tabela 3).
Picos de produção de óxido nitroso geralmente ocorrem imediatamente após
a adição de nitrato. Em poucas horas ou dias, a produção de N2O decresce e
incrementa a produção de N2, nos microsítios anaeróbicos, mesmo quando
concentrações relativamente altas de nitrato ainda estão presentes (ROLSTON et al.
1978).
No Organossolo drenado do presente estudo a elevação na emissão de
óxido nitroso não ocorreu em função da adição de nitrato, mas possivelmente devido
ao aumento nos teores de NO3- produzidos por nitrificação com a retirada da água.
TABELA 2 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE OS FLUXOS DE N2O, PELO MODELO ADITIVO GENERALIZADO - GAM.
Efeitos paramétricos p valor Tratamento (solo natural e drenado) < 2 x 10-16 *** Temperatura do ar (Componente principal 1)(1) 0,7196 ns Temperatura do ar (Componente principal 2)(2) 0,0607 ns Temperatura do ar+precipitação acumulada (Componente principal 3)(3) 0,9867 ns Efeitos não paramétricos p valor Tempo(4) (solo drenado) 7,7 x 10-7 *** Tempo(5) (solo natural) 0,493 ns
*** - altamente significativo (p < 0,001); ns - não significativo; (1) Temperatura mínima do ar das 9 às 12 horas no momento da coleta + temperatura mínima do ar durante o período de 1 dia que antecedeu as coletas. (2) Temperatura mínima do ar das 9 às 12 horas no momento da coleta + temperatura mínima do ar durante o período de 1 dia que antecedeu as coletas + temperatura média do ar nos períodos de 3 e de 5 dias que antecederam as coletas. (3) Temperatura mínima do ar das 9 às 12 horas no momento da coleta + temperatura mínima do ar durante o período de 1 dia que antecedeu as coletas + temperatura média do ar nos períodos de 3 e de 5 dias que antecederam as coletas + precipitação acumulada durante o período de 1 dia que antecedeu as coletas. (4) Número de dias a partir da drenagem. (5) Número de dias a partir do início do experimento.
Os fluxos ocorreram em vários pulsos de emissão, sendo que praticamente
cada elevação foi seguida de uma redução brusca (Figura 3). Esta alternância de
emissões altas e baixas sugere que houve alternância entre os processos de
nitrificação e desnitrificação. O primeiro processo teria originado as emissões
menores e o segundo os valores maiores.
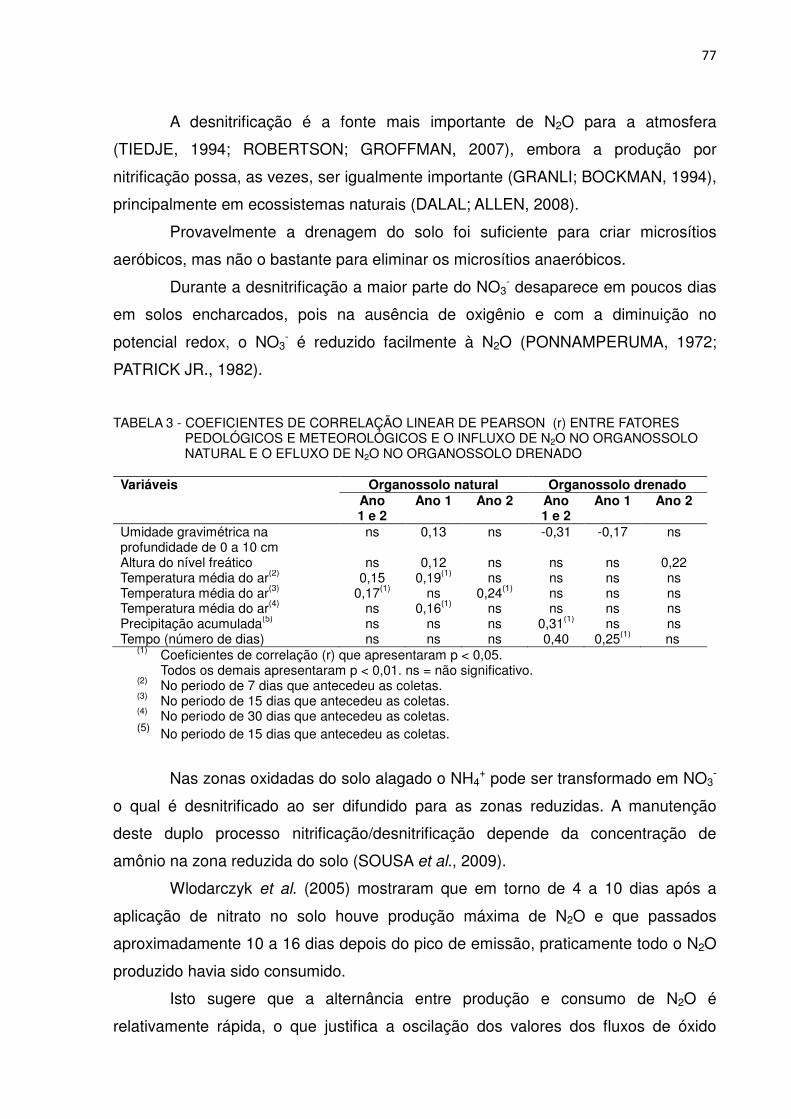
77
A desnitrificação é a fonte mais importante de N2O para a atmosfera
(TIEDJE, 1994; ROBERTSON; GROFFMAN, 2007), embora a produção por
nitrificação possa, as vezes, ser igualmente importante (GRANLI; BOCKMAN, 1994),
principalmente em ecossistemas naturais (DALAL; ALLEN, 2008).
Provavelmente a drenagem do solo foi suficiente para criar microsítios
aeróbicos, mas não o bastante para eliminar os microsítios anaeróbicos.
Durante a desnitrificação a maior parte do NO3- desaparece em poucos dias
em solos encharcados, pois na ausência de oxigênio e com a diminuição no
potencial redox, o NO3- é reduzido facilmente à N2O (PONNAMPERUMA, 1972;
PATRICK JR., 1982).
TABELA 3 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE FATORES PEDOLÓGICOS E METEOROLÓGICOS E O INFLUXO DE N2O NO ORGANOSSOLO NATURAL E O EFLUXO DE N2O NO ORGANOSSOLO DRENADO
Variáveis Organossolo natural Organossolo drenado
Ano 1 e 2
Ano 1 Ano 2 Ano 1 e 2
Ano 1 Ano 2
Umidade gravimétrica na profundidade de 0 a 10 cm
ns 0,13 ns -0,31 -0,17 ns
Altura do nível freático ns 0,12 ns ns ns 0,22 Temperatura média do ar(2) 0,15 0,19(1) ns ns ns ns Temperatura média do ar(3) 0,17(1) ns 0,24(1) ns ns ns Temperatura média do ar(4) ns 0,16(1) ns ns ns ns Precipitação acumulada(5) ns ns ns 0,31(1) ns ns Tempo (número de dias) ns ns ns 0,40 0,25(1) ns
(1) Coeficientes de correlação (r) que apresentaram p < 0,05. Todos os demais apresentaram p < 0,01. ns = não significativo.
(2) No periodo de 7 dias que antecedeu as coletas. (3) No periodo de 15 dias que antecedeu as coletas. (4) No periodo de 30 dias que antecedeu as coletas. (5) No periodo de 15 dias que antecedeu as coletas.
Nas zonas oxidadas do solo alagado o NH4+ pode ser transformado em NO3
-
o qual é desnitrificado ao ser difundido para as zonas reduzidas. A manutenção
deste duplo processo nitrificação/desnitrificação depende da concentração de
amônio na zona reduzida do solo (SOUSA et al., 2009).
Wlodarczyk et al. (2005) mostraram que em torno de 4 a 10 dias após a
aplicação de nitrato no solo houve produção máxima de N2O e que passados
aproximadamente 10 a 16 dias depois do pico de emissão, praticamente todo o N2O
produzido havia sido consumido.
Isto sugere que a alternância entre produção e consumo de N2O é
relativamente rápida, o que justifica a oscilação dos valores dos fluxos de óxido
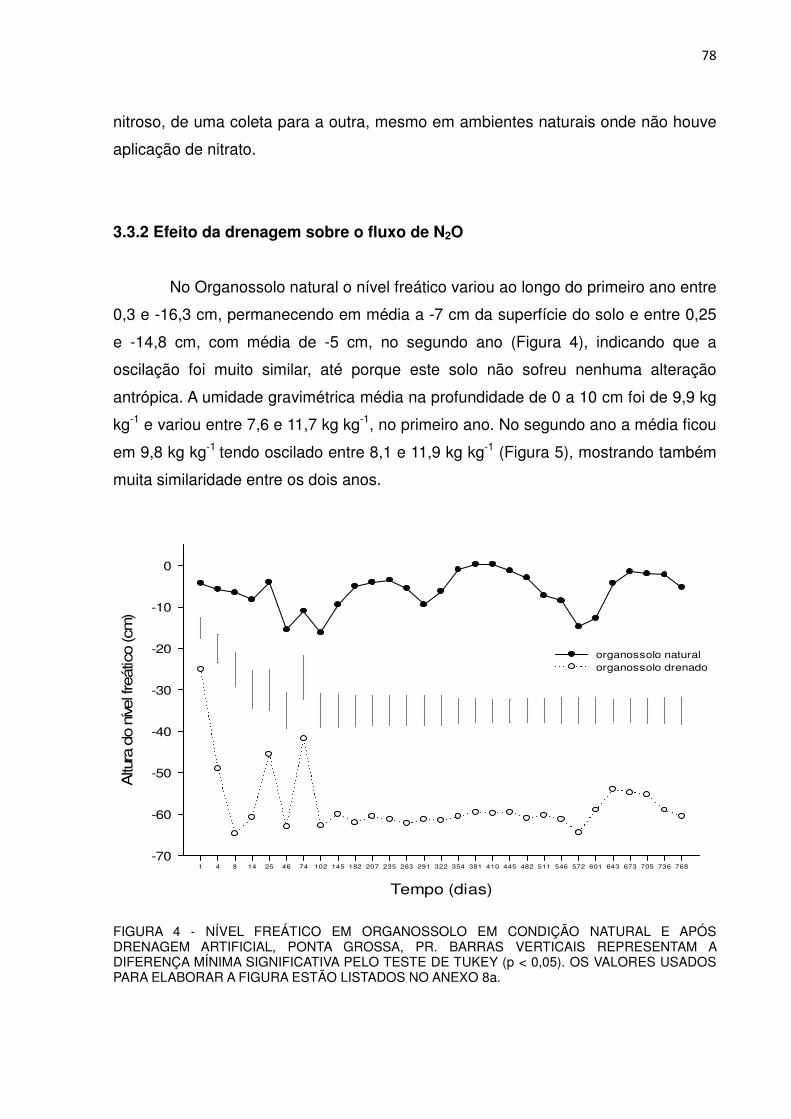
78
nitroso, de uma coleta para a outra, mesmo em ambientes naturais onde não houve
aplicação de nitrato.
3.3.2 Efeito da drenagem sobre o fluxo de N2O
No Organossolo natural o nível freático variou ao longo do primeiro ano entre
0,3 e -16,3 cm, permanecendo em média a -7 cm da superfície do solo e entre 0,25
e -14,8 cm, com média de -5 cm, no segundo ano (Figura 4), indicando que a
oscilação foi muito similar, até porque este solo não sofreu nenhuma alteração
antrópica. A umidade gravimétrica média na profundidade de 0 a 10 cm foi de 9,9 kg
kg-1 e variou entre 7,6 e 11,7 kg kg-1, no primeiro ano. No segundo ano a média ficou
em 9,8 kg kg-1 tendo oscilado entre 8,1 e 11,9 kg kg-1 (Figura 5), mostrando também
muita similaridade entre os dois anos.
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Altu
ra d
o ní
vel f
reát
ico
(cm
)
-70
-60
-50
-40
-30
-20
-10
0
organossolo naturalorganossolo drenado
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA ELABORAR A FIGURA ESTÃO LISTADOS NO ANEXO 8a.
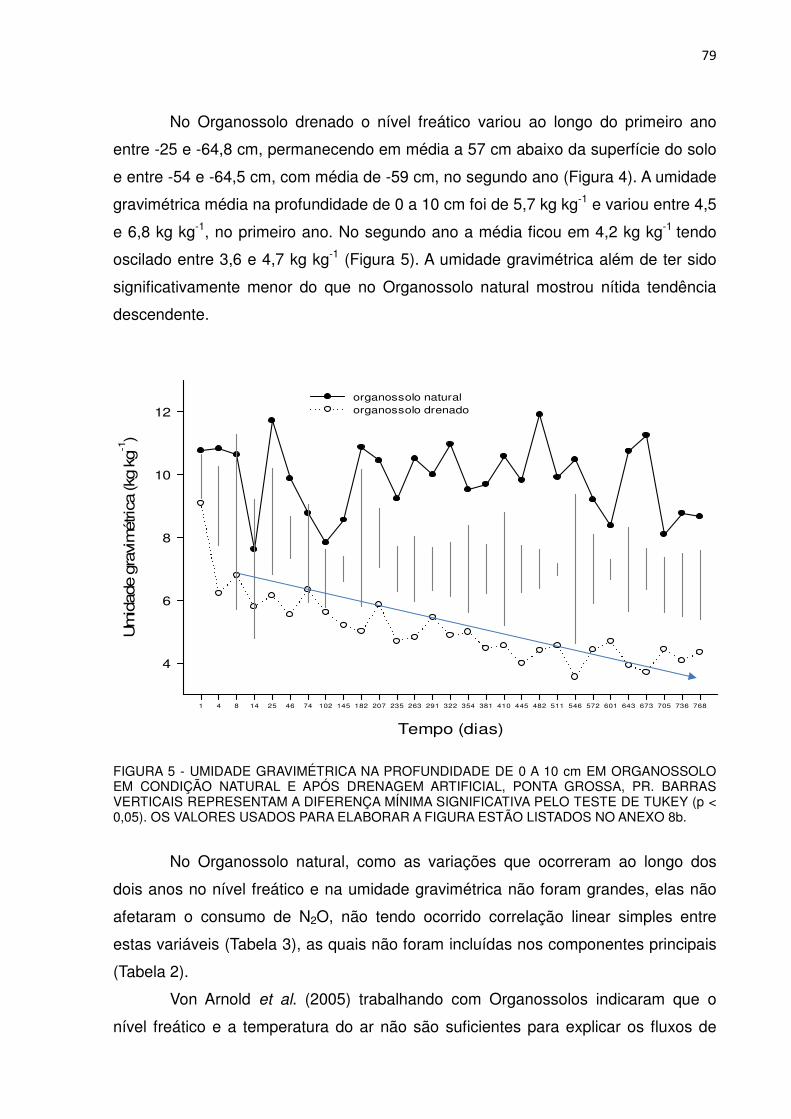
79
No Organossolo drenado o nível freático variou ao longo do primeiro ano
entre -25 e -64,8 cm, permanecendo em média a 57 cm abaixo da superfície do solo
e entre -54 e -64,5 cm, com média de -59 cm, no segundo ano (Figura 4). A umidade
gravimétrica média na profundidade de 0 a 10 cm foi de 5,7 kg kg-1 e variou entre 4,5
e 6,8 kg kg-1, no primeiro ano. No segundo ano a média ficou em 4,2 kg kg-1 tendo
oscilado entre 3,6 e 4,7 kg kg-1 (Figura 5). A umidade gravimétrica além de ter sido
significativamente menor do que no Organossolo natural mostrou nítida tendência
descendente.
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Um
idad
e gr
avim
étric
a (k
g kg
-1)
4
6
8
10
12organossolo naturalorganossolo drenado
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA ELABORAR A FIGURA ESTÃO LISTADOS NO ANEXO 8b.
No Organossolo natural, como as variações que ocorreram ao longo dos
dois anos no nível freático e na umidade gravimétrica não foram grandes, elas não
afetaram o consumo de N2O, não tendo ocorrido correlação linear simples entre
estas variáveis (Tabela 3), as quais não foram incluídas nos componentes principais
(Tabela 2).
Von Arnold et al. (2005) trabalhando com Organossolos indicaram que o
nível freático e a temperatura do ar não são suficientes para explicar os fluxos de

80
N2O, sendo necessárias variáveis adicionais. Estes mesmos autores afirmaram que
em solos com nível freático superficial os fluxos de óxido nitroso não são
linearmente relacionados com a temperatura do ar, havendo uma correlação múltipla
com a temperatura do ar e o nível freático.
No presente trabalho não houve correlação entre o consumo de N2O e a
umidade gravimétrica na profundidade de 0 a 10 cm, no organosolo natural (Tabela
3), pois o consumo do mesmo ocorre a profundidades maiores.
Van Groenigen et al. (2005) trabalhando em solo arenoso, identificaram que
na profundidade de 90 cm localizava-se a fonte principal de óxido nitroso, o qual
posteriormente foi consumido em seu caminho rumo à superfície do solo. Arah et al.
(1991) também se referem ao consumo de N2O em camadas mais profundas.
A emissão de N2O a partir da superfície do solo é função do tempo e da
profundidade específica onde se dá a produção e o consumo, no perfil (HOSEN et
al., 2000; CHAPUIS-LARDY et al., 2007). A maioria do N2O produzido no topo do
solo facilmente escapa para a atmosfera com curto período de permanência no solo,
enquanto o óxido nitroso produzido em profundidade pode nunca atingir a atmosfera
(YOH et al., 1997; NEFTEL et al., 2000). Segundo Metay (2005) há mesmo uma
defasagem de tempo entre a produção de N2O e sua difusão até a superfície.
O excesso de água no Organossolo natural do presente estudo ocasionou o
consumo de óxido nitroso, pois do mesmo modo observado por Robertson e
Groffman (2007), Farquharson e Baldock (2008) e Kogel-Knabner et al. (2010) pode
ter ocorrido desnitrificação completa do N2O com emissão de N2. Quanto mais
tempo o óxido nitroso permanece no solo, por ser produzido em camadas profundas
ou devido à redução na difusividade, mais ele é consumido (CHAPUIS-LARDY et al.,
2007).
Van Groenigen et al. (2005) relataram que pode ocorrer consumo de N2O
quando ele é difundido no sentido ascendente no perfil. Farquharson e Baldock
(2008) citam organismos desnitrificadores atuando neste processo.
No Organossolo natural a possível redução do N2O à N2 aconteceu em
profundidades maiores do que 15 cm, as quais apresentaram menores potenciais de
óxido-redução, como será visto adiante. Além disto o maior tempo de permanência
em condição de excesso de umidade deve ter contribuído.
O consumo de N2O, o qual segundo Inubushi et al. (2003) é altamente
solúvel em água, pode estar muito mais associado a dificuldade de difusão do

81
mesmo através do solo submerso do que ao próprio potencial de redução,
aumentando o tempo de permanência e induzindo os microrganismos a consumirem
o mesmo após a sua emissão (RYDEN, 1981; CHAPUIS-LARDY et al., 2007).
Isto sugere que mesmo em situações em que o potencial redox não se
encontrava muito baixo, como no caso das coletas 24 a 26 (Figura 6),
provavelmente o óxido nitroso tenha sido reduzido à N2, devido ao longo tempo de
submersão.
Na maioria das situações as quantidades de N2 emitidas durante a
desnitrificação são muito superiores às de N2O (CANTARELLA, 2007). Altos índices
de decomposição de matéria orgânica consomem muito oxigênio criando microsítios
anaeróbicos, o que favorece a desnitrificação (SMITH et al., 2003), sendo esperada
alta emissão de N2O (MALJANEN et al., 2003).
A emissão de óxido nitroso no Organossolo drenado foi atribuída a retirada
da água e a entrada de oxigênio em função do rebaixamento do nível freático, o que
estimulou a decomposição da matéria orgânica.
No primeiro ano, já a partir do primeiro dia após a drenagem, quando o nível
freático encontrava-se a 25 cm abaixo da superfície, o solo drenado apresentou
emissão de óxido nitroso, o que se prologou até o final do estudo. A partir de uma
semana após a drenagem o nível freático manteve-se constante a aproximadamente
60 cm abaixo da superfície na maior parte do tempo.
No Organossolo drenado, a umidade gravimétrica diminuiu ao longo do
tempo e a emissão de óxido nitroso aumentou, indicando correlação linear negativa
entre as duas variáveis. O nível freático, que se manteve praticamente constante,
não teve influência na emissão de N2O ao longo do tempo (Tabela 3).
Furukawa et al. (2005), no entanto, constataram que não houve relação
entre o nível freático e o fluxo de óxido nitroso, embora o pico de emissão máxima
tenha ocorrido quando o nível freático encontrava-se a apenas -20 cm. Para estes
autores o efeito da drenagem sobre o fluxo de N2O não foi claro pois a variação do
nível freático entre -10 e -30 cm possivelmente promoveu, tanto a nitrificação quanto
a desnitrificação.
Inúmeros autores observaram elevadas emissões de óxido nitroso quando o
nível freático foi rebaixado, nos processos de flutuação cíclica natural, criando
ambientes óxicos e anóxicos. Nestas condições o N2O é formado em grandes
quantidades durante a nitrificação e parte do nitrato produzido, pode mais tarde ser

82
reduzido a óxido nitroso, quando o potencial redox cai e as condições se tornam
favoráveis para a desnitrificação (BALDWIN; MITCHELL, 2000; HERNADEZ;
MITSCH, 2006; PAGE; DALAL, 2011), quando o solo torna-se novamente
encharcado.
O nitrato também pode ser transferido dos sítios aeróbicos para os
anaeróbicos nos quais pode ser reduzido.
A presença simultânea de sítios aeróbicos e anaeróbicos cria condições
favoráveis para a produção de N2O (DALAL; ALLEN, 2008), porque o nitrato formado
nos primeiros pode liberar N2O durante a nitritação (FARQUHARSON; BALDOCK,
2008) durante a transformação de NH4+ em NO3
- por bactérias quimioautotróficas
(GRANLI; BOCKMAN, 1994). No entanto a maior emissão se origina na
desnitrificação, na qual o nitrato anteriormente produzido é reduzido biologicamente
à N2O (TIEDJE, 1994; SMITH et al., 2003; ROBERTSON; GROFFMAN, 2007).
Possivelmente a emissão de N2O no Organossolo drenado da presente
pesquisa tenha ocorrido principalmente por desnitrificação, não descartando a
contribuição do processo de nitrificação. Provavelmente, a nitrificação tenha sido
mais intensa na superfície e nas zonas aeróbicas, enquanto a desnitrificação tenha
ocorrido com maior intensidade em profundidades maiores, nas zonas anaeróbicas.
Após a drenagem, um grande volume de solo tornou-se livre do excesso de
água e com maior disponibilidade de oxigênio, condição favorável para nitrificação e
liberação de óxido nitroso.
Por outro lado, embora o solo tenha se livrado do excesso de água, a
umidade ainda manteve-se relativamente elevada, o que continuou sendo propício
para a desnitrificação, principalmente nos microsítios anaeróbicos.
Wrage et al. (2001) referem-se a ambientes que reúnem condições para que
os dois processos ocorram ao mesmo tempo, desde que condições favoráveis tanto
para a nitrificação (anaerobismo), como para a desnitrificação (aerobismo), estejam
presentes em microhabitats vizinhos. Nas camadas mais profundas do solo estaria
ocorrendo desnitrificação e nas mais superficiais nitrificação (CANTARELLA, 2007)
Infelizmente não se determinou separadamente o volume de óxido nitroso
proveniente dos processos de nitrificação e de desnitrificação.
Durante os dois anos de estudo a redução da umidade em todo o perfil do
solo devido à drenagem foi o fator decisivo no aumento da emissão de óxido nitroso

83
ao longo do tempo (Figura 3), tendo o aumento do número de dias a partir da
drenagem apresentado correlação linear positiva com a emissão de N2O (Tabela 3).
No segundo ano aumentaram as diferenças nos fluxos de óxido nitroso entre
o solo natural e o drenado, pois devido ao maior tempo de drenagem, o solo
drenado passou a emitir, de modo crescente, mais N2O. É evidente a tendência
ascendente das emissões de óxido nitroso, com o tempo, no solo drenado (Figura
3).
A análise estatística efetuada pelo modelo GAM (Tabela 2), com todas as
variáveis em conjunto, permitiu concluir que o fluxo de N2O diferiu entre o solo
natural e o drenado devido à grande influência da drenagem, a qual superou o efeito
da temperatura média do ar, da precipitação acumulada, do nível freático e da
umidade gravimétrica de 0 a 10 cm. Houve diferença significativa nos fluxos de óxido
nitroso ao longo do tempo apenas no Organossolo drenado, pois a elevação da
emissão foi proporcional ao aumento no tempo de drenagem, ao longo do qual
houve incremento de oxigênio no solo.
3.3.3 Efeito do potencial de oxi-redução sobre o fluxo de N2O
No Organossolo natural o potencial redox nas profundidades de 5 a 40 cm
oscilou entre 267 e -178 mV. A variação foi grande com valores positivos e negativos
em todas as profundidades, de modo que em todas as determinações o potencial foi
diminuindo em profundidade, conforme esperado (Figura 6).
Na coleta 28 na qual o nível freático médio era de -2,3 cm, o segundo mais
elevado (Figura 4), não houve diferença significativa no potencial redox entre as
profundidades. Nas outras coletas as camadas mais profundas mostraram valores
significativamente menores.
Mesmo mostrando valores positivos em algumas coletas e profundidades o
potencial redox do solo encontrava-se moderadamente e fortemente reduzido na
maior parte do tempo (Figura 6). O estado fracamente reduzido ocorreu apenas nas
profundidades de 5 e 10 cm e somente na coleta 24, quando o nível freático (Figura
4) e a umidade gravimétrica (Figura 5), apresentaram os valores mais baixos.
O potencial redox mostrou correlação negativa com o fluxo de óxido nitroso
(Tabela 4) nas profundidades de 5, 10, 15, 20, 30 e 40 cm, de modo que quanto
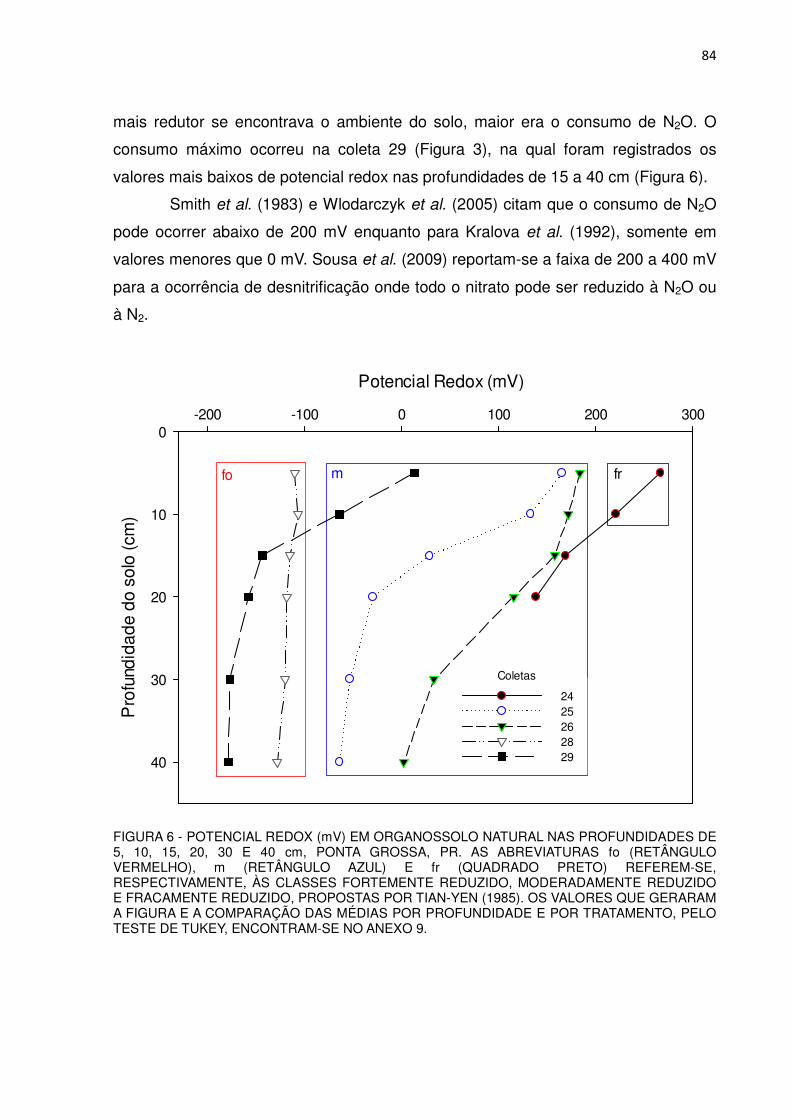
84
mais redutor se encontrava o ambiente do solo, maior era o consumo de N2O. O
consumo máximo ocorreu na coleta 29 (Figura 3), na qual foram registrados os
valores mais baixos de potencial redox nas profundidades de 15 a 40 cm (Figura 6).
Smith et al. (1983) e Wlodarczyk et al. (2005) citam que o consumo de N2O
pode ocorrer abaixo de 200 mV enquanto para Kralova et al. (1992), somente em
valores menores que 0 mV. Sousa et al. (2009) reportam-se a faixa de 200 a 400 mV
para a ocorrência de desnitrificação onde todo o nitrato pode ser reduzido à N2O ou
à N2.
Potencial Redox (mV)
-200 -100 0 100 200 300
Pro
fund
idad
e do
sol
o (c
m)
0
10
20
30
40
242526 28 29
Coletas
frmfo
FIGURA 6 - POTENCIAL REDOX (mV) EM ORGANOSSOLO NATURAL NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR. AS ABREVIATURAS fo (RETÂNGULO VERMELHO), m (RETÂNGULO AZUL) E fr (QUADRADO PRETO) REFEREM-SE, RESPECTIVAMENTE, ÀS CLASSES FORTEMENTE REDUZIDO, MODERADAMENTE REDUZIDO E FRACAMENTE REDUZIDO, PROPOSTAS POR TIAN-YEN (1985). OS VALORES QUE GERARAM A FIGURA E A COMPARAÇÃO DAS MÉDIAS POR PROFUNDIDADE E POR TRATAMENTO, PELO TESTE DE TUKEY, ENCONTRAM-SE NO ANEXO 9.

85
Como os valores de potencial redox encontrados nas camadas abaixo de 15
cm foram sempre menores do que 200 mV e muitas vezes inferiores a 0 mV, conclui-
se que o ambiente moderadamente e fortemente reduzido provavelmente foi
favorável para redução completa do N2O à N2.
Kralova et al. (1992) perceberam que os índices de desnitrificação e de
emissão de N2 continuavam aumentando quando o potencial redox atingia valores
mais baixos do que 0 mV.
No Organossolo drenado os valores de potencial redox oscilaram entre 368
e 120 mV, não tendo ocorrido nenhum valor negativo em nenhuma coleta e em
nenhuma profundidade (Figura 7), sendo a variação em profundidade muito menor
do que no Organossolo natural.
Conforme esperado o potencial redox no Organossolo drenado foi
significativamente maior do que no Organossolo natural, em todas as
determinações, indicando maior quantidade de oxigênio, com exceção da
profundidade de 5 cm na coleta 24, a única ocasião em que não houve diferença
significativa entre os dois tratamentos.
TABELA 4 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE POTENCIAL REDOX, NAS PROFUNDIDADE DE 5, 10, 15, 20, 30 E 40 cm E O INFLUXO DE N2O EM ORGANOSSOLO NATURAL E O EFLUXO DE N2O EM ORGANOSSOLO DRENADO
Solo Profundidades (cm)
5 10 15 20 30 40 Natural -0,21
(p<0,05) -0,29
(p<0,01) -0,25
(p<0,01) -0,26
(p<0,01) -0,29
(p<0,01) -0,30
(p<0,01)
Drenado 0,07 (p<0,01)
0,18 (p<0,01)
0,26 (p<0,01)
0,17 (p<0,01)
0,17 (p<0,01)
0,09 (p<0,01)
Valores de r obtidos com os dados dos dois anos de avaliação.
Também não houve diferença significativa no potencial redox entre as
profundidades em nenhuma das avaliações. Em todas as coletas e profundidades os
valores de potencial redox indicaram que o solo drenado permaneceu nos estados
de redução fracamente reduzido, principalmente, e moderadamente reduzido. Não
ocorreu a classe fortemente reduzido, conforme observado no solo natural (Figura
7).
O potencial redox apresentou correlação positiva com o fluxo de óxido
nitroso (Tabela 5) nas profundidades de 5, 10, 15, 20, 30 e 40 cm.
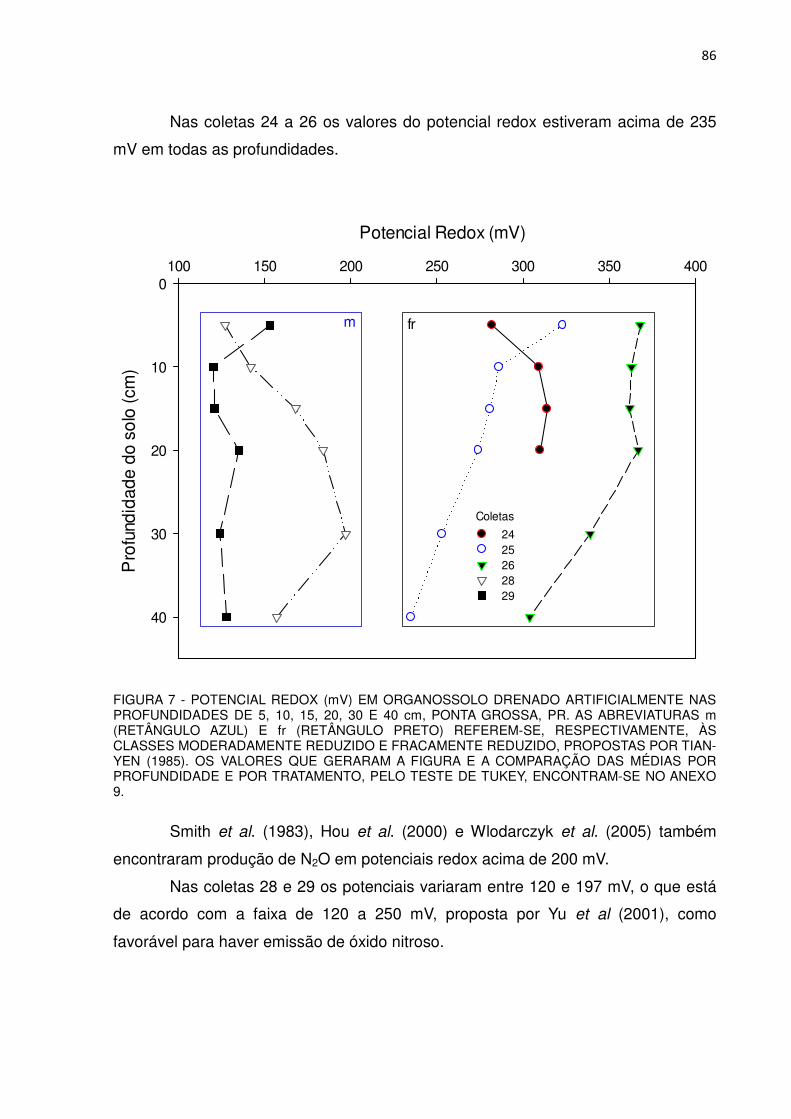
86
Nas coletas 24 a 26 os valores do potencial redox estiveram acima de 235
mV em todas as profundidades.
Potencial Redox (mV)
100 150 200 250 300 350 400
Pro
fund
idad
e do
sol
o (c
m)
0
10
20
30
40
242526 28 29
Coletas
frm
FIGURA 7 - POTENCIAL REDOX (mV) EM ORGANOSSOLO DRENADO ARTIFICIALMENTE NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, PONTA GROSSA, PR. AS ABREVIATURAS m (RETÂNGULO AZUL) E fr (RETÂNGULO PRETO) REFEREM-SE, RESPECTIVAMENTE, ÀS CLASSES MODERADAMENTE REDUZIDO E FRACAMENTE REDUZIDO, PROPOSTAS POR TIAN-YEN (1985). OS VALORES QUE GERARAM A FIGURA E A COMPARAÇÃO DAS MÉDIAS POR PROFUNDIDADE E POR TRATAMENTO, PELO TESTE DE TUKEY, ENCONTRAM-SE NO ANEXO 9.
Smith et al. (1983), Hou et al. (2000) e Wlodarczyk et al. (2005) também
encontraram produção de N2O em potenciais redox acima de 200 mV.
Nas coletas 28 e 29 os potenciais variaram entre 120 e 197 mV, o que está
de acordo com a faixa de 120 a 250 mV, proposta por Yu et al (2001), como
favorável para haver emissão de óxido nitroso.
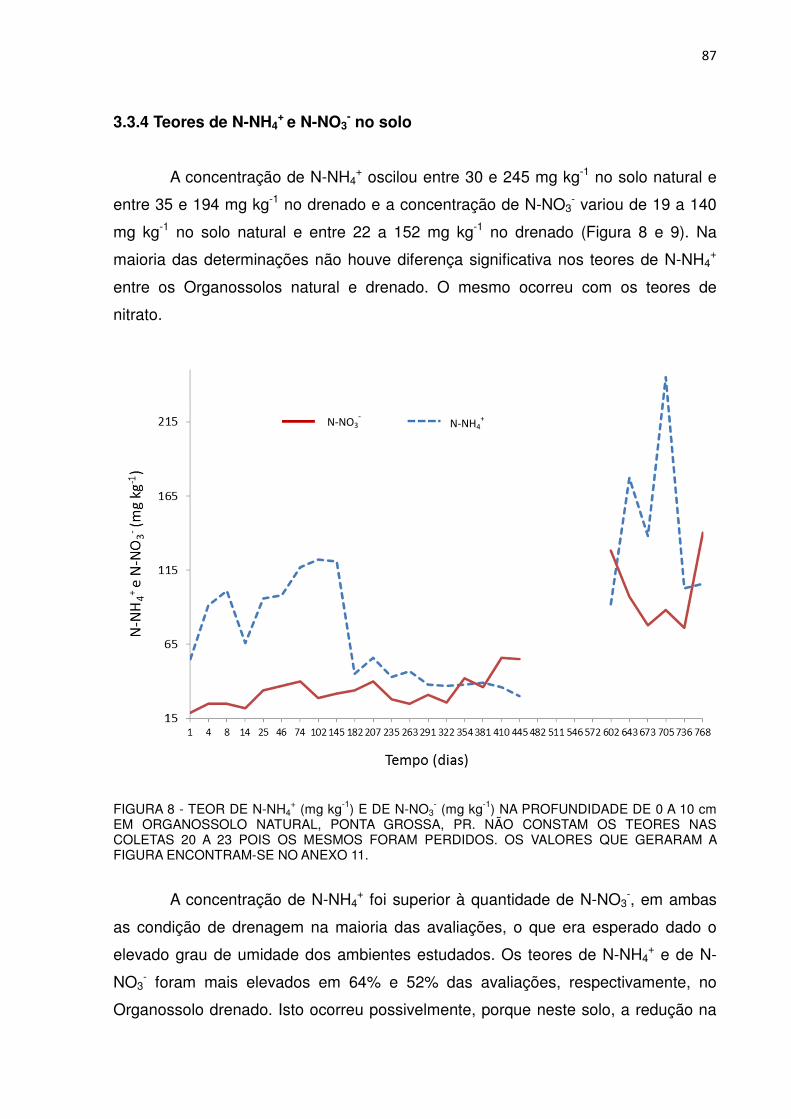
87
3.3.4 Teores de N-NH4+ e N-NO3
- no solo
A concentração de N-NH4+ oscilou entre 30 e 245 mg kg-1 no solo natural e
entre 35 e 194 mg kg-1 no drenado e a concentração de N-NO3- variou de 19 a 140
mg kg-1 no solo natural e entre 22 a 152 mg kg-1 no drenado (Figura 8 e 9). Na
maioria das determinações não houve diferença significativa nos teores de N-NH4+
entre os Organossolos natural e drenado. O mesmo ocorreu com os teores de
nitrato.
FIGURA 8 - TEOR DE N-NH4+ (mg kg-1) E DE N-NO3
- (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO NATURAL, PONTA GROSSA, PR. NÃO CONSTAM OS TEORES NAS COLETAS 20 A 23 POIS OS MESMOS FORAM PERDIDOS. OS VALORES QUE GERARAM A FIGURA ENCONTRAM-SE NO ANEXO 11.
A concentração de N-NH4+ foi superior à quantidade de N-NO3
-, em ambas
as condição de drenagem na maioria das avaliações, o que era esperado dado o
elevado grau de umidade dos ambientes estudados. Os teores de N-NH4+ e de N-
NO3- foram mais elevados em 64% e 52% das avaliações, respectivamente, no
Organossolo drenado. Isto ocorreu possivelmente, porque neste solo, a redução na
N-NO3
- N-NH4
+

88
umidade foi favorável ao aumento na decomposição da matéria orgânica do solo e
de resíduos de plantas. O aumento do nitrato sugere também a elevação da
atividade dos microrganismos nitrificadores, favorecidos pela drenagem.
Os valores observados para ambas as condições de drenagem avaliadas
indicam um elevado teor de N mineral, sendo provavelmente uma característica
intrínseca destes Organossolos. Corroborando com este estudo, Inubushi et al.
(2003) registraram em média teores de amônio de 857 mg kg-1 e 24 mg kg-1de
nitrato até a profundidade 30 cm em Organossolo cultivado com arroz irrigado. Em
áreas de lavouras de sequeiro abandonadas, também sobre Organossolos, o teor
médio de amônio reduziu-se para 246 mg kg-1 e o de nitrato se manteve (23 mg kg-
1).
Os valores de nitrato observados na presente pesquisa encontram-se muito
acima dos valores críticos para crescimento do milho, de 23 a 26 e 16 a 19 mg N kg-
1, na camada de 0-30 e 30-60 cm, respectivamente, obtidos por Binford et al. (1992).
Valores elevados como 400 mg N kg-1 de nitrato foram observados em áreas que
receberam elevadas doses de esterco (COOPER et al., 1984; ZEBARTH et al.,
1996).
Tanto no solo natural como no drenado as quantidades de N-NH4+ e de N-
NO3- mostraram variações específicas ao longo de três períodos bem diferenciados
(Figura 8 e 9).
No primeiro período, compreendido entre 1 e 145 dias (coletas 1 a 9),
correspondente ao final do inverno, à primavera de 2010 e ao início do verão de
2011, houve liberação de amônio devido a maior decomposição da matéria orgânica
nos Organossolos natural e drenado. Na Figura 3 do capítulo 3, percebe-se
nitidamente a elevação da emissão de dióxido de carbono durante este período.
Justifica-se a maior decomposição da matéria orgânica, em ambos os solos, pois a
umidade gravimétrica na profundidade de 0 a 10 cm e a altura do nível freático foram
diminuindo e a temperatura sofreu elevação ao longo do tempo. Como houve
aumento na emissão de CO2 e também na quantidade de amônio ao longo do
período, supõem-se que o aumento na decomposição da matéria orgânica do solo
tenha sido responsável pela elevação nos teores de N-NH4+ na profundidade de 0 a
10 cm, e que parte deste amônio tenha sido nitrificado. As correlações positivas
existentes entre a emissão de CO2 e o teor de amônio e de nitrato, entre amônio e
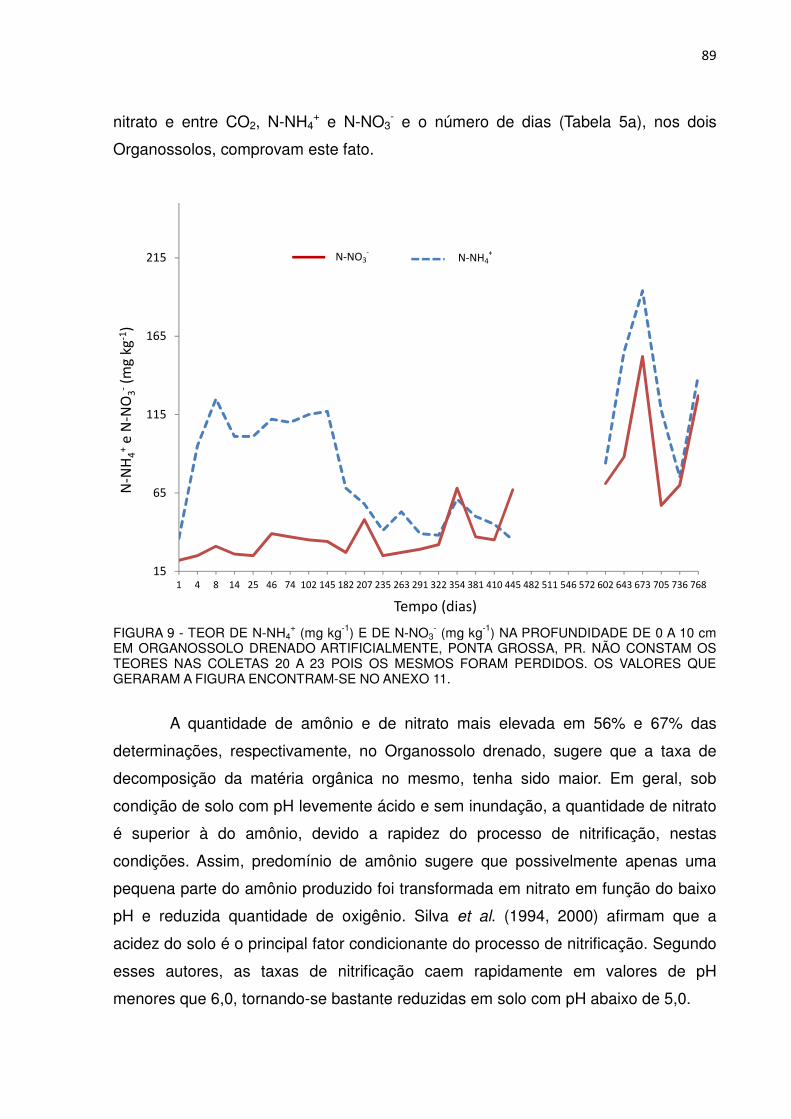
89
nitrato e entre CO2, N-NH4+ e N-NO3
- e o número de dias (Tabela 5a), nos dois
Organossolos, comprovam este fato.
FIGURA 9 - TEOR DE N-NH4+ (mg kg-1) E DE N-NO3
- (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR. NÃO CONSTAM OS TEORES NAS COLETAS 20 A 23 POIS OS MESMOS FORAM PERDIDOS. OS VALORES QUE GERARAM A FIGURA ENCONTRAM-SE NO ANEXO 11.
A quantidade de amônio e de nitrato mais elevada em 56% e 67% das
determinações, respectivamente, no Organossolo drenado, sugere que a taxa de
decomposição da matéria orgânica no mesmo, tenha sido maior. Em geral, sob
condição de solo com pH levemente ácido e sem inundação, a quantidade de nitrato
é superior à do amônio, devido a rapidez do processo de nitrificação, nestas
condições. Assim, predomínio de amônio sugere que possivelmente apenas uma
pequena parte do amônio produzido foi transformada em nitrato em função do baixo
pH e reduzida quantidade de oxigênio. Silva et al. (1994, 2000) afirmam que a
acidez do solo é o principal fator condicionante do processo de nitrificação. Segundo
esses autores, as taxas de nitrificação caem rapidamente em valores de pH
menores que 6,0, tornando-se bastante reduzidas em solo com pH abaixo de 5,0.
15
65
115
165
215
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 602 643 673 705 736 768
N-N
H4
+e
N-N
O3
-(m
g k
g-1
)
Tempo (dias)
N-NH4+ N-NO3
-

90
No segundo período, compreendido entre 182 e 445 dias (coletas 10 a 19),
referente ao final do verão e ao outono, inverno e primavera de 2011, houve redução
brusca na quantidade de N-NH4+ e elevação nos teores de N-NO3
-, na camada de 0
a 10 cm, em ambos os solos, ao longo do tempo. A correlação negativa e positiva,
respectivamente, entre a quantidade de amônio e de nitrato e o número de dias, nos
dois Organosolos, confirma o ocorrido (Tabela 5b). A inexistência de correlação
significativa e o baixo coeficiente de correlação entre CO2 e N-NH4+ no Organossolo
natural e drenado, respectivamente, sugerem que, os processos de decomposição
da matéria orgânica e a quantidade de N-NH4+ foram independentes, ou sofreram
efeitos de variáveis não monitoradas neste estudo. A ausência de correlação
significativa entre N-NH4+ e N-NO3
- em ambos os solos, a correlação positiva entre
N-NO3- e CO2 e a inexistência de correlação entre CO2 e o número de dias, também
em ambos os Organossolos, indica que seriam necessários estudos mais
aprofundados para explicar as complexas relações entre os teores de amônio, de
nitrato e as emissões de dióxido de carbono, que ocorreram no segundo período.
No entanto, os maiores teores de amônio no Organossolo drenado, em
relação ao natural, na grande maioria das avaliações do segundo período (Figura 9
e 8), bem como as maiores taxas de emissão de CO2 (Figura 3, capítulo 3), sugerem
que neste solo, em função da menor umidade, a taxa de decomposição da matéria
orgânica, foi ligeiramente maior do que no solo natural. Basta perceber que o teor de
amônio e de nitrato, no Organossolo drenado, foi superior em 90% e em 60% das
coletas, respectivamente.
Finalmente, no último período, compreendido entre 602 e 768 dias (coletas
24 a 29), em grande parte das avaliações, foram registrados os valores mais
elevados de amônio e de nitrato em ambos os Organossolos. Entretanto, ao
contrário dos períodos anteriores, o teor de amônio e de nitrato foi maior em apenas
33% e 17% das avaliações, respectivamente, no Organossolo drenado.
Considerando que, em ambos os solos, houve elevação significativa na
emissão de CO2 ao longo do tempo, mas ausência de correlação entre CO2 e N-
NH4+ e N-NO3
-, além de inexistência de correlação entre N-NH4+ e N-NO3
-, bem
como entre teor de nitrato e de amônio com o aumento no número de dias (Tabela
5c), conclui-se que também seriam necessários estudos mais aprofundados, para
justificar as relações entre os teores de amônio, de nitrato e as emissões de dióxido
de carbono, que ocorreram no terceiro período.

91
Ao analisar a evolução dos teores de amônio e de nitrato, ao longo de todo o
experimento, sem subdividí-lo nos três períodos, constata-se que também não
houve correlação significativa entre a emissão de CO2 e a quantidade de amônio,
em ambos os Organossolos (Tabela 5d). A despeito disto, em ambos os
Organossolos houve aumento significativo nos teores de amônio e de nitrato ao
longo do tempo, bem como correlação positiva entre NH4+ e NO3
- .
Segundo Wrage et al. (2001) há ambientes que reúnem condições para que
os dois processos ocorram ao mesmo tempo, desde que condições favoráveis tanto
para a nitrificação (aerobismo), como para a desnitrificação (anaerobismo), estejam
presentes em micro habitats vizinhos. O período menos úmido referente ao
rebaixamento do freático durante o qual o nitrato é produzido por nitrificação é
seguido por um período de maior umidade no solo, quando o freático torna a se
elevar e ocorre a desnitrificação (VON ARNOLD et al., 2005).
De qualquer maneira, seriam necessários outros estudos para compreender
de que forma ocorreram as inter-relações entre as emissões de dióxido de carbono e
as quantidades de amônio e de nitrato no solo.

92
TABELA 5 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE N-NH4+ E N-NO3
- NA PROFUNDIDADE DE 0 A 10 CM, EFLUXO DE CO2, E TEMPO, EM ORGANOSSOLO NATURAL E DRENADO, POR PERÍODO.
Periodo 1 (a)
Organossolo natural Organossolo drenado
Tempo(1) NH4+ NO3
- Tempo(1) NH4+ NO3
-
CO2 0,71
(p < 0,01)
0,53
(p < 0,01)
0,28
(p < 0,01)
0,59
(p < 0,01)
0,20
(p < 0,01)
0,32
(p < 0,01)
NO3- 0,52
(p < 0,01)
0,71
(p < 0,01)
- 0,62
(p < 0,01)
0,87
(p < 0,01)
-
NH4+ 0,78
(p < 0,01)
- - 0,46
(p < 0,01)
- -
Periodo 2 (b)
Organossolo natural Organossolo drenado
Tempo(1) NH4+ NO3
- Tempo(1) NH4+ NO3
-
CO2 ns ns 0,54
(p < 0,01)
ns 0,21
(p < 0,05)
0,29
(p < 0,01)
NO3- 0,65
(p < 0,01)
ns - 0,53
(p < 0,01)
ns -
NH4+ -0,83
(p < 0,01)
- - -0,52
(p < 0,01)
- -
Periodo 3 (c)
Organossolo natural Organossolo drenado
Tempo(1) NH4+ NO3
- Tempo(1) NH4+ NO3
-
CO2 0,41
(p < 0,01)
ns ns 0,22
(p < 0,01)
ns ns
NO3- ns ns - ns ns -
NH4+ ns - - ns - -
Periodo 1 + 2 + 3 (d)
Organossolo natural Organossolo drenado
Tempo(1) NH4+ NO3
- Tempo(1) NH4+ NO3
-
CO2 0,02
(p < 0,01)
ns 0,06
(p < 0,05)
-0,26
(p < 0,05)
ns -0,25
(p < 0,05)
NO3- 0,84
(p < 0,01)
0,47
(p < 0,01)
- 0,78
(p < 0,01)
0,60
(p < 0,01)
-
NH4+ 0,35
(p < 0,01)
- - 0,22
(p < 0,01)
- -
(1) Tempo em número de dias

93
3.3.5 Influência dos teores de N-NH4+ e N-NO3
- no solo sobre o fluxo de N2O
No Organossolo natural não houve correlação entre o consumo de N2O e a
quantidade de nitrato e de amônio na profundidade de 0 a 10 cm (Tabela 6a e 6c),
provavelmente porque o consumo ocorreu a maiores profundidades, em ambiente
mais reduzido.
A tendência do consumo de óxido nitroso representada por uma linha
praticamente paralela ao eixo X, a quantidade crescente de nitrato com linha de
tendência claramente ascendente (Figura 10) e a diminuição da quantidade de NH4
até 445 dias (coleta 19), seguida de brusca elevação (Figura 8), justificam a
afirmação anterior.
No Organossolo drenado houve correlação positiva entre os teores de nitrato
e a emissão de N2O (Tabela 6b), comprovando a tendência ascendente de ambos ao
longo do tempo (Figura 11). Isto pode estar associado à liberação de N2O que ocorre
durante a nitritação, conforme citado por Farquharson e Baldock (2008).
TABELA 6 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE N-NO3- E N-
NH4+ NA PROFUNDIDADE DE 0 A 10 cm E O INFLUXO DE N2O EM ORGANOSSOLO
NATURAL E O EFLUXO DE N2O NO ORGANOSSOLO DRENADO. Variáveis NO3
- NH4+
Natural (a) Drenado (b) Natural (c) Drenado (d) N2O ns 0,32 (p < 0,05) ns ns
Valores de r obtidos com os dados dos dois anos de avaliação. (1) Tempo contado em número de dias.
Neste tratamento também não houve correlação entre a quantidade de
amônio e a emissão de N2O (Tabela 6d).
No entanto, a correlação positiva que houve entre a quantidade de nitrato e
de amônio, em ambos os solos (Tabela 5d), reforça a importância da presença de
amônio como substrato para a ocorrência da nitrificação, bem como a
interdependência entre os dois processos.
Segundo Robertson e Groffman (2007) o fator mais importante que regula a
nitrificação, na maioria dos solos é o suprimento de NH4+. Porém, as quantidades de
nitrato e de amônio associadas ao teor de umidade do solo definem o rumo do ciclo
do N, orientando os processos de nitrificação e desnitrificação e os fluxos de N2O.
Geralmente o índice de desnitrificação aumenta com o aumento na concentração de
nitrato se a umidade do solo for alta o suficiente para favorecer a desnitrificação
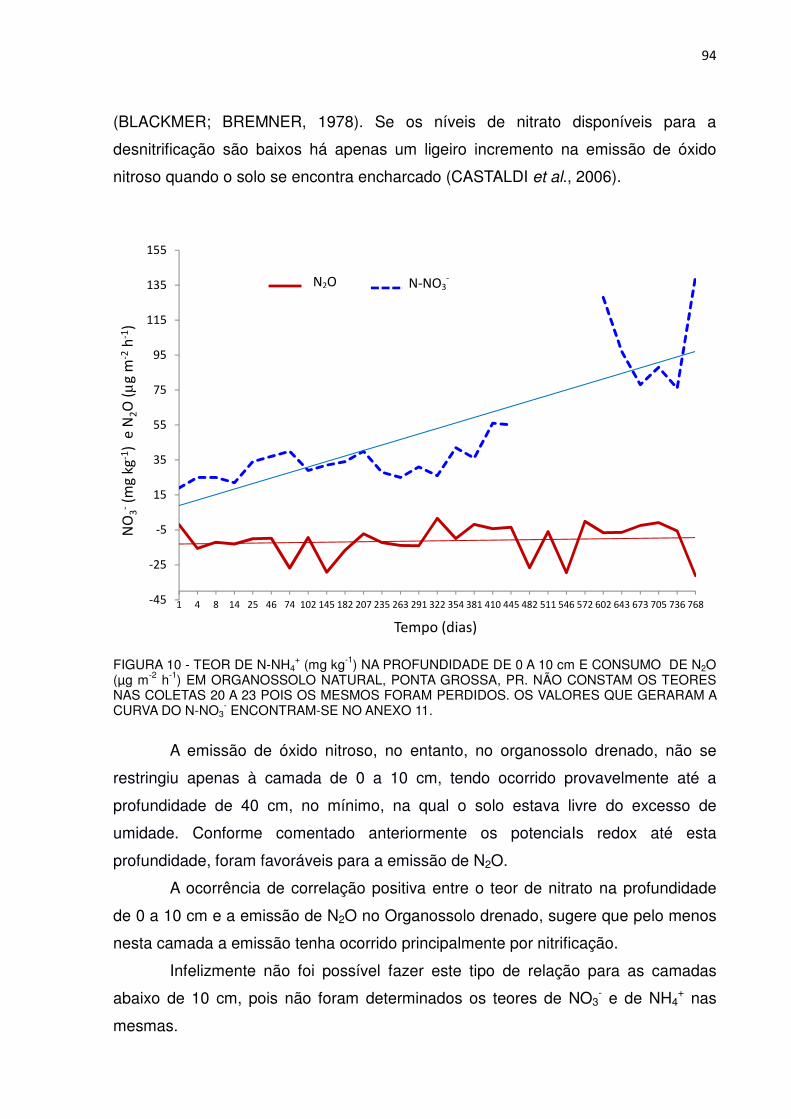
94
(BLACKMER; BREMNER, 1978). Se os níveis de nitrato disponíveis para a
desnitrificação são baixos há apenas um ligeiro incremento na emissão de óxido
nitroso quando o solo se encontra encharcado (CASTALDI et al., 2006).
FIGURA 10 - TEOR DE N-NH4
+ (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm E CONSUMO DE N2O (µg m-2 h-1) EM ORGANOSSOLO NATURAL, PONTA GROSSA, PR. NÃO CONSTAM OS TEORES NAS COLETAS 20 A 23 POIS OS MESMOS FORAM PERDIDOS. OS VALORES QUE GERARAM A CURVA DO N-NO3
- ENCONTRAM-SE NO ANEXO 11.
A emissão de óxido nitroso, no entanto, no organossolo drenado, não se
restringiu apenas à camada de 0 a 10 cm, tendo ocorrido provavelmente até a
profundidade de 40 cm, no mínimo, na qual o solo estava livre do excesso de
umidade. Conforme comentado anteriormente os potenciaIs redox até esta
profundidade, foram favoráveis para a emissão de N2O.
A ocorrência de correlação positiva entre o teor de nitrato na profundidade
de 0 a 10 cm e a emissão de N2O no Organossolo drenado, sugere que pelo menos
nesta camada a emissão tenha ocorrido principalmente por nitrificação.
Infelizmente não foi possível fazer este tipo de relação para as camadas
abaixo de 10 cm, pois não foram determinados os teores de NO3- e de NH4
+ nas
mesmas.
-45
-25
-5
15
35
55
75
95
115
135
155
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 602 643 673 705 736 768
NO
3-(m
g k
g-1
) e
N2O
(µ
g m
-2h
-1)
Tempo (dias)
N-NO3- N2O
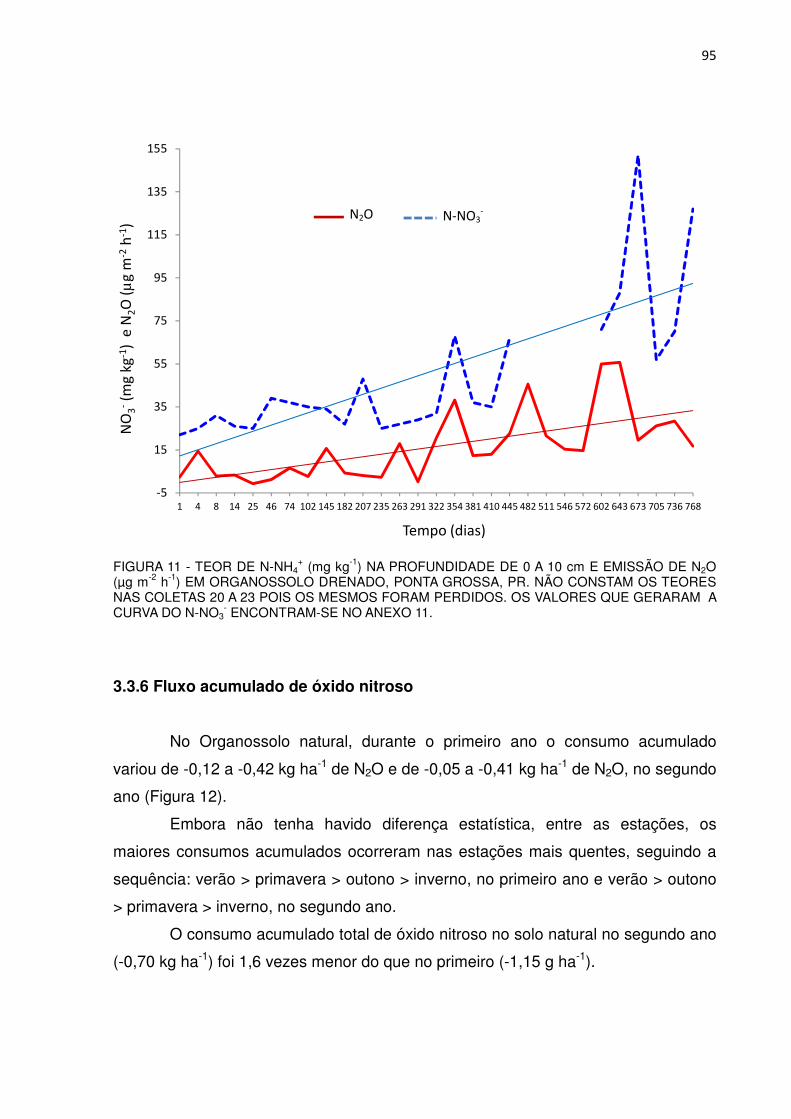
95
FIGURA 11 - TEOR DE N-NH4
+ (mg kg-1) NA PROFUNDIDADE DE 0 A 10 cm E EMISSÃO DE N2O (µg m-2 h-1) EM ORGANOSSOLO DRENADO, PONTA GROSSA, PR. NÃO CONSTAM OS TEORES NAS COLETAS 20 A 23 POIS OS MESMOS FORAM PERDIDOS. OS VALORES QUE GERARAM A CURVA DO N-NO3
- ENCONTRAM-SE NO ANEXO 11.
3.3.6 Fluxo acumulado de óxido nitroso
No Organossolo natural, durante o primeiro ano o consumo acumulado
variou de -0,12 a -0,42 kg ha-1 de N2O e de -0,05 a -0,41 kg ha-1 de N2O, no segundo
ano (Figura 12).
Embora não tenha havido diferença estatística, entre as estações, os
maiores consumos acumulados ocorreram nas estações mais quentes, seguindo a
sequência: verão > primavera > outono > inverno, no primeiro ano e verão > outono
> primavera > inverno, no segundo ano.
O consumo acumulado total de óxido nitroso no solo natural no segundo ano
(-0,70 kg ha-1) foi 1,6 vezes menor do que no primeiro (-1,15 g ha-1).
-5
15
35
55
75
95
115
135
155
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 602 643 673 705 736 768
NO
3-(m
g k
g-1
) e
N2O
(µ
g m
-2h
-1)
Tempo (dias)
N2O N-NO3-
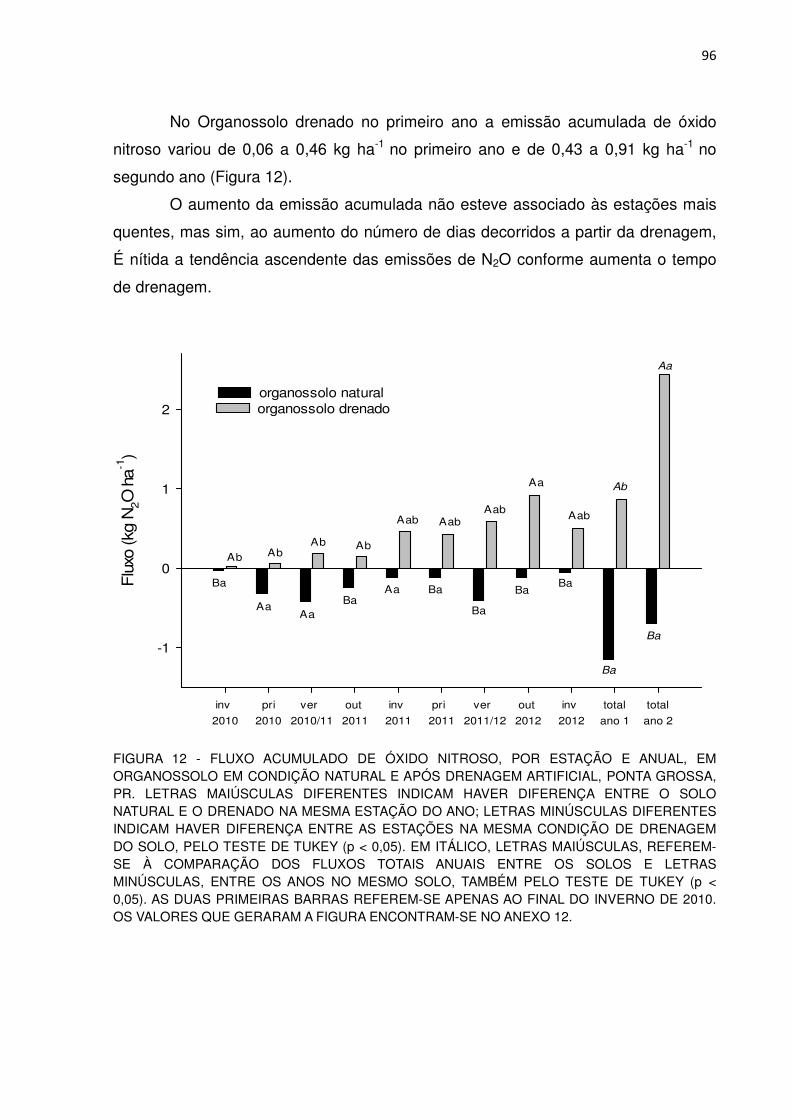
96
No Organossolo drenado no primeiro ano a emissão acumulada de óxido
nitroso variou de 0,06 a 0,46 kg ha-1 no primeiro ano e de 0,43 a 0,91 kg ha-1 no
segundo ano (Figura 12).
O aumento da emissão acumulada não esteve associado às estações mais
quentes, mas sim, ao aumento do número de dias decorridos a partir da drenagem,
É nítida a tendência ascendente das emissões de N2O conforme aumenta o tempo
de drenagem.
inv
2010
pri
2010
ver
2010/11
out
2011
inv
2011
pri
2011
ver
2011/12
out
2012
inv
2012
total
ano 1
total
ano 2
Flu
xo (k
g N
2O h
a-1)
-1
0
1
2
Aa
Ab
Ab AbAb Ab
Aab AabAab
Aa
Aab
organossolo natural organossolo drenado
Ba
AaAa
BaAa Ba
Ba
BaBa
Ba
Ba
FIGURA 12 - FLUXO ACUMULADO DE ÓXIDO NITROSO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. LETRAS MAIÚSCULAS DIFERENTES INDICAM HAVER DIFERENÇA ENTRE O SOLO NATURAL E O DRENADO NA MESMA ESTAÇÃO DO ANO; LETRAS MINÚSCULAS DIFERENTES INDICAM HAVER DIFERENÇA ENTRE AS ESTAÇÕES NA MESMA CONDIÇÃO DE DRENAGEM DO SOLO, PELO TESTE DE TUKEY (p < 0,05). EM ITÁLICO, LETRAS MAIÚSCULAS, REFEREM-SE À COMPARAÇÃO DOS FLUXOS TOTAIS ANUAIS ENTRE OS SOLOS E LETRAS MINÚSCULAS, ENTRE OS ANOS NO MESMO SOLO, TAMBÉM PELO TESTE DE TUKEY (p < 0,05). AS DUAS PRIMEIRAS BARRAS REFEREM-SE APENAS AO FINAL DO INVERNO DE 2010. OS VALORES QUE GERARAM A FIGURA ENCONTRAM-SE NO ANEXO 12.

97
A temperatura teve pouca influência sobre as emissões acumuladas, pois no
próprio inverno de 2011 a emissão de N2O foi a mais alta e em 2012 a emissão
acumulada no outono foi bem superior à do verão.
O consumo total anual no segundo ano (2,43 kg ha-1) foi 2,8 vezes superior
do que no primeiro ano (0,88 kg ha-1), o que reforça o efeito decisivo do maior tempo
de drenagem, comentado anteriormente.
3.3.7 Considerações adicionais
O Organossolo natural consumiu em média 0,92 kg de N2O ha-1 ano-1, o que
equivale a 0,27 Mg ha-1 ano-1 de CO2-eq, comprovando sua atuação como
sumidouro de óxido nitroso, além de todas as funções ecológicas comentadas no
Capítulo anterior.
Embora a quantidade de óxido nitroso retirada da atmosfera possa parecer
pequena ela é muito significativa, uma vez que se trata de um gás com alto potencial
de aquecimento global.
Além disto, estes Organossolos, em condições naturais, podem contribuir na
diminuição da quantidade de nitrato proveniente de atividades agro-silvo-pastoris
praticadas à montante. Em função do baixo potencial redox presente nestes
Organossolos, pode haver redução completa do nitrato à N2, o que também colabora
para atenuar a eutrofização dos corpos de água à jusante.
O Organossolo drenado, por sua vez, além de emitir grandes quantidades de
CO2, (conforme será visto no Capítulo seguinte) e reduzir o volume de água
armazenado, emitiu em média 1,65 kg de N2O ha-1 ano-1, o que equivale a 0,49 Mg
ha-1 ano-1 de CO2-eq.
A drenagem, além de causar outros prejuízos ambientais, cessa o consumo
de óxido nitroso e conforme será discutido no Capítulo 3, reduz o estoque de
carbono do solo.
Levando em conta as funções que os Organossolos desempenham no
ecossistema, os mesmos devem ser preservados e para isso não devem ser
drenados.

98
3.4 CONCLUSÕES
a) No Organossolo natural, devido ao excesso de água provocado pelo nível
freático elevado, houve consumo de N2O.
b) No Organossolo drenado, devido a redução do grau de umidade
ocasionada pelo rebaixamento do nível freático, houve emissão de N2O.
c) A elevação da temperatura média do ar favoreceu o consumo de N2O no
Organossolo natural.
d) O aumento na precipitação estimulou a emissão de N2O no Organossolo
drenado.
e) O potencial redox teve influência decisiva sobre os fluxos de óxido nitroso
nos Organossolos natural e drenado. No ambiente mais reduzido do Organossolo
natural ocorreu consumo de N2O e no Organossolo drenado, com potencial redox
bem mais elevado, houve emissão.

99
REFERÊNCIAS
ARAH, J. R. M.; SMITH, K. A.; CRICHTON, I. J.; LI, H. S. Nitrous oxide production and denitrification in Scottish arable soil. Journal of Soil Science, Oxford, v. 42, p. 351-367, 1991. BALDWIN, D. S.; MITCHELL, A. M. The effects of drying and re-flooding on the sediment and soil nutrient dynamics of lowland river-floodplain systems: a synthesis. Regulated Rivers: Research and Management, Sussex, v. 16, p. 457-467, 2000. BINFORD, G. D.; BLACKMER, A. M.; CERRATO, M. E. Relationships between corn yields and soil nitrate in late spring. Agronomy Journal, v. 84, p. 53-59, 1992. BLAIS, A.; LORRAIN, S.; TREMBLAY, A. Greenhouse gas fluxes (CO2, CH4 and N2O) in forests and wetlands of boreal, temperate and tropical regions. In: TREMBLAY, A.; VARFALVY, L.; ROEHM, C.; GARNEAU, M. (Ed.). Greenhouse gas emissions – fluxes and processes. Berlin: Springer-Verlag, 2005. p. 87-127. BLACKMER, A. M.; BREMNER, J. M. Inhibitory effects of nitrate on reduction of nitrous oxide to molecular nitrogen by soil microorganisms. Soil Biology & Biochemistry, Oxford, v. 10, p. 187-191, 1978. BREMNER, J. M. Inorganic forms of nitrogen. In: BLACK, C. A. (Ed.). Methods of Soil Analysis: chemical and microbiological properties. Madison: American Society of Agronomy, 1965. Pt. 2, p. 1179-1237. CANTARELLA, H. Nitrogênio. In: NOVAIS, R. F.; ALVAREZ V. V. H.; BARROS, N. F.; FONTES, R. L. F.; CANTARUTTI, R. B.; NEVES, J. C. L. (Ed.). Fertilidade do solo. Viçosa, MG: Sociedade Brasileira de Ciência do Solo, 2007. p. 373-470. CASTALDI, S.; ERMICE, A.; STRUMIA, S. Fluxes of N2O and CH4 from soils of savannas and seasonally-dry ecosystems. Journal of Biogeography, Oxford, v. 33, p. 401-415, 2006. CHAPUIS-LARDY, L.; WRAGE, N.; METAY, A.; CHOTTE, J. L.; BERNOUX, M. Soil, a sink for N2O?: a review. Global Change Biology, West Sussex, v. 13, p. 1-17, 2007. CLAESSEN, M. E. C. (Org.). Manual de métodos de análises de solo. 2. ed. rev. atual. Rio de Janeiro: EMBRAPA-CNPS, 1997. (Documentos, 1).

100
COOPER, J. R.; RENEAU, R. B.; KROOTJE, W.; JONES, G. D. Distribution of nitrogenous compounds in a Rhodic Paleudult following heavy manure application. Journal of Environment Quality, Madison, v. 13, n. 2, p. 189-193, 1984. CURI, N; KAMPF, N. Caracterização do solo. In: KER, J. C.; CURI, N.; SCHAEFER, C. E.; VIDAL-TORRADO, P. (Ed.). Pedologia: fundamentos. Viçosa, MG: Sociedade Brasileira de Ciência do Solo, 2012. p. 147-169. DALAL, R. C.; WANG, W. J.; ROBERTSON, G. P.; PARTON, W. J. Nitrous oxide emissions from Australian agricultural lands and mitigation options: a review. Australian Journal of Soil Research, Collingwood, v. 41, p. 165-195, 2003. DALAL, R. C.; ALLEN, D. E. Greenhouse gas fluxes from natural ecosystems. Australian Journal of Botany, Collingwood, v. 56, p. 369-407, 2008. DAVIDSON, E. A. Fluxes of nitrous oxide and nitric oxide from terrestrial ecosystems. In: ROGERS, J. E.; WHITMAN, W. B. (Ed.). Microbial production and consumption of greenhouse gases: methane, nitrogen oxides and halomethanes. Washington, DC: American Society for Microbiology, 1991. p. 219-235. ELDER, J. W.; LAL, R. Tillage effects on gaseous emissions from na intensively farmed organic soil in North Central Ohio. Soil and Tillage Research, Amsterdam, v. 98, p. 45-55, 2008. EPSTEIN, E.; BLOOM, A. J. Mineral nutrition of plants: principles and perspective. 2. ed. Sunderland: Sinauer Associates, 2005. FARQUHARSON, R.; BALDOCK, J. A. Concepts in modelling N2O emisions from land use. Plant and Soil, Dordrecht, v. 309, n. 1-2, p. 147-167, 2008. FENN, M. E.; POTH, M. A.; JOHNSON, D. W. Evidence for nitrogen saturation in the San Bernardino Mountains in sourthern California. Forest Ecology and Management, Amsterdam, v. 82, p. 211-230, 1996. FORSTER, P.; RAMASWAMY, V.; ARTAXO, P.; BERNSTEN, T.; BETTS, R.; FAHEY, D. W.; HAYWOOD, J.; LEAN, J.; LOWE, D. C.; MYHRE, G.; NGANGA, J.; PRINN, R.; RAGA, G.; SCHUTZ, M.; VAN DORLAND, R. Changes in atmospheric constituents and in radiative forcing. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z; MARQUIS, M.; AVERYT, K. B.; TIGNOR, M.; MILLER, H. L. (Ed.). Climate

101
change 2007: the physical science basis: contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge, UK: University Press, 2007. p. 129-234. FURUKAWA, Y.; INUBUSHI, K.; ALI, M.; ITANG, A. M.; TSURUTA, H. Effect of changing groundwater levels caused by land-use changes on greenhouse gas fluxes from tropical peat lands. Nutrient Cycling in Agroecosystems, Dordrecht, v. 71, p. 81-91, 2005. GLINSKI, J.; STEPNIEWSKA, Z. An evolution of soil resistance to reduction processes. Polish Journal of Soil Science, Lublin, v. 19, p. 15-19, 1986. GRANLI, T.; BOCKMAN, O. C. Nitrous oxide from agriculture. Norwegian Journal of Agricultural Sciences, Norway, v. 12 (Suppl.), p. 7-128, 1994. GROFMAN, P. M. Nitrogen in the environment. In: SUMNER, M. E. (Ed.). Handbook of Soil Science. Boca Raton: CRC Press, 2000. p. C190-C200. GUTKNECHT, J. L. M.; GOODMAN, R. M.; BALSER, T. C. Linking soil process and microbial ecology in freshwater wetlands ecossystems. Plant and Soil, The Hague, v. 289, p. 17-34, 2006. HADI, A.; INUBUSHI, K.; PURNOMO, E.; RAZIE, F.; YAMAKAWA, K.; TSURUTA, H. Effects of land-use changes on nitrous oxide (N2O) emissions from tropical peatlands. Chemosphere Global Change Science, Oxford, v. 2, p. 347-358, 2000. HADI, A.; HARIDI, M.; INUBUSHI, K.; PURNOMO, E.; RAZIE, F.; TSURUTA, H. Effect of land-use change in tropical peat soil on the microbial population and emission of greenhouse gases. Microbes and Environments, Miyagi, v. 16, p. 79-86, 2001. HADI, A.; INUBUSHI, K.; FURUKAWA, Y.; RASMALDI, M.; TSURUTA, H. Greenhouse gas emissions from tropical peat lands of Kalimantan, Indonesia. Nutrient Cycling in Agroecosystems, Dordrecht, v. 71, p. 73-80, 2005. HERNADEZ, M. E.; MITSCH, W. J. Influence of hidrologyc pulses, flooding frequency and vegetation on nitrous oxide emissions from created riparian marshes. Wetlands, Dordrecht, v. 26, n. 3, p. 862-877, 2006. HOSEN, Y.; TSURUTA, H.; MINAMI, K. Effects of the depth of NO and N2O

102
productions in soil on their emission rates to the atmosphere: analysis by a simulation model. Nutrient Cycling in Agroecosystems, Dordrecht, v. 57, p. 83-98, 2000. HOU, A. X.; CHEN, G. X.; WANG, Z. P.; VAN CLEEMPUT, O.; PATRICK, W. H. Methane and nitroux oxide emissions from a rice field in relation to soil redox and microbiological processes. Soil Science Society of American Journal, Madison, v. 64, n. 6, p. 2180-2186, 2000. HUANG, Y.; SUN, W. J.; ZHANG, W.; YU, Y. Q.; SU, Y. H.; SONG, C. C. Marshland conversion to cropland in northeast China from 1950 to 2000 reduced the greenhouse effect. Global Change Biology, West Sussex, v. 16, p. 680-695, 2010. INUBUSHI, K.; FURUKAWA, Y.; HADI, A.; PURNOMO, E.; TSURUTA, H. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use changes in tropical peatlands located in coastal area of South Kalimantan. Chemosphere, Oxford, v. 52, p. 603-608. 2003. INTERGOVERNMENTAL PANEL ON CLIMATE CHANGE (IPCC). Climate Change 2007: the physical science basis. Cambridge: Cambridge University 2007. JANTALIA, C. P.; SANTOS, H. P.; URQUIAGA, S.; BODDEY, R. M.; ALVES, B. J. R. Fluxes of nitrous oxide from soil under different crop rotations and tillage systems in the South of Brazil. Nutrient Cycling in Agroecosystems, Dordrecht, v. 82, p. 161-173, 2008. JIANG, C.; WANG, Y.; HAO, Q.; SONG, C. Effect of land-use change on CH4 and N2O emissions from freshwater marsh in Northeast China. Atmospheric Environment, Oxford, v. 43, p. 3305-3309, 2009. KÖGEL-KNABNER, I.; AMELUNG, W.; CAO, Z. H.; FIEDLER, S.; FRENZEL, P.; JAHN, R.; KALBITZ, K.; KOLBL, A.; SCHLOTER, M. Biogeochemistry of paddy soils. Geoderma, Amsterdam, v. 157, p. 1-14, 2010. KRALOVA, M.; MASSCHELEYN, P. H.; LINDAU, C. W.; PATRICK JR., W. H. Production of dinitrogen and nitrous oxide in soil suspensions as affected by redox potencial. Water, Air and Soil Pollution, Dordrecht, v. 61, p. 37-45, 1992. LONGORIA RAMÍREZ, R.; CARBAJAL BENÍTEZ, G.; MAR MORALES, B. E.; RUÍZ SUÁREZ, L. G. Nitrous oxide flux in maize and wheat cropped soils in the central region of Mexico during ‘El Niño’ year 1998. Atmósfera, Mexico, v. 16, p. 231-244,

103
2003. MALJANEN, M.; LIIKANEN, A.; SILVOLA, J.; MARTIKAINEN, P. J. Nitrous oxide emissions from boreal organic soil under different land-use. Soil Biology & Biochemistry, Oxford, v. 35, p. 1-12, 2003. MATHIEU, O.; LEVEQUE, J.; HENAULT, C.; MILLOUX, M. J.; BIZOUARD, F.; ANDREUX, F. Emissions and spatial variability of N2O, N2 and nitrous oxide mole fraction at the field scale, revealed with 15N isotopic techniques. Soil Biology and Biochemistry, Oxford, v. 38, p. 941-951, 2006. METAY, A. Carbon sequestration and greenhouse effect gases fluxes. Comparison between no-tillage system and conventional system in the Brazilian Cerrados. 231 p. Tese (PhD) - INA-PG, Paris, 2005. NEFTEL, A.; BLATTER, A.; SCHMID, M.; LEHMANN, B.; TARAKANOV, S. V. An experimental determination of the scale length of N2O in the soil of a grassland. Journal of Geophysical Research Atmosphere, Hoboken, v. 105, p. 12095-12103, 2000. PAGE, K. L.; DALAL, R. C. Contribution of natural and drained wetland systems to carbon stocks, CO2, N2O and CH4 fluxes: an Australian perspective. Soil Research, Rome, v. 49, p. 377-388, 2011. PATRICK JR., W. H. Nitrogen transformations in submerged soils. In: STEVENSON, F. J.; BREMNER, J. M.; HAUCK, R. D.; KEENEY, D. R. (Ed). Nitrogen in agricultural soils. Madison: American Society of Agronomy, 1982. p. 465-499. (Agronomy Series, 22) PATRICK, W. H.; JUGSUJINDA, A. Sequencial reduction and oxidation of inorganic nitrogen, manganese and iron in flooded soil, Soil Science Society of American Journal, Madison, v. 56, p.1071-1073, 1992. PONNAMPERUMA, F. N. The chemistry of submerged soils. Advances in Agronomy, Maryland Heights, v. 24, p. 29-96, 1972. R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing, 2011. Disponível em: <http://www.R-project.org/>. Acesso em: 25/05/2012.

104
ROBERTSON, G. P.; GROFMAN, P. M. Nitrogens transformation. In: PAUL, E. A. (Ed.). Soil Microbiology, Ecology and Biochemistry. San Diego: Academi Press, 2007. p. 342-364. ROLSTON, D. E.; HOFFMAN, D. L.; TOY, D. W. Field measurement of denitrification. 1. Fluxes of nitrogen and nitrous oxide. Soil Science Society of American Journal, Madison, v. 42, p. 863-869, 1978. ROSENKRANZ, P.; BRÜGGEMANN, N.; PAPEN, H.; XU, Z.; SEUFERT, G.; BUTTERBACH-BAHL, K. NO2, NO and CH4 exchange and microbial N turnover over a Mediterranean pine forest soil. Biogeosciences Discussions, Goettingen, v. 2, p. 673-702, 2005. RYDEN, J. C. N2O exchange between a grassland soil and the atmosphere. Nature, London, v. 292, p. 235-237, 1981. SANTOS, H. G. dos; JACOMINE, P. K. T.; ANJOS, L. H. C. dos; OLIVEIRA, V. A. de; OLIVEIRA, J. B. de; COELHO, M. R.; LUMBRERAS, J. F.; CUNHA, T. J. F. (Ed.). Sistema Brasileiro de Classificação de Solos. 2. ed. Rio de Janeiro: Embrapa Solos, 2006. SILVA, C. A.; VALE, F. R.; GUILHERME, L. R. G. Nitrificação em latossolos da região sul de Minas Gerais: efeito da acidez do solo. Ciência e Prática, Lavras, v. 18, p. 388-394,1994. SILVA, C. A.; VALE, F. R. Disponibilidade de nitrato em solos brasileiros sob efeito da calagem e de fontes e doses de nitrogênio. Pesquisa Agropecuária Brasileira, Brasília, v. 35, n. 12, p. 2461-2471, 2000. SMITH, C. J.; WRIGHT, M. F.; PATRICK JR., W. H. The effect of soil redox potential and pH on the reduction and production of nitrous oxide. Journal of Environmental Quality, Madison, v. 12, p. 186-188, 1983. SMITH, K. A.; BALL, T.; CONEN, F.; DOBBIE, K. E.; MASSHEDER, J.; REY, A. Exchange of greenhouse gases between soil and atmosphere: interactions of soil physical factors and biological processes. European Journal of Soil Science, West Sussex, v. 54, p. 779-791, 2003. SOIL SURVEY STAFF. Soil survey manual. Washington, DC: U.S. Department of. Agriculture, 1951. (Handbook, No. 18).

105
SOUSA, R. O.; VAHL, L. C.; OTERO, X. L. Química de solos alagados. In: MELO, V. F.; ALLEONI, L. R. F. (Ed.). Química e mineralogia do solo. Viçosa: Sociedade Brasileira de Ciência do Solo, 2009. p. 485-528. TIAN-YEN, Y. Soil and plants. In: TIAN-YEN, Y. Physical chemistry of paddy soils. Berlin: Springer-Verlag, 1985. p. 197-214. TIEDJE, J .M. Denitrifiers. In: WEAVER, J. S.; ANGELS, J. S.; BOTTOMLEY, P. S. (Ed.). Methods of soil analysis: microbilogical and biochemical properties. Madison: Soil Science Society of American, 1994. Pt. 2, p. 245-267. UNITED STATES ENVIRONMENTAL PROTECTION AGENCY (USEPA). Inventory of U.S. greenhouse gas emissions and sinks: 1990-2005. Washington, DC., 2007. (EPA 430-R-07-002). VAN GROENIGEN, J. W.; ZWART, K. B.; HARRIS, D.; VAN KESSEL, C. Vertical gradientes of δ15N and δ18O in soil atmosphere N2O – temporal dynamics in a sandy soil. Rapid Communications in Mass Spectrometry, West Sussex, v. 19, p. 1289-1295, 2005. VON ARNOLD, K.; NILSSON, M.; HANELL, B.; WESLIEN, P.; KLEMEDTSSON, L. Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biology & Biochemistry, Oxford, v. 37, p. 1059-1071, 2005. WLODARCZYK, T.; STEPNIEWSKI, W.; BRZEZINSKA, M. Nitrous oxide production and consumption in Calcaric Regosols as relacted to soil redox and texture. International Agrophysics, Lublin, v. 19, p. 263-271, 2005. WRAGE, N.; VELTHOF, G. L.; van BEUSICHEM, M. L. Role of nitrifier denitrification in the production of nitrous oxide. Soil Biology and Biochemistry, Oxford, v. 33, p. 1723-1732, 2001. YOH, M.; TODA, H.; KANDA, K-I.; TSURUTA, H. Diffusion analysis of N2O cycling in a fertilized soil. Nutrient Cycling in Agroecosystems, Amsterdam, v. 49, p. 29-33, 1997. YU, K. W.; WANG, Z. P.; VERMOESEN, A.; PATRICK, W. H.; VAN CLEMPUT, O. Nitrous oxide and methane emissions from diferente soil suspensions: effect of soil redox status. Biology and Fertility of Soils, Berlin, v. 34, p. 25-30, 2001.

106
ZEBARTH, B. J.; PAUL, J. W.; SCHMIDT, O.; McDOUGALL, R. Influence of time and rate of liquid-manure application on yield and nitrogen utilization of silage corn in south coastal British Columbia. Canadian Journal of Soil Science, v. 76, p. 153-164, 1996.

107
4 CAPÍTULO III
FLUXO DE CO2 E ESTOQUE DE CARBONO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM - PARANÁ - BR
RESUMO
Características pedológicas, variáveis climáticas e práticas de manejo, influenciam os fluxos de CO2 no solo. Organossolos estocam quantidades expressivas de carbono orgânico, podendo ser uma das formas mais eficientes de sequestrar e reter carbono. Condições de hidromorfia plena provocam baixos índices de decomposição de matéria orgânica, menor emissão de CO2 e grande armazenamento de carbono em comparação aos solos não hidromórficos. A drenagem aumenta o aporte de oxigênio e induz o solo a perder carbono. O objetivo deste estudo foi avaliar os fluxos de dióxido de carbono em Organossolo Háplico sáprico típico sob estepe hidrófila de altitude, em condição natural e após drenagem artificial. Variáveis de solo e de clima foram avaliadas para auxiliar a interpretação dos fluxos. Um experimento foi conduzido na nascente do rio Tibagi (Ponta Grossa, PR), onde foram avaliados os fluxos de CO2 em área natural e em área drenada. Seis câmaras estáticas foram instaladas em cada área, e no decorrer de 26 meses foram realizadas 29 coletas de amostras de ar, sendo a concentração de CO2 nas mesmas determinada por cromatografia gasosa. Em ambos os solos ocorreu emissão de dióxido de carbono em taxas similares. No solo sob condição natural as taxas de emissão variaram entre 244 e 870 mg de CO2 m
-2 h-1. Na área drenada artificialmente o fluxo oscilou de 238 a 1018 mg de CO2 m
-2 h-1. Embora não tenha ocorrido diferença significativa nas taxas de emissão entre o Organossolo natural e o drenado, este elevou em 17,9 Mg C-CO2-eq ha-1 ano-1 o potencial de aquecimento global, em função da perda de 6,5 % no estoque de carbono do solo até a profundidade de um metro. Organossolos sob estepe hidrófila possuem importantes funções no ecossistema, como acumular carbono orgânico, armazenar água e reduzir nitrato, não devendo portanto ser drenados, mas efetivamente preservados. A drenagem em Organossolos promove grandes perdas de carbono e contribui intensamente para aumentar o potencial de aquecimento global.
Palavras-chave: Gás de efeito estufa. Nascentes. Nível freático. Umidade gravimétrica. Temperatura do ar e precipitação. Perda de carbono. Potencial de aquecimento global.

108
FLUXES OF CO2 AND CARBON STOCK IN A WETLAND HISTOSOL IN NATURAL
AND DRAINAGE CONDITIONS - PARANA – BRAZIL
ABSTRACT
Pedological attributes and climatic factors, as well as management practices, influenced the flow of CO2 in the soil. Histosol stores significant amount of organic carbon, which may be one of the most efficient ways to sequester and retain carbon. Full conditions of hydromorphy causes low rates of decomposition of organic matter, lower CO2 emissions and high carbon storage compared to non-hydromorphic soils. Drainage increases the supply of oxygen and induces the soil to lose carbon. The purpose of this study was to evaluate the flow of carbon dioxide in a typical Haplic Histosol of a high altitude wetland in natural condition and under artificial drainage. Variables soil and climate were evaluated to assist in the interpretation of the flows. An experiment was conducted in the spring of the river Tibagi (Ponta Grossa, PR) were evaluated in CO2 fluxes in natural area and drained. Six static chambers were installed in each area, and in the course of 26 months were taken 29 samples of the air, and the CO2 concentration in the samples determined by gas chromatography. In both soils occurred emission of carbon dioxide at similar rates. In soil under natural condition emission rates varied between 244 and 870 mg CO2 m-2 h-1. In the artificially drained area the flow has ranged from 238 to 1018 mg CO2 m-2 h-1. Although there was no significant difference in emission rates between natural and drained Histosol, the drainage resulted in an increase of 17,9 Mg C- CO2 eq ha-1 yr-1, in the global warming potential due to the loss of 6,5 % in carbon stock soil. Histosols steppe hydrophilic play important roles in the ecosystem, such accumulate organic carbon, to store water and reduce nitrate, and therefore should not be drained, but effectively preserved. The histosol drainage promotes large carbon losses and contributes heavily to increase the global warming potential.
Keywords: Greenhouse gas. Springs. Groundwater level. Gravimetric moisture. Air temperature. Precipitation. Carbon loss and global warming potential.

109
4.1 INTRODUÇÃO
O dióxido de carbono é o mais importante gás do efeito estufa de origem
antrópica contribuindo em aproximadamente 60% com o aquecimento global (IPCC,
2007b). Durante o período de 1750 a 2005 o CO2 teve um incremento de
aproximadamente 280 ppm para 380 ppm, principalmente, devido ao uso de
combustíveis fósseis e mudanças do uso da terra (FORSTER et al., 2007).
Os ecossistemas naturais são sumidouros de CO2 com exceção da tundra e
principalmente as terras úmidas que são grandes fontes de CH4 (DALAL; ALLEN,
2008). Entretanto, Organossolos tropicais geralmente apresentam altas taxas de
emissão de CO2 (JAUHIAINEN et al., 2008). Em contrapartida tem sido considerado
que a emissão e a captação de CO2 em ecossistemas naturais varia muito pouco e
que o incremento no potencial de aquecimento global proveniente de fontes naturais
é oriundo das emissões de CH4 e de N2O (DENMAN et al., 2007). No entanto, a
função do fluxo de CO2 em solos hidromórficos ainda não está clara, tendo os
mesmos atuado tanto como fonte quanto como sumidouro em Organossolos do
hemisfério norte e também em áreas alagadas tropicais (BLAIS et al., 2005).
Em solos hidromórficos os índices de decomposição da matéria orgânica
são mais baixos, ocorrendo menor produção de CO2 e grande armazenamento de
carbono em comparação com solos não hidromórficos. Os primeiros, quando
vegetados, geralmente apresentam altos índices de produção primária tornando-se
sumidouros de CO2 (RAICH; POTTER, 1995; ALONGI et al., 2001; WHITING;
CHANTON, 2001; REDDY; DELAUNE, 2008). Organossolos podem ser umas das
formas mais eficientes de sequestrar e armazenar carbono, pois combinam grande
capacidade de produção e de conservação de biomassa em ambientes saturados
hidricamente (TAKAHASHI et al., 2002 e 2003).
Múltiplos fatores ambientais afetam a emissão de CO2 em solos
hidromórficos. A temperatura e a umidade do solo regulam a emissão de dióxido de
carbono (DAVIDSON et al., 1998; MOORE; DALVA, 1993), sendo grande a variação
nos fluxos de dióxido de carbono ao longo do ano. Inubushi et al. (2003) trabalhando
na Indonésia com turfeiras sob floresta secundária, arroz irrigado e culturas de
sequeiro, encontraram os maiores valores de emissão de dióxido de carbono nos
meses de março, abril e maio, o que corresponde à primavera, e em dezembro, que
se refere ao início do inverno.

110
Os valores de fluxos de dióxido de carbono encontrados na literatura para
Organossolos são menos variáveis do que os de óxido nitroso. Inubushi et al. (2003)
registraram variação entre 40 e 380 mg C m-2 h-1, Furukawa et al. (2005) entre 10 e
500 mg C m-2 h-1, Jauhiainen et al. (2008) entre 818 e 1151 mg CO2 m-2 h-1 e Koh et
al. (2009) de 67 a 908 mg CO2 m-2 h-1.
Inúmeros autores ressaltam a influência da oscilação do nível freático e da
drenagem sobre os fluxos de CO2. A emissão de CO2 em turfeiras mostra correlação
linear positiva com a profundidade do lençol freático (MOORE; DALVA, 1993 e 1997;
FURUKAWA et al., 2005). Furukawa et al. (2005) efetuando uma análise de
regressão de emissões de gases em Organossolos drenados cultivados com arroz
irrigado e mandioca, concluíram que o rebaixamento do nível freático à 10 cm da
superfície aumentou em 50% a emissão de CO2. Em Organossolos drenados, cujo
nível freático ficou a 15 e 18 cm, a emissão de CO2 foi praticamente o dobro da
ocorrida no Organossolo não drenado (VON ARNOLD et al., 2005). No entanto, os
fluxos de CO2 obtidos por Hadi et al. (2000,) não mostraram interrelação clara com o
nível freático. No caso de turfeiras tropicais a chuva induz à elevação do nível
freático durante a estação úmida, de modo que o solo torna-se anaeróbico, mas sem
alterar os fluxos de dióxido de carbono (INUBUSHI et al., 2003).
A drenagem aumenta a área de oxidação no perfil de solo e promove
incremento nas perdas de carbono (FURUKAWA et al., 2005). A emissão de CO2 a
partir de Organossolos drenados e cultivados pode ser de 2 a 4 vezes maior do que
em solos minerais, transformando-os de sumidouros para fontes de dióxido de
carbono (LOHILA et al., 2003; MALJANEN et al., 2004).
Entretanto, a remoção de carbono na forma de CO2 não é o único processo
de redução dos teores de carbono no solo. Podem ocorrer perdas de formas
solúveis de carbono por lixiviação. O transporte de carbono orgânico dissolvido é
determinado pelo regime do fluxo hídrico no solo e pela presença de
macroporosidade (KALBITZ et al., 2000). Easthouse et al. (1992) propuseram a
ocorrência de lixiviação de considerável quantidade de carbono orgânico dissolvido,
em horizontes orgânicos, devido a precipitação. A drenagem de organossolos
geralmente aumenta o fluxo de carbono orgânico dissolvido porque aumenta o
escorrimento superficial (MOORE, 1998), o qual incrementa a lixiviação.
Grandes áreas ocupadas por Organossolos tropicais tem sido drenadas
resultando em mudanças abruptas e permanentes no balanço ecossistêmico do

111
carbono, fazendo com que deixem de ser sumidouros e passem a funcionar como
fontes de CO2 (CANADELL et al., 2007; HIRANO et al., 2007). Furukawa et al.
(2005) e Von Arnold et al. (2005) também se referem a elevação no potencial de
aquecimento global em função da drenagem de Organossolos.
Neste contexto, concebeu-se a hipótese de que a drenagem elevaria
consideravelmente os fluxos de dióxido de carbono e que a temperatura do ar e a
precipitação também contribuíram para alterar os fluxos de CO2.
Este estudo objetivou avaliar os fluxos de CO2 em Organossolo Háplico
sáprico típico em estepe hidrófila de altitude, em condições naturais e após a
drenagem artificial, considerando a influência da temperatura, da precipitação, da
altura do nível freático e da umidade gravimétrica do solo.

112
4.2 MATERIAL E MÉTODOS
4.2.1 Experimento de campo
Foi implantado um experimento em área de estepe hidrófila com dimensão
de 6,5 ha (25o09´59´´S; 50o04´45´´O), a 860 m de altitude, situada na fazenda da
Embrapa SPM-EPGA (Serviço de Produtos e Mercado - Escritório de Ponta Grossa,
PR), segundo planalto paranaense, região dos Campos Gerais (Figura 1). A área de
estudo está localizada em uma nascente do rio Cará-Cará, afluente do rio Tibagi. O
clima local é do tipo Cfb, mesotérmico úmido, sem estação seca, temperatura média
do mês mais quente inferior a 22oC, temperatura média do mês mais frio menor do
que 18oC e precipitação entre 1100 e 2000 mm. A paisagem está inserida em
reverso distal de cuesta, sobre folhelho argiloso da Formação Ponta Grossa,
pertencente ao Grupo Paraná, Paleozóico. Pelo Sistema Brasileiro de Classificação
de Solos (SANTOS et al., 2006), o solo é um ORGANOSSOLO HÁPLICO sáprico
típico álico relevo plano fase campo subtropical hidrófilo de altitude, com 100 cm de
espessura, cujas principais características químicas e físicas encontram-se na
Tabela 1.
Foram testados dois tratamentos (solo em condição natural, e solo drenado).
Em ambos, houve 29 eventos de coleta e a amostragem foi realizada em 6 câmaras
estáticas, correspondentes a 6 repetições. Foi efetuada a drenagem de uma área de
30 m × 10 m, nos dias 08 e 09/09/2010. Usando uma retroescavadeira ligada a um
trator de esteira, foram abertos drenos trapezoidais com 1,5 m de base superior, 1,2
m de base inferior e 0,65 m de profundidade (Anexo 3). No Organossolo natural as
câmaras foram fixadas em uma área de 15 x 15 m, na mesma cota altimétrica, a 45
m da área drenada.
4.2.2 Coleta de amostras de ar
As primeiras coletas para avaliação do fluxo de CO2 foram realizadas a 1, 4,
8, 14 e 25 dias após a implantação do experimento. Depois destas, seguiram-se
mais 24 coletas mensais (Anexo 4).
113
Para apoiar as câmaras coletoras de gases, seis bases circulares de metal
foram fixadas no solo em cada tratamento. Estas bases tinham 38 cm de diâmetro
por 25 cm de altura, dos quais 15 cm eram cravados no solo (Anexo 5b). Para
remover o excesso de água da chuva, no período entre as coletas, na parte inferior
destas bases, próximo à superfície do solo, foram abertos quatro orifícios com dois
cm de diâmetro, os quais eram tampados com rolhas de borracha na hora das
coletas.
FIGURA 1 - LOCALIZAÇÃO DA ÁREA DE ESTUDO.
Câmaras estáticas de PVC com 40 cm de altura, colocadas sobre as bases,
foram usadas para coletar amostras de ar (Anexo 5d). Para impedir o vazamento
dos gases, entre a parte superior da base e a câmara havia uma canaleta de 4 cm
de altura por 6 cm de largura, a qual era preenchida com água no momento das
coletas. As câmaras foram equipadas com termômetro, ventilador para
homogeneizar o ar e torneira de três vias acoplada a uma mangueirinha para
transportar o ar para seringas de polipropileno de 20 ml. As amostras de ar eram
coletadas entre 9:00 e 11:00 horas, imediatamente após o fechamento das câmaras,
a 0, 15, 30 e 45 minutos.
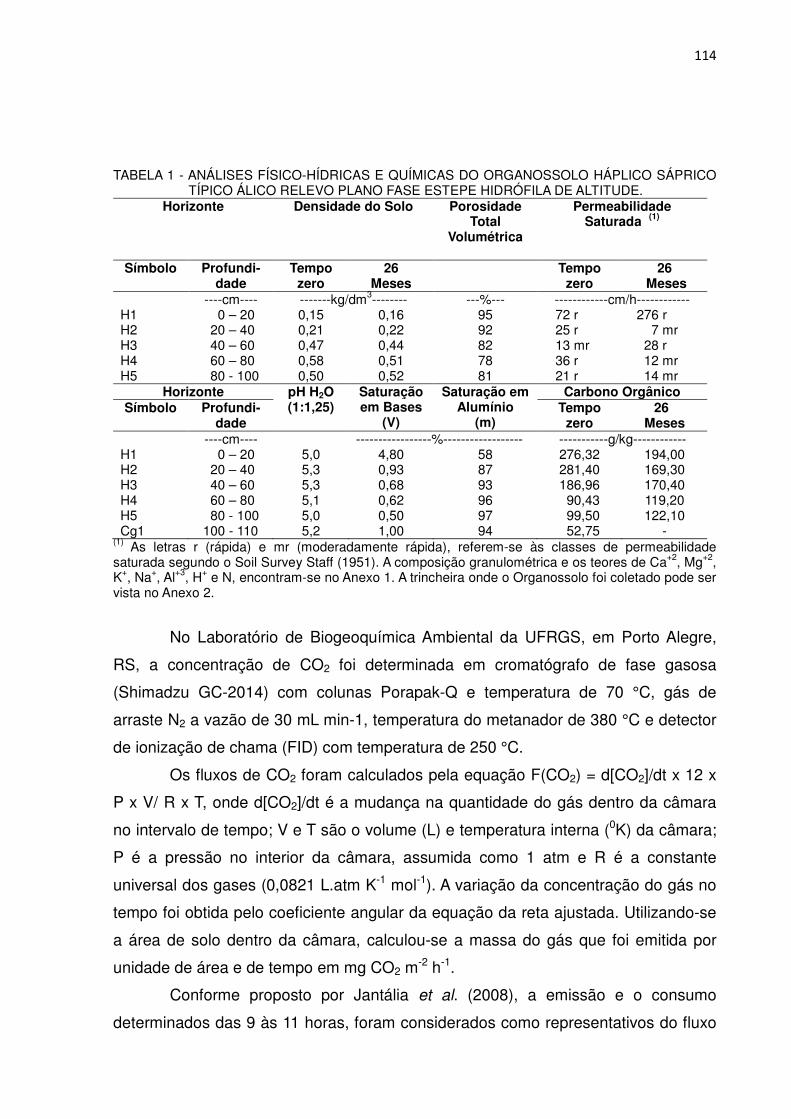
114
TABELA 1 - ANÁLISES FÍSICO-HÍDRICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO ÁLICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE.
Horizonte Densidade do Solo Porosidade Total
Volumétrica
Permeabilidade Saturada (1)
Símbolo Profundi-dade
Tempo zero
26 Meses
Tempo zero
26 Meses
----cm---- -------kg/dm3-------- ---%--- ------------cm/h------------ H1 0 – 20 0,15 0,16 95 72 r 276 r H2 20 – 40 0,21 0,22 92 25 r 7 mr H3 40 – 60 0,47 0,44 82 13 mr 28 r H4 60 – 80 0,58 0,51 78 36 r 12 mr H5 80 - 100 0,50 0,52 81 21 r 14 mr
Horizonte pH H2O (1:1,25)
Saturação em Bases
(V)
Saturação em Alumínio
(m)
Carbono Orgânico Símbolo Profundi-
dade Tempo
zero 26
Meses ----cm---- -----------------%------------------ -----------g/kg------------
H1 0 – 20 5,0 4,80 58 276,32 194,00 H2 20 – 40 5,3 0,93 87 281,40 169,30 H3 40 – 60 5,3 0,68 93 186,96 170,40 H4 60 – 80 5,1 0,62 96 90,43 119,20 H5 80 - 100 5,0 0,50 97 99,50 122,10 Cg1 100 - 110 5,2 1,00 94 52,75 -
(1) As letras r (rápida) e mr (moderadamente rápida), referem-se às classes de permeabilidade saturada segundo o Soil Survey Staff (1951). A composição granulométrica e os teores de Ca+2, Mg+2, K+, Na+, Al+3, H+ e N, encontram-se no Anexo 1. A trincheira onde o Organossolo foi coletado pode ser vista no Anexo 2.
No Laboratório de Biogeoquímica Ambiental da UFRGS, em Porto Alegre,
RS, a concentração de CO2 foi determinada em cromatógrafo de fase gasosa
(Shimadzu GC-2014) com colunas Porapak-Q e temperatura de 70 °C, gás de
arraste N2 a vazão de 30 mL min-1, temperatura do metanador de 380 °C e detector
de ionização de chama (FID) com temperatura de 250 °C.
Os fluxos de CO2 foram calculados pela equação F(CO2) = d[CO2]/dt x 12 x
P x V/ R x T, onde d[CO2]/dt é a mudança na quantidade do gás dentro da câmara
no intervalo de tempo; V e T são o volume (L) e temperatura interna (0K) da câmara;
P é a pressão no interior da câmara, assumida como 1 atm e R é a constante
universal dos gases (0,0821 L.atm K-1 mol-1). A variação da concentração do gás no
tempo foi obtida pelo coeficiente angular da equação da reta ajustada. Utilizando-se
a área de solo dentro da câmara, calculou-se a massa do gás que foi emitida por
unidade de área e de tempo em mg CO2 m-2 h-1.
Conforme proposto por Jantália et al. (2008), a emissão e o consumo
determinados das 9 às 11 horas, foram considerados como representativos do fluxo

115
médio diário. Para calcular os fluxos acumulados, integrou-se as áreas dos fluxos
diários de cada estação, com auxílio do programa Sigmaplot.
4.2.3 Parâmetros pedológicos
Com exceção do teor de carbono orgânico, o qual foi determinado por
oxidação sob atmosfera de gás oxigênio em determinador automático Analytik Jena,
as demais análises físicas e químicas do solo foram realizadas segundo a
metodologia proposta por Claessen (1997). Três amostras de solo de 0 a 10 cm
foram coletadas por parcela para determinação da umidade gravimétrica, em cada
coleta de ar.
Foram instalados 4 poços hídricos em cada tratamento, para determinar a
altura do nível freático. Para a confecção dos poços hídricos utilizou-se tubos de
PVC com 1,20 m de comprimento e 7,5 cm de diâmetro. Estes tubos foram
penetrados no solo até atingir o horizonte mineral. Para facilitar o escoamento da
água foram abertos 4 orifícios circulares com 1 cm de diâmetro, espaçados de 10
em 10 cm, do início até a base dos tubos. Cada poço hídrico foi posicionado entre
duas, câmaras. Em todas as coletas registrava-se a altura do nível freático com uma
trena.
O volume de água armazenado por hectare em cada horizonte do solo foi
obtido pela fórmula: volume de água = [porosidade total (cm3/cm3)/1000] x espessura
do horizonte (cm) x 100.000.000 (cm3). O somatório dos cinco horizontes foi
convertido em milhões de litros/ha, representando o volume total de água
armazenado no solo até a profundidade de 1 metro.
O estoque inicial de carbono no solo foi obtido pela fórmula: estoque de C =
teor de C (%) x densidade do solo (g/cm3) x espessura do horizonte (cm). O
somatório do carbono estocado em todos os horizontes, transformado em Mg ha-1,
indicou o estoque total de carbono até 1 m de profundidade. O estoque final de
carbono no solo foi determinado considerando a massa equivalente do solo no perfil
de referência, utilizando-se o cálculo proposto por Sisti et al. (2004), no qual foi
computada a taxa média de subsidência de 5,6 cm, no horizonte H1, o mais
suscetível ao processo.

116
Os fluxos anuais de metano e de óxido nitroso foram transformados em CO2-
eq, multiplicando-se os valores originais por 25 e 298, respectivamente, conforme
proposto pelo IPCC (2007a).
4.2.4 Variáveis meteorológicas
Para obtenção da precipitação e da temperatura do ar, a cerca de 1 km da
área experimental, foi instalada uma estação meteorológica portátil da marca Davis,
modelo Vantage Vue. Estas variáveis formam determinadas durante 24 meses.
4.2.5 Análise estatística
As diferenças na umidade gravimétrica do solo na profundidade de 0 a 10
cm, no nível freático e nos fluxos de CO2, entre o Organossolo natural e o drenado,
além da alteração nos teores de carbono nos horizontes H1 a H5, no Organossolo
drenado, em função do tempo de drenagem, foram comparadas pelo teste de Tukey
(p < 0,05).
Foram determinados os coeficientes de correlação linear de Pearson (r)
entre os fluxos de CO2 e a altura do nível freático, a umidade gravimétrica na
profundidade de 0 a 10 cm, a temperatura média do ar e a precipitação acumulada,
entre às 9 e 12 horas na ocasião das coletas e durante o período de 1, 3, 5, 7, 15 e
30 dias que antederam as coletas.
Para determinar os efeitos dos tratamentos (com e sem drenagem), dos
fatores pedológicos e climáticos e, do tempo, sobre os fluxos de CO2, utilizou-se o
Modelo Aditivo Generalizado (GAM) o qual além de analisar todas as variáveis
conjuntamente, pressupõe que os fluxos determinados em cada coleta são
interdependentes.
O Software R (R DEVELOPMENT CORE TEAM, 2011), foi utilizado para
realizar as análises estatísticas.

117
4.3 RESULTADOS E DISCUSSÃO
4.3.1 Efeito dos fatores meteorológicos sobre o fluxo de CO2
No primeiro ano de estudo a temperatura média anual foi 17,5 ºC e os
valores médios semanais variaram entre 22,2 e 10,3ºC. As maiores temperaturas
médias semanais ocorreram no final da primavera (21,8ºC), no verão (22,2ºC) e no
início do outono (20,9ºC) e as menores (10,3ºC) foram registradas no inverno. No
segundo ano de estudo a temperatura média anual foi 18,4 ºC e os valores médios
semanais oscilaram entre 24,7 e 10,3 ºC. As maiores temperaturas médias semanais
ocorreram no final da primavera (21,5 ºC), em meados do verão (24,7 ºC) e no início
do outono (20,3 ºC) e as menores (10,3 ºC) foram registradas no inverno (Figura 2).
Não houve diferenças marcantes na temperatura média semanal do ar nas
estações entre os dois anos. As temperaturas médias na primavera, verão, outono e
inverno foram de 18,5, 21,0, 16,2 e 14,8 0C, respectivamente, no primeiro ano e de
18,4, 21,3, 16,3 e 16,5 0C, respectivamente no segundo ano. No inverno de 2012 a
temperatura média semanal foi apenas 1,7 0C mais elevada do que no inverno de
2011.
A precipitação total acumulada ao longo do primeiro ano foi de 1837 mm
(Figura 2). As estações mais chuvosas foram a primavera (442 mm), o verão (543
mm) e, atipicamente, o inverno (580 mm). No outono a precipitação acumulada foi
254 mm. A precipitação total acumulada ao longo do segundo ano foi de 1518 mm.
As estações mais chuvosas foram o verão (446 mm) e atipicamente o outono (615
mm), seguidos pela primavera (298 mm) e inverno (114 mm). A precipitação anual no
segundo ano de estudo foi 319 mm menor do que no primeiro, sendo que com
exceção do outono, as outras estações foram menos chuvosas.
No Organossolo natural os fluxos variaram entre 244 e 850 mg m-2 h-1 de
CO2, com média de 528 mg m-2 h-1 no primeiro ano e de 292 a 870 mg m-2 h-1 com
média de 660 mg m-2 h-1 no segundo (Figura 3).
As emissões mínimas ocorreram nos meses mais frios e os picos nos meses
com temperaturas mais elevadas. Houve correlação positiva entre a entre a emissão
de dióxido de carbono e a temperatura do ar e a precipitação acumulada, embora a
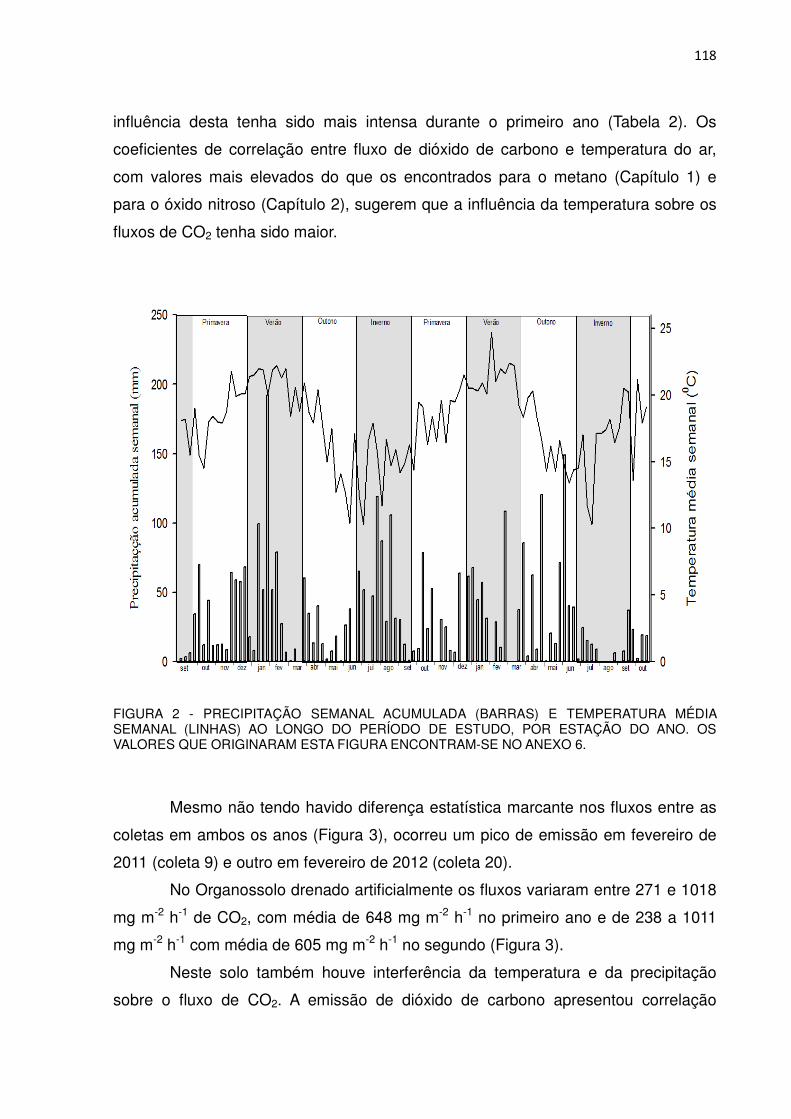
118
influência desta tenha sido mais intensa durante o primeiro ano (Tabela 2). Os
coeficientes de correlação entre fluxo de dióxido de carbono e temperatura do ar,
com valores mais elevados do que os encontrados para o metano (Capítulo 1) e
para o óxido nitroso (Capítulo 2), sugerem que a influência da temperatura sobre os
fluxos de CO2 tenha sido maior.
FIGURA 2 - PRECIPITAÇÃO SEMANAL ACUMULADA (BARRAS) E TEMPERATURA MÉDIA SEMANAL (LINHAS) AO LONGO DO PERÍODO DE ESTUDO, POR ESTAÇÃO DO ANO. OS VALORES QUE ORIGINARAM ESTA FIGURA ENCONTRAM-SE NO ANEXO 6.
Mesmo não tendo havido diferença estatística marcante nos fluxos entre as
coletas em ambos os anos (Figura 3), ocorreu um pico de emissão em fevereiro de
2011 (coleta 9) e outro em fevereiro de 2012 (coleta 20).
No Organossolo drenado artificialmente os fluxos variaram entre 271 e 1018
mg m-2 h-1 de CO2, com média de 648 mg m-2 h-1 no primeiro ano e de 238 a 1011
mg m-2 h-1 com média de 605 mg m-2 h-1 no segundo (Figura 3).
Neste solo também houve interferência da temperatura e da precipitação
sobre o fluxo de CO2. A emissão de dióxido de carbono apresentou correlação
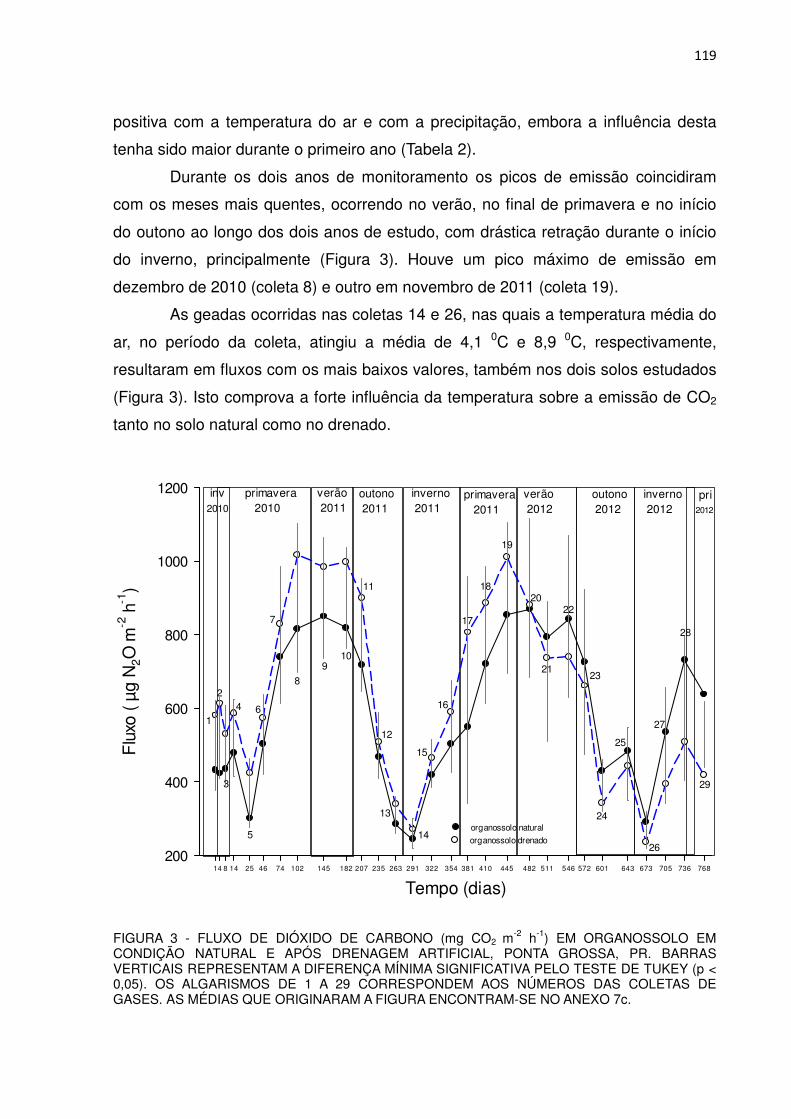
119
positiva com a temperatura do ar e com a precipitação, embora a influência desta
tenha sido maior durante o primeiro ano (Tabela 2).
Durante os dois anos de monitoramento os picos de emissão coincidiram
com os meses mais quentes, ocorrendo no verão, no final de primavera e no início
do outono ao longo dos dois anos de estudo, com drástica retração durante o início
do inverno, principalmente (Figura 3). Houve um pico máximo de emissão em
dezembro de 2010 (coleta 8) e outro em novembro de 2011 (coleta 19).
As geadas ocorridas nas coletas 14 e 26, nas quais a temperatura média do
ar, no período da coleta, atingiu a média de 4,1 0C e 8,9 0C, respectivamente,
resultaram em fluxos com os mais baixos valores, também nos dois solos estudados
(Figura 3). Isto comprova a forte influência da temperatura sobre a emissão de CO2
tanto no solo natural como no drenado.
organossolo natural
Tempo (dias)14 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Flu
xo (
µg N
2O m
-2 h
-1)
200
400
600
800
1000
1200 inv 2010
primavera 2010
verão 2011
outono 2011
inverno 2011
primavera 2011
verão 2012
outono 2012
inverno 2012
pri 2012
organossolo drenado
1
2
3
4
5
6
7
8
910
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
FIGURA 3 - FLUXO DE DIÓXIDO DE CARBONO (mg CO2 m-2 h-1) EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS ALGARISMOS DE 1 A 29 CORRESPONDEM AOS NÚMEROS DAS COLETAS DE GASES. AS MÉDIAS QUE ORIGINARAM A FIGURA ENCONTRAM-SE NO ANEXO 7c.
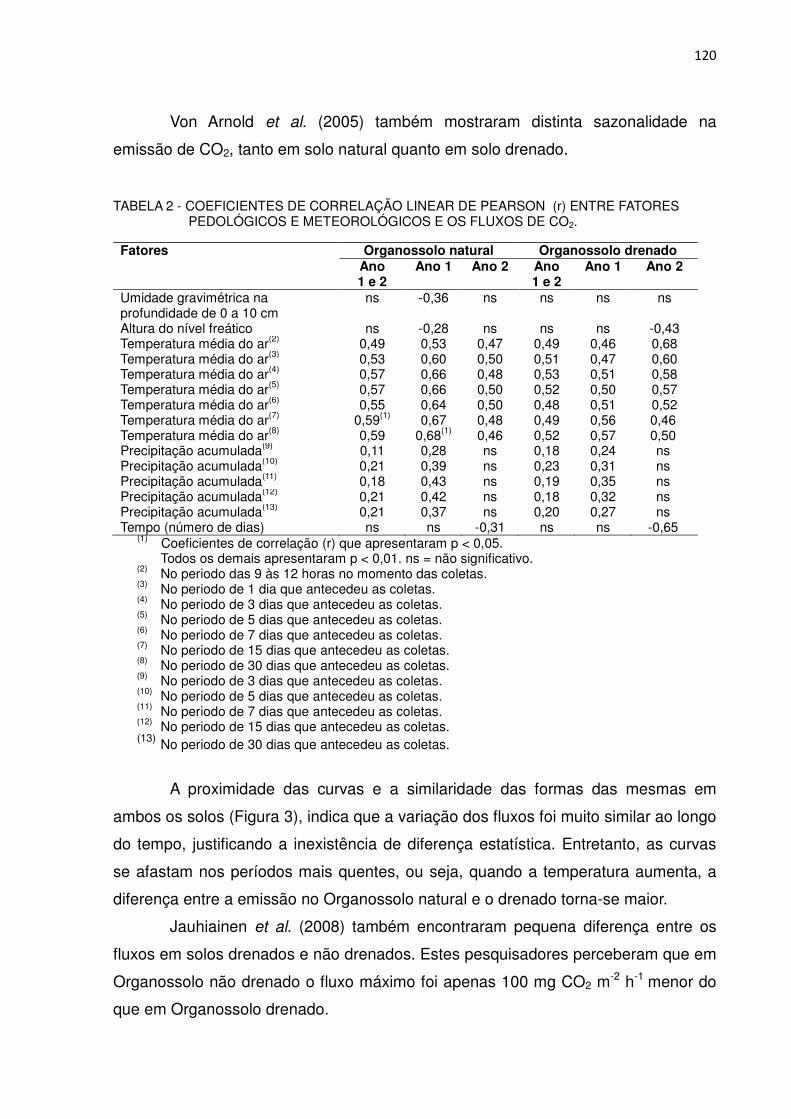
120
Von Arnold et al. (2005) também mostraram distinta sazonalidade na
emissão de CO2, tanto em solo natural quanto em solo drenado.
TABELA 2 - COEFICIENTES DE CORRELAÇÃO LINEAR DE PEARSON (r) ENTRE FATORES PEDOLÓGICOS E METEOROLÓGICOS E OS FLUXOS DE CO2.
Fatores Organossolo natural Organossolo drenado Ano 1 e 2
Ano 1 Ano 2 Ano 1 e 2
Ano 1 Ano 2
Umidade gravimétrica na profundidade de 0 a 10 cm
ns -0,36 ns ns ns ns
Altura do nível freático ns -0,28 ns ns ns -0,43 Temperatura média do ar(2) 0,49 0,53 0,47 0,49 0,46 0,68 Temperatura média do ar(3) 0,53 0,60 0,50 0,51 0,47 0,60 Temperatura média do ar(4) 0,57 0,66 0,48 0,53 0,51 0,58 Temperatura média do ar(5) 0,57 0,66 0,50 0,52 0,50 0,57 Temperatura média do ar(6) 0,55 0,64 0,50 0,48 0,51 0,52 Temperatura média do ar(7) 0,59(1) 0,67 0,48 0,49 0,56 0,46 Temperatura média do ar(8) 0,59 0,68(1) 0,46 0,52 0,57 0,50 Precipitação acumulada(9) 0,11 0,28 ns 0,18 0,24 ns Precipitação acumulada(10) 0,21 0,39 ns 0,23 0,31 ns Precipitação acumulada(11) 0,18 0,43 ns 0,19 0,35 ns Precipitação acumulada(12) 0,21 0,42 ns 0,18 0,32 ns Precipitação acumulada(13) 0,21 0,37 ns 0,20 0,27 ns Tempo (número de dias) ns ns -0,31 ns ns -0,65
(1) Coeficientes de correlação (r) que apresentaram p < 0,05. Todos os demais apresentaram p < 0,01. ns = não significativo.
(2) No periodo das 9 às 12 horas no momento das coletas. (3) No periodo de 1 dia que antecedeu as coletas. (4) No periodo de 3 dias que antecedeu as coletas. (5) No periodo de 5 dias que antecedeu as coletas. (6) No periodo de 7 dias que antecedeu as coletas. (7) No periodo de 15 dias que antecedeu as coletas. (8) No periodo de 30 dias que antecedeu as coletas. (9) No periodo de 3 dias que antecedeu as coletas. (10) No periodo de 5 dias que antecedeu as coletas. (11) No periodo de 7 dias que antecedeu as coletas. (12) No periodo de 15 dias que antecedeu as coletas. (13) No periodo de 30 dias que antecedeu as coletas.
A proximidade das curvas e a similaridade das formas das mesmas em
ambos os solos (Figura 3), indica que a variação dos fluxos foi muito similar ao longo
do tempo, justificando a inexistência de diferença estatística. Entretanto, as curvas
se afastam nos períodos mais quentes, ou seja, quando a temperatura aumenta, a
diferença entre a emissão no Organossolo natural e o drenado torna-se maior.
Jauhiainen et al. (2008) também encontraram pequena diferença entre os
fluxos em solos drenados e não drenados. Estes pesquisadores perceberam que em
Organossolo não drenado o fluxo máximo foi apenas 100 mg CO2 m-2 h-1 menor do
que em Organossolo drenado.

121
No presente estudo, a pequena diferença entre as emissões de CO2 nos
Organosolos natural e drenado, pode estar associada ao fato de que na camada de
0 a 10 cm, em ambos os solos, o acesso de oxigênio não foi restritivo, uma vez que,
segundo Maljanen et al. (2002), a emissão de CO2 ocorre justamente nas camadas
de 5 a 10 cm.
Por outro lado, Koh et al. (2009) analisando fluxos de dióxido de carbono em
Organossolos do Mississipi concluíram que nos solos ocasionalmente inundados e
não inundados a emissão oscilou entre 180 e 908 mg CO2 m-2 h-1 e nos solos
permanentemente inundados as emissões foram muito menores, variando entre 67 e
472 mg CO2 m-2 h-1. Von Arnold et al. (2005) perceberam que um Organossolo
drenado cujo nível freático oscilou entre -13 e -23 cm emitiu 1,6 vezes mais dióxido
de carbono do que o não drenado, onde o lençol freático variou de -1 a 3 cm, mas
permaneceu acima da superfície durante grande parte do tempo.
4.3.2 Efeito da drenagem sobre o fluxo de CO2
No Organossolo natural o nível freático variou ao longo do primeiro ano entre
0,3 e -16,3 cm, permanecendo em média a -7 cm da superfície do solo e entre 0,25
e -14,8 cm, com média de -5 cm, no segundo ano (Figura 4), indicando que a
oscilação foi muito similar, até porque este solo não sofreu nenhuma alteração
antrópica. O nível freático se manteve acima da superfície do solo em apenas duas
das 29 coletas. A umidade gravimétrica média na profundidade de 0 a 10 cm foi de
9,9 kg kg-1 e variou entre 7,6 e 11,7 kg kg-1, no primeiro ano. No segundo ano a
média ficou em 9,8 kg kg-1 tendo oscilado entre 8,1 e 11,9 kg kg-1 (Figura 5),
mostrando também muita similaridade entre os dois anos.
No Organossolo drenado o nível freático variou ao longo do primeiro ano
entre -25 e -64,8 cm, permanecendo em média a 57 cm abaixo da superfície do solo
e entre -54 e -64,5 cm, com média de -59 cm, no segundo ano (Figura 4), o que
também demonstra grande similaridade. A umidade gravimétrica média na
profundidade de 0 a 10 cm foi de 5,7 kg kg-1 e variou entre 4,5 e 6,8 kg kg-1, no
primeiro ano. No segundo ano a média ficou em 4,2 kg kg-1 tendo oscilado entre 3,6
e 4,7 kg kg-1 (Figura 5). A umidade gravimétrica além de ter sido significativamente
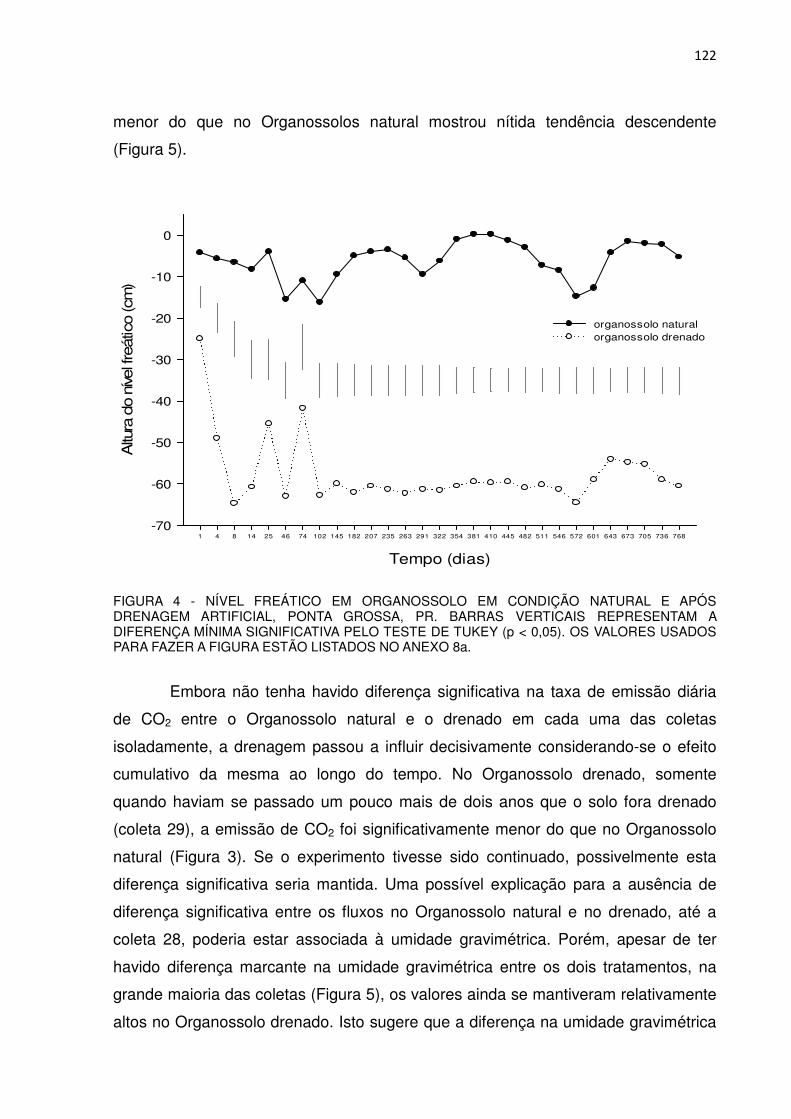
122
menor do que no Organossolos natural mostrou nítida tendência descendente
(Figura 5).
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Altu
ra d
o ní
vel f
reát
ico
(cm
)
-70
-60
-50
-40
-30
-20
-10
0
organossolo naturalorganossolo drenado
FIGURA 4 - NÍVEL FREÁTICO EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA FAZER A FIGURA ESTÃO LISTADOS NO ANEXO 8a.
Embora não tenha havido diferença significativa na taxa de emissão diária
de CO2 entre o Organossolo natural e o drenado em cada uma das coletas
isoladamente, a drenagem passou a influir decisivamente considerando-se o efeito
cumulativo da mesma ao longo do tempo. No Organossolo drenado, somente
quando haviam se passado um pouco mais de dois anos que o solo fora drenado
(coleta 29), a emissão de CO2 foi significativamente menor do que no Organossolo
natural (Figura 3). Se o experimento tivesse sido continuado, possivelmente esta
diferença significativa seria mantida. Uma possível explicação para a ausência de
diferença significativa entre os fluxos no Organossolo natural e no drenado, até a
coleta 28, poderia estar associada à umidade gravimétrica. Porém, apesar de ter
havido diferença marcante na umidade gravimétrica entre os dois tratamentos, na
grande maioria das coletas (Figura 5), os valores ainda se mantiveram relativamente
altos no Organossolo drenado. Isto sugere que a diferença na umidade gravimétrica
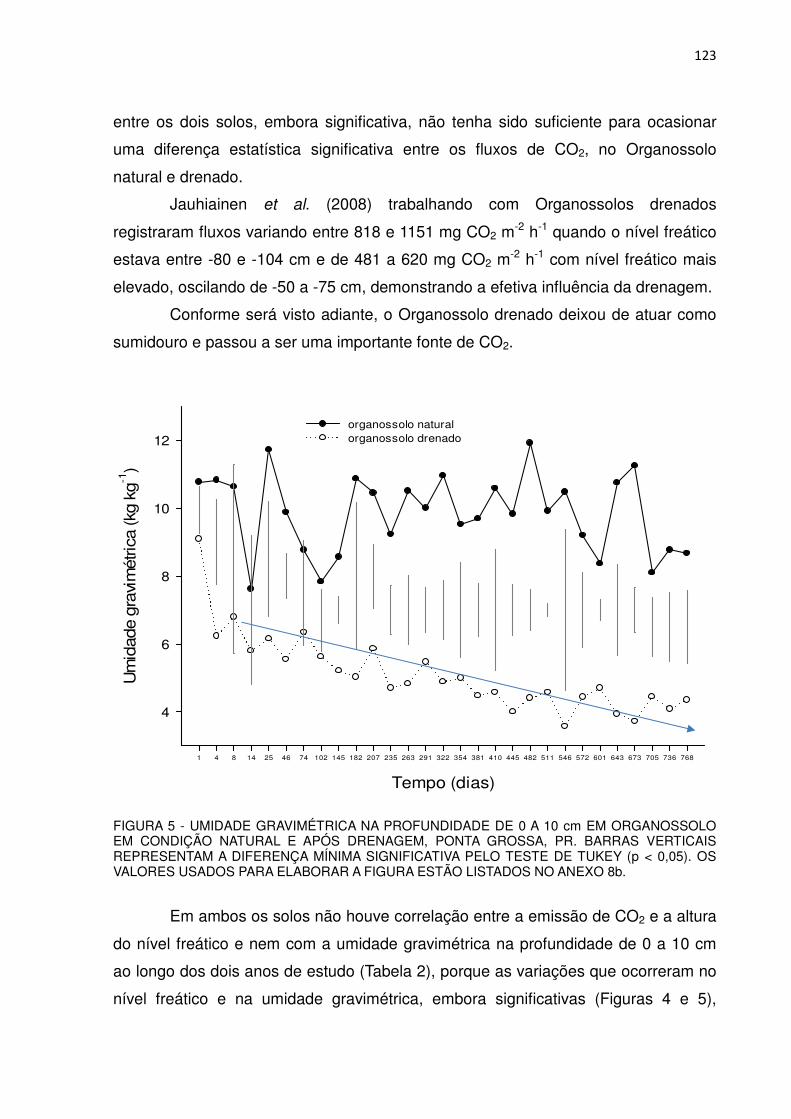
123
entre os dois solos, embora significativa, não tenha sido suficiente para ocasionar
uma diferença estatística significativa entre os fluxos de CO2, no Organossolo
natural e drenado.
Jauhiainen et al. (2008) trabalhando com Organossolos drenados
registraram fluxos variando entre 818 e 1151 mg CO2 m-2 h-1 quando o nível freático
estava entre -80 e -104 cm e de 481 a 620 mg CO2 m-2 h-1 com nível freático mais
elevado, oscilando de -50 a -75 cm, demonstrando a efetiva influência da drenagem.
Conforme será visto adiante, o Organossolo drenado deixou de atuar como
sumidouro e passou a ser uma importante fonte de CO2.
Tempo (dias)
1 4 8 14 25 46 74 102 145 182 207 235 263 291 322 354 381 410 445 482 511 546 572 601 643 673 705 736 768
Um
idad
e gr
avim
étric
a (k
g kg
-1)
4
6
8
10
12organossolo naturalorganossolo drenado
FIGURA 5 - UMIDADE GRAVIMÉTRICA NA PROFUNDIDADE DE 0 A 10 cm EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM, PONTA GROSSA, PR. BARRAS VERTICAIS REPRESENTAM A DIFERENÇA MÍNIMA SIGNIFICATIVA PELO TESTE DE TUKEY (p < 0,05). OS VALORES USADOS PARA ELABORAR A FIGURA ESTÃO LISTADOS NO ANEXO 8b.
Em ambos os solos não houve correlação entre a emissão de CO2 e a altura
do nível freático e nem com a umidade gravimétrica na profundidade de 0 a 10 cm
ao longo dos dois anos de estudo (Tabela 2), porque as variações que ocorreram no
nível freático e na umidade gravimétrica, embora significativas (Figuras 4 e 5),

124
possivelmente não foram suficientes para diferenciar de maneira mais intensa as
emissões de dióxido de carbono entre os dois solos estudados.
Vale enfatizar que apesar de no Organossolo drenado o excesso de água ter
sido removido, a umidade gravimétrica manteve-se relativamente elevada, pelo
menos na profundidade de 0 a 10 cm. Nesta profundidade apesar da umidade
gravimétrica ter sido significativamente inferior à do Organossolo natural, a mesma
ainda manteve-se bastante elevada em comparação a solos oxidados, nos quais
não há excesso de umidade. Certamente nas camadas abaixo de 10 cm a diferença
na umidade entre os dois solos tenha sido menor.
Chamou a atenção que a partir da coleta 20, no início do verão de 2012,
quando haviam se passado 16 meses que este Organossolo havia sido drenado, o
mesmo começou a emitir menos dióxido de carbono do que o natural, ao contrário
do que vinha ocorrendo (Figura 3).
Esta redução na emissão de CO2 possivelmente tenha ocorrido em função
da redução nos teores de carbono total do solo, devido à drenagem.
Entre 14 e 20 meses de drenagem os horizontes H1 e H3 sofreram uma leve
redução no teor de carbono de 7 e 8%, respectivamente (Figura 6).
Isto pode estar associado à redução que houve na emissão de CO2 no
organossolo drenado a partir da coleta 20, na qual haviam se passado 16 meses
que o solo havia sido drenado, pois se diminuiu a fonte de carbono no solo, era
esperado que ocorresse uma retração na emissão de dióxido de carbono.
Infelizmente o teor de carbono no solo não foi determinado aos 16 meses de
drenagem. Porém, se houve redução aos 14 e aos 20 meses, possivelmente tenha
ocorrido também aos 16 meses.
Aos 26 meses de drenagem a redução nos teores de carbono do solo se
acentuou, tendo sido de 30%, 40% e 9%, nos horizontes H1, H2, H3,
respectivamente, o que equivale a uma redução média de 26% no teor de carbono
no solo até a profundidade de 60 cm.
Por outro lado, ocorreu um incremento de 24% e 18% no teor de carbono do
horizonte H4 e H5, respectivamente, o que é um forte indício de que tenha ocorrido
remoção e transporte de formas solúveis de carbono dos horizontes superficiais para
os subsuperficiais.
A considerável redução no teor de carbono nos horizontes H1 a H3
possivelmente contribuiu para aumentar a diferença entre a emissão de dióxido de

125
carbono no solo natural e drenado após 26 meses de drenagem (coleta 29), ocasião
em que a emissão no Organossolo drenado foi significativamente menor do que no
natural (Figura 3).
Além da redução nos teores de carbono, duas suposições puderam
contribuir para justificar a redução da emissão de CO2 no Organossolo drenado após
482 dias de drenagem (coleta 20). Primeiramente, cogitou-se a hipótese de que
tenha ocorrido redução na quantidade de matéria orgânica lábil no solo, devido à
drenagem. Infelizmente esta hipótese não pode ser comprovada, pois não foi
avaliada a quantidade de matéria orgânica facilmente decomponível presente no
solo, antes e depois da drenagem. Reddy et al. (2006) citam que a matéria orgânica
lábil, presente no solo até a profundidade de 10 cm é facilmente oxidada após a
drenagem. Em segundo lugar supôs-se que a drenagem tenha alterado a fisiologia
das plantas nativas adaptadas ao excesso de água, fazendo com que as mesmas
passassem a emitir menos dióxido de carbono. Crow e Wieder (2005) perceberam
redução na emissão de CO2 em organossolos sob ciperáceas e arbustivas quando
se reduziu a umidade do solo. Rowson et al. (2013) afirmam que a alta produção de
CO2 estimulada por exsudatos radiculares só ocorre quando a planta se encontra em
pleno crescimento vegetativo e não durante períodos de senescência, como no
inverno. No Organossolo drenado do presente estudo, a redução na emissão de CO2
não teria ocorrido em função da senescência da vegetação, mas por alteração do
metabolismo, na ausência do excesso de água no solo.
Com exceção da redução do teor de carbono, não houve diferenças
marcantes nas características químicas e físicas do solo durante os dois anos de
drenagem. Embora a taxa média de subsidência tenha sido de 2,5 e de 5,6 cm no
primeiro e no segundo ano, respectivamente, a densidade do solo não foi alterada
(Tabela 1). Esta taxa de subsidência foi considerada no cálculo do estoque de
carbono do solo no final do experimento, tendo-se utilizado a espessura de 14,4 cm
para o horizonte H1.
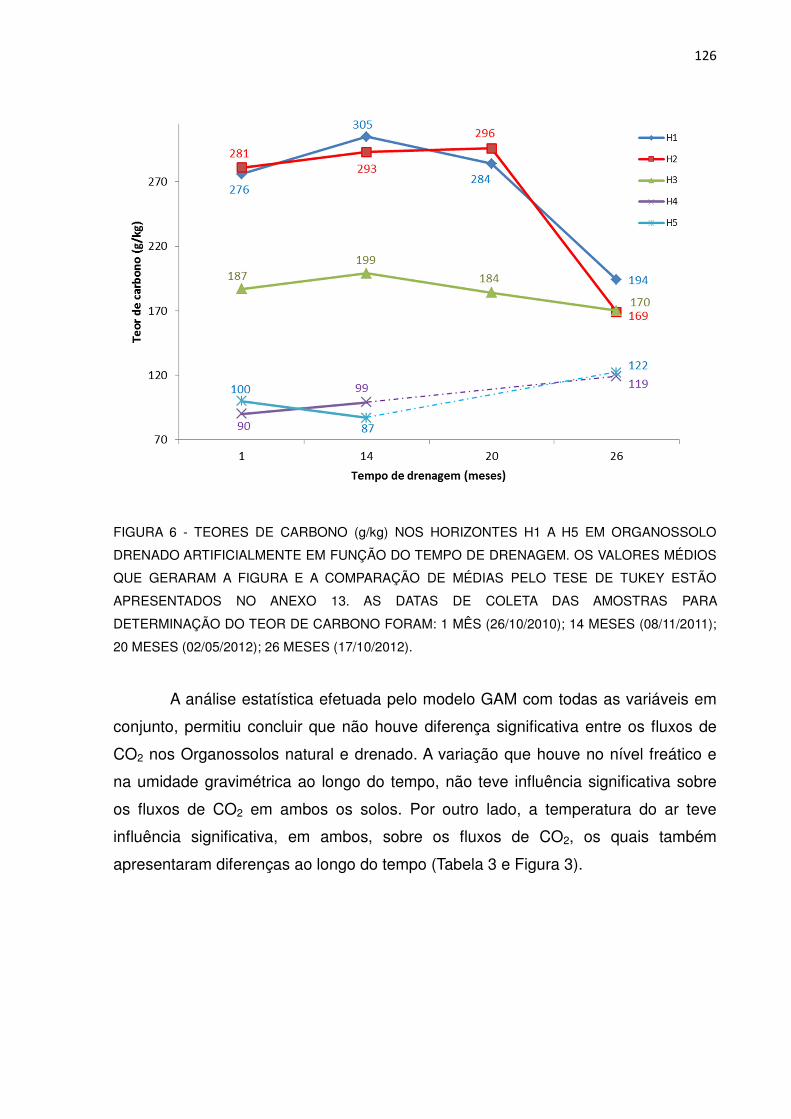
126
FIGURA 6 - TEORES DE CARBONO (g/kg) NOS HORIZONTES H1 A H5 EM ORGANOSSOLO
DRENADO ARTIFICIALMENTE EM FUNÇÃO DO TEMPO DE DRENAGEM. OS VALORES MÉDIOS
QUE GERARAM A FIGURA E A COMPARAÇÃO DE MÉDIAS PELO TESE DE TUKEY ESTÃO
APRESENTADOS NO ANEXO 13. AS DATAS DE COLETA DAS AMOSTRAS PARA
DETERMINAÇÃO DO TEOR DE CARBONO FORAM: 1 MÊS (26/10/2010); 14 MESES (08/11/2011);
20 MESES (02/05/2012); 26 MESES (17/10/2012).
A análise estatística efetuada pelo modelo GAM com todas as variáveis em
conjunto, permitiu concluir que não houve diferença significativa entre os fluxos de
CO2 nos Organossolos natural e drenado. A variação que houve no nível freático e
na umidade gravimétrica ao longo do tempo, não teve influência significativa sobre
os fluxos de CO2 em ambos os solos. Por outro lado, a temperatura do ar teve
influência significativa, em ambos, sobre os fluxos de CO2, os quais também
apresentaram diferenças ao longo do tempo (Tabela 3 e Figura 3).

127
TABELA 3 - ANÁLISE ESTATÍSTICA DO EFEITO DA DRENAGEM E DOS FATORES PEDOLÓGICOS E METEOROLÓGICOS SOBRE O EFLUXO DE CO2, PELO MODELO ADITIVO GENERALIZADO – GAM
Efeitos paramétricos p valor Tratamento (solo natural e drenado) 0,248 ns Nível freático (Componente principal 1) 0,492 ns Temperatura do ar (Componente principal 2)(1) 1,77 x 10-7 *** Temperatura do ar + precipitação acumulada (Componente principal 3)(2) 0,093 ns Efeitos não paramétricos p valor Tempo(3) (solo drenado) 8,35 x 10-10 *** Tempo(4) (solo natural) 0,00014 ***
*** - altamente significativo (p < 0,001); ** - significativo (p < 0,01); ns - não significativo; (1) Temperatura máxima e mínima do ar das 9 às 12 horas no momento da coleta + temperatura média e mínima do ar durante o período de 1, 3 e 5 dias que antecederam as coletas. (2) Temperatura máxima e mínima do ar das 9 às 12 horas no momento da coleta + temperatura média e mínima do ar durante o período de 3 e 5 dias que antecederam as coletas + precipitação acumulada no período das 9 às 12 horas no dia das coletas. (3) Número de dias a partir da drenagem. (4) Número de dias a partir do início do experimento.
4.3.3 Fluxo acumulado de dióxido de carbono
No Organossolo natural, no primeiro ano, a emissão acumulada variou de
8.678 kg CO2 ha-1 ano-1 no inverno a 17.334 kg CO2 ha-1 ano-1 no verão e de 9618 a
18.535 kg CO2 ha-1 ano-1, respectivamente, no inverno e verão do segundo ano
(Figura 7), reforçando a influência da temperatura.
A diferença estatística significativa nos fluxos acumulados de dióxido de
carbono ocorreu exatamente entre o verão e o inverno em ambos os anos.
A emissão acumulada total anual de dióxido de carbono foi de 49,4 Mg ha-1 e
de 56,2 Mg ha-1, no primeiro e segundo ano, respectivamente.
No Organossolo drenado, no primeiro ano a emissão acumulada de dióxido
de carbono variou de 11.096 kg ha-1 no inverno a 20.806 kg ha-1 no verão e de 7.207
kg ha-1 no inverno a 20.427 kg ha-1 na primavera do segundo ano (Figura 7).
No primeiro ano o fluxo acumulado de CO2 no verão foi estatisticamente
igual ao fluxo de primavera e diferente dos fluxos de outono e de inverno. No
segundo ano, embora o fluxo de primavera tenha sido superior ao do verão, não
houve diferença significativa entre eles. Neste ano o fluxo de primavera é que diferiu
dos fluxos de outono e de inverno.
A emissão total acumulada anual no primeiro e no segundo ano foi de 59,7
Mg CO2 ha-1 e de 55,6 Mg CO2 ha-1, respectivamente, (Figura 7), ou seja, houve
redução na emissão total no segundo ano.
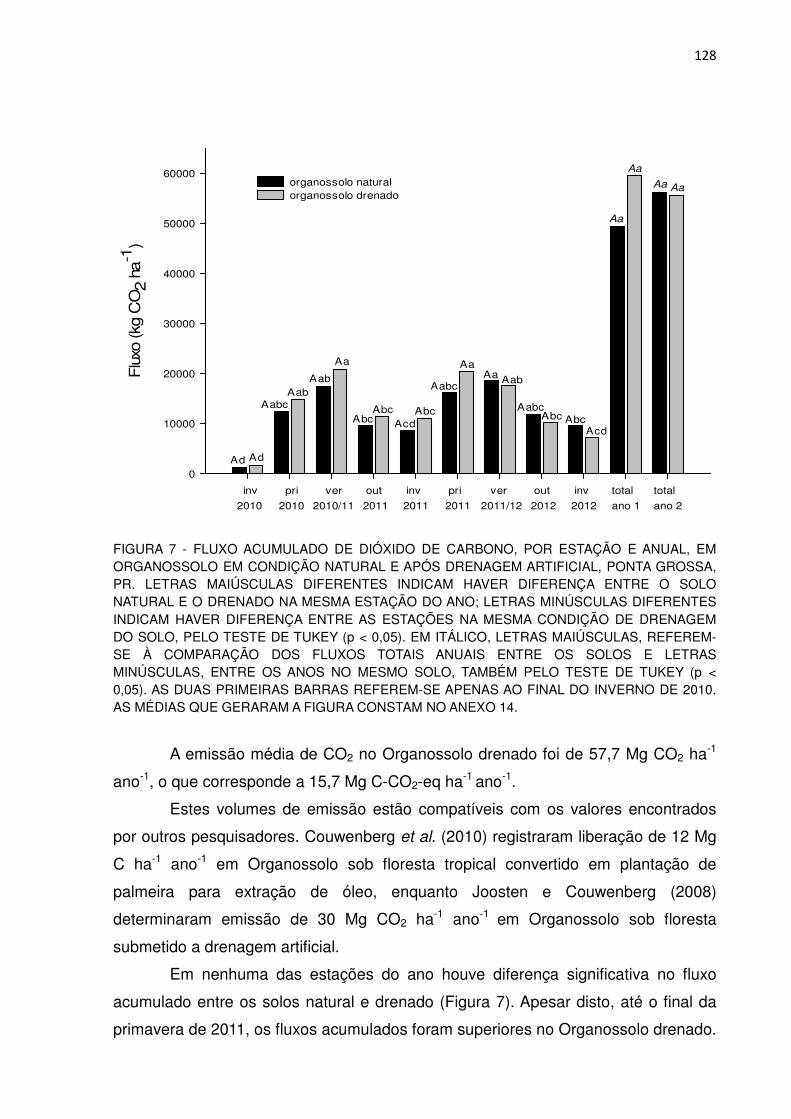
128
Aa
Aa
Aa Aa
inv
2010
pri
2010
ver
2010/11
out
2011
inv
2011
pri
2011
ver
2011/12
out
2012
inv
2012
total
ano 1
total
ano 2
Flu
xo (k
g C
O2
ha-1
)
0
10000
20000
30000
40000
50000
60000organossolo natural organossolo drenado
Ad Ad
AabcAab
Aab
Aa
AbcAbc
AcdAbc
Aabc
AaAa Aab
AabcAbc Abc
Acd
Aa
Aa
Aa Aa
FIGURA 7 - FLUXO ACUMULADO DE DIÓXIDO DE CARBONO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO EM CONDIÇÃO NATURAL E APÓS DRENAGEM ARTIFICIAL, PONTA GROSSA, PR. LETRAS MAIÚSCULAS DIFERENTES INDICAM HAVER DIFERENÇA ENTRE O SOLO NATURAL E O DRENADO NA MESMA ESTAÇÃO DO ANO; LETRAS MINÚSCULAS DIFERENTES INDICAM HAVER DIFERENÇA ENTRE AS ESTAÇÕES NA MESMA CONDIÇÃO DE DRENAGEM DO SOLO, PELO TESTE DE TUKEY (p < 0,05). EM ITÁLICO, LETRAS MAIÚSCULAS, REFEREM-SE À COMPARAÇÃO DOS FLUXOS TOTAIS ANUAIS ENTRE OS SOLOS E LETRAS MINÚSCULAS, ENTRE OS ANOS NO MESMO SOLO, TAMBÉM PELO TESTE DE TUKEY (p < 0,05). AS DUAS PRIMEIRAS BARRAS REFEREM-SE APENAS AO FINAL DO INVERNO DE 2010. AS MÉDIAS QUE GERARAM A FIGURA CONSTAM NO ANEXO 14.
A emissão média de CO2 no Organossolo drenado foi de 57,7 Mg CO2 ha-1
ano-1, o que corresponde a 15,7 Mg C-CO2-eq ha-1 ano-1.
Estes volumes de emissão estão compatíveis com os valores encontrados
por outros pesquisadores. Couwenberg et al. (2010) registraram liberação de 12 Mg
C ha-1 ano-1 em Organossolo sob floresta tropical convertido em plantação de
palmeira para extração de óleo, enquanto Joosten e Couwenberg (2008)
determinaram emissão de 30 Mg CO2 ha-1 ano-1 em Organossolo sob floresta
submetido a drenagem artificial.
Em nenhuma das estações do ano houve diferença significativa no fluxo
acumulado entre os solos natural e drenado (Figura 7). Apesar disto, até o final da
primavera de 2011, os fluxos acumulados foram superiores no Organossolo drenado.

129
A partir do verão de 2012, no entanto, os fluxos acumulados no Organossolo
drenado foram menores do que no natural, provavelmente em função da redução
nos teores de carbono no solo (Figura 6), a partir do início do verão de 2012.
4.3.4 Efeito da drenagem sobre o estoque de carbono no solo
Embora as emissões acumuladas de CO2 no Organossolo natural e no
drenado tenham sido muito próximas, o drenado sofreu redução no estoque de
carbono porque a decomposição da matéria orgânica foi maior do que a
acumulação, enquanto no natural o estoque se manteve constante, pois não houve
perdas.
A comparação dos estoques de carbono do solo no início do experimento
(581,13 Mg C ha-1) e após dois anos de drenagem (543,35 Mg C ha-1) indicou que
houve uma redução de 37,8 Mg C ha-1 (6,5%), o que corresponde a perda de 18,9
Mg C ha-1 ano-1.
Considerando que a emissão média de CO2 no Organossolo drenado foi de
15,7 Mg C-CO2-eq ha-1 ano-1, e que neste valor não está computado apenas o
dióxido de carbono proveniente da decomposição da matéria orgânica do solo,
deduz-se que parte do carbono do estoque do solo foi perdido de outras formas.
O aumento do teor de carbono nos horizontes H4 e H5 (Figura 6) legitima a
lixiviação de carbono dos horizontes H1, H2 e H3. A elevação da quantidade de
carbono no horizonte H4 de 90 para 119 g kg-1 e de 100 para 122 g kg-1 no H5,
comprova que parte do carbono foi removido e transportado da superfície e
depositado nos mesmos. Deve-se ter em conta que uma parte do carbono eluviado
possivelmente foi perdida para o freático, saindo do sistema.
A elevada porosidade total, a baixa densidade e a rápida permeabilidade do
solo nos horizontes H1, principalmente, e H2 (Tabela 1), associada a considerável
precipitação hídrica que ocorreu ao longo do experimento, justificam a remoção de
formas solúveis de carbono dos horizontes superficiais para os subsuperficiais e
para fora do sistema.
Buffam et al. (2001) e Marques et al. (2012) também sugeriram exportação
de carbono via solução do solo para as áreas de drenagem natural. O rápido
escorrimento superficial que ocorre durante a precipitação induz o transporte lateral

130
de carbono e a liberação de matéria orgânica dissolvida nos canais de drenagem
natural (WOHLFART et al., 2012; COOPER et al., 2007).
Por outro lado, a maior densidade, a menor porosidade total e a menor
permeabilidade nos horizontes H4 e H5, pelo menos em relação ao horizonte H1,
além do maior conteúdo de argila no horizonte H4, facilitaram a deposição de
carbono nos mesmos.
Marques et al. (2012) comprovaram que quanto maior é a porosidade total,
menor a densidade e mais rápida a permeabilidade do solo, maior é a quantidade de
carbono dissolvido na solução do solo, pois o aumento na macroporosidade do solo
reduz o contato de solutos orgânicos com a matriz do solo, predispondo a maiores
perdas de matéria orgânica dissolvida. Em contarpartida, Johnson et al. (2000)
confirmaram que em profundidade ocorre aumento na adsorção do carbono orgânico
dissolvido, devido a maior sorção do mesmo pela matriz do solo. Marques et al.
(2012) referiram-se à redução do transporte de carbono orgânico dissolvido em
profundidade, em função do aumento da densidade e da diminuição da
macroporosidade do solo.
Kalbitz et al. (2000) afirmaram que o maior conteúdo de matéria orgânica no
solo e o aumento no fluxo de água elevam a quantidade de carbono orgânico
dissolvido. Wohlfart et al. (2012) e Moore e Turunen (2004), trabalhando com
Organossolo, também encontraram relação positiva entre o teor de carbono orgânico
total e a quantidade de carbono orgânico dissolvido. Fiedler et al. (2008) afirmam
que em Organossolos com 65 cm de espessura e teor de carbono entre 397 e 498 g
kg-1, a quantidade de carbono orgânico dissolvido representa 35% do carbono
orgânico total.
Supôs-se que no Organossolo da presente pesquisa ocorreram processos
similares aos expostos acima.
4.3.5 Efeito da drenagem sobre potencial de aquecimento global
No Organossolo natural do presente trabalho houve consumo de 75,0 kg C-
CO2-eq ha-1 ano-1 de N2O, emissão de 1.136,8 kg C-CO2-eq ha-1 ano-1 de CH4 e
emissão de 14.391,4 kg C-CO2-eq ha-1 ano-1, resultando em uma emissão líquida de
15,5 Mg de C-CO2-eq ha-1 ano-1.

131
No Organossolo drenado houve emissão de 134,2 kg C-CO2-eq ha-1 ano-1 de
N2O, consumo de 70,0 kg C-CO2-eq ha-1 ano-1 de CH4 e emissão de 15.712,1 kg C-
CO2-eq ha-1 ano-1, o que resultou em uma emissão líquida de 15,78 Mg C-CO2-eq
ha-1 ano-1.
Entretanto, embora as emissões de CO2 do Organossolo natural e do
drenado tenham sido muito próximas, apenas o drenado sofreu redução no estoque
de carbono no solo.
No início do experimento o estoque de carbono no solo até a profundidade
de um metro foi de 581,13 Mg ha-1. Dentro de um período de 26 meses após a
drenagem o estoque reduziu-se para 543,35 Mg ha-1, o que significa que foram
perdidos 37,78 Mg C ha-1. Fazendo-se o balanço entre as entradas e as saídas de
carbono conclui-se que a drenagem aumentou o potencial de aquecimento global
em 17,89 Mg C-CO2-eq ha-1 ano-1 (Tabela 4).
TABELA 4 - BALANÇO DE CARBONO EM ORGANOSSOLO DRENADO SUA E RELAÇÃO COM O POTENCIAL DE AQUECIMENTO GLOBAL - PAG
Alterações devidas à drenagem Quantidade de carbono Redução do estoque de carbono no solo 18,89 Mg C ha-1 ano-1 Emissão de N2O 0,21 Mg C-CO2-eq ha-1 ano-1 Consumo de CH4 1,21 Mg C-CO2-eq ha-1 ano-1 Potencial de Aquecimento Global - PAG 17,89 Mg C-CO2-eq ha-1 ano-1
Percebe-se que a emissão de dióxido de carbono, originada pela
decomposição da matéria orgânica do solo, foi a principal causa da elevação do
potencial de aquecimento global. Portanto, é um grande equívoco drenar
Organossolos visando reduzir a emissão de metano, e/ou aumentar o consumo do
mesmo, uma vez que, em Organossolo drenado a emissão anual de dióxido de
carbono foi 224 vezes maior do que o consumo de CH4 expresso em CO2-eq.
Vários autores registraram elevação no potencial de aquecimento global em
função da retirada do excesso de água. A drenagem incrementou o potencial de
aquecimento de um Organossolo no sul da Suécia transformando-o em uma fonte
de 0,4 kg CO2-eq m-2 ano-1 (VON ARNOLD et al., 2005), enquanto Furukawa et al
(2005) trabalhando com Organossolos constataram que o rebaixamento do freático a
apenas 10 cm da superfície incrementou em 51 mg CO2-eq m-2 h-1 o potencial de
aquecimento global (FURUKAWA et al., 2005). A drenagem também provocou um
incremento de 4,3 Mg CO2-eq ha-1 ano-1 no potencial de aquecimento global de um
Organossolo na Escócia (YAMULKI et al., 2012).

132
Outros estudos mostram a influência decisiva do dióxido de carbono no
potencial de aquecimento global. A emissão de CO2 determinou o potencial de
aquecimento global contribuindo com 75 a 98%, conforme o manejo adotado em
Organossolos na região central da Escócia (YAMULKI et al., 2012). Em comparação
ao metano e ao óxido nitroso a emissão de CO2 em Organossolos e Espodossolos
teve um papel dominante sobre o potencial de aquecimento global, cuja participação
oscilou entre 71 e 96% (CARTER et al., 2012).
4.3.6 Considerações adicionais
No Organossolo natural a concentração de carbono orgânico nos diversos
horizontes entre 0 e 100 cm variou de 90 a 281 g kg-1 (Tabela 1), conferindo-lhe um
estoque total de 581 Mg ha-1, até a profundidade de um metro. Este montante, torna
irrelevante a quantidade de metano por ele emitida, de acordo com o discutido no
Capítulo 1. A baixa densidade, a alta porosidade e a localização do solo na
paisagem atribuíram ao mesmo uma capacidade de armazenar 8,6 milhões de litros
de água por hectare até a profundidade de 1 metro, o que equivale a uma lâmina de
860 mm, demonstrando a relevante contribuição deste solo como reserva hídrica
para recarga de córregos e rios.
A drenagem, na intensidade em que foi praticada, provocou uma perda de
37,8 Mg C ha-1 no estoque de carbono do solo, em apenas dois anos, e embora
tenha ocasionado o consumo de um pequeno volume de metano, causou a emissão
de um volume muito maior de dióxido de carbono e uma pequena quantidade de
N2O. Entretanto, não se deve subestimar o reduzido volume de óxido nitroso emitido,
pois, conforme indicado no Capítulo 2, a tendência é que ocorra aumento nas
emissões do mesmo, com o passar do tempo, em função da drenagem.
Se a drenagem tivesse sido mais drástica, com drenos mais profundos, por
exemplo, e as emissões tivessem sido determinadas por um período maior do que
26 meses, possivelmente a redução do estoque de carbono do solo, bem como a
elevação do potencial de aquecimento global, teriam sido superiores.
Estas informações dispensam maiores argumentações para justificar que
Organossolos sob estepe hidrófila não devam ser drenados, mas sim mantidos com
a vegetação natural.

133
4.4 CONCLUSÕES
a) Em dois anos o estoque de carbono do Organossolo drenado reduziu em
37,8 Mg C ha-1, o que elevou o potencial de aquecimento global em 17,9 Mg C-CO2-
eq ha-1 ano-1.
b) A temperatura do ar apresentou correlação positiva com os fluxos de CO2
no Organossolo natural e no drenado, tendo as maiores emissões ocorrido durante
os períodos mais quentes.
c) A precipitação teve menor efeito sobre os fluxos de CO2 do que a
temperatura do ar em ambos os solos. Isto se deve à constante saturação de água
no Organossolo natural e à alta permeabilidade no Organossolo drenado.

134
REFERÊNCIAS
ALONGI, D. M.; WATTAYAKORN, G.; PFITZNER, J.: TIRENDI, F.; ZAGORSKIS, I.; BRUNSKILL, G. J.; DAVIDSON, A.; CLOUGH, B. F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in Southern Thailand. Marine Geology, Amsterdam, v. 179, p. 85-103, 2001. BLAIS, A.; LORRAIN, S.; TREMBLAY, A. Greenhouse gas fluxes (CO2, CH4 and N2O) in forests and wetlands of boreal, temperate and tropical regions. In: TREMBLAY, A.; VARFALVY, L.; ROEHM, C.; GARNEAU, M. (Ed.). Greenhouse gas emissions: fluxes and processes. Berlin: Springer-Verlag, 2005. p. 87-127. BUFFAM, I.; GALLOWAY, J. N.; BLUM, L. K.; MCGLATHERY, K. J. A Stormflow/baseflow comparison of dissolved organic matter concentrations and bioavailability in an Appalachian stream. Biogeochemistry, Dordrecht, v. 53, p. 269-306, 2001. CANADELL, J. G.; PATAKI, D. E.; GIFFORD, R.; HOUGHTON, R. A.; LU0, Y.; RAUPACH, R. A.; SMITH, P.; STEFFEN, W. Saturation of the terrestrial carbono sink. In: CANADELL, J. G. D.; PATAKI, D.; PITELKA, L. (Ed.). Terrestrial ecosystems in a changing world. Berlin: Spring-Verlag, 2007. p. 59-78. (IGBP Series). CARTER. M. S.; LARSEN, K. S.; EMMETT, B.; ESTIARTE, M.; FIELD, C.; LEITH, I. D.; LUND, M.; MEIJIDE, A.; MILLS, R. T. E.; NIINEMETS, Ü.; PEÑUELAS, J.; PORTILLO-ESTRADA, M.; SCHMIDT, I. K.; SELSTED, M. B.; SHEPPARD, L. J.; SOWERBY, A.; TIETEMA, A.; BEIER, C. Synthesizing greenhouse gas fluxes across nine European peatlands and shrublands – responses to climatic and environmental changes. Biogeosciences, Goettingen, v. 9, p. 3739-3755, 2012. CLAESSEN, M. E. C. (Org.). Manual de métodos de análises de solo. 2. ed. rev. atual. Rio de Janeiro: EMBRAPA-CNPS, 1997. (Documentos, 1). COOPER, R.; THOSS, V.; WATSON, H. Factors influencing the release of dissolved organic carbon and dissolved forms of nitrogen from a small upland headwater during autumn runoff events. Hydrological Processes, West Sussex, v. 21, p. 622-633, 2007. COUWENBERG, J., DOMMAIN, R.; JOOSTEN, H. Greenhouse gas fluxes from tropical peatlands in South-East Asia. Global Change Biology, West Sussex, v. 16, p. 1715-1732, 2010.

135
CROW, S. E.; WIEDER, R. K. Sources of CO2 emission from a northern peatland: root respiration, exudation and decomposition. Ecology, Ithaca, v. 86, n. 7, p. 1825-1834, 2005. DALAL, R. C.; ALLEN, D E. Greenhouse gas fluxes from natural ecosystems. Australian Journal of Botany, Collingwood, v. 56, p. 369-407, 2008. DAVIDSON, E. A.; BELK, E.; BOONE, R. D. Soil water contente and temperature as independente or confounded factors conttolling soil respiration in a temperate mixed hardwood forest. Global Change Biology, West Sussex, v. 4, p. 217-227, 1998. DENMAN, K. L.; BRASSEUR, G.; CHIDTHAISONG, A.; CIAIS, P.; COX, P. M. Couplings between changes in the climate system and biogeochemistry. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z; MARQUIS, M.; AVERYT, K. B.; TIGNOR, M.; MILLER, H. L. (Ed.). Climate change 2007: the physical science basis: contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge, UK: University Press, 2007. p. 499-587. EASTHOUSE, K. B.; MULDER, J.; CHRISTOPHERSEN, N.; SEIP, H. M. Dissolved organic carbon fractions in soil and stream water during variable hydrological conditions at Birkenes, Southern Norway. Water Resources Research, Hoboken, v. 28, p. 1585-1596, 1992. FIEDLER, S.; HÖLL, B. S.; FREIBAUER, A.; STAHR, K.; DRÖSTER, M. SCHLOTER, M.; JUNGKUNST, H. F. Particulate organic carbon (POC) in relation to other pore water carbono fractions in drained and rewetted fens in Southern Germany. Biogeosciences, Goettingen, v. 5, p. 1615-1623, 2008. FORSTER, P.; RAMASWAMY, V.; ARTAXO, P.; BERNSTEN, T.; BETTS, R.; FAHEY, D. W.; HAYWOOD, J.; LEAN, J.; LOWE, D. C.; MYHRE, G.; NGANGA, J.; PRINN, R.; RAGA, G.; SCHUTZ, M.; Van DORLAND, R. Changes in atmospheric constituents and in radiative forcing. In: SOLOMON, S.; QIN, D.; MANNING, M.; CHEN, Z; MARQUIS, M.; AVERYT, K. B.; TIGNOR, M.; MILLER, H. L. (Ed.). Climate change 2007: the physical science basis: contribution of Working Group 1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Changes. Cambridge, UK: University Press, 2007. p. 129-234. FURUKAWA, Y.; INUBUSHI, K.; ALI, M.; ITANG, A. M.; TSURUTA, H. Effect of changing groundwater levels caused by land-use changes on greenhouse gas fluxes from tropical peat lands. Nutrient Cycling in Agroecosystems, Dordrecht, v. 71, p. 81-91, 2005.

136
HADI, A.; INUBUSHI, K.; PURNOMO, E.; RAZIE, F.; YAMAKAWA, K.; TSURUTA, H. Effects of land-use changes on nitrous oxide (N2O) emissions from tropical peatlands. Chemosphere Global Change Science, Oxford, v. 2, p. 347-358. 2000. HIRANO, T.; SEGAH, H.; HARADA, T.; LIMIN, S.; JUNE, T.; HIRATA, R.; OSAKI, M. Carbon dioxide balance of a tropical peat swamp forest in Kalimantan, Indonesia. Global Change Biology, West Sussex, v. 13, n. 2, p. 412-425, 2007. INUBUSHI, K.; FURUKAWA, Y.; HADI, A.; PURNOMO, E.; TSURUTA, H. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use changes in tropical peatlands located in coastal area of South Kalimantan. Chemosphere, Oxford, v. 52, p. 603-608, 2003. INTERGOVERNMENTAL PANEL OF CLIMATE CHANGE (IPCC). Climate Change 2007: the physical science basis. Cambridge: Cambridge University Press, 2007a. INTERGOVERNMENTAL PANEL OF CLIMATE CHANGE (IPCC). Climate Change 2007: synthesis. Geneve: IPCC, 2007b. (Report IPCC Plenary XXVII, Valencia, Spain, 12-17 November 2007, Working Group contributions to the Fourth Assessment Report). JANTALIA, C. P., SANTOS, H. P., URQUIAGA, S., BODDEY, R. M.; ALVES, B. J. R. Fluxes of nitrous oxide from soil under different crop rotations and tillage systems in the South of Brazil. Nutrient Cycling in Agroecosystems, Dordrecht, v. 82, p. 161-173, 2008. JAUHIAINEN, J.; LIMIN, S.; SILVENNOINEN, H.; VASANDER, H. Carbon dioxide and methane fluxes in drained tropical peat before and after hydrological restoration. Ecology, Ithaca, v. 89, n. 12, p. 3503-3514, 2008. JOHNSON, C. E.; DRISCOLL, C. T.; SICCAMA, T. G.; LIKENS, G. E. Element fluxes and landscape position in a northern hardwood forest watershed ecosystem. Ecosystems, New York, v. 3, p. 159-184, 2000. JOOSTEN, H.; COUWENBERG, J. Peatlands and carbon. In: PARISH, F.; SIRIN, A.; CHARMAN, D.; JOOSTEN, H.; MINAYEVA, T.; SILVIUS, M.; STRINGER, L. (Ed.). Assessment on peatlands, biodiversity and climate change. Kuala Lumpur: Global Environment Centre, 2008. p. 99-117.

137
KALBITZ, K.; SOLINGER, S.; PARK, J. H.; MICHALZIK, B.; MATZNER, E. Controls on the dynamics of dissolved organic matter in soils: a review. Soil Science, Philadelphia, v. 165, n. 4, p. 277-307, 2000. KOH, H. S.; OCHS, C. A.; YU, K. W. Hydrologic gradient and vegetation controls on CH4 and CO2 fluxes in a spring-fed forested wetland. Hydrobiologia, Dordrecht, v. 630, p. 271-286, 2009. LOHILA, A.; AURELA, M.; REGINA, K.; LAURILA, T. Soil and total ecosystem respiration in agricultural fields: effects of soil and crop type. Plant Soil, The Hague, v. 251, p. 303-317, 2003. MALJANEN, M.; MARTIKAINEN, P. J.; AALTONEN, H.; SILVOLA, J. Short-term variation in fluxes of carbon dioxide, nitrous oxide and methane in cultivated and forested organic boreal soils. Soil Biology & Biochemistry, Oxford, v. 34, p. 577-584, 2002. MALJANEN, M.; KOMULAINEN, V. M.; HYTONEN, J.; MARTIKAINEN, P. J.; LAINE, J. Carbon dioxide, nitrous oxide and methane dynamics in boreal organic agricultural soils with different soil characteristics. Soil Biology & Biochemistry, Oxford, v. 36, p. 1801-1808, 2004. MARQUES, J. D. O.; LUIZÃO, F. J.; TEIXEIRA, W. G.; FERREIRA, S. J. F. Variações do carbono orgânico dissolvido e de atributos físicos do solo sob diferentes sistemas de uso da terra na Amazônia Central. Revista Brasileira de Ciência do Solo, Campinas, v. 36, p. 611-622, 2012. MOORE, T. R. Dissolved organic carbon: sources, sinks and fluxes and role in the soil carbono cycle. In: LAL, R.; KIMBLE, J. M.; FOLLETT, R. F.; STEWART, B. A. (Ed.). Soil Processes Carbon Cycle. Boca Raton: CRC press, 1998. p. 281-292. MOORE, T. R.; DALVA, M. The influence of temperature and water-table position on carbon-dioxide and methane emission from laboratory columns of peat land soils. Journal of Soil Science, Oxford, v. 44, p. 641-664, 1993. MOORE, T. R.; DALVA, M. Methane and carbon dioxide exchange potentials of peat soils in aerobic and anaerobic laboratory incubations. Soil Biology & Biochemistry, Oxford, v. 29, p. 1157-1164, 1997. MOORE, T. R.; TURUNEN, J. Carbon accumulation and storage in mineral subsoil beneath peat. Soil Science Society of American Journal, Madison, v. 68, p. 690-

138
696, 2004. R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing, 2011. Disponível em: <http://www.R-project.org/>. Acesso em: 25/05/2012. RAICH, J. W.; POTTER, C. S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles, Hoboken, v. 9, p. 23-36, 1995. REDDY, K. R.; OSBORNE, T. Z.; INGLETT, K. S.; CORSTANJE, R. Soil Organic Matter Loss Rates as a Function of Water Level Relative to Soil Surface. In: REDDY, K. R.; OSBORNE, T. Z.; INGLETT, K. S.; CORSTANJE, R. (Ed.). Influence of Water Levels on Subsidence of Organic Soils in the Upper St. Johns River Basin. Gainesville: Wetland Biogeochemistry Laboratory Soil and Water Science Department University of Florida-IFAS, 2006. p. 36-77. REDDY, K. R.; DELAUNE, R. D. Biogeochemistry of wetlands: science and applications. London: Taylor & Francis Group, 2008. ROWSON, J. G.; WORRALL, F.; EVANS, M. G. Predicting soil respiration from peatlands. Science of the Total Environment, Amsterdam, v. 442, p. 397-404, 2013. SANTOS, H. G. dos; JACOMINE, P. K. T.; ANJOS, L. H. C. dos; OLIVEIRA, V. A. de; OLIVEIRA, J. B. de; COELHO, M. R.; LUMBRERAS, J. F.; CUNHA, T. J. F. (Ed.). Sistema Brasileiro de Classificação de Solos. 2. ed. Rio de Janeiro: Embrapa Solos, 2006. SISTI, C. P. J.; SANTOS, H. P.; KOHHANN, R.; ALVES, B. J. R.; URQUIAGA, S.; BODDEY, R. M. Change in carbon and nitrogen stocks in soil under 13 years of conventional or zero tillage in southern Brazil. Soil and Tillage Research, Amsterdam, v. 76, p. 39-58, 2004. SOIL SURVEY STAFF. Soil survey manual. Washington, DC: U.S. Department of. Agriculture, 1951. (Handbook, No. 18). TAKAHASHI, H.; SHIMADA, S.; IBIE, B.I.; USUP, A.; YUDHA; LIMIN, S.H. Annual changes of water balance and a drought index in a tropical peat swamp forest of Central Kalimantan, Indonesia. In: RIELEY, J. O.; PAGE, S. E.; SETIADI, B. (Ed.). Proceedings of the International Symposium on Tropical Peatland: Peatlands for people: natural resource functions and sustainable management. Jakarta: BPPT,

139
2002. p. 63-67. TAKAHASHI, H.; USUP, A.; HAYASAKA, H.; LIMIN, S. H. Estimation of ground water level in a peat swamp forest as an index of peat/forest fire. In: OSAKI, M. et al. (Ed.). Proceedings of the International Symposium on Land Management and Biodiversity in Southeast Asia. Sapporo: Hokkaiko University. 2003. p. 311-314. VON ARNOLD, K.; NILSSON, M.; HANELL, B.; WESLIEN, P.; KLEMEDTSSON, L. Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biology & Biochemistry, Oxford, v. 37, p. 1059-1071, 2005. WHITING, G. J.; CHANTON, J. P. Greenhouse carbon balance of wetlands: methane emission versus carbon sequestration. Tellus. Series B, Chemical Physical Meteorology, Hoboken, v. 53, p. 521-528, 2001. WOHLFART, T.; EXBRAYAT, J. F.; SCHELDE, K.; CHRISTEN, B.; DALGAARD, T.; FREDE, H. G.; BREUER, L. Spatial distribution of soils determines export of nitrogen and dissolved organic carbon from an intensively managed agricultural landscape. Biogeosciences, Goettingen, v. 9, p. 4513-4525, 2012. YAMULKI, S.; ANDERSON, R.; PEACE, A.; MORISON, J. I. L. Soil CO2, CH4, and N2O fluxes from an afforested lowland raised peatbog in Scotland: implications for drainage and restoration. Biogeosciences Discussions, Goettingen, v. 9, p. 7313-7351, 2012.

140
5 CONSIDERAÇÕES FINAIS
Organossolos, sob condições naturais, a despeito de constituírem-se em
fontes perenes de emissão de metano, possuem elevado potencial de estocar
grandes quantidades de carbono, bem como consumir óxido nitroso, funcionalidades
ecológicas de elevada importância. Abrigam inúmeras espécies da flora e fauna,
algumas endêmicas, armazenam/filtram expressivos volumes de água, contribuindo
para a regularização de fluxos fluviais, além de contribuírem significativamente na
redução de nitrato.
A drenagem dos mesmos, fato não incomum na região, transforma-os em
fontes consideráveis de CO2 e de N2O, o que sabidamente aumenta o potencial de
aquecimento global.
Portanto, estas áreas estão plenamente protegidas na proposição atual da
lei ambiental, onde as mesmas constituem APPs – Áreas de Preservação
Permanente, não devendo ser drenados.
6 RECOMENDAÇÕES
Para futuros trabalhos recomenda-se:
- determinar o potencial redox do solo por um período mínimo de 12 meses,
devido às possíveis pequenas oscilações do freático durante as diferentes estações
do ano;
- determinar a umidade gravimétrica e os teores de nitrato e de amônio, não
só entre 0 a 10 cm, mas em profundidades maiores, que sejam compatíveis com as
espessuras dos horizontes hísticos, devido à variação no conteúdo de carbono e no
teor de água dos mesmos;
- caracterizar a matéria orgânica do solo, relacionando-a com
especificidades fisionômicas e florísticas para melhor compreensão dos fluxos de
gases;
- monitorar a redução do estoque de carbono do solo com maior frequência,
coletando amostras em intervalos de tempo mais curtos;

141
- identificar e caracterizar as comunidades microbianas relacionadas com os
fluxos de CO2, CH4 e de N2O;
- utilizar passarelas para evitar a degradação do solo e da vegetação
durante o esforço amostral intenso e recorrente.

142
ANEXOS
ANEXO 1 - ANÁLISES FÍSICAS E QUÍMICAS DO ORGANOSSOLO
HÁPLICO SÁPRICO TÍPICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE..........................................................
144
ANEXO 2 - PERFIL DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE....
145
ANEXO 3 - CONSTRUÇÃO DOS DRENOS. a) COLOCAÇÃO DE PRANCHÕES DE EUCALIPTO SOBRE O SOLO PARA QUE O TRATOR DE ESTEIRA NÃO ATOLASSE; b) ABERTURA DOS DRENOS COM A RETROESCAVADEIRA; c) DIMENSÕES DOS DRENOS; E d) VISTA DA ÁREA DRENADA......................
145
ANEXO 4 - DATA DAS COLETAS DOS GASES, ESTAÇÃO DO ANO, NÚMERO DE DIAS A PARTIR DO INÍCIO DO EXPERIMENTO, INTERVALO DE DIAS ENTRE AS COLETAS, NÚMERO DA SEMANA DE CADA MÊS E ANO DE ESTUDO...........................
146
ANEXO 5 - CÂMARAS COLETORAS DE AMOSTRAS DE AR. a) BASE DA CÃMARA COM CANALETA PREENCHIDA COM ÁGUA; b) PORÇÃO DA BASE QUE FICA ACIMA (10 cm) E ABAIXO (15 cm) DO SOLO; c) VENTILADOR (PEÇA BRANCA) PARA HOMOGEINIZAR A AMOSTRA DE AR NO INSTANTE DA COLETA; E d) CÂMARA SOBRE A BASE NO MOMENTO DAS COLETAS DE AMOSTRAS DE AR..............................................
147
ANEXO 6 - TEMPERATURA MÉDIA SEMANAL E PRECIPITAÇÃO ACUMULADA SEMANAL ENTRE SETEMBRO DE 2010 E OUTUBRO DE 2012....................................................................
148
ANEXO 7 - FLUXOS DE CH4, N2O E CO2 EM ORGANOSSOLO DRENADO E NATURAL, PONTA GROSSA, PR.........................
150
ANEXO 8 - ALTURA DO NÍVEL FREÁTICO E UMIDADE GRAVIMÉTRICA DO SOLO (NA PROFUNDIDADE DE 0 A 10 cm) EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR................................
151
ANEXO 9 - POTENCIAL REDOX (mV) POR COLETA, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, EM ORGANOSSOLO DRENADO ARTIFICIALMENTE E NATURAL
152
ANEXO 10 - FLUXO ACUMULADO DE METANO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR................................
153
ANEXO 11 - TEOR DE N-NH4+ (a) E N-NO3
- (b) NA PROFUNDIDADE DE 0 A 10 cm, EM CADA COLETA, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICISLMRNTE...............................................
154
ANEXO 12 - FLUXO ACUMULADO DE ÓXIDO NITROSO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR................................
155
ANEXO 13 - TEORES DE CARBONO (g/kg) NOS HORIZONTES H1 A H5 NO ORGANOSSOLO DRENADO ARTIFICIALMENTE, EM FUNÇÃO DO TEMPO DE DRENAGEM......................................
155
ANEXO 14 - FLUXO ACUMULADO DE DIÓXIDO DE CARBONO, POR

143
ESTAÇÃO E ANUAL, EM ORGANOSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR.............
155

144
ANEXO 1 - ANÁLISES FÍSICAS E QUÍMICAS DO ORGANOSSOLO HÁPLICO SÁPRICO TÍPICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE
Horizonte Composição Granulométrica g/kg
Densidade do Solo - kg/dm3
Porosidade Volumétrica Total - %
Sím-bolo
Profundidade (cm)
Areia Silte Argila Tempo zero 26 meses
H1 0-20 19 (4-46) 3 854(834-892)3 127(100-160)3 0,15(0,10-0,17)9 0,16(0,15-0,17)4 95(93-99)9 H2 20-40 104(10-324)4 776(516-896)4 120(80-160) 4 0,21(0,20-0,22)9 0,22(0,22-0,23)4 92(91-94)9 H3 40-60 251(80-464)4 630(456-800)4 120(80-180) 4 0,47(0,39-0,53)9 0,44(0,42-0,45)4 82(79-87)9 H4 60-80 353(260-469)4 475(324-592)4 165(120-180)4 0,58(0,55-0,61)9 0,51(0,51-0,52)4 78(77-80)9 H5 80-100 214 (1) 666 (1) 120 (1) 0,50(0,48-0,52)3 0,52(0,50-0,61)4 81(81)3 Cg1 100-110 114 (1) 646 (1) 240 (1) - - - Hori-zonte
Permeabilidade Saturada - cm/h
pH H2O (1:2,5)
Complexo Sortivo - cmolc/kg
Tempo zero 26 meses Ca+2 + Mg+2 K+ Na+ H1 72 (4-384) 9 276(204-399)4 5,0 (4,7-5,3) 5 1,92 (1,0-2,6) 5 0,39 (0,13-0,49) 5 0,27 (0,12-0,52) 5 H2 25 (2-53) 9 7 (3-12) 4 5,3 (5,2-5,5) 5 0,48 (0,1-1,0) 5 0,07 (0,01-0,13) 5 0,03 (0,01-0,09) 5 H3 13 (1-27) 9 28 (0,9-64) 4 5,3 (5,0-5,6) 5 0,24 (0,0-0,6) 5 0,02 (0,01-0,03) 5 0,01 (0,01-0,01) 5 H4 36 (0,5-112) 9 12 (2-18) 4 5,1 (5,0-5,3) 4 0,17 (0,0-0,2) 4 0,02 (0,01-0,02) 4 0,01 (0,01-0,01) 4 H5 21 (0,5-60) 3 14 (2-30) 4 5,0 (4,9-5,0) 2 0,10 (0,0-0,2) 2 0,04 (0,01-0,06) 2 0,01 (0,01-0,01) 2 Cg1 - 5,2 (5,0-5,3) 2 0,15 (0,1-0,2) 2 0,03 (0,02-0,04) 2 0,01 (0,01-0,01) 2 Hori-zonte
Complexo Sortivo - cmolc/kg Valor V sat. por Bases - %
100.Al+3/S + Al+3 - % Valor S Al+3 H+ Valor T
H1 2,56 (1,2-3,2)5 3,48 (2,7-4,1)5 46,2 (39,7-50,0)5 52,2 (46,2-55,3)5 4,80 (2,0-6,0) 5 58 (48-77) 5 H2 0,56 (0,1-1,1)5 3,34 (2,6-4,2)5 49,9 (35,0-62,3)5 53,8 (37,7-66,1)5 0,93 (0,27-2,0) 5 87 (77-96) 5 H3 0,24 (0,1-0,6)5 2,90 (2,3-3,5)5 37,2 (24,1-57,9)5 40,3 (24,7-62,0)5 0,68 (0,0-1,0) 5 93 (85-100)5 H4 0,17 (0,0-0,2)4 2,75 (2,3-3,5)4 22,6 (18,7-27,2)4 25,4 (21,1-29,9)4 0,62 (0,0-1,0) 4 96 (93-100)4 H5 0,15 (0,0-0,3)2 3,05 (2,2-3,9)2 25,0 (24,6-25,4)2 28,2 (27,6-28,8)2 0,50 (0,0-1,0) 2 97 (93-100)2 Cg1 0,15 (0,1-0,2)2 2,15 (1,8-2,5)2 16,7 (16,0-17,3)2 19,0 (18,7-19,2) 2 1,00 (1,00-1,00) 2 94 (93-95) 2 Hori-zonte
Carbono (orgânico) - g/kg N - g/kg C/N Tempo zero 26 meses
H1 276,32 (266,60 - 285,20) 5 194,00 (179,20 - 204,00) 3 20,40 (18,10 - 22,40) 5 14 (13 - 15) 5 H2 281,40 (235,90 - 307,40) 5 169,30 (100,70 - 207,90) 3 19,06 (14,70 - 21,60) 5 15 (14 - 16) 5 H3 186,96 (103,40 - 270,30) 5 170,40 (101,20 - 205,40) 3 11,32 (5,10 - 18,30) 5 18 (15 - 21) 5 H4 90,43 (85,50 - 97,40) 4 119,20 (99,80 - 131,30) 3 3,95 (3,30 - 4,90) 4 23 (19 - 26) 4 H5 99,50 (81,4 - 117,6) 2 122,10 (103,40 - 134,80) 3 3,90 (3,60 - 4,20) 2 26 (23 - 28) 2 Cg1 52,75 (45,5 - 60,0) 2 - 2,50 (2,30 - 2,70) 2 21 (20 - 22) 2
O primeiro número antes do parêntese refere-se a média; os números entre parênteses correspondem aos valores mínimos e máximos, respectivamente e o número após o parêntese indica o número de observações que compôs a média. Valor S = soma de bases. Valor T = capacidade de troca de cátions.

145
ANEXO 2 - PERFIL DO ORGANOSSOLO HÁPLICO SÁPRICO RELEVO PLANO FASE ESTEPE HIDRÓFILA DE ALTITUDE
ANEXO 3 - CONSTRUÇÃO DOS DRENOS. a) COLOCAÇÃO DE PRANCHÕES DE EUCALIPTO SOBRE O SOLO PARA QUE O TRATOR DE ESTEIRA NÃO ATOLASSE; b) ABERTURA DOS DRENOS COM A RETROESCAVADEIRA; c) DIMENSÕES DOS DRENOS; E d) VISTA DA ÁREA DRENADA
d c
b a
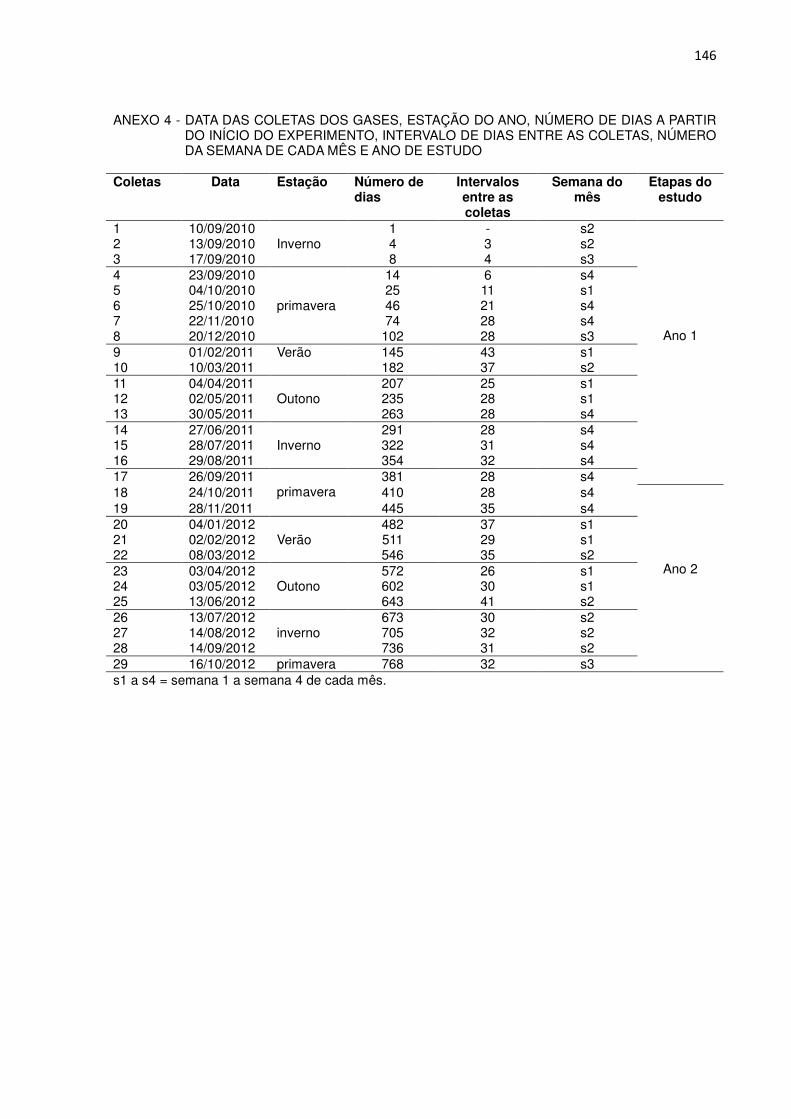
146
ANEXO 4 - DATA DAS COLETAS DOS GASES, ESTAÇÃO DO ANO, NÚMERO DE DIAS A PARTIR DO INÍCIO DO EXPERIMENTO, INTERVALO DE DIAS ENTRE AS COLETAS, NÚMERO DA SEMANA DE CADA MÊS E ANO DE ESTUDO
Coletas Data Estação Número de
dias Intervalos entre as coletas
Semana do mês
Etapas do estudo
1 10/09/2010 Inverno
1 - s2
Ano 1
2 13/09/2010 4 3 s2 3 17/09/2010 8 4 s3 4 23/09/2010
primavera
14 6 s4 5 04/10/2010 25 11 s1 6 25/10/2010 46 21 s4 7 22/11/2010 74 28 s4 8 20/12/2010 102 28 s3 9 01/02/2011 Verão 145 43 s1 10 10/03/2011 182 37 s2 11 04/04/2011
Outono 207 25 s1
12 02/05/2011 235 28 s1 13 30/05/2011 263 28 s4 14 27/06/2011
Inverno 291 28 s4
15 28/07/2011 322 31 s4 16 29/08/2011 354 32 s4 17 26/09/2011
primavera 381 28 s4
18 24/10/2011 410 28 s4
Ano 2
19 28/11/2011 445 35 s4 20 04/01/2012
Verão 482 37 s1
21 02/02/2012 511 29 s1 22 08/03/2012 546 35 s2 23 03/04/2012
Outono 572 26 s1
24 03/05/2012 602 30 s1 25 13/06/2012 643 41 s2 26 13/07/2012
inverno 673 30 s2
27 14/08/2012 705 32 s2 28 14/09/2012 736 31 s2 29 16/10/2012 primavera 768 32 s3 s1 a s4 = semana 1 a semana 4 de cada mês.

147
ANEXO 5 - CÂMARAS COLETORAS DE AMOSTRAS DE AR. a) BASE DA CÂMARA COM CANALETA PREENCHIDA COM ÁGUA; a) PORÇÃO DA BASE QUE FICA ACIMA (10 cm) E ABAIXO (15 cm) DO SOLO; c) VENTILADOR (PEÇA BRANCA) PARA HOMOGENEIZAR A AMOSTRA DE AR NO INSTANTE DA COLETA; E d) CÂMARA SOBRE A BASE NO MOMENTO DAS COLETAS DE AMOSTRAS DE AR
d
b a
c
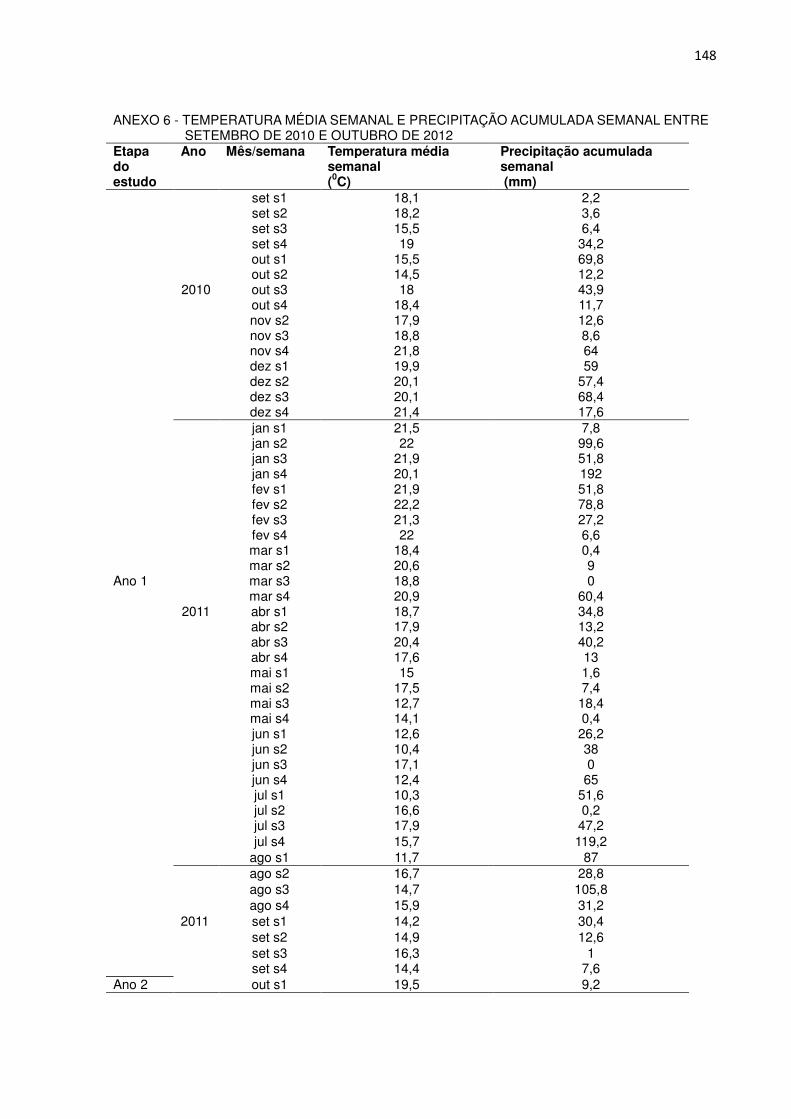
148
ANEXO 6 - TEMPERATURA MÉDIA SEMANAL E PRECIPITAÇÃO ACUMULADA SEMANAL ENTRE SETEMBRO DE 2010 E OUTUBRO DE 2012
Etapa do estudo
Ano Mês/semana Temperatura média semanal (0C)
Precipitação acumulada semanal (mm)
set s1 18,1 2,2 set s2 18,2 3,6 set s3 15,5 6,4 set s4 19 34,2 out s1 15,5 69,8 out s2 14,5 12,2 2010 out s3 18 43,9 out s4 18,4 11,7 nov s2 17,9 12,6 nov s3 18,8 8,6 nov s4 21,8 64 dez s1 19,9 59 dez s2 20,1 57,4 dez s3 20,1 68,4 dez s4 21,4 17,6 jan s1 21,5 7,8 jan s2 22 99,6 jan s3 21,9 51,8 jan s4 20,1 192 fev s1 21,9 51,8 fev s2 22,2 78,8 fev s3 21,3 27,2 fev s4 22 6,6 mar s1 18,4 0,4 mar s2 20,6 9 Ano 1 mar s3 18,8 0 mar s4 20,9 60,4 2011 abr s1 18,7 34,8 abr s2 17,9 13,2 abr s3 20,4 40,2 abr s4 17,6 13 mai s1 15 1,6 mai s2 17,5 7,4 mai s3 12,7 18,4 mai s4 14,1 0,4 jun s1 12,6 26,2 jun s2 10,4 38 jun s3 17,1 0 jun s4 12,4 65 jul s1 10,3 51,6 jul s2 16,6 0,2 jul s3 17,9 47,2
jul s4 15,7 119,2 ago s1 11,7 87 ago s2 16,7 28,8 ago s3 14,7 105,8 ago s4 15,9 31,2 2011 set s1 14,2 30,4 set s2 14,9 12,6 set s3 16,3 1
set s4 14,4 7,6 Ano 2 out s1 19,5 9,2
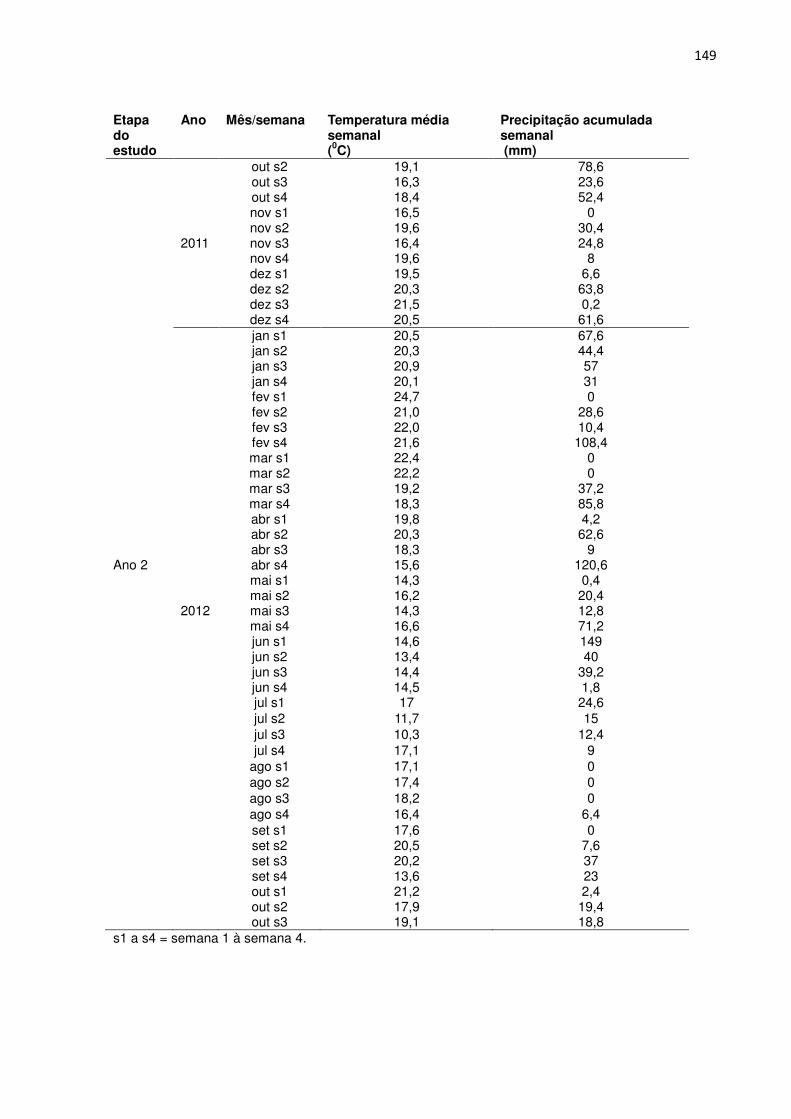
149
Etapa do estudo
Ano Mês/semana Temperatura média semanal (0C)
Precipitação acumulada semanal (mm)
out s2 19,1 78,6 out s3 16,3 23,6 out s4 18,4 52,4 nov s1 16,5 0 nov s2 19,6 30,4 2011 nov s3 16,4 24,8 nov s4 19,6 8 dez s1 19,5 6,6 dez s2 20,3 63,8 dez s3 21,5 0,2 dez s4 20,5 61,6 jan s1 20,5 67,6 jan s2 20,3 44,4 jan s3 20,9 57 jan s4 20,1 31 fev s1 24,7 0 fev s2 21,0 28,6 fev s3 22,0 10,4 fev s4 21,6 108,4 mar s1 22,4 0 mar s2 22,2 0 mar s3 19,2 37,2 mar s4 18,3 85,8 abr s1 19,8 4,2 abr s2 20,3 62,6 abr s3 18,3 9 Ano 2 abr s4 15,6 120,6 mai s1 14,3 0,4 mai s2 16,2 20,4 2012 mai s3 14,3 12,8 mai s4 16,6 71,2 jun s1 14,6 149 jun s2 13,4 40 jun s3 14,4 39,2 jun s4 14,5 1,8 jul s1 17 24,6 jul s2 11,7 15
jul s3 10,3 12,4 jul s4 17,1 9
ago s1 17,1 0 ago s2 17,4 0 ago s3 18,2 0 ago s4 16,4 6,4 set s1 17,6 0
set s2 20,5 7,6 set s3 20,2 37 set s4 13,6 23 out s1 21,2 2,4 out s2 17,9 19,4 out s3 19,1 18,8 s1 a s4 = semana 1 à semana 4.
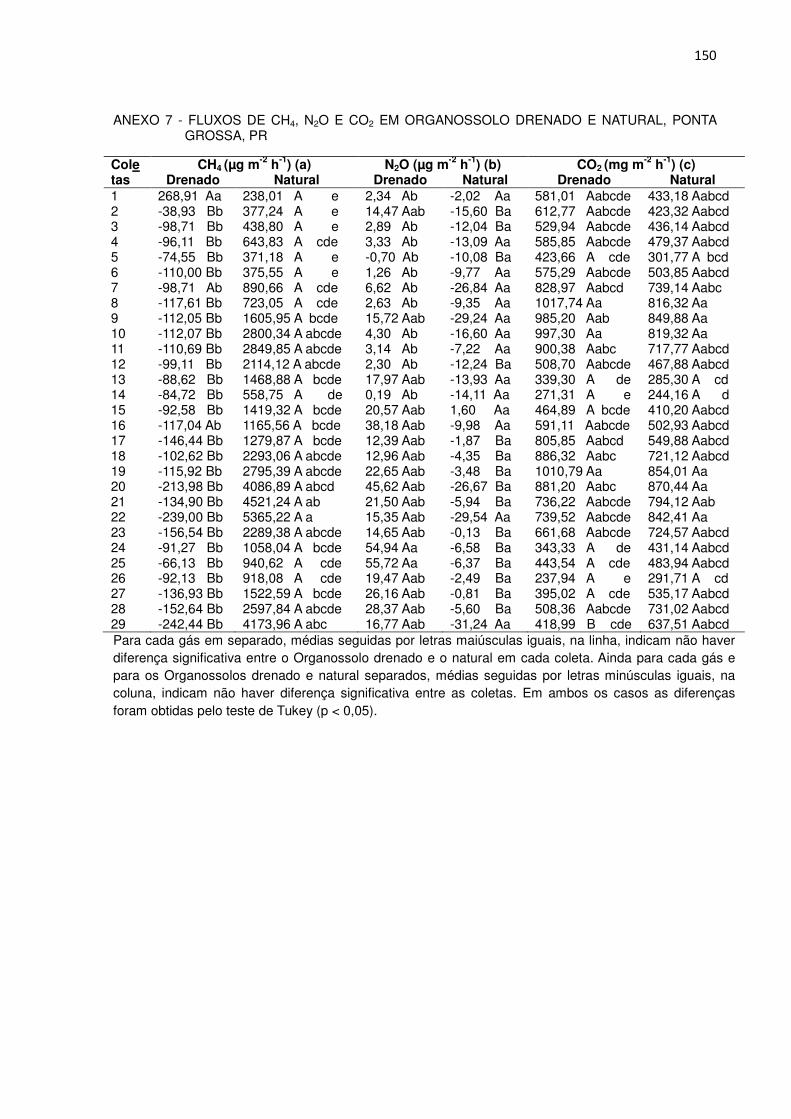
150
ANEXO 7 - FLUXOS DE CH4, N2O E CO2 EM ORGANOSSOLO DRENADO E NATURAL, PONTA GROSSA, PR
Cole tas
CH4 (µg m-2 h-1) (a) N2O (µg m-2 h-1) (b) CO2 (mg m-2 h-1) (c) Drenado Natural Drenado Natural Drenado Natural
1 268,91 Aa 238,01 A e 2,34 Ab -2,02 Aa 581,01 Aabcde 433,18 Aabcd 2 -38,93 Bb 377,24 A e 14,47 Aab -15,60 Ba 612,77 Aabcde 423,32 Aabcd 3 -98,71 Bb 438,80 A e 2,89 Ab -12,04 Ba 529,94 Aabcde 436,14 Aabcd 4 -96,11 Bb 643,83 A cde 3,33 Ab -13,09 Aa 585,85 Aabcde 479,37 Aabcd 5 -74,55 Bb 371,18 A e -0,70 Ab -10,08 Ba 423,66 A cde 301,77 A bcd 6 -110,00 Bb 375,55 A e 1,26 Ab -9,77 Aa 575,29 Aabcde 503,85 Aabcd 7 -98,71 Ab 890,66 A cde 6,62 Ab -26,84 Aa 828,97 Aabcd 739,14 Aabc 8 -117,61 Bb 723,05 A cde 2,63 Ab -9,35 Aa 1017,74 Aa 816,32 Aa 9 -112,05 Bb 1605,95 A bcde 15,72 Aab -29,24 Aa 985,20 Aab 849,88 Aa 10 -112,07 Bb 2800,34 A abcde 4,30 Ab -16,60 Aa 997,30 Aa 819,32 Aa 11 -110,69 Bb 2849,85 A abcde 3,14 Ab -7,22 Aa 900,38 Aabc 717,77 Aabcd 12 -99,11 Bb 2114,12 A abcde 2,30 Ab -12,24 Ba 508,70 Aabcde 467,88 Aabcd 13 -88,62 Bb 1468,88 A bcde 17,97 Aab -13,93 Aa 339,30 A de 285,30 A cd 14 -84,72 Bb 558,75 A de 0,19 Ab -14,11 Aa 271,31 A e 244,16 A d 15 -92,58 Bb 1419,32 A bcde 20,57 Aab 1,60 Aa 464,89 A bcde 410,20 Aabcd 16 -117,04 Ab 1165,56 A bcde 38,18 Aab -9,98 Aa 591,11 Aabcde 502,93 Aabcd 17 -146,44 Bb 1279,87 A bcde 12,39 Aab -1,87 Ba 805,85 Aabcd 549,88 Aabcd 18 -102,62 Bb 2293,06 A abcde 12,96 Aab -4,35 Ba 886,32 Aabc 721,12 Aabcd 19 -115,92 Bb 2795,39 A abcde 22,65 Aab -3,48 Ba 1010,79 Aa 854,01 Aa 20 -213,98 Bb 4086,89 A abcd 45,62 Aab -26,67 Ba 881,20 Aabc 870,44 Aa 21 -134,90 Bb 4521,24 A ab 21,50 Aab -5,94 Ba 736,22 Aabcde 794,12 Aab 22 -239,00 Bb 5365,22 A a 15,35 Aab -29,54 Aa 739,52 Aabcde 842,41 Aa 23 -156,54 Bb 2289,38 A abcde 14,65 Aab -0,13 Ba 661,68 Aabcde 724,57 Aabcd 24 -91,27 Bb 1058,04 A bcde 54,94 Aa -6,58 Ba 343,33 A de 431,14 Aabcd 25 -66,13 Bb 940,62 A cde 55,72 Aa -6,37 Ba 443,54 A cde 483,94 Aabcd 26 -92,13 Bb 918,08 A cde 19,47 Aab -2,49 Ba 237,94 A e 291,71 A cd 27 -136,93 Bb 1522,59 A bcde 26,16 Aab -0,81 Ba 395,02 A cde 535,17 Aabcd 28 -152,64 Bb 2597,84 A abcde 28,37 Aab -5,60 Ba 508,36 Aabcde 731,02 Aabcd 29 -242,44 Bb 4173,96 A abc 16,77 Aab -31,24 Aa 418,99 B cde 637,51 Aabcd Para cada gás em separado, médias seguidas por letras maiúsculas iguais, na linha, indicam não haver diferença significativa entre o Organossolo drenado e o natural em cada coleta. Ainda para cada gás e para os Organossolos drenado e natural separados, médias seguidas por letras minúsculas iguais, na coluna, indicam não haver diferença significativa entre as coletas. Em ambos os casos as diferenças foram obtidas pelo teste de Tukey (p < 0,05).
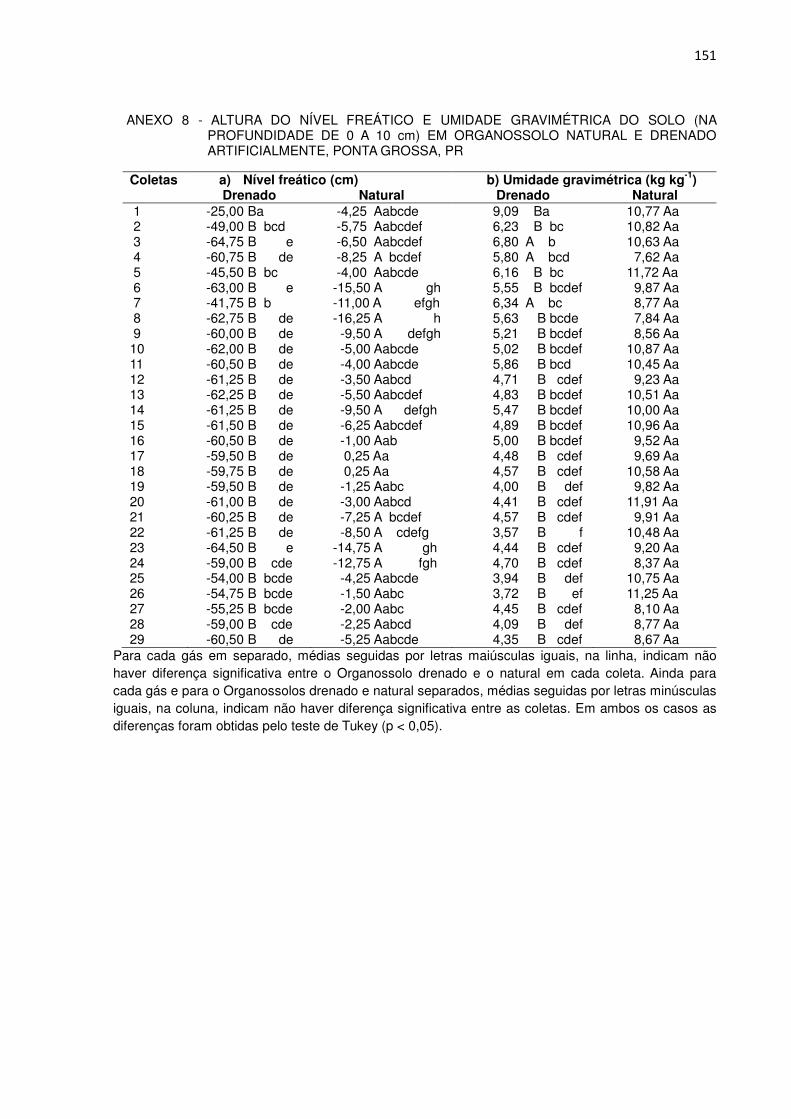
151
ANEXO 8 - ALTURA DO NÍVEL FREÁTICO E UMIDADE GRAVIMÉTRICA DO SOLO (NA PROFUNDIDADE DE 0 A 10 cm) EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR
Coletas a) Nível freático (cm) b) Umidade gravimétrica (kg kg-1) Drenado Natural Drenado Natural
1 -25,00 Ba -4,25 Aabcde 9,09 Ba 10,77 Aa 2 -49,00 B bcd -5,75 Aabcdef 6,23 B bc 10,82 Aa 3 -64,75 B e -6,50 Aabcdef 6,80 A b 10,63 Aa 4 -60,75 B de -8,25 A bcdef 5,80 A bcd 7,62 Aa 5 -45,50 B bc -4,00 Aabcde 6,16 B bc 11,72 Aa 6 -63,00 B e -15,50 A gh 5,55 B bcdef 9,87 Aa 7 -41,75 B b -11,00 A efgh 6,34 A bc 8,77 Aa 8 -62,75 B de -16,25 A h 5,63 B bcde 7,84 Aa 9 -60,00 B de -9,50 A defgh 5,21 B bcdef 8,56 Aa 10 -62,00 B de -5,00 Aabcde 5,02 B bcdef 10,87 Aa 11 -60,50 B de -4,00 Aabcde 5,86 B bcd 10,45 Aa 12 -61,25 B de -3,50 Aabcd 4,71 B cdef 9,23 Aa 13 -62,25 B de -5,50 Aabcdef 4,83 B bcdef 10,51 Aa 14 -61,25 B de -9,50 A defgh 5,47 B bcdef 10,00 Aa 15 -61,50 B de -6,25 Aabcdef 4,89 B bcdef 10,96 Aa 16 -60,50 B de -1,00 Aab 5,00 B bcdef 9,52 Aa 17 -59,50 B de 0,25 Aa 4,48 B cdef 9,69 Aa 18 -59,75 B de 0,25 Aa 4,57 B cdef 10,58 Aa 19 -59,50 B de -1,25 Aabc 4,00 B def 9,82 Aa 20 -61,00 B de -3,00 Aabcd 4,41 B cdef 11,91 Aa 21 -60,25 B de -7,25 A bcdef 4,57 B cdef 9,91 Aa 22 -61,25 B de -8,50 A cdefg 3,57 B f 10,48 Aa 23 -64,50 B e -14,75 A gh 4,44 B cdef 9,20 Aa 24 -59,00 B cde -12,75 A fgh 4,70 B cdef 8,37 Aa 25 -54,00 B bcde -4,25 Aabcde 3,94 B def 10,75 Aa 26 -54,75 B bcde -1,50 Aabc 3,72 B ef 11,25 Aa 27 -55,25 B bcde -2,00 Aabc 4,45 B cdef 8,10 Aa 28 -59,00 B cde -2,25 Aabcd 4,09 B def 8,77 Aa 29 -60,50 B de -5,25 Aabcde 4,35 B cdef 8,67 Aa
Para cada gás em separado, médias seguidas por letras maiúsculas iguais, na linha, indicam não haver diferença significativa entre o Organossolo drenado e o natural em cada coleta. Ainda para cada gás e para o Organossolos drenado e natural separados, médias seguidas por letras minúsculas iguais, na coluna, indicam não haver diferença significativa entre as coletas. Em ambos os casos as diferenças foram obtidas pelo teste de Tukey (p < 0,05).
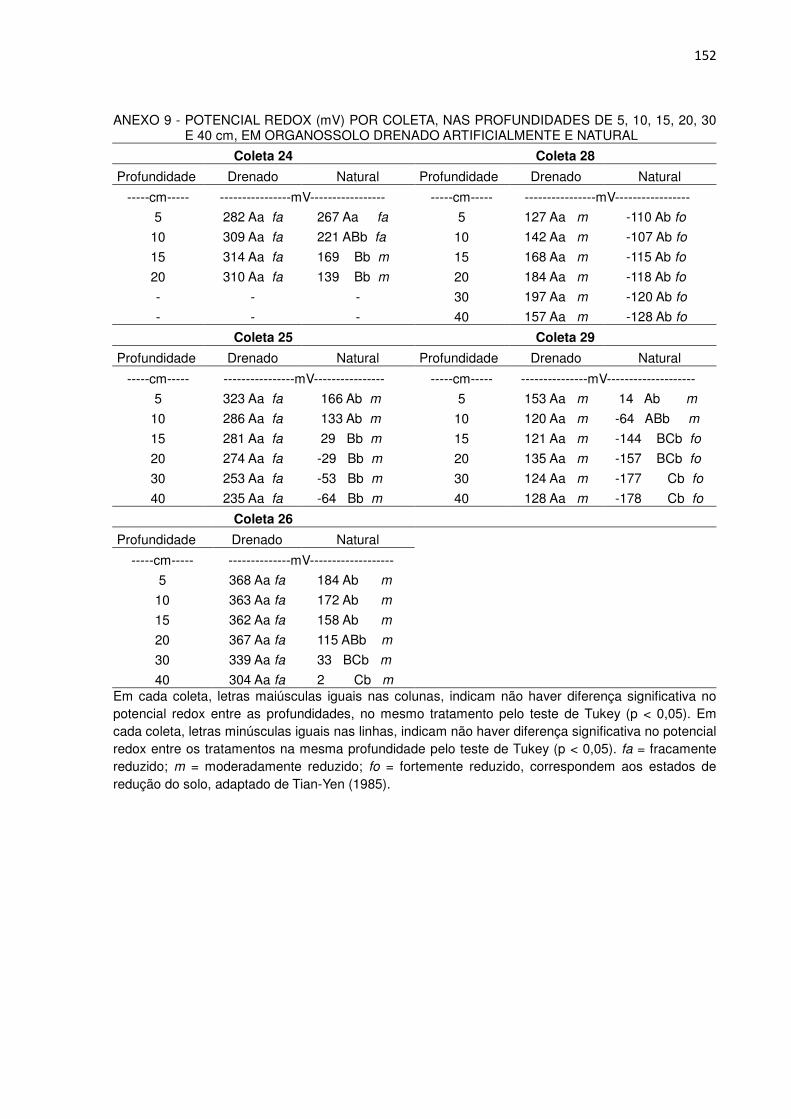
152
ANEXO 9 - POTENCIAL REDOX (mV) POR COLETA, NAS PROFUNDIDADES DE 5, 10, 15, 20, 30 E 40 cm, EM ORGANOSSOLO DRENADO ARTIFICIALMENTE E NATURAL
Coleta 24 Coleta 28
Profundidade Drenado Natural Profundidade Drenado Natural
-----cm----- ----------------mV----------------- -----cm----- ----------------mV-----------------
5 282 Aa fa 267 Aa fa 5 127 Aa m -110 Ab fo
10 309 Aa fa 221 ABb fa 10 142 Aa m -107 Ab fo
15 314 Aa fa 169 Bb m 15 168 Aa m -115 Ab fo
20 310 Aa fa 139 Bb m 20 184 Aa m -118 Ab fo
- - - 30 197 Aa m -120 Ab fo
- - - 40 157 Aa m -128 Ab fo
Coleta 25 Coleta 29
Profundidade Drenado Natural Profundidade Drenado Natural
-----cm----- ----------------mV---------------- -----cm----- ---------------mV--------------------
5 323 Aa fa 166 Ab m 5 153 Aa m 14 Ab m
10 286 Aa fa 133 Ab m 10 120 Aa m -64 ABb m
15 281 Aa fa 29 Bb m 15 121 Aa m -144 BCb fo
20 274 Aa fa -29 Bb m 20 135 Aa m -157 BCb fo
30 253 Aa fa -53 Bb m 30 124 Aa m -177 Cb fo
40 235 Aa fa -64 Bb m 40 128 Aa m -178 Cb fo
Coleta 26
Profundidade Drenado Natural
-----cm----- --------------mV-------------------
5 368 Aa fa 184 Ab m
10 363 Aa fa 172 Ab m
15 362 Aa fa 158 Ab m
20 367 Aa fa 115 ABb m
30 339 Aa fa 33 BCb m
40 304 Aa fa 2 Cb m Em cada coleta, letras maiúsculas iguais nas colunas, indicam não haver diferença significativa no potencial redox entre as profundidades, no mesmo tratamento pelo teste de Tukey (p < 0,05). Em cada coleta, letras minúsculas iguais nas linhas, indicam não haver diferença significativa no potencial redox entre os tratamentos na mesma profundidade pelo teste de Tukey (p < 0,05). fa = fracamente reduzido; m = moderadamente reduzido; fo = fortemente reduzido, correspondem aos estados de redução do solo, adaptado de Tian-Yen (1985).

153
ANEXO 10 - FLUXO ACUMULADO DE METANO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR
Estação do ano Organosolo natural (kg ha-1) Organossolo drenado (kg ha-1) inverno 2010 1,28 A b -0,11 Ba primavera 2010 12,90 A b -2,12 Bab verão 2010/11 36,81 A b -2,37 B b outono 2011 40,21 A b -2,05 Bab inverno 2011 24,66 A b -2,27 Bab primavera 2011 54,60 Aab -2,70 B b verão 2011/12 100,1 Aa -4,20 B b outono 2012 33,20 A b -2,30 Bab inverno 2012 29,80 A b -2,40 B b Total total ano 1 115,86 A b -8,90 Ba total ano 2 217,60 Aa -11,70 Ba
Letras maiúsculas iguais na linha indicam não haver diferença significativa entre o Organossolo natural e o drenado em cada estação; Letras minúsculas iguais na coluna indicam não haver diferença significativa entre as estações dentro de cada tratamento. Em ambos os casos, a comparação entre médias foi realizada pelo teste de Tukey (p < 0,05).
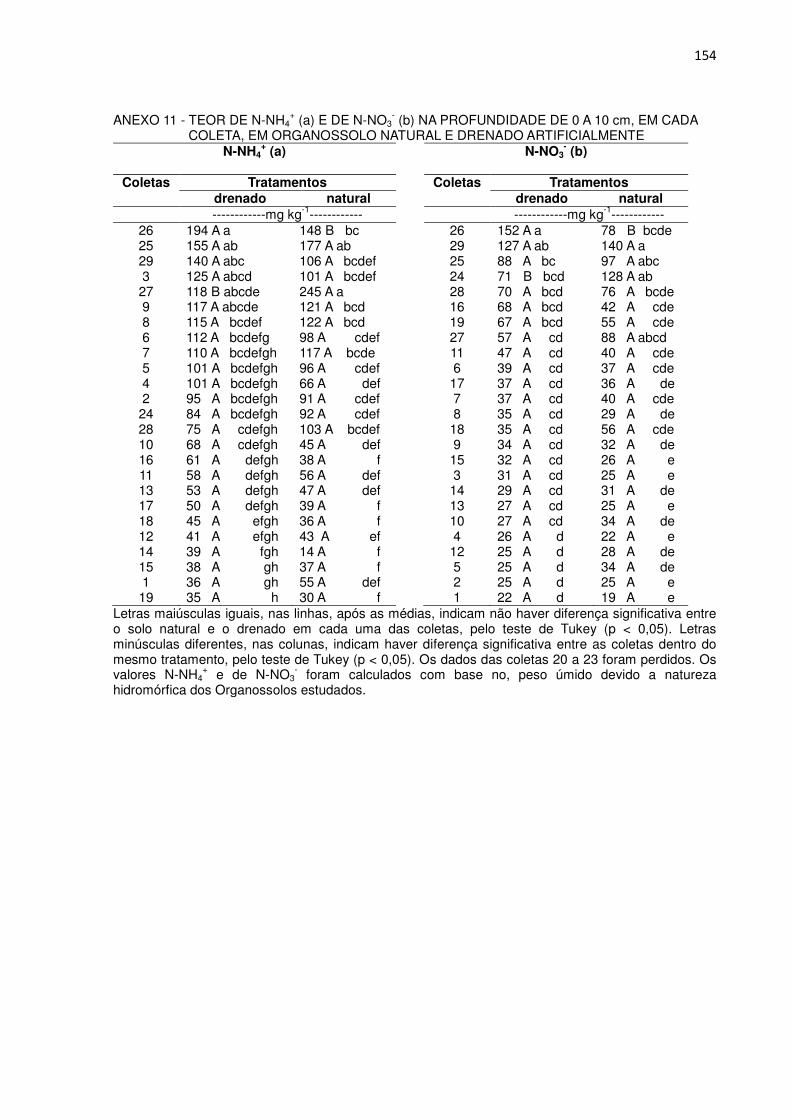
154
ANEXO 11 - TEOR DE N-NH4+ (a) E DE N-NO3
- (b) NA PROFUNDIDADE DE 0 A 10 cm, EM CADA COLETA, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE
N-NH4+ (a)
N-NO3
- (b)
Coletas Tratamentos Coletas Tratamentos drenado natural drenado natural
------------mg kg-1------------ ------------mg kg-1------------ 26 194 A a 148 B bc 26 152 A a 78 B bcde 25 155 A ab 177 A ab 29 127 A ab 140 A a 29 140 A abc 106 A bcdef 25 88 A bc 97 A abc 3 125 A abcd 101 A bcdef 24 71 B bcd 128 A ab 27 118 B abcde 245 A a 28 70 A bcd 76 A bcde 9 117 A abcde 121 A bcd 16 68 A bcd 42 A cde 8 115 A bcdef 122 A bcd 19 67 A bcd 55 A cde 6 112 A bcdefg 98 A cdef 27 57 A cd 88 A abcd 7 110 A bcdefgh 117 A bcde 11 47 A cd 40 A cde 5 101 A bcdefgh 96 A cdef 6 39 A cd 37 A cde 4 101 A bcdefgh 66 A def 17 37 A cd 36 A de 2 95 A bcdefgh 91 A cdef 7 37 A cd 40 A cde 24 84 A bcdefgh 92 A cdef 8 35 A cd 29 A de 28 75 A cdefgh 103 A bcdef 18 35 A cd 56 A cde 10 68 A cdefgh 45 A def 9 34 A cd 32 A de 16 61 A defgh 38 A f 15 32 A cd 26 A e 11 58 A defgh 56 A def 3 31 A cd 25 A e 13 53 A defgh 47 A def 14 29 A cd 31 A de 17 50 A defgh 39 A f 13 27 A cd 25 A e 18 45 A efgh 36 A f 10 27 A cd 34 A de 12 41 A efgh 43 A ef 4 26 A d 22 A e 14 39 A fgh 14 A f 12 25 A d 28 A de 15 38 A gh 37 A f 5 25 A d 34 A de 1 36 A gh 55 A def 2 25 A d 25 A e 19 35 A h 30 A f 1 22 A d 19 A e
Letras maiúsculas iguais, nas linhas, após as médias, indicam não haver diferença significativa entre o solo natural e o drenado em cada uma das coletas, pelo teste de Tukey (p < 0,05). Letras minúsculas diferentes, nas colunas, indicam haver diferença significativa entre as coletas dentro do mesmo tratamento, pelo teste de Tukey (p < 0,05). Os dados das coletas 20 a 23 foram perdidos. Os valores N-NH4
+ e de N-NO3- foram calculados com base no, peso úmido devido a natureza
hidromórfica dos Organossolos estudados.
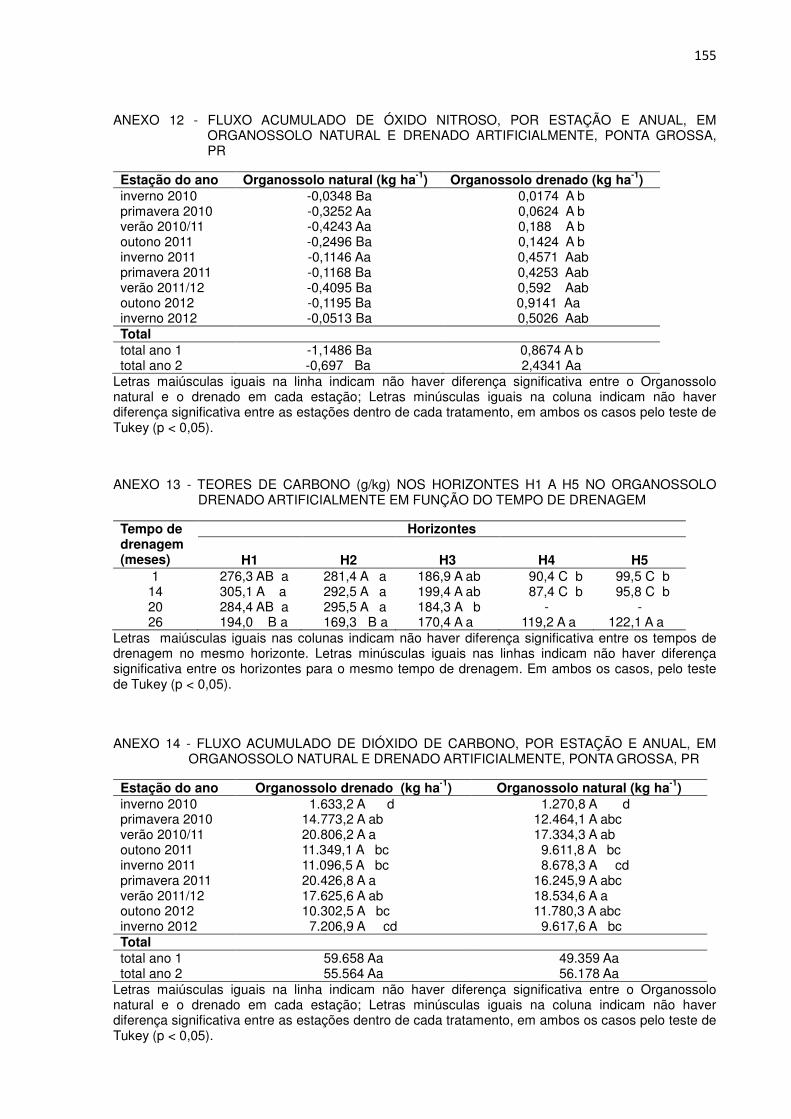
155
ANEXO 12 - FLUXO ACUMULADO DE ÓXIDO NITROSO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR
Estação do ano Organossolo natural (kg ha-1) Organossolo drenado (kg ha-1) inverno 2010 -0,0348 Ba 0,0174 A b primavera 2010 -0,3252 Aa 0,0624 A b verão 2010/11 -0,4243 Aa 0,188 A b outono 2011 -0,2496 Ba 0,1424 A b inverno 2011 -0,1146 Aa 0,4571 Aab primavera 2011 -0,1168 Ba 0,4253 Aab verão 2011/12 -0,4095 Ba 0,592 Aab outono 2012 -0,1195 Ba 0,9141 Aa inverno 2012 -0,0513 Ba 0,5026 Aab Total total ano 1 -1,1486 Ba 0,8674 A b total ano 2 -0,697 Ba 2,4341 Aa
Letras maiúsculas iguais na linha indicam não haver diferença significativa entre o Organossolo natural e o drenado em cada estação; Letras minúsculas iguais na coluna indicam não haver diferença significativa entre as estações dentro de cada tratamento, em ambos os casos pelo teste de Tukey (p < 0,05).
ANEXO 13 - TEORES DE CARBONO (g/kg) NOS HORIZONTES H1 A H5 NO ORGANOSSOLO DRENADO ARTIFICIALMENTE EM FUNÇÃO DO TEMPO DE DRENAGEM
Tempo de drenagem (meses)
Horizontes
H1
H2
H3
H4
H5 1 276,3 AB a 281,4 A a 186,9 A ab 90,4 C b 99,5 C b 14 305,1 A a 292,5 A a 199,4 A ab 87,4 C b 95,8 C b 20 284,4 AB a 295,5 A a 184,3 A b - - 26 194,0 B a 169,3 B a 170,4 A a 119,2 A a 122,1 A a
Letras maiúsculas iguais nas colunas indicam não haver diferença significativa entre os tempos de drenagem no mesmo horizonte. Letras minúsculas iguais nas linhas indicam não haver diferença significativa entre os horizontes para o mesmo tempo de drenagem. Em ambos os casos, pelo teste de Tukey (p < 0,05).
ANEXO 14 - FLUXO ACUMULADO DE DIÓXIDO DE CARBONO, POR ESTAÇÃO E ANUAL, EM ORGANOSSOLO NATURAL E DRENADO ARTIFICIALMENTE, PONTA GROSSA, PR
Estação do ano Organossolo drenado (kg ha-1) Organossolo natural (kg ha-1) inverno 2010 1.633,2 A d 1.270,8 A d primavera 2010 14.773,2 A ab 12.464,1 A abc verão 2010/11 20.806,2 A a 17.334,3 A ab outono 2011 11.349,1 A bc 9.611,8 A bc inverno 2011 11.096,5 A bc 8.678,3 A cd primavera 2011 20.426,8 A a 16.245,9 A abc verão 2011/12 17.625,6 A ab 18.534,6 A a outono 2012 10.302,5 A bc 11.780,3 A abc inverno 2012 7.206,9 A cd 9.617,6 A bc Total total ano 1 59.658 Aa 49.359 Aa total ano 2 55.564 Aa 56.178 Aa
Letras maiúsculas iguais na linha indicam não haver diferença significativa entre o Organossolo natural e o drenado em cada estação; Letras minúsculas iguais na coluna indicam não haver diferença significativa entre as estações dentro de cada tratamento, em ambos os casos pelo teste de Tukey (p < 0,05).


















