
Universidade Federal de Minas Gerais Instituto de Ciências...
79
Universidade Federal de Minas Gerais Instituto de Ciências Biológicas Pós Graduação em Ecologia, Conservação e Manejo de Vida Silvestre Comunidade Zooplanctônica da Região Litorânea das lagoas Carioca e Gambazinho, (Parque Estadual do Rio Doce,MG) JANAÍNA HORTA DE AZEVEDO LOPES Belo Horizonte- MG Dissertação apresentada ao Curso de Ecologia Conservação e Manejo de Vida Silvestre da Universidade Federal de Minas Gerais como requisito à obtenção do título de Mestre em Ecologia.
Transcript of Universidade Federal de Minas Gerais Instituto de Ciências...

1
Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Pós Graduação em Ecologia, Conservação e Manejo de Vida Silvestre
Comunidade Zooplanctônica da Região Litorânea das lagoas Carioca e Gambazinho,
(Parque Estadual do Rio Doce,MG)
JANAÍNA HORTA DE AZEVEDO LOPES
Belo Horizonte- MG
Dissertação apresentada ao Curso de Ecologia
Conservação e Manejo de Vida Silvestre da
Universidade Federal de Minas Gerais como requisito
à obtenção do título de Mestre em Ecologia.

2
Julhho/2014
JANAÍNA HORTA DE AZEVEDO LOPES
Comunidade Zooplanctônica da Região Litorânea das Lagoas Carioca e Gambazinho,
(Parque Estadual do Rio Doce, MG)
Dissertação apresentada ao Curso de
Ecologia, Conservação e Manejo da Vida
Silvestre da Universidade Federal de Minas
Gerais como requisito à obtenção de título de
Mestre em Ecologia.
Orientadora: Profa. Dra. Paulina Maria Maia Barbosa

3
Belo Horizonte- MG, Julho/2014
Dissertação defendida em __/__/____ e aprovada pela banca examinadora
constituída pelos professores:

4
Agradecimentos
Sou imensamente grata ao Espírito Santo, aos meus guias espirituais e todas formas
de vida, por minha existência e aprendizados tristes ou alegres, mas passos a mais no
despertar da minha consciência. Tão grande quanto é meu sentimento de gratidão para com
meus amados Arthur de Azevedo Lopes Ferreira, Maria Josely Horta de Azevedo e Geraldo
Elísio Machado Lopes, filho e pais preciosos. Grata também sou a toda minha família, que
me acolhe e alimenta com sua alegria, especialmente às minhas doces vovós, Dulce (in
memoriam) e Cândida, mulheres de força e fé.
Sou enormemente grata à Paulina Maria Maia Barbosa, minha orientadora, por seu
carinho, paciência e dedicação e à todos os professores do Programa de Pós Graduação em
Ecologia Conservação e Manejo de Vida Silvestre. Sou grata, especialmente, aos professores
Arnola Rietzler, Eneida Eskinazi e Ricardo Mota Pinto Coelho, por tão gentilmente aceitarem
ser da minha banca e contribuírem para a elaboração deste trabalho. Grata ao professor e
curador do herbário da UFV, Pedro B. Schuwartsburd, pela gentileza em identificar as
macrófitas. Manifesto ainda minha gratidão aos professores Reginaldo da Silva Romeiro. (in
memoriam) e Walter Yoshizo Okano, por me iniciarem no caminho da ciência.
À Rosinha grata por todos os ensinamentos e paciência em me ensinar a identificar as
espécies, e ser amiga tão gentil; ao Marcelo pelo carinho e disponibilidade em me instruir
em tantas coisas; aos queridos colegas do LIMNEA pelo companheirismo, solidariedade e
boas risadas, em especial Cínthia Tavares, Luisa Betim, Isabela Vas, Thécia Alfenas, Diego
Pujoni, Daniel Maronesi e Mirla. Aos queridos Cris e Fred, pela amabilidade e boa disposição!
Sou muito grata à Vinícius, Graziela, Xanda, Cláudio, Satyva, Tião e Canela, por me
apoiarem no Parque Estadual do Rio Doce com tanto desprendimento.
Grata à todos os amigos que me apoiaram durante o mestrado, especialmente dona
Efigênia, seu Luis, Virgínia, Rafaela, Rogério, Natália (Tchê) Fernanda Teixera e Michele; grata
também a minha querida psicóloga Claudia Prates e ao Dr. Adauto. Grata à Universidade
Federal de Pernambuco e aos amigos do curso de Ecologia da Caatinga, especialmente
Zezinho, Fernanda, Felipe, Xavier e aos professores Inara Leal, Marcelo Tabarelli e Alan
Andersen, pela gentileza e capacidade de doação, além de momentos divertidos.

5
Grata à Universidade Federal de Minas Gerais, ao Programa de Pós Graduação em
Ecologia Conservação e Manejo de Vida Silvestre, ao PELD, ao IEF, pela concessão de licença
de coleta e à CAPES, pela concessão da bolsa. Muito obrigada!

6
Índice
I. Resumo 7
II. Abstract 8
III. Introdução 9
IV. Material e métodos 15
V. Resultados 25
VI. Discussão 45
VII. Conclusão 62
VIII. Referências bibliográficas 64
IX. Anexos 76

7
Resumo
Pesquisas em ecologia sobre a comunidade zooplanctônica na região litorânea de corpos
de água dulcícolas são escassas na região tropical, quando em comparação com
pesquisas realizadas na região pelágica de lagos. Considerando a importância da região
litorânea, pela elevada riqueza e diversidade de espécies que esta apresenta, em
especial quando colonizada por macrófita, este estudo teve como objetivo caracterizar a
comunidade de Amoeba Testacea, Rotifera, Cladocera e Copepoda quanto à abundância
(org.m-3), riqueza Diversidade (Índice de Shannon) e distribuição de espécies na região
marginal com e sem plantas aquáticas das lagoas Carioca (com peixes não nativos) e
Gambazinho (com peixes nativos e exóticos), localizadas no Parque Estadual do Rio Doce,
MG. Para tanto, coletas bimensais de zooplâncton foram realizadas de junho de 2011 à
12 de 2012 nas lagoas contempladas nesta pesquisa. As diferenças observadas na
abundância de rotíferos e riqueza de espécies zooplanctônicas entre as lagoas é causada,
possivelmente, pela maior disponibilidade de recursos alimentares na lagoa Carioca, que
suportaria populações maiores e também um maior número de táxons quando
comparada à lagoa Gambazinho. Não houve diferenças quanto à riqueza, Diversidade e
abundância de espécies entre os habitas litorâneos com e sem plantas aquáticas, em
ambas as lagoas, provavelmente pelo fato destes habitats apresentarem condições
físico-químicas semelhantes e mesma disponibilidade de recursos algais (alimento para o
zooplâncton herbívoro), além da provável dispersão de espécies por correntes na água
no eixo horizontal, que promoveriam o intercâmbio de animais planctônicos nas áreas
com e sem macrófita.
Palavras-chave: zooplâncton, região litorânea, diversidade de espécies

8
Abstract
Ecological researches on zooplankton community in lake shoreline are scarce in tropics
when compaired to the ones conducted at pelagic region. Taking in to account the
importance of lakes shoreline due to its elevated species richness and Diversity, specially
when colonized by macropHytes, This study means to characterize the Amoeba Testacea,
Rotifera, Cladocera and Copepoda communities as for its abundance (org.m-3), Diversity
(Shannon Index) and species distribution in lake shoreline colonized and not colonized by
macropHytes. The research was conducted in lake Carioca (with exotic and native fish
fauna) and in lake Gambazinho (with native fish fauna, merely), both based in Parque
Estadual do Rio Doce, Minas Gerais State, Southest Brazil. To this end, bimensal
zooplankton sampling were donne from june/2011 up to December/2012 in these lakes.
Observed differences on Rotifera abundance and specie reachness among lake Carioca
and lake Gambazinho are probably due to more food disponibility, like algae and
detritus, in lake Carioca, what could sustain bigger populations and a greater number of
taxa than could do lake Gambazinho. There were no differences in relation to reachness,
Diversity and species abundance between litoral habitats with and without macropHytes,
what happened perhaps by the fact taht this environments presente the same pHisical-
chemical conditions and food resources disponibility, or by species dispertion on water
flow in horizontal axis from Wind, that could serve for plankton animal Exchange
between areas with and without aquatic vegetation.
Key words: zooplankton, lake shoreline, Species diversity

9
Introdução
Lagos são corpos de água continentais lênticos, sem ligação imediata com o oceano,
com baixas concentrações de íons em solução e curto tempo de vida, considerando a escala
de tempo geológica, devido a sedimentação de material orgânico e inorgânico advindos da
bacia de drenagem (Esteves, 1998). O ecossistema lacustre é dependente do ecossistema
terrestre adjacente e pode ser impactado pelas atividades ocorrentes no meio terrestre
circundante (Likens, 1992).
A maior parte dos lagos brasileiros é originária dos processos de erosão e
sedimentação de rios, do mar e de corais, o que explica a localização de muitos lagos
brasileiros em antigas áreas de drenagem de rios, cujos leitos se deslocaram, ou em regiões
litorâneas. Visto que a maioria dos lagos permanentes brasileiros estão em áreas costeiras,
são escassos aqueles oriundos exclusivamente de forças fluviais, como os lagos amazônicos,
do Pantanal e do Rio Doce (Trindade, 1984).
Os corpos d’água continentais apresentam diversidade faunística elevada, com
grupos exclusivos destes ambientes. Apesar disso, a riqueza de espécies contida em lagos,
rios e riachos ainda não é totalmente conhecida (menos de 30%), sendo que a maioria dos
artigos sobre a biota dulcícola trata da ictiofauna ou de espécies economicamente
importantes (Frouin e Lacobellis, 2003).
Os invertebrados dulcícolas compõe um conjunto diversificado, que abrange vários
filos, com participação ativa nas funções do ecossistema aquático (Purvis e Hector, 2000). No
Brasil, 3.134 espécies foram já descritas, porém, segundo Rocha (2003), estima-se a
existência de 8000 espécies ainda não registradas. Provavelmente, as lacunas no
conhecimento da fauna planctônica brasileira são decorrentes do fato da maioria dos
estudos focarem certas bacias hidrográficas, grupos taxonômicos específicos e serem
realizados nos habitats límnicos de ambiente lênticos, desconhecendo a biota dos ambientes
lóticos e região litorânea dos lagos (Eskinazi- Sant’ Anna et al, 2005).
Em um plano horizontal, o ecossistema lacustre pode ser dividido em dois
compartimentos com morfologia, condições ambientais e comunidades biológicas distintas:
região litorânea e limnética (ou pelágica). A região limnética abrange a porção interior do

10
lago, com profundidades que, frequentemente, determinam gradientes na distribuição de
luz, temperatura, concentração de oxigênio dissolvido e outras variáveis físicas e químicas
(Moretto, 2001). Os organismos característicos dessa região são representados pelo nécton,
notoriamente os peixes, e pelo plâncton (bacterio, fito e zooplâncton). As características
destas comunidades, e as variações saregiãois e espaciais nos parâmetros físicos e químicos
da região limnética tem sido relativamente bem estudadas (Esteves,1998 ).
A região marginal de lagos é o ecótono entre o corpo d’água e o ambiente terrestre
adjacente (Wetzel, 1983; Pieczynska, 1990). É uma região geralmente pouco profunda que
abriga uma riqueza enorme de espécies, especialmente quando coberta por macrófitas, pela
diversidade de habitats, abrigos e recursos alimentares disponíveis (Green, 1986; Maia-
Barbosa, 2008). Além disso, a região litorânea funciona como área de refugio para peixes
jovens contra seus predadores (Straskraba et al, 1993). Estas características tornam estas
áreas importantes para a preservação da biodiversidade.
De acordo com Straskraba et al (1993), são funções das margens de lagos: impedir a
dissipação de matéria alóctone para a região limnética, atuando como região tampão;
dispersar espécies para outros compartimentos lacustres; disponibilizar alimento e locais de
reprodução para peixes e aves paludícolas, além de participar ativamente da ciclagem de
nutrientes. A interação entre as regiões litorânea e limnética é dependente, do grau de
sinuosidade da margem do lago, representado pela sua fragmentação e seu contorno, e
mesurado pelo índice de desenvolvimento de margem (Kolasa e Zaleuski, 1995). Assim, lagos
mais irregulares, ou com mais reentrâncias, normalmente apresentarão maior riqueza de
espécies.
Estudos comparativos entre as regiões pelágica e litorânea de lagos (Pennak, 1966;
Serrano e Toja, 1998; Lima et al, 2003), apontam maior diversidade zooplanctônica no
habitats marginais, especialmente quando colonizados por macrófitas, devido à maior
heterogeneidade ambiental, além de maior disponibilidade de alimento e nichos ecológicos.
Esteves (1998) classifica as cadeias tróficas do região litorânea como de dois tipos:
cadeia trófica de herbivoria, onde os heterótrofos têm como fonte de energia a biomassa do
fitoplâncton, do perifíton e das macrófitas, e a cadeia trófica de detritivoria, onde
organismos heterótrofos têm como fonte energética a biomassa morta, alóctone (folhas,
galhos e sedimento do ambiente terrestre adjacente) e autóctone (organismos aquáticos

11
mortos, como algas, animais zooplanctônicos, macrófitas, bacterioplâncton), sendo esta a
principal cadeia na região marginal. Em ambos os tipos de cadeia participam invertebrados
zooplanctônicos, perifíticos e bentônicos, como Crustacea, Oligochaeta e estágios larvais de
Insecta (Moretto, 2001)
A região marginal, por sua menor profundidade, geralmente recebe radiação solar da
superfície ao fundo, favorecendo a fotossíntese e o desenvolvimento de várias espécies de
algas e plantas aquáticas em toda a coluna d’água, sendo um pólo de retenção de energia e
produção de matéria orgânica (Moretto, 2001). De acordo com as características da
vegetação e a profundidade, o região litorânea é dividido em duas regiãos: eulitoral, onde se
encontram macrófitas emergentes, e infralitoral, onde se encontram macrófitas submersas
(Wetzel e Likens, 1991). Além dessa regiãos, nem sempre facilmente distinguíveis, existe
uma terceira na qual as macrófitas não estão presentes, ou se estão não são abundantes.
Áreas litorâneas sem macrófitas ocorrem geralmente onde a vegetação terrestre
circundante provoca sombreamento na água, impedindo o desenvolvimento de plantas
aquáticas (Esteves, 1998).
Por serem geralmente pequenos e rasos, lagos tropicais são susceptíveis à
colonização por macrófitas de diversas espécies. A elevada concentração de nutrientes,
originários do meio terrestre adjacente e do sedimento, possibilita o desenvolvimento de
grandes bancos de macrófitas (Wetzel e Likens, 1991; Moretto, 2001) que servem de
substrato e oferecem habitat para muitos organismos, sustentando populações
consumidoras, de metabolismo rápido, que também contribuem para acelerar a ciclagem de
nutrientes. Além disso, as plantas aquáticas enraizadas são eficientes como rede de
transporte de nutrientes e energia entre o sedimento e a água, sendo mediadoras na
alocação de nutrientes entre esses compartimentos (Granáli e Solander, 1988). Funcionam
ainda como filtros importantes de retenção de detritos e poluentes das áreas adjacentes
(Engelhardt e Ritchie 2001).
Variações na região litorânea podem ocorrer por flutuações no nível da água,
alterações nas características físicas e químicas da água entre os períodos de chuva e seca,
ventos, ondas, topografia e declividade da região (Wetzel e Likens, 1991; Henry, 2003). Estas
variações determinam a eliminação e/ou substituição de espécies, alterando a estruturação
das comunidades que ali vivem (Carvalho, 2003; Henry, 2003; Nogueira, 2003; Smith et al

12
2003). Lagos naturais não conectados à rios, como aqueles do complexo lacustre do Médio
Rio Doce, tendem a apresentar maior estabilidade temporal e espacial, tanto nas variáveis
ambientais, como na diversidade de organismos e a relação destes entre si e com o meio.
Esta estabilidade relativa é decorrente do maior tempo de residência e baixa turbulência da
água, além de maior estabilidade do sedimento nas margens (Tundisi, 1990).
Dentre os fatores mais influentes no estabelecimento das espécies em uma
comunidade, seja em ambiente lênticos ou lóticos, está a qualidade do habitat. A biota do
ecossistema aquático é influenciada pelo meio físico do corpo: geomorfologia, velocidade da
corrente, vazão de água, tempo de retenção (Tate e Heiny,1995).
No ecossistema lacustre é comum a predominância, em abundância, de uma ou
poucas populações dentre um grupo taxonômico ou dentre a comunidade. Tal dominância
pode ser dependente dos parâmetros físicos da água, visto que a eficiência na utilização de
recursos difere entre os organismos sendo uns favorecidos por condições ambientais
específicas, bióticas, como a ausência de competidores e predadores, e abióticas, como a
concentração de oxigênio dissolvido na água e o pH (Moretto, 2001). Alterações nas
variáveis limnológicas podem, então, influenciar não só na qualidade e quantidade do
alimento disponível, como nas condições do habitat “ideais” para as diferentes espécies,
explicando a sucessão entre elas (Chapman et al, 1985).
As macrófitas podem determinar a estrutura e a dinâmica das comunidades
alterando padrões de riqueza, abundância e distribuição de espécies zooplanctônicas. Sua
presença no meio aumenta a heterogeneidade ambiental, fornecendo alimento pela
produção de matéria orgânica, substrato para o perifíton e refúgio para os animais
zooplanctônicos (Greco e Dabés, 2001; Maia-Barbosa et al, 2008; Pinto-Coelho et al, 2008;
Maia-Barbosa et al, 2010).
O conhecimento a respeito do funcionamento da região litorânea em ambientes
tropicais e sua importância ecológica é ainda pequeno, embora recentemente, algumas
pesquisas tenham revelado o papel crucial desta área na manutenção da diversidade
biológica (Thomaz, 1998; Moretto, 2001, Brito, 2003; Maia-Barbosa, 2008, Mormul et al,
2010). No Brasil dois sistemas têm sido estudados sistematicamente, principalmente após a
implantação dos Programas de Pesquisas Ecológicas de Longa Duração (PELD), incluindo
levantamentos em áreas litorâneas: a bacia de inundação do Rio Paraná (ex: Lansac-Tôha et

13
al, 1997; Velho et al, 1999; Rossa et al, 2001; Lima et al, 2003; Fulone, 2008) e o complexo
lacustre do Rio Doce (Moretto, 2001; Maia Barbosa et al, 2008; Maia-Barbosa et al, 2010;
Aoki, 2010).
Maia-Barbosa e colaboradores (2008) trabalhando em bancos de macrófitas no lago
Dom Helvécio no Parque Estadual do Rio Doce (PERD) registraram 130 novas ocorrências de
espécies zooplanctônicas para a região, demonstrando a importância de estudos que
contemplem a região litorânea para o conhecimento da biodiversidade de um lago. Outro
estudo que também comparou a diversidade de espécies da comunidade zooplanctônica em
bancos de macrófitas e na região limnética foi desenvolvido na lagoa dos Patos (PERD) e
registrou 228 e 51 espécies respectivamente, demostrando mais uma vez, a importância das
macrófitas para a maior riqueza de espécies (Aoki, 2010).
Para melhor conhecimento da biota e compreensão do funcionamento do
ecossistema lacustre as pesquisas devem incluir todos os compartimentos do lago, visto que
a biodiversidade geralmente tem grande variação entre os mesmos (Moretto, 2001). A
variabilidade temporal da comunidade zooplanctônica é caracterizada por alterações na
riqueza, abundância, diversidade, composição e densidade das espécies e seu estudo é
essencial para a compreensão do funcionamento do ecossistema. O acompanhamento das
variações nas comunidades é um meio eficaz de compreender como os sistemas naturais
respondem aos distúrbios, naturais ou antrópicos (Simões, 2010), como por exemplo, a
introdução de espécies exóticas.
A introdução de espécies exóticas é um problema global e tem sido considerada a
segunda causa de extinção de espécies, perdendo somente para a destruição de habitat
(Primack e Rodrigues, 2001).
Em lagos, o estabelecimento de espécies não nativas pode ter como consequências,
em consonância com a biologia da espécie introduzida e os processos ecológicos
característicos do local de introdução, alterações nas cadeias alimentares, diminuição ou
mesmo extinção de populações nativas, causadas por alterações estruturais dos habitats,
mudanças nas taxas de predação e competição entre as espécies, além do estabelecimento
de parasitas e patógenos alóctones (Welcomme, 1988).
No PERD a introdução de espécies é também um problema, e dificulta muito o
cumprimento de um dos principais objetivos de uma unidade de conservação, que é a

14
preservação da biodiversidade local e regional. No parque, um dos grupos que se destaca
pelo maior número de espécies introduzidas é o de peixes: Pygocentrus nattereri (piranha),
Cichla kelberi (tucunaré), Clarias gariepinus (bagre africano), Astronotus ocellatus (apaiari)
e Hoplosternum littorale (tamboatá) (Rebouças et al, 2006). Estudos têm demonstrado que,
pelo menos na lagoa Carioca, a presença de algumas destas espécies pode ser considerada
a principal causa de extinção local de peixes nativos (Godinho et al., 1994; Godinho, 1996,
Latini e Petrere, 2004 ). Da mesma forma, alterações na cadeia trófica decorrentes da
introdução de peixes não nativos foram apontadas por Pinto-Coelho et al (2008).
O objetivo desta pesquisa foi caracterizar e comparar a estrutura da comunidade
zooplanctônica quanto à riqueza, diversidade, abundância e distribuição de organismos, na
região litorânea, colonizada e não colonizada por macrófitas em duas lagoas tropicais
sediadas no Parque Estadual do Rio Doce (PERD, MG), uma com ictiofauna nativa (lagoa
Gambazinho) e a outra com ictiofauna nativa e exótica (lagoa Carioca).
Objetivos específicos
Caracterizar a comunidade zooplanctônica (Amoeba Testacea, Rotifera, Cladocera e
Copepoda) das lagoas Carioca e Gambazinho (PERD, MG), considerando a riqueza,
diversidade, abundância e distribuição das espécies;
Comparar a comunidade zooplanctônica entre as lagoas Carioca e Gambazinho
(PERD, MG) quanto aos mesmos parâmetros citados acima;Comparar, na região
litorânea de cada lagoa, a comunidade zooplanctônica entre os habitats colonizados
e não colonizados por macrófitas Comparar a comunidade zooplanctônica na região
litorânea (colonizada e não colonizada por macrófita) entre os meses chuvosos e
secos.
Predições
Haverá diferenças quanto à riqueza, diversidade, abundância e distribuição das
espécies de Amoeba Testacea, Rotifera, Cladocera e Copepoda entre as lagoas
Carioca e Gambazinho (PERD, MG)
Haverá diferenças quanto à riqueza, diversidade, abundância e distribuição das
espécies de Amoeba Testacea, Rotifera, Cladocera e Copepoda entre os habitats
colonizados e não colonizados por macrófitas na região litorânea.

15
Haverá diferenças quanto à riqueza, diversidade, abundância e distribuição das
espécies de Amoeba Testacea, Rotifera, Cladocera e Copepoda entre os períodos de
chuva e seca nos habitats litorâneos, colonizado e não colonizado por macrófitas.
Hipóteses
A lagoa Gambazinho apresentará maior riqueza, diversidade e abundância de
espécies zooplanctônicas do que a lagoa Carioca devido à menor pressão de
predação por peixes e invertebrados sobre o zooplâncton.
Na região litorânea das duas lagoas os valores de riqueza, diversidade e abundância
de espécies zooplanctônicas serão mais elevados, principalmente quando
colonizadas por macrófitas, devido à maior disponibilidade de habitats e recursos
alimentares para o zooplâncton
No período chuvoso, os valores de riqueza, diversidade e abundância de espécies
serão maiores nas duas lagoas, devido à maior disponibilidade de nutrientes
carreados pela chuva, maiores temperaturas e consequentemente maior produção
algal.
Material e Métodos
Área de Estudo
O vale do médio Rio Doce (MG) abriga mais de 60% da biodiversidade da Mata
Atlântica, incluindo uma porcentagem ainda maior das espécies endêmicas desse bioma,
que é um dos mais ameaçados do Brasil e de maior diversidade do mundo. A Mata Atlântica
do vale do Rio Doce foi reduzida a 4-5% de sua extensão original na região, e seu maior
remanescente, no Estado de Minas Gerais, encontra-se preservado no Parque Estadual do
Rio Doce (PERD) - 36.000 ha de florestas, predominantemente secundárias, entremeadas
por um sistema lacustre com 153 lagos nos mais variados estágios de evolução (Barbosa e
Moreno, 2002).
Segundo Setti (1996) os principais problemas decorrentes de atividades antrópicas na
Bacia do Rio Doce são: baixa qualidade da água, devido ao seu desvio e input de poluentes e

16
contaminação por substâncias inorgânicas, emitidas pela atividade siderúrgica e descarga de
efluentes domésticos de grandes cidades, como Governador Valadares. Além desses
problemas, outras fontes degradantes da Bacia do Rio Doce são as indústrias de celulose e
carvão, que substituíram grande parte da vegetação nativa por eucaliptais (Tundisi et al,
1997).
A bacia hidrográfica do Rio Doce situa-se no sudeste de Minas Gerais e abrange uma
área de drenagem de 83.400 km2, sendo 86% da área pertencente ao Estado de Minas
Gerais e o restante ao Estado do Espírito Santo. A Região é ocupada por 3,1 milhões de
habitantes em 222 municípios (Mello, 1997). Os principais tributários do Rio Doce são os rios
Santo Antônio, Piracicaba, Matipó e Casca (Moretto, 2001).
Um complexo hídrico com aproximadamente 160 lagoas ocorre na região entre o Rio
Piracicaba e o Rio Doce (Mello, 1997). Esses corpos d’água se originaram no período
Pleistoceno – Holoceno pela deposição de material carreado pelo Rio Doce, que acabou por
formar barragens que represavam a água de seus afluentes (Tundisi et al, 1997). Visto que
este rio teve seu leito deslocado diversas vezes, a maioria das lagoas encontradas em sua
bacia não estão próximas do leito atual do rio. Nos meses chuvosos é possível que ocorra
comunicação entre as lagoas entre si e também com rios tributários, favorecendo o
intercâmbio de espécies, inclusive aquelas alóctones (Godinho, 1996).
O sistema lacustre do vale do Rio Doce é utilizado pela população de cidades vizinhas
para abastecimento, pesca e lazer. A vegetação nativa, Floresta Tropical Úmida, do tipo
Mata Atlântica foi, em grande parte, substituída por cultivo de eucalipto destinado à
produção de carvão vegetal para as indústrias siderúrgicas do Vale do Aço. Além disso, há no
entorno do parque grandes áreas de secagem de arroz, cultivado nas áreas alagadas e lagoas
assoreadas (Moretto, 2001).
O Parque Estadual do Rio Doce PERD (19º 29’24’’- 19º 48’ 18’’ S; 42º 28’18’’- 42º
38’30’’ W, fig. 1), maior reserva de Mata Atlântica do Estado de Minas Gerais, abriga um dos
três grandes sistemas lacustres do Brasil. As temperaturas médias anuais oscilam entre 20 e
22º C. As chuvas concentram-se nos meses de verão e atingem, em média, 1.250mm,
podendo chegar até 1.500mm (De Meis, 1977), com meses de déficit hídrico de maio à
setembro (Minas Gerais, 1980). A cobertura vegetal nesta área é de floresta tropical úmida,
do tipo “Mata Atlântica”. O relevo é do tipo ondulado a montanhoso sendo o latossolo

17
vermelho-amarelo o tipo de solo predominante (Camargo et al., 1966). O PERD abrange os
municípios de Mariléia, Timóteo e Dionísio, na área denominada como Vale do Aço. O
parque é administrado pelo Instituto Estadual de Floresta de Minas Gerais (IEF-MG) e abriga
cerca de 50 lagoas, ou seja, 6% de sua área (Moretto, 2001).
As lagoas do PERD, sediadas entre morros, formam um mosaico crucial para a
preservação da biodiversidade regional, sendo que cada lagoa pode ser considerada uma
ilha em termos de biodiversidade, sendo um dos fatores responsáveis por este fato as
diferentes profundidades dos corpos de água, ou seja, características topográficas dos
mesmos (Meis e Tundisi, 1997). A tabela 1 resume as principais características das lagoas
Carioca e Gambazinho, segundo Barros (2010), protegidas pelo PERD e contempladas neste
estudo.
Figura 1: Mapa do Parque Estadual do Rio Doce, com algumas de suas lagoas preservadas por esta Unidade de Conservação, e a inserção do parque no Estado de Minas Gerais.

18
A lagoa Carioca (19° 45’ 26,0”S e 0,42° 37’ 06,2”W, figs. 2 e 3), classificada como
mesotrófica, apresenta um curto período de circulação durante o inverno (junho a agosto),
permanecendo estratificada durante os outros meses, o que a caracteriza como um lago
monomítico quente (Barbosa e Tundisi, 1989). Sua ictiofauna apresentou grandes alterações
desde a introdução do tucunaré (Cichla ocelaris). Atualmente predominam o tucunaré
(Cichla ocelaris) e a piranha (Pygocentrus nattereri) (Brito, 2003). Esta lagoa apresenta parte
das margens colonizadas por macrófitas emersas, submersas, e flutuantes sendo as
principais) Eleocharis interstincta (EupHorbiaceae); Mayaca fluviatilis Aubl. (Mayaceae);
NympHaea elegans (NympHaeaceae) e NympHoides indica (Menyanthaeceae) (Tavares,
2003)
As primeiras informações sobre a comunidade zooplanctônica desta lagoa foram
levantadas por Santos (1980), que comparou a composição e flutuação saregiãol de
cladóceros em cinco lagos em diferentes estágios de evolução, dentre eles a lagoa Carioca, e
Matsumura-Tundisi e Tundisi (1980), que apresentaram dados sobre a biomassa e
abundância de algumas espécies de cladóceros e copépodes. Apesar do volume grande de
informações sobre esta lagoa, coletados deste então, a região litorânea ainda foi pouco
explorada.
Altitude (m)
Profundidade (m)
DL
Área (km2)
Área da bacia (km2)
AB/AL
1,28
0,13
0,67
5,1
10
2,9
0,09
0,11
1,31
Lagoa Carioca Lagoa Gambazinho
270 260
10
Tabela 1: Altitude, profundidade, índice de desenvolvimento de margem (DL, segundo Hakanson, 2004) área (km2), área da bacia (km2) e AB/AL das lagoas Carioca e Gambazinho, PERD, MG, segundo Barros, 2010.

19
A lagoa Gambazinho (19° 47’ 07,7”S e 042° 34’ 45,5” W, figs. 4 e 5) não apresenta
estratificação térmica e é considerada oligotrófica. Segundo Ferreira et al, 2010, esta lagoa,
já a 6m da margem alcança profundidade de 1,3m, causando instabilidade do sedimento e
Figura 3: Foto da lagoa Carioca (PERD, MG)
1
2
3 4
5
6
Figura 2: Mapa da lagoa Carioca (PERD, MG) e os pontos de coleta: 1,3 e5 na região litorânea colonizada por macrófita 2,4 e 6 na região litorânea não colonizada por macrófita.
Foto: Cínthia Tavares

20
dificuldades para a permanência de macrófitas a longo prazo, principalmente as plantas
enraizadas, forma biológica mais frequente.
De acordo com os autores, nesta lagoa foram registradas 18 espécies de macrófitas
pertencentes à 14 famílias, a saber: (1) anfíbias: Eupatorium sp (Asteraceae), Andropogon
bicornis L., Panicum helobium Mej ex Henrard (Poaceae) e Borreria sp (Rubiaceae).(2)
Emersas: Eleocharis interstincta (Vahl) Roem. e Schult, Oxycoryum cubense (Popp. E Kunth)
Palla, Scleria melaleuca Rchb. Ex Schltr e Cham. (Cyperaeae), Caperonia castaneifolia (L.)a.
St. Hil (EupHorbiaceae), Hyptis lorentziana O. Hoffm.(Lamiaceae), Lycopodiella camporum B.
Ollg e P.G. Windisch (Lycopodiaceae), Rhynchanthera novemneria DC., Tococa sp
(Melastomataceae), Saweagesia erecta L. (Ochnaceae), Ludwigia sp (Onagraceae), Xyris
jupicae (Xyridaceae). (3) Submersa livre: Utricularia gibba L. (Lentibulariaceae). (4) Submersa
fixa: Mayaca fluviatilis Aubl. (Mayaceae) e (5) Flutuante fixa: NympHaea caerulea Sawigny
(NympHaeaceae). As figuras 6 (a, b, c, d, e, f) são registros fotográficos de macrófitas
observadas, durante as coletas, nas lagoas Carioca e Gambazinho e gentilmente
identificadas, através de fotos, pelo professor Pedro B. Schuartsburd, também curador do
Herbário da Universidade Federal de Viçosa. Para identificação das plantas, também foi
consultado o livro Plantas aquáticas do Pantanal (Pott e Pott, 2000).
Dentre os 42 lagos avaliados por Latini e colaboradores (2004) apenas 3 não
apresentavam espécies exóticas de peixes, sendo um deles a lagoa Gambazinho. Esta lagoa
mantém entre sua fauna íctica espécies como Oligosarcus solitarius, Moenkausia doceana,
Astyanax taeniatus, Cichla somafacetum e GeopHagus brasiliensis (Latini e Petrere, 2004),
possíveis predadoras de zooplâncton, além de Hoplias malabaricus e Leporinus
steindachneri. As figuras 7 (a, b, c, d, e, f) são fotografias dos peixes nativos e exóticos
observados nas lagoas estudadas.

21
Cínthia Tavares
1
2
3
4
5
6
Figura 4: Mapa da lagoa Gambazinho (PERD, MG) e os pontos de coleta: 1, 3 e 5 na região litorânea colonizada por macrófita; 2, 4 e 6 na região litorânea não colonizada por macrófitas.
Figura 5: Foto da lagoa Gambazinho (PERD, MG).
foto: Cíntjha Tavares

22
Figura 6a: Ludwigia sedoides (H.B.K.)Hara. Nome popular: Cruz de Malta. Família: Onagraceae. Macrófita observada na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Figura 6b: Eleocharis interstincta (Vahl) Roem. e Schult. Nome popular: junco. Família: Cyperaceae. Macrófita observada na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Figura 6c: Nymphoides sp. Nome popular: lagartixa. Família: Menyanthaceae. Macrófita observada na região litorânea da lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Figura 6d: Rhynchospora sp. Nome popular: capim navalha. Família: Cyperaceae. Macrófita observada na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Figura 6e: Salvinia auriculata. Aubl. Nome popular: orelha de onça. Família: Salvinaceae. Macrófita observada na região litorânea da lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Figura 6f: Nymphaea caerulea.Sawigyni. Nome popular: ninféia. Família: Nymphaeaceae. Macrófita observada na região litorânea da lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Foto: Janaína Lopes Foto: Janaína Lopes
Foto: Janaína Lopes Foto: Janaína Lopes
Foto: Janaína Lopes Foto: Janaína Lopes

23
Foto: Cínthia Tavares
a
Figuras 7 (a e b): Pygocentrus nattereri (Piranha) e Cichla kelberi (Tucunaré,) espécies introduzidas e registradas na região litorânea da lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Foto: Cínthia Tavares Foto: Cínthia Tavares
b
d c
Figuras 7 (c e d): Hoplosternum littorale (Tamboatá) espécie introduzida e registrada na região litorânea da lagoa Carioca (PERD, MG) e Astyanax aff. bimaculatus (Lambari), espécie nativa registrada nas lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
Foto: Cínthia Tavares
Foto: Cínthia Tavares Foto: Cínthia Tavares
Figuras 7 (e e f): Hoplias malabaricus (Traíra) e Moenkhausia doceana(Lambari), espécies nativas registradas na região litorânea da lagoa Carioca (PERD, MG) e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
e f
Foto: Cínthia Tavares

24
Coleta de dados
As coletas (pH, condutividade, temperatura da água, clorofila-a e concentração de
oxigênio dissolvido) foram realizadas bimensalmente, de junho de 2011 à dezembro de
2012, pela manhã, em seis pontos da região litorânea das lagoas Carioca e Gambazinho (3
em margem colonizada por macrófita, 3 em margem não colonizada por macrófita). Os
pontos de coleta foram estabelecidos na região litorânea, em profundidade de até 1,5 m.
Para a coleta de zooplâncton foram filtrados, em cada ponto, 100L de água da sub-
superfície com o auxílio de um balde e rede de plâncton de 45µc de abertura de malha. As
amostras foram fixadas em formalina 8 % neutralizada e coradas com Rosa de Bengala. Em
laboratório, os indivíduos foram identificados e contados sob microscópio óptico, em
câmara Sedgewik-Rafter, até que 200 indivíduos da espécie dominante fossem identificados.
Amostras com baixa densidade de organismos foram contadas na totalidade. As seguintes
chaves para identificação das espécies foram utilizadas: Edmondson (1959), Ruttner-Kolisko
(1974), Rocha e Matsumura-Tundisi (1976), Koste (1978), Sendacz e Kubo (1982), Reid
(1985), Paggi (1995), Segers (1995), El Moor-Loureiro (1997) e Souza (2008).
Amostras de água foram coletadas na sub-superfície para medição, em laboratório,
da condutividade elétrica (condutivímetro Tecnal, modelo TEC4MP) e pH (peagâmetro
Tecnal, modelo TEC5). A concentração de oxigênio dissolvido na água (mg.L-1) foi
determinada segundo Winckler (1888) e a concentração de clorofila-a segundo metodologia
proposta por Lorenzen (1967). A temperatura da água (ºC) foi medida in situ com um
termômetro manual (Incoterm).
Análises estatísticas
Para cálculo da diversidade de espécies foi utilizado o índice de Shannon para cada
ponto de amostragem em cada um dos meses em que foram realizadas as coletas. Também
foram calculados o índice de Equitabilidade, ambos pelo software PAST, e a riqueza de
2
4
5
1

25
espécies (somatório do número de táxons identificados nos pontos de amostragem nos
meses em que as coletas foram feitas). Para comparação da fauna zooplanctônica registrada
nas regiões litorâneas colonizada e não colonizada por macrófitas foi feita uma análise de
agrupamento (Cluster), por meio da abundância relativa das espécies, sendo as diferenças
expressas por meio da distância euclidiana. Para esta comparação também foram utilizados
os teste T e de Wilcoxon, sendo o teste paramétrico aplicado quando os dados
apresentavam distribuição normal, e o teste não paramétrico usado quando a distribuição
dos dados não se enquadrava à distribuição normal. A normalidade dos dados foi avaliada
pelo teste de Shapiro-Wilk. Os testes foram realizados no software R e para todos adotou-se
nível de significância de 5%.
Resultados
Variáveis abióticas
Lagoa Carioca X Lagoa Gambazinho
A tabela 2 apresenta, resumidamente, os valores máximo, médio e mínimo
registrados para as variáveis limnológicas mesuradas na região litorânea das lagoas Carioca e
Gambazinho no período compreendido entre 6/2011 à 12/2012. Dentre tais variáveis, as que
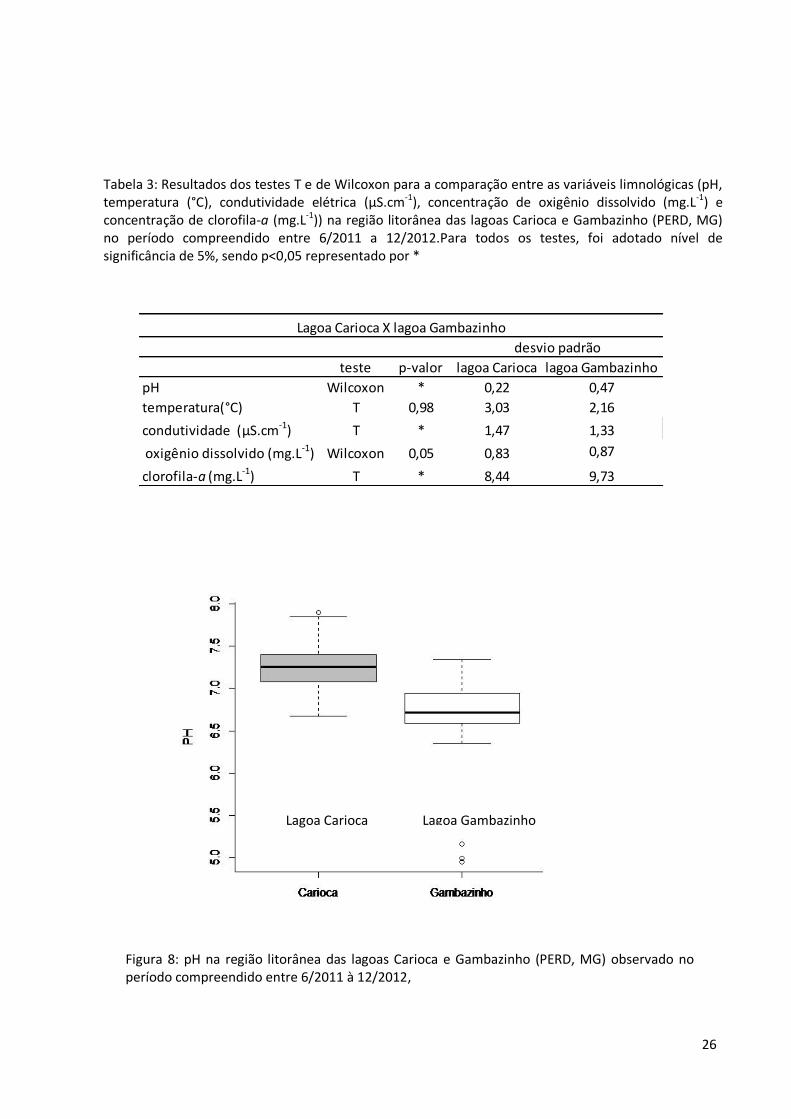
apresentaram diferenças estatisticamente significativas entre as lagoas foram: pH (fig. 8),
condutividade elétrica (fig.9) e concentração de clorofila-a (fig. 10), maiores na lagoa
Carioca. Os resultados dos testes T e de Wilcoxon estão resumidos na tabela 3.
Tabela 2: pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1) registrados na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 a 12/2012. As variáveis limnológicas marcadas com * apresentaram diferença estatisticamente significativa entre as lagoas
variáveis limnológicas valores mês lagoa Carioca mês lagoa Gambazinho
pH * máximo out/12 7,85 ago/11 7,34
médio 6,95 6,68
mínimo fev/12 6,67 jun/11 4,95
temperatura (°C) máximo out/12 35 dez/12 31,5
médio 28,13 28,12
mínimo jun/11 22,1 jun/11 23,4
condutividade (µS.cm-1) * máximo out/12 33,3 out/12 21,87
médio 27,75 14,09
mínimo dez/11 24,33 ago/12 12,31
oxigênio (mg.L-1) máximo ago/12 9,4 ago/12 9,1
médio 7,7 7,37
mínimo abr,12 6,16 jun/11 5,02
clorofila-a (mg.L-1) * máximo abr,12 48,65 jun/12 50,79
médio 22,03 17,96
mínimo out/12 14,43 out/11 3,2
Variáveis limnológica na região litorânea das lagoas Carioca e Gambazinho(PERD, MG)
26
Tabela 3: Resultados dos testes T e de Wilcoxon para a comparação entre as variáveis limnológicas (pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1)) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 a 12/2012.Para todos os testes, foi adotado nível de significância de 5%, sendo p<0,05 representado por *
Figura 8: pH na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) observado no período compreendido entre 6/2011 à 12/2012,
pH nas lagoas Carioca e Gambazinho
Lagoa Carioca Lagoa Gambazinho
teste p-valor lagoa Carioca lagoa Gambazinho
pH Wilcoxon * 0,22 0,47
temperatura(°C) T 0,98 3,03 2,16
condutividade (µS.cm-1) T * 1,47 1,33
oxigênio dissolvido (mg.L-1) Wilcoxon 0,05 0,83 0,87
clorofila-a (mg.L-1) T * 8,44 9,73
Lagoa Carioca X lagoa Gambazinho
desvio padrão

27
Figura 9: Condutividade elétrica (µS.cm-1) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) observada no período compreendido entre 6/2011 à 12/2012,
Condutividade elétrica nas lagoas Carioca e Gambazinho (PERD, MG)
Lagoa Carioca Lagoa Gambazinho
Co
nd
uti
vid
ade
elét
rica
(µ
S*cm
-1)
Concentração de clorofila-a nas lagoas Carioca e Gambazinho, (PERD, MG)
Lagoa Carioca Lagoa Gambazinho
Co
nce
ntr
ação
de
clo
rofi
la-a
(m
g.L-1
)
Figura 10: Concentração de clorofila-a (mg.L-1)na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) observada no período compreendido entre 6/2011 à 12/2012,
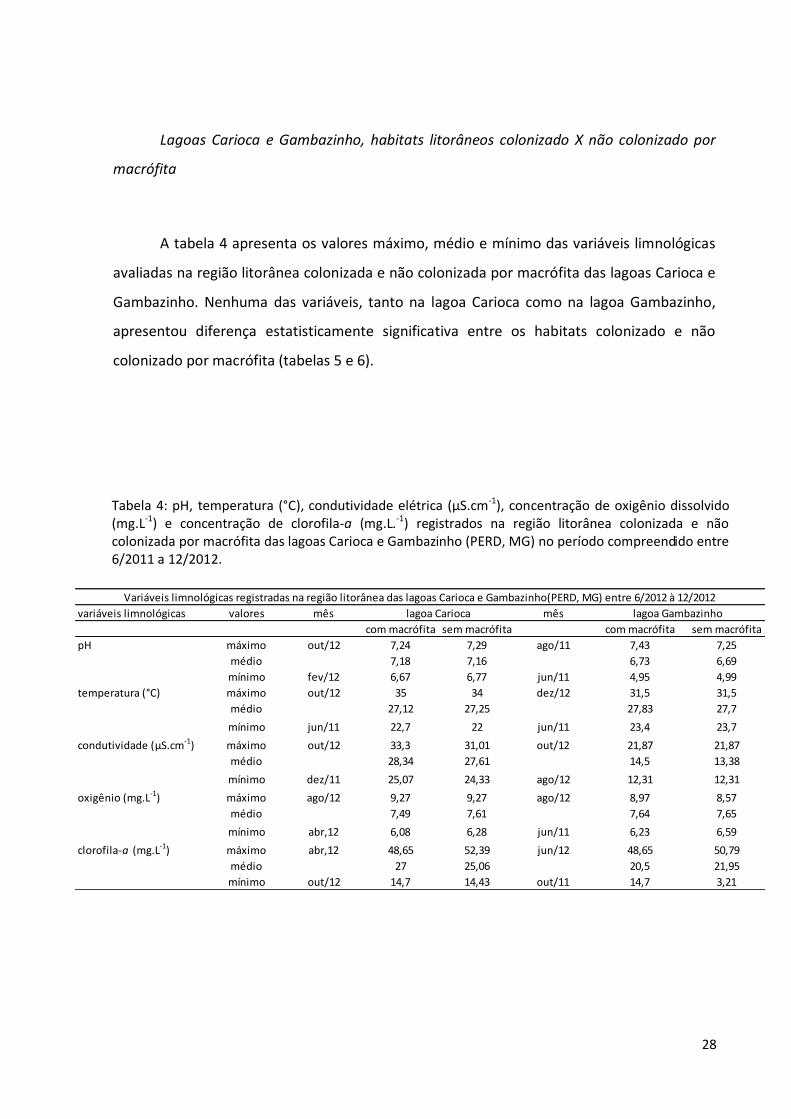
28
Lagoas Carioca e Gambazinho, habitats litorâneos colonizado X não colonizado por
macrófita
A tabela 4 apresenta os valores máximo, médio e mínimo das variáveis limnológicas
avaliadas na região litorânea colonizada e não colonizada por macrófita das lagoas Carioca e
Gambazinho. Nenhuma das variáveis, tanto na lagoa Carioca como na lagoa Gambazinho,
apresentou diferença estatisticamente significativa entre os habitats colonizado e não
colonizado por macrófita (tabelas 5 e 6).
Tabela 4: pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L.-1) registrados na região litorânea colonizada e não colonizada por macrófita das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 a 12/2012.
variáveis limnológicas valores mês mês
com macrófita sem macrófita com macrófita sem macrófita
pH máximo out/12 7,24 7,29 ago/11 7,43 7,25
médio 7,18 7,16 6,73 6,69
mínimo fev/12 6,67 6,77 jun/11 4,95 4,99
temperatura (°C) máximo out/12 35 34 dez/12 31,5 31,5
médio 27,12 27,25 27,83 27,7
mínimo jun/11 22,7 22 jun/11 23,4 23,7
condutividade (µS.cm-1) máximo out/12 33,3 31,01 out/12 21,87 21,87
médio 28,34 27,61 14,5 13,38
mínimo dez/11 25,07 24,33 ago/12 12,31 12,31
oxigênio (mg.L-1) máximo ago/12 9,27 9,27 ago/12 8,97 8,57
médio 7,49 7,61 7,64 7,65
mínimo abr,12 6,08 6,28 jun/11 6,23 6,59
clorofila-a (mg.L-1) máximo abr,12 48,65 52,39 jun/12 48,65 50,79
médio 27 25,06 20,5 21,95
mínimo out/12 14,7 14,43 out/11 14,7 3,21
lagoa Carioca lagoa Gambazinho
Variáveis limnológicas registradas na região litorânea das lagoas Carioca e Gambazinho(PERD, MG) entre 6/2012 à 12/2012
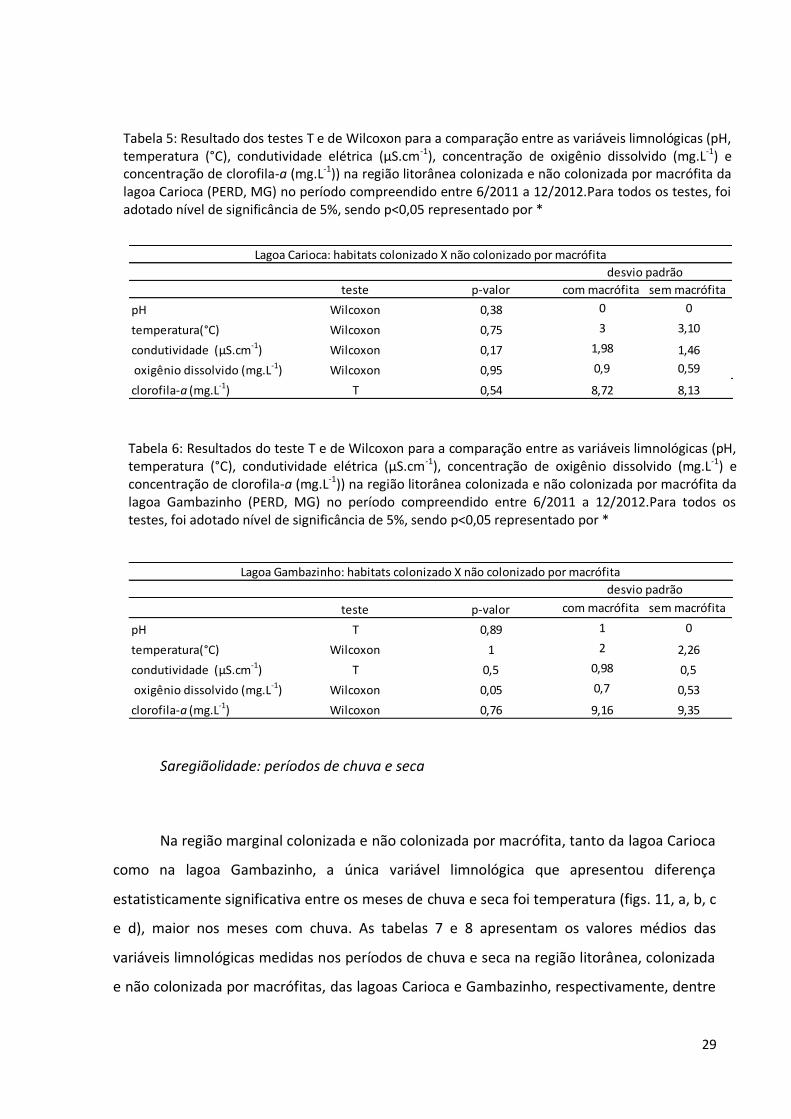
29
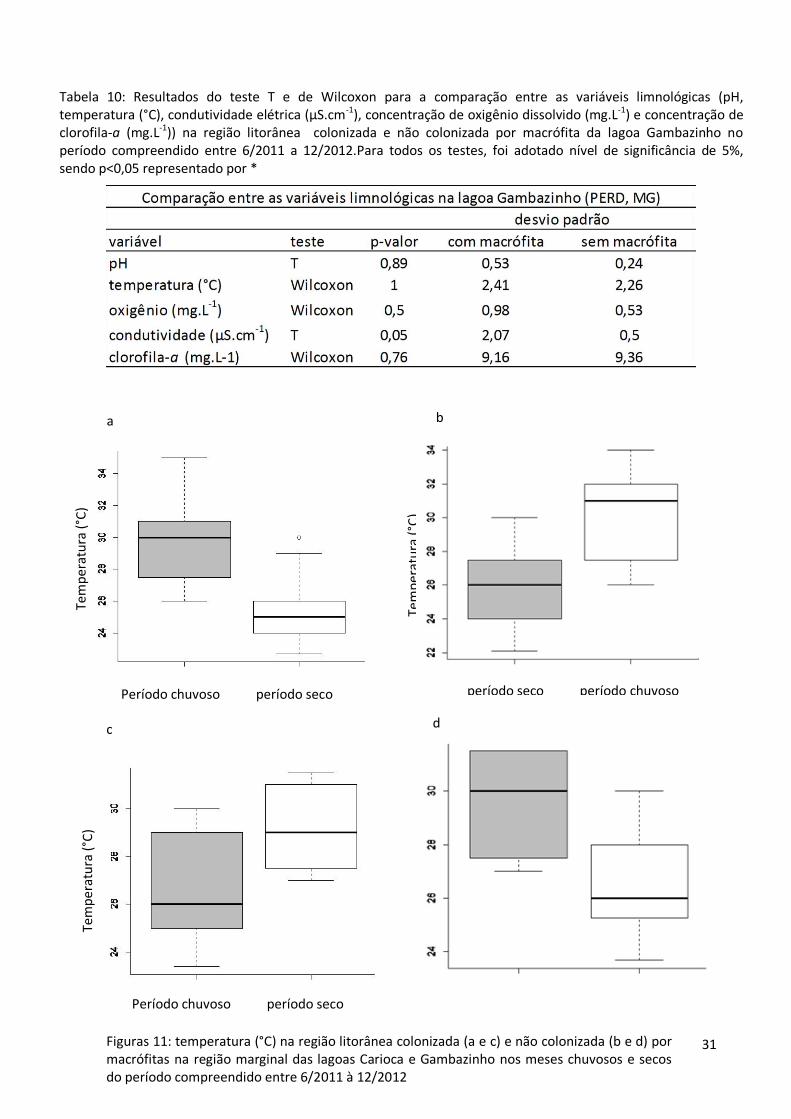
Saregiãolidade: períodos de chuva e seca
Na região marginal colonizada e não colonizada por macrófita, tanto da lagoa Carioca
como na lagoa Gambazinho, a única variável limnológica que apresentou diferença
estatisticamente significativa entre os meses de chuva e seca foi temperatura (figs. 11, a, b, c
e d), maior nos meses com chuva. As tabelas 7 e 8 apresentam os valores médios das
variáveis limnológicas medidas nos períodos de chuva e seca na região litorânea, colonizada
e não colonizada por macrófitas, das lagoas Carioca e Gambazinho, respectivamente, dentre
Tabela 5: Resultado dos testes T e de Wilcoxon para a comparação entre as variáveis limnológicas (pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1)) na região litorânea colonizada e não colonizada por macrófita da lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 a 12/2012.Para todos os testes, foi adotado nível de significância de 5%, sendo p<0,05 representado por *
Tabela 6: Resultados do teste T e de Wilcoxon para a comparação entre as variáveis limnológicas (pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1)) na região litorânea colonizada e não colonizada por macrófita da lagoa Gambazinho (PERD, MG) no período compreendido entre 6/2011 a 12/2012.Para todos os testes, foi adotado nível de significância de 5%, sendo p<0,05 representado por *
teste p-valor com macrófita sem macrófita
pH T 0,89 1 0
temperatura(°C) Wilcoxon 1 2 2,26
condutividade (µS.cm-1) T 0,5 0,98 0,5
oxigênio dissolvido (mg.L-1) Wilcoxon 0,05 0,7 0,53
clorofila-a (mg.L-1) Wilcoxon 0,76 9,16 9,35
Lagoa Gambazinho: habitats colonizado X não colonizado por macrófita
desvio padrão
teste p-valor com macrófita sem macrófita
pH Wilcoxon 0,38 0 0
temperatura(°C) Wilcoxon 0,75 3 3,10
condutividade (µS.cm-1) Wilcoxon 0,17 1,98 1,46
oxigênio dissolvido (mg.L-1) Wilcoxon 0,95 0,9 0,59
clorofila-a (mg.L-1) T 0,54 8,72 8,13
desvio padrão
Lagoa Carioca: habitats colonizado X não colonizado por macrófita
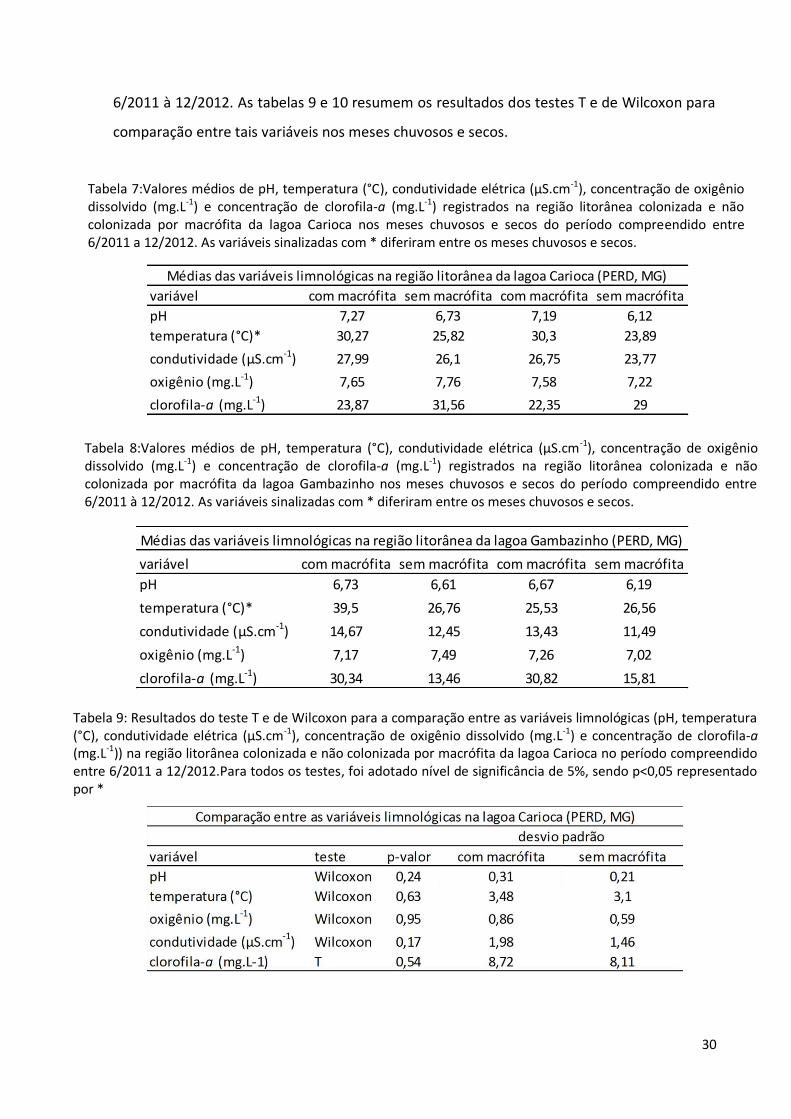
30
6/2011 à 12/2012. As tabelas 9 e 10 resumem os resultados dos testes T e de Wilcoxon para
comparação entre tais variáveis nos meses chuvosos e secos.
Tabela 7:Valores médios de pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1) registrados na região litorânea colonizada e não colonizada por macrófita da lagoa Carioca nos meses chuvosos e secos do período compreendido entre 6/2011 a 12/2012. As variáveis sinalizadas com * diferiram entre os meses chuvosos e secos.
Tabela 8:Valores médios de pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1) registrados na região litorânea colonizada e não colonizada por macrófita da lagoa Gambazinho nos meses chuvosos e secos do período compreendido entre 6/2011 à 12/2012. As variáveis sinalizadas com * diferiram entre os meses chuvosos e secos.
Tabela 9: Resultados do teste T e de Wilcoxon para a comparação entre as variáveis limnológicas (pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1)) na região litorânea colonizada e não colonizada por macrófita da lagoa Carioca no período compreendido entre 6/2011 a 12/2012.Para todos os testes, foi adotado nível de significância de 5%, sendo p<0,05 representado por *
variável com macrófita sem macrófita com macrófita sem macrófita
pH 7,27 6,73 7,19 6,12
temperatura (°C)* 30,27 25,82 30,3 23,89
condutividade (µS.cm-1) 27,99 26,1 26,75 23,77
oxigênio (mg.L-1) 7,65 7,76 7,58 7,22
clorofila-a (mg.L-1) * 23,87 31,56 22,35 29
Médias das variáveis limnológicas na região litorânea da lagoa Carioca (PERD, MG)
variável com macrófita sem macrófita com macrófita sem macrófita
pH 6,73 6,61 6,67 6,19
temperatura (°C)* 39,5 26,76 25,53 26,56
condutividade (µS.cm-1) 14,67 12,45 13,43 11,49
oxigênio (mg.L-1) 7,17 7,49 7,26 7,02
clorofila-a (mg.L-1) * 30,34 13,46 30,82 15,81
Médias das variáveis limnológicas na região litorânea da lagoa Gambazinho (PERD, MG)
31
Tabela 10: Resultados do teste T e de Wilcoxon para a comparação entre as variáveis limnológicas (pH, temperatura (°C), condutividade elétrica (µS.cm-1), concentração de oxigênio dissolvido (mg.L-1) e concentração de clorofila-a (mg.L-1)) na região litorânea colonizada e não colonizada por macrófita da lagoa Gambazinho no período compreendido entre 6/2011 a 12/2012.Para todos os testes, foi adotado nível de significância de 5%, sendo p<0,05 representado por *
b a
Tem
pe
ratu
ra (
°C)
Período chuvoso período seco
Temperatura na região litorânea colonizada por macrófitas da lagoa Gambazinho
Figuras 11: temperatura (°C) na região litorânea colonizada (a e c) e não colonizada (b e d) por macrófitas na região marginal das lagoas Carioca e Gambazinho nos meses chuvosos e secos do período compreendido entre 6/2011 à 12/2012
c d
Tem
per
atu
ra (
°C)
Temperatura na região litorânea não colonizada por macrófitas da lagoa Gambazinho
Tem
per
atu
ra (
°C)
Período chuvoso período seco período seco período chuvoso
Tem
per
atu
ra (
°C)
Período chuvoso período seco
d
32
Zooplâncton
Lagoa Carioca X Lagoa Gambazinho
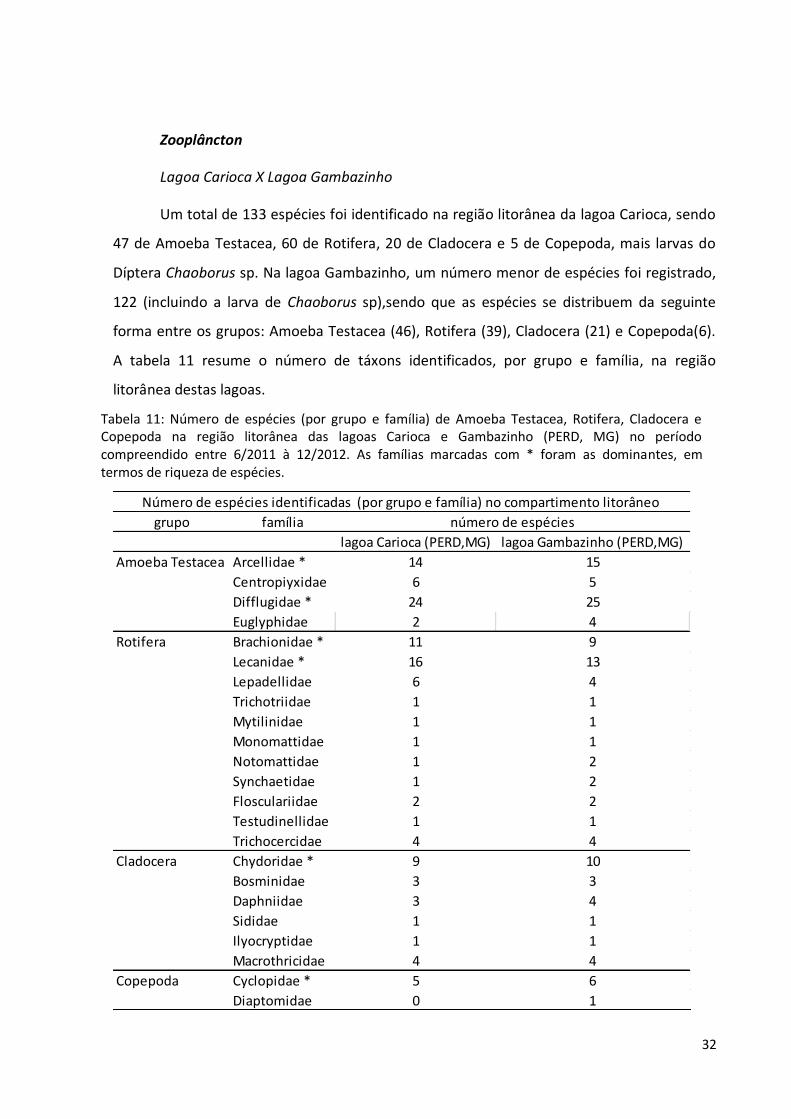
Um total de 133 espécies foi identificado na região litorânea da lagoa Carioca, sendo
47 de Amoeba Testacea, 60 de Rotifera, 20 de Cladocera e 5 de Copepoda, mais larvas do
Díptera Chaoborus sp. Na lagoa Gambazinho, um número menor de espécies foi registrado,
122 (incluindo a larva de Chaoborus sp),sendo que as espécies se distribuem da seguinte
forma entre os grupos: Amoeba Testacea (46), Rotifera (39), Cladocera (21) e Copepoda(6).
A tabela 11 resume o número de táxons identificados, por grupo e família, na região
litorânea destas lagoas.
grupo família
lagoa Carioca (PERD,MG) lagoa Gambazinho (PERD,MG)
Amoeba Testacea Arcellidae * 14 15
Centropiyxidae 6 5
Difflugidae * 24 25
Euglyphidae 2 4
Rotifera Brachionidae * 11 9
Lecanidae * 16 13
Lepadellidae 6 4
Trichotriidae 1 1
Mytilinidae 1 1
Monomattidae 1 1
Notomattidae 1 2
Synchaetidae 1 2
Flosculariidae 2 2
Testudinellidae 1 1
Trichocercidae 4 4
Cladocera Chydoridae * 9 10
Bosminidae 3 3
Daphniidae 3 4
Sididae 1 1
Ilyocryptidae 1 1
Macrothricidae 4 4
Copepoda Cyclopidae * 5 6
Diaptomidae 0 1
número de espécies
Número de espécies identificadas (por grupo e família) no compartimento litorâneo
Tabela 11: Número de espécies (por grupo e família) de Amoeba Testacea, Rotifera, Cladocera e Copepoda na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012. As famílias marcadas com * foram as dominantes, em termos de riqueza de espécies.
Co
nce
ntr
ação
de
clo
rofi
la-a
(m
g.L-1
)
c
33
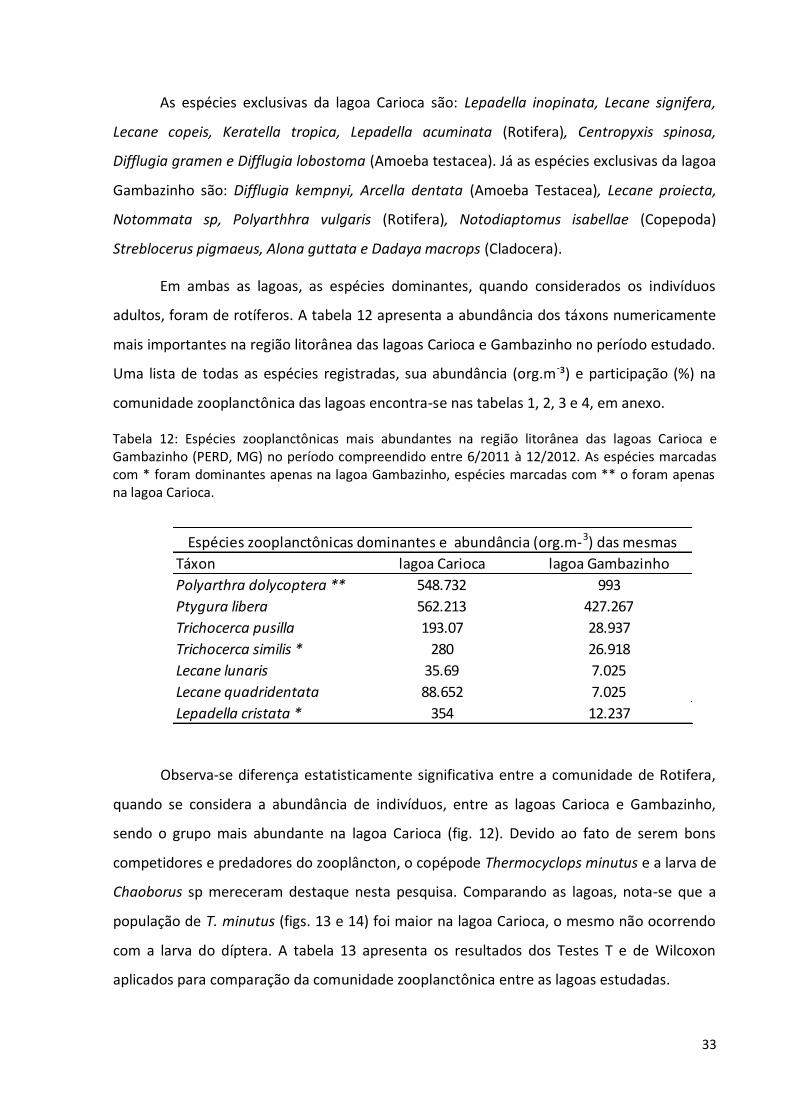
As espécies exclusivas da lagoa Carioca são: Lepadella inopinata, Lecane signifera,
Lecane copeis, Keratella tropica, Lepadella acuminata (Rotifera), Centropyxis spinosa,
Difflugia gramen e Difflugia lobostoma (Amoeba testacea). Já as espécies exclusivas da lagoa
Gambazinho são: Difflugia kempnyi, Arcella dentata (Amoeba Testacea), Lecane proiecta,
Notommata sp, Polyarthhra vulgaris (Rotifera), Notodiaptomus isabellae (Copepoda)
Streblocerus pigmaeus, Alona guttata e Dadaya macrops (Cladocera).
Em ambas as lagoas, as espécies dominantes, quando considerados os indivíduos
adultos, foram de rotíferos. A tabela 12 apresenta a abundância dos táxons numericamente
mais importantes na região litorânea das lagoas Carioca e Gambazinho no período estudado.
Uma lista de todas as espécies registradas, sua abundância (org.m-³) e participação (%) na
comunidade zooplanctônica das lagoas encontra-se nas tabelas 1, 2, 3 e 4, em anexo.
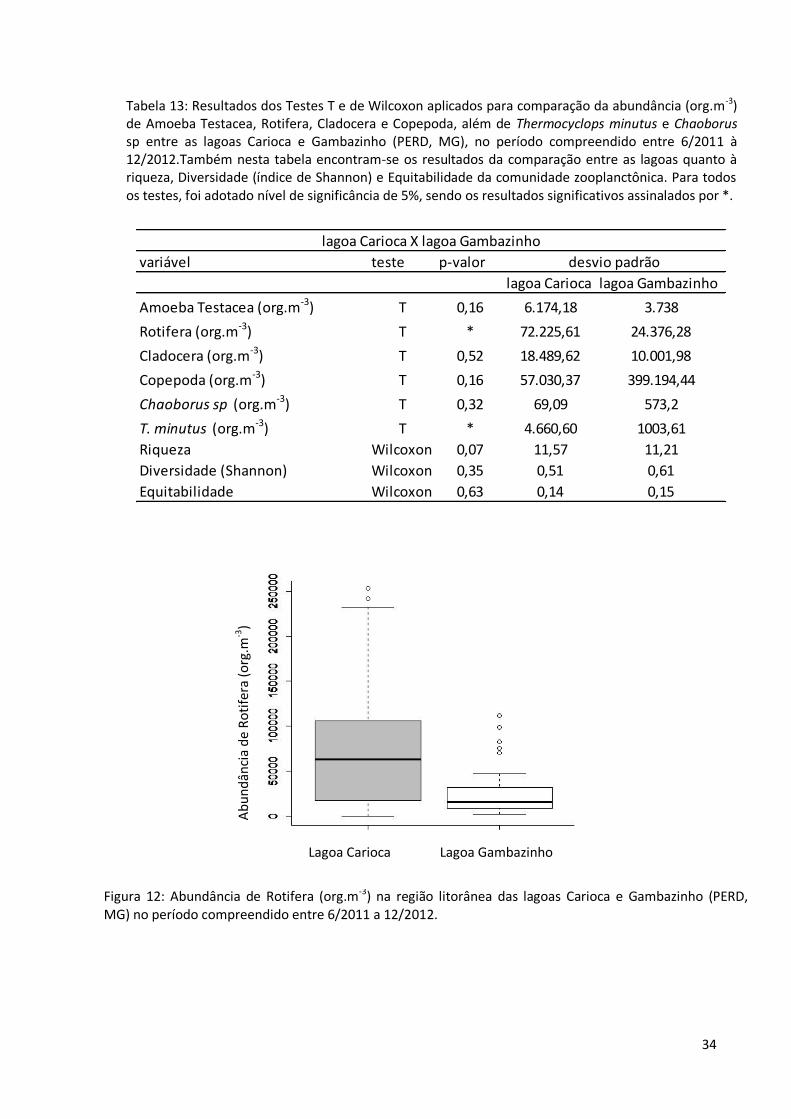
Observa-se diferença estatisticamente significativa entre a comunidade de Rotifera,
quando se considera a abundância de indivíduos, entre as lagoas Carioca e Gambazinho,
sendo o grupo mais abundante na lagoa Carioca (fig. 12). Devido ao fato de serem bons
competidores e predadores do zooplâncton, o copépode Thermocyclops minutus e a larva de
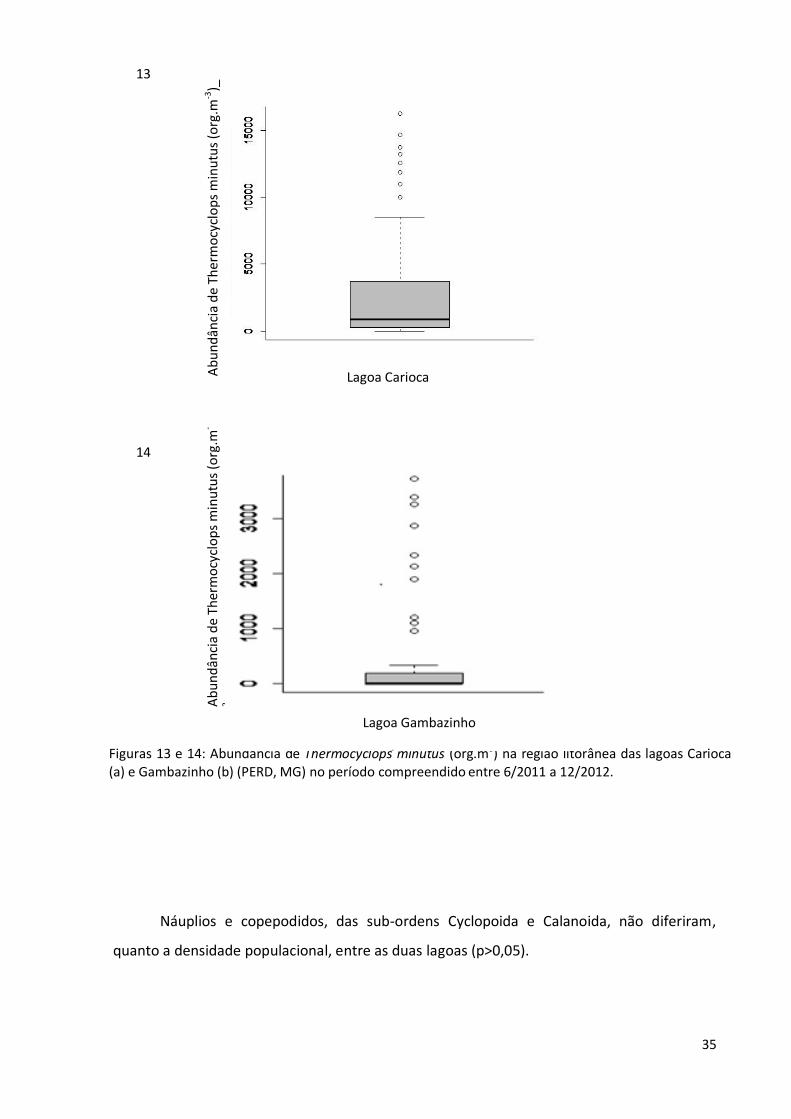
Chaoborus sp mereceram destaque nesta pesquisa. Comparando as lagoas, nota-se que a
população de T. minutus (figs. 13 e 14) foi maior na lagoa Carioca, o mesmo não ocorrendo
com a larva do díptera. A tabela 13 apresenta os resultados dos Testes T e de Wilcoxon
aplicados para comparação da comunidade zooplanctônica entre as lagoas estudadas.
Tabela 12: Espécies zooplanctônicas mais abundantes na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012. As espécies marcadas com * foram dominantes apenas na lagoa Gambazinho, espécies marcadas com ** o foram apenas na lagoa Carioca.
Táxon lagoa Carioca lagoa Gambazinho
Polyarthra dolycoptera ** 548.732 993
Ptygura libera 562.213 427.267
Trichocerca pusilla 193.07 28.937
Trichocerca similis * 280 26.918
Lecane lunaris 35.69 7.025
Lecane quadridentata 88.652 7.025
Lepadella cristata * 354 12.237
Espécies zooplanctônicas dominantes e abundância (org.m-3) das mesmas
34
Tabela 13: Resultados dos Testes T e de Wilcoxon aplicados para comparação da abundância (org.m-3) de Amoeba Testacea, Rotifera, Cladocera e Copepoda, além de Thermocyclops minutus e Chaoborus sp entre as lagoas Carioca e Gambazinho (PERD, MG), no período compreendido entre 6/2011 à 12/2012.Também nesta tabela encontram-se os resultados da comparação entre as lagoas quanto à riqueza, Diversidade (índice de Shannon) e Equitabilidade da comunidade zooplanctônica. Para todos os testes, foi adotado nível de significância de 5%, sendo os resultados significativos assinalados por *.
Figura 12: Abundância de Rotifera (org.m-3) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 a 12/2012.
variável teste p-valor
lagoa Carioca lagoa Gambazinho
Amoeba Testacea (org.m-3) T 0,16 6.174,18 3.738
Rotifera (org.m-3) T * 72.225,61 24.376,28
Cladocera (org.m-3) T 0,52 18.489,62 10.001,98
Copepoda (org.m-3) T 0,16 57.030,37 399.194,44
Chaoborus sp (org.m-3) T 0,32 69,09 573,2
T. minutus (org.m-3) T * 4.660,60 1003,61
Riqueza Wilcoxon 0,07 11,57 11,21
Diversidade (Shannon) Wilcoxon 0,35 0,51 0,61
Equitabilidade Wilcoxon 0,63 0,14 0,15
desvio padrão
lagoa Carioca X lagoa Gambazinho
Lagoa Carioca Lagoa Gambazinho
Ab
un
dân
cia
de
Ro
tife
ra (
org
.m-3
)
35
Náuplios e copepodidos, das sub-ordens Cyclopoida e Calanoida, não diferiram,
quanto a densidade populacional, entre as duas lagoas (p>0,05).
Figuras 13 e 14: Abundância de Thermocyclops minutus (org.m3) na região litorânea das lagoas Carioca (a) e Gambazinho (b) (PERD, MG) no período compreendido entre 6/2011 a 12/2012.
Lagoa Carioca Ab
un
dân
cia
de
Ther
mo
cycl
op
s m
inu
tus
(org
.m-3
)_ 13
14
Ab
un
dân
cia
de
Ther
mo
cycl
op
s m
inu
tus
(org
.m-
3)_
Lagoa Gambazinho

36
Habitats litorâneos colonizados X não colonizados por macrófitas
Não houve diferença estatisticamente significativa quanto à abundância de Amoeba
Testacea, Rotifera, Cladocera e Copepoda entre os habitats litorâneos colonizado e não
colonizado por macrófitas, tanto na lagoa Carioca, como na lagoa Gambazinho, porém foi
observada diferença quanto à riqueza de espécies zooplanctônicas entre estes habitats na
lagoa Carioca, o mesmo não acontecendo com Diversidade e Equitabilidade entre as
comunidades zooplanctônicas das regiões marginal com e sem plantas aquáticas desta
lagoa. Na lagoa Gambazinho, riqueza, Diversidade e Equitabilidade não diferiram entre as
regiões litorânea colonizada e não colonizada por macrófita.
Em relação aos náuplios e copepodidos Cyclopoida, tanto na lagoa Carioca como na
lagoa Gambazinho, não houve diferenças estatisticamente significativas entre os habitats
litorâneos com e sem macrófitas, o mesmo ocorrendo com os indivíduos jovens de
Calanoida (tabela 14). Considerando a abundância das populações de T. minutus e
Chaoborus sp nos habitats litorâneos em ambas as lagoas, também não se vê diferenças
entre as regiões com e sem vegetação marginal. Nas tabelas 15 e 16 estão os resultados dos
testes T e de Wilcoxon, usados para comparação da comunidade zooplanctônica da região
litorânea com e sem plantas aquáticas de ambas as lagoas.
Tabela 14: Abundância de náuplios e copepodidos, Cyclopoida e Calanoida, na região litorânea colonizada e não colonizada por macrófita das lagoas Carioca e Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012.
com macrófita sem macrófita com macrófita sem macrófita
náuplio Cyclopoida 1.429.496 1.170.432 286.243.456 1.244.880
copepodido Cyclopoida 321.898 334.343 451.259 286.353
náuplio Calanoida 25.815 0 615.330 590.018
copepodido Calanoida 7.833 130 44.045 25.118
Abundância (org.m-3) de copépodes jovens no compartimento litorâneo das lagoas Carioca e Gambazinho (PERD, MG)
lagoa Carioca lagoa Gambazinho
37
Tabela 15: resultados dos testes T e de wilcoxon aplicados para comparação da abundância de Amoeba Testacea, Rotifera, Cladocera, Copepoda, além de riqueza, Diversidade e Equitabilidade na comunidade zooplanctônica entre os habitats litorâneos colonizado e não colonizado por macrófita na lagoa Carioca (PERD, MG) no período compreendido entre 6/2011 à 12/2012. A tabela também mostra os resultados do mesmo teste para comparação entre a abundância de Thermocyclops minutus, Chaoborus sp, náuplios e copepodidos entre a região marginal com e sem plantas aquáticas.
teste p-valor com macrófita sem macrófita
Amoeba Testacea (org.m-3) Wilcoxon 0,55 5.050 654
Rotifera (org.m-3) T 0,19 28.606 19.157
Cladocera (org.m-3) T 0,5 7.623 12.161,10
Copepoda (org.m-3) Wilcoxon 0,41 55.908.816 46.434,40
Chaoborus sp (org.m-3) T 0,39 629,91 519,69
Thermocyclops minutus (org.m-3) T 0,67 918,02 1.100,78
Riqueza Wilcoxon 0,75 12,36 10,07
Diversidade (Shannon) Wilcoxon 0,2 0,54 0,56
Equitabilidade Wilcoxon 0,13 0,16 0,13
náuplio Cyclopoida (org.m-3) T 0,16 55.869.973 29.126,06
copepodido Cyclopoida (org.m-3) T 0,13 24.880,54 7.532,14
náuplio Calanoida (org.m-3) T 0,49 23.915,19 16.178,52
copepodido Calanoida (org.m-3) T 0,12 2.601,44 992,62
Lagoa Gambazinho: habitats colonizado X não colonizado por macrófita
desvio padrão
Tabela 16: resultados dos testes T e de Wilcoxon aplicados para comparação da abundância de Amoeba Testacea, Rotifera, Cladocera, Copepoda, além de riqueza, Diversidade e Equitabilidade na comunidade zooplanctônica entre os habitats litorâneos colonizado e não colonizado por macrófita na lagoa Gambazinho (PERD, MG) no período compreendido entre 6/2011 à 12/2012. A tabela também mostra os resultados do mesmo teste para comparação entre a abundância de Thermocyclops minutus, Chaoborus sp, náuplios e copepodidos entre a região marginal com e sem plantas aquáticas.
teste p-valor com macrófita sem macrófita
Amoeba Testacea (org.m-3) T 0,62 3.732 4.036
Rotifera (org.m-3) T 0,61 71.773 59.407
Cladocera (org.m-3) T 0,23 24.247 4.768,57
Copepoda (org.m-3) T 0,27 61.615 40.995,04
Chaoborus sp (org.m-3) T 0,69 448,96 373,70
Thermocyclops minutus (org.m-3) T 0,39 4.994,17 3.347,61
Riqueza Wilcoxon * 8,56 10,12
Diversidade (Shannon) Wilcoxon 0,21 0,51 0,54
Equitabilidade Wilcoxon 0,43 0,13 0,14
náuplio Cyclopoida (org.m-3) T 0,27 54.916,23 28.571,09
copepodido Cyclopoida (org.m-3) T 0,55 13.756,75 16.045,41
náuplio Calanoida (org.m-3) T 0,11 3.992,24 0
copepodido Calanoida (org.m-3) T 0,18 579,36 9,98
Lagoa Carioca: habitats colonizado X não colonizado por macrófita
desvio padrão

38
As espécies exclusivas da região litorânea colonizada por macrófita, não colonizada
por plantas aquáticas e comuns a ambos os habitats litorâneos nas lagoas Carioca e
Gambazinho, e suas respectivas abundâncias, estão listadas nas tabelas 1, 2, 3 e 4. em
anexo.
As figuras 15 (a, b, c, d, e, f) são os resultados das análises de cluster, expressos como
dendrogramas, para a comparação entre a abundância de espécies dos grupos Amoeba
Testacea, Rotifera, Cladocera, Copepoda, além da abundância de larvas de Chaoborus e
Thermocyclops minutus entre os habitats litorâneos colonizado e não colonizado por
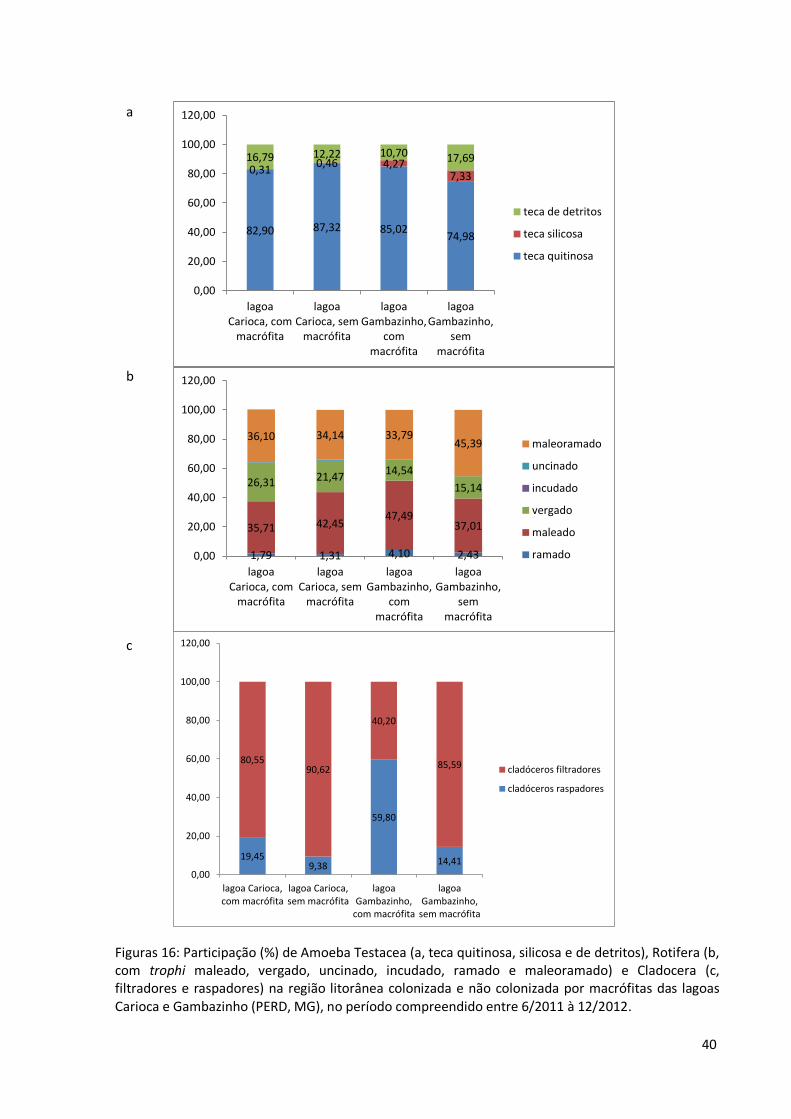
macrófitas nas lagoas estudadas. As figuras 16 (a, b, c e d) são referentes à participação (em
porcentagem) dos animais zooplanctônicos dentre guildas, cujos critérios para inclusão em
determinado grupo foram: tipo de teca para Amoeba Testacea (quitinosa, silicosa e de
detritos), morfologia do trofe para Rotifera (vergado, uncinado, maleoramado, incudado,
ramado), modo de alimentação para Cladocera (raspadores e filtradores) e hábito alimentar
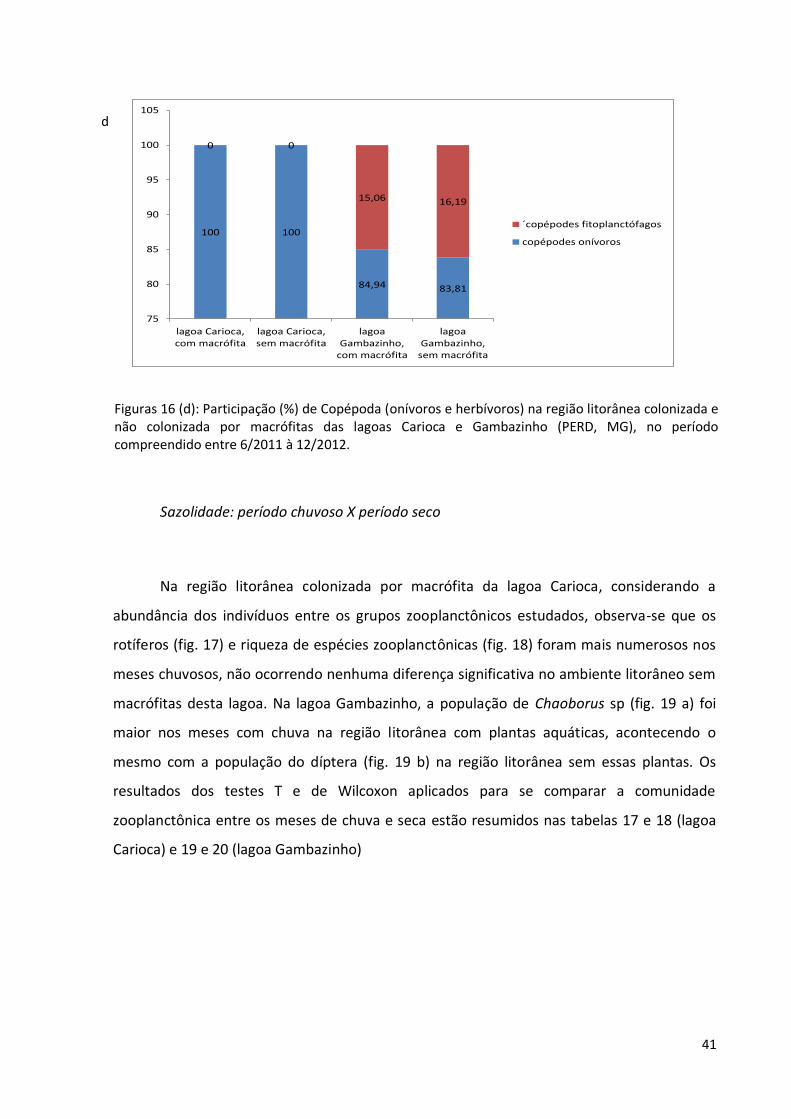
para Copepoda (onívoros e fitoplanctônicos)
Amoeba Testacea (org.m-3) Rotifera (org.m-3) a b
Dis
tân
cia
Eucl
idia
na
Dis
tân
cia
Eucl
idia
na
Figuras 15: comparação entre as comunidades de Amoeba Testacea (a) e Rotifera (b) entre os habitats litorâneos colonizados e não colonizados por macrófitas das lagoas Carioca e Gambazinho (PERD, MG), entre 6/2011 à 12/2012.

39
Dis
tân
cia
eucl
idia
na
Cladocera (org.m-3) Copepoda (org.m-3) c d
Dis
tân
cia
Eucl
idia
na
Dis
tân
cia
Eucl
idia
na
Figuras 15: comparação entre as comunidades de Cladocera(c) Copepoda (d), Chaoborus sp (e) e Thermocyclops minutus (f) entre os habitats litorâneos colonizados e não colonizados por macrófitas das lagoas Carioca e Gambazinho (PERD, MG), entre 6/2011 à 12/2012.
Chaoborus sp (org.m-3) Thermocyclops minutus (org.m-3) e f
Dis
tân
cia
Eucl
idia
na
Dis
tân
cia
Eucl
idia
na
40
82,90 87,32 85,0274,98
0,310,46 4,27
7,33
16,79 12,22 10,70 17,69
0,00
20,00
40,00
60,00
80,00
100,00
120,00
lagoaCarioca, com
macrófita
lagoaCarioca, sem
macrófita
lagoaGambazinho,
commacrófita
lagoaGambazinho,
semmacrófita
teca de detritos
teca silicosa
teca quitinosa
1,79 1,31 4,10 2,43
35,71 42,4547,49
37,01
26,31 21,4714,54
15,14
36,10 34,14 33,7945,39
0,00
20,00
40,00
60,00
80,00
100,00
120,00
lagoaCarioca, com
macrófita
lagoaCarioca, sem
macrófita
lagoaGambazinho,
commacrófita
lagoaGambazinho,
semmacrófita
maleoramado
uncinado
incudado
vergado
maleado
ramado
19,459,38
59,80
14,41
80,5590,62
40,20
85,59
0,00
20,00
40,00
60,00
80,00
100,00
120,00
lagoa Carioca,com macrófita
lagoa Carioca,sem macrófita
lagoaGambazinho,
com macrófita
lagoaGambazinho,
sem macrófita
cladóceros filtradores
cladóceros raspadores
Figuras 16: Participação (%) de Amoeba Testacea (a, teca quitinosa, silicosa e de detritos), Rotifera (b, com trophi maleado, vergado, uncinado, incudado, ramado e maleoramado) e Cladocera (c, filtradores e raspadores) na região litorânea colonizada e não colonizada por macrófitas das lagoas Carioca e Gambazinho (PERD, MG), no período compreendido entre 6/2011 à 12/2012.
a
b
c
41
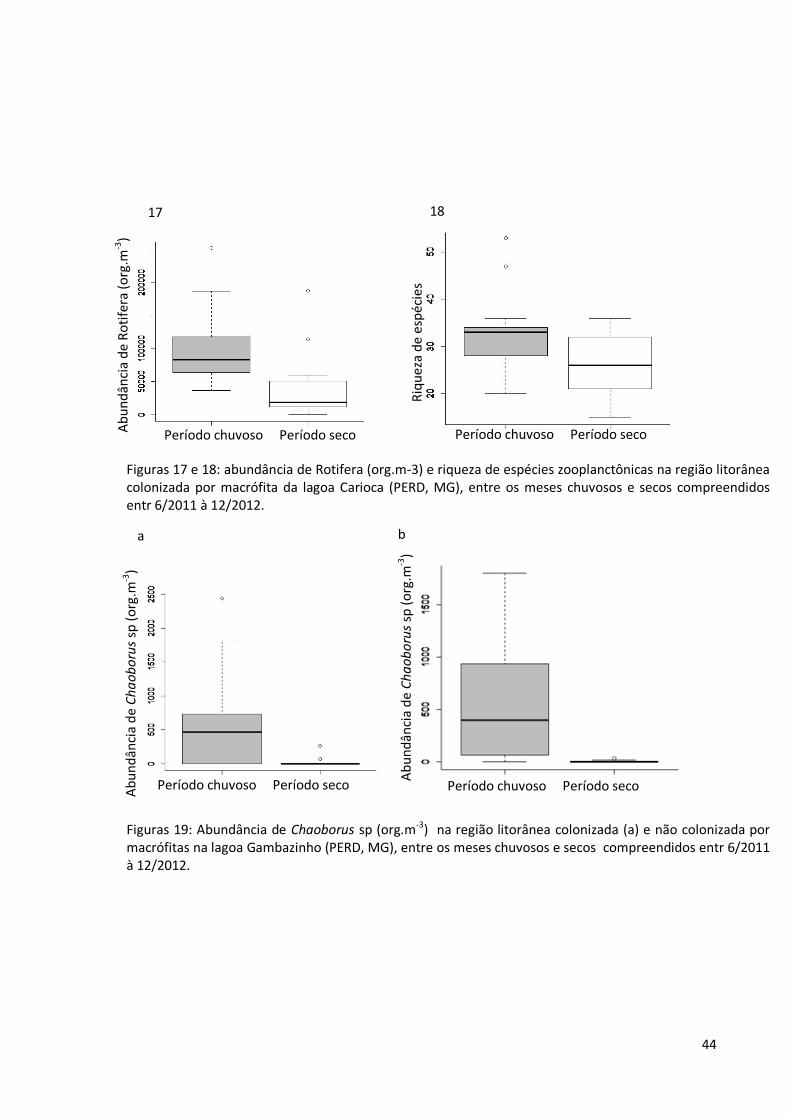
Sazolidade: período chuvoso X período seco
Na região litorânea colonizada por macrófita da lagoa Carioca, considerando a
abundância dos indivíduos entre os grupos zooplanctônicos estudados, observa-se que os
rotíferos (fig. 17) e riqueza de espécies zooplanctônicas (fig. 18) foram mais numerosos nos
meses chuvosos, não ocorrendo nenhuma diferença significativa no ambiente litorâneo sem
macrófitas desta lagoa. Na lagoa Gambazinho, a população de Chaoborus sp (fig. 19 a) foi
maior nos meses com chuva na região litorânea com plantas aquáticas, acontecendo o
mesmo com a população do díptera (fig. 19 b) na região litorânea sem essas plantas. Os
resultados dos testes T e de Wilcoxon aplicados para se comparar a comunidade
zooplanctônica entre os meses de chuva e seca estão resumidos nas tabelas 17 e 18 (lagoa
Carioca) e 19 e 20 (lagoa Gambazinho)
100 100
84,94 83,81
0 0
15,06 16,19
75
80
85
90
95
100
105
lagoa Carioca,com macrófita
lagoa Carioca,sem macrófita
lagoaGambazinho,
com macrófita
lagoaGambazinho,
sem macrófita
´copépodes fitoplanctófagos
copépodes onívoros
Figuras 16 (d): Participação (%) de Copépoda (onívoros e herbívoros) na região litorânea colonizada e não colonizada por macrófitas das lagoas Carioca e Gambazinho (PERD, MG), no período compreendido entre 6/2011 à 12/2012.
d
42
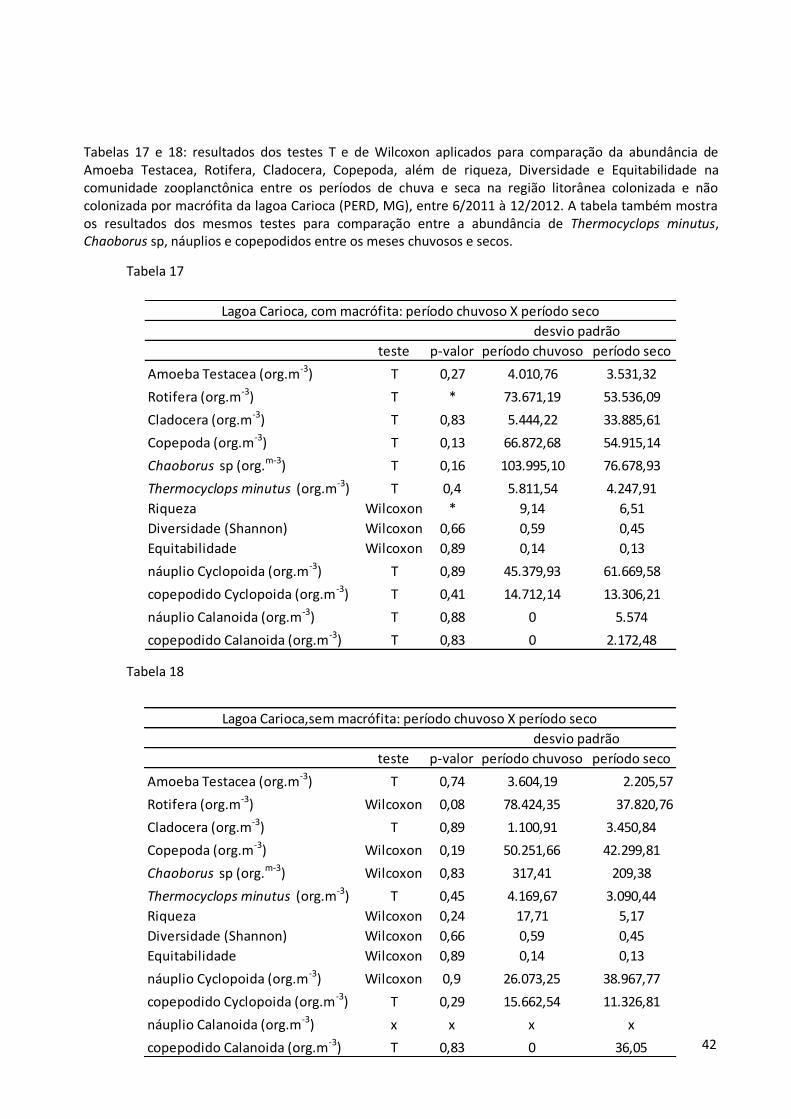
Tabelas 17 e 18: resultados dos testes T e de Wilcoxon aplicados para comparação da abundância de Amoeba Testacea, Rotifera, Cladocera, Copepoda, além de riqueza, Diversidade e Equitabilidade na comunidade zooplanctônica entre os períodos de chuva e seca na região litorânea colonizada e não colonizada por macrófita da lagoa Carioca (PERD, MG), entre 6/2011 à 12/2012. A tabela também mostra os resultados dos mesmos testes para comparação entre a abundância de Thermocyclops minutus, Chaoborus sp, náuplios e copepodidos entre os meses chuvosos e secos.
Tabela 17
Tabela 18
teste p-valor período chuvoso período seco
Amoeba Testacea (org.m-3) T 0,74 3.604,19 2.205,57
Rotifera (org.m-3) Wilcoxon 0,08 78.424,35 37.820,76
Cladocera (org.m-3) T 0,89 1.100,91 3.450,84
Copepoda (org.m-3) Wilcoxon 0,19 50.251,66 42.299,81
Chaoborus sp (org.m-3) Wilcoxon 0,83 317,41 209,38
Thermocyclops minutus (org.m-3) T 0,45 4.169,67 3.090,44
Riqueza Wilcoxon 0,24 17,71 5,17
Diversidade (Shannon) Wilcoxon 0,66 0,59 0,45
Equitabilidade Wilcoxon 0,89 0,14 0,13
náuplio Cyclopoida (org.m-3) Wilcoxon 0,9 26.073,25 38.967,77
copepodido Cyclopoida (org.m-3) T 0,29 15.662,54 11.326,81
náuplio Calanoida (org.m-3) x x x x
copepodido Calanoida (org.m-3) T 0,83 0 36,05
Lagoa Carioca,sem macrófita: período chuvoso X período seco
desvio padrão
teste p-valor período chuvoso período seco
Amoeba Testacea (org.m-3) T 0,27 4.010,76 3.531,32
Rotifera (org.m-3) T * 73.671,19 53.536,09
Cladocera (org.m-3) T 0,83 5.444,22 33.885,61
Copepoda (org.m-3) T 0,13 66.872,68 54.915,14
Chaoborus sp (org.m-3) T 0,16 103.995,10 76.678,93
Thermocyclops minutus (org.m-3) T 0,4 5.811,54 4.247,91
Riqueza Wilcoxon * 9,14 6,51
Diversidade (Shannon) Wilcoxon 0,66 0,59 0,45
Equitabilidade Wilcoxon 0,89 0,14 0,13
náuplio Cyclopoida (org.m-3) T 0,89 45.379,93 61.669,58
copepodido Cyclopoida (org.m-3) T 0,41 14.712,14 13.306,21
náuplio Calanoida (org.m-3) T 0,88 0 5.574
copepodido Calanoida (org.m-3) T 0,83 0 2.172,48
desvio padrão
Lagoa Carioca, com macrófita: período chuvoso X período seco
43
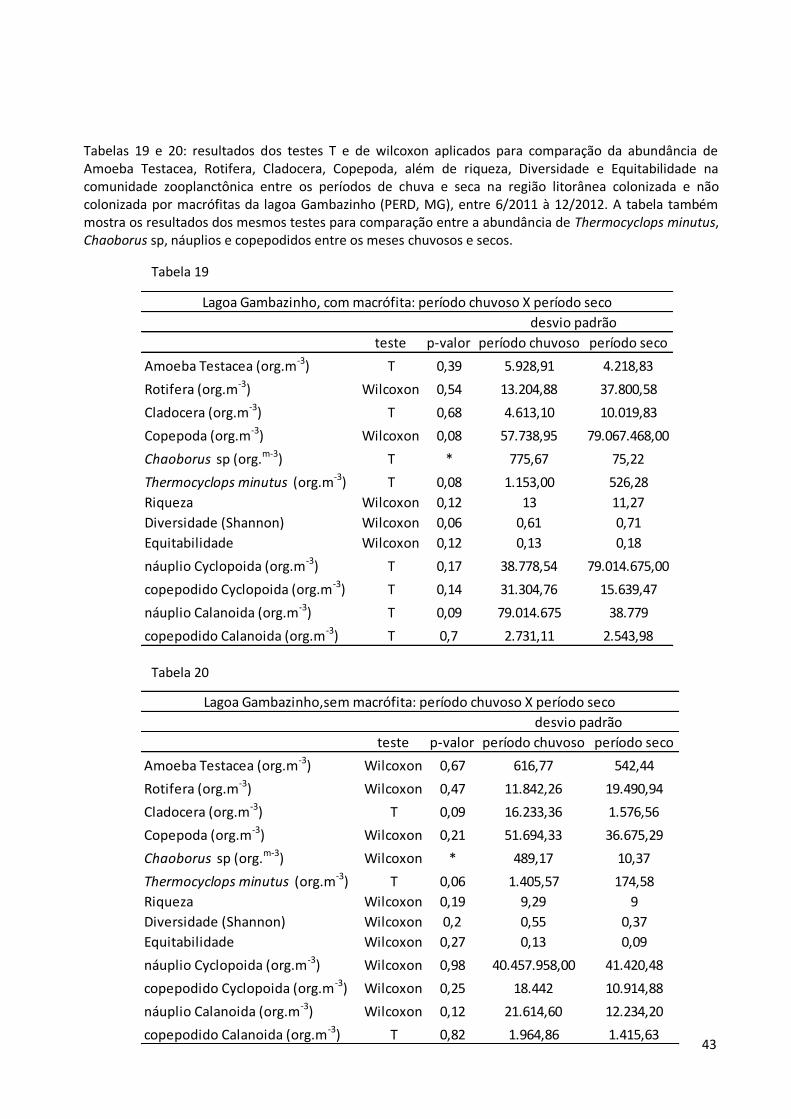
Tabelas 19 e 20: resultados dos testes T e de wilcoxon aplicados para comparação da abundância de Amoeba Testacea, Rotifera, Cladocera, Copepoda, além de riqueza, Diversidade e Equitabilidade na comunidade zooplanctônica entre os períodos de chuva e seca na região litorânea colonizada e não colonizada por macrófitas da lagoa Gambazinho (PERD, MG), entre 6/2011 à 12/2012. A tabela também mostra os resultados dos mesmos testes para comparação entre a abundância de Thermocyclops minutus, Chaoborus sp, náuplios e copepodidos entre os meses chuvosos e secos.
teste p-valor período chuvoso período seco
Amoeba Testacea (org.m-3) T 0,39 5.928,91 4.218,83
Rotifera (org.m-3) Wilcoxon 0,54 13.204,88 37.800,58
Cladocera (org.m-3) T 0,68 4.613,10 10.019,83
Copepoda (org.m-3) Wilcoxon 0,08 57.738,95 79.067.468,00
Chaoborus sp (org.m-3) T * 775,67 75,22
Thermocyclops minutus (org.m-3) T 0,08 1.153,00 526,28
Riqueza Wilcoxon 0,12 13 11,27
Diversidade (Shannon) Wilcoxon 0,06 0,61 0,71
Equitabilidade Wilcoxon 0,12 0,13 0,18
náuplio Cyclopoida (org.m-3) T 0,17 38.778,54 79.014.675,00
copepodido Cyclopoida (org.m-3) T 0,14 31.304,76 15.639,47
náuplio Calanoida (org.m-3) T 0,09 79.014.675 38.779
copepodido Calanoida (org.m-3) T 0,7 2.731,11 2.543,98
Lagoa Gambazinho, com macrófita: período chuvoso X período seco
desvio padrão
Tabela 19
Tabela 20
teste p-valor período chuvoso período seco
Amoeba Testacea (org.m-3) Wilcoxon 0,67 616,77 542,44
Rotifera (org.m-3) Wilcoxon 0,47 11.842,26 19.490,94
Cladocera (org.m-3) T 0,09 16.233,36 1.576,56
Copepoda (org.m-3) Wilcoxon 0,21 51.694,33 36.675,29
Chaoborus sp (org.m-3) Wilcoxon * 489,17 10,37
Thermocyclops minutus (org.m-3) T 0,06 1.405,57 174,58
Riqueza Wilcoxon 0,19 9,29 9
Diversidade (Shannon) Wilcoxon 0,2 0,55 0,37
Equitabilidade Wilcoxon 0,27 0,13 0,09
náuplio Cyclopoida (org.m-3) Wilcoxon 0,98 40.457.958,00 41.420,48
copepodido Cyclopoida (org.m-3) Wilcoxon 0,25 18.442 10.914,88
náuplio Calanoida (org.m-3) Wilcoxon 0,12 21.614,60 12.234,20
copepodido Calanoida (org.m-3) T 0,82 1.964,86 1.415,63
Lagoa Gambazinho,sem macrófita: período chuvoso X período seco
desvio padrão
44
Período chuvoso Período seco
Riq
uez
a d
e es
péc
ies
Período chuvoso Período seco Ab
un
dân
cia
de
Ro
tife
ra (
org
.m-3
)
Figuras 17 e 18: abundância de Rotifera (org.m-3) e riqueza de espécies zooplanctônicas na região litorânea colonizada por macrófita da lagoa Carioca (PERD, MG), entre os meses chuvosos e secos compreendidos entr 6/2011 à 12/2012.
Ab
un
dân
cia
de
Ch
aob
oru
s sp
(o
rg.m
-3)
Período chuvoso Período seco Período chuvoso Período seco
Ab
un
dân
cia
de
Ch
aob
oru
s sp
(o
rg.m
-3)
Figuras 19: Abundância de Chaoborus sp (org.m-3) na região litorânea colonizada (a) e não colonizada por macrófitas na lagoa Gambazinho (PERD, MG), entre os meses chuvosos e secos compreendidos entr 6/2011 à 12/2012.
a b
18 17

45
Discussão
Variáveis ambientais
Lagoa Carioca X lagoa Gambazinho
A lagoa Gambazinho apresentou temperatura média (28°C) maior que a temperatura
observada nesta lagoa durante o monitoramento PELD (24,71°C), em agosto de 2011, tal
diferença pode ser justificada pelo fato de que as coletas PELD são realizadas na região
limnética das lagoas, levando em consideração que a região limnética da lagoa Gambazinho
apresenta maior profundidade que a região litorânea, e portanto, maior massa de água por
unidade de volume, pode-se inferir que o aquecimento da água na região pelágica deste lago
é menor quando comparado ao aquecimento da água na região litorânea, mesmo sob a
mesma quantidade e tempo de incidência de luz. Evento semelhante ocorreu na lagoa
Carioca, provavelmente pelas mesmas causas inferidas para a lagoa Gambazinho. Em
comparação com o valor de temperatura observado no monitoramento do PELD, a lagoa
Carioca apresentou, nesta pesquisa, 4,5°C a mais que em agosto de 2011, cujo valor foi de
23,5°C.Comparando as lagoas entre si, vê-se que não há diferença entre as temperaturas
observadas em cada uma delas, provavelmente por ambas estarem submetidas ao mesmo
clima. Segundo Godinho (i996) e Moretto (2001), o clima da região caracteriza-se por
apresentar um período seco, de maio a agosto, e um período úmido, com temperaturas que
variam de 28 a 29°C nos meses chuvosos, e 7 a 20°C nos meses secos.
A concentração de oxigênio dissolvido na água é determinada por fatores diversos:
absorção do oxigênio atmosférico, atividade fotossintética e reações de oxi-redução, bióticas
ou abióticas. A solubilidade deste gás em meio aquoso é tanto menor quanto maior for a
temperatura, acontecendo o inverso em relação à pressão atmosférica. A solubilidade do
oxigênio também diminui na medida em que se aumenta a concentração de sais em solução
(Wetzel, 1983).
Comparando as concentrações de oxigênio dissolvido nas lagoas Carioca e
Gambazinho observadas entre junho de 2011 a dezembro de 2012 com aquelas observadas
durante o monitoramento PELD, observa-se menores concentrações de oxigênio dissolvido
nesta pesquisa (média de 7,3 mg.L-1, na lagoa Gambazinho; 7,7 mg.L-1na lagoa Carioca) do

46
que aquelas observadas em agosto de 2011 (média de 11,98 mg.L-1 na lagoa Gambazinho,
12,75 mg.L-1 na lagoa Carioca). Possivelmente essa diferença também se explica por serem as
coletas PELD feitas na região pelágica, onde é a região trofogênica, onde há produção de
oxigênio gasoso é mais distante do fundo, região trofolítica, onde o consumo deste gás é
elevado, além de haver liberação, pelo metabolismo microbiano, de gases, como dióxido de
carbono e gás sulfídrico, que competem com o oxigênio. Na região litorânea, a proximidade
com o sedimento, além da maior abundância e diversidade de organismos pode fazer com
que sejam menores a concentração do oxigênio dissolvido na água.
A maior temperatura observada na região marginal das lagoas Carioca e Gambazinho,
nesta pesquisa, poderia também ter determinado os menores valores de concentração de
oxigênio dissolvido, quando em comparação com o monitoramento PELD, visto que a
solubilidade deste gás diminui com a elevação da temperatura (Wetzel, 1983).
Utilizando apenas os valores de condutividade elétrica e concentração de clorofila-a a
lagoa Carioca mostrou maior grau de trofia do que a lagoa Gambazinho. Este resultado está
em concordância com os dados obtidos pelo monitoramento realizado pelo Programa PELD,
que utiliza além destas variáveis, as concentrações de fósforo e nitrogênio das águas. Águas
mais eutrofizadas, segundo Wetzel (1983) e Esteves (1996), apresentam maiores
concentrações de solutos originados da decomposição do plâncton e das macrófitas, o que
implica em maior condutividade, além disso, tais solutos, como íons nitrogenados e
fosfatados, fomentam a produtividade algal, elevando a concentração de pigmentos
fotossintetizantes na água.
Os maiores valores de pH observados na lagoa Carioca , quando em comparação com
aqueles registrados na lagoa Gambazinho podem ser decorrentes da natureza básica dos
compostos dissolvidos na água da lagoa Carioca, ou também da maior atividade
fotossintética nesta lagoa, realizada tanto por algas como por macrófitas, que consome o
gás carbônico do meio, evitando a formação de ácido carbônico, com consequente aumento
do pH da água.

47
Habitats litorâneos colonizado X não colonizado por macrófitas
Não houve diferença entre os parâmetros limnológicos observados nos habitats
colonizado e não colonizado por plantas aquáticas, tanto na lagoa Carioca como na lagoa
Gambazinho, possivelmente devido a ação de correntes horizontais na água, provocadas por
vento, e consequente homogeneização do pH, da temperatura, da concentração de oxigênio
dissolvido, condutividade elétrica e concentração de clorofila-a entre os ambientes
litorâneos. Maia- Barbosa et al (2008) também apontaram o vento e a mistura da água no
eixo horizontal como um possível fator determinante da semelhança entre as variáveis
limnológicas entre a região litorânea colonizada e não colonizada por macrófita na lagoa
Dom Helvécio (PERD, MG). O fato de serem as lagoas Carioca e Gambazinho corpos d’água
pequenos, também poderia ser um fator determinante da semelhança físico-química
observada na água entre a região marginal com e sem plantas aquáticas, pois torna menores
as distâncias percorridas por partículas dissolvidas ou em suspensão na água, quando
movimentadas por correntes.
Sazolidade: período chuvoso X período seco
As maiores temperaturas observadas nos meses chuvosos na região marginal das
lagoas Carioca e Gambazinho, tanto nos habitats colonizados como não colonizados por
macrófitas, são resultantes do aquecimento provocado pela incidência dos raios solares, e
também pela absorção de calor atmosférico, visto que nos meses chuvosos, na região
tropical, é maior a temperatura do ar.
Contrariando Esteves (1998), que afirma haver aumento da concentração de
clorofila-a nos corpos d’água durante o período chuvoso, devido ao input de nutrientes
advindos do meio terrestre adjacente, carreados pela água pluvial, na região litorânea com e
sem plantas aquáticas das lagoas Carioca e Gambazinho não houve diferença na
concentração de clorofila-a na água entre os meses chuvosos e secos. Possíveis causas para
este evento seria a maior eficiência de absorção de tais nutriente por macrófitas aquáticas,
especialmente as enraizadas, tornando o excedente de nutrientes advindos com a água
pluvial indisponível para a comunidade fitoplanctônica.
As semelhantes concentrações de clorofila-a observadas nos meses chuvosos,
quando comparadas às concentrações medidas nos meses secos, na margem não colonizada

48
por macrófitas em ambas as lagoas podem também ser resultantes do efeito diluidor:
diminuição das concentrações de solutos, como exemplo nitratos, amônio e fosfatos, por
aumento da quantidade de solvente (água). A diluição dos íons nitrogenados e fosfatados
reduziria a produtividade algal e por conseguinte, a concentração de pigmentos
fotossintetizantes no meio.
Comunidade zooplanctônica
A região marginal das lagoas Carioca e Gambazinho destaca-se pela elevada
diversidade de organismos zooplanctônicos, sendo um reservatório em potencial de
espécies para as lagoas como um todo. Os dados de riqueza de espécies levantados até o
momento para estas duas lagoas, referem-se a estudos realizados apenas na região
limnética. Estudos realizados na região litorânea de 2 outras lagoas do mesmo sistema, lagoa
Dom Helvécio e lagoa dos Patos, mostraram o acréscimo de 130 novas ocorrências de
espécies zooplanctônicas para a região, demonstrando a importância de estudos que
contemplem a região litorânea para o conhecimento da biodiversidade de um lago ( Maia-
Barbosa et al, 2008).
A predação, comumente aceita como fator determinante da riqueza zooplanctônica
pode manter baixa a abundância de determinadas espécies, diminuindo também a
competição interespecífica e permitindo a coexistência de numerosos táxons. A maioria dos
peixes faz uso do zooplâncton como alimento nas fases iniciais da vida, sendo que algumas
espécies o fazem também durante a fase adulta. Os estágios mais jovens predam
organismos menores, como protozoários, rotíferos e náuplios, enquanto que estágios
adultos podem se alimentar de zooplâncton de maior porte, como cladóceros, copépodos e
larvas de inseto (Santeiro e Pinto Coelho, 2000). Estudos sobre alimentação de jovens de
peixes estão sendo conduzidos nestas duas lagoas e nas mesmas regiões amostradas e têm
mostrado a utilização do zooplâncton como fonte alimentar (dados ainda não publicados).
Segundo Smith e colaboradores (2003), o ecótono água-terra, com suas plantas,
troncos, folhas, pedras e variedade de sedimento fornece aos peixes uma gama variada de
microhabitats e habitats para reprodução, deposição de ovos, forrageamento e abrigo
contra predadores. Durante as coletas foram avistados na margem da lagoa Carioca

49
cardumes numerosos de piranha (Pygocentrus nattereri) e tucunaré (Cichla ocelaris),
formados por indivíduos jovens, potenciais predadores do zooplâncton.
De acordo com Pinto-Coelho et al (2008), a introdução de piranha e tucunaré na
lagoa Carioca na década de 70 teve efeitos drásticos sobre a estrutura das comunidades
desta lagoa. Com a introdução destas espécies carnívoras, houve a redução das densidades
de peixes nativos, inclusive com a possível extinção local de seis espécies de peixes nativos
(Oporto, 2013). Nesta situação, larvas de Chaoborus sp. expandiram sua população
(Ecological Release) e utilizaram espécies da comunidade zooplanctônica como alimento,
reduzindo muito suas densidades. Isso talvez explique as baixas densidades de cladóceros na
lagoa Carioca. O tamboatá (Hoplosternum littorale), espécie bentônica também introduzida
nesta lagoa pode modificar a comunidade zooplanctônica ao ingerir espécies presentes no
sedimento. Hahn e colaboradores (1997) analisando o conteúdo estomacal de H. littorale
encontraram cladóceros (9,84%), amebas testáceas (12%), copépodes (5,91%) e rotíferos
(1,57%). Os microcrustáceos que se destacaram no estudo citado eram numerosos e
corriqueiros no ambiente onde foi feita a pesquisa. Segundo a autora, em muitos dos
estômagos analisados foram encontrados apenas cladóceros, sem vestígio de sedimento ou
detritos.
Comunidade zooplanctônica: Lagoa Carioca X Lagoa Gambazinho
Rotifera
Dentre os grupos zooplanctônicos o mais conhecido é o dos rotíferos, presentes em
quase todos os meios dulcícolas. No Brasil, 457 espécies foram descritas, segundo Oliveira-
Neto e Moreno (1999). Segers e Dumond (1994) afirmam que a maioria das espécies é
litorânea, devido a múltiplas causas: biogeografia, temperatura, concentração de oxigênio
dissolvido e pH.
Rotiferos filtram material em suspensão, incluindo bactérias e algas filamentosas,
sendo a maioria das espécies generalistas. Esses organismos têm elevadas taxas
reprodutivas e toleram condições adversas, (Bonecker et al , 2009; Lima et al, 2012)
características que podem determinar a predominância dos mesmos nos lagos tropicais
(Sendacz et al, 2006; Catunda, 2007; Lima et al, 2012). As famílias Lecanidae, Brachionidae e

50
Trichocercidae são as mais representativas, tanto em riqueza quanto abundância nesses
ambientes. Dentre estas famílias destaca-se a Lecanidae por apresentar gêneros
principalmente litorâneos (Panarelli, 2003; Aoyagui, 2008).
Lima et al, 2012, encontraram em lagoas marginais do Rio Cuiabá espécies das
famílias Lecanidae, Brachionidae e Trichocercidae, predominantemente dos gêneros Lecane,
Brachionus, Keratella e Trichocerca. Também nas lagoas Carioca e Gambazinho em áreas
com e sem macrófita, a abundância e riqueza de rotíferos foi grande sendo a família
Lecanidae a mais representativa e Brachionus o gênero dominante na lagoa Carioca e o
segundo em importância na lagoa Gambazinho.
A abundância de espécies dos gêneros Brachionus, Lecane, Trichocerca, Hexarthra e
Filinia estão diretamente relacionadas às concentrações de oxigênio dissolvido e clorofila-a
na coluna de água (Aoyagui, 2008). Isso talvez explique a abundância maior de B. mirus e H.
intermedia na lagoa Carioca (mesotrófica) do que na lagoa Gambazinho (oligotrófica). Tais
espécies podem ser mais tolerantes a menores concentrações de oxigênio dissolvido na
água, além de encontrarem alimento farto, devido à maior abundância de algas em meios
mais eutrofizados.
O predomínio de Rotifera em corpos d’água brasileiros, independentemente do grau
de trofia dos mesmos, é decorrente de seu ciclo de vida curto e reprodução
partenogenética, com elevadas taxas reprodutivas, além de seu comportamento oportunista
e maior resistência em ambientes instáveis e eutrofizados, quando comparados à outros
grupos zooplanctônicos (Matsumura-Tundisi et al, 1990; Sampaio et al, 2002). Muitos
rotíferos alimentam-se de matéria em suspensão de diversos tamanhos, como bactérias e
algas filamentosas, havendo também gêneros carnívoros (Lansac-Tôha, 1997). A
versatilidade dos hábitos alimentares deste grupo favorece também sua dominância nos
meios dulcícolas tornando-os elo importante de conversão do bacterio e fitoplâncton em
energia disponível aos consumidores secundários (Bonecker et al, 2005).
Em trabalho realizado no rio Cuiabá e lagoas marginais foram encontrados 114
táxons zooplanctônicos, sendo 72% de rotíferos, 16% de tecamebas, 9% de cladóceros e
1,5% de copépodos (Lima et al, 2012 ). Também no região litorânea das lagoas Carioca e
Gambazinho, Rotífera, quando considerados apenas indivíduos adultos, foi dominante. A
maior diversidade de Rotifera da lagoa Carioca, quando comparada à da lagoa Gambazinho

51
pode ser decorrente das condições tróficas das mesmas, sendo a primeira mesotrófica e a
última oligotrófica (Barbosa, 1979). Vanni (1987) relatou correlação positiva entre
enriquecimento nutricional e biomassa fito e zooplanctônica.
Pejler (1983) afirma que o aumento do grau de trofia da água fomenta o
desenvolvimento populacional de espécies especialistas no consumo de partículas
alimentares de pequenas dimensões, não favorecendo espécies filtradoras que tem
partículas de maior tamanho como recurso trófico. Berzins e Bertilsson (1988) observaram
relação entre táxons bentônicos, perifíticos e planctônicos com o estado trófico e os valores
de condutividade elétrica da água.
Segundo o mesmo autor, os hábitos alimentares destes animais são determinados
principalmente pela estrutura e função de dois órgãos: a coroa ciliar e o mástax. Espécies
com coroa ciliar bem desenvolvida e trophi dos tipos maleado e ramado, geralmente são
filtradores e selecionam partículas pequenas. O batimento dos cílios provoca uma corrente
na água que conduz partículas em suspensão até a boca do indivíduo. Rotíferos com mástax
do tipo incudado ou forcipado são macrófagos e predadores, como aqueles da família
Asplanchnidae e DicranopHoridae. As presas são capturadas acidentalmente, quando os
predadores encontram suas presas. Mástax uncinados, como nas espécies da família
Collothecacea, determinam a captura da presa (flagelados, ciliados) numa espécie de
alçapão, com cerdas no topo que impedem a fuga da presa. Indivíduos com mástax vergado
apresentam diversos comportamentos tróficos de acordo coma função da estrutura: sucção
ou “pumping”, algumas dessas espécies, como as do gênero AscomorpHa, perfuram o
tegumento do alimento e sugam seu conteúdo (Pourriot, 1977).
O tamanho das partículas alimentares ingeridas por rotíferos não filtradores vincula-
se ao tamanho do mástax, está por sua vez ligado ao tamanho corporal. Indivíduos de maior
porte se alimentam de presas e partículas de maiores dimensões (macrófagas), enquanto
que indivíduos menores se alimentam de partículas pequenas (micrófagas) (Pourriot, 1977).
Comparando a região litorânea das lagoas Carioca e Gambazinho observa-se que as
espécies detritívoras, bacteriófagas e/ou fitoplanctófagas são significativamente mais
abundantes na primeira lagoa, devido a maior disponibilidade de recursos alimentares para
essas espécies.

52
A decomposição parcial de macrófitas e algas tem como produto detritos (Wetzel,
1983) e favorece o desenvolvimento populacional de animais planctônicos detritívoros. Sua
decomposição total, libera na água nutrientes (cuja presença pode ser indicada também pela
condutividade elétrica, segundo o mesmo autor), como fósforo e nitrogênio, absorvido pelas
algas e bactérias, favorecendo as espécies fitoplanctófagas e bacteriófagas, como aquelas
dominantes na lagoa Carioca: Polyarthra vulgaris, P. dolycoptera e Trichocerca pusilla (fito e
zooplantófagas), Lecane quadridentata e L. lunaris (bacteriófagas e detritívoras). Tais
rotíferos, na lagoa Gambazinho, apresentam populações bem menores quando comparadas
as populações da lagoa Carioca. Outras espécies, cujas populações foram maiores na lagoa
Carioca, possivelmente por causa da maior disponibilidade de recursos alimentares na região
litorânea desta lagoa são: Hexarthra intermedia, Lecane hornemanii, Lecane lunaris, L.
quadridentata, Anuraeopsis fissa (bacteriófagas e detritívoras), Brachionus angularis, B.
mirus, B. quadridentatus (fitoplanctófagas, zooplanctófagas, bacteriófagas e detritívoras),
CepHalodella auriculata, CepHalodella gibba, Trichocerca pusilla (fito e zooplantófagas),
além de Euchlanis parva (fitoplanctófaga).
As espécies dominantes na lagoa Gambazinho, a saber: Trichocerca pusilla, T. similis
(fito e zooplanctófagas), Lepadella cristata, Ptygura libera,e Lecane lunaris (bacteriófagas e
detritívoras) apresentaram elevadas densidades populacionais quando comparadas a outras
espécies por encontrarem alimento farto na região litorânea desta lagoa e talvez explorarem
com mais eficiência que outros rotíferos. Estas espécies, excetuando-se T. pusilla,
apresentaram populações mais numerosas na lagoa Gambazinho do que na lagoa Carioca,
apesar da maior disponibilidade de alimentos nesta, provavelmente devido às menores
pressão de predação por alevinos e peixes jovens, visto que na lagoa Carioca, além de
alevinos e juvenis da ictiofauna nativa, há também alevinos e juvenis de peixes exóticos.
Em relação ao aparato bucal das espécies (segundo classificação de Stemberger,
1979), observa-se que espécies com tropHi dos tipos maleado, vergado e maleoramado são
mais abundantes na região litorânea da lagoa Carioca, sendo os últimos registrados
exclusivamente nesta lagoa. A maioria destas espécies se alimentam de partículas pequenas,
que segundo Wetzel (1983), tem maior ocorrência em ambientes com maior grau de trofia.
As espécies da família Trichocercidae, segundo Pejler e Berzins (1993), são
encontradas em diversos tipos de substratos, sedimento, vegetação submersa, macrófitas

53
flutuantes livres e também na coluna d’água, dentre o plâncton, onde depositam seus ovos e
se alimentam. Pesquisando em diversos lagos e riachos suecos, os autores constataram a
ocorrência de Trichocerca bicristata, T. pusilla e T. similis dentre organismos planctônicos e
plantas aquáticas (submersas e flutuantes), sendo a última espécie mais frequente no
plâncton. Também nas lagoas Carioca e Gambazinho foram observadas essas espécies,
sendo que apenas T. pusilla apresentou diferença, quanto à abundância entre as lagoas,
sendo sua população maior na lagoa Carioca.
De acordo com os mesmos autores, os recursos alimentares são influentes sobre a
presença e abundância dos tricocercídeos, sendo que espécies zooplanctófagas, como
Trichocerca pusilla são mais abundantes na ocorrência de outros animais zooplanctônicos,
visto que se alimentam, por sucção, dos ovos desses animais, e espécies como T. similis,
fitoplanctônicas, mais abundantes em “blooms” algais e dentre o perifiton. Além disso,
táxons como T.pusilla são indicadores de meios eutrofizados, podendo ser esta uma das
causas da maior abundância desta espécie na lagoa Carioca do que na lagoa Gambazinho. A
maior abundância de rotíferos na lagoa Carioca e o elevado número de náuplios e
copepodidos (possível fonte alimentar para T. pusilla), também poderiam ser uma das
causas determinantes do maior tamanho populacional de T. pusilla nesta lagoa, quando
comparada à mesma população na lagoa Gambazinho.
Microcrustáceos
Cladocera
Os microcrustáceos de água doce são representados principalmente pelos copépodes
e cladóceros, sendo que este grupo tem elevado grau de endemismo em águas interiores.
Nos meios dulcícolas brasileiros já foram descritas 76 espécies de Copepoda (Matsumura-
Tundisi e Silva, 1999) e 86 espécies de Cladocera (Rocha e Guntzel, 1999).
Segundo Rocha et al (1995), muitos estudos consideram pobre a fauna de cladóceros
em águas interiores tropicais, quando comparados aos corpos d’água doce de regiões
temperadas, provavelmente porque são apresentados resultados apenas de estudos
realizados na região limnética dos lagos, desconsiderando a fauna do plâncton típica da

54
região litorânea, subestimando a riqueza e abundância de espécies, como por exemplo, das
famílias Chydoridae e Macrothricidae.
A ocorrência Chydorus eurinotus, Macrothrix elegans e Ilyocriptus spinifer na região
litorânea das lagoas estudadas pode ser explicada porque estes organismos apresentam
características morfológicas e biológicas, ex. menor porte, carapaças mais rígidas e dotadas
de muitas cerdas e pelos, que permitem a sobrevivência de forma satisfatória nestes
ambientes (El Moor-Loureiro, 1997; Esteves, 1998; Nogueira, 2003; Panarelli, 2003)
Nas lagoas Carioca e Gambazinho as baixas riqueza e densidade de cladóceros nas
margens podem ter diversas causas: menor capacidade do grupo em explorar recursos
alimentares e habitats disponíveis no litoral quando comparados às amebas testáceas e aos
rotíferos (Fulone, 2008), maior pressão de predação (vertebrados e invertebrados) sobre
este grupo, principalmente pelo maior tamanho corporal (Matsumura-Tundisi e Okano,
1983). Especificamente para a lagoa Carioca, Pinto-Coelho et al (2008) propôs a teoria da
Cascata trófica invertida: com a introdução da ictiofauna exótica (majoritariamente
carnívora) a densidade das espécies de peixes nativos forrageiros (onívoros e planctófagos)
foi consideravelmente reduzida, o que levou ao relaxamento do nicho de Chaoborus
(ecological release) e aumento considerável de suas densidades. Com isso, aumentou a
pressão de predação sobre o zooplâncton de maior tamanho (cladóceros), explicando as
baixas densidades deste grupo naquela lagoa.
Segundo Castilho-Noll e Arcifa (2007), no lago Monte Alegre, Chaoborus apresentou
comportamento onívoro, sendo a microalga Peridinium ingerida principalmente pelos
instares I e II, náuplios por larvas do instar II e o cladócero Bosmina tubicen, copépodes e
rotíferos preferidos dos instares III e IV. Diferentemente do lago Monte Alegre, B. tubicen
apresentou maiores densidades na lagoa Carioca, justamente onde são mais abundantes as
larvas do chaoborídeo, enquanto que na lagoa Gambazinho este bosminídeo ocorreu em
densidades baixas, apesar da população do Chaoborus ser pequena quando comparada à
primeira lagoa. Segundo Pastorok (1981), a larva de Chaoborus geralmente utiliza as presas
mais abundantes, pois a frequência de encontros com o indivíduo é maior, e de maior porte,
visto que oferecem mais energia e compensam a energia desprendida na captura da presa.
Provavelmente na lagoa Carioca, a larva de Chaoborus sp esteja utilizando organismos como
copépodes do gênero Microcyclops e Mesocyclops, ou mesmo de náuplios, copepodidos e

55
rotíferos, organismos abundantes nesta lagoa. Um estudo sobre alimentação de larvas de
Chaoborus na lagoa Carioca está sendo desenvolvido como parte do Programa PELD/site4 e,
com certeza, esclarecerá o verdadeiro papel destas larvas naquele ambiente.
Diferentemente da lagoa Carioca, a lagoa Gambazinho não apresentou diferentes
abundâncias de cladóceros raspadores e filtradores, porém o mesmo não ocorreu em
relação ao número de táxons dentre cada guilda, sendo que os cladóceros raspadores foram
mais numerosos devido à ocorrência de Alona gutatta, Dadaya macrops e Streblocerus
pigmaeus exclusivos na lagoa Gambazinho.
Copepoda
Os copépodes colonizam todos os compartimentos lacustres e podem ser
classificados em três sub-ordens: Calanoida, tipicamente limnéticos, Cyclopoida,
predominantemente litorâneos e/ou bentônicos, com poucas espécies pelágicas, e
Harpacticoidae, com espécies litorâneas e bentônicas, sendo algumas delas semi- terrestres
(Reid, 1985).
Segundo Le Cren e Lowe-McConnell (1980), as populações de microcrustáceos
crescem com sucesso quando as condições alimentares são adequadas. Alterações na
composição, concentração e qualidade do alimento favorecem determinadas espécies,
eliminado ou reduzindo populações de outras. Grande parte das espécies de Cladocera e
Calanoida são herbívoros, sendo que os primeiros podem fazer uso também do
bacterioplâncton como suplemento alimentar. Cyclopoida são onívoros raptoriais, ingerindo
detritos, bactéria, fitoplâncton, microzooplâncton, incluindo seus ovos e náuplios (Melão,
1999).
As maiores densidades de Notodiaptomus isabellae registradas na lagoa Gambazinho
podem ser decorrentes do grau de trofia, oligotrófico, desta lagoa. Matsumura-Tundisi e
Tundisi (1997) trabalhando em lagoas do PERD, observaram que espécies de Calanoida
predominavam em ambientes mais oligotróficos, enquanto Cyclopoida em ambientes mais
eutroficos. Além disso, pelo maior tamanho corporal, a pressão de predação seria maior
sobre espécies de Calanoida na lagoa Carioca, fazendo com que sua abundância nesta lagoa
fosse menor. Na região litorânea da lagoa Gambazinho a ocorrência de N. isabellae pode

56
também estar vinculada a condições adversas no compartimento limnético da lagoa, como
predação exercida por peixes planctófagos e Chaoborus sp, além de competição exercida por
T. minutus.
Apesar da vegetação aquática não ser alimento direto para copépodes, a presença da
mesma pode contribuir com recursos importantes, como perifíton, detritos orgânicos,
protozoários, fungos e bactérias (Peixoto, 2007). A maior disponibilidade de recursos
poderia explicar a elevada abundância de náuplios e copepoditos Cyclopoida na região
marginal das lagoas Carioca e Gambazinho, Entretanto deve-se considerar que a dominância
de estágios jovens de copépodes é comum em lagos tropicais (Matsumura-Tundisi e Okano,
1983).
Na região litorânea das lagoas Carioca e Gambazinho, tanto em termos de riqueza de
espécies, quanto de abundância, os copépodes Cyclopoida foram dominantes em relação
aos Calanoida, provavelmente porque estes são organismos limnéticos e aqueles litorâneos,
de forma geral (Wetzel, 1983, Esteves, 1998). Também em ambas as lagoas predominaram
indivíduos jovens (náuplios e copepodidos, notoriamente os primeiros) de Cyclopoida, como
é comum nos ecossistemas lênticos, segundo Matsumura Tundisi e Okano (1983). A
abundância elevada de náuplios de Cyclopoida é reflexo da estratégia reprodutiva destes
animais, que garantem sua permanência no ambiente pela produção de muitos
descendentes, visto que apenas uma parcela pequena da nova geração alcançará a fase
adulta (Panarelli et al, 2003)
A subordem Calanoida foi representada por uma única família (Diaptomidae) apenas
na lagoa Gambazinho, onde houve ocorrência de indivíduos adultos de Notodiaptomus
isabellae. Na lagoa Carioca não foram observados indivíduos adultos da subordem
Calanoida, apenas náuplios e copepodidos. Corroborando Tundisi e Okano (1983), a
presença de N. isabellae apenas na lagoa Gambazinho pode vincular-se ao grau de trofia dos
corpos d’água estudados, visto que Calanoida são copépodes encontrados
preferencialmente em ambientes oligotróficos.
As espécies Thermocyclops minutus e Tropocyclops prasinus apresentaram diferenças
quanto suas respectivas abundâncias entre as lagoas, sendo a primeira mais abundante na
lagoa Carioca e a segunda na lagoa Gambazinho. Tundisi e Okano (1983) trabalhando na
lagoa Dom Helvécio (PERD), observaram o desaparecimento de T prasinus e um “boom”

57
populacional de T. minutus nesta lagoa após a introdução de peixes não nativos (carnívoros),
podendo ser a ictiofauna exótica na lagoa Carioca também causa determinante do tamanho
populacional destas espécies.
Amoeba Testacea
As amebas testáceas são organismos unicelulares de vida livre que apresentam uma
carapaça protetora. São heterotróficas e ocorrem em diversos ambientes, litoral marinho,
rios riachos, lagos, bromélias, poças e no solo. Caracterizam-se pela estrutura locomotora
denominada pseudópodo, sendo a forma típica de movimentação desses organismos
denominada ameboide (Souza, 2008).
A história de vida das espécies deve ser considerada quando se busca entender a
estrutura da comunidade zooplanctônica de uma área. As tecamebas são, geralmente, mais
dependentes do substrato do que os rotíferos e microcrustáceos, por isso podem responder,
de forma positiva, a um incremento na complexidade da região marginal, aumentando a
riqueza e abundância de espécies (Fulone, 2008). Nas lagoas estudadas amebas testáceas
não apresentaram aumento nas densidades, quando comparadas a rotíferos, cladóceros e
copépodos, talvez porque nenhuma coleta específica para este grupo tenha sido realizada.
A maioria das ambas testáceas é dulcícola e ocorre nas margens entre a vegetação
aquática, no sedimento, sobre rochas e detritos e também na coluna d’água. Em relação ao
comportamento alimentar, tais amebas podem ser consumidoras primárias,
(fitoplanctófagas), decompositoras (detritívoras), bacteriófagas ou micófagas. Ao se
alimentarem de detritos, as tecamebas disponibilizam, para as algas e as macrófitas,
nutrientes, sendo importantes na ciclagem de material orgânico no meio aquático. Algumas
espécies são carnívoras e predam outras amebas, rotíferos e microcrustáceos. Por outro
lado, estes protozoários são o recurso alimentar de invertebrados, como aqueles dos Filos
Rotifera e Arthropoda, e vertebrados, como peixes (Souza, 2008).
Na lagoa Carioca, de acordo com a classificação de Souza, (2008), as amebas com
tecas quitinosas ou construídas com partículas sedimentares detritos e proteínas foram
dominantes, tanto em termos de abundância quanto ao número de táxons, em relação às
amebas com tecas silicosas. Na lagoa Gambazinho, houve dominância de espécies com teca

58
quitinosa, sendo as espécies com carapaça silicosa escassa e aquelas com envoltório
proteico e sedimentar intermediárias. Quando se compara a abundância de organismos
dentre cada guilda entre estas lagoas, observa-se que as espécies com tecas quitinosas ou
construídas com detritos e proteínas apresentaram populações maiores na lagoa Carioca do
que na lagoa Gambazinho, o inverso ocorrendo com as espécies de teca sillicosa. A maior
abundância de amebas com tecas de sílica foi devida à ocorrência exclusiva de EuglypHa
denticulata e E. tuberculata nesta lagoa, além das populações de E. filifera e E.
acanthopHora serem menores na lagoa Carioca.A raridade das amebas testáceas com
carapaça silicosa (família EuglypHidae), em especial na lagoa Carioca, pode ser causada pela
escassez de sílica, essencial para a construção das tecas ou competição por este elemento,
exercida por algas, a exemplo das bacilariófitas.
Habitats litorâneos colonizados X não colonizados por macrófitas
Macrófitas aquáticas incrementam a heterogeneidade do habitat e favorecem
elevada diversidade biológica, sendo importantes na estrutura e funcionamento de
ecossistemas aquáticos (Esteves, 1998; Fulone, 2008). Na vegetação marginal a composição
da comunidade e a abundância das populações se alteram ao longo do tempo e refletem as
mudanças no meio físico, no que diz respeito à composição, distribuição e dinâmica dos
vegetais, além do ciclo biológico particular de cada espécie (Nogueira, 2003; Peixoto, 2008).
Em ambientes onde são escassas as macrófitas e menos abundantes os recursos é possível o
aumento da competição interespecífica e da exclusão competitiva na comunidade
zooplanctônica. Nos locais onde a vegetação marginal é densa permitindo uma variedade e
quantidade maior de recursos, é possível um relaxamento da competição interespecífica,
permitindo a coexistência de um maior número de taxa (Ribas et al, 2003)
Diferentemente do citado no parágrafo anterior, a comunidade zooplanctônica
habitante de banco de macrófita, tanto na lagoa Carioca como na lagoa Gambazinho, não foi
mais diversa que a comunidade habitante do região litorânea sem vegetação marginal. A
mesma disponibilidade de algas (medida indiretamente como concentração de clorofila- a),
recurso alimentar para muitos zooplanctontes, aliada a valores próximos de pH,
condutividade, temperatura e concentração de oxigênio dissolvido nestas regiões, pode
justificar a semelhança entre as comunidades.

59
Outra possível causa para que nestas lagoas a comunidade zooplanctônica
encontrada nos bancos de macrófita não tenha apresentado maior diversidade que aquela
presente na margem sem macrófita seria a presença de alevinos e peixes juvenis
zooplanctófagos na vegetação aquática, que eliminariam ou reduziriam as populações dos
animais de maior porte, como cladóceros e copépodes, e mesmo de rotíferos, resultando em
uma diversidade próxima entre as duas regiões analisadas. Em um estudo com a Tilápia do
Nilo (Oreochromis niloticus), espécie planctófaga nas fases iniciais do seu desenvolvimento e
filtradora na fase adulta, Santos (2010) observou que os alevinos davam preferência para
rotíferos, enquanto que peixes adultos predavam aleatoriamente Cladocera, Copepoda e
Rotifera
Pejler e Berzins (1993) afirmam que a predileção dos tricocercídeos por substratos
vegetais é decorrente do hábito de suas espécies de fixarem-se em outros corpos pela
produção de muco. Por não ser forte a ligação entre o rotífero e o substrato, é comum o
desprendimento do tricocercídeo, fazendo com que o mesmo se encontre na coluna d’água.
A inexistência de diferença estatisticamente significativa na abundância dos indivíduos, das
três espécies do gênero Trichocerca nos habitats colonizado e não colonizado por macrófita
nas lagoas Carioca e Gambazinho pode justificar-se talvez pela ineficiência da adesão do
muco produzido pelo indivíduo às plantas.
Comparando as espécies de Rotifera entre as áreas litorâneas com e sem plantas
aquáticas da lagoa Gambazinho, independentemente da guilda a qual pertencem, observa-
se que as populações de espécies habitantes dos bancos de macrófitas são mais numerosas
do que aquelas habitantes da margem não colonizada por macrófitas. Provavelmente,
dentre a vegetação aquática, há mais recursos alimentares para as espécies detritívoras,
reduzindo a competição interespecífica, permitindo a coexistência entre um maior número
de táxons.
Os indivíduos das famílias Chydoridae, Ilyocryptidae e Macrothricidae são raspadoras,
vivendo sobre plantas aquáticas (epifíticas) ou sobre o sedimento (bentônicas), segundo
ElMoor-Loureiro (1997). Espécies pertencentes a essas famílias foram registradas na região
litorânea colonizada e não colonizada por macrófita da lagoa Carioca. A única espécie
bentônica ou epifítica, de hábito alimentar raspador, exclusiva da região marginal com
plantas aquáticas foi Streblocerus pigmaeus, sendo Dunhevedia odontoplax e Karualona

60
muelleri espécies com mesmo comportamento alimentar e habitacional exclusivas do região
litorânea não colonizado por plantas aquáticas.
As espécies da família Sididae, Bosminidae e DapHnidae, segundo a mesma autora,
são filtradoras, como Bosmina hagmanni, B. tubicen, B. freyi, CeriodapHnia silvestrii,
SimocepHalus latirostris e ScapHoleberis armata, registradas tanto em áreas colonizadas
como não colonizadas por macrófita na lagoa Carioca, além de DiapHanosoma birgei e
ScapHoleberis spinifera, exclusivas da margem sem plantas aquáticas. Nos habitats
litorâneos com e sem macrófita da lagoa Carioca, os cladóceros filtradores foram mais
abundantes quando comparados aos cladóceros raspadores, devido à elevada
disponibilidade de recursos alimentares, como fitoplâncton e partículas em suspensão.
Os cladóceros raspadores, epifíticos ou bentônicos, também foram observados na
região litorânea da lagoa Gambazinho, a exemplo de Alona gutatta, Disparalona dadayi,
Chydorus eurinotus e Streblocerus pigmaeus. Táxons filtradores, livres na coluna d’água,
também foram registrados nos habitas litorâneos com e sem plantas aquáticas desta lagoa,
como aqueles do gênero Bosmina. Na lagoa Carioca, a margem colonizada por plantas
aquáticas apresentou maior abundância de cladóceros raspadores, ao contrário da margem
não colonizada por plantas aquáticas, onde dominaram as espécies filtradoras.
Possivelmente, as espécies filtradoras, de maior porte, são as presas preferenciais de
alevinos e peixes juvenis, que se abrigam dentre as plantas aquáticas.
Fulone (2008) trabalhando na região litorânea coberta por macrófitas em lagoas
marginais do rio Paraná registrou maior riqueza de amebas testáceas associando este fato à
heterogeneidade ambiental, o mesmo não acontecendo para microcrustáceos. Também
para os lagos estudados. As amebas testáceas foram dominantes em relação aos Copepoda
e Cladocera, talvez por explorarem com mais eficiência os microhabitats ofertados pela
vegetação aquática.
Os copépodes Cyclopoida observados na lagoa Carioca (Mesocyclops meridianus,
Microcyclops anceps, M finitimus, T. minutus e T. prasinus) foram mais abundantes nos
habitats litorâneos colonizados por macrófitas, o mesmo ocorrendo na lagoa Gambazinho,
exceto para T. minutus, cuja população foi maior em áreas sem vegetação aquática.
Possivelmente, dentre as macrófitas há maior disponibilidade de recursos alimentares (algas,
animais planctônicos e detritos) para estas espécies, majoritariamente onívoras.

61
Sazonalidade: período chuvoso X período seco
Algumas espécies apresentam maior abundância nos meses chuvosos, outras
desenvolvem-se melhor nos meses secos, ainda há espécies que apresentam dinâmica
populacional errática, desenvolvendo populações transitórias durante todo o ano, como
pequenos cladóceros e rotíferos (Allan, 1976).
A variabilidade temporal de uma comunidade é caracterizada por alterações na
diversidade, composição e densidade relativa das espécies e é essencial para pesquisas em
ecologia, visto que aponta os efeitos de fatores intrínsecos e extrínsecos sobre o
ecossistema e promove compreensão sobre o funcionamento do mesmo (Simões, 2010).
Comparando os períodos de chuva e seca na lagoa Carioca, os rotíferos apresentaram
variação em sua diversidade na margem coberta por macrófita, talvez por uma maior
susceptibilidade do grupo às variações de temperatura. Borges e Pedroso (2009)
encontraram correlação positiva de Polyarthra sp e Keratella sp com a temperatura, o
mesmo relatado por Landa e Mourguês Schurter (2000) para P. vulgaris A riqueza de
espécies da comunidade zooplanctônica da lagoa Carioca, determinada em grande parte por
Rotifera, foi maior no período chuvoso, onde normalmente predominam temperaturas mais
elevadas. Rocha (1978) trabalhando na mesma lagoa encontrou resultados semelhantes.
Na lagoa Gambazinho, considerada como oligotrófica (Barbosa, 1979), maiores
densidades populacionais foram registradas para Chaoborus sp em bancos de macrófitas
durante o período chuvoso, talvez por uma maior disponibilidade de detritos orgânicos
advindos da decomposição da vegetação marginal, que favoreceria o desenvolvimento das
populações zooplanctônicas que se alimentam deste recurso e por sua vez são o alimento do
chaoborídeo. Outra possível causa para este fato pode ser o efeito das temperaturas mais
elevadas registradas nesse período, que aceleram o metabolismo, a reprodução e de
crescimento da larva do Diptera (Melão, 1999). Na margem sem macrófita desta lagoa a
população de Chaoborus sp também apresentou diferentes abundâncias dentre os meses
chuvosos e secos, sendo as larvas mais abundantes na época de chuva, possivelmente pelas
mesmas causas descritas para a região litorânea colonizada por macrófita.

62
Na lagoa Carioca (mesotrófica) não foi observada diminuição da diversidade dos
grupos Tecameba, Cladocera e Copepoda na região marginal com vegetação aquática, por
efeito diluidor, durante o período de chuvas, conforme observado por Sendacz et al (2006)
nos reservatórios de Ponte nova e Guarapiranga, SP. Provavelmente uma entrada maior de
nutrientes, associada a maiores taxas de decomposição de macrófitas devido a temperaturas
mais elevadas, permitiu uma maior oferta de recursos alimentares conservando a
diversidade.
Conclusão
A região litorânea das lagoas Carioca e Gambazinho apresentam elevada riqueza e
diversidade de espécies zooplanctônicas devido à disponibilidade de hábitats e microhábitas,
além de recursos alimentares (algas e detritos) ofertados pelas macrófitas e pelo material
alóctone depositado na margem da lagoa, como folhas, troncos, seixos e sedimento. O
zooplâncton habitante da região litorânea tem papel crucial na cadeia trófica nas lagoas,
atuando como elo de transferência de energia e matéria entre os consumidores primários e
os níveis tróficos superiores.
Os rotíferos apresentam maior diversidade e riqueza na lagoa Carioca,
provavelmente pelo fato desta lagoa, quando comparada à lagoa Gambazinho, ter maior
grau de trofia, disponibilizando mais alimento para as espécies, sejam elas detritívoras,
bacteriófagas, fitopanctófagas, zooplanctófagas ou onívoras.
Por serem, em termos de número de táxons, os organismos dominantes em ambas as
lagoas, os rotíferos determinam a riqueza total de espécies zooplanctônicas, o que justifica o
valor mais elevado de riqueza de espécies ser observado na lagoa Carioca.
A ausência de Notodiaptomus isabellae adultos na lagoa Carioca pode estar vinculada
à predação exercida por alevinos e juvenis, tanto de peixes nativos quanto de peixes
exóticos, que buscam presas de maior tamanho corporal, como as espécies da família
Diaptomidae. Outra possibilidade para justificar a ausência dessa espécie na lagoa Carioca e
a sua presença na lagoa Gambazinho seria o grau de trofia destes corpos d’água,
considerando que Calanoida são adaptados a meios oligotróficos, como a lagoa Gambazinho.
A comunidade zooplanctônica habitante dos bancos de macrófita não diferiu daquela
habitante da região marginal não colonizada por macrófita, em ambas as lagoas,

63
provavelmente pelas condições ambientais (pH, temperatura, condutividade elétrica,
concentração de oxigênio dissolvido e concentração ce clorofila-a) semelhantes entre esses
hábitats. Além disso, há também a possibilidade de que os animais zooplanctônicos sejam
transportados por correntes de água no eixo horizontal, provocadas pelo vento, fazendo
com que a fauna planctônica entre as áreas com e sem plantas aquáticas seja parecida.
O aumento da temperatura favoreceu o crescimento populacional dos rotíferos, dos
copépodes (Thermocyclops minutus) e das larvas de Chaoborus nas lagoas Carioca e
Gambazinho, possivelmente por acelerar o crescimento e o desenvolvimento dos indivíduos,
fazendo com que se reproduzam mais cedo e produzam ninhada mais numerosa.

64
Referências Bibliográficas
ADRIAN, R.; FROST, T.M. Comparative feeding ecology of Tropocyclops prasinus mexicanus
(Copepoda, Cyclopoida). Journal of plankton Research, v. 14. n 10, p. 1369-1382. 1992.
ALLAN, J. DAVID. Stream Ecology – Structure and function of running waters. School of Natural
Resources and Environment, University of Michigan, USA. Chapman e Hall, 388p. 1995.
AOKI, A.Influência da Diversidade de Macrófitas na Composição e Abundância da
Comunidade Zooplanctônica na Lagoa dos Patos-Parque Estadual do Rio Doce(MG).
Dissertação. Belo Horizonte. UFMG. 52p. 2010.
AOYAGUI, ANDERSON SETSUO MIYASHIRO. Rotíferos em diferentes ambientes da Planície de
inundação do Alto Rio Paraná-PR/MS: riqueza, abundância e conectividade. dissertação.
Maringá. Universidade Estadual de Maringá. 32 p. 2008
BARBOSA, F.A.R. Produção primária e fatores ambientais na Lagoa Carioca, Parque Florestal
do Rio Doce-MG. Dissertação. São Carlos. UFSCAR . 1979.
BARBOSA, F. A. R. e TUNDISI, J. G., The influence of temperature variation on the distribution
of dissolved oxygen and nutrients. Arch. Hydrobiol., 116(3): 333-349. 1989
BARBOSA, F.A.R., AND P. MORENO. Mata Atlântica e Sistema Lacustre do Rio Doce. In:
Seeliger, U.; Cordazzo, C.; Barbosa, F. (Eds.). Os Sites e o Programa Brasileiro de
Pesquisas Ecológicas de Longa Duração. pp.69-81. 2002.
BEISNER.B.E.; IVES, A.R. e CARPENTER, S.R. The effects of an exotic fish invasion on the prey
communities of two lakes. Journal of Animal Ecology 72: 331-342. 2003.
BERZINS, B.e BERTILSSON, J. On limnic micro-crustaceans and tropHic degree.
Hydrobiologia. Vol. 185. P. 95-100. 1988.
BEZERRA-NETO, J.F. Migração vertical Diária e Cascata Trófica em Corpos Aquáticos
Tropicais: Influência da Larva do Díptera Chaoborus. 153 p. Tese. Belo Horizonte UFMG.
2007.
BONECKER, C. C.; COSTA, C.L., VELHO, L.F.M., LANSAC-TÔHA, F.A. Diversity and abundance of the
planktonic rotifers in different environments of the upper paraná river floodplain (Paraná State -
Mato Grosso do Sul State, Brazil). Hydrobiologia 546: 405- 414, 2005.

65
BONECKER, C. C; AOYGUI, A.S.M.; SANTOS, R.M. The impact of impoundment on the rotifers
communities of two tropical floodplain environments: interannual pulse variatons. Braz.
J. Biol., vol. 69 (2 suppl.), p:629:537. 2009.
BORGES, M.G. e PEDROSO, C.S. Zooplankton (Cladocera, Copepoda and Rotifera) richness,
diversity and abundance variations in the Jacuí Delta, RS, Brazil, in response to the
fluviometric level. Acta. Limnol. Bras. V.21. n. 1. P. 101-110.2009.
BRITO, S. L. Composição, distribuição espacial e biomassa do zooplâncton total do Lago Dom
Helvécio, Parque Estadual do Rio Doce-MG. Monografia. Belo Horizonte. UFMG. 2003.
BRITO, S. L. Interações bióticas e abióticas das populaçõeszooplanctônicas das lagoas Carioca
e Gambazinho (Parque Estadual do Rio Doce, Minas Gerais) e suas implicações na
estrutura do tamanho do corpo.dissertação.80p. Belo Horizonte. UFMG. 2005.
BRITO, S. L.. Caracterização limnológica e produtividade secundária das principais espécies
de microcrustáceos em dois braços dos reservatórios de Três Marias e Furnas, Minas Gerais,
Brasil. Tese. 112p. Belo Horizonte. UFMG 2010.
CARPENTER, S. R., J. K. KITCHELL, AND J. R. HODGSON. Cascading tropHic interactions and
lake productivity. BioScience 35:634-639. 1985.
CARVALHO, EDMIR DANIEL; CASTRO, REINALDO JOSÉ DE; DA SILVA, VALÉRIA FLÁVIA
BATISTA; VIDOTTO, ANA PAULA. A Estrutura das Assembleias de Peixes nas Regiãos de
Ecótono da Represa de Jurumirim (Alto do Rio Paranapanema, SP), pags: 249-278,
Ecótonos nas interfaces dos Ecossistemas Aquáticos, HENRY RAOUL, São Carlos: RiMa, ,
349p. 2003.
CASTILHO-NOLL, M. S. M.; ARCIFA M. S. Chaoborus diet in a tropical lake and predation of
microcrustaceans in laboratory experiments, Acta Limnol.Bras , v. 19 (2) p: 163-174.
2007.
CATUNDA-MARCELINO, SÉRGIO. Zooplâncton como bioindicador do estado trófico na
seleção de áreas aquícolas para piscicultura em tanque-rede no reservatório da UHE
Pedra no rio de Contas, Jequié, BA, 59fil. 2007.
CHAPMAN, M.A.; GREEN, J.D.; JOLLY, V.H. Relationships between zooplankton abundance
and tropHic state in seven New Zeland Lakes. Hydrobiologia. Vol. 123. P. 119-136. 1985.

66
DANTAS, E. W. ET AL. Efeito das variáveis abióticas e do fitoplâncton sobre a comunidade
zooplanctônica em um reservatório do Nordeste brasileiro, Iheringia, Porto Alegre, v.
99, n. 2, p. 132 – 141. 2009.
DE MEIS, MRM. and MONTEIRO, AMF. Upper quaternary ―rampas‖, Rio Doce Valley: S.E.
brazilian plateau. Z. GeomorpH., vol. 23, p. 132-151. 1979.
DUNSON, W. A. e TRAVIS, J. The role of abiotic factors in community organization.The American
Naturalist, 138:1067-1091; 1991.
EDMODSON, W.T. Fresh Water Biology. 2nd ed..John Wiley e Sons. Inc. 1959.
ELMOOR –LOUREIRO, L.M.A. Manual de identificação de cladóceros límnicos do Brasil. Ed.
Universa, Brasília.156p. 1997.
ENGELHARDT, K.A.M. e RITCHIE, M.E. Effects of macropHyte species richness on wetland ecosystem
functioning and services. Nature 411: 687-689. 2001.
ESKINAZI SANT’ANNA. EM.; MAIA-BARBOSA, PM; BRITO, SL e RIETZLER, AC. Zooplankton biodiversity
of Minas Gerais State: a prelimnary synthesis of presente knowledge. Acta Limnol. Bras vol. 17. N. 2.
P. 199-218. 2005.
ESTEVES, F. A. Fundamentos em limnologia. 2 ed. Rio de Janeiro: Interciência. 602p. 1998.
FERREIRA, FERNANDO ALVES; MORMUL, ROGER PAULO; PEDRALLI, GILBERTO;
POTT, VALI JOANA E POTT, ARNILDO. Estrutura da comunidade de macrófitas aquáticas em
três lagoas do Parque Estadual do Rio Doce, Minas Gerais, Brasil. Hoehnea 37(1): 43-52.
2010.
FULONE, LEANDRO JUNIO. Influência da complexidade estrutural de macrófitas aquáticas
sobre a diversidade de organismos perifíticos.. Dissertação. Maringá. Universidade
Estadual de Maringá.. 23 p. 2008.
FROIN, R. e LACOBELLIS, S. The absorbtion power of plankton. Annual report of Screpps
Institution of OceanograpHy. Vol. 4. P. 2-3. 2003.
GODINHO, A.L.,FONSECA, M.T., ARAÚJO, L.M. the ecology of predator fish introductions: the
case of Rio Doce valley lakes. In: COELHO, R.M.P., GIANI, A. SPERLING, E.V. eds. Ecology
and human impact on lake and reservoirs in Minas Gerais with special reference to
future development and management strategies. Belo horizonte, Segrac. P.77-83.1994.

67
GODINHO, A.L. Peixes do Parque Estadual do Rio Doce- Belo horizonte/Instituto Estadual de
Florestas/Universidade federal de Minas Gerais.48p. 1996
GRANÁLI, W. e SOLANDER, D. Influence of aquatic macropHytes on pHospHorus cycling in
lakes. Hydrobiologia. Vol. 170. P 245-266. 1988.
GREEN, J. 1986. Associations of zooplankton in six crater lakes in Ariregião, Mexico and New
Mexico. J. Zool., vol. 208, p. 138- 159.
GRECO, M.K.B. E DABÉS M. B.G.S. Comparação da microfauna, com e sem a presença da
macrófitas Mayaca sellowiana (Mayacaceae), Ribeirão Passageiro, MG. Bios. V. 9(9):p: 9-
14. 2001.
GLIWICZ, Z. M., AND A. PREJS. Can planktivorous fish keep in check planktonic crustacean
populations? A test of size-efficiency hypothesis in typical Polish lakes. Ekologia Polska v.
25: p.567-591. 1977.
GOPHEN, M. The impact of zooplankton status on the management of Lake Kinneret (Israel).
Hydrobiologia 113: 249-258. 1984.
HAHN, S.N. et al. Dieta e atividade alimentar de peixes do reservatório de Segredo. In
AGOSTINHO, A.A.; GOMES, L.C. (Ed.) Reservatório de Segredo: bases ecológicas para o
manejo. Maringá: Eduem, p.141-162. 1997.
HENRY, RAOUL. Os Ecótonos nas Interfaces dos Ecossistemas Aquáticos: Conceitos, Tipos,
Processos e Importância. Estudo de Aplicação em Lagoas Marginais ao Rio
Paranapanema na Região de Sua Desembocadura na Represa de Jurumirim, pags:1-28,
Ecótonos nas interfaces dos Ecossistemas Aquáticos, HENRY RAOUL, São Carlos: RiMa,
349p. 2003.
HORNE, A.J., GOLDMAN, C.R. Limnology ed. Mcgraw-Hill. New York. 576p.1994.
KEDDY, P.A. Plant region on lakeshores in Nova Scotia: a test of the resource specialization
hypothesis. Journal of Ecology 72: 797-808. 1984.
KOSTE, W. Rotatória. Die Rädertiere Mitteleuropas. Ein Bestimmungswerk begr. Von Max
Voigt. Überordnung Monogononta. V., 1-2, 673 p. 234pl. 1978.
KOLASA, J.e SALEWSKI, M. Notes on ecotone atributes and functions. Hydrobiologia. Vol.
303. P. 1-7. 1995.

68
KURATA e KIRA. Aspectos de calidad del agua. In: JORGENSEN, S.E.; LOFFLER, H. eds.
Directrizes para la gestion de lagos. Volumen 3. La gestion de la costa del lago. ILEC-
PNUMA. P.17-33. 1995.
LANDA G.G. E MOURGÊS-SCHURTER, L.R. Caracterização da comunidade zooplanctônica de
um sistema artificial (Represa Zootecnia), no Campus da Universidade Federal de Lavras
– MG. Acta Limnológica Brasiliensia, São Paulo, v. 12, n. 2, p. 69-83, 2000.
LANSAC-TÔHA,F.A.; BONECKER,C.C.; VELHO, L.F.M. E LIMA, A.F. Composição, distribuição e
abundância da comunidade zooplanctônica. In Vazzoler, A.E.A.M.; Agostinho, A.A. e
Hahn, N.S. (ed.) A planície de inundação do Alto Rio Paraná: aspectos físicos, biológicos
e socioeconômicos. EDUEM, Maringá.cap.2.p. 117-155. 1997.
LATINI, A.O. O efeito da introdução de peixes exóticos nas populações nativas de lagoas do
Parque Estadual do Rio Doce, MG. Dissertação. Belo Horizonte, UFMG. 62p. 2001.
LATINI, A.O. E PETRERE. M. Reduction of a native fish fauna by alien species: an example
from Brazilian freshwater tropical lakes. Fisheries Management and Ecology v. 11. P.: 71-
79. 2004.
LATINI, A.O.; PEREIRA, T.L.; LATINI, R.O.; GIACOMINI, H.C.; LIMA-JUNIOR, D.P.; OPORTO, L.T.
E ESPÍRITO-SANTO, H.M.V. Distribuição e efeitos de peixes exóticos sobre a ictiofauna
nativa dos lagos do médio rio Doce, MG, Brasil. In: ROCHA, O; ESPÍNDOLA, E.L.G.;
FENERICH-VERANI, N.; VERANI, J.R. E RIETZLER, A.C. (Orgs.). Espécies invasoras em águas
doces: Estudos de caso e propostas de manejo. São Carlos EDUFSCar. p.99-118. 2005.
LE CREN, E.D., LOWE-MCCONNELL, R.H., The functioning of freshwater ecosystems.
Cambridge: Cambridge University Press, 588p. (IBP-Handbook, 22). 1980.
LIKENS, G.E. The ecosystem approach: its use and abuse. Ecology Institute. Oldenhorf/ Lehe.
Germany. 1992.
LIMA, A.F.; LANSAC-TÔHA, F.A.; VELHO, L.F.M., BINI, L.M. E TAKEDA, A. M. Composition and
abundance of Cladocera (Crustacea) assembleges associated with Eichornia azurea(
Swartz) Kunth stands in the Upper Paraná River floodplain. Acta Scientiarium: Biological
Sciences. V.. 25(1). P.: 41-48. 2003.
LIMA, PLACIANO VIANA; LOVERDE–OLIVEIRA, SIMONI MARIA; SILVA, MATHEUS DE
CARVALHO; OLIVEIRA, VALDECI ANTÔNIO. Variação na riqueza de espécies

69
zooplanctônicas em lagoas marginais do Rio Cuiabá (Pantanal, MT). Biodiversidade. V.
11, n1, p.57. 2012.
LORENZEN, C.J. Determination of chloropHyll and pHeopigments: SpectropHotometric
equations. Limnol. Oceanogr. V. 12:p. 343-346. 1967.
MAIA-BARBOSA, P.M., PEIXOTO, R.S. E GUIMARÃES, A.S. Zooplankton in litoral Waters of a
tropical lake: a revisited biodiversity. Braz. J. Biol.v.68 (4 suppl.): 1069-1078. 2008.
MATSUZAKI, M.; MUCCI, J. L. N.; ROCHA, A. A. PHytoplankton community in a recreational
fishing lake, Brazil. Revista de Saúde Pública, v. 38, n. 5, p. 679-686, Oct.2004.
MARGALEF, R. .Limnologia. Barcelona. Ediciones Omega. 1983.
MATSUMURA-TUNDISI, T. and TUNDISI, JG. Biomass and zooplankton comunity struture of
three lakes of river Doce Valley (MG-Brazil); Fifth Japan-Brasil symposium on science and
technology, October Tokio, Japan. 1980.
MATSUMURA-TUNDISI, T. and OKANO, W.Y., Seasonal fluctuation of copepod population in
lake Dom Helvécio (Parque Florestal do Rio Doce, MG, Brazil). Rev. Hydrobiol. Trop., v. 6,
no. 1, p. 35-39. 1983.
MATSUMURA-TUNDISI, T.; LEITÃO, N. S.; AGUENA, S. L.; MIYAHARA, J. Eutrofização da represa
de Barra Bonita. Rev Bras Biol, Rio de Janeiro, v. 50, n. 4, p. 923-935, 1990.
MATSUMURA-TUNDISI, T. e SILVA, W.M. Crustáceos Copédodos Planctônicos. In: Joly, C.A.;.
Bicudo, C.E.M. org. Biodiversidade do Estado de São Paulo, Brasil: síntese do conhecimento do
final do século XX, 4: invertebrados de água doce/ Ismael, D.; Valenti, W.C.; Matsumura-Tundisi,
T.; Rocha, O. –São Paulo, FAPESP. P. 93 – 100. 1999.
MCQUEEN, D. J., AND J. R. POST. Enclosure experiments: the effects of planktivorous fish. in
Lake and reservoir management. Proceedings of the Fifth International Symposium on
Applied Lake and Watershed Management (NALMS), November 1985, Lake Geneva,
Wisconsin. American Lake Management Society, Washington, D.C., USA. V.2. P. 313-318.
1986.
MELÃO, M.G.G. Desenvolvimento e aspectos reprodutivos de cladóceros e copépodos de
águas continentais brasileiras. In: POMPÊO, M.L.M.(ed) perspectiva da limnologia no
Brasil. São Luiz do maranhão. Pag. 45-57.1999.

70
MELLO. Sedimentação de tectônica senozóicas no Médio Vale do Rio Doce (MG, sudeste do
Brasil) e suas implicações na evolução de um sistema de lagos. São Paulo, SP. Tese.
Instituto de Geociências. Universidade de São Paulo. 1997.
MEIS, M.R.M. e TUNDISI, J.G. GeomorpHologycal and limnological processes as a basis for lake
typology. The middle Rio Doce lake system. In: J.G. TUNDISI e Y. SAIJO (eds.) Limnological Studies on
the Rio Doce Valley Lakes, Brazil. Brasilian Academy of Sciences, Rio de Janeiro. 1997.
MILLS, E. L., J. L. FORNEY, AND K. J. WAGNER. Fish predation and its cascading effect on the
Oneida Lake food chain. P.118-131 in W. C. KERFOOT and A. SIH, editors. Predation.
Direct and indirect impacts on aquatic com- munities. University Press of New England,
Hanover, New Hampshire, USA. 1987.
MINAS GERAIS. Zoneamento agroclimático do Estado de Minas Gerais. Secretaria do Estado da
Agricultura. Belo Horizonte. 1980.
MORETTO,E.M. Diversidade zooplanctônica e variáveis limnológicas das regiões limnética e
litorânea de cinco lagoas do Rio Doce e sua relação com o entorno. 208 p. Dissertação,
São carlos. USP. 2001.
MORMUL, K.P., THOMAZ, S.M., HIGUTI, J.; MARTENS,K. Ostracod (Crustacea) colonization of
a native and a non native macropHyte species of Hydrocharitaceae in the Upper Paraná
Floodplain (Brazil): an experimental evaluation. Hydrobiologia 644: 185-193. 2010.
MUIRHEAD, RJ; EJSMONT-KARABIN e MACISAAC, JH. Quantyfing rotifer species richness in
temperate lakes. Freshwater Biology. Vol. 51. N. 9. P. 1696-1709. 2006.
NETO e MORENO. 1999.
NOGUEIRA, MARCOS GOMES; GEORGE, DAVID GLEN, JORCIN, ADRIANA. Estudo do
zoolâncton em Regiãos Litorâneas Lacustres: Um enfoque Metodológico. p.: 83-127,
Ecótonos nas interfaces dos Ecossistemas Aquáticos. HENRY RAOUL. São Carlos: RiMa, ,
349p. 2003.
OPORTO, LORENA TORRES. Aspectos ecológicos e sociais da invasão de peixes em lagos
naturais do médio Rio Doce, MG, Brasil Tese.127 pp. Belo Horizonte. UFMG.2013.
PANARELLI, E.; CASANOVA, S. M. C.; NOGUEIRA, M. G.; MITSUKA, P. M.; HENRY, R. A
Comunidade Zooplanctônica ao Longo de Gradientes longitudinais no Rio 89
Paranapanema/Represa de Jurumirim (São Paulo, Brasil). In: HENRY, R. Ecótonos nas Interfaces
dos Ecossistemas Aquáticos. São Carlos, Editora RiMa: 129-160, 2003.

71
PANARELLI, E.A. Flutuações mensais da comunidade zooplanctônica e dinâmica
daspopulações de Cladocera em lagoas marginais, na região de transição Rio
Paranapanema - Represa de Jurumirim. Tese. Botucatu.Universidade Estadual Paulista.
2004.
PAGGI, J. C. Crustacea Cladocera Ecossistemas de aguas continentales. Metodologias para su
studio III. Ediciones Sur, Plata. PEJLER, B.; BERZINS, B. On the ecology of Trichocercidae
(Rotifera). Hydrobiologia, v. 263, p. 55-59. 1995.
PASTOROK,. ROBERT A. Prey Vulnerability and Size Selection by Chaoborus Larvae. Ecology,
Ecological Society of America V. 62, No. 5 p. 1311-1324. 1981.
PATERSON, M. The distribution of microcrustacea in the litttoral zone of a freshwater lake.
Hydrobiologia. 263.pp.173-183. 1993.
PEIXOTO, R.S. Estudos ecológicos sobre as assembleias de microcrustáceos ( Copepoda e
Cladocera) na região litorânea do Lago Dom Helvécio, Minas gerais, Brasil. Dissertação.
Belo Horizonte. UFMG. 2007.
PEJLER, B. e BERZINS, B. On the ecology of Trichocercidae (Rotifera). Hydrobiologia. Vol. 263.
P: 55-59. 1993.
PEJLER, B. Relation to habitats in rotifers. Hydrobiologia. Vol. 313-314. P. 267-278. 1995.
PENNAK, PW. Structure of zooplankton populations in the litoral macropHytes zone of some
Colorado lakes. Trans. Am. Microsc. Soc. Vol. 85. N. 3. P.329-349. 1966.
PEREIRA, D. F. Estudo da comunidade zooplanctônica em um reservatório recémconstruído
(Reservatório do Lajeado), UHE Luis Eduardo Magalhães – TO. São Paulo: USP. 114p. Tese
(Doutorado em Ciências da Engenharia Ambiental). Escola de Engenharia de São Carlos,
Departamento de Hidráulica e Saneamento, Universidade de São Paulo. 2005.
PIECZYNSKA, P. Littoral habitats and communities. In JORGENSEN, SE., and LÖFFLER, H.
(Eds.). Guidelines of Lake Management. Lake shore management. International Lake
Environment Committee. United Nations Environment Programme. p. 39-72. 1990.
PINTO-COELHO, R.M.; BEZERRA-NETO, J.F.; MIRANDA, F.; MOTA, T.G.;RESCK, R.; SANTOS,
A.M.; MAIA-BARBOSA, P.M.; MELLO, N.A.S.T.; MARQUES, M.M.; CAMPOS, M.O.;
BARBOSA, F.A.R. The inverted tropHic cascade in tropical plankton communities:

72
impacts of exotic fish in the Middle Rio Doce lake district, Minas Gerais, Brazil. Brazilian
Journal of Biology, v.68, n.4 (suppl.). p.631-637. 2008.
POTT, VALI JOANA e POTT, ARNALDO. Plantas aquáticas do Pantanal. 404 p. Embrapa:
comunicação para transferência de tecnologia. Brasília, DF. 2000.
POURRIOT, R. Food and feeding habits of Rotifera. Hydrobiology. Vol 8 p: 243-260. 1977.
PRIMACK, R.B. E RODRIGUES, E. Biologia da Conservação. Ed. Planta, Londrina, PR. 328p.
2001.
PURVIS, A. e HECTOR, A. Getting the measure of biodiversity. Nature. Vol.405. p. 212-219.
2000.
REBOUÇAS, A.C.; BRAGA. B.; TUNDISI, J. G. Águas Doce no Brasil: capital ecológico, uso e
conservação,ed.3, Escrituras. 2006.
REID, J.W. Chave de identificação e lista de referências bibliográficas para as espécies
continentais sul-americanas de vida livre da Ordem Cyclopoida (crustacean, Copepoda).
Bolm.Zool. vol. 9. P.:17:143. 1985.
RIETZLER, A.C., Alimentação, ciclo de vida e análise da coexistência de espécies de na
represa de Barra Bonita,São Paulo.Tese. São Carlos: EESC/USP, 385p. 1995.
RIETZLER, A. C. Tempo de desenvolvimento, reprodução e longevidade de DiapHanosoma
birgei Korinek e CeriodapHnia silvestrii Daday em condições naturais de alimentação.
An. VIII Sem. Reg. Ecol., 8: 1159-1171, 1998.
ROCHA, O. E MATSUMURA-TUNDISI, T. Atlas do Zooplâncton Volume 1. UFSCAR. 1976.
ROCHA, O; SENDACZ, S.e mATSUMURA-TUNDISI, T. composition, biomass, and productivity
of zooplankton in natural lakes and reservoirs in Brazil In TUNDISI, J.G.; BICUDO, CEM. E
MATSUMURA-TUNDISI, T. (Eds).Limnology in Brazil. Rio de janeiro. Academia Brasileira
de Ciência. P. 151-166. 1995.
ROCHA, O. e GUNTZEL, A. Crustáceos Branchiópodos. In: Joly, C.A.;. Bicudo, C.E.M. org.
Biodiversidade do Estado de São Paulo, Brasil: síntese do conhecimento do final do século XX, 4:
invertebrados de água doce/ Ismael, D.; Valenti, W.C.; Matsumura-Tundisi, T.; Rocha, O. –São
Paulo, FAPESP. P. 109 - 120. 1999.

73
ROJAS, N. E. T.; MARINS, M. A. E ROCHA, O. The effect of abiotic factors on the hatching of
Moina micrura Kurz, 1874 (Crustacea: Cladocera) epHippial eggs. Brazilian Journal of
Biology, São Carlos, v.61, n.3, p. 371-376, 2001.
ROSSA,D.C.; LANSAC-TÔHA, F.A. e BONECKER, C.C. Abundance of cladocerans in the litoral
regions of two enviroments of the upper Paraná River floodplain, Mato Grosso do Sul,
Brazil. Revista Brasileira de Biologia 61(1): 45-53. 2001.
RUTTNER-KOLISKO, A. Plankton Rotifers. Biology and Taxonomy. Die Binnengewässer. Vol. 26
(1), Suppl., 146p. 1974.
SAMPAIO, E.V.; ROCHA, O.; MATSUMURA-TUNDISI, T.; TUNDISI, J.G. Composition and abundance
of zooplankton in the limnetic zone of seven reservoirs of the Paranapanema River, Brazil.
Brazilian journal of biology, 62(3): 525-545, 2002.
SANTEIRO, RACHEL MAGALHÃES E PINTO-COELHO, RICARDO MOTTA. Efeito de fertilização
na biomassa e qualidade nutricional do zooplâncton utilizado para alimentação de
alevinos na estação de hidrobiologia e piscicultura de Furnas, MG. Acta Scientiarium 22
(3):707, 716.2000.
SANTOS, L.C. Estudos das populações de Cladocera em cinco lagoas naturais (Parque
Florestal do Rio Doce-MG), que se encontram em diferentes estágios de
evolução.dissertação. São Carlos, UFSCAR, 260p. 1980.
SANTOS. PABLO LUCIO RUBIM COSTA. Respostas funcionais de diferentes tamanhos de
tilápia do Nilo, Oreochromis niloticus, predando zooplâncton.dissertação.Natal. UFRN.
2010.
SEGERS, H.; DUMONT, H.J. 102+ Rotifers species (Rotifera- Monogononta) in Broa Reservoir
(SP, Brazil) on 26 August 1994, with the description of three new species. Hydrobiologia.
Vol. 316. P. 183-197. 1994.
SEGERS, H. Rotifera. The lecanidae (Monogononta) .Guides to the identification of the
Microinvetebrates of the Continetal Waters of the World. V. 6 (2), 1995.
SENDACZ, S., CALEFFI, S. E SANTOS-SOARES. Zooplankton biomass of reservoirs in different
tropHic conditions in the state of São Paulo, Brazil. Brazilian Journal of Biology. V..
66(1B). p. 337-350. 2006.
SENDACZ, S., KUBO, E.. Copepoda (Calanoida e Cyclopoida) de Reservatórios do Estado de
São Paulo. B. Inst. Pesca v.. 9. P. 51-89. 1982

74
SERAFIM J.R. M.; LANSAC-TÔHA, F.A.; PAGGI, J.C.; VELHO, L.F.M.; ROBERTSON, B. Cladocera
fauna composition in a river-lagoon system of the uUpper 100 Paraná River floodplain, with a
new record for Brazil. Brazilian Journal of Biology, 63(2): 349- 356, 2003.
SERRANO, L. e TOJA, J. Interannual variability in the zooplanlton community of a shallow
temporary pond. Verh. Int. Ver. Limnol vol 26. P 1575-1581. 1998.
SETTI, A.A. A necessidade do uso sustentável dos recursos hídricos. Brasília, iBAMA. 1996.
SHAPIRO, J., B. FORSBERG, V. LAMARRA, G. LINDMARK, M. LYNCH, E. SMELTZER, AND G.
ZOTO.. Experiments and experiences in biomanipulation-studies of biological ways to
reduce algal abundance and eliminate blue-greens. EPA- 600/3-82-096. Corvallis
Environmental Research Labo- ratory. Environmental Protection Agency, Corvallis,
Oregon, USA. 1982.
SIMÕES, NADSON RESSYÉ. Variabilidade da comunidade zooplanctônica: testando a relação
diversidade estabilidade e identificando os efeitos de distúrbios hidrológicos ( secas
extremas) sobre a estabilidade das comunidades. Tese. Maringá.Universidade Estadual
de Maringá. 57 p. 2010
SMITH, WELBER SENTEIO;PEREIRA, CAMILA CRISTINA GALVÃO FRANCISCO; ESPÍNDOLA,
EVALDO LUIZ GAETA; ROCHA, ODETE. A Importância da Região Litoral para a
Disponibilidade de Recursos Alimentares à Comunidade de Peixes em Reservatórios, p.
233-248, Ecótonos nas interfaces dos Ecossistemas Aquáticos. HENRY RAOUL, São
Carlos: RiMa, 349p. 2003.
SOUZA, MARIA BEATRIZ GOMEZ. Guia das Tecamebas Bacia do Rio Peruaçu-Minas gerais-
Subsídio para conservação e monitoramento da bacia do Rio São Francisco. Editora
UFMG. 159p. 2008
STRASKRABA,M.; TUNDISI, J.G. e DUNCAN, A. Comparative reservoir limnology and water
quality management.Dordrech.Kluver academic Publisher. The Netherlands. 1993.
TATE, C. M. e HEINY, J. S. The ordination of benthic invertebrate communities in the South
Platte River Basin in relation to environmental factors. Freshwater Biology, 33: 439-454.
1995.
TAVARES, K. S. A comunidade de macrófitas aquáticas em reservatórios do médio e baixo Rio
Tietê (SP) e em lagos da bacia do médio Rio Doce. Dissertação de Mestrado em ecologia e
Recursos Naturais- Universidade federal de são carlos, 2003.

75
TRINDADE, M. Lagos: origem, classificação e distribuição geográfica. Departamento de
Ciências Biológicas. São Carlos. Universidade Federal de São Carlos. 1984.
TUDISI, J.G. Distribuição espacial, sequência temporal e ciclo saregiãol do fitoplâncton em
represas: fatores limitantes e controladores. Ver. Brasil, Biol. Vol. 50. N. 4. P. 937-955. 1990.
VANNI, MICHAEL J. Effects of nutrients and zooplankton size on the structure of
pHytoplankton communitie. Ecology. V. 63. N. 3. Pp. 624-635.1987.
VELHO,L.F.M.; LANSAC-TÔHA, F.A. e BINI, L.M. Spatial and temporal variation in densities of
testate amoebae in the plankton of the upper Paraná River floodplain, Brazil.
Hydrobiologia 411:103-113. 1999.
VIJVERBERG, J., AND W. L. T. VAN DENSEN. The role of the fish in the foodweb of
Tjeukemeer, The Netherlands. Internationale Vereinigung fur Theoretische und Ange-
wandte Limnologie, Verhandlungen 22:891-896. 1984.
WELCOMME, R.L. International introductions of inland aquatic species. FAO Fish. Tec. Pap.
Vol. 294. 1988.
WETZEL, R. G. E LIKENS, G. E. Limnological Analyses, segunda edição, New York: Springer-
Verlag. 1991.
WETZEL, R.G. Limnologia. Editora Fundação Calouste Gulbenkian. 915p . 1983.
WINKLER, L. W. Die Bertimmung des im wassergelösten Sauer-stoffs. Ber. dtsch. chem. Ges.,
21: 2843-2854. 1888.
76
Anexos
Tecameba autor família com macrófita sem macrófita com macrófita sem macrófita
Arcella costata * Ehrenberg, 1847 Arcellidae 8.026 5.921 2.559 1.417
Arcella braziliensis * Cunha, 1913 Arcellidae 480 72 0 25
Arcella conica* (Playfair, 1918) Deflandre, 1926 Arcellidae 1.130 200 161 153
Arcella costata angulosa * Ehrenberg, 1847 Arcellidae 5.833 2.733 1.178 367
Arcella crenulata * Deflandre,1928 Arcellidae 4.857 31.488 1.433 635
Arcella dentata * Ehrenberg,1938 Arcellidae 0 0 650 50
Arcella discoides * Ehrenberg,1871 Arcellidae 8.786 8.517 8.670 2.127
Arcella gibbosa * Pénard, 1890 Arcellidae 429 770 1.344 1.234
Arcella hemisphaerica undulata* Deflandre, 1928 Arcellidae 696 0 150 75
Arcella megastoma * Pénard,1913 Arcellidae 334 673 214 115
Arcella rota* Arcellidae 0 200 1.367 342
Arcella rotunda alta* Playfair, 1918 Arcellidae 173 910 0 33
Arcella sp.* Arcellidae 5.400 277 734 742
Arcella vulgaris* Ehrenberg,1930 Arcellidae 12.178 5.722 2.071 2.047
Arcella vulgaris undulata* Deflandre, 1928 Arcellidae 1.383 894 720 228
Centropyxis aculeata * Ehrenberg,1930 Centropiyxidae 4.568 9.487 12.824 4.358
Centropyxis aculeata oblonga * Deflandre, 1929 Centropiyxidae 1.435 738 15.319 2.945
Centropyxis discoides * Pénard,1890 Centropiyxidae 200 40 666 67
Centropyxis gibba * Deflandre, 1929 Centropiyxidae 1.286 1.130 6.479 1.809
Centropyxis sp* Centropiyxidae 50 0 5.650 425
Centropyxis spinosa * Cash, 1905 Centropiyxidae 0 134 0 0
Difflugia achlora** Pénard, 1902 Difflugidae 254 205 167 33
Difflugia corona ** Wallich, 1864 Difflugidae 392 33 40 105
Difflugia difficilis** Difflugidae 100 67 250 25
Difflugia gramen ** Pénard, 1902 Difflugidae 134 67 0 0
Difflugia kempnyi** Stepánek, 1953 Difflugidae 0 0 50 40
Difflugia limnetica** Pénard, 1912 Difflugidae 533 92 133 825
Difflugia litophila ** Pénard, 1902 Difflugidae 10 67 0 65
Difflugia lobostoma ** Leidy, 1877 Difflugidae 67 67 0 0
Difflugia muriformis** Gauthier-Lièvre e Thomas, 1958 Difflugidae 1.130 787 567 0
Difflugia penardi ** Hopkinson, 1909 Difflugidae 0 67 50 0
Difflugia pseudogramen** Gauthier-Lièvre e Thomas, 1960 Difflugidae 454 515 107 105
Difflugia sp** Difflugidae 200 210 650 50
Difflugia sp1** Difflugidae 0 67 0 58
Difflugia brevicola** Cash e Hopkinson, 1909 Difflugidae 200 0 100 75
Difflugia sp2** Difflugidae 0 0 100 0
Euglypha acanthophora*** Ehrenberg, 1841 Euglyphidae 100 267 737 988
Euglypha filifera*** Pénard, 1890 Euglyphidae 117 100 1.750 709
Euglypha denticulata*** Brown, 1912 Euglyphidae 0 0 40 96
Euglypha tuberculata *** Dujardin, 1841 Euglyphidae 0 0 599 84
Lesquereusia extranea ** Awerintzew, 1906 Difflugidae 133 0 0 0
Lesquereusia globulosa** Gauthier-Lièvre e Thomas, 1959 Difflugidae 915 317 200 240
Lesquereusia modesta minima ** Van oye, 1953 Difflugidae 0 200 267 50
Lesquereusia sp** Difflugidae 0 0 0 40
Lesquereusia spiralis ** (Ehrenberg) Butschli, 1880 Difflugidae 1.017 263 344 152
Lesquereusia spiralis caudata** Playfair, 1917 Difflugidae 0 0 283 234
Lesquereusia spiralis declotrei** Van oye, 1959 Difflugidae 67 0 67 0
Lesquereusia spiralis hirsuta** Gauthier-Lièvre e Thomas, 1959 Difflugidae 250 0 67 25
Netzelia labeosa ** Beyens e Chardez, 1997 Difflugidae 2.403 2.332 859 671
Netzelia oviformis ** Ogden, 1979 Difflugidae 2.512 1.803 1.939 1.023
Netzelia sp** Difflugidae 20 1.000 400 84
Netzelia sp.1** Difflugidae 434 1.383 972 360
Netzelia tuberculata** (Wallich, 1864) Netzel, 1983 Difflugidae 300 38 217 268
Protocucurbitela coriniformis ** Gauthier-Lièvre e Thomas, 1960 Difflugidae 67 200 0 0
Amoeba Testacea (org.m-3) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), de6/2011 à 12/2012.
lagoa Carioca lagoa Gambazinho
Tabela 19:Abundância (org.m-3) de Amoeba Testacea, por espécie, na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), entre 6/2011 à 12/2012. Espécies exclusivas da lagoa Gambazinho foram destacadas com a cor azul e as espécies exclusivas da lagoa Carioca o foram com a cor rosa.*=ameba com teca quitinosa, **=ameba com teca de detritos, ***=ameba com teca silicosa.
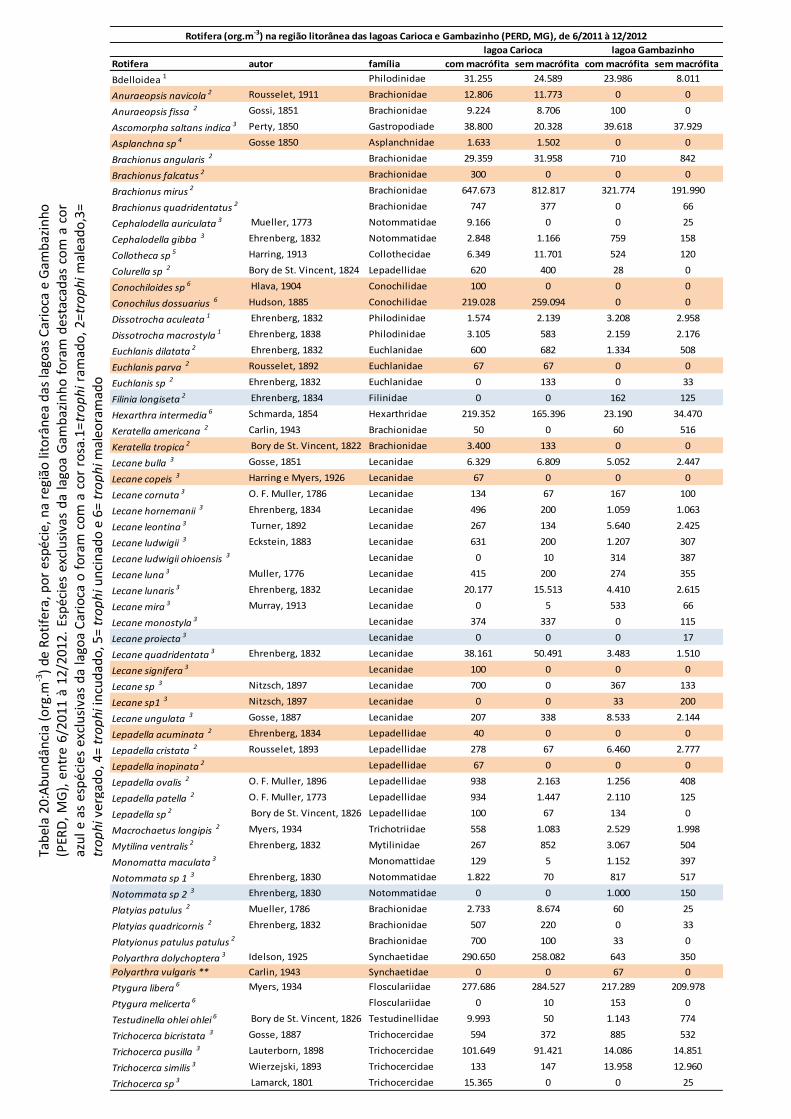
77
Tab
ela
20:A
bu
nd
ânci
a (o
rg.m
-3)
de R
oti
fera
, po
r es
péc
ie, n
a re
gião
lito
rân
ea d
as la
goas
Car
ioca
e G
amb
azin
ho
(P
ERD
, M
G),
en
tre
6/20
11 à
12
/20
12
. Es
péc
ies
excl
usi
vas
da
lago
a G
amb
azin
ho
fo
ram
des
taca
das
co
m a
co
r az
ul e
as
esp
écie
s ex
clu
siva
s d
a la
goa
Car
ioca
o f
ora
m c
om
a c
or
rosa
.1=
tro
ph
i ra
mad
o, 2
=tr
op
hi
mal
ead
o,3
=
tro
phi
ver
gad
o, 4
= tr
op
hi in
cud
ado
, 5=
tro
ph
i un
cin
ado
e 6
= tr
op
hi m
aleo
ram
ado
Rotifera autor família com macrófita sem macrófita com macrófita sem macrófita
Bdelloidea 1 Philodinidae 31.255 24.589 23.986 8.011
Anuraeopsis navicola 2 Rousselet, 1911 Brachionidae 12.806 11.773 0 0
Anuraeopsis fissa 2 Gossi, 1851 Brachionidae 9.224 8.706 100 0
Ascomorpha saltans indica 3 Perty, 1850 Gastropodiade 38.800 20.328 39.618 37.929
Asplanchna sp 4 Gosse 1850 Asplanchnidae 1.633 1.502 0 0
Brachionus angularis 2 Brachionidae 29.359 31.958 710 842
Brachionus falcatus 2 Brachionidae 300 0 0 0
Brachionus mirus 2 Brachionidae 647.673 812.817 321.774 191.990
Brachionus quadridentatus 2 Brachionidae 747 377 0 66
Cephalodella auriculata 3 Mueller, 1773 Notommatidae 9.166 0 0 25
Cephalodella gibba 3 Ehrenberg, 1832 Notommatidae 2.848 1.166 759 158
Collotheca sp 5 Harring, 1913 Collothecidae 6.349 11.701 524 120
Colurella sp 2 Bory de St. Vincent, 1824 Lepadellidae 620 400 28 0
Conochiloides sp 6 Hlava, 1904 Conochilidae 100 0 0 0
Conochilus dossuarius 6 Hudson, 1885 Conochilidae 219.028 259.094 0 0
Dissotrocha aculeata 1 Ehrenberg, 1832 Philodinidae 1.574 2.139 3.208 2.958
Dissotrocha macrostyla 1 Ehrenberg, 1838 Philodinidae 3.105 583 2.159 2.176
Euchlanis dilatata 2 Ehrenberg, 1832 Euchlanidae 600 682 1.334 508
Euchlanis parva 2 Rousselet, 1892 Euchlanidae 67 67 0 0
Euchlanis sp 2 Ehrenberg, 1832 Euchlanidae 0 133 0 33
Filinia longiseta 2 Ehrenberg, 1834 Filinidae 0 0 162 125
Hexarthra intermedia 6 Schmarda, 1854 Hexarthridae 219.352 165.396 23.190 34.470
Keratella americana 2 Carlin, 1943 Brachionidae 50 0 60 516
Keratella tropica 2 Bory de St. Vincent, 1822 Brachionidae 3.400 133 0 0
Lecane bulla 3 Gosse, 1851 Lecanidae 6.329 6.809 5.052 2.447
Lecane copeis 3 Harring e Myers, 1926 Lecanidae 67 0 0 0
Lecane cornuta 3 O. F. Muller, 1786 Lecanidae 134 67 167 100
Lecane hornemanii 3 Ehrenberg, 1834 Lecanidae 496 200 1.059 1.063
Lecane leontina 3 Turner, 1892 Lecanidae 267 134 5.640 2.425
Lecane ludwigii 3 Eckstein, 1883 Lecanidae 631 200 1.207 307
Lecane ludwigii ohioensis 3 Lecanidae 0 10 314 387
Lecane luna 3 Muller, 1776 Lecanidae 415 200 274 355
Lecane lunaris 3 Ehrenberg, 1832 Lecanidae 20.177 15.513 4.410 2.615
Lecane mira 3 Murray, 1913 Lecanidae 0 5 533 66
Lecane monostyla 3 Lecanidae 374 337 0 115
Lecane proiecta 3 Lecanidae 0 0 0 17
Lecane quadridentata 3 Ehrenberg, 1832 Lecanidae 38.161 50.491 3.483 1.510
Lecane signifera 3 Lecanidae 100 0 0 0
Lecane sp 3 Nitzsch, 1897 Lecanidae 700 0 367 133
Lecane sp1 3 Nitzsch, 1897 Lecanidae 0 0 33 200
Lecane ungulata 3 Gosse, 1887 Lecanidae 207 338 8.533 2.144
Lepadella acuminata 2 Ehrenberg, 1834 Lepadellidae 40 0 0 0
Lepadella cristata 2 Rousselet, 1893 Lepadellidae 278 67 6.460 2.777
Lepadella inopinata 2 Lepadellidae 67 0 0 0
Lepadella ovalis 2 O. F. Muller, 1896 Lepadellidae 938 2.163 1.256 408
Lepadella patella 2 O. F. Muller, 1773 Lepadellidae 934 1.447 2.110 125
Lepadella sp 2 Bory de St. Vincent, 1826 Lepadellidae 100 67 134 0
Macrochaetus longipis 2 Myers, 1934 Trichotriidae 558 1.083 2.529 1.998
Mytilina ventralis 2 Ehrenberg, 1832 Mytilinidae 267 852 3.067 504
Monomatta maculata 3 Monomattidae 129 5 1.152 397
Notommata sp 1 3 Ehrenberg, 1830 Notommatidae 1.822 70 817 517
Notommata sp 2 3 Ehrenberg, 1830 Notommatidae 0 0 1.000 150
Platyias patulus 2 Mueller, 1786 Brachionidae 2.733 8.674 60 25
Platyias quadricornis 2 Ehrenberg, 1832 Brachionidae 507 220 0 33
Platyionus patulus patulus 2 Brachionidae 700 100 33 0
Polyarthra dolychoptera 3 Idelson, 1925 Synchaetidae 290.650 258.082 643 350
Polyarthra vulgaris ** Carlin, 1943 Synchaetidae 0 0 67 0
Ptygura libera 6 Myers, 1934 Flosculariidae 277.686 284.527 217.289 209.978
Ptygura melicerta 6 Flosculariidae 0 10 153 0
Testudinella ohlei ohlei 6 Bory de St. Vincent, 1826 Testudinellidae 9.993 50 1.143 774
Trichocerca bicristata 3 Gosse, 1887 Trichocercidae 594 372 885 532
Trichocerca pusilla 3 Lauterborn, 1898 Trichocercidae 101.649 91.421 14.086 14.851
Trichocerca similis 3 Wierzejski, 1893 Trichocercidae 133 147 13.958 12.960
Trichocerca sp 3 Lamarck, 1801 Trichocercidae 15.365 0 0 25
lagoa Carioca lagoa Gambazinho
Rotifera (org.m-3) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), de 6/2011 à 12/2012

78
Tabela 21:Abundância (org.m-3) de Cladocera, por espécie, na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), entre 6/2011 à 12/2012. Espécies exclusivas da lagoa Gambazinho foram destacadas com a cor azul e as espécies exclusivas da lagoa Carioca o foram com a cor rosa.*=raspadores **=filtradores.
Cladocera autor família com macrófita sem macrófita com macrófita sem macrófita
Alona guttata * G. O. Sars, 1862 Chydoridae 0 0 150 0
Disparalona dadayi* Birge, 1910 Chydoridae 1.437 535 716 806
Bosmina hagmanni** Stingelin, 1904 Bosminidae 93.201 35.805 333 1.698
Bosmina tubicen ** Brehm, 1953 Bosminidae 38.982 29.272 266 2.409
Bosmina freyi ** De Melo e Hebert, 1994 Bosminidae 233 133 33 138
Ceriodaphnia silvestrii ** Daday, 1902 Daphniidae 399 77 0 33
Chydorus eurynotus* Sars, 1901 Chydoridae 495 201 67 0
Chydorus pubecens * Sars, 1901 Chydoridae 100 0 0 0
Coronatella poppei * Richard, 1897 Chydoridae 1.708 1.230 43.400 14.381
Dadaya macrops* Daday, 1898 Chydoridae 0 0 26.478 1.034
Diaphanosoma birgei ** Korinek, 1981 Sididae 0 67 50.031 101.046
Dunhevedia odontoplax* Sars, 1901 Chydoridae 0 5 0 400
Ephemeroporus barroisi * Richard, 1894 Chydoridae 13.098 2.625 4.116 572
Ephemeroporus hybridus* Daday, 1905 Chydoridae 11.385 1.160 543 131
Ephemeroporus tridentatus * Bergamin, 1939 Chydoridae 1.604 164 383 199
Ilyocriptus spinifer * Herrick, 1882 Ilyocryptidae 1.082 407 0 157
Karualona muelleri* Chydoridae 0 50 0 0
Macrothrix elegans* Brady, 1886 Macrothricidae 67 0 200 0
Macrothrix sp* Macrothricidae 67 133 100 0
Macrothrix spinosa* King, 1853 Macrothricidae 1.067 254 100 100
Simocephalus latirostris** Stingelin, 1906 Daphniidae 29 0 0 0
Scapholeberis armata ** Herrick e Turner, 1895 Daphniidae 116 0 528 233
Scapholeberis spinifera ** Nicolet, 1849 Daphniidae 0 0 150 58
Streblocerus pigmaeus* Sars, 1901 Macrothricidae 0 0 133 0
lagoa Carioca lagoa Gambazinho
Cladocera(org.m-3) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), de 6/2011 à 12/2012
Tabela 22:Abundância (org.m-3) de Copepoda, por espécie, na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), entre 6/2011 à 12/2012. Espécies exclusivas da lagoa Gambazinho foram destacadas com a cor azul e as espécies exclusivas da lagoa Carioca o foram com a cor rosa.*=onívoros **=fitoplanctófagos.
Copepoda autor família com macrófita sem macrófita com macrófita sem macrófita
Mesocyclops meridianus * Kiefer, 1926 Cyclopidae 6.041 1.163 67 25
Microcyclops anceps* Richard, 1897 Cyclopidae 33 0 50 0
Microcyclops finitimus* Cyclopidae 16.828 12.225 6.967 1.706
Notodiaptomus isabelae** Diaptomidae 0 0 24.273 18.543
Thermocyclops minutus* Cyclopidae 89.036 76.241 10.693 13.507
Tropocyclops prasinus* Fischer, 1860 Cyclopidae 4.720 3.406 119.106 80.743
lagoa Carioca lagoa Gambazinho
Copepoda (org.m-3) na região litorânea das lagoas Carioca e Gambazinho (PERD, MG), de 6/2011 à 12/2012

79


















