
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE …§ões-Teses... · JOSILAINE GONÇALVES DA SILVA...
72
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E ZOOTECNIA Programa de Pós-Graduação em Agricultura Tropical ASPECTOS FISIOLÓGICOS E PRODUTIVOS DO AMARANTO SUBMETIDO A DIFERENTES PERIODOS DE ESTRESSE HIDRICO JOSILAINE GONÇALVES DA SILVA CUIABÁ – MT 2015
-
Upload
nguyenminh -
Category
Documents
-
view
218 -
download
0
Transcript of UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE …§ões-Teses... · JOSILAINE GONÇALVES DA SILVA...

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
Programa de Pós-Graduação em Agricultura Tropical
ASPECTOS FISIOLÓGICOS E PRODUTIVOS DO AMARANTO
SUBMETIDO A DIFERENTES PERIODOS DE ESTRESSE HIDRICO
JOSILAINE GONÇALVES DA SILVA
CUIABÁ – MT
2015

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
Programa de Pós-Graduação em Agricultura Tropical
ASPECTOS FISIOLÓGICOS E PRODUTIVOS DO AMARANTO
SUBMETIDO A DIFERENTES PERIODOS DE ESTRESSE HIDRICO
JOSILAINE GONÇALVES DA SILVA
Engenheira Agrônoma
Orientador: Prof. Dr. ALOISIO BIANCHINI
Coorientador: Prof Dr. FRANCISCO DE ALMEIDA LOBO
Dissertação apresentada à Faculdade de
Agronomia, Medicina Veterinária e
Zootecnia da Universidade Federal de
Mato Grosso, para obtenção do título de
Mestre em Agricultura Tropical.
CUIABÁ–MT
2015

Dados Internacionais de Catalogação na F o n t e .
Ficha catalográfica elaborada automaticamente de acordo com os dados fornecidos pelo(a) autor(a).
Permitida a reprodução parcial ou total, desde que citada a fonte.
S586a SILVA, JOSILAINE GONÇALVES DA. ASPECTOS FISIOLÓGICOS E PRODUTIVOS DO AMARANTO
SUBMETIDO A DIFERENTES PERÍODOS DE ESTRESSE HÍDRICO / JOSILAINE GONÇALVES DA SILVA. -- 2015
69 f. ; 30 cm.
Orientador: ALOISIO BIANCHINI. Co-orientador: FRANCISCO DE ALMEIDA LOBO. Dissertação (mestrado) - Universidade Federal de Mato Grosso,
Faculdade de Agronomia e Medicina Veterinária, Programa de Pós-Graduação em Agricultura Tropical, Cuiabá, 2015.
Inclui bibliografia.
1. Amaranthus, crescimento vegetal, déficit hídrico, intercâmbio gasoso, relações hídricas. I. Título.


Aos meus pais José Gonçalves da Silva (in memoriam) e Nerli Terezinha do
Nascimento e Silva, que sempre foram meus exemplos de dignidade e apesar de
todas dificuldades que passamos me ensinaram que a educação é o caminho, a
eles que sempre me honraram com respeito e amor.
Dedico
Aos meus orientadores Prof Dr. Aloisio Bianchini e Prof. Dr. Francisco de
Almeida Lobo pela confiança, estímulo, paciência e, principalmente, pela
disposição em ensinar, a eles que contribuíram de forma significante para meu
crescimento profissional.
Ofereço

AGRADECIMENTOS
A Deus pela minha existência, pela força e sabedoria concedida, por abrir as portas
para a realização dos meus objetivos.
À Universidade Federal de Mato Grosso (UFMT), Programa de Pós-Graduação em
Agricultura Tropical (PPGAT), pela realização do curso de mestrado.
Ao Conselho Nacional (CNPq), pela concessão da bolsa durante o período do curso.
Á banca examinadora profª Drª Elisangela Clarete Camili e prof. Dr. Marco Antônio
Camilo de Carvalho por contribuírem nas correções e pelas sugestões neste trabalho.
Ao amigo Wininton Mendes pelos inúmeros esclarecimentos, pela valiosa
contribuição, revisões e, é claro, paciência e sugestões sempre que necessárias.
Aos professores do Programa de Pós-Graduação em Agricultura Tropical pelos
ensinamentos indispensáveis concedidos.
Á profª Drª Sânia Camargos, pela amizade, incentivo e por ser minha segunda mãe,
me apoiando sempre nos momentos mais difíceis.
Á Karoline Barros e Pâmela Palhâno pela amizade e dedicação e por estarem sempre
comigo.
Aos colegas da pós-graduação em especial aos meus amigos que aqui fiz: Adilson
Brandão, Alexandra de Paiva, Eduardo Vaz, Glauce Portela, João Rufino, Milton
Rodrigues e Tamires Brandão, pelo convívio e apoio durante o curso, a vocês que não
mediram esforços para me ajudar e estiveram sempre presentes quando eu precisei,
agradeço imensamente.
A todos que contribuíram para a realização deste trabalho meus sinceros
agradecimentos.

ASPECTOS FISIOLÓGICOS E PRODUTIVOS DO AMARANTO SUBMETIDO A
DIFERENTES PERIODOS DE ESTRESSE HIDRICO
RESUMO - O amaranto cultivado apresenta características nutricionais e agronômicas desejáveis de produção de biomassa e grãos com potencial para se tornar opção para compor sistemas produtivos no Cerrado. Entretanto, há carência de informações sobre o manejo da planta e o impacto da restrição hídrica para a produção comercial. O objetivo do presente trabalho foi avaliar os aspectos de crescimento, produtivos e as relações hídricas de plantas de amaranto sob diferentes regimes hídricos. Esta pesquisa foi realizada na Fazenda Experimental da Universidade Federal de Mato Grosso, no município de Santo Antônio do Leverger entre 30 de junho a 23 de setembro de 2014. O delineamento experimental utilizado foi em blocos casualizados em arranjo fatorial (2x3) com três repetições, sendo o primeiro fator as variedades de amaranto (‘BRS Alegria’ e ‘INCA’) e o segundo fator os níveis de estresse hídrico (plantas irrigadas até 45 dias após a emergência, plantas irrigadas até 60 dias após a emergência e irrigação o ciclo todo). Para análise quantitativa do crescimento foram avaliadas semanalmente: altura de plantas, diâmetro do caule, largura e comprimento da inflorescência, massa da matéria seca e massa seca da raiz. Ao final do período experimental foram avaliados os aspectos produtivos (produtividade total, índice de colheita e produtividade por unidade de água disponibilizada). As trocas gasosas (fotossíntese líquida, condutância estomática, transpiração e eficiência no uso da água) e relações hídricas foram avaliadas antes da imposição do estresse e na colheita (aos 63 dias após a emergência na variedade Inca e aos 84 dias após a emergência na variedade BRS Alegria). O déficit hídrico afetou o crescimento das plantas quanto à altura, diâmetro do caule e produção de matéria seca para os diversos órgãos da planta de amaranto exceto para a raiz que teve a massa acrescentada quando a imposição o estresse hídrico foi severo. A variedade Inca não apresentou diferença significativa entre os tratamentos quanto à produtividade. Na variedade BRS Alegria a melhor produtividade total e produtividade por unidade de agua disponibilizada foi obtida no tratamento com irrigação plena. O estresse severo provocou decréscimo na condutância estomática e fotossíntese. Para as variáveis das relações hídricas não foi possível verificar qualquer efeito dos tratamentos avaliados.
Palavras chave: Amaranthus, crescimento vegetal, déficit hídrico, intercâmbio
gasoso, relações hídricas.

PHYSIOLOGICAL ASPECTS AND PRODUCTION OF AMARANTH SUBMITTED
TO WATER STRESS
ABSTRACT-The cultivated amaranth presents nutritional and agronomic
characteristics desirable for the production of biomass and grain with potential to
become option to compoud productive systems in the Brazilian Savannah. But, there
is deficiency of information about the plant management and the impact of water
restriction for commercial production. The objective of this work was to evaluate
aspects of growth, production and water relations of amaranth plants under different
water regimes.This research was conducted at the Experimental Farm of the Federal
University of Mato Grosso, in the municipality of Santo Antônio do Leverger from 30
June to 23 September 2014. he experimental design was that of randomized block in
factorial arrangement (2x3) with three repettions, being the first factor the varieties of
amaranth (BRS Alegria 'and' INCA ') and the second factor the water stress levels
(irrigated plants until 45 days after emergence, irrigated plants until 60 days after
emergence and irrigation all cycle ).For quantitative growth analysis were evaluated
weekly: plant height, stem diameter, width and length of inflorescence, dry matter and
root dry mass. At the end of the trial period were evaluated productive aspects (grain
yield, harvest index and grain yield per available unit of water). The parameters of gas
exchange (net photosynthesis rate, stomatal conductance, transpiration and efficient
use of water) and water relations were evaluated before the stress and harvesting (at
63 days after emergence in the variety Inca and at 84 days after emergence in variety
BRS Alegria) The water deficit affected the growth of plants for height, stem diameter
and dry matter production for the various organs of the amaranth plant except for the
root that had the added mass when the water stress imposed was severe. The Inca
variety showed no significant difference between treatments for grain yield. In the
variety BRS Alegria better overall the grain yield and grain yield per available unit of
water was obtained in the treatment with full irrigation. The severe water deficit caused
a decrease in stomatal conductance and photosynthesis. For the variables of water
relations could not be verified any effect of the treatments evaluated.
Keywords: Amaranthus, plant growth, water deficit, gas exchange, water relations.

LISTA DE FIGURAS
Pág
Figura 1: Semeadura do amaranto nas linhas de plantio em junho de 2014, Santo
Antônio do Leverger-MT...................................................................................................
25
.Figura 2. Plântulas de Amaranthus cruentus com um par de folhas cotiledonares aos
dois dias após a emergência (A); Inicio do aparecimento da primeira folha verdadeira
aos cinco dias após a emergência (B) Santo Antônio do
Leverger,2014................................................................................................................
34
Figura 3. Primórdios da inflorescência A. caudatus cv Inca aos 35 dias após a
emergência (A) e primórdios da inflorescência na “BRS Alegria” aos 42 dias após a
emergência (B) Santo Antônio do Leverger,2014...........................................................
35
Figura 4 Plantas de A. caudatus cv ‘Inca’ em plena antese aos 39 dias após a
emergencia(A) e A. cruentus cv. ‘BRS Alegria’ aos 54 dias após a emergência em BRS
Alegria (B), Santo Antônio do Leverger,2014
.........................................................................................................................................
36
Figura 5. Ponto de colheita de A. caudatus ‘Inca’ (A) e A. cruentus ‘BRS Alegria’ (B),
Santo Antônio do
Leverger,2014..................................................................................................................
37
Figura 6: Dinâmica de crescimento do caule em plantas de amaranto variedade Inca
(A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60 dias (T2) e
durante o ciclo todo
(T3)..................................................................................................................................
38
Figura 7 Dinâmica de crescimento em altura de plantas de amaranto variedade Inca
(A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60 dias (T2) e
durante o ciclo todo (T3).).................................................................................................
40
Figura 8 Dinâmica de crescimento em massa seca de amaranto variedade Inca (A) e
BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60 dias (T2) e durante
o ciclo todo (T3).) ............................................................................................................
42
Figura 9. Dinâmica de crescimento em comprimento da panicula de plantas de
amaranto variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1)
até 60 dias (T2) e durante o ciclo todo (T3).)..............................................................
44
Figura 10. Dinâmica de crescimento largura de panícula de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60
dias (T2) e durante o ciclo todo (T3)........................................................................
45

Figura 11: : Dinâmica de crescimento em massa da panícula de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60
dias (T2) e durante o ciclo todo (T3)......................................
46
Figura 12. Dinâmica de crescimento em massa da panícula de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60
dias (T2) e durante o ciclo todo (T3)......................................
47

LISTAS DE TABELAS
Pág
Tabela 1. Características físico-químicas do solo da área experimental, Santo Antônio do
Leverger-MT,2014..................................................................................................................
24
Tabela 2. Precipitação e água fornecida na irrigação (em mm) durante o período
experimental, que foi de 26/06/2014 à 23/09/2014 em Santo Antônio de Leverger-
MT,2014.................................................................................................................................
32
Tabela 3. Duração dos estádios fenológicos do amaranto ‘BRS Alegria’ e ‘Inca’ cultivado
em Santo Antônio do Leverger-MT, no período de Julho a Setembro,
2014..................................................................................................................
33
Tabela 4. . Correlação de Pearson entre os parâmetros estudados na variedade INCA
(Amaranthus caudatus), agrupando-se os regimes hídricos
estudados...............................................................................................................................
49
Tabela 5. Correlação de Pearson entre os parâmetros estudados na variedade BRS Alegria
(Amaranthus cruentus), agrupando-se os regimes hídricos estudados.................................
50
Tabela 6. Taxas fotossintéticas líquidas para o desdobramento da interação entre o
momento da realização das medições do intercâmbio gasoso e o regime
hídrico...................................................................................................................................
52
Tabela 7. Taxas transpiratórias para o momento da realização das medições do intercâmbio
gasoso...................................................................................................................................
52
Tabela 8. Prova de médias das eficiências instantâneas no uso de água para o momento
da realização das medições do intercâmbio
gasoso..................................................................................................................................
54
Tabela 9 Produtividade (kg ha-1) de A. caudatus cv Inca e A. cruentus cv BRS Alegria em
diferentes regimes hídricos,Santo Antônio do Leverger-
MT,2009................................................................................................................................
57
Tabela 10. . Produtividade por unidade de água disponibilizada( kg ha.mm-1 (PUAD) para
desdobramento da interação em A. caudatus cv Inca e A. cruentus cv BRS Alegria em
diferentes regimes hídricos Santo Antônio do Leverger-
MT,2009................................................................................................................................
58
Tabela 11: Índice de colheita para A. Cruentus cv “BRS Alegria” e A. Caudatus cv
‘Inca’.......................................................................................................................................
59

Sumário
Página
1. INTRODUÇÃO ............................................................................................................................ 12
1.1 OBJETIVO GERAL ....................................................................................................... 14
1.1.1 Objetivos específicos ........................................................................................................ 14
1.1.2 Hipóteses ....................................................................................................................... 14
2. REVISÃO DE LITERATURA .................................................................................................... 15
2.1 ORIGEM E IMPORTÂNCIA DO AMARANTO ................................................................... 15
2. 2 CLASSIFICAÇÃO BOTÂNICA E DESCRIÇÃO DA PLANTA ....................................... 16
2.4 ECOFISIOLOGIA DA CULTURA DO AMARANTO .......................................................... 17
2.5 FENOLOGIA DA CULTURA DO AMARANTO .................................................................. 18
2.6 ESTRESSE HÍDRICO ............................................................................................................. 19
2.6.1 EFEITOS DO ESTRESSE HÍDRICO NO CRESCIMENTO VEGETAL ....................... 20
2.6.2 EFEITOS DO ESTRESSE HÍDRICO NAS RELAÇÕES HÍDRICAS ........................... 21
2.6.3 INDICADORES DE DÉFICIT HÍDRICO ............................................................................ 22
3. MATERIAL E MÉTODOS ......................................................................................................... 23
3.1 DELINEAMENTO EXPERIMENTAL .................................................................................... 23
3.2 ÁREA EXPERIMENTAL ....................................................................................................... 24
3. 4. AVALIAÇÕES ........................................................................................................................ 26
3.4. 1 Análise quantitativa do crescimento e analise dos estádios fenológicos: ....... 26
3.4.5 Medidas de intercâmbio gasoso .................................................................................... 27
3.4.6.Relações hídricas ............................................................................................................... 28
3.4.7 Aspectos produtivos................................................................................................... 30
3. 5 Análises estatísticas ............................................................................................................ 31
4. RESULTADOS E DISCUSSÃO ............................................................................................... 32
4.1 FENOLOGIA ............................................................................................................................ 33
4.2 ANALISE QUANTITATIVA DO CRESCIMENTO .............................................................. 37
4.2.1 Correlações de Pearson entre a análise quantitativa de crescimento e os
aspectos produtivos ................................................................................................................ 48
4.3 Intercâmbio Gasoso .................................................................................................... 51

4.4 Relações Hídricas ........................................................................................................ 55
4.5 ASPECTOS PRODUTIVOS ................................................................................................... 56
5. CONSIDERAÇÕES FINAIS ..................................................................................................... 60
6. CONCLUSÕES ........................................................................................................................... 61
7. REFERÊNCIAIS BIBLIOGRÁFICAS ...................................................................................... 63

12
1. INTRODUÇÃO
A utilização de novas espécies vegetais é importante para compor a
diversificação de cultivos. O amaranto desponta como alternativa ao criar
oportunidade para novos alimentos e gerando renda para o agricultor
No Brasil o cultivo de amaranto ainda é incipiente, no entanto, apresenta
perspectivas de se estabelecer como uma alternativa para a rotação de culturas
criando oportunidades para dietas alimentares mais saudáveis (Spehar et al., 2007).
Desde sua introdução no Brasil, a partir dos anos 1990, tem crescido a informação
sobre o cultivo, criando a necessidade de se ampliar o conhecimento e a tecnologia
de produção (Teixeira et al., 2003).
No Brasil, a depender da variedade e das condições ambientais, tem-se obtido
produtividades entre 1,000 e 2,359 kg ha-1 (Domingos et al., 2005; Spehar et al., 2003),
em condições de solo de Cerrado. No sul do país, tem se obtido produtividades de até
4,500 kg ha-1 (Brambilla et al., 2008) e no Distrito Federal, em solos anteriormente
cultivados com soja, obtiveram-se produtividades entre 1,000 a 3,000 kg ha-1 (Spehar
1998)
O amaranto tem múltiplos usos, tanto na alimentação humana, animal, na
indústria, na medicina e na ornamentação. Para o consumo humano o grão pode ser
utilizado inteiro ou moído na forma de farinha, assado, ou cozido. As folhas são usadas
da mesma forma a que folha outros legumes. Com grãos inteiros ou moídos pode
preparar pequenos-almoços, sopas, sobremesas, mingau, bolos, pudins, refrigerantes

13
e outros; também cozinhados, os grãos misturados com mel, melado ou chocolate,
dando-lhe diferentes formas em moldes de madeira ou metálicos conhecidos como
nougat de amaranto no Peru, "alegria" no México e "tadoos" na Índia (SinghaL;
Kulkarni, 1988).
Com a crescente utilização do amaranto para os mais diversos fins, faz-se
necessário um maior número de informações sobre o cultivo desta espécie, inclusive
sobre suas respostas a situações de deficiência hídrica. Do ponto de vista
ecofisiológico, os fatores externos por si só não proporcionam conhecimento suficiente
para se concluir sobre o grau de seca, devendo, portanto, ser a planta o objeto de
estudo principal, capaz de mostrar se a água é ou está sendo fator de estresse
(Larcher, 2006).
A adaptação de uma cultura aos sistemas de produção agropecuários depende
de uma série de características agronômicas. A seca é o estresse ambiental que mais
limita o crescimento e rendimento das culturas agrícolas (Guzmán et al., 2010). A
capacidade de se desenvolver e frutificar em restrição hídrica constitui uma valiosa
característica quando se fala em cultivos no Cerrado.
A resposta da planta à seca é caracterizada por mudanças fundamentais na
relação da célula com a água, nos seus processos fisiológicos, na estrutura de
membranas e de organelas, além das mudanças morfológicas e fenológicas da planta,
alterando a relação do seu dossel com o ambiente. Na planta, o efeito do estresse é
geralmente percebido como um decréscimo na fotossíntese líquida e crescimento e
está associado com alterações no metabolismo do carbono e nitrogênio (Yordanov et
al., 2000). As respostas vegetais à escassez hídrica são complexas, envolvendo
mudanças adaptativas e/ou efeitos deletérios (Chaves et al. 2002). Essas respostas
precisam ser investigadas para que se tenha informações necessárias para produção
em pequena ou larga escala da planta por inteiro ou de alguns de seus órgãos.
O estresse hídrico causa diferentes efeitos no vegetal dependendo da
intensidade, da velocidade de imposição do estresse e do estádio de desenvolvimento
da planta no momento em que o déficit hídrico ocorre (Pimentel, 2004).
O cultivo do amaranto surge com uma nova alternativa tanto na produção de
um alimento promissor, por ser uma fonte de nutrientes oriundos tanto do grão quanto
das partes vegetativas e não conter glúten, o que torna este produto de grande

14
importância na ampliação da oferta de alimentos nutritivos para portadores de doença
celíaca, quanto na introdução de uma nova espécie no sistema de produção. Apesar
disso, ainda é preciso levar em consideração que na produção do amaranto também
pode correr os mesmos riscos de redução na produção devido ao efeito dos veranicos.
Mediante a escassez de informações sobre efeito do estresse hídrico no
desenvolvimento do amaranto neste trabalho se teve o objetivo de avaliar o efeito de
diferentes períodos de restrição hídrica sobre o crescimento, a produção, o
intercâmbio gasoso e as relações hídricas em duas espécies de amaranto.
1.1 OBJETIVO GERAL
Estudar o efeito de diferentes períodos de restrição hídrica sobre o crescimento,
a produção, o intercâmbio gasoso e as relações hídricas em duas espécies de
amaranto
1.1.1 Objetivos específicos
1. Avaliar a produtividade de duas espécies de amaranto sob diferentes regimes
de estresse hídrico.
2. Descrever os estádios fenológicos do amaranto e sua relação com estresse
hídrico.
3. Avaliar o índice de colheita de duas variedades de amaranto.
4. Avaliar o intercâmbio gasoso e as relações hídricas de plantas de amaranto
antes da submissão do estresse e no pico do estresse hídrico (colheita).
5. Avaliar a análise quantitativa do crescimento e os estádios de desenvolvimento
do amaranto com e sem restrição hídrica.
6. Correlacionar os aspectos produtivos do amaranto com a análise quantitativa
do amaranto.
1.1.2 Hipóteses
1. O amaranto responde aos regimes de estresse hídrico, refletindo na fenologia,
reduzindo seu ciclo, com antecipação da floração e maturação dos frutos.
2. O déficit hídrico deve reduzir acúmulo de matéria seca na parte aérea, a relação
entre a parte aérea e a raiz e o índice de colheita.
3. A reposta do amaranto ao estresse hídrico depende da espécie, que influencia
na produtividade, produção e seus componentes

15
2. REVISÃO DE LITERATURA
2.1 ORIGEM E IMPORTÂNCIA DO AMARANTO
O amaranto é originário provavelmente das Américas do Sul e Central e
representou a base de diversas civilizações Pré colombianas, dentre elas os Maias,
Incas e Astecas (Capriles, 2012). Desde a pré história, antes mesmo do processo de
domesticação, inúmeras espécies de amaranto foram utilizadas em várias partes do
mundo, sob forma de hortaliças folhosas (Mújica-Sanchez et al., 1997).
A planta integra a lista das 36 culturas mais promissoras para alimentar a
humanidade e seu cultivo e consumo poderão aumentar a segurança alimentar de
populações vulneráveis (Ferreira et al.,2007). Embora tenha havido uma diminuição
de esforços para a produção desse vegetal nos Estados Unidos, em anos recentes,
as pesquisas de produção e utilização têm sido continuadas (Costa e Borges, 2005).
Nas duas últimas décadas, muitos cientistas dos Estados Unidos, da China, do México
e alguns poucos de outros países têm destinado grandes esforços no sentido de
melhorar os métodos de produção, caracterização e uso de sementes do amaranto.
As espécies cultivadas contêm óleo e proteína em quantidade e qualidade
superior à dos cereais (Spehar et al., 2003). Sob o ponto de vista agronômico, o
amaranto destaca-se por constituir opção para diversificar o cultivo de grãos no
cerrado. Ainda que a região tenha apresentado crescimento vertiginoso na agricultura,
esta baseia-se, principalmente na soja, no algodão, no milho e em menor escala no
feijão. Esse pequeno número de espécies torna a agricultura regional vulnerável,
ameaçando os avanços da pesquisa (Spehar,2009).

16
Por não ser autóctone no Brasil a planta tem sido muito pouco estudada e seu
consumo chega a ser praticamente desconhecido (Amaya-Farfan et al., 2005). As
duas espécies de amaranto mais comumente cultivadas são Amaranthus cruentus e
Amaranthus caudatus. No Brasil, os estudos concentram-se na espécie A. cruentus
cv. BRS Alegria (Gonçalves et al., 2006; Brambilla et al., 2008; Bianchini et al., 2010;
Piffer et al., 2010; Spehar & Trecentini, 2011; Nobre et al., 2013; Bianchini et al.,
2014; Ferreira, 2014).
2. 2 CLASSIFICAÇÃO BOTÂNICA E DESCRIÇÃO DA PLANTA
O amaranto é uma planta da família Amarantaceae, que se divide em mais de
70 gêneros e 850 espécies. É uma planta dicotiledônea, de porte herbáceo, com
inflorescência tipo capítulo, cacho ou panícula, com folhas inteiras, de disposição
espiralada ou oposta cruzada, sem estípulas (Amaya-Farfan et, al.,2005).
A raiz é pivotante, com abundantes ramificações e múltiplas radicelas finas que
se desenvolvem rapidamente. A raiz principal serve de suporte à planta, permitindo
manter o peso da panícula, principalmente quando os grãos se encontram cheios
antes da maturidade fisiológica (Spehar, 2007).
A planta, nos primeiros dias após a emergência, lembra o amaranto da espécie
daninha (Amaranthus viridis), também conhecido vulgarmente como caruru.
Entretanto, as plantas se desenvolvem vigorosas, com inflorescências (panículas ou
cachos) coloridas, com variações do amarelo ao roxo. Após 30 dias da emergência,
apresentam rápido crescimento e podem atingir até 2,0 m, com inflorescência de até
0,6 m, dependendo do manejo e suprimento de água. As sementes são muito
pequenas e arredondadas. Quando amadurecem, ao cair no solo, germinam
rapidamente na presença de umidade (Spehar et, al., 2003).
O caule é cilíndrico e anguloso, com grossas estrias longitudinais que lhe
confere aparência canelada com comprimento variável e espessura que diminui da
base ao ápice. As plantas são monóicas na maior parte das espécies de amaranto
(Brenner et al., 2000), porém, são predominantemente autógamas com variações no
percentual de polinização cruzada nas diferentes variedades.
As flores são unissexuais muito pequenas estaminadas no ápice e pistiladas.
O androceu é formado na base por cinco estames coloridos que mantém as anteras
por um ponto máximo da base, tornam-se bastante visíveis por ocasião da antese. O

17
gineceu apresenta ovário esférico, súpero, coroado por três estigmas filiformes e
pilosos que aloja uma semente (Tapia,1997). O fruto é uma cápsula pequena que se
denomina pixídio unilocular. Na maturidade essa cápsula abre de forma transversal
deixando cair a parte superior, o opérculo, descobrindo a parte inferior ou a urna onde
se encontra a semente. Em geral é deiscente, deixando cair a semente (Sanchez,
1980). A semente é pequena lenticular podendo alcançar de 1 a 1,5 mm ligeiramente
aplanada de cor bege ou branca, amarelada, rosada, roxa, marrom ou vermelha
(Nieto, 1990).
2.4 ECOFISIOLOGIA DA CULTURA DO AMARANTO
Para uma mesma região, o número de dias entre a emergência e a maturidade
pode variar com a latitude e a época de semeadura, indicando haver variabilidade
genética em A. cruentus para resposta ao fotoperíodo (Gimplinger et al., 2007; Earley
et al., 2005).
A luz corresponde a um dos principais aspectos da interação das plantas com
o ambiente, controlando o desenvolvimento, por influenciar processos como a
floração, germinação de sementes, crescimento de caules e folhas, formação de
órgãos de reservas e partição de assimilados. Fotoperiodismo é um termo usado na
botânica para descrever os efeitos e adaptações de plantas ao fotoperíodo, que
representa o comprimento de um dia e consiste na duração do período de luz de um
determinado lugar, dependendo da latitude e da estação do ano. Incluído no
fotoperíodo está o período de luz útil, que designa a duração, da qual a intensidade
luminosa é maior que o limiar de compensação fotossintética. É importante saber que
existem três tipos de plantas; plantas de dias curtos que florescem com fotoperíodos
inferiores ao fotoperíodo crítico, plantas de dias longos que florescem com
fotoperíodos superiores ou iguais ao fotoperíodo crítico, e plantas neutras ou
indiferentes. (Montenegro.,2010).
Estimar a data de início de florescimento é importante para o manejo da cultura,
pois, o intervalo de tempo de crescimento vegetativo afeta diretamente o tamanho da
planta e o índice de área foliar e, consequentemente a produção de massa de matéria
seca e produtividade (Rodrigues et al., 2001)

18
2.5 FENOLOGIA DA CULTURA DO AMARANTO
A caracterização do desenvolvimento das culturas é uma ferramenta
importante, pois possibilita identificar os diferentes estádios morfológicos da planta,
permitindo o estabelecimento de estratégias de manejo e consequentemente a
obtenção de rendimentos satisfatórios (Fancelli & Dourado Neto, 2000).
A descrição dos estádios fenológicos do amaranto segundo Mujica-Sanchez e
Quillahuamám,1989; Henderson,1993, são:
Emergência: As plântulas emergem do solo exibindo as duas folhas cotiledonares
com pelo menos 50% da densidade esperada no sulco. As folhas verdadeiras
apresentam tamanho menor que 2 cm de comprimento. Dura cerca de 8 dias a 21 dias
dependendo das condições agroclimáticas.
Vegetativa: Nesta fase, define-se o número de nós no caule ou haste principal onde
se encontram as folhas expandidas com pelo menos 2 cm de comprimento. O primeiro
nó é classificado como V1. A planta começa a ramificar no estádio V4.
Reprodutiva:
Início da formação de panícula (R1): ápice da inflorescência (panícula) torna-se
visível no extremo do caule entre 45 e 70 dias após a emergência.
Intermediária (R2): A panícula apresenta pelo menos 2 cm de comprimento.
Término (R3): A panícula apresenta pelo menos 5 cm de comprimento. Se a antese
teve início, a planta será classificada na etapa seguinte.
Antese (R4): Pelo menos uma flor por planta encontra-se aberta, mostrando os
estames separados e o estigma completamente visível. Em A. caudatus, as flores
hermafroditas são as primeiras a abrir. Em geral a antese começa pelo ponto
intermediário entre o eixo da panícula e as ramificações laterais dela. Nessa fase, a
planta se encontra mais sensível à falta de água e geadas.
Enchimento de grãos (R5): A antese se completou em pelo menos 95% do eixo
central da panícula. Essa etapa pode ser ainda subdividida em grão leitoso, quando
as sementes ao serem pressionadas entre os dedos deixam sair um liquido leitoso,
grão pastoso, quando as sementes ao serem pressionadas entre os dedos,
apresentam consistência pastosa, de cor esbranquiçada.

19
Maturidade fisiológica (R6): A maturação fisiológica ocorre quando a panícula muda
de coloração. Quando verdes, passam a exibir a coloração amarelo-dourada, as
vermelhas mudam para café avermelhado. As sementes são duras e, quando se agita
a panícula, elas se desprendem.
Ponto de colheita (R7): As folhas senescem e caem, a planta apresenta aspecto
seco, de coloração café. Nos cultivos sujeitos à geada, esta contribui para a seca do
talo.
2.6 ESTRESSE HÍDRICO
Atualmente, aproximadamente 70% da água potável disponível no mundo é
utilizada na agricultura e 40% dos alimentos são produzidas em solos irrigados. Nas
ultimas décadas, as bases moleculares e fisiológicas das respostas vegetais à seca
têm sido o foco principal de pesquisas intensas (Sadras & Milroy 1996; Yordanov et
al., 2000; Griffiths & Parry, 2002; Tyermann et al., 2002; Luu & Maurel, 2005; Jones,
2007).
O estudo da deficiência hídrica tem merecido especial atenção por parte dos
fisiologistas e agrônomos, devido a importância no crescimento e produção das
plantas superiores (Paulus et al., 2010).
Estresse é um desvio das condições ótimas de desenvolvimento do vegetal, no
qual podem ocorrer mudanças e respostas em parte do organismo ou nele como um
todo (Larcher, 2006). No início essas alterações podem ser reversíveis, mas,
dependendo da duração e da situação de estresse, podem ser irreversíveis, levando
a danos permanentes ou até à morte (Plaut, 1995).
As variedades tolerantes à seca podem reduzir a quantidade de água
consumida, e/ou produzirem mais grãos para uma com a mesma quantidade de água
(Sayed, 1983). O desenvolvimento de variedades tolerantes à seca deve ser um dos
objetivos primordiais nos programas de melhoramento genético (Bruckener e
Frohberg, 1987), e requer a identificação de mecanismos fisiológicos importantes para
serem usados como critério de seleção (Blum et al., 1981).

20
A capacidade da planta em responder e sobreviver ao estresse hídrico depende
de mecanismos que integram respostas celulares, sendo que tais respostas podem
ocorrer em alguns segundos ou em poucos minutos ou horas (Bray, 1997).
O déficit hídrico, afeta vários aspectos do desenvolvimento das plantas
destacando-se a redução do tamanho das plantas, de sua área foliar e do seu
rendimento físico (Farias et al., 2008). Inman-Bamber (2004) ressalta que o tempo de
exposição à seca, ou seja, a duração do período de déficit hídrico afeta negativamente
o crescimento da parte aérea e a taxa de fotossíntese das plantas. Assim a
produtividade das culturas é altamente dependente das interações entre as fases
fenológicas e as variações inter e intranuais da disponibilidade de água no solo, que
por sua vez, são altamente dependentes da distribuição temporal e espacial das
chuvas.
A disponibilidade hídrica é considerada um dos fatores de maior efeito sobre a
produtividade agrícola, sendo o fator que rege a distribuição das espécies nas
diferentes zonas climáticas do globo (Pimentel, 2004). Larcher (2006) considera que
a seca é um estado ambiental em que a demanda de energia pela planta é maior que
a produção, o que leva a problemas fisiológicos, seguidos por aclimatação e
estratégias de adaptação nas gerações seguintes. Aí reside a importância de estudos
que permitam compreender os efeitos do déficit hídrico sobre o desenvolvimento
vegetal (Martins, 2008)
2.6.1 EFEITOS DO ESTRESSE HÍDRICO NO CRESCIMENTO VEGETAL
Todos os aspectos de crescimento e desenvolvimento das plantas são afetados
pela deficiência hídrica nos tecidos, causada pela excessiva demanda evaporativa ou
limitado suprimento de água. Como consequência da deficiência hídrica, verifica-se
uma desidratação do protoplasto com consequente diminuição do volume celular e
aumento na concentração de solutos. Aliado a isto, a diminuição da turgescência pode
ser admitida como consequência do estresse hídrico, e o processo de crescimento,
principalmente em extensão, é o primeiro afetado (Nogueira et al., 2005).
A análise de crescimento é de fundamental importância para o entendimento
das reações do vegetal em consequência a fatores ambientais (Benincasa, 1988).
Expansão celular, síntese da parede celular e síntese protéica nos tecidos de
crescimento estão entre os processos mais sensíveis ao déficit hídrico, podendo se

21
concluir que a redução da expansão foliar é uma das primeiras respostas a pouca
água disponível (Sadras & Milroy, 1996).
Para espécies herbáceas, numerosos estudos documentam que, sob estresse
hídrico, há um maior investimento no desenvolvimento das raízes, resultando em
aumento do comprimento (Fitter & Hay 1987) e de matéria seca da raiz, redução da
matéria seca total (parte aérea + raiz), diminuição da expansão foliar e, em alguns
casos, há diminuição da produção de folhas (Steinberg et al. 1990). A deficiência de
água ocorrida em plantas durante a diferenciação celular das folhas determina
redução irreversível da área e do espessamento foliar e aumento da densidade
estomática (Zagdanska & Kosdoj, 1994).
2.6.2 EFEITOS DO ESTRESSE HÍDRICO NAS RELAÇÕES HÍDRICAS
Segundo Pimentel (2004), não há como saber se determinada espécie é
tolerante ou não à seca buscando essa resposta em apenas uma única variável. Há
que se fazer um estudo mais completo, no qual sejam analisados vários parâmetros
que levem a uma conclusão mais definitiva. Aliada à análise de crescimento, existem
outras variáveis que levam a uma visão mais completa do quadro do estresse no
vegetal.
A avaliação do potencial hídrico torna-se interessante porque é uma medida
fácil de ser obtida através de uma câmara de pressão (Scholander et al., 1965).
Por outro lado o estresse hídrico é apontado como o principal redutor da
capacidade fotossintética (Flexas e Medrano, 2002). Uma vez que tem efeito sobre
vários componentes estomáticos e não estomáticos, incluindo pigmentos
fotossintéticos, sistema de transporte de elétrons e vias de redução de CO2
(Rahanama et al., 2010).
Sob condições de baixa disponibilidade de água, o fechamento estomático,
promovido por altos níveis de ácido abscísico (ABA), constitui uma das primeiras
estratégias utilizadas pelas plantas para diminuir a taxa de transpiração e manter a
turgescência celular (Melcher et al., 2009). O fechamento estomático promove
redução da condutância estomática e limita a assimilação de CO2 (Asharaf et al.,
2013). Sob severo estresse hídrico, ocorre a desidratação das células do mesofilo e
inibição da base metabólica do processo de fotossíntese (Anjun et al., 2011).

22
2.6.3 INDICADORES DE DÉFICIT HÍDRICO
Vários indicadores de déficit hídrico são descritos na literatura, sendo os
principais: temperatura foliar, condutância estomática, conteúdo relativo de água nas
folhas (Morales, 2012). Quando o suprimento de água não atende as necessidades
da planta, desenvolve-se o estresse hídrico, o que pode eventualmente ocasionar
redução de fotossíntese, afetando adversamente o crescimento da cultura e, portanto
a produtividade (Pereira et al., 2002).
Durante períodos de deficiência hídrica, a manutenção do crescimento da
planta depende de sua capacidade de conservar a turgescência das células, através
do fenômeno conhecido como ajustamento osmótico (Jones et al., 1981). Através da
utilização da técnica conhecida como "Curvas pressão-volume", os componentes
osmótico (Ys) e pressão (Yp) do potencial total da água (Ya) podem ser estimados em
uma mesma folha, bem como as propriedades elásticas do tecido e os volumes de
água simplástica e apoplástica (Marur, 1999). Assim as curvas pressão-volume (PV)
consistem um método indireto para investigação de possível ajustamento osmótico
por plantas submetidas ao estresse hídrico (Warren et al., 2007).

23
3. MATERIAL E MÉTODOS
3.1 DELINEAMENTO EXPERIMENTAL
O experimento foi conduzido no período seco do ano, entre os meses de junho
e agosto de 2014, em área pertencente a Fazenda Experimental da Universidade
Federal de Mato Grosso, em Santo Antônio de Leverger-MT, localizada a 33 km de
Cuiabá, delimitada pelas coordenadas geográficas 15°51’S; 56°04’W e 140 m de
altitude
O delineamento experimental utilizado foi em blocos casualizados em arranjo
fatorial (2x3x2) com três repetições, sendo o primeiro fator as variedades de amaranto
(‘BRS Alegria’ e ‘INCA ') e o segundo fator os níveis de estresse hídrico (plantas
irrigadas até 45 dias após a emergência, plantas irrigadas até 60 dias após a
emergência e irrigação o ciclo todo). E o terceiro fator o momento da medição para o
intercâmbio gasoso.
O experimento para avaliação do intercâmbio gasoso consistiu da aplicação de
três fatores. O primeiro fator foram duas variedades, a Inca e a BRS Alegria. O
segundo fator foram os regimes hídricos, sendo um a irrigação por até 45 dias após a
emergência, outro com irrigação até os 60 dias após a emergência e um terceiro
irrigado permanentemente até o momento da colheita. O terceiro fator foi o momento
da medição do intercâmbio gasoso. O experimento foi conduzido em blocos
casualizados, com três repetições.

24
3.2 ÁREA EXPERIMENTAL
A área do experimento havia sido cultivada, anteriormente à instalação do
experimento, com milho e capim Tanzânia na sequência. O solo da unidade
experimental é definido como Cambissolo Háplico (EMBRAPA, 2006) cujas
características químicas encontram-se na Tabela 1.
Tabela 1: Características químicas do solo da área experimental, Santo Antônio do
Leverger-MT,2014.
Profundidade pH M.O P K+ Ca+2+Mg+2 Al+3 H+Al CTC V
(m) % mg dcm-3 cmolc dm3 %
0-0,20 4,9 2,34 9,2 36 2,5 0 3,5 6 42,9
0,20-0,40 5,2 1,62 3,7 27 2,3 0 2,4 4,8 49,7
O clima da região, segundo a classificação de Köppen, é Aw. A temperatura
média mensal varia de 22,0° a 27,2°C e a precipitação média anual é de 1.320 mm.
O período em que aparece déficit hídrico inicia em maio e termina em outubro, sendo
que, do início de junho ao final de agosto, a precipitação média mensal é menor do
que 15 mm, e a evapotranspiração potencial média mensal ultrapassa os 75 mm
(Pacheco e Campelo Júnior, 2001).
3.3 SEMEADURA E TRATOS CULTURAIS
A área experimental recebeu calagem adicional com calcário filler
(PRNT=99,9%) para elevar a saturação de bases (V%) ao nível de 70%, 8 dias antes
da semeadura. Na adubação a lanço, 5 dias antes da semeadura, utilizou-se 500 kg
ha-1 da fórmula 4-14-08 (N-P2O5-K2O) acrescido de enxofre e zinco.
O solo foi sulcado com enxada no espaçamento entre linhas de 0,5 m segundo
recomendação de Spehar et al (2003). Após o sulcamento realizou-se a semeadura
por meio de uma semeadora manual (Figura 1), nas linhas, em 26 de junho de 2014,
utilizando-se sementes das variedades BRS Alegria e Inca, produzidas em 2011 e
mantidas em câmara refrigerada, com 94 e 98 % de germinação, respectivamente.
Cada parcela foi composta por seis linhas, com 12 metros de comprimento cada
perfazendo 36 m² (12m x 0,5m x 6 linhas). A densidade de semeadura foi elevada
para permitir o desbaste e obtenção de um estande 200 000 plantas ha-1. O desbaste,

25
para obtenção da densidade de plantas desejada, foi realizada manualmente, aos 20
dias após a emergência, quando as plantas possuíam, aproximadamente, 15 cm de
altura na Inca e 20 cm na BRS Alegria.
Figura 1: Semeadura do amaranto (Amaranthus spp) nas linhas de plantio em junho de 2014, Santo Antônio do Leverger-MT
A adubação de cobertura foi realizada 36 dias após a emergência, onde se
aplicou 60 kg ha-1 de N e 40 kg ha-1 K2O, seguindo a recomendação de Brambilla et
al. (2008).
A irrigação foi realizada sempre pela manhã, mantendo-se teor de umidade do
solo próximo a capacidade de campo, que foi monitorada por meio de bateria de
sondas instaladas nas camadas de 0 a 200 mm e de 200 a 400 mm de profundidade.
A aplicação da água foi realizada por um sistema de irrigação tipo aspersão
convencional, utilizando aspersores setoriais para evitar a aplicação de água nas
parcelas adjacentes. Para a avaliação da uniformidade de distribuição de água foram
instalados pluviômetros, equidistantes, entre quatro aspersores de todas as parcelas

26
experimentais. O volume coletado em cada pluviômetro foi determinado, utilizando-se
de proveta graduada e posteriormente foi calculado o Coeficiente de Uniformidade
Christiansen.
O coeficiente de coeficiente de Christiansen foi calculado de acordo com a
Equação 1:
𝐶𝑈𝐶(%) = 100 (1 −∑ [ 𝑋𝐼−𝑋]𝑁
𝐼=1
𝑋) (Equação 1)
Em que:
CUC = coeficiente de uniformidade de Christiansen, %;
Xi = precipitação no pluviômetro de ordem i, mm;
X= média aritmética das precipitações, mm; e
n = número de pluviômetros
3. 4. AVALIAÇÕES
3.4. 1 Análise quantitativa do crescimento e analise dos estádios fenológicos:
A análise quantitativa do crescimento e avaliação dos estádios fenológicos foi
realizada semanalmente, em cada parcela experimento tomando-se três plantas, que
totalizou 27 plantas por espécie. A seguir se encontram as características avaliadas:
1) Fenologia: a partir de observações diárias foram determinados os estádios
de desenvolvimento das plantas de amaranto, sendo que só foi considerado
que as espécies de amaranto estavam nas fases correspondentes ao
descrito por Mujica-Sanchez e Quillahuamám,1989; Henderson,1993,
quando pelo menos 50% das plantas das parcelas tivessem na mesma
condição que a descrita por esses autores.
i. Emergência (VE): as plântulas foram classificadas nessa fase
quando pelo menos 50% destas haviam emergido, exibindo duas
folhas cotiledonares;
ii. Fase vegetativa (Vn): foram classificados de acordo com a
ocorrência de nós no caule, sendo o primeiro nó classificado como
V1 e assim sucessivamente até o aparecimento da inflorescência;

27
iii. Fase reprodutiva: quando a inflorescência tornou visível no ápice as
plantas passaram a ser classificadas nessa fase, sendo que até R3
a inflorescência apresentou 5 cm de comprimento. A antese ocorreu
quando as flores estavam abertas;
iv. Fase de maturidade fisiológica: a panícula apresentava-se com
coloração característica, na ‘BRS Alegria’ a panícula passou a exibir
uma coloração vermelho intenso, e na ‘Inca’ passou de verde para
amarelada;
v. Ponto de colheita: o ponto de colheita ocorreu quando as folhas
entraram em senescência e tinham aspecto seco e os grãos se
desprendiam da panícula com facilidade;
2) Diâmetro do caule: medido a 5 cm do solo com paquímetro digital;
3) Altura de plantas: efetuada com trena;
4) Comprimento e largura da inflorescência: realizada com trena;
5) Massa de matéria seca: foram colhidas plantas inteiras, separando-se
parte aérea e raiz e acondicionados e sacos de papel levados à estufa a
65ºC, por 72 horas, até atingir massa constante. No início da fase
reprodutiva foi incluído as medidas referentes à largura, comprimento e
peso da panícula;
3.4.5 Medidas de intercâmbio gasoso
O intercâmbio gasoso (taxa fotossintética líquida, A; taxa transpiratória, E e
condutância estomática, gs) foi determinado por meio do sistema portátil de medição
de fotossíntese, modelo LI-6400XT (LI-COR Bioscience Inc., Lincoln, NE, USA)
ajustado para uma temperatura do bloco de 28,0°C, umidade relativa do ar de análise
de 60 %, densidade de fluxo fotossinteticamente ativo de 1.000 µmol m-2 s-1 e
concentração de CO2 do ar de referência de 400 µmol mol-1. A partir dos valores de A
e de E, a razão A/E foi calculada para expressar o valor da eficiência instantânea no
uso de água (EUA).
As medições foram sempre realizadas na primeira ou segunda folha totalmente
expandida, a partir do ápice caulinar (geralmente a quarta ou quinta folha), exposta à
radiação solar e sem comprometimento fitossanitário. Todas as medições foram
realizadas no período diurno, entre 07:30 e 09:30 h (horário local).
28
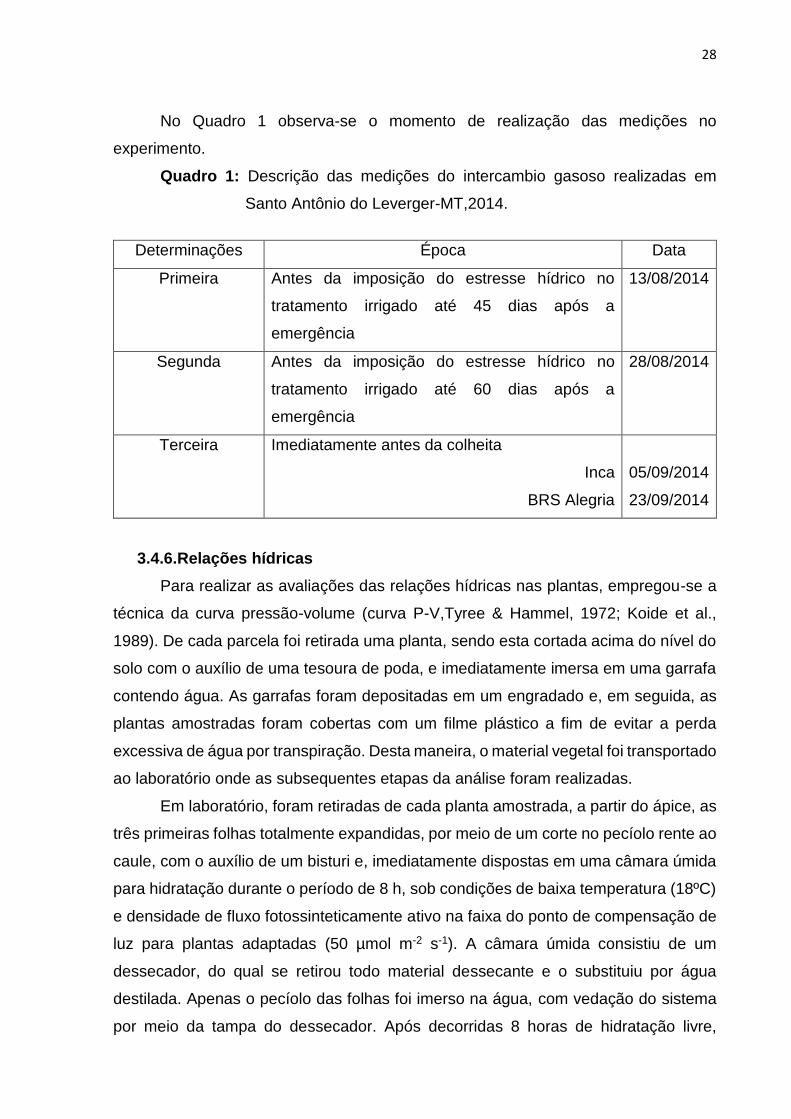
No Quadro 1 observa-se o momento de realização das medições no
experimento.
Quadro 1: Descrição das medições do intercambio gasoso realizadas em
Santo Antônio do Leverger-MT,2014.
Determinações Época Data
Primeira Antes da imposição do estresse hídrico no
tratamento irrigado até 45 dias após a
emergência
13/08/2014
Segunda Antes da imposição do estresse hídrico no
tratamento irrigado até 60 dias após a
emergência
28/08/2014
Terceira Imediatamente antes da colheita
Inca
BRS Alegria
05/09/2014
23/09/2014
3.4.6.Relações hídricas
Para realizar as avaliações das relações hídricas nas plantas, empregou-se a
técnica da curva pressão-volume (curva P-V,Tyree & Hammel, 1972; Koide et al.,
1989). De cada parcela foi retirada uma planta, sendo esta cortada acima do nível do
solo com o auxílio de uma tesoura de poda, e imediatamente imersa em uma garrafa
contendo água. As garrafas foram depositadas em um engradado e, em seguida, as
plantas amostradas foram cobertas com um filme plástico a fim de evitar a perda
excessiva de água por transpiração. Desta maneira, o material vegetal foi transportado
ao laboratório onde as subsequentes etapas da análise foram realizadas.
Em laboratório, foram retiradas de cada planta amostrada, a partir do ápice, as
três primeiras folhas totalmente expandidas, por meio de um corte no pecíolo rente ao
caule, com o auxílio de um bisturi e, imediatamente dispostas em uma câmara úmida
para hidratação durante o período de 8 h, sob condições de baixa temperatura (18ºC)
e densidade de fluxo fotossinteticamente ativo na faixa do ponto de compensação de
luz para plantas adaptadas (50 µmol m-2 s-1). A câmara úmida consistiu de um
dessecador, do qual se retirou todo material dessecante e o substituiu por água
destilada. Apenas o pecíolo das folhas foi imerso na água, com vedação do sistema
por meio da tampa do dessecador. Após decorridas 8 horas de hidratação livre,

29
procedeu-se a hidratação forçada por meio da imposição de vácuo na câmara úmida,
empregando-se uma bomba de vácuo, por um período de 15 min. Somente após a
hidratação forçada determinou-se as curvas P-V.
As curvas foram iniciadas com a pesagem inicial das folhas túrgidas e imediata
medição do potencial hídrico por meio de uma bomba de Scholander (Scholander et
al., 1965) de fabricação própria. Após a medição do potencial hídrico, as folhas foram
novamente pesadas e novamente submetidas à medição do novo potencial hídrico,
numa sequência contínua até que se verificasse que existiam pelo menos cinco
pontos dentro da fase linear (fase osmótica) da curva P-V, ou até que fosse possível,
porque algumas folhas se danificavam durante o processo e a partir desse momento
já não prestaram para novas medições.
Ao final das medições da massa da matéria fresca das folhas e respectivos
potenciais hídricos, estas foram submetidas à secagem em estufa de circulação
forçada à temperatura de 65ºC durante o período de 72 horas, para posterior
determinação da massa da matéria seca das mesmas.
As massas da matéria fresca e seca das folhas foram empregadas para a
determinação do conteúdo relativo de água e os pares de dados de potencial hídrico
e conteúdo relativo de água foram utilizados na construção das curvas P-V. Das
curvas, foram obtidos o conteúdo relativo de água no momento da perda de turgência
(CRAo), o potencial osmótico à plena turgência (100) a turgência incipiente (0) e o
módulo volumétrico de elasticidade da parede celular ().
O conteúdo relativo de água foi calculado de acordo com a Equação 2.
100
sfsat
sf
WW
WWCRA (Equação 2)
Em que: CRA = conteúdo relativo de água (%),
Wf = massa da matéria fresca atual da folha (g),
Ws = massa da matéria seca da folha (g),
Wfsat = massa da matéria fresca da folha saturada (g).

30
Ao se verificar que a massa da matéria fresca inicial obtida na primeira
pesagem logo após a hidratação não fosse correspondente à massa de matéria fresca
da folha saturada, essa última variável era estimada por meio da relação linear entre
a massa da matéria fresca e o potencial hídrico (w) verificada na primeira fase da
curva P-V, incluindo-se e o ponto de perda de turgor (0) e todos aqueles superiores
a este, obtendo-se o seu valor para um w igual a zero, de acordo com a aproximação
empregada por Kubiske & Abrams (1991).
O módulo volumétrico de elasticidade da parede celular foi obtido de acordo
com a Equação 2, para a primeira fase da curva P-V, incluindo-se o ponto de perda
de turgor (o) e todos aqueles superiores a este.
100
CRA
ψε
p
[Equação2]
Em que: = módulo volumétrico de elasticidade da parede celular (MPa),
p = variação do potencial de pressão (MPa),
CRA = variação do conteúdo relativo de água (%).
3.4.7 Aspectos produtivos
Na colheita foram avaliados nas duas linhas centrais excluindo a bordadura as
seguintes características.
a) Produtividade, obtida em gramas por parcela e, transformada para kg ha-1, com a
correção da umidade para 13%.
b) Ciclo da cultura, dimensionado por meio de contagem dos dias que decorreram,
desde a emergência até a data da colheita.
c) Produtividade por unidade de água disponibilizada (PUAD), é um valor tomado
a partir da Equação 3:
𝑃𝑈𝐴𝐷 =𝑃𝑅𝑂𝐷
𝐿𝑇𝐷.............................................................................................(Equação 3)
Em que: PUAD: produtividade por unidade de água disponibilizada (kg ha mm-1),
PROD: rendimento de grãos, em kg ha-1,

31
LTD: lâmina total disponibilizada referente à quantidade de água
disponibilizada na forma de irrigação e chuva durante o ciclo da cultura,
em mm.
d) Teor de água: Volumes iguais de sementes foram pesados e dispostos em
recipientes de metal. O material pesado foi submetido a secagem em estufa durante
24 horas á 105ºC. Depois foram transferidas para dessecador, até o resfriamento para
posterior pesagem em balança de precisão (BRASIL,2009).
e) Índice de Colheita: Foi calculado levando em conta a produção de grãos em
relação a matéria seca total da planta.
3. 5 Análises estatísticas
As curvas PV foram ajustadas ao modelo de Schulte & Hickley (1985) através
de uma rotina desenvolvida em planilha do Microsoft Excel.
Os parâmetros derivados das curvas PV foram analisados por meio de análise
de variância (ANOVA) e teste de F (α= 0,05%).
Os parâmetros derivados da análise de crescimento foram submetidos a
análise de regressão. Além disso, foi feita a correlação de Pearson, utilizando-se os
dados individuais das variáveis dos componentes de produção em relação as
variáveis de crescimento.
Os aspectos produtivos foram submetidos à análise individual e conjunta de
variância, com aplicação do teste de F. Para as comparações de médias de
tratamentos, foi utilizado o teste de médias de Tukey (P<0,05).
Os valores originais de A, E, gs e EUA foram submetidos à análise de variância
para medidas repetidas no tempo, com caracterização das probabilidades críticas (p)
para o teste de F e prova de médias por Tukey, considerando o erro alfa de 5%.

32
4. RESULTADOS E DISCUSSÃO
A uniformidade de aplicação de água do sistema, estimada através do
Coeficiente de Uniformidade de Christiansen (CUC), obteve valores de CUC de 87%,
sendo que o valor mínimo do CUC adotado como referência para sistemas de irrigação
por aspersão é de 80% (Merriam et al., 1973).
Na Tabela 2 encontra-se a precipitação e água fornecida na irrigação durante
o período experimental, no tratamento com irrigação plena (T3) forneceu-se em média
28,72 % de água a mais que no tratamento com irrigado até 45 dias (T1). A
precipitação foi maior no período de 6 de junho à 28 de julho de 2014,tendo valores
de 50 mm, sendo que para o mês de agosto não foi registrado precipitação na área
experimental e para o mês de setembro a precipitação foi de 37,7 mm.
Tabela 2. Precipitação e água fornecida na irrigação (em mm) durante o período
experimental, que foi de 26/06/2014 a 23/09/2014 em Santo Antônio de
Leverger.
T1 T2 T3
Irrigação
Precipitação
171,53
87,7
200,58
87,7
275,98
87,7
Total 259,23 288,28 363,68

33
4.1 FENOLOGIA
A semeadura foi realizada dia 26 de junho de 2014, sob dias curtos. A variedade
Inca apresentou ciclo de 63 dias e a BRS Alegria de 85 dias.
Na Tabela 3 observa-se a duração das fases fenológicas das variedades
cultivadas. O período vegetativo teve duração de 28 dias na Inca e 39 dias na ‘BRS
Alegria’. E o período reprodutivo durou 35 dias na variedade Inca e 45 dias BRS
Alegria. A colheita foi realizada no estádio R7, quando os grãos apresentavam-se
duros e se desprendiam com facilidade da panícula, o que ocorreu no dia 05 de
setembro de 2014 na Inca e 23 de setembro de 2014 na BRS Alegria
Spehar (2007) relata que entre a emergência e a maturidade fisiológica o ciclo
é de 90 dias. No presente experimento o ciclo foi menor que o citado na literatura. Tais
variações, provavelmente, ocorreram por causa do efeito do fotoperíodo ou
temperatura (Williams & Brenner, 1995). Teixeira et al. (2003) também observaram
efeito do fotoperíodo em acessos de amaranto (Amaranthus caudatus, A. cruentus e
A. hypochondriacus) e Mujica et al. (2001) e Duncan (2000) relatam que o amaranto
é uma planta de dias curtos.
Tabela 3. Duração dos estádios fenológicos do amaranto ‘BRS Alegria’ e ‘Inca’
cultivado em Santo Antônio do Leverger-MT, no período de Julho a
Setembro, 2014.
INCA BRS Alegria
Fases Inicio Término Inicio Término
Emergência 01/07/2014 01/07/2014
Vegetativo 06/07/2014 27/07/2014 06/07/2014 08/08/2014
Reprodutivo 28/07/2014 04/09/2014 09/08/2014 23/09/2014
Colheita 04/09/2014 23/09/2014

34
As plântulas emergiram cinco dias após a semeadura (Figura 2A), sendo este estádio
caracterizado pelo aparecimento de duas folhas cotiledonares e pelo menos 50% da
densidade de plantas esperada no sulco de semeadura. Na figura 2B, observa-se o
aparecimento da primeira folha verdadeira, que ocorreu no dia 6 de julho de 2014.
Figura 2 : Plântulas de Amaranthus cruentus com um par de folhas cotiledonares aos
dois dias após a semeadura (A); Inicio do aparecimento da primeira folha
verdadeira aos cinco dias após a emergência(B). Santo Antônio de
Leverger-MT,2014

35
Na Figura 3A observa-se início da formação da panícula em A. caudatus cv
Inca, que ocorreu aos 35 DAE, na “BRS Alegria’ o ápice da inflorescência se tornou
visível aos 42 DAE (Figura 3B).
Figura 3: Primórdios da inflorescência de A. caudatus cv Inca aos 35 dias após a
emergência(A) e primórdios da inflorescência na “BRS Alegria” aos 42 dias
após a emergência (B), Santo Antônio de Leverger-MT,2014

36
O início da antese é caracterizado pela abertura das flores na panícula, em ‘Inca’
ocorreu aos 39 DAE como pode ser observado na figura 4A e em ‘BRS Alegria’ aos
54 DAE (Figura 4B). A antese teve início pela parte intermediaria da panícula.
Figura 4: Plantas de A. caudatus cv ‘Inca’ em plena antese aos 39 dias após a (A) e
A. cruentus cv. ‘BRS Alegria’ aos 54 dias após a emergência em BRS
Alegria (B), Santo Antônio do Leverger-MT,2014.

37
Na Figura 5 observa se o ponto de colheita nas espécies estudadas em A.
cruentus a panícula apresentou coloração vermelho intenso e em A. caudatus a
inflorescência passou da coloração verde para um amarelado, com aspecto seco.
Figura 5: Ponto de colheita de A. caudatus ‘Inca’ (A) e A. cruentus ‘BRS Alegria’ (B),
Santo Antônio do Leverger-MT,2014.
4.2 ANALISE QUANTITATIVA DO CRESCIMENTO
Para avaliação do crescimento considerou T1,T2,T3, com sendo os regimes
hídrico com irrigação até 45, 60 dias após a emergência e irrigação o ciclo todo,
respectivamente. A variedade Inca foi classificada como V1 e a BRS Alegria como V2.
Os resultados permitem afirmar que as variações nos caracteres agronômicos
da planta, associados aos diferentes níveis de estresse hídrico, definiram um padrão
de efeito comportamental, o que auxilia na previsão e no adequado manejo da planta
de amaranto.
As variações no diâmetro do caule da variedade INCA seguiram uma função
potência e mostram que até os 28 dias após a emergência houve pequena variação
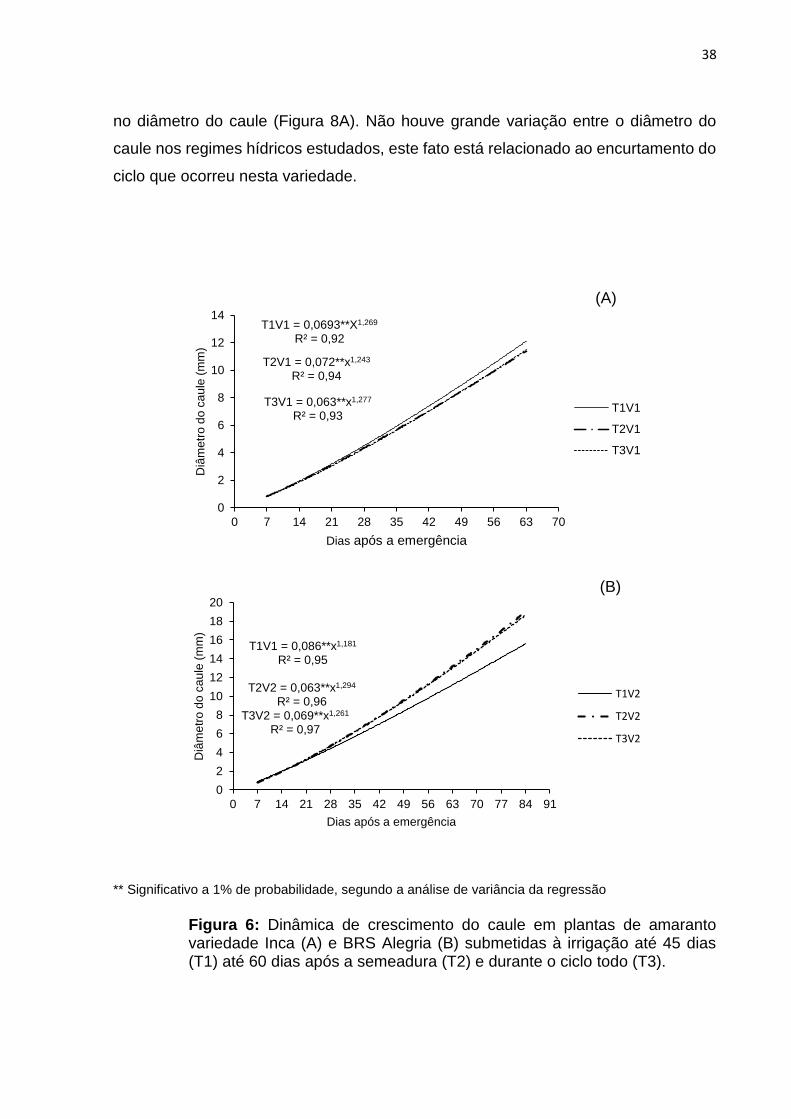
38
no diâmetro do caule (Figura 8A). Não houve grande variação entre o diâmetro do
caule nos regimes hídricos estudados, este fato está relacionado ao encurtamento do
ciclo que ocorreu nesta variedade.
** Significativo a 1% de probabilidade, segundo a análise de variância da regressão
Figura 6: Dinâmica de crescimento do caule em plantas de amaranto variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60 dias após a semeadura (T2) e durante o ciclo todo (T3).
T1V1 = 0,0693**X1,269
R² = 0,92
T2V1 = 0,072**x1,243
R² = 0,94
T3V1 = 0,063**x1,277
R² = 0,93
0
2
4
6
8
10
12
14
0 7 14 21 28 35 42 49 56 63 70
Diâ
me
tro
do
ca
ule
(m
m)
Dias após a emergência
(A)
T1V1
T2V1
T3V1
T1V1 = 0,086**x1,181
R² = 0,95
T2V2 = 0,063**x1,294
R² = 0,96
T3V2 = 0,069**x1,261
R² = 0,97
0
2
4
6
8
10
12
14
16
18
20
0 7 14 21 28 35 42 49 56 63 70 77 84 91
Diâ
me
tro
do
ca
ule
(m
m)
Dias após a emergência
(B)
T1V2
T2V2
T3V2

39
Na variedade BRS Alegria o diâmetro máximo médio do caule do tratamento
com regime hídrico pleno apresentou comportamento semelhante ao tratamento onde
houve suspensão aos 60 dias. No tratamento onde houve suspensão da irrigação aos
45 DAE, obteve o menor R2 e foi observada uma acentuada redução do diâmetro do
caule nas plantas sob o déficit hídrico (Figura 6B). Essa diminuição é relacionada,
possivelmente, à maior competição entre plantas por água e nutrientes. (Ferreira et
al., 2014)
O diâmetro do caule é importante por relacionar-se com o tombamento das
plantas, isso porque a panícula é dominante no ápice da planta. Essa panícula é
relativamente grande para o tamanho da planta tornando-a susceptível ao
tombamento. No presente experimento, ocorreu o tombamento de algumas plantas
nas parcelas experimentais na variedade INCA, sem haver prejuízos a coleta de
dados.
Uma explicação simplificada dessa contenção de crescimento pode ser feita a
partir da relação caule/parte aérea. Como o estresse hídrico causa diminuição da área
foliar e da massa da copa, o diâmetro caulinar da planta é contido ou diminuído, pois
a massa da parte aérea está sendo reduzido, ou seja, a planta está se adaptando às
novas condições. Além do mais, nesse momento a outra função básica do caule o
transporte de nutrientes, está em menor atividade, não havendo necessidade de
expansão, também devido ao menor fluxo de assimilados e de íons absorvidos
(Kerbauy, 2004).
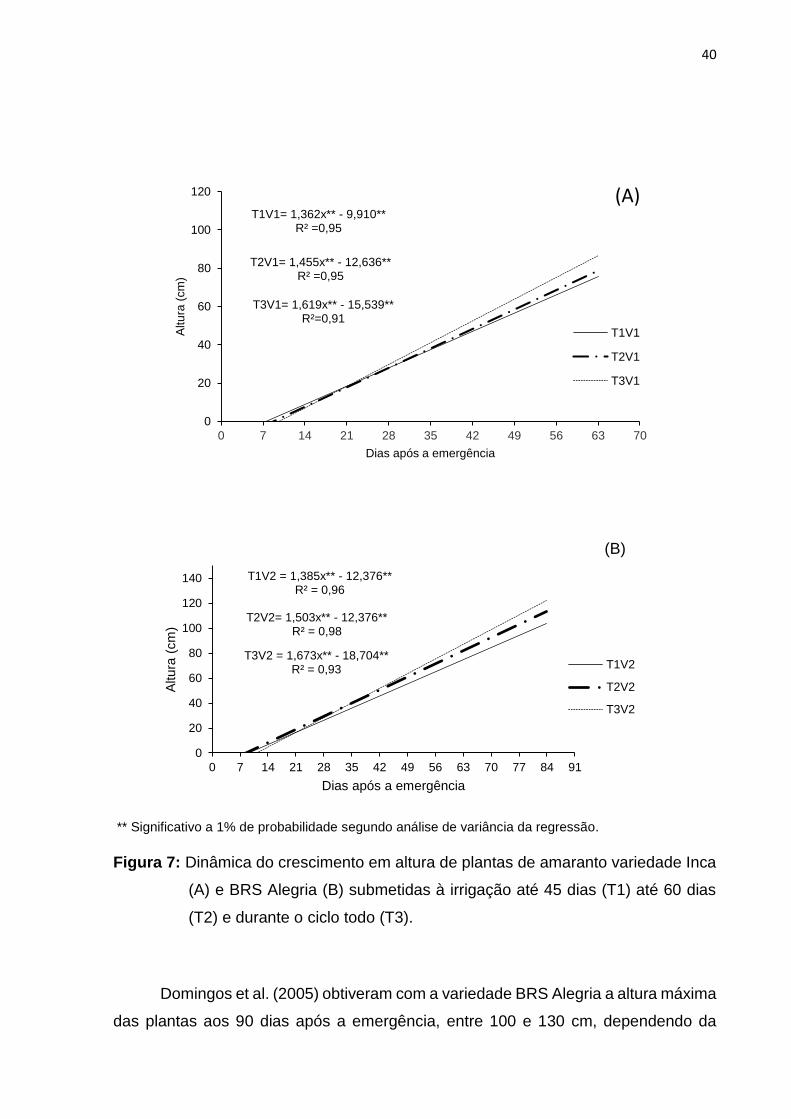
A altura de plantas apresentou resposta linear (Figura 7). A imposição do
estresse hídrico aos 45 dias após a emergência (período de florescimento) reduziu a
altura de plantas mostrando o efeito do déficit hídrico sobre a produção do amaranto,
confirmando a maior necessidade de água nas fases de floração e frutificação.
Queiroz et al. (2002) em experimento com a cultura da berinjela também obtiveram
menores valores de altura da planta quando submetida ao estresse hídrico no início
da frutifi cação. A primeira resposta fisiológica ao déficit hídrico é a diminuição da
turgescência e, associada a esse evento, a diminuição do crescimento da planta
porque as fases de floração e frutificação, para a maioria das culturas, são as que
apresentam maiores consumos de água, sendo, portanto, consideradas como fases
críticas (Taiz & Zeiger, 2004; Larcher, 2006).
40
** Significativo a 1% de probabilidade segundo análise de variância da regressão.
Figura 7: Dinâmica do crescimento em altura de plantas de amaranto variedade Inca
(A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60 dias
(T2) e durante o ciclo todo (T3).
Domingos et al. (2005) obtiveram com a variedade BRS Alegria a altura máxima
das plantas aos 90 dias após a emergência, entre 100 e 130 cm, dependendo da
T1V1= 1,362x** - 9,910**R² =0,95
T2V1= 1,455x** - 12,636**R² =0,95
T3V1= 1,619x** - 15,539**R²=0,91
0
20
40
60
80
100
120
0 7 14 21 28 35 42 49 56 63 70
Altu
ra (
cm
)
Dias após a emergência
T1V1
T2V1
T3V1
(A)
T1V2 = 1,385x** - 12,376**R² = 0,96
T2V2= 1,503x** - 12,376**R² = 0,98
T3V2 = 1,673x** - 18,704**R² = 0,93
0
20
40
60
80
100
120
140
0 7 14 21 28 35 42 49 56 63 70 77 84 91
Altura
(cm
)
Dias após a emergência
(B)
T1V2
T2V2
T3V2

41
quantidade de nutrientes aplicado. No presente trabalho a altura máxima encontrada
foi de 120 cm no tratamento sem déficit hídrico. A altura de plantas é um importante
parâmetro para a colheita mecanizada desta cultura com o uso da plataforma para
colheita de milho (Erasmo et al., 2004).
O A. cruentus cv ‘BRS Alegria’ possui maior altura de planta e diâmetro do caule
em relação ao A caudatus cv ‘Inca’. Esse fato leva a inferir que e A. cruentus é grande
produtora de matéria seca (Carmo & Aguiar, 2000), conforme pode ser observado na
Figura 8.
A cultura do amaranto, pela biomassa produzida, poderia ser utilizada em
sistema de plantio direto na região dos cerrados, por sua estabilidade na produção de
grãos e biomassa, principalmente na safrinha (Erasmo et al., 2004).
O estresse hídrico aplicado provocou reduções no acumulo de matéria seca
nas variedades estudadas sendo que o tratamento irrigado até 45 dias após a
emergência proporcionou menor acúmulo de matéria seca (Figura 8) o que se deve
ao fato de que em plantas estressadas ocorre a diminuição de folhas no caule (Souza
& Amorim, 2009), ou ainda o déficit hídrico pode reduzir a emissão de novas folhas. A
área celular reduzida se explica por uma menor taxa de divisão celular, reduzindo
assim o aparecimento de novas folhas, proporcionando um menor acúmulo de matéria
seca ao final do ciclo.
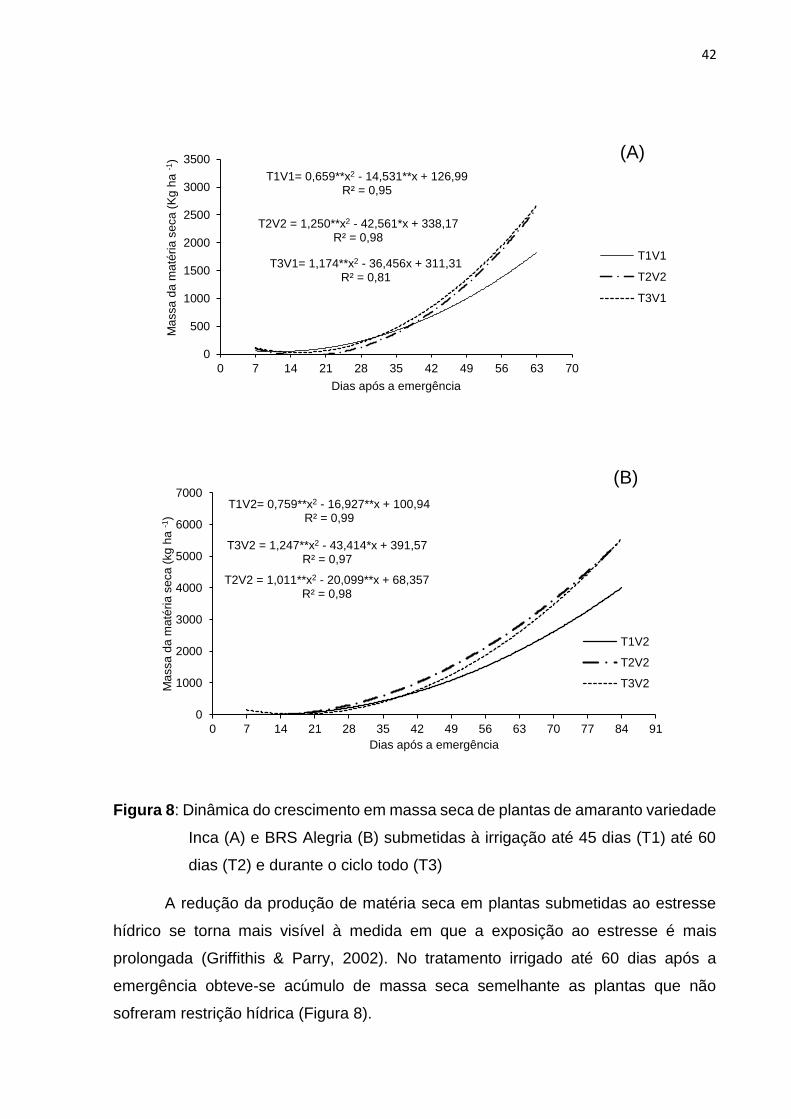
42
Figura 8: Dinâmica do crescimento em massa seca de plantas de amaranto variedade
Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1) até 60
dias (T2) e durante o ciclo todo (T3)
A redução da produção de matéria seca em plantas submetidas ao estresse
hídrico se torna mais visível à medida em que a exposição ao estresse é mais
prolongada (Griffithis & Parry, 2002). No tratamento irrigado até 60 dias após a
emergência obteve-se acúmulo de massa seca semelhante as plantas que não
sofreram restrição hídrica (Figura 8).
T1V1= 0,659**x2 - 14,531**x + 126,99R² = 0,95
T2V2 = 1,250**x2 - 42,561*x + 338,17R² = 0,98
T3V1= 1,174**x2 - 36,456x + 311,31R² = 0,81
0
500
1000
1500
2000
2500
3000
3500
0 7 14 21 28 35 42 49 56 63 70
Ma
ssa
da
ma
téri
a s
eca
(K
g h
a -1
)
Dias após a emergência
(A)
T1V1
T2V2
T3V1
T1V2= 0,759**x2 - 16,927**x + 100,94R² = 0,99
T2V2 = 1,011**x2 - 20,099**x + 68,357R² = 0,98
T3V2 = 1,247**x2 - 43,414*x + 391,57R² = 0,97
0
1000
2000
3000
4000
5000
6000
7000
0 7 14 21 28 35 42 49 56 63 70 77 84 91
Ma
ssa
da
ma
téri
a s
eca
(kg
ha
-1)
Dias após a emergência
(B)
T1V2
T2V2
T3V2

43
A primeira consequência fisiológica para plantas submetidas ao estresse
hídrico é a redução ou interrupção dos ritmos de crescimento, devido à redução na
diferenciação foliar influenciado pela restrição hídrica do tratamento adotado (Silva &
Nogueira, 2003).
O comprimento da panícula, em geral na variedade INCA não mostrou grande
variação (Figura 9A), uma vez que o estresse hídrico teve início aos 45 dias após a
emergência quando as plantas já se encontravam em plena antese. E aos 60 dias as
mesmas já se encontravam na maturidade fisiológica. Na variedade BRS Alegria
(Figura 9B) a restrição hídrica aos 45 dias após a emergência afetou o crescimento,
sendo que a curva de crescimento deste tratamento obteve os menores valores.
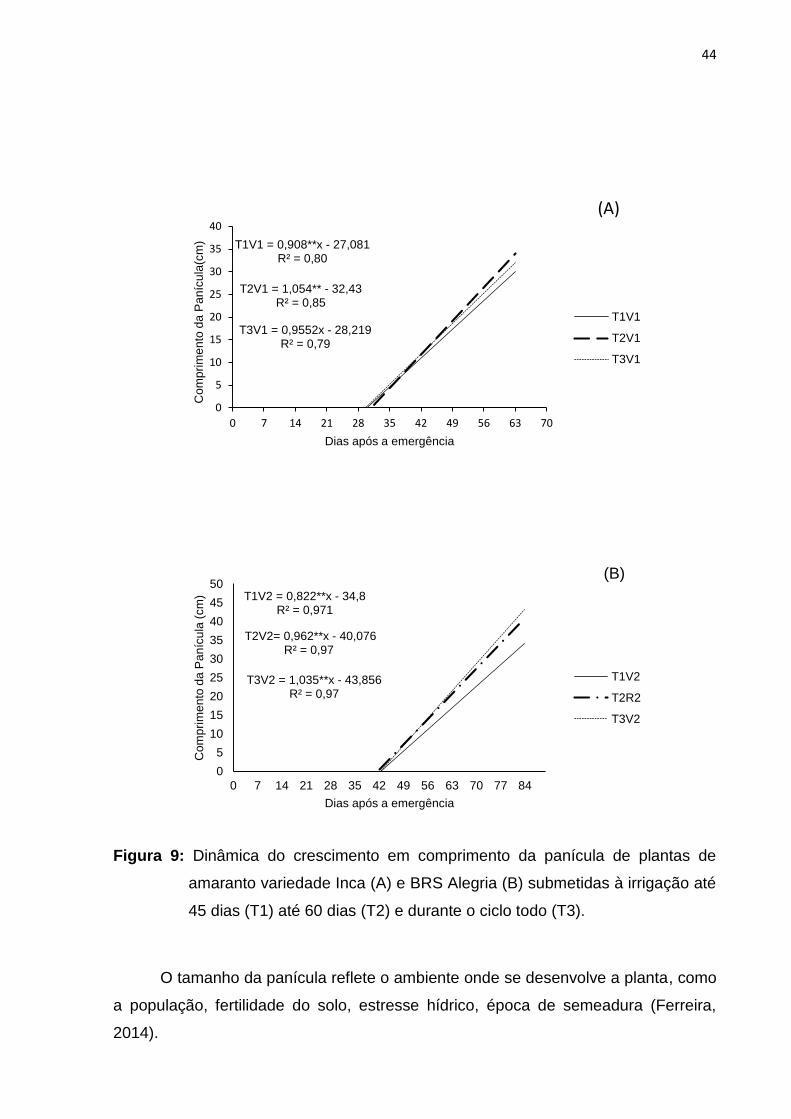
44
Figura 9: Dinâmica do crescimento em comprimento da panícula de plantas de
amaranto variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até
45 dias (T1) até 60 dias (T2) e durante o ciclo todo (T3).
O tamanho da panícula reflete o ambiente onde se desenvolve a planta, como
a população, fertilidade do solo, estresse hídrico, época de semeadura (Ferreira,
2014).
T1V1 = 0,908**x - 27,081R² = 0,80
T2V1 = 1,054** - 32,43R² = 0,85
T3V1 = 0,9552x - 28,219R² = 0,79
0
5
10
15
20
25
30
35
40
0 7 14 21 28 35 42 49 56 63 70
Co
mp
rim
en
to d
a P
an
ícu
la(c
m)
Dias após a emergência
(A)
T1V1
T2V1
T3V1
T1V2 = 0,822**x - 34,8R² = 0,971
T2V2= 0,962**x - 40,076R² = 0,97
T3V2 = 1,035**x - 43,856R² = 0,97
0
5
10
15
20
25
30
35
40
45
50
0 7 14 21 28 35 42 49 56 63 70 77 84
Com
pri
me
nto
da
Pa
níc
ula
(cm
)
Dias após a emergência
(B)
T1V2
T2R2
T3V2
45
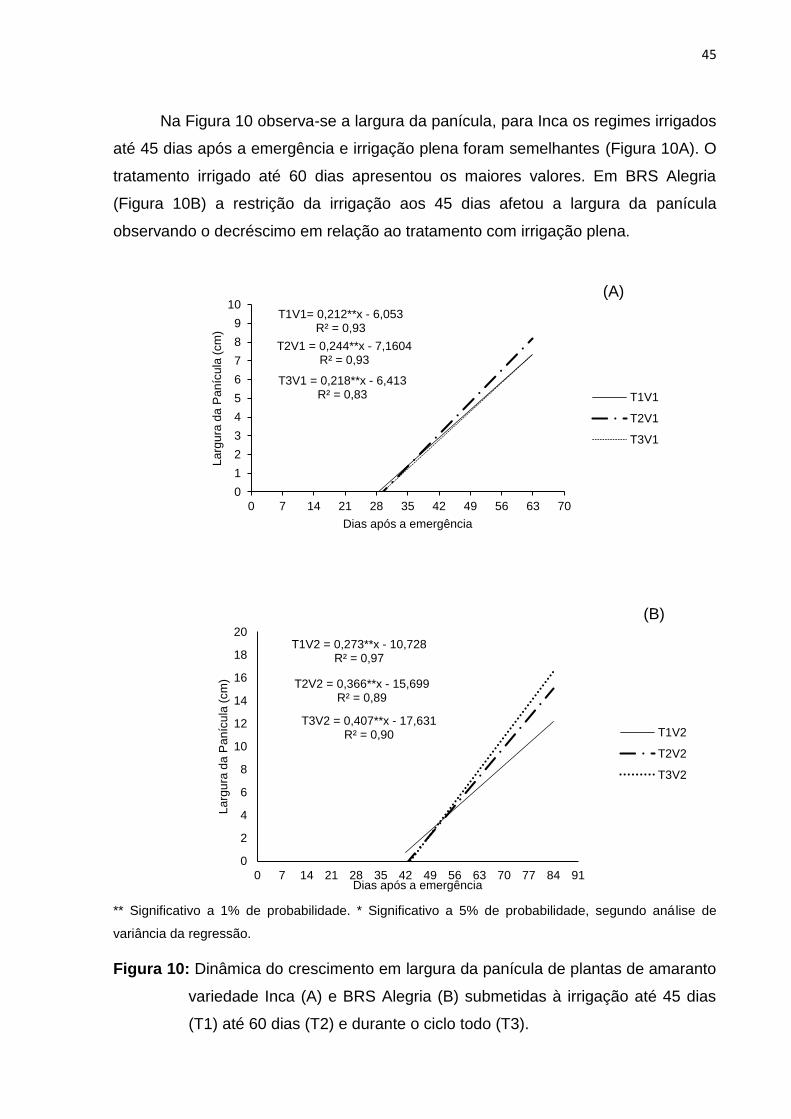
Na Figura 10 observa-se a largura da panícula, para Inca os regimes irrigados
até 45 dias após a emergência e irrigação plena foram semelhantes (Figura 10A). O
tratamento irrigado até 60 dias apresentou os maiores valores. Em BRS Alegria
(Figura 10B) a restrição da irrigação aos 45 dias afetou a largura da panícula
observando o decréscimo em relação ao tratamento com irrigação plena.
** Significativo a 1% de probabilidade. * Significativo a 5% de probabilidade, segundo análise de
variância da regressão.
Figura 10: Dinâmica do crescimento em largura da panícula de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias
(T1) até 60 dias (T2) e durante o ciclo todo (T3).
T1V1= 0,212**x - 6,053R² = 0,93
T2V1 = 0,244**x - 7,1604R² = 0,93
T3V1 = 0,218**x - 6,413R² = 0,83
0
1
2
3
4
5
6
7
8
9
10
0 7 14 21 28 35 42 49 56 63 70
La
rgu
ra d
a P
an
ícu
la (
cm
)
Dias após a emergência
(A)
T1V1
T2V1
T3V1
T1V2 = 0,273**x - 10,728R² = 0,97
T2V2 = 0,366**x - 15,699R² = 0,89
T3V2 = 0,407**x - 17,631R² = 0,90
0
2
4
6
8
10
12
14
16
18
20
0 7 14 21 28 35 42 49 56 63 70 77 84 91
La
rgu
ra d
a P
an
ícu
la (
cm
)
Dias após a emergência
(B)
T1V2
T2V2
T3V2
46
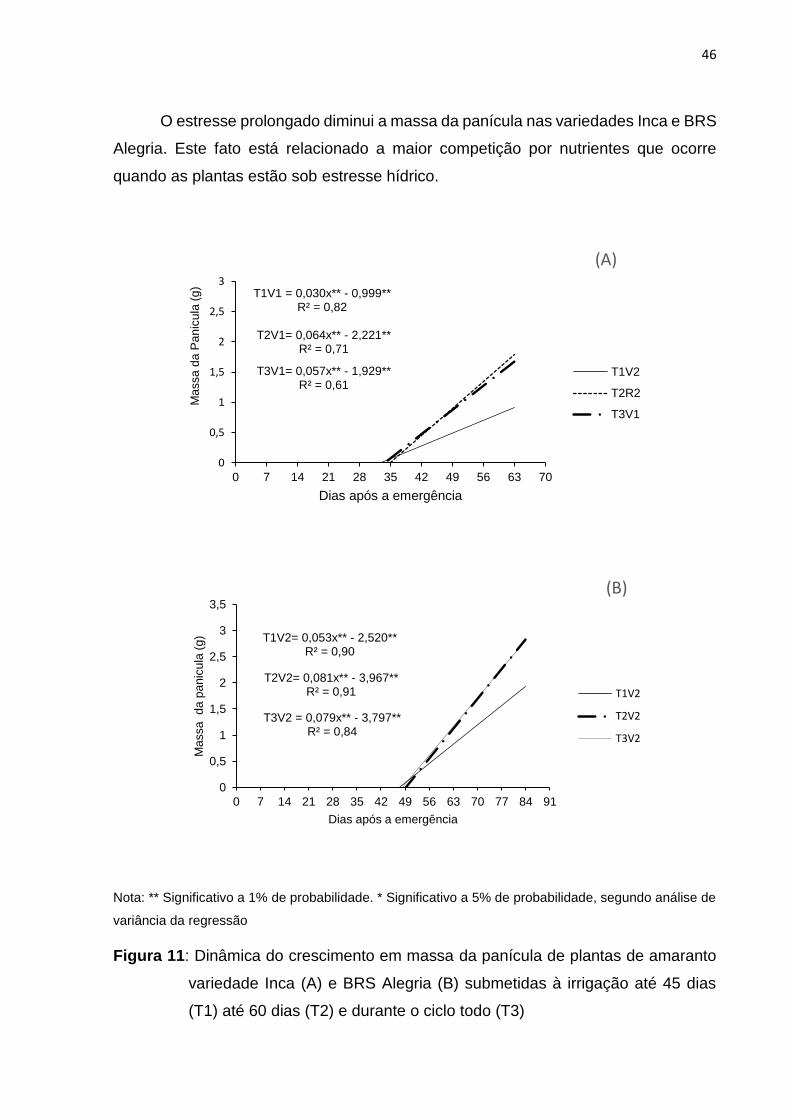
O estresse prolongado diminui a massa da panícula nas variedades Inca e BRS
Alegria. Este fato está relacionado a maior competição por nutrientes que ocorre
quando as plantas estão sob estresse hídrico.
Nota: ** Significativo a 1% de probabilidade. * Significativo a 5% de probabilidade, segundo análise de
variância da regressão
Figura 11: Dinâmica do crescimento em massa da panícula de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias
(T1) até 60 dias (T2) e durante o ciclo todo (T3)
T1V1 = 0,030x** - 0,999**R² = 0,82
T2V1= 0,064x** - 2,221**R² = 0,71
T3V1= 0,057x** - 1,929**R² = 0,61
0
0,5
1
1,5
2
2,5
3
0 7 14 21 28 35 42 49 56 63 70
Ma
ssa
da
Pa
nic
ula
(g
)
Dias após a emergência
(A)
T1V2
T2R2
T3V1
T1V2= 0,053x** - 2,520**R² = 0,90
T2V2= 0,081x** - 3,967**R² = 0,91
T3V2 = 0,079x** - 3,797**R² = 0,84
0
0,5
1
1,5
2
2,5
3
3,5
0 7 14 21 28 35 42 49 56 63 70 77 84 91
Ma
ssa
d
a p
an
icu
la (
g)
Dias após a emergência
(B)
T1V2
T2V2
T3V2
47
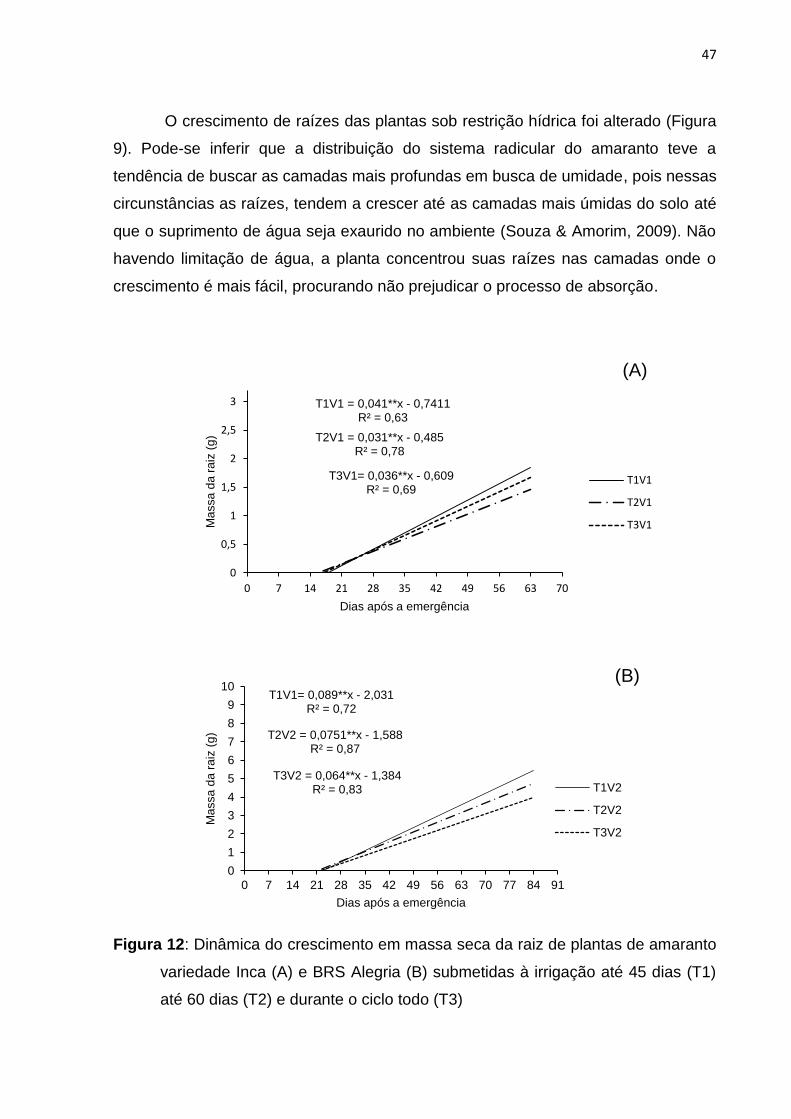
O crescimento de raízes das plantas sob restrição hídrica foi alterado (Figura
9). Pode-se inferir que a distribuição do sistema radicular do amaranto teve a
tendência de buscar as camadas mais profundas em busca de umidade, pois nessas
circunstâncias as raízes, tendem a crescer até as camadas mais úmidas do solo até
que o suprimento de água seja exaurido no ambiente (Souza & Amorim, 2009). Não
havendo limitação de água, a planta concentrou suas raízes nas camadas onde o
crescimento é mais fácil, procurando não prejudicar o processo de absorção.
Figura 12: Dinâmica do crescimento em massa seca da raiz de plantas de amaranto
variedade Inca (A) e BRS Alegria (B) submetidas à irrigação até 45 dias (T1)
até 60 dias (T2) e durante o ciclo todo (T3)
T1V1 = 0,041**x - 0,7411R² = 0,63
T2V1 = 0,031**x - 0,485R² = 0,78
T3V1= 0,036**x - 0,609R² = 0,69
0
0,5
1
1,5
2
2,5
3
0 7 14 21 28 35 42 49 56 63 70
Ma
ssa
da
ra
iz (
g)
Dias após a emergência
(A)
T1V1
T2V1
T3V1
T1V1= 0,089**x - 2,031R² = 0,72
T2V2 = 0,0751**x - 1,588R² = 0,87
T3V2 = 0,064**x - 1,384R² = 0,83
0
1
2
3
4
5
6
7
8
9
10
0 7 14 21 28 35 42 49 56 63 70 77 84 91
Ma
ssa
da
ra
iz (
g)
Dias após a emergência
(B)
T1V2
T2V2
T3V2

48
Ludlow e Muchow (1990) explicam que o aumento da quantidade de raiz
durante a seca ajuda a planta a obter água a níveis mais profundos no perfil do solo,
assim como ajuda a evitar déficits hídricos nas camadas mais superficiais do solo.
4.2.1 Correlações de Pearson entre a análise quantitativa de crescimento e os
aspectos produtivos
As variáveis produtivas (produtividade e índice de colheita) e as variáveis da
análise quantitativa de crescimento (massa da matéria seca total, massa da raiz,
largura da panícula, comprimento da panícula, massa da panícula, diâmetro do
caule e altura de plantas e relação raiz/parte aérea) foram submetidos a análise de
correlação de Pearson.
De acordo com esta análise (Tabela 4) não houve correlação entre diâmetro do
caule e as demais variáveis estudadas na variedade Inca. Na Tabela 5 observa se a
correlação entre diâmetro do caule e índice de colheita na variedade BRS Alegria.
Apaza-Gutierrez et al. (2002) consideram que o diâmetro do caule tem grande efeito
na produtividade de grãos, o que não ocorreu neste trabalho. Há poucos trabalhos
correlacionando características agronômicas em amaranto, mas Teixeira et al. (2002)
observaram correlação entre a altura de plantas e diâmetro do caule, porém não
obtiveram correlação entre os parâmetros de biometria com a produtividade da
cultura.
A variedade Inca obteve correlação positiva entre a matéria seca total e os
dados da panícula,a massa, a largura e comprimento da panícula significando que a
panícula tem grande influência no massa da matéria seca, uma vez que A. caudatus
a panícula é relativamente grande.
O massa da raiz em A. cruentus cv ‘BRS Alegria’ correlacionou se
positivamente com a altura de plantas (Tabela 5), uma vez que essa variedade
apresenta porte maior podendo chegar a 2 metros de altura (Spehar, 2009), dessa
forma é necessário um maior investimento em raiz para fixar a planta.
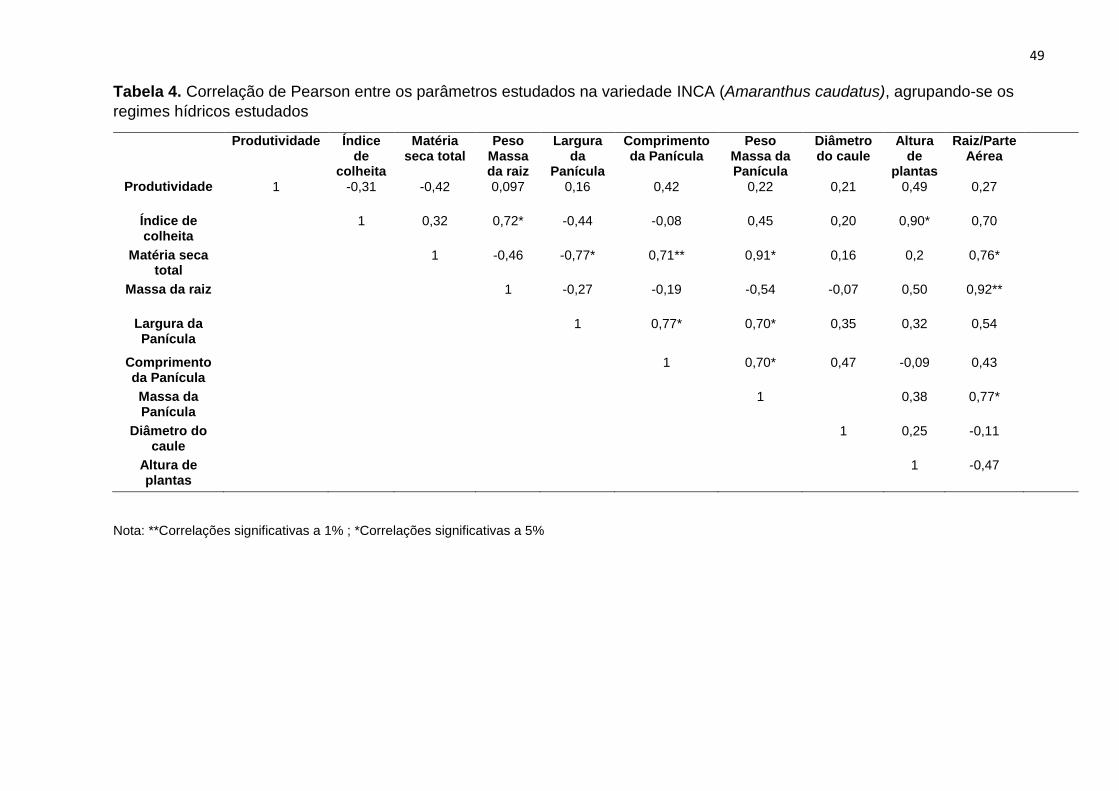
49
Tabela 4. Correlação de Pearson entre os parâmetros estudados na variedade INCA (Amaranthus caudatus), agrupando-se os
regimes hídricos estudados
Produtividade Índice de
colheita
Matéria seca total
Peso Massa da raiz
Largura da
Panícula
Comprimento da Panícula
Peso Massa da Panícula
Diâmetro do caule
Altura de
plantas
Raiz/Parte Aérea
Produtividade 1 -0,31 -0,42 0,097 0,16 0,42 0,22 0,21 0,49 0,27
Índice de colheita
1 0,32 0,72* -0,44 -0,08 0,45 0,20 0,90* 0,70
Matéria seca total
1 -0,46 -0,77* 0,71** 0,91* 0,16 0,2 0,76*
Massa da raiz 1 -0,27 -0,19 -0,54 -0,07 0,50 0,92**
Largura da Panícula
1 0,77* 0,70* 0,35 0,32 0,54
Comprimento da Panícula
1 0,70* 0,47 -0,09 0,43
Massa da Panícula
1 0,38 0,77*
Diâmetro do caule
1 0,25 -0,11
Altura de plantas
1 -0,47
Nota: **Correlações significativas a 1% ; *Correlações significativas a 5%
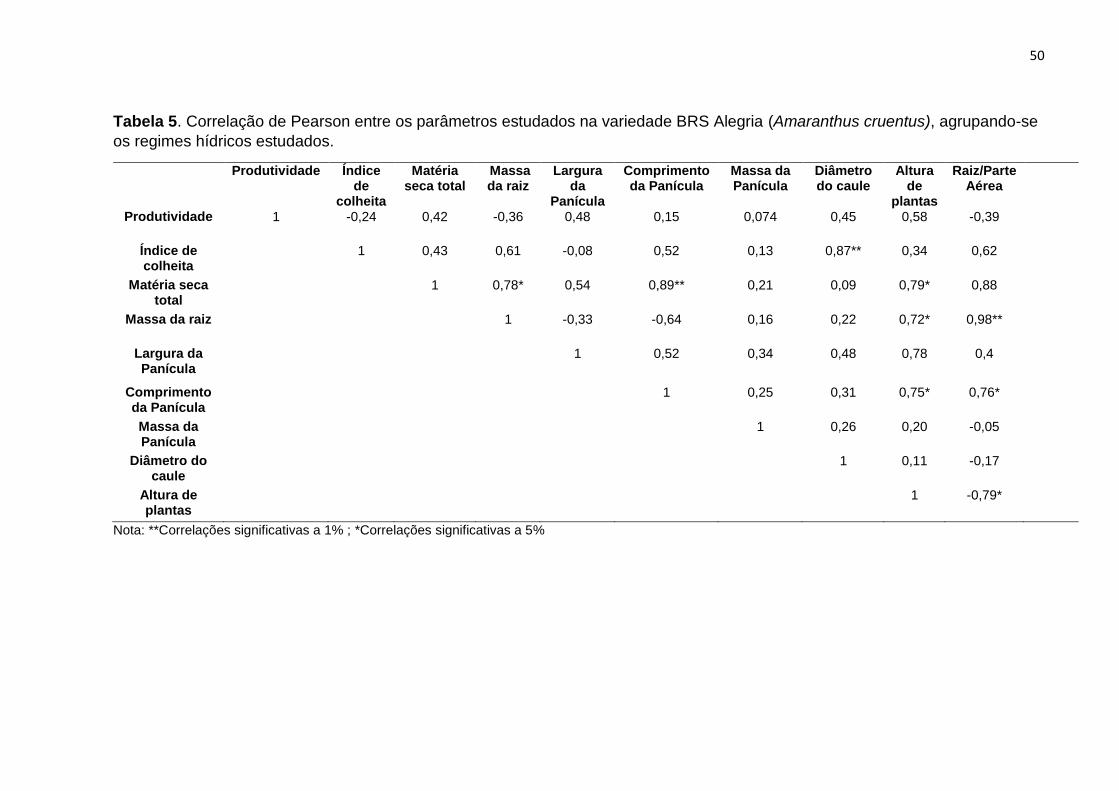
50
Tabela 5. Correlação de Pearson entre os parâmetros estudados na variedade BRS Alegria (Amaranthus cruentus), agrupando-se
os regimes hídricos estudados.
Produtividade Índice de
colheita
Matéria seca total
Massa da raiz
Largura da
Panícula
Comprimento da Panícula
Massa da Panícula
Diâmetro do caule
Altura de
plantas
Raiz/Parte Aérea
Produtividade 1 -0,24 0,42 -0,36 0,48 0,15 0,074 0,45 0,58 -0,39
Índice de colheita
1 0,43 0,61 -0,08 0,52 0,13 0,87** 0,34 0,62
Matéria seca total
1 0,78* 0,54 0,89** 0,21 0,09 0,79* 0,88
Massa da raiz 1 -0,33 -0,64 0,16 0,22 0,72* 0,98**
Largura da Panícula
1 0,52 0,34 0,48 0,78 0,4
Comprimento da Panícula
1 0,25 0,31 0,75* 0,76*
Massa da Panícula
1 0,26 0,20 -0,05
Diâmetro do caule
1 0,11 -0,17
Altura de plantas
1 -0,79*
Nota: **Correlações significativas a 1% ; *Correlações significativas a 5%

51
4.3 Intercâmbio Gasoso
Todas as variáveis do intercâmbio gasoso, assim como a eficiência do uso da
água (EUA), apresentaram pelo menos uma causa de variação significativa.
Para taxa fotossintética liquida (A), houve efeito significativo da época em que
se realizou a medição do intercâmbio gasoso (p < 0,0001), assim como do regime
hídrico (p = 0,0069) e da interação entre essas causas de variação (p = 0,0053). A
prova de médias para o desdobramento dessa interação pode ser observada na
Tabela 6.
No momento inicial não houve diferença entre os tratamentos (Tabela 6). A
maior abertura estomática encontrada na primeira avaliação, aos 45 dias após a
emergência das plantas (Quadro 2) pode ter contribuído para os maiores valores de
A encontrado nessa ocasião, pois nessa época as condições de umidade no solo eram
ideais, disponibilizando água suficiente para as plantas (Kozlowski e Pallardy,1996),
uma vez que as condições eram similares em todos os tratamentos. No momento
intermediário (aos 60 dias após a emergência das plantas) houve efeito do déficit
hídrico, sendo que tratamento irrigado até 45 dias após a emergência foi 44,2% inferior
que o tratamento com irrigação plena. Sabe-se que o estresse hídrico severo é um
dos causadores da diminuição da fotossíntese (Lauer e Boyer 1992). No momento
intermediário a atividade fotossintética foi superior nos tratamentos com suspensão
da irrigação aos 60 dias e no tratamento irrigado o ciclo todo, isso se deve ao fato de
que durante o desenvolvimento das plantas a atividade fotossintética aumenta quando
não há restrição hídrica.
Ao final do ciclo produtivo (colheita) não houve diferença significativa entre os
regimes hídricos, isso porque na colheita as espécies apresentam um decréscimo na
atividade fotossintética até sua senescência (Pimentel e Rossielo, 1995).
Comparando o momento de realização das medições dentro do regime hidrico
não se observou diferença significativa no tratamento irrigado até 45 dias após a
emergência. Para os tratamentos irrigado até 60 dias após a emergência e com
irrigação plena, a taxa fotossintética mostrou um acréscimo aos 60 dias após
emergência (momento intermediário) e posterior decréscimo na colheita (Tabela 6).
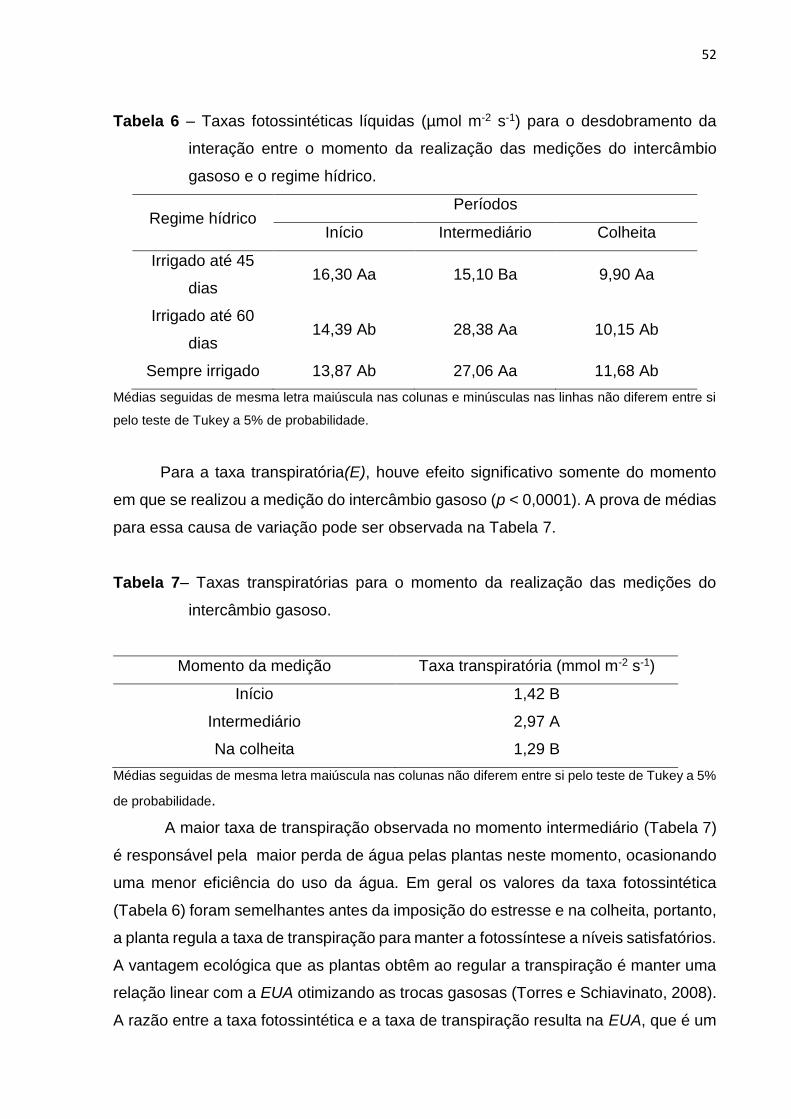
52
Tabela 6 – Taxas fotossintéticas líquidas (µmol m-2 s-1) para o desdobramento da
interação entre o momento da realização das medições do intercâmbio
gasoso e o regime hídrico.
Regime hídrico Períodos
Início Intermediário Colheita
Irrigado até 45
dias 16,30 Aa 15,10 Ba 9,90 Aa
Irrigado até 60
dias 14,39 Ab 28,38 Aa 10,15 Ab
Sempre irrigado 13,87 Ab 27,06 Aa 11,68 Ab
Médias seguidas de mesma letra maiúscula nas colunas e minúsculas nas linhas não diferem entre si
pelo teste de Tukey a 5% de probabilidade.
Para a taxa transpiratória(E), houve efeito significativo somente do momento
em que se realizou a medição do intercâmbio gasoso (p < 0,0001). A prova de médias
para essa causa de variação pode ser observada na Tabela 7.
Tabela 7– Taxas transpiratórias para o momento da realização das medições do
intercâmbio gasoso.
Momento da medição Taxa transpiratória (mmol m-2 s-1)
Início 1,42 B
Intermediário 2,97 A
Na colheita 1,29 B
Médias seguidas de mesma letra maiúscula nas colunas não diferem entre si pelo teste de Tukey a 5%
de probabilidade.
A maior taxa de transpiração observada no momento intermediário (Tabela 7)
é responsável pela maior perda de água pelas plantas neste momento, ocasionando
uma menor eficiência do uso da água. Em geral os valores da taxa fotossintética
(Tabela 6) foram semelhantes antes da imposição do estresse e na colheita, portanto,
a planta regula a taxa de transpiração para manter a fotossíntese a níveis satisfatórios.
A vantagem ecológica que as plantas obtêm ao regular a transpiração é manter uma
relação linear com a EUA otimizando as trocas gasosas (Torres e Schiavinato, 2008).
A razão entre a taxa fotossintética e a taxa de transpiração resulta na EUA, que é um
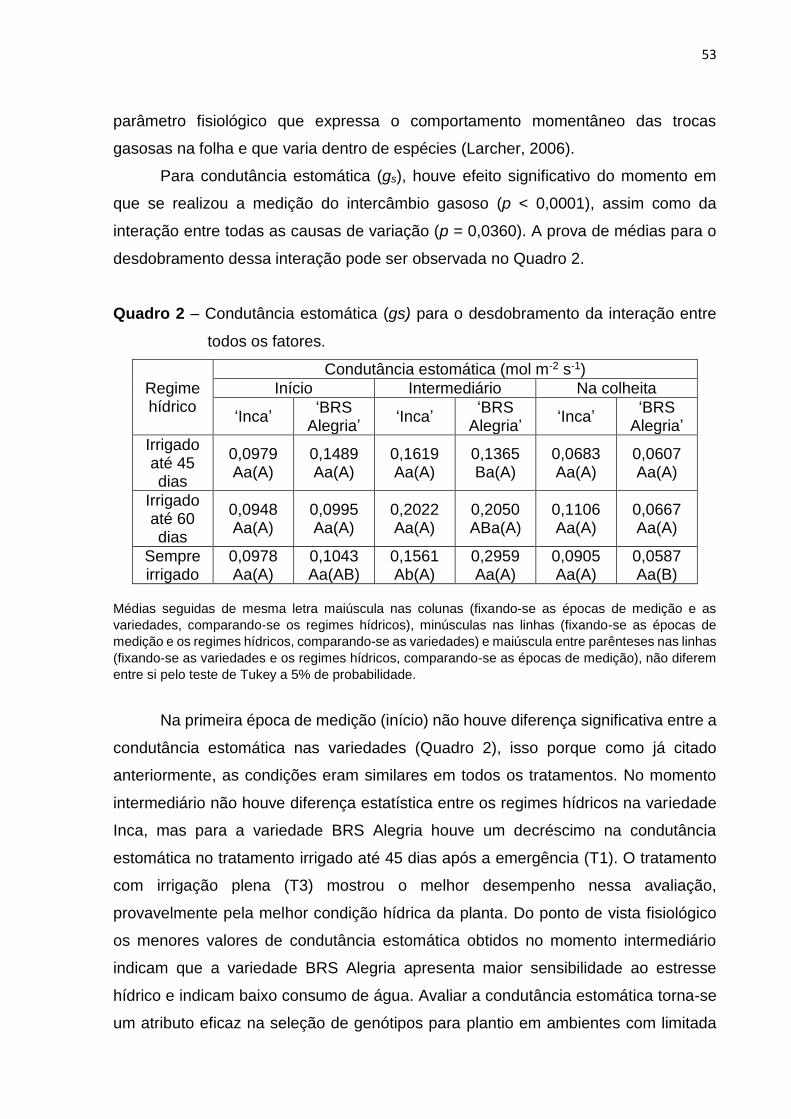
53
parâmetro fisiológico que expressa o comportamento momentâneo das trocas
gasosas na folha e que varia dentro de espécies (Larcher, 2006).
Para condutância estomática (gs), houve efeito significativo do momento em
que se realizou a medição do intercâmbio gasoso (p < 0,0001), assim como da
interação entre todas as causas de variação (p = 0,0360). A prova de médias para o
desdobramento dessa interação pode ser observada no Quadro 2.
Quadro 2 – Condutância estomática (gs) para o desdobramento da interação entre
todos os fatores.
Regime hídrico
Condutância estomática (mol m-2 s-1)
Início Intermediário Na colheita
‘Inca’ ‘BRS
Alegria’ ‘Inca’
‘BRS Alegria’
‘Inca’ ‘BRS
Alegria’
Irrigado até 45 dias
0,0979 Aa(A)
0,1489 Aa(A)
0,1619 Aa(A)
0,1365 Ba(A)
0,0683 Aa(A)
0,0607 Aa(A)
Irrigado até 60 dias
0,0948 Aa(A)
0,0995 Aa(A)
0,2022 Aa(A)
0,2050 ABa(A)
0,1106 Aa(A)
0,0667 Aa(A)
Sempre irrigado
0,0978 Aa(A)
0,1043 Aa(AB)
0,1561 Ab(A)
0,2959 Aa(A)
0,0905 Aa(A)
0,0587 Aa(B)
Médias seguidas de mesma letra maiúscula nas colunas (fixando-se as épocas de medição e as
variedades, comparando-se os regimes hídricos), minúsculas nas linhas (fixando-se as épocas de
medição e os regimes hídricos, comparando-se as variedades) e maiúscula entre parênteses nas linhas
(fixando-se as variedades e os regimes hídricos, comparando-se as épocas de medição), não diferem
entre si pelo teste de Tukey a 5% de probabilidade.
Na primeira época de medição (início) não houve diferença significativa entre a
condutância estomática nas variedades (Quadro 2), isso porque como já citado
anteriormente, as condições eram similares em todos os tratamentos. No momento
intermediário não houve diferença estatística entre os regimes hídricos na variedade
Inca, mas para a variedade BRS Alegria houve um decréscimo na condutância
estomática no tratamento irrigado até 45 dias após a emergência (T1). O tratamento
com irrigação plena (T3) mostrou o melhor desempenho nessa avaliação,
provavelmente pela melhor condição hídrica da planta. Do ponto de vista fisiológico
os menores valores de condutância estomática obtidos no momento intermediário
indicam que a variedade BRS Alegria apresenta maior sensibilidade ao estresse
hídrico e indicam baixo consumo de água. Avaliar a condutância estomática torna-se
um atributo eficaz na seleção de genótipos para plantio em ambientes com limitada

54
disponibilidade de água no solo. Segundo Krieg (1993) o fechamento dos estômatos
é uma das primeiras linhas de defesa contra a dessecação, uma tentativa favorável
de manter o conteúdo hídrico nos tecidos vegetais por mais tempo, contudo, essa
redução de abertura dos poros estomáticos restringe a troca gasosa entre o interior
da folha e a atmosfera causando diminuição na assimilação de CO2, utilizado no
processo de fotossíntese.
Comparando se os valores da condutância estomática entre as variedades e
os regimes hídricos, nas épocas de medição, houve diferença significativa apenas no
tratamento com regime hídrico de irrigação plena.
Para EUA foi verificada a mesma tendência observada para E. Houve efeito
significativo somente do momento em que se realizou a medição do intercâmbio
gasoso (p = 0,0033). A prova de médias para essa causa de variação pode ser
observada na Tabela 8.
Tabela 8 – Prova de médias das eficiências instantâneas no uso de água(EUA) para
o momento da realização das medições do intercâmbio gasoso.
Momento da medição Eficiência instantânea no uso de
água (mol mol-1)
Início 0,0106 A Intermediário 0,0080 B Na colheita 0,0079 B
Médias seguidas de mesma letra maiúscula nas colunas não diferem entre si pelo teste de Tukey a 5%
de probabilidade.
De modo geral os valores da EUA das plantas de amaranto sob estresse hídrico
diminuem com avanço do ciclo, indicando que a planta estaria sob uma restrição
hídrica mais severa. Isso ocorre em função de um maior aumento da resistência em
relação à difusão de CO2 no mesófilo diminuindo a eficiência de carboxilação (Lawlor
e Cornic, 2002). A maior eficiência do uso de água observada antes da imposição do
estresse hídrico (aos 45 dias após a emergência) é decorrente de que a taxa
transpiratória no início foi 52,1% menor que na fase intermediária, explicando dessa
forma porque na fase inicial a EUA foi maior que na fase intermediária. Firmano et al.,
(2009) relatam que plantas sem restrição hídrica apresentam um aumento no EUA
uma vez que a redução apenas parcial da abertura estomática limita mais fortemente
a transpiração do que a entrada de CO2 aumentando a eficiência do uso da água.

55
4.4 Relações Hídricas
Não foi possível verificar qualquer efeito das causas de variação (variedade,
regime hídrico, momento em que se realizaram as medições e interações) sobre o
conteúdo relativo de água no momento da perda de turgência (CRAo), o potencial
osmótico à plena turgência (100) à turgência incipiente (0) e o módulo volumétrico
de elasticidade da parede celular ().Os valores médios globais dessas variáveis
foram de 87,60% para CRAo, -0,530 MPa para 100, -0,892 MPa para 0 e 5,054 MPa
para .
Para cada 1,0 MPa de variação no potencial de pressão, há uma respectiva
variação em 2% no volume de água no interior das células. Isso significa uma parede
celular elástica e que se ajusta às variações de água dentro da célula, mantendo assim
o potencial de pressão positivo, fator decisivo na manutenção da capacidade de
reprodução celular. Por isso o amaranto não necessita de ajustamento osmótico. Há
casos em que plantas resistentes à seca apresentam parede celular rígida (elevados
valores de ), e para não perder o potencial de turgor, evita a variação de volume
celular (perda de água) por meio de ajustamento osmótico (potenciais osmóticos que
podem chegar à -2,0 MPa). O resultado observado indica que, do ponto de vista das
relações hídricas, o amaranto controla sua condição hídrica interna por meio de
controle estomático. Por ser uma planta C4 o amaranto pode fazer fotossíntese com
baixa ou até mesmo valores nulos de condutância estomática.
Os valores de encontrados para o amaranto é baixo, uma vez que valores
inferiores a 10 MPa são relativamente baixos e indicam parede celular elástica. Na
natureza, se encontram valores de entre 1 e 50 MPa. Meyer et al. (1992) concluíram
que os efeitos do estresse de água sobre a elasticidade do tecido não são
consistentes, sendo observados aumentos em algumas espécies e diminuições em
outras.

56
4.5 ASPECTOS PRODUTIVOS
As variáveis produtividade e índice de colheita apresentaram pelo menos uma
causa de variação significativa. O desdobramento da interação entre regime hídrico e
variedade para produtividade encontra-se na Tabela 9.
Comparando as variedades dentro do regime hídrico observa que na
suspensão da irrigação aos 45 dias a variedade Inca foi superior, demonstrando maior
resistência quando o estresse hídrico ocorre nessa fase, entretanto, na suspensão
aos 60 dias não houve diferença significativa entre as variedades.
Considerando este resultado quando a janela de semeadura for pequena é
recomendável a utilização da variedade Inca, uma vez que esta apresentou
produtividade superior e menor ciclo, ou seja, se o período de chuvas for pequeno, é
recomendável a semeadura da variedade Inca como alternativa de safrinha. Na
condição de irrigação plena, ou seja, disponibilidade de água durante todo o ciclo da
cultura, a variedade BRS Alegria apresentou a melhor produtividade, sendo 25 %
superior à variedade Inca para essa condição (Tabela 9).
Os diferentes regimes hídricos não afetaram significativamente a produtividade
de grãos na variedade Inca, mostrando que essa variedade é tolerante ao estresse
não havendo prejuízos a produção de grãos, outro fator importante é que a planta
pode economizar água para uso em períodos posteriores (Saha et al., 2008), como
por exemplo para atingir a produção de grãos. Assim os efeitos na produção de grãos
foram atenuados não havendo diferença significativa entre os tratamentos de déficit
hídrico na variedade Inca. Outro fator importante é o estádio de desenvolvimento que
o estresse hídrico ocorreu, aos 45 dias após a emergência (início da restrição hídrica
no tratamento T1) a variedade já estava em plena antese. De acordo com Kelling
(1995), o efeito do déficit hídrico sobre a produção das culturas está vinculado ao
período de ocorrência durante o desenvolvimento das plantas. Entretanto é importante
ressaltar que apesar de não haver diferença significativa entre os tratamentos T1 e
T2, a produtividade no segundo tratamento obteve diferença no valor médio de 23,8
%, respectivamente em relação ao tratamento irrigado até 45 dias após a emergência,
embora não significativo. Do ponto de vista prático, se esta diferença persistir para
ensaios com menores coeficientes de variação, esta diferença poderá vir a ser
significativa, o que representa um acréscimo de aproximadamente 305 kg por hectare
na produtividade.

57
Tabela 9. Produtividade (kg ha-1) de A. caudatus cv Inca e A. cruentus cv BRS
Alegria em diferentes regimes hídricos,Santo Antônio do Leverger-
MT,2009.
Variedades
Regime Hídrico Inca BRS Alegria
Irrigado até 45 dias 1.285,69 Aa 702,15 Cb Irrigado até 60 dias 1.591,01 Aa 1.544,92 Ba
Sempre irrigado 1.510,54 Ab 2.008,62 Aa
CV: 12,22% Médias seguidas pela uma mesma letra maiúscula nas colunas e minúsculas nas linhas não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Na variedade BRS Alegria o estresse teve efeito significativo na produção de
grãos. O déficit hídrico severo (início do estresse aos 45 dias após emergência)
reduziu em 65 % a produtividade de grãos nesta variedade. Uma das explicações para
a redução da produtividade de grãos em decorrência do déficit hídrico é que nessas
condições a planta sofre redução da condutância e da transpiração foliar e como
consequência há um aumento da temperatura foliar e redução na produção de
fotoassimilados, causando redução na produtividade de grãos (Taiz & Zeiger, 2004).
Os menores valores de produtividade encontrados nos tratamentos que tiveram
suspensão da irrigação aos 45 e 60 dias após a emergência comparado com o
tratamento que teve irrigação plena, estão relacionados ao fato de que nos primeiros
dois tratamentos o deficit hídrico ocorreu no período crítico, ou seja, da pré-floração
ao início do enchimento de grãos, assim a recuperação da capacidade produtiva da
cultura não ocorreu de forma satisfatória, uma vez que os eventos reprodutivos são
mais rápidos do que os verificados durante o crescimento vegetativo (Morizet &
Togola, 1984).
Na Tabela 10 observa-se a produtividade por unidade de água disponibilizada
(PUAD) nas variedades e a interação com os regimes hídricos. Para a variedade Inca
o regime hídrico com irrigação plena obteve o menor valor, sendo diferente
estatisticamente de T1 e T2. Em BRS Alegria o tratamento irrigado até 45 dias após a
emergência obteve a menor produtividade por unidade de água disponibilizada, ou
seja, o acréscimo na lâmina total disponibilizada promoveu um aumento na
produtividade desta variedade.
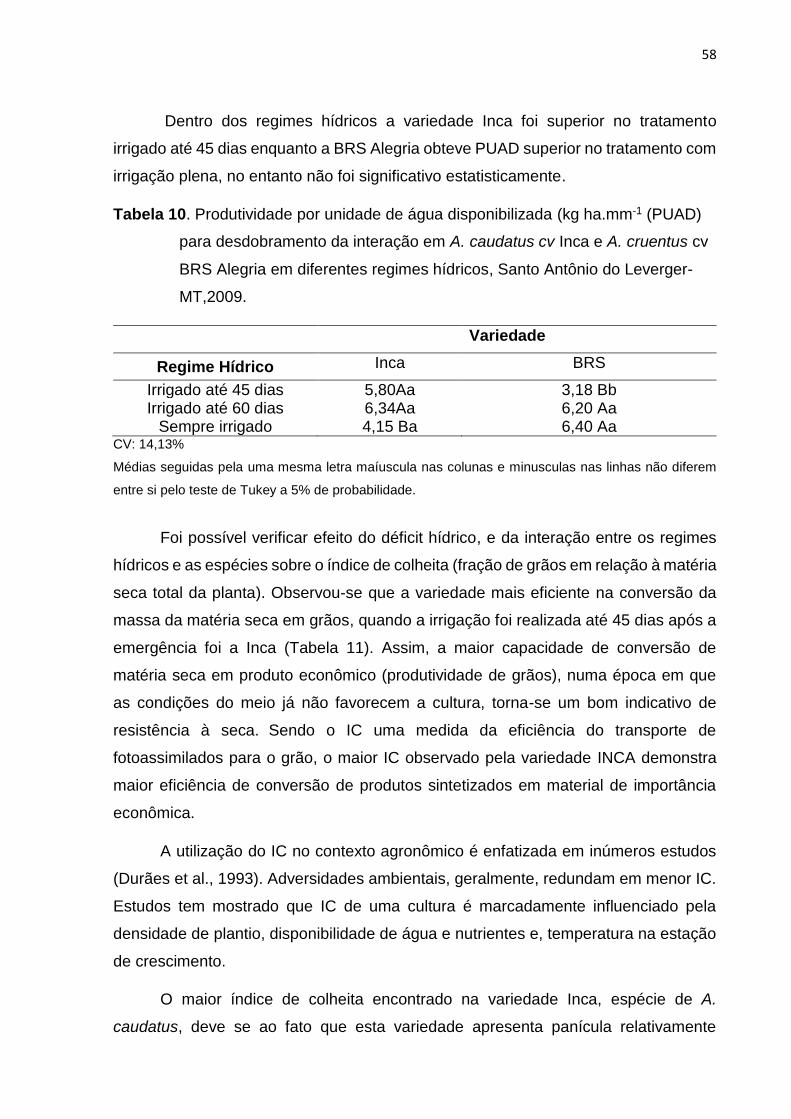
58
Dentro dos regimes hídricos a variedade Inca foi superior no tratamento
irrigado até 45 dias enquanto a BRS Alegria obteve PUAD superior no tratamento com
irrigação plena, no entanto não foi significativo estatisticamente.
Tabela 10. Produtividade por unidade de água disponibilizada (kg ha.mm-1 (PUAD)
para desdobramento da interação em A. caudatus cv Inca e A. cruentus cv
BRS Alegria em diferentes regimes hídricos, Santo Antônio do Leverger-
MT,2009.
Variedade
Regime Hídrico Inca BRS
Irrigado até 45 dias 5,80Aa 3,18 Bb Irrigado até 60 dias 6,34Aa 6,20 Aa
Sempre irrigado 4,15 Ba 6,40 Aa CV: 14,13%
Médias seguidas pela uma mesma letra maíuscula nas colunas e minusculas nas linhas não diferem
entre si pelo teste de Tukey a 5% de probabilidade.
Foi possível verificar efeito do déficit hídrico, e da interação entre os regimes
hídricos e as espécies sobre o índice de colheita (fração de grãos em relação à matéria
seca total da planta). Observou-se que a variedade mais eficiente na conversão da
massa da matéria seca em grãos, quando a irrigação foi realizada até 45 dias após a
emergência foi a Inca (Tabela 11). Assim, a maior capacidade de conversão de
matéria seca em produto econômico (produtividade de grãos), numa época em que
as condições do meio já não favorecem a cultura, torna-se um bom indicativo de
resistência à seca. Sendo o IC uma medida da eficiência do transporte de
fotoassimilados para o grão, o maior IC observado pela variedade INCA demonstra
maior eficiência de conversão de produtos sintetizados em material de importância
econômica.
A utilização do IC no contexto agronômico é enfatizada em inúmeros estudos
(Durães et al., 1993). Adversidades ambientais, geralmente, redundam em menor IC.
Estudos tem mostrado que IC de uma cultura é marcadamente influenciado pela
densidade de plantio, disponibilidade de água e nutrientes e, temperatura na estação
de crescimento.
O maior índice de colheita encontrado na variedade Inca, espécie de A.
caudatus, deve se ao fato que esta variedade apresenta panícula relativamente

59
extensa e menor tamanho da planta, sendo assim a fração de grãos em relação a
matéria seca total da planta é maior ocasionando maior índice de colheita para esta
variedade. Outro fator importante é que resultados experimentais têm mostrado que
plantas pequenas, adaptadas para condições de estresse, resultam em altos, ou mais
altos, IC que plantas maiores (Durães et al., 1998).
Para Irrigação até 60 dias variedade BRS Alegria se destacou obtendo IC 51%
superior a variedade Inca.
Tabela 11. Índice de colheita para A. cruentus cv “BRS Alegria” e A. caudatus cv
Inca. Santo Antônio do Leverger-MT,2009.
Variedade
Regime Hídrico Inca BRS
Irrigado até 45 dias 0.60 Aa 0.21 Bb Irrigado até 60 dias 0.27 Bb 0.53 Aa
Sempre irrigado 0,49 Aa 0,32 Bb CV = 18,66%
Médias seguidas pela uma mesma letra maiúscula na coluna e minúscula na linha não diferem entre si pelo teste de Tukey a 5% de probabilidade.

60
5. CONSIDERAÇÕES FINAIS
O déficit hídrico é uma situação comum à produção de muitas culturas,
podendo apresentar um impacto negativo e substancial no crescimento e
desenvolvimento das plantas. A necessidade em se resolver esta condição leva a
planta a desenvolver mecanismos morfofisiológicos, que as conduzem a economizar
água para uso em períodos posteriores.
Os estudos com amaranto ainda são incipientes, no entanto a cultura apresenta
perspectivas promissoras, dessa forma é importante que se realize mais pesquisas
no sentido de estabelecer quais processos fisiológicos, mudanças morfológicas e
fenológicas da planta se alteram em função do déficit hídrico.
Além disso faz se necessário um melhoramento das variedades de amaranto,
uma vez que o material genético ainda não apresenta estabilidade, apresentando
diferenças dentro do mesmo material genético.

61
6. CONCLUSÕES
Nas condições que o experimento foi conduzido foi possível concluir que:
I. A variedade de amaranto Inca (Amaranthus caudatus) mostrou capacidade de
produzir seu potencial granífero com apenas 45 dias de precipitação
pluviométrica.
II. A variedade de amaranto BRS Alegria (Amaranthus cruentus) apresentou
maior sensibilidade à restrição hídrica o que proporcionou decréscimos
significativos na produtividade e menor índice de colheita.
III. Uma vez que não se verificou efeito de espécies sobre o processo
fotossintético, a menor produtividade final de grãos no Inca em relação à BRS
Alegria pode ter sido devido à associação dos seguintes fatores: (a)
característica própria das variedades. (b) na Inca , menor tempo no período
vegetativo (10 dias menos), (c) menor tempo no período reprodutivo (10 dias
menos), (d) na INCA, menor tamanho da planta (possivelmente menor índice
de área foliar).
IV. O diâmetro de caule, a massa da matéria seca total, a massa da matéria seca
da parte aérea e o peso de raízes são variáveis indicadas para o estudo do
estresse hídrico em amaranto.
V. O déficit hídrico causou acréscimo na massa de raízes do amaranto, enquanto
as demais variáveis quantitativas de crescimento sofreram declínio.

62
VI. A irrigação até 45 dias após emergência proporcionou os menores valores de
matéria seca em A. caudatus e A. cruentus.
VII. O estresse hídrico provocou reduções significativas nas medidas de
intercambio gasoso das variedades estudadas.
VIII. Para as variáveis das relações hídricas não foi possível verificar os efeitos das
espécies de amaranto, do regime hídrico e da interação entre esses fatores
IX. As espécies aparentemente controlam eficientemente o uso de água por meio
de controle estomático. Isto porque não se verifica potenciais osmóticos e
módulo volumétrico de elasticidade da parede próprios de plantas resistentes
à seca.

63
7. REFERÊNCIAIS BIBLIOGRÁFICAS
AMAYA-FARFAN, J.; MARCÍLIO, R.; SPEHAR, C.R. Deveria o Brasil investir em novos grãos para a sua alimentação? A proposta do amaranto (Amaranthus sp.). Segurança Alimentar e Nutricional, v.12, p.47-56, 2005.
ANJUN, S.A.; XIE,X; WANG,L.; SALEEM, F.; MAN, C.; LEIL,W.; Morphological, physiological and biochemical responses of plants to drought stress. African Journal of Agricultural Research v.6, p.2026-2032, 2011.
APAZA-GUTIERREZ, V., A. Romero-Saravia, F.R. Guillen Portal, D.D. Baltensperger. 2002. Response of grain amaranth production to density and fertilization in Tarija, Bolivia. p. 107 -109. In: J. Janick and A. Whipkey (Eds.), Trends in new crops and new uses. ASHS Press, Alexandria, VA.
ASHARAF,M.; HARRIS,P.J.C.; Photosynthesis under stressful environments: An overview. Photosynthetica. v.51, 163-190, 2013.
BENINCASA, M.M.P. Análise de crescimento de plantas. 1ª ed. Jaboticabal: FUNEP,1988. 42 p
BIANCHINI, M.G.A.; BELÉIA, A.D.P. Umidade e congelamento de grãos de amaranto e sua capacidade de expansão térmica. Pesquisa Agropecuária Brasileira, v.45, n.8, p.917-924, 2010.
BIANCHINI, M.G.A.; BELÉIA, A.D.P.; BIANCHNI, A. Modificação da composição química de farinhas integrais de grãos de amaranto após a aplicação de diferentes tratamentos térmicos. Ciência Rural, v.44, p.167-173, 2014.

64
BRAMBILLA, T.R.; CONSTANTINO, A.P.; OLIVEIRA, P.S. Efeito da adubação nitrogenada na produção de amaranto. Semina: Ciências Agrárias, Londrina, v.29, n.4, p.761-768, 2008.
BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para Análise de Sementes. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Brasília, DF: Mapa/ACS, 2009. 395p.
BRAY, E.A . Plants responses to water deficit. Trends in Plant Science. Kidlington v.2, n.2 p.48-54, Feb,1997.
BRENNER, D.; WILLIAMS, J.T. Grain amaranth (amaranthus species). In: WILLIAMS, J.T. (Ed.). Underutilized Crops: cereals and pseudocereals. London: Chapman & Hall, 1995. p.128-186
CARMO, R.A.C; AGUIAR,L.G. Adaptação de espécies para a produção de grãos, proteção de solo e diversificação do sistema produtivo. Uberlandia.UFU.2000
CAPRILES, V.D.,;ARÊAS, J.A.G. Avaliação da qualidade tecnológica de snacks obtidos por extrusão de grão integral de amaranto ou de farinha de amaranto desengordurada e suas misturas com fubá de milho, Brazilian Journal of Food Technology. vol.15 no.1, p. 21-29, jan./mar. 2012 Campinas.
CHAVES, M. M.; PEREIRA, J. S.; MAROCO, J.; RODRIGUES, M. L.; RICARDO, C. P. P.; OSÓRIO, M. L.; CARVALHO, I.; FARIA, T.; PINHEIRO, C. How plants cope with water stress in the field. Photosynthesis and growth. Annals of Botany, v. 89, p. 907-916. 2002.
COSTA, D.J.A.; BORGES, A.S. AVALIAÇÃO DA PRODUÇÃO AGRÍCOLA DO AMARANTO (Amaranthus hypochondriacus), Holos, Ano 21, v.3, n.1, p.97-111, 2005.
DENADAI I.A.M; KLAR A.E; Resistência à seca em quatro variedadees de trigo: parâmetros fisiológicos. Scientia Agricola, 52:274-281. 1995
DOMINGOS, V.D.; ERASMO, E.A.L.; SILVA, J.I.C.; CAVALCANTE, G.D.; SPEHAR, C.R. Crescimento, produção de grãos e biomassa de variedades de amaranto (Amaranthus cruentus) em função de adubação com NPK. Bioscience Journal, v.21, p.29-39, 2005

65
DURÃES,F.O.M.; MAGALHÃES,P.C.; OLIVEIRA,A . C.; FANCELLI,A.L.; COSTA,J.D. Partição de fitomassa e limitações do rendimento de milho (Zea mays L.) relacionadas com a fonte dreno. In: CONGRESSO BRASILEIRO DE FISIOLOGIA VEGETAL, 4., Fortaleza, 1993. Resumos. Fortaleza, SBFV;UFCE, 1993. Revista Brasileira de Fisiologia Vegetal, São Carlos, 5(1):1-120, jan./jun. 1993.
EARLEY, G.S.; KAUL, H.P. KRUSE, M; AUFHAMMER, W. Yield and nitrogen utilization efficiency of the pseudocereals amaranth, quinoa, and buckwheat under differinf nitrogen fertilization. European Journal of Agronomy, n.22, p.95-100, 2005.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA –EMBRAPA. Centro Nacional de Pesquisa de Solos. Sistema brasileiro de classificação de solos. Rio de Janeiro, 2006. 306p
ERASMO, E.A.L.; DOMINGOS, V.D.; SPEHAR, C.R.; DIDONET, J.; SARMENTO, R.A.; CUNHA, A.M. Avaliação de variedadees de amaranto (Amaranthus spp.) em sistema plantio direto no sul de Tocantins. Bioscience Journal, v.20, p.171-176, 2004.
FANCELLI, A.L. & D. DOURADO-NETO. Produção de Milho. Ed. Agropecuária, Guaíba. 360 p., 2000
FARIAS, C. H. de A.; FERNANDES, P. D.;AZEVEDO, H. M.; DANTAS NETO, J. Índices de crescimento da cana-de-açúcar irrigada e de sequeiro no Estado da Paraíba. Revista Brasileira de Engenharia Agrícola e Ambiental, Campina Grande, v. 12, n. 4, p. 356-362, 2008
FAGERIA, N.K.; BALIGAR, V.C.; CLARK, R.B. Physiology of crop production. New York: Haworth Press, 2006. 345p
FERREIRA, T. A. P. C.; GUERRA-MATIAS, A. C.; ARÊAS, J. A. G. Características nutricionais e funcionais do Amaranto (Amaranthus sp.). Nutrire, São Paulo, v. 32, n. 2, p. 91-116, 2007
FERREIRA, C. C. Efeito da densidade de semeadura e doses de nitrogênio sobre a produtividade, e biometria do amaranto, no Cerrado do Planalto Central. Bioscience Journal. Uberlandia, v. 30, supplement 2, p. 534-546, Oct./14
FIRMANO, R. S.; KUWARA, F. A.; SOUZA, G. M. Relação entre adubação fosfatada e deficiência hídrica em soja. Ciência Rural, Santa Maria v.39, n.7, p.1967-1973, 2009.

66
FITTER, A.H. & HAY, R.K.M. 1987. Environmental Physiology of Plants. USA, Academic Press. 422p
FLEXAS, J.; MEDRANO, H. Drought-inhibition of photosynthesis in C3 plants: stomatal and non-stomatal limitations revisited. Annals of Botany. v.89, p.183-189, 2002.
GIMPLINGER, D.M.; DOBOS, G.; SCHÕNLECHNER, R.; KAUL, H.P. Yield and quality of grain amaranth (Amaranthus sp.) in Eastern Austria. Plant Soil Enviroment, v.53, n.3, p.105-112, 2007.
GONÇALVES, W.G.; JIMENEZ, R.L.; ARAÚJO FILHO, J.V.; ASSIS, R.L.; SILVA, G.P.; PIRES, F.R. Sistema radicular de plantas de cobertura sob compactação do solo. Engenharia Agrícola, v.26, p.67-75, 2006
GRIFFITHS, H.; PARRY, M. A. J. Plant responses to water stress. Annals of Botany, v. 89,p. 801-802, 2002.
HENDERSON, T.L. Agronomiv evaluation of grain amaranth in North Dakota. Tesis Ph. D. North Dakota State, North Dakota, USA. 1993
INMAN-BAMBER. Sugarcane water stress criteria for irrigation and drying off. Fields Crops Research, Australia, v.89, p.107-122. 2004
JONES, H. G. Monitoring plant and soil water status: established and novel methods revisitedand their relevance to studies of drought tolerance. Journal of Experimental Botany, v. 58, n. 2, p. 119-130. 2007
KERBAUY, G. B. Fisiologia vegetal. São Paulo: Guanabara Koogan, 2004. 452 p.
KOIDE, R.T; ROBICHAUX, R.H.; MORSE, S.R.; SMITH, C.M. 1989. Plant water status, hydraulic resistance and capacitance. In: Plant Physiological Ecology. Field Methods and Instrumentation (R.W. Pearcy, J.R. Ehleringer, H.A. Mooney, P.W. Rundel, Eds.). Chapman and Hall. London, New York. pp. 161-183.
KUBISKE, M.E.; ABRAMS, M.D. . Reydration effects on pressure-volume relationships in four temperate woody species: variability with site, time of season and drought conditions. Oecologia. 85: 537-542.1991

67
LARCHER, W. Ecofisiologia Vegetal. Tradução: Prado, C. H. B. A. São Carlos: Rima, 2006. 531 p.
LAUER, M. J. & BOYER, J. S. Internal CO2 measured directly in leaves. Abscisic acid and low leaf water potential cause opposing effects. Plant Physiology. v.98, p.1310-1316, 1992.
LAWLOR, D.W. Carbon and nitrogen assimilation in relation to yield: mechanisms are the key to understanding production systems. Journal of Experimental Botany, Oxford, v.53, n.370, p.773-
787, 2002
LUU, D. T.; MAUREL, C. Aquaporins in a challenging environment: molecular gears for adjusting plant water status. Plant, Cell and Environment, v. 28, p. 85-96. 2005
PIFFER, C.R. BENEZ, S.H.; BERTOLINI, E.V.; COMINETTI, F.R.; SILVA, P.R.A. Crescimento radicular de três espécies de cobertura vegetal em camadas de solo compactadas artificialmente. Varia Scientia Agrárias, v.01, p.31-43, 2010.
MARTINS, M. O.; Aspectos fisiológicos do nim indiano sob déficit hídrico em condições de casa de vegetação. Dissertação (Mestrado em Botânica) – Universidade Federal Rural de Pernambuco. Departamento de Biologia.84 f. 2008
MARUR, C.J. 1999. Curvas pressão-volume e expansão foliar em variedadees de algodoeiro submetidos a déficit hídrico. Scientia Agricola 56:563-569.
MELCHER, K., NG, L.M., ZHOU, X.E.; SOON , F.F.; XU, Y.; , Y.; SUINO- POWELL, K.M.; PARK, S.; WEINWE, J.J.; FUJII, H.; CHINNUSAMY, V.; KOVACH, A.; LI, J.; WANG, Y.; LI, J.; PETERSON, F.C.; JENSEN, D.R.; YOUNG, E. VOLKMAN, B.F.; CUTLER, S.R.; ZHU, J.; XU, H.E,. A gate-latch-lock mechanism for hormone signalling by abscisic acid receptors. – Nature v.462, p.602-608, 2009.
MERRIAM,J.L.;KELLER,J.;ALFARO,J. Irrigation: system evaluation and improvement Logan: Utah State University, 1973.
MORALES, R.G.F. Resistência ao déficit hídrico em famílias de tomateiros derivados de sollanum pennellii. Tese (DOUTORADO) Universidade Federal de Lavras,2012. 93p

68
MORIZET, J.; TOGOLA, D. Effect et arrière-effect de la sécheresse sur la croissance de plusieurs génotipes de maïs. In: CONFÉRENCE INTERNATIONALE DES IRRIGATIONS ET DU DRAINAGE, 1984, Versailles. Les bésoins en eau des cultures. Paris: Inra, 1984. p.351-360.
MUJICA-SÁNCHEZ,A.; BERTI-DIAZ,M.; IZQUIERDO,J. El Cultivo del Amaranto(Amaranthus spp); producción, mejoramineto genético y utilización. FAO. Oficina regional de la FAO para América Latina y el caribe: Santiago, Chile, 1997.
MUJICA-SÁNCHEZ, A.; QUILLAHUAMAN, A. Fenologia del cultivo de la kiwicha(Amaranthus caudatus l.) In; CURSO TALLER FENOLOGIA DE CULTIVOS ANDINOS Y USO DE LA INFORMACIONA AGROMEEREOLÓGICA..Puno, 7-10 agosto. INIA, Pica.Peru. p. 29-31. 1989.
MUJICA-SANCHEZ, A. et al. Quinua (Chenopodium quinoa Willd.): ancestral cultivo andino, alimento del presente y del futuro. Santiago: FAO, 2001.
NOBRE, D.A.C.; DAVID, A.M.S.S.; SOUZA, V.N.R.; OLIVEIRA, D.; GOMES, A.A.M.; AGUIAR, P.M.; MOTA, W.F. Influência do ambiente de armazenamento na qualidade fisiológica de sementes de amaranto. Comunicata Scientiae, v.4, p 216-219, 2013.
NOGUEIRA, R. J. M. C.; ALBUQUERQUE, M. B.; SILVA, E. C. Aspectos ecofisiológicos da tolerância à seca em plantas da caatinga. In: NOGUEIRA, R. J. M. C.; ARAÚJO, E. L.; WILLADINO, L. G.; CAVALCANTE, U. M. T. Estresses ambientais: danos e benefícios em plantas. Recife: UFRPE, Imprensa Universitária, p.22-31, 2005.
PAIVA, A. S.; FERNANDES, E. J.; RODRIGUES, T. J. D. TURCO J. E. P. Condutância estomática em folhas de feijoeiro submetido a diferentes regimes de irrigação. Engenharia. Agrícola. vol.25 no.1 Jaboticabal Jan./Apr. 2005
PAULUS D; DOURADO NETO D.; FRIZZONE J. A.; SOARES T. M. Produção e indicadores fisiológicos de alface sob hidroponia com água salina. Horticultura Brasileira, v.28, n. 29-35. 2010.
PIMENTEL, C. A relação da planta com a água. Seropédica, RJ: Edur. 2004. 191 p.
PLAUT, Z. Photosynthesis in plant crop under water and salt stress. In- PESSARAKLI, M. Handbook of plant crop physiology, Israel, cap. 27, p.587-601,

69
1995.
QUEIROZ, C.G.S.; GARCIA, Q.S.; LEMOS FILHO, J.P. 2002. Atividade fotossintética e peroxidação de lipídios de membrana em plantas de aroeira-do-sertão sob estresse hídrico e após reidratação. Brazilian Journal of Plant Physiology, 14: 59-63.
RAHANAMA, A.; POUSTINI, K.; MUNNS, R.; JAMES, R.A. Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Functional Plant Biology v.37.p.255–263, 2010 SADRAS, V. O.; MILROY, S. P. Soil-water thresholds for the responses of leaf expansion and gas exchange: A review. Field Crops Research, v. 47, p. 253-266, 1996. SAIBO, N.J.M., LOURENÇO, T., OLIVEIRA, M.M.: Transcription factors and regulation of photosynthetic and related metabolism under environmental stresses. – Annals of Botany. v.103, p. 609-623, 2009. SCHOLANDER, P.F.; HAMMEL, H.T.; HEMMINGSEN, E.A.; BRADSTREET, E.D. Sap pressure in vascular plants. Science, Washington, v. 148, p. 339-346, 1965. SPEHAR, C.R.; BRENNER, D. MUJICA-SANCHES, A. Origem e importância do Amaranto In: SPEHAR, C.R. (Ed.). Amaranto: Opção para diversificar a agricultura e os alimentos, Embrapa Cerrados, 2007, p.23-42. SPEHAR, C.R.; CABEZAS, W.A.R.L. Introdução e seleção de espécies para a diversificação do sistema produtivo nos cerrados. In: CABEZAS, W.A.R.L.; FREITAS, P.L. (Ed.). Plantio direto na integração lavoura pecuária. Uberlândia: Ed. da Universidade Federal de Uberlândia. 2001. p.179-189. SPEHAR, C.R.; SANTOS, R.L.B.; SOUZA, P.I.M. Novas plantas de cobertura para o sistema de produção de grãos. In: SEMINÁRIO INTERNACIONAL SOBRE O SISTEMA DE PLANTIO DIRETO, 2., 1997, Passo Fundo. Anais... Passo Fundo: Embrapa/CNPT, 1997. p.69-172, 1997. SPEHAR, C.R.; TEIXEIRA, D.L. Diferenças entre o pseudocereal amaranto e espécies de planta daninha, Amaranthaceae. Planaltina: EMBRAPA Cerrados, 2002. (Comunicado Técnico, 69). SPEHAR, C.R.; TEIXEIRA, D.L.; LARA CABEZAS, W.A.R.; ERASMO, E.A.L. Amaranto BRS Alegria: alternativa para diversificar os sistemas de produção. Pesquisa Agropecuária Brasileira, v.38, p.659-663, 2003.
SPEHAR, C.R. & TRECENTINI, R. Desempenho agronômico de espécies tradicionais e inovadoras da agricultura em semeadura de sucessão e entressafra no Cerrado do planalto central brasileiro. Bioscience Journal, v.27, p.102-111, 2011.

70
STEINBERG, S.L.; MILLER, J.C. & MCFARLAND, M.J. 1990. Dry matter partitioning and vegetative growth of young peach trees under water stress. Aust. J. Plant Physiol., 17: 23-36.
TAPIA,M. Cultivos Andinos Subexplotados y sua Aporte a la Alimentación. Oficina Regional de la FAO para ala América Latina y Caribe, Santiago, Chile. 217p. 1997.
TAIZ, L. e ZEIGER, E. Fisiologia Vegetal. Porto Alegre: Artmed Editora. 2004. 719p.
TEIXEIRA, D. L.; SPEHAR, C. R. Diferenças entre o pseudocereal amaranto e espécies de planta daninha, Amaranthaceae. Planaltina: EMBRAPA Cerrados, 2002. (Comunicado Técnico, 69).
TORRES, J. A.P; SCHIAVINATO, M. A. Crescimento, eficiência fotossintética e eficiência do uso da água em quatro espécies de leguminosas arbóreas tropicais. Hoehnea, v. 35, n. 3, p. 395-404, 2008.
TYERMANN, S. D.; NIEMIETZ, C. M.; BRAMLEY, H. Plant aquaporins: multifunctional
water and solute channels with expanding roles. Plant, Cell and Environment, v. 25, p. 173-194. 2002.
TYREE, M.T.; Hammel, H.T. 1972. The measurement of turgor pressure and the water relations of plants by the pressure-bomb technique. Journal of Experimental Botany. 23: 267-282.
YORDANOV, I.; VELIKOVA, V.; TSONEV, T. Plant responses to drought, acclimation and stress tolerance. Photosynth v. 38, n. 1, p. 171-186, 2000.


















