
UNIVERSIDADE FEDERAL DE JUIZ DE FORA FACULDADE DE ... · parcial para obtenção do título de...
51
UNIVERSIDADE FEDERAL DE JUIZ DE FORA FACULDADE DE FARMÁCIA TRABALHO DE CONCLUSÃO DE CURSO Douglas Massote Pestana AVALIAÇÃO COMPARATIVA DA ESTRUTURA DA COMUNIDADE BACTERIANA INTESTINAL, PELO USO DA TÉCNICA DE PCR-DGGE, EM INDIVÍDUOS EUTRÓFICOS E COM EXCESSO DE PESO Juiz de Fora 2016
Transcript of UNIVERSIDADE FEDERAL DE JUIZ DE FORA FACULDADE DE ... · parcial para obtenção do título de...

UNIVERSIDADE FEDERAL DE JUIZ DE FORA
FACULDADE DE FARMÁCIA
TRABALHO DE CONCLUSÃO DE CURSO
Douglas Massote Pestana
AVALIAÇÃO COMPARATIVA DA ESTRUTURA DA COMUNIDADE
BACTERIANA INTESTINAL, PELO USO DA TÉCNICA DE PCR-DGGE, EM
INDIVÍDUOS EUTRÓFICOS E COM EXCESSO DE PESO
Juiz de Fora
2016

Douglas Massote Pestana
Avaliação comparativa da estrutura da comunidade bacteriana intestinal, pelo
uso da técnica de PCR-DGGE, em indivíduos eutróficos e com excesso de
peso
Trabalho de Conclusão de Curso
apresentado ao corpo docente do Instituto
de Ciências Biológicas da Universidade
Federal de Juiz de Fora, como requisito
parcial para a obtenção do título de
Bacharel em Farmácia.
Orientador: Prof. Dr. Cláudio Galuppo Diniz
Juiz de Fora
2016

DOUGLAS MASSOTE PESTANA
Avaliação comparativa da estrutura da comunidade bacteriana intestinal, pelo
uso da técnica de PCR-DGGE, em indivíduos eutróficos e com excesso de
peso
Trabalho de Conclusão de Curso
apresentado ao corpo docente do Instituto
de Ciências Biológicas da Universidade
Federal de Juiz de Fora, como requisito
parcial para obtenção do título de Bacharel
em Farmácia.
Aprovado em: __/__/__
BANCA EXAMINADORA
_________________________________________
Prof. Dr. Cláudio Galuppo Diniz (Orientador)
Instituto de Ciências Biológicas (UFJF)
_________________________________________
Profa. Dra. Alessandra Barbosa Ferreira-Machado
Instituto de Ciências Biológicas (UFJF)
_________________________________________
MSc. Thaís Oliveira de Paula
Instituto de Ciências Biológicas (UFJF)


AGRADECIMENTOS
Agradeço primeiramente a Deus pela oportunidade de concluir a faculdade e
iniciar a vida profissional. Foram cinco anos de muita luta e garra, mas ao olhar
para trás, vejo que todo o esforço valeu a pena.
Agradeço ao meu orientador Prof. Dr. Cláudio Galuppo Diniz e à Profa. Dra.
Vânia Lúcia da Silva, por me concederem a oportunidade de trabalhar no
laboratório de microbiologia e por todos os ensinamentos que me transmitiram.
Agradeço à mestra Thaís Oliveira de Paula pela oportunidade de poder
trabalhar em seu projeto grandioso, que me fez despertar um interesse maior pela
ciência.
Agradeço a Alessandra, Juliana, Michele, Francis e Dionéia por todos os
conhecimentos que me passaram e pelo auxílio prestado durante os experimentos.
Agradeço a todos os colegas de laboratório que me ajudaram durante os
últimos três anos e me proporcionaram dias mais alegres. Foi uma experiência
incrível fazer parte da história de cada um.
Agradeço aos meus pais Paulo e Marli, à minha irmã Ana Paula e à minha
namorada Ohanna por todo o apoio que me deram durante esse período, além de
sempre me incentivarem a chegar até o final sem desistir de lutar.

RESUMO
A obesidade é um crescente problema de saúde pública no mundo inteiro.
Grande parte das despesas econômicas no sistema de saúde do mundo está
relacionada com as comorbidades decorrentes da obesidade, como diabetes,
hipertensão e câncer. Sua etiologia é multifatorial, envolvendo fatores genéticos,
endócrinos e ambientais. Estudos recentes classificam a obesidade como um
estado de inflamação crônica de baixo grau, enquanto outros estudos mostram que
a microbiota intestinal humana exibe um papel importante na manutenção da
homeostase energética. Uma das técnicas mais utilizadas para monitorar o
comportamento da microbiota intestinal é a Reação em Cadeia da Polimerase
seguida de Eletroforese em Gel com Gradiente Desnaturante (PCR-DGGE), uma
vez que esta fornece um fingerprinting da comunidade microbiana avaliada. O
presente estudo teve como objetivo avaliar comparativamente a estrutura da
comunidade bacteriana em indivíduos eutróficos e com excesso de peso, pelo
método de PCR-DGGE com primers para o domínio Bacteria. Foi utilizado um
grupo amostral de 72 indivíduos, classificados em obesos, com sobrepeso e
eutróficos, de acordo com o Índice de Massa Corporal (IMC) e as medidas
antropométricas. Por meio da similaridade dos perfis de bandeamento fornecidos
pelo DGGE, foi possível agrupar os indivíduos em três clusters diferentes. Tal fato
sugere que a comunidade bacteriana de indivíduos obesos se diferencia dos
indivíduos eutróficos, enquanto que os indivíduos com sobrepeso apresentam-se
em estágio de transição. Os resultados corroboram as evidências encontradas na
literatura e espera-se que auxiliem no desenvolvimento de estudos futuros, com a
finalidade de modular a microbiota intestinal e compreender melhor a patogenia da
obesidade.
Palavras-chave: Obesidade, Microbiota intestinal, Reação em Cadeia da
Polimerase-Eletroforese em Gel com Gradiente Desnaturante (PCR-DGGE).

ABSTRACT
Obestiy is a growing public health problem worldwide. Most of the economic
burden in the global health system is related to co-morbidities resulting from
obesity, such as diabetes, hypertension and cancer. Its etiology is multifactorial,
involving genetic, endocrine and environmental factors. In recent studies, obesity is
classified as a state of chronic low-grade inflammation, while other approaches
complements that the human intestinal microbiota exhibits an important role in
maintaining energy homeostasis. One of the most widely used techniques to
monitor the behavior of intestinal microbiota is the Polymerase Chain Reaction
followed by Denaturing Gradient Gel Electrophoresis (PCR-DGGE), since it is
capable to evaluate microbial communities through fingerprinting. This study aimed
at comparing the structure of the bacterial community in normal weight and
overweight individuals, by PCR-DGGE with universal primers for Bacteria. We
sampled a group of 72 individuals and classified them as normal weight, overweight
and obese, according to the Body Mass Index (BMI) and anthropometric
measurements. Through the similarity of banding profiles provided by DGGE, it was
possible to group individuals in three different clusters. This fact suggests that the
bacterial community of obese is different from normal weight individuals, while
overweight individuals are in the transition stage. These results support the
evidence found in the literature and are expected to assist in the development of
future studies, in order to modulate the gut microbiota and best understanding the
pathogenesis of obesity.
Keywords: Obesity, Intestinal microbiota, Polymerase Chain Reaction-Denaturant
Gradient Gel Electrophoresis (PCR-DGGE).

LISTA DE ILUSTRAÇÕES
Figura 1. DGGE fingerprint dos produtos de amplificação do DNA bacteriano
correspondente à região V6 do gene codificador para o rRNA 16S.......................33
Figura 2. Diagrama de Venn com a distribuição das bandas correspondentes à
região V6 do rRNA 16S bacteriano.........................................................................34
Figura 3. Agrupamento dos indivíduos participantes desse estudo (eutróficos, com
sobrepeso e obesos) de acordo com o perfil de bandeamento..............................35
Figura 4. Matriz de similaridade resultante da análise de agrupamento UPGMA com
coeficiente de Jaccard, do fingerprint dos produtos de amplificação do DNA
bacteriano, separados em DGGE...........................................................................36

LISTA DE TABELAS
Tabela 1. Classificação de peso de acordo com IMC e risco de comorbidades......15
Tabela 2. Sexo e idade dos participantes deste estudo...........................................30
Tabela 3. Características antropométricas dos participantes do estudo..................31
Tabela 4. Estatística descritiva dos resultados obtidos para o domínio Bacteria e
valores de p entre os grupos estudados..................................................................32

SUMÁRIO
1 INTRODUÇÃO E JUSTIFICATIVA....................................................... 12
2 REVISÃO DE LITERATURA................................................................. 14
2.1 Aspectos gerais da obesidade............................................................... 14
2.2 Obesidade e o sistema imunológico...................................................... 16
2.3 Tecido adiposo e sua função endócrina................................................ 17
2.4 Microbiota intestinal humana................................................................. 18
2.5 Alterações na microbiota intestinal........................................................ 20
2.6 Abordagens de técnicas independentes de cultivo................................ 21
3 OBJETIVOS.......................................................................................... 24
3.1 Objetivo geral......................................................................................... 24
3.2 Objetivos específicos............................................................................. 24
4 MATERIAIS E MÉTODOS..................................................................... 25
4.1 Seleção dos participantes do estudo..................................................... 25
4.2 Caracterização dos indivíduos............................................................... 25
4.3 Coleta de amostras fecais e preparação para análises microbiológicas......................................................................................
26
4.4 Extração de DNA a partir dos espécimes fecais.................................... 26
4.5 Eletroforese em Gel com Gradiente Desnaturante (DGGE).................. 27
4.5.1 Amplificação de rRNA 16S de representantes do domínio Bacteria..... 27
4.5.2 Condições da corrida eletroforética....................................................... 28
4.6 Análise estatística.................................................................................. 29
5 RESULTADOS...................................................................................... 30
5.1 Características epidemiológicas e clínicas dos participantes................ 30
5.2 Avaliação da estrutura da comunidade bacteriana................................ 32
6 DISCUSSÃO.......................................................................................... 38
7 CONCLUSÕES...................................................................................... 40
REFERÊNCIAS.................................................................................................... 41

ANEXO A - Parecer do comitê de ética em pesquisa – UFJF.............. 47
ANEXO B - Termo de consentimento livre e esclarecido ................. 49
ANEXO C - Ficha para dados antropométricos................................. 51

12
1 INTRODUÇÃO E JUSTIFICATIVA
A obesidade é um sério problema de saúde pública na atualidade.
Estimativas sugerem que cerca de 50% da população mundial está acima do peso,
o que a configura como uma pandemia. O maior problema relacionado à obesidade
se refere às comorbidades, sendo hipertensão arterial, diabetes mellitus tipo 2 e
dislipidemias as principais doenças crônicas não transmissíveis relacionadas ao
excesso de peso.
A etiopatologia da obesidade ainda não é totalmente esclarecida. Fatores
extrínsecos como alimentação rica em calorias vazias, ausência de exercícios
físicos e estresse elevado contribuem para que haja o desequilíbrio energético, o
qual culmina no ganho excessivo de peso. Muitas linhas de pesquisa procuram
elucidar os fatores intrínsecos relacionados às causas da obesidade. Ensaios
imunológicos demonstram que a obesidade comporta-se como uma inflamação
crônica de baixo grau, relacionada com a liberação contínua de determinadas
citocinas que induzem o acúmulo de gordura. Outra vertente de pesquisadores
correlaciona a comunidade microbiana intestinal, denominada microbiota intestinal,
com a elevada absorção de ácidos graxos e o balanço energético positivo.
Diversos estudos relatam que a microbiota intestinal apresenta um importante
papel na homeostase energética do indivíduo.
Os grandes avanços da Biologia Molecular permitiram o desenvolvimento de
técnicas apuradas, as quais já são utilizadas em certos laboratórios de rotina. A
técnica da Reação em Cadeia da Polimerase (PCR), inicialmente desenvolvida na
década de 80, representou um marco na história da pesquisa contemporânea. Com
o avançar do tempo, o aperfeiçoamento dessa técnica possibilitou o surgimento de
novas variantes, as quais são capazes de fornecer resultados com superior
qualidade e quantidade de informações.
A técnica de PCR-DGGE (Reação em Cadeia da Polimerase-Eletroforese
em Gel com Gradiente Desnaturante) é usada com o intuito de avaliar grandes
grupos genômicos. Em estudos de avaliação da diversidade microbiana em
determinado ambiente, esse método se destaca devido a sua capacidade de
organizar perfis genéticos semelhantes em agrupamentos, sendo bastante
consolidado na literatura.

13
O presente trabalho é uma fração de um projeto maior, realizado no
Laboratório de Fisiologia e Genética Molecular Bacteriana da Universidade Federal
de Juiz de Fora, o qual buscou avaliar a correlação entre a diversidade bacteriana
intestinal e a obesidade.

14
2 REVISÃO DE LITERATURA
2.1 Aspectos gerais da obesidade
A obesidade é descrita como uma doença crônica, com alta prevalência e
incidência no mundo inteiro, sendo considerada uma pandemia nos dias atuais. É
caracterizada pelo excessivo acúmulo de gordura no tecido adiposo, em
decorrência do rompimento da homeostase energética, que resulta em prejuízos
importantes na qualidade de vida do indivíduo. Preferencialmente, a energia em
excesso é armazenada na forma de gordura, a qual é metabolizada na ausência de
carboidratos para manter o funcionamento do organismo de maneira estável
(BORGES, 2011).
A etiologia da obesidade é multifatorial, envolvendo interações complexas
entre fatores genéticos, endócrinos e diversas condições ambientais desfavoráveis
à saúde, como hábitos alimentares inadequados e o sedentarismo. (MARTINEZ et
al., 2008) A obesidade é classificada como doença de caráter plurimetabólico por
coexistir com agravos como a hipertensão, dislipidemia e diabetes. Além disso,
está associada a fatores culturais e ambientais, apresentando impacto significativo
na saúde, no bem-estar psicossocial, na longevidade e na qualidade de vida.
(ESKINAZI et al., 2011)
O excesso de peso é um fator de risco conhecido para o desenvolvimento
de enfermidades crônicas não transmissíveis, como câncer, diabetes mellitus e
doenças cardiovasculares. Tais condições são responsáveis por grande parte das
despesas econômicas no sistema de saúde do mundo. De acordo com Bahia et al.
(2012), os custos de uma doença podem ser mensurados pelo impacto gerado no
sistema de saúde com os tratamentos (custos diretos) e por meio da queda na
qualidade de vida e no absenteísmo (custos indiretos). Estima-se que cerca de 2 a
6% dos gastos com saúde no mundo sejam decorrentes das comorbidades
envolvidas na obesidade. (BAHIA et al., 2012; TRAYHURN, 2013).
Atualmente, cerca de 80 milhões de brasileiros em idade adulta estão acima
do peso e um quinto se encontra obesa. 72% dos óbitos no país se referem às
doenças crônicas relacionadas à obesidade. A população com idade entre 35 a 64
anos é a que apresenta maior prevalência de obesidade no Brasil, e desse escopo,
o excesso de peso é maior observado nas mulheres. No mundo, atualmente 614

15
milhões de pessoas estão obesas e as estimativas sugerem que um quinto da
população global apresentará quadro de obesidade até o ano de 2025. (BRASIL,
2015; EZZATI, 2016)
A Organização Mundial da Saúde (OMS) classifica a obesidade de acordo
com o Índice de Massa Corporal (IMC), cujo cálculo se dá pelo valor do peso
corporal, em quilogramas, dividido pelo quadrado da altura, em metros quadrados,
conforme demonstrado na Tabela 1 (WHO, 2000).
Tabela 1. Classificação de peso de acordo com IMC e risco de comorbidades.
Classificação IMC (Kg/m2) Risco de comorbidades
Baixo peso < 18,5 Baixo
Peso normal 18,5 a 24,9 Médio
Pré-obeso 25,0 a 29,9 Aumentado
Obesidade grau I 30,0 a 34,9 Moderado
Obesidade grau II 35,0 a 39,0 Grave
Obesidade grau III ou mórbida ≥ 40,0 Muito grave
Fonte: Adaptado de WHO, 2000.
Apesar de ser um bom indicador, o IMC isolado não é totalmente
correlacionado com a gordura corporal. Dentre as suas limitações estão o fato de
ser incapaz de diferenciar massa gordurosa de massa magra, podendo ser pouco
estimado em indivíduos mais velhos e superestimado em indivíduos musculosos.
Além disso, este indicador não reflete a distribuição da gordura corporal, uma
variável importante na avaliação da obesidade devido ao fator de risco gerado pela
presença da gordura visceral. Dessa maneira, indivíduos com mesmo valor de IMC
podem apresentar proporções diferentes de gordura visceral, não se enquadrando,
portanto, no mesmo grau de risco. Para reduzir as limitações do indicador isolado,
a OMS sugere que o IMC seja utilizado de forma combinada com outras formas de
avaliação, como a medida da circunferência abdominal e a bioimpedância
(GODOY-MATOS et al., 2009; WHO, 2000).

16
2.2 Obesidade e o sistema imunológico
O estado de obesidade é caracterizado como uma “inflamação sistêmica de
baixo grau”, induzida por diversos mediadores inflamatórios, os quais foram
demonstrados pela primeira vez por Hotamisligil e seus colaboradores em 1993. O
tecido adiposo é considerado atualmente como um órgão endócrino, capaz de
mediar efeitos biológicos no metabolismo e inflamação, contribuir para a
manutenção da homeostase energética e, provavelmente, está relacionado com a
patogenia de disfunções metabólicas e inflamatórias relacionadas à obesidade
(HOTAMISLIGIL et al., 1993; WOZNIAK et al., 2009).
As causas e o mecanismo envolvido no estado inflamatório induzido na
obesidade ainda não são totalmente compreendidos. Grande parte dos estudos
estão centrados nas adipocinas, as quais são citocinas produzidas pelo tecido
adiposo com função imunológica, tais como a interleucina 6 (IL-6), o fator de
necrose tumoral α (TNF-α) e os fatores B, C3 e D (adipsina) do sistema
complemento. Algumas dessas moléculas possuem ação autócrina ou parácrina,
no entanto, podem também contribuir de forma significativa para a inflamação
sistêmica (BERG e SCHERER, 2005).
O TNF-α é uma citocina pró-inflamatória, produzida pelos tecidos muscular,
adiposo e linfoide. É capaz de reduzir a resposta à insulina por meio da diminuição
da expressão dos transportadores de glicose GLUT-4 na superfície celular,
fosforilação do substrato 1 nos receptores de insulina (IRS-1) e fosforilação
específica do receptor de insulina. Também é proposto que essa citocina possui
ação regulatória da massa de tecido adiposo, diminuindo a diferenciação de pré-
adipócitos pela indução da apoptose (in vitro) e da lipólise (in vitro e in vivo). O
TNF-α é comumente associado à insulino-resistência, sendo detectado em valores
elevados na obesidade, e reduzidos à medida que ocorre a perda de peso
(HOOPER et al., 2012).
O tecido adiposo (especialmente a gordura visceral), é a principal fonte de
IL-6 circulante nos estados não-inflamatórios. Há evidências de que a secreção de
IL-6 também ocorre na região do hipotálamo, o que denota um papel na regulação
do apetite e no gasto energético. Da mesma maneira que o TNF-α, a IL-6 está
relacionada com o metabolismo de lipídeos e da glicose, sendo também um
marcador para a resistência à insulina. Dentre suas principais funções, estão

17
envolvidas a inibição da lipoproteína lipase, a indução da lipólise e o aumento da
captação de glicose. Os níveis séricos de IL-6 estão aumentados na obesidade,
mas com a perda de peso, se encontram reduzidos (HERDER et al., 2013).
As moléculas do sistema complemento produzidas no tecido adiposo são o
fator B, fator C3 e fator D. Para que ocorra a síntese do fator C3a proveniente de
C3, é necessária a presença dos fatores B e D. O C3a é clivado posteriormente em
ASP (proteína estimulante de acilação). Tal proteína está diretamente relacionada
à síntese e armazenamento de triglicerídeos (TG). Experimentos em ratos
demonstraram que a sua deficiência está associada à diminuição da gordura
corporal e ao aumento da sensibilidade à insulina (HAVEL, 2004; XU et al., 2003).
2.3 Tecido adiposo e sua função endócrina
A leptina é um peptídeo fundamental na regulação do balanço energético.
Produzida quase que exclusivamente pelo tecido adiposo, é regulada por
alterações induzidas pela insulina no adipócito e seus níveis são estritamente
correlacionados com a massa desse tecido. Apresenta papel importante na
fertilidade, relacionada à liberação de gonadotrofinas e GnRH (hormônio liberador
de gonadotrofina), além de possuir ação moduladora da imunidade e da resposta
inflamatória (HOTAMISLIGIL, 2006; MANCO et al., 2007).
No hipotálamo, a leptina é capaz de controlar a liberação de neuropeptídios
anorexígenos. Como consequência, a resposta desencadeada pode determinar a
redução da energia adquirida, ou por uma via oposta, o aumento do gasto
energético. Perifericamente, essa molécula apresenta uma função importante na
redução da síntese e secreção de insulina (GREGOR e HOTAMISLIGIL, 2011).
É observado na obesidade que os níveis séricos elevados de leptina não
promovem a resposta esperada. É sugerido que a resistência à ação desse
hormônio esteja relacionada com a limitação do seu transporte a nível de barreira
hemato-encefálica. Tal fato poderá resultar na interrupção do eixo adipo-insular,
conduzindo ao hiperinsulinismo e ao diabetes mellitus tipo 2 associado à obesidade
(MITCHELL, et al., 2005).
A leptina não é considerada um fator de saciedade, no entanto, está ligada à
adaptação sob condições de baixa disponibilidade energética. Em humanos, a

18
mutação do gene da leptina ou de seu receptor está associado à obesidade
hiperfágica e à infertilidade (SMITH et al., 2007; ZHANG et al., 2009).
2.4 Microbiota intestinal humana
O intestino de uma pessoa adulta é colonizado principalmente por bactérias,
com um total de aproximadamente 100 trilhões delas e várias centenas de
espécies diferentes (RAJILIC-STOJANOVIC, 2013). A microbiota intestinal está
envolvida em diversos fatores biológicos intestinais, tais como a defesa contra
patógenos, imunidade, desenvolvimento dos microvilos intestinais e a degradação
de polissacarídeos complexos não-digeríveis. (HAN e LIN, 2014). Apesar de cada
indivíduo possuir uma microbiota diferente, o conjunto de genes expressos pelos
microrganismos intestinais (microbioma) é funcionalmente o mesmo para a grande
maioria dos indivíduos. As bactérias mais comuns que compõem a microbiota
intestinal são membros de dois filos: Bacteroidetes (Gram-negativos) e Firmicutes
(Gram-positivos), que juntos compõem mais de 90% do total das bactérias,
seguidos por Actinobacteria (Gram-positivos) e Proteobacteria (Gram-negativos)
(ESTEVE et al., 2011; MUSSO et al., 2010; SHEN et al., 2013).
A formação da microbiota se inicia ao nascimento e o padrão de colonização
será diferente de acordo com o modo de nascimento, uma vez que o bebê será
exposto primeiro à microbiota vaginal (como Lactobacillus e Prevotella) da mãe, se
for realizado um parto normal, ou à microbiota da pele (como Staphylococcus,
Corynebacterium, Propionibacterium), se for um parto por cesárea. O modo como o
recém-nascido é alimentado também influencia a composição da microbiota; com a
amamentação, observa-se grande quantidade de Bifidobacterium e pequena
quantidade de Clostridium e Bacteroides, enquanto que na ausência de
amamentação observa-se número elevado de Clostridium e Bacteroides. Durante a
infância, a microbiota será moldada de acordo com eventos como infecções,
exposição a antibióticos e, principalmente, alterações na dieta. Na primeira
infância, a microbiota do indivíduo é caracterizada por instabilidade e pouca
diversidade. Com o avançar da idade, ela se estabiliza e apresenta grande
diversidade (ALBENBERG e WU, 2014).
Boa parte da atividade metabólica microbiana é de grande relevância para o
hospedeiro. Seus efeitos benéficos envolvem o auxílio da digestão, síntese de

19
vitaminas, inibição de crescimento de patógenos e estímulo do sistema
imunológico. No entanto, também podem ser prejudiciais, como na síntese de
toxinas e compostos carcinogênicos, além da contribuição no desenvolvimento de
diarreia, constipação e infecções intestinais (RAJILIC-STOJANOVIC, 2013).
Vários estudos sugerem que a microbiota intestinal exibe um papel
importante no desenvolvimento de massa gorda e na homeostase energética
alterada. É importante para a compreensão do papel da microbiota intestinal na
manutenção da saúde do hospedeiro, o conceito de disbiose, o qual denota a
ruptura do equilíbrio entre as bactérias intestinais que promovem a saúde e
aquelas que não proporcionam benefícios ou que são prejudiciais para o
hospedeiro (SHEN et al., 2013).
As evidências a favor de um elo causal entre a microbiota e a obesidade
provêm a partir de modelos experimentais realizados em camundongos germ-free,
os quais demonstraram que os mesmos são resistentes à obesidade induzida por
dieta hipercalórica, principalmente devido à ausência de bactérias fermentadoras
capazes de processar polissacarídeos complexos, além da reduzida produção de
ácidos graxos de cadeia curta. A colonização de camundongos germ-free com
bactérias fermentadoras, juntamente com organismos que promovem processos
fermentativos, resultou em aumento de peso e obesidade. Tal estudo revelou que a
microbiota intestinal é um fator ambiental modulável, capaz de controlar o acúmulo
de gordura. Um dos mecanismos propostos sugere que a microbiota intestinal
otimiza o processo de extração da energia adquirida na dieta. Dessa maneira, a
habilidade de modular vias sinalizadoras do hospedeiro poderia influenciar no
balanço energético e no metabolismo (NICHOLSON, 2012).
Em condições patológicas, tais como obesidade e diabetes mellitus tipo 2, a
microbiota intestinal pode controlar o metabolismo do hospedeiro e contribuir para
o desenvolvimento de uma inflamação de baixo grau. É proposto que o
lipopolissacarídeo (LPS), derivado da microbiota intestinal, apresenta-se como uma
molécula crucial envolvida no desenvolvimento inicial da inflamação e das doenças
metabólicas. O LPS é uma poderosa molécula pró-inflamatória proveniente da
membrana externa de bactérias Gram-negativas e é continuamente liberada no
intestino do hospedeiro após a morte das mesmas (CANI et al., 2008; MUCCIOLI et
al.; 2010). O conceito de endotoxemia metabólica (níveis aumentados de LPS no
plasma) foi definido inicialmente em uma série de experimentos em camundongos.

20
A relação entre dieta hipercalórica, consumo de gorduras, obesidade, diabetes
mellitus e LPS foi posteriormente confirmada em diversos estudos realizados em
indivíduos humanos (AMAR et al., 2008; GHANIM et al., 2009).
2.5 Alterações na microbiota intestinal
Atualmente, a patologia da obesidade é reconhecida por estar associada a
alterações na diversidade e na composição da microbiota. Os primeiros estudos
que demonstraram os efeitos dessas mudanças foram realizados em camundongos
geneticamente obesos (genótipo ob/ob), os quais revelaram um aumento na
quantidade de Firmicutes e um decréscimo na proporção de Bacteroidetes, dois
filos dominantes da microbiota intestinal (LEY et al., 2005; TURNBAUGH et al.,
2006). Desde esses experimentos pioneiros, vários estudos caracterizaram a
microbiota intestinal em modelos animais obesos, com a maioria dos resultados
revelando um aumento na proporção de Firmicutes e decréscimo em Bacteroidetes
associado com a obesidade (HILDEBRANDT et al., 2009; MURPHY et al., 2010).
Em estudos que envolveram humanos, o aumento da razão
Firmicutes/Bacteroidetes em pacientes obesos ainda permanece em debate e
merece ser aprofundado. Um estudo foi capaz de indicar como a composição da
microbiota intestinal durante a infância é capaz de predizer a tendência para o
ganho de peso. Os autores identificaram níveis elevados de Staphylococcus aureus
e uma redução na quantidade de bifidobactérias nas amostras fecais de crianças
que estavam com sobrepeso (GEURTS et al., 2011; KALLIOMAKI et al, 2008;
TURNBAUGH et al., 2009)
É bem consolidado na literatura que alterações na composição e na
funcionalidade da microbiota intestinal geram um impacto na homeostase do
indivíduo. No entanto, diversas questões ainda necessitam de respostas e devem
ser debatidas. Ainda não há evidências claras se as alterações da microbiota
intestinal estão associadas com a dieta ou com a própria patologia da obesidade.
Além disso, muito se discute se as alterações da microbiota intestinal são a causa
ou a consequência da patogênese da obesidade (BACKHED, 2009)
Uma das principais funções do intestino é permitir a relação simbiótica entre
a microbiota intestinal e o hospedeiro, enquanto previne a invasão no tecido
hospedeiro. No entanto, disfunções na barreira intestinal podem representar uma

21
porta de entrada para microrganismos ou para moléculas derivadas de bactérias
oriundas do lúmen intestinal, como LPS, peptideoglicanos e flagelina. É sugerido
que alterações na permeabilidade da barreira intestinal poderiam ser responsáveis
pela endotoxemia metabólica presente em obesos (EVERARD et al., 2011). O
mecanismo envolvido nessas alterações está intimamente ligado à permeabilidade
paracelular, a qual é regulada por complexos multi-proteicos capazes de promover
a ligação entre as células epiteliais. Estudos envolvendo camundongos obesos
demonstraram que o aumento na permeabilidade intestinal pode estar associado
com a alteração na expressão, localização e distribuição de duas proteínas
específicas, ocludina e zonula occludens 1 (ZO-1), no intestino delgado. Além
disso, é relatado que alterações na microbiota intestinal pelo uso de antibióticos
aumentou a expressão de mRNA da ZO-1, reduzindo a permeabilidade intestinal e,
consequentemente, a endotoxemia metabólica em camundongos com obesidade
induzida por dieta hipercalórica (CANI et al., 2009; DE LA SERRE et al., 2010;
EVERARD et al., 2011).
Foi proposto em estudos recentes que o sistema endocanabinóide estaria
envolvido na regulação da barreira intestinal durante a obesidade. Esse sistema é
composto por lipídios endógenos bioativos que exercem a maioria das suas
funções pela ativação de dois receptores acoplados à proteína G, denominados
receptores canabinóides 1 (CB1) e 2 (CB2). Ambos são expressos através do trato
gastrointestinal em vários níveis, de acordo com o segmento. Antagonistas de CB1
diminuem a permeabilidade intestinal e a endotoxemia metabólica em
camundongos obesos através do aumento da distribuição e localização das
proteínas de junção ZO-1 e ocludina (ALHOUAYEK e MUCCIOLI, 2012). Em
estudos com animais, demonstrou-se que a microbiota intestinal também pode
regular o CB1, pois sua alteração com uso de prebióticos resultou em redução da
permeabilidade intestinal (EVERARD e CANI, 2013; HAN e LIN, 2014).
2.6 Abordagens de técnicas independentes de cultivo
A descoberta da sub-unidade do RNA ribossomal, o rRNA 16S, foi um marco
decisivo na evolução das metodologias de análise das comunidades bacterianas.
Essa molécula contém regiões de sequências de bases de nucleotídeos altamente
conservadas no domínio bacteriano, intercaladas por regiões variáveis e altamente

22
variáveis. A partir desse conceito, métodos moleculares para avaliação da
microbiota fecal foram desenvolvidos, nos quais os microrganismos constituintes de
uma amostra fecal são identificados a partir de seu material genético, independente
de serem cultiváveis ou não (TANNOCK, 2005).
Podem ser empregados diversos métodos para a análise filogenética da
microbiota intestinal. O padrão-ouro para análises de filogenia da microbiota
intestinal humana é baseado na construção de bibliotecas genômicas. Para tal, é
necessária a amplificação do gene rRNA 16S, seguindo-se da clonagem desse
produto de amplificação e posterior sequenciamento junto à análise dos clones
resultantes (ZOETENDAL et al., 2011).
O FISH (Hibridização Fluorescente In Situ) é um método no qual são
utilizadas sondas de oligonucleotídeos marcadas com um componente fluorescente
que hibridizam com o material genético presente na amostra. Dessa maneira,
microrganismos marcados na amostra podem ser identificados e quantificados
(FAVIER et al., 2002; ZOETENDAL et al., 2011).
Uma técnica que também tem sido utilizada para a quantificação de
diferentes grupos filogenéticos é a qPCR (ou Reação em Cadeia da Polimerase
quantitativa). Tal método utiliza iniciadores e sondas grupo-específicos, o que
garante uma alta sensibilidade, capaz de identificar a presença de microrganismos
em quantidades inferiores às necessárias em outras metodologias convencionais
(ZOETENDAL et al., 2011).
Com a finalidade de elucidar a dinâmica da microbiota intestinal, foram
introduzidas algumas variações das técnicas moleculares. Uma delas é a
eletroforese em gel com gradiente desnaturante (DGGE), uma técnica capaz de
determinar, descrever e monitorar a diversidade da comunidade microbiana de
ambientes complexos, tais como solo, água e o intestino humano. Essa técnica tem
sido empregada para análises da estrutura e da evolução de comunidades
bacterianas, a fim de determinar sua dinâmica em resposta a variações ambientais
(ERCOLINI, 2004).
A técnica de DGGE é um método de fingerprinting recente, no qual
fragmentos de DNA amplificados por PCR (amplicons) são separados de acordo
com suas sequências de pares de bases. Tem sido amplamente empregada em
estudos de microbiologia ambiental, microbiologia de alimentos e na análise de
comunidades microbianas que habitam o corpo humano. Anteriormente, estudos de

23
comunidade eram realizados primariamente por técnicas clássicas como cultura, ou
clonagem por PCR. A clonagem e o sequenciamento de amostras é um processo
altamente trabalhoso e caro de ser realizado, especialmente devido à grande
diversidade envolvida nesse tipo de estudo. Dessa maneira, técnicas de
fingerprinting genético como o DGGE, demonstram-se ideais para esta finalidade.
(BRAGA DA CRUZ, 2010; CARVALHO, 2012).
Quando aplicado para a análise de comunidades microbianas, o DGGE
assume o princípio de que moléculas de DNA dupla-fita de mesmo tamanho, mas
com diferentes sequências em pares de bases, podem ser parcialmente separadas
enquanto migram em um gel de poliacrilamida, contendo gradiente desnaturante
em concentração crescente e linear. Dessa forma, cada banda do DGGE
corresponde, teoricamente, a uma unidade taxonômica operacional (OTU), em que
o padrão de bandeamento total é capaz de refletir a riqueza e a diversidade das
espécies de uma comunidade. Quando combinada ao sequenciamento de clones
de rRNA 16S, a técnica de DGGE permite a determinação da taxonomia dos
membros de uma comunidade microbiana (FAVIER et al, 2003; CARVALHO,
2012).

24
3 OBJETIVOS
3.1 Objetivo geral
Avaliar comparativamente a diversidade bacteriana no ecossistema intestinal
de indivíduos eutróficos, com sobrepeso e obesos, os quais são
participantes do projeto “Diversidade bacteriana intestinal e parâmetros
nutricionais de indivíduos obesos, com sobrepeso e eutróficos” (DE PAULA,
2016).
3.2 Objetivos específicos
Obter o DNA representativo do metagenoma intestinal a partir de espécimes
fecais dos indivíduos participantes;
Avaliar a estrutura da comunidade bacteriana intestinal dos indivíduos pela
técnica de PCR seguida de eletroforese em gel com gradiente desnaturante
(PCR-DGGE) e agrupá-los de acordo com o perfil obtido;
Correlacionar os perfis eletroforéticos com as condições de indivíduos
eutróficos, com sobrepeso e obesos.

25
4. MATERIAIS E MÉTODOS
4.1 Seleção dos participantes do estudo
O presente estudo é parte de um projeto maior, denominado “Diversidade
bacteriana intestinal e parâmetros nutricionais de indivíduos obesos, com
sobrepeso e eutróficos”, o qual foi realizado nos anos de 2015 e 2016 no
Laboratório de Fisiologia e Genética Molecular Bacteriana da Universidade Federal
de Juiz de Fora. Tal projeto foi um estudo de caráter transversal, descritivo e
observacional, realizado com indivíduos de idade entre 18 e 60 anos e IMC a partir
de 18,5 kg/m², que foram selecionados na comunidade e no ambulatório do serviço
de nutrição do Hospital Universitário da Universidade Federal de Juiz de Fora.
Foram adotados os seguintes critérios de exclusão: histórico de doenças
intestinais, uso de antibióticos no último mês e diagnóstico confirmado de diabetes.
O estudo foi aprovado pelo Comitê de Ética em Pesquisa com Seres
Humanos da Universidade Federal de Juiz de Fora (Anexo A). Todos os voluntários
foram informados a respeito dos objetivos do estudo e assinaram o termo de
consentimento livre esclarecido (Anexo B).
4.2 Caracterização dos indivíduos
Para a avaliação antropométrica, foram utilizados peso atual (Kg), altura
(cm), medidas de índice de massa corporal (IMC), circunferências da cintura (CC),
abdominal (CA) e quadril (CQ) (Anexo C). O peso dos participantes foi aferido em
balança digital com o indivíduo em posição central, ereto, descalço, com os pés
juntos, e usando o mínimo de vestimentas possível. A altura foi verificada
utilizando-se um estadiômetro vertical fixo à balança, também descalço e ereto,
com calcanhares unidos, com a cabeça livre de adereços e olhando para o
horizonte. O IMC foi calculado a partir da relação peso (kg)/altura (m2) e avaliado
de acordo com o proposto pela Organização Mundial de Saúde (WHO, 2000).
Foram realizadas aferições das circunferências com fita métrica flexível e
inextensível de 150 cm de comprimento, estando no plano horizontal e no mesmo
nível em todas as partes, com o individuo em pé, ereto, braços estendidos ao longo
do corpo e sem comprimir os tecidos. A circunferência da cintura foi aferida a 2

26
dedos acima da cicatriz umbilical e com a roupa afastada, sendo analisada de
acordo com os pontos da Organização Mundial da Saúde (WHO, 2000), onde CC ≥
94 cm e ≥ 80 cm indica risco cardiovascular para homens e mulheres,
respectivamente. A circunferência abdominal foi medida na altura da cicatriz
umbilical, também com a roupa afastada e a circunferência do quadril foi aferida na
área de maior proeminência da região glútea. A relação cintura quadril (RCQ) foi
obtida por meio da divisão da circunferência da cintura pela circunferência do
quadril, sendo usado para análise com valores de corte considerados pela WHO
(1995), onde RCQ > 0,95 para homens e > 0,85 para mulheres são indicativos de
obesidade androide e risco aumentado de doenças metabólicas relacionadas com
a obesidade.
4.3 Coleta de amostras fecais e preparação para análises microbiológicas
As amostras fecais dos participantes foram coletadas em domicílio no início
da manhã, por demanda espontânea pelos participantes, em recipientes coletores
universais esterilizados e fornecidos pelos pesquisadores. Os recipientes com as
amostras fecais foram enviadas para o Laboratório de Fisiologia e Genética
Molecular Bacteriana do ICB/UFJF, onde foram aliquotadas. Para extração de DNA
metagenômico, foram pesados 200 mg de fezes, e armazenados em freezer à
temperatura de -20°C.
4.4 Extração de DNA a partir dos espécimes fecais
O DNA total foi extraído com a utilização do kit comercial QIAamp™ DNA
Stool Mini Kit e a plataforma automatizada Qiacube (Qiagen, Hilden, Alemanha), de
acordo com as instruções do fabricante e algumas alterações para aumento no
rendimento do DNA metagenômico. Inicialmente, três pérolas de vidro foram
adicionadas por amostra na etapa de lise celular e homogeneizadas em vortex
durante 2 minutos. Em seguida as amostras foram incubadas a 95°C, por 15
minutos, e subsequentemente agitadas em vortex, vigorosamente, por mais 2
minutos adicionais. Após esta etapa, 300 mL de tampão InhibiTex (fornecido no kit)
foram adicionados às suspensões fecais, seguido de homogeneização, incubação
e centrifugação. Posteriormente, o procedimento de obtenção dos extratos de DNA

27
prosseguiu na plataforma automatizada Qiacube, de acordo com o programa
específico para o QIAamp™ DNA Stool Mini Kit. O DNA representativo do
metagenoma intestinal foi eluído em volume de 200 µL e mantido em freezer a
-70ºC para estudos posteriores.
Para determinar a concentração e a pureza do DNA metagenômico, usou-se
o fluorímetro Qubit™ 2.0®, com o kit Qubit™ dsDNA HS Assay (Life Technologies,
California, USA), de acordo com as instruções do fabricante. A integridade do DNA
foi avaliada por eletroforese em gel de agarose a 0,8% em tampão TBE (Tris-HCl-
Borato-EDTA). O gel foi corado com brometo de etídio e analisado em
transiluminador de luz ultravioleta (GE Healthcare, Reino Unido). O DNA extraído
foi aliquotado e armazenado em freezer a -70°C.
4.5 Eletroforese em Gel com Gradiente Desnaturante (DGGE)
Com o intuito de avaliar a estrutura da comunidade bacteriana intestinal de
participantes obesos, com sobrepeso e eutróficos, foi usada a técnica de
eletroforese em gel com gradiente desnaturante.
4.5.1 Amplificação de rRNA 16S de representantes do domínio Bacteria
Os fragmentos do gene codificador do rRNA 16S de grupos microbianos
específicos representantes do domínio Bacteria foram amplificados por PCR.
Utilizou-se o DNA metagenômico extraído das amostras de fezes como molde para
as reações de amplificação.
Os primers (oligonucleotídeos iniciadores) utilizados nesta reação foram
F984GC (5’-gc.- AACGCGAAGAACCTTAC-3’) (gc = sequência rica em Guanina +
Citosina) (HEUER et al., 1997) e R1378 (5’-ACGGGCGGTGTGTACA -3’)
(BLACKWOOD et al., 2005) para amplificação da região V6 do DNA
correspondente ao rRNA 16S bacteriano. A reação de PCR apresentou
aproximadamente 20 ng/μL de DNA total, 0,5 μL de cada primer (10 μM) e como
mistura da reação foi utilizado o PCR Master Mix® (Promega, USA).
A PCR foi realizada contemplando-se as seguintes condições: temperatura
inicial de desnaturação a 94ºC por 5 minutos, seguida de 20 ciclos de 94ºC por 1

28
minuto para a desnaturação, 53ºC por 1 minuto para anelamento e 72ºC por 2
minutos para a extensão. O ciclo de amplificação foi seguido por uma extensão
final a 72ºC por 10 minutos (GELSOMINO; CACCO, 2006). Foi utilizado controle
negativo sem DNA molde. As reações de PCR foram realizadas no termociclador
Techne® TC-412. Os amplicons obtidos em cada reação foram visualizados em gel
de agarose (Sigma-Aldrich, USA) 1,2% em TBE (Tris-base, ácido bórico e EDTA), e
corados com brometo de etídio (Promega, USA). Para estimar o tamanho dos
amplicons, foi empregado o padrão de peso molecular de 100bp plus DNA ladder
ready-to-use (Bioron).
4.5.2 Condições da corrida eletroforética
Foi aplicado o volume de 20 μL dos produtos de PCR em gel de
poliacrilamida (acrilamida:N,N’-metilenobisacrilamida 37,5:1) vertical a 8% (p/v) em
tampão TAE (tris-acetato-EDTA) 1X. O gradiente desnaturante apresentou variação
linear de 30% a 70% utilizando-se como agentes desnaturantes ureia 7M, e
formamida deionizada 40% v/v, a partir de duas soluções estoque de poliacrilamida
a 0% (solução acrilamida/bisacrilamida 20% v/v, TAE 50X 2% v/v, água q.s.p) e
100% (solução acrilamida/bisacrilamida 20% v/v, TAE 50X 2% v/v, ureia 7M, e
formamida desionizada 40% v/v), as quais foram dispensadas pelo formador de
gradiente (Modelo 475 Gradient Delivery System – BIO-Rad Califórnia, USA). Além
das soluções estoque para a formação do gradiente, foram utilizados 0,03% (p/v)
de persulfato de amônio (polimerizador), 0,17% (v/v) de TEMED (N,N,N’,N’-
tetrametiletilenodiamino) (catalisador) e 50μL de corante (azul de bromofenol 0,5%,
xileno cianol 0,5% e TAE 1X) para caracterização visual do gradiente.
A eletroforese foi realizada em temperatura constante de 60ºC e voltagem
de 50V, durante 16 horas. O gel foi corado por 20 minutos com solução de SYBR®
Gold (Invitrogen™), de acordo com as recomendações do fabricante. A imagem do
gel foi visualizada no transiluminador ultravioleta ImageQuant 100 (GE Healthcare,
Reino Unido) e capturada com o auxílio da câmera ED LENS SP-500 UZ
(Olympus).

29
4.6 Análise estatística
Para análise dos dados foram utilizados os softwares de estatística SPSS
20.0, XLstat 2014 e PAST 3.0. Para as variáveis contínuas foram utilizadas
medidas de tendência central (média) e de dispersão (desvio padrão); para as
variáveis categóricas foi utilizada a distribuição percentual. Os dados obtidos foram
avaliados quanto ao tipo de distribuição, usando-se o teste de Kolmogorov-
Smirnov. Para comparações dos dados antropométricos, foi utilizado teste t de
Student, em que valores de p menores que 5% (p<0,05) foram considerados
significativos.
Por meio dos eletroforegramas obtidos pela técnica de PCR-DGGE e com
auxilio do programa pyElph 1.4 para a delimitação das bandas, foi construída uma
matriz binária, na qual a presença de banda correspondente a cada unidade
taxonômica operacional (OTU) foi codificada como “1” e ausência como “0”. A partir
disso, realizou-se uma análise de agrupamento. A estrutura da comunidade
microbiana foi avaliada de acordo com o cálculo do coeficiente de Jaccard de
similaridade e pela técnica Unweighted Pair Group Method with Arithmetic mean
(UPGMA) na análise de agrupamentos.

30
5 RESULTADOS
5.1 Características epidemiológicas e clínicas dos participantes
Após classificação dos indivíduos estudados nos grupos correspondentes
(eutrófico, com sobrepeso e obeso), foram avaliadas as características
demográficas. Dos 72 indivíduos amostrados, 63,9% eram do sexo feminino e
36,1% do sexo masculino, com uma média de idade de 39,6 anos, representativo
de indivíduos adultos. Os dados de gênero e idade por grupo são apresentados na
Tabela 2.
Tabela 2. Sexo e idade dos participantes deste estudo.
Eutrófico Sobrepeso Obeso
Masculino (%) 20,8 50,0 37.5
Feminino (%) 79,2 50,0 62.5
Média Idade 37,9 38,1 42,8
Os dados antropométricos e valores de p para teste de normalidade e
comparação de médias encontram-se na Tabela 3. Os valores de circunferência da
cintura, circunferência abdominal, circunferência do quadril e relação cintura quadril
foram analisados separadamente por sexo dentro dos grupos, uma vez que os
valores de referência são diferentes. Houve diferença estatística significativa em
todos os parâmetros analisados entre os três grupos em indivíduos do sexo
feminino, com um aumento nos valores proporcionalmente ao IMC. Já nos
indivíduos do sexo masculino não houve diferença estatisticamente significativa
nas medidas de circunferência da cintura e relação cintura–quadril entre nenhum
dos três grupos.
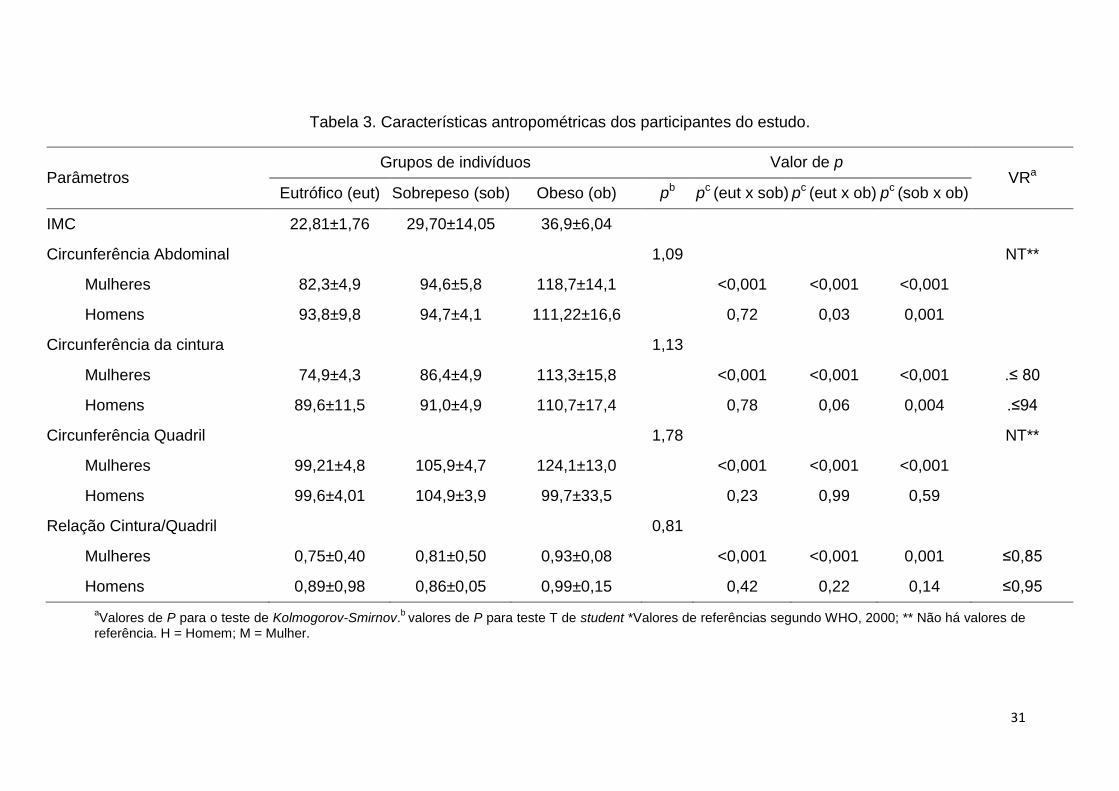
31
Tabela 3. Características antropométricas dos participantes do estudo.
Parâmetros Grupos de indivíduos Valor de p
VRa Eutrófico (eut) Sobrepeso (sob) Obeso (ob) pb pc (eut x sob) pc (eut x ob) pc (sob x ob)
IMC 22,81±1,76 29,70±14,05 36,9±6,04
Circunferência Abdominal 1,09 NT**
Mulheres 82,3±4,9 94,6±5,8 118,7±14,1 <0,001 <0,001 <0,001
Homens 93,8±9,8 94,7±4,1 111,22±16,6 0,72 0,03 0,001
Circunferência da cintura 1,13
Mulheres 74,9±4,3 86,4±4,9 113,3±15,8 <0,001 <0,001 <0,001 .≤ 80
Homens 89,6±11,5 91,0±4,9 110,7±17,4 0,78 0,06 0,004 .≤94
Circunferência Quadril 1,78 NT**
Mulheres 99,21±4,8 105,9±4,7 124,1±13,0 <0,001 <0,001 <0,001
Homens 99,6±4,01 104,9±3,9 99,7±33,5 0,23 0,99 0,59
Relação Cintura/Quadril 0,81
Mulheres 0,75±0,40 0,81±0,50 0,93±0,08 <0,001 <0,001 0,001 ≤0,85
Homens 0,89±0,98 0,86±0,05 0,99±0,15 0,42 0,22 0,14 ≤0,95
aValores de P para o teste de Kolmogorov-Smirnov.
b valores de P para teste T de student *Valores de referências segundo WHO, 2000; ** Não há valores de
referência. H = Homem; M = Mulher.

32
5.2 Avaliação da estrutura da comunidade bacteriana
Para amostrar a estrutura da comunidade bacteriana das amostras fecais
dos indivíduos, foi utilizado o perfil de bandeamento obtido pelo PCR-DGGE
(Figura 1). Considerou-se como OTU cada uma das bandas identificadas nos géis
e a estatística descritiva das mesmas pode ser observada na Tabela 4.
Tabela 4. Estatística descritiva dos resultados obtidos para o domínio Bacteria e
valores de p entre os grupos estudados.
Parâmetros
Domínio Bacteria Valor de p
Eutrófico
(eu)
Sobrepeso
(sob)
Obeso
(ob)
p (eu x sob)
p (eu x ob)
p (sob X ob)
Nº de Observações 24 24 24
Mínimo de bandas 1 3 1
Máximo de bandas 14 12 13
Média das bandas 5,96 5,41 6,87 0,53 0,36 0,07
Desvio Padrão 3,65 2,12 3,31

33
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Grupo de indivíduos eutróficos Grupo de indivíduos com sobrepeso Grupo de indivíduos obesosC C
Figura 1. DGGE fingerprint dos produtos de amplificação do DNA bacteriano correspondente à região V6 do gene codificador para o rRNA 16S a partir do
metagenoma fecal de indivíduos eutróficos, com sobrepeso e obesos. C = controle da eletroforese em gradiente de desnaturação contendo DNA de 4 espécies
bacterianas.

34
Foram identificadas 26 bandas distintas no total. Conforme demonstrado na
Figura 2, 15 dessas bandas estavam presentes nos três grupos de indivíduos
avaliados. 6 bandas pertenciam, simultaneamente, aos grupos eutrófico e com
sobrepeso, enquanto que 3 bandas foram detectadas exclusivamente no grupo
eutrófico e apenas 2 bandas únicas eram pertencentes ao grupo obeso.
Figura 2. Diagrama de Venn com a distribuição das bandas correspondentes à região V6 do rRNA
16S bacteriano, amplificadas a partir do DNA metagenômico extraído de amostras fecais de
indivíduos obesos, com sobrepeso e eutróficos.
Com a utilização de uma matriz de presença e ausência das bandas
detectadas, foi possível aplicar a técnica UPGMA, a qual agrupou os 72 indivíduos
participantes do estudo em 3 clusters: C1, C2 e C3, de acordo com o perfil de
bandeamento do DGGE (Figura 3). Um dendrograma de similaridade foi gerado
para visualizar a distribuição hierárquica dos 3 grupos (Figura 4).

35
C3: n = 20 indivíduos; 27,8%
C2: n = 8 indivíduos; 11,0%
C1: n = 44 indivíduos; 61,2%
C1
C2
C3
•Eutróficos: n=8; 18,5%•Com sobrepeso: n=16; 36,0%•Obesos: n=20; 45,5%
•Eutróficos: n=4; 50,0%•Com sobrepeso: n=3; 37,5%•Obesos: n=1; 12,5%
•Eutróficos: n=12; 60,0%•Com sobrepeso: n=5; 25,0%•Obesos: n=3; 15,0%
C1 + C2 + C3 => 72 indivíduos n = 24 Eutróficos n = 24 Com sobrepeson = 24 Obesos
Figura 3. Agrupamento dos indivíduos participantes desse estudo (eutróficos, com sobrepeso e
obesos) de acordo com o perfil de bandeamento correspondente à região V6 do rRNA 16S
bacteriano, amplificadas a partir do DNA metagenômico extraído de amostras fecais separadas por
DGGE.

36
C1
C2
C3
Figura 4. Matriz de similaridade resultante da análise de agrupamento UPGMA com coeficiente de
Jaccard, do fingerprint dos produtos de amplificação do DNA bacteriano, separados em DGGE,
correspondente à região V6 do gene codificador para o rRNA 16S a partir do metagenoma fecal de
indivíduos eutróficos, com sobrepeso e obesos. Bootstrap de 1000 réplicas.

37
Considerando-se o total de indivíduos em cada grupo, n=24, C1 é composto
por 83,3% dos indivíduos obesos avaliados, 66,6% dos indivíduos com sobrepeso
e 33,3% dos indivíduos eutróficos; C2 possui 4,2% dos obesos, 12,5% dos
indivíduos com sobrepeso e 16,7% dos eutróficos; enquanto que C3 apresenta
12,5% dos obesos, 20,8% dos indivíduos com sobrepeso e 50% dos eutróficos.
O cluster C1 é formado pela maioria dos participantes (44 indivíduos), sendo
20 obesos (45,5%), 16 com sobrepeso (36%) e 8 eutróficos (18,5%). Já o grupo
C2, de menor representatividade, é composto por 8 indivíduos, sendo 1 obeso
(12,5%), 3 com sobrepeso (37,5%) e 4 eutróficos (50%). O grupo C3 é composto
por 20 indivíduos, dos quais 3 são obesos (15%), 5 apresentam sobrepeso (25%) e
12 são eutróficos (60%).

38
6 DISCUSSÃO
A obesidade é uma pandemia associada a diversas comorbidades que
geram grande impacto no sistema de saúde. Recentemente, a relação entre a
microbiota intestinal humana e a obesidade tem sido abordada de diversas
maneiras em uma tentativa de elucidar as possíveis causas dessa enfermidade. Já
é consolidado na literatura o quão importante é o papel da microbiota intestinal na
influência sobre o processo saúde-doença.
No presente estudo, foi utilizada a técnica de PCR-DGGE para avaliar a
diversidade bacteriana da microbiota intestinal de 72 indivíduos, classificados como
obesos, com sobrepeso e eutróficos, de acordo com o IMC e as medidas
antropométricas. Nessa metodologia, a presença de bandas detectadas nos géis
permite inferir unidades taxonômicas operacionais (OTUs). No entanto, essa
técnica possui algumas limitações, como por exemplo, o tamanho dos fragmentos
não deve exceder 500pb para que possam ser bem separados na trama no gel de
poliacrilamida nas condições experimentais. Além disso, o DGGE dificilmente é
capaz de separar regiões caracterizadas apenas por pequenas variações de
sequência de aminoácidos (YU; MORRISON, 2004). A técnica detecta, a princípio,
apenas grupos ou espécies dominantes, porém, alguns relatos da literatura
demonstraram que a população detectada no gel pode não chegar a 1% da
população total. (FROMIN et al., 2002). A seleção de determinada região variável
(V) utilizada na amplificação e o método de extração do DNA, também são capazes
de influenciar na detecção das bandas (YU; MORRISON, 2004). Ainda assim, a
técnica de DGGE pode ser aplicada com êxito a uma grande variedade de estudos
de ecossistemas microbianos devido ao fato de permitir a visualização e a
comparação de perfis de comunidades microbianas com alta reprodutibilidade
(FROMIN et al., 2002; WU et al., 2010)
A análise estatística do perfil de bandeamento fornecido pelo PCR-DGGE
permitiu que os indivíduos fossem agrupados em três clusters, de acordo com a
similaridade entre os mesmos. A presença de um cluster dominante, composto em
sua maioria por indivíduos obesos e com sobrepeso, sugere um perfil de OTUs
predominantes na microbiota intestinal desses indivíduos. Da mesma maneira, um
segundo cluster formado majoritariamente por indivíduos eutróficos associados a
indivíduos com sobrepeso, indicam um outro perfil de OTUs característicos. No

39
terceiro grupamento, de menor representatividade, foi observada uma estrutura de
comunidade bacteriana intermediária quando comparada aos outros dois clusters.
Tal fato pode sugerir que os indivíduos com sobrepeso apresentam uma microbiota
intestinal em estágio de transição.
Quanto ao número de bandas avaliadas, não foi observada diferença
estatística significativa entre os três grupos, indicando que a obesidade está
relacionada com alterações na microbiota intestinal como um todo, e não com a
ação de um determinado grupo microbiano específico, conforme já demonstrado
por Wu e seus colaboradores (2010). A técnica de PCR-DGGE não apresenta a
sensibilidade necessária para determinar a diversidade total de uma amostra, no
entanto, a detecção de grupos dominantes é uma medida relativa que pode ser
utilizada para fins de comparação da diversidade entre ecossistemas microbianos.
Nesse estudo foi possível demonstrar que a microbiota intestinal de pessoas
obesas é diferenciada de indivíduos eutróficos, o que corrobora os achados da
literatura. No entanto, para descrever a maneira como ocorre essa diferença e a
forma como a microbiota pode ser modulada, novas abordagens seriam
necessárias, como o uso de metodologias complementares e a ampliação do
universo amostral.

40
7 CONCLUSÕES
A obesidade é uma enfermidade de relevância clínica mundial e a microbiota
intestinal humana é um importante alvo de estudos, principalmente devido
ao seu papel crucial na manutenção do equilíbrio energético;
Indivíduos obesos apresentam uma microbiota intestinal característica, que
se diferencia da microbiota de indivíduos eutróficos na população estudada.
Já os indivíduos com sobrepeso, no entanto, revelam uma microbiota em
caráter de transição entre eutróficos e obesos;
A técnica de PCR-DGGE apresenta limitações, como o tamanho limite de
500pb dos amplicons, o que dificultaria a sua resolução. O método também
se limita à detecção de apenas grupos dominantes, no entanto, pode ser
utilizada para comparação entre diferentes ecossistemas com alto grau de
reprodutibilidade. Sendo assim, novas abordagens metodológicas são
necessárias para que seja possível elucidar a correlação entre as alterações
nas comunidades microbianas intestinais e os mecanismos envolvidos no
processo da obesidade.

41
REFERÊNCIAS
ALBENBERG, L. G.; WU, G. D. Diet and the intestinal microbiome: associations,
functions, and implications for health and disease. Gastroenterology, v. 146, p.
1564–1572, 2014.
ALHOUAYEK, M.; MUCCIOLI, G. G. The endocannabinoid system in inflammatory
bowel diseases: from pathophysiology to therapeutic opportunity. Trends Mol Med,
v. 18, p. 615-25, 2012.
AMAR, J.; BURCELIN, R.; RUIDAVETS, J. B.; CANI, P. D.; FAUVEL, J.; ALESSI,
M. C.; Energy intake is associated with endotoxemia in apparently healthy men. Am
J Clin nutr, v. 87, p. 1219–23, 2008.
BACKHED, F.; Changes in intestinal microflora in obesity: cause or consequence?
J Pediatr Gastroenterol Nutr, v. 48, p. 56–7, 2009.
BAHIA, L.; COUTINHO, E. S. F.; BARUFALDI, L. A. ABREU, G. A. The costs of
overweight and obesity related diseases in the Brazilian public health system:
cross-sectional study. BMC Public Health. v. 12, n. 440, 2012.
BERG, A. H.; SCHERER, P. E. Adipose tissue, inflammation, and cardiovascular
disease. Circ Res, v. 96, n. 9, p. 939-949, 2005.
BLACKWOOD, C. B.; OAKS, A.; BUYER, J. S. Phylum- and class-specific PCR
primers for general microbial community analysis. Applied and Environmental
Microbiology, v. 71, n. 10, p. 6193-6198, out. 2005.
BORGES, N. Determinantes da epidemiologia mundial da obesidde. Cadernos de
saúde- número especial obesidade, v. 4, p.11-13, Universidade católica
Portuguesa, 2011.
BRASIL, Ministério da saúde. Vigitel Brasil 2014: vigilância de fatores de risco e
proteção para doenças crônicas por inquérito telefônico. Brasília, 2015.
CANI, P.D.; BIBILONI, R.; KNAUF, C.; WAGET, A.; NEYRINCK, A. M.;
DELZENNE, N. M. et al. Changes in gut microbiota control metabolic endotoxemia-
induced inflammation in high-fat diet-induced obesity and diabetes in mice.
Diabetes, v. 57, p. 1470–81, 2008.
CANI, P.D.; POSSEMIERS, S.; VAN DE, W. T.; GUIOT, Y.; EVERARD, A.;
ROTTIER, O. et al. Changes in gut microbiota control inflammation in obese mice
through a mechanism involving GLP-2-driven improvement of gut permeability. Gut,
v. 58, p. 1091-103, 2009.

42
CARVALHO, I. I. R. Análise da diversidade da microbiota fecal de crianças de
zero a doze meses de idade usando o método de eletroforese em gel com
gradiente desnaturante. 2012. Tese de Doutorado. Universidade de São Paulo.
Faculdade de Ciências Farmacêuticas
DE LA SERRE, C. B.; ELLIS, C. L.; LEE, J.; HARTMAN, A. L.; RUTLEDGE, J. C.;
RAYBOULD, H. E. Propensity to high-fat diet-induced obesity in rats is associated
with changes in the gut microbiota and gut inflammation. Am J Physiol
Gastrointest, Liver Physiol, v. 299, p. 440-8, 2010.
DE PAULA, T. O. Diversidade bacteriana intestinal e parâmetros nutricionais
de indivíduos obesos, com sobrepeso e eutróficos. Dissertação de Mestrado.
Universidade Federal de Juiz de Fora. Instituto de Ciências Biológicas. 2016
ERCOLINI, D. PCR-DGGE fingerprinting: novel strategies for detection of microbes
in food. Journal of microbiological methods, v. 56, n. 3, p. 297-314, mar. 2004.
ESKINAZI, F. M. V.; MARQUES, A. P. O.; LEAL, M. C. C.; DUQUE, A. M.
Envelhecimento e a epidemia da obesidade. Ciênc Biol Saúde, v. 13, p. 295-8,
2011.
ESTEVE, E.; RICART, W.; FERNANDEZ-REAL, J-M. Gut microbiota interactions
with obesity, insulin resistance and type 2 diabetes: did gut microbiote co-evolve
with insulin resistance? Current Opinion in Clinical Nutrition and Metabolic
Care, v.14, p. 483–490, 2011.
EVERARD, A.; CANI, P. D. Diabetes, obesity and gut microbiota. Best Practice &
Research Clinical Gastroenterology, v. 27, p. 73–83, 2013.
EVERARD, A.; LAZAREVIC, V.; DERRIEN, M.; GIRARD, M.; MUCCIOLI, G. M.;
NEYRINCK, A. M. et al. Responses of gut microbiota and glucose and lipid
metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice.
Diabetes, v. 60, p. 2775–86, 2011.
EZZATI, M. Trends in adult body-mass index in 200 countries from 1975 to 2014: a
pooled analysis of 1696 population-based measurement studies with 19,2 million
participants. Lancet , v. 387, p. 1377-96, 2016.
FAVIER, C. F.; DE VOS, W. M.; AKKERMANS, A. D. L. Development of bacterial
and bifidobacterial communities in feces of newborn babies. Anaerobe v. 9, n. 5, p.
219-229, 2003.
FAVIER, C. F.; VAUGHAN, E. E.; DE VOS, W. M.; AKKEMANS, A. D. L. Molecular
monitoring of succession of bacterial communities in humans neonates. Applied
and Enviromental Microbiology, v. 48, p. 219-226, 2002.

43
FROMIN, N.; HAMELIN, J.; TARNAWSKI, S.; ROESTI, D.; JOURDAIN‐MISEREZ,
K.; FORESTIER, N.; TEYSSIER‐CUVELLE, S.; GILLET, F.; ARAGNO, M.; ROSSI,
P. Statistical analysis of denaturing gel electrophoresis (DGGE) fingerprinting
patterns. Environmental Microbiology, v. 11, n. 4, p. 634-643, 2002.
GELSOMINO, A.; CACCO, G. Compositional shifts of bacterial groups in a
solarized and amended soil as determined by denaturing gradient gel
electrophoresis. Soil Biology & Biochemistry, v. 38, p. 91-102, jan. 2006.
GEURTS, L.; LAZAREVIC, V.; DERRIEN, M.; EVERARD, A.; VAN-ROYE, M.;
KNAUF, C. et al. Altered gut microbiota and endocannabinoid system tone in obese
and diabetic leptin-resistant mice: impact on apelin regulation in adipose tissue.
Front Microbiol, v. 2, p. 149, 2011.
GHANIM, H.; ABUAYSHEH, S.; SIA, C. L.; KORZENIEWSKI, K.; CHAUDHURI, A.;
FERNANDEZ-REAL, J. M. et al. Increase in plasma endotoxin concentrations and
the expression of Toll-like receptors and suppressor of cytokine signaling-3 in
mononuclear cells after a high-fat, high-carbohydrate meal: implications for insulin
resistance. Diabetes Care, v. 32, p. 2281–7, 2009.
GODOY-MATOS, A. F.; OLIVEIRA, J.; GUEDES, E. P.; CARRARO, L. Diretrizes
brasileiras de obesidade. Associação brasileira para o estudo da obesidade e
da síndrome metabólica, 3ª ed. São Paulo, 2009.
GREGOR, M. F.; HOTAMISLIGIL, G. S. Inflammatory Mechanisms in Obesity.
Annu. Reviews Immunology, v. 29, p. 415-445, jan. 2011.
HAN, J-L. H.; LIN, H-L. Intestinal microbiota and type 2 diabetes: from mechanism
insights to therapeutic perspective. World J Gastroenterol, v. 20, n. 47, p. 17737–
17745, 2014.
HAVEL, P.J.; Update on adipocyte hormones: regulation of energy balance and
carbohydrate/lipid metabolism. Diabetes, v. 53, p.43-51, 2004.
HERDER, C.; SCHNEITLER, S.; RATHMANN, W.; HAASTERT, B.; SCHNEITLER,
H.; WINKLER, H.; BREDAHL, R.; HAHNLOSER, E.; MARTIN, S. Low-grade
inflammation, obesity, and insulin resistance in adolescents. The Journal of
Clinical and Endocrinoly & Metabolism, v. 92, p. 4569-4574, jul. 2013.
HILDEBRANDT, M. A.; HOFFMANN, C.; SHERRILL-MIX, S. A.; KEILBAUGH, S.
A.; HAMADY, M.; CHEN, Y. Y. et al. High-fat diet determines the composition of the
murine gut microbiome independently of obesity. Gastroenterology, v. 137, p.
1716–24, 2009.

44
HOOPER, L. V.; LITTMAN, D. R.; MACPHERSON, A. J. Interactions between the
microbiota and the immune system. Science, v. 336, 2012.
HOTAMISLIGIL, G. S. Inflammation and metabolic disorders. Nature, v. 444, n.
1721, p. 860-867, dec. 2006.
HOTAMISLIGIL, G. S.; SHARGILL, N. S.; SPIEGELMAN, B. M. Adipose expression
of tumor necrosis factor-α: direct role in obesity-linked insulin resistance. Science,
v. 259, n. 5091, p. 87–91, 1993.
KALLIOMAKI, M.; COLLADO, M. C.; SALMINEN, S.; ISOLAURI, E. Early
differences in fecal microbiota composition in children may predict overweight. Am
J Clin Nutr, v. 87, p. 534–8, 2008.
LEY, R. E.; BACKHED, F.; TURNBAUGH, P.; LOZUPONE, C. A.; KNIGHT, R. D.;
Gordon, J. I. Obesity alters gut microbial ecology. Proc Natl Acad Sci USA, v. 102,
p. 11070–5, 2005.
MANCO, M.; FERNANDEZ-REAL, J. M.; EQUITANI, F.; VENDRELL, J.; VALERA-
MORA, M. E.; NANNI, G.; TONDOLO, V.; CALVANI, M.; RICART, W.;
CASTAGNET, M.; MINGRONE, G. Effect of massive weight loss on inflammatory
adipocytokines and the innate immune system in morbidly obese women. The
Journal of Clinical Endocrinology & Metabolism, v. 92, n. 2, p. 483-490, jul.
2007.
MARTINEZ, J. A.; PARRA, M. D.; SANTOS, J. L.; MORENO-ALIAGA, M. J.;
MARTI, A.; MARTINEZ-GONZALEZ, M. A. Genotype-dependent response to
energy-restricted diets in obese subjects: towards personalized nutrition. Asia Pac
J Clin Nutr, v. 17, p. 119-122, 2008.
MITCHELL, M.; ARMSTRONG, D. T.; ROBKER, R. L.; NORMAN, R.J. Adipokines:
implications for female fertility and obesity. Re-production, v. 130, n. 5, p. 583-597,
2005.
MUCCIOLI, G. G.; NASLAIN, D.; BACKHED, F.; REIGSTAD, C. S.; LAMBERT, D.
M.; DELZENNE, N.M. et al. The endocannabinoid system links gut microbiota to
adipogenesis. Mol Syst Biol, v. 6, p. 392, 2010.
MURPHY, E. F.; COTTER, P. D.; HEALY, S.; MARQUES, T. M.; O’SULLIVAN, O.;
FOUHY, F. et al. Composition and energy harvesting capacity of the gut microbiota:
relationship to diet, obesity and time in mouse models. Gut, v. 59, p. 1635-42, 2010.
MUSSO, G.; GAMBINO, R.; CASSADER, M. Obesity, Diabetes, and Gut
Microbiota. The hygiene hypothesis expanded? Diabetes Care, v. 33, p. 2277-
2284, 2010.

45
NICHOLSON, J. K.; HOLMES, E.; KINROSS, J.; BURCELIN, R.; GIBSON, G.; JIA,
W.; PETTERSSON, S. Host-gut microbiota metabolic interactions. Science, v. 336,
p. 1262–1267, 2012.
RAJILIC-STOJANOVIC, M. Function of the microbiota. Best Practice & Research
Clinical Gastroenterology, v. 27, p. 5–16, 2013.
SHEN, J.; OBIN, M. S.; ZHAO, L. The gut microbiota, obesity and insulin resistance.
Molecular Aspects of Medicine, v. 34, p. 39–58, 2013.
SMITH, A. G.; SHERIDAN, P. A.; HARP, J. B.; BECK, M. A. Diet-induced obese
mice have increased mortality and altered immune responses when infected with
influenza virus. Journal of Nutrition, v. 137, p. 1236-1243, may. 2007.
TANNOCK, G. W. New perspectives on the gut microbiota: implications for future
research. Gastroenterology clinics Am, v. 34, p. 361-382, 2005.
TRAYHURN, P. Hypoxia and adipose tissue function and dysfunction in obesity.
Physiology Reviews, v. 93. p. 1–21, Jan. 2013.
TURNBAUGH, P. J.; HAMADY, M.; YATSUNENKO, T.; CANTAREL, B. L.;
DUNCAN, A.; LEY, R. E. et al. A core gut microbiome in obese and lean twins.
Nature, v. 457, p. 480–4, 2009.
TURNBAUGH, P. J.; LEY, R. E.; MAHOWALD, M. A.; MAGRINI, V.; MARDIS, E.
R.; GORDON, J. I. An obesity-associated gut microbiome with increased capacity
for energy harvest. Nature, v. 444, p. 1027–31, 2006.
WORLD HEALTH ORGANIZATION. Obesity: preventing and managing the
global epidemic. Report of a WHO consultation on obesity. (WHO Technical
Report Series n. 894). Geneva, Switzerland: WHO, 2000.
WORLD HEALTH ORGANIZATION. WHO Expert Committee on Physical Status.
Physical status: the use and interpretation of anthropometry. Geneva: World
Health Organization, 1995.
WOZNIAK, S. E.; GEE, L. L.; WACHTEL, M. S.; FREZZA, E. E. Adipose tissue: the
new endocrine organ? A review article. Digestive Diseases and Sciences, v. 54,
n. 9, p. 1847–1856, 2009.
XU, H.; BARNES, G. T.; YANG, Q. et al. Chronic infIammation in fat plays a crucial
role in the development of obesity-related insulin resistance. J Clin Invest, v. 112,
n. 12, p. 1821-30, 2003.
YU, Z. MORRISON, M. Comparisons of different hypervariable regions of rrs genes
for use in fingerprinting of microbial communities by PCR-denaturing gradient gel

46
electrophoresis. Applied and environmental microbiology, v. 70, n. 8, p. 4800-
4806, apr. 2004.
ZHANG, N.; HUAN, Y.; HUANG, H.; SONG, G. M.; SUN, S. J.; SHEN, Z. F.
Atorvastatin improves insulin sensitivity in mice with obesity induced by
monosodium glutamate. Acta Pharmacologyca Sinica, v. 31, p. 35-42, Dec. 2009.
ZOETENDAL, E. G.; RAJILIC-STOJANOVIC, M.; DE VOS, W. M., High-throughput
diversity and functionality analysis of the gastrointestinal tract microbiota. Gut, v.
57, p. 1605-16, 2011.

47
ANEXO A - Parecer do comitê de ética em pesquisa - UFJF

48

49
ANEXO B - Termo de consentimento livre e esclarecido

50

51
ANEXO C – Ficha para dados antropométricos
Data
Peso atual
Altura
IMC/Classificação
CC
CA
CQ
Relação C/Q
IC







![2. Microbiologia - Aula 01 [Introdução à Microbiologia]](https://static.fdocumentos.tips/doc/165x107/55cf9336550346f57b9cd187/2-microbiologia-aula-01-introduaao-a-microbiologia.jpg)










