
UNIVERSIDADE FEDERAL DE GOIÁS PROGRAMA DE ......Entretanto este composto químico apresenta...
123
UNIVERSIDADE FEDERAL DE GOIÁS PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DA RELAÇÃO PARASITO-HOSPEDEIRO Amanda de Oliveira Melo Avaliação da toxicidade do cloridrato polihexametileno biguanida (PHMB) em Biomphalaria glabrata (Say 1818) Goiânia 2018
Transcript of UNIVERSIDADE FEDERAL DE GOIÁS PROGRAMA DE ......Entretanto este composto químico apresenta...

UNIVERSIDADE FEDERAL DE GOIÁS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DA
RELAÇÃO PARASITO-HOSPEDEIRO
Amanda de Oliveira Melo
Avaliação da toxicidade do cloridrato polihexametileno biguanida
(PHMB) em Biomphalaria glabrata (Say 1818)
Goiânia
2018


Amanda de Oliveira Melo
Avaliação da toxicidade do cloridrato polihexametileno biguanida
(PHMB) em Biomphalaria glabrata (Say 1818)
Dissertação de Mestrado apresentado ao
Programa de Pós-Graduação em Biologia da
Relação Parasito-Hospedeiro da Universidade
Federal de Goiás para obtenção do Título de
Mestre
Orientador: Prof. Dr. José Clecildo Barreto
Bezerra
Coorientador: Prof. Dr. Thiago Lopes
Rocha
Goiânia
2018



Programa de Pós-Graduação em Biologia da Relação Parasito-Hospedeiro da
Universidade Federal de Goiás
BANCA EXAMINADORA DA DISSERTAÇÃO DE MESTRADO
Aluna: Amanda de Oliveira Melo
Orientador: Prof. Dr. José Clecildo Barreto Bezerra
Coorientador: Prof. Dr. Thiago Lopes Rocha
Membros:
1. Prof. Dr. José Clecildo Barreto - UFG
2. Dra. Daniella de Sousa Mendes Moreira Alves - UFG
3. Profa. Dra. Luciana Damacena Silva - UEG
Data: 14/09/2018

A minha amada família, em
especial aos meus pais, minha
irmã e meu namorado.

AGRADECIMENTOS
Agradeço, primeiramente, a Deus por todas as conquistas concedidas a mim e a
toda minha família para o desenvolvimento deste trabalho ao qual eu me propus realizar.
Agradeço aos meus pais, Wilmar e Luciana, pelo carinho e amor. Vocês foram
meus exemplos de persistência, coragem, força e determinação, sempre me incentivando
a continuar e a buscar melhores resultados. A vocês todo o meu amor.
Agradeço a minha irmã Natália, uma pessoa iluminada que sempre me mostrou
que eu posso ir além.
Agradeço ao meu namorado Luiz Claudio, que me incentivou e me apoiou em
todas as minhas decisões, sempre me fazendo sorrir com sua alegria.
Agradeço a todos os meus familiares, em especial aos meus avós que me
motivaram com seus ensinamentos.
Agradeço ao meu orientador Prof. Dr. José Clecildo, pela confiança depositada
em meu trabalho, e por abrir as portas do seu laboratório a mim. Seus ensinamentos
colaboraram para minha formação durante o mestrado e eu os levarei para a vida toda.
Meus agradecimentos também se direcionam ao meu coorientador Prof. Dr.
Thiago, por ter me recebido tão bem, sempre disposto a me ajudar e a colaborar com a
minha pesquisa. Suas correções e instruções para o desenvolvimento deste trabalho foram
de grande valia. Obrigada pela dedicação!
Agradeço imensamente aos colaboradores deste trabalho, Profa. Dra. Luciana
Damacena Silva, Profa. Dra. Daniela Melo, Dra. Juliana Avelar, Ms. Fabrício Moreira e
Me. Daniela Braz por toda a ajuda para o desenvolvimento deste trabalho e pelos ricos
ensinamentos que recebi.
Agradeço também aos amigos do Laboratório de Estudos da Relação Parasito
Hospedeiro da Universidade Federal de Goiás, Ms. Guaraciara, Ms. Nayana, Geisa, Dra.
Daniella Sousa, Me. Thaynara, Ms. Heloísa, Pedro Eugênio, Paulo, Ms. Francesca, Ms.
Jaqueline, Dra. Caroline, Dr. Hanstter, Jade, Antônio, Daiane, Guiliane e Ms. Jéssica
pelos momentos de aprendizado.

Agradeço aos amigos e colegas do Laboratório de Biotecnologia Ambiental e
Ecotoxicologia da Universidade Federal de Goiás.
Agradeço aos pesquisadores e estudantes do Laboratório de Mutagênese da
Universidade Federal de Goiás, por me receberam tão bem, Dra. Fernanda Godoy, Dra
Wanessa Carvalho, Dr. Hugo Freire, Ms. Jhennefer Ramos, Ms. Thaís Alves e Ms. Alice
Tâmara.
Agradeço aos meus amigos da graduação Cárita Ribeiro, Leonardo Martins,
Déborah Carolina, Caroline Gusmão e Pedro Porto pelo apoio e incentivo durante o
mestrado.
Agradeço a secretaria e a coordenação do Programa de Pós-Graduação em
Biologia da Relação Parasito Hospedeiro.
Agradeço a Coordenação de Aperfeiçoamento de Pessoal de Nível Superior pela
concessão da bolsa, ao Instituto de Patologia Tropical e Saúde Pública e a Universidade
Federal de Goiás, por possibilitarem a realização deste trabalho.

SUMÁRIO
TABELAS E FIGURAS ............................................................................................... 12
SÍMBOLOS, SIGLAS E ABREVIATURAS ............................................................. 14
RESUMO ....................................................................................................................... 16
CAPÍTULO I .................................................................................................................. 1
1. INTRODUÇÃO / REVISÃO DE LITERATURA ................................................... 1
1.1 Histórico da esquistossomose ............................................................................ 1
1.1 Ciclo biológico de Schistosoma mansoni .......................................................... 4
1.2 Moluscos do gênero Biomphalaria .................................................................... 7
1.3 Características biológicas de Biomphalaria glabrata ........................................ 9
1.4 Distribuição geográfica de Biomphalaria glabrata no Brasil ......................... 14
1.5 Controle da esquistossomose ........................................................................... 15
1.6 Utilização de moluscicidas no controle da esquistossomose ........................... 16
1.7 Niclosamida ..................................................................................................... 18
1.8 Cloridrato polihexametileno biguanida (PHMB) ............................................ 20
1.8.1 Estrutura ................................................................................................... 20
1.8.2 Aplicações e benefícios ............................................................................ 21
1.9 Testes genotóxicos .......................................................................................... 22
1.9.1 O Ensaio cometa na avaliação de danos do DNA .................................... 23
1.9.2 Fundamentos do ensaio cometa ................................................................ 24
1.9.3 Ensaio cometa em Biomphalaria glabrata ............................................... 25
1.10 Teste do micronúcleo ....................................................................................... 26
2. JUSTIFICATIVA ..................................................................................................... 28
3. OBJETIVOS ............................................................................................................. 30
3.1 Objetivo Geral ...................................................................................................... 30
3.2 Objetivo Específico .............................................................................................. 30
CAPÍTULO II – ARTIGO I ........................................................................................ 32
1. Introduction ................................................................................................................. 35
2. Materials and methods ................................................................................................ 36
2.1 Test substance .................................................................................................. 36
2.2. Snails ................................................................................................................ 37
2.3. Snail embryotoxicity test ................................................................................. 37
2.4. Acute toxicity tests with new-borns and adult snails ....................................... 38

2.5. Behavioural assessment ................................................................................... 38
2.6 Statistical analysis ............................................................................................ 39
3. Results and discussion ................................................................................................ 39
3.1 Embryotoxicity ................................................................................................ 39
3.2 Toxicity to new-borns and adult snails ............................................................ 44
4. Conclusion .................................................................................................................. 48
References ...................................................................................................................... 50
CAPÍTULO III – ARTIGO II ..................................................................................... 58
1. Introduction ................................................................................................................. 61
2. Materials and Methods ............................................................................................... 62
2.1 Snails ................................................................................................................ 62
2.2 Substance test ................................................................................................... 63
2.3 Exposure .......................................................................................................... 63
2.4 Genotoxicity ..................................................................................................... 63
2.5 Mutagenicity .................................................................................................... 64
2.6 Statistical analysis ............................................................................................ 65
3. Results and discussion ................................................................................................ 65
3.1 Comet assay ..................................................................................................... 65
3.2 Micronucleus Test ............................................................................................ 69
4. Conclusion .................................................................................................................. 72
References ...................................................................................................................... 74
CAPÍTULO IV – CONSIDERAÇÕES FINAIS ........................................................ 82
ANEXOS ....................................................................................................................... 84
REFERÊNCIAS ........................................................................................................... 85

TABELAS E FIGURAS
Capítulo I
Figura 1 - Países e áreas de risco da esquistossomose no mundo ................................... 2
Figura 2 - Vermes adultos de Schistosoma mansoni ...................................................... 5
Figura 3 - Ciclo biológico das espécies de Schistosoma mansoni, Schistosoma japonicum
e Schitosoma haematobium ............................................................................................. 6
Figura 4 - Exemplar da espécie de Biomphalaria glabrata ........................................... 9
Figura 5 - Esquema geral do corpo de Biomphalaria glabrata ..................................... 10
Figura 6 - Massa ovígera de Biomphalaria glabrata com 12 horas de desenvolvimento
embrionário .................................................................................................................... 12
Figura 7 - Distribuição de Biomphalaria glabrata no Brasil ....................................... 14
Figura 8 - Países que utilizaram o controle de hospedeiros intermediários de Schistosoma
spp ................................................................................................................................. 17
Figura 9 - Fórmula molecular do composto 5-cloro-N-(2-cloro-4-nitrofenil)-
2hidroxibenzamida, Niclosamida, comercialmente chamada de Bayslucida® ............. 18
Figura 10 - Fórmula molecular do composto cloridrato polihexametileno biguanida
....................................................................................................................................... 20
Figura 11 - Imagem de nucleóide formando cometa com dano ao DNA ................... 24
Capitulo II
Figura 1 - Estádios iniciais de desenvolvimento de Biomphalaria glabrata ............. 38
Figura 2 - Frequência (%) dos estádios iniciais de desenvolvimento de Biomphalaria
glabrata após exposição ao cloridrato polihexametileno biguanida (PHMB) por 144 h
..................................................................................................................................... 40
Figura 3 - Taxa de eclosão (%) de Biomphalaria glabrata após exposição a cloridrato
polihexametileno biguanida (PHMB) por 144 h ....................................................... 41

Figura 4 - Alterações morfológicas em embriões de Biomphalaria glabrata após
exposição ao cloridrato polihexametileno biguanida (PHMB) por 144 h .................. 42
Figura 5 - Taxa de mortalidade (%) de embriões (A) de recém-nascidos (B) e adultos (C)
de Biomphalaria glabrata após exposição ao cloridrato polihexametileno biguanida
(PHMB) por 144 horas para embriões e 96 horas para recém-nascidos e adultos
..................................................................................................................................... 45
Figura 6 - Proporção de alterações no comportamento de Biomphalaria glabrata após a
exposição ao cloridrato polihexametileno biguanida (PHMB) por 96 horas
..................................................................................................................................... 47
Tabela 1 - Frequência (%) de alterações morfológicas nos estádios iniciais de
desenvolvimento de Biomphalaria glabrata após exposição ao cloridrato
polihexametileno biguanida (PHMB) por 144 h ....................................................... 43
Tabela 2 - Efeitos tóxicos do cloridrato polihexametileno biguanida (PHMB) nos
estádios iniciais de desenvolvimento, embriões, recém-nascidos e adultos de
Biomphalaria glabrata. Os resultados são apresentados como médias (limite inferior -
limites superiores) ..................................................................................................... 44
Capítulo III
Figura 1 - Danos no DNA expressos como % de DNA da cauda (média ± DP) em
Biomphalaria glabrata do controle e expostos ao cloridrato polihexametileno biguanida
(PHMB) durante 48 horas ......................................................................................... 66
Figura 2 - Imagem de ensaio cometa representativo em hemócitos de Biomphalaria
glabrata do controle e expostos ao cloridrato polihexametileno biguanida (PHMB), grupo
controle (A), 0,5 mg L-1 (B), 1,0 mg L-1 (C), 1,5 mg L-1 (D) ................................... 67
Figura 3 - Alterações nucleares de hemócitos de Biomphalaria glabrata do controle e
exposto ao cloridrato polihexametileno biguanida (PHMB) por 48 horas ............... 70
Figura 4 - Alterações nucleares de hemócitos observadas em Biomphalaria glabrata após
exposição ao PHMB ................................................................................................. 71
Tabela 1 - Frequência de hemócitos (%) distribuídos pelo grau de dano ao DNA em
Biomphalaria glabrata do controle e expostos ao cloridrato polihexametileno biguanida
(PHMB) por 48 horas ............................................................................................... 68

SÍMBOLOS, SIGLAS E ABREVIATURAS
ANOVA - Análise de Variância Multidirecional
AU - Unidade Arbitrária
BB - Núcleo Blebbed
BE - Núcleo Broken-eggs
BN - Binucleado
CL50 - Concentração Letal Mediana
DNA - Ácido Desoxirribonucleico
HNA - Alterações Nucleares Hemocitárias
IDH - Índice de Desenvolvimento Humano
IL - Interleucina
KN - Núcleo Kidney-shaped
LN - Núcleo Lobado
LOEC - Concentração do Menor Efeito Observado
MDA - Malondialdeído
MN - Micronúcleo
MOA - Mecanismo de Ação
NFKβ - Fator de Transcrição Nuclear β
NOEC - Concentração de Efeito Não Observado
NTD - Doenças Tropicais Negligenciadas
OCDE - Organização para a Cooperação e Desenvolvimento Econômico

OMS - Organização Mundial da Saúde
OTM - Moment Olive Tail
PHMB - Cloridrato Polihexametileno Biguanida
RNA - Ácido Ribonucleico
TL - Comprimento da Cauda
TNFα - Fator de Necrose Tumoral
WHO - World Health Organization

RESUMO
Os caramujos do gênero Biomphalaria atuam como primeiro hospedeiro no ciclo do
parasito Schistosoma mansoni, causador da esquistossomose no Brasil. O controle dessa
parasitose pode ser feito através do controle químico dos caramujos hospedeiros. A
Organização Mundial da Saúde (OMS), recomenda a niclosamida como agente
moluscicida. Entretanto este composto químico apresenta toxicidade à fauna e a flora
aquática, baixa solubilidade e alto custo, os quais demonstram a necessidade do
surgimento de novos compostos moluscicidas. O presente estudo propõe avaliar a
atividade moluscicida do cloridrato polihexametileno biguanida (PHMB) para o caramujo
B. glabrata. Os bioensaios foram realizados em três fases distintas do caramujo, sendo
elas embriões, recém-eclodidos e adultos, com as mesmas concentrações de PHMB
avaliadas: 0,6; 1,0; 1,6; 2,0; 2,6; 3,0 e 3,6 mg L-1, durante 144 e 96 horas respectivamente.
A avaliação da genotoxicidade e da mutagenicidade do PHMB foi realizada em hemócitos
de caramujos adultos através do ensaio cometa e do teste de micronúcleo e HNA
respectivamente, com as concentrações de 0,5; 1,0; 1,5 mg L-1 durante 48 horas de
exposição. O PHMB não impediu o desenvolvimento embrionário, mas impediu a eclosão
dos caramujos em concentrações acima de 1,6 mg L-1, resultando em mortalidade. A CL50
do PHMB para os embriões foi 0,98 mg L-1. Em caramujos recém-eclodidos e adultos, o
PHMB se demonstrou bastante tóxico, com CL50 de 1,43 e 1,49 mg L-1, respectivamente.
Os resultados apresentados pela avaliação da genotoxicidade e da mutagenicidade do
PHMB demonstraram que apenas as concentrações de 1,0 e 1,5 mg L-1 causaram efeito
genotóxico. E o PHMB na concentração de 1,0 mg L-1 durante 48 horas induziu a
formação de micronúcleos e alterações nucleares no hemócitos de B. glabrata. Este é o
primeiro estudo que avalia a atividade moluscicida do PHMB em B. glabrata. A
toxicidade do PHMB foi confirmada em todas as fases de B. glabrata analisadas, sendo
os embriões a fase mais sensível. Além disso o PHMB induziu genotoxicidade no
caramujo de forma não dependente da concentração, e apenas nas concentrações mais
altas este composto se mostrou mutagênico.
Palavras chave: esquistossomose, caramujo, moluscicida, ensaio cometa, teste do
micronúcleo, embriotoxicidade, biomarcadores.

ABSTRACT
The snails of the genus Biomphalaria act as the first host in the cycle of the parasite
Schistosoma mansoni, which causes schistosomiasis in Brazil. The control of this
parasitosis can be done through the chemical control of host snails. The World Health
Organization (WHO) recommends niclosamide as a molluscicidal agent. However, this
chemical presents toxicity to aquatic fauna and flora, low solubility and high cost, which
demonstrate the need for the appearance of new molluscicidal compounds. The present
study proposes to evaluate the molluscicidal activity of the polyhexamethylene biguanide
hydrochloride (PHMB) for the B. glabrata snail. The bioassays were performed in three
distinct stages of the snail, being embryos, new hatchlings and adults, with the same
PHMB concentrations evaluated: 0.6; 1.0; 1.6; 2.0; 2.6; 3.0 and 3.6 mg L-1, for 144 and
96 hours respectively. The genomic and mutagenicity evaluation of PHMB was
performed on adult sperm hemocytes through the comet assay and the micronucleus and
HNA test respectively, with concentrations of 0.5; 1.0; 1.5 mg L-1 for 48 hours of
exposure. PHMB did not prevent embryonic development, but prevented hatching of
snails at concentrations above 1.6 mg L-1, resulting in mortality. The LC50 of the PHMB
for the embryos was 0.98 mg L-1. In newly hatched snails and adults, PHMB was shown
to be quite toxic, with LC50 of 1.43 and 1.49 mg L-1, respectively. The results presented
by the PHMB genotoxicity and mutagenicity evaluation showed that only concentrations
of 1.0 and 1.5 mg L-1 caused genotoxic effect. The PHMB at the concentration of 1.0 mg
L-1 for 48 hours induced the formation of micronuclei and nuclear alterations in B.
glabrata hemocytes. This is the first study evaluating the molluscicidal activity of PHMB
in B. glabrata. The toxicity of PHMB was confirmed in all phases of B. glabrata
analyzed, the embryos being the most sensitive phase. In addition, PHMB induced
genotoxicity in the snail in a non-concentration-dependent manner, and only at the highest
concentrations did this compound prove to be mutagenic.
Key-words: schistosomiasis, snail, molluscicide, comet assay, micronucleus test,
embryotoxicity, biomarkers.

Capítulo I Revisão de Literatura

1
CAPÍTULO I
1. INTRODUÇÃO / REVISÃO DE LITERATURA
1.1 Histórico da esquistossomose
As esquistossomoses, popularmente conhecidas como bilharzíases ou barriga
d’água, são doenças parasitárias causadas por trematódeos do gênero Schistosoma que,
para o homem, tem como principais agentes etiológicos as espécies Schistosoma
haematobium, S. japonicum, S. mansoni. Na esquistossomose haematóbica, também
conhecida como esquistossomose urinária, ou geniturinária, o parasito S. haematobium
localiza-se de preferência no plexo vesical. Sua distribuição é predominante africana,
estendendo-se também a outras áreas da bacia do mediterrâneo. Os moluscos vetores são
espécies do gênero Bulinus. Já a S. japonicum é responsável pela esquistossomose
intestinal que ocorre ao Extremo Oriente e Pacífico Ocidental, onde se encontram os
hospedeiros intermediários do gênero Oncomelania. A espécie S. mansoni ocorre na
África, nas Antilhas e na América do Sul, onde determina uma infecção denominada
esquistossomose mansônica ou intestinal, pela localização dos parasitos nas vênulas da
parede do intestino grosso, sigmoide e reto. Sua distribuição geográfica está condicionada
pela distribuição de algumas espécies de moluscos de água doce, do gênero
Biomphalaria, hospedeiros intermediários de S. mansoni (Rey, 2008; Colley et al. 2014).
As primeiras observações sobre o agente etiológico da esquistossomose, foram
feitas em 1851 no Egito, pelo médico patologista alemão Theodor Bilharz. No Brasil a
história da esquistossomose teve início em 1866, quando em Berlim, o pesquisador
Griesinger enviou uma carta a Otto Wucherer, sugerindo-lhe verificar ovos de
Schistosoma haematobium, em pacientes com hematúria na urina (Paraense 2008).
Em 1902, Manson encontrou ovos de bilharzia, com espícula lateral por toda
massa fecal de alguns pacientes, e acreditou que havia duas espécies de bilharzia. No ano
de 1907 Sambon propôs a criação de uma nova espécie do parasito no homem, e
denominou-a de Schistosoma mansoni (Paraense 2008).

2
No ano de 1908, no Brasil, o cientista baiano Pirajá da Silva, encontrou um ovo
muito semelhante ao de S. mansoni, no sangue fresco de um adolescente nativo de
Salvador. Mas pouco se sabia sobre o ciclo do parasito e o hospedeiro intermediário ainda
era desconhecido. Porém Pirajá da Silva avançou em seus estudos, e concordou com
Manson e Sambon, que os ovos com espinho lateral indicavam uma terceira espécie de S.
mansoni.
Em 1916 Lutz, confirmou todos os resultados a que chegou Leiper, no Egito, sobre
o ciclo da esquistossomose, ou seja, confirmou a caracterização dos vermes adultos, além
disso reconheceu como hospedeiros intermediários a Biomphalaria glabrata (então
denominada de Planorbis divaceus e P. guadalculpensis) e a B. straminea (então P.
centimetralis), descrevendo as lesões principais nelas produzidas pelo parasito.
Descreveu a cercária, observando as condições de sua libertação do corpo do caramujo e
sua penetração em animais de laboratório. Confirmou a caracterização dos vermes adultos
feita por Pirajá da Silva e Leiper. E ocupou-se finalmente da infecção humana e
experimental sob aspectos sintomatológicos, patogênico, anatomopatológico, terapêutico
e profilática. (Paraense 2008).
Schistosoma mansoni foi introduzido as África para o hemisfério ocidental,
durante o tráfico de escravos. Quanto aos caramujos hospedeiros intermediários,
investigações sistemáticas moleculares têm indicado origem americana do gênero
Biomphalaria. Sobre a doença, a esquistossomose afeta cerca de 200 milhões de pessoas,
e mais de 600 milhões vivem em áreas sob o rico de contrair a doença (Figura 1). Além
disso cerca de 120 milhões de pessoas apresentam os sintomas da doença e 20 milhões
de pessoas apresentam a forma grave da doença. Atualmente a esquistossomose é
endêmica em 74 países, com destaque para África, América Latina e Ásia (WHO 2018).
Estima-se que pelo menos 91,4 % das pessoas que necessitam de tratamento para a doença
vivem na África (Tlamçani & Er-Rami 2014). Estudos recentes mostraram que que mais
de 200.000 mortes por ano são devidas à esquistossomose neste continente. Em países
como a Nigéria, a Tanzânia, Gana e Moçambique cerca de 76 milhões de pessoas estão
infectadas pela doença, sendo que 70 % dos casos apresentam as formas graves da doença,
com fibrose periportal, hepatomegalia e esplenomegalia (Adenowo et al. 2015).

3
Figura 1 – Países e áreas de risco da esquistossomose no mundo. Fonte World Health Organization 2014.
A esquistossomose se estabeleceu no Brasil através do tráfico de escravos vindos
do continente africano pela região litoral, mas expandiu-se amplamente dada as condições
climáticas favoráveis ao estabelecimento e reprodução do hospedeiro intermediário, bem
como as condições precárias de saneamento básico (Neves 2005). Abrange 19 Unidades
Federadas, com áreas endêmicas na região nordeste. Estima-se que, aproximadamente,
25 milhões de pessoas estejam expostas ao risco de contrair a doença, e que 2,5 a 6
milhões encontrem-se infectadas (Brasil 2010). A esquistossomose ocorre de forma
endêmica nos Estados: Alagoas, Bahia, Espirito Santo, Maranhão, Minas Gerais, Paraíba,
Pernambuco, Rio Grande do Norte e Sergipe. No Ceará, Distrito Federal, Goiás, Pará,
Paraná, Piauí, Rio de Janeiro, Rio Grande do Sul, Santa Catarina e São Paulo a
transmissão é focal (Brasil 2010; Domingues 2013). Vários estudos demonstraram que a
distribuição da esquistossomose não se faz de forma homogênea no Brasil, e nas regiões
consideradas endêmicas prevalece condições precárias e até mesmo inexistentes de
saneamento básico, pobreza e baixos níveis de escolaridade (Nascimento & Oliveira
2013).
A Organização Mundial da Saúde (OMS) caracteriza a esquistossomose com uma
Doença Tropical Negligenciada (NTD), uma classe de doenças que são consideradas
endêmicas em populações de baixa renda. Sua incidência está diretamente relacionada

4
com a fragilidade social e econômica da população, sendo associada com o baixo nível
de saneamento básico e acesso à educação sanitária desta população, comprovando a
estreita relação desta doença com o índice de desenvolvimento humano (IDH) (Lindoso
& Lindoso 2009; Abou-El-Naga 2015; Madinga et al. 2015).
Estudos demonstram que o financiamento mundial de inovação em doenças
negligenciadas não chega a 5 %, ainda que estas doenças estejam presentes em 149 países.
Isto porque a pesquisa desenvolvida em grandes indústrias farmacêuticas do setor privado
é sempre direcionada para a obtenção de lucro, e sendo esta categoria de doenças
endêmica em países subdesenvolvidos, o baixo poder aquisitivo dos pacientes não é capaz
de proporcionar um retorno financeiro exigido pelas grandes empresas (Brasil 2010;
Virmond 2010).
A esquistossomose é uma doença crônica e debilitante, pois compromete diversos
órgãos do corpo. Esta doença pode incapacitar os indivíduos que a manifestam e assim
como outras NTD, ela carrega um estigma social, fator este que dificulta ainda mais o
tratamento e a recuperação dos pacientes. A endemicidade da esquistossomose em uma
determinada região causa diversos impactos na população, já que esta doença
compromete a capacidade cognitiva, pode causar anemia, nanismo e atrapalha o
desenvolvimento social, em crianças e adultos jovens (Brasil 2010; Kealey 2010).
1.1 Ciclo biológico de Schistosoma mansoni
Os trematódeos do gênero Schistosoma são platelmintos, da classe Digenea e da
família Schistosomatidae. Algumas características tornam os representantes desse gênero
diferentes quando são comparados com os demais trematódeos. Primeiramente eles
apresentam sexos separados e bem definidos (dióicos), diferentemente dos outros
helmintos da classe que são hermafroditas. O macho adulto possui o corpo achatado e
largo, que é revestido por projeções em forma de espinhos, e apresenta uma fenda na
região anterior denominada um canal ginecóforo, onde a fêmea adulta se aloja. A fêmea
possui o corpo delgado e quase cilíndrico e de comprimento maior do que o macho
(Figura 2). Outra característica que os diferenciam, é que as fases larvais desse parasito,
completam parte do seu desenvolvimento em moluscos de água doce. Os ovos que
possuem formato elíptico, são liberados pelo ambiente externo embrionados (miracídios),
não são operculados e possuem um espículo lateral (Coley et al. 2014; Sah et al. 2015).

5
Figura 2 - Vermes adultos de Schistosoma mansoni. Fonte: Carvalho et al. 2008. Adaptado.
O ciclo biológico de S. mansoni é heteróxeno, composto por uma fase assexuada
que ocorre no hospedeiro intermediário, e outra sexuada que ocorre no hospedeiro
definitivo (Figura 3). O ciclo se inicia quando os ovos contendo miracídios são eliminados
pelo hospedeiro definitivo juntamente com as fezes. Esses ovos, ao entrarem em contato
com a água e em condições ambientais favoráveis de temperatura, luminosidade,
oxigenação e hipotonicidade, promovem a eclosão dos miracídios (fase livre nadante do
parasito), que por quimiotaxia, encontram caramujos do gênero Biomphalaria e penetram
pelos tecidos moles. Após 48 horas no interior do caramujo, os miracídios se transformam
em esporocistos primários, que através da poliembrionia se tornam esporocistos
secundários e depois cercárias. As cercárias (forma infectante para o hospedeiro
definitivo) possuem a cauda bifurcada e saem do caramujo através de estímulos de luz e
calor. As cercárias nadam e penetram ativamente na pele ou por qualquer região da pele
da conjuntiva ou da mucosa orofaringiana (regiões adequadas como porta de entrada) do
hospedeiro definitivo (homem, ou qualquer vertebrado que esteja na água). A infecção
via mucosas, pode ocorrer através dá ingestão de água que contém cercárias; estas não

6
chegam ao estômago, penetrando pelo esôfago (Barsoum et al. 2013; Colley et al. 2014;
Siqueira et al. 2017).
Figura 3 - Ciclo biológico de Schistosoma mansoni. Os vermes adultos que vivem nas vênulas de um
hospedeiro humano depositam os ovos (1), alguns dos quais ficam presos nos tecidos enquanto outros são
eliminados pelas fezes. Em contato com a água doce, eclodem os miracídios (2) presentes nos ovos livres
que infectam os caramujos do gênero Biomphalaria (3). Dentro do caramujo, os miracídios diferenciam-se
em esporocistos primários (4), posteriormente em esporocistos secundários (5). Cada esporocisto
secundário finalmente diferencia-se em cercárias (6) (forma infectante). As cercárias perdem suas caudas
durante a penetração da pele do hospedeiro definitivo (7) e tornam-se esquistossômulos (8). Estes migram
através da circulação sanguínea para as veias portais do fígado, onde se diferenciam e amadurecem em
vermes adultos (9). Os vermes machos e fêmeas adultos acasalam-se para completar o ciclo de vida. Fonte:
Pila et al. 2017, adaptado.
A penetração das cercárias é resultado da atividade lítica de suas glândulas de
penetração e pela ação mecânica promovida pelos seus movimentos vibratórios intensos
(Neves 2005; Caldas et al. 2008). Simultaneamente ao processo de penetração, ocorre a
perda da cauda bifurcada, e as larvas resultantes (esquistossômulos), migram através dos
tecidos subcutâneos, penetram em vasos sanguíneos e são levadas pela circulação para o
coração lado direito, depois pulmões, seguem pelas arteríolas pulmonares, pelos capilares
alveolares, chegando as veias pulmonares, e em seguida chegam ao lado esquerdo do
coração (Wynn & Cheever 1995; Barsoum et al. 2013). Seguindo o fluxo sanguíneo, os
esquistossômulos que chegam até o sistema porta intra-hepático onde podem completar

7
o seu desenvolvimento em aproximadamente 25-28 dias após a infecção, se diferenciando
em vermes adultos, machos e fêmeas. Há se então, o amadurecimento sexual dos machos
e o acasalamento, que é indispensável para que as fêmeas completem rapidamente eu
próprio desenvolvimento. A manutenção da fêmea no canal ginecóforo do macho, auxilia
na digestão do sangue e no transporte para os sítios de postura dos ovos. No entanto, como
observado em infecções unissexuais, o amadurecimento sexual do macho – com a
produção de espermatozoides – ocorre na ausência da fêmea (Lenzi et al. 2008).
Os vermes adultos migram, acasalados, para a veia mesentérica inferior e seus
ramos, alcançando muitos dos casais o plexo hemorroidário superior e áreas vizinhas onde
as fêmeas realizam a postura de, aproximadamente, 400 ovos por dia. Os ovos
depositados demoram cerca de 8 dias para se tornarem maduros, e aparecem nas fezes
após passarem pelos tecidos da mucosa intestinal. Os casais que mantém na circulação,
podem migrar para os espaços porta, levando à formação de granulomas, que são
caracterizados pela reação inflamatória aos antígenos do ovo, seguida da produção de
colágeno, podendo evoluir para um quadro de fibrose periportal hepática. A espícula
dorsal nos ovos de S. mansoni, provavelmente ajudam em sua retenção nos vasos. Uma
enzima produzida pelo miracídio difunde-se através da casca do ovo e ajuda a digerir o
tecido subjacente. A ação desta enzima, juntamente com necrose do tecido causado por
pressão e pelo efeito do espinho, ajuda a liberar o ovo dos tecidos p/ a luz do intestino
(Baptista & Andrade 2005; Wynn 2008; Andrade 2009; Colley et al. 2014; Souza 2015).
Os primeiros sintomas apresentados pela esquistossomose na fase aguda incluem
febre, distúrbios gastrointestinais e complicações pulmonares. Na fase crônica a doença
é caracterizada principalmente por inflamações hepáticas e esplênicas, com distensão
abdominal (Ross et al. 2007; Cantanhede et al. 2010).
1.2 Moluscos do gênero Biomphalaria
No Brasil, somente três, das onze espécies do gênero Biomphalaria são
encontradas naturalmente infectadas por S. mansoni, sendo elas Biomphalaria glabrata
(Say 1818), Biomphalaria straminea (Dunker 1848), Biomphalaria tenagophila
(D’orbigny 1835).
Os moluscos do gênero Biomphalaria encontram-se no filo Mollusca, classe
Gastropoda, subclasse Pulmonata, Ordem Basommatophora e família Planorbidae. Os

8
caramujos desse gênero possuem uma concha discoidal enrolada em espiral plana, a qual
apresenta um aprofundamento do giro central de ambos os lados, com coloração castanha
escuro, que pode variar dependendo de condições ambientais (Paraense 2008; Sanogo et
al. 2018).
O habitat desses moluscos são geralmente margens dos rios, lagos, lagoas, açudes,
pântanos, bueiros, brejos, canais de irrigação ou corpos d’água parados ou com baixa
correnteza e de pequena profundidade sobre temperatura de 20 º a 26 °C e pH de 7 a 8.
Com o passar dos anos observou-se uma ocorrência frequente deste hospedeiro
intermediário em áreas urbanas, que sofreram alguma alteração ambiental para satisfazer
as necessidades de consumo e lazer do ser humano. Em muitas regiões, principalmente
em cidades que possuem alguns bairros periféricos, encontra-se uma grande quantidade
de criadouros de caramujos em valas de hortas destinadas ao cultivo e provenientes de
drenagens fluviais (Tibiriça et al. 2011; Monteiro 2017). A alimentação desses
gastrópodes pode variar de acordo com o local onde se encontram e com a disponibilidade
deste alimento, apesar de serem considerados herbívoros, filtradores de plâncton,
predadores e até carnívoros em alguns casos, sua alimentação é quase que exclusivamente
vegetal (Barbosa 1995; Guimarães et al. 2009; Monteiro 2017).
Mesmo sendo hermafroditas, os caramujos desse gênero preferem a reprodução
por fecundação cruzada. A maturidade sexual é atingida em aproximadamente 35 a 50
dias. Os ovos são protegidos por uma substância gelatinosa e glicoproteica e depositados
em uma superfície, geralmente folhas de plantas, pedras ou até na concha de outros
caramujos. A eclosão embrionária ocorre em aproximadamente 7 – 9 dias (Paraense 1955;
Vianey-Liaud & Dussart 2002; Palasio et al. 2015). Dentre as três espécies do gênero
Biomphalaria, a espécie B. glabrata (Figura 4) é a principal espécie transmissora da
esquistossomose no Brasil, devido à sua ampla distribuição, altos índices de infecção,
maior susceptibilidade à infecção por S. mansoni (Boffi 1979). Este fato pode ser
comprovado ao analisar a distribuição de B. glabrata com as ocorrências da
esquistossomose, que coincidem sempre nas mesmas áreas (Giovanelli et al. 2001; Grault
2013; Filho 2016; Monteiro 2017).

9
Figura 4 - Biomphalaria glabrata. (ch) – concha; (gi) – giro interno ou apical; (pe) – pé; (te) – tentáculo.
Fonte: Laboratório de Estudos da Relação Parasito Hospedeiro – LAERPH/IPTSP/UFG.
1.3 Características biológicas de Biomphalaria glabrata
O caramujo B. glabrata adulto pode apresentar conchas com até 40 mm de
diâmetro, 11 mm de largura e com seis a sete giros, sinistrógiras aumentando
gradativamente o seu tamanho, enrolados em espiral plana sem opérculo (Souza &
Andrade 2006). A concha desses gastrópodes, que pode atingir o maior diâmetro e largura
dentre os representantes da família Planorbidae, em sua maioria das vezes possui
coloração marrom escuro, e é formada por várias camadas. A camada mais externa,
denominada de perióstraco, é constituída de material orgânico proteico, associado à
quinonas. As camadas internas são constituídas de cristais de carbonato de cálcio (Barnes
& Ruppert 1996; Bezerra 2005). Este órgão serve de proteção para as por partes moles,
divididas anatomicamente em: massa cefalopediosa e a massa visceral (Figura 4). Na
parte superior da massa cefalopediosa existem dois tentáculos longos e filiformes com os
olhos situados junto à base, que desenvolvem funções tátil. Abaixo dos tentáculos e acima
da boca encontra-se a mufla, que possui dois palpos labiais em forma de “T” quando vista
pela frente, sendo contornada pela mandíbula. Na parte inferior estão localizados os pés
alongados que são responsáveis pela locomoção, com o auxílio das glândulas podais que
secretam muco sobre o qual o caramujo prende-se ao substrato. É sobre essa estrutura que
ocorre o dobramento da epiderme, conhecida como manto, estrutura responsável pela

10
secreção de carbonato de cálcio, contribuindo assim para a formação da concha (Souza
& Andrade 2006; Paraense et al. 2008).
O sistema digestivo dos caramujos da espécie B. glabrata é formado pela boca,
cavidade bucal, esôfago, estômago, intestino, reto e ânus (Figura 5). A boca possui
espessamentos cuticulares denominados mandíbula e rádula. A rádula é uma fita alongada
e revestida de numerosos dentículos quitinosos em forma de gancho, direcionados para o
interior da cavidade bucal. A rádula sendo um órgão para alimentação, possui função
primordial de raspagem e maceração dos alimentos. A digestão é extracelular ocorre no
estômago, como resultado da ação de enzimas provenientes das bolsas esofágicas e do
ceco digestivo. O intestino estende-se desde a extremidade anterior do estômago
atravessando a massa visceral, para se abrir via ânus, do lado direito do manto. O sistema
digestório se finaliza no reto, que tem por função a formação, compactação e
armazenamento das fezes (Barnes & Ruppert 1996; Moore 2003; Brasil 2014).
Figura 5 - Esquema geral do corpo de B. glabrata. (ms) Massa cefalopediosa; (p) pé; (mf) mufla; (te)
tentáculo; (om) abertura genital mascula; (cm) colar do manto; (ps) pseudobrânquias, (an) ânus; (pn)
pneumóstoma; (mc) músculo columelar; (cl) crista lateral (cl), (ct) crista retal; (rt) reto; (tr) tubo renal; (cp)
cavidade pulmonar; (ga) glándula do albumen; (ia) intestino anterior; (et) estômago; (im) intestino médio;
(ip) intestino posterior; (gd) glándula digestiva (ot) ovoteste. Fonte: Paraense 2008.

11
O sistema genital desses moluscos hermafroditas é formado pelo ovoteste, órgão
em forma de cachos que produz os gametas masculinos e femininos. Depois de formados,
os gametas saem dessa estrutura e são transportados através do ovispermiduto passando
pelas vesículas seminais onde ocorre a maturação de espermatozóides e posteriormente
para a glândula do álbumem que tem a função de envolver os ovos produzidos. A partir
desta glândula, o ovispermiduto se bifurca, diferenciando-se em partes masculina e
feminina, até desembocar em uma pequena bolsa denominada de carrefour, que recebe
secreção da glândula de albume, destinada a envolver o ovo (Brasil 2007; Amaral et al.
2008; Paraense 2008; Cantinha 2008).
Os ovos são envoltos por uma substância gelatinosa, rica em proteínas,
glicoproteínas, galactogênio, glicogênio e lipídeos que são produzidos pela glândula de
albúmen e são agrupados por uma membrana protetora que é denominada massa ovígera
(Figura 6) (Ribeiro-Paes et al. 1994; Rosa 2008; Baron et al. 2013).
O desenvolvimento embrionário do caramujo B. glabrata ocorre através de
sucessivas clivagens espirais (Camey & Verdonk 1970; Carvalho 2008). Um ovo maduro
de B. glabrata tem 100 µm de comprimento. O primeiro estádio do desenvolvimento é
conhecido como blástula, que ocorre aproximadamente entre a 10a e 23a hora após a
primeira clivagem. Neste estádio ocorrem apenas mitoses sem aumento de volume celular
(Kawano 1983; Kawano et al. 2008).
Cerca de 24 a 39 horas após a primeira clivagem do ovo, inicia a gastrulação, que
pode ser caracteriza pelo fim da clivagem e início do crescimento, diferenciação e
movimentação celular, dando origem ao segundo estádio do desenvolvimento conhecido
como gástrula. O tipo de gastrulação no B. glabrata ocorre por invaginação ou endobolia.
Na região onde se encontram os corpúsculos polares está localizado o polo animal e
vegetal, que começam a se transformar e o embrião modifica sua forma de arredondada
para achatada, quando observado em perfil (Kawano et al. 1992; Kawano et al. 2008).
O estádio trocófora ocorre entre 40a e 89a após a 1a clivagem é identificada como
a primeira fase larval de B. glabrata, que se caracteriza pela formação do prototroco
(região do corpo que separa em duas partes: a região pré-trocal e pós-trocal), que é
formado de duas fileiras de células situadas acima da boca, com cílios recobrindo toda
sua superfície. É nesta fase que ocorre o início da movimentação larval, que de início é

12
muito lenta e com o passar do tempo vai se tornando mais rápida (Camey & Verdonk
1970; Kawano et al. 2008).
Os estádios de véliger (120 horas) e hipoestádio (144 horas de idade) em B.
glabrata ocorrem ainda dentro da cápsula do ovo. O estádio de véliger se apresenta
caracterizado pela formação da concha, que começa a cobrir uma parte do corpo. O
prototroco evolui para o velum, que é um órgão responsável pela movimentação intensa
da larva no interior da cápsula. Há um desenvolvimento maior do pé, formação dos olhos,
aumento do tamanho dos tentáculos e início do enrolamento da concha sobre o corpo. O
hipoestádio caracteriza-se pela formação completa do embrião e início da eclosão
(Tallarico et al. 2004; Kawano et al. 2008)
Figura 6 – Massa ovígera de Biomphalaria glabrata com aproximadamente 12 horas após a postura. (me)
– membrana externa; (ov) – ovo; (em) – embrião; (sc) – substância coloidal; (sp) – substância perivitelínica.
Fonte: Laboratório de Estudos da Relação Parasito Hospedeiro – LAERPH/IPTSP/UFG.
Os caramujos da espécie B. glabrata possuem um sistema circulatório composto
por coração com duas cavidades: aurícula e ventrículo. O fluido que preenche o corpo do
caramujo é chamado de hemolinfa, que se compõe de células denominadas hemócitos,
plasma rico em água, bicarbonatos, cloreto de sódio e hemoglobina. A hemoglobina pode
utilizar o oxigênio à baixa tensão porque contém ferro dissolvido. A hemoglobina

13
constitui 97 % das proteínas presentes na hemolinfa de B. glabrata. (Verrengia-Guerrero
et al. 1997; Pessôa & Martins 2011). Os hemócitos são as principais células do sistema
de defesa do caramujo, podem estar livres, circulantes na hemolinfa ou nos tecidos. A
hemolinfa dos caramujos circula em um sistema do tipo semiaberto. Ela é impulsionada
pelo coração, de onde parte a artéria aorta, que se ramifica para diversos tecidos, drenando
nos seios venosos e retornando ao coração pelas veias pulmonar e renal, após ser
reoxigenada na parede pulmonar (Negrão-Corrêa et al. 2008). Os hemócitos, também
chamados de amebócitos, podem ser produzidos por vários tecidos hematopoiéticos. Em
B. glabrata, estas células provavelmente são produzidas por uma região bem definida
localizada entre o pericárdio e o epitélio posterior da cavidade do manto, também
designado de amebocyte producing organ (APO). Outros estudos demonstraram que a
porção sacular dos túbulos renais, margeando o pericárdio e, ocasionalmente, a região
dorsal do reto, também é capaz de produzir hemócitos (Knaap & Loker 1990; Negrão-
Corrêa et al. 2008; Pila et al. 2017).
Os hemócitos são considerados as principais células efetoras do sistema de defesa
dos moluscos, participando da fagocitose de partículas como, por exemplo, bactérias e
protozoários, e do processo de encapsulação de patógenos maiores, como larvas de
helmintos (Loker et al. 1986; Adema & Loker, 1997). Além disso, os hemócitos também
produzem fatores solúveis, que são lançados na hemolinfa ou estão expressos em sua
membrana, que participam da ativação dos próprios hemócitos e na destruição de agentes
patogênicos (Negrão-Corrêa et al. 2008).
Alguns autores consideram a existência de subpopulações de hemócitos
circulantes na hemolinfa de Gastropoda. Em geral, os pesquisadores se baseiam nas
características morfológicas, ultra estruturais e no conteúdo enzimático destas células
para fazer essa classificação. Em B. glabrata a maioria dos autores distingue duas
subpopulações de hemócitos circulantes: os granulócitos e os hialinócitos. Os
granulócitos são células que medem aproximadamente 7 a 11 µm de diâmetro,
apresentam um núcleo excêntrico, esférico, rico em grumos de cromatina fortemente
condensada (heterocromatina) e um nucléolo proeminente. O citoplasma é granulado e é
envolto por uma membrana bem definida, e apresenta uma delimitação acentuada entre o
endoplasma, que é rico em organelas, e o ectoplasma homogêneo. Estas células possuem
ainda numerosos prolongamentos citoplasmáticos, delgados e longos denominados
filopódios. Bayne et al. (1980) consideram que os granulócitos de Gastropoda, em geral,

14
são células com funções semelhantes aos macrófagos de mamíferos. Os hialinócitos são
menores, medindo de 4 µm a 8 µm de diâmetro, apresentam contorno circular, citoplasma
contendo raras organelas e pouco ou nenhum lisossomo e ausência de pseudópodes
extensos (Harris, 1975; Yoshino, 1976; Lie et al. 1987; Barraco et al. 1993; Borges &
Andrade, 2003; Negrão-Corrêa et al. 2008)
1.4 Distribuição geográfica de Biomphalaria glabrata no Brasil
A espécie B. glabrata é considerada a espécie de molusco mais importante na
transmissão da esquistossomose mansônica, em vista de sua ampla distribuição
geográfica, grande compatibilidade com o parasito e os altos índices de infecção (Scholte
et al. 2012; Souza et al. 2017). Alguns estudos já relataram a presença dessa espécie em
17 estados do país, compreendendo 801 municípios (Alagoas, Bahia, Espírito Santo,
Distrito Federal, Goiás, Maranhão, Minas Gerais, Pará, Paraíba, Paraná, Pernambuco,
Piauí, Rio de Janeiro, Rio Grande do Norte, Rio Grande do Sul, São Paulo e Sergipe)
(Figura 7) (Carvalho et al. 2008; Brasil 2014; Silva 2018).
Figura 7 - Distribuição de Biomphalaria glabrata no Brasil. Fonte: Brasil 2014.
O estabelecimento e a expansão territorial de populações de Biomphalaria spp são
comumente relacionados a regiões de bacias hidrográficas (Carvalho et al. 1985; Telles
1996; Fernandez et al. 2001; Telles 2005; Lofty 2009; El-Wakeil et al. 2013; Abou-el-
Naga 2015; Leite et al. 2016). Entretanto, outros estudos sugerem que a presença de uma

15
bacia hidrográfica não é o principal determinante da frequência de locais de reprodução,
uma vez que os estados da região sul apresentam baixa frequência de criadouros, apesar
de possuir um número considerável de bacias hidrográficas. Nestes locais a ocupação dos
caramujos está fortemente relacionada às características da ocupação humana na área
(David et al. 2018).
Outro fator que contribui significativamente para a distribuição geográfica do B.
glabrata são as chuvas que ocorrem nestas regiões citadas. Alguns estudos têm relatado
que um aumento na precipitação faz com que a água flua para fora dos locais de
reprodução, provocando um efeito de arrasto e dispersão destes moluscos, criando novos
focos destes hospedeiros em outras localidades (Giovanelli et al. 2001; Araújo et al. 2007;
Souza et al. 2010; Barbosa et al. 2015).
Este cenário demonstra que a distribuição de B. glabrata é extremamente
relacionada à ocorrência de esquistossomose no Brasil (Scholte et al. 2012; Filho 2016).
O conhecimento da distribuição das espécies de Biomphalaria hospedeiras naturais de S.
mansoni podem servir como uma importante ferramenta para o planejamento e seleção
das intervenções de controle da esquistossomose, direcionando ações e economizando
recursos (Stensgaard et al. 2013).
1.5 Controle da esquistossomose
Através da percepção de que a esquistossomose é um problema de saúde pública
que depende de vários fatores, controle desta doença sempre foi uma pauta importante
para a OMS que adotou diversas estratégias desde 1950. Ao longo da história o combate
a esquistossomose se baseou em duas medidas principais: tratamento do hospedeiro
definitivo (humano), com ações sobre o parasito e medidas visando o hospedeiro
intermediário (moluscos) para impedir a morbidade e reduzir a transmissão,
respectivamente (WHO 2013). Estas estratégias empregadas sempre variaram de acordo
com os recursos técnicos disponíveis bem como com as necessidades e disponibilidades
financeira de cada país (Aagaard-Hansen & Bruun 2008).
Na década de 50 um comitê de especialistas em esquistossomose criado pela OMS
recomendou a realização de estudos nas áreas de taxonomia, morfologia, suscetibilidade,
ecologia e fisiologia dos moluscos hospedeiros, e outros fatores socioeconômicos
associados à transmissão da doença (WHO 1953). Como resultado desses estudos, esse

16
mesmo comitê determinou que os programas de controle da esquistossomose agissem
principalmente no combate do hospedeiro intermediário, através do uso de moluscicidas,
já que os estudos demonstraram que os fármacos disponíveis não eram promissores no
combate de S. mansoni (WHO 1965a; WHO 1965b; Barbosa et al. 2008).
Em 1979, foi lançado a oxamniquina (Pfizer) e o praziquantel, fármacos que
possuíam alta eficácia, poucos efeitos colaterais e custo. Com isso impulsionou-se o uso
de quimioterápicos associados a agente moluscicidas, com a comprovação que o uso
simultâneo desses novos fármacos permitia resultados mais rápidos sobre a incidência,
prevalência e transmissão da doença (WHO 2002; Carvalho et al. 2008; Rapado 2013;
Filho 2016).
Portanto a OMS estabeleceu algumas diretrizes para o combate a
esquistossomose. O tratamento de grupos de risco com o Praziquantel se tornou a medida
principal (WHO 2006). Mas sozinha essa medida não é eficaz, por isso o controle da
doença deve se basear na cooperação de diversos setores pare integrar abordagens
preventivas e direcionadas à população. Dentre essas abordagens se destacam: estrutura
de saneamento básico, educação sanitária e combate ao caramujo hospedeiro
intermediário do ciclo do parasito (WHO 2006; Grimes et al. 2015; Othman & Soliman
2015).
1.6 Utilização de moluscicidas no controle da esquistossomose
Visto a necessidade de controle dos moluscos hospedeiros intermediários de S.
mansoni muitas metodologias foram utilizadas para a eliminação desses caramujos
durante os anos (Figura 8). As principais medidas adotadas se dividem em três categorias:
alterações no habitat, controle biológico e uso de agentes químicos (Teles & Carvalho
2008; Moraes 2011; King & Bertsch 2015).

17
Figura 8 - Países que utilizaram o controle de hospedeiros intermediários de Schistosoma spp. para o
controle da esquistossomose. Os destaques em verde escuro indicam as áreas onde foi feito o controle dos
hospedeiros intermediários de Schistosoma spp. A parte destacada com um aumento mostra os países da
bacia do Caribe (adaptado de King & Bertsch 2015).
Os primeiros registros do uso de agentes químicos como moluscicidas para
controle do caramujo hospedeiro intermediário da esquistossomose foi datado em 1918
no Japão, com o óxido de cálcio (Jordan & Rosenfield 1983). Dois anos depois, o sulfato
de cobre começou a ser utilizado no Egito, e em 1940 um composto denominado
cianamida cálcica começou a ser utilizada. Com o avanço dos estudos nesta área, vários
agentes químicos com potencial moluscicida surgiram como o pentaclorofenato de sódio
e a tritilmorfolina. Em 1959 a Niclosamida teve suas propriedades moluscicidas
descobertas e com os resultados satisfatórios apresentados, passou a ser o fármaco
recomendado pela OMS (Jordan & Rosenfield 1983; Tanaka & Tsuji 1997; Inobaya et al.
2014; Filho 2016).
Em 1960 o trifenmorph, um fármaco com muitos efeitos tóxicos, teve sua ação
moluscicida comprovada, porém seu uso era limitado. Depois disso outros agentes
químicos foram descobertos, como o yurimin e o phebrol (Duncan 1981; Jordan &
Rosenfield 1983; Tanaka & Tsuji 1997; Inobaya et al. 2014). Paralelo aos estudos de
agentes químicos, estudos utilizando moluscicidas de origem vegetal também foram
realizados (Santos et al. 2000; Cantanhede et al. 2010).

18
1.7 Niclosamida
Desde a década de 60 até os dias atuais, a niclosamida (Bayluscida®) é o único
moluscicida recomendado pela OMS, por ser comprovada sua alta atividade contra todas
as fases de vida do caramujo em concentrações inferiores a 1 mg L-1 após 8 horas de
exposição. Além de moluscicida, a niclosamida possui grande toxicidade sobre helmintos
da classe Cestoda e está contida nas preparações de Yomesan®, Mansonil® e Lintex ®,
utilizados na medicina humana e veterinária no tratamento da teníase. Sua apresentação
comercial tem peso molecular de 327,1 g/mol e fórmula química C13H8Cl2N2O4 (2′,5-
dichloro-4′nitrosalicylanilide) (Figura 9) (Andrews et al. 1983; Filho 2016).
Figura 9 - Fórmula molecular do composto 5-cloro-N-(2-cloro-4-nitrofenil)-2hidroxibenzamida,
Niclosamida, comercialmente chamada de Bayluscida® (adaptado de
https://pubchem.ncbi.nlm.nih.gov/compound/4477#section=Top).
O mecanismo de ação da niclosamida ainda não foi totalmente elucidado.
Entretanto, sabe-se que ele é baseado no desacoplamento da fosforilação oxidativa
mitocondrial. Assim como outros desacopladores, a niclosamida estimula a respiração
mitocondrial em baixíssimas concentrações, porém inibe a respiração à medida que a
concentração é aumentada (Hollingworth et al. 2001). A niclosamida também diminui a
captação de água mesmo em concentrações abaixo da CL50 (0,08 mg L-1) (Duncan et al.
1977; Oliveira-Filho et al. 2010).

19
O fato de a niclosamida interferir nos processos osmorregulatórios já foi
mencionado por de Villiers & Mackenzie (1963). Aparentemente a niclosamida não é
irritante para os caramujos. Eles não evitam o contato com ela, tentando escapar da água,
um comportamento que é observado com outros moluscicidas (Etges & Gilbertson 1966).
Outro efeito estudado da niclosamida, foi nos tecidos dos caramujos. Sabe-se que
este composto inibe a oxidação de succinato em concentrações acima de 0,03 mg L-1.
Além disso também há a oxidação do glutamato e da redução da tetrametil-p-fenileno
diamina pelos tecidos de Biomphalaria sp (Ishak et al. 1970; Andrews et al. 1983). A
inibição de processos oxidativo também foi observada em B. alexandrina após exposição
a niclosamida (Ishak et al. 1972). Essa inibição pode ser parcialmente explicada pelo
acúmulo de oxaloacetato, um inibidor metabólico natural da enzima succinato
desidrogenase. Adição de ATP, que permite a remoção do oxaloacetato na forma de
piruvato de fosfoenol, preveniu completamente a inibição da oxidação do succinato. A
reação de transaminação do glutamato também pareceu ser sensível à niclosamida.
Entretanto, apesar de ser eficaz, a niclosamida possui desvantagens como a
toxicidade a outros animais aquáticos e plantas, apresenta dificuldade para ser
solubilizada tanto em solventes orgânicos quanto em água, possui alto custo, se decompõe
sob luz solar e provoca irritação em pele e mucosas (Yuan et al. 2005; Rapado et al. 2013;
Dai et al. 2014; King & Bertsch 2015).
Coura-Filho et al. (1992) demonstraram que o uso contínuo de niclosamida como
medida única não diminui a população de planorbídeos, já que após o uso do moluscicida
a população de caramujos se refazia em até três meses, garantindo assim a possibilidade
de transmissão de esquistossomose na área. Além disto, deve ser considerar que o uso
contínuo de um mesmo moluscicida químico pode provocar uma seleção de moluscos
resistentes ao produto químico aplicado. Todos esses fatores demonstram a necessidade
de investimento em pesquisas que busquem novas alternativas mais bem-sucedidas no
controle desses caramujos (Brasil 2010).

20
1.8 Cloridrato polihexametileno biguanida (PHMB)
1.8.1 Estrutura
O uso das biguanida como agentes microbianos para a preservação de produtos
industrializados tem sido bastante difundido ao longo dos anos. Por este motivo o
interesse no desenvolvimento de biguanida poliméricas como agentes antimicrobianos e
antissépticos tem despertado o interesse de diversos pesquisadores (Cunha et al. 2001).
A estrutura do cloridrato polihexametileno biguanida (PHMB) (Figura 10)
consiste em um polímero caracterizado por repetição de grupos biguanida ligados a
terminais que podem ser aminas, guanidinas ou ciano-guanidinas. A atividade
antimicrobiana do PHMB é conferida pelas moléculas da cadeia principal, que possuem
como característica elevada basicidade (Afonso 2013).
Figura 10 – Fórmula molecular do cloridrato de polihexametileno biguanida (PHMB) – (Adaptado de:
https://pubchem.ncbi.nlm.nih.gov/compound/52987646#section=Top.)
O PHMB forma soluções incolores, inodoras, de natureza catiônica,
quimicamente estável, não volátil, de alta atividade contra uma série de microrganismos,
principalmente bactérias Gram-negativas, e de baixa toxicidade para mamíferos
(Muangman et al. 2011). Muitos estudos têm sido realizados para elucidar o mecanismo
de ação antimicrobiana do PHMB, mas o atual entendimento sobre a molécula mostra
que este composto é um agente citoplasmático de membrana (Wessels & Ingmer 2013).
O mecanismo de ação deste composto se inicia quando o PHMB é adsorvido na superfície
da bicamada fosfatidilglicerol (PG) fosfolipídica ácida, via interação com os grupos de

21
cabeças polares dos lipídios com os grupos biguanida. Essa interação provoca uma
desorganização na bicamada de fosfatidilglicerol que por sua vez leva a maior fluidez,
expansão lateral e maior permeabilidade (Ikeda et al. 1984). Essas alterações causam uma
falha no mecanismo de defesa da célula e a ruptura da parede da mesma. O PHMB
provoca na célula bacteriana a perda de substâncias de baixo peso molecular, como íons
de potássio, cálcio e a inibição de enzimas responsáveis pela união da membrana, como
por exemplo a ATPase. Subsequentemente há a ruptura da membrana plasmática que
pode induzir à perda de substâncias macromoleculares e à precipitação das substâncias
celulares (Gilbert et al. 1990; Santos & Fernandes 2010; Wessels & Ingmer 2013).
1.8.2 Aplicações e benefícios
O PHMB tem sido alvo de vários estudos para uso como um composto em
formulações de desinfetantes para indústria alimentícia, farmacêutica, têxtil e bem como
a medicina possuindo uma excelente atividade no controle de microrganismos
patogênicos, como Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosas,
Salmonella cholerasuis, bem como endósporos de bactérias termo resistentes, alguns
fungos (Aspergillus niger, Candida albicans, Saccharomyces cerevisiae, Trichophyton
mentagrophytes), protozoários (Acanthamoeba sp.) e até alguns vírus (Herpes Vírus,
Fowlpox Vírus, Swine Vírus) (Allen et al. 2004; Santos & Fernandes 2010; Afonso 2013).
No que diz respeito à não toxicidade em seres humanos, o PHMB tem sido
referenciado como um composto bastante tolerado quando em terapia tópica nos olhos,
epitélio nasal e feridas (Afonso 2013). Por isso, durante muitos anos, esta substância tem
sido usada como antimicrobiano em cosméticos (Kramer, 2010; Frenzel et al. 2011).
Além disto o PHMB tem sido bastante usado em soluções de lente de contato já que se
demonstrou eficaz contra a ceratite causada por Acanthamoeba em concentrações baixa
como 0,025% (250 g / ml) como substância única, bem como em combinação com
propamidina e neomicina (Behrens-Baumann & Kramer 2002).
O custo benefício do PHMB é confirmado quando este é comparado aos
tradicionais desinfetantes, devido ao seu amplo espectro de ação na presença de matéria
orgânica aliado ao baixo índice tóxico tanto para a saúde pública como para o meio
ambiente, baixa corrosividade e alta solubilidade em água (Santos & Fernandes 2010).

22
Fumarola (2011) afirma que as propriedades anti-inflamatórias do PHMB estão
associadas a um registro insignificante de resistências, tornando-o um dos antissépticos
mais relevantes da atualidade. Além disto, este composto promove um aumento
significativo da cicatrização de feridas, sendo bem tolerado pela pele e mantendo uma
baixa toxicidade sistêmica. Os resultados satisfatórios dos estudos do PHMB como agente
antimicrobiano têm proporcionado a sua utilização em uma ampla variedade de
dispositivos médicos, assim como em soluções de desinfecção ocular (Afonso 2013).
Firdessa et al. (2015) demonstraram que o PHMB possui diversos potenciais
efeitos contra a leishmaniose. Entre eles destaca-se a aceleração do processo de
fechamento da ferida, melhorando assim as implicações provenientes da Leishmaniose
Cutânea e prevenindo infecções bacterianas e / ou fúngicas secundárias que muitas vezes
retardam a cicatrização de feridas em pacientes com Leishmaniose Cutânea. Os mesmos
autores também comprovaram a eficácia do PHMB em eliminar diretamente os parasitos.
1.9 Testes genotóxicos
Os testes de genotoxicidade são realizados com o intuito de esclarecer os efeitos
de agentes xenobióticos sobre o material genético e alterações nos produtos gênicos. Estes
agentes podem provocar quebras em fita simples ou dupla do DNA. Essas alterações na
molécula do DNA, quando não reparadas podem ocasionar mutações e morte celular
(Grisolia et al. 2004; Carvalho 2018). Os danos causados por esses agentes químicos
dependem de diversos fatores, concentração e duração da exposição, natureza química do
composto, a associação que este composto pode realizar com outros compostos químicos,
e de características da espécie estudada e do ambiente (Bolognesi 2003).
Os danos genéticos podem ser diferenciados em duas categorias, aneugênicos e
clastogênicos. Danos aneugênicos provocam alterações na distribuição dos cromossomos
durante o processo de divisão celular, dando origem as aneuploidias. Os danos
clastogênicos, induzem quebras e produzem alterações na estrutura do cromossomo
(Rabello-Gay et al. 1991; Collins 2004).
Em grande parte dos casos, os danos genéticos são reparados pelo próprio
organismo, mas essa capacidade de reparo do DNA diminui com o passar do tempo
(Grisolia 2004). Quando o reparo não ocorre ou não é feito de forma correta, as alterações
podem ser transmitidas para as células filhas, sendo então chamadas de mutações
hereditárias (Obe et al. 2004). A identificação dessas alterações pelos agentes

23
mutagênicos, em fases iniciais, é imprescindível para evitar danos mais extremos sobre o
comportamento da população estudada (Sarkar et al. 2006).
1.9.1 O Ensaio cometa na avaliação de danos do DNA
Ao longo dos anos o ensaio cometa, também conhecido como eletroforese em gel
de célula única, tornou-se um dos métodos referência para avaliar o dano do DNA. Suas
diversas modificações podem ser aplicadas em vários campos da genotoxicologia. Dentre
as diversas vantagens que a técnica apresenta destacam-se a simplicidade, sensibilidade
para detectar danos no DNA, possibilidade de análise de dados ao nível de células
individuais, uso de amostras de células extremamente pequenas, possibilidade de ser
realizado em praticamente qualquer célula eucariota, velocidade e o baixo custo. Embora
seja usado na maioria das vezes somente para demonstrar danos no DNA, o ensaio cometa
possui outras aplicações pouco exploradas (Collins 2004; Speit & Rothfuss 2012).
Atualmente existe quatro grandes áreas de pesquisa em que o ensaio cometa é
bastante adotado. A primeira é a avaliação de segurança de novas drogas, cosméticos ou
outros produtos químicos (Hartman et al. 2003). Normalmente é usado para avaliar
quebra no DNA, mas quando acrescido de outras técnicas como o aumento da
sensibilidade e a inclusão de endonucleases de reparo podem medir tipos de lesões
específicas (Asqueta & Collins 2013). A ecogenotoxicologia é a segunda área de
aplicação. A relevância da compreensão dos efeitos da genotoxicidade induzida por
poluentes gerados por atividades industriais, agrícolas e domésticas, e do risco imposto a
espécies ambientais, bem como para a saúde humana através da cadeia alimentar tornou
a técnica do ensaio cometa bastante utilizada para avaliar o nível de contaminação em
ambientes aquáticos e terrestres, através de organismos biomarcadores presentes nestes
locais (Grazeffe et al. 2008; Jah 2008; Martins & Costa 2015). Outra área de igual
importância é o biomonitoramento humano. Entre as aplicações na saúde humana, o
ensaio cometa é bastante empregado para o monitoramento da exposição ocupacional a
produtos químicos genotóxicos como inseticidas e pesticidas, na avaliação do estresse
oxidativo vinculado à algumas doenças, na detecção de danos causado pelo tabagismo e
dietas que diferem no conteúdo lipídico (Somorovská 1999; Jenkinson 2001; Izzotti et al.
2001; Bolognesi et al. 2004; Franco et al. 2016). Por fim, mas não menos importante, o
ensaio cometa também tem sido bastante utilizado para estudar os mecanismos de reparo
do DNA lesado (Asqueta & Collins 2013). Uma das abordagens amplamente utilizada

24
nesse campo de estudo é monitorar uma remoção dependente do tempo de lesões após o
tratamento com um agente prejudicial ao DNA.
1.9.2 Fundamentos do ensaio cometa
Na década de 70, Peter Cook e colaboradores desenvolveram uma metodologia
para estudar a estrutura nuclear através da lise de células com detergente não iônico e
cloreto de sódio de alta molaridade. Esses reagentes com alto teor de sal promovem a
remoção de vários componentes celulares, como membranas, citoplasma, nucleoplasma
e proteínas histonas. O que resta é o nucleóide, constituído de ácido ribonucleico e
proteínas, juntamente com o DNA condensado. A adição de brometo de etídio promoveu
o desenrolamento do DNA, permitindo a rotação livre do DNA (Collins 2004). Mais tarde
Ostling e Johanson (1984) fizeram o primeiro ensaio cometa em condições alcalinas,
referindo ao modelo de nucleóide de Cook e colaboradores. Esse modelo permitia a
detecção de quebras de fita dupla de DNA. Posteriormente Singh et al. (1988)
apresentaram a versão alcalina do teste, que além das detecções apresentadas pelo modelo
de Ostling e Johanson, ainda permitiam a detecção de quebras de fita simples, sítios álcali-
lábeis do DNA, dano oxidativo do DNA, DNA-DNA ou DNA-proteína e ligações
cruzadas (Pavanello & Clonfero 2000, Collins et al. 2008, Dhawan et al. 2009 Kumar &
Dhawan 2013; Gonçalves 2015).
Nos dias de hoje na técnica de eletroforese de microgel, ensaio cometa, um
pequeno número de células suspensas em um gel de agarose fino em uma lâmina de
microscópio, são lisadas, submetidas a eletroforese e coradas com um corante
fluorescente de ligação ao DNA. Como resultado da corrida eletroforética, os fragmentos
resultantes de DNA com lesões migram em velocidade diferente da cabeça do nucleóide,
formando a típica figura de um cometa (Figura 11), e quanto mais lesões apresentadas,
maior a cauda desse cometa e maior quantidade de DNA na cauda. Caso as células não
apresentem dano no DNA, este migrará em conjunto com o nucleóide, formando um
círculo (Olive et al. 1990; Collins et al. 1997; Speit & Rothfuss 2012).

25
Figura 11 – Imagem de nucleóide em hemócitos de Biomphalaria glabrata, formando cometa com
dano ao DNA. Fonte: Laboratório de Estudos da Relação Parasito Hospedeiro – LAERPH/IPTSP/UFG.
Existem vários programas de análise de imagem que podem medir os danos no
DNA e quantificar a intensidade de DNA na cauda. Os programas oferecem parâmetros
quantitativos que correspondem aos danos ocorridos no DNA em cada uma das células
analisadas. Os parâmetros mais utilizados são o comprimento da cauda, intensidade
relativa de fluorescência da cabeça e da cauda (normalmente expressa como porcentagem
do DNA na cauda) e momento da cauda olive (OTM) (Hartman et al. 2003; Collins 2004).
1.9.3 Ensaio cometa em Biomphalaria glabrata
Há algum tempo, pesquisadores vinham estudando a possibilidade da
padronização da técnica do ensaio cometa em caramujos de água doce. Essa escolha se
justificou pelo fato dos invertebrados representarem mais de 90 % das espécies aquáticas,
estando em contato com todos os resíduos que são descartados nos rios e lagoas. (Silva
2010; Monteiro 2017).
Foi demonstrado que o ensaio cometa é uma ferramenta rápida, sensível e útil para
detectar danos no DNA de caramujos da espécie B. glabrata. Como foi comprovado por
Grazeffe et al. (2008), que utilizou a técnica para detectar danos no DNA de hemócitos
circulantes do caramujo adulto B. glabrata após a exposição à radiação gama com doses

26
de 2,5; 5; 10 e 20 Gy de radiação gama Co60 a 2,82 KGy/h. Várias espécies de moluscos
aquáticos têm sido utilizadas para acompanhar a disseminação de mutagênicos ambientais
que são descartados no meio ambiente de forma descontrolada e sem acompanhamento
dos órgãos de fiscalização (Klobucar et al. 2003; Vilela et al. 2006; Bineli et al. 2008).
1.10 Teste do micronúcleo
Micronúcleos (MN) são pequenos fragmentos que se formam sempre que um
cromossomo ou um fragmento de um cromossomo não é incorporado em um dos núcleos
da célula filha após a divisão celular, devido à falta de centrômeros, defeitos de
centrímeros ou defeitos de citoquineses (Vincent-Hubert et al. 2011; Touahri et al. 2016).
O teste que avalia a presença de MN e consequentemente estima danos cromossômicos
em diferentes populações é denominado de teste do micronúcleo (MN) (Toosi et al. 2017).
O teste do MN in vitro é um método que fornece uma base abrangente para investigar o
potencial prejudicial do cromossomo porque tanto os danos de origem aneugênicos
quanto clastogênicos podem ser detectados em células que foram submetidos a divisão
celular durante ou após à exposição ao produto químico (OECD 2014). A primeira vez
que o MN foi utilizado como marcadores para danos citogenéticos foi em 1959, por Evans
e colaboradores. Mais tarde, Boller & Schimid (1970) comprovaram o avanço decisivo
dos MNs como sistema de ensaio para o potencial genotóxico de agentes externos,
sugerindo pela primeira vez o termo teste de MN (Kirsch-Volders 2003).
Para análise em nível celular de agentes xenobióticos o teste do MN pode ser
utilizado como bioindicador de genotoxicidade. Os xenobióticos podem agir de duas
formas no núcleo: diretamente sobre o cromossomo ou sobre o fuso mitótico, levando a
perda de parte dos cromossomos, ou de todo cromossomo (Arias 2007). Estes, durante a
citocinese, quando na formação do envoltório nuclear, se não forem incorporados pelo
núcleo principal, formam seu próprio envoltório sendo considerados MNs (Villela et al.
2006). Em alguns casos, observam-se pontes nucleoplásmicas entre núcleos em uma
célula binucleada. Estes cromossomos são chamados de dicêntricos, quando os dois
centrômeros foram puxados para polos opostos da célula e o DNA na ponte resultante é
coberto por membrana nuclear. Essas pontes nucleoplásmicas em células binucleadas,
também são alterações nucleares importantes na avaliação de dano cromossômico e
fornecem uma medida adicional de rearranjo, que pode ser pontuado juntamente com a
contagem de MN (Fenech & Morley 1985; Fenech 2000).

27
Os MNs são facilmente distinguidos do núcleo, já que para serem introduzidos
nesta categoria devem ter um tamanho de até 1/3 do núcleo principal. Além disso devem
possuir bordas distinguíveis e com a mesma refringência do núcleo principal (Al-Sabti &
Metcalfe 1995; Al-Sabti 2000; Gustavino et al. 2001). Para realizar a contagem desses
MNs são analisadas entre 1000 e 3000 células, que devem possuir membranas
citoplasmáticas e nucleares intactas, não sendo analisadas aquelas que estejam
sobrepostas ou danificadas (Al-Sabti & Metcalfe 1995).
Assim como o ensaio cometa, o MN também é recomendado para avaliação do
potencial mutagênico de agentes químicos e físicos, sendo bastante empregado em
estudos ecotoxicológicos. Além disso o MN também é utilizado para registro de novos
produtos químicos que serão liberados para comercialização (Gautier et al. 1999; Chung
et al. 2002; Ding et al. 2003; Ribeiro 2003).
A análise de MN só pode ser realizada em células que tenham a capacidade de
entrarem em divisão (Fenech 2000). Entretanto a técnica já foi padronizada em diversos
tipos celulares, como células de medula óssea, células de epitélio, células de brônquios,
da bexiga urinária e de sangue periférico (Nepomuceno et al. 1997). O MN também foi
padronizado em diversas espécies de animais: peixes (Al-Sabati 2000), anfíbios (Cabagna
et al. 2006), répteis (Lopéz-González et al. 2013) e mamíferos (Rodrigues 2013). Em
moluscos, o teste do MN foi utilizado por Filippi et al. (2018) para detectar o efeito
mutagênico causado pela poluição de uma usina de carvão na Itália no caracol terrestre
Helix aspersa. Neste experimento foi observado que após uma exposição de 13 dias em
locais de diferentes proximidades da usina de carvão, a frequência de micronúcleos foi
significativamente maior nos locais mais próximos a usina. Em outro estudo, o teste de
micronúcleo demonstrou aumento na frequência de micronúcleos em mexilhões Mytilus
galloprovincialis após a exposição a água contaminada de um porto comercial, em relação
ao não exposto (Touahri et al. 2016).
A importância de padronização do teste de micronúcleos e HNA no caramujo,
advém do fato de que o presente estudo é pioneiro na avaliação do teste de micronúcleos
e HNA em B. glabrata, uma espécie considerada como bioindicador aquático.

28
2. JUSTIFICATIVA
A esquistossomose é considerada um problema de saúde pública mundial, por ser
uma doença crônica, altamente debilitante para os acometidos e apresentar uma alta taxa
de morbidade em todo o mundo. Esta doença também causa um impacto negativo na
saúde, como consequência de manifestações inespecíficas, como anemia, intolerância ao
exercício, desnutrição e dificuldades de aprendizagem (Gazzinelli et al. 2015). Além
disso, as taxas de reinfecção apresentadas pela esquistossomose são bastante altas,
provocando graves consequências na economia dos países endêmicos (King 2010;
Barsoum 2013; Siqueira et al. 2017).
Diversos estudos demonstraram que a integração entre medidas profiláticas, como
o tratamento de pessoas infectadas, quimioterapia preventiva, controle dos moluscos
hospedeiros intermediários, educação em saúde, o fornecimento de água potável e o
saneamento básico podem controlar a esquistossomose no mundo. Atualmente o único
moluscicida recomendado para uso controlado pela OMS é a niclosamida. Entretanto este
composto químico provoca acumulo em órgãos de outros animais aquáticos, além do seu
alto custo impedir o seu uso em países subdesenvolvidos (Dai et al. 2014; King & Bertsch
2015). A niclosamida também possui baixa solubilidade em água e outros solventes
orgânicos, e o uso indiscriminado contribui para o aumento de casos de linhagens de
caramujos resistentes (Yuan et al. 2005; Dai et al. 2014; King & Bertsch 2015).
Os programas com foco no tratamento dos doentes são elaborados com base em
um único medicamento, o praziquantel (PQZ). O PQZ não apresenta eficácia quanto as
formas imaturas do parasito, e alguns estudos relataram resistência a essa droga (Song et
al. 2012; Cioli et al. 2014; King & Bertsch 2015; Othman e Soliman 2015). No Brasil
apenas 50,3 % da população tem acesso ao saneamento básico, e são poucos os programas
de educação sanitária com foco na esquistossomose (Brasil 2016; Snis 2015).
Na tentativa de busca de novos compostos químicos com potencial moluscicida,
o alvo do presente estudo é a molécula do PHMB. Este composto possui alto potencial
biológico, e a sua alta atividade antimicrobiana é conferida através da cadeia principal de
sua molécula, sendo considerado uma das substâncias mais promissoras disponíveis
atualmente do ponto de vista clínico (Hubner & Kramer 2010; Afonso 2013). Dentre
outras vantagens do uso desse composto químico destaca-se a boa solubilidade, o que

29
propicia o seu uso no ambiente natural do caramujo B. glabrata. Além disso, já foi
demonstrado que este composto possui baixa toxicidade para outros organismos não alvo,
comprovando a segurança do uso deste composto no meio ambiente (Christen et al. 2017).
Devido à sua interação forte e inespecífica com fosfolipídios de membrana, o PHMB tem
amplo espectro antimicrobiano incluindo bactérias gram-positivas e gram-negativas,
bactérias formadoras de placas e de formação de biofilme, bactérias formadoras de
esporos, bactérias intracelulares como clamídias e micoplasmas e fungos incluindo
Candida spp. bem como Aspergillus spp. A ruptura da parede bacteriana juntamente com
a inibição do metabolismo energético desses microrganismos torna o desenvolvimento de
resistência ao PHMB improvável (Hubner & Kramer 2010).
Levando em consideração todos os problemas da esquistossomose que ainda
precisam ser enfrentados, e a necessidade de novas buscas e identificação de compostos
moluscicidas que sejam altamente seletivos e não tóxicos para outras espécies de animais
e plantas aquáticas, este estudo objetiva encontrar possíveis alternativas para o controle
desta doença através da diminuição do B. glabrata, que é o hospedeiro intermediário de
S. mansoni mais importante (Knight et al. 2000). Este é o primeiro estudo que pretende
avaliar a atividade moluscicida do composto PHMB através de testes genotóxicos e
mutagênicos.

30
3. OBJETIVOS
3.1 Objetivo Geral
Avaliar a atividade moluscicida do cloridrato polihexametileno biguanida
(PHMB) em Biomphalaria glabrata por meio de testes de toxicidade.
3.2 Objetivo Específico
Avaliar a atividade moluscicida do PHMB em embriões, recém-eclodidos e
adultos do caramujo B. glabrata.
Determinar a CL50-144h do PHMB para embriões de B. glabrata, e a CL50-96h para
caramujos recém-eclodidos e adultos de B. glabrata.
Avaliar a taxa de eclosão de embriões de B. glabrata expostos ao PHMB.
Identificar alterações morfológicas em embriões de B. glabrata após a exposição
ao PHMB.
Avaliar alterações no comportamento do caramujo adulto de B. glabrata após a
exposição do PHMB.
Avaliar os efeitos genotóxicos da exposição do PHMB em adultos de B. glabrata
via ensaio cometa.
Identificar efeitos mutagênicos no caramujo adulto de B. glabrata causadas pela
exposição ao PHMB, através do teste do micronúcleo.

31
Capítulo II Artigo I
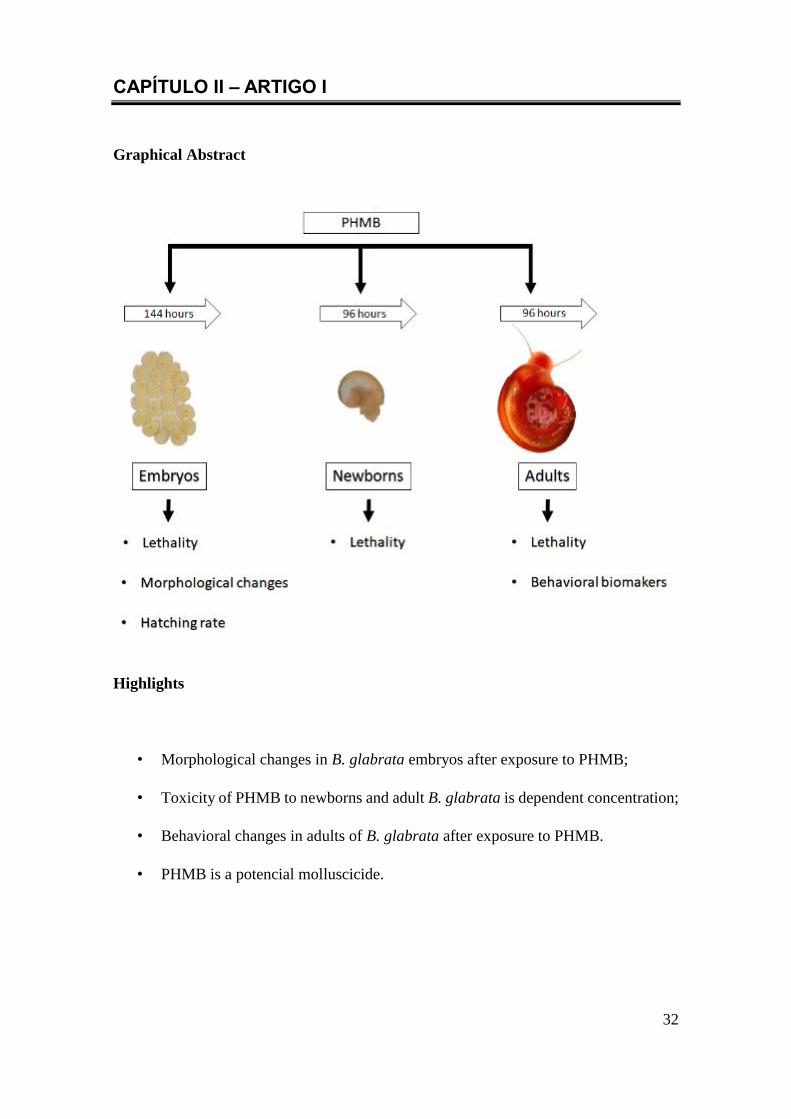
32
CAPÍTULO II – ARTIGO I
Graphical Abstract
Highlights
• Morphological changes in B. glabrata embryos after exposure to PHMB;
• Toxicity of PHMB to newborns and adult B. glabrata is dependent concentration;
• Behavioral changes in adults of B. glabrata after exposure to PHMB.
• PHMB is a potencial molluscicide.

33
Molluscicidal activity of polyhexamethylene biguanide hydrochloride on the early-
life stages and adults of the Biomphalaria glabrata (Say, 1818)
Atividade moluscicida do cloridrato polihexametileno biguanida nos estágios iniciais
de vida e adultos de Biomphalaria glabrata (Say, 1818)
Amanda de Oliveira Meloa; Daniela Braz Santosa; Luciana Damacena Silvab; Thiago Lopes
Rochac*; José Clecildo Barreto Bezerraa
a Laboratory of Studies of the Host-parasite Relationship, Institute of Tropical Pathology and
Public Health, Federal University of Goiás, Goiânia, Goiás.
b Laboratory of Host-Parasite Interactions, State University of Goiás, Anápolis, Goiás, Brazil.
c Laboratory of Environmental Biotechnology and Ecotoxicology, Institute of Tropical
Pathology and Public Health, Federal University of Goiás, Goiânia, Goiás.

34
ABSTRACT
Schistosomiasis is a disease that affects tropical and subtropical areas and is
considered the second most prevalent parasitic disease in the world. One of the ways of
combating this disease is the use of molluscicidal agents to eliminate or reduce the
population of intermediate host snails. Polyhexamethylene biguanide hydrochloride
(PHMB) is a chemical biocide commonly used as a disinfectant and antiseptic in the food
industry and very successfully for the disinfection of swimming pools. The US
Environmental Protection Agency (USEPA) indicated the PHMB as low environmental
risk. The present study aimed to evaluate the molluscicidal activity of the PHMB in
freshwater snail (Biomphalaria glabrata), intermediate host of Schistosoma mansoni. The
PHMB showed high toxicity against all stages of the snail B. glabrata: embryos, new-
borns and adults. The LC50 estimated was 0.98 mg L-1; 1.43 mg L-1 and 1.49 mg L-1,
respectively, after exposure of 144 hours for embryos and 96 hours for new-borns and
adults. PHMB did not prevent the development of embryos within the egg mass, since at
all concentrations evaluated 80 % of the embryos managed to develop until the hypo-
stage, which is the last stage of development before hatching. However, PHMB inhibited
the hatching of embryos by 100 % at all concentrations above 1.6 mg L-1. PHMB proved
to be a promising substance in the fight against schistosomiasis by eliminating the
intermediate host (B. glabrata). This was the first study that makes an experimental
observation of the molluscicidal activity of PHMB.
Keywords: schistosomiasis; snail; intermediate host; embryotoxicity; Schistosoma
mansoni; PHMB.

35
1. Introduction
Schistosomiasis, also known as bilharziosis, is a Neglected Tropical Disease
(NTD) considered by the World Health Organization (WHO) as the second most
widespread parasitic disease in more than 70 countries in the Americas, Africa and Asia.
It is estimated that some 390-600 million people are infected worldwide, while 800
million remain in areas at risk of infection (Hotez et al. 2014; Grimes et al. 2015). In
Brazil, it is estimated that more than 25 million people live in areas at risk for contracting
this disease. The disease reflects socioeconomic problems in endemic countries, and the
main approach to combating disease transmission is related to the periodic treatment of
people living in areas at risk of anti-schistosomiasis drugs (Lambertucci, 2010; Grimes et
al. 2015; Hotez et al. 2014).
Freshwater snails of the genus Biomphalaria act as the intermediate host of
schistosomiasis. Among the three freshwater snail species naturally infected by
Schistosoma mansoni in Brazil, Biomphalaria glabrata (Say, 1818) is more relevant due
to its wide geographic distribution, greater susceptibility to infection and efficacy in the
transmission of schistosomiasis (Costeau et al. 2015; Scholte et al. 2014). In addition to
the presence of an intermediate host with easy adaptation throughout the Brazilian
territory, the economic situation of the population, basic sanitation systems for only 50.3
% of the population and low level of health education further aggravate the transmission
of schistosomiasis in Brazil according reports (Brazil, 2010; Snis 2015).
Prophylactic public health measures in endemic regions for schistosomiasis
consist of treatment of infected persons, basic sanitation, health education, and control of
snails. However, it is recognized that the most effective schistosomiasis control method
to interrupt the cycle of S. mansoni is eliminating or reducing the number of intermediate
hosts in endemic areas (Pile et al, 2002; Brazil, 2008; Bruun & Aagaard-Hansen, 2008;
WHO, 2013; Sokolow et al. 2016).
The WHO recommends the use of synthetic molluscicide niclosamide
(Bayluscide®) as one of the strategies to combat schistosomiasis in the world (WHO,
2002). However, the niclosamide has high production value and can induced several
secondary effects, such as bioaccumulation and high toxicity in non-target animals
(Oliveira-Filho et al. 2010; King & Bertsch, 2015). In addition, the re-colonization of
snail populations is observed after a few months of application of the niclosamide,
demonstrating the niclosamide-resistant snail populations following repeated, extensive
use of this molluscicide (Coura-Filho et al. 1992; Martins et al. 2014). In this context, the

36
discovery of new molluscicides, which could be more selective to Biomphalaria and less
harmful to the aquatic environment is of urgent need for the combat of the schistosomiasis
(Albuquerque et al. 2014; Coelho & Caldeira, 2016; Silva et al. 2018; Rocha-Filho et al.
2015).
Polyhexamethylene biguanide hydrochloride (PHMB) is a chemical biocide and
a member of the polymeric guanidine family commonly used as a disinfectant and
antiseptic in the food industry and very successfully for the disinfection of swimming
pools (Mashat, 2016). The US Environmental Protection Agency (USEPA) indicated the
PHMB as low environmental risk (United States Enviromental Protection Agency, 2002).
Toxic effects of PHMB have been reported for gram-negative and gram-positive
bacteria (Staphylococcus aureus and Pseudomonas aeruginosa) (Fjeld & Lingaas, 2016)
and leishmaniosis with direct effect on the parasite (Firdessa et al. 2015). The mechanism
of action (MoA) and toxicity of the PHMB in gram-negative and gram-positive bacteria
was associated with binds to negatively charged phosphate groups in the cell wall, causing
greater stiffness, rupturing and altering the permeability of the cytoplasmic membrane,
and promoting the extravasation of cytoplasm that culminates in cell death. On the other
hand, no PHMB-resistant bacteria have been previously reported (Kaehn, 2010).
However, to the best of our knowledge, no data is available on the PHMB toxicity in the
snail Biomphalaria.
Accordingly, the aim of this study was to analyse the toxic effects of PHMB on
the early-life stages and adults of the snail B. glabrata. Therefore, the effects of PHMB
on the embryonic development, hatching rate, survival and morphological alterations of
embryos, new-borns and adult snails were determined. The hypothesis of this study was
that the PHMB has molluscicidal activity on B. glabrata, the snail intermediate host of S.
mansoni.
2. Materials and methods
2.1 Test substance
The polyhexamethylene biguanide hydrochlori (PHMB) (Vantocil IB®) (CAS nº
32289-58-0; 20 % w/w) was donated from Arch Química do Brasil (São Paulo, Brazil).
The PHMB stock solution (100 mg L-1) was made with milli-Q water. The Niclosamide
(Bayluscide®, Bayer AG Leverkusen, Germany) (CAS nº 50-65-7) was purchased from

37
Sigma-Aldrich (São Paulo, Brazil) and the stock solution (0.1 mg mL-1) was made in
milli-Q water.
2.2. Snails
Adult B. glabrata snails (shell diameter: 10 ± 2 mm; total weight: 0.40 ± 0.10 g)
were maintained in the Institute of Tropical Pathology and Public Health from Federal
University of Goiás, such as recommended by OECD guideline nº 243 (OECD, 2016).
Snails were kept in aerated glass tanks (24 × 16 × 9 cm) with 40 L of dechlorinated water
under 12:12 h light/dark cycles, temperature (25 ± 1 °C) and pH (7,0 ± 1). Snails were
fed ad libitum three times per week with lettuce leaves (Lactuca sativa).
2.3. Snail embryotoxicity test
The developmental toxicity of PHMB on B. glabrata was analyzed by
embryotoxicity test according to Ducrot & Charles (2015) and OECD guideline nº 243
(OECD, 2016). Snail egg-clutches with 25 ± 10 embryos in the blastula stage (~ 12 h)
were placed into 6-wells cell culture plates and exposed to PHMB (0.60, 1.0, 1.6, 2.0, 2.6,
3.0 and 3.6 mg L-1) during 144 h, jointly with a negative control group kept in
reconstituted water (OECD, 2016) and a positive control exposed to niclosamide
(Bayslucida®) at 0.08 mg L-1 (median lethal concentration – LC90-144h according to
Oliveira-Filho et al. 2010). All experimental procedures were conducted in a static
condition (5 mL per well) and a triplicate design (3 plates with 3 egg-clutches per
condition) under 12:12 h light/dark cycles and environmental temperature of 25 ± 1 °C.
During the exposure period (24, 48, 72, 96, 120 and 144 h), the frequency (%) of
viable and unviable embryos, embryo development stages and abnormal development
were analyzed using a microscope (Leica DM 750) associated with the LEICA ICC50
HD camera and the LAS EZ software. Embryos were considered as unviable according
to Oliveira-Filho et al. 2010: disaggregated dead embryos, embryos with no rotational
movements, no foot movements or a heartbeat. The early development stages of B.
glabrata were classified in five stages (blastulae, gastrulae, trochophore, veliger and
hippo-stage) (Fig. 1), such as recommended by Rapado et al. (2013). The morphological
changes during embryonic development of snails (shell malformations, poliophtalmia,
reduced size and colour changes) were analyzed according to Oliveira-Filho et al. (2010)
and Silva et al. (2018).

38
Figure 1. Early developmental stages of Biomphalaria glabrata. (A) Blastulae. (B) Gastrulae. (C)
Trocophore. (D) Veliger. (E) Hypo-stage. Magnification 40x; bar scale = 500 µm.
At the end of exposure period (144 h), the hatching rate (%) was determined using
the equation (1) described by OECD 243:
𝑁𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 ℎ𝑎𝑡𝑐ℎ𝑒𝑑 𝑒𝑚𝑏𝑟𝑦𝑜𝑠
𝑇𝑜𝑡𝑎𝑙 𝑛𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑒𝑚𝑏𝑟𝑦𝑜𝑠 𝑥 100 (1)
2.4. Acute toxicity tests with new-borns and adult snails
The acute toxicity tests (96 h) were conducted with new-borns (shell diameter: 2
± 2 mm) and adult snails (shell diameter: 10 ± 2; total weight: 0.45 ± 0.10 g). Ten new-
borns or adult snails were exposed to PHMB (0.60, 1.0, 1.6, 2.0, 2.6, 3.0 and 3.6 mg L-1)
during 96 h, jointly with a negative control group kept in reconstituted water (OECD,
2016) and a positive control exposed to niclosamide (Bayluscide®) at 0.10 mg mL-1 for
new-borns and adults (LC90-96h according to Ribeiro et al. 2009). Experiments were
conducted in a static condition were placed into 6-wells culture cells plates (5 mL per
well) for new-borns snail and glass aquariums (3 L) for adults, a triplicate design under
12:12 h light/dark cycles and environmental temperature of 25 ± 1 °C. The snails were
no fed during the exposure period (OECD, 2016). The animals were considered dead
according to the parameters described by Miyasato et al. (2012) and Tallarico et al.
(2014): immobility, discolouration of shells, exposure of visceral mass, release of
hemolymph and lack of heart beats.
2.5. Behavioural assessment
After the adult snail exposure to PHMB for 96 h, the frequency (%) of the
following behavioural alterations was determined: total retraction into its shell; lethargy

39
when stimulated in the cephalopodal mass with forceps; displacement to the periphery
and edges of the aquariums, production and secretion of mucus in the opening of the shell.
2.6 Statistical analysis
Statistical analyses were carried out using the Project R software (R Core Team,
2016). The results were compared using parametric tests (two-way ANOVA, followed by
Turkeys’ test). The median letal concentration (LC50) including its associated lower and
upper confidence limits, were estimated using an appropriated statistical method based
on a regression analysis. Furthermore, the No Observed Effect Concentration (NOEC)
and the Lowest Observed Effect Concentration (LOEC) were evaluated by one-way
analyses of variance (ANOVA) followed by multiple comparison procedure. Results
were considered significant when p < 0.05.
3. Results and discussion
3.1 Embryotoxicity
The unexposed snail embryos showed a low mortality rate (15 ± 5 %) and high
hatching rate (85 ± 5 %) after 144 h. Moreover, the embryos exposed to niclosamide
(positive control group) showed the effects described in the literature generating a high
mortality rate (95 ± 5 %), while complete inhibition of hatching (100 %) was observed
throughout the experimental period, such as reported in previous studies (Oliveira-Filho
et al. 2010). On the other hand, the results that showed that the embryos exposed to PHMB
at all concentrations (0.6 to 3.6) were able to develop to the last stage of development,
known as hypo-stage (Fig. 2), different from that of the positive control (Niclosamide)
when embryos can only achieve reach the trochophore.
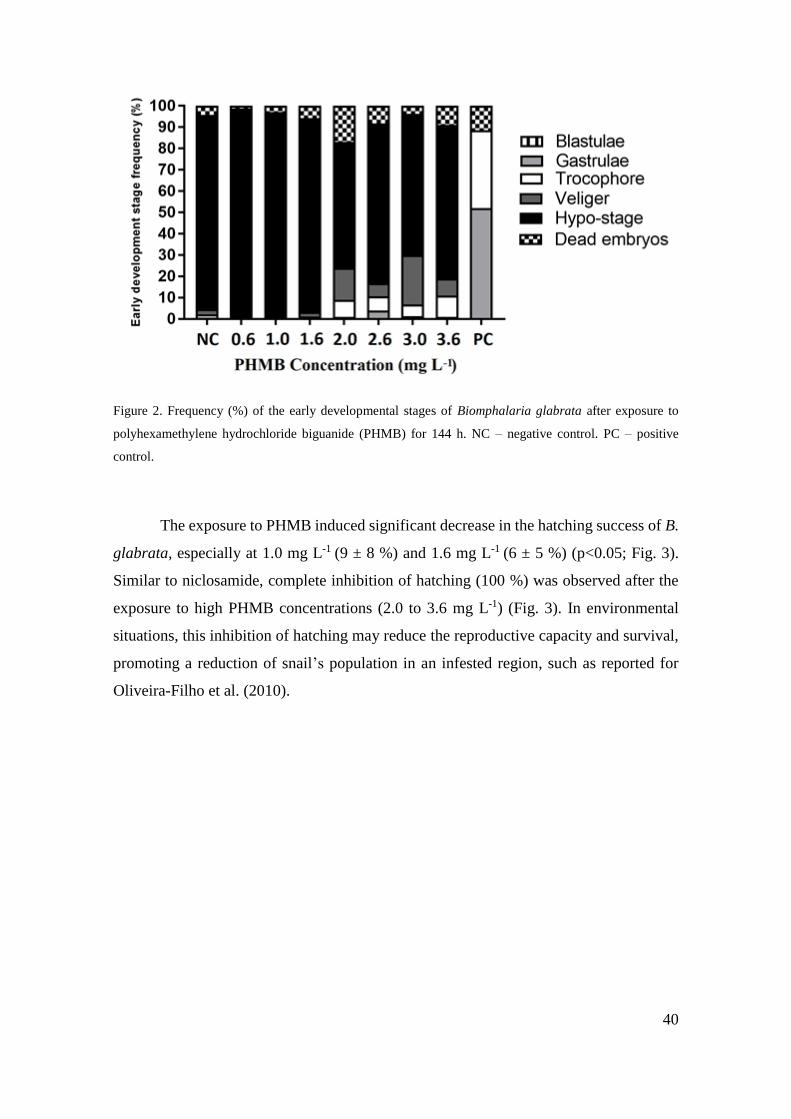
40
Figure 2. Frequency (%) of the early developmental stages of Biomphalaria glabrata after exposure to
polyhexamethylene hydrochloride biguanide (PHMB) for 144 h. NC – negative control. PC – positive
control.
The exposure to PHMB induced significant decrease in the hatching success of B.
glabrata, especially at 1.0 mg L-1 (9 ± 8 %) and 1.6 mg L-1 (6 ± 5 %) (p<0.05; Fig. 3).
Similar to niclosamide, complete inhibition of hatching (100 %) was observed after the
exposure to high PHMB concentrations (2.0 to 3.6 mg L-1) (Fig. 3). In environmental
situations, this inhibition of hatching may reduce the reproductive capacity and survival,
promoting a reduction of snail’s population in an infested region, such as reported for
Oliveira-Filho et al. (2010).
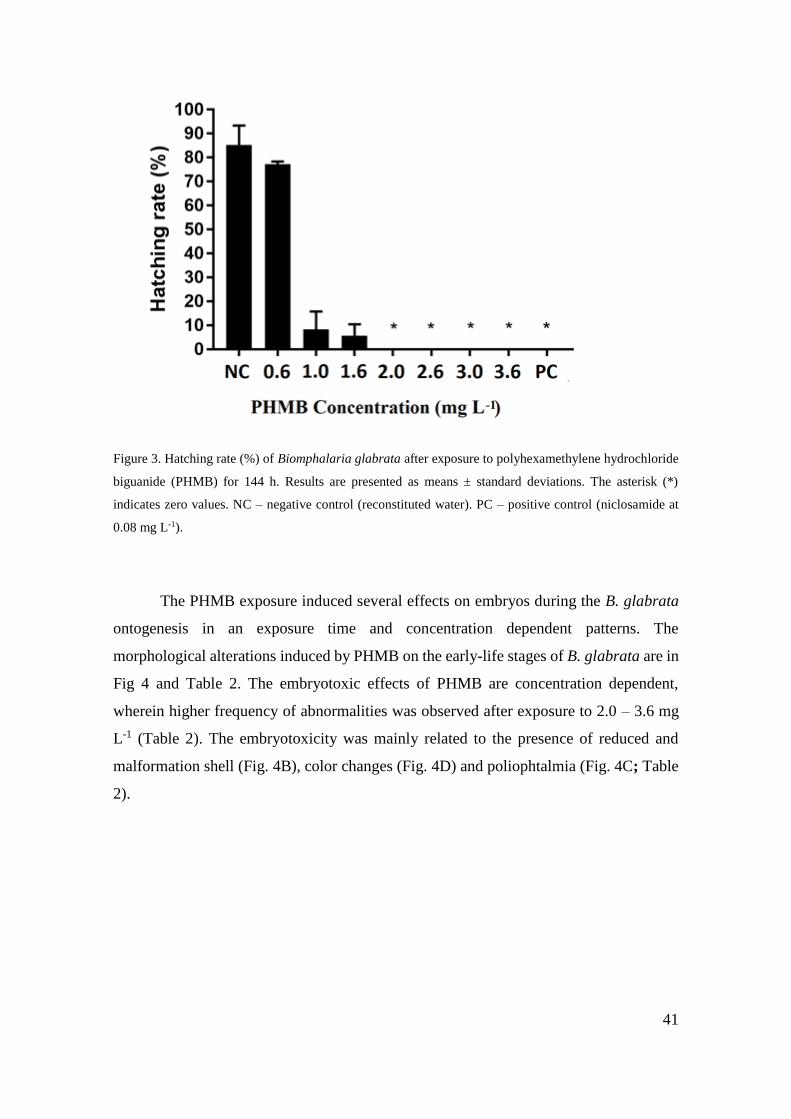
41
Figure 3. Hatching rate (%) of Biomphalaria glabrata after exposure to polyhexamethylene hydrochloride
biguanide (PHMB) for 144 h. Results are presented as means ± standard deviations. The asterisk (*)
indicates zero values. NC – negative control (reconstituted water). PC – positive control (niclosamide at
0.08 mg L-1).
The PHMB exposure induced several effects on embryos during the B. glabrata
ontogenesis in an exposure time and concentration dependent patterns. The
morphological alterations induced by PHMB on the early-life stages of B. glabrata are in
Fig 4 and Table 2. The embryotoxic effects of PHMB are concentration dependent,
wherein higher frequency of abnormalities was observed after exposure to 2.0 – 3.6 mg
L-1 (Table 2). The embryotoxicity was mainly related to the presence of reduced and
malformation shell (Fig. 4B), color changes (Fig. 4D) and poliophtalmia (Fig. 4C; Table
2).
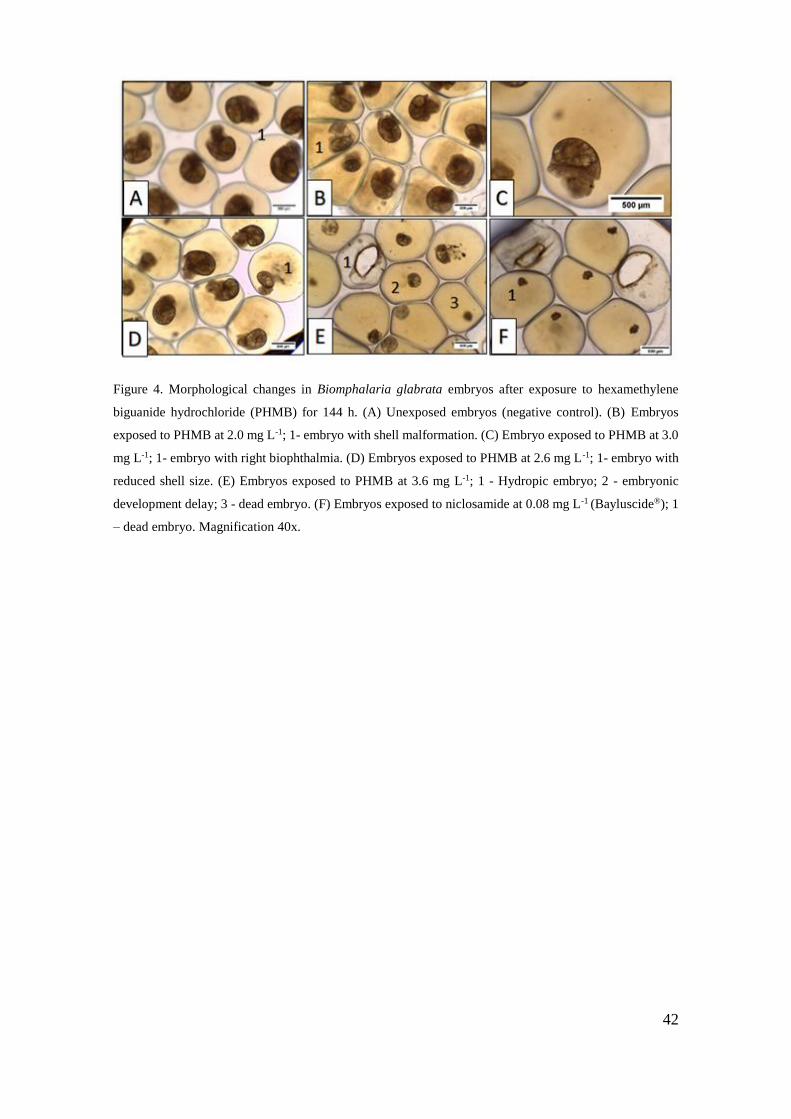
42
Figure 4. Morphological changes in Biomphalaria glabrata embryos after exposure to hexamethylene
biguanide hydrochloride (PHMB) for 144 h. (A) Unexposed embryos (negative control). (B) Embryos
exposed to PHMB at 2.0 mg L-1; 1- embryo with shell malformation. (C) Embryo exposed to PHMB at 3.0
mg L-1; 1- embryo with right biophthalmia. (D) Embryos exposed to PHMB at 2.6 mg L-1; 1- embryo with
reduced shell size. (E) Embryos exposed to PHMB at 3.6 mg L-1; 1 - Hydropic embryo; 2 - embryonic
development delay; 3 - dead embryo. (F) Embryos exposed to niclosamide at 0.08 mg L-1 (Bayluscide®); 1
– dead embryo. Magnification 40x.
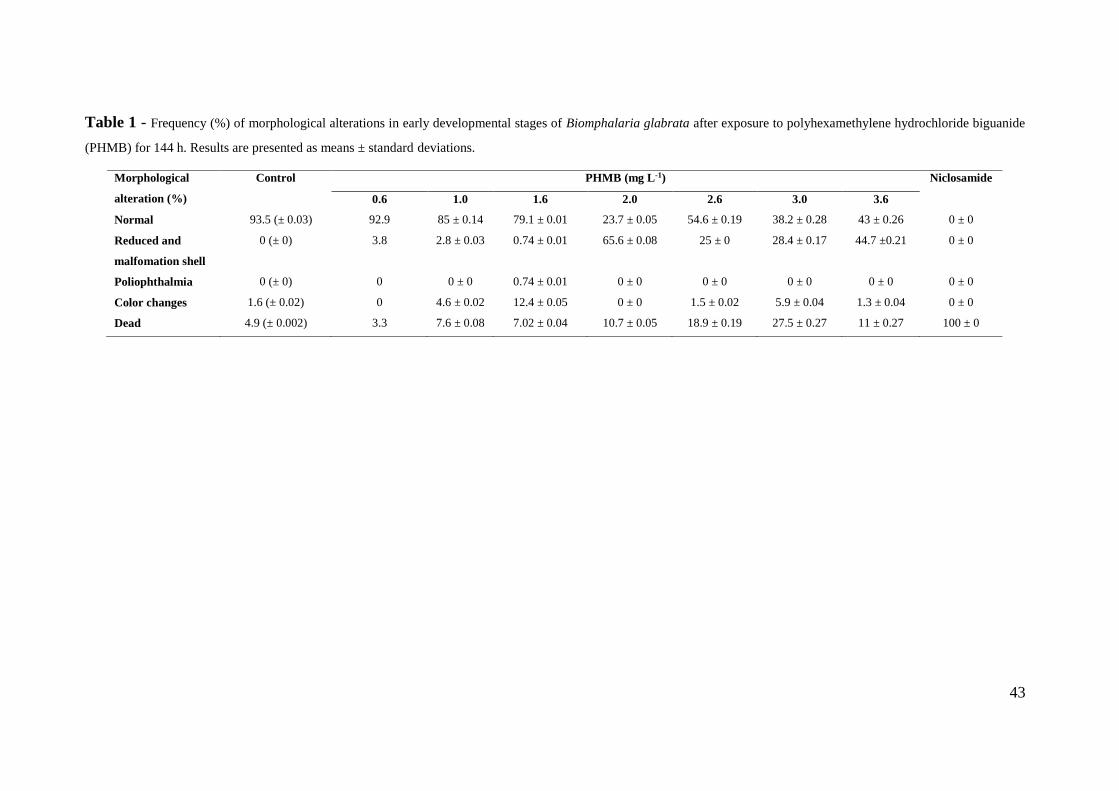
43
Table 1 - Frequency (%) of morphological alterations in early developmental stages of Biomphalaria glabrata after exposure to polyhexamethylene hydrochloride biguanide
(PHMB) for 144 h. Results are presented as means ± standard deviations.
Morphological
alteration (%)
Control PHMB (mg L-1) Niclosamide
0.6 1.0 1.6 2.0 2.6 3.0 3.6
Normal 93.5 (± 0.03) 92.9 85 ± 0.14 79.1 ± 0.01 23.7 ± 0.05 54.6 ± 0.19 38.2 ± 0.28 43 ± 0.26 0 ± 0
Reduced and
malfomation shell
0 (± 0) 3.8 2.8 ± 0.03 0.74 ± 0.01 65.6 ± 0.08 25 ± 0 28.4 ± 0.17 44.7 ±0.21 0 ± 0
Poliophthalmia 0 (± 0) 0 0 ± 0 0.74 ± 0.01 0 ± 0 0 ± 0 0 ± 0 0 ± 0 0 ± 0
Color changes 1.6 (± 0.02) 0 4.6 ± 0.02 12.4 ± 0.05 0 ± 0 1.5 ± 0.02 5.9 ± 0.04 1.3 ± 0.04 0 ± 0
Dead 4.9 (± 0.002) 3.3 7.6 ± 0.08 7.02 ± 0.04 10.7 ± 0.05 18.9 ± 0.19 27.5 ± 0.27 11 ± 0.27 100 ± 0

44
In addition to the sublethal effects, the embryos exposed to PHMB showed a linear
increase of the mortality during the exposure period (Y = 25.618X + 27.498, r2 = 0.7555,
p < 0.01). After the 144 h of exposure, IC50 of PHMB to snail embryos were 0.98 mg L-1
(upper limit: 0.97 – inferior limit: 1.0) (Table 1). The LC50 values of PHMB to B. glabrata
embryos were lower than the minimum inhibitory concentration (MIC: 0.5 to 4.0 mg L-
1) and minimum bactericidal concentration (MBC: 1.0 to 32 mg L-1) reported for gram-
positive and gram-negative bacteria (Koburger et al., 2010) and the LC50 of latex
Euphorbia milii (LC50-96h: 34.03 mg L-1) (Oliveira-Filho, 2010), confirming their toxicity
to early developmental stages of B. glabrata. On the other hand, the LC50 of PHMB for
B. glabrata embryos is higher than those reported for divaric acid (LC50, 24h: 0.01029 mg
L-1 (Silva et al. 2018) and potassium salt (LC50-24h 0.00577 μg ml-1) (Martins et al. 2014).
Table 2 - LC50-144 hours and LC50-96 hours values (in mg/L-1), based on embryos, new-borns and adult snails of
B. glabrata survival data analyses. Lowest Observed Effect Concentration (LOEC); No Observed Effect
Concentration (NOEC).
Parameters Embryos New-borns snail Adult snail
LC50 0.98
[0.97 – 1.0]
1.43
[ 0,44 – 2,00]
1.49
[ 0.78 – 1.64]
NOEC - - 0.6
LOEC 0.6 0.6 1.0
The toxicity of a compound to B. glabrata embryos depends on several factors
and particularities of the molecule studied. These organoleptic characteristics are
fundamental to determine the ability of the compound to penetrate the gelatinous capsule
of the egg, formed of a perivitelinic substance rich in lipids, proteins and glycogen, which
protects the embryo (Duarte et al., 2015; Hathaway et al., 2010; Miyasato et al., 2012;
Ribeiro-Paes et al., 1994).
3.2 Toxicity to new-borns and adult snails
The unexposed new-borns snail showed low mortality rate (5 ± 5 %) after 96 h,
while no mortality was observed in the and unexposed adult snails (Fig. 5). In addition,
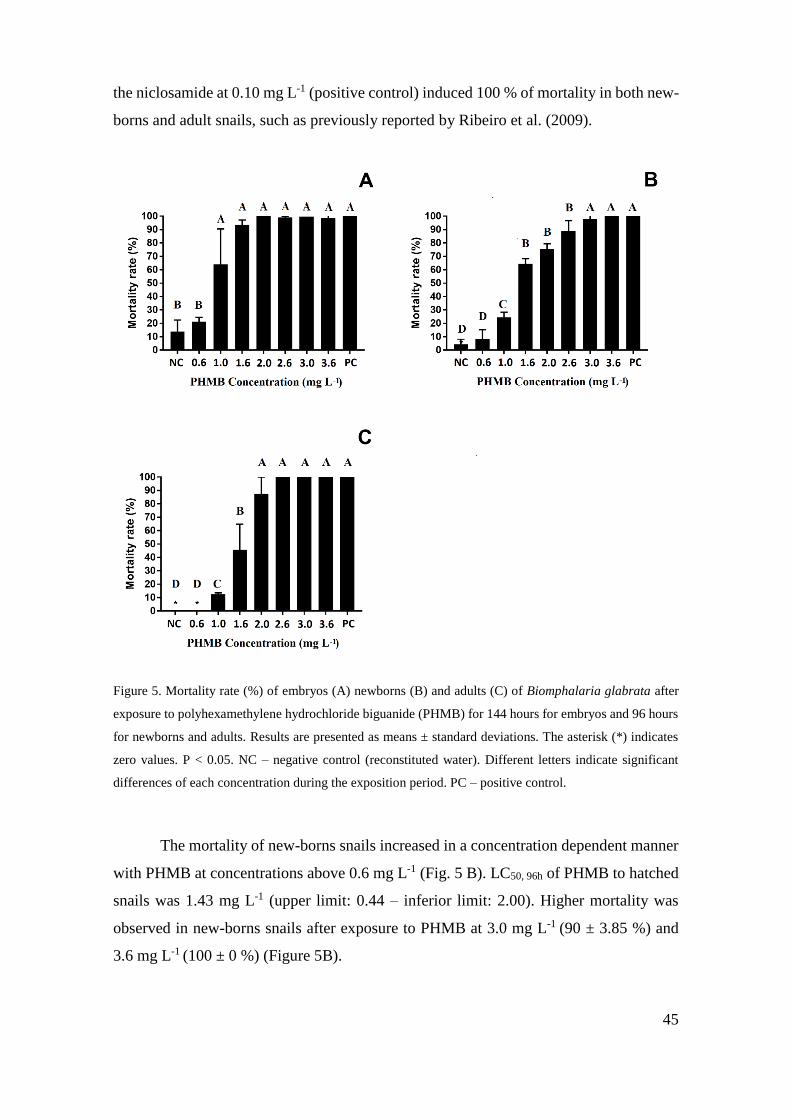
45
the niclosamide at 0.10 mg L-1 (positive control) induced 100 % of mortality in both new-
borns and adult snails, such as previously reported by Ribeiro et al. (2009).
Figure 5. Mortality rate (%) of embryos (A) newborns (B) and adults (C) of Biomphalaria glabrata after
exposure to polyhexamethylene hydrochloride biguanide (PHMB) for 144 hours for embryos and 96 hours
for newborns and adults. Results are presented as means ± standard deviations. The asterisk (*) indicates
zero values. P < 0.05. NC – negative control (reconstituted water). Different letters indicate significant
differences of each concentration during the exposition period. PC – positive control.
The mortality of new-borns snails increased in a concentration dependent manner
with PHMB at concentrations above 0.6 mg L-1 (Fig. 5 B). LC50, 96h of PHMB to hatched
snails was 1.43 mg L-1 (upper limit: 0.44 – inferior limit: 2.00). Higher mortality was
observed in new-borns snails after exposure to PHMB at 3.0 mg L-1 (90 ± 3.85 %) and
3.6 mg L-1 (100 ± 0 %) (Figure 5B).

46
Similar to new-borns snails, the toxicity of PHMB to adult snail was concentration
dependent (Table 1). The LC50 of PHMB for adult snail was 1.49 mg L-1 (0.78 – 1.64).
After 96 hours exposure, a higher mortality was observed after the exposure at 2.0 mg L-
1 (87.5 ± 12.5 %) and 2.6 mg L-1 (100 ± 0 %) (Figure 5C). The concentrations tested
showed statistically significant results in relation to the control (p <0.05).
The LC50 values for adult snail of B. glabrata obtained in this study were lower
than the values obtained in other older studies, such as the evaluation of molluscicidal
activity of Synadenium carinatum boiss (LC50-24h: 0.5 mg L-1); of lectins from Cratylia
floribunda (LC50-24h: 25.5 mg L-1); Dioclea guianensis (LC50-24h: 23.1 mg L-1);
Amphimedon viridis (LC50-24h: 20 mg L-1); Liagora farinosa (LC50-24h: 120 mg L-1); Piper
tuberculatum (LC50-24h: 4.19 mg L-1) and usnic acid potassium salt (LC50-24h: 5.77 mg L-
1) (Moreira et al. 2010; Santos et al. 2010; Miyassato et al. 2012; Rapado et al. 2013).
These results confirm that the PHMB has LC50 for embryos of B. glabrata smaller than
the studies mentioned.
The mortality of the B. glabrata embryos may be associated with PHMB
interference in the energy metabolism of the animals, as well as in the loss of calcium
ions and enzymes important for the cellular membrane, because in bacteria the MoA of
PHMB begins with the rapid attraction of the molecule, which has a cationic nature, on
the negatively charged bacterial surface, causing a failure in the defence mechanism of
the cell and rupture of the cell wall. PHMB is then attracted to the cytoplasmic membrane,
causing the loss of low molecular weight substances such as potassium ions, calcium and
inhibition of enzymes responsible for membrane junction, such as ATPase. Subsequently,
there is a rupture of the cytoplasmic membrane that can induce the loss of macromolecular
substances and the precipitation of cellular substances (Kaenh, 2010). This action
promoted by PHMB can justify the rupture of the cellular membrane of the cells of the
integument of B. glabrata snails, causing the hemolymph to be released. In addition, the
interaction between the compound and the egg mass can alter the gas exchanges
preventing embryo hatching. However, further studies are need to understand the MoA
of PHMB to snails and their effects on energy metabolism, calcium metabolism and
membrane integrity.
Previous studies with B. glabrata have confirmed that it is common for some
compounds to be more toxic to adult snails than embryos, in addition the evaluated

47
compound may be lethal to adult individuals without impeding embryo development
(Oliveira-Filho et al. 2010; Rapado et al. 2012). However, our data presented different
results, since the LC50 for embryos was lower than the LC50 for adult snails, and although
it does not impede the embryonic development, the PHMB prevents embryos from
hatching after 144 hours of analysis. Through the results found, we can state that the
PHMB can act in all the studied phases (embryos, new-borns and adults), being the
embryonic phase the most sensitive to the compound.
After 96 hours of exposure, no behavioural changes were observed in adult snails
from negative control, while 100 % of snails died after exposure to niclosamide (positive
control), inmates into their shells and displaced to the periphery of the aquariums. The
exposure to PHMB at 1.6 mg L-1 induced lethargy (26 ± 0.02 %), reclusion into the shell
(43 ± 0.02 %) and mucus secretion (63 ± 0.001 %), while 100 % of snails moved to the
edges of the aquarium. On the other hand, after exposure to PHMB at concentrations
above 2.6 mg L-1, all snails were already dead with reclusion into their shells and shifting
to the periphery of the test aquarium (Fig. 6). The behavior of the snail against exposure
of this compound, such as reclusion into the shell and mucus secretion demonstrate that
the molluscicidal action of PHMB is the result of several actions that affect more than a
system (WHO, 1983). Changes in behavior observed after exposure to compost can be
considered as a defence mechanism of the animal, the secretion of mucus in the opening
of the shell for example, can be understood as a way to block the entrance of the compost.
Figure 6. Proportion of changes in behaviour adults of Biomphalaria glabrata after the exposure to
polyhexamethylene hydrochloride biguanide (PHMB) for 96 hours. NC – negative control. PC – positive
control (niclosamide at 0.08 mg L-1).

48
Among the MoA of the PHMB, the breakdown of the cytoplasmic membrane due
to its electrostatic interaction with phospholipids present in it is outstanding (Kaehn,
2010; Mashat, 2016). In our study, all experimental aquaria with concentrations above
1.6 mg L-1 contained hemolymph scattered in the test solution. This fact may explain the
release of hemolymph from snails when in contact with the evaluated compound, a
behavior that was also observed in other studies evaluating the molluscicidal potential of
other substances (El-Beshbishi et al., 2015, Rapado et al., 2012).
PHMB demonstrated low toxicity to human liver cells when compared to
quaternary ammonium compounds such as BAC (composition: C12-C14- alkyl
(ethylbenzyl) dimethylammonium chloride), Barquat (composition: C14 alkyl 50 %, C16
dimethyl benzyl chloride 10 % ammonia, 40 % C12) and benzalkonium chloride. In
addition, PHMB also showed the highest LC50 and consequently low toxicity for
zebrafish fish cells when compared to BAC, Barquat, benzalkonium chloride and
glutaraldehyde (Christen et al. 2017), indicating that the PHMB has fewer side effects
such as toxicity and bioaccumulation, in non-target animals, than niclosamide.
4. Conclusion
The results of the present study demonstrated that the PHMB failed to interrupt
the development of the embryos until the last stage, a fact that can be explained by the
protection conferred by the gelatinous capsule around the embryos. However, even with
this result, and with the non-significant induction of malformations, PHMB was able to
inhibit embryonic hatching of B. glabrata, an effect that certainly contributes to the fight
against schistosomiasis.
All stages of B. glabrata snail evaluated in this study were susceptible to the toxic
effects of PHMB, the embryonic phase being the most sensitive. In new-borns snails the
LC50 was estimated to be 1.43 mg L-1. In adult snails PHMB was able to cause changes
in the snail behavior, and LC50 was estimated to be 1.49 mg L-1. These data demonstrate
that these results are within the parameters established by the WHO, which argues that
the molluscicidal activity of a compound is recognized at concentrations below 100 ppm
(WHO, 1993).

49
Acknowledgements
This work was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES, demanda social, edital 002/2016) and by Fundação de Amparo à
Pesquisa do Estado de Goiás (FAPEG, edital nº 04/17 – Programa Pesquisa para o SUS:
Gestão Compartilhada em Saúde ‒ FAPEG/SES-GO/CNPq/MS-
DECIT/2017– PPSUS/UFG/GO), Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq/MS/DECIT n. 37/2014. Projeto 470298/2014-6).
The authors thank the company Arch Quimica do Brasil for the donation of the
compound evaluated in the present study.

50
References
Albuquerque, L.P., Pontual, E.V., Santana, G.M.S., Silva, L.R.S., Aguiar, J.S., Coelho,
L.C.B.B, Rêgo, M.J.B.M., Pitta, M.G.R., Silva, T.G., Melo, A.M.M.A., Napoleão, T.H.,
Paiva, P.M.G., 2014. Toxic effects of Microgramma vacciniifolia rhizome lectin on
Artemia salina, human cells, and the schistosomiasis vector Biomphalaria glabrata. Acta
Tropica, 138. http://dx.doi.org/10.1016/j.actatropica.2014.06.005.
Brasil. Ministério da Saúde, 2008. Vigilância e Controle de Moluscos de Importância
Epidemiológica. Brasília. Editora MS.
Brasil. Ministério da Saúde, 2010. Guia de Vigilância Epidemiológica. 7a ed. Brasília.
Cad 10: 19–29. 2010.
Bruun, B., & Aagaard-Hansen, J., 2008. The social context of schistosomiasis and its
control: an introduction and annotated bibliography. World Health Organization, Geneva.
Christen, V., Faltermann, S., Brun, N.R., Kunz, P.Y., & Fent, K., 2017. Cytotoxicity and
molecular effects of biocidal disinfectants (quaternary ammonia, glutaraldehyde,
poly(hexamethylene biguanide) hydrochloride PHMB) and their mixtures in vitro and in
zebrafish eleuthero-embryos. Science of the Total Environment, 586.
https://doi.org/10.1016/j.scitotenv.2017.02.114.
Coelho, P.M.Z. & Caldeira, R.L., 2016. Critical analysis of molluscicide application in
schistosomiasis control programs in Brazil. Infectious Diseases of Poverty, 5.
http://doi.org/10.1186/s40249-016-0153-6.
Coura-Filho, P., Rocha, R.S., Lima e Costa, M.F.F.D., Katz, N., 1992. A municipal level
approach to the management of schistosomiasis control in Peri-Peri, MG, Brazil. Revista
do Instituto de Medicina Tropical de São Paulo, 34. http://dx.doi.org/10.1590/S0036-
46651992000600008.
Coustau, C., Gourbal, B., Duval, D., Yoshino, T.P., Adema, C.M., & Mitta, G., 2015.
Advances in gastropod immunity from the study of the interaction between the snail
Biomphalaria glabrata and its parasites: A review of research progress over the last
decade. Fish and Shellfish Immunology, 46. https://doi.org/10.1016/j.fsi.2015.01.036.

51
Tallarico, L.F., Borrely, S.I., Hamada, N., Grazeffe, V.S., Ohlweiler, F.P., Okazaki, K.,
Granatelli, A.T., Pereira, I.W., Pereira, C.A.B., Nakano, E., 2014. Developmental
toxicity, acute toxicity and mutagenicity testing in freshwater snails Biomphalaria
glabrata (Mollusca: Gastropoda) exposed to chromium and water samples.
Ecotoxicology and Environmental Safety, 110.
https://doi.org/10.1016/j.ecoenv.2014.09.005.
Duarte, G.F., Rodrigues, J., Fernandes, E.K.K., Humber, R.A., Luz, C., 2015. New
insights into the amphibious life of Biomphalaria glabrata and susceptibility of its egg
masses to fungal infection. Journal of Invertebrate Pathology.
https://doi.org/10.1016/j.jip.2014.12.013.
Ducrot, V., Charles, S., 2015. Development and validation of guidelines for mollusc
reproductive toxicity test. Regulatory Toxicology and Pharmacology,
https://doi.org/10.1016/j.yrtph.2014.09.004.
Firdessa, R., Good, L., Amstalden, M.C., Chindera, K., Kamaruzzaman, N.F., Schultheis,
M., Roger, B., Hecht, N., Oelschlaeger T.A., Meinel, L., Luhmann T., Moll, H., 2015.
Pathogen- and Host-Directed Antileishmanial Effects Mediated by Polyhexanide
(PHMB). PLoS Neglected Tropical Diseases, 9.
https://doi.org/10.1371/journal.pntd.0004041.
Fjeld, H., & Lingaas, E., 2016. Polyheksanid - Sikkerhet og effekt som antiseptikum.
Tidsskrift for Den Norske Laegeforening, 136. https://doi.org/10.4045/tidsskr.14.1041.
Grimes, J.E., Croll, D., Harrison, W.E., Utzinger, J., Freeman, M.C., Templeton, M.R.,
2015. The roles of water, sanitation and hygiene in reducing schistosomiasis: a review.
Parasites & Vectors, 8. https://doi.org/10.1186/s13071-015-0766-9.
Guimarães R.J.P.S., Freitas C.C., Dutra L.V., Felgueiras C.A., Moura A.C.M., Amaral
R.S., Drumond S.C., Scholte R.G.C., Oliveira G., Carvalho O.S., 2009. Spatial
distribution of Biomphalaria mollusks at São Francisco River Basin, Minas Gerais,
Brazil, using geostatistical procedures. Rev. Acta Tropica.
https://doi.org/10.1016/j.actatropica.2008.10.012.

52
Hartmann, D.B., Marim, R.A., Silva, Y.L., Zardeto G., Silva, I.A., Mattos, D.A., Júnior-
Laverde, A., 2011. Letalidade do extrato de Synadenium grantii hook. F. (Euphorbiaceae)
frente a caramujos Biomphalaria glabrata Say, 1818 (Gastropoda, Planorbidae).
Arquivos de Ciência Veterinária.
Zoológica. https://doi.org/10.25110/arqvet.v14i1.2011.3779.
Hathaway, J.J., Adema, C.M., Stout, B.A., Mobarak, C.D., Loker, E.S, 2010.
Identification of protein components of egg masses indicates parental investment in
immunoprotection of offspring by Biomphalaria glabrata (Gastropoda, Mollusca).
Developmental & Comparative Immunology. https://doi.org/10.1016/j.dci.2009.12.001.
Hotez, P.J., Alvarado, M., Basanez, M.G., Bolliger, I., Bourne, R., Boussinesq, M.,
Brooker, S.J., Brown, A.S., Buckle, G., Budke, C.M., Carabin, H., Coffeng, L.E., Fèvre,
E.M., Fürst, M., Halasa, Y.A., Jasrasaria, R., Johns, N.E., King, C.H., Lozano, L.,
Murdoch, M.E., O'Hanlon, O.S., Pion, S.D.S., Pullan, R.L., Ramaiah, K.D., Roberts, T.,
Shepard, D.S., Smith, J.L., Stolk, W.A., Undurraga, E.A, Utzinger, J., Wang, M., Murray,
C.J.L., Naghavi, M., 2014. The Global Burden of Disease study 2010: interpretation and
implications for the neglected tropical diseases. PLoS Neglected Tropical Diseases, 8.
https://doi.org/10.1371/journal.pntd.0002865.
Kaehn, K., 2010. Polihexanide: A safe and highly effective biocide. Skin Pharmacology
and Physiology, 23. https://doi.org/10.1159/000318237.
King, C.H. & Bertsch, D., 2015. Historical Perspective: Snail Control to Prevent
Schistosomiasis. PLoS Neglected Tropical Diseases, 9,
https://doi.org/10.1371/journal.pntd.0003657.
Koburger, T., Hübner, N.O., Braun, M., Siebert, J., & Kramer, A., 2010. Standardized
comparison of antiseptic efficacy of triclosan, PVP-iodine, octenidine dihydrochloride,
polyhexanide and chlorhexidine digluconate. Journal of Antimicrobial Chemotherapy,
65. https://doi.org/10.1093/jac/dkq212.
Lambertucci, J.R., 2010. Acute schistosomiasis mansoni: Revisited and reconsidered.
Memórias do Instituto Oswaldo Cruz, 105. http://dx.doi.org/10.1590/S0074-
02762010000400012.

53
Martins, M.C.B., Silva, M.C., Silva, L.R.S., Lima, V.L.M., Pereira, E.C., Falcão, E.P.S.,
Melo, A.M.M.A, Da Silva, N.H., 2014. Usnic acid potassium salt: An alternative for the
control of Biomphalaria glabrata (Say, 1818). PLoS ONE, 9.
https://doi.org/10.1371/journal.pone.0111102.
Mashat, B.H., 2016. Polyhexamethylene Biguanide Hydrochloride: Features and
Applications. British Journal of Environmental Sciences, 4.
http://www.eajournals.org/wp-content/uploads/Polyhexamethylene-Biguanide-
Hydrochloride-Features-and-Applications1.pdf.
Miyasato, P.A., Kawano, T., Freitas, J.C., Berlinck, R.G.S., Nakano, E., & Tallarico, L.F.,
2012. Molluscicidal activity of some marine substances against the snail Biomphalaria
glabrata (Mollusca, Planorbidae). Parasitology Research, 110.
https://doi.org/10.1007/s00436-011-2712-x.
Nacif-Pimenta, R., de Mattos, A.C.A., da Orfanó, A.S., Barbosa, L., Pimenta, P.F.P., &
Coelho, P.M.Z., 2012. Schistosoma mansoni in Susceptible and Resistant Snail Strains
Biomphalaria tenagophila: In Vivo Tissue Response and In Vitro Hemocyte Interactions.
PLoS ONE, 7. https://doi.org/10.1371/journal.pone.0045637.
OECD, 2016, Test No. 243: Lymnaea stagnalis Reproduction Test, OECD Guidelines for
the Testing of Chemicals, Section 2, OECD Publishing, Paris.
https://doi.org/10.1787/9789264264335-en.
Oliveira-Filho, E.C., Geraldino, B.R., Coelho, D.R., De-Carvalho, R.R., & Paumgartten,
F.J.R., 2010. Comparative toxicity of Euphorbia milii latex and synthetic molluscicides
to Biomphalaria glabrata embryos. Chemosphere, 81.
https://doi.org/10.1016/j.chemosphere.2010.06.038.
Moreira, CPS., Zani, C.L., Alves, T.M.A., 2010. Molluscicidal activity of Synadenium
Carinatum Boiss. (Euphorbiaceae) latex against Biomphalaria Glabrata and asolation of
aajor aompound, VII(3). https://doi.org/10.5216/ref.v7i3.12892.
Pereira, L.P.L.A., Dias, C.N., Miranda, M.V., Firmo, W.C.A., Rosa, C.S., Santos, P.F.,
Brito, M.C.A., Araruna, F.O.S., Araruna, F.B., Silva-Souza, N., Coutinho, D.F., 2017.
Molluscicidal effect of Euphorbia umbellata (Pax) Bruyns latex on Biomphalaria
glabrata, Schistosoma mansoni host snail. Revista do Instituto de Medicina Tropical de
São Paulo, https://dx.doi.org/10.1590/s1678-9946201759085.

54
Pile, E.A.M., Pastor, N., Santos, J.A., Barros, J.S.L., 2002. Aspectos histopátológicos de
Biophalaria glabrata Say, 1817 hospedeiro intermediário de Schistosoma mansoni, submetida a
Niclosamida. Brazilian Journal of Veterinary Research and Animal Science.
http://dx.doi.org/10.1590/S1413-95962002000400010.
R Core Team, 2016. R: A language and environmental for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria.
Rapado, L.N., Lopes, P.O.D.M., Yamaguchi, L.F., & Nakano, E., 2013 a. Ovicidal effect
of Piperaceae species on Biomphalaria glabrata, Schistosoma mansoni host. Revista Do
Instituto de Medicina Tropical de São Paulo, 55. https://doi.org/10.1590/S0036-
46652013000600009.
Rapado, L.N., Pinheiro, A. de S., Lopes, P.O. de M.V., Fokoue, H.H., Scotti, M.T.,
Marques, J.V., Ohlweiler, F.P., Borrely, S.I., Pereira, C.A.B., Kato, M.J., Nakano, E.,
Yamaguchi, L.F., 2013 b. Schistosomiasis Control Using Piplartine against Biomphalaria
glabrata at Different Developmental Stages. PLoS Neglected Tropical Diseases, 7.
https://doi.org/10.1371/journal.pntd.0002251.
Rassi, C., Kajungu D., Martin, S., Arroz, J., Tallant, J., Beyl, C.Z., Counihan, H., Newell,
J.N., Phillips, A., Muloliwa, A.M., Graham, K., 2016. Have You Heard of
Schistosomiasis? Knowledge, Attitudes and Practices in Nampula Province,
Mozambique. Neglected Tropical Diseases.
https://doi.org/10.1371/journal.pntd.0004504.
Ribeiro, K.A., de Carvalho, C.M., Molina, M.T., Lima, E.P., López-Montero, E., Reys,
J.R.M., Oliveira, M.B.F., Pinto, A.V., Santana, A.E.G., Goulart, M.O., 2009. Activities
of naphthoquinones against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae), vector
of dengue and Biomphalaria glabrata (Say, 1818), intermediate host of Schistosoma
mansoni. Acta Tropica. https://doi.org/10.1016/j.actatropica.2009.02.008.
Ribeiro-Paes, J.T., Costa P.I., Silva D., Barth L.R., Rodrigues V., 1994. Aspectos
imunológicos do sistema enzimático fenoloxidase de Schistosoma mansoni. Revista do
Instituto de Medicina Tropical. http://dx.doi.org/10.1590/S0036-46651994000500002.

55
Rocha-Filho, C.A.A., Albuquerque, L.P., Silva, L.R.S., Silva P.C.B., Coelho, L.C.C.B.,
Navarro, D.M.A.F., Albuquerque, M.C.P.A., Melo, A.M.M.A., Napoleão, T.H., Pontual,
V., Paiva, P.M.G., 2015. Assessment of toxicity of Moringa oleífera flower extract to
Biomphalaria glabrata, Schistosoma mansoni and Artemia salina. Chemosphere.
http://dx.doi.org/10.1016/j.chemosphere.2015.03.041.
Santos, A. F. dos, Cavada, B. S., Rocha, B. A. M. da, Nascimento, K. S. do, & Sant’Ana,
A. E. G., 2010. Toxicity of some glucose/mannose-binding lectins to Biomphalaria
glabrata and Artemia salina. Bioresource Technology, 101.
https://doi.org/10.1016/j.biortech.2009.07.062.
Say, T. 1818. Account of two new genera, and several new species, of fresh water and
land shells. J. Acad. Nat. Sci. Phila. 1: 276-284.
Schall, V.T, Vasconcellos, M.C., Rocha, S.R., Souza, C.P., Mendes, N.M., 2001. The
control of the schistosome-transmitting snail Biomphalaria glabrata by the plant
molluscicide Euphorbia splendens var. hislopii (syn milli Des. Moul): a longitudinal field
study in an endemic area in Brazil. Acta Tropica. https://doi.org/10.1016/S0001-
706X(01)00126-7.
Scholte, R.G.C., Gosoniu, L., Malone, J.B., Chammartin, F., Utzinger, J., Vounatsou, P.,
2014. Predictive risk mapping of schistosomiasis in Brazil using Bayesian geostatistical
models. Acta Tropica, 132: 57–63. https://doi.org/10.1016/j.actatropica.2013.12.007.
Silva, H. A. M. F., Siqueira, W. N., Sá, J. L. F., Silva, L. R. S., Martins, M. C. B., Aires,
A. L., Silva, N. H., 2018. Laboratory assessment of divaricatic acid against Biomphalaria
glabrata and Schistosoma mansoni cercariae. Acta Trop., 178.
https://doi.org/10.1016/j.actatropica.2017.09.019.
Sokolow, S.H., Wood, C.L., Jones, I.J., Swartz, S.J., Lopez, M., Hsieh, M.H., Lafferty,
K.D., Kuris A.M., Rickards C., De Leo G.A., 2016. Global Assessment of
Schistosomiasis Control Over the Past Century Shows Targeting the Snail Intermediate
Host Works Best. PLoS Neglcted Tropical Diseases, 10.
https://doi.org/10.1371/journal.pntd.0004794.
United States Environmental Protection Agency, 2002. Short-term Methods for
Estimating the Chronic Toxicity of Effluents and Receiving Waters to Freshwater

56
Organisms, 1. https://doi.org/http://www.dep.state.fl.us/water/wastewater/docs/ctf.pdf.
WHO. World Health Organization., 1983. Reports of the scientific working group on
plant molluscicides. Bulletin World Health Organization, 61(6).
https://doi.org/http://apps.who.int/iris/handle/10665/60086.
WHO. World Health Organization., 1993. The Control of Schistosomiasis, Technical
Report Series.
WHO. World Health Organization., 2002. World Health Organization. Niclosamide (2(,
5-dichloro-4(nitrosalicylanilide). WHO Specifications and Evaluations for Public Health
Pesticides. World Health Organization, Geneva. World Health Organization, 2015. Fact
Sheet 115.
WHO. World Health Organization, 2013. Schistosomiasis: progress report 2001 - 2011,
strategic plan 2012 - 2020. Available in: http://www.who.int/iris/handle/10665/78074.

57
Capítulo III
Artigo II
58
CAPÍTULO III – ARTIGO II
Graphical Abstract
Highlights
PHMB induces genotoxicity in snails B. glabrata in a non-concentration-
dependent manner.
Higher mutagenic effects in B. glabrata after at higher concentrations.
Hemocytes of B. glabrata are important target for the toxicity of PHMB.

59
Genotoxic and mutagenic effects of Polyhexamethylene biguanide
hydrochloride to Biomphalaria glabrata (Say, 1818)
Efeitos genotóxicos e mutagênicos do cloridrato polihexametileno
biguanida para Biomphalaria glabrata (Say, 1818)
Amanda de Oliveira Meloa; Daniela Braz dos? Santosa; Juliana Boaventura Avelara;
Daniela de Melo e Silvab; Thiago Lopes Rochac*; José Clecildo Barreto Bezerraa
aLaboratory of Studies of the Host-parasite Relationship, Institute of Tropical Pathology
and Public Health, Federal University of Goiás, Goiânia, Goiás.
b Laboratory of Mutagenese, Institute of Biological Sciences, Federal University of Goiás,
Goiânia, Goiás.
c Laboratory of Environmental Biotechnology and Ecotoxicology, Institute of Tropical
Pathology and Public Health, Federal University of Goiás, Goiânia, Goiás.
*Corresponding author at: T. L. Rocha, Universidade Federal de Goiás, Instituto de
Patologia Tropical e Saúde Pública, Rua 235, Setor Universitário, Goiânia, Goiás, Brasil.
CEP: 74605050, Goiânia, Goiás, Brasil. Tel.: +55 (62) 3209-6109; Fax: +55 (62) 3209-
6363. E-mail address: [email protected]

60
ABSTRACT
Schistosomiasis is a disease that is still prevalent in Africa, Asia and South America.
Considered the second most prevalent parasitic disease in the world, it affects more than
250 million people. The most affected patients live in conditions of poverty and little or
no sanitation, where there are few resources for research. The consensus on the best
control strategy for schistosomiasis has varied greatly over the years, but the World
Heatlh Organization (WHO) since 1940 advises on the use of integrated measures
including access to clean water and sanitation, health education, patient care and snail
control host of the parasite. Since then the efforts to discover new molluscicidal agents
have grown considerably. The present study aimed to evaluate the genotoxicity and
mutagenicity of polyhexamethylene biguanide hydrochloride (PHMB) in the
intermediate host of Schistosoma mansoni, the freshwater snail Biomphalaria glabrata.
Snails were exposed to PHMB (0.5, 1.0 and 1.5 mg L-1) for 48 h and the genotoxicity
(Comet assay – DNA damage) and mutagenicity (mincronucleus test and hemocyte
nuclear abnormalities assay) were analyzed in the circulating hemocytes. The results
showed that PHMB induced greater genotoxicity at 1.0 and 1.5 mg L-1 after 48 hours of
exposure. As for mutagenicity, PHMB induced the formation of micronuclei and nuclear
alterations at 1.0 mg L-1 after 48 hours of exposure. The results indicate that B. glabrata
hemocytes are immune cells suitable for evaluating the genotoxic and mutagenic effects
of potential molluscicidal compounds. Further investigations are needed to determine the
mode of action of PHMB on the snail and to explain the molluscicidal activity that this
compound has on B. glabrata.
Keywords: schistosomiasis; snail; micronucleus test, comet assay, intermediate host, S.
mansoni.

61
1. Introduction
According to the World Health Organization (WHO), about 240 million people
worldwide are affected by schistosomiasis, and another 700 million people live in areas
at risk of contracting the disease. The schistosomiasis belongs to the class of Neglected
Tropical Diseases (NTD) that cause great morbidity to the patients. This disease occurs
in 78 countries across South America, Asia, and Africa, and are originates from infection
with parasitic trematode worms of the genus Schistosoma. Among the damages caused
by this disease, can cite the decreases both the growth and intellectual development of
children and the production and working capacity of adults (Collaborators, 2017; Colley
et al., 2014; Sanogo et al., 2018).
Currently, three forms are used to control the transmission of schistosomiasis. The
first is periodical treatment of people living in risk areas with anti-schistosomicidal drugs.
The second method involves the elimination of the intermediate host from the biological
cycle of Schistosoma mansoni, such as the freshwater snail of the genus Biomphalaria
Last but not least, health education is the third form of disease control (Faria et al., 2018;
Othman & Soliman, 2015; WHO, 2010).
It is known that the distribution of the diseases caused by snailborne trematodes
especially schistosomiasis is focal. Therefore, the parasites distribution is strongly
dependente on the intermediate snail hosts distribution (Abe et al., 2018; Gordy et al.,
2016; Hanington et al., 2012). Among the intermediate host of schistosomiasis, the B.
glabrata snails play an integral role in the transmission of S. mansoni (Chistsulo et al.,
2000; Katz et al., 2018; Noya et al., 2015).
The WHO recommends utilization of niclosamide (Bayluscide®) to control of the
intermediate host of Schistosomiasis (WHO, 1992). However, the niclosamide has high
production value and can induced several secondary effects, such as photodegradation,
bioaccumulation and high toxicity in non-target animals (Faria et al., 2018; Hartmann et
al., 2011; Oliveira-Filho et al., 2010; Pereira et al., 2017). In addition, the re-colonization
of snail populations is observed after a few months of application of the niclosamide,
demonstrating the niclosamide-resistant snail populations following repeated, extensive
use of this molluscicide (Coura-Filho et al., 1992; Martins et al., 2014). Therefore, the
importance for the search of new compounds with potential molluscicidal activity.
Genotoxicity studies analyse the effects of xenobiotic agents on genetic material,
which may result in single or double strand breaks of DNA, also promoting the formation
of adducts. These changes occurring in the DNA molecule, when unrepaired may lead to

62
mutations and/or cell death, as well as decrease the fertility and their ability to respond to
environmental changes (Collins, 2004; Gunasekarana et al., 2015). In terms of
genotoxicity biomarkers, the comet assay has been indicated as a sensitive technique for
detection of DNA damage in hemocytes of several species of the molluscs (Ibrahim et al.
2018; Rocha-Filho et al., 2015). On the other hand, the micronucleus (MN) and hemocyte
abnormalities assay (HNA) are suitable approaches to assess the chromosomal alterations
(Filippi et al., 2018; Touahri et al. 2016).
The guanidines family, the Polyhexamethylene Hydrochloride Biguanide
(PHMB) is originally used as disinfectant and antiseptic (Mashat, 2016). PHMB acts on
gram positive and negative bacteria through the negatively charged phosphate groups
present in the wall of these microorganisms, promoting the extravasation of cytoplasm
that causes cell death (Kaehn, 2010). PHMB has been described as an inhibitory agent of
bacteria, fungi and parasites (Firdessa et al., 2015; Fjeld and Lingaas, 2016). The
mechanism of toxicity of PHMB has not yet been described for the snails of the genus
Biomphalaria sp.
In this study the genotoxic (DNA damage) and mutagenic effects (chromossal
alterations) of PHMB were analysed in hemolymph of in the adult snails B. glabrata
during acute exposure (48 h) in order to verify its mechanism of action (MoA) associated
to its molluscicidal activity.
2. Materials and Methods
2.1 Snails
Adult snails B. glabrata with shell diameter of 10 ± 2 mm and total weight of 0.40
± 0.10 g were kept at the Tropical Pathology and Public Health Institute of the Federal
University of Goiás, as recommended by the guideline of OECD nº 243 (OECD, 2016).
The snails were kept in aerated glass tanks of 24 × 16 × 9 cm diameter with a capacity of
40 L of dechlorinated water in light / dark cycles of 12:12 h, temperature of 25 ± 1 ° C
and pH around 7.0 ± 1. The snails were fed ad libitum three times a week with lettuce
leaves (Lactuca sativa).

63
2.2 Substance test
The Polyhexamethylene biguanide hydrochloride (PHMB), commercially known as
Vantocil IB® (CAS No. 32289-58-0; 20 % w / w), was donated from Arch Química do
Brasil (São Paulo, Brazil). The stock solution of PHMB (100 mg L-1) was made in Milli-
Q water.
2.3 Exposure
Adult snails were exposed to PHMB (0.5, 1.0, 1.5 mg L-1) during 48 h, jointly
with a negative control group kept in reconstituted water (OECD, 2016) in glass aquarium
(3 L final water volume in the aquarium). Experiments were conducted a triplicate design
under 12:12 h light/dark cycles and environmental temperature of 25 ± 1 °C. The snails
were no fed during the exposure period (OECD, 2016). The mortality was analyzed daily
(0, 24 and 48 h), while the cumulative mortality rate (%) was obtained at the end of the
experiment (48 h) was determined using the equation:
𝑀𝑜𝑟𝑡𝑎𝑙𝑖𝑡𝑦 𝑟𝑎𝑡𝑒 (%) = 𝑁𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑑𝑒𝑎𝑑 𝑠𝑛𝑎𝑖𝑙𝑠
𝑇𝑜𝑡𝑎𝑙 𝑛𝑢𝑚𝑏𝑒𝑟 𝑜𝑓 𝑠𝑛𝑎𝑖𝑙𝑠 ∗ 100
The animals were considered dead according to the parameters described by
Miyasato et al. (2011) and Tallarico et al. (2014): immobility, discoloration of shells,
exposure of visceral mass, release of hemolymph and lack of heart beats.
After the exposure period, the collection of hemolymph followed the procedures
described by Grazeffe et al. (2008). All the snails were carefully cleaned with absorbent
paper and 100 µL of hemolymph was collected by puncturing the foot with a
micropipette. The hemolymph samples were stored into 1.5 ml centrifuge microtube with
100 µL of EDTA.
2.4 Genotoxicity
The genotoxicity of PHMB in B. glabrata was evaluated by comet assay in
circulating hemocytes, according to Sing et al. (1988) and Grazeffe et al. (2008) with
modifications. A volume of 100 µL of hemolymph with EDTA was dissolved in 500 µL

64
of 0.5 % low melting agarose, and 100 µL of this solution was placed on microscope slide
covered with 1.5 % normal melting agarose (Sigma). After the solidification at 4°C (10 -
15 min), the slides were placed in lysis solution (1 % Triton X-100, 10 % DMSO, Stock
Lysis Solution pH = 10) at 4 °C during 2 h. The electrophoresis was conducted alkaline
conditions (300 mM NaOH, 1 mM EDTA, pH > 13, 30 min unwinding, 30 min
electrophoresis at 300 mA and 25V, at 4 °C). The slides were then neutralized with 0.4
M Tris buffer, pH 7.5, fixed with absolute ethanol for 10 min, and stained with 50 µL
SYBR green Staining solution S (430 Sigma Aldrich). The comets (n = ??? per animal; n
= ??? per experimental condition) were analyzed under a fluorescence microscope (Carl
Zeiss) associated to ISIS® software (MetaSystems, Altlußheim, Germany) at 400×, with
an exciting filter of 515–560 nm and a barrier filter of 590 nm. Slides were coded
independently and scored blindly.
For the evaluation of the DNA damage, used the TriTek Comet ScoreTM
program, version 1.5. This software evaluated pixel intensity to provide corresponding
values to estimate genomic damage, as arbitrary units (AU). DNA damages was
quantified in terms of the tail length (TL), the percentage of DNA in the tail (% DNA),
and the Olive tail moment (OTM) (Collins 2004). To classify the DNA degree in the tail,
the cells were classified according to the parameters of Almeida et al. (2011): zero or
minimal 10 % tail, low damage 10 e 25 %, mid damage 25 e 50 %, high damage 50 e 75
%, and extreme damage > 75 %.
2.5 Mutagenicity
Mutagenicity of PMBH to snails was analyzed by micronucleus (MN) test and
hemocyte abnormalities assay (HNA) according to the protocol described by Carrasco et
al. (1990) with modifications. The procedures used to prepare the slides in the MN test
were performed according to Silva (2010). A volume of 100 μl of hemolymph was
collected from each snail (n = 5 per experimental condition), which was centrifuged at
3000 rpm for 5 minutes at 4 °C. A volume of 20 μL of the centrifuged hemolymph was
then spread on a slide, which was placed in a moist chamber for 30 minutes. The slides
were fixed with methanol (10 %) for 10 minutes and stained with Pantype solution
(NewProv) for 10 minutes. An average of 3000 intact cells with nuclear and cellular
distinction were counted per slide (n = 3000 per animal; n = 3000 per experimental

65
condition). The MN test and HNA were measured by determining the frequency of
hemocytes with micronucleus (MN), binucleated nucleus (BN), lobed nucleus (LN),
kidney-shaped nucleus (KN), blebbed nucleus (BB) and nucleus type broken-eggs (BE),
according to Fenech et al. (2003); Pavlica et al. (2000) and Vignardi et al. (2015).
2.6 Statistical analysis
The results were compared using parametric tests (two-way ANOVA, followed
by the Tukey’s test) and/or non-parametric test (Kruskall-Wallis), depending on the
distribution of the data and homogeneity of variance (Shapiro-Wilk and Levene's tests).
Results were considered significant when p<0.05. Statistical analyses were carried out
using the Statistica 7.0 software (Statsoft Inc., 2005, Tulsa, OK, USA).
3. Results and discussion
3.1 Comet assay
DNA damage in B. glabrata adult hemocytes was analyzed by the comet assay,
and the results were expressed as percentage (%) DNA in the tail of the comet (Figure 1
and Figure 2). The unexposed snails (control group) showed no significant DNA damage,
and no significant changes were detected in any of the parameters during the experiment
(p> 0.05; Fig. 1). After 48 hours of exposure, early DNA damage was detected in
hemocytes of snails exposed to PHMB at 1.0 mg L-1, 2.9-fold greater than DNA damage
caused by the non-exposed group (p < 0.05; Figure 1), while the PHMB at 1.5 mg L-1
induced higher DNA damage (2.4-fold) when compared to control group. But this
damage was lower than the damage caused by concentration of 1.0 mg L-1 in the same
evaluated period (p <0.05).
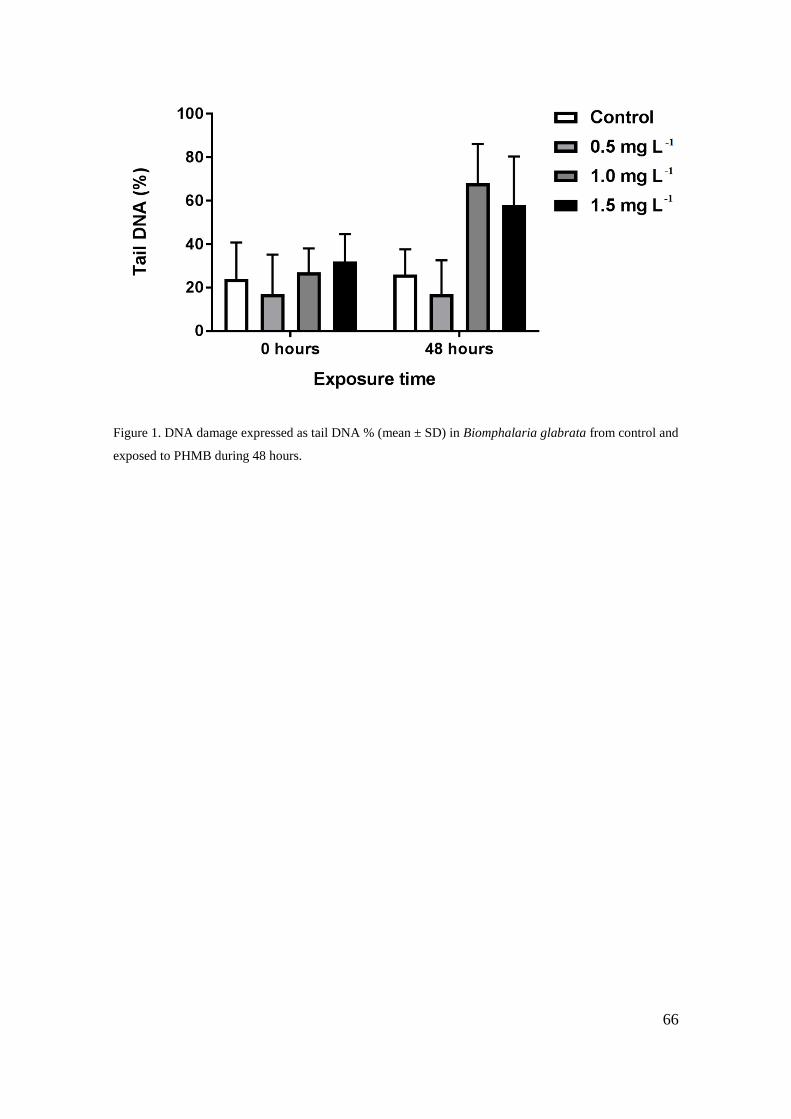
66
Figure 1. DNA damage expressed as tail DNA % (mean ± SD) in Biomphalaria glabrata from control and
exposed to PHMB during 48 hours.
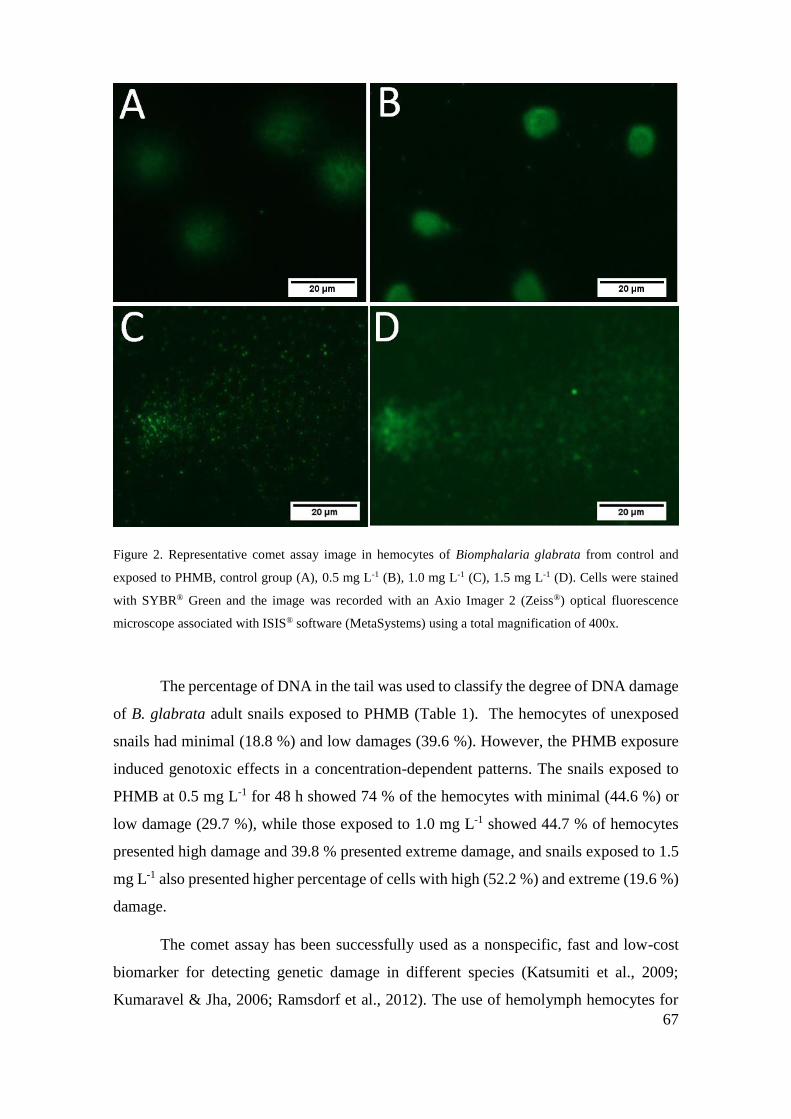
67
Figure 2. Representative comet assay image in hemocytes of Biomphalaria glabrata from control and
exposed to PHMB, control group (A), 0.5 mg L-1 (B), 1.0 mg L-1 (C), 1.5 mg L-1 (D). Cells were stained
with SYBR® Green and the image was recorded with an Axio Imager 2 (Zeiss®) optical fluorescence
microscope associated with ISIS® software (MetaSystems) using a total magnification of 400x.
The percentage of DNA in the tail was used to classify the degree of DNA damage
of B. glabrata adult snails exposed to PHMB (Table 1). The hemocytes of unexposed
snails had minimal (18.8 %) and low damages (39.6 %). However, the PHMB exposure
induced genotoxic effects in a concentration-dependent patterns. The snails exposed to
PHMB at 0.5 mg L-1 for 48 h showed 74 % of the hemocytes with minimal (44.6 %) or
low damage (29.7 %), while those exposed to 1.0 mg L-1 showed 44.7 % of hemocytes
presented high damage and 39.8 % presented extreme damage, and snails exposed to 1.5
mg L-1 also presented higher percentage of cells with high (52.2 %) and extreme (19.6 %)
damage.
The comet assay has been successfully used as a nonspecific, fast and low-cost
biomarker for detecting genetic damage in different species (Katsumiti et al., 2009;
Kumaravel & Jha, 2006; Ramsdorf et al., 2012). The use of hemolymph hemocytes for

68
genotoxic evaluation through the comet assay has already been described in other species
of molluscs (Al-Fanharawi et al., 2018; Bhattacharya et al., 2016; Sarkar et al., 2013),
and the positive result of the use of these cells is due to the fact that hemocytes play a
fundamental role in the transport of substances toxins that enter the body as well as the
defence mechanisms of animals (Bolognesi et al., 2004; Grazeffe et al., 2008), such as
observed in the B. glabrata exposed to PHMB (Figure 2).
Table 1 - Frequency of hemocytes (%) distributed by grade of DNA damage in Biomphalaria glabrata
from control and exposed to PHMB for 48 and 96 hours.
Control
Time (h)
0
Minimal
damage
18.8
Low
damage
39.6
Mid
damage
33.7
Hight
damage
6.9
Extreme
damage
1.0
48 22.5 41.9 29.9 4.8 0.9
0.5 mg L-1 48 44.6 29.7 21.8 3.5 0.5
1.0 mg L-1 48 0.0 2.9 12.6 44.7 39.8
1.5 mg L-1 48 4.3 6.5 17.4 52.2 19.6
PHMB penetrates the nucleus of the hemocytes and interacts with the DNA or
nuclear proteins indicating that this compound has genotoxic action and causes DNA
fragmentation in the hemocytes of the B. glabrata snail, since other authors have
demonstrated that PHMB does not interact indirectly with the molecule in mammalian
cells (Muller et al. 2013; Creppy et al., 2014).
Since there are no studies evaluating the genotoxic and mutagenic effects of the
exposure of other chemical compounds on the B. glabrata snail, it can’t be said that the
damage caused is a consequence of secondary non-nucleus lesions, such as through
interaction with other nuclear structures.
According to previous studies (OECD 2002a, OECD 2002b, OECD 2002c), the
PHMB was not classified as genotoxic for bacteria and mammalian cell. However, the
results of the present study demonstrated that PHMB is genotoxic for B. glabrata snail,
inducing DNA damage, as can be observed by the results of the comet assay.

69
In addition, PHMB does not induce significant oxidative stress in mammalian
cells, with MDA (malondialdehyde) production or lipoperoxidation, nor does it induce
DNA hydroxylation and / or hypermethylation. PHMB also failed to induce significant
production of mitogenic cytokines, such as TNF-α (tumor necrosis factor), interleukins
(IL-1 alpha) and transcription factor kappa nuclear factor B (NF-κB) that can cause
apoptosis or stimulate the growth of transformed cells or tumors, indicating that there was
no DNA cleavage, as demonstrated by Creppy et al. (2014). Therefore, this leads us to
believe that the damage caused by this compound is the result of direct interaction with
the DNA molecule.
The mechanism by which PHMB kills cells from different organisms is still the
subject of much research. It is known that mammalian cells treated with PHMB had no
or very weak activation of Caspase-3, indicating that the cells are not undergoing
apoptosis (Creppy et al., 2014). In our results, we observed that the hemocytes of the
snails after exposure to the concentration of 1.5 mg L-1 underwent apoptosis, which
decreased the amount of cells per slide and prevented the evaluation of this concentration
in the MN test.
3.2 Micronucleus Test
Six types of HNA induced by PHMB exposure were found in the present study
(Figure 4). The mutagenic effects induced by PHMB were concentration dependent, as
higher HNA were found at higher concentrations. It was not possible to observe nuclear
alterations in hemocytes of snails in the concentration of 1.5 mg L-1. The frequency of
MN and HNA in the control group remained unchanged throughout the bioassay. After
exposure to PHMB at 1.0 mg L-1, it was possible to observe 12, 86% of cells with some
kind of nuclear alteration. Moreover, in this same concentration, 6.16% had micronuclei,
and about 1.6% had 2 nuclei (binucleated haemocytes), corresponding to 35% and 13%,
respectively, of cells with this type of nuclear alteration of all the changes found. Figures
3 and 4 show some of the cellular changes that were analyzed in adult B. glabrata snails.
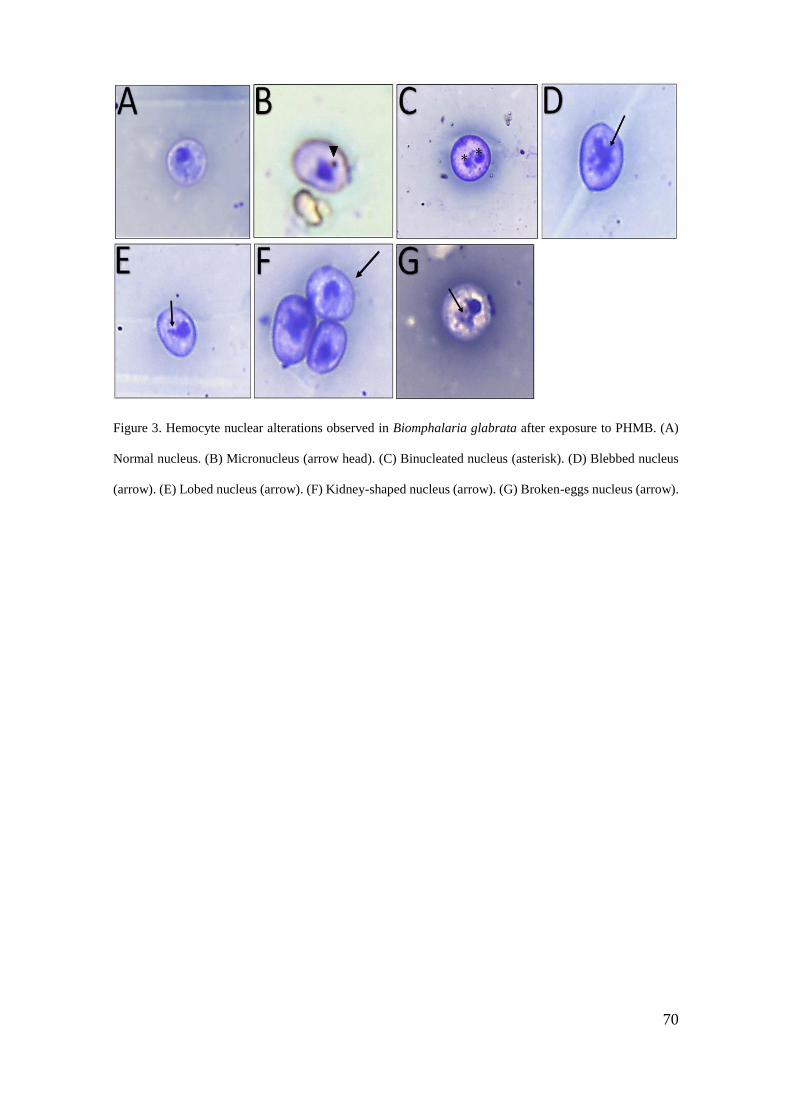
70
Figure 3. Hemocyte nuclear alterations observed in Biomphalaria glabrata after exposure to PHMB. (A)
Normal nucleus. (B) Micronucleus (arrow head). (C) Binucleated nucleus (asterisk). (D) Blebbed nucleus
(arrow). (E) Lobed nucleus (arrow). (F) Kidney-shaped nucleus (arrow). (G) Broken-eggs nucleus (arrow).
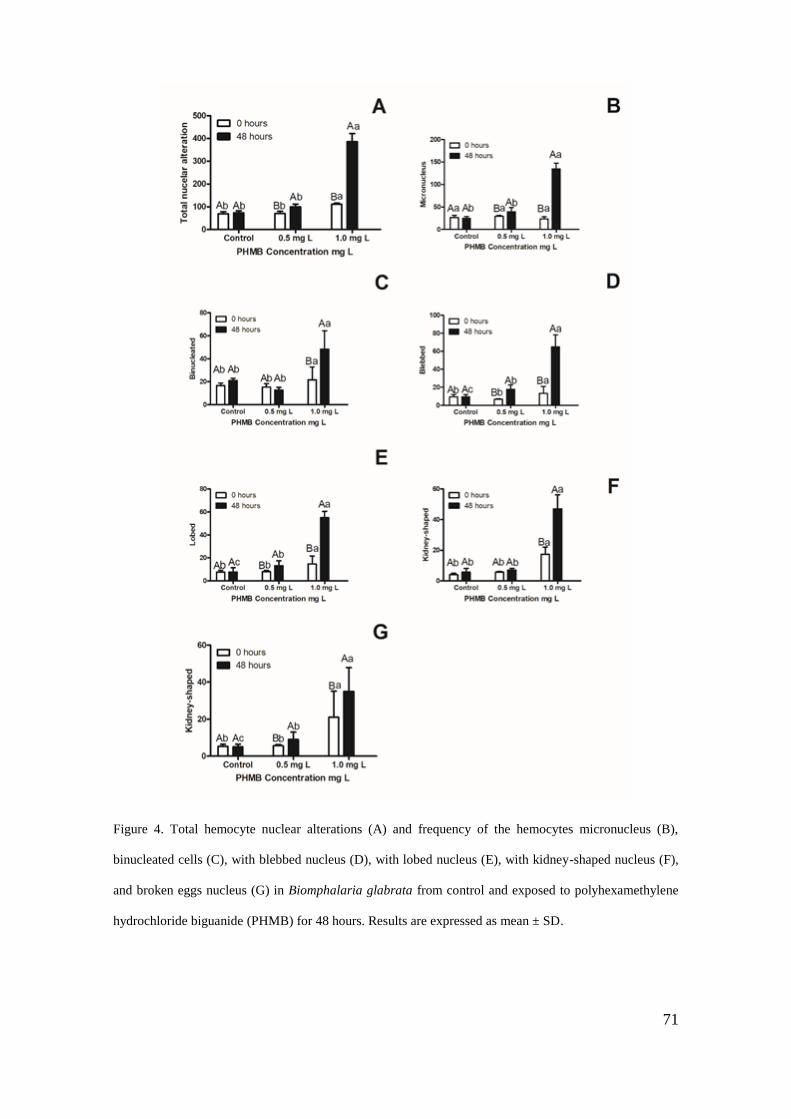
71
Figure 4. Total hemocyte nuclear alterations (A) and frequency of the hemocytes micronucleus (B),
binucleated cells (C), with blebbed nucleus (D), with lobed nucleus (E), with kidney-shaped nucleus (F),
and broken eggs nucleus (G) in Biomphalaria glabrata from control and exposed to polyhexamethylene
hydrochloride biguanide (PHMB) for 48 hours. Results are expressed as mean ± SD.

72
Genotoxicity tests are performed to clarify the effects of xenobiotic agents on
genetic material and changes in gene products (polypeptides and proteins). These agents
can cause single or double strand breaks of DNA. These changes in the DNA molecule,
when unrepaired, can cause mutations (Grisolia 2005).
The MN test is a method that provides a comprehensive basis for investigating the
harmful potential of the chromosome because both aneugenic and clastogenic origin
damage can be detected in cells that have been subjected to cell division during or after
exposure to the chemical (OECD 2014).
Micronuclei (MN) are small fragments that form whenever a chromosome or a
fragment of a chromosome is not embedded in one of the daughter cell nuclei after cell
division due to lack of centromere, centromere defects, or cytokine defects (Touahri et
al., 2016; Vincent- Hubert et al., 2011). As previously described, PHMB does not cause
indirect DNA damage, ie through mechanisms such as oxidative stress, lipoperoxidation
in mammalian cells (Creppy et al., 2014). In this context the results of the HNA frequency
of the short-term exposure of PHMB leads us to infer that the mechanisms that lead to the
formation of these nuclear alterations are derived from the direct interaction of the
compound with the DNA molecules. In addition, blebbed and fragmented nuclei, and
binucleate cells have been associated with failure of tubulin polymerization and mitotic
spindles caused by aneugenic actions (Qualhato et al., 2017; Sadiqul et al., 2016).
The binding between PHMB with single stranded DNA and double stranded DNA
and RNA molecules has been described (Allen et al., 2004). Taking into account that
PHMB interacts with the genetic code, and the fact that transcriptional regulation is a key
factor in the ability to adapt to hostile environments, after this interaction of the compound
with the DNA molecule, the mechanism of cell repair is probably acting to correct this
damaged DNA, since a large amount of damage is not observed through the micronucleus
test.
4. Conclusion
The present study evaluated the genotoxic and mutagenic effects of the PHMB,
which has been indicated as potential a molluscicide against S. mansoni intermediate host
snail. This is the first study to evaluate the genotoxicity and mutagenicity of this

73
compound in B. glabrata snail. The hemocytes of B. glabrata were cells of the immune
system suitable for analysis of DNA damage and nuclear alterations induced by
compounds with potential molluscicidal action.
The results indicated that the comet assay was an effective technique to assess
early DNA damage induced by exposure of PHMB in B. glabrata hemocytes. Other
studies indicated that PHMB is not genotoxic for other species, but the same compound
has been shown to be genotoxic to the snail, demonstrating a specific toxicity to B.
glabrata at the concentration of 1.0 mg L-1. The results presented by the MN test and
HNA showed that the PHMB at the concentration of 1.0 mg L-1 and the exposure time of
48 hours induced the formation of MN and nuclear alterations in hemocytes of B. glabrata.
The results indicated that the PHMB has genotoxic action on the genetic material of B.
glabrata.
Acknowledgements
This work was funded by the Foundation for Research Support of the State of
Goiás (FAPEG, edital nº 04/17 - Program for Research on SUS: Shared Health
Management - FAPEG / SES-GO / CNPq / MS-DECIT / 2017- PPSUS / UFG / GO) and
by the Coordination for the Improvement of Higher Education Personnel (CAPES, social
issue 002/2016), and the National Council for Scientific and Technological Development
(CNPq / MS / DECIT 37/2014 Project 470298/2014 -6 ]). We would like to thank the
Mutagenic Laboratory of the Federal University of Goiás (Labmut - UFG).
The authors thank the company Arch Quimica do Brasil for the donation of the
compound evaluated in the present study.

74
References
Abe, E.M., Guan, W., Guo, Y.H., Kassegne, K., Qin, Z.Q., Xu, J., Chen, J.H., Ekpo, U.F.,
Li, S.Z., Zhou, X.N., 2018. Differentiating snail intermediate hosts of Schistosoma spp.
Using molecular approaches: fundamental to successful integrated control mechanism in
Africa. Infectious diseases poverty, 7. https://doi.org/10.1186/s40249-018-0401-z.
Al-Fanharawi, A.A., Rabee, A.M., Al-Mamoori, A.M.J., 2018. Biochemical and
molecular alterations in freshwater mollusks as biomarkers for petroleum product,
domestic heating oil. Ecotoxicology and Environmental Safety, 158, 69-77,
https://doi.org/10.1016/j.ecoenv.2018.04.006.
Allen, M.J., Morby, A.P., White, G.F., 2004. Cooperativity in the binding of the cationic
biocide polyhexamethylene biguanide to nucleic acids. Biochemical and Biophysical
Research Communications, 318, 397-404, https://doi:10.1016/j.bbrc.2004.04.043.
Almeida, C., Pereira, C., Gomes, T., Bebianno, M.J. Cravo, A., 2011. DNA damage as a
biomarker of genotoxic contamination in Mytilus galloprovincialis from the south coast
of Portugal. Journal Enviromental Monitoring, 13, https://doi:10.1039/c1em10190k.
Bolognesi, C., Buschini, A., Branchi, E., Carboni, P., Furlini, M., Martino, A.,
Monteverde, M., Poli, P., Rossi, C., 2004. Comet and micronucleus assay in zebra mussel
cells for genotoxicity assessment of surface drinking water treated with three different
disinfectants. The Science of the total enviroment, 333, 127-
136, https://doi:10.1016/j.scitotenv.2004.05.018.
Bhattacharya, P., Swarnakar, S., Mukhopadhyay, A., Ghosh, S., 2016. Exposure of
composite tannery effluent on snail, Pila globosa: A comparative assessment of toxic
impacts of the untreated and membrane treated effluents. Ecotoxicology and
Environmental Safety, 126, 45-55, http://dx.doi.org/10.1016/j.ecoenv.2015.12.021.
Carrasco, K.R., Tilbury, K.L., & Myers, M.S., 1990. Assessment of the piscine
micronucleus test as an in situ biological indicator of chemical contaminant
effects. Canadian Journal of Fisheries and Aquatic Sciences, 47, 219-224,
https://doi.org/10.1139/f90-237.

75
Centres for Disease Control and Prevention: Schistosomiasis Fact Sheet, 2010.
https://www.cdc.gov/globalhealth/ntd/diseases/schisto_burden. html Weblink. Accessed
14 April, 2018.
Chitsulo, L., Engels, D., Montresor, A., Savioli, L., 2000. The global status of
schistosomiasis and its control. Acta Tropical, 77, 41-51, https://doi.org/10.1016/S0001-
706X(00)00122-4.
Collaborators, G.C.o.D., 2017. Global, regional, and national age-sex specific mortality
for 264 causes of death, 1980- 2016: a systematic analysis for the Global Burden of
Disease Study 2016. Lancet, 390. https://doi.org/10.1016/S0140-6736(17)32152-9.
Colley, D.G., Bustinduy, A.L., Secor, W.E., King, C.H., 2014. Human schistosomiasis.
Lancet, 383, 1151-1210, https://doi.org/10.1016/S0140-6736(13)61949-2.
Collins, A.R., 2004. The Comet Assay for DNA Damage and Repair. Molecular
Biotechnology, https://doi.org/10.1385/MB:26:3:249.
Collins, A.R., 2014. Measuring oxidative damage to DNA and its repair with the comet
assay. Biochimica et Biophysica Acta, 1840, 794-800,
http://dx.doi.org/10.1016/j.bbagen.2013.04.022.
Coura-Filho P., Rocha R.S., Lima e Costa M.F.F.D., Katz N., 1992. A municipal level
approach to the management of schistosomiasis control in Peri-Peri, MG, Brazil. Revista
do Instituto de Medicina Tropical de São Paulo, 34, 543-548,
http://dx.doi.org/10.1590/S0036-46651992000600008.
Creppy, E.E., Diallo, A., Moukha S., Eklu-Gadegbeku, C., Cros, D., 2014. Study of
Epigenetic Properties of Poly(HexaMethylene Biguanide) Hydrochloride (PHMB).
International Journal of Environmental Research and Public Health, 11, 8069-8092,
http://doi:10.3390/ijerph110808069.
Faria, R.X., Rocha, L.M., Souza, E.P.B.S.S., Almeida, F.B., Fernandes, C.P., Santos,
J.A.A., 2018. Molluscicidal activity of Manilkara subsericea (Mart.) dubard on
Biomphalaria glabrata (Say, 1818). Acta tropica, 178, 163-168,
http://doi.org/10.1016/j.actatropica.2017.11.012.

76
Fenech, M., 2003. The in vitro micronucleus technique. Mutation reserch, 455, 81-90,
https://doi.org/10.1016/S0027-5107(00)00065-8.
Firdessa, R., Good, L., Amstalden, M.C., Chindera, K., Kamaruzzaman, N.F., Schultheis,
M., Roger, B., Hecht, N., Oelschlaeger T.A., Meinel, L., Luhmann T., Moll, H., 2015.
Pathogen- and Host-Directed Antileishmanial Effects Mediated by Polyhexanide
(PHMB). PLoS Neglected Tropical Diseases, 9,
https://doi.org/10.1371/journal.pntd.0004041.
Fjeld, H., & Lingaas, E., 2016. Polyheksanid - Polyhexanide - safety and efficacy as an
antiseptic. Tidsskrift for Den Norske Laegeforening, 136, 707-711,
https://doi.org/10.4045/tidsskr.14.1041.
Gunasekarana, V., Raj, G.V., Chand, P., 2015. A Comprehensive Review on Clinical
Applications of Comet Assay. Journal of Clinical & Diagnostic Research, 9,
https://doi.org/ 10.7860/JCDR/2015/12062.5622.
Grisolia, C.K., 2005. Agrotóxicos- Mutações, Câncer e Reprodução. Brasília.
Grazeffe, V.S., de Freitas Tallarico, L., de Sa Pinheiro, A., Kawano, T., Suzuki, M.F.,
Okazaki, K., Pereira, C.A.B., Nakano, E., 2008. Establishment of the comet assay in the
freshwater snail Biomphalaria glabrata (Say, 1818). Mutation Research/Genetic
Toxicology and Environmental Mutagenesis, 654, 58-63,
https://doi.org/10.1016/j.mrgentox.2008.05.007.
Gordy, M.A., Kish, L., Tarrabain, M., Hanington, P.C., 2016. A comprehensive survey
of larval digenean trematodes and their snail hosts in central Alberta, Canada.
Parasitology Research, 115, 3867-3880, https://doi:
https://doi.org/10.1007/s0043601651529.
Hanington, P.C., Forys, M.A., Loker, E.S., 2012. A somatically diversified defense
factor, FREP3, is a determinant of snail resistance to schistosome infection. PLoS Negl
Trop Dis. https://doi.org/10.1371/journal.pntd.0001591.2012.
Hartmann, D.B., Marim, R.A., Silva, Y.L., Zardeto, G., Silva, I.A., Mattos, D.A., Júnior-
Laverde, A., 2011. Letalidade do extrato de Synadenium grantii hook. F. (Euphorbiaceae)
frente a caramujos Biomphalaria glabrata Say, 1818 (Gastropoda, Planorbidae).

77
Arquivos de Ciência Veterinária Zoológica, 14, 5-
11, https://doi.org/10.25110/arqvet.v14i1.2011.3779.
Kaehn, K., 2010. Polihexanide: A safe and highly effective biocide. Skin Pharmacology
and Physiology, 23. https://doi.org/10.1159/000318237.
Katsumiti, A., Valdez Domingos, F.X., Azevedo , M., da Silva, M.D., Damian , R.C.,
Almeida , M.I.M., Silva de Assis, H.C., Cestari, M.M., Randi , M.A.F., Ribeiro, O.C.A.,
Freire, C.A., 2009. An assessment of acute biomarker responses in the demersal catfish
Cathorops spixii after the Vicuña Oil Spill in a harbour estuarine area in Southern Brazil.
Environmental Monitoring Assessement 152. https://doi.org/10.1007/s10661-008-0309-
3.
Katz, N., 2018. Inquérito Nacional de Prevalência da Esquistossomose mansoni e Geo-
helmintoses. Avaliable in: https://www.arca.fiocruz.br/handle/icict/25662.
Knight, M., Ongele, E., Lewis, F.A., 2000. Molecular studies of Biomphalaria glabrata,
an intermediate host of Schistosoma mansoni. International Journal for Parasitology, 30,
535-540, https://doi.org/10.1016/S0020-7519(99)00182-4.
Kumaravel, T.S. & Jha, A.N., 2006. Reliable comet assay measurements for detecting
dna damage induced by ionising radiation and chemicals. Mutation research, 605, 7-16,
https://doi.org/10.1016/j.mrgentox.2006.03.002.
Lee, R.F. & Steinert, S., 2003. Use of the single cell gel electrophoresis/comet assay for
detecting DNA damage in aquatic (marine and freshwater) animals. Mutation
Research/Reviews in Mutation Research 544, 43-64, https://doi.org/10.1016/S1383-
5742(03)00017-6.
Martins, M.C.B., Silva, M.C., Silva, L.R.S., Lima, V.L.M., Pereira, E.C., Falcão, E.P.S.,
Melo, A.M.M.A., Silva, N.H., 2014. Usnic Acid Potassium Salt: An Alternative for the
Control of Biomphalaria glabrata (Say, 1818). PLoS ONE.
http://doi:10.1371/journal.pone.0111102.
Mashat, B.H., 2016. Polyhexamethylene Biguanide Hydrochloride: Features and
Applications. British Journal of Environmental Sciences, 4, 49-55,
http://www.eajournals.org/wp-content/uploads/Polyhexamethylene-Biguanide-
Hydrochloride-Features-and-Applications1.pdf.

78
Miyasato, P.A., Kawano, T., Freitas, J.C., Berlinck, R.G.S., Nakano, E., & Tallarico, L.F.,
2011. Molluscicidal activity of some marine substances against the snail Biomphalaria
glabrata (Mollusca, Planorbidae). Parasitology Research, 110, 1873-1879,
https://doi.org/10.1007/s00436-011-2712-x.
Noya, O., Katz, N., Pointier, J. P., Theron, A., & de Noya, B. A., 2015. Schistosomiasis
in America. Neglected Tropical Diseases-Latin America and the Caribbean, 11-43,
https://doi.org/10.1007/978-3-7091-1422-3_2.
OECD, 2016, Test No. 243: Lymnaea stagnalis Reproduction Test, OECD Guidelines for
the Testing of Chemicals, Section 2, OECD Publishing, Paris.
https://doi.org/10.1787/9789264264335-en.
Oliveira-Filho, E.C., Geraldino, B.R., Coelho, D.R., De-Carvalho, R.R., & Paumgartten,
F.J.R., 2010. Comparative toxicity of Euphorbia milii latex and synthetic molluscicides
to Biomphalaria glabrata embryos. Chemosphere, 81, 218-227,
https://doi.org/10.1016/j.chemosphere.2010.06.038.
Othman, A.A. & Soliman, R.H., 2015. Schistosomiasis in Egypt: A never-ending story?
Acta Tropica, 148, 179-190, https://doi.org/10.1016/j.actatropica.2015.04.016.
OECD 471. “Reverse Mutation Assay on Salmonella typhimurium his- and Escherichia
coli on PHMB P20D” according to OECD Guideline N°471; GLP Study N°2002-
DKY543-2 of September 5th, 2002. Unpublished work, 2002.
OECD 473. “In vitro Mammalian Chromosome Aberration Test Using Chinese Hamster
Ovary Cells (CHO) on PHMB P20D” according to OECD Guideline N°473; GLP Study
N°2002-DKY543-1 of September 5th, 2002. Unpublished work, 2002.
OECD 476. “In vitro Mammalian Cell Gene Mutation Test on PHMB P20D” according
to OECD Guideline N°476; GLP Study N°2002-DKY543-3 of September 5th, 2002.
Unpublished work, 2002.
Pereira, L.P.L.A., Dias, C.N., Miranda, M.V., Firmo, W.C.A., Rosa, C.S., Santos, P.F.,
Brito, M.C.A., Araruna, F.O.S., Araruna, F.B., Silva-Souza, N., Coutinho, D.F., 2017.
Molluscicidal effect of Euphorbia umbellata (Pax) Bruyns latex on Biomphalaria
glabrata, Schistosoma mansoni host snail. Revista do Instituto de Medicina Tropical de
São Paulo, 59, https://dx.doi.org/10.1590/s1678-9946201759085.

79
Qualhato, G., Lopes, T.R., Lima, E.C.O., Silva, D.M., Cardoso, J.R., Grisolia, C.K.,
Sabóia-Moraes, S.M.T., 2017. Genotoxic and mutagenic assessement of iron oxide
(maghemite-ɤ-Fe2O3) nanoparticule in the guppy Poecilia reticulata. Chemosphere, 183,
305-314, https://doi.org/10.1016/j.chemosphere.2017.05.061.
Ramsdorf, W.A., Vicari, T., Almeida, M.I.M., Artoni, R.F., Cestari, M.M., 2012.
Handling of Astyanax sp. for biomonitoring in Cangüiri Farm within a fountainhead (Iraí
River Environment Preservation Area) through the use of genetic biomarkers.
Environmental Monitoring Assessement, 184, 5841-5849
https://doi/org/10.1007/s10661-012-2752-4.
Pavlica, M, Klobucar, GIVM, Vetma, N, Erben, R, Papes, D. (2000). Detection of
micronuclei in haemocytes of zebra mussel and great ramshorn snail exposed to
pentachlorophenol. Mutation Research, 465, 145-150, https://doi.org/10.1016/S1383-
5718(99)00222-3.
Sadiqul, I.M., Ferdous, Z., Nannu, M.T.A., Mostakim, G.M., Rahman, M.K., 2016. Acute
exposure to a quinalphos containing insecticide (convoy) causes genetic damage and
nuclear changes in peripheral erythrocytes of silver barb, Barbonymus gonionotus.
Environ. Pollut. 219, 949-956. http://dx.doi.org/10.1016j.envpol.2016.09.066.
Sanogo, B., Yuan, D., Zeng, X., Zhang, Y., Wu, Z., 2018. Diversity and Compatibility of
Human Schistosomes and Their Intermediate Snail Hosts. Trends in Parasitology, 34,
493-510, https://doi.org/10.1016/j.pt.2018.03.004 1.
Sarkar, A., Bhagat, J., Ingole, B.S., Rao, D.P., Markad, V.L., 2013. Genotoxicity of
cadmium chloride in the marine gastropod Nerita chamaeleon using comet assay and
alkaline unwinding assay. Environmental Toxicology,
https://doi.org/10.1002/tox.21883.
Singh, N.P., McCoy, M.T., Tice, R.R., Schneider, E.L., 1988. A simple technique for
quantitation of low levels of DNA damage in individual cells. Experimental Cell
Research, 175, http://dx.doi.org/10.1016/0014-4827(88)90265-0.
Tallarico, L.F., Borrely, S.I., Hamada, N., Grazeffe, V.S., Ohlweiler, F.P., Okazaki, K.,
Granatelli, A.T., Pereira, I.W., Pereira, C.A.B., Nakano, E., 2014. Developmental
toxicity, acute toxicity and mutagenicity testing in freshwater snails Biomphalaria
glabrata (Mollusca: Gastropoda) exposed to chromium and water samples.

80
Ecotoxicology and Environmental Safety, 110, 208-215,
https://doi.org/10.1016/j.ecoenv.2014.09.005.
Torgerson, P.R., Devleesschauwer, B., Praet, N., Speybroeck, N., Willingham, A.L.,
Kasuga, F., 2015. World Health Organization. Estimates of the Global and Regional
Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS
Medice. 12. https://doi.org/10.1371/journal.pmed.1001920.
Touahri, H.G., Boutiba, Z., Benguedda, W., Shaposhnikov, S., 2016. Active
biomonitoring of mussels Mytilus galloprovincialis with integrated use of micronucleus
assay and physiological indices to assess harbor pollution. Marine Pollution Bulletin, 110,
52-64, http://dx.doi.org/10.1016/j.marpolbul.2016.06.029.
Vignardi, C.P., Hasue, F.M., Sartorio, P.V., Cardoso, C.M., Machado, A.S.D., Passos,
M.J.A.C.R., Santos, T.C.A., Nucci, J.M., Hewer, T.L.R., Watanabe, I.S., Gomes, V.,
Phan, N.V., 2015. Genotoxicity, potential cytotoxicity and cell uptake of titanium dioxide
nanoparticles in the marine fish Trachinotus carolinus (Linnaeus, 1766). Aquatic
Toxicology, 158, 2218-229, http://dx.doi.org/10.1016/ j.aquatox.2014.11.008.
Vincent-Hubert, F., Arini, A., Gourlay-Francé, C., 2011. Early genotoxic effects in gill
cells and haemocytes of Dreissena polymorpha exposed to cadmium, B[a]P and a
combination of B[a]P and Cd. Mutatation Research, 723, 23-35,
https://doi.org/10.1016/j.mrgentox.2011.03.008.
WHO. World Health Organization. (1992). The Role of Mollusciciding in
Schistosomiasis Control. WHO, Geneva (WHO/SCHISTO/92.107;1992). 1992.
WHO. World Health Organization (2010). Working to Overcome the Global Impact of
Neglected Tropical Diseases. First WHO Report on Neglected Tropical Diseases. WHO,
Geneva (WHO/HTM/NTD/2010.1).

81
Capítulo IV Considerações finais

82
CAPÍTULO IV – CONSIDERAÇÕES FINAIS
Este foi o primeiro trabalho realizado por meio de bioensaios, que avaliou a
atividade moluscicida do PHMB em todas as fases de vida do caramujo B. glabrata,
hospedeiro intermediário de S. mansoni. Também foi possível avaliar as alterações
comportamentais nos caramujos adultos, a genotoxicidade e a mutagenicidade através dos
danos genéticos causados pela exposição deste composto.
Os resultados demonstraram que o PHMB não interrompeu o desenvolvimento
embrionário, fato que pode ser explicado pela proteção conferida pela cápsula gelatinosa
ao redor dos embriões. No entanto, mesmo com este resultado, e com a indução não
significativa de malformações, o PHMB foi capaz de inibir a eclosão de B. glabrata, um
efeito que certamente contribui para a redução da população de caramujos de uma região
infestada.
Todas as fases do caramujo B. glabrata avaliadas neste estudo foram suscetíveis
aos efeitos tóxicos do PHMB, sendo a fase embrionária a mais sensível (CL50 0,98 mg L-
1). Nos caramujos recém-eclodidos a CL50 foi estimada em 1,43 mg L-1. Nos caramujos
adultos, o PHMB foi capaz de causar alterações no comportamento do caramujo e a CL50
foi estimada em 1,49 mg L-1.
Esses dados demonstram que esses resultados estão dentro dos parâmetros
estabelecidos pela OMS, que argumentam que a atividade moluscicida de um composto
é reconhecida em concentrações abaixo de 100 ppm (WHO 1993).
Os resultados dissertados confirmam que os hemócitos de B. glabrata foram
células do sistema imune adequadas para análise de danos no DNA e alterações nucleares
induzidas por compostos com potencial ação moluscicida. Os resultados também
indicaram que o ensaio cometa foi uma técnica eficaz para avaliar danos precoces no
DNA induzidos pela exposição de PHMB a hemócitos de B. glabrata. O PHMB nas
concentrações de 1,0 e 1,5 mg L-1 após 48 horas de exposição foi genotóxico sem efeito
concentração-resposta.
Os resultados apresentados pelo teste do MN mostraram que o PHMB na
concentração de 1,0 mg L-1 e no tempo de exposição de 48 horas induz a formação de
micronúcleos e alterações nucleares no hemócitos de B. glabrata. Os resultados

83
apresentados neste teste, juntamente com os resultados do ensaio cometa, podem sugerir
que o PHMB tem ação genotóxica e mutagênica nos hemócitos de B. glabrata.

84
ANEXOS
Produções científicas
Resumos em congressos e seminários
XXIV Congresso Latino-americano de Parasitologia:
Avaliação do efeito moluscicida do cloridrato polihexametileno biguanida em
Biomphalaria glabrata, hospedeiro intermediário de Schistosoma mansoni.
Alternativa de intervenção na esquistossomose por triagem in silico de novos
compostos moluscicidas para Biomphalaria glabrata (MOLLUSCA,
PLANORBIDAE)
Avaliação in silico da atividade do Tiabendazol, Albendazol, Alendronato e
Metronidazol contra enzimas do metabolismo energético do Schistosoma mansoni
Embriotoxicidade de nanopartículas de óxido de zinco em caramujos
Biomphalaria glabrata, hospedeiro intermediário de Schistosoma mansoni
Estímulo ao estudo da parasitologia pela educação empreendedora em
biotecnologia e formação de força de trabalho
XIII Seminário de Patologia Tropical e Saúde Pública e VIII Semana da
Biotecnologia:
Formação de força de trabalho em biotecnologia: resultados da educação
empreendedora no processo de formação, motivação e iniciativas profissionais na
última década no IPTSP/UFG.
Participação em cursos, estágios e congressos
Proponente da palestra “Iniciativa Biotec - Movimento Biotec Brasil” -
NÚCLEO’17 - III Encontro Nacional de Estudantes de Biotecnologia.
Proponente do minicurso “Gastrópodes de Importância Médica” - 28a Semana
do ICB.
Comissão organizadora do II Simpósio de Nanobiotecnologia e Saúde Ambiental.
Artigos submetidos em revistas científicas
Molluscicidal activity of polyhexamethylene biguanide hydrochloride on the
early-life stages and adults of the Biomphalaria glabrata (Say, 1818)

85
REFERÊNCIAS
Aagaard-Hansen J, Bruun B. The social context of schistosomiasis and its control: An
introduction and annotated bibliography. Disponível em:
http://www,who,int/tdr/publications/documents/social-contextschistosomiasis,pdf.
Acesso em: 15/06/2016, 2008.
Abreu FC, Goulart MOF, Oliveira Brett AM. Detection of the damage caused to DNA by
niclosamide using an electrochemical DNA-biosensor, Biosensors and Bioelectronics,
17, 913 – 919, 2002.
Abou-El-Naga IF. Demographic, socioeconomic and environmental changes affecting
circulation of neglected tropical diseases in Egypt, Asian Pac J Trop Med, 8, 881-888,
2015.
Adema CM & Loker ES. Specificity and immunobiology of larval digenean-sanil
associations. In: Fried B. & Graczyk TK. (Eds.) Advances in Trematode Biology. Boca
Raton: CRC Press, 1997.
Adenowo AF, Oyinloye BE, Ogunyinka BI, Kappo AP. Impact of human schistosomiasis
in sub-Saharan Africa, The Brazilian Journal of Infectious Diseases, 19, 196 – 205, 2015.
Afonso FR. PHMB como desinfetante e antisséptico, Portugal [Dissertação de Mestrado
em Ciências Farmacêuticas – ISCSEM] 2013.
Alford RA, Richards SJ. Global Amphibian Declines: A Problem in Applied Ecology,
Annual Review of Ecology and Systematics 30(1):133–165, 1999.
Al-Fanharawi AA, Rabee AM, Al-Mamoori AMJ, 2. Biochemical and molecular
alterations in freshwater mollusks as biomarkers for petroleum product, domestic heating
oil. Ecotoxicology and Environmental Safety, 158, 69-77. 2018.
Al-Sabti K. Chlorotriazine reactive Azo Red 120 textile dye induces micronuclei in fish,
Ecotoxicology and Environmental Safety, 47,149–155, doi: 10,1006/eesa,2000,1931,
2000.
Al-Sabti K, Metcalfe CD. Fish micronuclei for assessing genotoxicity in water, Mutat
Res, 343,121-35, 1995.

86
Anderson MJ, Scholz MT, Parks PJ. Ex vivo porcine vaginal mucosal model of infection
for determining effectiveness and toxicity of antiseptics, J Appl Microbiol 2013; 115: 679
– 88,2013.
Andrews P, Thyssen J, Lorke D. The biology and toxicology of molluscicides,
bayluscide®, Pharmacology & Therapeutics,19, 245 – 295, 1983.
Amaral RS, Thiengo SC, Pieri OS. Vigilância e controle de moluscos de importância
epidemiológica: Diretrizes técnicas, Programa de Vigilância e Controle da
Esquistossomose (PCE), Ministério da Saúde, 2008.
Andrade ZA. Schistosomiasis and liver fibrosis, Parasite Immunology, 31, 656-63, 2009.
Araújo KCGM, Resendes APC, Souza-Santos R, Júnior JCS, Barbosa CS. Análise
espacial dos focos de Biomphalaria glabrata e de casos humanos de esquistossomose
mansônica em Porto de Galinhas, Pernambuco, Brasil, no ano 2000. Cad Saude Publica,
23, 409–417, 2007.
Arias ARL, Buss DF, Albuquerque C de, Inácio AF, Freire MM, Egler M, Mugnai R,
Baptista DF. Utilização de bioindicadores na avaliação de impacto e no monitoramento
da contaminação de rios e córregos por agrotóxicos, Ciência & Saúde, 12, 61-72, 2007.
Attwood SW, Upathan ES. Observations on Neotricula aperta (Gastropoda:
Pomatiopsidae) population densities in Thailand and central Laos: implications for the
spread of Mekong schistosomiasis, Parasites & Vectors, 5, 2012.
Azqueta A, Collins AR. The essential comet assay: a comprehensive guide to measuring
DNA damage and repair, Archives of Toxicology, 87, 949-968, 2013.
Baptista AP & Andrade ZA. Angiogenesis and schistosomal granuloma formation,
Memórias Instituto Oswaldo Cruz, 100, 183-5, 2005.
Barbosa FS. Tópicos em malacologia médica, Fiocruz, 314, ISBN 85-85676-13-2,
Available from SciELO Books http://books,scielo,org, 1995.
Barbosa CS, Favre TC, Amaral RS, Pieri OS. Epidemiologia e Controle da
Esquistossomose Mansoni, In: Schistosoma mansoni e esquistossomose: uma visão
multidisciplinar, Rio de Janeiro, FIOCRUZ, 2008.
Barbosa CS, Santos RS, Gomes ES, Araújo K, Albuquerque J, Melo F, Guimarães RJ.
Epidemiologia da esquistossomose no litoral de Pernambuco. Rev Patologia Tropical,
43:436–445, 2015.
Barnes RD & Ruppert EE. Zoologia dos Invertebrados, 6, 1996.

87
Barraco MA, Steil AA, Gargioni R. Morphological characterization of the hemocytes of
the pulmonate snail Biomphalaria tenagophila. Memórias do Instituto Oswaldo Cruz,
88,73-83, 1993.
Barsoum RS, Esmat G, El-Baz T. Human Schistosomiasis: Clinical Perspective: Review,
Journal of Advanced Research, 4, 433 – 444, 2013.
Bayne CJ, Buckley PM, Dewan PC. Macrophage-like hemocytes of resistant
Biomphalaria glabrata are cytotoxic for sporocysts of Schistosoma mansoni in vitro. The
Journal of Parasitology, 66, 413-419, 1980.
Bezerra FSM. Moluscos transmissores da esquistossomose mansoni, In: Neves, D, P,
Parasitologia humana, 10, 213-221, 2005.
Borges CMC & Andrade ZA. Extra-cellular matriz changes in Schistosoma mansoni-
infected Biomphalaria glabrata. Memórias do Instituto Oswaldo Cruz, 98: 135-139,
2003.
Colaborators - Berriman M; Haas BJ, Loverde PT, Wilson RA, Dillon GP, Cerqueira GC,
Mashiyama ST, Al-Lazikani B, Andrade LF, Ashton PD, Aslett MA, Bartholomeu DC,
Blandin G, Caffrey CR, Coghlan A, Coulson R, Day TA, Delcher A, Demarco R, Djikeng
A, Eyre T, Gamble JÁ, Ghedin E, Gu Y, Hertz-Fowler C, Hirai H, Hirai Y, Houston R,
Ivens A, Johnston DA, Lacerda D, Macedo CD, Mcveigh P, Ning Z, Oliveira G,
Overington JP, Parkhill J, Pertea M, Pierce RJ, Protasio AV, Quail MA, Rajandream MA,
Rogers J, Sajid M, Salzberg SL, Stanke M, Tivey AR, White O, Williams DL , Wortman
J, Wu W, Zamanian M, Zerlotini A. Fraser-Liggett CM, Barrell BG, El-Sayed NM, The
genome of the blood fluke schistosoma mansoni, Nature, 460, 352-358, 2009.
Bhattacharya P, Swarnakar S, Mukhopadhyay A, Ghosh S. Exposure of composite
tannery effluent on snail, Pila globosa: A comparative assessment of toxic impacts of the
untreated and membrane treated effluents. Ecotoxicology and Environmental Safety, 126,
45-55, 2016.
Binelli A, Riva C, Cogni D, Provini A. Assessment of the genotoxic potential of
benzo(a)pyrene and pp’-dichlorodiphenyldichloroethylene in Zebra mussel (Dreissena
polymorpha), Mutation Research, 649, 135–145, 2008.
Boller K, Schmid W. Chemical mutagenesis in mammals, The Chinese hamster bone
marrow as an in vivo test system, Hematological findings after treatment with trenimon,
Humangenetik 11, 35–54, 1970.

88
Bolognesi C, Buschini A, Branchi E, Carboni P, Furlini M, Martino A, Monteverde M,
Poli P, Rossi C. Comet and micronucleus assays in zebra mussel cells for genotoxicity
assessment of surface drinking water treated with three different disinfectants, The
Science of the Total Environment, 333, 2003.
Bolognesi C, Buschini A, Branchi E, Carboni P, Furlini M, Martino A, Monteverde M,
Poli P, Rossi C. Comet and micronucleus assays in zebra mussel cells for genotoxicity
assessment of surface drinking water treated with three different disinfectants. The
Science of the Total Environment, 333, 127-136, 2004.Brasil, Ministério da Saúde.
Vigilância e Controle de Moluscos de Importância Epidemiológica, 2 ed, Brasília,
Secretaria de Vigilância em Saúde, 2010.
Brasil, Ministério da Saúde. Vigilância da esquistossomose mansoni, 4 ed, Brasília,
Secretaria de Vigilância em Saúde, 2014.
Cabagna MC, Lajmanovich RC, Peltzer PM, Attademo AM, Ale E. Induction of
micronuclei in tadpoles of Odontophrynus americanus (Amphibia: Leptodactylidae) by
the pyrethroid insecticide cypermethrin, Toxicological Environmental Chemistry 88,
729–737, doi: 10,1080/02772240600903805, 2006.
Caldas IR, Campi-Azevedo AC, Oliveira LF, Silveira AM, Oliveira RC, Grazzinelli G.
Human schistosomiasis mansoni: immune responses during acute and chronic phases of
the infection, Acta Tropica, 108, 109-17, 2008.
Cantanhede SPD, Marques AM, Silva-Souza N, Valverde AL. Atividade moluscicida de
plantas: uma alternativa profilática, Revista Brasileira de Farmacognosia, 20, 282 – 288,
2010.
Camey T, Verdonk NH. The early development of the snail Biomphalaria glabrata (Say)
and the origino f the head organs. Netherlands Journal of Zoology, 20, 93-121, 1970.
Carvalho OS, Guimarães CT, Massara CL, Bonesio JER. Situação atual da
esquistossomose mansoni no lago da Pampulha, Belo Horizonte, MG, Brasil. Rev Saúde
Pública, 19,270–277, 1985.
Carvalho OS, Amaral RS, Dutra LV, Scholte RGC, Guerra MAM. Distribuição Espacial
de Biomphalaria glabrata, B, straminea e B, tenagophila, Hospedeiros Intermediários do
Schistosoma mansoni no Brasil, In: Carvalho OS, Coelho PMZ, Lenzi HL, Schistosoma
mansoni e Esquistossomose: Uma visão multidisciplinar, 1ª ed, Rio de Janeiro: Editora
Fiocruz, 393 – 418, 2008.

89
Carvalho WF, [Tese de Doutorado apresentada ao Programa de Ciências Ambientais do
Instituto de Ciências Biológicas da Universidade Federal de Goiás] Goiãnia, 2013.
Christen V, Faltermann S, Brun NR, Kunz PY, Fent K. Cytotoxicity and molecular effects
of biocidal disinfectants (quaternary ammonia, glutaraldehyde, poly(hexamethylene
biguanide) hydrochloride PHMB) and their mixtures in vitro and in zebrafish eleuthero-
embryos. Science of the Total Environment, 586, 2017.
Cioli D, Pica-Mattoccia L, Basso A, Guidi A. Schistosomiasis control: praziquantel
forever? Molecular and Biochemical Parasitology, 195, 23–29, 2014.
Colley DG, Bustinduy AL, Secor WE, King CH. Human schistosomiasis, The Lancet,
383, 2253 – 2264, 2014.
Collins AR, Dobson VL, Dušinská M. The comet assay: what can it really tell us?
Mutation Research /Fundamental Molecular Mechanisms of Mutagenisis, 375, 183–193,
doi:10,1016/S0027-5107(97)00013-4, 1997.
Collins AR. The comet assay for DNA damage and repair, Molecular Biotechnology,
1073–6085,2004.
Collins AR, Amaia AO, Brunborg G, Gaivão I, Giovannelli L, Kruszewski M, C, Smith
CC, Stetina R. The Comet Assay: Topical Issues, Mutagenesis, 23, 143–51, 2008.
Coura-Filho P, Mendes NM, Souza CP, Pereira JP. The prolonged use of niclosamide as
a molluscicide for the control of Schistosoma mansoni, Revista do Instituto de Medicina
Tropical, 34, 427-431, 1992.
Creppy EE, Diallo A, Moukha S, Eklu-Gadegbeku C, Cros, D. Study of Epigenetic
Properties of Poly(HexaMethylene Biguanide) Hydrochloride (PHMB). International
Journal of Environmental Research and Public Health, 11, 8069-8092, 2014.
Cunha PHJ, Silva LAF, Mesquita AJ, Borges NC, Fioravante MCS, Moraes RR, Santana
AP. Avaliação da estabilidadedo cloridrato de polihexametileno biguanida em pedilúvio
para bovinos, Ciência Animal Brasileira, 2, 41-50, 2001.
Dai J, Li Y, Wang W, Xing Y, Qu G, Liang Y. Sensitivity of Oncomelania hupensis to
niclosamide: a nation-wide survey in China, Interntional Journal Environmental Research
Public Health, 11,3086–95, 2014.

90
David NF, Cantanhede SPD, Monroe NB, Pereira LPLA, Souza NS, Abreu-Silva ALA,
Oliveira VM, Tchaicka L. Spatial distribution and seasonality of Biomphalaria spp. in
São Luís (Maranhão, Brazil). Parasitology Research, 2018.
Dhawan A, Bajpayee M, Parmar D. Comet assay: a reliable tool for the assessment of
DNA damage in different models, Cell Biological Toxicology, 25, 5–32,
doi:10,1007/s10565008-9072-z, 2009.
Domingues EP, Lopes CN. Association between Schistosomiasis mansoni and hepatitis
C: systematic review, Revista de Saúde Pública, 47, 414-24, 2013.
Duarte GF, Rodrigues J, Fernandes EKK, Humber RA, Luz C. New insights into the
amphibious life of Biomphalaria glabrata and susceptibility of its egg masses to fungal
infection. Journal of Invertebrate Pathology, 2015.
Duncan J, Brown N, Dunlop RW. The uptake of the molluscicide, 4'-chloronicotinanilide
into Biomphalaria glabrata (Say) in a flowing water system, Pesticide Science banner, 8,
345-353, 1977.
Duncan J. The Toxicology of Molluscicides - Trifenmorph, Pharmacology Therapeutics,
14, 67–88, 1981.
Duthie SJ, Dobson V L. Dietary flavonoids protect human colonocyte DNA from
oxidative attack in vitro, Eur, J, Nutr, 38, 28–34, 1999.
Eberlein T, Haemmerle G, Signer M. Comparison of PHMB-containing dressing and
silver dressings in patients with critically colonised or locally infected wounds, J Wound
Care, 21: 12 – 20, 14 – 6, 18 – 20, 2012.
El-Beshbishi SN, El Bardicy S, Tadros M, Ayoub M, Taman A. Spotlight on the in vitro
effect of artemisinin-naphthoquine phosphate on Schistosoma mansoni and its snail host
Biomphalaria alexandrina, Acta Tropica, 14, 37-45, doi:
10,1016/j,actatropica,2014,09,018, 2015.
Elzinga G, van Doorn J, Wiersema AM. Clinical evaluation of a PHMB-impregnated
biocellulose dressing on paediatric lacerations, J Wound Care, 20: 280 – 4, 2011.
El-Wakeil KFA, Obuid-Allah AH, Mohamed AH, El FEZAA. Community structure of
molluscans in River Nile and its branches in Assiut governorate, Egypt. Egypt J Aquat
Res, 39,93–198, 2013.

91
Evans HJ, Neary GJ, Williamson FS. The relative biological efficiency of single doses of
fast neutrons and gamma rays in Vicia faba roots and the effect of oxygen, Part II,
Chromosome damage; the production of micronuclei, International Journal Radiation
Biology, 1, 230–240, 1959.
Etges FJ, Gilbertson DE. Repellent action of some chemical molluscicides on
schistosome vector snails, Americal Journal Tropical Medicine and Hygiene, 9, 618-624,
1966.
Fenech M, Morley AA. Solutions to the kinetic problem in the micronucleus assay,
Cytobios, 43, 233–246, 1985.
Fenech M. The in vitro micronucleus technique, Mutation Research/Fundamental and
Molecular Mechanisms of Mutagenesis, 455, 81-95, 2000.
Fernandez MA, Thiengo SC, Boaventura MF.Gastrópodes límnicos do Campus de
Manguinhos, Fundação Oswaldo Cruz, Rio de Janeiro, RJ. Rev Soc Bras Med Trop,
34,279–282, 2001.
Filippi S, Meschini R, Spognardi S, Papetti P, Angeletti, D. Genetic effects in Helix
aspersa near a coal plant revealed by the micronucleus test, Ecotoxicology, 27(2), 234-
243, 2018.
Filho JTM. Planejamento e Identificação de Novos Compostos Moluscicidas para
Biomphalaria glabrata (Mollusca, Planorbidae), [Dissertação de Mestrado em Biologia
da Relação Parasito Hospedeiro – Universidade de Federalde Goiás], Goiânia, 2016,
Firdessa R, Good L, Amstalden MC, Chindera K, Kamaruzzaman NF, Schultheis M,
Roger B, Hecht N, Oelschlaeger TA, Meinel L, Luhmann T, Moll H. Pathogen- and Host-
Directed Antileishmanial Effects Mediated by Polyhexanide (PHMB), PLoS Neglected
Tropical Diseases, 9, 2015.
Franco FC, Alves AA, Godoy FR, Avelar JB, Rodrigues DD, Pedroso TMA, Cruz AD,
Nomura F, Silva DM. Evaluating genotoxic risks in Brazilian public health agentes
occupationally exposed to pesticides: a multi-biomarker approach, Enviromental Science
and Pollution Research, 23, 19723-19734, 2016.
Fumarola S. Polyhexamethylene biguanide dressings in wound management Rev, Eletr,
Nursing Standart 25, 46 2011, Disponível em: https://rcni,com/nursing-
standard/search?search_api_views_fulltext=Polyhexamethylene%20biguanide%20dress

92
ings%20in%20wound%20management&sort_by=search_api_relevance, Acesso em:
10/10/2017.
Gazzinelli MF, Lobato L, Andrade G, Matoso LF, Diemert DJ, Gazzinelli A. Improving
the understanding of schistosomiasis among adolescents in endemic areas in Brazil: a
comparison of educational methods, Patient Education Counseling, 99, 1657–1662, 2015.
Giovanelli A, Soares MS, D’andréa SP, Gonçalves MML, Rey L. Abundância e infecção
do molusco Biomphalaria glabrata pelo Schistosoma mansoni no estado do Rio de
Janeiro. Rev Saúde Pública, 35,523–530, 2001.
Girardello F, Leite CC, Villela IV, da Silva Machado M, Juchem ALM, Roesch-Ely M,
Fernandes NA, Salvador M, Henriques JAP. Titanium dioxide nanoparticles induce
genotoxicity but not mutagenicity in golden mussel Limnoperna fortunei, Aquatic
Toxicology, 170, 223-228, 2016.
Giovanelli A, Soares MS, D'Andrea PS, Goncalves MM, Rey L. Abundance and infection
of Biomphalaria glabrata with Schistosoma mansoni in the state of Rio de Janeiro, Brazil,
Revista de Saúde Pública, 35, 523-30, 2001.
Gonçalves MW. Alterações genômicas e mutagênicas em duas espécies de anfíbios
anuros, Goiânia [Dissertação de Mestrado do Programa de Genética e Biologia Molecular
do Instituto de Ciências Biológicas– UFG], 2015.
Grault CE. Estudo de laboratório e de campo, sobre dinâmica populacional de
Biomphalaria spp,, hospedeiros intermediários do Schistosoma mansoni, no território
Brasileiro, [Tese de Doutorado do Programa de Pós-Graduação em Ciências - Instituto
Oswaldo Cruz] – Rio de Janeiro, 2013.
Grazeffe VS, de Freitas Tallarico L, de Sa Pinheiro A, Kawano T, Suzuki MF, Okazaki
K, Pereira CAB, Nakano E. Establishment of the comet assay in the freshwater snail
Biomphalaria glabrata (Say, 1818), Mutation Research/Genetic Toxicology and
Environmental Mutagenesis, 654, 2008.
Grimes JET, Croll D, Harrison, WE, Utzinger J, Freeman MC, Templeton MR. The roles
of water, sanitation and hygiene in reducing schistosomiasis: a review, Parasites &
Vectors, 8, 1 – 16, 2015.
Grisolia CK, Bilich MR, Formigli LM. A comparative toxicologic and genotoxic study
of the herbicide arsenal, its active ingredient imazapyr, and the surfactant nonylphenol
ethoxylate, Ecotoxicology Environmental Safety, 59, 123-126, 2004.

93
Grisolia CK. Agrotóxicos- Mutações, Câncer e Reprodução. Brasília.2005.
Guimarães RJPS, Freitas CC, Dutra LV, Felgueiras CA, Moura ACM, Amaral RS,
Drumond SC, Scholte RGC, Oliveira G, Carvalho OS. Spatial distribution of
Biomphalaria mollusks at São Francisco River Basin, Minas Gerais, Brazil, using
geostatistical procedures, Revista Acta Tropica, 109, 181-186, 2009.
Gustavino B, Scornajenghi KA, Minissi S, Ciccotti E. Micronuclei induced in
erythrocytes of Cyprinus carpio(teleostei, pisces) by X-rays and colchicine, Mutation
Research/Genetic Toxicology and Environmental Mutagenesis, 494, 151-159, 2001.
Harris KR. The Fine Structure of encapsulation in Biomphalaria glabrata. Annals of the
New York Academy of Sciences, 266, 446-464, 1975.
Hartmann A, Agurell E, Beevers C, Burlinson B, Clay P, Collins A, Smith A, Speit G,
Thybaud V, Tice RR. Recommendations for conducting the in vivo alkaline Comet assay,
Mutagenesis 18, 45–51, 2003.
Hathaway JJ, Adema CM, Stout BA, Mobarak CD, Loker ES. Identification of protein
components of egg masses indicates parental investment in immunoprotection of
offspring by Biomphalaria glabrata (Gastropoda, Mollusca). Developmental &
Comparative Immunology, 2010.
Hollingworth RM. Inhibitors and uncouplers of mitochondrial oxidative phosphorylation,
In: Krieger RI, Krieger WC, Handbook of Pesticide Toxicology, Elsevier-Academic
Press Inc, 1169 – 1261, 2001.
Inobaya MT, Olveda RM, Chau TN, Olveda DU, Ross AGP. Prevention and control of
schistosomiasis: a current perspective, Research and Report Tropical Medicine, 5, 65–
75, 2014.
Jah NA. Ecotoxicological applications and significance of the comet assay, Mutagenesis,
23, 207–221, 2008.
Ishak MM, Sharaf AA, Mohamed AM, Mousa AH, Mode of action of some molluscicides
on the snail Biomphalaria alexandrina, I. Effect of bayluscide, sodium
pentachlorophenate, and copper sulfate on succinate, glutamate and reduced TMPD
(Tetramethyl-p-phenylene diamine) oxidation, Comparative and General Pharmacology,
201-208, 1970.

94
Ishak MM, Sharaf AA, Mohamed AH. Studies on the mode of action of some
molluscicides on the snail, Biomphalaria alexandrina: II, Inhibition of succinate
oxidation by bayluscide, sodium pentachlorphenate, and copper sulphate, Comparative
and General Pharmacology, 3, 385-390, 1972.
Jenkinson AM, Collins AR, Duthie SJ, Wahle KWJ, Duthie GG. The effect of increased
intakes of polyunsaturated fatty acids and vitamin E on DNA damage in human
lymphocytes, FASEB Journal, 13, 2138–2142, 2001.
Jordan P, Rosenfield PL. Schistosomiasis Control: Past, Present, and Future, Annual
Review Public Health, 4, 311–34, 1983.
Kaehn K. Polihexanide: A safe and highly effective biocide. Skin Pharmacology and
Physiology, 23, 2010.
Katsumiti A, Valdez Domingos FX, Azevedo M, da Silva MD, Damian RC, Almeida
MIM, de Assis HCS, Cestari MM, Randi MAF, Ribeiro CAO, Freire CA. An assessment
of acute biomarker responses in the demersal catfish Cathorops spixii after the Vicuña
Oil Spill in a harbour estuarine area in Southern Brazil. Environ Monit Assess, 152, 209–
222, 2009.
Kawano T. Agentes que atuam sobre a morfogênese da região cefálica de alguns
moluscos. Ciência e Cultura, 116-166, 1983.
Kawano T, Okazaki K, Ré L. Embryonic development of Biomphalari glabrata (Say,
1818) (Mollusca, Gastropoda, Planorbidae): A pratical guide to the main stages.
Malacologia, 34, 25-32, 1992.
Kawano T, Nakano E, Watanabe LC. Estudo do desenvolvimento embrionário de
Biomphalaria glabrata (Mollusca, Planorbidae) e suas aplicações. In: Carvakho OS,
Coelho PMZ, Lenzi HL. Schitosoma mansoni e esquistossomose: uma visão
multidisciplinar. Rio de Janeiro: Editora Fiocruz, pp. 347-391, 2008.
Kealey A. Neglected Tropical Diseases: Infection, Modeling, and Control, Journal of
Health Care for the Poor and Underserved, 21, 53 – 69, 2010.
King CH. Parasites and poverty: the case of schistosomiasis, Acta Tropica, 113, 95–104,
2010.

95
King CH. Bertsch D, Historical Perspective: Snail Control to Prevent Schistosomiasis,
PLoS Neglected Tropical Diseases, 9, 2015.
Kirsch-Volders M, Sofuni T, Aardema M, Albertini S, Eastmond D, Fenech M, Jr I M,
Kirchner S, Lorge E, Morita T, Norppa H, Surrallés J, Vanhauwaert A, Wakata A. Report
from the in vitro micronucleus assay working group, Genetic Toxicology and
Environmental Muganenesis, 540, 153-163, 2003.
Knaap VWPW, Loker ES. Immune mechanism in trematode-snail interaction,
Parasitology Today, 6, 175 – 182, 1990.
Koblucar GIV, Pavlica M, Erben R, Papes D. Application of the micronucleus and comet
assays to mussel Dreissena polymorpha haemocytes for genotoxicity monitoring of
freshwater environments, Aquatic Toxicology, 64, 15-23, 2003.
Koburger T, Hübner NO, Braun M et al. Standardized comparison of antiseptic efficacy
of triclosan, PVP-iodine, octenidine dihydrochloride, polyhexanide and chlorhexidine
digluconate, Journal Antimicrobial Chemother, 65, 1712 – 9, 2010.
Koppen G, Angelis K J. Repair of X-ray induced DNA damage measured by the comet
assay in roots of Vicia faba, Enviromental Molecular Mutagenesis, 32, 281– 285, 1998.
Kumar A, Dhawan A. Genotoxic and carcinogenic potential of engineered nanoparticles:
an update, Archives of Toxicology, 87, 1883–1900, 2013.
Kumaravel TS & Jha AN. Reliable comet assay measurements for detecting DNA damage
induced by ionising radiation and chemicals. Mutation research, 605, 7-16, 2006.
Lee CK, Chua YP, Saw A. Antimicrobial gauze as a dressing reduces pin site infection:
a randomized controlled trial, Clin Orthop Relat Res 2012; 470: 610 – 5, 2012.
Leite MGP, Pimenta EC, Fujaco MAG, Eskinazi-Sant’Anna EM. Irrigation canals in
Melo creek basin (Rio Espera and Capela Nova municipalities, Minas Gerais, Brazil):
habitats to Biomphalaria (Gastropoda: Planorbidae) and potential spread of
schistosomiasis. Braz J Biol, 76,638–644, 2016.
Lenselink E, Andriessen A. A cohort study on the efficacy of a polyhexanide-containing
biocellulose dressing in the treatment of biofilms in wounds. Journal Wound Care, 9, 536
– 9, 2011.

96
Lenzi HL, Jurberg AD, Coelho PMZ, Lenzi JA. Migração e desenvolvimento de
Schistosoma mansoni no hospedeiro definitivo. In: Carvalho OS, Coelho PMZ, Lenzi HL.
Schitosoma mansoni e esquistossomose: uma visão multidisciplinar. Rio de Janeiro:
Editora Fiocruz, 2008.
Lie KJ, Jeong KH, Heyneman D. Molluscan host reactions to helminthic infection. In:
Soulsby EJL. Protozoa, Arthropods and Invertebrates, 1987.
Lindoso JAL, Lindoso AABP. Neglected Tropical Diseases in Brazil, Revista do Instituto
de Medicina Tropical de São Paulo, 51, 247 – 253, 2009.
Lotfy WM. Human schistosomiasis in Egypt: historical review, assessment of the current
picture and prediction of the future trends. J Med Res Inst, 30,1–7, 2009.
Loker ES, Environmental Assessment. Use of the Molluscicide Bayluscide in a Pilot
Program to Control Schistosomiasis in Kaele Division, Extreme North Province,
Cameroon, U, S, Agency for International Development, 81133, 1990.
López González EC, Latorre MA, Larriera A, Siroski PA, Poletta GL. Induction of
micronuclei in broad snouted caiman (Caiman latirostris) hatchlings exposed in vivo to
Roundup® (glyphosate) concentrations used in agriculture, Pesticide Biochemistry and
Physiology, 105, 131–134, 2013.
Madinga J, Linsuke S, Mpabanzi L, Meurs L, Kanobana K, Speybroeck N, Lutumba P,
Polman K. Schistosomiasis in the Democratic Republic of Congo: a literature review,
Parasitoloy Vectors, 8, 601, 2015.
Malhado M, Pinto DP, Silva ACA, Silveira GPE, Pereira HM, Santos JGF, Guilarducci-
Ferraz CVV, Viçosa AL, Nele M, Foncesa LB, Pinto JCCS, Calil ES. Preclinical
pharmacokinetic evaluation of praziquantel loaded in poly (methylmethacrylate)
nanoparticleusing a HPLC–MS/MS, Jounal Pharmacology Biomedicine Anal, 117, 405–
412, 2016.
Martins MCB, Silva MC, Silva LRS, Lima VLM, Pereira EC, Falcão EPS, Melo AMMA,
Da Silva NH. Usnic acid potassium salt: An alternative for the control of Biomphalaria
glabrata (Say, 1818). PLoS ONE, 9, 2014.
Martins M, Costa PM. The comet assay in Environmental Risk Assessment of marine
pollutants: applications, assets and handicaps of surveying genotoxicity in non-model
organisms, Mutagenesis, 30, 89–106, 2015.
Miyasato PA, Kawano T, Freitas JC, Berlinck RGS, Nakano E, Tallarico L.F.

97
Molluscicidal activity of some marine substances against the snail Biomphalaria glabrata
(Mollusca, Planorbidae). Parasitology Research, 110, 2012.
Monteiro AC, Revisão sistemática de marcadores de resistência e/ou susceptibilidade de
Biomphalaria sp, à infecção por Schistosoma mansoni, Alfenas [Dissertação de Mestrado
do Programa de Ciências Biológicas UFA], 2017.
Moore J. Uma introdução aos invertebrados, 1ª ed, São Paulo: Editora Santos, 2003,
Moraes J, Efeito in vitro de extratos de compostos naturais em Schistosoma mansoni,
[Tese Doutorado em Biotecnologia – Universidade de São Paulo], São Paulo, SP, 2011.
Moreira CPS, Zani CL, Alves TMA. Molluscicidal activity of Synadenium Carinatum
Boiss. (Euphorbiaceae) latex against Biomphalaria Glabrata and asolation of aajor
aompound, VII(3), 2010.
Muangman P, Nitimonton S, Aramwit P. Comparative Clinical Study of Bactigras and
Telfa AMD for Skin Graft Donor-Site Dressing, International journal of molecular
sciences, 12, 5031–8, 2011.
Nascimento GL, Oliveira MRF. Severe forms of schistosomiasis mansoni: epidemiologic
and economic impact in Brazil, 2010, Transactions of the Royal Society of Tropical
Medicine and Hygiene, 108, 29 – 36, 2013.
Negrão-Corrêa DA, Coelho PMZ, Andrade ZA, Martins-Souza RL, Pereira CAJ.
Biomphalaria e Schistosoma mansoni: papel do sistema interno de defesa do molusco na
susceptibilidade ao parasito. In: Carvalho OS, Coelho PMZ, Lenzi HL. Schitosoma
mansoni e esquistossomose: uma visão multidisciplinar. Rio de Janeiro: Editora Fiocruz,
2008, pp. 485-509.
Nepomuceno J,C,, Ferrari Í,, Spanó M,A,, Centeno A,J. Detection of micronuclei in
peripheral erythrocytes of Cyprinus carpio exposed to metallic mercury, Environmental
and Molecular Mutagenesis, 30, 293–297, 1997.
Obe G, Pfeiffer P, Savage JRK, Johannes C, Goedecke W, Jeppesen P, Natarajan AT,
Mart´ınez-López W, Folle GA, Drets ME. Chromosomal aberrations: formation,
identification and distribution, Mutation Research, 504,17–36, 2004.

98
OECD 471. “Reverse Mutation Assay on Salmonella typhimurium his- and Escherichia
coli on PHMB P20D” according to OECD Guideline N°471; GLP Study N°2002-
DKY543-2 of September 5th, 2002. Unpublished work, 2002 a.
OECD 473. “In vitro Mammalian Chromosome Aberration Test Using Chinese Hamster
Ovary Cells (CHO) on PHMB P20D” according to OECD Guideline N°473; GLP Study
N°2002-DKY543-1 of September 5th, 2002. Unpublished work, 2002 b.
OECD 476. “In vitro Mammalian Cell Gene Mutation Test on PHMB P20D” according
to OECD Guideline N°476; GLP Study N°2002-DKY543-3 of September 5th, 2002.
Unpublished work, 2002 c.
OECD. OECD guideline for the testing of chemicals, TG 487, Disponível em:
https://ntp,niehs,nih,gov/iccvam/suppdocs/feddocs/oecd/oecd-tg487-2014-508,pdf,
Acessado em: 11/03/2018, 2014.
Olive PL, Banath JP, Durand RE. Heterogeneity in Radiation Rnduced DNA Damage and
Repair in Tumor and Normal Cells Measured Using the “Comet” Assay, Radiation
Research 122, 86–94, 1990.
Oliveira-Filho EC, Geraldino BR, Coelho DR, De-Carvalho RR, Paumgartten FJR.
Comparative toxicity of Euphorbia milii latex and synthetic molluscicides to
Biomphalaria glabrata embryos, Chemosphere, 81, 2010.
Ostling O, Johanson KJ. Microelectrophoretic study of radiation-induced DNA damages
in individual mammalian cells, Biochemical and Biophysical Research Communication,
123, 291–298, 1984.
Othman AA, Soliman RH. Schistosomiasis in Egypt: A never-ending story? Acta
Tropica, 148, 179 – 190, 2015.
Palasio RGS, Casotti MO, Rodrigues TC, Menezez RMT, Zanotti-Magalhães EM, Tuan
R. The current distribution pattern of Biomphalaria tenagophila and Biomphalaria
straminea in the northern and southern regions of the coastal fluvial plain in the state of
São Paulo, Biota Neotropica, 15, 2015.
Paraense WL. Histórico do Gênero Biomphalaria, Morfologia e Sistemática Morfológica,
In: Carvalho OS, Coelho PMZ, Lenzi HL, Schistosoma mansoni e esquistossomose: uma
visão multidisciplinar, Fiocruz, Rio de Janeiro,285-308, 2008.

99
Pavanello S, Clonfero E. Biological indicators of genotoxic risk and metabolic
polymorphisms, Mutation Research, 463, 285–308, 2000.
Pavlica M, Klobučar GI, Vetma N, Erben R, Papeš D. Detection of micronuclei in
haemocytes of zebra mussel and great ramshorn snail exposed to
pentachlorophenol, Mutation Research/Genetic Toxicology and Environmental
Mutagenesis, 465, 145-150, 2000.
Pessôa SB & Martins AV. Parasitologia médica, ed, 11ª, Rio de Janeiro: Guanabara
Koogan S,A,, 2011.
Pila EA, Li H, Hambrook JR, Wu X, Hanington PC. Schistosomiasis from a Snail’s
Perspective: Advances in Snail Immunity, Trends in parasitology, 16, 2017.
Rabello-Gay MN, Rodrigues MALR, Monteleone-Neto R. Mutagênese, carcinogênese e
teratogênese: métodos e critérios de avaliação, Sociedade Brasileira de Genetica Revista
Brasileira de Genetica, 1991.
Qualhato G, Lopes TR, Lima ECO, Silva DM, Cardoso JR, Grisolia CK, Sabóia-Moraes
SMT. Genotoxic and mutagenic assessement of iron oxide (maghemite-ɤ-Fe2O3)
nanoparticule in the guppy Poecilia reticulata. Chemosphere, 183, 305-314, 2017.
Ramsdorf WA, Vicari T, Almeida MIM, Artoni RF, Cestari MM. Handling of Astyanax
sp. for biomonitoring in Cangüiri Farm within a fountainhead (Iraí River Environment
Preservation Area) through the use of genetic biomarkers. Environmental Monitoring
Assessement, 184, 5841-5849. 2012.
Rapado LN, Pinheiro AS, Lopes POMV, Fokoue HH, Scotti MT, Marques JV, Ohlweiler
FP, Borrely SI, Pereira CAB, Kato MJ, Nakano E, Yamaguchi LF. Schistosomiasis
control using pirplatine against Biomphalaria glabrata at different developmental stages,
PLoS Neglected Tropical Diseases, 7, 2251, 2013.
Ribeiro-Paes JT, Costa PI, Silva D, Barth LR, Rodrigues V. Aspectos imunológicos do
sistema enzimático fenoloxidase de Schistosoma mansoni, Revista do Instituto de
Medicina Tropical, 36, 396-412, 1994.
Ribeiro KA, de Carvalho CM, Molina MT, Lima EP, López-Montero E, Reys JRM,
Oliveira MBF, Pinto AV, Santana AEG, Goulart MO. Activities of naphthoquinones
against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae), vector of dengue and

100
Biomphalaria glabrata (Say, 1818), intermediate host of Schistosoma mansoni. Acta
Tropica. 2009.
Rizk MZ, Metwally NS, Hamed MA, Mohamed AM. Correlation between steroid sex
hormones, egg laying capacity and cercarial shedding in Biomphalaria alexandrina snails
after treatment with Haplophyllum tuberculatum, Exp, Parasitol, 132 (2012) 171-179,
doi: 10,1016/j,exppara,2012,06,011, 2012.
Rodrigues HG, Penha-Silva N, de Araújo MFP, Nishijo H, Aversi-Ferreira TA. Effects
of Roundup pesticide on the stability of human erytrocyte membranes and micronuclei
frequency in bone marrow cells of Swiss mice, The Open Biology Journal, 4, 54-59, 2011.
Ross AG, Vickers D, Oldes GR, Shah SM, McManus DP. Katayama syndrome, Lancet
Infect Dis, 7, 218-24, 2007.
Paraense WL. Histórico do Gênero Biomphalaria, Morfologia e Sistemática Morfológica,
In: Carvalho OS, Coelho PMZ, Lenzi HL, Schistosoma mansoni e esquistossomose: uma
visão multidisciplinar, Fiocruz, Rio de Janeiro, p, 285-308, 2008.
Sadiqul IM, Ferdous Z, Nannu MTA, Mostakim GM, Rahman MK. Acute exposure to a
quinalphos containing insecticide (convoy) causes genetic damage and nuclear changes
in peripheral erythrocytes of silver barb, Barbonymus gonionotus. Environ. Pollut. 219,
949-956, 2016.
Sah VK, Wang L, Min X, Rizal R, Feng Z, Ke Z, Deng M, Li L, Li H. Human
schistosomiasis: a diagnostic imaging focused review of a neglected disease, Radiology
Infections Disease, 2, 150–157, 2015.
Sambon LW. Descripitions of some new species of animal parasites. Proc. zool. Soc.
London. 19, 282-283, 1907.
Sanogo B, Yuan D, Zeng X, Zhang Y, Zhongdao W. Diversity and Compatibility of
Human Schistosomes and Their Intermediate Snail Hosts, Trends in Parasitology, 1745,
2018.
Say, T. Account of two new genera, and several new species, of fresh water and land
shells, Journal Academic National Science of Philadelphia, 1, 276-284, 1818.

101
Santos AF, Ferraz P, Pinto V, Pinto M do C, Goulart MO, Sant’Ana E. Molluscicidal
activity of 2-hydroxy-3-alkyl-1,4naphthoquinones and derivatives, International Journal
Parasitology, 30, 1199–202, 2000.
Santos PD, Fernandes PHS. Utilização de Cloridrato de Polihexametileno Biguanida
(PHMB) na desinfecção de indústrias cervejeiras, Teccen, 3, 2010.
Santos AF dos, Cavada BS, Rocha BAM. da, Nascimento KS do, Sant’Ana AEG.
Toxicity of some glucose/mannose-binding lectins to Biomphalaria glabrata and Artemia
salina. Bioresource Technology, 101, 2010.
Sarkar A, Bhagat J, Ingole BS, Rao DP, Markad VL. Genotoxicity of cadmium chloride
in the marine gastropod Nerita chamaeleon using comet assay and alkaline unwinding
assay. Environmental Toxicology, 2013.
Scholte RGC, Carvalho OS, Malone JB, Utzinger J, Vounatsou P. Spatial distribution of
Biomphalaria spp,, the intermediate host snails of Schistosoma mansoni, in Brazil,
Geospatial Health, 6, S95 – S101, 2012.
Sibbald RG, Coutts P, Woo KY. Reduction of bacterial burden and pain in chronic
wounds using a new polyhexamethylene biguanide antimicrobial foam dressing-clinical
trial results, Adv Skin Wound Care 2011; 24: 78 – 84, 2011.
Silva, HAMF. Atividade moluscicida e cercaricida do ácido divaricático extraído de
Ramalina áspera, [Dissertação de Mestrado do Programa de Bioquímica e Fisiologia do
Centro de Biociências – UFP], 2017.
Silva HAMF, Siqueira WN, Sá JLF, Silva LRS, Martins MCB, Aires AL, Silva NH.
Laboratory assessment of divaricatic acid against Biomphalaria glabrata and
Schistosoma mansoni cercariae. Acta Trop., 178, 2018.
Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of
low levels of DNA damage in individual cells, Experimental Cell Research, 175, 184–
191, 1988.
Siqueira LP, Fontes DAF, Aguileira CSB, Timóteo TRR, Ângelos MA, Silva LCPBB,
Melo CG, Rolim LA, Silva RMF, Neto PJR. Schistosomiasis: Drugs used and treatment
strategies, Acta Tropica, 176, 179–187, 2017.
Somorovská M, Tulinska J, Barancokova M, Zamecnicova M, Coliins A, Liskova A,
Vallova B, Petrovska H, Jahnova E, Vodicka P, Fuortes L, Dusinska M. The comet assay

102
in biomonitoring of occupational exposure in rubber factory and plastic lamination plant,
Comparison with cytogenetic and imune biomarkers, Neoplasma, 46, 23-25, 1999.
Song L, Li J, Xie S, Qian C, Wang J, Zhang W. Thioredoxin glutathione reductase as a
novel drug target: Evidence from schistosoma japonicum, PLoS One, 7, 2012.
Souza MAA, Barbosa VS, Albuquerque JO, Bocanegra S, Souza-Santo SR, Paredes H,
Barbosa CS. Aspectos ecológicos e levantamento malacológico para identificação de
áreas de risco para transmissão da esquistossomose mansoni no litoral norte de
Pernambuco, Brasil. Iheringia Ser Zool,100,19–24, 2010.
Souza MR, da Silva FR, de Souza CT, Niekraszewicz L, Dias JF, Premoli S, Corrêa DS,
Soares MC, Marroni NP, Morgam-Martins MI, da Silva, J. Evaluation of the genotoxic
potential of soil contaminated with mineral coal tailings on snail Helix
aspersa, Chemosphere, 139, 512-517, 2015.
Souza RP, Fatores imunológicos associados à resistência a infecção pelo Schistosoma
mansoni em uma área endêmica da Bahia, Brasil, [Tese de Doutorado apresentada ao
Programa de Pós Graduação em Imunologia, da Universidade Federal da Bahia], Bahia,
2015.
Speit G, Rothfuss A. The comet assay: a sensitive genotoxicity test for the detection of
DNA damage and repair, Methods Molecular Biology, 920, 79–90, 2012.
Stensgaard AS, Utzinger J, Vounatsou P, Hürlimann E, Schur N, Saarnak CFL, Simoonga
C, Mubita P, Kabatereine NB, Tchuem Tchuenté LA, Rahbek C, Kristensen TK. Large-
scale determinants of intestinal schistosomiasis and intermediate host snail distribution
across Africa: Does climate matter? Acta Tropica, 128, 378–90, 2013.
Tanaka H, Tsuji M. From discovery to eradication of schistosomiasis in Japan: 1847–
1996, Internatinal Journal Parasitology, 27, 1465–80, 1997.
Telles HMS. Distribuição de Biomphalaria straminea ao Sul da Região Neotropical. Rev
Saúde Pública, 30,341–349, 1996.
Telles HMS. Distribuição geográfica das espécies dos caramujos transmissores de
Schistosoma mansoni no Estado de São Paulo. Rev Soc Bras Med Trop, 38,426–432,
2005.
Teles HMS, Carvalho O dos S. Implicações da Biologia de Biomphalaria no Controle da
Esquistossomose, In: Schistosoma mansoni e esquistossomose: uma visão
multidisciplinar, Rio de Janeiro, Fiocruz, 2008.

103
Tibiriça SHC, Guimarães FB, Teixeira MTB. A esquistossomose mansoni no contexto da
política de saúde brasileira, Ciência e Saúde Coletiva, 16, 1375-1381, 2011.
Tlamçani Z, Er-Rami M. Schistosomiasis control: moroccan experience compared to
other endemic countries, Asian Pacific Journal of Tropical Disease, 4, 329 – 332, 2014.
Toossi MTB, Azimian H, Sarrafzadeh O, Mohebbi S, Soleymanifard S. Automatic
detection of micronuclei by cell microscopic image processing, Mutation Research, 806,
9-18 2017.
Touahri HG, Boutiba Z, Benguedda W, Shaposhnikov S. Active biomonitoring of
mussels Mytilus galloprovincialis with integrated use of micronucleus assay and
physiological indices to assess harbor pollution, Marine Pollution Bulletin, 110, 2016.
Verrengia-Guerrero NRV, Mozzarelli MN, Giancarlo H, Nahabedian D, Wider E.
Biomphalaria glabrata: Relevance of albino organisms as useful tool for environmental
lead monitoring, Bulletin of Environmental Contamination and toxicology, v, 59, p, 822-
827, 1997.
Vianey- Liaud M, Dussart G. Aspects of pairing and reproduction in the hermaphrodite
freshwater snail biomphalaria glabrata (gastropoda: pulmonata), Journal of Molluscan
Studies, 68, 243-248, 2002.
Villela IV, Oliveira IM, Silva J, Henriques JAP. DNA damage and repair in haemolymph
cells of golden mussel (Limnoperna fortunei) exposed to environmental contaminants,
Mutation Research, 605, 78– 86, 2006.
Villier JP & Mackenzie JG. Structure and activity in molluscicides: the phenacylhalides,
a group of potentially useful molluscicides, Bulletin of the World Health Organization,
29, 424-429, 1963.
Vincent-Hubert F, Arini A, Gourlay-Francé C. Early genotoxic effects in gill cells and
haemocytes of Dreissena polymorpha exposed to cadmium, B[a]P and a combination of
B[a]P and Cd, Mutation Research, 723, 26–35, 2011.
Virmond MCL. Pesquisa em doenças negligenciadas: um desafio presente, Salusvita, v,
29, p, 3 – 5, 2010.

104
Wynn TA & Cheever AW. Cytokine regulation of granuloma formation in
schistosomiasis, Current Opinion Immunology, 7, 505-11, 1995.
Wynn TA. Cellular and molecular mechanisms of fibrosis, Journal Pathology, 214, 199-
210, 2008.
WHO. Expert Committee on Bilharziasis: First Report. Disponível em:
https://extranet,who,int/iris/restricted/handle/10665/86986, Acesso em:
25/11/2016.1953.
WHO. Molluscicide screening and evaluation. Disponível em:
http://www,ncbi,nlm,nih,gov/pmc/articles/PMC2475882/pdf/bullwho0062 0-0132,pdf,
Acesso em: 25/06/2016. 1965 a.
WHO. Who expert committe on bilharziasis: Third Report. Disponível em:
https://extranet,who,int/iris/restricted/handle/10665/40617, Acesso em: 11/11/2016.
1965b.
WHO. World Health Organization. Reports of the scientific working group on plant
molluscicides. Bulletin World Health Organization, 61(6).
https://doi.org/http://apps.who.int/iris/handle/10665/60086.1983.
WHO. World Health Organization.. The Control of Schistosomiasis, Technical Report
Series.1993.
WHO. Preventive chemotherapy in human helminthiasis Preventive chemotherapy in
human helminthiasis. Disponível em:
http://apps,who,int/iris/bitstream/10665/43545/1/9241547103_eng,pdf, Acesso em:
11/11/2016. 2006.
WHO. Schistosomiasis: Progress report 2001–2011 and strategic plan 2012–2020, 2013,
Disponível em: http://apps,who,int/iris/handle/10665/78074, Acesso em: 05/12/2016.
2011.
Yoshino TP. The ultrasructure of circulating hemolymph cells of the marine snail
Cerithidea californica (Gastropoda: Prosobranchiata). Journal of Morphology, 150, 48,
1976.
Young ND, Jex AR, Li B, Liu S, Yang L, Xiong Z, Li Y, Cantacessi C, Hall RS, Xu X,
Chen F, Wu X, Zerlotini A, Oliveira G, Hofmann A, Zhang G, Fang X, Kang Y, Campbell
BE, Loukas A, Ranganathan S, Rollinson D, Rinaldi G, Brindley PJ, Yang H, Wang J,

105
Wang J, Gasser RB. Whole-genome sequence of schistosoma haematobium, Nature
genetics, 44, 221 – 225, 2012.
Yousef AAA, El-Kassas NB. Ultrastructure and histopathological effects of some plant
extracts on digestive gland of Biomphalaria alexandrina and Bulinus truncatus, J, Basic
Appl, Zool, 66 (2013) 27-33, 2013.
Yuan Y, Xu X-J, Dong H-F, Jiang M-S, Zhu H-G. Transmission control of
schistosomiasis japonica: implementation and evaluation of different snail control
interventions, Acta Tropica, 96, 191–7, 2005.


















