UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE … · problema de saúde pública, e o Brasil...
68
UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA FACULDADE DE VETERINÁRIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS BELARMINO EUGÊNIO LOPES NETO AVALIAÇÃO DO ESTRESSE OXIDATIVO NA LEISHMANIOSE VISCERAL CANINA FORTALEZA-CEARÁ 2012
Transcript of UNIVERSIDADE ESTADUAL DO CEARÁ PRÓ-REITORIA DE … · problema de saúde pública, e o Brasil...

UNIVERSIDADE ESTADUAL DO CEARÁ
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
BELARMINO EUGÊNIO LOPES NETO
AVALIAÇÃO DO ESTRESSE OXIDATIVO NA LEISHMANIOSE VISCERAL CANINA
FORTALEZA-CEARÁ 2012

BELARMINO EUGÊNIO LOPES NETO
AVALIAÇÃO DO ESTRESSE OXIDATIVO NA LEISHMANIOSE VISCERAL CANINA
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Veterinárias da Faculdade de Veterinária da Universidade Estadual do Ceará, como requisito parcial para a obtenção do grau de Mestre em Ciências Veterinárias. Área de Concentração: Reprodução e Sanidade Animal. Linha de Pesquisa: Reprodução e sanidade de carnívoros, onívoros, herbívoros e aves. Orientadora: Profa. Dra. Diana Célia Sousa Nunes Pinheiro.
FORTALEZA-CEARÁ 2012

Dados Internacionais de Catalogação na Publicação Universidade Estadual do Ceará
Biblioteca Central Prof. Antônio Martins Filho
L864a Lopes Neto, Belarmino Eugênio
Avaliação do estresse oxidativo na leishmaniose visceral canina / Belarmino Eugênio Lopes Neto. — 2012.
00 f. : enc. ; 30 cm. Dissertação (mestrado) – Universidade Estadual do Ceará,
Faculdade de Veterinária, Programa de Pós-Graduação em Ciências Veterinárias, Fortaleza, 2012.
Área de Concentração: Reprodução e Sanidade Animal. Orientação: Profa. Dra. Diana Célia Sousa Nunes Pinheiro.
1. Leishmaniose visceral canina. 2. Estresse oxidativo. 3. Catalase. 4. Glutationa peroxidade. 5. Biomarcador. I. Título.
CDD: 616.9364

BELARMINO EUGÊNIO LOPES NETO
AVALIAÇÃO DO ESTRESSE OXIDATIVO NA LEISHMANIOSE CANINA
Dissertação apresentada ao Programa de Pós- Graduação em Ciências Veterinárias da Faculdade de Veterinária da Universidade Estadual do Ceará, como requisito parcial para a obtenção do grau de Mestre em Ciências Veterinárias.
Aprovado em: _____/_____/_____
BANCA EXAMINADORA
_______________________________________________ Profa. Dra. Diana Célia Sousa Nunes Pinheiro
Universidade Estadual do Ceará
Orientadora
_________________________________ __________________________________ Profa. Dra. Erika Freitas Mota Profa. Dra. Virgínia Claudia Carneiro Girão
Universidade Federal do Ceará Universidade Federal do Ceará
Examinadora Examinadora
________________________________ Dr. José Claudio Carneiro de Freitas
Prefeitura Municipal de Fortaleza
Examinador

Aos meus pais,
Francisco Barroso Pinto e Maria Celina,
Dedico.

AGRADECIMENTOS
À Deus, pai supremo e fidedigno, por ter me concedido o dom da vida e da
sabedoria, me guiando e dando forças em mais esta etapa da minha vida.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pelo
apoio financeiro concedido durante os dois anos de Mestrado.
À Profa. Dra. Diana Célia Sousa Nunes Pinheiro, por sempre ter acreditado no
meu potencial, por sua inestimável orientação, colaboração, compreensão, incentivo e
acima de tudo amizade, meus sinceros agradecimentos.
À Profa. Dra. Romélia Gonçalves Pinheiro pela colaboração junto ao Laboratório
de Pesquisa em Hemoglobinopatias Genéticas das Doenças da UFC, permitindo a
realização de etapas fundamentais na execução deste trabalho.
Aos colegas do Laboratório de Imunologia e Bioquímica de Animais (LIBA), em
especial Glauco Jonas Lemos Santos, Adam Leal Lima e Tiago Cunha, cujas ajudas
foram de fundamental importância para a realização desse trabalho.
A todos os funcionários da Faculdade de Veterinária (FAVET), em especial aos
professores do Programa de Pós-Graduação em Ciências Veterinárias (PPGCV) da
UECE, pelos conhecimentos e experiências compartilhados.
Aos meus pais, Francisco Barroso Pinto e Maria Celina Lopes Barroso, que são
base da minha vida, por todo amor, carinho e dedicação, e por estarem sempre ao meu
lado, me apoiando em qualquer situação.
A minha noiva, Larissa Silva Barros, que teve paciência e me incentivou a sempre
seguir este caminho de dedicação e por todo o amor que existe nós.

RESUMO
A leishmaniose visceral canina (LVC) é uma doença que pode comprometer
diversos órgãos e tecidos. Estudos recentes têm sugerido a participação de espécies
reativas do oxigênio (ERO) na patogenia de diversas doenças infecciosas e parasitárias
em cães, incluindo a LVC. Neste trabalho objetivou-se avaliar o papel do estresse
oxidativo na LVC. Para tanto, foram utilizados 27 cães de ambos os sexos, com peso,
idade e raças variadas, oriundos do Centro de Controle de Zoonoses de Fortaleza e de
um canil particular. Os animais foram considerados com leishmaniose quando
apresentaram título ≥ 1:40 para Reação de Imunofluorescência Indireta (RIFI) e Ensaio
Imunoenzimático (ELISA), associado a três ou mais sinais característicos da doença. Os
animais foram divididos em dois grupos, Grupo A (n=12), animais sadios e o Grupo B
(n=15), animais positivos para LVC. Amostras de sangue foram coletadas por
venopunção jugular, distribuídas em tubos contendo anticoagulante (EDTA) e tubos
com gel de separação, para determinação dos parâmetros leucocitários, hematológicos,
bioquímicos séricos e as atividades das enzimas glutationa peroxidase (GPx; mU/mg
Hb) e catalase (CAT; mU/mg Hb) eritrocitárias. Os resultados foram expressos em
média e desvio padrão e a análise dos dados foi realizada através do teste t de Student
sendo as diferenças entre os grupos, consideradas significativas quando p < 0,05.
Também foi aplicado o teste de correlação de Pearson entre a variável GPx com He e
Hb e entre variável CAT com He e Hb. O protocolo experimental foi aprovado pelo
CEUA/UECE, n° 11516675-0/65. Dentre os parâmetros bioquímicos séricos, somente
as proteínas totais e a fosfatase alcalina encontravam-se elevadas no Grupo B. Para os
achados hematológicos, a contagem total de hemácia (He; p<0,001), hematócrito (Ht; p
< 0,05) e hemoglobina (Hb; p < 0,01) foi menor no Grupo B. As atividades de CAT e
GPx apresentaram-se inferiores (p < 0,001) no grupo B (189,358 ± 90,385; 3609,659 ±
1569,078) em relação ao grupo A (326,671±104,528; 5055,622±1569,078),
respectivamente. Houve correlação positiva entre hemácias e CAT, contudo, não foi
significativa. O estresse oxidativo está estabelecido na LC, além disso, a atividade da
enzima GPx e CAT pode ser utilizado como marcador da patogenia da doença associada
a outros resultados.
Palavras-chave: Estresse oxidativo, Catalase, Glutationa peroxidade, hemácias, leishmaniose visceral canina.

ABSTRACT
Canine visceral leishmaniasis (CVL) is a disease which affects many tissues and organs. Recent
studies suggested the involvement of reactive oxygen species (ROS) in the pathogenesis of
various infectious and parasitic diseases in dogs, including CVL. The aim of this work was to
evaluate the role of oxidative stress in the LVC. Therefore, we used 27 dogs, both sexes, weight,
age and varied breeds, from the Fortaleza Center of Zoonosis Control and also from a private
kennel. The animals were considered when presented with leishmaniasis titer ≥ 1:40 for
Immunofluorescence Assay (IFA) and enzyme linked immunosorbent assay (ELISA),
associated with three or more characteristic signs of the disease. The animals were divided into
two groups, Group A (n = 12), healthy animals, and Group B (n = 15), animals positive for
CVL. Blood samples were collected by jugular venipuncture, distributed into tubes containing
anticoagulant (EDTA) or separating gel for determination of leukocyte parameters,
hematological, serum biochemical and glutathione peroxidase (GPx, mU / mg Hb) and catalase
(CAT; mU/mg Hb) erythrocyte enzymes activities. Results were expressed as mean and
standard deviation. Data analysis was performed using the Student t test and the differences
between groups considered significant at p <0.05. It was also used Pearson correlation test
between the variable GPx with He and Hb and between variable CAT with He and Hb. The
experimental protocol was approved by CEUA / UECE, No. 11516675-0/65. Among the serum
biochemical parameters, only the total protein and alkaline phosphatase were higher in Group B.
Concerning the hematological findings, the total count of red blood cells (RBC, p <0.001),
hematocrit (Ht, p <0.05) and hemoglobin (Hb, p <0.01) were lower in Group B. The activities of
CAT and GPx showed up lower (p <0.001) in group B (189.358 ± 90.385; 3609.659 ±
1569.078) than in group A (326.671 ± 104.528, 5055.622 ± 1569.078) respectively. A positive
correlation between RBC and CAT, however, was not significant. These data suggest that
oxidative stress is laid on the CVL, and also that GPx and CAT enzyme can be used as a marker
of pathogenesis of disease associated with other results.
Keywords: Oxidative stress, catalase, glutathione peroxidase, erythrocytes, canine visceral
leishmaniasis.

LISTA DE FIGURAS
Figura 1. Formas amastigotas (a) e promastigotas (b) de Leishmania........................... 17
Figura 2. Fêmea de flebotomíneo ingurgitada............................................................. 17
Figura 3. Cão soropositivo para Leishmania chagasi sintomático apresentando severa emaciação.......................................................................................................... 22
Figura 4. Cão soropositivo para Leishmania chagasi apresentando Ceratoconjuntivite Seca............................................................................................................................ 22
Figura 5. Cão soropositivo para Leishmania chagasi apresentando lesão no focinho............. 22
Figura 6. Cão soropositivo para Leishmania chagasi apresentado onicogrifose...................... 23

9
LISTA DE TABELAS
Tabela 1. Estudos que investigaram o estresse oxidativo associado ao estado fisiológico canino.
34
Tabela 2. Estudos que investigaram o estresse oxidativo associado às infecções caninas. 35
CAPÍTULO I
Tabela 1. Parâmetros hematológicos e bioquímicos de cães saudáveis e com LVC. 47
Tabela 2. Parâmetros hematológicos e marcadores antioxidantes de cães saudáveis e de cães com LVC.
47

10
LISTA DE ABREVIATURAS
CAT Catalase
CAnT Capacidade antioxidante total
CDV Vírus da cinomose canina
CCZ Centro de Controle de Zoonoses
CD4+ Células T auxiliares
CD8+ Células T citotóxicas
CIC Complexos Imunes Circulantes
CPV Vírus da parvovirose canina
CREA Creatinina
CVL Canine Visceral Leishmaniasis
Cu+ Cobre
EAT Estado antioxidante total
EDTA Anticoagulante
ELISA Ácido etilenodiaminotetracético
ERO Espécie reativa ao oxigênio
ERN Espécie reativa ao nitrogênio
FA Fosfatase alcalina
Fe2+ Ferro
GPx Glutationa peroxidase
GSH Glutationa reduzida
GSH-Rd Glutationa-redutase
GSSH Glutationa oxidada
Hb Hemoglobina
HNO2 Ácido nitroso
Ht Hematócrito
HO•2- Radical hidroperoxil
H2O2 Peróxido de hidrogênio
IFN-γ Interferon gama
IgG Imunoglobulina G
IgM Imunoglobulina M

11
IL-2 Interleucina-2
IL-4 Interleucina-4
IL-10 Interleucina-10
IL-12 Interleucina-12
LC Leishmaniose canina
LC Leishmaniose canina
LO• Radical alcoxila
LOO• Radical peroxila
LOOH Hidroperóxido
LPO Lipoperoxidação
LT Leucócitos totais
LV Leishmaniose visceral
LVC Leishmaniose visceral canina
Ml Mililitros
MPO Mieloperoxidase
nm Nanômetros
NADPH Nicotinamida adenina dinucleotídeo fosfato
NO• Óxido nítrico
NOS Óxido nítrico sintetase
NOX NADPH oxidase
NO2− Nitritos
NO3−, Nitratos
N2O3 Óxido nitroso
O2 Oxigênio
O•2- Ânion superóxido
OCl- Hipoclorito
OH• Radical hidroxila
ONOO- Peroxinitro
ONOOH Forma protonada do peroxinitro
Prx Perorredoxinas
PT Proteínas totais
RBC Red blood cel
RIFI Reação de Imunofluorescência Indireta
RPM Rotações por minuto

12
SOD Superóxido dismutase
SRD Sem padrão de raça definida
TBARS Substâncias reativas ao ácido tiobarbitúrico
TGO Transaminase glutâmico-oxaloacética
TGP Transaminase glutâmico-pirúvica
TNF-α Fator de necrose tumoral alfa
Trx Tioredoxina
U/L Unidades por litro
µL Microlitros
U Uréia

13
SUMÁRIO
RESUMO ................................................................................................................ 6
ABSTRACT 7
LISTA DE FIGURAS ............................................................................................ 8
LISTA DE TABELAS ........................................................................................... 9
LISTA DE ABREVIATURAS E SIGLAS ........................................................... 10
1 INTRODUÇÃO ................................................................................................... 14
2 REVISÃO DE LITERATURA........................................................................... 16
2.1. Leishmaniose................................................................................... 16
2.1.1 Agente etiológico, vetor e reservatório........................................ 16
2.1.2 Resposta imunológica e patogênese à infecção por Leishmania.......
18
2.1.3 Apresentações Clínicas.....................................................................
21
2.2 O estresse oxidativo................................................................................. 25
2.2.1 Oxidantes........... .............................................................................. 25
2.2.2 Antioxidantes .................................................................................. 29
2.3 Estresse oxidativo em condições fisiológicas e patológicas................... 32
3 JUSTIFICATIVA ................................................................................................ 35
4 HIPÓTESE CIENTÍFICA ................................................................................. 36
5 OBJETIVOS ........................................................................................................ 37
6 CAPÍTULO I ....................................................................................................... 38
8 CONCLUSÕES ................................................................................................... 55
9 PERSPECTIVAS ................................................................................................ 56
10 REFERÊNCIAS BIBLIOGRÁFICAS ............................................................ 57
ANEXO 1................................................................................................................. 65
ANEXO 2................................................................................................................. 66

14
1. INTRODUÇÃO
As leishmanioses são zoonoses causadas pela infecção de um dos diferentes
protozoários das espécies do gênero Leishmania spp. A transmissão do parasito ocorre
através da picada de insetos dípteros, da família Psychodidae, gênero Phlebotomus e
Lutzomyia (GONTIJO; MELO, 2004). O complexo das leishmanioses representa grave
problema de saúde pública, e o Brasil está entre os países da América Latina que
apresenta maior número de casos humanos, cerca de 90% dos casos anuais
(MONTEIRO et al., 2005).
Leishmania spp. possui ciclo biológico heteroxeno, desenvolvendo-se no
aparelho digestivo do inseto flebotomíneo fêmea, atravessando estágios morfológicos
de diferenciação até se transformar na forma promastigota metacíclica infectante
(CAMARGO-NEVES et al., 2006). Após a picada do inseto infectado, o protozoário
continua seu desenvolvimento no hospedeiro mamífero no qual a forma promastigota
dentro do fagossomo transforma-se em amastigota, que persiste e prolifera dentro de
várias células do sistema imune (MORDASKI et al., 2009; REILING et al., 2010;
KAYE; SCOTT, 2011).
As diferentes espécies de Leishmania spp. são responsáveis por um amplo
espectro de doenças que acometem os mamíferos. Vale salientar que, embora o homem
possa atuar tanto como hospedeiro quanto reservatório do agente, outros mamíferos
podem atuar da mesma forma, incluindo os canídeos, roedores e marsupiais, sendo estes
apontados como responsáveis pela manutenção por longo prazo da Leishmania na
natureza. Diferente dos demais hospedeiros que estão localizados nas regiões rurais ou
silvestres, o cão se encontra adaptado às zonas urbanas, o que lhe é atribuído como um
dos principais responsáveis pela manutenção da cadeia epidemiológica urbana da
leishmaniose (RIBEIRO, 2007).
A leishmaniose visceral canina (LVC) é caracterizada por diferentes
apresentações, desde uma doença não aparente até casos severos, podendo levar o cão à
morte (BRASIL, 2006). Estes diferentes quadros clínicos vem sendo atribuídos à
resposta imune do cão e à cepa do parasito inoculado pela picada do vetor (SILVA,
2007). Portanto, a participação do sistema imune é determinante para a progressão ou
resolução da infecção (FREITAS; PINHEIRO, 2010).
Leishmania spp. são parasitos intracelulares obrigatórios e infectam células
fagocíticas as quais utilizam mecanismos químicos e celulares de resposta rápida para
controlar o crescimento ou eliminação do agente invasor (ABBAS, 2008). A eliminação

15
do material fagocitado pode ocorrer por mecanismos oxidativos e não oxidativos. O
mecanismo oxidativo é gerado através de processos que utilizam um elevado consumo
de oxigênio plasmático (explosão respiratória), sendo produzido quando um complexo
enzimático, NADPH oxidase (NOX), na membrana dos fagócitos é formado, resultando
na geração de produtos microbicidas, como intermediários reativos do oxigênio, do
nitrogênio e hipoaletos produzidos pela mieloperoxidase (SEGAL, 2005). Contudo, o
acúmulo dos produtos microbicidas tem como contrapartida a lesão de biomoléculas e,
se não controlados pelo sistema antioxidante tornam-se excessivos desenvolvendo-se o
estresse oxidativo. Nesse contexto, a leishmaniose canina vem sendo estudada quanto à
participação do status antioxidativo enzimático e não enzimático (BILDIK et al., 2004;
BRITTI et al., 2008).

16
2. REVISÃO DE LITERATURA
2.1. Leishmaniose
As leishmanioses são zoonoses que acometem o homem quando ele entra em
contato com o ciclo de transmissão do parasito (BRASIL, 2006). Existem várias formas
da apresentação, quais sejam as formas tegumentar, cutânea e mucocutânea, e visceral.
A doença visceral é caracterizada por uma evolução crônica com comprometimento
sistêmico e, se não tratada, pode resultar na morte em até 90% dos casos (MAIA-
ELKHOURY et al., 2008). Além do homem, alguns mamíferos podem ser acometidos
de leishmaniose, como as raposas (Dusicyonvetuluse cerdocyonthous), os marsupiais
(Didelphis albiventris), os saguis (Callithrix penicillata), o cão (Canis lupus familiares),
o gato (Felis catus), os quais podem servir como reservatório (BRASIL, 2006;
COELHO et al., 2010).
2.1.1 Agente etiológico, vetor e reservatórios
Os agentes causadores da leishmaniose canina (LC) são protozoários
pertencentes ao Reino Protista, da família Trypanosomatidae, gênero Leishmania, e no
Brasil são encontradas diversas espécies, como Leishmania amazonensis, L. braziliensis
e L. guyanensis, responsáveis pela leishmaniose tegumentar; e L. chagasi, responsável
pela forma visceral. Embora se discuta sobre a presença de outra espécie como agente
etiológico da leishmaniose visceral canina (LVC), Leishmania (Leishmania) infantum,
estudos com base nos perfis isoenzimáticos demonstram que os dois protozoários, na
realidade, sejam um só (BRASIL, 2006).
Em seu ciclo biológico, Leishmania spp. apresenta três formas morfológicas, a
forma amastigota (Figura 1a), medindo 2 a 5 µm, de corpo oval contendo um núcleo
excêntrico e flagelo não visível, em que a forma é intracelular obrigatória, sendo
encontrada nas células do sistema fagocítico mononuclear (SFM) do hospedeiro
vertebrado, a outra é a forma promastigota (Figura 1b), medindo de 10 a 15 µm, com
um flagelo longo de até 20 µm, encontrada no tubo digestivo do inseto vetor e no
hospedeiro vertebrado, quando este acaba de ser picado pelo inseto flebotomíneo e a
outra forma é a paramastigota, encontrada no hospedeiro invertebrado, que possui um
flagelo mais curto e mais robusto. Estes parasitos apresentam uma estrutura
característica contendo DNA mitocondrial (cinetoplasto), estando ligada a um corpo

17
basal, que dá origem a um único flagelo (CIARAMELLA; CORONA, 2003;
CAMARGO-NEVES et al., 2006).
Figura 1. Formas amastigotas (a) e promastigotas (b) de Leishmania spp.
(Fonte: http://www.fcfrp.usp.br/dactb/Parasitologia/Arquivos/Genero_Leishmania.htm)
O vetor biológico responsável pela transmissão das formas promastigotas da
Leishmania spp. são insetos dípteros da família Psychodidae, que pertencem ao gênero
Lutzomya encontrado nas Américas. No Ceará, o único flebotomíneo até hoje apontado
como transmissor da LVC é o Lutzomya longipalpis (BRASIL, 2006).
Figura 2. Fêmea de flebotomíneo ingurgitada (Fonte: BRASIL, 2006).

18
A infecção do vetor ocorre quando a fêmea do flebotomíneo (Figura 2) ao sugar
o sangue de mamíferos infectados, ingere células do sistema imune fagocítico
parasitadas com a forma amastigota da Leishmania. No trato digestivo do flebotomíneo
ocorre o rompimento dos macrófagos infectados liberando essa forma, que se reproduz
por divisão binária e se diferencia rapidamente em promastigota, que também se
reproduz rapidamente por divisão binária para a forma paramastigota que colonizam o
esôfago e a faringe do vetor, onde ficam aderidas ao epitélio pelo flagelo e,
posteriormente, se diferencia na forma infectante promastigota metacíclica
(MORDASKI et al., 2009).
Os animais que podem se contaminar com a Leishmania spp. através da picada
do flebótomo são mamíferos selvagens, como as raposas (Dusicyonvetuluse
Cerdocyonthous) e os marsupiais (Didelphisalbiventris), além do próprio homem que
também pode ser um reservatório (BRASIL, 2006). Com o incremento do processo de
domiciliação do ciclo zoonótico de transmissão das leishmanioses, animais
sinantrópicos e domésticos tem assumido importância relevante na infecção (DANTAS-
TORRES, 2007).
O papel do cão (Canis lupus familiaris) como reservatório da Leishmania foi
relatado pela primeira vez por Nicolle, em 1908, na Tunísia, quando experimentalmente
foi comprovada a infecção deste animal. Posteriormente, em inquérito realizado naquele
país, foi comprovada a transmissão natural em cães e assim registrado o primeiro foco
de LVC no mundo. No Brasil, a primeira evidência de transmissão da LVC foi em
Abaeté (PA), como resultado dos trabalhos desenvolvidos por comissão instituída pelo
Instituto Oswaldo Cruz para a avaliação dessa problemática. Porém, apenas em 1955 é
que foi estabelecido por Deane e Deane o papel do cão como reservatório da LVC,
quando constatada a transmissão em cães residentes em zona urbana do município de
Sobral (CE), verificando-se elevada carga parasitária nesses animais. Brener, em 1957,
concluiu frente aos achados de outros autores, que o cão tinha papel fundamental na
epidemiologia da leishmaniose visceral no Brasil (CAMARGO-NEVES et al., 2006).
2.1.2Resposta imunológica e a patogênese da infecção por Leishmania spp.
A infecção por diferentes espécies de Leishmania, em grande parte, depende do
tipo de resposta imunológica que o organismo do hospedeiro irá desenvolver (KAYE;
SCOTT, 2011). Nas infecções parasitárias, o sistema imune pode controlar tanto o

19
número de parasitos presentes no organismo quanto à resistência a infecção (ABBAS,
2008).
O estabelecimento da infecção implica que o parasito tem de entrar nas células
fagocitárias, monócitos, macrófagos residentes, células dendríticas e polimorfonucleares
neutrófilos através de um processo denominado fagocitose e, uma vez no interior da
célula, resistir à ação microbicida. Imediatamente após a infecção, um processo
inflamatório local é iniciado, e nas primeiras horas pós-infecção, as primeiras células a
serem recrutadas ao local são os polimorfonucleares neutrófilos, seguidos pelos
monócitos/macrófagos dois ou três dias depois (AGA et al., 2002). Após a fagocitose do
protozoário, mecanismos intrínsecos podem ser ativados fazendo com que o parasito se
torne resistente à ação das enzimas hidrolíticas e das espécies reativas ao oxigênio
(ASSCHE et al., 2011).
A disseminação e a localização estável do parasito no organismo é um requisito
prévio para a progressão do processo patogênico e, consequente manifestação clínica da
doença. Desta forma, o elemento patogênico primário na LVC é a infecção,
sobrevivência e multiplicação do parasito no interior das células do sistema
mononuclear fagocitário (MARQUES, 2008). Depois de se multiplicarem dentro dos
macrófagos no local da infecção após a picada do inseto, os parasitos podem deixar a
pele e se disseminar para os fagócitos mononucleares do sistema reticuloendotelial,
incluindo órgãos como baço, fígado e medula óssea, levando a uma doença crônica, e às
vezes fatal (GOTO; PRIANTI, 2009).
O sistema imunológico, além de agir inicialmente através dos mecanismos inatos
de defesa, também aciona os mecanismos específicos de defesa. Os linfócitos T
desempenham função no controle da infecção, tanto diretamente, por mediarem
respostas celulares e moleculares, quanto indiretamente, na regulação e produção de
anticorpos produzidos e secretados por linfócitos B ativados especificamente (ABBAS
et al., 2008).
A sintomatologia clínica da LVC tem sido associada às alterações imunológicas
com a participação das subpopulações de Linfócitos T efetores. Estas alterações incluem
a ausência de hipersensibilidade do Tipo IV aos antígenos da Leishmania, decréscimo
de Linfócitos T no sangue periférico, ausência de produção de interferon gama (IFN-γ)
e de Interleucina-2 (IL-2) pelas células mononucleares do sangue periférico in vitro.
Além disso, altos títulos de anticorpos anti-Leishmania foram detectados em animais
sintomáticos, principalmente da classe de Imunoglobulina G (IgG) (BARBIÉRI, 2006).

20
As principais subpopulações de linfócitos T incluem os Linfócitos T CD4+ (LT
CD4+, auxiliares, Th) e os Linfócitos T CD8+ (LT CD8+, citotóxicos). Em resposta aos
antígenos protéicos dos microorganismos, LT CD4+ podem se diferenciar em células
efetoras, denominadas Th1, Th2, que produzem e secretam distintos grupos de
citocinas, além dos Linfócitos T regulatórios, Treg (MCMAHON-PRATT;
ALEXANDER, 2004; GLAICHENHAUS et al., 2011). Várias pesquisas demonstraram
a participação de células efetoras LTCD4+ com a resistência ou susceptibilidade à
infecção por Leishmania (ALEXANDER; BRYSON, 2005).
Um perfil de aparente resistência a LVC tem sido observado quando a resposta
imune celular é mediada predominantemente por Th1, através da participação das
citocinas Interferon gama (IFN-γ), fator de necrose tumoral alfa (TNF-α) e Interleucina
2 (IL-2). Esta resistência é atribuída à ativação dos macrófagos pelas citocinas, IFN-γ e
TNF-α, capazes de ativar mecanismos que envolvem a participação do óxido nítrico
para eliminar formas amastigotas intracelulares (BARBIÉRI, 2006). Por outro lado, tem
sido relatada a participação de Treg que medeiam respostas supressoras através das
citocinas Interleucina 10 (IL-10) e fator de crescimento de transformação beta (TGF-β),
interferindo no controle da infecção por L. infantum em humanos e camundongos
(GLAICHENHAUS et al., 2011). A participação de Th2, através das citocinas
Interleucina 4 (IL-4) e IL-10, neste processo vem sendo relacionada a doença
progressiva, pois estão implicados mecanismos que elevam a produção de anticorpos
específicos anti-Leishmania. LTCD8+ podem também estar envolvidos no processo de
resistência ao parasito (KAYE; SCOTT, 2011).
Alguns estudos demonstraram que a formação das diferentes subclasses de
Imunoglobulinas da classe G (IgG) pode servir de indicador da evolução da LVC.
Assim, em cães assintomáticos foi observado um aumento de anticorpos anti-
Leishmania do tipo IgG1, e em animais clinicamente doentes um predomínio de
anticorpos anti-Leishmania do tipo IgG2 (INIESTA et al., 2005; FREITAS et al.,
2012b). Entretanto, Silva (2005) não conseguiu estabelecer uma correlação entre os
níveis de imunoglobulinas IgG, IgG1 e IgG2 com a classificação clínica dos cães
infectados por L. chagasi.
Outro fator importante no componente imunológico da resposta humoral é a
formação de complexos imunes circulantes (CIC), que se formam pela união de
antígenos de Leishmania, com anticorpos específicos das classes IgG ou IgM e que
possuem sítios de ligação para frações do complemento (principalmente a fração C3), os

21
quais podem ser depositados em órgãos e tecidos estratégicos, promovendo diversas
alterações patológicas, como vasculite, glomerulonefrite, artrite, entre outras
(MARQUES, 2008).
Além dos fatores imunológicos, as parasitoses, infecções secundárias, doenças
crônicas e alguns medicamentos podem levar ao aparecimento da LVC secundariamente
no indivíduo. Isto pode estar relacionado ao fato de alterarem o tipo de resposta imune
que o animal desenvolve, levando a ruptura do equilíbrio existente entre hospedeiro e
parasito, traduzindo-se na manifestação clínica da LVC (SILVA, 2007).
2.1.3Apresentações Clínicas
Como foi anteriormente relatado, cães com leishmaniose podem apresentar
diferentes apresentações sintomatológicas. Os animais infectados podem desenvolver
sintomas acentuados da infecção, podendo resultar em morte, enquanto outros
permanecem assintomáticos, ou ainda desenvolvem um ou mais sintomas brandos,
sendo classificados como oligossintomáticos (BRASIL, 2006).
A LVC, sendo uma doença de caráter crônica, pode apresentar sinais clínicos
com manifestacão de três meses a sete anos após a infecção. A apresentação clínica
inicial corresponde a uma alteração do estado orgânico do animal, com perda de peso
progressiva acompanhada de astenia, apatia e em casos extremos anorexia e febre, que
caracterizam uma síndrome geral inespecífica, de instalação lenta e evolução
progressiva (MARQUES, 2008).
Na leishmaniose, o estado geral do animal pode estar comprometido por diversas
manifestações clinicas, como emagrecimento, caquexia, apatia, prostração,
desidratação, febre, mucosas hipocoradas, epistaxe, esplenomegalia e linfoadenopatia
(SANTOS, 2006; ALBUQUERQUE et al., 2007; ABREU-SILVA et al., 2008;
MARQUES, 2008; PINHÃO, 2009; FREITAS et al., 2012a). A linfoadenomegalia é
um sinal característico da LVC, e esse aumento no tamanho dos linfonodos ocorre
devido à proliferação linfoplasmohistiocitário, podendo conter lesões hipertróficas nas
regiões corticais e medulares com formas amastigotas dentro de macrófagos medulares
(SILVA, 2007).

22
Figura 3. Cão soropositivo para Leishmania chagasi sintomático apresentando severa
emaciação.
O emagrecimento do animal (Figura 3) pode ser causado pela competição
parasitária por nutrientes, por mediadores químicos que induzem ao estado de caquexia.
A sintomatologia renal pode levar a anorexia, a anemia, tornando-o progressivamente
mais fraco e apático, podendo até ficar letárgico (PINHÃO, 2009).
Figura 4. Cão soropositivo para
Leishmania chagasi apresentando
Ceratoconjuntivite Seca.
Figura 5. Cão soropositivo para Leishmania chagasi
apresentando lesão no focinho.

23

24
Figura 6. Cão soropositivo para Leishmania chagasi apresentado onicogrifose.
As principais alterações cutâneas observadas são as dermatites crônicas,
alopécia, principalmente na zona periocular, pregas de pele e articulações, anomalias de
cornificação, seborréia seca, hiperqueratose, paroníquia, onicogrifose (Fig. 6), úlceras e
crostas; lesões no focinho (Fig. 5) e ponta da orelha (SANTOS, 2006; ABREU-SILVA
et al., 2008; MARQUES, 2008; PINHÃO, 2009; FREITAS et al., 2012a) .
As alterações dermatológicas são bastante relatadas em estudo clínico em cães
com leishmaniose, podendo chegar a 90% dos cães infectados (ALBUQUERQUE et al.,
2007). Este fato pode ser explicado por um processo inflamatório no local da picada do
vetor, migração de novas células para o sítio da infecção, gerando um infiltrado
inflamatório composto de neutrófilos, macrófagos e linfócitos que levam à formação de
granuloma. Neste ocorre a multiplicação do parasito em novas células, liberando
mediadores do processo inflamatório, ativação de células que culminam com a
ulceração superficial da pele (RIBEIRO, 2004). Os animais podem ainda apresentar
alopécia, que geralmente tem início na cabeça e orelhas, atingindo depois o tronco e os
membros. Esta pode também localizar-se ao redor dos olhos, dando origem a uma
imagem característica de “óculos” (PINHÃO, 2009).
As alterações oculares são bastante relatadas nos cães com leishmaniose, e
dentre elas a ceratoconjuntivite (Fig. 4), blefarite, inflamação mononuclear-
plasmocitário do trato uveal, conjuntivite, exoftalmia, corrimento e panoftalmia
(BRITO et al., 2004; COUTINHO, 2005; FREITAS et al., 2012a).
As lesões renais são a principal causa de óbito por LVC (RIBEIRO, 2004). Nos
rins, a deposição de imunocomplexos nos glomérulos pode acarretar em
glomerulonefrite membranoproliferativa e nefrite intersticial com comprometimento da
função renal (ALBUQUERQUE et al., 2005), muitas vezes sendo a principal causa da
morte dos cães. A nefropatia pode ser causada pelo infiltrado de LTCD4+ detectados na
região glomerular e intersticial dos rins de cães naturalmente infectados com L. chagasi
(COSTA et al., 2000). A insuficiência renal pode estar presente em cães sem os sinais
clássicos sistêmicos de LVC. A presença persistente de proteína na urina pode levar ao
aparecimento de sinais clínicos compatíveis com síndrome nefrótica, tais como
hipoalbuminemia, ascite, edema periférico e hipercolesterolemia (MARQUES, 2008).
No fígado, ocorre um infiltrado de células mononucleares devido à presença de
antígenos de Leishmania. Sant’ana et al. (2007) observaram que ocorre uma intensa

25
formação de granulomas hepáticos na LVC, e cães assintomáticos apresentam maior
número de granulomas do que os animais oligossintomáticos e sintomáticos.
Outro achado frequente na LVC é a esplenomegalia. As alterações patológicas
apresentam graus variados de intensidade, e o aspecto macroscópico das lesões está
relacionado com a evolução da doença (LUVIZOTTO, 2006). No baço ocorre uma
reação inflamatória crônica e difusa, com macrófagos organizados em granulomas e
repletos de amastigotas resultando em hipertrofia do órgão (XAVIER et al., 2006).
O exame citológico e histológico da medula óssea mostra padrão variável de
acordo com a fase da doença, podendo ser observado hipocelularidade das células da
linhagem eritrocitária e leucocitária, macrófagos repletos de hemossiderina, indicativo
de acúmulo de ferro não utilizado na produção de hemácias, sendo esse achado
relacionado à anemia profunda que, frequentemente, ocorre no desenvolvimento da
leishmaniose visceral, e que pode ser detectado ao exame necroscópico (LUVIZOTTO,
2006).
Outros sinais como complicação cardiorrespiratória e digestiva têm sido
associados aos animais com LVC. As alterações articulares também podem ser notadas,
como claudicação, dor, restrição de movimento e aumento de temperatura intra-articular
(EUGÊNIO et al., 2008).

26
2.2. Estresse oxidativo
2.2.1 Oxidantes
O oxigênio (O2) é uma molécula altamente requerida na formação de energia
pelos organismos aeróbicos, cerca de 85% a 90% do O2 é consumido na mitocôndria,
por meio da cadeia transportadora de elétrons. O restante é utilizado por diversas
enzimas oxidases e oxigenases e, ainda, por reações químicas de oxidação direta
(BARBOSAet al., 2010). Esta molécula participa na formação de vários agentes
oxidante por sua característica de ser um aceptor de elétrons, ou seja, possui elétrons
desemparelhados na sua última camada eletrônica pronta para receber outros elétrons
(IMLAY, 2003).
Os agentes oxidantes podem ser divididos em vários grupos, dependendo da sua
natureza ou sua reatividade. Uma classificação amplamente utilizada é se as moléculas
oxidantes são ou não radicais livre, ou seja, contém elétron não pareado na sua estrutura
química, levando a instabilidade da molécula (LYKKESFELDT; SVENDSEN, 2007).
A ação dos agentes oxidantes nas moléculas biológicas pode ocorrer através da
abstração de uma molécula de hidrogênio, abstração de um elétron, ou pela adição de
oxigênio. Por originarem-se bastante do metabolismo do oxigênio, as moléculas
oxidantes geralmente são classificadas como espécie reativa do oxigênio (ERO) como,
por exemplo, o ânion superóxido (O•2-) e peróxido de hidrogênio (H2O2). Os radicais
derivados do oxigênio representam a classe mais importante das espécies de radicais
gerados em sistemas vivos, principalmente por ter considerado grau de reatividade
(VALKO et al., 2007).
EROs são produzidas e liberadas no organismo por diferentes vias, como na
mitocôndria, pelas quais o oxigênio que escapa da cadeia de transporte de elétrons e
pode extravasar para o sarcoplasma. Também pode ter origem no endotélio capilar, em
que o processo de isquemia e reperfusão acontecem durante o exercício, além da
formação dos agentes oxidantes através de células inflamatórias mobilizadas para
reparar o dano tecidual e a auto-oxidação das catecolaminas, mioglobinas e
hemoglobina (KOHEN; NYSKA, 2002; CAMPOS; YOSHIDA, 2004). A formação da
grande maioria das EROs origina-se da redução de um único elétron de O2 que gera o
ânion superóxido, uma espécie de radical pouco reativo que tem dificuldade de
atravessar membranas biológicas. No entanto, o O•2- na sua forma protonada, isto é,
quando recebe um átomo de hidrogênio, forma o radical hidroperoxil (HO•2-). O radical
HO•2- tem maior facilidade em penetrar estruturas biológicas, embora o superóxido se

27
encontre mais na sua forma carregada sob pH fisiológico. Quando o radical O•2- sofre
dismutação, produz peróxido de hidrogênio, uma molécula altamente difusível no
organismo (UNDERHILL; OZINSKY, 2002).
Ao contrário dos radicais livres, o H2O2 tem meia vida longa no organismo e é
capaz de atravessar membranas celulares desencadeando mecanismos potencialmente
tóxicos para as células. Esta toxicidade pode ser aumentada em dez mil vezes pela
presença de ferro (DEATON; MARLIN, 2003). Além de ter ação deletéria em
moléculas orgânicas, o H2O2é utilizado como fonte para outros agentes oxidantes,
principalmente o radical hidroxila (OH•), formado via oxidação de metais de transição,
como os íons de ferro (Fe2+) e cobre (Cu+). O ferro é o metal pesado mais abundante no
organismo e está biologicamente mais capacitado para catalisar as reações de oxidação
de biomoléculas. Desta forma, através das reações de oxirredução dos íons de ferro pela
reação de Fenton (equação 1) ou Haber-Weiss (equação 2 e 3) é formado o radical
peróxido de hidrogênio (KOHEN; NYSKA, 2002).Radical OH• é um dos mais reativos
oxidantes das biomoléculas capaz de retirar átomos de hidrogênio do grupo metileno de
ácidos graxos poliinsaturados, dando início à peroxidação lipídica que acaba
provocando a lise de membranas(CAMPOS; YOSHIDA, 2004). A sua formação está
associada com danos celulares em diferentes níveis, incluindo danos a proteínas, ao
DNA e a própria membrana celular (POULSEN, 2005).
Fe2++ H2O2 → OH• + OH- + Fe3+ (1)
Fe3++ O•2- → Fe2++ O2 (2)
O2 + OH• +OH- → O•
2- + H2O2 (3)
Todas as membranas celulares são especialmente vulneráveis à oxidação devido
à sua alta concentração de ácidos graxos poliinsaturados. O processo de peroxidação
lipídica resulta na formação de três importantes EROs a partir dos lipídios insaturados:
o radical alcoxila (LO•), radical peroxila (LOO•) e o hidroperóxido (LOOH). Como no
caso do H2O2, o LOOH não é um radical livre, mas ele é uma molécula bastante
instáveis na presença de metais de transição. Já o LOO•e o LO• são radicais livres muito
reativas que participam diretamente do processo de propagação da lipoperoxidação
(HALLIWELL; GUITTERIDGE, 2007). As ações deletérias dos peróxidos lipídicos
mais frequentes são a ruptura da membrana celular, mutação do DNA, oxidação dos
lipídios insaturados, formação de resíduos químicos como o malondialdeído (MDA) e o

28
comprometimento dos componentes da matriz extracelular, proteoglicanos, colágeno e
elastina (LIMA; ABDALLA, 2001).
A formação dessas espécies ocorre através de uma reação em cadeia iniciada por
radicais livres (equação 4, 5 e 6) (LIMA; ABDALLA, 2001). Além disso, íons de
metais de transição, como Fe2+e Fe3+podem participar do processo catalisando a
formação de radicais lipídicos alcoxila, peroxila e hidroxila a partir dos hidroperóxidos
(equação 7 e 8).
LH+ OH• (ou LO•)→ L•+ H2O (ou LOH) (4)
L•+ O2 → LOO• (5)
LOO• + LH → LOOH + L• (6)
LOOH + Fe2+→ LO• + OH• + Fe3+ (7)
Fe3+ + LOOH → LOO• H+ + Fe2+ (8)
O mais importante complexo enzimático desencadeador da geração das EROs
nos processos inflamatórios e infecciosos é a NADPH oxidase (NOX). O NOX é um
conjunto de enzimas espalhadas na membrana e citoplasma das células de defesa, como
neutrófilos e macrófagos. Quando o fagócito é ativado, este complexo enzimático se
rearranja devido a diversas interações iônicas com suas proteínas, dando início a um
fenômeno chamado explosão respiratória. A molécula de nicotinamida adenina
dinucleotídeo fosfato (NADPH) que é gerado no ciclo das pentoses é oxidada pelo NOX
e uma molécula de oxigênio aceita um elétron doado, levando a geração do ânion
superóxido (equação 9). A interação de duas moléculas de O•2-são catalisadas pela
enzima superóxido dismutase (SOD), gerando uma molécula de peróxido de hidrogênio
(equação 10) (BABIOR, 2004).
NADPH + 2O2NOX
→NADP+ + H+ + 2O•2- (9)
2O•2- + 2H+SOD-Zn→H2O2 + O2 (10)
O peróxido de hidrogênio serve como substrato para formação de compostos
bactericidas, através da ação da mieloperoxidase (MPO). A MPO é uma das mais
importantes enzimas da explosão respiratória, pois catalisa a reação entre o H2O2e os
íons haletos (equação 11), produzindo hipoaletos (OCl-, OBr-, OI-, OSCN-). O
hipoclorito é o principal hipoaleto utilizado pelos neutrófilos devido a sua propriedade

29
de não se acumular nos sistemas biológicos, dissipando-se em múltiplas reações. A
formação do OCl- contribui para destruição das bactérias por meio da oxidação de suas
proteínas e lipídios, além de aumentar a atividade bactericida das enzimas lisossômicas
(SEGAL, 2005; van der VEEN et al., 2009).
H2O2 + Cl-MPO
→ H2O + OCl- (11)
Outra forma de eliminar os organismos invasores é através da produção de óxido
nítrico (NO•). O NO• é gerado quando ocorre a oxidação do aminoácido ʟ-arginina para
ʟ-citrulina na presença da molécula de oxigênio (equação 12), sendo esta reação
catalisada pela enzima óxido nítrico sintetase (NOS) (FRITZSCHE et al, 2010). A NOS
é uma enzima encontrada no organismo em diferentes isoformas, sendo duas
constitutiva geradas no endotélio e no tecido neuronal (eNOS e nNOS) e uma com
expressão induzida por uma ampla variedade de células e tecidos (iNOS) (BABIOR,
2004).
ʟ-arginina + NADPH + O2 → ʟ-citrulina + NO• + NADP+
(12)
O aumento da produção do óxido nítrico não necessariamente pode estar
relacionado com o estresse oxidativo. NO• pode ser observado desenvolvendo o papel
de mensageiro indutor da vasodilatação sanguínea, além de sua função como
mensageiro neuronal (LYKKESFELDT; SVENDSEN, 2007). Assim, o aumento da
produção de NO• por si só não é um indicador do estresse oxidativo, embora elevadas
concentrações desta molécula possam levar ao aumento da formação de oxidantes
deletérios denominados espécies reativas do nitrogênio (ERN). Podemos observar a
formação de ERN a partir do óxido nítrico formando o peroxinitro (ONOO-, equação
12), a forma protonada do peroxinitro (ONOOH) que é altamente reativo ao radical
hidroxila (equação 15), além do óxido nitroso (N2O3), ácido nitroso (HNO2), nitritos
(NO2−) e nitratos (NO3
−, equação 16) (VALKO et al., 2006).
NO• + O•2- → ONOO- (13)
ONOO- + H+ → ONOOH (14)

30
ONOOH → OH• + NO2 (15)
OH• + NO2 → NO3-+ H (16)
ERN são muito tóxicas, podendo causar danos diversos, como a peroxidação
lipídica através do processo de nitração, além de nitrar aminoácidos aromáticos, por
exemplo, a tirosina gerando nitrotirosina e as bases de DNA como a guanina se
transformando em 8-nitroguanina (BARREIROS et al., 2006).
2.2.2 Antioxidantes
EROs são encontradas em baixas concentrações nas células e tecidos biológicos.
Sua concentração é determinada pelo equilíbrio entre a taxa de produção e sua taxa de
remoção efetivada por várias enzimas e composta antioxidantes (DRÖGE, 2002). O
efeito prejudicial dos agentes oxidante ocorre quando eles estão em quantidade
excessiva no organismo, ultrapassando a capacidade do organismo de neutralizá-los
com os seus sistemas naturais. Para proteger-se, as células possuem sistema de defesa
que pode atuar em duas linhas. Uma delas atua como detoxificadora dos agentes antes
que eles causem lesões. Esta linha é constituída pelas enzimas glutationa reduzida
(GSH), superóxido dismutase (SOD), catalase (CAT), glutationa peroxidase (GPx) e
vitamina E. A outra linha de defesa tem a função de reparar a lesão ocorrida, sendo
constituída pelo ácido ascórbico, pela glutationa-redutase (GSH-Rd) e pela GPx, entre
outros. Com exceção da vitamina E (α-tocoferol), que é um antioxidante estrutural da
membrana, a maior parte dos agentes antioxidantes está no meio intracelular e no
organismo (CAMPOS; YOSHIDA, 2004; NAITO et al., 2010).
Todos os componentes que auxiliam na defesa do organismo contra o
desequilíbrio da homeostase provocado por agentes oxidantes formam uma rede de
moléculas antioxidantes. Esta rede pode atuar evitando as fontes de espécies reativas ou
fazendo a varredura dos mesmos, além de favorecer o reparo e a reconstituição de
estruturas biológicas (NAITO et al., 2010). Os principais componentes enzimáticos da
rede antioxidante são a SOD, CAT e GPx, visto que sua atividade catalítica em
transformar o ânion superóxido em peróxido de hidrogênio e água, podendo inativar
relevantes quantidades de oxidantes. Oligoelementos como selênio (Se), zinco (Zn),
cobre (Cu) e manganês (Mn) desempenham importante papel catalisador da atividade
enzimática de GPx com Se e SOD com Zn/Cu/Mn (KLOTZ et al., 2003;
KIRSCHVINK et al., 2008).

31
A primeira linha de frente do organismo contra agentes oxidantes são os
antioxidantes enzimáticos. A SOD participa do início da prevenção dos oxidantes e tem
como ação converter através do processo de dismutação o ânion superóxido em
peróxido de hidrogênio e oxigênio na presença do próton H+ (equação 10) (BATINIC-
HABERLE et al., 2010). Existem duas formas de SOD no organismo, a primeira
contém Cu2+ e Zn2+ como cofatores e ocorre no citosol, sendo que sua atividade não é
afetada pelo estresse oxidativo. A segunda contém Mn2+ como cofator, ocorrendo na
mitocôndria e sua atividade aumenta com o estresse oxidativo.
O produto final da dismutação do peróxido de hidrogênio pode ser removido
pela atividade da CAT e por membros da família das peroxidases e a GPx
(BARREIROS et al., 2006).
2H2O2CAT
�2H2O + O2 (17)
2GSH + H2O2GPx-Se
� GSSH + 2H2O (18)
GSSH + NADPH + H+GSH-Rd� 2GSH + NADP+ (19)
2H2O2+ NADPH + H+GSH
� 2H2O + NADP+ (20)
O peróxido de hidrogênio é uma das espécies reativas ao oxigênio mais comum
na biosfera, assim tanto o H2O2 como o produto de sua decomposição, o radical OH•
devem ser controladas, pois podem ser prejudiciais a quase todos os componentes
celulares (ZAMOCKY et al., 2008). A catalase é uma hemeproteína citoplasmática que
catalisa a redução de H2O2 em oxigênio e água utilizando o NADPH (equação 17),
encontrada em organelas intracelulares, como nos peroxissomas e em baixa
concentração na mitocôndria e reticulo endoplasmático. Dessa forma, o H2O2
intracelular não pode ser eliminado, a menos que se difunda para os peroxissomas
(LIMÓN-PACHECO; GONSEBATT, 2009). A atividade catalítica da CAT está
presente em todos os tecidos, porém com atividade enzimática elevada nos eritrócitos,
fígado, rim e tecido adiposo e uma baixa atividade no tecido nervoso (LIMA, 2004).
H2O2 também é alvo de compostos tióis que são potentes agentes redutores
graças ao grupamento sulfidrila (-SH), sendo o principal componente a glutationa na sua
forma reduzida (GSH). Para exercer sua atividade antioxidante o GSH liga-se a GPx
ligada ao cofator selênio, transformando o H2O2 em H2O mais glutationa oxidada
(GSSG) (equação 18). Para manter o nível de GSH basal é necessário que a enzima

32
GSH-Rd dependente de NADPH adicione elétrons na GSSG (equação 19). Além disso,
a GPx reduz tanto o peróxido de hidrogênio como hidroperóxidos orgânicos. Existem
várias isoformas de GPx as quais são específicas para fase lipofílica ou hidrofílica
(LIMA, 2004; LUBOS et al., 2011).
Outro antioxidante enzimático é a tioredoxina (Trx) e as perorredoxinas (Prx).
Trx são enzimas que reparam grupamentos -SH, removem H2O2 e removem radicais
livres e podem estar presentes em diversos compartimentos celulares como mitocôndria,
núcleo e citossol. Estas enzimas também estão envolvidas na regeneração do ácido
ascórbico oxidado (NAKAMURA, 2005). As enzimas antioxidantes Prx, também
chamadas de tioredoxina peroxidase pertencem à família de proteínas multifuncionais
cujas principais ações são a proteção celular contra o estresse oxidativo e a modulação
da cascata de sinalização que regulam a proliferação e diferenciação celular e a
expressão gênica e apoptose (IMMENSCHUH; VOGT, 2005).
Os antioxidantes que também têm suas atividades reconhecidas são os não-
enzimáticos. O ácido ascórbico (vitamina C) é encontrado no organismo na forma de
ascorbato, uma molécula bastante solúvel em água e de baixo peso molecular,
localizada nos compartimentos aquosos e nos tecidos orgânicos. O ascorbato atua como
agente redutor, reduzindo metais de transição, principalmente Fe3+ e Cu2+ presente nos
sítios ativos das enzimas ou em suas formas livres no organismo. Outros antioxidantes
não-enzimáticos que agem intensamente nas membranas prevenindo sua
lipoperoxidação são os tocoferóis (vitamina E). O α-tocoferol é um dos principais
antioxidantes na fase lipofílica do organismo e podem agir como doadora de prótons H+
para o radical peroxila, interrompendo o processo radicalar em cadeia da peroxidação
lipídica. Os carotenóides, principalmente o β-caroteno age como desativadores do
oxigênio singleto ou como sequestradores dos radicais peroxila, reduzindo a oxidação
do DNA e lipídios (SONG, 2004; BARREIROS et al., 2006).
O estresse oxidativo é conhecido como uma ruptura no equilíbrio dinâmico entre
moléculas oxidantes e antioxidantes, favorecendo assim a viabilidade dos agentes
oxidantes no organismo. No entanto, a detecção direta da produção destes oxidantes
livres no organismo é muito difícil, uma vez que estas moléculas têm uma meia vida
curta e por serem altamente reativos. Desta forma, os danos oxidativos são geralmente
analisados através de seus produtos secundários, incluindo os derivados de
aminoácidos, ácido nucléico e a peroxidação lipídica (KOHEN; NYSKA, 2002).

33
Pode-se medir o estresse oxidativo através de diversos marcadores. Glutationa
reduzida e glutationa oxidada (GSH/GSSH) são ótimos indicadores do processo
oxidativo no organismo, pois a GSH é um antioxidante primário das células, e
geralmente sua concentração apresenta-se baixa nas doenças crônicas. Outros
compostos químicos inflamatórios que são medidos no sangue ou urina para confirmar
o estresse oxidativo excessivo são o óxido nítrico (NO) e os produtos da peroxidação
lipídica, como o malondialdeído (MDA) (MANDELKER; VAJDOVICH, 2011).
A avaliação da peroxidação lipídica é amplamente utilizada como um indicador
do estresse oxidativo, já que a maioria dos EROs e ERNs reage com os ácidos graxos
poliinsaturados presentes na membrana celular, podendo levar a destruição estrutural da
célula, à falência dos mecanismos de troca metabólica e à morte celular. Desta forma, a
avaliação dos níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) é útil na
investigação da peroxidação lipídica e é expresso como resultado da lipoperoxidação
(LPO) (LIMA; ABDALLA, 2001).
O perfil antioxidante também é utilizado para avaliação do equilíbrio de
oxirredução no organismo. Para tanto, são avaliados os componentes enzimáticos
(SOD,CAT, GSH, GSH-Rd, GPx) e não enzimáticos (α-tocoferol, β-caroteno, ácido
ascórbico), entre outros (MANDELKER; VAJDOVICH, 2011).
Também outros marcadores do perfil antioxidante podem ser citados, como a
coenzima Q (CoQ), que é uma ubiquinona lipossolúvel, sendo este o único lipídio
sintetizado endogenamente no organismo que apresenta função redox auxiliando na
inibição da lipoperoxidação das membranas (BARREIRAS et al., 2006). As proteínas
de transporte de metais de transição como a transferrina e a ceruloplasmina são
indicadas para demonstrar a concentração de íons metálicos que são agentes pro-
oxidantes, como o Fe2+ e o Cu+ (VANSCONCELOS et al., 2007).
Alguns testes conseguem estimar a quantidade e a capacidade antioxidante total
do organismo, como os testes da capacidade antioxidante total (CAnT), do índice de
estresse oxidativo (IEO) e estado antioxidante total (EAT). CAnT tem como principal
vantagem, medir a capacidade antioxidante de virtualmente todos os componentes de
uma amostra biológica (sangue, urina, fezes) e não somente de um ou outro composto
(FERRARI, 2008). Esses tipos de análises são bastante úteis na visão geral do
organismo, mas o decréscimo destes marcadores não significa necessariamente que o
dano oxidativo ocorreu, pois pode ser resultado simplesmente de que os mecanismos de
defesa cumpriram sua função habitual (VASCONCELOS, 2007).

34
2.3. Estresse oxidativo em condições fisiológicas e patológicas
A formação de EROs ocorre naturalmente no organismo dos mamíferos, pois
muitos mecanismos fisiológicos estão associados a formação desses compostos (Tabela
1). Alguns estudos têm tentado padronizar valores de referência para a espécie canina
dos biomarcadores do estresse oxidativo, dentre eles os produtos do metabolismo
oxidativo e as moléculas antioxidantes (TODOVA et al., 2005; PASQUINE et al.,
2008).
Estados fisiológicos comumentes relacionados ao estresse oxidativo na espécie
canina já foram relatados por alguns estudos, como em cadelas prenhes (VANNUCCHI
et al., 2007) e em cães submetidos a diversos tipos de exercícios (Tabela 1) .
Tabela 1. Estudos que investigaram o estresse oxidativo associado ao estado fisiológico canino. Estado fisiológico Alteração apresentadas Referências
Gestação Diminuição do Retinol, Tocoferol e Magnésio
(VANNUCCHI et al., 2007)
Exercício de caça Aumento do d-ROM e diminuição do NPBA
(PASQUINI et al., 2010)
Exercício de corrida Diminuição de Tocoferol e aumento de Isoprostanos
(HINCHCLIFF, 2000)
Exercício com trenós Aumento CAnT (DUNLAP et al., 2006)
d-ROM: metabólitos reativos derivados do oxigênio; NPBA: nível do potencial biológico antioxidante; Retinol: vitamina A; Tocoferol: vitamina E; Isoprostanos: indicador da peroxidação lipídica; CAnT: capacidade antioxidante total
As alterações observadas no estresse oxidativo podem ser encontradas em
indivíduos saudáveis, contudo podem ser controladas pelos mecanismos antioxidantes.
Por outro lado, indivíduos que se encontram doentes, em sua grande maioria,
apresentam um desequilíbrio nos mecanismos de oxirredução, favorecendo um intenso
estresse oxidativo no organismo (DRÖGE, 2002). Estudos têm relacionado à
participação das desordens oxidativas nas diferentes doenças infecciosas nos animais
domésticos, incluindo equinos, bovinos, caprinos, ovinos, caninos e felinos
35
(LYKKESFELDT; SVENDSEN, 2007; WEBB et al., 2008; CELI, 2010;
PALTRINIERI et al., 2010).
Na espécie canina, a formação de EROs em condições patológicas pode estar
presente na atopia (KAPUN et al., 2011), no diabetes (CHANSAISAKORN et al.,
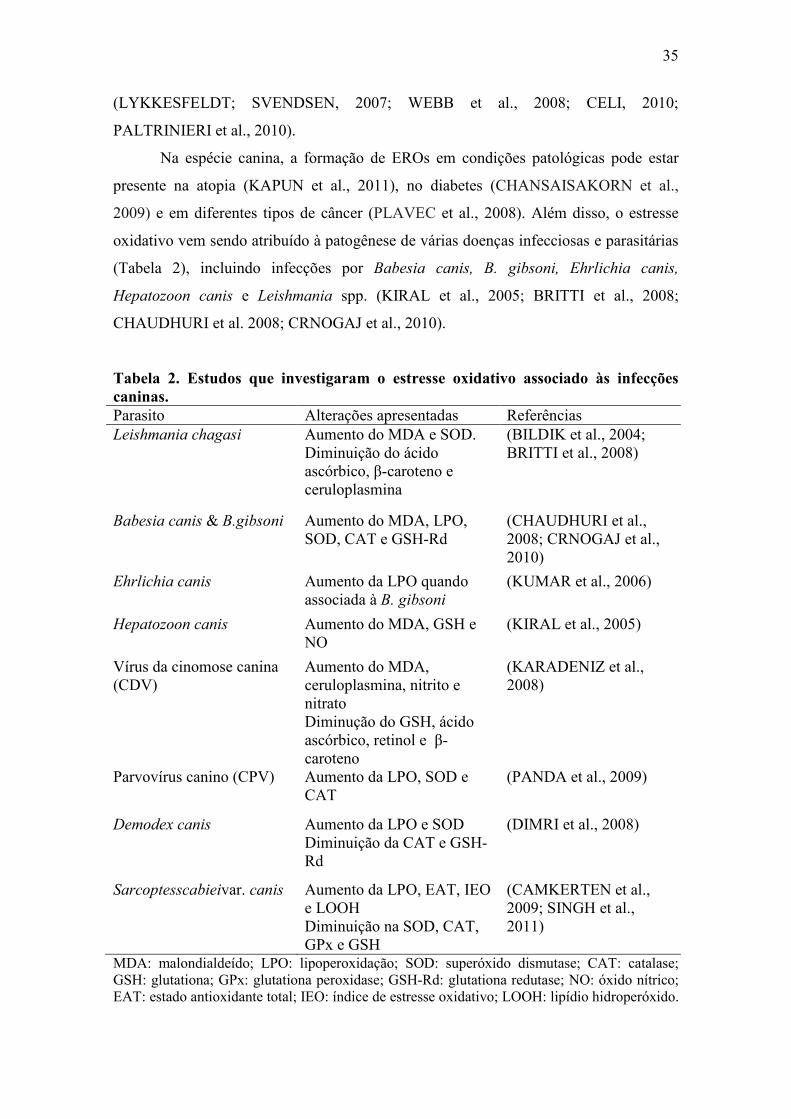
2009) e em diferentes tipos de câncer (PLAVEC et al., 2008). Além disso, o estresse
oxidativo vem sendo atribuído à patogênese de várias doenças infecciosas e parasitárias
(Tabela 2), incluindo infecções por Babesia canis, B. gibsoni, Ehrlichia canis,
Hepatozoon canis e Leishmania spp. (KIRAL et al., 2005; BRITTI et al., 2008;
CHAUDHURI et al. 2008; CRNOGAJ et al., 2010).
Tabela 2. Estudos que investigaram o estresse oxidativo associado às infecções caninas. Parasito Alterações apresentadas Referências Leishmania chagasi Aumento do MDA e SOD.
Diminuição do ácido ascórbico, β-caroteno e ceruloplasmina
(BILDIK et al., 2004; BRITTI et al., 2008)
Babesia canis & B.gibsoni Aumento do MDA, LPO, SOD, CAT e GSH-Rd
(CHAUDHURI et al., 2008; CRNOGAJ et al., 2010)
Ehrlichia canis Aumento da LPO quando associada à B. gibsoni
(KUMAR et al., 2006)
Hepatozoon canis Aumento do MDA, GSH e NO
(KIRAL et al., 2005)
Vírus da cinomose canina (CDV)
Aumento do MDA, ceruloplasmina, nitrito e nitrato Diminução do GSH, ácido ascórbico, retinol e β-caroteno
(KARADENIZ et al., 2008)
Parvovírus canino (CPV) Aumento da LPO, SOD e CAT
(PANDA et al., 2009)
Demodex canis Aumento da LPO e SOD Diminuição da CAT e GSH-Rd
(DIMRI et al., 2008)
Sarcoptesscabieivar. canis Aumento da LPO, EAT, IEO e LOOH Diminuição na SOD, CAT, GPx e GSH
(CAMKERTEN et al., 2009; SINGH et al., 2011)
MDA: malondialdeído; LPO: lipoperoxidação; SOD: superóxido dismutase; CAT: catalase; GSH: glutationa; GPx: glutationa peroxidase; GSH-Rd: glutationa redutase; NO: óxido nítrico; EAT: estado antioxidante total; IEO: índice de estresse oxidativo; LOOH: lipídio hidroperóxido.

36
3 JUSTIFICATIVA
O estresse oxidativo é conhecido como uma ruptura no equilíbrio dinâmico entre
moléculas oxidantes e antioxidantes. As alterações observadas no estresse oxidativo
podem ser encontradas em indivíduos saudáveis, contudo podem ser controladas pelos
mecanismos antioxidantes. Por outro lado, indivíduos que se encontram doentes, em sua
grande maioria, apresentam um desequilíbrio nos mecanismos de oxirredução,
favorecendo um intenso estresse oxidativo no organismo. O estresse oxidativo vem
sendo atribuído à patogênese de várias doenças infecciosas e parasitárias. O papel da
imunidade inata na defesa contra as diferentes espécies de Leishmania spp vem sendo
estudado, embora a função dos radicais livres na patogênese e na progressão de várias
doenças tenha sido discutida. A formação de espécies reativas ao oxigênio (ERO) de
maneira exagerada nas doenças infecciosas pode levar o organismo a um desequilíbrio
entre os agentes oxidantes e antioxidantes, proporcionando o estabelecimento do
estresse oxidativo. Este processo pode conduzir à oxidação de biomoléculas com
consequente perda de suas funções biológicas ou a um desequilíbrio homeostático que é
manifestado no organismo pela modificação de macromoléculas celulares, morte celular
por apoptose ou necrose, além de danos estruturais nos tecidos. Este assunto não tem
sido amplamente investigado na leishmaniose, o que torna a determinação de oxidantes
e antioxidantes no soro de grande valor e que podem funcionar como marcadores da
evolução na doença canina.

37
4. HIPÓTESE CIENTÍFICA
As enzimas antioxidantes podem ser utilizadas como marcadores do estresse
oxidativo na leishmaniose visceral canina.

38
5. OBJETIVOS
5.1. Objetivo geral
Avaliar o estresse oxidativo em cães naturalmente infectados com Leishmania chagasi. 5.2 Objetivos específicos
5.2.1
Avaliar os parâmetros leucocitários de cães naturalmente infectados com L. chagasi.
5.2.2 Avaliar os parâmetros hemáticos de cães naturalmente infectados com L.
chagasi. 5.2.3 Avaliar os parâmetros bioquímicos séricos de cães naturalmente infectados com
L. chagasi. 5.2.4 Determinar as atividades das enzimas glutationa peroxidase (GPx) e catalase
(CAT) eritrocitária em cães naturalmente infectados com L. chagasi. 5.2.5 Correlacionar os parâmetros hemáticos com a atividade das enzimas
antioxidantes.

39
6. Capítulo 1
Atividade das enzimas Catalase e Glutationa peroxidase em cães naturalmente
infectados por Leishmania chagasi
Catalase and glutathione peroxidase in dogs naturally infected with Leishmania
chagasi
Junho, 2012

40
Catalase and glutathione peroxidase in dogs naturally infected with Leishmania
chagasi
Belarmino Eugênio Lopes-Netoa; Glauco Jonas Lemos Santosa; Adam Leal Limaa;
Maritza Cavalcante Barbosab; Talyta Ellen Jesus dos Santosb; Daniel Couto Uchoac;
Evanisa Alves Venturad; Romélia Gonçalves Pinheirob; Diana Célia Sousa Nunes-
Pinheiroa*
aPrograma de Pós-Graduação em Ciências Veterinárias da Faculdade de
Veterinária/UECE; bLaboratório de Pesquisa em Hemoglobinopatias Genéticas das Doenças
Hematológicas/UFC; cCanil Grande Canafistula;
dCentro de Controle de Zoonoses de Fortaleza-CE
Corresponding author:
Universidade Estadual do Ceará/Laboratório de Imunologia e Bioquímica de
Animais/PPGCV/UECE, Av. Paranjana, 1700, CEP 60740-903 Fortaleza, Ceará, Brasil.
Fone. 55. 85.31019853; fax 55.85.31019860
e-mail: [email protected] ou [email protected]

41
Resumo: A leishmaniose visceral canina (LVC) é provocada por protozoário intracelular
obrigatório do gênero Leishmania e compromete diversos órgãos e tecidos. Estas
alterações podem resultar na geração de espécies reativas ao oxigênio (ERO). Estudos
recentes têm sugerido a participação de ERO na patogenia de diversas doenças. Neste
trabalho objetivou-se avaliar as atividades das enzimas antioxidantes na LC. Para tanto,
foram utilizados cães de ambos os sexos, peso, idade e raças variadas, oriundos do
Centro de Controle de Zoonoses de Fortaleza e de um canil particular de Fortaleza. Foi
considerada LVC animais com título ≥ 1:40 para Reação de Imunofluorescência
Indireta e para o Ensaio Imunoenzimático, associado ao sinais e sintomas característicos
da doença. Os animais foram divididos em dois grupos, Grupo A (n=12), animais sadios
e o Grupo B (n=15), animais com LVC. Foram determinados os parâmetros
leucocitários, hematológicos e bioquímicos séricos e as atividades das enzimas
glutationa peroxidase (GPx) e catalase (CAT) eritrocitárias. O estudo foi aprovado pelo
CEUA/UECE, n° 11516675-0/65. Dentre os parâmetros bioquímicos somente as
proteínas totais e a fosfatase alcalina encontravam-se elevados no grupo B. Os
parâmetros hemáticos do grupo B encontravam-se inferiores (p<0,05) aos do grupo A.
CAT e GPx apresentaram-se inferiores (p<0,001) no grupo B (189,358±90,385;
3609,659±1569,078) em relação ao grupo A (326,671±104,528; 5055,622±1569,078),
respectivamente. Houve correlação positiva entre hemácias e CAT, contudo, não foi
significativa. Conclui-se que CAT e GPx estão envolvidas no estresse oxidativo da
LVC.
Palavras-chaves: Estresse oxidativo, Catalase, Glutationa peroxidade, hemácias, Leihsmaniose canina.

42
Abstract
Canine visceral leishmaniasis (CVL) is caused by an obligatory intracellular parasite of
the Leishmania gender and undertakes several organs and tissues. These changes can
result in generation of reactive oxygen species (ROS). Recent studies address the role of
in the pathogenesis of many diseases. In this study, we aimed to access the activity of
the antioxidant enzymes in CVL. Therefore, were used dogs of both sexes, and varied
weight, ages and breeds, obtained in accordance with the Zoonosis Control Center in
Fortaleza city, Ceará, Brasil. Dogs were considered infected with ≥ 1:40 sera titration
for indirect immunofluorescent assay and ELISA, associated with classical symptoms of
the disease. The animals were divided in two groups, Group A (n=12), healthy animals
and group B (n=15), animals infected with CVL. The leukocytes, hematological and
biochemical parameters were assessed, along with the enzyme activity glutathione
peroxidase (GPx) and catalase (CAT) at erythrocyte. The present study was approved
by animal experimentation committee CEUA/UECE, nº 11516675-0/65. Among the
biochemical parameters only the total protein and alkaline phosphatase were increased
in group B. The hematological parameters in group B were decreased (p < 0.05) when
compared with group A. CAT and GPx also exhibited lower values (p < 0.001) in group
B (189.358±90.385; 3,609.659±1,569.078) when compared with group A
(326.671±104.528; 5,055.622±1,569.078), respectively. There was a positive
correlation between erythrocytes and CAT; however, it was not significant. We
conclude that CAT and GPx are involved in oxidative stress in CVL.
Keywords: Oxidative stress, erythrocyte, antioxidants enzymes, canine visceral
leishmaniasis.

43
1. Introdução
A leishmaniose é uma antropozoonose causada pela infecção de um dos
diferentes protozoários das espécies do gênero Leishmania. Sua transmissão ocorre
através da picada de insetos dípteros, da família Psychodidae, gênero Phlebotomus e
Lutzomyia (GONTIJO; MELO, 2004). Os diferentes tipos de Leishmania spp. são
responsáveis por um amplo espectro de doenças que acometem os mamíferos, e embora
o homem também possa ser hospedeiro e reservatório do agente, outros mamíferos
podem atuar da mesma forma, incluindo os canídeos, roedores e marsupiais, sendo estes
apontados como responsáveis pela manutenção a longo prazo da Leishmania na
natureza. Diferente dos demais hospedeiros que estão localizados nas regiões rurais ou
silvestres, o cão se encontra adaptado às zonas urbanas, o que lhe é atribuído como um
dos principais responsáveis pela manutenção da cadeia epidemiológica urbana da
leishmaniose (RIBEIRO, 2007).
A leishmaniose visceral canina (LVC) é caracterizada pelas diferentes
apresentações, podendo ser uma doença não aparente até casos severos, podendo levar à
morte. Estes diferentes quadros clínicos dependem da resposta imune do cão e da cepa
do parasito inoculado pela picada do vetor (SILVA, 2007). A participação do sistema
imunológico na disseminação da Leishmania no hospedeiro vertebrado é determinante
para a progressão ou resolução da infecção. LVC é uma doença sistêmica capaz de
comprometer diversos órgãos e tecidos, proporcionando alterações clinicopatológicas
nos animais como onicogrifose, hepatoesplenomegalia, linfadenomegalia, alopecia,
anemia (FREITAS et al., 2012b).
Estudos recentes têm sugerido a participação de espécies reativas ao oxigênio
(ERO) na patogenia de diversas doenças infecciosas e parasitárias em cães, incluindo a
cinomose (KARADENIZ et al., 2008), parvovirose (PANDA et al., 2009), babesiose
(CHAUDHURI et al., 2008), infecção por Hepatozoon canis (KIRAL et al., 2005),
sarna (SINGH et al 2011) e a própria LVC (BILDIK, 2004; BRITTI et al., 2008). A
formação de espécies reativas ao oxigênio (ERO) de maneira exagerada nas doenças
infecciosas pode levar o organismo a um desequilíbrio entre os agentes oxidantes e
antioxidantes, proporcionando o estabelecimento do estresse oxidativo. Este processo
pode conduzir à oxidação de biomoléculas com consequente perda de suas funções
biológicas ou a um desequilíbrio homeostático que é manifestado no organismo pela
modificação de macromoléculas celulares, morte celular por apoptose ou necrose, além

44
de danos estruturais nos tecidos (LYKKESFELDT E SVENDSEN, 2007; BARBOSA et
al., 2010).
Apesar do envolvimento do estresse oxidativo nos mecanismos fisiológicos e
nos mecanismos patológicos, poucos estudos foram realizados na leishmaniose canina.
Baseado no exposto, este estudo teve como objetivo avaliar atividade das enzimas
antioxidantes na leishmaniose canina.
2. Material e Métodos
2.1. Animais
Foram utilizados 27 cães adultos, ambos os sexos, com peso e idade variados,
sem raça definida. Os animais provenientes do Centro de Controle de Zoonoses de
Fortaleza (CCZ), recolhidos em domicílio através do programa SOS Cão, após
identificação de cães sorologicamente positivos para Leishmania chagasi, compuseram
o grupo teste, soropositivos. Os animais provenientes de Canil particular,
sorologicamente negativos para L. chagasi, compuseram o grupo controle,
soronegativos. O protocolo experimental foi aprovado pelo Comitê de Ética para o Uso
de Animais da Universidade Estadual do Ceará (CEUA/UECE), n° 11516675-0/65.
2.2. Sorologia para Leishmaniose
A sorologia foi realizada no CCZ, através da reação de Imunofluorescência
Indireta utilizando-se o kit IFI-leishmaniose-canina-Bio-Manguinhos (IFI-LC) e do
teste ELISA com o kit EIE-leishmaniose-canina-Bio-Manguinhos (EIE-LC), cujos
antígenos foram obtidos das formas promastigotas de L. chagasi. Os kits utilizados são
produzidos por Bio-Manguinhos/FIOCRUZ, Rio de Janeiro, Brasil, e todos os
procedimentos foram realizados de acordo com as instruções do manual técnico (Lira et
al., 2006). Foi considerado positivo os animais com títulos de IFI e ELISA ≥ 1:40.
2.3. Grupos experimentais
Os animais foram divididos em dois grupos, Grupo A (n=12), controle,
soronegativo e o Grupo B (n=15), teste, com LVC. O Grupo B, além de conter animais
soropositivos, apresentava dois ou mais sinais fortemente associados à doença incluindo
caquexia, onicogrifose, hepatoesplenomegalia, linfadenomegalia, alopecia, lesões
cutâneas, lesões perioculares, epistaxe, além de outros sinais relacionados à doença.

45
Foram considerados controles os cães sorologicamente negativos e sem sinais clínicos
característicos ou específicos de leishmaniose.
2.4. Amostras de sangue
Foram coletados 8 mL de sangue por venopunção jugular dos cães soropositivos
e soronegativos. Para tanto, utilizou-se uma seringa estéril, distribuindo-se 4 mL de
sangue em tubo contendo anticoagulante EDTA e 4 mL de sangue em tubo contendo gel
de separação, sem anticoagulante. As amostras foram devidamente transportadas em
caixas térmicas rapidamente para os laboratórios no intuito de prevenir a oxidação in
vitro do material. As análises laboratoriais foram realizadas no Laboratório de
Bioquímica e Imunologia Animal - LIBA/FAVET/UECE e no Laboratório de Pesquisa
em Hemoglobinopatias Genéticas das Doenças Hematológicas - UFC.
2.5. Avaliação hematológica e bioquímica sérica
As amostras de sangue em EDTA foram homogeneizadas e submetidas ao
aparelho de automação (HemaScreen® 18) para realização do hemograma completo. Os
resultados do hemograma foram realizados tendo-se como valores de referência
FELDMAN et al. (2000). Os parâmetros hematológicos avaliados da série branca
incluem a contagem de leucócitos totais (LT, x103/dL) e diferenciais: neutrófilos,
eosinófilos, monócitos, basófilos e linfócitos, e da série vermelha, a contagem de
hemácias (He, mm3), a dosagem de hemoglobina (Hb, g/dL) e a determinação do
hematócrito (Ht, %) além da contagem total de plaquetas.
Nas amostras de soros foram dosados os teores de uréia (U, mg/dL), creatinina
(CREA, mg/dL), proteínas totais (PT, g/dL) e a atividade das enzimas transaminase
glutâmico oxalacética (TGO, U/L), transaminase glutâmico pirúvica (TGP, U/L) e
fosfatase alcalina (FA, U/L) e proteínas totais em aparelho de automação (Metrolab®
2300 Plus) com Kits específicos utilizando-se as metodologias dos fabricantes.
2.6. Avaliação da atividade das enzimas antioxidantes
A atividade da enzima catalase (CAT) foi detectada seguindo o método descrito por
Aebi (1984), no qual a atividade da enzima é feita pela mensuração do decaimento de
um preparado de peróxido de hidrogênio em espectrofotometria. O método consiste na
preparação de um solução contendo o tampão da catalase (TRIS-HCl 1M e EDTA mM)
pH 8,0 acrescido de peróxido de hidrogênio a 30%. Previamente, as amostras de sangue

46
em EDTA foram centrifugadas a 4000 rpm por 15 minutos para separação das hemácias
do plasma e leucócitos. Em seguida foram realizadas três lavagens das hemácias com
solução fisiológica 0,9% sempre descartando o sobrenadante. Após a última lavagem as
hemácias foram hemolisadas com água destilada. Para avaliar CAT, 20µL da amostra
de sangue hemolisado foi adicionada a 2 mL da solução tampão e lida a uma
absorbância de 240 nm. O resultado foi expresso em U/mg Hb.
A medida da glutationa peroxidas (GPx) nas hemácias foi determinada
utilizando-se o kit Ransel® RS505 (Randox), para a determinação indireta da enzima. A
análise é baseada na oxidação da glutationa reduzida (GSH) para glutationa oxidada
(GSSG), catalizada pela GPx utilizando a glutationa redutase (GSH-Rd) e fosfato de
nicotinamida adenina dinucleotídeo (NADPH). A diminuição na absorbância do
NADPH medida a 340 nm, durante a oxidação de NADPH para NADP+ é indicativo da
atividade de GPx.Os resultados foram expressos em mU/mg Hb.
2.7. Análise estatística
Os resultados foram expressos em média e desvio padrão. O teste t de Student
não pareado foi utilizado para determinar as diferenças entre os grupos (p< 0,05). O
teste de correlação de Pearson foi realizado entre as variáveis GPx, hemácias e
hemoglobina e entre CAT, hemácias e hemoglobina. Todos os resultados foram
calculados através do programa GraphPadPrism® 5.04.
3. Resultados
Para a avaliação laboratorial dos animais dos Grupos A e B foram feitas
determinações hematológicas e análises bioquímicas cujos resultados encontram-se na
tabela 1. Os valores médios obtidos para os diferentes parâmetros nos Grupos A e B não
diferiram significativamente entre os grupos e todos se encontram dentro dos valores de
referência para a espécie canina. Contudo, os teores de proteínas totais e de fosfatase
alcalina encontram-se elevados no grupo B em relação ao grupo A. Estes dados
implicam em um aumento na produção de imunoglobulinas associadas à resposta imune
a L. chagasi. A fosfatase alcalina elevada juntamente com a gama glutamil transferase
(não determinada) pode estar associada a danos hepáticos.

47
Tabela 1. Parâmetros hematológicos e bioquímicos de cães saudáveis e com LVC.
Parâmetros Grupo A
(Controle, n=12) Grupo B
(Teste, n=15) Valores de Referência*
Leucócitos (x10³) 12,934 ± 1,777 12,47 ±2,429 6.000-17.000 Neutrófilos (%) 60,5 ± 17,753 68 ± 5,527 66-77 Linfócitos (%) 35,8 ± 19,949 30,25 ± 5,154 12-30 Monócitos (%) 3,3 ± 2,908 1,667 ± 1,435 3-10 Plaquetas (x10³) 297,667 ± 82,226 254,334 ± 213,213 200-500 Ureia (mg/dL) 33,334± 10,039 16,857 ± 6,212 21,4-59,92 Creatinina (mg/dL) 1,322 ± 0,188 0,996 ± 0,807 0,5-1,5 Proteínas totais (g/dL) 6,47 ± 1,11a 8,23 ± 1,22b 6-8 TGO (U/L) 30,084 ±13,419 24,143 ± 11,753 23-66 TGP (U/L) 42,834 ± 12,890 27,143 ± 13,518 21-102 FA (U/L) 109,083 ± 38,313a 165,4 ± 46,402b 20-156 * FELDMAN et al., 2000. Letras diferentes na mesma linha representam diferenças significativas ao nível de 1%.
Como a atividade das enzimas antioxidantes foi realizada nas hemácias, os
parâmetros da série vermelha foram colocados juntamente com os resultados da
avaliação do estresse oxidativo demonstrados na tabela 2.
As médias da contagem de hemácias (p < 0,001), a concentração de hematócrito
(p < 0,05) e o teor de hemoglobina (p < 0,01) do grupo B encontravam-se inferiores aos
do grupo A. Estes dados diferiram significativamente (p < 0,001) entre os grupos.
Tabela 2. Parâmetros hematológicos e marcadores antioxidantes de cães saudáveis e de cães com LVC.
Marcadores
Grupo A (Controle, n=12)
Grupo B (Teste, n=15)
Valor de p
Hemácias (x106/dL) 7,012 ± 0,656 5,559 ± 0,451 p<0,001 Hematócrito (%) 52,125 ± 2,439 48,155 ± 5,692 p<0,05 Hemoglobina (g/dL) 16,492 ± 1,308 14,744 ± 1,797 p< 0,01 CAT (U/g Hb) 326,6712 ± 104,528 189,358 ± 90,385 p <0,001 GPx (mU/mg Hb) 5055,622 ±1569,078 3609,659 ± 1569,078 p <0,001
Os valores médios da atividade das enzimas CAT e GPx apresentaram-se
inferiores nos animais do grupo B (189,358 ± 90,385; 3609,659 ± 1569,078) em relação
aos animais do grupo A (326,6712 ± 104,528; 5055,622 ±1569,078), respectivamente.
Estes dados diferiram significativamente (p < 0,001) entre os grupos.
Na tentativa de se verificar uma possível correlação entre os parâmetros
hematológicos e os marcadores do estresse oxidativo, verificou-se uma correlação

48
positiva entre a contagem de hemácias e CAT, contudo esta correlação não foi
significativa.
4. Discussão
As alterações relacionadas ao estresse oxidativo podem ser encontradas
fisiologicamente em indivíduos saudáveis, porém controlada por mecanismos que
levam a homeostase. Por outro lado, indivíduos que se encontram doentes, em sua
grande maioria, apresentam um desequilíbrio nos mecanismos redox, favorecendo um
intenso estresse oxidativo (DRÖGE, 2002).
O indivíduo necessita de um bom sistema de defesa para conseguir reverter a
multiplicação dos radicais livres e consequente injúria provocada por eles. Isto é
possível através dos agentes antioxidantes que atuam na prevenção oxidativa e na
regeneração tecidual e celular, tendo como linha de frente as enzimas glutationa
reduzida (GSH), superóxido dismutase (SOD), catalase (CAT) e glutationa peroxidase
(GPx) (CAMPOS; YOSHIDA, 2004; NAITO et al., 2010).
A leishmaniose canina é uma doença causada por um parasito intracelular
obrigatório caracterizada por apresentar evolução crônica com envolvimento sistêmico
em diversos órgãos e tecidos do hospedeiro (MAIA-ELKHOURY et al., 2008). A
sintomatologia clínica da LVC tem sido associada às alterações imunológicas
envolvendo células do sistema fagocítico mononuclear e diferentes populações de
linfócitos T efetores dos tipos Th1, Th2 e Treg, caracterizados pela secreção das
citocinas específicas das subpopulações celulares, bem como a produção de anticorpos
específicos através da produção de diferentes subclasses de imunoglobulinas
(BARBIÉRI, 2006; GLAICHENHAUS et al., 2011; KAYE; SCOTT, 2011; FREITAS
et al., 2012).
O desequilíbrio dos agentes oxidantes e antioxidantes está presente nas
diferentes espécies de animais domésticos em diversas situações patológicas, incluindo
as doenças infecciosas e parasitárias (LYKKESFELDT; SVENDSEN, 2007; WEBB et
al., 2008; CELI, 2010; PALTRINIERI et al, 2010). Neste trabalho é dado ênfase aos
marcadores do estresse oxidativo através da avaliação das atividades das enzimas CAT
e GPx (Tabela 2), as quais foram detectadas em hemácias. CAT é uma hemeproteína
citoplasmática que transforma o peróxido de hidrogênio (H2O2) em oxigênio e água
(PACHECO; GONSEBATT, 2009). A atividade enzimática da CAT está presente em
todos os tecidos, porém com sua função elevada nos eritrócitos, fígado, rim e tecido

49
adiposo e uma baixa atividade no tecido nervoso (ZAMOCKY et al., 2008). A alta
produção do H2O2 deve ser controlada pelo organismo através das enzimas
antioxidantes GPx e CAT, o que poderia levar a um grande exigência catalítica destas
enzimas. GPx é uma enzima que cataliza a redução do peróxido de hidrogênio (H2O2)
utilizando a glutationa como substrato, além de outros hidroperóxidos orgânicos
(HALLIWELL; GUTTERIDGE, 1999).
Quanto aos parâmetros hemáticos, neste estudo, a contagem de hemácias, a
concentração do hematócrito e o nível de hemoglobina de cães naturalmente infectados
com L. chagasi estavam reduzidos (p<0,05) em relação ao controle. Resultados
semelhantes foram encontrados por Britti et al. (2008). Tem sido relatado que a anemia
é um dos achados mais comuns na leishmaniose canina, afetando cerca de 60% dos
animais infectados (CIARAMELLA; CORONA, 2003), e em algumas situações é
caracterizada como severa somente em cães sintomáticos (REIS et al., 2006). As
alterações hematimétricas observadas nos cães com leishmaniose sugerem uma possível
correlação da anemia com o aparecimento dos sinais clínicos, que nem sempre é
observada (COSTA-VAL et al., 2007).
Os eritrócitos contém abundantemente as enzimas SOD, CAT e GPx, portanto,
alterações nestas células refletirão nas atividades destas enzimas. Sendo assim, neste
trabalho as mudanças no metabolismo de EROs na LVC foram determinadas através de
CAT e GPx nos eritrócitos. Em nosso estudo, somente CAT apresentou correlação
positiva com a contagem total de hemácias, contudo não foi significativa. Britti et al.
(2008) estudando LVC encontraram correlação positiva (p< 0,01) entre a atividade da
enzima GPx tanto com hemácias quanto com hemoglobina.
No presente estudo foi observado que GPx estava com atividade reduzida. Estes
resultados corroboram com o estudo realizado por Britti et al. (2008), que
demonstraram uma diminuição da GPx na LVC, embora não tenha sido significativa.
Contudo, estes mesmos autores encontraram uma correlação positiva entre a contagem
de hemácias e a atividade da GPx e entre os níveis de hemoglobina e a atividade da GPx
em animais com LVC. Estes resultados sugerem que a diminuição da atividade
detoxificante pode estar relacionada com a diminuição da concentração de eritrócitos e
que a atividade da GPx depende das alterações dos mecanismos de controle do sistema
antioxidante na LVC. Além disso, este resultado poderia estar associado com a
diminuição do número de células sanguíneas nos animais com LVC, pois a GPx é uma

50
enzima eritrocitária que desenvolve importante papel na proteção da hemoglobina
contra os danos oxidativos (LUBOS et al., 2011).
Bildik et al. (2004) estudando o estresse oxidativo e a atividade antioxidante não
enzimática em cães com leishmaniose visceral relataram que os níveis de ácido
ascórbico, β-caroteno e ceruloplasmina estavam reduzidos nos animais doentes, porém
os mesmos autores encontraram alto nível de malondialdeído (MDA). Estes dados
indicam a ocorrência da peroxidação lipídica na LVC. Em nosso trabalho a FA
encontrava-se aumentada no Grupo B, corroborando com o que foi encontrado em cães
com leishmaniose apresentando hepatite crônica (RALLIS et al., 2005).
Estudos avaliando o estresse oxidativo em cães infestados com Sarcoptes scabiei
var. canis relataram uma menor atividade da GPx nos animais com escabiose (SINGH
et al., 2011) o que levou os autores a conclusão de que a diminuição da enzima pode
estar relacionada coma produção excessiva dos agentes oxidante gerados nas células
inflamatórias recrutadas para combater o parasito, o que acarretaria na exaustão do
sistema antioxidante.
Outra enzima antioxidante importante identificada no presente estudo foi a
catalase. Animais com LVC apresentaram menor (p <0,001) atividade da enzima CAT
em relação aos animais controles. Resultados semelhantes foram encontrados na
leishmaniose cutânea em humanos (EREL et al., 1999). Cães com sarna demodécica
(Demodex canis) apresentaram uma atividade reduzida da enzima CAT (DIMRI et al.,
2008). Outros autores verificaram um aumento na atividade da CAT em cães com
parvovirose (PANDA et al., 2009) e babesiose (CHAUDHURI et al., 2008).
Este estudo torna-se relevante, pois existem poucos dados na literatura que
avaliam o perfil oxidativo na leishmaniose visceral canina.
6. Conclusão
Conclui-se que CAT e GPx estão envolvidas no estresse oxidativo da
leishmaniose canina, as quais podem funcionar como biomarcadores. Outros estudos
são necessários para avaliar o envolvimento completo do estresse oxidativo na
participação da patogenia da LVC nas diferentes formas clínicas, além de ser útil no
acompanhamento do tratamento desta doença.
8. Referências
Aebi, H., 1984. Catalase in vitro. Methods in Enzymology. 105, 121-128.

51
Barbiéri,C.L., 2006. Immunology of Canine leishmaniosis. Par. Immu. 28, 329-337.
Barbosa, K.B.F., Costa, N.M.B., Alfenas, R.C.G., de Paula, S.O., Minim, V.P.R.,
Bressan, J., 2010. Estresse oxidativo: conceito, implicações e fatores modulatórios. Rev.
Nut. 23, 629-643.
Bildik, A., Kargın, F., Seyrek, K., Pasa, S., Özensoy, S., 2004. Oxidative stress and
non-enzymatic antioxidative status in dogs with visceral Leishmaniasis. Res. Vet. Sci.
77, 63-66.
Britti, D., Sconza, S., Morittu, V.M., Santori, D., Boari, A., 2008. Superoxide dismutase
and Glutathione peroxidase in the blood of dogs with Leishmaniasis. Vet. Res.
Commun. 32, 251-254.
Campos, E.B.P., Yoshida, W.B., 2004. O papel dos radicais livres na fisiopatologia da
isquemia e reperfusão em retalhos cutâneos: modelos experimentais e estratégias de
tratamento. J. Vasc. Br. 3, 357-66.
Celi, P. 2010. The role of oxidative stress in small ruminants’ health and production. R.
Bras. Zootec. 39, 348-363.
Ciaramella, P., Corona, M., 2003. Canine leishmaniasis: clinical and diagnostic aspects.
Vet. Learn. 25, 358-368.
Chaudhuri, S., Varshney, J.P., Patra, R.C., 2008. .Erythrocytic antioxidant defense, lipid
peroxides level and blood iron, zinc and copper concentrations in dogs naturally
infected with Babesia gibsoni. Res. Vet. Sci. 85, 120–124.
Costa-Val, A.P., Cavalcanti, R.R., Gontijo, N.F., Michalick, M.S.M., Alexander, B.,
Williams, P., Melo, M.N., 2007. Canine visceral leishmaniasis: Relationships between
clinical status, humoral immune response, haematology and Lutzomyia (Lutzomyia)
longipalpis infectivity. The Vet. Journal. 174, 636-643.

52
Dimri, U., Ranjan, R., Kumar, N., Sharma, M.C., Swarup, D., Sharma, B., Kataria, M.
2008. Changes in oxidative stress indices, zinc and copper concentrations in blood in
canine demodicosis. Vet. Parasitol. 154, 98–102.
Dröge, W., 2002. Free radical in the physiological control of cell function. Physiol Rev.
82, 47-95.
Erel, O., Kocyigit, A., Bulut, V., Gurel, M.S., 1999. Free Radical Metabolisms in
Cutaneous Leishmaniasis. Mem. Inst. Oswaldo Cruz, 94, 179-183.
Freitas, J.C.C., Pinheiro, D.C.S.N., 2010. Aspectos celulares e moleculares da resposta
imunitária a Leishmania spp. RPCV. 109, 11-20.
Freitas, J.C.C., Pinheiro, D.C.S.N., Lopes-Neto, B.E., Santos, G.J., Abreu, C.R., Braga,
R.R., Campos, R.M., Oliveira, L.F., 2012a. Clinical and laboratory alterations in dogs
naturally infected by Leishmania chagasi. Rev.Soc. Bras. Med. Trop. 45, 24-29.
Freitas, J.C.C., Lopes-Neto, B.E., , Abreu, C.R., Coura-Vital, W., Braga, S. L., Reis, A.
B., Pinheiro, D.C.S.N., 2012b. Profile of anti-Leishmania antibodies related to clinical
picture in canine visceral leishmaniasis. Res. Vet. Sci. 93, 705-709.
Glaichenhaus, N., Mougneau, E., Bihl, F., 2011. Cell biology and immunology of
Leishmania. Immunol. Ver. 240, 286-296.
Gontijo, C.M.F., Melo, M.N., 2004. Leishmaniose Visceral no Brasil: quadro atual,
desafios e perspectivas. Ver. Bras. Epidemiol. 7, 338-349.
Halliwell, B., Guitteridge, J.M.C., Free Radicals in Biology and Medicine.4.ed. New
York: Clarendon Press, 2007, p.880.
Jain, N.C., 2000. Schalm’s, Veterinary Hematology. In: Feldman, B.F., Zinkl J.G vol. 1,
5th ed. Lippincott Williams & Wilkin,Philadelphia, pp. 1344.

53
Karadeniz, A., Hanedan, B., Cemek, M., Börküm, K., 2008. Relationship between
Canine Distemper and Oxidative Stress in dogs. Revue Méd. Vét.159, 462 – 467.
Kaye, P., Scott, P., 2011. Leishmaniasis: complexity at the host–pathogen interface.
Nature Rev. Microbiol., 9, 604-615.
Kiral F., Karagenc, T., Pasa, S., Yenisey, C., Seyrek, K., 2005. Dogs with Hepatozoon
canis respond to the oxidative stress by increased production of glutathione and nitric
oxide. Vet. Parasitol. 131, 15- 21.
Lima, M.H., 2004. Oxygen in biology and biochemistry: role of free radicals. In:
Storey, K.B., Functional Metabolism: Regulation and Adaptation, 4th ed. Wiley-
Iiss,New Jersey, pp. 319-366.
Lubos, E., Loscalzo, J., Handy, D.E., 2011. Glutathione Peroxidase-1 in Health and
Disease: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox
Signal. 15, 1958-1969.
Lykkesfeldt, J., Svendsen, O., 2007. Oxidants and antioxidants in disease: Oxidative
stress in farm animals. The Vet. Journal. 173, 502–511.
Maia-Elkhoury, A.N.S., Alves, W.A., Sousa-Gomes, M.L., Sena, J.M., Luna, E.A.,
2008. Leishmaníase visceral no Brasil: evolução e desafios. Cad. Saúde Pública. 24,
2941-2947.
Mancianti, F., Gramiccia, M., Gradoni, L., Pieri, S., 1988. Studies on canine
leishmaniasis control. Evolution of infection of different clinical forms of canine
leishmaniasis following antimonial treatment. Trans. R. Soc. Trop. Med. Hyg, 82, 566–
567.
Martinez, F.O., Helming, L., Gordon, S., 2009. Alternative activation of macrophages:
An immunologic functional perspective. Ann. Rev. Immunol. 27, 451-483.

54
Naito, Y., Lee, M.C., Kato, Y., Nagai, R., Yonei, Y., 2010. Oxidative stress markers.
Anti-Aging Medicine. 7, 36-44.
Pacheco, J.L., Gonsebatt, M.E., 2009. The role of antioxidants and antioxidant-related
enzymes in protective responses to environmentally induced oxidative stress. Mutat.
Res. 674, 137–147.
Panda, D., Patra, R.C., Nandi, S., Swarup, D., 2009. Oxidative stress indices in
gastroenteritis in dogs with canine parvoviral infection. Res. Vet. Sci. 86, 36–42.
Rallis, T., Day, M.J., Saridomichelakis, M.N., Adamama-Moraitou, K.K., Papazoglou,
L., Fytianou, A., Koutinas, A.F., 2005. Chronic hepatitis associated with canine
leishmaniosis (Leishmania infantum): a clinico pathological study of 26 Cases.
J.Comp.Path. 132, 145–152.
Reis, A.B., Martins-Filho, O.A., Teixeira-Carvalho, A., Carvalho, W.M., Mayrink, W.,
França-Silva, J.C., Giunchetti, R.C., Genaro, O., Corrêa-Oliveira, R., 2006. Parasite
density and impaired biochemical/hematological status are associated with severe
clinical aspects of canine visceral leishmaniasis. Res. Vet. Sci. 81, 68-75.
Ribeiro, V.M., 2007. Leishmaniose visceral canina: aspectos de tratamento e controle.
Clínica Vet., 71, 66-76.
Silva, F.S., 2007. Patologia e patogênese da leishmaniose visceral canina. RT-CAB. 1,
20-31.
Singh, S.K., Dimri, U., Sharma, M.C., Swarup, D., Sharma, B., 2011. Determination of
oxidative status and apoptosis in peripheral blood of dogs with sarcoptic mange. Vet.
Parasitol. 178, 330–338.
Webb, C., Lehman, T., Mccord, K., Avery, P., Dow, S. 2008. Oxidative stress during
acute FIV infection in cats. Vet. Immunol. Immunopathol. 122, 16-24.

55
Zamocky, M., Furtmüller, P.G., Obinger, C., 2008. Evolution of Catalases from
Bacteria to Humans. Antiox. Redox Signal. 10, 1527-1547.

56
7. CONCLUSÕES
As atividades das enzimas glutationa peroxidase (GPx) e catalase (CAT)
eritrocitária apresentaram-se reduzidas em cães naturalmente infectados com L. chagasi
e houve correlação positiva moderada entre CAT e hemoglobina (Hb).
Estes dados nos levam a concluir que CAT e GPx estão envolvidas no estresse
oxidativo da LVC e que podem ser utilizadas como biomarcadores. Contudo, outros
estudos são necessários para avaliar outros marcadores do estresse oxidativo na
patogenia da LVC em suas diferentes formas clínicas.

57
8. PERSPECTIVAS
A leishmaniose visceral canina no Brasil é uma doença presente em quase todas
as regiões do país e por ser uma zoonose, representa grave problema de saúde pública
levando vários desafios para os profissionais da saúde. Desta forma, há uma necessidade
cada vez maior de ações voltadas para o controle da leishmaniose visceral canina, já que
as infecções nestes animais precedem a infecção humana.
Pela grande relevância desta zoonose, todos os estudos que forneçam dados para
melhor compreensão da leishmaniose tornam-se importantes, pois muitos fatores ainda
não são bem esclarecidos na patogenia da doença. Assim, os resultados encontrados
neste trabalho são os primeiros descritos na literatura na leishmaniose visceral canina no
Brasil. Novos estudos devem ser realizados com o objetivo de avaliar o estresse
oxidativo de maneira sistêmica e nos diferentes órgãos e tecidos dos animais infectados
por L. chagasi.

58
9. REFERÊNCIAS
ABREU-SILVA, A.L.; LIMA, T.B; MACEDO, A.A.; DIAS, E.; BATISTA, Z.S.;
CALABRESE, K.S.; MORAES, J.L.; RABÊLO, J.M.; GUERRA, R.M.
Soroprevalência, aspectos clínicos e bioquímicos da infecção por Leishmania em cães
naturalmente infectados e fauna de flebotomíneos em uma área endêmica na ilha de São
Luís, Maranhão, Brasil. Revista Brasileira de Parasitologia Veterinária, v. 17, supl. 1,
p. 197-203, 2008.
ABBAS, A. B. ; LITCHMAN A. H. ; PILLAI, S. Imunologia celular e molecular. 6 ed.
Rio de Janeiro: Elsevier, 2008. 564p.
AGA, E.; KATSCHINSKI, D.M.; van ZANDBERGEN, G.; LAUFS, H.; HANSEN, B.;
MULLER, K.; SOLBACH, W.; LASKAY, T. . Inhibition of the spontaneous apoptosis
of neutrophil granulocytes by the intracellular parasite Leishmania major. The Journal
of Immunology, v. 169, p. 898-905, 2002.
ALBUQUERQUE, A.R.; ARAGÃO, F.R.; FAUSTINO, M.A.G.; GOMES, Y.M.; LIRA, R.A.; NAKASAWA, M.; ALVES, L.C. Aspectos clínicos de cães naturalmente infectados Leishmania
(Leishmania) chagasi na região metropolitana de Recife. Clínica Veterinária, v. 71, p.
78-84, 2007.
ALEXANDER, J.; BRYSON, K. T helper (h)1/Th2 and Leishmania: paradox rather
than paradigm. Immunology Letters, v. 99, p. 17–23, 2005.
ARTHER, R.G. Mites and Lice: Biology and Control. Veterinary Clinics Small Animal
Practice, v. 39, p. 1159–1171, 2009.
ASSCHE, T.V.; DESCHACHT, M.; DA LUZ, R.A.I.; MAES, L.; COS, P.Leishmania–
macrophage interactions: insights into the redox biology. Free Radical Biology &
Medicine, v.51, p. 337-351, 2011.
BABIOR, B.M. NADPH oxidase. Current Opinion in Immunology, v. 16, p. 42–47,
2004.
BARBIÉRI C.L. Immunology of canine leishmaniosis. Parasite Immunology, v. 28, p.
329-337, 2006.
BARREIROS, L.B.S.; DAVID, J.M.; DAVID, J.P. Estresse oxidativo: relação entre
geração de espécies reativas e defesas do organismo. Química Nova, São Paulo, v.
29, n. 1, 2006.

59
BATINIC-HABERLE, I.; REBOUÇAS, J.S.; SPASOJEVIC, I. Superoxide dismutase
mimics: chemistry, pharmacology, and therapeutic potential. Antioxidants & Redox
Signaling, v. 13, n. 6, p. 879-918, 2010.
BILDIK, A; KARGIN, F; SEYREK, K.; PASA, S.; ÖZENSOY, S. Oxidative stress
and non-enzymatic antioxidative status in dogs with visceral Leishmaniasis. Research in
Veterinary Science, v. 77, n.1, p.63-66, 2004.
BRASIL, Ministério da Saúde. Manual de Vigilância e Controle da Leishmaniose
Visceral. Brasília: Ministério da Saúde, 2006, 120 p.
BRITO, F. L. C ; et al. Uveitis associated to the infection by Leishmania chagasi in dog
from Olinda city, Pernambuco, Brazil, Ciência Rural, n.34, p. 925-929, 2004.
BRITTI, D; SCONZA, S; MORITTU, V. M.; SANTORI, D; BOARI, A. Superoxide
dismutase and Glutathione peroxidase in the blood of dogs with Leishmaniasis.
Veterinary Research Communication, v. 32, n. 1, p. 251-254, 2008.
CAMARGO-NEVES V. L. F. et al. Manual de Vigilância e Controle da Leishmaniose
Visceral Americana do Estado de São Paulo.São Paulo: Secretaria de Estado da Saúde,
2006, 145 p.
CAMKERTEN, I.; SAHIN, T.; BORAZAN, G.; GOKCEN, A.; EREL, O.; DAS. A.
Evaluation of blood oxidant/antioxidant balance in dogs with sarcoptic mange.
Veterinary Parasitology, v. 161, p. 106–109, 2009.
CAMPOS , E.B.P.; YOSHIDA, W.B. O papel dos radicais livres na fisiopatologia da
isquemia e reperfusão em retalhos cutâneos: modelos experimentais e estratégias de
tratamento. Jornal Vascular Brasileiro, v. 3, n. 4, 2004.
CHANSAISAKORN, W.; SRIPHAVATSARAKORN, P.; SOPAKDITTAPONG, P.;
TRISIRIROJ, M.; PONDEENANA, S.; BURANAKARL, C. Oxidative stress and
intraerythrocytic concentrations of sodium and potassium in diabetic dogs. Veterinary
Research Communication, v. 33, p. 67–75, 2009.
CHAUDHURI, S.; VARSHNEY, J.P.; PATRA, R.C. Erythrocytic antioxidant defense,
lipid peroxides level and blood iron, zinc and copper concentrations in dogs naturally
infected with Babesia gibsoni. Research in Veterinary Science, v. 85, p. 120–124, 2008.
CIARAMELLA, P.; CORONA, M. Canine leishmaniasis: clinical and diagnostic
aspects. VetLearn, v. 25, n.5, p. 358-368, 2003.
COELHO, W.M.D.; LIMA, V.M.F.; AMARANTE, A.F.T.; LANGONI, H.; PEREIRA,
V.B.R.; ABDELNOUR, A.; BRESCIANE, K.D.S. Occurrence of Leishmania
(Leishmania) chagasi in a domestic cat (Felis catus) in Andradina, São Paulo, Brazil:

60
case report. Revista Brasileira de Parasitologia Veterinária, v. 19, n. 4, p. 256-258,
2010.
COSTA, F. A. I. et al. CD4+ T cells participate in the nephropathy of canine viceral
leishmaniasis. Brazilian Journal of Medical and Biological Research, v. 33, p. 1455-
1458, 2000.
COUTINHO, J. F. V. Estudo clínico-laboratorial e histopatológico de cães naturalmente
infectados por Leishmania chagasi com diferentes graus de manifestação clínica. 2005.
p. 102. Dissertação (Mestrado em Bioquímica) – Universidade Federal do Rio Grande
do Norte, Natal. 2005.
CRNOGAJ, M.; PETLEVSKI, R.; MRLJAK, V.; KIS, I.; TORTI, M.; KUCER, N.;
MATIJATKO, V.; SACER, I.; STOKOVIC, I. Malondialdehyde levels in serum of
dogs infected with Babesia canis. Veterinary Medicine, v. 55, n. 4, p. 163–171, 2010.
DANTAS-TORRES, F. The role of dogs as reservoirs of Leishmania parasites with
emphasis on Leishmania (Leishmania) infantum and Leishmania (Viannia) brazileinsis.
Veterinay Parasitology, v. 149, p. 139-146, 2007.
DEATON, C.M.; MARLIN, D.J. Exercise-associated oxidative stress. Clinical
Techniques in Equine Practice, v. 2, n. 3, 2003.
DIMRI, U.; RANJAN, R.; KUMAR, N.; SHARMA, M. C.; SWARUP, D.; SHARMA,
B.; KATARIA, M. Changes in oxidative stress indices, zinc and copper concentrations
in blood in canine demodicosis. Veterinary Parasitology, v. 154, p. 98–102, 2008.
DRAPER, H.H.; HADLEY, M. Malondialdehyde determination as index of lipid
peroxidation. Methods in Enzymology, v. 186, p. 421–431, 1990.
DRÖGE W. Free radical in the physiological control of cell function. Physiological
Reviews, v.82, n. 1, 2002.
DUNLAP, K.L.; REYNOLDS, A.J.; DUFFY, L.K. Total antioxidant power in sled
dogs supplemented with blueberries and the comparison of blood parameters associated
with exercise. Comparative Biochemistry and Physiology, v. 143, p. 429 – 434, 2006.
EUGÊNIO, F.R.; ANDRADE, A.L.; BABO-TERRA, V.J. Estudo clínico-laboratorial
das articulações na leishmaniose experimental em cães. Veterinária e Zootecnia, v. 15,
n. 2, p. 278-287, 2008.
FERRARI, C.K.B. Capacidade antioxidante total (CAT) em estudos clínicos,
experimentais e nutricionais. Journal of the Health Sciences Institute, v. 28, p. 307-
310, 2010.

61
FREITAS, J. C. C.; PINHEIRO, D. C. S. N. Aspectos celulares e moleculares da
resposta imunitária a Leishmania spp. Revista Portuguesa de Ciências Veterinárias, v.
109, p. 11-20, 2010.
FREITAS, J. C. C.; PINHEIRO, D. N.; LOPES-NETO, B. E.; SANTOS, G. J.;
ABREU, C. R.; BRAGA, R. R.; CAMPOS, R. M.; OLIVEIRA, L. F. Clinical and
laboratory alterations in dogs naturally infected by Leishmania chagasi. Revista da
Sociedade Brasileira de Medicina Tropical, v. 45, n.1, p. 24-29, 2012a.
FREITAS, J. C. C. ; LOPES NETO, B. E.; ABREU, C. R. A.; COURA-VITAL, W.;
BRAGA, S. L.; REIS, A. B.; PINHEIRO, D. C. S. N. Profile of anti-Leishmania
antibodies related to clinical picture in canine visceral leishmaniasis. Research in
Veterinary Science, v. 93, p. 705-709, 2012b.
CONTIJO, C.M.F.; MELO, M.N. Leishmaniose visceral no Brasil: quadro atual,
desafios e perspectivas. Revista Brasileira de Epidemiologia, v. 7, n. 3, p. 338-349,
2004.
GOTO, H.; PRIANTI, M.G. Immunoactivation and immunopathogeny during active
visceral leishmaniasis. Revista do Instituto de Medicina tropical de São Paulo,v. 51, n.
5, p. 241-246, 2009.
GLAICHENHAUS, N.; MOUGNEAU, E.; BIHL, F. Cell biology and immunology of
Leishmania. Immunological Reviews, v. 240, p. 286-296, 2011.
HALLIWELL, B.; GUITTERIDGE, J. M. C. Free Radicals in Biology and Medicine.
4.ed. New York: Clarendon Press, 2007, p.880.
HINCHCLIFF, K.W.; REINHART, G.A.; DISILVESTRO, R.; REYNOLDS, A.
BLOSTEIN-FUJII, A. SWENSON, R.A. Oxidant stress in sled dogs subjected to
repetitive endurance exercise. American Journal of Veterinary Research, v. 61, n. 5,
p. 512-517, 2000.
IMLAY, J.A. Pathways of oxidative damage. Annual Review of Microbiology, v. 57, p.
395–418, 2003.
IMMENSCHUH, S.; VOGT, B. PEROXIREDOXINS, oxidative stress, and cell
proliferation. Antioxidant & Redox Signaling, v. 7, p. 768–777, 2005.
INIESTA, L.; GÁLLEGO, M.; PORTÚS, M. Immunoglobulin G and E responses in
various stages of canine leishmaniosis. Veterinary Immunol and Immunopathology, v.
103, n. 1/2, p. 77–81, 2005.

62
KAPUN, A.P.; SALOBIR, J.; LEVART, A.; KOTNIK, T.; SVETE, A.N. Oxidative
stress markers in canine atopic dermatitis. Research in Veterinary Science, v. 92, p.
469-470, 2012.
KARADENIZ, A.; HANEDAN, B.; CEMEK, M.; BÖRKÜM. K. Relationship between
Canine Distemper and oxidative stress in dogs. Revue Médicine Véterinaire, v.159, n. 8-
9, p. 462 – 467, 2008.
KAYE, P.; SCOTT, P. Leishmaniasis: complexity at the host–pathogen interface.
Nature Reviews: Microbiology, v. 9, p. 604-615, 2011.
KIRAL F, KARAGENC T, PASA S, YENISEY C, SEYREK K, , Dogs with
Hepatozoon canis respond to the oxidative stress by increased production of glutathione
and nitric oxide. Veterinary Parasitology, v. 131, p. 15- 21, 2005.
KIRSCHVINK, N.; MOFFARTS, B.; LEKEUX, P. The oxidant/antioxidant
equilibrium in horses. The Veterinary Journal, v. 177, p. 178–191, 2008.
KLOTZ, L.O.; KRONCKE, K.D; BUCHCZYK, D.P.; SIES, H. Role of copper, zinc,
selenium and tellurium in the cellular defense against oxidative and nitrosative stress.
The Journal of Nutrition, v. 133, p. 1448-1451, 2003.
KOHEN, R.; NYSKA, A. Oxidation of biological systems: Oxidative stress
phenomena, antioxidants, redox reactions, and methods for their quantification.
Toxicologic Pathology, v. 30, n. 6, p. 620-650, 2002.
KUMAR, A.; VARSHNEY, J.P.;. PATRA A .R. Comparative study on oxidative stress
in dogs infected with Ehrlichia canis with or without concurrent infection with Babesia
gibsoni. Veterinary Research Communications, v. 30, p. 917–920, 2006.
LIMA, E.S; ABDALLA, D.S.P. Peroxidação lipídica: mecanismos e avaliação em
amostras biológicas. Revista Brasileira de Ciências Farmacêuticas, v. 37, n. 3, p. 293-
303, 2001.
LIMA, M. H. Functional metabolism: regulation and adaptation, edited by Kenneth B.
Storey. 2004. 594 p. Ontario Canada. Editado por Wiley-Iiss, Inc., Hoboken, New
Jersey.
LIMÓN-PACHECO, J. L.; GONSEBATT, M. E. The role of antioxidants and
antioxidant-related enzymes in protective responses to environmentally induced
oxidative stress. Mutation Research, v. 674, p. 137–147, 2009.
LUBOS, E.; LOSCALZO, J.; HANDY, D.E. Glutathione peroxidase-1 in health and
disease: from molecular mechanisms to therapeutic opportunities. Antioxidants & Redox
Signaling, v. 15, n. 7, p. 1958-1969, 2011.

63
LUVIZOTTO, M.C.R. Alterações patológicas em animais naturalmente infectados. IN:
1º Fórum sobre Leishmaniose Visceral Canina. Faculdade de Ciências Agrárias e
Veterinárias, Universidade Estadual Paulista, Jaboticabal, março, 2006.
LYKKESFELDT, J.; SVENDSEN, O. Oxidants and antioxidants in disease: oxidative
stress in farm animals. The Veterinary Journal, n. 173, p. 502–511, 2007.
MCMAHON-PRATT D.; ALEXANDER, J. Does the Leishmania major paradigm of
pathogenesis and protection hold for new world cutaneous leishmaniases or the visceral
disease? Immunological Reviews, v. 201, p. 206-224, 2004.
MANDELKER, L.; VAJDOVICH, P. Oxidative stress in applied basic research and
clinical practice. 1ed. New York: Humana Press, 2011, 260p.
MARQUES, M. I. L. M. Leishmaniose canina. 2008. p. 131. Dissertação (Mestrado) –
Faculdade de Medicina Veterinária, Universidade Técnica de Lisboa, Lisboa. 2008.
MONTEIRO, E.M.; SILVA, J.C.; COSTA, R.T.; BARATA, R.A., PAULA, E.V.;
LINS, G.L.; ROCHA, M.F.; DIAS, E.S. Leishmaniose visceral: estudo de
flebotomíneos e infecção canina em Monte Claros, Minas Gerais. Revista da Sociedade
Brasileira de Medicina Tropical,v.38, n.2, p. 147-152, 2005.
MORDASKI, R.Y.; SOUSA, J.C.; COSTA, J.F.; FREITAS, K.C.; PASSONI, C.R.
Sousa. Leishmaniose visceral: ciclo biológico. Cadernos da Escola de Saúde, n. 03, p.
1-2, 2009.
NAITO, Y.; LEE, M.C.; KATO, Y.; NAGAI, R.; YONEI, Y. Oxidative stress markers.
Anti-Aging Medicine, v. 7, n. 5, p. 36-44, 2010.
NAKAMURA, H. Thioredoxin and its related molecules: Update 2005. Antioxidant &
Redox Signaling, v. 7, n.5, p. 823-828, 2005.
PALTRINIERI, S.; RAVICINI, S.; ROSSI, G.; ROURA, R. Serum concentrations of
the derivatives of reactive oxygen metabolites (d-ROMs) in dogs with leishmaniosis.
The Veterinary Journal, v. 186, p. 393–395, 2010.
PANDA, D.; PATRA, R. C.; NANDI, S.; SWARUP, D. Oxidative stress indices in
gastroenteritis in dogs with canine parvoviral infection. Research in Veterinary Science,
n. 86, p. 36–42, 2009.
PASQUINI A.; LUCHETTI, E.; MARCHETTI, V.; CARDINI, G.; IORIO, E.L.
Analytical performances of d-ROMs test and BAP test in canine plasma. Definition of
the normal range in healthy Labrador dogs. Veterinary Research Communication, v. 32,
p. 137–143, 2008.

64
PASQUINI, A.; LUCHETTI, E.; CARDINI, G. Evaluation of oxidative stress in
hunting dogs during exercise. Research in Veterinary Science , v.89, p. 120–123, 2010.
PIN, D.; BENSIGNOR, E.; CARLOTTI, D.N.; CADIERGUES, M.C. 2006.
Localisedsarcoptic mange in dogs: a retrospective study of 10 cases. The Journal of
Small Animal Practice, v. 47, n. 10, p. 611-614, 2006.
PINHÃO, C. P. R. Leishmaniose canina: estudo de 158 casos da região de Lisboa.
2009. p. 54. Dissertação (Mestrado) – Faculdade de Medicina Veterinária, Universidade
Técnica de Lisboa, Lisboa. 2009.
POULSEN, H.E. Oxidative DNA modifications. Experimental and Toxicologic
Pathology, v. 57, p. 1161-1169, 2005.
RIBEIRO V.M. 2004. Leishmaniose visceral canina. Manual técnico – Leishmaniose
Visceral Canina. Fort Dodge. 52p.
RIBEIRO, V. M. Leishmaniose visceral canina: aspectos de tratamento e controle.
ClínicaVeterinária, n.71, p. 66-76, 2007.
SANT’ANA, J. A .P. et al. Hepatic granulomas in canine visceral leishmaniasis and
clinical status. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.59, n.5,
p.1137-1144, 2007.
SANTOS, C. A. C. Percepção, epidemiologia e aspectos clínicos da leishmaniose
visceral canina em área urbana do Estado de Pernambuco. 2006. p. 61. Dissertação
(Mestrado em Ciência Veterinária) – Departamento de Medicina Veterinária,
Universidade Federal Rural de Pernambuco, Recife. 2006.
SEGAL, A.W. How neutrophils kill microbes. Annual Review Immunology, v. 23, p.
197-223, 2005.
SILVA, M. V. Avaliação de testes sorológicos para leishmaniose visceral canina
utilizando coleta de amostra sanguínea em papel filtro. 2005. p. 163. Dissertação
(Mestrado) – Universidade Federal de Ouro Preto, Minas Gerais, 2005.
SILVA, F. S. Patologia e patogênese da leishmaniose visceral canina. Revista tropica –
Ciências Agrárias e Biológicas.v. 1, n. 1, p. 20-31, 2007.
SINGH, S. K. ; DIMRI, U. ; SHARMA, M. C. ; SWARUP, D. ; SHARMA, B.
Determination of oxidative status and apoptosis in peripheral blood of dogs with
sarcoptic mange. Veterinary Parasitology, v. 178, p. 330–338, 2011.
SONG, O. Oxidative stress: a theoretical model or a biological reality? Comptes Rendus
Biologies, v. 327, p. 649–662, 2004.

65
TODOROVA, I.; SIMEONOVA, G.; KYUCHUKOVA, D.; DINEV, D.; GADJEVA,
V. Reference values of oxidative stress parameters (MDA, SOD, CAT) in dogs and cats.
Comparative Clinical Pathology, v. 13, p. 190–194, 2005.
UNDERHILL, D.M.; OZINSKY, A. Phagocytosis of microbes: complexity in action.
Annual Review Immunology, v. 20, p. 825-852, 2002.
VALKO, M.; LEIBFRITZ, D.; MONCOL, J.; CRONIN, M.T.D.; MAZUR M.;
TELSER, J. Free radicals and antioxidants in normal physiological functions and human
disease. The International Journal of Biochemistry & Cell Biology, v.39, p. 44-84, 2007.
van der VEEN, B.S.; MENNO, P.J.; de WINTHER.; HEERINGA, P. Myeloperoxidase:
molecular mechanisms of action and their relevance to human health and disease.
Antioxidant & Redox Signaling, v. 11, n. 11, p. 2900-2937, 2009.
VANNUCCHI, C.I.; JORDAO, A.A.; VANNUCCHI H. Antioxidant compounds and
oxidative stress in female dogs during pregnancy. Research in Veterinary Science, v.
83, p. 188-193, 2007.
VASCONCELOS, M. F. Aves observadas no Parque Paredão da Serra do Curral, Belo
Horizonte, Minas Gerais, Brasil. Atualidades Ornitológicas, v. 136, p. 6–11, 2007.
WEBB, C.; LEHMAN, T.; MCCORD, K.; AVERY, P.; DOW, S.Oxidative stress
during acute FIV infection in cats. Veterinary Immunology and Immunopathology, v.
122, p. 16-24, 2008.
XAVIER, S.C. et al. Canine visceral leishmaniasis: a remarkable histopathological
picture of one asymptomatic animal reported from Belo Horizonte, Minas Gerais,
Brazil. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.58, p.994-1000,
2006.
ZAMOCKY, M.; FURTMÜLLER, P. G.; OBINGER, C. Evolution of catalases from
bacteria to humans. Antioxidants & Redox Signaling, v. 10, n. 9, p. 1527-1547, 2008.

66
Anexo 1

67
Anexo 2
Comprovante de submissão