
UNIVERSIDADE ESTADUAL DE FEIRA DE … discente....ya UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA...
85
.ya UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INGRID ESTEFANIA MANCIA DE GUTIÉRREZ MICROPROPAGAÇÃO DE Bauhinia cheilantha (BONG.) STEUD. (FABACEAE) Feira de Santana, BA 2010
-
Upload
nguyenliem -
Category
Documents
-
view
215 -
download
0
Transcript of UNIVERSIDADE ESTADUAL DE FEIRA DE … discente....ya UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA...

.ya
UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
INGRID ESTEFANIA MANCIA DE GUTIÉRREZ
MICROPROPAGAÇÃO DE Bauhinia cheilantha (BONG.)
STEUD. (FABACEAE)
Feira de Santana, BA
2010

INGRID ESTEFANIA MANCIA DE GUTIÉRREZ
MICROPROPAGAÇÃO DE Bauhinia cheilantha (BONG.)
STEUD. (FABACEAE)
Dissertação apresentada ao Programa de Pós-graduação em
Biotecnologia, da Universidade Estadual de Feira de Santana como
requisito para o título de Mestre.
Orientador: Prof. Dr. José Raniere Ferreira de Santana
Feira de Santana, BA
2010

AGRADECIMENTOS
À Deus por ter me dado os pais e companheiro que tenho, pois deles recebi sempre apoio,
cuidado, carinho e compreensão durante toda essa trajetória para mais uma conquista.
À Deus por ter me aproximado de todas as pessoas que me orientaram e compartilharam seus
conhecimentos e experiências comigo, contribuindo para a conclusão de mais uma etapa na
minha formação profissional.
E mais uma vez à Ele, por ter conquistado todas as amizades que fiz, nas quais pude dividir
muitos momentos de alegria e de dificuldade, durante o período do mestrado.
Obrigada mais umas vez!

RESUMO
Bauhinia cheilantha (Bong.) Steud. é uma leguminosa típica do semiárido com relevante
importância sócio-econômica e medicinal. Suas sementes apresentam dormência o que
dificulta a obtenção de plantas uniformes e em curto período de tempo, diante disso, a
micropropagação promete ser uma alternativa a propagação da espécie. Objetivou-se nesse
trabalho estabelecer um protocolo de micropropagação para B. cheilantha. Avaliou-se os
efeitos de diferentes reguladores vegetais (BAP, KIN, TDZ e ANA) sobre a morfogênese in
vitro de diversos explantes (hipocótilo, segmento cotiledonar, cotilédone, epicótilo, segmento
nodal e internodal), obtidos de plântulas in vitro com 20 dias de idade. Na fase de
enraizamento, foram testadas diferentes auxinas (AIB, ANA e AIA) sob a influência do
carvão ativado. Avaliou-se a interferência do fechamento dos tubos de ensaio no percentual
de sobrevivência das mudas, durante a aclimatização. O meio de cultura utilizado em todos os
experimentos foi o WPM acrescido de 30,0 g L-1
de sacarose e 6,0 g L-1
de ágar. O segmento
nodal apresentou uma capacidade organogênica 2,4 vezes maior que o segmento cotiledonar.
O maior número de brotos (2,84) foi obtido em 2,0 mg L-1
de BAP. A utilização de ANA não
favoreceu a taxa de multiplicação. A utilização de 0,5 mg L-1
de AIA ou 1,0 mg L-1
de ANA
ou 0,25 mg L-1
de AIB promoveu o maior percentual de enraizamento (cerca de 65,0%). Em
relação ao carvão ativado, na concentração de 0,5 g L-1
houve efeito benéfico na maioria das
características analisadas. Os brotos enraizados foram transferidos para casa de vegetação e
aclimatizadas com sucesso.
Palavras-chave: Lenhosa medicinal. Cultivo in vitro. Organogênese. Reguladores vegetais.

ABSTRACT
Bauhinia cheilantha (Bong.) Steud. is a legume typical of the semi-arid region with the socio-
economic and medical relevance. Its seeds have a dormancy period which makes it difficult to
obtain uniform plants in a short period of time. As such, micropropagation is a promising
alternative for the propagation of the species. The objective of this work was to establish a
micropropagation protocol for B. cheilantha. This was undertaken through an evaluation of
the effects of different plant growth regulators (BAP, KIN, TDZ and NAA) on the
morphogenesis in vitro of different explants (hypocotyls, cotyledonary segment, cotyledon,
epicotyl, nodal and internodal segment), from 20 day old seedlings grown in vitro. In the
rooting phase, different auxins (IBA, NAA and IAA) were tested under the influence of
activated charcoal. The interference of the culture tube closure type in the percentage of
seedling survival during the acclimatization was also evaluated. The culture medium used in
all experiments was WPM plus 30.0 g L-1
sucrose and 6.0 g L-1
agar. The nodal segments
showed an organogenic capacity 2.4 times greater than the cotyledonary segment. The highest
number of shoots (2.84) was obtained at 2.0 mg L-1
BAP. The use of NAA did not favor the
multiplication rate. The use of 0.5 mg L-1
IAA or 1.0 mg L-1
NAA or 0.25 mg L-1
IBA
promoted the highest percentage of rooting (about 65.0%). As for the activated carbon at a
concentration of 0.5 g L-1
, there was a beneficial effect in most of the analyzed characteristics.
The rooted shoots were transferred to greenhouse and successfully acclimatized.
Keywords: Medicinal Woody. In vitro culture. Organogenesis. Plant growth regulators

LISTA DE FIGURAS
Figura 1 Distribuição das patentes identificadas com a palavra-chave
Bauhinia, nos campos de título e resumo, nos bancos de patentes
gratuitos (Fonte: INPI, EPO, USPTO, nov. 2009).
4
Figura 2 B. cheilantha: (A) Aspecto da casca; (B) Folha; (C) Sementes
maduras; (D) Inflorescência; (E) Fruto maduro (Fonte: Autora,
2010).
5
Figura 3 Distribuição geográfica de B. cheilantha () no Brasil. (Fonte:
Autora, adaptado de Vaz e Tozzi, 2003).
6
Figura 4 Número de Patentes depositadas com as palavras-chaves Cultura de
Tecidos Vegetais, Micropropagação, Embriogênese, Organogênese,
Enraizamento in vitro e Aclimatização, relacionada à área de
Reprodução de plantas por meio de técnicas de Cultura de Tecidos
(A01H 4/00) (Fonte: INPI, EPO e USPTO, nov., 2009).
9
Figura 5 Sementes de B. cheilantha, sob ocular 2X de esterescópico, seta
apontando o hilo (A) e sementes escarificadas (B) (Barra = 0,5 cm).
Feira de Santana, BA, 2010.
14
Figura 6 Esquema do seccionamento realizado para obtenção dos explantes
utilizados na etapa de multiplicação in vitro: (A) plântula de B.
cheilantha germinada in vitro em meio WPM; (B) segmento
internodal; (C) segmento nodal; (D) epicótilo; (E) cotilédone; (F)
segmento cotiledonar e (G) hipocótilo. Feira de Santana, BA, 2010.
15
Figura 7 Esquema da aclimatização das plantas enraizadas in vitro. (A) 1º dia
na casa de vegetação – garrafa pet tampada; (B) 7º dia – garrafa pet
desenroscada; (C) 16º dia – tampa retirada; (D) 30º dia – garrafa pet
retirada.
21
Figura 8 Crescimento da plântula de B. cheilantha germinada em meio de
cultura WPM durante 20 dias.
22
Figura 9 Porcentagem de explantes responsivos de B. cheilantha, aos 30 dias
de inoculados em meio de cultura WPM (A- Fator ANA; B- Fator
Explante; SC- segmento cotiledonar e SN- segmento nodal). Feira
de Santana, BA, 2010.
24
Figura 10 Aspecto dos calos nos explantes de hipocótilo (A), cotilédone (B),
epicótilo (C) e segmento internodal (D) aos 30 dias de inoculados
em meio de cultura WPM acrescido de 2,0 mg L-1
de BAP e 0,5 mg
L-1
de ANA. Feira de Santana, BA, 2010.
25
Figura 11 Número de brotos obtidos a partir do segmento nodal (A) e do
segmento cotiledonar (B) de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM com diferentes concentrações
de ANA e BAP ( 0,0 mg L-1
ANA; 0,25 mg L-1
ANA; 0,5
27

mg L-1
ANA). Feira de Santana, BA, 2010.
Figura 12 Número de gemas dos maiores brotos obtidos a partir de segmento
cotiledonar () e de segmento nodal () de B. cheilantha, aos 30
dias de inoculados em meio de cultura WPM suplementado com
diferentes concentrações de BAP (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
).
Feira de Santana, BA, 2010.
29
Figura 13 Comprimento de parte aérea (cm) obtida dos brotos de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM com
diferentes concentrações de ANA e BAP (** Significativo ao nível
de 1% de probabilidade pelo Teste F; * Significativo ao nível de 5%
de probabilidade pelo teste F; ns – não significativo) (A – Interação
BAP X ANA, () 0,0 mg L-1
, () 0,25 mg L-1
e () 0,5 mg L-1
de
ANA ; B- Interação BAP X EXP, segmento cotiledonar () e de
segmento nodal () ). Feira de Santana, BA, 2010.
32
Figura 14 Número de folhas obtido dos brotos a partir do segmento cotiledonar
() e segmento nodal () de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM em função de BAP (*
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não
significativo). Feira de Santana, BA, 2010.
33
Figura 15 Matéria seca da parte aérea advindas do segmento cotiledonar (SC)
e segmento nodal (SN) na fase de multiplicação de B. cheilantha,
aos 30 dias de inoculados em meio de cultura WPM em função de
BAP (* Significativo ao nível de 5% de probabilidade pelo teste F;
ns- não significativo). Feira de Santana, BA, 2010.
34
Figura 16 Brotos de B. cheilantha em meio de multiplicação WPM
suplementado com 2,0 mg L-1
de regulador vegetal, aos 30 dias de
inoculados. (A) Segmento cotilenodar em BAP; (B) Segmento nodal
em KIN; (C) Segmento nodal em BAP; (D) segmento cotiledonar
em TDZ; (E) segmento nodal em TDZ. (Barra = 1,0 cm) Feira de
Santana, BA, 2010.
38
Figura 17 Número de gemas obtidas dos brotos advindos de segmento
cotiledonar () e segmento nodal () e de B. cheilantha, aos 30
dias de inoculados em meio de cultura WPM com diferentes
concentrações de reguladores vegetais (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-
1) (* Significativo ao nível de 5% de probabilidade pelo teste F).
Feira de Santana, BA, 2010.
39
Figura 18 Comprimento dos brotos advindos de segmento cotiledonar () e
segmento nodal () de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM com diferentes concentrações de reguladores
vegetais (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) (* Significativo ao nível de
5% de probabilidade pelo teste F; ** Significativo ao nível 1% de
probabilidade pelo teste F). Feira de Santana, BA, 2010.
41

Figura 19 Porcentagem de enraizamento dos brotos de B. cheilantha, aos 45
dias de inoculados em meio de cultura WPM com diferentes
concentrações de AIB e acrescidos com diferentes concentrações de
carvão ativado (CA) ( 0,0 g L-1
; 1,0g L-1
e 2,0 g L-1
CA) (**
Significativo ao nível de 1% de probabilidade pelo teste F; ns – não
singificativo). Feira de Santana, BA, 2010.
44
Figura 20 Número de raízes adventícias nos brotos de B. cheilantha, aos 45
dias de inoculados em meio de cultura WPM com diferentes
concentrações de AIB e acrescidos em diferentes concentrações de
carvão ativado (CA) ( 0,0 g L-1
; 1,0g L-1
e 2,0 g L-1
CA) (**
Significativo ao nível de 1% de probabilidade pelo teste F; ns – não
significativo). Feira de Santana, BA, 2010.
45
Figura 21 Brotos de B. cheilantha em meio de enraizamento WPM, aos 45 dias
de inoculados em (A) 0,0 mg L-1
de AIB; (B) 0,0 mg L-1
de AIB e
2,0 g L-1
de carvão ativado; (C) 1,0 mg L-1
de AIB e (D) 2,0 mg L-1
de AIB. (Barra = 1,0 cm) Feira de Santana, 2010.
46
Figura 22 Número de raízes secundárias nos brotos de B. cheilantha, aos 45
dias de inoculados em meio de cultura WPM com diferentes
concentrações de AIB (0,0; 0,5; 1,0 e 2,0 mg L-1
) e acrescidos em
diferentes concentrações de carvão ativado (CA) ( 0,0 g L-1
; 1,0
g L-1
e 2,0 g L-1
CA) (** Significativo ao nível de 1% de
probabilidade pelo teste F; ns – não significativo). Feira de Santana,
BA, 2010.
47
Figura 23 Porcentagem de enraizamento dos brotos de B. cheilantha, aos 45
dias de inoculados em meio de cultura WPM com diferentes
concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (A-
AIB; B- ANA e C- AIA), acrescidos com diferentes concentrações
de carvão ativado (CA) ( 0,0 g L-1
; 0,5 g L-1
e 1,0 g L-1
CA)
(** Significativo ao nível de 1% de probabilidade pelo teste F; *
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não
singificativo). Feira de Santana, BA, 2010.
50
Figura 24 Número de raízes adventícias de B. cheilantha, aos 45 dias de
inoculados em meio de cultura WPM com diferentes concentrações
(0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (A- AIB; B- ANA e
C- AIA), acrescidos com diferentes concentrações de carvão ativado
(CA) ( 0,0 g L-1
; 0,5 g L-1
e 1,0 g L-1
CA) (** Significativo
ao nível de 1% de probabilidade pelo teste F;* Significativo ao nível
de %% de probabilidade pelo teste F; ns – não singificativo). Feira
de Santana, BA, 2010.
53
Figura 25 Brotos de B. cheilantha em meio de enraizamento WPM, aos 45 dias
de inoculados em (A) 0,0 mg L-1
de regulador; (B) 0,0 mg L-1
de
regulador e 0,5 g L-1
de carvão ativado; (C) 1,0 mg L-1
de AIB; (D)
1,0 mg L-1
de AIB + 0,5 g L-1
de carvão ativado; (E) 1,0 mg L-1
de
ANA; (F) 1,0 mg L-1
de ANA + 0,5 g L-1
de carvão ativado; (G) 0,5
54

mg L-1
de AIA; (H) 0,5 mg L-1
de AIA + 0,5 g L-1
de carvão ativado.
(Barra = 1,0 cm) Feira de Santana, BA, 2010.
Figura 26 Número de raízes secundárias nos brotos de B. cheilantha, aos 45
dias de inoculados em meio de cultura WPM com diferentes
concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (A-
AIB; B- ANA e C- AIA), acrescidos em diferentes concentrações de
carvão ativado (CA) ( 0,0 g L-1
; 0,5 g L-1
e 1,0 g L-1
CA) (**
Significativo ao nível de 1% de probabilidade pelo teste F; *
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não
significativo). Feira de Santana, BA, 2010.
56
Figura 27 Comprimento da maior raiz de B. cheilantha, aos 45 dias de
inoculados em meio de cultura WPM com diferentes concentrações
(0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (A- AIB; B- ANA e
C- AIA), acrescidos em diferentes concentrações de carvão ativado
(CA) ( 0,0 g L-1
; 0,5 g L-1
e 1,0 g L-1
CA) (** Significativo
ao nível de 1% de probabilidade pelo teste F; * Significativo ao
nível de 5% de probabilidade pelo teste F; ns – não significativo).
Feira de Santana, BA, 2010.
57
Figura 28 Percentual de sobrevivência das mudas de B. cheilantha, aos 30 dias
de aclimatização, referentes às condições de tamponamento durante
a etapa de enraizamento in vitro: filme de PVC, tampa plástica sem
PVC e tampa de algodão. Feira de Santana, BA, 2010.
60
Figura 29 Mudas de B. cheilantha (A) durante a aclimatização em casa de
vegetação e (B) após 30 dias de aclimatização. Feira de Santana,
BA, 2010.
60

LISTA DE TABELAS
Tabela 1 Estado Biotecnológico a respeito do Gênero Bauhinia (Fonte: INPI,
EPO, USPTO, nov. 2009).
4
Tabela 2 Atividades biológicas testadas para a espécie B. cheilantha.
7
Tabela 3 Tratamentos utilizados na indução de brotos em hipocótilo,
segmento cotiledonar, cotilédone, epicótilo, segmento nodal e
internodal de B. cheilantha, inoculados em meio WPM, acrescido de
diferentes concentrações de 6-benzilaminopurina (BAP) e ácido
naftaleno acético (ANA). Feira de Santana, BA, 2010.
16
Tabela 4 Tratamentos utilizados na indução de brotos em segmento
cotiledonar e segmento nodal de B. cheilantha, inoculados em meio
WPM, acrescido de diferentes concentrações de 6-
benzilaminopurina (BAP) ou cinetina (KIN) ou tidiazuron (TDZ).
Feira de Santana, BA, 2010.
17
Tabela 5 Tratamentos utilizados no enraizamento in vitro de B. cheilantha,
inoculados em meio WPM, acrescido de diferentes concentrações de
ácido indolbutírico (AIB) e acrescido de carvão ativado. Feira de
Santana, BA, 2010.
18
Tabela 6 Tratamentos utilizados no enraizamento in vitro de B. cheilantha,
inoculados em meio WPM, acrescido de diferentes concentrações de
ácido indolbutírico (AIB), ácido naftaleno acético (ANA) e ácido
indolacético (AIA). Feira de Santana, BA, 2010.
20
Tabela 7 Resumo da análise de variância para porcentagem de explantes
responsivos (% ER), número de brotos (NB), número de gemas do
maior broto (NG), número de folhas do maior broto (NF),
comprimento da parte aérea (CPA) (cm) e matéria seca da parte
aérea (MSPA) (mg) de plântulas de B. cheilantha, advindos de
segmento cotiledonar e nodal em diferentes concentrações de BAP
(0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e ANA (0,0; 0,25 e 0,5 mg L-1
) em
meio WPM aos 30 dias de inoculação in vitro. UEFS. Feira de
Santana, BA, 2010.
23
Tabela 8 Número de gemas obtidas do maior broto na fase de multiplicação
de B. cheilantha, aos 30 dias de inoculados em meio de cultura
WPM em função de ANA e explante. Feira de Santana, BA, 2010.
30
Tabela 9 Número de gemas a obtidas do maior broto na fase de multiplicação
de B. cheilantha, aos 30 dias de inoculados em meio de cultura
WPM em função de BAP e ANA. Feira de Santana, BA, 2010.
30
Tabela 10 Comprimento da parte aérea na fase de multiplicação de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM em
31

função de ANA e Explante (SC- segmento cotiledonar e SN-
segmento nodal). Feira de Santana, BA, 2010.
Tabela 11 Matéria seca da parte aérea na fase de multiplicação de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM em
função de BAP e ANA. Feira de Santana, BA, 2010.
34
Tabela 12 Resumo da análise de variância para a porcentagem de explantes
responsivos (% ER), número de brotos (NB), número de gemas do
maior broto (NG), número de folhas do maior broto (NF),
comprimento da parte aérea (CPA) (cm) e matéria seca da parte
aérea (MSPA) (mg) de plântulas de B. cheilantha, advindos de
segmento cotiledonar e nodal em diferentes concentrações de KIN,
BAP ou TDZ (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) em meio WPM aos 30
dias de inoculação in vitro. UEFS. Feira de Santana, BA, 2010.
36
Tabela 13 Porcentagem de explantes responsivos de B. cheilantha, aos 30 dias
de inoculados em meio de cultura WPM em função da concentração
das citocininas testadas (KIN; BAP e TDZ) e do tipo de explante
(SC-segmento cotiledonar e SN- segmento nodal). Feira de Santana,
BA, 2010.
37
Tabela 14 Número de brotos obtidos a partir de segmento cotiledonar (SC) e
segmento nodal (SN) de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM com diferentes concentrações de citocininas
(0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
). Feira de Santana, BA, 2010.
37
Tabela 15 Número de gemas obtidas dos brotos advindos de segmento
cotiledonar (SC) e segmento nodal (SN) de B. cheilantha, aos 30
dias de inoculados em meio de cultura WPM com diferentes
citocininas (KIN; BAP e TDZ). Feira de Santana, BA, 2010.
40
Tabela 16 Comprimento dos brotos advindos de segmento cotiledonar (SC) e
segmento nodal (SN) de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM suplementado com diferentes citocininas
(KIN; BAP e TDZ). Feira de Santana, BA, 2010.
40
Tabela 17 Comprimento dos brotos de B. cheilantha, aos 30 dias de inoculados
em meio de cultura WPM suplementado com diferentes
concentrações (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e tipos de citocininas
(KIN; BAP e TDZ). Feira de Santana, BA, 2010.
41
Tabela 18 Matéria seca da parte aérea (mg) de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM suplementado com diferentes
concentrações (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e tipos de citocininas
(KIN; BAP e TDZ). Feira de Santana, BA, 2010.
42
Tabela 19 Matéria seca da parte aérea (mg) de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM em função da concentração
dos reguladores testados e do tipo de explante. Feira de Santana,
42

BA, 2010.
Tabela 20 Resumo da análise de variância para % de enraizamento dos brotos
(%ENR), número de raízes adventícias por broto (NRA), número de
raízes secundárias da maior raiz (NRS), comprimento da maior raiz
(CMR) (cm) e % de sobrevivência dos brotos (%SOBR) de B.
cheilantha, em diferentes concentrações de AIB (0,0; 0,5; 1,0; e 2,0
mg L-1
) e carvão ativado (CA) (0,0; 1,0 e 2,0 g L-1
) em meio de
cultura WPM aos 45 dias de inoculação in vitro. UEFS. Feira de
Santana, BA, 2010.
43
Tabela 21 Comprimento da maior raiz (cm) de B. cheilantha, aos 45 dias de
inoculados em meio de cultura WPM suplementado com diferentes
concentrações de AIB (0,0; 0,5; 1,0; e 2,0 mg L-1
) e carvão ativado
(CA) (0,0; 1,0 e 2,0 g L-1
). Feira de Santana, BA, 2010.
48
Tabela 22 Resumo da análise de variância para % de enraizamento dos brotos
(%ENR), número de raízes adventícias por broto (NRA), número de
raízes secundárias da maior raiz (NRS), comprimento da maior raiz
(CMR) (cm) e % de sobrevivência dos brotos (%SOBR) de B.
cheilantha, em diferentes concentrações (C) (0,0; 0,25; 0,5 e 1,0 mg
L-1
) e tipos de auxinas (A) (AIB; ANA e AIA), em diferentes
concentrações de carvão ativado (CA) (0,0; 0,5 e 1,0 g L-1
) em meio
de cultura WPM aos 45 dias de inoculação in vitro. UEFS. Feira de
Santana, BA, 2010.
49
Tabela 23 Resumo da análise de variância para porcentagem de sobrevivência
das mudas (%SOBR) de B. cheilantha após um período de 30 dias
de aclimatização, pré-acondicionadas em diferentes condições de
tamponamentos (filme de PVC; tampa plástica sem filme de PVC e
tampão de algodão) durante o enraizamento in vitro. Feira de
Santana, BA, 2010.
59

SUMÁRIO
1 INTRODUÇÃO 1
2 REVISÃO DA LITERATURA 2
2.1 CONSIDERAÇÕES SOBRE A ESPÉCIE Bauhinia cheilantha 5
2.2 CULTURA DE TECIDOS VEGETAIS 7
2.2.1 Micropropagação 9
3 MATERIAL E MÉTODOS 13
3.1 MEIO DE CULTURA E CONDIÇÕES DE CULTIVO 13
3.2 ESTABELECIMENTO IN VITRO 14
3.3 MULTIPLICAÇÃO IN VITRO 15
3.3.1 Experimento 1 - Efeito da citocinina 6-benzilaminopurina (BAP) e da
auxina ácido naftaleno acético (ANA) na indução de brotações em
diferentes tipos de explantes de B. cheilantha.
15
3.3.2 Experimento 2 - Efeito do tipo de citocininas na indução de brotações
em diferentes tipos de explantes de B. cheilantha.
16
3.4
3.4.1
ENRAIZAMENTO IN VITRO
Experimento 3 – Efeito da auxina ácido indolbutírico (AIB) e carvão
ativado no enraizamento in vitro de brotos de B. cheilantha.
17
18
3.4.2 Experimento 4 - Efeito das auxinas ácido indolbutírico (AIB) ou ácido
naftaleno acético (ANA) ou ácido indolacético (AIA) e carvão ativado
no enraizamento in vitro de brotos de B. cheilantha.
18
3.5
3.5.1
ACLIMATIZAÇÃO
Experimento 5 – Efeito da aeração in vitro sob a aclimatização
19
19
3.6 ANÁLISE ESTATÍSTICA
21
4
4.1
4.1.1
RESULTADOS E DISCUSSÃO
MULTIPLICAÇÃO IN VITRO
Experimento 1 - Efeito da citocinina 6-benzilaminopurina (BAP) e da
auxina ácido naftaleno acético (ANA) na indução de brotações em
diferentes tipos de explantes de B. cheilantha.
21
22
22
4.1.2
4.2
4.2.1
4.2.2
4.3
4.3.1
5
Experimento 2 - Efeito do tipo de citocininas na indução de brotações
em diferentes tipos de explantes de B. cheilantha.
ENRAIZAMENTO IN VITRO
Experimento 3 - Efeito da auxina ácido indolbutírico (AIB) e carvão
ativado no enraizamento in vitro de brotos de B. cheilantha.
Experimento 4 - Efeito das auxinas ácido indolbutírico (AIB) ou ácido
naftaleno acético (ANA) ou ácido indolacético (AIA) e carvão ativado
no enraizamento in vitro de brotos de B. cheilantha. ACLIMATIZAÇÃO
Experimento 5 – Efeito da aeração in vitro sob a aclimatização
CONCLUSÃO
REFERÊNCIAS
35
43
43
48
59
59
61

1 INTRODUÇÃO
Bauhinia cheilantha (Bong.) Steud., conhecida popularmente como mororó ou pata-
de-vaca, é uma leguminosa típica da Caatinga, principal ecossistema existente no semiárido
brasileiro. Estima-se que cerca de 40% da área desse ecossistema nunca tenha sido coletada e
80% dela subamostrada (TABARELLI; VICENTE, 2004). Segundo Leal et al. (2005), a
Caatinga é, dentre todos os biomas brasileiros, o que registra menor número e menor extensão
de área protegida.
Para o nordeste brasileiro, a espécie em estudo tem uma importância sócio-econômica
relevante, devido ao seu vasto potencial como lenhosa forrageira sendo fonte alternativa de
renda para as comunidades, além do seu uso etnofarmacológico, para a produção de remédios
caseiros. São atribuídas as suas folhas ação antiinflamatória, antidiabética, sedativa,
antiparasitária, digestiva e expectorante, sendo comprovada cientificamente sua atividade
antioxidante, antinociceptiva e hipoglicemiante (LORENZI; MATOS, 2008).
Diversos países agregam esforços para investigar plantas medicinais com potente ação
hipoglicemiante, pois é sabido que a prevalência da diabetes em 2010 irá atingir 6,4% da
população mundial (285 milhões de adultos) e essa estimativa irá aumentar para 7,7% (439
milhões de adultos) em 2030 (SHAW; SICREE; ZIMMET, 2010). Sendo assim, a espécie é
uma planta de potente valor medicinal, utilizada pela população do semiárido brasileiro e
portanto, também pode ser utilizada no tratamento da doença em outras partes do mundo. A
espécie já possui uma metodologia de controle de qualidade para produção de fitoterápicos
que poderão ser desenvolvidos a partir de suas folhas (PEIXOTO-SOBRINHO et al., 2007,
2008), o que é de grande valia para o incremento do arsenal terapêutico no combate a
diabetes, uma vez que os medicamentos disponíveis hoje apresentam diversos efeitos
indesejáveis aos pacientes (HAMZA et al., 2010).
Nesse contexto, a produção de mudas de B. cheilantha é de relevante importância para
a indústria farmacêutica, pois o cultivo de plantas medicinais permite um controle maior dos
fatores extrínsecos (climáticos; edáficos-climáticos e edáficos) à cultura, fornecendo assim
uma matéria-prima vegetal de maior qualidade para a obtenção de fitoterápicos,
diferentemente da matéria-prima advinda de vegetais silvestres.
Desse modo, a Cultura de Tecidos Vegetais, através da micropropagação, permite a
multiplicação rápida de genótipos selecionados com maior produtividade, uniformidade e
desempenho no campo. A micropropagação ou propagação in vitro constitui uma alternativa
para a propagação de mudas sadias, em tempo e espaço físico reduzido, com alta fidelidade

genética (GEORGE, 2008). A aplicação da técnica pode ser vantajosa para a propagação da B.
cheilantha, pois suas sementes apresentam dormência tegumentar, o que dificulta a obtenção
de plantas uniformes e em curto período de tempo.
A micropropagação de espécies lenhosas tem se mostrado um método viável para a
produção de mudas em escala comercial, o que é de grande valia para a conservação de
espécies nativas e recuperação de áreas degradadas (MERKLE; NAIRN, 2005). Contudo,
diversas dificuldades durante o processo de cultivo in vitro tem sido relatadas, sendo comum a
contaminação por microorganismos endófíticos não removidos durante a desinfestação
superficial do tecido (HARRY; THORPE, 1994), a oxidação fenólica do meio de cultura e
explante (OLIVEIRA; NINO, 2009), a baixa taxa de multiplicação dos brotos (NHUT et al.,
2008), a abscisão foliar e o alongamento excessivo dos entrenós, o que reduz o número de
explantes para a fase de multiplicação (NEPOMUCENO et al., 2009) e a dificuldade no
enraizamento de partes aéreas (SOUZA; PEREIRA, 2007), o que pode interferir na
sobrevivência das mudas durante a fase de aclimatização.
Considerando a necessidade de produção de mudas de B.cheilantha para o
fornecimento de matéria-prima de qualidade para a produção de fitoterápicos e visto que até o
momento não há relatos com a espécie na área de propagação in vitro, objetivou-se com a
realização desse trabalho estabelecer um protocolo de micropropagação e aclimatização para
B. cheilantha.
2 REVISÃO DA LITERATURA
O gênero Bauhinia é composto por, aproximadamente, 300 espécies (VAZ ; TOZZI,
2005). Em 2008, a International Union for Conservation of Nature (IUCN) apontou 12
espécies do gênero em sua lista vermelha, nas seguintes categorias: 2 CR - Critically
Endangered – criticamente em perigo, 5 EN – Endangered- Perigo, 4 VU –Vulnerable-
vulneráveis, 1 LC - Least Concern (inclui LR/lc - Lower Risk, least concern)- baixo risco.
Sendo que destas espécies, 10 são nativas da América Latina e uma delas é nativa da Bahia, a
B. interregima. No Brasil ocorrem 91 espécies do gênero (VAZ; TOZZI, 2003), sendo três
destas consideradas raras: B. candelabriformis, B. leptantha e B. malacotrichoides
(QUEIROZ et al., 2009a).
Espécies desse gênero são classificadas na escala de sucessão vegetal como pioneiras
tardias, pois apresentam crescimento moderadamente rápido (LORENZI, 1992). Contudo, foi
demonstrado que suas sementes apresentam dormência tegumentar (ALVES et al., 2000,

2004). Seus exemplares são utilizados na recomposição vegetal, no reflorestamento de áreas
degradadas e também na arborização de ruas (ARAÚJO et al., 2009).
As plantas do gênero são comumente chamadas, no Brasil, de „mororó‟ (palavra de
origem tupi), „unha-de-vaca‟ ou „pata-de-vaca‟, devido as suas folhas fendidas lembrarem o
rastro da pata dos bovinos. Algumas espécies de Bauhina são utilizadas pela população para
fins medicinais e ritualísticos (ALONSO, 2004; MACEDO; FERREIRA, 2004; LORENZI;
MATOS, 2008; PIRES, 2009).
Os usos tradicionais dos „mororós‟ têm encontrado respaldo na literatura. Atividades
antifúngicas, antibacterianas, analgésicas, antiinflamatórias, antidiarréicas e especialmente
hipoglicemiante são relatadas em vários modelos experimentais, comprovando e justificando
o uso destas espécies na medicina popular (OLIVEIRA et al., 2001; SILVA; CECHINEL-
FILHO, 2002; SOSA et al., 2002; LINO et al., 2004; BARBOSA-FILHO et al., 2005).
Concomitantemente a esses estudos, foram isoladas diferentes classes de compostos orgânicos
de interesse medicinal: lactonas, flavonóides, terpenóides, esteróides, triterpenos, taninos e
quinonas (SILVA; CECHINEL-FILHO, 2002; DUARTE-ALMEIDA; NEGRI; SALATINO,
2004).
O gênero Bauhinia apresenta relevante potencial biotecnológico, uma vez que foram
encontradas diversas patentes depositadas no Instituto Nacional de Propriedade Intelectual
(INPI), European Patent Office (EPO) e United States Patent and Trademark Office (USPTO)
classificadas nas seguintes seções: Necessidades Humanas (A), Químicas e Metalúrgicas (C),
Têxteis e Papel (D) e Físicas (G) (Tabela 1), segundo a Classificação Internacional de
Patentes (CIP).
Foram identificadas 18 classificações para as patentes depositadas com a palavra-
chave Bauhinia, distribuídas em 564 grupos, sendo que a maior parte das patentes (52%) está
na seção de preparações com finalidades médicas, odontológicas ou higiênicas. Na seção de
microorganismos ou enzimas; propagação, preservação ou manutenção de tecidos e
engenharia genética, apenas sete patentes (1%) são encontradas (Figura 1).
Através de levantamento de patentes é possível visualizar as áreas de pesquisa que
estão incipientes em estudos. No que diz respeito ao gênero em estudo, é percebido que as
informações abrangendo o cultivo e a propagação de Bauhinia estão aquém da real
potencialidade do gênero.
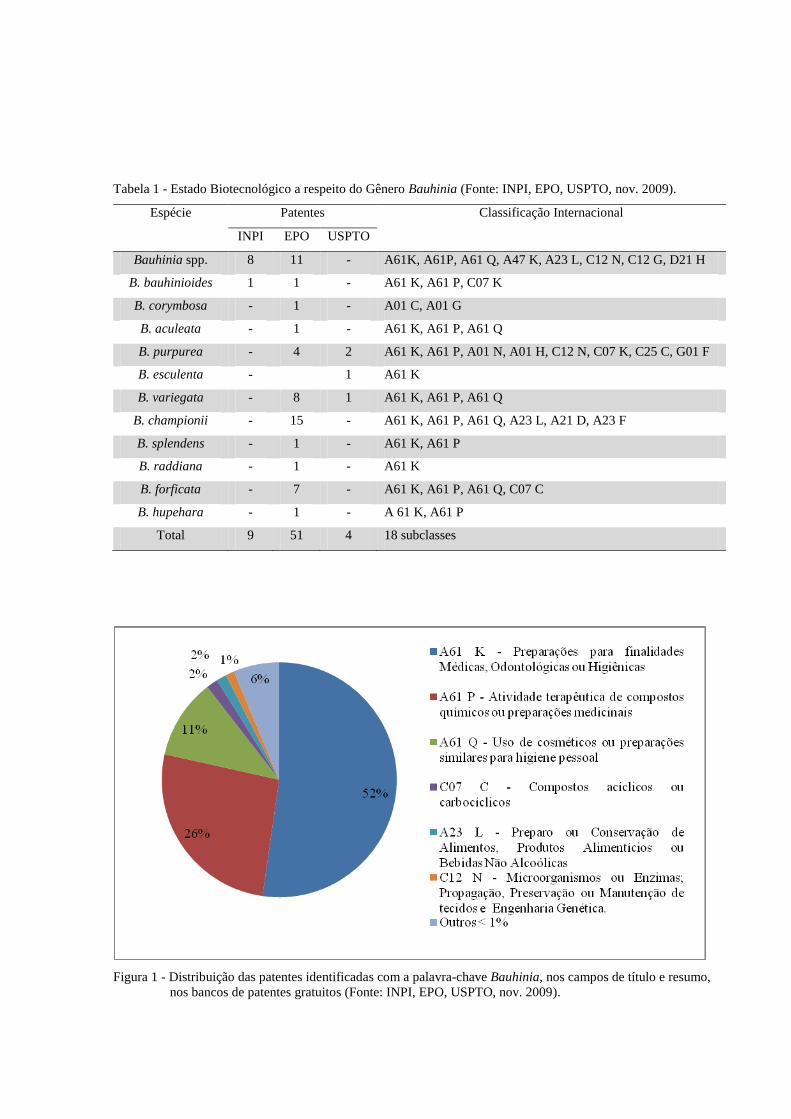
Tabela 1 - Estado Biotecnológico a respeito do Gênero Bauhinia (Fonte: INPI, EPO, USPTO, nov. 2009).
Espécie Patentes Classificação Internacional
INPI EPO USPTO
Bauhinia spp. 8 11 - A61K, A61P, A61 Q, A47 K, A23 L, C12 N, C12 G, D21 H
B. bauhinioides 1 1 - A61 K, A61 P, C07 K
B. corymbosa - 1 - A01 C, A01 G
B. aculeata - 1 - A61 K, A61 P, A61 Q
B. purpurea - 4 2 A61 K, A61 P, A01 N, A01 H, C12 N, C07 K, C25 C, G01 F
B. esculenta - 1 A61 K
B. variegata - 8 1 A61 K, A61 P, A61 Q
B. championii - 15 - A61 K, A61 P, A61 Q, A23 L, A21 D, A23 F
B. splendens - 1 - A61 K, A61 P
B. raddiana - 1 - A61 K
B. forficata - 7 - A61 K, A61 P, A61 Q, C07 C
B. hupehara - 1 - A 61 K, A61 P
Total 9 51 4 18 subclasses
Figura 1 - Distribuição das patentes identificadas com a palavra-chave Bauhinia, nos campos de título e resumo,
nos bancos de patentes gratuitos (Fonte: INPI, EPO, USPTO, nov. 2009).

2.1 CONSIDERAÇÕES SOBRE A ESPÉCIE Bauhinia cheilantha
Bauhinia cheilantha (Fam. Fabaceae ou Leguminosae) é uma árvore de caule duro,
com cascas fibrosas e com ausência de espinhos (LORENZI; MATOS, 2008). Possui um
porte pequeno, chegando atingir até 3,5 m de altura, folhas com lóbulos arredondados,
cartáceas a subcoriáceas, apresentando pecíolo com 2-3 cm de comprimento, suas
inflorescências chegam a atingir cerca de 5,0 cm de comprimento, apresentando pétalas
brancas e os frutos são do tipo legume deiscente (QUEIROZ, 2009b) (Figura 2).
Figura 2 – B. cheilantha: (A) Aspecto da casca; (B) Folha; (C) Sementes maduras; (D) Inflorescência; (E) Fruto
maduro (Fonte: Autora, 2010).
A B. cheilantha é típica da Caatinga do semiárido brasileiro, podendo ser encontrada
com menos freqüência em áreas de Cerrado, na área centro-oeste/Paraguai e também na
vegetação arbustiva próxima a Cuiabá (VAZ; TOZZI, 2003) (Figura 3).
A
B
D E
C

A espécie é usada na produção de remédios caseiros com ação antiinflamatória,
antidiabética, sedativa, antiparasitária, para distúrbios digestivos, asma e tosse (ALMEIDA et
al., 2005; ALBUQUERQUE et al., 2005, 2007; AGRA et al., 2007), e para uso alimentar
como bebida, a partir de suas sementes torradas (ALBUQUERQUE; ANDRADE, 2002;
LUCENA et al., 2007). Suas folhas são consumidas por bovinos, caprinos e ovinos, em
pastejo direto ou na forma de feno, pelo seu alto teor de proteínas (BATISTA; AMORIM;
NASCIMENTO, 2005; GUIMARÃES-BELEEN et al., 2006a, 2006b). Enquanto sua madeira
é utilizada em construção e como combustível (LUCENA et al., 2007).
Figura 3 - Distribuição geográfica de B. cheilantha () no Brasil. (Fonte: Autora, adaptado de Vaz e Tozzi,
2003).
Da espécie B. cheilantha foram isoladas do extrato etanólico diferentes classes de
compostos orgânicos de interesse medicinal: alcalóides, antraquinonas, esteróides livres,
flavonas, flavonóides e xantonas (folhas, madeira e raízes), fenóis (caule, folha), flavononas
(caule, raiz) (LUNA et al., 2005). Da fração metanólica da solução aquosa liofilizada das
folhas isolou-se dois flavonóides: quercitrina e afzelina, além da detecção de constituintes
voláteis nas folhas, como o óxido de cariofileno e trans-cariofileno. O óleo de suas sementes
apresentou 57,7 % de constituintes insaturados e 42,3 % de ácido linoléico (BRÍGIDO, 2001).
A atividade hipoglicemiante e antioxidante tem sido verificada em diversos extratos de
suas folhas (ALMEIDA et al., 1997; OLIVEIRA et al., 2002, 2008a, 2008b). O extrato
aquoso da entre-casca apresentou atividade antinociceptiva não sendo detectado toxicidade
aguda até a dose de 3g/Kg para a espécie (SILVA et al., 2005). O extrato etanólico de suas
raízes (100 ppm) apresentou alta toxicidade para Artemia salina (ca. 100% mortalidade),
enquanto o extrato etanólico de sua madeira (500 ppm) mostrou potente atividade larvicida
contra Aedes aegypti (LUNA et al., 2005) (Tabela 2).

Tabela 2 - Atividades biológicas testadas para a espécie B. cheilantha.
Parte da planta / extrato Atividade biológica
testada
Referência
Folhas / ext. metanólico Hipoglicemiante ALMEIDA; XAVIER; GUEDES, 1997.
Folhas / ext. bruto e fração
aquosa
Hipoglicemiante
Antioxidante
OLIVEIRA et al., 2002, 2008a, 2008b.
Folhas / ext. hidroalcoólico Anticolinesterásica TREVISAN; MACEDO, 2003.
Entre-casca / ext. aquoso Antinociceptiva SILVA et al., 2005.
Raiz / ext. etanólico Toxicidade LUNA et al., 2005.
Madeira / ext. etanólico Larvicida LUNA et al., 2005.
No que se refere a propagação da espécie, ela só ocorre a partir de suas sementes ou do
transplante de plantas jovens. Contudo suas sementes apresentam dormência tegumentar
(MAIA, 2004; SEIFFERT, 2006; LORENZI, MATOS, 2008). Diante disso, a Cultura de
Tecidos Vegetais, através da micropropagação, promete ser uma alternativa para a propagação
da espécie, possibilitando a obtenção de plantas uniformes e em curto período de tempo que
poderão ser utilizadas como matéria-prima para a produção de fitoterápicos.
2.2 CULTURA DE TECIDOS VEGETAIS
O termo Cultura de Tecidos Vegetais é geralmente utilizado para designar a cultura de
células, tecidos e órgãos isolados sob condições específicas in vitro. O conceito é baseado na
capacidade latente da célula vegetal poder regenerar um indivíduo completo, ou seja, a
totipotencialidade das células. As técnicas que envolvem cultura de tecidos, seja ela de tecidos
desorganizados ou organizados, são uma das mais importantes contribuições para a
propagação de plantas, possibilitando uma multiplicação clonal de indivíduos superiores com
rápido crescimento (GEORGE, 2008).
A micropropagação, uma das diversas técnicas de cultura de tecidos vegetais pode ser
dividas em dois grupos: organogênese e embriogênese somática (PASQUAL et al., 1997).
Este último, que consiste do desenvolvimento de embriões a partir de células somáticas, não
será abordado neste trabalho.
A organogênese é definida como a formação de estruturas a partir de tecidos e/ou
células, podendo ocorrer por duas vias: via direta, através da regeneração de plantas
provenientes de tecidos não meristemáticos, sem passar pela fase de calo, apresentando alta
fidelidade genética; e, por via indireta, quando o processo de regeneração é precedido pela

formação de calo (GRATTAPAGLIA; MACHADO, 1998). Entretanto, a regeneração via
calo, às vezes, resulta em aparecimento de variação somaclonal, tornando essa estratégia
questionável para multiplicação clonal (GEORGE, 2008).
A cultura de tecidos possui significativa aplicabilidade em diversas áreas, tais como a
indústria farmacêutica, com a produção de metabólitos de interesse a partir do cultivo de
suspensões celulares ou órgãos isolados (JEDINÁK et al., 2004); a horticultura e a
silvicultura, com o uso do DNA recombinante para o melhoramento genético e a propagação
vegetativa in vitro (SOUZA; COSTA; SILVA-NETO, 2006). Além das contribuições para a
indústria, essa técnica pode permitir uma melhor compreensão da fisiologia e do metabolismo
vegetal (DORAN, 2009).
Muitos protocolos de regeneração vegetal utilizando técnicas de cultura de tecidos
passaram a ser patenteados, ou seja, os resultados de investimentos em P&D foram
privatizados, passando então a serem comercializados dentro dos diferentes segmentos na área
dos agronegócios, tais como florestal, ornamental e hortigranjeijos, assim como da indústria
farmacêutica (CID, 2000).
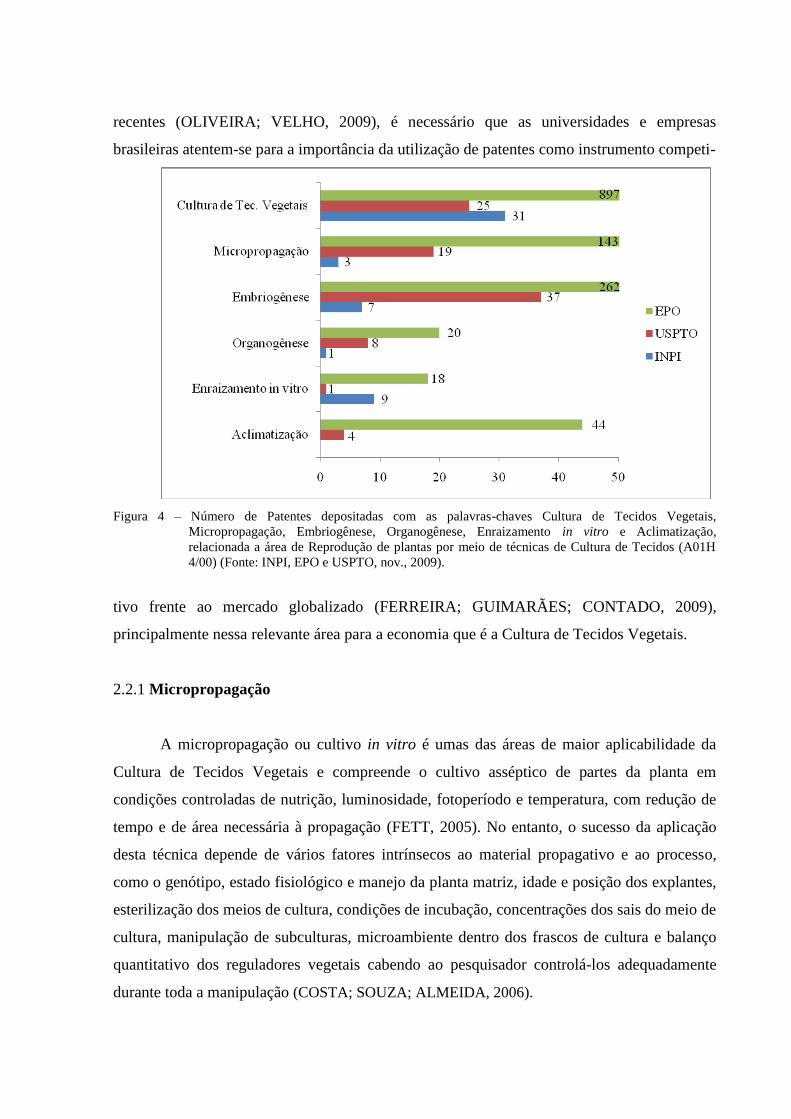
A análise exploratória das patentes, abrangendo a área de reprodução de plantas por
meio de técnicas de cultura de tecidos (Seção A01H 4/00, segundo a CIP), revela uma
quantidade inexpressiva de 97 patentes nacionais (INPI) comparando-se com as quantidades
encontradas no exterior, USPTO com 892 depósitos e EPO com 9.277. Dentro dessa seção
(A01H 4/00) foi realizada uma busca intercalada com as seguintes palavras chaves: Cultura de
Tecidos Vegetais, Micropropagação, Embriogênese, Organogênese, Enraizamento in vitro e
Aclimatização (Figura 4). O baixo número de concessões de patentes no país se contrapõe ao
expressivo número de publicações nacionais reportadas com igual temática. Diante do
exposto, é notória a negligência dos pesquisadores, empresas e universidades em proteger o
conhecimento gerado nas suas pesquisas.
No país, apenas uma única patente registra o protocolo de regeneração vegetal, sendo
esta para Acacia mangium (Fabaceae), sob o número BR0009112 (A), as demais patentes são
relacionadas a sistemas que facilitam o desenvolvimento da técnica. Enquanto que no exterior
diversos protocolos de propagação vegetativa in vitro são concedidos.
O fato é que, o Brasil possui grande parte da sua biodiversidade inexplorada e
enorme potencial na geração de produtos e processos que podem ser patenteáveis na área que
tange as técnicas de Cultura de Tecidos Vegetais. Apesar do processo de proteção e
comercialização das criações de pesquisas nacionais terem sido estimuladas por políticas
recentes (OLIVEIRA; VELHO, 2009), é necessário que as universidades e empresas
brasileiras atentem-se para a importância da utilização de patentes como instrumento competi-
Figura 4 – Número de Patentes depositadas com as palavras-chaves Cultura de Tecidos Vegetais,
Micropropagação, Embriogênese, Organogênese, Enraizamento in vitro e Aclimatização,
relacionada a área de Reprodução de plantas por meio de técnicas de Cultura de Tecidos (A01H
4/00) (Fonte: INPI, EPO e USPTO, nov., 2009).
tivo frente ao mercado globalizado (FERREIRA; GUIMARÃES; CONTADO, 2009),
principalmente nessa relevante área para a economia que é a Cultura de Tecidos Vegetais.
2.2.1 Micropropagação
A micropropagação ou cultivo in vitro é umas das áreas de maior aplicabilidade da
Cultura de Tecidos Vegetais e compreende o cultivo asséptico de partes da planta em
condições controladas de nutrição, luminosidade, fotoperíodo e temperatura, com redução de
tempo e de área necessária à propagação (FETT, 2005). No entanto, o sucesso da aplicação
desta técnica depende de vários fatores intrínsecos ao material propagativo e ao processo,
como o genótipo, estado fisiológico e manejo da planta matriz, idade e posição dos explantes,
esterilização dos meios de cultura, condições de incubação, concentrações dos sais do meio de
cultura, manipulação de subculturas, microambiente dentro dos frascos de cultura e balanço
quantitativo dos reguladores vegetais cabendo ao pesquisador controlá-los adequadamente
durante toda a manipulação (COSTA; SOUZA; ALMEIDA, 2006).

As células vegetais respondem aos estímulos físico, químico ou biológico, o que torna
possível o controle da morfogênese in vitro na cultura de tecidos. A depender das
interferências no ambiente, as células podem ser induzidas a tornar-se competentes, isto é, de
responder aos estímulos extra-celulares diferenciando ou regenerando-se, caso contrário são
chamadas de recalcitrantes. Uma vez competentes, inicia-se o processo da indução, onde uma
série de eventos, regulatórios do processo morfogenético celular, estabelece a expressão de
genes pré existentes. Posterior à etapa de indução vem a determinação, onde, em resposta aos
estímulos, a célula torna-se comprometida com uma rota específica, podendo acarretar a
organogênese. Sendo assim, a determinação precede aos eventos da diferenciação, onde uma
célula comprometida expressa suas características especializadas. Este último evento, nada
mais é que o processo de mudança bioquímica, anatômica, morfológica e fisiológica no qual
as células atingem sua função e forma final (HARTMANN et al., 1997; ALBERTS et al.,
1997; KLUG; CUMMINGS, 2000; PERES, 2002; GAHAN; GEORGE, 2008). No caso do
tecido vegetal possuir células meristemáticas, as células ganham novas competências se
tornando diferenciadas, apresentando alta fidelidade genética; e com células não
meristemáticas pode ocorrer a desdiferenciação do tecido, com a conseqüente formação de
calo, dando capacidade as células de iniciar uma resposta particular de diferenciação, ou então
ocorrer a rediferenciação que envolve a mudança da competência das células, onde em
condições apropriadas irá formar células competentes capazes de desenvolver novas
estruturas (GRATTAPAGLIA; MACHADO, 1998; PERES, 2002).
Os meios de cultura utilizados na micropropagação fornecem as substâncias essenciais
para o crescimento dos tecidos, suprindo as necessidades metabólicas, energéticas e
estruturais das células (CALDAS, HARIDASAN; ELIAS, 1998). O meio de cultura WPM
(Wood Plant Medium), elaborado por Lloyd e McCown em 1981 para a cultura de Kalmia
latifolia (Ericaceae), tem sido utilizado com sucesso para muitas lenhosas, em razão da sua
menor força iônica total em relação a outros meios empregados no cultivo in vitro
(PASQUAL, 2001; JHA; GHOSH, 2006). Na tentativa de otimizar o crescimento in vitro dos
tecidos vegetais, diversos estudos propõem a redução ou incremento de alguns macro e/ou
micronutrientes que compõem o meio de cultura, suprindo melhor as exigências nutricionais
de cada espécie, à exemplo dos meios modificados para amoreira-preta (Rubus sp.) e videira
(Vitis sp.) (VILLA et al., 2008, 2009).
A suplementação do meio de cultura com reguladores vegetais (substâncias sintéticas)
supre prováveis deficiências das quantidades de fitormônios (substâncias naturais) nos
explantes da planta matriz, induzindo os processos de desdiferenciação e rediferenciação, com

conseqüente formação de tecidos e órgãos (TERMIGNONI, 2005). Os reguladores vegetais
destacam-se como os principais controladores da morfogênese in vitro (TAIZ; ZEIGER,
2009). No campo biotecnológico, na ausência dessas substâncias chaves não haveria
viabilidade comercial para as biofábricas ou laboratórios comerciais que utilizassem os
protocolos de regeneração de plantas oriundas de condições in vitro.
As citocininas e auxinas são conhecidas por estimular ou inibir inúmeros processos
fisiológicos. Dentre as citocininas, está o 6-benzilaminopurina (BAP), que tem sido o mais
indicado para promover a proliferação de partes aéreas e indução de gemas adventícias in
vitro (GRATTAPAGLIA; MACHADO, 1998), mostrando-se bem eficiente na formação de
brotos em explantes de diversas espécies lenhosas. Outras substâncias com atividade
citocinica são a cinetina (KIN) e o tidiazuron (TDZ). Embora tenha sido desenvolvido para
ser utilizado como desfoliante para o algodoeiro, existem indicações de que o TDZ seria mais
efetivo para induzir multibrotações de gemas axilares de plantas lenhosas, quando utilizado
em pequenas concentrações. Entretanto, seu uso requer cautela, já que problemas como
hiperidricidade de brotações, brotações curtas e compactas, folhas com morfologia anormal e
dificuldades no alongamento e enraizamento têm sido observados na sua presença (CID,
2000).
Enquanto as citocininas promovem divisão celular nas partes aéreas e raízes, as
auxinas atuam no alongamento celular, sobretudo na promoção de raízes laterais e adventícias
(TAIZ; ZEIGER, 2009). Dentre as auxinas, o ácido indolbutírico (AIB) é o mais empregado
na indução de raízes em brotos originários da fase de multiplicação, por possuir maior
fotoestabilidade e menor degradação oxidativa que o ácido indolacético (AIA) e menor
conversão enzimática nos tecidos da planta que o ácido naftaleno acético (ANA)
(MACHAKOVA; ZAZIMALOVA; GEORGE, 2008). Segundo Gahan e George (2008), os
reguladores vegetais influenciam as diferentes fases do ciclo celular, as auxinas exercendo
efeito na replicação do DNA, enquanto as citocininas controlam os eventos da mitose. Desse
modo um balanço auxina/citocinina favorável às citocininas induz a formação in vitro de
brotos adventícios (PERES; KERBAUY, 2008).
Outra substância frequentemente adicionada ao meio de cultura é o carvão ativado,
com o intuito de otimizar o crescimento e desenvolvimento celular nas diferentes fases da
cultura de tecidos (VIEITEZ et al., 2009). O efeito benéfico do carvão se deve, em parte, a
sua grande área de superfície e volume o que lhe fornece a capacidade de adsorção. Desse
modo, ele modifica a composição e o pH do meio de cultura podendo melhorar ou regular o
crescimento in vitro (THOMAS, 2008). Além do efeito químico, o carvão fornece uma

condição de escuro favorável ao desenvolvimento das raízes adventícias, simulando as
condições do solo (MOSHKOV et al., 2008). Segundo Santos-Serejo et al. (2006), a
dificuldade no enraizamento pode estar ligada ao efeito residual de citocininas requeridas para
induzir a proliferação de brotações, na etapa de multiplicação, podendo o carvão ativado ser
utilizado, em etapa posterior, pela sua capacidade de adsorver concentrações excessivas
desses reguladores e composto tóxicos que inibem a morfogênese.
Na micropropagação convencional, as plantas crescem sob condições heterotróficas ou
mixotróficas devido à presença de açúcar no meio e ao uso de recipientes fechados. Nessas
circunstâncias, há pouca funcionalidade dos estômatos e uma deficiente formação de cera
cuticular nas plantas, além do reduzido desenvolvimento do mesofilo foliar, especialmente os
parênquimas clorofilianos e feixes vasculares (ZIV; CHEN, 2008). Desse modo, os órgãos
fotossintetizantes não operam normalmente e as plantas não desenvolvem capacidade
fotoautrotófica, o que facilita a rápida desidratação da planta quando transferida para o
ambiente normal, causando a morte de mudas durante a fase de aclimatização (NGUYEN;
KOZAI, 2001).
A aclimatização pode ser considerada a fase final da micropropagação e sua
importância é tal, que pode significar a limitação de todo processo de multiplicação in vitro de
plantas (SOUZA; COSTA; SILVA-NETO, 2006). Contudo, é possível estimular a planta
propagada in vitro à nutrição autotrófica, através da redução de sacarose no meio de cultura
e/ou do aumento da aeração dos recipientes, utilizando-se tamponamentos alternativos,
originando plantas anátomo-fisiológicas mais adaptadas ao processo de aclimatização
(THORPE et al., 2008).
Apesar das crescentes pesquisas abordando lenhosas, essas espécies enfrentam
diversas dificuldades durante várias etapas do processo de cultivo in vitro, tais como:
contaminação (HANDA; SAMPAIO; QUISEN, 2005), devido aos microorganismos
endófíticos desses vegetais não serem removidos com a desinfestação superficial do tecido
(HARRY; THORPE, 1994); oxidação fenólica do meio de cultura e explante (XAVIER,
OTONI, PENCHEL, 2007; OLIVEIRA; NINO, 2009), devido a produção frequente de
polifenóis e taninos, escurecendo rapidamente os tecidos, seguido da deterioração celular com
consequente necrose do explante; dificuldade no enraizamento de partes aéreas, que está
relacionado a uma gama de fatores endógenos e exógenos que ainda não estão completamente
elucidados (SOUZA; PEREIRA, 2007); a baixa taxa de multiplicação dos brotos (NHUT et
al., 2008); a abscisão foliar, causada pelo acúmulo de etileno nos frascos onde são mantidos o

material vegetal e o alongamento excessivo dos entrenós, o que reduz o número de explantes
para a fase de multiplicação (NEPOMUCENO et al., 2009).
No gênero Bauhinia, a B. purpurea foi a primeira espécie a ser regenerada in vitro via
brotos axilares (KUMAR, 1987) e posteriormente por organogênese indireta (KUMAR,
1992). B. variegata (MATHUR; MUKUNTHAKUMAR, 1992) e B. vahlii (DHAR; UPRETI,
1999; BHATT; DHAR, 2000) também tem sido propagadas via brotos axilares. No Brasil, a
B. forficata é a única representante do gênero com registro na área de cultivo in vitro
(CARVALHO, 1998), inclusive com o cultivo de células em suspensão (MELLO et al., 2001;
APPEZZATO-DA-GLÓRIA; MACHADO, 2004).
Com o intuito de superar as dificuldades encontradas no cultivo in vitro de lenhosas e
a insuficiência de protocolos para espécies de Bauhinia, é necessário a realização de estudos
mais específicos para a micropropagação e o desenvolvimento de metodologias que atendam
às exigências de cada espécie.
3 MATERIAL E MÉTODOS
Os experimentos foram realizados no Laboratório de Cultura de Tecidos Vegetais
(LCTV), pertencente à Unidade Experimental Horto Florestal (UEHF) da Universidade
Estadual de Feira de Santana (UEFS), localizado no município de Feira de Santana, região do
semiárido da Bahia.
Foram utilizadas sementes de B. cheilantha, que estavam armazenadas em sacos de
plásticos, no Banco de Sementes do Horto Florestal - UEFS, sob temperatura ambiente. Os
frutos maduros da espécie em estudo foram coletados manualmente no Campo Experimental
da Caatinga, da EMBRAPA Semiárido, localizado no submédio São Francisco, no município
de Petrolina – PE e fornecidos por esta instituição.
3.1 MEIO DE CULTURA E CONDIÇÕES DE CULTIVO
O meio de cultura utilizado foi o WPM (LLOYD; McCOWN, 1980), solidificado
com 0,6% de ágar (Himedia®). A sacarose foi utilizada como fonte de carbono na
concentração de 3%. O pH do meio de cultura foi ajustado para 5,7 ± 0,1 com hidróxido de
sódio (NaOH) ou ácido clorídrico (HCl) a 0,1 N antes da autoclavagem. Os reguladores
vegetais e o carvão ativado utilizados foram adicionados ao meio antes da sua esterilização.

O meio de cultura foi distribuído em tubos de ensaio (25x150 mm) vedados com tampa
plástica e esterilizados por autoclavagem durante 15 min à temperatura de 121 ºC e pressão de
1 atm.
As inoculações foram realizadas em câmara de fluxo laminar para a manutenção de
condição asséptica e os recipientes fechados com filme de Poli Cloreto de Vinila (PVC). As
culturas foram mantidas em sala de crescimento com temperatura de 25 ± 2ºC, sob
fotoperíodo de 16 horas, com umidade relativa de 60% e radiação fotossintética ativa de 60
µmol.m-2
.s-1
, fornecida por lâmpadas fluorescentes branco-frias.
3.2 ESTABELECIMENTO IN VITRO
Inicialmente as sementes de B. cheilanta foram submetidas à escarificação mecânica,
utilizando lixa de acabamento (nº 03), danificando o tegumento até o aparecimento do
endosperma no lado oposto ao hilo (Figura 5). Logo em seguida, as sementes escarificadas
foram lavadas com água estéril autoclavada e detergente e levadas para câmara de fluxo
laminar.
Figura 5 – Sementes de B. cheilantha, sob ocular 2X de esterescópico, seta apontando o hilo (A) e sementes após
escarificação (B) (Barra = 0,5 cm). Feira de Santana, BA, 2010.
Em câmara de fluxo laminar foram desinfestadas com imersão em etanol a 70% por 1
minuto, seguido de solução de hipoclorito de sódio - NaOCl [água sanitária comercial (Qboa
®) - 2,5% de cloro ativo] com 2 gotas de detergente neutro (Ypê ®) por 15 minutos e lavadas
8 vezes com água estéril. Em seguida foram inoculadas 10 sementes por placa de Petri
contendo papel germtest, previamente esterilizados, umedecidos com água estéril e então
vedadas com filme de PVC.
Após 72 horas de semeadura, quando as sementes já haviam emitido a radícula (com
aproximadamente 1,0 cm de comprimento), as plântulas foram transferidas para tubos de
ensaio, contendo 15 mL do meio de cultura WPM.

3.3 MULTIPLICAÇÃO IN VITRO
Para a multiplicação in vitro foram utilizadas plântulas oriundas da germinação in
vitro (Figura 6A). A indução de brotos foi realizada em seis tipos diferentes de explantes:
hipocótilo, segmento cotiledonar, cotilédone, epicótilo, segmento nodal e internodal (Figura
6B-6G).
Figura 6 - Esquema do seccionamento realizado para obtenção dos explantes utilizados na etapa de multiplicação
in vitro: (A) plântula de B. cheilantha germinada in vitro em meio WPM, com 15 dias de idade; (B)
segmento internodal; (C) segmento nodal; (D) epicótilo; (E) cotilédone; (F) segmento cotiledonar e
(G) hipocótilo. Feira de Santana, BA, 2010.
3.3.1 Experimento 1 - Efeito da citocinina 6-benzilaminopurina (BAP) e da auxina ácido
naftaleno acético (ANA) na indução de brotações em diferentes tipos de explantes de B.
cheilantha.
Neste experimento foram utilizadas plântulas de B. cheilantha com 15 a 20 dias de
idade. A indução de brotos foi realizada em hipocótilo, segmento cotiledonar, cotilédone,

epicótilo, segmento nodal e internodal, em diferentes concentrações de BAP (0,0; 0,5; 1,0;
1,5; 2,0 mg L-1
) combinado com ANA (0,0; 0,25; 0,5 mgL-1
) (Tabela 3).
Os explantes foram inoculados verticalmente em tubos contendo 5 mL de meio de
cultura por tubo. O delineamento experimental foi o inteiramente casualizado em esquema
fatorial 6 x 5 x 3 (seis tipos de explantes x cinco doses de citocinina x três dose de auxina),
totalizando 90 tratamentos. Cada tratamento foi constituído de cinco repetições, sendo cada
repetição composta por cinco tubos contendo um explante cada.
Tabela 3 – Tratamentos utilizados na indução de brotos em hipocótilo, segmento cotiledonar, cotilédone,
epicótilo, segmento nodal e internodal de B. cheilantha, inoculados em meio WPM, acrescido de
diferentes concentrações de 6-benzilaminopurina (BAP) e ácido naftaleno acético (ANA). Feira de
Santana, BA, 2010.
3.3.2 Experimento 2 - Efeito do tipo de citocininas na indução de brotações em diferentes
tipos de explantes de B. cheilantha.
Neste experimento foram utilizadas plântulas de B. cheilantha com 35 a 40 dias de
idade. A indução de brotos foi realizada em segmento cotiledonar e nodal, em diferentes
concentrações de 6-benzilaminopurina (BAP) ou cinetina (KIN) ou tidiazuron (TDZ) (0,0;
0,5; 1,0; 1,5 e 2,0 mg L-1
) (Tabela 4).
Tratamentos Concentrações dos Reguladores (mg L-1
)
BAP ANA
T 1 0,00 0,00
T 2 0,50 0,00
T 3 1,00 0,00
T 4 1,50 0,00
T 5 2,00 0,00
T 6 0,00 0,25
T 7 0,50 0,25
T 8 1,00 0,25
T 9 1,50 0,25
T 10 2,00 0,25
T 11 0,00 0,50
T 12 0,50 0,50
T 13 1,00 0,50
T 14 1,50 0,50
T 15 2,00 0,50

Os explantes foram inoculados verticalmente em tubos contendo 7 mL de meio de
cultura por tubo de ensaio. O delineamento experimental foi o inteiramente casualizado em
esquema fatorial 2 x 3 x 5 (dois tipos de explantes x três tipos de citocininas x cinco
concentrações de citocinina), totalizando 30 tratamentos. Cada tratamento foi constituído de
cinco repetições, sendo cada repetição composta por cinco tubos de ensaio contendo um
explante cada.
Tabela 4 – Tratamentos utilizados na indução de brotos em segmento cotiledonar e segmento nodal de B.
cheilantha, inoculados em meio WPM, acrescido de diferentes concentrações de 6-
benzilaminopurina (BAP) ou cinetina (KIN) ou tidiazuron (TDZ). Feira de Santana, BA, 2010.
Aos 30 dias da inoculação, para os dois experimentos de multiplicação realizados, os
explantes foram avaliados, sendo observadas as seguintes variáveis: frequência de explantes
responsivos, isto é, a porcentagem de explantes que formaram brotos, número de brotos por
explante, número de gemas do maior broto, número de folhas do maior broto, comprimento da
parte aérea e matéria seca da parte aérea.
3.4 ENRAIZAMENTO IN VITRO
Tratamentos Citocinina Concentração (mg L-1
)
T 1
BAP
0,0
T 2 0,5
T 3 1,0
T 4 1,5
T 5 2,0
T 6
KIN
0,0
T 7 0,5
T 8 1,0
T 9 1,5
T 10 2,0
T 11
TDZ
0,0
T 12 0,5
T 13 1,0
T 14 1,5
T 15 2,0

Para a indução de raízes, foram utilizados brotações provenientes do meio de cultura
suplementado com 2,0 mg L-1
de BAP, baseados nos resultados do experimento anterior,
medindo entre 1,5 a 5,0 cm de comprimento.
3.4.1 Experimento 3 – Efeito da auxina ácido indolbutírico (AIB) e carvão ativado no
enraizamento in vitro de brotos de B. cheilantha.
Os brotos foram transferidos para meio de cultura WPM suplementado com diferentes
concentrações de ácido indolbutírico (AIB) (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e acrescido de
carvão ativado (0,0; 1,0 e 2,0 g L-1
), sendo que cada tubo continha 14 mL de meio de cultura
(Tabela 5).
O delineamento experimental foi o inteiramente casualizado em esquema fatorial 4 x 3
(quatro concentrações de auxinas x três concentrações de carvão ativado) totalizando 12
tratamentos. Cada tratamento foi constituído de quinze repetições, sendo cada repetição
composta por sete tubos contendo um broto cada.
Tabela 5 – Tratamentos utilizados no enraizamento in vitro de B. cheilantha, inoculados em meio WPM,
acrescido de diferentes concentrações de ácido indolbutírico (AIB) e acrescido de carvão ativado.
Feira de Santana, BA, 2010.
Tratamentos
Concentração de AIB
(mg L-1
)
Carvão Ativado
(g L-1
)
T 1 0,0 0,0
T 2 0,0 1,0
T 3 0,0 2,0
T 4 0,5 0,0
T 5 0,5 1,0
T 6 0,5 2,0
T 7 1,0 0,0
T 8 1,0 1,0
T 9 1,0 2,0
T 10 2,0 0,0
T 11 2,0 1,0
T 12 2,0 2,0
3.4.2 Experimento 4 - Efeito das auxinas ácido indolbutírico (AIB) ou ácido naftaleno
acético (ANA) ou ácido indolacético (AIA) e carvão ativado no enraizamento in vitro de
brotos de B. cheilantha.

Os brotos foram transferidos para meio de cultura WPM suplementado com diferentes
tipos de auxinas: ácido indolbutírico (AIB), ácido naftaleno acético (ANA) e ácido
indolacético (AIA) em diferentes concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) e acrescido de
carvão ativado (0,0; 0,5 e 1,0 g L-1
), sendo que cada tubo continha 14 mL de meio de cultura
(Tabela 6).
O delineamento experimental foi o inteiramente casualizado em esquema fatorial 3 x 4 x
3 (três tipos de auxinas x quatro concentrações de auxinas x três concentrações de carvão
ativado) totalizando 36 tratamentos. Cada tratamento foi constituído de seis repetições, sendo
cada repetição composta por cinco tubos contendo um broto cada.
Aos 45 dias de inoculação, para os dois experimentos de enraizamento in vitro
realizados, os brotos foram avaliados, sendo observadas as seguintes variáveis: a porcentagem
de enraizamento dos brotos, o número de raízes adventícias por broto, o número de raízes
secundárias da maior raiz, o comprimento da maior raiz e a taxa de sobrevivência dos brotos.
3.5 ACLIMATIZAÇÃO
3.5.1 Experimento 5 – Efeito da aeração in vitro sob a aclimatização
No período de enraizamento in vitro, as plantas foram, concomitantemente, mantidas
em diferentes condições de tamponamento (filme de PVC; tampa plástica sem filme de PVC e
tampão de algodão) durante 45 dias e em seguidas transferidas para sacos de polietileno
contendo terra vegetal e mantidas em casa de vegetação. As plantas foram cobertas com uma
garrafa tipo pet com tampa para a manutenção da umidade relativa no microambiente e
mantida em casa de vegetação a 70% de luminosidade durante a aclimatização. Durante esse
período, a tampa da garrafa pet foi desenroscada no 7º dia para a redução da umidade relativa
e retirada no 16º dia, sendo a garrafa retirada no 30º dia (Figura 7).
Aos 30 dias de experimento foi avaliado o percentual de sobrevivência das mudas nos
três tratamentos, sendo cada tratamento constituído de dez repetições e cada repetição
composta por seis sacos contendo uma muda cada.
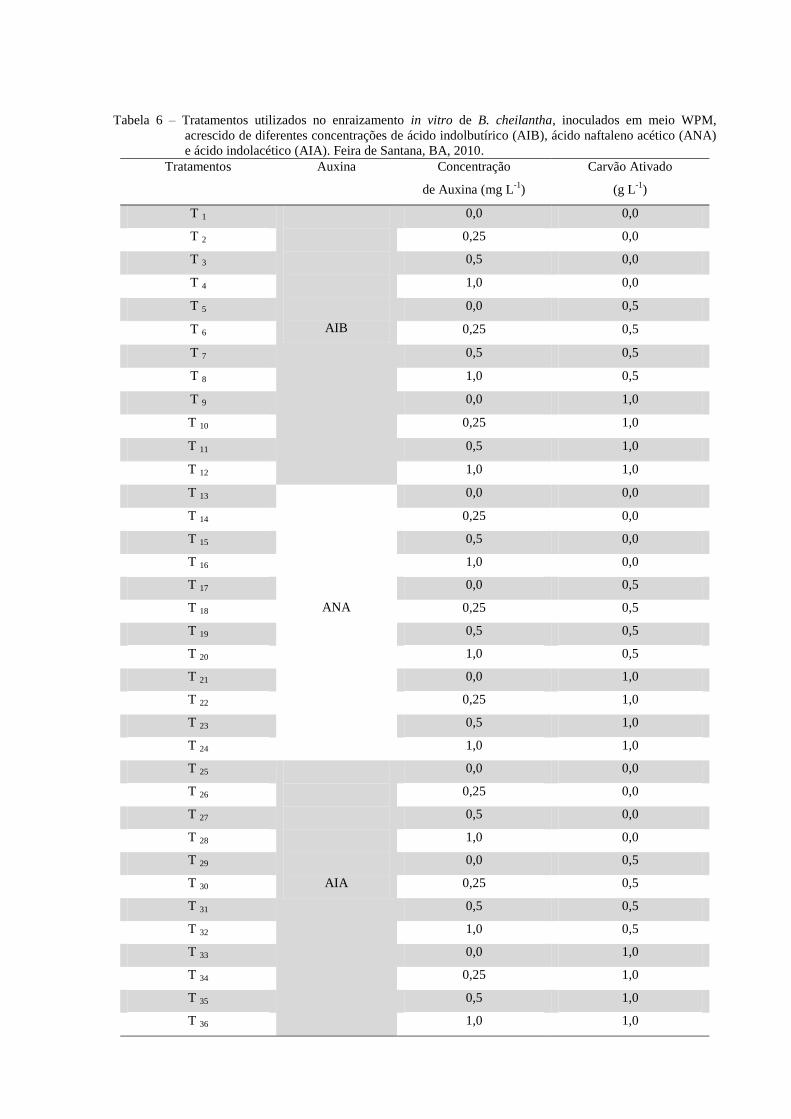
Tabela 6 – Tratamentos utilizados no enraizamento in vitro de B. cheilantha, inoculados em meio WPM,
acrescido de diferentes concentrações de ácido indolbutírico (AIB), ácido naftaleno acético (ANA)
e ácido indolacético (AIA). Feira de Santana, BA, 2010.
Tratamentos
Auxina Concentração
de Auxina (mg L-1
)
Carvão Ativado
(g L-1
)
T 1
AIB
0,0 0,0
T 2 0,25 0,0
T 3 0,5 0,0
T 4 1,0 0,0
T 5 0,0 0,5
T 6 0,25 0,5
T 7 0,5 0,5
T 8 1,0 0,5
T 9 0,0 1,0
T 10 0,25 1,0
T 11 0,5 1,0
T 12 1,0 1,0
T 13
ANA
0,0 0,0
T 14 0,25 0,0
T 15 0,5 0,0
T 16 1,0 0,0
T 17 0,0 0,5
T 18 0,25 0,5
T 19 0,5 0,5
T 20 1,0 0,5
T 21 0,0 1,0
T 22 0,25 1,0
T 23 0,5 1,0
T 24 1,0 1,0
T 25
AIA
0,0 0,0
T 26 0,25 0,0
T 27 0,5 0,0
T 28 1,0 0,0
T 29 0,0 0,5
T 30 0,25 0,5
T 31 0,5 0,5
T 32 1,0 0,5
T 33 0,0 1,0
T 34 0,25 1,0
T 35 0,5 1,0
T 36 1,0 1,0

A B C D
Figura 7 - Esquema da aclimatização das plantas enraizadas in vitro. (A) 1º dia na casa de vegetação – garrafa pet
tampada; (B) 7º dia – garrafa pet desenroscada; (C) 16º dia – tampa retirada; (D) 30º dia – garrafa pet
retirada.
3.6 ANÁLISE ESTATÍSTICA
Os dados obtidos das variáveis analisadas, nos diferentes experimentos, foram
submetidos à análise de variância (ANAVA) testando-se as médias pelo teste de Tukey em
nível de 5% de significância para os fatores qualitativos e ajuste de equações de regressão
polinomial para os fatores quantitativos, utilizando o software SISVAR (FERREIRA, 2008).
4 RESULTADOS E DISCUSSÃO
A fase de estabelecimento in vitro da espécie não foi influenciada pela
contaminação fúngica e bacteriana do material vegetal utilizado, sendo essa taxa irrelevante
(< 2,0%) (Figura 8). A superfície lisa do tegumento da semente pode ter favorecido a baixa
taxa de contaminação. Diferentemente de outros trabalhos com lenhosas, em que o índice de
contaminação bacteriana e fúngica foi alto, como o caso da oliveira (Olea europaea) com
93% (DONINI et al., 2008), do cultivo de Quercus rubra 46% (VIEITEZ et al., 2009), ou das
sementes de mogno (Swietenia macrophylla) com 31% de contaminação (COUTO et al.,
2004).

Figura 8 – Crescimento da plântula de B. cheilantha germinada em meio de cultura WPM durante 20 dias.
4.1 MULTIPLICAÇÃO IN VITRO
4.1.1 Experimento 1 - Efeito da citocinina 6-benzilaminopurina (BAP) e da auxina ácido
naftaleno acético (ANA) na indução de brotações em diferentes tipos de explantes de B.
cheilantha.
Observou-se efeito altamente significativo (p < 0,01) e significativo (p < 0,05) do fator
explante e do fator ANA para a variável explante responsivo, respectivamente. Para a variável
número de brotos por explante foi observado efeito altamente significativo (p < 0,01) da
interação tripla (tipo de explante x concentração de BAP x concentração de ANA), enquanto
que para a variável número de gemas do maior broto, todas as interações duplas (tipo de
explante x concentração de BAP; tipo de explante x concentração de ANA; concentração de
BAP x concentração de ANA) foram altamente significativas (p < 0,01). Foi observado efeito
significativo (p < 0,05) da interação dupla (tipo de explante x concentração de BAP) para a
variável número de folhas do maior broto e efeito altamente significativo (p < 0,01) de todas
as interações duplas para a variável comprimento da parte aérea. Para a variável matéria seca
da parte aérea as interações duplas (concentração de BAP x concentração de ANA; tipo de
explante x concentração de BAP) apresentaram efeito significativo (p < 0,05) (Tabela 7).
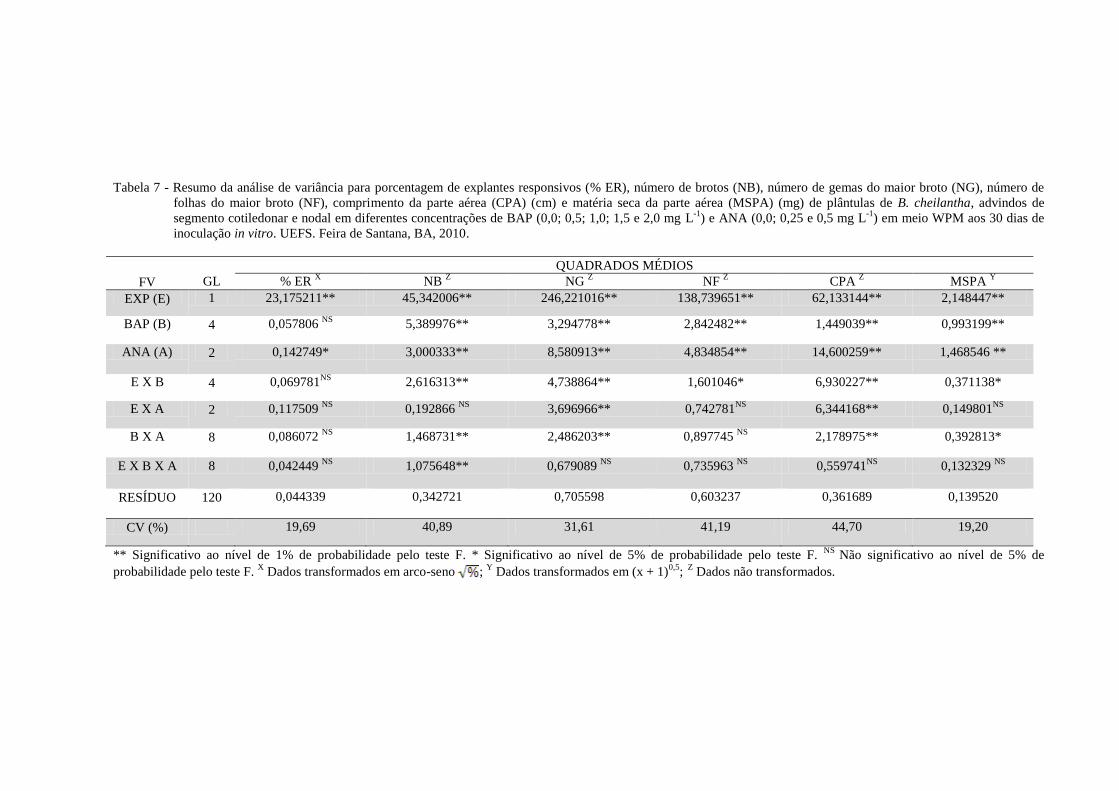
Tabela 7 - Resumo da análise de variância para porcentagem de explantes responsivos (% ER), número de brotos (NB), número de gemas do maior broto (NG), número de
folhas do maior broto (NF), comprimento da parte aérea (CPA) (cm) e matéria seca da parte aérea (MSPA) (mg) de plântulas de B. cheilantha, advindos de
segmento cotiledonar e nodal em diferentes concentrações de BAP (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e ANA (0,0; 0,25 e 0,5 mg L-1
) em meio WPM aos 30 dias de
inoculação in vitro. UEFS. Feira de Santana, BA, 2010.
QUADRADOS MÉDIOS
FV GL % ER X NB
Z NG
Z NF
Z CPA
Z MSPA
Y
EXP (E) 1 23,175211** 45,342006** 246,221016** 138,739651** 62,133144** 2,148447**
BAP (B) 4 0,057806 NS
5,389976** 3,294778** 2,842482** 1,449039** 0,993199**
ANA (A) 2 0,142749* 3,000333** 8,580913** 4,834854** 14,600259** 1,468546 **
E X B 4 0,069781NS
2,616313** 4,738864** 1,601046* 6,930227** 0,371138*
E X A 2 0,117509 NS
0,192866 NS
3,696966** 0,742781NS
6,344168** 0,149801NS
B X A 8 0,086072 NS
1,468731** 2,486203** 0,897745 NS
2,178975** 0,392813*
E X B X A 8 0,042449 NS
1,075648** 0,679089 NS
0,735963 NS
0,559741NS
0,132329 NS
RESÍDUO 120 0,044339 0,342721 0,705598 0,603237 0,361689 0,139520
CV (%) 19,69 40,89 31,61 41,19 44,70 19,20
** Significativo ao nível de 1% de probabilidade pelo teste F. * Significativo ao nível de 5% de probabilidade pelo teste F. NS
Não significativo ao nível de 5% de
probabilidade pelo teste F. X Dados transformados em arco-seno ;
Y Dados transformados em (x + 1)
0,5; Z
Dados não transformados.

Os explantes segmento cotiledonar e segmento nodal foram responsivos à formação de
brotos em todas as concentrações de BAP e ANA testadas (dados não mostrados) (Figura 9A).
O segmento nodal apresentou maior capacidade morfogenética (94,6%) que o
segmento cotiledonar (39,87%) (Figura 9B). Esse resultado difere do encontrado para angico-
vermelho (Parapiptadenia rigida), em que a porcentagem de explante responsivo, quando
utilizado segmento cotiledonar foi superior (98,8%) às taxas obtidas com o explante segmento
nodal (80%) (KIELSE et al., 2009). Os valores observados para B. cheilantha são superiores
aos reportados por Borchetia et al. (2009) em Camellia sinensis, em que a porcentagem
máxima de regeneração de brotos a partir de segmento nodal foi igual a 32,8% em meio de
cultura WPM suplementado com 3,0 mg L-1
de BAP, ou ainda a taxa de 84% de segmentos
nodais responsivos de Searsia dentata independente da dose de BAP acrescida no meio de
cultura MS (PRAKASH; STADEN, 2008).
Figura 9 – Porcentagem de explantes responsivos de B. cheilantha, aos 30 dias de inoculados em meio de
cultura WPM (A- Fator ANA; B- Fator Explante) (SC- segmento cotiledonar e SN- segmento
nodal) (Médias seguidas pela mesma letra, não diferem estatisticamente entre si ao nível de 5%, em
A, e 1%, em B, de probabilidade pelo teste de Tukey). Feira de Santana, BA, 2010.
A

Em B. cheilantha não foi possível observar, diante dos tratamentos testados, o
desencadeamento do processo morfogenético formando brotos nos explantes hipocótilo,
cotilédone, epicótilo e segmento internodal. Contudo, a formação de calo nos explantes foi
verificada (Figuras 10), mostrando ser diretamente proporcional à adição de BAP no meio de
cultura (dados não mostrados).
Figura 10 – Aspecto dos calos nos explantes de hipocótilo (A), cotilédone (B), epicótilo (C) e segmento
internodal (D) aos 30 dias de inoculados em meio de cultura WPM acrescido de 2,0 mg L-1
de
BAP e 0,5 mg L-1
de ANA. Feira de Santana, BA, 2010.
Segundo Ezura et al. (2000), a regeneração in vitro de muitas leguminosas é
considerada difícil. Provavelmente as combinações e/ou tipos de reguladores vegetais
utilizados no cultivo in vitro de B. cheilantha não foram suficientes para sinalizar essa
resposta específica de desenvolvimento ao nível celular. Na maioria das vezes, inúmeros
estudos laboriosos são necessários para que as células dos tecidos diferenciados adquiram
competência, como é o caso de Prunus salicina cv. Bruce, no qual uma taxa de regeneração
de brotos de 60% foi obtida para os cotilédones em meio de cultura QL (QUOIRIN;
LEPOIVRE, 1977) suplementado com 2,6 mg L-1
de TDZ e 0,5 mg L-1
de AIB, com
incubação de 10 dias no escuro com posterior transferência para fotoperíodo (CANLI; TIAN,
2009). Já em culturas de Psoralea coryfolia, os segmentos de hipocótilo mostraram-se com
alta capacidade organogênica (48,5 brotos/explante) quando inoculados em meio de cultura
combinando-se 0,66 mg L-1
de TDZ, 0,11 mg L-1
de BAP, 0,11 mg L-1
de KIN e 0,37 mg L-1
de ANA (BASKARAN; JAYABALAN 2009).
Em contra partida, há espécies lenhosas em que a alta freqüência de regeneração de
brotos a partir de tecidos não meristemáticos é facilmente obtida em meio suplementado com
uma única citocinina, a exemplo de Citrus spp. (MOURA et al., 2001; MOREIRA-DIAS et al.
2001; COSTA et al., 2004; ZENG et al., 2009). Essa maior capacidade morfogenética de
alguns tecidos pode ser causada pela variabilidade genética de cada cultura (GEORGE;
DEBERGH, 2008). Em Citrus reticulata é reportado uma taxa de regeneração de brotos de
72% a partir de epicótilo inoculado meio MT (MURASHIGE; TUCKER 1969) suplementado

com 2,0 mg L-1
de BAP (ZENG et al., 2009) e 93% a partir de epicótilo de citrange „Carizo‟
(Poncirus trifoliata x C. sinensis) inoculados no mesmo meio de cultura com 1,0 mg L-1
BAP
(SCHINOR et al., 2006). Para C. volkameriana, uma taxa de 77% de regeneração de brotos
foi registrada a partir de hipocótilo inoculado em meio de cultura EME (GROSSER;
GMITTER, 1990) suplementado com 1,0 mg L-1
de BAP, incubados por 30 dias no escuro e
em seguida transferidos por 15 dias em fotoperíodo (TAVANO et al., 2009).
A análise de regressão indicou um modelo quadrático (p < 0,01) das concentrações de
BAP para a variável número de brotos oriundos do explante segmento nodal, quando o meio
foi suplementado com 0,5 mg L-1
de ANA, ao passo que com 0,25 mg L-1
de ANA ou na sua
ausência, a tendência das brotações para segmento nodal apresentou um comportamento
linear (p < 0,01) (Figura 11 A).
A curva de resposta em 0,5 mg L-1
de ANA indica que a utilização de BAP na
concentração estimada de 1,23 mg L-1
atinge o maior valor estimado (3,11 brotos/explante)
para número de brotos. Incrementos de BAP superior a concentração estimada (1,23 mg L-1
)
tendem a desfavorecer a formação das brotações. Provavelmente, a presença de ANA na
maior dosagem testada sugere ter provocado uma diminuição nos níveis de substâncias
endógenas naturais que promoveriam a divisão celular. Em contrapartida, em uma
concentração menor de ANA (0,25 mg L-1
) ou na sua ausência, a indução dos brotos foi
crescente em função do aumento na concentração de BAP (Figura 11A).
O maior número de brotos observados (2,84) para o segmento nodal foi registrado
quando inoculado em meio de cultura WPM suplementado com 2,0 mg L-1
de BAP, tanto na
presença de 0,25 mg L-1
de ANA ou na sua ausência (Figura 11A). Como houve incremento
linear para esta variável, concentrações acima de 2,0 mg L-1
de BAP ainda podem ser testadas,
o que pode ser corroborado por Tamta et al. (2008) em cultivo in vitro de Quercus
semecarpifolia, e em S. dentata (PRAKASH; STADEN, 2008), que obtiveram uma média de
5,3 brotações a partir de segmento nodal em meio de cultura acrescido de 4,50 e 2,25 mg L-1
de BAP, respectivamente. Os resultados obtidos em B. cheilantha são semelhantes aos obtidos
por Mathur e Mukunthakumar (1992), quando obtiveram a partir de segmento nodal de B.
variegata uma média de 3,2 brotos em meio de cultura MS suplementado com 3,2 mg L-1
de
BAP. Já para B. vahlii, apenas um broto por segmento nodal foi observado na presença de
BAP (0,56 mg L-1
) (DHAR; UPRETI, 1999).
Os resultados obtidos para B. cheilantha são superiores aos reportados para outras
espécies lenhosas. Em goiaba-preta (Alibertia edulis), uma média máxima de 2,0 brotos foram
obtidos do segmento nodal inoculado em meio de cultura MS ¼ suplementado com 0,5 mg L-1
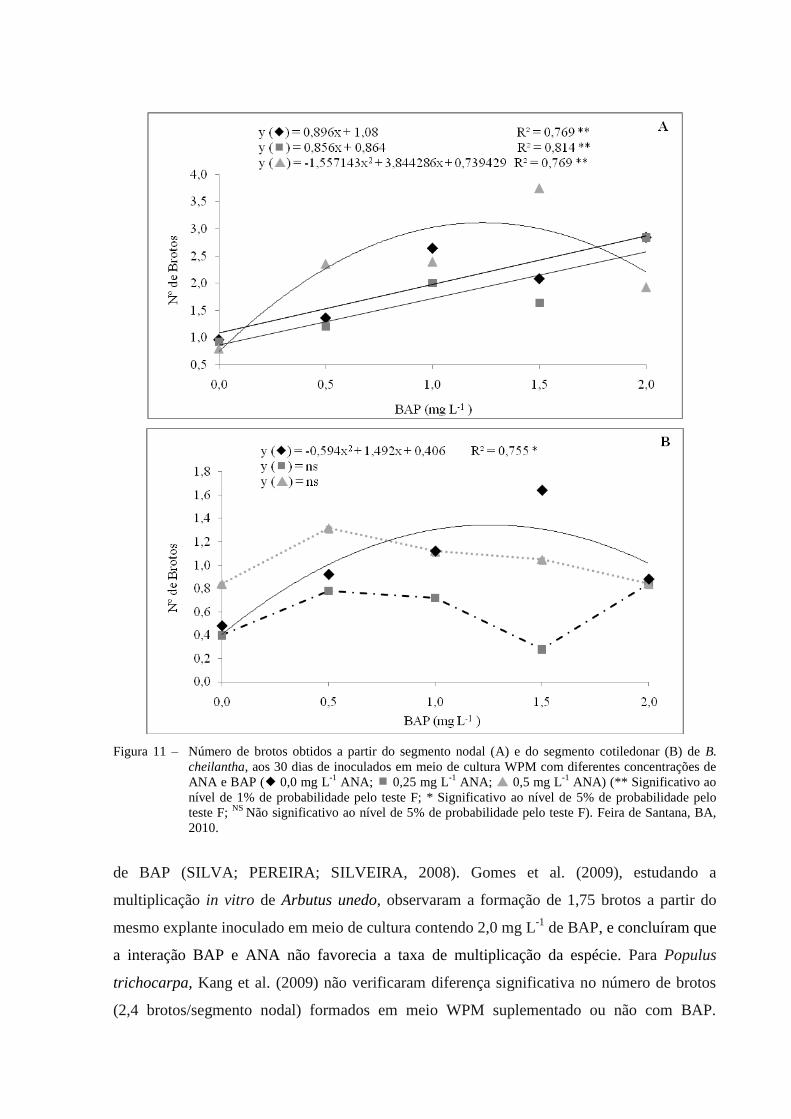
Figura 11 – Número de brotos obtidos a partir do segmento nodal (A) e do segmento cotiledonar (B) de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM com diferentes concentrações de
ANA e BAP ( 0,0 mg L-1
ANA; 0,25 mg L-1
ANA; 0,5 mg L-1
ANA) (** Significativo ao
nível de 1% de probabilidade pelo teste F; * Significativo ao nível de 5% de probabilidade pelo
teste F; NS
Não significativo ao nível de 5% de probabilidade pelo teste F). Feira de Santana, BA,
2010.
de BAP (SILVA; PEREIRA; SILVEIRA, 2008). Gomes et al. (2009), estudando a
multiplicação in vitro de Arbutus unedo, observaram a formação de 1,75 brotos a partir do
mesmo explante inoculado em meio de cultura contendo 2,0 mg L-1
de BAP, e concluíram que
a interação BAP e ANA não favorecia a taxa de multiplicação da espécie. Para Populus
trichocarpa, Kang et al. (2009) não verificaram diferença significativa no número de brotos
(2,4 brotos/segmento nodal) formados em meio WPM suplementado ou não com BAP.

Bopana e Saxena (2008), reportaram uma média máxima de 1,45 brotos em segmento nodal
de Asparagus racemosus, utilizando meio MS suplementado com 0,5 mg L-1
de BAP. Já
Iapichino e Airò (2008), concluíram que o segmento nodal de Metrosideros excelsa
inoculados em meio de cultura MS suplementado com 2ip (0,40 mg L-1
) e AIA (0,20 mg L-1
)
registraram uma maior taxa de multiplicação de brotos (2,2 brotos/segmento nodal) que os
observados com BAP (0,8 mg L-1
) e AIA (0,20 mg L-1
).
Todavia, os resultados encontrados para B. cheilantha são inferiores aqueles
reportados por Balaraju et al. (2008), que obtiveram média de 4,7 brotações por segmento
nodal de Vitex agnus-castus em meio de cultura MS enriquecido com 1,5 mg L-1
de BAP e 0,1
mg L-1
de ANA. Segundo esses mesmos autores, uma média superior para número de brotos
de V. agnus-castus foi obtida combinando-se BAP com KIN (5,7 brotos/segmento nodal) ao
invés da interação BAP e ANA. Em Camellia assamica, quando o segmento nodal foi
inoculado em meio de cultura MS ½, suplementado com BAP (6,0 mg L-1
), GA3 (1,0 mg L-1
)
e AIB (0,5 mg L-1
), a formação de brotos foi 8,5 vezes superior àquela encontrada em meio
WPM suplementado apenas com BAP (2,2 brotos/explante) (BORCHETIA et al., 2009).
Diante disso, percebe-se que é difícil predizer como a interação de dois ou mais reguladores
vegetais irão afetar o sistema fisiológico da planta. Segundo Staden, Zazimalova e George
(2008), a interação pode atuar sinérgica ou antagonicamente a depender de inúmeras
circunstâncias, uma vez que a presença de um determinado regulador pode afetar a biosíntese
ou metabolismo de um outro regulador, alterando os níveis das substâncias endógenas, ou
ainda os fatores ambientais inerentes ao processo podem modificar as respostas dos
reguladores ou até mesmo inativá-los.
Para o explante segmento cotiledonar, registrou-se comportamento quadrático (p <
0,05) das concentrações de BAP para a variável número de brotos na ausência de ANA. Essa
curva de resposta atingiu um valor máximo estimado de brotos (1,34) quando o meio foi
acrescido 1,26 mg L-1
de BAP. A partir dessa concentração houve uma tendência à diminuição
na formação de brotos para o explante segmento cotiledonar. Nos meios de cultura
suplementados com ANA, nas duas concentrações testadas, não foi possível o ajuste de uma
equação significativa com significado biológico, porém, é observado em 0,5 mg L-1
de BAP
uma tendência maior na formação das brotações (1,32 brotos/explante) quando o meio foi
suprido com igual concentração de ANA (Figura 11B).
Para as culturas de Dalbergia sissoo e Prosopis laevigata, ambas leguminosas, o
explante segmento cotiledonar da primeira obteve uma média de 2,8 brotos/explante em meio
de cultura MS acrescido de 1,0 mg L-1
de BAP (PRADHAN et al., 1998), enquanto que o
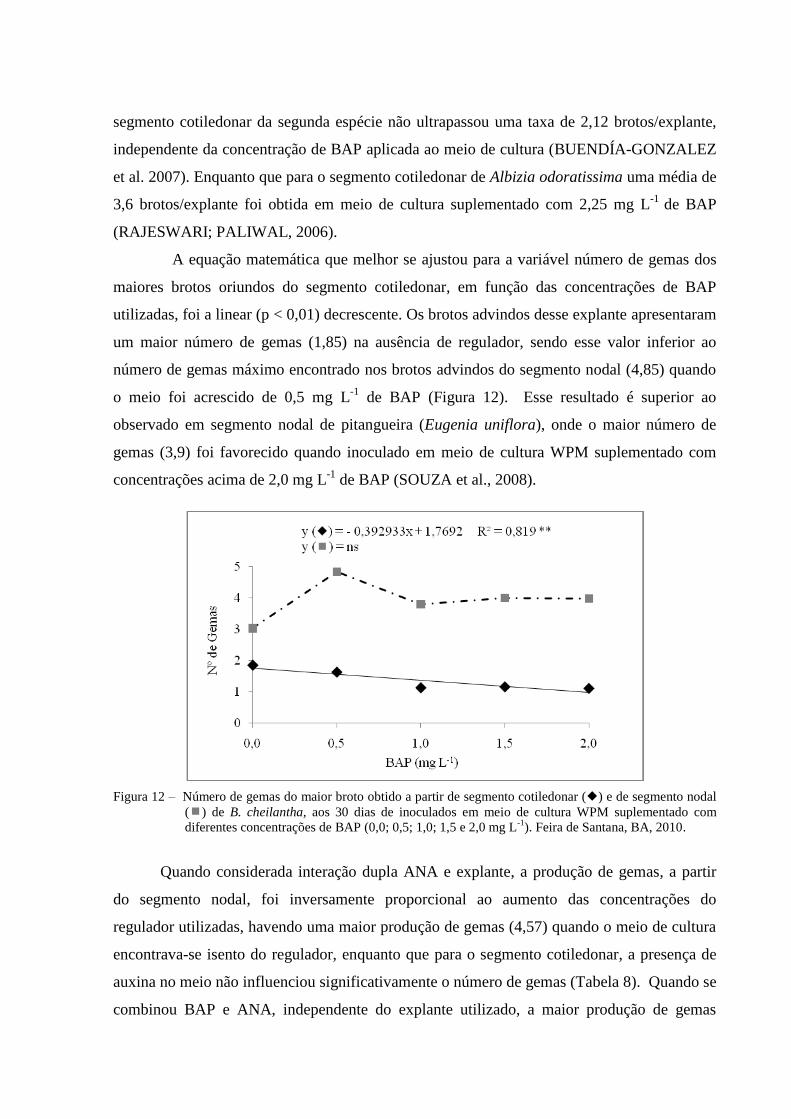
segmento cotiledonar da segunda espécie não ultrapassou uma taxa de 2,12 brotos/explante,
independente da concentração de BAP aplicada ao meio de cultura (BUENDÍA-GONZALEZ
et al. 2007). Enquanto que para o segmento cotiledonar de Albizia odoratissima uma média de
3,6 brotos/explante foi obtida em meio de cultura suplementado com 2,25 mg L-1
de BAP
(RAJESWARI; PALIWAL, 2006).
A equação matemática que melhor se ajustou para a variável número de gemas dos
maiores brotos oriundos do segmento cotiledonar, em função das concentrações de BAP
utilizadas, foi a linear (p < 0,01) decrescente. Os brotos advindos desse explante apresentaram
um maior número de gemas (1,85) na ausência de regulador, sendo esse valor inferior ao
número de gemas máximo encontrado nos brotos advindos do segmento nodal (4,85) quando
o meio foi acrescido de 0,5 mg L-1
de BAP (Figura 12). Esse resultado é superior ao
observado em segmento nodal de pitangueira (Eugenia uniflora), onde o maior número de
gemas (3,9) foi favorecido quando inoculado em meio de cultura WPM suplementado com
concentrações acima de 2,0 mg L-1
de BAP (SOUZA et al., 2008).
Figura 12 – Número de gemas do maior broto obtido a partir de segmento cotiledonar () e de segmento nodal
() de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM suplementado com
diferentes concentrações de BAP (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
). Feira de Santana, BA, 2010.
Quando considerada interação dupla ANA e explante, a produção de gemas, a partir
do segmento nodal, foi inversamente proporcional ao aumento das concentrações do
regulador utilizadas, havendo uma maior produção de gemas (4,57) quando o meio de cultura
encontrava-se isento do regulador, enquanto que para o segmento cotiledonar, a presença de
auxina no meio não influenciou significativamente o número de gemas (Tabela 8). Quando se
combinou BAP e ANA, independente do explante utilizado, a maior produção de gemas
(3,73) foi encontrada na presença de 0,5 mg L-1
de BAP e igual concentração de ANA (Tabela
9).
Tabela 8 – Número de gemas obtidas do maior broto na fase de multiplicação de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM em função de ANA e explante. Feira de Santana, BA, 2009.
ANA (mg L-1
) EXPLANTE
SC SN
0,0 1,64 A b 4,57 A a
0,25 1,17 A b 3,99 B a
0,5 1,32 A b 3,26 C a
Médias seguidas pela mesma letra maiúscula, na mesma coluna, ou minúscula, na mesma linha, não diferem
estatisticamente entre si ao nível de 1% de probabilidade pelo teste de Tukey.
Tabela 9 – Número de gemas a obtidas do maior broto na fase de multiplicação de B. cheilantha, aos 30 dias de
inoculados em meio de cultura WPM em função de BAP e ANA. Feira de Santana, BA, 2010.
BAP (mg L-1
) ANA (mg L-1
)
0,0 0,25 0,5
0,0 2,78 a 2,06 a 2,5 a
0,5 3,52 a 3,73 a 2,48 b
1,0 2,69 ab 2,86 a 1,86 b
1,5 3,68 a 1,92 b 2,15 b
2,0 2,86 a 2,34 a 2,45 a
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
As concentrações de BAP mostraram tendência quadrática (p < 0,05) ascendente para
a variável comprimento da parte aérea na ausência de ANA e comportamento quadrático
descendente (p < 0,05) quando o meio foi suplementado com 0,5 mg L-1
de ANA. Em meio
com 0,5 mg L-1
de ANA, estima-se que o comprimento dos brotos tende a ser desfavorecido
com o uso crescente de concentrações até 1,18 mg L-1
de BAP, em que atingi-se o valor
mínimo estimado (0,6 cm) e em concentrações superiores a estimada, o comprimento das
brotações tende a ser favorecido. Em meio de cultura isento de ANA, o maior comprimento
dos brotos estimado (2,28 cm) tende a ocorrer em meio suplementado com 1,08 mg L-1
de
BAP. O aumento da concentração a partir do valor estimado (1,08 mg L-1
de BAP), tende a
desfavorecer o comprimento das brotações (Figura 13A). Os brotos obtidos a partir de
segmento nodal apresentaram os maiores comprimentos da parte aérea em função das
concentrações de BAP utilizadas, sendo observado um valor máximo (2,47 cm) em 0,5 mg L-1
de BAP, enquanto que para o segmento cotiledonar o maior valor (1,38 cm) é observado na

ausência de regulador (Figura 13B). As citocininas podem favorecer o alongamento de brotos,
contudo, é necessário que se adicione ao meio a concentração adequada, sendo esta específica
para cada espécie (GRATTAPAGLIA; MACHADO, 1998).
Esses resultados são semelhantes aqueles reportados por Tilkat et al. (2009) em que
brotações de Pistacia vera atingem a 2,11 cm de comprimento no meio de cultura MS com
1,0 mg L-1
de BAP, porém, em P. trichocarpa, as brotações com semelhante comprimento
(2,2 cm) só foram obtidas quando inoculadas em meio MS na ausência de reguladores
vegetais (KANG et al., 2009). As médias de comprimento das brotações para B. cheilantha
foram superiores quando comparadas aos 0,43 cm obtidos por Kielse et al. (2009) em angico
vermelho (P. rigida), ou 0,39 cm obtidos por Souza et al. (2008) em pitangueira (E. uniflora),
ou 1,0 cm obtidos por Raghu et al. (2006), em guduchi (Tinospora cordifolia),
independentemente da concentração de citocinina utilizada no meio de cultura. Bopana e
Saxena (2008), estudando a multiplicação in vitro de A. racemosus reportaram uma média de
1,8 cm de comprimento das brotações mantidas em meio MS acrescido de 1,0 mg L-1
de BAP
e Vieitez et al. (2009), em Quercus Alba, obtiveram o maior comprimento dos brotos (1,7 cm)
em meio de cultura GD (GRESSHOFF; DOY 1972) suplementado com 0,2 mg L-1
de BAP.
Observou-se diferença significativa nos comprimentos dos brotos advindos do
segmento nodal em todas as concentrações de ANA utilizadas em relação aos do segmento
cotiledonar. Em meio de cultura isento de ANA foi detectada a maior altura dos brotos (2,92
cm) para o segmento nodal e com o aumento das concentrações de ANA utilizadas o
comprimento das brotações foi desfavorecido (Tabela 10).
Tabela 10 – Comprimento da parte aérea na fase de multiplicação de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM em função de ANA e Explante (SC- segmento cotiledonar e SN- segmento
nodal). Feira de Santana, BA, 2010.
Médias seguidas pela mesma letra maiúscula, na mesma coluna, ou minúscula, na mesma linha, não diferem
estatisticamente entre si ao nível de 1% de probabilidade pelo teste de Tukey.
ANA (mg L-1
) Explante
SC SN
0,0 0,88 A b 2,92 A a
0,25 0,71 A b 1,91 B a
0,5 0,51 A b 1,14 C a
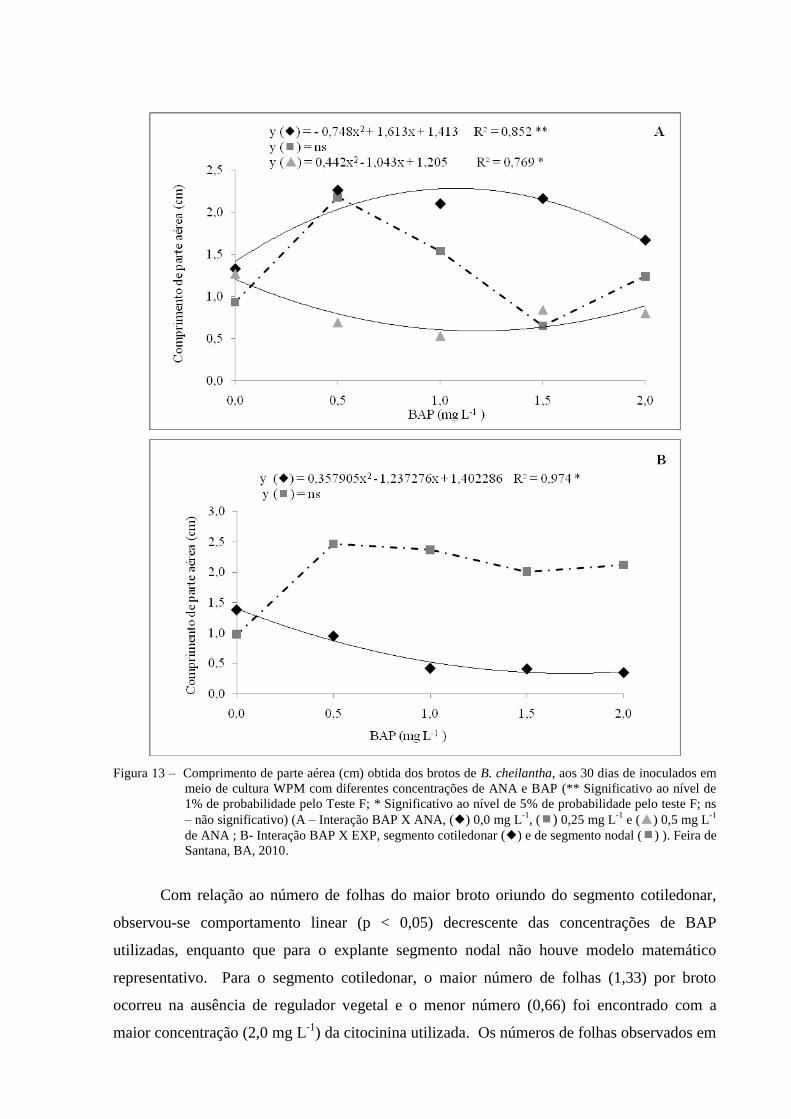
Figura 13 – Comprimento de parte aérea (cm) obtida dos brotos de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM com diferentes concentrações de ANA e BAP (** Significativo ao nível de
1% de probabilidade pelo Teste F; * Significativo ao nível de 5% de probabilidade pelo teste F; ns
– não significativo) (A – Interação BAP X ANA, () 0,0 mg L-1
, () 0,25 mg L-1
e () 0,5 mg L-1
de ANA ; B- Interação BAP X EXP, segmento cotiledonar () e de segmento nodal () ). Feira de
Santana, BA, 2010.
Com relação ao número de folhas do maior broto oriundo do segmento cotiledonar,
observou-se comportamento linear (p < 0,05) decrescente das concentrações de BAP
utilizadas, enquanto que para o explante segmento nodal não houve modelo matemático
representativo. Para o segmento cotiledonar, o maior número de folhas (1,33) por broto
ocorreu na ausência de regulador vegetal e o menor número (0,66) foi encontrado com a
maior concentração (2,0 mg L-1
) da citocinina utilizada. Os números de folhas observados em
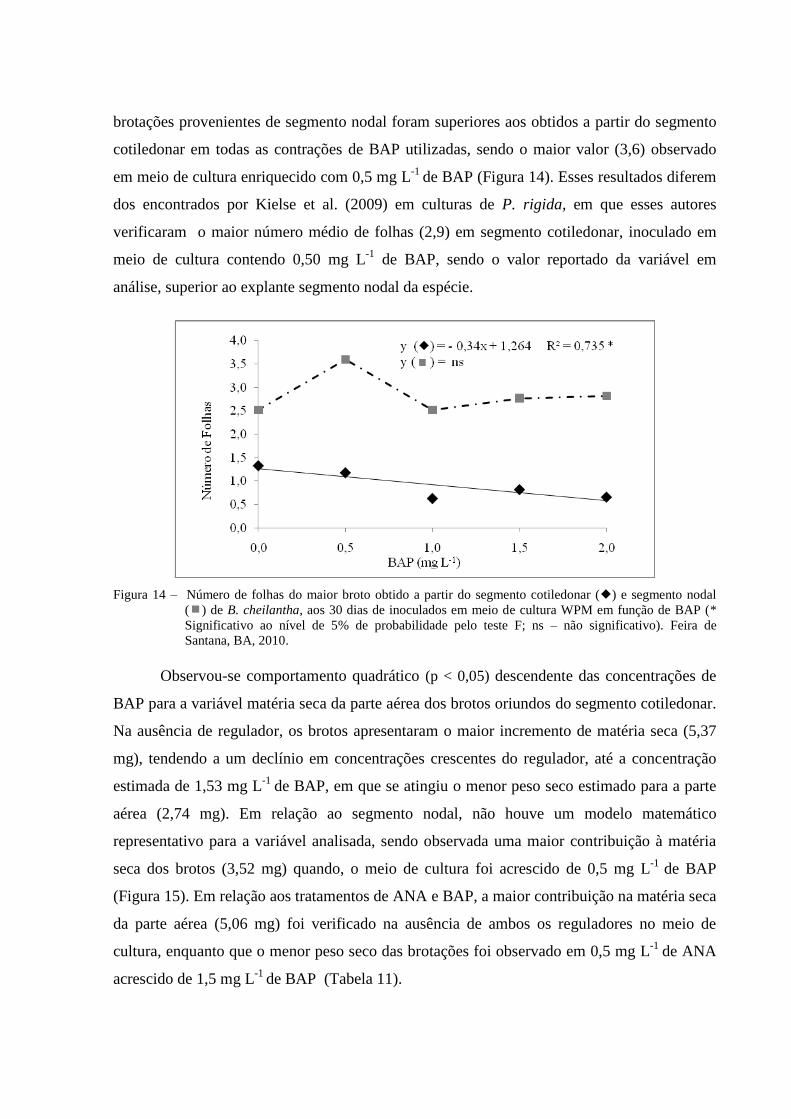
brotações provenientes de segmento nodal foram superiores aos obtidos a partir do segmento
cotiledonar em todas as contrações de BAP utilizadas, sendo o maior valor (3,6) observado
em meio de cultura enriquecido com 0,5 mg L-1
de BAP (Figura 14). Esses resultados diferem
dos encontrados por Kielse et al. (2009) em culturas de P. rigida, em que esses autores
verificaram o maior número médio de folhas (2,9) em segmento cotiledonar, inoculado em
meio de cultura contendo 0,50 mg L-1
de BAP, sendo o valor reportado da variável em
análise, superior ao explante segmento nodal da espécie.
Figura 14 – Número de folhas do maior broto obtido a partir do segmento cotiledonar () e segmento nodal
() de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM em função de BAP (*
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não significativo). Feira de
Santana, BA, 2010.
Observou-se comportamento quadrático (p < 0,05) descendente das concentrações de
BAP para a variável matéria seca da parte aérea dos brotos oriundos do segmento cotiledonar.
Na ausência de regulador, os brotos apresentaram o maior incremento de matéria seca (5,37
mg), tendendo a um declínio em concentrações crescentes do regulador, até a concentração
estimada de 1,53 mg L-1
de BAP, em que se atingiu o menor peso seco estimado para a parte
aérea (2,74 mg). Em relação ao segmento nodal, não houve um modelo matemático
representativo para a variável analisada, sendo observada uma maior contribuição à matéria
seca dos brotos (3,52 mg) quando, o meio de cultura foi acrescido de 0,5 mg L-1
de BAP
(Figura 15). Em relação aos tratamentos de ANA e BAP, a maior contribuição na matéria seca
da parte aérea (5,06 mg) foi verificado na ausência de ambos os reguladores no meio de
cultura, enquanto que o menor peso seco das brotações foi observado em 0,5 mg L-1
de ANA
acrescido de 1,5 mg L-1
de BAP (Tabela 11).
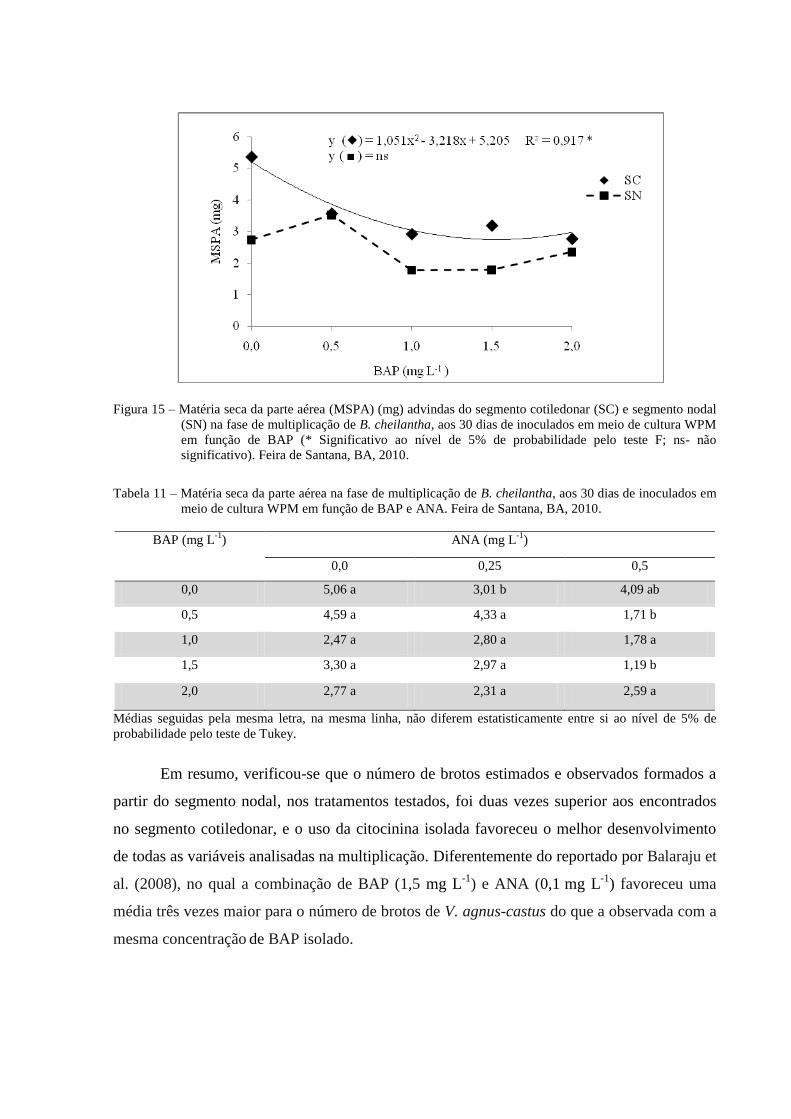
Figura 15 – Matéria seca da parte aérea (MSPA) (mg) advindas do segmento cotiledonar (SC) e segmento nodal
(SN) na fase de multiplicação de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM
em função de BAP (* Significativo ao nível de 5% de probabilidade pelo teste F; ns- não
significativo). Feira de Santana, BA, 2010.
Tabela 11 – Matéria seca da parte aérea na fase de multiplicação de B. cheilantha, aos 30 dias de inoculados em
meio de cultura WPM em função de BAP e ANA. Feira de Santana, BA, 2010.
BAP (mg L-1
) ANA (mg L-1
)
0,0 0,25 0,5
0,0 5,06 a 3,01 b 4,09 ab
0,5 4,59 a 4,33 a 1,71 b
1,0 2,47 a 2,80 a 1,78 a
1,5 3,30 a 2,97 a 1,19 b
2,0 2,77 a 2,31 a 2,59 a
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 5% de
probabilidade pelo teste de Tukey.
Em resumo, verificou-se que o número de brotos estimados e observados formados a
partir do segmento nodal, nos tratamentos testados, foi duas vezes superior aos encontrados
no segmento cotiledonar, e o uso da citocinina isolada favoreceu o melhor desenvolvimento
de todas as variáveis analisadas na multiplicação. Diferentemente do reportado por Balaraju et
al. (2008), no qual a combinação de BAP (1,5 mg L-1
) e ANA (0,1 mg L
-1) favoreceu uma
média três vezes maior para o número de brotos de V. agnus-castus do que a observada com a
mesma concentração de BAP isolado.

4.1.2 Experimento 2 - Efeito do tipo de citocininas na indução de brotações em diferentes
tipos de explantes de B. cheilantha.
Observou-se efeito altamente significativo (p < 0,01) da interação dupla (tipo de
explante x concentração de citocinina) para a variável porcentagem de explantes responsivos
e efeito altamente significativo (p < 0,01) da interação tripla (tipo de explante x tipo de
citocinina x concentração de citocinina) para a variável número de brotos por explante. Efeito
significativo (p < 0,05) da interação dupla (tipo de explante x tipo de citocinina) foi observado
para a variável número de gemas do maior broto e efeito altamente significativo (p < 0,01) da
interação dupla (tipo de explante x concentração de citocinina) para a mesma variável e para a
variável número de folhas do maior broto. Todas as interações duplas (tipo de explante x tipo
de citocinina; tipo de explante x concentração de citocinina; tipo de citocinina x concentração
de citocinina) foram altamente significativas (p < 0,01) para a variável comprimento da parte
aérea, enquanto que, apenas duas interações duplas (tipo de citocinina x concentração; tipo de
explante x concentração de citocinina) influenciaram de modo altamente significativo (p <
0,01) a variável matéria seca de parte aérea (Tabela 12).
Com relação à porcentagem de explantes responsivos, mais uma vez o segmento nodal
apresentou uma taxa superior ao segmento cotiledonar, independente do tipo e concentração
dos reguladores testados. A maior capacidade organogênica observada (97,33%) ocorre
quando o segmento nodal foi inoculado em meio de cultura acrescido da maior concentração
dos reguladores testados (2,0 mg L-1
) (Tabela 13). A taxa de explantes responsivos encontrado
para o segmento cotilenodar de B. cheilanta, é semelhante ao reportado por Nayak et al.
(2007), no qual uma média inferior a 56% foi observada para o segmento cotiledonar de
Aegles marmelos, independente da concentração de BAP ou KIN adicionada ao meio de
cultura MS, ou ainda as taxas inferiores a 30% encontradas para o segmento cotiledonar da
leguminosa Pterocarpus marsupium, independente da concentração de KIN utilizada no meio
de cultura (CHAND; SINGH, 2004). Contudo, Buendía-González et al. (2007) registraram
uma média acima de 90% de explantes responsivos para o segmento cotiledonar de P.
laevigata (Fabaceae), independente da concentração de BAP adicionada ao meio de cultura.
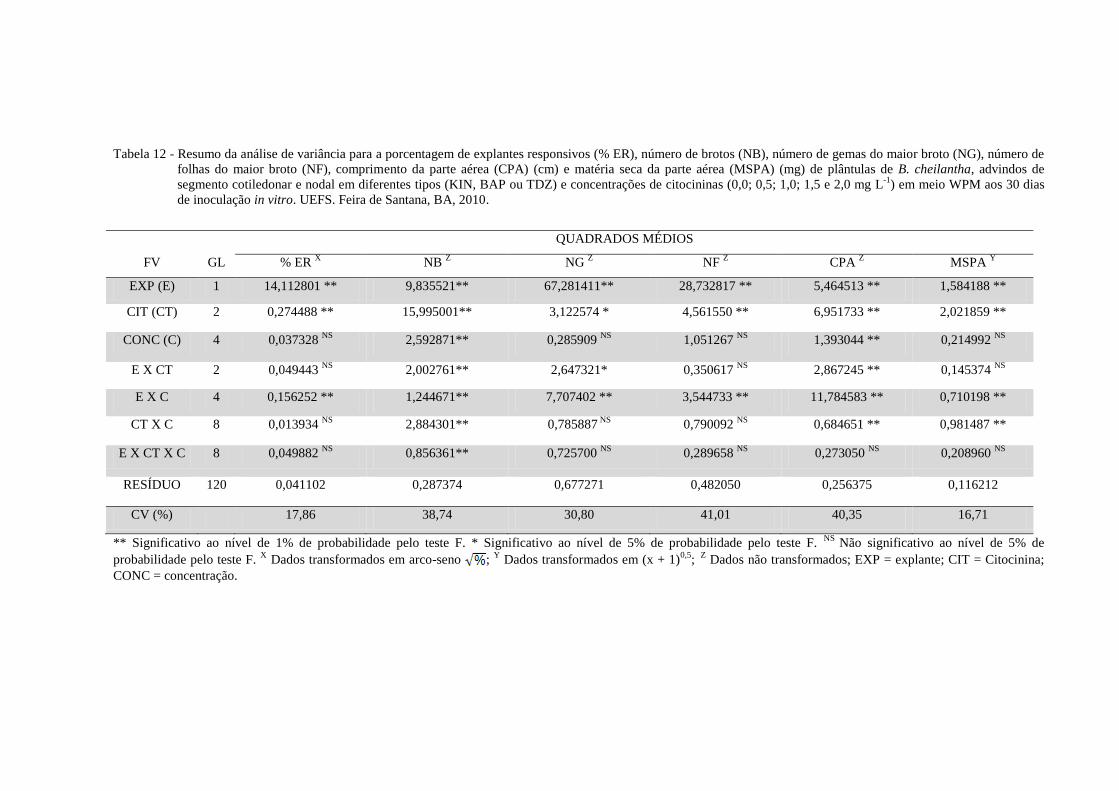
Tabela 12 - Resumo da análise de variância para a porcentagem de explantes responsivos (% ER), número de brotos (NB), número de gemas do maior broto (NG), número de
folhas do maior broto (NF), comprimento da parte aérea (CPA) (cm) e matéria seca da parte aérea (MSPA) (mg) de plântulas de B. cheilantha, advindos de
segmento cotiledonar e nodal em diferentes tipos (KIN, BAP ou TDZ) e concentrações de citocininas (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) em meio WPM aos 30 dias
de inoculação in vitro. UEFS. Feira de Santana, BA, 2010.
QUADRADOS MÉDIOS
FV GL % ER X NB
Z NG
Z NF
Z CPA
Z MSPA
Y
EXP (E) 1 14,112801 ** 9,835521** 67,281411** 28,732817 ** 5,464513 ** 1,584188 **
CIT (CT) 2 0,274488 ** 15,995001** 3,122574 * 4,561550 ** 6,951733 ** 2,021859 **
CONC (C) 4 0,037328 NS
2,592871** 0,285909 NS
1,051267 NS
1,393044 ** 0,214992 NS
E X CT 2 0,049443 NS
2,002761** 2,647321* 0,350617 NS
2,867245 ** 0,145374 NS
E X C 4 0,156252 ** 1,244671** 7,707402 ** 3,544733 ** 11,784583 ** 0,710198 **
CT X C 8 0,013934 NS
2,884301** 0,785887 NS
0,790092 NS
0,684651 ** 0,981487 **
E X CT X C 8 0,049882 NS
0,856361** 0,725700 NS
0,289658 NS
0,273050 NS
0,208960 NS
RESÍDUO 120 0,041102 0,287374 0,677271 0,482050 0,256375 0,116212
CV (%) 17,86 38,74 30,80 41,01 40,35 16,71
** Significativo ao nível de 1% de probabilidade pelo teste F. * Significativo ao nível de 5% de probabilidade pelo teste F. NS
Não significativo ao nível de 5% de
probabilidade pelo teste F. X Dados transformados em arco-seno ;
Y Dados transformados em (x + 1)
0,5; Z
Dados não transformados; EXP = explante; CIT = Citocinina;
CONC = concentração.
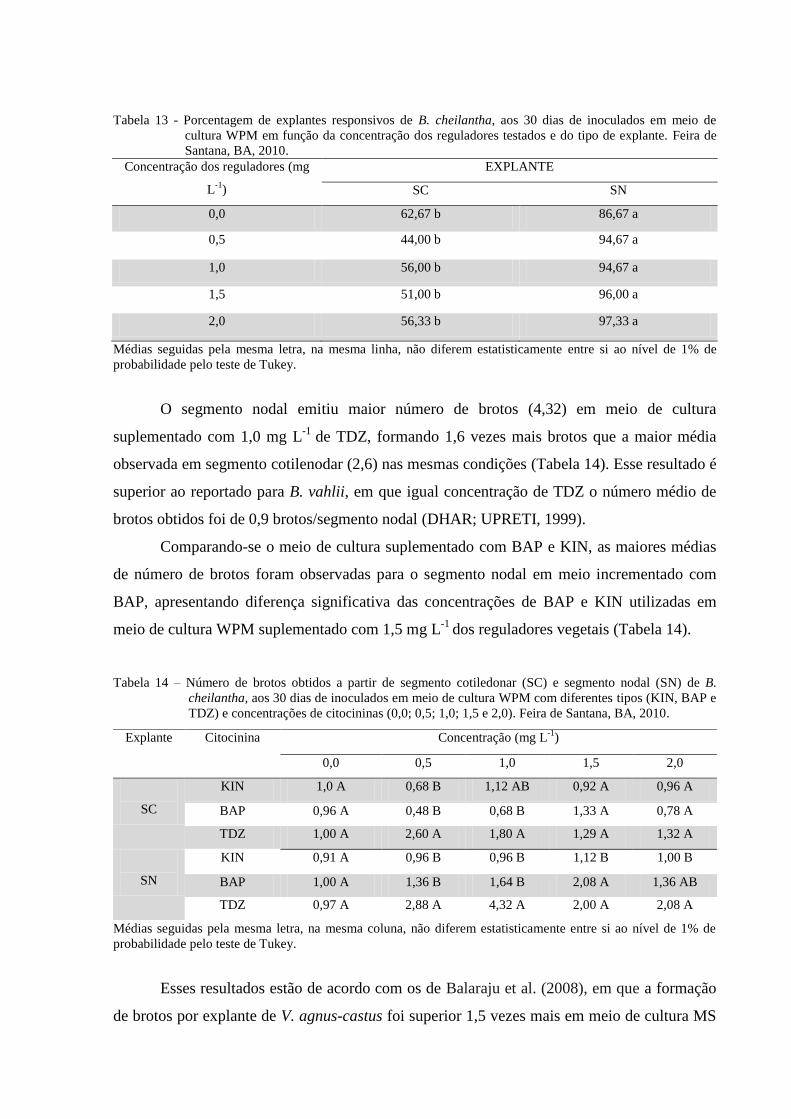
Tabela 13 - Porcentagem de explantes responsivos de B. cheilantha, aos 30 dias de inoculados em meio de
cultura WPM em função da concentração dos reguladores testados e do tipo de explante. Feira de
Santana, BA, 2010. Concentração dos reguladores (mg
L-1
)
EXPLANTE
SC SN
0,0 62,67 b 86,67 a
0,5 44,00 b 94,67 a
1,0 56,00 b 94,67 a
1,5 51,00 b 96,00 a
2,0 56,33 b 97,33 a
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
O segmento nodal emitiu maior número de brotos (4,32) em meio de cultura
suplementado com 1,0 mg L-1
de TDZ, formando 1,6 vezes mais brotos que a maior média
observada em segmento cotilenodar (2,6) nas mesmas condições (Tabela 14). Esse resultado é
superior ao reportado para B. vahlii, em que igual concentração de TDZ o número médio de
brotos obtidos foi de 0,9 brotos/segmento nodal (DHAR; UPRETI, 1999).
Comparando-se o meio de cultura suplementado com BAP e KIN, as maiores médias
de número de brotos foram observadas para o segmento nodal em meio incrementado com
BAP, apresentando diferença significativa das concentrações de BAP e KIN utilizadas em
meio de cultura WPM suplementado com 1,5 mg L-1
dos reguladores vegetais (Tabela 14).
Tabela 14 – Número de brotos obtidos a partir de segmento cotiledonar (SC) e segmento nodal (SN) de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM com diferentes tipos (KIN, BAP e
TDZ) e concentrações de citocininas (0,0; 0,5; 1,0; 1,5 e 2,0). Feira de Santana, BA, 2010.
Médias seguidas pela mesma letra, na mesma coluna, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
Esses resultados estão de acordo com os de Balaraju et al. (2008), em que a formação
de brotos por explante de V. agnus-castus foi superior 1,5 vezes mais em meio de cultura MS
Explante Citocinina Concentração (mg L-1
)
0,0 0,5 1,0 1,5 2,0
SC
KIN 1,0 A 0,68 B 1,12 AB 0,92 A 0,96 A
BAP 0,96 A 0,48 B 0,68 B 1,33 A 0,78 A
TDZ 1,00 A 2,60 A 1,80 A 1,29 A 1,32 A
SN
KIN 0,91 A 0,96 B 0,96 B 1,12 B 1,00 B
BAP 1,00 A 1,36 B 1,64 B 2,08 A 1,36 AB
TDZ 0,97 A 2,88 A 4,32 A 2,00 A 2,08 A

suplementado com BAP ao invés do incremento com KIN. Em M. excelsa (IAPICHINO;
AIRÒ, 2008) e em T. cordifolia (RAGHU et al., 2006), a melhor resposta da multiplicação in
vitro também foi relatada em meio de cultura suplementado com BAP em comparação a KIN.
Ao passo que em figueira “Roxo de Valinhos”, uma média superior a 2,0 brotos/explante foi
obtida em segmento nodal inoculado em meio de cultura WPM suplementado com 1,0 e 2,0
mg L-1
de KIN (FERREIRA; PASQUAL, 2008). Já em Stryphnodendron adstringens, à
medida que se aumentou a concentração de cinetina no meio de cultura, o número de brotos
formados a partir do segmento nodal foi favorecido, sendo registrada uma melhor resposta
(3,0 brotos/explante) quando se utilizou 5,0 mg L-1
de cinetina (NICIOLI et al., 2008). Para as
culturas de Rauvolfia tetraphylla, um elevado número de brotos (9,2) foi observado quando se
inoculou o segmento nodal em meio de cultura suplementado com 1,1 mg L-1
de TDZ
(FAISAL; AHMAD; ANIS, 2005).
Pode-se observar que na base dos explantes ocorreu formação de calo, que mostrou ser
diretamente proporcional ao aumento das concentrações dos reguladores vegetais no meio de
cultura, exceto para o TDZ, no qual independente da concentração utilizada, 100% dos
explantes apresentaram calos na sua base (dados não mostrados). O uso do TDZ revelou uma
ação tóxica nos brotos, apresentando-se hiperídricos e com brotações curtas, o mesmo sendo
detectado . Já os brotos cultivados em KIN possuíam uma coloração menos intensa de verde
que as brotações cultivadas em BAP, aparentando estarem menos vigorosos em meio de
cultura acrescido com KIN (Figura 16).
Figura 16 - Brotos de B. cheilantha em meio de multiplicação WPM suplementado com 2,0 mg L-1
de regulador
vegetal, aos 30 dias de inoculados. (A) Segmento cotilenodar em BAP; (B) Segmento nodal em KIN;
(C) Segmento nodal em BAP; (D) segmento cotiledonar em TDZ; (E) segmento nodal em TDZ.
(Barra = 1,0 cm). Feira de Santana, BA, 2010.
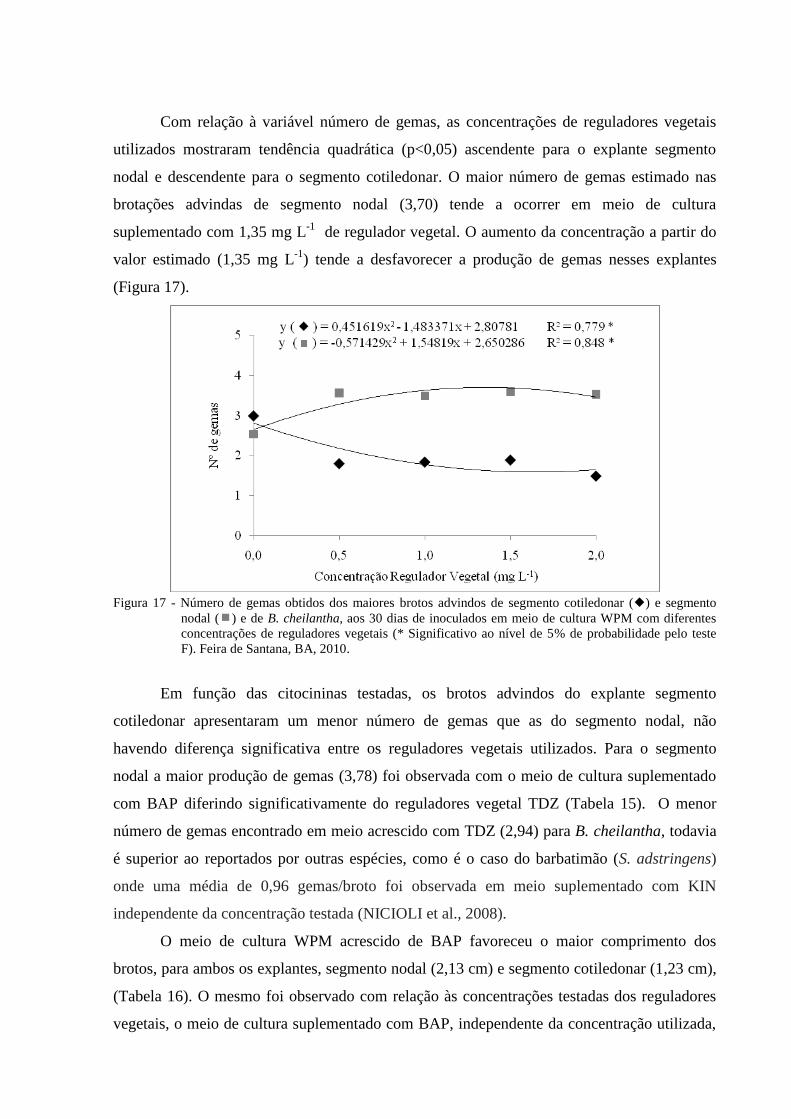
Com relação à variável número de gemas, as concentrações de reguladores vegetais
utilizados mostraram tendência quadrática (p<0,05) ascendente para o explante segmento
nodal e descendente para o segmento cotiledonar. O maior número de gemas estimado nas
brotações advindas de segmento nodal (3,70) tende a ocorrer em meio de cultura
suplementado com 1,35 mg L-1
de regulador vegetal. O aumento da concentração a partir do
valor estimado (1,35 mg L-1
) tende a desfavorecer a produção de gemas nesses explantes
(Figura 17).
Figura 17 - Número de gemas obtidos dos maiores brotos advindos de segmento cotiledonar () e segmento
nodal () e de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM com diferentes
concentrações de reguladores vegetais (* Significativo ao nível de 5% de probabilidade pelo teste
F). Feira de Santana, BA, 2010.
Em função das citocininas testadas, os brotos advindos do explante segmento
cotiledonar apresentaram um menor número de gemas que as do segmento nodal, não
havendo diferença significativa entre os reguladores vegetais utilizados. Para o segmento
nodal a maior produção de gemas (3,78) foi observada com o meio de cultura suplementado
com BAP diferindo significativamente do reguladores vegetal TDZ (Tabela 15). O menor
número de gemas encontrado em meio acrescido com TDZ (2,94) para B. cheilantha, todavia
é superior ao reportados por outras espécies, como é o caso do barbatimão (S. adstringens)
onde uma média de 0,96 gemas/broto foi observada em meio suplementado com KIN
independente da concentração testada (NICIOLI et al., 2008).
O meio de cultura WPM acrescido de BAP favoreceu o maior comprimento dos
brotos, para ambos os explantes, segmento nodal (2,13 cm) e segmento cotiledonar (1,23 cm),
(Tabela 16). O mesmo foi observado com relação às concentrações testadas dos reguladores
vegetais, o meio de cultura suplementado com BAP, independente da concentração utilizada,
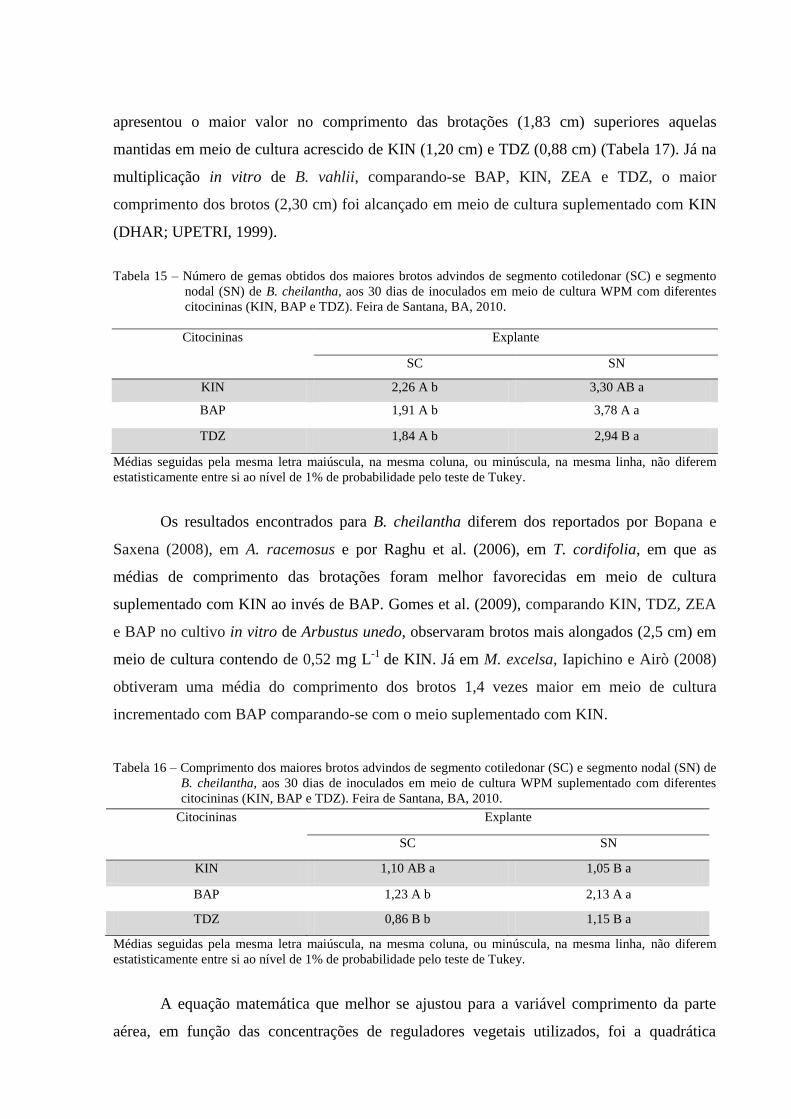
apresentou o maior valor no comprimento das brotações (1,83 cm) superiores aquelas
mantidas em meio de cultura acrescido de KIN (1,20 cm) e TDZ (0,88 cm) (Tabela 17). Já na
multiplicação in vitro de B. vahlii, comparando-se BAP, KIN, ZEA e TDZ, o maior
comprimento dos brotos (2,30 cm) foi alcançado em meio de cultura suplementado com KIN
(DHAR; UPETRI, 1999).
Tabela 15 – Número de gemas obtidos dos maiores brotos advindos de segmento cotiledonar (SC) e segmento
nodal (SN) de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM com diferentes
citocininas (KIN, BAP e TDZ). Feira de Santana, BA, 2010.
Médias seguidas pela mesma letra maiúscula, na mesma coluna, ou minúscula, na mesma linha, não diferem
estatisticamente entre si ao nível de 1% de probabilidade pelo teste de Tukey.
Os resultados encontrados para B. cheilantha diferem dos reportados por Bopana e
Saxena (2008), em A. racemosus e por Raghu et al. (2006), em T. cordifolia, em que as
médias de comprimento das brotações foram melhor favorecidas em meio de cultura
suplementado com KIN ao invés de BAP. Gomes et al. (2009), comparando KIN, TDZ, ZEA
e BAP no cultivo in vitro de Arbustus unedo, observaram brotos mais alongados (2,5 cm) em
meio de cultura contendo de 0,52 mg L-1
de KIN. Já em M. excelsa, Iapichino e Airò (2008)
obtiveram uma média do comprimento dos brotos 1,4 vezes maior em meio de cultura
incrementado com BAP comparando-se com o meio suplementado com KIN.
Tabela 16 – Comprimento dos maiores brotos advindos de segmento cotiledonar (SC) e segmento nodal (SN) de
B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM suplementado com diferentes
citocininas (KIN, BAP e TDZ). Feira de Santana, BA, 2010.
Médias seguidas pela mesma letra maiúscula, na mesma coluna, ou minúscula, na mesma linha, não diferem
estatisticamente entre si ao nível de 1% de probabilidade pelo teste de Tukey.
A equação matemática que melhor se ajustou para a variável comprimento da parte
aérea, em função das concentrações de reguladores vegetais utilizados, foi a quadrática
Citocininas Explante
SC SN
KIN 2,26 A b 3,30 AB a
BAP 1,91 A b 3,78 A a
TDZ 1,84 A b 2,94 B a
Citocininas Explante
SC SN
KIN 1,10 AB a 1,05 B a
BAP 1,23 A b 2,13 A a
TDZ 0,86 B b 1,15 B a
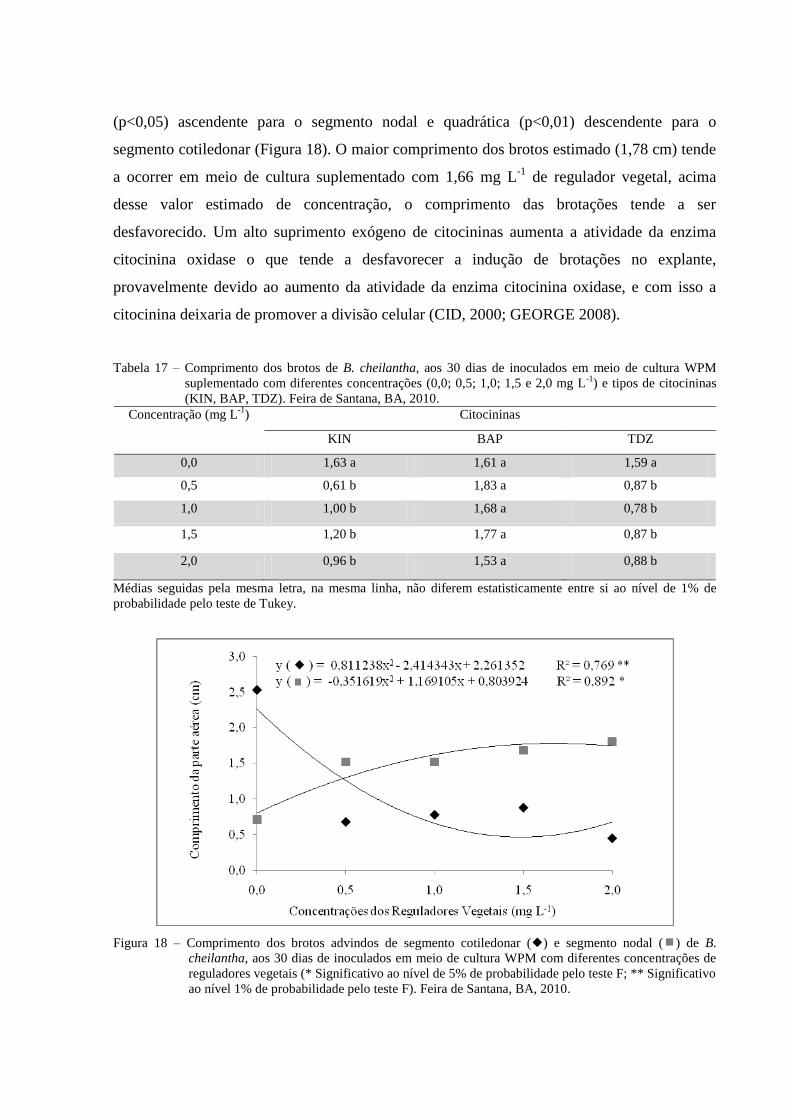
(p<0,05) ascendente para o segmento nodal e quadrática (p<0,01) descendente para o
segmento cotiledonar (Figura 18). O maior comprimento dos brotos estimado (1,78 cm) tende
a ocorrer em meio de cultura suplementado com 1,66 mg L-1
de regulador vegetal, acima
desse valor estimado de concentração, o comprimento das brotações tende a ser
desfavorecido. Um alto suprimento exógeno de citocininas aumenta a atividade da enzima
citocinina oxidase o que tende a desfavorecer a indução de brotações no explante,
provavelmente devido ao aumento da atividade da enzima citocinina oxidase, e com isso a
citocinina deixaria de promover a divisão celular (CID, 2000; GEORGE 2008).
Tabela 17 – Comprimento dos brotos de B. cheilantha, aos 30 dias de inoculados em meio de cultura WPM
suplementado com diferentes concentrações (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e tipos de citocininas
(KIN, BAP, TDZ). Feira de Santana, BA, 2010.
Concentração (mg L-1
) Citocininas
KIN BAP TDZ
0,0 1,63 a 1,61 a 1,59 a
0,5 0,61 b 1,83 a 0,87 b
1,0 1,00 b 1,68 a 0,78 b
1,5 1,20 b 1,77 a 0,87 b
2,0 0,96 b 1,53 a 0,88 b
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
Figura 18 – Comprimento dos brotos advindos de segmento cotiledonar () e segmento nodal () de B.
cheilantha, aos 30 dias de inoculados em meio de cultura WPM com diferentes concentrações de
reguladores vegetais (* Significativo ao nível de 5% de probabilidade pelo teste F; ** Significativo
ao nível 1% de probabilidade pelo teste F). Feira de Santana, BA, 2010.
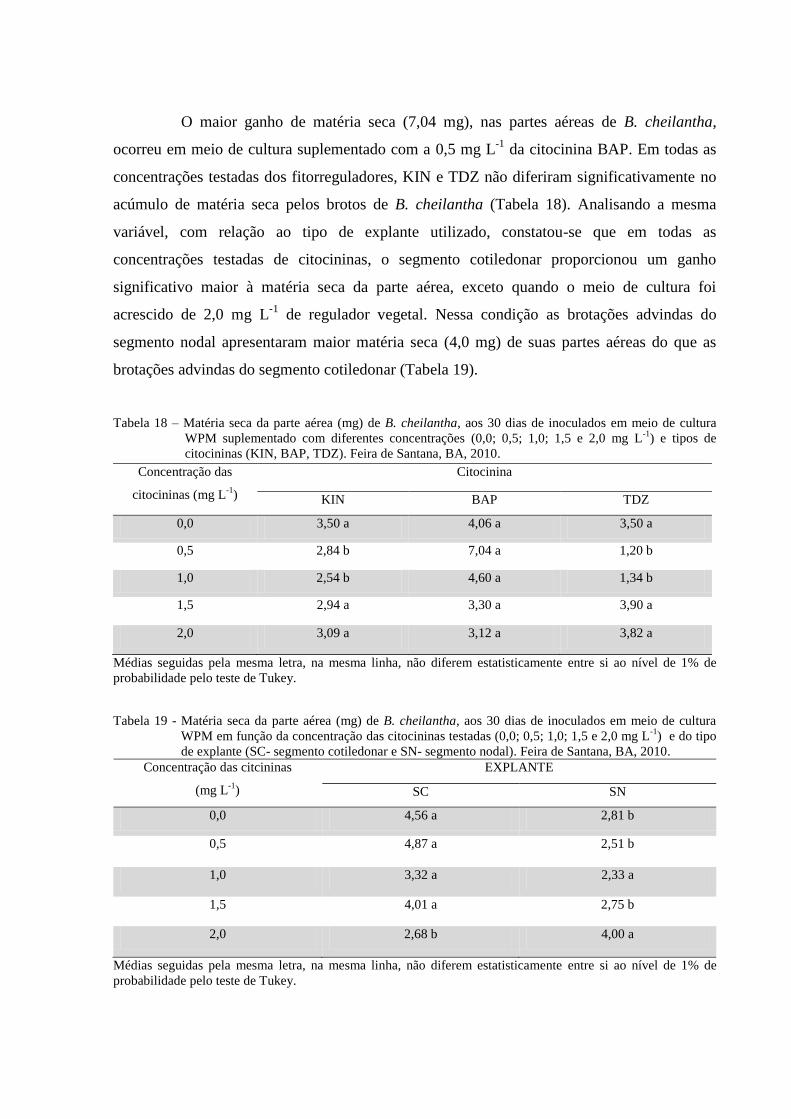
O maior ganho de matéria seca (7,04 mg), nas partes aéreas de B. cheilantha,
ocorreu em meio de cultura suplementado com a 0,5 mg L-1
da citocinina BAP. Em todas as
concentrações testadas dos fitorreguladores, KIN e TDZ não diferiram significativamente no
acúmulo de matéria seca pelos brotos de B. cheilantha (Tabela 18). Analisando a mesma
variável, com relação ao tipo de explante utilizado, constatou-se que em todas as
concentrações testadas de citocininas, o segmento cotiledonar proporcionou um ganho
significativo maior à matéria seca da parte aérea, exceto quando o meio de cultura foi
acrescido de 2,0 mg L-1
de regulador vegetal. Nessa condição as brotações advindas do
segmento nodal apresentaram maior matéria seca (4,0 mg) de suas partes aéreas do que as
brotações advindas do segmento cotiledonar (Tabela 19).
Tabela 18 – Matéria seca da parte aérea (mg) de B. cheilantha, aos 30 dias de inoculados em meio de cultura
WPM suplementado com diferentes concentrações (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e tipos de
citocininas (KIN, BAP, TDZ). Feira de Santana, BA, 2010.
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
Tabela 19 - Matéria seca da parte aérea (mg) de B. cheilantha, aos 30 dias de inoculados em meio de cultura
WPM em função da concentração das citocininas testadas (0,0; 0,5; 1,0; 1,5 e 2,0 mg L-1
) e do tipo
de explante (SC- segmento cotiledonar e SN- segmento nodal). Feira de Santana, BA, 2010.
Concentração das citcininas
(mg L-1
)
EXPLANTE
SC SN
0,0 4,56 a 2,81 b
0,5 4,87 a 2,51 b
1,0 3,32 a 2,33 a
1,5 4,01 a 2,75 b
2,0 2,68 b 4,00 a
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
Concentração das
citocininas (mg L-1
)
Citocinina
KIN BAP TDZ
0,0 3,50 a 4,06 a 3,50 a
0,5 2,84 b 7,04 a 1,20 b
1,0 2,54 b 4,60 a 1,34 b
1,5 2,94 a 3,30 a 3,90 a
2,0 3,09 a 3,12 a 3,82 a

4.2 ENRAIZAMENTO IN VITRO
4.2.1 Experimento 3 – Efeito da auxina ácido indolbutírico (AIB) e carvão ativado no
enraizamento in vitro de brotos de B. cheilantha.
De acordo com a análise de variância, observou-se efeito altamente significativo (p <
0,01) da interação dupla (concentrações de AIB e de carvão ativado) para todas as variáveis
analisadas: porcentagem de enraizamento dos brotos, número de raízes adventícias por broto,
número de raízes secundárias da maior raiz e comprimento da maior raiz, exceto para a
variável taxa de sobrevivência dos brotos, em que apenas o fator AIB influenciou de modo
significativo (p < 0,05) (Tabela 20).
Tabela 20 - Resumo da análise de variância para porcentagem de enraizamento dos brotos (%ENR), número de
raízes adventícias por broto (NRA), número de raízes secundárias da maior raiz (NRS),
comprimento da maior raiz (CMR) (cm) e porcentagem de sobrevivência dos brotos (%SOBR) de
B. cheilantha, em diferentes concentrações de AIB (0,0; 0,5; 1,0; e 2,0 mg L-1
) e carvão ativado
(CA) (0,0; 1,0 e 2,0 g L-1
) em meio de cultura WPM aos 45 dias de inoculação in vitro. UEFS. Feira
de Santana, BA, 2010.
Quadrados Médios
FV GL % ENR Z
NRA Y NRS
Y CMR
Y % SOBR
Z
AIB 3 0,297464** 0,601023** 0,331966 NS
0,056688 NS
0,083471*
CA 2 1,507754** 3,783546** 17,826969** 1,461211** 0,057741 NS
AIB X CA 6 0,313392** 0,580219** 3,077362** 0,458864** 0,049540 NS
RESÍDUO 120 0,005723 0,016251 0,447915 0,073132 0,026423
CV (%) 13,70 10,15 33,32 18,31 10,83
** Significativo ao nível de 1% de probabilidade pelo teste F. * Significativo ao nível de 5% de probabilidade
pelo teste F. NS
Não significativo ao nível de 5% de probabilidade pelo teste F. Y Dados transformados em arco-
seno ; Z Dados transformados em (x + 1)
0,5.
Para porcentagem de enraizamento dos brotos, a curva de regressão indicou um
comportamento quadrático (p < 0,01) ascendente em função das concentrações de AIB
utilizadas, na ausência de carvão ativado. Esse modelo matemático explica 98,02% de toda a
variação dos pontos no experimento. O valor estimado máximo (63,71%) da taxa de
enraizamento tende a ocorrer quando se adiciona 1,68 mg L-1
de AIB ao meio de cultura.
Concentrações do regulador superior a esta tendem a desfavorecer a formação de raízes em B.
cheilantha. Com a adição de carvão ativado ao meio de cultura, não foi possível o ajuste de
um modelo matemático significativo para a variável em análise, sendo observado que todos os
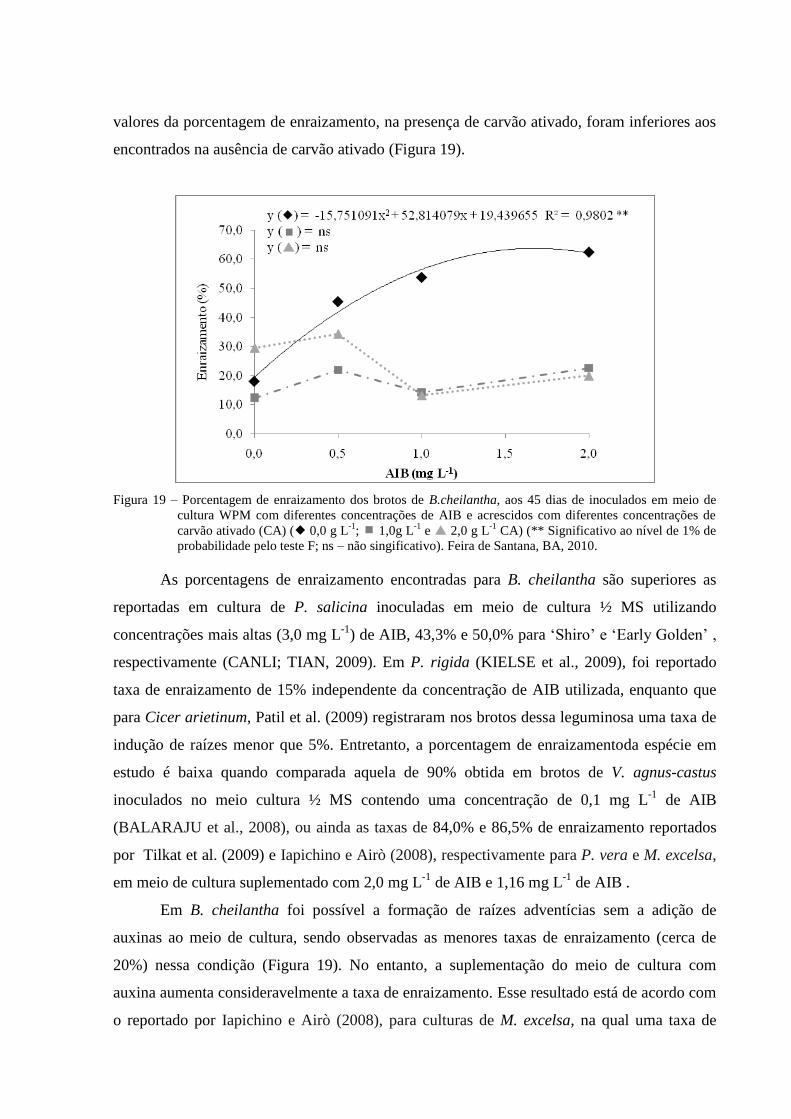
valores da porcentagem de enraizamento, na presença de carvão ativado, foram inferiores aos
encontrados na ausência de carvão ativado (Figura 19).
Figura 19 – Porcentagem de enraizamento dos brotos de B.cheilantha, aos 45 dias de inoculados em meio de
cultura WPM com diferentes concentrações de AIB e acrescidos com diferentes concentrações de
carvão ativado (CA) ( 0,0 g L-1
; 1,0g L-1
e 2,0 g L-1
CA) (** Significativo ao nível de 1% de
probabilidade pelo teste F; ns – não singificativo). Feira de Santana, BA, 2010.
As porcentagens de enraizamento encontradas para B. cheilantha são superiores as
reportadas em cultura de P. salicina inoculadas em meio de cultura ½ MS utilizando
concentrações mais altas (3,0 mg L-1
) de AIB, 43,3% e 50,0% para „Shiro‟ e „Early Golden‟ ,
respectivamente (CANLI; TIAN, 2009). Em P. rigida (KIELSE et al., 2009), foi reportado
taxa de enraizamento de 15% independente da concentração de AIB utilizada, enquanto que
para Cicer arietinum, Patil et al. (2009) registraram nos brotos dessa leguminosa uma taxa de
indução de raízes menor que 5%. Entretanto, a porcentagem de enraizamentoda espécie em
estudo é baixa quando comparada aquela de 90% obtida em brotos de V. agnus-castus
inoculados no meio cultura ½ MS contendo uma concentração de 0,1 mg L-1
de AIB
(BALARAJU et al., 2008), ou ainda as taxas de 84,0% e 86,5% de enraizamento reportados
por Tilkat et al. (2009) e Iapichino e Airò (2008), respectivamente para P. vera e M. excelsa,
em meio de cultura suplementado com 2,0 mg L-1
de AIB e 1,16 mg L-1
de AIB .
Em B. cheilantha foi possível a formação de raízes adventícias sem a adição de
auxinas ao meio de cultura, sendo observadas as menores taxas de enraizamento (cerca de
20%) nessa condição (Figura 19). No entanto, a suplementação do meio de cultura com
auxina aumenta consideravelmente a taxa de enraizamento. Esse resultado está de acordo com
o reportado por Iapichino e Airò (2008), para culturas de M. excelsa, na qual uma taxa de
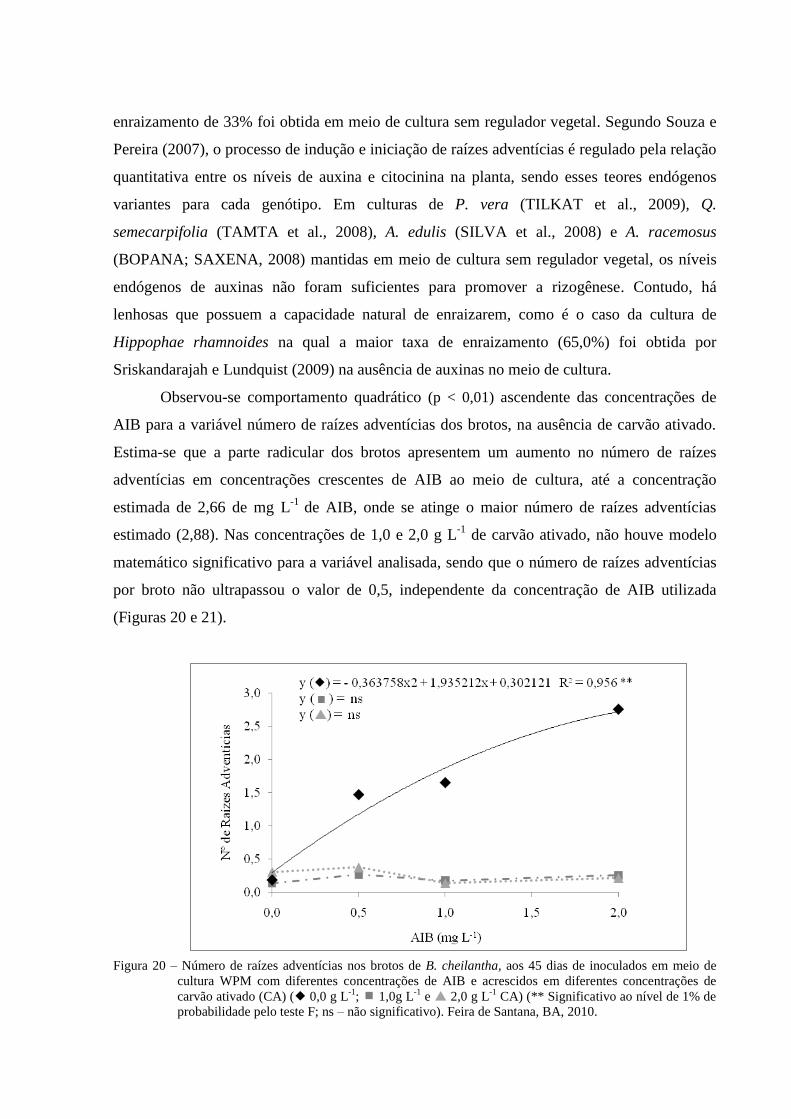
enraizamento de 33% foi obtida em meio de cultura sem regulador vegetal. Segundo Souza e
Pereira (2007), o processo de indução e iniciação de raízes adventícias é regulado pela relação
quantitativa entre os níveis de auxina e citocinina na planta, sendo esses teores endógenos
variantes para cada genótipo. Em culturas de P. vera (TILKAT et al., 2009), Q.
semecarpifolia (TAMTA et al., 2008), A. edulis (SILVA et al., 2008) e A. racemosus
(BOPANA; SAXENA, 2008) mantidas em meio de cultura sem regulador vegetal, os níveis
endógenos de auxinas não foram suficientes para promover a rizogênese. Contudo, há
lenhosas que possuem a capacidade natural de enraizarem, como é o caso da cultura de
Hippophae rhamnoides na qual a maior taxa de enraizamento (65,0%) foi obtida por
Sriskandarajah e Lundquist (2009) na ausência de auxinas no meio de cultura.
Observou-se comportamento quadrático (p < 0,01) ascendente das concentrações de
AIB para a variável número de raízes adventícias dos brotos, na ausência de carvão ativado.
Estima-se que a parte radicular dos brotos apresentem um aumento no número de raízes
adventícias em concentrações crescentes de AIB ao meio de cultura, até a concentração
estimada de 2,66 de mg L-1
de AIB, onde se atinge o maior número de raízes adventícias
estimado (2,88). Nas concentrações de 1,0 e 2,0 g L-1
de carvão ativado, não houve modelo
matemático significativo para a variável analisada, sendo que o número de raízes adventícias
por broto não ultrapassou o valor de 0,5, independente da concentração de AIB utilizada
(Figuras 20 e 21).
Figura 20 – Número de raízes adventícias nos brotos de B. cheilantha, aos 45 dias de inoculados em meio de
cultura WPM com diferentes concentrações de AIB e acrescidos em diferentes concentrações de
carvão ativado (CA) ( 0,0 g L-1
; 1,0g L-1
e 2,0 g L-1
CA) (** Significativo ao nível de 1% de
probabilidade pelo teste F; ns – não significativo). Feira de Santana, BA, 2010.

Figura 21 - Brotos de B. cheilantha em meio de enraizamento WPM, aos 45 dias de inoculados em (A) 0,0 mg L
-
1 de AIB; (B) 0,0 mg L
-1 de AIB e 2,0 g L
-1 de carvão ativado; (C) 1,0 mg L
-1 de AIB e (D) 2,0 mg L
-
1 de AIB. (Barra = 1,0 cm). Feira de Santana, 2009.
Esses resultados são semelhantes aos encontrados por Iapichino e Airò (2008), em M.
excelsa, e por Du e Pijut (2008), em Fraxinus pennsylvanica, em que constataram uma média
de 2,8 raízes/broto quando inoculados em meio de cultura contendo cerca de 1,0 mg L-1
de
AIB. Já para framboeseira „Batum‟ uma média de 1,82 raízes/broto foi reportada por Leitzke,
Damiani e Schuch (2009), quando este explante foi inoculado em meio WPM independente
da concentração de AIB utilizada. O número de raízes adventícias por broto de B. cheilantha é
superior quando comparada a média de 0,5 em culturas de P. rigida obtidas por Kielse et al.
(2009), independente da concentração de AIB adicionada ao meio de cultura, e inferior
quando comparada a média de 4,3 raízes adventícias/broto em culturas de S. dentata obtidas
por Prakash e Staden (2009), em meio acrescido de 2,0 mg L-1
de AIB, ou ainda a média de
9,2 em Rubus híbrido cv. Black Satin em meio de cultura ½ MS com 0,5 mg L-1
de AIB
(GUPTA; MAHALAXMI, 2009).
A equação matemática que melhor se ajustou para a variável número de raízes
secundárias da maior raiz em função das concentrações de AIB utilizadas foi a quadrática
(p<0,01) ascendente, na ausência de carvão ativado (Figura 22). O maior número de raízes
secundárias estimado (9,12) tende a ocorrer em meio de cultura suplementado com 1,16 mg L-
1 de AIB, acima desse valor estimado de concentração, o número de raízes secundárias tende a
ser desfavorecido. Já em F. pennsylvanica, o maior número de raízes secundárias (19,1) foi
observado em meio de cultura suplementado de 1,0 mg L-1
de AIB (DU; PIJUT, 2008).
Com o incremento de 2,0 g L-1
de carvão ativado ao meio de cultura, observou-se
comportamento linear (p < 0,01) decrescente das concentrações de AIB utilizadas para a
mesma variável. Na presença de carvão ativado o maior número de raízes (4,81) foi
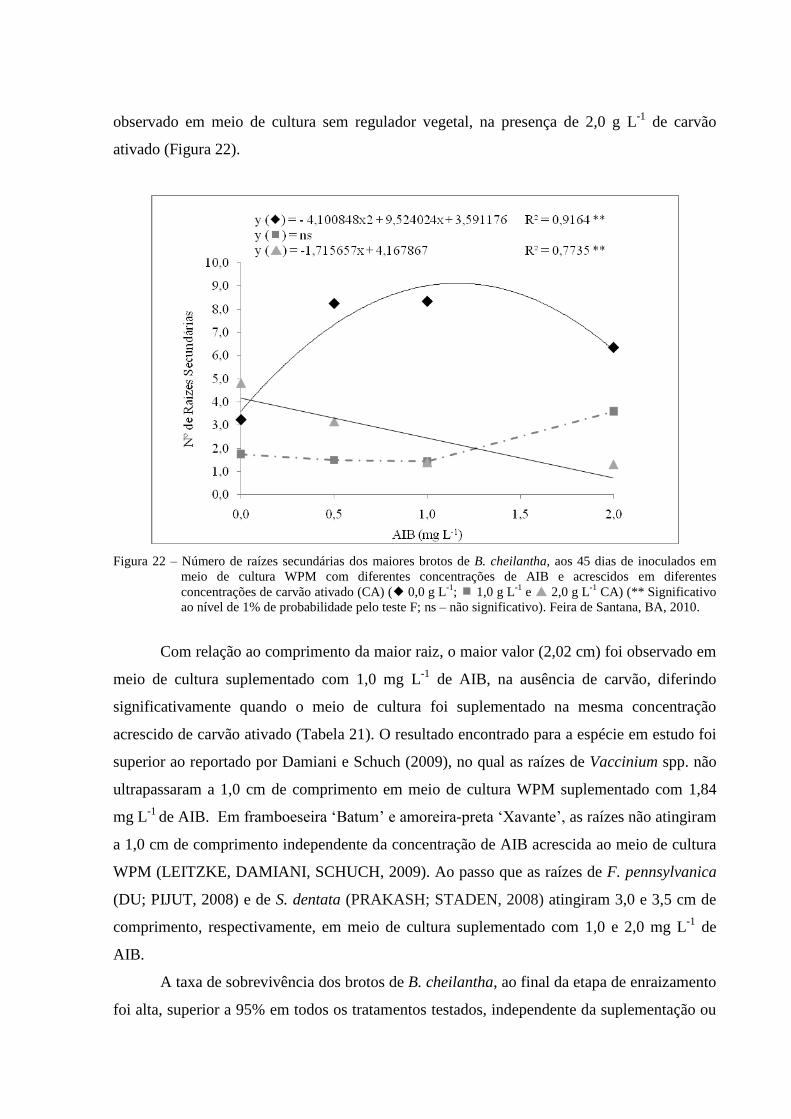
observado em meio de cultura sem regulador vegetal, na presença de 2,0 g L-1
de carvão
ativado (Figura 22).
Figura 22 – Número de raízes secundárias dos maiores brotos de B. cheilantha, aos 45 dias de inoculados em
meio de cultura WPM com diferentes concentrações de AIB e acrescidos em diferentes
concentrações de carvão ativado (CA) ( 0,0 g L-1
; 1,0 g L-1
e 2,0 g L-1
CA) (** Significativo
ao nível de 1% de probabilidade pelo teste F; ns – não significativo). Feira de Santana, BA, 2010.
Com relação ao comprimento da maior raiz, o maior valor (2,02 cm) foi observado em
meio de cultura suplementado com 1,0 mg L-1
de AIB, na ausência de carvão, diferindo
significativamente quando o meio de cultura foi suplementado na mesma concentração
acrescido de carvão ativado (Tabela 21). O resultado encontrado para a espécie em estudo foi
superior ao reportado por Damiani e Schuch (2009), no qual as raízes de Vaccinium spp. não
ultrapassaram a 1,0 cm de comprimento em meio de cultura WPM suplementado com 1,84
mg L-1
de AIB. Em framboeseira „Batum‟ e amoreira-preta „Xavante‟, as raízes não atingiram
a 1,0 cm de comprimento independente da concentração de AIB acrescida ao meio de cultura
WPM (LEITZKE, DAMIANI, SCHUCH, 2009). Ao passo que as raízes de F. pennsylvanica
(DU; PIJUT, 2008) e de S. dentata (PRAKASH; STADEN, 2008) atingiram 3,0 e 3,5 cm de
comprimento, respectivamente, em meio de cultura suplementado com 1,0 e 2,0 mg L-1
de
AIB.
A taxa de sobrevivência dos brotos de B. cheilantha, ao final da etapa de enraizamento
foi alta, superior a 95% em todos os tratamentos testados, independente da suplementação ou

não de carvão ativado ao meio de cultura (dados não mostrados). O que difere do observado
por Vieitez et al. (2009), em que a taxa de sobrevivência dos brotos de Quercus spp. foi maior
com a adição de carvão ativado ao meio de enraizamento. Contudo, o resultado observado
para B. cheilantha é superior ao reportado por Sudha et al. (2005), no qual taxas de
sobrevivência dos brotos de Decalepis arayalpathra não ultrapassam a 28%, independente da
concentração de AIB adiconada ao meio de cultura ½ MS na ausência de carvão ativado.
Tabela 21 – Comprimento da maior raiz (cm) nos brotos de B. cheilantha, aos 45 dias de inoculados em meio de
cultura WPM suplementado com diferentes concentrações de AIB e carvão ativado. Feira de
Santana, BA, 2010.
Concentração AIB
(mg L-1
)
Carvão ativado (g L-1
)
0,0 1,0 2,0
0,0 1,35 ab 0,50 b 1,95 a
0,5 1,72 a 1,14 a 1,50 a
1,0 2,02 a 0,71 b 0,67 b
2,0 1,88 a 1,28 ab 0,68 b
Médias seguidas pela mesma letra, na mesma linha, não diferem estatisticamente entre si ao nível de 1% de
probabilidade pelo teste de Tukey.
Diante do experimento realizado, pode-se inferir que a suplementação de carvão
ativado ao meio de cultura não foi eficiente para melhorar a etapa de enraizamento de B.
cheilantha. Provavelmente, a presença do carvão ativado deve ter alterado significativamente
o pH e a composição do meio de cultura, devido ao seu poder de adsorção de cátions,
nutrientes e reguladores vegetais, essenciais ao enraizamento dos brotos.
4.2.2 Experimento 4 - Efeito das auxinas ácido indolbutírico (AIB) ou ácido naftaleno
acético (ANA) ou ácido indolacético (AIA) e carvão ativado no enraizamento in vitro de
brotos de B. cheilantha.
De acordo com a análise de variância, a interação tripla (tipos, concentrações de
auxinas e carvão ativado) influenciou de modo altamente significativo (p < 0,01) as variáveis
porcentagem de enraizamento dos brotos, número de raízes adventícias por broto e número de
raízes secundárias da maior raiz, e de modo significativo (p < 0,05) as variáveis comprimento
da parte radicular e taxa de sobrevivência dos brotos (Tabela 22).
Com relação à porcentagem de enraizamento dos brotos, a curva de regressão indicou
comportamento quadrático (p < 0,01) descendente em função das concentrações de AIB e

ANA utilizadas na presença de 0,5 g L-1
(Figura 23 A e B) e uma tendência quadrática (p <
0,01) ascendente para as concentrações de AIA testadas (Figura 23 C). É interessante ressaltar
que o modelo matemático encontrado em função das concentrações de ANA utilizadas
explica 100% da variação da resposta analisada. Na ausência e na presença da maior
concentração de carvão ativado no meio de cultura, não foi possível o ajuste de um modelo
matemático significativo para a variável em análise independente da auxina utilizada, com
exceção da auxina AIB na presença de 1,0 g L-1
de carvão ativado no meio de cultura, em que
se observou uma tendência linear (p < 0,01) descendente para a porcentagem de enraizamento
dos brotos (Figura 23 A-C).
Tabela 22 - Resumo da análise de variância para porcentagem de enraizamento dos brotos (%ENR), número de
raízes adventícias por broto (NRA), número de raízes secundárias da maior raiz (NRS), comprimento
parte radicular (CPR) (cm) e porcentagem de sobrevivência dos brotos (%SOBR) de B. cheilantha,
em diferentes concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (AIB, ANA e AIA), em
diferentes concentrações de carvão ativado (CA) (0,0; 0,5 e 1,0 g L-1
) em meio de cultura WPM aos
45 dias de inoculação in vitro. UEFS. Feira de Santana, BA, 2010.
Quadrados Médios
FV GL % ENR Z
NRA Y NRS
Y CPR
Y % SOBR
Z
AUX (A) 2 0,185474** 0,068964** 1,989495 * 1,502961** 0,136439 NS
CON (C) 3 0,161453** 0,154996** 0,652367 NS
0,059545 NS
0,237147 *
CA 2 0,067159 * 0,166241** 0,486994 NS
0,366622* 0,243731 *
A X C 6 0,243038** 0,077885** 3,724064** 0,523805** 0,087544 NS
A X CA 4 0,282404** 0,208279** 6,646924** 0,707690** 0,174409 *
C X CA 6 0,370795** 0,168774** 2,733871** 1,212440** 0,156011 *
A X C X CA 12 0,044556** 0,035688** 1,519747** 0,246035* 0,133319 *
RESÍDUO 180 0,017699 0,011801 0,614182 0,112370 0,069082
CV (%) 20,81 8,63 30,95 19,23 18,66
** Significativo ao nível de 1% de probabilidade pelo teste F; * Significativo ao nível de 5% de probabilidade
pelo teste F; NS
Não significativo ao nível de 5% de probabilidade pelo teste F; Y Dados transformados em arco-
seno e Z Dados transformados em (x + 1)
0,5.
A taxa de enraizamento máxima no meio de cultura suplementado com AIB foi de
63,33%, observada na ausência de carvão ativado e 0,25 mg L-1
do regulador, porém
concentrações superiores a 1,0 mg L-1
de AIB na presença de 0,5 mg L-1
de carvão ativado
podem ser testadas, já que é estimado que concentrações superiores a 0,31 mg L-1
do
regulador tendem a favorecer o aumento da taxa de enraizamento nos brotos. Na presença de
1,0 mg L-1
de carvão ativado, o aumento das concentrações de AIB no meio de cultura não

Figura 23 – Porcentagem de enraizamento dos brotos de B. cheilantha, aos 45 dias de inoculados em meio de
cultura WPM com diferentes concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) e tipos de auxinas (A-
AIB; B- ANA e C- AIA) e acrescidos com diferentes concentrações de carvão ativado (CA) ( 0,0
g L-1
; 0,5 g L-1
e 1,0 g L-1
CA) (** Significativo ao nível de 1% de probabilidade pelo teste F;
* Significativo ao nível de 5% de probabilidade pelo teste F; ns – não singificativo). Feira de
Santana, BA, 2010.

favoreceu a formação de raízes adventícias nos brotos de B. cheilantha (Figura 23A). Com
relação às concentrações de ANA testadas, a maior taxa de enraizamento dos brotos foi de
66,67% quando o meio de cultura foi suplementado e 0,5 mg L-1
de carvão ativado e 1,0 mg
L-1
de ANA, sendo possível a adição de concentrações maiores do regulador, uma vez que a
taxa de enraizamento tende a ser favorecida em concentrações superiores a 0,25 mg L-1
de
ANA (Figura 23B). Na presença de AIA, estima-se que a maior porcentagem de enraizamento
dos brotos 65,7% tende a ocorrer em meio de cultura suplementado com 0,83 mg L-1
do
regulador e 0,5 mg L-1
de carvão ativado (Figura 23C). Diante desses resultados, inferi-se que
a taxa de enraizamento para B. cheilantha não foi significativamente influenciada pelos
diferentes reguladores vegetais, contrariamente, em V. agnus-castus, a porcentagem de
enraizamento dos brotos mantidos em meio de cultura incrementado com AIB foi superior
2,25 vezes a taxa obtida em meio suplementado com ANA ou AIA, independente da
concentração utilizada desses reguladores (BALARAJU et al. 2008).
As porcentagens de enraizamento obtidas para B. cheilantha são semelhantes aquelas
reportadas por Peternel et al. (2009), em culturas de Populus tremula, na qual a taxa de 60%
foi obtida em meio de cultura ½ MS suplementado com 1,0 mg L-1
de AIA na ausência de
carvão ativado, ou ainda a taxa de 65,5% de enraizamento em P. salicina cv. Bruce cultivado
em meio de cultura ½ MS suplementado com 3,0 mg L-1
de AIB (CANLI; TIAN, 2009).
Os valores encontrados para B. cheilantha são superiores as taxas de 38,8 e 30,4% de
enraizamento observada para B. vahlii em meio de cultura suplementado com 1,0 mg L-1
de
ANA e AIB, respectivamente (DHAR; UPRETI, 1999), ou ainda às taxas de 0,0-25,0%
obtidas em culturas de A. racemosus, independente da concentração de regulador (AIB, ANA
e AIA), individualmente ou em combinação, acrescido ao meio de cultura (BOPANA;
SAXENA, 2008). Para a leguminosa P. marsupium, a melhor taxa de enraizamento 30,0% e
21,6% foi registrada, respectivamente, em meio de cultura ½ MS suplementado com 2,0 mg
L-1
de AIB ou com 4,0 mg L-1
de AIA (CHAND; SINGH, 2004b). Já em culturas de Cornus
canadendis (FENG et al., 2009) e de Persea americana (NHUT et al., 2008), independente da
concentração de ANA adicionada ao meio de cultura, não foi possível induzir a formação de
raízes adventícias, contudo a taxa de enraizamento de 48% foi possível em brotos de C.
canadensis mantidos em meio de cultura suplementado com 1,0 mg L-1
de AIB na ausência de
carvão ativado. Para P. salicina, Canli e Tian (2009) não observaram enraizamento nos brotos
de „Shiro‟ e „Bruce‟ quando inoculados em meio de cultura acrescido de ANA,
diferentemente de „Early Golden‟ e „Gladstone‟ na qual a taxa de enraizamento foi de 40,0 e

46,7%, respectivamente, em meio de cultura suplementado com ANA, ou ainda a taxa de
67,2% obtida por Shan et al. (2009), em brotos de A. hypogaea mantidos em meio de cultura
suplementado com 0,5 mg L-1
de ANA.
Resultado superior foi registrado por Oliveira e Nino (2009), em que todos os brotos
de R. ideaus foram enraizados in vitro, quando inoculados em meio de cultura semissólido
MS com metade da concentração dos sais e 0,1 mg L-1
de ANA. Em B. variegata, 96% de
enraizamento foi reportado por Mathur e Mukunthakumar (1992), quando seus brotos foram
inoculados em meio MS suplementado com 1,0 mg L-1
de AIB, enquanto que para B. vahlii a
maior taxa (83,3 %) foi observada em meio de cultura MS com a concentração do seus sais
reduzidas à quarta parte acrescida de 0,2 mg L-1
de ANA (BHATT; DHAR, 2000).
Um comportamento quadrático (p < 0,05) descendente das concentrações de AIB e
ANA foi observado para a variável número de raízes adventícias dos brotos, na presença de
0,5 g L-1
de carvão ativado e tendência quadrática (p < 0,01) ascendente nas concentrações de
AIA. O maior valor (1,47) observado para o número de raízes adventícias ocorreu em meio de
cultura suplementado com 0,25 mg L-1
de AIB na ausência de carvão ativado, enquanto que
para as concentrações de ANA testadas o maior valor (1,10) observado foi verificado em 1,0
mg L-1
do regulador e 0,5 mg L-1
de carvão ativado. Estima-se que o sistema radicular dos
brotos apresentem um aumento no número de raízes adventícias em concentrações crescentes
acima de 0,25 mg L-1
de AIB e ANA, na presença de 0,5 mg L-1
de carvão ativado (Figura 24
A e B). Para as concentrações utilizadas do regulador AIA, o maior número de raízes
adventícias (1,19) estimado tende a ocorrer em meio de cultura suplementado com 0,74 mg L-
1 do regulador e 0,5 mg L
-1 de carvão ativado (Figura 24C). Machakova, Zazimalova e
George (2008) afirmaram que concentrações altas de auxinas in vitro tendem a desfavorecer a
formação das raízes adventícias, o que pode ser devido ao aumento da oxidação e da atividade
da peroxidase, reduzindo as concentrações de ácido indolacético endógeno.
Na ausência ou presença da maior concentração de carvão ativado, não foi possível o
ajuste de um modelo matemático significativo para a variável analisada, independente da
auxina utilizada, com exceção de AIB na presença de 1,0 g L-1
de carvão ativado no meio de
cultura, em que se observou uma tendência linear (p < 0,01) descendente para a variável
analisada (Figura 24C).
A rizogênese de B. cheilantha não induziu a formação de calos na base dos brotos
(Figura 25), sendo esse fato também relatado para outra espécie do gênero, B. variegata
(MATHUR E MUKUNTHAKUMAR, 1992). Esse comportamento pode ter facilitado a
formação de raízes nos brotos mesmo na ausência de auxinas exógenas (Figuras 23 e 24).
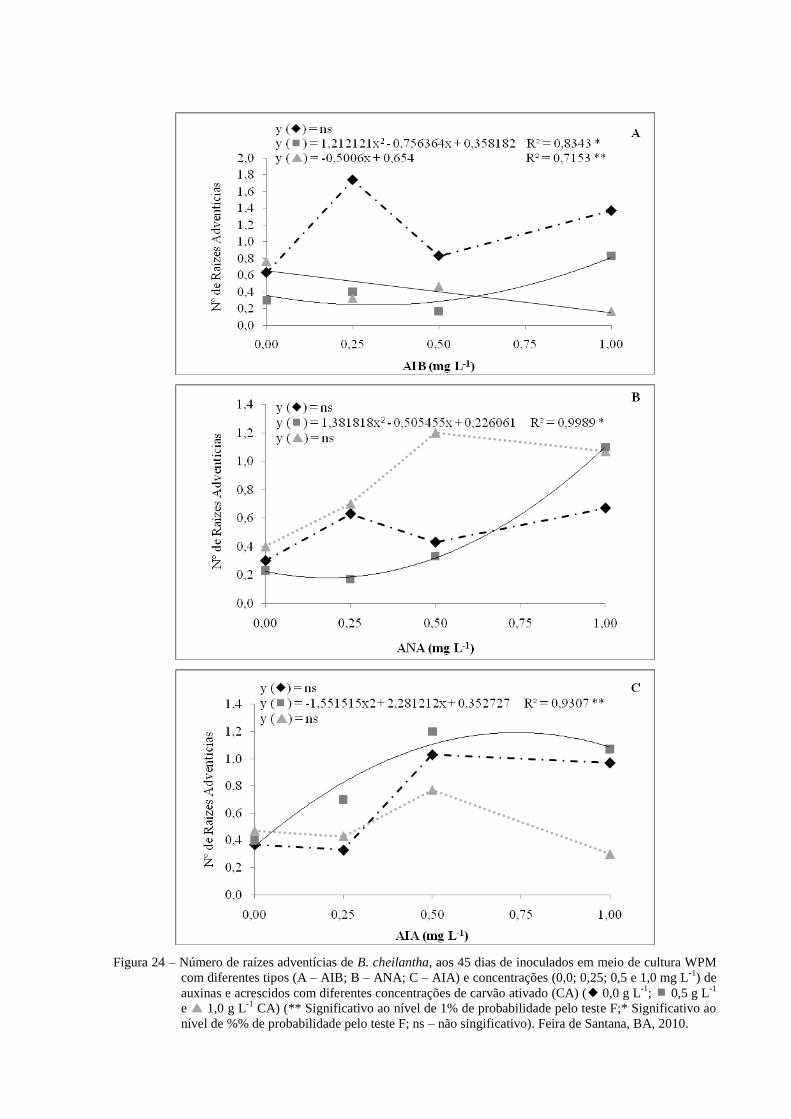
Figura 24 – Número de raízes adventícias de B. cheilantha, aos 45 dias de inoculados em meio de cultura WPM
com diferentes tipos (A – AIB; B – ANA; C – AIA) e concentrações (0,0; 0,25; 0,5 e 1,0 mg L-1
) de
auxinas e acrescidos com diferentes concentrações de carvão ativado (CA) ( 0,0 g L-1
; 0,5 g L-1
e 1,0 g L-1
CA) (** Significativo ao nível de 1% de probabilidade pelo teste F;* Significativo ao
nível de %% de probabilidade pelo teste F; ns – não singificativo). Feira de Santana, BA, 2010.

Figura 25 - Brotos de B. cheilantha em meio de enraizamento WPM, aos 45 dias de inoculados em (A) 0,0 mg L
-
1 de regulador; (B) 0,0 mg L
-1 de regulador e 0,5 g L
-1 de carvão ativado; (C) 1,0 mg L
-1 de AIB; (D)
1,0 mg L-1
de AIB + 0,5 g L-1
de carvão ativado; (E) 1,0 mg L-1
de ANA; (F) 1,0 mg L-1
de ANA +
0,5 g L-1
de carvão ativado; (G) 0,5 mg L-1
de AIA; (H) 0,5 mg L-1
de AIA + 0,5 g L-1
de carvão
ativado (Barra = 1,0 cm). Feira de Santana, BA, 2010.
No entanto, os teores endógenos de auxinas, presentes nas brotações de B. cheilantha,
não foram suficientes para promover um enraizamento em níveis satisfatórios para a
aclimatização.
Em cultura de H. rhamnoides o número de raízes adventícias foi superior ao
encontrado para B. cheilantha, 1,6 raízes/broto no meio de cultura sem regulador e 5,1
raízes/broto em meio suplementado com AIB (SRISKANDARAJAH; LUNDQUIST, 2009),
assim como a média de 3,15 e 2,50 raízes/broto reportada por Tilkat et al. (2009), em P. vera
na presença de 2,0 mg L-1
de AIB e por Bopana e Saxena (2008), em A. racemosus, na
presença de 0,3 mg L-1
de ANA, ambos na ausência de carvão ativado. Comparando-se com
outras espécies do gênero em estudo, culturas de B. vahlii, mantidas em meio de cultura ½
MS suplementado com 0,2 mg L-1
de ANA, obtiveram uma média de 2,6 (UPRETI; DHAR,
1996) e 3,0 raízes/broto (BHATT; DHAR, 2000). Para a mesma espécie, em estudo reportado
por Dhar e Upreti (1999), 1,86 mg L-1
do regulador ANA teve uma influência significativa
maior sob a variável em análise (4,5 raízes/broto) do que igual concentração do regulador AIB
(4,0 raízes/broto). Já para B. variegata o maior número de raízes adventícias (4,6) foi
observado em meio MS suplementado com 1,0 mg L-1
de AIB. Contudo, os resultados
encontrados para B. cheilantha são semelhantes aos reportados por Canli e Tian (2009), para

P. salicina „Early Golden‟ e „Gladstone‟, no qual uma média de 1,4 raízes/broto foi obtida na
presença do regulador ANA.
A equação matemática que melhor se ajustou para a variável número de raízes
secundárias da maior raiz foi a linear (p < 0,05) descendente em função das concentrações de
AIB utilizadas na presença de 1,0 mg L-1
de carvão ativado (Figura 26 A) , quadrática (p <
0,05) descendente para as concentrações de ANA (Figura 26 B) e quadrática (p < 0,01) ascen-
te para as concentrações de AIA (Figura 26 C), ambas na presença de 0,5 mg L-1
de carvão
ativado. O maior número de raízes secundárias (15,83) foi observado em meio de cultura
suplementado com 0,25 mg L-1
de AIB, na ausência de carvão ativado (Figura 26 A).
Enquanto que para F. pennsylvanica o maior número de raízes secundárias (19,0) foi
registrado em meio de cultura suplementado com maior concentração de AIB (1,0 mg L-1
)
(DU; PIJUT, 2008). No meio de cultura suplementado com 1,0 mg L-1
de carvão ativado, a
variável analisada foi desfavorecida com o aumento da concentração de AIB. Assim
concentrações mais baixas ou a ausência deste regulador são fundamentais para promover
uma maior emissão de raízes secundárias nessa condição. Nas concentrações de ANA
testadas, estima-se que concentrações superiores à 0,25 mg L-1
de ANA, na presença de 0,5
mg L-1
de carvão ativado, tendem a favorecer a variável em análise, sendo observado o maior
valor (12,23) para o número de raízes secundárias na maior concentração de ANA utilizada.
Para o regulador AIA, estima-se que a maior produção de raízes secundárias (13,07) ocorra
em meio de cultura WPM suplementado com 0,64 mg L-1
de AIA e 0,5 mg L-1
de carvão
ativado (Figura 26 A-C).
Percebe-se que a suplementação de baixas concentrações de carvão ativado ao meio de
cultura foi eficiente na promoção do enraizamento de B. cheilantha.
Com relação ao comprimento da maior raiz, observou-se comportamento quadrático (p
< 0,05) descendente para as concentrações de AIB e ANA utilizadas na presença de 0,5 mg L-
1 de carvão ativado. Para essas auxinas testadas, na ausência ou presença de 1,0 mg L
-1 de
carvão ativado no meio de cultura, não foi possível o ajuste de uma equação matemática
significativa. Sendo que em 0,5 mg L-1
de ANA e ausência de carvão ativado foi registrado o
menor comprimento (0,35 cm) das raízes para B. cheilantha (Figura 27 A e B). Na presença
de 0,5 e de 1,0 mg L-1
de carvão ativado, observou-se um modelo quadrático ascendente nas
concentrações de AIA utilizadas (Figura 27 C).
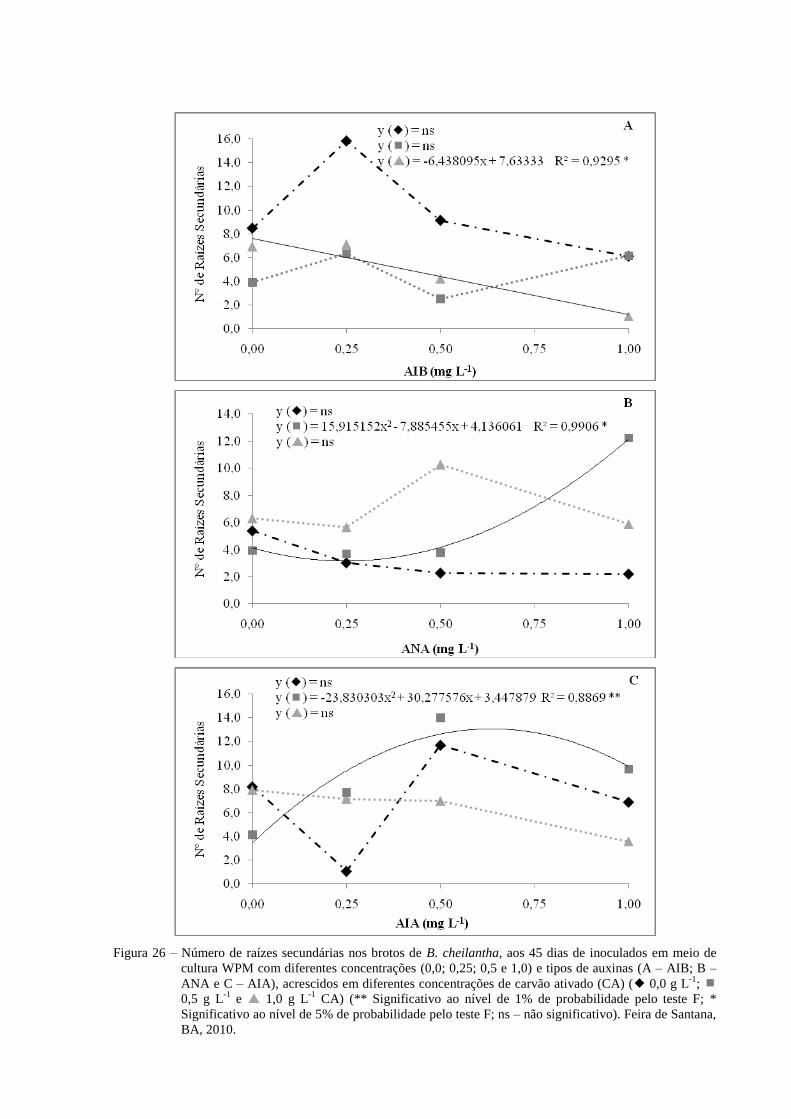
Figura 26 – Número de raízes secundárias nos brotos de B. cheilantha, aos 45 dias de inoculados em meio de
cultura WPM com diferentes concentrações (0,0; 0,25; 0,5 e 1,0) e tipos de auxinas (A – AIB; B –
ANA e C – AIA), acrescidos em diferentes concentrações de carvão ativado (CA) ( 0,0 g L-1
;
0,5 g L-1
e 1,0 g L-1
CA) (** Significativo ao nível de 1% de probabilidade pelo teste F; *
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não significativo). Feira de Santana,
BA, 2010.
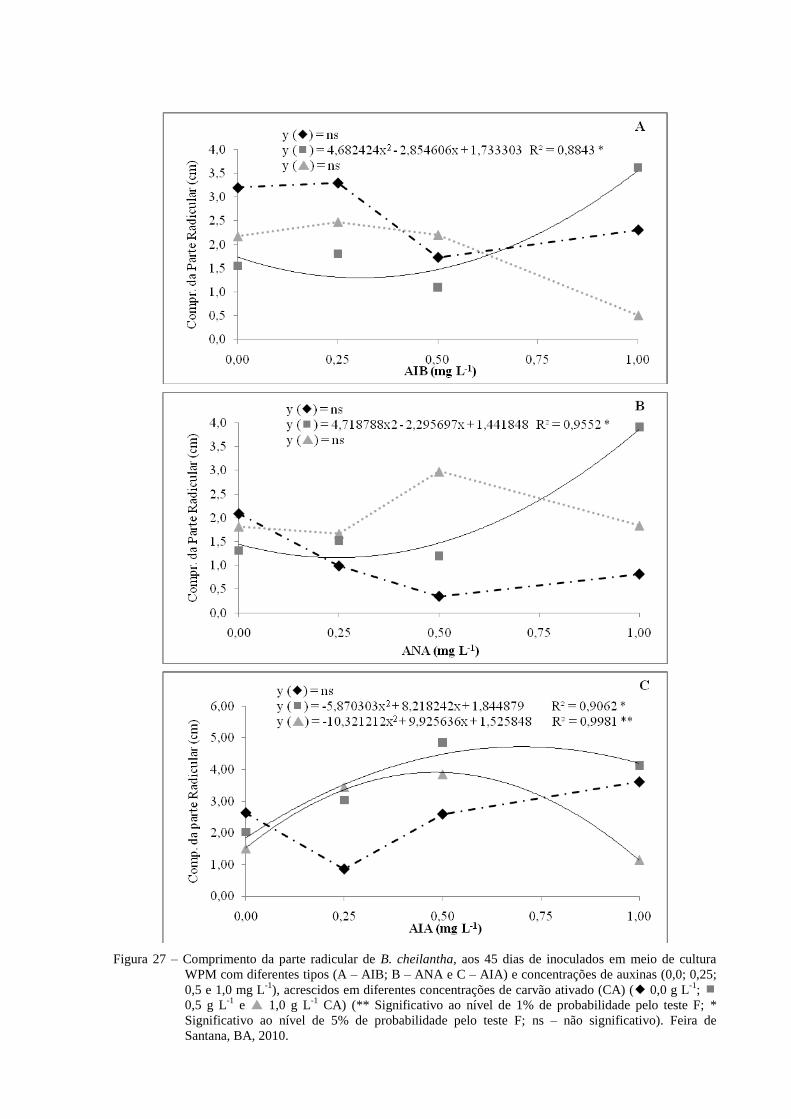
Figura 27 – Comprimento da parte radicular de B. cheilantha, aos 45 dias de inoculados em meio de cultura
WPM com diferentes tipos (A – AIB; B – ANA e C – AIA) e concentrações de auxinas (0,0; 0,25;
0,5 e 1,0 mg L-1
), acrescidos em diferentes concentrações de carvão ativado (CA) ( 0,0 g L-1
;
0,5 g L-1
e 1,0 g L-1
CA) (** Significativo ao nível de 1% de probabilidade pelo teste F; *
Significativo ao nível de 5% de probabilidade pelo teste F; ns – não significativo). Feira de
Santana, BA, 2010.

O maior valor (4,72 cm) estimado tende a ocorrer em meio de cultura suplementado
com 0,70 mg L-1
de AIA, na presença de 0,5 mg L-1
de carvão, concentrações do regulador
superior a estimada tendem a desfavorecer o comprimento das raízes (Figura 27 C). Efeito
inibitório exercido por altas concentrações de auxinas pode estar correlacionado a indução da
produção de etileno, inibidor do crescimento de raiz (MOSHKOV et al., 2008; TAIZ;
ZEIGER, 2009).
Em igual concentração de ANA testada para B. cheilantha (1,0 mg L-1
), na ausência de
carvão ativado, um comprimento de 0,04 cm das raízes de S. adstringens foi obtido por
Nicioli et al. (2008). Já em culturas de A. racemosus o comprimento de 0,20 cm de suas raízes
foi obtido em meio suplementado com 0,3 mg L-1
de ANA (BOPANA; SAXENA, 2008).
Enquanto que Iapichino e Airò (2008) reportaram os maiores comprimentos (1,3 e 1,0 cm)
para as raízes de M. excelsa, em meio de cultura suplementado com 0,58 mg L-1
de AIB e 1,0
mg L-1
de AIA, respectivamente. Para V. agnus-castus, o regulador AIB favoreceu o maior
comprimento das raízes (4,4 cm) com a suplementação de 0,1 mg L-1
de AIB no meio de
cultura, sendo 11,0 e 8,8 vezes maior o seu comprimento do que aquele obtido em igual
concentração de AIA e ANA, respectivamente (BALARAJU et al. 2008).
O enraizamento de B. cheilantha em meio de cultura suplementado com a menor
concentração de carvão ativado influenciou positivamente todas as variáveis analisadas. Silva
et al. (2008) constataram que a adição de carvão ativado era requerida para induzir o
enraizamento em brotações de A. edulis. Esse fato pode ser devido ao escurecimento do meio
de cultura favorecendo a indução e crescimento das raízes de B. cheilantha, pois na ausência
de luz há a promoção de reações fisiológicas que só ocorrem no escuro (MOSHKOV et al.,
2008). Além do que, a luz contribui para a diminuição dos teores endógenos de AIA, assim
como o acúmulo de fenóis e seus subprodutos, que inibem o enraizamento
(GRATTAPAGLIA; MACHADO, 1998). Contudo, o efeito da maior concentração de carvão,
adicionado ao meio de cultura, não favoreceu o processo de enraizamento de B. cheilantha, o
que pode ser explicado pela sua capacidade de adsorver parte de todos os elementos que
compõem o meio, o que pode ter acarretado deficiência nutricional e/ou uma menor
disponibilidade de auxina livre para os brotos (GRATTAPAGLIA; MACHADO, 1998),
prejudicando o enraizamento da espécie.
A taxa de sobrevivência dos brotos de B. cheilantha ao final da etapa de enraizamento,
foi alta, superior a 70% em todos os tipos de auxinas testadas, independente da suplementação
ou não de carvão ativado ao meio de cultura (dados não mostrados). Em D. arayalpathra, a

taxa de sobrevivência dos brotos não ultrapassou 46%, independente da concentração de AIB
e AIA utilizada ao meio de cultura, enquanto que a adição de ANA permitiu uma taxa de
sobrevivência superior a 70% dos brotos (SUDHA et al., 2005).
Inúmeros fatores podem ter influenciado as diferentes respostas no enraizamento dos
brotos de B. cheilantha. Segundo Machakova, Zazimalova e George (2008), a depender da
concentração e do tipo de auxina aplicada, esta pode sofrer influência da inativação no meio
de cultura ou dentro do explante, da sensibilidade dos tecidos, da sua taxa de transporte até o
tecido alvo, da síntese dos reguladores endógenos dentro do explante, da interação dos
reguladores exógenos e os níveis naturais das substâncias endógenas, ou até mesmo as
influências das condições ambientais, inerentes ao processo, sob a resposta e estabilidade das
mesmas.
4.5 ACLIMATIZAÇÃO
4.5.1 Experimento 5 – Efeito da aeração in vitro sob a aclimatização
Aos 30 dias de aclimatização, o percentual de sobrevivência das plantas pré-
acondicionadas, durante o enraizamento in vitro, em tubos fechados com tampa plástica e
algodão, não diferiram significativamente entre si, porém diferiram significativamente (p <
0,05) daquelas fechadas com filme de PVC (Tabela 23). As maiores taxas de sobrevivência
(86,5% e 78,33%) na aclimatização ocorreram quando os tubos de ensaio foram fechados com
tampa de plásticos e de algodão, respectivamente. Enquanto a menor taxa (61,0%) foi
observada nos tubos velados com filme de PVC (Figura 28).
Tabela 23 - Resumo da análise de variância para porcentagem de sobrevivência das mudas (%SOBR) de B.
cheilantha após um período de 30 dias de aclimatização, pré acondicionadas em diferentes
condições de tamponamentos (filme de PVC; tampa plástica sem filme de PVC e tampão de
algodão) durante o enraizamento in vitro. Feira de Santana, BA, 2010. FV GL Quadrado Médio
% SOBR Z
TRAT 2 1635,140163 *
RESÍDUO 27 438,771221
CV (%) 27,77
* Significativo ao nível de 5% de probabilidade pelo teste F e Z Dados não transformados.

A maior aeração promovida pelas tampas de plástico e de algodão pode ter favorecido
uma maior funcionalidade dos estômatos, formação de cera cuticular e/ou um
desenvolvimento mais acentuado do mesofilo foliar (Ziv; Chen, 2008), propiciando um
crescimento mais vigoroso, tanto da parte aérea quanto da parte radicular do que aquelas
cultivadas em recipiente fechado (PVC). Sendo assim, as plantas mantidas em condições
fotoautrotóficas testadas durante o enraizamento in vitro podem ter sofrido uma desidratação
menos intensa, quando transferidas para o ambiente ex vitro, promovendo uma maior taxa de
sobrevivência dessas mudas durante a fase de aclimatização.
Figura 28– Percentual de sobrevivência das mudas de B. cheilantha, aos 30 dias de aclimatização, referentes às
condições de tamponamento durante a etapa de enraizamento in vitro: filme de PVC, tampa plástica
sem PVC e tampa de algodão (Médias seguidas pela mesma letra não diferem estatisticamente entre
si ao nível de 5% de probabilidade pelo teste de Scott-Knot). Feira de Santana, BA, 2010.
Figura 29 - Mudas de B. cheilantha (A) durante a aclimatização em casa de vegetação e (B) após 30 dias de
aclimatização. Feira de Santana, BA, 2010.
Diferentemente do reportado por Bhatt e Dhar (2000), a taxa de sobrevivência das
mudas de B. vahlii não apresentou diferença significativa quando os brotos enraizados foram
pré-acondicionados em condições fotoautrotóficas (diferentes concentrações de sacarose: 0,0;
10,0; 20,0; 30,0; 40,0 e 50,0 g L-1
). Os resultados obtidos para B. cheilantha são semelhantes

aos obtidos em B. variegata, no qual o percentual de sobrevivência foi maior que 80% quando
as mudas foram transferidas para terra vegetal e regadas com os nutrientes do meio de cultura
MS diluídos 10 vezes (MATHUR; MUKUNTHAKUMAR, 1992). Já para as mudas de B.
purpurea uma taxa entre 33 a 60% de sobrevivência foi observada nos brotos enraizados
mantidos em substrato terra vegetal:areia (1:1) (KUMAR, 1992). Segundo Ziv e Chen (2008),
a baixa taxa de sobrevivência de algumas espécies, durante a aclimatização, deve-se a
deficiente vascularização entre a parte aérea e radicular em tecidos de plantas regeneradas in
vitro.
5 CONCLUSÃO
A propagação in vitro de B. cheilantha é possível utilizando-se o segmento nodal
como fonte de explante, em meio de cultura WPM suplementado com 2,0 mg L-1
de BAP. A
citocinina BAP demonstrou resposta superior à KIN e TDZ, na etapa de multiplicação da
espécie. O TDZ, apesar de ter proporcionado um maior número de brotos, estes se
apresentaram hiperídricos.
O enraizamento in vitro é possível na ausência de auxinas no meio de cultura. No
entanto, os níveis endógenos de auxinas não foram suficientes para promover uma rizogênese
adequada para a aclimatização. As maiores taxas de enraizamento, obtidas nos brotos de B.
cheilantha, foram semelhantes nas diferentes auxinas: AIA, ANA e AIB. No entanto, a
presença de ANA e AIA melhoraram alguns aspectos do enraizamento, como o número de
raízes adventícias, secundárias e comprimento da parte radicular. A presença de baixa
concentração de carvão ativado proporcionou maior porcentagem de enraizamento dos brotos,
assim como maior número e comprimento das raízes adventícias, independente do tipo de
auxinas adicionada ao meio de cultura.
É possível a aclimatização de B. cheilantha multiplicada e enraizada in vitro. A
aeração dos frascos de cultivo, durante o enraizamento in vitro, melhora a taxa de
sobrevivência durante a sua aclimatização.

REFERÊNCIAS
AGRA, Maria de Fátima et al. Medicinal and poisonous diversity of the flora of “Cariri
Paraibano”, Brazil. Journal of Ethnopharmacology, v. 111, p. 383–395, 2007.
ALBERTS, Bruce. Biologia molecular da celula. 3.ed Porto Alegre: Artes Médicas, 1997.
1294 p.
ALBUQUERQUE, U. P.; ANDRADE, L. H. C.; SILVA, A. C. O. Use of plant resources in a
seasonal dry forest (Northeastern Brazil). Acta Botanica Brasilica, v. 19, n. 1, p. 27-38, 2005.
ALBUQUERQUE, Ulisses Paulino et al. Medicinal plants of the caatinga (semi-arid)
vegetation of NE Brazil: A quantitative approach. Journal of Ethnopharmacology, v. 114, p.
325–354, 2007.
ALMEIDA, E. R.; XAVIER, H.; GUEDES, M. C. Estudo preliminar da ação hipoglicemiante
do extrato meatólico da Bauhinia cheilantha Steud (Fabaceae) em ratos. In: III JORNADA
PAULISTA DE PLANTAS MEDICINAIS, 1997. Campinas, SP, Anais... Campinas, 1997.
ALONSO, Jorge R. Tratado de Fitofármacos y Nutracéuticos. 1.ed. Rosario, Argentina:
Corpus Libros, 2004. 1359p.
ALVES, M. C. S. et al. Superação da dormência em sementes de Bauhinia monandra Britt. e
Bauhinia ungulata L. – Caesalpinoideae. Revista Brasileira de Sementes, v. 22, n. 2, p. 139-
144, 2000.
ALVES, A. U. et al. Superação da dormência em sementes de Bauhinia divaricata L. Acta
Botanica Brasilica, Paraíba, Brasil, v. 18, n. 4, p. 871-879, 2004.
APPEZZATO-DA-GLÓRIA, B.; MACHADO, S. R. Ultrastructural analysis of in vitro direct
and indirect organogenesis. Revista Brasileira de Botânica, São Paulo, SP, v. 27, n. 3,
jul./set. 2004.
ARAÚJO, A. C. et al.Análise quali-quantitativa da arborização no bairro Presidente Médici,
Campina Grande-PB. Revista da Sociedade Brasileira de Arborização Urbana, Piracicaba,
SP, v. 4, n. 1, p. 133-144, 2009.
BALARAJU, K. et al. Micropropagation of Vitex agnus-castus, (Verbenaceae)—a valuable
medicinal plant. In Vitro Cellular and Developmental Biology. Plant, Wallingford, UK, v. 44,
p. 436-441, 2008.
BARBOSA-FILHO, J. M. et al. Plants and their active constituents from South, Central, and
North America with hypoglycemic activity. Revista Brasileira de Farmacognosia/Braz J.
Pharmacogn., João Pessoa, PB, v. 15, n. 4, out./dez. 2005.

BASKARAN, P.; JAYABALAN, N. An improved protocol for adventitious shoot
regeneration and plant formation in Psoralea corylifolia L. Scientia Horticulturae,
Amsterdam, v. 123, n. 2, p. 283-286, dez. 2009.
BATISTA, A. M. V.; AMORIM, G. L.; NASCIMENTO, M. DO S. B. Forrageiras. 2005. In:
SAMPAIO, E. V. S. B. et al. Espécies da flora nordestina de importância econômica
potencial. Recife: Associação Plantas do Nordeste, 2005. 331 p.
BHATT, I. D.; DHAR, U. Combined effect of cytokinins on multiple shoot production from
cotyledonary node explants of Bauhinia vahlii. Plant Cell, Tissue and Organ Culture,
Amsterdam, v. 62, p. 79–83, 2000.
BOPANA, N.; SAXENA, S. In vitro propagation of a high value medicinal plant: Asparagus
racemosus Willd. In Vitro Cellular and Developmental Biology. Plant, Wallingford, UK, v.
44, p. 525-532, 2008.
BORCHETIA, S. et al. High multiplication frequency and genetic stability for
commercialization of the three varieties of micropropagated tea plants (Camellia spp.).
Scientia Horticulturae, Amsterdam, v. 120, p. 544–550, 2009.
BRÍGIDO, Cleiton Lisboa. Estudo dos Constituintes Químicos de Bauhinia cheilantha
(Bongard) Steudel. 2001. 1v. 148f. Dissertação (Mestrado em Química Orgânica) – Faculdade
de Química, Universidade Federal do Ceará, Fortaleza.
BUENDÍA-GONZÁLEZ, L. et al. Clonal propagation of mesquite tree (Prosopis laevigata
Humb. & Bonpl. ex Willd. M.C. Johnston). I. Via cotyledonary nodes. In vitro Cell
Development Biology—Plant, Wallingford, UK, v. 43, n. 3, p. 260–266, 2007.
CALDAS, L. S.; HARIDASAN, P. F.; ELIAS, M. Meios nutritivos. In: CALDAS, L. S.;
TORRES, A. C.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas.
Brasília: EMBRAPA-SPI/EMBRAPA - CNPH, v. 1, 1998. 509 p.
CANLI, F. A.; TIAN, L. Regeneration of adventitious shoots from mature stored cotyledons
of Japanese plum (Prunus salicina Lind.). Scientia Horticulturae, Amsterdam, v. 120, p. 64–
69, 2009.
CARVALHO, Ricardo Nevares. Cultivo in vitro de Bauhinia forficata Link. 1998. 1v. 52f.
Dissertação (Mestrado em Fisiologia Bioquímica de Plantas) – Escola Superior de Agricultura
Luiz de Queiroz, Universidade de São Paulo, Piracicaba.
CHAND, S.; SINGH, A. K. In vitro shhot regeneration from cotyledonary node explants of a
multipurpose leguminous tree, Pterocarpus marsupium Roxb. In Vitro Cell. Development
Biology. Plant, Wallingford, UK, v. 40, n. 5, p. 464-466, set./out. 2004.
CID, L. P. B. Citocininas. In: CID, L. P. B. (Ed.) Introdução aos Hormônios Vegetais.
Brasília: Embrapa Recursos Genéticos e Biotecnologia, 2000. p. 55-81.
COUTO, J. M. F. et al. Desinfestação e germinação in vitro de sementes de mogno (Swietenia
macrophylla King). Revista Árvore, Viçosa, MG, v. 28, n. 5, p. 633-642, 2004.

COSTA, M. A. P. C.; SOUZA, A. S.; ALMEIDA, W. A. B. Morfogênege in vitro. In:
SOUZA, A. S.; JUNGHANS, T. G. Introdução à micropropagação de plantas. Cruz das
Almas: Embrapa Mandioca e Fruticultura, Tropical, 2006. p. 115-130.
COSTA, M. G. C. et al. Morphogenic gradients of adventitious bud and shoot regeneration in
epicotyl explants of Citrus. Scientia Horticulturae, Amsterdam, v. 100, p. 63-74, 2004.
DAMIANI, C. R.; SCHUCH, M. W. Enraizamento in vitro de mirtilo em condições
fotoautotróficas. Ciência Rural, Santa Maria, v. 39, n. 4, p. 1012-1017, mar. 2009.
DHAR, U.; UPRETI. J. In vitro regeneration of a mature leguminous liana (Bauhinia vahlii
Wight & Arnott). Plant Cell Reports, v. 18, n. 664-669, 1999.
DONINI, L. P. et al. Estabelecimento in vitro de oliveira cv. Arbequina para início da
micropropagação. Ciência Rural, Santa Maria, v. 38, n. 6, set. 2008.
DORAN, P. M. Application of Plant Tissue Cultures in Phytoremediation Research:
Incentives and Limitations. Biotechnology and Bioengineering, v. 103, n. 1, p. 60-76, maio
2009.
DU, N.; PIJUT, P. M. Regeneration of plants from Fraxinus pennsylvanica hypocotyls and
cotyledons. Scientia Horticulturae, Amsterdam, v. 118, p. 74–79, 2008.
DUARTE-ALMEIDA, J. M.; NEGRI, G.; SALATINO, A. Volatile oils in leaves of Bauhinia
(Fabaceae-Caesalpinioideae). Biochemical Systematics and Ecology, v. 32, p. 747-753, 2004.
EPO. European Patent Office. Disponível em: < http://ep.espacenet.com/> Acesso dia: 29
nov. 2009.
EZURA, H. et al. In vitro plant regeneration in Macroptilium atropurpureum, a legume with
a broad symbiont range for nodulation. Plant Science, v. 159, p. 21-27, 2000.
FAISAL, M.; AHMAD, N.; ANIS, M. Shoot multiplication in Rauvolfia tetraphylla L. using
thidiazuron. Plant Cell, Tissue and Organ Culture, Amsterdam, v. 80, p. 187-190, 2005.
FENG, C.-M. et al. Shoot regeneration of dwarf dogwood (Cornus canadensis L.) and
morphological characterization of the regenerated plants. Plant Cell Tiss Organ Culture,
Amsterdam, v. 97, p. 27–37, 2009.
FERREIRA, A. A.; GUIMARÃES, E. R.; CONTADO, J. C. Patente como instrumento
competitivo e como fonte de informação tecnológica. Gestão & Produção, São Carlos, SP, v.
16, n. 2, abr./jun. 2009.
FERREIRA, D. F. Sisvar: um programa para análises e ensino de estatística. Revista
Científica Symposium, Lavras, v. 6, p. 36-4, 2008
FERREIRA, E. A.; PASQUAL, M. Otimização de protocolo para micropropagação da
figueira “Roxo de Valinhos”. Ciência Rural, Santa Maria, v. 38, n. 4, p. 1149-1153, jul. 2008.

FETT, M. S. Como produzir mudas de Eucalipto clonado? Serviço Brasileiro de Respostas
Técnicas. Piracicaba: SENAI, 2005. Disponível em:
<http://sbrt.ibict.br/upload/sbrt1352.pdf>. Acesso em: 10 maio 2009.
GAHAN, P. B.; GEORGE, E. F. Adventitious Regeneration. In: GEORGE, E. F. et al. (Ed.),
Plant Propagation by Tissue Culture. v.1 The Background. 3.ed. Dordrecht: Springer 2008. p.
355–401.
GEORGE, E. F. Plant Tissue Culture Procedure – Background. In: GEORGE, E. F. et al.
(Ed.), Plant Propagation by Tissue Culture. v.1 The Background. 3.ed. Dordrecht: Springer,
2008. p. 1–28.
GEORGE, E. F.; DEBERGH, P. C. Micropropagation: Uses and Methods. In: GEORGE, E.
F. et al. (Ed.), Plant Propagation by Tissue Culture, v.1 The Background. 3.ed. Dordrecht:
Springer, 2008. p. 29-64.
GOMES, F. et al. Effect of PGRs and genotype on the multiplication of micropropagated
cultures of Arbutus unedo L. (Strawberry Tree) adult selected trees. New Biotechnology, v.
25, suplem. 1, p. S311, set. 2009.
GRATTAPAGLIA, D.; MACHADO, M. A. Micropropagação. In: TORRES, A. C.;
CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas.
Brasília: Embrapa – SPI / Embrapa – CNPH, v. 1, 1998. p. 183-260.
GRESSHOFF P. M.; DOY C. H. Development and differentiation of haploid Lycopersicon
esculentum. Planta, v. 107, p. 161–170, 1972.
GUIMARÃES-BEELEN, P. M. et al. Characterization of condensed tannins from native
legumes of the Brazilian northeastern semi-arid. Sci. Agric., Piracicaba, SP, Rio Largo-AL, v.
63, n. 6, p. 522-528, nov./dez. 2006 (2006a).
GUIMARÃES-BEELEN, P. M. et al. Efeito dos taninos condensados de forrageiras nativas
do semi-árido nordestino sobre o crescimento e atividade celulolítica de Ruminococcus
flavefaciens FD1. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Rio Largo-AL, v.
58, n. 5, p. 910-917, 2006 (2006b).
GUPTA, S.; MAHALAXMI, V. In vitro high frequency direct plant regeneration from whole
leaves of blackberry. Scientia Horticulturae, Amsterdam, v. 120, p. 22-26, 2009.
HAMZA, N.; et al. Prevention of type 2 diabetes induced by high fat diet in the C57BL/6J
mouse by two medicinal plants used in traditional treatment of diabetes in the east of Algeria.
Journal of Ethnopharmacology, on line, 2010.
HANDA, L.; SAMPAIO, P. T. B.; QUISEN, R. C. Cultura in vitro de embriões e de gemas de
mudas de pau-rosa (Aniba rosaeodora Ducke). Acta Amazonica, Manaus, AM, v. 35, n. 1, p.
29-33, 2005.
HARRY, I. S.; THORPE, T. A. In vitro culture of forest trees. In: VASIL, I. K.; THORPE, T.
A. (Ed.), Plant cell and tissue culture. Dordrecht: Kluwer Academic Publishers, 1994. p. 539-
560. Disponível em:

<http://books.google.com.br/books?id=y4Z1l0ewvr4C&pg=PA559&lpg=PA559&dq=Plant+c
ell+and+tissue+culture.HARRY,+I.+S.%3B+THORPE&source=bl&ots=T4qqFZCRm9&sig=
9n01SVXHNJqRLgqRtnnxZOrh6vo&hl=pt-BR&ei=Yg4ES4qQE4-
muAeozYCmCA&sa=X&oi=book_result&ct=result&resnum=1&ved=0CAgQ6AEwAA#v=o
nepage&q=&f=false>. Acessado em: 18 nov. 2009.
HARTMANN, H. T. et al. Plant propagation: principles and practices. 6.ed. New Jersey:
Prentice Hall International, 1997. 770 p.
IAPICHINO, G.; AIRÒ, M. Micropropagation of Metrosideros excelsa. In Vitro Cellular and
Developmental Biology. Plant, Wallingford, UK, v. 44, p. 330-337, 2008.
INPI. Instituto Nacional de Propriedade Intelectual. Disponível em: <
http://www.inpi.gov.br>. Acesso dia: 29 nov. 2009.
IUCN 2008. Standards and Petitions Working Group. 2008. Guidelines for using the IUCN
International Union for Conservation of Nature, Red List of Threatened Species.Version 7.0.
Prepared by the Standards and Petitions Working Group of the IUCN SSC Biodiversity
Assessments Sub-Committee in August 2008. Disponível em: <www.iucnredlist.org>.
Acessado em: 19 nov. 2009.
JEDINÁK, A. et al. Approaches to flavonoid production in plant tissue cultures. Biologia, v.
59, n. 6, p. 697-710, 2004.
JHA, T. B.; GHOSH, B. Plant Tissue Culture: Basic and Applied. India: Universities Press
(India) Private Limited. 2006. 206p.
KANG, B. G. et al. Micropropagation of Populus trichocarpa „Nisqually-1‟: the genotype
deriving the Populus reference genome. Plant Cell Tiss Organ Culture, Amsterdam, v. 99, n.
3, p. 251-257, dez. 2009.
KIELSE, P. et al. Regeneração in vitro de Parapiptadenia rigida. Ciência Rural, Santa Maria,
v. 39, n. 4, jul. 2009.
KLUG, W. S.; CUMMINGS, M. R. Concepts of Genetics. 6. ed. Editora Prentice Hall. 2000
KUMAR, A. Pharmacognostic evaluation of certain forest trees with special reference to
tissue culture, anatomy, histochemistry and histoenzymological studies. Ph.D. Thesis, Visva-
Bharati University, Santiniketan, India, 1987.
KUMAR, A. Micropropagation of a mature leguminous tree - Bauhinia purpurea. Plant Cell,
Tissue and Organ Culture, Amsterdam, v. 31, p. 257-259, 1992.
LEAL, I. R. et al. Mudando o curso da conservação da biodiversidade na Caatinga do
Nordeste do Brasil. Megadiversidade, Belo Horizonte, MG, v. 1, n. 1, jul. 2005.
LEITZKE, L. N.; DAMIANI, C. R.; SCHUCH, M. W. Meio de cultura, concentração de AIB
e tempo de cultivo no enraizamento in vitro de amoreira-preta e framboeseira. Rev. Bras.
Frutic., Jaboticabal, SP, v. 31, n. 2, p. 582-587, jun. 2009.

LINO, C. S. de. et al. Antidiabetic activity of Bauhinia forficata extracts in alloxan-
diabetic rats. Biological and Pharmaceutical Bulletin, Japão, v. 27, n. 1, p. 125-127. 2004.
LLOYD, G.; McCOWN, B. Use of microculture for production and improvement of
Rhododendron spp. HortScience, Alexandria, v.15, p.415, 1980. (Abst. 321)
LORENZI, H. Árvores brasileiras. Manual de identificação e cultivo de plantas arbóreas
nativas do Brasil. Nova Odessa: 1992. 352p.
LORENZI, H.; MATOS, F. J. de A. Plantas Medicinais no Brasil: Nativas e exóticas. 2.ed.
Nova Odessa, SP: Instituto Plantarum, 2008. 544 p.
LUCENA, R. F. P. et al. Useful Plants of the Semi-Arid Northeastern Region of Brazil – A
Look at their Conservation and Sustainable Use. Environ Monit Assess, v. 125, p. 281-290,
2007.
LUNA, J. DE S. et al. A study of the larvicidal and molluscicidal activities of some medicinal
plants from northeast Brazil. Journal of Ethnopharmacology, v. 97, n. 2, p. 199-206, fev.
2005.
MACEDO, M.; FERREIRA, A. R. Plantas hipoglicemiantes utilizadas por comunidades
tradicionais na Bacia do Alto Paraguai e Vale do Guaporé, Mato Grosso-Brasil. Revista
Brasileira de Farmacogonosia, João Pessoa, PB, v. 14, supl. 01, p. 45-47, 2004.
MACHAKOVA, I.; ZAZIMALOVA, E.; GEORGE E. F. Plant Growth Regulators I:
Introduction: Auxins, their Analogues and Inhibitors. In: GEORGE, E. F. et al. (Ed.), Plant
Propagation by Tissue Culture. v. 1, 3.ed. Dordrecht: Springer, 2008. p. 175-204.
MAIA, G. N. Caatinga: árvores e arbustos e suas utilidades. São Paulo: D&Z Computação
grafica e Editora, 2004. 380 p.
MATHUR, J.; MUKUNTHAKUMAR, S. Micropropagation of Bauhinia variegata and
Parkinsonia aculeata from nodal explants of mature trees. Plant Cell, Tissue and Organ
Culture, Amsterdam, v. 28, n. 119-121, 1992.
MELLO, M. O. et al. Growth of Bauhinia forficata Link, Curcuma zedoaria Roscoe and
Phaseolus vulgaris L. cell suspension cultures with carbon sources. Scientia Agricola,
Piracicaba, SP, v. 58, n. 3, p. 481-485, jul./set. 2001.
MERKLE, S. A.; NAIRN, C. J. Hardwood tree biotechnology. In Vitro Cellular and
Developmental Biology. Plant, Wallingford, UK, v. 41, p. 602-619, set./out. 2005.
MOREIRA-DIAS, J. M. et al. Day length and photon flux density influence the growth
regulator effects on morphogenesis in epicotyl segments of Troyer citrange. Sciencia
Horticulturae, v. 87, p. 275-290, 2001.
MOSHKOV, I. E. et al. Plant Growth Regulators III: Gibberellins, Ethylene, Abscisic Acid,
their Analogues and Inhibitors; Miscellaneous Compounds. In: GEORGE, E. F. et al. (Ed.),
Plant Propagation by Tissue Culture, v. 1, 3.ed. Dordrecht: Springer, 2008. p. 277-282.

MOURA, T. L. et al. Citrus in vitro organogenesis related to BAP concentrations and explant
section. Revista Brasileira de Fruticultura, Jaboticabal, SP, v. 23, p. 240-245, 2001.
MURASHIGE T.; TUCKER D. P. H. Growth factor requirements of citrus tissue culture.
Proc First Int Citrus Symp, v. 3, p. 1155–1161, 1969.
NAYAK, P. et al. High frequency plantlet regeneration from cotyledonary node cultures of
Aegle marmelos (L.) Corr. In vitro Cell Developmental Biology–Plant, Wallingford, UK, v.
43, n. 3, p.231-236, maio 2007.
NEPOMUCENO, C. F. et al. Respostas morfofisiológicas in vitro de plântulas de
Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb) Altschul. Revista Árvore, Viçosa,
MG, v. 33, n. 3, p. 481-490, 2009.
NGUYEN, Q. T.; KOZAI, T. Photoautotrophic micropropagation of tropical and subtropical
woody plants. In: MOROHOSHI, N.; KOMAMINE, A. (Ed.). Molecular Breeding of Woody
Plants. Elsevier Science, 2001. p. 335-344
NHUT, D. T. et al. Peptone stimulates in vitro shoot and root regeneration of avocado
(Persea americana Mill.). Scientia Horticulturae, Amsterdam, v. 115, p. 124–128, 2008.
NICIOLI, P. M. et al. Ajuste do processo de micropropagação de barbatimão. Ciência Rural,
Santa Maria, v. 38, n. 3, p.685-689, maio/jun. 2008.
OLIVEIRA, A. M. F. et al. Atividade Hipoglicemiante de Bauhinia cheilantha (Bong.)
Steudel. In: 25ª REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE QUÍMICA (SBQ),
2002, Poços de Caldas-MG. Anais ... Poços de Caldas-MG: Sociedade Brasileira de Química,
2002. Disponivel em: <http://sec.sbq.org.br/cdrom/30ra/resumos/T1127-1.pdf> Acessado em:
15 abr. 2009.
OLIVEIRA, A. M. F. et al. Estudo Sazonal da Atividade Hipoglicemiante e Antioxidante de
Bauhinia cheilantha. In: XX SIMPÓSIO DE PLANTAS MEDICINAIS DO BRASIL / X
CONGRESSO INTERNACIONAL DE ETNOFARMACOLOGIA, 2008, São Paulo, Centro
de Convenções Frei Caneca CD Room, resumo 06.365 (2008a)
OLIVEIRA, A. M. F. Estudo Químico e Avaliação da Atividade Hipoglicemiante e
Antioxidante de Bauhinia cheilantha (Bong.) Steudel. 2008. 1v. 131f. Dissertação (Mestrado
em produtos naturais e sintéticos bioativos) - Universidade Federal da Paraíba, João Pessoa.
(2008b)
OLIVEIRA, C. de.; SANTANA, L. A.; CARMONA, A. K.; CEZARI, M. H.; SAMPAIO, M.
U.; SAMPAIO, C. A. M.; OLIVA, M. L. V. Structure of cruzipain/cruzain inhibitors isolated
from Bauhinia bauhinioides seeds. Biology Chemistry, Berlim, v. 383, p. 847-852, 2001.
OLIVEIRA, R. M. DE; VELHO, L. Benefícios e riscos da proteção e comercialização da
pesquisa acadêmica: uma discussão necessária. Ensaio: aval. pol. públ. Educ., Rio de Janeiro,
RJ, v. 17, n. 62, p. 25-54, jan./mar. 2009.

OLIVEIRA, R. P.; NINO, A. F. P. Potencial de Multiplicação in vitro de cultivares de
framboeseira. Revista Brasileira de Fruticultura, Jaboticabal, SP, v. 31, n. 1, p. 280-284, mar.
2009.
PASQUAL, M. Textos acadêmicos: meios de cultura. Lavras: FAEPE/UFLA, 2001. 127p.
PASQUAL, M.; HOFFMANN, A.; RAMOS, J. D. Cultura de tecidos vegetais: Tecnologia e aplicações: introdução: fundamentos básicos. Lavras: UFLA/FAEP. p. 7-45. 1997.
PATIL, G. et al. Glutamine improves shoot morphogenesis in chickpea (Cicer arietinum L.).
Acta Physiology Plant, v. 31, p.1077–1084, 2009.
PEIXOTO SOBRINHO, T. J. S. et al. Processos Extrativos para Flavonóides de Bauhinia
Cheilantha (Bong) Steudel (Pata-De-Vaca). In: 47º CBQ, 2007, Natal- RN. Anais ... Natal –
RN: Realização ABQ-RN, 2007. Disponível em:
<http://www.abq.org.br/cbq/2007/trabalhos/7/7-83-218.htm>. Acesso em: 07 abr. 2009.
PEIXOTO SOBRINHO, T. J. S. et al. Validação de metodologia espectrofotométrica para
quantificação dos flavonóides de Bauhinia cheilantha (Bongard) Steudel. Revista Brasileira
de Ciências Farmacêuticas, São Paulo, SP, v. 44, n. 4, out./dez., 2008.
PERES, L. E. P. Bases Fisiológicas e Genéticas da regeneração de plantas in vitro.
Biotecnologia Ciência & Desenvolvimento, n. 25, mar./abr. 2002.
PERES, L. E. P.; KERBAUY, G. B. Citocininas. In: KERBAUY, G. B. Fisiologia Vegetal.
2.ed. Rio de Janeiro: Guanabara Koogan, 2008. p. 212-234.
PETERNEL, S. et al. In vitro propagation of European aspen (Populus tremula L.) from
axillary buds via organogenesis. Scientia Horticulturae, Amsterdam, v. 121, n. 1, p. 109-112,
2009.
PIRES, M. V. et al. Etnobotânica de terreiros de candomblé nos municípios de Ilhéus e
Itabuna, Bahia, Brasil. Revista Brasileira de Biociência, Porto Alegre, v. 7, n. 1, p. 3-8,
jan./mar. 2009.
PRADHAN, C. et al. Propagation of Dalbergia sissoo Roxb. Through in vitro shoot
proliferation from cotyledonary nodes. Plant Cell Reports, v. 18, p. 122-126, 1998.
PRAKASH, S.; STADEN, J. V. Micropropagation of Searsia dentata. In Vitro Cellular and
Developmental Biology Plant, Wallingford, UK, v. 44, p. 338-341, 2008.
QUEIROZ, L. P. DE et al. Leguminosae. In: GIULIETTI, A. M. et al. (Org.), Co-editora:
Universidade Estadual de Feira de Santana. Plantas Raras do Brasil, Belo Horizonte, MG:
Conservação Internacional, 2009. p. 212-237. (2009a)
QUEIROZ, L. P. Leguminosas da Caatinga. Feira de Santana: Universidade Estadual de Feira
de Santana, 2009, 467p. (2009b)
QUOIRIN, M.; LEPOIVRE, P. Improved media for in vitro culture of Prunus sp. Acta Hort.,
v. 78, p. 437–442, 1977.

RAJESWARI, V.; PALIWAL, K. In vitro propagation of Albizia odoratissima L.F. (Benth.)
from cotyledonary node and leaf nodal explants. In Vitro Cellular and Developmental Biology
– Plant, Wallingford, UK, v. 42, n. 5, p. 399-404, set. 2006.
RAGHU, A. V. et al. In vitro clonal propagation through mature nodes of Tinospora
cordifolia (Willd.) Hook. F. & Thoms.: an important Ayurvedic Medicinal Plant. In Vitro
Cellular and Developmental Biology – Plant, Wallingford, UK, v. 42, p. 584–588, nov./dez.
2006.
SANTOS-SEREJO, J. A. et al. Meios Nutritivos para Micropropagação de Plantas. In:
SOUZA, A. S.; JUNGHANS, T. G. Introdução à micropropagação de plantas. Cruz das
Almas: Embrapa Mandioca e Fruticultura, Tropical, 2006. p. 80-97.
SCHINOR, E. H. et al. Organogênese in vitro a partir de diferentes regiões do epicótilo de
Citrus sp. Revista Brasileira Fruticultura, Jaboticabal, SP, v. 28, n. 3, p. 463-466, dez. 2006.
SEIFFERT, M. Estudos de alguns aspectos de germinação e bioquímicos de sementes de
Bauhinia cheiltanha (Bong.) Steud., sob diferentes condições de armazenamento. 2006. 1v.
69f. Tese (Doutorado em Botânica) – Faculdade de Ciências Biológicas, Universidade
Estadual Paulista Júlio Mesquita Filho, Botucatu.
SHAN, L. et al. High efficiency in vitro plant regeneration from epicotyls explants of Chinese
peanut cultivars. In Vitro Cellular and Developmental Biology – Plant, Wallingford, UK, v.
45, p. 525–531, 2009.
SHAW, J. E.; SICREE, R. A.; ZIMMET, P. Z. Global estimates of the prevalence of diabetes
for 2010 and 2030. Diabetes Research and Clinical Practice, Irlanda, v. 87, p. 4-14, 2010.
SILVA, A. M. O. et al. Antinociceptive activity of the aqueous extract of Bauhinia cheilantha
(Bong.) Steud. (Leguminosae: Caesalpinioideae). Biologia Geral e Experimental, São
Cristovão, SE, v. 5, n. 2, p. 10-15, 2005.
SILVA, F. A. B. DA; PEREIRA, L. A. R.; SILVEIRA, C. E. DOS S. Micropropagation of
Alibertia edulis Rich. Brazilian Archives of Biology and Technology, Curitiba, PR, v. 51, n. 6,
p.1103-1114, nov./dez. 2008.
SILVA, K. L.; CECHINEL-FILHO, V. Plantas do gênero Bauhinia: composição química e
potencial farmacológico. Química Nova, São Paulo, SP, v. 25, n. 3, p.449-454, 2002.
SOSA, S. et al. Topical anti-inflammatory activity of Bauhinia tarapotensis leaves.
Phytomedicine, v. 9, n. 7, p. 646-653, 2002.
SOUZA, A. V.; PEREIRA, A. M. S. Enraizamento de plantas cultivadas in vitro. Revista de
Brasileira de Plantas Medicinais, Botucatu, SP, v. 9, n. 4, p. 103-117, 2007.
SOUZA, F. V. D.; COSTA, M. A. P.; SILVA-NETO, H. P. Aclimatização. In: SOUZA, A.
S.; JUNGHANS, T. G. Introdução à micropropagação de plantas. Cruz das Almas: Embrapa
Mandioca e Fruticultura, Tropical, 2006. p. 131-140.

SOUZA, J. A. DE et al. Tipos e concentrações de citocinina na multiplicação in vitro de
pitangueira. Ciência Rural, Santa Maria, v. 38, n. 7, out. 2008.
SRISKANDARAJAH, S.; LUNDQUIST, P.-O. High frequency shoot organogenesis and
somatic embryogenesis in juvenile and adult tissues of seabuckthorn (Hippophae rhamnoides
L.). Plant Cell Tiss Organ Culture, Amsterdam, v. 99, p. 259-268, 2009.
STADEN, J. V.; ZAZIMALOVA, E.; GEORGE, E. F. Plant Growth Regulators II:
Cytokinins, their Analogues and Antagonists. In: GEORGE, E. F. et al. (Ed.), Plant
Propagation by Tissue Culture, v. 1, 3.ed. Dordrecht: Springer, 2008. p. 205-226.
SUDHA, C. G. et al. In vitro propagation of Decalepis arayalpathra, a critically endangered
ethnomedicinal plant. In Vitro Cellular and Developmental Biology – Plant, Wallingford, UK,
v. 41, n. 5, p. 648-654, set. 2005.
TABARELLI, M.; VICENTE, A. Conhecimento sobre plantas lenhosas da Caatinga: lacunas
geográficas e ecológicas. In: SILVA, J. M. C. et al. (Ed.) Biodiversidade da Caatinga: áreas e
ações prioritárias para a conservação. Brasília, Ministério do Meio Ambiente, 2004. p. 101-
110.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 4. ed. Porto Alegre: Artmed, 2009. 819 p.
TAMTA, S. et al. In vitro propagation of brown oak (Quercus semecarpifolia Sm.) from
seedling explants. In Vitro Cellular and Developmental Biology. Plant, Wallingford, UK, v.
44, p. 136-141, 2008.
TAVANO, E. C. R. et al. In vitro organogenesis of Citrus volkameriana and Citrus
aurantium. Biologia Plantarum, República Tcheca, v. 53, n. 2, p. 395-399, 2009.
TERMIGNONI, R. R. Cultura de tecidos vegetais. Porto Alegre: Ed. UFRGS, 2005, 182 p.
THOMAS, T. D. The role of activated charcoal in plant tissue culture. Biotechnology
Advances, v. 26, p. 618-631, 2008.
THORPE, T. et al. The Components of Plant Tissue Culture Media II : Organic Additions,
Osmotic and pH Effects, and Support Systems. In: GEORGE, E. F. et al. (Ed.), Plant
Propagation by Tissue Culture, v. 1, 3.ed. Springer, 2008. p. 115-174.
TILKAT, E. et al. Direct plant regeneration from mature leaf explants of pistachio, Pistacia
vera L. Scientia Horticulturae, Amsterdam, v. 121, p. 361–365, 2009.
TREVISAN, M. T. S.; MACEDO, F. V. V. Seleção de plantas com atividade anticolinesterase
para tratamento da doença de Alzheimer. Química Nova, São Paulo, SP, v. 26, n. 3, 301-304,
2003.
UPRETI, J.; DHAR, U. Micropropagation of Bauhinia valii Wight & Arnott – a leguminous
liana. Plant Cell Report, v. 16, p. 250–254, 1996.
USPTO. United States Patent and Trademark Office. Disponível em: <
http://www.uspto.gov/> Acesso em: 29 nov. 2009.

VAZ, A. M. S. F.; TOZZI, A. M. G. A. Bauhinia ser. Cansenia (Leguminosae:
Caesalpinioideae no Brasil). Rodriguésia, Rio de Janeiro, RJ, v. 54, p. 55-143, 2003.
VAZ, A. M. S. F; TOZZI, A. M. G. A. Sinopse de Bauhinia sect. Pauletia (Cav.) DC.
(Leguminosae: Caesalpinioideae: Cercideae) no Brasil. Revista Brasileira de Botânica, São
Paulo, SP, v. 28, n. 3, jul./set. 2005.
VIEITEZ, A. M. et al. In vitro regeneration of the important North American oak species
Quercus alba, Quercus bicolor and Quercus rubra. Plant Cell Tiss Organ Culture,
Amsterdam, v. 98, p. 135–145, 2009.
VILLA, F. et al. Cloreto de potássio e fosfato de sódio na multiplicação in vitro de amoreira-
preta cv. Tupy. Ciência e Agrotecnologia, Lavras, v. 32, n. 1, p. 37-41, 2008.
VILLA, F. et al. Micropropagação de duas espécies frutíferas, em meio de cultura DSD1,
modificado com fontes de boro e zinco. Ciência e Agrotecnologia, Lavras, v. 33, n. 2, p. 468-
472, mar./abr., 2009.
XAVIER, A.; OTONI, W. C.; PENCHEL, R. M. Micropropagação e enxertia in vitro de
espécies florestais. In: BORÉM, A. (Ed.). Biotecnologia Florestal. Suprema Gráfica e
Editora: Viçosa, 2007, p. 53-74.
ZENG, L. et al. High efficiency in vitro plant regeneration from epicotyls explants of Ponkan
Mandarin (Citrus reticulata Blanco). In Vitro Cellular and Developmental Biology. Plant,
Wallingford, UK, v. 45, p. 559–564, 2009.
ZIV, M.; CHEN, J. The Anatomy and Morphology of Tissue Cultured Plants. In: In:
GEORGE, E. F. et al. (Ed.), Plant Propagation by Tissue Culture, v. 1, 3.ed. Dordrecht:
Springer, 2008. p. 465- 478.


















