
UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE … · axonopodis pv. citri para sua futura...
96
UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE QUÍMICA VERÔNICA LEITE QUEIROZ CARACTERIZAÇÃO DE CELULASE RECOMBINANTE DE XANTHOMONAS AXONOPODIS PV. CITRI PARA APLICAÇÃO À HIDRÓLISE ENZIMÁTICA DO BAGAÇO DE LARANJA CAMPINAS 2017
-
Upload
nguyencong -
Category
Documents
-
view
213 -
download
0
Transcript of UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE … · axonopodis pv. citri para sua futura...

UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE QUÍMICA
VERÔNICA LEITE QUEIROZ
CARACTERIZAÇÃO DE CELULASE RECOMBINANTE DE XANTHOMONAS
AXONOPODIS PV. CITRI PARA APLICAÇÃO À HIDRÓLISE ENZIMÁTICA DO BAGAÇO DE LARANJA
CAMPINAS 2017

VERÔNICA LEITE QUEIROZ
CARACTERIZAÇÃO DE CELULASE RECOMBINANTE DE XANTHOMONAS
AXONOPODIS PV. CITRI PARA APLICAÇÃO À HIDRÓLISE ENZIMÁTICA DO BAGAÇO DE LARANJA
Dissertação de Mestrado apresentada ao Instituto de
Química da Universidade Estadual de Campinas como
parte dos requisitos exigidos para a obtenção do título
de Mestra em Química na área de Química Orgânica
Orientadora: Profa. Dra. Ljubica Tasic
ESTE EXEMPLAR CORRESPONDE À VERSÃO FINAL DA DISSERTAÇÃO DEFENDIDA PELA ALUNA VERÔNICA LEITE QUEIROZ, E ORIENTADA PELA PROFA. DRA. LJUBICA TASIC.
CAMPINAS 2017


BANCA EXAMINADORA
Profa. Dra. Ljubica Tasic (Orientadora)
Profa. Dra. Anete Pereira de Souza (IB-UNICAMP)
Profa. Dra. Luciana Gonzaga de Oliveira (IQ-UNICAMP)
A Ata da defesa com as respectivas assinaturas dos membros encontra-se no processo
de vida acadêmica do(a) aluno(a).
Este exemplar corresponde à redação final da
Dissertação de Mestrado defendida pelo aluno
VERÔNICA LEITE QUEIROZ, aprovada pela Comissão
Julgadora em 22 de junho de 2017.

À minha família, por todo apoio, carinho e amor.

AGRADECIMENTOS
Eu agradeço primeiramente à minha orientadora, Profª. Drª. Ljubica Tasic, pela
oportunidade de realizar esse trabalho, pela sua orientação e ensinamentos, pela amizade e
paciência, que permitiram a realização e conclusão dessa tarefa. Tenho orgulho de ter feito
parte desse grupo de pesquisa diversificado, que possibilitou que eu ampliasse minha visão
com relação a bioquímica e suas aplicações e que enriqueceu o meu aprendizado.
Agradeço à minha família que me incentivou e me apoiou durante o
desenvolvimento desse trabalho. Ao meu pai por sempre ter me encorajado a continuar com
a minha formação, e a minha mãe por todo o amor e carinho. À minha irmã Giovanna, pelas
conversas sempre cheias de atenção e amor. Agradeço ao meu marido Gabriel, por fazer parte
da minha vida, por todo o amor, compreensão, paciência e por nunca me deixar desistir. Todos
vocês são muito importantes na minha vida!
Aos meus líderes e colegas da Braskem, muito obrigada pelas sugestões,
discussões e apoio no desenvolvimento desse trabalho. Agradeço especialmente a minha
colega de trabalho e amiga Débora, que tive a sorte de conhecer na Braskem, obrigada por
toda a ajuda, ensinamentos e apoio.
Agradeço também às minhas melhores amigas, Isabela, Carol, Marilene e Thaís,
vocês fazem minha vida melhor e mais divertida, além de serem exemplos para mim.
Agradeço a minha banca de qualificação pelas sugestões e comentários.
Aos meus colegas do grupo do Laboratório de Química Biológica, obrigada pelas
contribuições, participações e apoio.
Agradeço também aos funcionários do Instituto de Química da Unicamp por todo
o auxílio e dedicação.

“Dans la vie rien n’est à craindre, tous est à comprendre”
Marie Curie

RESUMO
Palavras-chave: bagaço de laranja, Xanthomonas axonopodis pv. citri, hidrólise, enzimas,
celulase, etanol-2G.
O bagaço da laranja possui baixo custo, elevados níveis de carboidratos, alta susceptibilidade
à hidrólise enzimática, e sua utilização não compete com a produção de alimentos; tais
características tornam o bagaço uma promissora fonte de carbono para a produção de etanol
de segunda geração. Somam-se a isto o fato do Brasil ser o maior produtor de laranja do
mundo, e de apenas 50% da fruta ser utilizada para a produção de suco, o restante (bagaço)
é subaproveitado, sendo o principal destino a produção de pellets para alimentação de gado.
Nesse projeto foram realizados estudos com três diferentes tipos de biomassa semissólida
provenientes do processo de produção do suco de laranja e, também, do bagaço in natura
obtido localmente. Com esse projeto buscou-se a melhora do rendimento da hidrólise do
bagaço de laranja para a produção de etanol de segunda geração. Como objetivo visou-se a
caracterização bioquímica e biofísica de uma celulase recombinante de Xanthomonas
axonopodis pv. citri para sua futura aplicação à hidrólise enzimática do bagaço de laranja. A
celulase de Xanthomonas axonopodis pv. citri teve seus parâmetros cinéticos determinados,
mostrando-se ativa frente a hidrólise de carboximetil celulose, além disso foram
determinados a temperatura e pH ótimo de atividade. O estudo de sua estrutura secundária
através de dicroísmo circular e de estrutura terciária via fluorescência, também, foram
conduzidos com sucesso.

ABSTRACT
Key words: orange bagasse, Xanthomonas axonopodis pv. citri, hydrolysis, enzymes, cellulase,
ethanol-2G.
Orange bagasse is a low cost agro-industrial residue that possesses high carbohydrate levels,
high susceptibility to enzymatic hydrolysis, and it does not compete with food production;
such characteristics make bagasse a promising source of carbon for the production of second-
generation ethanol. In addition to the fact that Brazil is the greatest orange producer in the
world, and that only 50% of the fruit is used in a juice production, orange fruit residue
(bagasse) is underutilized, whose main destination is pellets production and use for livestock
feed. In this research, we have carried out experiments on three types of semi-solid biomass
from the orange juice production process, and on in natura bagasse obtained locally. Our aims
were to improve the yields of the hydrolysis of orange bagasse and posterior fermentation to
a second-generation ethanol. Our goals also included biochemical and biophysical
characterization of a recombinant cellulase from the Xanthomonas axonopodis pv. citri (Xac)
and its application in enzymatic hydrolysis of orange bagasse. This Xac cellulase had its kinetic
parameters determined, and it showed to be active in carboxymethyl cellulose hydrolysis. In
addition, its optimum working pH and temperature conditions were determined. Its
secondary structure features through circular dichroism and tertiary structure features via
fluorescence were also successfully evaluated.

LISTA DE ILUSTRAÇÕES
Figura 1: Gráfico construído com os dados obtidos do relatório da USDA de 2017 de produção
mundial de laranjas no ano de 20167.
Figura 2: Estrutura morfológica da laranja.
Figura 3: Composição do bagaço de laranja seco14,15.
Figura 4: Representação da estrutura da parede celular e suas subunidades.
Figura 5: Representação esquemática de uma molécula de celulose24.
Figura 6: Representação das ligações de hidrogênio supramolecular da celulose26.
Figura 7: Representação da molécula de hemicelulose29.
Figura 8: Representação de uma estrutura interna de lignina31.
Figura 9: Representação de uma molécula de pectina.
Figura 10: Representação da molécula de (R)-(+)-Limoneno e (S)-(-)-limoneno.
Figura 11: A) Laranja infectada pela bactéria Xanthomonas axonopodis pv citri (Xac). B)
Microscopia eletrônica de varredura da Xac (cepa 306).
Figura 12: Representação esquemática da atuação das enzimas envolvidas na hidrólise da
celulose.
Figura 13: Representação da luz polarizada circularmente utilizada na técnica de dicroísmo
circular.
Figura 14: Exemplo de dados obtidos por dicroísmo circular para diferentes tipos de proteínas.
A curva em preto foi obtida da poli-L-lisina em pH 11,1 na conformação de α-hélice; a curva
vermelha corresponde ao espectro da mesma proteína, porém na conformação de folha-β
antiparalela nas condições de pH 5,7. Na curva em ciano trata-se do colágeno desnaturado
em forma de uma proteína randômica, e nas curvas azul e verde o colágeno na conformação
estendida (Figura obtida dos espectros fornecidos por Dr. W. C. Johnson58).

Figura 15: Espectro de absorção e emissão dos aminoácidos fenilalanina, tirosina e triptofano
em pH 7 61.
Figura 16: Amostra de bagaço de laranja industrial contendo folhas e fibras antes e depois de
ser triturado.
Figura 17: Fluxograma do processo de produção da laranja e seus derivados.
Figura 18: A) Amostra de dorna industrial - licor. B) Amostra de pellet de bagaço de laranja
(Citrosuco, Matão, SP).
Figura 19: Isolamento de colônias a partir do bagaço da laranja em meio sólido YPD.
Figura 20: Gráfico de crescimento de colônias em YPD líquido contendo 0,5 % de d-limoneno
após 24 h a 30 °C com agitação.
Figura 21: Gráfico de crescimento das colônias em 24 h de cultivo a 30 °C com agitação de 200
rpm, medidas realizadas a 600 nm.
Figura 22: Gráfico de consumo de glicose medido ao longo de 24 h por HPLC.
Figura 23: Gráfico de produção de etanol durante 24 h medida realizada por HPLC.
Figura 24: Árvore filogenética demonstrando as relações evolutivas entre a sequência parcial
da região D1/D2 da amostra CPQBA 1332-16 DRM 01, correspondente a cepa 9, e as
sequencias de linhagens de micro-organismos relacionados presentes nas bases de dados CBS
e GenBank.
Figura 25: Gel de agarose 1% com marcador 1 kb e amostra de DNA genômico de Xac.
Figura 26: Gel obtido de um dos testes de amplificação. Nele é possível observar a amplificação
pouco especifica e também o efeito da temperatura de anelamento, que quanto mais alta
mais específica.
Figura 27: Predição de peptídeo sinal obtida através do servidor SignalP 4.1. C-score
corresponde a pontuação “raw cleavage site”; S-score a pontuação de peptídeo sinal e Y-score
a pontuação combinada de sítio de clivagem.
Figura 28: Domínios preditos utilizando a base de dados Pfam.

Figura 29: Mecanismos propostos para glicosil hidrolases com inversão de configuração (A) e
com retenção da configuração (B)81.
Figura 30: Foto do gel (12%) de acrilamida contendo as amostras de pET28 sem o gene e de
pET28 contendo a celulase em diferentes condições de expressão: 18 °C por 24 h, 28 °C por 4
h e 37 °C por 3 h. As setas indicam a presença da celulase alvo no Gel.
Figura 31: Gel 12% de acrilamida contendo as amostras de pET28 sem o gene e de pET28
contendo a celulase em diferentes concentrações de IPTG durante a expressão: 0,1 mmol L-1,
0,5 mmol L-1 e 1 mmol L-1.
Figura 32: SDS-PAGE das amostras de purificação da celulase recombinante. O gel contém
amostra da célula antes da indução com IPTG (não induzido) e após a indução (induzido – ME).
Também são mostrados as frações do início da purificação que não correspondem a proteína
de interesse (frações de 1 a 12) e as frações contendo a celulase recombinante em alto grau
de pureza nas frações de 15 e 16.
Figura 33: Cromatograma da purificação da celulase heteróloga em Origami 2 (DE3). O
segundo pico corresponde ao sinal da proteína de interesse.
Figura 34: Curva de calibração do fator k, calculado através da equação I, versus o logaritmo
da massa molar. A equação linear também é mostrada abaixo do gráfico.
Figura 35: Cromatograma de filtração em gel analítica da amostra de celulase previamente
purificada por afinidade. O pico correspondente a amostra apresenta tempo de eluição em
10,45 mL.
Figura 36: Predição da estrutura terciária da celulase de Xac obtida através da ferramenta I-
Tasser e visualizada através da ferramenta PyMol. Na primeira imagem estão as medidas de
comprimento e largura aproximados da proteína e na segunda imagem a proteína vista com
giros de 90°.
Figura 37: Solução 1% de CMC incubada com 200 nmol L-1 de celulase purificada por 30 min a
35 °C. É possível observar a alteração na viscosidade do tubo contendo a enzima versus o tubo
controle.
Figura 38: Produção de açúcares redutores em diferentes tempos de incubação, ensaio
realizado com 200 nmol L-1 de enzima, em tampão 50 mmol L-1 Na2HPO4/ácido cítrico pH 6 e
temperatura de 35 °C.

Figura 39: Influência do pH na atividade frente a CMC da celulase recombinante de Xac, ensaio
realizado com 200 nmol L-1 de enzima, 35 °C, 30 min de incubação e em diferentes valores de
pH do tampão 50 mmol L-1 Na2HPO4/ácido cítrico.
Figura 40: Influência da temperatura na atividade frente a CMC da celulase recombinante de
Xac, ensaio realizado em tampão 5 mmol L-1 Na2HPO4/ácido cítrico pH 6,5, 30 min de
incubação e 200 nmol L-1 de enzima.
Figura 41: Gráfico de velocidade inicial para a celulase recombinante de Xac, ensaio realizado
em tampão 50 mmol L-1 Na2HPO4/ácido cítrico pH 6,5, 30 °C, tempo de incubação de 30 min e
200 nmol L-1 da celulase recombinante de Xac.
Figura 42: Gráfico duplo-recíproco de Lineweaver-Burk. Efeito da concentração de substrato
(CMC) na velocidade da reação.
Figura 43: Espectro de dicroísmo circular obtido para a amostra purificada de celulase em
tampão 50 mmol L-1 ácido cítrico/Na2HPO4 pH 6,5 a 25 °C e 200 nmol L-1 de enzima.
Figura 44: Estabilidade térmica monitorada por dicroísmo circular a 222 nm de 20 a 90°C. Os
círculos fechados são os pontos referentes a curva de desnaturação e os círculos abertos os
pontos referentes a tentativa de enovelamento com diminuição da temperatura. O ensaio foi
realizado em tampão 50 mmol L-1 Na2HPO4/ácido cítrico, pH 6,5 com 200 nmol L-1 da enzima.
Figura 45: Estabilidade térmica monitorada por dicroísmo circular a 222 nm de 20 a 60°C. Os
círculos fechados são os pontos referentes a curva de desnaturação e os círculos abertos os
pontos referentes a tentativa de enovelamento com diminuição da temperatura. O ensaio foi
realizado em tampão 50 mmol L-1 Na2HPO4/ácido cítrico, pH 6,5 com 200 nmol L-1 da enzima.
Figura 46: Espectro de dicroísmo circular obtido para a amostra purificada de celulase (200
nmol L-1) em tampão 50 mmol L-1 ácido cítrico/Na2HPO4 em diferentes valores de pH a 25 °C.
Figura 47: Modelagem realizada no I-Tasser para a celulase recombinante de Xac. Em rosa
estão destacados os resíduos de triptofano da proteína recombinante de Xac.
Figura 48: Espectro de excitação para a proteína recombinante de Xac no tampão 50 mmol L-
1 Na2HPO4 a 25°C e pH 6,5.
Figura 49: Espectro de emissão para a proteína recombinante de Xac obtido em diferentes
valores de pH no tampão 50 mmol L-1 Na2HPO4/ácido cítrico e temperatura de 25 °C.

LISTA DE TABELAS
Tabela 1: Tabela contendo os dados obtidos da caracterização do bagaço de laranja comercial
e industrial
Tabela 2: Período de colheita por variedade de laranja e percentual da produção, tabela
retirada de “O retrato da Citricultura brasileira, 2010” 6
Tabela 3: Tabela contendo os dados obtidos da caracterização da amostra de licor industrial
(amostra Citrosuco, Matão, SP)
Tabela 4: Tabela contendo o resumo das informações das proteínas de Xac escolhidas para
serem amplificadas e clonadas em E. coli
Tabela 5: Tabela contendo os oligos diretos (primer forward – PF) e oligos reversos (primer
reverse – PR) usados na amplificação de cada sequência
Tabela 6: Valores encontrados para o cálculo da massa molar da celulase recombinante
Tabela 7: Parâmetros cinéticos da celulase recombinante de Xac utilizando CMC como
substrato
Tabela 8: Parâmetros cinéticos de celulases de diferentes microrganismos utilizando CMC
como substrato
Tabela 9: Comparação dos dados obtidos de estrutura secundária da celulase de Xac

SUMÁRIO
1. Introdução 17
1.3. Fibras Lignocelulósicas 19
1.3.1. Celulose 20
1.3.2. Hemicelulose 21
1.3.3. Lignina 22
1.3.4. Pectina 23
1.4. Óleo Essencial 24
1.5. Xanthomonas axonopodis pv. citri 24
1.6. Aplicação de enzimas na indústria 26
1.7. Tipos de pré-tratamento para hidrólise de biomassa 26
1.8. Hidrolases 28
1.9. Tipos de hidrólise 28
1.10. Hidrólise enzimática 29
1.10.1. Celulases 29
1.10.2. Pectinases 29
1.12. Caracterização estrutural de proteínas 31
1.12.1. Dicroísmo circular 31
1.12.2. Fluorescência 33
2. Objetivos 35
3. Materiais e métodos 36
3.1. Processamento do bagaço da laranja 36
3.2. Caracterização primária do bagaço de laranja 36
3.2.1. Determinação de sólidos totais e umidade 36
3.2.2. Determinação do conteúdo de cinzas 37
3.3. Determinação de fibras 37
3.3.1. Determinação de fibras em detergente neutro (FDN) 37
3.3.2. Determinação das fibras em detergente ácido (FDA) 38
3.3.3. Determinação de lignina em detergente ácido (LDA) 39
3.4. Determinação de Pectina 40
3.5. Extração do óleo essencial 40
3.6. Determinação de hesperidina 41
3.7. Isolamento de microrganismos do bagaço de laranja 42
3.8. Expressão do gene sintético, lise e purificação 43
3.9. Determinação das condições ótimas de ensaio de atividade 44
3.10. Cinética 44
3.11. Dicroísmo circular 45
3.12. Fluorescência 46
4. Resultados e Discussão 47
Parte I – Caracterização do Bagaço de Laranja e Isolamento de Microrganismo de Laranja 47

4.1. Caracterização do Bagaço de Laranja 47
4.2. Caracterização da amostra de licor e pellet industrial 50
4.3. Isolamento de Colônias de Bagaço de Laranja 52
Parte II – Hidrolase recombinante de Xanthomonas axonopodis pv. citri 57
4.4. Hidrolases de Xanthomonas axonopodis pv. citri 57
4.5. Celulase Q8PJK9 – Gene sintético 60
4.6. Expressão e purificação da celulase recombinante 63
4.6.1. Filtração em gel analítica 67
4.7. Resultados de otimização do ensaio de hidrólise enzimática 70
4.8. Cinética enzimática 73
4.9. Resultados do Dicroísmo Circular 76
4.10. Fluorescência 80
5. Conclusões 83
6. Referências 85
7. Anexos 90
Enzima 1 – Celulase Q8PJK9 90
Enzima 2 – Celulase Q8PPS3 91
Enzima 3 – Beta-Glicosidase Q8PFV3 92
Enzima 4 – Beta-Glicosidase Q8PMI0 93
Enzima 5 – Pectato Liase Q8PGQ4 94
Enzima 6 – Pectato Liase Q8PIB2 95

17
1. Introdução
1.1. Motivação
A busca por fontes renováveis de energia se torna cada vez mais importante como
alternativa sustentável às oriundas do petróleo, especialmente em um cenário mundial onde
as principais potencias e países em desenvolvimento assinaram um acordo, a COP21, no qual
se comprometem a um conjunto de ações visando manter o aumento da temperatura
limitado a 2 °C até o final deste século1.
Atualmente, o biocombustível mais presente no cotidiano é o etanol, que tem 80%
de sua produção vinda de fermentação2,3. A produção mundial de biocombustíveis aumentou
consideravelmente nos últimos anos passando de 18,2 bilhões de litros em 2000 para 60,6
bilhões de litros em 2007, sendo que desse total 85% corresponde ao bioetanol4. No entanto,
a maior parte deste biocombustível é produzida a partir de glicose, sendo nomeado etanol de
primeira geração, que compete com a produção de alimentos5. Como alternativa buscam-se
soluções através de fermentação de substratos chamados de biomassas para produção de
etanol de segunda geração, também conhecidos por substratos lignocelulósicos5.
O bagaço de laranja é uma alternativa para a fermentação e obtenção de etanol
de segunda geração, uma vez que essa biomassa tem baixo custo, não é utilizada para
alimentação direta de humanos e possui alto teor de carboidratos. Além disso, o Brasil é o
maior produtor mundial de laranjas6,7 (Figura 1) com produção em 2016 de aproximadamente
14,3 milhões de toneladas de acordo com o USDA (United States Department of Agriculture –
Departamento de Agricultura dos Estados Unidos)7.
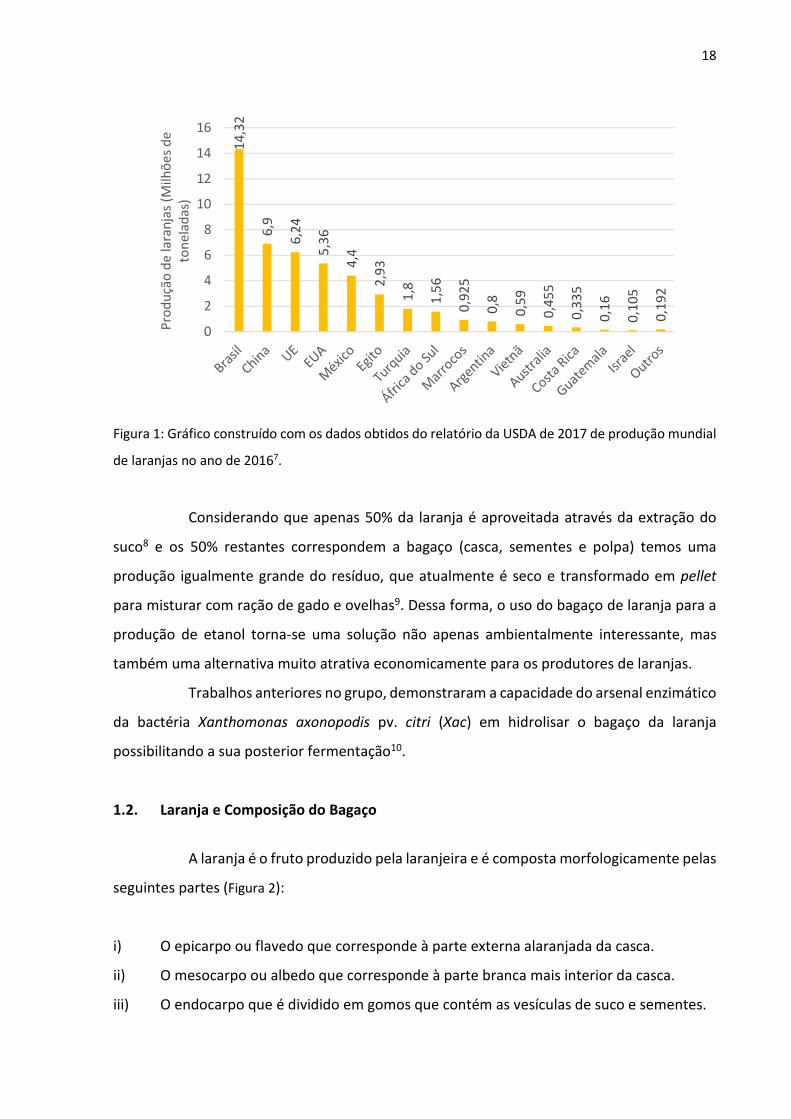
18
Figura 1: Gráfico construído com os dados obtidos do relatório da USDA de 2017 de produção mundial
de laranjas no ano de 20167.
Considerando que apenas 50% da laranja é aproveitada através da extração do
suco8 e os 50% restantes correspondem a bagaço (casca, sementes e polpa) temos uma
produção igualmente grande do resíduo, que atualmente é seco e transformado em pellet
para misturar com ração de gado e ovelhas9. Dessa forma, o uso do bagaço de laranja para a
produção de etanol torna-se uma solução não apenas ambientalmente interessante, mas
também uma alternativa muito atrativa economicamente para os produtores de laranjas.
Trabalhos anteriores no grupo, demonstraram a capacidade do arsenal enzimático
da bactéria Xanthomonas axonopodis pv. citri (Xac) em hidrolisar o bagaço da laranja
possibilitando a sua posterior fermentação10.
1.2. Laranja e Composição do Bagaço
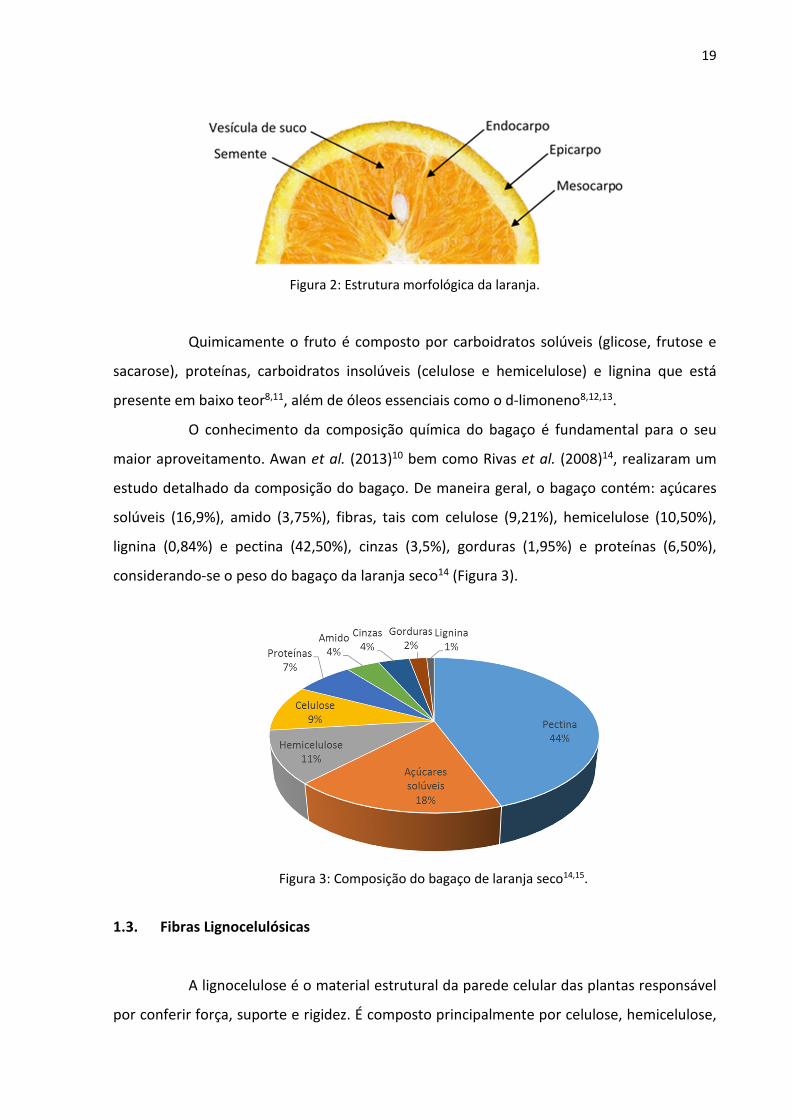
A laranja é o fruto produzido pela laranjeira e é composta morfologicamente pelas
seguintes partes (Figura 2):
i) O epicarpo ou flavedo que corresponde à parte externa alaranjada da casca.
ii) O mesocarpo ou albedo que corresponde à parte branca mais interior da casca.
iii) O endocarpo que é dividido em gomos que contém as vesículas de suco e sementes.
14
,32
6,9
6,2
4
5,3
6
4,4
2,9
3
1,8
1,5
6
0,9
25
0,8
0,5
9
0,4
55
0,3
35
0,1
6
0,1
05
0,1
92
0
2
4
6
8
10
12
14
16
Pro
du
ção
de
lara
nja
s (M
ilhõ
es d
e to
nel
adas
)
19
Figura 2: Estrutura morfológica da laranja.
Quimicamente o fruto é composto por carboidratos solúveis (glicose, frutose e
sacarose), proteínas, carboidratos insolúveis (celulose e hemicelulose) e lignina que está
presente em baixo teor8,11, além de óleos essenciais como o d-limoneno8,12,13.
O conhecimento da composição química do bagaço é fundamental para o seu
maior aproveitamento. Awan et al. (2013)10 bem como Rivas et al. (2008)14, realizaram um
estudo detalhado da composição do bagaço. De maneira geral, o bagaço contém: açúcares
solúveis (16,9%), amido (3,75%), fibras, tais com celulose (9,21%), hemicelulose (10,50%),
lignina (0,84%) e pectina (42,50%), cinzas (3,5%), gorduras (1,95%) e proteínas (6,50%),
considerando-se o peso do bagaço da laranja seco14 (Figura 3).
Figura 3: Composição do bagaço de laranja seco14,15.
1.3. Fibras Lignocelulósicas
A lignocelulose é o material estrutural da parede celular das plantas responsável
por conferir força, suporte e rigidez. É composto principalmente por celulose, hemicelulose,

20
lignina e pectina, tais polissacarídeos não são encontrados de maneira uniforme nas plantas,
sendo que sua composição difere de uma região morfológica a outra16,17.
Basicamente a parede celular é composta por uma camada primária (P) mais
externa e uma camada secundária (S), que por sua vez é composta por três subunidades que
se diferenciam pela orientação das microfibrilas: S1, S2 e S3 e uma camada chamada de lamela
mediana (LM) localizada entre células adjacentes17 (Figura 4).
Figura 4: Representação da estrutura da parede celular e suas subunidades (retirada de
http://www.sobiologia.com.br/conteudos/Citologia/cito4.php - acessado no dia 08/05/2016).
A camada primária (P) é composta principalmente por lignina, é a camada mais é
fina e flexível, o que permite o crescimento celular. Já a camada secundária é muito mais
espessa e rígida e sua composição é basicamente de carboidratos18, formada pelas
subunidades S1 e S3 que são normalmente finas e suas microfibrilas de celulose são
orientadas em uma hélice plana com relação ao eixo de elongação da célula19, a subunidade
S2, por sua vez é mais espessa possui de 75 a 85% das microfibrilas de celulose totais da
espessura da parede celular20. É importante ressaltar que hemicelulose e lignina estão
presentes em todas as camadas3,20
1.3.1. Celulose
A celulose é o material orgânico mais abundante da terra, já que é o principal
componente das plantas, tendo como função dar estrutura a plantas, árvores e gramíneas17,21.
Além disso, a celulose é um material biodegradável e renovável, e pode ser aplicada de
diversas formas devido a propriedades como hidrofobicidade, quiralidade, atoxicidade, alta

21
susceptibilidade a modificações químicas, no entanto boa parte da celulose produzida através
da agricultura e atividades industriais ainda é vista como descarte. Insolúvel na maioria dos
solventes devido as ligações de hidrogênio e cristalinidade22,23, já que é um polímero linear de
unidades de D-glicose ligadas através de ligações 1,4-β-glicosídicas (Figura 5). Com a fórmula
molecular (C6H10O5)n, duas unidades de glicose formam o dímero conhecido como celobiose
as longas cadeias de celulose cristalina fazem ligações de hidrogênio intermoleculares e
intramoleculares, o que confere maior rigidez e torna o ataque enzimático mais difícil22,24
(Figura 6), é interessante observar que os grupos hidroxilas das pontas da cadeia de celulose
possuem propriedades diferentes, o C1 tem propriedades redutoras, enquanto o C4 da
mesma cadeia é não redutor22,25.
Figura 5: Representação esquemática de uma molécula de celulose24.
Figura 6: Representação das ligações de hidrogênio supramolecular da celulose26.
1.3.2. Hemicelulose
Diferente da celulose, a hemicelulose não é quimicamente homogênea27, além de
majoritariamente amorfa e ramificada, trata-se de um heteropolissacarídeo cujo esqueleto é
formado por ligações β-1,4 com configuração equatorial em C1 e C428 composto por pentoses
(β-D-xilose, α-L-arabinose), hexoses (β-D-glicose, β-D-manose, α-L-galactose) e/ou ácidos de

22
açúcares (ácido glicurônico, ácido metilgalacturônico e ácido galacturônico), além disso, a
hemicelulose também pode conter em menores teores outros açúcares como ramnose e
fucose, os grupos hidroxílicos dos açucares também podem estar substituídos parcialmente
por grupos acetil27 (Figura 7). Além disso, os polissacarídeos de hemicelulose podem ser muito
diferentes uns dos outros em termos de estrutura e propriedades físicas e físico-químicas28.
O tipo mais abundante de hemicelulose são as xilanas, sendo a maior componente
da parede secundária celular27. Hemicelulose, em geral, representa de 15 a 35% da
composição da biomassa27 e pode ser facilmente hidrolisadas com o uso de soluções diluídas
de ácidos ou bases e são muito solúveis em álcalis28. Dependendo do processo e condições
usadas no tratamento para lise da hemicelulose, os açúcares podem se degradar em ácidos
fracos e furanos, que normalmente atuam como inibidores nos processos de fermentação27.
Figura 7: Representação da molécula de hemicelulose29.
1.3.3. Lignina
A lignina é um heteropolímero amorfo altamente ramificado presente na parede
celular normalmente encontrado adjacente às fibras de celulose formando o complexo
lignocelulósico30. A lignina é composta majoritariamente por três unidades de compostos
fenólicos: álcool p-cumarílico, álcool coniferílico e álcool sinapílico30 – sua função é conferir
rigidez, impermeabilidade e resistência aos ataques mecânicos e microbiológicos nos tecidos
vegetais (Figura 8).

23
Figura 8: Representação de uma estrutura interna de lignina31.
1.3.4. Pectina
A pectina é um polissacarídeo heterogêneo28 ramificado constituído
principalmente de unidades de ácido galacturônico (Figura 9). A pectina é um dos
componentes mais abundantes da parede celular primária de plantas que florescem, a grande
exceção é a família Poaceae (cana de açúcar, milho, trigo, arroz, por exemplo) que possuem
quantidades bem menores.
Figura 9: Representação de uma molécula de pectina.

24
1.4. Óleo Essencial
Óleos essências são conhecidos por sua ação antimicrobiana, sendo que a
atividade antimicrobiana irá depender da composição do óleo essencial32. Óleos essências são
misturas complexas formados por cerca de 400 compostos, mas sua composição irá depender
principalmente da forma de cultivo da laranja33. A composição do óleo essencial de laranja é
majoritariamente de d-limoneno correspondendo a 97%.
d-limoneno (C10H16) é um hidrocarboneto classificado como terpeno cíclico (Figura
10). É um líquido incolor a temperatura ambiente com um odor muito característico de
laranja.
Na laranja o óleo essencial é encontrado do epicardo (flavedo) em glândulas de
óleo.
Figura 10: Representação da molécula de (R)-(+)-limoneno e (S)-(-)-limoneno.
1.5. Xanthomonas axonopodis pv. citri
A Xanthomonas axonopodis pv. citri (Xac) é uma bactéria Gram-negativa34
patógena que infecta frutas cítricas, a doença é conhecida por cancro asiático, cancro tipo A
ou cancro cítrico35 (Figura 11) e é responsável por uma das doenças mais sérias que atingem
cítricos causando perda significativa na qualidade e produção de laranjas36,37.

25
Figura 11: A) Laranja infectada pela bactéria Xanthomonas axonopodis pv citri (Xac) (Figura obtida de
http://www.fundecitrus.com.br/doencas/cancro/7 - acessado 05/07/2014). B) Microscopia eletrônica
de varredura da Xac (cepa 306).
A bactéria entra no tecido hospedeiro através dos estômatos e feridas. A
temperatura ótima para a infecção é entre 20 e 30 °C, além disso a bactéria se multiplica de
3-4 unidades log por lesão em condições ótimas37, a forma mais eficiente para conter a
dispersão da doença é através da remoção das plantas recentemente infectadas e também
daquelas ao seu redor, o que resulta em programas de erradicação em massa36.
O cancro cítrico pode ser causado por outros variantes da bactéria X. axonopodis,
os sintomas são similares e a diferenciação é realizada através da análise do hospedeiro e de
característica fenotípicas e genotípicas das cepas37. O cancro cítrico tipo A causado pela
Xanthomonas axonopodis pv. citri é a forma mais difundida e severa da doença e por esse
motivo é uma das cepas mais estudadas35,37.
Devido a sua importância econômica a Xac (cepa 306) teve seu genoma
sequenciado em 2002 por Silva et al. 38. A bactéria possui um cromossomo circular com
5.175.554 pares de base e dois plasmídeos pXAC 33 (33.699 pares de base) e pXAC64 (64.920
pares de base), que codificam para proteínas em 85,59% das regiões de seu cromossomo,
sendo encontrados 2710 genes com função definida, 1272 de região conservada com função
hipotética, 331 genes hipotéticos, totalizando 4313 genes que codificam para a expressão de
proteínas38.
A Xac também possui um arsenal de genes relacionados à degradação de parede
celular, seu genoma possui ao menos seis Orfs (Open Reading Frames – fase de leitura aberta)
para enzimas com atividades pectinolíticas, doze Orfs com para enzimas celulolíticas, e doze

26
Orfs para enzimas hemicelulolíticas, totalizando ao menos 30 enzimas relacionadas à
degradação de parede celular38.
1.6. Aplicação de enzimas na indústria
Enzimas catalisam reações com grande especificidade e altas taxas de conversão,
sendo assim são ótimas candidatas para indústrias químicas e farmacêuticas preocupadas com
sustentabilidade, energia limpa, biomassa e com o meio ambiente39. A aplicação de enzimas
na indústria já ocorre em grande extensão e em diferentes áreas como na indústria
alimentícia, agricultura, indústria de papel, couro e de tecidos, sendo que seu uso resulta em
consideráveis ganhos econômicos39.
Estima-se que a produção de etanol oriundo de primeira geração, fermentação
dos açúcares livres que compete com a produção de alimentos, aumente para ao menos 100
bilhões de litros até 20224,40, consequentemente aumentando a necessidade de áreas
cultivadas e gerando preocupação com relação a biodiversidade e desmatamento4. Do ponto
de vista biotecnológico existe uma grande variedade de recursos de biomassa lignocelulósica
para serem convertidos em produtos, como o bioetanol e biocombustíveis41.
Nos últimos anos houve um grande desenvolvimento em biotecnologia para
tornar essas tecnologias viáveis, tendo em vista a poluição do meio-ambiente e aquecimento
global. Uma das possibilidades de se produzir o bioetanol é através do uso de biomassa
lignocelulósica, o processo de produção de bioetanol envolve as etapas de pré-tratamento
(mecânico, químico ou biológico), hidrólise (química ou enzimática) e fermentação42. No
entanto, o custo dessa produção é relativamente alto baseado nas tecnologias atuais e o
principal desafio é diminuir o custo do processo de hidrólise, tanto com o objetivo de otimizar
o pré-tratamento para a remoção de lignina e hemicelulose facilitando a hidrólise, quanto na
otimização das enzimas e do processo de hidrólise em si43,44.
1.7. Tipos de pré-tratamento para hidrólise de biomassa
A principal função do pré-tratamento do material lignocelulósico é aumentar a
acessibilidade das enzimas melhorando assim sua ação na celulose através da quebra da
lignina e das estruturas cristalinas de celulose. No entanto, cada tipo de pré-tratamento possui

27
um efeito na celulose, na hemicelulose e também na lignina, por isso diferentes tipos de pré-
tratamento devem ser aplicados dependendo do processo envolvido e também do tipo de
biomassa45.
Com relação aos tipos de pré-tratamento podemos dividi-los nos seguintes
grupos: mecânicos, físico-químico e químicos.
O mecânico tem por objetivo diminuir o tamanho da partícula aumentando sua
área superficial e diminuindo o grau de polimerização. O tratamento pode ser realizado
através de trituração ou moagem45.
O tratamento físico-químico mais utilizado é conhecido como explosão a vapor,
nele o material é exposto a vapor de água pressurizada no período que vai de segundos a
minutos e então bruscamente despressurizados. Nesse tipo de processo ocorre a auto
hidrólise e solubilização do grupo acetila presente na hemicelulose, além disso a variação de
pressão resulta na separação das fibras devido a rápida descompressão45. A explosão a vapor
apresenta como vantagem o uso da biomassa sem estar granulada, o baixo impacto ambiental
e o fato de não usar aditivos químicos, apesar da adição de ácido ser usada em alguns casos
melhorando a hidrólise43,45. É importante ressaltar que nesse tipo de pré-tratamento o
aumento da temperatura melhora a auto hidrólise da hemicelulose, mas também resulta no
aumento da degradação de pentoses e hexoses, assim como o uso de ácido em conjunto com
a explosão a vapor45.
Já o pré-tratamento químico pode ser ácido ou básico. O tratamento básico é
eficiente para a solubilização da lignina, mas apresenta menor solubilização para celulose e
hemicelulose do que os tratamentos ácidos ou de explosão a vapor, no entanto a vantagem
do tratamento básico é que pode ser feito a temperatura ambiente e aumenta a
digestibilidade da celulose, outro ponto favorável é que também resulta em menor
degradação de açúcares do que o processo ácido. Como reagentes podem ser usados
hidróxidos de sódio, cálcio, potássio e amônio, por exemplo. O pré-tratamento com hidróxido
de cálcio apresenta menores custos e menores requisitos de segurança do que NaOH ou KOH,
além de ser facilmente removido do hidrolisado através de uma reação com CO245–47.
Com relação ao tratamento ácido, o principal objetivo é remover e solubilizar a
hemicelulose tornando a celulose mais acessível ao ataque enzimático. O pré-tratamento
pode ser tanto com ácido diluído quanto concentrado, no entanto esse último apresenta
como desvantagem a formação de compostos que inibem a fermentação alcoólica, além da

28
desvantagem de corrosão e recuperação do ácido após a hidrólise, por esses motivos a uso do
ácido diluído é mais interessante do ponto de vista industrial. Soluções diluídas de ácidos
como sulfúrico, clorídrico, fosfórico e nítrico podem ser utilizadas nesse tipo de pré-
tratamento45,47.
1.8. Hidrolases
Antes da etapa de fermentação e após o pré-tratamento, quando já existe a maior
exposição das fibras de celulose, a celulose da biomassa precisa ser hidrolisada para então ser
fermentada para a produção de etanol. O processo pode ser realizado através de uma
hidrólise química ou enzimática. Tratamentos químicos como a hidrólise ácida geram mais
resíduos e os subprodutos dessas reações são tóxicos para a levedura prejudicando a etapa
seguinte de fermentação. Sendo assim, a hidrólise enzimática torna-se uma alternativa mais
verde e branda5,48.
1.9. Tipos de hidrólise
A hidrólise ácida pode ocorrer a partir do uso de ácidos concentrados ou diluídos.
Por exemplo, o uso de soluções diluídas (0,7–3,0%) necessita de temperaturas altas (220 –
240°C), enquanto as soluções concentradas apresentam as mesmas desvantagens expostas
para pré-tratamento com ácido concentrado. Tanto a hidrólise com soluções diluídas quanto
com ácido concentrado podem gerar subprodutos inibitórios para a fermentação49.
Já a hidrólise enzimática utiliza um conjunto de enzimas, tais como celulases,
coquetéis enzimáticos, e não produz inibidores de fermentação, além de exigir aplicação de
condições brandas de pH e temperatura44,49. As enzimas de fungos ou bactérias ou várias
enzimas como celulases, xilanases, ligninases, pectinases entre outras50 são necessárias para
a hidrólise completa da biomassa.
A classe de enzimas mais utilizada na hidrólise de polissacarídeos é EC 3.2 das
hidrolases, que catalisam a reação de hidrólise de polissacarídeos. As principais hidrolases
usadas em biotecnologia para a conversão dos polissacarídeos em monômeros são as
celulases, as β-glicosidases e as pectinases51.

29
1.10. Hidrólise enzimática
1.10.1. Celulases
As celulases são classificadas em três grupos principais: (i) endoglucanases ou 1,4-
β-D-glucano-4-glucano-hidrolase (EC 3.2.1.4), (ii) exoglucanases (EC 3.2.1.74) e 1,4-β-D-
glucano-celobio-hidrolases (celobio-hidrolases) (EC 3.2.1.91) e (iii) β-glicosidases (EC
3.2.1.21)51,52.
As endoglucanases hidrolisam de maneira randômica os sítios amorfos internos da
cadeia de polissacarídeos da celulose, gerando oligossacarídeos de vários tamanhos e com
novas extremidades51. Exoglucanases atuam nas extremidades da cadeia de polissacarídeos
da celulose liberando glicose ou celobiose como produtos principais (Figura 12).
As β-glicosidases (EC 3.2.1.21) hidrolisam celodextrinas solúveis e celobiose
liberando glicoses51. A quebra enzimática de uma ligação 1,4-β-glicosídica na celulose ocorre
através de um mecanismo de hidrólise ácida utilizando um doador de prótons e um nucleófilo
ou base.
Figura 12: Representação esquemática da atuação das enzimas envolvidas na hidrólise da celulose.
1.10.2. Pectinases
As pectinases, por sua vez, são as responsáveis pela hidrólise da pectina. As
pectinases estão entre as enzimas mais amplamente distribuídas em bactérias, fungos e
plantas. Protopectinases, poligalacturonases, liases e esterases pectina estão entre enzimas

30
pectinolíticas extensivamente estudadas. Protopectinases catalisam a solubilização de
protopectina. Poligalacturonases hidrolisam a cadeia do ácido poligalacturônico por adição de
água e as liases catalisam a clivagem trans-eliminativa do polímero de ácido galacturônico53.
1.11. Atividade enzimática – Fatores que afetam a atividade enzimática (pH, temperatura, agitação e inibição por produto)
Vários fatores podem afetar o rendimento de açúcares gerados a partir da
hidrólise de biomassa lignocelulósica, como o acesso das enzimas para a hidrólise,
concentração de enzima colocada, concentração de substrato, inibição de atividade pelo
acúmulo de glicose, e fatores diretamente ligados a atividade enzimática como pH,
temperatura e agitação43,44.
A escolha do pré-tratamento é fundamental para diminuir a cristalinidade da
celulose e remover a lignina, dessa forma menores concentrações de celulase podem ser
utilizadas para a hidrólise, além de um menor tempo de reação44,54.
Além disso, a inibição da atividade por acúmulo de produto é um problema
comum na hidrólise enzimática, mas pode ser resolvido através da sacarificação e
fermentação simultânea da biomassa54. Outro problema pode ser a alta concentração de
substrato, que também pode provocar a inibição enzimática resultando em uma taxa de
hidrólise muito lenta inicialmente, esse efeito está relacionado ao ajuste da proporção entre
substrato/enzima utilizados no processo43, problemas com a agitação e transferência de
massa também estão relacionados a uma alta concentração inicial de substrato54.
Valores ótimos de pH e temperatura dependem da enzima utilizada, do substrato
e da duração da hidrólise. Normalmente os valores de pH estão entre 4-5 e de temperatura
entre 40 – 50 °C44,54.

31
1.12. Caracterização estrutural de proteínas
1.12.1. Dicroísmo circular
Dicroísmo circular é uma técnica espectroscópica não destrutiva, amplamente
utilizada para o estudo de conformação e estabilidade de proteínas frente a diferentes
condições como temperatura, força iônica e presença de solutos ou moléculas pequenas55.
A técnica utiliza uma fonte de luz polarizada circularmente, na qual o vetor
rotacional oscila para a direita ou esquerda formando uma hélice ao redor do eixo de
propagação (Figura 13). Quando a luz polarizada passa por uma amostra opticamente ativa, a
amostra absorve de maneira diferente os componentes de luz polarizada a esquerda e a
direita, seguindo a lei de Beer-Lambert55,56.
Figura 13: Representação da luz polarizada circularmente utilizada na técnica de dicroísmo circular
(https://en.wikipedia.org/wiki/Circular_dichroism - acessado em 12/06/2016).
A técnica permite o estudo de estrutura secundária de proteínas, pois as ligações
peptídicas são assimétricas e moléculas que não possuem plano de simetria apresentam
dicroísmo circular. Os resultados são normalmente expressos como a média da elipticidade
molar do resíduo, [θ], dada pela equação abaixo:
��� �� � 100 �
� � � �
Onde θ é a elipticidade em graus, l é o caminho ótico em cm, C é a concentração
em mg mL-1, M é a massa molecular e n é o número de resíduos da proteína. A unidade de [θ]
é deg.cm2.dmol-1, tal unidade permite a comparação de dados obtidos em diferentes
bateladas de amostras ou diferentes laboratórios55.
32
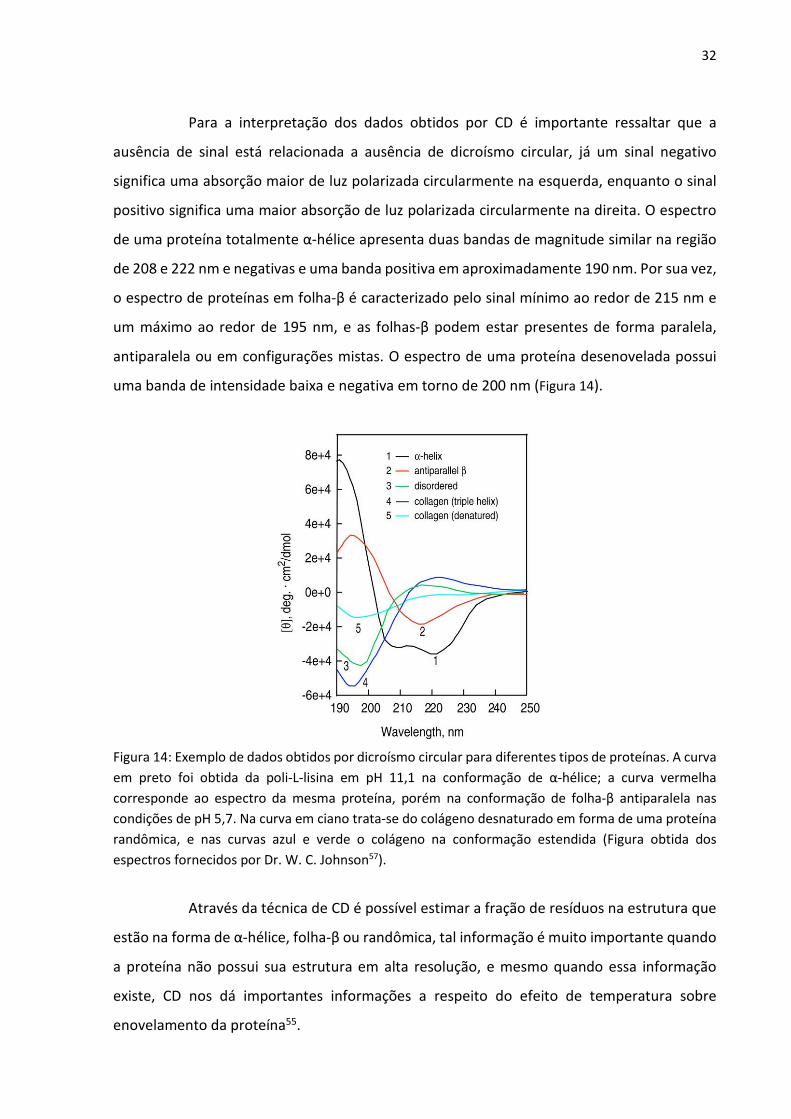
Para a interpretação dos dados obtidos por CD é importante ressaltar que a
ausência de sinal está relacionada a ausência de dicroísmo circular, já um sinal negativo
significa uma absorção maior de luz polarizada circularmente na esquerda, enquanto o sinal
positivo significa uma maior absorção de luz polarizada circularmente na direita. O espectro
de uma proteína totalmente α-hélice apresenta duas bandas de magnitude similar na região
de 208 e 222 nm e negativas e uma banda positiva em aproximadamente 190 nm. Por sua vez,
o espectro de proteínas em folha-β é caracterizado pelo sinal mínimo ao redor de 215 nm e
um máximo ao redor de 195 nm, e as folhas-β podem estar presentes de forma paralela,
antiparalela ou em configurações mistas. O espectro de uma proteína desenovelada possui
uma banda de intensidade baixa e negativa em torno de 200 nm (Figura 14).
Figura 14: Exemplo de dados obtidos por dicroísmo circular para diferentes tipos de proteínas. A curva
em preto foi obtida da poli-L-lisina em pH 11,1 na conformação de α-hélice; a curva vermelha
corresponde ao espectro da mesma proteína, porém na conformação de folha-β antiparalela nas
condições de pH 5,7. Na curva em ciano trata-se do colágeno desnaturado em forma de uma proteína
randômica, e nas curvas azul e verde o colágeno na conformação estendida (Figura obtida dos
espectros fornecidos por Dr. W. C. Johnson57).
Através da técnica de CD é possível estimar a fração de resíduos na estrutura que
estão na forma de α-hélice, folha-β ou randômica, tal informação é muito importante quando
a proteína não possui sua estrutura em alta resolução, e mesmo quando essa informação
existe, CD nos dá importantes informações a respeito do efeito de temperatura sobre
enovelamento da proteína55.
33
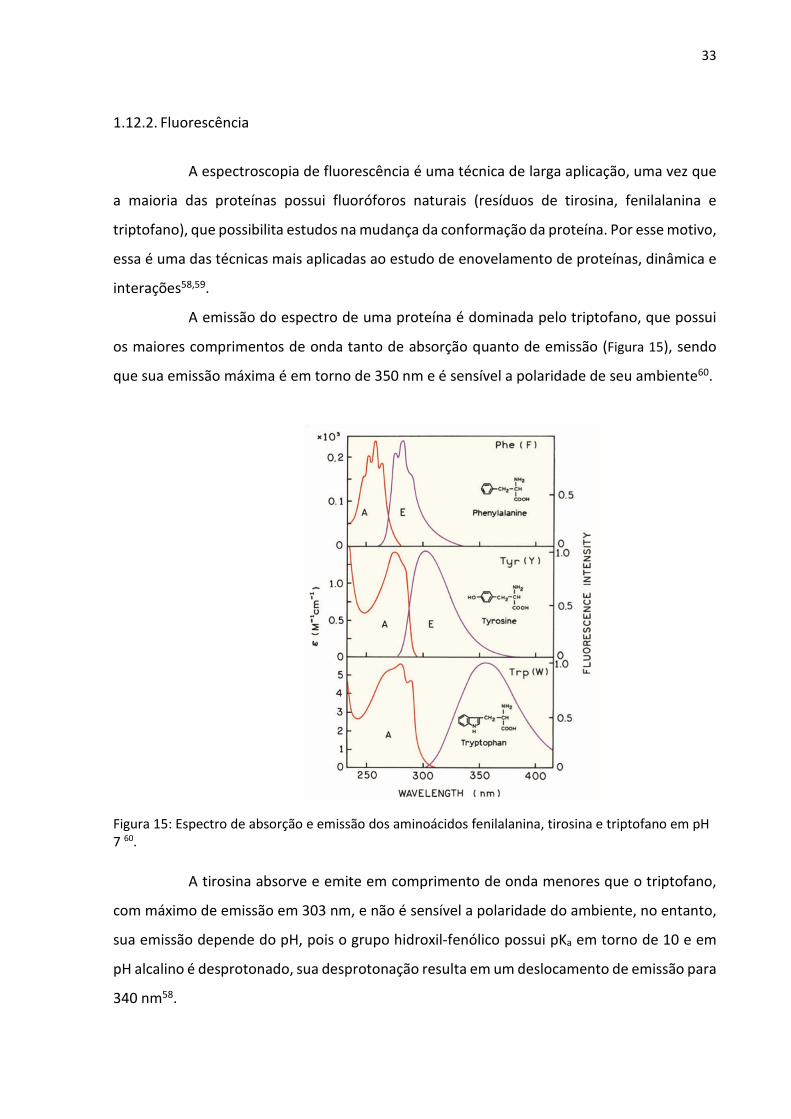
1.12.2. Fluorescência
A espectroscopia de fluorescência é uma técnica de larga aplicação, uma vez que
a maioria das proteínas possui fluoróforos naturais (resíduos de tirosina, fenilalanina e
triptofano), que possibilita estudos na mudança da conformação da proteína. Por esse motivo,
essa é uma das técnicas mais aplicadas ao estudo de enovelamento de proteínas, dinâmica e
interações58,59.
A emissão do espectro de uma proteína é dominada pelo triptofano, que possui
os maiores comprimentos de onda tanto de absorção quanto de emissão (Figura 15), sendo
que sua emissão máxima é em torno de 350 nm e é sensível a polaridade de seu ambiente60.
Figura 15: Espectro de absorção e emissão dos aminoácidos fenilalanina, tirosina e triptofano em pH 7 60.
A tirosina absorve e emite em comprimento de onda menores que o triptofano,
com máximo de emissão em 303 nm, e não é sensível a polaridade do ambiente, no entanto,
sua emissão depende do pH, pois o grupo hidroxil-fenólico possui pKa em torno de 10 e em
pH alcalino é desprotonado, sua desprotonação resulta em um deslocamento de emissão para
340 nm58.

34
A fenilalanina é o aminoácido que possui o menor comprimento de onda, tanto de
absorção quanto de emissão, com máximo de emissão próximo a 282 nm, no entanto quando
próximo a tirosina ou ao triptofano pode afetar o rendimento quântico através de
transferência de energia58,60.

35
2. Objetivos
O presente projeto teve por objetivo o estudo e caracterização de uma celulase
recombinante de Xanthomonas axonopodis pv. citri para posterior aplicação na hidrólise do
bagaço de laranja e produção de etanol de segunda geração.
Como objetivos específicos destacam-se:
• Caracterização de diferentes tipos de biomassa semissólida provenientes do
processo de produção do suco de laranja e bagaço in natura obtido localmente;
• Isolamento de microrganismo de bagaço de laranja com melhor rendimento
para fermentação de hidrolisados de bagaço;
• Amplificação, expressão e purificação de hidrolases recombinantes de Xac;
• Caracterização bioquímica e biofísica de uma celulase recombinante de Xac
através da determinação dos parâmetros cinéticos e estudos de dicroísmo circular
e fluorescência;

36
3. Materiais e métodos
3.1. Processamento do bagaço da laranja
O bagaço in natura da laranja foi recolhido de um restaurante em Campinas (São
Paulo, Brasil). Além disso, amostras in natura da empresa Citrosuco (Matão, SP – Brasil)
também foram analisadas, as amostras correspondiam ao bagaço logo após a extração do
suco (continha não apenas bagaço, mas também folhas e galhos).
Ambas as amostras foram processadas em um multiprocessador de cozinha
(Philco multipro, 800W, China), até que o tamanho da partícula ficasse entre 2-33 mm de
diâmetro, logo após as amostras foram colocadas em sacos plásticos e armazenadas a -20 °C.
3.2. Caracterização primária do bagaço de laranja
3.2.1. Determinação de sólidos totais e umidade
Para a determinação dos sólidos usou-se o protocolo descrito pelo NREL
(Laboratório Nacional de Energia Renovável - National Renewable Energy Laboratory)61.
Três placas de Petri previamente secas em estufa foram pesadas em uma balança
analítica e suas massas anotadas. Então colocou-se cerca de 17 g de bagaço de laranja
processado e homogeneizado em cada uma das placas que foram novamente pesadas. As
placas com bagaço foram levadas a estufa a 105 °C por 5 h. Passadas as 5 h as placas de Petri
com bagaço foram resfriadas em um dessecador até que atingissem a temperatura ambiente
para serem novamente pesadas.
O cálculo utilizado é mostrado abaixo:
% ����� �� �ó����� =3 − 1
2 − 1∙ 100
M1 = Placa de Petri vazia
M2 = Placa de Petri com bagaço de laranja úmido
M3 = Placa de Petri com bagaço de laranja seco
I)

37
Para o cálculo da porcentagem de umidade foi utilizada a equação abaixo:
% �� ������� = 100 − % ����� �� �ó�����
3.2.2. Determinação do conteúdo de cinzas
A determinação do conteúdo de cinzas foi realizada com as amostras de bagaço
previamente secas conforme descrição do item 2.2.1.
Em seguida, as amostras de bagaço seco foram colocadas em cadinhos de
porcelana previamente secos e de massas conhecidas, que foram novamente pesados em
balança analítica. Os cadinhos foram levados à mufla a 525 °C por 5 h, então em um
dessecador foram deixados resfriar a temperatura ambiente para serem pesados e o
conteúdo de cinzas calculado pela equação abaixo:
% ������ =3 − 1
2 − 1∙ 100
M1 = Cadinho de porcelana vazio
M2 = Cadinho de porcelana com bagaço de laranja seco
M3 = Cadinho com cinzas de BL
3.3. Determinação de fibras
3.3.1. Determinação de fibras em detergente neutro (FDN)
Cinco bolsas de filtros (F57, ANKOM Technology) foram pesadas em uma balança
analítica. Cerca de 0,50 g de bagaço de laranja previamente seco em estufa a 105 °C por 5 h
foi colocado em três das bolsas de filtro, as outras duas foram usadas como fator de correção.
Todas as bolsas de filtro foram fechadas selando as extremidades com calor.
A solução detergente foi preparada em um béquer de 1 L contendo: 15 g de
dodecil sulfato de sódio, 9,31 g de ácido etilenodiamino tetra-acético (EDTA), 3,40 g de borato
de sódio, 2,28 g de fosfato de sódio bibásico (anidro), 5 mL de trietilenoglicol em 500 mL de
II)

38
água destilada, para homogeneizar a solução foi aquecida por 5 minutos. Além disso,
adicionou-se 5 g de sulfito de sódio e 1 mL de α-amilase.
As bolsas de filtro foram mergulhadas nessa solução e deixadas em agitação por
75 min a 100 °C. Em seguida os filtros foram lavados com água destilada quente (80 °C), o
excesso de água foi removido e os filtros foram colocados em um béquer contendo acetona
por 5 min.
Posteriormente os filtros foram colocados em estufa a 105 °C até que estivessem
completamente secos para serem pesados em uma balança analítica.
A porcentagem de fibras neutras detergentes foi calculada de acordo com a
equação abaixo:
% �� =3 − (1 ∙ 1)
2∙ 100
M1 = Massa da bolsa de filtro
M2 = Massa do bagaço
M3 = Massa da bolsa de filtro com bagaço após a extração
C1 = Fator de correção (média das bolsas vazias após extração/média das bolsas
vazias antes da extração)
3.3.2. Determinação das fibras em detergente ácido (FDA)
Cinco bolsas de filtros (F57, ANKOM Technology) foram pesadas em uma balança
analítica. Cerca de 0,50 g de bagaço de laranja previamente seco em estufa a 105 °C por 5 h
foi colocado em três das bolsas de filtro, as outras duas foram usadas como fator de correção.
Todas as bolsas de filtro foram fechadas selando as extremidades com calor.
A solução detergente ácida foi preparada em um béquer de 1 L contendo: 10 g de
brometo de cetiltrimetilamônio (CTAB) em 500 mL de solução 1 mol.L-1 de H2SO4.
As bolsas de filtro foram mergulhadas nessa solução e deixadas em agitação por
1h a 100 °C. Em seguida os filtros foram lavados com água destilada quente (80 °C), o excesso
de água foi removido e os filtros foram colocados em um béquer contendo acetona por 5 min.

39
Posteriormente os filtros foram colocados em estufa a 105 °C até que estivessem
completamente secos para serem pesados em uma balança analítica.
A porcentagem de fibras neutras detergentes foi calculada de acordo com a
equação abaixo:
% ��# =3 − (1 ∙ 1)
2∙ 100
M1 = Massa da bolsa de filtro
M2 = Massa do bagaço
M3 = Massa da bolsa de filtro com bagaço após a extração
C1 = Fator de correção (média das bolsas vazias após extração/média das bolsas
vazias antes da extração)
3.3.3. Determinação de lignina em detergente ácido (LDA)
Depois da determinação de fibras em detergente ácido (FDA) descrita no tópico
2.3.2, as bolsas de filtro previamente pesadas foram colocadas em um béquer contendo uma
solução 72% de H2SO4, as bolsas de filtro foram mantidas submergidas e agitadas a cada 30
min.
Após 3 h as amostras foram lavadas até que todo o ácido tivesse sido removido, o
excesso de água foi removido e os filtros foram colocados em um béquer contendo acetona
por 5 min.
Posteriormente os filtros foram colocados em estufa a 105 °C até que estivessem
completamente secos para serem pesados em uma balança analítica.
A porcentagem de fibras neutras detergentes foi calculada de acordo com a
equação abaixo:
% $�# =3 − (1 ∙ 1)
2∙ 100
M1 = Massa da bolsa de filtro
M2 = Massa do bagaço
M3 = Massa da bolsa de filtro com bagaço após a extração

40
C1 = Fator de correção (média das bolsas vazias após extração/média das bolsas
vazias antes da extração).
3.4. Determinação de Pectina
A determinação de pectina foi feita em triplicata. 17 g de bagaço de laranja
processado e homogeneizado foram fervidos em 50 mL de uma solução de 0,05 mol.L-1 de HCl
por 1 h a 100 °C. Então o extrato foi filtrado com o auxílio de um funil de Büchner, kitassato,
bomba a vácuo e papel de filtro. O procedimento de extração e filtração foi repetido para o
resíduo retido no papel de filtro. Os dois filtrados foram misturados e deixados resfriar a
temperatura ambiente.
A pectina foi precipitada adicionando-se ao filtrado dois volumes de uma solução
95% de etanol acidificada com 0,05 mol.L-1 de HCl. A pectina resultante foi filtrada em papel
de filtro previamente pesado e lavada com uma solução de 65% de etanol acidificada e depois
com a mesma solução que foi utilizada na precipitação. O papel de filtro contendo a pectina
foi seco em estufa a 40 °C até que estivesse completamente seco.
A equação utilizada para o cálculo da porcentagem de pectina segue abaixo.
% %������ =2 − 1
3∙ 100
M1 = Massa do papel de filtro
M2 = Massa do papel de filtro com pectina seca
M3 = Massa do bagaço de laranja
3.5. Extração do óleo essencial
O óleo essencial do bagaço de laranja foi extraído por destilação a vapor. 50 g do
bagaço de laranja processado e homogeneizado foi colocado no balão de fundo redondo e o
vapor produzido em um frasco de destilação foi passado pelo bagaço por cerca de 40 min.
Nessa técnica o óleo é carregado pelo vapor e condensa no condensador caindo em um balão.

41
O óleo foi extraído com o auxílio de um funil de separação. No funil de separação
foi colocado éter dietílico e o destilado, então foi vigorosamente agitado e deixado que as
fases se separassem. A fase de aquosa foi removida e a fase orgânica, que contém o óleo, foi
coletada. A extração foi repetida, e a fase orgânica resultante das duas extrações foi seca com
sulfato de sódio anidro.
No rota-evaporador, todo o éter dietílico foi removido e o frasco contendo o óleo
foi pesado.
O teor de óleo essencial presente na amostra foi calculado de acordo com o a
equação mostrada abaixo:
% ó��� ��������� ����� =2 − 1
3∙ 100
M1 = Massa do balão de coleta
M2 = Massa do balão de coleta mais óleo
M3 = Massa de bagaço de laranja
3.6. Determinação de hesperidina
A hesperidina foi extraída a partir de 20 g do pellet triturado, usando um extrator
Soxhlet. Para isso o material triturado foi colocado em uma cápsula de celulose, e o extrator
Soxhlet foi acoplado a um balão de 500 mL contendo 250 mL de éter de petróleo como
solvente de extração e a um condensador. A mistura foi aquecida por 4 h sob forte refluxo.
Então o extrato de éter de petróleo foi removido. O conteúdo da cápsula foi removido do
extrator para que todo o éter evaporasse antes de se prosseguir com a extração.
Em seguida, realizou-se uma segunda etapa de extração, dessa vez com 250 mL de
metanol. O refluxo de metanol durou cerca de 4 h, até que o solvente, que inicialmente saía
de coloração marrom quando em contato com a amostra, passou a sair incolor.
O extrato do pellet em metanol foi evaporado em um rotaevaporador até a
consistência de um xarope. Ao xarope foram adicionados 50 mL de solução de ácido acético
6% para a formação de um precipitado. A suspensão foi centrifugada a 5000 rpm e o

42
sobrenadante descartado. Então o precipitado (hesperidina bruta) foi lavado com solução de
ácido acético 6% e seco a 60 °C.
A recristalização da hesperidina foi realizada dissolvendo-a em solução 5% de
dimetilsulfóxido sob agitação e aquecimento de 70 °C. Então desligou-se o aquecimento e
adicionou-se o mesmo volume de água destilada gota-a-gota sob constante agitação. A
solução foi deixada esfriar até atingir a temperatura ambiente quando se formaram os cristais
de hesperidina, que foram filtrados e deixados secar a 60 °C.
3.7. Isolamento de microrganismos do bagaço de laranja
O isolamento de culturas a partir do bagaço de laranja industrial foi realizado como
descrito por Tsukamoto et al. (2009)8.
Em meio sólido colocando-se pequenos pedaços de bagaço em placas de Petri
contendo meio sólido YPD (20 g L-1 de glicose, 10 g L-1 extrato de levedura, 20 g L-1 peptona,
15 g L-1 ágar e 30 mg mL-1 cloranfenicol) as placas foram então incubadas de 24-48 h a 30 °C.
Depois da incubação as colônias que cresceram isoladas foram selecionadas e
transferidas para outra placa de Petri contendo meio sólido YPD e novamente incubadas a 30
°C por 24 h. O procedimento foi repetido mais quatro vezes a fim de se garantir que a colônia
fosse de apenas um microrganismo.
Após o isolamento as colônias resultantes foram testadas frente ao crescimento
em presença de d-limoneno 0,5% em meio líquido de YPD. As colônias que apresentaram
maior densidade ótica em 600 nm (DO600) foram selecionadas para fermentação.
Para a fermentação, foi crescido um pré-inóculo a partir de uma colônia em 5 mL
de YPD e incubado por 16 h a 30 °C, então a cultura foi utilizada para inocular erlenmeyers de
250 mL contendo 50 mL de YPD com DO600 inicial de 0,2. Os inóculos foram incubados a 30 °C
por 24 h e amostras foram retiradas nos seguintes tempos: 0, 3,5, 6,5, 8,5 e 24 h para análise
por HPLC do consumo de glicose e produção de etanol, já o crescimento foi acompanhado
através da leitura da densidade ótica em 600 nm durante 24 h. As melhores culturas foram
armazenadas em glicerol 20% para posterior teste com hidrolisado.

43
3.8. Expressão do gene sintético, lise e purificação
O gene sintético clonado em pET28 foi transformado por protocolo
quimiocompetente em células competentes de Origami 2 (DE3), o cultivo cresceu em placa de
Petri contendo meio LB e as seleções de canamicina e tetraciclina por 16 h.
Então de uma colônia isolada foi realizado um pré-cultivo em 25 mL de meio de
cultura LB contendo canamicina e tetraciclina incubado a 37 °C com agitação de 220 rpm. Após
o crescimento de aproximadamente 4 h o pré-cultivo foi utilizado para inocular 300 mL de
meio LB contendo os mesmos antibióticos de seleção em uma A600 inicial = 0,1, o cultivo foi
novamente incubado a 37 °C e 220 rpm, e quando este atingiu A600 de 0,8-1,0 foi induzido
adicionando-se IPTG na concentração final de 1 mmol L-1 e incubado por 16 h a 18 °C com
agitação de 220 rpm.
A cultura foi centrifugada a 6000 rpm por 15 min e 4 °C. O sobrenadante foi
descartado e as células foram armazenadas a -80°C por ao menos 2 h. As mesmas foram
suspensas em 20 mL de tampão 50 mmol L-1 Tris-HCl, pH 7,5 e a lise foi realizada por sonicação
utilizando-se cinco ciclos de 10 s/10 s (sempre em gelo). Em seguida a amostra foi centrifugada
a 6000 rpm por 30 min a 4 °C.
A fração solúvel das proteínas foi filtrada em filtro 0,45 µm e então purificada. A
purificação foi realizada utilizando-se uma coluna de afinidade FF Crude 1 mL (GE). Para a
purificação utilizou-se um gradiente do tampão A (50 mmol L-1 fosfato de sódio pH 7,5, 300
mmol L-1 cloreto de sódio, PMSF, β-mercaptoetanol, 5 mmol L-1 imidazol) e do tampão B (50
mmol L-1 fosfato de sódio pH 7,5, 300 mmol L-1 cloreto de sódio, PMSF, β-mercaptoetanol, 500
mmol L-1 imidazol). As amostras da purificação foram recolhidas e armazenadas a 4°C.
Para a filtração em gel utilizou-se uma coluna Sephadex 75 (GE) com volume de
coluna de 24 mL. Injetou-se 100 µL da padrão BlueDextran para a determinação do volume
morto, em seguida foi realizada uma curva de calibração com os padrões de conoalbumina,
ovoalbumina, anidrase carbônica, ribonuclease e aprotinina.
Finalmente injetou-se a amostra e através do volume de eluição do pico foi
possível a determinação da massa molar da mesma.

44
3.9. Determinação das condições ótimas de ensaio de atividade
As condições ótimas de atuação da celulase frente a hidrólise de carboximetil
celulose (CMC) foram encontradas primeiramente variando o tempo de incubação da proteína
com o substrato. O ensaio foi realizado em pH 6 e temperatura de 35 °C. Após determinado
essa condição foi feito o teste de pH, no qual o pH foi variado a uma temperatura fixa, e em
seguida para se determinar a temperatura ótima do ensaio variou-se a temperatura no melhor
pH encontrado62.
Para isso utilizou-se o tampão Na2HPO4/Ácido cítrico a concentração de 50 mmol
L-1 que permite uma larga faixa de pH sem alteração da força iônica, os seguintes pHs foram
testados: 2,5; 3,5; 4,5; 5,5; 6,5 e 7,5. O ensaio foi realizado com 200 nmol L-1 da enzima
purificada em 1 mL do tampão contendo 0,5% de carboximetil celulose. A reação foi incubada
por 30 min a 50°C.
Com o objetivo de parar a reação e também quantificar a produção de açúcares
redutores adicionou-se 1 mL de reagente de ácido dinitrosalicílico (DNS) que foi então
incubado a 100 °C por 5 min. A absorbância a 540 nm foi lida em espectrofotômetro e a
concentração determinada a partir de uma curva de calibração de glicose63.
Após determinado o pH ótimo de atuação da enzima variou-se a temperatura. A
reação foi preparada conforme descrito anteriormente, porém com o tampão de pH
adequado. As temperaturas testadas foram as seguintes: 25, 30, 35, 40, 45, 50, 55, 60, 65, 70,
75 e 80 °C. Da mesma forma a reação foi parada e quantificada por reagente DNS62.
3.10. Cinética
Os ensaios de atividade foram realizados utilizando 200 nmol L-1 de enzima e
variando-se a concentração do substrato (CMC) no tampão 50 mmol L-1 ácido citríco/Na2HPO4
pH 6,5 a 35°C incubada por 30 min. As concentrações utilizadas foram as seguintes: 0,1, 0,5,
1,0, 1,5, 2,0, 2,5, 3,0, 3,5 e 4,0% (m/V) de CMC.
A concentração de açúcares redutores gerados foi monitorada pela reação de DNS
conforme descrito anteriormente.
Utilizando a equação de Michaelis-Menten64 (equação VII) e a equação de
Lineweaver-Burk (equação VIII) foi possível determinar a velocidade inicial (V0) velocidade

45
máxima (Vmáx), a constante de Michaelis-Menten (Km) e número de renovação (kcat)
enzimático.
&' =&(á*[+]
,( + [+] (&..)
1
&'
=,(
&(á*[+]+
1
&(á*
(&...)
3.11. Dicroísmo circular
Para o ensaio de dicroísmo circular realizou-se uma troca de tampão da fração da
purificação da celulase, para isso utilizou-se uma coluna de dessalinização PD-10 (GE), coluna
de gel filtração empacotada com Sephadex G-25. A coluna PD-10 foi equilibrada passando-se
15 mL de tampão 50 mmol L-1 Na2HPO4/ácido cítrico pH 6,5, então 2,5 mL de amostra foram
adicionados à coluna e eluída em 3,5 mL do tampão de equilíbrio65.
A concentração da proteína após a troca do tampão foi medida a 280 nm em
cubeta de quartzo de 1 cm. Utilizando-se o coeficiente de extinção molar (valor calculado de
121365 mol-1 L cm-1) foi possível calcular a concentração a partir da lei de Beer.
No equipamento Jasco J-720 foram realizadas medidas do tampão ácido
cítrico/Na2HPO4 fosfato 50 mmol L-1 (branco da amostra). As aquisições foram realizadas de
207 a 260 nm, em cubeta de 0,1 cm de caminho óptico, sendo realizadas 8 aquisições da
cubeta contendo apenas o tampão no qual a proteína foi eluída e, finalmente, foram
realizadas 16 aquisições da amostra no mesmo tampão.
Também foram realizadas leituras aumentando a temperatura da amostra de 25
a 90 °C e então esfriando-se lentamente a amostra de 90 a 25 °C, e uma outra análise
aumentando de 25 a 60 °C e depois de 60 a 25 °C.
Por fim, diferentes pH do tampão foram analisados. Para isso a proteína foi eluída
em tampão 50 mmol L-1 Na2HPO4/ácido cítrico nos seguintes pH: 2,5, 4,5, 5,5, 6,5 e 7,5 e
incubada por 16 h antes da análise de dicroísmo circular.

46
3.12. Fluorescência
As análises de fluorescência foram realizadas com as amostras em diferentes
valores de pH preparadas para o dicroísmo circular.
Primeiro foi realizado um espectro de excitação de 200 a 450 nm para encontrar
o máximo de excitação da amostra. Em seguida, foi realizado o espectro de emissão de 300 a
450 nm da amostra excitada a 278 nm.

47
4. Resultados e Discussão
Parte I – Caracterização do Bagaço de Laranja e Isolamento de Microrganismo
de Laranja
4.1. Caracterização do Bagaço de Laranja
Com o objetivo de conhecer melhor a natureza da matéria prima, e também
facilitar a interpretação dos resultados obtidos com os diferentes tipos de bagaço utilizados,
foi feita a caracterização dos mesmos para a determinação dos teores de sólidos totais e
umidade, cinzas e determinação de fibras (celulose, hemicelulose e lignina).
No método de determinação de Fibras em Detergente Neutro (FDN), a amostra de
bagaço é tratada com uma solução de detergente, após o tratamento, as fibras restantes são
compostas predominantemente por celulose, hemicelulose e lignina. No método de Fibras em
Detergente Ácido (FDA) o resíduo que sobra da digestão da biomassa com ácido sulfúrico e
CTAB é principalmente celulose e lignina. Já no método de Lignina em Detergente Ácido (LDA),
após o tratamento com ácido sulfúrico a celulose é removida e o objetivo é calcular a
porcentagem de lignina presente na amostra.
A Tabela 1 resume os principais resultados para a caracterização do bagaço de
laranja obtido do restaurante local (amostra comercial) e também os resultados obtidos para
a caracterização do bagaço industrial obtido da empresa Citrosuco (amostra industrial).
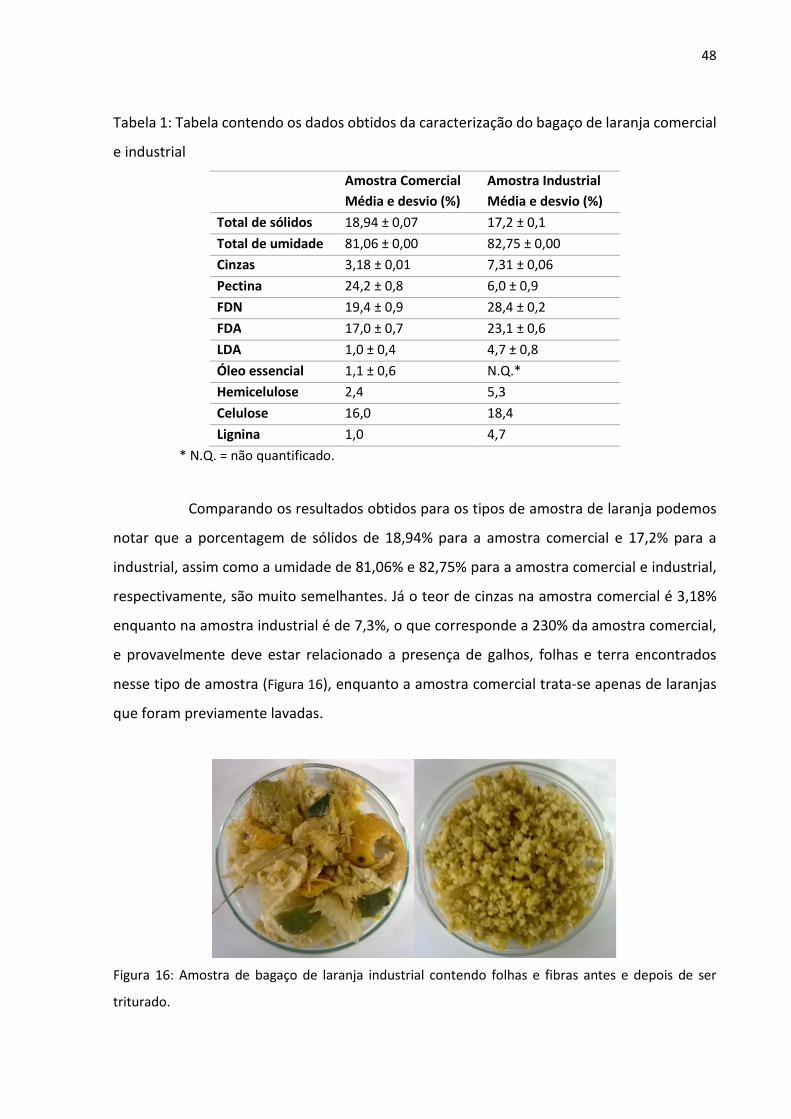
48
Tabela 1: Tabela contendo os dados obtidos da caracterização do bagaço de laranja comercial
e industrial
Amostra Comercial
Média e desvio (%)
Amostra Industrial
Média e desvio (%)
Total de sólidos 18,94 ± 0,07 17,2 ± 0,1
Total de umidade 81,06 ± 0,00 82,75 ± 0,00
Cinzas 3,18 ± 0,01 7,31 ± 0,06
Pectina 24,2 ± 0,8 6,0 ± 0,9
FDN 19,4 ± 0,9 28,4 ± 0,2
FDA 17,0 ± 0,7 23,1 ± 0,6
LDA 1,0 ± 0,4 4,7 ± 0,8
Óleo essencial 1,1 ± 0,6 N.Q.*
Hemicelulose 2,4 5,3
Celulose 16,0 18,4
Lignina 1,0 4,7
* N.Q. = não quantificado.
Comparando os resultados obtidos para os tipos de amostra de laranja podemos
notar que a porcentagem de sólidos de 18,94% para a amostra comercial e 17,2% para a
industrial, assim como a umidade de 81,06% e 82,75% para a amostra comercial e industrial,
respectivamente, são muito semelhantes. Já o teor de cinzas na amostra comercial é 3,18%
enquanto na amostra industrial é de 7,3%, o que corresponde a 230% da amostra comercial,
e provavelmente deve estar relacionado a presença de galhos, folhas e terra encontrados
nesse tipo de amostra (Figura 16), enquanto a amostra comercial trata-se apenas de laranjas
que foram previamente lavadas.
Figura 16: Amostra de bagaço de laranja industrial contendo folhas e fibras antes e depois de ser
triturado.

49
Por outro lado, a amostra comercial apresenta uma porcentagem muito menor de
pectina (6,0%) do que a amostra comercial (24,2%) para explicar essa diferença é importante
levar em consideração quando comparamos os dois tipos de bagaço que suas colheitas foram
realizadas em diferentes épocas do ano, a amostra comercial é do mês de agosto (inverno),
enquanto a amostra industrial é do mês de novembro (primavera), sabe-se que diferentes
variedades de laranjas são colhidas ao longo do ano para garantir o suprimento6, como pode
ser observado na Tabela 2, que mostra o período de colheita de cada variedade de laranja e o
percentual da produção. Portanto, é provável que a laranja comercial seja de uma variedade
de laranja precoce, enquanto a laranja industrial de variedade tardia, que corresponde ao
maior percentual de produção. Além disso, a mesma variedade de laranja pode apresentar
variação em sua composição dependendo da época em que foi colhida e devido alterações
das condições climáticas a que foram submetidas10.
Tabela 2: Período de colheita por variedade de laranja e percentual da produção, tabela retirada de “O
retrato da Citricultura brasileira, 2010” 6.
A quantidade de celulose, hemicelulose e lignina (Tabela 1) presentes na amostra
foram calculadas através dos testes de determinação de Fibras em Detergente Neutro (FDN),
Fibras em Detergente Ácido (FDA) e Lignina em Detergente Ácido (LDA) utilizando-se as
equações mostradas a seguir.

50
FDN - FDA = hemicelulose
FDA - LDA = celulose
LDA = lignina
Comparando-se as duas amostras percebemos que de maneira geral a amostra
industrial possui maior teor de hemicelulose, celulose e lignina. A maior porcentagem de
hemicelulose e celulose torna o bagaço industrial mais interessante do ponto de vista
econômico, já que a hidrólise resultará em maior concentração de açúcares fermentáveis para
a produção de etanol de segunda geração. Por sua vez o conteúdo de lignina apesar de ser
maior na amostra industrial (4,7%) é muito menor do que outras fontes de biomassa, como
por exemplo o bagaço de cana de açúcar que possui de 10,7 a 22,7% ou a palha de milho com
11% de lignina do conteúdo da biomassa28, o que torna o bagaço de laranja uma fonte muito
interessante para o tratamento enzimático, já que sua hidrólise produzirá menor quantidade
de compostos fenólicos e terá maior rendimento.
4.2. Caracterização da amostra de licor e pellet industrial
Abaixo segue um fluxograma do processo de produção industrial de laranja e seus
derivados (Figura 17), primeiramente é extraído o suco, então o bagaço é prensado e
centrifugado, dessa etapa temos três produtos: o licor, o óleo essencial e o resíduo
semissólido66. A amostra de licor, portanto, possui baixo teor de sólidos, bastante açúcar
dissolvido e não possui o óleo essencial da laranja, por esse motivo pode ser fermentada para
a produção de etanol 1G, mas além disso ainda é possível aproveitar o resíduo para uma etapa
de hidrólise e fermentação (Figura 18 A). O pellet por sua vez é o resíduo semissólido desse
processo que é seco e transformado em mistura para ração de gado9 (Figura 18 B).

51
Figura 17: Fluxograma do processo de produção da laranja e seus derivados.
Figura 18: A) Amostra de dorna industrial - licor. B) Amostra de pellet de bagaço de laranja (Citrosuco,
Matão, SP).
A Tabela 3 resume os principais resultados obtidos para a caracterização da
amostra de licor industrial.

52
Tabela 3: Tabela contendo os dados obtidos da caracterização da amostra de licor industrial (amostra
Citrosuco, Matão, SP)
Amostra Licor
Total de sólidos (%) 0,71 ± 0,05
Total de umidade (%) 99,29
FDN (%) 21,8 ± 0,7
FDA (%) 17,8 ± 0,8
LDA (%) 1,6 ± 0,4
Açúcares redutores totais (DNS) (g L-1) 79,4
Hemicelulose (%) 3,9
Celulose (%) 16,3
Lignina (%) 1,6
Observando-se os resultados podemos notar o grande conteúdo de açúcares
redutores totais (79,4 g L-1), o que justifica o uso da amostra como substrato de fermentação
para a produção de etanol de primeira geração.
Apesar do baixo conteúdo de sólidos presente na amostra, cerca de 0,7%, ainda
existe potencial para uma hidrólise enzimática e posterior fermentação para a produção de
etanol de segunda geração, principalmente devido ao teor de celulose (16,3%), hemicelulose
(3,9%) e baixo teor de lignina (1,6%), além disso, esse resíduo não compete com a produção
de ração de gado.
A amostra do pellet, que consiste no bagaço de laranja seco após prensagem, foi
utilizada para a extração da hesperidina, um flavonóide encontrado nas frutas cítricas de alto
interesse na indústria farmacêutica e cosmética devido as suas propriedades como agente
antioxidante, anti-inflamatória, analgésica, anticâncer e hipolipidêmico67–69. O teor de
hesperidina encontrado na amostra sólida de pellet foi de 1,47%.
4.3. Isolamento de Colônias de Bagaço de Laranja
Com o objetivo de buscar um microrganismo que pudesse fermentar melhor o
hidrolisado de bagaço de laranja frente a uma possível resistência à presença de óleo essencial
e outros compostos naturalmente presentes no bagaço foi feito o isolamento de colônias
nativas do bagaço de laranja em meio sólido (Figura 19). O resultado do primeiro isolamento
foram 32 colônias testadas frente a sua resistência ao crescimento em meio líquido contendo
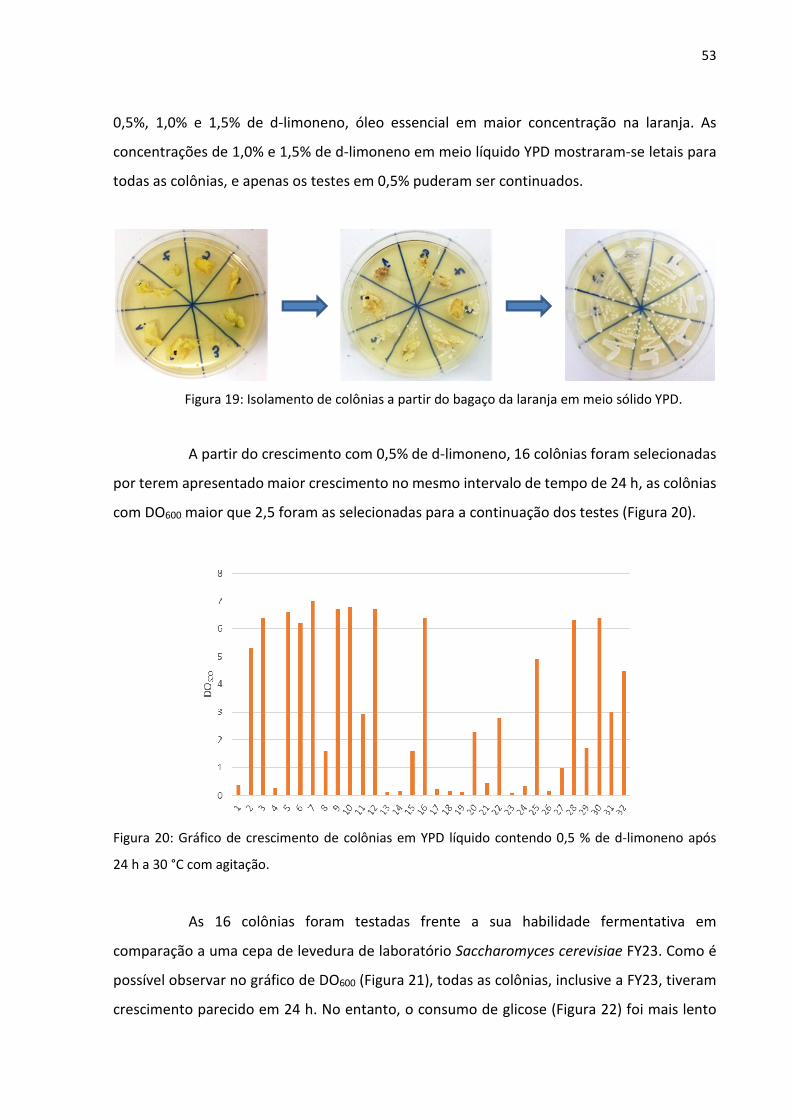
53
0,5%, 1,0% e 1,5% de d-limoneno, óleo essencial em maior concentração na laranja. As
concentrações de 1,0% e 1,5% de d-limoneno em meio líquido YPD mostraram-se letais para
todas as colônias, e apenas os testes em 0,5% puderam ser continuados.
Figura 19: Isolamento de colônias a partir do bagaço da laranja em meio sólido YPD.
A partir do crescimento com 0,5% de d-limoneno, 16 colônias foram selecionadas
por terem apresentado maior crescimento no mesmo intervalo de tempo de 24 h, as colônias
com DO600 maior que 2,5 foram as selecionadas para a continuação dos testes (Figura 20).
Figura 20: Gráfico de crescimento de colônias em YPD líquido contendo 0,5 % de d-limoneno após
24 h a 30 °C com agitação.
As 16 colônias foram testadas frente a sua habilidade fermentativa em
comparação a uma cepa de levedura de laboratório Saccharomyces cerevisiae FY23. Como é
possível observar no gráfico de DO600 (Figura 21), todas as colônias, inclusive a FY23, tiveram
crescimento parecido em 24 h. No entanto, o consumo de glicose (Figura 22) foi mais lento
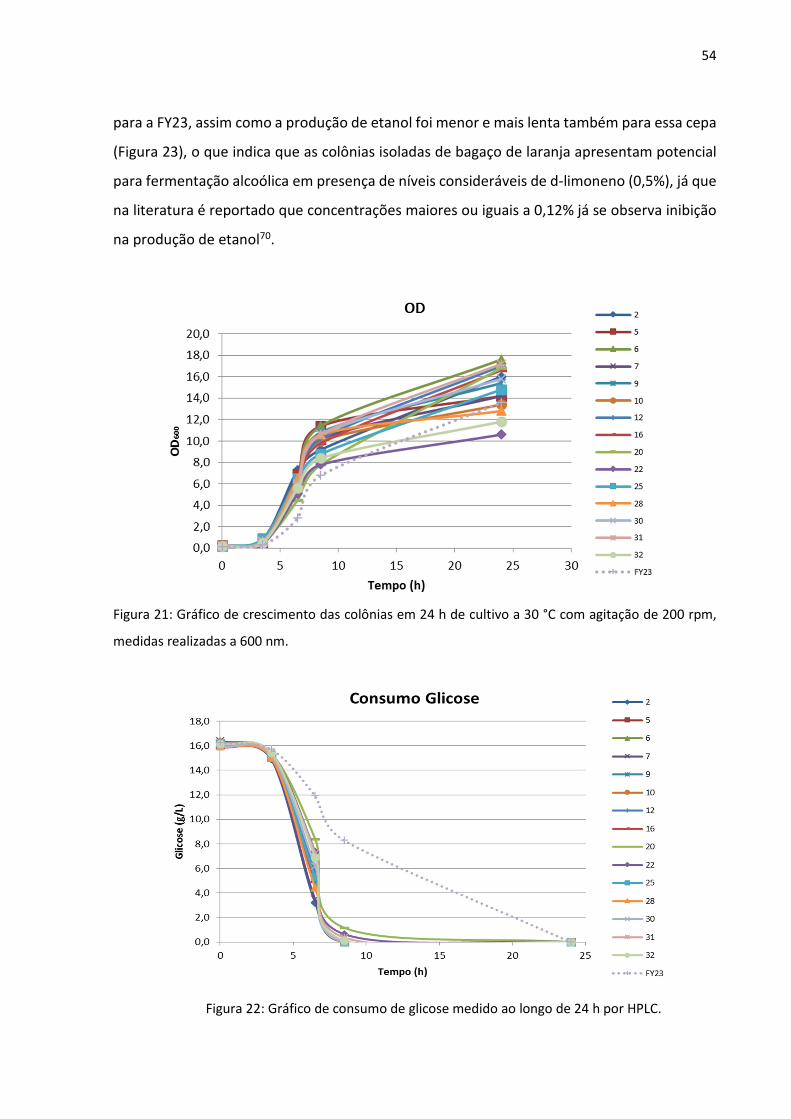
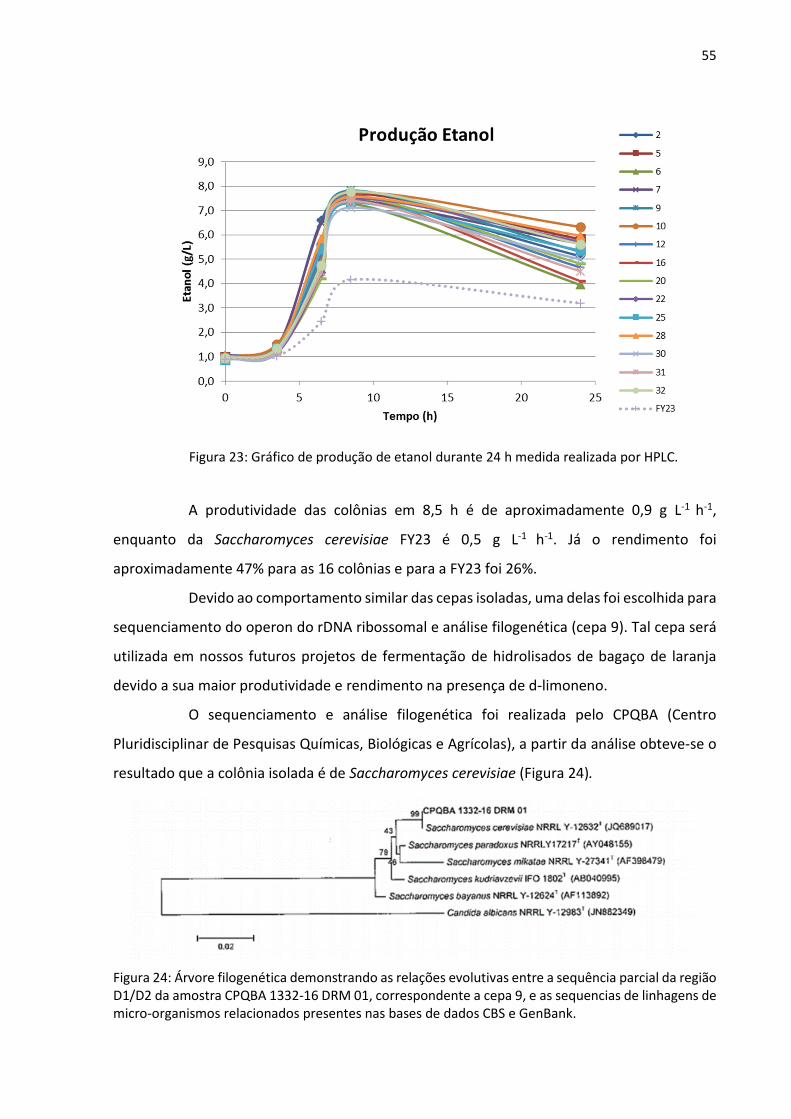
54
para a FY23, assim como a produção de etanol foi menor e mais lenta também para essa cepa
(Figura 23), o que indica que as colônias isoladas de bagaço de laranja apresentam potencial
para fermentação alcoólica em presença de níveis consideráveis de d-limoneno (0,5%), já que
na literatura é reportado que concentrações maiores ou iguais a 0,12% já se observa inibição
na produção de etanol70.
Figura 21: Gráfico de crescimento das colônias em 24 h de cultivo a 30 °C com agitação de 200 rpm,
medidas realizadas a 600 nm.
Figura 22: Gráfico de consumo de glicose medido ao longo de 24 h por HPLC.
55
Figura 23: Gráfico de produção de etanol durante 24 h medida realizada por HPLC.
A produtividade das colônias em 8,5 h é de aproximadamente 0,9 g L-1 h-1,
enquanto da Saccharomyces cerevisiae FY23 é 0,5 g L-1 h-1. Já o rendimento foi
aproximadamente 47% para as 16 colônias e para a FY23 foi 26%.
Devido ao comportamento similar das cepas isoladas, uma delas foi escolhida para
sequenciamento do operon do rDNA ribossomal e análise filogenética (cepa 9). Tal cepa será
utilizada em nossos futuros projetos de fermentação de hidrolisados de bagaço de laranja
devido a sua maior produtividade e rendimento na presença de d-limoneno.
O sequenciamento e análise filogenética foi realizada pelo CPQBA (Centro
Pluridisciplinar de Pesquisas Químicas, Biológicas e Agrícolas), a partir da análise obteve-se o
resultado que a colônia isolada é de Saccharomyces cerevisiae (Figura 24).
Figura 24: Árvore filogenética demonstrando as relações evolutivas entre a sequência parcial da região D1/D2 da amostra CPQBA 1332-16 DRM 01, correspondente a cepa 9, e as sequencias de linhagens de micro-organismos relacionados presentes nas bases de dados CBS e GenBank.

56
A melhor performance frente a presença de d-limoneno provavelmente está
relacionada a questões de adaptação da cepa, uma vez que esta foi isolada do bagaço de
laranja. Além disso, sabe-se que diferentes linhagens de um mesmo microrganismo podem
apresentar diferentes resistências frente a determinados compostos, como por exemplo HMF,
furfurais e mesmo maiores concentrações de etanol71.
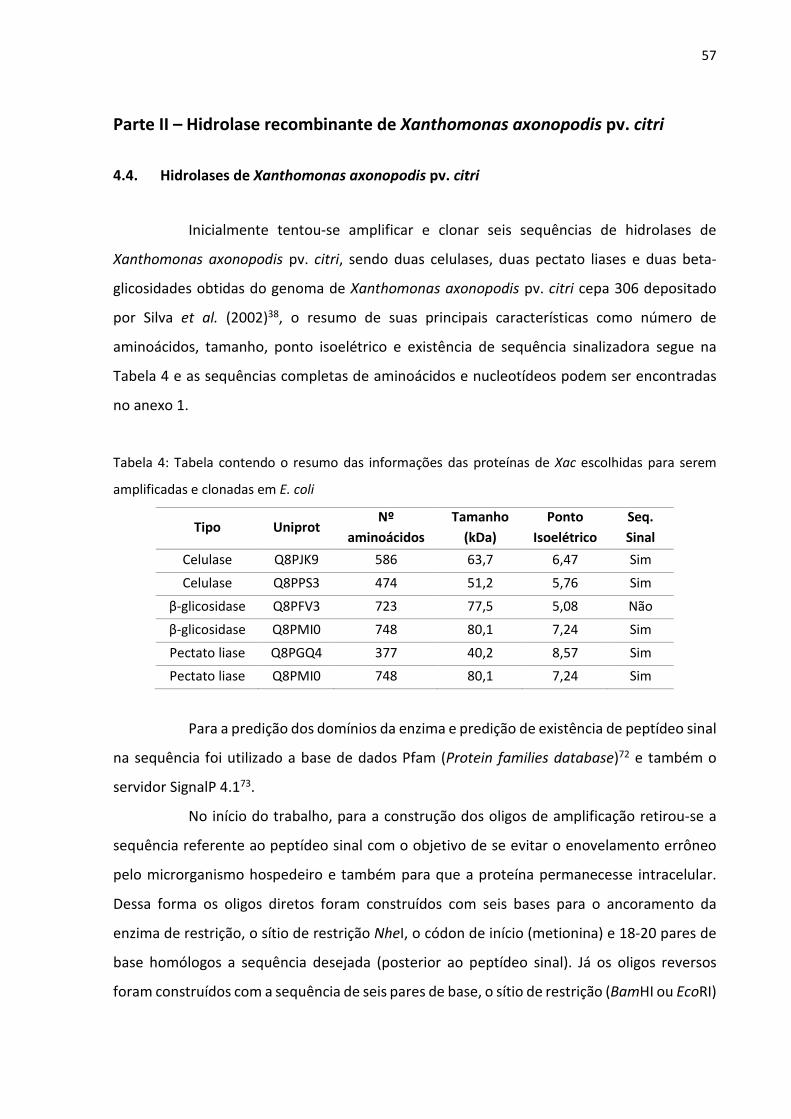
57
Parte II – Hidrolase recombinante de Xanthomonas axonopodis pv. citri
4.4. Hidrolases de Xanthomonas axonopodis pv. citri
Inicialmente tentou-se amplificar e clonar seis sequências de hidrolases de
Xanthomonas axonopodis pv. citri, sendo duas celulases, duas pectato liases e duas beta-
glicosidades obtidas do genoma de Xanthomonas axonopodis pv. citri cepa 306 depositado
por Silva et al. (2002)38, o resumo de suas principais características como número de
aminoácidos, tamanho, ponto isoelétrico e existência de sequência sinalizadora segue na
Tabela 4 e as sequências completas de aminoácidos e nucleotídeos podem ser encontradas
no anexo 1.
Tabela 4: Tabela contendo o resumo das informações das proteínas de Xac escolhidas para serem
amplificadas e clonadas em E. coli
Tipo Uniprot Nº
aminoácidos
Tamanho
(kDa)
Ponto
Isoelétrico
Seq.
Sinal
Celulase Q8PJK9 586 63,7 6,47 Sim
Celulase Q8PPS3 474 51,2 5,76 Sim
β-glicosidase Q8PFV3 723 77,5 5,08 Não
β-glicosidase Q8PMI0 748 80,1 7,24 Sim
Pectato liase Q8PGQ4 377 40,2 8,57 Sim
Pectato liase Q8PMI0 748 80,1 7,24 Sim
Para a predição dos domínios da enzima e predição de existência de peptídeo sinal
na sequência foi utilizado a base de dados Pfam (Protein families database)72 e também o
servidor SignalP 4.173.
No início do trabalho, para a construção dos oligos de amplificação retirou-se a
sequência referente ao peptídeo sinal com o objetivo de se evitar o enovelamento errôneo
pelo microrganismo hospedeiro e também para que a proteína permanecesse intracelular.
Dessa forma os oligos diretos foram construídos com seis bases para o ancoramento da
enzima de restrição, o sítio de restrição NheI, o códon de início (metionina) e 18-20 pares de
base homólogos a sequência desejada (posterior ao peptídeo sinal). Já os oligos reversos
foram construídos com a sequência de seis pares de base, o sítio de restrição (BamHI ou EcoRI)

58
e de 18-20 pares de base do complemento reverso da sequência desejada. Os oligos foram
sintetizados pela empresa IDT e seguem na Tabela 5 abaixo.
Tabela 5: Tabela contendo os oligos diretos (primer forward – PF) e oligos reversos (primer reverse –
PR) usados na amplificação de cada sequência
Enzimas Oligos
Enzima 1 PF: ACAATAGCTAGCATGGCCGAGACGCCCGGCAGGCAG (NheI)
PR: ATACTGGGATCCTCAGCGCGTCGACGCCTCGA (BamHI)
Enzima 2 PF: ATTGCCGCTAGCATGTATTCGGTCAGCAATAACAAG (NheI)
PR: TATACTGGATCCTCAACCTGCTGCGCAGAAGC (BamHI)
Enzima 3 PF: ACTTACGCTAGCATGGCTGCTGATCGCATC (NheI)
PR: TACACTGAATTCTCAGCGTTCCAGCAGCTCG (EcoRI)
Enzima 4 PF: ACATTAGCTAGCATGGCACCGCTGGCAGTCGCGCAG (NheI)
PR: AGATCTGAATTCTCACGGCAACTGTGCCGCCT (EcoRI)
Enzima 5 PF: ATATTAGCTAGCATGGATGCGGCGTTGGAGGTTGC (NheI)
PR: TATATCGGATCCTTACAGCTTGCCGGCGCCTG (BamHI)
Enzima 6 PF: ATATAAGCTAGCATGGGCCCGGTCGGCTACGGTGC (NheI)
PR: TGCACTGGATCCTCACTCCGCCAGGTTCTTGC (BamHI)
A extração do DNA genômico de Xac foi feito seguindo as instruções fornecidas
pelo kit de extração de DNA genômico da Invitrogen, o total de DNA obtido foi 100 µL com
concentração de 62,5 ng µL-1 (Figura 25).
Figura 25: Gel de agarose 1% com marcador 1 kb e amostra de DNA genômico de Xac.
Como o DNA de Xac (cepa 306) possui alto conteúdo GC (65%) foi importante
testar diferentes condições de reações de PCR na tentativa de se otimizar a amplificação das

59
ORFs desejadas. Os seguintes parâmetros foram testados: temperatura e tempo de
anelamento dos oligos, tempo de extensão, uso de diferentes DNA polimerases como de alta
fidelidade (Phusion High Fidelity, AccuPrime High GC e Kapa High fidelity) e Taq polimerase,
adição de diferentes concentrações de DMSO, adição de MgCl2. A visualização das
amplificações foi feita através de gel de agarose 1%, o tamanho dos fragmentos esperados
para cada enzima amplificada segue na tabela 5.
Dentre as enzimas testadas, as que geraram melhores resultados foram as de alta
fidelidade, sendo que a Taq polimerase foi incapaz de se ancorar ao DNA e amplificar qualquer
região, mesmo que não especificamente. Observou-se também que quanto menor a
temperatura de anelamento usada no ciclo de PCR maior o número de bandas inespecíficas,
isso ocorre pois em temperaturas mais baixas o primer consegue parear de forma errônea na
sequência gerando fragmentos não desejados (Figura 26).
Além disso, também se observou que o aumento da temperatura de anelamento
do primer apenas prejudicou a amplificação, sendo que menos bandas foram observadas
nesta condição. Acredita-se que o aumento do tempo de anelamento tenha provocado a
formação da dupla hélice o que impossibilitou a atuação da DNA polimerase.
No entanto, devido à falta de sucesso na clonagem decidiu-se por comprar o gene
de uma das celulases para a continuação dos estudos, uma vez que o objetivo do trabalho era
a caracterização de uma enzima hidrolítica de Xanthomonas axonopodis pv. citri.
Tabela 5: Tamanho dos fragmentos esperados para cada uma das sequencias de nucleotídeos
referente as enzimas de 1 a 6
Enzima 1 Enzima 2 Enzima 3 Enzima 4 Enzima 5 Enzima 6
1698 pb 1350 pb 2172 pb 2166 pb 1056 pb 993 pb
60
Figura 26: Gel obtido de um dos testes de amplificação. Nele é possível observar a amplificação pouco especifica e também o efeito da temperatura de anelamento, que quanto mais alta mais específica.
4.5. Celulase Q8PJK9 – Gene sintético
A celulase Q8PJK9 (identificação Uniprot) possui 586 aminoácidos e é codificada
por 1761 pares de base, além disso possui peso molecular de 63715 Da e ponto isoelétrico
teórico de 6,47, os parâmetros foram calculados pela ferramenta ProtParam do ExPASy74.
A sequência de aminoácidos segue abaixo (Accession: NP_642837.1 GI:21243255)
juntamente com os resultados obtidos no Pfam72 para domínios conservados e pelo SignalP73
para predição de sequência sinalizadora. Na sequência foram realçadas em amarelo a região
referente ao peptídeo sinal (Figura 27), em magenta o domínio semelhante a imunoglobulina
(Ig-like) no N-terminal e em ciano a família 9 de hidrolase glicosídica (Figura 28).
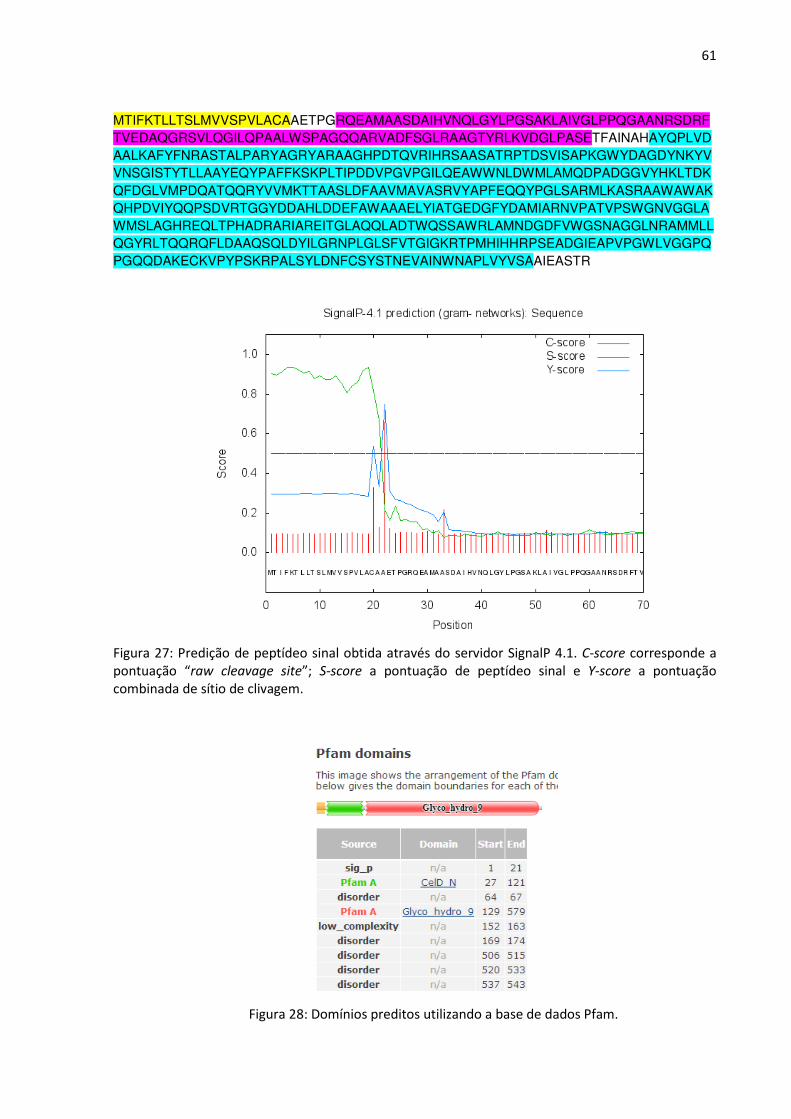
61
MTIFKTLLTSLMVVSPVLACAAETPGRQEAMAASDAIHVNQLGYLPGSAKLAIVGLPPQGAANRSDRF
TVEDAQGRSVLQGILQPAALWSPAGQQARVADFSGLRAAGTYRLKVDGLPASETFAINAHAYQPLVD
AALKAFYFNRASTALPARYAGRYARAAGHPDTQVRIHRSAASATRPTDSVISAPKGWYDAGDYNKYV
VNSGISTYTLLAAYEQYPAFFKSKPLTIPDDVPGVPGILQEAWWNLDWMLAMQDPADGGVYHKLTDK
QFDGLVMPDQATQQRYVVMKTTAASLDFAAVMAVASRVYAPFEQQYPGLSARMLKASRAAWAWAK
QHPDVIYQQPSDVRTGGYDDAHLDDEFAWAAAELYIATGEDGFYDAMIARNVPATVPSWGNVGGLA
WMSLAGHREQLTPHADRARIAREITGLAQQLADTWQSSAWRLAMNDGDFVWGSNAGGLNRAMMLL
QGYRLTQQRQFLDAAQSQLDYILGRNPLGLSFVTGIGKRTPMHIHHRPSEADGIEAPVPGWLVGGPQ
PGQQDAKECKVPYPSKRPALSYLDNFCSYSTNEVAINWNAPLVYVSAAIEASTR
Figura 27: Predição de peptídeo sinal obtida através do servidor SignalP 4.1. C-score corresponde a pontuação “raw cleavage site”; S-score a pontuação de peptídeo sinal e Y-score a pontuação combinada de sítio de clivagem.
Figura 28: Domínios preditos utilizando a base de dados Pfam.

62
Hidrolases glicosídicas (E.C. 3.2.1.-) catalisam a hidrólise das ligações glicosídicas
em oligos ou polissacarídeos, o último número E.C. depende da estrutura do substrato e do
produto. De acordo com a classificação do banco de dados Carbohydrate Active Enzyme (CAZy)
(ww.cazy.org) tais enzimas estão distribuídas em 135 famílias baseadas na similaridade de
sua sequência de aminoácidos75, no entanto novas famílias são adicionadas conforme novas
sequências de enzimas e padrões de atividade são descobertos76,77. Pelo motivo das enzimas
classificadas dentro da mesma família possuírem similaridades estruturais e mecanísticas,
podemos inferir algumas características de enzimas ainda não cristalizadas e caracterizadas,
dessa forma a classificação da base de dados CASy permite que predições do mecanismo de
reação sejam extrapoladas para suas similares77,78.
A família 9 de hidrolases glicosídicas possui 2321 sequências associadas, das quais
140 foram caracterizadas e 15 tiveram suas estruturas resolvidas. É comum encontrar nas
enzimas pertencentes a essa família módulos auxiliares de ligação à carboidratos (CBM –
carbohydrate-binding module), ou módulos de funções desconhecidas como o domínio similar
ao de imonoglobulina (Ig-like), como no caso da celulase estudada nesse trabalho76. Apesar
da função do domínio Ig-like não ser muito conhecida, a literatura mostra que sua deleção
resultou na perda total de atividade da enzima, e que o domínio pode estar relacionado a
estabilidade da proteína e ao seu enovelamento correto79.
Com relação ao sítio catalítico e mecanismo de reação de hidrólise é conhecido
que as glicosil hidrolases podem ser divididas em duas classes, a primeira classe que, durante
a hidrólise, causa a inversão da configuração anomérica e, a outra, que durante a hidrólise
mantém a configuração (Figura 29)80, sendo que, as ambas utilizam um par de ácidos
carboxílicos no sítio ativo em um mecanismo de catálise ácida que necessita de um doador de
prótons e um nucleófilo81. No mecanismo de inversão um dos resíduos de ácido carboxílico
atua como ácido e o outro como base, enquanto no mecanismo com retenção de configuração
um dos resíduos atua tanto como ácido quanto base, enquanto o outro atua como nucleófilo
e grupo de saída80.

63
Figura 29: Mecanismos propostos para glicosil hidrolases com inversão de configuração (A) e com retenção da configuração (B)80.
Na família 9 o mecanismo ocorre com inversão da configuração e o resíduo
catalítico doador de próton é um ácido glutâmico, enquanto o resíduo catalítico básico ou
nucleófilo é um ácido aspártico80–82.
A celulase Q8PJK9 possui uma sequência sinalizadora (peptídeo sinal), no entanto,
o gene foi sintetizado sem o peptídeo sinal para garantir uma expressão intracelular e dessa
forma maior facilidade para trabalhar com a proteína.
Além disso, o gene sintetizado teve seus pares de base otimizados para a
expressão em E. coli. A sequência sem o peptídeo sinal possui 566 aminoácidos, peso
molecular de 61625,3 Da e ponto isoelétrico teórico de 6,32, também calculados utilizando a
ferramenta ProtParam74.
4.6. Expressão e purificação da celulase recombinante
Para a otimização da superexpressão da celulase de interesse na fração solúvel
foram testadas diferentes condições de temperatura e tempos de indução após a adição de
IPTG quando o cultivo atingiu A600 ~ 0,9, as temperatura e tempos de indução testados foram:
18 °C durante 16 h, 28 °C durante 4 h e 37 °C durante 3 h. Além disso, testou-se diferentes
concentrações de IPTG: 1 mmol L-1, 0,5 mmol L-1 e 0,1 mmol L-1, os testes foram realizados
inicialmente na cepa de E. coli BL21(DE3). O gel, ilustrado na Figura 30 contém as amostras
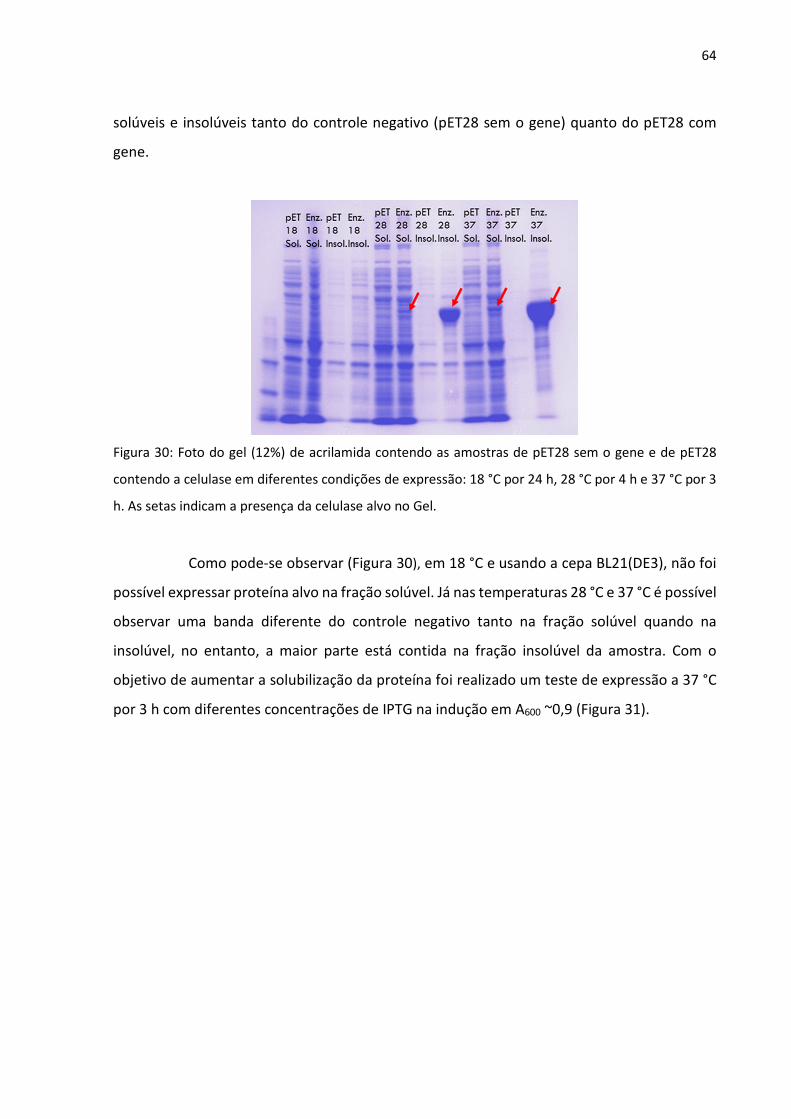
64
solúveis e insolúveis tanto do controle negativo (pET28 sem o gene) quanto do pET28 com
gene.
Figura 30: Foto do gel (12%) de acrilamida contendo as amostras de pET28 sem o gene e de pET28
contendo a celulase em diferentes condições de expressão: 18 °C por 24 h, 28 °C por 4 h e 37 °C por 3
h. As setas indicam a presença da celulase alvo no Gel.
Como pode-se observar (Figura 30), em 18 °C e usando a cepa BL21(DE3), não foi
possível expressar proteína alvo na fração solúvel. Já nas temperaturas 28 °C e 37 °C é possível
observar uma banda diferente do controle negativo tanto na fração solúvel quando na
insolúvel, no entanto, a maior parte está contida na fração insolúvel da amostra. Com o
objetivo de aumentar a solubilização da proteína foi realizado um teste de expressão a 37 °C
por 3 h com diferentes concentrações de IPTG na indução em A600 ~0,9 (Figura 31).
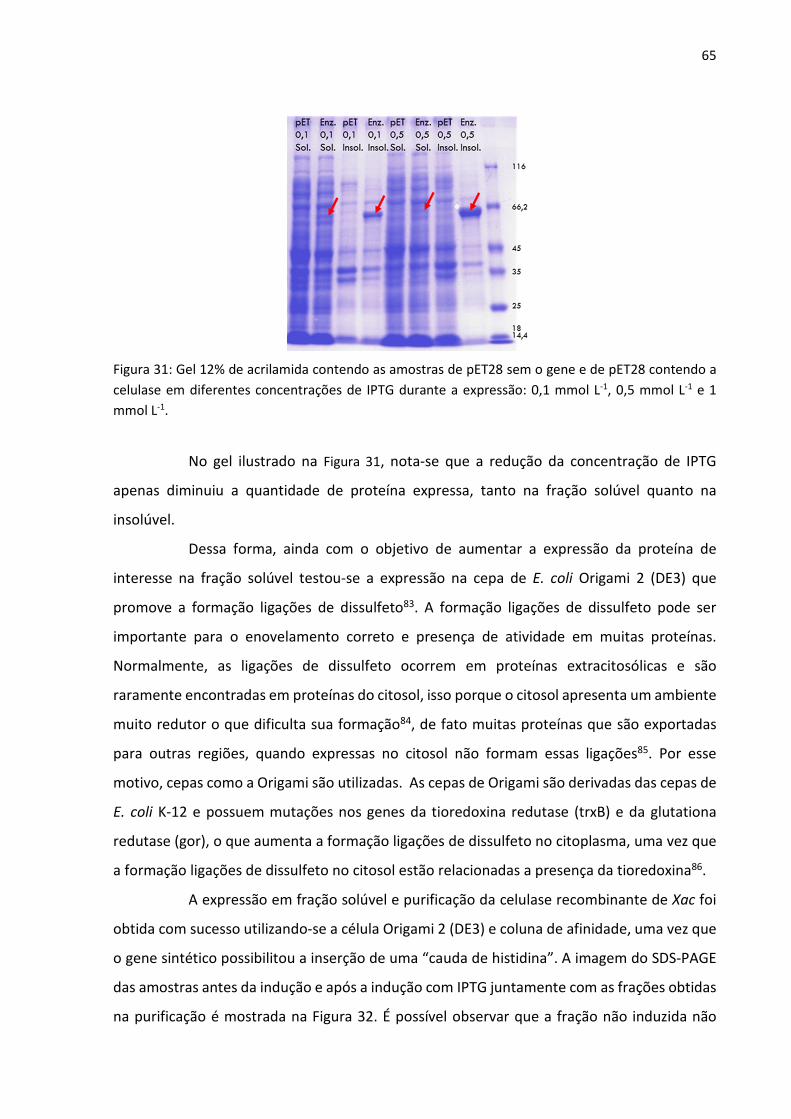
65
Figura 31: Gel 12% de acrilamida contendo as amostras de pET28 sem o gene e de pET28 contendo a
celulase em diferentes concentrações de IPTG durante a expressão: 0,1 mmol L-1, 0,5 mmol L-1 e 1
mmol L-1.
No gel ilustrado na Figura 31, nota-se que a redução da concentração de IPTG
apenas diminuiu a quantidade de proteína expressa, tanto na fração solúvel quanto na
insolúvel.
Dessa forma, ainda com o objetivo de aumentar a expressão da proteína de
interesse na fração solúvel testou-se a expressão na cepa de E. coli Origami 2 (DE3) que
promove a formação ligações de dissulfeto83. A formação ligações de dissulfeto pode ser
importante para o enovelamento correto e presença de atividade em muitas proteínas.
Normalmente, as ligações de dissulfeto ocorrem em proteínas extracitosólicas e são
raramente encontradas em proteínas do citosol, isso porque o citosol apresenta um ambiente
muito redutor o que dificulta sua formação84, de fato muitas proteínas que são exportadas
para outras regiões, quando expressas no citosol não formam essas ligações85. Por esse
motivo, cepas como a Origami são utilizadas. As cepas de Origami são derivadas das cepas de
E. coli K-12 e possuem mutações nos genes da tioredoxina redutase (trxB) e da glutationa
redutase (gor), o que aumenta a formação ligações de dissulfeto no citoplasma, uma vez que
a formação ligações de dissulfeto no citosol estão relacionadas a presença da tioredoxina86.
A expressão em fração solúvel e purificação da celulase recombinante de Xac foi
obtida com sucesso utilizando-se a célula Origami 2 (DE3) e coluna de afinidade, uma vez que
o gene sintético possibilitou a inserção de uma “cauda de histidina”. A imagem do SDS-PAGE
das amostras antes da indução e após a indução com IPTG juntamente com as frações obtidas
na purificação é mostrada na Figura 32. É possível observar que a fração não induzida não
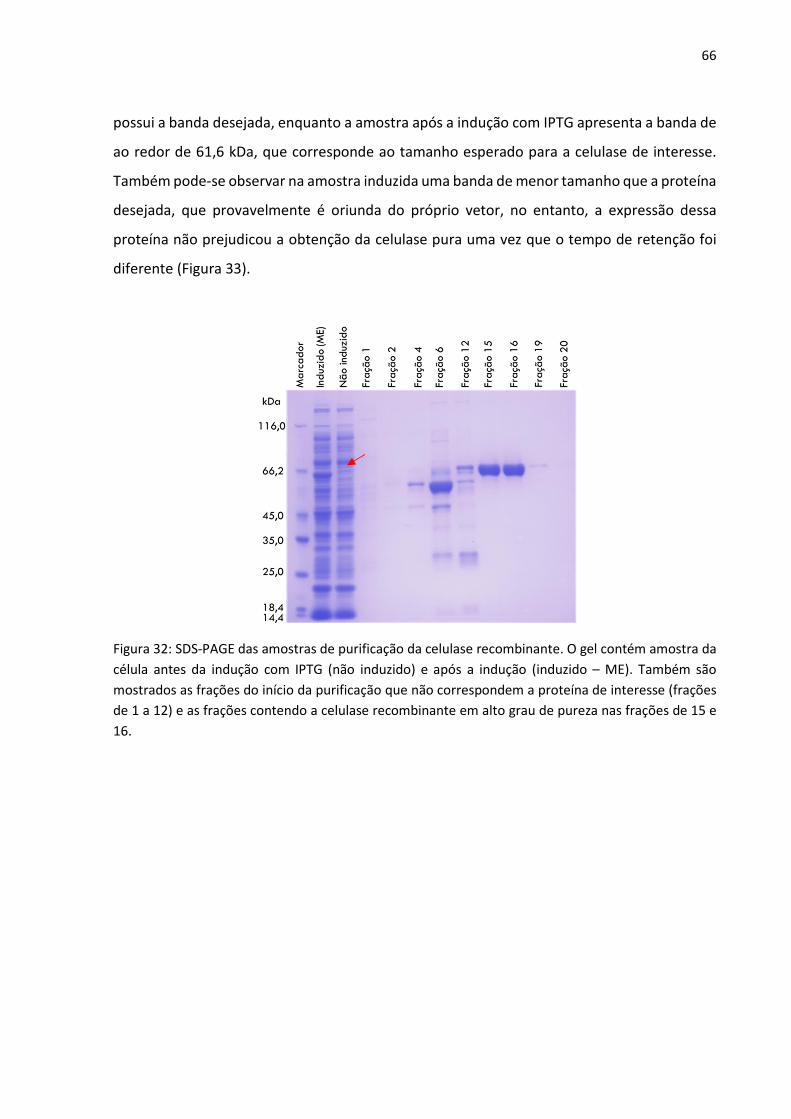
66
possui a banda desejada, enquanto a amostra após a indução com IPTG apresenta a banda de
ao redor de 61,6 kDa, que corresponde ao tamanho esperado para a celulase de interesse.
Também pode-se observar na amostra induzida uma banda de menor tamanho que a proteína
desejada, que provavelmente é oriunda do próprio vetor, no entanto, a expressão dessa
proteína não prejudicou a obtenção da celulase pura uma vez que o tempo de retenção foi
diferente (Figura 33).
Figura 32: SDS-PAGE das amostras de purificação da celulase recombinante. O gel contém amostra da
célula antes da indução com IPTG (não induzido) e após a indução (induzido – ME). Também são
mostrados as frações do início da purificação que não correspondem a proteína de interesse (frações
de 1 a 12) e as frações contendo a celulase recombinante em alto grau de pureza nas frações de 15 e
16.
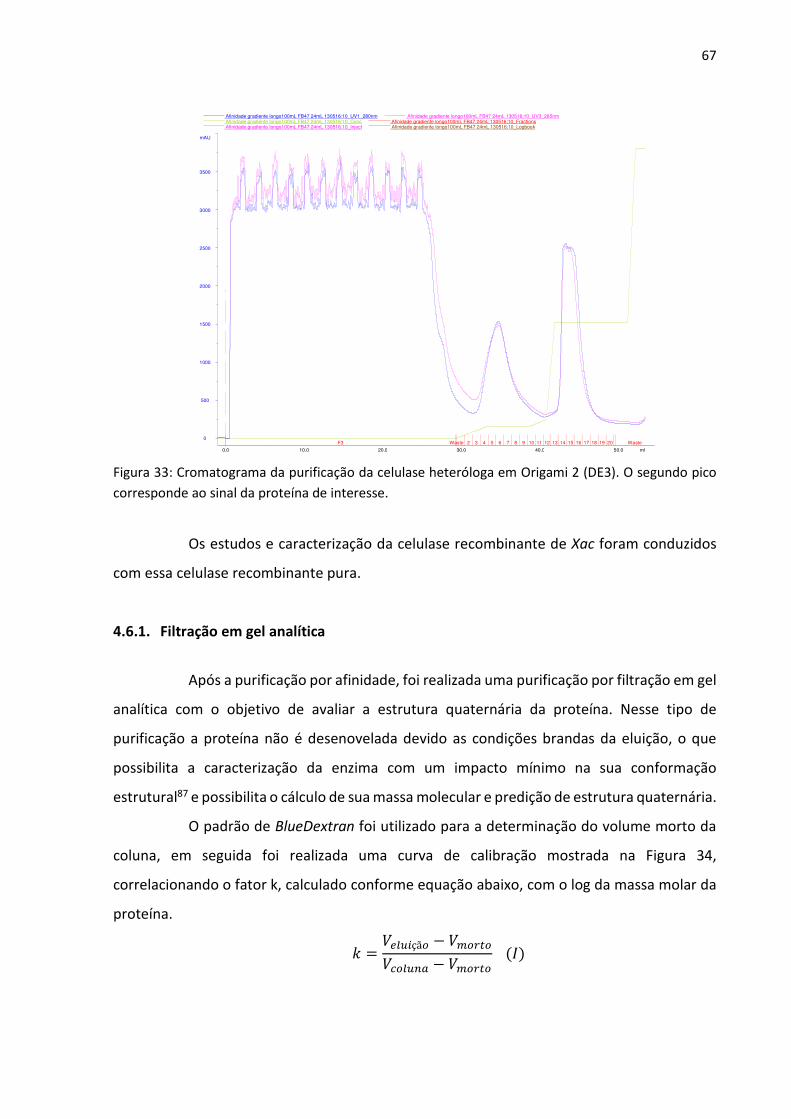
67
Figura 33: Cromatograma da purificação da celulase heteróloga em Origami 2 (DE3). O segundo pico
corresponde ao sinal da proteína de interesse.
Os estudos e caracterização da celulase recombinante de Xac foram conduzidos
com essa celulase recombinante pura.
4.6.1. Filtração em gel analítica
Após a purificação por afinidade, foi realizada uma purificação por filtração em gel
analítica com o objetivo de avaliar a estrutura quaternária da proteína. Nesse tipo de
purificação a proteína não é desenovelada devido as condições brandas da eluição, o que
possibilita a caracterização da enzima com um impacto mínimo na sua conformação
estrutural87 e possibilita o cálculo de sua massa molecular e predição de estrutura quaternária.
O padrão de BlueDextran foi utilizado para a determinação do volume morto da
coluna, em seguida foi realizada uma curva de calibração mostrada na Figura 34,
correlacionando o fator k, calculado conforme equação abaixo, com o log da massa molar da
proteína.
/ =&0123çã6 − &(6786
&9612:; − &(6786
(.)
Afinidade gradiente longo100mL FB47 24mL 130516:10_UV1_280nm Afinidade gradiente longo100mL FB47 24mL 130516:10_UV3_265nm Afinidade gradiente longo100mL FB47 24mL 130516:10_Conc Afinidade gradiente longo100mL FB47 24mL 130516:10_Fractions Afinidade gradiente longo100mL FB47 24mL 130516:10_Inject Afinidade gradiente longo100mL FB47 24mL 130516:10_Logbook
0
500
1000
1500
2000
2500
3000
3500
mAU
0.0 10.0 20.0 30.0 40.0 50.0 ml
F3 Waste 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Waste
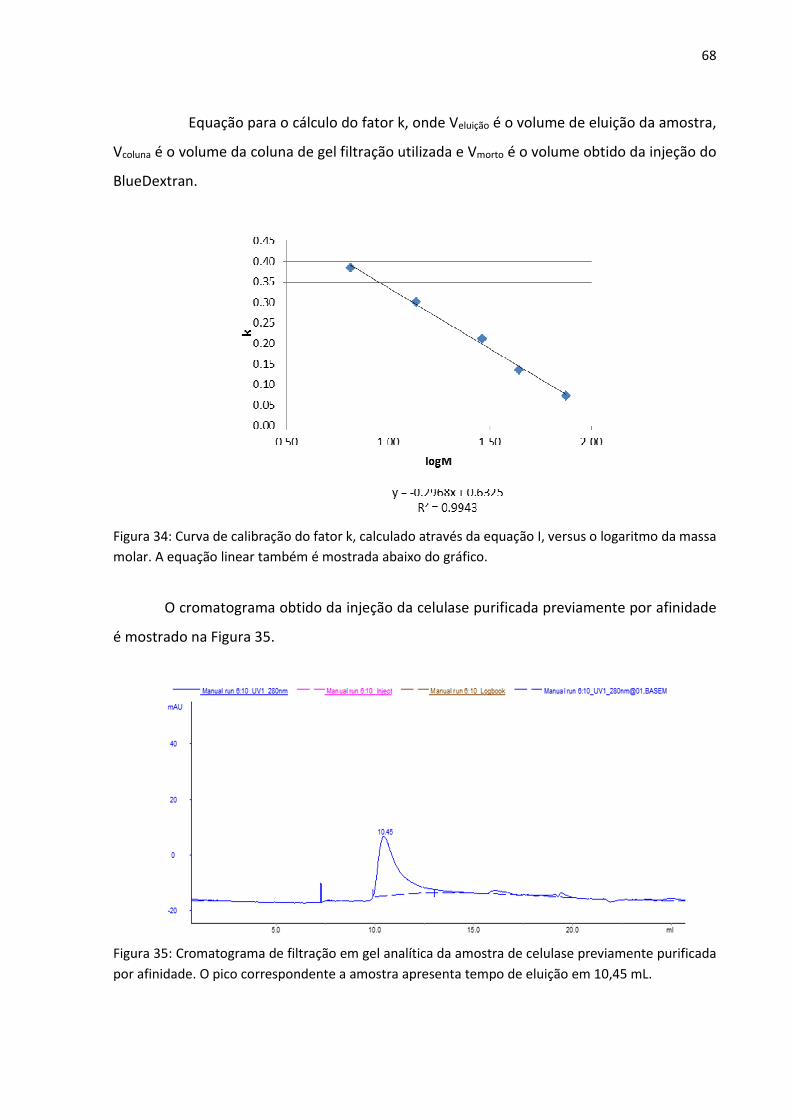
68
Equação para o cálculo do fator k, onde Veluição é o volume de eluição da amostra,
Vcoluna é o volume da coluna de gel filtração utilizada e Vmorto é o volume obtido da injeção do
BlueDextran.
Figura 34: Curva de calibração do fator k, calculado através da equação I, versus o logaritmo da massa
molar. A equação linear também é mostrada abaixo do gráfico.
O cromatograma obtido da injeção da celulase purificada previamente por afinidade
é mostrado na Figura 35.
Figura 35: Cromatograma de filtração em gel analítica da amostra de celulase previamente purificada
por afinidade. O pico correspondente a amostra apresenta tempo de eluição em 10,45 mL.

69
A partir da Figura 35 podemos observar apenas um pico com volume de eluição
de 10,45 mL referente a celulase. Através do volume de eluição da celulase foi possível
calcular o fator k, e utilizando a curva de calibração foi possível determinar a sua massa molar.
A Tabela 6, abaixo, apresenta os valores obtidos.
Tabela 6: Valores encontrados para o cálculo da massa molar da celulase recombinante
Volume de eluição (mL) 10,45
Fator k 0,135
logM 1,707
M (kDa) 50,99
O valor obtido de aproximadamente 51 kDa indica que a enzima se encontra na
forma monomérica, no entanto difere da massa estimada através da sequência primária de
aminoácidos de 61 kDa. Tal efeito pode ser explicado por não se tratar de uma proteína com
formato globular, e dessa forma ficar menos tempo retida na coluna. Para investigar essa
hipótese foi feita uma modelagem utilizando-se o software i-Tasser88–90, que utiliza em seus
modelos proteínas já cristalizadas e com alta identidade para gerar as estruturas propostas. O
modelo gerado pelo software é apresentado na Figura 36. Através da modelagem da estrutura
terciária fica evidente o formato alongado para proteína, apresentando 7,61 nm por 4,08 nm,
medidas realizadas utilizando-se o software PyMol91, que justificaria sua menor massa molar
estimada pela técnica de filtração em gel analítica.
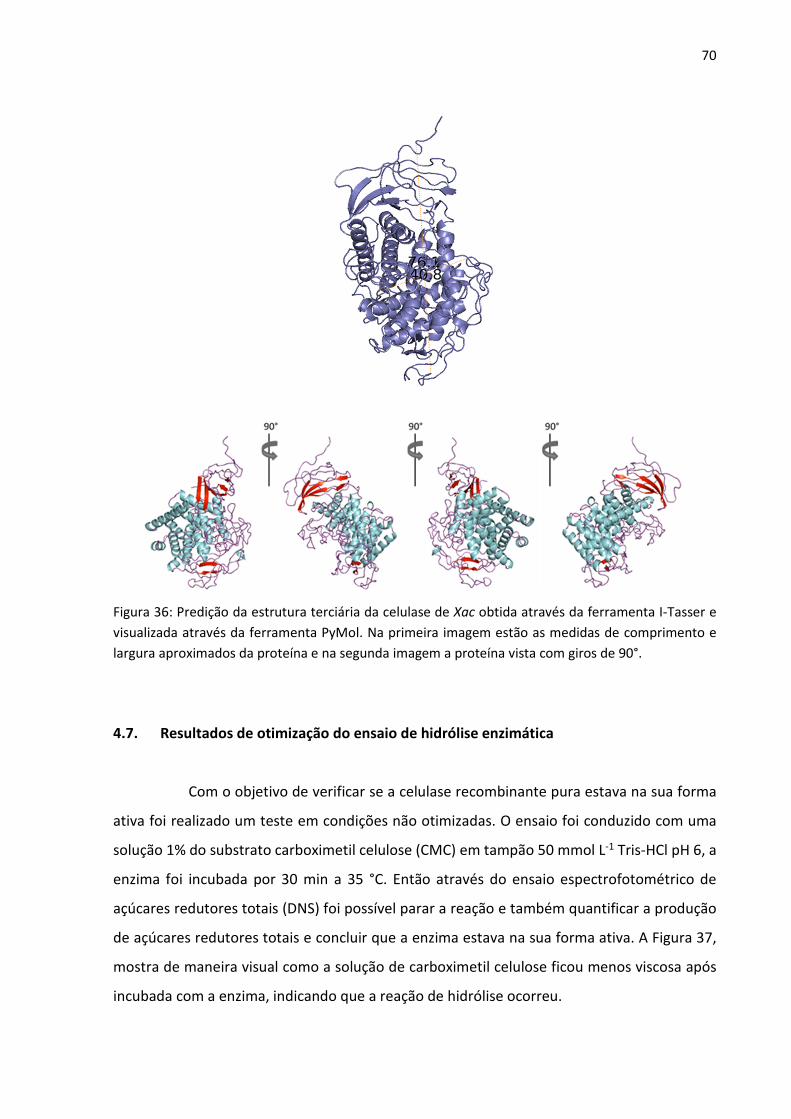
70
Figura 36: Predição da estrutura terciária da celulase de Xac obtida através da ferramenta I-Tasser e
visualizada através da ferramenta PyMol. Na primeira imagem estão as medidas de comprimento e
largura aproximados da proteína e na segunda imagem a proteína vista com giros de 90°.
4.7. Resultados de otimização do ensaio de hidrólise enzimática
Com o objetivo de verificar se a celulase recombinante pura estava na sua forma
ativa foi realizado um teste em condições não otimizadas. O ensaio foi conduzido com uma
solução 1% do substrato carboximetil celulose (CMC) em tampão 50 mmol L-1 Tris-HCl pH 6, a
enzima foi incubada por 30 min a 35 °C. Então através do ensaio espectrofotométrico de
açúcares redutores totais (DNS) foi possível parar a reação e também quantificar a produção
de açúcares redutores totais e concluir que a enzima estava na sua forma ativa. A Figura 37,
mostra de maneira visual como a solução de carboximetil celulose ficou menos viscosa após
incubada com a enzima, indicando que a reação de hidrólise ocorreu.

71
Figura 37: Solução 1% de CMC incubada com 200 nmol L-1 de celulase purificada por 30 min a 35 °C. É
possível observar a alteração na viscosidade do tubo contendo a enzima versus o tubo controle.
Em seguida, os parâmetros de tempo de reação, pH e temperatura foram
analisados com o objetivo de se encontrar as condições ótimas de atuação da celulase.
A Figura 38 mostra a variação da produção de açúcares redutores em diferentes
tempos de incubação. A reação foi incubada nos tempos: 0, 10, 20, 30, 40, 50 e 60 min.
Figura 38: Produção de açúcares redutores em diferentes tempos de incubação, ensaio realizado com
200 nmol L-1 de enzima, em tampão 50 mmol L-1 Na2HPO4/ácido cítrico pH 6 e temperatura de 35 °C.
Analisando o gráfico da Figura 38, determinou-se o tempo ótimo de reação como
30 min, já que a partir desse intervalo de incubação pequenas variações no tempo de reação
não representam variações significativas de produção de açúcares redutores.
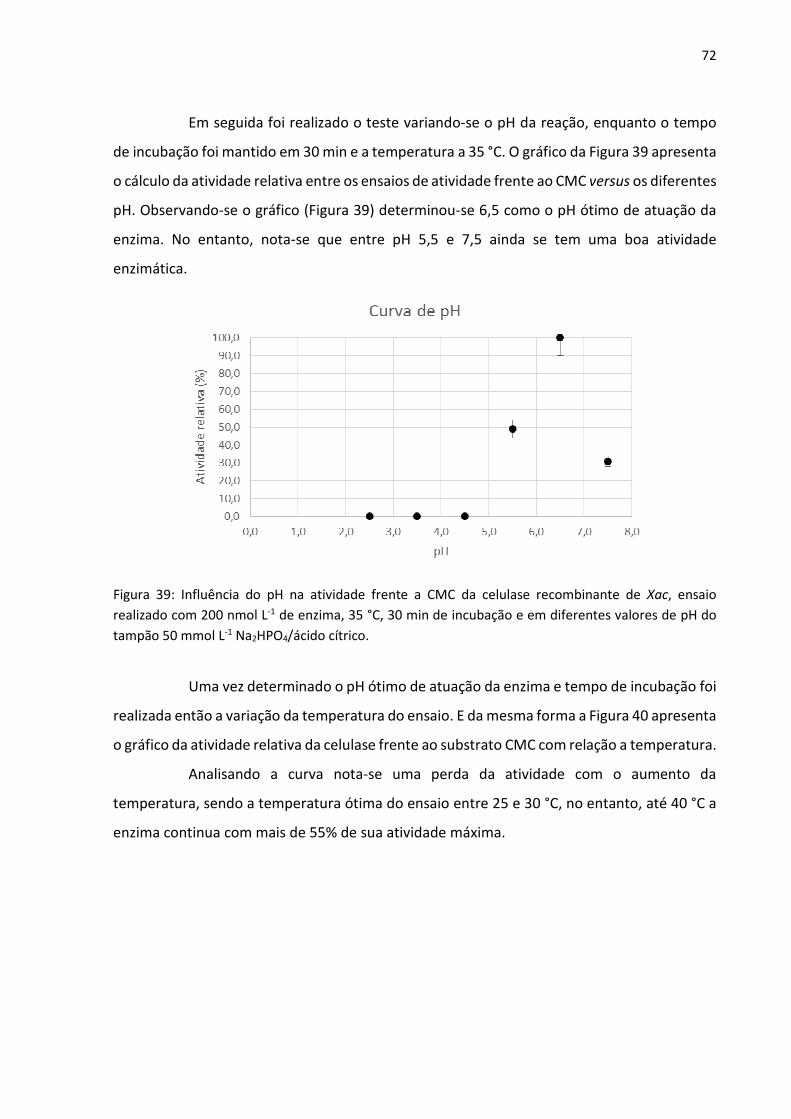
72
Em seguida foi realizado o teste variando-se o pH da reação, enquanto o tempo
de incubação foi mantido em 30 min e a temperatura a 35 °C. O gráfico da Figura 39 apresenta
o cálculo da atividade relativa entre os ensaios de atividade frente ao CMC versus os diferentes
pH. Observando-se o gráfico (Figura 39) determinou-se 6,5 como o pH ótimo de atuação da
enzima. No entanto, nota-se que entre pH 5,5 e 7,5 ainda se tem uma boa atividade
enzimática.
Figura 39: Influência do pH na atividade frente a CMC da celulase recombinante de Xac, ensaio
realizado com 200 nmol L-1 de enzima, 35 °C, 30 min de incubação e em diferentes valores de pH do
tampão 50 mmol L-1 Na2HPO4/ácido cítrico.
Uma vez determinado o pH ótimo de atuação da enzima e tempo de incubação foi
realizada então a variação da temperatura do ensaio. E da mesma forma a Figura 40 apresenta
o gráfico da atividade relativa da celulase frente ao substrato CMC com relação a temperatura.
Analisando a curva nota-se uma perda da atividade com o aumento da
temperatura, sendo a temperatura ótima do ensaio entre 25 e 30 °C, no entanto, até 40 °C a
enzima continua com mais de 55% de sua atividade máxima.
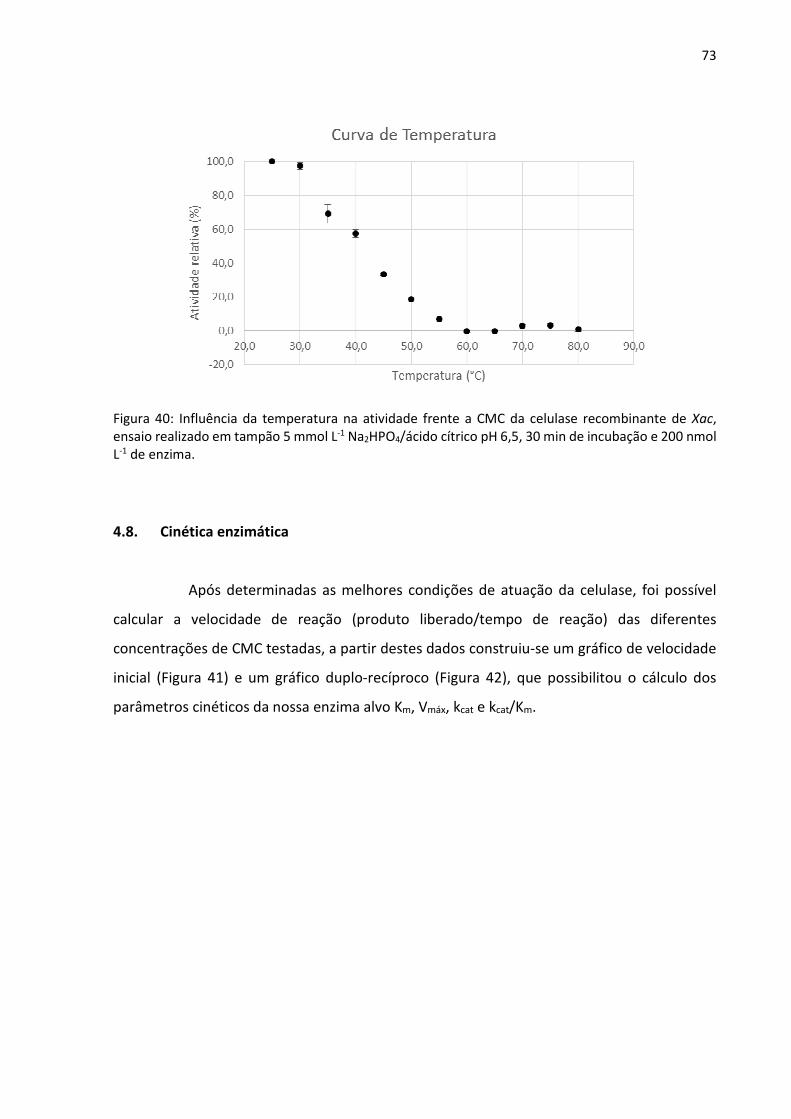
73
Figura 40: Influência da temperatura na atividade frente a CMC da celulase recombinante de Xac, ensaio realizado em tampão 5 mmol L-1 Na2HPO4/ácido cítrico pH 6,5, 30 min de incubação e 200 nmol L-1 de enzima.
4.8. Cinética enzimática
Após determinadas as melhores condições de atuação da celulase, foi possível
calcular a velocidade de reação (produto liberado/tempo de reação) das diferentes
concentrações de CMC testadas, a partir destes dados construiu-se um gráfico de velocidade
inicial (Figura 41) e um gráfico duplo-recíproco (Figura 42), que possibilitou o cálculo dos
parâmetros cinéticos da nossa enzima alvo Km, Vmáx, kcat e kcat/Km.
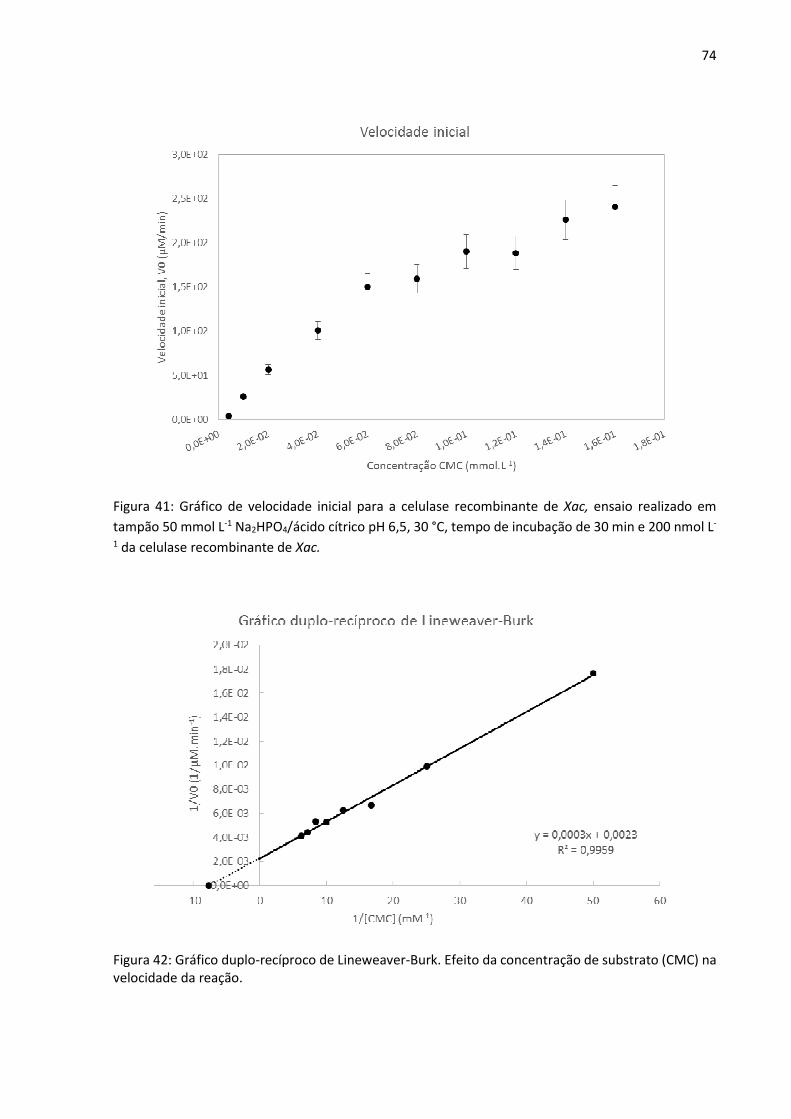
74
Figura 41: Gráfico de velocidade inicial para a celulase recombinante de Xac, ensaio realizado em
tampão 50 mmol L-1 Na2HPO4/ácido cítrico pH 6,5, 30 °C, tempo de incubação de 30 min e 200 nmol L-
1 da celulase recombinante de Xac.
Figura 42: Gráfico duplo-recíproco de Lineweaver-Burk. Efeito da concentração de substrato (CMC) na velocidade da reação.
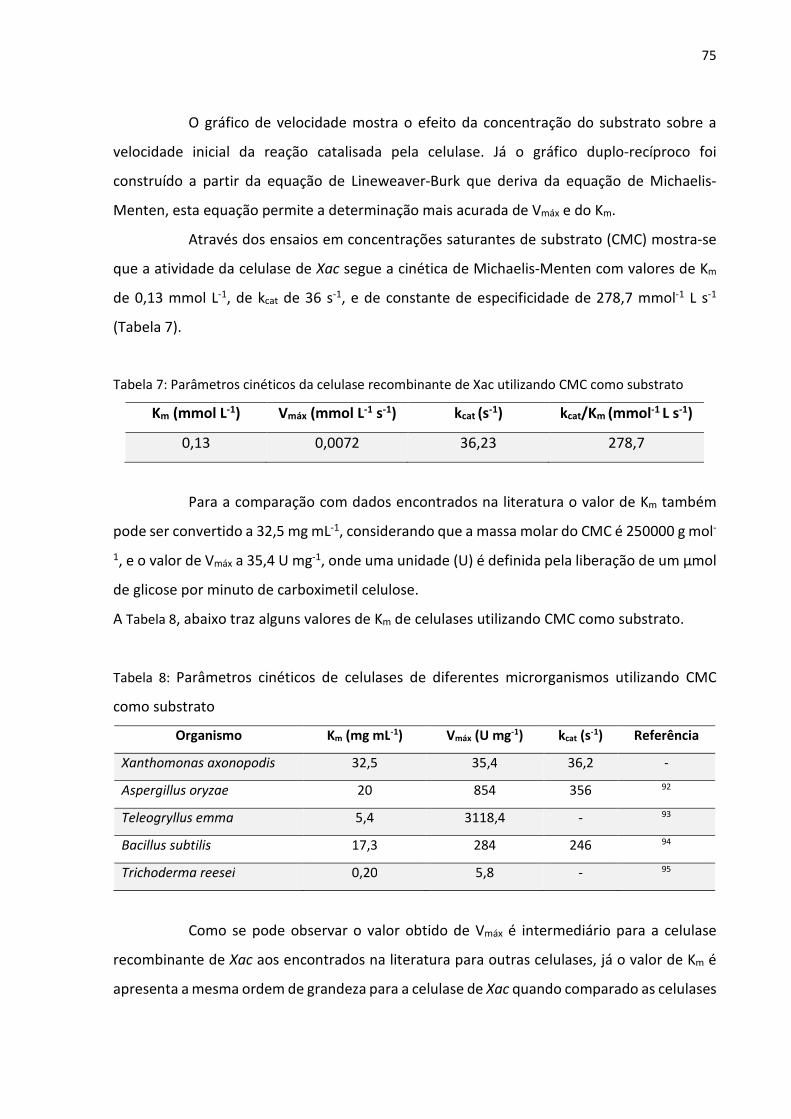
75
O gráfico de velocidade mostra o efeito da concentração do substrato sobre a
velocidade inicial da reação catalisada pela celulase. Já o gráfico duplo-recíproco foi
construído a partir da equação de Lineweaver-Burk que deriva da equação de Michaelis-
Menten, esta equação permite a determinação mais acurada de Vmáx e do Km.
Através dos ensaios em concentrações saturantes de substrato (CMC) mostra-se
que a atividade da celulase de Xac segue a cinética de Michaelis-Menten com valores de Km
de 0,13 mmol L-1, de kcat de 36 s-1, e de constante de especificidade de 278,7 mmol-1 L s-1
(Tabela 7).
Tabela 7: Parâmetros cinéticos da celulase recombinante de Xac utilizando CMC como substrato
Km (mmol L-1) Vmáx (mmol L-1 s-1) kcat (s-1) kcat/Km (mmol-1 L s-1)
0,13 0,0072 36,23 278,7
Para a comparação com dados encontrados na literatura o valor de Km também
pode ser convertido a 32,5 mg mL-1, considerando que a massa molar do CMC é 250000 g mol-
1, e o valor de Vmáx a 35,4 U mg-1, onde uma unidade (U) é definida pela liberação de um µmol
de glicose por minuto de carboximetil celulose.
A Tabela 8, abaixo traz alguns valores de Km de celulases utilizando CMC como substrato.
Tabela 8: Parâmetros cinéticos de celulases de diferentes microrganismos utilizando CMC
como substrato
Organismo Km (mg mL-1) Vmáx (U mg-1) kcat (s-1) Referência
Xanthomonas axonopodis 32,5 35,4 36,2 -
Aspergillus oryzae 20 854 356 92
Teleogryllus emma 5,4 3118,4 - 93
Bacillus subtilis 17,3 284 246 94
Trichoderma reesei 0,20 5,8 - 95
Como se pode observar o valor obtido de Vmáx é intermediário para a celulase
recombinante de Xac aos encontrados na literatura para outras celulases, já o valor de Km é
apresenta a mesma ordem de grandeza para a celulase de Xac quando comparado as celulases

76
de A. oryzae e B. subtilis, no entanto o valor obtido de kcat é mais baixo, indicando um turnover
menor para a enzima recombinante de Xac quando comparado às outras.
4.9. Resultados do Dicroísmo Circular
Inicialmente foi realizada uma previsão da estrutura secundária da celulase
utilizando-se a ferramenta GOR4 (https://npsa-prabi.ibcp.fr/cgi-bin/secpred_gor4.pl)96. De
acordo com a ferramenta, a proteína apresenta 40,6% de α-hélice, 14,13% de folha-β e 45,23%
de cadeia aleatória. Também foi realizada uma predição através do software I-Tasser88–90
(Tabela 4).
A proteína possui 566 resíduos e massa molar 61625,3 Da e a concentração de
proteína utilizada foi de 0,17 mg mL-1. A partir dos dados de elipticidade, θ, foi calculada [θ],
a média da elipticidade molar do resíduo utilizando-se a equação abaixo:
[�] =� × 100 ×
× � × �
Onde θ é a elipticidade em graus, l é o caminho ótico em cm, C é a concentração
em mg mL-1, M é a massa molecular e n é o número de resíduos da proteína. A unidade de [θ]
é deg cm2 dmol-1. O espectro de dicroísmo circular obtido para a amostra é ilustrado na Figura
43.
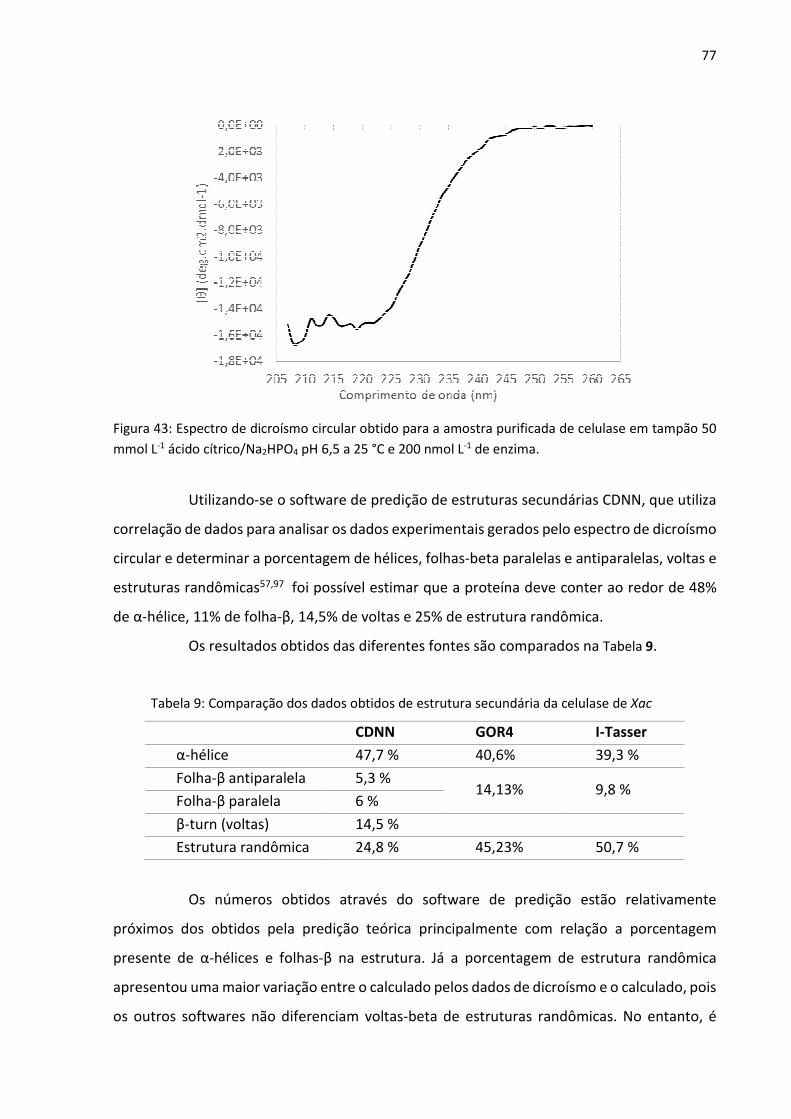
77
Figura 43: Espectro de dicroísmo circular obtido para a amostra purificada de celulase em tampão 50
mmol L-1 ácido cítrico/Na2HPO4 pH 6,5 a 25 °C e 200 nmol L-1 de enzima.
Utilizando-se o software de predição de estruturas secundárias CDNN, que utiliza
correlação de dados para analisar os dados experimentais gerados pelo espectro de dicroísmo
circular e determinar a porcentagem de hélices, folhas-beta paralelas e antiparalelas, voltas e
estruturas randômicas57,97 foi possível estimar que a proteína deve conter ao redor de 48%
de α-hélice, 11% de folha-β, 14,5% de voltas e 25% de estrutura randômica.
Os resultados obtidos das diferentes fontes são comparados na Tabela 9.
Tabela 9: Comparação dos dados obtidos de estrutura secundária da celulase de Xac
CDNN GOR4 I-Tasser
α-hélice 47,7 % 40,6% 39,3 %
Folha-β antiparalela 5,3 % 14,13% 9,8 %
Folha-β paralela 6 %
β-turn (voltas) 14,5 %
Estrutura randômica 24,8 % 45,23% 50,7 %
Os números obtidos através do software de predição estão relativamente
próximos dos obtidos pela predição teórica principalmente com relação a porcentagem
presente de α-hélices e folhas-β na estrutura. Já a porcentagem de estrutura randômica
apresentou uma maior variação entre o calculado pelos dados de dicroísmo e o calculado, pois
os outros softwares não diferenciam voltas-beta de estruturas randômicas. No entanto, é

78
importante notar que o espectro foi obtido na região de 207 a 265, o que compromete a
deconvolução dos dados de dicroísmo circular97.
Apesar das diferentes formas de predição de estrutura secundária, também deve
se ter em mente que a técnica de CD é mais precisa para a previsão de alfa-hélice. A predição
de folhas-beta, principalmente de sua orientação, paralela ou antiparalela, provou-se ser mais
difícil e tendenciosa devido a sua variedade espectral e menores amplitudes, o que é na
verdade uma limitação da técnica de dicroísmo circular98.
Com o objetivo de se determinar a temperatura de transição do estado de
enovelamento da proteína (Tm) foi feito o dicroísmo circular da celulase variando-se a
temperatura de 20 a 90 °C enquanto o comprimento de onda 222 nm foi monitorado, e então
voltou-se lentamente à temperatura inicial (Figura 44).
Analisando o gráfico (Figura 44) pôde-se calcular a Tm de 38 °C, que é coerente
com o ensaio de temperatura ótima realizado, onde observou-se que na temperatura de 40
°C menos de 60% da atividade foi vista. Além disso, pode-se notar que a celulase se desnatura
completamente e não retorna a seu estado nativo quando aquecida até 90 °C, indicando que
a desnaturação térmica seria irreversível. No entanto, como para o estado nativo a Tm foi de
38°C, um novo teste foi realizado subindo a temperatura apenas até 60 °C para observar se a
desnaturação continuaria irreversível até essa temperatura. O resultado dessa segunda
tentativa de desnaturação e re-enovelamento segue na Figura 45, que também apresentou
perda total de estrutura e desnaturação irreversível, além disso em ambos os casos se
observou formação de agregado durante o processo de desnaturação.
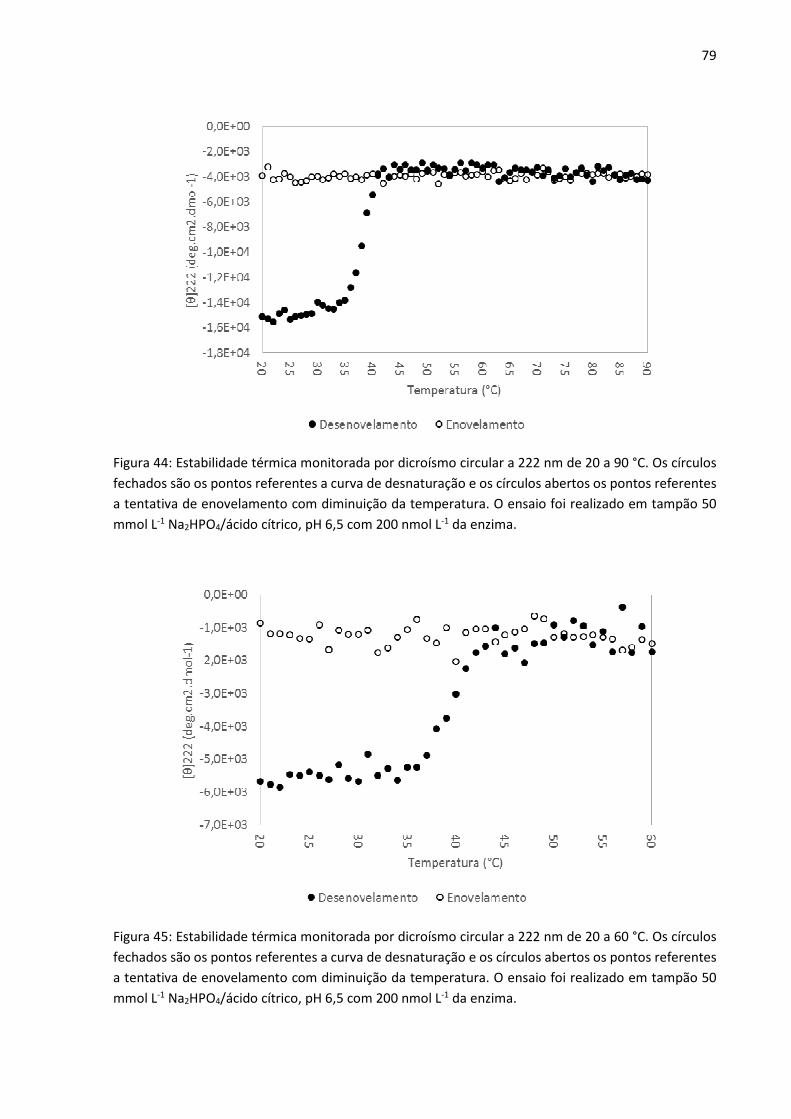
79
Figura 44: Estabilidade térmica monitorada por dicroísmo circular a 222 nm de 20 a 90 °C. Os círculos
fechados são os pontos referentes a curva de desnaturação e os círculos abertos os pontos referentes
a tentativa de enovelamento com diminuição da temperatura. O ensaio foi realizado em tampão 50
mmol L-1 Na2HPO4/ácido cítrico, pH 6,5 com 200 nmol L-1 da enzima.
Figura 45: Estabilidade térmica monitorada por dicroísmo circular a 222 nm de 20 a 60 °C. Os círculos
fechados são os pontos referentes a curva de desnaturação e os círculos abertos os pontos referentes
a tentativa de enovelamento com diminuição da temperatura. O ensaio foi realizado em tampão 50
mmol L-1 Na2HPO4/ácido cítrico, pH 6,5 com 200 nmol L-1 da enzima.
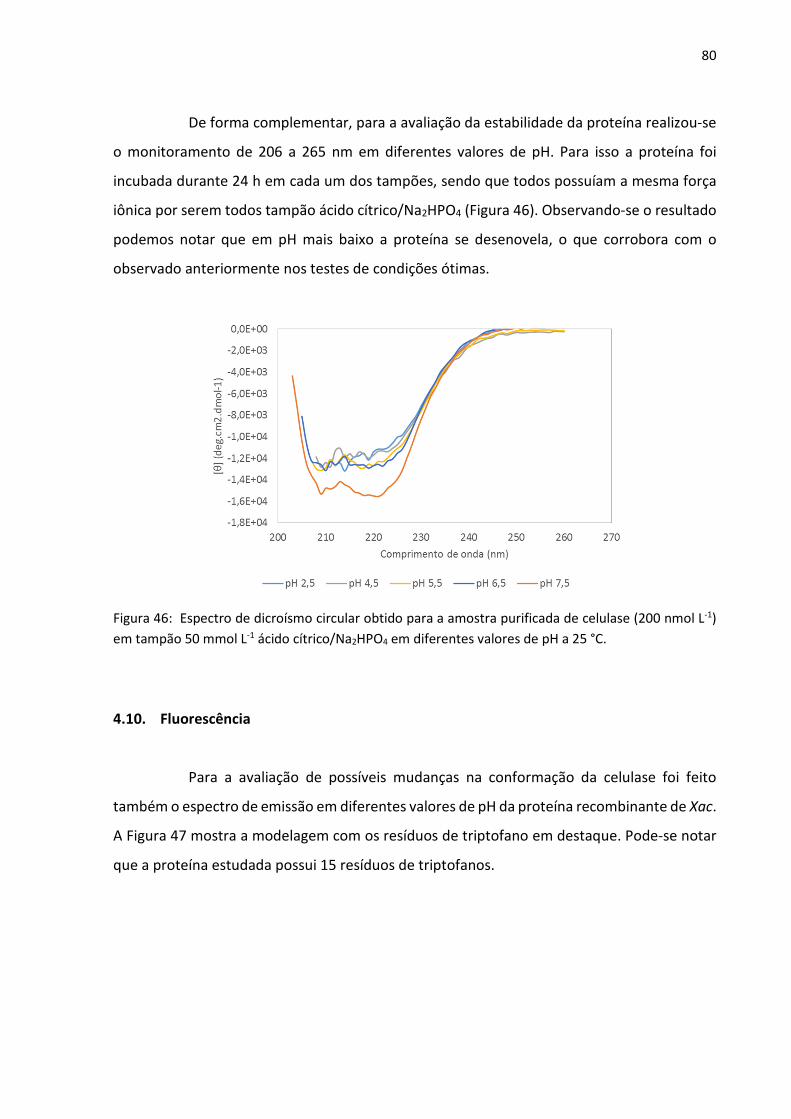
80
De forma complementar, para a avaliação da estabilidade da proteína realizou-se
o monitoramento de 206 a 265 nm em diferentes valores de pH. Para isso a proteína foi
incubada durante 24 h em cada um dos tampões, sendo que todos possuíam a mesma força
iônica por serem todos tampão ácido cítrico/Na2HPO4 (Figura 46). Observando-se o resultado
podemos notar que em pH mais baixo a proteína se desenovela, o que corrobora com o
observado anteriormente nos testes de condições ótimas.
Figura 46: Espectro de dicroísmo circular obtido para a amostra purificada de celulase (200 nmol L-1)
em tampão 50 mmol L-1 ácido cítrico/Na2HPO4 em diferentes valores de pH a 25 °C.
4.10. Fluorescência
Para a avaliação de possíveis mudanças na conformação da celulase foi feito
também o espectro de emissão em diferentes valores de pH da proteína recombinante de Xac.
A Figura 47 mostra a modelagem com os resíduos de triptofano em destaque. Pode-se notar
que a proteína estudada possui 15 resíduos de triptofanos.

81
Figura 47: Modelagem realizada no I-Tasser para a celulase recombinante de Xac. Em rosa estão destacados os resíduos de triptofano da proteína recombinante de Xac.
Um dos principais fatores que afetam a emissão do triptofano é a polaridade do
ambiente em que o resíduo se encontra, uma vez que o grupo indol pode formar ligações de
hidrogênio em ambientes polares, o que resulta em variações do máximo de emissão e
rendimento quântico do triptofano, tais alterações podem ser relacionadas com o
enovelamento da proteína e sua estrutura terciária. Observa-se que quando a proteína está
em sua conformação enovelada os resíduos de triptofano possuem menor exposição à
solventes polares e apresentam emissão em menores comprimentos de onda e com maior
rendimento quântico, logo as variações no espectro de emissão podem ser usadas para
acompanhar o desenovelamento de uma proteína60.
Na Figura 48 é mostrado o espectro de excitação da amostra e na Figura 49 o
espectro de emissão da mesma em diferentes valores de pH.
No espectro de emissão realizado o que se observa é uma variação do
comprimento de onda de emissão e intensidade com a variação do pH. Em pH ácido observa-
se comprimentos maiores e sinais menos intensos do que em pH neutro, o que sugere maior
exposição dos resíduos de triptofano e consequente mudança da estrutura terciária da
celulase, que corrobora com os dados de dicroísmo circular e dos parâmetros otimizados, tais
resultados indicam que as mudanças no comportamento enzimático e estabilidade da
proteína estão relacionados com a perda da conformação de estrutura terciária.
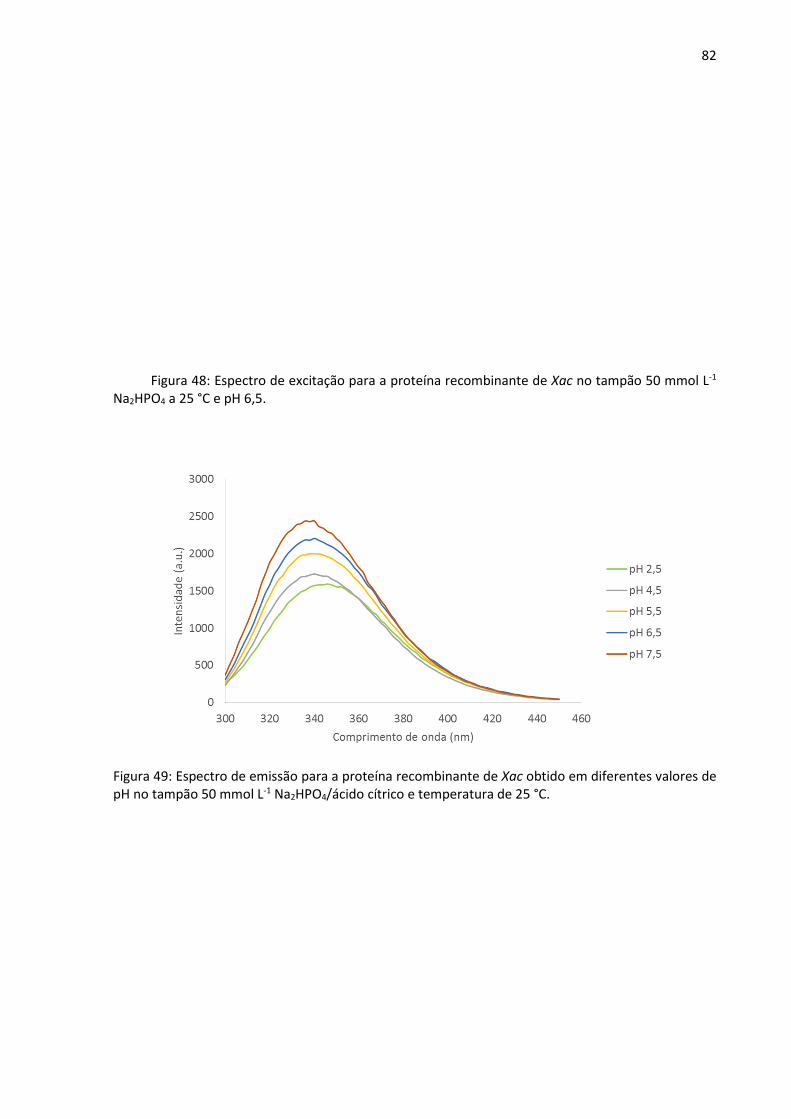
82
Figura 48: Espectro de excitação para a proteína recombinante de Xac no tampão 50 mmol L-1 Na2HPO4 a 25 °C e pH 6,5.
Figura 49: Espectro de emissão para a proteína recombinante de Xac obtido em diferentes valores de pH no tampão 50 mmol L-1 Na2HPO4/ácido cítrico e temperatura de 25 °C.

83
5. Conclusões
Como apresentado nas sessões anteriores a pesquisa foi bem-sucedida em várias
frentes, primeiramente frente à caracterização de diferentes tipos de bagaço de laranja
(comercial, industrial, amostra de dorna industrial e pellet), o que possibilitou uma melhor
compreensão dos tipos de bagaço e suas composições apresentando possibilidades para o uso
do resíduo da indústria de suco de laranja para a extração da hesperidina, óleo essencial e
fermentação de etanol de segunda geração agregando mais valor a cadeia de produção de
suco de laranja.
Além disso, foi possível isolar uma cepa de Saccharomyces cerevisiae proveniente
do bagaço de laranja industrial que resistiu a presença de 0,5% de concentração de d-
limoneno no meio de cultura, o que representa uma vantagem do ponto de vista de aplicação,
tendo em vista que concentrações baixas de d-limoneno já apresentam efeito inibitório na
fermentação, a cepa isolada apresentou maior rendimento (47%) e produtividade (0,9 g L-1 h-
1) em comparação com a cepa de levedura controle FY23 (26% de rendimento e 0,5 g L-1 h-1).
Na segunda parte dessa pesquisa foi realizado um estudo acerca de enzimas
recombinantes, infelizmente não se teve êxito na amplificação das mesmas. No entanto,
obteve-se um resultado inédito na caracterização da enzima recombinante de Xac
confirmando a proteína hipotética. A celulase foi expressa com sucesso na fração solúvel em
altas concentrações e obtida em alto nível de pureza. Além disso, foram determinadas suas
condições ótimas de atuação de tempo de incubação, pH e temperatura de atividade, nestas
condições foram obtidos seus parâmetros cinéticos (Km de 0,130 mmol L-1, kcat de 36 s-1, e
constante de especificidade de 278,7 mmol-1 L s-1), também foram determinamos que nas
condições do ensaio a proteína é monomérica. A partir do dicroísmo circular observou-se que
a enzima possui Tm de 38 °C, e não retorna a sua conformação original depois de desenovelada
formando agregados.
Os estudos de cinética, dicroísmo circular e fluorescência sugerem que as
mudanças no comportamento enzimático e estabilidade da proteína estão relacionados com
a perda da conformação de estrutura terciária em pH ácido.
Por final destaca-se que os estudos de avaliação de resíduos de suco de laranja,
busca por uma cepa resistente a d-limoneno e ensaio de caracterização da celulase
recombinante de Xac são promissores para o desenvolvimento de novos coquetéis

84
enzimáticos para o uso em hidrólise de biomassa, cepas com maior produtividade frente aos
resíduos de laranja e diferentes aplicações dos resíduos da indústria de suco de laranja, como
por exemplo para a produção de hesperidina e etanol de segunda geração.

85
6. Referências
1. Bagheri, H. C. & Del Amo, B. The Past and Future of Renewable Energy Deployment in Light of the COP21 Agreement: The Cases of Germany and the USA. (2016).
2. Chaudhary, N. & Qazi, J. Lignocellulose for ethanol production: A review of issues relating to bagasse as a source material. African Journal of Biotechnology 10, 1270–1274 (2013).
3. Zhao, X., Zhang, L. & Liu, D. Biomass recalcitrance. Part II: Fundamentals of different pre-treatments to increase the enzymatic digestibility of lignocellulose. Biofuels, Bioprod.
Biorefining 6, 561–579 (2012).
4. Saini, J. K., Saini, R. & Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: concepts and recent developments. 3 Biotech 5, 337–353 (2015).
5. Maitan-Alfenas, G. P., Visser, E. M. & Guimarães, V. M. Enzymatic hydrolysis of lignocellulosic biomass: converting food waste in valuable products. Curr. Opin. Food Sci. 1, 44–49 (2015).
6. Neves, M. F. et al. O retrato da citricultura brasileira. Markestrat (2010).
7. Citrus: World Markets and Trade. Foreign Agricultural Service (2017). Available at: http://usda.mannlib.cornell.edu/usda/fas/citruswm//2010s/2017/citruswm-01-25-2017.pdf. (Accessed: 25th February 2017)
8. Tsukamoto, J., Durán, N. & Tasic, L. Nanocellulose and Bioethanol Production from Orange Waste using Isolated Microorganisms. J. Braz. Chem. Soc. 24, 1537–1543 (2013).
9. Bampidis, V. A. & Robinson, P. H. Citrus by-products as ruminant feeds: A review. Anim. Feed
Sci. Technol. 128, 175–217 (2006).
10. Awan, A. T., Tsukamoto, J. & Tasic, L. Orange waste as a biomass for 2G-ethanol production using low cost enzymes and co-culture fermentation. RSC Adv. 3, 25071 (2013).
11. Grohmann, K., Cameron, R. G. & Buslig, B. S. Fractionation and pretreatment of orange peel by dilute acid hydrolysis. Bioresour. Technol. 54, 129–141 (1995).
12. Lohrasbi, M., Pourbafrani, M., Niklasson, C. & Taherzadeh, M. J. Process design and economic analysis of a citrus waste biorefinery with biofuels and limonene as products. Bioresour.
Technol. 101, 7382–7388 (2010).
13. Pourbafrani, M., Forgács, G., Horváth, I. S., Niklasson, C. & Taherzadeh, M. J. Production of biofuels, limonene and pectin from citrus wastes. Bioresour. Technol. 101, 4246–4250 (2010).
14. Rivas, B., Torrado, A., Torre, P., Converti, A. & Domínguez, J. M. Submerged citric acid fermentation on orange peel autohydrolysate. J. Agric. Food Chem. 56, 2380–7 (2008).
15. Queiroz, V. L., Awan, A. T. & Tasic, L. in Agro-Industrial Wastes as Feedstock for Enzyme
Production 111–131 (2016). doi:10.1016/B978-0-12-802392-1.00005-8
16. Wyman, C. Handbook on Bioethanol: Production and Utilization. (1996).
17. Hon, D. N.-S. & Shiraishi, N. Wood and Cellulosic Chemistry, Second Edition, Revised, and
Expanded. (2000).
18. Willats, W. T., McCartney, L., Mackie, W. & Knox, J. P. in Plant Cell Walls SE - 2 (eds. Carpita, N. C., Campbell, M. & Tierney, M.) 9–27 (Springer Netherlands, 2001). doi:10.1007/978-94-010-0668-2_2
19. Zhong, R., Ye, Z.-H., Zhong, R. & Ye, Z. Secondary Cell Walls. Encyclopedia of Life Sciences (John Wiley & Sons, Ltd, 2009). doi:10.1002/9780470015902.a0021256

86
20. Plomion, C., Leprovost, G. & Stokes, A. Wood Formation in Trees. PLANT Physiol. 127, 1513–1523 (2001).
21. O’Sullivan, A. C. Cellulose: the structure slowly unravels. Cellulose 4, 173–207 (1997).
22. Suhas et al. Cellulose: A review as natural, modified and activated carbon adsorbent. Bioresour.
Technol. 216, 1066–1076 (2016).
23. Klemm, D., Heublein, B., Fink, H.-P. & Bohn, A. Cellulose: Fascinating Biopolymer and Sustainable Raw Material. Angew. Chemie Int. Ed. 44, 3358–3393 (2005).
24. Santos, F. A. et al. Potential of sugarcane straw for ethanol production. Quim. Nova 35, 1004–1010 (2012).
25. Klemm, D., Philipp, B., Heinze, T., Heinze, U. & Wagenknecht, W. Comprehensive Cellulose
Chemistry. 1, (Wiley-VCH Verlag GmbH & Co. KGaA, 1998).
26. Lu, B. et al. Cation does matter: how cationic structure affects the dissolution of cellulose in ionic liquids. Green Chem. 16, 1326–1335 (2014).
27. Gírio, F. M. et al. Hemicelluloses for fuel ethanol: A review. Bioresour. Technol. 101, 4775–4800 (2010).
28. Zhou, X., Li, W., Mabon, R. & Broadbelt, L. J. A Critical Review on Hemicellulose Pyrolysis. Energy
Technol. 5, 52–79 (2017).
29. Sun, J., Tian, C., Diamond, S. & Glass, N. L. Deciphering transcriptional regulatory mechanisms associated with hemicellulose degradation in Neurospora crassa. Eukaryot. Cell 11, 482–93 (2012).
30. Zabed, H., Sahu, J. N., Boyce, A. N. & Faruq, G. Fuel ethanol production from lignocellulosic biomass: An overview on feedstocks and technological approaches. Renew. Sustain. Energy Rev. 66, 751–774 (2016).
31. Glazer, A. N. & Nikaido, H. Microbial Biotechnology: Fundamentals of Applied Microbiology. (Cambridge University Press, 2007).
32. Espina, L. et al. Chemical composition of commercial citrus fruit essential oils and evaluation of their antimicrobial activity acting alone or in combined processes. Food Control 22, 896–902 (2011).
33. Nannapaneni, R. et al. Campylobacter and Arcobacter species sensitivity to commercial orange oil fractions. Int. J. Food Microbiol. 129, 43–9 (2009).
34. Tasic, L., Borin, P. F. L., Khater, L. C. & Ramos, C. H. I. Cloning and characterization of three hypothetical secretion chaperone proteins from Xanthomonas axonopodis pv. citri. Protein
Expr. Purif. 53, 363–9 (2007).
35. Ferreira, C. B. et al. Identification of New Genes Related to Virulence of Xanthomonas axonopodis Pv. Citri during Citrus Host Interactions. Adv. Microbiol. 7, 22–46 (2017).
36. Di Lorenzo, F. et al. Xanthomonas citri pv. citri pathotypes: LPS structure and function as microbe-associated molecular patterns. ChemBioChem (2017). doi:10.1002/cbic.201600671
37. Graham, J. H., Gottwald, T. R., Cubero, J. & Achor, D. S. Xanthomonas axonopodis pv. citri: factors affecting successful eradication of citrus canker. Mol. Plant Pathol. 5, 1–15 (2004).
38. da Silva, A. C. R. et al. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 417, 459–63 (2002).
39. Beilen, J. B. va. & Li, Z. Enzyme technology: an overview. Curr. Opin. Biotechnol. 13, 338–344 (2002).
40. Goldemberg, J. & Guardabassi, P. The potential for first-generation ethanol production from

87
sugarcane. Biofuels, Bioprod. Biorefining 4, 17–24 (2010).
41. Anwar, Z., Gulfraz, M. & Irshad, M. Agro-industrial lignocellulosic biomass a key to unlock the future bio-energy: A brief review. J. Radiat. Res. Appl. Sci. 7, 163–173 (2014).
42. Wyman, C. E. Biomass Ethanol: Technical Progress, Opportunities, and Commercial Challenges. Annu. Rev. Energy Environ. 24, 189–226 (1999).
43. Sun, Y. & Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresour. Technol. 83, 1–11 (2002).
44. Sarkar, N., Ghosh, S. K., Bannerjee, S. & Aikat, K. Bioethanol production from agricultural wastes: An overview. Renew. Energy 37, 19–27 (2012).
45. Alvira, P., Tomás-Pejó, E., Ballesteros, M. & Negro, M. J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour.
Technol. 101, 4851–61 (2010).
46. Mosier, N. et al. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 96, 673–86 (2005).
47. Toquero, C. & Bolado, S. Effect of four pretreatments on enzymatic hydrolysis and ethanol fermentation of wheat straw. Influence of inhibitors and washing. Bioresour. Technol. 157, 68–76 (2014).
48. Brummer, V., Skryja, P., Jurena, T., Hlavacek, V. & Stehlik, P. Suitable Technological Conditions for Enzymatic Hydrolysis of Waste Paper by Novozymes® Enzymes NS50013 and NS50010. Appl.
Biochem. Biotechnol. 174, 1299–1308 (2014).
49. El-Zawawy, W. K., Ibrahim, M. M., Abdel-Fattah, Y. R., Soliman, N. A. & Mahmoud, M. M. Acid and enzyme hydrolysis to convert pretreated lignocellulosic materials into glucose for ethanol production. Carbohydr. Polym. 84, 865–871 (2011).
50. Singhania, R. R., Patel, A. K., Sukumaran, R. K., Larroche, C. & Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour.
Technol. 127, 500–507 (2013).
51. Lynd, L. R., Weimer, P. J., van Zyl, W. H. & Pretorius, I. S. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Microbiol. Mol. Biol. Rev. 66, 506–577 (2002).
52. Xia, T. et al. Identification of an Extracellular Endoglucanase That Is Required for Full Virulence in Xanthomonas citri subsp. citri. PLoS One 11, e0151017 (2016).
53. Jayani, R. S., Saxena, S. & Gupta, R. Microbial pectinolytic enzymes: A review. Process Biochem. 40, 2931–2944 (2005).
54. Taherzadeh, M. J. & Karimi, K. Enzymatic-based hydrolysis processes for ethanol from lignocellulosic materials : A review. BioResources 2, 707–738 (2007).
55. Corrêa, D. H. A. & Ramos, C. H. I. The use of circular dichroism spectroscopy to study protein folding, form and function. African J. Biochem. Res. 3, 164–173 (2009).
56. Schellman, J. A. Circular dichroism and optical rotation. Chem. Rev. 75, 323–331 (1975).
57. Greenfield, N. J. Using circular dichroism spectra to estimate protein secondary structure. Nat.
Protoc. 1, 2876–90 (2006).
58. Munishkina, L. A. & Fink, A. L. Fluorescence as a method to reveal structures and membrane-interactions of amyloidogenic proteins. Biochim. Biophys. Acta 1768, 1862–85 (2007).
59. Vivian, J. T. & Callis, P. R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 80, 2093–109 (2001).
60. Lakowicz, J. R. in Principles of Fluorescence Spectroscopy 529–575 (Springer US, 2006).

88
doi:10.1007/978-0-387-46312-4
61. Sluiter, A. et al. Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples Biomass and Total Dissolved Solids in Liquid Process Samples. in National
Renewable Energy Laboratory (2008).
62. Rosseto, F. R. Biochemical, Biophysical and Structural Studies of the Major Endoglucanase Secreted by Xanthomonas campestris pv. Campestris ATCC33913. (Universidade de São Paulo, 2011).
63. Miller, G. L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal.
Chem. 31, 426–428 (1959).
64. Johnson, K. A. & Goody, R. S. The Original Michaelis Constant: Translation of the 1913 Michaelis–Menten Paper. Biochemistry 50, 8264–8269 (2011).
65. Mahmood, R. T. et al. Production, purification, and characterization of exoglucanase by Aspergillus fumigatus. Appl. Biochem. Biotechnol. 170, 895–908 (2013).
66. Munhoz, J. R. & Morabito, R. Optimization approaches to support decision making in the production planning of a citrus company: A Brazilian case study. Comput. Electron. Agric. 107, 45–57 (2014).
67. Garg, A., Garg, S., Zaneveld, L. J. D. & Singla, A. K. Chemistry and pharmacology of the citrus bioflavonoid hesperidin. Phyther. Res. 15, 655–669 (2001).
68. Akdemir, F. N. E., Gülçin, İ., Karagöz, B., Soslu, R. & Alwasel, S. H. A comparative study on the antioxidant effects of hesperidin and ellagic acid against skeletal muscle ischemia/reperfusion injury. J. Enzyme Inhib. Med. Chem. 31, 114–118 (2016).
69. Cao, R., Yang, X., Strappe, P., Blanchard, C. & Zhou, Z. Natural products derived from tea on the solubility of hesperidin by LC-TOF/MS and NMR. Int. J. Food Prop. 1–9 (2017). doi:10.1080/10942912.2017.1295258
70. Wilkins, M. R., Widmer, W. W. & Grohmann, K. Simultaneous saccharification and fermentation of citrus peel waste by Saccharomyces cerevisiae to produce ethanol. Process Biochem. 42, 1614–1619 (2007).
71. Almeida, J. R. et al. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates bySaccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 82, 340–349 (2007).
72. Finn, R. D. et al. Pfam: the protein families database. Nucleic Acids Res. 42, D222-30 (2014).
73. Petersen, T. N., Brunak, S., von Heijne, G. & Nielsen, H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 8, 785–6 (2011).
74. Gasteiger, E. et al. in The Proteomics Protocols Handbook 571–607 (Humana Press, 2005). doi:10.1385/1-59259-890-0:571
75. Cantarel, B. L. et al. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics. Nucleic Acids Res. 37, D233–D238 (2009).
76. Ravachol, J., Borne, R., Tardif, C., de Philip, P. & Fierobe, H.-P. Characterization of all family-9 glycoside hydrolases synthesized by the cellulosome-producing bacterium Clostridium cellulolyticum. J. Biol. Chem. 289, 7335–48 (2014).
77. Henrissat, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 309–16 (1991).
78. Wyk, N. van, Drancourt, M., Henrissat, B. & Kremer, L. Current perspectives on the families of glycoside hydrolases of Mycobacterium tuberculosis: their importance and prospects for assigning function to unknowns. Glycobiology 27, 112–122 (2017).
79. Kataeva, I. A. et al. Interactions between immunoglobulin-like and catalytic modules in

89
Clostridium thermocellum cellulosomal cellobiohydrolase CbhA. Protein Eng. Des. Sel. 17, 759–769 (2004).
80. McCarter, J. D. & Stephen Withers, G. Mechanisms of enzymatic glycoside hydrolysis. Curr.
Opin. Struct. Biol. 4, 885–892 (1994).
81. Davies, G. & Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 3, 853–859 (1995).
82. CAZy - GH9. Available at: http://www.cazy.org/GH9_all.html. (Accessed: 1st March 2017)
83. Sørensen, H. P. & Mortensen, K. K. Advanced genetic strategies for recombinant protein expression in Escherichia coli. J. Biotechnol. 115, 113–28 (2005).
84. Bessette, P. H., Aslund, F., Beckwith, J. & Georgiou, G. Efficient folding of proteins with multiple disulfide bonds in the Escherichia coli cytoplasm. Proc. Natl. Acad. Sci. 96, 13703–13708 (1999).
85. Prinz, W. A., Aslund, F., Holmgren, A. & Beckwith, J. The Role of the Thioredoxin and Glutaredoxin Pathways in Reducing Protein Disulfide Bonds in the Escherichia coliCytoplasm. J. Biol. Chem. 272, 15661–15667 (1997).
86. Stewart, E. J., Aslund, F. & Beckwith, J. Disulfide bond formation in the Escherichia coli cytoplasm: an in vivo role reversal for the thioredoxins. EMBO J. 17, 5543–50 (1998).
87. Fekete, S., Beck, A., Veuthey, J.-L. & Guillarme, D. Theory and practice of size exclusion chromatography for the analysis of protein aggregates. J. Pharm. Biomed. Anal. 101, 161–173 (2014).
88. Roy, A., Kucukural, A. & Zhang, Y. I-TASSER: a unified platform for automated protein structure and function prediction. Nat. Protoc. 5, 725–38 (2010).
89. Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 9, 40 (2008).
90. Yang, J. et al. The I-TASSER Suite: protein structure and function prediction. Nat. Methods 12, 7–8 (2015).
91. Schrodinger LLC. The PyMOL Molecular Graphics System, Version 1.8. (2015).
92. Javed, M. R., Rashid, M. H., Nadeem, H., Riaz, M. & Perveen, R. Catalytic and thermodynamic characterization of endoglucanase (CMCase) from Aspergillus oryzae cmc-1. Appl. Biochem.
Biotechnol. 157, 483–97 (2009).
93. Kim, N. et al. Molecular cloning and characterization of a glycosyl hydrolase family 9 cellulase distributed throughout the digestive tract of the cricket Teleogryllus emma. Comp. Biochem.
Physiol. B. Biochem. Mol. Biol. 150, 368–76 (2008).
94. Wang, Y., Yuan, H., Wang, J. & Yu, Z. Truncation of the cellulose binding domain improved thermal stability of endo-beta-1,4-glucanase from Bacillus subtilis JA18. Bioresour. Technol. 100, 345–9 (2009).
95. Kupski, L., Pagnussatt, F. A., Buffon, J. G. & Furlong, E. B. Endoglucanase and total cellulase from newly isolated Rhizopus oryzae and Trichoderma reesei: production, characterization, and thermal stability. Appl. Biochem. Biotechnol. 172, 458–68 (2014).
96. Combet, C., Blanchet, C., Geourjon, C. & Deléage, G. NPS@: network protein sequence analysis. Trends Biochem. Sci. 25, 147–50 (2000).
97. Böhm, G., Muhr, R. & Jaenicke, R. Quantitative analysis of protein far UV circular dichroism spectra by neural networks. "Protein Eng. Des. Sel. 5, 191–195 (1992).
98. Micsonai, A. et al. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. U. S. A. 112, E3095-103 (2015).

90
7. Anexos
Todas as sequências utilizadas foram obtidas de Xanthomonas axonopodis pv. citri cepa 306
depositadas por Silva et al. (2002)38. Para a predição dos domínios da enzima e predição de
existência de peptídeo sinal na sequência foi utilizado a base de dados Pfam (Protein families
database)72 e também o servidor SignalP 4.173.
Enzima 1 – Celulase Q8PJK9
A celulase Q8PJK9 (identificação Uniprot) possui 586 aminoácidos e é codificada por 1761
pares de base. Além disso, possui peso molecular de 63715,0 Da e seu ponto isoelétrico
teórico é de 6,47. A sequência de aminoácidos segue abaixo (Accession: NP_642837.1
GI:21243255; XAC2522 gene egl2). Os resultados obtidos no Pfam e SignalP foram realçados
na sequência: em amarelo a região de peptídeo sinal, em magenta o domínio Ig-like N-
terminal e em ciano a família 9 de glicosil hidrolase.
MTIFKTLLTSLMVVSPVLACAAETPGRQEAMAASDAIHVNQLGYLPGSAKLAIVGLPPQGAANRSDRFTV
EDAQGRSVLQGILQPAALWSPAGQQARVADFSGLRAAGTYRLKVDGLPASETFAINAHAYQPLVDAALKA
FYFNRASTALPARYAGRYARAAGHPDTQVRIHRSAASATRPTDSVISAPKGWYDAGDYNKYVVNSGISTY
TLLAAYEQYPAFFKSKPLTIPDDVPGVPGILQEAWWNLDWMLAMQDPADGGVYHKLTDKQFDGLVMPDQA
TQQRYVVMKTTAASLDFAAVMAVASRVYAPFEQQYPGLSARMLKASRAAWAWAKQHPDVIYQQPSDVRTG
GYDDAHLDDEFAWAAAELYIATGEDGFYDAMIARNVPATVPSWGNVGGLAWMSLAGHREQLTPHADRARI
AREITGLAQQLADTWQSSAWRLAMNDGDFVWGSNAGGLNRAMMLLQGYRLTQQRQFLDAAQSQLDYILGR
NPLGLSFVTGIGKRTPMHIHHRPSEADGIEAPVPGWLVGGPQPGQQDAKECKVPYPSKRPALSYLDNFCS
YSTNEVAINWNAPLVYVSAAIEASTR
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PJK9,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:2954354-2956114 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome
ATGACTATCTTCAAGACTCTGCTCACCAGCCTGATGGTCGTTTCGCCAGTGCTCGCCTGCGCTGCCGAGACGCCC
GGCAGGCAGGAGGCCATGGCGGCCAGCGACGCCATCCACGTCAACCAGCTGGGCTATCTGCCGGGCTCGGCAAAA
CTGGCGATCGTCGGGCTGCCGCCGCAGGGCGCCGCGAACCGTTCGGACCGCTTCACGGTCGAAGATGCACAGGGA
CGCAGCGTGTTGCAGGGAATCCTGCAGCCGGCCGCGCTGTGGTCGCCGGCCGGTCAGCAGGCGCGGGTGGCGGAT
TTTTCCGGGCTGCGTGCGGCCGGGACCTATCGGTTGAAGGTCGACGGGCTGCCGGCATCGGAAACCTTCGCCATC
AACGCGCATGCCTACCAGCCCCTGGTCGATGCAGCGTTGAAGGCGTTCTACTTCAACCGTGCCAGCACCGCACTG
CCGGCACGCTACGCCGGGCGCTATGCGCGCGCGGCCGGGCACCCGGATACGCAGGTGCGCATCCATCGGTCCGCA
GCGTCTGCGACGCGTCCAACCGATAGCGTGATCAGCGCGCCCAAAGGTTGGTACGACGCGGGCGACTACAACAAG
TACGTCGTCAATTCCGGCATCAGCACCTACACCTTGCTGGCGGCCTACGAGCAGTATCCGGCGTTCTTCAAATCC
AAACCGTTGACCATCCCCGACGATGTGCCCGGCGTGCCCGGCATCCTGCAGGAGGCCTGGTGGAATCTGGACTGG

91
ATGCTCGCCATGCAGGACCCGGCCGATGGCGGCGTGTACCACAAGCTCACCGACAAGCAGTTCGATGGCTTGGTG
ATGCCGGATCAGGCCACGCAGCAGCGCTACGTGGTGATGAAGACGACTGCCGCGAGCTTGGACTTCGCTGCGGTG
ATGGCCGTGGCCAGCCGCGTGTATGCGCCGTTTGAACAGCAGTATCCGGGTCTGTCGGCGCGCATGTTGAAAGCC
TCGCGTGCTGCGTGGGCGTGGGCGAAGCAGCATCCGGATGTGATCTACCAGCAGCCCAGCGACGTGCGCACCGGC
GGTTACGACGATGCGCACCTGGACGACGAATTCGCCTGGGCGGCGGCCGAGTTGTACATCGCCACCGGCGAGGAT
GGGTTCTACGATGCGATGATCGCGCGCAACGTGCCGGCCACCGTGCCGTCGTGGGGCAATGTCGGTGGCCTGGCC
TGGATGTCGTTGGCCGGGCATCGCGAGCAACTCACGCCGCATGCCGATCGCGCGCGGATCGCACGGGAGATCACC
GGCCTGGCACAGCAGTTGGCCGACACCTGGCAATCATCGGCATGGCGGCTGGCGATGAACGATGGCGATTTCGTG
TGGGGCAGCAACGCAGGGGGGCTCAACCGCGCGATGATGCTGCTACAGGGCTACCGGCTGACGCAGCAGCGGCAA
TTTCTGGATGCGGCGCAATCGCAGCTGGACTACATCCTGGGCCGCAATCCGCTGGGGCTGTCCTTCGTTACGGGA
ATAGGCAAACGCACCCCGATGCATATCCATCACCGCCCCTCCGAGGCCGATGGTATCGAAGCGCCGGTACCGGGT
TGGCTGGTCGGCGGCCCGCAGCCGGGGCAGCAGGATGCCAAGGAGTGCAAGGTGCCGTATCCCTCCAAACGGCCG
GCGTTGTCGTACCTGGACAACTTCTGCAGCTACTCCACCAACGAGGTGGCGATCAACTGGAATGCGCCGCTGGTG
TATGTGAGCGCGGCGATCGAGGCGTCGACGCGCTGA
Enzima 2 – Celulase Q8PPS3
A celulase Q8PPS3 (identificação Uniprot) possui 474 aminoácidos e é codificada por 1425
pares de base, além disso possui peso molecular de 51294,4 Da e ponto isoelétrico teórico de
5,76. A sequência de aminoácidos segue abaixo (gi|21241383|ref|NP_640965.1). Os
resultados obtidos no Pfam e SignalP foram realçados: em amarelo a região de peptídeo sinal,
em magenta a família 5 de celulase glicosil hidrolase e em ciano o domínio de ligação de
celulose.
MSIFRAASTLALATMLALTSAPAFSYSVSNNKIVDDNGNVVQLKGVNVFGFETGNHVMHGLWARNWKEMI
NQMQGLGFNAVRLPFCPATLRSGQMPSSIDYSRNADLQGLTSLQILDKVINEFNARGMYVLLDHHTPDCG
AISELWYTSSYSEAQWLDDLRFVANRYKNVPSVIGVDLKNEPHGAATWGTGNAATDWNTAAERGSAAVLA
VAPKWIIAVEGITDNPVCSTNGGIFWGGNLQPLACTPLNIPANRLLLAPHVYGPDVFVQSYFNDSNFPNN
MPAIWDRHFGQFAGSHALLLGEFGGKYGEGDARDKVWQDALVKYLRSKGINEGFYWSWNPNSGDTGGILR
DDWTSVREDKMTLLRTLWGTVSNSTPTPTPTPTPTPTPTPSAGFGTKVIPDSTWNGGYCNRVQVTNTGTA
SGNWSISLTVTGTVNNAWNVTWSQSGTTLQASGVDYNRTLAPGATAEFGFCAAG
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PPS3,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:724505-725929 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome
ATGTCTATTTTCAGGGCTGCAAGTACGCTCGCATTGGCAACGATGTTAGCGCTCACCTCCGCACCGGCTTTCAGC
TATTCGGTCAGCAATAACAAGATCGTCGACGACAACGGCAATGTGGTTCAGCTCAAGGGCGTCAACGTGTTCGGC
TTCGAGACCGGCAACCATGTCATGCATGGTCTGTGGGCACGCAACTGGAAAGAGATGATCAACCAGATGCAGGGC
CTGGGCTTCAACGCGGTGCGCCTGCCATTCTGCCCGGCCACGCTGCGTAGCGGCCAGATGCCCAGCAGCATCGAC
TACAGCCGTAACGCCGATCTGCAGGGCCTGACCTCGCTGCAGATCCTCGACAAGGTGATCAACGAATTCAACGCA
CGCGGCATGTATGTGCTGCTGGATCACCACACGCCCGATTGCGGCGCGATTTCCGAGCTGTGGTACACCAGTTCG
TATTCCGAAGCGCAATGGCTGGATGATCTGCGCTTTGTCGCCAATCGTTACAAGAACGTGCCGAGCGTGATCGGC
GTGGATCTGAAGAACGAGCCGCACGGCGCCGCGACCTGGGGCACGGGTAACGCCGCCACCGACTGGAACACGGCC

92
GCCGAGCGCGGTTCTGCCGCTGTGCTTGCGGTGGCGCCGAAATGGATCATCGCGGTGGAAGGCATCACCGACAAC
CCGGTATGTTCGACCAACGGCGGCATCTTCTGGGGTGGCAACCTGCAGCCGCTGGCCTGCACCCCATTGAACATT
CCGGCCAACCGCCTGTTGCTGGCACCGCATGTGTACGGCCCGGACGTGTTCGTGCAGTCGTACTTCAACGACAGC
AACTTCCCCAACAACATGCCCGCGATCTGGGACCGCCATTTCGGCCAGTTCGCCGGCAGTCACGCGCTGCTGCTG
GGCGAATTCGGCGGCAAATACGGCGAAGGCGATGCGCGCGACAAGGTGTGGCAGGACGCGCTGGTGAAGTACCTG
CGCAGCAAGGGCATCAACGAAGGATTCTATTGGTCGTGGAACCCCAATAGCGGCGATACCGGCGGCATCCTGCGC
GACGACTGGACCAGCGTACGCGAGGACAAGATGACCTTGTTGCGGACGCTGTGGGGAACGGTGAGCAACAGCACA
CCGACGCCAACGCCGACCCCCACGCCGACGCCAACCCCAACACCAAGCGCGGGCTTCGGTACCAAGGTCATCCCG
GACAGCACCTGGAACGGTGGCTACTGCAACCGCGTGCAGGTCACCAACACCGGCACCGCCAGCGGCAATTGGTCG
ATCTCGCTGACAGTCACCGGCACCGTCAACAACGCCTGGAACGTCACCTGGTCACAGAGCGGCACCACGCTCCAG
GCCAGCGGCGTGGACTACAACCGCACGCTGGCGCCCGGTGCAACTGCCGAGTTCGGCTTCTGCGCAGCAGGTTGA
Enzima 3 – Beta-Glicosidase Q8PFV3
A celulase Q8PFV3 (identificação Uniprot) possui 723 aminoácidos e é codificada por 2172
pares de base, além disso possui peso molecular de 77499,9 Da e ponto isoelétrico teórico de
5,08.
A sequência de aminoácidos segue abaixo (gi|21244593|ref|NP_644175.1) juntamente com
os resultados obtidos no Pfam e SignalP. Na sequência foram realçadas em magenta o domínio
da família 3 de glicosil hidrolase N-terminal, em ciano o domínio da família 3 de glicosil
hidrolase C-terminal e em cinza o domínio tipo III de fibronectina, essa enzima não possui
peptídeo sinal.
>gi|21244593|ref|NP_644175.1| beta-glucosidase [Xanthomonas axonopodis pv. citri str.
306] MAADRIETLIARMTVEEKVGQLGVFADMVRPFAPDVNPEANVLNADEVLQQVRLGRVGSLFNGVGAALGV
QIQKVAVEESRLGIPVILAADVIHGMRTVFPIPLGEAASFEPALAERTARATAIEATAAGLHWTYAPAVD
IARDQRWGRGAEGAGEDVMLGAAFAAARVRGFQGSDLTAHDCLLATPKHFAAYGAVAAGMEYNTVDIAPQ
TLRDVHLPPFKAAFDAGALTVMSSFNDINGVPASANHELLTEILRGEWQFPGVVISDYTADMELIAHGYA
ADERDATKKAFLAGLDLSMQSGFYAAHLPSLVESGEVPMATLDASVRRILQLKEAIGLFDNPYRSLDPAR
EADTTHLPAHDALSRDAARRSIVLLKNEGGVLPLKKSGQRIALIGPFVQDRENIEGCWTLFGDKERYVTL
EQGVRAVVGTEDLSVVAGCGLEEALPGGISAAIDAAQAADVVVLALGEPQRFSGEAQSRTEITLPPAQQA
LAEAIAATGTPMVVLLRNGRALALGGAVRDADAIAVTWYLGTQTGTGVADVLFGDYNPSARLPISFPQAT
GQQPYFYNHLRTGRPELPTLSEYKARWREMPNEPLYPFGHGLSYTTFAYAQPQLSAAQLGWDEALTITTR
VSNTGKVAGEEVVQLYVHDRVASRVRPVRELKGFRKVLLQPGESQDVVFTLERDALAFTNHKGVFGAEPG
LFDVWVCASAKSGEAVTFELLER
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PFV3,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:4552011-4554182 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome

93
ATGGCTGCTGATCGCATCGAAACCCTGATTGCCCGGATGACCGTCGAAGAGAAGGTCGGACAGTTGGGTGTGTTC
GCCGACATGGTGCGGCCGTTCGCGCCGGACGTGAATCCCGAGGCCAACGTGCTCAATGCCGACGAGGTGTTGCAG
CAGGTGCGCTTGGGTCGGGTCGGCTCGCTGTTCAATGGCGTGGGCGCCGCGCTGGGCGTGCAGATCCAGAAAGTC
GCCGTGGAAGAGAGTCGCCTGGGGATCCCGGTGATCCTGGCCGCCGACGTGATCCACGGCATGCGCACGGTGTTC
CCGATCCCGCTGGGCGAGGCGGCCAGTTTCGAGCCGGCGTTGGCCGAGCGCACCGCGCGCGCCACCGCGATCGAG
GCCACCGCTGCCGGCCTGCACTGGACCTACGCGCCGGCCGTGGACATCGCGCGCGACCAGCGCTGGGGCCGTGGC
GCCGAGGGTGCCGGCGAAGACGTGATGCTGGGCGCGGCCTTTGCCGCTGCCCGTGTGCGCGGGTTCCAGGGCAGC
GACCTGACCGCCCACGATTGCCTGCTGGCCACGCCCAAGCATTTCGCCGCCTATGGCGCAGTGGCGGCAGGCATG
GAATACAACACCGTCGACATTGCGCCGCAGACGCTGCGCGATGTGCATTTGCCGCCGTTCAAGGCCGCCTTCGAT
GCCGGCGCATTGACCGTGATGTCGTCCTTCAACGACATCAACGGCGTACCGGCCAGCGCCAACCATGAGTTGCTG
ACCGAGATCCTGCGCGGGGAATGGCAGTTCCCCGGCGTGGTGATTTCCGATTACACCGCCGACATGGAATTGATC
GCCCATGGCTATGCCGCCGACGAGCGCGACGCGACCAAGAAGGCCTTCCTGGCCGGGTTGGACCTGAGCATGCAA
AGCGGCTTTTATGCCGCGCATCTGCCGTCGCTGGTGGAAAGCGGCGAGGTGCCGATGGCCACGTTGGATGCCAGC
GTGCGGCGCATCCTGCAGCTCAAAGAAGCGATCGGGCTGTTCGACAATCCGTATCGCTCGCTGGACCCGGCGCGC
GAAGCCGACACCACGCATCTGCCGGCGCACGATGCGCTGTCGCGCGACGCGGCGCGGCGTTCGATCGTGCTGTTG
AAAAACGAAGGCGGCGTGTTGCCTCTGAAGAAGAGCGGCCAGCGCATCGCGTTGATCGGCCCGTTCGTGCAGGAC
CGCGAAAACATCGAGGGCTGCTGGACCCTGTTCGGCGACAAGGAACGCTACGTGACCCTGGAGCAGGGCGTGCGT
GCGGTGGTCGGCACCGAAGATCTGAGCGTGGTGGCGGGCTGCGGACTGGAAGAGGCGCTGCCGGGTGGCATTTCC
GCTGCGATCGATGCGGCGCAGGCGGCCGATGTGGTGGTGCTGGCGTTGGGCGAGCCGCAGCGTTTCAGTGGCGAG
GCGCAATCGCGCACCGAGATCACCTTGCCGCCGGCGCAACAGGCACTGGCTGAAGCGATTGCAGCGACCGGCACC
CCGATGGTGGTGTTGTTGCGCAATGGTCGCGCGCTGGCCTTGGGTGGCGCGGTGCGCGATGCCGACGCGATTGCC
GTGACCTGGTACCTGGGCACGCAGACCGGTACCGGTGTGGCCGATGTGTTGTTCGGCGACTACAACCCGTCGGCG
CGTCTGCCGATCAGCTTCCCGCAGGCGACCGGGCAGCAACCTTATTTCTATAACCATCTGCGCACCGGGCGCCCG
GAATTGCCGACCCTGTCCGAGTACAAGGCGCGTTGGCGCGAGATGCCAAACGAGCCGTTGTATCCGTTCGGGCAT
GGCTTGAGCTACACCACGTTTGCGTATGCGCAGCCGCAGTTGAGCGCGGCGCAACTCGGCTGGGATGAGGCGCTG
ACCATCACCACGCGCGTCAGCAACACCGGCAAGGTCGCCGGCGAGGAAGTGGTGCAGTTGTATGTGCACGATCGC
GTGGCCAGTCGCGTGCGTCCGGTGCGCGAACTCAAGGGGTTTCGCAAGGTATTGCTGCAGCCCGGCGAGAGCCAG
GACGTGGTGTTCACGCTGGAGCGCGATGCGCTGGCATTCACCAACCACAAGGGCGTCTTCGGTGCGGAGCCGGGT
CTGTTCGATGTATGGGTGTGCGCGTCGGCCAAGAGCGGCGAGGCAGTCACCTTCGAGCTGCTGGAACGCTGA
Enzima 4 – Beta-Glicosidase Q8PMI0
A celulase Q8PMI0 (identificação Uniprot) possui 748 aminoácidos e é codificada por 2247
pares de base, além disso possui peso molecular de 80098,0 Da e ponto isoelétrico teórico de
7,24.
A sequência de aminoácidos segue abaixo (gi|21242200|ref|NP_641782.1) juntamente com
os resultados obtidos no Pfam e SignalP. Na sequência foram realçadas em amarelo a região
de peptídeo sinal, em magenta do domínio glicosil hidrolase N-terminal da família 3, em ciano
o domínio glicosil hidrolase C-terminal da família 3 e em cinza o domínio tipo III de
fibronectina.
>gi|21242200|ref|NP_641782.1| beta-glucosidase [Xanthomonas axonopodis pv. citri str.
306] MKPFTAHQNRPRPLVLAVVMGLGVAFAAPLAVAQGAPSSQPWMDTALTPDQRADRLLAQMSEDEKFQMLR
SYFGLGTDKIPKPEGALGSAGYVPGIPRLGIPAQQLADAGVGVTNPGGIRKGDYATAMPSGPSTASSWNR
QLAYTGGKTMGRESWQQGFNVLLAGSVNLQRDPRNGRNFEYAGEDPLLAGTMVGESIRGVQSEHVLSTMK
HFALNDMETRRNFHSAQIGEQAMHESDLLAFEIALKIGDPASVMCSYNKINGIYGCEHDYLLNQVLKQEW
KYPGYVMSDWGGVHSGSKAALAGLDQQSAGEVFDAAVFFDAPLRMAVSAGVVPRARFDDMVKRVLRSLFA
HGAFDHPTQRQPIDGKAGLLAAQRVAEEGSVLLRNEQAALPLSKDVRRIAVIGGYADKGVMSGGGSSRVD

94
YTINGGNAVPGITPTTWPGPVIIHPSSPLQALRAALPNVQIDYLDGTDRAAAARAAKAADVAIVFATQWA
AESVDLPDMRLPDNQDALIETVAKANPKTTVVLETNGPVRMPWAERVPAVLQAWYPGIGGGEAIANLLTG
AVNPSGHLPVTWPVDESQLPRPSIPGLGFKPAKPGEDSIDYAIEGANVGYKWFAARKLTPRYPFGHGLSY
TQFRMGGLQVEARGSQLTASFEVENTGPREGAAVPQLYVALPDGHATPLRLIGWQKLTLKPGEKRSVQVV
AEPKTLADFDAKARRWTIAAGTYRVQLARSTSEPVQTAEVTLQAAQLP
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PMI0,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:1671154-1673400 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome
ATGAAACCGTTTACAGCCCATCAGAATCGTCCGCGCCCCTTGGTTCTGGCGGTCGTGATGGGCCTTGGCGTCGCC
TTCGCCGCACCGCTGGCAGTCGCGCAGGGCGCGCCATCTTCGCAGCCGTGGATGGACACCGCACTGACCCCGGAC
CAGCGCGCCGACCGCCTGCTGGCGCAGATGAGCGAGGATGAAAAATTCCAGATGCTGCGCAGCTATTTCGGCCTG
GGCACCGACAAGATTCCCAAGCCCGAAGGCGCGCTCGGTTCGGCCGGCTACGTTCCCGGTATCCCGCGTCTGGGC
ATTCCCGCCCAGCAACTGGCCGATGCCGGCGTGGGCGTGACCAACCCCGGCGGCATCCGCAAGGGCGATTATGCC
ACCGCGATGCCGTCGGGACCGTCGACCGCATCGAGCTGGAATCGGCAACTGGCGTATACCGGCGGCAAGACCATG
GGCCGCGAATCCTGGCAGCAGGGCTTCAACGTGTTGCTCGCCGGCAGCGTCAATCTGCAGCGCGACCCGCGTAAT
GGGCGCAATTTCGAATACGCGGGTGAAGATCCGTTGCTGGCCGGCACCATGGTCGGCGAATCGATTCGCGGCGTG
CAAAGCGAACATGTGCTTTCCACGATGAAGCACTTCGCACTCAACGACATGGAAACGCGGCGCAACTTTCATAGC
GCGCAGATCGGCGAGCAGGCGATGCACGAATCGGACCTGCTGGCCTTCGAGATTGCGCTGAAGATCGGCGACCCG
GCCTCGGTGATGTGTTCGTACAACAAGATCAACGGTATCTACGGTTGCGAGCACGATTACCTGCTCAACCAGGTG
CTCAAGCAGGAATGGAAATACCCCGGTTACGTGATGTCCGATTGGGGCGGCGTGCATAGCGGATCCAAGGCGGCG
TTGGCCGGGCTGGATCAACAATCGGCCGGCGAAGTGTTCGACGCGGCAGTGTTTTTCGATGCGCCGTTACGCATG
GCGGTGTCTGCCGGCGTGGTGCCGCGCGCGCGTTTCGACGACATGGTCAAGCGCGTGCTGCGCAGCCTGTTCGCG
CACGGCGCCTTCGATCATCCTACCCAGCGTCAACCCATCGATGGCAAGGCCGGTCTTCTGGCTGCGCAGCGCGTG
GCCGAAGAAGGCAGCGTGTTGCTGCGTAACGAACAGGCCGCGTTGCCGCTGTCCAAGGACGTGCGCCGCATCGCG
GTGATCGGCGGCTACGCCGACAAGGGCGTGATGTCCGGCGGCGGCTCTTCGCGCGTGGACTACACCATCAACGGC
GGCAACGCGGTACCGGGCATCACGCCGACCACCTGGCCCGGCCCGGTGATCATCCATCCGTCCTCGCCGCTGCAG
GCACTGCGTGCGGCACTGCCGAACGTGCAGATCGATTACCTGGACGGCACCGACCGCGCCGCGGCCGCACGCGCC
GCCAAGGCGGCCGATGTGGCGATCGTGTTCGCCACCCAGTGGGCCGCCGAATCGGTGGATCTGCCGGACATGCGC
TTGCCCGACAACCAGGACGCGTTGATCGAGACCGTGGCCAAGGCCAACCCGAAGACCACCGTGGTGCTGGAAACC
AATGGACCGGTGCGCATGCCGTGGGCCGAACGCGTGCCGGCGGTGTTGCAGGCGTGGTATCCGGGCATCGGCGGC
GGCGAGGCGATCGCCAACCTGCTGACCGGCGCGGTGAATCCTTCCGGCCACCTGCCGGTGACCTGGCCGGTGGAC
GAGTCGCAGCTGCCGCGCCCGTCGATTCCGGGGCTGGGCTTCAAGCCGGCCAAACCGGGCGAAGACAGCATCGAC
TACGCCATCGAAGGCGCCAATGTCGGTTACAAGTGGTTCGCCGCGCGCAAGCTGACACCACGCTACCCGTTCGGC
CATGGCCTGTCGTACACGCAGTTCCGCATGGGCGGGCTGCAGGTGGAGGCACGCGGCAGCCAGCTCACCGCCAGC
TTCGAGGTGGAGAATACCGGCCCGCGCGAAGGCGCCGCCGTGCCGCAGCTCTACGTCGCCCTGCCCGACGGCCAC
GCCACCCCGCTGCGGCTGATCGGCTGGCAAAAGCTCACGCTAAAGCCTGGCGAAAAACGCAGCGTGCAGGTCGTC
GCAGAACCCAAGACCTTGGCCGATTTCGATGCCAAGGCACGTCGCTGGACGATCGCCGCGGGCACTTACCGCGTG
CAACTGGCGCGTTCGACGAGCGAGCCGGTGCAAACTGCGGAGGTCACGCTGCAGGCGGCACAGTTGCCGTGA
Enzima 5 – Pectato Liase Q8PGQ4
A celulase Q8PGQ4 (identificação Uniprot) possui 377 aminoácidos e é codificada por 1134
pares de base, além disso possui peso molecular de 40193,7 Da e ponto isoelétrico teórico de
8,57.

95
A sequência de aminoácidos segue abaixo (gi|21244287|ref|NP_643869.1) juntamente com
os resultados obtidos no Pfam e SignalP. Na sequência foram realçadas em amarelo a região
de peptídeo sinal, em magenta do domínio de pectato liase.
>gi|21244287|ref|NP_643869.1| pectate lyase [Xanthomonas axonopodis pv. citri str. 306] MKPKFSTAAAASLFVGSLLLTGVAYADAALEVATTGWATQNGGTKGGSKAAAANIYVVKNAAELKAALSA
SVGSNGRIIKVSGIIDISEGKPYTKTSDMKTRARLDVPTKTTLIGITSNAEIREGYFYVKANDVIIRNIT
IENPWDPEPVWDPDDGSAGNWNAEYDGLTVEGASNVWVDHVTFTDGRRTDDQNGTANGRPKQHHDGAMDI
KKGANFVTVSYSAFKSHEKNNLIGSSDSASSTDSGKLKVTIHNTLFENISARAPRVRFGQVHLYNNYHVG
STSNAVYPFSYAHGVGKESKIFSERNVFEISGVSSCDKIAADYGGSVYRDQGSLLNGKALSCSWNTNIGW
TPPYTYNLLSADKVAADVKAKAGAGKL
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PGQ4,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:4226043-4227176 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome
ATGAAGCCGAAATTTTCGACGGCGGCCGCCGCGTCGCTGTTCGTTGGGTCGTTGCTGCTCACCGGCGTTG
CCTATGCCGATGCGGCGTTGGAGGTTGCAACCACGGGTTGGGCGACCCAGAACGGCGGCACCAAGGGCGGCTCCA
AAGCAGCTGCCGCCAATATCTACGTCGTCAAGAACGCCGCCGAGTTGAAAGCAGCATTGAGCGCAAGCGTCGGGA
GCAACGGGCGCATCATCAAGGTCAGCGGCATCATCGATATCAGCGAAGGAAAGCCTTACACCAAAACCTCGGACA
TGAAGACGCGCGCGCGTCTGGACGTGCCCACCAAGACCACGCTGATCGGCATCACCAGCAACGCTGAAATTCGCG
AAGGCTATTTTTACGTCAAGGCCAACGACGTCATCATCCGCAATATCACCATCGAAAATCCATGGGACCCGGAGC
CGGTCTGGGACCCGGATGATGGCAGCGCCGGCAACTGGAATGCGGAGTACGACGGCCTGACCGTGGAAGGCGCCA
GCAACGTGTGGGTGGACCATGTCACCTTCACCGATGGCCGCCGTACCGACGATCAGAATGGCACTGCAAATGGTC
GTCCGAAACAGCACCACGATGGCGCGATGGACATCAAGAAAGGCGCCAATTTCGTCACCGTCTCGTACTCGGCCT
TCAAGTCGCACGAAAAGAACAACCTGATCGGCTCCAGCGACAGCGCGTCCAGTACCGACAGCGGCAAGCTCAAGG
TCACCATTCACAACACCCTGTTCGAAAACATTTCTGCGCGCGCCCCGCGCGTGCGCTTCGGTCAGGTGCACCTGT
ACAACAACTACCACGTCGGCAGCACCAGCAATGCGGTGTATCCGTTCTCCTACGCGCATGGCGTGGGCAAGGAGT
CGAAGATCTTCTCCGAGCGCAACGTGTTCGAGATCAGCGGTGTCAGCAGCTGCGACAAGATCGCCGCCGACTACG
GTGGCAGCGTGTATCGCGATCAGGGTTCGCTGCTCAACGGCAAGGCGCTGTCGTGCTCGTGGAATACCAACATCG
GCTGGACACCGCCCTATACCTACAATCTGCTATCGGCAGACAAGGTCGCTGCAGACGTCAAGGCGAAAGCAGGCG
CCGGCAAGCTGTAA
Enzima 6 – Pectato Liase Q8PIB2
A celulase Q8PIB2 (identificação Uniprot) possui 353 aminoácidos e é codificada por 1062
pares de base, além disso possui peso molecular de 37456,8 Da e ponto isoelétrico teórico de
8,24.
A sequência de aminoácidos segue abaixo (gi|21243713|ref|NP_643295.1) juntamente com
os resultados obtidos no Pfam e SignalP. Na sequência foram realçadas em amarelo a região
de peptídeo sinal, em magenta do domínio de pectato liase.

96
>gi|21243713|ref|NP_643295.1| pectate lyase II [Xanthomonas axonopodis pv. citri str. 306] MIIKTLQGALALVLSACAAGAIAGPVGYGAATTGGGNKTAVNVATFEAVQAAIDNYSGSGGLVLNYTGQF
DFGTIKDVCAQWKLPAKTVQIKNKSDITIKGANGSSANFGVRVVGDAHNVIVQNMTIGLLQGGEDADSIS
LEGNSSGEPSKIWIDHNTIFASLTKCSGAGDASFDGGIDMKKGVHHVTVSYNYVYNYQKVALNGYSDSDT
KNSAARTTYHHNRFENVESRLPLQRRGLSHVYNNYFNNVFTSGINVRMGGVAKIESNYFENIKNPVTSRD
SSEIGYWDLINNYVGSGITWSTPDGSKPYANATNWVSSKVFLESLGYIYTVTPAAQVKAKVIATAGAGKN
LAE
Além disso, segue abaixo a sequência de pares de base, que codifica para a proteína Q8PIB2,
que foi utilizada para a construção dos oligos utilizados na clonagem. A sequência
correspondente ao peptídeo sinal foi realçada em amarelo e em verde a região utilizada para
a construção dos oligos.
>gi|21240774:3490628-3491689 Xanthomonas axonopodis pv. citri str. 306 chromosome,
complete genome
ATGATCATCAAGACGCTCCAGGGCGCACTCGCGCTTGTACTGTCCGCTTGCGCCGCGGGCGCGATTGCCGGCCCG
GTCGGCTACGGTGCCGCCACCACCGGCGGCGGCAACAAGACAGCGGTCAACGTGGCGACGTTCGAAGCGGTGCAG
GCCGCCATCGACAACTATTCCGGTAGCGGCGGGCTGGTGCTCAACTACACCGGCCAGTTCGACTTCGGCACCATC
AAGGACGTGTGCGCGCAATGGAAGCTGCCGGCCAAGACAGTGCAGATCAAGAACAAGAGCGATATCACGATCAAG
GGCGCCAATGGCTCGTCGGCGAACTTCGGCGTGCGTGTGGTGGGCGATGCGCACAACGTGATCGTCCAGAACATG
ACCATCGGTCTGCTGCAAGGCGGTGAAGATGCCGATTCGATTTCGCTGGAAGGCAACTCCAGCGGCGAGCCGTCC
AAGATCTGGATCGACCACAACACCATCTTCGCCTCGCTCACCAAGTGTTCCGGTGCGGGCGATGCCTCCTTCGAT
GGCGGCATCGATATGAAGAAGGGCGTGCACCATGTCACCGTGTCCTACAACTATGTCTACAACTATCAAAAGGTC
GCGCTCAATGGCTACAGCGACAGCGACACCAAGAACTCGGCTGCACGCACCACGTATCACCATAACCGCTTCGAG
AACGTGGAATCGCGCTTGCCATTGCAGCGGCGCGGCTTGAGCCACGTCTACAACAACTACTTCAACAATGTGTTC
ACCTCAGGCATCAACGTGCGCATGGGTGGGGTGGCGAAGATCGAGTCGAACTACTTCGAGAACATCAAGAACCCG
GTGACCTCGCGCGACAGCAGCGAGATCGGCTACTGGGACCTGATCAACAACTACGTTGGCAGCGGCATCACCTGG
AGCACGCCGGACGGCAGCAAACCCTACGCCAATGCCACCAACTGGGTGAGCTCCAAGGTGTTCCTGGAGTCGCTG
GGCTACATCTACACCGTGACACCGGCGGCACAGGTCAAGGCCAAGGTGATCGCCACGGCGGGTGCGGGCAAGAAC
CTGGCGGAGTGA


















