
Isótopos estáveis de carbono e oxigênio em ostracodes do Cretáceo

441
Rev. bras. paleontol. 16(3):441-464, Setembro/Dezembro 2013© 2013 by the Sociedade Brasileira de Paleontologiadoi: 10.4072/rbp.2013.3.06
INTRODUÇÃO
A atual posição geográfi ca da Antártica, e as sucessões expostas nas áreas livres de gelo, apoiam as propostas mais recentes de sua ligação pretérita com outros continentes austrais (Lawver et al., 2009). Para suportar estas reconstituições, por seu caráter eminentemente continental, os fósseis de plantas das áreas antárticas constituem uma importante ferramenta, servindo ainda para a compreensão da moderna distribuição
das fl oras do Hemisfério Sul (Bose et al., 1990), a origem de seus elementos, e na discussão sobre o momento de chegada das angiospermas às altas latitudes (Cantrill & Nichols, 1996; Cantrill & Poole, 2012).
A tafofl ora dos níveis superiores do afl oramento do pontal Rip, uma exposição junto à costa norte da ilha Nelson, ilhas Shetland do Sul (Figura 1), é aqui discutida por seu caráter inédito e por reunir elementos macro e microfl orísticos, uma particularidade comum em depósitos lacustres proximais à
UMA FLORA DO FINAL DO CRETÁCEO NA ILHA NELSON, ILHAS SHETLAND DO SUL, PENÍNSULA ANTÁRTICA
BIBIANA LIGUORI BASTOS, TÂNIA LINDNER DUTRA, THIÈRS PORFÍRIO WILBERGER & CRISTINE TREVISAN
Programa de Pós-Graduação em Geologia, UNISINOS, Av. Unisinos, 950, 93.022-000, São Leopoldo, RS, Brasil. [email protected], [email protected], [email protected], [email protected]
ABSTRACT – A LATE CRETACEOUS FLORA FROM NELSON ISLAND, SOUTH SHETLAND ISLANDS, ANTARCTIC PENINSULA. A plant fossil assemblage (micro-and macrofl ora) from the northern sector of Nelson Island (Rip Point) is abborded on its composition and age. The macrofl ora shows few ferns and a conifer remain (cf. Papuacedrus) and dominant angiosperm fossil leaves represented by laurophyll (probable Elaeocarpaceae/Anacardiaceae affi nity), acrodromophyll (Melastomaceae?) and protophyll (Nothofagaceae) morphotypes. The absence of marginal characters in great part of the leaf remains incentivate their treatment by morphological groups, trying to guarantees at least the stablishment of its relation with other extant and fossil assemblages and the reconstructions of the geological process implied in its preservation. The microfl ora shows a distinct composition, with many fern and fungal spores and pollen grains of gymnosperms, with a minor component in angiosperms (Proteacea, Aquifoliacea and primitive Nothofagus). The presence of Classopollis and “ancestral” forms of Nothofagidites pollen grains indicate a Campanian-Lower Maastrichtian age to the Rip Point succession, where the occurrence of a probable Papuacedrus and Melastomataceae related forms have a strong evolutive value and phyllogenetic implications, could being its anciest know record. Leaves of deciduous Nothofagus dominate the macrofl ora and probably include juvenile forms. Fungi spores and foraminifera indicate a near-coast lake environment to the deposit, and the fl oristic composition, the remobilization of the leaf remains from a vegetation growing on the slopes of the proximal volcanoes. Comparable extant analogues grows today in rain forests of southern South America and New Zealand, and allows to infer a wet temperate to subtropical climate to the north of Antarctic Peninsula in the end of Cretaceous.
Key words: Nothofagus, Classopollis, Upper Cretaceous, Nelson Island, Antarctic Peninsula.
RESUMO – Uma fl ora inédita (macrorrestos e palinomorfos) do norte da ilha Nelson (pontal Rip), é caracterizada em sua composição e idade. Na macrofl ora foram identifi cadas raras pteridófi tas, uma conífera (cf. Papuacedrus) e angiospermas laurofílicas (exibindo afi nidade com Anacardiaceae, Elaeocarpaceae/Aquifoliaceae e Melastomataceae) e protofílicas, relacionadas a tipos primitivos de Nothofagus. A falta de caracteres marginais em grande parte dos elementos presentes levou a sua abordagem por grupos morfológicos (e morfotipos) buscando garantir, pelo menos, sua correlação com outras fl oras fósseis e modernas do Hemisfério Sul e suas aplicações paleoclimática e paleogeográfi ca. A microfl ora associada mostra uma representação proporcionalmente distinta, com domínio de fungos e pteridófi tas, muitas gimnospermas (Araucariacites, Cycadopites e Classopollis) e angiospermas restritas a proteáceas, aquifoliáceas e grãos de pólen primitivos de Nothofagus (Nothofagidites). A presença de Nothofagidites e Classopollis apoia uma idade Campaniano ou Maastrichtiano basal para a deposição do pontal Rip. Neste contexto, a ocorrência associada de prováveis representantes de Papuacedrus e Melastomataceae é de grande valor para a compreensão dos processos evolutivos destes grupos de plantas já que, se confi rmados, constituem seu mais antigo registro. Formas de Nothofagus são os únicos elementos constantes na macro e microfl ora e com proporções comparáveis e indicam formas primitivas de hábito decíduo. A composição distinta da macro e microfl ora sugere a prévia remobilização e o transporte dos macrorrestos, a partir de uma vegetação que crescia na encosta de vulcões próximos e, a ocorrência conjunta de testas de foraminíferos e fungos, uma deposição em corpos lacustres próximos à costa. Vegetações análogas modernas crescem hoje em áreas de clima oceânico do sul da América do Sul e da Nova Zelândia e sugerem condições temperadas a subtropicais úmidas para o clima nas áreas do norte da Península Antártica no fi nal do Cretáceo.
Palavras-chave: Nothofagus, Classopollis, Cretáceo Superior, ilha Nelson, Península Antártica.

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013442
ambientes vulcânicos (Leroy & Roiron, 1996; Ollivier et al., 2010). Um primeiro registro desta ocorrência havia sido feito por Dutra et al. (1998), mas sem descrições formais, e o estudo prévio, e ainda inédito, de um nível mais basal (Figura 2), mostrou grãos de pólen de Classopollis associados à macrorrestos de ramos e lenhos carbonizados e frondes de Coniopteris (Trevisan, 2011).
Como parte das áreas ocidentais da Península Antártica, a ilha Nelson parece ter sido submetida aos mesmos processos de subducção que afetaram a península como um todo, inserindo-a num contexto de antearco até, pelo menos, o fi nal do Cretáceo Médio (Smellie, 1981; Thomson et al., 1983; Tokarski, 1991). A partir daí, processos superimpostos de formação de um arco de ilhas (ou hot spot), são atestados pelo rejuvenescimento das litologias de sul para norte, nas ilhas do arquipélago das Shetland do Sul (Pankhurst, 1982; Birkenmajer et al., 1986). Refl etindo estes eventos, a ilha Livingston contém depósitos distribuídos entre o Jurássico e o Cretáceo inferior (Griffi ths & Oglethorpe, 1998; Cantrill & Nagalingum, 2005), enquanto King George (KG), no extremo norte, exibe litologias e fósseis distribuídos entre o Cretáceo Superior e o Mioceno (Birkenmajer, 1980; Smellie et al., 1984; Shen, 1994; Birkenmajer, 2001). Neste contexto, o estudo dos fósseis na ilha Nelson pode complementar o conhecimento sobre fl oras potencialmente cretácicas no norte da Península e sua transição para aquelas que caracterizarão o Paleógeno.
Aspectos geológicos e registro fóssilA sucessão do pontal Rip, com cerca de 10 m de espessura,
é composta em sua maior parte por lavas andesíticas e corpos intrusivos e se inicia no nível do mar (Fontoura et al., 1988; Dutra et al., 1998). Lentes restritas de piroclásticas (tufos e lápili) e vulcanoclásticas de cores cinza escuro ocorrem entre as lavas e aglomerados e concentram a macro e microfl ora. As impressões de folha e os palinomorfos aqui discutidos provêm da intercalação superior composta por tufi tos cinza-amarronados (Figura 2). O caráter alterado das litologias e as cores escuras difi cultaram a visualização dos fósseis, algumas vezes só possível pelo caráter vítreo dos minerais que recobrem as impressões.
Na vizinha ilha KG esses tipos litológicos, caracterizados ainda por conter intercalações finas de carvão (pontais Half Three e Price), haviam sido atribuídos ao Jurássico e considerados os mais antigos na sucessão (Hawkes, 1961). Dados absolutos de idade e estudos posteriores com os restos de plantas permitiram reavaliar a idade e propor uma deposição correspondente ao fi nal do Cretáceo (Birkenmajer, 1981; Smellie et al., 1984; Shen, 1994; Dutra & Batten, 2000). Para o pontal Half Three, sul da ilha KG, exposição mais próxima de Rip, dados absolutos de idade (Rb-Sr) forneceram valores entre 71-77 Ma para os basaltos cálcico-alcalinos que se sobrepõem às camadas de tufos com palinomorfos e restos de folhas carbonizadas (Li et al., 1989; Cao, 1994). Outras deposições na ilha KG, com macro e microfl ora comparável,
Figura 1. Mapa das ilhas King George e Nelson, e a localização do pontal Rip (estrela). Os círculos claros apontam os locais com estratos atribuídos ao Cretáceo Superior e os de cor cinza, os referentes ao fi nal do Paleoceno e/ou Eoceno Inferior (modifi cado de Reguero et al., 2013). Idades isotópicas de Birkenmajer et al. (1986), Li et al. (1989) e Shen (1994). 1, Half Three Point; 2, Skua Bay (Wickle Point); 3, Price Point; 4, Fossil Hill; 5, Rocky Bay; 6, Collins Glacier; 7, Barton Peninsula; 8, Potter Peninsula; 9, Paradise Cove; 10, Block Point; 11, Zamek Hill; 12, Admiralen Peak; 13, Dufayel Island; 14, Point Thomas; 15, Keller Peninsula; 16, Point Hennequin.
Figure 1. Location map of King George and Nelson islands detaching the studied area at Rip Point (star). White circles marks the places of Upper Cretaceous age proposed, and the grey ones those where the levels are considered to correspond to Late Paleocene and/or Early Eocene (modifi ed from Reguero et al., 2013). Isotopic data from Birkenmajer et al. (1986), Li et al. (1989) and Shen (1994). For numbers 1-16 see Portuguese legend above.

443BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
são as do pontal Price (Dutra et al., 1996), da Península Barton (Tokarski et al., 1987) e as de Stacek Cove (Birkenmajer, 1980, 2001). No conjunto, contêm fl oras mais empobrecidas que aquelas do Paleógeno e caracterizadas pelo domínio de Nothofagus, pteridófi tas e raras coníferas (Dutra & Batten, 2000; Dutra, 2001, 2004).
MATERIAL E MÉTODOS
O material aqui estudado foi coletado durante as atividades do Programa Antártico Brasileiro e está depositado no Laboratório de História da Vida e da Terra (LaViGæa), seção Antártica, da Universidade do Vale do Rio dos Sinos, UNISINOS, sob a sigla ANTF 34. Foram analisadas 109 amostras, das quais 45 mostraram folhas que, embora incompletas, possuiam elementos morfológicos e da venação preservados. Foram preparadas para estudo com a remoção do material superfi cial que recobria as impressões, e desenhadas em microscópio estereoscópico Olympus, com câmara clara acoplada, sob iluminação de baixo ângulo, para destacar seus
caracteres. Os desenhos foram digitalizados e redesenhados no programa Corel Draw e as amostras fotografadas com câmera digital Canon EOS REBEL, com 12 Mpx de resolução.
Os exemplares referentes à pteridófi tas e o único resto de conífera presente foram comparados com os já descritos para as bacias austrais e com formas modernas herborizadas do herbário do Instituto Anchietano (IAC) e do herbário-folheário do LaViGæa, ambos da UNISINOS. Quando possível, a inserção taxonômica baseou-se, respectivamente, em Smith et al. (2006) e Eckenwalder (2009).
Na descrição dos macrorrestos de angiospermas foi utilizada a terminologia proposta por Hickey (1973) e Hickey & Wolfe (1975), atualizada, quando necessário, pelo Manual of Leaf Arquitecture (Ellis et al., 2009). A preservação parcial dos restos de folha e a presença de grãos de pólen de grupos primitivos, que sugeriam uma idade para a deposição anterior ao início do Paleógeno, incentivaram uma abordagem preferencial por grupos morfológicos (morfotipos), buscando evitar o uso de nomenclaturas que remetessem a associações genéticas com as formas modernas, que poderiam mascarar os processos evolutivos ocorridos. Além disto, a constatação de que muitos restos de folha mostravam aspectos morfológicos de mescla, hoje presentes em distintos grupos de angiospermas, ou um padrão desorganizado de venação apoiando o caráter primitivo da tafofl ora do pontal Rip, incentivaram um destaque àqueles parâmetros presentes na venação.
A ausência de caracteres foliares de margem, base, ou ápice, é uma realidade comum às macrofl oras da porção ocidental da Península Antártica e está provavelmente relacionada à proximidade com áreas submetidas a vulcanismo ativo, por grande parte de sua história evolutiva (Dusén, 1908; Tokarski, 1991; Zastawniak, 1981, 1994; Dutra & Batten, 2000). No estudo destas tafofl oras, mesmo a abordagem por paratáxons é de difícil aplicação, e não contribui para as correlações com outras assembleias previamente estudadas.
Neste trabalho e, após o estabelecimento das principais arquiteturas foliares presentes, procurou-se agrupar os distintos morfotipos em grupos de morfologia e, para tanto, foram utilizadas e, por vezes, modifi cadas, as propostas prévias feitas para outras fl oras austrais do fi nal do Cretáceo (Pole, 1992; Zastawniak, 1994; Cantrill & Nichols, 1996, Iglesias et al., 2007). Quando pertinentes, os grupos propostos por Hickey (1984) e Crabtree (1987) para fl oras setentrionais de mesma idade foram igualmente empregados.
Três grupos morfológicos se destacaram na assembleia do pontal Rip: (i) os tipos pinados broquidódromos (magnoliofílicos; laurofílicos ou cinamomofílicos); (ii) os acródomos (mais raros), hoje presentes em distintas ordens de eudicotiledoneas (Canellales, Piperales, Laurales -”malvids”- e Mirtales, incluindo as Melastomataceae) e monocotiledoneas e, (iii) os pinados craspedódromos (protofílicos), das Proteales, Canellales, “fabids” e, por sua importância nas fl oras austrais, nas Fagales. Nesta última categoria, entre o material estudado, todos possuem, além disto, uma venação comparável à das Nothofagaceae (ou Fagaceae). Diante desta expressiva representatividade, é aqui proposto o grupo informal Nothofagus. Este reúne além do caráter pinado craspedódromas sua venação caracteriza-se
Figura 2. Perfi l geológico da sucessão vulcânica e vulcanoclástica do pontal Rip, Ilha Nelson, evidenciando os níveis com plantas (modifi cado de Dutra et al., 1998).
Figure 2. Geological section of the volcanic and volcanoclastic succession from the Rip Point, Nelson Island, showing the levels with plant fossil (modifi ed from Dutra et al., 1998).

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013444
pela primária destacada, secundárias rectipinadas e terciárias percorrentes. A adoção deste epíteto não pretende refl etir um parentesco biológico com o gênero moderno, mas reunir um tipo arquitetural de folha capaz de ser comparado com outros presentes, especialmente em floras do final do Cretáceo e Cenozoico basal, nas fl oras fósseis dos distintos continentes austrais. Na Península Antártica morfotipos comparáveis ocorrem desde o Cretáceo Inferior (Albiano) e desde altas latitudes (70ºS), como demonstraram Cantrill & Nichols (1996) em seu estudo da tafofl ora da ilha Alexander.
Na designação dos morfotipos incluídos nos diferentes grupos morfológicos utilizou-se a sigla RIP, referente ao local de ocorrência, buscando uniformizar com as análises prévias de Pole (1992), Cantrill & Nichols (1996) e Iglesias et al. (2007) em outras fl oras cretácicas. Afi nidades propostas a nível familiar e genérico com outros tipos fósseis, quando possível, foram baseadas em Stewart & Rothwell (1993). E a provável afi nidade às famílias modernas segue o APG III (Angiosperm Phylogeny Group, 2009).
Lâminas palinológicas foram processadas com a metodologia usual, através da eliminação dos carbonatos e silicatos, seguida de oxidação (Jones & Rowe, 1999), nos laboratórios de palinologia da Universidade Federal do Rio Grande do Sul (UFRGS) e do Instituto Tecnológico de Micropaleontologia (ITT Fossil, UNISINOS), utilizando para isto, as mesmas amostras de onde provinham os macrofósseis. As lâminas foram fotografadas em aumento de 400 x e estão armazenadas no LaViGæa, sob a sigla ANTF 34, adicionada da sigla P (palinomorfos), número 9528 (a-d). Sua abordagem é aqui feita de modo qualitativo, sem descrições taxonômicas mais formais, uma vez que o objetivo almejado era a confi rmação da idade dos depósitos e a identifi cação de elementos que auxiliassem no estabelecimento da diversidade da tafofl ora e suas afi nidades. Uma amostra foi analisada no laboratório da Companhia de Pesquisas de Recursos Minerais - CPRM, buscando informações de caráter petrográfi co e a determinação de sua origem vulcânica ou vulcanoclástica.
SISTEMÁTICA PALEOBOTÂNICA
Divisão MONILOPHYTA Classe POLYPODIOPSIDA (= PTERIDOPSIDA)
Ordem POLYPODIALES(Figura 3A)
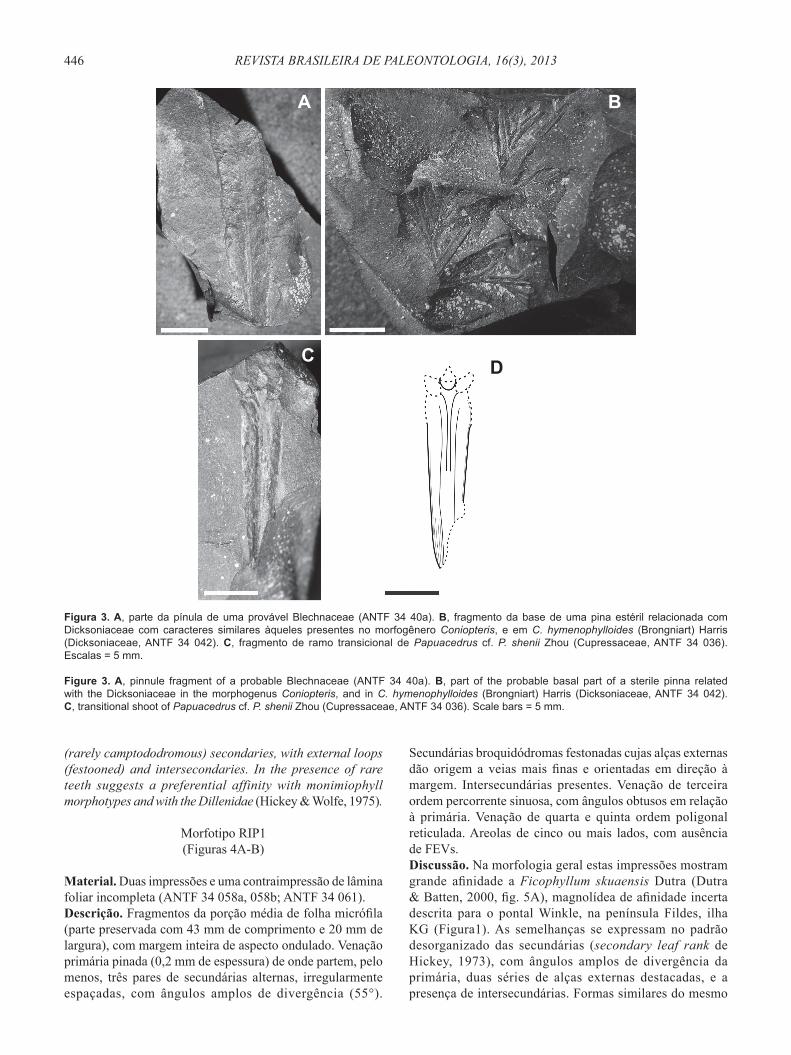
Material. Impressões parciais de pinas (ANTF 34 40a, 40b) e ráquis (ANT 34 40c, 40d, 40e).Descrição. Fragmentos incompletos de pinas de lados paralelos (pecopteroides/aletopteroides), coriáceas, de base ampla e parcialmente adpressa à ráquis (25 mm de comprimento e 5 mm de largura na porção preservada). Margem inteira e destacada, levemente ondulada, ápice ausente. Veia média proeminente. Veias laterais de curso reto, divergindo do eixo em ângulos amplos e homogêneos e para ambos os lados da pínula, bifurcadas admedialmente e terminando na margem da pínula. Não são visíveis anastomoses.Discussão. Os caracteres vistos aproximam estas impressões
dos representantes modernos tanto de Blechnaceae como de Aspleniaceae. Contudo, os lados paralelos, margens onduladas e lisas e a venação central bem marcada, com laterais bifurcadas, se assemelham mais ao que é comum na primeira família e, entre seus representantes, com as formas do gênero Blechnum. Na comparação com tipos modernos, no entanto, distingue-se pela menor densidade de veias laterais. Atualmente Blechnum caracteriza as fl orestas temperadas dos continentes austrais e, na Nova Zelândia, é o segundo gênero mais abundante de pteridófi tas (Brownsey & Smith-Dodsworth, 2000).
Na ilha KG próxima, as Blechnaceae são registradas entre os palinomorfos desde o fi nal do Cretáceo (Dutra et al., 1996) e como macrofósseis a partir do Paleógeno (Troncoso, 1986). Na América do Sul, formas de Blechnum são reconhecidas em níveis do Eoceno Inferior de RioTurbio, Argentina (Baldoni et al., 1985), onde se associa a Nothofagus, e na fl ora de Quinamavida, no Chile, onde foi associada a Blechnum sp. (Troncoso, 1992). Entre os tipos previamente descritos, a forma chilena é a que compartilha o maior número de caracteres com a aqui descrita, pela pínula com veia média destacada e laterais bifurcadas. Possui, contudo, a densidade maior de veias e margem crenada que caracteriza os tipos modernos da família, como por exemplo, B. chilensis (Kaulf.) Mett (Tryon & Tryon, 1982).
Ordem CYATHEALESFamília ?DICKSONIACEAE Bower, 1908
?Coniopteris (Brong.) Harris, 1961(Figura 3B)
Material. Parte e contraparte de fragmentos, aparentemente de uma única pina (ANTF 34 042).Descrição. Pina de ordem alta, coriácea, pinada, com segmentos cuneados (wedge) e bifurcados. A parte basal parece sugerir uma inserção catadrômica das pínulas. Pínulas de lados paralelos, divergentes, com uma veia central destacada (cenopteroidea) para cada segmento e raras ramifi cações livres.Discussão. O material escasso e incompleto impede uma maior afi nidade além daquela que, pelo caráter coriáceo da pina e pínulas bifurcadas oblongas de venação simples e livre, pode ser feita com algumas frondes estéreis do gênero Coniopteris (Dicksoniaceae), como C. bujerensis (pínula simples e linear), C. hymenophylloides (Harris, 1961) e C. bella. Das duas últimas espécies, o material do pontal Rip se distingue pela inserção alterna das pínulas e o tipo simples de venação, sem muitas veias laterais. A presença do esporo Cyatheacidites minor Couper nos mesmos níveis e de outras frondes de Coniopteris nos níveis basais do mesmo afl oramento (Trevisan, 2011), apoia esta afi nidade.
Os restos aqui descritos ainda compartilham caracteres, como a fi sionomia coriácea e a dicotomia das pinas, com as Polipodiales da família Davalliaceae (e.g. Davallia Sm.), fetos epifíticos comuns nos ambientes insulares modernos do Hemisfério Sul, em especial na Austrália e Nova Zelândia

445BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
(Tryon & Tryon, 1982). A forma moderna de Davallia, no entanto, possui frondes em geral anadrômicas.
Classe GYMNOSPERMOPSIDAOrdem CONIFERALES
Família CUPRESSACEAE Gray, 1822
Papuacedrus H.L. Li., 1953
cf. Papuacedrus sp.(Figura 3C)
Material. Impressão de parte de um ramo vegetativo (amostra ANTF 34 036). Descrição. Fragmento de ramo vegetativo como eixo recoberto por duas folhas laterais alongadas (21 mm de comprimento e 5 mm de largura) e três folhas faciais pequenas (3 mm de comprimento e 2-1 mm de largura). As folhas laterais possuem base convexa, e contornos levemente curvos, estando fundidas pela margem em uma cicatriz espessa. Os ápices são agudos, mas pouco fl etidos (spreading), e parcialmente preservados. A superfície é marcada por linhas fi nas e longitudinais, que provavelmente correspondem às múltiplas bandas de estômatos, e por um canal resinífero proeminente, com orientação paralela à margem. As folhas faciais, visíveis na porção distal do par de laterais, são quilhadas, de forma romboide a ovada e com ápices levemente acuminados.Discussão. Apesar da preservação parcial, este único fragmento de ramo mostra características que o inserem entre os presentes nos representantes da família Cupressaceae. A afi nidade é sugerida pela presença de folhas dimórfi cas, de inserção oposta decussada, com folhas laterais grandes e alongadas, e faciais pequenas (Farjon, 2005; Eckenwalder, 2009). A presença de folhas dimórfi cas os insere, além disto, no grupo E de Eckenwalder (2009), que inclui Austrocedrus Florin & Boutelje (endêmico do Chile), Libocedrus Endl. (exclusivo da Nova Zelândia e Nova Caledônia) e Papuacedrus H.-L.Li (endêmico de Papua-Nova Guiné). Papuacedrus se distingue pelo canal resinífero proeminente, pelas bandas longitudinais de estômatos e pela fusão apical das folhas laterais com as faciais (Wilf et al., 2009, Tabela 2), caracteres aparentemente aqui presentes. O caráter pouco acuminado e sem spreading do ápice das folhas laterais, e as bifaciais acuminadas, sugerem que se trate de parte de um ramo transicional (Wilf et al., 2009).
Entre os morfotipos previamente descritos em áreas próximas, a maior afi nidade é com os ramos de Papuacedrus shenii Zhou, descritos por Zhou & Li (1994, pl. 2:5-12; pl. 3:4) e apontados por Dutra (1997a, sem descrição formal, ambos em níveis do Eoceno Inferior da ilha KG, Formação Fossil Hill). As semelhanças se manifestam no heteromorfi smo das laterais e faciais, as últimas emergindo aparentemente soldadas, da porção apical das laterais. Entre os tipos fi gurados por estes autores destacam-se ainda pela maior semelhança, as folhas ilustradas para ramos jovens ou transicionais (os adultos possuem folhas equidimensionais). O material foi
associado por Zhou & Li (1994) a forma atual P. papuana (F. Mueller) Lie e, junto com a forma aqui descrita, atesta a presença das Cupressaceae nas sucessões do norte da Península Antártica, entre o Cretáceo e o Eoceno.
Tipos cupressoides haviam sido pioneiramente identificados por Berry (1938) para o Eoceno de Rio Pichileufú, Argentina, mas foram associados ao gênero Libocedrus (L. prechilensis). Posteriormente Wilf et al. (2009), ao abordarem os fósseis de Laguna Del Hunco, de idade correlata, incluíram o material de Berry (1938) no gênero Papuacedrus, baseando-se no caráter fusionado das folhas laterais. Para os autores e pela mesma razão, Austrocedrus australis, do Oligoceno da Tasmânia, deveria ser também aí incluída. Na Nova Zelândia, Papuacedrus só é registrado após o início do Mioceno, a partir de ramos vegetativos com cutículas preservadas (Pole, 2007).
Ao analisar a fi logenia das Cupressaceae, Mao et al. (2012) propuseram uma origem triássica para a família e que as relações entre as espécies e gêneros teriam se conformado entre o fi m do Jurássico e início do Cretáceo. A partir daí (153 Ma), as formas cupressoides setentrionais teriam se separado das caliptroides gondwânicas.
O registro fóssil de Papuacedrus e sua moderna distribuição constituem um bom argumento para uma origem e distribuição das Cupressaceae profundamente ligada à quebra do Gondwana (Mao et al., 2012). Mesmo parcialmente preservado e raro, as feições presentes no pequeno ramo de Papuacedrus aqui descrito, atestam sua provável condição de constituir o mais antigo registro conhecido para o gênero e, junto com as formas eocênicas da própria Península e Patagônia, uma origem das Cupressaceae Callitroideae em áreas ocidentais do sul do Gondwana, com posterior migração para o setor oriental. As análises fi logenéticas de Quinn & Price (2003), além disto, suportam que Papuacedrus teria sido a forma Callitroidea que primeiro divergiu, em um tempo que se seguiu à individualização entre as Taxodiaceae (Sciadopityaceae) e Cupressaceae.
A manutenção de Papuacedrus até o inicio do Eoceno no Gondwana Ocidental, demonstra ainda seu importante papel na evolução das Cupressaceae austrais, e para os modelos biogeográfi cos, atestando que terras contínuas existiam até esse momento entre Antártica/América do Sul e Australásia.
Papuacedrus cresce hoje nas altitudes médias as subalpinas das encostas submetidas a clima oceânico, em Papua-Nova Guiné (Hill, 2001).
Divisão MAGNOLIOPHYTA
Grupo Laurof í l ico (~pinado broquidódromo/cinamomofílicode Crabtree, 1987)
Folhas ovadas a elípticas, margem inteira, venação primária pinada, secundárias broquidódromas (raras vezes camptódromas), com alças externas e intersecundárias. A presença de raros dentes indica uma afi nidade preferencial, mais que com as Magnolidae, aos tipos monimiofílicos e às Dillenidae (Hickey & Wolfe, 1975)/ovate to elliptic leaves, entire margin, pinnate primary, brochidodromous
REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013446
(rarely camptododromous) secondaries, with external loops (festooned) and intersecondaries. In the presence of rare teeth suggests a preferential affinity with monimiophyll morphotypes and with the Dillenidae (Hickey & Wolfe, 1975).
Morfotipo RIP1(Figuras 4A-B)
Material. Duas impressões e uma contraimpressão de lâmina foliar incompleta (ANTF 34 058a, 058b; ANTF 34 061).Descrição. Fragmentos da porção média de folha micrófi la (parte preservada com 43 mm de comprimento e 20 mm de largura), com margem inteira de aspecto ondulado. Venação primária pinada (0,2 mm de espessura) de onde partem, pelo menos, três pares de secundárias alternas, irregularmente espaçadas, com ângulos amplos de divergência (55°).
Secundárias broquidódromas festonadas cujas alças externas dão origem a veias mais fi nas e orientadas em direção à margem. Intersecundárias presentes. Venação de terceira ordem percorrente sinuosa, com ângulos obtusos em relação à primária. Venação de quarta e quinta ordem poligonal reticulada. Areolas de cinco ou mais lados, com ausência de FEVs.Discussão. Na morfologia geral estas impressões mostram grande afi nidade a Ficophyllum skuaensis Dutra (Dutra & Batten, 2000, fi g. 5A), magnolídea de afi nidade incerta descrita para o pontal Winkle, na península Fildes, ilha KG (Figura1). As semelhanças se expressam no padrão desorganizado das secundárias (secondary leaf rank de Hickey, 1973), com ângulos amplos de divergência da primária, duas séries de alças externas destacadas, e a presença de intersecundárias. Formas similares do mesmo
Figura 3. A, parte da pínula de uma provável Blechnaceae (ANTF 34 40a). B, fragmento da base de uma pina estéril relacionada com Dicksoniaceae com caracteres similares àqueles presentes no morfogênero Coniopteris, e em C. hymenophylloides (Brongniart) Harris (Dicksoniaceae, ANTF 34 042). C, fragmento de ramo transicional de Papuacedrus cf. P. shenii Zhou (Cupressaceae, ANTF 34 036). Escalas = 5 mm.
Figure 3. A, pinnule fragment of a probable Blechnaceae (ANTF 34 40a). B, part of the probable basal part of a sterile pinna related with the Dicksoniaceae in the morphogenus Coniopteris, and in C. hymenophylloides (Brongniart) Harris (Dicksoniaceae, ANTF 34 042). C, transitional shoot of Papuacedrus cf. P. shenii Zhou (Cupressaceae, ANTF 34 036). Scale bars = 5 mm.
A
C
B
D

447BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
gênero haviam sido descritas para o Albiano da ilha Alexander como F. palustris Cantrill (Cantrill & Nichols, 1996), mas mostram venação ainda mais desorganizada (embora mantenham o caráter broquidódromo festonado) e irregularidades marginais (ou dentes), estas impossíveis de avaliar no material de Rip. Numerosos morfotipos foliares de venação comparável, com margens ainda mais fortemente onduladas e raros dentes distanciados, foram descritos para o fi nal do Paleoceno e início do Eoceno na ilha KG (Zastawniak, 1981; Birkenmajer, 1985; Czajkowski & Rösler, 1986; Li, 1994, fi g. 5; pl. 4:1-2) e associados às Anacardiaceae. Em sua maior parte e pela presença dos poucos dentes, um caráter restrito às formas temperadas de Anacardiaceae modernas, foi associado ao gênero Rhoophyllum (R. nordenskjoeldii Dusén,1899). Contudo, os tipos descritos por Hünicken (1995, pl. E: 25-26) para o Cretáceo de Rio Guillermo (América do Sul) possuem, como aqui, bordas aparentemente lisas e veias secundárias broquidódromas, com ângulos amplos de divergência. É possível, portanto, que a presença de dentes marginais nas formas de Rhoophyllum do sul da América, refl ita apenas uma distinta adaptação ao clima e latitude.
Berry (1932) e Romero (1980), ao abordarem a coleção de plantas de Chubut (Cañadon Hondo), Argentina, também associaram morfotipos similares a Rhoophyllum nordenskjoeldii e às Anacardiaceae. Embora reconhecessem a falta de caracteres diagnósticos, esta afi nidade familiar foi mantida pelos autores mais pela presença da família nas fl oras modernas da América do Sul. Czajkowski & Rösler (1986, est. III, fi g. 17-18), utilizando material da sucessão de Fossil Hill, e Fisch (2009, fi g. 20) com impressões da ilha Dufayel, também salientaram o caráter problemático da afi nidade destes restos com Anacardiaceae, para o que a preservação incompleta do material aqui descrito não fornece novos elementos.
Na incerteza de afinidades mais próximas, uma associação preferencial, mesmo a nível familiar é de difícil estabelecimento para estes exemplares, até que lâminas foliares melhor preservadas sejam identifi cadas.
Morfotipo RIP2(Figuras 4C-D)
Material. Impressão e contraimpressão de uma folha isolada (ANTF 34 066a, 066b).Descrição. Folha microfílica parcialmente preservada (32 mm de comprimento e 15 mm de largura), simétrica, base arredondada a cuneada, pecíolo espesso de base alargada (7 mm de comprimento, 1 mm de largura), margem inteira e ápice ausente. Primária pinada (0,5 mm de espessura), destacada e reta. Secundárias broquidódromas festonadas, de inserção oposta e ângulos amplos (70°), formando alças externas de contorno quadrangular. Intersecundárias presentes. Venação de terceira ordem percorrente, de ângulo obtuso em relação à primária. Raros setores mostram venação de quarta ordem do tipo reticulada regular poligonal. Discussão. O escasso material e a preservação parcial desta impressão impossibilita o estabelecimento de afi nidade mais
precisa. Embora o padrão de venação seja similar ao visto para RIP 1 (broquidódroma festonada), esta forma possui veias menos destacadas e alças mais regulares, e um menor número de alças externas. Os ângulos de divergência da primária são também mais amplos, a textura aparentemente mais coriácea (pela margem mais ressaltada do sedimento) e preserva um pecíolo do tipo “pulvinar”. Um aspecto distintivo importante é a presença ocasional de uma alça broquidódroma extra que une duas secundárias principais em alguns setores e próximo à margem.
Entre as famílias modernas de eudicotiledôneas, padrões similares são encontrados entre as Elaeocarpaceae (Eurosidae I, Oxilalidales), como em Sloanea L. (especialmente, S. hirsuta Planch. et Benth., hoje sinonimizada em S. monosperma Vell), forma endêmica do Brasil distribuída ao longo da costa atlântica e até os 29ºS de latitude, quando passa a fazer parte da fl oresta com Araucária (Silveira, 2009). Uma nova espécie do gênero (S. lasiocoma K. Schum) foi recentemente registrada pela primeira vez, para o norte da Argentina (Keller et al., 2012). Além disto, Sloanea sect. Antholoma é acompanhante comum de Nothofagus na vegetação de altitude da Nova Guiné e Nova Caledônia (Heads, 2006). Seu registro mais antigo no setor do Gondwana oriental data da base do Eoceno da Austrália, onde hoje é representado exclusivamente por S. australis (Coode, 1983). Pole (1993a) descreve o morfotipo MANU-5 para o Mioceno da Nova Zelândia, que associa a Sloanea/Elaeocarpus, mas que difere de Rip 2, pela presença de dentes glandulares.
Nas sucessões das ilhas KG e Seymour, as Elaeocarpaceae são bem representadas tanto na macroflora, como nos microfósseis, desde o Cretáceo (Zastawniak, 1994) até o Eoceno médio (Shen, 1994; Askin, 1997). As impressões nos níveis mais antigos tem venação eucamptódroma a broquidódroma irregular, e como aqui, veias intersecundárias (Dicotylophyllum sp. 7, fi g. 8:7, e Dicotylophyllum sp. 10, pl.4:3 e fi g. 8:9, Zastawniak, 1994), constituindo, portanto, bons elementos comparativos.
Folhas broquidódromas simples e inteiras caracterizam igualmente as Monimiaceae (e.g. Peumus sp.), Proteaceae e Aquifoliaceae, as duas últimas também presentes na microfl ora aqui associada. Folhas simples e coriáceas, pecíoladas, de venação pouco destacada, e com alças externas ocasionais são comuns, por exemplo, a algumas espécies de Ilex, como I. theezans Mart. ex Reissek., outro elemento da fl oresta com Araucária do sul do Brasil.Tipos relacionados a Ilex ocorrem desde o Turoniano nos depósitos das áreas de abrangência da Província Weddelliana, ou seja, Austrália-Nova Zelândia e sul da América do Sul, passando pela costa ocidental da Antártica (Cantrill & Poole, 2012). Assim, constituem igualmente uma afi nidade possível para o morfotipo aqui descrito.
Morfotipo RIP3(Figuras 4E-F)
Material. Impressão de folha isolada (amostra ANTF 34 097).Descrição. Folha microfílica (30 mm de comprimento, 17 mm de largura), com parte do ápice e base presentes. Margem inteira, parcialmente preservada. Primária pinada e destacada (1 mm de largura), de onde partem pelo menos quatro pares

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013448
de secundárias broquidódromas, com um conjunto externo adicional de alças pouco visíveis, das quais emergem no setor basal da lâmina, veias de calibre mais fi no que alcançam a margem. As secundárias divergem da primária em ângulos retos, inicialmente mais amplos e reduzindo para topo. Intersecundárias presentes, venação terciária fracamente preservada e percorrente.Discussão. As bordas lisas, ângulo amplo de inserção das secundárias, de tipo geral broquidódromo a broquidódromo festonado, se assemelham ao dos tipos anteriormente descritos, em especial com RIP1, razão de sua inserção no agrupamento laurofílico. RIP 3 difere, no entanto, no aspecto menos coriáceo, secundárias mais destacadas e no maior desenvolvimento de veias externas às alças das secundárias (festoons), especialmente junto à base. Mas a falta de alguns elementos fi sionômicos, como da porção basal em RIP 1, e da margem com secundárias externas em RIP 2, torna desaconselhável reuní-los em um único morfótipo, até que materiais mais completos sejam identifi cados. Por esta mesma razão foi evitada a proposta de afi nidades familiares.
Tipos como RIP 1, 2 e 3 têm sido tradicionalmente incluídos entre os morfotipos laurofílicos e caracterizam as litologias mais antigas na ilha KG (Czajkowski & Rösler, 1986; Zastawniak, 1994; Dutra & Batten, 2000) e Seymour (Lauriphyllum nordenskjoldii Dusén, 1908). No Eoceno, ou se tornam mais escassas, confi rmando sua relação com grupos primitivos de angiospermas, ou passam a apresentar uma venação mais organizada que propicia sua mais adequada inserção taxonômica (Berry, 1925, 1938; Troncoso, 1992).
Alguma comparação ainda pode ser feita com o morfotipo fi gurado por Tokarski et al. (1987, fi g. 8-h) para a Península Barton, na Ilha KG, em níveis de provável idade cretácica. Igualmente de difícil associação a grupos modernos, foi atribuído apenas a dicotyledonous leaves of uncertain affi nity e mostra uma forma geral da lâmina similar e secundárias, com amplos ângulos de divergência.
Grupo acrodomofílico (acrodomophyll morphotype)Folhas de venação acródroma, com três ou mais primárias, ou secundárias laterais destacadas, convergindo para o ápice (Zastawniak, 1994; Cantrill & Nichols, 1996; Iglesias et al., 2007). Para Pole (1992) podem ter intramarginais./Leaf with acrodromous venation, three or more primaries or detached lateral secondaries, converging to the apex. To Pole (1992) could have intramarginal veins.
Morfotipo RIP4(Figuras 4G-H)
Material. Impressão da porção basal e média de uma folha (ANTF 34 106).Descrição. Folha microfílica (44 mm de comprimento, 15 mm de largura na porção preservada), de lâmina simétrica e margem lisa. Base cuneada, de ângulo agudo, ápice ausente. Venação do tipo acródroma, com uma veia central de curso retilíneo e calibre destacado (1 mm de espessura), sugerindo a presença de um pecíolo largo, e duas laterais que parecem
convergir superiormente formando arcos amplos e paralelos a margem. Veias de menor calibre fortemente opostas e percorrentes divergem da central em ângulos retos e uniformes e unem as duas laterais à principal originando pregas ou saliências em toda a extensão do limbo. Um sistema de veias mais fi nas e igualmente ortogonais distribui-se entre o padrão principal de venação (intersecundárias?). Veias intramarginais presentes, divergindo em ângulos retos do par lateral. Uma veia de menor calibre, de curso similar ao das primárias, acompanha a borda. Discussão. A falta do ápice, que permitiria confi rmar a confl uência apical das veias laterais, difi culta signifi cativamente o estabelecimento das relações de afinidade. Contudo, os caracteres da base e porção média presentes – lâmina com saliências entre as veias, margens inteiras, três veias acródomas, a mais central de maior calibre na base, unidas por secundárias de curso ortogonal e regularmente distribuídas sobre a lâmina – mostram uma relação muito próxima com a arquitetura das folhas modernas de Melastomataceae e, especialmente, com o gênero Miconia Ruiz & Pav., hoje de distribuição tropical a subtropical na América do Sul, desde a Colômbia (M. acutipetala Sprague) e Peru (M. amplexicaulis Naudin), até o Uruguai e Argentina, incluindo o Brasil (Miconia affi ni D.C.), segundo Cogniaux (1891) e Michelangeli et al. (2004).
Por extensão, apoia igualmente uma afi nidade com a espécie fóssil Miconiiphyllum australe, também referente a uma folha incompleta, e descrita por Dusén (1908, pl. I:14), para o “Terciário” da ilha Seymour. M. australe difere, no entanto, na venação mais delicada e numerosa, com intramarginais de ângulos agudos de divergência que sugerem a presença de alças broquidódromas externas, caracteres que não são típicos das formas de Miconia modernas e incentivam uma rediscussão do epíteto genérico de Dusén (1908).
Outros gêneros de Melastomataceae, considerados derivados de Miconia (Michelangeli et al., 2004), como Clidemia D. Don e Leandra Raddi, do setor nor-oriental da Cordilheira dos Andes e sudeste do Brasil, possuem folhas trinervadas similares, mas de maior tamanho e mais largas (Reynel & Honorio, 2004; Baumgratz et al., 2013), um aspecto que pode dever-se apenas à melhores condições de calor e umidade.
Já Melastoma e Tibouchina Aubl. (tribo Melastomae), possuem folhas pentanervadas, de veias menos destacadas e as de ordem mais alta com padrão reticulado e ângulos agudos de divergência. O mesmo tipo geral caracteriza Huberia D.C. (tribo Merianieae), embora se assemelhe a forma de Rip pelo caráter trinervado (Baumgratz, 2004).
Cantrill & Nichols (1996) descreveram uma forma acródroma primitiva e trinervada, com veias de segunda ordem broquidódromas (Timothyia trinervis Cantrill), para níveis do Cretáceo basal da ilha Alexander na Península Antártica, e que poderia estar relacionada com a forma aqui descrita. A ausência do ápice nas formas de RIP previne uma melhor comparação, mas de qualquer modo, o material de Rip possui uma veia central duas vezes mais espessa, e ângulos mais agudos de inserção das primárias laterais e secundárias.

449BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
Estes morfotipos, no entanto, atestam a presença pioneira dos tipos acródromos nas altas latitudes do sul e sua inclusão no estoque ancestral de angiospermas da Península Antártica.
Zastawniak (1994, fig. 10:5), estudando morfotipos acrodromofílicos do fi nal do Cretáceo (Formação Zamek) da ilha KG, chamou a atenção para a diversidade de familias que podem conter tipos acródromos (Menispermaceae, Melastomataceae, Lauraceae, Epacridaceae, Proteaceae, Piperaceae, Rhamnaceae e Smilacaceae), mas sugere que, em fl oras austrais, eles têm sido tradicionalmente associados às Melastomataceae. E é importante destacar, que quando presentes os caracteres apicais, muitas destas famílias podem ser eliminadas.
Entre os materiais descritos por Czajkowski & Röesler (1986, pl. I, fi gs. 3 e 5) para o Eoceno basal da mesma ilha KG (Formação Fossil Hill), uma associação preferencial com Miconiiphyllum australe Dusén foi proposta para os tipos
acródromos, apesar de se tratarem de lâminas pentanervadas. Já Zastawniak (Birkenmajer & Zastawniak, 1989, fi g. 4-A e pl. 5-A), descreveu uma forma praticamente idêntica, sem nominar o trabalho de Czajkowski & Rösler (1986) e propôs a nova espécie, Dicotylophyllum duseni Zastawniak, posteriormente emendada como Pentaneurum duseni por Li (1994), para a mesma Formação Fossil Hill. No conjunto, estas impressões pentanervadas e com veias de menor calibre desorganizadas, são todas distintas da aqui designada a RIP 4.
Levando em conta a preservação parcial e escassa do tipo aqui descrito, e até que mais elementos sejam encontrados, no momento e pela presença de três veias principais, RIP 4 se aproxima de Miconiiphyllum australe (com as ressalvas feitas a respeito de sua relação com Miconia), da ilha Seymour, e de Timothyia trinervis Cantrill & Nichols, do Albiano da ilha Alexander.
Figura 4. Grupos Magnolifílico e Laurofílico (pinado broquidódromo correspondente ao cinamomofílico de Crabtree, 1987) (A-F) e Grupo Acrodromofílico com venação acrodomica (G-H). A-B, Morfotipo RIP 1 (ANTF 34 058a); C-D, Morfotipo RIP 2 (ANTF 34 066a); E-F, Morfotipo RIP 3 (ANTF 34 097); G-H, Morfotipo RIP 4 (ANTF 34 106). Escalas = 10 mm.
Figure 4. Angiosperm morphotypes from the Magnoliaephyll and Laurophyll groups (or Cinamomophyll from Crabtree, 1987) (A-F) and Acrodomophyll Group, with acrodomous venation (G-H). A-B, Morphotype RIP 1 (ANTF 34 058a); C-D, Morphotype RIP 2 (ANTF 34 066a); E-F, Morphotype RIP 3 (ANTF 34 097); G-H, Morphotype RIP 4 (ANTF 34 106). Scale bars = 10 mm.
A
E
B
F
C
G
D
H

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013450
Entre os representantes modernos, a relação mais próxima é com aqueles incluídos na tribo Miconiae de Melastomataceae. Formas associadas à Miconia são registradas na macro e microfl ora do Brasil desde o Eoceno (Duarte, 1956; Lima & Salard-Cheboldaeff, 1981), uma área hoje considerada, junto com Equador e Peru, como centros de diversidade das Melastomataceae (Jaramillo & Dilcher, 2001; Baumgratz, 2004). No Chile, o registro é ainda mais antigo, datando do fi nal do Paleoceno, e mostra que o gênero era parte de uma associação tropical-subtropical de climas úmidos (Hinojosa et al., 2006). As Melastomataceae irão se manter no registro deste último país até o Mioceno médio, mas representadas por formas pentanervadas (Gutiérrez et al., 2013, fi g. 4).
Deste modo, a presença de tipos acródromos para o fi nal do Cretáceo na ilha Nelson e sua manutenção até o Paleoceno?-Eoceno da ilha KG, é de especial interesse. Se confi rmada a afi nidade com as Melastomataceae, inserem as áreas da Península como um dos prováveis centros de origem de mais um dos elementos hoje característicos dos biomas do Hemisfério Sul, como ocorre com Nothofagus, e muitos tipos modernos de Araucariaceae e Podocarpaceae. No caso da família Melastomataceae, contraria inclusive os dados oriundos das análises fi logenéticas, que propõem uma origem para a família no limite Paleoceno-Eoceno da Laurásia (Renner et al., 2001), ou África Central (Michelangeli et al., 2004), e sua irradiação a partir daí, através das áreas tropicais do Gondwana oriental. Para Morley & Dick (2003), esta controvérsia é causada pelo, em geral, desconhecimento do registro meridional e da difi culdade em distinguir os grãos de pólen da família, daquele das Combretaceae. Para estes autores o padrão biogeográfi co de distribuição das melastomatáceas seria derivado de uma linhagem gondwânica, à semelhança do proposto por Burnham & Johnson (2004), para as formações tropicais.
Grupo Nothofagus (protofílico)Folha pinada (margem se presente, não inteira, com 1 a 3 dentes por intervalo de secundária), primária destacada (razão 2:1 a 3:1), de curso reto, curvo ou sinuoso, secundárias craspedódromas, opostas a alternas, regularmente espaçadas, rectipinadas ou descrevendo uma curva levemente ascendente, com ângulos agudos e homogêneos de divergência (40º-45º), mais amplos na base (60-50º), podendo bifurcar exmedialmente. Ramifi cações do par basal de secundárias podem estar presentes. Lâminas pregueadas com secundárias destacadas e terciárias percorrentes indicam formas decíduas, enquanto as planas, com veias mais delicadas e que se reduzem em calibre para o ápice num padrão bifurcado-reticulado, sugerem formas perenes (exceto N. moorei)./Pinnately craspedodromous leaf, with non-entire margin, one to three theeths per secondaires, 1ª straight, detached, curved or sinuous, 2ª regularlly spaced, straight in course or slight arcuate, opposed or alternate and inserted in acute and homogeneous angles, broader in basal, sometimes pectinal ones. Folded lamina with sharp main venation and percurrent tertiaries indicate a deciduous habit (except N. moorei), the smooth and more coriaceous ones, with bifurcate or reticulate faintly visible secondaries, the perennial forms.
Morfotipo RIP 5(Figuras 5A-D)
Material. Impressões da porção apical (ANTF 34 001), de parte e contraparte da porção média da folha (ANTF 34 046a, 046b), e outras impressões isoladas (ANTF 34 014a,017, 037, 097B, 097C, 101).Descrição. Folhas microfílicas (larguras mínimas entre 17 mm, na parte média, e 10 mm na porção apical), Margem serrada com duas ordens de dentes, de ápice saliente e arredondado (mucronado), reto-concavo/convexo (rt-cc/cv), os de primeira ordem recebendo centralmente a fi nalização de uma secundária. Primária pinada espessa (0,25-0,3 mm), de curso sinuoso. Pelo menos 6 pares de secundárias craspedódromas destacadas, de inserção oposta a suboposta, em ângulos agudos (43°-45°, em média), e curso levemente arqueado apicalmente. Terciárias fracamente preservadas, percorrentes.Discussão. o caráter pinado craspedódromo da venação secundária, a presença de dentes duplos do tipo platanoide ou rosoide, e secundárias de curso ascendente, podem ser encontrados em distintas formas primitivas de angiospermas, como entre as Hammamelididae e/ou Rosidae, e Dillenidae pinadas (Hickey, 1973; Hickey & Wolfe, 1975). Na maior parte destes aspectos são também característicos, contudo, das formas do gênero austral Nothofagus e, entre elas, daquelas reunidas por Tanai (1986) em seu Grupo II, que inclui folhas pequenas, como Nothofagus antarctica e N. pumilio, da América do Sul e N. gunnii, da Nova Zelândia. Associados ao caráter pregueado da lâmina, a veia media sinuosa, as secundárias com ângulos agudos de inserção, e as terciárias percorrentes, assemelham-se a outros restos atribuídos às Nothofagaceae nos depósitos das bacias austrais (Thomson & Burn, 1977; Romero & Dibbern, 1985; Tanai, 1986).
Rip 5 compartilha características em sua porção preservada com um grande número de elementos presentes no registro fóssil das bacias austrais. Com Nothofagus magelhaenica (Engelh.) Dusén, 1899, do Eoceno-Oligoceno de Rio Las Minas (Formação Loreto), Punta Arenas, Chile (Tanai, 1986, text. fi g. 7G; pl. 10:4, 6-8 e 12), se assemelha pelos dentes duplos e mucronados, com sinus profundos. Entre as fl oras cretáceas revisadas por Dutra & Batten (2000, fi g. 12 A-B) para a ilha KG, é comparável a N. glaucifolia Dutra (similar a forma moderna N. glauca (Phil.) Kraser, da secção Lophozonia Turcz., hoje endêmica do Chile central), pela primária sinuosa, margem serrada, secundárias alternas a subopostas, com dois a três dentes por intervalo de secundária principal.
A presença de bordas serrilhadas e dentes pronunciados caracteriza igualmente Nothofagus lanceolata Dusén (Tanai, 1986) e N. serrulata Dusén, tipos eocênicos (Formação Loreto, Dusén, 1899, pl. IX: 4 e 5), a primeira também presente em KG (Torres et al., 2009). Segundo Hünicken (1966), N. serrulata ocorre também nos níveis de Rio Turbio, de mesma idade. Alguma semelhança ainda existe com N. plicata (Scriven et al., 1995, pl. 1:2) dos níveis do fi nal do Eoceno

451BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
do sul da Austrália. Como RIP 5, esta forma possui margem serrada, dentes de ápice arredondado (neste caso, glandulares), folha fortemente plicada, venação secundária rectipinada com ângulos agudos de inserção e terciárias percorrentes. Para os autores constitui o mais antigo macrofóssil de um tipo decíduo do gênero para aquele continente.
É possível ainda, como visto acima, que RIP 5 se refi ra a uma forma decídua juvenil (fortemente pregueada), similar às encontradas por Dutra & Batten (2000, fi g. 11) nos níveis cretácicos da ilha KG. De qualquer modo, comparada com os outros elementos da assembleia do pontal RIP, é um morfotipo exclusivo pelo acentuado pregueamento da lâmina, tamanho reduzido e pequeno número de secundárias, o que reforça seu provável caráter juvenil.
O grande número de formas com esta arquitetura previamente descritas para as bacias austrais atesta: o caráter em geral homogêneo e conservador da morfologia foliar em Nothofagus, a possibilidade de que eventuais distinções morfológicas possam ser devidas a distintos estágios ontogenéticos, e a necessidade de uma revisão destes elementos à luz das novas análises fi logenéticas.
Morfotipo RIP 6(Figuras 5E-F)
Material. Impressão de folha (ANTF 34 070).Descrição. Folha pinada, simétrica, microfílica (42 mm de comprimento na porção preservada), aparentemente oblongo-ovalada, levemente pregueada na base e plana na porção média e superior. Veia primária espessa na base da lâmina (1,2 mm de largura), afi nando em direção ao ápice (0,25 mm). Dez pares de veias secundárias são visíveis, igualmente destacadas (0,8 mm), de curso reto e paralelo, distanciadas entre 3-3,5 mm, de inserção oposta a suboposta, e em ângulos agudos maiores na base (60°), que no ápice (50º-45º). Caracteres da margem, base, ápice, e de venação de ordem mais alta ausentes.Discussão. Em que pese a preservação muito incompleta e a escassez do material, impedindo afi nidades mais específi cas, a arquitetura geral da lâmina, aparentemente alongada, levemente pregueada, e com numerosas secundárias rectipinadas (proporcionalmente ao tamanho da lâmina), de inserção próxima e paralela, com pares basais com ângulos mais amplos de inserção, sugere sua relação com Nothofagus, e em especial, com a espécie moderna N. alpina (syn. Nothofagus procera), secção Lophozonia, da Argentina e Chile.
Ao levantar aspectos distintivos entre as formas modernas de Nothofagus Dutra (1997b) propôs para N. alpina uma maior densidade comparativa de secundárias, menor largura relativa, e valores comparativos mais altos de razão comprimento-largura (C/L= 3/2).
Entre as formas do registro esta morfologia é comum a Fagus subferruginea (Dusén, 1899, apenas os tipos da plate VIII, fi gs. 2, 3), posteriormente transferida para Nothofagus por Tanai (1986). A semelhança se manifesta no aspecto geral e tamanho da lâmina, e no número, proximidade e regularidade das secundárias. N. subferruginea, como
Figura 5. Morfotipos de angiospermas do Grupo Nothofagus. A-D, Morfotipo RIP 5 (ANTF 34 001); E-F, Morfotipo RIP 6 (ANTF 34 070). Escalas: A-B = 5 mm; C-F = 10 mm.
Figure 5. Angiosperm Nothofagus Group morphotype. A-D, Morphotype RIP 5 (ANTF 34 001); E-F, Morphotype RIP 6 (ANTF 34 070). Scale bars: A-B = 5 mm; C-F = 10 mm.
veremos a seguir, é a forma mais frequente nos depósitos do K-Paleógeno da ilha KG (Zastawniak et al., 1985; Shen, 1994; Dutra & Batten, 2000).
Morfotipo RIP 7(Figuras 6A-B)
Material. Impressão da parte basal e média de uma folha (ANTF 34 023) e outras impressões incompletas (ANTF 34 005bB, 002, 007, 013a, 013c, 033, 047a, 047d, 047g, 052a, 052b, 077a, 077b, 080, 098).
A
C
E
B
D
F

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013452
Descrição. Folha pinada, microfílica (35 mm de comprimento e 19 mm de largura na porção preservada), aparentemente elíptica ou ovada, lâmina fortemente pregueada. Margem lisa, apenas visível na porção basal. Veia primária destacada (1 mm de espessura) de onde partem oito secundárias de curso reto (0,2 mm de espessura), alternas a subopostas, e regularmente espaçadas (3,5 mm). Divergem da primária em ângulos agudos (40°) e sem ramifi cações externas visíveis. Venação terciária percorrente oposta e sinuosa, fortemente marcada, de disposição ortogonal às secundárias, e em ângulos obtusos em relação à primária.Discussão. Embora o mais abundante entre as restos de folha do pontal Rip, a ausência de uma melhor preservação das margens previne o estabelecimento de afi nidades mais específi cas para este morfotipo. Os caracteres do limbo, no entanto, mais uma vez atestam sua relação com formas decíduas de Nothofagus, no caráter pregueado e número expressivo de secundárias. Formas modernas do gênero com esta morfologia restringem-se às latitudes mais baixas da distribuição e às altitudes médias da Cordilheira dos Andes (Veblen et al., 1996).
Entre as espécies aí presentes, RIP 7 compartilha caracteres, tanto com Nothofagus alessandrii (sect. Fuscospora), como com N. macrocarpa (A.D.C.) Vásquez & Rodr. (sect. Lophozonia), no maior tamanho da folha, dentes pouco desenvolvidos e, secundárias apicais arqueadas superiormente. As diferenças entre estas duas espécies concentram-se nos caracteres marginais e, em sua ausência, como aqui, são indistinguíveis.
No registro fóssil da ilha KG, tipos comparáveis são encontrados em Nothofagus cretaceae Zastawniak, dos níveis do Campaniano da Formação Zamek (Zastawniak, 1994). Nos exemplares desta espécie faltam também as margens, e o curso percorrente das secundárias e terciárias é muito similar. Já entre as muitas formas fi guradas por Zastawniak et al. (1985), para níveis do Eoceno basal, a maior parte sem margens, também estão formas comparáveis, nas folhas grandes, de secundárias destacadas, separadas por dobras da lâmina. Estes autores propuseram a associação de cinco dos seus morfotipos com a espécie Nothofagus alessandrii moderna e, entre eles, o da fi g. 10:8, pelo tipo percorrente e destacado da venação terciária, é a que mais se aproxima de RIP 7. Também para a ilha Seymour, Zastawniak (in Doktor et al., 1996) fi gurou e descreveu brevemente formas de Nothofagus sp., similares em tamanho e tipo de venação, destacando sua semelhança à N. subferruginea.
Li (1994, pls. 1:1; 2:2) ilustrou para o Eoceno basal de KG (Formação Fossil Hill) tipos de folhas grandes e muitas secundárias, sem bifurcações externas aparentes, que também associou a Nothofagus subferruginea (Dusén) Tanai (cuja ilustração original reúne distintas morfologias). O exame conjunto das formas de N. subferruginea ilustradas por Dusén (1899, fi g. 8:1-2), e por Tanai (1986, fi gs. 9:1 e 4), espécie mantida por Romero & Dibbern (1985) e Romero (1980), mostra muitos tipos sem bordos e comparáveis com RIP 7, na forma da lâmina, número e curso das secundárias.
Finalmente, pelo caráter da venação, tamanho e forma da base, a forma aqui descrita é igualmente comparável a Nothofagus ulmifolia (Ett.) Oliver, do Cretáceo da Nova Zelândia (Pole, 1992, fi gs. 46-52).
Morfotipo RIP8(Figuras 6C-D)
Material. impressão de parte da porção mediana de uma folha (ANTF 34 005a) e outras impressões foliares igualmente incompletas (ANTF 34 004aB, 004bB, 005bA, 014A, 029, 079a).Descrição. Folha larga (pelo menos 3,5 cm de largura), primária pinada (0,8 mm de largura) e pelo menos 6 pares de secundárias alternas a subopostas (0,4 mm de espessura), paralelas entre si e uniformemente espaçadas, com ângulos amplos e uniformes de divergência (50-55°), de curso reto e paralelo. Pregas suaves entre as secundarias. Discussão. As folhas amplas, com primária destacada e secundárias equidistantes de curso reto e ângulos amplos de divergência, caracterizam mais uma vez formas relacionadas aos subgêneros Fuscospora e Lophozonia de Nothofagus, cujos análogos modernos crescem nas latitudes mais baixas da distribuição do gênero na América do Sul (N. alessandrii e N. alpina). RIP 8 no entanto, distingue-se dos outros morfotipos presentes, pela maior largura da lâmina, o caráter menos pregueado do limbo e pelas secundárias numerosas e delicadas, de curso reto (sem recurvar exmedialmente).
Na deposição da ilha KG, lâminas largas, com numerosas secundárias de curso reto e paralelo, ocorrem como visto, tanto nas tafofloras do final do Cretáceo (Formação Zamek), como do Eoceno basal (formações Fossil Hill e Mount Wawel), reduzindo-se a partir daí e dando lugar a formas de folhas pequenas. Nas associações cretácicas são representadas, por exemplo, por Nothofagus hillii Dutra (Dutra, 2000) e N. zastawniakiae Dutra (Dutra & Batten, 2000), esta última incluindo parte das formas referidas a N. cretaceae por Zastawniak (1994, fi gs. 9.2, 11-12) e N. subferruginea (Dusén) Tanai, de Li (1994, pl.2:2). Nas ilustrações originais de N. subferruginea de Tanai (1986), coincide com a representada na pl. 9:10, e em Dusén (1899), aos fi gurados na pl. VIII:6 e 8, caracterizadas por folhas mais largas, de venação secundária delicada, numerosa e regularmente espaçada.
Romero & Dibbern (1985), para a América do Sul, e Oliver (1936), trabalhando com material cenozoico da Nova Zelândia, identifi caram morfotipos “fagaleanos” similares e os atribuíram às formas setentrionais de Fagus e Carpinus, com o que concordava Dusén (1907). Tanai (1986), contudo, sem comentar as afi nidades propostas, mantém a associação apenas a Nothogafus.
Nothofagus novaezealandiae (Oliver) Pole, descrita para o Grupo Manuherikia, Mioceno da Nova Zelândia, é igualmente comparável em tamanho, número e curso da venação principal (Pole, 1993b). O autor comenta que esta forma compartilha características com a forma decídua sul-americana N. alessandrii, e com N. moorei (F. Muell.) Krasser,
453BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
a única forma moderna perene de folhas grandes (subgênero Lophozonia) e também relictual das áreas altas do leste da Austrália. Não é possível descartar que RIP 8 se refi ra a uma forma perene do gênero (ou a uma forma adulta decídua) dada a ausência de pregas no limbo.
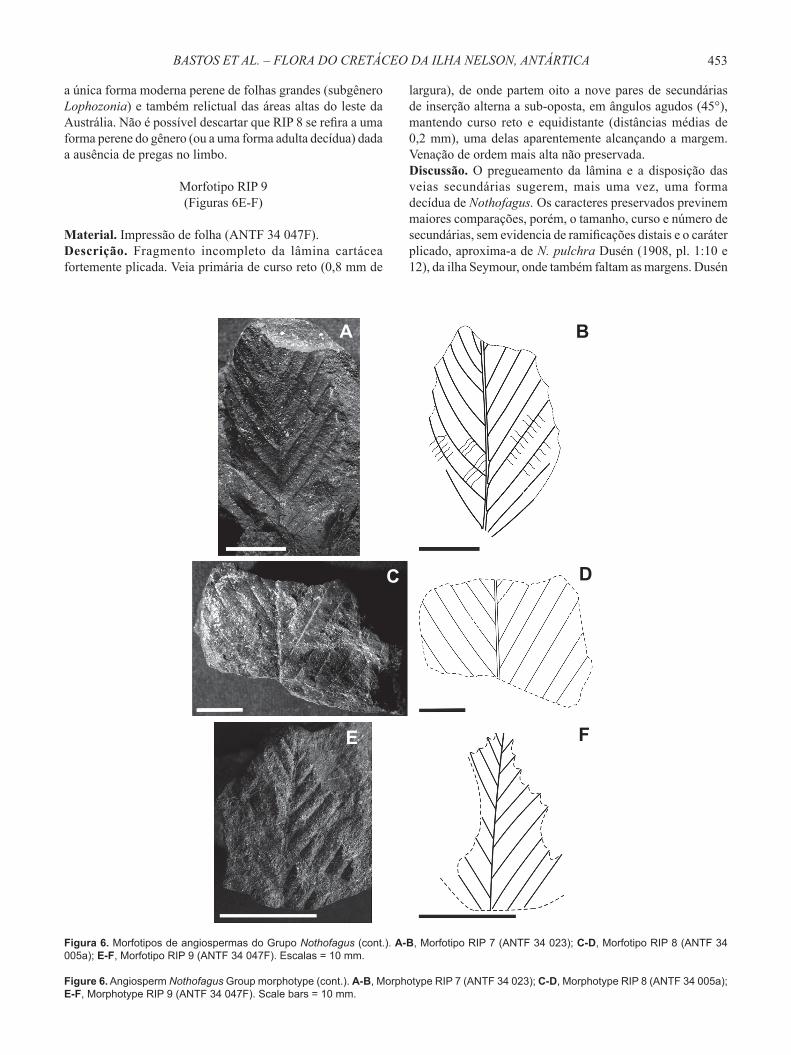
Morfotipo RIP 9(Figuras 6E-F)
Material. Impressão de folha (ANTF 34 047F).Descrição. Fragmento incompleto da lâmina cartácea fortemente plicada. Veia primária de curso reto (0,8 mm de
Figura 6. Morfotipos de angiospermas do Grupo Nothofagus (cont.). A-B, Morfotipo RIP 7 (ANTF 34 023); C-D, Morfotipo RIP 8 (ANTF 34 005a); E-F, Morfotipo RIP 9 (ANTF 34 047F). Escalas = 10 mm.
Figure 6. Angiosperm Nothofagus Group morphotype (cont.). A-B, Morphotype RIP 7 (ANTF 34 023); C-D, Morphotype RIP 8 (ANTF 34 005a); E-F, Morphotype RIP 9 (ANTF 34 047F). Scale bars = 10 mm.
largura), de onde partem oito a nove pares de secundárias de inserção alterna a sub-oposta, em ângulos agudos (45°), mantendo curso reto e equidistante (distâncias médias de 0,2 mm), uma delas aparentemente alcançando a margem. Venação de ordem mais alta não preservada. Discussão. O pregueamento da lâmina e a disposição das veias secundárias sugerem, mais uma vez, uma forma decídua de Nothofagus. Os caracteres preservados previnem maiores comparações, porém, o tamanho, curso e número de secundárias, sem evidencia de ramifi cações distais e o caráter plicado, aproxima-a de N. pulchra Dusén (1908, pl. 1:10 e 12), da ilha Seymour, onde também faltam as margens. Dusén
A
C
E
B
D
F

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013454
(1908) apontou a semelhança de N. pulchra com os tipos por ele descritos para o Rio Condor, na Terra do Fogo (Chile), que havia atribuído a N. densinervosa (Dusén, 1899, tab. 9:14-19). Embora tenha validado esta espécie, Tanai (1986) fi gura tipos de tamanho muito variável (Tanai, 1986, pl. 12:1-12), mas igualmente chama a atenção para a semelhança com N. pulchra, caracterizando-a como dotada de margens crenadas, impossíveis de avaliar no morfotipo aqui descrito.
O número expressivo de secundárias (8-10), em uma lâmina de tamanho restrito, aproxima RIP 9 igualmente de outro elemento de Rio Condor, Nothofagus simplicidens Dusén, forma comum nos depósitos de sul do Chile (Punta Arenas, Carmen Silva, e rios Beta e Guillermo), segundo Tanai (1986). Na revisão de Romero & Dibbern (1985), não referida por Tanai (1986), N. pulchra e N. densinervosa foram consideradas inválidas por seu caráter incompleto e por representarem provavelmente formas decíduas juvenis.
MICROFÓSSEIS
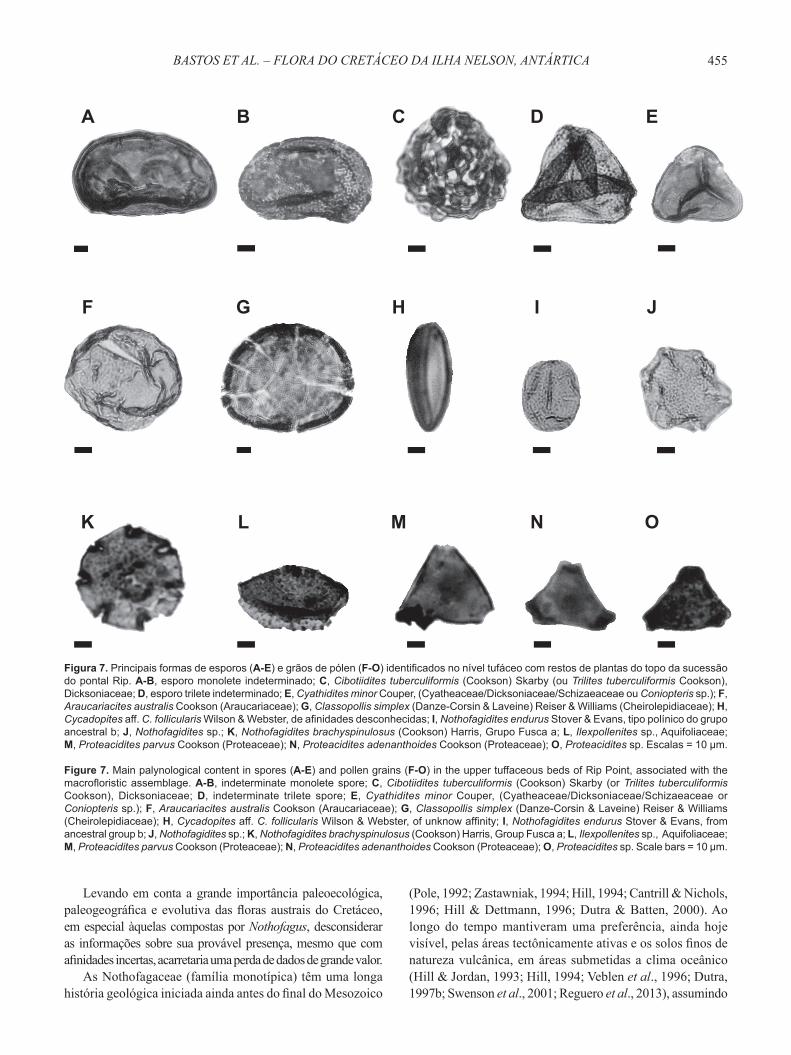
Uma análise preliminar e qualitativa dos palinomorfos presentes nos mesmos níveis de onde provêm as impressões (Figura 7) mostra que a microfl ora possui uma composição distinta. Entre os palinomorfos dominam os esporos (61%), seguidos de grãos de pólen de gimnospermas (22%) e angiospermas (12%). Esporos de fungos (4%) e testas de foraminíferos (visualizadas em lâmina petrográfi ca) estão igualmente presentes e sugerem ambientes subaquosos e costeiros para a deposição.
As pteridófitas estão representadas por Trilites parvallatus Krutzsch, 1959 (associada à Dicksonia, segundo Volkheimer et al., 2007), Cyathidites minor Couper (Cyatheaceae/Dicksoniaceae/Schizaeaceae ou Coniopteris sp.) e Rugulatisporites mallatus Stover (Stover & Partridge, 1973), relacionado com Thyrsopteridaceae ou Osmundaceae. Entre as gimnospermas, Araucariacites australis Cookson é a forma mais abundante, com 50% de representação, seguido de Classopollis simplex (Danze-Corsin & Laveine) Reiser & Williams 1969 (Cheirolepidiaceae, para Sirivastava, 1976), e Cycadopites aff. C. follicularis Wilson & Webster, de afinidade indeterminada e restrita ao Cretáceo (Raine et al., 2011).
Um dos poucos aspectos comuns entre a macro e a microfl ora é a presença signifi cativa dos grãos de pólen de Nothofagidites, a forma mais abundante entre as angiospermas. Está representado pelos tipos primitivos N. asperus (Cookson, 1959) Romero, 1973, do grupo ancestral a de Dettmann et al. (1990), e N. endurus Stover & Evans e N. kaitangataensis (Te Punga) Romero (ou N. kaitangata Raine et al., 2011), do grupo ancestral b (Dettmann et al., 1990). Estas formas compartilham uma distribuição cronológica restrita a parte mais alta do Cretáceo e um primeiro aparecimento nas altas latitudes do Gondwana (Dettmann et al., 1990; Hill & Dettmann, 1996).
Em menor proporção estão presentes tipos associados às Proteaceae (Proteacidites spp.) e Aquifoliaceae (Ilexpolenites sp.), esta última também com provável presença na
macrofl ora, como visto. As Proteaceae, em que pese seu registro abundante entre os grãos de pólen, não foram identifi cadas entre os macrofósseis, embora algum dos tipos laurofílicos possam indicar sua presença. Folhas relacionadas a esta família são pouco comuns igualmente nas associações do fi nal do Cretáceo de outras áreas da Península Antártica e do lado oriental da América do Sul, embora ocorram signifi cativamente nas fl oras da Austrália e da Nova Zelândia (Tabela 1).
No conjunto, os palinomorfos apoiam uma idade campaniana para a tafocenose do pontal Rip, pela presença de tipos primitivos de Nothofagidites, associados à gimnospermas restritas ao Mesozoico (Classopollis simplex e Cycadopites aff. C. follicularis). Este tipo de associação microfl orística é comum nos níveis do fi nal do Cretáceo das ilhas orientais da Península (Seymour e James Ross), associados à pteridófi tas (Dicksoniaceae, Cyatheaceae e Osmundaceae), coníferas, proteáceas, aquifoliáceas, monimiáceas e magnolídeas indeterminadas (Askin, 1997; Dettmann et al., 1990; Dutra & Batten, 2000).
CONSIDERAÇÕES FINAIS
Características do Grupo Morfológico Nothofagus (pro-tofílico), suas relações com Nothofagus Bl. e a aplicação à paleoecologia e paleobiogeografi a
Quando integralmente preservadas, as folhas de Nothofagus se caracterizam por margens não inteiras, com sinus e dentes simples ou compostos, mais ou menos salientes (menos visíveis ou arredondados nas formas perenes), que recebem uma ou mais terminações das secundárias e suas ramifi cações exmediais (venação craspedódroma, com raras exceções, Romero & Dibbern, 1985; Pole, 1993b, 1994; Dutra, 1997b). Para Tanai (1986), as formas modernas do gênero ainda seriam caracterizadas por uma veia fi mbrial, sem a qual não haveria garantia sobre a presença do gênero no registro.
A presença, contudo, de folhas inteiras é muito reduzida nas associações tafofl orísticas do K-Paleógeno do setor ocidental da Península Antártica, e em outras localidades austrais que compartilham uma gênese em ambientes de arco tectônico, onde eventos catastrófi cos e de vulcanismo podem gerar tefras em altas temperaturas que queimam a vegetação ainda antes de seu soterramento. Assim, associações compostas por um grande número de restos foliares, desprovidos de caracteres marginais ou ápice, ou de expansões da lâmina foliar de maior contato com o ambiente, são comuns nestes ambientes. A tafofl ora do pontal Rip, preservada em tufos entre níveis de lava, que atestam a proximidade com o edifício vulcânico, constitui um bom exemplo deste processo e os restos foliares incompletos que a compõem ampliam as difi culdades, já comuns às tafofl oras compostas exclusivamente por impressões foliares, no estabelecimento das afi nidades. Soma-se a isto a maior antiguidade dos níveis, sugerida pelas assembleias polínicas. Diante disto, optou-se por seu tratamento por grupos morfológicos e, quando possível, apontando as relações específi cas ou genéricas mais prováveis.
455BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
Figura 7. Principais formas de esporos (A-E) e grãos de pólen (F-O) identifi cados no nível tufáceo com restos de plantas do topo da sucessão do pontal Rip. A-B, esporo monolete indeterminado; C, Cibotiidites tuberculiformis (Cookson) Skarby (ou Trilites tuberculiformis Cookson), Dicksoniaceae; D, esporo trilete indeterminado; E, Cyathidites minor Couper, (Cyatheaceae/Dicksoniaceae/Schizaeaceae ou Coniopteris sp.); F, Araucariacites australis Cookson (Araucariaceae); G, Classopollis simplex (Danze-Corsin & Laveine) Reiser & Williams (Cheirolepidiaceae); H, Cycadopites aff. C. follicularis Wilson & Webster, de afi nidades desconhecidas; I, Nothofagidites endurus Stover & Evans, tipo polínico do grupo ancestral b; J, Nothofagidites sp.; K, Nothofagidites brachyspinulosus (Cookson) Harris, Grupo Fusca a; L, Ilexpollenites sp., Aquifoliaceae; M, Proteacidites parvus Cookson (Proteaceae); N, Proteacidites adenanthoides Cookson (Proteaceae); O, Proteacidites sp. Escalas = 10 μm.
Figure 7. Main palynological content in spores (A-E) and pollen grains (F-O) in the upper tuffaceous beds of Rip Point, associated with the macrofl oristic assemblage. A-B, indeterminate monolete spore; C, Cibotiidites tuberculiformis (Cookson) Skarby (or Trilites tuberculiformis Cookson), Dicksoniaceae; D, indeterminate trilete spore; E, Cyathidites minor Couper, (Cyatheaceae/Dicksoniaceae/Schizaeaceae or Coniopteris sp.); F, Araucariacites australis Cookson (Araucariaceae); G, Classopollis simplex (Danze-Corsin & Laveine) Reiser & Williams (Cheirolepidiaceae); H, Cycadopites aff. C. follicularis Wilson & Webster, of unknow affi nity; I, Nothofagidites endurus Stover & Evans, from ancestral group b; J, Nothofagidites sp.; K, Nothofagidites brachyspinulosus (Cookson) Harris, Group Fusca a; L, Ilexpollenites sp., Aquifoliaceae; M, Proteacidites parvus Cookson (Proteaceae); N, Proteacidites adenanthoides Cookson (Proteaceae); O, Proteacidites sp. Scale bars = 10 μm.
Levando em conta a grande importância paleoecológica, paleogeográfi ca e evolutiva das fl oras austrais do Cretáceo, em especial àquelas compostas por Nothofagus, desconsiderar as informações sobre sua provável presença, mesmo que com afi nidades incertas, acarretaria uma perda de dados de grande valor.
As Nothofagaceae (família monotípica) têm uma longa história geológica iniciada ainda antes do fi nal do Mesozoico
(Pole, 1992; Zastawniak, 1994; Hill, 1994; Cantrill & Nichols, 1996; Hill & Dettmann, 1996; Dutra & Batten, 2000). Ao longo do tempo mantiveram uma preferência, ainda hoje visível, pelas áreas tectônicamente ativas e os solos fi nos de natureza vulcânica, em áreas submetidas a clima oceânico (Hill & Jordan, 1993; Hill, 1994; Veblen et al., 1996; Dutra, 1997b; Swenson et al., 2001; Reguero et al., 2013), assumindo
A
F
K
B
G
L
C
H
M
D
I
N
E
J
O

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013456
assim o papel de um verdadeito ecotipo (Margalef, 1968). Embora o contexto ambiental instável possa eventualmente favorecer a preservação, mantendo a diversidade original e detalhes morfológicos mais fi nos (Burnham & Spicer, 1986), os processos tafonômicos aí envolvidos podem levar a perda daqueles caracteres que possuem de maior contato com o ambiente, em geral mais afetados pelo calor (Dutra, 1997a). No caso de Nothofagus, esta difi culdade de preservação é, em parte compensada, pela grande durabilidade post-mortem de suas folhas, resistentes aos meios ácidos, e pelo hábito decíduo de parte de seus representantes, que lhes garante expressiva participação nas serapilheiras (Remmert, 1988; Steart et al., 2005).
Mas suas lâminas foliares, como visto, possuem um padrão básico e “conservador”, mantido por, pelo menos, 50 milhões de anos (Swenson et al., 2001). Se isto em parte facilita seu reconhecimento no registro e a avaliação de suas adaptações, por outro, é mais um aspecto que perturba sua mais precisa inserção taxonomica. Estes aspectos são tão evidentes no registro, que o uso do epíteto genérico Nothofagus é admitido mesmo para associações pré-cenozoicas.
Graças a esta condição as modifi cações de sua fi sionomia refl etem suas distintas adaptações ao clima e latitude e são capazes de ser percebidas mesmo em formas incompletas. Phillipson & Phillipson (1988) e Pole (1994), por exemplo, demonstraram que as folhas dos tipos decíduos de Nothofagus expressam em suas lâminas jovens e maduras, o caráter plicado de sua vernação, exibindo dobras ou arqueamentos entre as veias secundárias. A possibilidade de sua abcissão as torna igualmente mais delicadas em textura e dotadas de expansões marginais mais expressivas (dentes), já que podem ser eliminadas nas fases mais drásticas do clima. Nos indivíduos perenes, as lâminas costumam ser menores e aplainadas, com menos expansões marginais e de textura coriácea, e ao contrário do esperado, são as formas modernas que crescem sob os climas mais frios (Veblen et al., 1996; Dutra, 1997b). Esta qualidade de Nothofagus o torna um elemento independente e muito útil ao trabalho geológico, permitindo testar nos ambientes continentais, as inferências paleoclimáticas provenientes das faunas marinhas (Alves et al., 2004).
Em termos paleogeográfi cos, a moderna distribuição disjunta de Nothofagus em áreas que preteritamente compunham o sul do Gondwana, faz dele ainda um elemento chave para os modelos paleobiogeográfi cos de vicariância e dispersão (van Steenis, 1972; Heads, 2006), e auxilia a testar as hipóteses derivadas das análises cladísticas. Para Ming Li (2011), por exemplo, os processos de dispersão seriam mais frequentes para as formas do gênero que aqueles resultantes da vicariância, um aspecto ainda discutível quando se examina o registro fóssil. Por outro lado, também com este tipo de análise fi logenética, Sauquet et al. (2012) obtiveram dados de idade para o crown group de Nothofagus que variaram enormemente, entre 13 até 113 Ma.
Na assembleia do pontal Rip, tipos morfológicos relacionados a Nothofagus compõem 25% da tafofl ora. Entre eles, a maior parte se enquadra no Grupo I de Tanai (1986),
que reúne formas exclusivas da América do Sul e espécies modernas decíduas (N. alessandrii, N. alpina, N. glauca e N. obliqua), portadoras de grãos de pólen do tipo Fusca a (subgênero Fuscospora) ou Menziesii (subgênero Lophozonia, de Hill & Read, 1991). N. alessandrii, a forma moderna mais relacionada aos tipos fósseis é hoje endêmica das áreas subtropicais (latitudes mais baixas) úmidas (Veblen et al., 1996), um bom modelo para as reconstruções paleoambientais das áreas da Península ocidental no fi nal do Mesozoico.
Quando se analisa o registro do norte da Península Antártica como um todo, formas morfologicamente análogas estão presentes desde o fi nal do Campaniano (Tabela 1) e compartilham a arquitetura que levou à caracterização, principalmente, das morfoespécies Nothofagus subferruginea (Dusén) Tanai, e N. magelhaenica (Engelhardt) Dusén, também incluídas no grupo I de Tanai (1986). Estes dados demonstram que indivíduos de folhas simples, com numerosas secundárias rectipinadas, e lâminas pregueadas (decíduas), caracterizaram estas áreas e os tipos mais primitivos. Nothofagus e seus relacionados se manterão aí até o fi nal do Eoceno, migrando para as áreas mais ao norte com a chegada dos climas frios (Birkenmajer et al., 2005).
Deste modo, o registro aqui discutido para a ilha Nelson, junto com as áreas do centro-sul da ilha KG (Dutra & Batten, 2000) correspondem ao mais antigo registro conhecido de macro e microfósseis do gênero, já capazes de ser associadas a forma moderna e confi rma o papel das áreas do norte da Península Antártica como seu centro de origem. Na falta de dados absolutos de idade não é possível precisar o exato momento em que isto ocorreu, mas assembleias polínicas da bacia de James Ross (Dolding, 1992), sugerem que este evento evolutivo coincide com o início do Campaniano (Cantrill & Poole, 2012).
A tafofl ora do pontal Rip: composição, paleoambiente e idade
A presença da tafoflora aqui estudada em níveis de tufos e vulcanoclásticas atesta uma gênese próxima a áreas submetidas a vulcanismo ativo (Dutra et al., 1998). O caráter seletivo da assembleia (macro e microfl ora de composição distinta), por outro lado, aponta para processos que afetaram os elementos que cresciam nas áreas de encosta, como Nothofagus, fetos arborescentes e coníferas, seguidos de um transporte dos macrorrestos, muitos previamente carbonizados, até as áreas baixas e corpos de água costeiros. Estas inferências são apoiadas pela presença nas mesmas camadas, de fragmentos devitrifi cados de material vulcânico e charcoal, e testas de foraminíferos (Figura 8).
Composicionalmente, a união dos dados da macro e microfl ora permite sugerir o fi nal do Campaniano como o tempo de deposição dos níveis do pontal Rip, apoiada pela presença conjunta de gimnospermas Cheirolepidiaceae e formas primitivas de Nothofagus, e nas correlações com as microfloras da bacia de James Ross (Dolding, 1992; Dettmann et al., 1990). Esta idade é ainda apoiada por vários outros elementos. Na associação de Rip estão ausentes as Bennettitales e Cycadales, comuns nas deposições juro-

457BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICATa
bela
1. G
rupo
s m
orfo
lógi
cos
de fo
lha
de a
ngio
sper
mas
par
a o fi n
al d
o C
retá
ceo
nas
alta
s la
titud
es d
o H
emis
fério
Sul
, com
ênf
ase
nos
cara
cter
es d
a ve
naçã
o (g
rupo
s se
gund
o C
rabt
ree,
198
7; P
ole,
19
92; Z
asta
wni
ak, 1
994;
Igle
sias
et a
l., 2
007;
Hay
es e
t al.,
200
6) e
sua
s po
ssív
eis
afi n
idad
es. I
lha
King
Geo
rge
(Zas
taw
niak
, 199
4; C
ao, 1
994;
Dut
ra &
Bat
ten,
200
0), S
eym
our/J
ames
Ros
s (D
usén
, 19
08; U
pchu
rch
& As
kin,
198
9; H
ayes
et a
l., 2
006;
Can
trill &
Poo
le, 2
012)
; Nov
a Ze
lând
ia (P
ole,
199
2) e
Aus
trália
(McL
ough
lin e
t al.,
1995
); Ba
cia
Aust
ral, A
rgen
tina
(Fre
ngue
lli, 1
953;
Men
énde
z, 1
969;
H
ünic
ken,
199
5; P
assa
lia e
t al.,
200
1; Ig
lesi
as e
t al.,
200
7); P
unta
Are
nas,
Chi
le (T
ronc
oso
& R
omer
o, 1
998;
Yab
e et
al.,
200
6; L
eppe
et a
l., 2
011)
. Abr
evia
ções
. ~, r
epre
sent
a po
ssív
eis
afi n
idad
es
fam
iliare
s ou
gen
éric
as; 1
, pal
eola
titud
e no
Cre
táce
o (P
ole,
199
2); 2
, par
a Po
le (1
992)
tipo
s lo
bado
s de
mar
gem
não
inte
ira, c
om v
enaç
ão c
rasp
edód
rom
a, p
odem
est
ar re
laci
onad
os a
Not
hofa
gus;
3,
as d
istin
tas
form
as d
e St
ercu
lia d
e Be
rry (1
927,
193
8) e
Men
énde
z (1
969)
são
man
tidas
nes
te g
rupo
por
seu
car
áter
pla
tano
fílic
o (e
.g. S
. was
hbur
ni B
erry
), m
as a
pres
enta
m 2
ª cra
sped
ódro
mas
e lo
bos
agud
os, s
imila
res
a Br
achy
chito
n Sc
hott
& En
dl. (
Ster
culo
idea
, Mal
vace
ae),
da A
ustrá
lia/N
ova
Gui
né, e
Acr
opog
on d
a N
ova
Cal
edôn
ia. J
á as
Ste
rcul
ia m
oder
nas
poss
uem
folh
as s
impl
es o
u pa
lmad
o-co
mpo
stas
(ven
ação
cam
ptód
rom
a a
broq
uidó
drom
a). E
S. s
ehue
nsis
Ber
ry é
trilo
bada
. Est
es a
spec
tos
dem
onst
ram
a n
eces
sida
de d
e um
a re
visã
o da
s fo
rmas
ass
ocia
das
a St
ercu
lia n
o re
gist
ro.
Tabl
e 1.
Lat
e C
reta
ceou
s m
orph
olog
ical
gro
ups
of a
ngio
sper
m le
aves
to th
e so
uthe
rnm
ost S
outh
Hem
isph
ere
(sug
gest
ed a
ffi ni
ties
emph
asis
ing
the
vena
tion
appe
als)
. Gro
up n
ames
com
pile
s th
ose
used
to fl
oras
from
this
tim
e by
Cra
btre
e (1
987)
, Pol
e (1
992)
, Zas
taw
niak
(199
4), I
gles
ias
et a
l. (2
007)
and
Hay
es e
t al.
(200
6). K
ing
Geo
rge
Isla
nd (Z
asta
wni
ak, 1
994;
Cao
, 199
4; D
utra
& B
atte
n,
2000
), S
eym
our/J
ames
Ros
s (D
usén
, 190
8; U
pchu
rch
& A
skin
, 198
9; H
ayes
et a
l., 2
006;
Can
trill
& P
oole
, 201
2); N
. Zea
land
(Pol
e, 1
992)
and
Aus
tralia
(McL
ough
lin e
t al.,
1995
); A
ustra
l Bas
in,
Arg
entin
a (F
reng
uelli
, 195
3; M
enén
dez,
196
9; H
ünic
ken,
199
5; P
assa
lia e
t al.,
200
1; Ig
lesi
as e
t al.,
200
7); P
unta
Are
nas,
Chi
le (T
ronc
oso
& R
omer
o, 1
998;
Yab
e et
al.,
200
6; L
eppe
et a
l., 2
011)
. A
bbre
viat
ions
. ~, s
ugge
st p
ossi
ble
fam
iliar
or g
ener
ic a
ffi ni
ties;
1, C
reta
ceou
s pa
leol
atitu
de (P
ole,
199
2); 2
, to
Pol
e (1
992)
cra
sped
odro
mou
s le
aves
with
non
ent
ire m
argi
ns c
ould
be
rela
ted
with
N
otho
fagu
s; 3
, the
impo
rtant
K-P
aleo
gene
mor
phog
enus
Ste
rcul
ia fr
om B
erry
(192
7; 1
938)
is h
ere
inse
rted
in g
roup
5 b
y its
“pla
tano
id” g
ener
al m
orph
olog
y (e
.g. S
. was
hbur
ni B
erry
). B
ut th
e hi
gh
orde
r ven
atio
n ch
arac
ters
in th
e pr
eser
ved
foss
ils a
re m
ore
sim
ilar t
o th
ose
foun
d in
Bra
chyc
hito
n (A
ustra
lia/N
ew G
uine
a), a
nd A
ndro
pogo
n (N
ew C
aled
onia
) gen
era.
The
ext
ant s
peci
es o
f Ste
rcul
ia
have
, in
gene
ral,
sim
ple
or p
alm
ate
com
poun
d le
aves
, whi
ch s
egm
ents
hav
e br
ochi
drod
omou
s or
cam
ptod
rom
ous
seco
ndar
y ve
natio
n. Y
et, t
he fo
ssil
spec
ies
S. s
ehue
nsis
Ber
ry is
a tr
iloba
te le
af.
This
cal
ls th
e at
tent
ion
to th
e ne
ed o
f a c
aref
ul re
visi
on o
f thi
s m
orph
otax
a, ta
king
in a
ccou
nt it
s ro
le in
the
Cre
tace
ous fl o
ras
and
the
new
phy
loge
netic
stu
dies
.
MO
RFO
TIPO
S FO
LIA
RES
(m
orph
otyp
es)/
A
FIN
IDA
DES
(aff.
)
Nel
son-
65ºS
(est
e tra
balh
o/th
is w
ork)
(Cam
pan.
-Maa
stric
h.?)
PEN
. AN
T
Kin
g G
eorg
e-62
o SA
dmira
lty B
ay(Z
amek
Fm
., C
ampa
nian
?) ty
pes
from
Zas
taw
niak
(1
994)
PEN
.AN
T.
Kin
g G
eorg
e-62
o S(H
alf T
hree
Poi
nt
Fm.)
(Cam
pan.
-Maa
str.)
PEN
. AN
T.
Jam
es R
oss/
Seym
our-6
4º S
Hid
den
Lake
(C
onia
cian
) &
Sant
a M
arta
(C
ampa
nian
) Fm
.ty
pes f
rom
Hay
es
et a
l.(2
006)
PE
N. A
NT.
New
Zea
land
/A
ustr.
45º
S (8
5o S,
1) to
25º
SM
atak
aen
Gr.
(Cam
pani
an)
type
s fro
m P
ole
(199
2)N
.ZEL
./AU
ST
Punt
a Are
nas-
41ºS
Cer
ro T
oro,
Tre
s Pa
sos,
Dor
otea
fm
s.(C
amp.
-Maa
st.)
CH
ILE
Bac
ia A
ustra
l-50º
SM
. Am
arill
a Fo
rm.
(Con
iac.
/San
ton.
)(m
orph
. gro
ups f
rom
Ig
lesi
as e
t al.,
200
7)A
RG
ENTI
NA
1.M
AG
NO
LIO
FÍLI
CO
(m
agno
liaep
hyll
)“m
agno
liid
com
plex
”[M
agno
liale
s/C
anel
lale
s]
RIP
1?
Fico
phyl
lum
skua
ensi
s Dut
ra“M
agno
liaep
hyll
mor
phot
ype”
Mag
nolii
daep
hyllu
m
birk
enm
ajer
i Zas
taw
.D
icot
ylop
hyllu
m sp
.1
Dic
otyl
ophy
llum
sp.1
1 (~
Win
tera
ceae
)
“Mag
nolia
ephy
ll m
orph
otyp
e”
Dic
otyl
ophy
llum
sp
.
Mag
nolia
les
(dom
inan
t. H
.Lak
e Fm
.)La
urip
hyllu
m
nord
ensk
ioel
dii
Dus
énLa
urel
ia in
sula
ris
Dus
én
MAT
A 1
(ins
.sedi
s)M
ATA
5
MAT
A 4
?TA
RA
25?
Laur
ophy
llum
spp.
Laur
elia
sp.
Peum
us sp
.
2.LA
UR
OFÍ
LIC
O (l
auro
phyl
l or c
inam
omop
hyll
from
C
rabt
ree,
198
7)“m
agno
liids
”[L
aura
les]
“f
abid
s”[O
xida
lale
s]“a
ster
ids”
(in
part)
RIP
1?
Roop
hyllu
m n
orde
nskj
oeld
ii D
usén
RIP
2(~
Elae
ocar
pace
ae/
Aqu
ifolia
ceae
/ /M
onim
iace
a w
ith e
ntire
mar
gin)
R
IP 3
“Lau
roph
yll
mor
phot
ype”
~ La
urop
hyllu
m
nord
ensk
jold
ii D
usén
~ N
ecta
ndra
pro
lific
a B
erry
Dic
otyl
ophy
llum
sp.2
D
icot
ylop
hyllu
m sp
.3~
Nec
tand
ra sp
p.
Dic
otyl
ophy
llum
sp.7
D
icot
ylop
hyllu
m sp
.10
(~El
aeoc
arpa
ceae
)
Fico
phyl
lum
sk
uaen
sis D
utra
D
icot
ylop
hyllu
m
sp. 3
(Z
asta
wni
ak, 1
994)
cf
. Ore
opan
ax
guin
azui
Ber
ry
aff.
Laur
elia
spp.
(B
erry
, 193
8)
Fico
phyl
lum
pa
lust
ris C
antri
ll
Myr
ica
nord
ensk
joel
dii
Dus
énC
inna
mom
oide
s sp.
Pr
oteo
phyl
lum
sp.
Mol
lined
ia
seym
oure
nsis
D
usén
Laur
ales
: Mor
phot
. 2,
3,11
and
flow
ers
(Hay
es e
t al.,
200
6)
TAR
A 1
4M
ATA
4(“
Cin
nam
omum
inte
rmed
ium
” Et
t.)
“dic
ot w
ith si
mpl
y se
rrat
ed m
argi
n”C
inna
mon
um sp
. Sc
haef
fcf
. Mon
imia
ceae
Gru
po 1
(2nd
. cou
ld b
e ca
mpt
odro
mou
s)M
A10
0 (~
Peum
us c
lark
i,La
urop
hyllu
m
prot
eafo
lium
&L.
kur
tzi B
erry
(193
7)

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013458
3.AC
RO
DO
MO
FÍLICO
(acrodomophyll) “m
agnollids”[C
anellales, Piperales]“m
alvids”[M
irtales, Melastom
at.] “m
onocots” [Liliaceae]
RIP 4
aff. tribo Miconieae
(Melastom
ataceae)
“Acrodrom
ophyll”D
icotylophyllum sp.5
Miconiiphyllum
austral D
usén (aff. M
iconia)
Bignonites ovatus B
erryM
enispermites sp.
Lesq.M
enispermites sp.
cf. M. piatnitzkyi
Berry
Grupo 2
MA
101 & M
A102
(~Bignonites chalianus B
erry &
Menisperm
ites piatnitzkyi B
erry)
4.MIRTO
FÍLICO
(myrtophilll)
“malvids”
[Mirtales: M
yrtaceae] “rosids”
“Myrtophill”
Myrciophyllum
santacruzansis (B
erry) Zastaw
niak
Myrtophill
cf. Cassia B
erry M
yrtaceae (flowers
and cuticles, Hayes
et al., 2006)Phyllites sp. 6 D
usén (1908)
Grupo 3
MA 103, 104,105
(~Myrcia acutifolia)
Myrtoidea patagonica
Passalia et al.( 2011)5. PEN
TALO
BO
FÍLICO
/AR
ALIA
EFÍLICO
(3)(pentalobophyll/ araliaephyll)“m
agnolids” [Proteales] “m
alvids”(=Palm
ate Dillenidae)]
“asterids”[Araliaceae]
“Pentalobophyll/ A
raliaephyllM
orphotype” (3), D
icotylophyllum sp. 4
Araliaephyllum
sp. aff. A. quinquelobatus C
antrill Sterculiaephyllum
australis D
utra (3)“platanoid m
orphotypes”
Araliaephyllum sp.
Araliopsoides sp. Sassafras sp.TA
RA
22 (Aralia tasm
ani Ett.) TA
RA
23,26,28
Sterculia sehuensis B
erry, (trilobate)Sterculia spp. (3)
Grupo 4
MA
106 (~Sterculia w
ashburni,~Sterculia sehuensis, ~C
issus guidoensis H
ünicken 95), ~C
issites Menéndez
696. PA
LMA
DO
CR
ASPED
ÓD
RO
MO
(palm
ate craspedrosomous)
“fabids”[Proteales]“m
alvids”
only in Early C
retaceousM
orphotype 2 (H
ayes et al., 2006): Sterculiaceae
MATA
3TA
RA
5TA
RA
21, 18TA
RA
16 (2)TA
RA
15 (2)A
ngiosperm sp.
C (M
cLoughlin et al. 1995)
“Dicot w
ith lobed m
argin” C
issites parvifolius M
enéndez
Grupo 5
MA
107 (~TAR
A 15),
MA
108 (~TAR
A 16 &
21 from
Pole, 1992)Thorhallenia dentata Passalia, 2007
7.CU
NO
NIO
FÍLICO
(cunoniophyll)“fabids”[C
unoniaceae]“m
agnollids”,[Proteales,Proteaceae]“m
alvids”[A
nacard/Sapindaceae]M
onimiales
RIP 1?
Roophyllum nordenskjoeldii
Dusén?
(Anacardiaceae?)
RIP 5?
“Cunoniophyll”
Dicotylophyllum
sp. 6“sapindophyll”D
icotylophyllum
elegans Li, 1994 M
onimiophyllum
sp. aff. antarcticum
Zastaw
niak
Knithia andreae
Dusén?
Lomatia
angustiloba Dusén
Caldcluvia
mirabilis D
usénLaurelia am
arillana B
erry
TAR
A 29
Ribes spp.C
upania spp.Ribes extincta (M
enendez, 1969)
Grupo 6
MA
109 to MA
111 m
orphotypes (cunonioid or clorantoid teeths) Laurelia am
arillana B
erry (1928)
8.PRO
TOFÍLIC
O(protophyll) “m
agnoliids”[Proteales: Proteaceae][C
anellales:Winteraceae]
“fabids”[Fagales]
Knithia andreae
Dusén?
Drym
is antarctica (w
eak 2rs.)
Proteophyllum sp.
Winteraceae (H
ayes et al., 2006)
MATA
2 (N
othofagus?)TA
RA
18 (N
. obliqua?)
Schinopsis sp.
9. Gr. N
OTH
OFA
GU
S (2)(protophyll)“fabids”[Fagales:N
othofagaceae]
RIP 5 (~N
. glaucifolia Dutra)
RIP 6?
RIP 7 cf. N
. subferruginea (Li, 1994; Zastaw
niak et al., 86 &N
. ulmifolia from
Pole, 92)R
IP 8(~ N
. zastawniakae D
utra)R
IP 9 (Nothofagus sp.)
“Protophyll m
orphotype”N
othofagus cretaceae Zast.N
othofagus sp.
N. cretacea Zast.
N. zastaw
niakiae D
utraN
. glaucifolia Dutra
Fagus dicksoni D
usénF. obscura D
usénN
. magellaenica
Dus.
N. pulchra D
us.
TAR
A 15
N. m
elanoides Pole (2).TA
RA
16N
. praequercifolia (Ett.) Pole (2)TA
RA
17N
. ulmifolia (Ett.)
Oliver
TAR
A 19
TAR
A 24
Nothofagus ?
Dusén, 1899
(Troncoso &
Rom
ero, 98)M
. Leppe (inf. verbal)
Thorhallenia dentata Passalia 2007? (2)
Tabela 1. Cont.
Table 1. Cont.

459BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
eocretáceas mais antigas da Península (Birkenmajer & Ociepa, 2008) e, dos níveis do Coniaciano da Formação Hidden Lake, ilha James Ross. Faltam aqui também os tipos primitivos do Complexo Fagaleano, identifi cados para o Albiano da ilha Alexander (Cantrill & Nichols, 1996) e elementos comparáveis aos presentes nas fl oras de angiospermas (sem Nothofagus), que caracterizam o Albiano na ilha Livingston (Cantrill & Nagalingum, 2005).
No conjunto e comparativamente, três dos cinco tipos morfológicos que caracterizam o Albiano na Península (Cantrill & Nichols, 1996), já têm arquiteturas de venação comparáveis com as impressões de RIP, e demonstram que angiospermas ligadas às Laurales, Magnoliales, Piperales e Fagales já haviam chegado às altas latitudes do sul. São representados pelos morfotipos craspedódromos, broquidódromos (Ficophyllum palustris Cantrill, similar em morfologia a forma RIP 1), e acródromos trinervados (Timothyia trinervis Cantrill), com venação secundária broquidódroma e semelhante a RIP 4. Já os tipos exclusivos do Albiano de Alexander, são formas pecioladas fl abeladas, de margem crenada e venação actinodrômica (das Saururaceae, Piperaceae e Aristolochiaceae) e palinactinodrômica (folhas palmadas pentalobadas, com 3 a 5 veias decorrentes), representado por Araliaephyllum quinquelobatus Cantrill, de afi nidades lauraleanas.
Entre as tafofl oras do fi nal do Cretáceo conhecidas (Tabela 1), a maior identidade se expressa com aquelas do flanco ocidental da própria Península Antártica (ilha KG), também gerada em um contexto de antearco. Em menor grau, e com uma diversidade comparativamente menor, estão as fl oras da Bacia de James Ross, na região de retroarco (Dettmann & Jarzen, 1990; Truswell, 1990; Askin, 1997; Hayes et al., 2006; Reguero et al., 2013). E, entre as fl oras de KG, a maior similaridade é com aquelas dos pontais Half Three e Price (Cao, 1994; Dutra & Batten, 2000), na Península Fildes, locais exclusivos ainda por conter macro e microfósseis, a presença
de camadas de carvão, e evidências da infl uência marinha. A presença de coníferas Cupressaceae, é contudo, exclusiva do pontal Rip, e só registrada no Eoceno basal de KG. Seguem-se as áreas próximas e de menor paleolatitude, do sul da América e Austrália-Nova Zelândia. Mas falta nas assembleias de Rip e KG, os tipos lobados simples e protofílicos comuns nestas áreas e, em troca, Nothofagus está ausente nestas latitudes mais baixas.
Estas comparações atestam que havia um livre trânsito das fl oras através da província Weddelliana (Zinsmeister, 1979), o conjunto de terra emersas proposto para o fi nal do Cretáceo nas altas latitudes do sul (Hill & Dettmann, 1996; Cantrill et al., 2012; Reguero et al., 2013).
Hayes et al. (2006), utilizando análises fi sionômicas das folhas das formações Hidden Lake (Santoniano), com 30 morfotipos, e Santa Marta (Campaniano), com 32 morfotipos, obtiveram níveis médios de temperatura entre 17ºC e 19ºC, respectivamente, e consideraram que os períodos de chuva eram provavelmente sazonais (embora a ausência também sazonal da luz tenha aí um papel importante). Em termos comparativos, a reduzida diversidade da fl ora do pontal Rip (nove morfotipos), com pteridófi tas arborescentes, angiospermas laurofílicas e o domínio das Nothofagaceae, parece sugerir condições ainda mais frias para às áreas do arco, menos protegidas. Mas é igualmente provável, dado o condicionamento tectônico, que refl ita uma vegetação de caráter pioneiro, similar à que costuma colonizar os ambientes pós-eruptivos. Deste modo, um clima regional de caráter temperado a subtropical, com abundante umidade é sugerido para as áreas da Península ocidental neste intervalo. Um análogo moderno adequado, pela composição de suas fl oras, são as áreas montanhosas da costa chilena da América do Sul, entre 30º e 35ºS de latitude.
AGRADECIMENTOS
Às equipes dos laboratórios de palinologia da UFRGS (Laboratório Marleni Marques Toigo) e da UNISINOS (ITT Fóssil). A A. Sander pela confecção e análise da lâmina petrográfi ca e a D.A. Spiazzi, A.B. Duarte e R. Barboni pelas fotografi as dos macrofósseis. Ao CNPq pela bolsa de mestrado da primeira autora e bolsa de produtividade da segunda autora. Ao PROANTAR-CNPq, pelo suporte durante as expedições Antárticas.
REFERÊNCIAS
Alves, L.S.R.; Guerra-Sommer, M. & Dutra, T.L. 2004. Paleobotany and Paleoclimatology: grothw rings in fossil woods and leaf assemblages. In: E.A.M Koutsoukos (ed.) Applied Stratigraphy, Springer, p. 179-202. doi: 10.1007/1-4020-2763-X_9
Angiosperm Phylogeny Group (AGP). 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of fl owering plants: APG III. Botanical Journal of the Linnean Society, 161:105-121. doi:10.1111/j.1095-8339.2009.00996.x
Askin, R.A. 1997. Eocene–?earliest Oligocene terrestrial palynology of Seymour Island. In: C.A. Ricci (ed.) The Antarctic Region: Geological Evolution and Processes. Terra Antarctica Publication, p. 993-996.
Figura 8. Sessão delgada de uma das amostras com impressões de folha, evidenciando a presença de fragmentos de carvão e de uma testa de foraminífero (círculo). Escala = 0,3 mm.
Figure 8. A thin section from one sample containing leaf impressions, attesting the associate occurrence of charcoal and foraminifers (circle). Scale bar = 0,3 mm.

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013460
Baldoni, A.M.; Dibbern, M.& Genini, A. 1985. Nota sobre una especie de Blechnum (Blechnaceae-Pteridophyta) e impronta de Fagus subferruginea Dusen en el Terciario de la Provincia de Chubut, Argentina. Ameghiniana, 22:229-232.
Baumgratz, J.F.A. 2004. Synopsis of Huberia D.C. (Melastomataceae: Merianieae). Revista Brasileira de Botânica, 27:545-561.
Baumgratz, J.F.A. et al. 2013. Melastomataceae. Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. Disponível em http://fl oradobrasil.jbrj.gov.br/jabot/fl oradobrasil/FB161; acessado em 11/12/2013.
Berry, E.W. 1925.Fossil plants from the Tertiary of Patagonia and their signifi cance. Proceedings of the National Academy of Sciences, 11:404-405.
Berry, E.W. 1932. Fossil Plants from Chubut Territory collected by the Scarritt Patagonian Expedition. American Museum Novitates, 536:1-10.
Berry, E.W. 1938. Tertiary fl ora from the Rio Pichileufú, Argentina. Geological Society of America Special Papers, 12:1-149.
Birkenmajer, K. 1980. Tertiary volcanic-sedimentary succession at Admiralty Bay, King George Island (South Shetland Islands, Antarctica). Studia Geologica Polonica, 64:7-65.
Birkenmajer, K. 1981. Lithostratigraphy of the Point Hennequin Group (Miocene vulcanics and sediments) at King George Islands, Antarctica. Studia Geologica Polonica, 72:59-73.
Birkenmajer, K. 1985. Oligocene-Miocene glacio-marine sequences of King George Island (South Shetland Islands), Antarctica. Palaeontologia Polonica, 49:9-36.
Birkenmajer, K. 2001. Mesozoic and Cenozoic stratigraphic units in parts of the South Shetland Islands and northern Antarctic Peninsula. Studia Geologica Polonica, 188 p. (Geological Results of the Polish Antarctic Expeditions, 118).
Birkenmajer, K. & Ociepa, A.M. 2008. Plant-bearing Jurassic strata at Hope Bay, Antarctic Peninsula (West Antarctica): geology and fossil-plant description. Studia Geologica Polonica, 128:5-96.
Birkenmajer, K. & Zastawniak, E. 1989. Late Cretaceous-Early Tertiary fl oras of King George Island, West Antarctic: their stratigraphic distribution and palaeoclimatic signifi cance. In: J.A. Crame (ed.) Origins and Evolution of the Antarctic Biota. The Geological Society, p. 227-240 (Special Publication 47).
Birkenmajer, K.; Delitala, M.C.; Narebski, W.; Nicoletti, M. & Petrucciani, C. 1986. Geochronology and migration of Cretaceous through Tertiary plutonic centers, South Shetland Islands (West Antarctica): subduction and hot spot magmatism. Bulletin of the Polish Academy of Sciences, Earth Sciences, 34:243-255.
Birkenmajer, K.; Gaździcki, A.;Krajewski, K.P.;Przybycin, A.; Solecki, A.; Tatur, A. & Ho Il Yoon, 2005. First Cenozoic Glaciers in West Antarctica. Polish Polar Research, 26:3-12.
Bose, M.N.; Taylor, E.L. & Taylor, T.N. 1990. Gondwana Floras of India and Antarctica. A survey and appraisal. In: T.N. Taylor & E.L. Taylor (eds.) Antarctic Paleobiology, Springer Verlag, p. 118-148.
Brownsey, P.J. & Smith-Dodsworth, J.C. 2000. New Zealand ferns and allied plants. 2nd ed. Albany, David Bateman Ltd., 168 p.
Burnham, R.J. & Johnson, K.R. 2004. South American paleobotany and the origins of neotropical rainforests. Philosophical Transactions of the Royal Society of London B, 359:1595-1610. doi: 10.1098/rstb.2004.1531
Burnham, R.J. & Spicer, R.A. 1986. Forest Litter Preserved by volcanic activity at El Chichón, Mexico: a potentially
accurate record of the Pre-Eruption Vegetation. Palaios, 1:158-161. doi: 10.2307/3514509
Cantrill, D.J. & Nagalingum, N.S. 2005. Ferns from the Cretaceous of Alexander Island, Antarctica: implications for Cretaceous phytogeography of the Southern Hemisphere. Review of Palaeobotany and Palynology, 137:83-103. doi:10.1016/j.revpalbo.2005.08.004
Cantrill, D.J. & Nichols, G.J. 1996.Taxonomy and paleoecology of Early Cretaceous (Late Albian) angiosperm leaves from Alexander Island, Antarctica. Review of Palaeobotany and Palynology, 92:1-28. doi:10.1016/0034-6667(95)00105-0
Cantrill, D.J. & Poole, I. 2012. The vegetation of Antarctica through geological time. New York, Cambridge University Press, 480 p.
Cao, L. 1994. Late Cretaceous palynofl ora in King George Island of Antarctic with references to its paleoclimatic signifi cance.In: Shen Yanbin (ed.) Stratigraphy and Palaeontology of Fildes Peninsula, King George Island, Antarctic, Science Press, p. 51-83 (Monograph 3).
Cogniaux, A. 1891. Melastomataceae. In: A. de Candolle & C. de Candolle (eds.) Monographiae Phanerogamarum 7, G. Masson, p. 1-1256.
Coode, M.J.E. 1983. A conspectus of Sloanea (Elaeocarpaceae) in the Old World. Kew Bulletin, 38:347-428.
Crabtree, D.R. 1987. Angiosperms of the Northern Rocky Mountains: Albian to Campanian (Cretaceous) megafossil fl oras. Annals of Missouri Botanical Garden, 74:707-747. doi:10.2307/2399448
Czajkowski, S. & Rösler O. 1986. Plantas fósseis da Península Fildes, Ilha King George (Shetland do Sul): morfografi a das impressões foliares. Anais da Academia Brasileira de Ciências, 58:99-100.
Dettmann, M.E. & Jarzen, D.M. 1990. The Antarctic/Australian rift valley: Late Cretaceous cradle of northeastern Australasian relicts? Review of Palaeobotany and Palynology, 65:131-144.doi:10.1016/0034-6667(90)90064-P
Dettmann, M.E.; Pocknall, D.T.; Romero, E.J. & Zamaloa, M.C. 1990. Nothofagidites Erdtman ex Potonie, 1960; a catalogue of species with notes on the palaeogeographic distribution of Nothofagus Bl. (Southern Beech). Paleontological Bulletin of the New Zealand Geological Survey, 60:1-79. doi:10.1016/0034-6667(92)90024-B
Doktor, M.; Gazdzicki, A.; Jerzmanska, A.; Porebski, S.J. & Zastawniak, E. 1996. A plant-and-Fish assemblage from the Eocene La Meseta Formation of Seymour Island (Antarctic Peninsula) and its environmental implications. Palaeontologia Polonica, 55:127-146.
Dolding, P.J.D. 1992. Palynology of the Marambio Group (Upper Cretaceous) of northern Humps Island. Antarctic Science, 4:311-326. doi:10.1017/S0954102092000476
Duarte, L. 1956. Melastomataceae fósseis da bacia terciária de Fonseca, Minas Gerais. Rio de Janeiro, Departamento Nacional de Produção Mineral, p. 7-32 (Boletim 161).
Dusén, P. 1899. Über die tertiäre Flora der Magellans-länder. Wissennschaftliche Ergebnisse der Schwedischen Expedition nach den Magellandändern 1895-1897, 1:87-107.
Dusén, P. 1907. Über dir tertiäre flora der Magellansländer. Wissennschaftliche Ergebnisse der Schwedischen Expedition nach den Magellandändern 1895-1897, 1:241-248.
Dusén, P. 1908. Über die tertiäre Flora der Seymour-Insel. Wissenschaftliche Ergebnisse der Schwedischen Sudpolar-Expedition 1901-1903. Lithographisches Institut des Generalstabs, 1-3:1-27.

461BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
Dutra, T.L. 1997a. Composição e história da vegetação do Cretáceo e Terciário da ilha Rei George, Península Antártica. Curso de Pós Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, Tese de Doutoramento, 488 p.
Dutra, T.L. 1997b. Nothofagus leaf architecture, an old design to a new Gondwana: the use of the modern subtropical and temperate foliar character of the genus in paleoecology. In: SOUTHERN CONNECTION CONGRESS, 2, 1997. Noticeros de Biologia, Valdivia e Santiago, Universidad de Chile, p. 148-149.
Dutra, T.L. 2000. Nothofagus no norte da Península Antártica (ilha King George, ilhas Shetland do Sul). II. Paleoceno Superior-Eoceno Inferior. Revista da Universidade Guarulhos, Geociências, 5:131-136.
Dutra, T.L. 2001. Paleofl ora da ilha 25 de Mayo, Península Antártica: contribuição à paleogeografi a, paleoclima e para a evolução de Nothofagus. Asociación Paleontologica Argentina, p. 29-37 (Publicación Especial 8).
Dutra, T.L. 2004. Paleofl oras da Antártica e sua relação com os eventos tectônicos e paleoclimáticos nas altas latitudes do sul. Revista Brasileira de Geociências, 34:401-410.
Dutra, T.L. & Batten, D.J. 2000. Upper Cretaceous fl oras of King George Island, West Antarctica, and their palaeoenvironmental and phytogeographic implications. Cretaceous Research, 21:181-209. doi:10.1006/cres.2000.0221
Dutra, T.L.; Leipnitz, B.; Faccini, U.F. & Lindenmayer, Z. 1996. A non marine Upper Cretaceous interval in West Antarctica (King George Island, Northern Antarctic Peninsula). SAMC (South Atlantic Mesozoic Correlations) News, 5:21-22.
Dutra, T.L.; Hansen, M.A.F. & Fleck, A. 1998. New evidences of wet and mild climates in Northern Antarctic Peninsula at the end of Cretaceous. In: ANNUAL CONFERENCE OF IGCP PROJECT 381, 3, 1998. Expanded Abstracts, Buenos Aires, p. 10-13.
Eckenwalder, J.E. 2009. Conifers of the world: the complete reference. Portland, Timber Press, 720 p.
Ellis, B.; Daly D.C.; Hickey L.J.; Johnson K.R.; Mitchell J.D.; Wilf P. &Wing S.L. 2009. Manual of Leaf Architecture. New York, Cornell University Press, 190 p.
Farjon, A. 2005. A monograph of Cupressaceae and Sciadopitys. United Kingdom, Royal Botanic Gardens, 643 p.
Fisch, F. 2009. Taxonomia e contexto geológico da tafofl ora da ilha Dufayel, ilha King George, Península Antártica. Programa de Pós-Graduação em Geologia, Universidade do Vale do Rio dos Sinos, Dissertação de Mestrado, 140 p.
Fontoura, M.A.; Fensterseifer, H.C. & Troian, F.L. 1988. Geology and petrography of intrusive bodies of Stansbury Peninsula, Nelson Island, South Shetland Islands, Antarctica. Serie Científi ca INACH, 38:45-58.
Frenguelli , J. 1953. La Flora fósil de la región del Alto Río Chalía en Santa Cruz (Patagonia). Notas del Museo de la Plata, 16:239-257.
Griffi ths, C.J. & Oglethorpe, R.D.J. 1998. The stratigraphy and geochronology of Adelaide Island. Antarctic Science, 10:462-475. doi:10.1017/S095410209800056X
Gutiérrez, N.M.; Hinojosa, L.F.; Le Roux, J.P. & Pedroza, V. 2013. Evidence for an Early-Middle Miocene age of the Navidad Formation (central Chile): paleontological, paleoclimatic and tectonic implications. Andean Geology, 40:66-78. doi:10.5027/andgeoV40n1-a03
Harris, T.M. 1961. The Yorkshire Jurassic floraI. Tallophyta, Pteridophyta. London, British Museum (Natural History), 212 p.
Hawkes, D.D. 1961.The Geology of the South Shetland Islands. I. The petrology of the King George Island. Science Reports of the Falkland Island Dependencies, 26:1-28.
Hayes, P.A.; Francis, J.E.; Cantrill, D.J. & Crame, J.A. 2006. Palaeoclimate analysis of Late Cretaceous angiosperm leaf floras, James Ross Island, Antarctica. In: J.E. Francis; D. Pirrie & J.A. Crame (eds.) Cretaceous-Tertiary high-latitude palaeoenvironments, James Ross Basin, Antarctica. Geological Society of London, p. 49-62 (Special Publications 258).
Heads, M. 2006. Panbiogeography of Nothofagus (Nothofagaceae): analysis of the main species massings. Journal of Biogeography, 33:1066-1075. doi:10.1111/j.1365-2699.2006.01479.x
Hickey, L.J. 1973.Classifi cation of the architecture of dicotyledoneous leaves. American Journal of Botany, 60:17-33.
Hickey, L.J. 1984.Changes in the angiosperm fl ora across the Cretaceous-Tertiary boundary. In: W.A. Berggren & J. Van Couvering (eds.) Cathastrophes and Earth History - The New Uniformitarism, Princeton University Press, p. 279- 314.
Hickey, L.J. & Wolfe, J.A. 1975. The bases of angiosperm phylogeny: vegetative morphology. Annals of the Missouri Botanical Garden, 62:538-589. doi: 10.2307/2395267
Hill, R.S. 1994. The history of selected Australian taxa.In: R.S. Hill (ed.) History of the Australian Vegetation: Cretaceous to Recent. Cambridge University Press, p. 390-420.
Hill, R.S. & Dettmann, M.E. 1996. Origin and diversifi cation of the genus Nothofagus. In: T.T. Veblen; R.S. Hill & J. Read (eds.) The Ecology and Biogeography of Nothofagus Forests, New Haven, p. 11-24.
Hill, R. & Jordan, G.J. 1993. The evolutionary history of Nothofagus, Nothofagaceae. Australian Systematic Botanical, 6:111-126.
Hill, R. 2001. The Cenozoic macrofossil record of the Cupressaceae in the Southern Hemisphere. Acta Palaeobotanica, 41:123-132.
Hill, R.S. & Read, J. 1991.A revised infrageneric classifi cation of Nothofagus (Fagaceae). Botanical Journal of the Linnean Society, 105:37-72. doi:10.1111/j.1095-8339.1991.tb00199.x
Hinojosa, L.F.; Pesce1, O.; Yabe, A.; Uemura, K. & Nishida, H. 2006. Physiognomical analysis and paleoclimate of the Ligorio Márquez fossil fl ora, Ligorio Márquez Formation, 46o 45´S, Chile. In: H. Nishida (ed.) Post-Cretaceous fl oristic changes in Southern Patagonia, Chile, Chuo University, p. 45-55.
Hünicken, M. 1966. Flora Terciaria de los estratos Rio Turbio, Santa Cruz (niveles plantíferos del Arroyo Santa Flavia). Revista de la Facultad de Ciencias Exactas, Fisicas y Naturales, Serie Ciencias Naturales, 56:139-260.
Hünicken, M.A. 1995. Floras cretácicas y terciarias. In: P.N. Stipanicic & M.A. Hünicken (eds.) Revisión y actualización de la obra paleobotánica de Kurtz en la República Argentina, Atlas de la Academia Nacional de Ciencias de Córdoba, p. 199-219 (Número 10).
Iglesias, A.; Zamuner, A.B.; Poiré, D.G. & Larriestra, F. 2007. Diversity, taphonomy and palaeoecology of an angiosperm fl ora from the Cretaceous (Cenomanian-Coniacian) in Southern Patagonia, Argentina. Paleontology, 50:445-466. doi:10.1111/j.1475-4983.2007.00639.x
Jaramillo, C.A. & Dilcher, D.L. 2001. Middle Paleogene palynology of central Colombia, South America: a study of pollen and spores from tropical latitudes. Palaeontographica Abt B, 258:87-213.
Jones, T.P. & Rowe, N.P. 1999. Fossil plants and spores: modern techniques. Londres, The Geological Society, p. 71-75.

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013462
Keller, H.A.; Sampaio, D. & Tressens, S.G. 2012.Primer registro de Sloanea (Elaeocarpaceae) para la Argentina. Darwiniana, 50:154-156.
Lawver, L.A.; Dalziel, I.W.D.; Norton, I.O. & Gahagan, L.M. 2009. The PLATES 2009 Atlas of Plate Reconstructions (750 Ma to Present Day), PLATES Progress Report No. 325-0509 . University of Texas, 157 p. (Technical Report 196).
Leppe, M.; Mihoc, M.; Varela, N.; Stinnesbeck, W.; Mansilla, H.; Bierma, H.; Cisterna, K.; Frey, E. & Jujihara, T. 2011. Evolution of the Austral-Antarctic fl ora during the Cretaceous: new insights from a paleobiogeographic perspective. Revista Chilena de Historia Natural, 85:369-392.
Leroy, S.A.G. & Roiron, P. 1996. Latest Pliocene pollen and leaf fl oras from Bernasso palaeolake (Escandorgue Massif, Hérault, France). Review of Palaeobotany and Palynology, 94:295-328. doi:10.1016/S0034-6667(96)00016-4
Li Haonai. 1994. Early Tertiary fossil fl ora from Fildes Peninsula of King George Island, Antarctica. In: Y. Shen (ed.) Stratigraphy and Palaeontology of Fildes Peninsula, King George Island, Antarctica, Science Press, p. 133-171 (Monograph 3).
Lima, R. L. & Salard-Cheboldaeff, M. 1981. Palynologie des Bassins de Gandarela et Fonseca (Eocene de l’etat de Minas Gerais, Bresil). Boletin do Instituto de Geociências da USP, 12:33-54.
Li, Z.; Lui, X.; Zheng, X.; Jin, Q. & Li, G. 1989.Tertiary volcanism and formation of volcanic rocks in the Fildes Peninsula, King George Island, Antartic. In: K. Guo (ed.) Proceedings of the International Symposium on Antarctic Research, Ocean, p. 114-118.
Mao, K.; Milne, R.I.; Libing Zhangd; Yanling Penga; Jianquan Liua; Thomas, P.; Mill, R.R. & Renner, S.S. 2012. Distribution of living Cupressaceae refl ects the breakup of Pangea. Proceedings of the National Academy of Sciences of the United States of America. Early Edition, 1-6. doi:10.1073/pnas.1114319109
Margalef, R. 1968. Perspectives in ecological theory. Chicago, University of Chicago, 111 p.
McLoughlin, S.; Drinnan, A.N. & Rozefelds, A.C. 1995. The Cenomanian flora of the Winton Formation, Eromanga Basin, Queensland, Australia. Memoirs of the Queensland Museum, 38:273-313.
Michelangeli, F.A.; Penneys, D.S.; Giza, J.; Soltis, D.; Hills, M.H.& Skean, J.D. Jr. 2004. A preliminary phylogeny of the tribe Miconieae (Melastomataceae) based on nrITS sequence data and its implications on infl orescence position. Taxon, 53:279-290. doi:10.2307/4135608
Menéndez, C.A. 1969: Die fossilen fl oren Südamerikas. In: E.J. Fittkau; J. Illies; H. Klinge; G.H. Schwabe & H. Siol (eds.) Biogeography and Ecology in South America, The Hague, p. 519-558.
MingLi Z. 2011. A cladistic scenario of Southern Pacific biogeographical history based on Nothofagus dispersal and vicariance analysis. Journal of Arid Land, 3:104-113. doi: 10.3724/SP.J.1227.2011.00104
Morley, R.J. & Dick, C.W. 2003. Missing fossils, molecular clocks, and the origin of the Melastomataceae. American Journal of Botany, 90:1638-1644. doi:10.3732/ajb.90.11.1638
Oliver, W.R.B. 1936. The Tertiary fl ora of the Kaikorai Valley, Otago, New Zealand. Transactions of the Royal Society of New Zealand, p. 284-304.
Ollivier, V. et al. 2010. Quaternary volcano-lacustrine patterns and palaeobotanical data in southern Armenia.
Quaternary International, 223-224:312-326. doi:10.1016/j.quaint.2010.02.008
Pankhurst, R.J. 1982. Rb-Sr geochronology of Graham Land, Antarctica. Journal of the Geological Society, 139:701-711. doi:10.1144/gsjgs.139.6.0701
Passalia, M.G.; Romero, E.J. & Panza, J.L. 2001. Improntas foliares del Cretácico de la Provincia de Santa Cruz, Argentina. Ameghiniana, 38:73-84.
Philipson, W.R. & Philipson, M.N. 1988. A classifi cation of the genus Nothofagus (Fagaceae). Botanical Journal of the Linnean Society, 98:27-36.
Pole, M. 1992. Cretaceous macrofl oras of eastern Otago, New Zealand: angiosperms. Australian Journal of Botany, 40:169-206.
Pole, M.S. 1993a. Early Miocene flora of the Manuherikia Group, New Zealand. 5. Smilacaceae, Polygonaceae, and Elaeocarpaceae. Journal of the Royal Society of New Zealand, 23:289-302.
Pole, M.S. 1993b. Early Miocene fl ora of the Manuherikia Group, New Zealand. 8. Nothofagus. Journal of the Royal Society of New Zealand, 23:329-344.
Pole, M.S. 1994. Deciduous Nothofagus leaves from the Miocene of Cornish Head, New Zealand. Alcheringa, 18:79-83. doi:10.1080/03115518.1994.9638766
Pole, M.S. 2007. Conifer and cycad distribution in the Miocene of southern New Zealand.Australian Journal of Botany, 55:143-164. doi:10.1071/BT06056
Pole, M.S.; Hill, B.D. & Harwood, D.M. 2000. Eocene plant macrofossils from erratics, McMurdo Sound, Antartica. In: J.D. Stilwell & R.M. Feldmann (eds.) Paleobiology and Paleoenvironments of Eocene Rocks, McMurdo Sound, East Antarctica, Antarctic Research Series, 76:243-251. doi: 10.1029/AR076
Quinn, C.J. & Price, R.A. 2003. Phylogeny of the Southern Hemisphere Conifers. Acta Horticulturae, 615:129-136.
Raine, J.I.; Mildenhall, D.C. & Kennedy, E.M. 2011. New Zealand fossil spores and pollen: an illustrated catalogue. 4th ed., GNS Science Miscellaneous Series. Disponível online em http://data.gns.cri.nz/sporepollen/index.htm; acessado em 25/03/2013.
Reguero, M.; Goin, F.J.; Hospitaleche, C.A.; Dutra, T. & Marenssi, S.A. 2013. Late Cretaceous/Paleogene West Antarctica terrestrial biota and its intercontinental affi nities. London, SpringerBriefs in Earth System Sciences, 120 p. doi 10.1007/978-94-007-5491-1
Remmert, H. 1988. Autoecologia, Ecologia de Populações e Estudo de Ecossistemas. Barcelona, Editorial Blume, 304 p.
Renner, S.S.; Clausing, G. & Meyer, K. 2001. Historical biogeography of Melastomataceae: the roles of tertiary migration and long-distance dispersal. American Journal of Botany, 88:1290-1300. doi: 10.2307/3558340
Reynel, C. & Honorio, E. 2004. Diversidad y composición de la fl ora arbórea en un área de Ladera de Bosque Montano: Pichita, Valle de Chanchamayo, 2000-2500 msnm. In: D. Antón & C. Reynel (eds.) Relictos de bosques de excepcional diversidad en los Andes Centrales del Perú, Herbario de la Facultad de Ciencias Forestales de la Universidad Nacional Agraria La Molina, p. 45-98.
Romero, E.J. 1973. Polen fósil de Nothofagus (Nothofagidites) del Cretácico y Paleoceno de Patagonia. Revista del Museo de la Plata, 7:291-303.

463BASTOS ET AL. – FLORA DO CRETÁCEO DA ILHA NELSON, ANTÁRTICA
Romero, E.J. 1980. Arquitectura foliar de las especies sudamericanas de Nothofagus Blume. Boletín da la Sociedad Argentina de Botanica, 19:289-308.
Romero, E.J. & Dibbern M.C. 1985. A review of the species described as Fagus and Nothofagus by Dusen. Palaeontographica Abt B, 197:123-137.
Silveira, D.S. 2009. Revisão taxonômica das espécies neotropicais extra-amazônicas de Sloanea L. (Elaeocarpaceae) na América do Sul. Programa de Pós-Graduação em Biologia Vegetal, Universidade de Campinas, Tese de Doutoramento, 168 p.
Sauquet, H. et al. 2012. Testing the impact of calibration on molecular divergence times using a fossil-rich group: the case of Nothofagus (Fagales). Systematic Biology, 61:289-313. doi:10.1093/sysbio/syr116
Scriven, L.J.; McLoughlin, S. & Hill, R.S. 1995. Nothofagus plicata (Nothofagaceae), a new deciduos Eocene macrofossil species, from southern continental Australia. Review of Palaeobotany and Palynology, 86:199-209. doi: 10.1016/0034-6667(94)00145-A
Shen Yanbin. 1994. Cretaceous to Paleogene biostratigraphy in Antarctic peninsula and its signifi cance in the reconstruction of Gondwanaland. In: Y. Yanbin (ed.) Stratigraphy and Palaeontology of Fildes Peninsula, King George Island, Antarctica, Science Press, p. 329-348 (Monograph 3).
Smellie, J.J. 1981. A complete arc-trench system recognized in Gondwana sequences of the Antarctic Peninsula region. Geological Magazine, 118:139-159.
Smellie, J.L.; Pankhurst, R.J.; Thomson, M.R.A. & Davies, R.E.S. 1984.The geology of the South Shetland Islands: VI. Stratigraphy, geochemistry and evolution. British Antarctic Survey Scientifi c Reports, 87:1-85.
Smith, A.R.; Pryer, K.M.; Schuettpelz, E.; Korall, P.; Schneider, H. & Wolf, P.G. 2006. A classifi cation of extant ferns. Taxon, 3:705-731. doi:10.2307/25065646
Srivastava, S.K. 1976. The fossil pollen genus Classopollis. Lethaia, 9:437-457.
Steart, D.C.; Greenwood, D.R. & Boon, P.I. 2005. Paleoecological implications of differential biomass and litter production in canopy trees in Australian Nothofagus and Eucalyptus Forests. Palaios, 20:452-462. doi:0883-1351/05/0020-0452
Stewart, W.N. & Rothwell, G.W. 1993. Paleobotany and the evolution of plants. 2nded., Cambridge, Cambridge University Press, 535 p.
Stover, L.E. & Partridge, A.D. 1973. Tertiary and late Cretaceous spores and pollen from the Gippsland Basin, southeastern Australia. Proceedings of the Royal Society of Victoria, 85:237-286.
Swenson, U.; Hill, R.S. & McLoughlin, S. 2001. Biogeography of Nothofagus supports the sequence of Gondwana Break-up. Taxon, 50:1025-1041. doi:10.2307/i252380
Tanai, T. 1986. Phytogeography and phylogenetic history of the genus Nothofagus Bl. (Fagaceae) in the Southern Hemisphere. Journal of Faculty Science Hokkaido University, 21:505-582.
Thomson, M.R.A. & Burn, W. 1977. Angiosperm fossils from latitude 70” S. Nature, 269:139-141. doi:10.1038/269139a0
Thomson, M.R.A.; Pankhurst, R.J. & Clarkson, P.D. 1983. The Antarctic Peninsula - a Late Mesozoic-Cenozoic arc (review). In: R.L. Oliver; P.R. James & J.B. Jago (eds.) Antarctic Earth Sciences, Australian Academy of Sciences, p. 289-294.
Tokarski, A.K. 1991. The Late Cretaceous-Cenozoic structural history of King George Island, South Shetland Islands, and its
plate-tectonic setting. In: M.R.A. Thomson; J.A. Crame & J.W. Thomson (eds.) Geological Evolution of Antarctica. Cambridge University Press, p. 493-498.
Tokarski, A.K.; Danowski, W. & Zastawniak, E. 1987. On the age of fossil fl ora from Barton Peninsula, King George Island, West Antarctica. Polish Polar Research, 8:293-302.
Torres, T.; Méon, H.; Otero, R. & Galleguillos, H. 2009. Palinofl ora y macrofl ora de la Formación Loreto, Punta Arenas, Región de Magallanes, Chile. In: CONGRESO GEOLÓGICO CHILENO, 12, 2009. Resumen expandido, Santiago, p. 1-4.
Trevisan, C. 2011. Fósseis de pteridófi tas das ilhas King George e Nelson: afi nidade taxonômica e importância no contexto deposicional e do clima nas áreas setentrionais da Península Antártica. Programa de Pós-Graduação em Geologia, Universidade do Vale do Rio dos Sinos, Dissertação de Mestrado, 44 p.
Troncoso, A. 1986. Nuevas órganos-especies en la tafofl ora terciaria inferior de Peninsula Fildes, Isla Rey Jorge, Antártica. Série Científi ca del Instituto Antártico Chileno, 34:23-46.
Troncoso, A. 1992. La tafofl ora terciaria de Quinamavida (VII Región, Chile). Boletin del Museo Nacional de Historia Natural, 43:155-178.
Troncoso, A. & Romero, E.J., 1998. Evolución de las comunidades fl orísticas en el extremo sur de sudamérica durante el cenofítico. In: CONGRESO LATINOAMERICANO DE BOTÁNICA, 6, 1998. Proceedings, Mar del Plata, p. 149-172.
Truswell, E.M. 1990. Cretaceous and Tertiary vegetation of Antarctica: a palynological perspective. In: T.N. Taylor & E.L. Taylor (eds.) Antartic Paleobiology, its role in the reconstruction of Gondwana, Springer-Verlag, p. 71-88.
Tryon, R.M. & Tryon, A.F. 1982. Ferns and allied plants, with special reference to tropical America. New York, Springer Verlag, 857 p.
Upchurch, G.R. & Askin, A.R. 1989. Latest Cretaceous and earliest Tertiary dispersed plant cuticles from Seymour Island. Antarctic Journal of the United States, 24:7-10.
van Steenis, C.G.G.J. 1972. Nothofagus, key genus of plant geography. In: D.H. Valentine (ed.) Taxonomy, Phytogeography and Evolution, Academic Press, p. 275-288.
Veblen, T.T.; Hill, R.S. & Read, J. 1996. The Ecology and Biogeography of Nothofagus Forests. New Haven, Yale University Press, 403 p.
Volkheimer, W.; Scafati, L. & Melendi, D.L. 2007. Palynology of a Danian warm climatic wetland in Central Northern Patagonia, Argentina. Revista Española de Micropaleontología, 39:117-134.
Wilf, P.; Little, S.A.; Iglesias A.; Zamaloa, M.C.; Gandolfo, M.A.; Cúneo, N.R. & Johnson, K.R. 2009. Papuacedrus (Cupressaceae) in Eocene Patagonia, a new fossil link to Australasian rainforests. American Journal of Botany, 96:2031-2047. doi:10.3732/ajb.0900085
Yabe, A.; Uemura, K.; Nishida, H. & Yamada, T. 2006. Geological notes on plant megafossil localitiesat Cerro Guido, Ultima Esperanza, Magallanes (XII) region, Chile. In: H. Nishida (ed.) Post-Cretaceous Floristic Changes in Southern Patagonia, Chile, Chuo University, p. 5-10.
Zastawniak, E. 1981.Tertiary leaf fl ora from the Point Hennequin Group of King George Island (South Shetland Island, Antarctica). Preliminary Report. Studia Geologica Polonica, 72:97-108.

REVISTA BRASILEIRA DE PALEONTOLOGIA, 16(3), 2013464
Zastawniak, E. 1994. Upper Cretaceous leaf flora from the Blaszyk Moraine (Zamek Formation), King George Island, South Shetland Islands, West Antarctica. Acta Paleobotanica, 34:119-163.
Zastawniak, E.; Wrona, R.; Gazdzicki, A. & Birkenmajer, K. 1985. Plant remains from the top part of the Point Hennequin Group (Upper Oligocene), King George Island (South Shetland Islands, Antarctica). Studia Geologica Polonica, 81:143-164.
Zinsmeister, W.J. 1979. Biogeographic signifi cance of the Late Mesosoic and Early Tertiary molluscan faunas of Seymour Island (Antarctic Peninsula) to the fi nal breakup of Gondwanaland.
In: J. Gray & A.J. Boucot (eds.) Historical Biogeography, Plate Tectonics, and the Changing Environment, Oregon State University, p. 349-355.
Zhou Zhiyan & Li Haomin. 1994. Early tertiary gimnosperms from Fildes peninsula, King George island, Antarctica. In: Shen Yanbin (ed.) Stratigraphy and palaeontology of Fildes Peninsula, King George Island, Antarctica, Science Press, p. 208-219 (Monograph 3).
Received in July, 2012; accepted in August, 2013.


















