
UFRRJ INSTITUTO DE VETERINÁRIA CURSO DE PÓS-GRADUAÇÃO EM...
69
UFRRJ INSTITUTO DE VETERINÁRIA CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS TESE ANÁLISES GENOTÍPICA E FILOGENÉTICA DO GENE 18S RNAr DE ISOLADOS BRASILEIROS DE Cytauxzoon felis (APICOMPLEXA: THEILERIIDAE) ALESSANDRA SCOFIELD AMARAL 2006
Transcript of UFRRJ INSTITUTO DE VETERINÁRIA CURSO DE PÓS-GRADUAÇÃO EM...

UFRRJ
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
TESE
ANÁLISES GENOTÍPICA E FILOGENÉTICA DO GENE 18S RNAr
DE ISOLADOS BRASILEIROS DE Cytauxzoon felis
(APICOMPLEXA: THEILERIIDAE)
ALESSANDRA SCOFIELD AMARAL
2006

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
ANÁLISES GENOTÍPICA E FILOGENÉTICA DO GENE 18S RNAr
DE ISOLADOS BRASILEIROS DE Cytauxzoon felis
(APICOMPLEXA: THEILERIIDAE)
ALESSANDRA SCOFIELD AMARAL
Sob a Orientação do Professor Dr. Adivaldo Henrique da Fonseca
Co-orientação do Professor
Dr. Paulo Fernando Vargas Peixoto
e do Dr. Cleber Oliveira Soares
Tese submetida ao Curso de Pós-
Graduação em Ciências Veterinárias,
área de Concentração em Parasitologia
Veterinária como requisito parcial para
obtenção do grau de Doutor em
Ciências.
Seropédica, RJ
Fevereiro de 2006

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
ALESSANDRA SCOFIELD AMARAL
Tese submetida ao curso de Pós-Graduação em Ciências Veterinárias, área de
concentração em Parasitologia Veterinária, como requisito parcial para obtenção do
grau de Doutor em Ciências.
TESE APROVADA EM 22/02/06
__________________________________________
Adivaldo Henrique da Fonseca (Ph.D.)
UFRRJ (Orientador)
___________________________________________
Cleber Oliveira Soares (Ph.D.)
Embrapa Gado de Corte
___________________________________________
Carina Elisei de Oliveira (Ph.D.)
Embrapa Gado de Corte
_________________________________________
Kátia Regina dos Santos Teixeira (Ph.D.)
Embrapa Agrobiologia
__________________________________________
Carlos Luiz Massard (Ph.D.)
UFRRJ

A Deus pelo auxílio e proteção constantes em minha vida.
À minha Mãe Geralda Janeth Scofield Amaral
e Irmãs Vivianne Scofield Amaral
e Carolline Scofield Amaral, pelo amor, carinho,
apoio, estímulo, conselhos e por compreenderem a
minha ausência física em busca dos meus ideais.
Vocês são fundamentais em minha vida!
Ao meu saudosoPai Aroldo Amaral Costa,
pelo amor, carinho e educação.
Amo vocês!

Ao Alessandro Antônio da Silva pelo carinho, paciência, apoio, estímulo e companheirismo.
Aos meus irmãos de espírito Franziska Huber, Jadier C. de Oliveira Júnior, Maria Dalila
Forlano, Marcos Antônio Vargas, Carina Elisei, Daniele Ramos e Astrid Barbosa
pela amizade, carinho, apoio e maravilhosa convivência durante todos esses anos.

AGRADECIMENTOS
Ao Prof. Dr. Adivaldo Henrique da Fonseca, pela amizade, orientação e apoio
durante os dez anos de trabalho em conjunto. Obrigada pela confiança depositada!
Agradeço também à Profa. Marília pela amizade e carinho.
Ao Pesquisador Dr. Cleber Oliveira Soares da Embrapa Gado de Corte, pelo
apoio incondicional, ensinamentos e por me apresentar o mundo fascinante da pesquisa
científica. Obrigada, meu Amigo! Você contribuiu muito para a minha formação!
Ao Prof. Dr. Carlos Luiz Massard, pelos valiosos ensinamentos, amizade e amor
pela parasitologia.
Ao Dr. W. O. Neitz por ser constante fonte de inspiração.
Ao Prof. Dr. Paulo Vargas Peixoto, pelos conselhos, apoio, incentivo e amizade.
Ao Prof. Dr. Carlos Hubinger Tokarnia e à Profa. Dra. Marilene de Farias Brito
Queiroz, pela amizade e importante apoio.
À Pesquisadora Dra. Kátia Regina dos Santos Teixeira da Embrapa
Agrobiologia e a Profa. Dra. Teresa Cristina Bérgamo do Bomfim, pela amizade e
incentivo.
Ao Pesquisador Dr. Flábio Ribeiro Araújo da Embrapa Gado de Corte, pela
amizade e orientação durante o processamento das amostras.
Ao Pesquisador Dr. Cláudio Roberto Madruga da Embrapa Gado de Corte, pela
contribuição durante a execução deste trabalho.
À Professora Dra. Karla S. C. Yotoko da Universidade Federal de Juiz de Fora,
pela amizade e orientação durante as análises filogenéticas.
Aos Pesquisadores Dr. Stênio Perdigão Fragoso e Paulo R. C. Arauco do
Instituto de Biologia Molecular do Paraná, pelo apoio e realização do seqüenciamento
das amostras estudadas.
A todos os Professores, Funcionários e Amigos do Curso de Pós-graduação em
Ciências Veterinárias, pelos ensinamentos, incentivo e amizade.
A todos os Professores e Funcionários do Curso de Graduação em Medicina
Veterinária da Universidade Federal Rural do Rio de Janeiro, o meu muito obrigada!
A todos os Funcionários do Projeto Sanidade Animal-Embrapa, pela amizade e
apoio.
À equipe do Laboratório de Doenças Parasitárias - Projeto Sanidade Animal/
Embrapa: Cátia Marques da Costa, Charles Rangel, Jania Rezende, Fábio Jorge Moreira
da Silva, Genevaldo Barbosa da Silva, Rafaela Câmara Teixeira, Marlone Martins
Coelho, Victor Dantas Whittmann, Fabíola do Nascimento Corrêa, Raquel Lisbôa,
Nathalie Costa da Cunha, Renata Cunha Madureira, Daniel da Silva Guedes Júnior e
Luciana Rodrigues de Almeida pela amizade, importante apoio e por compartilharem de
muitos momentos na realização deste trabalho.
À equipe da área de Sanidade Animal, Embrapa Gado de Corte: Carlos Ramos,
Gisele Bacanelli, Elaine Melo, Renata, Maristeli B. Oliveira e Midori Miguita pelo
apoio e amizade.
A todos os meus familiares, pelo carinho, orações e em especial ao meu tio,
padrinho e amigo Wagner Rachid Scofield, pelo importante apoio durante todos esses
anos. Obrigada por tudo! Amo vocês!
Aos meus queridos cunhados Adriano Mendonça Joaquim e Robério Farias Reis
pela amizade e apoio.

À D. Maria José Gomes Alexandre e Alessandra Maria Gomes da Silva, pelo
apoio e carinho.
À querida Tia Célia Elisei e ao amigo Thiago Elisei, pelo carinho e apoio.
Aos amigos Carlo José Freire de Oliveira e Jaime da Silva Pena, pelo importante
auxílio no início da fase experimental.
Aos animais que tanto contribuíram para a realização deste estudo.
Aos amigos da Universidade Federal do Pará: Valíria Duarte Cerqueira, Gabriela
Riet Correa, Vivina Monteiro e Gustavo Góes Cavalcante, pelo importante apoio nesta
etapa final.
A todos que de alguma forma auxiliaram na realização deste trabalho e me
incentivaram no decorrer desta jornada.
Ao CNPq, Capes, Faperj, Curso de Pós-graduação em Ciências Veterinárias –
UFRRJ e Embrapa Gado de Corte pelo apoio e auxílio financeiro.

BIOGRAFIA
Alessandra Scofield Amaral, filha de Aroldo Amaral Costa e Geralda Janeth
Scofield Amaral, é natural da cidade de Itambacuri, Estado de Minas Gerais (M.G.),
onde cursou o ensino primário na Escola Estadual Dr. Tristão da Cunha e o ensino
fundamental na Escola Estadual Madre Serafina de Jesus. O ensino médio foi cursado
no Colégio Ibituruna da cidade de Governador Valadares – M.G., no período de 1990 a
1993.
No ano de 1995, ingressou no Curso de Medicina Veterinária da Universidade
Federal Rural do Rio de Janeiro (UFRRJ), colando grau e obtendo o título de Médica
Veterinária em 25 de novembro de 2000.
Foi bolsista de Iniciação Científica pelo Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) no período de fevereiro de 1997 a novembro de 2000,
junto a projetos de pesquisa na área de hemoparasitologia. Foi bolsista de
Aperfeiçoamento Científico do CNPq de dezembro de 2000 a fevereiro de 2001 na
mesma linha de pesquisa.
Durante o período acadêmico participou de projetos de pesquisa no
Departamento de Epidemiologia e Saúde Pública e no Departamento de Parasitologia
Animal da UFRRJ. Colaborou no desenvolvimento de projetos de pesquisa em conjunto
com a Universidade de São Paulo, Universidade Federal Fluminense, Ministério da
Agricultura, Pecuária e Abastecimento e com a Embrapa Gado de Corte. Participou de
cerca de 50 publicações científicas entre artigos em periódicos indexados, anais de
congressos e eventos científicos nacionais e internacionais.
Em março de 2001 ingressou no Curso de Pós-graduação em Ciências
Veterinárias – Área de concentração Parasitologia Veterinária da UFRRJ em nível de
Mestrado, onde foi bolsista da Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES) entre março de 2001 a fevereiro de 2002. Foi contemplada, entre
seus pares, como Bolsista de Mestardo do Programa “Bolsa Nota 10” da Fundação de
Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) entre março de 2002 a
outubro de 2002.
Em outubro de 2002 defendeu a dissertação intitulada “Caracterizações
morfológica, morfmétrica e ultraestrutural de formas intraeritrocíticas de Cytauxzoon
felis simile” obtendo o título de Magister Science.
Em junho de 2005 prestou concurso público de provas e títulos na Universidade
Federal do Pará (UFPA) - Campus Castanhal, para o cargo de Profesoor Assistente I da
Disciplina Zoologia/Parasitologia, sendo aprovada em primeiro lugar e contratada em
agosto do mesmo ano.
Em novembro de 2002 ingressou no Curso de Pós-graduação em Ciências
Veterinárias - Parasitologia Veterinária da UFRRJ em nível de Doutorado. E nesta data,
apresenta e defende a presente tese como requisito parcial para a obtenção do grau de
Doutor em Ciências.

SUMÁRIO
Páginas
1. INTRODUÇÃO................................................................................................ 1
2. REVISÃO DE LITERATURA.......................................................................
3
2.1. Breve Histórico e Aspectos Gerais sobre a Citauxzoonose............................
3
2.2. Gênero Cytauxzoon.........................................................................................
3
2.2.1. Citauxzoonose felina...................................................................................
4
2.3. Cytauxzoon felis..............................................................................................
5
2.3.1. Biologia, morfologia e ultraestrutura...........................................................
5
2.3.1.1. Formas intrahistiocíticas............................................................................
5
2.3.1.2. Formas intraeritrocíticas............................................................................
7
2.3.1.3. Gametogonia e esporogonia......................................................................
8
2.3.2. Vetores.........................................................................................................
8
2.3.3. Hospedeiros..................................................................................................
9
2.3.4. Diagnóstico...................................................................................................
12
2.3.4.1. Diagnóstico direto.....................................................................................
12
2.3.4.2. Diagnóstico indireto e exames auxiliares..................................................
13
2.3.4.3. Diagnóstico diferencial..............................................................................
13
2.3.5. Tratamento.................................................................................................... 14
2.3.6. Distribuição.................................................................................................. 14
2.3.7. Potencial zoonótico...................................................................................... 15
2.4. Filogenia e evolução.......................................................................................
15
2.4.1. Posição sistemática e status taxonômico.....................................................
16
2.5. Análises Moleculares...................................................................................... 17
2.5.1. PCR (Polymerase chain reaction)............................................................... 17

2.5.2. RNA ribossômico (RNAr)........................................................................... 18
2.5.3. Análises moleculares e filogenéticas........................................................... 19
3. MATERIAL E MÉTODOS............................................................................ 20
3.1. Origem das Amostras de Cytauxzoon felis simile........................................... 20
3.2. Manutenção do Isolado de Panthera onca em Gatos Domésticos................. 20
3.2.1. Detecção da parasitemia e coleta das amostras do isolado de Panthera
onca........................................................................................................................
20
3.3. Escolha do Gene............................................................................................. 21
3.4. Desenho dos Oligonucleotídeos Iniciadores................................................... 21
3.5. Extração de DNA ...................................................................................... ..... 21
3.6. Amplificação das Amostras de DNA de Cytauxzoon felis simile................... 22
3.7. Análise dos Produtos de Amplificação........................................................... 22
3.8. Clonagem dos Produtos de PCR..................................................................... 23
3.9. Lise e Análise de Colônias............................................................................. 24
3.10. PCR de Colônia............................................................................................ 24
3.11. Reação de Minipreparação de Plasmídeos (Miniprep)................................. 24
3.12. Sequenciamento............................................................................................ 25
3.13. Análise das Seqüências (in silico)................................................................ 25
3.14. Alinhamento e Análise Filogenética............................................................. 26
4. RESULTADOS................................................................................................
29
4.1. Desenho dos Oligonucleotídeos Iniciadores..................................................
29
4.2. Amostras Analisadas e Extração de DNA......................................................
29
4.3. Amplificação das Amostras de DNA de Cytauxzoon felis ............................
30
4.4. Clonagem e PCR de Colônia........................................................................... 32
4.5. Sequenciamento.............................................................................................. 33

4.5.1. Análise de identidade entre as seqüências................................................... 33
4.6. Análise Filogenética........................................................................................ 36
5. DISCUSSÃO.................................................................................................... 38
CONCLUSÕES.................................................................................................... 41
REFERÊNCIAS BIBLIOGRÁFICAS............................................................... 42

ÍNDICE DE TABELAS
Páginas
Tabela 1. Hospedeiros naturais e experimentais e distribuição geográfica da
infecção causada por Cytauxzoon felis....................................................................
11
Tabela 2. Espécies de parasitos, número de acesso no GenBank, hospedeiros e
país de origem usados na construção da árvore filogenética e a análise de
agrupamento das seqüências de 18S RNAr de Cytauxzoon felis............................
27

ÍNDICE DE FIGURAS
Páginas
Figura 1. Representação esquemática da unidade de repetição do DNA dos
eucariontes. A proporção entre as diversas regiões dentro da unidade não está
necessariamente representada na figura. ETS: espaçador externo; 18S região que
codifica para RNAr 18S; ITS: espaçador interno; 25-28S: região que codifica
para RNAr 25-28S; NTS: espaçador externo ou espaçador intergênico (GORAB,
2001 adaptado)..........................................................................................................
19
Figura 2. Esquema do plasmídeo pTrcHis – TOPO (INVITROGEN®) utilizado
na clonagem dos produtos de PCR dos isolados de Cytauxzoon felis de Panthera
onca e Leopardus tigrinus........................................................................................
23
Figura 3. A-B. Esquema dos dois pares de iniciadores (Cfe304F e Cfe1159R e
Cfe843F e Cfe1680R) utilizados na amplificação do gene 18S RNAr de
Cytauxzoon felis e seus respectivos sítios de anelamento; C. Representação dos
sítios de anelamento dos iniciadores na seqüência consenso de Cytauxzoon felis
(AF399930)...............................................................................................................
29
Figura. 4. Trofozoítas de Cytauxzoon felis () em esfregaço sangüíneo de gato
doméstico infectado experimentalmente com isolado de Panthera onca. Giemsa (1000x)......................................................................................................................
30
Figura 5. Eletroforese dos produtos de amplificação do gene 18S RNAr de
Cytauxzoon felis do isolado de Panthera onca, em gel de agarose a 1%. M)
Marcador de pares de base; 1) DNA total de extrato de sangue de um gato
doméstico negativo com iniciadores Cfe843F e Cfe1680R; 2) DNA total de
extrato de sangue do gato-doméstico positivo com iniciadores Cfe843F e
Cfe1680R; 3) DNA total de extrato de sangue de um gato doméstico negativo
com iniciadores Cfe304F e Cfe1159R, e 4) DNA total de extrato de sangue do
gato doméstico positivo com iniciadores Cfe304F e Cfe1159R...............................
31
Figura 6. Eletroforese dos produtos de amplificação do gene 18S RNAr de
Cytauxzoon felis da amostra de Leopardus tigrinus, em gel de agarose a 1%. M)
Marcador de pares de base; 1) DNA total de extrato de sangue de um gato
doméstico negativo com iniciadores Cfe843F e Cfe1680R 2) DNA total de
extrato de sangue de Leopardus tigrinus com iniciadores Cfe843F e Cfe1680R;
3) DNA total de extrato de sangue de um gato doméstico negativo com
iniciadores Cfe304F e Cfe1159R, e 4) DNA total de extrato de sangue do
Leopardus tigrinus com iniciadores Cfe304F e Cfe1159R......................................
32

Figura 7. Eletroforese dos produtos de amplificação da PCR de colônia do gene
18S RNAr de Cytauxzoon felis da amostra de Leopardus tigrinus, em gel de
agarose a 1%. M) Marcador de pares de base; 1) Produto de amplificação da
colônia contendo inserto amplificado com iniciadores Cfe843F e Cfe1680R; 2)
Colônia sem inserto amplificado com iniciadores Cfe304F e Cfe1159R; 3-4)
Produto de amplificação das colônias contendo inserto amplificado com
iniciadores Cfe304F e Cfe1159R..............................................................................
33
Figura 8. Seqüência consenso do gene 18S de Cytauxzoon felis obtida do isolado
de Panthera onca: em azul está marcada a seqüência do iniciador Cfe843F e em
amarelo está a seqüência do iniciador Cfe1159R......................................................
34
Figura 9. Seqüência consenso do gene 18S de Cytauxzoon felis obtida da amostra
de Leopardus tigrinus: em azul está marcada a seqüência do iniciador Cfe843F e
em amarelo a seqüência do iniciador Cfe1159R.......................................................
34
Figura 10. Estruturas secundárias do gene 18S RNAr de espécies de Cytauxzoon.
A) C. felis isolado de Panthera onca (DQ382276), Brasil. B) C. felis de amostra
de Leopardus tigrinus (DQ382277), Brasil. C) C. felis de gato doméstico
(AF399930), Estados Unidos. D) C. manul de gato Pallas (AY485690),
Mongólia...................................................................................................................
35
Figura 11. Árvore filogenética com as duas seqüências de Cytauxzoon felis
obtidas neste trabalho indicadas por setas (). Em rosa estão identificadas as
espécies do gênero Cytauxzoon, em vermelho as espécies do gênero Babesia, em
azul claro as espécies do gênero Theileria, em azul escuro os Piroplasmidas não
identificados e em preto o grupo externo representado pelas espécies Neospora
sp. e Toxoplasma gondii. (*) Espécies que recebem sinonímia de Gonderia e (**)
espécie que recebe sinonímia de Cytauxzoon............................................................
37

RESUMO
AMARAL, Alessandra Scofield. Análises genotípica e filogenética do gene 18S
RNAr de isolados brasileiros de Cytauxzoon felis (Apicomplexa: Theileriidae).
Seropédica: UFRRJ, 2006. 53p. (Tese, Doutorado em Ciências Veterinárias).
O presente trabalho teve como objetivos determinar a espécie de Cytauxzoon que
infectou onça-pintada (Panthera onca) e gato-do-mato (Leopardus tigrinus) do
Zoológico Municipal de Volta Redonda, Estado do Rio de Janeiro, utilizando técnicas
de diagnóstico molecular, comparar as seqüências consenso obtidas neste trabalho com
seqüências de Cytauxzoon, Theileria, Babesia e de Piroplasmida depositadas em bancos
de dados (GenBank), e realizar análises filogenéticas com a espécie estudada. Para
caracterizar os isolados de Cytauxzoon diagnosticados em Panthera onca e Leopardus
tigrinus, foi realizada a reação em cadeia da polimerase (PCR) com dois pares de
iniciadores universais do gene codificador 18S RNAr desenhados a partir da seqüência
de Cytauxzoon felis (n° de acesso no GenBank AF399930). Dois gatos domésticos
foram inoculados experimentalmente com sangue de P. onca infectado com
Cytauxzoon. Amostras de sangue desses animais e de L. tigrinus naturalmente infectado
foram utilizadas para extração de DNA. DNA extraído de sangue de um gato doméstico
não infectado foi utilizado como controle negativo. Os produtos da PCR das duas
amostras foram clonados e seqüenciados. As seqüências consenso foram obtidas com o
auxílio do programa computacional Clustal-W e alinhadas com outras seqüências do
gene 18S RNAr de espécies de Cytauxzoon depositadas no GenBank com o auxílio do
programa computacional Blastn. Posteriormente, algumas seqüências homólogas foram
selecionadas para o alinhamento e análises filogenéticas segundo os programas
computacionais MEGA 3, GeneBee e PAUP. A seqüência consenso do isolado da P.
onca apresentou 947 pares de bases (n° de acesso no GenBank DQ382276) e identidade
de 99% com a espécie C. felis originária dos EUA, 98% com Cytauxzoon sp. e
Cytauxzoon manul originário da Mongólia e com Cytauxzoon sp. diagnosticado em
lince da Espanha, e 97% com Cytauxzoon sp. diagnosticado em gato doméstico da
Espanha, segundo o programa computacional Blastn. A seqüência consenso da amostra
de L. tigrinus apresentou 1204 pares de bases (n° de acesso no GenBank DQ382277) e
identidade de 99% com a espécie C. felis originária dos EUA e de 96% com Cytauxzoon
sp. e C. manul originário da Mongólia, e Cytauxzoon sp. isolado na Espanha. As
análises filogenéticas demonstraram que as seqüências estudadas agruparam com outras
seqüências de Cytauxzoon felis que infecta felídeos nos EUA formando um grupo irmão
das espécies C. manul que infecta gatos Pallas da Mongólia e Cytauxzoon sp. que
infecta lince e gato doméstico da Espanha. Pôde-se observar também o monofiletismo
das espécies do gênero Cytauxzoon e a ausência deste nos grupos de Babesia e
Theileria. Os resultados obtidos neste trabalho confirmam que o agente da
citauxzoonose felina em P. onca e L. tigrinus é C. felis.
Palavras-chave: Cytauxzoon felis 18S RNAr-PCR, sequenciamento, análise filogenética.

ABSTRACT
AMARAL, Alessandra Scofield. Genotypic and phylogenetic analysis of the 18S
rRNA gene to brazilian Cytauxzoon felis (Apicomplexa: Theileriidae) isolates. Seropédica: UFRRJ, 2006. 53p. (Thesis, Doctor in Veterinary Sciences).
The objectives of this work were to determine the species of Cytauxzoon that infected one
jaguar (Panthera onca) and one wild cat (Leopardus tigrinus) of the Volta Redonda
Municipal Zoo, Rio de Janeiro State, using molecular diagnostic techniques; compare the
consensus sequences obtained in this study with the sequences of Cytauxzoon, Theileria,
Babesia and Piroplasmida deposited in data banks (GenBank); and to perform
phylogenetic analysis of the studied species. In order to characterize the species of
Cytauxzoon diagnosticated in Panthera onca e Leopardus tigrinus, polymerase chain
reaction (PCR) assays were done using two pairs of universal primers of the coding gene
18S rRNA designed from Cytauxzoon felis (AF399930) deposited in GenBank. Two
domestic cat were experimentally infected with blood of the Cytauxzoon infected P. onca.
Blood samples of these cats and from the naturally infected L. tigrinus were used for
DNA extraction. DNA extracted from a non infected domestic cat was used as negative
control. The products of the PCR were cloned and sequenced. The consensus sequences
were obtained using the computer program Clustal-W; they were aligned with other
sequences of the 18S rRNA gene of Cytauxzoon species deposited in GenBank using the
computer program Blastn. Afterwards, some homologous sequences were selected for the
alignment and phylogenetic analysis based on the computer programs MEGA3, GeneBee
and PAUP. The consensus sequence of the jaguar isolate showed 947 pairs of basis
(GenBank acess nr. DQ382276); identity of 99% with the species C. felis originated from
the USA, 98% with the Cytauxzoon sp. and C. manul originated from Mongolia;
Cytauxzoon sp. diagnosed in Lynx from Spain and 97% with the Cytauxzoon sp.
diagnosticated from domestic cats in Spain. The consensus sequence of the wild cat
sample showed 1204 pairs of basis (GenBank acess nr. DQ382277), identity of 99% to
the species C. felis originated from the USA and 96% to Cytauxzoon sp. and C. manul
originated from Mongolia and Cytauxzoon sp. from Spain. The phylogenetic analysis
showed that the studied sequences clustered with other sequences of Cytauxzoon felis that
infects felids in the USA, forming a related group of the C. manul species that infects
Pallas cats of Mongolia and Cytauxzoon sp. that infects lynx and domestic cats of Spain.
The monophyletic characteristic of the species of the genus Cytauxzoon, in contrast with
the poliphyletic structure of Babesia and Theileria groups could also be observed. The
results obtained in this study confirmed that the agent of feline cytauxzoonosis in P. onca
and L. tigrinus is C. felis.
Keywords: Cytauxzoon felis18S rRNA-PCR, sequencing, phylogenetic analysis.

1
1. INTRODUÇÃO
Os membros do gênero Cytauxzoon Neitz e Thomas, 1948 (Apicomplexa:
Theileriidae) são hemoparasitos transmitidos por carrapatos que causam enfermidades
principalmente em ruminantes e felinos. Esses protozoários caracterizam-se por realizar
esquizogonia em histiócitos, nos quais um único grande esquizonte é formado, e por
apresentar formas intraeritrocíticas diminutas com cromatina nuclear compacta e
citoplasma hialino, típicas dos teilerídeos (NEITZ e THOMAS, 1948; NEITZ, 1956;
SIMPSON et al., 1985a; KOCAN et al., 1992; SOARES, 2002).
A citauxzoonose foi primeiramente descrita em ruminantes silvestres no
continente africano (Neitz e Thomas, 1948; NEITZ, 1956; 1957). Mais tarde uma
doença fatal foi reportada em felídeos domésticos na América do Norte (WAGNER,
1976; KIER et al., 1982a;b, 1987). Atualmente são conhecidas cinco espécies deste
gênero: Cytauxzoon silvicaprae, Cytauxzoon strepsicerosi e Cytauxzoon taurotragi, os
quais infectam ungulados na África (NEITZ, 1956; BROCKLESBY, 1962;
MEHLHORN et al., 1994), Cytauxzoon felis, que parasita, principalmente, felídeos nos
Estados Unidos da América (EUA) (WAGNER et al., 1976; 1980; GARNER et al.,
1996; MEIER e MOORE, 2000) e Cytauxzoon manul que infecta gatos Pallas oriundos
da Mongólia (REICHARD et al., 2005).
Na América Latina os primeiros registros sobre a citauxzoonose felina natural
foram feitos em leões (Panthera leo) (PEIXOTO et al., 1999), em onça-pintada
(Panthera onca) (SOARES et al., 1999a) e em gato-do-mato pequeno (Leopardus
tigrinus) (SOARES et al., 1999b), todos no Estado do Rio de Janeiro. Mais
recentemente, formas parasitárias intraeritrocíticas semelhantes a C. felis foram
diagnosticadas no esfregaço sangüíneo de uma onça-parda (Puma concolor) em Mato
Grosso do Sul. Embora não tenham sido observadas formas esquizogônicas, o parasito
foi denominado de Cytauxzoon felis-like (JULIANO et al., 2004).
A citauxzoonose felina cursa, de forma severa e fatal, em gatos domésticos,
linces da Flórida, em tigre branco, nos EUA e em tigre de bengala na Alemanha
(WAGNER et al., 1976; FERRIS, 1979; KIER et al., 1982a; GARNER et al., 1996;
JAKOB e WESEMEIER, 1996). Em lince americano, puma do Texas, puma da Flórida,
guepardo e experimentalmente em ovinos a doença é assintomática e de curso longo
(ZINKL et al., 1981; BUTT et al., 1991; WEHINGER et al., 1995; ROTSTEIN et al.,
1999). Um agente morfologicamente semelhante a C. felis determinou doença fatal em
macaco rhesus no Panamá (GARDINER, 1998 – comunicação pessoal). No Brasil, a
enfermidade causada por C. felis simile cursa de forma fatal em leões (PEIXOTO et al.,
1999) e com quadro mais brando em onça-pintada (SOARES, 2002); em gato-do-mato
pequeno, experimentalmente em gato doméstico (SCOFIELD et al., 1999; SOARES,
2002), e em onça-parda (JULIANO et al., 2004) a infecção pode ser inaparente.
Até o momento, o único transmissor de C. felis comprovado experimentalmente
é o carrapato Dermacentor variabilis (BLOUIN et al., 1984; 1987; KOCAN et al.,
1992), todavia, para as espécies de Cytauxzoon que ocorrem no continente africano,
diversas espécies de ixodídeos têm sido incriminadas como transmissores (NEITZ,
1956; BROCKLESBY, 1962; YOUNG et al., 1980; JARDINE, 1992; MEHLHORN et
al., 1994).
A citauxzoonose vem ganhando importância não só pela sua dispersão e
patogenicidade para felinos de companhia, mas também por ser capaz de infectar
bovinos, ovinos, e até mesmo primata (ALLSOPP et al., 1994; COX, 1994;

2
GARDINER, 1998; KJEMTRUP et al., 2000a,b; SOARES, 2002). Não obstante, tem-se
pouco conhecimento a respeito de Cytauxzoon spp. fora da América do Norte. Ademais,
muitas dúvidas ainda persistem quanto a sua biologia, transmissão e seus aspectos
morfológicos e ultraestruturais. O diagnóstico de citauxzoonose é baseado,
principalmente, na observação de formas intraeritrocíticas e/ou formas esquizogônicas
teciduais típicas do agente. Entretanto, na ausência das formas esquizogônicas, torna-se
difícil o diagnóstico específico.
A dificuldade de diagnóstico baseado em parâmetros fenotípicos e o
desconhecimento de muitos fatores biológicos de diferentes grupos de parasitos têm
incentivado pesquisadores a empregar técnicas moleculares e análises filogenéticas no
diagnóstico e na classificação e/ou reclassificação de diferentes grupos. Dentre as
seqüências de DNA mais utilizadas neste tipo de abordagem, destaca-se a que codifica a
subunidade 18S RNAr, que constitue a subunidade menor dos ribossomas, em estudos
filogenéticos para um grande número de espécies de protozoários. O sequenciamento de
regiões do DNA que codificam esta subunidade pode incrementar ainda mais o
conhecimento a respeito das espécies, subespécies ou cepas e ainda auxiliar na
identificação das mesmas facilitando o diagnóstico da doença.
No Brasil o agente diagnosticado em Panthera onca e Leopardus tigrinus foi
denominado de Cytauxzoon felis simile, não sendo determinada a espécie (SOARES,
2002).
O presente estudo teve como objetivos determinar a espécie de Cytauxzoon que
infectou onça-pintada (Panthera onca) e gato-do-mato (Leopardus tigrinus) do
Zoológico Municipal de Volta Redonda, Estado do Rio de Janeiro utilizando técnicas de
diagnóstico molecular, comparar as seqüências consenso obtidas neste trabalho com
seqüências de Cytauxzoon, Theileria, Babesia e Piroplasmida depositadas em bancos de dados (GenBank), e realizar análises filogenéticas com a espécie estudada.

3
2. REVISÃO DE LITERATURA
2.1. Breve Histórico e Aspectos Gerais sobre a Citauxzoonose
A citauxzoonose é uma hemoparasitose determinada por protozoários do gênero
Cytauxzoon Neitz e Thomas, 1948, que acomete ruminantes domésticos, ruminantes
silvestres e felídeos (NEITZ, 1956; 1957; BROCKLESBY, 1962; KIER et al., 1987;
SOARES, 2002). Foi descrita pela primeira vez na África do Sul, causando uma doença
aguda em duiquer (Sylvicapra grimmia) (NEITZ e THOMAS, 1948), logo em seguida
foi reportada em kudus (Strepsiceros strepsiceros) (NEITZ e de LANGE, 1956) e em
elande (Taurotragus oryx) (MARTIN e BROCKLESBY, 1960). Nos Estados Unidos da
América (EUA), em 1976, uma citauxzoonose fatal foi descrita em gato doméstico
(WAGNER, 1976).
Embora os gêneros Cytauxzoon e Theileria possuam algumas semelhanças,
Cytauxzoon spp apresentam características biológicas e morfológicas que podem ser
consideradas apomórficas deste gênero, principalmente quanto à esquizogonia tecidual,
que pode ocorrer em diversos tipos de células mononucleares (NEITZ, 1957; KIER et
al., 1987), enquanto que Theileria spp realizam esquizogonia exclusivamente em
linfócitos. A despeito do conhecimento biológico destes Theileriidae, alguns
taxonomistas consideram os dois gêneros sinônimos e os agrupam em um único gênero,
Theileria (GROOTENHUIS et al., 1979). Outros preferem mantê-los separados
(NEITZ, 1957; BARNETT, 1977; ALLSOPP et al., 1994; UILENBERG, 1995).
Na maioria dos hospedeiros susceptíveis a Cytauxzoon sp., a enfermidade cursa
com o quadro severo, geralmente agudo, de hipertermia, anorexia, perda de peso,
anemia, icterícia, dispnéia, letargia e fraqueza, com evolução para coma e morte
(NEITZ, 1957; BROCKLESBY, 1960; GLENN e STAIR, 1984; KIER et al., 1987;
GARNER et al., 1996; MEINKOTH et al., 2000).
Uma ampla variedade de mamíferos, principalmente ruminantes e carnívoros, tem
sido descrita como hospedeiros de Cytauxzoon sp. no continente africano e nos EUA
(McCULLY et al., 1970; YOUNG et al., 1980; GLENN et al., 1982; KIER et al., 1982a;
GARNER et al., 1996; JAKOB e WESEMEIER, 1996). Recentemente, foi verificada
citauxzoonose em primatas não-humanos no Panamá (GARDINER, 1998).
No Brasil, Estado do Rio de Janeiro, a citauxzoonose foi diagnosticada
primeiramente em leões (PEIXOTO et al., 1999) e logo em seguida em onça-pintada
(SOARES et al., 1999a) e gato-do-mato pequeno (SOARES et al., 1999b). Estudos
experimentais dessa enfermidade em gatos domésticos também foram realizados
avaliando os aspectos clínicos, patológicos e parasitológicos (SCOFIELD et al., 1999;
SOARES, 2002; SCOFIELD, 2002). Em 2004, formas parasitárias intraeritrocíticas
semelhantes a C. felis foram observadas em esfregaço sangüíneo de uma onça-parda em
Campo Grande-M.S. e o agente identificado como C. felis like (JULIANO et al., 2004).
2.2. Gênero Cytauxzoon
O gênero Cytauxzoon Neitz e Thomas, 1948 foi assim nominado para referir-se a
dezenas ou centenas de diminutos (0,5 a 0,75 mm) corpos circulares regularmente
distribuídos, que caracterizavam a esquizogonia desse parasito no citoplasma de
histiócitos. Este parasita se multiplica por fissão binária simples em eritrócitos e por

4
esquizogonia em histiócitos (NEITZ e THOMAS, 1948; NEITZ, 1956; 1957) e tem
Cytauxzoon silvicaprae Neitz e Thomas, 1948 como espécie-tipo.
Até o momento, são reconhecidas cinco espécies pertencentes ao gênero
Cytauxzoon: Cytauxzoon silvicaprae Neitz e Thomas, 1948; C. strepsicerosi Neitz e de
Lange, 1956; C. taurotragi Martin e Brocklesby, 1960; C. felis Kier, 1979 e C. manul
Reichard et al., 2005, porém existem suspeitas de novas espécies (LOPEZ-REBOLLAR
et al., 1999; SOARES, 2002).
2.2.1. Citauxzoonose felina
Uma doença fatal foi descrita em quatro gatos domésticos (Felis catus) no
Sudoeste do Estado de Missouri, EUA, entre setembro de 1973 e junho de 1975. Esses
animais apresentavam quadro grave que, em princípio, foi diagnosticado como anemia
infecciosa felina (= hemobartonelose) com base na sintomatologia. No terceiro caso,
contudo, detectaram-se formas intraeritrocíticas com aspecto de anel de rubi no
esfregaço sangüíneo, sendo estabelecido então o diagnóstico ante-mortem de
citauxzoonose (WAGNER, 1976).
Pelo acompanhamento clínico dos quatro gatos, verificou-se uma doença aguda e
fatal caracterizada por hipertermia, anorexia, fraqueza, letargia, palidez de mucosas,
desidratação, anemia, icterícia e morte. Observaram-se trofozoítas intraeritrocíticos com
média de parasitemia oscilando entre 1 e 4 % e formas predominantemente em anel de
rubi, alongadas ou puntiformes, medindo 1 a 1,2 mm (WAGNER, 1976).
À necropsia havia icterícia generalizada, petéquias e equimoses na superfície do
coração e dos pulmões e distensão do saco pericárdico com líquido seroso amarelo-
claro. O exame microscópico revelou formas esquizogônicas em diferentes fases de
desenvolvimento, parasitando histiócitos e células retículo-endoteliais de diversos
órgãos (pulmão, fígado, baço, coração, rins e vesícula biliar). Observaram-se oclusão
parcial ou total de vasos sangüíneos pelos esquizontes e marcada eritrofagocitose
tecidual. Os esquizontes pleomórficos continham centenas de pequenos corpos medindo
de 0,1 a 0,2 mm (= merozoítas), enquanto outros esquizontes evidenciavam dezenas de
corpos maiores (citômeros), com muitos dos pequenos corpos (WAGNER, 1976).
Com base nestas observações, a enfermidade foi primariamente descrita como
uma citauxzoonose simile fatal em gatos (WAGNER, 1976). Mais tarde verificou-se
que a doença era endêmica no Sudoeste de Missouri, pois muitos casos de infecção
natural surgiram com os mesmos aspectos clínico-patológicos (WAGNER, 1976;
WAGNER et al., 1976). A enfermidade foi reproduzida em outros gatos domésticos
pela inoculação de sangue e fragmentos de baço, por passagens consecutivas; todos os
animais inoculados apresentaram doença aguda e morreram entre sete e 12 dias após
inoculação (WAGNER et al., 1976). Os estudos iniciais evidenciaram que o gato
doméstico é o hospedeiro definitivo terminal para esta citauxzoonose (WAGNER, 1976;
WAGNER et al., 1976).
Inicialmente pensou-se que esta nova doença de gatos norte americanos estava
restrita ao Sudoeste de Missouri. A enfermidade, porém, foi posteriormente registrada
nos Estados do Texas, Arkansas e Geórgia. Em todos os casos, os animais estavam
parasitados por carrapatos ou co-habitavam locais com bovinos, eqüinos, caprinos,
ovinos ou com cães parasitados por carrapatos (WAGNER, 1976; WAGNER et al.,
1976; WIGHTMAN et al., 1977; FERRIS, 1979).
Estes fatos despertaram o interesse na investigação da doença em outros
mamíferos, especialmente animais de produção, bem como no seu possível potencial

5
zoonótico, principalmente pelos seguintes motivos: i) a citauxzoonose só havia sido
descrita em ungulados na África (NEITZ, 1956); ii) felinos ocorrem em todos os
continentes e iii) animais silvestres são continuamente transportados de um continente
para outro (WAGNER, 1976; WAGNER et al., 1976; WIGHTMAN et al., 1977;
FERRIS, 1979).
Adicionalmente, o Departamento de Agricultura dos Estados Unidos concentrou
esforços para melhor compreender esta enfermidade, descobrir sua relação com as
citauxzoonoses africanas e verificar o potencial de infecção para animais de produção.
Mais de 500 gatos foram estudados sob condições experimentais e outros sob condições
naturais e mais de 100 passagens seriadas do agente foram feitas no intuito de
compreender as doenças natural e experimental, o diagnóstico diferencial, o prognóstico
e a epizootiologia (FERRIS, 1979).
Ainda assim, muitas dúvidas permaneceram como por exemplo: existe esta doença
em gatos em outros continentes ou ilhas? A doença tem potencial para se dispersar? É
cosmopolita ou regional? Quais os hospedeiros e reservatórios? Qual é realmente o
agente etiológico (espécie)? (WAGNER et al; 1976; FERRIS, 1979).
2.3. Cytauxzoon felis
O agente etiológico da doença fatal para gato doméstico, semelhante à
citauxzoonose dos ungulados africanos foi inicialmente diagnosticado, nos EUA, com
base nos caracteres morfológicos e biológicos, especialmente das formas
esquizogônicas, sendo tentativamente classificado como pertencente ao gênero
Cytauxzoon Neitz e Thomas, 1948, família Theileriidae (WAGNER, 1976; WAGNER
et al., 1976).
Posteriormente, com base em estudos de ultraestrutura, histológicos, tipos de
célula infectada e modo de multiplicação, o protozoário foi nominado Cytauxzoon felis.
Esta nominação, no entanto, não atende aos requisitos do Código Internacional de
Nomenclatura Zoológica (CINZ) (UILENBERG, 1995), pois foi proposta no trabalho
de tese de Ann B. Kier, o qual não é um documento aceito pelo CINZ. Logo em
seguida, contudo, foram publicados os estudos clínico-patológicos e da transmissão
experimental do agente para gatos domésticos e para 30 espécies de mamíferos
domésticos e silvestres, nos quais se considerou como agente etiológico C. felis Kier,
1979 (KIER et al., 1982a,b). Pelo fato da literatura internacional reconhecer até o
presente momento os estudos e a nominação dada por Ann B. Kier, trataremos a espécie
como C. felis Kier, 1979.
Recentemente foram diagnosticadas formas parasitárias intraeritrocíticas
semelhantes a C. felis em gatos Pallas (Otocolobus manul) oriundos da Mongólia e
importados pelos EUA. Ketz-Riley et al. (2003) sugeriram uma nova espécie para esse
agente e Reichard et al. (2005) classificaram-no como Cytauxzoon manul após estudos
morfométricos, moleculares e filogenéticos.
2.3.1. Biologia, morfologia e ultraestrutura
2.3.1.1. Formas intrahistiocíticas
O carrapato inocula no hospedeiro esporozoítas que infectam histiócitos para
formar esquizontes (KOCAN et al., 1992). Para C. felis apenas um esporozoíta infecta

6
um histiócito. O desenvolvimento do parasito sob observação ultraestrutural, começa
como uma indistinta massa dentro do citoplasma da célula hospedeira, que
progressivamente torna-se mais densa. Com o desenvolvimento, a divisão nuclear
ocorre e vários núcleos individualizados tornam-se mais evidentes com o aumento da
densidade do nucleoplasma e o aparecimento da membrana nuclear (SIMPSON et al.,
1985a).
Neste estágio, o parasito inicia um processo de multilobulação, cujos lóbulos são
interconectados por numerosas pontes citoplasmáticas. Com o aumento de tamanho e
complexidade do parasito, organelas são formadas ao longo das margens da membrana
do parasito. O citóstoma torna-se visível no plasmalema, inicia-se o aparecimento do
retículo endoplasmático, do anel polar, da mitocôndria não-pregueada e o núcleo torna-
se mais denso que o citoplasma (SIMPSON et al., 1985a). Com o desenvolvimento,
aumentam as lobulações e as róptrias aparecem no ápice das lobulações. Os merozoítas
surgem da fissão multinucleada de um único esporonte existente nesta fase. Os
citômeros são porções desenvolvidas deste único esporonte interconectadas por
numerosas pequenas pontes citoplasmáticas (KOCAN et al., 1992).
A massa do parasito diminui de tamanho, formando o corpo residual e os
merozoítas permanecem associados ao corpo residual que contém o núcleo do parasito e
organelas (SOARES, 2002). Os merozoítas são produzidos a partir de um arranjo com
aspecto de roseta em torno do pequeno corpo residual (SIMPSON et al., 1985a;
KOCAN et al., 1992). O merozoíta formado contém um único núcleo em posição oposta
às róptrias e aos micronemas. Assim, diversos merozoítas em desenvolvimento
permanecem nutrindo-se no citoplasma da célula hospedeira até sua completa
maturação (SIMPSON et al., 1985a).
Aparentemente, a ruptura da célula hospedeira só ocorre após a completa
maturação dos merozoítas (SIMPSON et al., 1985a).
As marcantes diferenças na esquizogonia entre Theileria sp. e Cytauxzoon sp.
fundamentam-se no fato de que, no primeiro gênero, este processo ocorre apenas em
linfócitos e no segundo, em histiócitos. No gênero Cytauxzoon, apenas um único
esporozoíta invade a célula hospedeira para desenvolver o esquizonte, enquanto que em
Theileria muitos esporozoítas invadem linfócitos, dando origem a vários esquizontes
(MEHLHORN e SCHEIN, 1984; KOCAN et al., 1992; MEHLHORN et al., 1994).
Os esquizontes de C. felis são pleomórficos, com formas predominantemente
arredondadas com dimensão variando entre 15 e 250 µm de diâmetro (WAGNER, 1976;
WAGNER et al., 1976; KIER et al., 1987; PEIXOTO et al., 1999; SOARES, 2002). A
esquizogonia em C. felis parece ser um processo de curta duração e limitado (BLOUIN
et al., 1987). Experimentalmente observou-se que após inoculação de esporozoítas por
adultos de Dermacentor variabilis, em gatos domésticos, os esquizontes foram
visualizados nos tecidos aos 11 dias após inoculação. Aos 30 dias pós-inoculação, via
carrapato, não mais havia esquizontes, permanecendo os animais-portadores de formas
intraeritrocíticas, com baixa parasitemia, por longo período (BLOUIN et al., 1987). Por
outro lado, o inóculo de fragmentos de tecidos de felídeos (gato doméstico ou linces)
com esquizontes em gatos domésticos e/ou linces resultou na formação de esquizontes,
baixa parasitemia de formas eritrocíticas e doença grave, com morte entre sete e 20 dias
(WAGNER et al., 1976; WIGHTMAN et al., 1977; FERRIS, 1979; WAGNER et al.,
1980; KIER et al., 1982a,b; 1987).

7
2.3.1.2. Formas intraeritrocíticas
O merozoíta maduro livre possui uma camada enrugada sobre a membrana
plasmática e entra no eritrócito circulante por um processo semelhante à endocitose.
Quando o merozoíto entra em contato com o eritrócito, a membrana deste inicia um
processo de modificação no ponto de contato com o merozoíto, invaginando-se. À
proporção que o merozoíto entra na célula, a invaginação alarga-se e aprofunda-se para
acomodá-lo. A margem da membrana invaginada fica intimamente aderida ao
merozoíto. A camada enrugada do merozoíto desaparece quando o mesmo está
parcialmente internalizado. Uma vez que o merozoíto se encontra dentro do eritrócito, a
membrana invaginada fusiona-se, abraçando o merozoíto e formando um vacúolo. A
entrada do merozoíto ocorre sem ruptura da membrana do eritrócito, porém o merozoíto
sofre fissão binária, dando origem à trofozoítas, com capacidade de invadir outros
eritrócitos (KOCAN et al., 1992).
Sob exame ultraestrutural, as formas intraeritrocíticas possuem uma dupla
membrana citoplasmática, núcleo pobremente definido contendo discreta cromatina,
retículo endoplasmático, ribossomos livres, mitocôndrias não-pregueadas, vacúolo
alimentar e citóstoma na membrana plasmática que circunda o parasito (SIMPSON et
al., 1985b). No Brasil estudos ultraestruturais de trofozoítas de C. felis simile isolado de
Panhtera onca revelaram a presença de membrana citoplasmática dupla e delgada,
núcleo excêntrico, circundado por uma membrana também dupla, mitocôndria atípica
(não-pregueada), com dupla membrana e localizada no pólo oposto ao núcleo e
vacúolos digestivos limitados por dupla membrana. Microtúbulos extranucleares,
ribossomos, retículo endoplasmático e lisossomos foram observados dispersos pelo
citoplasma do trofozoíto. Foram visualizados também citóstoma com dupla membrana e
pregas citoplasmáticas interligando formas parasitárias durante o processo de divisão
binária (SCOFIELD, 2002).
Nos eritrócitos, encontram-se usualmente um ou dois trofozoítas, eventualmente
três ou quatro (em cruz-de-malta). As formas intraeritrocíticas podem ser ovais,
arredondadas, puntiformes, piriformes, em anel de rubi, alongadas ou em tétrades
(WAGNER, 1976; KIER et al., 1982a,b; 1987; SIMPSON et al., 1985b; SCOFIELD,
2002). A cromatina nuclear, geralmente periférica, cora-se basofílicamente e o
citoplasma cora-se azul-hialino ou cora-se fracamente. Esta forma pode medir de 0,5 a
2,5 µm de diâmetro, mas usualmente atinge 0,5 a 1,5 µm de diâmetro, e encontra-se
freqüentemente no centro ou excêntrico quanto à localização no eritrócito (WAGNER,
1976; KIER et al., 1987; SIMPSON et al., 1985b; GLENN e STAIR, 1984; SOARES,
2002; SCOFIELD, 2002).
Normalmente a parasitemia por C. felis é baixa, entre 0,5 e 5% e, na maioria dos
casos de infecção natural está em torno de 1% (WAGNER, 1976; WAGNER et al.,
1976; 1980; FERRIS, 1979; SIMPSON et al., 1985b; BLOUIN et al., 1987; KIER et al.,
1987; SCOFIELD et al., 1999; SOARES, 2002; SCOFIELD, 2002). Os trofozoítas
permanecem infectando eritrócitos e pelo repasto do carrapato, dá-se continuidade ao
ciclo biológico. Os trofozoítas são infectantes para outros hospedeiros, via transmissão
sangüínea iatrogênica, mas aparentemente dão origem apenas a novas formas
eritrocíticas (BLOUIN et al., 1984; 1987; SIMPSON et al., 1985b).

8
2.3.1.3. Gametogonia e esporogonia
O ciclo de C. felis no carrapato ainda é desconhecido. Com base no ciclo de C.
taurotragi em R. appendiculatus (YOUNG et al., 1980) e nos fenômenos de
gametogonia, singamia e esporogonia que ocorrem em todos os Piroplasmida (NEITZ,
1956; 1957; MEHLHORN e SCHEIN, 1984; KAKOMA e MEHLHORN, 1994;
MEHLHORN et al., 1994), poder-se-ia comentar resumidamente um possível ciclo:
alguns trofozoítas nos eritrócitos sofrem diferenciação e quando ingeridos pelo
carrapato, formam microgametócitos ou macrogametócitos que, posteriormente, dão
origem a micro ou macrogametas, respectivamente. Estes formam o zigoto, por
singamia, que ao penetrar em células intestinais diferencia-se em cineto. O cineto, via
hemolinfa, invade os ácinos de glândulas salivares, realiza a esporogonia, com liberação
de esporozoítas que são inoculados pelo carrapato (MEHLHORN e SCHEIN, 1984;
KAKOMA e MEHLHORN, 1994; MEHLHORN et al., 1994).
2.3.2. Vetores
Os Piroplasmida são transmitidos biologicamente apenas por carrapatos (NEITZ,
1956; HOOGSTRAAL, 1985; SONESHINE, 1991; COX, 1994). Os teilerídeos são
transmitidos essencialmente por carrapatos heteroxenos (NEITZ, 1956; MEHLHORN e
SCHEIN, 1984; UILENBERG, 1995), apesar de existir um relato sobre Boophilus
annulatus transmitindo Theileria mutans para bovinos exportados dos EUA para a
Alemanha (REICHENOW, 1935, citado por NEITZ, 1956).
O carrapato transmissor natural de C. felis não é conhecido. Experimentalmente,
foi verificado que Dermacentor variabilis alimentado no estágio de ninfa em lince
(Lynx rufus) parasitado por C. felis, adquire a infecção e transmite para gato doméstico
quando adulto (BLOUIN et al., 1984). Esta constatação foi confirmada posteriormente
com outros experimentos similares (BLOUIN et al., 1987; KOCAN et al., 1992).
De forma geral, a literatura considera D. variabilis como vetor natural de C. felis
(KIER et al., 1987; BUTT et al., 1991; GARNER et al., 1996; JAKOB e WESEMEIER,
1996). Por outro lado, vários autores têm observado desde a descrição original da
enfermidade (WAGNER, 1976) que nas áreas onde ocorre a doença em gato doméstico
e lince, o carrapato mais prevalente é Amblyomma americanum (WAGNER, 1976;
WAGNER et al., 1976; 1980; WEHINGER et al., 1995; GARNER et al., 1996).
Recentemente, Bondy Jr. et al. (2005) realizaram análises moleculares para
detecção de DNA de C. felis em 1362 exemplares de carrapatos das espécies A.
americanum, D. variabilis e Rhipicephalus sanguineus. Esses carrapatos foram
coletados em diferentes cidades do Estado de Missouri onde já havia sido diagnosticada
citauxzoonose felina. Entre os exemplares analisados, apenas três ninfas de A.
americanum foram positivas e essa espécie foi considerada como potencial vetor de C.
felis.
Adicionalmente, em um estudo conduzido entre 1974 e 1991, na Flórida, para
determinar a incidência de carrapatos em lince (Lynx rufus floridanus) e em puma ou
pantera da Flórida (Felis concolor coryi), observou-se que estes animais (n= 189) eram
parasitados principalmente por adultos de Ixodes scapularis, I. affinis, D. variabilis, A.
americanum e A. maculatum (WEHINGER et al., 1995), sendo a espécie I. scapularis a
mais incidente em ambos os hospedeiros. Assim, estes autores consideram que
principalmente I. scapularis pode ser considerado como um potencial vetor de C. felis
para estes felídeos (WEHINGER et al., 1995).

9
As espécies de Theileria são transmitidas, principalmente, por carrapatos dos
gêneros Rhipicephalus e Hyalomma (NEITZ, 1956; SCHEIN, 1975; MEHLHORN e
SCHEIN, 1984; UILENBERG, 1995), enquanto que as espécies do gênero Cytauxzoon
que acometem ungulados são transmitidas por Rhipicephalus spp. As espécies Theileria
mutans e T. velifera, que ocorrem tanto na África quanto no Caribe, são transmitidas por
diversas espécies do gênero Amblyomma: Amblyomma variegatum, Amblyomma
hebraeum, Amblyomma gemma, Amblyomma lepidum, Amblyomma coherens entre
outras (NEITZ, 1956; MEHLHORN et al., 1994; UILENBERG, 1995). Theileria cervi,
que ocorre na América do Norte, é transmitida principalmente por A. americanum
(HAZEN-KARR et al., 1987; MEHLHORN et al., 1994).
No Brasil, foi verificada a presença de Amblyomma cajennense no recinto de leões
mantidos em cativeiro que morreram em decorrência de citauxzoonose (PEIXOTO et
al., 1999). Na mesma localidade, outros felinos como onça-pintada e gato-do-mato
pequeno também foram encontrados naturalmente parasitados por C. felis simile
(SOARES et al., 1999a;b). No entanto, ainda não é conhecido o transmissor deste
teilerídeo no País.
2.3.3. Hospedeiros
Cytauxzoon felis foi originalmente descrito parasitando gato doméstico (Felis
catus) (WAGNER, 1976; WAGNER et al., 1976; WIGHTMAN et al., 1977), no qual
determinava invariavelmente uma doença fatal (FERRIS, 1979; WAGNER et al., 1980;
KIER et al., 1987) e de curso rápido, o que levou os pesquisadores a considerarem ser o
gato doméstico um hospedeiro acidental e/ou final para o parasito (KIER et al., 1982a,b;
1987; SIMPSON et al., 1985a,b; BLOUIN et al., 1987).
Posteriormente, verificou-se que a inoculação de esquizontes e trofozoítas foi bem
sucedida em duas espécies de linces, sendo que o lince americano (L. rufus rufus)
apresentou apenas formas sangüíneas e não desenvolveu a doença, enquanto o lince da
Flórida (L. rufus floridanus) apresentou trofozoítas e esquizontes e desenvolveu uma
doença fatal idêntica à observada em gatos domésticos (KIER et al., 1982a,b). Nos
EUA, diversos casos foram descritos de linces americanos parasitados por C. felis. Na
maioria deles, os animais apresentavam apenas formas intraeritrocíticas do parasito.
Pelo inóculo de sangue desses animais em gatos, porém desenvolveu-se uma doença
fatal com formação de esquizontes teciduais (GLENN et al., 1982, 1983; KIER et al.,
1982a,b; BLOUIN et al., 1984) (Tabela 1). Acredita-se, portanto, que o lince é o
principal reservatório de C. felis na América do Norte (GLENN et al., 1982, 1983;
BLOUIN et al., 1987; WEHINGER et al., 1995).
Experimentalmente foi demonstrado que ovinos são susceptíveis à C. felis
(Tabela 1). Estes animais não apresentam sintomas aparentes, mas desenvolvem uma
baixa e persistente parasitemia (KIER et al., 1982a). Ao se avaliar experimentalmente o
potencial de transmissão interespécies de C. felis, proveniente de gato doméstico, em
quatro espécies de animais domésticos de produção, nove espécies de animais de
laboratório e 17 espécies de animais silvestres, observou-se que somente lince
americano, lince da Flórida e ovino foram infectados (KIER et al., 1982a).
As infecções naturais por C. felis em guepardo (ZINKL et al., 1981), em puma ou
pantera da Flórida (BUTT et al., 1991; ROTSTEIN et al., 1999) e em puma ou pantera
do Texas (ROTSTEIN et al., 1999) têm sido descritas com formas eritrocíticas e quadro
assintomático, e, em tigre branco (GARNER et al., 1996) e em tigre de bengala
(JAKOB e WESEMEIER, 1996) com formas eritrocítica e esquizogônicas e doença

10
fatal. Recentemente verificou-se, em macaco rhesus, uma típica citauxzoonose, com
formas esquizogônicas semelhantes à C. felis (GARDINER, 1998) (Tabela 1).
No Brasil, C. felis simile foi observado primeiramente em leões causando uma
enfermidade severa e lesões patognomônicas, caracterizadas pela oclusão parcial ou
total de vasos sangüíneos (PEIXOTO et al., 1999). Em seguida foi diagnosticado o
parasitismo por formas intraeritrocíticas típicas deste Theileriidae em onça-pintada
(SOARES et al., 1999a) e em gato-do-mato pequeno (SOARES et al., 1999b).
Posteriormente, estudos experimentais avaliaram os aspectos clínicos, patológicos e
parasitológicos em gatos domésticos (SCOFIELD et al., 1999; SOARES, 2002;
SCOFIELD, 2002). Em 2004, JULIANO et al. diagnosticaram em um esfregaço
sangüíneo de uma onça-parda capturada no estado do Mato Grosso do Sul formas
parasitárias intraeritrocíticas semelhantes a C. felis. Embora não tenham sido observadas
formas esquizogônicas, o parasito foi denominado de Cytauxzoon felis-like.

11

12
2.3.4. Diagnóstico
2.3.4.1. Diagnóstico direto
O diagnóstico direto de citauxzoonose é baseado, principalmente, na observação
de formas intraeritrocíticas e/ou formas esquizogônicas teciduais, típicas do agente. O
diagnóstico de trofozoíta é feito pela confecção de esfregaço a partir de sangue
periférico, obtido por punção de vasos marginais. Estes esfregaços são fixados em
metanol absoluto e podem ser corados por derivados do Romanowsky como Giemsa,
Leishman, May-Grünwald-Giemsa etc, ou por laranja de acridina (BOBADE e NASH,
1987; MacWILLIAMS, 1987; FOREYT, 1989; SOARES, 2002).
O exame sob microscopia fotônica deve basear-se na verificação de formas
intraeritrocíticas de pequeno porte (0,5–1,5 m x 0,8–2,0 m), predominantemente
arredondadas e em forma de anel de rubi. Estes tipos morfológicos encontram-se em
posição central no eritrócito. Formas em tétrade também ocorrem, porém em menor
freqüência. Os trofozoítas possuem cromatina nuclear basofílica e de localização
periférica e citoplasma azul-hialino fracamente corado (WAGNER, 1976;
WIGHTMAN et al., 1977; GLENN e STAIR, 1984; SIMPSON et al., 1985a,b; KIER et
al., 1987; MEIER e MOORE, 2000; SOARES, 2002; SCOFIELD, 2002).
As formas intrahistiocíticas podem ser diagnosticadas por citologia de
fragmentos de tecidos e órgãos. Na observação citológica de aposições ou “arrastes” de
tecidos e órgãos, processados como descrito para esfregaços de sangue, pode-se
verificar histiócitos contendo no citoplasma esquizontes com citômeros e inúmeros
merozoítas. Na observação de tecidos processados por técnicas histológicas e corados
por hematoxilina-eosina ou por Giemsa, verificam-se histiócitos repletos de esquizontes
com citômeros e merozoítas aderidos ao endotélio de vasos sangüíneos de diversos
órgãos (FERRIS, 1979; WAGNER et al., 1980; KIER et al., 1982b; 1987; GLENN e
STAIR, 1984; GARNER et al., 1996; JAKOB e WESEMEIER, 1996; MEIER e
MOORE, 2000; SOARES, 2002).
Cabe salientar, porém, que as formas intraeritrocíticas de C. felis, quando da
infecção via carrapato, aparecem na corrente sangüínea geralmente entre um e três dias
após o surgimento de esquizontes em histiócitos, período no qual surgem os sintomas
(HAUCK et al., 1982; SIMPSON et al., 1985a,b; BLOUIN et al., 1987; KIER et al.,
1987; MEIER e MOORE, 2000).
A parasitemia por C. felis usualmente é baixa (0,5 a 3%) persistente por um
longo período, e o animal também pode permanecer portador crônico assintomático
(WAGNER et al., 1980; KIER et al., 1987; HOOVER et al., 1994; WALKER e
COWELL, 1995; MEINKOTH et al., 2000; SCOFIELD, 1999; SOARES, 2002;
SCOFIELD, 2002). E pelo fato da esquizogonia ser limitada e aparentemente
transitória, o animal pode resistir à infecção e não apresentar esquizontes, persistindo
apenas parasitos intraeritrocíticos (SIMPSON et al., 1985b; BLOUIN et al., 1987; KIER
et al., 1987; KOCAN et al., 1992).
Pode-se proceder, ainda, o diagnóstico molecular, realizado principalmente por
meio da técnica da reação de polimerase em cadeia (PCR), a qual amplifica fragmentos
de gene do parasito utilizando iniciadores. A PCR, seguida do seqüenciamento,
possibilita a comparação de seqüências do gene 18S RNA ribossomal de C. felis com
outras espécies de Cytauxzoon, Theileria e Babesia disponíveis no GenBank
(www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=Nucleotide) (ALLSOPP et al., 1994;
MEINKOTH et al., 2000; REICHARD et al., 2005).

13
2.3.4.2. Diagnóstico indireto e exames auxiliares
O diagnóstico sorológico tem sido feito pela detecção de anticorpos da classe
IgG por meio da técnica de imunofluorescência indireta e pelo imunoensaio
microfluorimétrico. Ambas as provas utilizam formas esquizogônicas como antígeno
(SHINDEL et al., 1978; UILENBERG et al., 1987; COWELL et al., 1988). Estes
ensaios, além de serem um meio indireto de diagnóstico, contribuem para o
conhecimento de possíveis animais portadores em estado subclínico e para a
epidemiologia, determinando a prevalência e distribuição da enfermidade.
Os exames hematológico e bioquímico são auxiliares no diagnóstico da
citauxzoonose. As alterações mais freqüentes e importantes são anemia normocítica
normocrômica, leucopenia, trombocitopenia, decréscimo do volume globular e da
concentração de albumina, aumento da concentração de proteínas plasmáticas totais, de
glicose e de alanina transaminase (KOCAN e BLOUIN, 1985; GARNER et al., 1996;
ROTSTEIN et al., 1999).
Apesar das drogas derivadas das diamidinas e das naftoquinonas terem algum
efeito contra Cytauxzoon spp, o diagnóstico terapêutico não é empregado para a
citauxzoonose felina devido à baixa eficácia das drogas (MOTZEL e WAGNER, 1990;
WALKER e COWELL, 1995; GREENE et al., 1999; MEINKOTH et al., 2000).
2.3.4.3. Diagnóstico diferencial
Os aspectos clínicos das hemoparasitoses em geral são muito semelhantes, e
freqüentemente contribuem para falsos diagnósticos de citauxzoonose felina, quando na
verdade trata-se, em muitos casos, de babesiose ou hemobartonelose (SOARES, 2002).
A qualidade dos preparados citológicos ou teciduais é um fator imprescindível para o
correto diagnóstico de qualquer hemoparasito, pois simples artefatos de técnica e
sujidades freqüentemente confundem profissionais durante a observação microscópica.
Assim, a acuidade e a cautela na diferenciação de hemoparasitos, especialmente no que
diz respeito aos aspectos morfológicos e biológicos do possível agente em questão,
devem ser considerados antes da definição diagnóstica (SOARES, 2002).
Diversos hemoparasitos têm sido objeto de dúvidas em relação a C. felis.
Haemobartonella felis Clark, 1942 é um microrganismo da Ordem Rickettsiales parasita
de eritrócitos de felídeos (CLARK, 1942; BOBADE e NASH, 1987; NEIMAK et al.,
2001). Esta riquétsia é considerada por muitos autores como o principal agente a ser
diferenciado de C. felis (WAGNER, 1976; KIER et al., 1987; FOREYT, 1989; MEIER
e MOORE, 2000; SOARES, 2002). Morfologicamente, todavia, esse agente apresenta
predominantemente formas cocóides e em cadeia, de coloração basofílica densa e
homogênea, tamanho diminuto (0,2–0,9 m) e localizadas perifericamente na superfície
do eritrócito, portanto, formas completamente distinguíveis das de C. felis (WAGNER
et al., 1976; WIGHTMAN et al., 1977; MacWILLIAMS, 1987; COWELL et al., 1988;
HOOVER et al., 1994).
Embora ocorra, aparentemente, com baixa prevalência em felídeos, Babesia spp
pode ser considerada mais importante na diferenciação com C. felis que H. felis. Os
Babesiidae que acometem felídeos são: Babesia felis Davies, 1929; Babesia cati
Mudaliar, Achiary e Alwar, 1950; Babesia herpailuri Dennig, 1967; Babesia pantherae
Dennig e Brocklesby, 1972 e Babesia sp. Lopez-Rebollar, Penzhorn, de Wall e Lewis,
1999 (espécie não-nominada; previamente codificada como pequeno piroplasma KNP).

14
Apesar de haver alguma semelhança entre determinados tipos morfológicos de
Babesia spp com C. felis, não foram verificadas reações sorológicas cruzadas entre estes
dois agentes (UILENBERG et al., 1987; LOPEZ-REBOLLAR et al., 1999).
As formas esquizogônicas de C. felis presentes em tecidos podem ser
consideradas patognomônicas para citauxzoonose, entretanto cabe salientar que
esquizontes de Hepatozoon spp. em felídeos também ocorrem em tecidos de diversos
órgãos e, eventualmente, no endotélio de vasos (SOARES, 2002). Pseudocistos de
Toxoplasma gondii podem ser visualizados em diversos órgãos e tecidos (FOREYT,
1989). Embora em menor freqüência, ninho leishmanióide contendo formas amastigotas
de Trypanosoma cruzi são observados também em felídeos (MacWILLIAMS, 1987;
FOREYT, 1989).
2.3.5. Tratamento
O tratamento eficiente para a citauxzoonose ainda está por ser estabelecido. O
uso de terapia de suporte com solução de Ringer lactato e analgésico associada a
diamidinas, parece apresentar melhor resultado (HOOVER et al., 1994; GREENE et al.,
1999; MEHINKOTH et al., 2000). Contudo, cabe salientar que tanto o aceturato de
diaminazeno quanto o dipropionato de imidocarb possuem moderada ação tóxica para
felinos (GREENE et al., 1999). Ambas as drogas são tóxicas também para outras
espécies animais, inclusive para bovinos (PEREGRINE, 1994). Quando a doença for
aguda, as drogas diamidinas devem ser administradas juntamente com heparina,
mantendo-se a fluidoterapia (MEIER e MOORE, 2000).
A administração de aceturato de diaminazeno e de dipropionato de imidocarb, na
dose única de 2mg/Kg, apresentou eficácia de 50% em gatos com infecção natural por
C. felis (GREENE et al., 1999). O tratamento, com dipropionato de imidocarb, em gatos
com citauxzoonose experimental induzida, via carrapatos, foi ineficiente (MEINKOTH
et al., 2000).
Apesar das naftoquinonas serem consideradas as melhores drogas teilericidas,
essas não têm mostrado eficiência sobre Cytauxzoon spp. A avaliação de parvaquone na
dose única de 20 ou 30 mg/Kg e de buparvaquone na dose de 5 ou 10 mg/Kg, revelou
que ambas são ineficientes no tratamento da citauxzoonose felina experimental
(MOTZEL e WAGNER, 1990).
2.3.6. Distribuição
A citauxzoonose felina foi observada primeiramente em gato doméstico no
sudoeste do Estado de Missouri, EUA (WAGNER, 1976). Posteriormente foi
diagnosticada na região de fronteira entre os Estados de Missouri e Arkansas
(WAGNER et al., 1976) e, em seguida, a enfermidade foi registrada nos Estados do
Texas e Georgia (WIGHTMAN et al., 1977).
Cytauxzoon simile foi reportado infectando naturalmente guepardos cativos (A.
jubatus) no Estado da Califórnia (ZINKL et al., 1981). Só em 1982, no Estado de
Oklahoma, foi diagnosticada a infecção por Cytauxzoon sp. em linces (Tabela 1)
naturalmente parasitados (GLENN et al., 1982).
Posteriormente, a citauxzoonose foi registrada acometendo felinos domésticos e
silvestres experimental e/ou naturalmente infectados em diversos estados americanos
(Tabela 1), principalmente, nas regiões Sul, Sudeste, Centro-oeste e ao longo da costa

15
do Golfo do México (FERRIS, 1979; HAUCK et al., 1982; GLENN et al., 1983; BUTT
et al., 1991; HOOVER et al., 1994; WALKER e COWELL, 1995; GARNER et al.,
1996; ROTSTEIN et al., 1999; MEINKOTH et al., 2000). Experimentalmente, foi
descrita a infecção em ovinos no Estado de Missouri (KIER et al., 1982a).
A maioria dos casos de citauxzoonose felina foi observada nos EUA,
adicionalmente, foi relatada citauxzoonose fatal em tigre de bengala (P. tigris) na
Alemanha (JAKOB e WESEMEIER, 1996) e em macaco rhesus no Panamá
(GARDINER, 1998). Em ambos os casos, o diagnóstico foi estabelecido pelo estudo
histopatológico. Não são conhecidos nestes países quaisquer dados adicionais sobre a
enfermidade e seu agente (Tabela 1).
Na América Latina, os registros de citauxzoonose felina foram realizados no
Brasil, Estado do Rio de Janeiro, em leões (PEIXOTO et al., 1999), em onça-pintada
(SOARES et al., 1999a) em gato-do-mato pequeno (SOARES et al., 1999b) e
experimentalmente em gatos domésticos (SCOFIELD et al., 1999; SOARES, 2002;
SCOFIELD, 2002); no Estado do Mato Grosso do Sul, formas parasitárias
intraeritrocíticas de C. felis like foram diagnosticadas em onça parda (JULIANO et al.,
2002) (Tabela 1).
2.3.7. Potencial zoonótico
O potencial zoonótico de C. felis e outras espécies do gênero Cytauxzoon não é
conhecido, no entanto, pelo fato de ter sido diagnosticada infecção natural e fatal por C.
felis simile em macaco rhesus (GARDINER, 1998) e, por outras características
biológicas e moleculares, não se pode descartar a possibilidade deste hematozoário
infectar humanos.
Curiosamente, as principais espécies de Babesia que infectam humanos, B.
microti, B. rodhaini e B. equi, possuem uma relação molecular e filogenética estreita
com C. felis (ALLSOPP et al., 1994; BRONSDON et al., 1999; KJEMTRUP et al.,
2000b).
Diversas espécies de Babesia têm sido descritas infectando e determinando
doença em seres humanos e entre elas Babesia divergens, B. bovis, B. microti, B. canis,
B. rodhaini e B. equi (TELFORD III et al., 1993; ESKOW et al., 1999). Além disso,
existem cinco espécies descritas e outras ainda não-nominadas em primatas não-
humanos, com potencial zoonótico (TELFORD III et al., 1993; BRONSDON et al.,
1999; ESKOW et al., 1999).
2.4. Filogenia e evolução
A Ordem Piroplasmida foi incluída como grupo próximo aos Amoebae, porém, os
primeiros realizam reprodução sexuada (COX, 1994). Alguns autores colocaram-na
sistematicamente próximo aos Coccidia, enquanto outros a relacionaram aos
Haemosporida (LEVINE, 1988; MEHLHORN, 1988). Como se vê, a classificação dos
Piroplasmida não está bem clara e não satisfaz ao que se conhece a respeito deste grupo
(COX, 1994). Essa ordem possui duas famílias de importância, Babesiidae e
Theileriidae. A primeira família, com o gênero Babesia, contém cerca de 90 espécies
(LEVINE, 1988), a segunda, com o gênero Theileria, contém cerca de 40 espécies
(LEVINE, 1988); o gênero Cytauxzoon, até o momento, apresenta cinco espécies
reconhecidas (SOARES, 2002; REICHARD et al., 2005).

16
A expansão dos Piroplasmida também tem relação com movimentação de seus
hospedeiros, a exemplo do que ocorreu com Babesia bovis e Babesia bigemina para
bovinos (KUTTLER, 1988), mas, principalmente, com a dispersão dos vetores, como
exemplo Boophilus sp. para babésias de bovinos, Amblyomma sp. para Theileria mutans
e Theileria velifera e Rhipicephalus sanguineus para Babesia canis, dentre outros
(NEITZ, 1956; 1957; KUTTLER, 1988; KAKOMA e MEHLHORN, 1994;
MEHLHORN et al., 1994).
Estudos realizados por Allsopp et al. (1994), Cox (1994) e Reichard et al. (2005)
com análises de seqüências de RNA ribossomal 18S revelaram que os membros da
família Theileriidae constituem grupos monofiléticos; assim como alguns membros da
família Babesiidae; ambas as famílias são grupos-irmãos, derivados de um ancestral
parafilético comum (ALLSOPP et al.,1994).
Para alguns autores, o grupo parafilético que originou os taxa monofiléticos
Babesia stricto sensu e Theileria stricto sensu também deram origem a outro taxa
monofilético, o qual contém as espécies Babesia equi, Babesia rodhaini e C. felis
(ALLSOPP et al., 1994). Para estes autores, os três piroplasmídeos encontram-se em
uma posição taxonômica incerta, apesar de possuírem seqüências de RNA similares aos
Theileriidae e Babesiidae. Dessa forma, foi sugerido, com base em similaridades
fenotípicas e filogenéticas, que as três espécies, juntamente com as espécies de Babesia
de aves, formassem uma nova família. Pelo fato de ter sido proposta a reclassificação de
Babesia equi para Nicollia equi (KRYLOV, 1981, citado por ALLSOPP et al., 1994), o
nome apropriado para esta nova família seria Nicolliidae (ALLSOPP et al., 1994).
Mais recentemente, Reichard et al. (2005) realizaram novas análises filogenéticas
com membros da ordem Piroplasmida revelando que o agrupamento das espécies de
Babesia é parafilético, encontrando-se dividido em diferentes clados não-irmãos. Essas
análises também propuseram a reclassificação de Babesia equi e Theileria equi em um
gênero próprio, assim como foi proposto por Allsopp et al. (1994).
Como as mutações no DNA ocorrem de forma aleatória e relativamente constante,
teoricamente é possível medir a divergência entre duas espécies (unidades
reprodutivamente isoladas, ou seja, que não trocam genes) a partir da estimativa das
diferenças encontradas em um determinado fragmento de DNA ou RNA. Os resultados
dessas análises tendem ser mais regulares que a avaliação de caracteres morfológicos
(ALLSOPP et al., 1994; COX, 1994; KJEMTRUP et al., 2000a,b). No entanto, a
associação das análises moleculares às morfológicas e biológicas fornece hipóteses mais
robustas acerca da evolução de organismos (WOESE, 1981; STEWART, 1993; COX,
1994).
2.4.1. Posição sistemática e status taxonômico
Até o momento, foram propostas diversas classificações taxonômicas para os
protozoários, muitas delas baseadas apenas em caracteres morfológicos ou por simples
agrupamento combinatório de caracteres. Assim, a posição sistemática do gênero
Cytauxzoon seria melhor definida com base na associação de diversos parâmetros
morfológicos, biológicos, fisiológicos e patológicos, definidos pelos trabalhos de Neitz
(1956; 1957), Wagner (1976) e Cox (1994).
O status taxonômico de Cytauxzoon spp ainda encontra-se incerto. Para Neitz
(1956), na família Theileriidae du Toit, 1918 encontram-se protozoários que se
multiplicam por esquizogonia em linfócitos e finalmente invadem eritrócitos. As formas
eritrocíticas não se reproduzem e são possíveis gametócitos ou gametas. Esta família é

17
representada por um único gênero Theileria, cuja principal espécie seria Theileria
parva.
Na família Gonderidae Neitz e Jansen, 1956, encontram-se parasitos que se
multiplicam por esquizogonia em linfócitos (Gonderia sp.) ou em histiócitos
(Cytauxzoon sp.) e também em eritrócitos. Nas hemácias se reproduzem por divisão
binária, originando dois ou quatro indivíduos-filhos, que posteriormente assumem a
característica forma de cruz-de-malta. O estágio final do parasito no eritrócito é
possivelmente um gametócito ou gameta. Esta família é representada por dois gêneros,
Gonderia e Cytauxzoon (NEITZ, 1956).
Os membros do gênero Gonderia se multiplicam por esquizogonia em linfócitos e
por fissão em eritrócitos. Possuem diversas espécies: Gonderia annulata, Gonderia
mutans, Gonderia hirci, Gonderia ovis, Gonderia lawrencei, entre outras (NEITZ,
1956). Posteriormente, esse gênero foi considerado sinonímia de Theileria (NEITZ,
1957; LEVINE, 1971; 1985) e todos os piroplasmídeos que realizam esquizogonia em
linfócitos foram classificados como Theileria spp. Alguns autores, todavia, consideram
o gênero Cytauxzoon como Theileria (LEVINE, 1985; GROOTENHUIS et al., 1979) e
outros, baseados em diversos parâmetros citados anteriormente, o mantém na condição
original (WAGNER, 1976; KIER et al., 1982a,b; ALLSOPP et al.,1994). Em
concordância com essa última colocação, será adotada a nomenclatura original do
gênero.
Desta forma, de acordo com Soares (2002), a posição sistemática do parasita em
estudo pode ser assim definida: Domínio Eucarya, Reino Protozoa, Sub-reino Alveolata,
Super-filo Apicomplexa, Filo Sporozoa, Classe Piroplasmea, Ordem Piroplasmida,
Família Theileriidae, Gênero Cytauxzoon e provável espécie Cytauxzoon felis.
2.5. Análises Moleculares
O controle das doenças parasitárias de importância veterinária depende do
diagnóstico rápido e exato dos agentes etiológicos, bem como da habilidade para
caracterizar os agentes parasitários baseando-se na epidemiologia, incluindo
especificidade do hospedeiro, potencial zoonótico, virulência e sensibilidade às drogas.
As técnicas de diagnóstico tradicionais que envolvem microscopia óptica têm sido
complementadas com uma variedade de ferramentas moleculares. A aplicação destas
ferramentas também tem ajudado a resolver problemas taxonômicos que no passado
foram motivo de controvérsia quando novas espécies eram descobertas e descritas
baseadas nas características fenotípicas ou observações epidemiológicas, principalmente
em áreas endêmicas. Recentemente, o enfoque tem sido dirigido em definir e
caracterizar regiões específicas do DNA, que podem ser usados como marcadores para
detectar variações entre diferentes taxons. Isto requer a eleição da ferramenta molecular
capaz de diferenciar as variantes genéticas nos diferentes níveis hierárquicos. Estas
técnicas têm-se mostrado úteis e apropriadas no desenvolvimento e avanço da
taxonomia em diversas áreas da biologia, inclusive na parasitologia (ALLSOPP et al.,
1994; MATHEW et al., 2000).
2.5.1. PCR (Polymerase chain reaction)
A técnica denominada de reação em cadeia da polimerase (PCR) é um método
que envolve a síntese enzimática “in vitro” de bilhões de cópias de um segmento

18
específico de DNA na presença da enzima termoestável, a Taq DNA polimerase. A
reação de PCR se baseia no pareamento de oligonucleotídeos iniciadores ou
“primers”(pequenas seqüências de ácidos nucléicos de fita simples) que delimitam a
região do DNA alvo que será multiplicada. Esses iniciadores são sintetizados
artificialmente de maneira que suas seqüências de nucleotídeos sejam complementares
às seqüências específicas de cada fita do DNA que flanqueiam a região alvo
(ALBERTS et al., 1997).
O princípio da técnica de PCR se baseia em três etapas: desnaturação,
pareamento ou “annealing” e extensão. Estas etapas são organizadas em ciclos, sendo
que cada ciclo da reação é caracterizado pela separação das fitas da dupla-hélice de
DNA genômico durante a etapa da desnaturação. O resfriamento da mistura de PCR na
etapa de pareamento permite que se formem ligações entre os iniciadores com as
seqüências genômicas complementares. Após o pareamento, a DNA-polimerase, na
presença dos desoxirribonucleosídeos trifosfatados (dNTP’s), inicia a etapa de extensão
a partir das regiões extremas de cada uma das fitas de DNA pareadas com os
iniciadores. A cada ciclo, o procedimento de duplicação é repetido e os novos
fragmentos sintetizados servem como moldes para a ligação de outros iniciadores, o
produto final predominante dessa reação de amplificação exponencial é um fragmento
de DNA delimitado pela região onde os iniciadores parearam. Normalmente são
necessários de 20 a 30 ciclos de reação para que amplificação do DNA seja efetiva.
Cada ciclo dobra a quantidade de DNA sintetizada no ciclo anterior resultando em
multiplicação exponencial do número de moléculas do DNA alvo (SAMBROOK et al.,
1989).
2.5.2. RNA ribossômico (RNAr)
Os RNAs ribossômicos são seqüências essenciais na fisiologia celular. Estes
componentes interagem de modo específico com as proteínas ribossômicas para formar
as subunidades dos ribossomos que atuam no processo de síntese de proteínas. Os
RNAs ribossomais são os principais produtos da transcrição em qualquer célula,
constituindo geralmente de 80 a 90% da massa de RNA total dos procariontes e
eucariontes (GORAB, 2001).
As seqüências que codificam para o RNAr são altamente conservadas e podem
ocorrer várias cópias nos diversos organismos estudados. Nos eucariontes, essas cópias
de DNAr estão organizadas em tandem em uma ou mais regiões cromossômicas. Os
RNAs ribossômicos são classificados conforme seu coeficiente de sedimentação em
campo gravitacional de centrifugação de acordo com tamanho utilizando-se como
medida as unidades Svedberg (S). De forma geral, cada unidade de repetição do DNAr
eucarionte possui uma organização conservada, constituída por: um espaçador externo
(external transcribed spacer-ETS), transcrito em uma seqüência que contém a
extremidade 5´da molécula precursora do RNAr 18S com alto coeficiente de
sedimentação (até 45S); região que codifica para RNAr 18S; um espaçador interno que
é transcrito (internal transcribed spacer-ITS) e que contém a região que codifica o
RNAr 25-28S e um espaçador externo (non transcribed spacer NTS), que não é
transcrito (GORAB, 2001).

19
Figura 1. Representação esquemática da unidade de repetição do DNAr dos
eucariontes. A proporção entre as diversas regiões dentro da unidade não está
necessariamente representada na figura. ETS: espaçador externo; 18S região que
codifica para RNAr 18S; ITS: espaçador interno; 25-28S: região que codifica para
RNAr 25-28S; NTS: espaçador externo ou espaçador intergênico (GORAB, 2001
adaptado).
2.5.3. Análises moleculares e filogenéticas
A classificação taxonômica dos parasitos baseava-se anteriormante nas
características morfológicas, ecológicas, epidemiológicas e clínicas. Atualmente a
dificuldade de diagnóstico específico baseado em parâmetros fenotípicos e o
desconhecimento de muitos fatores biológicos de diferentes grupos de parasitos têm
incentivado pesquisadores a empregar técnicas moleculares e análises filogenéticas no
diagnóstico, na classificação e/ou reclassificação de diferentes grupos (ALLSOPP et al.,
1994; KJEMTRUP et al., 2001; REICHARD et al., 2005). O gene 18S tem sido
frequentemente utilizado em análises filogenéticas de diferentes espécies de
protozoários por ser conservado e por se tratar de gene efetivo na fisiologia dos
organismos (ALBERTS et al; 1997; GORAB, 2001; REICHARD et al., 2005).
Para o gênero Cytauxzoon a utilização dessas técnicas tem auxiliado na
identificação de espécies e seqüenciamento de subunidades do gene. Reichard et al.
(2005), utilizando caracterísicas morfológicas, PCR e análises filogenéticas do gene 18S
descreveram a espécie C. manul, que infecta gatos Pallas (Odocoelus manul) oriundos
da Mongólia e importados pelos EUA. Recentemente Bondy Jr. et al. (2005)
identificaram e sequenciaram o gene ITS-1 de C. felis.
ITS
ETS
18S
NTS
25S/28S

20
3. MATERIAL E MÉTODOS
3.1. Origem das Amostras de Cytauxzoon felis simile
Foram utilizadas três amostras de C. felis simile, uma obtida a partir de órgãos
de uma leoa (Panthera leo) de oito anos e sete meses, outra a partir de onça-pintada
(Panthera onca) macho, com cinco anos, e uma terceira a partir de gato-do-mato,
pequeno (Leopardus tigrinus) macho, com quatro anos, procedentes do Zoológico
Municipal de Volta Redonda (altitude = 390m, latitude sul = 22° 31’ 23” e longitude
oeste = 44° 06’ 15”) (CIDE, 1997), Estado do Rio de Janeiro (SOARES, 2002).
As análises moleculares de C. felis simile do isolado da P. onca foram realizadas
a partir de sangue de gato doméstico inoculado experimentalmente (Item 3.2) e para a
amostra do L. tigrinus foi utilizado uma alíquota de sangue criopreservada em
nitrogênio líquido, mantida no Laboratório de Doenças Parasitárias desde o ano de
1998. Fragmentos de órgãos contendo esquizontes de C. felis simile, conservados em
formalina a 10%, desde 1998, foram doados gentilmente pelo Professor Paulo Peixoto
para a realização das análises moleculares.
3.2. Manutenção do Isolado de Panthera onca em Gatos Domésticos
Em 1998, amostras de sangue de P. onca contendo trofozoítas de C. felis simile
(SOARES et al., 1999a; 2002) foram inoculadas experimentalmente em gatos
domésticos (Felis catus) para manutenção e estudos clínico-biológicos (SCOFIELD et
al., 1999; SOARES, 2002; SCOFIELD, 2002). Em 2003, dois gatos domésticos, com
idade de aproximadamente cinco meses, saudáveis e negativos para hemoparasitos em
esfregaço sangüíneo, foram utilizados para multiplicar o isolado da P. onca. Cada
animal recebeu três inóculos de sangue (1,0 ml) de um gato doméstico doador inoculado
em 1998. As inoculações foram realizadas por via subcutânea, com intervalo de 30 dias
entre cada inóculo. Entre o segundo e terceiro inóculo foi realizada imunossupressão em
cada animal, sendo administrado dexametasona (1mg/Kg) via oral, durante sete dias.
Esses animais foram doados gentilmente por um médico veterinário logo após o
desmame.
Os animais foram mantidos em ambiente isolado, segundo recomendações do
Serviço de Sanidade Animal/Delegacia Federal de Agricultura/RJ–Ministério da
Agricultura, Pecuária e Abastecimento, no biotério do Laboratório de Doenças
Parasitárias–Projeto Sanidade Animal, Município de Seropédica (altitude = 26m,
latitude sul = 22° 44’ 38” e longitude oeste = 43° 42’ 27”) (CIDE, 1997), Estado do Rio
de Janeiro. Esses foram alimentados com água e ração comercial para gatos (Top Cat e
Leroy), ad libitum e acompanhados pelo período de dois anos e meio.
3.2.1. Detecção da parasitemia e coleta das amostras do isolado de P. onca
Esfregaços sangüíneos foram confeccionados, a cada três dias a partir do
primeiro inóculo, por veno-punção de vasos marginais da orelha dos dois animais até a
detecção da parasitemia. Os esfregaços sangüíneos periféricos foram secados ao ar,

21
fixados em álcool metílico absoluto, corados pelo Giemsa diluído em tampão Sorensen
pH 6,8 (LUBINSKY, 1960) e observados em microscópio óptico (Dialux 20 ES, Leitz).
A partir da detecção de trofozoítas circulantes, quinze dias após o terceiro
inóculo, amostras de sangue foram coletadas assepticamente, por venopunção radial
utilizando tubos de ensaio do tipo “vacuntainer” contendo anticoagulante citrato-
fosfato-dextrose. As amostras de sangue foram levadas ao Laboratório de Doenças
Parasitárias, Projeto Sanidade Animal/Embrapa onde foi adicionado glicerol 80%.
Posteriormente, essas amostras foram mantidas em nitrogênio líquido até o momento de
serem transportadas para o Laboratório de Biologia Molecular, da área de Sanidade
Animal, Embrapa - Centro Nacional de Pesquisa de Gado de Corte (CNPGC), no
Município de Campo Grande-MS, onde foram processadas para análise genotípica.
3.3. Escolha do Gene
A escolha do gene foi baseada nos trabalhos de Allsopp et al. (1994) e Meinkhot
et al. (2000) que utilizaram o gene 18S (região Ssu-rDNA, menor sub-unidade do
ribossomo nuclear) para discutir a evolução da Ordem Piroplasmida e realizar análises
moleculares e filogenéticas de C. felis. Durante as análises deste estudo, este gene
também foi o único que permitiu a comparação com outras seqüências de Cytauxzoon
depositadas em bases de dados do GenBank (http://www.ncbi.nlm.nih.gov), pois até o
momento das análises havia apenas seqüências do gene 18S para C. felis. Além disso,
em estudos de filogenia o gene 18S tem sido frequentemente utilizado por ser
conservado e por se tratar de gene efetivo na fisiologia dos organismos.
3.4. Desenho dos Oligonucleotídeos Iniciadores
Devido ao escasso número de trabalhos referentes a estudos moleculares para o
gênero Cytauxzoon, foram desenhados dez pares de oligonucleotídeos iniciadores
utilizando o programa computacional DNAStar (http://www.dnastar.com/web/r6.php)
tendo como base a seqüência parcial do gene 18S da subunidade ribossomal de C. felis
(n° de acesso AF399930) depositada no GenBank. Dois pares de iniciadores foram
escolhidos baseando-se na temperatura média, sítio de anelamento e tamanho do
produto amplificado. A seqüência consenso gerada pelo sequenciamento dos produtos
de PCR amplificados com os pares de iniciadores desenhados poderia apresentar
aproximadamente 1445 pares de base (pb).
3.5. Extração de DNA
O DNA nuclear de C. felis simile do isolado de P. onca e da amostra de L.
tigrinus foram obtidos a partir de amostras de 350,0 μl de sangue total de gato
doméstico (Item 3.2.1) e de L. tigrinus (Item 3.1) respectivamente, utilizando o Kit
Easy-DNATM (Genomic Isolation-INVITROGEN ®), conforme especificação do
fabricante para extração de amostras de sangue. O DNA nuclear de C. felis presente nas
amostras de tecido de P. leo (Item 3.1) foi extraído utilizando o mesmo Kit, conforme
especificação do fabricante para extração de amostras de tecido.
Como controle negativo, foi utilizada amostra de sangue de um gato doméstico
saudável e negativo para hemoparasitos em esfregaço sangüíneo. O DNA genômico foi

22
extraído utilizando o Kit Easy-DNATM (Genomic Isolation-INVITROGEN®), conforme
especificação do fabricante para extração de amostras de sangue.
A concentração do DNA extraído de cada amostra foi determinada por
espectrofotômetro (Gene Quant®, Amersham Pharmacia, EUA), sob absorbância de
280 m.
3.6. Amplificação das Amostras de DNA de Cytauxzoon felis simile
A amplificação do gene 18S de Cytauxzoon felis simile foi realizada por meio da
PCR (Polymerase Chain Reaction), com os dois pares de oligonucleotídeos iniciadores
desenhados.
Realizou-se a padronização da PCR, utilizando a amostra extraída de sangue de
gato doméstico (isolado de Panthera onca). Para cada reação foram testados dois
volumes de cada par de iniciadores 100ηg/µL (0,5 e 1,5 µL), três volumes de MgCl2
50Mm (1,5, 2,5 e 3,0 µL), três volumes de DNA (0,5, 1,0 e 2,0 µL) e dois volumes de
Taq DNA polimerase 5 U/L (CENBIOT®) (0,25 e 0,5 µL). Para completar a reação
utilizou-se 2,5 µL tampão de reação 10x (10mM de tris-HCl pH 8,3; 50mM de KCl,
3,0), 0,5 µL de uma mistura contendo 250 M de cada dNTPs, e volume de água
comercial para PCR livre de DNAse (INVITROGEN®), perfazendo o volume final de
25,0 µL de reação.
Após a padronização da reação acima foi amplificado o DNA do isolado de L.
tigrinus. Para essa amostra, utlizou-se também o Kit PCR Master Mix (PROMEGA®),
com a seguinte reação de PCR para cada par de iniciadores: 12,5 µL do PCRMasterMix
(tampão pH 8,5; 3,0 mM de MgCl2; 400 µM de cada dNTPs; Taq DNA polimerase 50
U/mL), 0,5 µL de cada iniciador (100 ηg/µL), 2,0 µL DNA total e 9,5 µL de água
comercial para PCR livre de DNAse (INVITROGEN®), perfazendo um volume final de
25,0 µL de reação.
Para a PCR da amostra de fígado e rim de P. leo, foi utilizado uma reação para
cada par de iniciadores com 2,0 µL tampão de reação 10x (10mM de tris-HCl pH 8,3;
50mM de KCl, 3,0), 1,2µL de MgCl2 50Mm, 0,4 µL de uma mistura contendo 250 µM
de cada dNTPs, 0,4 µL de cada iniciador (100ηg/µL), 0,4 µL de Taq DNA polimerase 5
U/µL (CENBIOT®). Dois volumes de DNA foram testados: 4,0 e 5,0µL. Cada reação
foi completada para um volume final de 20,0 L com água comercial para PCR livre de
DNAse (INVITROGEN®).
As reações de amplificação de DNA de Cytauxzoon felis simile isolado de P.
onca foram realizadas em um termociclador (MJ – RESEARCH® - modelo PTC – 200)
programado da seguinte maneira: um ciclo de um minuto a 94C; 35 ciclos de 94C por
um minuto, gradiente de temperatura de anelamento com as seguintes variações 47, 53,
55, 57, 57,7, 60 e 61C por um minuto e 72C por um minuto. Um ciclo final de
extensão a 72 C por sete minutos foi realizado ao término dos ciclos e as reações foram
resfriadas a 4 °C para interromper a amplificação. Após a padronização, todas as
reações de amplificação de DNA de Cytauxzoon felis simile foram realizadas conforme
descrito acima, alterando apenas a temperatura de anelamento que foi de 60°C.
3.7. Análise dos Produtos de Amplificação
Os produto de PCR (5,0 μL de cada) (Item 3.6) foram adicionados ao tampão de
amostra (1,0 μL), distribuídos separadamente em géis de agarose (1,0%) com Tampão

23
TAE (Tris-acetato-EDTA pH 8,0) e submetidos à eletroforese por uma hora a 60 V.
Para estimar o peso molecular dos produtos de PCR foi utilizado o marcador 1Kb plus
de 100 a 12000 pares de base (INVITROGEN ®) como padrão. Os géis foram corados
em solução de brometo de etídio (5 g/L) por 15 minutos. As bandas obtidas foram
visualizadas em transiluminador (SPECTROLINE® - modelo TL-312A – 312 m
ultravioleta) e fotografadas usando máquina fotográfica digital interligada ao sistema
AlphaDigidoc® (Gel Documentatation e Imagem Analysis System - Alpha Innotech
Corporation).
3.8. Clonagem dos Produtos de PCR
Para a clonagem dos produtos de PCR dos isolados de C. felis de P. onca e de L.
tigrinus, foi realizada uma reação de ligação, catalisada pela enzima topoisomerase I,
adicionando em microtubos de 1,5 mL, 0,5 μL de plasmídeo pTrcHis – TOPO
(INVITROGEN®) e 4,5 μL do produto de PCR (Item 3.7) (Figura 2). Realizou-se
também, para a clonagem dos produtos de PCR do isolado de C. felis de L. tigrinus uma
reação de ligação com 0,5 μL de plasmídeo pTrcHis – TOPO (INVITROGEN®) e 6,0
μL do produto de PCR. As amostras foram homogeneizadas levemente e incubadas por
cinco minutos em temperatura ambiente (Figura 2).
Figura 2. Esquema do plasmídeo pTrcHis – TOPO (INVITROGEN®) utilizado na
clonagem dos produtos de PCR dos isolados de Panthera onca e Leopardus tigrinus.
Os produtos de ligação foram utilizados na transformação de células E. coli TOP
10 produzidas segundo protocolo do Laboratório de Biologia Molecular da Embrapa
Gado de Corte; esses produtos foram adicionados às células homogeneizando levemente
e incubando em gelo por 30 minutos. Posteriormente os tubos foram colocados por 30
segundos em banho-maria à 42°C sem agitação. Em seguida estes foram transferidos
imediatamente para o gelo, onde permaneceram por um período suficiente para serem
transportados até a câmara de fluxo laminar, onde adicionou-se em cada tubo 250,0 μL
de meio SOC. Os tubos foram vedados e submetidos à agitação horizontal a 37°C por
30 minutos. Os produtos transformados foram semeados com auxílio de uma alça de
vidro em placas de Petri 20 x 24 cm devidamente identificadas, contendo meio Luria –
Bertani (LB) ágar com 100 μg/mL de ampicilina. As placas foram incubadas por 12
horas à 37°C para o crescimento das colônias bacterianas.

24
3.9. Lise e Análise de Colônias
As colônias bacterianas contendo o inserto de DNA de C. felis foram
selecionadas. Com auxílio de palitos de dente estéreis, foram removidas doze colônias
das placas de Petri do isolado de Panthera onca e quatro colônias das placas de Petri da
amostra de Leopardus tigrinus; todas as colônias foram distribuídas separadamente em
microtubos de 1,5 mL. Com os mesmos palitos usados para distribuir as colônias,
preparou-se placas “mãe” com meio LB àgar com 100 μg/mL de ampicilina que foram
incubadas durante 12 horas à 37°C para o desenvolvimento das colônias bacterianas.
Nos microtubos contendo as bactérias adicionou-se 15,0 μL de água comercial
para PCR livre de DNAse (INVITROGEN®), homogeneizou-se em vortex por 10
segundos e incubou-se por 10 minutos em banho-maria a 100°C. As amostras (5,0 μL)
mais 1,0 μL de tampão de amostra foram aplicadas em gel de agarose a 1% (utilizando
pentes com dentes finos), em tampão tris – EDTA – acetato (TAE) pH 8,0 e separadas
por eletroforese submarina horizontal em gel sob corrente de 60 V. O gel foi corado em
solução de brometo de etídio (5 g/L) por 15 minutos e as bandas foram visualizadas
sob luz ultravioleta em transiluminador (SPECTROLINE® - modelo TL-312A – 312
m ultravioleta).
Após essa etapa as amostras foram centrifugadas e o sobrenadante utilizado para
a PCR de colônia.
3.10. PCR de Colônia
A PCR de colônia foi realizada para identificar os clones bacterianos que
continham os insertos de interesse. Assim como na PCR (Item 3.6), foram utilizados os
dois pares de oligonucleotídeos iniciadores desenhados.
A reação de PCR para cada par de iniciadores foi realizada com 2,0 µL tampão
de reação 10x (10 mM de tris-HCl pH 8,3; 50 mM de KCl, 3,0), 1,2 µL de MgCl2 50
Mm, 0,4 µL de uma mistura contendo 250 M de cada dNTPs, 0,4 µL de cada iniciador
(100 ηg/µL), 0,4 µL de Taq DNA polimerase 5 U/µL (CENBIOT®) e 2,0 µL do
sobrenadante resultante da lise das colônias (Item 3.9). Cada reação foi completada para
um volume final de 20,0 L com água comercial para PCR livre de DNAse
(INVITROGEN®).
As reações de amplificação foram realizadas em termociclador (MJ –
RESEARCH® - modelo PTC – 200) programado da seguinte maneira: um ciclo de um
minuto a 94C; 35 ciclos de 94C por um minuto, 60C por um minuto e 72C por um
minuto. Um ciclo final de extensão a 72C por sete minutos foi realizado ao término dos
ciclos e as reações foram resfriadas a 4°C para interromper a amplificação.
3.11. Reação de Minipreparação de Plasmídeos (Miniprep)
Para purificação do DNA plasmidial das amostras do gene 18S de C. felis a
partir de cultura de E. coli, utilizou-se o Kit Ultra Clean TM (MOBIO Laboratories®),
conforme especificação do fabricante.
As células transformadas (Item 3.8), previamente selecionadas pela técnica de
PCR de colônia, foram transferidas para tubo “Falcon” de 15,0 mL contendo 2,0 mL de
LB líquido com 100 μg/mL de ampicilina, e incubadas overnight à 37°C sob agitação.

25
Em seguida, 2,0 mL das células cultivadas foram transferidos para tubos de
microcentrífuga, devidamente identificados e centrifugados por um minuto a 10.000 xg.
Descartou-se o sobrenadante por inversão dos tubos e o restante foi retirado com auxílio
de uma pipeta. Ressuspendeu-se o pellet adicionando 50,0 μL da solução de suspenção
(solução I), homogeneizou-se no vortex por um minuto, sendo dez segundos no vortex e
um segundo fora. Adicionou-se 100,0 μL da solução de desnaturação de proteína e de
DNA bacteriano (solução II), homogeneizou-se os tubos invertendo-os por uma vez.
Adicionou-se 325,0 μL da solução de precipitação de proteína e de renaturação de DNA
plasmidial (solução III), homogeneizou-se os tubos invertendo-os por uma vez.
Centrifugou-se os tubos por dois minutos à 10.000 xg. Os sobrenadantes com o DNA
plasmidial dos diferentes isolados foram transferidos para novos microtubos, contendo
colunas com membrana de sílica. Centrifugou-se o conjunto por 30 segundos à 10.000
xg. Adicionou-se 300,0 μL da solução de lavagem. Centrifugou-se o conjunto por 30
segundos à 10.000xg e descartou-se o líquido da solução de lavagem (solução IV).
Finalmente adicionou-se 50,0 μL da solução de eluição (solução V), centrifugou-se o
conjunto por 30 segundos à 10.000 xg e descartou-se as colunas.
Para analisar a qualidade do DNA plasmidial obtido dos diferentes isolados,
aplicou-se 5,0 μL da solução e 1,0 μL de tampão de amostra em gel de agarose a 1%,
em tampão tris – EDTA – acetato (TAE) pH 8,0 e separou-se por eletroforese
submarina horizontal em gel sob corrente de 60V. O gel foi corado em solução de
brometo de etídio (5 g/L) por 15 minutos, as bandas foram visualizadas sob luz
ultravioleta em transluminador e documentado usando o sistema AlphaDigidoc® (Gel
Documentatation e Imagem Analysis System – Alpha Innotech Corporation).
3.12. Sequenciamento
Os produtos obtidos na minipreparação de plasmídios e placas de Petri contendo
as colônias das diferentes amostras foram enviadas para o Instituto de Biologia
Molecular do Paraná – Curitiba, PR, onde foram seqüenciadas (BigDy®Terminatorv
1.1 Cicle Sequencing Kit - Apllied Biosystems), em seqüenciador automático ABI 3100
de 16 capilares (Apllied Biosystems) pelo método dideoxi (SANGER et al., 1977),
segundo as recomendações do fabricante. As amostras foram seqüenciadas com os
iniciadores do gene 18S de Cytauxzoon felis e com os iniciadores do vetor pTrcHis –
TOPO (XpressF e PtrcHisR).
3.13. Análise das Seqüências (in silico)
Para avaliar a qualidade das seqüências foram utilizados três pacotes
computacionais Phred, Phrap, Cross-match (http://www.phrap.org) e Blast
(http://www.ncbi.nlm.nih.gov/blast/); essas análises foram realizadas na Embrapa Gado
de Corte.
O programa Phred faz a leitura dos cromatogramas, traduz a seqüência das bases
em letras e atribui valores de qualidade para cada uma das bases. O valor de qualidade
está relacionado com a probabilidade de erro na leitura da base. Quanto menor a
probabilidade de erro, maior é o valor da qualidade. O valor da qualidade é calculado
pela fórmula:

26
VQ=-10 x log (PE)
VQ = Valor de qualidade
PE = probabilidade de erro
Log = logaritmo na base 10
O programa Phrap faz a detecção de sobreposições e alinhamento das
seqüências e o consenso destas. O Cross-match é um programa de aplicação geral para
comparação de dois conjuntos de seqüências, este programa é utilizado para verificar se
a seqüência obtida não é do plasmídeo utilizado na clonagem.
O Blast consiste em uma busca heurística desenvolvida para ser uma alternativa
mais rápida de comparar seqüências. Esse programa busca, em um banco contendo
milhares de seqüências, aquelas que são similares aos fragmentos de interesse (neste
caso, verifica se os fragmentos recém seqüenciados são realmente do RNAr 18S de
espécies de Cytauxzoon).
3.14. Alinhamento e Análise Filogenética
Os programas computacionais ClustalW (http://www.ebi.ac.uk/clustalw) e Blast
foram utilizados para gerar a seqüência consenso de C. felis de cada amostra estudada e
para compará-las com seqüências de espécies de Cytauxzoon depositadas no GenBank.
O alinhamento das seqüências estudadas com outras seqüências homólogas depositadas
no GenBank (Tabela 2) foi realizado usando o programa MEGA 3
(http://www.megasoftware.net/) e GeneBee
(http://www.genebee.msu.su/services/rna2reduced.html) que leva em conta a estrutura
secundária do RNA ribossomal. As espécies Neospora sp. e Toxoplasma gondii foram
utilizadas como grupo externo com base nos trabalhos de Ketz-Riley et al. (2003) e
Reichard et al. (2005).
A árvore filogenética foi inferida através do método de Máxima
Verossimilhança (MV) (CAVALLI-SFORZA e EDWARDS, 1967; FELSENSTEIN,
1981) com o uso do programa PAUP* (SWOFFORD, 1998). Neste tipo de abordagem,
a seleção cautelosa do modelo de substituição de nucleotídeos é fundamental para
aumentar a confiabilidade da estimativa. Assim, foi utilizado o programa ModelTest
3.06 para estabelecer o modelo de substituição que melhor se ajusta aos nossos dados
(HUELSENBECK e CRANDALL, 1997; POSADA e CRANDALL, 1998). O modelo
escolhido foi o TrN (TAMURA e NEI, 1993), com 44% de sítios invariáveis e o valor
do parâmetro gama em 0,41. Depois de escolhido o modelo, foi construída uma árvore
preliminar através do método de Neighbor Joining (NJ) (SAITOU e NEI, 1987) que foi
otimizada pelo algoritmo TBR (tree-bissection-reconnection) para obter a árvore de
Máxima Verossimilhança. A análise Bayesiana foi utilizada para obter a sustentação dos
nós internos através do programa Mr. Bayes 3.1 (HUELSENBECK e RONQUIST,
2001). Foram realizadas 1.000.000 de gerações através da cadeia de Monte Carlo,
amostradas de 100 em 100 para gerar a árvore consenso.

27
Tabela 2. Espécies de parasitos, número de acesso no GenBank, hospedeiros e país de
origem usados na construção da árvore filogenética e a análise de agrupamento das
seqüências de 18S RNAr de Cytauxzoon felis (continua).
Parasito Nº de acesso no GenBank Hospedeiro Origem
Babesia microti AB032434 Humano Japão
B. microti AY144698 Gambá EUA
B. microti AY144700 Cão Espanha
B.microti AB197940 Quati Japão
B. microti AY144701 Quati EUA
B. microti AY144702 Raposa EUA
B. gibsoni AF158702 Cão EUA
B. rhodaini AB049999 Roedor -
B. canis canis AY649326 Carrapato Rússia
B. canis vogelli AY371198 Cão EUA
Babesia sp. AF188001 Cão Espanha
B. crassa AY260176 Ovino Irã
B. ovata AY603401 Ovino China
B. motasi AY533147 Ovino Espanha
B. bovis AY150059 Bovino Portugal
B. bigemina AY603402 Bovino China
B. odocoilei AY144689 Veado EUA
Babesia sp. WA1 AY027816 Humano EUA
Babesia sp. WA1 AY027815 Humano EUA
B. felis AY542707 Gato África do Sul
B. felis AF244912 - -
B. leo AF244911 Leão África do Sul
B. caballi AY534883 Equino Espanha
B. equi AY150062 Equino Espanha
B. equi AY150063 Equino Espanha
B. equi AY150064 Cão Espanha
B. equi Z15105 Equino África do Sul
Theileria equi AY534882 Equino Espanha
Theileria sp. AB012194 Cervo Japão
Theileria sp. AB012195 Cervo Japão
T. cervi AF086804 Cervo EUA
Theileria sp. AF162432 Veado -
Theileria sp. AY421708 Veado -
Theileria sp. AF162433 Veado -
Theileria sp. U97054 Veado EUA
Theileria sp. AF529273 Cervo Japão
Theileria sp. AF529271 Cervo Japão
T. ovis AY260171 Ovino Sudão
T. ovis AY260172 Ovino Turkia
T. ovis AY533144 Ovino Espanha
T. separata AY260175 Ovino África do Sul
Theileria sp. AY262115 Ovino China
Theileria sp. AY262116 Ovino China
Theileria sp. AY533143 Ovino Espanha
Theileria sp. AY533145 Ovino Espanha
T. cf. ovis AY260174 Caprino Tanzânia
28
Tabela 2. Continuação.
Parasito Nº de acesso GenBank Hospedeiro Origem
T. cf. ovis AY260173 Caprino Tanzania
Theileria sp. AB012200 Caprino Japão
Theileria sp. AB012201 Caprino Japão
Theileria sp. AF078816 Bovino África do Sul
T. sergenti AF081137 Bovino China
Theileria sp. AF162431 Bovino EUA
Theileria sp. AF036336 Bovino China
T. mutans AF078815 Bovino Kenya
T. annulata AY524666 Bovino Turkia
Theileria sp. U97051 Bovino Korea
T. buffeli Z15106 Bovino -
T. buffeli AB000272 - -
T. parva AF013418 Búfalo Kenya
T. parva L02366 - -
T. velifera AF097993 Bovino Tanzânia
T. youngi AF245279 Roedor EUA
T. bicornis AF499604 Rinoceronte preto África do Sul
T. lestoquardi AJ006446 - -
T. taurotragi L19082 - -
Cytauxzoon felis AF399930 Gato EUA
C. felis L19080 Gato -
Cytauxzoon sp. AY309956 Gato Espanha
Cytauxzoon sp. AY496273 Lince Espanha
Cytauxzoon sp. AF531418 Gato Pallas Mongólia
C. manul AY485690 Gato Pallas Mongólia
C. manul AY485691 Gato Pallas Mongólia
Piroplasmida gen. sp. AF158701 Hamster EUA
Piroplasmida gen. sp. AF158706 Cervo EUA
Piroplasmida gen. sp. AF158708 Ovino EUA
Piroplasmida gen. sp. AF158709 Ovino EUA
Neospora sp. U17345 Bovino -
Toxoplasma gondii L37415 - -

29
4. RESULTADOS
4.1. Desenho dos Oligonucleotídeos Iniciadores
Foram utilizados dois pares de iniciadores para cada amostra estudada de DNA
de C. felis simile com o objetivo de obter uma seqüência maior com menor
probabilidade de erros. O primeiro par de iniciadores (Cfe304F 5’ -
ATTGCTTTATGCTGGCGATGTATCA - 3’ e Cfe1159R 5’ -
ACCTGGTAAGTTTTCCCGTGTTGAGTC - 3’) favoreceu a amplificação de um
fragmento de aproximadamente 922 pares de base (pb), enquanto que o segundo par
(Cfe843F 5’ - TTGGGGGCATTCGTATTTAACTGTCAGAG - 3’ e Cfe1680R 5’ -
TTCACAAAACTTTCCATGGCGTATCTAAC - 3’) favoreceu a amplificação de um
fragmento de aproximadamente 866pb (Figura 3). A região de sobreposição entre os
fragmentos amplificados pelos dois pares de oligonucleotídeos iniciadores foi de
aproximadamente 343 pb.
Figura 3. A-B. Esquema dos dois pares de iniciadores (Cfe304F e Cfe1159R e Cfe843F
e Cfe1680R) utilizados na amplificação do gene 18S RNAr de Cytauxzoon felis e seus
respectivos sítios de anelamento; C. Representação dos sítios de anelamento dos
iniciadores na seqüência consenso de Cytauxzoon felis (AF399930).
4.2. Amostras Analisadas e Extração de DNA
As amostras de sangue e órgãos dos animais estudados foram transportadas ao
Laboratório de Biologia Molecular da Embrapa Gado de Corte onde foram processadas
para análises moleculares. É importante ressaltar que o gato doméstico doador
inoculado com isolado de P. onca viveu por aproximadamente cinco anos com baixa e
persistente parasitemia, e que os animais utilizados na multiplicação desse isolado
viveram por mais de dois anos e meio não apresentando sintomatologia de
citauxzoonose.
Em relação à extração de DNA total, a utilização do protocolo do Kit Easy-
DNATM (Genomic Isolation-INVITROGEN®) promoveu um bom resultado nas
Cfe304F
Cfe1159R
Cfe843F
Cfe1680R
Cfe1680R
A
B
C
Cfe843F
Cfe304F
Cfe1159R

30
amostras de sangue de gato doméstico (isolado de P. onca) com parasitemia média de
0,8% (Figura 4), de L. tigrinus com parasitemia média de 0,4% (SOARES, 2002) e do
controle negativo (gato doméstico não infectado por C. felis simile). A concentração de
DNA total das amostras de sangue foi de 115,8 ηg/µL, 55,6 ηg/µL e 70,4 ηg/µL,
respectivamente. Vale ressaltar que após a utilização do DNA extraído dessas amostras
em algumas PCR, este se degradava e novas extrações eram realizadas.
Em relação à extração de DNA total das amostras de fígado e rim de P. leo, foi
observada uma baixa concentração, 3,6 ηg/µL e 2,5 ηg/µL, respectivamente.
Figura. 4. Trofozoítas de Cytauxzoon felis () em esfregaço sangüíneo de gato
doméstico infectado experimentalmente com isolado de Panthera onca. Giemsa (1000x).
4.3. Amplificação das Amostras de DNA de Cytauxzoon felis
A padronização da PCR utilizada para amplificação das amostras de DNA de
Cytauxzoon felis foi realizada durante, aproximadamente, nove meses de análises.
Foram testados diferentes volumes para os reagentes, sendo escolhidos os seguintes:
para cada par de iniciadores 0,5 µL (100 ηg/µL); para o MgCl2 50Mm 1,5 µL; para cada
amostra de DNA 2,0 µL, e para a Taq DNA polimerase 5 U/L (CENBIOT®) 0,5 µL,
em uma reação de PCR com volume final de 25,0 µL. Os demais componentes da
reação foram utilizados segundo protocolo padrão do Laboratório de Biologia
Molecular da Embrapa Gado de Corte, previamente padronizadas para outras espécies
de hemoparasitos de modo a diminuir a quantidade de testes a serem realizados.
O DNA de C. felis extraído das amostras de sangue de gato doméstico (isolado
da P. onca) e do L. tigrinus (Item 3.2) foi amplificado na presença dos iniciadores
Cfe304F e Cfe1159R apresentando um perfil de banda de 922 pb (Figuras 5 e 6). Essas
amostras também foram amplificadas na presença dos iniciadores Cfe843F e Cfe1680R

31
apresentando um perfil de banda de 866 pb (Figuras 5 e 6). O DNA do controle
negativo não amplificou com nenhum par de iniciadores (Figuras 5 e 6).
O DNA total extraído das amostras esquizogônicas de C. felis de fígado e rim de
P. leo (Item 3.2) não amplificou com nenhum par de iniciadores utilizados.
Figura 5. Eletroforese dos produtos de amplificação do gene 18S RNAr de Cytauxzoon
felis do isolado de Panthera onca, em gel de agarose a 1%. M) Marcador de pares de
base; 1) DNA total de extrato de sangue de um gato doméstico negativo com iniciadores
Cfe843F e Cfe1680R; 2) DNA total de extrato de sangue do gato-doméstico positivo
com iniciadores Cfe843F e Cfe1680R; 3) DNA total de extrato de sangue de um gato
doméstico negativo com iniciadores Cfe304F e Cfe1159R, e 4) DNA total de extrato de
sangue do gato doméstico positivo com iniciadores Cfe304F e Cfe1159R.
Pb
12.000
5.000
2000
1650
1000
850
650
500
400
300
200
100
M 1 2 3 4
922pb 866pb
Cytauxzoon felis
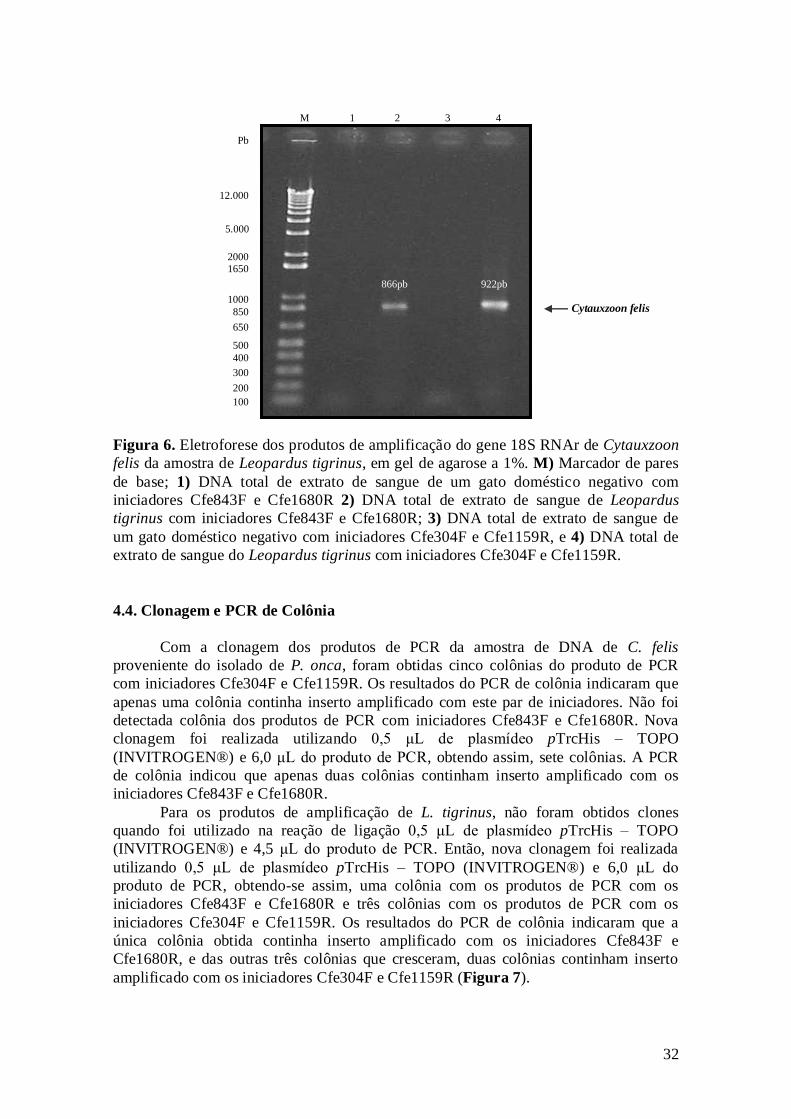
32
M 1 2 3 4
Figura 6. Eletroforese dos produtos de amplificação do gene 18S RNAr de Cytauxzoon
felis da amostra de Leopardus tigrinus, em gel de agarose a 1%. M) Marcador de pares
de base; 1) DNA total de extrato de sangue de um gato doméstico negativo com
iniciadores Cfe843F e Cfe1680R 2) DNA total de extrato de sangue de Leopardus
tigrinus com iniciadores Cfe843F e Cfe1680R; 3) DNA total de extrato de sangue de
um gato doméstico negativo com iniciadores Cfe304F e Cfe1159R, e 4) DNA total de
extrato de sangue do Leopardus tigrinus com iniciadores Cfe304F e Cfe1159R.
4.4. Clonagem e PCR de Colônia
Com a clonagem dos produtos de PCR da amostra de DNA de C. felis
proveniente do isolado de P. onca, foram obtidas cinco colônias do produto de PCR
com iniciadores Cfe304F e Cfe1159R. Os resultados do PCR de colônia indicaram que
apenas uma colônia continha inserto amplificado com este par de iniciadores. Não foi
detectada colônia dos produtos de PCR com iniciadores Cfe843F e Cfe1680R. Nova
clonagem foi realizada utilizando 0,5 μL de plasmídeo pTrcHis – TOPO
(INVITROGEN®) e 6,0 μL do produto de PCR, obtendo assim, sete colônias. A PCR
de colônia indicou que apenas duas colônias continham inserto amplificado com os
iniciadores Cfe843F e Cfe1680R.
Para os produtos de amplificação de L. tigrinus, não foram obtidos clones
quando foi utilizado na reação de ligação 0,5 μL de plasmídeo pTrcHis – TOPO
(INVITROGEN®) e 4,5 μL do produto de PCR. Então, nova clonagem foi realizada
utilizando 0,5 μL de plasmídeo pTrcHis – TOPO (INVITROGEN®) e 6,0 μL do
produto de PCR, obtendo-se assim, uma colônia com os produtos de PCR com os
iniciadores Cfe843F e Cfe1680R e três colônias com os produtos de PCR com os
iniciadores Cfe304F e Cfe1159R. Os resultados do PCR de colônia indicaram que a
única colônia obtida continha inserto amplificado com os iniciadores Cfe843F e
Cfe1680R, e das outras três colônias que cresceram, duas colônias continham inserto
amplificado com os iniciadores Cfe304F e Cfe1159R (Figura 7).
Pb
12.000
5.000
2000
1650
1000
850
650
500
400
300
200
100
922pb 866pb
Cytauxzoon felis
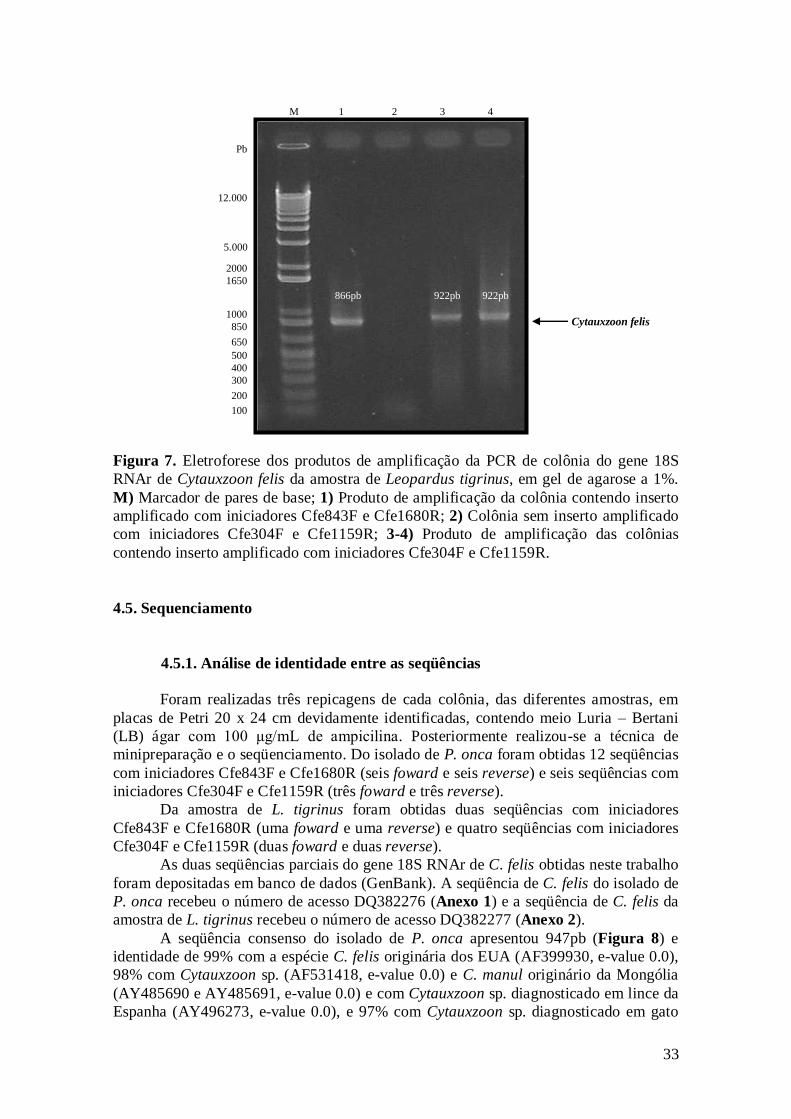
33
M 1 2 3 4
Figura 7. Eletroforese dos produtos de amplificação da PCR de colônia do gene 18S
RNAr de Cytauxzoon felis da amostra de Leopardus tigrinus, em gel de agarose a 1%.
M) Marcador de pares de base; 1) Produto de amplificação da colônia contendo inserto
amplificado com iniciadores Cfe843F e Cfe1680R; 2) Colônia sem inserto amplificado
com iniciadores Cfe304F e Cfe1159R; 3-4) Produto de amplificação das colônias
contendo inserto amplificado com iniciadores Cfe304F e Cfe1159R.
4.5. Sequenciamento
4.5.1. Análise de identidade entre as seqüências
Foram realizadas três repicagens de cada colônia, das diferentes amostras, em
placas de Petri 20 x 24 cm devidamente identificadas, contendo meio Luria – Bertani
(LB) ágar com 100 μg/mL de ampicilina. Posteriormente realizou-se a técnica de
minipreparação e o seqüenciamento. Do isolado de P. onca foram obtidas 12 seqüências
com iniciadores Cfe843F e Cfe1680R (seis foward e seis reverse) e seis seqüências com
iniciadores Cfe304F e Cfe1159R (três foward e três reverse).
Da amostra de L. tigrinus foram obtidas duas seqüências com iniciadores
Cfe843F e Cfe1680R (uma foward e uma reverse) e quatro seqüências com iniciadores
Cfe304F e Cfe1159R (duas foward e duas reverse).
As duas seqüências parciais do gene 18S RNAr de C. felis obtidas neste trabalho
foram depositadas em banco de dados (GenBank). A seqüência de C. felis do isolado de
P. onca recebeu o número de acesso DQ382276 (Anexo 1) e a seqüência de C. felis da
amostra de L. tigrinus recebeu o número de acesso DQ382277 (Anexo 2).
A seqüência consenso do isolado de P. onca apresentou 947pb (Figura 8) e
identidade de 99% com a espécie C. felis originária dos EUA (AF399930, e-value 0.0),
98% com Cytauxzoon sp. (AF531418, e-value 0.0) e C. manul originário da Mongólia
(AY485690 e AY485691, e-value 0.0) e com Cytauxzoon sp. diagnosticado em lince da
Espanha (AY496273, e-value 0.0), e 97% com Cytauxzoon sp. diagnosticado em gato
Pb
12.000
5.000
2000
1650
1000
850
650
500
400
300
200
100
922pb 922pb
866pb
Cytauxzoon felis

34
doméstico da Espanha (AY309956, e-value 0.0), segundo o programa computacional
Blastn (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi).
A seqüência consenso da amostra de L. tigrinus apresentou 1204 pb (Figura 9) e
identidade de 99% com a espécie C. felis originária dos EUA (AF399930, e-value 0.0) e
de 96% com Cytauxzoon sp. (AF531418, e-value 0.0) e C. manul originário da
Mongólia (AY485690, AY485691, e-value 0.0), e com Cytauxzoon sp. isolado da
Espanha (AY496273; AY309956, e-value 0.0), segundo o programa computacional
Blastn (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi).
1 tatttatgtt gtggcttttt ctggtgatta tatttcggta tgattatcca gattgttact
61 ttgagaaaat tagagtgctt aaagcaggca tttgccttga atactttagc atggaataac
121 taagtaggac tttggttcta ttttgttggt ttaagagcca aagtaatgat taataggaac
181 agttgggggc attcgtattt aactgtcaga ggtgaaattc ttagatttgt taaagacgaa
241 ctactgagaa agcatttgcc gaggatgttt tcattaatca agaacgaaag ttaggggatc
301 gaagacgatc agataccgtc gtagtcctaa ccataaacta tgccgactag agattggagg
361 tcgtcagttt gaacgactcc ttcagcacct tgagagaaat caaagtcttt gggttctggg
421 gggagtatgg tcgcaaggct gaaacttaaa ggaattgacg gtagggcacc accaggcgtg
481 gagcctgcgg cttaatttga ctcaacacgg gaaaacttac caggtccaga cagaggaagg
541 attgacagat tgatagctct ttcttgattc tttgggtggt ggtgcatggc cgttcttagt
601 tggtggagtg atttgtctgg ttaattccgt taacgaacga gaccttaacc tgctaaatag
661 gatctgagaa taaactttat gttgtctcag catcgcttct tagagggact ttgcggntat
721 aaatcgcaag gaagtttaag gcaataacag gtctgtgatg cccttanatg tcctgggctg
781 cacgcgcgct acactgatgc attcatcgag tatatccttg gccgagaggc ttgggtaatc
841 tttagtatgc atcgtgatgg ggattgatta ttgcaattat taatcatgaa cgaggaatgc
901 ctagtagacg cgagtcatca gctcgtgtcg attacgtccc tgccctt
Figura 8. Seqüência consenso do gene 18S de Cytauxzoon felis obtida do isolado de
Panthera onca: em azul está marcada a seqüência do iniciador Cfe843F e em amarelo
está a seqüência do iniciador Cfe1159R.
Figura 9. Seqüência consenso do gene 18S de Cytauxzoon felis obtida da amostra de
Leopardus tigrinus: em azul está marcada a seqüência do iniciador Cfe843F e em
amarelo a seqüência do iniciador Cfe1159R.
1 aacggggaat tagggttcga ttccggagag ggagcctgag aaatggctac cacatctaag
61 gaaggcagca ggcgcgtaaa ttacccaatc ctaacacagg gaggtagtga caagaaataa
121 caatacgagg cttaaagtct tgtaattgga atgacggaaa tttaagctct ttccggagta
181 tcaattggag ggcaagtctg gtgccagcag ccgcggtaat tccagctcca atagcgtata
241 ttaaacttgt tgcagttaaa aagctcgtag ttgaatttct gctgcatcat ttatattcct
301 taatcggttt attcatgttg tggctttttc tggtgattat acttcggtat tattatccag
361 attgttactt tgagaaaatt agagtgctta aagcaggcat ttgccttgaa tactttagca
421 tggaataact aagtaggact ttggttctat tttgttggtt taagagccaa agtaatgatt
481 aataggaaca gttgggggca ttcgtattta actgtcagag gtgaaattct tagatttgtt
541 aaagacgaac tactgcgaaa gcatttgcca aggatgtttt cattaatcaa gaacgaaagt
601 taggggatcg aagacgatca gataccgtcg tagtcctaac cataaactat gccgactaga
661 gattggaggt cgtcagtttg aacgactcct tcagcacctt gagagaaatc aaagtctttg
721 ggttctgggg ggagtatggt cgcaaggctg aaacttaaag gaattgacgg aagggcacca
781 ccaggtgtgg agcctgcggc ttaatttgac tcaacacggg aaaacttacc aggcccagac
841 agaggaagga ttgacagatt gatagctctt tcttgattct ttgggtggtg gtgcatggcc
901 gttcttagtt ggtggagtga tttgtctggt taattccgtt aacgaacgag accttaacct
961 gctaaatagg atctgagaat aaactttatg ttgtctcagc atcgcttctt agagggactt
1021 tgcggttata aatcgcaagg aagtttaagg caataacagg tctgtgatgc ccttagatgt
1081 cctgggctgc acgcgcgcta cactgatgca ttcatcgagt atatccttgg ccgagaggct
1141 tgggtaatct ttagtatgca tcgtgatggg gattgattat tgcaattatt aatcatgaac
1201 gagg
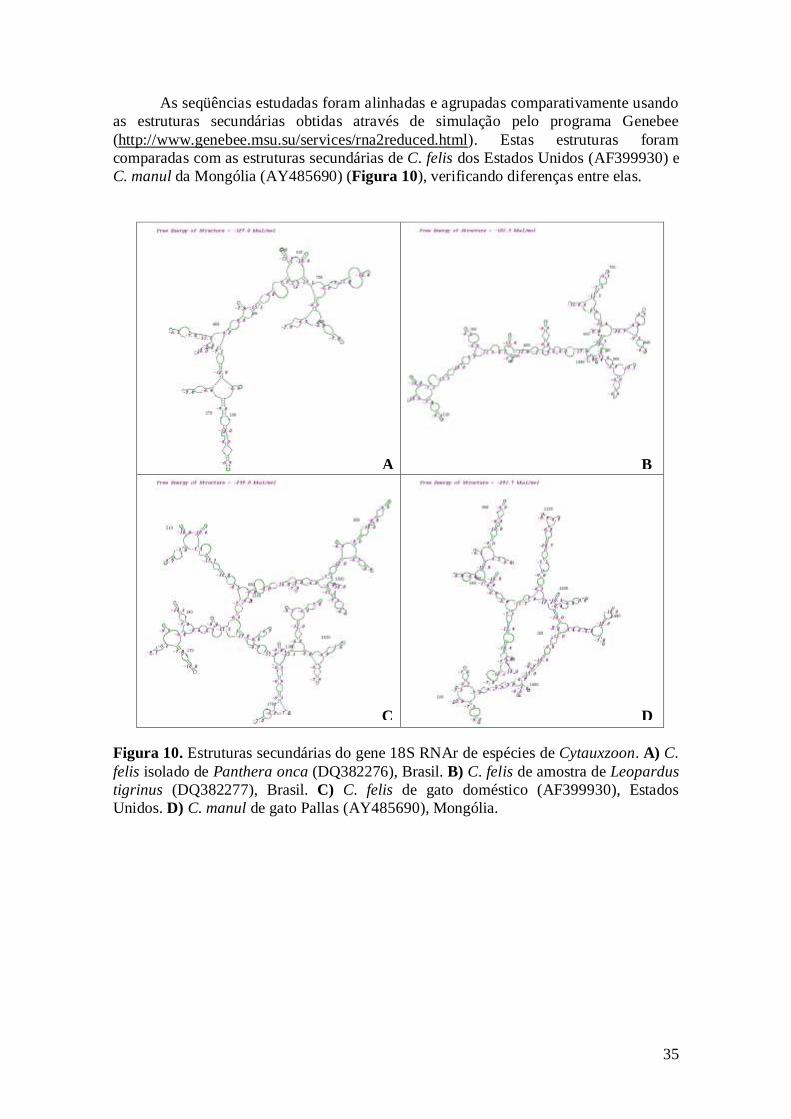
35
As seqüências estudadas foram alinhadas e agrupadas comparativamente usando
as estruturas secundárias obtidas através de simulação pelo programa Genebee
(http://www.genebee.msu.su/services/rna2reduced.html). Estas estruturas foram
comparadas com as estruturas secundárias de C. felis dos Estados Unidos (AF399930) e
C. manul da Mongólia (AY485690) (Figura 10), verificando diferenças entre elas.
Figura 10. Estruturas secundárias do gene 18S RNAr de espécies de Cytauxzoon. A) C.
felis isolado de Panthera onca (DQ382276), Brasil. B) C. felis de amostra de Leopardus
tigrinus (DQ382277), Brasil. C) C. felis de gato doméstico (AF399930), Estados
Unidos. D) C. manul de gato Pallas (AY485690), Mongólia.
B
C D
A

36
4.6. Análise Filogenética
As seqüências consenso obtidas pela amplificação, clonagem e sequenciamento
de DNA de C. felis do isolado de P. onca e da amostra do L. tigrinus foram submetidas
à análise filogenética para testar a identidade das amostras e a distância entre estas
seqüências com as outras seqüências de Cytauxzoon, Babesia, Theileria e Piroplasmidas
não identificados depositadas no GenBank. A Tabela 2 (Item 3.14) mostra todas as
espécies utilizadas neste estudo, seus números de acesso no GenBank, o hospedeiro que
parasitam e o país de origem.
A árvore de MV foi construída com sete seqüências do gene 18S de Cytauxzoon,
27 de Babesia, 38 de Theileria, quatro de Piroplasmidas não identificados e duas do
grupo externo (Toxoplasma gondii e Neospora sp.). Os valores representados nos nós
internos da árvore correspondem à freqüência de árvores que recuperaram cada um
deles nas 1.000.000 árvores geradas durante a análise bayesiana. Essa árvore (Figura
11) mostra a distribuição das espécies do gênero Cytauxzoon que infectam felídeos,
indicando que as amostras estudadas provavelmente são representantes da espécie C.
felis.
Após a separação basal de Toxoplasma gondii e Neospora sp. (espécies
utilizadas como grupo externo), as demais espécies de Piroplasmida dividiram-se em
dois grandes grupos. O primeiro (de cima para baixo) reúne espécies de Babesia que
infectam roedores, marsupiais, humanos e felinos. O segundo grande grupo se subdivide
em três grupamentos de espécies de Babesia, um grupamento de Piroplasmidas não
identificados, dois grupamentos de espécies de Theileria e um grupamento de espécies
de Cytauxzoon. Esse último grupamento foi dividido em dois grupos discretos, o
primeiro contendo apenas seqüências identificadas como C. felis originárias dos EUA,
onde estão também inseridas as duas seqüências estudadas, e o segundo grupo onde
estão inseridas seqüências de Cytauxzoon sp. e Cytauxzoon manul oriundas da
Mongólia, e Cytauxzoon sp. diagnosticado na Espanha.
Pôde-se observar que as espécies de Cytauxzoon que infectam felídeos formam
um grupo monofilético, diferindo das espécies do gênero Babesia que formaram quatro
grupos distintos e das espécies de Theileria que formaram dois grupos distintos. Vale
ressaltar que o grupo das espécies de Babesia que infectam felídeos, principal
diagnóstico diferencial para a espécie C. felis, ficou distante do grupo das espécies de
Cytauxzoon.
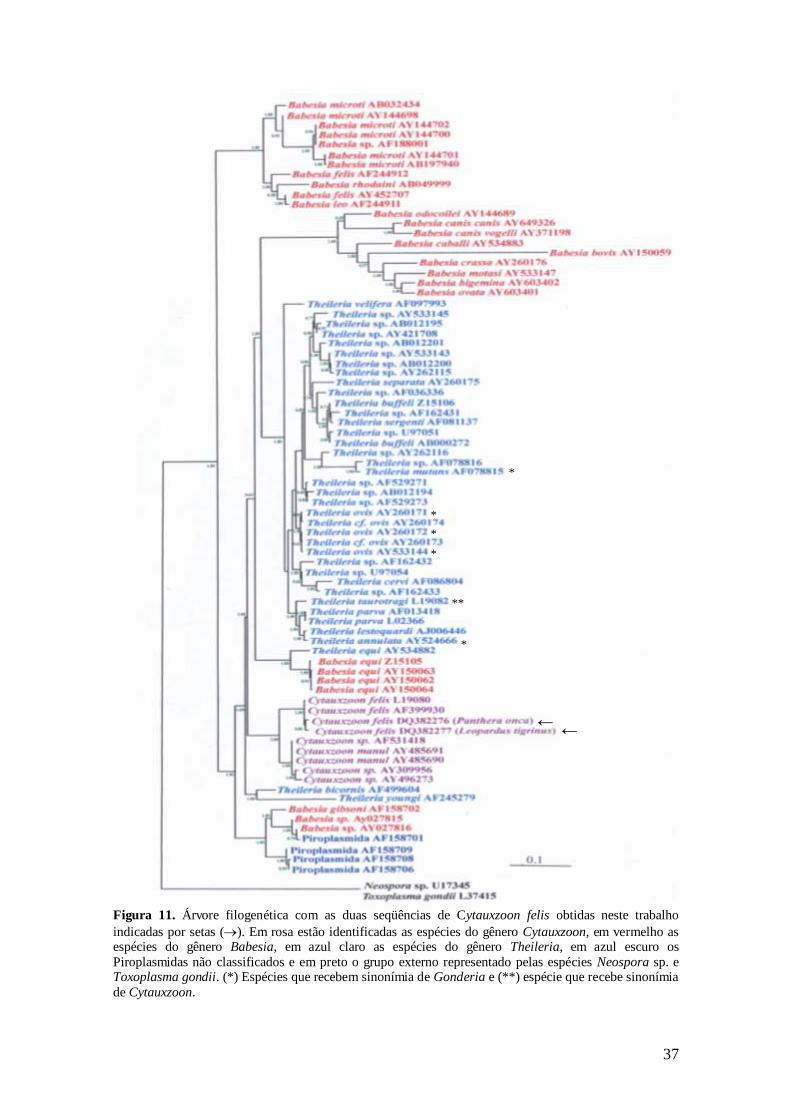
37
Figura 11. Árvore filogenética com as duas seqüências de Cytauxzoon felis obtidas neste trabalho
indicadas por setas (). Em rosa estão identificadas as espécies do gênero Cytauxzoon, em vermelho as espécies do gênero Babesia, em azul claro as espécies do gênero Theileria, em azul escuro os
Piroplasmidas não classificados e em preto o grupo externo representado pelas espécies Neospora sp. e Toxoplasma gondii. (*) Espécies que recebem sinonímia de Gonderia e (**) espécie que recebe sinonímia
de Cytauxzoon.
←
←
*
*
*
*
**
*

38
5. DISCUSSÃO
O presente estudo relata a caracterização molecular de isolados de C. felis
diagnosticados em Panthera onca e Leopardus tigrinus no Brasil. A amplificação e o
sequenciamento do gene 18S RNAr de C. felis obtido a partir de sangue de gato
doméstico (isolado de P. onca) e de L. tigrinus revelaram a sensibilidade do diagnóstico
molecular, confirmando o diagnóstico microscópico de trofozoítas intraeritrocíticos de
C. felis simile realizado por Soares et al. (1999a,b). Vale ressaltar que a parasitemia de
C. felis do isolado de P. onca obtida neste estudo foi em média de 0,8% e da amostra de
L. tigrinus foi de 0,4% segundo Soares (2002). Meinkoth et al. (2000) também
relataram a sensibilidade do diagnóstico molecular, confirmando com PCR casos
suspeitos de cytauxzoonose felina em animais que sobreviveram à infecção.
A não amplificação do gene 18S RNAr de C. felis das amostras de rim e fígado
de Panthera leo pode ter ocorrido devido a baixa concentração de DNA em decorrência
de degradação promovida pelo longo tempo de conservação em formalina 10%. Estudos
revelaram que o processo de fixação por formalina promove a degradação de ácidos
nucléicos e proteínas, modificando também as ligações covalentes do RNA, dificultando
a extração. A presença de endonucleases nas amostras teciduais também pode atuar
como fator inibitório de amplificação de DNA (SHI et al, 2002; KÖRBLER at al.,
2003).
O desenho dos iniciadores consistiu em uma etapa importante neste estudo, uma
vez que os iniciadores utilizados nos trabalhos de Allsopp et al. (1994), Meinkoth et al.
(2000) e Ketz-Riley et al. (2003) amplificaram muitas espécies da Ordem Piroplasmida.
Os dois pares de iniciadores utilizados, Cfe304F e Cfe1159R e Cfe843F e Cfe1680R,
além de amplificarem um número menor de espécies dessa Ordem, permitiram também
a obtenção de uma seqüência com maior número de pares de base e menor número de
erros. As duas seqüências parciais do gene 18S RNAr de C. felis obtidas neste trabalho
foram depositadas em banco de dados GenBank. A seqüência consenso do gene 18S
RNAr de C. felis obtido a partir de sangue de gato doméstico (isolado de P. onca)
resultou em 947 pb (DQ382276), e a seqüência consenso obtida a partir de sangue de L.
tigrinus apresentou 1204 pb (DQ382277). Os dois pares de iniciadores não
amplificaram o DNA do controle negativo (DNA extraído do sangue de dato doméstico
não infectado). Em 2005, Bondy Jr. et al. identificaram e sequenciaram o gene ITS-1 de
C. felis, desenhando um iniciador reverse específico. Os dados obtidos no presente
trabalho com o gene 18S de C. felis não foram comparados com os dados destes autores,
pois, até o momento das análises realizadas, todas as seqüências de Cytauxzoon
depositadas no GenBank eram do gene 18S.
Em relação às análises das estruturas secundárias das duas seqüências de C. felis
estudadas (DQ382276 e DQ382277), da seqüência de C. felis dos EUA (AF399930) e
da seqüência de C. manul (AY485690) foi possível observar as diferenças estruturais
existentes. Apesar da estrutura secundária ser resultante da simulação a partir de uma
pequena região codificadora para 18S RNAr, suas formas estão associadas à diferença
entre algumas bases das seqüências dessa região nestas espécies.
As análises filogenéticas das duas seqüências estudadas foram realizadas com
outras 78 depositadas no GenBank. Duas espécies, Toxoplasma gondii e Neospora sp.,
foram utilizadas como grupo externo com base nos estudos filogenéticos realizados por
Ketz-Rilley et al. (2003) e Reichard et al. (2005). As análises realizadas revelaram um
agrupamento irmão entre os grupos de espécie de C. felis que parasita felinos nos

39
Estados Unidos, com as duas seqüências obtidas neste trabalho e o grupo de Cytauxzoon
manul oriundo da Mongólia e Cytauxzoon sp. da Espanha. Pôde-se observar o
monofiletismo entre as espécies de Cytauxzoon que infectam felídeos, concordando com
os resultados obtidos por Ketz-Riley et al. (2003) e Reichard te al. (2005). Já Allsopp et
al. (1994), com base em similaridades fenotípicas e filogenéticas, propuseram a
reclassificação de C. felis, Babesia equi e B. rodhaini em uma nova família denominada
Nicolliidae.
Quanto ao grupo das espécies de Theileria, as análises realizadas revelaram que
esse foi dividido em dois subgrupos distintos: um pequeno com as espécies T. youngi e
T. bicornis, e outro com todas as outras espécies de Theileria, não sendo um grupo
monofilético. Esses resultados discordam daqueles apresentados por Reichard et al.
(2005) que defendem o monofiletismo desse grupo, propondo uma nova classificação
para a espécie Theileria youngi; a seqüência de Theileria bicornis não foi analisada por
esses autores. As duas seqüências de C. felis obtidas neste estudo apresentaram 96% de
identidade com T. youngi e T. bicornis. Existe a possibilidade destas espécies
pertencerem ao gênero Cytauxzoon, mas outros marcadores devem ser usados e mais
informações sobre a biologia dessas espécies devem ser estudadas para confirmar esta
reclassificação. O presente trabalho também confirma a sinonímia de Theileria annulata
(= Gonderia annulata), T. mutans (= G. mutans) e T. ovis (= G. ovis, G. hirci) citada por
Neitz (1957) e Levine (1988); essas espécies ficaram agrupadas com outras espécies de
Theileria.
Outra seqüência depositada no GenBank e utilizada nas análises filogenéticas do
presente estudo foi Theileria taurotragi que ficou agrupada com outras espécies do
gênero Theileria. Essa espécie foi descrita inicialmente como Cytauxzoon taurotragi
Martin e Brocklesby, 1960 (BROCKLESBY, 1962), mas outros autores como YOUNG
et al. (1980) classificaram-na como T. taurotragi. Os resultados obtidos concordam com
YOUNG et al. (1980) classificando esta espécie como membro do gênero Theileria.
Em relação ao agrupamento das espécies de Babesia, pôde-se observar a
formação de quatro grupos distintos: um agrupando as espécies que infectam felinos,
roedores, humanos e marsupiais; um segundo com as espécies que infectam ruminantes,
cães e eqüinos, um terceiro grupo representado pela espécie B. equi (=T. equi) e um
quarto representado por espécies diagnosticadas em cães, humanos, ovinos, cervos e
hamsters. Torna-se evidente o não monofiletismo desse grupo, corroborando com os
resultados apresentados por Allsopp et al. (1994) e Reichard te al. (2005). Foi observado
também que as seqüências de B. equi formaram um grupo mais próximo do
agrupamento das espécies de Theileria, do que das espécies de Babesia, corroborando
com os resultados obtidos por Reichard et al. (2005). A classificação dessa espécie,
ainda está incerta, pois alguns autores classificam-na como Nicollia equi (KRYLOV,
1981, citado por ALLSOPP et al.,1994), outros como Babesia equi (GUIMARÃES et
al., 1998) e ainda como Theileria equi (MELHORN e SCHEIN, 1998). Neste estudo foi
adotada a classificação da espécie como B. equi, sugerindo que mais estudos devam ser
realizados com o objetivo de definir a correta classificação taxonômica desse agente.
Neste estudo foram identificados dois novos hospedeiros naturais de C. felis,
onça-pintada (P. onca) e gato-do-mato (L. tigrinus) no Brasil. Anteriormente infecção
natural por C. felis tinha sido relatada em guepardo (ZINKL et al., 1981), em puma ou
pantera da Flórida (BUTT et al., 1991; ROTSTEIN et al., 1999), em puma ou pantera do
Texas (ROTSTEIN et al., 1999), em tigre branco (GARNER et al., 1996), em tigre de
bengala (JAKOB e WESEMEIER, 1996) e em leões (PEIXOTO et al., 1999).
As informações referentes ao ciclo biológico, aspectos morfológicos,
patogenicidade e dados epidemiológicos de um determinado agente parasitário,

40
associadas às análises moleculares e filogenéticas são de fundamental importância, uma
vez que as combinações dessas informações garantem um diagnóstico mais seguro e de
maior sensibilidade, além de contribuir para a taxonomia do agente estudado.

41
CONCLUSÕES
1. Com base nos estudos moleculares do gene 18S RNAr e análises filogenéticas
das amostras estudadas, a espécie de teilerídeo diagnosticada em Panthera onca e
Leopardus tigrinus é Cytauxzoon felis;
2. A PCR do gene 18S RNAr é indicada como método de diagnóstico para C.
felis.
3. O grupo das espécies de Cytauxzoon que parasitam felídeos é monofilético,
sendo que o grupo de C. felis onde estão inseridas as seqüências do presente trabalho é
irmão do grupo de C. manul da Mongólia e Cytauxzoon sp. isolado da Espanha.
4. Duas novas espécies de hospedeiros naturais de C. felis, Panthera onca e
Leopardus tigrinus, são relatadas ampliando a faixa de hospedeiros conhecidos.

42
REFERÊNCIAS BIBLIOGRÁFICAS
ALBERTS, B.; BRAY, D.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WATSONN, J. D.
Biologia molecular da célula. 3º Edição, Artes Médicas Porto Alegre, 2983p. 1997.
ALLSOPP, M. T. E. P.; CAVALIER-SMITH, T.; DE WAAL, D. T.; ALLSOPP, B. A.
Phylogeny and evolution of the piroplasms. Parasitology, v. 108, p. 147-152. 1994.
BARNETT, S. F. Theileria. In: KREIER, J. P. Parasitic Protozoa. Vol. 4. Academic
Press, New York. p. 77-114. 1977.
BLOUIN, E. F.; KOCAN, A. A.; GLENN, B. L.; KOCAN, K. M. Transmission of
Cytauxzoon felis Kier, 1979 from bobcats, Felis rufus (Schreber), to domestic cats by
Dermacentor variabilis (Say). Journal Wildlife Disease, v. 20, n. 3, p. 241-242. 1984.
BLOUIN, E. F.; KOCAN, A. A.; KOCAN, K. M. Evidence of a limited schizogonous
cycle for Cytauxzoon felis in bobcats following exposure to infected ticks. Journal
Wildlife Disease, v. 23, n.3, p. 499-501. 1987.
BOBADE, P. A.; NASH, A. S. A comparative study of the efficiency of acridine orange
and some Romanowsky staining procedures in the demostration of Haemobartonella
felis in feline blood. Veterinary Parasitology, v. 26, p. 169-172. 1987.
BONDY Jr., P. J.; COHN, L. A.; TYLER, J. W.; MARSH, A. E. Polymerase chain
reaction detection of Cytauxzoon felis from field-collected ticks and sequence analysis
of the Small subunit and Internal transcribed spacer 1 region of the ribosomal RNA
gene. Journal of Parasitology, v. 91, n. 2, p. 458-461. 2005.
BOURDEAU, P. Les babésioses félines. Le Point Vétérinaire, v. 27, n.173, p. 947-953.
1996.
BROCKLESBY, D. W. Cytauxzoon taurotragi Martin e Brocklesby, 1960 a piroplasm
of the eland (Taurotragus oryx pattersonianus Lydekker, 1906). Research in Veterinary
Science, v. 3, p. 334-348. 1962.
BUTT, M. T.; BOWMAN, D.; BARR, M. C.; ROELKE, M. E. Iatrogenic transmission
of Cytauxzoon felis from a Florida panther (Felis concolor coryi) to a domestic cat.
Journal of Wildlife Disease, v. 27, n. 2, p. 342-347. 1991.
CASADEVALL, A.; PIROFSKI, L-A. Host-pathogen interactions: Basic concepts of
microbial commensalism, colonization, infection, and disease. Infection and Immunity,
v. 68, n. 12, p. 6511-6518. 2000.
CAVALLI-SFORZA, L. L.; EDWARDS, A. W. F. Phylogenetic analysis: models and
estimation procedures. American Journal Human Genetics, v. 19, p. 122-257. 1967.

43
CIDE. Território, Rio de Janeiro. Fundação de Informação e Dados do Rio de Janeiro.
80p. 1997.
CLARK, R. Eperythrozoon felis (Sp. Nov.) in a cat. Journal of South African
Veterinary Medical Association, XIII, p. 15-16. 1942.
COWELL, R. L.; FOX, J. C.; PANCIERA, R. J.; TYLER, R. D. Detection of
anticytauxzoon antibodies in cats infected with a Cytauxzoon organism from bobcats.
Veterinary Parasitology, v. 28, p. 43-52. 1988.
COX, F. E. G. The evolutionary expansion of the sporozoa. International Journal for
Parasitology, v. 24, p.1301-1316. 1994.
DAVIS, L. J. On a piroplasm of the Sudanese wild cat (Felis ocreata). Transaction of
the Royal Society of Tropical Medicine and Hygiene, v. 22, p. 523-534. 1929.
DENNIG, H. K. Eine unbekannte babesienart beim jaguarundi (Herpailurus
yaguarondi). Die Kleintierpraxis, v. 12, p. 146-152. 1967.
DENNIG, H. K.; BROCKLESBY, D. W. Babesia pantherae sp. nov., a piroplasm of
the leopard (Panthera pardus). Parasitology, v. 64, p. 525-532. 1972.
ESKOW, E.; KRAUSE, P. J.; SPIELMAN, A.; FREEMAN, K.; ASLANZADEH, J.
Southern extension of the range of human babesiosis in the Eastern United States.
Jounal of Clinical Microbiology, v. 37, n. 6, p. 2051-2052. 1999.
FELSENSTEIN, J. Evolutionary trees from DNA sequences: a maximum likelihood
approach. Journal of Molecular Evolution, v. 17, p. 368-376. 1981.
FERRIS, D. H. A progress report on the status of a new disease of American cats:
Cytauxzoonosis. Comparative Immunology Microbiology and Infectious Diseases, v. 1,
p. 269-276. 1979.
FOREYT, W. J. Diagnostic Parasitology. Veterinary Clinical North American Small
Animal Practice, v. 19, n. 5, p. 979-1001. 1989.
GARDINER, C. H. Comunicação pessoal. (Armed Forces Institute of Pathology,
Washington, DC, USA). Seminário apresentado no Departamento de Parasitologia
Animal da UFRRJ, Brasil, em 19 de dezembro de 1998.
GARNER, M. M.; LUNG, N. P.; CITINO, S.; GREINER, E. C.; HARVEY, J. W.;
HOMER, B. L. Fatal cytauxzoonosis in a captive-reared write tiger (Panthera tigris).
Veterinary Pathology, v. 33, p. 82-86. 1996.
GLENN, B. L.; ROLLEY, R. E.; KOCAN, A. A. Cytauxzoon-like piroplasms in
erythrocytes of wild-trapped bocats in Oklhoma. Journal of the American Veterinary
Medical Association, v. 181, n. 11, p. 1251-1253. 1982.
GLENN, B. L.; KOCAN, A. A.; BLOUIN, E. F. Cytauxzoonosis in bobcats. Journal of
the American Veterinary Medical Association, v. 183, n. 11, p. 1155-1158. 1983.

44
GLENN, B. L; STAIR, E. L. Cytauxzoonosis in domestic cats: report of two cases in
Oklahoma, with a review and discussion of the disease. Journal of the American
Veterinary Medical Association, v. 184, n. 7, p. 822-825. 1984.
GORAB, E. Evolução dos genes nucleares de RNA ribossômico. In: MATIOLI, S.R.
Biologia Molecular e Evolução. Holos Editora. Ribeirão Preto, p. 202. 2001.
GREENE, C. E.; LATIMEER, K.; HOPPER, E.; SHOEFFLER, G.; LOWER, K.;
CULLENS, F. Administration of diminazene aceturate or imidocarb dipropionate for
treatment of cytauxzoonosis in cats. Journal of the American Veterinary Medical
Association, v. 215, n. 4, p. 497-500. 1999.
GROOTENHUIS, J. G.; YOUNG, A. S.; DOLAN, T. T.; STAGG, D. A. The
characteristics of Theileria species (eland) infections in eland and cattle. Research in
Veterinary Science, v. 27, p. 59-68. 1979.
GUIMARÃES, A.M.; LIMA, J.D.; RIBEIRO, M.F.B.; CAMARGOS, E.R.S.; BOZZI,
I.A. Ultrastructure of sporogony in Babesia equi in salivary glands of adult female
Boophilus microplus ticks. Parasitology Research, v. 84, p. 69-74. 1998.
HAUCK, W. N.; SNIDER, T. G.; LAWRENCE, J. E. Cytauxzoonose in a native
Louisiana cat. Journal of the American Veterinary Medical Association, v. 180, n. 12, p.
1472-1474. 1982.
HAZEN-KARR, C. G.; KOCAN, A. A.; KOCAN, K. M.; HAIR, A. The ultrastructure
of sporogony in Theileria cervi (Bettencourt et al., 1907) in salivary glands of female
Amblyomma americanum (L.) ticks. Journal of Parasitology, v. 73, n. 6, p. 1182-1188.
1987.
HOOVER, J. P.; WALKER, D. B.; HEDGES, J. D. Cytauxzoonosis in cats: Eight cases
(1985-1992). Journal of the American Veterinary Medical Association, v. 205, n. 3, p.
455-460. 1994.
HOOGSTRAAL, H. Argasid and Nuttalliellid ticks as parasites and vectors. Advances
in Parasitology, v. 24, p. 135-238. 1985.
HUELSENBECK, J. P.; CRANDALL, K. A. Phylogeny estimations and hypothesis
testing using maximum likelihood, Annual Review Ecology Systematic, v. 28, p. 437-
466. 1997.
HUELSENBECK, J. P.; RONQUIST, F. Mr. Bayes: Bayesian inference of phylogenetic
tree, Bioinformatics Applications Note, v. 17, p. 754-755. 2001.
INVITROGEN. Invitrogen life technologies - Instruction Manual. Printe in USA, p. 6-
7. 2003.
JAKOB, W.; WESEMEIER, H. H. A fatal infection in a bengal tiger resembling
cytauxzoonosis in domestic cats. Journal of Comparative Pathology, v. 114, p. 439-444.
1996.

45
JARDINE, J. E. The pathology of cytauxzoonosis in a tsessebe (Damaliscus lunatus).
Journal of South African Veterinary Association, v. 63, n. 1, p. 49-51. 1992.
JULIANO, R. S.; SOUZA, A. I.; TÉRCIO NETTO, N.; SCHEIDE, R.; SCOFIELD, A.
Cytauxzoon felis like in the Mountain Lion (Puma concolor): A Case Report. Journal of
Animal and Veterinary Advances, v. 3, n. 12, p. 820-823. 2004.
KAKOMA, I.; MEHLHORN, H. Babesia of domestic animals. In: KREIER, J. P.
Parasitic Protozoa. 2ª edition, Vol. 7. Academic Press, New York. 1994. p. 141-216.
KETZ-RILEY, C. J.; REICHARD, M. V.; BUSSCHE, R. A. V. D.; HOOVER, J. P.;
MEINKOTH, J.; KOCAN, A. A. An intraerythrocytic Small piroplasm in wild-caugth
Pallas’s cats (Otocolobus manul) from Mongolia. Journal of Wildlife Disease, v. 39, n.
2, p. 424-430. 2003.
KIER, A. B.; WIGHTMAN, S. R.; WAGNER, J. E. Interspecies transmission of
Cytauxzoon felis. American Journal of Veterinary Research, v. 43, n. 1, p. 102-105.
1982a.
KIER, A. B.; WAGNER, J. E.; MOREHOUSE, L. G. Experimental transmission of
Cytauxzoon felis from bobcats (Lynx rufus) to domestic cats (Felis domesticus).
American Journal of Veterinary Research, v. 43, n. 1, p. 97-101. 1982b.
KIER, A. B.; WAGNER, J. E.; KINDEN, D. A. The pathology of experimental
cytauxzoonosis. Journal of Comparative Pathology, v. 97, p. 415-432. 1987.
KJEMTRUP, A. M.; THOMFORD, J.; ROBINSON, T.; CONRAD, P. A. Phylogenetic
relationships of human and wildlife piroplasm isolates in the Western United States
inferred from the 18S nuclear small subunit RNA gene. Parasitology, v. 120, p. 487-
493. 2000a.
KJEMTRUP, A. M.; KOCAN, A. A.; WHITWORTH, L.; MEINKOTH, J.;
BIRKENHEUER, A. J.; CUMMINGS, J.; BOUDREAUX, M. K.; STOCKHAM, S. L.;
IRIZARRY-ROVIRA, A.; CONRAD, P. A. There are at least three genetically distinct
small piroplasms from dogs. International Journal for Parasitology, v. 30, p. 1501-
1505. 2000b.
KOCAN, A. A.; BLOUIN, E. F. Hematologic and serum chemical values for free-
ranging bobcats, Felis rufus (Schreber), with reference to animals with natural
infections of Cytauxzoon felis Kier, 1979. Journal of Wildlife Disease, v. 21, n. 2, p.
190-192. 1985.
KOCAN, A. A.; KOCAN, K. M.; BLOUIN, E. F.; MUKOLWE, S. W. A redescription
of schizogony of Cytauxzoon felis in the domestic cat. Annals New York Academy of
Sciences, v. 653, p. 161-167. 1992.
KÖRBLER, T.; GRSKOVIÉ, M.; DOMINIS, M.; ANTICA, M. A simple method for
RNA isolation from formalin-fixed and paraffin-embedded lymphatic tissues.
Experimental and Molecular Pathology, v. 74, p. 336-340. 2003.

46
KUTTLER, K. L. World-wide impact of babesiosis. In: RISTIC, M. Babesiosis of
Domestic Animals and Man. CRC Press, Boca Raton. 1988. p. 1-22.
LEVINE, N. D. Taxonomy of the piroplasms. Transactions of the American
Microscopical Society, v. 90, p. 2-33. 1971.
LEVINE, N. D. Veterinary Protozoology. Iowa State University Press, Ames. 1985. 414
p.
LEVINE, N. D. The Protozoan Phylum Apicomplexa. Vol. 2. CRC Press, Boca Raton.
1988. 154 p.
LOPEZ-REBOLLAR, L. M.; PENZHORN, B. L.; DE WAAL, D. T.; LEWIS, B. D. A
possible new piroplasm in lions from the Republic of South Africa. Journal of Wildlife
Disease, v. 35, n. 1, p. 82-85. 1999.
LUBINSKY, G. Note of the use of McLrvaine’s buffer solutions in staining blood
Protozoa. Canadian Journal of Zoology, v. 38, p. 437-438. 1960.
MACWILLIAMS, P. S. Erythrocytic rickettsia and protozoa of the dog and cat
Veterinary Clinical North American Small Animal Practice, v. 17, n. 6, p. 1443-1461.
1987.
MARTIN, H.; BROCKLESBY, D. W. A new parasite of eland. Veterinary Record, v.
72, p. 331-332. 1960.
MATHEW, J. S.; VAN DEN BUSSCHE, R. A., EWING, S. A. Phylogenetic
relationships of Hepatozoon (Apicomplexa: Adeleorina) based on molecular,
morphologic, and life-cicle characters. The Journal of Parasitology, v. 86, p. 366-372.
2000.
McCULLY, R. M.; KEEP, M. E.; BASSON, P. A. Cytauxzoonosis in a giraffe (Giraffa
camelopardalis (Linneaus, 1758) in Zululand. Onderstepoort Journal of Veterinary
Research, v. 37, p. 7-10. 1970.
MEINKOTH, A.; KOCAN, A. A.; WHITWORTH, L.; MURPHY, G.; FOX, J. C.;
WOODS, J. P. Cats surviving natural infection with Cytauxzoon felis: 18 cases (1997-
1998). Journal of Veterinary Internal Medicine, v. 14, p. 521-525. 2000.
MEHLHORN, H.; SCHEIN, E. The piroplasms: life cycle and sexual stages. Advances
in Parasitology, v. 23, p. 37-103. 1984.
MEHLHORN, H.; SCHEIN, E. Redescription the Babesia equi Laveran,1901 as
Theileria equi Mehlhorn e Schein, 1998. Parasitology Research, v. 84, p. 467-475.
1998.
MEHLHORN, H.; SCHEIN, E.; AHMED, J. S. Theileria. In: KREIER, J. P. Parasitic
Protozoa. 2ª edition, Vol. 7. Academic Press, New York. 1994. p. 217-304.

47
MEIER, H. T.; MOORE, L. E. Feline cytauxzoonosis: a case report and literature
review. Journal of the American Animal Hospital Association, v. 36, p. 493-496. 2000.
MOTZEL, S. L.; WAGNER, J. E. Treatment of experimentally induced cytauxzoonosis
in cats with parvaquone and buparvaquone. Veterinary Parasitology, v. 35, p. 131-138.
1990.
MUDALIAR, S. V.; ACHARY, G. R.; ALWAR, V. S. On a species of Babesia in a
Indian wild cat (Felis catus). Indian Veterinary Jounal, v. 26, p. 391-395. 1950.
NAGORE, D.; GARCIA-SANMARTIN, J.; GARCIA-PEREZ, A.L.; JUSTE, R.A.;
HURTADO, A. Detection and identification of equine Theileria and Babesia species by
reverse line blotting: epidemiological survey and phylogenetic analysis. Veterinary
Parasitology, v. 123, n. 1-2, p. 41-54. 2004.
NEIMARK, H.; JOHANSSON, K. E.; RIKIHISA, Y.; TULLY, J. G. Proposal to
transfer some members of the genera Haemobartonella and Eperythrozoon to genus
Mycoplasma with descriptions of Candidatus Mycoplasma haemofelis, Candidatus
Mycoplasma haemomuris, Candidatus Mycoplasma haemosuis and Candidatus
Mycoplasma wenyonii. International Journal of Systematic and Evolutionary
Microbiology, v. 51, p. 891-899. 2001.
NEITZ, W. O.; THOMAS, A. D. Cytauxzoon sylvicaprae gen. nov., spec. nov., a
protozoan responsible for a hitherto undescribed disease in the duiker Sylvicapra
grimmia (Linné). Onderstepoort Journal of Veterinary Science Animal Industry, v. 23,
p. 63-76. 1948.
NEITZ, W. O. Classification, transmission and biology of piroplasms of domestic
animals. Annals New York Academy of Sciences, v. 64, p. 56-111. 1956.
NEITZ, W. O.; de LANGE, M. Onderstepoort, Experimental observations. (citado em
NEITZ, 1957). 1956.
NEITZ, W. O. Theileriosis, gonderiosis and cytauxzoonosis: a review. Onderstepoort
Journal of Veterinary Research, v. 27, p. 275-430. 1957.
PEIXOTO, P. V.; SOARES, C. O.; SANTIAGO, C. D.; BARROS, S. S.; BARROS, C.
L. S.; CUNHA, B. M. Citauxzoonose fatal em leões (Panthera leo) no Brasil. In: XXVI
CONGRESSO BRASILEIRO DE MEDICINA VETERINÁRIA, 1999, Campo Grande,
M.S. Anais eletrônico... Campo Grande, M.S: CONBRAVET, 1999. CD-ROM.
PEREGRINE, A. S. Chemotherapy and delivery systems: haemoparasites. Veterinary
Parasitology, v. 54, p. 223-248. 1994.
POSADA, D.; CRANDALL, K. A. Modeltest: testing the model DNA substitution.
Bioinformatics, v. 14, p. 817-818. 1998.
PROMEGA. Promega protocols and applications guide. Second edition. Printe in USA,
p, 52-53. 1991.

48
REICHARD, M. V.; BUSSCHE, R. A. V. D.; MEINKOTH, J. H.; HOOVER, J. P.;
KOCAN, A. A. A new species of Cytauxzoon from Pallas’ cats caught in Mongolia and
comments on the systematics and taxonomy of Piroplasmids. Journal of Parasitology,
v. 91, n. 2, p. 420-426. 2005.
RZHETSKY, A.; NEI, M. A simple method for estimating and testing minimum-
evolution trees. Molecular Biology Evolution, v. 9, p. 945-967. 1992.
ROTSTEIN, D. S.; TAYLOR, S. K.; HARVEY, J. W.; BEAN, J. 1999. Hematologic
effects of cytauxzoonosis in Florida panthers and Texas cougars in Florida. Journal of
Wildlife Disease, v. 35, n. 3, p. 613-617. 1999.
SAITOU, N.; NEI, M. The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Molecular Biology Evolution, v. 4, p. 406-425. 1987.
SANGER, F.; NICKLEN, S.; COULSON, A. R. DNA sequencing with chain-
terminating inhibitors. Proceedings of the National Academy of Sciences, v.74, p. 5463-
5467. 1977.
SAMBROOCK, J.; FRITSCH, E.F.; MANIATIS, T. Molecular Cloning. A Laboratory
Manual. Cold Spring Harbor Laboratory Press, New.York. 1989.
SCHEIN, E. On the life cycle of Theileria annulata (Dschunkowsky and Lunhs, 1904)
in the midgut and hemolymph of Hyalomma anatolicum excavatum (Kock, 1844).
Zeitschrift für Parazitenkunde, v. 47, p. 165-167. 1975.
SIMPSON, C. F.; HARVEY, J. W.; LAWMAN, M. J. P.; MURRAY, J.; KOCAN, A.
A.; CARLISLE, J. W. Ultrastructure of schizonts in the liver of cats with
experimentally induced cytauxzoonosis. American Journal of Veterinary Research, v.
46, n. 2, p. 384-390. 1985a.
SIMPSON, C. F.; HARVEY, J. W.; CARLISLE, J. W. Ultrastructure of the
intraerythrocytic stage of Cytauxzoon felis. American Journal of Veterinary Research,
v. 46, n. 5, p. 1178-1180. 1985b.
SHI, S. R.; COTE, R. J.; WU, L.; LIU, C.; DATAR, R.; SHI, Y.; LIU, D.; LIM, H.;
TAYLOR, C. R. DNA extraction from archival formalin-fixed, paraffin-embedded
tissue sections base don the antigen retrieval principle: heating under the influence of
pH. The Journal of Histochemistry and Cytochemistry, v. 50, n. 8, p.1005-1011. 2002.
SHINDEL, N.; DARDIRI, A. H.; FERRIS, D. H. An indirect fluorescent antibody test
for the detection of Cytauxzoon-like organisms in experimentally infected cats.
Canadian Journal of Comparative Medicine, v. 42, p. 460-465. 1978.
SCOFIELD, A.; SOARES, C. O.; MANERA, G. B.; FONSECA, A. H.; PEIXOTO, P.
V.; GONZALEZ, A. A P. Citauxzoonose felina experimental em gato doméstico (Felis
catus). In: XI SEMINÁRIO BRASILEIRO DE PARASITOLOGIA VETERINÁRIA,
1999a, Salvador, B.A. Anais... Salvador: SBPV, 1999a. p. 236.

49
SCOFIELD, A. Caracterizações morfológica, morfmétrica e ultraestrutural de formas
intraeritrocíticas de Cytauxzoon felis simile. 2002. 56f. Dissertação (Mestrado em
Ciências Veterinárias - Parasitologia Veterinária) - Curso de Pós-graduação em Ciências
Veterinárias da Universidade Federal Rural do Rio de Janeiro, Seropédica, 2002.
SOARES, C. O.; PEIXOTO, P. V.; SCOFIELD, A.; FONSECA, A.H.; MASSARD, C.
L.; SANTIAGO, C. D. Cytauxzoon sp. Neitz e Thomas, 1948 (Apicomplexa:
Theileriidae) em Panthera onca. In: XI SEMINÁRIO BRASILEIRO DE
PARASITOLOGIA VETERINÁRIA, 1999a, Salvador, B.A. Anais... Salvador: SBPV,
1999a. p.235.
SOARES, C. O.; PEIXOTO, P. V.; SCOFIELD, A.; FONSECA, A.H.; SANTIAGO, C.
D. Cytauxzoon sp. Neitz e Thomas, 1948 (Apicomplexa: Theileriidae) em Felis tigrina
(Leopardus tigrinus) no Brasil. In: XXVI CONGRESSO BRASILEIRO DE
MEDICINA VETERINÁRIA, 1999b, Campo Grande, M.S. Anais eletrônico... Campo
Grande, M.S: CONBRAVET, 1999b. CD-ROM.
SOARES, C. O. Estudos biológicos e clínico-patológico da citauxzoonose causada por
Cytauxzoon felis simile (Apicomplexa: Theileriidae) no Brasil. 2002. 150f. Tese
(Doutorado em Ciências Veterinárias - Parasitologia Veterinária) - Curso de Pós-
graduação em Ciências Veterinárias da Universidade Federal Rural do Rio de Janeiro,
Seropédica, 2002.
SONENSHINE, D. E. Biology of Ticks. Vol. 2. Oxford University Press, New York.
1991. 464 p.
SUAREZ, C. E.; FLORIN-CHRISTENSEN, M.; HINES, S. A.; PALMER, G. H.;
BROWN, W. C.; McELWAIN. Characterization of allelic variation in the Babesia bovis
merozoite surface antigen 1 (MSA-1) locus and identification of a cross-reactive
inhibition-sensitive MSA-1 epitope. Infection and Immunity, v. 68, n. 12, p. 6865-6870.
2000.
SWOFFORD, D. L. PAUP* – A computer program for phylogenetic inference using
Maximum Parsimony (*and other methods). Version 4. Sinauer Associates, Sunderland,
Massachusetts. 1998.
TAMURA, K.; NEI, M. Estimation of the number of nucleotide substitutions in the
control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology
and Evolution, v. 10, p.512-526. 1993.
TELFORD III, S. R.; GORENFLOT, A.; BRASSEUR, P.; SPIELMAN, A. Babesial
infections in man and wildlife. In: KREIER, J. P. Parasitic Protozoa. 2ª edition, Vol. 5.
Academic Press, New York. 1993. p. 1-47.
UILENBERG, G. International collaborative research: Significance of tick-borne
hemoparasitic diseases to world animal health. Veterinary Parasitology, v. 57, p. 19-41.
1995.

50
UILENBERG, G.; FRANSSEN, F. F. J.; PERIÉ, N. M. Relationships between
Cytauxzoon felis and African piroplasmids. Veterinary Parasitology, v. 26, p. 21-28.
1987.
WALKER, D. B.; COWELL, R. L. Survival of a domestic cat with naturally acquired
cytauxzoonosis. Journal of the American Veterinary Medical Association, v. 206, n. 9,
p. 1363-1365. 1995.
WAGNER, J. E. A fatal cytauxzoonosis-like disease in cats. Journal of the American
Veterinary Medical Association, v. 168, n. 7, p. 585-588. 1976.
WAGNER, J. E.; KIER, A. B.; MOREHOUSE, L. G. Feline cytauxzoonosis: A newly
reported blood protozoan disease from Southwestern Missouri. Missouri Veterinary, v.
26, n. 2, p. 12-13. 1976.
WAGNER, J. E.; FERRIS, D. H.; KIER, A. B.; WIGHTMAN, S. R.; MARING, E.;
MOREHOUSE, L. G.; HANSEN, R. D. Experimentally induced cytauxzoonosis-like
disease in domestics cats. Veterinary Parasitology, v. 6, p. 305-311. 1980.
WEHINGER, K. A.; ROELKE, M. E.; GREINER, E. C. Ixodid ticks from panthers and
bobcats in Florida. Journal of Wildlife Disease, v. 31, n. 4, p. 480-485. 1995.
WIGHTMAN, S. R.; KIER, A. B.; WAGNER, J. E. Feline cytauxzoonosis: Clinical
features of a newly described blood parasite disease. Feline Practice, May, p. 23-26.
1977.
WOLTERS, K. The troublesome parasites: molecular and morphological evidence that
Apicomplexa belong to the dinoflagellate-ciliate clade. BioSystems, v. 25, p. 75-83.
1991.
WOESE, C. R. Archaebacteria. Scientific American, v. 244, n. 6, p. 98-122. 1981.
XIAO, L.; ESCALANTE, L.; YANG, C.; SULAIMAN, I.; ESCALANTE, A. A.;
MONTAL, R. J.; FAYER, R.; LAL, A. A. Phylogenetic analysis of Cryptosporidium
parasites based on the small-subunit rRNA gene locus. Applied and Environmental
Microbiology, v. 65, n. 4, p. 1578-1583. 1999.
ZINKL, J. G.; McDONALD, S. E. Cytauxzoon-like organisms in erythrocytes of two
cheetahs. Journal of the American Veterinary Medical Association, v. 179, n. 11, p.
1261-1262. 1981.
YOUNG, A. S.; GROOTENHUIS, J. G.; LEITCH, B. L.; SCHEIN, E. The
development of Theileria = Cytauxzoon taurotragi (Martin and Brocklesby, 1960) from
eland in its tick vector Rhipicephalus appendiculatus. Parasitology, v. 81, p. 129-144.
1980.

51
ANEXOS
Páginas
Anexo 1. Informações sobre o depósito da seqüência de Cytauxzoon felis isolado
de Panthera onca (DQ382276) no Brasil................................................................
52
Anexo 2. Informações sobre o depósito da seqüência de Cytauxzoon felis isolado
de Leopardus tigrinus (DQ382277) no Brasil.........................................................
53

52
Anexo 1. Informações sobre o depósito da seqüência de Cytauxzoon felis isolado de
Panthera onca (DQ382276) no Brasil.
LOCUS DQ382276 947 bp DNA linear INV 16-FEB-2006
DEFINITION Cytauxzoon felis from Panthera onca small subunit ribosomal RNA gene, partial sequence.
ACCESSION DQ382276
VERSION DQ382276
KEYWORDS .
SOURCE Cytauxzoon felis
ORGANISM Cytauxzoon felis
Eukaryota; Alveolata; Apicomplexa; Piroplasmida; Theileriidae;Cytauxzoon.
REFERENCE 1 (bases 1 to 947)
AUTHORS Scofield, A.
TITLE Characterization of genotype and phylogeny of Cytauxzoon felis in Brazil
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 947)
AUTHORS Scofield, A., Araujo, F.R., Soares, C.O., Elisei, C., Fragoso, S.P.,Arauco, P.R.C. and
Oliveira, C.J.
TITLE Direct Submission JOURNAL Submitted (30-JAN-2006) Animal Parasitology, Veterinary, Br 465, Rio de Janeiro, RJ,
Brazil
FEATURES Location/Qualifiers
source 1..947
/organism="Cytauxzoon felis"
/mol_type="genomic DNA"
/isolation_source="blood"
/specific_host="Panthera onca"
/db_xref="taxon:27996"
/country="Brazil"
rRNA <1..>947
/product="small subunit ribosomal RNA"
1 tatttatgtt gtggcttttt ctggtgatta tatttcggta tgattatcca gattgttact
61 ttgagaaaat tagagtgctt aaagcaggca tttgccttga atactttagc atggaataac
121 taagtaggac tttggttcta ttttgttggt ttaagagcca aagtaatgat taataggaac
181 agttgggggc attcgtattt aactgtcaga ggtgaaattc ttagatttgt taaagacgaa
241 ctactgagaa agcatttgcc gaggatgttt tcattaatca agaacgaaag ttaggggatc
301 gaagacgatc agataccgtc gtagtcctaa ccataaacta tgccgactag agattggagg
361 tcgtcagttt gaacgactcc ttcagcacct tgagagaaat caaagtcttt gggttctggg
421 gggagtatgg tcgcaaggct gaaacttaaa ggaattgacg gtagggcacc accaggcgtg
481 gagcctgcgg cttaatttga ctcaacacgg gaaaacttac caggtccaga cagaggaagg
541 attgacagat tgatagctct ttcttgattc tttgggtggt ggtgcatggc cgttcttagt
601 tggtggagtg atttgtctgg ttaattccgt taacgaacga gaccttaacc tgctaaatag
661 gatctgagaa taaactttat gttgtctcag catcgcttct tagagggact ttgcggntat
721 aaatcgcaag gaagtttaag gcaataacag gtctgtgatg cccttanatg tcctgggctg
781 cacgcgcgct acactgatgc attcatcgag tatatccttg gccgagaggc ttgggtaatc
841 tttagtatgc atcgtgatgg ggattgatta ttgcaattat taatcatgaa cgaggaatgc
901 ctagtagacg cgagtcatca gctcgtgtcg attacgtccc tgccctt

53
Anexo 2. Informações sobre o depósito da seqüência de Cytauxzoon felis isolado de
Leopardus tigrinus (DQ382277) no Brasil.
LOCUS DQ382277 1204 bp DNA linear INV 16-FEB-2006
DEFINITION Cytauxzoon felis from Leopardus tigrinus small subunit ribosomal RNA gene, partial sequence.
ACCESSION DQ382277
VERSION DQ382277
KEYWORDS
SOURCE Cytauxzoon felis
ORGANISM Cytauxzoon felis
Eukaryota; Alveolata; Apicomplexa; Piroplasmida; Theileriidae; Cytauxzoon.
REFERENCE 1 (bases 1 to 1204)
AUTHORS Scofield, A.
TITLE Characterization of genotype and phylogeny of Cytauxzoon felis in Brazil
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 1204)
AUTHORS Scofield, A., Soares, C.O., Araujo, F.R., Elisei, C., Fragoso, S.P., Arauco, P.R.C. and
Rangel, C.P.
TITLE Direct Submission
JOURNAL Submitted (30-JAN-2006) Animal Parasitology, Veterinary, Br 465, Rio de Janeiro, RJ,
Brazil FEATURES Location/Qualifiers
source 1..1204
/organism="Cytauxzoon felis"
/mol_type="genomic DNA"
/isolation_source="blood"
/specific_host="Leopardus tigrinus"
/db_xref="taxon:27996"
/country="Brazil"
rRNA <1..>1204
/product="small subunit ribosomal RNA"
1 aacggggaat tagggttcga ttccggagag ggagcctgag aaatggctac cacatctaag
61 gaaggcagca ggcgcgtaaa ttacccaatc ctaacacagg gaggtagtga caagaaataa
121 caatacgagg cttaaagtct tgtaattgga atgacggaaa tttaagctct ttccggagta
181 tcaattggag ggcaagtctg gtgccagcag ccgcggtaat tccagctcca atagcgtata
241 ttaaacttgt tgcagttaaa aagctcgtag ttgaatttct gctgcatcat ttatattcct
301 taatcggttt attcatgttg tggctttttc tggtgattat acttcggtat tattatccag
361 attgttactt tgagaaaatt agagtgctta aagcaggcat ttgccttgaa tactttagca
421 tggaataact aagtaggact ttggttctat tttgttggtt taagagccaa agtaatgatt
481 aataggaaca gttgggggca ttcgtattta actgtcagag gtgaaattct tagatttgtt
541 aaagacgaac tactgcgaaa gcatttgcca aggatgtttt cattaatcaa gaacgaaagt
601 taggggatcg aagacgatca gataccgtcg tagtcctaac cataaactat gccgactaga
661 gattggaggt cgtcagtttg aacgactcct tcagcacctt gagagaaatc aaagtctttg
721 ggttctgggg ggagtatggt cgcaaggctg aaacttaaag gaattgacgg aagggcacca
781 ccaggtgtgg agcctgcggc ttaatttgac tcaacacggg aaaacttacc aggcccagac
841 agaggaagga ttgacagatt gatagctctt tcttgattct ttgggtggtg gtgcatggcc
901 gttcttagtt ggtggagtga tttgtctggt taattccgtt aacgaacgag accttaacct
961 gctaaatagg atctgagaat aaactttatg ttgtctcagc atcgcttctt agagggactt
1021 tgcggttata aatcgcaagg aagtttaagg caataacagg tctgtgatgc ccttagatgt
1081 cctgggctgc acgcgcgcta cactgatgca ttcatcgagt atatccttgg ccgagaggct
1141 tgggtaatct ttagtatgca tcgtgatggg gattgattat tgcaattatt aatcatgaac
1201 gagg


















